Embed Size (px)
Citation preview

UNIVERSIDADE DE SÃO PAULO
FACULDADE DE MEDICINA DE RIBEIRÃO PRETO
DEPARTAMENTO DE BIOQUÍMICA E IMUNOLOGIA
Contribuição da microbiota para o colapso dos mecanismos reguladores da homeostase
intestinal pós-infecção por Toxoplasma gondii
Murilo Solano Dias
RIBEIRÃO PRETO-SP
2015
Dissertação apresentada ao Programa de Pós-
Graduação em Imunologia Básica e Aplicada da
Faculdade de Medicina de Ribeirão Preto da
Universidade de São Paulo como parte das
exigências para obtenção do título de Mestre em
Ciências. Área de Concentração: Imunologia
Básica e Aplicada.

UNIVERSIDADE DE SÃO PAULO
FACULDADE DE MEDICINA DE RIBEIRÃO PRETO
DEPARTAMENTO DE BIOQUÍMICA E IMUNOLOGIA
Contribuição da microbiota para o colapso dos mecanismos reguladores da homeostase
intestinal pós-infecção por Toxoplasma gondii.
Murilo Solano Dias
Nome do Orientador: Prof. Dr. João Santana da Silva
RIBEIRÃO PRETO-SP
2015
Dissertação apresentada ao Programa de Pós-
Graduação em Imunologia Básica e Aplicada da
Faculdade de Medicina de Ribeirão Preto da
Universidade de São Paulo como parte das
exigências para obtenção do título de Mestre em
Ciências. Área de Concentração: Imunologia
Básica e Aplicada.

FOLHA DE APROVAÇÃO Nome: SOLANO DIAS, Murilo Título: Contribuição da microbiota para o colapso dos mecanismos reguladores da homeostase intestinal pós-infecção por Toxoplasma gondii . Aprovado em: ___/___/___
Banca Examinadora Prof. Dr.: João Santana Silva Instituição: Faculdade de Medicina de Ribeirão Preto - USP Julgamento: __________________Assinatura:___________ Prof. Dr.: Angelica Thomaz Vieira Instituição: Instituto de Ciências Biológicas - UFMG Julgamento: __________________Assinatura:___________ Prof. Dr.: Ademilson Panunto Castelo Instituição: Faculdade de Filosofia, Ciências e Letras de Ribeirão Preto – USP Julgamento: __________________Assinatura:___________
Dissertação apresentada à Faculdade de Medicina de Ribeirão Preto da Universidade de São Paulo para obtenção do título de Mestre em Ciências

Autorizo a divulgação total ou parcial deste trabalho, por qualquer meio convencional
ou eletrônico para fins de estudo e pesquisa desde que citada a fonte.
Solano Dias, M.
Contribuição da microbiota para o colapso dos mecanismos reguladores da
homeostase intestinal pós-infecção por Toxoplasma gondii.90p: iL ; 30cm
Dissertação apresentada ao Programa de Pós-Graduação em
Imunologia Básica e Aplicada da Faculdade de Medicina de Ribeirão Preto da
Universidade de São Paulo como parte das exigências para obtenção do título
de Mestre em Ciências.
Orientador: João Santana da Silva
1.Células T reguladoras. 2. Toxoplasma gondii . 3.Microbiota Intestinal. 4. Inflamação Intestinal. 5. Receptores da Imunidade Inata.

Dedicatória...
Aos meus pais, Silvia Cristina Solano Dias e Irios Aparecido Dias pelo apoio
incondicional e fundamental para meu desenvolvimento pessoal e intelectual. A eles, meu
muito obrigado.

Agradecimentos
Ao Professor Dr. João Santana da Silva, pelos ensinamentos, orientação e por sempre
proporcionar aos seus alunos o melhor, dentro do possível. Ao senhor, toda minha
admiração.
À Dra. Denise Morais da Fonseca, pela excelente orientação e por todos os ensinamentos
técnicos, científicos e pessoais. A ela devo todas as oportunidades que me surgiram nos
quatro anos (iniciação científica e mestrado) dedicados à Imunologia. Meus mais sinceros
agradecimentos.
À Dra. Maria do Carmo de Souza e à Dra. Gabriela Gonçalves Oliveira pela parceria
científica, amizade e carinho quase maternal.
Ao grande amigo Msc. Rodrigo Pereira de Almeida Rodrigues, pessoa brilhante e que esteve
presente na execução da maioria dos experimentos realizados durante este trabalho. Muito
obrigado.
À Msc. Marcela Davoli Ferreira, pela imensa ajuda prestada no desenvolvimento desse
trabalho e também pela amizade. Muito obrigado.
À grande amiga cubana, Malena Martinez, pela amizade sem igual e pela imensa ajuda
prestada na fase final de conclusão desse trabalho! Muchas gracias!
Aos amigos de laboratório, Jéssica Lobo (eterna caloura), Larissa Marfori, Gretel Rodriguez,
Havier Ricardo Rubio Peña, Pedro Alexandre Sampaio, Verônica Saltarelli, Mariana
Thomaz, Msc. Alynne Karen e Dra. Carla Duque Lopes, pelo companheirismo na ciência e
na amizade. Muito obrigado por tudo!
Aos demais companheiros de laboratório, Msc. Laís Sacramento, Mikael Haruro, Ketelut-
Carneiro, Msc. Gustavo Quirino, Dra. Sandra, Msc. Maria Cláudia Silva, Msc. Frederico

Ribeiro, Dr. Giuliano Bonfá, Dra. Grace Kely da Silva, Dra. Daniela Carlos e Dra. Luciana
Benevides, por toda ajuda prestada.
Aos funcionários Wander Cosme Ribeiro, Cristiane Milanezi, Denise Ferraz, Rubilan
Quioneiro, Edinelson, Ana Cristine e Júlio Anselmo Siqueira, pela eficiência e destreza nos
serviços prestados.
Aos demais docentes do Programa de Pós-Graduação em Imunologia Básica e Aplicada, em
especial, à Professora Dra. Vânia Luiza Deperon Bonato e Professora Dra. Beatriz Ferreira,
pelos ensinamentos durante o desenvolvimento do Projeto de Extensão Jovem Imunologista.
À Dra Yasmine Belkaid, pela oportunidade de estágio no Laboratório de Doenças
Parasitárias (NIH/Bethesda/EUA).
Aos colegas do Laboratório de Doenças Parasitárias (NIH/Bethesda/EUA), em especial
Mike Askenase, Seong Ji Han, Samira Tamoutounour e Timoty Hand, pela ajuda científica e
amizade.
Às secretárias do Professor João, do passado e do presente, em especial, Isadora Lima e
Letícia Alves Ferreira, pela ajuda indispensável com assuntos burocráticos.
Aos amigos de graduação sempre presentes, em especial, Rafael Gil de Castro, Luanne
Caires Cruz Souza, Sophia Araujo do Val da Silva e Carolina Corrocher.
Aos docentes do Departamento de Biologia da Faculdade de Filosofia, Ciências e Letras da
Universidade de São Paulo, pela formação científica inicial.
À Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP), pelo financiamento
deste trabalho - Processos 2013/15123-0; 2014/03249-2.

APOIO E SUPORTE FINANCEIRO
Este projeto foi desenvolvido no Laboratório de Imunoparasitologia, coordenado pelo
Professor João Santana da Silva - Departamento de Bioquímica e Imunologia da Faculdade
de Medicina de Ribeirão Preto - Universidade de São Paulo. Parte do projeto também foi
desenvolvido no Laboratório de Doenças Parasitárias, coordenado pela Dra Yasmine Belkaid
- National Institute of Alergy and Infeccious Diseases - National Institutes of Health,
Bethesda, EUA, com apoio das seguintes agências de fomento e instituições:
- Fundação de Amparo à pesquisa do Estado de São Paulo (FAPESP) – Processos
2013/15123-0; 2014/03249-2; 2012/14524-9.
- Conselho Nacional de Desenvolvimento Científico e Tecnológico - Processo 477751/2010-
5

"Happy is the person who is able to discern the causes of things."
(Virgil 37 A.C.)

SUMÁRIO
Lista de figuras........................................................................................................................12
Lista de Abreviações e símbolos.............................................................................................13
Resumo....................................................................................................................................15
Abstract...................................................................................................................................16
1 - INTRODUÇÃO................................................................................................................18
2 - OBJETIVOS......................................................................................................................28
2.1 - Objetivos gerais..............................................................................................................28
2.2 - Objetivos específicos......................................................................................................28
3 - MATERIAL E MÉTODOS...............................................................................................30
3.1- Animais............................................................................................................................30
3.2- Parasito e infecção...........................................................................................................30
3.3- Obtenção e purificação de anticorpo monoclonal contra CD25......................................31
3.4- Diferenciação de Macrófagos e de Células Dendríticas..................................................32
3.5- Preparo das amostras de macerado intestinal...................................................................33
3.6- Detecção de citocinas por ensaio imunoenzimático........................................................33
3.7- Avaliação da translocação bacteriana..............................................................................34
3.8- Extração de células da Lâmina Própria intestinal............................................................34
3.9- Marcações de moléculas de superfície e intranucleares para citometria de fluxo...........35
3.10- Extração de RNA e reação por qPCR............................................................................35
3.11-Tratamento dos camundongos para depleção da flora intestinal....................................37
3.12- Detecção de células produtoras de citocinas por citometria de fluxo............................37
3.13 – Análise Histopatológica...............................................................................................38
3.14- Análise de citometria de fluxo.......................................................................................39
3.15- Análises estatísticas dos resultados................................................................................41
4 – RESULTADOS.................................................................................................................43
4.1- Resposta imune adaptativa após infecção oral por T. gondii...........................................43
4.2 - Papel de IL-6 na patogênese da doença inflamatória induzida no intestino de animais
infectados com T. gondii.........................................................................................................45
4.3-Produção in vitro de IL-6 por células dendríticas e macrófagos em resposta a T.
gondii.......................................................................................................................................47
4.4-Translocação de bactérias da microbiota intestinal e produção diferencial de IL-6 entre

camundongos resistentes e susceptíveis..................................................................................48
4.5-Neutrófilos e monócitos inflamatórios são majoritariamente parasitados durante infecção
por T. gondii............................................................................................................................51
4.6-Produção de IL-6 por neutrófilos e monócitos inflamatórios in vivo durante infecção por
T. gondii..................................................................................................................................52
4.7 Contribuição de células dendríticas para produção de IL-6 durante infecção por T.
gondii.......................................................................................................................................56
4.8 A via da molécula adaptadora Myd88 está associada à produção de IL-6 após infecção
por T. gondii............................................................................................................................58
4.9 O colapso de células Treg durante a infecção por T. gondii não é dependente do
reconhecimento do parasito por receptores da via de TLR, NOD1 e NOD2..........................60
4.10 Translocação de bactérias da microbiota intestinal em camundongos deficientes para
Myd88, TLR2, TLR4 e TLR9.................................................................................................62
4.11 Receptores TLR11 e TLR12 não influenciam no colapso de Tregs durante infecção por
T. gondii..................................................................................................................................64
4.12 Envolvimento da microbiota intestinal na produção de IL-6 e inflamação tecidual
durante a infecção por T. gondii..............................................................................................67
5- DISCUSSAO......................................................................................................................72
6 - CONCLUSÃO...................................................................................................................82
7 – REFERÊNCIAS................................................................................................................84

LISTA DE FIGURAS
Figura 1: Caracterização das populações de linfócitos T CD4+ durante infecção oral por T.
gondii.......................................................................................................................................44
Figura 2: Papel IL-6 durante a infecção com T. gondii.. ....................................................... 46
Figura 3: Produção de IL-6 por células dendríticas e macrófagos ........................................ 48
Figura 4: Translocação de bactérias da microbiota intestinal e produção de citocinas entre
camundongos resistentes e susceptíveis ................................................................................. 50
Figura 5: Populações de células parasitadas durante infecção por T.gondii.. ....................... 52
Figura 6: Dot plot representativo das populações de monócitos inflamatórios, neutrófilos e células produtoras de IL-6 durante a infecção por T. gondii...................................................53
Figura 7: Produção de IL-6 por monócitos inflamatórios durante a infecção por T. gondii...
................................................................................................................................................ 54
Figura 8: Produção de IL-6 por neutrófilos durante a infecção por T.
gondii.......................................................................................................................................55
Figura 9: Avaliação das subpopulações de células dendríticas, APCs e produção de IL-6
durante a infecção oral por T.gondii. ...................................................................................... 57
Figura 10: Rastreamento de receptores da imunidade inata possivelmente envolvidos na via
de produção da citocina IL-6 por macrófagos ........................................................................ 59
Figura 11: Rastreamento de receptores da imunidade inata possivelmente envolvidos na via
de produção da citocina IL-6 por células dendríticas ............................................................. 60
Figura 12: Avaliação do papel receptores da imunidade inata no colapso de Tregs e
produção de IL-6..................................................................................................................... 61
Figura 13: Avaliação da translocação de bactérias da microbiota intestinal e análise de
sobrevivência em animais selvagens e deficientes em receptores TLRs durante infecção por
T.gondii. .................................................................................................................................. 63
Figura 14: Avaliação da população de Tregs e da resposta imunológica durante infecção
oral por T. gondi em camundongos deficientes para TLR11 e 12.......................................... 65
Figura 15: Relação entre translocação da microbiota intestinal e produção de IL-6 em
camundongos susceptíveis. ..................................................................................................... 68
Figura 16: Papel da microbiota na inflamação intestinal mediante infecção por T.gondii..................................................................................................................................70

13
LISTA DE ABREVIAÇÕES E SÍMBOLOS
APC do inglês, Antigen-presenting cells, “Células apresentadoras de antígeno”
CD do inglês, Cluster of differentiation, “Cluster de diferenciação”
cDNA do inglês, Complementary DNA, “DNA complementar”
DNA do inglês, Deoxyribonucleic Acid, “Ácido Desoxirribonucleico”.
DPI Dias pós-infecção
ELISA do inglês, Enzyme-Linked Immunosorbent Assay, “Ensaio imunoenzimático
de fase sólida”
Foxp3 Fator de transcrição forkhead Box P3
H&E/HE Hematoxilina e Eosina
IFN Interferon
IL- Interleucina
iNOS do inglês Inducible Nitric Oxide Synthase, “Óxido nítrico sintase
induzível”.
i.p Intraperitoneal
KO do inglês, knockout, “nocaute”.
ME-49 Cepa de baixa virulência de T. gondii tipo II
NK do inglês, Natural Killer cell, “Célula Naturalmente citotóxica”.
NO do inglês, Nitric oxide, “Óxido Nítrico”.
PBS do inglês, Phosphate buffered saline, “Solução salina isotônica tamponada
com fosfato”.
PMA do inglês, Phorbol 12-myristate 13-acetate, “12-O- tetradecanoilforbol-13-
acetato ou acetate de miristato de forbol”
qPCR do inglês, Real-time quantitative polymerase chain reaction, “Reação em
Cadeia da Polimerase quantitativa em Tempo Real”

14
RFP do inglês, Red fluorescent protein, “Proteína Vermelho Fluorescente”
RPMI Meio de cultura celular 1640
SBF Soro Bovino Fetal
TGF- do inglês, Transforming growth factor, “Fator de crescimento e
transformação”
TNFdo inglês, Tumor necrosis factor α, “Fator de necrose tumoral”
Tregs Células T reguladoras
UFC Unidades formadoras de colônias
WT do inglês Wild type, “Tipo Selvagem”

15
SOLANO-DIAS, M. Contribuição da microbiota para o colapso dos mecanismos reguladores da homeostase intestinal pós-infecção por Toxoplasma gondii. Dissertação de mestrado. Faculdade de Medicina de Ribeirão Preto, Universidade de São Paulo, Ribeirão Preto, SP, 2015, 89 pg.
RESUMO
Neste trabalho, nós avaliamos a influência da microbiota intestinal na produção diferencial da citocina IL-6 e a consequente redução das células T reguladoras (Tregs) durante a inflamação intestinal induzida por Toxoplasma gondii. Nesse contexto, sítios de alta exposição antigênica, como o trato gastrointestinal desenvolvem mecanismos sofisticados de manutenção da homeostase imunológica tecidual. Quando tais mecanismos falham, a tolerância imunológica local é quebrada e ocorre o desenvolvimento de lesão tecidual, resultando em um conjunto de doenças conhecidas como doenças inflamatórias intestinais. O modelo murino de infecção oral por T. gondii
tem sido utilizado para o estudo da patogênese de tais doenças. O protozoário induz inflamação intestinal em camundongos susceptíveis C57BL/6, que sucumbem durante a fase aguda da infecção devido à indução de uma forte resposta Th1 dirigida contra o parasito e também contra componentes da microbiota intestinal. Por outro lado, animais da linhagem BALB/c são resistentes à doença. Nossos dados demonstram que a progressão dessa resposta inflamatória em camundongos susceptíveis está associada à redução da frequência de Tregs. Animais C57BL/6 apresentam maior produção de IL-6 em relação a camundongos BALB/c. Este fenômeno está associado ao colapso das Tregs, visto que animais C57BL/6 deficientes para a produção de IL-6 são resistentes à infecção pelo parasito. Tais resultados sugerem que o aumento da produção de IL-6 durante a infecção por T. gondii influencia negativamente a manutenção de células Treg, favorecendo o desenvolvimento da inflamação intestinal. Nesse sentido, observamos que durante a infecção, ocorre a translocação de bactérias da microbiota intestinal em paralelo à produção de citocinas como IL-12, IL-10 e IL-6 de maneira exacerbada nos animais susceptíveis C57BL/6. A produção de IL-6 é desencadeada tanto pelo parasito quanto por produtos da microbiota, visto que macrófagos e células dendríticas produzem IL-6 in vitro em resposta a antígenos do parasito e da microbiota. In vivo, verificamos que as populações majoritariamente infectadas pelo parasito são populações de neutrófilos e monócitos inflamatórios, as quais também atuam de maneira preponderante na produção diferencial de IL-6. Curiosamente, monócitos inflamatórios e neutrófilos não parasitados, mas provenientes de animais infectados, produzem quantidades maiores de IL-6 quando comparadas às células infectadas pelo parasito, reforçando o papel de produtos microbianos no desencadeamento da inflamação intestinal. Desse modo, receptores NOD1, NOD2 e TLR2, TLR4, TLR9 e especificamente TLR11 e TLR12, associados ao reconhecimento inato de T. gondii, não estão envolvidos na produção diferencial de IL-6 e redução das populações de Tregs. Paralelamente, a depleção da microbiota intestinal leva à recuperação da população de Tregs nos animais susceptíveis, além de diminuir os níveis de IL-6 secretados. Em conjunto, os dados demonstram que a microbiota intestinal, juntamente com o parasito, colabora para a produção diferencial de IL-6 e para o consequente comprometimento de Tregs em animais susceptíveis.

16
SOLANO-DIAS, M. “The microbiota-induced IL-6 production mediates Treg impairment during Toxoplasma gondii infection”. Dissertation, School of Medicine of Ribeirão Preto, University of São Paulo, Ribeirão Preto SP, 2015, 89 pg.
ABSTRACT
During this work, we evaluated the influence of microbiota in the IL-6 differential production and the Treg impairment during gut inflammation induced by T. gondii infection. In this context, sites of high antigenic exposure, such as the gastrointestinal tract requires sophisticated mechanisms for maintaining the tissue immune homeostasis. When such mechanisms fail, the local immune tolerance is broken and tissue injury may develop, resulting in the development of inflammatory intestinal disorders. The murine model of oral infection with T. gondii has been used to study the pathogenesis of such diseases. The C57BL/6 susceptible mice succumb during the acute phase of infection due to a strong Th1 response against the parasite and also against components of gut microbiota. On the other hand, BALB/c mice are resistant to T. gondii oral infection. Our data demonstrate that the progression of the inflammatory response in susceptible mice is associated to the impairment of regulatory T cells (Tregs). C57BL/6 mice produced high levels of IL-6 as compared to BALB/c mice. This phenomenon is associated with the collapse of Tregs, since IL-6-deficient C57BL/6 mice are resistant to infection, suggesting that the increased IL-6 production during T. gondii infection leads to Treg collapse. Accordingly, gut bacteria translocates from the gut to mesenteric lymph nodes during the infection. In parallel, we observed the over production of IL-12, IL-10, IL-6 in C57BL/6 mice. Indeed, in vitro experiments showed that bone marrow-derived macrophages and dendritic cells produced high levels of IL-6 after stimulation with the parasite or microbiota antigens. In vivo, during T. gondii infection, the major cell population infected by T. gondii was not only macrophages, but also, neutrophils and inflammatory monocytes, which produce high amounts of IL-6. Interestingly, after in vivo infection, bystander uninfected inflammatory monocytes and neutrophils are the major IL-6-producing population when compared to the parasite-infected cells. These data suggests that during T. gondii infection, these cell population might be primed other antigens rather than the parasite, such as the microbiota products. In agreement, NOD1, NOD2, TLR2, TLR4, TL9, TLR11 and TLR12 receptors, which are involved in parasite recognition, were not involved in Treg collapse and IL-6 over production. In this context, when the gut microbiota is depleted with antibiotics, the Tregs are restored and the IL-6 production is reduced. Taken together, the data show that during T. gondii
infection, the gut microbiota collaborates for the IL-6 production and Treg collapse.

