Embed Size (px)
Citation preview


REVISTA THEOBROMA
Janeiro-março 1974
AnoiV N?l
Publicação rrimesual dedicada à divulgação de in· vestigação científica relacionada com problemas agro· nõmicos e sócio-econômicos de áreas cacaueiras. Editada pelo Centro de Pesquisas do Cacau (CEPEC), DePa"amenlo da Comissão Executiva do Plano da Lavoura Cacaueira (CEPLAC). f
COMISSÃO EDITORIAL
Alúm, Paulo d"' T. Cruz, Luiz C. Sales, José C. de Vello, Fernando
ASSESSORES CIENTÍFICOS
Ai<ken, William M. Alencar, Maria H. Alvim, Ronald Barroco, H.O,Jio E. Barros, Raimundo S. Cabala R .• F. Percy Carletto, Geraldo A. Loureiro, Edmar S. Maravalhas, Nelson Mariano, Antônio H. Medeiros, Arnaldo G. Menezes, José A. S. Miranda, Emo Ruy de Pereira, Clovis P. Rocha, Hermínio M. Silva, Luiz F. da Silva, Pedriro Ventocilla, José A.
Endereço para correspondência (Adreu for cnrre· spondence):
Revisra Theobroma Cenrro de Pesquisas do Cacau (CEPEC). Caixa Posral 7 4~-600- habuna-Bahia
Brasil
Tiragem 3.000 exemplares
CONTEÚDO
l. tdentidade e caracteriz<lção taxonômica prelimintJt das mosq uinbas Forcipomyia (Diptera, Ceratopogonidae) associadas com a polinização do cacaueiro na B<lhia. S. de J. S01ia e W.W. Wirth . .............. _
Identification and preliminary taxonomic characterizatíon of Forcipomyia midges (Diptera, Cera topo· gonidae) related to the pollination of cacao in Bahia. (Summary)p.l2.
2, Zoneamento agrícola da região cacaueira baiana. L. F. da Silva, A. H. Mariano e A.C. da C.P. Dias, .....
Agricultura! zoning of the Bahian cocoa region. (Summary) p. 27.
3. Cuatro especies de trips en el cscao de Tabasco, Mexico. R.M. Johan· sen N ...... .
Four species of thrips on cocoa in Tabasco, Mexico. (Summary) p. 38.
4, Nemutodes o/ the cocoa region of Bahia, Brazil. 111. Plant parasitic and free-living nematodes in tbe rizospheres o/ six di!ferent plant species. R.D. Sharma e P.A.A. Loof. ...................... . ................ .
Nematôdios da região cacaueira da Bahia, Brasil. III. Nematódios parasitos e não parasitos na rizosfera de seis diferentes espécies de plantas. {Resumo) _p. 43.
NOTA
Segunda reunião regional americana do
3
13
29
"
grupo de Phytophthora palmivora. 4-4
Second American regional meeting of PhytophthOTa palmívora group. p. 48.

IDENTIDADE E CARACTERIZAÇÃO TAXONOMICA PRELIMINAR
DAS MOSQUINHAS Forcipomyia
(DIPTERA, CERATOPOGONIDAE) ASSOCIADAS COM A POLINIZAÇÃO
DO CACAUEIRO NA BAHIA•
Saulo de Jesus Soria•• Willis w. Wirtb •u
A caracterização taxonômica das mesquinhas Forcipomyia spp. tem por objetivo o seu reconhecimento nos estudos de caráter econômico, que estão sendo conduzidos na região cacaueira da Bahia. Essa caracterização é considerada preliminar, porquanto foi baseada em um número reduzido de elementos taxonômicos, facilmente medidos em adultos de ambos os sexos. Concomitantemente são descritos os hábitos das mesquinhas em função da polinização do cacaueiro.
A literatura sobre a taxonomia de Forcipomyia é rica devido à grande diversidade de espécies existentes neste gênero. Saunders (10) dividiu inicialmente o gênero Forcipomyia em oito subgêneros
com base nos caracteres dos estados imaturos. Novos subgêneros foram sucessivamente adicionados~ à medida que o estudo do gênero Forcipomyia avançava, Dessart (2, 3) elaborou uma chave para a identificação das espécies polinizadoras do cacaueiro e outras do gênero Forc;_pomyia, que ocorrem no Congo, Africa. Saunders (9), Wirth (16) e Dow e Wirth (4) desenvolveram chaves para as espécies neotropicais e as neárticas dos subgêneros Tbyridomyia e Syntbrydomyia. Wirth (17) descreveu espécies neotropicais do subgênero Microbelea.
Winder e Silva (13, 14), em colaboração com W.W. Wirth, pesquisaram a complexa fauna de dípteros nos cacauais da Bahia, pro-
'*Recebido para publicação em novembro, 1973. ** Ph.D., Divisão de Entomologia, CEPEC.
'*"'"'Ph.D., U. S, Department of Agriculture, e/o U. S. National Museum, Washington D.C. 20560, U,S.A.
Revista Theobroma, CEPEC, Ilhéus, BITisil, 4 (I J: 3. 12. }an.-mar. 1974.
3

curando principalmente as mosquinhas Forripomyia, em criadouros naturais e nas flores do cacaueiro.. Vârios substratos orgânicos, presentes nos cacauais, demonstraram o;er criadouros de Forcipom) ia spp., particularmente as Bromeliaceae epífitas, das quais emergiram exemplares de F. (Pro
jordpomria), reconhecidos como polinizadores importantes em outras áreas da América.
"''o presente trabalho, não se pretende fazer a revisão completa do assunto, uma vez que estudos mais aprofundados, baseados nas descrições dos estados imaturos correspondentes ao presente material, estão sendo conduzidos sob a liderança de W. W. W irth do La. boratório de Entomologi~ Sistemâtica, do Departamento de Agricultura dos Estados Unidos da América (USDA).
:\IATERIAL E :\1ÉTODOS
A determinação qualitativa e quantitativa, dinâmica populacional e polinização pelas mosquinhas Porcipomyia spp. foram estudadas em quadras experimentais do CEPEC, município de Ilhéus, BA, nos anos de 1972 e 1973.
Na terminologia usada, a denominação F.uforcipomyia Malloch (7) para o subgênero Proforcipomyia Saunders (10) volta a ter validez por prioridade de descrição, segundo a opinião de Wirth (18). No presente trabalho, a relação tarsal (T.R.) é o resultado da di-
4
visão do comprimento do primeiro tarsômero pelo comprimento do segundo (7). A relação antena! (A.R.), nas fêmeas, é o comprimento combinado dos cinco flagelômeros distais dividido pelo comprimento combi9ado dos oito flagelômeros precedentes (3). Nos machos, a A.R. é o comprimento combinado dos cinco flagelômeros distais dividido pelo comprimento combinado dos oito flagelômeros precedentes (3). O comprimento da asa, de acordo com a terminologia adotada por Wirth (15), é medido a partir do arculus basal até o extremo distai da asa; a relação costal (C. R.), também de acordo com Wirth (16), é o comprimento da costa medido a partir do arculus basal até o extremo distai da célula radial secundária dividido pelo comprimento da asa.
Para determinar qualitativa e quantitativamente as mosquinhas Porcipomyia spp., três operários treinados em reconhecê-las nos cacauais efetuaram coletas diárias sistemáticas em 108 árvores etiquetadas. As coletas foram feitas das 8:00 às 10:00 horas, no período de 15 de abril a 28 de novembro de 1972.
Quando descoberta numa flor, a mosquinha era capturada e a flor imediata'll.ente encerrada em um vidro com 2,5 em de diâmetro e 11 em de comprimento. As mosquinhas capturadas eram conduzidas ao laboratório, onde eram mortas e conservadas e'll álcool a 70% para estudos taxonômicos.
I (
{
'
1.
••
Cada exemplar fêmea era dissecado com o auxilio de um estereomicroscópio, segundo a metodologia de Saunders (11), separando-se e orientando-se a cabeça, uma asa e o abdômem. A genitalia ma~culina foi separada com os três últimos segmentos abdominais para permitir a observação das suas estruturas internas e externas; do mesmo modo, procedeu-se com a cabeça, tórax e asa para medição das antenas, tarsos, asas e outras estruturas. As estruturas foram cobertas com lamínulas para serem estudadas posteriormente ao microscópio. As medições das asas e demais estruturas foram feitas sob aumentos de 20x e 80x, respectivamente, usando-se escala micrométrica.
RESUL T !\DOS E DISCUSSÃO
F. (Euforcipomyia)
Quarenta e seis mosquinhas foram coletadas individualmente em 46 flores no ato da polinização. O número total de flores observadas foi de 327.000 durante o período de amostragem (101 dias). O número total de mosquinhas coletadas em atitude de descanso nas flores, assim como em botões florais e pontas secas de folha, foi de 487 fêmeas e 158 machos. A relação entre o número de flores disponíveis e o de mosquinhas polinizadoras foi de 7.122:1; no entanto, a taxa de flores disponíveis sobre o total de mosquinhas Forcipomyia presentes nos cacauais foi de 508:1.
5
As mosquinhas fêmeas coletadas no ato da polinização foram todas enquadradas no subgênero F. (Euforcipomyia) Malloch e se apresentaram em dois grupos principais, separados entre si pelo tamanho das espermatecas, a saber:
F. (Euforcipomyia) sp. 1. Caracteriza-se por ter espermatecas piriformes, marcadamente desiguais (Quadro 1 e Figura 18). A espermateca maior tem comprimento médio de 70 micra e largura de 50 micra. A espermateca me· nor tem comprimento de 57 micra e largura de 40 micra, A T .R. média foi de 1,97 e a A.R. foi de 1,13. O comprimento médio da asa, a partir do arculus, foi de 0,80 mm; a largura média foi de 0,36 mm na sua maior expansão e a C.R. foi 0,54.
F. (Euforcipomyia) sp. 2. Caracteriza-se por ter espermatecas quase iguais (Quadro 1 e Figura lC). Cada espermateca tem com~ primento médio de 64 micra e largura média de 49 micra. A diferença dimensional entre as espermatecas é de apenas 3 micra em comprimento. Outras características subgenéricas foram T.R. média de 1,63; A.R. média de 1,03; comprimento da asa de 1,04 mm; largura 0,42 mm e C.R. de 0,52. Como se vê, existe uma notável diferença entre os tamanhos das asas de F. (Euforcipomyia) sp. 2 e F. (Euforcipomyia) sp, 1, sendo a primeira maior que a segunda. Esta espécie foi recentemente identificada como F. (Euforcipomyia) spatulifera Saunders.

Quadro 1 - ( flrnc teristicas de c spc'cies poli111zadoras do subgê11em Euforcipomyia, Jcimeas (' machos. 1/l>éus , Babia. 19"'2 73 .
Fêmeas Machos Elementos taxonômtcos
sp. l sp. 2 (não diferenciados)
Comprimento da asa (mm) 0,80 1,04 1,05 Largura da a~a (mm) 0,36 0,42 0,34
Espermateca maior D1âmetro ma10r (mícron) 70 64 Diâmetro menor (mícron) 50
Espermatcc~ m<'nor Diâmetro mawr (mícron) 57 Diâmetro menor (mícron) 40
Relação tarsal (T.R.) 1,97 Relação antena) (A.R.) 1,13 Relação costal (C.R.) 0,54
N~ mdivíduos estudados
O exame dos machos mostrou os dois upodemas bas istilares unidos em ângulo agudo, característica da F. (/:'ujorcipom)ia) sens. lat. (Quadro 1 e Figura lD). A medição dos elementos morfológicos externos permitiu a ca racter ização de um grupo subgenérico único com T.R. média de 1,62 (Quadro 1), A. R. média de 0,98, comprimento e largura méd10s da asa de 1 ,OS mm e 0,35 mm, respectivamente, c C.R . de 0,48. Este grupo prov«velmentc será subd ivid ido em vá ri <Js espéc 1es, q uand o um número m<ns represcn (<.~tivo de caructer íst1C<JS for medido para su<1 curactcr 17-<~Çilo.
Ohscr va<..ôes f<•JIHs soh o mi e roscf,pw nns 4f, flores visitad;,::;
lO
49
1,68 1,62 1,03 0,98 0,52 0,48
14 23
pelas mosquinhas, no ato de poltnização, indicmam que somente em 52% dos casos o pólen foi deposita do no es l!lo e estigma florais. A ocorrência limitada de polinizações (52%) na presente amostragem parece ter sido 1 nfluenciada pe lo pouco te mpo que as mosquinhas pe rmaneceram n<ls flores . I sto teve lugar enquanto a mosquinha andava e prova\'<J a super f ície inter ior dos estaminóides . Nessa ocasião, eventual mente, a mosqu in h a entr<.~ra n;:Js conch::is das péta l as, onde o pólen ader i u ao seu dorso. O csp.u; o ent re o estigma e os es taminóides pa reciél pcrm1 tlf a pássagem do inseto, mas nn1) da SUU Cé1rga de gr~os de pólen, que ficava dcpositHdil no cst 1gma nu sua passa -
..
Figura 1 - A. Fêmea de Forcipomyia (Euforcipomyia) sp., 30x: B. Espermatecas de F. (Euforc ipomyia) sp. 1: C. Espermatecas de F. (Euforcipomyia) sp. 2; D. Genitalia masculina de F. (Euforcipomyia) sens. lat. 11/Jéus, Bahia, 1972/ 73.
gem. Esta mecamca de polinização concorda com as observações feitas por Billes (1) e Posnette (8), em Trinidad, e Hernandez (5), na Costa Rica. Neste mesmo país, Soria (12) regis trou um período de 10 a 45 minutos necessários para que as mosquinhas efetuassem uma polinização típica.
7
Forcipomyia sens. lat.
O gênero Forcipomyia, na Bahia, apresenta grande variabilidade, de acordo com as determinações feitas por Wirth (Quadro 2). Com base nas variações da T .R. para o gênero Forcipomyia, medi das em exemplares co letados no

Quadro 2 - Mosqui71has Forcipomyia coletadas nas flores do cacaueiro. Ilhéus, Bahia, 197P.
Mosquinhas -Fêmeas Machos ·~ (número) (número) I F. (Hujorcipomyia) sp. 1 139 71 ' }:'
F. (EMforcipom_via} spp. 18 2 F. (Microhf.'/ea) fuliRinosa (Meigen) 12 27 F. (Mir:rohelea) erioph"ora (Williston) 3 4 F, (Mir:rohelea) squamosa Lub; 8 2 F. (Microhdea) sp. 2 F. (Fwcipomyia) genualis (Loew) 16 9 F. (Forcípomyia) pictoni Macfie 2 F, (Forc:ipomyia) sp. 1 I 13 F, (Forc:ipomvia) sp. 2 4 2 F, (Rhvnchoforcipomyia) brachyrhynchus
Wirth & Dow I F, (Lasiohelea} stylifera Lutz 6 F. (Lasioht'/ea) sp. 1 3 I Dasyhelea sp. B 4 I
*Determinadas por W. W. Wirth, ARS-USDA.
CEPEC, foi comprovada a maior freqüência das mosquinhas Euforcipomyia (T.R. 1,3-2,0) nas flores (Quadro 3). Somente F. (Euforâpomyia) Malloch comportou-se como polinizadora das flores do cacaueiro. O índice elevado de polinização observado na região cacaueira da Bahia, no período de abril a agosto de 1973, esteve aparentemente relacionado com a freqüência das mosquinhas Euiorcipomyia nos cacauais.
Os estudos baseados na conformação dos parâmeros e apodemas internos da genitalia mascu-
8
na indicaram, até hoje, a presença dos subgêneros Fuforcipomyia, Lasiobelea, Forcipomyia e Microhelea e das mosquinhas Dasyhelea como categorias predominantes nos cacauais da Bahia (Figura 2). Machos de Euforcipomyia com parâmeros fusionados como arco gótico (10) não foram encontrados polinizando as flores do cacaueiro na Bahia. Vale salientar que Kaufmann (6) registrou, na África, a ação polinizadora dos machos Forcipomyia squamipennis Ingram & Macfie, razão porque o seu papel nessa operação deve ser estudado também na Bahia.
I f
•
r'
' I
Quadro 3 - Freqüência de exemplares de Forcipomyia co/etados em cacaueiros, agrupados de acordo com os valores de relação tarsal (T.R.) de Saunders (1956). Ilhéus, Bahía, 1973.
;p ....,. Exemplares 4es~tqs.
"faixa 1' .R. pelas _espermatecas Freqüência Proporçolo
relativa <\J Critério m · 0,50 :.. I ,29 Iguais 47 (PfirtipotJ~:yiaJ Desieuais 21 68 I
1,30-,;;;:,2,00 Iguais 147 /fl!fEÚfon:ff,omyi4) Desi~is 72 219 3
~OI- 3,()() Iguais 9 Desiguais 7 16 '< I ___: aforcipo,.yial- ... ... •
Subgênero E •f"n:if>o"'1"' Forci{XJIIIJill Forcip.>•yi11 ~icrohlet> Uasyhl~a
h .... ~ ~ ~ f:ró 6mJ f/0 Gnitt>litl
. .
A.R. 0.98 "' 0.81 LOó l.20
T.R. 1.62 0.-18 0.52 0.43 2.50
C. R. 0.48 0.41 "' 0.43 o.so
Figura 2 - Sinopse comparativa dos quatro elementos morfol6gicos que caracterizam quatro subgêneros de Forcipomyia e um de Dasyhelea, Ilhéus, Bahia, 1972/73.
AGRADECIMENTOS
Ao Engenheiro Agrônomo Pedrito Silva, por sua entusiástica dedicação à revisão do trabalho. Aos Eng~ Agr~ João Manuel Abreu e José A. Ventocilla, pelas críticas, Ao B.S. William Martin Aitken, pela revi-
9

s:w rh1 rl'sumo ('1\1 inl',l<'s, (' ao Sr. Florisvaldo A. Galvão, pela ajuda na rt'<l<l~',hl d(' .1rt igo.
LITER:\TVR1\ CITADA
L BILLES, D. J. Pollination of Tl>eobruma cncao L in Trinidad. B. W.L Tropical Agriculture (Trinidad) 18(8):151-156. 1941.
2. DESSART, P. Contribution a l'etude des Ceratopogonidae (Diptera); TI. Revision des Fordpomria congolais descrits par le Dr. Geotghebuer, Bulletin Annuel de Societé Royale Belge 97: 315-376. 1961.
3. ____ Contribution a l'étude des Ceratopogonidae (Diptera); VII. Bruxelles, Institut Royal de Sciences Naturelles de Belgique 72(2):1-151. 1%3.
4. DOW, M. I. e WIRTH, W. W. Studies on the genus Forcipomyia sp. 2. The Neartic species of the subgenera Thyridomyia and Synthridomyia (Diptera, Ceratopogonidae), Annals of the Entomological Society of America 65(1):177-201. 1972.
5. HERNANDEZ, J. lnsect pollination of cacao (Theobroma cacao L.) in Costa Rica. Ph.D. Thesis. Madison, University of Wisconsin, 167 p. 1965.
6. KAUFMANN, T. Correspondência particular. 1973.
7. MALLOCH, J.R. Some aditional records of Chiromidae Illinois. Bulletin of Illinois State Laboratory of Natural History 11:305-363. 1915.
8. POSNETTE, A.F. Pollination of cacao in Trinidad. Tropical Agriculture (Trinidad) 21:115-118. 1944.
9. SAUNDERS, L.G. On the life history, morphology and sistematic position of Apelma Kieffer and Thyridomyia n. gen. (Diptera Nemt. Ceratopogonidae). Parasitology 17:252-277. 1925.
10. Revision of the Genus Forcipomyia based on characteres of all stages. Canadian Journal of Zoology 34:657-705. 1956.
11. Methods for studying Forcipomyia midges with special reference to cacao pollination species {Diptera, Ceratopogonidae). Canadian Journal of Zoology 37:33-51. 1959.
12. SORIA, S. Studies on Forcipomyia spp, midges (Diptera, Ceratopogonidae) related to the pollination of Theobroma cacao L. Ph.D. Thesis. Madison, University of Wisconsin, 142 p. 1970.
lO
•
/
•
'
13. WINDER, J.A. e SILVA, P. Cacao pollination: Microdiptera of cacao plantations and some of their breeding places. Bulletin of Entomological Research 61:651-655. 1972.
14. ____ e Pesquisas sobre a polinização do cacaueiro por insetos na Bahia, Revista Theobroma (Brasil) 2(3):36-46. 1972.
15. WIRTH, W.W. The Heleidae of California, Publications Entomology 9:95-266.
University !952.
of California
16. ____ The Neotropical Forcipomyia midges of the subgenus Thyridomyia Saunders (Diptera, Ceratopogonidae). Studia Entomologica 13(1-4):429-440. 1970.
17. ____ The Neotropical Forcipomyia (Microhelea) species related to the caterpillar parasite F. fuliginosa {Diptera, Ceratopogonidae). Annals of Entomological Society of America 65{3): 564-577. 1972.
18. __ _ Correspondência particular. 1973.
RESUMO
São descritos vários caracteres taxonômicos de mosquinhas Forcipomyia spp. adultas e os seus hábitos de polinização no cacaueiro. Foram medidos alguns caracteres morfológicos em adultos machos e fê~
meas, para duas espécies de F. (F.uforcipomyia), preliminarmente denominadas sp. 1 e sp. 2. Sugere-se que o nome subgenérico F.uforcipomyia Malloch (1915) seja usado no lugar de Proforcipomyia Saunders {1956). F, (F.uforcipomyia) spp., F. (Lasiohelea) spp. e F, (Microhelea) spp. foram identificados de acordo com a genitalia dos machos. Estes taxa e as mosquinhas Dasyhelea spp. são freqüentemente encontrados nas flores do cacaueiro, nunca, porém, efetuando qualquer atividade polinizadora. Após examinar uma amostra de 327.000 flores, concluiu-se que as mosquinhas F. (F.uforcipomyia) são as principais responsáveis pela polinização do cacaueiro na Bahia. Tal qual ocorre em outras regiões, observaram-se depósitos maciços de pólen quando a mosquinha Euforcipomyia caminha nas superfícies interiores dos estaminóides e prova o tecido epidérmico vermelho. Ao que parece, o espaço entre os estaminóides e o estigma é suficientemente grande para que o inseto passe sozinho, mas não os grãos de pólen, que ficam acumulados no estigma, caracterizando a polinização típica da Euforcipomyia no cacaueiro.
11

IDENTIFICATION AND PRELIMINARY TAXONOMIC CHARACTERIZATION
OF Forcipomyia MIDGES (DIPTERA, CERATOPOGONIDAE) RELATED TO THE POLLINATION
OF CACAO IN BAHIA
SU:\'IM.-\RY
Severa! taxonomic characters of adult Forcipomyia spp. midges and their pollinating habits in cocoa are described. Morphological characters,. based on adult males and females for two species of Forcipomyia (Euforcipomyia) were measured. It is suggested that the subgenus of Euforcipomyia Malloch (1915) be used in preference to its present name Pro/orcipom)'ia Saunders (1956). Forcipomyia (Euforcipomyia) spp., F, (Lasiohelea) spp. and F. (lllicrohelea) spp., were identified according to their male genitalia, These and the closely related Dasyhel(:a spp. midges were frequently found in cocoa flowers but no pollinating activity was observed. From over 327,000 flowers examined it is concluded that it is the F. (Euforcipomyia) midges which are mainly responsible for pollination of cocoa in Bahia. Similar to descriptions from other regions it was found that massive pollen deposits ocurred when the Euforcipo· myia midge was walking and probing on the inner surfaces of the staminodes, lt appears that the space between the staminodes and the stigma is large enough for only the midge to pass through, the pollen grains being left on the stigma as a massive deposit, considered characteristic of Eufordpomyia pollination of cocoa.
12
I ..
•
ZONEAMENTO AGRÍCOLA DA REGIÃO CACAUEIRA BAIANA'
Luiz F erre ira da Silva** Antônio Henrique Mariano*** Antônio C. da C.P. Dias ****
A região cacaueira baiana, aqui entendida, circunscreve uma área de aproximadamente 91.000 km 2 e é constituída de 89 municípios, 73 dos quais produtores de cacau (1) e os demais estreitamente ligados a estes por relações de natureza sócio-econômica. Trata-se de uma área de economia eminentemente cacaueira, responsável por quase 95% da produção brasileira de cacau, cujo cultivo se distribui em 400.000 ha. Estende-se do vale do Baixo Jequiriçá, ao Norte, ao vale do Baixo Mucuri, ao Sul, e comporta uma população estimada em 250 milhões de cacaueiros (3).
Apesar da existência de outras atividades agrícolas (seringueira, coco-da-Bahia, dendê e piaçava) e pecuária, a Região é tida como e· conomicamente monocultora. C are-
ce, portanto, de aproveitamento racional de seus recursos naturais, haja vista ser dotada de características peculiares ao desenvolvimento econômico: vias de acesso aos grandes centros de consumo (Rio de Janeiro, Salvador, Vitória e Belo Horizonte), facilidades de assistência técnica e aquisição de insumos, porto inte•rnacional e potencialidade agro-pastoril.
Efetivamente, há necessidade de se proceder o zoneamento agrícola dessa região, que permitirá diagnosticar as possibilidades de aproveitamento agrícola dos seus solos. Através de estudos de solos, clima e topografia (fisiografia), o presente trabalho objetiva, em primeira aproximação, fornecer informações para fins de planejamentos diversos,
* Recebido para publicação em março, 1974.
** Eng.-Agr~, Divisão de Solos, CEPEC.
*** M.S., Divisão de Genética, CEPEC.
**** M.S., Divisão de Solos, CEPEC.
Revista Theobroma. CEPEC,Jlhéus, Brasil, 4(1): 13-28. Jan.-mar. 1974.
13

ASPECTOS PF.DOLÓGICOS
Do ponto de vista pedológico, a região cacaueira da Bahia se constitui em uma área complexa, cuja variação de solos se relaciona com a diversidade geológica. Apresenta 30 unidades cartográficas (Quadro 1), as quais se encontram perfeitamente caracterizadas e classificadas (11).
Há uma dominância de solos velhos (Oxisols), em conseqÜência pobres, de maior ocorrência na faixa costeira úmida, derivados de sedimentos desgastados do Terciário e outros materiais pseudo-autóctones do Pré-Cambriano (9).
Por outro lado, dentro desta faixa tropical úmida, ainda se encontram solos relativamente jovens,
Qúadro 1 - Legenda de identificação dos solos da região cacaueira baiana.
.~pameato• """ Ordens
<tJJ:/ • <!' MolUsols "zi"-.
I,,
Sol'os Ultisols <<>m
8 textl;lral
Alfiscls
Solos
''"" 8 Oltisols
Iat:~óJ.ieo
Solos eom B incipiente
lnceptisols
Solos com B Spodosols podzólico
• ) •'
Cortr;éituaçiq
t:utt6neos •
Meso!tóÚeos h
1Hstrôlicos <
Endo-distróficos d
Eqtro?ficoS
Distróficos
Eutrófi.cos
Eutróficol'l
Mesotróficos
Distrófkos
14
Unidades ca'rtográfkase
2 3 4 5
6 7
• • lO
·'13 ,14
', :: 17 18
19
20
22
Continua
. ..
•
• •
Quadro 1 - Continuação
Solos pouco Entisols Distróficos 23 desenvolvidos 24
Solos orgânicos Histosols Mesotróficos 25
Solos Aridisols Eutróricos 26
Indi!eretlci&dOB Inceptisols I 27 Entisols Mesottóficos 28
Tipos de 29 Jerrellos ~ 30
a - Saturação de bases 1n3ior que 50% em todo o perfil b- " " entre 30 e 50% " " '' " c - " " " menor que 30'l"b " " " " d- Horizonte A mesotrófico e horizonte 8 distrófico e - Unidades çartográficas :
1. Argiustolls 2. Tropudults variação Il:llbuna 3. Camaçlln 4 . .. .. Morro Redondo S. .. .. São Paulinho 6. Nazaré 7. Cururupe 8. São Ma teus 9. Vargito
!0. Itagimirim !1. Tropudalfs !2. Haplustalfs !3. Hllplortho>ts variação T11buleiro !4 . Haplustoxs Tabuleiro !5 . Haplortho>ts Cristalino
ricos em bases trocáveis, oriundos de rochas intermediárias ou básicas, nos quais o cacaueiro se desenvolve bem.
A oeste, já em clima de transição, na zona pastoril, grandes faixas de solos férteis se derivam de rochas gnaissicas do complexo cristalino, cuja pedogênese está estreitamente relacionada com a mudança climática.
Nos grandes rios - notadamente Pardo, Jequitinhonha e Mucuri -
15
16. Haplustoxs vllriação Cristalino 17. .. .. Chapada 18. Umbriorthoxs 19. Eutrortho>tS 20 . Eutropepts 21. Dystropepts 22. Tropaquods 23. Quartzipsamments variação Catru 24. Quart:<:ipsamments var. Areias Costeiras 25. Troposaprists 26. Solos semi-áridos 27. Solos Aluviais 28. Solos Hidromórficos 29. Mangues 30. Ch11rcos.
são encontrados solos de aluvião, ricos, devido sobretudo ao processo de colmatagem desses rios que, periodicamente, lançam um lodo fértil sobre as camadas sedimen~ tadas anteriormente (10).
Finalmente, a noroeste, surgem os solos férteis das caatingas e, em posição de altiplano, os terrenos velhos do planalto sul-baiano.
ASPECTOS CLIMÁTICOS
Com base em estudos de Frota (5), Santos (8) e Domingues e Keller

(4), o clima da Região é caracterizado a seguir em suas diversas feições
O elemento mais importante na caracterização do clima dessa região parece ser a precipitação, contribuindo os diversos regimes pluviométricos para imprimir à paisagem nuanças fisiográficas.
De um modo geral, pode-se distinguir duas regiões climáticas marcantes: uma úmida, na faixa costeira, cuja precipitação anual é superior a 1.000 mm - chegando até 2.297 mm (Valença)- e outra mais seca, mais para o interior, comprecipitação inferior a 750 mm.
As abundantes chuvas no litoral são devidas à ação dos ventos alísios que desempenham importante papel na génese das chuvas na Região. São ventos muito úmidos, devido ao longo trajeto sobre o oceano, que, penetrando no Continente, sofrem súbita ascensão e, conseqÜentemente, descompressão adiabática que provoca um abaixamento da temperatura, especialmente no inverno.
Avançando em direção ao oeste, observa-se que a umidade vai diminuindo sensivelmente, de forma gradual, começando a se fazer sentir o regime pluviométrico de uma estação seca que, mais para o interior, torna -se bem marcante. Tudo indica que, aí, existe menor influência dos ventos alísios, predominando as chuvas de verão.
A freqüência das chuvas, independentemente da sua quantidade,
!6
varia bastante desde 55 dias em Poções até 225 em Gandu, ocorrendo a maior freqÜência na faixa c oste ira.
A:.: temperaturas se mantêm com relativa uniformidade, apresentando médias elevadas e pequenas oscilações no decorrer do ano. As médias anuais são de aproximadamente 24°C, com índices médi<JS mais elevados (26°C) nos meses de verão e mais baixos nos de inverno, nunca atingindo, porém, valores médios inferiores a l8°C.
Quanto à umidade relativa do ar, a Região apresenta uma média anual de 78%, com valores médios superiores a 80% na costa, bastante correlacionados com o regime pluviométrico.
Para fins de interpretação das condições agrícolas, dividiu-se a Região em três zonas climáticas, descritas a seguir.
Clima úmido. Faixa próxima ao litoral, com uma largura média de 50 km, precipitação superior a 1.000 mm, temperatura em torno de 24°C e umidade relativa acima de 80%, ocorrendo as máximas pluviométricas de fevereiro a julho. Não possui estação seca definida, apesar de, na faixa mais afastada do litoral (últimos 10 km), já se evidenciar uma tendência (dados dos últimos 6 anos) de período seco (agosto a setembro /"utubro).
Encontram-se nesu. faixa os municípios de Valença, C.amamu,
... ,.
'
Ubaitaba, Taperoá, Cairu, ltuberá, Ibirapitanga, Maraú, ltacaré, Uruçuca, ltajuípe, ltabuna, Ilhéus, Buerarema, Una, Canavieiras, Belmonte, Porto Seguro, Itamaraju, Mucuri, Caravelas e outras, responsáveis por mais de 90% da produção de cacau da Bahia.
Clima de transição. Zona de estação seca definida (3 a 4 meses), com vegetação típica semi-caducifólia. Caracteriza-se pela precipitação mediana (800 a 1.100 mm) e temperaturas elevadas, Abrange toda a zona pastoril representada pelos seguintes municípios mais importantes: Itapetinga, Itororó, Itambé, Potiraguá, ltarantim, Maiquinique, ltaju do Colônia, Ipiaú, Guaratinga, Medeiros Neto, Ibirapuã, ltagibá, ltagi e Jtanhém.
Clima seco. Faixa oeste da Região, de precipitação aquém de 800 mm e estação seca de mais de 5 meses (abril a setembro). A vegetação é xerófila (acatingada) nas áreas de altitude inferior a 600 m, e caducifólia (mata-cipó) no planalto sul-baiano (mais de 750 m de altitude). Compreende os municípios de Jequié, Manoel Vitorino, Anagé, Vitória da Conquista, Poções, Cândido Sales etc.
ASPECTOS TOPOGRÁFICOS
Utilizando-se imagens de radar na escala de 1-250.(X)Q, elaborou-se o mapa esquemático dos principais tipos de relevo da Região (Figura 1), descritos a seguir:
17
Relevo praticamente plano. Compreende as restingas, formadas pela sedimentação marinha regressiva, os deltas, resultantes da deposição de materiais na embocadura dos rios, em ambiente salobro, e as planícies aluviais oriundas da colmatagem dos rios (Pardo, Jequi· tinhonha e Mucuri).
Relevo suave ondulado. Área dos tabuleiros típicos, caracterizados por suas formas aplainadas de topo achatado e cortados por vales de seção transversal em forma de "U" (7). Inclui-se também o planalto sul-baiano de superfície peneplanizada e altitude superior a 750 m.
Relevo ondulado. Área essencialmente integrada por seql.iência de colinas arredondadas de modelado suave, muitas vezes associadas a uma topografia ondulada de topos concordantes (tabuleiros).
Relevo forte ondulado, Compreende as cristas baixas e os patamares nivelados do Pré-Cambriano, geralmente de direção SW-NE. Topografia formada por outeiros e morros (elevações de 100 a 200 m de altitude relativa) com declives fortes.
Relevo movimentado, Constituídodas cristas altas do Pré-Cambriano e encostas ou áreas dissectadas do planalto sul-baiano. Superfície de topografia vigorosa, com predominância de formas acidentadas, apresentando desnivelamentos relativamente grandes e declives fortes ou muito fortes.

nc.ou•dnu •• 40 •• • ....
-I!H4-
HABORAÇIO OO E•~uf?'rt:RRI!:IRA OA SILVA
J,.OCALIZAcÃO
~ IE:m Rol•"' ,..,.., 011<1•/G~o
li!II-R....,.o on~lllool<l
-Rolo"" fo<lo Mtl<l
liiDilD - "'o•!..,onlollo
O.oenhlolo corlo't<IJf•·~-~ Pf'OOLOG!A -- CEPI!:C-- CI!:PJ..AC
...
Figura 1 - Principais tipos de relevo da região cacaueiro baiana.
18
•
•
'
'
Em síntese, como é mostrada no Quadro 2, trata-se de uma região com mais de 42% de topografia acentuada, sem possibilidades, portanto, de utilização com máquinas agrícolas. Mais de 30% de suas áreas não apresentam quaisquer impedimentos (relevo suave, sem pedregosidade e boas condições físicas do solo) à mecanização e o restante, ligeiros impedimentos, sobretudo pela presença de pedras superficiais e/ou más condições físicas do solo.
ZONEAMENTO AGRÍCOLA
As condições edafo-climáticas e a topografia constituem os fatores mais importantes para a seleção dos cultivos de uma região. Os aspectos edafo-climáticos, chamados também de aspectos ecológi-
cos, tem relação com o uso agrícola da terra e com a vegetação natural (6), enquanto a topografia, aspecto agrícola, possui maior identidade com a agricultura em si.
O clima, sobretudo pela presença e duração de um período seco, o solo, pela sua fertilidade, textura e profundidade efetiva, e a topografia, pelas suas formas de relevo (altitude relativa, declividade e pedregosidade) permitem, através da integração de seus parâmetros, estabelecer zonas de distintas potencialidades agro-pastoris .
Isto posto, procedeu-se, em primeira aproximação, ao zoneamento agrícola da Região (Quadro 3 e Figura 2). Para tanto, foi ela dividida em oito macrozonas (fatores edafo-climáticos) e 23 áreas agrícolas (adição do 3? fator, a topografia), descritas a seguir.
Quadro 2 - Distribuição das classes de relevo na região cacaueira baiana .
Áreas (km) Possibilidade de mecanizaçilo
Fonnas de relevo agrícola
Praticamente plano 4.880 5,40 Ligeiros problemas (condições físicas dos solos).
Suave ondulado 27.800 30,79 Área ideal, sem quaisquer impedimentos.
' Ondulado 18.960 21,01 Problemas com pedregosidade
' em grande parte da área.
Forte ondulado 21.200 23,48 Sem possibilidades de mecanização.
Movimentado 17.440 19,31 Sem possibilid.ades de mec.ani~.açAo.
19

Clima tímido
(I) Solos fértei.s medi.anamPnte profundos. Correspondem aos Alfi-
sois, Ultisols mesotróficos, Dystropepts, Aluviais e Hidromórficos. São caracterizados pelo médio e alto índice de saturação de bases,
Quadro 3- Alternativas de utilização agro-pastoril e zoneamento agríc:ola da rr:gião cacaueira baiana.
C~ima
11
Úmido
lU
!V
v
Trensiçio
VI
• Fádes topogtá4co
I
2
3
4
5
6
7
8
9
to
11
12
13
14
15
16
17
18
20
Área (tm1
1.320
520
1.280
$.240
490
3.560
320
13.120
4.840
4.120
1.040
3.960
4.840
5.680
4.800
6.400
7.160
4.520
• PoaaibUidadn agrícolas
cacau (solos argilosos • bem drenadm~) e c:ul· tivos de subsistência.
Cacau e frutleulturfl,
Cà~u e pntagens.
Cacau e reflorestamento
Pastagen&_:(regitmt in-tensivo) e"cuttivotJ anuais.
Coco-da-Baia, caju e abacaxi.
Serin.gueiJ"a, dendê, cana-de-açúcar, pimen· t.-c:lo·teino, cravo-dalndia e cultivos anua.is..
Seringueira, dendê, pi· menta--do-reino, cravoda-índia
Pastagens
Refl(IJ'estarnento
Cultivos anuais
Fruticultura
Pastagem;
Pastagens e refloresta· mento
Cultivos anuais
Pastagens
Pastagens e reservas norestais {topos).
Continua
•
'
f
' .
vn
VIU
19
20
21
22
23
920
2.640
3.840
9.800
1.920
Palnl& rorrageira, tn.11·
mona, sisal, algodio perene e girassol.
Pastagens
Reserva florestal
Olericultura
Fruticultura subtropical, café e pastagens (regime de conCinamento).
I -Solos férteis de profundidade mediana II - Solos férteis rasos
V -Solos pobres VI -Solos férteis
VII -Solos férteis VIII - Solos pobres
III - Solos pobres e arenosos IV - Solos pobres argilosos
I. Praticamente plano 2. Ondulado 3. Forte ondulado 4. Movimentado 5. Ondulado 6. Praticamente plano 7. Forte ondulado 8. Suave ondulado 9. Ondulado
10. Forte ondulado 11. Movimentado 12. Sua v e ondulado
pH moderadamente ácido, boa capacidade de retenção de umidade e presença de minerais primários em quase todo o perfil.
Área de pouco mais de 6.000 km', com vocação agrícola para a cacauicultura em sua maior parte. Dado a ocorrência de manchas cuja reduzida profundidade ou deficiência de drenagem torna o solo limitante para a agricultura, as fazendas não possuem, de modo geral, toda a sua área ocupada com esse cultivo (2).
A diversificação agrícola das fazendas poderá possibilitar o to-
21
13. Ondulado 14. Forte ondulado 15. Movimentado 16. Ondulado 17. Forte ondulado 18. Movimentado 19. Sua v e ondulado 20. Forte ondulado 21. Movimentado 22. Suave ondulado 23. Ondulado
tal aproveitamento dessas áreas, com grandes benefícios sociais para a população rural. Entretanto, sendo esparsas e reduzidas as áreas disponíveis para essa prática, não se pode esperar uma diversificação com amplos reflexos na economia local. Objetivando-se uma diversificação de efeito principalmente social, poder-se-iam utilizar as áreas planas de baixadas com cultivos de subsistência, as onduladas com fruticultura e as demais com pastagens e reflorestamento.
(11) Solos férteis rasos. Terrenos de características quase idên-

