Embed Size (px)
Citation preview

UNIVERSIDADE DE SÃO PAULO
FFCLRP - DEPARTAMENTO DE BIOLOGIA
PROGRAMA DE PÓS-GRADUAÇÃO EM ENTOMOLOGIA
Produção “in vitro” de rainhas e ocorrência natural de machos de Frieseomelitta varia
(Apidae: Meliponina)
Ana Rita Tavares de Oliveira Baptistella
Dissertação apresentada à Faculdade de
Filosofia, Ciências e Letras de Ribeirão Preto
da USP, como parte das exigências para a
obtenção do título de Mestre em Ciências,
Área: Entomologia.
RIBEIRÃO PRETO -SP
Ano 2009

ANA RITA TAVARES DE OLIVEIRA BAPTISTELLA
Produção “in vitro” de rainhas e ocorrência natural de machos de Frieseomelitta varia
(Apidae: Meliponina)
Dissertação apresentada à Faculdade de
Filosofia, Ciências e Letras de Ribeirão Preto
da USP, como parte das exigências para a
obtenção do título de Mestre em Ciências.
Área de Concentração: Entomologia.
Orientador: Prof. Dr. Ademilson Espencer
Egea Soares.
RIBEIRÃO PRETO -SP
Ano 2009

FOLHA DE APROVAÇÃO
BAPTISTELLA, Ana Rita Tavares de Oliveira
Título: Produção “in vitro” de rainhas e ocorrência natural de machos de Frieseomelitta
varia (Apidae: Meliponina)
Dissertação apresentada à Faculdade de
Filosofia, Ciências e Letras de Ribeirão Preto
da USP, como parte das exigências para a
obtenção do título de Mestre em Ciências.
Área: Entomologia.
Aprovado em: ___/___/_____
Banca Examinadora
Prof. Dr. ___________________________________________________________________
Instituição: ____________________________ Assinatura: ___________________________
Prof. Dr. ___________________________________________________________________
Instituição: ____________________________ Assinatura: ___________________________
Prof. Dr. ___________________________________________________________________
Instituição: ____________________________ Assinatura: ___________________________

Aos meus pais: fonte constante
de inspiração, apoio e ensino
diário.
Dedico

AGRADECIMENTOS
É muito bom dizer obrigada a tantas pessoas que, neste período, em que se é acometido de
tantos surtos de tristeza, euforia, incerteza, cansaço, alegria, conseguiram se manter
simplesmente presente.
Ao bom Deus, que iluminou meu caminho e me amparou nos momentos de fraqueza.
Aos meus pais, Paulinho e Ana, e meu irmão Nando pelo imenso amor, constante estímulo,
enorme compreensão, confortável força e pelas orações.
Ao meu orientador, sempre amigo e professor, Ademilson Espencer Egea Soares, pelo apoio,
entusiasmo e por me apresentar ao fantástico mundo das abelhas.
A CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior), que forneceu
todas as condições para que esta pesquisa fosse realizada.
À Weyder Santana, que me ensinou os primeiros passos da pesquisa científica. Obrigada pelo
carinho, eterna paciência e pelas valiosas orientações que foram essenciais para a conclusão
deste trabalho.
À Camila Calixto, que esteve presente em cada instante. Obrigada por ser amiga, irmã e mãe
e, por me fazer acreditar que na terra existem “anjos”.
À Amanda e Aline Makert, duas amigas imprescindíveis na vida de qualquer ser humano.
Obrigada por fazerem meu coração sorrir.
À Camila Maia, Vanessa, Marina e Daiana, pela amizade e pelos ótimos momentos
compartilhados.
À Ivan Akatsu, que por muitas vezes além de amigo foi professor
À Mauro Prato, pelo companheirismo e pelas risadas compartilhadas em nossas viagens.
À Geusa Simone Freitas, pela simpatia, apoio e ajuda inestimável.
À Professora Luci Bego, pela atenção oferecida e pelas valiosas sugestões.
À Professora Márcia Salomão por todo apoio e zelo.
Aos colegas do Laboratório de Biologia e Genética de Abelhas, Aline Simoneti, Omar, Paulo
Emílio, Ivan, Jairo, Marcela, Vera, pelo carinho e pelos momentos de descontração.
Às encantadoras abelhinhas Frieseomelitta varia, pois sem elas nenhuma dessas páginas
estaria completa.
Meus sinceros agradecimentos a todos que contribuíram para a conclusão desse trabalho.

RESUMO
BAPTISTELLA, A. R. T. O. Produção “in vitro” de rainhas e ocorrência natural de
machos de Frieseomelitta varia (Apidae, Meliponina). 2009. 70 f. Dissertação de Mestrado
(Entomologia) – Faculdade de Filosofia Ciências e Letras, Universidade de São Paulo,
Ribeirão Preto.
Nas abelhas da espécie Frieseomelitta varia, as castas são determinadas pela quantidade de
alimento ingerida pela larva em desenvolvimento, sendo que os imaturos destinados a
tornarem-se rainha ingerem quantidades maiores de alimento larval do que aquelas destinadas
a ser operária. Os machos por sua vez, são determinados geneticamente, independente do
volume de alimento ingerido pela larva. Sabendo da importância ecológica das abelhas sem
ferrão que atuam na manutenção da diversidade de plantas nativas e como agentes
polinizadores, o conhecimento da biologia e o aprimoramento de técnicas de produção que
visem ampliar o número de colônias destas abelhas são necessários para a preservação destas
espécies e da biodiversidade a ela associada, portanto este trabalho teve por objetivos gerais:
aperfeiçoar a técnica de “produção in vitro” de rainhas de F. varia a partir da manipulação de
diferentes quantidades de alimento larval; verificar a eficiência de Frieseomelitta varia em
formar colônias diminutas; verificar a (s) época (s) de ocorrência natural de machos. O
alimento larval utilizado para produção de rainhas foi coletado por sucção à vácuo de células
de cria recém operculadas e posteriormente, diferentes quantidades de alimento (25, 30, 35,
40 e 50 µl) foram oferecidos às larvas em fase pré-defecante. As rainhas nascidas foram
introduzidas em colônias órfãs com algumas operárias jovens, alimento e pólen e foram
acompanhadas diariamente para verificar seu desenvolvimento. A produção natural de
machos foi verificada através de coletas mensais de pupas, analisando seu aparelho reprodutor
sob lente de aumento. Encontramos um volume médio de 26,70 ± 3,55 µl de alimento por
célula de operária de Frieseomelitta varia. A estimativa da taxa de crescimento da cápsula
cefálica das larvas, K=1,37 e o coeficiente de determinação (R2) de 0,98, resultou na presença
de quatro ínstares larvais, assim as 339 larvas transferidas apresentavam cápsula entre 0,866 e
1,143 mm. Nos experimentos em que foram fornecidas às larvas quantidades de alimento de
30 e 40 µl (dezembro/fevereiro e abril/julho), houve uma maior freqüência de larvas
diferenciadas em rainha. Os experimentos de maio e outubro (35 µl), apesar da taxa de
mortalidade das larvas ter sido elevada, todas as larvas nascidas se diferenciaram em rainha,
já no experimento realizado em agosto (50µl), houve mortalidade total das larvas (n=32). As
56 rainhas produzidas foram introduzidas em mini-colônias e verificamos uma alta taxa de
mortalidade das rainhas pelas operárias (77%). Quanto à ocorrência de machos, a freqüência
de células com machos prestes a nascer variou de 4% em outubro e setembro a 46 % em
janeiro e 45 % em dezembro e julho. Verificamos que o volume total médio de alimento
necessário para a diferenciação de uma larva jovem de F. varia em rainha é de 59,20 ± 5,00
µl, ou seja, a rainha consome 2,23 vezes mais alimento que a operária. Alguns elementos
interferiram no sucesso das rainhas produzidas, no entanto, algumas rainhas (n=6) se
estabeleceram como fêmea dominante. Os dados de ocorrência de machos mostram que a
produção de machos nas colônias, embora ocorra durante todo ano, ocorre com maior ou
menor intensidade em determinadas épocas.
Palavras-chave: abelha sem ferrão, produção de rainha, machos, alimento larval.

ABSTRACT
BAPTISTELLA, A. R. T. O. Production “in vitro” of queens and natural occurrence of
males of Frieseomelitta varia (Apidae, Meliponina). 2009. 70 f. Dissertação de Mestrado
(Entomologia) – Faculdade de Filosofia Ciências e Letras, Universidade de São Paulo,
Ribeirão Preto.
In Frieseomelitta varia bees species, the castes are determined by the amount of food ingested
by the larvae in development, being the immature ones destined to become queen, ingest
larger amount of larval food than those destined to be workers. The males ones, in their turn,
are genetically determined independently from the food volume ingested by the larvae.
Knowing about the ecological importance of the stingless bees that act in the maintenance of
the diversity of native plants and as pollinating agents, the knowledge of the biology and the
improvement of production techniques that seek to enlarge the number of colonies of these
bees are necessary for the preservation of these species and the biodiversity associated to it.
Therefore, the general aim of this work were: to improve the technique of "in vitro
production" of F. varia queens starting from the manipulation of different amounts of larval
food; to verify the efficiency of Frieseomelitta varia to form tiny colonies and verify the
period (s) of natural occurrence of males. The larval food used for production of queens was
collected by vacuum suction of cells recently covered, and later different amount of food (25,
30, 35, 40 and 50 µl) were offered to the larvae in pre-defecating phase. The born queens
were introduced in orphan colonies with some young workers, food and pollen and were
accompanied daily to verify their development. The natural production of males was verified
through monthly collections of pupae, analyzing its reproductive system under magnifying
glass. We found an average volume of 26.70 ± 3.55 µl of food per worker cell of
Frieseomelitta varia. The estimate rate growth of the cephalic capsule of the larvae, K=1.37
and the determination coefficient (R2) of 0.98, resulted in the presence of four instar larvae,
thus, the 339 transferred larvae, presented capsule between 0.866 and 1.143 mm. In the
experiments in which the larvae amount of food of 30 and 40 µl (december/february and
april/july) were supplied, there was a larger frequency of larvae differentiated into queen. The
experiments of may and october (35 µl), in spite of the mortality rate of the larvae being
raised, all of the born larvae differentiated into queen, whereas in the experiment carried out
in august (50µl), there was total mortality of the larvae (n=32). The 56 produced queens were
introduced in colonies and it was recorded a high mortality rate of the queens by the workers
(77%). As for the occurrence of males, the frequency of cells with ready to be born males
varied from 4% in october and september to 46% in January and 45% in december and july.
We verified that the total average volume of necessary food for the differentiation of a young
larva of F. varia in queen is of 59.20 ± 5,00 µl, in other words, the queen larvae consumes
2.23 times more food than the workers one. Some elements interfered in the success of the
queens produced, but, some queens (n=6) established themselves as dominant female in the
new colonies. The data of occurrence of males show that the production of males in colonies,
although it happens during the whole year, happens with larger or smaller intensity in certain
periods.
Key-words: stingless bee, queen production, males, larval food.

LISTA DE ILUSTRAÇÕES
FIGURA 1 - Mecanismos de determinação das castas nas abelhas da subtribo
Meliponina. a) espécies do gênero Melipona; b) abelhas dos demais
gêneros; c) algumas espécies dos gêneros Leurotrigona, Frieseomelitta e
Plebeia........................................................................................................15
FIGURA 2 - Células de cria das espécies Frieseomelitta varia (A) e Plebeia lucii (B)
agrupadas na forma de cacho......................................................................16
FIGURA 3 - Célula típica de rainha de Frieseomelitta varia (círculo): célula auxiliar
com alimento extra (a); célula com a larva jovem
(b)................................................................................................................16
FIGURA 4 - Vista interna de uma das colônias de Frieseomelitta varia utilizada no
experimento. Observar a quantidade de células de cria (A), de potes de mel
(B) e potes de pólen (C)..............................................................................22
FIGURA 5 - (A) Instrumentação para coleta de alimento larval por sucção, I – bomba de
sucção a vácuo, II - recipiente de vidro para coleta de resíduos, III – tubo
Falcon mantido em recipiente com gelo, IV – ponteira de micropipeta
acoplada à mangueira para a sucção do alimento, V – tampa da placa de
Petri para acomodar as células de cria. (B) Detalhe do tubo Falcon coletor
de alimento; (C) Detalhe da ponteira de micropipeta e tampa de placa de
Petri.............................................................................................................23
FIGURA 6 - Alimento larval de células de cria de Frieseomelitta varia sendo transferido
para a placa de Elisa....................................................................................24
FIGURA 7 - (A) Recipiente plástico protegido por filme de PVC, contendo a placa de
Elisa com alimento e as larvas transferidas, solução saturada de água e
NaCl (seta); (B) Todo conjunto foi envolto em papel laminado.................25
FIGURA 8 - Rainha de Frieseomelitta varia produzida in vitro após o nascimento, com
marcação no mesonoto indicada pela seta..................................................25
FIGURA 9 - Variação do volume de alimento larval em células de cria de Frieseomelitta
varia, durante os meses de coleta................................................................29
FIGURA 10 - Variação do volume total de alimento larval nos alvéolos de Frieseomelitta
varia nas diferentes épocas de coleta dos dados (época
seca/chuvosa)..............................................................................................30
FIGURA 11 - Curva de distribuição de freqüência da largura das cápsulas cefálicas (mm)
de larvas de Frieseomelitta varia (n=494)..................................................32

FIGURA 12 - Detalhe do último par de pernas dos adultos nascidos do experimento
realizado em dezembro (30µl de alimento larval). A: Operária, B: Rainha,
C: indivíduo com características intermediárias entre rainha e
operária........................................................................................................35
FIGURA 13 - Fatores responsáveis pela alta mortalidade das rainhas de Frieseomelitta
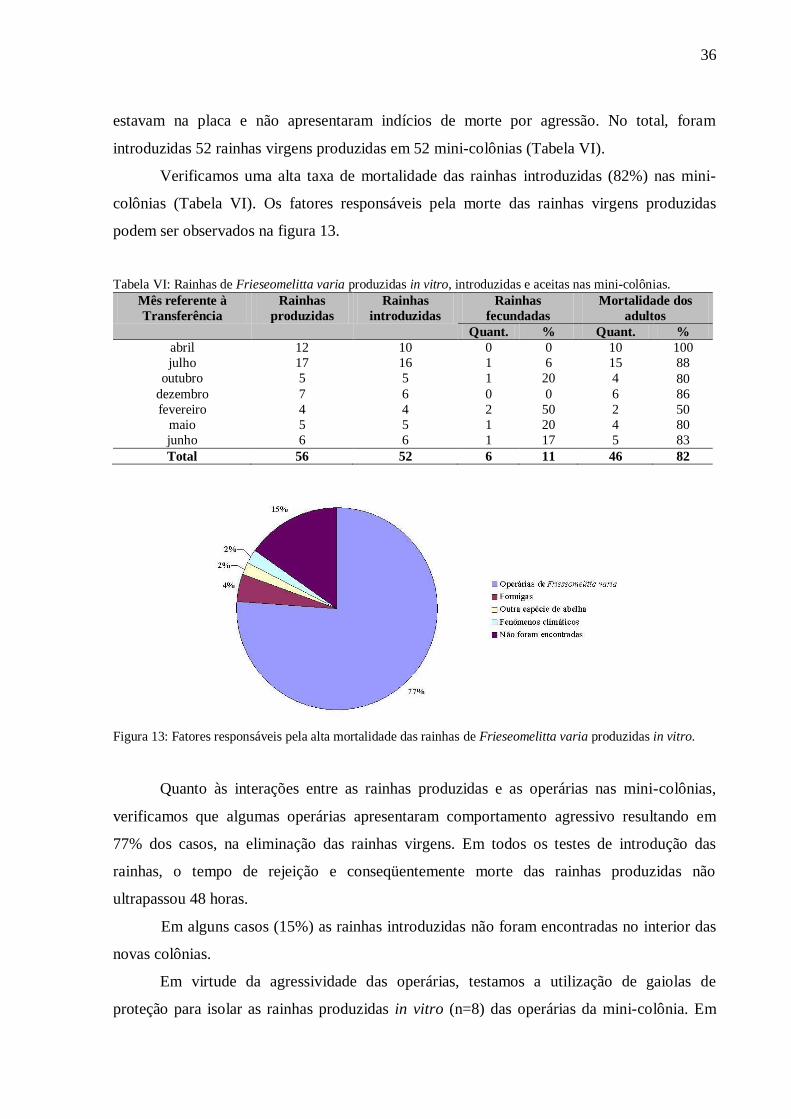
varia produzidas in vitro.............................................................................36
FIGURA 14 - Gaiola dentro da qual as rainhas produzidas foram mantidas isoladas das
operárias da mini-colônia. Detalhe da tela com fragmentos de cerume que a
cobriam.......................................................................................................37
FIGURA 15 - A) Mini-colônia com rainha produzida “in vitro” (círculo), B) detalhe da
rainha marcada e fecundada. A seta evidencia o abdômen desenvolvido da
rainha marcada............................................................................................38
FIGURA 16 - Célula de aprisionamento com rainha virgem de Frieseomelitta
varia............................................................................................................39
FIGURA 17 - Colônia experimental para Frieseomelitta varia, construída em módulos de
madeira e laterais de vidro. A) Vista lateral; B) Vista frontal; C) Vista
interna da colônia, D) Detalhe interno dos módulos preenchidos..............39
FIGURA 18 - Freqüência relativa (%) de machos e operárias de Frieseomelitta varia
coletados em 1784 células de cria em três colônias distintas ao longo de 12
meses..........................................................................................................43
FIGURA 19 - Freqüência relativa (%) de machos (VERMELHO) e operárias (PRETO)
por mês de amostragem em cada uma das três colônias de Frieseomelitta
varia. Colônia 1 (A), Colônia 2 (B) e Colônia 3 (C)..................................44
FIGURA 20 - Freqüência de machos de Frieseomelitta varia, em estágio de pupa, nos
meses de coleta dos dados (12 meses). As marcações em vermelho indicam
os meses em que as rainhas produzidas foram fecundadas
(n=6)............................................................................................................55

