Embed Size (px)
Citation preview

UNIVERSIDADE DE LISBOA
FACULDADE DE CIÊNCIAS
DEPARTAMENTO DE BIOLOGIA ANIMAL
MOVIMENTOS E ACTIVIDADES DOS GOLFINHOS-ROAZES
(TURSIOPS TRUNCATUS) NA ZONA TERMINAL DO ESTUÁRIO DO SADO
E POSSÍVEIS INFLUÊNCIAS ECOLÓGICAS
Maria Cecília Vilhena Ferreira
MESTRADO EM ECOLOGIA MARINHA
2010

UNIVERSIDADE DE LISBOA
FACULDADE DE CIÊNCIAS
DEPARTAMENTO DE BIOLOGIA ANIMAL
MOVIMENTOS E ACTIVIDADES DOS GOLFINHOS-ROAZES
(TURSIOPS TRUNCATUS) NA ZONA TERMINAL DO ESTUÁRIO DO SADO
E POSSÍVEIS INFLUÊNCIAS ECOLÓGICAS
Maria Cecília Vilhena Ferreira
Dissertação orientada por:
Prof. Doutor Francisco Andrade – DBA / FCUL
Prof. Doutor Manuel Eduardo dos Santos – ISPA - Instituto Universitário
MESTRADO EM ECOLOGIA MARINHA
2010

ÍNDICE
Índice
Agradecimentos ............................................................................................................... i
Resumo .......................................................................................................................... iii
Abstract ...........................................................................................................................iv
1. Introdução ................................................................................................................... 1
1.1. Caracterização geral da espécie .......................................................................... 1
1.1.1. A comunidade residente de golfinhos-roazes do estuário do Sado .............. 3
1.2. Padrões de movimento e utilização do habitat .................................................... 4
1.2.1. Ciclo de marés ............................................................................................... 5
1.2.2. Hora do dia e estações do ano ...................................................................... 6
1.2.3. Estudos sobre movimentos e utilização do habitat na comunidade de
golfinhos-roazes do estuário do Sado ..................................................................... 8
1.3. Objectivos ............................................................................................................ 9
2. Materiais e métodos ................................................................................................. 10
2.1. Área de estudo ................................................................................................... 10
2.1.1. Marés ........................................................................................................... 11
2.1.2. Caracterização hidrológica do estuário do Sado ......................................... 12
2.2. Metodologia de recolha de dados ...................................................................... 12
2.3. Análise de dados ................................................................................................ 18
2.3.1. Utilização do habitat .................................................................................... 18
2.3.2. Número de golfinhos em relação à actividade dominante aparente ........... 19
2.3.3. Influência do estado da maré e hora do dia no número de golfinhos .......... 19
2.3.4. Influência da fase da maré e da hora do dia no tipo de movimento e na
actividade dominante aparente ............................................................................. 19
2.3.5. Influência de variáveis ambientais no movimento e na actividade dos
golfinhos ................................................................................................................ 20
3. Resultados ................................................................................................................ 21
3.1. Esforço de amostragem ..................................................................................... 21
3.2. Distribuição de actividades ................................................................................ 23

ÍNDICE
3.2.1. Número de golfinhos em relação à actividade dominante aparente ........... 23
3.3. Utilização do habitat ........................................................................................... 24
3.4. Ciclo de marés ................................................................................................... 26
3.4.1. Número de golfinhos em relação ao estado da maré .................................. 26
3.4.2. Tipo de movimento em relação à fase da maré .......................................... 27
3.4.3. Actividade dominante aparente em relação à fase da maré ....................... 28
3.5. Hora do dia ......................................................................................................... 29
3.5.1. Número de golfinhos em relação à hora do dia ........................................... 29
3.5.2. Tipo de movimento em relação à hora do dia ............................................. 30
3.5.3. Actividade dominante aparente em relação à hora do dia .......................... 31
3.6. Tipo de movimento em relação à fase da maré por hora do dia ........................ 32
3.7. Influência de variáveis ambientais no movimento e actividade dos golfinhos ... 33
4. Discussão ................................................................................................................. 36
4.1. Utilização do habitat ........................................................................................... 36
4.2. Actividade dominante aparente .......................................................................... 37
4.3. Movimento .......................................................................................................... 39
4.4. Limitações deste estudo .................................................................................... 41
4.5. Sugestões para estudos futuros ........................................................................ 41
5. Bibliografia ................................................................................................................ 43
ANEXO ......................................................................................................................... 53

AGRADECIMENTOS
Agradecimentos
Durante a realização deste trabalho, foram várias as pessoas que, de uma forma ou
de outra, deram o seu contributo e às quais apresento o meu agradecimento:
Ao Prof. Doutor Francisco Andrade, por ter aceite a orientação deste trabalho e pelo
interesse e disponibilidade demonstrados durante todo este percurso.
Ao Prof. Doutor Manuel Eduardo dos Santos, pela oportunidade que me deu de
trabalhar com estes golfinhos, há três anos e com um objectivo de estudo totalmente
diferente deste, mas que me permitiu chegar até aqui.
Ao Prof. Doutor Henrique Cabral, pela ajuda que me deu no tratamento estatístico dos
dados e pela disponibilidade que sempre demonstrou em ajudar-me.
Ao Prof. Doutor João Maroco, pela ajuda nas análises multivariadas realizadas.
À Tânia Ferreira, por todas as dicas de ArcMap e pela ajuda na construção dos
mapas.
A todas as pessoas que contribuíram com o envio de bibliografia, palavras de apoio e
incentivo e toda a ajuda prestada durante este percurso, e ao Jorge Forte Faria pela
imensa ajuda que me deu com o ROV (quando o tema de mestrado era outro…).
À Erica, pela amizade e incentivo e por todo o apoio nos últimos tempos.
À Patrícia, pela amizade e boa disposição nas nossas muitas horas no cubículo, e
pelas sugestões à versão (quase) final da tese; à Joana, pela amizade e ajuda,
mesmo à distância; e à Rita e ao Miguel, pelo apoio e amizade sempre demonstrados.
Obrigada a todos pelas longas horas no Sado a “fazer ciência”.
À Mariana, o meu obrigada do fundo do coração, pelas correcções e sugestões a um
manuscrito desta tese, pela amizade e pelo apoio, e pelas palavras certas nos
momentos certos.
A todos os meus amigos, por saberem sempre quando precisava de esquecer a tese
sem ser necessário dizer: obrigada por me desencaminharem.
Aos meus Pais, que são a minha fonte de inspiração, por terem possibilitado, em todos
os aspectos, a realização desta tese. Obrigada por me apoiarem de forma incansável,
por me incentivarem sempre a fazer aquilo que mais gosto e nunca me deixarem
desistir. Quaisquer palavras não serão suficientes para vos agradecer por tudo o que
Cecília V. Ferreira i

AGRADECIMENTOS
Cecília V. Ferreira ii
sempre fizeram por mim. Vocês são o espelho de quem eu quero ser, e este trabalho
é dedicado a vocês.
À minha Irmã, por todo o apoio e palavras de incentivo, sempre e desde sempre,
obrigada por acreditares sempre em mim; e ao Cali, pelos abraços e pela boa
disposição que tanto me anima.
À minha família, que me incentiva e me apoia incondicionalmente. Aos meus tios e
primos e à minha avó Lígia, obrigada pela preocupação, interesse e incentivo
constantes. Obrigada à minha avó Albertina, que nunca compreendeu muito bem este
meu fascínio pelos “peixes” mas nunca deixou de me apoiar na minha escolha.
E ao Nuno, por ter dado um sentido novo e especial a este ano. Obrigada pelo
carinho, apoio, amizade, paciência, compreensão e amor. Obrigada por todos os
momentos.

RESUMO
Resumo
A comunidade de golfinhos-roazes do estuário do Sado é uma das poucas
comunidades residentes de golfinhos na Europa, sendo a única em Portugal
Continental. Pretende-se neste trabalho estudar as possíveis influências ecológicas no
movimento e nas actividades destes animais na zona terminal do estuário.
A recolha de dados foi efectuada a partir de um ponto de observação em terra
ao longo de 18 dias entre Agosto e Novembro de 2009. Em 206.5 horas de
observação, os golfinhos-roazes foram avistados na área de estudo durante 34.7
horas, correspondentes a 71 scans positivos (scans com golfinhos presentes)
utilizados para tratamento estatístico. A análise dos dados foi realizada recorrendo a
testes de Kruskal-Wallis, estatística ACTUS2, regressão logística e análise
discriminante.
As actividades mais frequentemente observadas foram a deslocação (75%) e a
alimentação (19%). A regressão logística revelou que a altura da maré é a variável que
mais influencia o movimento dos golfinhos-roazes. Pelo menos nesta época do ano, os
golfinhos-roazes entram no estuário mais frequentemente no período enchente da
maré, abandonando a área de estudo ao final da tarde durante o período vazante. O
número mediano de golfinhos na zona terminal do estuário variou significativamente
com o ciclo de marés, atingindo o máximo 3 horas após a baixa-mar.
Estes resultados parecem sugerir que os golfinhos utilizam as correntes de
maré para se deslocarem entre as águas costeiras e o interior do estuário, poupando
assim energia com o movimento. Por outro lado, poderão aproveitar-se da
previsibilidade das marés e dos movimentos passivos das suas presas, que facilitarão
a sua captura. O canal Sul foi o mais frequentado pelos golfinhos, onde foram
observados episódios de alimentação, tendo evitado o canal Norte. Este estudo
denota assim a importância do estuário para estes animais, sendo essencial que se
implementem novas medidas de protecção deste habitat.
Palavras-chave: Golfinho-roaz; Tursiops truncatus; Sado; movimentos; actividades;
regressão logística.
Cecília V. Ferreira iii

ABSTRACT
Abstract
The bottlenose dolphin community of the Sado estuary is one of the few
resident communities of dolphins in Europe and the only one in mainland Portugal. The
aim of this work is to study the possible ecological influences on bottlenose dolphins’
movement and activities near the mouth of the estuary.
Observations were carried out from an observation point on land over 18 days
between August and November of 2009. During 206.5 hours of observation effort,
dolphins were present in the study area for 34.7 hours, and a total of 71 positive scans
were used for statistical analysis. Data analysis was done using Kruskal-Wallis tests,
ACTUS2, logistic regression and discriminant analysis.
Travelling (75%) and feeding (19%) were the most frequently observed
activities. Logistic regression results showed that tidal height was the variable that
influenced dolphins’ movements the most. At least in this time of year, bottlenose
dolphins enter the estuary more frequently during the flood tide, leaving the study area
in the afternoon during the ebb tide. The median number of dolphins varied significantly
with the tidal cycle, with more dolphins being sighted 3 hours after low water.
These results seem to suggest that bottlenose dolphins use the tidal currents to
move between the coastal waters and the estuary, thus saving energy with movement.
On the other hand, they may take advantage of the predictability of their prey’s
movements, which facilitates capture. The South channel was the most frequently used
by these dolphins, where several feeding episodes were observed; an avoidance of the
North channel was also observed. This study denotes the importance of the estuary to
these animals, and the implementation of new protection measures is essential.
Keywords: Bottlenose dolphin; Tursiops truncatus; Sado; movements; activities;
logistic regression.
Cecília V. Ferreira iv

INTRODUÇÃO
1. Introdução
1.1. Caracterização geral da espécie
O golfinho-roaz, Tursiops truncatus (Montagu, 1821), é uma espécie
pertencente à família Delphinidae e uma das espécies de cetáceos mais bem
estudada. Tem uma distribuição geográfica cosmopolita, podendo ocorrer em águas
temperadas e tropicais (Klinowska, 1991; Jefferson et al., 2008; Wells & Scott, 2009).
Os limites de distribuição desta espécie parecem estar relacionados com a
temperatura, de forma directa ou indirectamente devido à distribuição de presas (Wells
& Scott, 1999).
Conhecem-se para esta espécie dois ecótipos distintos: um costeiro e um
pelágico (Klinowska, 1991; Wells & Scott, 2009). As populações costeiras ocorrem em
habitats como baías, lagoas, estuários e rios (Klinowska, 1991; Hammond et al.,
2008), e as pelágicas em águas offshore, perto de ilhas oceânicas e ao longo da
plataforma continental (Wells & Scott, 2009); muitas das populações costeiras ocupam
de forma residente estuários ou faixas restritas da zona costeira devido à elevada
riqueza alimentar que aí se encontra disponível (Klinowska, 1991).
Apresenta um corpo robusto de tamanho médio com uma coloração dorsal e
lateral que pode variar entre cinzento e preto, sendo quase branca (por vezes rosada)
na zona ventral (Jefferson et al., 2008; Wells & Scott, 2009). O indivíduo adulto mede
entre 1.9 e 3.8 m, existindo no entanto uma grande variação entre populações. O peso
máximo ronda 650 kg (Jefferson et al., 2008). Apesar de os machos poderem ser
ligeiramente maiores do que as fêmeas, não existe um dimorfismo sexual evidente
(Connor et al., 2000; Jefferson et al., 2008).
Os golfinhos-roazes são predadores generalistas (dos Santos, 1998; Wells &
Scott, 2009). Alimentam-se de várias espécies de cefalópodes e peixes (Connor et al.,
2000) apesar de aparentarem ter uma preferência por presas pertencentes às famílias
Mugilidae, Scombridae e Sciaenidae (Wells & Scott, 2009). Também os peixes
produtores de sons parecem ser escolhidos pelos golfinhos, pensando-se que os sons
emitidos pelas presas podem auxiliá-los na sua localização (dos Santos, 1998).
Apresentam estratégias de caça bastante diversificadas: caçam individualmente ou em
grupo, e apresentam um vasto repertório de comportamentos durante a alimentação
que estará adaptado às estratégias que as próprias presas desenvolvem para
evitarem ser detectadas e capturadas (Connor et al., 2000).
Cecília V. Ferreira 1

INTRODUÇÃO
Relativamente à estrutura social, o golfinho-roaz apresenta um padrão de
associação fluido e dinâmico do tipo fissão-fusão (Connor et al., 2000; Jefferson et al.,
2008), em que os indivíduos se associam em pequenos grupos cuja composição e
comportamento mudam frequentemente, por vezes numa base temporal muito curta; a
composição dos grupos pode variar de acordo com factores como o sexo, grau de
parentesco e estado reprodutivo, entre outros (Connor et al., 2000).
A longevidade varia entre os sexos, atingindo os machos mais de 40 anos e as
fêmeas mais de 50 anos. A maturação sexual nas fêmeas surge entre os 5 e os 10
anos de idade, ocorrendo mais tardiamente nos machos, entre os 8 e os 12 anos
(Connor et al., 2000). Tipicamente existe um pico de nascimentos entre a Primavera e
o início do Verão, podendo ocorrer um segundo pico no Outono dependendo da
localização geográfica (Jefferson et al., 2008; Scott et al., 1990). Apresentam um
período de gestação de cerca de 12 meses, e a amamentação dura até a cria atingir
cerca de 1 ano (Wells & Scott, 2009), podendo prolongar-se por vários anos (Mann et
al., 2000; Wells & Scott, 2009); no entanto, já foi encontrado alimento sólido em
conteúdos estomacais de crias com 4 meses de idade (Wells & Scott, 2009). O maior
crescimento ocorre durante os dois primeiros anos de vida (Wells & Scott, 2009). As
crias permanecem com as mães num período que pode ir dos 3 aos 6 anos após o
nascimento (Wells & Scott, 2009), e podem continuar a ser acompanhadas durante a
gravidez seguinte (Mann et al., 2000; Wells & Scott, 2009).
Os golfinhos-roazes produzem sons que se podem agrupar em três categorias:
assobios (sons tonais de banda estreita que se pensa terem funções comunicativas),
estalidos (com banda larga e de curta duração utilizados na ecolocalização) e sons
pulsados (sons mais complexos e variáveis associados à expressão de emoções e à
comunicação). A ecolocalização é, no entanto, um dos mais especializados processos
sensoriais nos odontocetos, em que o animal, ao emitir um estalido, obtém informação
sobre o meio que o rodeia através da interpretação mental do eco que recebe (dos
Santos, 1998).
A União Internacional para a Conservação da Natureza (IUCN) atribui a esta
espécie a categoria Least Concern no Livro Vermelho de Espécies Ameaçadas,
estimando-se um total de 600.000 animais em todo o mundo (Hammond et al., 2008).
Esta espécie está também incluída no Apêndice II da Convenção CITES (Convenção
sobre o Comércio Internacional de Espécies da Fauna e da Flora Selvagem
Ameaçadas de Extinção) e no Anexo II da Directiva Habitats (Rede Natura 2000).
Cecília V. Ferreira 2