17
1111---- IntroduçãoIntroduçãoIntroduçãoIntrodução

18
1- INTRODUÇÃO
As respostas imunológicas adaptativas são mediadas por eventos envolvendo a
interação entre células efetoras, auxiliares e moléculas expressas e/ou secretadas por
essas células, direcionamento para os padrões de resposta especializada, como um
resultado da diferenciação de linfócitos T em células Th1, Th2, Th9, Th17 e Th22
(Mosmann, Cherwinski et al. 1986; Veldhoen, Hocking et al. 2006; Veldhoen,
Uyttenhove et al. 2008; Eyerich, Eyerich et al. 2009). Existem ainda subtipos de
linfócitos responsáveis pelo controle das respostas imunológicas como as células T
reguladoras (Tregs). Tais células, denominadas inicialmente como T supressoras são
linfócitos inibidores da reposta imunológica Inicialmente, as Tregs foram caracterizadas
pela expressão de moléculas de superfície, como a cadeia α do receptor de IL-2 (CD25),
essencial para sua diferenciação e sobrevida. Em modelo murino, a identificação das
Tregs tem sido feita pela expressão do fator de transcrição Foxp3 (do inglês, forkhead
transcription factor) (Hori, Nomura et al. 2003). Vários subtipos de Tregs têm sido
descritos com base na sua origem, indução e mecanismos efetores as quais são subdivididas
em células Tregs tímicas e Tregs periféricas (Abbas, Benoist et al. 2013). As primeiras
emergem do timo com fenótipo supressor enquanto que as Tregs periféricas diferenciam-se
na periferia a partir de células T convencionais após a exposição aos antígenos e a sinais
como citocinas reguladoras (IL-10 e TGF-β)
A capacidade das Tregs em controlar respostas imunológicas é derivada de
vários mecanismos supressores, os quais incluem a privação de outras populações
celulares em obter IL-2 do microambiente tecidual através da captura de IL-2 por CD25
(Sakaguchi, Sakaguchi et al. 1995) e a secreção de citocinas reguladoras, como IL-10
(Hawrylowicz and O'Garra 2005), IL-35 (Collison, Workman et al. 2007) e TGF-β
(Joetham, Takeda et al. 2007). Também são capazes de promover a lise celular por meio

19
da liberação de grânulos de granzima e perforina (Gondek, Lu et al. 2005). Podem
ainda modular a função ou maturação de células dendríticas através da interação
dependente de CTLA-4 (Oderup, Cederbom et al. 2006). As Tregs também podem
exercer seus mecanismos supressores através da expressão das ectoenzimas CD39 e
CD73, as quais clivam ATP em adenosina e suprimem as células efetoras através do
receptor da adenosina 2A (A2AR) (Borsellino, Kleinewietfeld et al. 2007; Deaglio,
Dwyer et al. 2007).
A efetuação de uma resposta adequada pelas Tregs é essencial em sítios de alta
exposição antigênica, como o trato gastrointestinal. A importância dessas células nesse
sítio é notória em fenômenos como o da tolerância oral, situação em que ocorre
ausência de resposta imunológica local e sistêmica contra um determinado antígeno, a
qual pode ser induzida por administração oral de antígenos inócuos, como antígenos
alimentares. Esse mesmo processo também regula respostas para bactérias comensais do
intestino e está associado à prevenção de desordens intestinais, tais como alergia
alimentar, doença celíaca, e doenças inflamatórias do intestino. Populações
especializadas de células dendríticas atuam na indução da tolerância imunológica. Nesse
contexto, células dendríticas induzem Tregs, as quais se expandem na região da lâmina
própria intestinal e atuam na inibição de hipersensibilidade contra antígenos inócuos
(Pabst and Mowat 2012).
Na mucosa intestinal, o sistema imunológico é capaz de estabelecer uma
complexa relação com a microbiota residente. Normalmente esta relação é não
patogênica, havendo assim manutenção da homeostase local. No entanto, em
determinados casos, bactérias comensais podem ativar a imunidade inata e adaptativa
levando ao desenvolvimento de respostas contra antígenos alimentares e comensais
(Elson and Cong 2012). Consequentemente pode haver o desenvolvimento de processos

20
inflamatórios conhecidos como “Doenças Inflamatórias Intestinais” (Inflammatory
Bowel Diseases -IBDs), sendo a doença de Crohn e colite ulcerativa os tipos mais
comuns (Brugman and Nieuwenhuis 2010; Hooper, Littman et al. 2012).
As IBDs são caracterizadas por resposta imunológicas crônicas que afetam
diferentes porções do trato gastrointestinal. Durante a fase inflamatória, a progressão de
uma IBD resulta no aumento acentuado de linfócitos e monócitos ativados, sendo que
nas áreas afetadas, é tipicamente observada a ocorrência de hiperplasia linfoide, edema
submucoso, lesões ulcerativas e fibrose (Brugman and Nieuwenhuis 2010).
Experimentalmente são descritos vários modelos de indução de colite que
demonstram a importância de Tregs na homeostase intestinal e no desenvolvimento das
IBDs. O principal modelo se baseia na transferência de células T efetoras CD4+
CD45RBhigh para animais linfopênicos, como camunodongos deficientes para enzima
RAG (do inglês, Recombination-Activating Genes) e camundongos SCID (do inglês,
Severe combined immunodeficiency) (Coombes, Robinson et al. 2005; Kanai,
Kawamura et al. 2006). A ausência de células Tregs dentre as células TCD4+
CD45RBhigh transferidas para esses animais está associada ao desenvolvimento da
inflamação intestinal. Nesse sentido, a transferência de Tregs juntamente com células
efetoras TCD4+ CD45RBhigh previne o desenvolvimento de colite, demonstrando assim
a importância das Tregs na regulação do desenvolvimento de processos inflamatórios na
mucosa intestinal (Yu, Saruta et al. 2007; Reardon, Wang et al. 2008).
No contexto da mucosa intestinal, é possível que a invasão de microrganismos
possa causar a quebra da tolerância contra a microbiota comensal, além de modificações
na frequência das populações de bactérias residentes, ou seja, disbiose intestinal (Raetz,
Hwang et al. 2013) e consequentemente, induzir um padrão de inflamação semelhante
ao que ocorre durante as IBDs. Desse modo, diferentes patógenos podem ser utilizados

21
como modelo na indução de inflamação intestinal, como o protozoário Toxoplasma
gondii. (Liesenfeld 2002).
Experimentalmente, diferentes linhagens de camundongos respondem de modo
distinto à infecção por esse parasito, sendo, portanto resistentes ou susceptíveis
(Johnson 1984). Dessa forma, ao se inocular 100 cistos da cepa ME-49 de T. gondii em
camundongos da linhagem C57BL/6, observa-se mortalidade desses animais após sete
dias de infecção, ao passo que camundongos BALB/c progridem para a fase crônica da
doença (Liesenfeld 2002).
No que se refere à fase aguda, a infecção oral leva ao desenvolvimento de
inflamação intestinal nos camundongos susceptíveis (C57BL/6), causada pela quebra da
tolerância à microbiota do intestino delgado (Heimesaat, Bereswill et al. 2006).
Camundongos C57BL/6 germ free (deficientes da microbiota intestinal), são resistentes
ao desenvolvimento dessa resposta inflamatória após infecção por T. gondii (Raetz,
Hwang et al. 2013). Por outro lado, animais C57BL/6 que possuem microbiota
desenvolvem a doença, a qual é caracterizada pelo desenvolvimento de necrose grave
no íleo, local no qual as vilosidades intestinais e mucosas tornam-se seriamente
comprometidas, ao passo que camundongos BALB/c não apresentam tais alterações
(Liesenfeld, Kosek et al. 1996). Dessa forma, a microbiota assume um papel crucial na
inflamação intestinal durante a infecção por T.gondii.
A patogênese da doença intestinal induzida pelo parasito está associada à
capacidade da infecção por T. gondii em modular fortemente a resposta imunológica
adaptativa, induzindo uma intensa resposta de padrão Th1 na lâmina própria, que se
caracteriza pela alta produção de IFN-γ por linfócitos TCD4+ e TCD8+ (Khan, Smith et
al. 1988). No entanto, deve-se ressaltar que a produção de IFN-γ é essencial para a
eliminação do parasita, visto que animais deficientes para a produção dessa citocina

22
sucumbem à infecção. Assim, a exacerbação da resposta Th1 é o fator associado ao
desenvolvimento da inflamação intestinal durante a infecção por T.gondii (Liesenfeld,
Kosek et al. 1996).
Durante a toxoplasmose experimental, a resposta imunológica inicia-se pela
ativação de células apresentadoras de antígenos (APCs), localizadas no sítio da infecção
(lâmina própria) que migram para os órgãos linfoides onde ativam células TCD4+ que
passam a produzir IFN-γ (Gazzinelli, Wysocka et al. 1994). IFN-γ juntamente com
TNF-α (do inglês, tumor necrosis fator) levam à ativação de fagócitos como
macrófagos e monócitos inflamatórios, os quais produzem altos níveis de intermediários
reativos de oxigênio, fator essencial para o controle do parasito (Kasper, Courret et al.
2004). A interação de taquizoítos com receptores da imunidade inata induz a produção
de IL-12, TNF-α e outras citocinas pró-inflamatórias por esses fagócitos. A citocina IL-
12 produzida pelas células da imunidade inata leva à diferenciação de linfócitos Th1
(Sher, Denkers et al. 1995). Ainda durante a infecção por T. gondii, ocorre a produção
de IFN-γ pelas células NK (do inglês, natural killers) (Sher, Oswald et al. 1993).
Para que ocorra a produção de IL-12 por meio de mecanismos associados à
imunidade inata, durante a toxoplasmose, há a necessidade de o parasito ser reconhecido
por receptores do hospedeiro. Desse modo, os TLRs (do inglês, Toll-Like Receptors)
tem um papel importante no processo. Assim, camundongos deficientes para a molécula
adaptadora MyD88 (do inglês, myeloid differentiation factor-88), necessária durante a
sinalização da maioria dos TLRs, são susceptíveis à fase aguda da toxoplasmose, pois
apresentam falha na produção de IL-12 e IFN-γ e consequentemente não sobrevivem à
infecção pela cepa ME-49 de T. gondii (Scanga, Aliberti et al. 2002).
Existem TLRs específicos implicados na resposta imunológica durante a
infecção com T. gondii, incluindo os TLR2, TLR4, TLR9, TLR11 e TLR12. TLR11 e

23
TLR12 estão envolvidos no reconhecimento e resposta a profilina (Yarovinsky, Zhang
et al. 2005; Jenkins, Tuo et al. 2010), enquanto TLR2 e TLR4 reconhecem glicosil
inositol fosfolipídios (GPIs) presentes na superfície do parasito (Debierre-Grockiego,
Campos et al. 2007). Além disso, em decorrência da infecção oral por T. gondii,
antígenos bacterianos do intestino ativam TLR2, TLR3 e TLR9 o que contribui para o
desenvolvimento da resposta imune Th1 (Benson, de Jongh et al. 2010). Nesse
contexto, camundongos deficientes para TLR2 sucumbem apenas quando infectados
com altas cargas do parasito (Scanga, Aliberti et al. 2002).
O receptor intracelular TLR11, tem um importante papel durante a infecção
induzida por T. gondii. Ele interage com UNC93B1, proteína residente no retículo
endoplasmático, a qual é essencial para o reconhecimento do parasito por células
dendríticas, via interação TLR11 (Pifer, Benson et al. 2011). Essa interação leva à
produção de IL-12 via sinalização por MyD88 (Pifer, Benson et al. 2011). Desse modo,
tais animais são capazes de sobreviver à fase aguda da doença, apresentando, porém,
elevado número de cistos cerebrais e resistência prejudicada durante a fase crônica
(Yarovinsky, Zhang et al. 2005). Além de TLR11, TLR12 também está implicado no
reconhecimento de profilina e e indulççao da produção de IL-12 e IFN-γ. Recentemente
relatou-se que homodímeros de TLR-12 ou heterodímeros de TLR-12/11 também são
capazes de reconhecer profilina associada à T. gondii e induzir assim, a produção de IL-
12 e IFN-γ (Koblansky, Jankovic et al. 2013).
Os receptores da família NLR (do inglês, NOD-like receptor) também são
importantes sinalizadores da imunidade inata e atuam no reconhecimento de PAMPS
(do inglês, pathogen-associated molecular pattern) intracelulares. Membros dessa
família, os receptores NOD1 e NOD2 reconhecem fragmentos de peptideoglicano de
bactérias. Desse modo, o ácido γ-D-glutamil-meso-diaminopimélico, característico de

24
bactérias Gram negativas estimula a sinalização por NOD1 (Girardin, Boneca et al.
2003). Já NOD2 é ativado por MDP (do inglês, muramyl dipeptide), um constituinte do
peptideoglicano presente tanto em bactérias Gram positivas quanto Gram negativas
(Girardin, Boneca et al. 2003). O papel dos NLR durante a infecção experimental por T.
gondii é controverso. Inicialmente foi demonstrado por Shaw e colaboradores (2009)
que o receptor NOD2 tem importante papel na defesa do hospedeiro contra o parasito.
Nesse trabalho, foi verificado que camundongos deficientes de NOD2 apresentam baixa
produção de IL-2 associada a pouca proliferação de células T auxiliares (Shaw, Reimer
et al. 2009). Em contrapartida, Caetano e colaboradores (2011) afirmam que NOD2 não
possui papel importante na defesa contra T. gondii, uma vez que animais deficientes em
NOD foram capazes de induzir resposta Th1 durante a infecção pelo parasito (Caetano,
Biswas et al. 2011).
Durante as respostas imunológicas nas regiões das mucosas intestinais, subtipos
de células dendríticas desempenham importante papel no combate às infecções e na
regulação e indução da resposta imunológica adaptativa. Entre os subtipos existentes,
ocorrem duas subpopulações principais de células dendríticas migratórias, ambas
caracterizadas pela expressão do marcador CD103 e diferenciadas pela expressão ou
não de CD11b, havendo portanto, células CD103+CD11b+ e CD103+CD11b-
(Grainger, Askenase et al. 2014). As células CD103+CD11b+ estão presentes em
tecidos mucosos não linfoides como a lâmina própria e possuem a capacidade de migrar
para o linfonodo mesentérico após a captura do antígeno (Varol, Vallon-Eberhard et al.
2009). Já as células CD103+CD11b- estão presentes em abundância nas placas de Peyer
e em folículos linfoides (Wang, Kim et al. 2005). No microambiente da mucosa
intestinal existe ainda uma população de APCs derivadas de monócitos e outros
precursores e que não expressam CD103, mas expressam CD11b (Bogunovic, Ginhoux

25
et al. 2009). Essa subpopulação contém células dendríticas e macrófagos, que podem ser
identificados pela expressão do receptor de quimiocina CX3CR1, ao contrário das
células dendríticas (Bogunovic, Ginhoux et al. 2009). No contexto das respostas
imunológicas da mucosa intestinal, as células CD103-CD11b+ estão relacionadas à
produção de IL-12 e indução do padrão Th1 de resposta (Merad et al, 2013; Denning et
al, 2011; Cerovic et al, 2013). Já as células CD103+CD11b+ e CD103+CD11b- são
importantes na manutenção da homeostase e tolerância intestinal (Coombes et al, 2007;
Grainger et al, 2014). Essas células migram para os linfonodos mesentéricos,
metabolizando ácido retinóico e TGF-β e participam da indução de células T
reguladoras. Desse modo, animais deficientes para IRF4, importante para o
desenvolvimento e manutenção das células CD103+CD11b+, apresentam falhas na
indução de tolerância oral, devido à redução de células Tregs. As células dendríticas
CD103+CD11b+ também atuam na produção de IL-6 e IL-23, e assim induzem células
do padrão Th17 no intestino (Kinnebrew, Buffie et al. 2012; Persson, Uronen-Hansson
et al. 2013).
A infecção oral por T. gondii é capaz de levar a um desbalanço na regulação da
resposta imunológica da mucosa intestinal por diferentes vias: induzindo recrutamento
de populações de células inflamatórias, alterando o balanço de subpopulações de células
dendríticas ou ainda, modulando a função e sobrevida das células Treg. Desse modo,
dados do nosso grupo e da literatura demonstram a redução significativa do número e da
função supressora da população de células Treg na placa de Peyer, linfonodo
mesentérico e lâmina própria de camundongo susceptível C57BL/6 (Oldenhove,
Bouladoux et al. 2009). O colapso da população de Tregs durante a infecção é um dos
fenômenos determinantes para a susceptibilidade à doença. Oldenhove e colaboradores

26
(2009) mostraram que tal fenômeno está associado à diminuição da disponibilidade de
IL-2 que ocorre no microambiente intestinal após a infecção pelo parasito (Oldenhove,
Bouladoux et al. 2009; Benson, Murray et al. 2012). Outras citocinas como IL-6
também podem exercer influência negativa na atividade das células Treg (Fujimoto,
Nakano et al. 2011). Desse modo, a atividade de Tregs periféricas fica prejudicada em
camundongos que produzem quantidades relativamente altas de IL-6 (Fujimoto, Nakano
et al. 2011). A citocina IL-6 também pode atuar na modificação do fenótipo das células
Tregs, inativando a expressão do gene Foxp3 em células Treg tímicas, resultando assim
na perda da capacidade supressora dessas células, e até mesmo na induzindo conversão
de Tregs em outras populações celulares (Kastner, Dwyer et al. 2010).
De uma maneira geral, a resposta inflamatória induzida pela interação do
parasito ou de componentes da microbiota com receptores da imunidade inata pode
colaborar tanto para o controle do parasito, quanto para a patogênese da doença
inflamatória intestinal.
Portanto, desvendar as vias de sinalização da imunidade inata envolvidas na
potencialização da produção da citocina IL-6 e comprometimento das Tregs, bem como
os mecanismos envolvidos no controle de manutenção das Tregs em sítios de alta
exposição antigênica durante a infecção por T. gondii, torna-se importante para o
entendimento geral das reposta imunológicas em mucosas.

27

28
2222---- OBJETIVOSOBJETIVOSOBJETIVOSOBJETIVOS
2 - OBJETIVOS
Objetivo geral
Determinar o papel da produção de IL-6 induzida via receptores da imunidade
inata sobre os mecanismos envolvidos na manutenção da população de células Treg
durante a inflamação intestinal desenvolvida após infecção por T. gondii.
Objetivos específicos
1) Caracterizar a dinâmica de produção de IL-6 e a disbiose da microbiota intestinal
em camundongos resistentes (BALB/c) e susceptíveis (C57BL/6) à infecção por T.
gondii e que apresentam manutenção ou eliminação de células Treg durante a
progressão da doença. Paralelamente, identificar se existe correlação entre
produção de IL-6 e redução de Tregs no intestino dos animais.
2) Identificar as células responsáveis pela produção de IL-6 durante a infecção in vitro
e in vivo por T. gondii em camundongos C57BL/6.
3) Determinar os mecanismos envolvidos na produção de IL-6 por macrófagos,
monócitos e células dendríticas: identificar os receptores da imunidade inata
envolvidos na produção de IL-6 induzida pelo parasito in vitro e in vivo.

