Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DA PARAÍBA
CENTRO DE CIÊNCIAS DA SAÚDE
LABORATÓRIO DE TECNOLOGIA FARMACÊUTICA – LFT
PROGRAMA DE PÓS-GRADUAÇÃO EM PRODUTOS NATURAIS E SINTÉTICOS BIOATIVOS
Rossana Miranda Pessoa Antunes
AÇÃO ANTIMICROBIANA E ANTIPLASMIDIAL DO LAPACHOL E SEUS DERIVADOS SOBRE BACTÉRIAS E FUNGOS LEVEDURIFORMES
João Pessoa – PB 2007

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

AÇÃO ANTIMICROBIANA E ANTIPLASMIDIAL DO LAPACHOL E SEUS
DERIVADOS SOBRE BACTÉRIAS E FUNGOS LEVEDURIFORMES
Rossana Miranda Pessoa Antunes
Orientadores:
Profª. Drª. Edeltrudes de Oliveira Lima
Prof. Dr. José Maria Barbosa Filho
Co-Orientadores:
Profª. Drª. Maria do Socorro Vieira Pereira
Prof° Dr° Celso Amorim Câmara
Tese apresentada ao Programa de Pós-graduação
em Produtos Naturais e Sintéticos Bioativos do
Centro de Ciências da Saúde – Laboratório de
Tecnologia Farmacêutica da Universidade Federal
da Paraíba, em cumprimento às exigências para
obtenção do título de Doutor em Produtos Naturais e
Sintéticos Bioativos (Farmacologia).

- II -
AÇÃO ANTIMICROBIANA E ANTIPLASMIDIAL DO LAPACHOL E SEUSDERIVADOS SOBRE BACTÉRIAS E FUNGOS LEVEDURIFORMES
Rossana Miranda Pessoa Antunes
Tese submetida ao corpo docente do Programa de Doutorado em Produtos
Naturais e Sintéticos Bioativos/LTF da Universidade Federal da Paraíba. Como parte
dos requisitos necessários à obtenção do seu grau de Doutora.
Aprovada por:
_______________________________________Profª. Drª. Edeltrudes de Oliveira Lima
Orientadora
_______________________________________Profª. Drª. Lindomar de Farias Belém
_______________________________________Profº Dr. Evandro Leite de Souza
_______________________________________Profª. Drª. Margareth de Fátima F. M. Diniz
_______________________________________Profº Dr. José Pinto Siqueira Júnior

- III -
D E D I C A T Ó R I A
À Deus
A meus pais e irmãos
A meu esposo Thúlio e aos meus filhos Beatriz e Daniel grandes companheiros.

- IV -
A G R A D E C I M E N T O S
A Deus, por ter me guiado nesta caminhada, me iluminado para eu concluir este
estudo.
Aos meus filhos, Beatriz e Daniel, pela compreensão das ausências, nas horas
necessárias.
Ao meu marido, Thúlio Antunes de Arruda, pela grande ajuda na elaboração deste
trabalho.
Aos meus familiares e amigos, que me incentivaram.
À professora Edeltrudes de Oliveira Lima, obrigada pelos seus valiosos
ensinamentos e correções que tanto enriqueceram esta tese.
Ao professor José Maria Barbosa Filho, pela acolhida inicial, pelos seus
ensinamentos e disponibilidade sempre presentes em todos os momentos.
Ao professor Celso Amorim Câmara, pelas substâncias alvo desta pesquisa e pelas
suas valiosas correções.
À professora Maria do Socorro Vieira Pereira, pela contribuição, muito obrigada.
À professora Bagnólia Araújo Costa, coordenadora do Programa de Qualificação
Institucional – PQI/ Universidade Federal da Paraíba-UFPB/ Laboratório de
Tecnologia Farmacêutica – LTF, agradeço pela disponibilidade e pelos seus
ensinamentos enquanto docente do programa de doutorado.
À professora Lindomar de Farias Belém, coordenadora do PQI/ Universidade
Estadual da paraíba – UEPB, por estar trilhando, com competência, este caminho.
À amiga Profª Raïssa Mayer Ramalho Catão, companheira do curso de doutorado,
com a qual trabalhei nesta pesquisa, agradeço pelos ensinamentos na área de
microbiologia, sem os quais teria sido mais difícil chegar até aqui.
Aos professores, amigos e companheiros do PQI/UEPB, com os quais compartilhei
esta importante experiência de estudo, o meu agradecimento.

- V -
A todos os demais professores, colegas e funcionários do programa de doutorado do
LTF.
A todos os professores e funcionários do Laboratório de Análises Clínicas – LAC/
UEPB, onde funciona o laboratório de pesquisa em microbiologia do PQI/UEPB,
meu agradecimento.
Ao Laboratório DIAGNOSE, na pessoa da profª Raïssa Mayer Ramalho Catão.
Aos professores e funcionários da Pró-Reitoria de Pós-graduação da UEPB.
À CAPES, pela bolsa de estudos a mim concedida, importante apoio financeiro,
muito obrigada.

- VI -
S U M Á R I O
LISTA DE FIGURAS............................................................................................... VIII LISTA DE TABELAS ................................................................................................ X
LISTA DE QUADROS............................................................................................... XI SIGLAS E ABREVIATURAS ................................................................................... XII RESUMO................................................................................................................. XIII ABSTRACT.............................................................................................................XIV 1. INTRODUÇÃO ..................................................................................................... 15 2. OBJETIVOS......................................................................................................... 21
2.1. Objetivo Geral................................................................................................ 22
2.2. Objetivos Específicos..................................................................................... 22
3. REVISÃO DE LITERATURA ............................................................................... 23
3.1. Microrganismos: generalidades ..................................................................... 24
3.2. Patogenese microbiana ................................................................................. 26
3.2.1. Gênero Staphylococcus....................................................................... 26
3.2.2. Gênero Escherichia ............................................................................. 28
3.2.3. Gênero Pseudomonas ......................................................................... 30
3.2.4. Gênero Candida .................................................................................. 31
3.3. Antimicrobianos ............................................................................................. 32
3.3.1. Antibacterianos.................................................................................... 32
3.3.2. Antifúngicos ......................................................................................... 33
3.3.3 Resistência aos antimicrobianos .......................................................... 34
3.3.4. Mecanismo genético de aquisição de resistência aos antimicrobianos 36
3.4. Plasmidios ..................................................................................................... 42
3.5. Transposons .................................................................................................. 44
3.6. Aspectos botânicos e fitoquimicos do lapachol e seus derivados semi-
sintéticos e nitrogenados ............................................................................... 45
4. MATERIAL E MÉTODOS .................................................................................... 52
4.1. Local da pesquisa.......................................................................................... 53
4.2. Produtos testados.......................................................................................... 53
4.3. Substâncias/ Antibióticos............................................................................... 55

- VII -
4.4. Espécies microbianas.................................................................................... 55
4.4.1. Isolamento e identificação dos microrganismos.................................. 56
4.4.2. Preparo da suspensão microbiana ..................................................... 56
4.5. Meios de cultura ............................................................................................ 57
4.6. Metodologia ................................................................................................... 57
4.6.1. Determinação da atividade antimicrobiana – triagem ......................... 57
4.6.2. Determinação da Concentração Inibitória Mínima (CIM),
Concentração Bactericida Mínima (CBM) e Concentração Fungicida
Mínima (CFM) ..................................................................................... 59
4.6.3. Determinação da cinética de crescimento bacteriano......................... 59
4.6.4. Determinação da cinética fúngica ....................................................... 61
4.6.5. Caracterização fenotípica dos padrões de resistência aos
antimicrobianos................................................................................... 63
4.6.6. Tratamento por substratos sintéticos obtidos a partir do lapachol e
seus derivados sintéticos – avaliação da atividade curagênica .......... 63
4.6.7. Estudo da relação estrutura-atividade.................................................. 65
5. RESULTADOS E DISCUSSÃO ........................................................................... 66
5.1. Resistência Antimicrobiana ........................................................................... 67
5.2. Avaliação da atividade antimicrobiana do lapachol e seus derivados ........... 69
5.3. Determinação da CIM e CBM........................................................................ 71
5.4. Cinética bacteriana das cepas de S. aureus ATCC 25923, S. aureus 319U
e S. aureus 122U, frente aos produtos derivados do lapachol ...................... 77
5.5. Eliminação de resistência a drogas pelo lapachol e seus derivados ............. 85
6. CONCLUSÕES .................................................................................................... 91 REFERÊNCIAS........................................................................................................ 94 ANEXOS................................................................................................................. 105

- VIII -
L I S T A D E F I G U R A S
Figura 1. Forma e arranjo celular de S. aureus ......................................................26
Figura 2. S. aureus MRSA......................................................................................27
Figura 3. Aspecto morfológico de E. coli ................................................................29
Figura 4. Forma e arranjo celular de P. aeruginosa ...............................................30
Figura 5. Forma e arranjo celular de C. albicans ....................................................31
Figura 6. Plasmídio.................................................................................................43
Figura 7. Ipê-Roxo..................................................................................................45
Figura 8. Estruturas químicas do lapachol e seus derivados semi-sintéticos .........46
Figura 9. Formação de enamina a partir do lapachol .............................................48
Figura 10. Estruturas químicas do lapachol e seus derivados..................................54
Figura 11. Técnica de difusão em meio sólido, processo cavidade-placa. ...............58
Figura 12. Fluxograma da cinética bacteriana..........................................................60
Figura 13. Fluxograma da cinética fungica...............................................................62
Figura 14. Fluxograma da Avaliação da Atividade Curagênica ................................64
Figura 15. Atividade antimicrobiana do lapachol (1), α-nor-lapachona (2); β-
lapachona (3); α-lapachona (4); β-nor-lapachona (5), frente a S.
aureus ATCC 25923................................................................................70
Figura 16. Avaliação da CIM e da CBM do lapachol (A), α-lapachona (B), β-
lapachona (C) e β-nor-lapachona (D) nas concentrações de 200 (1),
100 (2), 50 (3) e 25g/ mL (4) frente à cepa MSSA ATCC 25923...........72
Figura 17. Percentuais de inibição do lapachol e derivados frente as cepas de S.
aureus. ....................................................................................................76
Figura 18. Cinética bacteriana de S. aureus ATCC 25923 frente a lapachol, α-
lapachona, β-lapachona, α-nor-lapachona, β-nor-lapachona, α-I-
lapachona, β-I-lapachona. .......................................................................79

- IX -
Figura 19. Cinética bacteriana de S. aureus 319U frente aos produtos Lapachol,
α-lachona, β-lapachona, α-nor-lapachona, β-nor-lapachona, α-I-
lapachona, β-I-lapachona. .......................................................................81
Figura 20. Cinética bacteriana de S. aureus 122U frente ao lapachol, α-lachona,
β-lapachona, α-nor-lapachona, β-nor-lapachona, α-I-lapachona, β-I-
lapachona................................................................................................83
Figura 21. Cinética bacteriana do lapachol (1), α-lapachona (2), β-lapachona (3),
α-nor-lapachona (4) sobre a cepa S. aureus ATCC 25923 em 8h e
24h. .........................................................................................................84
Figura 22. Colônias possilvelmente curadas pelo β-I-lapachona para a marca de
resistência plasmidial à eritromicina para a cepa S. aureus 319U. .........87
Figura 23. Colônias possilvelmente curadas pelo lapachol para a marca de
resistência plamidial à penicilina para a cepa S. aureus 319U................88
Figura 24. Freqüência de cura da marca de resistência plasmidial à penicilina
pelo lapachol e derivados em S. aureus 319U (a) e S. aureus 122U (b).89
Figura 25. Freqüência de cura da marca de resistência plasmidial à eritromicina
pelo lapachol e derivados em S. aureus 319U (a) e S. aureus 122U (b)...89

- X -
L I S T A D E T A B E L A S
Tabela 1. Identificação e origem das cepas microbianas testadas .........................55
Tabela 2. Perfil de sensibilidade das cepas de S. aureus frente a
Antimicrobianos.......................................................................................68
Tabela 3. Avaliação da atividade antimicrobiana do lapachol e derivados, frente
a microrganismos de origem ambulatorial e cepas ATCC.......................69
Tabela 4. Avaliação da CIM e da CBM do lapachol e derivados frente a
microrganismos. ......................................................................................73
Tabela 5. Atividade antimicrobiana do lapachol e derivados frente as cepas de
S. aureus de origem humana, animal e ambiental x estrutura química. ..74
Tabela 6. Atividade antimicrobiana do lapachol e seus derivados frente à cepas
de E. coli..................................................................................................77
Tabela 7. Concentração Inibitória Mínima (CIM) e subinibitória (1/2 x CIM) para
a cepa de S. aureus 319U.......................................................................85
Tabela 8. Concentração Inibitória Mínima (CIM) e subinibitória (1/2 x CIM) para
a cepa de S. aureus 122U.......................................................................86
Tabela 9. Freqüência de cura da marca de resistência plasmidial à penicilina
pelo lapachol e seus derivados semi-sintéticos em linhagens de S.
aureus. ....................................................................................................87
Tabela 10. Freqüência de cura da marca de resistência plasmidial à eritromicina
pelo lapachol e seus derivados semi-sintéticos em linhagens de S.
aureus . ...................................................................................................88

- XI -
L I S T A D E Q U A D R O S
Quadro 1. Cinética bacteriana de S. aureus ATCC 25923 frente a lapachol, α-
lapachona, β-lapachona, α-nor-lapachona, β-nor-lapachona, α-I-
lapachona, β-I-lapachona. .......................................................................79
Quadro 2. Cinética bacteriana de S. aureus 319U frente ao lapachol, α-
lapachona, β-lapachona, α-nor-lapachona, β-nor-lapachona, α-I-
lapachona, β-I-lapachona. .......................................................................81
Quadro 3. Cinética bacteriana de S. aureus 122U frente ao lapachol, α-lachona,
β-lapachona, α-nor-lapachona, β-nor-lapachona, α-I-lapachona, β-I-
lapachona................................................................................................83

- XII -
S I G L A S E A B R E V I A T U R A S
AMH - Agar Mueller-Hinton AS - Agar Sangue ASD - Agar Sabourand Dextrose ATCC - American Type Culture Collection BAB - Blood Agar base BHI - Broth heart infusion BHI - Brain Heart Infusion BORSA - Boderline oxacilin – resistant S. aureus CBM - Concentração bactericida mínima CFM - Concentração fungicida mínima CIM - Concentração inibitória mínima CMH - Caldo Mueller Hinton DAEC - Escherichia coli que adere difusamente DMSO - Dimetil sulfórido EAggEC - Escherichia coli enteroagressiva EHEC - Escherichia coli enterohemorrágica EPEC - Escherichia coli enteropatogênica clássica ETEC - Escherichia coli enterotoxigênica LAFEP - Laboratório Farmacêutico do Estado de Pernambuco MAS - Agar Manitol Salgado MRSA - Staphylococcus aureus resistente a meticilina MSSA - Staphylococcus aureus sensível a meticilina NCCLS - National Committee for Chemical Laboratory Standands OMS - Organização Mundial de Saúde ORSA - Staphylococcus aureus resistente a oxacilina PCR - Reação em cadeia da polimerase PFGE - Pulsed-field gel eletrophoresis TSA - Teste de sensibilidade a antibióticos VRE - Enterococcus faecium vancomicina resistente

- XIII -
R E S U M O
ANTUNES, R. M. P. Ação antimicrobiana e antiplasmidial do lapachol e seus derivados sobre bactérias e fungos leveduriformes. [Tese-Doutorado], Programa de Pós-graduação em Sintéticos Bioativos/CCS/LTF, João Pessoa (PB): UFPB, Universidade Federal da Paraíba, 2007. Diante da problemática da resistência microbiana, as pesquisas apontam para a descoberta de novos antibióticos que sejam eficazes ante os patógenos emergentes. Neste contexto, estudou-se a ação antimicrobiana e antiplasmidial do lapachol e seus derivados semisintéticos e nitrogenados frente a S.aureus ATCC 25923, E. coli ATCC 25922, P.aeruginosa ATCC 27853, C.albicans ATCC 76643, 08 cepas de S. aureus de origem animal, 10 cepas de S. aureus de origem ambulatorial, 01 cepa MRSA ambulatorial, 02 cepas de S. aureus de origerm ambiental, 10 cepas de E. coli de origem ambulatorial e 06 cepas de C. albicans de origem ambulatorial. A Concentração inibitória mínima (CIM), a Concentração bactericida mínima (CBM) e a curva de morte foram realizadas para os microrganismos ATCC. Um estudo de estrutura-atividade frente às cepas de S. aureus também foi realizado bem como uma investigação sobre a atividade curagênica das bactérias com perfil plasmidial conhecido. Os resultados estimulam o aprofundamento das pesquisas em microbiologia como também para outras atividades biológicas dessas substâncias obtidas a partir de modificações estruturais da molécula do lapachol por semi-síntese, sendo elas as mais ativas frente à S. aureus ATCC, inclusive MRSA. As curvas de morte indicam que estes produtos têm ação bacteriostática. Os compostos -lapachona e nor-lapachona apresentaram menor CIM para todos os microrganismos testados podendo ser os mais eficazes da série. As mudanças na estrutura química do lapachol aumentaram consideravelmente sua atividade antimicrobiana diante de S. aureus. O produto -lapachona apresentou ação simultânea para a eliminação das marcas de resistência para penicilina e a eritromicina. O potencial promissor dessas substâncias as colocam como alvo de interesse farmacológico e químicos sintéticos devido à variedade de atividades biológicas que essas naftoquinonas apresentam. Palavras-chaves: Atividade antimicrobiana, atividade antiplasmidial, fitoconstituintes, produtos de síntese, lapachol e derivados

- XIV -
A B S T R A C T
ANTUNES, R. M. P. Ação antimicrobiana e antiplasmidial do lapachol e seus derivados sobre bactérias e fungos leveduriformes. [Tese-Doutorado], Programa de Pós-graduação em Sintéticos Bioativos/CCS/LTF, João Pessoa (PB): UFPB, Universidade Federal da Paraíba, 2007. Once there is the problematic with microbial resistance, the researches point to the use of new antibiotics that are efficient with the emerging pathogens. In this context, it were studied the antimicrobial and antiplasmidial actions of lapachol and its semi synthetic and nitrogenated derivates in S.aureus ATCC 25923, E. coli ATCC 25922, P.aeruginosa ATCC 27853, C.albicans ATCC 76643, 08 animal originated strains of S. aureus, 10 ambulatorial originated strains of S. aureus, 01 ambulatorial MRSA strain, 02 S. aureus originated from the ambient, 10 strains of E. coli originated from ambulatory and 06 strains of C. albicans originated from ambulatory. The MIC, the MBC and the death curve were done with the ATCC microorganisms. A study structure-activity with the strains of S. aureus was done and also an investigation of cure activity in bacteria with plasmidial profile known. The results stimulate the deepening of researches in microbiology and also to other biological activities of these substances obtained by structural modifications of molecule of lapachol by semi synthesis, being those more active in S. aureus ATCC, including MRSA. The death curves indicate that these products have bacteriostatic action. -lapachone and nor-lapachone presented lower MIC to all microorganisms tested, and can be the most potent in the series. The changes in chemical structure of lapachol enhanced considerably its antimicrobial activity with S. aureus. The product lapachone presented simultaneous action to elimination of resistance marks to penicillin and eritromicine. The promising potential of these substances make them a target of pharmacological and synthetic chemical interest due to the variety of biological activities that these naphtoquinones present. Key words: antimicrobial activity, antiplasmidial activity, phytoconstitutes, synthesis products, lapachol and derivates.

I N T R O D U Ç Ã O

16
1. INTRODUÇÃO
Com a notável descoberta dos medicamentos de síntese como os antibióticos
e antinflamatórios e com o avanço da indústria farmacêutica, os produtos naturais
caíram em desuso. Entretanto, o aparecimento de numerosos microrganismos
resistentes às drogas, os altos níveis de resíduos tóxicos nos alimentos, aliados ao
desequilíbrio ecológico causado pelo homem, fizeram com que se buscassem
alternativas compatíveis com o modo de vida humano, nas mais diversas facetas do
bem-estar bio-psico-social. A resistência aos antimicrobianos tem se tornado um fato
corriqueiro desde o início da era dos antibióticos quando a descoberta de potentes
agentes antimicrobianos foi considerada como uma das grandes contribuições da
medicina no Séc. XX (MOELLERING Jr., 2000).
Desde o início dos anos 80, o número de antimicrobianos em fase de
desenvolvimento diminuiu consideravelmente. Ao mesmo tempo, a resistência aos
antimicrobianos cresce de forma imensurável. Nos anos 70 e 80, as bactérias Gram-
negativas resistentes eram consideradas como o principal flagelo, entretanto no final
do Séc. XX, as bactérias Gram-positivas, resistentes, têm se tornado mais
importantes. Os microrganismos estão cada vez mais desenvolvendo uma série de
novos mecanismos de resistência. A resistência microbiana resulta em crescente
morbidade e mortalidade, como também eleva os custos de saúde e o
desenvolvimento de qualquer novo antimicrobiano vem sendo acompanhado pelo
aparecimento da resistência bacteriana. Segundo MOELLERING Jr. (2000);
MOREIRA (2004), este fato resulta de vários fatores, tais como: o uso indiscriminado
de antimicrobianos, o uso crescente de procedimentos invasivos, em grande número
de hospedeiros susceptíveis e falhas no controle de infecções, ocasionando
aumento da transmissão de organismos resistentes, principalmente em ambientes
hospitalares e em pacientes imunocomprometidos.
Vários são os mecanismos pelos quais os microrganismos podem escapar
dos efeitos dos antimicrobianos, entre eles incluem-se: alteração da estrutura
molecular de antimicrobianos ou produção de enzimas que inativam a droga a
exemplo das β-lactamases ou enzimas modificadoras de aminoglicosídeos,
alteração das proteínas ligantes da penicilina ou outros pontos-alvo nas paredes das
células, alvos modificados da DNA-girase, mutações de permeabilidade e
modificações ribossômicas (FILE Jr., 2000).

17
O uso cada vez maior de antimicrobianos na prática clínica, assim como
também a enorme quantidade de antibióticos utilizados na agricultura, criação de
peixes e criação de animais, possibilitam condições favoráveis para a seleção de
microrganismos resistentes (TOMAZ, 1994; PEREIRA, 2000; MOREIRA, 2004).
A resistência a antimicrobianos pode ser transferida entre bactérias pelos
plasmídios que são moléculas extracromossomiais de DNA, transposons que são
segmentos móveis de DNA, ou pelos mecanismos de inserção seqüencial. Os
plasmídios transferíveis podem possuir genes que apresentam código de
transferência em relação a uma ampla gama de drogas antimicrobianas. Assim, para
os microrganismos Gram-positivos e Gram-negativos, uma única transferência pode
resultar na aquisição de vários determinantes de resistência microbiana. A pressão
ambiental proveniente do uso excessivo de agentes antimicrobianos evidentemente
contribuiu para disseminar os determinantes de resistência. Praticamente todas as
bactérias patogênicas adquiriram genes de resistência antimicrobiana (FILE Jr.,
2000; PEREIRA, 2000; UENO; JORGE, 2001).
Entre as várias propriedades mediadas pelos plasmídios de maior relevância
médica, estão os relacionados com a resistência a antibióticos e aos fatores de
virulência. Existem muitas variedades de plasmídios, diferentes em tamanho,
composição genética e capacidade de transferência entre bactérias. Alguns
plasmídios realizam sua própria transferência entre bactérias da mesma espécie ou
de espécies diferentes, estes são chamados de plasmídios de conjugação. Muitos
plasmídios de resistência a antibióticos, também são plasmídios de conjugação.
Alguns apresentam capacidade de se replicar em diferentes hospedeiros e propagar
a resistência às drogas entre espécies bacterianas não-relacionadas. Esses
plasmídios contribuem para o drástico aumento da resistência antimicrobiana em
populações naturais (SCHAECHTER, et al., 2002). Os plasmídios podem ser
curados ou removidos da célula, depois de serem submetidos a diferentes condições
de stress, como mudanças na temperatura e presença de determinados substratos
(PEREIRA; SIQUEIRA-JUNIOR, 1995; TRABULSI, 2006; PEREIRA, 2000; UENO;
JORGE, 2001).
A resistência microbiana é um problema emergente na medicina humana e
exige uma mudança na terapia empírica e revisão das estratégias dos
procedimentos de testes. A velocidade na qual se desenvolvem nos microrganismos

18
está relacionada à sua exposição aos agentes antimicrobianos (TENOVER, 2000;
SOUZA; REIS; PIMENTA, 2005).
Grande parte da resistência que o gênero Staphylococcus adquire às drogas,
é de natureza extracromossômica, isto é, determinada por plasmídios portadores de
genes para resistência (LYON; SKURRAY, 1987). Uma variedade de compostos tais
como corantes de acridina, brometo de etídio, rifampicina, sal de bis-amônio e mais
tioridazina (uma fenotiazina), assim como também antibióticos inibidores da sub-
unidade β da DNA girase, novobiocina e coumermicina, têm sido reportadas como
eliminadores de plasmídios (HOOPER et al., 1984; WEISSER; WIEDMANN, 1985;
PEREIRA, 2000; PEREIRA; SIQUEIRA-JÚNIOR; TAKAKI; 2004).
A transferência de material genético entre organismos da mesma ou de
diferentes espécies desempenha um papel crucial na evolução da resistência a
drogas em bactérias. Em Staphylococcus aureus essa transferência pode ocorrer
por transformação, por transdução e por processos que envolvem contato físico
celular - conjugação, mediada por fagos ou conjugação propriamente dita. Alguns
fatores podem afetar a transferência conjugativa de plasmídios, como a composição
do meio, pH e temperatura. Antibióticos como gentamicina e vancomicina estimulam
a freqüência de transferência de 10 a 20 vezes enquanto meticilina e alguns
inibidores da síntese de proteínas reduzem essa freqüência (AL – MASSAUD; DAY;
RUSSEL, 1991).
A resistência mediada por plasmídios, pode ser simples, porém na maioria
das vezes é múltipla, tornando a bactéria resistente a dois ou mais antimicrobianos.
Isto se deve à presença de genes de resistência, para diferentes antimicrobianos,
num só plasmídio (TRABULSI, 2006, UENO; JORGE, 2001).
Dentre os vários microrganismos freqüentemente isolados em infecções
humanas que apresentam elevada resistência aos antimicrobianos, destacam-se:
Staphylococcus aureus e Escherichia coli. Entretanto outros microrganismos
considerados emergentes, como por exemplo, Listeria monocytogenes e fungos,
também têm sua importância devido à gravidade de seus processos infecciosos,
principalmente quando atinge pacientes imunossuprimidos. Entre os fungos
leveduriformes Candida aIbicans é a levedura mais freqüente envolvida na etiologia
de infecções fúngicas (MOREIRA, 2004).
Staphylococcus aureus é um coco Gram-positivo grande que cresce em
cachos. Trata-se de uma das mais resistentes bactérias não formadoras de esporos,

19
capaz de sobreviver por longos períodos em objetos inanimados secos. É também
relativamente termo-resistente. Essas propriedades permitem ao S. aureus
sobreviver em qualquer tipo de ambiente onde se encontram seres humanos. São
patógenos potentes, amplamente encontrados na biota humana, podendo causar
vários processos infecciosos, piogênicos algumas vezes levando a formação de
abscessos em tecidos profundos. Além disso, podem produzir enfermidades
mórbidas distintas por meio da produção de toxinas específicas. A vertente na
pesquisa de novos compostos vegetais com ação antimicrobiana, se apresenta
como um modelo ecologicamente correto, de produzir substâncias que sejam
eficazes e menos agressivas ao meio ambiente e aos homens, contribuindo para
uma melhor qualidade de vida, conforme estabelece a Carta Européia do Ambiente
e da Saúde, publicada pela OMS em 1989 (DEOUX; DEOUX, 1998).
A utilização de plantas medicinais na promoção da saúde e prevenção das
doenças foi sempre prática corrente na história da humanidade, sendo a importância
das plantas medicinais tão grande que até meados do século XX a grande maioria
dos medicamentos era de origem vegetal. Como exemplo, existem escritos que
remetem à época de Hipócrates, cerca de 400 a.C., sobre a utilização da casca e
das folhas do salgueiro pelas suas propriedades analgésicas. Já no século XX, esta
dimensão foi diminuindo com o aparecimento dos medicamentos de síntese,
notadamente os antibióticos e os antinflamatórios que evidenciaram bons
resultados terapêuticos (CARVALHO; SARTI, 1995; SALLÉ, 1996; MONTANARI,
2001; YUNES, 2001)
Na área farmacêutica, as plantas e os extratos vegetais foram e
continuam sendo de grande relevância, tendo em vista a utilização das substâncias
ativas como protótipos para o desenvolvimento de fármacos e como fonte de
matérias-primas de fármacos e adjuvantes, ou ainda, de medicamentos elaborados
exclusivamente à base de extratos vegetais, os fitoterápicos (DI STASI, 1996;
GOTTLIEB et al., 1996; YAMADA, 1998; MONTONARI, 2001; YUNES, 2001).
O vegetal, além da organização trófica conseguida através da
fotossíntese, possui um mecanismo regulador que o ajuda a desempenhar suas
funções vitais. Por meio desse mecanismo, o metabolismo de ordem secundária, a
planta consegue elaborar substâncias que desempenharão papéis específicos em
casos de stress, crescimento, reprodução, repelir organismos invasores, atrair
insetos ou pássaros necessários para a polinização e adaptação às mudanças

20
físicoquímicas do meio externo. Devido a estas características, são atribuídos aos
princípios químicos vegetais diversas ações biológicas constituindo a planta num
enorme laboratório de síntese orgânica, fruto de milhões de anos de evolução
(HARBONE, 1988; BEART; LILLEY; HASLAM, 1985; CARVALHO; SARTI, 1995;
MACHADO; PALMA; COSTA, 1995; DI STASI, 1996; SALLÉ, 1996; VON POSER;
MENTZ, 2000; DlAS, 2001; MONTONARI, 2001; YUNES, 2001).
Nesta pesquisa estudou-se o lapachol e seus derivados semi sintéticos, assim
como alguns derivados nitrogenados obtidos por semi-síntese, com o objetivo de
conhecer sua ação antimicrobiana e antiplasmidial sobre microrganismos. Além do
mais, estas substâncias já exibem alguma atividade biológica, seja ela
antinflamatória, antimalárica, antiviral, anticancerígena e antimicrobiana (FONSECA;
BRAGA; SANTANA, 2003; LIRA et al, 2004), moluscicida (SILVA et al, 2005) ou
atuação inibitória sobre sistemas reparadores (DA SILVA; FERREIRA; SOUZA,
2003).
O lapachol é uma hidroxi-naftoquinona substituída, 2-hidroxi-3-(3-metil-
2butenil) – 1,4-naftalenodiona, descoberto e estudado desde o século passado, e
isolado pela primeira vez do lenho da árvore argentina “Lapacho” Tabebuia
avellanedae, Lar (Bignoniaceae), por E. Paterno. Posteriormente, o lapachol também
foi encontrado em várias outras espécies como, por exemplo, no pau d’arco roxo
(Tabebuia sp).

