Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DO CEARÁ
CENTRO DE CIÊNCIAS
DEPARTAMENTO DE BIOQUÍMICA E BIOLOGIA MOLECULAR
PROGRAMA DE PÓS-GRADUAÇÃO EM BIOQUÍMICA
ALYSSON CHAVES ALMEIDA
CARACTERIZAÇÃO ESTRUTURAL E BIOLÓGICA DE UMA LECTINA DE
SEMENTES DE Centrolobium tomentosum GUILL. EX BENTH.
FORTALEZA
2016

ALYSSON CHAVES ALMEIDA
CARACTERIZAÇÃO ESTRUTURAL E BIOLÓGICA DE UMA LECTINA DE
SEMENTES DE Centrolobium tomentosum GUILL. EX BENTH.
Tese de Doutorado submetida à
Coordenação do Curso de Pós-Graduação
em Bioquímica da Universidade Federal do
Ceará como requisito parcial para obtenção
do grau de Doutor em Bioquímica.
Orientador: Prof. Dr. Benildo Sousa
Cavada.
FORTALEZA
2016



Aos meus pais Marta e Almeida, por terem me
oferecido o amor mais puro e terem feito de
seus sonhos os meus;
Aos meus avós Lurdinha e Zequinha (In
memorian) e à minha segunda mãe Mundinha,
por todo amor e carinho;
À minha amada esposa Rebecca, pelos
conselhos, apoio e muito amor;
Dedico.

AGRADECIMENTOS
Agradeço primeiramente a Deus pela proteção, pela presença constante em minha
vida e pela benção que agora me oferece;
Ao meu orientador, Prof. Dr. Benildo Sousa Cavada, que acreditou no meu trabalho
e me acolheu em seu grupo. Agradeço também pelo exemplo de grande ser humano, de grande
liderança;
À Prof.ª. Dr.ª Kyria Santiago do Nascimento, pela colaboração na realização deste
trabalho e pelos valiosos conhecimentos transmitidos durante o convívio no laboratório.
À Prof.ª Dr.ª Alana de Freitas Pires, por aceitar fazer parte da banca examinadora
dessa tese, pelos valiosos ensinamentos e pela colaboração nos testes de atividade biológica.
Ao Prof. Dr. Francisco Nascimento Pereira Junior, por compor a banca
examinadora desta tese, pelos ensinamentos, pela amizade e honestidade sempre constantes.
Ao Prof. Dr. João Batista Cajazeiras, por aceitar fazer parte da banca examinadora
dessa tese.
À Prof.ª Dr.ª Ana Maria Assreuy, pela disponibilidade e atenção, e toda a equipe do
LAFFIN pelo apoio e colaboração nos testes de atividade biológica.
Um agradecimento especial aos amigos Mayara Queiroz, Vinícius Silva e Vanir
Reis, três grandes amigos, que colaboraram diretamente não só com a redação desse trabalho
de tese, mas também com seu artigo científico. Além disso, agradeço os inúmeros ensinamentos
nessa nossa nova empreitada pela bioinformática. Obrigado pela amizade, pelo convívio sempre
alegre, pela consideração e pela compreensão. Serei eternamente grato a vocês.
Aos amigos Alfa, Ana Cecília, William, Jorge, Mayara Queiroz, Mayara Torquato,
Vanir e Vinícius. Obrigado por tantos momentos de alegria que vivemos juntos, pelos milhares
de ensinamentos que me passaram e pelo apoio nos momentos difíceis. Agradeço também por
toda a ajuda na redação deste trabalho;
Aos colegas do BioMol-Lab, Clareane, Cleane, Cláudia, Ivanice, David, Gleiciane,
Batista, Lucas, André, Ronniery, Simoni e a todos os demais que compõe o BioMol-Lab que
contribuíram durante esta caminhada e a quem aprendi a ter um grande respeito.
À todos os professores e funcionários do Departamento de Bioquímica e Biologia
Molecular da UFC, pelos valiosos ensinamentos que me proporcionaram ao longo do curso;
À Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) pelo
auxílio financeiro.

De maneira mais que especial, agradeço aos meus pais, Marta e Almeida. Mãe,
obrigado pelo amor e carinho mais verdadeiro que pode existir neste mundo, por estar ao meu
lado em todos os momentos, por nunca parar de orar por mim um só segundo. Pai, obrigado
pelo exemplo de homem trabalhador e pai amoroso que, mesmo em face à dificuldade, nunca
deixou de acreditar que a vida é uma dádiva de Deus. Se hoje sou um homem vitorioso, é porque
vocês me ensinaram a seguir o caminho correto. Amo vocês e nunca os abandonarei.
À toda a minha família, que sempre acreditou em mim. Em especial, aos meus avós
Lurdinha e Zequinha, que me ensinaram a ter amor pela natureza e por me receberem sempre
com um abraço apertado e um sorriso. À minha avó pelo carinho mais tenro e pelas orações
constantes; ao meu avô, pelo exemplo de homem simples, íntegro, dedicado à família, sempre
perseverante e alegre. Amo vocês.
À Mundinha, minha segunda mãe, por ter feito parte da minha vida desde o primeiro
minuto e cuidado de mim sempre com muito amor. Obrigado por me mostrar que não há
problema algum na vida que valha a pena perder o sorriso e a esperança num futuro melhor.
Amo você.
À minha amada esposa Rebecca, por estar ao meu lado quando mais precisei, por
ter me mostrado tanta coisa que não conhecia, por ter me ajudado a ser o homem que sou hoje,
acreditando no meu potencial e me incentivando a caminhar sempre em frente. Obrigado por
ter iluminado minha vida com seu lindo sorriso. Você é a mulher com quem quero compartilhar
todos os meus sonhos, minha eterna companheira, a futura mãe dos meus filhos. Você fez e faz
minha vida muito mais feliz. Eu te amo muito;
À todas as pessoas que me incentivaram e acreditaram na minha vitória.

“O mundo não é um mar de rosas, é um lugar duro e
maldoso e não importa o quanto você ache que é forte, em
algum momento ele vai te deixar de joelhos e te deixará
assim permanentemente se você permitir. Nem você nem
ninguém vai bater tão forte como a vida. Mas isso não é
sobre o quão forte você bate, é sobre o quão forte você
aguenta ser golpeado e seguir adiante. É assim que a vitória
é conquistada..."
Rocky Balboa

RESUMO
Uma lectina glicosilada (CTL) com especificidade a manose e glucose foi detectada e purificada
a partir de sementes de Centrolobium Tomentosum, uma leguminosa pertencente à tribo
Dalbergieae. CTL foi isolada por cromatografia de afinidade de Sepharose-Manose. A estrutura
primária foi determinada por espectrometria de massas e consiste em 245 aminoácidos e um
sitio de N-glicosilação, demonstrando similaridade com a lectina de Platypodium elegans
(PELa), Pterocarpus angolensis (PAL), dentre outras, oriundas da mesma tribo. Duas estruturas
cristalinas de CTL, de formas monoclínica e tetragonal, ambas complexadas com metil-
dimanosídeo, foram resolvidas a 2,25 e 1,9 Å, respectivamente, apresentando alta similaridade
entre si. A lectina mostrou adotar uma organização dimérica canônica típica de lectinas de
leguminosas. O domínio de reconhecimento de carboidratos (CRD), local de ligação do metal
e local de glicosilação foram caracterizados e a base estrutural para a interação com carboidratos
foi elucidado. CTL mostrou efeito inflamatório agudo em um modelo de edema de pata. A
estrutura da proteína foi submetida a uma análise de interações com dimanosídeos e
trimanosídeos por Docking Molecular, revelando sua maior afinidade por trimanosídeos e suas
interações foram comparadas com lectinas similares que possuam a mesma especificidade de
ligação. Esse é o primeiro relato de estrutura cristalina de uma lectina nativa manose/glucose
específica da tribo Dalbergieae com atividade pró-inflamatória.
Palavras chave: lectina; Centrolobium tomentosum; estrutura primária; estrutura cristalográfica;
atividade pró-inflamatória; Docking molecular.

ABSTRACT
A glycosylated lectin (CTL) with specificity for mannose and glucose has been detected and
purified from seeds of Centrolobium tomentosum, a legume plant from the Dalbergieae tribe.
CTL was isolated by mannose-Sepharose affinity chromatography. The primary structure was
determined by tandem mass spectrometry and consists of 245 amino acids and one N-
glycosylation site, possessing high similarity with the lectin Platypodium elegans (PELa) and
Pterocarpus angolensis (PAL) derived from the same tribe. Two crystal structures of CTL, with
monoclinic and tetragonal forms, both complexed with methyl dimanosídeo has been solved at
2.25 and 1.9 Å, respectively, with high similarity. The lectin adopts a typical canonical dimeric
organization of legume lectins. The carbohydrate recognition domain (CRD), metal binding
site, and glycosylation site have been characterized and the structural basis for interaction with
carbohydrate been elucidated. CTL showed acute inflammatory effect in a paw edema model.
The protein structure was subjected to ligand screening (dimannosides and trimannoside) by
molecular docking, revealing a higher affinity for trimannosides and their interactions were
compared with similar lectins, which have the same binding specificity. This is the first report
of a crystal structure of a native mannose lectin / specific glucose Dalbergieae tribe with pro-
inflammatory activity.
Keywords: Lectin; Centrolobium tomentosum; Primary structure; Crystal structure;
Proinflammatory activity; Molecular docking.

LISTA DE ILUSTRAÇÔES
Figura 1. Classificação estrutural de lectinas vegetais. ............................................................ 26
Figura 2. Classificação das lectinas de plantas em famílias evolutivamente relacionadas. ..... 28
Figura 3. Processamento pós-traducional durante a biossíntese de ConA. .............................. 31
Figura 4. Estrutura de monômero típica das lectinas de leguminosas. ..................................... 32
Figura 5. Estruturas tridimensionais de lectinas da tribo Dalbergieae. .................................... 35
Figura 6. Processamento pós-traducional proposto para a lectina da Vatairea macrocarpa. .... 37
Figura 7. Centrolobium tomentosum Guill. ex Benth. ............................................................. 39
Figura 8. Representação esquemática dos três métodos de difusão de vapor para cristalografia
de macromoléculas. .................................................................................................................. 46
Figura 9. Etapas da Cristalografia de raios X. .......................................................................... 48
Figura 10. Diagrama esquemático dos passos do processo de autofagia. ................................ 56
Figura 11. Sequência de aminoácidos da CTL. ........................................................................ 62
Figura 12 - Alinhamento da sequência de aminoácidos de Centrolobium tomentosum (CTL)
com lectinas da tribo Dalbergieae. ........................................................................................... 63
Figura 13. Cristais de CTL complexada com Metil-Dimanosídeo. .......................................... 69
Figura 14. Gráfico de Ramachandran das coordenadas de CTL. ............................................. 72
Figura 15. Estrutura geral do monômero de CTL. ................................................................... 73
Figura 16. Estrutura tridimensional de CTL complexada com metil-dimanosídeo. ................ 73
Figura 17. Sítio de N-Glicosilação de CTL. ............................................................................. 74
Figura 18. Sítio de ligação à metais de CTL. ........................................................................... 75
Figura 19. Sítio de ligação a carboidratos de CTL interagindo com metil-dimanosídeo. ........ 75
Figura 20. Sobreposição do sítio de ligação a carboidratos das formas monoclínica e tetragonal
de CTL. ..................................................................................................................................... 78
Figura 21. Sobreposição dos monômeros das formas monoclínica e tetragonal de CTL. ....... 79
Figura 22. Sobreposição de entre as estruturas de CTL, PELa e PAL complexadas com
trimanosídeos. ........................................................................................................................... 80
Figura 23. Indução de edema da pata por CTL. ....................................................................... 85
Figura 24. Avaliação do efeito vasorelaxante de CTL. ............................................................ 86
Figura 25. Ensaio de citotoxidade de CTL em células tumorais. ............................................. 87

LISTA DE TABELAS
Tabela 1. Lectinas da tribo Dalbergieae. ................................................................................. 34
Tabela 2. Estatísticas da coleta de dados de difração de raios X de CTL em complexo com
metil-dimanosídeo. ................................................................................................................... 71
Tabela 3. Interações entre CTL e metil-dimanosídeo. ............... Erro! Indicador não definido.

LISTA DE ABREVIATURAS E SIGLAS
AAL Lectina de Andira anthelmia
ABU Ácido α-aminobutírico
ACA Lectina de Amaranthus caudatus
ACh Acetilcolina
AUC Área sob a curva
cDNA DNA complementar
CEUA Comitê de Ética da Universidade Estadual do Ceará para Uso de Animais
CFL Lectina de Cratylia floribunda
CGL Lectina de Canavalia grandiflora
CID Dissociação induzida por colisão
ConA Lectina de Canavalia ensiformis
ConA Lectina de Canavalia ensiformis
ConBr Lectina de Canavalia brasiliensis
CRD Domínio de reconhecimento a carboidratos
CRLI Lectina I de Cymbosema roseum
CTL Lectina de Centrolobium tomentosum
CVL Lectina de Canavalia virosa
DDA Aquisição Dependente de Dados
DM Docking molecular
DMSO Dimetilsulfóxido
DRC Domínio de reconhecimento a carboidratos
DSL Lectina de Dioclea sclerocarpa
DVL Lectina de Dioclea violácea
ESI Ionização por Eletrospray
GMPc Monofosfato cíclico de guanosina
GNA Lectina de Galanthus nivalis
GTP Trifosfato de guanosina
HBSS Solução salina balanceada de Hank
LaL Lectina de Lonchocarpus araripensis
LC-MS/MS Cromatografia líquida acoplada a espectrometria de massas sequencial
LNLS Laboratório Nacional de Luz Síncrotron
MALDI Dessorção/Ionização a Laser Auxiliada por Matriz

MDM Metil-dimanosídeo
MS/MS Espectrometria de massas sequencial
MT1-MMP proteína de membrana - metaloproteinase tipo 1
MTT 3-(4,5-dimetiltiazol-2-il)-2,5-difeniltetrazólio
NAG N-acetil-D-glicosamina
NCBI National Center of Biotechnology Information
NO Óxido nítrico
PAL Lectina de Pterocarpus angolensis
PCT Teste de Pré-Cristalização
PDB Protein Data Bank
PELa Lectina de Platypodium elegans
PPL-1 Lectina 1 de Parkia platycephala
PPL-2 Lectina 2 de Parkia platycephala
RIP Proteínas inativadoras de ribossomos
RMSD Raiz quadrada do desvio médio (do inglês root mean square deviation)
SDS-PAGE Eletroforese em gel de poliacrilamida com de Sódio Dodecil Sulfato
TNF-α Fator de Necrose Tumoral α
ToF Tempo de vôo (do inglês Time off light)
TriMan Trimanosídeo
TxLCI Lectina do bulbo da tulipa
UFC Universidade Federal do Ceará
VGL Lectina de Vatairea guianensis
VML Lectina de Vatairea macrocarpa

SUMÁRIO
1 INTRODUÇÃO ........................................................................................................... 16
2 OBJETIVOS ............................................................................................................... 19
2.1 Objetivo Geral ............................................................................................................. 19
2.2 Objetivos Específicos .................................................................................................. 19
3 CAPÍTULO I: FUNDAMENTAÇÃO TEÓRICA ................................................... 21
3.1 Lectinas vegetais ......................................................................................................... 21
3.1.1 Aspectos históricos e definição de lectinas ................................................................. 21
3.1.2 Generalidades sobre lectinas vegetais ......................................................................... 23
3.1.3 Classificação estrutural de lectinas vegetais ............................................................... 25
3.1.4 Lectinas de leguminosas .............................................................................................. 30
3.1.5 Lectinas de Dalbergieae ............................................................................................... 33
3.1.6 Lectina de Centrolobium tomentosum ........................................................................ 38
3.2 Aspectos gerais da espectrometria de massas .......................................................... 40
3.3 Cristalografia de raios X ............................................................................................ 44
3.4 Docking molecular ...................................................................................................... 48
3.5 Atividades biológicas de lectinas vegetais ................................................................. 50
3.6 Aspectos gerais do processo inflamatório ................................................................. 51
3.7 Aspectos gerais da contratilidade .............................................................................. 52
3.8 Câncer, gliomas e autofagia ....................................................................................... 53
3.8.1 Câncer .......................................................................................................................... 53
3.8.2 Gliomas ......................................................................................................................... 54
3.8.3 Autofagia ...................................................................................................................... 55
4 CAPÍTULO II: DETERMINAÇÃO DA ESTRUTURA PRIMÁRIA DE CTL POR
ESPETROMETRIA DE MASSAS ....................................................................................... 59
4.1 Digestão proteolítica in gel e obtenção de peptídeos ................................................ 59
4.2 Sequenciamento de peptídeos por MS/MS ............................................................... 60
4.3 Análise da estrutura primária por bioinformática .................................................. 60
4.4 Resultados e Discussão ............................................................................................... 61
4.4.1 Estrutura primária de CTL .......................................................................................... 61
5 CAPÍTULO III: RESOLUÇÃO DA ESTRUTURA TRIDIMENSIONAL DE CTL
POR CRISTALOGRAFIA DE RAIOS X ............................................................................ 66
5.1 Ensaio de Cristalização .............................................................................................. 66
5.2 Coleta de dados e resolução da estrutura cristalográfica ....................................... 67

5.3 Docking Molecular ........................................................... Erro! Indicador não definido.
5.4 Resultados e Discussão ............................................................................................... 69
5.4.1 Cristalização e coleta de dados .................................................................................... 69
5.4.2 Coleta de dados e refinamento da estrutura ............................................................... 69
5.4.3 Estrutura cristalográfica de CTL em complexo com metil-dimanosídeo .................. 72
5.4.4 Comparação entre as formas monoclínica e tetragonal de CTL ............................... 77
5.4.5 Docking molecular ....................................................................................................... 79
6 CAPÍTULO IV: AVALIAÇÃO Do ATIVIDADES INFLAMATÓRIAS,
VASORELAXANTES E DA CITOTOXICIDADE EM CÉLULAS TUMORAIS .......... 82
6.1 Modelo de edema de pata ........................................................................................... 82
6.2 Teste de contratilidade em aortas isoladas ............................................................... 82
6.3 Análise estatística ........................................................................................................ 83
6.4 Avaliação da Citotoxicidade em Células Tumorais ................................................. 83
6.4.1 Análise estatística ......................................................................................................... 84
6.5 Resultados e Discussão ............................................................................................... 84
6.5.1 Atividade inflamatória de CTL .................................................................................... 84
6.5.2 Avaliação do efeito relaxante de CTL ......................................................................... 85
6.5.3 Avaliação da atividade citotóxica de CTL ................................................................... 87
7 CONCLUSÃO ............................................................................................................. 89
REFERÊNCIAS ..................................................................................................................... 90

16
1 INTRODUÇÃO
Os carboidratos são a maior fonte de biomassa do planeta Terra e sua complexidade,
especialmente em termos de variações estruturais, não tem igual em outras classes de
biomoléculas (NELSON; COX, 2014).
O enorme potencial biológico codificado por carboidratos resulta da sua capacidade
de criar estruturas complexas, incluindo pontos de ramificação a partir de monômeros simples
(lineares). Esta característica dos carboidratos é única entre todos os polímeros biológicos. Por
exemplo, dois aminoácidos diferentes podem se ligar apenas por uma de duas maneiras. Por
outro lado, um monossacarídeo simples, como a glicose, pode se ligar com o seu carbono
anomérico (α ou β) a cinco hidroxilas presentes em uma segunda glucose, dando origem a até
onze diferentes isômeros estruturais. É compreensível, portanto, porque o número de variações
estruturais fornecidas por este tipo de moléculas é enorme (SHARON; LIS, 1993).
A estrutura de glicanos (oligossacarídeos, glicoconjugados, polissacarídeos)
depende fortemente da sua composição e, além disso, a presença de ligações glicosídicas
introduz uma flexibilidade estrutural e conformacional considerável. A consequência direta
dessa variabilidade estrutural é uma grande diversidade no papel e localização de carboidratos
nos organismos vivos (TAYLOR; DRICKAMER, 2003).
Essa complexidade presente nos carboidratos permite que esses funcionem como
moléculas de sinalização, reconhecimento e adesão. Além disso, eles estão envolvidos em
várias funções fisiologicamente importantes que incluem: armazenamento de energia; defesa;
desenvolvimento embrionário normal; diferenciação; crescimento; inibição de contato;
reconhecimento célula-célula; interação hospedeiro-patógeno durante a infecção; resposta
imune do hospedeiro; desenvolvimento de doença; metástase; tráfego e localização intracelular
e controle de degradação e da rigidez da membrana celular (GHAZARIAN; IDONI;
OPPENHEIMER, 2011; TAYLOR; DRICKAMER, 2003).
No entanto, por mais de um século a pesquisa nas áreas de ácidos nucléicos,
proteínas e lipídios detiveram toda a atenção de pesquisadores em todo o mundo. Os
carboidratos, provavelmente por serem muito complexos e não codificados no genoma,
passaram a receber maior atenção algum tempo depois (GABIUS et al., 2011). No início do
século XX, os conhecimentos sobre a biologia dos carboidratos eram, ainda, limitados àqueles
com papéis estruturais, como a celulose nas plantas e a quitina dos exoesqueletos de insetos, ou
de acúmulo e transporte de energia, como a glicose e o amido. Apenas em 1968, os carboidratos
foram reconhecidos como moléculas biológicas extremamente importantes. Nessa época,

17
admitia-se que todas as informações biológicas eram realizadas pelas duas principais classes de
moléculas: proteínas e ácidos nucleicos (LIS; SHARON, 1998; SHARON; LIS, 1993).
Grandes descobertas que marcaram a elucidação do papel dos oligossacarídeos: o
estabelecimento da estrutura glicídica dos determinantes do sistema ABO; a demonstração da
aglutinação de eritrócitos pelo vírus influenza pela interação entre a hemaglutinina e ácido
siálico; a observação de que a defucosilação de linfócitos impedia sua migração pelos tecidos.
Estas descobertas foram a fonte de muita pesquisa que tem ligado o papel dos carboidratos nos
processos de comunicação e reconhecimento molecular (SHARON; LIS, 1993).
Os carboidratos localizados sobre a superfície das células contribuem para a maioria
das interações entre as células e o ambiente. Por formarem uma camada que recobre as células,
os carboidratos que constituem o glicocálice geralmente são as primeiras moléculas a serem
encontradas e reconhecidas por outras células, anticorpos e microrganismos invasores, como
vírus e bactérias. Moléculas secretadas, tais como hormônios e toxinas, também se ligam a
motivos de carboidratos que atuam como receptores para essas moléculas. Em função do já
conhecido papel da glicosilação na comunicação celular, não é surpreendente que padrões de
glicosilação anormais são conhecidos como marcadores ou, em alguns casos, até mesmo a causa
de certas doenças como, por exemplo, câncer (DENNIS et al., 1999).
Os avanços na pesquisa de carboidratos fizeram surgir o termo “Glicobiologia”,
definida como a ciência que estuda as estruturas, propriedades e funções dos carboidratos
presentes em diversidade nos seres vivos. Estes estudos não se restringem apenas aos
carboidratos, mas também a todas as outras classes de biomoléculas (proteínas, lipídios,
nucleotídeos) que interagem com eles. O objetivo do glicobiologia estrutural é, naturalmente,
determinar a estrutura tridimensional e estabelecer modelos para as interações envolvendo essas
moléculas (TAYLOR; DRICKAMER, 2003).
No entanto, não há como discutir glicobiologia sem referenciar as lectinas. Essas,
por serem moléculas capazes de “decifrar os glicocódigos” codificados na estrutura dos
glicoconjugados que compõem as membranas celulares, desempenham um papel fundamental
em muitos processos biológicos, tais como comunicação celular, resposta imunológica,
fertilização, desenvolvimento de infecções parasitárias e metástase de tumores (GABIUS;
GABIUS, 1997). Um dos maiores incentivos à pesquisa sobre lectinas, provavelmente, está
relacionado com sua capacidade única de decifrar as informações biológicas que são
codificadas na estrutura tridimensional de carboidratos. As lectinas são, de fato, os receptores
para as interações proteína-açúcar, tendo papel fundamental em vários processos de
reconhecimento molecular e sinalização celular (AMBROSI et al., 2005).

18
Lectinas estão largamente distribuídas na natureza, sendo encontradas em seres
unicelulares (IMBERTY et al., 2004), animais (MOURA et al., 2006) e vegetais (SILVA et al.,
2011). Fica evidente, então, que a descoberta, o isolamento e a caracterização química, físico-
química e biológica de novas lectinas se revestem de grande importância, na medida em que
novas lectinas com diferentes aplicabilidades podem ser encontradas. Além disso, tendo em
vista que, atualmente, é forte a corrente que defende o papel de moléculas de comunicação para
as lectinas, a descoberta de outras lectinas com especificidades diferentes se torna atrativa, pois
podem servir como novos modelos para o entendimento dos processos dinâmicos de
comunicação célula-célula, célula-microrganismo ou célula-vírus (PEUMANS; VAN
DAMME, 1995).
Os estudos estruturais de lectinas servem como ferramentas para a compreensão do
reconhecimento celular entre carboidratos da membrana celular e proteínas e qual a sua
importância em diversos processos fisiológicos. Dentre as várias técnicas utilizadas para a
elucidação estrutural de proteínas, a espectrometria de massas e a cristalografia de raios X se
destacam pela sua eficiência.
A Espectrometria de Massas (do inglês Mass Spectrometry, MS) é uma técnica
muito utilizada em química de proteínas para identificar, caracterizar modificações pós-
traducionais, sequenciar proteínas e peptídeos com importância biológica, caracterizar a
interação em proteínas e outras moléculas (HOFFMANN; STROOBANT, 2007; DUIJN et al.,
2009; SCARFF et al., 2009; RUOTOLO et al., 2005).
Por sua vez, a cristalografia de raios X é considerada a principal técnica de
resolução da estrutura tridimensional de proteínas, sendo capaz de elucidar a estrutura de
moléculas com alto peso molecular e de resolver estruturas oligoméricas (BLUNDELL;
JONHSON, 1976; DRENTH, 1994).

19
2 OBJETIVOS
2.1 Objetivo Geral
O presente trabalho tem por objetivo caracterizar estruturalmente a lectina de
sementes de Centrolobium tomentosum, CTL, avaliar sua atividade em modelos de inflamação
e testar sua citotoxicidade em células tumorais.
2.2 Objetivos Específicos
Determinar a estrutura primária da lectina utilizando espectrometria de massas;
Cristalizar a lectina em complexo com ligantes específicos;
Difratar os cristais obtidos e obter o conjunto dos padrões de difração;
Resolver a estrutura cristalográfica de CTL;
Simular interações por Docking molecular com CTL e carboidratos diversos;
Determinar o efeito inflamatório da lectina em modelo de edema de pata;
Avaliar o efeito vasorelaxante de CTL em segmentos isolados de aortas de ratos;
Avaliar a citotoxicidade da lectina em células tumorais.

