Embed Size (px)
Citation preview

SANTOS, F.R. et al. Desenvolvimento digestivo e aproveitamento energético em frangos de corte. PUBVET, Londrina, V. 6, N. 18, Ed. 205, Art. 1373, 2012.
PUBVET, Publicações em Medicina Veterinária e Zootecnia.
Desenvolvimento digestivo e aproveitamento energético em
frangos de corte
Fabiana Ramos dos Santos1, Paula Rodrigues Oliveira2, Cibele Silva Minafra1,
Eduardo Ferreira Duarte2, Regiane Rocha de Almeida2, Weslaine Justina da
Silva2
¹ Zootecnista, Profª. Me. Instituto Federal Goiano – Campus Rio Verde,
Rodovia Sul Goiana, Km 01, Zona Rural, Caixa Postal 66, CEP – 75.901-970,
Rio Verde – GO. Tel. (64) 3620-5618, e-mail: [email protected]
² Discente do curso de Zootecnia do Instituto Federal Goiano, Campus Rio
Verde, Rodovia Sul Goiana, Km 01, Zona Rural, Caixa Postal 66, CEP – 75.901-
970, Rio Verde – GO.
Resumo
Com este trabalho pretendeu-se descrever o processo de maturação do
sistema digestório em frangos de corte, de forma a fornecer subsídios para o
entendimento do aproveitamento energético em diferentes fases de
desenvolvimento. Após o nascimento, o sistema digestório da ave está
anatomicamente completo, porém, sua capacidade funcional ainda não permite
a adequada digestão e absorção de todos os nutrientes. Dessa forma, são de
suma importância o conhecimento dos processos de maturação dos órgãos e
das enzimas digestivas na tentativa de fornecer melhor nutrição aos pintos e,
consequentemente, obter melhor desempenho. Assim, pesquisas que

SANTOS, F.R. et al. Desenvolvimento digestivo e aproveitamento energético em frangos de corte. PUBVET, Londrina, V. 6, N. 18, Ed. 205, Art. 1373, 2012.
objetivam determinar os fatores que influenciam o desenvolvimento digestivo
e o aproveitamento nutricional nas diferentes fases de criação são relevantes
para garantir melhor eficiência nutricional e econômica para a indústria
avícola.
Palavras-chave: aves de corte, enzimas digestivas, nutrição
Digestive development and energy utilization in broiler
Abstract
This work was conducted to describe the process of maturation of the digestive
tract in broilers, and to provide a basis for knowledge the energy utilization at
different stages of development. When chickens are born, its digestive system
is anatomically complete, however, their functional capacity does
not allow proper digestion and absorption of nutrients. Thus, these are
extremely important to understand the processes of maturation of organs
and digestive enzymes in an attempt to provide better nutrition to chicks and
hence better performance. Thus, research which objective to determine the
factors that influence the development and digestive energy utilization at
different stages of creation are important to ensure better nutritional efficiency
and economical for the poultry industry.
Keywords: poultry farming, digestive enzymes, nutrition
1. INTRODUÇÃO
O bom desempenho das aves depende da obtenção adequada de energia
e compostos químicos pelo organismo. Para que isso ocorra é necessário que o
trato digestivo apresente características estruturais funcionais adequadas
desde a ingestão dos alimentos até sua absorção (BOLELI et al., 2002).
Embora o sistema digestório do pinto esteja anatomicamente completo
logo após o nascimento este se encontra imaturo do ponto de vista funcional.
Portanto, o sistema digestivo das aves, à semelhança do que ocorre com o

SANTOS, F.R. et al. Desenvolvimento digestivo e aproveitamento energético em frangos de corte. PUBVET, Londrina, V. 6, N. 18, Ed. 205, Art. 1373, 2012.
sistema termorregulador e o imunológico, sofre maturação no período pós-
eclosão.
Diversos fatores intrínsecos e extrínsecos á aves podem estimular este
processo entre eles pode-se citar a idade e peso das matrizes e pintos, o
período de jejum pós-eclosão, a quantidade de saco vitelino residual,
quantidade e qualidade de alimentos e água, nível de enzimas pancreáticas e
intestinais, área de superfície do trato gastrintestinal (TGI), transportadores de
nutrientes e a sua digestibilidade.
Dessa forma, o estudo das variáveis que afetam o processo de
maturação digestiva é de suma importância, principalmente, se for levado em
consideração que o aproveitamento energético depende em grande parte do
estágio de desenvolvimento anatomofisiológico do sistema digestório da ave.
Assim, a escolha do nutricionista sobre qual alimento utilizar na formulação da
dieta está intimamente ligada à forma como os nutrientes serão aproveitados
nas diferentes fases da vida dos frangos de corte.
Por isso, atualmente, em virtude da variação do aproveitamento
nutricional do frango de corte moderno nas diferentes fases de criação é
comum a utilização de programas alimentares múltiplos. De acordo com
CARVALHO, (2010) os programas de alimentação semanais levam em
consideração que a capacidade digestiva das aves varia em função da idade
ocorrendo um aumento no aproveitamento nutricional em função do
desenvolvimento dos órgãos acessórios e do próprio sistema digestório.
Portanto, objetivou-se, com este trabalho, descrever o processo de
maturação do sistema digestório em frangos de corte, de forma a fornecer
subsídios para o entendimento do aproveitamento energético em diferentes
fases de desenvolvimento.

SANTOS, F.R. et al. Desenvolvimento digestivo e aproveitamento energético em frangos de corte. PUBVET, Londrina, V. 6, N. 18, Ed. 205, Art. 1373, 2012.
2. REVISÃO DA LITERATURA
2.1 SISTEMA DIGESTÓRIO DE FRANGOS DE CORTE
2.1.1 Estrutura geral do trato digestório
O rápido e adequado ganho de peso por parte das aves estão
diretamente relacionados com a nutrição das mesmas e, portanto, é
imprescindível que se estabeleçam critérios de manejo para manter a
integridade do sistema digestório, de suas glândulas anexas e reduzam as
enfermidades entéricas que podem comprometer a eficiência funcional destes
órgãos (BOLELI et al., 2002). No entanto, para a correta aplicação das técnicas
de manejo e nutricionais é importante conhecer as características
morfofuncionais do sistema digestório.
Anatomicamente, o TGI das aves, segundo ARTONI (2004) e REECE
(2006), é um tubo oco e fibromusculoso que pode ser descrito, da região
anterior para a posterior, da seguinte maneira: cavidade oral, faringe, esôfago,
papo, proventrículo, moela, intestino delgado, cecos e cólon. A ele também
estão conectadas duas glândulas anexas, o fígado e o pâncreas (Figura 1).
FIGURA 1. Esquema geral do sistema digestório do frango de corte
Fonte: BOLELI et al. (2002).

