Embed Size (px)
Citation preview

UNIVERSIDADE ESTADUAL DE CAMPINAS
Instituto de Geociências
JULIANA SAMPAIO DA COSTA
ASSEMBLÉIA MACROFITOFÓSSIL DO RIO CAPIVARI: UM REGISTRO DA
FLORA PRÉ-GLOSSOPTERIS – TIETÊ (SP), BRASIL
CAMPINAS
2017

JULIANA SAMPAIO DA COSTA
ASSEMBLÉIA MACROFITOFÓSSIL DO RIO CAPIVARI: UM REGISTRO DA
FLORA PRÉ-GLOSSOPTERIS – TIETÊ (SP), BRASIL
DISSERTAÇÃO APRESENTADA AO INSTITUTO DE
GEOCIÊNCIAS DA UNIVERSIDADE ESTADUAL DE
CAMPINAS PARA OBTENÇÃO DO TÍTULO DE MESTRA
EM GEOCIÊNCIAS NA ÁREA DE GEOLOGIA E
RECUSROS NATURAIS
ORIENTADORA: PROFA. DRA. FRESIA SOLEDAD RICARDI TORRES BRANCO
ESTE EXEMPLAR CORRESPONDE À VERSÃO FINAL
DA DISSERTAÇÃO DEFENDIDA PELA ALUNA
JULIANA SAMPAIO DA COSTA E ORIENTADA PELA
PROFA. DRA. FRESIA SOLEDAD RICARDI TORRES
BRANCO.
CAMPINAS
2017

Agência(s) de fomento e nº(s) de processo(s): CNPq, 132015/2015-3
Ficha catalográficaUniversidade Estadual de CampinasBiblioteca do Instituto de GeociênciasCássia Raquel da Silva - CRB 8/5752
Costa, Juliana Sampaio da, 1991- C823a CosAssembléia macrofitofóssil do Rio Capivari : um registro da Flora Pré-
Glossopteris - Tietê (SP), Brasil / Juliana Sampaio da Costa. – Campinas, SP :[s.n.], 2017.
CosOrientador: Fresia Soledad Ricardi Torres Branco. CosDissertação (mestrado) – Universidade Estadual de Campinas, Instituto de
Geociências.
Cos1. Gangamopteris cyclopteroides. 2. Gondwana (Geologia). 3.
Glossopteris. 4. Bacias (Geologia). I. Ricardi-Branco, Fresia, 1963-. II.Universidade Estadual de Campinas. Instituto de Geociências. III. Título.
Informações para Biblioteca Digital
Título em outro idioma: Macrophitofossil assemblage of Capivari River's outcrop: : the Pre-Glossopteris Flora - state of São Paulo, BrazilPalavras-chave em inglês:Gangamopteris cyclopteroidesGondwana (Geology)GlossopterisBasins (Geology)Área de concentração: Geologia e Recursos NaturaisTitulação: Mestra em GeociênciasBanca examinadora:Fresia Soledad Ricardi Torres Branco [Orientador]Rafael Souza de FariaCarolina ZabiniData de defesa: 28-08-2017Programa de Pós-Graduação: Geociências
Powered by TCPDF (www.tcpdf.org)

UNIVERSIDADE ESTADUAL DE CAMPINAS
INSTITUTO DE GEOCIÊNCIAS
AUTORA: Juliana Sampaio da Costa
Assembléia Macrofitofóssil do Rio Capivari:
um registro da flora pré-glossopteris - Tieté (SP), Brasil
ORIENTADORA: Profa. Dra. Fresia Soledad Ricardi Torres Branco
Aprovado em: 28 / 08 / 2017
EXAMINADORES:
Profa. Dra. Fresia Soledad Ricardi Torres Branco - Presidente
Prof. Dr. Rafael Souza de Faria
Profa. Dra. Carolina Zabini
A Ata de Defesa assinada pelos membros da Comissão Examinadora,
consta no processo de vida acadêmica do aluno.
Campinas, 28 de agosto de 2017.

Aos meus pais, Cristiane e José,
por todo o apoio e amor incondicional.

AGRADECIMENTOS
- Agradeço a todos que participaram dessa jornada, direta e indiretamente, pois não há como
realizar algo sem ajuda neste mundo.
- Agradeço primeiramente aos meus pais por todo apoio, compreensão e amor incondicional.
- À minha irmã, Bárbara, pela amizade, paciência e união, apesar da distância geográfica na
maior parte do tempo.
- Aos meus avós, Ana e Carlos, pelo carinho, apoio e compreensão.
- À minha orientadora, Fresia, pelos anos dedicados de trabalho, companheirismo e amizade.
- Ao Instituto de Geociências da Unicamp e seus funcionários, por disponibilizarem todos os
recursos necessários para que este trabalho fosse realizado.
- Ao geólogo Fábio e ao técnico Cristiano, cuja ajuda foi imprescindível em muitas etapas
deste trabalho, especialmente nas etapas de campo.
- Às instituições FAPESP e CNPq pelos fundos fornecidos para esta pesquisa.
- Aos meus colegas de trabalho e bancada, Ariel, Adriana, Amanda, Isabela, Flávia e
Francisco, por compartilharem material, dúvidas e momentos de descontração. Em especial à
minha colega de trabalho e amiga Isabel, por toda sua disposição em me ajudar sempre.
- Aos meus amigos Mateus, Du, Deh, Jack, Biel, Camis, Mary, Oton, Felipe, Gabriel e
Vanessa por compartilharem experiências comigo. Em especial ao Rodrigo, que muito me
ajudou a realizar este trabalho; distância nenhuma separa.
- À Gabriela, pelos anos compartilhados, pelo apoio e por estar sempre ao meu lado.

SÚMULA
Juliana Sampaio da Costa é geóloga (2015) pela Universidade Estadual de Campinas. Tem
experiência na área de Geociências, especificamente Paleontologia com ênfase em
Paleobotânica, Tafonomia e reconstrução paleoambiental. Atuou como auxiliar didática na
disciplina de Paleontologia (de 2011 a 2016) do curso de Graduação em Geologia pela
UNICAMP e também colaborou no Programa de Formação Interdisciplinar Superior (2015 e
2016). Trabalhou no projeto FAPESP (2013/11563-6) “Integração de Dados Multifonte para
Análise Paleobotânica da Bacia do Paraná”, contribuindo com o estudo de Briófitas e
Megásporos do Grupo Itararé. Atualmente está finalizando o mestrado na área de Geociências
pela Universidade Estadual de Campinas, com a temática voltada para a descrição tafonômica
e paleoambiental de assembleias macrofitofossilíferas do afloramento Rio Capivari, na região
entre Piracicaba e Tietê, estado de São Paulo. Suas principais publicações incluem a co-
autoria em um artigo dedicado ao estudo das Paleofloras do Grupo Itararé, contribuindo com
o estudo de briófitas e megásporos supracitados, e publicações em congressos nacionais e
internacionais.

“se nem
for
terra
se
trans
for
mar”
(Paulo Leminski)

RESUMO
As assembleias fossilíferas permo-carboníferas da Bacia do Paraná exibem alta copiosidade em se
tratando de conteúdo vegetal, característica que se observa também na paleoflora do Afloramento Rio
Capivari, SP. Sua ocorrência sugere o início do estabelecimento da Província Paleoflorística de
Glossopteris no paleocontinente Gondwana antes mesmo do fim da glaciação permocarbonífera. O
conteúdo fóssil dos diferentes níveis estudados no Afloramento Rio Capivari atesta a influência de
climas episódicos mais amenos, em meio a pulsos glaciais, momento em que essas florestas puderam
se instaurar. A partir do estudo tafonômico das densidades e orientação das folhas in situ, foi possível
propor interpretações paleoclimáticas, bem como estabelecer relações paleoambientais em conjunto
com a análise do tamanho, forma e gênero de folhas, troncos e sementes carbonificadas ali
encontradas. A etapa de amostragem resultou em onze níveis descritos e orientados, 252 amostras
coletadas e cinco taxa identificados. No perfil de quatro metros, exposto na adjacência do Rio
Capivari, foi aberta uma seção de 150x100x50 centímetros do topo à base, exibindo a posição
estratigráfica e preservando a distribuição espacial dessas assembleias. Os resultados taxonômicos das
folhas apontam a presença apenas do gênero Gangamopteris, sendo a espécie G. obovata de maior
ocorrência. Esse gênero predominante é típico da Flora Pré-Glossopteris, e caracteriza-se por não
possuir nervura mediana em seu padrão de venação, configurando em uma vegetação típica do
Permiano do Gondwana Sul Ocidental em estágio inicial de instalação. Sua presença em meio a
estruturas glaciais encontradas no perfil, como diamictitos e seixos em flat iron, aponta estágios
cíclicos de clima mais brando, intercalados a momentos mais frios, revelando que o clima estava se
aquecendo regionalmente e permitindo a instalação dessa vegetação mais perene. Sua perenidade é
legitimada pela ocorrência de órgãos vegetais distintos na assembleia, especialmente sementes, cuja
produção ocorre sazonalmente. Dados palinológicos obtidos em estudos anteriores sugerem que estas
assembleias pertencem à zona Vitattina, subzona Protohaploxypinus goraiensis, no intervalo
Asseliano-Sakmariano. A orientação dos espécimes indica que não há paleocorrente evidente ou
sentido preferencial, caracterizando as assembleias como parautóctones. A litologia da coluna e
estruturas sedimentares encontradas, somada a distribuição espacial e taxonomia das assembleias,
evidencia que o paleoambiente pode estar relacionado às bordas de um lago ou planície de inundação.
Estes fitofósseis se localizam abaixo dos últimos diamictitos glaciais e representam prova veemente de
um período interglacial durante a deposição da assembleia.
Palavras-chave: Gangamopteris, Gondwana, Grupo Itararé, Pré-Glossopteris, Bacia do Paraná.

ABSTRACT
Permo-Carboniferous fossil assemblages on the Paraná Basin exhibit high copiousness on
plant content, which is also observed on the paleoflora of Rio Capivari (SP) outcrop. This
occurrence suggests the beginning of the Glossopteris Paleofloristic Province on the
Gondwana paleocontinent even before the Permo-Carboniferous glaciation. The fossil content
on its different layered levels testify the influence of episodic mild climates, between glacial
pulses, in which these forests bloomed. The in situ taphonomic study of leaves density and
orientation allowed tracing paleoclimate interpretations, as well as establishing
paleoenvironmental relationships, by analyzing the carbonized leaves, trunks, and seeds size,
form, and genus. Sampling resulted in eleven described levels, 252 collected samples and five
identified taxa. A 150x100x50 centimeters section was opened on the four-meter profile,
outcropped on Capivari River adjacencies, exhibiting the stratigraphic positioning and
preserving the spatial distribution of its assemblages. The taphonomic results showed
presence of the Gangamopteris genus, with higher occurrence of the G.abovata species,
typical from the Pre-Glossopteris flora. This genus is distinguished by the absence of midrib
on the venation pattern, which spatially and temporally allocates this vegetation at the South
Ocidental Gondwana, on its first stages of development, at the Permian. Its presence along
with the glacial evidences on the profile, such as dimictites and flat iron pebbles, suggests
cyclic stages of mild climates, interspersed by colder climates, revealing a regional warming
that allowed the installation of this perennial vegetation. This perenniality is confirmed by the
occurrence of distinct plants on the assembly, especially seeds, whose production occurs
seasonally. Palynological data of previously studies suggests these assemblages belongs to the
Vitattina zone, Protohaploxypinus goraiensis subzone, on the Asselian-Sakmarian interval.
The specimens’ orientation indicate the absence of evident paleocurrents or preferential sense,
characterizing a parautoctonous assembly. The profile lithologies and sedimentary structures,
as well as the assemblages special distribution and taphonomy, suggests the paleoenvironment
is related to the edges of a lake or a floodplain. These phitofossils are located below the
glacial diamictites and represent vehement proof of an interglacial period during the assembly
deposition.
Keywords: Gangamopteris, Gondwana, Itararé Group, Pré-Glossopteris, Paraná Basin.

LISTA DE FIGURAS
DISSERTAÇÃO
FIGURA 1: MAPA DE LOCALIZAÇÃO DO AFLORAMENTO RIO CAPIVARI, SP, PRÓXIMO À RODOVIA SP-
127. ................................................................................................................................................ 17
FIGURA 2: AFLORAMENTO ÀS MARGENS DO RIO CAPIVARI. A. VISTA PANORÂMICA DO
AFLORAMENTO A JUSANTE; B. VISTA A MONTANTE; C. SEÇÃO PARCIALMENTE DECAPEADA; D.
TRINCHEIRA ABERTA COM BANCADA EXPONDO OS NÍVEIS FÓSSEIS; E. DETALHE DA BANCADA
EXPONDO A ASSEMBLEIA. .............................................................................................................. 18
FIGURA 3: ESTRUTURAS ENCONTRADAS AO LONGO DA SEÇÃO ESTRATIGRÁFICA. A: INTERCALAÇÕES
DE LAMITO (ESCURA) E CAMADAS MAIS ARENOSAS (CLARAS) COM LAMINAÇÃO PLANO-
PARALELA; B: CAMADA ARENOSA INTERCALADA A FINAS CAMADAS DE SILTITO SOBREJACENTE A
CAMADA SILTOSA CONTENDO CONCREÇÕES; C: CAMADA CONTENDO GRETAS DE CONTRAÇÃO
PREENCHIDAS POR SEDIMENTO LIGEIRAMENTE MAIS GROSSO; D:LAMITO ARENOSO COM
ICNOFÓSSEIS E MARCAS DE RASTOS; E: DIAMICTITO COM ESTRUTURAS GLACIAIS (SEIXOS FLAT
IRON). ............................................................................................................................................. 20
FIGURA 4: COLUNA ESTRATIGRÁFICA DO CARBONÍFERO SUPERIOR AO PERMIANO INFERIOR DA
BACIA DO PARANÁ (MODIFICADO DE MILANI ET AL., 2007). ........................................................ 21
FIGURA 5: GONDWANA NO LIMITE PERMO-CARBONÍFERO QUANDO SE ENCONTRAVA SOB INFLUÊNCIA
GLACIAL (VEEVERS ET AL., 2004). ................................................................................................. 22
FIGURA 6: COLUNA ESTRATIGRÁFICA DO AFLORAMENTO RIO CAPIVARI, COM A INDICAÇÃO DOS
NÍVEIS DESCRITOS E PRINCIPAIS ESTRUTURAS (BASEADA EM ROHN ET AL., 2000). ...................... 25
FIGURA 7: PERFIL GEOLÓGICO DA ESTRADA NOVA TIETÊ-PIRACICABA (SOARES ET AL., 1977). ......... 26
FIGURA 8: MÉTODOS DE TRABALHO; A: MALHA QUADRICULADA EM USO PARA MEDIÇÃO DA
DENSIDADE; B: DECAPEAMENTO DENOTANDO O NÍVEL FÓSSIL A SER DESCRITO (NÍVEL 9); C:
TRINCHEIRA COM O NÍVEL FÓSSIL EXPOSTO. D: LOCAL DO LEVANTAMENTO ESTRATIGRÁFICO
PARCIALMENTE ABERTO, CONFORME O MÉTODO DE TRABALHO. ................................................. 30
ARTIGO
FIGURE 1: CAPIVARI RIVER OUTCROP LOCALIZATION MAP, AT TIETÊ, SÃO PAULO STATE. ...... 35
FIGURE 2: GEOLOGIC PROFILE OF THE NEW TIETÊ-PIRACICABA ROAD (SOARES ET AL., 1977). 36
FIGURE 3: WORKING METHODS; A. CHECKERED GRID USED FOR DENSITY MEASUREMENTS; B.
OUTCROOP DECAPPING AND CLEARING OF THE FOSSIL LEVEL (LEVEL 9); C. TRENCH WITH
EXPOSED FOSSIL LEVEL. D. PARTIALLY CLEARED OUTCROP, USED FOR THE STRATIGRAPHIC
SURVEY, ACCORDING TO THE WORK METHODS. .................................................................. 38
FIGURE 4: SPECIMENS STUDIED ON THE PALEOBOTANIC SYSTEMATICS. A: SAMPLE CP1/625B OF
GANGAMOPTERIS AFF. MOSESII (SCALEBAR = 1 CM); SAMPLE CP1/669 OF GANGAMOPTERIS AFF.
OBOVATA E SAMAROPSIS SP. AFF. S. RIGBYI (SCALEBAR = 1 CM); C: SAMPLE CP1/633 OF
SAMAROPSIS SP. AFF. S. RIGBYI (SCALEBAR = 2 MM); D: SAMPLE CP1/666 OF GANGAMOPTERIS
AFF. ANGUSTIFOLIA (SCALEBAR = 1 CM). ............................................................................ 45
FIGURE 5: DIFFERENTIATED SPECIMENS ON THE PALEOBOTANIC SYSTEMATICS. A. SAMPLE CP1/650A
OF GANGAMOPTERIS AFF. OBOVATA (SCALEBAR = 1 CM); B SAMPLE CP1/756 OF
GANGAMOPTERIS AFF. ANGUSTIFOLIA (SCALEBAR = 2 CM); C. CP1/626B, CORRESPONDS TO
LEVEL 6, VERY DENSE, ALMOST UNDIFFERENTIATED LEAF LAMINA (VISIBLE GANGAMOPTERIS
AFF. ANGUSTIFOLIA; SCALEBAR = 1 CM); D. CP1/627 CORRESPONDS TO LEVEL 5, VERY DENSE,

HOWEVER THE LEAF LAMINA IS CLEARLY DIFFERENTIATED. BLUE ARROW: GANGAMOPTERIS AFF.
OBOVATA, RED ARROW: GANGAMOPTERIS AFF. ANGUSTIFOLIA (SCALEBAR = 2 CM). ....... 46
FIGURE 6: ABSOLUTE DENSITY QUALITATIVE GRAPHICAL REPRESENTATIONS, FROM CAPIVARI RIVER
COLLECTED DATA, BASED ON DENSITY DATA OBTAINED USING THE POINT COUNTER METHOD
(SPICER, 1980 E 1988). TEN LEVELS OF GRIDS (1 TO 10) ILLUSTRATING THE IN SITU POSITIONING
OF THE PHYTOFOSSILS. THE OUTCROP CUT IS IN THE 242N DIRECTION, DISPLAYED ON THE
IMAGES. ............................................................................................................................... 48
FIGURE 7: LEVELS PRESENTING THE STUDIED ASSEMBLAGES: A. LEVEL 8; B. MUDCRACKS BETWEEN
LEVELS 6 AND 7; C. LEVEL 9; D AND E. LEVEL 6; F. COMPARISON BETWEEN LEVELS 9 (UPPER)
AND 8 (LOWER). ................................................................................................................... 49
FIGURE 8: PERCENTAGE-BASED DENSITY GRAPH COMPARING THE AVERAGE DENSITIES ON EACH OF
THE STUDIES LEVELS (GRIDS). ............................................................................................ 50
FIGURE 9: ORIENTATION ROSETTES WITH PREFERENTIAL ORIENTATION OF THE VEGETAL FOSSIL
ASSEMBLAGES. .................................................................................................................... 52
FIGURE 10: PLANT ORGANS PROPORTION ON EACH STUDIED LEVEL OF THE CAPIVARI RIVER
PALEOFLORISTIC ASSOCIATION. THE X AXIS SHOWS THE DENSITY OF THE 30X30 CENTIMETERS
GRID. .................................................................................................................................... 53
FIGURE 11: AVERAGE FRAGMENTATION OF THE RIO CAPIVARI’S ASSEMBLY PHYTOFOSSILS, IN
PERCENTAGE. TO VALUES ABOVE 50% THERE ARE WELL PRESERVED SPECIMENS; TO VALUES
EQUAL OR BELOW 50% THE SPECIMENS ARE POORLY PRESERVED. .................................... 54
FIGURE 12: STRATIGRAPHIC COLUMN ON THE OUTCROP SECTION, WITH MACROPHITOFOSSILS
ASSEMBLAGES, SPECIES, AND STRUCTURES INDICATED. ..................................................... 55
FIGURE 13: COMPARATIVE DENSITIES OF ALL LEVELS (LEAVES AND TWIGS) ON THE CAPIVARI RIVER
PALEOFLORISTIC ASSOCIATION. .......................................................................................... 63
FIGURE 14: RECONSTRUCTION OF THE CAPIVARI RIVER TAPHOFLORA ENVIRONMENT, BASED ON THE
TAPHONOMIC DESCRIPTION AND FOSSILS OF THE ITARARÉ GROUP, SÃO PAULO STATE. ... 64