29
3333---- MATERIAL E MATERIAL E MATERIAL E MATERIAL E MÉTODOSMÉTODOSMÉTODOSMÉTODOS

30
3- MATERIAL E MÉTODOS
3.1- Animais
Foram utilizados camundongos selvagens da linhagem BALB/c e C57BL/6, e
destituídos dos genes de RIP2 (RIP2-/-), NOD1 (NOD1-/-) e 2 (NOD2-/-), TLR2 (TLR2-
/-), 4 (TLR 4-/-), 9 (TLR 9-/-), 11/12 (TLR11/12-/-) e MyD88 (MyD88-/-), cujo
background era C57BL/6. Os animais tinham entre 6 e 8 semanas de idade, sendo os
selvagens obtidos do Biotério Central do Câmpus da USP de Ribeirão Preto e os
deficientes do Biotério de Animais Especiais da Faculdade de Medicina de Ribeirão
Preto (FMRP-USP). Foram mantidos no biotério do Departamento de Bioquímica e
Imunologia da FMRP-USP em estantes micro-isoladoras, providas de ar filtrado, com
livre acesso à água e ração, previamente esterilizados. Os camundongos TLR11/12-/-
foram doados pelo Dr. Alan Sher (National Institutes of Health - NIH, Bethesda, EUA),
e mantidos no biotério de camundongos do NIH. Todos os experimentos foram
desenvolvidos de acordo com os Princípios Éticos na Experimentação Animal adotado
pelo Colégio Brasileiro de Experimentação Animal (COBEA) – Protocolo para Uso de
Animais em Experimentação n°105/2013.
3.2- Parasito e infecção
Para a manutenção da cepa ME-49 selvagem ou RFP (do inglês, Red Fluoresent
Protein) de T. gondii, camundongos C57BL/6 foram inoculados com 10 cistos do
parasito no peritônio. Os cistos foram obtidos a partir do macerado de cérebro coletado
de animais com 30 a 60 dias de infecção, quantificados em microscópio de luz ou de
fluorescência (no caso da cepa ME-49 RFP) e diluídos para uma concentração de 20
cistos/200 µL de solução salina tamponada com fosfato a 10 mM PBS (do inglês,

31
Phosphate Buffered Saline). A infecção foi realizada pela inoculação de 200 µL por
gavagem.
3.3- Obtenção e purificação de anticorpo monoclonal contra CD25
Para obtenção do anticorpo contra a molécula de superfície CD25, células do
hibridoma PC61 foram cultivadas em meio RPMI (1640-Medium, Sigma, St. Louis,
EUA) completo com 5% de soro bovino fetal - SBF (GIBCO, Grand Island, USA), 1%
de L-glutamina, Sigma, St. Louis, 1% de penicilina -10,000 Unidades/mL,
estreptomicina 10000μg/mL, GIBCO, Grand Island, USA). Uma vez confluentes nas
garrafas de cultura, as células foram coletadas e 2,5 × 106 células foram inoculadas no
peritônio de camundongos nude da linhagem BALB/c, previamente tratados com 500
µL de óleo mineral. Dez dias após o inóculo do hibridoma, foi coletado o líquido
ascítico do peritônio dos camundongos. Após coletar o líquido ascítico, este foi
centrifugado por 10 minutos, 332 × g, 4ºC, para retirada das hemácias. O líquido foi
então congelado a -20ºC e mantido nessas condições até a realização do protocolo de
precipitação de proteínas para a obtenção dos anticorpos monoclonais contra CD25.
As amostras de líquido ascítico foram descongeladas e centrifugadas por 20
minutos, 332 × g, 4ºC, e toda a camada de lipídeos presente no sobrenadante foi
descartada. A amostra então foi incubada a 56ºC por 45 minutos para inativação de
proteínas do complemento. Ao líquido ascítico foi adicionada lentamente por
gotejamento a solução saturada de sulfato de amônio [(NH4)2SO4], num volume
suficiente para que a amostra ficasse com saturação final de 45%, segundo a fórmula
abaixo indicada:
Volume de sulfato de amônio saturado = 1 (Saturação Final – Saturação Inicial)
Volume de líquido ascítico 1 – Saturação Final

32
A solução permaneceu em agitação por 30 minutos no gelo e então foi incubada
a 4ºC permanecendo nesta temperatura até o dia seguinte, quando a amostra foi
novamente centrifugada por 30 minutos, a 332 × g, 4ºC O sobrenadante foi descartado e
o material precipitado foi diluído em 20 mL de PBS. A amostra então foi colocada em
sacos de membranas semipermeáveis, os quais permaneceram embebidos em 2L de
PBS (trocados por três vezes ao dia), sob agitação a 4ºC para realização do processo de
diálise. A comprovação da presença do anticorpo contra CD25 foi feita por meio de gel
de poliacrilamida SDS PAGE.
3.4- Diferenciação de Macrófagos e de Células Dendríticas
Para a obtenção de macrófagos, as células da medula óssea foram extraídas do
fêmur de camundongos C57BL/6, BALB/c e de nocautes e foram cultivadas na
presença de meio RPMI (Sigma) contendo 10% de SBF e 30% de sobrenadante de
cultura de células L929 (LCCM) em placas de Petri (BD Optilux, Franklin Lakes, EUA)
e incubadas a 37ºC em uma atmosfera de 5% de CO2 por 7 dias. Para a obtenção de
células dendríticas, as células da medula óssea foram cultivadas na presença de 10 mL
de meio RMPI completo contendo 10% de SBF, 1% de L-Glutamina e 1% de penicilina
e 20 μg/mL de GM-CSF (Pepro-Tech, Rocky Hill, EUA), em placas de Petri e incubadas
a 37ºC em uma atmosfera de 5% de CO2 por 7 dias.
As células diferenciadas foram infectadas ou não com 1 taquizoíto de T. gondii
para cada 10 células ou estimulados ou não com 10 µg/ml de STAg (do inglês, Soluble
T.gondii antigen) ou 10 µg/ml de lisado de Escherichia coli.

33
3.5- Preparo das amostras de macerado intestinal
Para avaliar a presença de citocinas no sítio da infecção por T. gondii, o intestino
delgado de camundongos infectados ou não foi coletado e incluído em 2 mL de inibidor
de protease (CompleteTM, Roche, Alemanha) e congelado imediatamente em nitrogênio
líquido. As amostras foram mantidas congeladas a -70°C até o processamento. Para o
processamento, as amostras foram descongeladas a 4°C e trituradas com homogenizador
mecânico. As amostras foram centrifugadas a 500xg, 4°C e o sobrenadante foi
armazenado a -70°C até a dosagem das citocinas.
3.6- Detecção de citocinas por ensaio imunoenzimático
As concentrações das citocinas IL-6, IL-10, e IL-12 foram mensuradas no
macerado intestinal e a citocina IL-6 no sobrenadante de macrófagos e células
dendríticas diferenciados a partir de medula óssea pelo método de ensaio
imunoenzimático (ELISA). As reações foram feitas seguindo as instruções do
fabricante, Duoset R&D (Minneapolis, EUA) e OpTEIA, BD Bioscience (San Diego,
EUA). Placas de poliestireno de 96 poços (Costar, Corning, EUA) foram sensibilizadas
com anticorpos de captura e incubadas overnight a 4°C. Após a incubação, as placas
foram lavadas com PBS/Tween 0,05% em lavadora automática (Immunowash157, Bio-
Rad Lab, EUA). Foi feito o bloqueio dos sítios inespecíficos na placa com tampão
indicado pelo fabricante (BSA a 1% em PBS). Posteriormente, as amostras e a curva
padrão da reação (50 µL/poço) foram adicionadas, seguido de um tempo de 2 horas de
incubação. Após incubação, adicionou-se o anticorpo de detecção biotinilado e a
peroxidase. Após a etapa de lavagem foi adicionado a peroxidase como substrato e o
indicador de reação tetrametilbenzidina (TMB, KPL, Gaithersburg, EUA). A reação foi
finalizada pela adição de H2SO2 2N e a densidade óptica das amostras determinadas a

34
450 nm em leitor de placa (EMax, Molecular Devices Corporation®, Sunnyvale, EUA).
Os resultados da leitura da densidade óptica das amostras foram interpolados na curva
padrão para a obtenção da concentração final da citocina.
3.7- Avaliação da translocação bacteriana
Para extração das bactérias possivelmente translocadas para baço e linfonodos
mesentéricos, foi feito macerado desses órgãos utilizando-se o êmbolo de uma seringa
de 3 mL e Cell Strainer (BD-Falcon, One River Front Plaza, EUA) e ressuspendendo-se
o material extraído em 1 mL de PBS estéril. Posteriormente, duas alíquotas (10 µL e
100 µL) da suspensão celular foram espalhadas ágar BHI (Brain Heart Infusion) e
acondicionadas em estufa a 37°C por 48 horas para a contagem das unidades
formadoras de colônia.
3.8- Extração de células da lâmina própria intestinal
As células da lâmina própria foram extraídas a partir da digestão do intestino
delgado coletado de camundongos infectados ou não com 20 cistos de T. gondii. Para
tal, foram removidas as placas de Peyer intestinais e as fezes. Posteriormente, os
intestinos foram fragmentados em segmentos de 2 centímetros e incubados sob
agitação, a 37°C, em 20 mL de meio RPMI contendo 3% de SBF, 5 mM de EDTA e
0,145 mg/mL de DTT (Dithiotheitol, Gaithersbug, USA), por 20 minutos. Após
incubação, os fragmentos foram agitados em meio RPMI incompleto (contendo 2 mM
de EDTA) e passados em peneira de metal por 3 vezes (descartando-se a cada vez o
meio) para a remoção dos linfócitos intra-epiteliais. Em seguida, os segmentos foram
fragmentados e transferidos para 3 mL de meio RPMI incompleto contendo 0,05% de
DNAse (Sigma, St. Louis, EUA), 0,2 mg/mL de Liberase TL (Roche, Indianápolis,

35
EUA) e incubados sob agitação, a 37°C, por 30 minutos. Na sequência, foram
adicionados 20 mL de meio RPMI completo contendo 3% de SFB para inibir a
atividade das enzimas de digestão. O material foi transferido para Cell Strainner de 70
µm e macerado. Após centrifugação (332 × g, 4°C, 10 minutos), o sobrenadante foi
descartado e adicionado 10 mL de meio RPMI completo contendo 3% de SBF, sendo o
material transferido e filtrado em Cell Strainer de 40 µm e centrifugado. O
sobrenadante foi descartado e as células ressuspensas em 1 mL de meio RPMI completo
contendo 5% de SBF e contadas.
3.9- Marcações de moléculas de superfície e intranucleares para citometria de
fluxo
As células obtidas a partir do linfonodo mesentérico e lâmina própria foram
marcadas com anticorpos monoclonais conjugados a fluorocromos para identificação
celular através de citometria de fluxo. Para marcação de moléculas de superfície celular,
1 × 106 células do linfonodo mesentérico ou da lâmina própria foram incubadas por 30
minutos em solução contendo 10% de soro de coelho inativado e anticorpos
monoclonais conjugados a fluorocromos, de acordo com a marcação desejada e
incubados por 30 minutos no escuro.
Para a marcação de moléculas intracelulares, como fator de transcrição, as células
obtidas dos órgãos de interesse foram fixadas e permeabilizadas com o auxílio dos
tampões providos pelo kit comercial (Foxp3/Transcription Factor
Fixation/Permeabilization Concentrate and Diluent e-Bioscience). Após
permeabilização, as células foram incubadas por 1 hora em solução contendo 10% de
soro de coelho inativado e anticorpos monoclonais conjugados a fluorocromos para
marcação intracelular e de superfície. Posteriormente, as células foram lavadas com

36
PBS. Para análise de citometria, foram adquiridos de 200.000 a 1.000.000 eventos de
cada amostra em citômetro de fluxo Canto II (BD – Biosciences) ou Fortesa (BD –
Biosciences). Para análise das populações de células T reguladoras utilizamos os
anticorpos anti-CD3 (clone 1452C11-BD- Biosciences), anti-CD4 (clone RM4-4-BD-
Biosciences), Foxp3 (clone MF23-BD-Bioscience). Os dados foram analisados com
auxílio do software FlowJo (TreeStar).
3.10- Extração de RNA e reação por qPCR
Para a análise da expressão gênica das citocinas IL-6 foi utilizada a reação em cadeia
da polimerase quantitativa em tempo real (qPCR, quantitative polymerase chain
reaction), foi feito o máster mix utilizando-se: SYBR Green (Promega, Madison, EUA),
IL-6 foward e IL-6 reverse em água livre de nuclease. A leitura foi feita por meio do
equipamento GeneAmp7700 (Applied Biosystems, EUA). Para tal, foram coletadas
amostras dos segmentos intestinais (duodeno, jejuno e íleo) de camundongos selvagens
e deficientes para os seguintes receptores da imunidade: NOD1, NOD2, Myd88, TLR2,
TLR4 e TLR9, infectados ou não. As amostras foram armazenadas e trituradas em
reagente de Trizol (Invitrogen, Carlsbad, EUA) e o RNA foi extraído utilizando-se kit
comercial “Total Isolation System” (Promega, Madison, EUA), de acordo com as
recomendações do fabricante. Após isolamento do RNA das amostras, foi sintetizado o
cDNA pela reação da enzima transcriptase reversa, de acordo com as recomendações do
kit “High Capacity cDNA Reverse Transcriptio Kit” (Applied Biosystems, Foster City,
EUA) . O cDNA foi construído a partir de 1µg do RNA obtido. Após a quantificação do
cDNA, a expressão dos genes de interesse foram determinadas por qPCR com as
seguintes condições de ciclagem: 2 minutos a 50°C, 10 minutos a 95°C e quarenta
ciclos de 15 segundos a 95°C e 1 minuto a 60°C e um último ciclo de vinte minutos

37
com temperatura crescente de 60° a 95°C. Os resultados foram analisados com base no
valor de CT (CT, do inglês, cycle threshold) ou ponto de corte, que compreende o
número de ciclos onde a amplificação atinge um limiar de detecção permitindo a análise
quantitativa do gene alvo. A expressão relativa do gene alvo foi estimada pela fórmula
2-∆∆CT onde o ∆CT = CT do gene alvo - CT do gene endógeno (β-actina) e o ∆∆ct =
∆CT da amostra - ∆CT da amostra controle.
3.11-Tratamento dos camundongos para depleção da microbiota intestinal
A depleção das bactérias da microbiota intestinal foi realizada como
previamente descrito (Hand, Dos Santos et al. 2012). De forma resumida, foi
administrada diariamente aos camundongos a seguinte combinação de antibióticos: 5
mg de trissulfato de neomicina (Sigma), 2,5 mg de vancomicina (Sigma), 5 mg de
metronidazol 5 mg/dia (Sigma) e 5 mg de ampicilina sódica (Sigma). A combinação de
antibióticos foi diluída em 200 μL de água por animal, a qual foi administrada via
gavage oral por 30 dias.
3.12- Detecção de células produtoras de citocinas por citometria de fluxo
Células dos linfonodos mesentéricos e lâmina própria foram isoladas e
cultivadas em meio RPMI 1640 suplementado com 5% de SBF, 1% de penicilina /
estreptomicina, HEPES (Sigma), 1% de L-glutamina (Sigma, St Louis, EUA),
aminoácidos não essenciais (GIBCO, Grand Sland EUA), e 50 mM de β-
mercaptoetanol na presença de 500 ng / mL de ionomicina (Sigma, St Louis, EUA), 50
ng / mL de forbol 12-myristrate 13 acetato (PMA - Sigma, St Louis, EUA) e 1 µg / mL
de Stop Golgi (TM - BD – Biociences, San Diego, EUA). Depois de 4 horas, as células
foram coletadas e marcadas para moléculas de superfície. Após a marcação de

38
superfície, as membranas celulares foram permeabilizadas para a marcação de citocinas
intracelulares. Para tal, as células estimuladas foram coletadas e primeiramente fixadas
com tampão de fixação (PBS / paraformoldeído 2%) por 10 minutos a temperatura
ambiente. Após esta etapa, as células foram lavadas com tampão (1% de SBF em PBS)
e, posteriormente, ressuspensas em 1 mL de tampão de permeabilização (SFB a 1%,
azida sódica a 0,1%, saponina a 0,5% em PBS). As células foram então centrifugadas,
lavadas novamente com tampão de permeabilização e ressuspensas em tampão de
bloqueio (tampão de permeabilização com 10% de soro de coelho inativado) e com
solução contendo anticorpos monoclonais conjugados a fluorocromos específicos para
as citocinas. Após incubação por 1 hora, as células foram novamente lavadas, e a
produção de citocinas por cada subtipo celular foi avaliada por citometria de fluxo. Para
a marcação de citocinas produzidas por monócitos, macrófagos e células dendríticas, as
amostras foram cultivadas por 2 horas na presença de Stop Golgi (TM - BD –
Biociences, San Diego, EUA), e as células foram marcadas com anticorpos
monoclonais específicos para CD11b (clone M1/70- BD -Biosciences), CD11c (clone
N418- BD - Biosciences), Ly6C (clone HK1.4- BD - Biosciences), Ly6G (clone RB6-
8C5-BD-Biosciences), moléculas do MHC classe II (clone M5/114.15.2 -BD -
Biosciences), CD103 (clone 2E7- BD - Biosciences), permeabilizadas e marcadas para
as citocinas, IL-6 (clone MP5-20F3- BD - Biosciences) e IL-12 (clone C15.6 - BD -
Biosciences). Para a marcação de IFN-γ produzidos por células TCD4, utilizamos
anticorpos monoclonais específicos para: CD3 (clone 1452C11-BD- Biosciences), CD4
(clone RM4-4-BD- Biosciences) e IFN-γ (clone XMG1.2 - BD - Biosciences).

39
3.13 – Análise Histopatológica
Para observação da morfologia e alterações histológicas no intestino dos animais
infectados e não infectados, segmentos do intestino delgado foram coletados após o 8°
dia de infecção. O intestino foi seccionado em três partes: duodeno, jejuno e íleo. O
tecido coletado foi fixado em formol 10% tamponado com fosfato dissódico de potássio
(Na2PO4) e fosfato monossódico hidratado (NaH2PO4H2O) e após 18 a 24h de fixação,
os tecidos foram submetidos à bateria de desidratação com o uso de concentrações
crescentes de álcool e posteriormente xilol. Os fragmentos foram incluídos em parafina
e cortes de 5 μm de espessura dos tecidos foram obtidos com o auxílio de um
micrótomo (Leica, Nussloch, Alemanha). Os cortes foram dispostos em lâminas e
depois incubados a 60°C para fixação em estufa (overnight), para inicio do processo de
retirada de parafina. Em seguida as lâminas foram passadas em xilol e reidratadas com
concentrações decrescentes de álcool (99% - 70%). Os cortes foram então corados com
hematoxilina e eosina.
3.14- Análise de citometria de fluxo.
As análises das populações celulares envolvidas na resposta imunológica durante
infecção por T. gondii foram feitas utilizando o software FlowJo V10. A determinação
dos tipos celulares foi baseada na marcação por anticorpos conjugados a fluorocromos.
Desse modo, após aquisição das amostras por meio do citômetro de fluxo, foi excluída,
por meio de parâmetros baseados em área (FSC-A) e altura (FSC-H), os dados
referentes à fragmentos de células mortas e grumos de células que possam de alguma
maneira interferir na análise. As análises relacionadas à expressão de Foxp3 (células
Treg) e IFN-γ (células Th1) foram feitas em relação às populações de linfócitos, as
quais foram delimitadas baseadas nos parâmetros de tamanho por granulosidade (FSC-

40
A/SSC-A), sendo tal população caracterizada por pequeno tamanho e pouca
granulosidade. Dentro da população de linfócitos, analisou-se a expressão das moléculas
CD3 e CD4, delimitando-se as células CD3+CD4+. Dentro dessas células duplo
positivas, analisou-se a expressão do fator de transcrição Foxp3 e da citocina IFN-γ,
obtendo-se uma porcentagem relativa à população CD3+CD4+ e um número baseado no
total de células extraídas dos órgãos coletados (lâmina própria intestinal e linfonodo
mesentérico) .
Com relação às análises de monócitos inflamatórios e neutrófilos, após a
exclusão dos detritos, delimitou-se uma população de células com maior tamanho e
maior granulosidade. Dentro dessa população analisou-se as células expressando as
moléculas de superfície Ly6C e CD11b, delimitando-se uma população nas células
duplo positivas (Ly6C+CD11b+). Dentro dessa população, analisou-se a expressão das
moléculas de MHC de classe II e Ly6G. As células MHCII+ Ly6G- foram delimitadas
como monócitos inflamatórios. Já a população MHCII-Ly6G+ foi delimitada como
neutrófilo. Também se analisou as populações de células apresentadoras de antígenos
(APCs). Essas células foram marcadas a partir da expressão das molécuças do MHC de
classe II e CD11c dentro da população de granulócitos, delimitando-se uma gate nas
células duplo positivas (MHCII+CD11c+). Dentro dessa população, analisou-se a
expressão das moléculas CD11b e CD103. A partir dessas moléculas pôde-se
estabelecer 3 populações celulares, sendo elas as células dendríticas CD103+CD11b-,
CD103+CD11b+ e APCs CD103-CD11b+, que contém tanto macrófagos intestinais,
quanto células dendríticas. Dentro de cada uma dessas populações, analisou-se a
produção das citocina IL-6, obtendo-se uma porcentagem relativa à produção dessas
citocinas por cada uma dessas populações e um número baseado no total de células
extraídas dos órgãos coletados (lâmina própria intestinal ou linfonodo mesentérico). A

41
produção de IL-6 por células infectadas ou não foi avaliada de acordo com a produção
de IL-6 em células RFP+ (infectadas) e RFP-(não infectadas).
3.15- Análises estatísticas dos resultados
Os dados foram expressos a partir da média ± desvio padrão e comparados
usando-se análise de variância (ANOVA) seguida pelo teste de Tukey em experimentos
com mais de dois grupos. No caso de experimentos com apenas dois grupos
experimentais, foi utilizado a análise com teste t de Student (não-pareado). O software
utilizado foi o Prisma 5.0 (GraphPad Software Inc., San Diego, CA, EUA). Os testes
Mantel-Cox e qui-quadrado foram utilizados para comparar a taxa de sobrevivência dos
grupos experimentais.