SANTOS, F.R. et al. Desenvolvimento digestivo e aproveitamento energético em frangos de corte. PUBVET, Londrina, V. 6, N. 18, Ed. 205, Art. 1373, 2012.
Segundo BOLELI et al. (2002), os órgãos tubulares do sistema digestório
das aves seguem um modelo estrutural básico, constituído por quatro túnicas
concêntricas, com características histológicas e funcionais distintas. Tais
estruturas são túnicas denominadas, da luz para a periferia do órgão, de
mucosa, submucosa, muscular e serosa (Figura 2).
FIGURA 2. Esquema Geral das túnicas que constituem a parede do Sistema
Digestório
Fonte: BOARO (2009).
BOARO (2009) descreve estas estruturas da seguinte forma:
- A túnica mucosa tem por função controlar os movimentos da mucosa
intestinal que são independentes dos da túnica muscular. Em alguns
segmentos do sistema digestório, pode aparecer glândulas chamadas de
mucosas nessa túnica.
- A túnica submucosa é constituída de tecido conjuntivo moderadamente
denso e tal como a mucosa, pode conter glândulas submucosas. Em ambos os
tipos de glândulas, os ductos se abrem na luz intestinal. Tanto na mucosa

SANTOS, F.R. et al. Desenvolvimento digestivo e aproveitamento energético em frangos de corte. PUBVET, Londrina, V. 6, N. 18, Ed. 205, Art. 1373, 2012.
como na submucosa, o tecido conjuntivo é rico em vasos sanguíneos e
linfáticos e pode conter nódulos linfóides.
- A túnica muscular possui, em geral, duas camadas de músculos lisos. A
camada interna é formada de fibras musculares dispostas de forma circular ao
trato digestório. Sua contração alonga e constringe o intestino. A camada
externa é constituída de fibras musculares dispostas longitudinalmente, que
servem para encurtar o sistema digestório. A ação coordenada dessas duas
camadas, da túnica muscular, promove a peristalse e a segmentação.
- A túnica serosa, por sua vez, é formada de tecido conjuntivo envolto
por mesentério, com exceção da porção cervical do esôfago.
De forma geral, todas as porções do TGI se encontram imaturas durante
a fase embrionária e, após a eclosão, passam a ter grande importância no
crescimento das aves. Nos primeiros dias de vida, os segmentos do TGI sofrem
sensíveis alterações para o consumo e utilização de alimentos, sendo
necessário um período de maturação até que o intestino delgado atinja o
tamanho relativo adequado e o pâncreas alcance níveis de produção
enzimática que não limitem a taxa de crescimento (NITSAN, 1995). Assim, é
de suma importância o conhecimento dos processos de maturação destes
órgãos na tentativa de fornecer melhor nutrição aos pintos e,
consequentemente, obter melhor desempenho.
2.1.2 Desenvolvimento anatômico do TGI em frangos de corte
Conforme anteriormente discutido, as aves recém nascidas têm seu
sistema gastrintestinal imaturo e sofrem processos adaptativos, buscando
maior eficiência nos processos digestivos. Na eclosão o sistema digestório da
ave está anatomicamente completo, porém, sua capacidade funcional ainda
não permite a adequada digestão e absorção de todos os nutrientes. Dessa
forma, segundo MURAROLLI, (2008) o TGI das aves passará por alterações
morfológicas que irão proporcionar um aumento na área de superfície de
digestão e absorção.

SANTOS, F.R. et al. Desenvolvimento digestivo e aproveitamento energético em frangos de corte. PUBVET, Londrina, V. 6, N. 18, Ed. 205, Art. 1373, 2012.
No TGI de frangos de corte, as alterações morfológicas podem ocorrer
principalmente pelo processo de hiperplasia, seguida de aumento no número
de células intestinais e crescimento de órgãos digestivos.
No intestino delgado, alterações morfológicas mais evidentes são: o
aumento no comprimento do intestino, na altura e densidade dos vilos, no
número e volume de suas células epiteliais denominadas enterócitos
(responsáveis pela absorção dos nutrientes), células caliciformes (produtoras
de mucina) e enteroendocrinas (produtoras de hormônios).
Estas alterações ocorrem marcadamente entre o 17° dia de incubação e
o sétimo dia pós-eclosão (UNI et al., 1998), podendo o tamanho das
vilosidades do duodeno aumentar em 200 a 300% no período final da
incubação (SKLAN, 2001). No jejuno e íleo, o crescimento continua até o 14º
dia, resultando em aumento no número de enterócitos por vilo (NOY e SKLAN,
1997). Durante esse período, a profundidade de cripta também aumenta,
sendo maior no duodeno e menor no íleo (NITSAN et al., 1991). Porém, a
maturação do trato gastrintestinal se estabelece aos 16 dias de idade em
frangos de corte (UNI et al., 1998).
Segundo SELL (1996), a capacidade do trato digestório do frango na
primeira semana de vida pode ser considerada um fator limitante tanto para o
consumo de alimentos quanto para a digestão e a absorção de nutrientes.
Sendo assim, quanto mais cedo o intestino atingir sua maturidade funcional
mais precocemente o pintainho poderá utilizar os nutrientes dietéticos e
expressar todo seu potencial genético para ganho de peso (UNI & FERKET,
2004).
O desenvolvimento da mucosa intestinal decorre primariamente de dois
eventos citológicos associados: renovação celular (proliferação e diferenciação
das células localizadas na cripta e ao longo dos vilos) e perda celular (extrusão
que ocorre normalmente no ápice dos vilos). Segundo BOARO (2009), o
equilíbrio entre esses dois processos determina um turnover (proliferação –
migração - extrusão) constante, ou seja, a manutenção do tamanho dos vilos.

SANTOS, F.R. et al. Desenvolvimento digestivo e aproveitamento energético em frangos de corte. PUBVET, Londrina, V. 6, N. 18, Ed. 205, Art. 1373, 2012.
O tempo de turnover celular é o tempo necessário para que uma célula
originada no processo mitótico entre cripta-vilo migre para a ponta do vilo e
descame para o lúmen intestinal. Em média, o turnover celular no intestino de
pintos de corte oscila entre 90 a 96 horas, ou seja, aproximadamente quatro
dias. Este período de tempo parece curto; contudo, considerando que o
período de criação do frango (aproximadamente 42 dias) representa nada
menos do que 10% do tempo de vida da ave (FURLAN, 2009).
Quando o intestino responde a algum agente (infecção com
microrganismos, inadequação ambiental ou nutricional) ocorre o desequilíbrio
neste turnover e desta forma há modificação na altura dos vilos. Assim,
segundo FURLAN (2009), se considerarmos a perda de 10% no sistema de
produção devido a distúrbios da mucosa intestinal (ineficiência no turnover
celular), em 20 mil aves haveria a queda de aproximadamente 5 toneladas de
peso vivo/lote.
O crescimento do trato gastrintestinal é importante no desenvolvimento
geral da ave. O intestino delgado aumenta seu peso a velocidade maior que o
peso corporal e de órgãos essenciais, como o pulmão. Por outro lado, outros
órgãos do trato digestório como a moela e o pâncreas não apresentam
desenvolvimento proporcional ao corpo (UNI et al., 1999). Segundo os
autores, logo após o nascimento, ocorre proliferação da mucosa em resultado
a hiperplasia celular, aumentando a superfície de absorção e o peso dos
intestinos. Este processo de crescimento relativo atinge o desenvolvimento
máximo entre o 6º e 10º dia de vida da ave.
Em acordo com estes resultados, KONDO (2003), em estudo comparativo
entre diferentes linhagens de frangos, observou que o maior desenvolvimento
do tamanho do intestino delgado ocorreu na primeira semana de vida dos
pintos, sendo que, o desenvolvimento gradual nas diferentes regiões
intestinais dos frangos de corte tenha se prolongado até 36 dias de idade.
De acordo com LONGO (2004), o aumento no comprimento do intestino
delgado e na área de superfície da mucosa com a idade é também uma
estratégia para compensar a perda de atividade enzimática por célula ou por