LISTA DE TABELAS
TABLE 1: CHART OF THE DESCRIBED SPECIMENS TO EACH LEVEL AND ITS CATALOGUE NUMBERING
ON THE HIDROGEOLOGY LABORATORY OF THE GEOSCIENCES INSTITUTE (UNICAMP),
CORRESPONDING TO THE PALEOBOTANY COLLECTION 1 (PC1). ........................................ 56
TABLE 2: SPECIES DISTRIBUTION PER LEVEL. ANALOGOUS TO THE FIGURE 12 COLUNM. ......... 62

SUMÁRIO
1. INTRODUÇÃO ................................................................................................................. 15
2. OBJETIVOS ...................................................................................................................... 16
3. ÁREA DE ESTUDO.......................................................................................................... 17
4. CONTEXTO GEOLÓGICO ............................................................................................ 21
4.1 AFLORAMENTO RIO CAPIVARI .......................................................................................... 23
5. MATERIAIS E MÉTODOS ................................................................................................ 28
5.1 MATERIAL DE ESTUDO ...................................................................................................... 28
5.2 MÉTODOS DE ESTUDO ....................................................................................................... 29
5.2.1 Estudos de Campo ...................................................................................................... 29
5.2.2 Estudos de Laboratório .............................................................................................. 31
6. ARTIGO: TAPHONOMIC CHARACTERIZATION OF THE CAPIVARI RIVER
MACROPHITOFOSSIL ASSEMBLY: A PRE-GLOSSOPTERIS RECORD – SÃO PAULO,
BRAZIL. .................................................................................................................................... 32
INTRODUCTION .................................................................................................................. 33
PARANÁ BASIN AND THE GONDWANA PALEOZOIC .................................................. 34
Capivari River outcrop........................................................................................................ 35
MATERIALS AND METHODS ............................................................................................ 37
RESULTS ............................................................................................................................... 39
Paleobotany systematic ....................................................................................................... 39
Density ................................................................................................................................. 47
Orientation .......................................................................................................................... 50
Distribution ......................................................................................................................... 53
CAPIVARI RIVER OUTCROP AND THE PRE-GLOSSOPTERIS FLORA ........................................ 57
Paleobiogeography ............................................................................................................. 57
Paleobotany systematics ..................................................................................................... 57
Biostratigraphy ................................................................................................................... 59
Taphonomy .......................................................................................................................... 59
CONCLUSIONS ....................................................................................................................... 65
ACKNOWLEDGEMENTS ...................................................................................................... 66
REFERENCES .......................................................................................................................... 66
ATTACHMENT I ................................................................................................................... 76
7. CONSIDERAÇÕES FINAIS ........................................................................................... 81
8. BIBLIOGRAFIA ............................................................................................................... 82

15
1. INTRODUÇÃO
O estudo das paleofloras da Bacia do Paraná desperta interesse desde 1908, quando
White descreveu exemplares de lenhos fósseis permineralizados em camadas de carvão,
oriundos do Permiano do Rio Grande do Sul. Desde então muito material foi estudado,
somando décadas de contribuição científica acerca dessas assembleias no Brasil.
As paleofloras pertencentes ao Grupo Itararé no estado de São Paulo se encontram
distribuídas estratigraficamente desde os níveis mais basais do grupo (briófitas do Município
de Campinas, SP – Bashkiano ao Moscoviano), até seu topo (Flora de Cerquilho – Asseliano
ao Sakmariano). Os fitofósseis mais abundantes do Grupo Itararé são Gynkgophytas e
Coniferophytas, embora também sejam encontradas Lycophyta e Sphenophyta e, em menor
número de espécies, Bryophyta (Christiano-de-Souza e Ricardi-Branco, 2013; Ricardi-Branco
et al., 2013).
Mune e Bernardes-de-Oliveira (2007) dividiram tais ocorrências dentro de nove floras
(Campinas, Buri, Itapeva, Araçoiaba da Serra, Monte Mor, Itu, Itaporanga, Tietê e Cerquilho),
posicionando-as estratigraficamente de acordo com sua idade e conteúdo fossilífero.
Posteriormente, Bernardes de Oliveira et al. (2016) reuniram as floras do Grupo Itararé dentro
de dois conjuntos. O mais antigo refere-se aos fitofósseis encontrados em depósitos da
Formação Lagoa Azul, que vai do Westphaliano ao Stephaniano e o mais recente, diz respeito
às paleofloras presentes na Formação Campo Mourão, do Stephaniano ao Sakmariano (Milani
et al., 1998; Milani et al., 2007).
Rohn et al. (2000) registraram em estudo preliminar a ocorrência dos vegetais fósseis
coletados para este trabalho, preservados como compressões carbonificadas. Esse tipo de
fossilização conserva estruturas anatômicas (e.g. cutículas) e morfológicas, possibilitando a
identificação dos gêneros dos macrofitofósseis.
A seção estudada do afloramento Rio Capivari é extremamente rica em termos de
conteúdo fóssil. Onze níveis com assembleias fitofósseis distintas foram descritas, adquirindo
informações sobre sua densidade, orientação e morfologia dos espécimes. A partir dos dados
obtidos, foi possível determinar quais condições da deposição e quais os principais eventos
até o soterramento desse material. O estudo do afloramento do Rio Capivari se justifica pelas
poucas publicações científicas a respeito da localidade. Além disso, o viés tafonômico do
estudo é inédito para exemplares da Bacia do Paraná, dada a dificuldade de encontrar
exposições com alto grau de preservação no Brasil, tornando este tipo de pesquisa muito raro
no país.

16
2. OBJETIVOS
O objetivo central deste estudo é analisar a tafonomia, as informações paleoambientais
e possíveis informações paleoecológicas, através do mapeamento estratigráfico e tafonômico
in situ, para as assembleias fitofossilíferas encontradas no afloramento Rio Capivari.
Como objetivos específicos se integrará a descrição morfológica e análise taxonômica
dos espécimes a nível de gênero, a fim de associá-las a espécies previamente descritas na
literatura. Também consta inferir o posicionamento estratigráfico do afloramento dentro das
biozonas descritas para o Grupo Itararé e comparar seus morfotipos a outras assembleias
inseridas na Bacia do Paraná. Dessa forma, contribuir-se-á com o estudo das palelofloras do
Grupo Itararé no Estado de São Paulo.
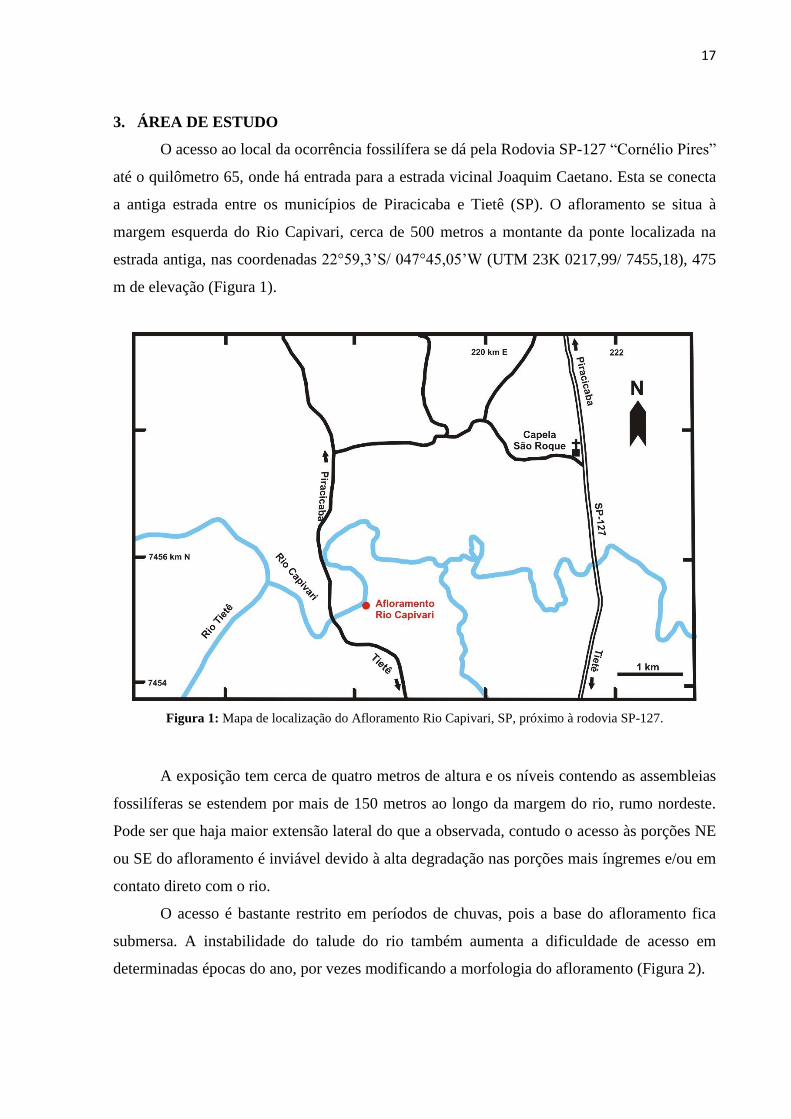
17
3. ÁREA DE ESTUDO
O acesso ao local da ocorrência fossilífera se dá pela Rodovia SP-127 “Cornélio Pires”
até o quilômetro 65, onde há entrada para a estrada vicinal Joaquim Caetano. Esta se conecta
a antiga estrada entre os municípios de Piracicaba e Tietê (SP). O afloramento se situa à
margem esquerda do Rio Capivari, cerca de 500 metros a montante da ponte localizada na
estrada antiga, nas coordenadas 22°59,3’S/ 047°45,05’W (UTM 23K 0217,99/ 7455,18), 475
m de elevação (Figura 1).
Figura 1: Mapa de localização do Afloramento Rio Capivari, SP, próximo à rodovia SP-127.
A exposição tem cerca de quatro metros de altura e os níveis contendo as assembleias
fossilíferas se estendem por mais de 150 metros ao longo da margem do rio, rumo nordeste.
Pode ser que haja maior extensão lateral do que a observada, contudo o acesso às porções NE
ou SE do afloramento é inviável devido à alta degradação nas porções mais íngremes e/ou em
contato direto com o rio.
O acesso é bastante restrito em períodos de chuvas, pois a base do afloramento fica
submersa. A instabilidade do talude do rio também aumenta a dificuldade de acesso em
determinadas épocas do ano, por vezes modificando a morfologia do afloramento (Figura 2).

18
Figura 2: Afloramento às margens do Rio Capivari. A. Vista panorâmica do afloramento a jusante; B. Vista a montante;
C. Seção parcialmente decapeada; D. Trincheira aberta com bancada expondo os níveis fósseis; E. Detalhe da bancada
expondo a assembleia.
A
B C
D
B
E
B

19
Níveis carbonosos aparecem por toda a base da área intercalados a níveis lamíticos e
arenosos de coloração mais clara. Essa variação quase cíclica é atribuída a sazonalidade do
ambiente deposicional. Nesse contexto, dentro dos níveis carbonificados, se encontram as
assembleias macrofitofossilíferas. Estas estão alocadas nas porções mais finas de níveis
carbonosos das intercalações, por todo o perfil, e bem distribuídas espacialmente, com boa
continuidade lateral (Rohn et al., 2000).
É possível observar diversas estruturas sedimentares distintas, como laminações
plano-paralelas a cruzadas, concreções, paleossolos, diamictitos de origem glacial,
icnofósseis, gretas de contração, pavimento estriado e flasers, as quais agregam evidências
para a reconstituição da história evolutiva da área e caracterização do paleoambiente (Figura
3).

20
Figura 3: Estruturas encontradas ao longo da seção estratigráfica. A: Intercalações de lamito (escura) e
camadas mais arenosas (claras) com laminação plano-paralela; B: Camada arenosa intercalada a finas
camadas de siltito sobrejacente a camada siltosa contendo concreções; C: Camada contendo gretas de
contração preenchidas por sedimento ligeiramente mais grosso; D:Lamito arenoso com icnofósseis e marcas
de rastos; E: Diamictito com estruturas glaciais (seixos flat iron).
E
B
D
B
C
B A

21
4. CONTEXTO GEOLÓGICO
A Bacia do Paraná é oriunda de uma grande sinéclise na porção central-sul do
continente Sul-Americano. Sua forma é ovalada e alongada no sentido NE-SW, com 1,5
milhão de quilômetros quadrados e profundidades de até 5000 m na região do depocentro.
Sua área estende-se a Argentina, Brasil, Paraguai e Uruguai (Bacia do Chaco-Paraná),
podendo também ser correlacionada a Bacia do Karoo (África do Sul) e Bacia de Huab
(Namíbia) (Ricardi-Branco et al., 2016; Milani e DeWitt, 2008). Foi dividida por Milani et al.
(2007) em seis grandes superciclos deposicionais, do Ordoviciano ao Cretáceo, a citar: Rio
Ivaí, Paraná, Gondwana I, Gondwana II, Gondwana III e Bauru.
Segundo Milani et al. (1998, 2007), a Supersequência Gondwana I (Carbonífero-
Triássico), possui espessuras que variam entre 1,5 a 2,4 km e perfaz o maior volume
sedimentar da Bacia do Paraná. Ela aflora como uma estreita e contínua faixa na borda leste
da bacia, direção nordeste-sudoeste, de São Paulo ao Rio Grande do Sul, estendendo-se ao
Uruguai (Bacia Chaco-Paraná). É subdividida nos Grupos Itararé, Guatá e Passa Dois,
unidades que refletem variadas condições deposicionais ao longo do tempo geológico (Figura
4).
Figura 4: Coluna estratigráfica do Carbonífero Superior ao Permiano Inferior da Bacia do Paraná (modificado
de Milani et al., 2007).
Cada Grupo registra um tipo de ambiente, desde marinho com influência
glacial/interglacial/pós-glacial, até continental árido com domínio de campos de dunas
eólicas. Essas características enfatizam um ciclo transgressivo-regressivo completo que
ocorreu no Gondwana Ocidental, onde a deglaciação da calota sul-americana ocasionou um
aumento no nível do mar e o fechamento da bacia às incursões marinhas vindas da porção

22
oeste. O fechamento foi ocasionado pelo soerguimento de arcos estruturais que circundam a
bacia devido à tectônica compressional nas margens do Gondwana (Daemon e Quadros,
1970; Schneider et al., 1974; Milani et al., 2007).
Durante o Paleozoico Superior a porção Sul Ocidental do paleocontinente Gondwana
estava sob influência da Glaciação do Paleozóico Superior (Late Paleozoic Ice Age – LPIA),
devido a sua posição geográfica (Figura 5). O Grupo Itararé corresponde ao mais expressivo
registro deste evento apresentando grande diversidade de litofácies, característica que
dificulta sua compartimentação e delimitação estratigráfica (Longhim, 2003; Veevers et al.,
2004).
Figura 5: Gondwana no limite Permo-Carbonífero quando se encontrava sob influência glacial (Veevers et al.,
2004).
A deposição do Grupo Itararé teve início no Moscoviano (Pensilvaniano) e prolongou-
se ao Sakmariano (Cisuraliano). Nesse período a bacia possuía uma entrada glacial
parcialmente isolada devido ao controle tectonossedimentar, ou um estuário com a salinidade
controlada pela variação do nível médio oceânico global, regulando um ciclo transgressivo-
regressivo (Rocha-Campos et al., 2008). Isto posto, a diversidade faciológica abrange
depósitos plataformais subaquosos (Caetano-Chang, 1984; Souza Filho, 1986; Santos, 1987),
glaciais terrestres (Rocha-Campos, 1967; Santos et al., 1996) e de ambientes marinhos
proximais (Saad, 1977; Santos, 1979) e distais (Gama Jr et al., 1992; Santos et al., 1996;
Canuto et al., 2001).

23
Algumas estruturas que compõem as litofácies, como diamictitos, conclomerados,
varvitos, turbiditos, arenitos, lamitos, estratificações cruzadas, laminação plano-paralela,
pavimentos estriados e camadas carbonosas, são comuns ao Itararé. Estas também
interpretadas como sequências de degelo, corroborada pelo recuo dos tratos de sistema
deposicionais, e breves registros do avanço da geleira intercalados (Vesely e Assine, 2006).
Assim como suas fácies, o conteúdo fossilífero do Grupo Itararé é bastante variado.
Em seu registro constam foraminíferos, braquiópodes, moluscos insetos, peixes e fósseis
vegetais (Veseely e Assine, 2006; Rocha-Campos et al., 2008). Os últimos geralmente estão
associados aos acamamentos carbonosos, salientando o desenvolvimento das paleofloras nos
intervalos das glaciações. São encontradas Bryophyta, Lycophyta, Sphenophyta, Filicophyta,
Pteridospermatophyta, Gynkgophyta, sendo que os intervalos mais recentes abrigam os
primeiros elementos da Flora de Glossopteris (Rohn et al., 2000).
Sobrejacente ao Itararé situa-se o Grupo Guatá, apresentando fácies de ambientes
fluviais, deltaicos, costeiros e marinhos resultantes de um momento sob influência do
aumento gradual da umidade e da temperatura (Souza, 2006). Possui também leitos de carvão,
os quais contém abundantes registros de macro e microfitofósseis da Flora de Glossopteris
(Ricardi-Branco, 1998).
4.1 Afloramento Rio Capivari
O Afloramento Rio Capivari está inserido no Grupo Itararé no estado de São Paulo e
corresponde aos registros glaciais/interglaciais encontrados na região. Situa-se em um talude à
margem esquerda do Rio Capivari, caracterizado por ser geomorfologicamente instável. O
acesso ao local é bastante restrito, especialmente em períodos de chuvas (outubro a abril)
quando a base do afloramento fica submersa. A caminhada até o local é de cerca de 500 m
dentro de mata ciliar fechada, com descida de nível próximo a zona do afloramento. A
exposição tem cerca de 150 m de extensão lateral, com diversas descontinuidades
morfológicas. Escolheu-se a área mais estável para fazer o perfil estratigráfico do
afloramento, cuja altura é de cerca de quatro metros.
A porção basal é composta de lamitos cinzas com grande concentração de folhas nos
níveis mais escuros e intercalações arenosas próximas ao topo. Dentre as estruturas
sedimentares encontradas podemos citar o acamamento plano-paralelo com laminação muito
fina por toda a secção, com intercalações de areia fina na porção superior. Essa unidade
contém os níveis 0 (corresponde ao chão e ao nível estudado por Rohn et al., 2000), 1, 2, 3, 4

24
e 5, é sobreposta por um siltito arenoso cinza bastante deteriorado, gerando uma
descontinuidade de 60 centímetros acima dessas assembleias.
Os mesmos lamitos cinza que compõem a porção basal da coluna aparecem na porção
superior, após a descontinuidade citada acima. Novamente eles estão repletos de intercalações
carbonosas contendo as assembleias deste estudo, em níveis reconhecidos por 6, 7, 8, 9 e 10.
Também se deve citar a presença de estruturas diferenciadas como gretas de contração,
paleossolo e icnofósseis. A coluna gerada a partir das descrições e medições de campo é a
apresentada na (Figura 6).