20
CÁPITULO I:
FUNDAMENTAÇÃO TEÓRICA

21
3 CAPÍTULO I: FUNDAMENTAÇÃO TEÓRICA
3.1 Lectinas vegetais
3.1.1 Aspectos históricos e definição de lectinas
As lectinas foram primeiramente denominadas de hemaglutininas ou
fitohemaglutininas, pelo fato de que essas proteínas foram detectadas pela primeira vez através
da aglutinação de eritrócitos e por terem sido encontradas quase que exclusivamente em plantas
(SHARON; LIS, 1993).
O primeiro relato de lectinas decorre do ano de 1888, tendo sido esse realizado por
Stillmark, quem pesquisava na época o princípio tóxico da Ricinus communis (Euphorbiaceae)
em sua tese de doutorado intitulada: “Ricina, um fermento tóxico de sementes de Ricinus
communis L. e algumas outras espécies de euforbiáceas”, publicada na Universidade de Dopart
(hoje Universidade de Tartu, Estônia). Stillmark descobriu que extratos de sementes de Ricinus
communis eram capazes de aglutinar células sanguíneas de diferentes animais. Ele denominou
essa substância de Ricina. (SHARON; LIS, 2004).
Na verdade, a ricina com que Stillmark (1888) trabalhou ainda era uma mistura
complexa de moléculas tóxicas de ricina com outras aglutininas não tóxicas. Mesmo assim, seu
trabalho pioneiro foi um marco na biologia, unindo a toxicidade da mamona à ocorrência de
um fator proteico hemaglutinante. Além disso, sua descoberta foi também um marco na
bioquímica de plantas, pois a ricina foi a primeira proteína vegetal cuja atividade biológica pôde
ser determinada (VAN DAMME et al., 1998, SHARON; LIS, 2004).
Substâncias tóxicas similares a ricina foram posteriormente identificadas em
sementes de Croton tiglium (crotina), Abrus precatorius (abrina) e em casca de Robinia
pseudoacacia (robina). Em 1898, Elfstrand introduziu pela primeira vez o termo
“hemaglutinina” referindo-se a proteínas hemaglutinantes vegetais como um nome comum a
todas as proteínas de plantas que causassem agrupamento de células. Esse novo termo foi
claramente inspirado na extrema similaridade entre a atividade macroscópica de algumas
proteínas vegetais e algumas aglutininas de soro animal e humano, essas últimas descritas
inicialmente por Landois (1875). (VAN DAMME et al., 1998).
Ricina e abrina foram amplamente utilizadas como modelos de antígenos em
estudos imunológicos. Isso permitiu a Paul Ehrlich, do Instituto Real de Terapia Experimental
(Frankfurt), estabelecer, no final da década de 1890, vários dos princípios fundamentais da

22
imunologia, dentre eles, a especificidade da relação antígeno/anticorpo, o fenômeno da
memória imunológica e a transferência da imunidade humoral de mãe para filho (SHARON;
LIS, 2004).
A ideia de que a toxicidade é uma propriedade intrínseca das lectinas foi
abandonada no início do século XX, depois que Ladsteiner e Raubitscheck, em 1907, relataram
pela primeira vez a presença de uma lectina não tóxica nas leguminosas Phaseolus vulgaris
(feijão), Pisum sativum (ervilha), Lens culinaris (lentilha) e Vicia sativa. Após esse trabalho,
muitas outras hemaglutininas vegetais não tóxicas foram descobertas. Tornou-se, a partir de
então, evidente que lectinas estão difundidas no reino vegetal e que a toxicidade atribuída às
mesmas é exceção, e não regra (VAN DAMME et al., 1998).
Em 1919, James Sumner, ao purificar a urease através de precipitação e
cristalização a partir de sementes de Canavalia ensiformis, co-purificou outra proteína que ele
próprio nomeou de Concanavalina A (Con A). Dessa forma, Sumner obteve a primeira lectina
pura. Contudo, somente em 1936, Sumner e Howell demonstraram que a Concanavalina A
aglutinava células tais como eritrócitos e, também, precipitava glicogênio de soluções. Os
mesmos demonstraram posteriormente que a hemaglutinação era inibida pela sacarose da cana-
de-açúcar, monstrando, pela primeira vez, a especificidade das lectinas por açúcares
(SHARON; LIS, 2004). No entanto, somente em 1952, foi demonstrado que as propriedades
hemaglutinantes de lectinas eram baseadas em uma atividade específica de ligação a
carboidratos (WATKINS; MORGAN, 1952).
Alguns anos depois, Renkonen (1948) e Boyd e Reguera (1949) obtiveram um novo
marco para a história das lectinas vegetais. Eles constataram que algumas hemaglutininas
exibiam uma preferência clara por hemácias de um tipo particular de grupo sanguíneo humano
do sistema ABO. Foi observado que algumas proteínas vegetais obtidas de sementes de planta
podiam reconhecer um grupo específico e aglutiná-lo. Hemácias do sistema ABO respondiam
ao contato com essas hemaglutininas de maneira distinta, umas aglutinando e outras não (VAN
DAMME et al., 1998). A habilidade das aglutininas de plantas em distinguir eritrócitos de
diferentes tipos sanguíneos levou Boyd e Shapleigh a propor, em 1954, o termo “lectina”
(legere, do latim, significando “selecionar” ou “escolher”) para nomear tal proteína (SHARON;
LIS, 2004).
O marco seguinte foi relevante não só para a história da lectinologia, mas também
teve impacto revolucionário sobre a imunologia. No ano de 1960, Peter C. Nowell demonstrou
que a lectina de Phaseolus vulgaris possui atividade mitogênica sobre linfócitos. Essa
descoberta foi marcante na imunologia pelo fato de que até aquele momento acreditava-se que

23
os linfócitos eram células incapazes de se dividirem ou se diferenciarem em outros tipos
celulares (SHARON; LIS, 2004).
Ao serem reconhecidas como proteínas ligantes de carboidratos, as lectinas
puderam ser distinguidas de outras proteínas com base em um critério funcional bem definido.
Goldstein e colaboradores (1980) conceituaram lectinas como sendo proteínas ou
glicoproteínas de origem não-imune ligantes a carboidratos que são capazes de aglutinar células
e/ou precipitar polissacarídeos ou glicoconjugados.
Atualmente, a definição mais aceita de lectinas de plantas e também mais utilizada
foi proposta por Peumans e Van Damme (1995), que definem lectinas como sendo proteínas de
origem não imune, capazes de se ligar reversível e especificamente a carboidratos e substâncias
que contenham açúcar, sem alterar a estrutura covalente de nenhum ligante glicosil.
3.1.2 Generalidades sobre lectinas vegetais
As lectinas representam um grupo heterogêneo de proteínas oligoméricas variando
em tamanho, estrutura, organização molecular e entre os sítios de ligação a carboidratos. Em
relação às suas cadeias polipeptídicas, as lectinas vegetais são caracteristicamente ricas em
aminoácidos ácidos e hidroxilados e, encontram-se associadas por interações hidrofóbicas,
pontes de hidrogênio e em alguns casos pontes de dissulfetos (KENNEDY et al., 1995;
RÜDIGER; GABIUS,1993).
Pelo número de cadeias polipeptídicas, as lectinas são denominadas monoméricas
quando apresentam apenas uma cadeia, como a lectina de Solanum tuberosum, com 50 kDa
(ALLEN; NEUBERGER, 1973), a heveína, com apenas 43 aminoácidos e 4 pontes de
dissulfeto (SOEDJANAATMADJA et al., 1995), e a arcelina de Phaseolus vulgaris (LORIS,
2002). Quando as lectinas apresentam duas cadeias polipeptídicas elas são denominadas
diméricas, com cadeias semelhantes ou diferentes entre si, ganhando respectivamente a
denominação de lectinas homodiméricas ou heterodiméricas. Erythrina speciosa possui uma
lectina homodimérica com 27,6 kDa por subunidade (KONOZY et al., 2003). Em Ricinus
communis a lectina é heterodimérica, com uma subunidade de 32 kDa e outra de 34 kDa
(FRIGERIO; ROBERTS, 1998), bem como a lectina de Erythrina indica com 65 kDa que é
heterodimérica, com uma cadeia polipeptídica de 30 kDa e outra de 35 kDa (KONOZY et al.,
2002).
Lectinas compostas por quatro subunidades são denominadas tetraméricas, como a
lectina JCA, também denominada jacalina, com 66 kDa (PRATAP et al., 2002; KABIR, 1998).

24
Também a SBA, com massa molecular de 120 kDa, é tetramérica e cada uma de suas
subunidades possui massa molecular de 30 kDa, (CARVALHO, 1990). Ocorrem outros tipos
estruturais de lectinas, como as triméricas, pentaméricas, hexaméricas ou poliméricas. Uma
lectina trimérica ocorre em Geodia cydonium com cadeias de massa molecular 12,2, 13,0 e 13,8
kDa, conforme Müller e colaboradores (1983). Em Araucaria angustifólia ocorre, nas
sementes, uma lectina com 10 subunidades de 20 kDa, totalizando 200 kDa, e uma segunda
lectina com massa molecular relativa de 204 kDa, hexamérica, com subunidades de 34 kDa,
conforme Datta e colaboradores (1991).
As lectinas são encontradas nos vegetais desde organismos simples como musgos
(MOLINA; VINCENTE, 1995) até os mais complexos como gimnospermas (SANTI-
GADELHA et al., 2006) e angiospermas (MORAES et al., 1996; WONG; NG, 2005). Desde o
início do estudo sobre lectinas centenas delas têm sido identificadas, isoladas e caracterizadas.
Graças a disponibilidade e diversidade de ligação a carboidratos, as lectinas de vegetais são o
grupo melhor estudado.
A grande maioria das lectinas vegetais já isoladas foi obtida de sementes,
principalmente em leguminosas, onde são acumuladas no período de maturação, desaparecendo
após a germinação. No entanto, lectinas também podem ser encontradas em folhas (COELHO;
SILVA, 2000; RATANAPO et al., 2001), frutos (SAMPIETRO et al., 2001; WU; WANG; TB,
2011), cascas (INA et al., 2 005), raízes (HE et al., 2011; PEUMANS et al., 1997), e
tubérculos (OHIZUMI et al., 2009; SUSEELAN et al., 2002).
Nas células vegetais, as lectinas podem ser encontradas em corpos proteicos (ou
vacúolos de estocagem) que são pequenos vacúolos que sofreram diferenciação para serem
capazes de estocar proteínas durante a maturação da semente e hidrolisá-las durante períodos
de crescimento (CHRISPEELS, 1984). Ainda no ambiente celular, já foram identificadas
lectinas presentes no citoplasma, no núcleo (VAN DAMME et al., 2004), além de também
ocorrerem no espaço intercelular (ETZLER et al., 1986).
As lectinas mais estudadas são da família Leguminosae. Entretanto, muitas lectinas
de outras famílias também têm sido frequentemente isoladas e caracterizadas como, por
exemplo, lectinas de Solanaceae (PEUMANS et al., 2003), Cucurbitaceae (SULTAN;
KAVITHA; SWAMY, 2009), Amaranthaceae (RINDERLE et al., 1989), Cactaceae
(ZENTENO et al., 1995), Euphorbiaceae (VAN PARIJS et al., 1991), Labiateae
(FERNÁNDEZ-ALONSO et al., 2003), Moraceae (MOREIRA et al., 1998) e Urticaceae
(PEUMANS; DOLEY; BRACKAERT, 1984), entre diversas outras.

25
Muitas funções têm sido propostas para as lectinas, tais como proteção contra
patógenos e insetos, transporte e armazenamento de carboidratos, reconhecimento celular
(dentro da célula, entre células ou entre organismos), proteínas de reserva ou reguladores de
crescimento (PUSZTAI, 1991).
Atualmente, essas proteínas têm mostrado aplicabilidades em diversas áreas
científicas, abrangendo áreas que vão da medicina à agricultura. Muitas dessas proteínas são
consideradas como ferramentas básicas para áreas de ponta, como a biotecnologia, tendo em
vista suas várias propriedades, tais como: atividade anti-inflamatória e pró-inflamatória (PIRES
et al.,2016; ROCHA et al., 2015; SANTIAGO et al., 2014; TEIXEIRA et al., 2014), efeito
vasorelaxante em aortas isoladas de ratos (BARROSO-NETO et al., 2016; ALVES et al., 2015),
efeito antidepressivo em ratos (RIEGER et al., 2014), efeito antiproliferativo em células
tumorais (SILVA et al., 2014), atividade inseticida (KAUR et al., 2015; MACEDO;
OLIVEIRA; OLIVEIRA, 2015), efeito tóxico para Artemia salina (ALMEIDA et al., 2014;
BARI et al., 2013), agentes em drug delivery (IKEMOTO et al., 2016; BOGOEVA;
PETROVA; TRIFONOV, 2014), indução do fenômeno de apoptose (BARBOSA et al., 2001),
dentre outros.
Embora lectinas apresentem a propriedade comum de se ligarem, reversivelmente,
a carboidratos (LIS; SHARON, 1998), elas apresentam características próprias, principalmente
no que diz respeito a aplicações biológicas. Assim, mesmo lectinas que apresentem homologia
de sequências entre si, mostram-se diferentes quanto às várias propriedades biológicas. Isso faz
com que, em regra, cada lectina tenha as suas potencialidades de aplicação, o que justifica que
cada uma delas, por mais semelhante que possa parecer com outra lectina, mereça ser estudada
isoladamente.
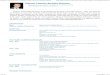
3.1.3 Classificação estrutural de lectinas vegetais
As lectinas formam um grupo heterogêneo de proteínas. Van Damme et al. (1998)
subdividiu as lectinas vegetais de acordo com a estrutura em quatro classes principais:
merolectinas, hololectinas, quimerolectinas e superlectinas (Figura 1).
As merolectinas são proteínas que consistem exclusivamente de um único domínio
de ligação a carboidrato. Devido seu caráter monovalente, as merolectinas são incapazes de
precipitar glicoconjugados ou aglutinar células. A heveína, uma proteína do látex de seringueira
(Hevea brasiliensis), que se liga à quitina, é uma típica merolectina (VAN PARIJS et al., 1991).

26
Figura 1. Classificação estrutural de lectinas vegetais.
Fonte: Adaptado de VAN DAMME et al., 1998. DRC: domínio de reconhecimento a carboidratos.
As hololectinas também são compostas exclusivamente de domínios de ligação a
carboidrato, mas com dois ou mais sítios ligantes que são idênticos ou parecidos. Devido ao
fato de hololectinas serem divalentes ou multivalentes, elas podem aglutinar células e/ou
precipitar glicoconjugados. As hololectinas comportam-se como verdadeiras aglutininas e
compreendem a maioria das lectinas de plantas, representando a classe de lectinas mais bem
estudada. São exemplos típicos de hololectinas as lectinas encontradas em sementes de plantas
pertencentes à subtribo Diocleinae (CAVADA et al., 2001).
As quimerolectinas são a fusão de proteínas compostas de um ou mais domínios de
ligação a carboidratos e de um domínio não relacionado com uma atividade catalítica bem
definida que age independentemente dos domínios de ligação a carboidratos. Dependendo do
número de sítios de ligação a carboidratos, as quimerolectinas comportam-se como
merolectinas ou hololectinas. Um representante desse grupo são as proteínas inativadoras de
ribossomos (RIP tipo 2), como por exemplo a Ricina (toxina da mamona), que possui dois
domínios de ligação para carboidratos comportando-se como uma hololectina e um domínio
para a inativação de ribossomo (PEUMANS; VAN DAMME, 1998). Um outro exemplo é a
PPL-2, uma lectina quitina-ligante isolada de sementes de Parkia platycephala que possui além
do sítio ligante a carboidrato um sítio catalítico com atividade endoquitinásica (CAVADA et
al., 2006).

27
As superlectinas consistem de no mínimo dois domínios de ligação para
carboidratos. Diferentemente das hololectinas, os domínios de ligação para carboidratos das
superlectinas reconhecem açúcares estruturalmente e funcionalmente diferentes. Como
exemplo, a lectina do bulbo de tulipa (TxLCI) que são formados por dois domínios de ligação
a carboidrato, que reconhecem manose e N-acetil-D-galactosamina, respectivamente (VAN
DAMME et al., 1996).
O avanço da biologia estrutural em conjunto com modernas técnicas de DNA
recombinante deu um salto significativo na elucidação das mais diversas moléculas presentes
em estruturas dos sistemas biológicos. Muitas já possuem seus sítios de reconhecimento a
carboidratos bem estabelecidos, apresentando semelhanças quanto a aspectos estruturais.
Outras características como sequência primária, reconhecimento a açúcares e até mesmo quanto
a função desempenhada, já são bem conhecidas e estudadas (VAN DAMME et al., 1998).
Isso ocorreu de uma forma que tornou possível classificar lectinas não somente
quanto aos aspectos estruturais vistos anteriormente, mas como também em famílias que
estejam evolutivamente relacionadas, ou seja, que não possuam apenas estruturas quaternárias
semelhantes, mas que compartilham outras características que possam estar diretamente
relacionadas com a função desempenhada por cada uma delas (VAN DAMME et al., 1998).
Van Damme e colaboradores (1998) classificaram as lectinas de plantas em sete
grupos, ou famílias, de acordo com as semelhanças evolutivas e estruturais. Esses grupos são
nomeados de: lectinas de leguminosas, lectinas ligantes a quitina contendo domínios
heveínicos, lectinas de monocotiledôneas que se ligam a manose, proteínas inativadoras de
ribossomo tipo II (RIPs tipo II), lectinas de floema de Cucurbitaceae, lectinas relacionadas à
jacalina e lectinas de Amaranthaceae (Figura 2).
As lectinas de leguminosas correspondem à família de lectinas mais estudada até o
presente, com dezenas de representantes já purificados e caracterizados. As proteínas desse
grupo caracterizam-se por possuírem uma alta similaridade estrutural, porém, com
especificidade por carboidratos e propriedades biológicas distintas. Enquadram-se nesse grupo
a Concanavalina A (ConA), extraída de sementes de Canavalia ensiformes, e as lectinas
extraídas de outras plantas da subtribo Diocleinae (CAVADA et al.,2001).
As lectinas que se ligam a quitina são capazes de se ligar à resíduos de N-acetil-D-
glicosamina (principal unidade monomérica constituinte da quitina) e têm como característica
principal o fato de possuir um “domínio heveínico” em suas moléculas (VAN DAMME et al.,
1998). O termo domínio heveínico refere-se a uma pequena proteína (uma merolectina) de 43
resíduos de aminoácidos, extraída do látex da Hevea brasiliensis, a popular seringueira

28
(WALJUNO et al., 1975). Lectinas pertencentes a esse grupo têm sido encontradas em várias
famílias de plantas não relacionadas taxonomicamente tais como Gramineae, Leguminosae,
Solanaceae, Urticaceae, Papaveraceae e Amaranthaceae. São também exemplos de lectinas
que se ligam a quitina as quitinases classe I (COLLINGE et al., 1993).
Figura 2. Classificação das lectinas de plantas em famílias evolutivamente relacionadas.
Fonte: Protein Data Bank (PDB). A: Lectinas de leguminosas (ConBr, código PDB 3JU9); B: Lectina ligante à
quitina composta por domínios heveínicos (Heveína, código PDB 1HEV); C: Lectinas de monocotiledôneas
ligantes à manose (GNA, código PDB 1NIV); D: RIP do tipo II (Abrina, código PDB 2ZR1); E: Lectinas
relacionadas à Jacalina (PPL-1, código PDB 1ZGR); F: Lectina da família da Amarantina (ACA, código PDB
1JLY).
O termo “lectinas de monocotiledôneas que se ligam a manose” refere-se a uma
família de lectinas estreitamente relacionadas que possuem especificidade para manose,
encontradas exclusivamente em plantas monocotiledôneas, como nas famílias Amaryllidaceae,
Alliaceae, Araceae, Liliaceae, Orchidaceae e Bromeliaceae. Estudos comparativos indicam

29
que essas proteínas compartilham um alto grau de similaridade de sequência primária, porém
com uma completa heterogeneidade de estrutura molecular (VAN DAMME et al., 1998).
As proteínas inativadoras de ribossomos tipo II (RIP’s tipo II) são lectinas do tipo
quimerolectina, formadas por duas cadeias polipeptídicas sintetizadas a partir de uma única
molécula precursora que passa por modificações pós traducionais através da excisão de uma
ligação entre as cadeias. A primeira dessas cadeias (cadeia A) é formada por um polipeptídeo
constituído de um domínio adenosina glicosidase e a outra cadeia (cadeia B) constituída por um
domínio de ligação a carboidratos, sendo que as cadeias permanecem unidas por pontes
dissulfeto. Ao penetrar nas células, esta ligação dissulfeto é quebrada e a cadeia A adquire uma
forte atividade N-glicosidase tornando-se capaz de inativar cataliticamente ribossomos
eucarióticos e procarióticos (VAN DAMME et al., 1998). A Ricina (Ricinus communis) e
Abrina (Abrus precatorius) são exemplos clássicos de RIP’s tipo 2 com potentes atividades
biológicas (PEUMANS et al., 2001).
As lectinas do floema de Cucurbitaceae são proteínas diméricas compostas de duas
subunidades idênticas de 24 kDa que se ligam a resíduos de N-acetil-D-glicosamina. Todas as
lectinas conhecidas dos floemas de Cucurbitaceae mostram um grau de similaridade
sequencial, mas não apresentam qualquer similaridade sequencial com outras lectinas de
plantas ou proteínas (VAN DAMME et al., 1998).
A expressão “lectinas relacionadas à Jacalina” é usada para nomear um grupo de
lectinas que são evolucionaria e estruturalmente relacionadas a Jacalina, uma lectina com
especificidade para D-galactose extraída de sementes de jaca (Artocarpus integrifolia). Esse
tipo de lectina é encontrado principalmente em espécies de Moreaceae e Convolvulaceae (VAN
DAMME et al., 1998).
A Amarantina, uma lectina extraída de Amaranthus caudatus, não se assemelha a
nenhuma outra lectina de plantas, tanto com relação à sequência de aminoácidos quanto com
relação à estrutura tridimensional. Baseado nessas informações, a Amarantina é considerada
um protótipo da família de lectinas das Amaranthaceae. Várias outras lectinas extraídas de
plantas do gênero Amaranthus, como por exemplo, Amaranthus leucocarpus, contêm lectinas
que são muito similares à Amarantina, sendo todas elas lectinas ligantes específicas por N-
acetil-D-galactosamina (VAN DAMME et al., 1998).
Apesar de parecer clara, esta classificação das lectinas vegetais tende a deixar de
ser baseada em aspectos evolutivos e passa a levar em consideração somente os aspectos
estruturais. Isso se dá pelo fato de que, à medida que novas lectinas vão sendo descobertas e
caracterizadas, deixa de existir um limite taxonômico bem definido entre as famílias de lectinas

30
vegetais. Um exemplo claro deste fato é a lectina específica para glicose e manose extraída de
sementes da leguminosa Parkia platycephala, PPL-1, cuja estrutura é composta por três
domínios repetidos relacionados à Jacalina. Outra lectina também purificada a partir de
sementes de Parkia platycephala, PPL-2, também é um exemplo de exceção, pois trata-se de
uma RIP tipo II (CAVADA et al., 2006; GALEGO DEL SOL et al., 2005; MANN et al., 2001).
3.1.4 Lectinas de leguminosas
Dentre as classificações em família evolutivamente relacionadas, as lectinas mais
bem estudadas pertencem ao grupo das leguminosas. Dezenas de lectinas deste grupo já foram
isoladas e caracterizadas, sendo a maioria dos estudos relacionados à lectinas de membros da
subfamília Papilionoideae, principalmente da tribo Phaseoleae (MANN et al., 2001;
SHARON; LIS, 1990). Em sementes, principal fonte de lectinas neste grupo, foi denotado que
o percentual de lectina pode variar de 1 a 10% do total de proteínas solúveis, embora algumas
espécies possam apresentar concentrações abaixo de 0,1% do total de proteínas solúveis
presentes nas sementes (SHARON; LIS, 1990; VAN DAMME, et al., 1998).
Sabe-se que algumas lectinas de leguminosas, especialmente as da subtribo
Diocleinae, são sintetizadas no retículo endoplasmático na forma de pré-pro-lectinas ou pro-
lectina glicosilada (Figura 3). A região correspondente ao cDNA contém 29 resíduos de uma
sequência sinal, que é removido posteriormente durante o transporte da pré-pro-lectina para o
lúmen do reticulo endoplasmático, originando uma pró-lectina. Em seguida encontra-se uma
região codificante referente aos aminoácidos 119-237 da Concanavalina A, e uma outra região
codificante correspondente aos aminoácidos 1-118 da lectina, seguida por último por uma
extensão C-terminal de 9 resíduos. Além disso, há entre os dois peptídeos uma unidade
oligomanose N-ligada a uma pentadecapeptídeo interno, o qual é excisado durante a maturação,
produzindo os dois peptídeos. Acredita-se que a presença da glicosilação iniba a atividade de
ligação a carboidrato da lectina. A lectina madura e ativa, portanto, é não-glicosilada. A maior
parte dos dois peptídeos formados são então covalentemente ligados por uma reação de
transpeptidação, concomitante com a clivagem da extensão C-terminal, formando a lectina
madura, onde o alinhamento dos resíduos 1-118 e 119-237 é o inverso de como se apresenta no
precursor. Esse processamento recebeu o nome de “Permutação Circular” (SHARON & LIS,
1990; SHARON, 2007).