INTRODUÇÃO
1.1.1. A comunidade residente de golfinhos-roazes do estuário do Sado
A comunidade de golfinhos-roazes do estuário do Sado é uma das poucas
comunidades residentes de golfinhos na Europa, sendo a única em Portugal
Continental. O tamanho desta comunidade tem vindo a decrescer ao longo das últimas
décadas: em 1987 estimavam-se pelo menos 40 indivíduos neste grupo (dos Santos &
Lacerda, 1987), número que decresceu até aos actuais 24 indivíduos, o mais baixo
registado até à data. Destes 24 animais, a maioria já é adulta, contando a comunidade
com apenas 2 crias e 3 subadultos (Augusto et al., in press). Esta comunidade habita
uma zona fortemente impactada devido à proximidade a um grande centro urbano,
identificando-se como principais ameaças à sua sobrevivência a degradação da
qualidade da água, a pesca, a poluição acústica e o tráfego marítimo (Silva, 2008).
A primeira referência bibliográfica a esta comunidade data de 1863 e é da
autoria do naturalista Bocage (dos Santos & Lacerda, 1987). No entanto, os primeiros
estudos realizados sobre esta comunidade só foram iniciados em 1980 com os
trabalhos de Teixeira & Duguy (1981), Hussenot (1982), dos Santos (1985) e dos
Santos & Lacerda (1987).
Mais recentemente, têm sido realizados estudos sobre comportamento
(Harzen, 1995; Carvalho, 2000; Gaspar, 2003; Augusto, 2007; dos Santos et al.,
2007), acústica (dos Santos, 1998; Couchinho, 1999; Brito, 2001; dos Santos et al.,
2005), utilização do habitat (Freitas, 1995; Harzen, 1998; Nunes, 2001; Cândido, 2003)
e avaliação de impactos antropogénicos (Cascão, 2001; Luís, 2008; Silva, 2008).
O estuário do Sado apresenta-se como um habitat de extrema importância para
estes golfinhos, que dele dependem do ponto de vista alimentar devido à sua elevada
riqueza biológica. Estão descritas 3 zonas preferenciais de alimentação: a extremidade
da península de Tróia, a zona junto à Caldeira de Tróia e o Canal da Comporta
(Freitas, 1995; Cândido, 2003). São suas presas confirmadas a enguia (Anguilla
anguilla [Linnaeus, 1758]), as tainhas (Mugil cephalus Linnaeus, 1758; Liza sp. e
Chelon labrosus [Risso, 1827]), o polvo-comum (Octopus vulgaris Cuvier, 1797) e o
choco (Sepia officinalis Linnaeus, 1758) (dos Santos et al., 2007).
Devido a esta dependência do estuário, os golfinhos apresentam um elevado
número e variedade de lesões cutâneas, desconhecendo-se a sua origem; pensa-se,
no entanto, que estas lesões possam estar relacionadas com a degradação do habitat
(Harzen & Brunnick, 1997).
Cecília V. Ferreira 3

INTRODUÇÃO
As ameaças à sobrevivência desta comunidade levaram a que, em 2009, se
procedesse à escrita e implementação de um Plano de Acção para a salvaguarda e
monitorização da população de Roazes do estuário do Sado (Sequeira et al., 2009).
Este Plano inclui um conjunto de acções e medidas que tentarão facilitar a recolha de
informação actualizada sobre esta comunidade e a interacção entre as várias
entidades que actuam no estuário, para que possam ser tomadas decisões que
melhorem a qualidade do estuário do Sado e, a longo prazo, protejam esta
comunidade ameaçada de golfinhos.
1.2. Padrões de movimento e utilização do habitat
O movimento dos animais é um processo ecológico que determina a estrutura
espacial e demográfica das populações (Wiens, 2000), tendo evoluído como uma
resposta às características bióticas e abióticas do ambiente em que vivem (Stevick
et al., 2002). As causas que condicionam o movimento dos mamíferos marinhos estão
relacionadas com a necessidade de realização de determinadas actividades em
determinado local e numa escala temporal específica, bem como a maximização da
utilização de recursos de um determinado habitat; por outro lado, relacionam-se
também com a necessidade de se deslocarem para longe de locais que possam impor
riscos à sua sobrevivência, como a existência de predadores, doenças ou locais com
condições físicas prejudiciais (Stevick et al., 2002).
Compreender o padrão de movimento dos animais no tempo e no espaço é
fundamental para o estudo da sua ecologia, permitindo o estabelecimento e
implementação de estratégias eficazes de conservação e gestão de recursos (Pittman
& McAlpine, 2003).
Na revisão da literatura feita por Klinowska (1986) relativamente a ritmos
diurnos de várias espécies da Ordem Cetacea, a autora menciona que estes animais,
que se caracterizam por tantas adaptações à vida no meio aquático, parecem
depender totalmente do ambiente que os rodeia na organização das suas rotinas
diárias, avançando com a explicação de que não ter uma rotina diária definida, como é
característico de outros grupos de animais, poderá ter sido a forma mais vantajosa de
sobreviver num ambiente aquático. No entanto, a autora encontrou referência a várias
“rotinas” diárias em espécies desta Ordem, diferentes consoante o local e a espécie
estudados.
Cecília V. Ferreira 4

INTRODUÇÃO
Desde então vários estudos têm sido realizados em populações de diversas
espécies de cetáceos que habitam ambientes costeiros, uma vez que existe uma
maior facilidade de encontrar e estudar animais que utilizam normalmente habitats
mais “confinados” e de mais fácil acesso, como é o caso de estuários e baías. Apesar
da maioria destes trabalhos ter como objecto de estudo o golfinho-roaz, existem
trabalhos sobre outras espécies de cetáceos.
Os padrões diurnos de movimento e uso do habitat parecem estar relacionados
com um número diverso de factores, dependentes da ecologia do habitat, que fazem
variar os padrões comportamentais dos golfinhos. Estes factores incluem o ciclo de
marés (e.g. Shane, 1990a; Irvine et al., 1981) bem como a hora do dia (e.g. Gregory &
Rowden, 2001) e a fisiografia do habitat (e.g. Ingram & Rogan, 2002).
1.2.1. Ciclo de marés
O movimento dos golfinhos tem uma associação variável com as correntes de
maré, dependendo da localização geográfica do habitat em que são estudados: podem
afectar directamente o comportamento, ao ajudar ou impedir a deslocação dos
animais, e indirectamente influenciando os movimentos das suas presas (Shane,
1990a). Os estudos já realizados sobre este assunto mostram efectivamente algum
tipo de associação, embora as razões sejam, em muitos casos, meramente
especulativas.
Em Sanibel Island, Florida, os golfinhos-roazes movimentam-se
significativamente mais contra do que a favor da maré (Shane, 1990a), tendo sido
observado um comportamento semelhante em Port Aransas, Texas (Shane, 1990b),
Ensenada De La Paz, México (Acevedo, 1991) e no estuário de Shannon, Irlanda
(Berrow et al., 1996). Em Sarasota, Florida (Irvine et al., 1981), os golfinhos foram
observados a movimentar-se maioritariamente a favor da maré, comportamento que
também foi observado em Cardigan Bay, West Wales (Gregory & Rowden, 2001) e em
New South Wales, Austrália (Fury, 2008).
Mendes et al. (2002) realizaram um estudo para perceber qual o efeito de uma
frente de maré existente em Moray Firth, Escócia, na abundância de golfinhos-roazes.
Realizaram para tal observações a partir de terra, em 1994 para investigar a relação
entre o ciclo de marés e a abundância de golfinhos e em 2000 para caracterizar
espacial e temporalmente a frente de maré. Os autores avistaram mais golfinhos
Cecília V. Ferreira 5

INTRODUÇÃO
durante o período enchente da maré, tendo sugerido que os animais se associam à
zona superficial da frente de maré e que esta pode influenciar o comportamento das
suas presas, que se poderão acumular perto dessa frente.
Outras espécies de cetáceos que apresentam uma associação com as
correntes de maré incluem o boto, Phocoena phocoena (Bay of Fundy, Canadá:
Johnston et al., 2005; North Devon, Reino Unido: Goodwin, 2008; e Wales, Reino
Unido: Pierpoint, 2008), o boto-cinza, Sotalia guianensis (Baía de Sepetiba, Brasil:
Flach et al., 2008; e Baía de Babitonga, Brasil: Cremer et al., 2009) e o golfinho-
comum, Delphinus delphis (Bay of Plenty, Nova Zelândia: Neumann, 2001).
Alguns destes estudos abordam as razões para esta associação com as
marés. Os autores associam o movimento dos animais essencialmente à alimentação
e à busca de presas. No caso em que os animais se movimentam contra a maré, foi
sugerido que este movimento contra-corrente se trata de uma estratégia de caça, na
medida em que, uma vez que as suas presas se movimentam essencialmente com a
maré, os golfinhos movimentam-se na direcção das suas presas (Shane, 1980;
Acevedo, 1991), apesar dos maiores custos energéticos que advêm de um movimento
contra uma forte corrente (Shane, 1980). Por outro lado, o movimento a favor da maré,
ou quando as correntes são mais fracas, permite a redução dos custos energéticos
associados à deslocação (Gregory & Rowden, 2001).
1.2.2. Hora do dia e estações do ano
Várias espécies de cetáceos apresentam padrões de movimento e
comportamento que se podem relacionar com a hora do dia (Klinowska, 1986). A hora
do dia, tal como as estações do ano, podem condicionar o movimento dos golfinhos ao
influenciarem directamente o movimento das suas presas.
Hansen (1990) registou, em dois locais na Califórnia, que o tamanho médio de
grupo de golfinhos-roazes atingiu o seu máximo durante o período da manhã. Em
Galveston Bay, Texas, durante os meses de Verão, Bräger (1993) evidenciou a
existência de um padrão diurno no comportamento dos golfinhos: a frequência de
alimentação apresentou-se com um pico principal durante a manhã e um pico
secundário mais baixo ao final da tarde; a frequência de socialização aumentou
quando a alimentação diminuiu, tendo apresentado um pico durante a tarde; os
animais foram observados em deslocação principalmente ao final da tarde,
Cecília V. Ferreira 6

INTRODUÇÃO
correspondente ao regresso à baía. Durante os meses de Outono em que estes
animais foram estudados, este padrão alterou-se, havendo uma diminuição da
frequência de socialização e deslocação, com a alimentação a ocorrer ao longo de
todo o dia. O autor refere semelhanças nestes padrões com aqueles observados
noutros locais no Texas, apesar dos diferentes métodos de recolha de dados. Em
contraste, um estudo realizado por Shane (1990a) em Sanibel Island, Florida, mostra
que os comportamentos relacionados com a alimentação foram mais comuns durante
a manhã, tendo a socialização apresentado um pico ao final do dia e a deslocação
durante a tarde. A autora detectou também o aumento da alimentação e a diminuição
da deslocação na transição do Verão para o Outono.
Algumas espécies apresentam um padrão circadiano bastante rígido e
previsível, como é o caso do golfinho-rotador (Stenella longirostris). Os animais desta
espécie estudados no Hawaii deslocam-se ao final do dia para zonas offshore para se
alimentarem, no período nocturno, a profundidades de cerca de 100 m; iniciam depois
um movimento contrário em direcção a baías protegidas onde permanecem durante o
dia, geralmente em repouso (Norris & Dohl, 1980). Comportamento semelhante é
observado nos golfinhos-rotadores estudados no Arquipélago de Fernando de
Noronha, Brasil (Silva & Silva-Jr, 2009).
O golfinho-de-Heaviside (Cephalorhynchus heavisidii) estudado em Agterbaai,
África do Sul, apresentou também um padrão marcado de movimento onshore-
offshore ao longo do dia. Estes animais permaneceram mais próximo de costa durante
a manhã, iniciando depois um movimento offshore em direcção a zonas de maior
profundidade (Elwen et al., 2006).
O boto-cinza (Sotalia guianensis) estudado na Baía Norte, Brasil, apresentou
uma variação no padrão do comportamento ao longo do dia, verificando-se uma
tendência inversa nas frequências de deslocação e na alimentação, com um
decréscimo progressivo da alimentação e um aumento progressivo da deslocação ao
longo do dia (Daura-Jorge et al., 2005). Na Baía de Sepetiba, Brasil, Flach et al. (2008)
estudaram também o boto-cinza, tendo detectado durante a manhã uma frequência
mais elevada de comportamentos relacionados com a alimentação, enquanto no
período da tarde houve uma maior frequência dos comportamentos de repouso e
socialização.
Cecília V. Ferreira 7

INTRODUÇÃO
1.2.3. Estudos sobre movimentos e utilização do habitat na comunidade de golfinhos-roazes do estuário do Sado
Os movimentos dos golfinhos-roazes residentes na região do Sado foram já
abordados no passado, de forma mais ou menos detalhada, tendo sido estudada a
relação com o ciclo de marés e com a hora do dia. Não foram, no entanto,
determinados os factores passíveis de influenciar o movimento e actividades dos
animais, nem exploradas a fundo as razões para tal influência.
Dos Santos & Lacerda (1987) não assinalaram qualquer relação entre o
movimento dos grupos de golfinhos e as marés a partir da análise dos seus dados.
Dois estudos realizados posteriormente por Harzen (1998, 2002), com dados
recolhidos em vários períodos entre Junho de 1986 e Dezembro de 1993, mostraram
que os golfinhos aparentam entrar no estuário durante a manhã e com a maré
enchente, alimentando-se nessa corrente de entrada; com a maré vazante e durante a
tarde, os animais movimentaram-se gradualmente para fora da área de estudo.
Nunes (2001), por sua vez, não detectou qualquer relação entre as marés e o
sentido de deslocação dos golfinhos, apesar de notar a aparente preferência pela
maré vazante para as actividades de alimentação.
No que diz respeito à utilização deste habitat, os golfinhos são observados
durante todo o ano no estuário do Sado e na zona marinha adjacente, utilizando mais
frequentemente o canal Sul e a foz (dos Santos & Lacerda, 1987; Harzen, 1998).
Trata-se de uma importante área de alimentação para estes animais (dos Santos &
Lacerda, 1987; Nunes, 2001), que utilizam uma grande variedade de técnicas de caça,
dependentes da zona do estuário em que se encontram (dos Santos & Lacerda, 1987).
Um estudo realizado por Cândido (2003) utilizou um sistema de informação
geográfica para realizar uma análise individual de comportamentos associados aos
movimentos e à utilização espacial do estuário e da zona marinha adjacente. Este
trabalho permitiu estimar áreas vitais dos indivíduos estudados, e determinou como
centros de actividade para a maioria dos animais observados a zona da Caldeira de
Tróia e a extremidade da península de Tróia.
Cecília V. Ferreira 8

INTRODUÇÃO
Cecília V. Ferreira 9
1.3. Objectivos
O objectivo principal deste trabalho é analisar a influência de factores
ecológicos no movimento dos golfinhos-roazes na zona terminal do estuário do Sado.
A utilização do habitat pelos golfinhos-roazes será analisada em relação à
actividade comportamental, e serão determinadas as actividades dominantes dos
golfinhos em relação ao ciclo de marés e à hora do dia, para além da quantificação do
tempo total despendido nas várias actividades que compõem o seu repertório
comportamental.
Serão aplicadas metodologias de análise multivariada (regressão logística e
análise discriminante) que permitam identificar as influências de diferentes variáveis
ambientais quasi-independentes no movimento e comportamento dos golfinhos-
roazes.