O B J E T I V O S

22
2. OBJETIVOS
2.1. Objetivo Geral
Submeter o lapachol e seus derivados semi-sintéticos e nitrogenados a
análise da atividade antimicrobiana verificando a que grupo farmacológico
pertencem: bactericida ou bacteriostático e estudar a ação dessas
substâncias sobre plasmídios de resistência em amostras bacterianas.
2.2. Objetivos Específicos
Verificar a atividade antimicrobiana para determinação da Concentração
Inibitória Mínima – CIM, Concentração Bactericida Mínima - CBM e
Concentração Fungicida Mínima - CFM do lapachol e seus derivados.
Analisar a relação estrutura/atividade dos derivados semi-sintéticos do
lapachol sobre Staphylococcus aureus.
Avaliar a interferência das substâncias sobre a curva de crescimento
microbiano - cinética bacteriana, cinética fúngica.
Detectar a influência das substâncias em teste sobre a cura de plasmídios de
resistência a antimicrobianos.

R E V I S Ã O D E L I T E R A T U R A

24
3. REVISÃO DE LITERATURA
3.1. Microrganismos: Generalidades
Estima-se que nos seres humanos o número de células bacterianas que
fazem parte da microbiota normal ultrapasse em 10 vezes o número de células do
próprio organismo. A microbiota humana apresenta uma série de atividades
benéficas para o homem e colabora com os mecanismos de proteção infecciosa. De
um modo geral, os microrganismos são capazes de sobreviver em ambientes de
diversas condições físicas. Entretanto, existe uma limitação da capacidade de
sobrevivência de determinado microrganismo, em um meio ambiente desfavorável,
de modo que este recurso foi aproveitado pelo homem para o controle de
microrganismos. Os investigadores logo descobriram que os microrganismos
desenvolvem resistência a antimicrobianos por um processo conhecido como
seleção natural. Quando uma população microbiana é exposta a um antibiótico,
organismos mais sensíveis são destruídos e somente sobreviverão aqueles mais
resistentes ao antimicrobiano. (RIBSON, 2002; BRANDILEONE et al., 2006).
A parede celular bacteriana é uma estrutura rígida que recobre a membrana
citoplasmática e confere forma às bactérias. Ela é constituída por ácido
diaminopimérico (DPA), ácido murâmico e ácido teicóico além de aminoácidos,
carboidratos e lipídeos. Todos esses compostos estão reunidos para formar
substâncias poliméricas complexas que por sua vez estruturam a parede celular.
Uma macromolécula complexa denominada peptideoglicano (mucopeptídeo ou
mureína) forma a estrutura rígida da parede. Além disso, a parede celular protege a
célula, mantém a pressão osmótica intrabacteriana, impedindo o rompimento da
célula devido à entrada de água, e funciona como suporte de antígenos somáticos
bacterianos. A divisão das bactérias Gram-positivas e Gram-negativas, de acordo
com sua resposta à coloração de Gram é decorrente das diferenças na composição
e estrutura da parede celular (SANTOS, 2004; SOUZA; REIS; PIMENTA, 2005).
As bactérias Gram-positivas possuem uma quantidade maior de
peptideoglicano em sua parede celular, o que torna a parede dessas bactérias mais
espessa e rígida do que a das bactérias Gram-negativas. Composta de proteínas,
lipídeos, peptideoglicano e ácidos teicóicos (cadeias de polifosfato com resíduos de
ribitol e glicerol), essas bactérias são sensíveis a lisozima e sua parede constitui o

25
local de ação de alguns antibióticos, além de apresentar elementos básicos para
identificação sorológica (SANTOS, 2004; SOUZA; REIS; PIMENTA, 2005).
A parede celular das bactérias Gram-negativas é menos espessa e elas são
mais complexas do que as Gram positivas por apresentarem uma membrana
externa cobrindo a fina camada de peptídeoglicano. A membrana externa é o que
distingue as bactérias Gram negativas, servindo como uma barreira seletiva para a
entrada e saída de algumas substâncias da célula e podendo ainda causar efeitos
tóxicos sérios em animais infectados. A estrutura da membrana externa é composta
por fosfolipídios, lipoproteínas e lipopolissacarídeos (LPSs). Os lipopolissacarídeos
estão localizados exclusivamente na camada externa da membrana, enquanto que
os fosfolipídeos estão presentes quase completamente na camada interna. Os LPSs
são compostos por três segmentos ligados covalentemente: (1) lipídeo A,
firmemente embebido na membrana; (2) cerne do polisssacarídeo, localizado na
superfície da membrana; e (3) antígenos O, que são polissacarídeos que se
estendem como pêlos a partir da superfície da membrana em direção ao meio
circundante. A porção lipídica do LPSs é também conhecida como endotoxina e
pode atuar como um veneno, causando febre, diarréia, destruição das células
vermelhas do sangue e choque potencialmente fatal (SANTOS, 2004; SOUZA;
REIS; PIMENTA, 2005).
Quanto aos fungos, incluídos no reino Fungi, são encontrados em vegetais,
em animais, no homem, em detritos e em abundância no solo, participando
ativamente do ciclo dos elementos. Sua dispersão na natureza é feita por várias
vias, principalmente pelo ar atmosférico através dos ventos (TRABULSI, 2006).
Todas as células fúngicas são eucarióticas, isto é, possuem núcleo com
membrana nuclear. Os fatores de virulência têm sido pouco estudados. Como
possíveis fatores citam-se a variabilidade fenotípica, aderência nos tecidos dos
hospedeiros, produção de toxinas e enzimas, a exemplo das proteinases, lípases e
fosfolipases. Estas enzimas hidrolíticas extracelulares são importantes na
patogenicidade dos fungos, causando danos às células do hospedeiro. A proteinase
ácida e fosfolipase de Candida albicans têm sido investigadas como fatores de
virulência. Entretanto, pouco se conhece sobre enzimas como condroitin-sulfatase e
hialuronidase nas demais espécies do gênero Candida e em outros fungos
patogênicos (FURLETTI, 2006; LOGUERCIO-LEITE et al., 2006).

26
3.2. Patogênese microbiana
3.2.1. Gênero Staphylococcus
Staphylococcus spp. são cocos Gram-positivos e catalase positivos (Figura 1)
que tendem a formar agrupamentos semelhantes a cachos de uva. São sensíveis a
lisostafina, característica que permite sua diferenciação do gênero Micrococcus.
Staphylococcus são amplamente distribuídos na natureza e fazem parte da
microbiota normal da pele e mucosas de mamíferos e aves. Atualmente, o gênero é
composto por cerca de 27 espécies, sendo que algumas são frequentemente
associadas a uma ampla variedade de infecções de caráter oportunista em seres
humanos e animais. Tradicionalmente são divididos em duas categorias: coagulase
positivos e coagulase negativos. Essa classificação é baseada na capacidade de
coagular o plasma que é uma propriedade considerada há muito tempo, como
importante marcador de patogenicidade do gênero Staphylococcus. Entre as
espécies coagulase positivas S. aureus representa a espécie geralmente envolvida
em infecções humanas, sendo a mais estudada, pois produz um amplo espectro de
doenças desde lesões superficiais até severas infecções sistêmicas (TRABULSI,
2006; PEREIRA, 2002).
Figura 1. Forma e arranjo celular de S. aureus Fonte: http://www.visualsunlimited.com/browse/vu350/vu350691.htm
S. aureus é um importante patógeno nosocomial, sendo descrito como agente
etiológico significativo em infecções hospitalares adquiridas, desde 1950. Ainda é o
microrganismo mais freqüente associado à mastite caprina e bovina (PEREIRA,
2000; FREITAS et al., 2003; PEREIRA, 2002).

27
A patogênese de S. aureus é devida a fatores de virulência na forma de
toxinas, enzimas e outras proteínas associadas a parede celular mediadas por
genes plasmidiais ou cromossômicos que combinados conduzem à doença. S.
aureus é o agente mais comum de infecções piogênicas podendo também causar
vários tipos de intoxicações importantes como a síndrome da pele escaldada,
intoxicação alimentar e síndrome do choque tóxico (TRABULSI, 2006).
Espécies de Staphylococcus utilizam extensivas estratégias para sobrepujar
as defesas microambientais do hospedeiro infectado e, potencialmente, colonizar os
tecidos. Evidências recentes sugerem que as exotoxinas estafilocóccicas são super
antígenos reconhecidos como proteínas multifuncionais que invariavelmente exibem
atividade letal, pirogenicidade e capacidade de induzir hipersensibilidade a
endotoxinas (PEREIRA, 2000; FREITAS et al., 2003;).
Staphylococcus aureus resistente a meticilina - MRSA
A espécis MRSA (Figura 2) representa uma ameaça por causa da sua
distribuição em comunidades e ambientes hospitalares, visto que apresenta
resistência a quase todos os antibióticos como os - lactâmicos, as fluorquinolonas,
cloranfenicol, clindamicina, tetraciclina, aminoglicosídeos e mais recentemente a
vancomicina. Associações na antibioticoterapia tem sido usadas para controlar as
infecções por MRSA, mas por um período curto devido sua possível alteração a
essa susceptibilidade (WICKENS; WADE, 2005).
Figura 2. S. aureus MRSA. Fonte: http://education.denniskunkel.com/catalog/product_
info.php?products_id=512

28
O uso abusivo e indiscriminado de agentes antimicrobianos na prática clínica
veterinária tem um efeito seletivo no surgimento e manutenção de resistência a
drogas. Particularmente, S. aureus, com reconhecida versatibilidade no
desenvolvimento de resistência contribui para sua sobrevivência em ambientes
hospitalares e difusão entre os pacientes (CLOUTIER, 1995; PEREIRA, 2000;
SOUZA; REIS; PIMENTA, 2005).
Os mecanismos pelos quais os genes de resistência se movimentam entre os
organismos são complexos e fascinantes. Em S. aureus a resistência múltipla resulta
principalmente da presença de plasmídios que geralmente se encontram em
múltiplas cópias, que permite sua transferência em uma freqüência mais elevada.
Outra estratégia é a aquisição de genes de resistência ao cromossomo, assim
produzindo uma multiresistência mais estável, a exemplo do MRSA (WICKENS;
WADE, 2005; GRUNDMANN et al., 2006).
MRSA tem sido descrito como o maior patógeno nosocomial em todo o
mundo, capaz de causar uma grande variedade de infecções. Para o tratamento,
embora tenha sido registrado recentemente casos de resistência a vancomicina,
este antibiótico ainda tem sido potencialmente usado, bem como o teicoplamina.
Diante deste panorama, o desenvolvimento de novos compostos anti-MRSA é de
grande e relevante importância. Novos compostos, tais como quinupristina –
dalfopristina e linesolida têm sido utilizados, sendo que linesolida é viável na forma
de administração oral (PEREIRA, 2000; RIVERON et al, 2003; WICKENS & WADE,
2005; GRUNDMANN et al, 2006).
3.2.2. Gênero Escherichia
O gênero Escherichia compreende as espécies E. coli, E. blatae, E.
fergusonii, E. hermannii e E. vulneris. Entretanto a única espécie de maior
importância prática é a E. coli (TRABULSI, 2006).
Escherichia coli A espécie E. coli (Figura 3), compreende grande número de grupos e tipos
sorológicos, identificados por meio de anti-soros preparados contra as três
variedades de antígenos que ocorrem na espécie, ou seja, os antígenos O, K e H.
No momento, devido a grande variedade de antígenos, a única identificação de
rotina do estudo sorológico de amostras de E. coli diz respeito ao diagnóstico de

29
infecções intestinais medidas por Escherichia coli enteropatogênica clássica (EPEC)
e enteroinvasora (EIEC), E. coli enteroxigênica (ETEC), E. coli enterohemorrágica
(EHEC), E. coli enteroagregativa (EAggEC) e E. coli que adere difusamente (DAEC)
(SILVA; DA SILVA, 2005).
Figura 3. Aspecto morfológico de E. coli Fonte: http://www.visualsunlimited.com/browse/vu214/vu214556.html
Quanto às infecções extra-intestinais, as cepas de E. coli mencionadas até
então, raramente estão envolvidas neste processo. E. coli é encontrada
normalmente nos intestinos e pode se localizar em qualquer órgão ou tecidos do
corpo humano. Entretanto, as infecções mais freqüentes são urinárias, meningite do
recém-nascido. É a causa mais comum de infecção urinária, sendo responsável por
90% ou mais, das infecções adquiridas na comunidade, sendo as cepas envolvidas
denominadas E. coli uropatogênicas. O diagnóstico das infecções extra-intestinais é
feito pelo isolamento e posterior identificação bioquímica. O antimicrobiano a ser
utilizado no tratamento deve ser selecionado tendo-se por base não apenas a
sensibilidade do agente etiológico, mas, também a concentração que atinge no local
da infecção (NATARO; KAPER, 1998; TRABULSI, 2006; BALLESTER; ESCOBAR;
GRISI, 2002; CAMARGO et al., 2002; ESMERINO; GONÇALVES; SHELESKY,
2003).

30
3.2.3. Gênero Pseudomonas
O gênero compreende grande número de espécies de bacilos Gram-
negativos, normalmente diferenciados por meio de provas bioquímicas, testes de
sensibilidade a antibióticos (TSA), formação de pigmentos, número e localização de
flagelos. Na realidade, 90% das amostras isoladas de material clínico correspondem
a Pseudomonas aeruginosa (TRABULSI, 2006).
Pseudomonas aeruginosa Uma das características de P. aeruginosa (Figura 4) é a capacidade de
produzir um pigmento azul-esverdeado, denominado piocianina. É por esta razão
que a espécie é também denominada bacilo piociânico. Entretanto, nem todas as
amostras produzem este pigmento.
Figura 4. Forma e arranjo celular de P. aeruginosa Fonte: http://www.biotox.cz/toxikon/bakterie/bakterie/obr/
pseudomonas_aeruginosa_1.htm
Pseudomonas aeruginosa é um germe tipicamente oportunista que pode
causar várias doenças, tais como bacteremias severas (processos cirúrgicos ou
queimaduras), infecções urinárias, pneumonia em casos de fibrose cística e em
pacientes que usam respiradores contaminados, ceratites oculares (cirurgias
oculares), meningites e endocardites (após punção e cirurgias cardíacas). É
produtora de toxina A, que é mais tóxica e seu mecanismo de ação é idêntico ao da
toxina diftérica. As amostras não produtoras de toxina A são menos virulentas,
naturalmente resistentes a vários antibióticos como beta-lactâmicos, às tetraciclinas
e cloranfenicol. Este microrganismo pode adquirir resistência a qualquer agente
terapêutico, com exceção da polimixina, através de mutações ou da aquisição de

31
plasmídios de resistência. Recomenda-se a realização de antibiograma para a
seleção do antibiótico a ser usado no tratamento (TRABULSI, 2006; LAMBERT,
2002, SCHWEIZER, 2003).
Em relação a sua epidemiologia, esta bactéria normalmente habita o solo,
água e vegetais. Pode ser encontrada na pele, fezes e garganta de indivíduos
sadios. Em pacientes hospitalizados, a taxa de portadores pode ser bastante
elevada. Atualmente é responsável por aproximadamente 15% dos casos de
bacteremia causadas por germes Gram-negativos. A elevada freqüência de P.
aeruginosa em hospitais é explicada, em parte, pela elevada resistência a muitos
antibióticos e anti-sépticos leves (TRABULSI, 2006; PAVIANI; STADNIK; HEINEK,
2004; ÁLVAREZ et al., 2005).
3.2.4. Gênero Candida
Figura 5. Forma e arranjo celular de C. albicans Fonte:http://education.denniskunkel.com/catalog/
product_info.php?products_id=817
O gênero Candida (Figura 5) é responsável por diversas manifestações
clínicas conhecidas como candidíase. O agente etiológico mais comum é C.
albicans, que tem sido isolada da boca, tubo digestivo, intestino, orofaringe, vagina e
pele de indivíduos sadios.
A maior parte das infecções causadas por C. albicans é de origem endógena.
A transmissão exógena, principalmente intra-hospitalar de C. albicans e outras
espécies do gênero, tem sido relatada como infecções oportunistas pelo seu poder
invasor em pacientes debilitados, sendo grave a candidíase sistêmica de diagnóstico
e isolamento difícil devido ao polimorfismo das lesões (TRABULSI, 2006). Infecções

32
oportunistas por Candida spp são de grande interesse e sua pesquisa tem
aumentado nas últimas décadas. Candidíase sistêmica é descrita em 20% de
pacientes com câncer e em aproximadamente 25% em pacientes com transplante
de medula. O diagnóstico é feito através de exame clínico, com a verificação da
forma invasiva, as hifas, através da análise de material de biópsia ou raspado das
lesões. Para tratamento das infecções micóticas estão incluídos desde os
antissépticos convencionais (tintura de iodo, violeta de genciana, ácido salicílico e
benzóico), derivados do anel benzênico (ácido benzóico, sulfamídicos, corantes,
quinonas), derivados triazíncos, derivados imidazólicos (miconazol, econazol,
cetoconazol) e derivados alilamínicos (nistatina, anfotericina B). O uso das drogas
antifúngicas necessita de critérios valiosos, principalmente quando aplicados por via
sistêmica devido aos efeitos colaterais e tóxicos produzidos pela maioria
(TRABULSI, 2006; SUDBERY, 2004; VUCKOVIC et al., 2004; BORIOLLO et al.,
2005; LOGUERCIO-LEITE, 2006; LIMA et al., 2006).
3.3. Antimicrobianos
Os antibióticos são substâncias químicas produzidas por organismos vivos
(fungos, bactérias, actinomicetos e plantas), ou seus equivalentes sintéticos. Estas
substâncias são capazes de destruir ou impedir o crescimento dos microrganismos
patogênicos (MOREIRA, 2004).
3.3.1. Antibacterianos
Dentre os grupos de antimicrobianos merecem destaque os -lactâmicos que
se caracterizam por apresentarem um anel -lactâmico essencial para sua atividade
anti-bacteriana. São antibióticos bactericidas que inibem a síntese da parede celular
através da interrupção do processo de transpeptidação. Podem atuar mais
facilmente nas bactérias Gram-positivas (MACEDO et al, 2005). São antibióticos -
lactâmicos as penicilinas naturais, penicilinas semi-sintéticas e as cefalosporinas, os
monolactâmicos e as carbapanemas (TRABULSI, 2006).
As quinolonas são um grupo de substâncias antibacterianas. Existem
quinolonas de primeira geração (ácido nalidíxico, flumequina e o ácido oxolínico) e
segunda geração (enrofloxacino, ciprofloxacino e perfloxacino) reconhecidas como
fluorquinolonas. As quinolonas são antimicrobianos bactericidas e sua atividade se

33
relaciona a inibição da enzima DNA girase, impedindo o enrolamento da fita de DNA.
O interesse nesses compostos reside na sua possível administração por via oral,
uma absorção rápida e boa difusão graças a sua lipossolubilidade (FREITAS et al.,
2003; PEREIRA et al., 2000; TRABULSI, 2006).
Os antibióticos macrolídeos são caracterizados pela presença de um anel
lactona macrolídico em sua estrutura química e apresentam um espectro de ação
limitado. Um dos principais macrolídeos de aplicação terapêutica é a eritromicina.
São ativos contra bactérias Gram-positivas e micoplasma e possuem boa atividade
contra bactérias anaeróbicas. Agem impedindo a síntese protéica, sendo
bacteriostáticos. São bastante lipossolúveis atravessando as barreiras celulares com
facilidade. São bem absorvidos, biotransformados e eliminados (SANTOS; SOUSA;
SANTANA, 2006).
3.3.2. Antifúngicos
O número de fármacos disponíveis para o tratamento de infecções fúngicas
sistêmicas é limitado. Nos últimos anos, a anfotericina B e os azóis, principalmente
cetoconazol, fluconazol e itraconazol, têm sido os fármacos de primeira escolha na
terapia antifúngica. Estas duas classes de medicamentos têm como alvo a
membrana celular dos fungos. Os polienos ligam-se a uma porção esterol,
basicamente ergosterol, presente na membrana de fungos sensíveis, formando
poros ou canais. O resultado é um aumento na permeabilidade da membrana que
permite o extravasamento de diversas pequenas moléculas, levando à morte celular.
A anfotericina B é um antibiótico fungicida de largo e potente espectro de ação, mas
seu uso é associado a efeitos adversos significantes, como nefrotoxicidade e febre
com calafrios, como reação aguda à infusão intravenosa, já que a farmacocinética
deste fármaco não permite a administração oral (GOODMAN; GILMAN, 1996).
Novas formulações da anfotericina B, na forma de lipossomas e de dispersão
coloidal, produzem menos efeitos colaterais, como resultado da redistribuição do
fármaco nos tecidos e da seletividade de liberação, mas o preço destas formulações
é muitas vezes maior que o das antigas (HARSTE; BOLARD, 1996; LOGUERCIO-
LEITE et al., 2006).
Os azóis são compostos totalmente sintéticos. O mecanismo de ação destes
fármacos baseia-se na inibição do esterol-14--demetilase, um sistema enzimático

34
microssomal dependente do citocromo P450, prejudicando a síntese do ergosterol
na membrana citoplasmática e levando ao acúmulo de 14--metilesteróis. Esses
metilesteróis não possuem a mesma forma e propriedades físicas que o ergosterol e
levam à formação da membrana com propriedades alteradas, que não desempenha
as funções básicas necessárias ao desenvolvimento do fungo. Os azóis causam
menos reações adversas que a anfotericina B, mas são menos potentes que a
mesma. Podem ter ação fungistática ou fungicida. O uso excessivo dos azóis levou
ao aparecimento de resistência em espécies suscetíveis. Além disso, os azóis ainda
apresentam a desvantagem da resistência cruzada (GOODMAN; GILMAN, 1996;
WILLIAMS; LEMKE, 2002; LOGUERCIO-LEITE et al., 2006).
Outro agente antifúngico sistêmico utilizado é o pró-fármaco flucitosina. Todos
os fungos sensíveis são capazes de desaminar a flucitosina em 5-fluorouracila, um
potente antimetabólito; como resultado final, a síntese de ácido desoxirribonucléico
(ADN) dos mesmos fica prejudicada. Embora as células dos mamíferos não
convertam a flucitosina em fluorouracila, o que é crucial para ação seletiva do
composto, alguns microrganismos da flora intestinal o fazem, causando certa
toxicidade aos humanos. A flucitosina tem espectro de ação restrito, tem atividade
clinicamente útil contra Cryptococcus neoformans, Candida spp. e os agentes da
cromomicose e a resistência medicamentosa que surge durante a terapia é causa
importante de fracasso terapêutico (GOODMAN; GILMAN, 1996; WILLIAMS;
LEMKE, 2002; LOGUERCIO-LEITE et al., 2006).
No tratamento das micoses superficiais a diversidade de fármacos disponíveis
para a terapia é maior. Além dos polienos anfotericina B e nistatina, da flucitosina e
de uma maior variedade de azóis (bifonazol, clotrimazol, econazol, isoconazol,
oxiconazol, sertaconazol, miconazol, terconazol e tioconazol), existem ainda outros
antifúngicos: o derivado da morfolina, amorolfina; os tiocarbamatos tolnaftato e
tolciclato; as alilaminas naftifina, terbinafina e butenafina e o composto ciclopirox,
além do antibiótico griseofulvina (GOODMAN; GILMAN, 1996; WILLIAMS; LEMKE,
2002; KOROLKOVAS, 2003; LOGUERCIO-LEITE et al., 2006).
3.3.3. Resistência aos antimicrobianos
Após a descoberta das moléculas antimicrobianas, seu uso tornou-se muito
disseminado e doenças que anteriormente eram tidas como de mau prognóstico,