""
'L/Mil ÚMIDO
(Jl SOLOS fÉRTEIS
OE PROFUNOIOAOE MEOIANA
I. Pro,comonto ~lano l'..Ondu>ado 3- F"''" or>dulado 4 Movimontodo
(ll)SOLOS HRTEIS RASOS
5 On4'""do
••'
"'
lllll SOLOS POBRES ARE,.OSOS '-'>r----G;;';I ,, 6.Prolioomonl< plano 7 fotto on<*Jiodo
[IV) SOLOS POBRES ARGILOSOS
8. Suo•• ondulado 9. Ondulado 12 F orlo ..,dulado II.M .. ImoMa<lo
CLIMA DE TRANSIÇ.lO
lVI SOLOS POBRES
12 Suo•• ondulado 13 Otlduiodo 14 Fone ondulodo 15. Mo•imoniMo
(VllSOLOS FERTEIS
16 O...dula4o 17 For lo ondulado 18 Movlmontodo
CliMA SECO (VII)SOLOS FERT~IS
19 Sua .. ondulado 20FO<to ..,dulodo 21.Movimentodo
(VJIUSOL.OS POBRES
22 Su••• ondulado 23 Ondulado
.........
T ..... -1 ,.
,,
,..
....•. ., . ELABORA~i,ÃO 00 [ftgllAtrt: L..U F"-LU-LL1l..o1.4. SiUG...
OoHo~loto cart<ÍQ..,Io:--wa..U:.o,P---M~
Figura 2- Zoneamento agrícola da região cacaueira baiana.
22
•
'
ticas às do grupo anterior, diferindo basicamente no tocante à profundidade efetiva, que não excede 50 em. Correspondem aos Eutropepts cuja pequena cubagem não fornece condições a um armazenamento de água capaz de satisfazer às exigências do cacaueiro.
Estes solos, que abrangem pequenas áreas e são de preço elevado, são excelentes para cultivos de pequeno porte. São recomendáveis para pastagens desde que explorados em regime intensivo. Podem igualmente ser utilizados com cultivos anuais de subsistência devido principalmente ao seu fácil acesso e proximidade às maiores cidades da Região (Itabuna, Ilhéus, etc.), onde o poder aquisitivo e população possibilitam preço compensador para os produtores. Área de 480 km1
•
(111) Solos pobres arenosos. Solos de textura excessivamente leve, tais como: Quartzipsaments e Tropaquods, cuja potencialidade agrícola se restringe a cultivos psamófilos como a piaçava (Attalea funifera Mart), coco-da-Bahia (Cocos nucifera L.), caju (Anacardium occidentale L.) e abacaxi (Ananas sativus Schult.). Área de 3.880 km'.
(IV) Solos pobres argilosos. Compreendem os Oxisols (Haplortoxs) e Ultisols (distróficos), cuja limitação principal de utilização agrícola é a baixa fertilidade, apesar de possuirem boas condições físicas. Trata-se de solos com saturação de bases inferior a 30%, alta saturação de alumínio, pH áci-
do e baixa reserva de minerais primários. Área de 23.080 km1 •
Possuem uma significativa área de topografia suave (13.000 km1).
que deve ser reservada para cultivos de seringueira (Hevea brasiliensis Muell Arg.), dendê (Elaeis guineensis Jacq.), cravo-da-Índia (Eugenia cariophyllata Thunb.), pimenta-do-reino (Piper nigrum L.), guaraná (Paullinia Cupana H.B.K.) e cana-de-açúcar (Saccharum officinarum L.). Muitos desses cultivos poderão se estender às áreas onduladas.
As pastagens e reflorestamento têm seu melhor aproveitamento nos solos de topografia forte ondulada e movimentada respectivamente.
Outros cultivos que poderão ser localizados também nos Oxisols de topografia suave são os de ciclo curto principalmente pela presença de estradas de escoamento, facilidade de mecanização e baixo preço das terras.
Clima de transição
(V) Solos pobres. Oxisols do grupo Haplustoxs, os quais, muitas vezes, apresentam o horizonte A (O - 30 em) com fertilidade mediana diferindo, portanto, dos Oxisols de clima úmido, cuja pobreza química é praticamente homogênea em todo o perfil. Área de 19.280 km'.
Desde que convenientemente manejados, podem ser utilizados com cultivos anuais, especialmen-
23

te nas áreas de topografia suave, fruticultura nos relevos ondulados e pastagem nas demais áreas, podendo ser associadas com reflorestamento.
(VI) Solos férteis, Pertencem a este grupo, principalmente, os Argiustolls e Haplustalfs, que se caracterizam pela alta saturação de bases, argila de atividade elevada, pH praticamente neutro, minerais primários em todo o perfil e profundidade efetiva entre 50-150cm.
Possuem problemas de deficiência de água (clima e profundidade do solo) e, em grande parte da área, impedimentos ao uso de maquinaria agrícola (topografia e/ou pedregosidade). Constituem-se nas melhores áreas de pastagem daRegião, sobretudo pela riqueza de seus solos. Área de 18.080 km'.
Possivelmente, o mais racional seria a utilizaçãodestes solos com pastagens nas áreas de relevo acentuado e cultivos anuais nas zonas intermontanas não pedregosas de relevo ondulado, aproveitandose as boas condições edafo-climáticas.
Clima seco
(VII) Solos férteis. Região semiárida, altitude em torno de 450 m, relevo variável e fertilidade alta, algumas vezes com problemas de salinidade (zonas abaciadas). Área de 7.400 km 2
•
Pelas condições climáticas adversas e pouco desenvolvimento do
24
solo são terrenos de baixa potencialidade agrícola, a não ser para cultivos adaptados, tais como a palma forrageira (Opuntia spp.), mamona (Ricinus communis L.). sisal (Aga!Je rigida Mill) e girassol (Helianthus annus L.), que devem ser reservados para as áreas de re
levo suave. Também podem ser utilizadas com pastagens as zonas de topografia forte ondulada, deixando as áreas movimentadas para reservas florestais.
(VIII) Solos pobres. Oxisols de altitude (relevo de altiplano acima de 750 m), dotados de excelentes condições físicas e baixa fertilidade natural. Área de 11.720 km'.
As áreas mais planas prestamse perfeitamente para a mecani:.;ação agrícola. As de posição abaciada e altitude acima de 900 m oferecem melhores condições para a exploração agrícola. Dado sua riqueza em matéria orgânica e condições desfavoráveis ao desenvolvimento de muitas enfermidades, devem ser reservados à olericultura. Além destas vantagens, a ligação dessas áreas com os centros consumidores, por estradas asfaltadas tornam-nas por demais promissoras às atividades aí desenvolvidas. Já as áreas com relevo ondulado, poderão ser aproveitadas com cafeicultura, fruticultura subtropical e pastagens (regime de confinamento).
OISCIJSS.o\.0 E CONCLUSÕES
Como se depreende, a região cacaueira baiana possui excelen-
•
'
tes condições agrícolas e suas peculiaridades permitem uma diversificação agro-pecuária bastante ampla, graças à variedade de seus solos, clima e topografia.
Economicamente, a área mais importante é a zuna cacaueira propriamente dita {solos férteis, medianamente profundos, clima úmido), caracterizada pela predominância de relevo acentuado e/ou pedregoso, cuja opção mais racional de utilizacão agrícola é o cacaueiro, pelas suas exigências e qualidades de planta preservadora das condições ambientais. É evidente a necessidade de uma cacauicultura implantada com material genético mais produtivo e conduzida com técnicas modernas. Simultaneamente, para aproveitamento de áreas impróprias para a cacauicultura, pode ser conduzida uma diversificação ao nível de propr;edade com finalidades predominantemente sociais.
Outra zona de importância para ~ Região é a pastoril {solos férteis, clima de transição), cujos so-
los férteis estão sendo subutilizados. Além das pastagens, que devem ser melhor manejadas, há condições de obtençao de altas produções em cultivos anuais, nas áreas de topografia ondulada (não pedregosa), cuja atividade agrícola deve ser considerada, haja vista a disponibiiidude de áreas e a carência de plantas alimentícias na Região.
De alta potencialidade agrícola, por outro lado, é a zona dos tabuleiros sul-baianos (solos pobres), cuja topografia e condições físicas permitem uma tecnologia agrícola avançada (insumos modernos e mecanização agrícola), com possibibilidades de aproveitamento com cultivos perenes dos trópicos úmidos, para a qual deve s~r dirigido um plano objetivo de diversificação.
Em síntese, trata-se de uma região com excelentes perspectivas de aproveitamento agrícola, cujas nuanças de solos, fatores climáticos e topografia, aliados às boas condições de infra-estrutura, permitem uma diversificação técnica e economicamente racional.
i\GRAOECDIENTOS
Aos Eng.-Agr'?s Kozen Igue e Paulo Frota, pelas sugestões na elaboração do texto.
LITl-:R.o\TLJR,\ CITADA
1. ÁLVARES-AFONSO, F.M. Características do sistema da produção: cacau, seringa, coco, dendê, piaçava, pecuária de corte, alimentícios. In Seminário de Desenvolvimento Regional Integra-
25

do, Itabuna, Bahia, Brasil, 8 a 13 de junho, 1970. Introdução à região cacaueira, Itabuna, Centro de Pesquisas do Cacau. v. 2. pp. 4-63
2. ALENCAR, M.H. Aspectos da concentração da produção de cacau e da estrutura fundiária na região cacaueira do Estado da Bahia. Itabuna, Brasil. Centro de Pesquisas do Cacau. Comunicação Técnica n~ 3 7. 1970. 27 p.
3. BARROCO, H.E. A importância econômica da reg1ao cacaueua para a Bahia e para o Brasil. A economia cacaueira: produção e tendência. Út Seminário de Desenvolvimento Regional Integrado, ltabuna, Bahia, Brasil, 8 a 13 de junho, 1970. Introdução à região cacaueira. ltabuna, Centro de Pesquisas do Cacau. V· 3. pp.4-11.
4. DOMINGUES, A.J .P. e KELLER, E.C. de S. Bahia. Rio de Janeiro, Conselho Nacional de Geografia, 1958. 310 p.
S. FROTA, P.C.E. Notas sobre o clima da região cacaueira baiana. Cacau Atualidades (Brasil) 9 (2): 17-24. 1972.
6. JAN BEEK, K. et al. Interpretação preliminar da carta de solos da região cacaueira. In Congresso Brasileiro de Ciência do Solo, 1~. Piracicaba, São Paulo, Brasil, 19 a 30 de julho, 1%5.
7. LEÃO, A.C. e SILVA, L.F. da. Fotointerpretação aplica~a a estudos de solos na região cacaueira baiana. In Congreft,.C! -Brasileiro de Cartografia, 6~. Rio de Janeiro, 15 a 22 de ju~, 1973.
8. SANTOS, R.F .A. dos. Chuvas na Bahia, máximas e mínimas. Salvador, Ministério da Viação e Obras Públicas, 1962. s.p.
9. SILVA, L.F. da. Possibilidades de aproveitamento agrícola dos solos da região cacaueira baiana. Cacau Atualidades (Brasil) 9 (4), 10-14. 1973.
10. et ai. Solos dos vales dos rios Pardo e Jequitinhonha da Bahia. ltabuna, Brasil. Centro de Pesquisas do Cacau. Comunicação Técnica n~ 16. 1%8. 30 p.
11. et al. Solos da região cacaueira baiana. Itabuna, Brasil, Centro de Pesquisas do Cacau, 276 p. (Datilografado).
26
>.
.. ,
RESUMO
Com base em fatores climáticos, pedológicos e topográficos, considerados fundamentais na seleção de cultivos, procedeu-se o zoneamento agrícola da região cacaueira baiana. Para tanto foi ela dividida em oito macro-zonas e 23 áreas agrícolas, as quais são descritas no presente trabalho e dadas suas mais viáveis possibilidades de aproveitamento agropastoril.
Dentre as zonas delimitadas, três são enfatizadas pela sua importância econômica e/ou geográfica: A. zona cacaueira propriamente dita, para a qual se recomenda uma cacauicultura melhor manejada e aproveitamento das áreas vizinhas com cultivos de subsistência a nível de fazenda; B. zona pastoril, de solos férteis subutilizados, para a qual recomenda-se aproveitar as áreas onduladas (não pedregosas) com cultivos anuais e ra· cionalizar o manejo das pastagens; e C. zona dos tabuleiros úmidos sulbaianos, considerada de alta potencialidade agrícola, com possibilidades de aproveitamento com cultivos perenes dos trópicos úmidos, tendo em vista as boas condições físicas (externas e internas) dos seus solos.
Finalmente, conclui-se que a Região tem excelentes perspectivas de aproveitamento agrícola, dado suas boas condições de infra-estrutura e variedade edafo-climática.
AGRI~.l(.RAL ZONING OF THE BAHIAN COCOA REGION
SUMMARY
From the various climatic, pedologic and topographic data available, the Bahian cocoa region has been divided up into eight micro-regions and 23 agricultura! areas. Each has been outlined giving the most favorable methods for their agricultura! utilization.
Of the different zones three are given special attention due to their economic and/or geographical importance: (i) The present cocoa zone for which a better management for the actual cocoa is recommended together with the use of neighboring areas for subsistence crops to be grown at the farm size levei; (ii) The grassland zone with its underutilized fertile soils, for which annual crops are recommended especially on the stone free undulating areas together with a better management of the existing pastures; (Ui) The Southern Bahian humid lowland plateau zone, which is considered to have great agricultura! potential in particular f't,b~N~ tropical tree
"~"t. C.-fo e ~'\~ - ~P. 27 vç ., !BLIOTECA 0•· c/ D ~ ... ,.., . . "

cwps due to the excellent physical charucteristics (both internai and exlt'rna!) of its soils.
In summury it is concluded that the overali region has good possibilities for the future as regards agricultura! utilization, given its advanced infrastructure and good edapho-climatic conditions.
28
CUATRO ESPECIES DE TRIPES EN EL CACA O
DE TABASCO, MEXICO *
Roberto M. ]ohansen Naime**
E! propósito dei presente trabajo es informar sobre los trips (Thysanoptera) que habitao en las plantas de cacao dei Estado de Tabasco, México. Muchas veces se considera y se generaliza en México que, cuando hay trips en el cacao, son necesariamente de la especie Selenolhrips rubrodnc:tus {Giard), dando lugar a errares, con esas afirmaciones. Las especies aqui consideradas pertenecen al suborden Terebrantia y son: Selenothrips ruhrocinctus (G i a rd), Prankliniella parvula Hood, Sericolhrips im)ersus Hood y /.eucothrips piercei V<li. theohromae Priesner.
La primera especie fue noticiada y estudiada como plaga dei cacao en México, aproximadamente desde 1952 (3, 4, 5, 7, 11). Actualmente su presencia ha sido com· probada fuera dei cacao, en el almendro (J'erminalia catappa) (8, 11, 12).
F. parvula, es una plaga común en e! plátano (Musa paradisíaca) donde lleva a cabo su ciclo vital, pero ha sido observada también causando manchas necróticas fo· liares en el cacao. Existen antecedentes de su presencia en el follaje dei cacao en Panamá e Indias Occidentales (10) y como polinizador en Colombia (1).
El S. int)ersus ha sido descrito como una especie frecuente en pastos (9). No hay datas que permitan tener ante.cedentes de la especie en México y menos en plantas de cacao.
Por lo que respecta a L. piercei, existe el antecedente de haber sido colectada en follaje joven de cacao en Surinám (9) y no hay mención sobre su presencia en México. Moulton (9) menciona que la especie l.eucothrips piercei Morgan, sí existe en México, pero no en la planta de cacao.
* Recibido para publicación en marzo de 1972. ** Hiólogo, Laboratorio de Entomologia, Colegio Superior de Agricultura
Tropical, Secretaría de Agricultura y Ganadería. Cárdenas, Tabasco, México.
R<'vista Theobroma. (fj'fl~. 1/hé,.s. Brasil, 4!1): 29-38. Jan.-mar. 1974.
29

MATERIALES Y METODOS
Las colectas fueron realizadas en las plantas de alimentación, mediante un pincel humedecido en una solución de alcohol 96°, ácido acético al 5% y agua, en la proporción 95:3:2, conservando los ejemplares en frascos, con la misma solución. Después fueron deshidratados con alcoholes de 70; 80; 90; 96 e 100°, aclarados con xilol y m~ntados entre porta y cubreobjetos con bálsamo dei Canadá, para su identificación. Las observaciones fueron hechas con microscopio compuesto de campo claro a menor aumento (60x) y mayor aumento (100x). Las microfotografías fueron hechas con película de grano fino, usándose además filtros azul, verde claro y amarillo para contrastar. Todos los ejemplares quedaron depositados en la colección dei autor.
RESULTADOS Y DISCUSION
Descripción de las especies
S. rubrocinctus
Los estados inmaduros, larva, prepupa y pupa (Figuras 1, 2, 3, 4 y 5) son de color amarillo transparente en algunos ejemplares, contrastando con anaranjado y además, con un característico cinturón de pigmento rojo, generalmente en los tres primeros segmentos abdominales. Las larvas están desprovistas de rudimentos alares y tienen en el extremo abdominal una característica corona de cer-
30
I
Figura 1 - Selenotrips rubrocinctus. Larva de primera fase, en vista verltral. 32x. Filtro azul.
das largas, negras; son muy activas y se les observa generalmente con el abdomen encorvado hacia arriba, transportando una esfera de líquido excremental de color obscuro. Abundao sobre todo cerca de las nervaduras central y secundarias de la hoja. La prepupa se caracteriza por la aparición de los rudimentos alares pequenos y transparentes, y las antenas dirigidas bacia adelante. La pupa se distingue dei estado anterior, por el mayor desarrollo de los sacos alares, la posición de las antenas dirigidas sobre el dorso y además, cerca de su madurez se pueden
Figura 2 - Selenotrips rubrocinctus. Larva en segunda fase en vista ventral. 32x. Filtro verde.
distinguir pupas masculinas y femeninas, por la aparición de los genitales externos en ambos casos. Los dos estados son virtualmente inmóviles y reposan en grupos generalmente cerca de los márgenes externos de las hojas.
Adulto
Hembra, macróptera. L a r g o 0,96-1,54 mm (Figura 6). De color café ambarino en algunos ejemplares especialmente los jóvenes; y en muchos ejemplares, esta coloración es casi negra especialmente en el abdomen, pero los úl-
31
Figura 3 - Selenotrips rubrocinctus. Prepupa en uista dorsal. 32x. Piltro verde }' azul.
timos segmentos abdominales son siempre más claros; la cabeza y el tórax son en general de un tono más claro que el abdomen; las coxas y fémures, café, igual ai tórax; tíbias i gua I mente cafés per o ligeramente amarillentas en los extremos distales; tarsos de coloración amarilla muv clara. Alas anteriores de color uniformemente café, con pubescencia. Alas posteriores con la mitad externa salpicada de café, la mitad basal de un tono más claro. La superfície ex
terna de) cuerpo presenta una ornamentación a base de surcos y crestas que formao áreas poligo-

Figura 4 - Selenotrips rubrocinctus. Pupa masc-ulina e11 z•ista dorsal. J.?x. Piltro azul. (,\'rjte~e
la plac-a gl'nital f'll e/ e>.lrt>IIIO de/ ahdomen ),
nales. J~s cua les dan un as pecto caracteris t i co de mallu : las antenas con el segmento I v 11, café, JII claro tru.,slúcido, hgeramente s alpicado con café, grtsáceo en la mitad distai : TV tgual ui antertor ; V mttad basul translúcida y obscurecido con café en la distai; VI café; VII y Vlll café grisáceo claro.
Macho, macróptero. Lurgo 1,31 mm. (Figuro 7). En general con la misma colornci6n que la hembra, pero de cucrpo mucho más delgado Las ah•s son relativumcnte más
32
Figura 5 Se lenotrí ps rubro-cinctus. Pufm lt menma en ris ta 1 entra/. 3.:?\ . l'iltm azul ) 1 Prde. (,\ 'ritese e/ 01 r,poçitor).
cortas; el abdomen achatado en el extremo y con tres pares de espinas cortas lanccolr.das en el dorso del novena segmento <tbdominal.
Localidad: Cárdenas y La Chonlolpa, en el Estado de Tabasco, y Ros11no·lzapa, en el Estado de Chiapas.
Hospcderas: cncao r almC'ndro.
1 ~. pan ufa
LüS lan·as jóvenes dcl pnmer eswdro son de calor blanquecino
'.
Figura 6 - Selenotrips rubrocinctus. t\dulto bembra t 11 l"Ísta
dorsal. 32x. Filtro azul.
transparente mientras que las del segundo estadia son de calor amari llento que en algunos casos es más fuerte (Figura 8). Son muy activas en las inflorescenctas del plátano donde abundao y causao manchas necróticas de color purduzco.
De los estados prepupa y pupa, no tenemos ejemplares, pero de acuerdo com De Leon (2) son estados de reposo en la tierra adyacente a la planta de alimentación.
Adulto
Hembra, macróptera. Lurgo 1,1-1,3 mm (Figura 9). De colar café
33
Figura 7 - Se!enotrips rubro· cinctus. trlulto mac-ho 1!11 riS/a nmlral. Jl.\. Filtro azul.
cla~o; abdomen con los segmentos apicales de un tono más obscuro; fêmures cafés, más claros apicalmente; tíbias y tarsos amaríllo claro. Las alas anteriores de calor café v las posteriores claras; antenas con los segmentos l-li y IV-VUT de calor café, el 11 es el más obscuro; lll amarillo grisáceo; IV-V más claros basalmente. La cabeza y protórax, con algunas estrías trnnsversa!es en su superCicie dorsal.
Macho, macróplero (Figura 10). De color amanllo claro, con una mancha parduzca en med1o de los terguitos 2-8. Alas anteuores ela-

Figura 8 - Frank liniella parvula. Lan·a en s<•gunda fase cn 1 ista dorsal. J2x. Fdtm amarillo.
ras; antenas cnsi blancas; los segmentos IV-V obscurecidos, con gris ap1calmente; VI-VIII grises.
Local1dad: Cárdenas v La Chontalpa, en el Estado d~ Tabasco: X1co, Texolo, Coatepcc \' jalapa, en el Estado de Veracruz.
Hospeder.1s: huevecillo v estadios l~trV<Jrios cn lus inflorescencias, tanto cn pie, como las tiradas en el suclo: los ndultos, tanto en inflorcsccncias como cn frutos de plátuno. En follaje llerno de cacao, fldultos solnmente.
34
Figura 9 - Frankliniella parvula. Adulto hembra en l'ista dorsal. 19x. Filtro azul ) amarillo.
S. im ersus
Hembra, macróptera (Figura 11). De color uniformemente amarillo claro, algo ambarino a los lados, extremo abdominal, tórax \' cono bucal; con algunas manchas cafés en el protórax, dos en el metascutum y una a cada lado de los terguitos abdominales 11-VIl. Las antenas con los segmentos 1-llf gn ses; lV-Vlll café negruzco. Al~s anteriores blanco transparente, con las venas de color anaraniado claro. La cabeza y et tórax· con arrugas transversales conspícuas. Las patas y abdomen presentan
Figura 10 - Frankliniella parvula. Adullo macl>o 1!11 tisla dorsal, J.h. f-'illro terde.
pubescenc1a, especialmente a ambos lados de este último. Ojos café obscuro; ocelos con el crecíente de color anaranjado rojizo.
Macho, macróptero (Figura 12). Virtualmente semejante a la hembra en coloración pero de menor tamaiio v más delgado. Destaca la fuerte coloracíón anaranjada de los testículos.
Localidad· Cárdenas, en el Estado de T~basco.
Hospederas: follaje tierno de cacao.
Figura 11 - Sericothrips inversus. ,\dullo l>embra en 1 isla dor· sal. 19x. Pi/lro 1 erde v amard/().
L~ piercei
Hembra, macróptera (Figura 13 ). Largo 0,49-0,58 mm. De color blanquecino amarillento en general, pero el tórax, cono bucal y O\'ipositor de un tono más fuene. Alas anteriores blanco transparente, con una conspícua mancha de color rejo vivo entre las bases de las antenas. Ojos negruzcos y los oce los con el crec1ente de color rojo \'ÍVO.
La superficie de! cuerpo lisa.
Localidad· Cárdenas, en el Estado de Tabasco.
Hospederas: follaje tierno de caca o.
35

Fi~ura 12 - Sericothrips inversus . ,\du/tr, marl•n ,.,, I 'Í~Ia dorsttl. J.h 1:iltro a::ul.
C 0'\Cl l <,lO' I .._
De las obsen:aciones realizad.ts se puede hacer el siguiente <.ln<~llsts por espccte:
ruhrl)cÍintu~. Las les10nes nc:-cróttcas que produce en los tejidos foliares v cn el fruto, son causadas pnncip.1lmente por los cstadtos larvari<>S. Los estados de prepupa y pupa son de rcposo. Los adultos caus.1n t;~mbtén daõos de consideración, siendo muy ucltvos en el follajc, llores v frutos. El ciclo de vida completo se lleva a cabo en la mtsma pl<~nt:.~ de ali-
36
Figura 13 - Leucothrips ptercei. t ar theobromae. ,\clultn l>«·mhra <'11 1 isla 1 entrai, J2,, I tltr() 1 c>rdf!.
mentación, tanto en c I cr.cao corno en el almendro. Es probable que el almendro sea una hospedem importante, sobre todo St el cacao est<t cercano.
f'. pan ufa. Esta especte ha estado b«jo estudio cn Tabasco para conocer su ciclo d(~ \'Jda (2) v la forma de combut irl;1 químicamente (6). Se sabe que Jus larvas se alimentao y des,trrullan en las flores dei plátano donde provocan lesiones como manchas parduzcas. lõ:t prepupa y la pupa pasan este ciclo en el suelo ;1dvacente a lu
planta de alímentación v los adull<·'i pululan en las flores y frutos. En la localidad de Tabasco, un alto porcentaje de las plantacíones de plátano están infestudas y
extste la circunstancia de que algunos cacaoteros son sombreados con estas plantas, de tal manera que esto podría explicar. a prime-
ra VtSta, la presencia de los adultos en el c&cao.
Por lo que respecta a 5. inver~IH y 1 •. piercei, no tenemos aún datos sobre sus ctclos de vida y podemos mencionar que ambas especies causan manchas necróticas foliares en el cacao, sin mayor importancta económka.
L I TERATURA CITADA
1. DE FRANCO, T.H. Catálogo provisional de los insectos encontrados en el cacaolero, en Colombia. In Conferencia Interamericana de Cacoo, 7a, Palm tra, Colombia, 1958. Palmira, Colombia, Ministerio de Agricultura, ,1958. pp. 231-239.
2. DE LEÓN, ] . Comunicaciones personales acerca dei ciclo de vida de FrallkliniPIItt tJart ula, Hood. Cárdenas, México, Coleg10 Superior de Agricultura Tropical, Secretaria de Agucultura y
Gaoadería. 1972.
3. EL CACAOTERO, cultivo y plagas. México, D.F. Secretaria de Agricultura ~ Ganadería, Dirección General de Agncultura en cooperación con Du-Pont, México. pp. 33-38. 1952.
4. EL CULTIVO del cacao, insectos dei cacao. México, D.F. Secretaria de Agrtculturã y Ganadería, Dirección General de Agucultura. pp. 62-70. 1953.
S. HECHT, O. Nota acerca de s~/~notJ.rips ruhrocinctus (Giard), plaga del cacaotero. Fitófilo (México) 6(5):33-42. 1952.
6. !BARRA, G.E., GARCIA, R. y F'AVELLA, A. Combate de trips de! fruto dei plátano con tratamtcntos de insecticida y bolsas de poltetilcno. VH Congreso Nacional de Entomología. Folia Entomológica de México n~ 23-24:64-65. 1972.
7. KNOKE, ] .K. lnsectos que atacan ai cacao en América v su com-bate. C',cuo (Costa Rica) 10(2)·1-7. 1965. ·
8. MEDINA G., S. The Thysanopteru o f Puerto Rico. UnJverstly o f Puerto Rtco, Agrtcullural Expenment Station, Technical Paper 32. 159 p. 1%1
9. MOULTON, D. The Th'" ... noptera of South America 11. Re\ista de Entomologio (P .... sil) 3(1):99-106. 1933.
37

10. MOULTON, O. The genus Frankliniella Karny,with keys for the determination of species (Thysanoptera). Revista de Entomologia (Brasil) 19(1-2): 55-114. 1948.
11. SILVA, P. Tripes do cacaueiro - causador do queima da folha e da ferrugem do fruto. Cacau Atualidades (Brasil) 1(9 -10): 1-4. 1964.
12. WOLCOTT, G.N. The insects of Puerto Rico. Thysanoptera. journal of Agriculture of University o{ Puerto Rico 32(1): 94-102. 1948.
RESUMEN
Frecuentemente se afirma que solamente el Selenothrips rubrocinctus (Giard) es el responsable por los daíios causados en los cacaotales de México, Sin embargo, se encontrá que los thrips Frankliniella parvula Hood, Sericothrips inversus Hood y Leucothrips piercei var. theobromae Priesner son también plagas dei cacaotero. Por la primera vez estas tres especies son noticiadas en cacaoteros de México. Los hábitos, ciclos de vida y distribución de las cuatro especies de thrips, son mencionadas en este estudio.
FOUR SPECIES OF THRIPS ON COCOA IN TABASCO, MEXICO
SUMMAR\'
lt is often said that only the thrips Selenothrips rubrocinrtus (Giard) is responsible for damage in the cocoa plantations of Mexico, However, it was found that the thrips Frankliniella parvula Hood, Sericothrip:-; inversu:-; Hood and Leucothrips piercei var. lheobromae Priesner are also pests of cocoa. This is the first time that these three species have been reported on cocoa in Mexico. Habits, life cycles and distribution of all the four thrips species are mentioned.
' 38
NEMATODES OF THE COCOA REGION OF BAHIA, BRAZIL
[[[ - PLANT PARASITIC AND FREE-LIVING NEMATODES IN THE RHIZOSPHERES OF SIX DIFFERENT PLANT SPECIES•
Ravi Datt Sharma** Pieter Aart Albertus Loof***
The occurrence and distribution of nematodes from the rhizospheres of different crops in the cocoa region of Bahia has already been widely reported (1, 2, 3, 4, 5, 6, 7). This paper presents information on the plant parasitic and free-living nematodes from six different plant species hitherto nnsampled for nematodes.
\IATJ.:RIALS ASD METHODS
As a part of general nematode survey started in May, 1971, 13 soil and root samples were taken from the following plant species: 1'heobroma grandiflora, T. bicolor, Erythrina glauca, (;/adiolus sp., Momordíca charantia and Psidium gua;al!a. Two samples were taken each from E, glauca, Gladiolus sp. and T. grandiflora, one from M. charantia and T. bicolor and five
from P. gua;al!a. The samples from T. grandiflora and T. bicolor were collected at CEPLAC's Junior School of Agriculture (EMARC), Uruçuca, the remainder being sampled at the Cocoa Research Center (CEPEC) in Ilhéus. The methods involved in the extraction of the nematodes, including their further permanent mounting, follow those previously described by Sharma and Loof (1).
RESUL TS ANO DISCUSSION
The plant parasitic and freeliving nematodes found in the different samples are listed in Tables 1 and 2 respectively.
Of all the plant parasitic nematodes Helicotvlenchus dihys· lera was the dominant species. Except for 11. californicus, Macro·
* Received for publication in August, 1973. ** D.Sc, (Agric), Division of Entomology, CEPEC.
*** Dr .. Department of Nematology, Binnenhaven·lS, Agticultural University, Wageningen, The Netherlands.
Revista Theobroma. CEPEC, Ilhéus, Brasil, 4 (l): 19-43. ]an .. mar. 1974 ·
39

Table 1 - Frequency of occurrence of planl parasilic nematodes associated with six different plant species in 13 samples in lhe cocoa region of Bahia.
Frequency of occunence t'Jematodes
Plant species ,., of samples .... 1Jasiri11 sp. l>o/ichodor11s sp, 1/e/icoty/encbus sp.
E, F o A
H. ctlliforniciiS H. áihysltra Hewticyc/iophora cf, /oofi \4acroposthoflia cf. or~tala M. onottnsis
c A,B,C,O,E,F c
3 2 1 l
11 I 1 4 1 3 1 2 1 1 6 6 3 1 s
c A,B
'tftloidog) ne sp. D .\f. incognita .\f. lha me si
C, O, F D
Paratri< hodor11s cbristiei Peltam;gratus sp.
C, O F
P. ho/de,aui Rol) /enchullts rf'uifor•is X íphinnna sp. X. bret icfJ/1,•
c A,D,E C, O, E A,B
X. ifacoiMIII X . I.,,,,,. o
A, C
A. Theobroma grandiflora; B. Theobroma bicolor: C. Erythrina glauca; D. Gladiolus sp., E. Mormordica charautia: e F. Psidium guajava.
posthonia cf. ornata and Xipl>inema brel'icolle, ali the plant parasitic nernatodes listed in Table 1 have already been reported from a wide range of plant species in Bahia (1, 2, 3, 4, 5, 6, 7). Out of 46 genera and 74 species of freeliving nematodes found in this survey, 34 genera and 44 species have already been recorded previouslv from Bahia (1, 2).
40
Bearing in mind the large number of nematodes found it is recommended that a systematic nernatode survey of ali the important crops of the cocoa region of Bahia be undertaken to determine their role as plant parasite both under laboratory and fie ld conditions. Such a study may reveal many unrecorded nematode species of economic importance.
Table 2 - Frequency of occurrem e offree·lit i11g nematotles associatedwith six different fJianf species ir1 lJ samples in the cocoa region o/ Bahia.
Nematodes
Achrot~~llllora sp. AcroiH/es sp. Aciitroltli,,s sp. Al•i•.s sp. A•l'ltitlelt~s sp. A. Ctlrulitl•s Á. tlol•ch•r•s A•~itiMyl•••as sp Apbeletrch•s IIVe•tle
porcd•i•.lhls sp. • ohiMSÍCti.JIIIIIS . cf, ptlrtlnlllroCBCII$ IITollu•oU/u sp. xcmcb••• sp. . ••pllcolle tiS rroty/e /JIIIS COTQIItlllls aslatritl sp. elontlir• sp. • cblvlctiJUitlllls • cf. fOIIIui • ten111tle•s eiontl~tellll leru
Carchmolllimus for,os•s C, piz11i
bitwootlr11s tr11nsr t~~~lensis obhonch11s sp. ryploncb•s tlbno,.is r pios Ct1J11•r coroul11s ipthero/'bortl sp. orylaitul111s sp. • clavat•s • cf. spical•s utlorylai,•s sp. • ettersbergensis • grtln111ijerus
•• ,.inutus eJerocephlllobus sp.
11. JongrcaMáatus • pulchrr
rouus lorzgtcaudatlls
Frequency of occurrence
Plant species
F o c A,B,C,O,E,F A, F E D o D.E,F O,F A,C,D c c B A, O c o A,B,F A,B A A c F O, F c C, O, F c D,F F C, O, F B F A,B,C,O,E,F F A,C,O,E,F C, O c C,D,F o F, E
41
N? of samples
1 2 6 6 2 1 1 1 4 4 a 1 1 1 2 2 2 5 3 1 1 1 1 3 1 s 1 2 1 5 1 1
10 1 9 3 1 4 2 2
Continued

Tahle ? - Continued
lotoncbus sp. I. c f. Ir i c l>ur145 Leptatlchus sp. l.. gramtlos rts
,\trsodo')·la(•us sp. \I. parasllhtilis .'tlt•r.rJrhabditis sp. ,\1, paucipapillala M iconchus sp. MCJnhp;tera sp. ,'tly/rmchulus sp. ,\1: index :\1. mírwr .''h golaimus sp. Oif!olaimelltJ sp. O.Yidirus sp. O. tropicus Poliouchulrts sp. Prirmchu/us rnuscorum PrismauAoimus inlermedius ProdrHvlaimus sp. PrtJII' ptonr hus sp. Psli<•11chu.s sp. P. cC. hilar~llus RhabdUis sp. .\pomnchulu.<; denlalus Thomenema sp. r. catalc:anlii '/. mauritianum '1. lririosum '/ rischisloma arenicola T) lencJm/ai,rllus sp. 'f. cf. "'agnide11s 1')·/uchus sp.
A, C 8 A, C D C, E, F D,F F D A, E, F D A,D,F F C, O D,F A A,B,C,F A F A A,D A,B,C,D,F B,C A E D B,D A, F A,C,D,E,F F, O F D D,F A,D.F D
3 1 3 2 3 3 1 1 3 1 5 1 2 2 1 5 1 1 1 2 7 2 1 1 1 2 2
10 2 1 2 3 ~ 1
1\. Theobroma gra,Jdif/ora; B. Theobroma hicolor; C. Erythrina glauca; D. (./adinlus sp.; E. Momordica charanlia; e F. Psidium guajava.
LITERATURE CITED
1. SHARMA, R.D. and LOOF, P.A.A. Nematodes associated with different plants at the Centro de Pesquisas do Cacau, Bahia. Revista Theobroma (Brasil) 2(4):38-43. 1972.
2. and Nematodes of the Cocoa Region of Bahia, Brazil. I - Plant -parasitic and frec-living nematodes associated
42
with rubber (1/<'I'C?O hrrl\ilieus is Muell.-Arg). Revi~tn Thcobrom<t (Brasil) 3(1):36-41. 1973.
3. and SHER, S./\. Nematodes associated with banunu in Ra-hia, Brazil. Ciêncic1 e Cultura (Brastl) 25(7):665-668. 1973.
4. and . Nematodes associated with c1trus 1n 8<th1a, Braz i L Ciência e Cultura (Brasil) 25(7):668-672. 1973.
S. and Nematodes of lhe Cocoa Region of Bahia, Braz i!. li - Occurrence and distribution of plant paras li ic nematodes associated with cocoa ( Theohroma cacao L.). Revista Theobroma (Brasil) 3(3):17- 24. 1973.
SUMMARY
Ten genera and 19 species of plant parasitic and 46 genera and 74 species of free-living nematodes were found in 13 samples collected from the rhizospheres of six different plant species: Tb(!ohmma J.!'OIIdt flora. T. bicolor, Er\1/lrina Rlauca. \lomordica cbora111ia and Pstdimn f!llajar·a. The most common plant parasitic species encountred in the samples was H elico/y/enchus dihrs ter a. The nematodes Rol\ lellcbulus rl!ntformis, Xipbinema sp., X. t•tdgare, Do/ichodorus sp., and t\facropostho1lia mzoe11sis were found in 30-50% of the samples.
NEMATÓDIOS DA REGIÃO CACAUEIRA DA BAHIA, BRASIL. 111 - NEMATÓDIOS PARASITOS E NÃO PARASITOS
NA RIZOSFERA DE SEIS DIFERENTES ESPÉCIES DE PLANTAS
R E S U ~tO
Em 13 amostras de solo coletadas na rizosfera de 1'beobroma grandiflora, T. bicolor, Erythrina glauca, Gladiolus sp., ,\fomordica charantia e Psidium guajava, foram encontrados 10 gêneros e 19 espécies de nematódios parasitos e 46 gêneros e 74 espécies de nematódios não parasitos (de vida livre). A espécie mais freqüente de parasitos de plantas encontrado nas amostras foi lfelicotylenchus dihystera. Rotv· [(!nchulus reniformis, Xiphin(!mf1 sp., X. t•uLKare, Dolichodoms sp. e ,\facroposthonia onoeTISis foram encontrados em 30-50% das amostras.
43