25
Figura 6: Coluna estratigráfica do afloramento Rio Capivari, com a indicação dos níveis descritos e principais
estruturas (baseada em Rohn et al., 2000).

26
Um grande corpo diamictítico com características glaciais sobrepõe todas as litologias
com contato erosivo, talvez denotando o fim do interglacial. Soares et al. (1977) apresentaram
um perfil geológico ao longo da nova estrada Tietê-Piracicaba (aproximadamente paralela a
estrada antiga) enfatizando a ocorrência de camadas delgadas de carvão logo abaixo dos
últimos diamictitos do Gr. Itararé (Figura 7). Também descreveram a associação litológica
que abriga tais camadas como fluvial/lacustre, de costa irregular, com existência de golfos
onde ocorreram episódios pontuais de transgressão marinha.
Figura 7: Perfil geológico da estrada nova Tietê-Piracicaba (Soares et al., 1977).
Foram documentados inúmeros níveis fósseis e exemplares além dos descritos por
Rohn et al. (2000). Os níveis das assembleias fósseis se situam estratigraficamente abaixo dos
últimos diamictitos descritos para o Gr. Itararé, estes em contato discordante com a Formação
Tatuí, aproximadamente na cota 500 m (Soares et al., 1977).
Análises palinológicas de amostras do Rio Capivari (Callegari, 2001; Souza e
Calegari, 2004) indicaram que os depósitos correspondem à Zona Vittatina de Souza e
Marques-Toigo (2001), Subzona Protohaploxypinus goraiensis, correlacionado aos sub-
intervalos H3 + I de Daemon e Quadros (1970), possivelmente do Asseliano-Sakmariano
(Souza, 2000). A subzona abrange depósitos da parte superior do Grupo Itararé e da parte
inferior da Formação Rio Bonito (ou equivalentes), não apresentando resolução para separar
as duas unidades. Na assembleia do Rio Capivari há elementos indicativos de condições com

27
certa influência marinha, por exemplo, algas e fungos. Contudo, predominam na palinoflora
pólens de Spermatophytas, além de esporos de Lycophyta e Sphenophyta (Callegari, 2001;
Souza e Callegari, 2004).

28
5. MATERIAIS E MÉTODOS
5.1 Material de Estudo
O material estudado provém do Afloramento Rio Capivari, situado no município de
Tietê (SP). Posteriormente foi catalogado e inserido na Coleção Paleobotânica (CP1) do
Laboratório de Paleohidrogeologia, no Departamento de Geologia e Recursos Naturais
(Instituto de Geociências – UNICAMP). Tratam-se de exemplares fósseis de restos vegetais,
preservados em camadas delgadas de lamito cinza e escuro, que se intercalam a camadas
arenosas, amareladas e mais claras.
Os fósseis são majoritariamente compostos por folhas de Glossopteridales e algumas
de suas sementes, sendo as primeiras o principal objeto deste estudo. Há também fragmentos
de caules de Sphenophyta e lenhos de Spermatophyta e gênero não identificado, que apesar
disso compõem a assembleia e complementam a interpretação tafonômica. Nas folhas, é
possível identificar marcas de danos causados por insetos (e.g.: herbivoria) e, por entre as
laminações, marcas carbonificadas indicando grande quantidade de carbono disponível,
assinalando a carbonificação como principal processo de fossilização.
As amostras coletadas totalizam 252 e compreendem de CP1/625-676 a CP1/680-881,
sendo que nem todas elas possuem exemplares capazes de serem individualizados para estudo
morfológico. São 153 espécimes estudados, correspondentes a CP1/625-634, 636, 638-642,
644, 661, 665-671, 673-676, 685-751, 753-815. Parte do material foi gentilmente cedido pela
Profa. Dra. Rosemarie Rohn (Unesp), o qual fora coletado em 2000 com a ajuda da Dra.
Márcia E. Longhim e a Profa. Dra. Mary E. C. Bernardes de Oliveira, durante um projeto para
reencontrar a localidade fossilífera descrita originalmente por Barbosa e Almeida (1949).
Ambos os materiais adquiridos para este estudo, tanto o coletado quanto o doado, são
concordantes em termos de conteúdo vegetal fóssil. Contudo, a necessidade de novas coletas
se deu com o intuito de executar um estudo tafonômico dessas assembleias. Com novas
amostras em mãos, foi descrita a morfologia afim de classificar os espécimes a nível de
gênero. Assim, o material cedido foi utilizado para fins comparativos entre os taxa
encontrados na localidade.
Todas as amostras utilizadas foram coletadas com orientação, exceto àquelas cedidas.
Porém, havia o conhecimento de sua localização estratigráfica e de que pertenciam a um
mesmo acamamento (nível zero), o qual corresponde ao pavimento da base, ou chão, quando
Rio Capivari está no seu no seu nível mais baixo durante o período de estiagem.

29
De posse dessas informações, este nível, ainda que não houvessem novas coletas, foi
posicionado na coluna estratigráfica do Afloramento Rio Capivari confeccionada para este
trabalho. Por conter a assembleia mais próxima ao rio é, consequentemente, aquela mais
degradada e intemperizada. Assim, dada a friabilidade do material, não foi possível realizar
novas coletas do nível base (zero), apenas um breve levantamento para fins comparativos.
Durante os trabalhos de campo, foi realizado o registro fotográfico do afloramento,
incluindo todos os níveis descritos. Na medida em que o decapeamento avançava, foram
tomadas fotos para posterior comparação com os dados coletados para cada malha. As
amostras foram retiradas cuidadosamente durante processo de abertura de trincheira,
atentando para manter sua orientação em relação ao afloramento preservada. Apenas deste
modo seria possível propor uma interpretação tafonômica e ambiental coerente e fiel a
realidade.
5.2 Métodos de Estudo
5.2.1 Estudos de Campo
Os levantamentos de campo ocorreram ao longo de 19 meses, começando em maio de
2015 e se estendendo até outubro de 2016. Durante esse período foram realizados 5 trabalhos
de campo no afloramento, onde se observou a presença de numerosas camadas com alta
densidade fóssil por toda região. Dessa forma, foi possível encontrar níveis diversificados de
assembleias vegetais fossilizadas e situá-los na coluna estratigráfica do afloramento,
estabelecendo comparações entre elas e uma evolução paleoflorística local.
Para melhorar a qualidade e precisão dos dados adquiridos, utilizou-se o método de
abertura de trincheira, atingindo assim camadas com conteúdo fóssil, sem que estas fossem
danificadas. Para isso, foram utilizadas pás, picaretas e martelos, abrindo pouco a pouco uma
trincheira que resultaria em mais de duas toneladas de rocha retirada e dimensões de
150x100x50 centímetros. Iniciou-se então, de cima para baixo, isto é, do mais novo para o
mais antigo, uma varredura minuciosa das camadas fossilíferas que pudessem ser
identificadas ou expostas o suficiente para realizar as medições necessárias para confecção
dos gráficos de orientação e densidade.
A partir do decapeamento das camadas sobrejacentes, foi possível posicionar uma
malha quadriculada transparente de 30x30 centímetros e obter a densidade para cada nível
através do método de Point Counter, utilizado por Spicer (1988) (Figura 8). Com o conteúdo
macrofóssil exposto, mediu-se a orientação através de bússola (Clar) de cada espécime visível

30
que pudesse ser distinto em meio a matéria orgânica acumulada. Por fim, as amostras
expostas em cada superfície foram coletadas e encaminhadas ao laboratório, a fim de estudar
os exemplares e classificá-los a nível de gênero.
Figura 8: Métodos de trabalho; A: Malha quadriculada em uso para medição da densidade; B:
Decapeamento denotando o nível fóssil a ser descrito (nível 9); C: Trincheira com o nível fóssil exposto. D:
Local do levantamento estratigráfico parcialmente aberto, conforme o método de trabalho.
A
B
B
B
C
B
D
B

31
5.2.2 Estudos de Laboratório
Apesar de bastante restrita, a área foi vasculhada a procura de informações litológicas
que pudessem corroborar com os estudos paleoambientais desenvolvidos. Com os dados
obtidos em campo e descritos previamente por Rohn et al. (2000), confeccionou-se uma
coluna estratigráfica detalhada do Afloramento Rio Capivari. A partir dela e da identificação
dos níveis das assembleias fossilíferas, foi determinada sua sucessão na coluna. Ademais, as
paleocorrentes puderam ser definidas por meio das análises das rosetas de orientação e da
densidade dos níveis medidas em campo.
O conteúdo fóssil das amostras foi minunciosamente analisado baseando-se nas
descrições da literatura pertinente. Os parâmetros utilizados para caracterizar as folhas
basearam-se nas descrições de McCoy (1860). Foram mensurados e observados na lâmina
foliar, quando possível, largura, comprimento, quantidade de nervuras por centímetro na
borda da margem, ângulo entre o eixo mediano da costa da folha e se o ângulo da nervação ao
atingir a margem é obtuso ou agudo, além do ápice e base, padrão de nervuras (se contém
anastomoses) e sua natureza. No caso dos exemplares vegetais da assembleia
macrofitofossilífera do afloramento Rio Capivari, não houve exemplar algum com nervura
média registrada. Já para caracterização de sementes baseou-se nas propostas de Taylor e
Taylor (1993), Millan (1994) e Archangeslky (2000) e foram observados largura e
comprimento do colpo, da asa e da semente no total, dimensão da abertura superior e também
a relação entre a largura da testa e a largura do núcleo para determinar o gênero.
Para maior detalhe da morfologia, foi necessário utilizar o estereomicroscópio Zeiss
combinado a ferramentas que pudessem desagregar a amostra com mais facilidade para exibir
mais aspectos. Também foi feito o registro fotográfico de todos exemplares estudados,
criando um banco de dados completo da assembleia. Por fim, o gênero dos espécimes
descritos foi incluído na coluna nível a nível, permitindo assim inferir pontos importantes
sobre a evolução paleoflorística local e a paleoflora alóctone.
Com base nos dados de densidade dos fitofósseis nos níveis individualizados em
campo, e nos de orientação dos espécimes in situ, foram confeccionadas tabelas no software
da Microsoft® Excel. Fundamentada nos dados de densidades de cada nível, foram calculadas
as concentrações de fitofósseis. Também com eles se produziu gráficos de forma a tornar
mais simples a compreensão da comparação entre as assembleias. Tais informações,
associadas aos resultados obtidos com o estudo sistemático, viabiliza a elaboração da
discussão e também as comparações com a literatura paleobotânica do final do Paleozoico no
Gondwana.

32
6. ARTIGO: TAPHONOMIC CHARACTERIZATION OF THE CAPIVARI RIVER
MACROPHITOFOSSIL ASSEMBLY: A PRE-GLOSSOPTERIS RECORD – SÃO
PAULO, BRAZIL.
Juliana Sampaio da Costaa, Fresia Ricardi-Branco
b, Rosemarie Rohn-Davies
c, Ariel Milani Martine
a, Rodrigo
César Teixeira de Gouvêad
a Geociences Institute Graduate Program, University of Campinas, Campinas, SP.
b Geociences Institute Geology and Natural Resources Department, University of Campinas, Campinas, SP.
c Geociences Institute Applied Geology Departament, Unesp, Rio Claro Campus, Rio Claro, SP.
d Geociences Institute Graduate Program, University of Pará, Belém, PA.
ABSTRACT: Permo-Carboniferous fossil assemblages on the Paraná Basin exhibit high copiousness
on plant content, which is also observed on the paleoflora of Rio Capivari (SP) outcrop. This
occurrence suggests the beginning of the Glossopteris Paleofloristic Province on the Gondwana
paleocontinent even before the Permo-Carboniferous glaciation. The fossil content on its different
layered levels testify the influence of episodic mild climates, between glacial pulses, in which these
forests bloomed. The in situ taphonomic study of leaves density and orientation allowed tracing
paleoclimate interpretations, as well as establishing paleoenvironmental relationships, by analyzing
the carbonized leaves, trunks, and seeds size, form, and genus. Sampling resulted in eleven described
levels, 252 collected samples and five identified taxa. A 150x100x50 centimeters section was opened
on the four-meter profile, outcropped on Capivari River adjacencies, exhibiting the stratigraphic
positioning and preserving the spatial distribution of its assemblages. The taphonomic results showed
presence of the Gangamopteris genus, with higher occurrence of the G.abovata species, typical from
the Pre-Glossopteris flora. This genus is distinguished by the absence of midrib on the venation
pattern, which spatially and temporally allocates this vegetation at the South Ocidental Gondwana, on
its first stages of development, at the Permian. Its presence along with the glacial evidences on the
profile, such as dimictites and flat iron pebbles, suggests cyclic stages of mild climates, interspersed
by colder climates, revealing a regional warming that allowed the installation of this perennial
vegetation. This perenniality is confirmed by the occurrence of distinct plants on the assembly,
especially seeds, whose production occurs seasonally. Palynological data of previously studies
suggests these assemblages belongs to the Vitattina zone, Protohaploxypinus goraiensis subzone, on
the Asselian-Sakmarian interval. The specimens’ orientation indicate the absence of evident
paleocurrents or preferential sense, characterizing a parautoctonous assembly. The profile lithologies
and sedimentary structures, as well as the assemblages special distribution and taphonomy, suggests
the paleoenvironment is related to the edges of a lake or a floodplain. These phitofossils are located
below the glacial diamictites and represent vehement proof of an interglacial period during the
assembly deposition.
Keywords: Gangamopteris, Gondwana, Itararé Group, Pré-Glossopteris, Paraná Basin.

33
INTRODUCTION
Gangamopterids has great paleoenvironmental importance as it registers the ascension
and establishment of the Glossopteris Flora on the Gondwana paleocontinent, under the
influence of glacial and interglacial periods (Limarino et al., 2013; Christiano-de-Souza and
Ricardi-Branco, 2015). These species are dominant on the paleoflora of the Lower Permian
and occurs singularly or together with protoglossopteridales (Rubidgea spp.) or primitive
glossopteridales species (Christiano-de-Souza and Ricardi-Branco, 2015). They differentiate
from glossopteridales from the absence of midrib, being restricted to densification of the
secondary veins, on the median portion of the leaf lamina, in some species.
Regardless the differences, both species are quite similar, with common taphonomic
features. These similarities and the species bioestratigraphic distribution suggests a feasible
ancestry of the Gangamopteris in relation to the Glossopteris (Bernardes-de-Oliveira, 1978;
Bernardes-de-Oliveira et al., 2016).
The recognized paleofloras on Itaraté Group (Permo-Carboniferous) were deposited
under glacial conditions on the Late Paleozoic (Late Paleozoic Ice Age – LPIA). This rock
assembly is one of the most expressive deposits with glacial/interglacial influence of the
Paraná Basin (Scotese, 2000; Veevers et al., 2004; Milani et al., 2007; Bernardes-de-Oliveira
et al., 2016).
In this scenery emerges the Capivari River taphoflora, whose record is characterized
by extremely rich fossil assemblages, preserved though the carbonification process.
Therefore, the morphological and anatomical structures are well preserved despite the
exposition to the riverbank weathering. It was possible to distinguish eleven levels, including
the basal, of which was acquired density, orientation and/or morphology of the specimens.
Barbosa and Almeida (1949) registered the first record about this fossil plants
occurrence, reporting a place on the Capivari River riverbank. Later, Soares et al. (1977)
reaffirmed its presence as thin carbonized layers along the Piracicaba-Tietê road profile.
Lastly, Rohn et al. (2000) ratified the existence of these assemblages in preliminary study,
describing specimens collected along the Capivari River riverbanks, highlighting its botanical
affinities to the Itararé Group paleofloras.
The main goal of this study is to propose taphonomical, paleoenvironmental and, at
times, paleoecological interpretations, through in situ mapping of the Capivari River outcrop
fitofossil assemblages. Particularly associate the morphological descriptions of the specimens
at the genus level to previously descripted species on literature. Which will also results in
stratigraphic positioning of the outcrop in the Itararé Grup biozones, in addition to compare

34
its morphotypes to other assemblages of the Paraná Basin, contributing to the understanding
of its paleofloras.
Understanding the Rio Capivari taphoflora is important firstly because of the scarcity
of detailed publications on this outcrop. Furthermore, it is fundamental to study the
conclusive proof of Glossopteris flora installation on the Gondwana paleocontinent before the
glaciation (Rohn et al., 2000). Finally, its taphonomic character is of major importance, due to
the scarcity of preserved outcrops on the Paraná Basin.
PARANÁ BASIN AND THE GONDWANA PALEOZOIC
The Paraná Basin is formed due to a major sineclisis on the central-south portion of
the South American continent. Its elongated and oval shape and is on the NE-SW direction,
with 1.5 milion square kilometers and up to 5000 meters depth on the depocenter region. Its
area includes Brazil, Paraguai and Uruguai (Chaco-Paraná Basin), and is correlated to the
Karoo Basin (South Africa) and Huab Basin (Namíbia) (Milani and DeWitt, 2008; Ricardi-
Branco et al., 2016).
Milani et al. (2007) categorize the basin in six depositional supercyles, from
Ordovician to Cretaceous. The Itararé Group is located on the Gondwana I Supersequence.
This Group deposition started at the Moscovian (Pennsylvanian) and lasted until the
Sakmarian (Cisuralian). At this period, the basin held a partially isolated glacial entrance due
to tecnosedimentary control or an estuary with salinity controlled by variations on the
medium global ocean level, regulating a regressive-trangressive cycle (Rocha-Campos et al.,
2008). There is a range of faciological diversity, with subaqueous platform deposits, glacial
terrestrial, as well as proximal and distal marine environments (Gama Jr. et al., 1992; Santos
et al., 1996; Canuto et al., 2001).
The Itarare Group paleofloras on São Paulo State are represented, on the basal level,
by the bryophytes of Campinas and Salto cities, to the Cerquilho flora, on the top. The most
abundant fitofossils of the Itararé Group are Ginkgophytas and Coniferophytas, and in less
proportion Lycophyta and Sphenophyta, with few species of Bryophyta (Christiano-de-Souza
and Ricardi-Branco, 2013; Ricardi-Branco et al., 2013).
Mune and Bernardes-de-Oliveira (2007) divided such occurences into nine
stratigraphically positioned floras (Campinas, Buri, Itapeva, Araçoiaba da Serra, Monte Mor,
Itu, Itaporanga, Tietê and Cerquilho). Finally, Bernardes-de-Oliveira et al. (2016), proposed a
new fitostratigraphic division of the Itararé Group paleofloras, inserting the Capivari River
outcrop in the Association Gangamopteris-Arberia-Stephanophyllites of Asselian-Sakmarian

35
age. Which allowed to correlate it to the Sítio Toca do Índio outcrop (Cerquilho, SP) and the
basal portion of the Rio Bonito Formation (Daemon, 1974; Rösler, 1978; Arai, 1980; Souza,
2006).
Capivari River outcrop
The outcrop is located at the left margin of the Capivari River, about 500 meters
upstream from the bridge on the old road, at the coordinates 22°59,3’S/ 047°45,05’W (UTM
23K 0217,99/ 7455,18), and at 475 meter high. The access is over the SP-127 “Cornélio
Pires” highway at the km 65, where is located the local road Joaquim Caetano, which is
connected to the old road that connects Piracicaba and Tietê cities (Figure 1).
Figure 1: Capivari River outcrop localization map, at Tietê, São Paulo state.
It is composed of grey mudstones, interleaved by lighter sandy and darker carbonous
levels, in which the eleven fossiliferous assemblages mapped on this study are hosted.
Distinctive features such as radial mudcracks, paleosoils, and ichnofossils are found in

36
between some of the levels. Overlaying all the lithologies, in an erosive contact, there is a big
diamictite body with glacial features, perhaps evidencing the end of the interglacial period.
Soares et al. (1977) presents a geological profile along the new Tietê-Piracicaba road
(somewhat parallel to the old road), emphasizing the occurrence of thin coal layers right
under the last diamictites on the Itararé Group (Figure 2). The authors also described the
lithological association hosting these layers as fluvial/lacustrine, on an irregular shore, with
gulfs that witnessed punctual episodes of marine transgression.
Altogether, it was documented 10 fossil levels and 153 fossil specimens, besides the
ones described by Rohn et al. (2000). The assemblies’ levels are stratigraphically below the
last diamictites described on the Itararé Group, which is in discordant contact with Tatuí
Formation, approximately at 500 m elevation (Soares et al., 1977).
Figure 2: Geologic profile of the new Tietê-Piracicaba road (Soares et al., 1977).