LISTA DE TABELAS
TABELA I - Volumes médios de alimento larval coletado em relação ao número de
células analisadas em Frieseomelitta varia................................................29
TABELA II - Volumes médios de alimento larval em células de cria de Frieseomelitta
varia em diferentes estações do ano e comparação das médias pelo teste
Z..................................................................................................................31
TABELA III - Número de ínstares larvais em Frieseomelitta varia, amplitude, média da
largura da cápsula cefálica (mm) e intervalo de confiança (IC).................32
TABELA IV - Desenvolvimento in vitro das larvas de Frieseomelitta varia....................33
TABELA V - Diferentes quantidades de alimento larval oferecidos às larvas para
produção de rainhas in vitro de Frieseomelitta varia.................................34
TABELA VI - Rainhas de Frieseomelitta varia produzidas in vitro, introduzidas e aceitas
nas mini-colônias........................................................................................36
TABELA VII - Peso (g) das rainhas produzidas em condições naturais e das rainhas
produzidas em laboratório que se estabeleceram nas mini-
colônias.......................................................................................................40
TABELA VIII - Teste-t: Comparação do peso ao nascer entre as rainhas naturais (N) e
rainhas produzidas em condições controladas (P) que se estabeleceram nas
mini-colônias..............................................................................................41
TABELA IX - Teste-t: Comparação do peso ao nascer entre as rainhas naturais (N) e
rainhas produzidas in vitro que morreram (Pm).........................................41
TABELA X - Freqüência de machos e operárias em três colônias naturais de
Frieseomelitta varia em diferentes épocas do ano......................................41
TABELA XI - Meses nos quais ocorreu a fecundação das rainhas de Frieomelitta varia
produzidas in vitro......................................................................................54

SUMÁRIO
I- INTRODUÇÃO ..................................................................................................................11
II- OBJETIVOS......................................................................................................................18
III- MATERIAL E MÉTODOS.............................................................................................20
3.1. Quantificação do volume de alimento larval em células de cria de Frieseomelitta
varia.....................................................................................................................21
3.2. Determinação do número de ínstares larvais em Frieseomelitta varia..................21
3.3. Produção in vitro de rainhas de Frieseomelitta varia............................................22
3.3.1. Coleta e estocagem do alimento larval....................................................22
3.3.2. Transferência das larvas de Frieseomelitta varia para placas de cultivo
celular......................................................................................................23
3.2. Montagem e desenvolvimento de mini-colônias de Frieseomelitta varia
utilizando-se rainhas produzidas in vitro..............................................................25
3.5. Ocorrência de machos em colônias naturais de Frieseomelitta
varia......................................................................................................................27
VI- RESULTADOS.................................................................................................................28
4.1. Quantificação do volume de alimento larval em células de cria de Frieseomelitta
varia.....................................................................................................................29
4.2. Determinação do número de ínstares larvais em Frieseomelitta varia..................31
4.3. Produção in vitro de rainhas de Frieseomelitta varia............................................33
4.3.1 Possível ocorrência de intercasta..............................................................34
4.4. Mini-colônias de Frieseomelitta varia utilizando as rainhas produzidas in
vitro.......................................................................................................................35
4.5. Ocorrência de machos em colônias naturais de Frieseomelitta varia....................40
V- DISCUSSÃO.......................................................................................................................45
VI- CONCLUSÕES................................................................................................................58
REFERÊNCIAS BIBLIOGRÁFICAS..................................................................................60

11
I- INTRODUÇÃO

12
As abelhas pertencem à família Apidae e à subfamília Apinae (Michener, 1974) e são
representadas por um grupo muito diversificado, apresentando 17 tribos dentre as quais 13
estão representadas no Brasil (Silveira et al., 2002). Todos os Apidae apresentam algum grau
de organização social, no entanto, os indivíduos das subtribos Apina e Meliponina (Silveira et
al., 2002) possuem o comportamento social mais elaborado entre as abelhas (Michener, 1974;
2000; Sakagami, 1982, Wille, 1983; Winston, 1991).
A socialidade pressupõe a existência de uma organização entre os indivíduos
pertencentes à mesma colônia, que cooperam com o cuidado da cria e dividem o trabalho no
ninho (Wilson, 1971).
Nos insetos sociais em geral, o desenvolvimento das castas está relacionado com um
amplo fenômeno de polimorfismo (Nijhout & Wheeler, 1982) que nas abelhas envolve a
população feminina (rainhas e operárias), apresentando diferenças acentuadas quanto à
divisão de trabalho dentro da colônia. O desenvolvimento de operárias e rainhas diverge do
ponto de vista morfológico e fisiológico, culminando em adultos que diferem também em
suas características comportamentais (Weaver, 1966; Engels & Imperatriz-Fonseca, 1990).
As rainhas possuem função exclusivamente reprodutiva, copulam com os machos e
passam o resto de seus dias realizando a postura de ovos. Sua presença na colônia é
fundamental, garantindo a integridade e funcionalidade da sociedade. Quanto às operárias,
elas são responsáveis pela manutenção da colônia realizando diversas atividades, tais como,
cuidado com a prole, coleta de alimento, construção e defesa do ninho, limpeza e
aprovisionamento das células de cria, dentre outras (Michener, 1974; Laidlaw, 1992; Page &
Peng, 2001).
Em uma colônia de abelha eussocial são produzidos além da rainha e das operárias,
indivíduos do sexo masculino. Normalmente, alguns dias após emergirem, os machos são
expulsos da colônia pelas operárias (Michener, 1948; Kerr et al.1996) e têm como função
principal fecundar a rainha virgem durante o vôo nupcial. Entretanto, alguns autores
observaram machos de espécies de meliponíneos realizando algumas tarefas dentro da colônia
envolvendo atividades de construção, trofaláxis, desidratação de néctar (Imperatriz-Fonseca,
1973; Cortopassi-Laurino, 1979) e algumas atividades relacionadas à defesa do ninho (Kerr,
1990).
As atividades desenvolvidas pelos machos é assunto a ser investigado com mais
afinco, uma vez que não há evidências suficientes que comprovem estes comportamentos.
Para alguns autores, estas atividades são meramente acidentais (Nogueira-Neto, 1997). Até o
momento, não há evidências de que a atividade de desidratação do néctar pelos machos

13
contribua de qualquer forma como uma vantagem para a colônia (Cortopassi-Laurino, 2007).
Assim, de acordo com Velthuis e colaboradores (2005), a produção de machos é um
investimento que está em conflito com o investimento no tamanho da colônia, pois ao
contrário das operárias, a maioria dos machos não participa das atividades regulares da
colônia.
Dentre as diferenças morfológicas entre rainhas e operárias podem-se observar, por
exemplo: sistema reprodutor desenvolvido com ovários com maior número de ovaríolos em
rainhas (Michener, 1974) e a presença de corbícula, estrutura encontrada na tíbia das pernas
posteriores de todos os Apidae (Michener, 1999), formada por uma cavidade envolvida por
várias cerdas ou pêlos (Snodgrass, 1956), que é mais elaborada nas operárias e está
relacionada com a coleta de pólen e resina (Thorpe, 1979).
Os insetos da subtribo Meliponina que compreendem as abelhas do gênero Melipona e
todos os demais gêneros (Silveira et al., 2002), são conhecidas popularmente como abelhas
sem ferrão, abelhas indígenas ou meliponíneos, por apresentarem seu acúleo (ferrão) atrofiado
(Nogueira-Neto, 1997). São insetos sociais de grande diversidade e de ampla distribuição
geográfica (Sakagami, 1982; Roubik, 1989), podem ser encontradas em áreas tropicais e
subtropicais do mundo, mas sua maior diversidade é observada nas regiões Neotropicais (com
mais de 300 espécies descritas) e na região Indo-Malaia (com cerca de 60 espécies), sendo
que certos táxons são restritos às regiões tropicais e subtropicais do hemisfério sul (Camargo,
1989; Camargo & Pedro, 1992). No Brasil, essa subtribo é bastante representativa e já foram
descritas aproximadamente 192 espécies pertencentes a 27 gêneros (Silveira et al., 2002).
Segundo Koedam (1995), na maioria dos insetos eussociais a seleção natural
favoreceu o surgimento de uma plasticidade comportamental, que permite às operárias serem
capazes de realizar qualquer comportamento inerente às fêmeas daquela espécie, exceto a
postura de ovos fecundados e, a adaptação dos ovários das rainhas, relacionada às altas taxas
de oviposição, é uma especialização que está acompanhada por uma mudança
morfológica/fisiológica, limitando o desempenho de outros comportamentos. Entretanto,
segundo Bourke (1988) esta especialização não transforma os outros grupos de fêmeas, como
as operárias, em uma casta completamente estéril.
As operárias do gênero Apis podem ter seus ovários desenvolvidos durante a ausência
da rainha ou quando a população é muito grande (Groot & Voogd, 1954). Nos meliponíneos a
situação é mais complexa. Em algumas espécies, por exemplo, em Leurotrigona muelleri,
assim como em Apis mellifera as operárias têm seus ovários diferenciados apenas na ausência
da rainha (Sakagami & Zucchi, 1966; Cruz-Landim, 2000), entretanto, em algumas espécies

14
as operárias apresentam ovários desenvolvidos (ativos) e produzem ovos mesmo na presença
da rainha (Sakagami et al., 1963). Estas operárias podem produzir dois tipos de ovos : 1)
reprodutivos, que contribuem com a produção de machos da colônia (Beig, 1972; Machado et
al., 1984; Imperatriz-Fonseca & Kleinert, 1998; Sommeijer et al., 1999; Koedam et al., 2001;
Tóth et al., 2002; Chinh et al., 2003; Paxton et al., 2003; Tóth et al., 2004); 2) ovos tróficos,
que são ingeridos pela rainha antes da postura de seus ovos (Silva et al., 1972; Contel, 1976;
Bego, 1982; 1990; Sommeijer & van Buren, 1992; Koedam et al, 2005).
De acordo com Zucchi et al. (1977;1999) as operárias de colônias sociais avançadas
geralmente se mantêm estéreis, como pode ser observado em algumas espécies tais como
Duckeola ghilianii (Sakagami & Zucchi, 1968), Tetragonula laeviceps (Sakagami et al.,
1983), Tetragonula minangkabau (Suka & Inoue, 1993), incluindo espécies do gênero
Frieseomelitta (Suka & Inoue, 1993; Zucchi et al., 1999; Boleli et al., 1999; 2000; Cruz-
Landim, 2000).
Os mecanismos de determinação de castas nas abelhas eussociais dependem, direta ou
indiretamente, da alimentação na fase larval, mesmo naquelas espécies em que os fatores
genéticos podem estar envolvidos (Hartfelder et al., 2006).
Em abelhas do gênero Apis a quantidade e a qualidade do alimento larval são
fundamentais para a diferenciação de rainhas e operárias (Beetsma, 1979). As larvas que
originarão rainhas recebem alimento diferenciado daquelas destinadas a se tornarem
operárias. As futuras rainhas são alimentadas com geléia real durante todo o seu
desenvolvimento e as operárias recebem uma mistura de mel e pólen após o terceiro dia de
alimentação larval (Rembold et al., 1974; Weaver, 1974; Asencot & Lensky, 1984). O
processo de alimentação das larvas nas abelhas melíferas é progressivo, em virtude do fato
das operárias nutridoras depositarem o alimento nas células durante visitas periódicas
(Michener, 1974).
Por outro lado, nas abelhas sem ferrão, não foram encontradas diferenças qualitativas
entre o alimento oferecido às larvas que originarão rainhas e aquele oferecido às larvas que
originarão operárias (Kerr et al., 1966; Camargo, 1972, Hartfelder & Engels, 1989). Os
meliponíneos possuem um mecanismo diferente de aprovisionamento do alimento larval, que
é constituído por uma secreção glandular produzida pelas glândulas hipofaríngeas das
operárias nutridoras e uma mistura de pólen e mel com aproximadamente, 16% de pólen e 8%
de mel (Laidlaw et al., 1956; Hartfelder & Engels,1989). Todo o alimento necessário para o
desenvolvimento do imaturo é depositado na célula pelas operárias antes da postura do ovo
pela rainha. Depois a célula é operculada pelas operárias e não ocorre contato entre elas e as

15
larvas em desenvolvimento. Este processo recebe o nome de aprovisionamento massal (Kerr,
1948; Michener, 1974; Nogueira-Neto,1997).
Nas abelhas da subtribo Meliponina, o sistema de produção de rainha difere entre as
espécies. Nas espécies do gênero Melipona o mecanismo de determinação das castas tem base
genética (Kerr, 1950 apud Kerr et al., 1996) modulado por influências ambientais (Kerr &
Nielsen, 1966; Kerr, 1974; Engel & Imperatriz-Fonseca, 1990; Velthuis e Sommeijer, 1991).
As rainhas se desenvolvem em células de mesmo tamanho das células de operárias e de
machos (Kerr et al., 1996) (Figura 1a). Neste gênero, a variação na quantidade de alimento
associado aos mecanismos genéticos é responsável por regular o sistema de diferenciação
entre rainhas e operárias (Kerr, 1950 apud Kerr et al., 1996).
Nos demais gêneros de abelhas indígenas, a quantidade de alimento larval é o fator
decisivo na determinação das castas (Camargo, 1972; Michener, 1974; Engels & Impertriz-
Fonseca, 1990). As rainhas emergem de células de tamanho maior que as células de operárias,
estas células reais ou realeiras contém uma quantidade de alimento larval maior que as demais
células (Figura1b). Deste modo, as larvas destinadas a se tornarem rainhas ingerem uma
quantidade de alimento larval maior que aquele ingerido pelas larvas que irão se desenvolver
em operárias (Kerr, 1948; Camargo, 1972).
No entanto, outro mecanismo diferencial de produção de rainhas pode ser observado
em algumas espécies dos gêneros Leurotrigona, Frieseomelitta e Plebeia que contróem
células de cria arranjadas na forma de cacho (Figura 2) (Michener, 1974; Nogueira-Neto,
1970; Moure, 2004, respectivamente). Nestas abelhas não ocorre construção de células reais
típicas (Figura 1c), as rainhas se desenvolvem em casulos reais formados a partir da junção de
duas células de cria de tamanho normal (Terada, 1974; Faustino et al., 2002; Teixeira, 2007),
Figura 1: Mecanismos de determinação das castas nas abelhas da subtribo Meliponina. a) espécies do gênero
Melipona; b) abelhas dos demais gêneros; c) algumas espécies dos gêneros Leurotrigona, Frieseomelitta e
Plebeia.Fonte: modificado de Ribeiro et al., 2006.

16
Figura 2: Células de cria das espécies Frieseomelitta varia (A) e Plebeia luciii (B) agrupadas na forma de cacho.
Fonte figura 1B: Moure, 2004.
Em Frieseomelitta varia, espécie estudada neste trabalho, este mecanismo de
produção de rainha foi estudado por Terada (1974) e Faustino e colaboradores (2002) que
demonstraram experimentalmente que as larvas que tem acesso somente ao alimento da sua
célula se desenvolvem em operárias, por outro lado, aquelas que se alimentam de uma
quantidade extra de alimento larval aprovisionado em uma célula adjacente, diferenciam-se
em rainha. Esta célula auxiliar contendo alimento, com o tamanho aproximado de uma célula
normal (Figura 3), é construída pelas operárias ao lado ou acima da célula contendo a larva
escolhida para tornar-se rainha (Terada, 1974; Faustino et al., 2002).
Figura 3: Célula típica de rainha de Frieseomelitta varia (círculo): célula auxiliar com alimento extra (a); célula
com a larva jovem (b). Fonte: Faustino, 2002.
Além destes mecanismos, existem ainda alguns gêneros de meliponíneos onde as
rainhas podem emergir tanto de uma célula de tamanho normal de operária quanto de células
maiores ou células reais (realeiras). As rainhas produzidas em células de mesmo tamanho que
as de operárias são menores que as rainhas emergidas de células reais típicas, estas rainhas
são usualmente chamadas de rainhas miniaturas ou rainhas anãs (Engels & Imperatriz-
B A

17
Fonseca, 1990; Imperatriz-Fonseca et al., 1997; Bourke & Ratnieks, 1999; Ribeiro, 2004;
Wenseelers et al., 2005; Ribeiro et al., 2006).
Atualmente, têm-se registro de ocorrência de rainhas miniaturas em nove espécies de
abelha sem ferrão, entre elas: Cephalotrigona capitata (Schwarz, 1948); Schwarziana
quadripunctata (Camargo, 1974; Imperatriz-Fonseca & Darakjian, 1993; Nogueira-Ferreira et
al., 2000; Wenseelers et al., 2005), Plebeia julianii (Juliani, 1962), Plebeia remota
(Imperatriz-Fonseca et al., 1975; Ribeiro et al. 2003; Ribeiro et al., 2006), Nannotrigona
testaceicornis (Imperatriz-Fonseca et al., 1997); Plebeia lucii (Teixeira & Campos, 2005).
Sabe-se que as abelhas sem ferrão constituem um grupo ecologicamente importante,
uma vez que, salvo algumas exceções, como Trigona crassipes (Fabricius, 1793) e Trigona
hypogea (Silvestri, 1902), que possuem hábitos necrófagos (Nogueira-Neto, 1997; Velthuis,
1997), apresentam uma estreita relação de dependência com as plantas, as quais fornecem o
alimento para as abelhas, principalmente pólen e néctar (fontes de proteína e energia,
respectivamente) e em troca recebem os benefícios da transferência de pólen (Kevan & Baker,
1983; Proctor et al., 1996) atuando portanto na manutenção da diversidade das plantas como
agentes polinizadores. Acredita-se, que no Brasil cerca de 40 a 90% das árvores nativas são
polinizadas por abelhas sem ferrão, dependendo do ecossistema (Kerr, 1996).
Assim, a grande necessidade de se conservar as espécies de meliponíneos brasileiros
reside principalmente, na ampla ação como polinizadores da nossa flora que, por sua vez,
garante a presença e desenvolvimento da fauna e de toda biodiversidade (Kerr et al., 1996).
Da mesma forma, a conservação da flora nativa também é necessária, uma vez que a
fragmentação e degradação dos ambientes naturais podem prejudicar as comunidades de
abelhas nativas (Kremen et al., 2002) causando a perda ou a dissociação de recursos
importantes para sua alimentação e locais de nidificação (Potts et al., 2005).
A importância das abelhas sem ferrão aumenta à medida que mais conhecimento se
tem sobre elas. O interesse cada vez maior na sua utilização como agentes polinizadores de
culturas agrícolas, na comercialização de seus produtos como o pólen, o mel, o própolis,
aumenta a procura por suas colônias. Estudar a produção de machos, rainhas e operárias é o
primeiro passo para possibilitar a multiplicação dos ninhos em larga escala, viabilizando sua
utilização tanto ecológica como economicamente, além de possibilitar estudos sobre a
biologia e a determinação de castas nos meliponíneos.