SANTOS, F.R. et al. Desenvolvimento digestivo e aproveitamento energético em frangos de corte. PUBVET, Londrina, V. 6, N. 18, Ed. 205, Art. 1373, 2012.
unidade de superfície da mucosa intestinal, que ocorre com o passar da idade
da ave. Em idades mais avançadas, as vilosidades se estendem para sustentar
uma alta atividade total das enzimas em relação aos pequenos vilos de aves
jovens (IJI et al., 2001).
Confirmando esta afirmativa, MARCHINI et al. (2009) avaliaram o
desenvolvimento da altura dos vilos (VI), profundidade das criptas (CR),
relação vilo/cripta (VI/CR) da mucosa duodenal de frangos de corte do
primeiro até o 42º dia de idade submetidos a temperaturas elevadas e criados
em ambiente termoneutro.
Os pesquisadores verificaram que á medida que as aves envelhecem
ocorre aumento na VI e CR independente da temperatura de criação. Este
aumento foi mais expressivo (em torno de duas vezes a altura de vilo e
profundidade de cripta inicial) nos primeiros 14 dias de vida da ave.
No entanto, em função do estresse fisiológico gerado pelas altas
temperaturas, a VI máxima foi obtida em 33 dias de criação pelas aves criadas
em ambiente termoneutro, enquanto que as aves criadas em temperaturas
elevadas atingiram em 41 dias de vida. Estes resultados sugerem que o
estresse calórico pode ser um agente inibidor do desenvolvimento digestivo e
consequentemente do desempenho de frangos de corte.
É importante ressaltar que o comprimento do intestino delgado aumenta
na primeira semana mesmo quando a ave não recebe alimento, mas, para o
desenvolvimento das vilosidades, a alimentação é essencial. Este fato salienta
a importância do fornecimento precoce de alimentos, pois, logo após a eclosão,
a maior parte da demanda de energia e proteína das aves é direcionada para o
desenvolvimento do trato digestório, principalmente do intestino (MAIORKA,
2001).
Com o estímulo do fornecimento de alimento tem sido verificado que o
peso do intestino delgado aumenta cerca de 600% dentro dos primeiros sete
dias (NOY et al., 2001) e o tamanho dos vilos e profundidade das criptas
intestinais aumentam consideravelmente dos 4 aos 21 dias de idade (BATAL &
PARSONS, 2002).

SANTOS, F.R. et al. Desenvolvimento digestivo e aproveitamento energético em frangos de corte. PUBVET, Londrina, V. 6, N. 18, Ed. 205, Art. 1373, 2012.
Isso ocorre porque após a eclosão, ocorre rápida transição no suprimento
da energia fornecida por nutrientes endógenos do saco vitelínico para
fornecimento pela ração exógena rica em carboidratos que passa a ser
absorvida pelo intestino.
Aproximadamente, 20% da proteína residual do saco vitelino é
representada pelas imunoglobulinas maternais e a gordura bruta residual é
constituída basicamente de triglicerídeos, fosfolipídios e colesterol. É evidente
que o uso destes compostos para fins nutricionais priva o neonato da proteção
de anticorpos. Além disso, fosfolipídios e colesterol não são uma eficiente via
de utilização da energia sendo mais eficientes quando utilizados como
componentes essenciais das membranas celulares. Se todo o triacilglicerol
presente no saco vitelino fosse metabolizado com 100% de eficiência, o
máximo de energia oferecida seria de 9,0 kcal, valor inferior à necessidade de
energia para mantença do neonato no primeiro dia, que é de 11,0 kcal
(DIBNER et al., 1998).
Este fato evidencia a importância do fornecimento do alimento exógeno
não somente para a manutenção das funções vitais, mas também pelo fato
que a presença do alimento sólido, rico em carboidratos e proteínas, estimula
o desenvolvimento do trato digestório e induz a produção de secreções
digestivas (SKLAN, 2003)
Neste contexto, ALMEIDA et al. (2006) avaliaram o efeito do período de
jejum (0, 24 e 48 horas) entre o nascimento e o alojamento sobre o
desempenho de frangos de corte provenientes de matrizes de diferentes idades
(30 e 60 semanas) e verificaram que aos 42 dias de idade, os frangos que não
foram submetidos ao jejum após o nascimento apresentaram maior ganho de
peso em relação aos demais (Tabela 1). Os autores atribuem o melhor
desempenho das aves não submetidas ao jejum ao maior consumo de ração,
associados a outros fatores fisiológicos, como produção enzimática,
desenvolvimento de enterócitos nas criptas, aumento na quantidade e
tamanho das vilosidades por área intestinal, favorecidas por um consumo
precoce de alimento.

SANTOS, F.R. et al. Desenvolvimento digestivo e aproveitamento energético em frangos de corte. PUBVET, Londrina, V. 6, N. 18, Ed. 205, Art. 1373, 2012.
Ainda neste ensaio, verificou-se que os pintos provenientes de matrizes
mais velhas apresentaram melhor desempenho em relação aos oriundos de
matrizes jovens. O trato digestório de pintainhos provenientes de matrizes
velhas apresenta-se mais desenvolvido no momento da eclosão, fato que pode
contribuir para uma melhor adaptação à alimentação exógena e para o
consequente melhor desempenho destes animais (HUDSON et al., 2004).
TABELA 1. Consumo de ração (CR), ganho de peso (GP) e conversão
alimentar (CA) de frangos de corte, proveniente de matrizes de
diferentes idades com diferentes tempos de alojamento.
Desempenho de 1 a 42 dias
CR (g) GP(g) CA (g) Viabilidade,%
Idade da matriz
30 semanas 3503 b 2244 b 1,699 a 95,31
60 semanas 3929 a 2394 a 1,749 b 96,15
Tempo de alojamento
0 3859 a 2347 a 1,746 93,27
24 3706 b 2346 a 1,720 97,11
48 3582 c 2264 b 1,706 96,81
CV,% 7,69 4,50 2,73 5,92
Letras distintas nas colunas diferem entre si (Tukey, 5%)
Adaptado de: ALMEIDA et al. (2006)
Segundo ROCHA et al. (2008), o peso do saco vitelino é correlacionado
ao peso do pinto, que, por sua vez, é correlacionado ao peso do ovo e a
porcentagem de gema. Portanto, os ovos de matrizes mais velhas por serem
maiores, proporcionam aos pintos maior aporte nutricional, além de permitir
que os mesmos adquiram elevada capacidade de formação de resposta
imunológica ativa, uma vez que cerca de 10 a 20% dos níveis médios de
imugoglobulinas das matrizes são transferidos à progênie através da gema