SEGUNDA REUNIÃO REGIONAL AMERICANA DO GRUPO DE Phytophthora palmivora
Guayaquil, Equador, 8 a 23 de junho de 1973
RESUMO DOS TRABALHOS APRESENTADOS
Foram apresentados oito trabalhos na seção destinada ao Phytophra palmivora, com discussões sobre podridão parda e cancro.
1. Uma nova espécie de Phytophthora, denominado P. megasperma, foi registrada em cacaueiro, na Venezuela, Este organismo causa uma doença denominada Mancha de Agua ou podridão aquosa e é encontrado numa região daquele país, A inoculação dos ramos, folhas e frutos ocasionou reprodução dos sintomas da enfermidade em vários cultivares. O fungo é bem diferente do P. palmivora, sendo homotálico, Com a aplicação de Cupravit, foi obtido algum controle em frutos afetados pela podridão. (Lilian C. de Reyes, Estación Experimental de Caucagua, Venezuela).
2. Também na Venezuela, quatro espécies do gênero Phytophthora foram registrados com isolados dos frutos do cacaueiro: P. palmivora, P. megasperma, P. parasitica e P. syringae. A inoculação de sementes, ramos e frutos ocasionou reprodução de sintomas com todas as espécies, A sua agressividade, entretanto, varia de espécie para espécie. (Lilian C. de Reyes e llumberto Reyes, Estación Experimental de Caucagua, Venezuela).
3. Estudos efetuados na Jamaica demonstraram que os frutos podem ser contaminados pelo Phytophthora proveniente de um cancro localizado no tronco da árvore. O cancro também pode ter início a partir de um fruto contaminado, pelo crescimento do fungo através do pedúnculo, atingindo o tronco. Durante a estação seca, os cancros atravessam um período de inatividade total. Práticas sanitárias e pulverização são importantes na redução da incidência de cancro e prodidão parda. (Caroll Henry, Jamaica).
4. O uso de suspensão de zoospórios para inocular frutos demonstrou que estes são altamente susceptíveis quando novos (2 meses);
44
esta susceptibilidade decresce quando os frutos tem 3 meses e aumenta novamente com a idade, até o seu amadurecimento. Em testes de inoculação de frutos, com base no diâmetro da lesão, os cultivares Catongo, CC-42, Sca-6 e UF-29 mostraram maior resistência. (j. Laurence, UCA, Turrialba, Costa Rica).
S. Experimentos de campo feitos na Estação Experimental La Lola, Costa Rica, durante 6 anos, demonstraram que os cultivares CC-42, UF-29 e CC41 tiveram baixa percentagem de frutos com podridão parda enquanto CC10 e UF-667 tiveram uma alta percentagem de frutos atacados. Os cultivares CC45, UF-668 e UF-221 deram uma percentagem intermediária. Escape, mais do que a resistência à doença, pode estar incluída em alguns dos resultados desses experimentos. A lesão dos frutos, 6 dias após a inoculação, demonstrou tendências semelhantes às encontradas com Catongo, que também mostrou resistência. (j. Soria, lfCA, Turrialba, Costa Rica).
6. Pesquisas feitas na Bahia, Brasil, deram informações sobre diversos aspectos da epidemiologia e controle da podridão parda. O período crítico para infecção de frutos na Bahia é de maio a setembro. Fatores climáticos relacionados à incidência da enfermidade incluem deficit de pressão de vapor, número de dias chuvosos e número de horas de chuva. A redução da sombra diminuiu o número de frutos contaminados. O uso de Kocide 101 e Cobre Sandoz deu bom resultado no controle da podridão parda, com duas ou quatro pulverizações anuais. Sca-6, Sca-12, alguns cultivares da série Catongo e seleções TSH e TSA mostraram resistência ao P. palmivora com a inoculação dos frutos. Medidas de saneamento são também recomendadas para um controle mais eficiente. (H. A1, Rocha, CEPEC, Ilhéus, Bahia, Brasil).
7. Em testes feitos com diversas culturas da coleção da Universidade da Califórnia, Riverside, inclusive isolados de diversos países e diferentes hospedeiros, isolados de P. palmivora mostraram variações quanto à morfologia e patogenicidade. Baseado nos cancros desenvolvidos nos caules de seedlings um isolado (Gana), por exemplo, era altamente patogênico em relação ao Sca-6; sua patogenicidade era intermediária quanto ao UF -29 e baixa em relação ao Catongo. A patogenicidade de um isolado da Costa Rica era intermediária em relação ao Sca-6, baixa em relação ao UF -29 e alta em relação ao Catongo. Outros isolados evidenciaram variações similares. (G. A. Zentmyer, Univer· sidade da Califórnia, Rir1erside, E.V,A.).
8 A resistência de sete cultivares de cacau em relação a quatro isolados do fungo (Costa Rica, Ceilão, Gana e Brasil) foi testada pelo método de inoculação em caule de seMlings. Sca-6 e UF -29 demons-
45

traram resistência a três dos quatro isolados. Pound 12 demonstrou ser muito susceptível a todos eles. Em geral, CC42, UF -613, Catongo e UF-221 mostraram reações intermediárias, sendo resistentes a alguns isolados e com resistência intermediária em relação a outros. (G, A, Zentmver, Universidade da Califórnia, RitJerside, E.U.A.).
RECOMENDAÇÕES
Gerais
É preciso aumentar os contactos e comunicações entre os pesquisadores de fitopatologia que trabalham com enfermidades do cacaueiro nos diversos países americanos.
As agências governamentais, bem como as indústrias, devem dar ênfase e publicidade à importância das enfermidades que afetam o cacaueiro. Maior cgoperação entre os países produtores de cacau deve ser encorajada. E necessário dar maior ênfase aos programas de extensão, de maneira que informações atualizadas sobre os problemas do cultivo do cacau sejam levadas rapidamente aos cacauicultores.
Aumentar os estudos básicos em relação às enfermidades que afetam o cacaueiro, especialmente :\lonilia e \'assoura-de-bruxa. (Marasmiuspernic:iosus), Estudos adicionais deste tipo também são necessários com relação à podridão parda, Ceratoc_ystis e Fusarium. Estes devem incluir pesquisas detalhadas sobre aspectos tais como fisiolo!'!:ia do fungo, esporulação, radiação solar e genética dos organismos causadores da enfermidade bem como sobre epidemiologia. Muitos desses aspectos podem ser enfatizados em diversos laboratórios dos Estados Unidos.
No campo, intensificar os estudos de epidemioi.ogia, relações do meio-ambiente, práticas agronômicas e de cultivo, inclusive nutrição, que podem ser usadas na redução da incidência de enfermidades.
Desenvolver uma metodologia padronizada e usá-la nos testes de resistência para avaliação dos diversos tipos de tratamento e controle das diferentes enfermidades. Condições do meio-ambiente, inoculação e plantas para material de pesquisas devem ser, tanto quanto possível, uniformes de maneira que se possa :..dequadamente avaliar os resultados obtidos nos diversos países.
Intensificar as pesquisas sobre o processo infeccioso, inclusive estudos de propagulos infectivos, relação entre quantidade de inóculo e infecção e fatores que influenciam a infecção.
46
Aumentar os estudos sobre controle com produtos químicos, especialmente podridão parda, .\lvnilia e vassoura-de-bruxa, inclusive pesquisas com novos fungic1das sistêmicos e métodos de aplicação. Problemas de resíduos também devem ser considerados.
Conduzir estudos detalhados, in \'i\'o, sobre esporulação e germinação dos diferentes pató~enos.
Comparar o comportamento dos diversos cultivares em relação aos diferentes patógenos e estudar a interrelação e interação das diversas enfermidades.
Para testes comparativos de resistência a enfermidades feitos em diferentes localidades, alguns clones comuns devem ser ~sados inclusive aqueles já conhecidos pela sua susceptibilidade generali~ada ao patógeno específico. Isto é particularmente significativo em relação à podridão parda devido à sua ampla distribuição pelo mundo.
Intensificar os estudos e fisiologia de maneira a aspecto do seu controle.
\'a ssoura -dP -bruxa
de ecologia, meio-ambiente, epidemiologia facilitar o escape à enfermidade como um
Específicas
Aumentar as pesquisas sobre a relação do meio-ambiente quanto à infecção. Intensificar os estudos sobre controle com produtos químicos por aspersão foliar e fungicidas sistêmicos.
.\lonilia
Aumentar as pesquisas sobre o ciclo de vida do fungo, incluindo fatores que envolvem a produção e germinação de esporos, identificação do estágio sexual e estudos fisiológicos e biológicos.
Podridão parda
Intensificar os estudos sobre efeitos do meio-ambiente (umidade, deficit de pressão de vapor, sombra e sol) e fatores de culti\'o, indusive fertilização, no desenvolvimento da enfermidade.
Selecionar diversos clones comuns para testar a resisto:'ncia ao 1'/,ylofJ/•Ihora em diferentes localidades, usando paw isto pe\1, menos um que Rej;~ conhecido por sua alta suRceptihilidade.
47

Estudar a relação entre quantidade e tipo de inóculo usado (esporângios, zoosporos, clamidosporos e oosporos) e infecção do fruto,
Estudar mais detalhadamente a relação entre Phytophthora palmiiJora e outras espécies de Phytupbthora (registradas na Venezuela) e pesquisar a existência destas espécies em outras áreas cacaueiras,
Intensificar os estudos sobre variações na patogenicidade do P. palmiiJora, isolados dos diferentes países e diferentes hospedeiros com o objetivo de identificar raças e também desenvolver uma série de clones diferenciais.
Continuar e intensificar as pesquisas sobre saneamento em relação à red~ção de enfermidades, incluindo ítens como tratamento do solo e do tronco com fungicidas para reduzir o inóculo, remoção de frutos e tratamento de casqueiro.
Fusarium
Maior número de trabalhos de v e ser feito com o objetivo de encontrar uma técnica para reproduzir a enfermidade em plantas adultas. Deve-se dar ênfase à determinação das condições do meio-ambiente que favorecem a infecção e desenvolvimento da enfermidade. Mais trabalhos devem ser feitos para determinar a relação entre os diferentes tipos de galhas produzidas pelo fungo.
SECOND AMERICAN REGIONAL MEETING OF Phytophthora palmivora GROUP Guayaquil-Ecuador, June 8-23-1973
SUMMARY OF PAPERS PRESENTF:D
Eight presentations were made in the session on black pod, with discussion of canker as well as black pod.
1. A m•w species of Phytophtbora on cocoa, designated as P. megasperma, was reported from Venezuela. This organism causes a disease termed Mancha de Agua or watery rot, in one region of Venezuela. Inoculations of stems, leaves and fruit resulted in disease production in severa! cultivars. The fungus is very distinct from P. palmivora,
48
being homothallic. Some contrai was obtained of fruit rot by application of Cupravit. (l.ilian C, de Reyes, Estación Exp<!rimental de CaucaRua. V ene zue la).
2. In another report on Phytophthora on cocoa in Venezuela, four species of the genus were iisted as isolated from pods: P. palmivora, P. mf'gasperma, P. parasitü:a, and P. syringae. Inoculations of seeds, stems, and pods resulted in disease production with all four species, with variation in aggressiveness in the different species. (Li/ian C. de Reyes and ltumberto Reyes, Estación Experimental d"' Caucagua, Venezuela.
3. Studies in Jamaica showed that pods can become infected with Phytopbthora from a canker on the trunk. Also P. palmivora can grow from an infected pod into the trunk to initiate a canker. Cankers undergo a period of dormancy during the dry season. Sanitation and spraying are both important in reducing the incidence of canker and black pod. (Caroll Henry, Jamaica).
4. Inoculations of pods with zoospore suspension showed that susceptibility is high when the pods are young (2 months old), then decreases when pods are 3 months old, and increases again with age until pods become mature. In inoculation tests with pods, Catongo, CC42, Sca-6, and UF29 were the most resistant, based on measurements of lesion diameter. (]. Laurence, IICA, 1'urrialba, Costa Rica).
5. In field trials at the La Lo la Experiment Station in Costa Rica over a 6-year period, the cultivars CC42, UF29 and CC41 had low percentages of deseased pods, whereas CCIO and UF667 had very high percentages of deseased pods, and the cultivars CC45, UF668, and UF221 were intermediate. Disease escape rather than resistance may be involved in some of these results of field trials. Pod lesions 6 days after inoculation showed similar trends with Catongo also showing resistance. (]. Soria, IICA, Turrialba, Costa Rica).
6. Research in Bahia, Brazil, provided information on a number of phases of black pod epidemiology and control. The criticai period for pod infection in Bahia is from May to September; climatic factors related to disease incidence included vapor presswe deficit, number of rainy days, and number of hours with rain. Reducing shade resulted in reduced pod rot. Kocide 101 and Copper Sandoz gave good contrai of black pod with either low volume or conventional spraying. Sca-6 and Sca-12, some Catongo cultivars and selections of TSH and TSA showed resistance to P. palmivora in pod inoculations, Sanitary measures also are recommended for control. (H.M.. Rocha, CI!PEC, Ilhéus, Brasil),
49

7. Isolates of P. palmivora were found to vary in morphology and pathogenicity, in tests with many cultures from the collection at the University of California, Riverside, involving isolates from various countries and different hosts. Based on cankers developed from stem inoculation of seedlings, an isolate from Ghana, for example, was highly pathogenic to Sca-6 intermediate in pathogenicity to UF29, and low in pathogenicity to Catongo. Pathogenicity of an isolate from Costa Rica was intermediate to Sca-6 low to UF29, and high to Catongo. Similar variation was evident with other isolates. (G. A. Zentmyer, Universíty of California, Riverside, V.S.A.).
8. Resistance of seven cultivars of cocoa to four isolates of the fungus (Costa Rica, Ceylon, Ghana, Brazil) was tested by the stem inoculation method. Sca-6 and UF29 were resistant to three of the four isolates. Pound 12 was very susceptible to ali four isolates. In general, CC42, UF612, Catongo, and UF221 showed intermediate reactions, being resistant to some isolates, and intermediate in resistance or susceptibility to others. (G. A. Zentmyer, University of California, Riverside, V.S.A.).
RECOMENDATIONS
General
lncre<.~sed contact and communication is needed between cocoa disease research workers in the various countries in the Americas.
Governmental agencies as well as industry should emphasize and publicize the importance of cocoa diseases. lncreased cooperation between the various cocoa producing countries should be encouraged. More extension emphasis is needed, so that current information on cocoa problems can become readily available to the growers.
Increase basic studies in relation to diseases of cocoa, especially on Monilia pod rot, and witches broom, caused by Marasmius perniciosus, Additional studies of this type are also needed on black pod, Ceratocystis, and Fusarium. This should include detailed investigations of such aspects as fungus physiology, sporulation, radiation and genetics of the causal organisms and epidemiology. These aspects could be emphasized in severallaboratories in the U.S.
Under field conditions, intensify studies of epidemiology, of environmental relations, agronomic and cultural practices including nutrition, that could be used to reduce disease incidence_
50
Develop and agree upon uniform methOOs to use in testing resistance and in evaluating different types of contrai treatments in relation to various diseases. Conditions of environment, inoculum and plant material should be as uniform as possible in order to adequately evaluate responses in different countries.
Intensify research on the infection process, including studies of infective propagules, relation between the amount of inoculum and infection, and factors influencing infection.
Increase studies of chemical control, especially of black pod, Monilia, and witches broom, including research on new systemic fungicides and methOOs of application. Residue problems must also be considered.
Conduct detailed studies in vivo on sporulation and germination in the severa! disease problems.
Compare responses of different cultivars to the different pathogens, and study interrelationships and interactions between the severa! diseases.
For comparative tests of resistance to the diseases in different localities a few common clones should be used, including some that are known to be generally susceptible to the particular pathogen. This is particularly significant in relation to black pOO, with its world-wide distribution.
Intensify ecological, environmental, epidemiological and physiological studies so that disease avoidance or escape can be facilitated as one aspect of controlling these diseases.
SPECIFIC
Witchs Broom
Increase research on environmental relationship in regard to infection. Inte~s~fy studies on chemical control by foliar sprays and systemic fungtctdes.
Monília
Increase research on the Iife cicle of the fungus, including studies of factors involved in spore production and germination, identification
51

of the sexual stage, and physiological and biological studies of the fungus.
IUack Pod
Intensify studies of the effect of environmental (moisture, vapor pressure deficit, shade and suo), and cultural factors including fertilization, on the development of black pod.
Select severa! common clones to test for resistance to Phytophthora in different localities, using at !east one that is known to be generally vety susceptible.
Study the relation between amount of inoculum and type of inoculum (sporangia, zoospores, chlamydospores, oospores), and infection of pods.
Study further the relationship between Phytophthora palmivora and the other species of Phytophthora reported from cocoa in Venezuela, and search for these species in other cocoa areas.
lncrease studies on variation in pathogenicity of P. palmivora isolates from different countries and different hosts with the aim of designating races and also developing a series of differential hosts.
Continue and increase research on sanitation in re!ation to disease reduction, including such items as treating the soil and the trunk with fungicides to reduce inoculum, pod remova!, and treatment of pod piles,
Pusarium
• More work should be conducted with the purpose of finding a
technique to reproduce the disease on fully grown plants.
Emphasis should be given to the determination of the environmental conditions that favor ínfection and disease development,
More work is needed to determine the relation between the different types of galls incited by this fungus,
52
REVISTA THEOBROMA
Abril-junho 1974
Ano IV N~ 2
Publicação uimesual dedicada ã divulgação de investigação científica relacionada com problema~ agronõmicos e sócio-econõmicos de áreas cacaueiras. Editada pelo Centro de Pesquisas do Cacau (CEPEC), Departamento da Comissão E:~ecutiva do Piano da Lavoura Cac:aueira (CEPLACl.
COMISSÃO EDITORIAL
A! vim, Paulo de T. Cruz, Luiz C. Sales, Josê C. de Vello, Fernando
ASSESSORES CIENTÍFICOS
Aitken, William M. Alencar, Maria H. Alvim, Ronald Barroco, Hêlio E. Barros, Raimundo S. Cabala R., F. Percy Carleuo, Geraldo A. Loureiro, Edmar S. Maravalhas, Nelson Mariano, Anrõnio H. Medeiros, Arnaldo G. Menezes, José A. S. Miranda, Emo Ruy de Pereira, Clovis p. Rocha, Hermínin M. Silva, Luiz F. da Silva, Pedrito Ventocilla, José A.
Endereço para correspondência {Adress for correspondente);
Revista Theobroma Centro de Pe~quisas do Cacau (CEPEC). Caiu Postal 7 4~.600 - lta~na-Bahia
Brasil
Tiragem
3.000 e:remplfres
CONTEÚDO
1, Indução da produção em cacaueiro com uso de atomizador motorizado portátil na Bahia, Brasil. S. de J. Soria. 3
Mechanical pollination of cocoa using motorised knapsack sprayers .in Bahia, Brazil. {Summary}. p. 12.
2. Um método colorímétrico simples para determinação dJl intensidade de contaminação com fumaça em amêndoas de cacau. P.R.F. Berbert, T.F. Bsqu.ivel e N. Marava-lhas. 14
A colorimetric method for determining the intensity of smoke contamination in cocoa beans. (Summary). p. 19.
3. Economicitkldedos insumos modernos em fazendas de cacau. ].A. de S. Meneses, B. Paniago, H.P. La-deira e A.L. Bandeira. 21
Economic aspeds of modero inputs used in Bahian cocoa farms. (Summary). p. 32.
4. Observações citológicas run células mães de polem de cacaueiros. G. A . Carletto. 34
Cytological aberrat.ions of lhe pollen mother cells in Theobroma cacao L. (Summary). p. 39.

•
INDUÇÃO DA PRODUÇÃO EM CACAUEIRO COM USO DE ATOMIZADOR MOTORIZADO PORTÁTIL
NA BAHIA, BRASIL*
Saulo de f esus Soria **
A colheita de cacau da Bahia é feita em duas épocas do ano. Uma delas, denominada regionalmente temporão, ocorre no primeiro semestre e a outra, denominada safra, no segundo.
Devido à escassez de insetos polinizadores no período novembro-janeiro, tem-se notado um deficit de polinização natural por ocasião do temporão. Este fato parece refletir-se diretamente sobre a produção, tanto assim que o temporão, nas lavouras comerciais do CEPEC, foi aproximadamente um terço da safra nos últimos 4 anos***. Na Bahia, em geral, analisando-se a média anual das colheitas nos últimos 16anos (152.844 toneladas), constata-se uma diferença de cerca de 10.000 em favor
da safra****· Por outro lado, nos principais países produtores da África, a produção concentra-se praticamente em um período, que corresponde à safra baiana. Esta ocorrência coloca o temporão da Bahia em posição relativamente privilegiada no âmbito internacional, uma vez que o produto é posto no mercado numa época em que os países competidores não dispõem de produção. Do exposto pode-se concluir que há uma vantagem evidente em se reforçar o temporão, seja através de polinização artificial, seja de outro método de eficiência técnica e econômica comprovada .
Com base nessa premissa, realizou-se o presente trabalho, com o objetivo de estimar a viabilidade
• Recebido para publicação em abril, 1974. ** Ph.D., Divisão de Fitoparasitologia, CEPEC .
••• ........ Dados fornecidos pelo Setor de Produção do CEPEC, 1973 .
Dados fornecidos pela Divisão de S6cio·Economía, CEPEC.
Revista Tbeobromu. CEPEC, IlMus, Brasil, 4 (2): 3 -13. Abr.-jun. 1974
3

da indução da polinização em cacaueiros mediante o uso de atomizadores. A economicidade do método é medida em termos do lucro proporcionado pelo adicional de produção devido ao tratamento.
REVISÃO DE LITERATURA
O uso de atomizadores para induzir a polinização de cacau tem sido um método bem sucedido nas variedades autocompatíveis. Knoke e Saunders (3), em Costa Rica, induziram um aumento do número de frutos com a aplicação de inseticidas por atomizadores. Soria (8), também em Costa Rica, incrementou 3 e 4 vezes o rendimento nos tratamentos pela atomização de água e vento, respectivamente, comparados com a testemunha. Soria e Cerdas (6) observaram aumentos significativos na produção de cacau, agitando flores de clones autocompatíveis com vassouras de sorgo. Posteriormente, os mesmos autores (7) conseguiram, com polinização manual, aumentar a produção até 10 vezes em relação à polinização natural.
Entre os vários métodos de polinização artificial avaliados por Arevalo (1), o manual foi o mais eficiente para elevar os rendimentos. Os métodos ensaiados de ordenho (despetalamento), pincelamento e atomização só elevaram os rendimentos e tornaram-se rentáveis quando aplicados sobre o clone autocompatível UF-221. A maior frutificação resultante da
4
aplicação dos tratamentos incrementou a perceritagem de frutos pecos e diminuiu o índice de sementes. Medina (5), no Equador, incrementou significativamente a produção usando a polinização manual, mas fracassou nesse propósito quando soprou flores do cacaueiro com ar puro e/ou inseticidas mediante atomizador motorizado portátil. A polinização manual foi considerada anti-econômica devido ao elevado custo da operação. O consenso geral na literatura é que a polinização mecânica com atomizadores e outros equipamentos é efetiva somente nas variedades autocompatíveis.
A disponibilidade de uma floração razoavelmente intensa durante os meses de novembro, dezembro e janeiro, na região cacaueira da Bahia (9), coincidente com uma baixa taxa de fertilização, estimulou a condução de estudos sobre polinização mecânica, como um artifício para suplantar a falta de insetos polinizadores nesse período.
MATERIAL E MÉTODOS
Cento e vinte cacaueiros de variedade Comum, com espaçamento irregular e idade aproximada de 50 anos, foram selecionados e etiquetados dentro de uma plantação na área experimental do CEPEC. Ilhéus, Bahia. Seis parcelas de 10 árvores cada foram comparadas com uma quantidade equivalente
de parcelas testemunhas, num desenho experimental de blocos <lO
acaso (parcelas pareadas).
Na mesm<~ áre<~ experimental, num bloco arbitrariamente denon'!Jnado testemunha adicional, foram selecionadas outras 20 árvores da mesma idade e tamanho menor, nas quais eram contados semanalmente todas as flores e bilros presentes para acompanhar a flutuação natural de floração. Este bloco não fez parte do desenho experiment<~l
mencionado, pois pretendeu apenas destacar um parâmetro adicional de floração em condições vegetativas diferentes.
Um atomizador FONTAN JLO, tipo L77L, de fabricação alemã, foi usado para soprar as flores. Em sua máxima aceleração, a máquina produz um jato de ar de intensidade equivalente a um vento de 51 quilômetros por hora, medido a 1,5 metros de distância a partir da boca do atomizador (distância média entre esta e as inflorescências). Nestas condições, as flores foram agitadas violentamente até o ponto em que aquelas não fertilizadas, e de mais de 2 dias de idade, eram arrancadas da árvore. As flores foram sopradas 6 vezes, em dias alternados, nas primeiras horas da manhã, durante a floração de janeiro de 1973.
' A eficiênci<~ do tratamento face
a testemunha foi determinada con-tando-se semanalmente as flores bilros e frutos maduros colhido~ por árvore e pesando-se as semen-
5
tes frescas. O peso seco foi calculado na base de 40'7o do peso úmido, de acordo com a recomendação feita pela Divisão de Genética do CEPEC.
Para o estudo dos custos operacionais, foram coletados os seguintes dados: horas totais e horas efetivas de trabalho dos operários no campo, manutenção e depreciação do atomizador, combustível consumido, transporte de pessoa\ e máquinas para o campo e preços do cacau no mercado local por ocasião da venda. O cálculo do custo operacional foi baseado em metodologia sugerida por Boss e Pond,citado por Brandão (2),que consiste em considerar os fatores de custo para manutenção da máquina, depreciação, juros e reparos.
A soma desses fatores constitui o custo anual que, dividido pelo número de dias trabalhados, representa o custo diário de operação da máquina, ao qual deverão ser adicionados os gastos variáveis de operação (combustível, lubrificante, mão-de-obra etc.).
A depreciação (O) constitui a relação entre o preço de aquisição da máquina (P) e sua duração provável (vida útil}, em anos (T}:
p D - -- sendo P =Cr$ 1.500,00
T T.:: 5 anos
Os juros são calculados a partir da taxa que incide sobre o capi-

tal investido (preço de aquisição + depreciação). Essa é arbitrária, mas eleve ser pelo menos igual à menor taxa que este capital obteria a prazo fixo nas entidades financeiras oficiais (12~ a. a.).
O valor pera reparos é também estabelecido através de uma taxa arbitrária (1~) sobre o preço de aquisição, de acordo com a análise de investimento e inflação feita por Machline (4).
Estabeleceu-se ainda um custo adicional relativo ao transporte que, na prática, ocorre, seja feito em veículo, animal ou pelo operá-
rio a pé. No (Weseafle caso, oade se utilize-.- pick-ap Fout F-75, o custo de baDSpolte foi estmado em C4 O.SO/ba. de acordo co. cálcaJos feitos pelo Setor de Tauaspca-tes da CEPLAC (1973).
RESI1TADOS E DfiCnssÃO
Fie« tã- e ___.,ele ele Oc:n- cio
Os resultados dos estudos de flutuação de floração no tratameato, coap;u:ados COla a testeCDUJJha, ao ~.t.ei.ro se.estre de 1973 (Figura 1). indicam que os cacaueiros
I • 7 J
F1gura 1 - Distribuição da [loTação, frrdifialfM:J ~ /IJrM•çMJ J~ CIICII•
nas áreas de sopTo e testl!munha. llhiiiS, Babi4. Br~il, l~s~•estr~. 197J.
6
mostraram três picos destacados de floração (janeiro, março- abril e junho). Dos três picos, somente
0 primeuo f01 aproveitado para induzir a frutificação. Os outros dois, particularmente o segundo, que foi o ma1S intenso, foram deixados para poliniZação hvre (aberta).
Verificou-se uma grande variação entre as intensidades de floração das árvores amostradas, independentemente da época do ano. A média das flores nas 60 árvores da testemunha experimental (aproximadamente 400 flores/planta) mostrou-se 10 vezes maior que a das 20 árvores da testemunha paralela (Quadro 1).
Flutuação de lrutificação e indução de produção
Temporão
A curva correspondente à colheita do primeiro semestre (Figura 1) apresentou uma elevação a partir de junho para tratamento e testemunhas, mas com intensidades s igmficativamente diferentes . A frutificação no tratamento incrementou após seis aplicaçõés de sopro, até um limite de 20 frutos por árvore. Este número permaneceu relativamente estável até ju· nho, época em que se iniciou a colheita do temporão. A tendência da cu1va correspondente à testemunha indicou um incremento lento
Quadro 1 - Média de [loTes por árvore nas áreas testemunha t!xperi· mental (a) e adicional (b). CEPEC, ilhéus, Bahia, Brasil, janeiTo·fevereiro, 1973.
Testemunha Testemunha Data experimental (a) adicional (b)
Jaa. 5 20 2 12 500 46 19 1000 118 26 600 44
Fev. 2 200 32 9 70 6
Média/árvore 398 41 Taxa aproximada a/b 10:1
(a) Média de 60 árvores (b) Média de 20 árvores
7
300+ (300 + 1.500) 12 + 150 100
de frutificação a partir de janeiro, que foi marcadamente acelerado a partir de maio (Figura 1). X: =
180
= Cr$ 3,70 /dia Os resultados relacionados com
indução de produção (Quadro 2) indicam que o sopro triplicou o rendimento no primeiro semestre, comparado com a testemunha.
onde: D = depreciação da máquina
Receita líquida e lucro = valor máximo
vida útil
Os resultados com relação ao cálculo de custos foram os seguintes:
J = Juros do investimento
Custo diário do trabalho da mãquina (X):
= (valor da máq. + deprec.) taxa 100
R = reparos (10% do custo inicial) X= D+J+R =
N N = dias de utilização
Quadro 2 - Cacau mole (Jeg) f!roduzido por parcela de lO cacaueiros nos tratamentos de sopro e testemunha. Ilhéus, Bahia, Brasil, 197).
I~ S...atre 2~ Semeatre Total
Sopro• Teatemunha Sopro Teatemunba Sopro Teatemunh.a
1,90 0 ,85 60,95 47, 10 62,85 47,95
11 ,05 1,40 34,45 39,05 45,50 40,45
13,10 1,75 43,75 28,00 56,85 29,75
9,65 2,75 69,15 63,28 78,80 66,03
7,15 0,90 48,80 59,25 55,95 60,15
3,25 8,85 50,85 64,25 54,10 73, 10
Total 46,10 16,50 307,95 300,93 354,05 317,43
Peao aeco (ki) .. 18,44 6,60
* O Jllto dt• nr rot aplicado somente no primeiro seme11tre. • • Daseodo em recomendação da Divisão de Genética, CEPEC (peso seco=
40% do pello do cacau mole).
8
Foram arbitrariamente tomados 180 dtas de utilização anual média da máquina, embora, no presente trabalho, tenham stdo uttltzados apenas 6 dtas, uma vez que, na prática,a máquinatemoutros usos.
Ao capttal de Cr$ 65,70, investido no tratamento sopro (Quadro 3), correspondeu uma receita líquida de Cr$ 5,34 (Quadro 4)- ou seja, 8% sobre esse capital - 6 meses após o início do processo. O lucro talvez pudesse ser awnentado intensificando-se o insumo sopro, particularmente no que se refere ao número de aplicações e número de operários ativos, pois o custo atribuído a transporte ficaria quase invariável dentro dos limites de capacidade de transporte do carro.
Safra
A safra, resultante da pohnização natural das flores entre março e junho, foi quantitativamente domtnante, comparada ao temporão (Quadro 2). A colheita das áreas experimentats ocorreu a partir de setembro, estendendo-se até princípios de janeiro de 1974 e foi parttcularmente e levada nesse período. Completada esta, procedeu-se a análise de variação da colheita total do ano para as áreas de tratamento e testemunha. A colheita total no tratamento sopro foi aproximadamente igual à encontrada na testemunha (Quadro 2). A média anual de rendimento foi de2,36 e 2,12 kg de cacau seco/árvore para as áreas de tratamento e testemunha, respectivamente.
Quadro 3 - Custo total atribuído ao msumo sopro. Ilhéus, Bahia. Bra · si/, 1973.
I B••••lftuton
Atoalndor (J-dM de t Hlea) 3 3,78• U,IG CoabQatl"l - paollaa (U.._) 24 .. , .. 11,60 Lalatlflc.ate - 6Jeo 2 T (lltrw) I ••• 6,00 lllo-de..tn (lHas)•• 24 1,10 24,00 -c.,._...._.._ ........ ~(A) 59.7G Traaporte ............... (lia) (8) 12 G,50 ••• c.. .......... ~~(A+., -
• Valor esUmado com base no custo duírao de trabalho da máquina (jornada de 8 horas).
*• Considerou-se um salário mínimo de Cr$ 240,00/mês, sem Incluir os encargos SOClQlS.
9

Quadro 4 - Receita líquida adicional proporcionada pelo insumo sopro, com base na produção de 60 cacaueiros. no pe,;odo do "temporão". tlhlus, Bahia, Brasil, 1913.
Áreas Produçlo Receita Cuato Rece1ta
peso aeco (lti) bruta (Cr$) • total (Cr$) líquida (Cr$)
Sopro 18,44 110,64 65,70 44,94
Te a temunha 6,60 39,60 39,60
ao sopro 11,&4 71,04 65,70 5,34
• Foi considerado o preço de Cr$ 90,00 por arroba (15 kg) de cacau seco.
Quando se usou como critério de comparação as produções semestrais, encontrou-se uma dtferença altamente signtficahva entre a produção do primeiro semestre frente à do segundo: a colhetta obtida no segundo semestre (safra) foi 12,5 vezes maior que a do pumeiro. Esta superioridade quantitativa na safra/1973 determinou que as diferenças altamente significativas entre tratamentos no primeiro semestre, ficassem insignificantes no segundo.
CONCLUSÕES
1. A prática de sopro, nas condições experimentais descutas, mostrou-se eficiente como método, triplicando a produção nas plantações comerciats de cacau Comum autocompatível trabalhadas, durante o primeiro semestre;
2. A técnica de sopro tncrementou o lucro no primeiro período
10
de produção do ano (temporão), dentro dos limites das condições ambientais reinantes de floração, custo de insumos e mercado de venda do produto na respectiva época de colhetta;
3. O rendtmento anual (temporão + safra) não foi significativamente incrementado pelo tratamento sopro;
4. Ensaios de avaliação de produtos químicos agrícolas que utilizam jato de ar como vetor, devem incluir uma testemunha adacional para isolar a vauável correspondente ao pegamento adicional devido ao jato de ar; e
S. Considerando-se que a aplicação de sopro foi feita somente em um período do ano, em uma população de cacaueiros com uma média de 400 flores/planta, há necessidade de se repetir o experimento sob diferentes condições de
intensadade de florac;-Jo e em meses d aferentes parii ~wa la ar ~H é
que ponto a aplicação do sopro é economacamente v1ável.
Aos Eng.-Agr?s Fernando Vello, Geraldo Carletto, Pedrito Silva, João l\t . de Abreu e ao 8. Se. Wdham Martin Aitken,1 pela revisão do artigo e sugestões, aos Eng.-Agr~ Maria Helena Alencar. Nilson de Matos Sab1• no. Ubaldo Santos e Economasta Áureo Brandão, pela onentação no cál culo de custos e receita líquada; aos Eng.-Agr?s José A. Ventocilla e Cláudto Miranda, pelos dados de produção da área, e ao Sr. Florisvaldo A. Galvão, pela ajuda na redação do trabalho.
LITERI\TURA CITADA
1. AREVALO, R.A . Evoluación de cuatro metodos de polinización artificial en cacao (I hf'obromo cacao L.J. Thesis Magister Science. Tumalba, Costa R1ca, Instituto Interamencano de Ciências Agrícolas, 39 p.l972.
2 BRANDÃO, E.D. Aulas de Administração Rural. Viçosa, MG, Escola Supenor de Agricultura, 105 p. 1958. (mimeografado).
3. KNOKE, J .K. e SAUNDE.RS, J. Induced frutt set of Theobroma cacao L. b} mastblower apphcations of insecticides. Journal of Economic Entomology 59(6) : 1427-1430. 1966.
4. MACHLINE, C. Anãhse de investimentos e inflação. Revista de Administração de Empresas 6(18) : 51-123. 1966.
S. MEDINA, L.M.F. Efecto de las polintzaciones artific1ales y entornofilas en el rendamtento de! cacao. Tesas de Ingeniero Agronomo. Faculdad de Agronomia y Vetennaria, Universidad de Guayaquil, Ecuador, 44p. 1973.
6. SORIA, J. e CERDAS, M Pohnizac10nes por movimientos de flores con escobilla de sorgo. Cacao (Costa Rica) 11(3):8-9. 1966.
7. ----e . Rendimiento de I cacao en relación con polma-
zacaón artifícaal y abonamiento. Cacao (Costa Rica) 15(3)·11-12. 1970.
11

8. SORIA, S. de J. Studies on Forcil'o-ric spp. aidges (Dipled.. Cen.topogonidae) related to tbe polliaatiom o( Tk._., c•uo L. Dissertation Abstracts IntematioaaU 31(5):2744-B. 19m.
9. VELLO, F. Fatores relacionados com a pol.iaüzação~ tertilizaçlio e produção do cacaueiro na Bahia. Il!léss. Bahia. Brasil, Centro de Pesquisas do Cacau, 26 p. (~)-
RESUMO
Na região cacaueira da Bahia, tem-se obsenr.ldocolbeitas aeoores durante a primeira metade do ano, motivadas aparentesaente pela falta de agentes polinizadores no período de floração que vai de aovemko a janeiro. Foi aventada e testada experimentalDeatte a hipótese de que a aplicação de um jato de ar proveniente de um atomõzadot motorizado portãtil poderia induzir polinização nas flores da variedade c-, autocompatível. As aplicações do jato de ar triplicaram o remlimeoto de cacau seco em relação com a testemunha, durante o pimeim seiik!She de 1973. A receita líquida obtida foi equivalente a 8% do capital investido oo t:Jatamento, 6 meses após o início do processo.
Por outro lado, o rendimento dumote todo o aoo não foi significativamente incrementado, pois uma safra excepciooa~me gmnde. equivalente a 12,5 vezes o temporão, aparentemente obscwecen os resultados obtidos no primeiro semestre. Sugere-se que a técnica de sap-o é válida unicamente nas plantações de variedades autocom.pativeis.
MECHANICAL POLLINATION OF COCOA USING MOTORISED KNAPSACK SPRA YERS
IN BAHIA, BRAZIL
SUMMAB.Y
In the Bahian cocoa region, the cocoa harvest dwing the first 6 months of tbe year is usually less than the secood, a fact« attributed to tbe low leveis of insect pollinators between November adl January. A trial carried out by blowing air, using a motorised lmapsack SpGiyet, over tbe flowers of the self-compatible variety Com- in ao aUempt to indnce mechanical pollination, tripled the yield compared to the untreated plots. However, the net profit computed of the 6 montht period was equivalent to only 8% of the treahnent costs.
12
Due to the very high yields of the second 6 month period for the two areas, the accumulated yields for whole year show little difference between the treatments. It is to be remembered that increasing pollination by blowing air is only valid for self-compatible cocoa varieties,
.. ..
13