31
Figura 3. Processamento pós-traducional durante a biossíntese de ConA.
Fonte: Santiago, 2013.
Do ponto de vista estrutural, as lectinas de leguminosas são geralmente compostas
de 2 ou 4 subunidades, iguais ou diferentes com massa molecular em torno de 25 a 30 kDa,
compostas geralmente de uma cadeia polipeptídica simples com cerca de 250 aminoácidos, cuja
união é estabelecida por forças não covalentes como ligações de hidrogênio, interações
hidrofóbicas e eletrostáticas, que formam dímeros canônicos e estabilizam o tetrâmero pela
união destes dímeros. Cada uma destas subunidades apresenta um único sítio de ligação a
carboidratos com a mesma especificidade, além de um ou mais sítio de ligação a íons, podendo
ainda apresentar um ou dois oligossacarídeos N-ligados. Entretanto, algumas lectinas possuem
subunidades formadas por duas cadeias polipeptídicas, entre elas as lectinas da tribo Vicieae,
gêneros Pisum, Vicia, Lathyrus e Lens. Estas lectinas são dímeros formados por duas
subunidades iguais e cada subunidade é constituída de uma cadeia α (5 a 7 kDa) e uma β (15 a
19 kDa) mantidas por ligações não-covalentes (SHARON; LIS, 1989).
As estruturas dos monômeros de diferentes lectinas de leguminosas são
extremamente similares (Figura 6). Aproximadamente 20% dos resíduos de aminoácidos são
invariantes em todas as lectinas de leguminosas e outros 20% são similares. Entre os
aminoácidos conservados incluem-se aqueles envolvidos na interação com o carboidrato e
quase todos os resíduos que coordenam íons metais (BANERJEE et al., 1996; HAMELRYCK
et al., 1998). A resolução de estruturas tridimensionais de lectinas de leguminosas tem mostrado

32
que cada subunidade é constituída de cerca de 60% de fitas-beta antiparalelas arranjadas em
duas ou mais folhas beta interconectadas por voltas, mais conhecido como enovelamento do
tipo “β-sanduíche”. Para todas as lectinas de leguminosas conhecidas, a estrutura terciária é
constituída de duas folhas-β antiparalelas, uma com seis fitas β (atrás) e outra com sete fitas β
(frente), conectadas por uma outra folha-β de 5 fitas (superior), gerando o conhecido motivo
"jellyroll', também referido como "lectin fold". (SRINIVAS et al., 1996; HAMELRYCK et al.,
1998) (Figura 4).
Figura 4. Estrutura de monômero típica das lectinas de leguminosas.
Fonte: Adaptado de Benevides, 2011. À esquerda, diagrama esquemático da estrutura terciária de um monômero
típico de lectinas de leguminosas. Do lado direito, estrutura terciária do monômero da lectina de Centrolobium
tomentosum (CTL) complexada com metil-dimanosídeo (código PDB:5EYX).
O sítio de ligação a carboidratos está localizado no lado côncavo do β-sanduíche,
próximo ao sítio de ligação a metais. O sítio de ligação a carboidratos consiste de diversos
“loops” com diferentes graus de variabilidade (SHARMA; SUROLIA, 1997). As
conformações destes loops são determinadas pela presença de íons de metais de transição, como
o cálcio e o manganês, na estrutura (BOUCKAERT et al., 2000; LORIS et al., 1998). Esses
metais se localizam a uma distância de aproximadamente 4,5 Å e ambos são coordenados por
cadeias laterais de pelo menos quatro aminoácidos e duas moléculas de água. O reconhecimento
das lectinas de leguminosas por íons é reversível e a remoção desses metais resulta em
importantes mudanças conformacionais. Em ConA, por exemplo, com a falta dos íons cálcio e
manganês, o peptídeo Ala207-Asp208, que para participar efetivamente do reconhecimento ao

33
seu carboidrato específico necessita estar na conformação trans, passa a assumir uma
conformação do tipo cis (BOUCKAERT et al., 1995). A ausência desses metais, portanto,
resulta em uma instabilidade local e na perda da capacidade de ligar-se a carboidratos (LORIS
et al., 2004).
Além de um sítio de ligação a carboidratos, altamente conservado, juntamente com
um sítio de ligação a metais, que é responsável pela estabilização do “loop” que compõe o sítio
de ligação a carboidratos, estudos recentes mostraram a capacidade dessas proteínas de
interagirem com compostos hidrofóbicos. Algumas lectinas de leguminosas podem interagir
com outras espécies de moléculas, que não carboidratos, tais como adenina (HAMELRYCK et
al., 1998) e o ácido α-aminobutírico (ABU) (DELATORRE et al., 2007). A ligação das lectinas
a essas moléculas se dá através de uma região estrutural hidrofóbica, altamente conservada em
muitas lectinas de leguminosas e que, devido a esse alto grau de conservação, pode estar
envolvida em algum papel biológico importante (DELATORRE et al., 2007).
Algumas lectinas de leguminosas, como as lectinas extraídas de espécies da
subtribo Diocleinae, exibem uma oligomerização dependente de pH, onde a proporção entre
proteínas no estado dimérico e no estado tetramérico pode ser alterada de acordo com o pH do
meio (CALVETE et al., 1999; NAGANO et al., 2008). A alteração desse equilíbrio
dímero/tetrâmero dependente de pH pode influenciar diretamente nas atividades biológicas
dessas lectinas, pelo fato dessas proteínas serem capazes de se ligar aos receptores
glicoconjugados da superfície das membranas com uma maior afinidade quando na forma
tetramérica (DELATORRE et al., 2006).
Muitas lectinas de leguminosas são expressas nas formas glicosiladas e não
glicosiladas. É o caso da lectina de Vatairea macrocarpa que apresenta cadeias ativas com
um único ponto ou dois pontos de glicosilação, além de fragmentos, também ativos com frações
glicosilados e não glicosiladas (CALVETE et al., 1998). Algumas variações também são
encontradas nas lectinas de sementes de Phaseolus vulgaris, Griffonia simplicifolia e Vicia
villosa (VAN DAMME et al., 1998). Alguns pesquisadores citam que a glicosilação pode ser
uma estratégia celular para que novas estruturas tridimensionais surjam devido a impedimento
espacial causado pela presença do glicano (SHAANAN et al., 1991). Outros relatam que a
glicosilação em lectinas pode estar envolvida em outras funções celulares, como o
enovelamento diferenciado (TURTON et al., 2004).
3.1.5 Lectinas de Dalbergieae

34
A tribo Dalbergieae, pertencente à família Leguminosae (Fabaceae), subfamília
Papilionoideae, compreende 48 gêneros e 1.200 espécies aproximadamente. Até o presente
momento, foram relatadas 16 lectinas pertencentes a esta tribo. A tabela 1 relata as espécies as
quais estas lectinas pertencem, além de suas especificidades a carboidratos.
Tabela 1. Lectinas da tribo Dalbergieae.
Espécie Especificidade Referência
Andira fraxinifolia Man/Glc RANGEL et al., 2009
Andira pisonis Man/Glc LÓSSIO et al., 2014
Andira surinamensis Man/Glc NOBRE, 2012
Andira anthelmia Man/Glc NASCIMENTO et al., 2015
Centrolobium microchaete Man/Glc VASCONCELOS, 2015
Centrolobium tomentosum Man/Glc ALMEIDA et al., 2014
Lonchocarpus araripensis GlcNac PIRES et al., 2008
Lonchocarpus capassa Gal/GalNAc JOUBERT et al., 1986
Lonchocarpus sericeus GlcNAc ALENCAR et al., 1999
Machaerium acutifolium GlcNAc BEZERRA et al., 2003
Platymiscium floribundum Man/Glc PEREIRA-JÚNIOR et al., 2012
Platypodium elegans Man/Glc BENEVIDES et al., 2012
Pterocarpus angolensis Man/Glc LORIS et al., 2003
Pterocarpus rotundifolius Man/Glc MARONDEDZE et al., 2004
Vatairea guianensis Gal/GalNAc SILVA et al., 2012
Vatairea macrocarpa Gal/GalNAc CAVADA et al., 1998
Fonte: Elaborado pelo autor.
Apesar da tribo Dalbergieae apresentar um número considerável de lectinas
caracterizadas, apenas três lectinas apresentam estruturas tridimensionais determinadas: a
lectina de Pterocarpus angolensis (PAL) e a lectina de Platypodium elegans (PELa)
(BENEVIDES et al., 2012; LORIS et al., 2003), ambas manose/glicose específicas; e a lectina
de Vatairea macrocarpa (VML), específica à galactose e seus derivados (SOUSA et al., 2015).
As estruturas tridimensionais das três lectinas estão ilustradas na figura 5.

35
Figura 5. Estruturas tridimensionais de lectinas da tribo Dalbergieae.
Fonte: Elaborado pelo autor. Estrutura geral das lectinas: A: PAL (código PDB: 1Q8S), B: PELa (código PDB:
3ZVX) e C: VML (código PDB: 4WV8). Os monômeros estão representados em cartoon, com seus monômeros
coloridos em diferentes tons de cinza.
A estrutura da PAL (Figura 5A) contém um loop clássico de especificidade a
manose, mas seu loop de ligação a metal assemelha-se a de lectinas com especificidades não
relacionadas como Ulex europeus e Maackia amurensis. As interações com manose no sítio de
ligação primário são conservadas, mas detalhes da extensão desse sítio de ligação a carboidratos
difere de outros vistos em complexos a carboidratos equivalentes em ConA (LORIS et al.,
2004).
A estrutura tridimensional da PELa (Figura 5B) possui uma organização dimérica
canônica típica de lectinas de leguminosas. Cada monômero de PELa adota o clássico fold β-
sanduíche, com a presença de um íon cálcio e um íon manganês. Os dois monômeros na unidade
assimétrica se associam através de um arranjo paralelo de suas folhas β (trás), resultando num
dímero canônico típico, classicamente observado lectinas de leguminosas diméricas e
tetraméricas (BENEVIDES et al., 2012).
A estrutura da VML, por sua vez, é caracterizada como um tetrâmero, contendo 240
aminoácidos por monômero (CALVETE et al., 1998). A estrutura monomérica da VML
apresenta o enovelamento característico de lectinas de leguminosas, consistindo em um

36
sanduíche β, formado por uma folha estendida de seis fitas e uma folha curvada de sete fitas
conectadas por alças de tamanhos variados (LORIS et al., 1998). O tetrâmero da VML é
composto por dois dímeros arranjados de forma oposta, formando uma grande cavidade central
(Figura 5C). A cavidade central do oligômero é formada por meio de interações entre as fitas
externas das folhas estendidas de cada monômero, originando uma interface canônica típica de
leguminosas. Os sítios de ligação a carboidrato e a metais apresentam-se conservados entre
lectinas de leguminosas (SOUSA et al., 2015).
As lectinas pertencentes à tribo Dalbergieae possuem características peculiares em
se tratando de seu processamento pós-traducional. Diferentemente das lectinas da tribo
Phaseoleae, subtribo Diocleinae, onde suas lectinas são processadas por permutação circular,
como citado anteriormente, as lectinas da tribo Dalbergieae podem apresentar um
processamento por uma asparaginil-endopeptidase, gerando cadeias que não sofrem religação,
mas que interagem entre si e são co-purificadas por cromatografia de afinidade. Um exemplo
de ocorrência desse processamento é a lectina de Vatairea macrocarpa (CALVETE et al.,
1998).
Calvete e colaboradores (1998) propuseram que o processamento pós-traducional
da VML funciona da seguinte forma: a cadeia α sofre uma clivagem pelo rompimento da ponte
entre Asn114 e Lys115. Essa clivagem é diretamente dependente da deglicosilação do resíduo de
Asn111. A partir dessa clivagem são gerados os fragmentos γ (N-terminal deglicosilado) e β (C-
terminal glicosilado). O fragmento β pode ainda sofrer ainda uma deglicosilação no resíduo
Asn183 seguido de processamento N- e C-terminal, gerando isoformas. Esses fragmentos são
reunidos por ligações não covalentes fazendo com que ela tenha estrutura tridimensional
praticamente idêntica àquela da cadeia alfa (não processada) (Figura 6).
Outro grupo que sofre o mesmo processo é a tribo Sophoreae. Exemplos de lectinas
pertencentes a esta tribo são a lectina de Luetzelburgia auriculata (OLIVEIRA et al., 2002) e a
lectina da casca de Sophora japonica (HANKINS; KINDINGER; SHANNON, 1988).

37
Figura 6. Processamento pós-traducional proposto para a lectina da Vatairea macrocarpa.
Fonte: CALVETE et al., 1998.
Mesmo com um número pequeno de lectinas isoladas e caracterizadas, várias
atividades biológicas já foram demonstradas pelas lectinas da tribo Dalbergieae. A lectina mais
bem caracterizada é a lectina de Vatairea macrocarpa. Seus efeitos biológicos já demonstrados
incluem indução de efeito depressivo e expressão de marcadores neuroinflamatórios em ratos
(GONÇALVES et al., 2013), indução da infiltração de leucócitos em edema de pata
(ALENCAR et al., 2004), liberação de mediadores quimiotáticos por macrófagos (ALENCAR
et al., 2007), aumento da resistência vascular renal, da taxa de filtração glomerular e do fluxo
urinário (MARTINS et al., 2005), migração de neutrófilos in vivo por mecanismo indireto
(ALENCAR et al., 2003) e apresenta especificidade para resíduos de antígeno do tipo Tn (DAM
et al., 2007).
A lectina de mesmo gênero, Vatairea guianensis (VGL), exibiu atividade anti-
inflamatória associada à inibição da migração de leucócitos e uma atividade antinociceptiva
que pode estar associada à inibição da dor inflamatória. Além disso, VGL também exibiu um
efeito vasorrelaxante in vitro em aortas isoladas de ratos, sendo esse efeito dependente da

38
presença de endotélio e envolvendo a participação do óxido nítrico e é mediado pelo sítio de
ligação a carboidratos da lectina (SILVA et al., 2012).
A lectina de Andira anthelmia (AAL) apresentou efeito analgésico envolvendo o
domínio de lectina por meio de mecanismos de nocicepção inflamatória periféricos
(NASCIMENTO et al., 2015).
Algumas atividades biológicas também foram evidenciadas em uma lectina extraída
de sementes de Lonchocarpus sericeus, como alta atividade anti-inflamatória em modelo de
edema de pata (ALENCAR et al., 1999), efeito anti-inflamatório e antibacteriano em um
modelo de peritonite infecciosa (ALENCAR et al., 2005), diminuição da migração leucocitária
e hipernocicepção mecânica pela inibição da produção de citocinas e quimiocinas
(NAPIMOGA et al., 2007).
Segundo Pires et al. 2008, a lectina de Lonchocarpus araripensis (LaL) possui ação
anti-inflamatória sobre o edema de pata induzido por carragenina, mas é desprovida de ação
pró-inflamatória quando injetada localmente. Recentemente, a lectina demonstrou atenuar a
inflamação aguda celular em ratos (PIRES et al., 2016).
Uma fração lectínica de Platypodium elegans (PELa) apresentou atividade
termiticida contra operários e soldados de Nasutitermis corniger, além de potencial fungicida
contra os fungos Fusarium solani, Fusarium oxysporium e Fusarium lateritium (BENEVIDES
et al., 2008). Essas duas atividades reforçam a participação de lectinas na defesa vegetal,
atuando contra fitopatógenos.
3.1.6 Lectina de Centrolobium tomentosum
A espécie Centrolobium tomentosum (Figura 7) é uma espécie exclusivamente
brasileira, ocorrendo na Bahia, Espírito Santo, Distrito Federal, Rio de Janeiro, São Paulo,
Minas Gerais, Paraná, Santa Catarina e Rio Grande do Sul. Habita preferencialmente matas
mesófilas semidecíduas e matas de galeria nas formações de Cerrado, podendo ocorrer também
em descampados, cerrados, matas mais secas e formações litorâneas, em solos de média e boa
fertilidade, rasos ou profundos.

39
Figura 7. Centrolobium tomentosum Guill. ex Benth.
Fonte: LORENZI, 1992. A: Visão geral da árvore. B: Ramos com flores. C: Sâmaras. D: Flor.
Filogenia:
Família Leguminosae (Fabaceae)
Subfamília Papilionoideae
Tribo Dalbergieae DC.
Gênero Centrolobium Mart. ex Benth
Centrolobium tomentosum Guill. ex Benth.
A lectina de sementes de C. tomentosum é uma proteína ligante à manose/glicose e
relacionados, purificada através de cromatografia de afinidade em Sepharose-manose.
Demonstrou ser uma glicoproteína (1,7% de carboidratos em sua composição). A lectina nativa
é formada por 2 subunidades de 27 kDa, que formam um dímero em solução, independente de

40
pH (ALMEIDA et al., 2014). Esse dado foi corroborado pela massa intacta obtida por
espectrometria de massas, que determinou que CTL possui uma massa de 27.452 ± 2 Da.
Quanto às suas atividades biológicas, CTL exibiu baixa toxicidade contra náuplios de Artemia
sp., sendo esse efeito mediado por seu domínio de reconhecimento a carboidratos com glicanos
presentes no trato digestórios dos náuplios (ALMEIDA et al., 2014).
3.2 Aspectos gerais da espectrometria de massas
A espectrometria de massas pode ser entendida como uma técnica analítica que
permite a identificação da composição química de um determinado composto isolado, ou de
diferentes compostos em misturas complexas, através da determinação de suas massas
moleculares na forma ionizada (ou seja, com carga elétrica líquida, positiva ou negativa) (VAN
BRAMER, 1998). Esta análise se dá através da determinação da relação entre a massa e a carga
(m/z) das espécies ionizadas em fase gasosa (AEBERSOLD; MANN, 2003).
A espectrometria de massas é uma das técnicas analíticas mais úteis e poderosas
para a investigação científica, tendo sido aplicada em várias áreas, como ciências biológicas,
biomédicas, geologia, produtos naturais, sequenciamento de proteínas, identificação de
marcadores biológicos entre outras. A espectrometria de massas pode ser utilizada em análises
quantitativas, mas é em análises qualitativas que ela tem se destacado, como na identificação
de compostos em misturas e, principalmente, na caracterização estrutural de compostos
desconhecidos (NANDAKUMAR et al., 2009).
Um espectrômetro de massas é constituído fundamentalmente por três partes: uma
fonte de ionização, que é fundamental para gerar os íons das moléculas de interesse a serem
analisadas; um analisador, que separa as moléculas pela sua m/z; um detector que converte os
íons em sinais eletrônicos; e um sistema de aquisição de dados, que são softwares que
interpretam os sinais elétricos emitidos pelo detector e os converte em dados para análise
(STEEN; MANN, 2004). Além disso, os íons podem chegar à fonte de ionização através de
insersor como um cromatógrafo gasoso ou líquido.
O princípio básico da espectrometria de massas consiste em gerar íons a partir de
compostos (orgânicos ou inorgânicos) através de um método de ionização apropriado, separá-
los através de sua m/z em um analisador de massa, e detectar quali e/ou quantitativamente os
compostos a partir da m/z e suas respectivas abundâncias por meio de um detector, que “conta”
os íons e transforma o sinal em corrente elétrica. A magnitude do sinal elétrico em função da

41
m/z é convertida por um processador de dados, que gera o espectro de massas correspondente
(GROSS, 2004).
Inicialmente, a aplicação da espectrometria de massas era restrita à análise de gases,
substâncias voláteis e termicamente estáveis, pois as primeiras técnicas de ionização
empregadas (impacto de elétrons e química) são processos constituídos por duas etapas, nas
quais o analito é vaporizado com aquecimento e a ionização ocorre uma vez que a molécula
esteja na fase gasosa (KINTER; SCHERMAN, 2000).
O desenvolvimento de novas técnicas de ionização foi fundamental, pois permitiu
a produção de íons a partir de compostos de alta massa molecular e não voláteis. A fonte de
ionização é um passo primordial, pois é nela onde o analito é convertido em íon e posto em fase
gasosa. Há diversos métodos de ionização, porém os principais métodos para análise de
biomoléculas utilizados são a Dessorção/Ionização a Laser Auxiliada por Matriz (MALDI)
(KARAS; HILLENKAMP,1988) e Ionização por Eletrospray (FENN et al., 1990).
Nestas técnicas de ionização, íons são obtidos através da protonação ou
desprotonação, ou ainda pela adição de outros íons, como Na+, K+, NH4+ e Cl-, formando
aductos. Enquanto na ionização por elétrons, formam-se íons com energia interna elevada, que
se fragmentam e fornecem informações sobre características estruturais das moléculas, ESI e
MALDI são técnicas brandas de ionização, que formam íons com baixa energia interna,
permitindo a observação do íon molecular, entretanto com muito pouca ou nenhuma
fragmentação (CHAPMAN, 1993).
A ionização por electrospray (ESI) é um método de ionização brando, muito
utilizado para análise de proteínas e diversas moléculas polares e possui capacidade de análise
de moléculas com menos de 100 Da até maiores de 1.000.000 Da e é desenvolvida a pressão
atmosférica. Neste método, a amostra é dissolvida em um solvente polar e volátil e bombeada
através de um capilar de aço inoxidável em um fluxo de 1 µL/min a 1 mL/min. Uma alta
voltagem é aplicada a ponta do capilar (3 a 4 kV) formando um alto campo elétrico, a amostra
sai da ponta do capilar dispersa em forma de aerossol com gotas altamente carregadas. Esse
processo é auxiliado por um fluxo de gás nebulizador (geralmente nitrogênio) por fora do
capilar. Este gás auxilia o direcionamento do spray para o espectrômetro de massas e a
evaporação do solvente (HOFFMANN; STROOTBART, 2007). À medida que o solvente é
evaporado e as gotas vão diminuindo de tamanho, a repulsão eletrostática entre as moléculas
carregadas vai se tornando altíssima. Nesse mecanismo vão ocorrendo sucessivas explosões
coulômbicas de modo que gotas contendo apenas um íon são formadas. Paralelo a isso a

42
diminuição da gota também faz com que a tensão superficial não suporte tantas cargas (limite
de Rayleigh) e induz a transferência dos íons para a fase gasosa (CANTU et.al., 2008).
A ionização por MALDI possui características muito importantes que a fazem uma
fonte de ionização importante na caracterização de proteínas como a alta sensibilidade
(amostras em baixíssima concentração e quantidade podem ser analisadas por esta técnica) e
tolerância a contaminação das amostras com sais, tampões, detergentes etc. (CHEN et al., 1998;
STUMP et al., 2002). Dentro da fonte e sob vácuo, a mistura de matriz e amostra será irradiada
por um laser pulsado durante um determinado período de tempo. O mecanismo pelo qual os
íons são formados no MALDI não é completamente elucidado, porém postula-se que a
irradiação pelo laser induz um rápido aquecimento do cristal formado pela matriz pela
acumulação de uma grande quantidade de energia na fase condensada pela excitação da matriz.
Este rápido aquecimento causa a sublimação dos cristais da matriz, ablação de uma porção da
superfície cristalina e expansão da matriz para a fase gasosa, levando o analito intacto na pluma
em expansão (DREISEWERD, 2003).
Posteriormente a ionização, os íons produzidos e em fase gasosa serão separados
de acordo com sua m/z. Na verdade o que é mensurado na espectrometria de massas é a m/z
para cada íon e esta é a função dos analisadores. Este papel é realizado pelos analisadores de
massas. Eles podem ser divididos em duas grandes classes baseados em suas propriedades.
Analisadores tipo varredura que transmitem os íons de diferente m/z sucessivamente em uma
escala de tempo. O analisador do tipo quadrupolo é um exemplo. Outro grupo é composto pelos
analisadores que transmitem simultaneamente todos os íons como o analisador por tempo de
vôo (Time of Flight - ToF) (HOFFMANN; STROOTBART, 2007).
Cada analisador tem características e aplicações distintas. Além disso, estes
analisadores podem ser utilizados de maneira individual ou em conjunto, acoplados entre si,
dando origem a equipamentos classificados como híbridos. Tais equipamentos permitem que
experimentos em sequência (tandem) sejam realizados, isto é, sendo possível detectar um
determinado íon e posteriormente submetê-lo a uma etapa de fragmentação. Uma vez
separados, esses íons são detectados por eletromultiplicadoras que constituem os detectores
mais largamente usados (CANTU et al., 2008).
A possibilidade da criação de espectrômetros de massas que contém mais de um
analisador de massas iguais ou diferentes foi um grande marco da evolução das tecnologias em
MS, dando início a geração de equipamentos híbridos. A análise de dados por mais de um
analisador de massas é chamada de espectrometria de massas sequencial. O primeiro aparelho

43
híbrido comercialmente disponível e mais popularmente utilizado foi o Triplo Quadrupolo, que
é constituído de três analisadores quadrupolo em sequência.
A utilização de analisadores de massas em sequência possibilita utilizá-los para
desvendar a estrutura química de diversos compostos como peptídeos e proteínas. Diversas
técnicas são utilizadas nestes equipamentos, porém a mais popular e de melhor resposta é a
dissociação induzida por colisão (CID). Essa técnica consiste em selecionar o íon a ser
fragmentado utilizando o primeiro analisador de massas e em seguida enviá-lo a uma célula de
colisão, onde este íon irá colidir com um gás inerte. Em seguida, a energia cinética irá ser
transferida parcialmente em energia vibracional e os fragmentos resultantes serão analisados
por um segundo analisador de massas (HOFFMANN; STROOTBART, 2007).
A inserção de reflectrons nos analisadores ToF foi um incremento significativo à
resolução desse analisador. O acoplamento de um analisador quadrupolo como filtro de íons e
um analisador do tipo ToF com reflectron (gerando os equipamentos do tipo Q-ToF) foi um
salto de qualidade nas análises de estrutura de peptídeos, possibilitando diferenciar aminoácidos
que possuem diferenças de milidaltons em sua massa monoisotópica (HOFFMANN;
STROOTBART, 2007)
Os programas mais comumente empregados para a identificação de proteínas em
bancos de dados a partir de dados de MS são o “Sequest” e o “Mascot”. Ambos os programas
correlacionam espectros de massas de fragmentação (não interpretados) de peptídeos com
sequências de aminoácidos de proteínas registradas em bancos de dados (CHAMRAD et al.,
2004; ELIAS et al., 2005). Além disso, esses softwares também têm a capacidade de usar
sequências de nucleotídeos para fazer tal correlação. Para tal, eles primeiramente simulam as
sequências primárias potenciais das proteínas correspondentes àquelas sequências de
nucleotídeos encontradas nos bancos de genes, utilizando-se do código genético universal.
Posteriormente, simulam a fragmentação destas sequências primárias. De forma geral, estes
programas têm como objetivo encontrar a sequência de aminoácidos, em um determinado banco
de dados, que melhor descreve os íons fragmentos encontrados em um espectro. As sequências
“candidatas” são procuradas nos bancos de dados de acordo com a massa do peptídeo intacto e
com o espectro de fragmentação obtido para cada peptídeo (CANTU et al., 2008).
Além disso, a interpretação manual de espectros é recomendada em todos os casos
e indispensável em algumas situações. Por fim, existem situações nas quais o genoma de uma
determinada espécie ainda não está completamente sequenciado ou disponível e, neste cenário,
é necessário derivar a sequência primária de aminoácidos de um determinado peptídeo baseado

44
única e exclusivamente nos dados obtidos por espectrometria de massas, isto é, sem recorrer a
banco de dados (sequenciamento “De novo”) (STEEN; MANN, 2004).
Atualmente a espectrometria de massas se tornou uma das ferramentas mais
utilizadas na caracterização de proteínas, devido a algumas características como a rapidez e
relativa simplicidade da amostra, a necessidade de quantidades pequenas de amostra e a
precisão dos dados obtidos. Esta técnica é capaz de determinar a massas molecular de uma
proteína, estados oligoméricos, localização de pontes de sulfeto, caracterização de modificações
pós traducionais (glicosilações e fosforilações) e identificação de proteínas provenientes da
separação de uma eletroforese bidimensional. A caracterização de proteínas e suas
modificações por MS tem se tornado uma técnica em constante expansão.
O potencial de aplicação de MS em estudos biológicos tem sido bastante estendido,
em razão dos impressionantes avanços observados nos últimos anos nas áreas de genômica, de
transcriptômica, de metabolômica, de proteômica, de lipidômica e de outras
plataformas “omicas”, e do desenvolvimento extraordinário dos equipamentos (FENG et al.,
2008; HOFFMANN; STROOTBART, 2007). A MS é atualmente uma técnica bastante
utilizada para identificação de proteínas e para estudo de modificações pós-traducionais em
diferentes condições fisiológicas. Além disso, a MS vem sendo utilizada no monitoramento e
na caracterização de diversos processos industriais, tais como processos fermentativos e até
mesmo análise de microrganismos intactos (CLAYDON et al., 1996; FENSELAU; DEMIREV,
2001).
3.3 Cristalografia de raios X
A cristalografia de raios X é provavelmente o método de análise estrutural mais
poderoso na elucidação de estruturas tridimensionais de moléculas e macromoléculas
biológicas. A aplicação da cristalografia de macromoléculas para o estudo estrutural,
especialmente de proteínas, deve o seu nascimento aos estudos de Max Perutz e John Kendrew,
que receberam o Prêmio Nobel em química (1962) pelo seu trabalho sobre a determinação da
estrutura tridimensional da hemoglobina e mioglobina. De 1959 até o presente, a biologia
estrutural tem avançado consideravelmente.
O principal objetivo da cristalografia é o estudo estrutural, por métodos de difração
de raios X. Através desta técnica pode-se determinar espacialmente a posição atômica de todos
os átomos que constituem uma macromolécula biológica como uma proteína. Esse estudo

45
estrutural é fundamental para o entendimento molecular de vários mecanismos biológicos
(DELATORRE, 2006).
A cristalização de proteínas é, basicamente, um procedimento de tentativa e erro.
Algumas características potencializam a cristalização de proteínas. De uma forma geral, quanto
mais pura estiver a proteína, maiores as chances de crescimento de cristais. Neste caso, técnicas
como a espectrometria de massas são importantes para uma análise prévia da pureza da
preparação. A proteína é dissolvida em uma solução tampão aquosa contendo aditivos e agentes
precipitantes como sulfato de amônio, dentre outros. Cada proteína cristaliza sob determinados
parâmetros. As proteínas são peculiares devido ao seu alto conteúdo de solvente, valores de pH
e força iônica definidas para a sua estabilidade e função, dentre outros. Logo, estas proteínas
têm de ser cristalizadas sob suas condições nativas, através da utilização de soluções complexas
(DRENTH, 2007).
Cristais são caracterizados por apresentarem alto grau de ordenação interna, ou seja,
são formados por repetições translacionais de moléculas ou átomos em todas as direções. A
periodicidade interna é descrita por uma pequena unidade de volume uniforme da qual o cristal
pode ser considerado um derivado, chamada de cela unitária. Cada cela unitária é caracterizada
por três vetores a, b e c que definem suas arestas e pelos ângulos α, β e γ existentes entre elas
(DELATORRE et al., 2006).
Para crescer cristais de qualquer composto, as moléculas ou íons têm de ser
deslocados a um estado de supersaturação, um estado que é termodinamicamente instável, no
qual pode se desenvolver numa fase amorfa ou cristalina quando este retornar ao equilíbrio. A
Supersaturação pode ser alcançada pela evaporação lenta do solvente ou pela variação de alguns
dos parâmetros como temperatura, pH e força iônica. Neste estado, a solução de proteína
alcança a zona de nucleação, onde podem se formar os núcleos (primeiros agregados ordenados
da proteína). Com a diminuição da solubilidade da proteína, estes núcleos atingem a zona
metaestável, onde ocorrerá o crescimento do cristal (DUCRUIX; GIEGÉ, 1992).
Existem muitas técnicas para a cristalização de macromoléculas biológicas, todas
tendo como objetivo trazer a solução a um estado de supersaturação. A fim de obter cristais de
proteínas, um método frequentemente utilizado é a difusão de vapor. (HAMPEL et al., 1968).
Nesta técnica, uma gota contendo poucos microlitros da macromolécula biológica a ser
cristalizada em um agente de cristalização, tampão e aditivos, é equilibrada contra um
reservatório contendo a solução de cristalização a uma concentração mais alta do que na gota e
um volume largamente maior. O equilíbrio prossegue por meio da difusão das espécies voláteis
(água e solventes orgânicos) até que a pressão de vapor na gota se iguale àquela do reservatório.