35
passaram a ser controladas com sucesso, ocasionando a redução drástica da
mortalidade causada por microrganismos (SILVEIRA et al, 2006). Por outro lado a
disseminação do uso de antibióticos fez com que as bactérias desenvolvessem
defesas relativas aos agentes antibacterianos, com o conseqüente aparecimento de
resistência a tais compostos. Este fato impõe sérias limitações às opções para o
tratamento de infecções bacterianas, representando uma ameaça para a saúde
pública. Esta resistência prolifera-se rapidamente através de transferência genética
atingindo algumas das principais bactérias Gram-positivas como Enterococcus,
Staphylococcus e Streptococcus (LIU, 1999; TAVARES, 2000; SILVEIRA et al.,
2006).
A última linha de defesa contra a ameaça do S. aureus surgiu a partir da
descoberta do antibiótico glicopeptídeo vancomicina isolado do fungo Streptomyces
orientalis, em 1956. A vancomicina tornou-se quase numa lenda devido a sua
excelente performance frente a MRSA, e outras bactérias Gram-positivas, mas não
apresenta atividade contra Gram-negativas. Em 1988, os primeiros casos de
resistência a espécie de Enterococcus resistentes a vancomicina conhecidos por
VRE, causou enorme alarme, pois estes microrganismos são capazes de infectar
pacientes imunodeficientes como os transplantados e vítimas da AIDS (SILVEIRA,
2006).
A resistência bacteriana aos antibióticos está associada a seu uso
inadequado. O mecanismo celular desta resistência deve-se a uma modificação
genética estável, transmissível através das gerações. Apesar da mutação ser
frequentemente a causa dessa resistência, outros mecanismos de transferência do
material genético estão envolvidos, como a transdução, transformação, conjugação,
elementos extracromossomicos (plasmídios) e DNA recombinante. A escolha
racional do antimicrobiano é um processo complexo que exige diagnóstico clínico
laboratorial e conhecimento farmacológico dos agentes infecciosos, representando
um desafio para os médicos e farmacêuticos (LIU, 1999; OLIVEIRA, et al., 2004;
MOREIRA, 2004; CALBO; GARAU, 2005). Desta forma, a comunidade médica e
científica vem procurando compreender os fenômenos responsáveis pelo
mecanismo adaptado de resistência. A demanda crescente por novas substâncias
capazes de inibir, em concentrações baixas, processos vitais de uma ou mais
espécies de microrganismos resistentes tem provocado uma verdadeira corrida em
busca de agentes antibacterianos, de origem natural, semissintética ou sintética,

36
cada vez mais eficientes (SMITH; COAST, 2002; ELLIS et al., 2004; SILVEIRA,
2006).
3.3.4. Mecanismo genético de aquisição de resistência aos antimicrobianos
O aparecimento de bactérias resistentes a antibióticos pode ser considerado
como uma manifestação natural regida pelo princípio evolutivo da adaptação
genética de organismos a mudanças no seu meio ambiente. Atualmente, a
resistência bacteriana adquirida é descrita em praticamente todas as espécies de
bactérias, conhecendo-se os mecanismos de aquisição de resistência e os
mecanismos moleculares da manifestação da resistência (CLOUTIER, 1995;
TAVARES, 2000; ANDERSON, 2004).
O DNA em bactérias é uma macromolécula em forma de uma dupla fita
circular altamente empacotado e dobrado para se manter dentro de uma DNA-
girase, a qual converte o DNA em diferentes configurações e se agrupa dentro das
topoisomerases que podem ser topoisomerase I, que corta o DNA em fita simples e
topoisomerase II que corta o DNA em duas fitas simultâneas no processo de
replicação (TRABULSI, 2006).
A resistência aos antimicrobianos é um fenômeno genético, relacionado a
existência de genes contidos no microrganismo que codificam diferentes
mecanismos bioquímicos que impedem a ação das drogas. A resistência pode ser
originada em mutações que ocorreram no microrganismo durante seu processo
reprodutivo e resultam de erros de cópia na seqüência de bases que formam o DNA
cromossômico responsáveis pelo código genético. A outra origem da resistência é a
importação dos genes causadores do fenômeno, consistindo na resistência
transferível. Esta resistência faz-se através dos mecanismos de transdução,
transformação e conjugação e, frequentemente, envolve genes situados em
plasmídios e transposons (TAVARES, 2000; SILVEIRA et al, 2006).
Em inúmeros microrganismos, o fenômeno da resistência é natural.
Considerando que os microrganismos produtores de antibióticos existentes no meio
ambiente apresentam mecanismos de proteção (ex.: Streptomyces cervicalae
produtor da eritromicina é resistente a eritromicina) codificados geneticamente,
admite-se que, além da mutação a origem da resistência adquirida em bactérias e
fungos causadores de infecção no homem e outros mamíferos esteja relacionada

37
principalmente, à transferência de genes de resistência contidas nestes
microrganismos presentes na natureza. Portanto, a transferência de genes de
resistência de bactérias não patogênicas ou de baixa patogenicidade para
microrganismos patogênicos provavelmente é um fenômeno comum. Este fenômeno
natural ganha importância, devido ao amplo emprego clínico, industrial (conservação
de alimentos), comercial (engorda de animais) e experimental dos antimicrobianos,
os quais podem desenvolver sua ação selecionadora de microrganismos resistentes
nos mais variados ambientes (RUSSEL, 1998; TAVARES, 2000; SILVEIRA et al.,
2006).
Dentre os microrganismos que sofreram grandes modificações na
sensibilidade aos antimicrobianos com o decorrer dos anos, destacam-se
Staphylococcus, Enterobacter, Pseudomonas, Acinetobacter e recentemente
Meningococcus, Enterococus e Pneumococcus. Na atualidade, é motivo de grande
preocupação entre os cientistas a resistência entre bactérias Gram-positivas, que
vêm se tornando um problema na terapêutica antiinfecciosa (SILVEIRA et al, 2006).
Staphylococcus aureus: mecanismo da resistência identificação
laboratorial e tratamento O isolamento de Staphylococcus resistentes às penicilinas pôde ser
observado logo após os primeiros experimentos com a introdução da penicilina G na
clínica em 1941. Em 1950, cerca de 80% desses microrganismos isolados em
hospitais mostravam-se resistentes às penicilinas, devido a produção de
penicilinases codificadas geneticamente em plasmídios transferíveis por transdução.
Para combater o gênero Staphylococcus foram descobertas as penicilinas
antiestafilocócicas (meticilina e a oxacilina e seus derivados) e as cefalosporinas de
primeira e segunda geração. Contudo, logo após, surgiram bactérias resistentes a
tais compostos em todas as partes do mundo. Na atualidade, espécies de
Staphylococcus vêm mostrando também crescente resistência a lactâmicos,
caracterizando-os como Staphylococcus aureus resistente a meticilina (MRSA) ou
Staphylococcus aureus resistente a oxacilina (ORSA) (TAVARES, 2000).
MRSA emergiu como patógeno nosocomial no início da década de 60. Há
basicamente dois mecanismos responsáveis pela resistência de S. aureus aos
agentes antimicrobianos que possuem anel -lactâmico. Um mecanismo é a
produção de -lactamases, que são enzimas responsáveis pelo desenvolvimento da

38
resistência à penicilina antiestafilocócicas e que levam ao desenvolvimento de
penicilinases anti-estafilocócicas, resistentes as -lactamases, como a meticilina.
Outro mecanismo é uma alteração nas proteínas fixadoras de penicilinas (PBPs). As
PBPs são enzimas que catalizam a etapa final da síntese da parede bacteriana,
além disso, a resistência pode ser devido a alteração na permeabilidade impedindo
o antibiótico de atingir o seu receptor. Todas as cepas de MRSA produzem uma
PBP (PBPza ou PBP2’) (UENO; JORGE, 2001).
As cepas de S. aureus com menor susceptibilidade à meticilina por causa da
alta produção de beta-lactamase são chamadas de BORSA para borderline oxacillin
– resistent S. aureus. Esta resistência, assim chamada borderline pode ser diferente
da resistência intrínseca de MRSA, porém no laboratório é difícil distinguir entre
ambas. Algumas linhagem de BORSA não contém PBP2a, apresentando portanto
outro mecanismo de resistência. A resistência borderline é considerada
primeiramente mediada por plasmídio (diferente da resistência mediado por
cromossomo, como no caso de MRSA) (UENO; JORGE, 2001; PEREIRA, 2002;
MACEDO et al., 2005; SOUSA; REIS; PIMENTA, 2005).
Desde a década de 70, Staphylococcus meticilina resistente tornou-se a
principal causa de infecções hospitalares em todo o mundo. Vancomicina era o
único antibiótico efetivo contra os mesmos, mas, em 1977, foram descritos S. aureus
com resistência à vancomicina e a teicoplamicina que receberam a sigla VRSA
(vancomycin resistant Staphylococcus aureus), estando seu mecanismo de
resistência também ligado as PBP2 e PBP2’, espessamento da parede celular e
aprisionamento de drogas (SOUSA; REIS; PIMENTA, 2005).
Os testes para avaliação laboratorial da susceptibilidade aos antimicrobianos
utilizados são: susceptibilidade aos antibióticos pelo teste de difusão em discos; E-
test, que contém um gradiente de concentração crescente de antibióticos,
fornecendo o valor da CIM diretamente. Reação em Cadeia da Polimerase, PCR,
utilizada na identificação de MRSA; eletroforese em campo pulsado (PFGE),
utilizado para a genotipagem de Staphylococcus, indicado para estudos
epidemiológicos (NCCLS, 2002).
As drogas utilizadas para o tratamento das estafilococcias são amoxicilina,
amoxicilina + clavulanato de potássio, gentamicina, vancomicina. Atualmente se
utilizam para esse tratamento: as cefalosporinas (resistentes a inativação por -
lactamases) cefalosporinas de quarta geração, a cefpiroma, grupos dos

39
glicopeptídeos (vancomicina e teicoplamina para tratar MRSA, as
isoxazolilpenicilinas e as cefalosporinas (para tratamento das pneumonias,
osteomielites, septicemias endocardites, meningoencefalites). Novos antibióticos
para o tratamento de estafilococos são ainda as estreptograminas que são formadas
por uma mistura de duas classes de componentes quimicamente distintos,
denominados de estreptograminas A e B – quinupristina – dalfopristina que é ativa
pelo efeito bactericida sinergístico, contra uma ampla variedade de gram-positivos
incluindo MRSA e VRE (Enterococcus faecium vancomicina resistentes). Outro
grupo de novos antibióticos utilizados para o tratamento de infecções causadas por
bactérias gram-positivas é a das oxazolidinonas que não apresentam ligações com
outras classes de antimicrobianos, a linezolida cujo mecanismo de ação está ligado
à síntese protéica, age inclusive contra MRSA e VRE e tem amplo espectro de
atividade contra microrganismos gram-positivos (SOUSA; REIS; PIMENTA, 2005).
Escherichia coli – Mecanismos de resistência
Escherichia coli enteropatogênica é importante causa de diarréias infantis, de
maneira perversa e está associada a fatores como subnutrição, condições precárias
de habitação e falta de saneamento básicos. Biópsias do intestino de pacientes
infectados com Escherichia coli enteropatogênica (EPEC), mostraram que as
bactérias aderem à superfície do epitélio intestinal, na forma de micro colônias
localizadas, e, na célula infectada causando lesões típicas destruindo as
microvilosidades. Essa aderência intima é mediada pela interação entre uma
proteína da membrana externa da bactéria, a intimina, e seu receptor TIR,
translocado da bactéria para a superfície das células epiteliais. A interação intimina –
TIR provoca rearranjo dos componentes do citoesqueleto, na região da adesão da
célula infectada (SILVA; DA SILVA, 2005).
EPEC produz vigorosa e persistente produção de anticorpos em seres
humanos que vivem em áreas endêmicas e em modelos animais. Essas
observações reforçam as recomendações de organismos ligados a saúde pública
sobre a importância da amamentação do recém-nascido com leite materno e o
desenvolvimento de vacinas e anticorpos protetores para a prevenção e o
tratamento de infecções causadas por EPEC (SILVA; DA SILVA, 2005).

40
Diarréias causadas por EPEC, em geral, são mais severas do que as
causadas por outros patógenos, com prevalência de óbito superior a 30%. É
importante causa de diarréias infantis nos países em desenvolvimento e nos
desenvolvidos em todo o mundo. Os principais sintomas clínicos da doença são
diarréia aquosa, febre, mal-estar e vômitos. A Organização Mundial de Saúde, OMS,
reconheceu, em 1980 doze sorogrupos O designados: O26, O55, O86, O111, O114, O119,
O125, O126, O127, O128, O142, O158. Nestes sorogrupos estão incluídas cepas de EPEC
típicas e atípicas (isoladas de diferentes espécies de animais), E.coli
enteroagregativas, além de outras variantes de E.coli causadoras de diarréias
(SILVA; DA SILVA, 2005).
E.coli é o bacilo gram-negativo mais comum isolado de pacientes sépticos e é
responsável por mais de 80% de todas as infecções das vias urinárias adquiridas na
comunidade e pela maioria das infecções hospitalares causadas por linhagens
multiresistentes a antibióticos, tornando as opções de tratamento limitadas. Até o
momento eram susceptíveis aos carbapanêmicos como imipinem e meropenem,
entretanto já tem sido isoladas linhagens que apresentaram uma resistência
adicional ao imipinem. Estas linhagens possuem plasmídios transmissíveis que
codificam beta-lactamases do tipo ampicilina (TURRI, 2003). Além disso, sistemas
ativos de efluxo em segmentos transmembranares que expulsam o antimicrobiano
de dentro da célula, também conferem resistência aumentando a CIM (BAMBEKE,
et al., 2003; MARTINS, 2005).
Pseudomonas aeruginosa
É o mais importante patógeno humano do gênero Pseudomonas. P.
aeruginosa faz parte da microbiota humana e raramente se torna a causa de
infecções comunitárias em indivíduos saudáveis. No entanto, em ambientes
hospitalares, esta bactéria torna-se um agente infeccioso importante, principalmente
em pacientes que apresentam quebra de barreira física e imunossupressão. Além
disso o número de fatores de virulência é significativamente maior em cepas
provindas de isolados clínicos, quando comparados a cepas ambientais (PAVIANI,
STADNIK; HEINEK, 2004). Muitas publicações têm mencionado um aumento
importante na freqüência e na resistência da P. aeruginosa aos antimicrobianos.
Pseudomonas aeruginosa é intrinsecamente resistente a vários antimicrobianos,

41
incluindo o cloranfenicol, tetraciclinas, algumas quinolonas e beta-lactâmicos. Entre
os antimicrobianos usados para o tratamento desta bactéria estão a penicilina
(piperacilina), cefalosporinas (ceftazidima, cefepima), carbapenêmicos (imipimem,
meropenem), monobactamico (aztreonam), aminoglicosídios (gentamicina,
tobramicina, amicacina) e fluorquinolonas (ciprofloxacino) (PAVIANI; STADNIK;
HEINEK, 2004; ALVAREZ et al., 2005).
A resistência aos -lactâmicos ocorre por mecanismos variados: produção de
-lactamase AmpC (ampicilinase), β-lactamases de espectro estendido,
carbapenemases, diminuição da permeabilidade celular e mecanismo de efluxo
estão entre os descritos. A perda da permeabilidade e o mecanismo ativo de efluxo
podem afetar a atividade de outros tipos de fármacos como aminoglicosídeos e
quinolonas. Esta atividade pode ser completada com alterações da DNA girase.
Atualmente tem sido relatado a incidência de P. aeruginosa multi-resistentes
(LAMBERT, 2002; BAMBEKE, 2003; PAVIANI; STADNIK; HEINEK, 2004;
SCHWEIZER, 2003).
Fungos Leveduriformes
Os fungos podem se apresentar como patógenos primários capazes de
causar infecção sem fatores predisponentes, como também, como patógenos
oportunistas, manifestando seu potencial patogênico em hospedeiros
imunologicamente ou fisiologicamente comprometido, neste último grupo estão
inseridas as leveduras de gênero Candida. O status fisiológico do hospedeiro tem
sido reconhecido então como o fator etiológico primário no desenvolvimento das
infecções fúngicas, mais que a virulência intrínseca do microrganismo oportunista.
Entretanto, apenas pequenas alterações neste estado podem modificar o
comportamento desses microrganismos que passa a manifestar seu potencial de
virulência e assim, de comensal inofensivo tornar-se um patógeno agressivo.
(FURLETTI, 2006).
C. albicans pode se apresentar sobre várias formas, chamadas de variações
adaptativas, sendo a forma, de hifa a mais virulenta e mais aderente em relação à
célula leveduriforme. Dessa forma, a capacidade dessa espécie de formar tubos
germinativos, parece contribuir para sua virulência no homem e nos animais
(FURLETTI, 2006; MOREIRA, 2003).

42
A patogenia de Candida spp. pode ser observada por algumas características
como a formação das hifas e pseudohifas com mais de 200 µm de comprimento,
podendo representar um obstáculo à fagocitose, que é o principal meio de defesa do
organismo contra a infecção fúngica. A produção de alguns metabólicos por cepas
de Candida, é capaz de desencadear manifestações alérgicas. Somente as espécies
capazes de crescer a 37ºC são potencialmente patogênicas ao homem. Os fatores
que estão relacionados ao microrganismo na instalação do processo infeccioso são
a presença de cepas com melhor aderência celular ao hospedeiro, produção de
pseudo-hifas e pseudomicélio, presença de toxinas e a produção de enzimas
proteolíticas (MOREIRA, 2003).
Essas enzimas proteolíticas podem degradar ou transformar constituintes da
membrana celular do hospedeiro induzindo uma disfunção ou destruição física,
sendo que a invasão das células dos tecidos do hospedeiro por tais microrganismos
implica na penetração e danos ao envelope externo celular sendo esse processo
mediado por meios físicos e enzimáticos ou pela combinação de ambos (FURLETTI,
2006).
Os fatores de virulência de C. albicans estão relacionados com a produção de
hialuronidase, condoitina sulfatase, proteinase e fosfolipases. os mesmos podem
atuar de maneira conjunta ou separada determinando a intensidade de virulência e
também são considerados essenciais para o microrganismo (FURLETTI, 2006).
3.4. Plasmídios
Os plasmídios (Figura 6), são moléculas extracromossomiais circulares de
DNA encontradas em muitas espécies bacterianas e em algumas espécies de
eucariotos. São moléculas que podem ser curadas ou removidas da célula depois de
serem submetidas a diferentes condições de estresse, como mudanças na
temperatura, presença de certos corantes ou carência de certos nutrientes (LYON,
1987; PEREIRA, 2000; UENO; JORGE, 2001).

43
Figura 6. Plasmídio. Fonte: http://www.libertaria.pro.br/d_ressurgentes_arquivo/plasmídio.gif
Os plasmidios podem conferir resistência a um antibiótico ou a um metal
pesado. A replicação do plasmídio pode ocorrer em dois momentos: primeiro quando
a célula bacteriana se divide, o DNA plasmidial também se divide assegurando que
cada célula filha receba uma cópia deste. Segundo, durante o processo de
conjugação, a molécula de DNA replicada pode entrar na célula receptora. O
plasmídio se replicará até alcançar seu número de cópias que é controlado pela
iniciação de síntese de DNA. Uns se apresentam em baixo número de cópias (1 a
10), outros o fazem em alto número (10 a 100). Supõe-se que o plasmídio codifica
inibidores que afetam a taxa de iniciação da própria síntese, controlando, portanto o
número de cópias (TRABULSI, 2006). A presença de plasmidios, geralmente em
múltiplas cópias, garante não somente sua distribuição durante a divisão celular,
mas, principalmente, permite a transferência em freqüência mais elevada, sem
causar na maioria das vezes um custo biológico para a célula bacteriana. Uma
variedade de compostos, tais como corantes de acridina, brometo de etídio,
rifampicina, sal de bis-amônio e mais a tioridazina, uma fenotiazina, assim como
antibióticos inibidores da subunidade B da DNA girase, novobiocina e coumermicina
tem sido reportados para eliminar plasmidios. No entanto, são raros os relatos a cura

44
de plasmidios por inibidores da subunidade A da DNA girase, particularmente em
linhagens de S. aureus de origem animal (PEREIRA, 2000).
Os plasmídios de uma célula normalmente perfazem menos de 5% do DNA
total. A análise de perfil plasmidial requer primeiramente a sua separação do DNA
cromossômico de modo que existem vários métodos de preparação rápida para
extração de plasmídios em pequena escala, as quais podem ser utilizadas com
propósito de identificação e estudos epidemiológicos de Staphylococcus. Uma das
técnicas amplamente utilizadas devido sua alta reprodutibilidade e simplicidade é a
técnica de análise de DNA plasmidial usando endonuclease de restrição (REAP)
(UENO; JORGE, 2001).
3.5. Transposons
São segmentos móveis de DNA em bactérias, que são movimentados em
baixa freqüência dentro do cromossomo. A freqüência de transposição tanto em
procariontes como em eucariontes é relativamente baixa (10-7 por geração)
dependendo do elemento em particular. Sendo o cromossomo uma molécula
contínua de DNA, a transposição dos elementos móveis é um processo de
intercâmbio de DNA caracterizado como um tipo de recombinação, (não sendo esta
a recombinação clássica homóloga). Muitos transposons de bactéria possuem genes
facilmente identificáveis, que podem ou não existir em outro lugar do genoma.
Genes de resistência a antibióticos são comuns, e transposons levando esses genes
são os mais frequentemente estudados (SHLAES et al., 1997; TRABULSI, 2006;
SEABRA et al., 2004; FERREIRA, 2005). Estes são designados Tn (ex: Tn1 amp),
quando não é reconhecida a marca, estes elementos são designados seqüenciais
de inserção ou elementos IS (ex: 1S1, 1S2 etc.). Os transposons estão
frequentemente localizados dentro de um gene particular gerando neste uma
mutação. Os transposons levam outros marcadores em adição à informação para a
transposição. A marca mais comum é a resistência a antibióticos, outros marcadores
são genes para fermentação de lactose, metabolismo de rafinose, formação de
enterotoxina em Escherichia coli e resistência a metais pesados. Existem
transposons que são capazes de se transferir de uma célula para outra sem auxilio
de plasmídios, num processo em que é necessário o contato direto entre a célula
doadora e a receptora. Este tipo de transposon está presente nos Streptococcus e é
conhecido como transposon conjugativos. Ele é importante na disseminação da

45
resistência múltipla a antibióticos nesse gênero bacteriano e possivelmente em
outras bactérias Gram-positivas (SHLAES et al., 1997; TRABULSI, 2006; SEABRA
et al., 2004; FERREIRA, 2005).
3.6. Aspectos botânicos e fitoquimicos do Lapachol e seus derivados semi-
sintéticos e nitrogenados
O lapachol foi isolado primeiramente do lenho da árvore argentina “Lapacho”
Tabebuia avellanedae lor. família Bignonaceae (Figura 07).
Figura 7. Ipê-roxo. Fonte: http://www.linkbrazil.com.br/nature/pantanal/flora/ipe_roxo/ipe_roxo_1.jpg
Em 1956, Gonçalves de Lima e coll. constataram no cerne do pau d’arco roxo,
Tabebuia spp., a existência de um composto de coloração amarela com atividade
antimicrobiana, principalmente para o gênero Brucella e que chegaram à conclusão
de que era lapachol. A presença do lapachol foi constatada posteriormente em
várias outras espécies e diferentes famílias tais como: Bignoniaceae (Tabebuia
flavesons Benth, & Hook, T. gnayacan Hems, Tavellanedae Lor Ex Griseb, T.
serratifolia (Vahl) Nichols, T. rosa, T. pentaphylla (Linn) Hemls, Paratecoma peroba
(recors) Kuhlm, Tecoma araliaceae DC, Stereospernum suaveolen DC, S.
kunthianum, Millingtonia hortensis Linn, Stereospermum tetragonum DC, Catalpa
longíssima, Macfadyena ungis – cati, Tabebuia bata (E. Mey) Sandw); Verbenaceae
(Tectona grandis L.fil, Avicennia tomentosa Joacq, Avicenia officinalis), Proteaceae
(Conospremum teretifolium R. Br.), Leguminosae (Diphysa latifólia) outras espécies
(Sterocarpus salignus, Haplophragma adenopphyllum, Kigelia pinnata, Markhamia

46
stipulata Wall, Phyllarthon conorense, Radermachera Xylocarpa, Avicenia alba)
(FONSECA et al., 2003).
O lapachol é uma hidroxi-naftoquinona substituída, [2 – hidroxi – 3 – (3 – metil
– 2 butemil) – 1,4 – naftalenodina]. Os estudos químicos foram desenvolvidos
inicialmente por S.C. Hooker em 1892 e 1896. Além do lapachol foram isolados na
espécie Tabebuia avellanedae Lor. ex giseb (Bignoniaceae) e em outras espécies,
seis naftoquinonas e nove antraquinonas. A α-lapachona e β-lapachona estão
citadas no primeiro grupo. A presença de filoquinona (vitamina K) foi registrada
apenas nas folhas desta planta. As substâncias mais estudadas nesta espécie são o
lapachol, os dois isômeros (derivados éter metílico do lapachol) α-lapachona e β-
lapachona e a xiloidona (desidro – α – lapachona), todos biologicamente ativos
(FONSECA et al., 2003; Lyra et al., 2004)
lapachol α-lapachona
β-lapachona xiloidona
Figura 8. Estruturas químicas do lapachol e seus derivados semi-sintéticos
O lapachol pode ser obtido por vários métodos de extração, sempre se
baseando na sua solubilidade em solvente orgânico e do seu sal em meio aquoso

47
alcalino, gerando, respectivamente, soluções amarelas e vermelhas. Os métodos de
extração desenvolvidos por Gonçalves et al., 1962, consistem na extração do
lapachol da serragem do cerne de Tabebuia spp. (pau d’arco roxo) com benzeno ou
com acetona. A forma de extração do lapachol mais empregada foi descrita por
Paterno em 1882. Este método propicia a obtenção de 80g de lapachol puro,
cristalizado para cada 1 kg de serragem do cerne do vegetal. Os principais
derivados do lapachol podem ser obtidos da seguinte forma: α–lapachona e β-
lapachona podem ser obtidas a partir do lapachol. A xiloidona pode ser obtida
quimicamente pelo método que consiste em refluxar o lapachol, e por um segundo
método que consiste em refluxar o lapachol em anidrido acético e piridina, seguindo-
se de uma hidrólise e oxidação lenta. O lapachol também pode ser transformado em
α–lapachona, β–lapachona e xiloidona por via seca (calor); e a xiloidona aquecida é
transformada em α–lapachona e β–lapachona. Fieser propôs uma rota para a
obtenção sintética do lapachol em 1927, seguido por Hooker, em 1936 e Pettit e
Houghton, em 1971 (FONSECA; BRAGA; SANTANA, 2003).
Em relação as propriedades físico-quimicas, o lapachol tem a forma de
cristais prismáticos ou folhas amarelas, de peso molecular 242,26, sendo sua
formula química C15H14O3, com ponto de fusão 139,5 – 140,2ºC. É bastante solúvel
em acetona, etanol, metanol, clorofórmio, benzeno e ácido acético; solúvel em éter
etílico, insolúvel em água a frio, sendo ligeiramente solúvel em água em ebulição
promovendo uma coloração alaranjada. A solução dissolve-se prontamente em
soluções alcalinas; na presença de hidróxido de sódio, forma um sal sódico que
confere às soluções aquosas uma coloração vermelha brilhante. Esta forma é
extremamente ativa como corante (FONSECA; BRAGA; SANTANA, 2003).
Derivados nitrogenados do lapachol
Atualmente, a exploração da reatividade química da cadeia lateral do
lapachol, tem sido realizada para aumentar a atividade biológica, a exemplo da
susbstituição da hidroxila do lapachol por aminas que vem sendo um dos alvos de
interesse dos pesquisadores. As substituições diretas feitas com simples adição de
aminas secundárias apropriadas à temperatura ambiente tem bons rendimentos,
resultam em um aumento da atividade citotóxica em relação ao lapachol. Estas
substâncias são identificadas neste trabalho com os códigos L-01 a L-08.