UM MÉTODO COLORIMÉTRICO SIMPLES PARA DETERMINAÇÃO DA INTENSIDADE DE CONTAMINAÇÃO
COM FUMAÇA EM AMÊNDOAS DE CACAU *
Paulo Romeu F. Berbert ** Terezinha F. Esquivei***
O cheiro de fumaça é considerado defeito grave em cacau porque amêndoas contaminadas, quando utilizadas na fabricação de chocolate, dão-lhe um gosto desagradável de fumaça, Tal tipo de contaminação ocorre quando as amêndoas entram em contato direto com fumaça durante a secagem artificial em secadores a lenha, ou mesmo durante a secagem natural, em barcaças. No primeiro caso, a contaminação é devida à existência de vazamento na chapa tubular do secador, ou ao fato de a chaminé do secador ser mais baixa que a cumeeira do mesmo. No segundo caso, o cacau é contaminado com fumaça proveniente de fogão a lenha ou de candeeiro de iluminação existentes em residências construídas sob as barcaças. Devido a este problema,
com a finalidade de fazer melhorar a qualidade do cacau brasileiro, o Governo proibiu a exportação de cacau contaminado com fumaça (1). Para melhor controle do cumprimento de tal resolução, a CEPLAC, Órgão do Governo Federal, tem aberto novos Postos de Classificação de cacau em vários locais das zonas produtoras e tem feito campanhas junto aos cacauicultores com a finalidade de torná-los mais esclarecidos sobre o problema.
Atualmente, a classificação do cacau, no que diz respeito à contaminação de fumaça, é feita cheirando-se, uma por uma, 300 amêndoas que foram cortadas longitudinalmente. O grau de contaminação é expresso em função da percentagem de amêndoas encontradas com
* Recebido para publicação em junho, 1974. ** Eng.-A.,-?, Divisão de Fitologia, CEPEC.
*** Farmacêutica Bioquímica, Divisão de Bioengenharia, CEPEC.
Revista Theobroma. CEPEC, Ilhéus, Brasil, 4 (2): 14-20. Abr.-;un. 1974.
14
cheiro de fumaça. Este método é insatisfatório uma vez que leva em conta apenas o número de amêndoas contaminadas, não considerando a intensidade de contaminação.
A existência de um método para detecção e quantificação de fumaça, menos subjetivo e mais preciso que o método em uso, seria de grande utilidade para um melhor controle da qualidade do cacau. Pensouse então em um método colorimétrico, no qual extratos de amostras quaisquer pudessem ser comparados visualmente ou espectrofotometricamente com extratos padrões provenientes de amostras com diferentes graus de contaminação de fumaça. Após diversas tentativas com diferentes solventes orgânicos, tais como álcool etílico, álcool metílico, benzo! etc., notou-se que álcool etílico a 96% é o que melhor extrai os componentes da fumaça, adquirindo uma coloração amarela mais intensa que no caso dos demais solventes. Além disso, o ãlcool etílico a 96%, quando comparado com solventes mais polares, como por exemplo o álcool metílico, apresenta a vantagem de ser um mau solvente para os pigmentos existentes na amêndoa de cacau.
Sendo a determinação da quantidade absoluta de fumaça um processo impraticável no caso, pelas dificuldades analíticas que apresenta, procurou-se um método capaz de quantificá-la de maneira relativa, comparando-se o extrato do lote de amêndoas do qual se deseja
conhecer o grau de contaminação de fumaça com extratos padrões provenientes de lotes de cacau com percentagens conhecidas de amêndoas uniformemente enfumaçadas.
A finalidade do presente trabalho é descrever um método simples e prático para determinar, de maneira mais precisa que a técnica em uso, a intensidade de contaminação de fumaça em amêndoas de cacau. Espera-se que tal método• possa ser utilizado nos Postos de Classificação e por outros interessados, facilitando dessa maneira o controle da qualidade do cacau brasileiro para exportação, no que se refere a cheiro de fumaça. Trabalho preliminar sobre o assunto foi publicado recentemente (5).
MATERIAIS E MÉTODOS
Para obtenção de amêndoas uniformemente contaminadas com fumaça a serem utilizadas no preparo de extratos padrões, colocou-se cerca de 20kg de cacau fermentado em bandeja de madeira com fundo de tiras separadas entre si cerca de 0,5 em, semelhante à bandeja de fermentação de Rohan (6). Em seguida, procedeu-se a secagem em fogo brando, produzido por lenha, com a bandeja a 1,5 m dosolo. Durante o dia, as amêndoas eram revolvidas freqüentemente e, à noite, a bandeja era coberta com lona, permanecendo o fogo aceso. Após 72 horas, o fogo foi retirado e a secagem continuou, ao sol, até
15

as amêndoas alcançarem cerca de 7% de umidade. Prepararam-se amostras de 100 g misturando-se 2; 5; 10; 25 e 50% de amêndoas contaminadas, com amêndoas livres de fumaça, todas elas inteiras e com casca. Cada amostra assim preparada foi colocada em erlenmeyer de 500 ml, ao qual se adicionaram 33,3 ml de álcool etílico a 96%. O conteúdo foi agitado por 5 minutos em um agitador rotativo de 60 a 260 ciclos por minuto, .de Eberbach Corp., Ann Arbor, Michigan, a 1/4 de volta abaixo da sua velocidade máxima. Para cada tipo de amostra, foram feitas três repetições, de cujos extratos filtrados foram lidas as respectivas absorvâncias num espectrofotômetro Zeiss, modelo PQM 11 - M4Q III, a 325 m11. Como blank, utilizou-se o extrato de um lote de 100g de amêndoas livres de fumaça. De cada uma das três repetições de cada tratamento, foram retirados 10 ml, os quais, depois de misturados, foram colocados em tubos de ensaio de 200/25 que, em seguida, foram fechados a fogo, obtendo-se assim os padrões para comparações visuais.
Procurou-se relacionar a coloração dos extratos obtidos com o gosto de fumaça detectável no produto final. Utilizando-se amêndoas do mesmo lote contaminado com fumaça, foram então confeccionadas barras de chocolate de amostras preparadas de maneira idêntica à descrita. Essas barras foram testadas organolepticamente,
•
utilizando-se um painel de degustação, para determinar o nível mínimo de contaminação de fumaça que pode ser detectado pelo paladar.
RESULTADOS E DISCUSSÃO
As leituras espectrofotométricas dos diversos extratos alcoólicos padrões, a 325 mp., forneceram as absorvâncias apresentadas no Quadro 1.
Quadro 1 - Absorvâncias a 325 m11 dos diversos extratos padrões contaminados com fumaça.
Porcentagem de amêndoas
contaminadas
2 5
lO 25 50
Absorvâncias
(325m")
0,110 0,200 0,380 0,910 1,800
Sugere-se que estes sejam considerados os extratos padrões. Quaisquer outros padrões com essas mesmas percentagens de contaminação que venham a ser preparados posteriormente devem possuir aquelas absorvâncias. Os padrões com as percentagens de contaminação acima mencionados dão margem a uma comparação visual conveniente para uso rotineiro na classificação de cacau. É possível, entretanto, a depender da conveni-
16
ência, prepararem-se extratos padrões com qualquer intensidade de contaminação em relação aos padrões preparados inicialmente. Para isto, basta se observar as absorvâncias na curva de calibração obtida a partir daqueles padrões (Figura 1). Por exemplo, caso haja necessidade de serem preparados extratos padrões com 3% e 6% de contaminação de fumaça, a curva de calibração mostra que suas absorvâncias deverão ser respectivamente 0,130 e 0,250.
A fumaça constitui-se principalmente de uma mistura de subs-
0,40
1,20
~ • 1,00 " " • 0,80 • o z •• > 0,60 • o • • • 0,40
0,20
4 8 " "
tâncias fenólicas (2, 3, 4), que podem sofrer leves alterações qualitativas conforme sua origem. Tal fato, apesar de não ser problema no caso da avaliação visual dos extratos, pode resultar em variações significativas na análise espectrofotométrica, caso as fontes de origem da fumaça sejam muito divt:rsas. Como o conhecimento das absorvâncias é importante no preparo dos extratos padrões,estes deverão então sempre ser preparados com amêndoas contaminadas com fumaça proveniente da queima de madeira. Somando-se este fato ao alto custo do equipamento e necessidade de mão-de-obra especia-
20 ,. ,. •• PERCENTAGENS OE AMÊNDOAS CONTAMINADAS
Figura 1 - Curva espectrofotogramétrica de calibração, apresentando a intensidade relativa de contaminação por fumaça, em percentagem de amêndoas contaminadas, e suas respectivas ahsorvâncias.
17

•
lizada, leituras feitas com espectrofotômetro ou colorímetro, apesar de mais sensíveis, não seriam convenientes no caso. Os padrões seriam preparados em laboratórios equipados com o instrumental necessário e distribuídos para os Postos de Classificação e demais interessados. Na utilização usual do método, a intensidade de contaminação de fumaça em um lote qualquer de amêndoas seria avaliada pela simples comparação visual de seu extrato com os estratos padrões.
A contaminação mínima passível de detecção pelo painel de 'degustação foi ao nível de 3,5% de amêndoas contaminadas. O teor máximo de contaminação por fumaça admissível em amêndoas de cacau, quando determinado pelo método proposto, deve estar, portanto, em torno de 3%.
Os extratos padrões não podem ser utilizados indefinidamente uma vez que suas colorações são alteradas pela luz e diferenças das intensidades de suas cores são ob-
servadas a partir do terceiro mês, Na prática, esse inconveniente pode ser contornado pelas seguintes possibilidades, dentre outras:
a, Manutenção dos extratos padrões na ausência de luz;
b. Renovaçãodos extratos padrões a cada 3 meses;
c, Formulação de extratos de colorações semelhantes aos padrões que não sejam alterados com o tempo ou pela ação de luz; e
d. Utilização de escalas de cores com base na coloração dos extratos padrões,
Ainda que não forneça os teores absolutos de fumaça, mas somente valores relativos de intensidade de contaminação, o presente método é mais sensível e bem mais exato que o utilizado atualmente no Brasil, pelo que se espera que o mesmo venha a ser utilizado nos Postos de Classificação de cacau e por outros interessados.
AGRADECIMENTO
Os autores expressa~ seu reconhecimento ao Eng. Químico Edmar Loureiro, pela colaboração prestada na execução dos testes organolépticos.
LITERATURA CITADA
1. BASTOS, G.A.C. Padronização e classificação do cacau. Cacau Atualidades (Brasil) 11(2): 9-18. 1974.
18
I·
í
2. FIDDLER, W., WASSERMAN, A.E. e DOERR, R.C. Smoke flavor fraction of a liquid smoke solution. Journal of Agricultura! Food Chemistry 18(5): 934-936. 1970.
3. ISSENBERG, P., KORNREICH, M.R. e LUSTRE, A.O. Recovery of phenolic wood smoke components from smoked foods and model systems. Journal of Food Science 36:107-109. 1971.
4. KORNREICH, M.R. e ISSENBERG, P. Determination of phenolic wood smoke components as trimethysilyl ethers. Journal of Agricultura! Food Chemistry 20(6): 1109-1112. 1972.
S. MARAVALHAS, N., BERBERT, P.R.F. e ESQUIVEL, T.F. Um método fácil para determinação de cheiro de fumaça. Cacau Atualidades (Brasil) 10(4): 2-4. 1973.
6. ROHAN, T.A. Processing of raw cocoa for the market. Rome, FAO, 1%3. p. 109.
RESUMO
Foi desenvolvido um método colorimétrico simples para determinar a intensidade de contaminação de fumaça em amêndoas de cacau. O método é baseado no fato de substâncias normalmente encontradas em material contaminado por fumaça serem francamente solúveis em álcool etílico a 96%, ao qual dão uma cor amarela característica. A intensidade dessa cor é proporcional à quantidade de fumaça e pode ser medida no comprimento de onda de 325 m11- Para fins práticos, o método permite uma determinação visual satisfatória, comparando-se os extratos dos lotes de cacau a serem classificados, com extratos padrões. Espera-se que este método possa ser utilizado nos Postos de Classificação de cacau com a finalidade de se detectar cacau contaminado com fumaça.
A COLORIMETRIC METHOD FOR DETERMINING THE INTENSITYOF SMOKE CONTAMINATION
IN COCOA BEANS
SUMMARY
A simple colorimetric method was developed for determining the intensity of smoke contamination in cocoa beans. The tar compounds typically
v':. DOCiJMENTAC.f to>,•" o h
~~- <; .14: 6 , 6 BIBLIOTECA ~oi;.
,~ q ' ~ ' .n1 .• c:, ·c;.

•
found with smoke contamination are freely soluble in ethyl alcohol %% giving a characteristicyellow color theintensity of which càn be assessed both visually and spectrophotometrically. For practical purposes the method permits an accurate visual determination by comparing prepared standards with samples made on the spot from any lot of cocoa beans. Such a method is expected to be used at the cocoa grading stations to detect smoke contaminated cocoa beans.
20
ECONOMICIDADE DOS INSUMOS MODERNOS EM FAZENDAS DE CACAU•
José Alexandre de Souza Menezes** Euter Paniago *** Hércio Pereira Ladeira**** Antonio Lima Bandeira****
A CEPLAC vem, desde 1%4, orientando e estimulando os cacauicultores no uso de melhores métodos e modernos fatores de produção e, ao mesmo tempo, proporcionando meios para a ampliação das áreas de cultivo (1). Espera-se que os acréscimos de produção, obtidos através do uso de recursos maio;; eficientes, possam diminuir os custos marginais de produção e que tal fato resulte na expansão da própria oferta do produto, em condições competitivas.
Aceita-se, em geral, que o objetivo do agricultor, ao adotar inovações, é basicamente obter uma vantagem econômica a qual depende, em princípio, de como se comportam as produtividades físi-
cas dos insumos modernos utilizados, das relações de preços desses fatores e do produto resultante.
Todavia, a introdução de inovações com a finalidade de modernizar a agricultura, nem sempre redunda em benefícios a nível de fazendas.
Com respeito a fertilizantes, Schuh e Tollini (3) opinam que os agricultores poderão incorrer em grandes perdas, se não tomarem decisões corretas acerca da quantidade e do tipo de fertilizante a usar.
A Teoria Econômica pressupõe que a firma opere no sentido de maximizar lucros sob condições em que cada recurso deva ser
*Recebido para publicação em agosto, 1973. * Resumo da tese inlitulada"Produtividade e Taxa Marginal de Retomo de
Insumos Modemos em Fazendas de Cacau, Região Cacaueira da Bahia, Ano Agrícola 1971 172~, apresentada pelo primeiro autor à Universidade Federal de Viçosa para obtenção do grau de ~Magister Scientiae".
** M.S., Divisão de Sócio-Economia, CEPEC. *** Ph.D., Departamento de Economia Rural, Universidade Federal de Viçosa
(DER-UFV), Viçosa, MG. **** M.S .. DER-UFV.
Revista Theobwrna. CBPBC, Ilhéus, Brasil, 4(2):21-33. Abr.-;r.m. 1974.
21

• . ,
usado no nível em que o valor do seu produto marginal iguale o preço do referido recurso.
Os insumos modernos, recomendados pela CEPLAC, requerem inversões adicionais cujos retornos dependem das suas produtividades marginais e do preço do produto obtido. O problema se afigura na incerteza de se saber se o valor da produtividade marginal dos recursos é suficientemente alto para cobrir tais inversões,- dada certas condições de preços.
A utilização, a nível de fazenda, dos insumos modernos recomendados pela CEPLAC conduz a duas importantes indagações: 1. É o valor da produtividade marginal desses insumos adequado no presente nível de uso? e 2. Qual a taxa marginal de retorno desses insumos?
Tais indagações motivaram o presente estudo cujo objetivo consiste em analisar a produtividflde e taxa marginal de retorno dos recursos utilizados na produção de cacau, principalmente insumos modernos recomendados pela CEPLAC, em fazendas de cacau da Bahia, no ano agrícola de 1971/72. Especificamente, .procurou-se:
1. Estimar e examinar o valor do produto marginal dos recursos e insumos modernos;
2. Estimar as elasticidades parciais e totais de produção dos recursos e insumos modernos; e
3. Estimar e analisar o retorno lí-
22
quido marginal e a taxa marginal de retorno dos recursos e insumos modernos.
MATERIAL E MÉTODO
Origem dos dados
Os dados utilizados no presente trabalho provêm de um projeto de contabilidade agrícola, conduzido em 80 fazendas de cacau -localizadas nos principais municípios produtores da Bahia - sob orientação do Departamento de Extensão da CEPLAC (DEPEX). Esse projeto tem o objetivo de oferecer aos~ cacauicultores participantes um processo simples e racional para medir os resultados alcançados em suas atividades agrícolas e desenvolver o hábito de utilizar a contabilidade agrícola como um instrumento de administração. Além disso, o projeto visa a coletar dados para determinar padrões e requerimentos de mão-de-obra para as práticas agrícolas, insumos modernos, dispêndios, margens de lucro da empresa por roça*, gastos e rendimentos de mão-de-obra sob regimes de trabalho por empreitada ou por diária. As unidades selecionadas para o projeto estavam enquadradas em critérios tais como: empresa de fácil acesso e com área entre 20 e 200 hectares; agricultores conscientes, dispostos a participarem do projeto, receptivos a mudanças tecnológicas, alfabetizados e com
* O termo "roça" refere-se às divisões, em áreas produtivas, das fazendas de cacau.
idade adequada para o desempenho da tarefa.
Modelo conceitual
Tendo em vista os objetjvos do estudo e o número de observações utilizáveis no que diz respeito a graus de liberdade e outros aspectos estatísticos, escolheu-se como instrumento de análise a função de produção tipo Cobb-Douglas, expressa em termos logarítmicos,
Avaliação e especificação das variáveis
De acordo com os objetivos do estudo, foram definidas as seguintes variáveis para a construção de equações que permitissem explicar a produção de cacau:
Y - Produção de cacau (arrobas). X, - Valor de uso da terra com
cacauais (cruzeiros). X
2 -Área com cacauais produti
vos (hectares). X1 - Investimento em animais de
serviço (cruzeiros). X4 - Investimento em benfeitorias
(cruzeiros). X5 -Investimento em equipamen
tos (cruzeiros). X6 - Mão-de-obra em tratos cultu
rais (cruzeiros). X7 - Mão-de-obra em beneficia
mento (cruzeiros), X8 - Despesas diversas (cruzei-
ros). x. -Inseticida (sacos). X,o- Inseticida (cruzeiros). X, - Fungicida (sacos). X11 - Fungicida (cruzeiros). X11 - Fertilizantes (sacos).
Xu- Fertilizantes (cruzeiros). X,. -Calcário dolomítico (sacos). X16 -Calcário dolomítico (cruzei-
ros). x,7 - X1 + x4 + x. X,.-X6 +X7 X19 - X, 0 + X,1
~o- X,4 + X,6
XH - x20 + X,o Xa- X2o + X, 2
X11 - X,9 + X20
X24 - X18 + X"
Renda bruta X,,-
Custos totais variável
auxiliar assumida para introduzir o efeito da administração na função de produção. Esta variável foi utilizada por Ladeira (2).
23
Sabe-se que a função de produção é um relacionamento entre o produto físico obtido e os recursos físicos utilizados, daí a conveniência de se mensurar esse relacionamento em unidades físicas apropriadas. Todavia, devido à necessidade de se agregar insumos, medidos em unidades heterogêneas e cuja medição só pode ser feita em unidades homogêneas, fêz-se a agregação usando-se termos de valor. Sabe-se que, se existir outra relação que não seja competição perfeita para os mercados de fatores e do produto, a medição das variáveis em termos de valor pode causar tendenciosidade (4). Foi assumido, neste estudo, para a medição das variáveis da função de prOO ução de CobbDouglas, especificadas em termos de valor, a existência de merca-

dos de competição perfeita. Esta pressuposição é feita com base em razões lógicas, desde que não é conhecida a natureza dos mercados de fatores e do produto, através de estudos empíricos, para a região estudada.
Na avaliação das vanaveis, considerou-se a produção de cacau (Y), no ano agrícola 1971/72. A variável terra com cacauais (X1), refere-se ao fluxo de serviços prestados na produção pelos cacauais produtivos, fluxo esse que foi calculado através de depreciação linear, pelo período de 50 anos. Os investimentos em animais de serviço (X,), benfeitorias (X 4 ) e equipamentos (X 5 )
referem-se ao fluxo de serviços prestados, o qual foi calculado através de sua depreciação além de se considerar os reparos e manutenção. As variáveis mão-deobra para tratos culturais (X,) e mão-de-obra para beneficiamento (X1) foram medidas em cruzeiros, dada a dificuldade de expressá-las em unidades físicas. As despesas diversas (X8 ) referem-se a várias despesas não enquadráveis nas categorias já descritas. Os insumos modernos foram medidos em termos de valor e em físicos. Os preços dessas variáveis foram estabelecidos com base no valor do dinheiro empregado em cada uma delas. O preço da variáve I dependente (Y) foi estimado em Cr$23,33 por arroba. Para as variáveis independentes, arbitrou-se uma taxa de juro$ para cada cruzeiro investido.
24
Procedeu-se ao ajustamento dos parâmetros das equações de regressão pelo processo de estimação de mínimos quadrados.
RESULTADOS E DISCUSS,\0
Resultados estatísticos
Os dados foram ajustados pelo método dos mínimos quadrados à equação tipo Cobb-Douglas, expressa em termos logarítmicos. Dentre as várias equações ajustadas, uma delas sobressaiu-se por apresentar coeficientes de regres são altamente significativos e outros indicadores estatísticos com características desejáveis para a análise econômica. As variáveis componentes desta equação estão expressas no Quadro 1, onde se observa que:
a. Todos os coeficientes de regressão da equação escolhida apresentam-se com o sinal esperado;
b. Todos os coeficientes de regressão apresentam estimativas confiáveis no que tange ao nível de significância estatística (teste t);
c. A equação escolhida apresenta alto coeficiente de determinação múltipla (R2
), sendo aproximadamente igual a 0,99, indicando que cerca de 99% da variação total de Y é explicada pela variação das variáveis independentes;
d. Não foi observada, na matriz de correlação simples, presença da multicolinearidade, entre as variáveis independentes;
Quadro 1 -Valores eslalísticos das lJariát•eis da equação, ajustada para a produção de cacau. Região cacaueira bai011a. ano agrícola 1971 /72 .
Variáveis
Xt -Terra com cacauais (Cr$) Xs - Despesas diversas (Cr$) X17- Benf., animais e equip. (Cr$) Xts- Mão-de-obra (Cr$) X23- Insumos modernos (Cr$) X2s- Administração
Constante de regressão (a) 0,546.
Coefic. de regressão
0,221 0,048 0,14Q 0,437 0,108 0,914
Erros padrão
0,021 + +
0,007 +. 0,014 •• 0,026-+ t
0,017 ~ +
0,033 + t-
Coeficiente de determinação múltipla (R2) • 0.99. Estatística de Durbin Watson (d) 2,149. + + Significante ao nível de 1%.
e. O teste de Durbin-Watson, para os resíduos, sugere ausência de correlação serial ou perturbação no termo do erro.
A representante da equação escolhida, para análise econômtca, em forma logarítmica é:
log y = 0,5460 + 0,2210 log xl + 0,481 log X, +0,1495 log X17 + 0,4368 log xl. + 0,1082 log XH + O, 9142 log X25 •
Análise econômica
O Quadro 2 apresenta o uso atual (médio) dos recursos nas fazendas estudadas e a produção média obtida. Dos valores expostos, calculou-se que, para se produzir 2.460 arrobas de cacau, valor considerado como média geométrica da produção das 80 fazendas, o custo total foi de aproximadamente Cr$34.420,00 (média
25
geométrica). Considerando-se o pre<,:o médio anual de cacau, no período de 1971/72 (Cr$23,33), para uso tota I dos recursos e para a produção de 2.460 arrobas, assinalou-se uma renda líquida aproXJmada de Cr$22.960,00. Fazendose o cálculo para 1.000 arrobas, tem-se o custo total igual a Cr$ 13.990,00, renda bruta de Cr$23.330,00 e renda líquida de Cr$9.340,00. Assim, a renda bruta auferida no ano considerado paga o fluxo dos serviços dos fatores terra , capital em benfeitorias, animais e equipamentos, mão-de-obra, despesas diversas e insumos modernos. E sobram Cr$9.340,00/ 1.000 arrobas para remunerar a capacidade empresarial da firma.
Para se observar como cada recurso influi no custo total, foi calculada a participação percentual de cada uma das principais categorias de recursos (Quadro 3).

Quadro 2 - I'rodu('ào e u<;o atual dos recursos r1as fazendas de carau estudada<;, Rf!git7o racauC'ira baiana, ano agrícola 1971 /72.
Especifícaçllo
Produção de cacau Área da fazenda Valor de uso da terra Valor de uso das benfeitorias Valor de uso dos equipamentos Capital investido em anim. de aerv. Mi<Hie-obra para tratos culturais MAo~e-obra pua beneficiamento Despesas diversas Inseticidas Fungicidas Fertilizantes Calcário dolomitico
*Média geométrica.
Unidade
arroba h a Cr$ Cr$ Cr$ Cr$ Cr$ Cr$ Cr$ saco saco saco saco
Quantidade*
2.460,00 49,50
5.532,00 3.748,00
334,00 1.097,00 5.319,00 6.092,00 2.356,00
23,00 3,00
136.00 6,00
Quadro 3 - Participação do.-; recursos utilizados pelas fazendas de cacau no custo totul tie protiuçilo. Região cacaueiro baiana. ano agríco · la 1971/72.
Recursos % do custo total*
Valor de uso da terra com cacauais Valor de uso de benfeit., animais e equipamentos Mão-de-obra
18 20 39 15 Insumos modernos
Despesas diversas
*Média geométrica.
As elasticidades parciais de produção, representadas pelos coeficientes de regressão do modelo selecionado, na média para cada recurso, mantido os outros constantes, são apresentadas no Quadro 4. Os resultados indicam que:
a. Um aumento de 10% no valor do capital do fluxo de serviços prestados por benfeitorias, an i-
26
8
mais e equipamentos (X 17 ) determinaria um aumento de 1,5"/o na produção de cacau.
b. Um aumento de 10"/o no recurso de mão-de-obra (X,.) determinaria um aumento de 4,4"/o na produção de cacau.
c. Um aumento de 10% no valor dos recursos insumos modernos (X,) acarretaria um aumento de 1,1% na produção de cacau.
d. Um aumento de 10% no fluxo de
Quadro 4 - Flasttcidade'> parciais de produção tios recursos, envoluitlos na produção tie cacau. Região cacaueiro baiana, ano agrícola 1971 /72.
Recursos
Terra com cacauais
Elasticidade parcial de produçlo {bi) *
Benfeitorias, animais e equipamentos Despesas diversas
0,221 0,149 0,048 0,437 0,108
Mão-de-obra Insumos modernos
* bi = Coeficientes das variáveis independentes da regressão e que no modelo escolhido equivale à elasticidade parcial de produção.
serviços prestados pelo recurso terra com cacauais (X,), resultaria em um acréscimo de 2,2% na produção de cacau.
e. Um aumento de 10% nas despesas diversas (X,) acarretaria um aumento de 0,5% na produção de cacau.
Observou-se que as elasticidades parciais de produção são positivas e menores que a unidade. Na média, 1sso sugere que os fazendeuos estão operando com os recursos, como considerados, na amphtude racional de produção.
Caso se aumentasse de 10% todos os fatores simultaneamente, a produção aumentaria de 9,63%.
O teste estatístico comprovou que a soma dos coeficientes de regressão é igual à unidade. Não foi incluído na soma o coeficsente da variável auxlliar adm1n1stração (XH ), pois poderia ser sugendo que o retorno, possivelmente crescente, era devido à administração, e não à escala.
A efíciênc1a do uso dos recursos na produção de cacau, com insumos modernos, foi avaliada através da relação do valor dos orodutos marginais dos fatores (VPM) e seus respectivos preços (Pxi) (Quadro 5). Notou-se que os VPM para os recursos considerados na média são sempre maiores que seus respectivos preços, o que sugere que os recursos, apesar de estarem sendo usados na amplitude racional de produção, não satisfazem à condição do emprego no nível óhmo. Para alcançar este nível, todos os recursos devem ser incrementados.
Pressupondo condições de competição perfeita para produto e fatores de produção, obteve-se a estimativa do retorno líquido marginal (RLM), subtraindo-se do valor do VPM o preço do recurso (Pxi). A TMR foi obtida divid1ndo-se o RLM pelo Pxi. É aqui admitido que, no ponto onde ocorre o nível de emprego ótimo de recurso, o RLM e a TMR são iguais a zero; 1sto é, ambos podem ser pos1tivos
27
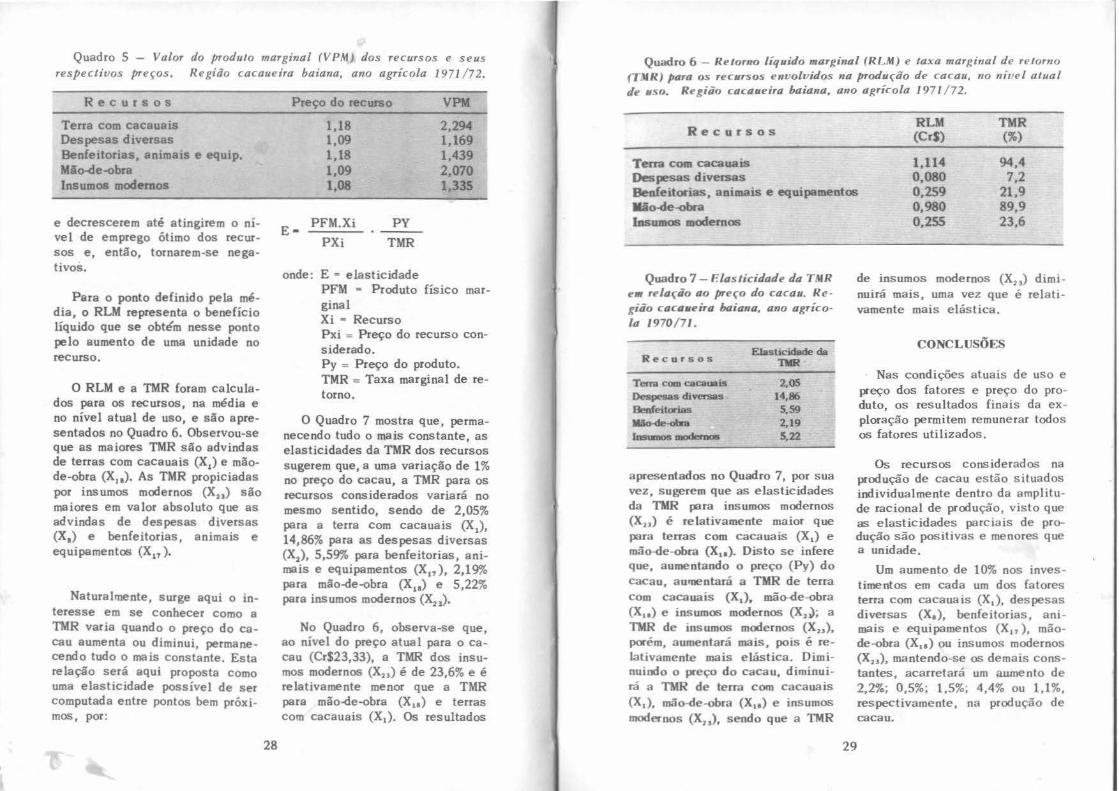
Quadro 5 - Valor do produto marginal (VPM) dos recursos e seus respectivos preços. Região cacaueiro baiana, ano agrícola 1971/72.
Recursos
Terra com cacauais Despesas diversas Benfeitorias, animais e equip. Mio-de-obra Insumos modernos
e decrescerem até atingirem o níveL de emprego ótimo dos recursos e, então, tornarem-se negativos.
Para o ponto definido pela média, o RLM representa o benefício líquido que se obtém nesse ponto pelo aumento de uma unidade no recurso.
O RLM e a TMR foram calculados para os recursos, na média e no nível atual de uso, e são apresentados no Quadro 6. Observou-se que as maiores TMR são advindas de terras com cacauais (X1) e mãode-obra (X,.). As TMR propiciadas por insumos modernos (X23) são maiores em valor absoluto que as advindas de despesas diversas (X,) e benfeitorias, animais e equipamentos (X17 ).
Naturalmente, surge aqui o interesse em se conhecer como a TMR varia quando o preço do cacau aumenta ou diminui, permanecendo tudo o mais constante. Esta relação será aqui proposta como uma elasticidade possível de ser computada entre pontos bem próximos, por:
28
Preço do recurso VPM
E•
1,18 1,09 1,18 1,09 1,08
PFM.Xi
PXi
py
TMR
2,29-t 1,169 1,439 2,070 1,335
onde: E = elasticidade PFM 5 Produto físico marginal Xi - Recurso Pxl = Preço do recurso considerado. Py = Preço do produto. TMR = Taxa marginal de retorno.
O Quadro 7 mostra que, permanecendo tudo o mais constante, as elasticidades da TMR dos recursos sugerem que, a uma variação de 1% no preço do cacau, a TMR para os recursos considerados variará no mesmo sentido, sendo de 2,05% para a terra com cacauais (X
1),
14,86% para as despesas diversas (X2), 5,59% para benfeitorias, animajs e equipamentos (XI?), 2,19% para mão-de-obra (X11) e 5,22% para insumos modernos (~ 1).
No Quadro 6, observa-se que, ao nível do preço atual para o cacau (Cr$23,33), a TMR dos insumos modernos (X~,) é de 23,6% e é relativamente menor que a TMR para mão-de-obra (Xu) e terras com cacauais (X1). Os resultados
Quadro 6- Retorno líq11ido marginal (RLM) e taxa mar~inal de retomo fT.URJ ptiTa os recursos envolv idos na produção de cacau. no nível atual tfe •so. Região caca•eira hnia11a, ano agrícola 197 1/72.
Recursos
Terra com cacauais Despesas diversas
RLM (Cr$)
TMR (%)
94,4
Benfeitorias, allimais e equipamentos lU o-de-obra
1,114 0,080 0,259 0,980 0,255
7,2 21,9 89,9 23,6 las1U1106 modernos
Quadro 7- 1:/asliciJaJe da TMR e• rt!lação ao fJreço do cacau. Região caca11eira baiana, ano agrícola 1970/71.
Recursos
Teml com caca .. ill Despesas diversas Benfeitorias lllo-de-obna .......,.....,_
2,05 14,86
5,59 2,19 5,22
apresentados no Quadro 7, por sua vez, sugerem que as elasticidades da TMR para insumos modernos (Xu) é relativamente maior que para terras com cacauais {X1 ) e mão-de-obra (X .. ). Disto se infere que, aumentando o preço (Py) do cacau, aumentam a TMR de terra com cacauais (X,), mão-de-obra (Xu) e insumos modernos (X2); a TMR de insumos modernos (X,), porém, aumentará mais, pois é relativamente mais elástica. Diminuindo o pr-eço do cacau, diminui rá a TMR de terra com cacauais (X,), mão-de-obra (X,.) e insumos modernos (X73), sendo que a TMR
de msumos modernos <Xn) diminuirá mais, uma vez que é relativamente mais elástica.
CONCLUSÕF.S
Nas condições atuais de uso e preço dos fatores e preço do produto, os resultados finais da exploração permitem remunerar todos os fatores utilizados.
Os recursos considerados na produção de cacau estão situados individualmente dentro da amplitude racional de produção, visto que as elasticidades parciais de produção são positivas e menores que a unidade.
Um aumento de 10% nos mvestimentos em cada um dos fatores terra com cacauais (X,), despesas diversas (Xa), benfeitorias, animais e equipamentos (X17 ), mãode-obra (X,.) ou insumos modernos (X21), mantendo-se os demais constantes, acarretará um aumento de 2,2%; 0,5%; 1,5%, 4,4% ou 1,1%, respectivamente, na produção de cacau.
29

'
Todos os recursos c-onsiderados no modelo podem ter seus usos incrementados, uma vez que o valor de suas produtividades marginais são maiores que seus respectivos preços.
A soma dos expoentes da função ajustada é, estatisticamente, igual à unidade e define um retorno constante à escala.
Os insumos modernos (X,.), assim como os demais recursos, se forem aumentados à margem, apresentam resposta econômica favorável. A taxa marginal de retorno para insumos rnodernos (X,) é de 24%.
Dentro das possibilidades de produção, deve-se incrementar os investimentos nos recursos usados na produção de cacau, segundo a ordem seguinte· 1. Terra com cacauais (X,); 2. Mão-de-obra (Xu); 3. Insumos modernos (X"); 4. Benfeitorias, animais e equipamentos (X11 ) e 5 Despesas diversas
(X.).
Aumentando o preço do cacau, (tudo o mais constante) aumentará a TMR de todos os recursos utilizados na produção. A TMR de insumos modernos (Xll), porém, aumentará mais que as de terra com cacauais (X,) e mão-de-obra (X,.), uma vez que é mais elástica.
AGRADECIME:\ITOS
Ao DEPEX, especialmente ao Eng'? Agr. Edmundo P. Mandarino e agrônomos participantes do projeto de contabilidade agrícola, pela coleta das informações sem as quais não seria possível a execução do presente trabalho, e à Eng? Agr. Maria l-lelena Alencar, pelas constantes sugestões.
LITERATURA CITADA
1. A L VIM, P. de T. e ROSÁRIO, M. Cacau ontem e hoje. ltabuna, CEPLAC, 1972. 83 p.
2. LADEIRA, H.P. Produtividade dos recursos na produção de cacau, região cacaueira, Bahia. Tese de M.S. Viçosa, MG, Universidade Federal. 1971. 75 p.
3. SCHUH, G.E. e TOLLINl, H. Análise econômica de ensaios de adubação. In Reunião Brasileira de Fertilidade de Solo, 7a., Itabuna, Bahia, Brasil, 1972.
4. TEIXEIRA FILHO, A.R. An economic evaluation of methodology employed in the estimation of farm levei production functions. Ph.D. Thesis. s.l. Purdue University. 1970. 199 p.
30
5. TOLLINI, H. Indústria de fertilizantes na América Latina: Preços relativos a requisitos de desenvolvimento. Viçosa, MG, Universidade Federal, 1971, 8 p. (mimeografado).
RESUMO
O presente estudo focaliza a utilização, a nível de fazenda de cacau, no Sul da Bahia, dos insumos modernos recomendados pela Comissão Executiva do Plano da Lavoura Cacaueira (CEPLAC), no ano agrícola 1971/72. Esses insumos demandam inversões adicionais e os retornos a esses investimentos dependem da produtividade marginal desses recursos e do preço do produto obtido. Tal fato não permite saber se 0 valor da produtividade marginal é suficiente para cobrir tais inversões.
Os objetivos específicos do estudo foram: 1. Estimar e analisar o valor do produto físico marginal dos recursos e insumos modernos; 2. Estimar as elasticidades parciais e totais de produção; e 3. Estimar e analisar o retorno líquido marginal e a taxa marginal de retorno.
Para estudar aspectos ligados à fase produtiva, usaram-se dados provenientes do projeto de Contabilidade Agrícola conduzido nas fazendas pelos produtores e orientado pela CEPLAC.
Foram ajustadas funções de produção do tipo Cobb-Douglas, por regressões, usando-se a técnica dos quadrados mínimos.
Entre as equações ajustadas, foram apresentadas quatro e, entre elas, escolhida uma, por se apresentar com coeficientes de regressão altamente significativos e outros indicadores estatísticos com características desejáveis do ponto-de-vista estatístico, com vistas à análise econômica.
O produto obtido e os recursos utilizados foram medidos em arrobas (15 kg) e pelo valor do fluxo de serviços prestados, respectivamente.
A elasticidade parcial de produção indicou que todos os fatores de produção, inclusive insumos modernos, estão positivamente relacionados com a produção do cacau. Um aumento de 10% em investimentos com insumos modernos aumentaria em 1,1% a produção de cacau. Os produtores de cacau estão atuando no estádio racional de produção sem, contudo, atingir ao ótimo econômico. É necessário aumentar o uso de todos os recursos considerados. Nas condições atuais de uso desses recursos, os resultados finais permitem remunerar todos os fatores de produção.
31

