Embed Size (px)
Citation preview

Universidade de São PauloInstituto de Química de São Carlos
Tema 17: Direcionamento Termodinâmico para a Síntese da Ligação Peptídica e Fosfodiéster
SQM0416- Bioquímica IIProf. Dr. Júlio César Borges
Daniel Luis do NascimentoGustavo Mancini
Murilo Gustavo Januário

Sumário1) Introdução.
a) Termodinâmica Geral;b) Ativação de Reações Químicas;c) Relação do equilíbrio químico reacional com o conceito de
deslocamento de reações.
2) Ligações Peptídicas.a) Custo energético da síntese de proteínas;b) Etapas da síntese de proteínas.
3) Ligação Fosfodiéster.a) Mecanismo;b) Balanço energético.
4) Cálculo aproximado de NTPs.

Conceitos termodinâmicos aplicados à bioquímica
• Energia livre de Gibbs: expressa a quantidadede energia capaz de realizar trabalho duranteuma reação à temperatura e pressãoconstantes.
• Entropia: é a desordem do sistema (caos).
• Entalpia: é o conteúdo de calor do sistemareagente.

Conceitos termodinâmicos aplicados à bioquímica
• 1a lei da termodinâmica: A energia é conservada, elanão pode ser criada ou destruida.
• 2a lei da termodinâmica: A entropia do universoaumenta.
• Processo espontâneo (liberação de energia ΔG).• Um processo pode ser espontâneo mesmo com ΔH > 0,
que se opõe ao processo,se a ΔS > 0 for suficiente.Ex.: Dissolução de NaCl em H2O.
• Processo não espontâneo (variação de ΔG positiva).• Estado padrão ΔG0 1 atm e 25 0C.

Equilíbrio e energia livre de Gibbs
• Onde K é a constante de equilíbrio.
• 𝐾𝐾 = [𝑝𝑝𝑝𝑝𝑝𝑝𝑝𝑝𝑝𝑝𝑝𝑝𝑝𝑝𝑝𝑝][𝑝𝑝𝑟𝑟𝑟𝑟𝑟𝑟𝑟𝑟𝑟𝑟𝑝𝑝𝑟𝑟𝑝𝑝]

Reversebilidade de reações biológicas
• Reações biológicas nem sempre sãoreversiveis se a liberação for muito grande deenergia. O processo é irreversível.
• Biologicamente, isso se deve principalmentepor não ter uma reação de acoplamentoenergético que consiga reverter a reação.

Ativação de reações
• Energia de barreira.• Acúmulo de energia para quebra da barreira
energética.
RTEAAek /−=

Papel enzimático
• A enzima diminui a energia de ativação dareação não mexendo com a termodinâmicadela, mas sim com a velocidade de formação.
• Ela interage com o substrato, ativando-o pelamudança de conformação do mesmo.

Formas de controle termodinâmico de reações
• Princípio de le Chatelier.• Excesso de produtos gera a reação reversa.• Controle de temperatura.• A temperatura define a constante de equilíbrio, ou
seja, ela define até que ponto a reação se propagaconsiderando reações rápidas.
• As reações de liberação de calor (quebra de ligação)são inibidas quando há aumento da temperatura.
• As reações endotérmicas são beneficidas com oaumento da temperatura ou acoplamentos de reações.

Formas de controle termodinâmico de reações
• Acoplamento de reações redox são as maisimportantes já que em reações de oxido-redução a oxidação do composto liberaenergia. Essa energia gerada pelo fluxo deelétrons é utilizada na formação de ligaçõesmais energéticas como a fosfodiéster porexemplo.