48
Figura 9. Formação de enamina a partir do lapachol (BARBOSA, 2006)
Atividades Biológicas
Atividade Anticarcinogênica - Vários estudos mostraram ser positiva a
atividade anticancerígena do lapachol. Atualmente no Brasil o lapachol é
comercializado sob a forma de cápsulas gelatinosas indicadas para pacientes
portadores de várias neoplasias malignas, como carcinoma epidermóide do colo
uterino, carcinoma epidermóide do assoalho da boca, adenocarcinoma do aparelho
digestivo, além de auxiliar na quimioterapia (FONSECA; BRAGA; SANTANA, 2003).
Os primeiros estudos antineoplásicos para o lapachol foram desenvolvidos
por Hatwell em 1967. Estes estudos mostraram uma alta atividade antineoplásica do
lapachol sobre o sarcoma de Yoshida (86% de inibição). Posteriormente, foram
pesquisadas as suas ações sobre o carcinoma de Walker 256 e contra o
linfosarcoma de Murphy Sturm. O sal sódico do lapachol é ativo contra leucemia
L1210 e o lapachol também mostrou atividade contra carcinoma ascitíco de Ehrlich.
Sabe-se que a concentração plasmática do lapachol necessária para atividade
contra alguns tipos de câncer (ex: carcinoma de Walker256) é de 30 a 25 µg/mg,
mas que nunca foi alcançada devido o surgimento de efeitos tóxicos (FONSECA;
BRAGA; SANTANA, 2003).
Objetivando o estudo apropriado da relação entre a estrutura química do
lapachol e sua atividade farmacológica, alguns autores observaram que a
substituição dos grupamentos metil por cloro ou bromo, resulta em compostos com
muito boa atividade anticancerígena. Ao contrário de outras drogas anticâncer do
tipo benzantraquinona, que estimulam a peroxidação lipídica dependente de
NADPH: citocromomo P450 redutase microssomal, o lapachol inibe por um
mecanismo potente e desconhecido. O lapachol, como um antagonista da vitamina
K (efeito hipoprotrobinêmico), deve usar todas as reações vitamina K-dependentes,
incluindo a interação 6as6 – AX1 e então bloquear as transduções dos sinais que

49
estimulam a produção em tumores onde o oncogene AX1 é expresso. O lapachol
pode ser um agente anticâncer oncogene – especifico. Isso reacende a possibilidade
do uso do lapachol como agente anticâncer (FONSECA; BRAGA; SANTANA, 2003).
Atividade antiinflamatória
Extratos hidroalcoóticos do líber de Tabebuia spp. foram utilizados para o
tratamento de cervicites e cervico-vaginites crônicas com ação antinfamatória e
cicatrizante. Também foram utilizados no tratamento de úlceras de perna, bursites,
tendinites, sinusites e otites agudas e crônicas. Formulações tópicas do lapachol
mantém suas propriedades antiinflamatórias e analgésicas (FONSECA; BRAGA;
SANTANA, 2003; LIRA et al., 2004).
Atividade antimicrobiana
Desde os primeiros trabalhos sobre o lapachol que sua atividade
antimicrobiana é estudada, observando-se uma predominância contra certos
microrganismos Gram-positivos e ácidos resistentes e uma baixa atuação contra
Gram-negativos, exceto contra o gênero Brucella. Verificou-se também que, a
medida que se eleva o grau de pureza do lapachol, diminui-se progressivamente a
sua atividade antimicrobiana, o que pode ser explicado pelo fato de que outras
naftoquinonas extraídas junto com o lapachol, também possuem essa atividade. Nos
testes de atividade fungicida observou-se que a β-lapachona tem uma maior
eficiência do que o lapachol. No total, tanto a β-lapachona quanto o lapachol foram
mais ativos que o cetoconazol. Também tem sido estudada sua atividade antibiótica
contra vírus e atividade tripanosomicida, antimalárica e moluscicida(FONSECA;
BRAGA; SANTANA, 2003; DA SILVA et al., 2003).
Atividade inibidora de sistemas celulares reparadores
Nesse processo as quinonas naturais, cujo principal destaque das
naftoquinonas é o lapachol, atuam de diferentes formas. Como exemplo, destaca-se
o estresse oxidativo que provocam ao induzirem a formação deletéria endógena de
espécies bioativas derivadas do oxigênio (O2-, OH e H2O2) como ocorre no
Trypanosoma cruzi, agente causador da doença de chagas. Outra atividade
marcante destas substâncias, descoberta um tanto recentemente, é a inibição do
complexo das topoisomerases, ação que provoca o desenvolvimento da apoptose
celular (suicídio celular). A interferência das quinonas na apoptose, constitui-se hoje
uma pesquisa interdisciplinar de fronteira na química medicinal, existindo grande

50
expectativa quanto a delineação de estratégias racionais, visando o combate de
neoplasias, principalmente as relacionadas ao câncer de próstata. A química das
quinonas, já há muito vem sendo descrita em vários livros publicados, em inúmeras
e excelentes revisões, estando assim bem documentada sua evolução ao longo do
tempo. Destaca-se no grupo das naftoquinonas a β-lapachona com acentuada
atuação tanto na química como na farmacologia e o lapachol, outrora comercializado
para o tratamento de certos tipos de câncer, como coadjuvante, fabricado pelo
LAFEP (Laboratório Farmacêutico do Estado de Pernambuco) não mais disponível
no mercado. Esta naftoquinona também já constou do catálogo da Aldrich Chemical
Co, em edição antiga (1989). Também não consta nas últimas edições do Índice
Merck (DA SILVA; FERREIRA; SOUZA, 2003).
O principal interesse no lapachol reside em sua capacidade de induzir o
estresse oxidativo através da formação intracelular de espécies reativas do oxigênio
como o peróxido de hidrogênio (H2O2), o ânion radical superóxido (O2-) e o radical
hidroxila (OH). Estas espécies podem danificar alguns componentes celulares
importantes, tanto de células normais como de malignas. Esta interferência
xenobiótica altera o balanço natural de sinais que interferem na divisão celular em
pontos específicos da evolução morfogênica natural (checkpoint ou ponto de
checagem). A alteração da normalidade pode induzir a apoptose como alternativa,
caso não se consiga eliminar por completo o estresse oxidativo. Estudos realizados
indicam que o estresse oxidativo induzido pelo Lapachol, ocorre na enzima P450
redutase. Neste processo espécies reativas do oxigênio, promovem a cisão do DNA.
Este tipo de mecanismo de ação é importante, pois alguns microrganismos
patogênicos são muito mais sensíveis ao estresse oxidativo que os hospedeiros
humanos (DA SILVA; FERREIRA; SOUZA, 2003).
A β-lapachona, exibe in vitro variados tipos de atividade contra diferentes
linhagens de células, principalmente células malignas dos cânceres de pulmão,
mama, colo retal, próstata, melanoma e leucemia. Apesar do amplo espectro de
bioatividades os mecanismos de ação da β-lapachona em modelos experimentais
ainda não estão bem delineados. Além do estresse oxidativo, também se observam
vias biorredutivas alquilantes sobre ácidos nucléicos e/ou protéicos. Entretanto, a
atuação inibitória da β-lapachona sobre sistemas reparadores parece não ser o seu
único modo de atuação uma vez que fungos que não expressam topoisomerase I
também são inibidos por essa quinona. Ao que parece a β-lapachona é um

51
xenobiótico de atuação critica sobre mais de um alvo intracelular que não só a
topoisomerase I. Com as informações disponíveis na literatura pode-se apontar para
uma ação de indução da apoptose através da inibição do complexo topoisomerase –
DNA. A possibilidade de uma ação direta da β-lapachona no ciclo catalítico das
enzimas topoisomerases I e II, assim como a geração endógena de O2־ e H2O2, são
sinais decisivos que desencadeiam a apoptose (DA SILVA; FERREIRA; SOUZA,
2003).
Efeitos colaterais e toxicidade
Apesar de sua baixa toxicidade, o lapachol apresenta alguns efeitos como a
anorexia, náusea e mais raramente vômitos, o que pode ser facilmente controlado
com antieméticos. Uma prolongação reversível no tempo de protrombina é
observada a doses elevadas o que é revertido com a administração parenteral de
vitamina K. Apesar da β-lapachona possuir atividades antineoplásica e antibiótica
superiores às do lapachol, bem como atividade antiviral, esse isômero apresenta-se
20 vezes mais tóxico que o Lapachol. A dose de 0,25g de Lapachol/kg administrada
diariamente, via oral durante 28 dias, é considerada a máxima dose tolerada em
macacos e cachorros. A administração de 100 mg/kg de Lapachol em ratas prenhes,
mostrou uma grande incidência de reabsorção do ovo por ação blastocistóxica –
antizigótica e uma atividade abortiva, levando a malformação (FONSECA; BRAGA;
SANTANA, 2003)
De todo modo, não se deve considerar a toxicidade elevada da β-lapachona
como um fator limitante ao uso clínico. Novas expectativas colocam-se nos estudos
futuros da relação atividade biológica versus estrutura química, uma área de atuação
aberta à química medicinal, que pode levar-se à solução de problemas de
toxicidade. (DA SILVA; FERREIRA; SOUZA, 2003).

M A T E R I A L E M É T O D O S

53
4. MATERIAL E MÉTODOS 4.1. Local da Pesquisa
As atividades de pesquisa foram desenvolvidas nos Laboratórios de Micologia
(DCF/CCS/UFPB), Genética Microbiana (CCEN/UFPB) e Microbiologia (LAC/UEPB),
onde funciona o Laboratório de Pesquisas em Atividades Antimicrobianas do
Programa de Qualificação Institucional – PQI da UEPB. 4.2. Produtos testados As substâncias testadas, lapachol e derivados, foram obtidas no Laboratório
de Tecnologia Farmacêutica (LTF/UFPB) e gentilmente cedidas pelo Profº Dr. Celso
Amorim Câmara. As mesmas foram sintetizadas conforme procedimentos descritos
na literatura (CÂMARA et al, 2001; CAMARA et al, 2002, BARBOSA et al, 2005;
SILVA et al, 2005; BARBOSA, 2006; BARBOSA-FILHO et al, 2006) São elas:
Lapachol e seus derivados semi-sintéticos -lapachona, -nor-lapachona, -I-
lapachona, -lapachona, nor-lapachona, -I-lapachona e seus derivados
nitrogenados obtidos por semisíntese: 2-amino-1,4-naftalenodiona (L-01), 2-(2,2-
dimetoxietilamino)-3-(3-metil-2-butemil)-1,4-naftalenodiona (L-02), 1-(2,2-dimetoxi-
etil)-2,2-dimetil-1,2-diidro-benzo[g]quinolina-5,10-diona (L-03), 1-benzil-2,2-dimetil-
1,2-diidro-benzo[g]quinolina-5,10-diona (L-04), 2-(2-hidroxietilamino)-3-(3-metil-2-
butemil)-1,4-naftalenodiona (L-05), 4-(1-hydroxi-1-metil-etil)-4,5-diidro-1H-nafto[2,3-
b]azepina-6,11-diona (L-06), N-[1,4-diidro-3-(3-metil-2-butenil)-1,4-dioxo-2-naftalenil]-
glicina (L-07), 2-amino-3-(3-metil-2-butemil)-1,4-naftalenodiona (L-08).
As soluções apresentando diferentes concentrações dos produtos foram
obtidas, da seguinte forma: Em um tubo de ensaio (15 x 16 mm) estéril foi colocado
12mg do produto; 0,04mL de Tween 80; 50µl de Dimetil sulfóxido (DMSO) e 6,0 mL
de água destilada estéril. Essa mistura foi homogeneizada por aproximadamente 5
minutos. A partir de tal procedimento, obteve-se uma solução com concentração
final de 200mg/mL (solução padrão). Seguindo-se o processo de diluições seriadas,
preparou-se uma série de 3 tubos de ensaio onde cada um continha 2,5 mL de água
destilada estéril. A partir da solução (padrão), as diluições seriadas foram realizadas,
transferindo-se 2,5 mL dessa diluição para o primeiro tubo de ensaio da série,
homogeneizou-se por 5 vezes e transferiu-se 2,5 ml para o tubo seguinte e assim
sucessivamente até o último tubo da série, obtendo-se soluções com as seguintes
concentrações: 100, 50 e 25 µg/mL.

54
O
O
OH
Lapachol
O
O
OH
Norlapachol
O
O
O
-Norlapachona
O
O
O
I
3-Iodo--lapachona
Der
ivad
os s
emi-s
inté
ticos
do
Lapa
chol
O
OI
O
3-Iodo--
lapachona
O
O
O
-Lapachona
O
O
O
-Lapachona
OO
O
-Norlapachona
O
O
NH2
L01
2-Amino-1,4- naftalenodiona
O
O
NHCH2CH(OCH3)2
L02
2-(2,2-Dimetoxietilamino)-3-(3-
metil-2-butenil)-1,4-naftalenodiona
O
O
N
CH2CH(OCH3)2
L03
1-(2,2-Dimetoxi-etil)-2,2-dimetil-1,2-diidro-
benzo[g]quinolina-5,10-diona
O
O
N
Ph
L04 1-Benzil-2,2-dimetil-
1,2-diidro-benzo[g]quinolina-
5,10-diona
Der
ivad
os n
itrog
enad
os d
o La
pach
ol
O
O
NHCH2CH2OH
L05
2-(2-Hidroxietilamino)- 3-(3-metil-2-butenil)- 1,4-naftalenodiona
O
O
NH
OH L06
4-(1-Hidroxi-1-metil-etil)-4,5-diidro-1H-
nafto[2,3-b]azepina-6,11-diona
O
O
NHCH2CO2H
L07
N-[1,4-diidro-3-(3-metil-2-butenil)-1,4-dioxo-2-naftalenil]-
glicina
O
O
NH2
L08
2-Amino-3-(3-metil-
2-butenil)-1,4-naftalenodiona
Figura 10. Estruturas químicas do lapachol e seus derivados

55
4.3. Substâncias
Tween 80 (Sigma Chemical)
DMSO – Dimetilsulfóxido (Merck)
Água Destilada
Solução salina (NaCl 0,85%)
Antibióticos
- Tetraciclina (Lote: 1112, Fornecedor DGE. Origem: China)
- Penicilina G (Laboratório de genética de microrganismos da UFPB)
- Estolato de eritomicina (Lote: 200592, Fornecedor: AMT do Brasil. Origem:
Índia)
- Sulfato de estreptocimina (Lote: 826.545 Fornecedor: INLAB – Brasil)
- Discos de Antibióticos (Cefar) - Antibiograma
4.4. Espécies microbianas
Foram utilizadas neste trabalho, amostras de Staphylococcus aureus,
meticilina-sensíveis (MSSA) e S.aureus meticilina-resistentes (MRSA), Escherichia
coli e Candida albicans, além de cepas padrão American Type Culture Collection
(ATCC), apresentadas na Tabela 1.
Tabela 1. Identificação e origem das cepas microbianas testadas
Cepas/Identificação Origem/Fornecedor S.aureus – MSSA: ATCC 25923 Cefar
S.aureus – MRSA: 171c Humana/ Lab. Genética - UFPB
S. aureus: orofaringe (8, 18, 43, 44, 45, 46, 47, 48, 49, 50) Humana/ Diagnose
S. aureus: fossas nasais (129 FN, 233 FN, 322 FN) Animal / Lab. Genética - UFPB
S.aureus: úbere (122U, 146U, 223U, 312U, 319U) Animal / Lab. Genética - UFPB
S. aureus: ambiente hospitalar (1A e 2A) Diagnose
E. colii ATCC 25922 Cefar
E.coli: infecção urinária (1, 2, 3, 4, 5, 6, 7, 8, 9, 10) Humana/ Diagnose
P.aeruginosa ATCC 27853 Cefar
C.albicans ATCC 76643 Cefar
C. albicans spp Humana / Lab. Micologia - UFPB
C. albicans: (1, 2, 3, 4, 5, 6) Humana / Lab. Micologia - UFPB

56
4.4.1. Isolamento e Identificação dos microrganismos
Para os ensaios foram utilizados os meios Agar Sangue, Agar Manitol
Salgado e Agar Müeller-Hinton (BBL-difco). O Agar Sangue foi utilizado para verificar
a pureza e a capacidade hemolítica das cepas bacterianas e o Agar Manitol Salgado
para verificação da capacidade de utilização do manitol pelas cepas de S.aureus. O
Agar Müeller-Hinton foi utilizado para a realização do Teste de Sensibilidade aos
Antimicrobianos-TSA, por ser o meio de cultura padrão de acordo com Bauer et al.
(1966) e NCCLS (2002).
As cepas de S. aureus de origem humana e ambiental, utilizadas nesse
trabalho foram adquiridas e identificadas por métodos microbiológicos tradicionais,
pelo Laboratório de Microbiologia da Universidade Estadual da Paraíba – UEPB e
pelo Setor de Bacteriologia da Diagnose-Clínica de Análises Especializadas – LTDA,
Campina Grande-Pb. As cepas de origem animal foram doadas da coleção do
Laboratório de Genética da UFPB e as cepas ATCC foram adquiridas através da
PROBAC do BRASIL.
Para a identificação bioquímica das cepas de S.aureus, foram realizados
testes rotineiros para detectar a produção das enzimas catalase, coagulase e
DNAse além de observar a utilização do manitol (fermentação e oxidação) e a
produção de hemólise em Ágar Sangue.
Para as leveduras foram selecionadas cepas de C. albicans, isoladas de
amostras clínicas de pacientes portadores de infecções superficiais e/ou profundas.
Os cultivos foram realizados em Agar Sabouraud Dextrose (ASD), incubados a 35°C
e observados por um período de 2 a 3 dias, a fim de evidenciar o crescimento
fúngico. As características macromorfológicas, microscópicas e bioquímicas dos
cultivos foram estudadas segundo a chave de identificação proposta por GUÉHO et
al. (1994).
4.4.2. Preparo da suspensão microbiana
Durante a realização do ensaio, as cepas foram mantidas em meio Agar
Müeller-Hinton (difco) e repicadas para caldo Brain Heart Infusion - BHI (BBL-difco) e
incubadas a 37ºC/24 horas. Após este período, procedeu-se o semeio pela técnica
de estrias (para obtenção de colônias isoladas) em placas de Agar Sangue, que
foram incubadas a 37ºC/24 horas. Com relação a obtenção do inóculo bacteriano,

57
foram selecionadas 3 a 5 colônias semelhantes as quais foram transferidas para
5,0mL de caldo BHI e incubadas a 37ºC durante 24 horas. Após este período, foi
feito o subcultivo, transferindo 50µl do inóculo inicial para 30mL de caldo Müeller-
Hinton, incubando-o à 37ºC/1h de modo a produzir uma leve turvação, de densidade
visualmente equivalente ao tubo 0,5 da escala de McFarland, obtendo-se assim um
inóculo de concentração final de 106 UFC/mL. Essa suspensão, foi semeada no
tempo máximo de 15 a 20 minutos após sua preparação.
Quanto as leveduras, as culturas foram mantidas em Agar Sabouraud
Dextrose (ASD) a 35°C durante 24-72 horas. A suspensão de levedura foi preparada
e padronizada em solução de cloreto de sódio a 0,85% estéril. Foi obtida uma
suspensão com turvação comparativa com a do tubo 0,5 da escala de McFarland. A
mesma foi ajustada no espectrofotômetro (Leitz-Photometer 340-800), para conter
aproximadamente uma concentração final de 106 UFC/mL (BAUER et al., 1966;
CASALS, 1979; FROMTLING; PUI-YU; SHADOMY, 1983; PLEMPEL et al., 1986;
DRUTZ, 1987; ODDS et al., 1989: NASSIS et al., 1989; CLEELAND; SQUIRES,
1991; ELLIS, 1994; NCCLS, 2002).
4.5. Meios de cultura
Para a manutenção dos microrganismos e ensaios de atividades biológicas,
foram usados os meios de cultura Blood Agar Base (BAB), Agar Sangue (AS), Agar
Manitol Salgado (MAS), Agar Mueller-Hinton (AMH), Caldo Brain Heart Infusion (BHI)
e Caldo Mueller-Hinton (CMH) para bactérias; e Agar e caldo Sabouraud Dextrose –
ASD para fungos. Os meios de cultura (difco), foram preparados conforme as
instruções do fabricante.
4.6. Metodologia
4.6.1. Determinação da atividade antimicrobiana – triagem
Com relação a determinação da sensibilidade dos microrgansmos frente ao
lapachol e seus derivados sobre microrganismos, foi realizado inicialmente a triagem
da atividade antimicrobiana pelo método de difusão em meio sólido, processo
cavidade-placa (BAUER et al., 1966; CLEELAND; SQUIRES, 1991; NCCLS, 2002).

58
Quanto a realização do teste de difusão, foram utilizadas placas de Petri (90 x
15 mm) descartáveis, estéreis, contendo 20mL de Agar Müeller-Hinton, que foram
inoculadas pela técnica de espalhamento em superfície (Figura 11). Com auxílio de
"swabs" estéreis, mergulhados na suspensão, foi retirado o inóculo, eliminando-se o
excesso do líquido por pressão nas paredes do tubo. O inóculo foi semeado em toda
a superfície do agar, de modo a se obter um crescimento uniforme e semi-
confluente.
As placas foram colocadas para secar, durante 3 a 5 minutos, antes de se
fazer as cavidades de 6 mm de diâmetro, com o auxílio de perfuradores descartáveis
estéreis. Em cada cavidade foi adicionado 50 l da solução de cada produto testado
nas concentrações de 200g/mL, 100g/mL, 50g/mL e 25g/mL, a fim de verificar
a presença ou não de atividade antimicrobiana. Foi utilizado como controle em uma
das cavidades, 50L da solução de DMSO a 2% usada como solvente. As placas
foram incubadas a 37°C por 24 horas e após este período, foram medidas, em
milímetros, as zonas (halos) de inibição de crescimento bacteriano. Os ensaios
foram realizados em triplicata e o resultado final foi determinado pela média
aritmética do tamanho dos halos de inibição (mm) dos valores obtidos nos ensaios.
Na triagem para a avaliação da atividade antimicrobiana, foi considerado como teste
positivo quando a média dos halos foi igual ou superior a 10 mm de diâmetro
(DROUHET; DUPONT, 1978; GUNDIDZA, 1986). E no caso das leveduras, 1 mL do
inoculo foi colocado nas placas de Petri (90x15 mm) estéreis e, em seguida, foi
adicionado 20 mL do agar Sabouraud dextrose liquefeito à 50°C. As placas foram
homogeneizadas e deixadas em repouso até a solidificação (WONG-LEUNG, 1988;
SAKAR, 1988).
Figura 11. Técnica de difusão em meio sólido, processo cavidade-placa.
Placa contendo meio de cultura Placa contendo
cavidades de 6mm de diâmetro +
50l dos produtos incubadas 24h/37°C "Swab" com o inóculo: 10
6 UFC/mL -
tubo 0,5 da escala de Mac Farland
Zonas de inibição indicam atividade
antimicrobiana