SANTOS, F.R. et al. Desenvolvimento digestivo e aproveitamento energético em frangos de corte. PUBVET, Londrina, V. 6, N. 18, Ed. 205, Art. 1373, 2012.
(GRINDSTAFF et al., 2005). A melhor nutrição e maior resposta imunológica,
entre outros fatores ambientais, contribuem para que os pintos de matrizes
mais velhas possam se desenvolver mais rapidamente.
2.1.3 Desenvolvimento de enzimas digestivas em frangos de corte
Além do desenvolvimento anatômico, as aves passam por alterações
fisiológicas que visam à melhor eficiência das funções digestivas. Estas
alterações fisiológicas estão relacionadas ao aumento da produção de enzimas
digestivas (MURAROLLI, 2008). As rápidas alterações do trato digestório
possibilitam aumento de consumo de ração e alteram a digestibilidade dos
nutrientes.
O desenvolvimento das enzimas digestivas exerce grande influência no
aproveitamento de nutrientes pelos frangos de corte. As atividades das
enzimas digestivas pancreáticas e intestinais aumentam com a idade do
frango. Segundo NOY & SKLAN (1995), a secreção de amilase, tripsina e lipase
é pequena até o quarto dia de vida da ave, porém, os níveis de secreção
destas enzimas aumentam em 100, 50 e 20 vezes respectivamente até os 21
dias de idade. NITSAN et al., (1991) afirmam que a atividade das enzimas
digestivas (unidade/kg de peso vivo) terão seus valores máximos obtidos no
pâncreas aos 80 dias para amilase e lipase e aos 110 dias de idade, para
tripsina e quimiotripsina.
SAKOMURA et al. (2004a) também avaliaram o efeito da idade dos
frangos sobre a atividade enzimática e digestibilidade dos nutrientes e
observaram que a atividade da amilase, tripsina e lipase pancreática aumentou
com a idade das aves, assim como o crescimento alométrico do pâncreas. Os
autores verificaram que a maior taxa de crescimento pancreático ocorreu na
segunda semana, coincidindo com a fase de maior aumento da atividade das
enzimas digestivas (Tabela 2).
Entretanto, embora o desenvolvimento enzimático seja altamente
dependente da idade da ave, a secreção de enzimas digestivas pode ser

SANTOS, F.R. et al. Desenvolvimento digestivo e aproveitamento energético em frangos de corte. PUBVET, Londrina, V. 6, N. 18, Ed. 205, Art. 1373, 2012.
estimulada pelo fornecimento do alimento sólido. SELL et al. (1991)
observaram que aves alimentadas com dietas ricas em carboidratos
apresentaram maiores atividades de dissacaridases. MORAN Jr. (1985) indicou
que a digestão e a absorção de carboidratos em aves não são fixas, mas são
altamente adaptáveis de acordo com sua presença na dieta.
TABELA 2. Crescimento alométrico do pâncreas1(CAP), atividade da amilase,
tripsina e lipase (U/mg de proteína) de acordo com a idade das
aves
Idade (dias) CV
% 1 a 7 8 a 14 15 a 21 22 a 28 35 a 42
CAP 1,98 1,62 1,39 1,12 0,39 15,15
Amilase 14,73 33,82 59,03 90,08 127,51 35,85
Tripsina 1,69 5,56 16,11 24,58 38,62 33,35
Lipase 42,24 128,59 257,94 397,74 417,83 26,54 1Valores expressos em peso absoluto: (Peso do pâncreas da ave na idade x/
peso do pâncreas da ave com um dia de idade)/( peso corporal da ave na
idade x/ peso corporal da ave com um dia de idade)
Adaptado de SAKOMURA et al. (2004a).
Da mesma maneira, a digestão de proteínas também sofre adaptações
marcantes no período pós-eclosão. As pesquisas indicam que esses processos
são influenciados pelo nível de alimentação e a composição da dieta.
STRINGHINI et al. (2009) estudaram os efeitos de níveis decrescentes de
proteína com suplementação de aminoácidos essenciais na ação das enzimas
amilase pancreática e tripsina de pintos de corte de 4 a 14 dias de vida. Os
autores constataram que a atividade das enzimas é absolutamente alta para
garantir sua ação nos substratos provenientes dos alimentos, especialmente
para a tripsina.

SANTOS, F.R. et al. Desenvolvimento digestivo e aproveitamento energético em frangos de corte. PUBVET, Londrina, V. 6, N. 18, Ed. 205, Art. 1373, 2012.
As principais fontes energéticas dos pintos antes da eclosão são as
proteínas e os ácidos graxos (principalmente triglicerídeos) provenientes do
saco vitelino. Após a eclosão, ocorre rápida transição no suprimento da energia
fornecida pelos nutrientes endógenos do saco vitelínico (50% lipídios) para
fornecimento pela ração exógena rica em carboidratos (SKLAN, 2001) que no
intestino delgado serve como estímulo ao crescimento e desenvolvimento do
trato gastrintestinal (TGI) e, conseqüentemente, das funções de digestão e
absorção.
Entretanto, no que se refere aos lipídeos dietéticos, é observado uma
ineficiência na digestão deste nutriente pelas aves nos primeiros dias de vida.
De acordo com MAIORKA et al. (2004), as aves jovens tem ineficiência na
produção da enzima lipase, além de possuírem imaturidade na circulação
entero-hepática afetando desta maneira a secreção de sais biliares e
resultando na menor digestibilidade de lipídeos da dieta.
Já VIEIRA & POPHAL (2000) afirmam que utilização da lipase pancreática
aumenta linearmente até 16 dias após a eclosão, estabilizando-se a seguir.
Esta é dependente da presença de lipídios na dieta, pois a ingestão de dietas
com altos níveis de gordura aumentou em torno de 10 vezes a sua
concentração quando comparado com dietas com baixo teor de gordura.
Portanto, uma vez os níveis de lipase se elevam com o teor de lipídeos da dieta
os autores creditam a menor digestibilidade das gorduras pelas aves jovens
apenas à imaturidade em sua circulação entero-hepática já que a adição de
sais biliares à dieta resultam na melhor utilização da gordura dos alimentos,
conforme demonstrado por POLIN & HUSSEIN (1982)
Segundo WISEMAN & SALVADOR (1989) as maiores diferenças na
capacidade das aves em utilizar gordura são mais proeminentes entre a
primeira e terceira semanas, sendo ausente ou mínima entre a quinta e sétima
semanas de vida. Estes fatos sugerem que os níveis de gordura nas primeiras
semanas de vida devam ser adequadamente controlados.