MATERIAIS E MÉTODOS
2. Materiais e métodos
2.1. Área de estudo
O estuário do Sado localiza-se na região ocidental de Portugal Continental
(figura 1), junto à cidade de Setúbal. É o segundo maior estuário português e constitui
uma das zonas húmidas mais importantes do país. Estende-se no sentido NW-SE ao
longo de cerca de 45 Km (Sequeira et al., 2009) e apresenta uma área aproximada de
180 Km2 (Cabral, 2000) desde a foz (38°29' N, 8°55' W) até ao limite de influência da
maré, a cerca de 25 Km a montante de Alcácer do Sal (Cândido, 2003). A principal
contribuição de água doce provém do rio Sado, com caudais que variam entre 1 m3 s-1
no Verão e 1800 m3 s-1 no Inverno (Martins et al., 2002).
Na zona a montante do estuário, junto a Alcácer do Sal, atinge-se a largura
máxima encontrada, cerca de 5 Km. Ocorre depois um estreitamento a jusante, na foz,
onde a largura é de cerca de 1,5 Km (Sequeira et al., 2009). O contacto com o mar
faz-se neste local, e a profundidade chega a atingir 50 m junto à margem do Outão
(dos Santos, 1998).
A zona intermédia do estuário apresenta um conjunto de bancos de areia
intertidais que divide esta zona em dois canais. O canal Norte localiza-se junto à
cidade de Setúbal, na margem norte, onde se situam as principais indústrias da região
e que funciona como canal de navegação de acesso ao Porto de Setúbal. Este canal
apresenta uma profundidade máxima de 15 m e os sedimentos são mais envasados
(Cabral, 2000). O canal Sul localiza-se junto à península de Tróia e atinge 25 m de
profundidade. Neste canal, os sedimentos são diversos e encontram-se distribuídos
em manchas, sendo a areia fina e a vasa os tipos predominantes (Cabral, 2000).
O estuário do Sado apresenta uma elevada importância ecológica devido à sua
riqueza biológica e de habitats, razão pela qual está incluído em vários documentos
legais que lhe conferem estatutos de conservação. Assim, parte do estuário do Sado
está definida desde 1980 como Reserva Natural (Decreto-Lei n.º 430/80, de 1 de
Outubro). No âmbito da Rede Natura 2000, foi definido o Sítio de Importância
Comunitária “Estuário do Sado” (PTCON0011) ao abrigo da Directiva Habitats
(Resolução do Conselho de Ministros n.º 142/97, de 28 de Agosto) e uma Zona de
Protecção Especial inserida na Directiva Aves (Decreto-Lei n.º 384-B/99, de 23 de
Setembro). O estuário está ainda definido como Zona Húmida de Importância
Internacional (PT023) na Lista de Sítios da Convenção de RAMSAR.
Cecília V. Ferreira 10
MATERIAIS E MÉTODOS
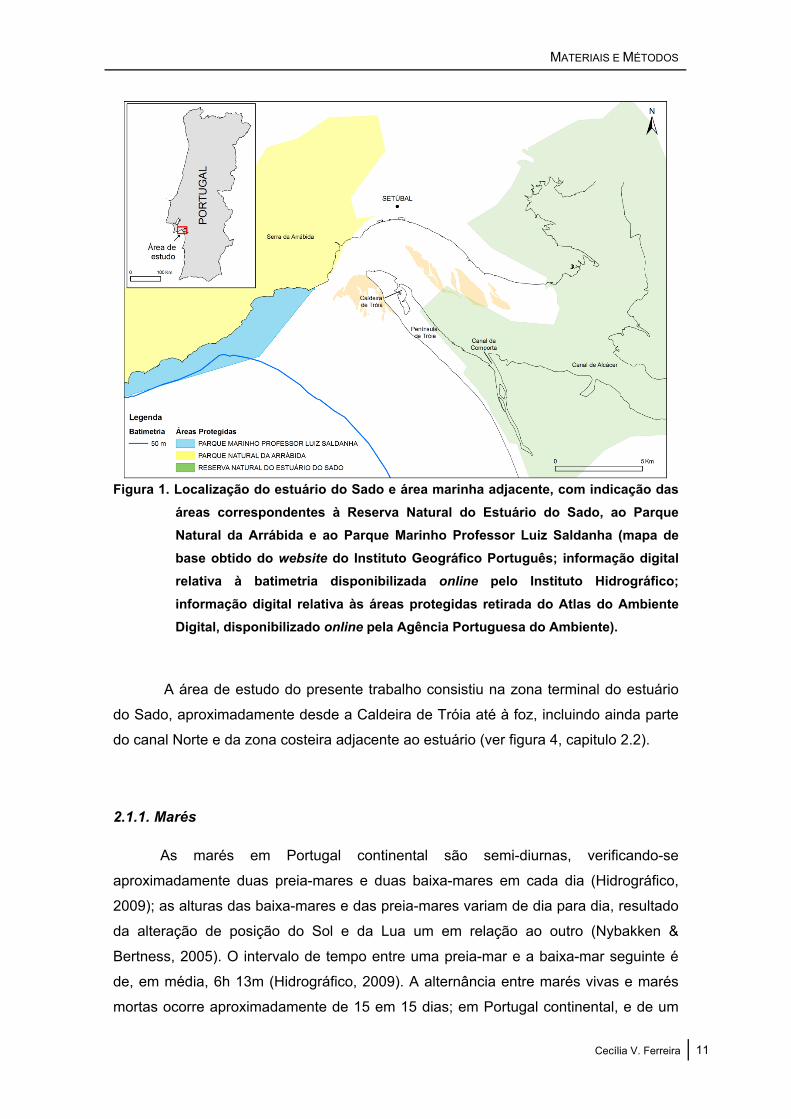
Figura 1. Localização do estuário do Sado e área marinha adjacente, com indicação das áreas correspondentes à Reserva Natural do Estuário do Sado, ao Parque Natural da Arrábida e ao Parque Marinho Professor Luiz Saldanha (mapa de base obtido do website do Instituto Geográfico Português; informação digital relativa à batimetria disponibilizada online pelo Instituto Hidrográfico; informação digital relativa às áreas protegidas retirada do Atlas do Ambiente Digital, disponibilizado online pela Agência Portuguesa do Ambiente).
A área de estudo do presente trabalho consistiu na zona terminal do estuário
do Sado, aproximadamente desde a Caldeira de Tróia até à foz, incluindo ainda parte
do canal Norte e da zona costeira adjacente ao estuário (ver figura 4, capitulo 2.2).
2.1.1. Marés
As marés em Portugal continental são semi-diurnas, verificando-se
aproximadamente duas preia-mares e duas baixa-mares em cada dia (Hidrográfico,
2009); as alturas das baixa-mares e das preia-mares variam de dia para dia, resultado
da alteração de posição do Sol e da Lua um em relação ao outro (Nybakken &
Bertness, 2005). O intervalo de tempo entre uma preia-mar e a baixa-mar seguinte é
de, em média, 6h 13m (Hidrográfico, 2009). A alternância entre marés vivas e marés
mortas ocorre aproximadamente de 15 em 15 dias; em Portugal continental, e de um
Cecília V. Ferreira 11

MATERIAIS E MÉTODOS
modo geral, a amplitude de maré varia entre 1.50 m em marés vivas e 0.70 m em
marés mortas em relação aos níveis médios (Hidrográfico, 2009). A previsão da hora e
altura das marés em Portugal está disponível para consulta online no website do
Instituto Hidrográfico.
2.1.2. Caracterização hidrológica do estuário do Sado
O estuário do Sado, em particular a sua foz, apresenta-se como uma zona de
forte influência das correntes de maré. No estuário existem dois vórtices principais: um
ciclónico, centrado no início dos bancos de areia, e um anti-ciclónico centrado mais a
montante do estuário, na região da Setenave. A corrente residual é mais forte junto da
foz e em particular na zona da Califórnia (junto à Caldeira de Tróia), onde os dois
vórtices se encontram. Os bancos de areia intertidais impedem que o mesmo aconteça
no canal Norte, que tem assim correntes mais fracas (Neves, 1985 in Rodrigues &
Quintino, 1993). No canal de navegação forma-se um jacto e dois vórtices adjacentes.
O vórtice ciclónico localiza-se na zona do Cambalhão e é de maiores dimensões,
apresentando também maiores velocidades. Estes dois vórtices põem em evidência a
entrada da água no estuário pelas zonas junto a terra (INAG/MARETEC, 2001). Estes
fluxos são importantes uma vez que os fenómenos de upwelling e downwelling
associados contribuem para a mistura das massas de água e o movimento de
nutrientes para a superfície (Robinson, 2010).
2.2. Metodologia de recolha de dados
A recolha de dados foi feita a partir de um ponto de observação em terra, na
margem Norte do estuário do Sado (Forte de S. Filipe: 38° 31.040'N, 8° 54.583'W), a
114 m de altitude. Este local foi escolhido por permitir um amplo campo de visão que
compreende os canais Norte e Sul do estuário bem como a foz e parte da área
marinha adjacente (figura 2).
Cecília V. Ferreira 12

MATERIAIS E MÉTODOS
Cecília V. Ferreira 13
Figura 2. A – Campo de visão da área de estudo a partir do local de observação; B – Recolha de dados com recurso a binóculos e telescópio e preenchimento
da ficha de amostragem.
O período de amostragem decorreu entre Agosto e Novembro de 2009,
abrangendo apenas o período diurno (o início e o fim da amostragem em cada dia
variou ao longo do período de amostragem, no entanto as observações iniciaram-se
entre as 6h30 e as 8h30 e terminaram entre as 17h e as 20h).
A metodologia de recolha de dados utilizada foi a amostragem de grupo focal
(Mann, 1999), tendo sido considerado como grupo o conjunto de animais envolvidos
na mesma actividade aparente num raio de 200 m (dos Santos et al., 2007).
Foram feitos scans à área de estudo com o auxílio de binóculos Helios
Naturesport 7x50 mm a cada 30 minutos, tendo cada scan a duração de 10 minutos.
Os scans em que não havia avistamento de golfinhos foram anotados como scans
negativos e o procedimento era repetido 30 minutos depois.
Quando era avistado um grupo de golfinhos-roazes, a observação tinha a
duração de 10 minutos, recorrendo a um telescópio Bresser Optics 25-75x100 mm. A
posição aproximada de cada avistamento foi marcada num mapa da área de estudo,
recorrendo para tal a bóias de navegação existentes no estuário, localização de
bancos de areia e marcos proeminentes em terra (como edifícios) facilmente
visualizados do ponto de observação. Esta posição aproximada apresenta um erro
associado, uma vez que não foi possível determinar exactamente a posição geográfica
dos animais; no entanto, tentou minimizar-se este erro utilizando diversas
A
B

MATERIAIS E MÉTODOS
conhecenças presentes no estuário. Durante estes scans positivos, se os animais se
movimentassem para locais não visíveis do ponto de observação, o observador
dirigia-se para um ponto secundário no Forte de S. Filipe de onde fosse possível
avistar o grupo. Após a marcação da posição do avistamento, a observação era
realizada durante 10 minutos, findos os quais se registava numa ficha de amostragem
(em anexo) a hora do avistamento, estado da maré, quadrante, direcção aparente de
deslocação, tamanho de grupo e actividade dominante aparente, entre outras. Os
dados recolhidos em cada dia foram introduzidos em folhas de cálculo no software
Microsoft® Excel© no final de cada dia de amostragem. De seguida explicita-se a forma
como foi caracterizada cada uma destas variáveis.
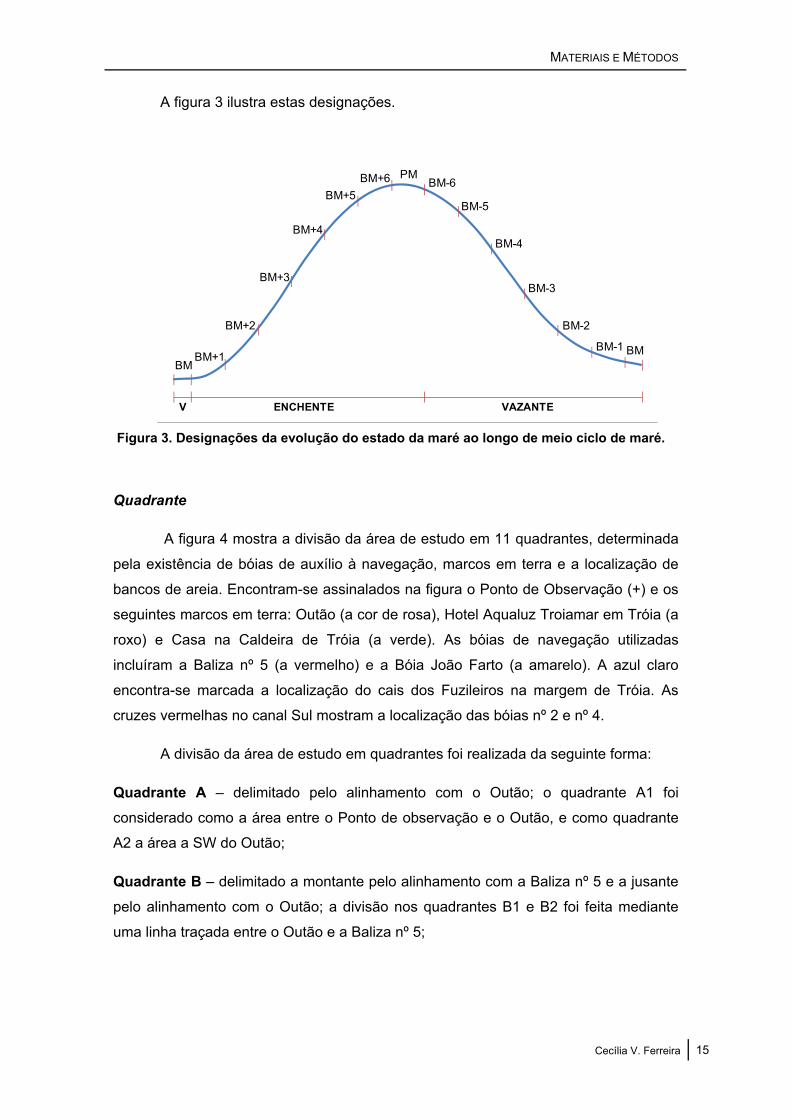
Estado da maré
De forma a acompanhar o movimento e actividade dos animais com a evolução
da maré, foi atribuída, a cada scan, uma categoria do estado da maré em função da
hora de baixa-mar anterior à primeira observação, recorrendo à designação BM (de
Baixa-Mar), e uma única categoria com a designação PM (de Preia-Mar). As
categorias BM e PM foram definidas como o intervalo de tempo compreendido entre
os 15 minutos anteriores e os 15 minutos posteriores à hora tabelada para cada uma
das marés. Ao contrário das outras categorias, estas tiveram a duração de 30 minutos,
correspondente à duração do período de estofo da maré, em que ocorre a inversão do
sentido da maré e a intensidade da corrente é praticamente nula, não se verificando
alteração na altura. Durante a maré enchente, as categorias seguintes à categoria BM,
com a duração de 1 hora e a designação BM+x (em que x corresponde ao número de
horas depois da baixa-mar), foram obtidas a partir da anterior tendo como limite
inferior o limite superior da categoria que a precede. Durante a maré vazante, e após a
categoria PM, atribuiu-se a designação BM-x, obtidas igualmente a partir da categoria
anterior utilizando como limite inferior o limite superior da categoria que a antecede.
Utilizaram-se ainda as designações “enchente”, quando os indivíduos foram
observados durante o período enchente da maré (período entre uma baixa-mar e a
preia-mar seguinte), e “vazante”, quando os indivíduos foram observados durante o
período vazante da maré (período entre uma preia-mar e a baixa-mar seguinte).
Incluíram-se na designação “enchente” as observações realizadas desde a hora
tabelada de baixa-mar (exclusive) até à hora de preia-mar, e na designação “vazante”
as observações realizadas desde a hora tabelada de preia-mar (exclusive) até à hora
de baixa-mar seguinte.
Cecília V. Ferreira 14
MATERIAIS E MÉTODOS
Cecília V. Ferreira 15
A figura 3 ilustra estas designações.
Figura 3. Designações da evolução do estado da maré ao longo de meio ciclo de maré.
Quadrante
A figura 4 mostra a divisão da área de estudo em 11 quadrantes, determinada
pela existência de bóias de auxílio à navegação, marcos em terra e a localização de
bancos de areia. Encontram-se assinalados na figura o Ponto de Observação (+) e os
seguintes marcos em terra: Outão (a cor de rosa), Hotel Aqualuz Troiamar em Tróia (a
roxo) e Casa na Caldeira de Tróia (a verde). As bóias de navegação utilizadas
incluíram a Baliza nº 5 (a vermelho) e a Bóia João Farto (a amarelo). A azul claro
encontra-se marcada a localização do cais dos Fuzileiros na margem de Tróia. As
cruzes vermelhas no canal Sul mostram a localização das bóias nº 2 e nº 4.
A divisão da área de estudo em quadrantes foi realizada da seguinte forma:
Quadrante A – delimitado pelo alinhamento com o Outão; o quadrante A1 foi
considerado como a área entre o Ponto de observação e o Outão, e como quadrante
A2 a área a SW do Outão;
Quadrante B – delimitado a montante pelo alinhamento com a Baliza nº 5 e a jusante
pelo alinhamento com o Outão; a divisão nos quadrantes B1 e B2 foi feita mediante
uma linha traçada entre o Outão e a Baliza nº 5;
BMBM+1
PM
BM+2
BM+3
BM+4
BM+5BM+6 BM-6
BM-5
BM-4
BM-3
BM-2
BM-1 BM
ENCHENTE VAZANTEV