37
MATERIALS AND METHODS
The studied material corresponds to 252 samples (CP1/625-676 e CP1/680-881), of
which CP1/625-634, 636, 638-642, 644, 661, 665-671, 673-676, 685-751, 753-815 could be
used for morphological studies. In addition to the material collected in the year 2000, and
kindly provided by Prof. Doc. Rosemarie Rohn (Unesp), included in this research to add to
the bibliographic database, which is scarce on the outcrop, as well as contribute to the
morphological and botanical descriptions previously performed on theses specimens.
The assemblages are mostly composed of Glossopteridales leaves, the focus of this
study, and some of its seeds. There is also Sphenophyta branches and unidentified trunks. On
the leaves, it is possible to identify insect-based damage (e.g. herbivoria) and, in between
laminations, carbonized marks suggesting a large amount of available carbon, pointing out
carbonification as the main fossilization process.
Based on the Point Counter method (Bateman, 1999; Spicer, 2010), a 150x100x50
centimeters trench was opened, allowing the use of a 30x30 centimeters checkered grid (with
1x1 centimeters squares). Thereby, orientation data was obtained (Clar measurements) for
each specimen on each baseline, as well as density data (using the checkered grid) to each of
the macrofitofossil assemblages (Figure 3). Lastly, the samples were morphologically
described to determine its paleobotanical affinities.

38
Figure 3: Working methods; A. Checkered grid used for density measurements; B. Outcroop decapping and clearing of
the fossil level (level 9); C. Trench with exposed fossil level. D. Partially cleared outcrop, used for the
stratigraphic survey, according to the work methods.
A
B
C D

39
RESULTS
The first stage of this study was a field trip survey, in order to collect samples and
perform the oriented mapping and density measurements of the plant assemblages. Data
collection from the outcrop allowed the elaboration of lithological descriptions, including a
stratigraphic column with the fossil levels localization. Posteriorly, in the laboratory, using
the basal level samples as comparative parameter, the taxonomic study was developed.
A defined ground zero on the outcrop basal level corresponds to the floor and to the
level previously described in Rohn et al. (2000), which is also studied on this work. Due to
difficulties in decapping the layers and the friability of the assemblages hosting material, the
density and orientation of this level was not measured. At this zone there is higher degree of
weathering because of the proximity to the Capivari River riverbed and the influence of the
flooding periods. The rest of the assemblages were numbered from one to ten, from the base
to the top.
Paleobotany systematic
The taxonomic study was based on the Plantӕ Kingdon by Taylor et al. (2009).
Kingdom: PLANTAE
Phylum: PTERIDOSPERMOPHYTA
Order: GLOSSOPTERIDALES
Genus: Gangamopteris, McCoy, 1860
Figure 4 and Figure 5
Species-type: Cyclopteris (?) angustifolia McCoy, 1847, Ann. & Mag. Nat. Hist., 20: 148, est.
9, figs. 3, 3ª depository collection Sedgwick Museum, Cambridge.
Gangamopteris angustifolia McCoy, 1875, Victoria Geol. Survey, Dec. 2: 11, est. 12, fig. 1;
est. 12, figs. 2, 2a, Mudgee. New South Wales, Australia.
The genus Gangamopteris was instituted based on the Cyclopteris (?) angustifolia
form, described by McCoy, in 1847, for Australian material, from Bacchus Marsh, in
Victoria. In 1860, the same author originally diagnosed the Gangamopteris genus as “frond
simple, entire, greatly varied in shape, elliptical, broadly and elongately obovate, broadly
lanceolate, sub-linear, etc., contracted towards base. Midrib absent. Median nerves more or
less parallel, anastomosing; lateral nerves arising by repeated dichotomy from the median
nerves or from the base, arched, bifurcating, and anastomosing to form a network”.

40
In 1875, the same author elected the Cyclopteris (?) angustifolia to the genus species-
type. Posteriorly Feistmantel (1879), Arber (1905), Maithy (1966) and Pant and Singh (1968)
modified this genus diagnose as follows: “leaf simple, sessile or petiolate, lamina linear or
broadly ovate, apex broadly rounded to acute or mucronate, base gradually tapering or
contracted; midrib never well defined, but has a number of parallel veins forming,
longitudinally elongated mesher or justordinary mesher, venation elsewhere reticulate”.
In this way, all the described samples of the Capivari River outcrop paleoflora belong
to this classification, being noticeable the presence of three different species. In turn, these
samples were classified according to the gondwanic species botanic affinity, which are
described next:
Division: Glossopteridophyta
Order: Glossopteridales
Gangamopteris aff. G. obovata (Carruthers) White (1908)
Figure 4 C; Figure 5 A and D
Collection: Paleohidrogeology Laboratory – DGRN/IG/UNICAMP, CP1/ 633B, 633D, 634,
639, 640, 641, 644, 647, 648A, 648B, 650A, 669, 672, 673, 695, 697A, 697B, 711, 724, 730,
731, 735, 736, 753, 755, 756, 769, 777, 778, 779, 782, 790, 792, 795, 797.
Provenance: Capivari River left margin, about 500 meters upstream from the bridge on the
old Piracicaba-Tietê road, coordinates 22°59,3’S/ 047°45,05’W (UTM 23K 0217,99/
7455,18), at 475 m elevation. Short provenance name: Capivari River Outcrop.
Lithostratigraphy: Itararé Group, Paraná Basin, São Paulo state.
Age: Upper Carboniferous – Lower Permian.
Description: Leaf fragments notophyll-obovate with flat margins, apex obtuse-rounded, and
base narrow, from acute to concave. Variable length between 3.12 and 5.74 cm, wider width
between 2.75 and 4.78 cm. Venation pattern varying from distinct to almost imperceptible
with subparallel veins near the base. Abundant presence of dichotomies and anastomoses,
forming elongated meshes. Veins caliber are indistinctive between the median and lateral
portions, but gradually smaller towards the apex. Approximately parallel central veins and

41
lateral veins slightly arched to the margins, forming 80º angle to the median axis of the leaf
(commonly around 60º).
Observations: G. obovata is frequently cited on literature and it is registered in a number of
Gondwana basins (Bernardes-de-Oliveira, 1978; Tybusch and Iannuzzi, 2008; Limarino et al.,
2013; Christiano-de-Souza and Ricardi-Branco, 2015), but widely varying in morphology.
For instance, the Argentinian (e.g. Archangelsky, 1958; Leguizamon, 1972) and the Rio
Grande do Sul shapes (e.g. Guerra-Sommer e Cazzullo-Klepzig, 1993; Tybusch e Iannuzzi,
2008) seem to approach the lanceolate pattern with slightly arched venation, besides
presenting clearer median bundle.
Comparing to the Brazilian records, the Capivari River Outcrop specimens resemble,
in shape and dimensions, one of the G. obovatta specimen of Cerquilho outcrop (Millan,
1989, est. 1, fig. 1). Distinguishing from others by denser venation and more arched to the
lateral margins, its shape and venation match the Criciúma and Rio Bonito Formation G.
obovate specimens (Bernardes-de-Oliveira et al., 2016), but in smaller dimensions.
Stratigraphic distribution:
Brazil: G. obovata, in a broader sense, occurs on the Itararé Group [Cerquilho (Millan, 1989),
Capivari River (this study), Acampamento Velho, Morro Papaleo (Tybusch and Iannuzzi,
2008)]; Rio Bonito Formation [Bainha, Bairro 20, Hospital, São Marcos, in Criciúma, Lauro
Müller, Cambuí, Arroio dos Ratos, Arroio dos Cachorros, Candiota, Faxinal (Bernardes-de-
Oliveira, 1977, Tybusch and Iannuzzi, 2008; Christiano-de-Souza and Ricardi-Branco, 2015;
Benardes-de-Oliveira et al., 2016)]. Argentina: Lubeckense A, B and “Bonetense” (medium
to inferior part of lower Permian); Índia: Talchir, Karharbari and Barakar Formations in local
stages (Lower Permian); South Africa: Dwyka to Lower Beaufort (Lower to Upper Permian);
Zimbabwe: Upper Wankie (Lower Permian, lower portion); Mozambique: Productive Series;
Australia: almost entire Permian; Antarctica: Medium-Upper Permian (Archangelsky et al.,
2013, Cariglino, 2013).
Gangamopteris aff. G. mosesi Dolianiti, 1954
Figure 4 A

42
Collection: Paleohidrogeology Laboratory – DGRN/IG/UNICAMP, CP1/ 625, 633C, 642,
655B, 656, 668, 670, 676, 693, 699, 701, 702, 705, 714, 722, 723, 724, 725, 729, 734, 741,
742, 748, 756, 762, 766, 798, 801, 809, 810.
Provenance: Capivari River left margin, about 500 meters upstream from the bridge on the
old Piracicaba-Tiete road, coordinates 22°59,3’S/ 047°45,05’W (UTM 23K 0217,99/
7455,18), at 475 m elevation. Short provenance name: Capivari River Outcrop.
Lithostratigraphy: Itararé Group, Paraná Basin, São Paulo state.
Age: Upper Carboniferous – Lower Permian.
Description: Leaf notophyll, oblong-spatulate, flat margins, base soft and apex slightly obtuse
and rounded. Variable length between 2.10 and 4.19 cm, wider width between 0.57 and 2.07
cm. Venation pattern characterized by dichotomies and anastomoses, the latter more frequent
near the margins, in the form of oblique connecting veins or by fusion of two veins. Veins
course fairly parallel and slightly divergent, gently arched to the margins, locally wavy due to
conservation problems. Veins caliber slightly variable from the leaf base to the apex.
Observations: Venation and shape of the Capivari River specimens resemble the G. mosesi
holotype (Dolianiti, 1954). According to the species original diagnosis, it presents lanceolate
form and acute or mucrunate apex. However, the holotype photography shows obovate form,
its apex, actually, is on the boundary of acute and obtuse (~90º), and the mucrunate feature is
not clearly identified. Other G. mosesi specimen was presented by Bernardes-de-Oliveira
(1977: fig. 68), with even more obtuse apex (~105º), whilst clearly mucrunate. Capivari River
specimens are smaller and its venation patter less arched to the margins, but it seems to
suitably match the species features. It was not possible to establish comparisons to the
Argentinian forms.

43
Stratigraphic distribution:
Brazil: Itararé Group, Capivari River; Rio Bonito Formation, Criciúma. Also occurring in
Argentina, Lubeckense B and “Bonetense” (medium part of the Lower Inferior) and in
Australia, on Newcastle Coal Measures and the Sydney Basin (Upper Permian), however the
morphological correspondence could not be confirmed (Guerra-Sommer and Cazzulo-
Klepzig, 2000; Tybusch and Iannuzzi, 2008, Cariglino, 2013).
Gangamopteris aff. G. angustifolia McCoy 1875
Figure 4 B; Figure 5 B, C e D
Collection: Paleohidrogeology Laboratory – DGRN/IG/UNICAMP, CP1/ 629, 628, 631, 636,
638, 648, 651, 654, 655, 662, 665, 674, 671, 676, 696, 700, 701, 702, 727, 728, 732, 740, 749,
751, 758, 761, 762, 763, 776, 787, 788, 791, 793, 798, 799, 802, 803, 811, 814, 815.
Provenance: Capivari River left margin, about 500 meters upstream from the bridge on the
old Piracicaba-Tietê road, coordinates 22°59,3’S/ 047°45,05’W (UTM 23K 0217,99/
7455,18), at 475 m elevation. Short provenance name: Capivari River Outcrop.
Lithostratigraphy: Itararé Group, Paraná Basin, São Paulo state.
Age: Upper Carboniferous – Lower Permian.
Description: Leaf fragments simple, linear, narrowly oblong and oblanceolate, full margins,
symmetric to slightly asymmetric, apex acuminate, base narrow. Variable length between
4.35 and 8.73 cm, wider width between 2.65 and 3.17 cm. Venation pattern composed of
parallel to subparallel veins dichotomizing distally, rare anastomoses, X connections, oblique
and transversal, forming narrow and elongated reticles, on the medium portion and towards
the margins. Divergence angles varying between 5º to 10º, with very thin veins gently arching
towards the margins.
Observations: The venation pattern resembles the McCoy (1875) described holotype, despite
differing in absence of well-defined midrib.

44
Stratigraphic distribution:
Brazil: Itararé Group in Acampamento Velho (RS) and, doubtfully, in Cerquilho (SP).
Argentina: Lower Member of La Golondrina Formation (Lower Permian “upper”). Australia:
Bacchus Marsh, Victoria (Lower Permian). Índia: andares Talchir, Karharbari (Lower
Permian), besides some unverified mentions on Africa (Mcloughlin et al., 2005; Tybusch and
Iannuzzi, 2008; Srivastava and Agnihotri, 2010; Cariglino, 2013; Benardes-de-Oliveira et al.,
2016; Tybusch et al., 2016).
GYMNOSPERMAE
Semina Incertae Sedis
Samaropsis Goeppert 1864
Samaropsis sp. aff. S. rigbyi Millan 1977
Figure 4 C and D
Species-type: Samaropsis Goeppert, 1864.
Collection: Paleohidrogeology Laboratory – DGRN/IG/UNICAMP, CP1/625, 630, 646, 661,
664, 669, 676B, 685, 689, 690, 741, 742, 750, 755, 757, 760, 765, 767, 768, 771, 774, 775,
784, 786, 793, 798, 806.
Provenance: Capivari River left margin, about 500 meters upstream from the bridge on the
old Piracicaba-Tietê road, coordinates 22°59,3’S/ 047°45,05’W (UTM 23K 0217,99/
7455,18), at 475 m elevation. Short provenance name: Capivari River Outcrop.
Lithostratigraphy: Itararé Group, Paraná Basin, São Paulo state.
Age: Upper Carboniferous – Lower Permian.
Description: Circular to subcircular seed, smooth, with bilateral symmetry and poorly defined
contour in most samples. Average length of 8.82 mm, width variable between 4.2 and 6.9
mm. Nucellus apex acute and base cordate, length of up to 5.9 mm and width between 2.04
and 3.75 mm. Wing of analogous general length of the seed and width between 1.31 and 2.77
mm. The specimens have differentiated testa, with sclerotesta of apex acute and round base
and well-developed sarcotesta.

45
Stratigraphic distribution: Itararé Group, Cerquilho, São Paulo state (Millan, 1989).
Figure 4: Specimens studied on the paleobotanic systematics. A: Sample CP1/625B of Gangamopteris aff. mosesii
(scalebar = 1 cm); Sample CP1/669 of Gangamopteris aff. obovata e Samaropsis sp. aff. S. rigbyi
(scalebar = 1 cm); C: Sample CP1/633 of Samaropsis sp. aff. S. rigbyi (scalebar = 2 mm); D: Sample
CP1/666 of Gangamopteris aff. angustifolia (scalebar = 1 cm).
A B
C
D

46
Figure 5: Differentiated specimens on the paleobotanic systematics. A. Sample CP1/650A of Gangamopteris aff.
obovata (scalebar = 1 cm); B Sample CP1/756 of Gangamopteris aff. angustifolia (scalebar = 2 cm); C.
CP1/626B, corresponds to level 6, very dense, almost undifferentiated leaf lamina (visible Gangamopteris
aff. angustifolia; scalebar = 1 cm); D. CP1/627 corresponds to level 5, very dense, however the leaf
lamina is clearly differentiated. Blue arrow: Gangamopteris aff. obovata, red arrow: Gangamopteris aff.
angustifolia (scalebar = 2 cm).
A B
C
D

47
Density
All ten levels of fossils observed on the Capivari River outcrop are relatively different
concerning the density of the assemblages. These levels are over the ground zero level,
numbered from one to ten, the first being stratigraphically older, and the tenth the youngest
and closest to the overlaying diamictite.
The qualitative graphical representations of the studied bioclasts are show next,
always looking to keel its original position in relation to the outcrop, trench and stratigraphic
positioning (Figure 6).
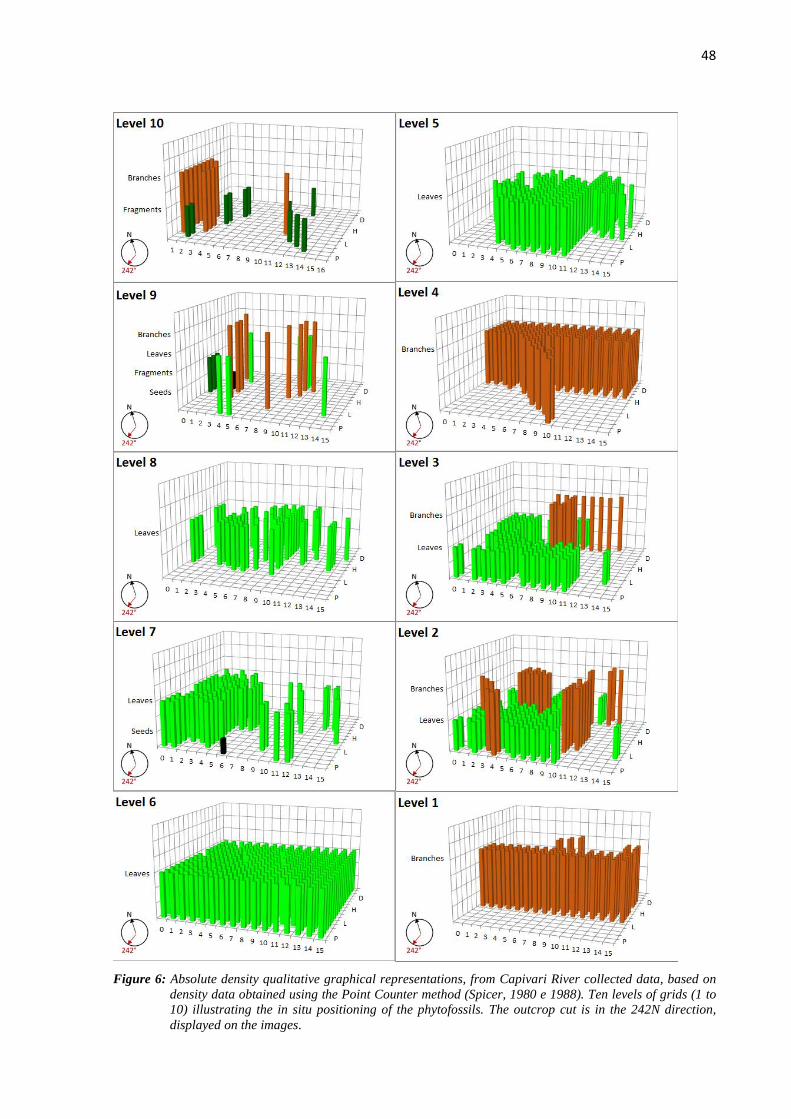
48
Figure 6: Absolute density qualitative graphical representations, from Capivari River collected data, based on
density data obtained using the Point Counter method (Spicer, 1980 e 1988). Ten levels of grids (1 to
10) illustrating the in situ positioning of the phytofossils. The outcrop cut is in the 242N direction,
displayed on the images.