46
Se o equilíbrio ocorre por meio de troca da água (da gota para o reservatório), isto leva a uma
diminuição do volume da gota, e consequentemente a um aumento na concentração de todos os
constituintes da gota de cristalização, inclusive da proteína. Para espécies químicas com pressão
de vapor maior que a pressão de vapor da água, a troca ocorre do reservatório para a gota. Os
mesmos princípios se aplicam para gotas suspensas (hanging drops), gotas sentadas (sitting
drops) e gotas sanduíches (sandwich drops) (Figura 8).
Figura 8. Representação esquemática dos três métodos de difusão de vapor para cristalografia de macromoléculas.
Fonte: PEREIRA-JUNIOR, 2014. A: Gota suspensa, B: Gota sentada e C: Gota sanduíche.
Com o aumento do número de macromoléculas biológicas cristalizadas com
sucesso, tornou-se óbvio que muitas das condições de cristalização se assemelhavam, ou seja,
havia uma concentração de resultados positivos de cristalização que usava um número limitado
de precipitantes, tampões e aditivos (CARTER; CARTER, 1979), levando à produção dos kits
comerciais contendo numerosas condições com diferentes tipos de agentes precipitantes,
concentrações de precipitantes, tampões, pHs e sais. Uma vez que as condições iniciais foram
encontradas, é possível otimizar as condições de cristalização para a obtenção de melhores
cristais quanto ao tamanho e forma (WOOH et al., 2003; JANCARIK; KIM, 1991).
O passo seguinte na obtenção da estrutura de uma proteína é a difração do cristal
obtido em raios X. O sucesso dessa etapa depende, principalmente, da qualidade dos cristais e
da fonte de A coleta e o processamento dos dados a partir de difração do cristal em raios X é o
próximo passo no objetivo de se obter a estrutura. O sucesso desta etapa depende da qualidade
dos cristais e da fonte de radiação utilizada. Nesta etapa, os cristais são caracterizados, ou seja,
seus parâmetros de rede e suas simetrias são determinados. O conhecimento das amplitudes e
das fases permite a reconstituição da densidade eletrônica do cristal.
Ao ser colocado em um feixe de raios X, a densidade eletrônica de cada átomo do
cristal interage com a radiação, dispersando-a em todas as direções do espaço. É a presença de
uma rede cristalina que amplifica a radiação somente em determinadas direções, dando origem
ao padrão de difração característico. O cristal é girado em torno de um eixo geralmente

47
perpendicular ao feixe de raios X e os raios difratados são coletados em um detector capaz de
gravar um grande número de pontos (spots) em um tempo de aquisição bem curto. Cada imagem
corresponde a uma rotação de 0,2-1º grau e pode ser considerada uma "fatia" do espaço
recíproco. A posição de uma reflexão contém as informações sobre a geometria do grupo
espacial e da rede cristalina, enquanto que a intensidade I medida depende da distribuição da
densidade eletrônica na unidade assimétrica. De posse das primeiras imagens, é possível
realizar a indexação e determinar o grupo espacial, dimensões e orientação da célula unitária,
além de estimar a mosaicidade. Esse passo é necessário para planejar uma estratégia de coleta
de dados eficiente, minimizando o tempo e maximizando os resultados (SMYTH; MARTIN,
2000; WLODAWER et al., 2013).
Durante o processo de difração é possível medir a amplitude de cada reflexão, mas
não sua fase correspondente, que é invariavelmente perdida durante o experimento de difração,
ou seja, toda a informação angular é perdida. Isso é chamado de “problema da fase” (LI; HE;
ZHANG, 2015; WLODAWER et al., 2013). Existem diferentes métodos criados para resolver
esse “problema”. A técnica mais utilizada para esse fim é a substituição molecular que se baseia
no princípio de que macromoléculas que apresentam alta homologia sequencial tendem a se
enovelar de forma similar, e desta forma, uma proteína já resolvida serve como modelo inicial
para a obtenção de um conjunto de fases que será subsequentemente refinado (ABERGEL,
2013; WLODAWER et al., 2013).
Uma vez que o modelo preliminar foi conseguido o próximo passo é o refinamento
das posições atômicas, ou seja, encontrar uma melhor concordância entre o modelo proposto
para a estrutura da molécula e sua estrutura real. O acompanhamento da qualidade do modelo
é analisado pelo fator de correlação Rfactor, que é um fator de correlação entre o modelo atual e
o modelo real (HOLTON et al., 1992; MURSHDOV et al., 2011). Uma estrutura com resolução
menor que 2,0 Å está considerada resolvida quando o valor do Rfactor atingir valores menores
eu igual a 20%. Um outro fator importante é o Rfree que é uma pequena porcentagem das
reflexões que são excluídas do refinamento e utilizadas como um teste para verificar se não
houve superrefinamento da estrutura, em geral a diferença entre o Rfactor e o Rfree não deve
ultrapassar 5% (KARPLUS; DIEDERICHS, 2015; MORRIS et al., 2014).
O último passo é a validação do modelo quanto aos parâmetros estruturais. Esses
programas realizam análises das as distâncias ideais entre as ligações, os ângulos de ligação, os
ângulos diédricos e os contatos de Van der Waals, usando para isso a comparação com proteínas
modelos. Uma das saídas destes programas é o diagrama de Ramachandran, que afere as
medidas dos ângulos diédricos Φ e Ψ e é um bom indicador da qualidade do modelo (COOPER

48
et al., 2012; HOLLINGSWORTH; WLODAWER et al., 2013). Um resumo do processo de
cristalografia de proteínas está ilustrado na figura 9.
Figura 9. Etapas da Cristalografia de raios X.
Fonte: Elaborado pelo autor. A: Cristalização. B: Difração de cristal e obtenção do padrão de difração. C:
Resolução da estrutura por bioinformática. D: Estrutura tridimensional (VML, código PDB: 4WV8).
3.4 Docking molecular
O docking molecular é uma ferramenta computacional muito aplicada em projetos
de drug discovery e em estudos de interações de proteínas com ligantes. Tradicionalmente, o
docking molecular é utilizado para abordar uma das seguintes aplicações: 1) dada uma molécula
de um ligante e um receptor proteico, fazer a predição do modo de ligação do ligante com o
receptor, 2) realizar uma varredura de uma coleção de moléculas pequenas com um receptor e
fazer um ranking de acordo com a probabilidade de ligação com o mesmo e 3) dada uma
molécula de um ligante e um receptor alvo, predizer a afinidade de ligação entre os dois. A
orientação assumida pelas moléculas em um complexo depende da complementaridade em
tamanho, forma, distribuição de cargas, polaridade e potencial de interações, e da afinidade

49
entre as moléculas (POLANSKI; KIMMEL, 2007). A simulação de docking pode ser realizada
utilizando-se um algoritmo capaz de prever o modo ideal de ligação entre o ligante e o sítio de
ligação de uma molécula alvo. No docking, usualmente a estrutura do receptor permanece rígida
e várias posições de possíveis para o ligante são identificadas pelo software (COLEMAN et al.,
2013; POLANSKI; KIMMEL, 2007).
Essa técnica permite realizar a triagem de grandes bibliotecas de compostos,
avaliando afinidade e a especificidade a partir de propriedades estruturais e químicas, como
tamanho, geometria, distribuição de cargas, polaridade e potencial de interações hidrofóbicas e
ligações de hidrogênio. O objetivo da triagem de bancos de possíveis ligantes é identificar
compostos que se ligam a uma proteína alvo. Dessa forma, a reação bioquímica que a proteína
alvo realiza pode ser alterada, permitindo a descoberta de possíveis ferramentas médicas e/ou
biotecnológicas (ROCHA, 2011). Atualmente, o acesso via internet a banco de dados de
estruturas de proteínas, como o PDB, e de grandes bibliotecas de pequenas moléculas
comercialmente acessíveis, como o ZINC database (IRWIN; SHOICHET, 2005) fornecem um
grande número de possíveis novos ligantes, uma vez que esses ligantes forem testados in silico
eles podem ser comprados ou sintetizados para teste in vitro e/ou in vivo, agilizando desta forma
a pesquisa farmacêutica e de biotecnologia (SHOICHET, 2004). Essa abordagem de se testar
uma grande quantidade de compostos por meio de docking molecular é chamada de screening
virtual.
Análises de DM podem permitir a localização de sítios de ligação em proteínas,
locais de ligação de drogas, análises de ligação entre lectinas e carboidratos, medida da força
das interações entre as moléculas. Além disso com o uso do docking é possível verificar como
alterações estruturais podem afetar a ligação da proteína com os ligantes, por meio análises
comparativas com moléculas mutadas (REGO, 2012).
O docking de moléculas com possíveis ligantes segue uma ordem. Primeiramente
se faz a separação dos prováveis sítios de ligação, depois disso se faz a sobreposição destas
regiões e por meio de análises estatísticas e funções de pontuação tenta-se obter os complexos
mais próximos do que ocorre experimentalmente. A análise dos encaixes é dada pontuando-se
a formação de cada complexo. A partir desta pontuação é realizada uma análise comparativa,
em que cada complexo formado é avaliado com uma pontuação, que ao final será posicionado
em um ranking (SCHNEIDMAN-DUHOVNY et al., 2012; SCHNEIDMAN-DUHOVNY et
al., 2005).
O docking molecular vem sendo bastante aplicado no estudo com lectinas no
objetivo de buscar estruturas glicídicas capazes de se ligar as lectinas de interesse. Alguns

50
exemplos de aplicação dessa técnica são: uso do DM na lectina de Dolichos lablab para análise
de interação entre a proteína e oligomanosídeos (TEIXEIRA et al., 2014) busca de ligantes para
lectinas de bactérias (TOPIN et al., 2013), uso de DM para análise da forma de interação das
lectinas de Yucca filamentosa e Arum maculatum com ácido siálico (XU et al., 2012).
3.5 Atividades biológicas de lectinas vegetais
Cada vez mais as lectinas têm demonstrado novas aplicabilidades em diversas
áreas, abrangendo desde a medicina até a agricultura, vários estudos têm demonstrado que as
lectinas, sejam de origem vegetal ou de outras fontes, exibem uma gama de atividades
biológicas desencadeadas pela interação dessas proteínas com a porção glicídica dos
glicoconjugados da superfície celular.
Demonstrou-se já que lectinas de especificidades semelhantes por carboidratos
podem, em alguns casos, apresentar afinidade pelo ligante específico de maneira diferenciada,
como se alguma pequena diferença estrutural entre as lectinas pudesse ocasionar diferenças de
afinidade em cada complexo estabelecido entre lectina-ligante, e isto se referir na potência das
atividades biológicas desempenhadas por estas proteínas (CAVADA et al., 2001).
Dentre as atividades biológicas podemos citar a indução de efeito depressivo e
expressão de marcadores neuroinflamatórios em ratos (GONÇALVES et al., 2013), liberação
de mediadores quimiotáticos por macrófagos (ALENCAR et al., 2007), aumento da resistência
vascular renal, da taxa de filtração glomerular e do fluxo urinário (MARTINS et al., 2005),
migração de neutrófilos in vivo por mecanismo indireto (ALENCAR et al., 2003) a proliferação
de linfócitos com produção de interferon γ (BARRAL-NETTO et al., 1992), produção de oxido
nítrico (ANDRADE et al., 1999) e indução de apoptose (BARBOSA et al., 2001),
reconhecimento específico e atividade antiproliferativa sobre células cancerígenas (WU et al.,
2004; LIU et al., 2009), atividade anti (PIRES et al.,2016; NASCIMENTO et al., 2015) ou pro-
inflamatória SANTIAGO et al., 2014, TEIXEIRA et al., 2014), atividade vasorrelaxante
(BARROSO-NETO et al., 2016; ALVES et al., 2015), atividade antinociceptica (ALMEIDA
et al., 2015; PIRES et al., 2014), atividade cicatrizante (NASCIMENTO-NETO, 2012), agentes
em drug delivery (BIES et al., 2004), atividade inseticida (ZHU et al., 1996; MACEDO et al.,
2007), atividade antifúngica (CIOPRAGA et al., 1999; SOUZA et al., 2011), atividade antiviral
(FAVACHO et al., 2007) e antibacteriana (COSTA et al., 2010).
As possibilidades do uso de lectinas vegetais como ferramentas biotecnológicas são
consideradas devido ao grande número de trabalhos científicos exibindo atividades biológicas

51
relevantes para essas moléculas (KITADA et al., 2010; KIMBLE et al., 2010). A propriedade
que muitas lectinas possuem de aglutinar células talvez seja seu aspecto imunológico mais
importante. Devido à multivalência de reconhecimento de carboidratos, muitas lectinas fazem
ligações cruzadas entre células e seus receptores (DUBOIS et al., 1998).
3.6 Aspectos gerais do processo inflamatório
O processo infamatório é uma sequência complexa de eventos que ocorre em
tecidos vascularizados, em resposta a agressão por agentes lesivos. Esta reação tem como
principal objetivo livrar o organismo do agente causador da injúria e também desencadear
processos que tendem a reparação do tecido lesado (McGEER; McGEER, 2000). Uma
característica importante deste processo é que, independente da natureza do estímulo, a resposta
inflamatória segue um padrão característico, podendo ser observadas modificações discretas no
padrão de resposta, inerente ao agente etiológico, ao tecido ou órgão lesado e ao estado
patológico do hospedeiro (RANG et al., 2001).
Os sinais clínicos da inflamação, eritema, calor, inchaço e dor, foram descritos por
Cornelius Celsus no início da era cristã (FANTONE; WARD, 1990). O quinto sinal da
inflamação, a lesão aguda dos tecidos, com perda da função dos órgãos foi mencionado mais
tarde por Virchow, no século XIX (ROCHA; SILVA, 1978).
A inflamação pode ser dividida em uma fase aguda e fase crônica. A primeira das
fases da reação inflamatória, fase aguda, depende de o estímulo ser ou não persistente, esta
reação pode tornar-se crônica, podendo, muitas vezes, ser prejudicial ao organismo.
Frequentemente, após a fase aguda, ocorre a resolução do processo, em razão da eliminação
dos agentes causadores.
Vários tipos de células estão envolvidos no processo inflamatório e estas podem ser
classificadas em três tipos: células endoteliais; células próprias do tecido (mastócitos,
fibroblastos e macrófagos residentes); células migratórias (neutrófilos, eosinófilos, basófilos,
linfócitos sanguíneos, plasmócitos e macrófagos livres).
A resposta inflamatória aguda envolve fenômenos vasculares (vasodilatação e
aumento da permeabilidade celular) e celulares (infiltração celular) decorrentes da liberação
local de mediadores químicos formados e liberados concomitantemente ou sequencialmente no
local da lesão. Estes mediadores podem ser divididos em nas seguintes categorias: Aminas
vasoativas, metabólitos do ácido araquidônico, fator ativador plaquetário, citocinas,

52
quimiocinas, óxido nítrico, radicais livres derivados de oxigênio e neuropeptídeos (FERENCÍK
& STVRTINOVÁ, 1996; ABBAS et al., 2010).
A vasodilatação é um fenômeno que corresponde a alterações no calibre vascular,
que conduz a um aumento do fluxo sanguíneo, decorrente da ação de mediadores
principalmente em arteríolas. O aumento da permeabilidade vascular se deve a ação de
mediadores inflamatórios sobre as células endoteliais venulares, induzindo a contração das
mesmas. Isto permite a passagem de proteínas plasmáticas para o interstício, as quais não seriam
filtradas em condições fisiológicas. O aumento da permeabilidade vascular somado ao aumento
da pressão de filtração, por consequência da vasodilatação, leva à formação do edema
inflamatório (ABBAS et al., 2010).
A migração de leucócitos da microcirculação e seu acúmulo no foco da agressão é
uma das etapas fundamentais para a defesa do organismo. Os leucócitos ingerem os agentes
agressores, destroem as bactérias e outros micróbios e degradam o tecido necrótico e os
antígenos estranhos. Porém, os leucócitos podem prolongar a inflamação e induzir lesão
tecidual mediante a liberação de enzimas, de mediadores químicos e de radicais livres do
oxigênio (ABBAS et al., 2010).
A mobilização adequada dos leucócitos, da microcirculação para o foco
inflamatório (tecido intersticial), é também uma etapa fundamental para a defesa do organismo
e é denominada de extravasamento. Pode ser dividida nas seguintes etapas: (1) intraluminais:
marginação, rolamento e adesão; (2) transmigração através do endotélio (também denominada
de diapedese); e (3) migração nos tecidos intersticiais na direção do estímulo quimiotático
(ABBAS et al., 2010).
3.7 Aspectos gerais da contratilidade
O mecanismo contrátil do músculo liso vascular é de grande importância para o
conhecimento dos papéis funcionais, fisiológicos e fisiopatológicos do mesmo. O músculo liso
vascular, geralmente, opera em um estado de tensão devido a pressão que o sangue exerce sobre
a parede dos vasos. Este estado é conhecido como tônus, a partir do qual ele pode contrair ou
relaxar, induzindo assim o aumento ou redução da pressão no interior dos vasos (MONCADA
et al., 1991; FURCHGOTT; ZAWADZKI, 1980).
O estado contrátil do músculo liso é regulado pela concentração de cálcio livre no
citosol. Uma variedade de estímulos que induzem a contração do músculo liso, como

53
despolarização da membrana, α-adrenárgicos, e agonistas muscarínicos ativam um aumento no
nível de Ca2+ (ALESSI et al., 1992; KARAKI et al., 1997).
Várias substâncias (fatores) que são produzidas pelas células endoteliais difundem-
se ou são liberadas, tanto para o sangue como para as outras camadas do vaso. Dessa forma,
elas podem controlar funções das células musculares lisas e das células circulantes que
participam das funções regulatórias. Dentre os fatores produzidos, destacam-se o óxido nítrico
(NO), a prostaciclina, o tromboxano, as endotelinas, a angiotensina II, o fator hiperpolarizante
do endotélio, a bradicinina, a serotonina, a histamina e os radicais livres de oxigênio, dentre
outros (VANE, et al., 1990; LUSHER; BARTON, 1997; VAPAATALO; MERVAALA, 2001).
Uma dessas funções regulatórias consiste no controle do tônus da musculatura lisa
vascular pela produção de mediadores que podem produzir vasodilatação ou vasoconstrição.
Os principais fatores relaxantes derivados do endotélio são o NO, o fator hiperpolarizante
derivado do endotélio e a prostaciclina (CARVALHO et al., 2001).
Dentre os fatores relaxantes, o NO é o mais potente e é produzido no endotélio a
partir da ação da enzima NO sintase sobre o substrato L-arginina, com formação de NO e L-
citrulina. O NO difunde-se para o interior das células musculares lisas, onde interage com o
átomo de ferro do grupo heme da molécula de guanilato ciclase, levando à ativação dessa
enzima; essa enzima ativada, por sua vez, atua sobre o trifosfato de guanosina (GTP),
transformando-o no composto ativado monofosfato cíclico de guanosina (GMPc). O aumento
da concentração de GMPc nas células musculares leva à diminuição da [Ca+2]i e ao consequente
relaxamento do vaso. Sob condições basais, o NO é continuamente liberado pelas células
endoteliais saudáveis (SILVA, 2006).
3.8 Câncer, gliomas e autofagia
3.8.1 Câncer
O termo câncer inclui diferentes patologias que tem como características comuns:
desregulação na divisão celular, perda da capacidade de diferenciação das células, aumento da
sobrevivência celular e aumento da capacidade de migração celular (CORNER; BAILEY,
2008). A tumorigênese em humanos é um processo complexo que reflete alterações genéticas
que direcionam uma transformação progressiva de células normais em derivativos altamente
malignos (TABASSUM; POLYAK, 2015).

54
O processo pelo qual células normais se transformam progressivamente em células
tumorais pode ser resultado de processos endógenos como erros na replicação do DNA, a
instabilidade química intrínseca de algumas bases do DNA, a perda de heterozigose ou um
ataque de radicais livres gerados durante o metabolismo. Danos ao DNA também podem ser
resultado de interações com agentes exógenos como radiações ionizantes, radiação UV e/ou
carcinogênicos químicos (BERTRAM, 2000). As células possuem mecanismos para reparar
tais danos, mas, por várias razões erros podem ocorrer e, com isso, mutações no genoma dessas
células (BERTRAM, 2000).
A transição de célula normal para tumoral envolve a participação de genes
envolvidos em mecanismos de homeostase que controlam a proliferação e morte celular. Se
essas mudanças e mutações induzem a ativação de genes que estimulam a proliferação ou
protegem a célula da morte, esses genes são referidos como protooncogenes. Se mutações
inativam genes que normalmente inibem a proliferação, esses genes são referidos como
supressores de tumor (CHIAL, 2008; ZHU et al., 2015).
As marcas do câncer são seis capacidades adquiridas durante o processo de
desenvolvimento dos tumores. Elas incluem a manutenção de sinalização proliferativa, evasão
dos supressores proliferativos, resistência a morte celular, ativação da imortalidade replicativa,
indução de angiogênese e capacidade de invasão e metástase (HANAHAN; WEINBERG,
2011).
3.8.2 Gliomas
Gliomas malignos são os tumores cerebrais primários mais comuns. Os gliomas
mais comuns incluem astrocitomas, oligodendrocitomas, ependimomas e glioblastomas.
Mesmo que relativamente raros (menos de 1 % de incidência), tipicamente representam 30 %
de todos os tumores cerebrais de adultos, e cerca 80 % de todos os tumores cerebrais malignos
de adultos (AMERICAN CANCER SOCIETY, 2014). Eles são caracterizados por uma
proliferação local, uma infiltração pelo parênquima cerebral e uma angiogênese robusta. O atual
tratamento convencional inclui cirurgia, radioterapia e quimioterapia. Muitos agentes
terapêuticos como a temozolomida (TZM), cisplatina, carmustina e lomustina vem sendo
usados para tornar mais lenta a progressão desses canceres incuráveis (JOHNSON; WHITE;
GRIMALDI, 2011). Um problema que ocorre é que as células de glioma frequentemente
adquirem resistência a esses agentes (HEGI et al., 2005). O tempo médio de vida para pacientes
diagnosticados com glioblastoma é de aproximadamente 12 meses, por essa razão, tem se feito

55
esforços para a busca de vias moleculares visando o desenvolvimento de novos agentes e
terapias contra este tipo de câncer (FURNARI et al., 2007).
Existem diversos tipos de tumores cerebrais, a eles são classificados pela
Organização Mundial de Saúde (OMS) baseando-se na sua origem celular e aparência
histopatológica.
Astrocitomas: a maioria dos tumores que se desenvolvem no cérebro começam
como células gliais chamadas astrócitos. Esses tumores são chamados astrocitomas. Dois de
cada dez tumores no cérebro são astrocitomas. A maioria desses pode se espalhar amplamente
pelo cérebro e se misturar com o tecido sadio, o que os tornam bastante difíceis de se remover
cirurgicamente. É muito raro que eles se espalhem para fora do cérebro ou medula espinhal.
Astrocitomas são geralmente classificados como de baixo grau, grau intermediário e de alto
grau baseado em como na aparência das células ao microscópio. O astrocitoma de mais alto
grau é conhecido como glioblastoma, sendo esse o de progressão mais rápida e o tipo de tumor
maligno cerebral mais comum em adultos (AMERICAN CANCER SOCIETY, 2014).
Oligodendrogliomas: esses tumores iniciam em célula do cérebro chamadas
olidendrócitos. Como astrocitomas, a maioria deles pode se infiltrar nos tecidos próximos e não
podem ser completamente removidos com cirurgia. Oligodendrogliomas raramente se
espalham para fora do cérebro ou medula espinhal. Formas muito agressivas desses tumores
são chamadas oligodendrogliomas anaplásticos. Apenas 2 % dos tumores cerebrais são
oligodendrogliomas (AMERICAN CANCER SOCIETY, 2014).
Ependimomas: esses tumores surgem a partir de células ependimais que delimitam
os ventrículos. Eles podem variar de tumores de grau muito baixo (menos agressivo) até
tumores de alto grau, chamados de ependimomas anaplásticos. Ependimomas não se espalham
para fora do cérebro ou medula espinhal. Diferentemente dos tipos anteriores esse tipo de tumor
não infiltra em tecidos saudáveis permitindo, na maioria das vezes, sua retirada por cirurgia
(AMERICAN CANCER SOCIETY, 2014).
3.8.3 Autofagia
Autofagia é um processo celular evolutivamente conservado de degradação e
reciclagem de proteínas e organelas. É um mecanismo induzido como resposta a diversos
estímulos como estresse oxidativo, hipóxia ou depleção de nutrientes. Sendo um processo
constitutivo, a autofagia basal tem importante função na manutenção na homeostase, através da
manutenção do controle de qualidade de proteínas e organelas. Mesmo que a maioria das

56
evidências suportem uma função de manutenção da sobrevivência da célula, paradoxalmente,
a morte celular resultando de um consumo de células progressivo tem sido atribuído a autofagia
irrestrita (MIZUCHIMA, 2007; REEF et al., 2006).
Os mecanismos que regulam as funções mutualmente-opostas de sobrevivência e
morte para a autofagia ainda não são conhecidos. A explicação mais plausível é que o
catabolismo por autofagia é predominantemente para manutenção da sobrevivência. Mas um
desbalanço no metabolismo celular onde o consumo celular autofágico excede a capacidade
celular de síntese, promove a morte celular (MATHEW; KARANTZA-WADSWORTH;
WHITE, 2007).
Existem três formas de autofagia: macroautofagia, autofagia mediada por
chaperonas e microautofagia. Sendo a mais observada a macroautofagia (RAVIKUMAR et al.,
2010).
O processo de macroautofagia ocorre da seguinte forma: fagóforos (também
chamados de estruturas pré-autafagossomais ou membranas de isolamento) elongam e se
fundem enquanto engolfam uma porção do citoplasma em vesículas, chamadas
autofagossomos. Esses autofagossomos primeiro se fundem com endossomos para formar
organelas híbridas chamadas anfissomos que posteriormente se fundem com lisossomos ácidos
onde o conteúdo citoplasmático é degradado (Figura 10). Os mecanismos moleculares
responsáveis pela regulação da autofagia ainda não foram completamente elucidados, o que se
sabe é que diversos genes de autofagia (ATG) participam da regulação desse processo celular
(RAVIKUMAR et al., 2010).
Figura 10. Diagrama esquemático dos passos do processo de autofagia.
Fonte: Adaptado de Melendez e Levine (2009).

57
Nesse contexto, Pratt e colaboradores (2012) mostraram a capacidade de ConA de
induzir morte autofágica de células de glioma na linhagem de células de glioma humano U87.
Foi mostrado que essa atividade se deve a regulação da proteína MT1-MMP (proteína de
membrana - metaloproteinase tipo 1), uma glicoproteína fortemente expressa em gliobastomas
radio- e quimioresistentes. Essa proteína é responsável por mediar a sinalização pró-apoptótica
e autofágica em células de câncer do cérebro. A lectina foi capaz de induzir a atividade da
proteína com consequente aumento de vacúolos ácidos característicos de processos autofágicos.