MATERIAIS E MÉTODOS
Quadrante C – delimitado a montante pelo alinhamento com o Hotel Aqualuz Troiamar
e a jusante pela Baliza nº 5; traçou-se uma linha entre a Baliza nº 5 e o Hotel Casino
para obter os quadrantes C1 e C2;
Quadrante D – delimitado a montante pelo alinhamento com a Casa na Caldeira e a
jusante com o Hotel Casino; a posterior delimitação nos quadrantes D1 e D2 foi feita
mediante uma linha traçada entre a Bóia João Farto e o pórtico azul na margem Norte
do estuário;
Quadrante E – quadrante a montante do quadrante D; o quadrante E1 é delimitado
por uma linha traçada entre a Bóia João Farto e o pórtico azul na margem Norte do
estuário; e os quadrantes E2N e E2S foram obtidos a partir de um eixo alinhado com
os bancos de areia que dividem o estuário no canal Norte (junto a Setúbal) e canal Sul
(junto à Península de Tróia).
Figura 4. Mapa da área de estudo com divisão nos quadrantes de amostragem.
Direcção aparente de deslocação
A direcção aparente de deslocação foi definida como “UP” quando o grupo se
encontrasse em deslocação para o interior do estuário (sentidos Nordeste e Sudeste),
“DOWN” quando o movimento fosse no sentido de saída do estuário (sentidos
Cecília V. Ferreira 16

MATERIAIS E MÉTODOS
Noroeste e Sudoeste), ou “IND” (indeterminado) se o grupo aparentava um movimento
sem direcção definida.
Tamanho de grupo
Durante um scan positivo, o número de indivíduos do grupo foi contabilizado,
sempre que os animais vinham à superfície, como o número de animais à superfície
num dado instante. Quando se obtinham várias contagens diferentes do número de
animais durante um scan, considerou-se, de entre todas as contagens, o número
mínimo de animais contabilizado à superfície.
Actividade dominante aparente
A definição no campo da actividade dominante aparente em que os golfinhos
se encontravam envolvidos foi baseada em comportamentos observados, tais como a
direcção e a velocidade de natação e a duração dos mergulhos, bem como o contacto
físico entre indivíduos e a presença de presas (Bearzi et al., 1999).
Com base na literatura, foram utilizadas quatro categorias comportamentais
para determinar a actividade dominante aparente (dos Santos, 1998; Cândido, 2003;
Constantine et al., 2004; Lusseau, 2006):
Deslocação – Os animais emergem próximos uns dos outros, num só grupo compacto
ou em várias unidades separadas dispostas, geralmente, ao longo de um eixo de
deslocação. A deslocação do grupo é rápida e a direcção bastante marcada. As
submersões são normalmente curtas, embora possa registar-se a ocorrência de
mergulhos prolongados, de duração relativamente constante (Lusseau, 2006), que
alternam com sequências de ventilação. São raros outros comportamentos à
superfície para além de saltos (dos Santos, 1998);
Alimentação – Os animais encontram-se envolvidos em actividades de captura e
consumo de presas, evidenciadas pela presença de presas à superfície e técnicas de
caça como natação em círculo (Constantine et al., 2004). Os mergulhos têm duração
variável e não são sincronizados (Cândido, 2003);
Socialização – Os animais encontram-se junto à superfície, não sendo detectada a
presença de presas. Podem ocorrer diversos tipos de interacções entre os indivíduos,
Cecília V. Ferreira 17

MATERIAIS E MÉTODOS
como saltos sincronizados e contacto físico (dos Santos 1998), e os intervalos de
mergulhos são de duração variável. Ocorrem pequenos sub-grupos espalhados
(Lusseau, 2006). Pode ter um papel social e/ou reprodutor (Constantine et al., 2004);
Repouso – Os animais deslocam-se muito próximos uns dos outros, com movimentos
lentos e formando uma só unidade que se desloca praticamente imóvel à superfície
(dos Santos, 1998).
2.3. Análise de dados
A estatística descritiva e os testes estatísticos foram realizados recorrendo aos
programas Microsoft® Excel©, PASW Statistics versão 18.0 e ACTUS2 (Estabrook &
Estabrook, 1989). O nível de significância (α) adoptado em todos os testes foi de 5%.
A normalidade dos dados foi testada, quando necessário, recorrendo ao teste
de Shapiro-Wilk (Zar, 2009), rejeitando-se a hipótese nula de normalidade dos dados
sempre que o valor de p obtido foi inferior ao nível de significância adoptado
(p < 0.05).
2.3.1. Utilização do habitat
A estatística de simulação ACTUS2 foi utilizada para comparar a utilização de
cada quadrante da área de estudo em relação à actividade dominante. Esta análise
utiliza tabelas de contingência que não poderiam ser analisadas recorrendo à
estatística de χ2 por não cumprirem os seus pressupostos (ou seja, tabelas com várias
células com frequências baixas (< 5) ou com valor nulo) (Almada & Oliveira, 1997).
Assim, o ACTUS2 simula 10000 tabelas de contingência de valores esperados e
devolve os valores de χ2 e de p associados, bem como as células em que os valores
observados diferem significativamente dos esperados, à semelhança do que se obteria
num teste a posteriori.
Foi analisada uma tabela de contingência que continha a frequência de scans
positivos registados durante o período de amostragem em cada quadrante para cada
uma das actividades consideradas. Não se utilizaram dados relativos ao repouso por
apresentarem uma frequência muito baixa.
Cecília V. Ferreira 18

MATERIAIS E MÉTODOS
2.3.2. Número de golfinhos em relação à actividade dominante aparente
A normalidade dos dados foi testada através do teste de Shapiro-Wilk. Uma
vez que os dados não seguem uma distribuição normal, foi realizado um teste não
paramétrico de Kruskal-Wallis para determinar a existência de diferenças
estatisticamente significativas no número mediano de golfinhos em relação à
actividade dominante em que os animais se encontravam envolvidos. Não se
utilizaram nesta análise os scans em que os animais foram registados como estando
em repouso, pela sua baixa frequência.
2.3.3. Influência do estado da maré e hora do dia no número de golfinhos
Uma vez que os dados não apresentam uma distribuição normal, foi aplicado
um teste não paramétrico de Kruskal-Wallis para determinar se existem diferenças
estatisticamente significativas no número mediano de indivíduos em relação ao estado
da maré bem como à hora do dia. Quando o resultado foi estatisticamente significativo,
aplicou-se um teste de comparações múltiplas de Dunn (Zar, 2009). Utilizou-se o
número mediano de golfinhos para comparabilidade com Mendes et al. (2002).
Utilizaram-se as 14 categorias adoptadas para o estado da maré (ver capítulo
2.2), tendo sido calculado o número mediano de golfinhos registado em cada estado
de forma a acompanhar a abundância de animais ao longo de meio ciclo de maré (de
uma baixa-mar à seguinte). Relativamente à hora do dia, os scans positivos foram
atribuídos a uma categoria horária, de duração igual a 1 hora, tendo sido depois
calculado o número mediano registado em cada hora.
2.3.4. Influência da fase da maré e da hora do dia no tipo de movimento e na actividade dominante aparente
Foram criadas tabelas de contingência que foram posteriormente analisadas
recorrendo ao programa ACTUS2.
Para determinar a influência da fase da maré no tipo de movimento e na
actividade dominante aparente, agruparam-se os scans positivos em quatro classes de
fase da maré: baixa-mar; enchente; preia-mar e vazante. A classe “baixa-mar” agrupou
as categorias BM-1, BM e BM+1; a classe “enchente” incluiu as categorias BM+2,
Cecília V. Ferreira 19

MATERIAIS E MÉTODOS
Cecília V. Ferreira 20
BM+3, BM+4 e BM+5; a classe “preia-mar” agrupou as categorias BM+6, PM e BM-6;
e por fim a classe “vazante” incluiu as categorias BM-5, BM-4, BM-3 e BM-2.
Para determinar a influência da hora do dia no tipo de movimento e na
actividade dominante aparente, os scans positivos registados ao longo do dia foram
agrupados nas seguintes classes: [8h – 12h[; [12h – 16h[; [16h – 20h].
2.3.5. Influência de variáveis ambientais no movimento e na actividade dos golfinhos
Realizou-se uma regressão logística, como descrito por Maroco (2010), para
analisar o efeito conjunto de diversas variáveis ambientais na direcção aparente de
deslocação dos golfinhos, codificada para cada categoria de direcção dominante. As
variáveis ambientais quasi-independentes incluíram hora do dia, estado da maré,
altura da maré e amplitude da maré.
A altura da maré foi calculada recorrendo a informação constante do website
do Serviço Hidrográfico e Oceanográfico da Marinha Francesa (SHOM) para o porto
de Setúbal (disponível no endereço http://www.shom.fr/ann_marees/cgi-
bin/predit_ext/choixp?opt=1&zone=1&port=SETUBAL).
A variável dependente – direcção dominante aparente – foi convertida para o
sistema binário, tendo sido atribuído 0 (zero) ao movimento de entrada no estuário
(UP) e 1 ao movimento de saída (DOWN). A categoria IND foi retirada da análise por
ser pouco representativa.
A regressão logística foi realizada no software PASW Statistics v.18.0.
Realizou-se também uma análise discriminante para identificar qual ou quais
das variáveis ambientais em estudo (hora do dia, estado da maré, altura da maré e
amplitude de maré) permitem diferençar significativamente as actividades dominantes
dos golfinhos (deslocação [D] e alimentação [A]).
Nesta análise, a variável dependente – actividade dominante aparente – foi
codificada da seguinte forma: D = 1; A = 2. Os dados relativos às actividades
Socialização e Repouso não foram utilizados por serem insuficientes para a realização
da análise.
Esta análise foi realizada no software PASW Statistics v.18.0.
RESULTADOS
3. Resultados
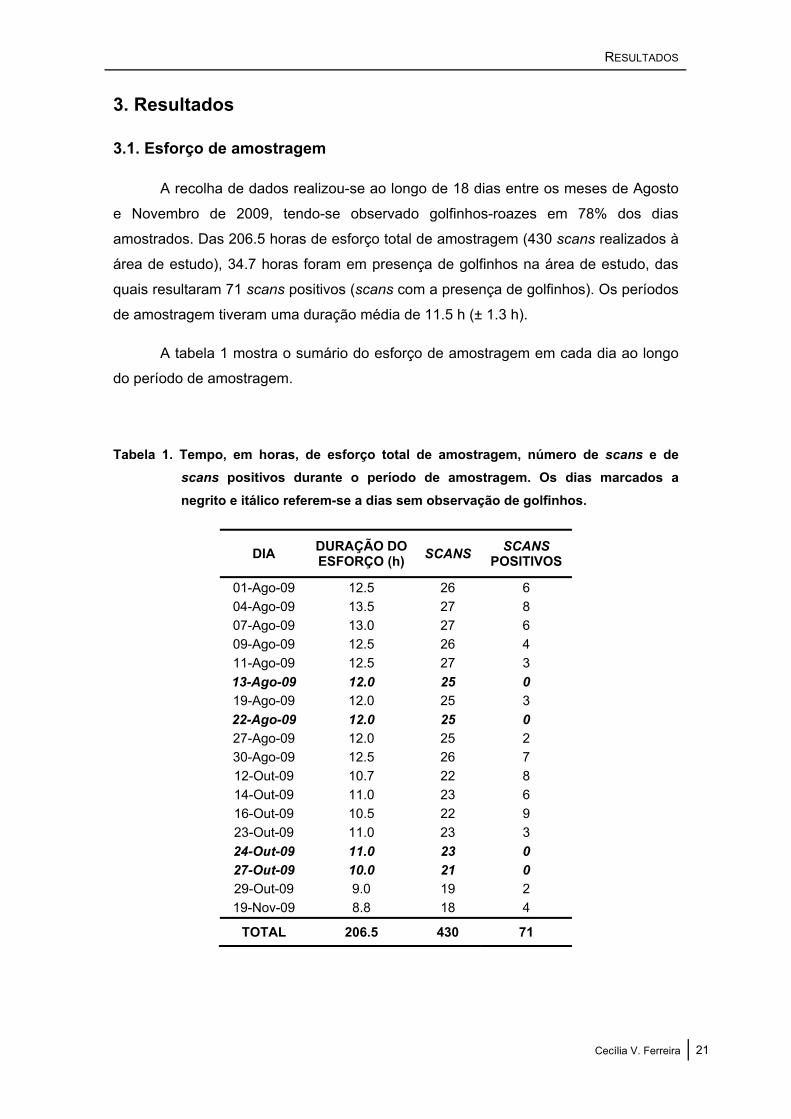
3.1. Esforço de amostragem
A recolha de dados realizou-se ao longo de 18 dias entre os meses de Agosto
e Novembro de 2009, tendo-se observado golfinhos-roazes em 78% dos dias
amostrados. Das 206.5 horas de esforço total de amostragem (430 scans realizados à
área de estudo), 34.7 horas foram em presença de golfinhos na área de estudo, das
quais resultaram 71 scans positivos (scans com a presença de golfinhos). Os períodos
de amostragem tiveram uma duração média de 11.5 h (± 1.3 h).
A tabela 1 mostra o sumário do esforço de amostragem em cada dia ao longo
do período de amostragem.
Tabela 1. Tempo, em horas, de esforço total de amostragem, número de scans e de scans positivos durante o período de amostragem. Os dias marcados a negrito e itálico referem-se a dias sem observação de golfinhos.
DIA DURAÇÃO DO ESFORÇO (h) SCANS SCANS
POSITIVOS
01-Ago-09 12.5 26 6 04-Ago-09 13.5 27 8 07-Ago-09 13.0 27 6 09-Ago-09 12.5 26 4 11-Ago-09 12.5 27 3 13-Ago-09 12.0 25 0 19-Ago-09 12.0 25 3 22-Ago-09 12.0 25 0 27-Ago-09 12.0 25 2 30-Ago-09 12.5 26 7 12-Out-09 10.7 22 8 14-Out-09 11.0 23 6 16-Out-09 10.5 22 9 23-Out-09 11.0 23 3 24-Out-09 11.0 23 0 27-Out-09 10.0 21 0 29-Out-09 9.0 19 2 19-Nov-09 8.8 18 4
TOTAL 206.5 430 71
Cecília V. Ferreira 21
RESULTADOS
Cecília V. Ferreira 22
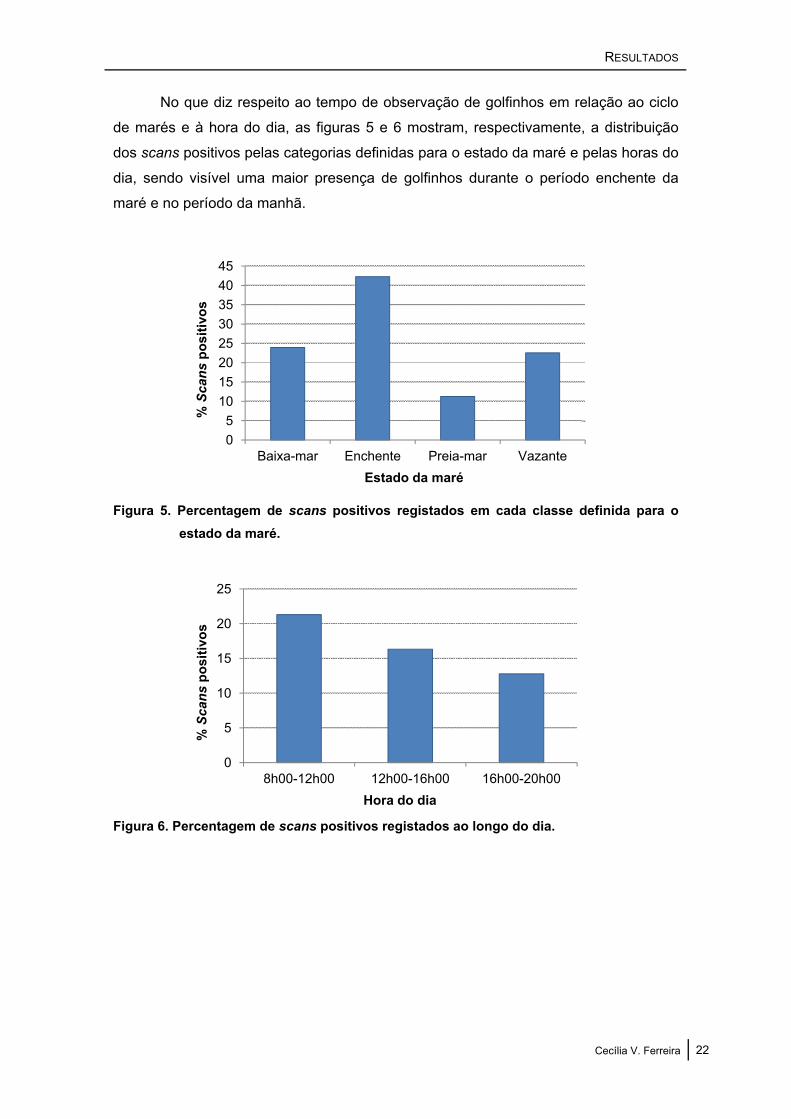
No que diz respeito ao tempo de observação de golfinhos em relação ao ciclo
de marés e à hora do dia, as figuras 5 e 6 mostram, respectivamente, a distribuição
dos scans positivos pelas categorias definidas para o estado da maré e pelas horas do
dia, sendo visível uma maior presença de golfinhos durante o período enchente da
maré e no período da manhã.
Figura 5. Percentagem de scans positivos registados em cada classe definida para o estado da maré.
Figura 6. Percentagem de scans positivos registados ao longo do dia.
05
1015202530354045
Baixa-mar Enchente Preia-mar Vazante
% S
cans
posi
tivos
Estado da maré
0
5
10
15
20
25
8h00-12h00 12h00-16h00 16h00-20h00
% S
cans
posi
tivos
Hora do dia