18
II- OBJETIVOS

19
O presente trabalho teve por objetivos:
1. Adaptar e aperfeiçoar a técnica de “produção artificial” ou “produção in vitro” de
rainhas de meliponíneos a partir da manipulação de quantidades diferenciais de alimento
larval;
2. Verificar a capacidade de Frieseomelitta varia em formar colônias diminutas a
partir dos indivíduos obtidos pelo método de produção in vitro, para a multiplicação de ninhos
dessa espécie;
3. Avaliar a aceitação das rainhas de Frieseomelitta varia produzidas em condições
controladas;
4. Verificar a (s) época (s) de ocorrência natural de machos, a fim de sincronizar com
a produção in vitro de rainhas;
5. Verificar a viabilidade reprodutiva das rainhas virgens produzidas in vitro
introduzidas através da fecundação e início da postura.

20
III- MATERIAL E MÉTODOS

21
1. Quantificação do volume de alimento larval em células de cria de Frieseomelitta varia
O material biológico utilizado neste estudo foi retirado de uma colônia de
Frieseomelitta varia, mantida em colônia racional no meliponário do Departamento de
Genética da FMRP/USP. A colônia apresentava excelentes condições e foi escolhida com
base na quantidade de potes de alimento, no número de células de cria e no número de
operárias em atividade na colônia.
Durante os meses de maio, junho, agosto, novembro (2007), fevereiro e abril (2008),
foram coletadas células de cria recém operculadas, todas contendo ovos não eclodidos em seu
interior, estas células são facilmente identificáveis, pois possuem coloração mais escura que
as demais e geralmente se concentram na porção superior da área de cria.
Em laboratório, sob condições assépticas e, com o auxílio de um estilete de ponta fina,
as células foram desoperculadas e os ovos removidos e descartados. O alimento larval de cada
célula foi coletado e quantificado volumetricamente utilizando-se uma micropipeta (10µl).
Após o processo de extração, o alimento foi acondicionado em tubo plástico tipo Falcon e
conservado em freezer à - 20ºC.
Após a coleta do alimento larval, as células restantes foram devolvidas à colônia de
origem para serem reutilizadas pelas operárias do ninho.
2. Determinação do número de ínstares larvais em Frieseomelitta varia
O número de ínstares larvais foi determinado a partir da medida da largura máxima da
cápsula cefálica de 494 larvas provenientes de três colônias distintas de Frieseomelitta varia.
Os indivíduos foram fixados inicialmente em solução de Bouin (150 ml de álcool 80%, 60 ml
de solução com formol, 15 ml ácido acético glacial, 1g ácido pícrico), após 48 horas foram
transferidos para álcool 70%, onde foram conservadas, para posterior análise.
As medições das cápsulas cefálicas foram feitas através de uma ocular graduada
acoplada a um microscópio estereoscópio (Leica Wild M3B). As larvas com as mesmas
medidas foram agrupadas em frascos tipo eppendorf contendo álcool 70% e etiquetados com
os seus respectivos valores.
Os dados foram plotados em gráfico de distribuição de freqüência e, utilizou-se o
modelo matemático com base na regra de Dyar, conforme preconizado por Parra e Haddad

22
(1989), para determinação do número real de ínstares. A partir deste modelo foram obtidos a
taxa de crescimento (K) e o coeficiente de determinação (R2).
3. Produção in vitro de rainhas de Frieseomelitta varia
3.1 Coleta e estocagem do alimento larval
Para a extração do alimento larval foram retiradas células de cria recém operculadas de
três colônias em condições estáveis, com grande disponibilidade de alimento e bom número
de postura pela rainha, observado através da quantidade de células recém operculadas (Figura
4).
As células retiradas do ninho foram colocadas em placa de Petri e conduzidas ao
laboratório, onde foram desoperculadas e os ovos removidos utilizando-se um estilete de
ponta fina.
Figura 4: Vista interna de uma das colônias de Frieseomelitta varia utilizada no experimento. Observar a
quantidade de células de cria (A), de potes de mel (B) e potes de pólen (C).
O alimento foi coletado por sucção através de um tubo de borracha de silicone de fino
calibre, com uma ponteira de pipeta na extremidade, conectado a um tubo do tipo Falcon e
ligado a uma bomba de vácuo (Figura 5). O tubo plástico foi mantido em uma caixa de isopor
com gelo a temperatura aproximada de 4ºC para não alterar as características físico/químicas
do alimento durante o processo de extração.
A
B
C

23
Após a coleta do alimento, o tubo Falcon foi mantido em freezer a – 20ºC, onde foi
mantido até a sua utilização. As células das quais os alimentos foram extraídos foram
devolvidas à colônia original para reaproveitamento da cera pelas operárias.
Figura 5: (A) Instrumentação para coleta de alimento larval por sucção, I – bomba de sucção a vácuo, II -
recipiente de vidro para coleta de resíduos, III – tubo Falcon mantido em recipiente com gelo, IV – ponteira de
micropipeta acoplada à mangueira para a sucção do alimento, V – tampa da placa de Petri para acomodar as
células de cria. (B) Detalhe do tubo Falcon coletor de alimento; (C) Detalhe da ponteira de micropipeta e tampa de placa de Petri.
3.2. Transferência das larvas de Frieseomelitta varia para placas de cultivo celular
Para a realização dos experimentos envolvendo a transferência das larvas, o alimento
previamente coletado (ver item 3.1) foi descongelado gradualmente em um recipiente de
isopor com gelo e posteriormente homogeneizado com o auxílio de um agitador elétrico.
Em uma placa de Elisa esterilizada (Enzyme-Linked ImmunoSorbent Assay) com 96
pocinhos, medindo: 0,7 mm de diâmetro, 1 cm de profundidade e volume total de 400 µl
(Figura 6), o alimento foi transferido com uma micropipeta Gilson em diferentes
concentrações (25, 30, 35, 40 e 50 µl), suficientes para que as larvas se desenvolvessem em
rainhas.
I
II
III
IV
V C
B
A

24
Figura 6: Alimento larval de células de cria de Frieseomeiltta varia sendo transferido para a placa de Elisa.
Das mesmas colônias das quais foram retiradas as células para extração do alimento
larval, foram selecionadas e coletadas células de cria velha, as quais foram abertas uma a uma
com o auxílio de pinça de ponta fina, para a seleção das larvas na fase de alimentação para
posterior transferência.
Foram selecionadas larvas no último estágio de alimentação (pré-defecante), ou seja,
haviam ingerido completamente o conteúdo de sua célula, porém não iniciaram o processo de
defecação, apresentando coloração branca-leitosa e o intestino repleto com alimento.
Tais larvas foram imediatamente transferidas para a placa de Elisa, depositadas
cuidadosamente sobre o alimento, utilizando-se uma pinça com ponta arredondada, tomando o
cuidado necessário para não perfurar a larva.
As placas tipo Elisa contendo as larvas transferidas foram acondicionadas no interior
de recipientes plásticos com tampa (Tupperware) com uma solução de NaCl saturada em seu
interior para manter a umidade relativa em torno de 75%. Os recipientes foram envolvidos
com filme de PVC mantendo as condições de umidade interna e com papel laminado a fim de
evitar a luminosidade externa (Figura 7).

25
Figura 7: (A) Recipiente plástico protegido por filme de PVC, contendo a placa de Elisa com alimento e as
larvas transferidas, solução saturada de água e NaCl (seta); (B) Todo conjunto foi envolto em papel laminado.
Os experimentos foram mantidos em estufa tipo BOD a 28ºC e aproximadamente 70%
UR (Camargo, 1972; Buschini & Campos, 1995). O desenvolvimento das larvas foi
acompanhado diariamente até o nascimento do adulto na placa de Elisa.
4. Montagem e desenvolvimento de mini-colônias de Frieseomelitta varia utilizando-se
rainhas produzidas in vitro
As rainhas virgens emergidas foram pesadas individualmente, em balança eletrônica
de precisão Metter (AE50). Após a pesagem, foram marcadas no mesonoto com caneta
Creative Maker Compactor atóxica e sem odor (Figura 8), para facilitar sua identificação na
colônia.
Figura 8: Rainha de Frieseomelitta varia produzida in vitro após o nascimento, com marcação no mesonoto
indicada pela seta.
As rainhas recém nascidas foram acondicionadas em placas de Petri de tamanho
grande, com aproximadamente 10 a 15 operárias jovens (de coloração clara), solução de
sacarose a 50% (50% açúcar e 50% água) e uma pequena quantidade de cerume retirada da
A
B

26
colônia da qual posteriormente, foram retiradas as células de cria e as operárias jovens para a
montagem das novas colônias.
As placas permaneceram na estufa por alguns dias para a observação do
comportamento de aceitabilidade das operárias com relação à nova rainha e, para que a rainha
introduzida adquirisse o odor da nova colônia, diminuindo desta forma o risco de rejeição
pelas operárias.
Após este período, a rainha juntamente com as operárias foi introduzida em uma mini-
colônia (caixa de madeira 16 x 14 x 16 cm com tampa de vidro) contendo xarope (solução de
sacarose 50%), pólen natural, mais 20 operárias novas e/ou nutridoras coletadas de colônias
naturais e células de cria velha contendo indivíduos na fase de pupa prestes a emergir,
garantindo dessa forma a manutenção e estabelecimento da nova colônia.
Em alguns casos, a fim de evitar o ataque às rainhas virgens produzidas in vitro,
utilizamos uma gaiola de proteção para abelhas formada por uma base de madeira e uma tela
de arame de pequeno diâmetro. Este teste consistia em isolar dentro da gaiola a rainha virgem
produzida, um pedaço de algodão embebido em xarope e em média 3 a 5 operárias jovens.
Inicialmente, as mini-colônias foram mantidas em estufa BOD a 28º C e 70% de
umidade, por sete dias em média, dependendo das condições climáticas. Após este período
elas foram transferidas para o meliponário e o orifício de entrada foi desobstruído, permitindo
o fluxo dos indivíduos e a saída da rainha virgem para o vôo nupcial.
As colônias foram monitoradas periodicamente, verificando a presença da rainha, a
população de operárias e a disponibilidade de alimento. Quando necessário, as colônias
iniciais recebiam reforços de cria e/ou operárias e alimento.
Durante os experimentos constatamos a construção de algumas celas de
aprisionamento de rainhas virgens em colônias naturais de Frieseomelitta varia. Estas
câmaras construídas com cerume foram retiradas das colônias, juntamente com as rainhas
virgens aprisionadas, com a ajuda de uma espátula. As rainhas foram acondicionadas em
placas de Petri e levadas ao laboratório onde foram expostas rapidamente a baixas
temperaturas em gelo (4ºC), apenas o suficiente para que adormecessem (+/- 1 minuto),
pesadas e marcadas no mesonoto com tinta atóxica. Passado algum tempo, quando as rainhas
despertaram por completo, foram devolvidas à colônia de origem.

27
5. Ocorrência de machos em colônias naturais de Frieseomelitta varia
Para verificar a(s) época(s) de produção de machos de Frieseomelitta varia, foram
utilizadas três colônias (colônia 1, 2 e 3) priorizando-se o tamanho populacional (número
grande de operárias) e disponibilidade de alimento estocado. Estas colônias foram utilizadas
apenas para esse propósito.
Os dados foram coletados durante um ano, a partir de setembro/2007 até agosto/2008.
Para registrar a proporção do sexo dos membros da colônia, foram coletadas mensalmente de
cada colônia, células de cria contendo indivíduos em estágio pupal. O número de células de
cria foi coletado de acordo com as condições e disponibilidade da colônia. Estas células foram
colocadas em placas de Petri contendo a identificação do ninho de origem e, levadas ao
laboratório onde foram desoperculadas com o auxílio de uma pinça de ponta fina.
As pupas retiradas das células, depois de verificado o sexo, foram colocadas em placas
de Petri identificadas com o número da colônia de origem e mantidas em estufa à 28º C para
completarem seu desenvolvimento. Depois de nascidas, as abelhas foram devolvidas aos seus
respectivos ninhos para que a sua ausência não interferisse na dinâmica da colônia.
Foram coletados 1784 indivíduos, para a identificação do sexo com o auxílio de um
estereoscópio (Leica Wild M3B), através da presença (macho) ou ausência (fêmea) do
gonóstilo que caracteriza o aparelho reprodutor masculino. Desta forma a razão sexual, a
freqüência de machos e fêmeas, de cada colônia foi registrada mensalmente.
Para a análise dos dados coletados, computamos o número total de indivíduos de ambos os
sexos em cada colônia e estabelecemos uma freqüência mensal. Para a análise de múltiplas
amostras independentes utilizou-se o teste de Kruskal-Wallis (ANOVA) considerando
significativos os valores de probabilidade menores que 5% (p<0,05). O programa Statistic 6.0
foi utilizado para realizar tais testes estatísticos.

28
IV- RESULTADOS
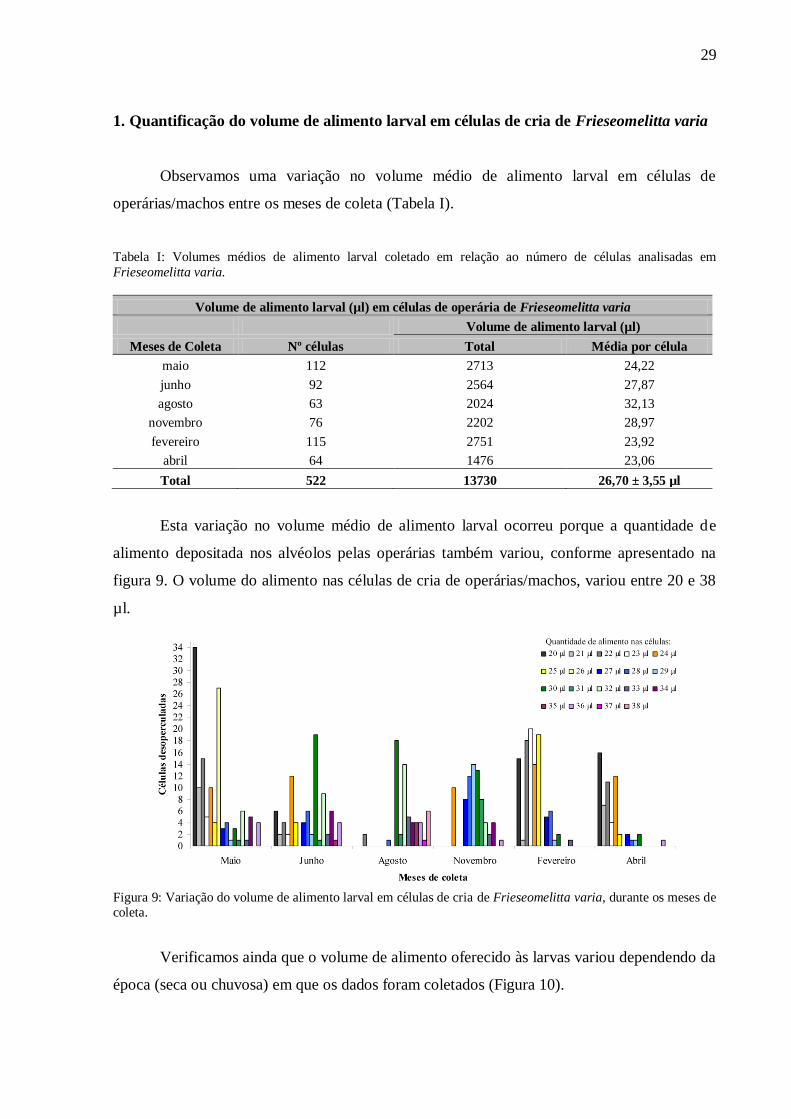
29
1. Quantificação do volume de alimento larval em células de cria de Frieseomelitta varia
Observamos uma variação no volume médio de alimento larval em células de
operárias/machos entre os meses de coleta (Tabela I).
Tabela I: Volumes médios de alimento larval coletado em relação ao número de células analisadas em
Frieseomelitta varia.
Volume de alimento larval (µl) em células de operária de Frieseomelitta varia
Volume de alimento larval (µl)
Meses de Coleta Nº células Total Média por célula
maio 112 2713 24,22
junho 92 2564 27,87
agosto 63 2024 32,13
novembro 76 2202 28,97
fevereiro 115 2751 23,92
abril 64 1476 23,06
Total 522 13730 26,70 ± 3,55 µl
Esta variação no volume médio de alimento larval ocorreu porque a quantidade de
alimento depositada nos alvéolos pelas operárias também variou, conforme apresentado na
figura 9. O volume do alimento nas células de cria de operárias/machos, variou entre 20 e 38
µl.
Figura 9: Variação do volume de alimento larval em células de cria de Frieseomelitta varia, durante os meses de coleta.
Verificamos ainda que o volume de alimento oferecido às larvas variou dependendo da
época (seca ou chuvosa) em que os dados foram coletados (Figura 10).