58
CAPÍTULO II:
DETERMINAÇÃO DA ESTRUTURA PRIMÁRIA DE
CTL POR ESPETROMETRIA DE MASSAS

59
4 CAPÍTULO II: DETERMINAÇÃO DA ESTRUTURA PRIMÁRIA DE CTL POR
ESPETROMETRIA DE MASSAS
4.1 Digestão proteolítica in gel e obtenção de peptídeos
A estrutura primária da lectina de sementes de C. tomentoseum foi determinada
através de sobreposição das sequencias de peptídeos oriundos de digestões proteolíticas
determinadas por espectrometria de massas sequencial (MS/MS).
O procedimento de digestão proteolítica da lectina para sequenciamento foi
realizado seguindo o protocolo descrito por Shevchenko e colaboradores (2006). A proteína
foi aplicada a um gel de 12% de poliacrilamida (SDS-PAGE), realizada segundo uma adaptação
ao método descrito por Laemmli (1970). A banda referente à cadeia alfa da proteína foi
recortada do gel, descorada por sucessivas lavagens utilizando uma solução de 50% de
acetonitrila contendo 25 mM de bicarbonato de amônio, desidratada em 100% de acetonitrila e
seca em Speedvac (LabConco).
Após a secagem, o gel contendo a lectina foi reidratado em uma solução contendo
uma das seguintes soluções contendo enzimas proteolíticas: 50 mM de bicarbonato de amônio
com tripsina (Sigma-Aldrich) ou quimiotripsina (Sigma-Aldrich); 25mM de bicarbonato de
amônio com endoproteinase Asp-N (Sigma-Aldrich); HCl 10 mM contendo Pepsina. A razão
enzima:substrato utilizada nos experimentos foi de 1:50 (m/m). As reações de digestão
permaneceram overnight à 37 ºC, sendo interrompidas com a adição de ácido fórmico a 2%. Os
peptídeos oriundos das digestões proteolíticas foram então extraídos do gel utilizando uma
solucao de 5% de ácido fórmico em 50%, concentrados em Speedvac e ressuspensos com 25
µL com ácido fórmico 0,1%.
Os peptídeos oriundos das digestões proteolíticas foram então extraídos do gel
utilizando uma solução de 5% de ácido fórmico em 50% de acetonitrila, sob agitação durante
15 minutos. Este procedimento foi repetido três vezes. O sobrenadante contendo os peptídeos
extraídos foram unidos, concentrados em Speedvac e ressuspensos com 25 μL com ácido
fórmico 0,1%.

60
4.2 Sequenciamento de peptídeos por MS/MS
Os peptídeos obtidos das diferentes digestões proteolíticas foram injetados em um
sistema de cromatografia liquida de ultra performance (NanoAcquity®, Waters Corp. Milford,
USA) diretamente conectado a uma fonte de nanoeletrospray de um espectrômetro de massas
(Synapt HDMS, Waters Corp., Milford, USA). Os peptídeos foram previamente separados por
cromatografia fase reversa em coluna BEH300 C18 (75 μm x 100 mm), eluída com um gradiente
linear partindo de 10% a 85% de acetonitrila contendo 0,1% de ácido fórmico à 60 µL/min.
O espectrômetro de massas foi operado em modo positivo, com a temperatura da
fonte de 90 ºC e a voltagem do capilar de 3,0 kV. Os experimentos de LC-MS/MS foram
realizados utilizando a função DDA (Data Dependent Acquisition – Aquisição Dependente de
Dados), onde os íons precursores com carga entre +2 e +4 foram selecionados para análise de
MS/MS sendo fragmentados através de CID (Collision Induced Dissociation - Dissociação
induzida por colisão), utilizando argônio como gás de fragmentação. O instrumento foi
previamente calibrado com fragmentos do íon duplamente carregado do [glu1]-fibrinopeptideo
B (m/z = 785,84) gerados por CID enquanto que a correção de massas realizadas durante os
experimentos foi realizada com este íon intacto.
Os dados foram coletados, processados e analisados utilizando o programa
MassLynx v 4.1 (Waters Corp) e ProteinLynx v 2.5 (Waters Corp). Os peptídeos com sequência
de aminoácidos comuns a outras proteínas serão identificados por buscas em banco de dados
utilizando as ferramentas de pesquisa por padrão de fragmentação dos peptídeos ProteinLynx
2.4 (Waters Corp) e MASCOT (Matrix Science). Os peptídeos que não forem identificados por
estes meios terão suas sequências determinadas através da interpretação manual dos espectros
de fragmentação (sequenciamento de novo).
4.3 Análise da estrutura primária por bioinformática
Uma vez obtida a sequência primaria, esta foi submetida a analise onde utilizados
programas de alinhamento para análise de similaridade e homologia entre as sequencias de
aminoácidos obtidas e todo o banco não redundante de proteínas depositadas no National Center
of Biotechnology Information (NCBI). Inicialmente, a sequência primaria de CTL foi
submetida ao programa BLAST (ALTSCHUL et al., 1997) e as proteínas com maior score
foram selecionados para os alinhamentos de sequencias utilizando o ESPript 3.0 (ROBERT;
GOUET, 2014). A informação de ponto isoelétrico (pI) e a massa molecular media teórica da

61
sequência de aminoácidos da lectina foram calculadas usando a ferramenta PeptideMass
(http://web.expasy.org/peptide_mass/) (GASTEIGER et al., 2009). A presença de sítios de N-
glicosilação foi avaliada usando o NetNGlyc (http://www.cbs.dtu.dk/services/NetNGlyc/)
(GUPTA; JUNG; BRUNAK, 2004).
4.4 Resultados e Discussão
4.4.1 Estrutura primária de CTL
A estrutura primária da lectina de sementes de C. tomentosum foi determinada
através da sobreposição das sequências de peptídeos, oriundos de digestões proteolíticas com
as enzimas tripsina, quimotripsina, pepsina e endoproteinase Asp-N, determinadas por
espectrometria de massas sequencial (MS/MS) (Figura 11). Como resultado, foram
sequenciados 15 peptídeos de tripsina, 10 de quimiotripsina, 6 de pepsina e 8 de endoproteinase
Asp-N, resultando em 245 aminoácidos, totalizando uma massa de 27.062,9 Da e um pI teórico
de 4,76. A sequência possui um sitio de N-glicosilação previsto (118QNE120).
Entre a sequência, seis resíduos (71QSQF74, 201VV202) não puderam ser identificados
por MS/MS, sendo determinados através do mapa de densidade eletrônica da estrutura
cristalográfica de CTL (descrito posteriormente). Os dados da estrutura primária foram
depositados no Uniprot Knowledgebase, um banco de dados de informações funcionais sobre
proteínas, sob o número de adesão C0HJX1.
A estrutura primária de CTL apresentou similaridade com outras lectinas de
leguminosas manose/glicose específicas pertencentes à tribo Dalbergieae, como a lectina de
Platypodium elegans (89% de identidade), Pterocarpus rotundifolius (84%), Pterocarpus
angolensis (77%) e Arachis hypogaea (71%) (Número de acesso: AEK69351, AAT57665,
CAD19804 e AAA74572, respectivamente). Além disso, CTL mostrou uma alta similaridade
(93%) com a lectina do mesmo gênero, Centrolobium microchaete (VASCONCELOS et al.,
2015). O alinhamento das sequencias estão exibidas na figura 12. Após alinhamento com
lectinas de Dalbergieae relacionadas e que já apresentam estruturas tridimensionais
determinadas, foi possível observar na sequência de da lectina de C. tomentosum regiões
bastante conservadas em relação ao sítio de ligação à metais e sítio de ligação á carboidratos,
bem como os cinco loops descritos como integrantes do sítio de ligação a carboidratos (LORIS
et al., 1998; SHARMA; SUROLIA, 1997).

62
Figura 11. Sequência de aminoácidos da CTL.
Fonte: Elaborado pelo autor. A sequência de aminoácidos de DrfL foi montada pela sobreposição das sequências
de aminoácido de peptídeos, oriundos de digestões proteolíticas com Tripsina (T), Quimiotripsina (Q), Pepsina (P)
e Endproteinase Asp-N (D), determinadas por espectrometria de massas sequencial (MS/MS).
Outras lectinas pertencentes à tribo Dalbergieae já foram sequenciadas, porém
demonstraram baixa similaridade com a estrutura primária de CTL. Um exemplo disso são as
lectinas isoladas do gênero Vatairea, VML e VGL, que são específicas à galactose e seus
derivados, apresentam dois sítios de N-glicosilação e possuem um complexo mecanismo pós-
traducional envolvendo seus sítios de N-glicosilação (CALVETE et al., 1998; SILVA et al.,
2012). Por outro lado, as lectinas de Dalbergieae manose/glicose específicas, incluindo CTL,
possuem apenas um sítio de N-glicosilação, um pequeno peptídeo sinal, mas nenhuma
evidência de qualquer processamento proteolítico em suas regiões internas, apresentando
apenas uma cadeia polipeptídica (PEREIRA-JUNIOR et al., 2012; LORIS et al., 2004;
ALMEIDA et al., 2014; BENEVIDES et al., 2012). Essas diferenças sugerem a existência de
pelo menos dois grupos distintos de lectinas na tribo Dalbergieae. Além disso, pode-se inferir

63
que as lectinas manose/glicose específicas de Dalbergieae devem exibir um mecanismo pós-
traducional diferente do descrito para as lectinas do gênero Vatairea (CALVETE et al., 1998).
Figura 12 - Alinhamento da sequência de aminoácidos de Centrolobium tomentosum (CTL) com lectinas da tribo
Dalbergieae.
Fonte: Elaborado pelo autor. Alinhamento da sequência de aminoácidos de CTL com lectinas da tribo Dalbergieae
usando o programa ESPript 3.0 (ROBERT; GOUET, 2014). Centrolobium microchaete (VASCONCELOS et al.,
2015), Platypodium elegans (Número de acesso: AEK69351), Pterocarpus rotundifolius (Número de acesso:
AAT57665), Pterocarpus angolensis (Número de acesso: CAD19804) e Arachis hypogaea (Número de acesso:
AAA74572). O alinhamento múltiplo e predições de estrutura secundária foram alcançados usando a ESPript 3.0
(ROBERT; GOUET, 2014). Os símbolos são como se segue: α-hélices (α), 310-hélices (η), folhas β (β), loops-β
(TT), resíduos do sítio de ligação a carboidratos (●), resíduos do sítio de ligação a metal (○) e sítio de N-
glicosilação predito (♦).

64
A região C-terminal das lectinas de leguminosas geralmente exibem processamento
proteolítico (YOUNG et al., 1995). Embora este processamento não seja claramente
compreendido, Chrispeels e Raikhel (1992) sugerem que as proteínas são direcionadas para os
vacúolos através de pequenos domínios peptídicos, situados nas regiões C- e N-terminal.
Lectinas de Dalbergieae que tiveram seu sequenciamento realizado por cDNA exibem uma
extensão nas em suas regiões terminais em comparação à sequência de CTL, o que sugere que
a ela também sofra um processamento proteolítico em ambas as regiões (BENEVIDES et al.,
2012; LORIS et al., 2004).

65
CAPÍTULO III:
RESOLUÇÃO DA ESTRUTURA TRIDMENSIONAL
DE CTL POR CRISTALOGRAFIA DE RAIOS X

66
5 CAPÍTULO III: RESOLUÇÃO DA ESTRUTURA TRIDIMENSIONAL DE CTL
POR CRISTALOGRAFIA DE RAIOS X
5.1 Ensaio de Cristalização
Para determinar a concentração adequada de lectina para o ensaio de cristalização,
foi realizado o teste de pré-cristalização segundo manual de seu fabricante (PCT® - Pre-
Crystallization Test, Hampton Research). O teste PCT propõe-se a minimizar ou prevenir
situações em que ensaio de cristalização produza uma abundância de precipitados amórficos ou
gotas claras.
O kit PCT contém 4 reagentes únicos, pré-formulados e estéreis, utilizados para
avaliar a concentração de proteína para o posterior ensaio de cristalização. Inicialmente, uma
amostra da proteína é misturada com dois dos reagentes para determinar se sua concentração é
adequada para o ensaio de cristalização. Se a proteína é muito sensível à concentração de sal
ou precipitante, com base nos resultados de PCT iniciais, a proteína pode ser avaliada utilizando
o segundo conjunto de reagentes de PCT. Os resultados posteriores fornecerão informações
tanto à concentração da amostra adequada ou indicar que outros testes de diagnóstico, tais como
eletroforese em gel nativo devem ser realizados para demonstrar a homogeneidade da amostra
apropriada para cristalização.
O teste de pré-cristalização foi realizado com diferentes concentrações de lectina,
onde o melhor resultado foi obtido com a lectina à concentração de 15 mg/mL. Portanto, esta
concentração foi utilizada no ensaio de cristalização
Para o ensaio de cristalização, a lectina liofilizada foi solubilizada em água
ultrapura (Milli-Q®) na concentração final de 15 mg/mL. A solução da proteína foi então
centrifugada à 4.000 g por 10 minutos e o sobrenadante foi utilizado para o ensaio. A proteína
em solução foi também incubada com três carboidratos específicos, o 5-bromo-6-chloro-3-
indolyl-α-D-mannopiranosídeo (X-Man), o dissacarídeo Metil-O3-(α-D-manose)-α-D-manose
(Metil-Dimanosídeo), e α-(1,3)-D-manose (Dimanosídeo) na concentração final de 5 mM
durante uma hora. O carboidrato em solução foi utilizado para caracterizar e/ou estabilizar o
domínio de reconhecimento à carboidrato (CRD). A proteína foi então submetida ao ensaio de
cristalização, utilizando o método da matriz esparsa inicialmente descrito por Jancarik e Kim
(1991). Os kits utilizados foram o “Crystallization Extension Kit For Proteins” (Sigma
Aldrich), bem como o “Crystal Screen I” e “Crystal Screen II” (Hampton Research).

67
O método utilizado foi no ensaio foi a difusão de vapor e gota suspensa com uso de
placas de microtitulação de 96 poços (fundo reto). As placas foram manuseadas no robô TTP
Labtech's Mosquito® Crystal. Foram adicionados 100 μL da condição de cristalização e a gota
foi composta por 100 nL da solução de proteína e 100 nL da condição de cristalização. Os poços
foram vedados com ViewDrop® II para placa de 96 poços para até 3 gotas suspensas. O poço
foi então vedado com silicone e deixado em repouso à temperatura de 18 ºC.
Após obtenção de cristais numa determinada condição, otimizações foram
realizadas, variando principalmente a concentração de concentração de sal, agente precipitante
e pH da solução, repetindo todo o ensaio de cristalização. Essa otimização visa melhorar a
condição de formação do cristal, produzindo assim, um cristal com características necessárias
para que ele possa ser mais bem difratado quando submetido aos raios X.
Diferentes condições foram manualmente preparadas para os ensaios de otimização
com base na formulação das condições originais onde obtiveram-se cristais. O ensaio utilizou
placas de Linbro de 24 poços. Foram colocados em cada poço da placa de cristalização 300 μL
da solução de otimização e a gota foi composta por 2 μL da solução de proteína à 5mg/mL e 2
μL da condição de cristalização. O poço foi então vedado com silicone e deixado em repouso à
temperatura de 18 ºC.
5.2 Coleta de dados e resolução da estrutura cristalográfica
Os dados de difração de raios X foram coletados a temperatura de -173,5 °C. Para
evitar formação de gelo, os cristais foram mergulhados em uma solução crioprotetora aquosa
de glicerol (30%). Os dados foram coletados em um comprimento de onda de 1,54 Å, utilizando
uma fonte de luz de radiação síncrotron (estação MX2 no Laboratório Nacional de Luz
Síncrotron - LNLS - Campinas, Brasil), utilizando um detector PILATUS2M® (Dectris,
Suíça). Foram coletadas 180 imagens com oscilação de 1º. Os conjuntos de dados foram
indexados, integrados e escalonados através do programa MOSFLM (LESLIE; POWELL,
2007) e SCALA (EVANS, 1993). Ambos fazem parte do pacote de CCP4 (WINN et al., 2011).
Para resolução do problema de fase foi utilizado o método de substituição molecular
utilizando o programa MOLREP (VARGIN; TAPLYAKOV, 1997). As coordenadas atômicas
usadas como modelo foram obtidas da lectina de sementes de Platypodium elegans complexada
com trimanosídeo (PDB 3ZVX) (BENEVIDES et al., 2012), que apresenta 89% de similaridade
com a estrutura primária de CTL. Após a utilização de operações de simetria de rotação e
translação, obteve-se a melhor solução de acordo com os parâmetros Rfactor e o coeficiente de

68
correlação, admitindo como modelo de qualidade a ser refinado os resultados que apresentaram
valores de Rfactor e coeficiente de correlação inferior a 40% e superior a 60%, respectivamente.
Após a substituição molecular, a estrutura obtida foi submetida a refinamentos de
corpo rígido e refinamentos posicionais para um melhor ajuste da molécula em sua unidade
assimétrica através do uso dos programas REFMAC5 (MURSHUDOV et al., 2011) e PHENIX
(ADAMS et al., 2010). A finalidade deste refinamento foi encontrar uma melhor concordância
entre o modelo proposto e a estrutura real da molécula, colocando o modelo na mesma posição
que a molécula real ocupa na sua rede cristalina. Para avaliar os resultados dos cálculos do
refinamento foram utilizados como referenciais os valores de Rfactor e Rfree (BRÜNGER, 1992).
Os mapas de densidade eletrônica foram gerados e visualizados através do
programa Coot (EMSLEY et al., 2010). Foram realizados ajustes manuais no modelo
(refinamento posicional), posicionadas corretamente as moléculas de metil-dimanosídeo, e
verificadas as moléculas de água. Todos os passos manuais foram seguidos de ciclos de
refinamentos posicionais e os valores do Rfactor e Rfree foram monitorados. Análises
estereoquímicas, dos ângulos de ligação, distâncias interatômicas e interações foram checadas
manualmente com ajuda do gráfico de Ramachandran e mapa de Fourier e a qualidade do
modelo foi avaliada através dos programas Molprobity (CHEN et al., 2010) e PDB Validation
Tool (READ et al., 2011).
As estruturas cristalográficas de CTL complexada com Metil-Dimanosídeo foi
visualizada através do programa Coot. Todas as figuras foram feitas utilizando os programas
PyMol (DELANO, 2002) e LigPlot (WALLACE et al., 1996). As coordenadas foram
depositadas no Protein Data Bank (PDB) nos códigos 5EYX e 5EYY.
5.3 Docking molecular
A capacidade de CTL em interagir com oligomanosídeos foi avaliado por Docking
Molecular. O Docking rígido foi realizado utilizando o software CLC Drug Discovery
Workbench Software (CLC Bio, Boston, MA, USA), uma plataforma integrada de predição de
interações de proteínas. O Raio selecionado foi de 11 Da ao redor do CRD e o máximo de
interações foi 5.000. A energia de interação proteína-ligante foi expressa na forma de escore
em unidades arbitrárias onde um valor mais negativo indica uma interação mais forte.

69
5.4 Resultados e Discussão
5.4.1 Cristalização e coleta de dados
CTL foi cristalizada a partir de uma solução de 15 mg/mL da lectina solúvel em
água ultrapura, previamente incubada com 5 mM de Metil-Dimanosídeo por uma hora antes do
ensaio de cristalização. Cristais foram obtidos através do método de difusão de vapor em gota
suspensa após três semanas, com a placa sendo deixada em repouso à temperatura constante de
20 °C. A condição que apresentou crescimento de cristais foi a nº 30 do Kit “Crystallization
Extension Kit For Proteins” (HEPES pH 7.5 0.1 M, 2-Methyl-2,4-pentanediol 5%, PEG 6000
10%). Foram realizadas otimizações desta condição em placas de 24 poços variando o pH e a
concentração do agente precipitante. Foram realizadas otimizações desta condição o pH e a
concentração do agente precipitante, obtendo-se cristais em duas condições de otimização: A:
HEPES pH 7.0 0.1 M, 2-Methyl-2,4-pentanediol 5%, PEG 8.000 10% e B: HEPES pH 8.0 0.1
M, 2-Methyl-2,4-pentanediol 5%, PEG 10.000 10% (Figura 13).
Figura 13. Cristais de CTL complexada com Metil-Dimanosídeo.
Fonte: Elaborado pelo autor. Cristais obtidos a partir da otimização da condição nº 30 do kit “Crystallization
Extension Kit For Proteins”. A: HEPES pH 7.0 0.1 M, 2-Methyl-2,4-pentanediol 5%, PEG 8.000 10%. B: HEPES
pH 8.0 0.1 M, 2-Methyl-2,4-pentanediol 5%, PEG 10.000 10%. As setas brancas apontam os cristais de cada
condição utilizados para a difração de raios X.
5.4.2 Coleta de dados e refinamento da estrutura
Após congelados em solução crioprotetora (condição de cristalização com glicerol
30% v/v), dados de difração de raios X de cristais de CTL com metil-dimanosídeo foram
coletados, utilizando como fonte de radiação. Os dados foram coletados em um comprimento

70
de onda de 1,54 Å, utilizando uma fonte de luz de radiação sincrotron a estação MX2 no
Laboratório Nacional de Luz Sincrotron (LNLS - Campinas, Brasil), utilizando um detector
PILATUS2M® (Dectris, Suíça). Foram coletadas 180 imagens de cada cristal, com um ângulo
de oscilação de 1º.
Após a indexação dos dados, determinou-se a existência de dois grupos espaciais
dentre os cristais difratados: um monoclínico C121 (α = 90º, β = 102.40º e γ = 90º) com
parâmetros de célula a = 145,84 Å, b = 41,55 Å, c = 94,64 Å; e um tetragonal P412121 (α = 90º,
β = 90º e γ = 90º) com parâmetros de célula a = 72.58 Å, b = 72.58 Å e c = 128.45Å. O
coeficiente de Matthew (2,59 Å3/Da) indicou a presença de um dímero na unidade assimétrica
do cristal monoclínico, com a presença de 52,60% de solvente. Já para o cristal tetragonal, seu
coeficiente de Matthew (3,38 Å3/Da) indicou a presença de um monômero na unidade
assimétrica, com a presença de 63,67 % de solvente. A tabela 2 demonstra as estatísticas
detalhadas da coleta e processamento de dados.
A estrutura cristalográfica da lectina de C. tomentosum foi obtida por substituição
molecular utilizando como modelo as coordenadas da estrutura da lectina de Platypodium
elegans (PDB 3ZVX) para o cristal pertencente ao grupo monoclínico. Para o grupo espacial
tetragonal, a substituição molecular utilizou a estrutura obtida no grupo espacial monoclínico.
As estruturas obtidas foram submetidas a refinamentos de corpo rígido e
posicionais, a fim de encontrar uma melhor concordância entre o modelo proposto e a estrutura
real da molécula, colocando o modelo na mesma posição que a molécula real ocupa em sua
rede cristalina. Após o último refinamento, a estrutura monoclínica obteve resolução máxima
de 2,25 Å e valores de Rfactor e Rfree de 17,98% e 23,45%. Já a estrutura tetragonal obteve
resolução máxima de 1,9 Å e valores de Rfactor e Rfree de 15,6% e 17,7%.
A qualidade estereoquímica da estrutura foi comprovada a partir do gráfico de
Ramachandran, gerado pelo programa Molprobity (CHEN et al., 2010), que analisa as torções
dos ângulos Φ e Ψ. O gráfico da forma monoclínica mostrou um nenhum resíduo em regiões
não permitidas e 2,5% dos resíduos presentes em regiões adicionalmente permitidas. A
tetragonal também não mostrou nenhum resíduo em regiões não permitidas e 3,4% dos resíduos
presentes em regiões adicionalmente permitidas (Figura 14).

71
Tabela 2. Estatísticas da coleta de dados de difração de raios X de CTL em complexo com metil-dimanosídeo.
Parâmetros Grupo Monoclínico Grupo Tetragonal
Dados de Coleta
Grupo especial C121 P412121
Parâmetros de célula unitária
a, b, c (Å) 145,84 Å, 41,55 Å, 94,64 Å 72,58 Å, 72,58 Å, 128,45Å
Ângulos de célula (α, β, γ) 90,00°, 102,40°, 90,00° 90,00º, 90,00º, 90,00º
Reflexões totais 91.499 219.670
Reflexões únicas 26.323 27.589
Moléculas por unidade assimétrica 2 1
Limite de resolução (Å) 35,92 - 2,25 (2,33 - 2,25) 32,46 - 1,90 (1,97 - 1,902)
Rmergea (%) 9,7 (41,9)d 6,7 (16,5) d
Completeza (%) 98,4 (99,6)d 99,0 (94,3) d
Multiplicidade 3,5 (3,4) 8 (7,8)
(I)/σ (média) 9,7 (3,7)d 19,1 (10,2)d
Substituição Molecular
wRfactor 0,458 0,452
Score 0,580 0,650
Refinamento
Faixa de resolução (Å) 35,92 - 2,25 32,46 - 1,90
Rfactorb (%) 17,98 15,6
Rfreec (%) 23,45 17,7
Resíduos na unidade assimétrica 490 245
Moléculas de água 323 226
R.M.S.D
Comprimento de ligação (Å) 0,008 0,0101
Ângulos de ligação (graus) 1,04 0,958
Fator de Temperatura
Média do fator B (Å2) 21,64 17,10
Gráfico de Ramachandran
Resíduos em regiões mais favoráveis (%) 97,5 97
Resíduos em regiões adic. permitidas (%) 2,5 3,4
Resíduos em regiões não permitidas (%) 0 0
Fonte: Elaborado pelo autor.
a where I(hkl) is the intensity of the measurement of the reflection h and I(hkl) is the
mean value of the I(hkl)i for all I measurements.
b
c Calculated with 5% of the reflections omitted from refinement. d Values in parentheses represent the high resolution shell.

72
Figura 14. Gráfico de Ramachandran das coordenadas de CTL.
Fonte: Elaborado pelo autor. A: Forma monoclínica de CTL. B: Forma Tetragonal de CTL. Os aminoácidos estão
representados em azul, as regiões mais favoráveis estão representadas em vermelho, as regiões adicionais mais
permissivas são representadas em marrom, as regiões permissivas aparecem em amarelo e as regiões não
permissivas correspondem à cor bege.
5.4.3 Estrutura cristalográfica de CTL em complexo com metil-dimanosídeo
A estrutura geral do monômero da lectina de Centrolobium tomentosum
complexada com metil-dimanosídeo apresenta 245 resíduos de aminoácidos, 323 moléculas de
água, 1 íon cálcio e 1 íon manganês. Uma molécula de N-acetil-D-glicosamina foi modelada no
seu sítio de N-glicosilação e a molécula de metil-dimanosídeo foi modelada no domínio de
reconhecimento a carboidratos (CRD). Dos 245 resíduos presentes na estrutura primária de
CTL, todos puderam ser identificados no mapa de densidade eletrônica, exceto pelos quatro
últimos aminoácidos da extremidade C-terminal (242AQKE245).
O monômero de CTL (Figura 15) apresenta o arranjo clássico de lectinas de
leguminosas (Sanduíche β), incluindo um íon cálcio e um íon manganês próximos a seu sítio
de ligação. Os monômeros são compostos por 14 fitas beta e 2 pequenas hélices alfa, sendo
interligadas por voltas (loops) e alças formando uma estrutura de forma achatada. Esse tipo de
enovelamento é conhecido como “β-sanduíche”, por ser composto majoritariamente de folhas
betas antiparalelas (cerca de 60%). A orientação antiparalela das folhas beta, e a mudança entre
uma face e outra, forma uma estrutura rígida forte, semelhante a um sanduíche (CHANDRA et
al., 2001).
CTL apresentou-se formando um dímero canônico, também observado em muitas
outras lectinas de leguminosas diméricas e tetraméricas (MANOJ; SUGUNA, 2001;
SRINIVAS et al., 2001). A estrutura tridimensional dimérica completa de CTL complexada

73
com metil-dimanosídeo está ilustrada na figura 16. É possível observar a molécula de metil-
dimanosídeo localizada no sítio de ligação a carboidrato, assim como os metais Ca2+ e Mn2+
situados no sítio de ligação a metais. Um sítio de N-glicosilação em Asn119 encontra-se situado
sobre uma pequena hélice-α na superfície da proteína, sendo, portanto susceptível a estar
glicosilado na planta (Figura 17).
Figura 15. Estrutura geral do monômero de CTL.
Fonte: Elaborado pelo autor. Monômero de CTL representado em verde, seus ligantes estão representados em
sticks na cor amarela, o íon cálcio está representado em branco e o manganês em violeta.
Figura 16. Estrutura tridimensional de CTL complexada com metil-dimanosídeo.
Fonte: Elaborado pelo autor. Visão geral da estrutura dimérica de CTL em complexo com metil-dimanosídeo,
formada pelos monômeros A em verde e monômero B em vermelho. Presença dos metais Ca2+ e Mn2+ (esferas
branca e violeta, respectivamente), do ligante metil-dimanosídeo (MDM) em amarelo, e duas moléculas de N-
acetil-D-glicosamina (NAG) em amarelo.