49
According to the acquired data, level 6 has the densest assembly of fossils. It is
enriched in plant based organic matter, which is undifferentiated given the large number of
overlapping leaves, densely packed on the surface, making it difficult to individualize leaf
laminas. Vegetal matter is quite dark, suggesting intense action of the carbonization process
comparing to the other levels (Figure 7).
Figure 7: Levels presenting the studied assemblages: A. Level 8; B. Mudcracks between levels 6 and 7;
C. Level 9; D and E. Level 6; F. Comparison between levels 9 (upper) and 8 (lower).
A B
C D
E F

50
Processing the acquired data at all studied levels (Attachment 1) generated a
comparative diagram of density (Figure 8). For this purpose, the density values were
calculated on binary tables, indicating presence or absence of phytofossils, in a 30x30
centimeters area. This percentage-based density can be defined as the frequency of plant
based organic matter over the standard grid area on this work (30x30 cm). These results
allowed comparing all levels, highlighting levels with high or low surface organic matter
content (Figure 8).
Figure 8: Percentage-based density graph comparing the average densities on each of the studies levels
(Grids).
Orientation
Direction and sense are distinct attributes, the first being the angle between the
measured direction and the NS direction, and the latter defined by the quadrant, indicating the
sense. In other words, every direction has two senses. To the Capivari River assemblages,
only direction measures were made. This is because sense indicators features were absent on
the outcrop, being impossible to establish the exact path of the fluid responsible for the plant
organs’ deposition. However, a subtle difference was observed on the denser levels (more
twigs and leaves), which indicate these detached organs are not reliable to demonstrate
paleocurrent sense, only to establish its activity on the past.
The fossil assembly orientation rosettes are presented on Figure 9. Such parameters
were established based on specimens’ alignment in relation to the magnetic north on the
compass, obtaining a direction measure for each recognizable fossil plant. Despite some
0% 20% 40% 60% 80% 100%
Level 10
Level 9
Level 8
Level 7
Level 6
Level 5
Level 4
Level 3
Level 2
Level 1

51
variability, the paleocurrents presented a NE-SW approximate direction on the denser levels
near the top, and NW-SE on the basal levels. However, a regional study, in another scale,
should be performed to interpret and refine this information.

52
Figure 9: Orientation rosettes with preferential orientation of the vegetal fossil assemblages.

53
Distribution
After laboratory description, the specimens were combined according to its
paleobotanic classification and stratigraphic level. Levels 1 and 4 showed total absence of leaf
records. In levels 5, 6, 7 and 8, it was identified dense organic matter, mainly composed of
leaves, suggesting paleoflora dominance during interglacial pulses. Sphenophytas were
identified on level 7 in tiny amounts, accreted to the plant matter, and more frequently on the
ground zero level, although fragmented. Figure 10 shows the proportion of plant organs on
each studied level.
Figure 10: Plant organs proportion on each studied level of the Capivari River paleofloristic
association. The X axis shows the density of the 30x30 centimeters grid.
Specimen fragmentation is also a parameter to interpret the local taphonomy. It is
closely related to material nature and its interaction with the environment. The Capivari River
paleoflora is mostly composed of twigs and leaves detached from the base, and whole seeds.
Twigs and seeds are more resistant and, therefore most conserved, their degradation being
related to post lithification processes. The leaves, in turn, are more fragile and easily
fragmented. Ultimately, every foliar material with less than 50% preservation is considered
fragmented (Figure 11).
0 0,05 0,1 0,15 0,2 0,25 0,3
Level 10
Level 9
Level 8
Level 7
Level 6
Level 5
Level 4
Level 3
Level 2
Level 1
Leaves Twigs Vegetal Fragment Seeds

54
Figure 11: Average fragmentation of the Rio Capivari’s assembly phytofossils, in percentage.
To values above 50% there are well preserved specimens; to values equal or below
50% the specimens are poorly preserved.
Even with three different species identified, it was not necessary to individualize each
species on the stratigraphic column, because all three species were found on each of the leaf-
composed levels. Besides the plant fossils content, it was also found layers with icnofossils
and sedimentary structures, which are important to the paleoenvironmental interpretations.
Bentonic organisms’ marks were found on level 10, revealing the shallow depth from the
water blade and a less energetic environment (Anderson, 1975; Lima et al., 2015). Levels 7
and 8 contain in situ root marks and, right under level 7 and above level 6, mudcracks were
present. These features suggests the environment drying process as well as less influence of
the waterbody. The stratigraphic column representation, with information about levels and
species founded on the fossil record of the Capivari River outcrop, is introduced on Figure 12.
0 0,2 0,4 0,6 0,8 1 1,2
Level 10
Level 9
Level 8
Level 7
Level 6
Level 5
Level 4
Level 3
Level 2
Level 1
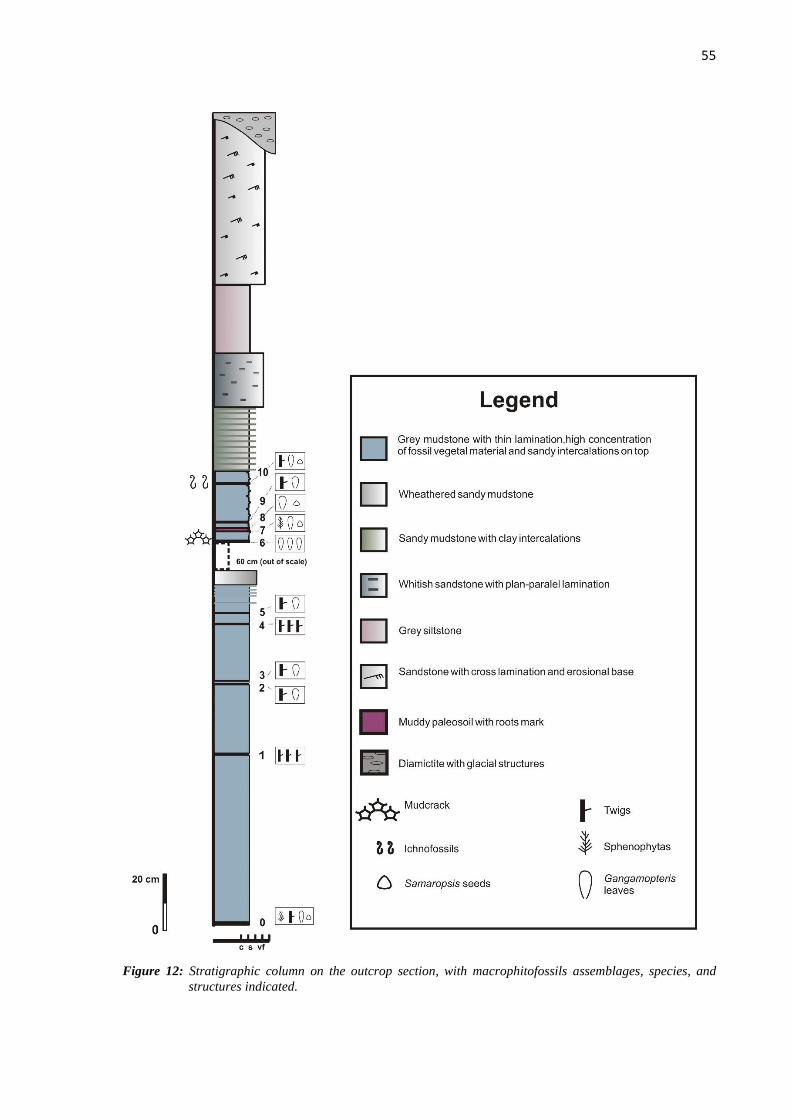
55
Figure 12: Stratigraphic column on the outcrop section, with macrophitofossils assemblages, species, and
structures indicated.

56
The stratigraphic column above (Figure 12), with its distribution on the stratus, genus
level classification and, when possible, proximity to the gondwanic plant species, was based
on Table 1 presented below. It is important to highlight some levels presenting high content of
plant organic matter, leaves and rests, mostly indistinctive, making it difficult to identify
genus or species in some samples. It should also be noted that not all observed and
documented organs (in situ) during the field activities could be sampled and transported to the
laboratory, due to the material’s friability. In this sense, a partial picture of all the observed
specimens is dicussed, which reveals the necessity for new methodological approaches in
order to explore its botanical affinities.
Table 1: Chart of the described specimens to each level and its catalogue numbering on the Hidrogeology Laboratory of the
Geosciences Institute (Unicamp), corresponding to the Paleobotany Collection 1 (PC1).
Table 1: Gangamopteris
sp.1 aff. Obovate
Gangamopteris
sp. 2 aff. mosesii Gangamopteris
sp.3 aff.
angustifolia
Samaropsis sp. 1 Sphenophyta
Branches
Twigs
N
Level 0
769, 777, 778, 779, 762, 766, 798, 801,
809, 810 777, 798, 809,
810
685, 689, 690, 741,
742, 750, 755, 757,
760, 765, 767, 768,
771, 774, 775, 784,
786, 793, 798, 806
757, 765, 768
Level 1 675A, 675B,
667
Level 2 736, 753, 755, 756 734, 741A, 742 736, 741B
Level 3 730, 731, 735 725, 729, 734, 748,
756
725, 756
Level 4 793, 798,
806
Level 5 711, 724, 627 722, 723, 724 627
Level 6 626A, 695, 697A,
697B, 782, 790, 792,
795, 797
693, 699, 701, 702,
705, 714
626B
Level 7 673 655B, 668, 676,
670
648, 655, 654,
662, 676
664, 661, 676B 664A, 676C
Level 8 669, 672, 648A,
648B, 647, 650A
656 651, 665, 654,
674, 671
669, 646
Level 9 634, 639, 641, 640,
644, 633B, 633D
642, 633C 636, 638 784, 786,
793
Level 10 625 629, 628, 631 630, 625 774, 775

57
Capivari River Outcrop and the Pre-Glossopteris Flora
Paleobiogeography
On the São Paulo state surroundings there is evidence for a glacial lobe called
“Kaokoveld”, which persisted until the end of Itararé Group deposition, advancing and
retreating according to glacial and interglacial periods (Santos et al., 1996). Retreat phases
were forthwith succeeded by deposition of thin marine layers. Those were followed by
deposition of clastic wedges, coastal to continental, due to isostatic lifting of the deglaciated
basin margins, with developing of paralic tuffs (Santos et al., 1996).
In this circunstances there was deposition of coals from Buri, Monte Mor and
Cerquilho. Coals are absent on the Capivari River outcrop, although it can be found thin
layers of shale, extremely enriched in leaf and other carbonized plant remains. Thus, Capivari
River fossiliferous interval is probably related to an interglacial period, not rejecting,
however, synchronicity between the vegetation accommodation and retraction of glaciers, as
observed nowadays in Alaska and Tierra del Fuego.
The Capivari River taphoflora can be considered vehement evidence that the Flora of
Glossopteris, or at least its precursor, settled in South Gondwana before the end of the Permo-
Carboniferous glaciation. The found Gangamopteris, still lacking differentiated median
venation suggests a very primitive stage of the Glossopteris paleoflora, perhaps one of the
oldest ever recorded. Its taxonomic features indicate this paleoflora to be older than the one
registered in Cerquilho and the most primitive until now.
This fossiliferous interval is stratigraphically superimposed by the latter glacial
diamictite, indicator of the cold climates in which these plants developed. The paleolatitude is
based on the apparent magnetic poles determined by Smith (1999) and it is estimated in ~50-
55º. Both Capivari River and Cerquilho outcrops may have been, at the beginning of Permian,
at equivalent latitudes to the Westfalian outcrops in Argentina (Iannuzzi and Rösler, 2000;
Bernardes-de-Oliveira et al., 2016)
Paleobotany systematics
Rio Capivari’s macrophitofossiliferous assemblies are entirely composed by
Glossopteridales of Gangampteris genus, since all collected samples present absence of
differentiated median venation. The absence implies in Pre-Glossopteris flora, mostly
composed of Gangamopteris, which possibly, due to its perenniality, reveals the first
warming records of an interglacial period.

58
The Cerquilho outcrop Gangamopteris, unlike the Capivari River samples, exhibit
medium venation denoted by anastomosis in the central portion of the leaf strand. Iannuzzi
and Rohn (1995) identified and classified the Cerquilho specimen as Glossopteris, since there
were prominent medial bundles without anastomoses. According to the evolutive proposal of
Bernardes-de-Oliveira (1978) and Srivastava (1991), the Permian
gangamopterids/glossopterids had median axis increasingly distinguished, with gradually
more anastomosed veins, until the appearance of a thick primary vein. The absence of such
venation features lead to believe Capivari River’s Gangamopteris are older than Cerquilho’s.
Nevertheless, other factors cannot be disregarded, which could have equally
influenced this character (e.g., light and humidity availability; Gulbranson et al., 2014), since
glossopterides with thin median bundles existed until the Lopingian (Rohn et al., 1984). The
Capivari River outcrop leaf venation density seems lower than the Rio Bonito Formation
specimens (above Itararé Group). The latter were deposited under warm and humid
conditions, as suggested by the size of its leaves (larger) and the charcoal around it (Ricardi-
Branco and Bernardes-de-Oliveira, 2000). Capivari River leaf assemblages are predominantly
microphyll and notophyll, typical from glacial periods.
In general, the assemblages’ specimens have leaves with apex wide, obovate and/or
spatulate (Gangamopteris obovata) and not as wide (Gangamopteris mosesii). Wide apex on
Glossopteridales specimens are common and widely spread over Gondwana during the
Permian, especially Glosspteris (Tybusch and Iannuzzi, 2008). Thereby, such feature is not
decisive for paleoecological interpretations.
Species diversity is low and homogeneous, shown by the identification of G. obovata,
G. mosesii and G. angustifolia specimens only. However, specimens differentiation on more
dense and organic matter enriched levels is hard (e.g. levels 5 and 6) due to foliar laminas
juxtaposition. It is also important to notice the host rock friability, under constant weathering,
which difficulted sampling and conservation.
Levels with the studied Gangamopteris can be interpreted as seasonal accumulation
(autumn), of short transported leaves. Leaves with more prominent medial bundles can mask
its abundance. Willis and McElwain (2002) define glossopterides: trees that could reach 10
meters high, deciduous, leaves abscission on autumn and winter, and typical of cold temperate
climates as pointed by its occurrences during the Permian. Such features are also observed on
most of the analyzed material of this study; therefore autumn leaves abscission could explain
the origin of Capivari River assemblages.

59
Biostratigraphy
This study paleoflora, as mentioned above, is similar to the one of Cerquilho (SP),
with more primitive venation pattern. Palynological data (Souza and Callegary, 2004) reveal
Upper Asselian to Lower Sakmarian ages. Macrofitofossils levels are below the glacial
diamictites of the Itararé Group, as observed on this work and by Rohn et al. (2000). Soares et
al. (1977) presented the Tietê-Piracicaba road geological profile, posteriorly revised by
Bernardes-de-Oliveira et al. (2016), on the new Tietê-Piracicaba road.
A plant remains occurrence is highlighted on this profile, approximately at the same
altitude of the Capivari River outcrop (around 475-480 m). Such occurrence would also be
overlapped by the Itararé Group diamictites, whose upper limit is with Tatuí Formation at 500
m altitude. The authors also reported these thin coal layers and interpreted it as
fluvial/lacustrine. The coast was probably irregular, with gulfs episodically allowing marine
invasion to continental areas.
Bernardes-de-Oliveira et al. (2016) suggests a new bioestratigraphical classification to
the Pensilvanian to Eocisuralian interglacial paleofloras of the Paraná Basin. According to
this classification, the Capivari River taphoflora is part of the Gangamopteris-Arberia-
Stephanophyllites Association (GASAs), with Cerquilho paleoflora. It is the oldest
association after the Dwykea-Sublagenicula-Calasmospora Association, reassuring the
transition from a glacial climate, resembling a tundra, to a milder climate where forests could
flourish.
Palinological analysis from Capivari River samples, performed by Souza and
Callegary (2004), suggested this assemblages belongs to the Vittatina Zone of Souza and
Marques-Toigo (2001), Protohaploxypinus goraiensis Subzone, on the H3 + I sub-intervals of
Daemon and Quadros (1970), possibly on the Asselian-Sakmarian intreval (Souza, 2000).
This subzone covers the upper part of the Itararé Group and lower part of the Rio Bonito
Formation (or equivalent), impossible to individualize due to low resolution.
There are indicative elements of marine conditions on the Capivari River assemblages
(Callegari, 2001). The correlation between these elements and the Itararé Group deposits in
São Paulo could not be confirmed, but this information is valuable, especially if combined
with palinological data from Souza (2000).
Taphonomy
Phytofossil assemblages of leaves are generally parautochthonous and accurately
picture the local vegetation, which enable clear paleoecological reconstructions (Boulter et
al., 1988; Burnham, 1989; Behrensmeyer et al., 2000; Greenwood, 1991, 1992; Ricardi-