A taxa marginal de retorno foi estimada em 94,4% para terras com cacauais; 7,?% para despesas diversas; 21,9% para fluxo de serviços em benfeitorias, animais de serviços e equipamentos; 89,9% para mão-de-obra e 23,6% para insumos modernos. O retorno líquido marginal indicou que, para o próximo cruzeiro investido em terras com cacauais, haverá lucro de Cr$ 1,11; Cr$ 0,08 para despesas diversas; Cr$ 0,26 para benfeitorias animais e equipamentos; Cr$ 0,98 para mão-de-obra e Cr$ 0,25 para insumos modernos.
Os resultados sugerem que os insumos modernos aplicados em cacauais baianos são rentáveis. Esta rentabilidade, porém, aproximadamente igual a 24%, pode ser aumentada mais rapidamente que a dos outros recursos, dado o comportamento do preço desses insumos, orientação da pesquisa agrícola e elasticidade da taxa marginal desses mesmos insumos ser mais sensível às variações de preço de cacau que a dos demais recursos.
ECONOMIC ASPECTS DF MODERN INPUTS USED IN BAHIAN COCO A F ARMS
SUMMARY
The present study considers the utilization of the modero inputs such as fertilizers, fungicides, insecticides, etc recommended by Comissão Executiva do Plano da Lavoura Cacaueira (CEPLAC) in cocoa production, at the farm levei in southern Bahia during the agricultura! year 1971/72. Considering that these inputs require additional investments and the return of these investments depends on the marginal productivity of the resources and the price of the product obtained, the problem seems to be the uncertainty of knowing whether the value of the marginal productivity is sufficient to cover that investment.
The specific objetives of this study were; 1) to estimate and analyze the value of the marginal physical product of the resources and modero inputs, 2) to estimate the partia! and total elasticities of production, and 3) to estima te and analyze the marginal net return and the marginal mte of return of investment.
The data used to study the relevant aspects of cocoa production were obtained from farm records from the Agricultura! Records Project conducted by CEPLAC. These farms, which comprised a distinct population, kept records in a uniform manner. All utilized fertilizers, herbicides and fungicides according to CEPLAC's recomendations.
32
Cobb-Douglas production functions were estimated using the ordinary least squares technique. Four regression equations were estimated, of which one was chosen for detailed analysis. Its regression coefficients were highly significant. Also included in the analysis were other indicators, such as the corre!ation matrices. These demonstrate desirable characteristics from a statistical point of view and also provide insights for economic analysis.
The production obtained was measured 10 units of arrobas (1 arroba '" 15 kg), and the resources utilizeJ were measured by the value of the flow of services rendered.
The partia} elasticities of production suggested that ali the factors, including modero inputs, made a positive contribution to the production of cocoa. An increase of 10% in investments in modero inputs causes a 1.1% increase in the physical production of cocoa. Constant returnsto-scale were implied by the analysis,
The results suggest that the cocoa producers studied are producing at the rational stage of production although the economic optimum has still not been reached. lncreases are implied in the levei of use of all resources considered.
Under current conditions of resource use, the conclusion is reached that the output obtained is adequate to remunerate ali the factores of production.
The results suggested that the modero inputs applied in Bahia cocoa production are profitable, but this profitability, approximately equal to 24%, could be increased more rapidly than that for other inputs, given the behavior of the prices of modern inputs, given the orientation of agricultura! research, and given that the elasticity of the marginal rate of return on investment for modem inputs is more sensitive to variations in the price of cocoa than is that for the other resources,
The marginal rate. of return for land in cocoa production was estimated at 94.4%; for miscellaneous expenditures, 7.2%; for flow for services of improvements, livestock and equipment, 21.9%; for manual labor, the estimated rate for return was 89.9% and for the modero inputs, 23.6%. The marginal net return on working capital indicates that for the next cruzeiro invested in land in cocoa production, there is a gain of Cr$ l.ll; for miscellaneous expenditures, Cr$ 0.08; for buíldings, livestock and equipment, Cr$ 0.26; for manual labor Cr$ 0.98, and for modero inputs, Cr$ 0.25.
® ® ®
33

OBSERVAÇÕES CITOLÓGICAS EM CÉLULAS MÃES DE POLEM
DE CACAUEIROS * Geraldo Adami Carletto **
Embora se constitua numa planta de grande importância econômica, o cacaueiro não tem recebido dos biologistas a devida atenção no que se refere aos aspectos citogenéticos relacionados com a produção.
Enriquez (1) observa que os trabalhos existentes sobre a estrutura cromossômica do cacaueiro são escassos e versam sobre o número, tamanho, forma e processo meiótico de divisão das células mães de polem (CMP).
Recentemente, Opeke e Jacob (4) mostra ram que o clone autoincompatível Na-32 apresenta associações cromossômicas mono, bi e tetravalentes, diferentes da associação teórica esperada para um di-
plóide. Por este motivo, formularam a hipótese de que o cacaueiro seria uma tetraplóide e que estas irregularidades no pareamento dos cromossomos poderiam servir para explicar o fenômeno de incompatibilidade.
A necessidade de maiores conhecimentos sobre os mecanismos citogeoéticos do cacaue1to motivou a presente investigação.
MA TERIA L E MÉTODO
O presente trabalho foi realizado no Centro Tropical de Ensino e Investigação do Instituto l nteramericano de Ciências Agrícolas (IICACTEI), Turrialba, Costa Rica, utilizando-se os clones autocompatí-
• Recebado pora publlcação em fevereiro, 1973. • Extraado do tese Ulhtulada "Estudo dos mecanismos detemanantes de graus
de autocom.,.tíb&lidade em Theobroma cacao L. •, apresentada ao UCACTEI, Turriatba, Costa Rica, para obtenção do título de Maglster Scientiae.
•• !11. S., Di•isio de Genética, CEPEC.
Ret•ísla TJ.cobroma. CE:.PEC., Ilhéus, Brasil, 4 (2) · 34 -40. Abr. -Jun. 1974.
34
veis ICS-1, UF-667, UF-29 e seus luõridos com os autoincompatíveis Sca-6 e DIC-67.
Foram coletados, em todos os cultivares, botões florais joveos e flcxes abertas no mesmo diâ, entre 7 e 12 horas da manhã. Os botões florais foram fixados em Cacnoy 6: 3 : 1, poc 36 horas, em baixa temperatura, para observação das CMP. As associações cromossômicas das CMP foram observadas em metafase I através de preparações temporárias coloridas com aceto-carmim.
A coleta das flores para observação da normalidade dos grãos-depolem foi feita em bolsas de polietileno revestido internamente com papel de filtro umedecido. Considerou-se normal todo grão-de-polem colorido pelo Cottoa bJue e que apresentasse .a forma esférica típica. Para efeito de uniformização de amostragem, somente foram considerados aqueles grãos que estavam dentro da área previamente estabelecida no campo visual do microscópio.
ltESUL TADOS
Nos Quadros 1 e 2, são apresentados os resultados de observações do pareamento de cromossomos de CMP de clones autocompatíveis e seus lu"bridos com um pai autoincompativeL
Os clones UF -667 e UF -29 apresentaram um número de associações cromossômicas bivalentes maior do
Quadro 1- Associações cromos sômicas das células mães de polem (CMP) em metáfase 1 dos clones ••tocom/)atíveis.
X!CMP
l 3 2 3 I I 1
11 2 I
9 J I 1
I
o 2 4 1 3 9 4
o I 4
• 2 .. •
Aaaocs.çaea•
B m ICI- 1
10 o 9 o I o I I 7 I 4 1 5 2
UF- 667
10 o I 1 5 2
UF- 29
lO o 9 o • o
' 2
• Associações: 1-mooovalentes; nbivalentes e m - trivalentes.
que a do ICS-1. Estes valores indicam que existe maior uniformidade no processo da divisão das CMP para formação dos grãos-de-polem nos dois primeiros cultivares. Nos híbridos com um pai autoincompatível observou-se uma baixa freqüência de associações bivalentes.
Partindo do pressuposto de que se deveria encontrar uma relação perfeita entre a freqüência de pareamento normal dos cromossomos e a normalidade dos grãos-de-polem, foram procedidas as observações
35

Quadro 2-A'>.'oocioções cromos· ~tJmíco~ dt> célula~ mde.s áe polem f(. MJ>J em metáloH' f dr~s híhridos.
N! CMP
4 3 2 2 2 1
.. 3 2 2 2
.. 4 2 I I I
3 5 3
3 5 I 3 I I
As•oc'-ç6e• •
11 DI
ICS - 1 aJIIC- 67
o 2 4 6 I 7
lO 9 8 7 8 5
o o o o I I
ICS - 1 x Sca -6
o .. 3 I 8
lO
• 1 8 3
o o I 1 l
UF - 667 • Sca-6
o 2 4 6 I 3 8
lO 9 8 7 8 7 3
o o o o I I 2
UF - 667 x IIIC - 67
o 4 2 I 5 2
lO 8 9 8 6 6
o o o I 1 l
UF -29 11 IIIC - 67
.. 2 8 ~ 7 o
8 9 6 6 5 7
o o o I 1 2
• As~ocaaçõe,:; · 1- monovalenlcs; 11 -bavah>nh.•s e 111 - lravalentes.
constantes dos Quadros 3 e 4. nos clones autocomp;itíveis e em seus híbndos com um dos pais autoin · compatíveis. As percentagens médias de pólens norma1s foram bas tante altas (91% a 99%), tanto nos clones como em seus hibrtdos.
DISC USS;\o t-; CONCLUSÕES
Tanto os clones uutocompatívei:; (JCS-1, UF -667 e UF -29) como os seus híbndos com os clones autoincompatíveis (Sco.~ -6 e IMC-67) apresentaram dSSOC1ações cromossômicas mono, bi e trivalentes na formação de gametas. Estes resultados divergem do reg1strado por Opeke e Jacob (4) que encontraram associações cromossômicas mono, bi e tetravalentes no cultivar Na-32. Da1 formularem a hspotese de que o cacaueuo seria um tetraplóide e que essas irregularid<.1des no p<.~re<J · mento de cromossomos poderiam servir de base, também, para expli car o fenômeno da 1ncompat iblll dade.
36
Os resultados da presente mves · t igação. juntamente com as mformações de Muõoz Orteg<~ (3}. que o número bás1co de cromossomos de todas as espécies do gênero Theohromo é 2n = 20, e Knight (2). que polípló1des Induzidos em cacaueiros têm o comportnmenlottplcodes sas plantilS, sugt>rem que a hipó· tese formulada por Opeke e Jacob (4)deva ser objeto de estudos ma1s profundos.
Quadro 3 - \"ti mero á e grãos-dt!-pol~m norma i\ (\' J. anormais ( ;\N J <'
pcrcentagt'l'l médio de normais f"ó ~) de algum, c fones .
j N~ da observaçio
l 2 3 4 5 6 7 8 9
10 11 12 13 14 15 16 17 18 19 20
I 20
~N
ICS- 1 N AN
43 49 27 56 34 54 47 44 45 72 56 44 92 86 44 56 77 28 31 87
7 6 8 4 2 3 3 1 7 6 7 8 8 4 2 5 1 6 4 8
1072 100
91,47
Considerou-se que a normalidade do polem poder ia estar relacionada com a alta freqüência de associações cromossômicas biva lentes no processo de div1são das CMP. Os valores encontrados para pólens norma1s (91 a 99%) indicam que não há relação entre os dois fenômenos, uma vez que não se cumpriu a expectação teórica tanto nos cultivares autocompatíve1s como nos
UF - 667
N AN
43 34 32 38 28 66 47 37 30 34 34 39 33 42 37 70 46 67 39 43
o o 2 o 1 o o o 2 o l 1 o o 1 2 o 1 o o
839 11
98,57
UF - 29 N AN
41 28 43 32 29 46 32 37 38 29 28 42 35 38 32 56 28 44 49 39
1 o 2 4 4 1 o 1 2 o 3 2 3 o o o o 3 1 o
746 27
96,50
seus híbrido.<; com clones amazônicos autoincompatíveis. Os va !ores de polem normais encontrados servem para determinar graus de fertshdade em cacaueiros e não aJudam à explicação do comportamento de cacaue1ros incompatíveis.
Admitem-se como preliminares os resultados obt1dos no presente trabalho, uma vez que as observa-
37

Quadro 4- Núm~ro de gri!os-de-polem normais (N), anormais (A .'l), e percentagem d~ normais i!f.N) de alguns cultivares híbridos.
UF-f67a UF-667 X JCS-1 X ICS-1 X UF-29 X I N~ da Scll-6 IIIC-67
!Pbservaçio N AN N AN
1 89 2 53 o 2 62 4 63 3 3 45 2 88 o 4 43 1 40 1 5 78 1 44 3 6 94 1 44 2 7 72 l 101 18 8 38 2 81 1 9 41 o 142 3
10 56 l 27 42 11 36 o 44 3 12 74 l 89 o 13 62 o 74 o 14 46 1 27 1 15 36 4 94 7 16 93 4 64 5 17 36 2 99 3 18
. 37 4 78 1
19 43 1 49 o 20 62 o 101 6
I. 20 1143 32 l4cr2 99
"N 9'1:rl 93.40
ções do pareamento de cromossomos não puderam ser feitas em prófase (paquiteno e diacnesis) e sim em metáfase I, face ao pequeno ta-
IIIC-67 Sca-6 IMC-67 N AN N AN N AN
33 o 32 o 36 1 45 o 38 2 83 o 38 o 39 1 42 o 49 o 51 o 60 1 63 1 41 o 39 o 64 o 32 1 46 o 49 o 37 4 118 o 67 o 43 4 51 o 42 1 32 6 46 o 34 o 34 6 67 9 31 5 32 1 63 o 40 o 43 2 56 o 39 o 30 1 75 o 48 o 32 o 60 2 66 o 37 2 74 8 38 4 42 1 83 4 64 o 39 2 60 o 32 1 56 o 44 3 41 o 51 1 43 2 58 3 44 2 58 o
941 15 786 36 1204 30 98",48 95,62 97,33
manho dos cromossomos e às dificuldades encontradas na coloração das células, onde substânc1as como mucilagens também se coloriam.
AGR·ADECIMENTO
Desejo expressar sinceros agradecimentos ao Dr. Jorge Soria e Dr. Carl Moh, pela ouentação dada.
38
.I
I
I
LITERATURA C ITADA
1. ENRIQUEZ C., G.A. Revisión de literatura de citogenetica en cacao. s.n.t. 9 p. (Mecanografado)
2. KNIGHT, R. Induced polyploidy in Tht>ohroma cacao L. and related spec1es. Journal of Horticultura! Sciences 32(1):1-8. 1957.
3. MuNOZ ORTEGA, J .M. Estudios cromosomicos en el genero Theo· bromo L. Tesis Mag. Agr. Turnalba, Costa R1ca, Instituto lnteramericano de Ciencias Agrícolas, 1948. 43 p.
4. OPEKE, L.K. e JACOB, V .J. Citological irregularities in Theobroma cacao L. In Conferência Internac10nal de Pesquisas em Cacau, 2a., Salvador e ltabuna, Bahia, Brasil, novembro 19-26, 1967. Memórias, Itabuna, Centro de Pesquisas do Cacau, 1969. pp. 114 - 115.
RESUMO
O estudo citológiCO das células mães de polem (CMP) de cacaueiros autocompatíveis e seus híbridos com um pa1 autoincompativel mostrou o aparecimento de assoc1ações cromossômicas mono, bi e trivalentes no processo de formação de grãos-de-polem.
Considerou-se que a normalidade do polem poderia estar relacionada com a freqüência de associação cromossômica bivalente no processo de divisão das CMP. Entretanto, os valores encontrados para pólens normais, tanto nos cultivares autocompatíveis como nos seus híbridos com um pai autoincompatível, ind1cam que não há relação entre esses fenômenos.
CYTOLOGICAL A BERRA TIONS OF THE POLLEN MOTHER CELLS
IN Theobroma cacao L.
S UMMARY
Mono, bi- and trivalent chromosomal pairing was observed 1n the pollen grain formation process for both self-compatible cocoa and cocoa hybrids
39

where one of the parents was self-compatible. It had been considered that pollen viability was related to the frequency of bivalent chromosomal pairing in the division of the pollen mother cell. However, values found for viable pollen grains for both self-compatible cultivars and cultivars with one self-incompatible parent indicate there is no relationship between viability and frequency of bivalent chromosomal pairing.
40
REVISTA THEOBROMA
Julho-serembro 1974
Ano IV N~ 3
Publicaçio nimesrrod dedicada à divulgaçio de investigação dendfica reJa· cionada com problemas agro· nômkos e sócio-econômicos de áreas cacaueiras. Editada pelo Cenuo de Pesquisas do Cacau (CEPEC), Departamen<o da Comissio E:ucutiva do Plano da Lavoura Cacaueira CCEPLAC).
COMISSÃO EDITORIAL
Alvim. Paulo de T.
Cruz, Luiz C. Sales, Josil' C. de Vello, Fernando
ASSESSORES CIENTÍFICOS
Aitken, 1Villiam M. Alencar, Maria H. Alvim. Ronald Barroco, Hil'lio E. Barros, Raimundo S. Cabala R., F. Percy Carteuo, Geraldo A. Loureiro, Edmar S. Maravalbas, Nelson Mariaoo, Antônio H. Medeiros, Arnaldo G. Menezes, José A. S. Miranda, Emo Ruy de Pereira, Clovis P. Rocha, Hermínio M. Silva, Luiz F. da SHva, Pedri<o Veotocilla, José A.
Endeteço para correspon• dênda (Adress for corre• spnndence):
Revista Tbeobroma Centro de Pesquisas do Cacau (CEPEC). Caixa Postal 7 45.600- ltabuna·Bahia
Brasil
Titagem 3.000 e:remplares
CONTEÚDO
I. The possible role of abscisic acid and cytokinins in growth rhythms of Theobroma cacao L. R. Aluim, P. de T. Aluim, R. LOf'etrzi e P. F. Satmders.
Fitormônios e ritmos de crescimento em Theobrorrza cacao L. (Resumo) p. 11.
2. Mortalidade dos politri.zadOf'es Forcipomyia spp, (Diptera, Ceralopogonidae) causada pela aplicm;do de inseticidas nos cacauais baianos. S. de]. Soria e]. M. A-breu, ----------·-·-
Mortality of Forcipomyia spp. midges (Diptera, Ceratopogonidae) following insecUcide applicaUons in Bahian cocoa plantations. (Summary) p. 25.
J, Nematodes of tbe cocoa regicm of Bahia, Brazil. IV - Ne-todes in the rhizospherr>s of pepper (Piper nigrum L.) and clave (Eugenia caryophyllata Thunb.), R. D. Shamra e P.A.A. Loof. _____ _
Nematódios da regilo cacaueira da Bahia, Brasil. IV - Nematódios na rü:osfera de pimenta-do-reioo (Piper nigr- L.) e cravo-da-India (Eugenia caryophyllala Thunb.). (Resumo) p. 31.
NOTA
Palogenicidade do Bacillus thuringiensis Berliner ccmlra o .. _ndarov4" da seringueira (Erinnyis ello l...) (Lepidoptera:Sphingidae). ]. M.
A breu. ·····---- --·-····-····-····-·----
Pathogenicity of Bacillu.s tburingietrsis Berliner against the rubber leaf caterpillar (Erinnys ello L.) (Lepidoptera: Sphingidae). p. 35.
l
ll
26
ll

THE POSS!BLE ROLE OF ABSCISIC ACID AND CYTOKININS
L .•
IN GROWTH RHYTHMS OF Theobroma cacao
R. Alvim ** P. de T. AI vim*** R. Lorenzi **** P. F. Saunders ****
In the State of Bahia, Brazil, cacao buds are usually dormant between May and August. From September to April the tree shows intermittent growth, when periods of rest alternate with periods of profuse bud activity, the latter comprising two main llushings in September;October and February/March, sometimes with two or three minor ones between October and February (3).
The influence that climatic factors exert upon growth periodicity of tropical species is not so easy to understand as in plants of temperate regions, where well defined changes in environmental conditions occur. Nevertheless, cycles of Ieaf flushing in the cacao tree are undoubtedly under the influence
of such factors. Temperatures below 28°C were reported to be unfavourable to flushing (8). A relatively small temperature rise during the rest period (from 21-23 to 24-27°C) has been suggested to be necessary for leaf bud burst in Bahia (3). Bud growth in Trinidad was also correlated with the occurrence of a relatively dry period after the rainy season or, conversely, with the beginning of rains after a dry period (11). The March/April flush of cacao in Ghana was associated with the onset of rains (9). Cacao in Bahia undergoes intensive flushing when rains follow a dry spell (4). Observations under controlled soil moisture conditions have shown that depletion of soil water results in abundant flushing almost immediatety after cacao has been wa-
* Received for publication in July, 1974. Portion of a Ph.D. thesis submitted to the University of Wales, Aberystwyth, Wales, U. K.
** Ph.D., Division of Plant Physiology, CEPEC. ... Ph.D., Division of Plant Physiology, CEPEC . Ph.D., Department of Botany and Microbiology, University of Wales. Aberystwyth, Wales, U. K.
Revi.~la Theobroma. CEPEC, Ilhéus, Brasil, 4 (3}: J -12. juf.-sel. J<J74.
3

tered, whilst plants continuously irrigated eventually cease flushing (14). The frequency and quantity of raios also affect leaf fali in cacao, which occurs in greater intensity during the dry periods. Leaf abscission in Bahia takes place throughout the year, but is particularly conspicuous during the major peaks of leaf flushing (3).
On the other hand, bud growth in cacao appears to be held in check simply by correlative inhibition exerted by the mature leaves of the shoot, since defoliation experiments have been shown effective in bringingabout flushing even from May to August, the response being proportional to the number of leaves removed (3).
In view of these facts, Alvim et al (4) proposed an explanation for the growth rhythm of cacao from September to April, when temperature in Bahia is not a limiting factor. The hypothesis postulates a hydroperiodic reaction to the environment, which would involve hormonal changes, starting with a possible increase in the abscisic
acid (ABA) content of the mature leaves, triggered by a dry spell, and the consequent formation of abscission layers in these !eaves. lt is well known that water stress can bring about large increases of ABA in leaves of many species (13, 16, 19) and that this inhibitor is involved in abscission (17). As a consequence of leaf abscission, the amount of ABA translocated
4
from the leaves to the shoot tips would decrease. Bud growth would be expected to occur soon after the abscission process starts or, in case of severe drought, with the onset of rains, as a result of a shift in thehormonalcontent of the terminal buds towards growth promoting substances. The defoliation experiments cited above have also been taken to indicate that promoters produced by the roots are involved in the contrai of bud growth in cacao. This possibility is supported by the observation that if some branches are girdled bud growth is promoted on intact branches.
The following account describes the identification of abscisic acid and the detection of cytokinin activity in extracts of cacao buds. Additionally, the hypothesis that short periods of water stress incresse the ABA content of cacao leaves and that defoliation is capable of modifying the inhibitor/ promoter balance of the terminal buds of this plant have been tested.
MATERIALS i\ND METHODS
Abscisic acid was identified in cacao huds by comhined gas-liquid chromatography /mass spectroscopy. Leaf buds were harvested in liquid nitrogen and freeze-dried at the Centro de Pesquisas do Cacau in Bahia, Brazil, and then sent by air mail to the University of Wales, in Aberystwyth, U. K., where they were extracted and purified f o r qualitative analysis by gas-liquid
chromatography (GLC) by the method of Lenton et al (10).
A severe drought (total rainfal! of approximately 60 mm) occurred in the area se!ected for the defoliation experiment durin-g t h e months of January and February and atl trees were dormant when the treatments were applied. The treatments given consisted of defoliating five equally illuminated shoots from each of 20 cacao trees on three separate occasions (14, 19 and 24/2/73). One hundred terminal buds were harvested from each treatment on the day follow· ing the !ast defoliation. At the same time an equal number of control buds were excised from shoots which had been left intact. On this date, des pite the little rainfa ll, bud growth was apparent in the shoots which had been defoliated on 14/2 and 19/2/73. Quantitative analysis of ABA by GLC was performed as described by Alvim (5). Brief!y, radioactive abscisic acid was added as internai standard and the tissues were homogenized and extracted in 80% aqueous methanol, using 100 ml of solvent per gram dry weight of buds. The methanol was evaporated and the resulting aqueous solutions centrifuged and acidified to pH 2.6. ABA was then extracted in three equal volumes of diethyl ether. The remaining aqueous fractions were kept frozen for cytokinin bioassays and the ether layers were combined, reduced to dryness, taken up in a suitable volume of 5% sodium bicarbonate and their pH was adjusted to 7.0.
5
The resulting solutions were slurried in water washed polyvinylpyrrolidone powder, filtered, acidified to pH 2.6 and re-extracted in ether as before. After evaporation of the ether the extracts were chromatographed on plates, methylated matographed.
thin-layer and gas-chro-
Aqueous phases from the first ether partition were taken to pH 8.0 and extracted in three equal volumes of water saturated normal butano!. The butano! was dried and the residues redissolved in a small volume of 80% ethanol for pape r chromatography in isopropanol: ammonia : water, 10: 1 : 1 (v/v/v). Soybean callus assays (12) were performed with an amount of extract equivalent to 1.54 grams dry weight of buds. These bioassays were repeated twice.
The effect of moisture stress on the ABA content of cacao leaves was determined by applying different irrigation regimes to 6 month old seedlings growing in 12 x 28 em pots. Water which had been applied daily was stopped on different days so that the plants were harvested after 8, 6 or 4 days of no water. Each treatment consisted of 10 seedlings from each of which five selected leaves wer~ harvested. The tissues were macerated, extracted and analysed as previously described. The water potential of the leaves was measured at harvest by means of a pressure chamber (15).

RESllL TS :\ND DISC'USSION
In Figure 1 the ABA contents of leaves subjected to various leveis of moisture stress are presented and in Figure 2 the variation in leaf water potential. As the period of imposed drought increased there was in increase in the amount of ABA found in the leaves. After 8 days of no water wilting had still not occurred in the test plants although this did start to develop 10 days after water began to be withheld in some of the border plants, when very low leaf water potentials were found for these plants (Figure 2).
r-100
i 80 ,---• • r o ~ 60 ,---• • -' 40 • o a • 20 • •
control 4 • 8 doys withouf wof.,.
Figwe 1 - Absr::isic acid con· tents o/ cacao leaves sub;ected to a varying uumher o/ days of ímposed drought.
6
-· . ! -· ' ! -10
• -: -IZ
" -· ' ' • • • " Do,. ••••u• •••n
Figure 2- Changes in leaf water potential in plants suh;ected to díf/erent leveis of moisture stress.
Figure 3 shows the effect of defoliation on the amount of ABA in the buds. In Figure 4 are shown the results for the soybean callus bioassays. Following defoliation ABA leveis decreased markedly and cytokinin activity increased, both effects occurring before bud out-growth.
Combined gas-liquid chromatographic/mass spectroscopy confirmed the identity of the A B A peak previously recorded by gasliquid chromatography alone .
The hypothesis that shoot growth is influenced by a hormone formed in the roots was first put forward by Went (18), who used the name caulocaline to refer to this hypothetical root promoter. More recently,Doorembos (6)sugested that
lO
8
o 6 S! ' ~ • 4
2
Day• aft•r defoliation
Figure 3 - The elfect of defoliation on the ABA content of cacao huds. Concentrations expressed as J1- ABA/g dry weight represented by hatched columns andas J1- ABA/100 buds by open columns.
growth periOOicity is under the influence of inhibitory substances prOOuced by the leaves and stimulating factors originating in the roots. The evidence obtained in the present work gives support to this view and is in agreement with suggestion by Alvim et ai (4) concerning the role of ABA in the contrai of shoot growth in cacao. During the inter-flush periods growth of cacao buds seems to be held in check by a direct inhibitory effect of ABA prOOuced by the foliage. The results indica te also t h a t
7
cytokinnins play an important role in the mechanism of flushing .
An analogy could be tentatively traced between the processes leading to an inter-flush period in the cacao tree and the gradual decrease of extension growth occurring during the preparative stages for dormancy in temperate species (5). In both cases, an inhibitor/promoter balance appears to be involved. rt• rould be suggested that, with the development of an increasing leaf area, more and more ABA is tmnsIocated from the leaves to the grow-

' "
o
Figure 4 - The effect of defoliation on the cytokinin activity of ca. cao buds, as assessed by tbe soybean callus bioassay. Kinetin contrais in p./l.
ing points and larger amounts ofroot cytokinins are diverted to the leaves. The inhibitor/promoter ratio in the shoot tips would consequently be gradually increased. A situation may be reached in which the leaf area attains a criticai value, at which the action of ABA totally overcomes the growth effects of cytokinins and other promoters. For growth to be resumed following this state of total cone la tive inhibition, the criticai leaf area would have to be somewhat reduced. Perhaps at this stage a dry spell, which appears to enhance or even to be obligatory for the renewal of growth in some species, would exert its role. A criticai number of water stressed leaves could increase ABA leveis to the point of inducing
3
the formation of abscission layers. These layers would act as barriers against both the diversion of root cytokinins to the leaves and leaf ABA to the shoot tips, thus triggering the mechanism of growth flushes.
It is noteworthy thut leaf abscission in cacao is highly correlated with soil moisture: when a dry petiod follows a wet or medium period, leaves of different ages are shed intensively. In plants continuously irrigated many of the original leaves still remain after nearly 600 days and these plants stop flushing (14). It is also relevant that young cacao plants do not show the seasonal growth periodicity of mature trees, flushing
episodically throughout the year, even during periods of lower temperatures and shorterdays(2). This may have ontogenic reasons, but could be equally due to the fact that the total leaf area of small plants is not large enough to maintain the apex inhibited through an adverse ABA/cytokinin balance when large supplies of cytokinins are available from a rapidly developing root system. A similar situation occurs in other intermittently growing species (1, 7). lt is known that cacao trees on the outskirts of a plantation flush earlier that those in the centre (8), this probably being the result of higher exposure to the factors which increase transpiration and could lead to an earlier formation of abscission layers. The correlation between flushing of cacao and maxi-
mum air temperatures reported by Humphries (8) could also be due to the indirect effect of high temperatures on transpiration. Alvim et ai (4) observed that when a drought occurred in June/July, 1971, which are normally the wettest months of the year in Bahia, an off-season leaf fali was followed by a conspicuous flushing in July/August. Flushing during this period of the year is rarely ohserved in Bahia.
The above mentioned observations together with the results reported in this work, lend support to the theory according to which internai moisture stress, apparently because of its effect on ABA/cytokinin balance within the vegetative buds, would be the primary stimulus for synchronous growth flushes in mature cacao trees as normally observed under field conditions.
LITERATVRE CITED
1. ALVIM, P. de T. Tree growth periodicity in tropical climates. In Zimmermann, N.H., ed. Formation of wood in forrest trees. New York, Academic Press, 1964. pp. 479-495.
2. . Eco-physiology of the cacao tree. In Conference Inter-nacionale sur les Recherches Agronomiques Cacaoyeres. Abdijan, 15-20 novembre, 1%5. Paris, Jouve, 1967. pp. 23-25.
3. , MACHADO, A.D. and GRANGIER Jr., A. Alguns estu-dos sobre as relações de água, solo e crescimento do cacaueiro. In Conferência Internacional de Pesquisas em Cacau, 2a., Salvador e Itabuna, Bahia, Brasil, 19 a 26 de novembro, 1967. Itabuna, Centro de Pesquisas do Cacau, 1%9. pp. 316-324.
4. , MACHADO, A.D. and VELLO, F. Physiological re-sponses of cacao to environmental factors. In International Cocoa Research Conference, 4th., Trinidad and Tobago, 1972.
9

~. ,\L\'Il\1, R. Studies on endogeneous hormones in relation to dormancy 1n wood plants. Ph.D. Thesis. Aberystwith, University of \\'ales, 158 p. 1973.
o. OOOREMBOS, j. Review of the literature on dormancy in buds of wood plants. Mededelingen van de Landbouwhogeschool te Wageningen (Nederland) 53: 1·24. 1953.
1 • HOLDSWORTH, M. Intermitent growth of the mango tree. Journal of Western Africa Science Association 7 : 163 -171. 1973.
8. !IUMPHRIES, E.C. A consideration of the factors controlling the opening of buds in the cacao tree (Theobroma cacao L.) Annals ofBotany(n.s.)8:259-267. 1944.
9. HURD, R.G. and CUNNINGHAM, R.K. A cocoa shade and manurial experiment at the West Africa Cocoa Research 1nstitute. 111. Physiological results. journal of Horticultura! Science 36:126 - 127. 1961.
10. LENTON, j.R., PERRY, V.M. and SAUNDERS, P.F. The identification and quantitative analysis of abscisic acid in plant extracts by gas-liquid chromatography. Planta (Germany) %:271-280. 1971.
11. McDONALD, J.A. An environmental study of the cacao tree. PartI. In Imperial College of Tropical Agriculture. Annual Report on Cacao Research 1:29-38. 1971.
12. MILLER, C.O. In Modern methods of plant analysis. Linkskens, H. F. and Tracey, M.V., eds. Berlin, Springer Vetlag, 1963. p.194.
13. MOST, B.H. ABA in immature apical tissue of sugar cane and in leaves of plants exposed to drought. Planta (Germany) 101:67-
75. 1971.
14. SALE, P.j.M. Growth, flowering and fruiting of cacao under controlled soil moisture conditions. Journal of Horticultura! Science 45:99-118. 1970.
.15. SCHOLANDER, P.F. et ai. Sap pressure in vascular plants. Science (U.S.A.) 148:339-346. 1965.
lO
16. SIMPSON, G.M. and SAUNDERS, P.F. Abscisic acid associated with wilting in dwarf and tall Pisum sathmm. Planta (Germany) 102: 272-276. 1972.
17. WAREING,P.F. and PHILLIPS, I.O.J. The contrai of growth and differentiation in. plants. Asprey, G.F. and Lyon, A.G., eds. Oxford, Pergamon Press, 1970. 303 p.
18. WENT, F.W. Specific factors other than auxins affecting growth and root formation. Plant Physiology 13: 55-80. 1938.
19. WRIGHT, S.T.C. and HIRON, R.W.P. (±) Abscisic acid, the growth inihibitor induced in detached wheat leaves by a period of wilting. Nature (England) 224:719-720, 1969.
SUMMARY
Abscisic acid was identified in extracts of cacao buds by combined gas-liquid chromatography/mass spectroscopy. Cytokinin activity was detected in these extracts by biological assay. Both hormones were found to vary in concentration in response to defoliation treatments. ABA leveis in the leaves increased following depletion of soil moisture. The possible role of these hormones in the control of bud growth is discussed.
lt is suggested that a feed-back inhibition may operate in cacao and other species of intermittent growth, in that shoot growth can eventually become limited by a criticai leaf area, According to this hypothesis, internai moisture stress, which appears to be a prerequisite for synchronous flushing of cacao under field conditions, would trigger the hormonal control of bud growth, thanks to its effect on leaf shedding or on the formation of abscission layers,
FITORMONIOS E RITMOS DE CRESCIMENTO EM Theobroma cacao L.
RESUMO
Através de cromatografia gas-líquida e espectroscopia de massa, identificou-se ácido abscísico em extratos de gemas vegetativas do cacaueiro . Constatou-se ainda a presença de citocininas nestes extratos, através de sua atividade no crescimento de calos cotiledonais de soja.
11