74
Já foi demonstrado em outras lectinas de Dalbergieae manose/glicose específicas o
mesmo padrão de glicosilação nas mesmas posições, quando comparadas as suas sequências de
aminoácidos. PELa apresenta seu sítio em Asn127 e PAL em Asn126 (BENEVIDES et al., 2012;
LORIS et al., 2004). Além disso, PELa apresenta também uma outra glicosilação próxima à
seu N-terminal (Asn7), sendo essa possivelmente relacionada ao mecanismo pós-traducional
ainda não esclarecido que as lectinas de Dalbergieae manose/glicose específicas realizam.
Figura 17. Sítio de N-Glicosilação de CTL.
Fonte: Elaborado pelo autor. Representação do sítio de glicosilação de CTL com mapa de omissão 2mFo-dFc (1σ)
para N-acetil-D-glicosamina em A) forma monoclínica e B) forma tetragonal.
A estrutura cristalográfica de CTL possui um sítio de ligação a metal contendo
resíduos conservados. Este sítio é semelhante aos presentes nas outras lectinas de Dalbergieae
que fazem a coordenação dos íons Ca2+ e Mn2+, bem como lectinas de leguminosas já
caracterizadas (BENEVIDES et al., 2012; DELATORRE et al., 2007; LORIS et al., 2004). O
sítio de ligação a metal presente na estrutura monomérica situa-se nas proximidades do CRD e
a ligação aos íons favorecem interações que auxiliam na estabilização do sítio de ligação a
carboidratos (LORIS et al., 2004). Quatro aminoácidos, principalmente através de suas cadeias
laterais, e duas moléculas de água coordenam cada íon metálico. O Ca2+ é coordenado pelos
resíduos Asp131, Phe133, Asn139 e Asp142. O íon Mn2+ é coordenado pelos resíduos Glu129, Asp131,
Asp142 e His147. A representação do sítio de ligação a metais de CTL está ilustrado na figura
18.

75
Figura 18. Sítio de ligação à metais de CTL.
Fonte: Elaborado pelo autor. Representação do sítio de ligação com mapa de densidade eletrônica diferencial
(mFo-dFc) contornada à 3σ para Ca2+ e Mn2+ em A) forma monoclínica, B) forma tetragonal.
Analisando o metil-dimanosídeo complexado no domínio de reconhecimento a
carboidrato (CRD) de CTL, a ligação entre eles foi estabilizada por uma rede de ligações de
hidrogênio conectando os aminoácidos Gly107, Ser138, Asn139, Gly221 Glu222 e Gln223 aos átomos
de oxigênio O1’, O2’, O3’, O4’, O5’ e O6’ do metil-dimanosídeo. A interação também é
estabilizada por interações de Van der Waals com o subsítio hidrofóbico, envolvendo os
resíduos Ala86, Asp87, Gly107, Phe133, Ser138, Gly221 e Gln223. A representação do domínio de
reconhecimento a carboidratos de CTL está ilustrado na figura 19. A tabela 3 descreve todas as
interações entre CTL e metil-dimanosídeo.
Figura 19. Sítio de ligação a carboidratos de CTL interagindo com metil-dimanosídeo.
Fonte: Elaborado pelo autor. Sítio de ligação à carboidratos interagindo com metil-dimanosídeo com representação
do mapa de omissão 2mFo-dFc de 1σ contornando MDM. Traços azuis representam os contatos polares. A) Forma
monoclínica, B) Forma tetragonal.
A estabilização da estrutura cristalina na presença de um ligante já foi previamente
demonstrada em outras lectinas da tribo Dalbergieae, como PELa (BENEVIDES et al., 2012)

76
e PAL (LORIS et al., 2004), bem como em lectinas da tribo Phaseoleae como CRLI (ROCHA
et al., 2015), DVL (BEZERRA et al., 2013) e CGL (BARROSO-NETO et al., 2014).
Tabela 3. Interações entre CTL e metil-dimanosídeo.
Aminoácido Metil-Dimanosídeo Forma Monoclínica (Å) Forma Tetragonal (Å)
Van der Waals
Gln223 CD C7’ 3,46 ̶
Gln223 OE1 C7’ 3,06 ̶
Gln223 NE2 C7’ 3,19 ̶
Gln223 NE2 C6 3,45 ̶
Phe133 CZ O2’ 3,43 3,25
Phe133 CD1 O4 3,30 3,44
Phe133 CE1 O4 3,41 ̶
Phe133 CE1 O2’ ̶ 3,45
Ser138 CB O2’ 3,22 ̶
Ser138 CB O3’ ̶ 3,48
Ala86 CB O6 3,45 ̶
Gly221 CA O6 3,33 3,28
Gly 221 C O6 3,43 3,44
Asp87 OD1 C4 3,42 3,45
Asp87 CG O4 ̶ 3,46
Gly107 N C4 3,47 3,50
Gly106 CA O3 ̶ 3,48
Ligações de Hidrogênio
Asp87 OD1 O4 ̶ 2,58
Asp87 OD2 O6 ̶ 2,73
Gln223 NE2 O1’ 3,05 ̶
Ser138 OG O2’ 2,85 2,86
Ser138 OG O3’ ̶ 3,35
Glu222 N O5 2,94 3,00
Gly221 N O6 3,16 3,20
Gln223 O O6 3,17 3,39
Gln223 N O6 2,98 3,08
Glu222 N O6 3,16 3,11
Asn139 ND2 O4 2,71 ̶
Asn139 OD1 O4 ̶ 2,89
Gly107 N O4 3,24 3,24
Gly107 N O3 2,69 2,68
Fonte: Elaborado pelo autor.
Comparando a estrutura tridimensional de CTL com as outras duas lectinas da tribo
Dalbergieae manose/glicose específicas que já tiveram suas estruturas resolvidas até o presente
momento, PELa e PAL, é possível notar algumas semelhanças importantes. Sharma e Surolia
(1997) descreveram que o sítio de ligação a carboidrato das lectinas de leguminosas consiste
em resíduos pertencentes a até cinco loops de cadeias polipeptídicas, nomeadas de A a E, as
quais variam em diferentes graus entre as lectinas com diferentes especificidades (Figura 12).
Os loops A e B, em geral, contêm um resíduo Asp essencial (invariavelmente
precedido por uma ligação cis-peptídica) e um grupo -NH suporte (usualmente de um resíduo
Gly), respectivamente. Em CTL, foi possível identificar o resíduo Asp87 no loop A e a Gly107

77
no loop B, concordando com essa arquitetura, típica do reconhecimento a manose. O mesmo
padrão foi encontrado em PELa e PAL.
O loop C é a de ligação a metal, que envolve os íons cálcio e manganês
estruturalmente importantes. Em estruturas cristalinas conhecidas, cinco diferentes tamanhos
de loop (12-16 resíduos) adotando oito diferentes conformações são observadas (LORIS et al.,
2004). Não existe uma relação simples entre tipo de especificidade a monossacarídeos e o
tamanho e conformação do loop C. Em CTL, esse loop tem o comprimento de 16 resíduos,
similar em comprimento e conformação à PELa e PAL. Cadeias laterais específicas nesse loop
influenciam na natureza do açúcar que pode ser acomodado no sítio de ligação (LORIS et al.,
2004). Entre os resíduos descritos no sítio de ligação a de CTL, encontram-se nesse loop os
resíduos Phe133, Asn139 e Asp142, participantes do sítio de ligação a metal. Ao mesmo tempo,
também encontramos os resíduos Ser138 e Asn139 participando do sítio de ligação ao metil-
dimanosídeo.
Em contraste aos loops A-C, o D não interage diretamente com o íon cálcio
estrutural. Em sua grande variabilidade de comprimento, conformação e sequência, é referido
comumente como o loop determinante da especificidade ao monossacarídeo, assim como do
oligossacarídeo (LORIS et al., 1998). Em CTL, a conformação adotada por esse loop de 9
resíduos é idêntica àquelas encontrados em estruturas de lectinas manose/glicose específicas da
tribo Dalbergieae como PELa e PAL (BENEVIDES et al., 2012; LORIS et al., 2004), bem
como na subtribo Diocleinae, como por exemplo a lectina de Canavalia brasiliensis (SANZ-
APARICIO et al., 1997) e Dioclea guianensis (WAH et al., 2001). Em CTL, nesse loop
encontramos os resíduos Gly221, Glu222 e Gln223, participantes do sitio de ligação ao
dimanosídeo.
Finalmente, e em poucos casos, o loop E é encontrado interagindo com o
carboidrato ligado, geralmente entre oligossacarídeos e lectinas, agindo como um subsítio,
como em PAL em complexo com Man(α1-2)Man (LORIS et al., 2004). Em CTL, esse loop não
se encontra envolvida na ligação ao dimanosídeo. Na lectina PELA, esse loop também não está
envolvido no sítio de ligação.
5.4.4 Comparação entre as formas monoclínica e tetragonal de CTL
Como descrito anteriormente, a difração de raios X da lectina de C. tomentosum
gerou dois grupos de dados de difração: um pertencente ao grupo espacial monoclonal e um
pertencente ao grupo espacial tetragonal. Ambas as estruturas tridimensionais correspondentes

78
foram resolvidas e suas coordenadas depositadas no Protein Data Bank (PDB) nos códigos
5EYX e 5EYY. Com relação à sua estrutura geral não há diferenças marcantes entre as
estruturas.
As estruturas diferem em apenas 1 aminoácido na extremidade C-terminal, Thr241,
presente na forma monoclínica e ausente na tetragonal. Essas diferenças podem se relacionar à
qualidade do mapa de densidade eletrônica gerada por cada difração. No entanto, quando
analisados os sítios de ligação à metal, à carboidrato, N-glicosilação, as estruturas são idênticas,
diferindo apenas pelo fato da mobilidade natural da molécula durante seu processo de
cristalização. Os aminoácidos que coordenam o sítio de ligação a carboidratos são os mesmos
em ambas as estruturas, da mesma forma que os aminoácidos que coordenam o sítio de ligação
a metais. Na figura 20 está ilustrado a sobreposição do sítio de ligação à carboidratos das duas
formas. Logo em seguida, a figura 21 apresenta a sobreposição dos monômeros de ambas as
formas da estrutura tridimensional de CTL.
Figura 20. Sobreposição do sítio de ligação a carboidratos das formas monoclínica e tetragonal de CTL.
Fonte: Elaborado pelo autor. Representação da interação do metil-dimanosídeo no sítio de ligação a carboidratos
das estruturas monoclínica e tetragonal de CTL. Os resíduos de aminoácidos da forma monoclínica estão ilustrados
em azul, os resíduos da forma tetragonal em violeta. As moléculas de metil-dimanosídeo estão ilustradas em
amarelo e verde.

79
Figura 21. Sobreposição dos monômeros das formas monoclínica e tetragonal de CTL.
Fonte: Elaborado pelo autor. A: Representação em cartoon das formas monoclínica (vermelho) e tetragonal
(verde) de CTL. B: Representações em superfície da forma monoclínica (vermelho) e tetragonal (verde) de CTL.
5.4.5 Docking molecular
A fim de analisar as interações entre CTL e diferentes manosídeos, foram testadas
algumas interações por docking molecular com sua estrutura cristalográfica resolvida (forma
tetragonal). Os testes mostraram que, entre os açúcares que interagiram com a lectina, CTL se
ligou mais fortemente ao trimanosídeo Man(α1-3)Man(α1-6)Man com um escore de -63,11
(Figura 22), seguido pelo dimanosídeo Man(α1-6)Man com escore de -54,34 e por outro
dimanosídeo Man(α1-2)Man com escore -54,32.
Resultados anteriores demonstraram a forte ligação das lectinas de Pterocarpus
angolensis e Platypodium elegans à trimanosídeos (LORIS et al., 2004; BENEVIDES et al.,
2012). A superposição de da estrutura cristalográfica CTL com as estruturas de PEla (código
PDB: 3ZVX) e PAL (código PDB: 1Q8V) complexados com trimanosídeos demonstraram uma
notável semelhança em seu modo de ligação, especialmente nas duas primeiras moléculas de
manose, como visualizado na figura 22. A terceira manose apresentou um alto nível de

80
variabilidade nas três estruturas, provavelmente devido ao menor número de interações com as
cadeias laterais de aminoácidos CRD.
Figura 22. Sobreposição de entre as estruturas de CTL, PELa e PAL complexadas com trimanosídeos.
Fonte: Elaborado pelo autor. Representação da sobreposição da estrutura de CTL complexada com o trimanosídeo
Man(α1-3)Man(α1-6)Man com as estruturas das lectinas de Platypodium elegans (PELa) e Pterocarpus angolensis
(PAL) complexadas com o mesmo ligante (trimanosídeo com carbonos amarelos: CTL; carbonos azuis: PELa;
carbonos laranja: PAL).
Trimanosídeos compõem o núcleo dos N-glicanos das células. Geralmente, glicanos
específicos estão relacionados a diferentes processos celulares (ROCHA et al., 2011). Baseado
nisso, a hipótese que o domínio lectínico e gliconjugados contendo resíduos relacionados com
manosídeos podem contribuir para atividades especiíficas, como o efeito inflamatório de CTL
(descrito posteriormente). Os N-glicanos de compostos manosídeos e os glicanos complexos de
manose estão envolvidos no reconhecimento molecular de células (MITOMA et al., 2007).

81
CAPÍTULO IV:
AVALIAÇÃO DAS ATIVIDADES
INFLAMATÓRIAS, VASORELAXANTES E
AVALIAÇÃO DA CITOTOXICIDADE EM
CÉLULAS TUMORAIS

82
6 CAPÍTULO IV: AVALIAÇÃO DO ATIVIDADES INFLAMATÓRIAS,
VASORELAXANTES E DA CITOTOXICIDADE EM CÉLULAS TUMORAIS
6.1 Modelo de edema de pata
Ratos Wistar fêmeas (150-200 g) foram criadas e mantidas (n = 6 por gaiola) em
salas com um ciclo controlado luz/escuro 12/12 h a 25 °C com comida e água ad libitum. Os
protocolos experimentais foram aprovados pelo Comitê de Ética Institucional da Universidade
Estadual do Ceará para uso de animais (CEUA nº 10130208-8/40).
O volume da pata foi medido imediatamente antes (tempo zero) da injeção por via
subcutânea (s.c.) de estímulos inflamatórios na pata traseira direita dos ratos (n = 30) e depois
no intervalo de tempo selecionado (0,5, 1, 2-7 e 24 h) através de hidropletismometro. Os
resultados foram calculados como a variação do volume da pata (mL) ou a área sob a curva do
curso-tempo (AUC, em unidades arbitrárias) em relação ao tempo zero (LANDUCCI et al.,
1995). CTL, testada aqui como estímulo inflamatório, foi solubilizada em solução salina estéril
(NaCl 0,15 M), filtrada e injetada na concentração de 30 a 300 µg/pata por injeção via
subcutânea (s.c.). Foi também investigado o envolvimento do domínio lectínico de CTL, a
lectina foi previamente incubada (60 minutos, 37 ºC) com seu açúcar específico, metil-
dimanosídeo (0,1 M) antes das injeções. CTL e o metil-dimanosídeo também foram incubados
em soluções separadas utilizando as mesmas condições, como controles.
6.2 Teste de contratilidade em aortas isoladas
Ratos Wistar machos (250-300 g) foram criados e mantidos (n = 6 por gaiola) em
salas com um ciclo controlado luz/escuro 12/12 h a 25 °C com comida e água ad libitum. Os
protocolos experimentais foram aprovados pelo Comitê de Ética Institucional da Universidade
Estadual do Ceará para uso de animais (CEUA nº 10130208-8/40).
Após o sacrifício dos animais por concussão cerebral, a aorta torácica foi
rapidamente removida e limpa e os segmentos em anel (3-5 mm) foram preparados para a
gravação da tensão (2 g) em banhos para órgãos de 10 ml preenchidos com solução de Tyrode
modificada (NaCl 136 mM, KCl 5 mM, MgCl2 0,98 mM, 2 CaCl2 mM, NaH2PO4 0,36 mM,
NaHCO3 11,9 mM) e 5,5 mM de glucose, pH 7,4 (a 37 °C, 95 % de O2 e 5 % de CO2). Em todos
experimentos a aorta foi testada com KCl (60 mM) após 45 minutos de equilíbrio para assegurar
a viabilidade do tecido. A resposta contrátil foi medida usando um transdutor de força ligado a

83
um pré-amplificador e um sistema de aquisição de dados computadorizado (Powerlab, AD
Instruments). O software Chart® (versão 4.1) foi utilizado para análise de dados.
Para avaliar o efeito relaxante de CTL, foram realizadas curvas de concentração
cumulativa de lectina (1-100 µg/ml) em um platô de concentração induzido pela fenilefrina
(Phe, 0,1 µM) em segmentos de aorta com endotélio intacto ou desnudo. Aorta de controle
recebeu o mesmo volume de Tyrode. A remoção do endotélio foi realizada por meio de fricção
mecânica da superfície da íntima da aorta. O endotélio intacto foi considerado para respostas
relaxantes a acetilcolina (ACh, 1 µM) 75% maior que induzido por Phe (FURCHGOTT;
ZAWADZKI, 1980).
6.3 Análise estatística
Os resultados foram apresentados como média ± erro padrão da média (EPM) e as
diferenças estatísticas entre os grupos foram obtidas através da análise de variância (ANOVA),
seguida de teste de Bonferroni. Valores de p < 0,05 foram considerados significantes.
6.4 Avaliação da Citotoxicidade em Células Tumorais
Para avaliação da citotoxicidade em células tumorais, o método utilizado foi o
ensaio colorimétrico com sal de tetrazólio, MTT (3-(4,5-dimetiltiazol-2-il)-2,5-
difeniltetrazólio), proposto por Mosmann (1983).
Para a realização do ensaio, suspensão células C6 (linhagem celular de glioma de
rato), com concentração de 50.000 células/mL, obtidas após dissociação química por tripsina,
foram distribuídas em placas de 96 cavidades (100μL/cavidade). As placas foram incubadas
por 24 h, a 37 ºC, em estufa de 5% de CO2 até a confluência. Após 24 h, os poços foram tratados,
em diferentes concentrações da lectina (10, 30, 50 e 100 µg/mL). Decorridas as 24 h de
incubação com a lectina, o meio de cada cavidade foi substituído por 100 μL de uma solução
de MTT a 0,5 mg/mL, diluído em solução salina balanceada de Hank, HBSS (137 mM de NaCl,
0,63 mM de Na2HPO4, 4,17 mM de NaHCO3, 5,36 mM de KCl, 0,44 mM de KH2PO4, 1,26
mM de CaCl2, 0,41 mM de MgSO4, 0,49 mM de MgCl2 e 5,55 mM de glicose).
As placas foram incubadas por 1 h em estufa a 37°C. Em seguida, o meio contendo
MTT foi retirado e se adicionou 100 μL de DMSO/cavidade para dissolução dos cristais de
formazan. A placa foi agitada por 5 minutos à temperatura ambiente para que toda o formazan
fosse dissolvido e a absorbância foi medida em espectrofotômetro (Labsystems Multiskan MS

84
352, Haverhill, MA, USA) a 540 nm. Os valores de absorbância, medidos para cada
concentração de cada amostra, foram transformados em porcentagens de viabilidade celular
(%) em relação à média dos controles celulares, considerados 100% viáveis.
6.4.1 Análise estatística
Os resultados foram apresentados como média ± erro padrão da média (EPM) e as
diferenças estatísticas entre os grupos foram obtidas através da análise de variância (ANOVA),
seguida de teste de Bonferroni. Valores de p < 0,05 foram considerados significantes.
6.5 Resultados e Discussão
6.5.1 Atividade inflamatória de CTL
A injeção subcutânea intraplantar de CTL nos ratos foi hábil em produzir edema
apenas a 300 µg/pata, atingindo seu pico de atividade a 120 minutos (0,6 ± 0,06 mL × 0,13 ±
0,06 mL), mantendo-se significativa até 300 minutos (Figura 24A). O aumento do volume da
pata promovido por CTL (AUC: 85 ± 9,93) foi aproximadamente 67% maior em comparação
com os animais injetados com o controle de solução salina (AUC: 28 ± 9,04) (Figura 24B).
Além disso, o efeito edematogênico de CTL (AUC: 56,25 ± 9,14) foi inibido em 66% pela
incubação prévia da lectina (300 µg/pata) com seu carboidrato específico: metil-dimanosídeo
(AUC: 18,9 ± 4,07). O carboidrato injetado sozinho não promoveu edema (Figura 24C).
Edema, um sinal clínico importante da inflamação aguda, tem sido amplamente
investigado no modelo experimental de edema da pata induzido por carragenina em ratos. Neste
modelo, vários mediadores inflamatórios estão envolvidos, incluindo a serotonina, a histamina
e a bradicinina (CRUNKHORN; MEACOCK, 1971; FERREIRA et al., 1974) na sua fase
inicial (0-2h); e prostaglandina E2, o óxido nítrico (SAUTEBIN et al., 1995) e citocinas na fase
tardia (2-5h) (FERREIRA et al., 1974), quer localmente ou sistemicamente (VAZQUEZ et al.,
2015).
O tempo de edema de pata induzido por CTL (2-4 h) sugere a participação de
mediadores da fase tardia. Assim como CTL, outra lectina da tribo Dalbergieae, VML, também
demonstrou atividade pró-inflamatória. VML induziu o edema da pata (ALENCAR et al., 2004)
e a migração de neutrófilos mediada pela liberação de TNF-α por macrófagos (ALENCAR et
al., 2003; ALENCAR et al., 2007). É notório salientar que o efeito edematogênico de CTL foi

85
inibido pela sua associação com seu açúcar específico, metil-dimanosídeo, sugerindo a
participação direta do domínio de reconhecimento a carboidrato no efeito edematogênico
encontrado. A atividade inflamatória de VML também demonstrou ser dependente do domínio
lectínico. Com base na seletividade demonstrada por CTL em exibir um curso temporal curto
de edema (2-4 h), CTL pode ser posteriormente investigado como uma nova ferramenta em
estudos de inflamação.
Figura 23. Indução de edema da pata por CTL.
Fonte: Elaborado pelo autor. Os ratos foram injetados s.c. com solucao salina, CTL (30, 300 µg/pata), metil-
dimanosídeo (0.1 M) ou CTL (300 µg/pata) + metil-dimanosídeo (após incubação por 1h, a 37 °C). O edema foi
medido antes (tempo zero) e a 0.5, 1, 2-7 e 24 h após a injeção e expresso como a média ± S.E.M. (n=6) da
diferença do volume da pata (mL) (A) área sob a curva do curso-tempo (AUC, em unidades arbitrárias) em relação
ao tempo zero (B, C). * p<0.05 em comparação com a solução salina.
6.5.2 Avaliação do efeito relaxante de CTL
CTL (2,4% ± 2,7) não foi hábil a promover o relaxamento ou contração em anéis
de aorta à 1 e 100 µg/ml, após 40 minutos de estabilização comparado com o grupo controle

86
(TN; 4,7 ± 5,2%) (Figura 25A). Em aortas pré-contraídas com fenilefrina (96,3% ± 5,2), a
adição cumulativa de CTL (107,7 ± 11,7%) não promoveu efeito relaxante em nenhuma das
doses testadas (Figura 25B).
Figura 24. Avaliação do efeito vasorelaxante de CTL.
Fonte: Elaborado pelo autor. A: CTL (1-100 µg/mL) não apresentou efeito sobre o tônus basal de aortas isoladas
de ratos. B: CTL não apresentou efeito relaxante em aortas pré-contraídas com fenilefrina (Phe). TN: solução de
Tyrode.
Os resultados demonstraram pela primeira vez a ausência de efeito relaxante de uma
lectina pertencente a tribo Dalbergieae sobre aortas isoladas de ratos. No entanto, lectinas que
possuem alta similaridade de estruturas, primária e terciária, e mesma especificidade a
carboidratos com CTL, tais como PAL, PFL e PELa (BENEVIDES et al, 2012; LORIS et al,
2004; PEREIRA- JUNIOR et al, 2012), não tiveram até o presente momentos seus efeitos
vasoativos avaliados in vitro.
Carboidratos localizados na superfície celular contribuem para a maioria das
interações entre as células e o ambiente externo. A lectina de sementes de Vatairea guianensis
(VGL), também pertencente a tribo Dalbergieae, porém com especificidade por galactosídeos,
apresentou o primeiro relato de atividade vasorelaxante em aortas isoladas de ratos (Silva et al.,
2012). No entanto, a ausência de atividade vasorelaxante em CTL não pode ser devido à sua
diferente especificidade em relação à VGL, pois outras lectinas glicose/manose específicas de
leguminosas já apresentaram efeito relaxante, como as lectinas de Dioclea sclerocarpa (DSL),
Canavalia grandiflora (CGL) e Canavalia virosa (CVL) (BARROSO-NETO et al., 2016;
BARROSO-NETO et al., 2014; OSTERNE et al., 2014).
Fica evidenciado a reavaliação dos parâmetros dos testes de contratilidade em
aortas para avaliação do efeito vasorelaxante da lectina de C. tomentosum, ou a necessidade de
um novo modelo para a validação ou não do efeito de CTL.

87
6.5.3 Avaliação da atividade citotóxica de CTL
A avaliação da atividade citotóxica de CTL em células tumorais utilizou o método
do MTT. Este é um teste amplamente utilizado para determinação da viabilidade de células
isoladas. No ensaio o MTT, um sal de tetrazólio solúvel em água, é convertido em cristais de
formazan de cor púrpura, insolúveis em água, após clivagem do anel de tetrazólio por
desidrogenases mitocondriais e outras enzimas lisossomais (LIU et al., 1997). Na verdade, o
MTT não interage diretamente com as desidrogenases e sim com os seus subprodutos, NADH
e NADPH (LIU et al., 1997). Os cristais de formazan são solubilizados, formando assim um
composto colorido cuja medição da densidade óptica é feita em espectrofotômetro.
A intensidade do produto colorido formado é diretamente proporcional ao número
de células viáveis presente na amostra, confirmando a capacidade redutora do sistema sobre o
MTT (LIU et al., 1997; HEINRICH et al, 2005). Outros estudos têm examinado a
correspondência entre os resultados obtidos durante contagem direta de células e confirmou-se
que a análise com MTT é um indicador válido do número de células viáveis (KIM et al., 2009).
Como resultado do ensaio, verificou-se que CTL não foi hábil em reduzir
significativamente a viabilidade celular dos gliomas de (Figura 26).
Figura 25. Ensaio de citotoxidade de CTL em células tumorais.
Fonte: Elaborado pelo autor. Gráfico de “Concentração × Viabilidade celular” de CTL. Média ± Média do erro
padrão (n = 3).
Estudos anteriores mostraram a capacidade de da lectina de Canavalia ensiformis
(ConA) de induzir morte autofágica de células de glioma na linhagem de células de glioma
humano U87. Foi mostrado que essa atividade se deve a regulação da proteína MT1-MMP
(proteína de membrana – metaloproteinase tipo 1), uma glicoproteína fortemente expressa em

88
gliobastomas radio- e quimioresistentes. Essa proteína é responsável por mediar a sinalização
pró-apoptótica e autofágica em células de câncer do cérebro. A lectina foi capaz de induzir a
atividade da proteína com consequente aumento de vacúolos ácidos característicos de processos
autofágicos (BELKAID et al., 2007; GINGER et al., 2000; PRATT; ROY; ANNABI, 2012).
Outros trabalhos também demonstraram que as lectinas de Canavalia ensiformis,
Canavalia brasiliensis e Cratylia floribunda foram citotóxicas, inibindo tanto a viabilidade
quanto a proliferação celular de diferentes linhagens tumorais murina (J774) e humanas (MCF-
7, HL-60 e MOLT-4) (FAHEINA-MARTINS, 2009; FAHEINA-MARTINS et al., 2012). Foi
possível observar no trabalho que houve uma variação na ação das lectinas em cada linhagem,
Sendo ConA, mais tóxica sobre as linhagens, pelo fato desta encontrar-se completamente
tetramérica em pH fisiológico, conseguindo exercer um efeito mais pronunciado que ConBr e
CFL, que se apresentam numa mistura de dímeros e tetrâmeros em pH fisiológico (SANS-
APARÍCIO et al., 1997; CALVETE et al., 1999).