SANTOS, F.R. et al. Desenvolvimento digestivo e aproveitamento energético em frangos de corte. PUBVET, Londrina, V. 6, N. 18, Ed. 205, Art. 1373, 2012.
2.2 APROVEITAMENTO NUTRICIONAL POR FRANGOS DE CORTE
2.2.1 Formas de aproveitamento da energia
Para a formulação de rações economicamente viáveis é importante
conhecer o valor nutritivo dos alimentos. Para isso, é necessário determinar a
composição química, a disponibilidade dos nutrientes, a concentração e o
aproveitamento de energia dos alimentos (GENEROSO et al., 2008).
Na literatura, os valores energéticos dos ingredientes de ração variam
muito. No Brasil, vários estudos já foram desenvolvidos para se determinar o
valor de energia metabolizável (EM) dos alimentos utilizados em rações para
aves (LANNA et al., 1979; ALBINO, 1981; COELHO, 1983; ALBINO, 1984,
ROSTAGNO et al., 2000). ROSTAGNO et al. (2005), com base em pesquisas
realizadas na Universidade Federal de Viçosa, elaboraram a tabela brasileira de
composição de alimentos, em que se observam variações nos valores de
composição de alimentos quando comparados com tabelas estrangeiras.
Isso ocorre porque os valores de EM dos ingredientes utilizados na
alimentação de aves são influenciados por diversos fatores, tais como:
deficiências de aminoácidos e vitaminas, níveis de cálcio e fósforo, níveis de
inclusão de gorduras, idade e peso das aves, sexo, metodologia utilizada,
componentes da ração, entre outros. Portanto, associando os fatores inerentes
aos alimentos à constante evolução genética da avicultura de corte, faz-se
necessário determinar os valores nutricionais dos alimentos utilizados na
formulação de rações para aves, proporcionando informações para atualização
das tabelas de composição de alimentos (SAKOMURA et al., 2004b).
A energia metabolizável é a forma normalmente utilizada para aves
sendo obtida pela diferença entre a EB do alimento e a EB das excretas (fezes,
urina e gases da digestão) (Figura 3).

SANTOS, F.R. et al. Desenvolvimento digestivo e aproveitamento energético em frangos de corte. PUBVET, Londrina, V. 6, N. 18, Ed. 205, Art. 1373, 2012.
FIGURA 3. Esquema da utilização da energia pelas aves conforme proposto
por SIBBALD (1982)
Fonte: Adaptado de SAKOMURA et al. (2004).
A energia perdida na forma de gases em aves é muito baixa sendo
desprezada nos cálculos da EM. Para aves, a EM pode ser determinada e
expressa como: energia metabolizável aparente (EMA), energia metabolizável
aparente corrigida para balanço de nitrogênio (EMAn), energia metabolizável
verdadeira (EMV) ou energia metabolizável verdadeira corrigida para balanço
de nitrogênio (EMVn).
A EMAn difere da EMA pela correção associada ao balanço de nitrogênio
(N). HILL & ANDERSON (1958) propuseram um fator de correção para o teor
de N retido ou excretado (8,22 kcal/g de N). Esta correção se baseia no fato de

SANTOS, F.R. et al. Desenvolvimento digestivo e aproveitamento energético em frangos de corte. PUBVET, Londrina, V. 6, N. 18, Ed. 205, Art. 1373, 2012.
que em aves em crescimento, a proteína retida no organismo da ave e,
conseqüentemente não catabolizada até os produtos de excreção nitrogenada,
não contribui para a energia das fezes e urina. Por outro lado, em aves
adultas, parte dos compostos nitrogenados são catabolizados e excretados
como ácido úrico (SIBBALD, 1982). Assim, aves com diferentes graus de
retenção nitrogenada proporcionam diferentes valores de energia excretada,
para a mesma digestibilidade do alimento. A correção pelo balanço de N
(ganhos ou perdas de nitrogênio corporal) tem como objetivo padronizar e
reduzir a variação nos valores de EM dos alimentos medidos em diferentes
condições que podem resultar em maior ou menor ganho de peso ou em perda
de peso dos animais (SAKOMURA & ROSTAGNO, 2007).
Entre as metodologias empregadas para avaliação dos alimentos para
aves, destacam-se os métodos tradicionais de coleta total de excretas
(SIBBALD & SLINGER, 1963), alimentação precisa (SIBBALD, 1976), e o
método indireto que utiliza as equações de predição (NRC,1994).
O método de coleta total de excretas com pintos de corte é o mais
utilizado, entretanto, apresenta alguns contratempos, como a interferência da
idade das aves nos valores de energia.
Segundo BRUMANO et al. (2006), aves mais jovens possuem menor
capacidade de digestão e absorção de nutrientes, por não possuírem sistema
digestivo completamente desenvolvido, em especial para as gorduras
presentes na dieta resultantes da baixa secreção de lipase e sais biliares. Em
contrapartida, aves mais velhas, apresentam sistema digestivo plenamente
desenvolvido, com maior tamanho do trato digestivo, o que possibilita maior
permanência do alimento em contato com as enzimas e secreções gástricas e,
portanto, melhor aproveitamento dos alimentos.
SAKOMURA et al. (2004b) apresentaram a relação entre EMA e EMAn
com dados obtidos em ensaios realizados com pintos e galos usando a
metodologia de coleta total por CAFÉ (1993). Conforme os dados
demonstrados as diferenças entre a EMA e EMAn foram maiores no ensaio com
pintos do aquelas obtidas com galos. Esta diferença ocorreu em função das