60
Branco et al., 2009; Ellis and Johnson, 2013). Capivari Rivers’ fossiliferous levels are
composed of plant organs, mostly Gangamopteris leaves and Spermatophyte twigs, which are
enough evidence to sustain and substantiate a reliable paleoenvironmental description.
According to Greenwood (1991) the distance to the burial and fossilization site on the
sedimentation process is analogous to the density of leaves on the assemblage. In other words,
the lower the leaves counting, the greater the distance covered in sedimentation. That is a
response to the foliar coating polymer’s fragility (cutin), which easily degrades in this
process. Otherwise, the lignin on the twigs is more resistant to mechanical degradation,
allowing them to travel long distances until final deposition and burial (Spicer, 1989, 1991).
Besides that, the twigs have a more hydrodynamic shape, in comparison to leaves, being
cylindrical while leaves are planar and blade-shaped.
On this concern, levels 5, 6, 7 and 8 have large amount of leaves, intensely
carbonified, especially on level 6, which presents the highest density of plant origin material.
Levels 1 and 4 present only lignified material, such as twigs and trunks. Levels 2 and 3
include both leaves and twigs, with lower proportion of the latter. The foliar blades are clearly
distinguished on these assemblages, due to even distribution and spacing among specimens on
these levels. In turn, levels 9 and 10, despite including both, present a very fragmented plant
origin material (less than 50% of preserved foliar blade). That evidences an intense
degradation process during fossildiagenesis, probably caused by transport.
The appearance of seeds on levels 6 (not punctuated during field mapping because of
its diffusion within organic matter), 7, 8 and 10 denote the large dispersion capacity of
gangamopterides. Its conservation state suggests little damage during transport and
deposition. Seeds also evidence the paleoflora’s perenniality, once it is reproductive organs
with seasonal character; in other words, they occur at a specific time of the year, suggesting
the paleoflora to be established for at least one reproductive cycle (Greenwood, 1991).
The higher density levels 5, 6 and 7 have assemblages consisting almost entirely of
juxtaposed leaves. This evidence indicates no sign of currents and, therefore, no transport and
low energy conditions, in which leaves would float until reaching a water saturation state and
then sink. That is also evidenced by low degradation observed on the leaves, once the organic
matter on the plant organs are susceptible to weathering, implying in additional degradation
according to increase in the source-to-sink distance (Benoit et al., 1968; Spicer, 1989, 1991).
Therefore, these assemblages can be considered autochthonous (Table 2).
Bentonic organisms’ marks are found on level 10, revealing higher depth from the
water blade and a less energetic environment (Anderson, 1975; Lima et al., 2015). Levels 7

61
and 8 contain in situ root marks and right under level 7 and above level 6 it was found
mudcracks. These features, along with its spatial distribution and leave density, suggest the
environment possibly to be a lake border, where leaves accumulate.
Despite this brief interpretation of the results, it is important to account for the rate of
plant organs production. For instance, leave abscission is more common than on twigs,
implying dominance of leaves over twigs. Seeds, on the other hand, have high rates of
production, although seasonal, varying in rate during the year (Spicer, 1989; Greenwood,
1991). The fossil assemblies are the result of this production rates combined. Therefore,
abundance of a certain type of plant organ does not account for its production rate on a
determined time.
The rosette diagram data shows no paleocurrent distinction, because it was not
possible to distinguish diagnostic characters on sediments or plant organs (e.g. imbrication,
morphology) to define its sense. Even on levels where there is alignment of plant organs, it
was not possible to distinguish paleocurrent sense, only its direction.
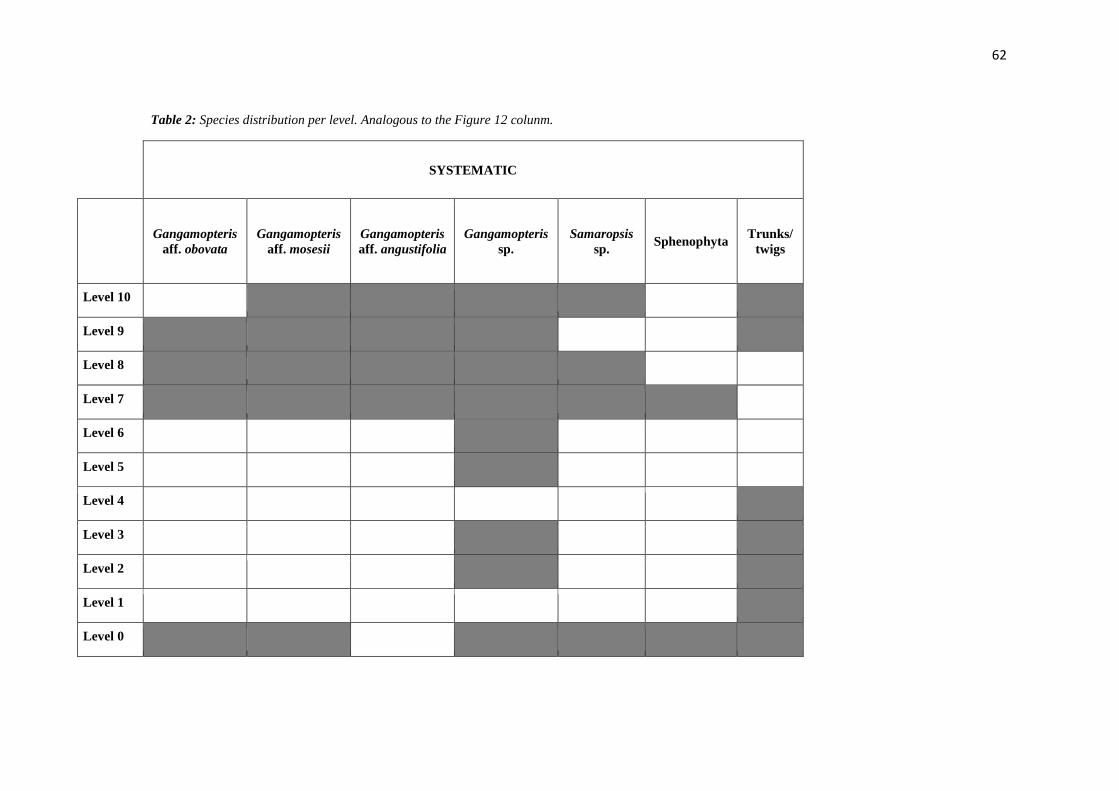
62
Table 2: Species distribution per level. Analogous to the Figure 12 colunm.
SYSTEMATIC
Gangamopteris
aff. obovata
Gangamopteris
aff. mosesii
Gangamopteris
aff. angustifolia
Gangamopteris
sp.
Samaropsis
sp. Sphenophyta
Trunks/
twigs
Level 10
Level 9
Level 8
Level 7
Level 6
Level 5
Level 4
Level 3
Level 2
Level 1
Level 0
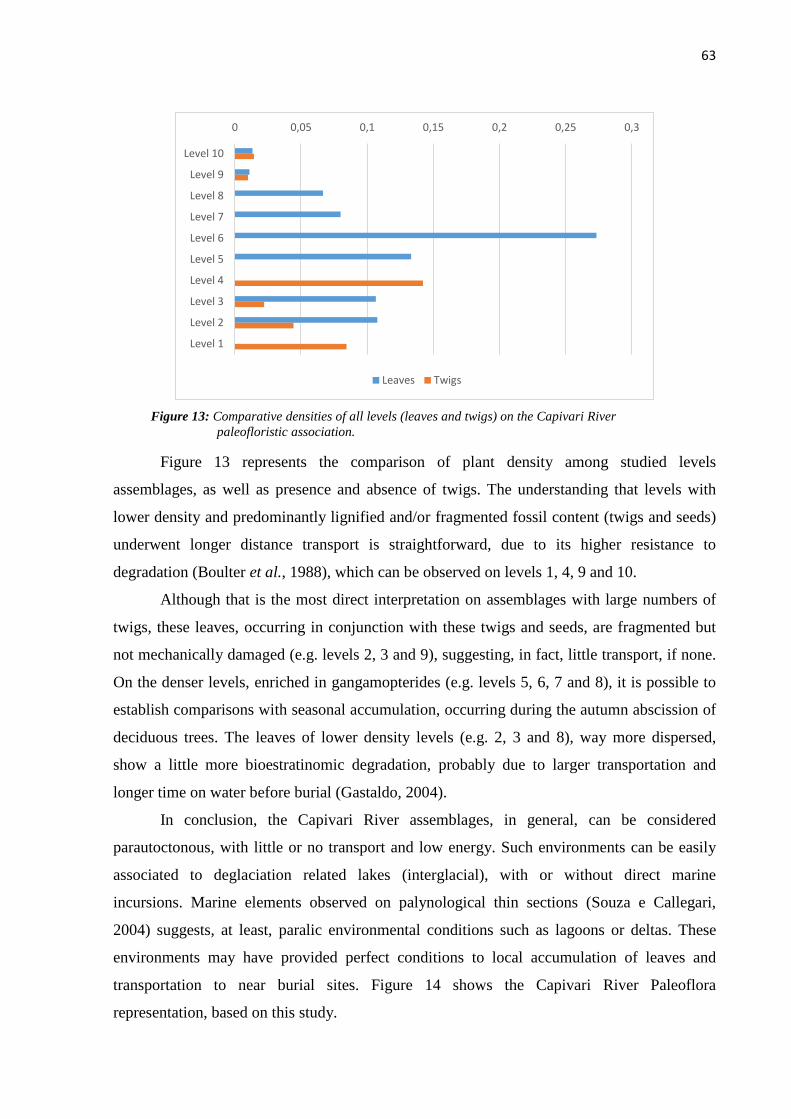
63
Figure 13: Comparative densities of all levels (leaves and twigs) on the Capivari River
paleofloristic association.
Figure 13 represents the comparison of plant density among studied levels
assemblages, as well as presence and absence of twigs. The understanding that levels with
lower density and predominantly lignified and/or fragmented fossil content (twigs and seeds)
underwent longer distance transport is straightforward, due to its higher resistance to
degradation (Boulter et al., 1988), which can be observed on levels 1, 4, 9 and 10.
Although that is the most direct interpretation on assemblages with large numbers of
twigs, these leaves, occurring in conjunction with these twigs and seeds, are fragmented but
not mechanically damaged (e.g. levels 2, 3 and 9), suggesting, in fact, little transport, if none.
On the denser levels, enriched in gangamopterides (e.g. levels 5, 6, 7 and 8), it is possible to
establish comparisons with seasonal accumulation, occurring during the autumn abscission of
deciduous trees. The leaves of lower density levels (e.g. 2, 3 and 8), way more dispersed,
show a little more bioestratinomic degradation, probably due to larger transportation and
longer time on water before burial (Gastaldo, 2004).
In conclusion, the Capivari River assemblages, in general, can be considered
parautoctonous, with little or no transport and low energy. Such environments can be easily
associated to deglaciation related lakes (interglacial), with or without direct marine
incursions. Marine elements observed on palynological thin sections (Souza e Callegari,
2004) suggests, at least, paralic environmental conditions such as lagoons or deltas. These
environments may have provided perfect conditions to local accumulation of leaves and
transportation to near burial sites. Figure 14 shows the Capivari River Paleoflora
representation, based on this study.
0 0,05 0,1 0,15 0,2 0,25 0,3
Level 10
Level 9
Level 8
Level 7
Level 6
Level 5
Level 4
Level 3
Level 2
Level 1
Leaves Twigs

64
Figure 14: Reconstruction of the Capivari River taphoflora environment, based on the taphonomic description and
fossils of the Itararé Group, São Paulo state.

65
CONCLUSIONS
This study contributes to the paleoenvironmental understanding of the Capivari River
Permian taphoflora on the São Paulo state. These assemblages’ taphonomical interpretations
suggest autochtonous to parauthoctonous origin. The first is related to seasonal accumulation
of leaves on low energy burial sites, possibly on lake borders or deltas, with marine
incursions. The latter suggests transportation, due to presence of lignified material such as
twigs, which are resistant to degradation and hydrodynamic, indicating transportation for
longer distances until final deposition.
Palinological data, upper Itararé Group paleofauna information and age estimates of
the Gondwana’ Permo-Carboniferous glaciation suggest that the Capivari River deposits to
have upper Asselian to lower Sakmarian age. The assemblages are similar to the Cerquilho
(SP) ones, but the venation pattern of the gangamopterides seem slightly primitive. The same
pattern is verified on other Gondwanic taphofloras, suggesting this to be one the oldest
registered occurrences of the Glossopteris Flora.
There are more common elements with the Andar Karharbari in India (Artinskian)
and, secondly, with the Ecca Médio Group in Africa (Artinskian) and the Bajo de Veliz
Formation in Argentina (probably lower Permian). However, the general characteristics of
these macrophitofossils assemblages, such as absence of Glossopteris, low diversity and small
size of leaves, are similar to the Andar Talchir in India (Sakmarian), which is located just
above the glacial deposits.
The most abundant species is Gangamopteris obovatta, but the preserved fossils do
not substantiate consistent paleoecological and paleoclimatic interpretations, due to low
diversity of the assemblage. Nevertheless, small sized leaves could evidence colder climates,
and the fact these occurrences are located right under the last glacial deposits (diamictites)
seems to constitute evidence for cold climatic conditions, even with vegetation development.

66
ACKNOWLEDGEMENTS
The authors would like to thank to FAPESP (Fundação de Amparo à Pesquisa do Estado de
São Paulo – the São Paulo Foundation of Support to Research Work) by the financial support.
Grant numbers 2013/11563-6: “Integration of multisource data for the paleobotanical analysis
of the Paraná Basin in the Carboniferous-Permian interval”; and 2016/20927-0: “Studies of
Modern and fossil bioclast accumulations related to continental and coastal environments”. It
was also funded by CNPQ (Conselho Nacional de Desenvolvimento Científico e Tecnológico
- National Council for Scientific and Technological Development) grants.
REFERENCES
Anderson, A.M., 1975. Turbidites and arthropod trackways in the Dwyka glacial deposits
(Early Permian) of Southern Africa. Transactions of the Geological Society of South Africa
78, 265-273.
Arai, M., 1980. Contribuição dos pólens estriados na bioestratigrafia neopaleozóica da parte
nordeste da Bacia do Paraná. São Paulo. Boletim IG-USP 11, 98-105.
Arber, E.A.N., 1905. Catalogue of the Fossil Plants of the Glossopteris Flora. Department of
Geology, British Museum, London, 295 p.
Archangelsky, S. 1958. Estudio geológico y paleontológico del Bajo de la Leona (Santa
Cruz). Acta Geol. Lilloana 2, 5-133.
Barbosa, O, Almeida, F.F.M., 1949. A Série Tubarão na Bacia do Rio Tietê, Estado de São
Paulo. Boletim DNPM/DGM 48, 1-16.
Behrensmeyer, A.K., Kidwell, S.M., Gastaldo, R.A., 2000. Taphonomy and paleobiology.
Paleobiology 26, 103-147.

67
Bateman, R.M., 1999. Particle orientation and palaeoenvironments. In: Jones, T.P., Rowe,
N.P. (Eds), Fossils plants and Spores: modern techniques. Geological Society, pp. 199-205.
Benoit, R.E., Starkey R.L., Basaraba, J., 1968. Effect of purified plant tannin on
decomposition of some organic compounds and plant materials. Soil Sciences, 105, 153–158
Bernardes-De-Oliveira, M.E.C. 1978. Ensaio sobre a utilização de caracteres biométricos das
Glossopteridófitas em estratigrafia. Boletim IG-USP 9, 57-152.
Bernardes-de-Oliveira, M.E.C., Kavali, P.S., Mune, S.E, Shivanna, M., Souza, P.A., Iannuzzi,
R., Jasper, A., Hoelzel, A., Boardman, D.R., Rohn, R., Ricardi-Branco, F., 2016.
Pennsylvanian-Early Cisuralian interglacial macrofloristic succession in Paraná Basin of the
State of São Paulo. Journal of South American Earth Sciences 72, 351-374.
Bernardes-de-Oliveira, M. E.C., Mune, S.E., Garcia, M.J., Iannuzzi, R., Guerra-Sommer, M.,
Jasper, A., Kavali, P.S., Shivanna, M., Pereira, K.G., 2016. The Artinskian Siderópolis
Member macroflora, Rio Bonito Formation and its stratigraphical correlation with other early
Permian macrofloras of Paraná Basin, Brazil. Geologia USP. Série Científica, 16, 65-85.
Boulter, M.C., Spicer, R.A., Thomas, B.A., 1988. Patterns of plant extinction from some
palaeobotanical evidence. In: Larwood GP, (Ed.), Extinction and survival in the fossil record.
Oxford University Press, Oxford, UK, pp. 1–36.
Burnham, R.J., 1989. Relationships between standing vegetation and leaf litter in a
paratropical forest: Implications for paleobotany. Review of Palaeobotany and Palynology 58,
5–32, doi:10.1016/0034-6667(89)90054-7.

68
Canuto, J.R.; Santos, P.R.; Rocha-Campos, A.C. 2001. Estratigrafia de Sequências do
Subgrupo Itararé (Neopaleozóico) no Leste da Bacia do Paraná, nas Regiões Sul do Paraná e
Norte de Santa Catarina, Brasil. Revista Brasileira de Geociências, 31, 107-116.
Cariglino, B., 2013. Fructifi cation diversity from the La Golondrina Formation (Permian),
Santa Cruz Province, Argentina. Geobios 46, 183-193. doi:10.1016/j.geobios.2012.10.017
Christiano-de-Souza, I. C.,Ricardi-Branco, F., 2013. Swamp floral changes through the
Pennsylvanian – Early Permian interval in the Gondwana Floristic Province of the Brazilian
portion of the Parana Basin. New Mexico Museum of Natural History and Science, Bulletin
60, 66-69.
Christiano-de-Souza, I.C., Ricardi-Branco, F.S., 2015. Study of the West Gondwana Floras
during the Late Paleozoic: A paleogeographic approach in the Paraná Basin–
Brazil. Palaeogeography, Palaeoclimatology, Palaeoecology, 426, 159-169.
Daemon, R.F., Quadros, L.P., 1970. Bioestratigrafia do Neopaleozóico da Bacia do Paraná.
Anais Congresso Brasileiro de Geologia, 24., Brasília, pp. 359-412.
Daemon, R. F.1974. Palinomorfos guias do Devoniano Superior e Carbonífero Inferior das
Bacias do Amazonas e Parnaíba. Anais da Academia Brasileira de Ciências 46, 549-807.
Dolianiti, E.,1954. A flora do Gondwana Inferior em Santa Catarina V: o gênero
Gangamopteris. Notas Preliminares e Estudos 89, 1-12
Ellis, B., Johnson, K.R., 2013. Comparison of leaf samples from mapped tropical and
temperate forests: implications for interpretations of the diversity of fossil assemblages.
PALAIOS 28, 163-177.

69
Feistmantel, O. 1879. Fossil Flora of the Gondwana System. I suppl.) - The flora of the
Talchir-Karharbari beds. Memoirs of the Geological Survey of India, Palaeontologia Indica
Series 12, 1-64.
Gama Jr, E.G., Perinotto, J.A.J., Ribeiro, H.J.P.S., Padula, E.K., 1992. Contribuição ao estudo
de ressedimentação do Subgrupo Itararé: um gruia temático de campo. Revista Brasileira de
Geociências 22, 237-247.
Gastaldo, R.A., Stevanovic-Walls, I., Ware, W.N., 2004. In situ, erect forests are evidence for
large-magnitude, coseismic base-level changes within Pennsylvanian cyclothems of the Black
Warrior Basin, USA: In: Pashin, J.C., Gastaldo, R.A., (Eds.), Sequence Stratigraphy,
Paleoclimate, and Tectonics of Coal-bearing Strata, American Association of Petroleum
Geologists Studies in Geology 51, 219-238.
Goeppert, H.P., 1864. Die fossile flora der Permischen Formation. Palaeontographica 12, 1-
244.
Greenwood, D.R., 1991. The taphonomy of plant macrofossils. In: Donovan, S.K. (Ed.), The
Processes of Fossilization. Columbia University Press, New York, pp. 145-169.
Greenwood, D.R., 1992, Taphonomic constraints on foliar physionomic interpretations of
Late Cretaceous and Tertiary palaeoclimates. Review of Palaeobotany and Palynology 71,
142-196.
Guerra-Sommer, M., Cazzulo-Klepzig, M., 1993. Bioestratigraphy of the souther Brasilian
Neopaleozoic Gondwana Sequence: a preliminar paleobotanical approach. Comptes Rednus
Int. Congres Stratigraphie et Geologie du Carbonifere et Permien, 12, Buenos Aires, 2, pp.
61-72.