89
7 CONCLUSÃO
Os resultados apresentados neste trabalho de tese forneceram dados importantes
sobre a caracterização estrutural e bioquímica da lectina de sementes da espécie pertencente a
tribo Dalbergieae, Centrolobium tomentosum. A lectina demonstrou alta similaridade de
estrutura primária com outras lectinas de sua tribo. Além disso, os resultados sobre sua
sequência corroboraram a possibilidade da existência de dois grupos na tribo Dalbergieae,
apresentando especificidade a carboidratos, sítios de glicosilações e, principalmente,
processamentos pós-traducionais diferentes.
A partir de dados de cristalografia de raios X foram obtidas duas estruturas de CTL,
uma de forma monoclínica e a outra tetragonal, sendo elucidados dados importantes sobre suas
interações proteína-ligante. Esses foram aprofundados com o estudo de Docking molecular. A
lectina xibiu forte afinidade para manosídeos, sendo essa característica demonstrada tanto por
co-cristalização, como por Docking molecular.
De posse desses resultados, a lectina pode ser utilizada para reconhecer glicanos
específicos envolvidos eventos biológicos, tanto para entendimento do evento, quanto para
aplicação biotecnológica. Um bom exemplo disso foi a indução do efeito edematogênico
demonstrado por CTL no modelo de edema da pata, mostrando uma resposta inflamatória
aguda, com a participação de seu domínio de reconhecimento a carboidratos. A elevada
afinidade por complexos de manose apresentada pela sua estrutura cristalográfica pode explicar
os resultados obtidos nos testes em modelos de inflamação.
Este estudo apresentou, pela primeira vez, a estrutura tridimensional de uma lectina
nativa, específica à manose, pertencente à tribo Dalbergieae.

90
REFERÊNCIAS
ABBAS, A. K.; FAUSTO, N.; KUMAR, V. Robbins e Cotran - Patologia: bases patológicas
das doenças. 8ªed. Rio de Janeiro: Elsevier. 2010.
ABERGEL, C. Molecular replacement: tricks and treats. Acta Crystallogr. D Biol.
Crystallogr., v. 69, p. 2167-2173, 2013.
ADAMS, P.D. et al. PHENIX: a comprehensive Python-based system for macromolecular
structure solution. Acta Cryst., v. 66, p. 213-221, 2010.
AEBERSOLD, R.; MANN, M. Mass spectrometry-based proteomics. Nature, v. 422, n. 6928,
p. 198-207, 2003.
ALENCAR, N. M. et al. The galactose binding lectin from Vatairea macrocarpa seeds induces
in vivo neutrophil migration by indirect mechanism. Int. J. Biochem. Cell Biol., v. 35, p. 1674-
1681, 2003.
ALENCAR, N. M. et al. Vatairea macrocarpa (Leguminosae) lectin activates cultured
macrophages to release chemotactic mediators. Naunyn Schmiedebergs Archives of
Pharmacology, v. 374, p. 275-282, 2007.
ALENCAR, N. M. et al. Vatairea macrocarpa lectin induces paw edema with leukocyte
infiltration. Prot. Pept. Lett., v. 11, p. 195-200, 2004.
ALENCAR, N. M. et al. Anti-inflammatory and antimicrobial effect of lectin from
Lonchocarpus sericeus seeds in an experimental rat model of infectious peritonitis. J. Pharm.
Pharmacol., v. 57, n. 7, p. 919-22, 2005.
ALENCAR, N. M. et al. Leguminous lectins as tools for studying the role of sugar residues in
leukocyte recruitment. Mediators. Inflamm., v. 8, p. 107-13, 1999.
ALESSI, D. et al. The control of protein phosphatase-1 by targetting subunits. The major
myosin phophastase in avian smooth muscle is a novel form of protein phosphatase-1. Eur. J.
Biochem., v. 210, p. 1023-1035, 1992.
ALLEN, A. K.; NEUBERGER, A. The purification and properties of the lectin from potato
tubers, a hydroxyproline-containing glycoprotein. Biochemical Journal, v.135, p. 307-324,
1973.
ALMEIDA, A. C. et al. Purification and partial characterization of a new mannose/glucose-
specific lectin from Centrolobium tomentosum Guill. ex Benth seeds exhibiting low toxicity on
Artemia sp. International Journal of Indigenous Medical Plants, v. 47, p. 1567-1577, 2014.
ALMEIDA, L.M. et al. Edematogenic and antinociceptive activities of the lectin isolated from
the albumin fraction of Acacia farnesiana. Ciencia Animal (UECE), v. 25, p. 19-22, 2015.
ALTSCHUL, S. F. et al. Gapped BLAST and PSI-BLAST: A new generation of protein
database search programs. Nucleic Acids Research, v. 25, p. 3389-3402, 1997.

91
ALVES, A. C. et al. A novel vasorelaxant lectin purified from seeds of Clathrotropis nitida:
partial characterization and immobilization in chitosan beads. Archives of Biochemistry and
Biophysics, v. 588, p. 33-40, 2015.
AMBROSI, M.; CAMERON, N. R.; DAVIS, B. G. Lectins: tools for the molecular
understanding of the glycocode. Org. Biomol. Chem., v. 3, p. 1593-1608, 2005.
ANDRADE, J. L. et al. Lectin-induced NO production. Cellular Immunology, v. 194, p. 98-
102, 1999.
BRÜNGER, A. T. Free R value: a novel statistical quantity for assessing the accuracy of crystal
structures. Nature, v. 355, p. 472-475, 1992.
BANERJEE, R. et al. Conformation, Protein-Carbohydrate Interactions and a Novel Subunit
Association in the Refined Structure of Peanut Lectin-Lactose Complex. J. Mol. Biol., v. 259,
p. 281-296, 1996.
BARBOSA, T. et al. In Vivo lymphocyte activation and apoptosis by lectins of The Diocleinae
Subtribe. Memórias do Instituto Oswaldo Cruz, v. 96, n. 5, p. 673-678, 2001.
BARBOSA, T. et al. In Vivo Lymphocyte Activation and Apoptosis by lectins of the Diocleinae
Subtribe. Memórias do Instituto Oswaldo Cruz., v. 96, n. 5, p. 673-678, 2001.
BARI, A. U. et al. Purification and partial characterization of a new mannose/glucose-specific
lectin from Dialium guineense Willd seeds that exhibits toxic effect. JMR. Journal of
Molecular Recognition, v. 26, p. 351-356, 2013.
BARRAL-NETTO, M. et al. Human lynphocyte stimulation by legume lectins from the
Diocleae tribe. Immunol. Invest., v. 21, p. 297-303, 1992.
BARROSO-NETO, I. L. et al. Structural analysis of a Dioclea sclerocarpa lectin: Study on the
vasorelaxant properties of Dioclea lectins. International Journal of Biological
Macromolecules, v. 82, p. 464-470, 2016.
BARROSO-NETO, I. L. et al. Vasorelaxant activity of Canavalia grandiflora seed lectin: A
structural analysis. Archiv. of Biochem. and Biophys., v. 543, p.31-39, 2014.
BELKAID, A. et al. Necrosis induction in glioblastoma cells reveals a new "bioswitch"
function for the MT1-MMP/G6PT signaling axis in proMMP-2 activation versus cell death
decision. Neoplasia, v. 9, p. 332-340, 2007.
BENEVIDES, R. G. et al. A Lectin from Platypodium elegans with Unusual Specificity and
Affinity for Asymmetric Complex N-Glycans. Journal of Biological Chemistry, v. 287, p.
26352-26364, 2012.
BENEVIDES, R.G. Avaliação do Potencial Fungicida e Termiticida de uma Fração Protéica
Lectínica de Sementes de Platypodium elegans Vogel e Obtenção da Lectina Purificada.
Dissertação (Mestrado em Bioquímica) - Centro de Ciências, Universidade Federal do Ceará,
Fortaleza, 151 p., 2008.

92
BENEVIDES, R.G. Caracterização bioquímica e estrutural de uma lectina recombinante de
sementes de Platypodium elegans Vogel, Tese (Doutorado em Bioquímica) – Universidade
Federal do Ceará, Fortaleza, 292 p. 2011.
BERMAN, H. M. et al. The Protein Data Bank And The Challenge Of Structural Genomics.
Nat. Struc. Biol., v. 7, p. 957-959, 2000.
BERTRAM, J. S. The molecular biology of cancer. Mol. Aspects Med., v. 21, p. 167-223,
2000.
BEZERRA, D. P. et al. Purificação de uma lectina presente em sementes de Machaerium
acutifolium. Anais da XXVI Reunião Nordestina de Botânica, In: XXVI Reunião Nordestina
de Botânica, Fortaleza, 2003.
BEZERRA, M. J. B. et al. Crystal structure of Dioclea violacea lectin and a comparative study
of vasorelaxant properties with Dioclea rostrata lectin. Internat. J. of Bioc. & Cell Biol., v.
45, p. 807-815, 2013.
BIES, C.; LEHR, C.; WOODLEY, J. F. Lectin-mediated drug targeting: history and
applications. Advanced Drug Delivery Reviews, v. 56, p. 425–435, 2004.
BLUNDELL, T.L.; JONHSON, L.N. Protein crystallography. Academic Press. USA, 1976.
BOGOEVA, V. P.; PETROVA, L. P.; TRIFONOV, A. A. New Activity of a Protein from
Canavalia ensiformis. Sci. Pharm., v. 82, n. 4, p. 825-34, 2014.
BOUCKAERT, J. et al. Crystallographic structure of metal-free concanavalin A at 2.5 Å
resolution. Proteins Struct. Funct. Genet., v. 23, p. 510-524, 1995.
BOUCKAERT, J.; DEWALLEF, Y.; POORTMANS, F.; WYNS, L.; LORIS, R. The
structural features of concanavalin A governing non-proline peptide isomerization. Journal of Biological Chemistry, v. 275, p. 19778-19787, 2000.
CALVETE, J. J. et al. Amino acid sequence, glycan structure, and proteolytic processing of the
lectin of Vatairea macrocarpa seeds. FEBS Letters, v. 425, p. 286-292, 1998.
CALVETE, J. J. et al. Molecular characterization and crystallization of Diocleinae lectins.
Biochimica et Biophysica Acta., v. 1430, p. 367-375, 1999.
CANTU, M. et al. Sequenciamento de peptídeos usando espectrometria de massas: um guia
prático. Quím. Nova, v. 31, n. 3, p. 669-675, 2008.
CARTER, C. W. JR.; CARTER, C. W. Protein crystallization using incomplete factorial
experiments. J. Biol. Chem., v. 254, p. 12219, 1979.
CARVALHO, H. F. Aspectos moleculares e biológicos das lectinas. Ciência e Cultura., v. 42,
p. 884-893, 1990.

93
CARVALHO, M.H.C. et al. Hipertensão arterial: o endotélio e suas múltiplas funções. Rev.
Bras. Hipertens., v. 8, n. 1, 2001.
CAVADA B.S. et al. Purification and characterization of a lectin from seeds of Vatairea
macrocarpa Duke. Phytochemistry, v. 49, n. 3, p. 675-80, 1998.
CAVADA, B. S. et al. Revisiting proteus: Do minor changes in lectin structure matter in
biological activity? Lessons from and potential biotechnological uses of the Diocleinae subtribe
lectins. Current Protein and Peptide Sciences, v. 2, p. 123-135, 2001.
CAVADA, B.S. et al. cDNA cloning and 1.75 Å crystal structure determination of PPL2, an
endochitinase and N-acetylglucosamine-binding hemagglutinin from Parkia platycephala
seeds. The FEBS Journal, v. 273, p. 3962-3974, 2006.
CHAMRAD, D. C. et al. Evaluation of algorithms for protein identification from sequence
databases using mass spectrometry data. Proteomics., v. 4, n. 3, p. 619-628, 2004.
CHANDRA, N. R. et al. Structural similarity and functional diversity in proteins containing the
legume lectin fold. Protein Eng., v. 14, p. 857-866, 2001.
CHAPMAN, J. R. Practical Organic Mass Spectrometry - A guide for chemical and
biochemical analysis. 2ª.Ed., Chinchester: John Wiley & Sons, 1993.
CHEN, X. J.; CARROLL, J. A.; BEAVIS R. C. Near-UV-induced matrix-assisted laser
desorption/ionization as a function of wavelength. J. Am. Soc. Mass. Spectrom., v. 9, n. 9, p.
885-91, 1998.
CHEN, V. B. et al. MolProbity: all-atom structure validation for macromolecular
crystallography. Acta Crystallographica D, v. 66, p. 12-21, 2010.
CHIAL, H. Proto-oncogenes to oncogenes to cancer. Nat. Education., v.1, 33 p., 2008.
CHRISPEELS, M. J. Biosynthesis, processing, and transport of storage proteins and lectins in
cotyledons of developing legume seeds. Philos. Trans. R. Soc. Lond., v. 304, p. 309-322,
1984.
CHRISPEELS, M. J.; RAIKHEL, N. V. Short peptide domains target proteins to plant vacuoles.
Cell, v. 68, p. 613-616, 1992.
CIOPRAGA, J. et al. Fusarium spp. growth inhibition by wheat germ agglutinin. Biochim.
Biophys. Acta., v. 1428, p. 424-432, 1999.
CLAYDON M. et al. The rapid identification of intact microorganisms using mass
spectrometry. Nat. Biotechnol., v. 14, n. 11, p. 1584-1586.
COELHO, L. C. B. B.; SILVA, M. B. R. Simple method to purify milligram quantities of the
galactose-specific lectin from the leaves of Bauhinia monandra. Phytochemical Analysis, v.
11, p. 295-300, 2000.

94
COLEMAN, R.G. et al. Ligand Pose and Orientational Sampling in Molecular Docking. Plos
One., v. 8, 2013.
COOLINGE, D. B. et al. Plant chitinases. Plant Journal, v. 3, p. 31-40, 1993.
COOPER, D.R. et al. X-ray crystallography: Assessment and validation of protein-small
molecule complexes for drug discovery. Expert. Opin. Drug. Discov., v. 6, p. 771-782, 2012.
CORNER, J.; BAILEY, C. Cancer nursing: care in context, 2ª ed. Oxford, Wiley-Blackwell,
2008.
COSTA, R. M. P. B., et al. A new mistletoe Phthirusa pyrifolia leaf lectin with antimicrobial
properties. Process Biochemistry, v. 45, p. 525-533, 2010.
CRUNKHORN, P.; MEACOCK, S.C.R. Mediators of the inflammation induced in the rat paw
by carrageenin. Br. J. Pharmac., v. 42, p. 392-402, 1971.
DAM, T. K. et al. Diocleinae lectins: a group of proteins with conserved binding sites for the
core trimannoside of asparagine-linked oligosaccharide and differential specificities for
complex carbohydrates. The Journal Of Biological Chemistry, v. 273, n. 20, p. 12082-12088,
1998.
DAM, T. K. et al. Thermodynamic Binding Studies of Lectins from the Diocleinae Subtribe to
Deoxy Analogs of the Core Trimannoside of Asparagine-Linded Oligosaccharides. The
Journal of Biological Chemistry, v. 275, n.21, p. 16119-16126, 2000.
DAM, T. K. et al. Binding studies of alpha-GalNAc-specific lectins to the alpha-GalNAc (Tn-
antigen) form of porcine submaxillary mucin and its smaller fragments. The Journal Of
Biological Chemistry., v. 282, p. 28256-28263, 2007.
DATTA, P. K.; FIGUEROA, M. O. R.; LAJOLO, F. M. Purification and characterization of
two major lectins from Araucaria brasiliensis syn Araucaria angustifolia seeds (pinhão). Plant
Physiology., v. 97, p. 856-862, 1991.
DELANO, W. L. The PyMOL Molecular Graphics System. DeLano Scientific, San Carlos,
CA, USA, 2002.
DELATORRE, P. et al. Structure of a lectin from Canavalia gladiata seeds: new structural
insights for old molecules, BMC Struct. Biol., v. 7, p. 52, 2007.
DELATORRE, P. Lectinas de leguminosas: novos enfoques estruturais para velhas
moléculas. Tese (Doutorado em Bioquímica) – Centro de Ciências, Universidade Federal do
Ceará, Fortaleza, 2006.
DENNIS, J. W.; GRANOVSKY, M.; WARREN, C. E. Protein glycosylation in development
and disease. BioEssay, v. 21, p. 412-21, 1999.
DREISEWERD K. (2003) The Desorption Process in MALDI. Chem Rev, v. 103, p. 395.
DRENTH, J. Principles of Protein X-Ray Crystallography. Springer Advanced Texts in
Chemistry, 3rd ed., 332 p. 2007.

95
DUBOIS B. et al. Regulation of gelatinase B (MMP-9) in leukocytes by plant lectins. FEBS
Letters, 427, 275-278,1998.
DUCRUIX, A.; GIEGE, R. Crystallization of nucleic acids and proteins. A practical approach.
Oxford University Press, 435 p., 1992.
DUIJN, E. et al. Chaperonin Complexes Monitored by Ion Mobility Mass Spectrometry.
Journal of the American Chemical Society, v. 131, p. 1452-1459, 2009.
ELIAS, J. E. et al. Comparative evaluation of mass spectrometry platforms used in large-scale
proteomics investigations. Nat. Methods, v. 2, p. 667-675. 2005.
Emsley, B. et al. Features and development of Coot. Acta Crystallogr. D, v. 66, p. 486-501,
2010.
ETZLER, M. E. Distribution and function of plant lectins. Lectins. Properties, Function, and
Applications in Biology and Medicine. Academic Press, p. 371-435, 1986.
EVANS, P. “Data reduction”, Proceedings of CCP4 Study Weekend. Data Collection &
Processing, p. 114-122, 1993.
FAHEINA-MARTINS, G. V. et al. Antiproliferative effects of lectins from Canavalia
ensiformis and Canavalia brasiliensis in human leukemia cell lines. Toxicology in Vitro, v.
26, n. 7, p. 1161-1169, 2012.
FANTONE, J. C. et al. Inflamação: Patologia. Rio de Janeiro, Brasil: Guanabara Koogan, p.
34-58, 1990.
FAVACHO, A. R. M. et al. In vitro activity evaluation of Parkia pendula seed lectin against
human cytomegalovirus and herpes virus 6. Biologicals., v. 35, p. 189-194, 2007.
FENG, H. et al. Clonal integration of a polyomavirus in human Merkel cell carcinoma. Science,
v. 319, n. 5866, p. 1096-1100, 2008.
FENN, J. B. et al. Electrospray ionization-principles and practice. Mass Spectrometry
Reviews, v. 9, p. 37-70, 1990.
FENSELAU C.; DEMIREV P. A. Characterization of intact microorganism by MALDI mass
spectrometry. Mass Spectrom. Rev., v. 20, n. 4, p. 157-171, 2001.
FERENCÍK, M.; STVRTINOVÁ, V. Endogenous control and modulation of inflammation.
Folia Biol. Praha., v. 42, p. 47-55, 1996.
FERNÁNDEZ-ALONSO, J. L.; PÉREZ, G.; VEGA, N. Lectin prospecting in Colombian
Labiatae. A systematic-ecological approach. Biochemical Systematics and Ecology, v. 31, n.
6, p. 617-633, 2003.
FERREIRA, S. H. et al. Proceedings: the concomitant release of bradykinin and prostaglandin
in the inflammatory response to carrageenin. Br. J. Pharmacol., v. 52, p. 108-109, 1974.

96
FRIGERIO, L.; ROBERTS, L. M. The enemy within: ricin and plant cells. Journal of
Experimental Botany, v. 49, n. 326, p. 1473-1480, 1998.
FURCHGOTT, R.; ZAWADZKI, Z. The obligatory role of the endothelial cells in relaxation
of arterial smooth muscle by acetycholine. Nature, v. 288, p. 373-376, 1980.
FURNARI, F. B. et al. Malignant astrocytic glioma: genetics, biology, and paths to treatment.
Genes Dev., v. 21, p. 2683-2710, 2007.
GABIUS, H. J. et al. From lectin structure to functional glycomics: principles of the sugar code.
Trends Biochem. Sci., v. 36, n. 6, p. 298-313, 2011.
GABIUS, H. J.; GABIUS, S. Glycoscience: status and perspectives. Chapman & Hall,
Weinheim, Germany, 1997.
GALEGO DEL SOL, F. et al. The First Crystal Structure of a Mimosoideae Lectin Reveals a
Novel Quaternary Arrangement of a Widespread Domain. J. Mol. Biol., v. 353, p. 574-583,
2005.
GASTEIGER, E. et al. Protein identification and analysis tools on the ExPASy Server. In:
Walker JM editor. The Proteomics Protocols Handbook Totowa: Humana Press., p. 571-
607, 2009.
GHAZARIAN, H.; IDONI, B.; OPPENHEIMER, S. B. A. Glycobiology Review:
Carbohydrates, Lectins, and Implications in Cancer Therapeutics. Acta Histochemica, v. 113,
p. 236, 2011.
GINGER, D. et al. Rapid activation of matrix metalloproteinase-2 by glioma cells occurs
through a posttranslational MT1-MMP-dependent mechanism. Biochim. Biophys. Acta., v.
1497, p. 341-350, 2000.
GINGRAS, D. et al. Rapid activation of matrix metalloproteinase-2 by glioma cells occurs
through a posttranslational MT1-MMP-dependent mechanism. Biochim. Biophys. Acta., v.
1497, p. 341-350, 2000.
GOLDSTEIN, I. J. et al. What should be called a lectin? Nature, v. 285, p. 66, 1980.
GONÇALVES, F. M. et al. Vatairea macrocarpa Lectin (VML) Induces Depressive-like
Behavior and Expression of Neuroinflammatory Markers in Mice. Neurochemical Research,
v. 38, p. 2375-2384, 2013.
NASCIMENTO-NETO, L. G. et al. Characterization of Isoforms of the Lectin Isolated from
the Red Algae Bryothamnion seaforthii and Its Pro-Healing Effect. Marine Drugs, v. 10, p.
1936-1954, 2012.
GROSS, J. H. Mass Spectrometry - A Textbook. Berlin, Springer, 2004.
GUPTA, R.; JUNG, E.; BRUNAK, S.; Prediction of N-glycosylation sites in human proteins.
In preparation. 2004.

97
HAMELRYCK, T. W. et al. Properties and structure of the legume lectin family. Trends in
Glycoscience and Glycobiology, v. 10, p. 349-404, 1998.
HAMPEL, A., LABANANSKAS, M., CORNNERS, P. G., KIRKEGARD, L.,
RAJBHANDARY, U. L., SIGLER, P. B., AND BOCK, R. M. Single crystals of transfer RNA
from formylmethionine and phenylalanine transfer RNA's. Science, 162, 1384, 1968.
HANAHAN, D.; WEINBERG, R. A. Hallmarks of cancer: the next generation. Cell., v. 144,
p. 646-674, 2011.
HANKINS, C. N.; KINDINGER, J. I.; SHANNON, L. M. The lectins of Sophora japonica II.
Purification, properties, and N-terminal amino acid sequences of five lectins from bark. Plant
Physiol., v. 86, p. 67-70, 1988.
HE, X. M. et al. Purification, characterization, and molecular cloning of a novel antifungal
lectin from the roots of Ophioglossum pedunculosum. Appl. Biochem. Biotechnol., v. 165, p.
1458-1472, 2011.
HEGI, M. E. et al., MGMT gene silencing and benefit from temozolomide in glioblastoma. N.
Engl. J. Med., v. 352, p. 997-1003, 2005.
HEINRICH, E. L. et al. Direct Targeting of Cancer Cells: A Multiparameter Approach. Acta
Histochem., v. 107, n. 5, p. 335-344, 2005.
HEINRICH, E. L. et al. Mass Spectrometry Principles and Applications. 3 ed. Wiley, England.
2007.
HOFFMAN, E. H.; STROOBANT, V. Mass Spectrometry. Principles and applications. Wiley,
502 p., 2007.
HOLLINGSWORTH, S.A.; KARPLUS, P.A. A fresh look at the Ramachandran plot and the
occurrence of standard structures in proteins. Biomol. Concepts., v. 1, p. 271-883, 2011.
HOLTON, J.M. et al. The R-factor gap in macromolecular crystallography: an untapped
potential for insights on accurate structures. Febs J., v. 281, p. 4046-4060, 2014.
IKEMOTO, K. et al. Bauhinia purpurea agglutinin-modified liposomes for human prostate
cancer treatment. Cancer Sci., v. 07, n. 1, p. 53-59, 2016.
IMBERTY, A. et al. Structures of the lectins from Pseudomonas aeruginosa: insights into the
molecular basis for host glycan recognition. Microbes and Infection, v. 6, n. 2, p. 221-228,
2004.
INA, C. et al. Screening for and purification of novel self-aggregatable lectins reveal a new
functional lectin group in the bark of leguminous trees. Biochimica et Biophysica Acta, v.
1726, n. 1, p. 21-27, 2005.
IRWIN, J.J.; SHOICHET, B.K. ZINC--a free database of commercially available
compounds for virtual screening. J. Chem. Inf. Model., v. 45, p. 177-182, 2005.

98
JANCARIK, J.; KIM, S. H. Sparse matrix sampling: a screening method for crystallization of
proteins. Journal of Applied Crystallography, v. 24, p. 409-411, 1991.
JOHNSON, G. G.; WHITE, M. C.; GRIMALDI, M. Stressed to death: targeting endoplasmic
reticulum stress response induced apoptosis in gliomas. Curr. Pharm. Des., v. 17, p. 284-292,
2011.
JOUBERT, F. J.; SHARON, N.; MERRIFIELD, E. H. Purification and properties of a lectin
from Lonchocarpus capassa (apple-leaf) seed. Phytochemistry, v. 25, n. 2, p. 323-327, 1986.
KABIR, S. (1998). Jacalin: a jackfruit (Artocarpus heterophyllus) seed-derived lectin of
versatile applications in immunobiological research. Journal of Immunological Methods. Nº
212, p. 193-211.
KARAKI, H. et al. Calcium movements, distribution, and functions in smooth muscle.
Pharmacol Rev., v. 49, p. 157-230, 1997.
KARAS, M.; HILLENKAMP, F. Laser desorption ionization of proteins with molecular masses
exceeding 10000 Daltons. Anal. Chem., v. 60, n. 20, p. 2299–2301, 1988.
KARPLUS, P.A.; DIEDERICHS, K. Acessing and maximizing data quality in macromolecular
crystallography. Curr. Opi. Struct. Biol., v. 34, p. 60-68, 2015.
KAUR, M. et al. Assessment of Sauromatum guttatum lectin toxicity against Bactrocera
cucurbitae. J. Environ. Biol., v. 36, n. 6, p. 1263-1268, 2015.
KENNEDY, J. F. et al. Lectins, versatile proteins of recognition: a review. Carbohydrate
Polymers, v. 26, p. 219-30, 1995.
KIM, H. et al. Discriminative cytotoxicity assessment based on various cellular damages.
Toxicol. Lett., v. 184, n. 1, p. 13-17, 2009.
KIMBLE, B., RAMIREZ NIETO, G., PEREZ, D. R. Characterization of influenza virus sialic
acid receptors in minor poultry species. Virol. J., v. 7, 365 p., 2010.
KINTER, M.; SCHERMAN, N.E. Protein Sequence and Identification using Tandem Mass
Spectrometry. New York: John Wiley & Sons, 2000.
KITADA, M., KURODA, Y., DEZAWA, M. Lectins as a Tool for Detecting Neural
Stem/Progenitor Cells in the Adult Mouse Brain. Anat. Rec. Hoboken., v. 1, p. 1-19, 2010.
KONOZY, E. H. E. et al. Isolation, purification, and physicochemical characterization of a D-
galactose-binding lectin from seeds of Erythrina speciosa. Archives of Biochemistry and
Biophysics, v. 410, p. 222-229, 2003.
KONOZY, E. H. E. et al. Purification, some properties of a D-galactose-binding leaf lectin
from Erythrina indica and further characterization of seed lectin. Biochimie, v. 84, p. 1035-
1043, 2002.