RESULTADOS
Cecília V. Ferreira 23
3.2. Distribuição de actividades
A figura 7 representa a percentagem de tempo empregue pelos animais em
cada uma das actividades consideradas durante o período de amostragem. Os
indivíduos despenderam a maioria do tempo em deslocação (75%), seguindo-se a
alimentação e a socialização. A categoria comportamental repouso foi apenas
observada em 3 ocasiões (durante a manhã, entre as 8h e as 10h), e por essa razão
não foi utilizada nas análises subsequentes.
Figura 7. Distribuição de actividades dos golfinhos-roazes durante o período de amostragem.
3.2.1. Número de golfinhos em relação à actividade dominante aparente
Realizou-se um teste de Kruskal-Wallis para avaliar a existência de diferenças
significativas no número mediano de golfinhos em relação à actividade em que os
indivíduos se encontravam envolvidos (figura 8). O teste não revelou diferenças
estatisticamente significativas (χKW2 (2) = 2.46; p > 0.05; n = 68).
75%
19%
6%
Deslocação
Alimentação
Socialização

RESULTADOS
Cecília V. Ferreira 24
Figura 8. Distribuição do número mediano de golfinhos em relação à actividade dominante aparente. As diferenças não foram estatisticamente significativas
(χKW2 (2) = 2.46; p > 0.05; n = 68). O marcador azul representa a mediana, e os
extremos inferior e superior da caixa representam, respectivamente, o 1º e o 3º Quartil. As barras inferiores e superiores representam os valores mínimo e máximo, respectivamente, não se observando outliers.
3.3. Utilização do habitat
Os quadrantes mais utilizados pelos golfinhos-roazes foram os quadrantes
E2S, C1 e B2 (E2S: 29.4%, C1 e B2: 16.2%), não tendo sido avistados animais nos
quadrantes A1 e A2 durante o período de amostragem. Nos quadrantes E1 e E2N, que
correspondem essencialmente ao canal Norte do estuário, foram registados golfinhos
em apenas 3% do total de scans positivos (tabela 2).
D A S
Actividade dominante aparente
0
2
4
6
8
10
12
Núm
ero
med
iano
de
golfi
nhos
Deslocação Alimentação Socialização

RESULTADOS
Tabela 2. Percentagem de scans positivos registados em cada quadrante.
QUADRANTE % SCANS POSITIVOS
A1 0 A2 0 B1 11.8 B2 16.2 C1 16.2 C2 7.4 D1 5.9 D2 10.3 E1 1.5
E2N 1.5 E2S 29.4
Todos os quadrantes da área de estudo foram utilizados durante a deslocação
(excepto os quadrantes A1 e A2), e foi nos quadrantes D1, D2 e E2S que se
registaram as maiores frequências de alimentação; a socialização foi apenas registada
na zona da foz do estuário, junto à baliza nº 5 (figura 9). A análise realizada no
ACTUS2 não mostrou diferenças estatisticamente significativas (embora próximo
disso) na distribuição das actividades pelos vários quadrantes (χ2 = 25.57; df = 20;
p = 0.058), apesar da frequência de alimentação observada no quadrante E2S se ter
revelado significativamente superior ao esperado (tabela 3).
Cecília V. Ferreira 25
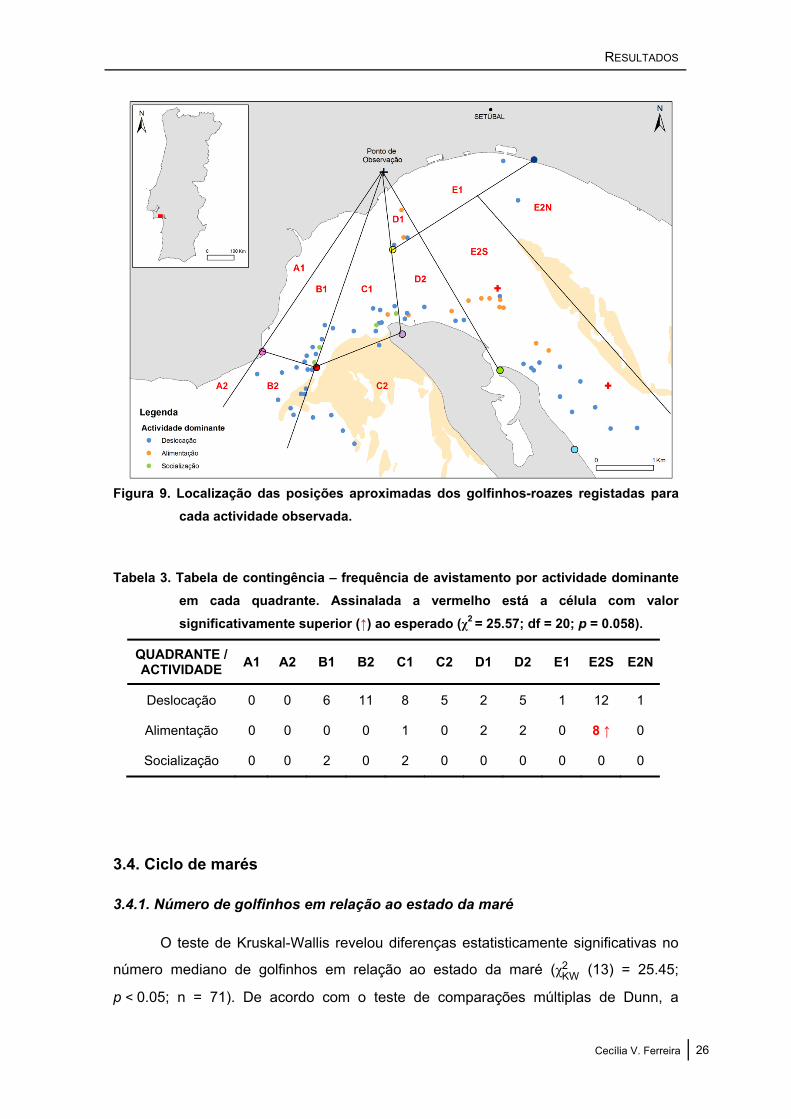
RESULTADOS
Figura 9. Localização das posições aproximadas dos golfinhos-roazes registadas para cada actividade observada.
Tabela 3. Tabela de contingência – frequência de avistamento por actividade dominante em cada quadrante. Assinalada a vermelho está a célula com valor significativamente superior (↑) ao esperado (χ2 = 25.57; df = 20; p = 0.058).
QUADRANTE / ACTIVIDADE A1 A2 B1 B2 C1 C2 D1 D2 E1 E2S E2N
Deslocação 0 0 6 11 8 5 2 5 1 12 1
Alimentação 0 0 0 0 1 0 2 2 0 8 ↑ 0
Socialização 0 0 2 0 2 0 0 0 0 0 0
3.4. Ciclo de marés
3.4.1. Número de golfinhos em relação ao estado da maré
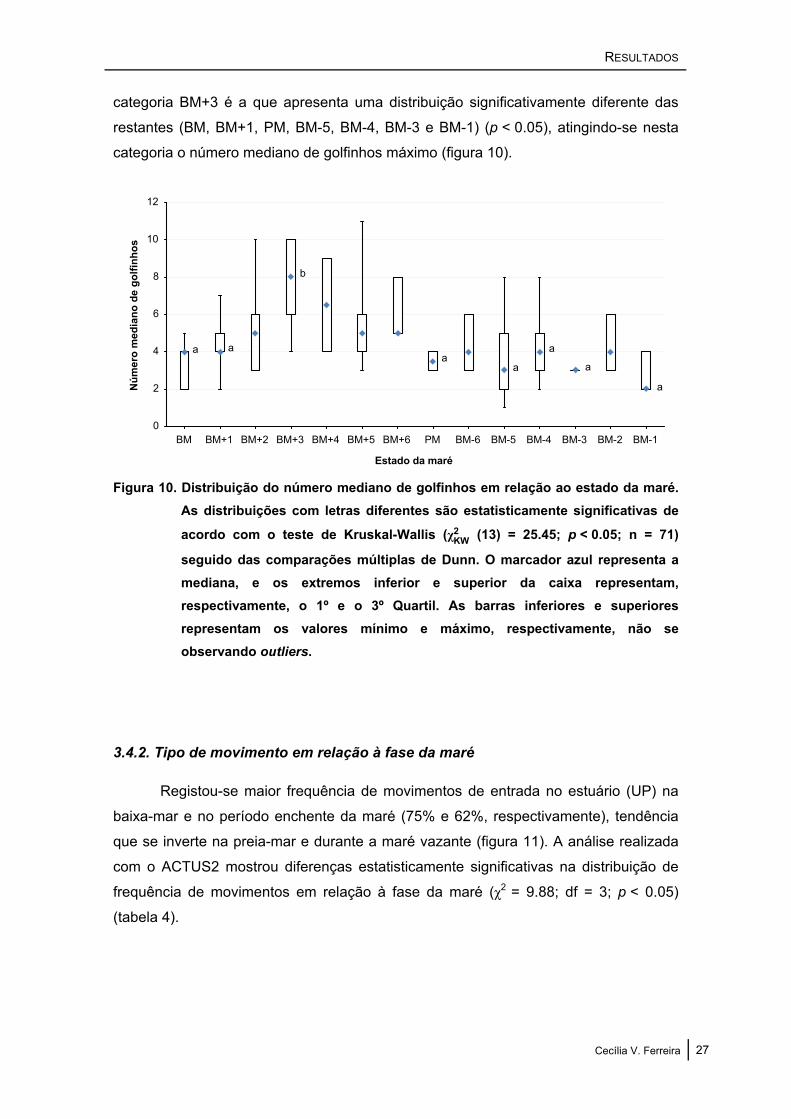
O teste de Kruskal-Wallis revelou diferenças estatisticamente significativas no
número mediano de golfinhos em relação ao estado da maré (χKW2 (13) = 25.45;
p < 0.05; n = 71). De acordo com o teste de comparações múltiplas de Dunn, a
Cecília V. Ferreira 26
RESULTADOS
Cecília V. Ferreira 27
categoria BM+3 é a que apresenta uma distribuição significativamente diferente das
restantes (BM, BM+1, PM, BM-5, BM-4, BM-3 e BM-1) (p < 0.05), atingindo-se nesta
categoria o número mediano de golfinhos máximo (figura 10).
Figura 10. Distribuição do número mediano de golfinhos em relação ao estado da maré. As distribuições com letras diferentes são estatisticamente significativas de
acordo com o teste de Kruskal-Wallis (χKW2 (13) = 25.45; p < 0.05; n = 71)
seguido das comparações múltiplas de Dunn. O marcador azul representa a mediana, e os extremos inferior e superior da caixa representam, respectivamente, o 1º e o 3º Quartil. As barras inferiores e superiores representam os valores mínimo e máximo, respectivamente, não se observando outliers.
3.4.2. Tipo de movimento em relação à fase da maré
Registou-se maior frequência de movimentos de entrada no estuário (UP) na
baixa-mar e no período enchente da maré (75% e 62%, respectivamente), tendência
que se inverte na preia-mar e durante a maré vazante (figura 11). A análise realizada
com o ACTUS2 mostrou diferenças estatisticamente significativas na distribuição de
frequência de movimentos em relação à fase da maré (χ2 = 9.88; df = 3; p < 0.05)
(tabela 4).
BM BM+1 BM+2 BM+3 BM+4 BM+5 BM+6 PM BM-6 BM-5 BM-4 BM-3 BM-2 BM-1
Estado da maré
0
2
4
6
8
10
12
Núm
ero
med
iano
de
golfi
nhos
b
a a
a
a
a
aa

RESULTADOS
Cecília V. Ferreira 28
Figura 11. Percentagem de scans positivos registados durante as quatro classes da fase da maré em relação aos dois tipos de movimento registados – UP e DOWN.
Tabela 4. Tabela de contingência – frequência de scans positivos por tipo de movimento em cada fase da maré. A análise desta tabela de contingência revela que a hipótese de independência entre linhas e colunas deve ser rejeitada (χ2 = 9.88; df = 3; p < 0.05).
TIPO DE MOVIMENTO / FASE DA MARÉ UP DOWN
Baixa-mar 12 4
Enchente 18 11
Preia-mar 3 5
Vazante 4 12
3.4.3. Actividade dominante aparente em relação à fase da maré
A figura 12 mostra que os golfinhos exibiram comportamentos de alimentação
mais frequentemente durante a baixa-mar e a preia-mar (cerca de 30% e 40%
respectivamente), tendo socializado ligeiramente mais durante o período enchente da
maré (cerca de 11%). Estas diferenças não foram, no entanto, significativas (χ2 = 9.10;
df = 6; p > 0.05).
0,0
20,0
40,0
60,0
80,0
100,0
Baixa-mar Enchente Preia-mar Vazante
% S
cans
posi
tivos
Fase da maré
UP
DOWN

RESULTADOS
Cecília V. Ferreira 29
Figura 12. Percentagem de scans positivos registados durante as quatro classes da fase da maré consideradas em relação à actividade dominante.
3.5. Hora do dia
3.5.1. Número de golfinhos em relação à hora do dia
O teste de Kruskal-Wallis não mostrou diferenças estatisticamente
significativas no número mediano de golfinhos em relação à hora do dia
(χKW2 (11) = 11.74; p > 0.05; n = 71) (figura 13). O número mediano de golfinhos
registado foi 4, não sendo assinalável um padrão de abundância ao longo do dia.
0,0
20,0
40,0
60,0
80,0
100,0
Baixa-mar Enchente Preia-mar Vazante
% S
cans
posi
tivos
Fase da maré
Deslocação
Alimentação
Socialização