Exemplo de acoplamento energético:
glicose + Pi -> glicose-6-fosfato + H2O ΔG0= + 13,8 kJ.mol-1
ATP + H20 -> ADP + Pi ΔG0 = -30,5 kJ.mol-1
_________________________________________________glicose + ATP -> glicose-6-fosfato + ADP ΔG0 = -16,7 kJ.mol-1

Custo Energético Na Síntese De Proteínas
• A síntese de uma proteína pelo seu respectivomRNA requer energia.
• A formação de cada aminoacil-tRNA utilizadois grupos fosfatos de alta energia.
Estrutura tridimensional do RNAt.
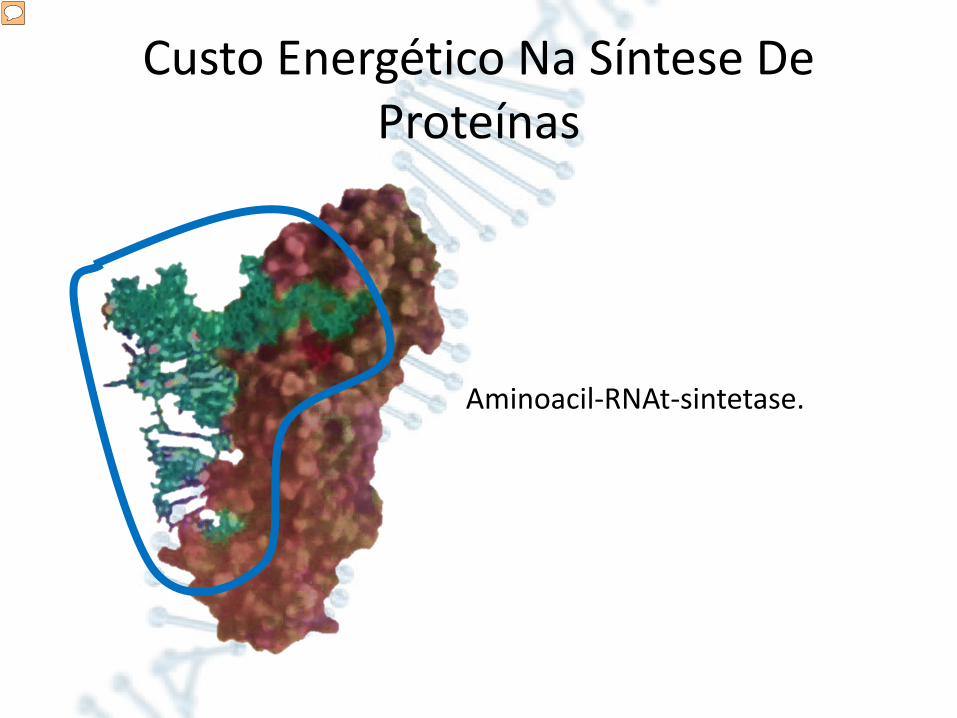
Custo Energético Na Síntese De Proteínas
Aminoacil-RNAt-sintetase.

Custo Energético Na Síntese De Proteínas
• Um GTP é clivado durante a primeira etapa dealongamento e outro durante a etapa detranslocação.
• Para a formação da ligação peptídica énecessária a energia derivada da hidrólise dequatro NTPs.

Custo Energético Na Síntese De Proteínas
• pelo menos 4 x 30,5 kJ.mol-1 = 122kJ.mol-1 deenergia de ligação fosfodiéster para gerar umaligação peptídica.
• Cada um dos compostos fosfatados gastosnesse processo têm papel fundamental.

Etapas da síntese de proteína
Ativação do aminoácido• Para que a síntese inicie é necessário que o
aminoácido seja ativado.
Ativação do aminoácido