99
SAUTEBIN, L. et al. Endogenous nitric oxide increases prostaglandin biosynthesis in
carrageenin rat paw edema. Eur. J. Pharmacol., v. 286, p. 219-222, 1995.
LAEMMLI, U. K. Cleavage of structural proteins during the assembly of the head of
bacteriophage T4. Nature, v. 227, p. 680-685, 1970.
LANDUCCI, E. C. et al. Inhibition of carrageenin-induced rat paw edema by crotapotin, a
polypeptide complexed with phospholipase A2. British Journal of Pharmacology, Londres,
v. 114, p. 578-583, 1995.
LESLIE, A.; POWELL, H. Processing Diffraction Data with Mosflm. Evolving Methods for
Macromolecular Crystallography, v. 245, p. 41-51, 2007.
LI, H.; HE, M.; ZHANG, Z. Image definition evaluation functions for X-ray crystallography: a
new perspective on the phase problem. Acta Cryst., v. 71, p.526-533, 2015.
LIS, H.; SHARON, N. Lectins: Carbohydrate-specific proteins that mediate cellular
recognition. Chem. Rev., v. 98, p. 637-674, 1998.
LIU, B. et al. Antiproliferative activity and apoptosis-inducing mechanism of Concanavalin A
on human melanoma A375 cells. Arch. Biochem. Biophys., v. 482, n. 1-2, p. 1-6, 2009.
LIU, Y. et al. Mechanism of cellular 3-(4,5-Dimethyazol-2-yl-2,5-Diphenyltetrazolium
bromide (MTT) reduction. Journal of Neurochemistry, v. 69, p 581-592, 1997.
LORENZI, H. Árvores brasileiras: manual de identificação e cultivo de plantas arbóreas nativas
do Brasil. Nova Odessa: Plantarum, 352 p., 1992.
LORIS R. Principles of structures of animal and plant lectins. Bioch. Bioph. Acta, v. 1572, p.
198-208, 2002.
LORIS, R. et al. Legume lectin structure. Biochimica et Biophysica Acta, v. 1383, p. 9-36,
1998.
LORIS, R. et al. Structural basis of oligomannose recognition by the Pterocarpus angolensis
seed lectin. Journal of Molecular Biology, v. 335, p. 1227-1240, 2004.
LORIS, R.; HAMELRYCK, T.; BOUCKAERT, J.; WYNS, L. Legume lectin structure.
Biochimica et Biophysica Acta, v. 1383, p. 9-36. 1998.
LORIS, R. et al. Crystal structure of Pterocarpus angolensis lectin in complex with glucose,
sucrose and turanose. J. Biol. Chem., v. 278, p. 16297-16303, 2003.
LOSSIO, C. F. et al. Isolation, purification and partial physicochemical characterization of a
lectin in Andira Pisonis. Mart seed. BMC Proc., v. 8, suppl. 4, p. 226, 2014.
LUSHER, T.F., BARTON, M. Biology of the endotellium. Clin Cardiol., v.20, n. 2, 310 p.,
1997.

100
MACEDO, M. L. R. et al. Insecticidal action of Bauhinia monandra leaf lectin (BmoLL)
against Anagasta kuehniella (Lepitoptera: Pyralidae), Zabrotes fubsasciatus and
Callosobruchus maculatus (Coleoptera: Pruchidae). Comp. Biochem. Physiol., v. 146, p. 486-
498, 2007.
MACEDO, M. L.; OLIVEIRA, C. F.; OLIVEIRA, C. T. Insecticidal activity of plant lectins
and potential application in crop protection. Molecules, v. 20, n 2, p. 2014-2033, 2015.
MANN, K. et al. The amino-acid sequence of the glucose/mannose-specific lectin isolated from
Parkia platycephala seeds reveal three tandemly arranged jacalin related domains. Eur. J.
Biochem., v. 218, p. 4414-4422, 2001.
MANOJ, N.; SUGUNA, K. Signature of quaternary structure in the sequences of legume
lectins. Protein Eng., v. 14, p. 735-745, 2001.
MARONDEDZE, C.; CHIDZWONDO, F.; SITHOLE-NIANG, I.; READ, J. S. Pterocarpus
rotundifolius lectin gene. Submitted to the EMBL/GenBank/DDBJ databases, 2004.
MARTINS, A.M. et al. Renal effects induced by the lectin from Vatairea macrocarpa seeds.
J. Pharm. Pharmacol., v. 57, p. 1329-1333, 2005.
MARTINS, G. V. F. Avaliação do potencial citotóxico das lectinas de Canavalia ensiformis,
Canavalia brasiliensis e Cratylia floribunda. Dissertação (Mestrado em Farmacologia) -
Universidade Federal da Paraíba, João Pessoa, 125 p., 2009.
MATHEW, R. et al. Role of autophagy in cancer. Nat. Rev. Cancer., v. 7, p. 961-967, 2007.
McGEER, P. L.; McGEER, E.G. Autotoxicity and Alsheimer Disease. Archieves of
Neurology, v. 57, p. 789-790, 2000.
MEAGHER, J. L. et al. Crystal structure of banana lectin reveals a novel second sugar binding
site. Glycobiology, v. 15, n. 10, p. 1033-42, 2005.
MELENDEZ, A.; LEVINE, B. Autophagy in C. elegans. The C. elegans Research
Community, 2009.
MITOMA, J. X. et al. Critical functions of N-glycans in L-selectin-mediated lymphocyte
homing and recruitment. Nat. Immunol., v. 8, p. 409-418, 2007.
MIZUSHIMA, N. Autophagy: process and function. Genes Dev., v. 21, p. 2861-2873, 2007.
MOLINA, M. C.; VINCENTE, C. Correlationships between enzymatic activity of lectins,
putrescine content and chloroplast damage in Xanthoria parietina phycobionts. Cell
Communication and Adhesion, v. 3, p. 1-12. 1995.
MONCADA, S. et al. Nitric oxide: Physiology, pathophysiology and pharmacology.
Pharmacol. Rev., v. 43, p. 109-142, 1991.

101
MORAES, S. M. D. et al. Purification, physicochemical characterization and biological
properties of a lectin from Erythrina velutina forma aurantiaca seeds. Braz. J. Med. Biol. Res.,
v. 29, n.8, p. 977-985, 1996.
MOREIRA, R. A. et al. Isolation and partial characterization of a lectin from Artocarpus incisa
L. seeds. Phytochemistry, v. 47, n. 7, p. 1183-1188, 1998.
MORRIS, D. M. et al. Extending the range of pentasubstituted cyclopentadienyl compounds:
The synthesis of a series of tetramethyl(alkyl or aryl) cyclopentadienes (Cp∗R), their iridium
complexes and their catalytic activity for asymmetric transfer hydrogenation. Polyhedron, v.
84, p. 120-135, 2014.
MOSMANN, T. Rapid colorimetric assay for cellular growth and survival: application to
proliferation and cytotoxicity assays. J. Immunol. Methods., v. 16, p. 55-63, 1983.
MOURA, R. M. et al. CvL, a lectin from the marine sponge Cliona varians: Isolation,
characterization and its effects on pathogenic bacteria and Leishmania promastigotes.
Comparative Biochemistry and Physiology A, v. 145, n. 4, p. 517-523, 2006.
MÜLLER, W. E. G.et al. Characterization of trimeric, self-recognizing Geodia cydonium lectin
I. European Journal Biochemistry., v. 133, p. 263-267, 1983.
MURSHUDOV, G et al. Refmac 5 for the refinement of macromolecular crystal structures.
Acta Crystallographica D, v. 67, p. 335-367, 2011.
NAGANO, C. S. et al. Insights in to the structural basis of the pH-dependent dimer-tetramer
equilibrium through crystallographic analysis of recombinant Diocleinae lectins. Biochem. J.,
v. 409, p. 417-428, 2008.
NANDAKUMAR, R.; MADAYIPUTHIYA, N.; FOUAD, A. F. Proteomic analysis of
endodontic infections by liquid chromatography-tandem mass spectrometry. Oral Microbiol.
Immunol., v. 24, p. 347-352, 2009.
NAPIMOGA, M. H. et al. Lonchocarpus sericeus lectin decreases leukocyte migration and
mechanical hypernociception by inhibiting cytokine and chemokines production. Int.
Immunopharmacol., v. 7, n. 6, p. 824-835, 2007.
NASCIMENTO, K. S. et al. Purification of a thermostable antinociceptive lectin isolated from
Andira anthelmia. J. Mol. Recognit., 2015. doi: 10.1002/jmr.2523 (Epub ahead of print).
NELSON, D. L.; COX, M. M. Princípios de bioquímica de Lehninger. 6. ed. Porto Alegre:
Artmed, 1328 p., 2014.
NOBRE, C.B. Caracterização físico-química e efeito sobre bactérias orais de uma lectina
de sementes de Andira surinamensis (Bondt) Splitg. ex Amshoff. Dissertação (Mestrado em
Bioquímica) - Centro de Ciências, Universidade Federal do Ceará, Fortaleza, 2012.
OHIZUMI, Y. et al. Mannose-binding lectin from yam (Dioscorea batatas) tubers with
insecticidal properties against Helicoverpa armigera (Lepidoptera: Noctuidae). J. Agric. Food
Chem., v. 57, n. 7, p. 2896-2902, 2009.

102
OLIVEIRA, J. T. A. et al. Purification and physicochemical characterization of a cotyledonary
lectin from Luetzelburgia auriculada. Phytochemistry, v. 61, p. 301-310, 2002.
OSTERNE, V. J. S. et al. Purification, Partial Characterization, and Cnbr-Sepharose
Immobilization of a Vasorelaxant Glucose/Mannose Lectin from Canavalia virosa Seeds.
Applied Biochemistry and Biotechnology, v. 172, p. 3342-3353, 2014.
PEREIRA-JUNIOR, F. N. et al. Purification and characterization of a mannose/N-acetyl- D-
glucosamine-specific lectin from the seeds of Platymiscium floribundum Vogel. Journal of
Molecular Recognition, v.25, p. 443-449, 2012.
PEUMANS, W. J. et al. Classification of plant lectins in families of structurally and
evolutionary related proteins. Adv. Exp. Med. Biol., v. 491, p. 27-54, 2001.
PEUMANS, W. J.; DOLEY, M.; BRACKAERT, W. F. An unusual lectin from stinging nettle
(Urtica dioica) rhizomes. FEBS Letters., v. 177, p. 99-103, 1984.
PEUMANS, W. J.; ROUGÉ, P.; VAN DAMME, E. J. M. The tomato lectin consists of two
homologous chitin-binding modules separated by an extensin-like linker. Biochemical
Journal, v. 376, p. 717-724, 2003.
PEUMANS, W. J.; VAN DAMME, E. J. M. Lectins as plant defense proteins. Plant Physiol.,
v. 109, p. 347-352, 1995.
PEUMANS, W. J.; VAN DAMME, E. J. M. Plant Lectins: versatile proteins with important
perspectives in biotechnology. Biotechnology and Genetic Engineering Reviews, v.15, p.
199-227, 1998.
PEUMANS, W. J. et al. Isolation of a novel plant lectin with and unusual specificity from
Calystegia sepium. Glycoconjugate Journal, v. 14, p. 259-265, 1997.
PIRES, A. F. et al. A novel N-acetyl-glucosamine lectin of Lonchocarpus araripensis
attenuates acute cellular inflammation in mice. Inflammation Research (Printed ed.), v. 65,
p. 43-52, 2016.
PIRES, A. F. et al. Atividade Antiinflamatória Da Lectina De Sementes De Lonchocarpus
araripensis. 3ª Reunião Regional FESBE, Fortaleza, CE, 2008.
PIRES, A. F. et al. Alternative pathways involved in the anti ncociceptive effect of Canavalia
brasiliensis lectin: nitric oxid and adennergic system. Current Topics in Pharmacology, v.
13, p. 45, 2014.
POLANSKY, A.; KIMMEL. M. Bioinformatics. Berlin, Springer-Verlag, 2007.
PRATAP, J. V. et al. Crystal structures of artocarpin, a Moraceae lectin with mannose
specificity, and its complex with methyl-a-D-mannose: Implications to the generation of
carbohydrate specificity. Journal of Molecular Biology., v. 317, p. 237-247, 2002.

103
PRATT, J.; ROY, R.; ANNABI, B. Concanavalin-A-induced autophagy biomarkers requires
membrane type-1 matrix metalloproteinase intracellular signaling in glioblastoma cells.
Glycobiol., v. 22, p.1245-1255, 2012.
PUSTZAI, A. Plant Lectins. Cambridge University Press, 1991.
READ, R. J. et al. New generation of crystallographic validation tools for the protein data bank.
Structur., v.19, p. 1395-1412, 2011.
RANG, H. P.; DALE, M. M.; RITTER, J.M. Pharmacology. New York, Churchill
Livingstone, 797 p. 2001.
RANGEL, T. B. A. et al. Isolamento e caracterização parcial de lectina proveniente de sementes
da espécie vegetal Andira fraxinofolia (Leguminosae-Fabaceae). Anais/Resumos da 61ª
Reunião Anual da SBPC, issn 2176-1221, 2009.
RATANAPO, S.; NGAMJUNYAPORN, W.; CHULAVATNATOL, M. Interaction of a
mulberry leaf lectin with a phytopathogenic bacterium, P. syringae pv Mori. Plant Science, v.
160, p. 739-744. 2001.
RAVIKUMAR, B. et al. Regulation of mammalian autophagy in physiology and
pathophysiology. Physiol. Rev., v. 90, p. 1383-1435, 2010.
READ, R. J. et al. New generation of crystallographic validation tools for the protein data bank.
Structure, v. 19, p. 1395-1412, 2011.
REEF, S. et al. A short mitochondrial form of p19ARF induces autophagy and caspase-
independent cell death. Mol. Cell., v. 22, p. 463-475, 2006.
REGO, F. O. R. Análise in silico de proteínas: modelo baseado na Apolipoproteína-e
relacionada à doença de Alzheimer. Monografia (Bacharelado em Ciências Biológicas) -
Universidade Federal da Bahia, 2012.
RIEGER, D. K. et al. Antidepressant-like effect of Canavalia brasiliensis (ConBr) lectin in
mice: Evidence for the involvement of the glutamatergic system. Pharmacology,
Biochemistry and Behavior, v. 122, p. 53-60, 2014.
RINDERLE, S. J. et al. Isolation and characterization of amaranthin, a lectin present in the
seeds of Amaranthus caudatus, that recognizes the T- (or cryptic T)-antigen. J. Biol. Chem., v.
264, n. 27, p. 16123-16131, 1989.
ROBERT, X.; GOUET, P. Deciphering key features in protein structures with the new
ENDscript server. Nucleic Acids Research, v. 42, p. 320-324, 2014.
ROCHA E SILVA, M. O. Brief history of inflammation. In: VANE, J. R.; FERREIRA, S. H.,
Handbook of experimental pharmacology. New York: Springer-Verlag, 65 p., 1978.
ROCHA, B. A. M. et al. CRLI induces vascular smooth muscle relaxation and suggests a dual
mechanism of eNOS activation by legume lectins via muscarinic receptors and shear stress.
Archiv. of Biochem. and Biophys., v. 565, p. 32-39, 2015.

104
ROCHA, B. A. M. et al. Structural basis for both pro- and anti-inflammatory response induced
by mannose-specific legume lectin from Cymbosema roseum. Biochimie, v. 93, p. 806-816,
2011.
ROCHA, B. A. M. et al. Structural basis of ConM binding with resveratrol, an anti-
inflammatory and antioxidant polyphenol. International Journal of Biological
Macromolecules, v. 72, p. 1136-1142, 2015.
RÜDIGER, H.; GABIUS, H. Lectinologie: Gestchichte, konzepte und pharmazeutische
bedeutung. Deutsche Apotheker Zeitung, V. 133, nº 26, p. 15-36, 1993.
RUOTOLO, B.T. et al. Evidence for Macromolecular Protein Rings in the Absence of Bulk
Water. Science, v. 310, p. 1658. 2005.
SAMPIETRO, A. R. et al. An N-acetylglucosamine oligomer binding agglutinin (lectin) from
ripe Cyphomandra betacea Sendt. Fruits Plant Science, v. 160, p. 659-667. 2001.
SANTIAGO, M. Q. et al. Purification, characterization and partial sequence of a pro-
inflammatory lectin from seeds of Canavalia oxyphylla Standl. & L. O. Williams. JMR.
Journal of Molecular Recognition, v. 27, p. 117-123, 2014.
SANTI-GADELHA, T. et al. Purification and biological effects of Araucaria angustifólia
(Araucariaceae) seed lectin. Biochem. Biophys. Res. Commun., v. 350, p. 1050-1055, 2006.
SANZ-APARICIO, J. et al. Endogenous nitric oxide increases prostaglandin biosynthesis in
carrageenin rat paw edema. Eur. J. Pharmacol., v. 286, p. 219-222, 1995.
SANZ-APARICIO, J. et al. The crystal structure of Canavalia brasiliensis lectin suggests a
correlation between its quaternary conformation and its distinct biological properties from
concanavalin A. FEBS Letters, v. 405, p. 114-118, 1997.
SCARFF, C.A. et al. Probing Hemoglobin Structure by Means of Traveling-Wave Ion Mobility
Mass Spectrometry. Journal of The American Society for Mass Spectrometry, v. 20, p. 625-
631, 2009.
SCHNEIDMAN-DUHOVNY, D. et al. A method for integrative structure determination of
protein-protein complexes. Bioinformatics., v. 28, p. 3282-3289, 2012.
SCHNEIDMAN-DUHOVNY, D. et al. PatchDock and SymmDock: servers for rigid and
symmetric docking. Nucleic Acids Res., v. 33, 2005.
SHAANAN, B.; LIS, H.; SHARON, N. Structure of a legume lectin with an ordered N-linked
carbohydrate in complex with lactose. Science, v. 254, p. 862-866, 1991.
SHARMA, V.; SUROLIA, A. Analyses of carbohydrate recognition by legume lectins: size of
the combining site loops and their primary specificity. J. Mol. Biol., v. 267, p. 433-445, 1997.
SHARON, N. Lectins: carbohydrate-specific reagents and biological recognition molecules. J.
Biol. Chem., v. 282, n. 5, p. 2753-2764, 2007.

105
SHARON, N.; LIS, H. Carbohydrates in cell recognition. Scientific American, v. 268, n. 3, 12
p., 1993.
SHARON, N.; LIS, H. History of lectins: from hemagglutinins to biological recognition
molecules. Glycobiology, v. 14, n. 11, p. 53-62, 2004.
SHARON, N.; LIS, H. Lectins. Chapman and Hall, London, v.126, 1989.
SHARON, N.; LIS, H. Legume Lectins - A large family of homologous proteins. FASEB
Journal, v. 4, p. 3198-3208, 1990.
SHEVCHENKO, A. et al. In-gel digestion for mass spectrometric characterization of proteins
and proteomes. Nature protocols, v. 1, n. 6, p. 2856-2860, 2006.
SHOICHET, B. K. Virtual screening of chemical libraries. Nature. v. 16, p. 862-865, 2004.
SILVA, F. O. et al. Antiproliferative effect of Canavalia brasiliensis lectin on B16F10 cells.
Research in Veterinary Science, v. 96, p. 276-282, 2014.
SILVA, H. C. et al. Purification and primary structure determination of a galactose-specific
lectin from Vatairea guianensis Aublet seeds that exhibits vasorelaxant effect. Process
Biochemistry, v. 47, p. 2347-2355, 2012.
SILVA, H.C. et al. Purification and Partial Characterization of a New Pro-Inflammatory Lectin
from Bauhinia bauhinioides Mart (Caesalpinoideae) Seeds. Protein and Peptide Letters, v.
18, p. 396-402, 2011.
SILVA, R.B.L. Endotélio: Um novo alvo terapêutico no tratamento das doenças
cardiovasculares. Revista Médica Ana Costa, v. 11, n. 1, 2006.
SMITH, M. S.; MARTIN, J. H. J. X Ray crystallography. Mol. Pathol., v. 53, n. 1, p. 8-14,
2000.
SOEDJANAATMADJA, U. M. S.; SUBROTO, T.; BEINTEMA, J. J. Processed products of
hevein precursor in the latex of the rubber tree (Hevea brasiliensis). Federation of European
Biochemical Societies, v. 363, p. 211-213, 1995.
SOUSA, B. L. et al. High-resolution structure of a new Tn antigen-binding lectin from Vatairea
macrocarpa and a comparative analysis of Tn-binding legume lectins. International Journal
of Biochemistry & Cell Biology, v. 59, p. 103-110, 2015.
SOUZA, J. D. et al. A new Bauhinia monandra galactose-specific lectin purified in milligram
quantities from secondary roots with antifungal and termiticidal activities. International
Biodeterioration & Biodegradation, v. 65, n. 5, p. 696-702, 2011.
SRINIVAS, V. R. et al. Legume lectin family, the ‘natural mutants of the quaternary state’,
provide insights into the relationship between protein stability and oligomerization. Biochim.
Biophys. Acta, v. 1527, p. 102-111, 2001.

106
SRINIVASAN, N. et al. A superfamily of proteins with the lectin fold. Chemrracrs-Biochem
Mol Biol., v. 6, p. 149-164, 1996.
STEEN, H.; MANN, M. The ABC's (and XYZ's) of peptide sequencing. Nat. Rev. Mol. Cell.
Biol., v. 5, p. 699-711, 2004.
STUMP M. J. et al. Matrix-assisted laser desorption mass spectrometry. Applied Spectroscopy
Reviews, v. 37, p. 275-303, 2002.
SULTAN, N. A.; KAVITHA, M.; SWAMY, M. J. Purification and physicochemical
characterization of two galactose-specific isolectins from the seeds of Trichosanthes cordata.
IUBMB Life, v. 61, n. 4, p. 457-469, 2009.
SUSEELAN, K. N. et al. Purification and characterization of a lectin from wild sunflower
(Helianthus tuberosus L.) tubers. Archives of Biochemistry and Biophysics, v. 407, p. 241-
247, 2002.
TABASSUM, D.P.; POLYAK, K. Tumorigenesis: it takes a village. Nat. Rev. Cancer., v. 15,
p. 473-483, 2015.
TAYLOR, M.; DRICKAMER, K. Introduction to Glycobiology. Oxford University Press,
Oxford, 207 p., 2003.
TEIXEIRA, C. S. et al. Mannose-specific legume lectin from the seeds of Dolichos lablab
(FRIL) stimulates inflammatory and hypernociceptive processes in mice. Process
Biochemistry, v. 49, p. 529-534, 2014.
TOPIN, J. et al. Deciphering the Glycan Preference of Bacterial Lectins by Glycan Array and
Molecular Docking with Validation by Microcalorimetry and Crystallography. PLOS One, v.
19, 2013.
TURTON, K. et al. Crystal structures of Erythrina cristagalli lectin with bound N-linked
oligosaccharide and lactose. Glycobiology, v. 14, n. 10, p. 923-929, 2004.
VAN BRAMER, S. E. An introduction to mass spectrometry. Widener University
Department of Chemistry One University Place Chester, PA, 1998.
VAN DAMME, E. J. et al. Potato lectin: an updated model of a unique chimeric plant protein.
Plant J., v. 37, n.1, p. 34-45, 2004.
VAN DAMME, E. J. et al. The bark of Robinia pseudoaccacia contains a complex mixture of
lectins. Plant Physiol., v. 107, p. 833-43, 1995.
VAN DAMME, E. J. M. et al. Molecular cloning of two different mannose-binding lectins from
tulip bulbs. European journal of biochemistry, v. 236, p. 419-427, 1996.
VAN DAMME, E. J. M. et al. Plant lectin: A composite of several distinct families of structural
and evolutionary related proteins with deverse biological roles. Crit. Ver. Plant Sciences, v.
17, n. 6, p. 575-692, 1998.

107
VAN PARIJS, J. et al. Hevein: An antifungal protein from rubber-tree (Hevea brasiliensis)
latex. Planta, v. 183, p. 258-264, 1991.
VANE, J. R.; ANGGARD, E. E.; BOTTING, R. M. Regulatory functions of the vascular
endothelium. N. Engl. J. Med., v. 323, p. 27-36, 1990.
VAPAATALO, H.; MERVAALA, E. Clinically important factors influencing endonthelial
function. Med. Sci. Monit., v. 7, p. 1075-85, 2001.
VARGIN, A.; TEPLYAKOV, A. MOLREP: an automated program for molecular replacement.
J. Appl. Crystallographica, v. 30, p. 1022-1025, 1997.
VASCONCELOS, M. A. et al. Purification and primary structure of a novel mannose-specific
lectin from Centrolobium microchaete Mart seeds. International Journal of Biological
Macromolecules, v. 81, p. 600-607, 2015.
VAZQUEZ, E. et al. Systemic changes following carrageenan-induced paw inflammation in
rats. Inflamm. Res., v. 64, p. 333-342, 2015.
W. Delano, The Pymol Molecular Graphics System, 2002.
WAH, D. A. et al. Crystal structure of native and Cd/Cd-substituted Dioclea guianensis seed
lectin. A novel manganese-binding site and structural basis of dimer-tetramer association. J.
Mol. Biol., v. 310, p. 885-894, 2001.
WALJUNO, K. et al. Amino acid sequence of hevein. Proc. Int. Rubber Conf., Kuala
Lumpur, v. 2, p. 518-531, 1975.
WALLACE, A. C.; LASKOWSKI, R. A.; THORNTON, J. M. LIGPLOT: A program to
generate schematic diagrams of protein-ligand interactions. Prot. Eng., v. 8, p. 127-134, 1996.
WATKINS, W. M.; MORGAN, W. T. J. Neutralization of the anti-H agglutinin in eel by simple
sugars. Nature, v. 169, p. 825-826, 1952.
WINN, M. D. et al. Overview of the CCP4 suite and current developments. Acta
Crystallographica D, v.67, p. 235-242, 2011.
WLODAWER, A. et al. Protein crystallography for aspiring crystallographers, or how to avoid
pitfalls and traps in macromolecular structure determination. FEBS J., v. 280, p. 5705-5736,
2013.
WONG, J. H.; NG, T. B. Isolation and characterization of a glucose/mannose-specific lectin
with stimulatory effect on nitric oxide production by macrophages from the emperor banana.
International Journal of Biochemistry and Cell Biology, v. 38, p. 234-43, 2005.
WOOH, J.W. et al. Comparison of three commercial sparse-matrix crystallization screens. Acta
Crystallogr. D Biol. Crystallogr., v. 59, p. 769-772, 2003.
WU, A.M. et al. Recognition profile of Bauhinia purpurea agglutinin (BPA). Life Sci., v. 74,
p. 1763–1779, 2004.

108
WU, Y.; WANG, H.; NG, T. B. Purification and characterization of a lectin with
antiproliferative activity toward cancer cells from the dried fruit bodies of Lactarius flavidulus.
Carbohydr. Res., v. 346, n. 16, p. 2576-2581, 2011.
XU, H.L. et al. Molecular modeling, docking and dynamics simulations of GNA-related lectins
for potential prevention of influenza virus (H1N1). J. Mol. Model., v. 18, n. 1, p. 27-37, 2012.
YOUNG, N. M. et al. C-terminal post-translational proteolysis of plant lectins and their
recombinant forms expressed in Escherichia coli. Characterization of “ragged ends” by mass
spectrometry, Journal of Biology and Chemistry, v. 270, p. 2563-2570, 1995.
ZENTENO, E. et al. Specificity of the isolectins from the plant cactus Machaerocereus eruca
for oligosaccharides from porcine stomach mucin. Glycoconjugate Journal, v. 12, p. 699-706,
1995.
ZHU, K. et al. Oncogenes and tumor suppressor genes: comparative genomics and network
perspectives. BMC Genom. v. 16, 2015.
ZHU, K. et al. An insecticidal N-acetylglucosamine-specific lectin gene from Griffonia
simplicifolia (Leguminosae). Plant Physiol., v. 110, p. 195-202, 1996.