30
Figura 10: Variação do volume total de alimento larval nos alvéolos de Frieseomelitta varia nas diferentes épocas de coleta dos dados (época seca/chuvosa).
Na época seca (maio, junho e agosto), a quantidade de alimento nas células de operária
variou consideravelmente, encontramos células com o volume mínimo de 20 µl até o volume
máximo de 38 µl, apesar de estas últimas aparecerem em menor número. Por outro lado, na
época chuvosa (novembro, fevereiro e abril), a quantidade de alimento concentrou-se entre 20
e 30 µl, havendo uma baixa freqüência de células analisadas com volume superior a 32 µl.
Analisando o número total de células desoperculadas e o volume total de alimento
coletado nos meses de maio, junho, agosto (Estação Seca) e novembro, fevereiro e abril
(Estação Chuvosa) (Tabela II), encontramos que o volume total médio de alimento larval
existente em uma célula de operária/macho de F. varia na estação seca e na estação chuvosa,
foram de 27,34 ± 5,32 µl e 25,21± 3,68 µl, respectivamente.
0
5
10
15
20
25
30
35
40
20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38
Volumes alimento larval (µl)
To
tal
de c
élu
las
deso
percu
lad
as Época seca
0
5
10
15
20
25
30
35
40
20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38
Volumes alimento larval (µl)
To
tal
de c
élu
las
deso
percu
lad
as
Época chuvosa

31
Tabela II: Volumes médios de alimento larval em células de cria de Frieseomelitta varia em diferentes estações
do ano e comparação das médias pelo teste Z.
Estações do ano
Seca Chuvosa
Tamanho amostral 267 255
Volume médio de alimento (µl) 27.34 25.21
Variância paramétrica 28.40 13.60
Desvio Padrão 5.32 3.68
(Z)= 5.3370 ---
(p)unilateral = < 0.0001 ---
Poder (0.05) 0.9999 ---
Diferença entre as médias = 2.1328 ---
IC 95% (Dif. entre Médias) = 2.129797 2.135812
Comparando as médias das duas estações (seca e chuvosa), por meio do teste Z,
encontramos que α = 0,0001, indicando que existe variação entre a média de alimento larval
nas células de operárias/machos, nos períodos seco e chuvoso.
Com base no intervalo de confiança entre as médias das amostras (±2,13), podemos
dizer que o volume de alimento larval em uma célula de cria de F. varia encontra-se, com
grau de confiança de 95%, entre 24,57 µl (limite inferior) e 28,83 µl (limite superior).
2. Determinação do número de ínstares larvais em Frieseomelitta varia
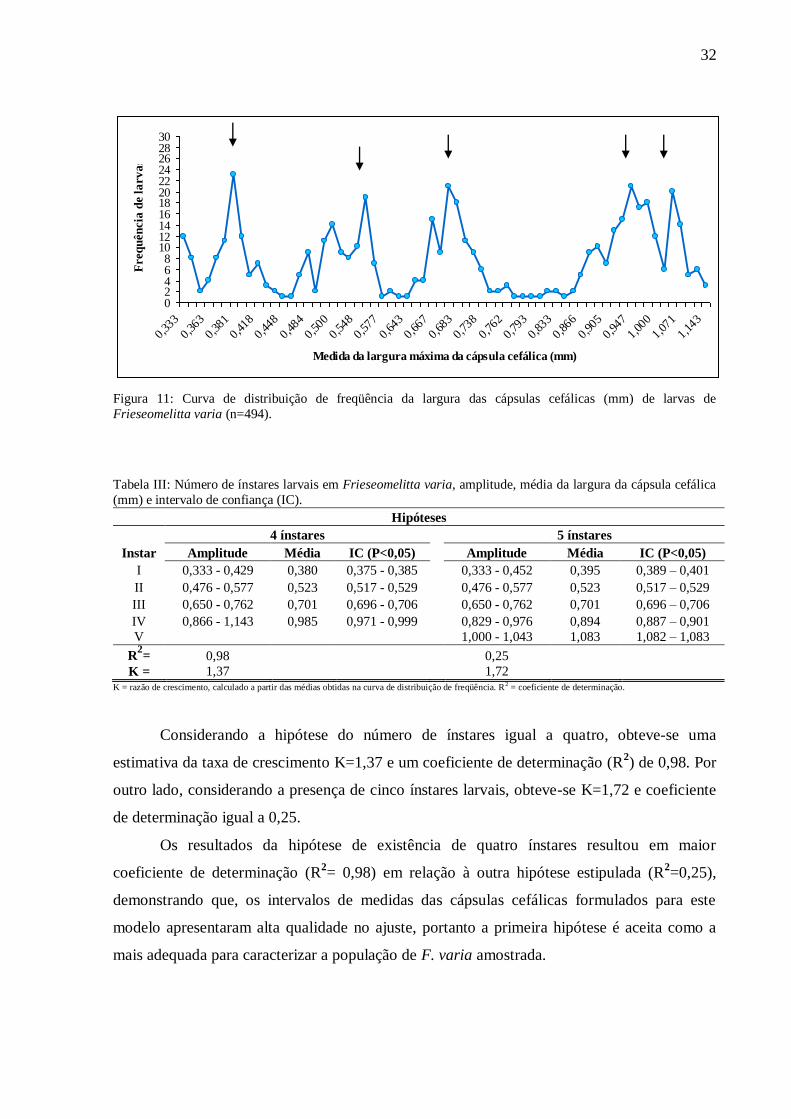
Através da análise da curva de distribuição de freqüências da largura da cápsula
cefálica (Figura 11) observamos a presença de cinco picos, sendo os três primeiros bem
distintos e os dois últimos bastante próximos. Os declives da curva de distribuição sugerem a
ocorrência de no mínimo quatro e, no máximo, cinco ínstares larvais.
Neste sentido, foram testadas duas hipóteses a fim de determinar o número de ínstares
larvais para a espécie Friesomelitta. varia: 1) a existência de quatro ínstares larvais; 2) a
existência de cinco ínstares, por meio do modelo de Dyar, obtendo-se os dados apresentados
na Tabela III.
32
02468
1012141618202224262830
0,333
0,363
0,381
0,418
0,448
0,484
0,500
0,548
0,577
0,643
0,667
0,683
0,738
0,762
0,793
0,833
0,866
0,905
0,947
1,000
1,071
1,143
Medida da largura máxima da cápsula cefálica (mm)
Freq
uên
cia
de l
arv
as
Figura 11: Curva de distribuição de freqüência da largura das cápsulas cefálicas (mm) de larvas de
Frieseomelitta varia (n=494).
Tabela III: Número de ínstares larvais em Frieseomelitta varia, amplitude, média da largura da cápsula cefálica
(mm) e intervalo de confiança (IC).
Hipóteses
Instar
4 ínstares 5 ínstares
Amplitude Média IC (P<0,05) Amplitude Média IC (P<0,05)
I 0,333 - 0,429 0,380 0,375 - 0,385 0,333 - 0,452 0,395 0,389 – 0,401
II 0,476 - 0,577 0,523 0,517 - 0,529 0,476 - 0,577 0,523 0,517 – 0,529
III 0,650 - 0,762 0,701 0,696 - 0,706 0,650 - 0,762 0,701 0,696 – 0,706
IV 0,866 - 1,143 0,985 0,971 - 0,999 0,829 - 0,976 0,894 0,887 – 0,901
V 1,000 - 1,043 1,083 1,082 – 1,083
R2= 0,98 0,25
K = 1,37 1,72 K = razão de crescimento, calculado a partir das médias obtidas na curva de distribuição de freqüência. R2 = coeficiente de determinação.
Considerando a hipótese do número de ínstares igual a quatro, obteve-se uma
estimativa da taxa de crescimento K=1,37 e um coeficiente de determinação (R2) de 0,98. Por
outro lado, considerando a presença de cinco ínstares larvais, obteve-se K=1,72 e coeficiente
de determinação igual a 0,25.
Os resultados da hipótese de existência de quatro ínstares resultou em maior
coeficiente de determinação (R2= 0,98) em relação à outra hipótese estipulada (R
2=0,25),
demonstrando que, os intervalos de medidas das cápsulas cefálicas formulados para este
modelo apresentaram alta qualidade no ajuste, portanto a primeira hipótese é aceita como a
mais adequada para caracterizar a população de F. varia amostrada.
33
3. Produção in vitro de rainhas de Frieseomelitta varia
Sabendo que nas abelhas desta espécie a diferenciação das castas ocorre por meio da
quantidade de alimento ingerido pela larva, sendo que os indivíduos diferenciados em rainha
ingerem uma quantidade maior de alimento do que aquelas que originam operárias e, tendo
encontrado um volume médio de alimento larval de 26,70 ± 3,55 µl por célula de operária,
tomamos como base este dado para dar início aos testes de manipulação de diferentes
quantidades de alimento oferecidas às larvas.
Os experimentos foram realizados a partir da transferência de larvas na fase pré-
defecante com cápsula cefálica medindo entre 0,866 e 1,143 mm.
Foram realizados nove experimentos de transferência de larvas de Frieseomelitta
varia, nos meses de fevereiro, abril, maio, junho, julho, agosto, outubro, novembro e
dezembro, totalizando 339 larvas transferidas e destas, 109 indivíduos (32%) completaram
seu desenvolvimento até a fase adulta (Tabela IV).
Tabela IV: Desenvolvimento in vitro das larvas de Frieseomelitta varia.
Meses de
Transferência
Número de larvas
transferidas
Natalidade Mortalidade
Quant. % Quant. %
abril 48 16 33 32 67
julho 35 23 66 12 34
outubro 38 5 13 33 87
dezembro 65 9 14 56 86
fevereiro 25 5 20 20 80
maio 36 5 14 31 86
junho 18 15 83 3 17 agosto* 32 0 0 32 100
novembro** 42 31 74 11 26
Total 339 109 32 230 68 * Mortalidade total das larvas transferidas
** Nascimento de machos
No experimento realizado durante o mês de novembro (n=42 larvas transferidas), após
o nascimento dos adultos, os indivíduos foram analisados sob lente de aumento (40x) e
verificamos que todas as abelhas deste grupo (n=31 indivíduos emergidos) pertenciam ao
sexo masculino. Fato este que despertou nosso interesse a respeito das possíveis épocas de
produção de machos para as abelhas desta espécie. Estes dados serão apresentados adiante.
Sabendo que a quantidade média de alimento em uma célula de cria de F. varia é de
aproximadamente 26,7 µl e, supondo que a célula auxiliar contenha o mesmo volume de
34
alimento, procedemos os experimentos com diferentes volumes de alimento a partir do
volume conhecido (ver item 1), fornecendo às larvas 25, 30, 35, 40 e 50 µl (Tabela V).
Tabela V: Diferentes quantidades de alimento larval oferecidos às larvas para produção de rainhas in vitro de
Frieseomelitta varia.
Meses referentes às
transferências
Vol. de
alimento
larval (µl)
Larvas
Transferidas Natalidade
Rainhas
Emergidas
Operárias
Emergidas
Número % Número % Número %
junho 25 18 15 83 6 40 9 60
dezembro/fevereiro 30 90 14 16 11 79 3 21
maio/outubro 35 74 10 14 10 100 0 0
abril/julho 40 83 39 47 29 74 10 26
agosto 50 32 0 0 0 0 0 0
Total 297 78 56 22
A partir dos dados sumarizados na tabela acima, podemos observar que no
experimento realizado em agosto (50µl de alimento larval), houve mortalidade total das larvas
transferidas (n=32). Por sua vez, nos experimentos em que foram fornecidas às larvas
quantidades menores de alimento, 30 e 40 µl (dezembro/fevereiro e abril/julho), houve uma
maior freqüência de larvas diferenciadas em rainha. Os experimentos realizados nos meses de
maio e outubro nos quais foi oferecido às larvas em desenvolvimento um volume de alimento
larval de 35 µl, apesar da taxa de mortalidade das larvas ter sido elevada, todas as larvas
nascidas (n=10) se diferenciaram em rainha.
3.1 Possível ocorrência de intercasta
No experimento realizado em dezembro (30 µl), nasceram nove indivíduos (Tabela
IV), entre eles seis rainhas e três operárias, curiosamente verificamos o nascimento de um
indivíduo com algumas características peculiares que, em princípio pensamos ser uma rainha,
pois apresentava o abdômen mais desenvolvido do que de uma operária e com coloração mais
clara, como o de uma rainha (Fig.12B). Entretanto, ao observar este indivíduo sob lente de
aumento do estereomicroscópio, observamos a presença da corbícula no último par de pernas,
semelhante à corbícula de uma operária (Fig. 12A) embora fosse menos aparente (Fig. 12C).
Esta abelha recebeu o mesmo tratamento dado às rainhas produzidas, a fim de
observarmos o seu comportamento junto às operárias e sua viabilidade reprodutiva.

35
Figura 12: Detalhe do último par de pernas dos adultos nascidos do experimento realizado em dezembro (30µl
de alimento larval). A: Operária, B: Rainha, C: indivíduo com características intermediárias entre rainha e
operária.
Ao introduzir a abelha marcada na mini-colônia, observamos que: ao perceber a
presença das operárias, sua primeira reação foi procurar refúgio atrás dos potes de pólen
localizados em um dos cantos da mini-colônia e ali ficou por aproximadamente 12 minutos;
quando saiu do esconderijo caminhou rapidamente para a tampa de vidro da colônia e ali
ficou passando constantemente as pernas posteriores sobre o abdômen. Após terem notado a
presença da nova rainha, três jovens operárias nutridoras permaneceram ao seu redor, por
vezes tocando-a com as antenas.
Durante o tempo de observação (3h e 20 minutos) não observamos comportamento
agressivo das operárias, entretanto a possível “intercasta” se apresentava agitada à
aproximação das operárias.
No terceiro dia após ter sido introduzida, encontramos a “intercasta” morta no
depósito de lixo da colônia.
4. Mini-colônias de Frieseomelitta varia utilizando-se rainhas produzidas in vitro
As rainhas produzidas sob condições controladas, logo após o nascimento, foram
pesadas e marcadas na região torácica (mesonoto) e transferidas para placas de Petri com
alimento (xarope) e algumas operárias jovens, para posteriormente serem introduzidas nas
mini-colônias. Dentre as rainhas produzidas (n=56), quatro fêmeas morreram quando ainda
A C B
36
estavam na placa e não apresentaram indícios de morte por agressão. No total, foram
introduzidas 52 rainhas virgens produzidas em 52 mini-colônias (Tabela VI).
Verificamos uma alta taxa de mortalidade das rainhas introduzidas (82%) nas mini-
colônias (Tabela VI). Os fatores responsáveis pela morte das rainhas virgens produzidas
podem ser observados na figura 13.
Tabela VI: Rainhas de Frieseomelitta varia produzidas in vitro, introduzidas e aceitas nas mini-colônias.
Mês referente à
Transferência
Rainhas
produzidas
Rainhas
introduzidas
Rainhas
fecundadas
Mortalidade dos
adultos
Quant. % Quant. %
abril 12 10 0 0 10 100
julho 17 16 1 6 15 88
outubro 5 5 1 20 4 80
dezembro 7 6 0 0 6 86
fevereiro 4 4 2 50 2 50
maio 5 5 1 20 4 80
junho 6 6 1 17 5 83
Total 56 52 6 11 46 82
Figura 13: Fatores responsáveis pela alta mortalidade das rainhas de Frieseomelitta varia produzidas in vitro.
Quanto às interações entre as rainhas produzidas e as operárias nas mini-colônias,
verificamos que algumas operárias apresentaram comportamento agressivo resultando em
77% dos casos, na eliminação das rainhas virgens. Em todos os testes de introdução das
rainhas, o tempo de rejeição e conseqüentemente morte das rainhas produzidas não
ultrapassou 48 horas.
Em alguns casos (15%) as rainhas introduzidas não foram encontradas no interior das
novas colônias.
Em virtude da agressividade das operárias, testamos a utilização de gaiolas de
proteção para isolar as rainhas produzidas in vitro (n=8) das operárias da mini-colônia. Em

37
75% dos casos (n=6), em dois dias (±48 horas) após a introdução das gaiolas de proteção nas
mini-colônias, as operárias cobriram completamente a tela das gaiolas com cerume (Figura
14) ocasionando a morte das rainhas. Este comportamento se repetiu nas duas mini-colônias
restantes em um período um pouco mais longo de 3 dias (±72 horas).
Figura 14: Gaiola dentro da qual as rainhas produzidas foram mantidas isoladas das operárias da mini-colônia. Detalhe da tela com fragmentos de cerume que a cobriam.
O estabelecimento da fisogastria pelas rainhas produzidas in vitro foi observado em
seis casos. Depois de passar pela aceitação das operárias, as rainhas produzidas nos
experimentos de julho, outubro, fevereiro, maio e junho (Tabela VI), realizaram o vôo nupcial
com sucesso. Embora não tenhamos observado diretamente este comportamento, verificamos
a construção, o aprisionamento e a oviposição de células de cria pela rainha, além disso,
observarmos a presença das rainhas marcadas com o abdômen dilatado, fisogástrico, nas
mini-colônias (Figuras 15A e 15B).

38
Figura 15: A) Mini-colônia com rainha produzida “in vitro” (círculo), B) detalhe da rainha marcada e fecundada.
A seta evidencia o abdômen desenvolvido da rainha marcada.
Durante as visitas de monitoramento dos ninhos no meliponário, observamos o
aparecimento de três rainhas virgens naturais aprisionadas em câmaras de cerume. A primeira
rainha foi encontrada em abril/2007 (Figura 16) presa em uma câmara de cerume construída
pelas operárias (1,8 cm de diâmetro e 2 cm de espessura da parede) na parede lateral de vidro
de uma colônia experimental, montada a partir de módulos de madeira e com parede externa
de vidro (Figura 17), elaborada por Jairo de Souza e Ademilson Espencer Egea Soares do
Departamento Genética da FMRP/USP com a finalidade de facilitar as observações diretas da
colônia. Acompanhamos esta rainha virgem por 10 dias, até verificarmos no décimo dia a
destruição da cela de aprisionamento e a antiga rainha fisogástrica caminhando pela colônia.
A B

39
Figura 16: Célula de aprisionamento com rainha virgem de Frieseomelitta varia. Foto: Weyder Santana
Figura 17: Colônia experimental para Frieseomelitta varia, construída em módulos de madeira e laterais de
vidro. A) Vista lateral; B) Vista frontal; C) Vista interna da colônia, D) Detalhe interno dos módulos
preenchidos. Fotos: Weyder Santana e Geusa Simone de Freitas.
Nos outros dois casos, encontramos as rainhas virgens aprisionadas nas câmaras de
cerume construídas na tampa de duas colônias racionais tradicionais, nos meses de maio/2007
e setembro/2008. A rainha encontrada no mês de maio/2007 (p=0,0223 g) foi eliminada pelas
operárias em menos de 24 horas após ter sido devolvida à colônia, seu corpo estava
totalmente coberto com cerume e grudado em uma das paredes do ninho.
A B
C D

40
No dia seguinte à reintrodução da rainha encontrada em setembro/2008 (p=0,0241 g),
observamos que ela foi novamente aprisionada pelas operárias. Nove dias depois, a câmara de
aprisionamento estava destruída e o corpo da rainha virgem marcada foi encontrado na área
reservada para depósito de lixo da colônia, estava apenas com uma das pernas anteriores.
Em meados de outubro/2007, notamos a presença de dois casulos reais em uma
colônia de Frieseomelitta varia. A colônia se encontrava muito populosa, com muitas células
de cria, estoque de mel e pólen e a rainha mãe foi vista caminhando agilmente pelos favos.
Em setembro/2008, outro casulo real foi encontrado em outra colônia localizada no
meliponário, entretanto, não foi observada a presença da rainha dominante nesta colônia.
Os três casulos reais foram retirados cuidadosamente de suas respectivas colônias e
mantidos em estufa a 28 ºC, dentro de pequenas placas de Petri até o seu nascimento, quando
foram pesadas, marcadas e devolvidas às colônias. Com exceção de uma rainha que morreu
acidentalmente durante o procedimento de pesagem.
O peso destes indivíduos produzidos em condições naturais foi comparado com o peso
das rainhas produzidas artificialmente que obtiveram sucesso ao se fixarem como fêmea
dominante nas mini-colônias (Tabela VII) e com o restante das rainhas nascidas (Tabela
VIII).
Tabela VII: Peso (g) das rainhas produzidas em condições naturais e das rainhas produzidas em laboratório que
se estabeleceram nas mini-colônias.
Peso das rainhas virgens após o nascimento (g)
Rainhas naturais Rainhas in vitro
Encontrada em: Peso Peso Produzida em:
maio 0,0223 0,0218 fevereiro
setembro 0,0241 0,0168 fevereiro
setembro 0,0198 0,0209 maio
outubro 0,0219 0,0181 junho
outubo 0,0269 0,0199 julho 0,0149 outubro
Média 0,0230 0,0187
Segundo o Teste-t para duas amostras independentes (Tabela VIII) as rainhas
produzidas “in vitro” (P) e as rainhas produzidas naturalmente nas colônias (N) não possuem
pesos médios diferentes estatisticamente entre si, uma vez que o valor-p foi menor que 0,05
(0,0128), adotando nível de significância de 5%.