59
4.6.2. Determinação da Concentração Inibitória Mínima (CIM), Concentração
Bactericida Mínima (CBM) e Concentração Fungicida Mínima (CFM)
Os testes para a determinação da CIM, da CBM e CFM foram realizados por
difusão em meio sólido, processo cavidade-placa conforme os protocolos de Bauer
et al (1966); CLEELAND; SQUIRES, 1991; HADACEK; GREGER (2000); NCCLS
(2002).
As placas de Petri (90x15 mm), contendo 20mL do meio de cultura ágar
Müeller-Hinton, foram inoculadas pela técnica de espalhamento em superfície, com
auxílio de "swabs" estéreis mergulhados na suspensão bacteriana para retirar o
inóculo. E no caso das leveduras, utilizou-se o meio de cultura agar Sabouraud
dextrose. Em cada placa foram feitas cavidades, de 6mm de diâmetro, com o auxílio
de perfuradores descartáveis estéreis. Nesses orifícios foram colocados 50l dos
produtos, para verificar a presença ou não de atividade antimicrobiana. As placas
foram incubadas à 35°C por 24 horas e após este período, foram medidas, as zonas
de inibição, em milímetros. Cada ensaio foi realizado em triplicata, para cada cepa
selecionada. O resultado final foi determinado pela média aritmética do tamanho dos
halos de inibição (mm) dos valores obtidos nos três ensaios.
A CIM foi considerada como a menor concentração do produto que inibiu o
crescimento visível dos microrganismos após 24h/37°C de incubação para bactérias
e 24-48h/35ºC, para as leveduras. A CBM e CFM foram consideradas como a
concentração do produto imediatamente superior à da CIM (NCCLS, 2002).
4.6.3. Determinação da cinética do crescimento bacteriano
Foram determinadas as curvas de crescimento do lapachol e seus derivados
sobre amostras selecionadas de S. aureus, estabelecendo-se a concentração
relacionada à CIM, em diferentes tempos de incubação, segundo o método proposto
por Peyret et al, (1990). As amostras foram inoculadas em caldo nutritivo (BHI),
incubadas a 37ºC por 18 à 24h. Após este período, foram cultivadas, transferindo-se
50 µL do inóculo inicial para 50 mL de caldo Mueller-Hinton, e incubando-o à
37ºC/1h de modo a produzir uma leve turvação, de densidade visualmente
equivalente ao tubo 0,5 da escala de McFarland, com concentração final de 10-6
UFC/mL. Transferiu-se 9,0mL desta cultura bacteriana e adicionou-se 1,0 mL do
produto a ser testado e no tubo controle foi adicionado 1,0 mL de água destilada
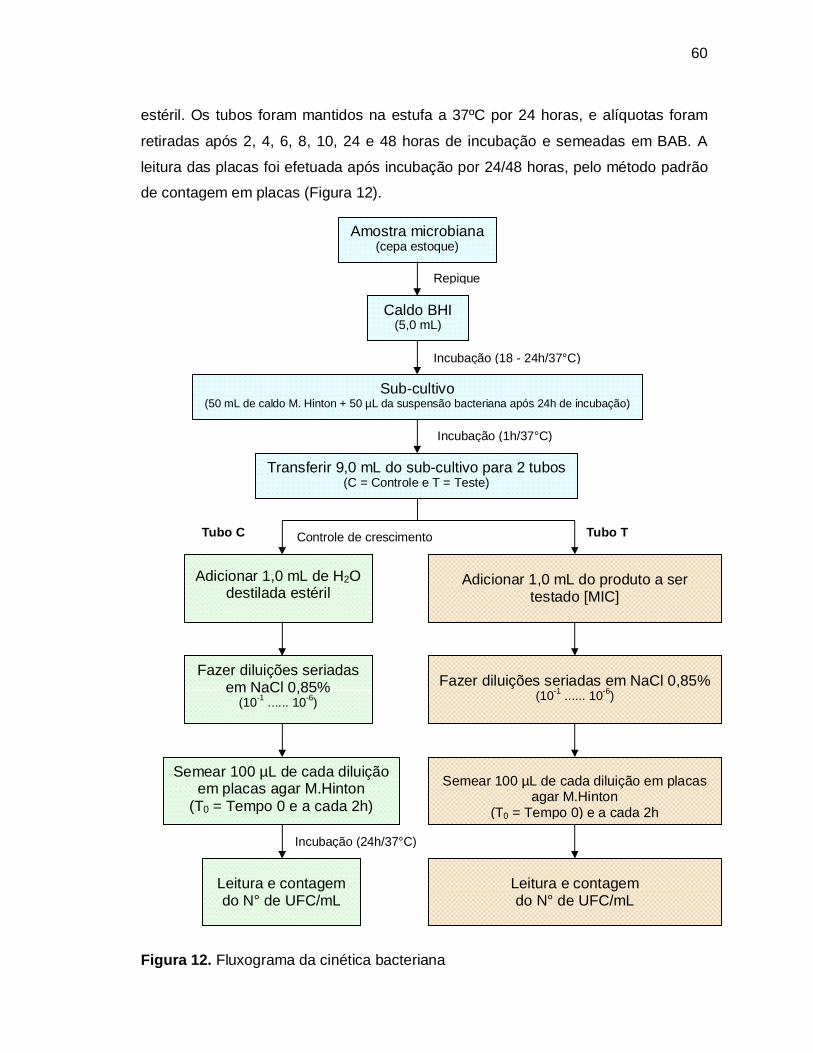
60
estéril. Os tubos foram mantidos na estufa a 37ºC por 24 horas, e alíquotas foram
retiradas após 2, 4, 6, 8, 10, 24 e 48 horas de incubação e semeadas em BAB. A
leitura das placas foi efetuada após incubação por 24/48 horas, pelo método padrão
de contagem em placas (Figura 12).
Figura 12. Fluxograma da cinética bacteriana
Amostra microbiana (cepa estoque)
Caldo BHI (5,0 mL)
Sub-cultivo (50 mL de caldo M. Hinton + 50 µL da suspensão bacteriana após 24h de incubação)
Transferir 9,0 mL do sub-cultivo para 2 tubos (C = Controle e T = Teste)
Adicionar 1,0 mL de H2O destilada estéril
Fazer diluições seriadas em NaCl 0,85%
(10-1 ...... 10-6)
Fazer diluições seriadas em NaCl 0,85% (10-1 ...... 10-6)
Semear 100 µL de cada diluição em placas agar M.Hinton
(T0 = Tempo 0 e a cada 2h)
Leitura e contagem do N° de UFC/mL
Semear 100 µL de cada diluição em placas agar M.Hinton
(T0 = Tempo 0) e a cada 2h
Leitura e contagem do N° de UFC/mL
Adicionar 1,0 mL do produto a ser testado [MIC]
Incubação (18 - 24h/37°C)
Tubo C
Repique
Incubação (1h/37°C)
Tubo T Controle de crescimento
Incubação (24h/37°C)

61
4.6.4. Determinação da cinética fúngica
Foram determinadas as curvas de morte microbiana após adição dos
produtos sobre a amostra selecionada, Candida albicans ATCC 76643,
estabelecendo-se concentração relacionada à concentração inibitória mínima (CIM),
em diferentes tempos de incubação, segundo o método proposto por Peyret et al,
(1990).
As amostras foram inoculadas em caldo Sabouraud e incubadas a
35ºC/24h. Após este período, foram subcultivadas, transferindo-se 50 µL do inóculo
inicial para 50 mL de caldo Sabouraud, incubando-o à 35ºC/1h de modo a produzir
uma leve turvação, de densidade visualmente equivalente ao tubo 0,5 da escala de
McFarland, com concentração final de 106 UFC/mL. Transferiu-se 9,0 mL desta
cultura fúngica para um tubo teste (T) e um tubo controle (C), adicionando-se 1.0mL
do produto a ser testado no tubo T e 1,0 mL de água destilada estéril, no tubo C.
Placas de agar Sabouraud dextrose (ASD), foram imediatamente semeadas com
0,1 mL dessas suspensões fúngicas, usando-se alças calibradas estéreis. As placas,
previamente identificadas em T e C (Tempo Oh) foram incubadas a 35ºC/24-48h. Os
tubos T e C foram mantidos na estufa a 35ºC por até 48 horas, e alíquotas foram
retiradas após 2, 4, 6, 8, 10, 24 e 48 horas de inoculação das substâncias e
semeadas em placas de ASD, as quais foram lidas após incubação por 35ºC/24-48
horas, pelo método padrão de contagem em placas (Figura 13).

62
Figura 13. Fluxograma da cinética fungica
Amostra microbiana (cepa estoque)
Caldo Sabouraud
(5,0 mL)
Sub-cultivo (50 mL de caldo Sabouraud + 50 µL da suspensão bacteriana após 24h de incubação)
Transferir 9,0 mL do sub-cultivo para cada tubo (C = Controle e T = Teste)
Adicionar 1,0 mL de H2O destilada estéril
Fazer diluições seriadas em NaCl 0,85%
(10-1 ...... 10-6) Fazer diluições seriadas em NaCl 0,85%
(10-1 ...... 10-6)
Semear 100 µL de cada diluição em placas ASD
(T0 = Tempo 0 e a cada 2h)
Leitura e contagem do N° de UFC/mL
Semear 100 µL de cada diluição em placas ASD
(T0 = Tempo 0) e a cada 2h
Leitura e contagem do N° de UFC/mL
Adicionar 1,0 mL do produto a ser testado [CIM]
Incubação (24h/35°C)
Tubo C
Repique
Incubação (1h/35°C)
Tubo T Controle de crescimento
Incubação (24h – 48h/35°C)

63
4.6.5. Caracterização fenotípica dos padrões de resistência aos antimicrobianos
A determinação do teste de sensibilidade aos antimicrobianos (Antibiograma)
foi realizada pelo método de disco-difusão (BAUER et al., 1966) usando-se discos
de antibióticos da Cefar, seguindo-se as recomendações do fabricante e do NCCLS
(2002).
4.6.6. Tratamento por substratos sintéticos obtidos a partir do lapachol e seus
derivados sintéticos – avaliação da atividade curagênica
A avaliação da atividade curagênica dos substratos sintéticos obtidos a partir
dos produtos testados sobre S.aureus resistentes a drogas foi determinada
utilizando-se o valor da metade da CIM. Todas as amostras bacterianas foram
inoculadas em caldo BHI (Brain Heart Infusion, DIFCO) a 37ºC por 18-24 horas e
diluídas 100 vezes no mesmo meio adicionado da concentração subinibitória (1/2
CIM) de cada produto ensaiado. Após incubação a 37ºC/24horas, sob agitação, as
culturas foram convenientemente diluídas em solução salina esterilizada (NaCl
0,85%) e alíquotas de 0,1mL semeadas em BAB (Blood Agar Base, difco) para
determinação do título (concentração em que cresceram nas placas 30 a 300
colônias). A perda da resistência a drogas foi determinada por réplica para meios de
cultura, acrescidos de cada antibiótico estudado (LEDEBERG; LEDEBERG, 1952;
PEREIRA, 2000). As confirmações da ocorrência de eliminação de plasmídios nas
variantes possivelmente curadas foram em placas com meio sólido (BAB) contendo
os antibióticos selecionados: Penicilina e Eritromicina.
Observa-se a cura quando a placa contendo o antibiótico apresenta a
ausência de colônias variantes anteriormente resistentes presentes na placa matriz,
sem antibiótico, indicando que as mesmas possivelmente perderam a característica
fenotípica de resistência quando tratada com os produtos testados (Figura 14).
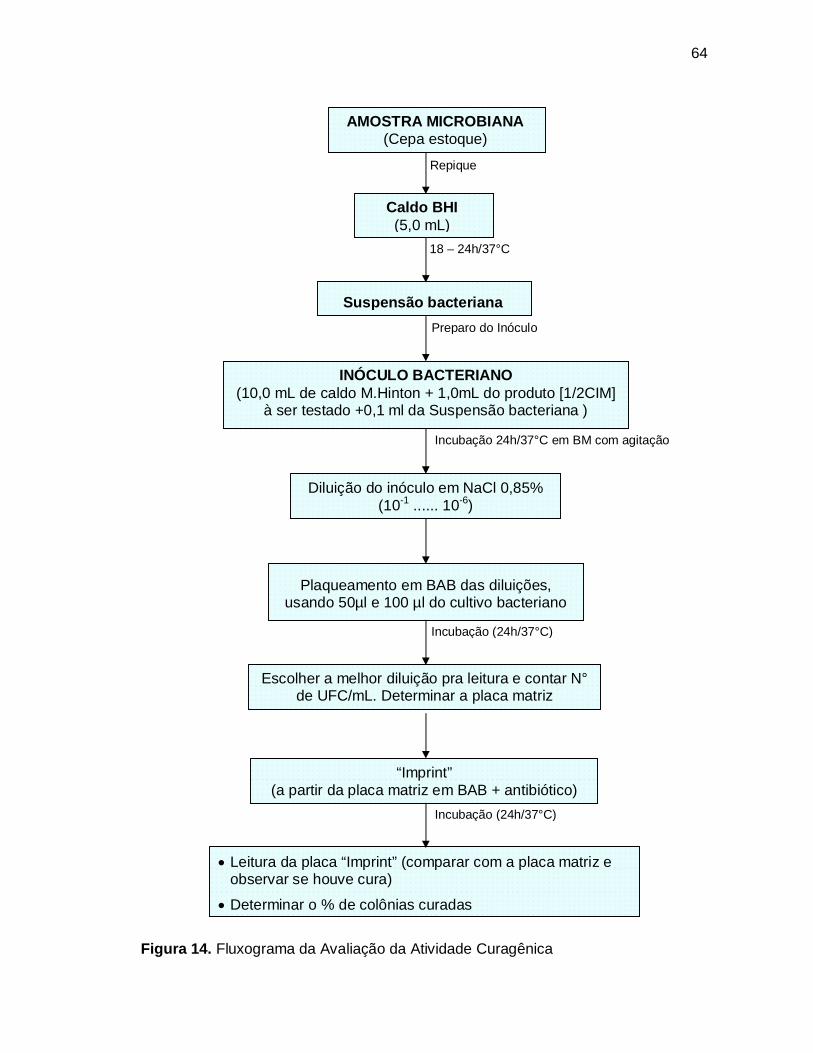
64
Figura 14. Fluxograma da Avaliação da Atividade Curagênica
Incubação (24h/37°C)
Incubação (24h/37°C)
Preparo do Inóculo
18 – 24h/37°C
Incubação 24h/37°C em BM com agitação
INÓCULO BACTERIANO (10,0 mL de caldo M.Hinton + 1,0mL do produto [1/2CIM]
à ser testado +0,1 ml da Suspensão bacteriana )
Diluição do inóculo em NaCl 0,85% (10-1 ...... 10-6)
Plaqueamento em BAB das diluições, usando 50µl e 100 µl do cultivo bacteriano
Escolher a melhor diluição pra leitura e contar N° de UFC/mL. Determinar a placa matriz
“Imprint” (a partir da placa matriz em BAB + antibiótico)
Leitura da placa “Imprint” (comparar com a placa matriz e observar se houve cura)
Determinar o % de colônias curadas
AMOSTRA MICROBIANA (Cepa estoque)
Repique
Caldo BHI (5,0 mL)
Suspensão bacteriana

65
Os experimentos controles para a determinação de eliminação espontânea de
plasmídios foram realizados anteriormente por Pereira & Siqueira (1995), utilizando
o método da diluição em placas caldo e por eletroforese em gel de agarose após lise
com lisostafina (GOERING; RUFF, 1983).
4.6.7. Relação estrutura-atividade
O estudo da estrutura-atividade foi realizado observando-se a relação da
mudança na cadeia do lapachol e a variação da sua atividade antimicrobiana, em
detrimento das mudanças estruturais (ANTUNES et al., 2006).

R E S U L T A D O S E D I S C U S S Ã O

67
5.0. RESULTADOS E DISCUSSÃO
5.1. Resistência Antimicrobiana
A resistência antimicrobiana é um dos principais problemas de saúde pública
mundial, despertando preocupação na busca contínua por novos antimicrobianos. A
potência efetiva dos antimicrobianos existentes vem sendo gradativamente reduzida
ou mesmo anulada pela emergência de cepas que se apresentam resistentes aos
mesmos. Essas cepas são freqüentemente envolvidas na etiologia de infecções
humanas e animais, além de serem também comumente encontradas em infecções
hospitalares (XU; LEE, 2001).
A versatilidade e flexibilidade genética das bactérias são fatores que
contribuem para eficiência do fenômeno de resistência que tem se disseminado
entre diferentes gêneros e espécies de bactérias (PONTES et al., 2004). É sobre
essa variabilidade genética que o homem vem, paradoxalmente, exercendo alta
pressão seletiva (hospital, comunidade, agropecuária, meio ambiente) favorecendo
os genótipos resistentes (CHARTONE-SOUZA, 2004). De modo que, os estudos de
produtos naturais e sintéticos bioativos têm-se mostrado como uma alternativa
terapêutica e várias pesquisas estão sendo realizadas no intuito de avaliar as
propriedades de atividade antimicrobianas de substâncias de origem vegetal e
sintéticas. Dentro dessa perspectiva, o interesse em produtos vegetais com
propriedades medicamentosas tem evoluído bastante em todo o mundo. Vários
trabalhos descritos na literatura buscam avaliar produtos de plantas que possuam
atividade antimicrobiana (THUILLE; FILLE; NAGL, 2003; VASCONCELOS, et al.,
2004; BRAGA et al., 2005).
Na Tabela 2 observa-se o perfil de sensibilidade das cepas de S. aureus
frente a 15 antimicrobianos usados rotineiramente na clínica médica humana e
veterinária. Observou-se ainda nesse estudo, que a maioria (15/22) das cepas
apresentou resistência a penicilina G e a ampicilina e que as cepas 129FN, 233FN,
322FN (origem animal) e as cepas 8 e 18 (origem humana) foram sensíveis a todos
os antimicrobianos testados. Apenas duas cepas 171c (humana) e 2A (ambiental)
apresentaram resistência a oxacilina, sendo classificadas como MRSA. Todas as
cepas foram sensíveis a vancomicina, cloranfenicol, ciprofloxacina e
amplicilina/sulbactan. Esse estudo prévio de avaliação do perfil de sensibilidade

68
permitiu mostrar que a diversidade fenotípica das cepas em relação aos
antimicrobianos tradicionais, independe da sua origem.
Das 22 cepas ensaiadas, 7 foram sensíveis a todos os antimicrobianos
testados, as demais apresentaram multirresistência que variou de 2 a 7
antimicrobianos, sendo 6 cepas resistentes apenas à penicilina e à ampicilina. As
cepas mais resistentes foram 171c, 46 e 2A, respectivamente, resistentes a 7, 6 e 5
tipos de antimicrobianos. As cepas resistentes à penicilina foram consideradas como
produtoras de plasmídio penicilinase (TRABULSI, 2006).
Tabela 2. Perfil de sensibilidade das cepas de S. aureus Frente a Antimicrobianos
ANTIMICROBIANOS TESTADOS RM
C
epas
PEN
ERI
TET
AM
P
VAN
OXA
AM
I
GEN
SFT
AM
P/S
UL
CIP
CLI
CLO
AZI
EST
n° a
ntib
iótic
os
122 U R R S R S S S S S S S S S I S 3 146 U R I S R S S S S S S S S S S S 2 223 U R S S R S S S S S S S S S S S 2 312 U R S S R S S S S S S S S S S S 2 319 U R I S R S S S S S S S S S S S 2
129 FN S S S S S S S S S S S S S S S 0 233 FN S S S S S S S S S S S S S S S 0 322 FN S S S S S S S S S S S S S S S 0
171c R R R R S R I S R S S S S R S 7 8 S S S S S S S S S S S S S S S 0
18 S S S S S S S S S S S S S S S 0 43 R S S R S S S S S S S S S S S 2 44 R I R R S S S S R S S S S S S 4 45 R I R R S S S S S S S R S S S 4 46 R I R R S S R R S S S S S S R 6 47 R I R R S S S S S S S S S S S 3 48 R S S R S S S S S S S S S S S 2 49 R R S R S S S S R S S S S S S 4 50 R R S R S S S S R S S S S S S 4
ATCC S S S S S S S S S S S S S S S 0 1A S S S S S S S S S S S S S S S 0 2 A R R R R S R S S S S S S S S S 5
Legenda: S=sensível, I=intermediário, R=resistente, PEN=Penicilina G, EST=Estreptomicina, ERI=Eritromicina, TET=Tetraciclina, AMP= Ampicilina, VAN= vancomicina, OXA= Oxacilina, AMI=Amicacina, GEN=Gentamicina, SFT=Sulfametoxazol-Trimetropim, AMP/SUL=Ampicilina/Sulbactam, CIP= Ciprofloxacim, CLI=Clindamicina, CLO=Cloranfenicol, AZI=azitomicina, U=úbere, FN=fossas nasais, c=clínica, A=ambiental, ATCC = American Type Culture Collection ; RM = Resistência Múltipla

69
5.2. Avaliação da atividade antimicrobiana do lapachol e seus derivados
A Tabela 3, apresenta os resultados da avaliação da atividade antimicrobiana
do lapachol e seus derivados. Esses produtos foram avaliados frente a cepas de
Candida albicans, Staphylococcus aureus, Escherichia coli, Pseudomonas
aeruginosa, usadas rotineiramente como padrão em estudos de atividade
antimicrobiana, de acordo com o NCCLS (2002).
Tabela 3. Avaliação da atividade antimicrobiana do lapachol e derivados, frente a microrganismos de origem ambulatorial e cepas ATCC
Derivados nitrogenados do lapachol (200µg/mL)
lapachol e derivados semi-sintéticos (200µg/mL)
L-01
L-03
L-04
L-05
L-06
L-07
L-08
Lapa
chol
α-la
pach
ona
β -n
or-
lapa
chon
a
β-la
pach
ona
α-no
r-la
pach
ona
β-I-
lapa
chon
a
α-I-
Lapa
chon
a
Microrganismos
Zonas de inibição (mm) Zonas de inibição (mm) P. aeruginosa ATCC
27853 0 0 0 0 0 0 0 11 16 19 14 0 12 13
E. coli ATCC 25922 0 0 0 0 0 0 0 0 0 0 0 0 0 0 S. aureus ATCC
25923 0 0 12 0 0 0 15 11 18 20 19 13 20 16 C. albicans ATCC
76645 0 0 0 0 0 0 0 0 0 0 0 0 0 0
C. guill LM 06 (03) 0 0 0 0 0 0 0 0 0 0 0 0 0 0
S. cerevisae (-) 0 0 0 0 0 0 0 0 0 13 10 0 0 0
C. tropicale (04) 0 0 0 0 0 0 0 0 0 0 0 0 0 0 C. tropicale LM 124
(05) 0 0 0 0 0 0 0 0 0 0 0 0 0 0
C. stella LM13 (6) 0 0 0 0 0 0 0 0 0 0 0 0 0 0 C. parapisi LM 1D
(07) 0 0 0 0 0 0 0 0 0 0 0 0 0 0
C. Stella LM 1 (08) 0 0 0 0 0 0 0 0 0 0 0 0 0 0 C. albicans LM 300
(01) 0 0 0 0 0 0 0 0 0 0 0 0 0 0
C. albicans 556 (02) 0 0 0 0 0 0 0 0 0 0 0 0 0 0
S. aureus 319U 0 0 0 0 0 0 0 0 12 0 26 0 25 0
S. aureus 122U 0 0 0 0 0 0 0 0 11 0 24 0 23 0
Os resultados demonstraram que o lapachol e seus derivados foram mais
ativos para S. aureus ATCC 25923 (Figura 15) e P. aeruginosa ATCC 27853,
corroborando as afirmações de Fonseca, Braga e Santana (2003). Estes autores
verificaram a predominância do lapachol com atividade antimicrobiana para
microrganismos G(+) e baixa atuação para os G(-), aqui confirmado com o teste para

70
a E. coli, onde os resultados dos ensaios foram negativos, ou seja, sem atividade
antimicrobiana.
Figura 15. Atividade antimicrobiana do lapachol (1), α-nor-lapachona(2); β-
lapachona (3); α-lapachona (4); β-nor-lapachona (5), frente a S. aureus ATCC 25923
A maioria das substâncias derivadas do lapachol mostrou potencial
antimicrobiano diante das cepas testadas. Para os derivados nitrogenados obtidos a
partir do lapachol, observou-se halos de inibição de crescimento apenas para L-04
(12 mm) e para L-08 (15 mm), frente a S. aureus ATCC 25923. As demais
substâncias desse grupo não apresentaram atividade antimicrobiana para nenhuma
das cepas testadas. Com relação às cepas de S. aureus 319U e S. aureus 122U,
apenas a -lapachona, -lapachona e -lapachona formaram halo de inibição.
Quanto às leveduras, apenas o -nor-lapachona e o ß-lapachona
apresentaram halo de inibição de crescimento respectivamente de 13 mm e 10 mm,
evidenciando atividade antifúngica frente a uma cepa de Saccharomices cerevisae.
De acordo com Fonseca, Braga e Santana (2003), esses resultados também se
mostram compatíveis uma vez que os autores supracitados afirmam ser a ß-
lapachona mais ativa que o próprio lapachol, dados também observados neste
ensaio. Os resultados dos testes preliminares realizados com as leveduras (Tabela
3) apontaram baixa atividade. Por esta razão, foram selecionadas apenas as
bactérias para os testes seguintes. Na Tabela 3, pode se observar que as cepas P.
aeruginosa ATCC 27853, S. aureus ATCC 25923, S. aureus 319U, S. aureus 122U e