70
Guerra-Sommer, M., Cazzulo-Klepzig, M., 2000. Permian Paleofloras in Southern Brazilian
Gondwana. Revista Brasileira de Geociências, Special Issue 30, 486-491.
Gulbranson, E.L., Ryberg, P.E., Decombeix, A.L., Taylor, E.L., Taylor, T.N., Isbell, J.L.,
2014. Leaf habit of Late Permian Glossopteris trees from high-palaeolatitude forests. Journal
of the Geological Society 171, 493-507.
Iannuzzi, R., Rohn, R., 1995. Discussão sobre a presença do gênero Glossopteris na tafoflora
Cerquilho (Grupo Tubarão, Bacia do Paraná, Estado de São Paulo) e seu possível significado
biocronoestratigráfico. Atas Congresso Brasileiro de Paleontologia, 14, Uberaba, pp. 62–63.
Iannuzzi, R., Rösler, O., 2000. Floristic migration in South America during the
Carboniferous: phytogeographic and biostratigraphic implications. Palaeogeography,
Palaeoclimatology, Palaeoecology 161, 71- 94.
Leguizamón, R.R., 1972. Estudio paleobotánico de la Formación Tasa Cuna, Pérmico Inferior
de la Provincia de Córdoba. Ameghiniana 9, 305–342.
Lima, J.H.D., Netto, R.G., Corrêa, C.G., Lavina, E.L.C., 2015. Ichnology of deglaciation
deposits from the Upper Carboniferous Rio do Sul Formation (Itararé Group, Paraná Basin) at
central-east Santa Catarina State (southern Brazil). Journal of South American Earth Sciences
63, 137-148.
Limarino, C.O., Césari, S., Spalletti, L., Taboada, A.C., Isbell, J., Geuna, S., Gulbranson, E.L.
2013. Paleoclimatic evolution of southern South America during the Late Paleozoic: a record
from Icehouse to Greenhouse conditions. Gondwana Research 25, 1396-1421.
Maithy, P.K. 1965. Studies in the Glossopteris flora of India – 26. Glossopteridales from the
Karharbari beds, Giridih coalfield, India. The Paleobotanist 13, 248-263.

71
McCoy, F., 1847. On the fossil Botany and Zoology of rocks associated with the coal of
Australia. Annual Magazine of Natural History Museum 20, 145-298.
McCoy, F., 1860. A commentary on “A communicaton made by the Rev. W. B. Clarke” & C.
Transaction of Royal Society 5, 98.
McCoy, F., 1875. Prodromus of the palaeontology of Victoria. Geological Survey of Victoria,
Melbourne, Decades I, p. 1-43, II, p. 1-37.McLoughlin, S., Larsson, K., Lindström, S., 2005.
Permian plant macrofossils from Fossilryggen, Vestfjella, Dronning Maud Land. Antarctic
Science 17, 73-86.
Milani, E.J., Dewitt, M.J., 2008. Correlations between the classic Paraná and Cape-Karoo
sequences of South America and southern Africa and their basin infills flanking the
Gondwanides: du Toit revisited. Geological Society, Special Publications 294, 319-342.
Milani, E.J., Melo, J.H.G., Souza, P.A., Fernandes, L.A., França, A.B., 2007. Bacia do
Paraná. Boletim de Geociências da Petrobrás 15, 265-287.
Millan, J.H., 1977. Sementes platispérmicas da flórula gondwanica de Monte Mor, estado de
São Paulo, Brasil. Ameghiniana 14, 33-44.
Millan, J.H., 1989. Sobre a presença de formas foliares mais primitivas de glossopterídeas
(Gênero Gangamopteris) no Município de Cerquilho, carvão do Eogonwana do Estado de São
Paulo. Anais da Academia brasileira de Ciências 61, 201-213.
Mune, S.E., Bernardes de Oliveira, M.E.C., 2007. Revisão da tafoflora interglacial
neocarbonífera de Monte Mor, SP (Subgrupo Itararé), nordeste da Bacia do Paraná. Revista
Brasileira de Geociências 37, 32-45.

72
Pant, D.D., Singh, K.B. 1968. On the Genus Gangmopteris McCoy. Palaeontographica Abt.
B 124, 83-101.
Ricardi-Branco, F., Branco, F.C, Garcia, R.F., Faria, F.S, Pereira, S.Y., Portugal, R, Pessenda,
L.C., Pereira, P.R.B., 2009. Features of plant accumulations along the Itanhaém river, on the
southern coast of the Brazilian state of São Paulo. PALAIOS 24, 416-424.
Ricardi-Branco, F., Rohn, R., Longhim, M.E., Costa, J.S., Martine, A.M., Christiano-De-
Souza, I.C. 2016. Rare Carboniferous and Permian glacial and non-glacial bryophytes and
associated lycophyte megaspores of the Paraná Basin, Brazil: A new occurrence and
paleoenvironmental considerations. Journal of South American Earth Sciences 72, 63-75.
Ricardi-Branco, F.S., Bernardes-de-Oliveira, M.E.C. 2000. Gangamopteris roesleri sp. nov.,
da Tafoflora eopermiana de Cerquilho (SP), Grupo Tubarão, Bacia do Paraná, Brasil. Rev.
Uni. Guar. Geo (special number), 44-48.
Ricardi-Branco, F.S., Costa, J.S., Christiano-de-Souza, I.C., Rohn, R., Longin, M., Faria,
R.S., 2013. Bryophytes associated with Pennsylvanian Periglacial Environments in Southern
Gondwana (São Paulo State, Itararé Group, Paraná Basin, Brazil). New Mexico Museum of
Natural History and Science 60, 343-347
Rocha-Campos, A.C., dos Santos, P.R., Canuto, J.R., 2008. Late Paleozoic glacial deposits of
Brazil: Paraná Basin. Geological Society of America Special Papers 441, 97-114.
Rohn, R.; Babinski, M.E.C.B.O.; Rosler, O., 1984. Glossopteris da Formação Rio do Rasto
no Sul do estado do Paraná. Anais Congresso Brasileiro de Geologia 32, Rio de Janeiro, pp.
1047-1061.

73
Rohn, R., Longhim, M.E., Bernardes-De-Oliveira, M.E., Navarro, G.R.B., 2000. Nova
ocorrência fitofossilífera neocarbonífera-eopermiana do Subgrupo Itararé, à margem esquerda
do rio Capivari, Município de Tietê, SP, Brasil. Revista Universidade de Guarulhos,
Geociências, 5, 57-61.
Rösler, O., 1978. The Brazilian eogondwanic floral successions. Boletim IG-USP 9, 85-91.
Santos, P.R., Rocha Campos, A.C., Canuto, J.R., 1996. Patterns of Late Paleozoic
deglaciation in the Paraná Basin, Brazil. Palaeogeography, Palaeoclimatology, Palaeoecology
125, 165-184.
Scotese, C.R. 2000. Paleomap project. http://www.scotese.co
Smith, A.G., 1999. Gondwana: its shape, size and position from Cambrian to Triassic times.
Journal of African Earth Sciences 28, 71-99.
Soares, P.C., Landim, P.M.B., Sineli, O, Wernick, E., Wu, F.T., Fiori, A.P., 1977.
Associações litológicas do Subgrupo Itararé e sua Interpretação Ambiental. Revista Brasileira
de Geociências 7, 131-149.
Souza, P.A., Marques-Toigo, M., 2001. Zona Vittatina: marco palinobioestratigráfico do
Permiano Inferior da Bacia do Paraná. Correlação de Sequências Paleozóicas Sul-
Americanas. Ciência, Técnica, Petróleo. Seção: Exploração de Petróleo 20, 153-159.
Souza, P.A., Callegari, L.M., 2004. An Early Permian palynoflora from the Itararé Subgroup,
Paraná Basin, Brazil. Revista Española de Micropaleontologia 36, 439-450.
Souza, P.A., 2006. Late Carboniferous palynostratigraphy of the Itararé Subgorup,
northeastern Paraná Basin, Brazil. Review of Paleobotany and Palynology 138, 9-29.

74
Spicer, R.A., 2010. Recording palaeontological information. In: Coe, A.L. (Ed.), Geological
Field Techniques. Open University, pp. 79-101.
Spicer, R.A., 1989. Physiological characteristics of land plants in relation to environment
through time. Trans R Soc Edinburgh (Earth Sc) 80, 321-329.
Spicer, R., 1991. Plant taphonomic processes. Taphonomy: Releasing the Data Locked in the
Fossil Record: Topics in Geobiology 9, 71-113.
Srivastava, A.K., 1991. Evolutionary tendency in the venation pattern of Glossopteridales.
Geobios 4, 383-386
Srivastava, A.K., Agnihotri, D., 2010. Morphological consequence of Gangamopteris McCoy
in Glossopteris flora. Journal of Asian Earth Sciences 39, 760-769.
Taylor, T.N., Taylor E.L., Krings, M., 2009. Paleobotany. The biology and evolution of fossil
plants. Elsevier/Academic Press Inc., Burlington
Tybusch, G.P., Iannuzzi, R., 2008. Reavaliação taxonômica dos gêneros Gangamopteris e
Rubidgea, Permiano Inferior da Bacia do Paraná, Brasil. Revista Brasileira de Paleontologia
11, 73-86.
Tybusch, G.P., Iannuzzi, R., Bernardes-de-Oliveira, M.E.C., Lopes, R.C., 2016. Revaluation
of the Glossopterids from the Lower Permian of Cambaí Grande Outcrop, Paraná Basin, RS.
Geologia USP. Série Científica, 16, 41-51.
Veevers, J.J. 2004. Gondwanaland from 600-570 Ma assembly through 320 Ma merger in
Pangea to 160 Ma breakup: supercontinental tectonics via stratigraphy and radiometric dating.
Earth Sciences Review 68, 1-132.

75
White, D., 1908. Fossil flora of the coal measures of Brazil. In: White I.C. (Ed) Comissão de
estudos das Minas de Carvão de Pedra do Brasil. Relatório Final. Pp. 558-568 (2a ed.).
Brasília: Departamento Nacional de Produção Mineral.
Willis, K.J., Mcelwain, J.C., 2002. The Evolution of Plants. Oxford University Press Oxford.

76
ATTACHMENT I LEVEL 1
0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1
0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1
0 0 0 0 0 0 1 0 0 0 0 0 0 0 1 1
0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0
LEVEL 2
1 0 1 0 1 0 1 1 1 1 1 0 0 0 0
1 1 1 1 1 1 1 1 1 1 1 0 0 0 0 0
0 1 1 1 1 1 1 1 1 0 0 0 0 0 0 0
0 1 1 1 1 1 1 1 1 0 1 0 0 0 0 1
1 1 1 1 1 1 1 1 0 0 1 1 0 0 0 1
1 1 0 1 1 1 1 1 1 0 1 1 0 0 0 0
1 1 1 1 1 1 1 1 1 0 1 1 0 0 0 0
1 1 0 1 1 1 0 1 1 0 0 1 0 0 0 0
0 1 0 1 1 1 0 0 0 0 1 1 0 0 0 0
0 0 0 1 1 1 1 1 1 0 0 0 0 0 0 0
0 0 0 1 1 1 1 0 1 0 0 0 0 0 0 0
0 0 0 1 1 1 1 1 1 0 0 0 0 0 0 0
0 1 1 1 1 1 1 1 1 0 1 0 0 0 0 0
1 1 1 1 1 0 1 1 0 0 1 1 1 0 0 0
1 1 1 1 1 0 1 0 0 0 0 1 0 1 0 0
0 1 0 0 0 0 1 0 0 0 0 1 1 0 0 0

77
LEVEL 3
1 0 1 1 1 1 0 1 1 1 1 1 0 0 0 0
1 0 1 1 1 1 0 1 1 1 1 1 0 0 0 0
0 0 0 0 1 1 1 1 1 1 1 1 0 0 0 0
0 0 0 1 1 1 1 1 1 1 1 1 0 0 1 0
0 0 0 1 1 1 1 1 1 1 1 1 0 0 1 0
0 1 1 1 1 1 0 1 0 1 1 0 0 0 0 0
0 1 1 1 1 0 0 0 0 0 0 0 0 0 0 0
0 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0
0 0 0 1 0 0 0 1 1 0 0 0 0 0 0 0
0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0
1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0
1 1 1 0 0 0 0 0 1 0 0 0 0 0 0 0
1 1 1 1 0 0 1 1 1 0 0 0 0 0 0 0
1 1 1 1 0 1 1 1 1 1 1 1 1 0 0 0
1 1 1 1 0 1 0 1 1 1 0 0 0 1 0 0
1 1 1 1 0 1 0 1 1 1 0 0 0 0 0 0
LEVEL 4
0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0
0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0
0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0
0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0
0 0 0 0 0 0 0 1 1 1 0 0 0 0 0 0
0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0
0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0
0 0 0 0 0 1 1 0 0 0 0 0 0 0 0 0
0 0 0 1 0 1 1 0 0 0 0 0 0 0 0 0
1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1

78
LEVEL 5
0 0 0 0 0 0 1 1 1 1 1 1 0 0 0 0
0 0 0 0 1 1 1 1 1 1 1 1 0 0 0 0
0 0 0 0 1 1 1 1 1 1 1 1 0 0 0 0
0 0 0 0 0 0 1 1 1 1 1 1 0 0 0 0
0 0 0 0 0 1 1 1 1 1 1 1 0 0 0 0
0 0 0 1 1 1 1 1 1 1 1 1 0 0 0 0
0 0 1 1 1 1 1 1 1 1 1 1 0 1 1 1
0 1 1 1 0 0 1 1 1 1 1 1 1 1 0 0
0 0 1 1 0 0 1 1 1 1 1 1 1 1 0 0
0 0 0 0 0 1 1 0 1 1 1 1 1 1 0 0
0 0 0 0 0 1 1 0 1 1 1 1 1 1 1 1
0 0 1 0 1 1 1 0 1 1 0 1 1 1 0 0
0 0 1 0 1 1 1 0 1 0 0 1 1 1 0 0
0 0 0 0 0 0 0 0 0 0 0 1 1 1 0 0
0 0 0 0 0 1 1 0 0 0 0 1 0 0 0 0
0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0
LEVEL 6
1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
1 1 1 1 1 1 1 1 1 1 1 0 0 1 1 1
1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
1 1 0 0 1 1 1 1 1 1 1 1 1 1 1 1
1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
1 1 1 1 1 0 1 1 1 0 1 1 1 1 1 1
0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1
1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 1
1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1

79
LEVEL 7
1 1 0 0 0 0 1 0 0 0 0 1 1 0 0 0
1 1 0 0 1 0 0 0 0 0 0 0 1 0 0 0
1 1 1 1 1 1 0 0 0 1 0 0 0 0 0 0
1 1 1 1 1 0 0 0 0 1 0 1 0 0 0 0
0 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0
0 0 1 1 1 0 0 0 0 0 0 0 0 0 0 0
0 0 0 1 1 0 0 0 0 0 0 0 0 0 0 1
1 0 0 1 1 1 1 0 0 0 0 0 0 0 0 0
1 1 0 0 1 1 1 0 0 0 0 1 0 0 0 0
1 0 1 1 0 0 1 0 0 0 0 0 0 0 0 0
1 0 1 1 1 0 1 0 0 0 0 0 0 1 1 0
1 1 1 1 1 0 0 0 0 1 1 0 0 1 1 0
1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0
0 1 1 1 1 0 0 0 0 0 0 0 0 0 0 0
0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0
1 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0
LEVEL 8 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
0 0 0 0 0 1 1 0 0 1 0 0 0 0 0 0
1 0 0 1 1 1 1 1 0 0 0 0 0 0 0 0
1 0 0 1 1 1 0 0 0 1 0 0 0 0 0 0
1 0 0 1 1 0 0 0 0 1 0 0 0 0 1 0
0 0 0 1 1 0 1 1 1 0 0 1 0 0 1 0
0 0 0 1 1 0 0 0 1 1 0 1 0 0 1 0
0 0 0 0 1 0 0 1 1 1 0 0 1 0 0 0
1 0 0 0 0 0 1 0 0 1 0 0 0 0 0 1
1 0 0 1 1 0 0 0 0 1 0 0 0 0 0 0
1 0 0 0 1 0 1 1 1 1 0 1 0 0 0 0
0 0 0 0 0 0 1 1 0 1 0 1 0 0 0 0
0 0 0 0 0 0 0 1 1 1 0 1 0 0 0 0

80
LEVEL 9
0 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0 1 0 0 0 0 0 1 0
0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0
1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
1 0 0 1 0 0 0 0 0 1 0 0 0 0 0 0
1 0 1 1 0 0 0 0 0 0 1 0 0 0 0 0
0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0 0 0 1 1 0 0 0 0
0 0 0 1 0 0 0 0 0 1 1 0 0 0 0 0
0 0 1 0 0 0 0 0 0 0 1 0 0 0 0 0
0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
LEVEL 10
0 0 0 0 0 0 0 0 0 0 0 0 0 1 0
1 0 0 0 0 0 0 0 0 0 0 0 1 0 0
1 0 0 0 0 0 0 0 0 0 0 1 0 0 0
0 1 0 0 0 0 0 0 0 0 1 0 0 0 0
0 1 0 0 0 0 0 0 0 0 0 0 0 0 0
0 1 0 0 0 0 0 0 0 1 0 0 0 0 0
0 0 1 0 0 0 0 0 0 1 0 0 0 0 0
0 0 1 0 0 0 0 0 0 0 0 0 0 0 0
1 0 0 0 0 0 0 0 0 0 0 0 0 0 0
0 0 0 1 0 0 0 0 0 0 0 0 0 0 0
0 0 0 1 0 0 0 0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0 0 0 1 0 0 0 0
0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0 0 0 0 0 0 0 0

81
7. CONSIDERAÇÕES FINAIS
O mapeamento tafonômico executado neste trabalho revelou a existência de
assembleias parautóctones no Afloramento Rio Capivari, Grupo Itararé, Bacia do Paraná (SP).
Estas, por sua vez, formaram-se a partir de acumulações sazonais em locais de baixa energia
de fluxo, apontada pela distribuição e orientação preferencial dos espécimes em cada patamar
fossilífero estudado na localidade. Os dados palinológicos obtidos por Callegari (2001)
sugerem influência marinha, possivelmente pela localização do sistema deposicional nas
bordas de um lago ou delta, e também sugerem a idade do final do Asseliano ao início do
Sakmariano.
A assembleia do Rio Capivari assemelha-se morfologicamente e estratigraficamente à
paleoflora encontrada em Cerquilho (SP), Afloramento Sítio Toca do Índio. No entanto, o
padrão de venação das gangamopterídeas do Rio Capivari parece ser ligeiramente mais
primitivo. O mesmo pode ser constatado em relação a outras tafofloras do Gondwana,
sugerindo-se que o Rio Capivari represente uma das ocorrências mais antigas da Flora de
Glossopteris. A ausência de Glossopteris, baixa diversidade e tamanho pequeno das folhas,
leva ao pressuposto de que a flora do Rio Capivari se assemelha mais àquelas do Andar
Talchir da Índia (Sakmariano), o qual está situado logo acima dos últimos depósitos glaciais.
A espécie mais abundante é Gangamopteris obovatta, porém os macrofitofósseis
preservados, em si, não parecem fundamentar interpretações paleoecológicas e
paleoclimáticas muito consistentes devido a diversidade relativamente baixa da assembleia
(Spicer, 1989; Greenwood, 1991). Contudo o tamanho pequeno das folhas poderiam ser
evidências de clima frio e o próprio fato da ocorrência estar localizada abaixo dos últimos
depósitos glaciais parece constituir uma prova de que as condições climáticas eram
provavelmente bastante frias, mesmo que a vegetação tenha se desenvolvido durante uma fase
interglacial.