SANTOS, F.R. et al. Desenvolvimento digestivo e aproveitamento energético em frangos de corte. PUBVET, Londrina, V. 6, N. 18, Ed. 205, Art. 1373, 2012.
aves em crescimento apresentarem maior retenção de N corporal em relação
aos galos. Entretanto, quando se compara a diferença entre a EMA obtida para
pintos e galos estas foram inferiores em relação às diferenças entre as EMAn,
evidenciando o efeito da idade da ave quando se considera a correção da EMA
pelo balanço de nitrogênio .
2.2.2 Determinação do valor energético dos alimentos com frangos de
corte em diferentes idades
De acordo com os estudos analisados anteriormente, a idade associada
ao grau de desenvolvimento digestivo da ave pode afetar o aproveitamento
nutricional.
De acordo com FREITAS et al. (2006), há aumento da EMAn dos
alimentos com a idade, e essas diferenças devem ser consideradas no cálculo
das rações. Nesse contexto, várias pesquisas são realizadas com o objetivo de
se determinar o aproveitamento energético das aves em diferentes idades
(RODRIGUES et al., 2001; RODRIGUES et al., 2002; BRUMANO et al., 2006;
FREITAS et al., 2006; GENEROSO et al., 2008; MELLO et al., 2009,
CARVALHO, 2010).
GENEROSO et al. (2008) determinaram os valores de EMA e EMAn de
alguns ingredientes com pintos de 21 a 30 e 41 a 50 dias de idade. Os autores
verificaram que os valores de EMA e EMAn de todos os alimentos obtidos em
aves mais novas foram inferiores aos encontrados em aves mais velhas,
atestando a maior capacidade digestiva destas últimas (Tabela 3).
Confirmando este resultado, MELLO et al. (2009) trabalharam com aves
de 10 a 17, 26 a 33, 40 a 47 dias de idade e galos adultos e observaram que
os valores de EMA do farelo de soja, do sorgo, do farelo de arroz integral, das
farinhas de penas e do plasma sangüíneo e os valores de EMAn desses
alimentos, assim como os do farelo de trigo, aumentam de acordo com a idade
das aves. Portanto, segundo os autores, ao formular rações para aves, deve-se
considerar que os valores energéticos dos alimentos diferem em cada idade.
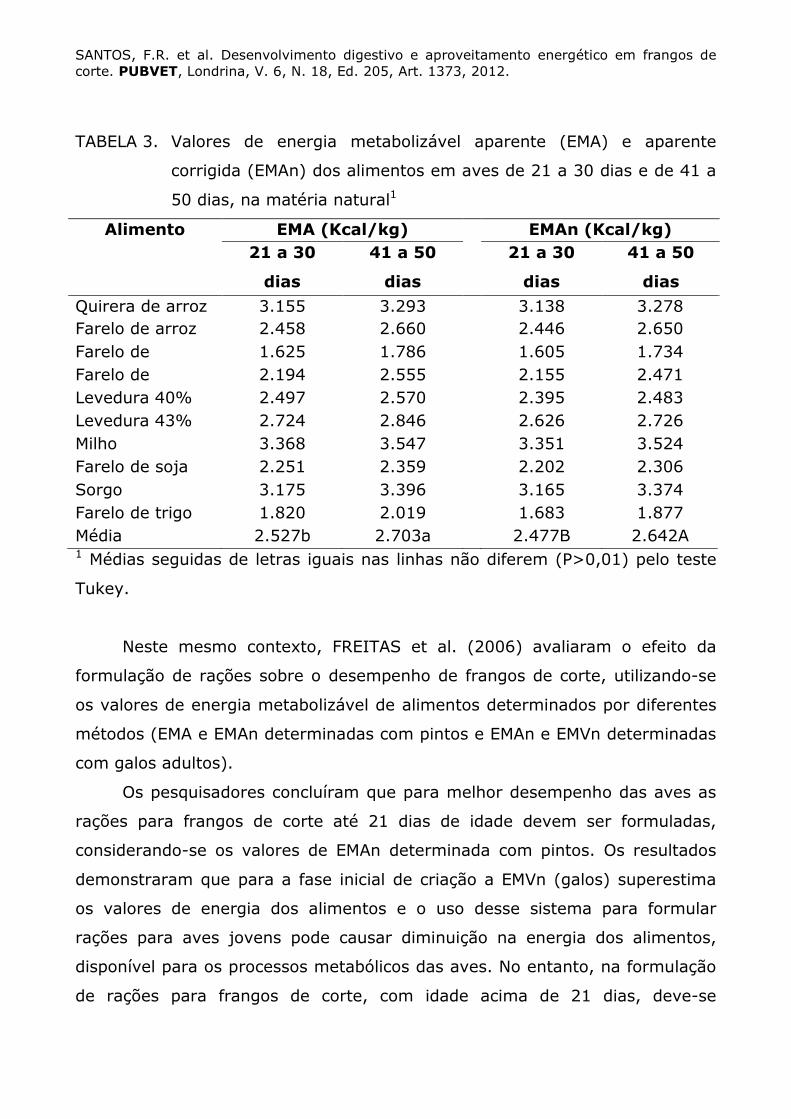
SANTOS, F.R. et al. Desenvolvimento digestivo e aproveitamento energético em frangos de corte. PUBVET, Londrina, V. 6, N. 18, Ed. 205, Art. 1373, 2012.
TABELA 3. Valores de energia metabolizável aparente (EMA) e aparente
corrigida (EMAn) dos alimentos em aves de 21 a 30 dias e de 41 a
50 dias, na matéria natural1
Alimento EMA (Kcal/kg) EMAn (Kcal/kg)
21 a 30
dias
41 a 50
dias
21 a 30
dias
41 a 50
dias
Quirera de arroz 3.155 3.293 3.138 3.278 Farelo de arroz 2.458 2.660 2.446 2.650 Farelo de 1.625 1.786 1.605 1.734 Farelo de 2.194 2.555 2.155 2.471 Levedura 40% 2.497 2.570 2.395 2.483 Levedura 43% 2.724 2.846 2.626 2.726 Milho 3.368 3.547 3.351 3.524 Farelo de soja 2.251 2.359 2.202 2.306 Sorgo 3.175 3.396 3.165 3.374 Farelo de trigo 1.820 2.019 1.683 1.877 Média 2.527b 2.703a 2.477B 2.642A 1 Médias seguidas de letras iguais nas linhas não diferem (P>0,01) pelo teste
Tukey.
Neste mesmo contexto, FREITAS et al. (2006) avaliaram o efeito da
formulação de rações sobre o desempenho de frangos de corte, utilizando-se
os valores de energia metabolizável de alimentos determinados por diferentes
métodos (EMA e EMAn determinadas com pintos e EMAn e EMVn determinadas
com galos adultos).
Os pesquisadores concluíram que para melhor desempenho das aves as
rações para frangos de corte até 21 dias de idade devem ser formuladas,
considerando-se os valores de EMAn determinada com pintos. Os resultados
demonstraram que para a fase inicial de criação a EMVn (galos) superestima
os valores de energia dos alimentos e o uso desse sistema para formular
rações para aves jovens pode causar diminuição na energia dos alimentos,
disponível para os processos metabólicos das aves. No entanto, na formulação
de rações para frangos de corte, com idade acima de 21 dias, deve-se

SANTOS, F.R. et al. Desenvolvimento digestivo e aproveitamento energético em frangos de corte. PUBVET, Londrina, V. 6, N. 18, Ed. 205, Art. 1373, 2012.
considerar o aumento na digestibilidade dos nutrientes, com a idade das aves,
e valorizar a contribuição energética dos alimentos, utilizando-se os valores de
EMAn (galos) ou EMVn (galos) para a formulação.
Este resultado confirma os dados observados nos ensaios de
digestibilidade, os quais sugerem que para maior eficiência produtiva dos
frangos de corte as tabelas de composição nutricional deveriam conter dados
do aproveitamento energético de acordo com a fase de desenvolvimento da
ave. No entanto, infelizmente, no Brasil a maioria das tabelas nutricionais para
a formulação de ração não adotam esta prática que poderia contribuir para
redução de custos com a formulação além de diminuir os problemas
metabólicos gerados pelo excesso de energia nas rações.
3 CONSIDERAÇÕES FINAIS
Os pintos jovens apresentam o sistema digestivo imaturo e por isso
necessitam de práticas de manejo que lhes garantam bom desenvolvimento
morfofisiológico do trato gastrintestinal.
É sabido que práticas nutricionais como o fornecimento precoce de
alimentos de qualidade e a adequação dos níveis nutricionais nos primeiros
dias de vida da ave podem favorecer desenvolvimento da TGI. No entanto, é
importante lembrar que a idade tem papel preponderante no processo de
maturação digestiva do frango e, portanto, as pesquisas que objetivam
determinar o aproveitamento nutricional nas diferentes fases de criação são de
suma importância para garantir melhor eficiência nutricional e econômica para
a indústria avícola.
4. REFERÊNCIAS
ALBINO, L.F.T.; FIALHO, E.T.; ROSA, P.S. Composição química e valores energéticos alguns alimentos para frangos de corte. Pesquisa Agropecuária Brasileira, v.19, n.7, p.897 -903, 1984.
ALBINO, L.F.T.; ROSTAGNO, H.S.; FONSECA, J.B. et al. Tabela de composição de alimentos concentrados. Valores de composição química e de energia determinados com aves em