41
Tabela VIII: Teste-t: Comparação do peso ao nascer entre as rainhas naturais (N) e rainhas produzidas em
condições controladas (P) que se estabeleceram como fêmeas dominantes nas mini-colônias.
N P
Média = 230.00 187.33
Variância = 709.00 685.86
Variância = 696.14 ---
T = 2.67 ---
Graus de liberdade = 9 ---
p (unilateral) = 0.0128 ---
Poder (0.05) 0.8469 ---
Diferença entre as médias = 42.66 ---
IC 95% (Dif. entre médias) = 6.52 a 78.80
Por sua vez, os dados de peso médio das rainhas produzidas in vitro que morreram
(Pm) comparado com os dados coletados das rainhas virgens naturais (N), sumarizados na
Tabela IX, demonstraram que a média do peso das duas amostras também não são diferentes
entre si, pois o valor-p encontrado é menor que 0,05 (p= 0,0023).
Tabela IX: Teste-t: Comparação do peso ao nascer entre as rainhas naturais (N) e rainhas produzidas in vitro que
morreram (Pm).
N Pm
Tamanho = 5 50
Média = 230 178
Variância = 709 1485
Variância = 1426.04 ---
T = 2.96 ---
Graus de liberdade = 53 ---
p (unilateral) = 0.0023 ---
Poder (0.05) 0.9583 ---
Diferença entre as médias = 52.44 ---
IC 95% (Dif. entre médias) = 16.93 a 87.94
5. Ocorrência de machos em colônias naturais de Frieseomelitta varia
A ocorrência de todos os indivíduos (machos e fêmeas) durante os meses de coleta
estão representados na Tabela X e Figura 18.
Tabela X: Freqüência de machos e operárias em três colônias naturais de Frieseomelitta varia em diferentes
épocas do ano.
Colônia
Frequência (Nº e %) células com pupas de machos e
operárias
Machos Operárias Total de células
Quant. Freq.(%) Quant. Freq.(%) n=
Setembro
Colônia 1 0 0 34 100 34
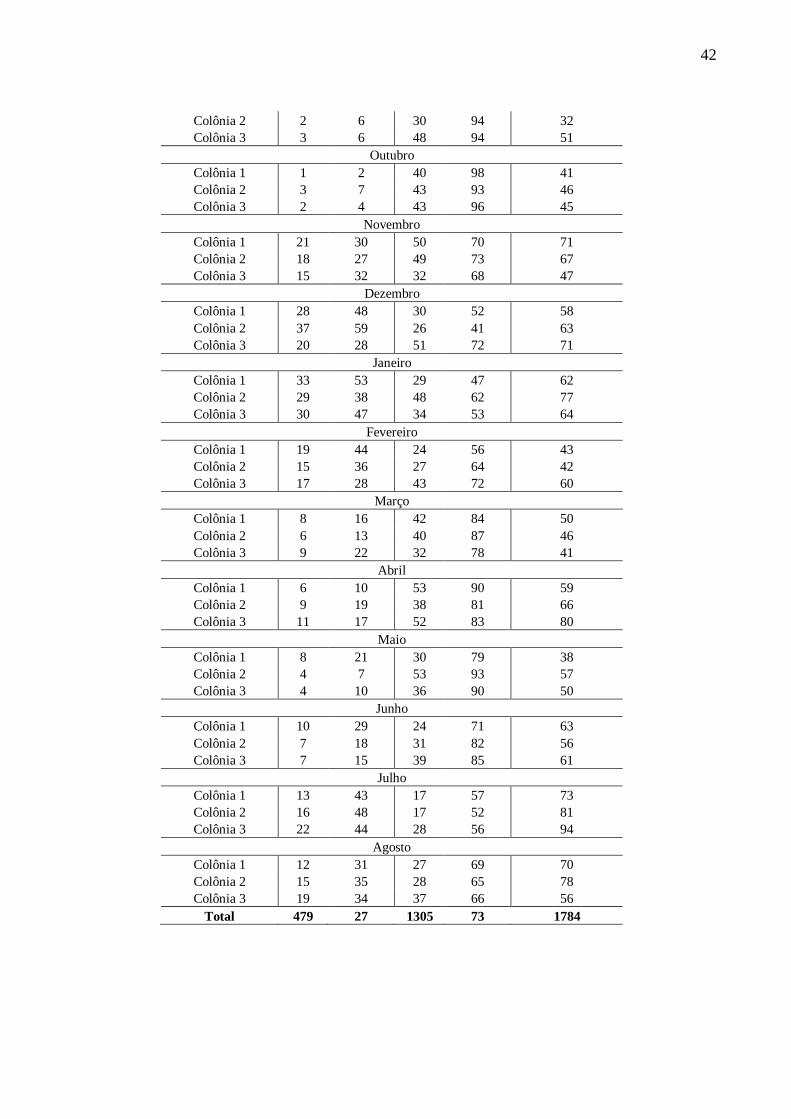
42
Colônia 2 2 6 30 94 32
Colônia 3 3 6 48 94 51
Outubro
Colônia 1 1 2 40 98 41
Colônia 2 3 7 43 93 46
Colônia 3 2 4 43 96 45
Novembro
Colônia 1 21 30 50 70 71
Colônia 2 18 27 49 73 67
Colônia 3 15 32 32 68 47
Dezembro
Colônia 1 28 48 30 52 58
Colônia 2 37 59 26 41 63
Colônia 3 20 28 51 72 71
Janeiro
Colônia 1 33 53 29 47 62
Colônia 2 29 38 48 62 77
Colônia 3 30 47 34 53 64
Fevereiro
Colônia 1 19 44 24 56 43
Colônia 2 15 36 27 64 42
Colônia 3 17 28 43 72 60
Março
Colônia 1 8 16 42 84 50
Colônia 2 6 13 40 87 46
Colônia 3 9 22 32 78 41
Abril
Colônia 1 6 10 53 90 59
Colônia 2 9 19 38 81 66
Colônia 3 11 17 52 83 80
Maio
Colônia 1 8 21 30 79 38
Colônia 2 4 7 53 93 57
Colônia 3 4 10 36 90 50
Junho
Colônia 1 10 29 24 71 63
Colônia 2 7 18 31 82 56
Colônia 3 7 15 39 85 61
Julho
Colônia 1 13 43 17 57 73
Colônia 2 16 48 17 52 81
Colônia 3 22 44 28 56 94
Agosto
Colônia 1 12 31 27 69 70
Colônia 2 15 35 28 65 78
Colônia 3 19 34 37 66 56
Total 479 27 1305 73 1784
43
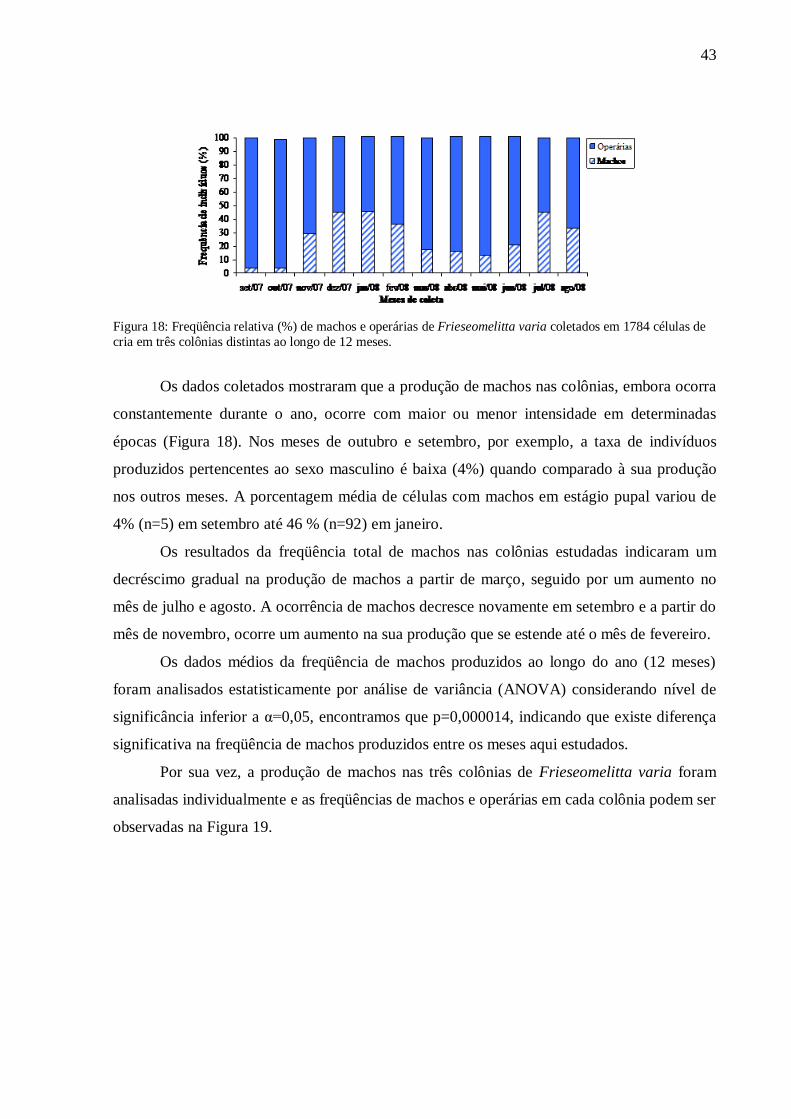
Figura 18: Freqüência relativa (%) de machos e operárias de Frieseomelitta varia coletados em 1784 células de
cria em três colônias distintas ao longo de 12 meses.
Os dados coletados mostraram que a produção de machos nas colônias, embora ocorra
constantemente durante o ano, ocorre com maior ou menor intensidade em determinadas
épocas (Figura 18). Nos meses de outubro e setembro, por exemplo, a taxa de indivíduos
produzidos pertencentes ao sexo masculino é baixa (4%) quando comparado à sua produção
nos outros meses. A porcentagem média de células com machos em estágio pupal variou de
4% (n=5) em setembro até 46 % (n=92) em janeiro.
Os resultados da freqüência total de machos nas colônias estudadas indicaram um
decréscimo gradual na produção de machos a partir de março, seguido por um aumento no
mês de julho e agosto. A ocorrência de machos decresce novamente em setembro e a partir do
mês de novembro, ocorre um aumento na sua produção que se estende até o mês de fevereiro.
Os dados médios da freqüência de machos produzidos ao longo do ano (12 meses)
foram analisados estatisticamente por análise de variância (ANOVA) considerando nível de
significância inferior a α=0,05, encontramos que p=0,000014, indicando que existe diferença
significativa na freqüência de machos produzidos entre os meses aqui estudados.
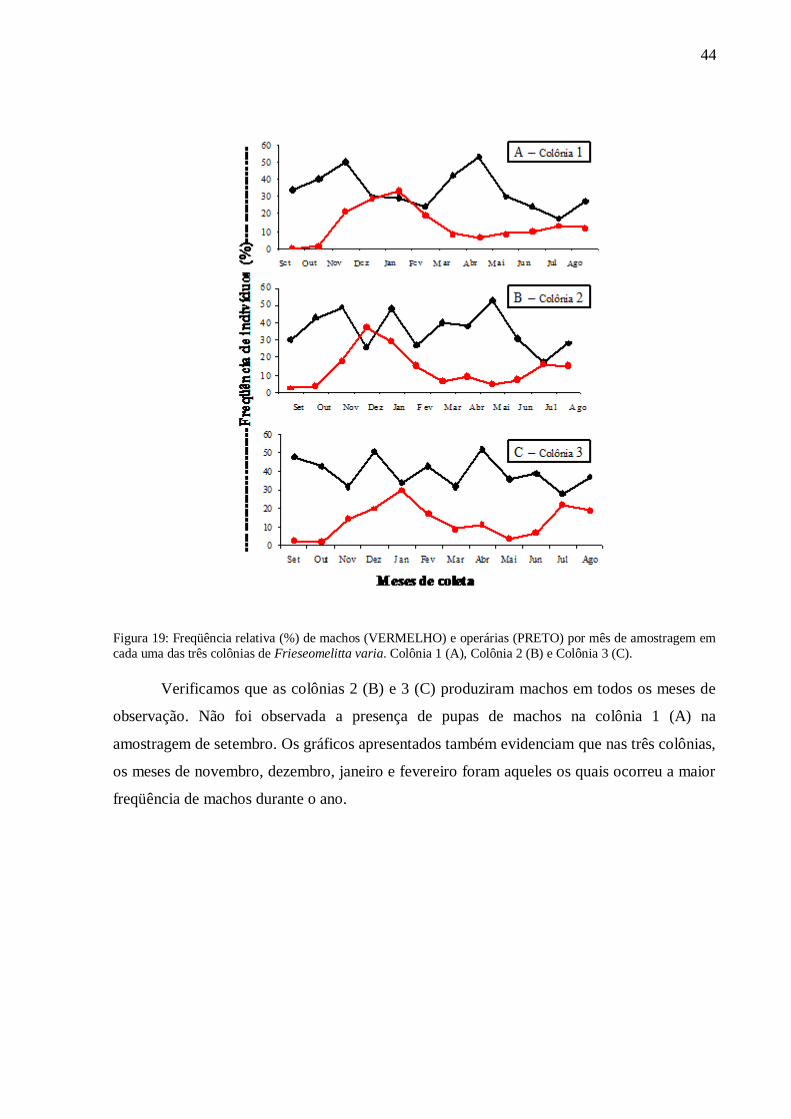
Por sua vez, a produção de machos nas três colônias de Frieseomelitta varia foram
analisadas individualmente e as freqüências de machos e operárias em cada colônia podem ser
observadas na Figura 19.
44
Figura 19: Freqüência relativa (%) de machos (VERMELHO) e operárias (PRETO) por mês de amostragem em
cada uma das três colônias de Frieseomelitta varia. Colônia 1 (A), Colônia 2 (B) e Colônia 3 (C).
Verificamos que as colônias 2 (B) e 3 (C) produziram machos em todos os meses de
observação. Não foi observada a presença de pupas de machos na colônia 1 (A) na
amostragem de setembro. Os gráficos apresentados também evidenciam que nas três colônias,
os meses de novembro, dezembro, janeiro e fevereiro foram aqueles os quais ocorreu a maior
freqüência de machos durante o ano.

45
V- DISCUSSÃO

46
1. Quantificação do volume de alimento larval em células de cria de Frieseomelitta varia
As abelhas sem ferrão seguem a estratégia de aprovisionamento massal de alimento
em suas células de cria (Kerr, 1948; Michener, 1974; Nogueira-Neto, 1997), entretanto, o
mecanismo de determinação das castas difere entre as abelhas do gênero Melipona e as
abelhas dos demais gêneros. Nos meliponíneos, exceto os do gênero Melipona, a indução da
casta parece ser determinada apenas pela quantidade de alimento ingerido pela larva durante
seu desenvolvimento (Hartfelder et al, 2006). Camargo (1972) demonstrou em prática esta
teoria, produzindo rainhas de Scaptotrigona postica a partir de quantidades excessivas de
alimento larval.
O passo inicial para a realização dos experimentos a cerca da teoria de determinação
das castas com base na quantidade de alimento fornecido à larva, é a quantificação do
alimento larval nas células de operária. Neste trabalho encontramos para Frieseomelitta varia,
um volume total médio de alimento larval em célula de operária igual a 26,7 ± 3,55 µl.
As operárias de F. varia participam ativamente do estágio de aprovisionamento das
células depositando alimento em seu interior, em média 2,0 a 4,0 vezes (Faustino, 2002) até
completar a quantidade suficiente de alimento necessária para o desenvolvimento da larva.
Verificamos a ocorrência de uma variação do volume de alimento larval contido nas células
de cria entre os meses de coleta: estação seca (maio, junho e agosto) de 20 µl à 38 µl; estação
chuvosa (novembro, fevereiro e abril) a maior freqüência de células ocorreu com quantidades
de alimento larval entre 20 e 30 µl.
A variação na quantidade de alimento aprovisionado nas células de cria está
relacionada à disponibilidade de alimento estocado e/ou à densidade populacional da colônia,
uma vez que as abelhas coletam quantidades maiores ou menores de recursos alimentares
(néctar e pólen) de acordo com a sua disponibilidade na natureza e os fornecem às larvas da
mesma forma. Nas épocas chuvosas do ano, por exemplo, embora tenha mais recursos
disponíveis, o investimento na produção de cria nas colônias também aumenta,
conseqüentemente, a demanda de alimento aumentaria e as operárias por sua vez investiriam
menos na alimentação das larvas, depositando menores quantidades de alimento por célula
para atender as necessidades da colônia.
Entretanto, não podemos afirmar com base nos dados apresentados que a situação
descrita acima ocorra desta forma, para isso é necessário verificar futuramente o efeito do

47
estoque de alimento e da densidade populacional sobre a quantidade de alimento fornecida às
larvas.
2. Determinação do número de ínstares larvais em Frieseomelitta varia
O processo de muda é de alto custo energético para o inseto e o conteúdo calórico e
nutricional de uma cutícula mudada pode representar mais de 20% da produção total de
biomassa larval (Parra, 1991). Assim, o inseto alcança um peso máximo em cada ínstar e
perde peso durante a muda, pois a cutícula mudada e a energia usada na ecdise contribuem
para perdas que chegam a ser da ordem de 45% (Waldbauer, 1962). A cutícula totalmente
esclerotizada não expande, portanto, o crescimento de partes duras somente ocorre quando
houver uma muda, quando então uma nova cutícula será produzida e expandida (Parra &
Haddad, 1989). Deste modo, a avaliação das medidas das cápsulas cefálicas é um bom
parâmetro para se avaliar o crescimento dos insetos, como previsto pelo modelo de DYAR.
O número de estágios larvais parece não ser constante nas abelhas. Para as espécies de
abelha da subtribo Meliponina, por exemplo, tem sido verificada a presença de três a cinco
ínstares larvais. Darchen & Delage-Darchen (1974) e Tambasco (1975), citado por Rossini
(1989), encontraram cinco ínstares larvais para Melipona bicolor, Melipona beecheii e
Melipona quadrifasciata anthidioides, respectivamente. Entretanto, Rossini e Bueno (1986) e
Rossini (1989) demonstraram através de observações diretas do desenvolvimento larval e pela
análise da cápsula cefálica das larvas, a existência de três ínstares para Melipona fasciculata
(K=1,42) e para Melipona quadrifasciata anthidioides (K=1,33).
Inicialmente, para as abelhas da espécie Scaptotrigona postica, Oliveira (1960)
encontrou cinco ínstares larvais, posteriormente Neves e Cruz-Landim (1984) caracterizaram
o desenvolvimento larval dessa espécie pela presença de quatro ínstares, separados por três
mudas e uma taxa de crescimento (K) de 1,38. Azevedo (1997) encontrou dificuldades em
confirmar a presença de quatro ou cinco ínstares em Partamona helleri, levando-se em conta
somente o modelo de Dyar. É possível que os diferentes resultados encontrados pelos autores
e as dificuldades em estabelecer precisamente o número de ínstares larvais, ocorram devido às
diferentes metodologias empregadas pelos pesquisadores, além disso, deve-se levar em conta
que as taxas de crescimento de um estágio para o outro pode ser alterado por vários fatores,
como as condições de temperatura e umidade dentro da colônia, quantidade e/ou qualidade do
alimento fornecido à larva.