RESULTADOS
Cecília V. Ferreira 30
Figura 13. Distribuição do número mediano de golfinhos em relação à hora do dia. As
diferenças não foram estatisticamente significativas (χKW2 (11) = 11.74;
p > 0.05; n = 71) O marcador azul representa a mediana, e os extremos inferior e superior da caixa representam, respectivamente, o 1º e o 3º Quartil. As barras inferiores e superiores representam os valores mínimo e máximo, respectivamente, não se observando outliers.
3.5.2. Tipo de movimento em relação à hora do dia
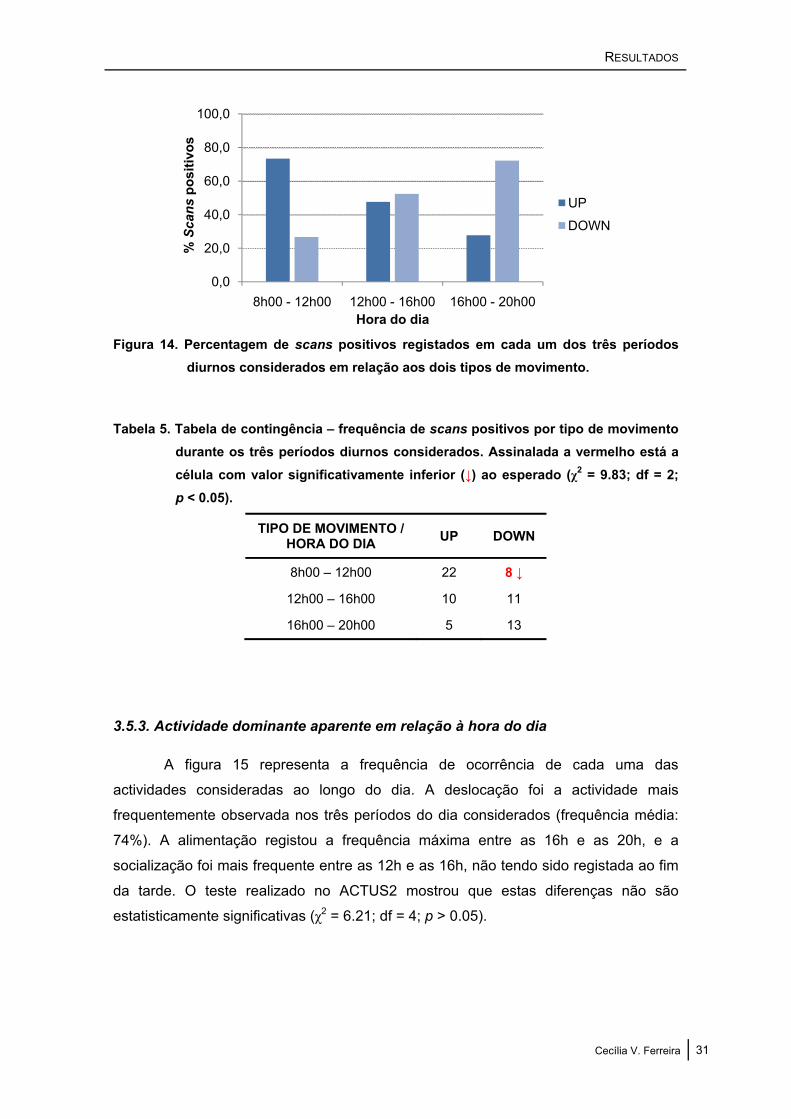
A figura 14 representa a frequência do tipo de movimento registado em cada
período diário considerado. Os animais foram observados a entrar no estuário (UP)
com maior frequência entre as 8h00 e as 12h00 (cerca de 70%), tendo-se observado o
movimento inverso entre as 16h00 e as 20h00 (DOWN: cerca de 70%). A análise no
ACTUS2 revelou que estas diferenças são estatisticamente significativas (χ2 = 9.83;
df = 2; p < 0.05) (tabela 5).
8h -
9h
9h -
10h
10h
- 11h
11h
- 12h
12h
- 13h
13h
- 14h
14h
- 15h
15h
- 16h
16h
- 17h
17h
- 18h
18h
- 19h
19h
- 20h
Hora do dia
0
2
4
6
8
10
12
Núm
ero
med
iano
de
golfi
nhos
RESULTADOS
Cecília V. Ferreira 31
Figura 14. Percentagem de scans positivos registados em cada um dos três períodos diurnos considerados em relação aos dois tipos de movimento.
Tabela 5. Tabela de contingência – frequência de scans positivos por tipo de movimento durante os três períodos diurnos considerados. Assinalada a vermelho está a célula com valor significativamente inferior (↓) ao esperado (χ2 = 9.83; df = 2; p < 0.05).
TIPO DE MOVIMENTO / HORA DO DIA UP DOWN
8h00 – 12h00 22 8 ↓
12h00 – 16h00 10 11
16h00 – 20h00 5 13
3.5.3. Actividade dominante aparente em relação à hora do dia
A figura 15 representa a frequência de ocorrência de cada uma das
actividades consideradas ao longo do dia. A deslocação foi a actividade mais
frequentemente observada nos três períodos do dia considerados (frequência média:
74%). A alimentação registou a frequência máxima entre as 16h e as 20h, e a
socialização foi mais frequente entre as 12h e as 16h, não tendo sido registada ao fim
da tarde. O teste realizado no ACTUS2 mostrou que estas diferenças não são
estatisticamente significativas (χ2 = 6.21; df = 4; p > 0.05).
0,0
20,0
40,0
60,0
80,0
100,0
8h00 - 12h00 12h00 - 16h00 16h00 - 20h00
% S
cans
posi
tivos
Hora do dia
UP
DOWN

RESULTADOS
Cecília V. Ferreira 32
Figura 15. Percentagem de scans positivos registados durante os três períodos diurnos considerados em relação à actividade dominante registada.
3.6. Tipo de movimento em relação à fase da maré por hora do dia
A figura 16 representa o resumo do tipo de movimento dos golfinhos na área
de estudo em função da hora do dia e da fase da maré. Destaca-se o movimento de
entrada a favor da maré (UP/enchente) durante os períodos da manhã (8h – 12h) e
meio-dia (12h – 16h), invertendo-se este padrão no período da tarde (16h – 20h),
período em que os animais são avistados mais frequentemente a sair do estuário
novamente a favor da maré (DOWN/vazante).
Figura 16. Percentagem de scans positivos registados para cada tipo de movimento em função da hora do dia e da fase da maré.
0,0
20,0
40,0
60,0
80,0
100,0
8h - 12h 12h - 16h 16h - 20h
% S
cans
posi
tivos
Hora do dia
Deslocação
Alimentação
Socialização
0,0
10,0
20,0
30,0
40,0
50,0
60,0
70,0
Enchente Vazante Enchente Vazante Enchente Vazante
8h - 12h 12h - 16h 16h - 20h
% S
cans
posi
tivos
Fase da maré / hora do dia
UP
DOWN

RESULTADOS
3.7. Influência de variáveis ambientais no movimento e actividade dos golfinhos
A regressão logística revelou que o estado da maré (χWald2 (1) = 3.378;
p = 0.066) e a amplitude da maré (χWald2 (1) = 0.343; p = 0.558) não apresentaram um
efeito estatisticamente significativo sobre o Logit da probabilidade de saída do
estuário. Em contraste, as variáveis hora do dia (βHora do dia = 0.323; χWald2 (1) = 9.814;
p = 0.002; OR = 1.381) e altura da maré (βAltura da maré = 1.523; χWald2 (1) = 8.833;
p = 0.003; OR = 4.585) apresentaram um efeito estatisticamente significativo. Da
análise do odds ratio conclui-se que a maior influência provém da altura da maré:
quando a maré sobe 1 m, a probabilidade dos golfinhos saírem do estuário aumenta
4.5 vezes. A tabela 6 resume os coeficientes de regressão logística, bem como a sua
significância no modelo.
Tabela 6. Coeficientes Logit do modelo de regressão logística da variável “Tipo de movimento” em função da Hora do dia, Altura da maré, Estado da maré e Amplitude da maré.
Parâmetro df β χWald2 p-value Odds ratio (OR)
Hora do dia 1 0.323 9.814 0.002 1.381
Altura da maré 1 1.523 8.833 0.003 4.585
Estado da maré 1 -0.185 3.378 0.066 0.831
Amplitude da maré 1 -0.333 0.343 0.558 0.717
Constante 1 -0.668 11.664 0.001 0.001
Cecília V. Ferreira 33
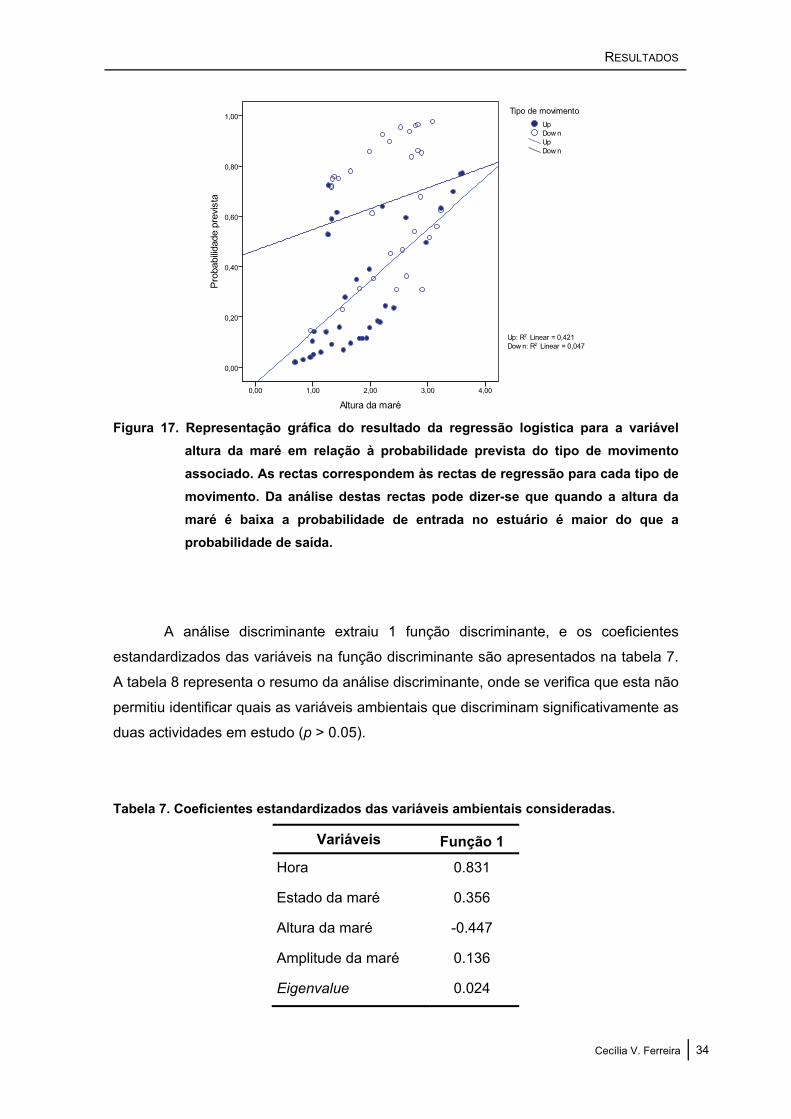
RESULTADOS
Figura 17. Representação gráfica do resultado da regressão logística para a variável altura da maré em relação à probabilidade prevista do tipo de movimento associado. As rectas correspondem às rectas de regressão para cada tipo de movimento. Da análise destas rectas pode dizer-se que quando a altura da maré é baixa a probabilidade de entrada no estuário é maior do que a probabilidade de saída.
A análise discriminante extraiu 1 função discriminante, e os coeficientes
estandardizados das variáveis na função discriminante são apresentados na tabela 7.
A tabela 8 representa o resumo da análise discriminante, onde se verifica que esta não
permitiu identificar quais as variáveis ambientais que discriminam significativamente as
duas actividades em estudo (p > 0.05).
Tabela 7. Coeficientes estandardizados das variáveis ambientais consideradas.
Variáveis Função 1
Hora 0.831
Estado da maré 0.356
Altura da maré -0.447
Amplitude da maré 0.136
Eigenvalue 0.024
Cecília V. Ferreira 34

RESULTADOS
Cecília V. Ferreira 35
Tabela 8. Tabela-resumo da análise discriminante.
Função Lambda de Wilks χ2 df p-value
1 0.977 1.373 4 0.849

DISCUSSÃO
4. Discussão
As observações realizadas a partir de terra permitem monitorizar os
movimentos e comportamento dos animais e têm como principais vantagens não
interferir com o comportamento dos animais (Würsig & Jefferson, 1990; Berrow et al.,
1996) e permitir um campo de visão alargado sobre a área de estudo (Berrow et al.,
1996), apresentando, por outro lado, custos de execução relativamente baixos (Würsig
& Jefferson, 1990) quando comparados com os da observação a partir de uma
embarcação. O principal inconveniente deste tipo de observação prende-se com a
distância do observador ao objecto de estudo. Esta distância deve ser tida em
consideração quando se determina a espécie, o número de indivíduos presente na
área e o padrão comportamental apresentado (Elwen et al., 2009), uma vez que pode
conduzir a determinações incorrectas destas variáveis; neste tipo de observação é
quase impossível o reconhecimento individual dos animais devido à distância entre o
observador e o animal (Würsig & Jefferson, 1990).
Os resultados deste trabalho mostram que o estuário do Sado é um importante
habitat para a comunidade de golfinhos-roazes. Durante o período de amostragem,
estes animais foram avistados em 78% dos dias amostrados, frequência que traduz
assim o carácter residente desta comunidade, que utiliza o estuário quase
diariamente. É preciso admitir que nos 22% dos dias remanescentes poderá ter
ocorrido alguma presença não detectada, por exemplo no período nocturno.
4.1. Utilização do habitat
Detectaram-se alguns padrões de utilização da área de estudo em relação às
diversas actividades observadas, apesar do teste estatístico realizado não ter
permitido encontrar diferenças estatisticamente significativas. Estes padrões vão de
encontro ao obtido noutros estudos de utilização deste habitat (Freitas, 1995; dos
Santos, 1998; Harzen, 1998; Nunes, 2001; Augusto, 2007).
De um modo geral, observou-se uma preferência pelo canal Sul e zona da foz
do estuário, enquanto no canal Norte foram observados golfinhos em apenas 3% dos
scans realizados, tratando-se estas observações de animais em deslocação. Apesar
da área de estudo deste trabalho não incluir a totalidade do estuário, este evitamento
do canal Norte foi já evidenciado em vários estudos realizados anteriormente (Harzen,
1998; Nunes, 2001; Cândido, 2003; Augusto, 2007).
Cecília V. Ferreira 36

DISCUSSÃO
O canal Norte, junto à cidade de Setúbal, sofre o impacto directo de diversas
actividades humanas que se estabeleceram nessa margem: é este o canal de acesso
ao porto de Setúbal, bem como aos portos de pesca e de recreio, apresentando assim
níveis bastante elevados de poluição acústica (Ferreira et al., 1996; dos Santos, 1998;
Luís, 2008), para além da poluição química da água decorrente da existência de
efluentes domésticos e industriais que descarregam neste canal. Apesar disso, este
canal apresenta uma elevada abundância de espécies ícticas (Lopes da Cunha, 1994),
nomeadamente as tainhas (uma das presas confirmadas destes golfinhos; dos Santos
et al., 2007), o que poderá explicar as pontuais incursões dos golfinhos a esta zona
para se alimentarem destes peixes. Outros estudos realizados apontam para uma
situação semelhante em anos recentes, com Augusto (2007) a detectar a presença de
menos grupos nas zonas correspondentes ao canal Norte do estuário.
Relativamente à restante área de estudo, a sua utilização foi também selectiva
no que diz respeito ao comportamento, tendo os golfinhos demonstrado uma
preferência pelo canal Sul para a alimentação; foram observados comportamentos de
socialização apenas na zona da foz do estuário.
4.2. Actividade dominante aparente
Os golfinhos residentes no estuário do Sado utilizam este habitat
extremamente produtivo nas suas rotinas diárias, nomeadamente para a alimentação.
Esta actividade foi a segunda mais frequentemente observada durante o período de
amostragem (19%), antecedida pela deslocação (75%). O número mediano de
golfinhos foi, no entanto, independente da actividade dominante, como foi também
observado por Augusto (2007).
A maior frequência de deslocação registada na área de estudo (75%) em
comparação com as restantes actividades pode ser explicada pelo facto da zona da
foz do estuário (a área de estudo) ser uma zona de ligação entre o estuário e a zona
marinha adjacente. Augusto (2007) registou um maior número de grupos em
deslocação em comparação com a alimentação e a socialização, o que está de acordo
com o cenário apresentado no presente estudo.
Como já foi referido, a actividade de alimentação foi essencialmente observada
no canal Sul do estuário; outros autores detectaram um padrão semelhante (Nunes,
2001; Augusto, 2007), sendo o canal Sul, e em particular a zona da Caldeira de Tróia,
Cecília V. Ferreira 37