71
S. cerevisae apresentaram-se sensíveis ao lapachol e derivados semi-sintéticos
(200g/ mL) cujos halos de inibição medem de 10 a 26 mm de diâmetro.
O lapachol e seus derivados semi-sintéticos apresentaram atividade
antimicrobiana significativa para as cepas S. aureus testadas. Uma possibilidade da
elucidação desse mecanismo, é a capacidade que essas substâncias possuem de
causar estresse oxidativo e apoptose. Segundo da Silva, Ferreira e Sousa (2003),
alguns microrganismos patogênicos são muito mais sensíveis ao estresse oxidativo
que os hospedeiros humanos. Da Silva, Ferreira e Sousa (2003) e Barbosa (2006),
afirmaram que lapachol e -lapachona podem danificar alguns componentes
celulares importantes por induzirem a formação deletéria endógena de ânion radical
superóxido (O2-), radical hidroxila (HO) e peróxido de hidrogênio (H2O2), os
chamados radicais livres, bem como a apoptose ao inibirem o complexo das
topoisomerases (lapachol e β-lapachona). Estes estudos podem direcionar novos
caminhos para continuidade das pesquisas nos alvos intracelulares que elucidariam
os mecanismos de ação dessas substâncias nas células microbianas.
5.3. Determinação da CIM e CBM
A CIM é a maior diluição do produto que conseguiu inibir o crescimento
microbiano, enquanto que a CBM é a diluição imediatamente anterior à CIM
representando a diluição bacteriostática do produto. Em relação à avaliação da
concentração inibitória mínima (CIM), foram utilizadas apenas as substâncias mais
ativas, que foram testadas frente a S. aureus ATCC 25923, conforme a Figura 16, E.
coli ATCC 25922 e P. aeruginosa ATCC 27853, cepas recomendadas pelo NCCLS
(2002), quando se avalia o perfil de atividade de antibióticos. Incluiu-se também uma
cepa de S. aureus multirresistente (S. aureus 171c MRSA) de origem humana, bem
como cepas com perfil plasmidial de resistência à antibióticos conhecido (S. aureus
319U e 122U) de origem animal.
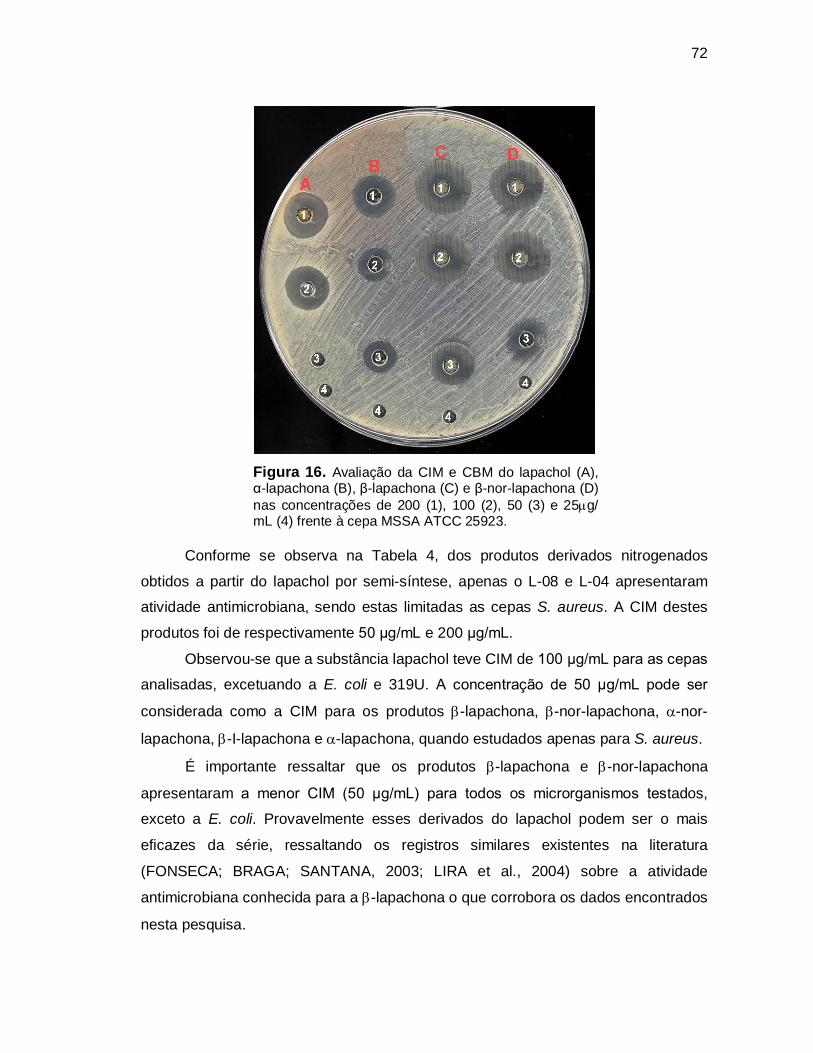
72
Figura 16. Avaliação da CIM e CBM do lapachol (A), α-lapachona (B), β-lapachona (C) e β-nor-lapachona (D) nas concentrações de 200 (1), 100 (2), 50 (3) e 25g/ mL (4) frente à cepa MSSA ATCC 25923.
Conforme se observa na Tabela 4, dos produtos derivados nitrogenados
obtidos a partir do lapachol por semi-síntese, apenas o L-08 e L-04 apresentaram
atividade antimicrobiana, sendo estas limitadas as cepas S. aureus. A CIM destes
produtos foi de respectivamente 50 µg/mL e 200 µg/mL.
Observou-se que a substância lapachol teve CIM de 100 µg/mL para as cepas
analisadas, excetuando a E. coli e 319U. A concentração de 50 µg/mL pode ser
considerada como a CIM para os produtos -lapachona, -nor-lapachona, -nor-
lapachona, -I-lapachona e -lapachona, quando estudados apenas para S. aureus.
É importante ressaltar que os produtos -lapachona e -nor-lapachona
apresentaram a menor CIM (50 µg/mL) para todos os microrganismos testados,
exceto a E. coli. Provavelmente esses derivados do lapachol podem ser o mais
eficazes da série, ressaltando os registros similares existentes na literatura
(FONSECA; BRAGA; SANTANA, 2003; LIRA et al., 2004) sobre a atividade
antimicrobiana conhecida para a -lapachona o que corrobora os dados encontrados
nesta pesquisa.
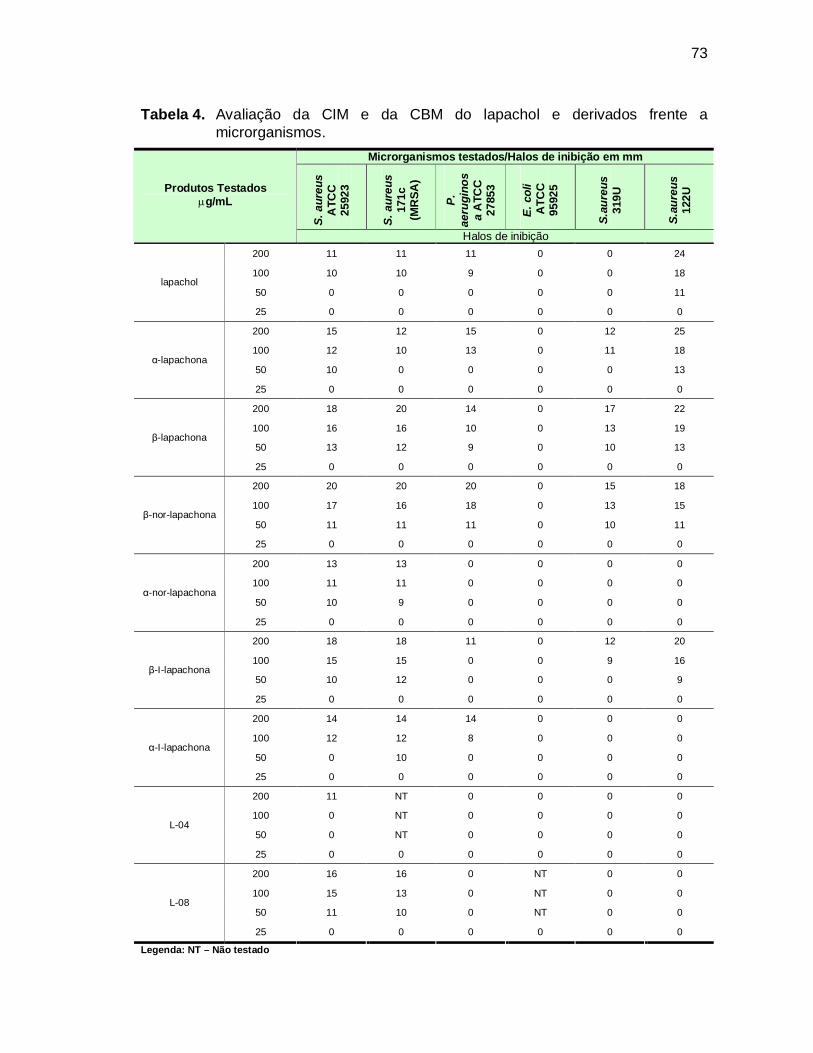
73
Tabela 4. Avaliação da CIM e da CBM do lapachol e derivados frente a microrganismos.
Microrganismos testados/Halos de inibição em mm
S. a
ureu
s A
TCC
25
923
S. a
ureu
s 17
1c
(MR
SA)
P.
aeru
gino
sa
ATC
C
2785
3
E. c
oli
ATC
C
9592
5
S.au
reus
31
9U
S.au
reus
12
2U Produtos Testados
g/mL
Halos de inibição 200 11 11 11 0 0 24
100 10 10 9 0 0 18
50 0 0 0 0 0 11 lapachol
25 0 0 0 0 0 0
200 15 12 15 0 12 25
100 12 10 13 0 11 18
50 10 0 0 0 0 13 α-lapachona
25 0 0 0 0 0 0
200 18 20 14 0 17 22
100 16 16 10 0 13 19
50 13 12 9 0 10 13 β-lapachona
25 0 0 0 0 0 0
200 20 20 20 0 15 18
100 17 16 18 0 13 15
50 11 11 11 0 10 11 β-nor-lapachona
25 0 0 0 0 0 0
200 13 13 0 0 0 0
100 11 11 0 0 0 0
50 10 9 0 0 0 0 α-nor-lapachona
25 0 0 0 0 0 0
200 18 18 11 0 12 20
100 15 15 0 0 9 16
50 10 12 0 0 0 9 β-I-lapachona
25 0 0 0 0 0 0
200 14 14 14 0 0 0
100 12 12 8 0 0 0
50 0 10 0 0 0 0 α-I-lapachona
25 0 0 0 0 0 0
200 11 NT 0 0 0 0
100 0 NT 0 0 0 0
50 0 NT 0 0 0 0 L-04
25 0 0 0 0 0 0
200 16 16 0 NT 0 0
100 15 13 0 NT 0 0
50 11 10 0 NT 0 0 L-08
25 0 0 0 0 0 0
Legenda: NT – Não testado
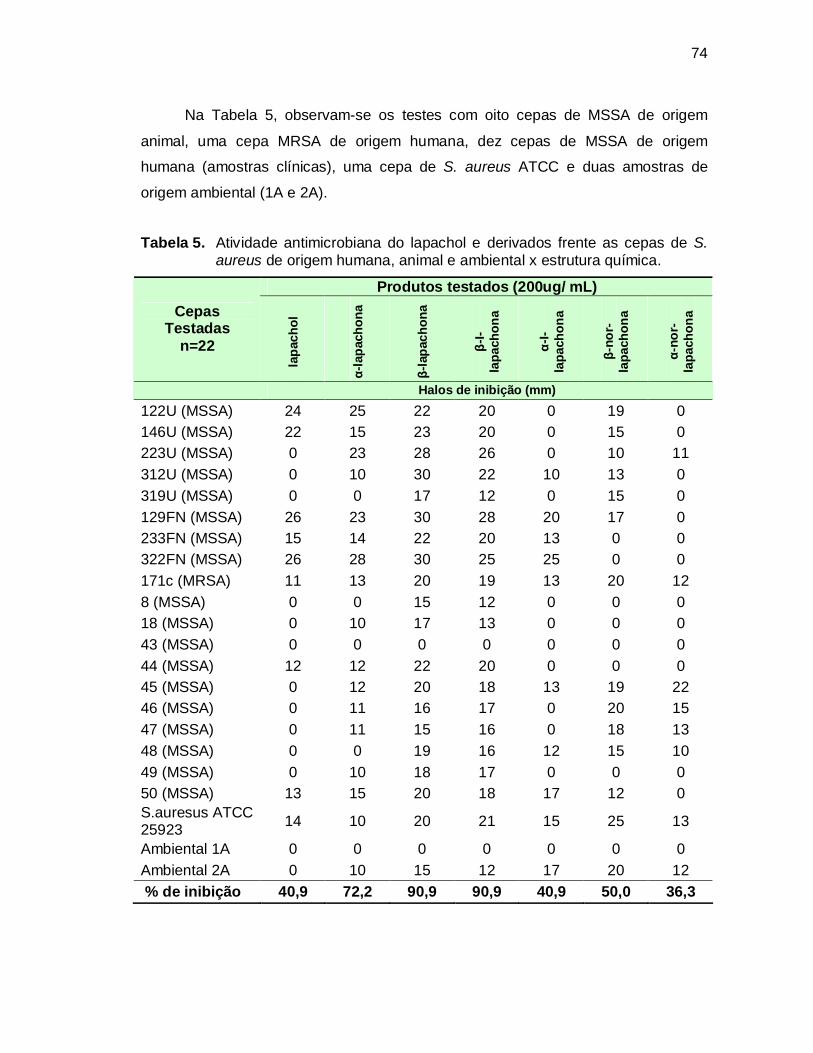
74
Na Tabela 5, observam-se os testes com oito cepas de MSSA de origem
animal, uma cepa MRSA de origem humana, dez cepas de MSSA de origem
humana (amostras clínicas), uma cepa de S. aureus ATCC e duas amostras de
origem ambiental (1A e 2A).
Tabela 5. Atividade antimicrobiana do lapachol e derivados frente as cepas de S.
aureus de origem humana, animal e ambiental x estrutura química.
Produtos testados (200ug/ mL) Cepas
Testadas n=22
lapa
chol
α-la
pach
ona
β-la
pach
ona
β-I-
lapa
chon
a
α-I-
lapa
chon
a
β-no
r-la
pach
ona
α-no
r-la
pach
ona
Halos de inibição (mm) 122U (MSSA) 24 25 22 20 0 19 0 146U (MSSA) 22 15 23 20 0 15 0 223U (MSSA) 0 23 28 26 0 10 11 312U (MSSA) 0 10 30 22 10 13 0 319U (MSSA) 0 0 17 12 0 15 0 129FN (MSSA) 26 23 30 28 20 17 0 233FN (MSSA) 15 14 22 20 13 0 0 322FN (MSSA) 26 28 30 25 25 0 0 171c (MRSA) 11 13 20 19 13 20 12 8 (MSSA) 0 0 15 12 0 0 0 18 (MSSA) 0 10 17 13 0 0 0 43 (MSSA) 0 0 0 0 0 0 0 44 (MSSA) 12 12 22 20 0 0 0 45 (MSSA) 0 12 20 18 13 19 22 46 (MSSA) 0 11 16 17 0 20 15 47 (MSSA) 0 11 15 16 0 18 13 48 (MSSA) 0 0 19 16 12 15 10 49 (MSSA) 0 10 18 17 0 0 0 50 (MSSA) 13 15 20 18 17 12 0 S.auresus ATCC 25923 14 10 20 21 15 25 13
Ambiental 1A 0 0 0 0 0 0 0 Ambiental 2A 0 10 15 12 17 20 12 % de inibição 40,9 72,2 90,9 90,9 40,9 50,0 36,3

75
As substâncias testadas apresentaram halo de inibição de crescimento para
quase todos os microrganismos com exceção de uma cepa de origem clínica a 43
(MSSA) e uma cepa de origem ambiental, 1A.
É importante observar que a cepa MRSA de origem humana foi inibida por
todas as substâncias testadas (Tabela 5), mostrando o potencial antimicrobiano do
lapachol e seus derivados diante de uma cepa resistente a quase todos os
antibióticos, segundo Wickens e Wade (2005). Nota-se ainda que, as cepas MSSA
43 e 1A cresceram diante das substâncias, enquanto que a cepa MRSA – 171C, foi
sensível sugerindo diferentes mecanismos de defesa das mesmas em relação aos
produtos. Estes resultados sugerem que, posteriormente, se investigue outras
possibilidades de atividade biológica dessas substâncias, devido a importância
epidemiológica dos S. aureus em todo o mundo, quando se envolvem em um amplo
espectro de doenças ou quando são vistos como importantes patógenos
nosocomiais causadores de infecções hospitalares. Ou ainda, quando se afirma que
são os mais freqüentes microrganismos associados às mastites de animais sem
mencionar-se as extensivas estratégias que estes microrganismos utilizam para
sobrepujarem as defesas microambientais do hospedeiro infectado e também os
mecanismos de resistência à drogas que desenvolvem. Estas afirmações podem ser
referendadas em Trabulsi (2006), Pereira (2002a), Pereira (2002b) e Freitas et al.
(2003).
Conforme se observa na Figura 17, os produtos -lapachona, -lapachona e
-I-lapachona inibiram as cepas testadas nos percentuais de 77,2%, 90,9% e 90,9%.
O produto -nor-lapachona, inibiu 50% das cepas e os produtos lapachol, -I-
lapachona e -nor-lapachona foram os que menos formaram halos de inibição,
embora tenham apresentado um percentual de 40,9%, 40,9% e 36,3%.

76
40,9%
72,2%
90,9% 90,9%
49,9% 50,0%
36,3%
0
10
20
30
40
50
60
70
80
90
100
% d
e In
ibiç
ão
Lapachol α-lapachona β-lapachona β-I-lapachona α-I-lapachona β-nor-lapachona α-nor-lapachona
Substâncias
Figura 17. Percentuais de inibição do lapachol e derivados frente as cepas de S. aureus.
Com relação às mudanças nas estruturas químicas dos compostos derivados
do lapachol e suas atividades antimicrobianas (estudo de atividade biológica x
estrutura química), observa-se que, os produtos -lapachona e -I-lapachona
apresentaram maior atividade antimicrobiana em relação aos demais, embora a
adição do Iodo à molécula do -lapachona não tenha alterado sua atividade
antimicrobiana neste evento, pois ambas as substâncias apresentaram iguais
percentuais de inibição (90,9%). Porém, a adição do Iodo na estrutura da -
lapachona diminuiu sua atividade frente aos microrganismos testados em 36,3
pontos percentuais. Já com relação aos compostos -nor-lapachona e -nor-
lapachona observa-se que o primeiro apresentou atividade antimicrobiana em um
percentual mais expressivo que o segundo (40,9% e 36,3%). Essas mudanças
estruturais mostram a importância da exploração da reatividade química, atualmente
aplicada aos compostos (SILVA; FERREIRA; SOUZA, 2003; OLIVEIRA et al., 2001;
BARBOSA, 2006), notadamente evidenciada nesta pesquisa quando o lapachol é o
composto que menos apresentou potencial antimicrobiano, superado apenas pela -
nor-lapachona com percentuais de inibição respectivamente de apenas 50% e
36,3%, enquanto que os demais derivados do lapachol apresentaram percentuais de
inibição bem superiores, aumentando sua atividade biológica.
Segundo Fonseca, Braga e Santana (2003), o lapachol e seus derivados
apresentam baixa atividade antimicrobiana frente a microrganismos Gram negativos.
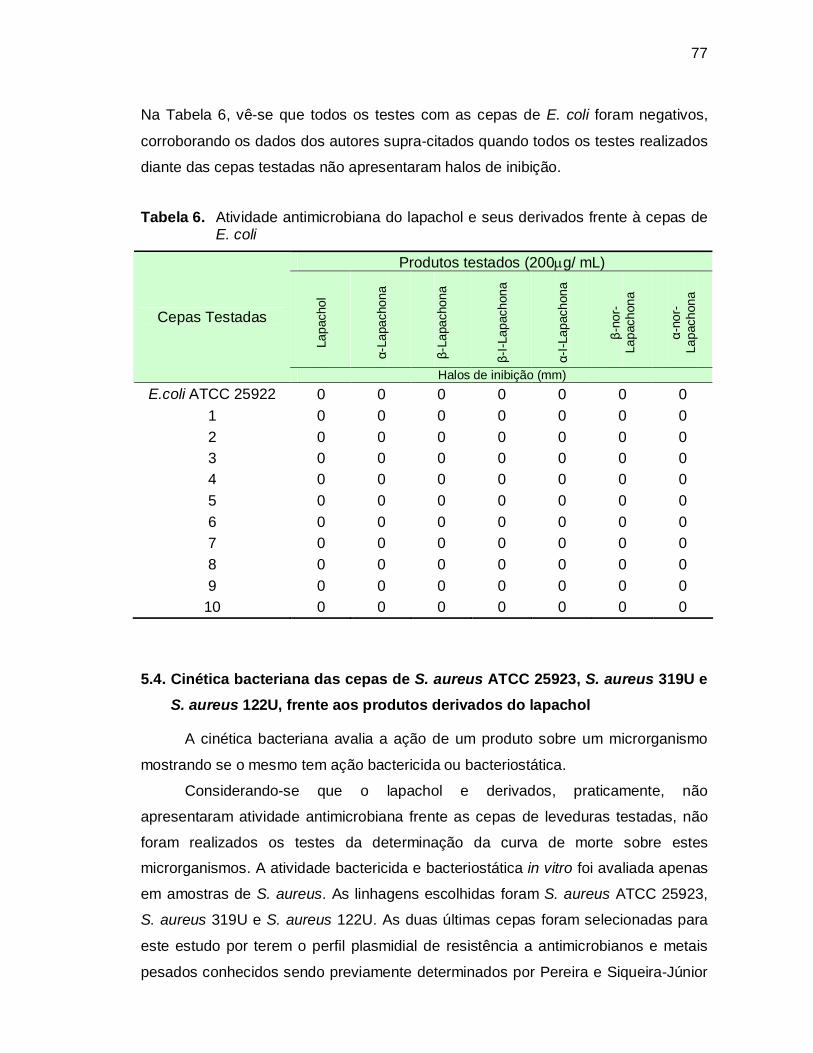
77
Na Tabela 6, vê-se que todos os testes com as cepas de E. coli foram negativos,
corroborando os dados dos autores supra-citados quando todos os testes realizados
diante das cepas testadas não apresentaram halos de inibição.
Tabela 6. Atividade antimicrobiana do lapachol e seus derivados frente à cepas de
E. coli
Produtos testados (200g/ mL)
Lapa
chol
α-La
pach
ona
β-La
pach
ona
β-I-L
apac
hona
α-I-L
apac
hona
β-no
r-La
pach
ona
α-no
r-La
pach
ona
Cepas Testadas
Halos de inibição (mm) E.coli ATCC 25922 0 0 0 0 0 0 0
1 0 0 0 0 0 0 0 2 0 0 0 0 0 0 0 3 0 0 0 0 0 0 0 4 0 0 0 0 0 0 0 5 0 0 0 0 0 0 0 6 0 0 0 0 0 0 0 7 0 0 0 0 0 0 0 8 0 0 0 0 0 0 0 9 0 0 0 0 0 0 0 10 0 0 0 0 0 0 0
5.4. Cinética bacteriana das cepas de S. aureus ATCC 25923, S. aureus 319U e
S. aureus 122U, frente aos produtos derivados do lapachol
A cinética bacteriana avalia a ação de um produto sobre um microrganismo
mostrando se o mesmo tem ação bactericida ou bacteriostática.
Considerando-se que o lapachol e derivados, praticamente, não
apresentaram atividade antimicrobiana frente as cepas de leveduras testadas, não
foram realizados os testes da determinação da curva de morte sobre estes
microrganismos. A atividade bactericida e bacteriostática in vitro foi avaliada apenas
em amostras de S. aureus. As linhagens escolhidas foram S. aureus ATCC 25923,
S. aureus 319U e S. aureus 122U. As duas últimas cepas foram selecionadas para
este estudo por terem o perfil plasmidial de resistência a antimicrobianos e metais
pesados conhecidos sendo previamente determinados por Pereira e Siqueira-Júnior

78
(1995), utilizadas neste experimento para a realização da cura de plasmídios. Os
resultados dos testes mostram que todos os compostos influenciaram o crescimento
bacteriano, pois todos os testes se mantiveram abaixo da curva de controle de
crescimento bacteriano.
O Quadro 1 mostra a cinética bacteriana de S. aureus ATCC 25923 frente aos
derivados do lapachol. Nota-se que todos os produtos até 24 horas apresentaram
uma ação bacteriostática, pois após seis horas praticamente, se mantiveram no
mesmo Log até 48 horas em relação ao Log inicial (Figura 18). Apenas -nor-
lapachona apresentou uma queda mais acentuada em relação aos demais
compostos após 10 horas, no entanto continua com ação bacteriostática, pois de
acordo com Jones; Anderegg; Deshpande (2002) e Shelburne et al. (2004), para
uma substância ter ação bactericida seu Log precisa cair pelo menos três vezes em
relação ao inóculo do Log inicial, o que não se observa em nenhum evento
registrado na Figura 18 até 24 horas.
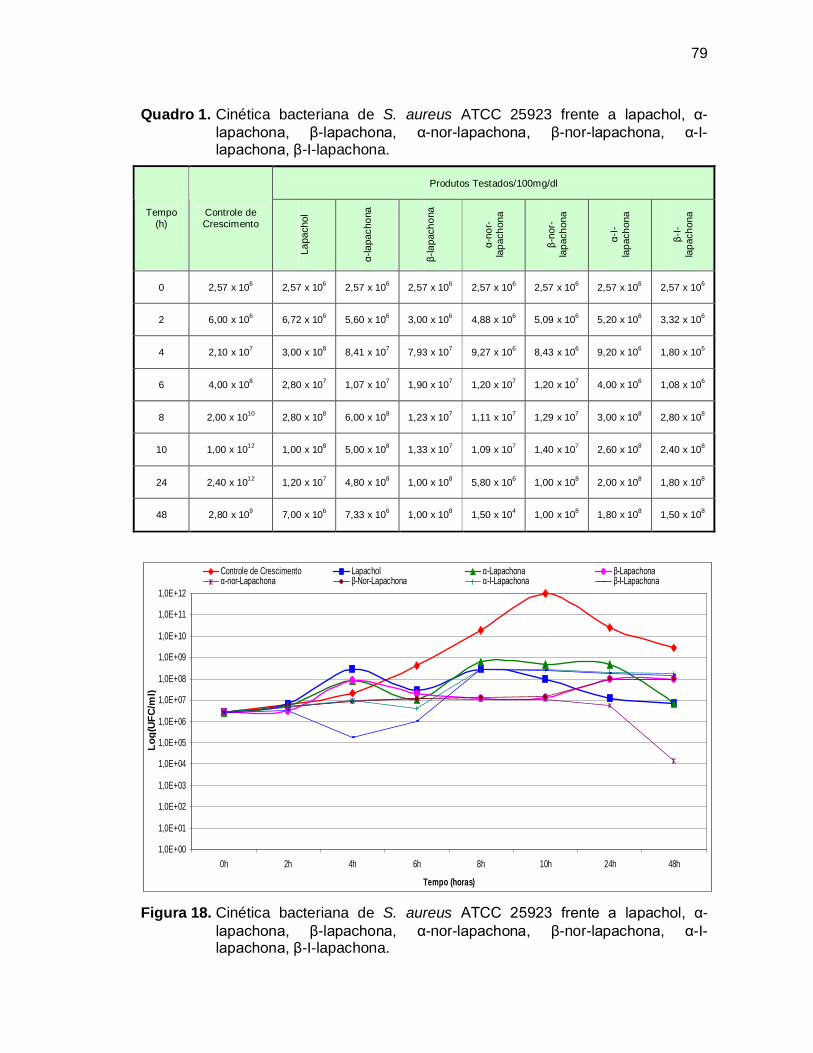
79
Quadro 1. Cinética bacteriana de S. aureus ATCC 25923 frente a lapachol, α-lapachona, β-lapachona, α-nor-lapachona, β-nor-lapachona, α-I-lapachona, β-I-lapachona.
Produtos Testados/100mg/dl
Tempo (h)
Controle de Crescimento
Lapa
chol
α-la
pach
ona
β-la
pach
ona
α-no
r-la
pach
ona
β-no
r-la
pach
ona
α-I-
lapa
chon
a
β-I-
lapa
chon
a
0 2,57 x 106 2,57 x 106 2,57 x 106 2,57 x 106 2,57 x 106 2,57 x 106 2,57 x 106 2,57 x 106
2 6,00 x 106 6,72 x 106 5,60 x 106 3,00 x 106 4,88 x 106 5,09 x 106 5,20 x 106 3,32 x 106
4 2,10 x 107 3,00 x 108 8,41 x 107 7,93 x 107 9,27 x 106 8,43 x 106 9,20 x 106 1,80 x 105
6 4,00 x 108 2,80 x 107 1,07 x 107 1,90 x 107 1,20 x 107 1,20 x 107 4,00 x 106 1,08 x 106
8 2,00 x 1010 2,80 x 108 6,00 x 108 1,23 x 107 1,11 x 107 1,29 x 107 3,00 x 108 2,80 x 108
10 1,00 x 1012 1,00 x 108 5,00 x 108 1,33 x 107 1,09 x 107 1,40 x 107 2,60 x 108 2,40 x 108
24 2,40 x 1012 1,20 x 107 4,80 x 108 1,00 x 108 5,80 x 106 1,00 x 108 2,00 x 108 1,80 x 108
48 2,80 x 109 7,00 x 106 7,33 x 106 1,00 x 108 1,50 x 104 1,00 x 108 1,80 x 108 1,50 x 108
1,0E+00
1,0E+01
1,0E+02
1,0E+03
1,0E+04
1,0E+05
1,0E+06
1,0E+07
1,0E+08
1,0E+09
1,0E+10
1,0E+11
1,0E+12
0h 2h 4h 6h 8h 10h 24h 48h
Tempo (horas)
Log(
UFC
/ml)
Controle de Crescimento Lapachol α-Lapachona β-Lapachonaα-nor-Lapachona β-Nor-Lapachona α-I-Lapachona β-I-Lapachona
Figura 18. Cinética bacteriana de S. aureus ATCC 25923 frente a lapachol, α-
lapachona, β-lapachona, α-nor-lapachona, β-nor-lapachona, α-I-lapachona, β-I-lapachona.

80
No Quadro 2 e Figura 19, observa-se a curva de morte do S. aureus 319U
frente ao lapachol e seus derivados. Nota-se também que, todos os compostos
tiveram ação bacteriostática frente a esse microrganismo até 24 horas. O Log do
lapachol caiu em quatro horas, se manteve estável até 48 horas e foi o mais baixo
do conjunto, sendo superado apenas após 24 horas pelo -lapachona que declina
após este horário e apresenta ação bactericida em 48 horas. Pode-se verificar que,
em oito horas, todas as substâncias apresentaram atividade antimicrobiana em
relação ao controle, caindo apenas após esta hora o Log do -lapachona e do -
lapachona. O Log dos demais compostos se manteve estável até 48 horas.