42
4444---- RESULTADOSRESULTADOSRESULTADOSRESULTADOS

43
4-RESULTADOS
4.1- Resposta imune adaptativa após infecção oral por T. gondii. Inicialmente, avaliamos a população de células Treg e o padrão de resposta
imunológica durante a fase aguda da infecção oral por T. gondii. Para tal, camundongos
C57BL/6 foram infectados com 20 cistos da cepa ME-49 e após 8 dias de infecção, as
populações de células T CD4+ expressando os fatores de transcrição Foxp3, T-bet e
GATA3 foram identificadas em células da lâmina própria do intestino delgado por
citometria de fluxo. Quando comparamos animais infectados com os controles não
infectados, observamos que no sítio da infecção houve uma redução do número total de
células Treg (Figura 1A), bem como o estabelecimento de um padrão de resposta Th1,
determinado pelo aumento do número absoluto de células T CD4+T-bet+ e de células T
CD4+ produtoras de IFN-γ (Figura 1B). Também verificamos redução do número
absoluto de células T CD4+ produtoras de IL-17 (Figura 1A, 1B) e de Tregs
expressando o fator de transcrição GATA3 no sítio infeccioso (Figura 1C). Portanto, a
infecção por T. gondii induz ativação de células Th1 e colapso das células Treg em
camundongos susceptíveis.
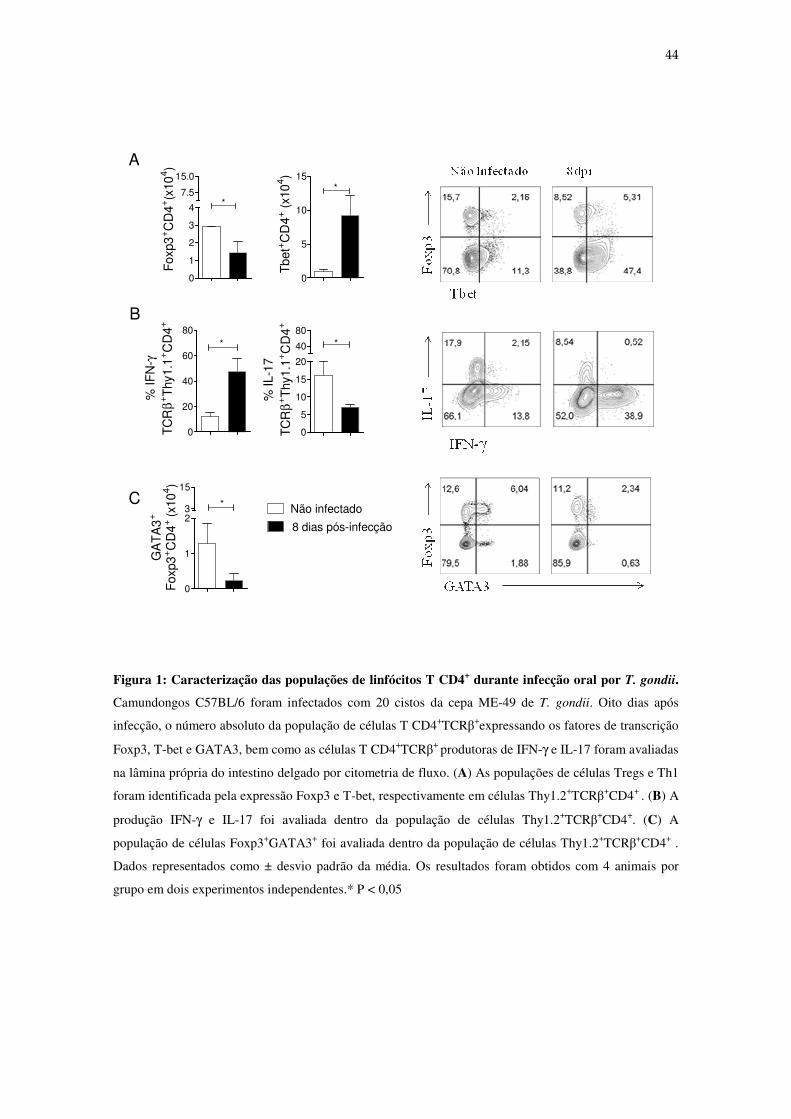
44
Figura 1: Caracterização das populações de linfócitos T CD4+ durante infecção oral por T. gondii.
Camundongos C57BL/6 foram infectados com 20 cistos da cepa ME-49 de T. gondii. Oito dias após
infecção, o número absoluto da população de células T CD4+TCRβ+expressando os fatores de transcrição
Foxp3, T-bet e GATA3, bem como as células T CD4+TCRβ+ produtoras de IFN-γ e IL-17 foram avaliadas
na lâmina própria do intestino delgado por citometria de fluxo. (A) As populações de células Tregs e Th1
foram identificada pela expressão Foxp3 e T-bet, respectivamente em células Thy1.2+TCRβ+CD4+ . (B) A
produção IFN-γ e IL-17 foi avaliada dentro da população de células Thy1.2+TCRβ+CD4+. (C) A
população de células Foxp3+GATA3+ foi avaliada dentro da população de células Thy1.2+TCRβ+CD4+ .
Dados representados como ± desvio padrão da média. Os resultados foram obtidos com 4 animais por
grupo em dois experimentos independentes.* P < 0,05
0
1
2
3
4
7.5
15.0 F
oxp3
+C
D4
+(x
10
4)
*
0
5
10
15
Tb
et+
CD
4+ (
x10
4)
*
0
20
40
60
80
% I
FN
-γ
TC
Rβ
+T
hy1.1
+C
D4
+
*
0
5
10
15
20
40
80
% I
L-1
7
TC
Rβ
+T
hy1
.1+C
D4
+
*
0
1
23
15
GA
TA
3+
Foxp3
+C
D4
+(x
10
4)
*
A
B
CNão infectado
8 dias pós-infecção

45
4.2- Papel de IL-6 na patogênese da doença inflamatória induzida no intestino de
animais infectados com T. gondii
Uma vez observado o impacto da infecção com o parasito T. gondii sobre as
células Tregs no sítio infeccioso, buscamos analisar quais citocinas poderiam estar
envolvidas no comprometimento da regulação da resposta imunológica durante o curso
da doença. Como a citocina IL-6 possui atividade e exerce influência sobre as células
Treg (Kastner, Dwyer et al. 2010; Fujimoto, Nakano et al. 2011), analisamos se essa
citocina estaria sendo produzida diferencialmente entre camundongos resistentes e
susceptíveis, uma vez que dados de nosso grupo demonstraram que camundongos
resistentes não apresentam colapso de Tregs (Fonseca, 2015, comunicação pessoal). Os
resultados mostraram que existe um aumento significativo na produção de IL-6 em
camundongos susceptíveis C57BL/6, quando comparados aos animais resistentes,
BALB/c (Figura 2A). Paralelamente, também verificamos que animais C57BL/6
deficientes para a produção de IL-6 (IL-6-/-) são resistentes à infecção por T. gondii e
apresentam um aumento significativo da frequência de Tregs no intestino em relação
aos animais C57BL/6 capazes de produzir IL-6 (Figuras 2B e 2C). Também
constatamos que animais IL-6-/- tornam-se susceptíveis à infecção por T. gondii
mediante depleção das células Tregs por meio de tratamento com anticorpo monoclonal
contra CD25 (Figura 2C).
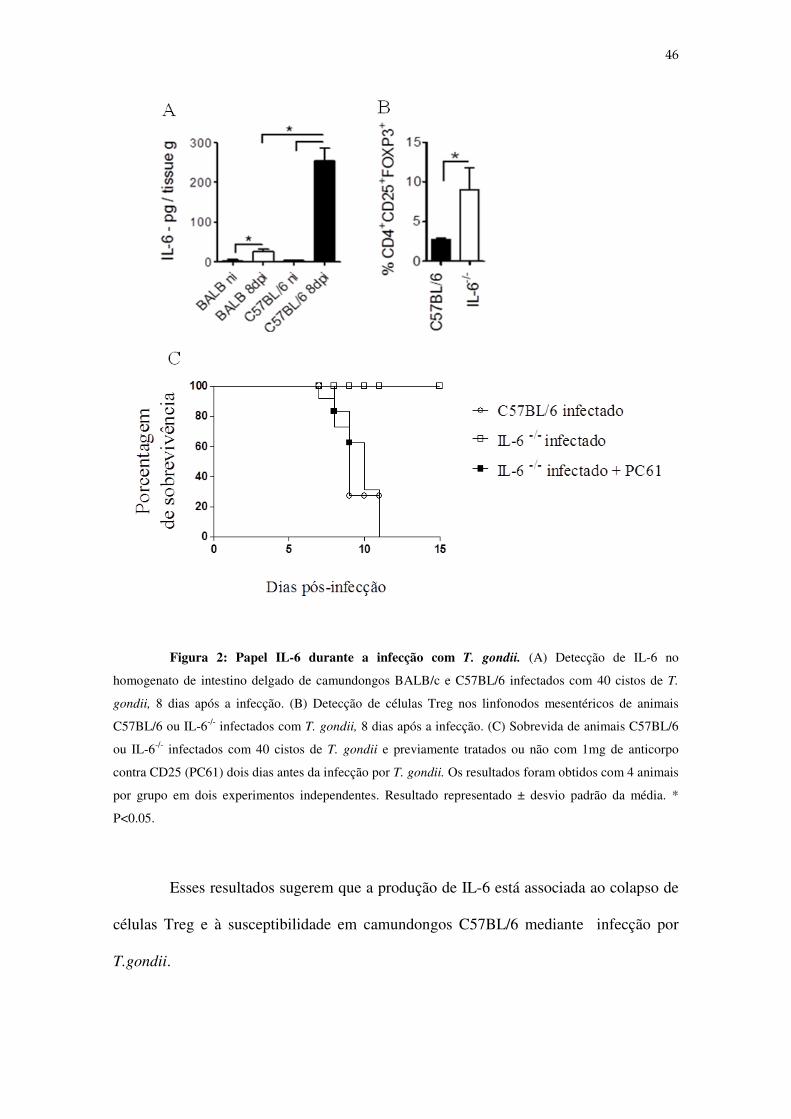
46
Figura 2: Papel IL-6 durante a infecção com T. gondii. (A) Detecção de IL-6 no
homogenato de intestino delgado de camundongos BALB/c e C57BL/6 infectados com 40 cistos de T.
gondii, 8 dias após a infecção. (B) Detecção de células Treg nos linfonodos mesentéricos de animais
C57BL/6 ou IL-6-/- infectados com T. gondii, 8 dias após a infecção. (C) Sobrevida de animais C57BL/6
ou IL-6-/- infectados com 40 cistos de T. gondii e previamente tratados ou não com 1mg de anticorpo
contra CD25 (PC61) dois dias antes da infecção por T. gondii. Os resultados foram obtidos com 4 animais
por grupo em dois experimentos independentes. Resultado representado ± desvio padrão da média. *
P<0.05.
Esses resultados sugerem que a produção de IL-6 está associada ao colapso de
células Treg e à susceptibilidade em camundongos C57BL/6 mediante infecção por
T.gondii.

47
4.3-Produção in vitro de IL-6 por células dendríticas e macrófagos em
resposta a T. gondii
Com o objetivo de avaliar a capacidade do parasito em induzir a produção de IL-
6 por macrófagos e células dendríticas, realizamos inicialmente um ensaio in vitro. Para
tal, células dendríticas e macrófagos diferenciados a partir de precursores da medula
óssea de camundongos BALB/c e C57BL/6 foram infectados com taquizoítos da cepa
ME-49 de T. gondii (1 parasito: 10 células) ou estimulados com 10 μg/mL de STAg.
Células não infectadas ou estimuladas com lisado de E. coli, foram utilizadas como
controle experimental negativo e positivo, respectivamente. Detectamos produção
significativa de IL-6 tanto por células dendríticas quanto por macrófagos, de ambas
linhagens de camundongos quando infectados com taquizoítos (Figura 3A). É
importante notar que antígenos do parasito também foram capazes de induzir a
produção de IL-6, visto que macrófagos e células dendríticas estimulados com STAg
produziram quantidades significativas de IL-6 em relação às células não estimuladas
(Figura 3B). Por fim, estímulos derivados de bactérias presentes na microbiota intestinal
também foram capazes de estimular a produção de IL-6, uma vez que macrófagos e
células dendríticas estimulados com lisado de E. coli produziram quantidades
significativas de IL-6 em relação às células não estimuladas (Figura 3C).

48
12 24 12 24 12 24 12 24 0
20
40
60
BALB/c C57BL6
Taquizoíto (1:10)
MeioTaquizoíto (1:10)
Meio
**
*
**
*
horas
IL-6
(ng
/mL
)
12 24 12 24 12 24 12 240
2
4
6
8
10
**
*
**
BALB/c C57BL6
Taquizoíto (1:10)
MeioTaquizoíto (1:10)
Meio
horas
IL-6
(ng
/mL
)
12 24 12 24 12 24 12 240
20
40
60
80
BALB/c C57BL6
STAGMeioSTAGsMeio
horas
**
**
IL-6
(ng/
mL
)
12 24 12 24 12 24 12 24 0
5
10
15
BALB/c C57BL6
STAGMeioSTAGMeio
horas
**
**
IL-6
(ng
/mL
)
12 24 12 24 12 24 12 240
10
20
30
40
50
BALB/c C57BL6
E. coliMeioE. coliMeio
**
*
**
horas
IL-6
(ng
/mL
)
12 24 12 24 12 24 12 240
5
10
15
20
**
*
**
*
*
BALB/c C57BL6
E. coliMeioE. coliMeio
horas
IL-6
(ng/
mL
)
Células Dendríticas MacrófagosA
B
C

49
Figura 3: Produção de IL-6 por células dendríticas e macrófagos. Células dendríticas e macrófagos
foram diferenciadas a partir de precursores da medula óssea de camundongos BALB/c e C57BL/6 e
infectadas ou não com taquizoítos de T. gondii (A) ou estimulados ou não com 10 µg/ml de STAg (B) ou
com 10 µg/ml de lisado de E. coli (C). A produção de IL-6 foi detectada por ELISA, 12 e 24 horas após a
infecção, conforme indicado no eixo X. Os resultados foram obtidos com 4 duplicatas por grupo.* P <
0,05.
De maneira interessante é válido ressaltar que macrófagos derivados de
camundongos susceptíveis produziram quantidades significativamente maiores de IL-6
em relação aos camundongos resistentes. Já o mesmo não aconteceu em relação às
células dendríticas derivadas da medula óssea de camundongos resistentes, as quais
produzem quantidades significativamente maiores de IL-6 em relação aos animais
susceptíveis.
4.4-Translocação de bactérias da microbiota intestinal e produção diferencial de
IL-6 entre camundongos resistentes e susceptíveis
Com o intuito de detectar fatores diferenciais que promovem a inflamação
intestinal e colapso de células Treg em animais susceptíveis, avaliamos a produção de
citocinas inflamatórias e a translocação bacteriana após a infecção por T. gondii em
camundongos C57BL/6 e BALB/c. Não constatamos translocação de bactérias para os
linfonodos mesentéricos de camundongos BALB/c após a infecção pelo parasito (Figura
4A). No entanto, a translocação bacteriana ocorreu nos linfonodos mesentéricos de
camundongos C57BL/6 infectados (Figura 4A). Paralelamente, verificamos um
aumento significativo da produção de IL-6 e IL-12 no intestino dos camundongos
C57BL/6 no oitavo dia de infecção em relação aos animais não infectados e a
camundongos BALB/c (Figura 2B e 2C). Também avaliamos a produção de IL-10 no
homogenato intestinal de ambas as linhagens de camundongos. A produção de IL-10

50
atingiu o pico de produção durante o oitavo dia após infecção nos animais susceptíveis
(Figura 4E). Portanto, esses resultados mostram que, in vivo, camundongos susceptíveis
C57BL/6 são mais propensos à produção local de IL-6 e translocação de bactérias
intestinais.
Figura 4: Translocação de bactérias da microbiota intestinal e produção de citocinas entre
camundongos resistentes e susceptíveis. Camundongos BALB/c e C57BL/6 foram infectados ou não
pela via oral com 20 cistos de T. gondii (ME-49), nos dias indicados no eixo X das figuras. Foram
avaliadas as unidades formadoras de colônia (UFC) translocadas para os linfonodos mesentéricos de
camundongos BALB/c e C57BL/6 através da cultura de homogenato tecidual (A). Paralelamente, 0, 2, 5 e
8 dias após a infecção foi realizada a detecção da produção de IL-6 (B), IL-12p40 (C), IL-10 (D) no
0 2 5 8 0 2 5 8 0
50
100
150
200
250 *
*
*
BALB/c C57BL/6
Dias pós-infecção
IL-6
(pg/
g de
teci
do)
0 2 5 8 0 2 5 80
1
2
3
BALB/c C57BL/6
Dias pós-infecção
*
**
IL-1
2 p4
0 (p
g/m
g de
teci
do)
0 2 5 8 0 2 5 80.0
0.2
0.4
0.6
BALB/c C57BL/6
Dias pós-infecção
*
**
IL-1
0 (p
g/m
g de
teci
do)
A B
C D
0 2 5 8 0 2 5 80
2
4
6
8
BALB/c C57BL/6
Dias pós-infecção
#
UFC
(Log
2)
51
homogenato intestinal dos animais BALB/c e C57BL/6. Os resultados foram obtidos com 4 animais por
grupo em dois experimentos independentes. Resultado representado como ± desvio padrão da média. *
P<0.05.
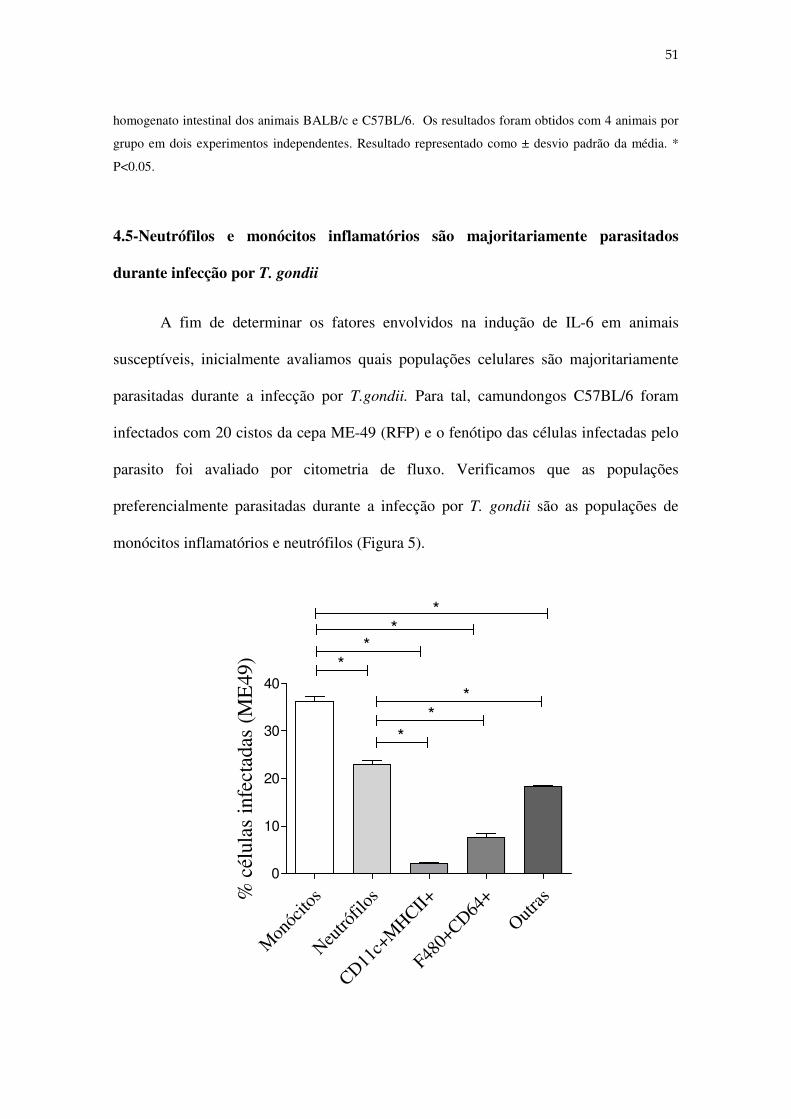
4.5-Neutrófilos e monócitos inflamatórios são majoritariamente parasitados
durante infecção por T. gondii
A fim de determinar os fatores envolvidos na indução de IL-6 em animais
susceptíveis, inicialmente avaliamos quais populações celulares são majoritariamente
parasitadas durante a infecção por T.gondii. Para tal, camundongos C57BL/6 foram
infectados com 20 cistos da cepa ME-49 (RFP) e o fenótipo das células infectadas pelo
parasito foi avaliado por citometria de fluxo. Verificamos que as populações
preferencialmente parasitadas durante a infecção por T. gondii são as populações de
monócitos inflamatórios e neutrófilos (Figura 5).
M
onóc
itos
Neutró
filos
CD11c+
MHCII+
F480+
CD64+
Outras
0
10
20
30
40
*
**
*
**
*
% c
élul
as in
fect
adas
(M
E49
)

52
Figura 5: Populações de células parasitadas durante infecção por T.gondii. Camundongos C57BL/6
foram infectados com 20 cistos da cepa ME-49 (RFP) de T. gondii. Oito dias após infecção, a frequência
de macrófagos (F480+CD64+), células dendríticas (CD11c+MHCII+), neutrófilos (Ly6G+MHCII-) e
monócitos inflamatórios (Ly6G-MHCII+) foi analisada dentro da população celular parasitada por T.
gondii (RFP+). Os resultados foram obtidos com 4 animais por grupo. Resultado representado como ±
desvio padrão da média. * P<0.05.
4.6-Produção de IL-6 por neutrófilos e monócitos inflamatórios in vivo durante
infecção por T. gondii.
Tendo-se em vista o aumento da produção de IL-6 mediante infecção por T.
gondii em paralelo à translocação de bactérias da microbiota intestinal, e parasitismo
predominante de monócitos inflamatórios e neutrófilos, avaliamos a produção de IL-6
por essas populações celulares. Para tal, camundongos susceptíveis foram infectados ou
não com a cepa ME-49 RFP e a produção da citocina IL-6 foi avaliada ex vivo em
monócitos inflamatórios e neutrófilos, na lâmina própria intestinal (Figura 6).