As concentrações desses hormônios nas gemas terminais variaram quando se retiraram as folhas de seus ramos. Mostrou-se ainda que os nÍ\'eis de ácido abscísico nas folhas aumentam à medida que diminui a disponibilidade de água para a planta.
Discutem-se os possíveis papéis desses hormônios na dormência da gema vegetativa do cacaueiro.
12
MORTALIDADE DOS POLINIZADORES Foccipomyia spp. (DIPTERA, CERATOPOGONIDAE)
CAUSADA PELA APLICAÇÃO DE INSETICIDAS NOS CACAU AIS BAIANOS*
Saulo de f esus Soria ** João Manuel Abreu***
O uso de inseticidas com amplo espectro de ação, como o BHC, para combater as pragas do cacaueiro na Bahia, tem originado controvérsias. Não se conhece em profundidade o grau de benefício dessa prática em confronto' com a sua influência sobre as populações de polinizadores.
A avaliação do efeito de inseticidas na entomofilia do cacaueiro tem sido feita por métodos indiretos, nos quais foram considerados a polinização, frutificação e produção, na pressuposição de que estes fenômenos são o resultado da interferência dos polinizadores.Cardona (1), na Colômbia, estudou a influência de sete inseticidas na polinização e frutificação do cacaueiro, tendo verificado que o BHC e o Dieldrin, nas dosagens comerciais foram os menos preju-
diciais. Stelzer e colaboradores (12), na Costa Rica, concluíram que aplicações de Dieldrin ou de DDT não tiveram efeito adverso na polinização do cacaueiro, tendo, pelo contrário, proporcionado maior produção. Ainda na Costa Rica, Knoke e Saunders (7), ao estudarem a influência doCarbaryl, Malathion, Dieldrin e Dimetoato no desenvolvimento da flor e produção de frutos, constataram que o Carbaryl aumentou a produção total de frutos, enquanto o Malathion, Dieldrin e Dimeotato, em ordem decrescente, tiveram efeito adverso, quando comparados com a pulverização com água.
Em Gana, Entwistle e Hurd (4) realizaram experimentos para verificar o efeito de pulverizações mensais com Lindane e Dieldrin sobre a polinização e fertiliZação
* Recebido para publicação em 11gosto, 1974. ** Ph.D., Divisão de Fitopan•sitologia, CEPEC.
*** M.S., Divisão de Fitopt~rasitologia, CEPEC.
Reuista Tbeob,oma. CEPEC, Ilhéus. B'asif, 4(3): IJ-25. ]ui-sei. 1974.
13

das fi<)res do cacaueiro e número de mosquinhas ceratopogonideas. Os resultados obtidos mostraram que estes inseticidas provocaram diminuição de população dos insetos polinizadores, não se encontrando diferenças aparentes na sua densidade e nos índices de fertilização. Sobre este experimento, Entwistle (3) opina que os resultados negativos do efeito do Lindane e Dieldrin sobre os níveis de polinização e frutificação podem ser intensificados quando são tratadas grandes áreas por um período muito prolongado. Estudos realizados por Ventocilla (14), na Bahia, indicam que o BHC 1% (pó), aplicado por meio de helicóptero provocou uma insignificante diminuição na polinização dos cacaueiros.
Num estudo preliminar efetuado pelos autores, em agosto de 1972, no CEPEC, Ilhéus, Bahia, foi verificado que o polvilhamento de cacauais com BHC 1 ,5% provocou um declínio de 90% na população de Forcipomyia spp. O estudo foi efetuado em área onde estavam sendo feitas amostragens diárias de polinizadores para determinação das curvas de flutuação populacional. A falta de um esquema apropriado de experimentação para determinar mortalidades, considerando a flutuação em área não polvilhada, não permitiu estabelecer conclusões sobre o efeito do inseticida nestas populações.
Com base nos trabalhos acima referidos, foi realizada a presente
pesquisa para avaliar o efeito de três inseticidas em termos de mortalidade de polinizadores. O nome Forcipomyia, neste trabalho,abrange as espécies Forcipomyia (For· cipomyia) genualis (Loew) e, em menor freqüência, F. (Forcipomyia) pictoni Macfie e F. (Fordpomyia) spp. (10). O nome Euforcipomyia abrange as espécies Forcipomyia (Euforcipomyia) sp. 1 e, em menor
freqüência, F. (Euforcipomyia) spatulifera Saunders (10).
MA.TERIA.L E MÉTODOS
Os ensaios que se descrevem a seguir foram conduzidos em áreas experimentais do CEPEC durante as épocas de maior intensidade de população de polinizadores (maioagosto)*, nos anos de 1973 e 1974,
Ensaio I.
Nesse ensaio, foi feita a avaliação da mortalidade de Euforcipomyia e Forcipomyia como resultado de polvilhamentos c o m BHC 1,5%, Carbaryl 7,5% e Parathion metílico 1,5%, aplicados à razão de 16 kg/ha. Para cada inseticida, foi destinada uma parcela de 625m2
, em cujo centro foram tomados, ao acaso, 10 cacaueiros, sob os quais foram estendidos lençóis de 4 x 4 m. A seguir, proce-
14
* Observação feít!l pelo putor principal em pesquisa pinda em andamento.
deu-se ao tratamento das parcelas, às primeiras horas da manhã. As coletas dos insetos foram feitas 2; 6 e 24 horas após o tratamt:nto. Ao final destas coletas, procedeu-se ao tratamento de choque das árvores amestradas com BHC 12%, de acordo com o método de Collyer (2). As coletas dos indivíduos mortos pelo tratamento de ch(Xjue foram feitas 2; 6 e 24 horas após a sua aplicação. Foi usada uma lupa manual com aumento de lOx para distinguir microdípteros dos demais microinsetos presentes nos lençóis. Os microdípteros de cada árvore foram coletados e acondicionados em frascos pequenos com álcool 70% e conduzidos ao laboratório para posterior identificação genérica e quantificação com o auxílio de estereomicroscópio. A população adulta total foi calculada somando-se o total de insetos mortos devido à ação de cada inseticida ao total de insetos mortos por aplicação do BHC 12%.
Ainda neste ensaio, foram utilizadas duas parcelas adicionais, com níveis de floração contrastantes, também com 625 m2
, em cujo centro foram tomados, ao acaso, 10 cacaueiros. Nestas parcelas, foram avaliadas, pelo método de choque, as populações dos polinizadores a fim de verificar a interferência da floração sobre a sua população.
Ensaio 11.
Efetuado 1 ano após o ensaio anterior, do qual diferiu pelo ta-
15
manho das parcelas ( 2.500 m'), método de contagem da população adulta total e por terem sido tomados, ao acaso, cinco cacaueiros no centro das parcelas. Como no ensaio anterior, foi empregado o método de choque, tendo sido somados os números de insetos mortos encc·ntrados nos lenç:'.iis 24 horas após a aplicação do inseticida (tratamento) e número de insetos mortos por aplicação do BHC 12%. A diferença de tamanho das parcelas se deveu a um dos objetivos do ensaio, ou seja, verificar os efeitos da bordadura sobre as avaliações de mortalidade.
Ensaio 111.
Neste ensaio, foi feita a avaliação da mortalidade de Forcipomyia spp., contando-se o número de insetos sobreviventes após polvilhamento com BHC 1,5% aplicado à razão de 16 kg/ha em duas parcelas de 2.500 m2 cada. No meio de cada uma delas, foram etiquetadas 36 plantas, nas quais era'll efetuadas amostragens semanais das populações.
As amostragens foram feitas pela observação visual dos polinizadores presentes nos botões florais, pétalas, estaminóides, sépa!as, ápices secos de folhas e frutos pecos até a altura de 1,80 m, entre as 8:00 e 10:00 horas, antes e de ... pois do polvilhamento, sendo os dados registrados em cartões individuais para cada árvore (9).
A avaliação foi feita comparando-se o número de indivíduos oh-

!:'ervados nas 36 árvores tratadas, em relação às 36 não tratadas. A percentagem de mortalidade dos polinizadores foi calculada pela fórmula de Henderson e Tilton (5).
% = 100 11 - AxB CxD
em que
A = número de insetos contados após o polvilhamento;
B = número de insetos contados na testemunha, antes do polvilhamento;
C = número de insetos contados antes do polvilhamento;
D = número de insetos contados na testemunha, depois do polvilhamento.
Os dados foram tomados semanalmente, por um período de 21 dias, para se observar se ocorreria restauração da população aos níveis anteriores ao polvilhamento.
RESULTADOS
Ensaio I.
A mortalidade de Buforcipomyia e Forcipomyia, após a aplicação do BHC 1,5%, foi de 75 e 36%, respectivamente. As mortalidades, após a aplicação do Parathion metílico, foram de 44 e 46% para Euforcipo· myia e Forcipomyia, respectivamente, enquanto que pela ação do Carbaryl foram de 68 e 69%, respectivamente (Quadro 1).
A tendência das curvas de mortalidade de Euforcipomyia e Forcipomyia por ação do BHC mostra que a mortalidade mais elevada ocorreu 2 horas após o tratamento, caindo a níveis mínimos 6 horas após o mesmo e, daí em diante, aumentou em graus variáveis mas não atingiu os níveis iniciais (Figuras 1 e 2). A tendência das curvas de mortalidade, sob a ação do Parathion metílico e do Carbaryl, para os mesmos polinizadores, foi simi-
Quadro 1 - Populações de Forcipomyia spp. e sua mortalidade (%) 24 horas após o polvilhamento com diversos inseticidas (avaliação feita pelo método de choque). CEPEC, julho, 1973.
Inseticida Bu(orcipomyia ForcitJomyia
N? % N? %
BHC 1,5% 740 75 993 36
Carbaryl 7,5% 34 68 63 69
Parathion metílico 1,5% 35 44 52 46
16
~ o ~ • o • ~ w • o o c N
z ~ o • w o
o • w • 'O z
400
200
" M. PARATHION l,S%
10
•
lO CARBARYL 7,S%
6
2L-.---~----~----r---~----~---2 6 10 14 10 22
HORAS APÓS TRATAMENTO
Figura 1- ,'tlortalidade de F. (Euforcipomyia) spp. após polvilhamentos com inseticidas em parcelas experimentais do CEPEC, Bahia, Brasil, agosto-setembro /73.
17

400 8HC 1,~%
200
~ o ~ « o 2 .. ~ w « 20 N. PARATHION I, 5 CJb o o c
" N
z
" 10 o • w • o
o « w 2 -~ z
20
CAR8ARYL 7,S% lO
10
• 2 • 10 14 10 22 24
HORAS APÓS TRATAMENTO
Figura 2 - Mortalidade de F. (Forcipomyia) spp. após polvilhamento com inseticidas em parcelas experimentais do CEPEC, Bahia, Brasil, agosto-sete mbro/7 3.
18
:<
lar à encontrada para o BHC (Figuras 1 e 2).
Os resultados relacionados com a abundância de polinizadores face a níveis contrastantes de floração (Quadro2) indicaram que as populações de polinizadores na parcela com densidade baixa de floração foi mais do que o dobro da encontrada na parcela com densidade alta de floração (32 vezes superior).
Ensaio 11.
Os resultados relacionados com a determinação de mortalidade pelo método de choque indicaram que as de Euforcipomyia e Forcipomyia, após aplicação de BHC 1,5% (Quadro 3), foram 79% para ambas as espécies. As mortalidades de Eu· forcipomyia, após aplicação de Carbaryl e Parathion metílico, foram de 70 e 83%, respectivamente,
enquanto para Porcipomyia foram de 66 e 87%, respectivamente.
Ensaio 111.
Os totais de mosquinhas encontradas nas 36 árvores amostradas na área tratada e testemunha podem ser observados no Quadro 4. Sobre os totais apresentados neste quadro, foi observada uma mortalidade de 91% de mosquinhas Porcipomyia spp.
Os resultados relacionados com a determinação do período d~ restauração das populações (QuadroS) na parcela polvilhada, face à testemunha, indicaram que o nível inicial de população, aproximadamente 166 mosquinhas por parcela, foi recuperado 1 semana após o polvilhamento, seguido de um incremento progressivo nas 2 semanas de observação seguintes.
Quadro 2- Número de flores e populações acumuladas de Forcipomyia spp. em duas parcelas de 10 cacaueiros cada, com densidade contrastante de floração. CEPEC, julho, 1973.
Floração
Densidade alta (A)
Densidade baixa (B)
Relação A/B
Número de flores '
2896
90
32/1
19
.._ Níveis de populações
Euforcipomyia Forcipomyia
294 422
740 983
1/2 1/2

Quadro 3 - Populações de Forcipomyia spp. e sua mortalidade (%), 24 horas após o polvilhamento com diversos inseticidas, em duas par. celas de cinco cacaueiros cada (avaliação feita pelo método de cho· que). CEPEC, julho, 1974.
Euforcipomyia Forcipomyia Média
BHC 1,5- Tratamento 31 64 Choque 8 17
% 79 79
Carbaryl 7,5% Tratamento 30 53 Choque 13 27
% 70 66 68
Parathion metílico 1,5% Tratamento 66 137 Choque 14 21
% 83 87 85
Quadro 4 - Totais de mosquinhas Forcipomyia spp. encontradas nas 36 árvores tratadas e não tratadas antes e depois da aplicação do BHC 1,5%. CEPEC, julho-agosto, 1974.
Tratamento Testemunha
Antes
Insetos 166
Mortalidade (%)
DISCUSSÃO
A avaliação de populações de polinizadores em parcelas com níveis contrastantes de floração per· mitiu comprovar que o método de choque utilizado nos dois primeiros
Depois Antes Depois
14 224 221
91
ensaios é o mais eficiente porque não depende da floração, mas sim da disponibilidade de insetos.
20
Comparando a mortalidade dos inseticidas em relação aos dois grupos de polinizadores (Quadro 3),
Quadro 5 - Flutuação dsa populações de Forcipomyia spp. amostra• das em 36 árvores em área polvilhada com BHC 1,5% face à área testemunha. CEPBC, julho-agosto, 1974.
Variãvel Data de contagem
29-7 31-7 6-8 13-8 20-8
Tratamento * 166 14 136 224 284
Testemunha 224 221 92 162 484
* As árvores foram polvilhadas em 30 de julho.
verificou-se ser o Parathion metílico o mais nocivo (85% de mortali· dade), seguido do BHC (79ro) e, p« último, do Carbaryl (68%).
A mortalidade mais elevada observada no segundo ensaio, quando comparada à do primeiro, foi aceita como a mais confiável, pois os seus resultados foram obtidos de árvores mais protegidas da afluência de insetos das áreas adjacentes às parcelas tratadas. A possibilidade de penetração de insetos até as cinco árvores etiquetadas, no centro das parcelas de 2.500 m2 , no segundo ensaio, é muito menor que a de atingirem as lO árvores marcadas no centro das parcelas de 625 m2 utilizadas no primeiro ensaio.
A mortalidade iniciai elevada pode ser atribuída à ação do inseticida aplicado. O decréscimo do número de insetos mortos, no in· tervalo de 2 a 6 horas após o tratamento, pode ser atribuído à dimi· nuição da população de adultos, eliminados pelos inseticidas. O aumento do número de insetos mor-
21
tos no intervalo entre 6 e 24 horas após o tratamento foi atribuído à emergência de adultos dos criadouros naturais bem como ao transporte casual pelas correntes aéreas de populações das áreas não tratadas.
A mortalidade calculada com base na contagem visual de insetos vivos nas diversas partes do cacaueiro foi elevada (91%), quando comparada com a mais alta encontrada pelo método de choque (79%). Esta última - que abrange a população total do ambiente - foi aceito como a mais conclusiva, tendo em vista que a contagem visual está mais sujeita a falhas humanas que o método de choque. Apesar disso, a amostragem visual permitiu acompanhar a flutuação das populações por um período determi· nado até se obter a restauração da população.
Considerando-se que as taxas de crescimento das populações de insetos no ecossistema do cacaual ainda são desconhecidas, é prema· turo discutir o que ocorreria com

essas populações após a aplicação eventual ou periódica de inseticidas. Estudos de Knipling (6) com outros insetos mostraram que um nível inferior a 80% de mortalidade manteria a população estável desde que ela apresentasse uma taxa de crescimento normal cinco vezes maior em cada geração. Tendo em vista que as mortalidades avaliadas pelo método de choque (admitidas como conclusivas) foram de 79; 68 e 85% para o BHC 1,5%, Carbaryl 7,5% e Parathion metílico 1,5%, respectivamente,e que a taxa de crescimento normal de Forcipomyia e Euforcipomyia é provavelmente superior à mencionada por Knipling (6), conclui-se que os níveis mencionados de mortalidade manteriam as populações estáveis.
O controle químico de inset~ é altamente eficiente em termos de número de insetos mortos quanào a população é alta, mas ineficiente quando ela é baixa (6). De acordo com os resultados do presente trabalho, sabe-se que os níveis populacionais originais são restabelecidos 1 semana após a aplicação do inseticida; donde se conclui que essas aplicações, em épociis de baixo nível populacional das mosquinhas, teriam pouco efeito na polinização do cacaueiro. Além disso, a disponibilidade de ovos, larvas e pupas dos polinizadores, protegidos nos criadouros naturais (11), provavelmente facilitou a rápida restauração das populações.
A persistência do poder residual do BHC nos solos de clima tempe-
22
rado é de 2 anos (8), a do Parathion metílico, 45 dias e a do Carbaryl, 9 dias (13). Estes inseticidas, em confronto com outros, de efeito residual mais prolongado, são mais seguros, evitando a acumulação de tóxicos no ambiente, bem como os efeitos nocivos aos polinizadores.
Os dados aqui apresentados podem ser usados como uma explicação para os resultados encontrados por Ventocilla (14), Cardona (1), Stelzer e colaboraàores (12), Knoke e Saunders (7) e Entwistle e Hurd (4), nos quais não foram encontradas mudanças significativas nos níveis de polinização e/ou frutificação do cacaueiro após ensaios com inseticidas no campo.
CONCLUSÕES
1. A mortalidade de Euforcipomyia após aplicação de BHC 1,5%, avaliada pelo método de choque, foi de 79% enquanto utilizando a fórmula de Henderson e Tilton foi de91%. As mortalidades deste inseto por aplicação do Parathion metílico e Carbaryl exclusivamente pelo método de choque foi de 83 e 70%, respectivamente.
2. A mortalidade de Forcipomyia por aplicação do BHC 1,5% avaliada pelos métodos de choque e de Henderson e Tilton atingiu níveis de 79 e 91% respectivamente. As mortalidades deste inseto por aplicação de Parathion metílico e Carbaryl, exclu-
sivamente pelo método de choque, foram de 87 e 66%, respectivamente.
3. O Parathion metílico foi o inseticida que causou, em conjunto, mortalidade mais elevada (85%), seguido do BHC (79%) e doCarbaryl (68%).
4. A abundância do Euforcipomyia e Forcipomyia não está relacionada com a disponibilidade de flores.
S. O método de choque (knock-down) para avaliar mortalidade de polinizadores mostrou-se o mais eficiente, pois a avaliação não depende da disponibilidade de fio-
res por árvore e sim da presença de insetos.
6. A restauração das populações de polinizadores é iniciada 24 horas após o tratamento, atingindo os níveis originais 1 semana após a aplicação dos inseticidas.
7. O combate às pragas do cacaueiro com BHC 1,5%, Parathion metílico 1,5% ou Carbaryl 7,5%, na dosagem de 16 kg/ha, aplicados nas épocas de renovação de folhas - quando as populações dos polinizadores se encontram em níveis mais baixos -, não compromete a sua sobrevivência e, conseqüentemente, íl polinização do cacaueiro.
AGRADECIMENTOS
Aos Eng.·Agr'?s Pedrito Silva e José A. Ventocilla, pelas sugestões apresentadas na elaboração deste artigo.
LITERATURA CITADA
1. CARDONA, E. Influencia de siete insecticidas en la polinización y fructificación dei cacao y breve estudio de los insectos polinizadores. Cacao en Colombia 2:41-61. 1953.
2. COLLYER, E. A method for the estimation of insect populations on fruit trees. Report E. Malling Research Station. 1949- 1950. pp. 148- 151. 1951.
3. ENTWISTLE, P.F. Pests of cacao. London, Longman,1972. 779 p.
4. ENTWISTLE, H.M. e HURD, J. The effects of insecticides on pollination, flower setting and numbers of pollinator insects. 17J West African Cocoa Research lnstitute. Annual Report 1957 · 58. Tafo, Ghana, 1959. pp. 42- 44.
23

S. HENDERSON, C.F. e TILTON, E.W. Tests with acaricides against the brown wheat mite. Journal of Economic Entomology 48 (2)' 157- 161. 1955.
6. KNIPLING, E.F. Some basic principies in insect population suppression. Bulletin of the Entomological Society of America 12(1),7-15. 1966.
7. KNOKE, S.K. e SAUNDERS, J.L. Induced fruit set of 1"heobroma cacao by mistblower applications of insecticides. Journal of Economic Entomology 59 (6): 1427- 1430. 1966.
8. MENZIE, C.M. Fate of pesticides in the environment. AnnualReview of Entomology 17: 199-222. 1972.
9. SORIA, S. de J. Locais de coleta e distribuição de Forcipomyia (Diptera, Ceratopogonidae) relacionadas com a floração e frutificação do cacaueiro na Bahia, Brasil. Revista Theobroma (Brasil) 3(2): 41-49. 1973.
10.
11.
-----, Identidade e caracterização taxonômica preliminar das mosquinhas Porcipomyia (Diptera, Ceratopogonidae) associadas com a polinização do cacaueiro na Bahia. Revista Theobroma (Brasil) 4(1): 3-12. 1974.
-----. e SILVA, P. A polinização do cacaueiro na Bahia e algumas implicações do uso de inseticidas e outras práticas culturais. Cacau Atualidades (Brasil) 11(1): 15-20. 1974.
12. STELZER, M.U. et al. Resultados de pruebas preliminares para determinar los efectos de Ias aspersiones de insecticida sobre la producción del cacao en Costa Rica, Cacao (Costa Rica) 6(2),1-14. 1961.
13. TECHNICAL INFORMATION on Sevin Carbaryl insecticide. Y ork, Union Carbide Coqxlration, Agricultura! Products, 1968.
New 56 p.
14. VENTOCILLA, J.A. Efeito do BHC sobre as mosquinhas Ceratopogonideas polinizadoras do cacaueiro. In Conferência Internacional de Pesquisas em Cacau, 2a., Salvador e Itabuna, Bahia Brasil, 19- 26 de novembro, 1967. Memórias. ltabuna, Centro de Pesquisas do Cacau, 1969. pp. 287-289.
RESUMO
O BHC 1,5%, Parathion metílico 1,5% e Carbaryl 7,5% foram usados em experimentos de campo na dosagem de 16 kg/ha para avaliar o seu
24
efeito sobre populações de F.uforcipomyia e Porr:ipomyia, os dois grupos de insetos mais importantes para a polinização do cacaueiro.
As contagens de mosquinhas mortas foram feitas 2; 6 e 24 h após a aplicação do inseticida. Após a última contagem, foi feito um tratamento de choque (knock down) com BHC 12%, para calcular a sua população total e as respectivas percentagens de mortalidade. Ambos os grupos de mosquinhas foram afetados pelos inseticidas. O Parathion metílico foi o mais tóxico (85% de mortalidade) seguido do BHC (79%) e Carbaryl (68%). A mortalidade mais elevada foi registrada no intervalo de 2 h após a aplicação dos inseticidas. A população das mesquinhas iniciou a re~
cuperação 24 h após o polvilhamento com o BHC e dentro de 1 semana ultrapassou os níveis originais, Após a aplicação de Carbaryl e Parathion metílico, precisa-se provavelmente do mesmo período para a recuperação dos níveis originais de população dos insetos. Isto sugere que, nas dosagens empregadas e com intervalos apropriados entre os polvilhamentos - períodos entre lançamentos de folhas -, esses inseticidas não são prejudiciais à sobrevivência das mesquinhas polinizadoras do cacaueiro.
MORTALITY OF F orcipomyia spp. MIDGES (DIPTERA, CERATOPOGON!DAE)
FOLLOWING INSECTICIDE APPLICATIONS IN BAHIAN COCOA PLANTATIONS
SUMMARl'
Three insecticides BHC 1.5%, Methyl Parathion 1.5% and Carbaryl 7.5% were used in field trials at the rate of 16 kg commercíal product per hectare to assess their relative toxicity on Euforcipomyía and Forcipomyia populations, the two midge groups chiefly responsible for the pollination of cocoa.
Dead midge counts were made 2, 6 and 24 hours after insecticide application. Following the 24 hours collection a 12% BHC knock-down application was made to calculate total adult population and thereby the respective mortality percentages. Both midge groups were equally affected by each insecticide. Of the three insecticides, Methyl Parathion was the most toxic - 85% mortality, followed by BHC - 79% and Carbaryl 68%. Highest mortality was recorded for the 2 hours collection. The midge population started to recover within 24 hours of the BHC treatment and within 1 week had surpassed the pre-application leveis. This suggests that at the given dosages and with sufficient time intervals between applications the above insecticides, particularly BHC are not prejudicial to the overall survival of either of the two cocoa pollinating midges •
25

NEMATODES OF THE COCOA REG!ON OF BAHIA, BRAZIL. IV - NEMATODES IN THE RHIZOSPHERES
OF PEPPER (Piper nigrum L.) ANO CLOVE (Eugenia caryophyllata Thunb.) *
R avi Datt Sharma ** Pieter Aart Albertus Loof***
8oth pepper (Piper nigrum L.) and clove (Eugenia caryophyllata Thunb,), used as spices or condiments in flavouring foodstuffs, are being increasingly planted in the rainforest zone of Bahia. The total area in pepper and clove is variously estimated as 268 ha and 588 ha respectively with corresponding yields of ahout 195 and 133 tonnes (1,6).
As part of a general nematode survey carried out between May, 1971 and May, 1974 in the southern part of the State of Bahia, soil and root samples were collected both from pepper and clove. Sampling involved both heal[hy and diseased plants.
The symptoms of the sick pepper vines were characterized by stunting, bronzing and interveinal yellowing of leaves, followed by
complete yellowing and defoliation, dieback and the gradual death of the vines. Root systems of such vines were severely galled and cracked due to what appeared to be heavy root-knot nematode infestatíon. The sick plants had very low fruit production or no fruit at ali. In one location the heavily galled system of diseased vines were also black and detetiorated, possibly due to combined attack by nematodes and Fusarium solani F. piper.
Some clove trees showed deteriorated root system and progressive drying of leaves resulting in death of the trees,
This paper deals with plant parasitic and free-living nematodes found in some rhizospheres of pepper and clove in Bahia.
* Received for publication in September, 1973. H Oivision of Phytoparasitology, CEPEC. ... R<•vi~ta 'l'ht•{)hrmna. C/:'1'/:'C, 1/hPus, fJ,a.~il. 4 (3}: 26. 32. ]ul-set. 1974.
26
MATERIALS AND METIIODS
Eighteen soil and root samples were taken, nine being from pepper plantations and nine from c I o v e plantations.
Of the pepper samples four were taken from one property in Una and other five from three properties in Ituberá. The clove samples carne from six farms in Ituberá.
The methods for extraction of the nematodes and subsequently mounting for permanent slides, are described by Sharma and Loof (9).
RESUL TS ANO DISCUSSION
The results of the survey are listed in Tables 1 and 2. The rootknot nematode, Meloidogyne incognita was the dominant species found associated with pepper yel· lows, a finding also observed by Sher et ai (13) in Thailand. From a literature search (2, 3, 5, 7, 12, 13, 14) it would appear that Aphelenchoides sp., Discocriconemella limitanea, Dolichodorus sp., Helicotylenchus sp., H. dihystera, Trichodorus sp. andXiphinema vulgare have been found for the first time in association with pepper in the field. Radopholus similis, the usual cause of serious yellows was not found in our samples, but some of the field symptoms were quite s i m i 1 a r to those described by Thorne (13)and Van derVecht (14). It is realized that identical symptoms may be produced by different root-infesting nematode species.
27
R. similis was recently reported associated with some banana plant importations in Bahia (8) and it must be hoped that measures will be taken to limit its spread.
The spiral nematode H. dihystera was the dominant species found associated with clove. Dolichodorus sp., H. dihystera, M. incognita, Rotylenchulus reniformis and Trichodorus sp., were common to both pepper and clove whereas Aphelenchoides sp., D. limitanea, Helicotylenchus sp. and X. vulgare were associated only with pepper. The root-knot nematode, Meloidogyne sp. reported by Ghesquiere (4) was not found in our samples. The association between clove and the five species listed in Table 1 is considered as the first of its kind.
Baseei on the results of previous surveys (9, 10, 11), out of the 29 genera and 38 species of free-living nematodes listed in Table 2 12 species are recorded for th~ first time in Bahia namely: Acrobeloides sp., Bastiania acarayensis, Cruznema lambdiense, Dorylaimoides parvus, lotonchus cf. risoceiae, Monohystera villosa, Mylonchulus brachiuris, Prismarolaimus sp. Prodorylaimus hamatus Sporonchulus sp. Trischistoma pel~ lucidum and Tylenchus leptosoma.
CONCLUSION
The root-knot nematode, Meloidogyne incognita is probably a serious pest of pepper in Bahia and praticai means of control should

>( Table 1 - Plant parasitic nematodes found associated with pepper
(Piper nigrum L.) and clave (Eugenia caryophyllata Thunb.) in the Cocoa Region of Bahia.
Genus I Species
Apbelencboídes sp.
Discocriconemella /imitanea
Oolichodorus minor
Helicotylenchus sp.
H. dihystera
Meloidogyne incognita
Rotylenchulus reniformis
Trichodorus sp.
Xiphinema vulgare
* Number of positive samples.
Pepper (9 samples)
2
1
2
1
6
9
2
1
1
Ocurrence * Clave
(9 samples)
o o 1
o 7
2
2
4
o
Table 2- Free·living nematodes found associated with pepper (Piper nigrum) and clove (Eugenia caryophyllata Thunb.) in the Cocoa Region of BaMa.
Genus I Species
Achromadora sp
Acrobeles sp. Acrobeloides sp. Alaimus sp. Aphelenchus avenae Aporcelaimellus sp.
A. obtusicaudatus A. cf. paracentrocercus Axoncbium amplicole
nas;imtyleptus sp.
+ == present :: absent
28
Occurrence * Pepper
(9 samples)
+ + + + +
+ + +
Clove (9 samples)
+ + + + +
+ +
Table 2 - Continued
Genus I Species
Rastiania acarayensis Cobbonchus sp. Cbronogasler sp. Cruznema lambdiense
Dorylaimellus sp. Dorylaimoides parvus Eudorylaimus sp. E. ettersbergensis E. granuliferus (group) E. cf. minutus Heterocephalobus longicaudatus lotonchus sp. I. cf, risoceiae Mesodorylaimus parasubitilis Mesorhabditis sp. M. cf. paucipapillata Monohystera villosa Mylonchulus minar ,\1, bracbyuris Prlsmatolaímus sp. Prodorylaimus hamatus Rhabditis sp. Sporonchulus sp. S. dentatus Thornenema sp. Trischistoma pellucidum Tylenchus leptosoma Zeldia sp.
+ :: present == absent
be considered. While the burrowing nematode Radopholus similis. well known from pepper, banana and
29
Occurrence * Pepper
(9 samples
+ +
+
+ + +
+ + + +
+
+ + + + + + + + + + +
Clove (9 samples)
+ + + + + + + + + + +
+ +
+
+
citrus, was not encountered in our samples, it is recommended that strict quarantine measures should

be taken to prevent its spread, es~ pecially bearing in mind that the CEPLAC Diversification Scheme
plans to ptomote increased clave and pepper plantings within the Bahian cocoa region.
LITERATURE CITED
1. CASTRO NETO, A. de, et al. O cravo-da-lndia na região cacaueAüCa, estudo preliminar. In Ilhéus, Bahia, Brasil. DEPEX/CEPL , Informe, July, 1972. 10 p.
2. CHRISTIE, J .R. The yellows disease of citrus. Plant disease Reporter 41 : 267- 268. 1957.
3. DELACROIX, H. Pathologie vegetale rnaladies vermiculaires.ITA-S~ une maladie du proivier (Piper nigrum L) en Cochichine. gnculture Pratique de Pays Chauds. 1:672-680. 1901-02.
4. GESQUIERE, J. Laboratoire d'Entomologie d'Eala (Equateur). Reports de l'entomologiste. Bulletin Agricole du Congo Belge 12:703- 732. 1921.
5. LUC, M. aitd GUIRAN, G. de. Les nematodes associés aux plantes de l'Ouest Africain. Liste preliminaire. Agronomie Tropicale 15(4): 434-449. 1960.
• 6. MANDARINO, E. Relatório sobre a pimenta-do-reino na Bahia. In
Ilhéus, Bahía, Brazil. CEPLAC/DEPEX, Informe, 1974.
7. MARTIN, G.C. The common root-knot nematode and crop rotation. Rhodesian Farmer (Reprint of a series of articles, 13th July, 1956 onwards) 20 p.
8. SHARMA, R. O. Nematóide cavernícola atacando a bananeira na Bahía. Cacau Atualidades (Brazil) 11 (1): 4-6. 1974.
9. and LOOF, P.A.A. Nematodes associated with different
10.
plants at the Centro de Pesquisas do Cacau, Bahia. Revista Theobroma (Braz i!) 2 (4): 38-43. 1972.
---;:-c-e- and Nematodes of the Cocoa Region of B~hi~. Hrazil. I - Plant parasitic and free-living nematodes ~ssociated wlth ruhber (llevta brasiliensis Muell. Arg.). Revist~ Theohroma (Erazil) 3(1):36-41. 1973.
30
11. SHARMA, R.D.and LOOF, P.A.A. Nematodes of theCocoaRegion of Bahia, Brazil. III - Plant parasitic and free-living nematodes in the rhizospheres of six different plant species. Revista Theobroma (Brazil) 4 (1): 39-43. 1974.
12. SHER, S.A., CHURAM, C. and PHOLCHARDEN, S. Pepper yellows disease and nematodes in Thailand. FAO Plant Protection Bulletin 17 (2): 33. 1%9.
13. THORNE, G. Principies of nematology. New York, McGrow-Hill, 1961. 553 p.
14. VECHT, ]. van der. Op planten parasiterende aaltjes (Tylenchidae). In Kalshoven, L.G.E., ed. De plagen van de cultuurgewassen in Indonesie. s'Gravenhage, W. van Hoeve, 1959. pp. 16-42.
SUMMARY
Eighteen soil and root samples collected from the rhizospheres of pepper (Piper nigrum L.) and clave (Eugenia caryophyllata Thunb.) revealed the presence of nine species of plant parasitic nematodes. Nematodes common to both pepper and clove were Dolichodorus sp., Helicotylenchus dihystera, Meloidogyne incognita, Rotylenchulus reniformis and Trichodorus sp. Besides the above mentioned nematode species, Aphelenchoides sp., Discocriconemella limitanea, Helicotylenchus sp . and Xiphinema vulgare were also associated with pepper. Out of 29 genera and 38 species of free-living nematodes recorded in this survey, 12 species are recorded for the first time in Bahia namely: Acrobeloides sp., Bastiania acarayensis, Cruznema lambdiense, Dorylaimoides parvus, lotonchus cf. risoceiae, Monohystera villosa, Mylonchulus brachyuri.s, Prismatolaimus sp., Prodorylaimus hamatus, Sporoncbulus sp., Trischistoma pellucidum and Tylenchus leptosoma.
NEMATÓDIOS DA REGIÃO,CACAUEIRA DA BAHIA, BRASIL IV - NEMATODIOS NA RIZOSFERA
DE PIMENTA-DO-REINO (Pipec nigrum L.) E CRA VO-DA-INDIA (Eugenia caryophy !lata Thunb.)
RESL'MO
Em 18 amostras de solo e raízes coletadas na rizosfera de pimenta-doreino (Piper nigrum L.) e cravo-da-lndia (Eugenia caryophyllata Thunb.)
31

foi detectada a presença de nove espécies de nematódios parasitas. Os nematódios associad"Js a ambos os cultivos são: Dolichodoms sp., llelicotylenchus dihystera, Me!oidogyne incognita, Rotylenchulus reniformis e Trichodorus sp. Além destes, Aphelenchoides sp., Disc::ocriconemella limitanea, Helicotylenchus sp. e Xiphir1ema z•l!_lf!.are foram encontrados associados somente à pimenteira, Dos 29 gêneros e 38 espécies de nematódios não parasitas, 12 novas espécies são encontradas pela primeira vez na Bahia, a saber: Acrobeloides sp., Bastiania acarayensis, Cruznema !ambdiense, Dorylaimoides partJUS, fotonchus cf, risoceiae, ,\tonohystera vil/asa, Mylonchulus brachyuris, Prismatolaimus sp., Prodorylaimus hamatus, Sporonchulus sp., Trischistoma pellucidum e Tylen
chus leptosoma.
32
NOTA
PATOGENICIDADE DO Bacillus thuringiensis BERLINER
CONTRA O "MANDAROVA" DA SERINGUEIRA ( E•innyis ello L.) (LEPIDOPTERA : SPHINGIDAE).*
João Manuel Abreu**
A lagarta da mariposa Erinnyis ello L. é a praga mais importante que ataca a seringueira (Hevea brasiliensis Muell. Arg.), o segundo cultivo mais importante da região cacaueira da Bahia. A lagarta se alimenta das folhas e, durante os surtos mais severos, causa a desfolhação completa de grandes plantações.
O sucesso alcançado nos últimos 20 anos com a aplicação do Bacillus thuringiensis Berliner contra mais de 137 espécies de insetos das ordens Lepidoptera, Hymenoptera, Diptera e Coleoptera (1) motivaram a realização de um ensaio explorativo para determinar a patogenicidade desta bactéria contra E. ello. O ensaio constou de cinco tratamentos e quatro repetições. As lagartas foram coletadas no campo e permaneceram 1 dia em repouso no laboratório. Cinco lagartas do 3? instar foram confinadas em cubas de vidro (14 em de diâmetro e 5 em de altura), cobertas com placas de Petri contendo papel de filtro umedecido. Duas cubas de vidro foram usadas para cada repetição. As lagartas foram alimentadas com folhas de seringueira previamente mergulhadas em diluições de esporos de 8. thuringiensís (Dipel, pó molhável-Abbott Laboratórios do Brasil) e secas ao ar. A partir do segundo dia, todas as lagartas receberam o mesmo alimento da testemunha. A percentagem de mortalidade foi calculada 1;3;5 e 7 dias após o início do ensaio. Ao final do ensaio, as fezes secas ao ar (11% de umidade) foram pesadas para determinar a quantidade relativa de alimento ingerida.
* Recebido para publicação em março, 1974.
* Realizado com recursos do Convênio SUDHEVEA /CEPLAC.
** M.S., Divisão de Fitoparasitologia, CEPEC.
Revista Theobroma, CBPEC, Ilhéus, Brasil, 4 (3}: 33-36. ]ul.- set. 1974.
33

O Quadro 1 mostra as percentagens de mortalidade e o peso das fezes das lagartas. De um modo geral, a mortalidade cresceu com o aumento da concentração de esporos e o númerv de dias após o tratamento, embora não tenha atingido a eliminação total das lagartas. A mortalidade na testemunha foi elevada, provavelmente por deficiência na manipulação das lagartas. Em termos de peso de fezes, a análise mostrou que as di· ferenças entre os tratamentos foram significativas ao nível de 1% de pro· habilidade. As lagartas doentes deixam de se alimentar.
Quadro 1 - Mortalidade e peso de fezes de lagarta de E. e !lo alimentadas com folhas de seringueira tratadas com o Bacillus thutingiensis. CBPEC, 1973.
Mortalidade (%) Peso de fezes
Diluições * Dias após o tratamento (g)
I 3 5 7
1: 1.000 5 33 48 70 11,9 2: 1.000 3 38 53 70 9,0 3: 1.000 lO 50 60 80 8,2 4: 1.000 8 70 80 88 3,0
Testemunha 3 13 23 45 23,9
* A diluição 1: 1.000 contém 25 x 10 6 esporos/ ml.
A conclusão provisória desse ensaio é que os esporos do 8. thurin· giensis, em todas as concentrações usadas afetam drasticamente a capacidade alimentar da lagarta E. ello provocando a sua morte. Novos ensaios deverão ser feitos com o aprimoramento da técnica de manipula· ção do materiaL É preciso registrar que as lagartas utilizadas no ensaio foram trazidas para o CEPEC de uma plantação distante 150 km e que, apenas 1 dia de repouso no laboratório não é suficiente para a sua recuperação. A adição de um espalhante adesivo à suspensão de esporos deverá também melhorar a eficiência do tratamento.
LITERATURA CITADA
1. HEIMPEL, A.M. A criticai review of Bacillus thuringiensis var. thuringiensis Berliner and other crystaliferous bacteria. Annual Review of Entomology 12: 287-322. 1967.
34
PATHOGENICITY OF Bacillus thuringiensis BERLINER AGAINST THE RUBBER LEAF CATERPILLAR
(Eriooyis ello L.) (LEPIDOPTERA : SPHINGIDAE).
The caterpillar of the hawkmoth Eri1myis ello L. is the most destructive insect pest of the rubber tree (Hevea brasiliensis Muell Arg.), the principal diversification crop grown in the Bahian cocoa region. The caterpillar feeds on the young leaves and in severe outbreaks can cause the complete defoliation of large areas.
Due to the success over the last 20 years of the crystal·forming bacteria against more than 137 insect species from the orders Lepidoptera, Hymenoptera, Diptera and Coleoptera (1), an explorative laboratory test was carried out with Bacillus thuringiensis Berliner to determine its pathogenicity against E. ello. The experiment consisted of four concen· tration treatments together with a control, with four replicates of each. Catterpillars were collected from a rubber estate and kept at rest for one day in the laboratory. Five caterpillars of the 3rd instar were placed in deep culture dishes (14 em diameter, 5 em high) with the inside of their loose fittinglids covered with moistened filter paper. Two culture dishes were used for each replicate. The test caterpillars received for 1 day air-dtied young fully expanded rubber leaves previously dipped in the various spore solutions of B. thuringiensis (Dipel (wettable powder), Abbott Labs). From the second day onwards all caterpillars were fed on undipped rubber leaves. Mortality (%) was calculated 1, 3, 5 and 7 days after the start of the treatment. At the end of 7 days the air-dried faeces (11% moisture) were weighed to detennine relative food consumption.
Mortality and faeces weight for the different treatments are shown in Table 1. Although mortality increased in relation to time and spore concentration, a 100'1c. kill was not achieved in any of the treatments. The unexpectedly high mortality among the control caterpillars is attributed to poor handling techniques. The differences in faeces weight, signifcant at the 1% levei, show that the bacteria caused the caterpillars to stop eating.
The preliminary conclusions which can be drawn from this experiment clear!y show that even at a concentration of 1 : 1000 the spores of H. thuringiensis drastically diminish the appetite of the E. e/lo caterpillar and can kill it. However further laboratory experiments are required in which the handling techniques are improved. lt is worth remembering that the caterpillars used in this trial carne from a rubber estate over 150 km from the CEPEC research station and the one day rest in the laboratory
35