Etapas da síntese de proteína
Ativação do aminoácido • Formação de umaminoacil-adenilato.
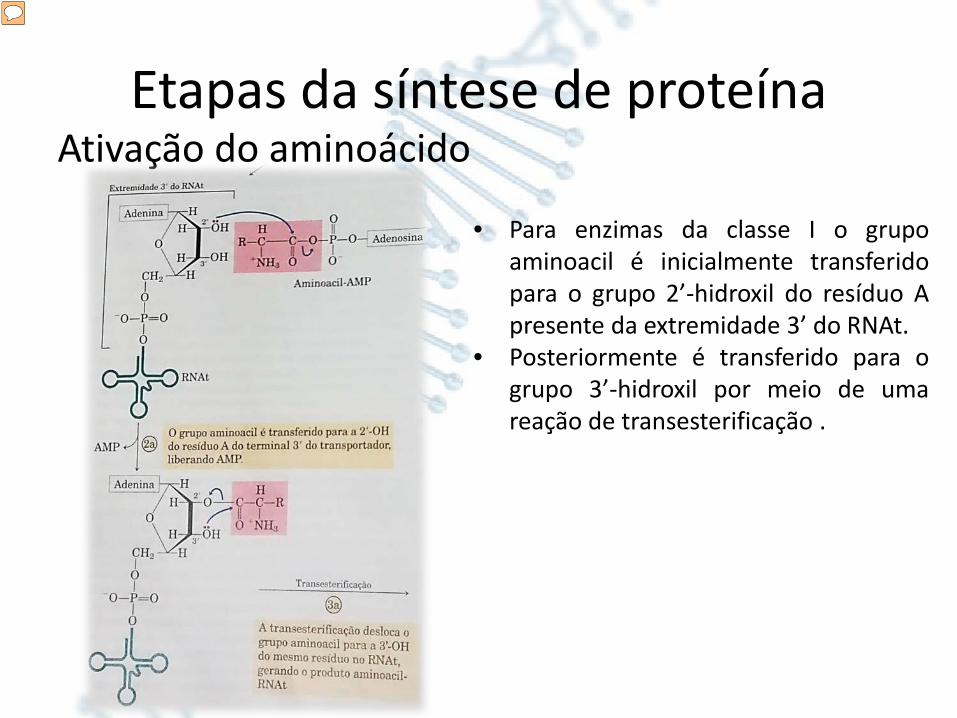
Etapas da síntese de proteínaAtivação do aminoácido
• Para enzimas da classe I o grupoaminoacil é inicialmente transferidopara o grupo 2’-hidroxil do resíduo Apresente da extremidade 3’ do RNAt.
• Posteriormente é transferido para ogrupo 3’-hidroxil por meio de umareação de transesterificação .

Etapas da síntese de proteínaAtivação do aminoácido
• Para enzimas da classe II o grupoaminoacil é transferido diretamentepara o grupo 3’-hidroxil do resíduo Apresente da extremidade 3’ do RNAt.

Etapas da síntese de proteínaIniciação
• aminoácido ativado pode sertransportado ao RNAm, onde ocódon e o anticódon se ligam.

Etapas da síntese de proteínaAlongamento: Ligação do segundo aminoacil-RNAt
• O segundo aminoacil-RNAt entra no sítio Ado ribossomo.
• Assim como o primeiro, o segundo AAtambém é ativado.

Etapas da síntese de proteínaAlongamento: Formação da ligação peptídica
• O grupo amino do aminoácido que entrou nosítio A do complexo do ribossomo, atacanucleofilicamente o grupo carboxil doprimeiro aminoácido.

Etapas da síntese de proteínaAlongamento: Translocação
• O RNAt que contem dois aminoácidos ligados transloca-se para este sítio desocupado.

Etapas da síntese de proteínaTerminação
• Esse processo apenas necessita de um doscódons de terminação.
• A reciclarem do ribossomo é necessários dofator de reciclagem e um GTP para que sedissociem os componentes envolvidos natradução.

Ligação Fosfodiéster
Relembrando:

Ligação Fosfodiéster

Ligação Fosfodiéster
Mecanismo:

Ligação Fosfodiéster
Balanço Energético:Em condições normais a hidrólise de
polinucleotídeos em nucleotídeos é umprocesso termodinamicamente favorecido.
(𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶 𝐶𝐶𝐶𝐶 𝑃𝑃𝑃𝑃𝑃𝑃𝐶𝐶𝑃𝑃𝑃𝑃𝑃𝑃𝑃𝑃𝐶𝐶𝑃𝑃𝑃𝑃𝑃𝐶𝐶𝐶𝐶𝑃𝑃)𝑁𝑁 + 𝑃𝑃𝑃𝑃𝑃𝑃𝑃𝑃𝐶𝐶𝑃𝑃𝑃𝑃𝑃𝐶𝐶𝐶𝐶𝑃𝑃 𝑚𝑚𝑃𝑃𝑃𝑃𝑃𝑃𝑚𝑚𝑃𝑃𝑚𝑚𝑚𝑚𝐶𝐶𝑃𝑃𝑃𝑃⇌ (𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶 𝐶𝐶𝐶𝐶 𝑃𝑃𝑃𝑃𝑃𝑃𝐶𝐶𝑃𝑃𝑃𝑃𝑃𝑃𝑃𝑃𝐶𝐶𝑃𝑃𝑃𝑃𝑃𝐶𝐶𝐶𝐶𝑃𝑃)𝑁𝑁+1 + 𝐻𝐻2𝑂𝑂
ΔGo= ~ +25,0 kJ.mol−1

Ligação Fosfodiéster
Então como de fato é possível a formação de polinucleotídeos ?
Acoplamento de reações !
𝑁𝑁𝑃𝑃𝑃𝑃𝑃𝑃𝐶𝐶𝑃𝑃𝑃𝑃𝑃𝐶𝐶𝐶𝐶𝑃𝑃 𝑃𝑃𝑡𝑡𝐶𝐶𝑚𝑚𝑃𝑃𝑚𝑚𝑚𝑚𝐶𝐶𝑃𝑃𝑃𝑃 + 𝐻𝐻2𝑂𝑂 ⇌ 𝑃𝑃𝑃𝑃𝑃𝑃𝑃𝑃𝐶𝐶𝑃𝑃𝑃𝑃𝑃𝐶𝐶𝐶𝐶𝑃𝑃 𝑚𝑚𝑃𝑃𝑃𝑃𝑃𝑃𝑚𝑚𝑃𝑃𝑚𝑚𝑚𝑚𝐶𝐶𝑃𝑃𝑃𝑃 + 𝑃𝑃𝐶𝐶𝑡𝑡𝑃𝑃𝑚𝑚𝑃𝑃𝑚𝑚𝑚𝑚𝐶𝐶𝑃𝑃𝑃𝑃 𝑃𝑃𝑃𝑃𝑖𝑖 ΔGo= −31,0 kJ.mol−1
(𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶 𝐶𝐶𝐶𝐶 𝑃𝑃𝑃𝑃𝑃𝑃𝐶𝐶𝑃𝑃𝑃𝑃𝑃𝑃𝑃𝑃𝐶𝐶𝑃𝑃𝑃𝑃𝑃𝐶𝐶𝐶𝐶𝑃𝑃)𝑁𝑁 + 𝑃𝑃𝑃𝑃𝑃𝑃𝑃𝑃𝐶𝐶𝑃𝑃𝑃𝑃𝑃𝐶𝐶𝐶𝐶𝑃𝑃 𝑚𝑚𝑃𝑃𝑃𝑃𝑃𝑃𝑚𝑚𝑃𝑃𝑚𝑚𝑚𝑚𝐶𝐶𝑃𝑃𝑃𝑃 ⇌ (𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶 𝐶𝐶𝐶𝐶 𝑃𝑃𝑃𝑃𝑃𝑃𝐶𝐶𝑃𝑃𝑃𝑃𝑃𝑃𝑃𝑃𝐶𝐶𝑃𝑃𝑃𝑃𝑃𝐶𝐶𝐶𝐶𝑃𝑃)𝑁𝑁+1 + 𝐻𝐻2𝑂𝑂 ΔGo= +25,0 kJ.mol−1
_________________________________________________________________________________________________________
(𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶 𝐶𝐶𝐶𝐶 𝑃𝑃𝑃𝑃𝑃𝑃𝐶𝐶𝑃𝑃𝑃𝑃𝑃𝑃𝑃𝑃𝐶𝐶𝑃𝑃𝑃𝑃𝑃𝐶𝐶𝐶𝐶𝑃𝑃)𝑁𝑁 + 𝑁𝑁𝑃𝑃𝑃𝑃𝑃𝑃𝐶𝐶𝑃𝑃𝑃𝑃𝑃𝐶𝐶𝐶𝐶𝑃𝑃 𝑃𝑃𝑡𝑡𝐶𝐶𝑚𝑚𝑃𝑃𝑚𝑚𝑚𝑚𝐶𝐶𝑃𝑃𝑃𝑃 ⇌ (𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶𝐶 𝐶𝐶𝐶𝐶 𝑃𝑃𝑃𝑃𝑃𝑃𝐶𝐶𝑃𝑃𝑃𝑃𝑃𝑃𝑃𝑃𝐶𝐶𝑃𝑃𝑃𝑃𝑃𝐶𝐶𝐶𝐶𝑃𝑃)𝑁𝑁+1 + 𝑃𝑃𝐶𝐶𝑡𝑡𝑃𝑃𝑚𝑚𝑃𝑃𝑚𝑚𝑚𝑚𝐶𝐶𝑃𝑃𝑃𝑃 𝑃𝑃𝑃𝑃𝑖𝑖 ΔGo= −6,0 kJ.mol−1