53
Figura 6: Countor plot representativo das populações de monócitos inflamatórios, neutrófilos e
células produtoras de IL-6 durante a infecção por T. gondii. Camundongos C57BL/6 foram infectados
ou não com 20 cistos da cepa ME-49 RFP de T. gondii. Oito dias após infecção, o número absoluto e a
frequência da população de monócitos inflamatórios, neutrófilos, bem como a produção de IL-6 por
monócitos parasitados (RFP+) ou não (RFP-) foram avaliados na lâmina própria do intestino delgado. Para
identificação dos monócitos inflamatórios, foi utilizada a expressão de MHC II dentro da população de
células Ly6ChiCD11b+. Para identificação dos neutrófilos, foi utilizada a expressão de Ly6G dentro da
população de células Ly6ChiCD11b+.
ME-49
IL-6
MHCII
Ly6G
CD11b
ME-49
IL-6
MHCII
Ly
6G
CD11b
Não infectado
8 dias pós-infecção
Ly6C
Ly6C
ME-49
IL-6
ME-49
IL-6
ME-49
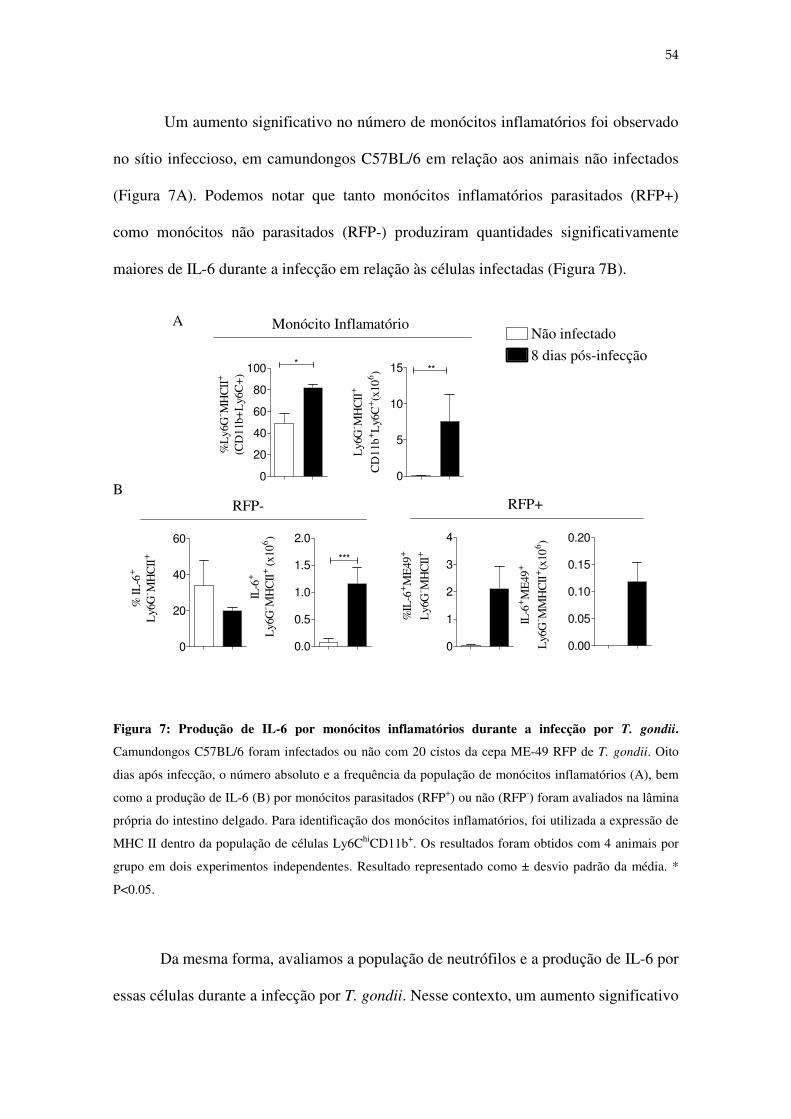
54
Um aumento significativo no número de monócitos inflamatórios foi observado
no sítio infeccioso, em camundongos C57BL/6 em relação aos animais não infectados
(Figura 7A). Podemos notar que tanto monócitos inflamatórios parasitados (RFP+)
como monócitos não parasitados (RFP-) produziram quantidades significativamente
maiores de IL-6 durante a infecção em relação às células infectadas (Figura 7B).
Figura 7: Produção de IL-6 por monócitos inflamatórios durante a infecção por T. gondii.
Camundongos C57BL/6 foram infectados ou não com 20 cistos da cepa ME-49 RFP de T. gondii. Oito
dias após infecção, o número absoluto e a frequência da população de monócitos inflamatórios (A), bem
como a produção de IL-6 (B) por monócitos parasitados (RFP+) ou não (RFP-) foram avaliados na lâmina
própria do intestino delgado. Para identificação dos monócitos inflamatórios, foi utilizada a expressão de
MHC II dentro da população de células Ly6ChiCD11b+. Os resultados foram obtidos com 4 animais por
grupo em dois experimentos independentes. Resultado representado como ± desvio padrão da média. *
P<0.05.
Da mesma forma, avaliamos a população de neutrófilos e a produção de IL-6 por
essas células durante a infecção por T. gondii. Nesse contexto, um aumento significativo
0
20
40
60
80
100
%L
y6G
- MH
CII
+
(CD
11b+
Ly6
C+
)
*
0
5
10
15L
y6G
- MH
CII
+
CD
11b+
Ly6
C+(x
106 ) **
0
20
40
60
% IL
-6+
Ly6
G- M
HC
II+
0.0
0.5
1.0
1.5
2.0
IL-6
+
Ly6
G- M
HC
II+
(x10
6 )
***
0
1
2
3
4
%IL
-6+M
E49
+
Ly6
G- M
HC
II+
0.00
0.05
0.10
0.15
0.20IL
-6+M
E49
+
Ly6
G- M
MH
CII
+(x
106 )
Monócito Inflamatório
RFP- RFP+
A
B
Não infectado
8 dias pós-infecção
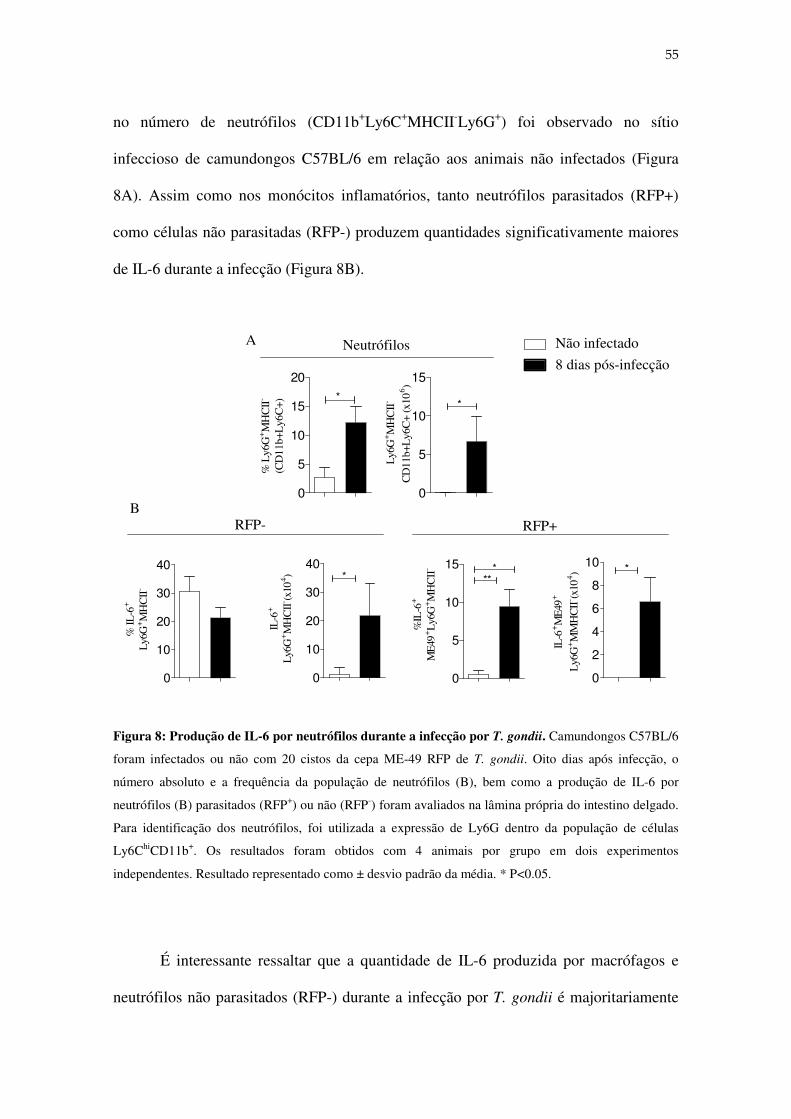
55
no número de neutrófilos (CD11b+Ly6C+MHCII-Ly6G+) foi observado no sítio
infeccioso de camundongos C57BL/6 em relação aos animais não infectados (Figura
8A). Assim como nos monócitos inflamatórios, tanto neutrófilos parasitados (RFP+)
como células não parasitadas (RFP-) produzem quantidades significativamente maiores
de IL-6 durante a infecção (Figura 8B).
Figura 8: Produção de IL-6 por neutrófilos durante a infecção por T. gondii. Camundongos C57BL/6
foram infectados ou não com 20 cistos da cepa ME-49 RFP de T. gondii. Oito dias após infecção, o
número absoluto e a frequência da população de neutrófilos (B), bem como a produção de IL-6 por
neutrófilos (B) parasitados (RFP+) ou não (RFP-) foram avaliados na lâmina própria do intestino delgado.
Para identificação dos neutrófilos, foi utilizada a expressão de Ly6G dentro da população de células
Ly6ChiCD11b+. Os resultados foram obtidos com 4 animais por grupo em dois experimentos
independentes. Resultado representado como ± desvio padrão da média. * P<0.05.
É interessante ressaltar que a quantidade de IL-6 produzida por macrófagos e
neutrófilos não parasitados (RFP-) durante a infecção por T. gondii é majoritariamente
0
5
10
15
20
% L
y6G
+M
HC
II-
(CD
11b+
Ly6
C+)
*
0
5
10
15
*
Ly6
G+M
HC
II-
CD
11b+
Ly6
C+
(x10
6 )
0
10
20
30
40
% IL
-6+
Ly6
G+M
HC
II-
0
10
20
30
40
IL-6
+
Ly6
G+M
HC
II- (x
104 ) *
0
5
10
15
%IL
-6+
ME4
9+L
y6G
+M
HC
II- *
**
0
2
4
6
8
10IL
-6+M
E49
+
Ly6
G+M
MH
CII
- (x10
4 ) *
Neutrófilos
RFP- RFP+
A
B
Não infectado
8 dias pós-infecção

56
maior em porcentagem e número absolutos quando comparados às mesmas populações
celulares parasitadas (RFP+), sugerindo que produtos da microbiota translocada
poderiam participar da indução de IL-6 por essas células (Figura 7B e Figura 8B).
4.7 Contribuição de células dendríticas para produção de IL-6 durante infecção por T. gondii
A produção de IL-6 também foi avaliada nas diferentes subpopulações de células
apresentadoras de antígenos. Para tal, camundongos C57BL/6 foram infectados ou não
com 20 cistos da cepa ME-49 RFP de T. gondii e a produção de IL-6 foi avaliada nas
populações celulares CD11c+MHCII+ que expressavam CD103 e/ou CD11b. Desse
modo, podem-se estabelecer as seguintes populações celulares: C103+CD11b-,
CD103+CD11b+ e CD103-CD11b+. Em relação às células CD103+CD11b-, foi possível
observar um aumento significativo do número total dessa população na lâmina própria
intestinal em relação aos animais não infectados (Figura 9A). Já a frequência de células
CD103+CD11b+, encontrava-se diminuída após a infecção (Figura 9B). Mediante a
infecção por T. gondii, ocorreu um aumento no número absoluto de células CD103-
CD11b+ em relação aos animais não infectados (Figura 9C).

57
Figura 9: Avaliação das subpopulações de células dendríticas e produção de IL-6 durante a infecção por T. gondii. Camundongos C57BL/6 foram infectados ou não com 20 cistos da cepa ME-49 RFP de T. gondii pela via oral. Oito dias após a infecção, a frequência e o número absoluto das populações de células dendríticas CD103+CD11b- (A), CD103+CD11b+ (B) e CD103-CD11b+ (C), foram avaliadas na lâmina própria do intestino delgado. As subpopulações de células dendríticas foram identificadas pela expressão de CD103 e/ou CD11b em células CD11c+MHCII+. A produção de IL-6 foi avaliada dentro das populações celulares parasitadas (RFP+) ou não (RFP-), colocadas em cultura por 2,5 horas em meio acrescido com inibidor de transporte intracelular. Resultado representado como ± desvio padrão da média. P < 0,05.
0
2
4
6
8
10C
D10
3+C
D11
b-
CD
11c+
MH
CII
+ (x
104 ) *
0
5
10
15
IL-6
+C
D10
3+C
D11
b-
CD
11c+
MH
CII
+ (
x104 )
*
0.0
0.2
0.4
0.6
0.8
1.0
IL-6
+ M
E49
+C
D10
3+C
D11
b-
CD
11c+
MH
CII
+(x
103 ) *
0
10
20
30
40
50
%C
D10
3+C
D11
b+
(CD
11c+
MH
CII
+)
*
0
2
4
6
8
IL
-6 +
CD
103+
CD
11b+
CD
11c+
MH
CII
+(x
104 )
*
0.0
0.1
0.2
0.3
0.4
IL-6
+M
E49
+C
D10
3+C
D11
b+
CD
11c+
MH
CII
+(x
104 ) *
0
2
4
6
CD
103- C
D11
b+
CD
11c+
MH
CII
+(x
105 ) *
0.0
0.5
1.0
1.5
IL-6
+C
D10
3- CD
11b+
CD
11c+
MH
CII
+(x
104) *
0.00
0.01
0.02
0.03
0.04
IL-6
+M
E49
+C
D10
3- CD
11b+
CD
11c+
MH
CII
+(x
104 ) *
CD103+
CD11b-
CD103+
CD11b+
CD103-
CD11b+
A
B
C
RFP+RFP-
Não infectado
8 dias pós-infecção

58
Em relação à produção de IL-6, as três populações celulares provenientes de
camundongos infectados apresentaram aumento do número absoluto de células
parasitadas (RFP+) ou não (RFP-) atuando na produção de IL-6 (Figura 8), embora a
célula não infectadas (RFP-) produzam quantidades maiores de IL-6 em relação às
células parasitadas (RFP+).
4.8 A via da molécula adaptadora MyD88 está associada à produção de IL-6 após
infecção por T. gondii
Uma vez estabelecido que as populações de monócitos inflamatórios e
neutrófilos estão envolvidas na produção local de IL-6 em camundongos susceptíveis,
procuramos identificar qual via da imunidade inata poderia mediar a indução dessa
citocina e se tal indução seria dependente do reconhecimento do parasito ou não. Para
tal, diferenciamos macrófagos e células dendríticas da medula óssea de camundongos
C57BL/6 e de camundongos deficientes na expressão de receptores da imunidade inata
e moléculas sinalizadoras envolvidas no reconhecimento de T. gondii. Desse modo,
foram utilizados camundongos deficientes para a expressão de NOD1, NOD2 e MyD88
(mesmo background genético de camundongos C57BL/6). As células diferenciadas
foram infectadas in vitro com taquizoítos de T. gondii ou estimuladas com STAg ou
lisado de E. coli para a detecção da produção de IL-6 nos sobrenadantes de cultura.
Concentrações semelhantes de IL-6 foram detectadas nos sobrenadantes de cultura de
macrófagos e células dendríticas diferenciadas a partir de células provenientes de
camundongos WT, NOD1-/-, NOD2-/- (Figuras 10 e 11). No entanto, tanto macrófagos,
quanto células dendríticas derivadas de animais MyD88-/- foram incapazes de secretar
IL-6 após a infecção in vitro com taquizoítos de T. gondii. Tais resultados mostram que
a produção de IL-6 induzida pelo parasito tanto em macrófagos, quanto em células