48
Os resultados encontrados neste trabalho demonstraram que o coeficiente de
determinação (R=0,98), considerando-se a hipótese de quatro ínstares representou melhor
ajuste dos dados, como preconizado por Parra e Haddad (1989).
Quanto à razão de crescimento (K), foram observados valores médios que obedecem a
uma progressão geométrica, como postulado por Dyar (1890). Considerando a presença de
quatro ínstares, a média da razão de crescimento igual a 1,37 se encontra dentro do intervalo
de variação preconizado pelo modelo de Dyar, que postula que as peças do exoesqueleto dos
insetos crescem de maneira mais ou menos constante a cada muda, por uma razão geométrica
próxima a 1,4 vezes, e aproximou-se da razão média de 1,52 registrada por Cole (1980) para
insetos holometábolos.
De acordo com os resultados encontrados, pode-se inferir que as larvas de
Frieseomelitta varia amostradas, apresentam quatro ínstares larvais, caracterizado pelos
intervalos de largura da cápsula cefálica entre 0,333 - 0,429 mm, como sendo o primeiro
ínstar, 0,476 - 0,577 mm, o segundo, 0,650 - 0,762 mm, o terceiro e, 0,866 - 1,143 mm, o
quarto e último ínstar larval.
3. Transferências de larvas de Frieseomelitta varia com diferentes quantidades de
alimento, visando a produção de rainhas “in vitro”
A determinação das castas nos Apidae é de origem trofogênica ou trofogenética. A
determinação trofogênica ocorre quando as diferenças entre as castas surgem somente devido
às diferenças alimentares, ou seja, na quantidade ou na qualidade do alimento ingerido pela
larva, ou ambas (Camargo, 1972; Hartfelder & Engels 1989; Buschini & Campo, 1995). E a
trofogenética se restringe, até o momento, às espécies do gênero Melipona, e envolve fatores
genéticos atuando em conjunto com fatores alimentares (Kerr et al., 1966).
A produção de rainhas nos meliponíneos com determinação trófica das castas é
condicionada pelas operárias, ao construírem células de cria de tamanho maior capazes de
receber mais alimento. No entanto, Terada (1974) e Faustino (2002) demonstraram que nas
abelhas da espécie Frieseomelitta varia não ocorre construção de células reais típicas
diferente das células de operária e macho. Colônias mantidas sob condições de orfandade são
capazes de produzir rainhas, construindo células auxiliares próximas às células que contém
larvas de operárias no fim do estágio larval, que ainda estejam se alimentando. Estas células
auxiliares, através de uma conexão com as células contendo larvas, fornecem um suprimento

49
extra de alimento para as mesmas. Ao final deste período de “superalimentação”, a larva dará
origem a uma rainha.
Testando o efeito nutricional na determinação das castas em Frieseomelitta varia,
através da manipulação de doses suplementares de alimento oferecidas às larvas no último
estágio de alimentação, pudemos observar que a quantidade de alimento consumido pelas
larvas foi o fator determinante na diferenciação final do adulto com características fenotípicas
bem definidas. O desenvolvimento de rainhas ocorreu a partir da administração de 25 até 40µl
de alimento larval extra. Volumes maiores de alimento, de 50µl, não foram completamente
ingeridos pelas larvas demonstrando que possivelmente, elas apresentem um limite de
ingestão de alimento necessário para seu desenvolvimento, neste caso representado pelo
volume máximo de 40 µl.
Os experimentos foram realizados a partir da transferência de larvas na fase pré-
defecante, com cápsula cefálica medindo entre 0,866 e 1,143 mm, estas larvas além de serem
mais resistentes à manipulação são encontradas com maior facilidade nos favos de cria. No
entanto não podemos afirmar a idade das larvas utilizadas nas transferências. Embora algumas
tentativas, infelizmente sem sucesso, tenham sido realizadas na esperança de obter estes
dados, alguns fatores como a própria disposição dos favos de cria (em forma de cacho), a
postura desordenada das células e a intolerância das operárias frente a estímulos externos
como o manuseio e marcação das células, impossibilitaram a coleta dos dados. O fato de não
sabermos exatamente a idade das larvas utilizadas nas transferências e a quantidade de
alimento ingerido por elas anteriormente aos experimentos, talvez expliquem o nascimento de
operárias nas transferências realizadas. Em estudos futuros pretendemos utilizar a colônia de
módulos desenvolvida por Souza e Soares para desenvolver um método de observação de
postura controlada da rainha, visando estudar o tempo de desenvolvimento dos imaturos dessa
espécie a fim de aperfeiçoar e padronizar a técnica de produção de rainhas em laboratório.
Considerando que as larvas de operárias de Frieseomelitta varia possam se diferenciar
em rainhas após consumir o alimento aprovisionado em sua célula, e adicionalmente o
alimento da célula auxiliar, ambas de tamanhos similares (Faustino & Zucchi, 2002), supõe-se
que as rainhas ingerem duas vezes mais alimento que as operárias durante o desenvolvimento
larval. Verificamos que a quantidade média de alimento larval nas células de operária/macho
é em média de 26,70 ± 3,55 µl e, supondo que as larvas utilizadas nos experimentos ingeriram
esta mesma quantidade anteriormente às transferências, verificamos que após o consumo de
volumes extras de alimento (32,50 ± 6,45 µl) as larvas se diferenciaram em rainhas. Assim, o
volume total médio de alimento necessário para a diferenciação de uma larva jovem de F.

50
varia em rainha é de aproximadamente 59,20 ± 5,00 µl, ou seja, a larva destinada a ser rainha
consome 2,23 vezes mais alimento que a futura operária.
Em suma, estes resultados reforçam a teoria de que a determinação das castas nas
abelhas da espécie Frieseomelitta varia depende exclusivamente da quantidade de alimento
fornecida à larva em desenvolvimento. Os dados encontrados em nosso trabalho são
semelhantes a outros estudos a respeito do efeito da nutrição larval na determinação das castas
das abelhas sem ferrão. De acordo com Prato (2007), as larvas de Tetragonisca angustula
destinadas a se tornarem rainhas são alimentadas em média, 6,62 vezes mais (53 µl) do que as
futuras operárias (8 µl); em Trigona spinipes as abelhas diferenciadas em rainhas consomem
10 vezes mais alimento que as operárias, a larva de rainha ingere em torno de 360 µl de
alimento, enquanto a larva de operária consome apenas 36 µl do mesmo alimento (Buschini &
Campos, 1995; Lisboa et al., 2005). Estudos realizados com Scaptotrigona quadripunctata
demonstraram que o fornecimento de doses extras de alimento às larvas de operárias, fez com
que se desenvolvessem em rainhas (Camargo, 1972), Campos & Costa (1989) e recentemente
Castilho-Hyodo (2001) verificaram ainda que, rainhas de Schwarziana quadripunctata podem
emergir de células de operárias sendo resultantes de larvas que receberam uma quantidade de
alimento maior, que aquela recebida pelas operárias.
No início dos experimentos, um dos fatores responsáveis pela grande mortalidade das
larvas (68%) foi o efeito negativo da manipulação dos imaturos durante o processo de
transferência da célula de cria para os poços da placa de Elisa contendo alimento larval, sendo
que estas larvas apresentam uma película bastante delicada e susceptível à lesões. Entretanto,
mesmo após adquirir habilidade considerável no manejo das larvas, a mortalidade dos
indivíduos transferidos permanecia elevada. Verificamos que as larvas não ingeriam o
alimento fornecido, pois esse havia solidificado, provavelmente devido à baixa umidade
relativa do ar, razão pela qual passamos a controlar a umidade dos experimentos
acondicionados em estufa, utilizando solução supersaturada de NaCl e água (1:1) nos
experimentos seguintes, a partir daí não constatou-se mais a solidificação do alimento larval.
Em relação ao tempo de desenvolvimento dos indivíduos obtidos nos experimentos,
verificamos que as rainhas (n= 56) levaram em média 29,25 ± 2,63 dias para emergir, o que
corrobora com as observações realizadas por Faustino (2002) que acompanhou o processo de
produção natural de duas rainhas de F. varia, desde a construção da célula auxiliar até a
emergência dos adultos, levando 25 e 28 dias para completar seu desenvolvimento. As
operárias por sua vez, apresentaram tempo menor de desenvolvimento, em média 22,56 ±
2,38 dias. O tempo total de desenvolvimento das rainhas foi maior do que o das operárias,

51
pois está relacionado com o maior consumo de alimento ingerido pelas larvas de rainha
(Camargo, 1972) que desencadeia tardiamente os processos de muda.
Quanto ao método empregado para a produção “in vitro” de rainhas, o sucesso no
nascimento das larvas transferidas em placa tipo Elisa foi menor (32%) que o esperado. No
experimento realizado em agosto, houve mortalidade total das larvas transferidas (n=32), isto
ocorreu porque o volume de alimento oferecido às larvas transferidas de 50µl foi superior à
quantidade máxima ingerida por elas. Observamos que algumas larvas se afogaram no grande
volume de alimento, e aquelas que conseguiram se alimentar não ingeriram todo o alimento.
O restante de alimento larval não consumido em conjunto com as condições dentro da estufa
tornou o ambiente propício ao desenvolvimento de fungos que proliferaram rapidamente
causando a morte das larvas.
No entanto, os dados relacionados à alta mortalidade das larvas podem auxiliar no
aprimoramento da técnica aqui utilizada em estudos futuros, quanto ao controle dos prováveis
fatores responsáveis pela mortandade dos imaturos, como: a contaminação por fungos e
bactérias, seja no alimento ou em qualquer instrumento empregado no processo; a
manutenção das condições de acondicionamento dos experimentos em estufa; o manuseio das
larvas durante as transferências e afogamento dos imaturos em demasiadas quantidades de
alimento.
Segundo Velthuis & Sommeijer (1991) em colônias naturais de abelhas sem ferrão não
ocorre intercasta, uma vez que o resultado final da diferenciação das castas nos insetos sociais
é o desenvolvimento de fêmeas muito diferenciadas, tanto em atributos morfológicos quanto
comportamentais, ou seja, rainhas (funções reprodutivas) ou operárias (funções mantenedoras
da colônia).
No entanto, a literatura afim registra escassos exemplos de ocorrência de intercastas
em colônias naturais. De fato, neste aspecto, têm-se apenas as informações de Cappas e Sousa
(1992) que encontrou intercastas em uma colônia de Melipona quadrifasciata e, Mateus e
colaboradores (2002) em colônia natural de Melipona seminigra mantida em laboratório.
Com relação à intercastas obtidas em experimentos realizados sob condições
controladas, também foram relatados alguns prováveis casos em espécies de meliponíneos,
como por exemplo, em Melipona beechei (Velthuis et al., 1991).
Em nosso estudo, não foi possível determinar com base em apenas um indivíduo
produzido se o adulto obtido, portador de características combinadas de rainha e operária
pertencia ou não a uma intercasta. Para tanto, seria necessário considerar outras características
relacionadas à sua morfologia externa e interna.

52
Assim, o caráter recente da descoberta em Frieseomelitta varia, não permite o
estabelecimento de bases sólidas para discutir adequadamente o problema das intercastas,
entretanto, afirmam a importância do efeito nutricional na determinação da casta real nas
abelhas desta espécie. Possivelmente, as operárias desta espécie sejam fêmeas subalimentadas
que uma vez ingerindo uma dose insuficiente de alimento durante seu desenvolvimento para
produzir uma rainha completamente diferenciada, originam fêmeas estéreis com ovaríolos
totalmente degenerados ainda durante o período pupal, como verificado por Boleli e
colaboradores (1999, 2000), demonstrando desta forma sua capacidade de totipotência.
4. Montagem e desenvolvimento de mini-colônias de Frieseomelitta varia utilizando as
rainhas produzidas in vitro
Vários fatores como aqueles relacionados ao ataque de inimigos naturais, como
formigas e algumas espécies de abelhas como Scaptotrigona depilis, bem como acidentes
naturais causados por fenômenos climáticos, contribuíram para a mortalidade das rainhas
introduzidas, e nestes casos em particular (8%) na morte de toda a população das mini-
colônias. Em 15% dos casos, as rainhas produzidas não foram encontradas no ninho,
provavelmente ao saírem para realizar o vôo nupcial, ou o vôo de reconhecimento da colônia
possam ter sido predadas ou então não conseguiram localizar a colônia de origem.
Entretanto é interessante notar que as operárias foram responsáveis pela morte das
rainhas introduzidas, em 77% dos casos, o que provavelmente está relacionado com a
capacidade de reconhecimento entre os indivíduos do ninho, precisamente entre as operárias e
a nova rainha virgem, uma vez que os insetos sociais exalam compostos que funcionam como
assinaturas químicas individuais carregando informações a respeito da casta, idade, sexo e
status reprodutivo (Howard & Blomquist, 2005).
O reconhecimento ocorre através de duas etapas principais: a liberação de uma pista
pelo animal e o reconhecimento dela por um segundo indivíduo (Beecher, 1982). Os insetos
sociais não são uma exceção à regra, o que pode ser facilmente entendido ao observarmos
que, após uma rápida “antenação” as operárias são capazes de identificar os companheiros do
ninho.
Diversos estudos comportamentais e químicos confirmam que as pistas utilizadas no
reconhecimento são representadas por compostos associados à cutícula dos insetos (Breed &
Stiller, 1992; Gamboa et al., 1996) que é recoberta por uma camada de ceras impermeáveis
constituída dos hidrocarbonetos (Lockey, 1988).

53
Nunes (2008) demonstrou que rainhas naturais recém emergidas de Frieseomelitta
varia não possuem 16 compostos característicos presentes na cutícula de rainhas fisogástricas,
em contrapartida apresentam 2 compostos que não são encontrados nos adultos. Os indivíduos
de F. varia reconhecem pistas indesejadas no perfil cuticular daqueles não companheiros do
ninho, e essas pistas são as responsáveis pela rejeição dos indivíduos. Nessa proposta, a
aceitação se dá não pela presença das pistas desejadas, mas sim pela ausência das indesejadas
(Couvillon e Ratnieks, 2008; Nunes, 2008).
Os diversos odores presentes em uma colônia são absorvidos pela cera, e redistribuída
por ela formando um odor homogêneo dentro do ninho. Breed et al (1995), estudando a cera
das abelhas melíferas, concluíram que ela é o componente com maior importância na
formação do padrão de hidrocarbonetos presentes na cutícula destas abelhas. Seguindo esta
mesma linha, acredita-se que o cerume em colônias de F. varia também atue como unificador
de compostos químicos presentes na colônia, por este motivo as rainhas produzidas em
laboratório foram expostas a pequenas quantidades de cerume para que adquirissem o odor
característico da colônia, na tentativa de auxiliar no processo de reconhecimento destas
operárias. Segundo Nunes (2008) em experimentos de reconhecimento e análises químicas
realizadas com indivíduos de F. varia, ocorre transferência de pistas utilizadas no
reconhecimento através de um breve contato com o cerume. As análises químicas para os
indivíduos que permaneceram por 10 minutos em contato com o cerume mostrou que a
concentração de alguns compostos mudou, e outros foram adquiridos.
Quanto às interações rainha virgem – operárias, verificamos uma grande inquietação
por parte das rainhas introduzidas que se movimentavam rapidamente no interior da colônia e
eram intensamente assediadas pelas operárias. Este comportamento está de acordo com
Imperatriz-Fonseca & Zucchi (1995) que observaram este comportamento em rainhas virgens
de abelhas sem ferrão não pertencentes ao gênero Melipona. Além deste comportamento,
observamos que a maioria das rainhas introduzidas apresentou outros três tipos de
comportamentos distintos, embora nem sempre com a mesma intensidade ou freqüência: 1)
movimentação constante das asas, mesmo quando a rainha se encontrava parada, 2)
agressividade contra as operárias e 3) as rainhas passavam as pernas posteriores sobre o
abdome, esticando-o, provavelmente para estimular as glândulas responsáveis pela liberação
do cheiro de atratividade da rainha.
Embora não tenhamos observado diretamente o vôo nupcial das rainhas produzidas
que se estabeleceram nas mini-colônias (n=6), consideramos que elas haviam sido fecundadas
a partir da observação da construção, aprovisionamento e presença de novas células