diferentes idades. Revista da Sociedade Brasileira de Zootecnia, v.10, n.1, p.133-146, 1981.
ALMEIDA, J.G.; DAHLKE, F.; MAIORKA, A.; MACARI, M.; FURLAN, R.L. efeito do jejum no intervalo entre o nascimento e o alojamento sobre o desempenho de frangos de corte provenientes de matrizes de diferentes idades. Archives of Veterinary Science, v. 11, n. 2, p. 50-54, 2006.
ARTONI, S. M. B. Anatomia do sistema digestório das aves. In: Curso de Fisiologia da Digestão e Metabolismo de Nutrientes em Aves, UNESP Jaboticabal, out. 2004. 1 CD-ROM.
BATAL, A. B.; PARSONS, C. M. Effects of age on nutrient digestibility in chicks fed different diets. Poultry Science, v. 81, p. 400-407, 2002.
BOARO, M. Morfofisiologia do trato intestinal. In: Conferência APINCO, 2009. FACTA, Porto Alegre, 261-274,2009.
BOLELI, I.C.; MAIORKA, A.; MACARI, M. Estrutura funcional do trato digestório. In: MACARI, M., FURLAN, R.L.; GONZALES, E. Fisiologia Aviária plicada a frangos de corte. Jaboticabal: /funep/UNESP, 2002. p. 75-95.
BRUMANO, G.; GOMES, P.C.; ALBINO, L.F.T.; ROSTAGNO, H.S.; GENEROSO, R.A.; SCHMIDT, M. Composição química e valores de energia metabolizável de alimentos protéicos determinados com frangos de corte em diferentes idades. Revista Brasileira de Zootecnia, v.35, n.6, p.2297-2302, 2006.
CAFÉ, M.B. Estudo do valor nutricional da soja integral processada para aves. 1993, 97f, Dissertação (Mestrado) Universidade Estadual Paulista, Jaboticabal.
CARVALHO, F.B. Efeito da temperatura ambiente e da idade do frango de corte sobre o valor energético do milho, farelo e óleo de soja. 2010, 115f. Universidade Estadual Paulista, Faculdade de Medicina Veterinária e Zootecnia, Botucatu.
COELHO, M.G.R. Valores energéticos e de triptofano metabolizável de alimentos para aves, utilizando duas metodologias. 1983. 87f. Dissertação (Mestrado em Zootecnia) – Universidade Federal de Viçosa, Viçosa.
DIBNER, J.; KNIGHT, C. D.; KITCHELL, M. L. et al. Early feeding and development of the immune system in neonatal poultry. Journal of Applied Poultry Research, v. 7, p. 425-436, 1998.
FREITAS, E.R., SAKOMURA, N.K., EZEQUIEL, J.M.B., NEME, R., MENDONÇA, M.O. Energia metabolizável de alimentos na formulação de ração para frangos de corte. Pesquisa agropecuária brasileira, v.41, n.1, p.107-115, 2006.
FURLAN, R. Uso de prebióticos, probióticos e flora de exclusão competitiva http://www.avisite.com.br/cet/img/20091105_probiotico.pdf 2009. Acessado em 11/11/2010.
GENEROSO, R.A.R. GOMES, P.C.; ROSTAGNO, H.S.; ALBINO, F.T.; BARRETO, L.T.; BRUMANO, G. Composição química e energética de alguns alimentos para frangos de corte em duas idades. Revista Brasileira de Zootecnia, Viçosa, v.37, n.7, p.1251-1256, 2008.

GRINDSTAFF, J.L., DEMAS, G.E. and KETTERSON, E.D. Diet quality affects egg size and number but does not reduce maternal antibody transmission in Japanese quail Coturnix japonica. Journal Animal and Ecology, v.74, p.1051-1058, 2005.
HILL, F.W.; ANDERSON, D.L. Comparation of metabolizable energy and productive energy determinations with growing chicks. Journal nutrition, v.64, p.587-603, 1958.
HUDSON, B.P.; DOZIER, W.A.; WILSON, J.L.; SANDER, J.E.; WARD, T.L. Reproductive performance and immune status of caged broiler breeder hens provided diets supplemented with either inorganic or organic sources of zinc from hatching to 65 wk of age. Journal of Applied Poultry Research, v.13, p.349-359, 2004.
IJI, P.A.; SAKI, A.; TIVEY, D.R. Body and intestinal growth of broiler chicks on a commercial starter diet. 2. Development and characteristics of intestinal enzymes. British Poultry Science, v. 42, p. 514-522, 2001
KONDO, N. Estudo das características morfométricas de diferentes regiões do intestino delgado e índices zootécnicos em quatro linhagens de frangos de corte. 2003. Tese (Doutorado em Zootecnia) Faculdade Medicina Veterinária e Zootecnia, Universidade Estadual Paulista, Botucatu.
LANNA, P.A.S.; ROSTAGNO, H.S.; SILVA, D.J. Tabela de composição de alimentos concentrados. I. Valores de composição química e de energia metabolizável determinados com pintos. Revista da Sociedade Brasileira de Zootecnia, v.8, n.3, p.516-523, 1979.
LONGO, F.A. Avaliação de fontes de carboidrato e proteína e sua utilização na dieta pré-inicial de frangos de corte. 2004. 110 f. Tese (Doutorado em Agronomia), Universidade de São Paulo, Piracicaba.
MAIORKA, A. Adaptações digestivas pós-eclosão. In: Conferência apinco de ciência e tecnologia avícolas, 2001, Santos, São Paulo. Anais... Campinas: FACTA, 2001. v.2, 141-151.
MAIORKA, A., SANTIN, E., SILVA, A.V. F., ROUTMAN, K. S., PIZAURO Jr., J. M., MACARI, M. Effect of Broiler Breeder Age on Pancreas Enzymes Activity and Digestive Tract Weight of Embryos and Chicks. Brazilian Journal Poultry. Sciece, v.6, n.1, p.19-22, 2004.
MARCHINI C.F.P.; SILVA, P.L.; NASCIMENTO, M.R.B.M.; BELETTI, M.E.; GUIMARÃES, E.C.; SOARES, H.L. Morfometria da mucosa duodenal em frangos de corte submetidos à temperatura ambiente cíclica elevada. Arq. Bras. Med. Vet. Zootec., v.61, n.2, p.491-497, 2009.
MELLO, H.H.C.; GOMES, P.C.; ROSTAGNO, H.S.; ALBINO, L.F.T.; SOUZA, R.M.; CALDERANO, A.A. Valores de energia metabolizável de alguns alimentos obtidos com aves de diferentes idades. Revista Brasileira de Zootecnia, v.38, n.5, p.863-868, 2009.
MORAN JR., E. T. Digestion and absortion of carbohydrates in fowl and events through perinatal development. Journal of Nutrition, v. 115, p. 665-674, 1985.
MURAROLLI, V.D.A. Efeito de prebiótico, probiótico e simbiótico sobre o desempenho, morfologia intestinal e imunidade de frangos de corte. 2008. 101 f. Dissertação (Mestrado em Medicina Veterinária) - Faculdade de Medicina Veterinária e Zootecnia, Universidade de São Paulo, Pirassununga.