59
dendríticas diferenciadas a partir de precursores da medula óssea é dependente de
sinalização via MyD88.
Figura 10- Rastreamento de receptores da imunidade inata envolvidos na via de produção de IL-6.
Macrófagos foram diferenciados a partir da medula óssea de camundongos C57BL/6 e camundongos
deficientes para os receptores NOD1, NOD2 e MyD88 e infectados ou não com 1 taquizoíto de T. gondii
para cada 10 células (A) ou estimulados ou não com 10 µg/ml de STAg (B) ou 10 µg/ml de lisado de E.
coli (C). A produção de IL-6 foi detectada por ELISA, 12 horas após a infecção. Os resultados foram
obtidos com 4 replicatas por grupo. Resultado representado como ± desvio padrão da média.* P < 0,05.
0
5000
10000
15000 *
WT NOD1-/-Myd88-/- NOD2-/-
IL-6
(pg/
mL
)
0
1000
2000
3000
4000
5000
WT NOD1-/-Myd88-/- NOD2-/-
*
IL-6
(pg/
mL
)
0
2000
4000
6000
8000
10000
*
WT NOD1-/-Myd88-/- NOD2-/-
IL-6
(pg
/mL
)
MeioEstímulo
Taquizoíto STAG
E. coli
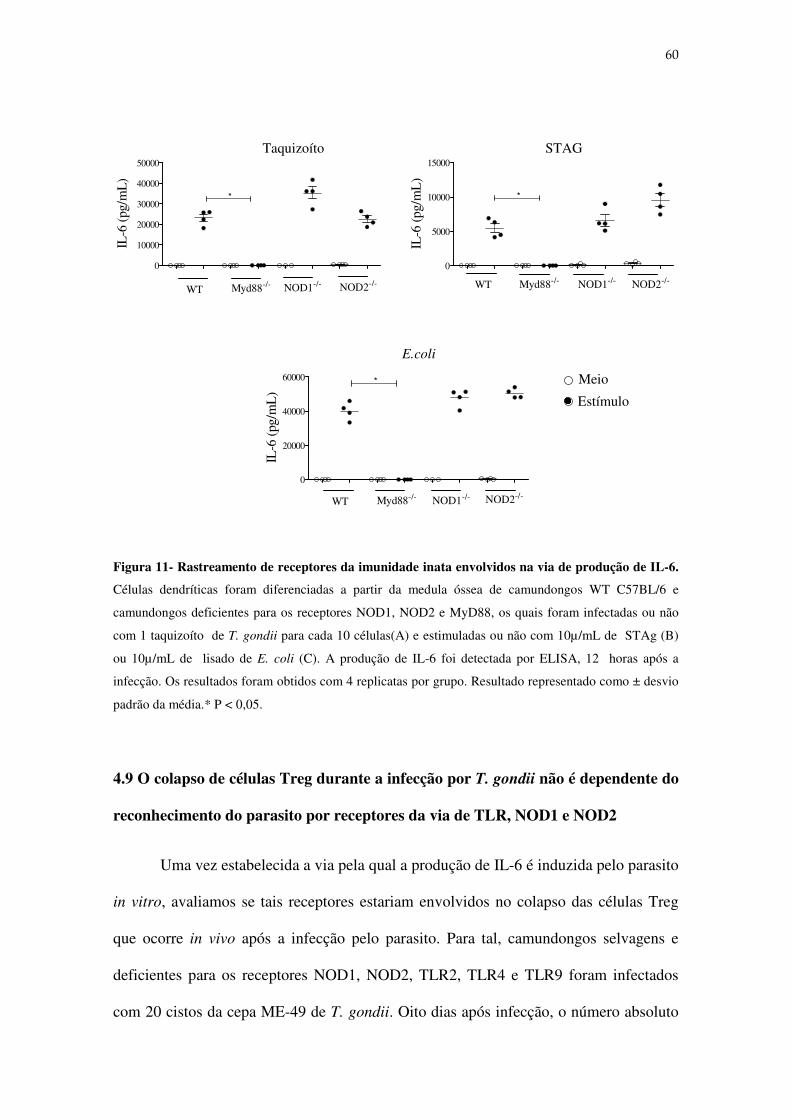
60
Figura 11- Rastreamento de receptores da imunidade inata envolvidos na via de produção de IL-6.
Células dendríticas foram diferenciadas a partir da medula óssea de camundongos WT C57BL/6 e
camundongos deficientes para os receptores NOD1, NOD2 e MyD88, os quais foram infectadas ou não
com 1 taquizoíto de T. gondii para cada 10 células(A) e estimuladas ou não com 10µ/mL de STAg (B)
ou 10µ/mL de lisado de E. coli (C). A produção de IL-6 foi detectada por ELISA, 12 horas após a
infecção. Os resultados foram obtidos com 4 replicatas por grupo. Resultado representado como ± desvio
padrão da média.* P < 0,05.
4.9 O colapso de células Treg durante a infecção por T. gondii não é dependente do
reconhecimento do parasito por receptores da via de TLR, NOD1 e NOD2
Uma vez estabelecida a via pela qual a produção de IL-6 é induzida pelo parasito
in vitro, avaliamos se tais receptores estariam envolvidos no colapso das células Treg
que ocorre in vivo após a infecção pelo parasito. Para tal, camundongos selvagens e
deficientes para os receptores NOD1, NOD2, TLR2, TLR4 e TLR9 foram infectados
com 20 cistos da cepa ME-49 de T. gondii. Oito dias após infecção, o número absoluto
0
10000
20000
30000
40000
50000
WT NOD1-/-Myd88-/- NOD2-/-
*
IL-6
(pg/
mL
)
0
20000
40000
60000
WT NOD1-/-Myd88-/- NOD2-/-
*
IL-6
(pg
/mL
)0
5000
10000
15000
WT NOD1-/-Myd88-/- NOD2-/-
*
IL-6
(pg/
mL
)
Taquizoíto STAG
E.coli
Meio
Estímulo

61
de Tregs foi avaliado no linfonodo mesentérico (Figura 12A e 12B). Também avaliamos
a expressão relativa de mRNA para IL-6 no intestino delgado desses animais. Como
resultado, observamos que nenhum dos receptores estudados estavam envolvidos no
colapso de Tregs, uma vez que mediante infecção, tais camundongos não apresentaram
WT ni
WT in
f
NOD1-/-
inf
NOD2-/-
inf
TLR2-/-
inf
TLR9-/-
inf
0
20
40
60 #
Foxp
3+C
D3+
CD
4+
(x10
4 )
ni inf inf-/-
TLR4
0
1
2
3
4
5
Foxp
3+C
D3+
CD
4+
(x10
5 )
**
WT ni
WT in
f
NOD1-/-
inf
NOD2-/-
inf
TLR2-/-
inf
TLR9-/-
inf
0
10
20
30
40
*
***
IL-6
RN
Am
expr
essã
o re
lativ
a ( β
-act
in)
A
B
C

62
Figura 12- Avaliação do papel receptores da imunidade inata no colapso de Tregs e produção de
IL-6. Camundongos C57BL/6, NOD-/-,NOD2-/-,TLR2-/-,TLR4-/- ,TLR9-/- e RIP2-/- foram infectados com
20 cistos da cepa ME-49 de T. gondii. A) e B) Oito dias após infecção, o número absoluto da população
de células CD3+CD4+ expressando o fator de transcrição Foxp3, foi avaliado no linfonodo mesentérico.
C) Oito dias após infecção, a expressão relativa de mRNA para IL-6 foi avaliada no intestino delgado. Os
resultados foram obtidos com 4 animais por grupo. Resultado representado como ± desvio padrão da
média.* P < 0,05. # P < 0,05 em relação aos demais grupos.
diferença no número absoluto de Tregs em relação ao camundongo selvagem infectado
(Figuras 12A, 12B). Paralelamente e de acordo com a nossa hipótese de que a produção
local de IL-6 exerce um papel relevante para o colapso das células Treg, os
camundongos deficiente para NOD1, NOD2, TLR2 e TLR9 não apresentaram alteração
na expressão de mRNA para IL-6 em relação aos camundongos selvagens infectados
(Figura 12C). Portanto, esses resultados demonstram que, mesmo na ausência de
receptores envolvidos no reconhecimento do parasito in vitro, observa-se produção
acentuada de IL-6 e colapso de Tregs.
4.10 Translocação de bactérias da microbiota intestinal em camundongos
deficientes para MyD88, TLR2, TLR4 e TLR9
Como constatamos que alguns dos receptores que podem atuar no
reconhecimento do parasito não estão envolvidos no colapso de Tregs e na produção
diferencial de IL-6 após a infecção por T. gondii, avaliamos a ocorrência ou não de
translocação de bactérias da microbiota intestinal para o linfonodo mesentérico em
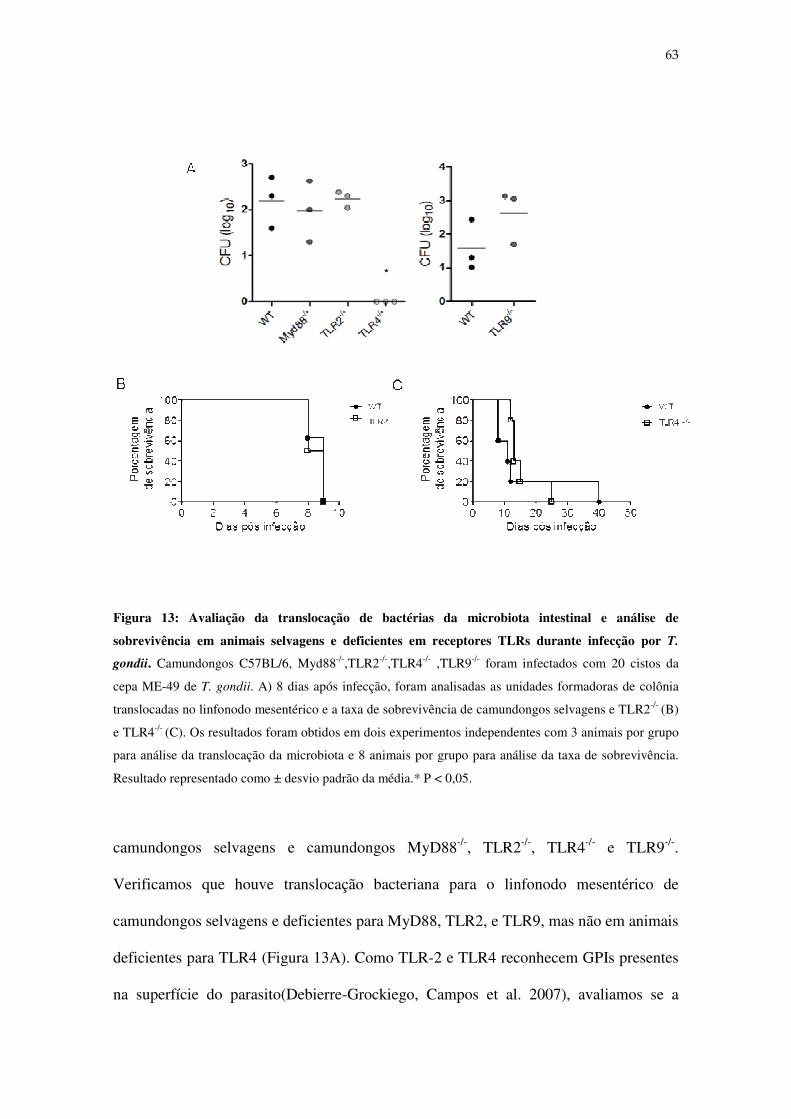
63
Figura 13: Avaliação da translocação de bactérias da microbiota intestinal e análise de
sobrevivência em animais selvagens e deficientes em receptores TLRs durante infecção por T.
gondii. Camundongos C57BL/6, Myd88-/-,TLR2-/-,TLR4-/- ,TLR9-/- foram infectados com 20 cistos da
cepa ME-49 de T. gondii. A) 8 dias após infecção, foram analisadas as unidades formadoras de colônia
translocadas no linfonodo mesentérico e a taxa de sobrevivência de camundongos selvagens e TLR2-/- (B)
e TLR4-/- (C). Os resultados foram obtidos em dois experimentos independentes com 3 animais por grupo
para análise da translocação da microbiota e 8 animais por grupo para análise da taxa de sobrevivência.
Resultado representado como ± desvio padrão da média.* P < 0,05.
camundongos selvagens e camundongos MyD88-/-, TLR2-/-, TLR4-/- e TLR9-/-.
Verificamos que houve translocação bacteriana para o linfonodo mesentérico de
camundongos selvagens e deficientes para MyD88, TLR2, e TLR9, mas não em animais
deficientes para TLR4 (Figura 13A). Como TLR-2 e TLR4 reconhecem GPIs presentes
na superfície do parasito(Debierre-Grockiego, Campos et al. 2007), avaliamos se a

64
ausência desses receptores teria algum impacto na taxa de sobrevivência de
camundongos susceptíveis mediante a infecção por T. gondii. Não observamos
diferenças nas taxas de sobrevida em camundongos deficientes para os receptores TLR2
e TLR4 (Figuras 13B, 13C) em relação ao camundongo selvagem infectado. Portanto o
reconhecimento do parasito por si só não é determinante na susceptibilidade mediante a
infecção por T.gondii.
4.11- Receptores TLR11 e TLR12 não influenciam no colapso de Tregs durante
infecção por T. gondii.
Até o momento, os dados coletados não demonstraram papel efetivo dos
receptores NOD1, NOD2, TLR2, TLR4 e TLR9 no colapso de Tregs e produção
diferencial de IL-6. Embora tais receptores tenham importância em diferentes aspectos
da resposta imunológica contra T. gondii, não reconhecem exclusivamente o parasito.
Nesse sentido, investigamos se receptores específicos para T. gondii estariam
envolvidos no colapso de Tregs e na produção diferencial de IL-6. Para tal,
camundongos selvagens e duplo deficientes para os receptores TLR11 e TLR12 foram
infectados com 20 cistos da cepa ME-49 de T. gondii. Oito dias após infecção, a carga
parasitária, o número absoluto de Tregs, células TCD4+T-bet+ e Tregs expressando o
fator de transcrição GATA3, bem como a produção diferencial de IL-6 por monócitos e
neutrófilos foram avaliados na lâmina própria intestinal (Figura 14).
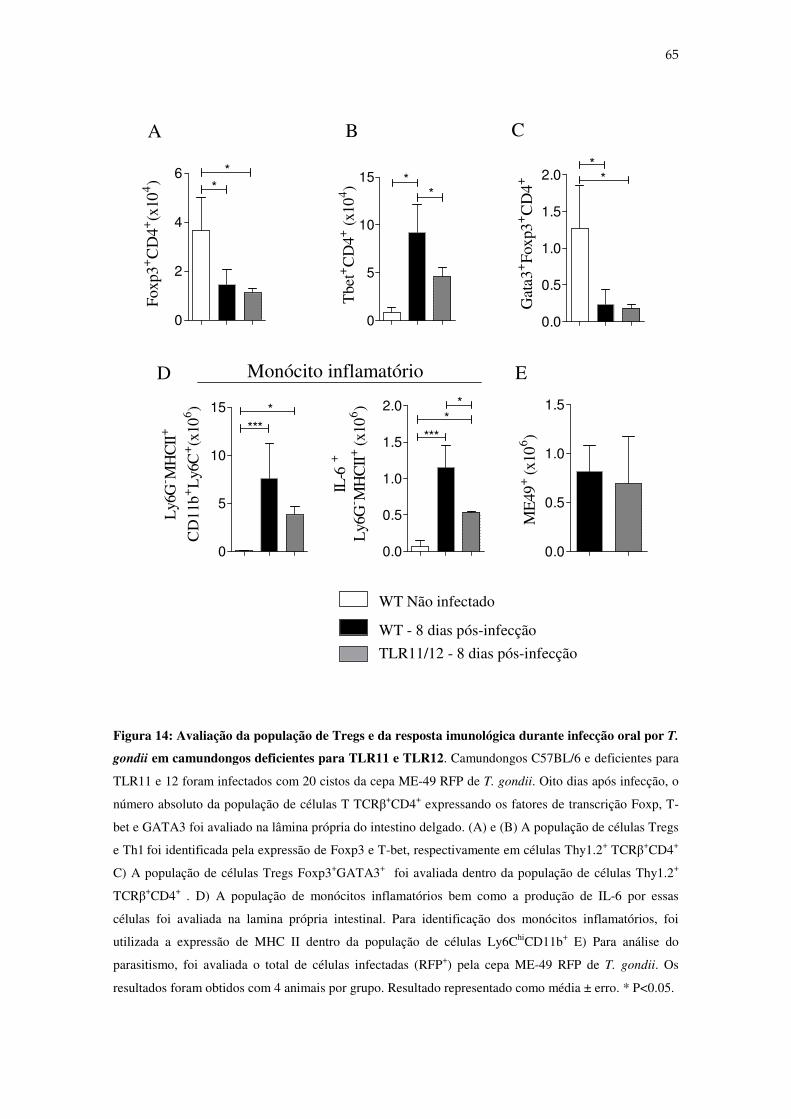
65
Figura 14: Avaliação da população de Tregs e da resposta imunológica durante infecção oral por T.
gondii em camundongos deficientes para TLR11 e TLR12. Camundongos C57BL/6 e deficientes para
TLR11 e 12 foram infectados com 20 cistos da cepa ME-49 RFP de T. gondii. Oito dias após infecção, o
número absoluto da população de células T TCRβ+CD4+ expressando os fatores de transcrição Foxp, T-
bet e GATA3 foi avaliado na lâmina própria do intestino delgado. (A) e (B) A população de células Tregs
e Th1 foi identificada pela expressão de Foxp3 e T-bet, respectivamente em células Thy1.2+ TCRβ+CD4+
C) A população de células Tregs Foxp3+GATA3+ foi avaliada dentro da população de células Thy1.2+
TCRβ+CD4+ . D) A população de monócitos inflamatórios bem como a produção de IL-6 por essas
células foi avaliada na lamina própria intestinal. Para identificação dos monócitos inflamatórios, foi
utilizada a expressão de MHC II dentro da população de células Ly6ChiCD11b+ E) Para análise do
parasitismo, foi avaliada o total de células infectadas (RFP+) pela cepa ME-49 RFP de T. gondii. Os
resultados foram obtidos com 4 animais por grupo. Resultado representado como média ± erro. * P<0.05.
0
2
4
6
Fox
p3+C
D4+
(x10
4 ) **
0
5
10
15 *
Tbe
t+C
D4+
(x1
04 )
*
0.0
0.5
1.0
1.5
2.0
Gat
a3+Fo
xp3+
CD
4+ **
0.0
0.5
1.0
1.5
ME
49+
(x10
6 )0
5
10
15
Ly6
G- M
HC
II+
CD
11b+
Ly6
C+(x
106 )
***
*
0.0
0.5
1.0
1.5
2.0
IL-6
+
Ly6
G- M
HC
II+
(x10
6 )
***
**
A B C
ED Monócito inflamatório
WT Não infectado
WT - 8 dias pós-infecção
TLR11/12 - 8 dias pós-infecção

66
Os resultados demonstraram que os camundongos deficientes para os receptores
TLR11 e 12 não apresentam diferenças nas populações de Tregs e na expressão de
GATA3 pelas Tregs quando comparados aos animais selvagens durante infecção por T.
gondii. Portanto, TLR11 e 12 não estão envolvidos no colapso de Tregs e na expressão
de GATA3 por esta população celular (Figura 14A e 14C). No entanto, foi possível
observar nos animais deficientes para os receptores TLR11 e 12 uma redução
significativa no número absoluto de células expressando o fator de transcrição T-bet
(Figura 14B) quando comparados aos animais selvagens, durante infecção por T. gondii.
Também observamos um menor número absoluto de monócitos inflamatórios e de
produção de IL-6 por essas células (Figura 14D). Entretanto, a produção de IL-6 por
animais nocautes infectados ainda foi significativamente maior em relação aos animais
WT não infectados. Não notamos diferença em relação à carga parasitária de
camundongos deficientes para TLR11 e 12 e animais selvagens (Figura 14E). Desse
modo, é possível inferir que, embora não estejam envolvidos no colapso de Tregs,
TLR11 e TLR12 são importantes no reconhecimento do parasito e geração de resposta
Th1 (Andrade, Souza Mdo et al. 2013) e que o parasito pode atuar na indução de IL-6 in
vivo. No entanto, essa citocina é produzida de forma diferencial em relação aos
camundongos selvagens não infectados, o que nos leva supor que componentes da
microbiota é que estariam majoritariamente envolvidos na produção de IL-6 e colapso
de Tregs.