Ligação Fosfodiéster
𝑃𝑃𝐶𝐶𝑡𝑡𝑃𝑃𝑚𝑚𝑃𝑃𝑚𝑚𝑚𝑚𝐶𝐶𝑃𝑃𝑃𝑃 𝑃𝑃𝑃𝑃𝐶𝐶 + 𝐻𝐻2𝑂𝑂 ⇌ 2 𝑃𝑃𝐶𝐶 ΔGo= −33,0 kJ.mol−1
Δ𝑮𝑮𝟎𝟎 = +25,0 kJ.mol−1
Δ𝑮𝑮𝟎𝟎 = −39,0 kJ.mol−1

Cálculo Energético
• Quantidade de equivalentes de ATP para aformação da ligação fosfodiéster em DNA eRNA:
𝐷𝐷𝑁𝑁𝐴𝐴(𝑁𝑁) + 𝑁𝑁𝑁𝑁𝑃𝑃 ⇌ 𝐷𝐷𝑁𝑁𝐴𝐴(𝑁𝑁+1) + 2𝑃𝑃𝑖𝑖
1lig.fosfodiéster : 2NTP

Cálculo Energético
• Exemplo: Para uma proteína de 100 aminoácidos, partindo do DNA e passando por um mRNA:– DNA: Na cadeia polinucleotídica de DNA, um
conjunto de 3 nucleotídeos (códon) corresponde a um aminoácido.2 NTPs x 300 nucleotídeos x 2 (fitas) = 1200 NTPs
– mRNA: Temos apenas uma fita:1200 + 600 = 1800 NTPs

Calculo EnergéticoNumero de NTPs usadosAtivação: 1 NTPIniciação: 1 NTPAlongamento: 2 NTPTerminação: 1 NTP1 NTP ≈ 30,5 kJ.mol-1
4 x 30,5 kJ/mol = 122kJ.mol-1
Para uma proteína de 100 amino-ácidos
100 x 122 kJ/mol + 1 NTP ≈ 12230,5 kJ.mol-1
=12,23MJ.mol-1

Calculo Energético
• Número total de NTPs usados:- DNA: 1200 NTPs- mRNA: 600 NTPs- Proteína com 100 aminoácidos: 400 NTPs
TOTAL = 2200 NTPs = 67,1 MJ.mol-1

Referências1) NELSON D. L., COX M. M., Princípios de Bioquímica de Lehninger.
Sarvier, 4ª edição, 2006.2) Disponível em: http://www.jbc.org/content/282/2/e1.full.pdf3) Disponível em: http://bitesizebio.com/10279/the-basics-how-
does-dna-ligation-work/4) Disponível em:
https://www.chem.wisc.edu/deptfiles/genchem/netorial/modules/biomolecules/modules/dna1/dna13.htm
5) Disponível em: https://www.youtube.com/watch?v=VfjCn3MPEDE6) Disponível em:
https://www.youtube.com/watch?v=EiPQMSuxH4Q7) Slides de aula do Professor Dr. Júlio Borges, Bioquímica I.

OBRIGADO !





