DISCUSSÃO
reconhecido como uma zona preferencial de alimentação para estes golfinhos
(Cândido, 2003), sendo uma zona extremamente rica em macroinvertebrados e
espécies ictícas (Lopes da Cunha, 1994; Cabral, 1999). É de notar que esta frequência
de alimentação apenas inclui a percentagem de scans em que os golfinhos foram
observados a alimentar-se à superfície ou exibiram comportamentos associados à
alimentação. Na eventualidade de um golfinho realizar um mergulho profundo e longo
para procurar uma presa, esse comportamento poderá ser interpretado como
deslocação se não existirem outros comportamentos que indiquem que se trata
efectivamente de um episódio de alimentação, o que poderá conduzir a uma incorrecta
atribuição da actividade dominante em que o animal se encontra. Por outras palavras,
o valor de 19% reportado é, muito provavelmente, uma estimativa por defeito.
Os períodos de baixa-mar e preia-mar foram aqueles em que se registou maior
frequência de alimentação, apesar da análise com o ACTUS2 não ter revelado
diferenças estatisticamente significativas na actividade dos animais em relação à fase
da maré. Este resultado contradiz o observado por Hanson & Defran (1993) na sua
área de estudo na costa da Califórnia, EUA, em que os golfinhos-roazes foram
observados a alimentar-se com maior frequência em períodos de enchente e vazante,
em que as correntes são mais fortes, podendo esta estratégia estar relacionada com o
movimento das suas presas nesse local, sendo várias as espécies que acompanham
as correntes para se movimentarem (Gibson, 2003).
A socialização foi apenas observada na zona da foz do estuário e em 6% dos
scans realizados, e esta baixa frequência é semelhante à registada por Augusto
(2007), que apenas observou comportamentos de socialização em 8% das suas
amostras.
Durante o período de amostragem, foram observados animais em repouso em
apenas 3 scans positivos, e estes foram registados durante o período da manhã (entre
as 8h e as 10h) e apenas em dias de semana. Este padrão pode ser explicado pelo
facto de serem alturas em que o estuário apresenta um menor tráfego marítimo
(principalmente no respeitante a embarcações de recreio), factor passível de perturbar
os golfinhos durante o repouso. Estudos anteriores revelam um padrão semelhante,
com Harzen (1998) a detectar uma frequência muito baixa de repouso nas suas
observações (apenas 0.2%), ao passo que Nunes (2001) e Augusto (2007) não
observaram animais nesta categoria comportamental.
Cecília V. Ferreira 38

DISCUSSÃO
4.3. Movimento
A abundância e o movimento de espécies de cetáceos parecem ser
influenciados principalmente pela disponibilidade temporal e espacial de alimento, que
pode ser secundariamente influenciada por características oceanográficas (Davis
et al., 1998). Será razoável considerar que, tendo em conta as características do
estuário do Sado que o tornam num habitat tão rico do ponto de vista ecológico e tão
importante para os golfinhos-roazes nas suas rotinas diárias, os movimentos destes
animais estarão principalmente relacionados com a eficaz obtenção de alimento, para
além da poupança de energia subjacente a um movimento a favor das correntes, em
que os animais não despendem energia enquanto se movimentam entre áreas. Por
outro lado, as correntes de maré são fenómenos mais ou menos previsíveis no tempo
e no espaço, e os golfinhos poderão aproveitar-se desse facto para maximizarem o
seu esforço de movimentação, regulando assim os seus movimentos de acordo com
as correntes.
As análises estatísticas realizadas neste trabalho mostraram que, durante o
período de amostragem, os golfinhos foram observados mais frequentemente a
deslocar-se a favor da maré (entrada no estuário durante o período enchente da maré
e saída durante a vazante). O número mediano de golfinhos atingiu um máximo
durante o período de maré enchente, à semelhança do observado por Mendes et al.
(2002) em Moray Firth, Escócia. Neste local, os autores caracterizaram uma frente de
maré e detectaram que os golfinhos são mais abundantes durante o período de maré
enchente, distribuindo-se preferencialmente na região frontal da frente de maré,
possivelmente devido à distribuição ou comportamento das suas potenciais presas. De
facto, fenómenos hidrográficos como frentes de maré e eddies apresentam elevada
produtividade biológica (Kimura et al., 1997), e em vários locais do mundo os
golfinhos-roazes distribuem-se mais abundantemente na entrada de estuários e baías
onde existem fortes correntes (e.g. Irvine et al., 1981; Ballance, 1992; Wilson et al.,
1997). No caso do estuário do Sado, que apresenta na sua foz dois vórtices, é
possível que também ocorra uma influência deste tipo de fenómeno no
comportamento dos golfinhos, mas sem conhecer melhor estes fenómenos e a sua
relação com as várias espécies que lá ocorrem, nomeadamente as presas dos
golfinhos e os próprios golfinhos, não é possível tirar qualquer conclusão sobre a sua
importância para os animais. Por outro lado, os movimentos e a distribuição dos
animais podem também estar ligados à batimetria, tendo sido evidenciado em Moray
Firth, Escócia, que a maior ocorrência de comportamentos de alimentação em zonas
com gradientes acentuados pode dever-se, por um lado, à maior concentração de
Cecília V. Ferreira 39

DISCUSSÃO
presas nessas zonas, e por outro à maior facilidade de captura dessas mesmas
presas (Hastie et al., 2004). A foz do estuário do Sado apresenta um acentuado
declive (Martins et al., 2002) que poderá ser importante para os processos de
enriquecimento orgânico e recirculação desta zona de ligação entre o interior do
estuário e as águas costeiras adjacentes.
À semelhança do que se observa noutros locais, os movimentos de entrada e
saída dos golfinhos do estuário do Sado são influenciados principalmente pela maré,
como mostram os resultados da regressão logística realizada. Neste trabalho, os
golfinhos foram observados a entrar no estuário com a enchente em cerca de 65% dos
scans positivos, invertendo-se este padrão durante a vazante, quando os golfinhos
foram observados a abandonar a área de estudo com maior frequência. Este padrão
foi também observado em locais como Sarasota, Florida (Irvine et al., 1981), Cardigan
Bay, West Wales (Gregory & Rowden, 2001) e New South Wales, Austrália (Fury,
2008).
Os movimentos destes golfinhos no estuário do Sado poderão ser cruzados
com os movimentos de algumas das suas presas. As tainhas (família Mugilidae) foram
confirmadas como presas dos golfinhos neste local (dos Santos et al., 2007), sendo,
aliás, um conjunto de espécies universalmente capturadas pelos golfinhos-roazes (e.g.
Irvine et al,. 1981; Shane, 1990a; Díaz López, 2006). No estuário do rio Mira, onde
foram estudadas por Almeida (1996), as tainhas (Liza ramada) movimentaram-se
essencialmente a favor das marés. A verificar-se o mesmo cenário no estuário do
Sado, os movimentos de entrada com a enchente e saída com a vazante dos
golfinhos-roazes poderão estar relacionados com este movimento previsível das
tainhas, o que poderá maximizar a sua captura.
Apesar de menos acentuada, a influência da hora do dia no movimento dos
golfinhos não pode ser negligenciada. De facto, os animais foram avistados a entrar no
estuário com maior frequência durante a manhã (cerca de 70% dos scans positivos),
tendo abandonado a área de estudo ao final da tarde. Este padrão pode estar também
relacionado com o movimento das tainhas, espécie que no estuário do rio Mira
pareceu ter uma maior taxa de consumo de alimento durante o dia, reduzindo durante
a noite (Almeida, 2003). Nestes períodos de alimentação, as tainhas poderão ser mais
facilmente vistas pelos golfinhos devido ao seu movimento junto à superfície, o que
poderá facilitar a sua captura.
O número mediano de golfinhos não apresentou diferenças estatisticamente
significativas em relação à hora do dia. Esta comunidade apresenta um efectivo
Cecília V. Ferreira 40

DISCUSSÃO
populacional bastante reduzido (Augusto et al., in press), o que, aliado ao facto de este
se tratar de um habitat com uma grande abundância de espécies das quais os
golfinhos se alimentam, contribui para uma diminuição da competição entre indivíduos,
favorecendo assim a existência de grupos maiores e de abundância não variável ao
longo do tempo e nas várias actividades (Augusto, 2007).
Por outro lado, ficam por explicar as razões que levam os golfinhos a
abandonar a área de estudo ao final do dia. O estuário do Sado, que oferece
protecção contra potenciais predadores e que no período nocturno se apresenta como
um habitat calmo em termos de ruído subaquático, é assim preterido em relação às
águas costeiras, onde os animais permaneceram à noite, pelo menos aparentemente,
durante o período de amostragem. É relevante assinalar que não há registos de
utilização nocturna regular de estuários por golfinhos residentes, talvez devido às
dificuldades de observação neste período.
4.4. Limitações deste estudo
As observações realizadas a partir de terra para trabalhos desta natureza
podem ser aperfeiçoadas recorrendo ao uso de um teodolito, um equipamento que
permite obter posições geográficas dos animais ou grupos de animais (Würsig et al.,
1991). Neste trabalho não foi possível utilizar esse equipamento nem foi possível fazer
observações com cruzamento de azimutes por vários observadores, pelo que as
localizações aqui apresentadas deverão ser consideradas com alguma reserva, não
devendo ser utilizadas em estudos em que se pretenda estabelecer relações entre a
ocorrência de golfinhos e a distribuição fina de parâmetros ambientais, uma vez que
as localizações terão um erro associado que, no caso desses estudos, não pode ser
negligenciado. Não existem também informações relativas a utilização do habitat
durante o período nocturno nem foi possível recolher dados ao longo das estações do
ano para uma análise sazonal dos movimentos e actividades.
4.5. Sugestões para estudos futuros
Tendo em conta as limitações deste estudo apresentadas anteriormente,
sugere-se que em trabalhos futuros seja incorporado um teodolito nos métodos de
amostragem, de forma a permitir determinar, com maior exactidão, a localização dos
animais e assim abrir portas para novos estudos de utilização do habitat,
Cecília V. Ferreira 41

DISCUSSÃO
Cecília V. Ferreira 42
nomeadamente relacionados com movimentos e utilização do espaço em relação a
variáveis ambientais como a profundidade, o tipo de sedimento, a batimetria, a
temperatura da água e correntes, entre outras.
Sugere-se a continuação deste trabalho de forma a obter dados ao longo de
todo o ano, o que poderá permitir a distinção de padrões de movimento e actividade
em relação à estação do ano. Trata-se de uma variável ambiental que noutros locais
influenciou o comportamento (e.g. Bräger, 1993; Hanson & Defran, 1993) e a utilização
do habitat (Fury, 2008), pelo que seria interessante verificar se no estuário do Sado
ocorre uma influência semelhante e procurar explicar as razões para tal relação. De
forma a melhor compreender os movimentos dos golfinhos no estuário do Sado, seria
muito útil obter informação detalhada sobre os padrões de distribuição e movimento
das suas presas (confirmadas e potenciais) e as suas migrações em relação ao ciclo
de marés. Esta informação poderia então ser cruzada com a dos movimentos dos
golfinhos, permitindo assim a comparação entre as distribuições das presas e dos
seus predadores.
Este tipo de investigação poderá também beneficiar imensamente se for
complementado por metodologias de monitorização acústica passiva, que poderiam
permitir ultrapassar as óbvias limitações causadas pelos períodos nocturno ou de
baixa visibilidade, expandindo no tempo e no espaço a detecção de golfinhos nesta
zona de entrada e saída do estuário.

BIBLIOGRAFIA
5. Bibliografia
Acevedo, A. (1991). Behaviour and movements of bottlenose dolphins, Tursiops
truncatus, in the entrance to Ensenada De La Paz, Mexico. Aquatic Mammals
17.3: 137-147.
Almada, V. C. & Oliveira, R. F. (1997). Sobre o uso de estatística de simulação em
estudos de comportamento. Análise Psicológica 15: 97–109.
Almeida, P. R. (1996). Estuarine movement patterns of adult thin-lipped grey mullet,
Liza ramada (Risso) (Pisces, Mugilidae), observed by ultrasonic tracking. Journal
of Experimental Marine Biology and Ecology 202: 137–150.
Almeida, P. R. (2003). Feeding ecology of Liza ramada (Risso, 1810) (Pisces,
Mugilidae) in a south-western estuary of Portugal. Estuarine, Coastal and Shelf
Science 57: 313-323.
Augusto, J. F., Lopes, P. R. & dos Santos, M. E. (in press). Social structure of the
declining resident community of common bottlenose dolphins in the Sado Estuary,
Portugal. Journal of the Marine Biological Association of the United Kingdom.
Augusto, J. F. F. (2007). Análise da estrutura social, da composição dos grupos e
associações nos golfinhos-roazes (Tursiops truncatus) residentes na região do
Sado. Dissertação de Mestrado em Ecologia Marinha, Faculdade de Ciências da
Universidade de Lisboa. 60 pp.
Ballance, L. T. (1992). Habitat Use Patterns and Ranges of the Bottlenose Dolphin in
the Gulf of California, Mexico. Marine Mammal Science 8(3): 262-274.
Bearzi, G., Politi, E. & Notarbartolo Di Sciara, G. (1999). Diurnal behavior of free-
ranging bottlenose dolphins in the Kvarneric (Northern Adriatic Sea). Marine
Mammal Science 15(4): 1065–1097.
Berrow, S. D., Holmes, B. & Kiely, O. R. (1996). Distribution and abundance of bottle-
nosed dolphins Tursiops truncatus (Montagu) in the Shannon estuary. Biology and
Environment: Proceedings of the Royal Irish Academy 96B(1): 1-9.
Brito, C. (2001). Estudo da possível influência de factores ecológicos e
comportamentais nas emissões acústicas dos Golfinhos-roazes Tursiops
Cecília V. Ferreira 43

BIBLIOGRAFIA
truncatus no estuário do Sado. Dissertação de Mestrado em Etologia, Instituto
Superior de Psicologia Aplicada. 94 pp.
Bräger, S. (1993). Diurnal and seasonal behavior patterns of bottlenose dolphins
(Tursiops truncatus). Marine Mammal Science 9(4): 434-438.
Cabral, H. N. (2000). Distribution and Abundance Patterns of Flatfishes in the Sado
Estuary, Portugal. Estuaries 23(3): 351-358.
Cabral, H. N. (1999). Ictiofauna do Estuário do Sado. Relatórios Científicos e Técnicos
Instituto de Investigação das Pescas e do Mar 47: 1-30.
Carvalho, I. (2000). Observação e análise dos padrões de comportamento dos
golfinhos-roazes, Tursiops truncatus (Montagu, 1821), no estuário do Sado.
Relatório de estágio da Licenciatura, Faculdade de Ciências da Universidade de
Lisboa. 46 pp.
Cascão, I. (2001). Measuring the impacts resulting from interactions between
approaching boats and resident bottlenose dolphins (Tursiops truncatus), in the
Sado estuary, Portugal. Relatório de estágio da Licenciatura, Faculdade de
Ciências da Universidade de Lisboa.
Connor, R. C., Wells, R. S., Mann, J. & Read, A. J. (2000). The Bottlenose Dolphin –
Social Relationships in a Fission-Fusion Society. In Cetacean Societies: Field
Studies of Whales and Dolphins. (eds J. Mann, R.C. Connor, P.L. Tyack & H.
Whitehead), pp. 91-126. The University of Chicago Press, Chicago, USA.
Constantine, R., Brunton, D. H. & Dennis, T. (2004). Dolphin-watching tour boats
change bottlenose dolphin (Tursiops truncatus) behaviour. Biological
Conservation 117: 299-307.
Couchinho, M. N. (1999). Padrões de emissão de assobios por golfinhos-roazes
(Tursiops truncatus) na região do Sado. Relatório de estágio da Licenciatura,
Faculdade de Ciências da Universidade de Lisboa. 50 pp.
Cremer, M. J., Simões-Lopes, P. C. & Pires, J. S. R. (2009). Occupation pattern of a
harbor inlet by the estuarine dolphin, Sotalia guianensis (P. J. Van Bénéden,
1864) (Cetacea, Delphinidae). Brazilian Archives of Biology and Technology
52(3): 765-774.
Cecília V. Ferreira 44

BIBLIOGRAFIA
Cândido, A. T. M. B. (2003). Utilização de um Sistema de Informação Geográfica (SIG)
para a Análise da Distribuição e Padrões de Movimentos da População de
Golfinhos-Roazes (Tursiops truncatus) do Estuário do Sado. Dissertação de
Mestrado em Etologia, Instituto Superior de Psicologia Aplicada. 64 pp.
Daura-Jorge, F. G., Wedekin, L. L., Piacentini, V. D. Q. & Simões-Lopes, P. C. (2005).
Seasonal and daily patterns of group size, cohesion and activity of the estuarine
dolphin, Sotalia guianensis (P.J. van Bénéden) (Cetacea, Delphinidae), in
southern Brazil. Revista Brasileira de Zoologia 22(4): 1014-1021.
Davis, R. W., Fargion, G. S., May, N., Leming, T. D., Baumgartner, M., Evans, W. E.,
Hansen, L. J. & Mullin, K. (1998). Physical habitat of cetaceans along the
continental slope in the north-central and western Gulf of Mexico. Marine Mammal
Science 14(3): 490-507.
Díaz López, B. (2006). Bottlenose Dolphin (Tursiops truncatus) Predation on a Marine
Fin Fish Farm: Some Underwater Observations. Aquatic Mammals 32(3): 305-310.
dos Santos, M. E. (1985). Estudo preliminar da população sedentária de roazes,
Tursiops truncatus (Montagu, 1891) (Cetacea Delphinidae) no Estuário do Sado.
Relatório de estágio da Licenciatura, Faculdade de Ciências da Universidade de
Lisboa. 185 pp.
dos Santos, M. E. (1998). Golfinhos-Roazes do Sado: Estudos de Sons e
Comportamento. Colecção Teses 4, Instituto Superior de Psicologia Aplicada,
Lisboa. 279 pp.
dos Santos, M. E., Coniglione, C. & Louro, S. (2007). Feeding behaviour of the
bottlenose dolphin, Tursiops truncatus (Montagu, 1821) in the Sado estuary,
Portugal, and a review of its prey species. Revista Brasileira de Zoociências 9(1): 31-39.
dos Santos, M. E. & Lacerda, M. (1987). Preliminary observations of the bottlenose
dolphin (Tursiops truncatus) in the Sado estuary (Portugal). Aquatic Mammals
13.2: 65-80.
dos Santos, M. E., Louro, S., Couchinho, M. & Brito, C. (2005). Whistles of Bottlenose
Dolphins (Tursiops truncatus) in the Sado Estuary, Portugal: Characteristics,
Cecília V. Ferreira 45