67
4.12 Envolvimento da microbiota intestinal na produção de IL-6 e inflamação
tecidual durante a infecção por T. gondii.
Os dados coletados indicam que a produção de IL-6 e o colapso de Tregs em
camundongos susceptíveis não são completamente dependentes do reconhecimento do
parasito por receptores da imunidade inata, sugerindo que tal fenômeno estaria
associado à detecção de bactérias da microbiota intestinal translocadas durante a
infecção. A fim de testar a validade desta hipótese, camundongos susceptíveis foram
tratados com coquetel de antibióticos (ampicilina, metronidazol, neomicina e
vancomicina) previamente à exposição ao parasito. Após 30 dias de tratamento, os
camundongos foram infectados com 20 cistos da cepa ME-49 de T. gondii e após 8 dias
de infecção, o número absoluto de Tregs foi avaliado na lâmina própria e no linfonodo
mesentérico de camundongos susceptíveis (Figura 15A e B). Também foi avaliada a
produção de IL-6 no homogenato intestinal de camundongos susceptíveis (Figura 15C),
além da população de monócitos inflamatórios (Figura 15D), bem como a produção de
IL-6 por essas células (figura 15E).
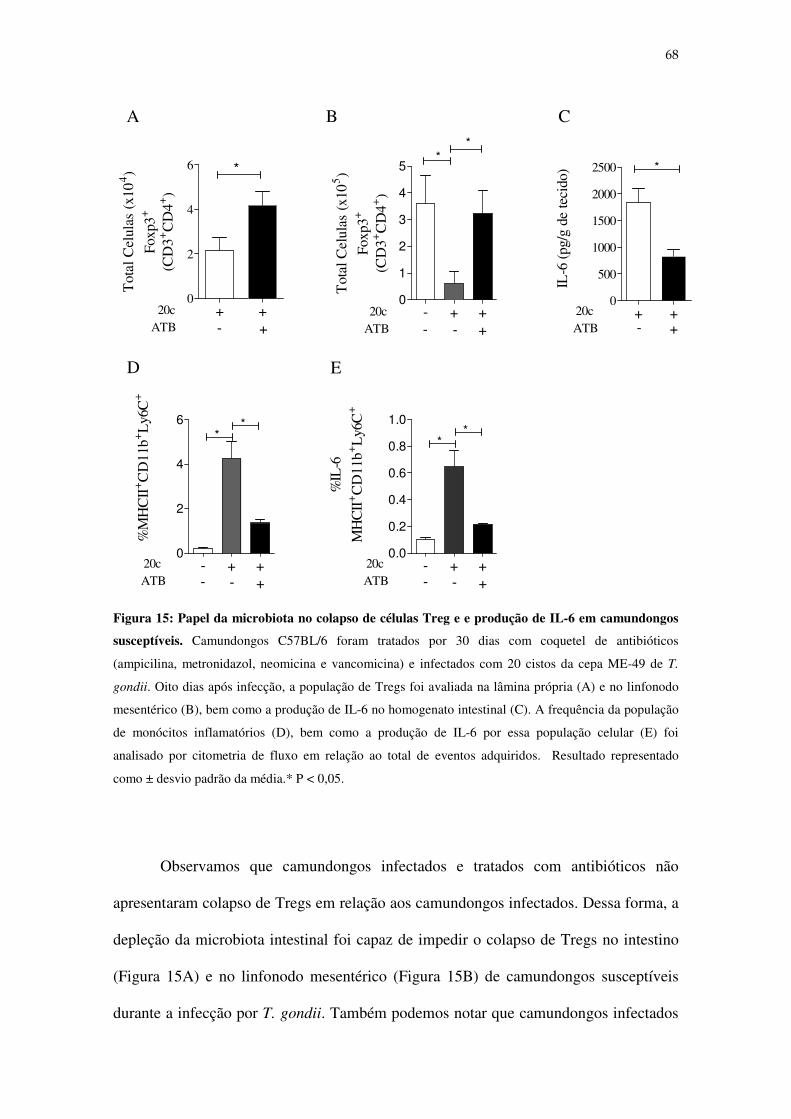
68
Figura 15: Papel da microbiota no colapso de células Treg e e produção de IL-6 em camundongos
susceptíveis. Camundongos C57BL/6 foram tratados por 30 dias com coquetel de antibióticos
(ampicilina, metronidazol, neomicina e vancomicina) e infectados com 20 cistos da cepa ME-49 de T.
gondii. Oito dias após infecção, a população de Tregs foi avaliada na lâmina própria (A) e no linfonodo
mesentérico (B), bem como a produção de IL-6 no homogenato intestinal (C). A frequência da população
de monócitos inflamatórios (D), bem como a produção de IL-6 por essa população celular (E) foi
analisado por citometria de fluxo em relação ao total de eventos adquiridos. Resultado representado
como ± desvio padrão da média.* P < 0,05.
Observamos que camundongos infectados e tratados com antibióticos não
apresentaram colapso de Tregs em relação aos camundongos infectados. Dessa forma, a
depleção da microbiota intestinal foi capaz de impedir o colapso de Tregs no intestino
(Figura 15A) e no linfonodo mesentérico (Figura 15B) de camundongos susceptíveis
durante a infecção por T. gondii. Também podemos notar que camundongos infectados
+ +0
2
4
6
Tot
al C
elul
as (
x104 )
Foxp
3+
(CD
3+C
D4+
)
- +
20cATB
*
- + +0
1
2
3
4
5
Tot
al C
elul
as (
x105 )
Foxp
3+
(CD
3+C
D4+
)
- +
20cATB -
**
+ +0
500
1000
1500
2000
2500 *
IL-6
(pg
/g d
e te
cido
)
- +
20cATB
- + +0
2
4
6
- +
20cATB -
**
%M
HC
II+C
D11
b+L
y6C
+
- + +0.0
0.2
0.4
0.6
0.8
1.0
- +
20cATB -
**
%IL
-6
MH
CII
+C
D11
b+L
y6C
+
A B C
D E

69
e tratados com o coquetel de antibióticos apresentaram menor concentração de IL-6 no
homogenato intestinal quando comparados aos animais infectados por T. gondii e que
não receberam antibiótico (Figura 15C). Soma-se a isso uma menor porcentagem de
monócitos inflamatórios e produção de IL-6 por essas células nos linfonodos
mesentéricos dos animais infectados e tratados com antibióticos em relação aos animais
infectados e não tratados (Figura 15D e 15E). Em conjunto, esses dados sugerem que a
microbiota intestinal tem importante papel na indução da produção exacerbada de IL-6,
a qual impacta na manutenção das populações de Tregs durante a infecção oral por T.
gondii.
Também avaliamos a influência da microbiota na lesão tecidual do intestino
delgado durante a infecção por T. gondii. Para tal, camundongos susceptíveis foram
tratados com coquetel de antibióticos. Após 30 dias de tratamento, os camundongos
foram infectados com 20 cistos da cepa ME-49 de T. gondii. 8 dias após infecção, a
região do íleo do intestino delgado de animais tratados ou não foi coletada para análise
histopatológica (Figura 16).
Como é classicamente descrito (Liesenfeld 2002), camundongos C57BL/6
desenvolveram uma intensa inflamação intestinal mediante infecção por T. gondii,
principalmente nas regiões distais do intestino delgado, como no Íleo (Figura 16).
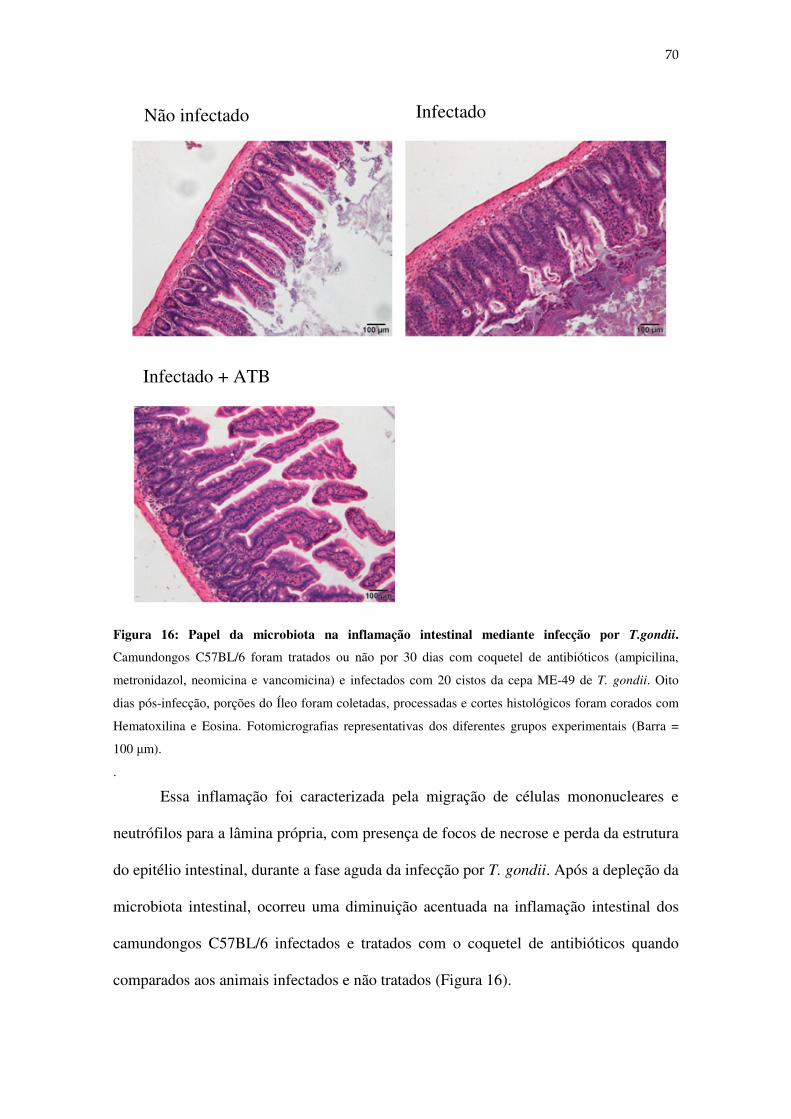
70
Figura 16: Papel da microbiota na inflamação intestinal mediante infecção por T.gondii.
Camundongos C57BL/6 foram tratados ou não por 30 dias com coquetel de antibióticos (ampicilina,
metronidazol, neomicina e vancomicina) e infectados com 20 cistos da cepa ME-49 de T. gondii. Oito
dias pós-infecção, porções do Íleo foram coletadas, processadas e cortes histológicos foram corados com
Hematoxilina e Eosina. Fotomicrografias representativas dos diferentes grupos experimentais (Barra =
100 μm).
.
Essa inflamação foi caracterizada pela migração de células mononucleares e
neutrófilos para a lâmina própria, com presença de focos de necrose e perda da estrutura
do epitélio intestinal, durante a fase aguda da infecção por T. gondii. Após a depleção da
microbiota intestinal, ocorreu uma diminuição acentuada na inflamação intestinal dos
camundongos C57BL/6 infectados e tratados com o coquetel de antibióticos quando
comparados aos animais infectados e não tratados (Figura 16).
Não infectado Infectado
Infectado + ATB

71
5555---- DISCDISCDISCDISCUSSÃOUSSÃOUSSÃOUSSÃO

72
5- DISCUSSÃO Nesse trabalho demonstramos que a inflamação intestinal observada durante a
infecção por T. gondii está relacionada à translocação de bactérias comensais do
intestino, produção diferencial de IL-6 e consequente colapso de Tregs.
Ao infectar as células do hospedeiro murino, o parasito intracelular T. gondii
interage intimamente com o sistema imunológico levando a uma série de condições
patológicas relacionadas à virulência da cepa, à via de infecção e à carga parasitária.
Nesse sentido, a infecção por T. gondii é bastante útil como modelo de encefalite,
inflamação ocular, infecção congênita (Subauste 2012) e no caso deste trabalho, como
modelo no estudo da inflamação e imunologia da mucosa intestinal. Dessa forma, dados
da literatura (Liesenfeld 2002) e de nosso grupo demonstraram que a infecção oral por
T. gondii é capaz de induzir inflamação intensa no intestino delgado de camundongos
C57BL/6, que são susceptíveis à infecção. Nesses animais, esse processo inflamatório é
acompanhado de intensa resposta Th1, o que pode ser verificado pelo aumento da
expressão de Tbet, aumento da produção de IL-12 presente no homogenato intestinal e
produção de IFN-γ por células T CD4+ (Liesenfeld 2002) em paralelo à diminuição das
populações de Tregs (Oldenhove, Bouladoux et al. 2009). É válido ressaltar que o
desenvolvimento de resposta Th1 é importante no controle do parasito, sendo a
exacerbação da resposta o motivo maior do desenvolvimento da inflamação intestinal.
Nesse sentido, foi demonstrado recentemente que células T e células NK são capazes de
produzir IFN-γ na ausência de T-bet e controlar a replicação do parasito no sítio
infeccioso. No entanto, em sítios secundários de infecção, esse controle falha devido à

73
falha migração de células T específicas contra o parasito (Harms Pritchard, Hall et al.
2015).
O comprometimento das células Tregs durante processos infecciosos e
inflamatórios tem sido associado ao desenvolvimento de doenças autoimunes e doenças
inflamatórias, evidenciando sua importância no controle dessas doenças (Vignali,
Collison et al. 2008). No contexto da inflamação intestinal induzida por T. gondii,
grandes esforços tem sido feitos a fim de desvendar os mecanismos associados ao
comprometimento de Tregs. Nesse sentido, Oldenhove e colaboradores (2009)
mostraram que a diminuição da disponibilidade de IL-2 no microambiente intestinal
após a infecção pelo parasito está associada ao comprometimento das Tregs
(Oldenhove, Bouladoux et al. 2009; Benson, Murray et al. 2012). Paralelamente, as
células Tregs dos animais infectados passam a expressar T-bet e secretar IFN-γ em
consequência do microambiente inflamatório assumindo assim, um fenótipo efetor
(Oldenhove, Bouladoux et al. 2009). Dados do nosso grupo mostram ainda que as Tregs
reminiscentes nos animais susceptíveis infectados com T. gondii apresentam um
comprometimento significativo na sua capacidade supressora (Fonseca, 2015,
comunicação pessoal). Nesse contexto, acreditamos que essas Tregs funcionalmente
comprometidas contribuam para a progressão da doença inflamatória intestinal durante
a infecção por T. gondii no animal susceptível. De fato, quando realizamos a depleção
de Tregs em camundongos resistentes, esses animais tornam-se susceptíveis à doença e
apresentam menor sobrevida e maior lesão tecidual no intestino delgado. De maneira
complementar, a transferência adotiva de Tregs para camundongos susceptíveis
C57BL/6 aumenta a sobrevida desses animais (Fonseca, 2015, comunicação pessoal).

74
O acúmulo e sobrevivência de células Treg no ambiente inflamatório está
relacionado à expressão de fatores de transcrição (Yu, Sharma et al. 2015). Dentre eles,
GATA3 e T-bet tem sua expressão regulada de maneira transitória nas Tregs, estando
sob influência das citocinas presentes no microambiente celular (Yu, Sharma et al.
2015). A ausência concomitante desses fatores está relacionada ao desenvolvimento de
doenças autoimunes devido ao aumento na expressão de RORγt e diminuição na
expressão de Foxp3 nas Tregs (Yu, Sharma et al. 2015). Nesse ambiente inflamatório,
GATA3, o qual tem sua expressão nas Tregs diminuída durante a infecção por T.
gondii, assume papel importante na fisiologia dessas células, principalmente em tecidos
de barreira como pele e mucosa intestinal. Sua expressão está associada ao acúmulo de
Tregs em sítios infecciosos e à expressão de Foxp3 por essas células (Wohlfert,
Grainger et al. 2011).
A fim de entendermos os outros fatores associados à diminuição de Tregs no
contexto da infecção por T. gondii, investigamos o papel da citocina pró-inflamatória
IL-6, a qual é relatado ter influência na atividade/plasticidade das células Treg
(Fujimoto, Nakano et al. 2011). Nesse sentido, é descrito que a função supressora de
células Treg periféricas fica prejudicada em camundongos que produzem quantidades
relativamente altas de IL-6 (Fujimoto, Nakano et al. 2011). A citocina IL-6 atuaria ainda
na modificação do fenótipo das células Treg, inativando a expressão do gene Foxp3 em
Tregs naturais, resultando na perda da capacidade supressora dessas células, e até
mesmo induzindo a conversão de Tregs em outras populações celulares (Kastner,
Dwyer et al. 2010). Nesse contexto, dados do grupo demonstraram que em culturas de
células de baço derivadas de camundongos C57BL/6, infectadas com taquizoítos de T.
gondii e tratadas com a citocina IL-6 recombinante, apresentaram uma redução bastante
acentuada na frequência das células Treg quando comparada com as culturas infectadas

75
e não tratadas (Fonseca, 2015, comunicação pessoal). É importante ressaltar que, o
efeito de IL-6 sobre a redução das Tregs ocorre apenas na presença do parasito, sendo
que o tratamento com IL-6 recombinante na ausência de infecção, não acarreta em
alteração na população de Tregs. Dessa forma, além da IL-6 o parasito pode induzir
outros mediadores ou ativar moléculas da imunidade inata que atuariam em sinergia
com essa citocina exercendo um efeito deletério sobre as Tregs.
Assim, supomos que IL-6 é capaz de potencializar a redução da frequência de
Tregs induzida por T. gondii. Soma-se a isso o fato de que camundongos deficientes
para a produção de IL-6 (background C57BL/6) são resistentes à infecção por T. gondii,
possuindo inclusive, uma quantidade maior de células Treg quando comparados aos
camundongos susceptíveis. Assim, quando depletamos as células Treg dos
camundongos IL-6-/-, estes se tornaram susceptíveis. A mortalidade dos camundongos
associada à alta produção de IL-6 também pode estar relacionada ao fato de que essa
citocina é capaz de aumentar a replicação intracelular de T. gondii em macrófagos,
favorecendo assim a progressão da inflamação tecidual e amplificando o colapso das
células Treg (Beaman, Hunter et al. 1994).
O papel dos micro-organismos comensais que habitam regiões como pele, trato
gastro intestinal e outros sítios mucosos tem sido evidenciado, principalmente no
contexto da indução de Tregs por bactérias. Em condições naturais, bactérias presentes
na microbiota intestinal secretam ácidos graxos de cadeia curta, derivados da
fermentação de fibras alimentares. Esses ácidos graxos, sendo o butirato o principal
deles, atuam na indução de células Tregs na região da lâmina própria do cólon intestinal
(Arpaia, Campbell et al. 2013). A microbiota intestinal é capaz ainda de atuar na
educação pós-tímica, uma vez que é observada a presença Tregs com TCR específico

76
para antígenos microbianos no contexto da mucosa intestinal, evento importante na
indução da tolerância do sistema imunológico a esses organismos (Lathrop, Bloom et al.
2011). No entanto, condições patológicas podem alterar a dinâmica protetora conferida
pelos organismos comensais, levando a uma modificação na composição populacional
da comunidade microbiana no trato gastrointestinal, e à translocação dessa microbiota
para locais secundários, como linfonodo mesentérico, baço e fígado (Hand, Dos Santos
et al. 2012). Nesse contexto, Hand e colaboradores (2012) mostraram que camundongos
infectados pela via oral com cistos de T. gondii desenvolvem uma inflamação intestinal
grave e um desbalanço na população da microbiota bacteriana comensal do intestino,
levando à diferenciação de células T específicas homeostáticas para componentes da
microbiota em células efetoras inflamatórias (Hand, Dos Santos et al. 2012). Sendo
assim, a disbiose, observada durante a infecção por T. gondii está associada à destruição
de células de Paneth, importantes na produção de peptídeos antimicrobianos que
controlam a diversidade populacional da microbiota. A perda dessas células leva à
expansão de bactérias da família das Enterobacteriaceae (Gram-negativas), fenômeno
estritamente associado ao desenvolvimento da inflamação intestinal induzida pelo
parasito, uma vez que camundongos germ-free (do inglês, livres de micro-organismos)
não apresentam essa inflamação intestinal (Raetz, Hwang et al. 2013). Todos esses
dados previamente descritos demonstram o profundo impacto da infecção por T. gondii
na dinâmica do sistema imunológico no sítio infeccioso e a relação dessas alterações
com a microbiota comensal residente. Nesse contexto, a hipótese de que o colapso
observado nas células Tregs estaria intimamente associado à translocação de bactérias
da microbiota intestinal parece bastante razoável, uma vez que a translocação da
microbiota intestinal é observada apenas no pico da fase aguda em animais susceptíveis
e ocorre em paralelo ao pico da produção de citocinas como IL-12, IL-10, IL-6 e ao

77
colapso de Tregs. A produção de IL-10 nos animais susceptíveis é reflexo do aumento
da inflamação tecidual relacionada ao controle da resposta inflamatória. Embora não
seja suficiente para impedir a progressão da inflamação tecidual em camundongos
C57BL/6, a deficiência na produção de IL-10 torna os animais significativamente
susceptíveis à infecção pelo parasito (Gazzinelli, Wysocka et al. 1996). Assumindo a
citocina IL-6 como determinante na redução das células Tregs, verificamos que tanto o
parasito em si quanto produtos microbianos são capazes de induzir a produção de IL-6
in vitro por macrófagos e células dendríticas derivadas de medula óssea de
camundongos resistentes e susceptíveis.
Na infecção intestinal por T. gondii foi demonstrado que neutrófilos e os
monócitos inflamatórios apresentam um papel importante na regulação da inflamação,
atuando na contenção de bactérias comensais (Dunay, Fuchs et al. 2010; Dunay and
Sibley 2010; Grainger, Wohlfert et al. 2013). Os monócitos inflamatórios possuem
importante papel na regulação da homeostase intestinal. Num contexto de inflamação,
em modelos de colite induzida por DSS (sulfate sodium salt), essas células são capazes
de induzir a produção de IL-6 e IL-23, mediante aumento da expressão de TLR2 e
NOD2, o que os tornam mais sensíveis aos produtos da microbiota (Zigmond, Varol et
al. 2012). Durante a infecção por T.gondii, o monócito inflamatório também pode
apresentar papel regulador através da produção de mediadores como prostaglandina E2
(PGE2) e IL-10 em resposta a organismos comensais (Grainger, Wohlfert et al. 2013).
PGE2 atua sobre neutrófilos, inibindo a ação destes e controlando a patogênese da
doença (Grainger, Wohlfert et al. 2013). Essas células são importantes no controle do
parasito e em modelos de infecção crônica por T. gondii, é demostrado que o
recrutamento de monócitos inflamatórios em resposta à profilina (ligante de TLR11/12)
é capaz de tornar camundongos resistentes à coinfecção de outros microorganismos,