81
Quadro 2. Cinética bacteriana de S. aureus 319U frente ao lapachol, α-lapachona, β-lapachona, α-nor-lapachona, β-nor-lapachona, α-I-lapachona, β-I-lapachona.
Produtos Testados/100mg/dl
Tempo (h)
Controle de Crescimento
Lapa
chol
α-la
pach
ona
β-la
pach
ona
α-no
r-la
pach
ona
β-no
r-la
pach
ona
α-I-
lapa
chon
a
β-I-
lapa
chon
a
0 2,27 x 106 2,27 x 106 2,27 x 106 2,27 x 106 2,27 x 106 2,27 x 106 2,27 x 106 2,27 x 106
2 1,60 x 108 2,70 x 108 8,00 x 107 6,50 x 106 1,60 x 108 2,76 x 106 2,42 x 106 3,00 x 106
4 4,00 x 108 1,00 x 107 7,00 x 108 8,10 x 108 2,40 x 108 6,40 x 106 5,60 x 106 4,80 x 106
6 7,00 x 108 1,80 x 106 6,00 x 108 7,60 x 108 3,00 x 108 7,20 x 107 6,70 x 106 5,70 x 107
8 2,40 x 1012 2,10 x 106 5,10 x 108 7,10 x 108 2,90 x 108 8,10 x 107 7,70 x 108 7,90 x 107
10 2,00 x 1011 3,20 x 106 4,80 x 108 3,80 x 105 2,70 x 108 9,20 x 107 8,10 x 108 8,90 x 108
24 1,00 x 1010 3,40 x 106 3,20 x 108 1,00 x 108 1,00 x 108 8,70 x 108 9,20 x 108 8,90 x 108
48 2,20 x 109 2,10 x 106 3,20 x 103 1,00 x 108 1,00 x 107 8,60 x 108 9,80 x 108 9,30 x 108
1,0E+00
1,0E+01
1,0E+02
1,0E+03
1,0E+04
1,0E+05
1,0E+06
1,0E+07
1,0E+08
1,0E+09
1,0E+10
1,0E+11
1,0E+12
1,0E+13
0h 2h 4h 6h 8h 10h 24h 48hTempo (horas)
Log
(UFC
/ml)
Controle de Crescimento Lapachol α-Lapachona β-Lapachonaα-nor-Lapachona β-Nor-Lapachona α-I-Lapachona β-I-Lapachona
Figura 19. Cinética bacteriana de S. aureus 319U frente aos produtos Lapachol, α-
lachona, β-lapachona, α-nor-lapachona, β-nor-lapachona, α-I-lapachona, β-I-lapachona.

No Quadro 3, está registrado os resultados da cinética bacteriana do S.
aureus 122U frente aos derivados do lapachol. Também frente a esta cepa estes
produtos tiveram ação bacteriostática até 24 horas, apresentando em média pico
máximo de crescimento em oito horas. Após esta hora o Log de todos os compostos
declina.
Manteve-se estável até 48 horas apenas a α-nor-lapachona em Log de 108. O
Log das demais substâncias declina até 48 horas e o Log do lapachol mantém-se
estável a partir de quatro horas, horário em que cai. A α-lapachona mantém um
comportamento parecido com o teste da cepa 319U, quando após 24 horas é o
produto que apresenta queda de três Log, sendo bactericida (Figura 20).
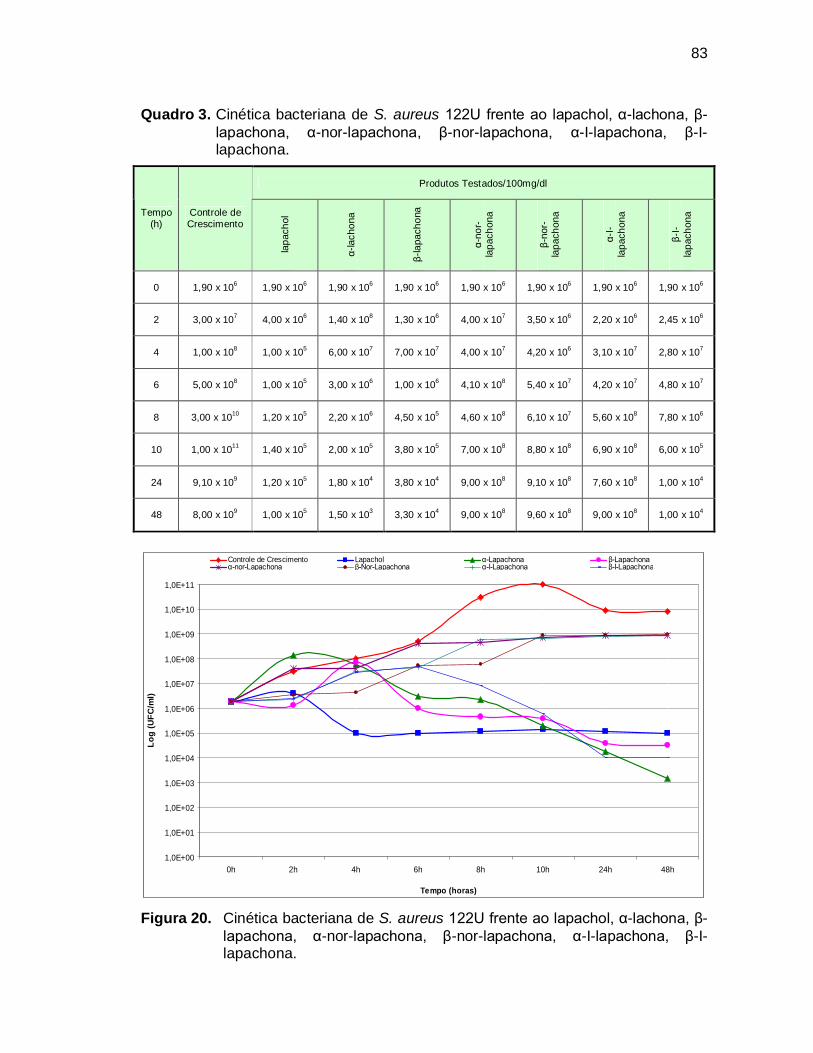
83
Quadro 3. Cinética bacteriana de S. aureus 122U frente ao lapachol, α-lachona, β-lapachona, α-nor-lapachona, β-nor-lapachona, α-I-lapachona, β-I-lapachona.
Produtos Testados/100mg/dl
Tempo (h)
Controle de Crescimento
lapa
chol
α-la
chon
a
β-la
pach
ona
α-no
r-la
pach
ona
β-no
r-la
pach
ona
α-I-
lapa
chon
a
β-I-
lapa
chon
a
0 1,90 x 106 1,90 x 106 1,90 x 106 1,90 x 106 1,90 x 106 1,90 x 106 1,90 x 106 1,90 x 106
2 3,00 x 107 4,00 x 106 1,40 x 108 1,30 x 106 4,00 x 107 3,50 x 106 2,20 x 106 2,45 x 106
4 1,00 x 108 1,00 x 105 6,00 x 107 7,00 x 107 4,00 x 107 4,20 x 106 3,10 x 107 2,80 x 107
6 5,00 x 108 1,00 x 105 3,00 x 106 1,00 x 106 4,10 x 108 5,40 x 107 4,20 x 107 4,80 x 107
8 3,00 x 1010 1,20 x 105 2,20 x 106 4,50 x 105 4,60 x 108 6,10 x 107 5,60 x 108 7,80 x 106
10 1,00 x 1011 1,40 x 105 2,00 x 105 3,80 x 105 7,00 x 108 8,80 x 108 6,90 x 108 6,00 x 105
24 9,10 x 109 1,20 x 105 1,80 x 104 3,80 x 104 9,00 x 108 9,10 x 108 7,60 x 108 1,00 x 104
48 8,00 x 109 1,00 x 105 1,50 x 103 3,30 x 104 9,00 x 108 9,60 x 108 9,00 x 108 1,00 x 104
1,0E+00
1,0E+01
1,0E+02
1,0E+03
1,0E+04
1,0E+05
1,0E+06
1,0E+07
1,0E+08
1,0E+09
1,0E+10
1,0E+11
0h 2h 4h 6h 8h 10h 24h 48h
Tempo (horas)
Log
(UFC
/ml)
Controle de Crescimento Lapachol α-Lapachona β-Lapachonaα-nor-Lapachona β-Nor-Lapachona α-I-Lapachona β-I-Lapachona
Figura 20. Cinética bacteriana de S. aureus 122U frente ao lapachol, α-lachona, β-
lapachona, α-nor-lapachona, β-nor-lapachona, α-I-lapachona, β-I-lapachona.

84
Na Figura 21 observa-se a curva de morte da bactéria S. aureus ATCC 25923
em 08 horas e em 24 horas, sob a ação do lapachol, α-lapachona, -lapachona e α-
Nor-lapachona.
(1) T8 – 10-3(1) T8 – 10-3
(1) T24 – 10-3(1) T24 – 10-3
(2) T8 – 10-3(2) T8 – 10-3 (2) T24 – 10-3(2) T24 – 10-3
(3) T8 – 10-3(3) T8 – 10-3 (3) T24 – 10-3(3) T24 – 10-3
(4) T8 – 10-3(4) T8 – 10-3 (4) T24 – 10-3(4) T24 – 10-3
(C) T8 – 10-3(C) T8 – 10-3 (C) T24 – 10-3(C) T24 – 10-3
Figura 21. Cinética bacteriana do lapachol (1), α-lapachona (2), β-lapachona (3), α-
nor-lapachona (4) sobre a cepa de S. aureus ATCC 25923 em 8h e 24h.
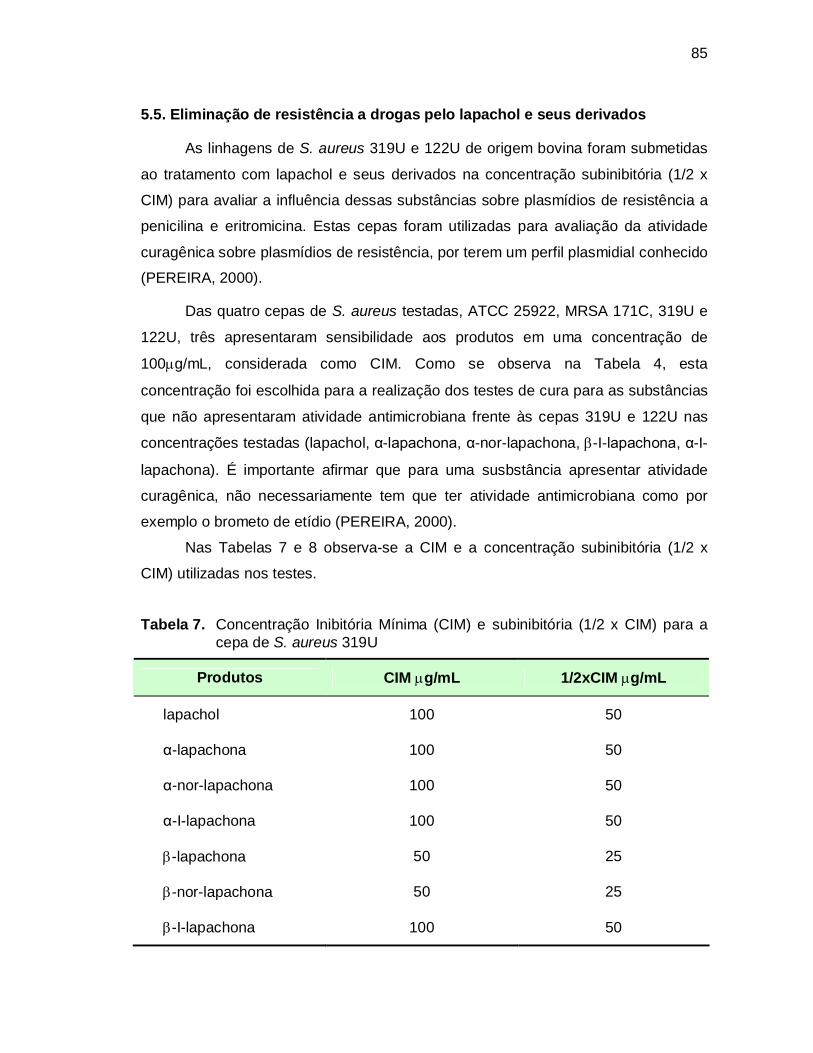
85
5.5. Eliminação de resistência a drogas pelo lapachol e seus derivados
As linhagens de S. aureus 319U e 122U de origem bovina foram submetidas
ao tratamento com lapachol e seus derivados na concentração subinibitória (1/2 x
CIM) para avaliar a influência dessas substâncias sobre plasmídios de resistência a
penicilina e eritromicina. Estas cepas foram utilizadas para avaliação da atividade
curagênica sobre plasmídios de resistência, por terem um perfil plasmidial conhecido
(PEREIRA, 2000).
Das quatro cepas de S. aureus testadas, ATCC 25922, MRSA 171C, 319U e
122U, três apresentaram sensibilidade aos produtos em uma concentração de
100g/mL, considerada como CIM. Como se observa na Tabela 4, esta
concentração foi escolhida para a realização dos testes de cura para as substâncias
que não apresentaram atividade antimicrobiana frente às cepas 319U e 122U nas
concentrações testadas (lapachol, α-lapachona, α-nor-lapachona, -I-lapachona, α-I-
lapachona). É importante afirmar que para uma susbstância apresentar atividade
curagênica, não necessariamente tem que ter atividade antimicrobiana como por
exemplo o brometo de etídio (PEREIRA, 2000).
Nas Tabelas 7 e 8 observa-se a CIM e a concentração subinibitória (1/2 x
CIM) utilizadas nos testes.
Tabela 7. Concentração Inibitória Mínima (CIM) e subinibitória (1/2 x CIM) para a cepa de S. aureus 319U
Produtos CIM g/mL 1/2xCIM g/mL
lapachol 100 50
α-lapachona 100 50
α-nor-lapachona 100 50
α-I-lapachona 100 50
-lapachona 50 25
-nor-lapachona 50 25
-I-lapachona 100 50

86
Tabela 8. Concentração Inibitória Mínima (CIM) e subinibitória (1/2 x CIM) para a cepa de S. aureus 122U
Produtos CIM g/mL 1/2xCIM g/mL lapachol 100 50 α-lapachona 50 25 α-nor-lapachona 100 50 α-I-lapachona 100 50
-lapachona 50 25 -nor-lapachona 50 25
-I-lapachona 50 25
Observa-se os resultados da freqüência da eliminação de plasmídios
resistentes à eritromicina (Figura 22) e penicilina (Figura 23), pelo lapachol e seus
derivados semi-sintéticos, nas Tabelas 9 e 10 e Figuras 24 e 25. Ao analisar estas
tabelas observa-se que os produtos testados não apresentaram efeito frente à cepa
S. aureus 122U para a marca de resistência à penicilina. Os produtos lapachol e α-
lapachona apresentaram, respectivamente, freqüência de cura de 0,78% e 0,23%
para a penicilina frente à cepa S. aureus 319U. Os produtos -lapachona, -I-
lapachona e -I-lapachona apresentaram respectivamente 0,61%, 1,06% e 2,45%
de freqüência de cura para a marca de resistência à Eritromicina diante da cepa
319U.
Apenas -I-lapachona apresentou freqüência de cura com 0,39% para a cepa
S. aureus 122U para a marca de resistência à eritromicina.
O produto mais eficaz para a eliminação da marca de resistência da
eritromicina foi o -I-lapachona por ter atuado para as cepas S. aureus 319U
(Figuras 22 e 25) e 122U com freqüência de cura respectivamente de 1,06% e
0,39%.
O produto -lapachona apresentou ação simultânea para a eliminação das
marcas de resistência à penicilina, com freqüência de cura de 0,61%; e à
eritromicina, cuja freqüência de cura foi de 0,23%, o que sugere a existência de um
plasmídio codificando ambas as propriedades fenotípicas (PEREIRA, 1992;

87
PEREIRA; SIQUEIRA-JUNIOR, 1995; PEREIRA, 2000; PEREIRA; SIQUEIRA-
JUNIOR; TAKAKI, 2004).
O lapachol não apresentou halo de inibição frente à cepa 319U, porém
apresentou freqüência de cura de 0,78% para a marca de resistência à penicilina
(Figuras 23 e 25), dados que corroboram as afirmações de Pereira (2000) quando
diz que uma substância não precisa ter necessariamente atividade antimicrobiana
para apresentar atividade curagênica, pois o produto pode interferir, possivelmente,
apenas na replicação plasmidial e não exercer uma ação antimicrobiana sobre a
cepa estudada.
Tabela 9. Freqüência de cura da marca de resistência plasmidial à penicilina pelo
lapachol e seus derivados semi-sintéticos em linhagens de S. aureus.
N° de colônias ensaiadas/Nº de variantes sensíveis Freqüência de cura % Produtos
319U 122U 319U 122U lapachol 384/3 299/0 0,78 0,0
-lapachona 433/1 90/0 0,23 0,0
-I-lapachona 163/0 101/0 0,0 0,0
-lapachona 515/0 162/0 0,0 0,0
-I-lapachona 748/0 254/0 0,0 0,0
Figura 22. Colônias possilvelmente curadas pelo β-I-lapachona para a marca de resistência plasmidial à eritromicina para a cepa S. aureus 319U.
a) b)
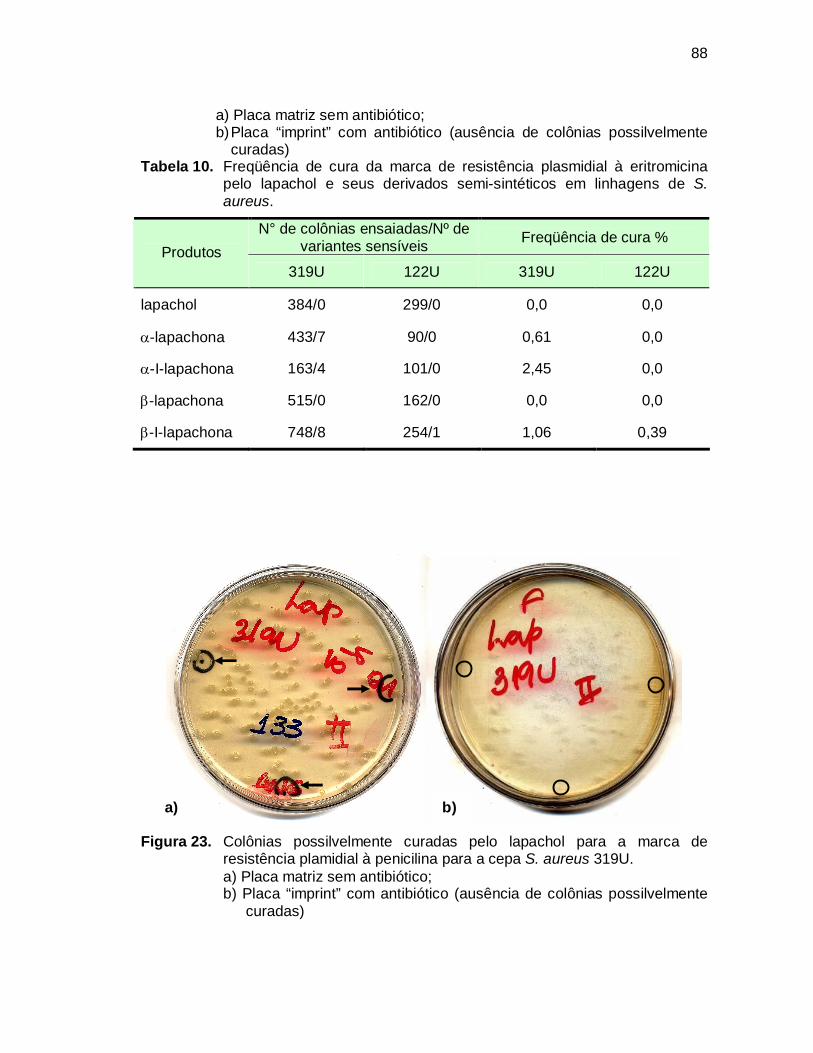
88
a) Placa matriz sem antibiótico; b) Placa “imprint” com antibiótico (ausência de colônias possilvelmente
curadas) Tabela 10. Freqüência de cura da marca de resistência plasmidial à eritromicina
pelo lapachol e seus derivados semi-sintéticos em linhagens de S. aureus.
N° de colônias ensaiadas/Nº de variantes sensíveis Freqüência de cura %
Produtos 319U 122U 319U 122U
lapachol 384/0 299/0 0,0 0,0
-lapachona 433/7 90/0 0,61 0,0
-I-lapachona 163/4 101/0 2,45 0,0
-lapachona 515/0 162/0 0,0 0,0
-I-lapachona 748/8 254/1 1,06 0,39
Figura 23. Colônias possilvelmente curadas pelo lapachol para a marca de
resistência plamidial à penicilina para a cepa S. aureus 319U. a) Placa matriz sem antibiótico; b) Placa “imprint” com antibiótico (ausência de colônias possilvelmente
curadas)
a) b)

89
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
Freq
üênc
ia d
e cu
ra
Lapachol a-lapachona ß-lapachona ß-I-lapachona a-I-lapachona
Substâncias 0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1
Freq
üênc
ia d
e cu
ra
Lapachol a-lapachona ß-lapachona ß-I-lapachona a-I-lapachona
Substâncias Figura 24. Freqüência de cura da marca de resistência plasmidial à penicilina pelo
lapachol e derivados em S. aureus 319U (a) e S. aureus 122U (b)
0
0,5
1
1,5
2
2,5
Freq
üênc
ia d
e cu
ra
Lapachol a -lapachona ß-lapachona ß-I-lapachona a-I-lapachona
Substâncias
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
Freq
üênc
ia d
e cu
raLapachol a -lapachona ß-lapachona ß-I-lapachona a-I-lapachona
Substâncias Figura 25. Freqüência de cura da marca de resistência plasmidial à eritromicina pelo
lapachol e derivados em S. aureus 319U (a) e S. aureus 122U (b)
A transferência de material genético entre organismos da mesma ou
diferentes espécies desempenha um papel crucial na evolução de resistência a
drogas em bactérias. Embora a existência de plasmídios em S. aureus de origem
bovina já tenha sido demonstrada por Pereira (2000), o seu papel na determinação
de resistência à drogas continua sendo de extrema importância, pois sendo este
apenas um dos mecanismos utilizados dentre outros, que os microrganismos lançam
maõs para se defenderem a exemplo dos mecanismos de defesa cromossomiais é
importante a pesquisa de novos compostos para a prevenção e/ou controle de
linhagens multiresistentes. Os resultados obtidos neste trabalho mostram a
importância do estudo de agentes com atividade antimicrobiana ou não na
eliminação de plasmídios, o que poderá contribuir quando associados a outros
estudos, para a prevenção dessas linhagens multiresistentes, notadamente
enfatizada em Pereira (2000) e Ferreira et al., (2006), quando afirmam que em geral
a capacidade de compostos para eliminar plasmídios tais como SDS (dodecil sulfato
de sódio), acriflavina ou acridina laranja, é muito tóxica para uso terapêutico.
Pesquisas de compostos não tóxicos têm sido realizadas a exemplo do ácido
ascórbico que elimina plasmídio penicilinase em S. aureus. Certamente, os estudos
a b
a b

90
de estrutura-atividade que estão sendo desenvolvidos na atualidade com as
modificações na estrutura molecular dos compostos com atividades biológicas já
conhecidas, a exemplo do lapachol e seus derivados, vem se tornando cada vez
mais significativos quando tentam diminuir a toxicidade destes compostos.
Os resultados dessa investigação sugerem que, posteriormente, se investigue
outras possibilidades de atividades biológicas, devido ao potencial promissor das
moléculas obtidas a partir das modificações estruturais do lapachol, pois tanto o
lapachol como as lapachonas têm sido alvo de constante interesse farmacológico,
devido a variedade de atividades biológicas que essas naftoquinonas apresentam.

C O N C L U S Õ E S

92
6. CONCLUSÕES
No estudo sobre a ação antimicrobiana e antiplasmidial do lapachol e
derivados sobre bactérias e fungos leveduriformes, conclui-se que:
As substâncias lapachol e seus derivados semi-sintéticos -lapachona, -nor-
lapachona, -I-lapachona, -lapachona, -nor-lapachona e -I-lapachona, foram
as mais ativas frente a S. aureus ATCC 25922, P. aeruginosa ATCC 27853,
MRSA 171C, S. aureus 319U e S. aureus 122U.
As substâncias lapachol e derivados não apresentaram atividade antimicrobiana
sobre as cepas de E. coli ATCC 95925 e Candida spp.
Somente S. aureus ATCC 25923, foi sensível frente as substâncias L04 e L08,
derivados nitrogenados do lapachol.
Todos os derivados semi-sintéticos obtidos a partir do lapachol tiveram atividade
antimicrobiana significativa frente as cepas de S. aureus inclusive a MRSA.
Os produtos -lapachona e -nor-lapachona apresentaram a menor CIM
(50g/mL) e menor CBM (100g/mL) para todos os microrganismos testados,
com exceção da E. coli, podendo ser considerados os mais eficazes da série.
Nos testes de estrutura-atividade, observa-se que as cepas MRSA 171C e 2ª
(com exceção do lapachol), foram inibidas por todas as substâncias testadas
mostrando o potencial antimicrobiano do lapachol e seus derivados semi-
sintéticos.
Os produtos mais ativos frente as cepas de Staphylococcus testadas foram -
lapachona e -I-lapachona, apresentando ambos os percentuais de inibição de
90,9%.
As mudanças na estrutura química do lapachol aumentaram consideravelmente
sua atividade antimicrobiana frente às cepas de Staphylococcus aureus.

93
As curvas de morte (cinética bacteriana) mostram que o lapachol e seus
derivados semi-sintéticos têm ação bacteriostática, com exceção dos produtos α-
lapachona e α-nor-lapachona que após 48h tem ação bactericida.
Apenas o produto -lapachona apresentou ação simultânea para a eliminação
das marcas de resistência à penicilina e à eritromicina.
O produto mais eficaz para eliminação da marca de resistência à eritromicina foi
o -I-lapachona.
O lapachol não apresentou atividade antimicrobiana frente a cepa de S. aureus
319U, porém apresentou uma freqüência de cura de 0,78% para a marca de
resistência à penicilina.