BIBLIOGRAFIA
Production Rates, and Long-Term Contour Stability. Aquatic Mammals 31.4:
453-462.
Elwen, S. H., Best, P. B., Reeb, D. & Thornton, M. (2009). Diurnal Movements and
Behaviour of Heaviside’s Dolphins, Cephalorhynchus heavisidii, with some
Comparative Data for Dusky Dolphins, Lagenorhynchus obscurus. South African
Journal of Wildlife Research 39(2): 143-154.
Elwen, S., Meÿer, M. A., Best, P. B., Kotze, P. G. H., Thornton, M. & Swanson, S.
(2006). Range and movements of female Heaviside’s dolphins (Cephalorhynchus
heavisidii), as determined by satellite-linked telemetry. Journal of Mammalogy
87(5): 866-877.
Estabrook, C. B. & Estabrook, G. F. (1989). ACTUS: a solution to the problem of small
samples in the analysis of two-way contingency tables. Historical Methods 22: 5-8.
Ferreira, A., Bento-Coelho, J. L. & dos Santos, M. E. (1996). Underwater noise in the
Sado estuary. Acustica – acta acustica 82: s225.
Flach, L., Flach, P. A. & Chiarello, A. G. (2008). Aspects of behavioral ecology of
Sotalia guianensis in Sepetiba Bay, southeast Brazil. Marine Mammal Science
24(3): 503-515.
Freitas, A. (1995). Estudo preliminar dos padrões de utilização do habitat pela
população de golfinhos-roazes (Tursiops truncatus) no estuário do Sado.
Relatório de estágio de Licenciatura, Faculdade de Ciências da Universidade de
Lisboa. 39 pp.
Fury, C. A. (2008). Abundance, behaviour and habitat use patterns of Indo-Pacific
bottlenose dolphins (Tursiops aduncus) in the Clarence and Richmond River
estuaries in northern New South Wales, Australia. Tese de Doutoramento,
Southern Cross University, Australia. 212 pp.
Gaspar, R. (2003). Status of the resident bottlenose dolphin population in the Sado
estuary: past, present and future. Tese de Doutoramento, University of St.
Andrews, Scotland. 194 pp.
Gibson, R. N. (2003). Go with the flow: tidal migration in marine animals. Hydrobiologia
503: 153-161.
Cecília V. Ferreira 46

BIBLIOGRAFIA
Goodwin, L. (2008). Diurnal and Tidal Variations in Habitat Use of the Harbour
Porpoise (Phocoena phocoena) in Southwest Britain. Aquatic Mammals 34(1): 44-53.
Gregory, P. R. & Rowden, A. A. (2001). Behaviour patterns of bottlenose dolphins
(Tursiops truncatus) relative to tidal state, time-of-day and boat traffic in Cardigan
Bay, West Wales. Aquatic Mammals 27(2): 105-113.
Hammond, P. S., Bearzi, G., Bjørge, A., Forney, K., Karczmarski, L., Kasuya, T.,
Perrin, W. F., Scott, M. D., Wang, J. Y., Wells, R. S. & Wilson, B. (2008). Tursiops
truncatus. IUCN 2010, IUCN Red List of Threatened Species - Version 2010.1.
Hansen, L. J. (1990). California Coastal Bottlenose Dolphins. The Bottlenose Dolphin.
(eds S. Leatherwood & R.R. Reeves), pp. 403-420. Academic Press, San Diego.
Hanson, M. T. & Defran, R. H. (1993). The behavior and feeding ecology of the Pacific
coast bottlenose dolphin, Tursiops truncatus. Aquatic Mammals 19.3: 127-142.
Harzen, S. (1995). Behaviour and Social Ecology of the Bottlenose Dolphin, Tursiops
truncatus, in the Sado Estuary, Portugal. Tese de Doutoramento, University of
Bielefeld, Germany. 259 pp.
Harzen, S. (1998). Habitat use by the bottlenose dolphin (Tursiops truncatus) in the
Sado estuary, Portugal. Aquatic Mammals 24.3: 117-128.
Harzen, S. (2002). Use of an electronic theodolite in the study of movements of the
bottlenose dolphin (Tursiops truncatus) in the Sado Estuary, Portugal. Aquatic
Mammals 28.3: 251-260.
Harzen, S. & Brunnick, B. J. (1997). Skin disorders in bottlenose dolphins (Tursiops
truncatus), resident in the Sado estuary, Portugal. Aquatic Mammals 23.1: 59-68.
Hastie, G. D., Wilson, B., Wilson, L. J., Parsons, K. M. & Thompson, P. M. (2004).
Functional mechanisms underlying cetacean distribution patterns: hotspots for
bottlenose dolphins are linked to foraging. Marine Biology 144: 397-403.
Instituto Hidrográfico (2009). Marés. Glossário científico.
Cecília V. Ferreira 47

BIBLIOGRAFIA
Hussenot, E. (1982). Premier recensement du grand dauphin (Tursiops truncatus)
dans l’estuaire du Sado au Portugal. Actes du VI Colloque de la Société Française
pour l’Etude et la Protection des Mammifères. pp. 65-73.
Ingram, S. N. & Rogan, E. (2002). Identifying critical areas and habitat preferences of
bottlenose dolphins (Tursiops truncatus). Marine Ecology Progress Series 244:
247-255.
Irvine, A. B., Scott, M. D., Wells, R. S. & Kaufmann, J. H. (1981). Movements and
activities of the atlantic bottlenose dolphin, Tursiops truncatus, near Sarasota,
Florida. Fishery Bulletin 79(4): 671-688.
Jefferson, T. A., Webber, M. A. & Pitman, R. L. (2008). Marine Mammals of the World –
A Comprehensive Guide to their Identification. Academic Press/Elsevier, San
Diego, California.
Johnston, D. W., Westgate, A. J. & Read, A. J. (2005). Effects of fine-scale
oceanographic features on the distribution and movements of harbour porpoises
Phocoena phocoena in the Bay of Fundy. Marine Ecology Progress Series 295:
279-293.
Kimura, S., Kasai, A., Nakata, H., Sugimoto, T., Simpson, J. H. & Cheok, J. V. S.
(1997). Biological productivity of meso-scale eddies caused by frontal
disturbances in the Kuroshio. ICES Journal of Marine Science 54: 179-192.
Klinowska, M. (1986). Diurnal rhythms in Cetacea - a review. Rep. Int. Whal. Commn.
(Special Issue 8): 75-88.
Klinowska, M. (1991). Dolphins, Porpoises and Whales of the World – The IUCN Red
Data Book. IUCN, Gland, Switzerland and Cambridge, UK.
Lopes da Cunha, P. (1994). Estrutura e dinâmica da ictiofauna do estuário do Sado.
Tese de Doutoramento, Faculdade de Ciências da Universidade de Lisboa.
397 pp.
Lusseau, D. (2006). The short-term behavioral reactions of bottlenose dolphins to
interactions with boats in Doubtful Sound, New Zealand. Marine Mammal Science
22(4): 802-818.
Cecília V. Ferreira 48

BIBLIOGRAFIA
Luís, A. R. F. (2008). Avaliação do impacto de construções portuárias no
comportamento e no ambiente acústico da população de golfinhos-roazes
(Tursiops truncatus) do estuário do Sado. Dissertação de Mestrado em Ecologia e
Gestão Ambiental, Faculdade de Ciências da Universidade de Lisboa. 110 pp.
Mann, J. (1999). Behavioral sampling methods for cetaceans: a review and critique.
Marine Mammal Science 15(1): 102-122.
Mann, J., Connor, R. C., Barre, L. M. & Heithaus, M. R. (2000). Female reproductive
success in bottlenose dolphins (Tursiops sp.): life history, habitat, provisioning,
and group-size effects. Behavioral Ecology 11(2): 210-219.
INAG/MARETEC, 2001. Limites de Jusante dos Estuários Portugueses. Lisboa.
Martins, F., Leitão, P. & Neves, R. (2002). Simulating vertical water mixing in
homogeneous estuaries: the SADO Estuary case. Hydrobiologia 475/476:
221-227.
Martins, F., Leitão, P., Silva, A. & Neves, R. (2001). 3D modelling in the Sado estuary
using a new generic vertical discretization approach. Oceanologica Acta 24: 1-12.
Maroco, J. (2010). Análise estatística com o PASW Statistics. ReportNumber, Pêro
Pinheiro. 953 pp.
Mendes, S., Turrell, W., Lütkebohle, T. & Thompson, P. (2002). Influence of the tidal
cycle and a tidal intrusion front on the spatio-temporal distribution of coastal
bottlenose dolphins. Marine Ecology Progress Series 239: 221-229.
Neumann, D. R. (2001). Activity budget of free-ranging common dolphins (Delphinus
delphis) in the northwestern Bay of Plenty, New Zealand. Aquatic Mammals 27.2:
121-136.
Norris, K. S. & Dohl, T. P. (1980). Behavior of the Hawaiian spinner dolphin, Stenella
longirostris. Fishery Bulletin 77(4): 821–849.
Nunes, S. M. P. D. C. (2001). Estudo da utilização do habitat pela população
sedentária de golfinhos-roazes, Tursiops truncatus (Montagu, 1821), no estuário
do Sado, Portugal. Relatório de Estágio de Licenciatura, Faculdade de Ciências
do Mar e do Ambiente da Universidade do Algarve. 49 pp.
Cecília V. Ferreira 49

BIBLIOGRAFIA
Nybakken, J. W. & Bertness, M. D. (2005). Marine Biology: An Ecological Approach (6ª
Edição Internacional). Pearson Education, Inc., San Francisco.
Pierpoint, C. (2008). Harbour porpoise (Phocoena phocoena) foraging strategy at a
high energy, near-shore site in south-west Wales, UK. Journal of the Marine
Biological Association of the United Kingdom 88(6): 1167 - 1173.
Pittman, S. J. & McAlpine, C. A. (2003). Movements of Marine Fish and Decapod
Crustaceans: Process, Theory and Application. Advances in Marine Biology 44:
205-294.
Robinson, I. S. (2010). Mesoscale ocean features: Eddies. Discovering the Oceans
from Space: The Unique Applications of Satellite Oceanography. pp. 69-114.
Springer.
Rodrigues, A. M. J., & Quintino, V. M. S. (1993). Horizontal biosedimentary gradients
across the Sado estuary, W. Portugal. Netherlands Journal of Aquatic Ecology
27(2-4): 449-464.
Scott, M. D., Wells, R. S. & Irvine, A. B. (1990). A Long-Term Study of Bottlenose
Dolphins on the West Coast of Florida. The Bottlenose Dolphin. (eds S.
Leatherwood & R.R. Reeves), pp. 235-244. Academic Press, San Diego.
Sequeira, M., Matias, S., Farinha, J. C., Gaspar, R., Silva, C., Augusto, J., Ferreira, C.
V., Fonseca, M. J., Narra, P. & Luís, A. R. (2009). Bases para o plano de acção
para a salvaguarda e monitorização da população de Roazes do estuário do
Sado. Instituto da Conservação da Natureza e da Biodiversidade. 80 pp.
Shane, S. H. (1980). Occurrence, movements, and distribution of bottlenose dolphin,
Tursiops truncatus, in southern Texas. Fishery Bulletin 78(3): 593-601.
Shane, S. H. (1990a). Behavior and Ecology of the Bottlenose Dolphin at Sanibel
Island, Florida. The Bottlenose Dolphin. (eds S. Leatherwood & R.R. Reeves), pp.
245-265. Academic Press, San Diego.
Shane, S. H. (1990b). Comparison of Bottlenose Dolphin Behavior in Texas and
Florida, with a Critique of Methods for Studying Dolphin Behavior. The Bottlenose
Dolphin. (eds S. Leatherwood & R.R. Reeves), pp. 541-558. Academic Press, San
Diego.
Cecília V. Ferreira 50

BIBLIOGRAFIA
Silva, A. C. V. (2008). A população residente de Tursiops truncatus (Montagu, 1821)
num quadro de gestão integrada do estuário do Sado: proposta de um acordo
voluntário. Dissertação de Mestrado em Ciências e Tecnologias do Ambiente,
Faculdade de Ciências da Universidade de Lisboa. 114 pp.
Silva, F. J. D. L. & Silva-Jr, J. M. D. (2009). Circadian and seasonal rhythms in the
behavior of spinner dolphins (Stenella longirostris). Marine Mammal Science
25(1): 176-186.
Stevick, P. T., McConnell, B. J. & Hammond, P. S. (2002). Patterns of Movement.
Marine Mammal Biology: An Evolutionary Approach. (ed A.R. Hoelzel),
pp. 185-211. Blackwell Science.
Teixeira, A. M. & Duguy, R. (1981). Observations de Delphinidés dans les eaux
côtières portugaises. Relatórios de Actividades do Aquário Vasco da Gama 9: 1-9.
Wells, R. S. & Scott, M. D. (1999). Bottlenose dolphin Tursiops truncatus (Montagu,
1821). Handbook of Marine Mammals – Volume 6: The Second Book of Dolphins
and the Porpoises. (eds S.H. Ridgway & S.R. Harrison), pp. 137-182. Academic
Press, San Diego, CA.
Wells, R. S. & Scott, M. D. (2009). Common Bottlenose Dolphin - Tursiops truncatus.
Encyclopedia of Marine Mammals, Second Edition. (eds W.F. Perrin, B. Würsig &
J.G. Thewissen), pp. 249-255. Academic Press/Elsevier, San Diego, CA, USA.
Wiens, J. A. (2000). Ecological heterogeneity: an ontogeny of concepts and
approaches. The ecological consequences of environmental heterogeneity. (eds
M.J. Hutchings, L.A. John & A.J.A. Stewart), pp. 9-31. Cambridge University
Press.
Wilson, B., Thompson, P. M. & Hammond, P. S. (1997). Habitat use by bottlenose
dolphins: seasonal distribution and stratified movement patterns in the Moray
Firth, Scotland. Journal of Applied Ecology 34: 1365-1374.
Würsig, B., Cipriano, F. & Würsig, M. (1991). Dolphin Movement Patterns: Information
From Radio And Theodolite Tracking Studies. Dolphin Societies: Discoveries And
Puzzles. (eds K. Pryor & K.S. Norris), pp. 79-112. University Of California Press,
Berkeley, CA.
Cecília V. Ferreira 51

BIBLIOGRAFIA
Cecília V. Ferreira 52
Würsig, B. & Jefferson, T. A. (1990). Methods of photo-identification for small
cetaceans. Rep. Int. Whal. Commn. (Special Issue 12): 43-52.
Zar, J. H. (2009). Biostatistical Analysis (5ª Edição Internacional). Pearson Education,
New Jersey, USA.

ANEXO
FICHA DE AMOSTRAGEM UTILIZADA PARA A RECOLHA DE DADOS.

I - Ficha de amostragem utilizada para a recolha de dados.
DATA BM METEO OBSERVADORES INÍCIO ESFORÇO PM AVISTAMENTOS
FIM
HORA ESTADO DA MARÉ QUADR. LOCAL
DIRECÇÃO APARENTE DE DESLOCAÇÃO
TAMANHO GRUPO
ACTIV. DOMINANTE APARENTE
PRESENÇA DE EMBARCAÇÕES OBSERVAÇÕES (BEAUFORT,
COMPORTAMENTO, ...) NÚMERO TIPO
:
:
:
:
:
:
:
:
:
:
Cecília V. Ferreira 54