54
operculadas. Segundo Ferreira (1993), a construção das primeiras células de cria está
relacionada com o estímulo do vôo nupcial da rainha, a partir do qual estas estruturas
começam a ser construídas após a fecundação da rainha.
Foi relatado anteriormente que alguns elementos, bióticos e abióticos, interferiram
negativamente no sucesso das rainhas produzidas in vitro, no entanto, algumas rainhas se
estabeleceram como fêmea dominante (n=6).
A disponibilidade de machos na natureza pode ter contribuído para o sucesso e
estabelecimento das rainhas produzidas em condições controladas como fêmeas dominantes
nas mini-colônias (Tabela XI).
Tabela XI: Meses nos quais ocorreu a fecundação das rainhas de Frieomelitta varia produzidas in vitro
Experimento de
transferência das
larvas
Mês referente à
introdução da rainha
virgem
Tempo (dias)
decorrente até à
fecundação
Mês referente à
fecundaçao da
rainha
julho agosto 19 setembro
outubro novembro 38 dezembro
fevereiro março 58 maio
fevereiro março 17 março
maio junho 72 agosto
junho Julho 27 agosto
Após a introdução das rainhas nas colônias órfãs, observamos que elas levaram em
média 38,5 ± 22,25 dias para fecundar, fato confirmado através de observações diretas sobre a
morfologia externa das rainhas, precisamente a visível dilatação da região abdominal.
Observando os dados acima, percebemos que em dois casos particulares: rainha
introduzida em março e rainha introduzida no mês de junho, a fecundação ocorreu
tardiamente, 58 e 72 dias após a introdução, respectivamente. As demais rainhas introduzidas
em agosto, novembro, março e julho, fecundaram em média três vezes mais rápido (22,75 ±
5,56 dias).
A relação entre os meses em que ocorreu a fecundação das rainhas virgens produzidas
e os meses de produção natural de machos em colônias de F. varia estão representados na
Figura 20.
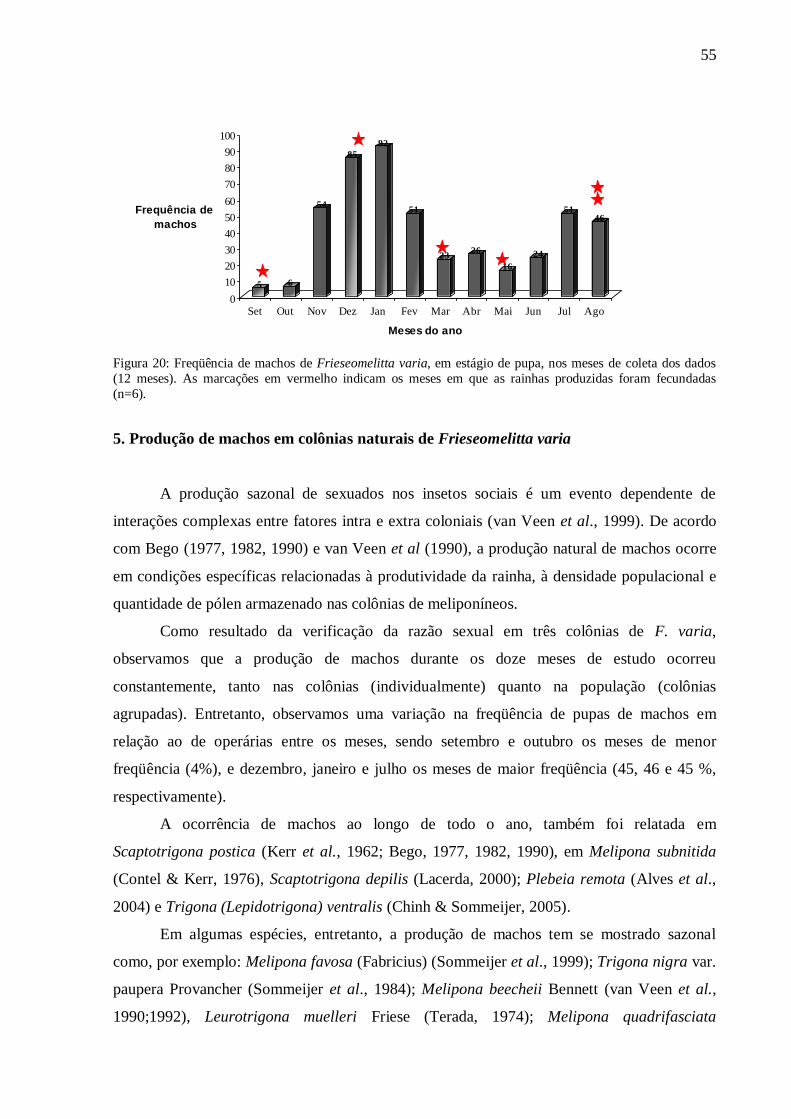
55
5 6
54
85
92
51
2326
16
24
5146
0
10
20
30
40
50
60
70
80
90
100
Frequência de
machos
Set Out Nov Dez Jan Fev Mar Abr Mai Jun Jul Ago
Meses do ano
Figura 20: Freqüência de machos de Frieseomelitta varia, em estágio de pupa, nos meses de coleta dos dados
(12 meses). As marcações em vermelho indicam os meses em que as rainhas produzidas foram fecundadas
(n=6).
5. Produção de machos em colônias naturais de Frieseomelitta varia
A produção sazonal de sexuados nos insetos sociais é um evento dependente de
interações complexas entre fatores intra e extra coloniais (van Veen et al., 1999). De acordo
com Bego (1977, 1982, 1990) e van Veen et al (1990), a produção natural de machos ocorre
em condições específicas relacionadas à produtividade da rainha, à densidade populacional e
quantidade de pólen armazenado nas colônias de meliponíneos.
Como resultado da verificação da razão sexual em três colônias de F. varia,
observamos que a produção de machos durante os doze meses de estudo ocorreu
constantemente, tanto nas colônias (individualmente) quanto na população (colônias
agrupadas). Entretanto, observamos uma variação na freqüência de pupas de machos em
relação ao de operárias entre os meses, sendo setembro e outubro os meses de menor
freqüência (4%), e dezembro, janeiro e julho os meses de maior freqüência (45, 46 e 45 %,
respectivamente).
A ocorrência de machos ao longo de todo o ano, também foi relatada em
Scaptotrigona postica (Kerr et al., 1962; Bego, 1977, 1982, 1990), em Melipona subnitida
(Contel & Kerr, 1976), Scaptotrigona depilis (Lacerda, 2000); Plebeia remota (Alves et al.,
2004) e Trigona (Lepidotrigona) ventralis (Chinh & Sommeijer, 2005).
Em algumas espécies, entretanto, a produção de machos tem se mostrado sazonal
como, por exemplo: Melipona favosa (Fabricius) (Sommeijer et al., 1999); Trigona nigra var.
paupera Provancher (Sommeijer et al., 1984); Melipona beecheii Bennett (van Veen et al.,
1990;1992), Leurotrigona muelleri Friese (Terada, 1974); Melipona quadrifasciata

56
(Camargo, 1976 e Bezerra, 1995), Plebeia (Friesella) schrotkyii Friese (Camillo-Atique,
1977) e Schwarziana quadripunctata (Alves & Imperatriz-Fonseca, 2005).
Velthuis et al. (2005) reviram os padrões de produção de machos em algumas espécies
de abelha sem ferrão e verificaram que os períodos cíclicos de produção de machos (MPP)
ocorrem em colônias individuais, também observado em Melipona favosa (Chinh et al., 2003)
e Melipona bicolor (Alves, 2004), embora isto não reflita no contexto populacional, ou seja,
geralmente a produção de machos ocorre continuamente ao longo do ano.
A respeito da influência de fatores externos na produção de machos, Bego (1982)
aponta que os fatores extrínsecos à colônia como a temperatura e precipitação, que
desempenham papel na disponibilidade de alimento no ambiente, não são os fatores
determinantes na produtividade de machos; segundo a autora, os fatores intrínsecos como a
densidade populacional da colônia, que pode ser conseqüência do estoque de pólen e mel, são
os fatores determinantes.
Em seu estudo com Melipona beecheii, Moo Valle et al ( 2001) testando o efeito da
nutrição na produção de sexuados, observaram que as colônias pouco alimentadas produziram
0,7% de machos na população, enquanto as colônias superalimentadas produziram 23,4%,
concluindo que as reservas de alimento tem forte influência na produção dos machos.
É válido ressaltar que, as células de cria coletadas mensalmente nas colônias de F.
varia eram compostas por células que continham abelhas nos estágios de pupa. Observamos
que as pupas de macho coletadas levaram em média 14 a 20 dias para completar seu
desenvolvimento e emergir do casulo. Portanto, devemos considerar que na natureza, os
indivíduos adultos que estão aptos a desempenharem seu papel reprodutivo, serão encontrados
no mês seguinte ao da coleta dos dados aqui apresentados
Os dados obtidos acerca da produção de machos em Frieseomelitta varia, são
importantes pois sabendo as épocas de maior disponibilidade de machos na natureza,
podemos sincronizar com a produção “in vitro” das rainhas, aumentando o sucesso no
estabelecimento das mini-colônias, haja vista que, algumas rainhas produzidas in vitro e que
foram introduzidas desapareceram do ninho, provavelmente não haviam machos próximos
para realizar a cópula e, ao saírem para realizar o vôo nupcial, por longo tempo se perderam
ou foram predadas.
Como mencionado anteriormente, a quantidade de alimento estocado nas colônias,
influenciam na produção de sexuados, entretanto, não observamos escassez de alimento nas
colônias de Frieseomelitta varia utilizadas na coleta dos dados, mesmo porque as abelhas
desta espécie são conhecidas por serem excelentes coletoras de pólen e as colônias foram

57
selecionadas para este estudo considerando suas condições internas de estoque de alimento.
Contudo, estudos mais detalhados a respeito de tal tema devem ser realizados, para que a
influência do alimento estocado na produção de sexuados nesta espécie possa ser esclarecida.

58
VI- CONCLUSÕES

59
De acordo com os resultados obtidos, podemos concluir que:
Em uma célula de operária/macho de Frieseomelitta varia, é depositado em média
26,7 ± 3,55 µl de alimento larval. Entretanto, a quantidade de alimento aprovisionado nas
células pode variar dependendo da estação do ano (seca ou chuvosa) e da disponibilidade de
recursos alimentares.
A hipótese da presença de quatro ínstares larvais para as abelhas da espécie
Frieseomelitta varia, representou o melhor ajuste dos dados, com coeficiente de determinação
(R2) igual a 0,98 e média da razão de crescimento (K) igual a 1,37. Assim, as larvas
amostradas neste estudo apresentaram intervalos de largura da cápsula cefálica entre 0,333 -
0,429 mm como sendo o primeiro instar; 0,476 - 0,577 mm o segundo; 0,650 - 0,762 mm o
terceiro e, 0,866 - 1,143 mm o quarto e último ínstar larval.
O desenvolvimento das larvas transferidas (em fase tardia de desenvolvimento) em
rainha ocorre a partir da ingestão de 25 até 40µl de alimento larval extra, desta forma as larvas
que darão origem a rainhas se alimentam em média 2,23 mais que as larvas que se
desenvolverão em operárias.
Em relação ao tempo de desenvolvimento dos indivíduos obtidos nos experimentos,
verificamos que as rainhas produzidas sob condições controladas (n= 56) levaram em média
29,25 ± 2,63 dias para emergir. As operárias por sua vez, apresentaram tempo menor de
desenvolvimento, em média 22,56 ± 2,38 dias.
Com base em apenas um indivíduo de F. varia produzido, portador de características
combinadas de rainha e operária, não é possível afirmar se pertencia ou não a uma intercasta.
Sugerimos que novos experimentos sejam realizados a fim de se produzir maiores
quantidades destes indivíduos para que sejam analisados detalhadamente.
A maioria das rainhas produzidas “in vitro” que foram introduzidas nas mini-colônias
apresentaram comportamentos semelhantes ao de rainhas virgens naturais quando se
encontravam na presença das operárias.
Após a introdução das rainhas virgens produzidas nas mini-colônias, elas levaram em
média 38,5 ± 22,25 dias para fecundar.
Machos de Frieseomelitta varia são produzidos constantemente ao longo do ano,
embora ocorra uma variação na sua produção natural em alguns meses. Encontramos uma
menor freqüência de pupas de machos nos meses de setembro e outubro, por outro lado nos
meses de dezembro, janeiro e julho a ocorrência destes indivíduos em relação as operárias foi
maior.

60
REFERÊNCIAS BIBLIOGRÁFICAS

61
ALVES, D. A. & IMPERATRIZ-FONSECA, V. L. One year-study on the production of
sexuals in a stingless bee species, Schwarziana quadripunctata (Lepeletier, 1836)
(Hymenoptera: Apidae: Meliponini). In: ABSTRACTS HANDBOOK OF ASSOCIATION
FOR TROPICAL BIOLOGY AND CONSERVATION, p. 103, 2005.
ALVES, D. A. Produção de operárias, rainhas e machos em Melipona bicolor Lepeletier,
1836 (Apidae, Meliponini). 2004. Trabalho de conclusão do curso, apresentada à
Universidade Presbit. Mackenzie, São Paulo, 2004.
ALVES, D. A.; RIBEIRO, M. F.; SANTOS-FILHO, P. S.; IMPERATRIZ-FONSECA V. L.
Production of gynes and males in Plebeia remota Holmberg, 1903 (Apidae, Meliponini), In:
PROC. 8TH IBRA INT. CONF. ON TROPICAL BEES AND VI ENCONTRO SOBRE
ABELHAS, Ribeirão Preto: Funpec, p. 753, 2004.
AZEVEDO, G. G. Atividade de vôo e determinação do número de ínstares larvais em
Partamona helleri (Friese) (Hymenoptera, Apidae, Meliponinae). 1997. 64 f. Dissertação
apresentada ao Departamento de Biologia, Universidade Federal de Viçosa, Viçosa, 1997.
BEECHER, M. D. Signature systems and kin recognition. American Zoologist, v. 22, p. 477-
490, 1982.
BEETSMA, J. The process of queen-worker differentiation in the honey bee. Bee World, v.
60, p. 24-39, 1979.
BEGO, L. R. On social regulation in Nannotrigona (Scaptotrigona) postica Latreille, with
special reference to productivity of colonies (Hymenoptera, Apidae, Meliponinae). Rev. Bras.
Entomol., v. 34, p. 721-738, 1990.
BEGO, L. R. Aspectos da regulação social em Nannotrigona (Scaptotrigona) postica
Latreille (Hymenoptera, Apidae, Meliponinae). 1977. 180 f. Tese apresentada à Faculdade
de Medicina de Ribeirão Preto, Universidade de São Paulo, Ribeirão Preto, 1977.
BEGO, L. R. On social regulation in Nannotrigona (Scaptotrigona) postica Latreille, with
special reference to male production cycles (Hym., Apidae, Meliponinae). Bol. Zool., São
Paulo, v. 7, p. 181–196, 1982.
BEIG, D. The production of male in queenright colonies of Trigona (Scaptotrigona) postica.
Jour. Apic. Res, v. 11, n. 1, p. 33-39, 1972.

62
BOLELI, I. C.; PAULINO-SIMÕES, Z. L; BITONDI, M. M. G. Cell death in ovarioles
causes permanent sterility in Frieseomelitta varia worker bees. J. Morphol, v. 242, n. 3, p.
271-82, 1999.
BOLELI, I. C.; PAULINO-SIMÕES, Z. L; BITONDI, M. M. G. Regression of the lateral
oviducts during the larval adult transformation of the reproductive system on Melipona
quadrifasciata and Frieseomelitta varia. J. Morphol. v. 243, p. 141 – 151, 2000.
BOURKE, A. F. G. Worker reproduction in the highly eussocial Hymenoptera. Quart. Rev.
Biol., v. 63, p. 291 – 311, 1988.
BREED, M. D., STILLER, T. M. Honey bee Apis mellifera, nestmate discrimination:
Hydrocarbon effects and the evolutionary implications of comb choice. Anim. Behav., v. 43,
p. 875-883, 1992.
BREED, M. D.; GARRY, M. F.; PEARCE, A. N.; HIBBARD, B. E.; BJOSTAD, L. B.;
PAGE., R. E. The role of wax comb in honey bee nestmate recognition. Anim. Behav., v. 50,
p. 489-496, 1995.
BUSCHINI, M. L. T., CAMPOS, L. A. O. Caste determination in Trigona spinipes
(Hymenoptera, Apidae): influence of the available food and the juvenile hormone. Rev.
Brasil. Biol., v. 55, p. 121 -129, 1995.
CAMARGO, C. A. Determinação de castas em Scaptotrigona postica Latreille
(Hymenoptera, Apidae). Rev. Bras. Biol., v. 32, p. 133-138, 1972.
CAMARGO, J. M. F.; PEDRO, S. R. M. Systematics, phylogeny and biogeography of the
Meliponinae: a mini review. Apidologie. v. 23, p. 509-522, 1992.
CAMILLO-ATIQUE, C. Estudo da variabilidade etológica de Friesella incluindo a
caracterização de espécies crípticas (Hym., Meliponinae). 1977. 203 f. Tese apresentada à
Faculdade de Medicina, Universidade de São Paulo, Ribeirão Preto, 1977.
CAMPOS, L. A. O. & COSTA, M. A. Determinação do sexo em abelhas: XXVIII.
Determinação das castas em Schwarziana quadripunctata (Hymenoptera, Apidae). Rev.
Bras. Biol., v. 49, n. 4, p. 999-1001, 1989.
CAPPAS E SOUSA, J. P. Os Meliponíneos em Portugal e na Europa. In: Anais do V
Congresso Ibérico de Entomologia. Lisboa, p. 53-68, 1992.