78
como L. monocytogenes mediante produção de IFN-γ (Neal and Knoll 2014). Já os
neutrófilos possuem um papel variado podendo, tanto contribuir com o aumento da
lesão tecidual através da produção de TNF-α e de citocinas inflamatórias (Fournier and
Parkos 2012), quanto conter a disseminação de bactérias ao evitar o contato destas com
o epitélio intestinal (Molloy, Grainger et al. 2013). Nossos dados demonstram que
monócitos inflamatórios e neutrófilos estariam contribuindo significativamente com a
exacerbação da resposta inflamatória mediante infecção por T. gondii. Durante a fase
aguda da infecção, ambas as populações são bastante recrutadas para a lâmina própria
intestinal, sendo as subpopulações mais infectadas pelo parasito. Essas populações
produzem quantidades significativamente altas da citocina IL-6. Na lâmina própria
Intestinal, subpopulações de células dendríticas atuam de maneira importante na
homeostase. Dentre as populações, observamos que além de produzirem quantidades
exacerbadas de IL-6, subpopulações com células CD103+CD11b+, importantes para a
indução de Tregs, encontram-se diminuídas após a infecção (Denning, Wang et al.
2007). Já a subpopulação de células CD103-CD11b+, dentre as quais é possível
encontrar, macrófagos, identificados pela expressão do receptor de quimiocina CX3CR1
(Bogunovic, Ginhoux et al. 2009) encontra-se aumentada. É importante destacar que a
produção de IL-6 por células dendríticas pode exercer uma influência mais acentuada
sobre as populações de Treg dos órgãos linfóides drenantes em relação a lâmina Própria,
visto que após ativação, tais células migram para os linfonodos drenantes.
Nesse contexto, a produção diferencial de IL-6 parece estar relacionada à via dos
receptores TLRs, uma vez que macrófagos e células dendríticas derivadas da medula
óssea de camundongos deficientes para a moléculas adaptadora MyD88 tem a produção
dessa citocina prejudicada. Quando testamos in vivo a relação de receptores mais
relacionados ao reconhecimento do parasito como TLR2, TLR4, TLR9, TLR11 e

79
TLR12, com o colapso de Tregs e produção de IL-6, não obtivemos resultados que
demonstrassem que uma dessas vias estivesse associada ao colapso das células Tregs.
Em conjunto, esses dados reforçam a hipótese de que a microbiota intestinal estaria
relacionada à exacerbação da inflamação intestinal durante a infecção experimental por
T. gondii. De fato, quando avaliamos a translocação bacteriana em camundongos
deficientes para TLR2, TLR4 e TLR9, observamos a ocorrência de translocação em
camundongos deficientes para TLR2 e TLR9. Durante a infecção por T. gondii, é
demonstrado que nos camundongos deficientes para o receptor TLR4, a população de E.
coli aumenta no íleo. No entanto, esses animais teriam a resistência à infecção
aumentada em decorrência do não reconhecimento das bactérias intestinais na ausência
de TLR4. Desse modo, ocorre menor produção de IFN-γ e óxido nítrico (NO) na região
terminal do íleo. Esses dados divergem de nossos resultados, uma vez que, embora não
apresentem translocação da microbiota intestinal durante a infecção por T. gondii, os
animais deficientes para o receptor TLR4 sucumbem à infecção ainda na fase aguda. Já
o receptor TLR9 não está envolvido na inflamação intestinal desencadeada durante a
infecção por T. gondii, uma vez que a patogênese observada nos animais deficientes
para TLR9 apresentam ileíte comparável aos animais selvagens infectados, havendo, no
entanto, maior produção de IFN-γ e maior carga parasitária (Bereswill, Kuhl et al.
2014). Nesse sentido, nossos dados demonstram que os animais deficientes para o
receptor TLR9 apresentam redução da população de Tregs comparável aos animais
selvagens, além de altos níveis de RNAm para IL-6 no intestino delgado.
Os dados demonstram que o reconhecimento da microbiota intestinal está mais
relacionado à produção diferencial de IL-6 e colapso de Tregs. Isso é reforçado pelo não
envolvimento dos receptores TLR11 e TLR12, específicos no reconhecimento da
profilina de T. gondii e desenvolvimento de resposta Th1 contra T. gondii (Raetz,

80
Kibardin et al. 2013). Outro dado que reforça a hipótese é a produção de IL-6 em maior
quantidade por monócitos inflamatórios e neutrófilos não infectados durante a infecção
por T. gondii. Assim, monócitos inflamatórios e neutrófilos seriam sensibilizados em
resposta às bactérias translocadas durante a fase aguda da infecção por T. gondii. Nesse
sentido, a depleção da microbiota intestinal, mediante tratamento com antibióticos de
amplo espectro leva à recuperação da população de Tregs nos animais susceptíveis,
além de diminuir os níveis de IL-6 secretados. A depleção da microbiota mediante
tratamento com antibióticos inibe ainda a patogênese tecidual observada nos animais
susceptíveis durante a infecção por T. gondii.
Portanto, a utilização da infecção experimental por T. gondii como modelo para
estudo de fenômenos associados às respostas imunológicas de mucosa é bastante válida.
Assim, utilizando-se dessa abordagem, pudemos demonstrar que o parasito é capaz de
induzir inflamação intestinal exacerbada devido à perda dos mecanismos reguladores da
resposta imune tecidual, mais especificamente, do intestino delgado. Isso leva à
produção diferencial de IL-6 em resposta não somente ao parasito, mas à microbiota
translocada, e ao consequente colapso das células Treg no sítio de infecção e órgãos
linfoides associados. Dessa forma, o modelo contribui para o entendimento dos
mecanismos envolvidos no desenvolvimento de grande diversidade de doenças
inflamatórias intestinais associadas à quebra da tolerância contra organismos comensais.

81
6666---- CONCLUSÃOCONCLUSÃOCONCLUSÃOCONCLUSÃO

82
6 – CONCLUSÃO
Os dados gerados durante o trabalho permitem concluir que a microbiota possui
importante papel na mediação da redução da população das células Tregs no contexto
da infecção oral por T.gondii. Bactérias comensais translocadas do intestino delgado
após a infecção estimulam a produção de IL-6 por monócitos inflamatórios e neutrófilos
não parasitados. Desse modo a infecção por T. gondii, ao desencadear a translocação da
microbiota intestinal, induz a produção exacerbada de IL-6 e a consequente redução das
células Tregs.

83
7777----REFERÊNCIASREFERÊNCIASREFERÊNCIASREFERÊNCIAS

84
7-REFERÊNCIAS
Abbas, A. K., C. Benoist, et al. (2013). "Regulatory T cells: recommendations to simplify the nomenclature." Nat Immunol 14(4): 307-308.
Andrade, W. A., C. Souza Mdo, et al. (2013). "Combined action of nucleic acid-sensing Toll-like receptors and TLR11/TLR12 heterodimers imparts resistance to Toxoplasma gondii in mice." Cell Host Microbe 13(1): 42-53.
Arpaia, N., C. Campbell, et al. (2013). "Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation." Nature 504(7480): 451-455.
Beaman, M. H., C. A. Hunter, et al. (1994). "Enhancement of intracellular replication of Toxoplasma gondii by IL-6. Interactions with IFN-gamma and TNF-alpha." J Immunol 153(10): 4583-4587.
Benson, A., S. Murray, et al. (2012). "Microbial infection-induced expansion of effector T cells overcomes the suppressive effects of regulatory T cells via an IL-2 deprivation mechanism." J Immunol 188(2): 800-810.
Benson, N., J. de Jongh, et al. (2010). "Pharmacokinetic-pharmacodynamic modeling of alpha interferon response induced by a Toll-like 7 receptor agonist in mice." Antimicrob Agents Chemother 54(3): 1179-1185.
Bereswill, S., A. A. Kuhl, et al. (2014). "The impact of Toll-like-receptor-9 on intestinal microbiota composition and extra-intestinal sequelae in experimental Toxoplasma gondii induced ileitis." Gut Pathog 6: 19.
Bogunovic, M., F. Ginhoux, et al. (2009). "Origin of the lamina propria dendritic cell network." Immunity 31(3): 513-525.
Borsellino, G., M. Kleinewietfeld, et al. (2007). "Expression of ectonucleotidase CD39 by Foxp3+ Treg cells: hydrolysis of extracellular ATP and immune suppression." Blood 110(4): 1225-1232.
Brugman, S. and E. E. Nieuwenhuis (2010). "Mucosal control of the intestinal microbial community." J Mol Med (Berl) 88(9): 881-888.
Caetano, B. C., A. Biswas, et al. (2011). "Intrinsic expression of Nod2 in CD4+ T lymphocytes is not necessary for the development of cell-mediated immunity and host resistance to Toxoplasma gondii." Eur J Immunol 41(12): 3627-3631.

85
Collison, L. W., C. J. Workman, et al. (2007). "The inhibitory cytokine IL-35 contributes to regulatory T-cell function." Nature 450(7169): 566-569.
Coombes, J. L., N. J. Robinson, et al. (2005). "Regulatory T cells and intestinal homeostasis." Immunol Rev 204: 184-194.
Deaglio, S., K. M. Dwyer, et al. (2007). "Adenosine generation catalyzed by CD39 and CD73 expressed on regulatory T cells mediates immune suppression." J Exp Med 204(6): 1257-1265.
Debierre-Grockiego, F., M. A. Campos, et al. (2007). "Activation of TLR2 and TLR4 by glycosylphosphatidylinositols derived from Toxoplasma gondii." J Immunol 179(2): 1129-1137.
Denning, T. L., Y. C. Wang, et al. (2007). "Lamina propria macrophages and dendritic cells differentially induce regulatory and interleukin 17-producing T cell responses." Nat Immunol 8(10): 1086-1094.
Dunay, I. R., A. Fuchs, et al. (2010). "Inflammatory monocytes but not neutrophils are necessary to control infection with Toxoplasma gondii in mice." Infect Immun 78(4): 1564-1570.
Dunay, I. R. and L. D. Sibley (2010). "Monocytes mediate mucosal immunity to Toxoplasma gondii." Curr Opin Immunol 22(4): 461-466.
Elson, C. O. and Y. Cong (2012). "Host-microbiota interactions in inflammatory bowel disease." Gut Microbes 3(4): 332-344.
Eyerich, S., K. Eyerich, et al. (2009). "Th22 cells represent a distinct human T cell subset involved in epidermal immunity and remodeling." J Clin Invest 119(12): 3573-3585.
Fournier, B. M. and C. A. Parkos (2012). "The role of neutrophils during intestinal inflammation." Mucosal Immunol 5(4): 354-366.
Fujimoto, M., M. Nakano, et al. (2011). "The influence of excessive IL-6 production in vivo on the development and function of Foxp3+ regulatory T cells." J Immunol 186(1): 32-40.
Gazzinelli, R. T., M. Wysocka, et al. (1994). "Parasite-induced IL-12 stimulates early IFN-gamma synthesis and resistance during acute infection with Toxoplasma gondii." J Immunol 153(6): 2533-2543.

86
Gazzinelli, R. T., M. Wysocka, et al. (1996). "In the absence of endogenous IL-10, mice acutely infected with Toxoplasma gondii succumb to a lethal immune response dependent on CD4+ T cells and accompanied by overproduction of IL-12, IFN-gamma and TNF-alpha." J Immunol 157(2): 798-805.
Girardin, S. E., I. G. Boneca, et al. (2003). "Nod1 detects a unique muropeptide from gram-negative bacterial peptidoglycan." Science 300(5625): 1584-1587.
Girardin, S. E., I. G. Boneca, et al. (2003). "Nod2 is a general sensor of peptidoglycan through muramyl dipeptide (MDP) detection." J Biol Chem 278(11): 8869-8872.
Gondek, D. C., L. F. Lu, et al. (2005). "Cutting edge: contact-mediated suppression by CD4+CD25+ regulatory cells involves a granzyme B-dependent, perforin-independent mechanism." J Immunol 174(4): 1783-1786.
Grainger, J. R., M. H. Askenase, et al. (2014). "Contextual functions of antigen-presenting cells in the gastrointestinal tract." Immunol Rev 259(1): 75-87.
Grainger, J. R., E. A. Wohlfert, et al. (2013). "Inflammatory monocytes regulate pathologic responses to commensals during acute gastrointestinal infection." Nat Med 19(6): 713-721.
Hand, T. W., L. M. Dos Santos, et al. (2012). "Acute gastrointestinal infection induces long-lived microbiota-specific T cell responses." Science 337(6101): 1553-1556.
Harms Pritchard, G., A. O. Hall, et al. (2015). "Diverse Roles for T-bet in the Effector Responses Required for Resistance to Infection." J Immunol 194(3): 1131-1140.
Hawrylowicz, C. M. and A. O'Garra (2005). "Potential role of interleukin-10-secreting regulatory T cells in allergy and asthma." Nat Rev Immunol 5(4): 271-283.
Heimesaat, M. M., S. Bereswill, et al. (2006). "Gram-negative bacteria aggravate murine small intestinal Th1-type immunopathology following oral infection with Toxoplasma gondii." J Immunol 177(12): 8785-8795.
Hooper, L. V., D. R. Littman, et al. (2012). "Interactions between the microbiota and the immune system." Science 336(6086): 1268-1273.
Hori, S., T. Nomura, et al. (2003). "Control of regulatory T cell development by the transcription factor Foxp3." Science 299(5609): 1057-1061.
Jenkins, M. C., W. Tuo, et al. (2010). "Neospora caninum: cloning and expression of a gene coding for cytokine-inducing profilin." Exp Parasitol 125(4): 357-362.

87
Joetham, A., K. Takeda, et al. (2007). "Naturally occurring lung CD4(+)CD25(+) T cell regulation of airway allergic responses depends on IL-10 induction of TGF-beta." J Immunol 178(3): 1433-1442.
Johnson, A. M. (1984). "Strain-dependent, route of challenge-dependent, murine susceptibility to toxoplasmosis." Z Parasitenkd 70(3): 303-309.
Kanai, T., T. Kawamura, et al. (2006). "TH1/TH2-mediated colitis induced by adoptive transfer of CD4+CD45RBhigh T lymphocytes into nude mice." Inflamm Bowel Dis 12(2): 89-99.
Kasper, L., N. Courret, et al. (2004). "Toxoplasma gondii and mucosal immunity." Int J Parasitol 34(3): 401-409.
Kastner, L., D. Dwyer, et al. (2010). "Synergistic effect of IL-6 and IL-4 in driving fate revision of natural Foxp3+ regulatory T cells." J Immunol 185(10): 5778-5786.
Khan, I. A., K. A. Smith, et al. (1988). "Induction of antigen-specific parasiticidal cytotoxic T cell splenocytes by a major membrane protein (P30) of Toxoplasma gondii." J Immunol 141(10): 3600-3605.
Kinnebrew, M. A., C. G. Buffie, et al. (2012). "Interleukin 23 production by intestinal CD103(+)CD11b(+) dendritic cells in response to bacterial flagellin enhances mucosal innate immune defense." Immunity 36(2): 276-287.
Koblansky, A. A., D. Jankovic, et al. (2013). "Recognition of profilin by Toll-like receptor 12 is critical for host resistance to Toxoplasma gondii." Immunity 38(1): 119-130.
Lathrop, S. K., S. M. Bloom, et al. (2011). "Peripheral education of the immune system by colonic commensal microbiota." Nature 478(7368): 250-254.
Liesenfeld, O. (2002). "Oral infection of C57BL/6 mice with Toxoplasma gondii: a new model of inflammatory bowel disease?" J Infect Dis 185 Suppl 1: S96-101.
Liesenfeld, O., J. Kosek, et al. (1996). "Association of CD4+ T cell-dependent, interferon-gamma-mediated necrosis of the small intestine with genetic susceptibility of mice to peroral infection with Toxoplasma gondii." J Exp Med 184(2): 597-607.
Molloy, M. J., J. R. Grainger, et al. (2013). "Intraluminal containment of commensal outgrowth in the gut during infection-induced dysbiosis." Cell Host Microbe 14(3): 318-328.

88
Mosmann, T. R., H. Cherwinski, et al. (1986). "Two types of murine helper T cell clone. I. Definition according to profiles of lymphokine activities and secreted proteins." J Immunol 136(7): 2348-2357.
Neal, L. M. and L. J. Knoll (2014). "Toxoplasma gondii profilin promotes recruitment of Ly6Chi CCR2+ inflammatory monocytes that can confer resistance to bacterial infection." PLoS Pathog 10(6): e1004203.
Oderup, C., L. Cederbom, et al. (2006). "Cytotoxic T lymphocyte antigen-4-dependent down-modulation of costimulatory molecules on dendritic cells in CD4+ CD25+ regulatory T-cell-mediated suppression." Immunology 118(2): 240-249.
Oldenhove, G., N. Bouladoux, et al. (2009). "Decrease of Foxp3+ Treg cell number and acquisition of effector cell phenotype during lethal infection." Immunity 31(5): 772-786.
Pabst, O. and A. M. Mowat (2012). "Oral tolerance to food protein." Mucosal Immunol 5(3): 232-239.
Persson, E. K., H. Uronen-Hansson, et al. (2013). "IRF4 transcription-factor-dependent CD103(+)CD11b(+) dendritic cells drive mucosal T helper 17 cell differentiation." Immunity 38(5): 958-969.
Pifer, R., A. Benson, et al. (2011). "UNC93B1 is essential for TLR11 activation and IL-12-dependent host resistance to Toxoplasma gondii." J Biol Chem 286(5): 3307-3314.
Raetz, M., S. H. Hwang, et al. (2013). "Parasite-induced TH1 cells and intestinal dysbiosis cooperate in IFN-gamma-dependent elimination of Paneth cells." Nat Immunol 14(2): 136-142.
Raetz, M., A. Kibardin, et al. (2013). "Cooperation of TLR12 and TLR11 in the IRF8-dependent IL-12 response to Toxoplasma gondii profilin." J Immunol 191(9): 4818-4827.
Reardon, C., A. Wang, et al. (2008). "Transient local depletion of Foxp3+ regulatory T cells during recovery from colitis via Fas/Fas ligand-induced death." J Immunol 180(12): 8316-8326.
Sakaguchi, S., N. Sakaguchi, et al. (1995). "Immunologic self-tolerance maintained by activated T cells expressing IL-2 receptor alpha-chains (CD25). Breakdown of a single mechanism of self-tolerance causes various autoimmune diseases." J Immunol 155(3): 1151-1164.
Scanga, C. A., J. Aliberti, et al. (2002). "Cutting edge: MyD88 is required for resistance to Toxoplasma gondii infection and regulates parasite-induced IL-12 production by dendritic cells." J Immunol 168(12): 5997-6001.

89
Shaw, M. H., T. Reimer, et al. (2009). "T cell-intrinsic role of Nod2 in promoting type 1 immunity to Toxoplasma gondii." Nat Immunol 10(12): 1267-1274.
Sher, A., E. Y. Denkers, et al. (1995). "Induction and regulation of host cell-mediated immunity by Toxoplasma gondii." Ciba Found Symp 195: 95-104; discussion 104-109.
Sher, A., I. P. Oswald, et al. (1993). "Toxoplasma gondii induces a T-independent IFN-gamma response in natural killer cells that requires both adherent accessory cells and tumor necrosis factor-alpha." J Immunol 150(9): 3982-3989.
Subauste, C. (2012). "Animal models for Toxoplasma gondii infection." Curr Protoc Immunol Chapter 19: Unit 19 13 11-23.
Varol, C., A. Vallon-Eberhard, et al. (2009). "Intestinal lamina propria dendritic cell subsets have different origin and functions." Immunity 31(3): 502-512.
Veldhoen, M., R. J. Hocking, et al. (2006). "TGFbeta in the context of an inflammatory cytokine milieu supports de novo differentiation of IL-17-producing T cells." Immunity 24(2): 179-189.
Veldhoen, M., C. Uyttenhove, et al. (2008). "Transforming growth factor-beta 'reprograms' the differentiation of T helper 2 cells and promotes an interleukin 9-producing subset." Nat Immunol 9(12): 1341-1346.
Vignali, D. A., L. W. Collison, et al. (2008). "How regulatory T cells work." Nat Rev Immunol 8(7): 523-532.
Wang, Y. G., K. D. Kim, et al. (2005). "Stimulating lymphotoxin beta receptor on the dendritic cells is critical for their homeostasis and expansion." J Immunol 175(10): 6997-7002.
Wohlfert, E. A., J. R. Grainger, et al. (2011). "GATA3 controls Foxp3(+) regulatory T cell fate during inflammation in mice." J Clin Invest 121(11): 4503-4515.
Yarovinsky, F., D. Zhang, et al. (2005). "TLR11 activation of dendritic cells by a protozoan profilin-like protein." Science 308(5728): 1626-1629.
Yu, F., S. Sharma, et al. (2015). "Dynamic expression of transcription factors T-bet and GATA-3 by regulatory T cells maintains immunotolerance." Nat Immunol 16(2): 197-206.

90
Yu, Q. T., M. Saruta, et al. (2007). "Expression and functional characterization of FOXP3+ CD4+ regulatory T cells in ulcerative colitis." Inflamm Bowel Dis 13(2): 191-199.
Zigmond, E., C. Varol, et al. (2012). "Ly6C hi monocytes in the inflamed colon give rise to proinflammatory effector cells and migratory antigen-presenting cells." Immunity 37(6): 1076-1090.





