Table 1. M.ortality and we;gbt offaeces of E. ello caterpWars fed with rubber leaves dipped in B. thuringiensis spore solutions at CBPBC, 1973.
Mortality (%) Weight of faeces
Concentration Days after Treatment (g)
I 3 5 7
1:1000• 5 33 48 70 11.9 2' 1000 3 38 53 70 9.0 3' 1000 lO 50 60 80 8.2 4: 1000 8 70 80 88 3.0
Control 3 13 23 45 23.9
• 1 : 1000 concentration contains 25 x 1 O ti esporos/ml
was not sufficient for the caterpillars to recuperate. The addition of a spreader would also improve the efficacy of the bacteria treatment.
36
REVISTA THEOBROMA
Outubro-Dezembro 1974
Ano IV N~ 4
publicação trimesnal dedicada à divulgação de investigação científica relacionada com problemas agronômicos e sócio-econômicos de iress cacaueiras. Editada pelo Centro de Pesquisas do Cacau (CEPEC), Departamento da Comissão Ezecutiva do Plano da Lavoura Cacaueira (CEPLAC).
COMISSÃO EDITORIAL
Ahim, Paulo de T.
Cruz, Luiz C. Sales, Jos~ C. de Vello, Fernando
ASSESSORES CIENTÍFICOS
Ailken, WiUiam M. Alencar, Maria H. Alvim, Ronald Barroco, H~lio E. Buros, Raimundo S. Cabala R., F. Percy Carleuo, G"raldo A. Loureiro, Edmar S. Maravalbas, Nelson Mariano, Antônio H. Medeiros, Arnaldo G. Menezes, José A. S. Miranda, Emo Ruy de Pereira, Clovis P. Rocha. Hermínio M. Silva. Luiz F- da Silva, Pedrito Ventocilla, JDS~ A.
Endereço para correspondência (Adress for corre· spondence):
Revista Tbeobroma Centro d., Pesquisas do Cacsu (CEPEC). Cai:u Postal 7 4~.600- ltabuna-Bahia
Brasil
Tiragem 3.000 exemplares
CONTEÚDO
1. Pbysiological responses of cacao lo environmental factors. P. de T. Alvim, A.D. Machado e F. Vello.
Influência de fatores Qmbientais sobre a fisiologia do cacaueiro. {Resu·
mo). p. 24
2. Nematódios da região cacaueira do Espírito Santo, Brasil - I - Nema· tódios associados a o cacaueiro (Theobroma cacao L.) R.D. Sharma e S.A. Sher.
Nematodes of the Cocoa Region of Espírito Santo, Brazil - I - Nematodes associated with c o c o a (Tbeobroma cacao L.). (Summary) p. 30
3. Fatores qut? influem na captura dt? Eri.nnyis ello L. (Lepdoptera: Sphin· gidae) por armadilhas luminosas.
].M. Abreu. --·-·-·····-········-···--
The influence of some factors on Hght trap catches of the bawkmoth Erinnyis ello L. {Lepdoptera: Sphingi
dae). (Summary). p. 43
3
26
•
32

PHYSIOLOGICAL RESPONSES OF CACAO TO ENVIRONMENTAL FACTORS *
Paulo de Tarso Alvim** Antonio Dantas Machado*** Fernando Vello ****
In a previous review on physiology and ecology of the cacao tree (6) an attempt was made to summarize and appraise available information on effects of environmental factors on shoot-growth rhytms,cambial activity, flowering, fruit growth, and cherelle wilt. A few papers dealing with growth and flowering rhythms of cacao havt" appeared since then (7, 9, 15, 21, 22, 23, 24, 25). In Brazil, additional research has been carried out during the last 6 years on physiological responses of cacao to environmental factors. Although most of these studies in Brazil are still in progress it is felt that some of the results so far obtained are worth discussing in the light
of recent findings reported in the literature. This paper will review studies on leaf water potential, growth rhythms (flushing and leaf fall), flowering, and fruit development.
LEAF WATER POTENTIAL
Field measurements of leaf water potential were made with the pressure chamber technique (26). The method consists of enclosing an excised leaf in a pressure chamber, with the petiole projecting through the lid and applying a gas pressure (nitrogen) until sap exudes from the cut surface. The pressure necessary to displace the sap is taken to be equivalent to
* Received for publication in December, 1974.
* Presented at the Intemational Cocoa Research Conference. Trinidad and Tobago, January 9-18, 1972.
** Ph.D .• Division of Phyto1ogy, CEPEC . ... ****
M.S., Division of Phytology, CEPEC. Present address: Conselho Nacional
de Desenvolvimento Científico e Tecnológico (CNPq), Rio de Janeiro. M.S .• Division of Genetics, CEPEC.
Revista Theobroma. CEPEC, Ilhéus, Brasil, 4(4): 3·25. Out.-dez. 1974.
3

the tension or water potential in the xylem before excising the leaf.
lnitial trials were performed by sampling Ieaves at random over the whole tree, a procedure that soon proved to be inadequate since wide variation was observed in water potential in leaves of the same tree, and even for the same branch. Subsequent measurements were oriented toward the influence of leaf age and position on its water balance.
Figure 1 shows diurna! variation in water potential of 4-month-old leaves taken from the upper part o f the tree, all facing in the same direction. The measurements were made hourly on a cloudy day in a
• "
- ' -. - , - ' - '
600 800 D 00 1200 "'00 1600 1800
H O UI!
"
" "
Figure 1 -Diurna/ variation in water potentíal o/ 4-month-old cacao leaves.
4
field where a leaf retention experiment was being conducted. This permitted an securate estimate of leaf age. The number of leaves sampled, which was never less than five, varied according to the degree of variability found in exploratory measurements.
Differences in leaf water potential within the hourly samples were small, especially in those taken in the early morning and at dusk. Water potential decreased (became more negative) progressively to midday, then increased during the afternoon.
Water potential of leaves of different age and with the same position in relation to the sun was evaluated using 4, 7 and lO monthold-leaves. The results (Figure 2) followed the pattern already described with respect to diurna! variation, with no clear differences shown in water potential of Ieaves of different ages.
Since leaves having the same position on the tree with uniform orientation in relation to the sun showed no significant differences in water potential, independently of leaf age, water potentials of shaded leaves and those exposed to the sun were determined. Tenmonth-old Ieaves were sampled from the upper part of the trees (leaves directly exposed to the sunlight) and compared with shaded Ieaves taken from the lower part of the tree.
í ~ ' ' • í
i
'
_,
. .
J
• " w ~ . ' " .
19 ~
.._ -10.~ ' ~....,.'"""'~=~~ 600 aOOIOOO 1200 .. 001600 '800
HO 011
Figure 2 - Diurna/ variation in water potential o/ leaves o/ different ages.
Substantial differences in water potential occured as illustrated in Figure 3. The three pairs of curves shown were obtained on alternate days and provide a good example of the effect of atmosphere humidity of leaf water potential in the absence of a soil moisture stress. The first pair of curves refers to a cloudy day, during which it rained between 10:30 and 11:30 a.m.; the middle pair to a brighter day with a rain shower around 9:20 a.m.; and the third pair to a clear and warm day without rain. The water potential of the upper exposed, leaves was appreciable lower than that of shaded leaves, and the difference was accentuated with decrease in rel-
OGTOO .. "•'9'' OOfOOEO >0, '"''
_, _,
-· _,
-" _,.
. \ !
~
0001000'>00•400 1000
HOUR
' . • '
28 ~
Figure 3 - Diurna! variations in leaf water as related to air temperalrae and relative humidity, for J dilferent days.
5

ative humidity. Since all data were taken during a relatively rainy period it is clear that the daily variation in water potential of cacao leaves is much more dependent on relative humidity of the air than on soil water availability. Natural~ ly, during periods of prolonged drought soil moisture supply becomes increasingly important in influencing leaf water balance.
In a recent experiment on effects of shade removal on pedodicity of leaf fali and leaf flushing of cacao, leaf water potentials were measured in an open atea and in a densely shaded ones. The selected areas were near each other and had similar slope and soil characteristics. The initial results (Table 1) showed that leaves in the shaded area were under considerably less water stress than the leaves in the open area. Differences in water potential between upper and lower leaves were minimal wheteas there was wide variation for the same leaves in the unshaded plot. In addition, tem-
perature of the upper sunlit leaves in the unshaded plot was 10° c higher than amhient temperature. As expected, such a difference was not observed with the lower and shaded leaves of both plots.
GROWTH RHYTHMS
In spite of growing in areas with little seasonal variation in climate, patticularly with regatd to temperature and day-lenght, cacao, like many tropical trees does not grow continuously thtoughout the year but behaves as an intermittently growing evergreen, exhibiting petiods of intensive leaf growth alternating with periods of vegetative rest. These growth ~ulses are generally recognized as "seasonal" in the sense that they are repeated at about the same time of year, varying according to region but within a region usually showing little change from Y.ear to year. Attention will be given in this discussion to the role played by environmental factors in control of shoot flushing and leaf abscission.
Table 1 - Ef!ect of shade and time of day on leaf water potential.
Time
10:00
12:00
14:00
Leal water potential (bars)
With shade No shade
Upper leaves Lower leaves Upper leaves Lower leaves
-6.3 ± 0.9 -4.3 ± 0.4 -13.7:!: 1.2 -5.3 ± 0.6
-5.0 ± 0.2 -4.3 ± 0.4 -12.4:!: 0.2 -7.3:!:1.1
-4.9 ± 0.1 -4.2 ± 0.1 -12.7 ± 0.4 -6.1±0.9
6
Age eUect
When studying growth rhythms of trees it is most important to recognize that juvenile plants behave very differently from_ adult plants. Greathouse, Laetsch and Phinney (15) recently studied growth rhythms of young cacao plants (about 1 year old) undd~r controUed environmental con I
ti.ons (29°C; 75-100% telative humidity; 600 ft C light intensity, and 12-hr photo-period). The growth cycle 0 { a shoot showed an average duratiort of about 65 days, and in their opinion, once this cycle was compteted a new flushing cycle automatically started, provided environmental conrlitions wete suitable for growth. Great~ house et ai (15) observed that plants unrler controlled conditions were not synchronous with each other with regard to shoot-growth "as it appears to be in the field" (sic). and concluded thttt their data "strongly suggested that ftushing was conttolled by an endogenous mechanism".
The fact that young c a c a o plants are asynchronous with regard to growth rhythms is w e ll known (5, 6). Equally well known is the fact that mature plants in the field exhibit ctose s-ynchrOf\iSm in growth behaviour. Jhe underlying physiological reasons for this difference between matute and juvenite plants are not well understood. There is little doubt, however that growth rhythm of m:ilture plants in the field is strongly con-
trotled by environmental factors and not by endogenous rhythms.
Internai factors
Whatever the nature of the externai stimulus which induces growth flushes in cacao, thete is Iittle doubt that internai factors -be they growth promoters, growth inhibitors, nutrients, ora balance of such internai requirements -a r e obviously involved in the mechanism of growth and rest. With certain species of temperate regions, some progress has been made on endogenous hormones controlling growth rhythms (8, 10, 13, 30). ln the case of cacao, good data are scarce but some indirect evidence illustrates the role of intern<tl factors in the mechanism of flushing,
Studies in Brazil (6, 7) and in the Camerouns (9) have shown tbat ftusbing and leaf fali are closely related phenomena. Intensive leaf fali invariably occurs simultaneously of just before or after an intensive flush of shoot growth. Ordinarily, the main flush .in Bahia occurs in September-October and is followed by two or three minot Hushes between November and April, with flushing in February-March usually second in importance. Maximum leaf fall as a rule t<:~kes place soon after the intensive growth flush of SeptemberOctober. From May to mid-September the plants remain in a state of
7

rest, with practically no f!ushing and very limited leaf-fall taking place (Figure 4).
Two hypothesis may be suggested to explain the synchronism between leaf-fall and flushing; a) the concentration of an inhibítor responsible for bud dormancy and originating in the leaves is reduced as a consequence of leaf abscissíon, thus triggering bud expansion; b) flushing imposes a strain for nutrients and/or hormones, which would acce!erate
•• ~ ! -r. . \~+i, .
1:PA,I~ ·~-
' ! ', I
'
I
senescence and abscission of old leaves. The first hypothesis is supported by the fact that flushing can be induced during any period of the year by means of heavy pruning or by various degrees of defoliation. Studies in Bahia have shown that leaf remova! caused intensive flushing even during a period when plants normally are in a state of rest (e.g. in June) (7).
In 1971, a drought ocurred during June/July, normally the wettest period of the year in Bahia,
. . o
·~ ' i
'
JASONOJF "'MJJASONOJ MAMJJASOND
I 9 6 9 I 9 7 0 ' 9 7 '
Figure 4- Bi-weekly data on flusbing of cacao in ]uçari, Bahia, from July 1969 to Novemher 1971, Data are expressed in% o/ hud break and number of newly formed [eavq, together with data on leaf fall/m' /week, temperature, rainfall, and cumulative soil moisture defícit and excess.
8
causing off-season leaf-drop which was immediately followed by intensive flushing in July-August (Figure 4). This was the first time in many years that flushing was
0 bserved in Bahia during this period.
As shown in Figure 4 1 leaf fall in October 1969, October 1970, and Decembet 1971 occurred 2-3 weeks after flushing. This does not necessary invalidate the first hypothesis, as the abscission }ayer is obviously formed before actual leaf shedding takes place and could therefore precede onset of flushing .
A growth prometer moving from the roots to the shoots through the phloem also appears to play an imporlant role in the mechanism of flushing. The indirect evidence which appears to justify the assertion is the following (7): a) when a complete ring of bark is removed from the trunk or from one of the main branches of a cacao tree thereby preventing upward phloem transport, flushing is inhibited in the wholc tree or in the ringed branch, respectively; h)
when all but one of the ma in branches are ring-barked, the ungirdled branch is stimulated to f 1 u s h off-season, whereas ali ringed branches do not flush (suggesnng that the growth prometer originating in the roots moved in higher concentrations to the ungirdled branch}; c) flushing is also inhibited when some lateral roots are "pruned" by opening a trench about 25 em deep with a circumference 2 m in diameter around the tree,
Competition for food between growing pods and shoots also appears to reduce flushing intensity. This was observed in the seed garden plots of the CacaoResearch Center, in ltabuna, Bahia, in 1970 and 1971, Plants of the Catongo cultivar about 4-5 years old, which had been intensively hand pollinated to produce hybrid seeds exhibited very little flushing when compared with otherCatongo plants in the same area which had not been hand pollinated. Heavy fruit setting on the hand pollinated plants also appeared to be responsible for die-back and even death of a few trees. Die-back symptoms disappeared and normal flushing was promptly resumed when the affected plants were stripped of ali fruits.
9
Environmental factors
Ear!ier theories about the effect of environmental factors on growth rhythms of cacao were reviewed in a previous paper ( 6 ). Research carried out between 1966 and 1972 (7, 24, 25} gives support to a theory f i r s t suggested by McDonald (19) which attributed particular importance to the P.ffect of soil moisture and/or air relative humidity on flushing. On the basis of field studies in Brazil, the following hypothesis was proposed to explainthe re\ationship between moisture availability and flushing (7): a) decreased rainfall (or increased moisture stress) induces leaf abscission; b) abscission breaks bud dormancy (because of

reduced production o{ ar. inhibitor or dormancy inducing substance originating in Jeaves); c) flushíng occurs soon after abscission or, under conditions of severe moisture stress, after the onset of rains.
Sale (25) demonstrated that each time cacao plants were watered following a period of soil moisture deficiency vigorous flushing occutted about 10 days after waterir.g. Many of the f!ushes were produced from axillary buds as well as from terminal buds. Plants which were irrigated frequently flushed less vigorous than droughted plants and only at the terminat buds, with peaks of flushing occurring every S-6 weeks. In spite of flushing Iess intensively, the total leaf area was higher for the ftequently watereO plants than for droughted ones because of reduced leaf shedding or longer Jeaf of the former. Sale (21) also demonstrated under controlled ccmditions that flushing was not induced by increase in temperature above 83°F, as previously advanced by Humphries (17) and Gteenwood and Posnette (16), or by increase in temperature range {thermo-periodicity) as suggested in earlier papers of Atviro(2). Sale (21) found however, that the intervais between flushes were reduced when temperature was raised, with plants growing at 74 ° F flushing approximately every 95 days and plants growing 80 o F and 86 a F flushing every 36 and 20 days, respectively.
10
Figure 4 shows data on flushing and leaf shedding injuçari, Bahia, from July 1969 to November 1971, together With data on r a in f a 11, roean tempetature, and cumulattve soil moisture deficit and excess. Flushing was recorded on 20 plants, about 12 years otd, by weekly counting of newly formed leaves on 200 weU exposed shoots (10 shoots per plant). The percentage of shoots initiating flushing (bud break) was also recorded at weekly intervals. Twenty, 1 sq.m wooden boxes were placed under the trees for weekly counting of leaf fall. For the sake of simplicity, dat<' on bud bteak and number of newly formed leaves are presented for 2-week intervals.
Cumulative soil moisture deficits were estimated foBowing the method of Smith (27), assuming a soil depth (root space) of 45 em, an apparent specific gravity of the soil of 1.4, a field capacity of 32% and a wílting point of 16%. These values give a ~K" value of 0.01, anrl a water retention capacity of lOOmm. The cumulative soil moisture excess was derived from the balance between tainfall and potential evapotranspiration. Data on solar radiation as measured with a solarigraph, relative humidity, and vapour pressure deficit, were <'liso analysed in this study but are not included in Figure 4 as they did not show any correlatíon with growth rhythms.
By comparing the data on flushing with that on cumulative soil
woisture deficit and excess - and to a lesser extent with rainfall at 10-day interva1s - it becomes apparent that periods of increased bud break and subsequent leaf expansion were always prec~ded by a peüodof soit moisture depletion. In most cases the beginning of a flushing cycle took place simultaneously with/or soon after the {irst rainy days following a relatively dry period. There were cases, however, (e.g. in Decembet 197 O) when flushing apparently started before the onset of rain. Further studies e.re needed to determine whether a dry-wet sequence is always necessary to initiate flushing or if moisture depletion. alone can trigger flushing (provided it does not reach the pointof checkinggrowth ot causing wilting).
The importance of moisture stress in breaking dormancy has been clearly demonstrated for coffee flower buds (3). Moistu~e stress was shown to be an essential prerequisite for breaking dormancy, but anthesis could take place only following irrigation 01 rain. A siroilat type of responSe to a relatively dry period followed by a wet one has been observed in citrus, in connection With Howering and flushing cycles (11) and probably is of common occurrence among tropical trees, The term "hydroperiodicity" has been suggested for this physiological response to a succession of dryness and wetness (S).
11
Special attention should be given to the fact tb.at between 1963 and 1970 (6, 7), f!ushing had neve r been reported in Bahia during the months of May to August, which cotresponds to the period of reta. tively frequent rainfall, reduced solar radiatíon and low tempetatute. In 1971 intenSive leaf fali followed by vigorous flushing C'i>
curred for the first time during the month of July, and this was clearly associated with an unusually severe drought which ocurred in June (Figure 4). Temperature and solar radiation showed no apparent relationship to this off season flush. These results seem to indi· cate that the absence of flushing normally observed ftom May to August in Bahia cannot be attributed to low temperature or low solar radiation, as previously believed (6, 7), but to normal absence of moistwe stress during the period.
In Sa1e's experiments(25) plants w h i c h were frequently watered (''wet treatment~) did not flush at all after they reached the age of about 2 years, whereas plants of the same age but previously submitted to soil moisture stress exhibited vigorous flushing. Sale carried out his e'ltpetiments with plants aged 12 to 30 months and it seems likely that the "juvenility effect" overshadowed responses to the water regime during the eatlier part of his experiments, i. e., when plants were younger. He agreed with the previous suggestion of Alvi:m, Machado and Gtangier (7) that watet stress caused the

buds to develop to a stage where they were ready to grow and appeared to be a "necessary requirement for the plants to continue to either initiate or expand new leaves ''. Sale recognize that this conclusion did not invalidate the suggestion of Cheesman (12) "that the absence of a dry season may be wholly favourable to cacao -for under most field conditions it is probable that soil water would be depleted occasionally to at least 50%, and thus provide any growth check that might be necessary for continued flushing".
1t is well known that c a c a 0
g r ow in g without shade flushes much more intensively than does shaded cacao. This probably is associated with higher internai moisture stress in unshaded plants, as shown by our studies on the effect of shade on leaf water potential.
Studies with species of temperate regions have shown that moisture stress increases the amount of abscisic acid in the leaves this apparently being the ma in fac~ tor controlling leaf shedding (30). Abscisic acid has been found also to impose bud dormancy in some species (8, 10, 13,29, 30). Our previous hypothesis to explain the effect of moisture depletion on flushing could be tentatively rewritten as follows:
a) Moisture stress increases the concentration of abscisic acid, thus accelerating senescense and leaf shedding;
12
h) Translocation of abscisic acid or another rest-imposing substance from the leaves to buds decreases as a result of formation of abscission layer or because of actual !eaf drop;
c) Buds start growing;
d) Shoots and leaves expand, provided water is available for growth.
FLOWERING
As in the case of flushing, age effects and internai factors deserve attention when studying the influence of environmental factors on the flowering cyc!es of cacao.
1\ge effect
The age at which cacao starts producing flowers in the field varied a great deal with genetic origin and cultural practices. Un
.der good management conditions most of the new hybrids now bein~ planted in Brazil start flowering at an age of about 18 months after transplanting to the field. An extreme case of genetically controlled premature flowering was observed by Soria (28) on some seedlings of pure criollo cacao from Mexico, which began to produce normal flowers at the age of 3 months when the cotyledons had not yet fallen from the plants. Plants growing in nutrient solution under greenhouse conditions in Bahia, usually start producing flowers at the age of about 12 months from seed. Unselected amelonado plants under traditional
cultural pract:ices used by farmers · Bahia take from 3 to 4 years, on m . the average, to start flowenng.
As previously reported (6), the flowering cycle of mature cacao
tants in Bahia follows a Tk.arly :stabilished seasonal pattern, with practically no flowers appearing during the months of July, August, September or October. In young plants flowering is reduced but not completely arrested during t h e same period. The difference be-
N D
tween mature and young p 1 a n t s with regarei to flowering has recently been confirmed in a study comparing the flowering cycle of the cultivar Catongo aged 5, 12 and 23 years old growing in neighboring areas, in Uruçuca, Bahia. To estimate flowering intensity, the soil surface under each tree was covered with a layer of sand a nd the number of unfertilized flowers falling on the sand was counted at intervals of 2or 3 days. Figure 5 shows the total number
I I o N D
'------------- MONTHS Figure 5- Montbly flower production in Catongo trees, aged 5, 12, and
23 years, in Uruçuca, Bahia, from November 1970 to Dece~ber 1971 (average for eight trees in each age group). Data for a stenle 12 year old tree which produced only one pod during the period of the study, are included for comparative purpose (see text).
13

of flowers produced per month (average ofeight trees for each group) from November 1970 to November 1971. Attention should be called to the fact that because of the very wide changes in the number of flowers during the course of this study - particularly in the case of older plants - a logarithmic scale was used. As shown in Figure 5 seasonal variability in flowering increased in direct proportion to plant age. The 23-yea,r-old plants produced about 10 times more flowers than the 5-yearold plants during the peak of flowering (February-April) and about 10 times less during the low flowering period (August-September). The figures for the 12 year old plants fell between the other two age groups.
lt may be possible that the microclimate around the plant rather than plant age is responsible for the difference between mature and young plants with regard to flowering and cropping patterns. Among other things, younger plants,
because of their lower leaf area index, receive more solar energy per unit-leaf area than older plants, and this might be one important factor associated with the difference in their response to variations in climate. Another possibility is that older plants, because of their larger proportion of nonphotosynthetic tissues (roots, trunks etc) or large "sink", would be more easily depleted of flowerforming substances than younger
14
plants. Further research is obviously needed on this point.
Internai factors
A chemical stimulus originating in the leaves and moving down through the inner bark (phloem) appears to play an important role in the mechanism of flowering. This can be demonstrated by removing a ring of bark from the trunk of mature cacao trees. Studies in Brazil have shown that following such stem girdling during the normal blooming season, flowering was very profuse on the portion of the trunk above the ring and was completely arrested bellOVI the ring, this response occurring within 2-3 weeks following treatment. Accumulation of photosynthates and/or flowering hormones is apparently responsible for the enhancement of flowering above the stem girdle. Similar responses to bark ringing has been reported for citrus (14).
The fruit load of the plant also affects flowering intensity. This was observed during the course of our study on the effect age. One of the 12 -year-old Catongo plants included by chance in the experiment has proven to be practically sterile, as it produced only one pod during the course of the study. The flowering cycle of this plant is also included in Figure 5 (12 years without fruits). lt not only bloomed nmch more heavily than the normal bearing 12-year-old plants, but showed a relatively
slight decrease in flowering ftom July to November when other plants, irrespective of age, showed much Iower flowering intensity. Together with our observations on the effect of ringing, these results seem to indicate that fruits com~ pete with flowers for a substance or substances (which could very well be carbohydrates) whose concentration is related to flowering intensity. Competition be t w e e n f r u i t s and flowers is known to play an important role in deter· mining flowering intensity of other trce crops, such as citrus (18).
Environmental factors
Accurate data on- flowering cycles of cacao have been obtained at the Experimental Station in Juçari, for a period of 38 months (January 1968 to February 1971), using 12 clonal plants which about 8-years-old at the beginning of the study. A circular nylon sheet about 3 m in diameter was placed under the canopy of each tree at a height of about 50 em from the ground, resembling an inverted umbrella with the trunk of the cacao tree as the central rod. The number of flowers which dropped into the umbrella was recorded every two days. The average number of flowers per day/plant is given in Figure 6 for 10-day intervals. Data on cumulative soil moisture deficit and excess, calculated by the method mentioned under growth rhythm as well as data on mean temperature, are also presented. In Figure 6 other climatological
data, such as solar radiation as measured by a solarigraph, celative humidity, and vapor pressure deficit, did not seem to show any relevant relationship with the flowering data and will not be included in this disc..ussion.
As shown in Figure 6, the period of intensive flowering usually occurred from November to June, whereas the months of least flowering were August to October. Pte· vious studies carried out at anoth· er station (Uruçuca) with older trees (6) showed minimum flowering from June to September. It is not known if this discrepancy is due to difference in climate, age of trees, varietal differences, or methods of study. It must be recognized, however, that the observations of Figure 6 as well as of Figure 5, were more accurately taken than those previously reported, and are thus probably more reliahle.
15
In previous studies on flowering carried out in Bahia (6) it was suggested that the decrease in flowering from June to September was a result of low temperature alone. Because temperature drop in Bahia starts in May, when flowering is still quite intensive, it was postulated that the effect of low temperature was indirect in the sense that it first affected vegetative growth which in turn affected flowering, this accounting for the time lag between supposed cause and effect. This the ory needs now to be revised in view

DATE Figure 6 - Flowering intensity of cacao as expressed in number of
flowers/pl~nt/day (mean figures for 12 clonal plants), in Juçari, Bahia, together wtth data on temperature and cumulative soil moisture deficit and excess, at 10 day intervals from January 1968 to March 1971.
of recent findings. As shown in our studies on flushing (Figure 4), temperature did not show any re~
lationship with growth rhythms of cacao. lntensive bud break and leaf expansion could take place even during period when tempera~ tures were relatively low (e.g., in July 1971). Sa!e (23) also did not find support for an "indirect" ef~ fect of temperature on flowering. He confirmed that low temperature decreased flowering, but the effect
16
seemed to be direct. We are in~
clined to think that reduced flow~ ering in Bahia from ]une ~July to September~October is caused by internai competition between fruits and flowers, with the effect of low temperature apparently being of lesser importance. As will be shown !ater (Figure 8) the peri.od of maximum fruit load in Bahia, as expressed in total dry weight of hanging pods, coincides with the period of minimum flowering.
The flowering season for plants in our studies occurred from No~ vember to )une. It is worth point~ ing out that the beginning of this season coincided every year with the period when plants had the lowest fruit load (Figure 8), This lends support to our hypothesis about the importance of internai competition between fruits a nd flowers. It appears from our re~
sults that, as longas this compe~ tition exists, flowering shows practically no res ponse to changes in environmental conditions. Only after the plants reach the period of minimum fruit !oad or decreased internai competi.tion between fruits and flowers, blooming becomes responsive to environmental con~ ditions, and exhibits wide flue~ tuation from time to time. lt is well established that flowering of cacao is inhibited during period of moisture stress (1, 4, 6, 20, 25). 1t has also been demonstrated that flowering becomes particularly pro~ fuse after the first heavy rains following an extended dry period (6),
By investigating the data on cumulative soil moisture deficit and excess with the number of flowers per plant (Figure 6) during the flowering season, it may be observed that periods of increased flowering were u;>ually preceded by a relatively dry period followed by a wet period and that decreased flowering was associated either with a dry spell (e.g. April 1970) or excessive soil moisture (e,g. December 1968, December 1969,
February 1970, and November 1970), In some instances, such as in December 1970 and january 1971, increased flowering appeared to result from decreased soil mois~ ture following a period of moisture excess.
Sale (25) obtained profuse flow~ ering in potted cacao plants v.~hen~ ever the soil was watered follow~ ing a period of moisture stress, and concluded that flower initiation had been enhanced during the dry period and only flower growth was inhibited by moisture deficiency. With regard to coffee, the flower buds remain in a state of complete rest for as long as the plants are frequently watered and kept under high atmospheric humidity, a "dry~ shock" being appatently needed for rest breaking (3), As pointed out by Sale(25)this does not seem to be the case with cacao, as the flower buds do not go through a rest period imposed by high water potential, some flowering always occurring even under conditions of frequent watering or high hu~ midity. Sale (24) also demonstrat~ ed, under controlled conditions, that profuse flowering can be in~ · duced by transfening the plants from a low (50-60%) and medium (70~80%) relative humidity to a high one (90~95% ).
17
It is worth pointing out that flowering of citrus in tropical a~ reas has been shown to be con· trolled by alternations between dry and rainy periods and that in irrigated citrus orchards it is pos~

sible to induce out of season flowering by irrigation following a drought stress (11).
From the foregoing discussion it may be concluded that the maio factors controlling flowering of cacao appear to be, internally, the competition between fruits and flowers, and externally, the alternation between dry and wet periods, ot hydroperiodicity of the environment.
FRUIT PHYSIOLOGY
Cacao harvesting in Bahia usually extends for about 8 or 9 months, Le. from mid-April to midJanuary. Plants up to 4-6 years old, tend to produce fruits throughout the year, hut the crop is usually much reduced from mid-January to mid-April. In August-September the crop from mature plants is also reduced but its production is not completely stopped. The harvest from Aptil to August is referred to as "temporão" or midcrop, and from September to midJanuary, •safra" or maio crop. The cropping pattern shows some variation from year to year but on the average there is practically no difference between "temporão" and "safra" in volume of production.
Temperature effect on fruit development
In a study carried out with the white seeded cultivar "Catongo", it was found that the period from
18
pollination to' ftuit ripening varied in Bahia from 140 to 205 days, the average being 167 ± 0,78 days. As previously reported (6) when pods of the "temporão" crop (pollinated in November-January and harvested in April-August) were compared with the "safra" crop (pollinated in March-June and harvested in September-January) it was found that the former matured in 140-175 days whereas the latter took 168-205 days reach rcaturity.
A highly significant negative corre lation was found between the number of days from pollination to harvest and the mean temperature during the period. Based on this correlation the following equation was developed to estimate the number of days from pollination to harvest for the Catongo cultivar in Bahia:
N = 2500
T-9 Where N is the number of days and T the daily mean temperature (0 C) during the period.
Figure 7 gives the estimated number of days from pollination to harvest for fruits set at different periods of the year calculated olf the basis of the above equation, using the mean temperature values for Uruçuca, Bahia.
Seasonal changes in fruit Ioad
In order to estimate changes in fruit load during the year, careful pod counts were made monthly on
"' ...
•••
i ,,, ...
"' !
~ ,,, "'
• ' '
.... O!' .... , • .,,..
the same 12 clonal plants used in the study on flowering cycles. Figure 8 shows the monthly numbers of harvested pods, developing pods, and wilted cherelles. On the basis of a previous study on changes in pod dry weight as re~
Figure 7 - Tbeoretical periods hetween fruil setling (pollination) and barvesl, expressed in days, as a function of date of pollina· tion. Dates on the curve were calculated on the assumption that pollination occurred on the first day of each month. In Bahia, as a rule, there is no crop between mid· january and mid-April.
lated to fruit age (6), and using the final number of harvested pods as a reference, the total dry weight of the hanging pods could be estimated. Wilted cherelles were not included in this estimation. As indicated in Figure 8, the period of maximum fruit load on a dry weight basis did not coincide with the period of maximum number of pods per tree. As a rule, the latter occurred from March to July with the peak in May/June,
Figure g _ Pod counts at monthly intervals for 12 clonal plants from }anuary 1968 to February 1971, in Juçari, Bahia, together witb estimated total dry weight of pods per plant/month.
19

whereas the former carne about 3 months later, with the peak in August/September. The period of minimum fruit load occurred from December to February, As previously stated, changes in flowering intensity appeared to be related to changes in fruit load.
Very likely, internai competition between fruit growth and vegetative growth (flushing, root a n d cambial growth) plays an important r o l e in determining changes in fruit load during the year. In other words, when externai conditions are favorable for growth (OctoberA p r i l) the vegeta tive sink incresses and fruit load decreases;
• . ~ . .
' . ' . . ~
•
.
•
.
.
li li [I 1/
~· (h 1 •
I'
·' i
• • ' • • •
' ;; I
I :1"1 li i I , I
' '
!
' I
I ••
!
'
• • •
when is arrested (May-September) the vegetative sink decreases and fruit Ioad increases, This hypothesis implies that vegetative tissue always are the strongest sinks and that changes in fruit Ioad during the year are primarily due to environmental effects on vegetative growth, this in turn affecting fruiting,
Seasonal changes in fruit setting
In our study on flowering and fruit Ioad at Juçari, data on fruit setting were also obtained by counting the number of n e w I y formed pods at monthly intervals, Results are presented in Figure 9 together with data on w i 1 te d
mm- -., ... , '""' "' ' I
,.-n H,, ..
li I I '"" ., , .. , . ' !
iT' ! '
u i • ' . ' •
Figure 9- Fruit setting, cheridle wilt, percentage of settingfno.newly formed pods x 100/total number o/flowers), and estimated total dry weight of pods per plant, at monthly intervals, in 12 clonal plants grown in Ju· çari, Bahia, from January 1968 to Fehruary 1971.
20 t
cherelles. Also included are data
011 the percentage of setting (nurnber of newly forrned pods x 100/ numberof flower)and the estimated fruit load on a dry weight basis.
B y comparing these results with Figure 6 it-will be noticed that the heaviest fruit setting periods occurred when the plants were flowering heavily but the percentage of setting reached its peak (up to 85.7%) during the lowflowering periods. The population of pollinating insects shows great variation during the year (31) but changes in the ratio of pollinating insects to nurnber of flowers appear to be related more to this
wide variation in percentage of setting.
In earlier papers (6) the absence of crop in Bahia from rnidJanuary to mid-April was attributed to reduced flowering in July-October, Results in Figure 9 indicate quite clearly that the actual cause was not absence of fruit setting in in July-October but greater loss of young cherelles overruling fruit se t ti n g during that period. As shown Figure 9 the incidence of cherelle wilt appears to be related to fruit load. This indicates that, under the conditions of this study cornpetition between fruits probably was the main cause of wilt.
LITERATURE CITED
1. ALVIM, P. de T. Correlação entre chuva, temperatura e produção do cacaueiro. In Conferência Interamericana de Cacau, 6a., Salvador, Bahia, Brasil, 20-27 de maio, 1956. Bahia, Brasil, Instituto de Cacau, 1957. pp.133-136.
2. . Fatores que controlam os lançamentos do cacaueiro. in Conferência Interamericana de Cacau, 6a., Salvador,Bahia, Brasil, 20-27 de maio, 1956. Bahia, Brasil, Instituto de Cacau, 1957. pp. 117- 125 .
3. ----- . Mo.isture stress as a requirement for flowering of coffee. Science 132 : 354. 1%0.
4.
S.
----- . Las necesidades de agua dei cacao. Turrialba 10: 6-16. 1960.
. Tree growth periodicity in tropical climates. In Zimmermann, N.H., ed. Forrnation of wood in forest trees. New York, Academic Press, 1%4. pp. 479-495.
21

6. ALVIM, P. de T. Eco-physiology of the cacao tree.ln Conference Intemationale sur les Recherches Agronomiques Cacaoyêres, Abidjan, 15-20 Nov., 1965. Paris, Jouve, 1967. pp. 23-25.
7. , MACHADO, A.D. and GRANGIER Jr., A. Alguns estu-dos sobre as relações de água, solo e crescimento do cacaueiro. In Conferência Internacional de Pesquisas em Cacau, 2a., Salvador e Itabuna, Bahia, Brasil, Novembro 19-26, 1967. Memórias. ltabuna, Centro de Pesquisas do Cacau, 1%9. pp. 316- 326.
8. BOWEN, M.R. and HOAD, G.V. ll)hibitor content of phloem and xylem sap obtained from willow (Salix viminalis L.) entering dormancy. Planta 81: 64-70. 1968.
9. BOYER, J. lnfluence des régimes hydriques, radiatif et thérmique du climat sur l'activité vegetative et la floraison de cacaoyers cultivés au Cameroun. Café, Cacao, Thé 14: 189-201. 1970.
10. BROWNING, G. and HOAD, G.V. Identification of abscisic acid in flower buds of Coffea arabica (L.) Planta 94: 213-219. 1970.
11. CASSIN, J. et al. The influence of climate upon the blooming of citrus in tropical areas. In International Citrus Symposium, 1969. Ptoceedings. V. l. pp. 315-323.
12. CHEESMAN, E.E. Report on the potentialities for the cultivation of cocoa in Malaya, Sarawak, and N. Borneo. London, H.M.S.O., 1948. 44 p. (Colonial Office n. 230).
13. CONFORTH, J.W., MILBORROW, B.V. and RYBACK, G. Identity of sycamore "'Dormin" with abscisin li. Nature 205: 1269-1270. 1965.
14. FURR, J.R., COOPER, W.C. and REECE, P.C. An investigation of flower formation in adult and juvenile citrus trees. American Journal of Botany 34: 1-8. 1947.
15. GREATHOUSE, D.C., LAETSCH, W.M. and PHINNEY, 8.0. The shoot-growth rhythm of a tropical tree, Theobroma cacao. American Journal of Botany 58: 281-286. 1971.
16. GREENWOOD, M. and POSNETTE, A, F. The growth flushes of cocoa. journal of Horticultura! Sc:ience 25: 164- 174. 1950.
22
17. HUMPHRIES, E.C. A consideration of the factors controlling the opening of buds in the cacao tfee (Theohroma cacao L.). Annals of Botany (n.s.) 8: 259-267. 1944.
18. LENZ, F. Effects of day lenght and temperature on the vegetative and reproductive growth of "'Washington navel" orange. In International Citrus Symposium, 1969. Proceedings v. 1., pp. 333-337.
19. McDONALD, ].A. An environmental study of the cacao tree. In Imperial College of Tropical Agriculture. Trinidad. Annual Report on Cacao Research 1: 29-38, 1932; 2: ii-iv, 1933.
20. MURRAY, D.B. Soil moisture regimes. In Imperial College of Tropical Agriculture. Trinidad. Report on Cacao Research, 1965. pp. 34-39. 1966.
21. SALE, P.J.M. Flushing and leaf growth of cacao under controlled temperature conditions. Journal of Agriculture Science 43: 475-489. 1968.
22.
23.
24.
25.
----- . Extension growth of cacao under controlled temperature conditions. Journal of Horticultura} Science 44 : 189-193. 1969.
Flowering of cacao under controlled temperature conditions. Journal of Horticultura} Science 44: 163-173. 1969.
----- . Growth and floweting of cacao under controlled atmospheric relative humidities. Journal of Horticultura} Science 45: 119-132. 1970.
----- . Growth, flowering and fruiting of cacao under controlled soil moisture conditions. Journal of Horticultura! Science 45: 99-118. 1970.
26. SCHOLANDER, P .F. et ai. Sap pressure in vascular plants. Science (U.S.A.) 148, 339-346. 1965.
27. SMITH, G.W. The relativity between rainfall, soil, water and yield of copra on a coconut estate in Trinidad. Journal of Applied Ecology 3: 117-125. 1966.
28. SORIA V., J. A note on cacao plants which flowered at the age of three months. Cacao (Costa Rica) 6: 11-12. 1961.
23