NATIONAL RESEARCH COUNCIL (NRC) - Nutrients requirements of poultry. 9th Rev. Edition Natl. Acad. Press, Washington, DC., 1994, 155p.
NITSAN, Z.; DUNNINGTON, E. A.; SIEGEL, P. B. Organ growth and digestiveenzymes levels to fifteen days of age in lines of chickens differing in body weight. Poultry Science, v. 70, n. 12, p. 2040-2048, 1991.
NITSAN, Z. The development of digestive tract in posthatched chicks. In: European symposium on Poultry Nutrition, 10., 1995, Antalya. Anais… Antalya: Europ. Poultry Science Assoc. p.21-28.
NOY, Y.; SKLAN, D. Digestion and absorption in the young chick. Poultry Science, v.74, p. 366-373, 1995.
NOY Y, SKLAN D. Posthatch development of poultry. Journal of Applied Poultry Research, v.6, n.3, p. 344-354, 1997.
NOY, Y.; SKLAN, D. Yolk and exogenous feed utilization in the posthatch chick. Poultry Science, v. 80, p. 1490-1495, 2001.
POLIN, D.; HUSSEIN, T.H. The effect of bile acid on lipid and carcass composition and dietary metabolizable energy in very young chicks. Poultry Science, v.61, p.1697–1707, 1982.
REECE, W. O. Dukes - Fisiologia dos Animais Domésticos. 12ª ed. Rio de Janeiro: Guanabara Koogan, 2006. 942p.
ROCHA, J.S.R.; . LARA, L.J.; BAIÃO, CN.C.; CANÇADO,S.V.; BAIÃO,L.E.C., SILVA,T.R. Efeito da classificação dos ovos sobre o rendimento de incubação e os pesos do pinto e do saco vitelino. Arq. Bras. Med. Vet. Zootec., v.60, n.4, p.979-986, 2008.
RODRIGUES, P.B.; ROSTAGNO, H.S.; ALBINO, L.F.T; GOMES, P.C.; BARBOZA, W.A.; SANTANA, W.A. valores energéticos do milheto, do milho e subprodutos do milho, determinados com frangos de corte e galos adultos. Revista Brasileira de Zootecnia, v.30, n.6, p.1767-1778, 2001.
RODRIGUES, P.B.; ROSTAGNO, H.S.; ALBINO, L.F.T; GOMES, P.C.; NUNES, R.V.; TOLEDO, R.S. Valores energéticos da soja e subprodutos da soja, determinados com frangos de corte e galos adultos. Revista Brasileira de Zootecnia, v.31, n.4, p.1771- 1782, 2002.
ROSTAGNO, H.S.; ALBINO, L.F.T.; DONZELE, J.L.; GOMES, P.C.; FERREIRA, A.S.; OLIVEIRA, R.F.M.; LOPES, D.C. Tabelas brasileiras para aves e suínos (composição de alimentos e exigências nutricionais). Viçosa: Universidade Federal de Viçosa, 2000.
ROSTAGNO, H.S.; ALBINO, L.F.T.; DONZELE, J.L.; GOMES, P.C.; OLIVEIRA, R.F.; LOPES, D.C.; FERREIRA, A.S.; BARRET, S.L.T. Tabelas brasileiras para aves e suínos - composição e alimentos e exigências nutricionais. 2. ed. p. 141. Viçosa: Universidade Federal de Viçosa, 2005.
SAKOMURA N.K.; DEL BIANCHI, M.; PIZAURO JR., J.M., CAFÉ, M.B.; FREITAS, E.R. Efeito da idade dos frangos de corte sobre a atividade enzimática e digestibilidade dos nutrientes do farelo de soja e da soja integral. Revista Brasileira de Zootecnia, v.33, n.4, p.924-935, 2004a.

SAKOMURA, N.K.; SÁ FORTES, C.M.L.; SANTOS, F.R. Determinação da digestibilidade dos alimentos para aves. In: Curso de Fisiologia da Digestão e Metabolismo de Nutrientes em Aves, UNESP Jaboticabal, out. 2004b. 1 CD-ROM.
SAKOMURA, N. K.; ROSTAGNO, H. S. Métodos de pesquisa em nutrição de monogástricos. Jaboticabal: Funep, 2007. 283 p.
SELL, J.; ANGEL, C.; PIQUER, F. et al. Development patterns of selected characteristics of the gastrointestinal tract of young turkeys. Poultry Science, v. 70, p. 1200-1205, 1991.
SELL, J.L. Physiological limitations anda potential for improvement in gastrointestinal tract function of poultry. Journal of Applied Poultru Reseach, v. 05, p. 96-101, 1996.
SIBBALD, I. R. A biossay for true metabolizable energy in feedingstuffs. Poultry Science, v. 55, v. 1, p. 303-308, 1976.
SIBBALD, I. R. Measurement of bioavailable energy in poultry feedingstuffs: review. Canadian Journal of Animal Science, 62: 983-1048, 1982.
SIBBALD, I. R.; SLINGER, S.J. A biological assay for metabolizable energy in poultry feed ingredients together with findings wich demostrate some of the problems associeted with the evaluation of fats. Poultry Science, v. 59, p. 1275-1279, 1963.
SKLAN, D. Development of the digestive tract of poultry. World's Poultry Science Journal, v.57, n.1, p 415-428, 2001.
SKLAN, D. Fat and carbohydrate use in posthatch chicks. Poultry Science, v. 82, p.117-122, 2003.
STRINGHINI, J.H. ; ANDRADE, M.L. ; XAVIER, S.A.G.; ANDRADE, M.A. ; MINAFRA, C.S.; CAFÉ, M.B. . Protein and Amino Acid Supplementation Levels for Broilers in Pre-Starter Ration. International Journal of Poultry Science, v. 8, p. 946-951, 2009.
UNI, Z., GANOT, S., SKALAN, D. Posthatch development of mucosa function in the broiler small intestine Poultry Science, v.77, n.1, 75-82, 1998.
UNI, Z.; NOY, Y.; SKLAN, D. Posthatch Development of Small Intestinal Function in the Poult. Poultry Science, v.78, n.1, p.215–222, 1999.
UNI, Z.; FERKET, P. R. Methods for early nutrition and their potential. World's Poultry Science Journal, v.60, n.1, p.101–111, 2004
VIEIRA, S.L.; POPHAL, S. Nutrição pós-eclosão de frangos de corte. Revista Brasileira de Ciência Avícola, v.2, n.3, p.189-1999, 2000.
WISEMAN, J.; SALVADOR, F. Influence of age, chemical composition and rate of inclusion on the apparent metabolisable energy of fats fed to broilers chicks. Britsh Poultry Science, v.30, n.3, p.653-662, 1989.





