R E F E R Ê N C I A S

95
REFERÊNCIAS
AL-MASAUDI, S. B.; DAY, M. J.; RUSSEL, A. D. Effect of some antibioticsand biocides on plasmids transfer in Staphylococcus aureus. J. Applied Bacter, v.71, p.239- 243, 1991.
ÁLVAREZ, C. A. G.; CASTRO, A. L. L.; GONZALEZ, M. J. P.; JIMENEZ, M. L. N. Mecanismos de resistência em Pseudomonas aeruginosa: entendiendo a um peligroso enemigo. Rev Fac Univ Nac Colomb, v.53, n.1, 2005.
ANDERSON, D.; The ways in which bacteria resist antibiotics, A multidisciplinary muting at the drags. Hammarskjöld Foundation, p. 5-7, 2004.
ANTUNES, R. M. P.; LIMA, E. O.; PEREIRA, M. S. V.; CAMARA, C. A.; ARRUDA, T. A.; CATÃO, R. M. R.; BARBOSA, T. P. B.; NUNES, X. P.; DIAS, C. S.; SILVA, T. M. S. Atividade antimicrobiana “in vitro” e determinação da concentração inibitória mínima (CIM) de fitoconstituintes e produtos sintéticos sobre bactérias e fungos leveduriformes. Rev Bras Farmacogn, v.16, n.4, p. 517-524, 2006.
BALLESTER, D.; ESCOBAR, A. M. U.; GRISI, S. J. F. E. Diarréia persistente: revisão dos principais aspectos fisiopatogênicos, fatores de risco e implicações terapêuticas. Pediatria, v.24, n.3/4, p.112-121, 2002.
BAMBEKE, F.V; GLUPEZYNSKI, Y; PLESIAT, P; PECHERE, J.C; TULKENS, P.M; Antibiotic eflux pumps in procariatic cels: occurrence, impacto n resistence and estrategies for the future of antimicrobial ferapy, Journal of Antimicrobial Chemotherapy, v.51, p.1055-1065, 2003.
BARBOSA – FILHO, J.M; CÂMARA, C.A; SILVA, T.M.S; GIOLIETTI, A.M.; BOTELHO, M.P; SANTOS, R.R. Processo de síntese da 3-iodo-alfa-lapachona e 3-iodo-beta-lapachona e uso como imunomodulardor, antimicrobiano e antiflamatório. Depósito de patente publicado na Revista da Propriedade Industrial 1843, de 02/05/2006, referente ao PI 0403686-7.
BARBOSA, T.P. Derivados nitrogenados do norlapachol: Síntese e atividade biológica. Dissertação de mestrado, LTF-UFPB, 152.pp.2006.
BARBOSA, T.P; CÂMARA, C.A.; SILVA, T.M.S; MARTINS, R.M; PINTO, A.C; VARGAS, M.D. New 1, 2, 3, 4 – tetrahydro-1-aza-anthraquinones and 2-aminoalkyl compounds from norlapachol with molluscicidae actvity, Biorganic & Medicinal Chemistry, v.13, p.6464-6469, 2005.
BAUER, A. N., KIRBY, W. M. M., SHERRIS, J. C., TURCK, M. Antibiotic susceptibility testing by a standardized single disc method. Am. J. Clin. Pathol, v.45, p.493-496, 1966.

96
BEART, J. E.; LILLEY, T. H.; HASLAM, E. Plant polyphenols - secondary metabolism and chemical defense: some observations. Phytochemestry, v.24, p.33-8, 1985.
BORIOLLO, M. F. G.; HÖFLING, J. F.; MENDES, A.; ROSA, E. A. R. Ferramentas moleculares para a caracterização de Candida albicans (Robin) Berkhout (1923) em estudos epidemiológicos. Estud Biolog, v.27, n.58, p.21-47, 2005.
BRAGA, L. C.; LEITE, A. A. M; XAVIER, K, G. S.; TAKAHASHI, J. A.; BEMQUERER; CHARTONE-SOUZA, E.; NASCIMENTO, A. M. A. Synergic interation between pomegrate extract an antibiotics against Staphylococcus aureus. Can J Microbiol, v.51, p.541-547, 2005.
BRANDILEONE, M. C.C.; CASAGRANDE, S. T.; GUERRA, M. L. L. S.; ZANELLA, R. C.; ANDRADE, A. L. S. S.; AND DI-FABIO, J. L. Increase in numbers of ß-lactam-resistant invasive Streptococcus pneumoniae in Brazil and the impact of conjugate vaccine coverage. J Med Microbiol, v.55, p.567-574, 2006.
BURGESS, D. S. Princípios Farmacodinâmicos dos Tratamentos Antimicrobianos na Prvenção da Resistência. In: PLE CHEST The Cardiopulmonary and Critical Care Journal (edição em português), Suplemento 2:1(19-23). 2000.
CALBO, E.; GARAU, J.; Aplication of pharma cokinetics and pharmaco dynamics to antimicrobial terapy of com munity acquired respiratory tract infections, Respiration, v.72, p.561-571, 2005.
CAMARA, C.A; PINTO, A.C; VARGAS, M.D; Azepines from the intramolecular prins cyclization of an aminoderivative of lapachol, Tetrahedron, v.58, p.6135-6140, 2002.
CAMARA, C.A; ROSA, M.A; PINTO, A.C; VARGAS, M.D; Secondary amines and unexpected 1-aza-antraquinones from 2-methoxy-lapachol. Tetrahedron, v.57, p.9569-9574, 2001.
CAMARGO, C. B. S.; PEDRO, C. C.; LOURENÇO, D. S.; GIRONI, R. H. A.; MARTINEZ, R. Infecção de vias urinárias na comunidade de Ribeirão Preto-SP: etiologia, sensibilidade bacteriana a antimicrobianos e implicações terapêuticas. Medicina, v.35: 173-178, 2002.
CARVALHO, J. C. T.; SARTI, S. J. Produção de extratos vegetais para uso terapêutico e cosmético. Ribeirão Preto: Núcleo de Pesquisa de Produtos Naturais de Ribeirão Preto 1995.
CASALS, J. B. Tablet sensibility testing of pathogenic fungi. J Clin Pathol, v.32, p.719-722, 1979.
CHARTONE-SOUZA, E. Genética de Resistência Bacteriana a Drogas. In: Anais XXIV Reunião de Genética de Microrganismos. Gramado. Rio Grande do Sul: Comunicação Impressa, 2004, p.52.

97
CLEELAND, R.; SQUIRES, E. Evaluation of new antimicrobials in vitro and in experimentalanimal infections In: LORIAN, V. Antibiotics in laboratory medicine. 3 ed. Baltimore: Willians & Wilkins, 739-788, 1991.
CLOUTIER, M.J.; Antibiotics, Mechanisms of action and the acquisition of resistance – When magic Bullets lose their magic, American Journal of Pharmaceutical Education, v.59, p.167-172, 1995.
DA SILVA, M.N; FERREIRA, V.F; de SOUZA, M.C.B.V. Um panorama atual da química e da farmacologia de naftoquinonas com ênfase na β-lapachona e derivados. Química nova, v.26, p.407-416, 2003.
DÉOUX, S.; DÉOUX, P. Ecologia é a saúde - o impacto da deteriorização do ambiente na saúde. Lisboa: Instituto Piaget, 1998.
Dl STASI, L. C. Arte, ciência e magia In: Dl STASI, L. C. (Org.) Plantas medicinais: arte e ciência - um guia de estudo interdisciplinar. São Paulo: Unesp, p.161-186, 1996.
DROUHET, E.; DUPONT, B. Sensitivity of fungi to antifungal compounds, Bull Soc Fr Mycol Med, v.7, p. 165-170, 1978.
DRUTZ, D. J. In vitro antifungical susceptibility testing and measurement of levels of antifungical agents in body fluids. Rev. Infect. Dis, v.9, n.2, p.411-15, 1987.
ELLIS, D. H. Clinical Mycology. The human opportunistic mycosis. Australia: Pfizer, 1994.
ESMERINO, L. A.; GONÇALVES, L. G.; SCHELESKY, M. E. Perfil de sensibilidade antimicrobiana de cepas de Escherichia coli isoladas de infecções urinárias comunitárias. Publ UEPG Biol Saúde, V.9, N.1, 31-39, 2003.
FERREIRA, L. L.; Estrutura clonal e multiresistência em Pseudomonas aeruginosa. [Dissertação – Mestrado]. INCQSP. Fundação Oswaldo Cruz. Rio de Janeiro: 2005, 98p.
FERREIRA, L. M.; NADER-FILHO, A.; OLIVEIRA, E.; ZAFALON, L. F.; SOUZA, V. Variabilidades fenotípica e genotípica de estirpes de S. aureus isoladas em casos de mastite subclinica bovina. Ciência Rural, v.36. n.4, p.1228-1234, 2006.
FILE Jr.,T. M. Visão Geral Sobre Resistência Bacteriana nos Anos 90. In: PLE CHEST The Cardiopulmonary and Critical Care Journal (edição em português). Suplemento 2:1(3-9). 2000.
FONSECA, S.G. da C.; BRAGA, R.M.C.; SANTANA, D.P. Lapacho Química, Farmacologia e Métodos de Dosagens, Revista Brasileira de Farmácia, v.84, n.1, p.9-16, 2003.

98
FREITAS, D.B.; SIQUEIRA-JUNIOR,.J.P. de; FREITAS, F.I. de S; PEREIRA, M. do S.V.; Atividade antimicrobiana de fluoquinolonas sobre Staphylococcus aureus hospitalares. Revista Brasileira de Ciências da Saúde, v.7, n.2, p.143-148, 2003.
FROMTLING, R. A.; PUI-YU, H.; SHADOMY, S. In vitro inhibitory activities of 2 new orally absorbable imidazole derivatives: BAYN 7133 and BAYN 913. Sabouraudia, v.21, p.179-184, 1983.
FURLETTI, V.F; Suscetibilidade de isolados orais de cândida ssp. Provenientes de pacientes com doença periodontal aos antifúngicos azólicos e a anfotericina B. Dissertação de Mestrado, Universidade Federal de Campinas, Fac. De Odontologia de Piracicaba, Piracicaba, 2006.
GOERING, R. V. & RUFF, E. A. Comparative analysis of conjugative plasmids mediating gentamicin resistance in Staphylococcus aureus. Antimicrobial Agents and Chemotherapy, v.24, p.450 – 452, 1983.
GOODMAN, L. S; GILMAN, A. As Bases Farmacológicas da Terapêutica. 9 ed. Rio de Janeiro: McGraw Hill, p. 1436, 1996.
GOTTLIEB, O. R.; KAPLAN, M. A. C.; BORIN, M. R. M. B. Biodiversidade: um enfoque químico-farmacoIógico. Rio de Janeiro: UFRJ, 1996.
GRUNDMANN, H.; SOUSA, M. A.; BOYCE, J.; TIEMERSMA, E. emergence and resurgence of meticillin-resistant Staphylococcus aureus as a public-health threat. Lancet, v.368, p.874-885, 2006.
GUÉHO, E; IMPROVISI, L.; HOOG, G. S. de; DUPONT, B. Trichosporon on humans: a pratical account. Mycoses, v.37, p.3-10, 1994.
GUNDIDZA, M. Screening of extracts from Zimbabwean higher plants II: antifungical properties. Fitoterapia, v.57, p.111-114, 1986.
HADACEK, F. & GREGER, H. Testing of antifungical natural products: methodologies, comparatibility of results and assay choise. Phytochem Anal, v.11, p.137-147, 2000.
HARBORNE J. B. Introduction to ecological biochemestry 3 ed., London, academic, 1988.
HARSTE, S.; BOLARD J. Amphotericin B: new life for an old drug. Trends Pharmac Sci, v.17, p.445-449. 1996.
HOOPER, D. C., WOLFSON, J. S., McHUGH, G. L., SWARTZ, M. D., TUNG, C. AND SWARTZ, M. N. Elimination of plasmid pMG110 from Escherichia coli by novobocin and other inibitors of DNA gyrase. Antimicrobial Agents Chemotherapy, v.25, p.586-590, 1984.

99
JONES,R. N.; ANDEREGG, T. R.; DESHPANDE, L.M. AZD 2563, A new oxazolidinone bactericidal activity and synergy studies combined with gentamicin or vancomycin against staphylococci and streptococcal stains. Diagn Microbiol Infec Dis, v.43, n.1, p.87-90,2002.
KOROLKOVAS, A. Dicionário Terapêutico Guanabara 2002/2003. ed. 2002/2003 Rio de Janeiro, Guanabara Koogan. 2003, p.300
LAMBERT, P.A; Mechanisms of antibiotic resistance in Pseudomonas aeruginosa, Journal of the Royal Society of Medicine Suplement, v.95, n.41, 2002.
LEDEBERG, J. & LEDEBERG, E. M. Replic plating and indirect selection of bacterial mutants. Journal of Bacteriology, v.63, p.399 – 406, 1952.
LEWIS, K.; SALYERS, A.A.; TABER, H.W.; WAX, R.G.; Bacterial resistance to antimicrobials. Marcel Dekker, v.5, p.97-99, New York, 2002.
LIMA, I. O.; OLIVEIRA, R. A. G.; LIMA, E. O.; FARIAS, N. M. P.; SOUZA, E. L. Atividade antifúngica de óleos essenciais sobre espécies de Candida. Rev Bras Farmacognosia, v.16, p.197-201, 2006.
LIRA, A.A.M.; SESTER, E. de A; ABREU, L.R.P.; SILVA, L.B.L.; WANDERLEY, A.G.; SANTANA, D.P. Desenvolvimento preliminar de gel de Lapachol: Estudo de permeação in vitro. Revista Brasileira de Ciência Farmacêutica, v.40, n.1, 2004.
LIU, H.H.; Antibiotic resistance in bacteria Rheumaderm, v.455, p.387-395, 1999.
LOGUERCIO-LEITE, C.; GROPOSO, C.; DRESCHLER-SANTOS, E. R.; FIGUEIREDO, N. F.; GODINHO, P. S.; ABRÃO, R. L. A particularidade de ser um fungo – I. constituintes celulares. Biotemas, v.19, n.2, p.17-27, 2006.
LYON, B. R. & SKURRAY, R., 1987. Antimicrobial resistance of Staphylococcus aureus: genetic basis. Microbiology Rewiews, v.51, p.88-134.
MACEDO, M. de L. de A. P.; CARTAXO, R. de S.; ALMEIDA, T.C. da C.; SOUSA, L.B.S.; SANTANA, W.J.; COUTINHO, H.D.M. Mecanismos de resistência e detecção das β-lactamases. Ciências Biol. Saúde, v.7,n.1, p.59-63, 2005.
MACHADO, V. L. L.; PALMA, M. S.; COSTA, O. M. Ação repelente das frações de óleos essenciais da folha de louro (Laurus nobilis) em ninfas e adultos de Periplaneta amaericana L. (Blataria: Blattidade). An. Soc. Entomol. Brasil, v.24, n.1, p.13-29, 1995.
MARTHINS, A.F; Caracterização de metalo-β-lactomases produzidas por amostras de Pseudômonas aeruginosa isoladas em dois hospitais de Porto Alegre. Dissertação de Mestrado, Universidade Federal de Rio Grande do Sul, Fac. De Farmácia, Porto Alegre, 2005.

100
McGINNIS, M. R. Laboratory handbook of medical mycology. New York: Academic Press, 1980, p.411-416.
MOELLERING, R. C. Jr. Novos desafios no campo das doenças infecciosas. In: Patógenos Emergentes nas Doenças Infecciosas: Relatório Especial Hospital Practice. Euromédice, p.5-7. 2000.
MONTANARI, C. A.; BOLZANI, V. da S.; Planejamento racional de fármacos baseado em produtos naturais, Quim. Nova, v.24, p.1, 105-111, 2001.
MOREIRA, L, B; Princípios para o uso racional de antimicrobianos, Revista AMRIGS, v.48, n.2, p.118 – 120, Porto Alegre, 2004.
MOREIRA, L.S; Estudo da atividade de drogas antifúngicas através de curvas de crescimento de candida albicans utilizando o sistema automatizado Bio-cell tracer . Dissertação de mestrado, Universidade Estadual de Campinas, 2003.
NASSIS, C. Z.; GIESBRECHT, A. M.; SAITO, M. L.;OLIVEIRA, F. Atividade antibiótica de extratos de espécies de Annonaceae. Cienc. Cult, v.41, n.7, p.527 1989.
NATARO ,J. P.; KAPER J. B. Diarrheagenic Escherichia coli. Clin Microb Rev, v.11, n.1, p.142-201, 1998.
NATTIONAL COMMITTEE FOR CLINICAL LABORATORY STANDART - NCCLS, Methods for diluition antimicrobial susceptibility tests for bacteria that grow aerobically. 2 ed. Tentative standart. NCCLS Document M7-T2, V. B. n. B. Villa Nova PA, National Commitee for Clinical Laboratory Standart, 2002.
ODDS, F. C. Antifungal activity of saperconazole (R. 66905) in vitro. J Antimicrob Chemother, v.24, p.533-537, 1989.
OLIVEIRA, A.O.T. de; PEDROSO, C.F.; MIGUEL, M.D.; MONTRUCCHIO, D.; ZANIN, S.M.W.; DORNELES, D.; Atenção farmacêutica na antibioticoterapia visão acadêmica, v.5, n.1, p.7-4, 2004.
OLIVEIRA, C. G. T.; MIRANDA, F. F.; FERREIRA, V. F.; FREITAS, C. C.; RABELLO, R. F.; CARBALLIDO, J. M.; CORRÊA, L. C. D. Synthesis and antimicrobial evaluation of 3-hydrazino-napfthoquinones as analogs of lapachol. J Bras Chem Soc, v.12, n.3, p. 339-345, 2001.
PAVIANI, E. R; STADNIK, C.B; HEINEK, J; Estudo da epidemiologia e perfil de sensibilidade da Pseudomonas aeruginosa. Infarma, v.15, n.11-12, 2004.
PEREIRA, M. do S.V; Atividade in vitro de fluorquinolonas e ação sobre plasmídios em amostra de staphylococcus aureus humanas e bovinas, Tese de doutorado UFPB, CCB, 2000.

101
__________. Staphylococcus aureus: o micro vilão da resistência a antibióticos, on line, janeiro de 2002a. http//WWW.biologianaweb/biomural/ divulga/ staph/socorro.html
__________. Staphylococcus aures: o micro vilão da resistência a antibióticos, on line, janeiro de 2002b. http//WWW.biologianaweb/biomural/ divulga/staphy/socorrol.html.
___________. Alguns aspectos genéticos da resistência a drogas em amostras bovinas de Staphylococcus aureus isoladas no estado da Paraíba (Dissertação de Mestrado. UFPB). 87p. 1992.
PEREIRA, M. do S.V.; SIQUEIRA Jr., J. P. Antimicrobial drugs resistance in Staphylococcus aureus isolated from cattle in Brasil. Letters in Applied Microbiology, v.20, p.391-395,1995.
PEREIRA, M. do S.V.; SIQUEIRA Jr. J.P; TAKAKI G.M.C. Eliminação de resistência a drogas por fluorquinolonas em Staphylococcus aureus de origem bovina. Pesq. Vet. Bras, v.24, p.11-14, 2004.
PEYRET, M.; CARRET, G.; CARRE, C.; FARDEL, G.; FLANDROIS, J. P. etude mathematique de la curve de mortalité d’Escherichia coli exposure aux polymixines. Pathologie Biologie, v.38, p.441-445, 1990.
PLEMPEL, M.; BERG, D.; BUCHEL, D. B.; ABBINK, D. Test methods for antifungalagents – a critical review. Mykosen, v.30, n.1, p.28-34, 1986.
PONTES, D.S. et al. Perfil de resistência a antibióticos e ao mercúrio no trecho médio da bacia do Rio Doce (Parque Estadual do Rio Doce). In: Anais XXIV Reunião de Genética de Microrganismos. Gramado. Rio Grande do Sul. Comunicação Impressa, 2004. p.211.
RIBSON, G. R. Human gut microbiology: the end of the food chain or the start of good health? Microbiol Tod, v.29, p.4-6, 2002.
RIVERÓN, F. F.; HERNANDÉZ, J. L.; MARTÍNEZ, L. M. P.; BETARTE, C. M. Resistencia bacteriana. Rev Cub Méd Milit, v.32, n.1, p.44-48, 2003.
RUSSEL, A.D. Bacterial resistance to disinfectants: Present know large anal future problems, Journal of Hospital infection, v.43, p.557-568, 1998.
SALLÉ, J. O totum em fitoterapia - abordagem de fito-bioterapia. São Paulo: Rose, 1996.
SAKAR, M. K. Antimicrobial activities of some Hypericum species growing in Turkey, Fitoterapia, v.59, p.49-51, 1988.

102
SANTOS dos J.C.A.; SOUSA de R.V.; SANTANA, G. de C. Princípios da antibioticoterapia em medicina veterinária. Disponível na internet. Acessado em 10/08/2006.
SANTOS, B. A. As infecções respiratórias em crianças e a resistência bacteriana: um desafio para o pediatra. Rev AMRIGS, v.48, n.2, p.135-138, 2004
SCHAECHTER, M.; ENGLEBERGI N. C.; EISENSTEIN, B. I.; MEDOFF, G. Microbiologia. 3ªEd. Guanabara Koogan:120-127. 642p. 2002.
SCHWEIZER, H.P; Efflux as a mechanism of resistance to antimicrobials in Pseudomonas aeruginosa and related bacteria: Unanswered questions.Genetic and Molecular Research, v.2, p.48-62, 2003.
SEABRA, F. R. G.; SEABRA, B. G. M.; SEABRA, E. G. Antibioticoterapia profilática em cirurgias periodontais. Odontol Clin Cient, v.3, n.1, p.11-16, 2004.
SHADOMY, S.; ESPINEL-INGROF, A.; GABHAR, R. J. In vitro studies with sf 86-327, a new orally active allylamione derivatives. J Med Vet Mycol, v.23, p.125-132, 1985.
SHELBURNE, S. A.; MUSHER, D. M.; HULTEN, K.; CEASAR, H.; LU,M. Y.; BHAILA, I., HAMILL, J. In vitro killing of community-associated methicillin-resistant Staphylococcus aureus with drugs combinations. Antimicrob Agents Chemother, v.48, n.10, p.4016-4019, 2004.
SHLAES, D. M.; GERDING, D. N.; CRAIG, W. A.; BORNSTEIN, D. L.; DUNCAN, R. A.; ECKMAN, M. R.; FARRER, W. D.; GREENE, W. H. Society for healthcare epidemiology of America and infections diseases society of America joint committee of the prevention of antimicrobial resistance: guidelines for the prevention of antimicrobial resistance in hospitals. SHEA, v.18, n.4, p.275-295, 1997.
SILVA, J.A; da SILVA, W.D. Escherichia coli Enteropatogênica (EPEC), ao contrário da Escherichia coli comensal, adere sinalisa e lesa enterócitos. Revista de Patologia Tropical, v.34, p.175-196, 2005.
SILVA, M. N.; FERREIRA, V. F.; SOUZA, M. C. B. V. Um panorama atual da química e da farmacologia de naftoquinonas, com ênfase na -lapachona e derivados. Rev Quim Nova, v.26, n.3, p.407-417, 2003.
SILVA, T.M.S; CÂMARA, C.A; BARBOSA, T.P; SOARES, A.Z; PINTO, A.C; VARGAS, M.D; Mollucicidae activity of syntetic Lapachol amino- and hydrogenated – derivatives. Bioorg Med Chem, v.13, p.193-196, 2005.
SILVEIRA, G.P. NOME, F.; GESSER, J.C.; AS, M. M.; TERENZI, H.; Estratégias utilizadas no combate a resistência bacteriana, Química Nova, v.29, n.4, p.844-855, 2006.

103
SMITH, R.; COAST, J. Antimicrobial resistance: a global response; Bulletin of the World Heath Organization, v.80, n.2, 2002.
SOUZA, M. V.; REIS, C.; PIMENTA, F. C. Revisão sobre a aquisição gradual de resistência de Staphylococcus aureus aos antimicrobianos. Rev Patol Trop, v.34, n.1, p.27-36, 2005.
SUDBERY, P.; GOW, N; BERMAN, J. The distinct morphogenic states of Candida albicans. Trends in Microbiol, v.12, n.7, p.217-324, 2004.
TAVARES, W.; Bactérias Gram-positivas problemas: resistência do estafilococo e do enterococo e do pneumococo dos antimicrobianos. Revista da Sociedade Brasileira de Medicina Tropical, v.33, n.3, p.281-301, 2000.
TENOVER, F. C. A melhor época, a pior época: a evolução do problema de resistência das bactérias aos agentes antimicrobianos. In: Patógenos emergentes nas doenças infecciosas: relatório Especial Hospital Practice. Euromédice. Ed. Médicas. 8-13, 2000.
THUILLE, N.; FILLE, M.; NAGL, M. Batericidal activity of herbal extracts. Int. J. Hyg. Environ. Health, v.206, n.3, p.217-221, 2003.
TOMAZ, A. Mulpiple-antibiotic-resistance pathogenic bacteria: a repot on Rockefeller - University Worksop. J. Med. n.330, p.1247-1251. 1994.
TRABULSI, L.R.; ALTERTHUM, F.; Microbiologia, Atheneu, 4ed., São Paulo, 718p., 2006.
TURRI, R. de J.G. Estudo de fatores de virulência em amostras de Klebsiella pneumonial e Escherichia coli β-lactanases espectro estendidas. Tese de Doutorado da Universidade Estadual de Campinas, 2003.
UENO, M.; JORGE, A. O.C. Caracterização de Staphylococcus aureus resistentes a meticilina, envolvidas em infecções nosocomiais, por meio de técnicas fenotípicas e análise de perfil plasmidial. Revista Biociências, v.7, p.15-22, 2001.
VASCONCELOS, L.C.S. et al., Use of Punica granatum Linn. As antifungal agent against candidioses associeted with denture stomatitis. Mycoses, v.46, n.6, p.192-196, 2004.
VON POSER.G. L.; MENTZ, L. A. Diversidade biológica e sistemas de classificação. In: SIMÕES, C. M. O.; SCHENKEL, E. P.; GOSMANN, G.; MELLO, J. C. P. de; MENTZ, L. A.; PETROVICK, P. R. (Orgs.). Farmacognosia: da planta ao medicamento. 2.ª ed. Porto Alegre/Florianópolis: UFRGS/URFSC, 2000, p.66-70.

104
VUCKOVIC, N.; BOKOR-BRATIC, M.; VUCKOVIC, D.; PICURIC, I.; Presence of Candida albicans in potentially malignant oral mucosal lesions. Arch Oncol, v.12, n.1, p.51-54, 2004.
WEISSER, J. & WIEDEMANN B. Elimination of plasmids by new 4-quinolones. Antimicrobial Agents and Chemotherapy, v.28, p.700 - 702. 1985.
WICKENS, H.; WADE, P. Understanding antibiotic resistance. Pharm J, v.274, p.501-505, 2005.
WILLIAMS, D. A.; LEMKE, T. L. Foye’s Principles of Medicinal Chemistry. 5 ed. Philadelphia, Lippincot Williams & Wilkins. p.1114. 2002.
WONG-LEUNG, Y. L. Antibacterial activities of some Hong-Kong plants used in Chinese medicine. Fitoterapia, v.69, n.1, p.11-16, 1988.
XU, HONG-XI: LEE, F. Song Activity of plant Flavonoids Against Antibiotic-Resistant Bacteria. Phytotherapy Research, v.15, p.39-43, 2001.
YAMADA, C. S. B. Fitoterapia: sua história e importância. Fitoterapia - Revista Racine, São Paulo, n. 43, p.50-51, 1998.
YUNES, R. A.; Fármacos e Fitoterápicos: A necessidade do desenvolvimento da indústria de fitoterápicos e fitofármacos no Brasil. Química Nova, v.24, n.1, p.147-152, 2001.
Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo





