29. VAN DER VEEN, R. Plant hormones and flowering in coffee. Acta Botanica Neerlandica 17: 373 -376. 1968.
30. WAREING, P.F. and SAUNDERS, P.F. Hormones and Annual Review of Plant Physiology 22: 261-288.
dormancy, 1971.
31. WINDER, J.A. and SILVA, P. Current research on insect pollination of cacao in Bahia. In InternationalCocoa Research Conference, 4th, Trinidad and Tobago, January, 1972. 14 p.
SUMMARY
Effects of environmental factors on leaf water potential, growth rhyÍ:hms, flowering, and fruit development of cacao are discussed in the Iight of recent research in Brazil and elsewhere. Some plant factors, such as age and internai competition between fruits and flowers must be considered when studying growth and flowering responses to environmental factors, Flushing and leaf abscission were found to be correlated with each other. Among various environmental factors studied in connection with flushing, moisture stress appeared to be the most criticai one, both for bud break (flushing initiation) and leaf abscission. Rain following a dry period apparently stimulates intensive flushing (including leaf expansion). No support has been found for earlier theories according to which flushing would &e controlled by changes in air temperature.
Flowering seems to be controlled internally by competition with fruits (fruit load) and externally by a succession of dry-wet periods. The term ,.hydroperiodicity~ is proposed to refer to growth and flowering responses to a dry wet sequence. Flowering is inhibited by extended dry periods as well as by excessive soil moisture,
Data on leaf water potential, fruit setting, rate of fruit development, and seasonal changes in fruit load, are also discussed in connection with environmental influence.
INFLUÊNCIA DE FATORES AMBIENTAIS SOBRE A FISIOLOGIA DO CACAUEIRO
RESUMO
São discutidos à luz de pesquisas recentes, efetuadas no Brasil e no exterior, os efeitos de fatores ambientais sobre o potPncial de água das
24
J folhas, o ritmo de crescimento, a floração e o desenvolvimento do fruto do cacaueiro. Alguns fatores da planta, tais como idade e competição interna entre frutos e flores devem ser considerados no estudo das respostas de crescimento e floração a fatores ambientais. Encontrou-se que o fluxo de crescimento e a abscisão das folhas se correlacionam mutuamente. Entre os vários fatores ambientais estudados em relação aos fluxos de crescimento, a deficiência hídrica parece ser o mais crítico, tanto para a abertura das gemas (iniciação do lançamento) como para a abscisão da folha. A chuva logo após período seco aparentemente estimula o fluxo foliar intensivo (inclusive desenvolvimento foliar)]Não se encontrou apoio para as teorias anteriormente formuladas segundo as quais os fluxos foliares seriam controlados por mudanças na temperatura ambiental.
A floraçãc parece ser controlada internamente pela competição com os frutos (carga de frutos) e externamente por uma sucessão de períodos secos e úmidos. O termo "hidroperiodicidade" é sugerido para designar a correlação entre crescimento e floração em resposta a uma seqüência de período seco-úmido. A floração é inibida por um período seco prolongado bem como por excesso de água no solo.
Dados sobre o potencial de água em folhas, frutificação, taxa de crescimento do fruto e mudanças sazonais na carga de frutos são tambem discutidos em relação à influência do ambiente.
.. .. ..
25

NEMA TÓDIOS DA REGIA O CACAUEIRA DO ESPÍRITO SANTO, BRASIL
I - NEMATÓDIOS ASSOCIADOS AO CACAITFIRO (Theobroma cacao L.)•
R avi Datt Sharma ** Samuel Alexis Sher***
Pouco se sabe a respeito de ,lematódios associados com o cacaueiro no Espírito Santo; as informações disponíveis sobre o assunto são de Sharma (4). Em vista disso, um levantamento nematológico sistemático foi efetuado na região cacaueira desse Estado com o objetivo de determinar a ocorrência e distribuição dos nematóides fitoparasitas associados com diversos cultivos de modo geral, e no cacaueiro, de modo particular.
M.-\TERIAIS E MÉTODOS
O levantamento foi conduzido em 27 propriedades, distribuídas em três municípios do Espírito Santo (Unhares, Colatina e São Mateus), no período de outubro de 1971 a março de 1974. Foram coletadas 89 amostras de solos e
raízes de cacaueiros sadios e doentes. A amostragem e extração dos nematódios foram realizadas segundo técnica de Sharma e Loof (5). Para a identificação e montagem permanentes desses nematódios, foi utilizado o processo descrito por Sharma e Sher (6). Os locais de amostragem bem como o número de amostras, sintomas apresentados pelas plantas e tipos de solos estão mencionados no Quadro 1. Os sintomas apresentados pelas plantas doentes foram anteriormente descritos por Sharma e Sher (6).
RESULTADOS
F oi detectada uma população mista de 20 gêneros e 26 espécies de nematóides fitoparasitas (Quadro 2), Os gêneros encontrados com
* Recebido para publicação em agosto, 1974. ** D.Sc. (Agric.), Divisão de Filoparasilologia, CEPEC.
*** Ph.D., Universidade da Califórnia, Riverside, Cal., E.U,A.
Revista Theobrom<l. CEPEC, llhtfus, Brasil, 4(4}: 26-31. Out.-dez. 1974.
26
Quadro 1- Levantamento nematologico do cacaueiro no Espírito Santo (locais de coleta, número de amostras, sintomas apresentados pelas plantas e tipo de solo), outubro/71 a março/74.
Locais de coleta (Fazendas e municípios)
Angélíca (Ll Batista (L) Boa Esperança (C) Brazilãndia (L) Brejo Grande (L) Bugrinha (L) Contendas (L) Esperança (C) Est. Exp. Minist, Agricultura (L) Est. Exp. Filogônio Peixoto (L) Goiana (L) Guanabara (L) Grafite Sossego (L) Havaí (L) lpanema (L) lpiranga (L) Paraíso (L) Piraquê (L) Relíquia (L) Santo Antônio (L) Santa Rita (L) Santa Terezinha Siio Jorge (SM) Sãc Pedro (SM) São Sebastião (L) Tris Marias (L) Vila Nova (L)
Municípios: (L) = Linh11res (C)= Co latina
(SM) = São Ma teus
mais freqüência foram: Helicoty· lenchus, Meloidogyne, Hemicriconemoides, Xiphinema, Criconemoides e Paratylenchus. Foram constatadas também em todas as amostras, espécies saprofíticas e predadoras.
Criconemoides mauritiana, Dolichodorus sp., Gracilacus aculen-
N~de
amostras Sintomas Tipos de solo
2 D,S Franco 3 SD Argila pesada
17 S,MS Franco 1 MS Franco 1 SD Argila pesada 4 S,SD,MS Franco 1 SD Argila pesada 1 SD Argila pesada 2 SD Argila pesada
11 SD Franco 1 SD Argila pesada 8 S,SD,MS Argila pesada 1 SD Franco 2 MS Franco 2 SD,MS Argila pesada 2 SD Argila pesada 1 SD Argila pesada 1 SD Argila pesada 4 s Argila pesada
11 S,MS Argila pesada 2 SD Argila pesada 1 SD Argila pesada 2 MS Argila pesada 1 MS Argila pesada 1 SD Franco 1 SD Argila pesada 5 SD Argila pesada
Sintomas: S = sadio SD = seca descendente MS = morte súbita
tus, Tylenchus davainei, Tylenchorhychus martini e Xiphinema setariae estavam associadas aos cacaueiros que apresentavam sintomas de seca descendente (dieback). Aphelencbus avenae, Aphelenchoides sp., Boleodorus sp., Helicotylenchus multicinctus, Longidorus sp., X iphinema americanum e um gênero não identificado
{ ~ uoC\IMENTAC.fo '1 ~ ~
27 -~~ ... "' .... (ç:- !TG'l"EC:: o-9 /f -' .I ~ ~ • ' ;..:~
0 L 0 ~ 'JJ ~

Quadro 2 - Freqiiência de ocorrência de nematóides parasitas de plantas associados ao cacaueiro (Theobroma cacao L.) em 89 amostras na regiao cacaueira do Espírito Santo.
Freqüência de ocorrência
Nematóides Locais de amostragens* %das 89 amostras
Apbelenchu.s avenae Y,Zo 4.5 Aph€knchoides sp. C,Zo 7.9 Boleodoru.s sp. M 1.1 Criconemoides sp, A,E,F,H,j,L,N,O,P,R,S,T,V,
W,Z,Zo 29.2 C. mauritiana u 1.1 Dolichodorus sp. N,U 3.4 Gracikzcus acu.lentus H,K 2.2 Helichotylenchu.s sp. A, B,C ,D,D,F, G,H,I,J, K,L,N ,0,
P,R,T,U, V,Z,Zo 56.2 H. dihystera C,J,j,S,V,Y,Zo 20.2 H. multicinctus j,P,Zo 3.4 Hemicycliophora sp, G,J,P.Zo 4.5 Hemicriconemoides sp, A,C,F ,G,J ,M,N, R,S, U,X,Zo 35.9 Longidoru.s sp, Zo 2.2 Meloidogynfi' incognita A,B,C,F,G,I,j,L,M,N,O,P,R,S,
T,V,W,X,Y,Z 58.4 Paratylencbus sp, A,F ,l,j,L,M,Q,S, T,U,Zo 25,8 Peltamigratu.s sp, B,H,L,N,S,U,Zo 19.1 Rotylenchulu.s reniformis C,j,L,S,X,Z,Zo 20.2 Trichodorus sp. C,Zo 3.4 T ropbwu.s s p. B,j,L,M,O,P,Q, Y,Z,Zo 18.0 Tylenchorhynchu.s martini B,Zo 2.2 Tylenchu.s sp. B,L,Z,Zo 6.7 T, daminei Zo 1.1 Tylenchulidae (familia) F,W,Zo 4.5 Xiphínema sp, A,C,D ,H, I,] ,L,N,O,P,R,S,,
U,V,X,Y,Z,Zo 34.5 X. americanu.m Zo 1.1 X, setariae y 1.1
* Fazendas ou estações experimentais.
A= Angélica; 8 = Batista; C= Boa Esperança; D = Brasilândia; E = Brejo Gran~ de; F= Bugrinha; G =Contendas; H= Esperança; I= Eatação Exp, Min. Agric,; J = Estação Exp. F. Peixoto; K =Goiana; L= Guanabara; M =Grafite Sossego; N = Havaí; O= Jp~~nema; P = Ipiranga; Q = Paraíso; R= Piraquê; S =Relíquia; T = Sta Rita; U = Sta Terezinha; V= Síío Jorge; W =São Pedro; X= São Sebastião; Y =Três Marias; Z =Vila Nova; Zo = St~ Antônio.
28
da família Tylenchulidae foram encontrados associados aos cacau~ eiros com sintomas de morte súbita. Os outros nematóides apre~ sentados no Quadro 2 estavam associados com os dois tipos de sin~ tomas acima mencionados.
DISCUSSÃO
Meloidogyne incognita foi a es~ pécie mais freqüente no levanta~ mento e tem sido registrada neste cultivo em diferentes partes do mundo (6, 7). A mesma espécie foi também encontrada parasitando cafeeiros no Espírito Santo, demonstrando sua ampla distribuição (1). M. e:âgua, parasita comum de cafeeiros (2), não foi encontrada no presente levantamento, assim como Rotylenchus robustus, detectado em cacaueiros em Trinidad (7). M. incognita foi encontrado em solos argilosos e pesados,
quando normalmente ocorrem em solos leves e arenosos (3).
Apbelenchoídes sp., Boleodorus sp.,Criconemoides mauritiana, Gracilacus aculentus e o gênero não identificado da família Tylenchulidae foram encontrados associados ao cacaueiro pela primeira vez, o mesmo acontecendo no Brasil com as espécies Helicotylenchus multicinctus, Tylenchorhynchus martini e Xiphinema americanum.
Os resultados deste levantamento indicam um grande número de espécies de nematóides fitoparasitas amplamente distribuídos e associados ao cacaueiro. Pelo ex~ posto, há necessidade de se definir esta associação e de se deter~ minar o papel desses nematóides nos processos patológicos das raí~ zes do cacaueiro e sua possível relação com a morte súbita e seca descendente.
LITERATURA CITADA
1. INSTITUTO BRASILEIRO do Café e GERCA, Informativo 8(1):1-8. 1974.
2. CHEBABI, A. e LORDELLO, L.G.E. Ckorrência de nematóides em cafezais do Espírito Santo. Revista de Agricultura (Brasil) 43: 140. 1%8.
3. SASSER, J.N. Identification and host~parasite relationships of certain root-knot nematodes (Meloidogyne spp.). Maryland Agricultura} Experimental Station Bulletin A77: 1954. 31 p.
4, SHARMA, R.D. Relatório Anual de Nematologia. Ilhéus, Bahia, Brasil, Centro de Pesquisas do Cacau, 1973. 25 p. (Datilografado).
29

S. SHARMA, R.D. e LOOF, P.A.A. Nematodes of the cocoa region of Bahia, Brazil. I - Plant parasitic and free·living nematodes associated with rubber (Hevea brasiliensis Muell. Arg.). Revis· ta Theobroma (Brasil) 3(1):36·41. 1973.
6. . e SHER, S.A. Nematodes of the Cocoa Region of Bahia, Brazil. Occurrence and distribution of plant parasitic nemato· des associated with cocoa (Theobroma cacao L.). Revista Theobroma (Braz i!) 3 (3): 17· 24. 1973.
7. SINGH, N.D. Preliminary report on plant parasitic nematodes asso. ciated with important crops in Trinidad. Nematropica 3(2):56. 61. 1973.
RESUMO
Vinte gêneros e 26 espécies de nematódios fitoparasitas foram encon· trados associados ao cacaueiro (Theobroma cacao L) em 89 amostras de solos e raízes provenientes de 27 fazendas localizadas em três muniCÍ· pio do Estado do Espírito Santo. Doze espécies estavam associadas a cacaueiros com sintomas de seca descendente (die·back) e morte súbita (seis de cada). Os gêneros mais comuns foram: Helicotylenchus, Meloido· gyne, Xiphinema, Criconemoides e Paratylenchus. As espécies identifj. cadas foram: Aphelenchus avenae, Aphelenchoides sp., Boleodorus sp., Criconemoides sp., C. mauritiana, Dolichodorus sp., Gracilacus aculentus, llelicotylenchus sp., H, dihystera, H. multicinctus, Hemicycliophora sp., llemicriconemoides sp., Longidorus sp., Meloidogyne incognita, Paraty· lenchus sp., Peltamigratus sp., Rotylenchulus reniformis, Trophurus sp., Trichodorus sp., Tylenchorhynchus martini, Tylenchus sp., T. davainei, Tylenchulidae (família), Xiphinema sp., X. americanum e X. setaria;;,
NEMATODES OF THE COCOA REGION
OF ESPIRITO SANTO, BRAZIL.
I- NEMATODES ASSOCIATED WITH COCOA (Theobroma cacao L.)
SUMMARY
A total of 89 soil and root samples ftom 27 farms, frorn three munlCl· palities in Espírito Santo (Linhares, Colatina and São Mateus) contained mixed populations of 20 genera and 26 species of plant parasitic nema·
30
todes. Twelve species associated with cocoa trees had syrnptoms of die· back and sudden death (six each). The most common nematode genera were: Meloidogyne, Helicotylenchus, Hemicriconemoides, Xiphinema, Criconemoides and Paratylenchus, The plant nematode species identified were: Aphe!enchus avenae, Aphelenchoides sp., Boleodorus sp., C rico· nemoides sp., C. mauritiana, Do/ichodorus sp., Gracilacus aculentus, H e· /icotylenchus sp., H. dihys"tera, H. multicinctus, Hemicycliophora sp., Hemicriconemoides sp., Longidorus sp., Meloidogyne incognita, Paraty· !enchus sp., Peltamigratus sp., Rotylenchulus reniformis, Trichodorus sp. Trophurus sp., Tylenchorhynchus martiní, Tylenchus sp., T. davainei, Tylenchulidae (family), Xiphinema sp., X, americanum and X. setariae.

FATORES QUE INFLUEM NA CAPTURA DE Ecinnyi' ello L. (LEP!DOPTERA: SPHING!DAE)
POR ARMADILHAS LUMINOSAS*
f oão Manuel A breu**
A lagarta da mariposa Erím1yis
ello L. é a praga mais importante da seringueira (Hevea brasiliensis M,.eJ!. Arg.) na Bahia. Alimenta-se das folhas, podendo provocar a desfolhação completa por ocasião de surtos violentos.
Tratando-se de um inseto de vôo noturno, fototrópico positivo, as amostragens da sua população são efetuadas com armadilhas luminosas. No entanto, é preciso considerar alguns fatores ambientais que interferem na sua captura por estas armadilhas.
Os fatores que influem na captura de msetos por armadilhas luminosas foram estudados por diversos pesquisadores (1, 2, 3, 5, 6, 9, 10, 14, 16, 17) com espécies de insetos associados aos mais diversos cultivos, tendo Hoiloway
(7) citado que a temperatura mínima noturna, chuva, vento, neblina, lua, periodicidade e nuvens são os mais importantes para as mariposas.
Na Bahia, Winder e Abreu (18), em experimentos preliminares com uma armadilha luminosa "omnidirecional~ instalada em um seringal adulto, no município de Ituberá, obtiveram resultados preliminares sobre o comportamento de vôo de Erinnyis ello e Eri1wyis alope.
Na Malásia, Rao (11) usou armadilhas luminosas para controlar o escarabeídeo Lechnostema bidelltata Burm., praga dos seringais em crescimento. Para tanto, estudou modelos de armadilhas, fontes luminosas, efeito de altura, habitas de vôo e proporção dos sexos desse besouro.
* Recebido para publicação em setembro, 1974.
* Realizado com recursos do Convênio SUDHEVEA/CEPLAC. ** MS, Divisão de Fitoparasitologia, CEPEC.
Revista Theobroma. CEPEC, llhéu~. Rrasil, 4(4): 32-43. Out. -dez. 1974.
32
O presente trabalho foi efetuado com o propósito de determinar o efeito da temperatura mínima noturna, chuva, lua, altura e periodicidade de vôo, proporção dos sexos e atração pela luz artificial na captura de E. ello, em seringais. A determinação destes fatores tem importância para estabelecer métodos mais seguros nos estudos básicos e aplicados, visando o controle dessa importante praga dos seringais.
MATERIAL E MÉTODOS
A captura das mariposas foi efetuada com armadilhas luminosas modelo "Luiz de Queiroz" (13), instaladas na Fazenda Marimbu, município de Ituberá (13° 40'S e 39° lO'W), em dois mastros de madeira com 13m de altura, distanciados 500 m um do outro (Figura 1). Os dois mastros foram fixados no topo de duas colinas em meio a seringais em produção. As alturas das seringueiras variavam entre 8 e 11 metros.
A determinação da periodicidade de vôo foi conduzida com uma armadilha colocada a 13m de altura, acima, portanto, do nível superior das copas mais altas, e provida de uma lâmpada fluorescente ultravioleta modelo Fl5 T8/BL (black-light). As coletas foram feitas durante a noite de hora em hora, pela substituição do saco coletor. A primeira série de amostragens cobriu o período de 25/01 a 05/06 /73, tendo a armadilha operado durante 10 noites consecutivas. A
33
••
Figura 1 - Armadilhas luminosas modelo "Luiz de Queiroz" instaladas em seringal. Fazenda Marimbu, ltuberá, Bahia, 1973.
segunda série cobriu o período de 23/10 a 25/11/73, operando a armadilha em 15 noites, alternadamente.
As determinações da atração pela luz e altura de vôo foram realizadas utilizando-se nas armadilhas lâmpadas fluorescentes modelos FlS T8/LD (luz do dia), com emissão máxima entre 550 e 600 nm, e F15 T8/BLB (black-light blue) com emissão máxima de 365 nm. As armadilhas operaram a 1; 5; 9 e 13m do nível do solo.
As demais determinações foram efetuadas operando-se as armadi-

lhas equipadas apenas com a lâmpada modelo FlS T8/BL, colocadas a 13m de altura. A pesquisa cobriu o período de janeiro de 1973 a março de 1974.
RESULTADOS E DISCUSSÃO
Temperatura mínima noturna - Nas áreas de clima com estações bem definidas, a temperatura é um fator que tem influência sobre a atividade de vôo dos insetos. Estes, em geral, só voam a partir de uma temperatura limiar, que permite tal
c; 23
'-
" 22
~ • • • • • 21 ~
20 ~ 80 ~ ~
~ h
70
' <l 60
~ ~ 50
"' a ~ 40 ~ ~ ~
li 30
a 20 ~ ~ ~
lO -~ ~
•
atividade (4, 15). É sabido também que a temperatura mínima noturna está diretamente relacionada C'Om as capturas noturnas das mariposas pelas armadilhas luminosas. As capturas diminuem com a queda de temperatura (7).
Considerando que nesta latitude não ocorrem mínimas muito baixas, o fato das capturas diminuirem com a queda de temperatura, não é aplicável para o caso de E. e/lo, conforme pode ser observado nas Figuras 2 e 3, onde as captu-
• • • /-
18.00 19.00 20.00 2100 22.00 23.00 24.00 01.00 02.00 03.00 04.00 0!100 06.00
HORAS
Figura 2 - Periodicidade de vôo de E. ello capturadas por armadilhas luminosas. ltuberá, Bahia, período maio-junho, 1973.
34
ras ainda são acentuadas após a meia-noite, quando ocorrem as mínimas, particularmente nas coletas de outubro-.wvembro (Figura 3). Em maio-junho, o máximo de capturas se concentrapróximo·da meia -noite.
Chuva - Para determinar a influência da chuva sobre a atividade de E. ello, foram consideradas as capturas das mariposas feitas em noites sem luar e as precipitações ocorridas durante a noite. A Figura 4 mostra que a precipitação, mesmo intensa, não interferiu so-
24 - ........ ~
'- 23 • • ~
~ 22
~
â 21
~ 7
' ~ 600 ~
;;j 500 a ~ ~ 400 q ~
~ 300
a ~ 200
·~ 100 ~
bre a atração das mariposas pelas armadilhas.
Lua - A lua é a fonte natural de luz que orienta o vôo de insetos noturnos. Ao voar em direção à lua, eles atingem uma zona de correntes aéreas acima das barreiras naturais, sendo favorecida, deste modo, a dispersão e possibilitando, inclusive, o entrecruzamento de populações (7, 13).
As amostragens para estudos da flutuação sazonal da população de E. e/lo, evidenciaram a influência da fase lunar na captura das
.......... • ~ • •
18.00 19.00 20.00 2100 2200 23.00 <>!-00 01.0002.00 O.WO 04.00 05.00 06DO
HORAS
Figura 3 - Periodicidade de vôo de E. ello capturadas por armadilhas luminosas. ltuberá, Bahiil, período de outubro-novembro, 1973.
35

" ~
" " " Í\ \~ ' . ' ' ""' ~ ' ' ,. lb:n
00
00
00 rnn- h . ri] cu - ' • " " " " " ' " " u ~ " " ' " " " SETE .. BRO OUTUBRO
23 25 27
NOVENBIIO
1 9 1' 3
" >.
' )~ ' /\~ '
' .A ' ' ' ' ' 00
~
wo
wo
~o
h-Dr. >00 n-rfll ~ ,n
" ' • tô!OZ22 l6
DtzUIBRO
1973
17 lt 21 23 25 27
J.o\l'tt:IRO
5 1 !f 11 ll "
I'IE:VIE:REIRO
'" Figura 4 - Influência da chuva na captura de Erinnyis ello por armadilhas luminosas em noite sem luar. ltuberá, BA, 1973/74.
mariposas. As fases da lua, com as diferentes intensidades luminosas, têm influência marcante na captura dessas mariposas pelas armadilhas luminosas conforme pode ser observado na Figura 5, onde se verifica uma relação pronunciada entre as capturas e a fase
36
lunar. Nas fases de quarto minguante e lua nova, o número de mariposas capturadas é mais abundante que no quarto crescente e lua cheia. Este fenômeno se repete todos os meses, independentemente da flutuação sazonal da população.
o •o
50
<O
" " " "
o
000
"o <00
,00
'o "
) • ct, .-- \
! ' : ' ' . ' '
·---.----. '
20 22 24 26 28
ogosto
) •
24 26
IIOY&mbro 1 9 7 3
'o '
/
··, ,· '
" "'
' '
'
5 ' ' " setembro
o o IYO cheio,
) rninQuonte
• novo
crescente
• ' ' deze!flbro
'o
Figura 5- Efeito da fase lunar sobre a captura de Erinnyis ello por armadilha luminosa. ltuberá, Bahia, 1973.
Williams, mencionado por Holloway (7), sugere que, além do efeito competitivo do luar em rela-
37
ção à fonte luminosa da armadilha, deve existir um fator fisiológico reduzindo a atividade de uma ma-

riposa devido à luminosidade provocada pela lua. Outro fator importante é a posição da armadilha, desde que se considere ser a lua responsável pela dispersão. Como as mariposas tendem sempre a subir em direção à fonte luminosa ou voar no mesmo plano, as armadilhas capturarão apenas aquelas que estiverem abaixo ou no mesmo plano da armadilha, conforme demonstra Silveira Neto (12), com Azochis gripusalis Walker, praga da figueira. Deste modo, no caso de E. e/lo, mesmo as armadilhas colocadas no nível superior da copa das seringueiras, onde as mariposas são mais abundantes, só atr2iriam aquelas que estivessem neste nível ou abaixo dele e na zona de influência da armadilha. As mariposas acima deste plano, segundo a hipótese, escapariam à atração da armadilha em noites de luar.
Periodicidade de vôo - Segundo Johnson {8), a periodicidade de vôo reflete os movimentos migratórios e é influenciada também pela atividade dos insetos para a alimentação, acasalamento e aviposição.
As curvas de periodicidade de vôo (Figuras 2 e 3) mostram para maio-junho atividade mais intensa entre 22:00 h e 5:00 h, e para outubro-novembro a partir das 20:00h, atingindo o máximo entre 1:00 h e 3:00 h. Considerando os sexos, no período maio-junho, a atividade do macho se concentra entre as 23:00 h e 4:00 h, enquanto a da fêmea apresenta dois picos: um entre
38
23:00 e 24:00 h e outro entre 2:00 e 3:00 h. No período de outubronovembro, há certa variação na periodicidade de vôo. Nesta época do ano, os machos intensificam a atividade entre 1:00 e 3:00 h e as fêmeas a partir das 20:00 h, atingindo o pico máximo entre as 2:00 e 3:00 h.
As curvas de periodicidade de vôo dos machos são unimodais, variando apenas o pico de acordo com a época do ano. Nos meses mais frios (maio-junho), a atividade é intensificada mais cedo(23:00 h) e cobre um período de 5 horas, enquanto nos meses mais quentes é intensificada mais tarde (1:00 h) e cobre um período de 3 horas. Estas diferenças podem estar relacionadas com as temperaturas mínimas noturnas, conforme discutido anteriormente, que atingem níveis mais baixos em maio-junho do que em outubro-novembro. Com as fêmeas, o fenômeno é similar: o maior pico de maio-junho ocorre antes da meia-noite e o de outubro -novembro após a meia-noite, possivelmente pelas mesmas razôes apresentadas para os machos. Cabe salientar que a população total é mais elevada nos meses quentes.
Atração pela luz artificial e altura de vôo - Os resultados do Quadro 1 mostram que a captura das mariposas pela armadilha equipada com a lâmpada Fl5 T8/BLB é 8 vezes superior à equipada com a lâmpada Fl5 T8/LD. Nos experimentos em que foram comparadas as lâmpadas FlS T8/BLB e Fl5 T8/BL não houve diferença significativa
Quadro 1 -Mariposas capturadas a diferentes alturas do solo, utilizando-se armadilhas luminosas equipadas com lâmpadas LD (luz do dia) e BLB (luz negra). ltuberá, BA. período de 10/01 a 01/02/73.
Altura da Lâmpada FlS TS/LD
armadilha Macho Fêmea
1m 6 5 Sm 18 17 9m 40 41
13m 149 108
Total 213 171
nas capturas (Quadros 2 e 3). As capturas foram mais abundantes nas armadilhas situadas a 13m, independentemente do tipo de luz (Quadros 1 e 2).
Quando foram comparadas as capturas nas armadilhas instaladas a 9 e 13m, as situadas a 9 m de altura efetuaram capturas mais abundantes (Quadro 3), Embora estas diferenças não sejam expres-
Lâmpada F15 T8/BLB Total
Macho Fêmea
98 72 181 156 140 331 457 372 910 993 771 2.021
1.704 1.355 3.443
sivas, elas foram mais evidentes nas armadilhas equipadas com a lâmpada F15 T8/BL. É provável que, para este caso, a maior captura nas armadilhas a 9 m se deva ao fato da lâmpada atrair as mariposas em atividade no plano ou abaixo do plano desta armadilha, considerando serem as lâmpadas acionadas ao mesmo tempo, no mesmo mastro, e também, como já foi dito anteriormente, pelo fato de
Quadro 2 - Mariposas capturadas a diversas alturas do solo, utilizando-se armadilhas luminosas equipadas com lâmpadas BL e BLB. ltuberá, BA, período de 03 a 09/04/73.
Altura da Lâmpada F15 T8/BL Lâmpada F15 T8/BLB Total
armadilha Macho Fêmea Macho Fêmea
1m 9 8 16 11 44 Sm 40 25 34 25 124 9m 63 46 64 48 221
13m 65 53 63 44 225
Total 177 132 177 128 614
39

Quadro 3 - Nmriposas capturadas em armadilhas luminosas, instaladas a 9 e 13m de altura do solo, equipadas com lâmpadas BL e BLR. ftuberá, BA, período de 10 a 20/03/73.
Altura da Lâmpada F15 T8/BL
armadilha Macho Fêmea
9m 1.081 1.092
13m 970 947
Total 2.051 2.039
qc~e a tendência de vôo das mariposas é sempre para cima, a fim de ultrapassarem as barreiras para dispersão.
Proporção dos sexos - Os dados apresentados nos Quadros 1, 2 e 3, mostram que a proporção dos sexos permaneceu praticamente inalterada em relação à altura de vôo. No período em que os dados foram obtidos (janeiro, março e abril de 1973), a proporção sexual foi de 1 : 1. A proporção dos sexos varia também com a flutuação sazonal da população, sendo que, na época dos surtos, a captura de fêmeas chega a atingir 80%, o que dá a proporção de 1 : 4.
Em relação à periodicidade de vôo, ocorre também variação na proporção dos sexos, conforme pode ser observado nas Figuras 2 e 2>. As fêmeas predominam durante toda a noite no período de outubronovembro, mas a proporção sexual é reduzida entre 1 e 3 horas da madrugada (Figura 3).
40
Lâmpada F15 T8/BLB Total
Macho Fêmea
912 937 4.022
914 927 3.758
1.826 1.864 7.780
CONCLUSÕES
1. Com a queda de temperatura, há uma tendência para aumentar a atividade das mariposas;
2. A chuva não influencia a atividade de vôo de E. ello;
3. O luar reduz a captura destas mariposas pelas armadilhas luminosas, criando, deste modo, uma variável adicional nas amostragens;
4. A atividade do macho é mais acentuada a partir da meia-noite. A da fêmea apresenta dois picos: um antes da meia-noite e outro pela madrugada;
S. A atividade das populações de E. e !lo está concentrada ao nível da copa das seringueiras;
6. A captura de fêmeas chega a atingir 80% nas épocas em que a população é mais abundante;
7. As fêmeas predominam durante toda a noite no período de outu-
bro-novembro, a proporção sexual porém, é reduzida entre 1 e 3 horas da madrugada; e
8. As armadilhas luminosas devem ser instaladas ao nível supe-
rior da copa das seringueiras e providas, de preferência, com lâmpadas do tipo F 15 T8/BL. Na falta desta, a FlS TS/BLB também oferece bons resultados.
AGRADECIMENTOS
Aos Drs. Saulo Sória, P. Silva e Arnaldo G. Medeiros, pelas sugestões apresentadas quando da elaboração do presente artigo. Aos Práticos Agrícolas Carlos Antonio Niella e Adão Gonçalves da Rocha, pela colaboração na execução dos trabalhos de campo.
LITERATURA CITADA
1. BIDLINGMAYER, W.L. The effect of moonlight on the flight activity of mosquitoes. Ecology 45 (1): 87-94. 1964.
2. BOWDEN, J. e CHURCH, B.M. The influence of moonlight on catches of insects in light-traps in Africa, 11. The effect of moon phase on light-trap catches. Bulletin of Entomological Research 63 : 129-142. 1973.
3. BROWN, E.S. e TAYLOR, L.R. Lunar cycles in the distribution and abundance of airbone insects in the equatorial highlands of East Africa. Journal of Animal Ecology 40 : 767-779. 1971.
4. DRY, W.M. e TAYLOR, L.R. Light and temperature thresholds for take-off by aphids. Journal of Animal Ecology 39 : 493-504. 1970.
5. FISK, F.W. e PEREZ PEREZ, R. h Fh1·~ht .actPueivity1 ~~~odsJof the
1 sugarca borer Díatraea sac:c ara ts, 1n r o o. ourna of Agril:lture ÍJniversity of Puerto Rico 53 (2) : 93-99 .. 1969,
6. FROST, S.W. Insects attracted to light p(14a)cedss"a-' dsis~ere1n~Sheights. Journal of Economic Entomology 51 : • 8.
7. HOLLOWAY, J.D. Studies and suggestionHs.o,n tshe be3h!a-4vi5ou<1o9f67moths at light. Proc. S. London Ent. Nat. 15 · oc. ' ·
8. JOHNSON, C.G. Migration and dispersai of insects by flight. London,
Methuen, 1969. 763 P•
41

9. PINCHIN, R,D, e ANDERSON, ]. On the nocturnal activity of tipulinae (Diptera) as measured by a light trap; 2. The influence of temperature, cloud and moonlight on the number of insects captured. Proceedings of the Royal Entomological Society of London (A) 11, PT - 3-5. 75-78. 1936.
10. PROVOST, M. W. The influence of moonlight on light trap catches of mosquitoes. Annals of the Entomological Society of America 52 (3) '261-271. 1959.
11. RAO, B.S. The use of light traps to contrai the cockchafer Lechnos terna bidentata Burm. in Malayan rubber plantations. Journal of Rubber Research lnstitute of Malaya 18:243-252. 1964.
12. SILVEIRA NETO, S. Flutuação da população e controle das principais pragas da família Pyraustidae, com emprego de armadilhas luminosas. Tese de doutoramento. Piracicaba, São Paulo, Brasil, Escola Superior de Agricultura "Luiz de Queiroz", 1969. 96 p.
13, . et al. Ecologia dos insetos. Piracicaba, São Paulo, Brasil, Escola Superior de Agricultura "Luiz de Queiroz" ,1973. 254 p.
14. STEWART, P.A. et ai. Activity of tobacco hornworm and corn earworm moths as determined by traps equipped with black-light lamps. Journal of Economic Entomology 60 (6) : 1520-1522. 1967.
15. TAYLOR, L.R. Analysis of the effect of temperature on insects in flight. Journal of Animal Ecology 32: 99-117. 1963.
16. , e CARTER, C.l. The analysis of numbers and disttibu-tion in an aerial population of macrolepidoptera. Transactions of the Royal Entomological Society of London 113: pt. 12. 369-386. 1961.
17. WILLIAMS, C.B. et ai. An investigation into the possible effects of moonlight on the activity of insects in the field. Proceedings of the Royal EntomologicalSociety of London(A) 31, PTS. 10-12. 135-144. 1956.
18, WINDER, J.A. e ABREU, J.M. Preliminary results on the flight behaviour of the Sphingid moths E'Tinnyis ello and Erinnyis alope using a light trap. (Inédito).
42
RESUMO
For~m r:al~ados experimentos de campo com a finalidade de determi-nar a Influencia da temperatura mínima noturna h lua altura e pe-·oo· 'd d d - , c uva, , per1 ICI a e. e v~o, atração pela luz artificial e proporção dos sexos na capt_ura de Er~nnyts ello L., praga da seringueira na Bahia, por meio de armadtlhas luminosas.
A chuva e temperatura mínima noturna não têm influência sobre a atividade de vôo de E. e !lo. As fases da lua têm influência marcante na captura de E. e/lo pelas armadilhas luminosas, ocorrendo coletas mais abundantes na ausência de luar. O macho apresenta atividade de vôo mais acentuada a partir da meia-noite, enquanto a fêmea apresenta dois picos: um antes da meia-noite e outro de madrugada. As armadilhas luminosas devem ser instaladas ao nível da copa das seringueiras e equipadas preferencialmente com lâmpadas modelo F15 T8/BL. As variações na proporção dos sexos são apresentadas em função da altura de vôo, época do ano e periodicidade de vôo.
THE INFLUENCE OF SOME FACTORS ON LIGHT TRAP CATCHES OF THE HAWKMOTH Erinnyis ello L. (LEPIDOPTERA' SPHINGIDAE)
SUMMARY
Field trials were carried out to find the influence of some physical and biological factors on light trap catches of the hawkmoth Erinnyis ello L., which is the major pest of rubber tree in Bahia, Brazil.
The results showed that the rainfall and the lowest nocturnal temperature has no inftuence on the flight activity of E. ello. The moon phases have strong influence on the light trap catches o_f the moth, and the most h ndant catches occurred in absence of moonhght. The male flies are :o~e active after midnight, while the female presents two flight peaks;
bef ·d · ht d ther at dawn These same results showed that one ore mt ntg an ano · the artificial light traps must be located a little above the canopy of the rubber trees, and must be equipped with F15 '!8~~L ~odel la~ps, Pre. · b . 1 t d to sex ratio vanabtltty tn funchon of the
hmmary o servahons re a e 'od' ·t 1 · 1 d height, seasonal fluctuation and flight pen tCt Y were a some u ed. . . ..
43

~ ESTf liVRO DEYf SER DUOUJOO
~ 11 OUIMI DAli ClftiMBIDI
. }fv:Ji ~~~%~ ····-··-·· ·······-···-···· t)Df7l~}o'3~ ':.9 -~~ .15 LJ/clN 1 ................. . 2,6.oq ,A' . {o . .. . ............ .
.23 ,o .5'-lt rdi!/Zco ··················· ········· vJ--
S·Df ..••••••.••..•• _ .• _._·_-._-_-._-.---.·.·---------······-······· '"'-oq-JJ~- ····················· ························· 1-~MAr\W
J< I ~GG 919 .. 29 sn 979 ...................... .
03/-:fuL ........ . ....................... . .J4/3/t ....... ············ ········- ······· ···········
JfjG~~9 m /·••••••••••• •••• m
CEf'LA,C- OU2.004H -

























![Theobroma grandiflorum(Willd. Ex Spreng.) Shum.]](https://img.document.onl/doc/110x75/61929208a34c6b42ce5de00a/theobroma-grandiflorumwilld-ex-spreng-shum.jpg)