63
CASTILHO-HYODO, V. C. C. Rainha ou operária? Um ensaio sobre a determinação de
castas em Schwarziana quadripunctata (Lepeletier, 1836) (Hymenoptera, Apidae,
Meliponini). 2001. 134 f. Tese (Doutorado em Ecologia), apresentada ao Departamento de
Ecologia, Universidade de São Paulo, São Paulo. 2001.
CHINH, T. X. & SOMMEIJER, M. J. Production of sexuals in the stingless bee Trigona
(Lepidotrigona) ventralis flavibasis Cockerell (Apidae, Meliponini) in northern Vietnam.
Apidologie. v. 36, p. 493-503, 2005.
CHINH, T. X.; GROB, G. B. J., MEEUWSEN, F. J. A. J.; SOMMEIJER, M. J. Patterns of
male production in the stingless bee Melipona favosa (Apidae, Meliponini). Apidologie, v.
34, p. 161–170, 2003.
COLE, J. B. Growth ratios in holometabolous and hemimetabolous insects. In: Annals of the
Entomological Society of America. v. 64, p. 540-544, 1980.
CONTEL, E. P. B.; KERR, W. E. (1976). Origin of males in Melipona subnitida estimated
from data of an isozymic polymorphic system. Genética, v.46, p. 271-277, 1976.
CORTOPASSI-LAURINO, M. et al. Global meliponiculture: challenges and opportunities.
Apidologie, v. 37, n. 2, p. 275-292, 2006.
CORTOPASSI-LAURINO, M. Observações sobre atividades de machos de Plebeia droryana
Friese (Apidae, Meliponinae). Rev. Bras. Entomol. v. 24, p. 177-191, 1979.
COUVILLON, M. J., RATNIEKS, F. L. W. Odour transfer between colonies of the stingless
bee Frieseomelitta varia demonstrates that entrance guards use an “undesirable-absent” cue
recognition system. Behavioural Ecology and Sociobiology, 2008 (accepted).
CRUZ–LANDIM, C. Ovarian development in Meliponinae bees (Hymenoptera: Apidae) the
effect of queen presence and food on worker ovary development and egg production.
Genetics and Molecular Biology. v. 23, n. 1, p. 83-88, 2000.
DARCHEN, R. & DELAGE-DARCHEN, B. Contribution à l’étude d’une abeille du Mexique
Melipona beecheii B. (Hymenoptère : Apide). Apidologie, v. 6, p. 295–339, 1975. apud
ROSSINI, S. A. Caracterização das mudas ontogenéticas e biometria dos corpora allata
de Melipona quadrifasciata anthidioides Lep. (Hymenoptera, Apidae). 1989. 82 f. Tese
apresentada à Universidade Estadual Paulista Júlio de Mesquita Filho, Rio Claro, 1989.
DYAR, H. G. The number of molts of Lepidopterous larvae. Psyche. v. 5, p. 420-422. 1980.

64
ENGELS, W.; IMPERATRIZ-FONSECA, V. L. Caste development, reproductive strategies
and control of fertility in honey bees and stingless bees. In: ENGELS, W. (ed.), Social
Insects: Springer-verlag, Berlin, p 167-230, 1990.
FAUSTINO, C. D.; SILVA-MATOS, E. V.; MATEUS, S.; ZUCCHI, R. First record of
emergency queen rearing in stingless bees. Insectes Soc. v. 49, p. 111-113, 2002.
FAUSTINO, C. D.; ZUCCHI, R. Mini-colônias de Frieseomelitta varia (Hymenoptera,
Apinae, Meliponini) e sua utilização na solução de problemas bionômicos. 2002. 67 f.
Dissertação apresentada à Faculdade de Filosofia, Ciências e Letras de Ribeirão Preto,
Universidade de São Paulo, Ribeirão Preto, 2002.
FERREIRA, F. H. N. Aspectos da estratégia reprodutiva em Tetragonisca angustula
angustula Latreille, 1811 (Hymenoptera, Apidae, Meliponinae).1993. 40 f. Dissertação
(Mestrado em Entomologia) apresentada à Faculdade de Filosofia, Ciências e Letras de
Ribeirão Preto, Universidade de São Paulo, Ribeirão Preto, 1993.
GAMBOA, J. G.; GRUDZIEN, T. A.; ESPELIE, K. E.; BURA, E. A. Kin recognition in
social wasps: combining chemicals and behavioral evidence. Animal Behaviour, v. 51, p.
625-629, 1996.
HARTFELDER, K. et al. Physiological and genetic mechanisms underlying caste
development, reproduction and division of labor in stingless bees. Apidologie. v. 37, n. 2, p.
144-163, 2006.
HARTFELDER, K.; ENGELS, W. The composition of larval food in stingless bees:
Evaluating nutritional balance by chemosystematic methods. Insectes Sociaux, v. 36, p. 1-14,
1989.
HOWARD, R. W., BLOMQUIST, G. J. Ecological, behavioral, and biochemical aspects of
arthropod hydrocarbons. Annual Review of Entomology, v. 50, p. 371-393, 2005.
IMPERATRIZ-FONSECA, V. L. & ZUCCHI, R. Virgin queens in stingless bees (Apidae,
Meliponinae) colonies: a review. Apidologie, v. 26, p. 231-244, 1995.
IMPERATRIZ-FONSECA, V. L.; CONTRERA, F. A. L.; KLEINERT, A. M. P. A
Meliponicultura e a iniciativa Brasileira dos polinizadores. In: ANAIS DO XV CONGRESSO
BRASILEIRO DE APICULTURA E 1º CONGRESSO BRASILEIRO DE
MELIPONICULTORES, Natal, 2004.

65
IMPERATRIZ-FONSECA, V. L.; GONÇALVES, L. S.; DE JONG, D.; FREITAS, B. F.;
CASTRO, M. S.; SANTOS, I. A.; VENTURIERI, G. C. Abelhas e desenvolvimento rural no
Brasil. Rev. Mensagem Doce, v. 80, p. 03-08, 2005.
KERR, W. E. Estudos sobre o gênero Melipona. In: An. Esc. Sup. Agr. “Luiz de Queiroz”.
v. 5, n. 2, p. 181-276, 1948.
KERR, W. E. Some aspects of the evolution of social bees (Apidae). Evolutionary Biology,
v. 3, p. 119-175, 1969.
KERR, W. E.; CARVALHO, G. A.; NASCIMENTO, V. A. Abelha Uruçu: Biologia, manejo
e conservação. Belo Horizonte: Fundação Acangaú, p. 143, 1996.
KERR, W. E.; NIELSEN, R. A. Evidence that genetically determined Melipona queens can
become workers. Genetic. v. 54, p. 859-865, 1966.
KERR, W. E.; STORT, A. C.; MONTENEGRO, M. J. Importância de alguns fatores
ambientais na determinação das castas do gênero Melipona. Acad. Bras. Ciênc., v. 38, p.
149-168, 1966.
KERR, W. E.; ZUCCHI, R.; NAKADAIRA, I. T.; BUTOLO, J. E. Reproduction in the social
bees. J. N. Y. Entomol. Soc. v. 70, p. 265-276, 1962.
KOEDAM, D. et al. Behavioural and physiological implications of queen dominance in
stingless bees. Utrecht: Universiteit Utrecht. p. 109, 1995.
KOEDAM, D. et al. The behaviour of laying workers and the morfology and viability of their
eggs in Melipona bicolor bicolor. Physiol. Entomol. v. 26, n. 3, p. 254-259, 2001.
KOEDAM, D., CONTRERA, F. A. L.; IMPERATRIZ-FONSECA, V. L. Clustered male
production by workers in the stingless bee Melipona subnitida Ducke (Apidae, Meliponinae).
Insectes Soc., v. 46, p. 387-391, 1999.
KOEDAM, D.; CONTRERA, F. A. L.; FIDALGO, A. O.; IMPERATRIZ-FONSECA,. V. L.
How queens and workers share in male production in the stingless bee Melipona subnitida
Ducke (Apidae, Meliponini). Insectes Soc. v. 52, p. 114-121, 2005.
LACERDA, L. M. Morfometria e morfologia de ovos e aspectos relacionados à
reprodução de Scaptotrigona aff. depilis (Moure, 1942) (Hymenoptera, Apidae,

66
Meliponinae). 2000. 73 f. Tese apresentada à Facul. Filos. Ciên. Letras de Ribeirão Preto,
Universidade de São Paulo, Ribeirão Preto, 2000.
LACERDA, M. L. & SIMÕES, L. P. Z. Ovos produzidos por rainhas e operárias de
Scaptotrigona depilis (Hymenoptera, Apidae, Meliponina): morfometria e aspectos
relacionados. Iheringia, v. 96, n. 1, p. 89-93, 2006.
LAIDLAW, H. H.; GOMES, F. P.; KERR, W. E. Estimation of the number of lethal alleles in
a panmitic population of Apis mellifera. L. Genetics, v. 41, p. 179-188, 1956.
LISBOA, L. C. O.; SERRÃO, J. E.; CRUZ-LANDIM, C.; CAMPOS, L. A. O. Effect of larval
food amount on ovariole development in queens of Trigona spinipes (Hymenoptera, Apinae).
Anat. Histol. Embriol., v. 34, p. 179-184, 2005.
LOCKEY, K. H. Lipids of the insect cuticle: origin, composition and function. Comparable
Biochemistry and Physiology, v. 89B, p. 595-645, 1988.
MATEUS, S.; HRNCIR, M.; FAUSTINO, C. D.; ZUCCHI, R. Ocorrência de intercastas
adultas em colônias de Melipona seminigra: aspectos morfo-etológicos preliminares
(Hymenoptera, Apidae, Meliponini). In: ANAIS DO V ENCONTRO SOBRE ABELHAS,
Ribeirão Preto, p. 211-216, 2002.
MICHENER, C. D. Observations on the nests and behavior of Trigona in Australia an New
Guinea. American Museum Novitates, v. 2026, p. 1-46, 1961.
MICHENER, C. D. The bees of the world. Baltimore: The Johns Hopkins University Press,
p. 913, 2000.
MICHENER, C. D. The social behavior of the bees: a comparative study. Cambridge,
Massachussetts: Belknap Press of Harvard Univ. Press, p. 404, 1974.
MOO-VALLE, H.; QUEZADA-EUÁN, J. J. G.; WENSELEERS, T. The effect of food
reserves on the production of sexual offspring in the stingless bee Melipona beecheii (Apidae,
Meliponini). Insectes Sociaux, v. 48, p. 398-403, 2001.
NEVES, N. A., CRUZ-LANDIM, C. Determinação do número de estágios de Scaptotrigona
postica (Hymenoptera, Apidae). Ciên. Cult., v. 36, p. 912, 1984.

67
NIJHOUT, H. F.; WHEELER, D. F. Juvenile hormone and the physiological basis of insect
polymorphisms. The Quartely Review of Biology, v. 57, n. 2,p. 109-133, 1982.
NOGUEIRA-NETO, P. Vida e criação de abelhas indígenas sem ferrão. São Paulo:
Nogueirapis, p. 445, 1997.
NUNES, T. M. Reconhecimento parental em abelhas eussociais Neotropicais
(Hymenoptera: Apinae, Meliponini): uma análise dos mediadores químicos e seus
determinantes em Frieseomelitta varia. 2008. 52 f. Dissertação apresentada à Facul. Filos.
Ciên. Letras de Ribeirão Preto, Universidade de São Paulo, Ribeirão Preto, 2008.
OLIVEIRA, B. L. Mudas ontogenéticas em larvas de Melipona nigra schenki Gribodo
(Hymenoptera, Apoidea). Boletim da Universidade do Paraná - Zoologia, v. 2, p. 1-16,
1960.
PARRA, J. R. P. Consumo e utilização de alimentos por insetos. In: PANIZZI, A. R. &
PARRA, J. R. P. (ed.). Ecologia nutricional de insetos e suas implicações no manejo de
pragas. São Paulo: Manole, p. 9-65, 1991.
PARRA, J. R. P.; HADDAD, M. L. Determinação do número de ínstares de insetos.
Piracicaba: FEALQ, p. 49, 1989.
PAXTON, R. J.; BEGO, L. R.; SHAH, M. M.; MATEUS, S. Low mating frequency of
queens in the stingless bee Scaptotrigona postica and worker maternity of males. Behav.
Ecol. Sociobiol. v. 53, p. 174-181, 2003.
PRATO, M. Produção artificial de rainhas em Tetragonisca angustula Latreille 1811
(Hymenoptera, Apidae). 2007. 57 f. Trabalho de conclusão do curso (Biologia) apresentada
à Facul. Filos. Ciên. Letras de Ribeirão Preto, Universidade de São Paulo, Ribeirão Preto,
2007.
ROSSINI, S. A. Caracterização das mudas ontogenéticas e biometria dos corpora allata
de Melipona quadrifasciata anthidioides Lep. (Hymenoptera, Apidae). 1989. 82 f. Tese
apresentada à Universidade Estadual Paulista Júlio de Mesquita Filho, Rio Claro, 1989.
ROSSINI, S. A., BUENO, O. D. Mudas ontogenéticas nos Meliponinae: Melipona
compressipes (Hymenoptera, Apidae). Ciên. Cult., v. 38, p. 845, 1986.
ROUBIK, D. W. Ecology and natural history of tropical bees. Cambridge: Cambridge
University Press, p. 514, 1989.

68
SAKAGAMI, S. F. & ZUCCHI, R. Estudo comparativo do comportamento de várias espécies
de abelhas sem ferrão, com especial referência ao processo de aprovisionamento e postura das
células. Ciência e Cultura, v. 18, p. 283-296, 1966.
SAKAGAMI, S. F. & ZUCCHI, R. Oviposition process in a stingless bee Trigona
(Scaptotrigona) postica (Hymenoptera). Studia Entomológica, v. 6, n. 1/4, p. 497-510, 1963.
SAKAGAMI, S. F. Stingless bees In: Hermann H.R. (Ed.), Academic Press, New York:
Social Insects, v. 3, p. 361-423, 1982.
SILVA, D. L. N.; ZUCCHI, R.; KERR, W. E. Biological and behavioral aspects of the
reproduction in some species of Melipona (Hymenoptera, Apidae, Meliponinae).
Anim.Behav., v. 20, p. 123-132, 1972.
SOMMEIJER, M. J.; BRUIJIN, L. L. M.; MEEUWSEN, F. J. A. M.; MARTENS, E. P.
Natural patterns of caste and sex allocation in the stingless bees Melipona favosa and
Melipona trinitatis related to worker behaviour. Insectes Soc., v.50, p. 38–44, 2003.
SOMMEIJER, M. J.; BUREN, N. J. M. VAN. Male production by laying workers in
queenright colonies of Melipona favosa (Apidae, Meliponinae), In: Billen J. (Ed.), Biology
and Evolution of Social Insects, Leuven Univ. Press, p. 89-97, 1992.
SOMMEIJER, M. J.; CHINH, T. X.; MEEUWSEN, F. J. A. J. Behavioural data on the
production of males by workers in the stingless bee Melipona favosa (Apidae, Meliponinae).
Insectes Soc., v. 46, p. 92-93, 1999.
SOMMEIJER, M. J.; VAN ZEIL, M. & DOHMEN, M. R. Morphological differences
between worker – laid eggs from a queenright colony and queenless colony of Melipona
rufiventris paraensis (Hymenoptera: Apidae). Entomologishe Berichichten, v. 44, n. 1, p.
91-95, 1984.
SUKA, T.; INOUE, T. Nestmates recognition of the stingless bee Trigona (Tetragonula)
minangkabau (Apidae: Meliponinae). Journal of Ethology, v. 11, p. 141-147, 1993.
TAMBASCO, A. J. Aspectos do processo reprodutivo e da citologia de fêmeas de
meliponíneos. 1975. Dissertação apresentada à Faculdade de Medicina de Ribeirão Preto,
Universidade de São Paulo, Ribeirão Preto, 1975.

69
TERADA, Y. (1974). Contribuição ao estudo da regulação social em Leurotrigona mulleri
e Frieseomelitta varia (Hymenoptera, Apidae). 1974. 96 f. Dissertação apresentada à
Faculdade de Medicina de Ribeirão Preto, Universidade de São Paulo, Ribeirão Preto, 1974.
TÓTH, E.; QUELLER, D. C.; IMPERATRIZ-FONSECA, V. L.; STRASSMANN, J. E.
Genetic and behavioral conflict over male production between workers and queens in the
stingless bee Paratrigona subnuda. Behav. Ecol. Sociobiol., v. 53, p. 1-8, 2002.
TÓTH, E.;QUELLER, D. C.; DOLLIN, A.; STRASSMANN, J. E. Conflict over male
parentage in stingless bees. Insectes Soc., v. 51, p. 1-11, 2004.
VEEN, J. W. VAN & SOMMEIJER, M. J. Tropical beekeeping: the production of males in
stingless bees (Melipona). Amsterdam: NEV, Proc. Exper. & Appl. Entomol., v. 1; p. 171-
176, 1990.
VEEN, J. W. VAN; ARCE, H. G. & SOMMEIJER, M. J. Brood production in Melipona
beecheii in relation to dry season foraging. In: Biology and Evolution of Social Insects, J.
Billen Editor, Leuven University Press, p. 81-88, 1992.
VEEN, J. W. VAN; SOMMEIJER, M. J. & ARCE, H. G. The role of colony development
and resource availability in the regulation of queen production in Melipona beecheii
(Apidae, Meliponini). In: Colony Reproduction in Stingless Bees, 1999. p.79-87. PhD.
Dissertation, Faculty of Biology, Utrecht University, 1999.
VEEN, J. W. VAN; SOMMEIJER, M. J.; MEEUWSEN, F. Behavior of drones in Melipona
(Apidae, Meliponinae). Insectes Soc., v. 44, p. 435-447, 1997.
VELTHUIS, H. H. W. & SOMMEIJER, M. J. Roles of morphogenetic hormones in caste
polymorphisms in stingless bees. In: Rutgers University Press, New Jersey: New
Brunswick, v. 9, p. 346-383, 1991.
VELTHUIS, H. H. W. Biologia das abelhas sem ferrão. São Paulo: Edusp, v. 33. 1997.
VELTHUIS, H. H. W.; KOEDAM, D. & IMPERATRIZ-FONSECA, V. L. Males of
Melipona and other stingless bees, and their mothers. Apidologie, v. 36, p. 169-185, 2005.
WAULDBAUER, G. P. The growth and reproduction of maxillectomizeds tobacco
hornworms feeding on normally reject non-solanaceous plants. Entomologia Experimentalis
et Applicata, v. 5, p. 147-158, 1962.

70
WEAVER, N. Physiology of caste determination. Ann Rec. Entomol., v. 11, p. 79-102,
1966.
WIRTZ, P.; BEETSMA, J. Introduction of caste differentiation in the honey bee (Apis
mellifera) by juvenile hormone. Entomology Experiment Application, v. 15, p. 517-520,
1972.
ZUCCHI, R. Aspectos etológicos evolutivos da bionomia de Meliponinae (Hymenoptera,
Apidae). 1977. 204 f. Tese apresentada à Faculdade de Filosofia, Ciências e Letras de
Ribeirão Preto, Universidade de São Paulo. Ribeirão Preto, 1977.
ZUCCHI, R.; SILVA-MATOS, E. V.; NOGUEIRA-FERREIRA, F. H.; AZEVEDO, G. G.
On the cell provisioning and oviposition process (POP) of the stingless bees, nomenclature
reappraisal and evolutionary considerations (Hymenoptera, Apidae, Meliponinae).
Sociobiology, v. 34, p. 65-86, 1999.




