82
8. BIBLIOGRAFIA
Alvin, K.L.. Fraser, C.J. & Spicer, R.A. 1981. Anatomy and palaeoecology of
Pseudofrenelopsis and associated conifers in the English Wealden. Paleontology,
24:759-778.
Anderson, A.M. 1975. Turbidites and arthropod trackways in the Dwyka glacial deposits
(Early Permian) of Southern Africa. Transactions of the Geological Society of South
Africa, 78:265-273.
Arai, M. 1980. Contribuição dos pólens estriados na bioestratigrafia neopaleozóica da parte
nodeste da Bacia do Paraná. São Paulo. Boletim IG-USP, 11:98-105.
Arber, E.A.N. 1905. Catalogue of the Fossil Plants of the Glossopteris Flora. Department of
Geology, British Museum, London, 295 p
Archangelsky S. 1958. Estudio geológico y paleontológico del Bajo de la Leona (Santa Cruz).
Acta Geol Lilloana, 2:5-133.
Barbosa, O & Almeida, F.F.M., 1949. A Série Tubarão na Bacia do Rio Tietê, Estado de São
Paulo, Rio de Janeiro, DNPM/DGM, Not. Prel. Boletim 48.
Behrensmeyer, A.K.; Kidwell, S.M. e Gastaldo, R.A. 2000. Taphonomy and paleobiology.
Paleobiology, 26:103-147.
Benoit RE, Starkey RL, Basaraba J. 1968. Effect of purified plant tannin on decomposition of
some organic compounds and plant materials. Soil Sciences, 105:153–158
Bernardes-de-Oliveira, M. E. C. 1977. Tafoflora Eogondvânica da camada Irapuá, Formação
Rio Bonito (Grupo Tubarão), SC. Tese (Doutorado). São Paulo: Instituto de
Geociências – USP.
Bernardes-de-Oliveira, M. E. C., Kavali, P. S., Mune, S. E, Shivanna, M., Souza, P. A.,
Iannuzzi, R., Jasper, A., Hoelzel, A., Boardman, D. R., Rohn, R., Ricardi-Branco, F.
2016. Pennsylvanian-Early Cisuralian interglacial macrofloristic succession in
Paraná Basin of the State of São Paulo. Journal of South American Earth Sciences,
72:351-374.
Bernardes-De-Oliveira, M.E.C. 1978. Ensaio sobre a utilização de caracteres biométricos das
Glossopteridófitas em estratigrafia. Advances in Palaeobotany and allied scienes in
Brazil. Boletim IG. Instituto de Geociências, USP, 9:57-152.
Boulter MC, Spicer RA, Thomas BA. 1988. Patterns of plant extinction from some
palaeobotanical evidence In: Larwood GP, editor. , ed. Extinction and survival in the
fossil record. Oxford, UK: Oxford University Press, 1–36.

83
Burnham, A.K., 1989, A simple kinetic model of petroleum formation and cracking:
Lawrence Livermore National Lab Report, USA, UCID- 21665, 11 p.
Caetano-Chang, M.R.. 1984. Análise Ambiental e Estratigráfica do Subgrupo Itararé (PC) no
Sudoeste do Estado de São Paulo. Tese de Doutoramento, Instituto de Geociências,
Universidade de São Paulo, São Paulo, 310 p.
Callegari, L. M. Palinologia da porção superior do Subgrupo Itararé na região de Jumirim e
Tietê, Estado de São Paulo. São Paulo, 2001. 52 p. Monografia (Trabalho de
Formatura)- Instituto de Geociências, Universidade de São Paulo
Canuto, J.R.; Santos, P.R.; Rocha-Campos, A.C.. 2001. Estratigrafia de Sequências do
Subgrupo Itararé (Neopaleozóico) no Leste da Bacia do Paraná, nas Regiões Sul do
Paraná e Norte de Santa Catarina, Brasil. Revista Brasileira de Geociências,
31(1):107-116.
Cariglino, B. 2013. Fructifi cation diversity from the La Golondrina Formation (Permian),
Santa Cruz Province, Argentina. Geobios, 46:183-193.
doi:10.1016/j.geobios.2012.10.017
Christiano-De-Souza, I.C. & Ricardi-Branco, F.S. 2015. Study of the West Gondwana Floras
during the Late Paleozoic: A paleogeographic approach in the Paraná Basin – Brazil.
Palaeogeography, Palaeoclimatology, Palaeoecology, 426:159-169.
Christiano-de-Souza, I.C., Ricardi-Branco, F.S., Silva, A.M., El-Dash, L.G. Faria, R.S., 2014.
New approach for the study of paleofloras using geographical information systems
applied to Glossopteris Flora. Brazilian Journal of Geology. 44:681-689.
Daemon, R. F.; Quadros, L. P. Bioestratigrafia do Neopaleozóico da Bacia do Paraná. In:
CONGRESSO BRASILEIRO DE GEOLOGIA, 24., Brasília, 1970.
Daemon, R. F.1974. Palinomorfos guias do Devonlano Superlor e Carbonífero Inferior das
Bacias do Amazonas e Parnaíba. Anais da Academia Brasileira de Ciências, 46:549-
807.
Ellis, B. Johnson, K.R. 2013. Comparison of leaf samples from mapped tropical and
temperate forests: implications for interpretations of the diversity of fossil
assemblages. PALAIO, 28:163-177.
Feistmantel, O. 1879. Fossil Flora of the Gondwana System. I suppl.) - The flora of the
Talchir-Karharbari beds. Memoirs of the Geological Survey of India, Palaeontologia
Indica series 12(3):1-64.

84
Gama Jr, E.G.; Perinotto, J.A.J.; Ribeiro, H.J.P.S.; Padula, E.K.. 1992. Contribuição ao estudo
de ressedimentação do Subgrupo Itararé: um gruia temático de campo. Revista
Brasileira de Geociências, 22(2):237-247.
Gastaldo, R.A., Stevanovic-Walls, I., and Ware, W.N., 2004. In situ, erect forests are
evidence for large-magnitude, coseismic base-level changes within Pennsylvanian
cyclothems of the Black Warrior Basin, USA: In: Pashin, J.C., and Gastaldo, R.A.,
eds., Sequence Stratigraphy, Paleoclimate, and Tectonics of Coal-bearing Strata:
American Association of Petroleum Geologists Studies in Geology, 51:219-238.
Greenwood, D. R. 1991. The taphonomy of plant macrofossils. In: Donovan, S. K. (Ed.), The
processes of fossilisation, pp.141-169. Belhaven Press.
Greenwood, D.R., 1991. The taphonomy of plant macrofossils: In: Donovan, S.K., ed., The
Processes of Fossilization: Columbia University Press, New York, p. 141-169.
Greenwood, D.R., 1992, Taphonomic constraints on foliar physionomic interpretations of
Late Cretaceous and Tertiary palaeoclimates: Review of Palaeobotany and
Palynology, 71:142-19
Guerra-Sommer, M.; Cazzulo-Klepzig, M. 1993. Bioestratigraphy of the souther Brasilian
Neopaleozoic Gondwana Sequence: a preliminar paleobotanical approach. In:
International Congres De La Stratigraphie Et Geologie Du Carbonifere Et Permien,
12, Buenos Aires, 1991. Comptes rednus…Buenos Aires, 2:61-72
Harris, T.M. 1958. Forest fire in the Mesozoic. Journal of Ecology 46:447–453.
Iannuzzi R., Rohn R., 1995. Discussão sobre a presença do gênero Glossopteris na tafoflora
Cerquilho (Grupo Tubarão, Bacia do Paraná, Estado de São Paulo) e seu possível
significado biocronoestratigráfico. In: CONGRESSO BRASILEIRO DE
PALEONTOLOGIA, 14, Atas, Uberaba, 62–63.
Iannuzzi, R. & Rösler, O. 2000. Floristic migration in South America during the
Carboniferous: phytogeographic and biostratigraphic implications.
Palaeogeography, Palaeoclimatology, Palaeoecology, 161:71- 94.
Iannuzzi, R.; Rösler, O. Floristic migration in South America during the Carboniferous:
phytogeographic and biostratigraphic implications. Palaeogeography,
Palaeoclimatology, Palaeoecology, 161: 71-94, 2000.
Leguizamón, R.R. 1972. Estudio paleobotánico de la Formación Tasa Cuna, Pérmico Inferior
de la Provincia de Córdoba. Ameghiniana 9:305–342.

85
Lermen, R. E. (2006). Assinaturas icnológicas em depósitos glacigênicos do Grupo Itararé no
RS. Dissertação de Mestrado: Universidade Vale do Rio dos Sinos. 2006. São
Leopoldo (RS).
Limarino, C.O.; Césari, S.; Spalletti, L.; Taboada, A.C.; Isbell, J.; Geuna, S.; Gulbranson,
E.L. 2013. Paleoclimatic evolution of southern South America during the Late
Paleozoic: a record from Icehouse to Greenhouse conditions. Gondwana Research
25(4):1396-1421.
Maheshwari, S. K., Gupta, P. C. and Gandhi, S. K. 1991. Evaluation of different
fungitoxicants against early blight of tomato. Agricultural Science Digest (Kamal),
11:201-202.
Maithy, P. K. 1965. Studies in the Glossopteris flora of India – 26. Glossopteridales from the
Karharbari beds, Giridih coalfield, India. The Paleobotanist, 13(1):248-263.
McCoy, F. 1847. On the fossil Botany and Zoology of rocks associated with the coal of
Australia. Annual Magazine of Natural History Museum, 20(1):145,226,298.
McCoy, F. 1860. A commentary on “A communicaton made by the Rev. W. B. Clarke” & C.
Transaction of Royal Society, Victoria 5, 98.
McLoughlin P. D., Dunford J. S., Boutin S. 2005. Relating predation mortality to broad-scale
habitat selection. J. Anim. Ecol.74, 701–707
Milani, E.J., Dewitt, M.J.. 2008. Correlations between the classic Paraná and Cape-Karoo
sequences of South America and southern Africa and their basin infills flanking the
Gondwanides: du Toit revisited. Geological Society, London, pp. 319-342. Special
Publications 294.
Milani, E.J.; Melo, J.H.G.; Souza, P.A.; Fernandes, L.A.; França, A.B.. 2007. Bacia do
Paraná. Boletim de Geociências da Petrobrás. 15: 265-287.
Millan, J.H. 1987. Os pisos florísticos do carvão do Subgrupo Itararé do Estado de São Paulo
e suas implicações. In: CONGRESSO BRASILEIRO DE PALEONTOLOGIA, 10,
Rio de Janeiro. Anais..., p.832-857.
Millan, J.H. 1989. Sobre a presença de formas foliares mais primitivas de glossopterídeas
(Gênero Gangamopteris) no Município de Cerquilho, carvão do Eogonwana do
Estado de São Paulo. Anais da Academia brasileira de Ciências. 61(2):201-213.
Mune, S.E., Bernardes de Oliveira, M.E.C., 2007. Revisão da tafoflora interglacial
neocarbonífera de Monte Mor, SP (Subgrupo Itararé), nordeste da Bacia do Paraná.
Revista Brasileira de Geociências, 37:32-45.

86
Nykvist, N. 1962. Leaching and decomposition of litter. V. Experiments on leaf litter of Alnus
glutinosa, Fagus and Quercus robur. Oikos, 13:232-48.
Pant, D. D. & Singh, K. B. 1968. On the Genus Gangmopteris McCoy. Palaeontographica
Abt. B, 124:83-101.
Ricardi-Branco, F. S. R. T. Tafoflora Gondvânica do Membro Triunfo, Formação Rio Bonito
(Eopermiano), no Município de Figueira. Tese de doutorado. Instituto de
Geociências, Universidade de São Paulo (USP), São Paulo. 1998.
Ricardi-Branco, F., Branco, F.C, Garcia, R.F., Faria, F.S, Pereira, S.Y., Portugal, R, Pessenda,
L.C., Pereira, P.R.B. 2009. Features of plant accumulations along the Itanhaém river,
on the southern coast of the Brazilian state of São Paulo. PALAIOS. 24:416-424
Ricardi-Branco, F.; Rohn, R.; Longhim, M.E.; Costa, J.S.; Martine, A.M.; Christiano-De-
Souza, I.C.. 2016. Rare Carboniferous and Permian glacial and non-glacial
bryophytes and associated lycophyte megaspores of the Paraná Basin, Brazil: A new
occurrence and paleoenvironmental considerations. Journal of South American Earth
Sciences. 72:63-75.
Ricardi-Branco, F.S. & Bernardes-de-Oliveira, M.E.C. 2000. Gangamopteris roesleri sp. nov.,
da Tafoflora eopermiana de Cerquilho (SP), Grupo Tubarão, Bacia do Paraná, Brasil.
Ver. Uni. Guar. Geo (special number) p.44-48.
Ricardi-Branco, F.S., Costa, J.S., Christiano-de-Souza, I.C., Rohn, R., Longin, M., Faria,
R.S., 2013. Bryophytes associated with Pennsylvanian Periglacial Environments in
Southern Gondwana (São Paulo State, Itararé Group, Paraná Basin, Brazil). New
Mexico Museum of Natural History and Science. 60:343-347
Rocha-Campos, A.C.. 1967. The Tubarão Group in the Brazilian portion of the Paraná Basin.
In: J.J. Bigarella; R.D. Becker; I.D. Pinto (Editors), Problems in Brazilian
Gondwana Geology, Curitiba, p. 27-102.
Rohn, R.; Babinski, M.E.C.B.O.; Rosler, O. Glossopteris da Formação Rio do Rasto no Sul
do estado do Paraná. In: XXXII CONGRESSO BRASILEIRO DE GEOLOGIA, Rio
de Janeiro, Anais..., p. 1047-1061, 1984.
Rohn, R.; Longhim, M.E.; Bernardes-De-Oliveira, M.E.; Navarro, G.R.B. 2000. Nova
ocorrência fitofossilífera neocarbonífera-eopermiana do Subgrupo Itararé, à margem
esquerda do rio Capivari, Município de Tietê, SP, Brasil. Revista Universidade de
Guarulhos, Geociências, 5 (nº especial): 57-61.
Rosler, O. 1978. The Brazilian eogondwanic floral successions. Boletim IG-USP, São Paulo,
9:85-91.

87
Saad, A.R.. 1977. Estratigrafia do Subgrupo Itararé no centro e sul do Estado de São Paulo.
Dissertação de Mertrado, Instituto de Geociências, Universidade de São Paulo, São
Paulo, 107 p.
Santos, P.R,; Rocha-Campos, A.C.; Canuto, J.R.. 1996. Patterns of Late Palaeozoic
Deglaciation in the Paraná Basin, Brazil. Palaeogeography, Palaeoclimatology,
Palaeocology, 125(1-4):165-184.
Santos, P.R. Distribuição Estratigráfica, Características, e Fácies de Diamictitos e Rochas
Associadas do Subgrupo Itararé no Centro e Sul do Estado de São Paulo. São Paulo,
1979, 135 p. Dissertação (Mestrado) – Instituto de Geociências, Universidade de São
Paulo.
Santos, P.R.. 1987. Fácies e Evolução Paleogeográfica do Subgrupo Itararé/Grupo
Aquidauana (Neopaleozóico) na Bacia do Paraná, Brasil. Tese de Doutoramento.
Instituto de Geociências, USP, 128p.
Santos, P.R.; Rocha Campos, A. C.; Canuto, J.R. 1996. Patterns of Late Paleozoic
deglaciation in the Paraná Basin, Brazil. Palaeogeography, Palaeoclimatology,
Palaeoecology., 123:165-184.
Scotese, C.R. 2000. Paleomap project. http://www.scotese.com
Smith, J.R. 1999. Late Quaternary glacial history of Lake Hazen Basin and eastern Hazen
Plateau, northen Ellesmere Island, Nunavut, Canada. Canadian Journal of Earth
Sciences, 36:1547-1565.
Soares, P. C.; Landim, P. M. B.; Sineli, O. Wernick, E. Wu, F. T.; Fiori, A. P. 1971.
Associações litológicas do Subgrupo Itararé e sua Interpretação Ambiental. Revista
Brasileira de Geociências, 7(2)131-149.
Soares, P. C.; Landim, P. M. B.; Sineli, O. Wernick, E. Wu, F. T.; Fiori, A. P. 1977
Associações litológicas do Subgrupo Itararé e sua Interpretação Ambiental. Revista
Brasileira de Geociências, 7(2):131-149.
Souza Filho, E.E.. 1986. Mapeamento Faciológico do Subgrupo Itararé na Quadrícula de
Campinas (SP). Dissertação de Mestrado. Instituto de Geociências, USP. São Paulo.
121 p.
Souza, P.A. & Marques-Toigo, M. 2001. Zona Vittatina: marco palinobioestratigráfico do
Permiano Inferior da Bacia do Paraná. In: Correlação de Sequências Paleozóicas Sul-
Americanas. Ciência, Técnica, Petróleo. Seção: Exploração de Petróleo, 20:153-159.
Souza, P.A. 2006. Late Carboniferous palynostratigraphy of the Itararé Subgorup,
northeastern Paraná Basin, Brazil. Review of Paleobotany and Palynology, 138:9-29.

88
Spicer RA. 1989. Physiological characteristics of land plants in relation to environment
through time. Trans R Soc Edinburgh (Earth Sc) 80: 321-329.
Spicer, R. A., 1975, The sorting of plant remains in a recent depositional environment, Ph.D.
thesis, University of London.
Srivastava, A.K. 1991. Evolutionary tendency in the venation pattern of Glossopteridales.
Geobios, 4(24):383-386
Srivastava, A.K.; Agnihotri, D. 2010. Morphological consequence of Gangamopteris McCoy
in Glossopteris flora. Journal of Asian Earth Sciences, 39:760-769.
Taylor TN, Taylor EL, Krings M. 2009.Paleobotany. The biology and evolution of fossil
plants. Elsevier/Academic Press Inc., Burlington MA, London, San Diego CA, New
York NY, USA
Tybusch, G. P. , Iannuzzi, R. , Bernardes-De-Oliveira, M. E. C. , Lopes, R. C. 2016.
Revaluation of the Glossopterids from the Lower Permian of Cambaí Grande
Outcrop, Paraná Basin, RS. Geologia USP. Série Científica, 16:41.
Tybusch, G.P., Iannuzzi, R. 2008. Reavaliação taxonômica dos gêneros Gangamopteris e
Rubidgea, Permiano Inferior da Bacia do Paraná, Brasil. Revista Brasileira de
Paleontologia 11(2):73-86.
Tybusch, G.P., Iannuzzi, R., 2008. Taxonomic reevaluation of the genera Gangamopteris and
Rubidgea, Lower Permian of the Paraná Basin, Brazil. Revista Brasileira de
Paleontologia, 11:73–86.
Veevers, J.J.. 2004. Gondwanaland from 600-570 Ma assembly through 320 Ma merger in
Pangea to 160 Ma breakup: supercontinental tectonics via stratigraphy and
radiometric dating. Earth Sciences Review, 68:1-132.
Willis, K.J.; Mcelwain, J.C. The Evolution of Plants. Oxford: Oxford University Press, 2002,
378 p.





























