Embed Size (px)
Citation preview

i
UNIVERSIDADE ESTADUAL DE CAMPINAS
INSTITUTO DE BIOLOGIA
MAURICIO BONESSO SAMPAIO
"ECOLOGIA, MANEJO E CONSERV A<;AO DO BURITI
(MAURITIA FLEXUOSA; ARECACEAE) NOS BREJOS DO
BRASIL CENTRAL"
' -1<~-:xemplar 00it3sponde e redavao final I :~ .. ~ese defendida pelo(a) candidate (a) I ~~~Ct'' p(JNJP f~·g . !
.. .. , ada pela Comi~ Julgado.;-- 1
Tese apresentada ao Institute de Biologia para obtenyao do Titulo de Doutor em Biologia Vegetal.
or. . fflEJ Orientador(a): Prof. Dr
1
Flavio Antonio Maes dos Santos
Campinas, 2012

ii

iii

iv
"O senhor estude: o buriti é das margens, ele cai seus
cocos na vereda – as águas levam – em beiras, o
coquinho as águas mesmas replantam; dai o buritizal, de
um lado e do outro se alinhando, acompanhando, que
nem que por um cálculo."
João Guimarães Rosa – Grande Sertão: Veredas

v
AGRADECIMENTOS
Ao Flavio Antonio Maës dos Santos por ter me incentivado na execução deste
estudo, por ter proporcionado as condições necessárias para que o trabalho se tornasse
viável, pelas conversas sempre repletas de informação e experiência, pelos conhecimentos
transmitidos nas suas ótimas disciplinas e por ter sugerido soluções inteligentes para os
problemas que surgiram durante o estudo.
À Isabel Belloni Schmidt, que me convidou para começar um estudo preliminar
com o buriti no Jalapão em 2005 (convite que surgiu a partir de uma demanda da
comunidade da Mumbuca), contribuiu com discussões sobre metodologia, com a
localização de algumas das áreas de estudo no Jalapão, com centenas de incentivos para
que eu continuasse o trabalho, me ajudou a fazer os aceiros, foi companheira nas viagens de
campo ao Jalapão, contribuiu com as análises e revisou boa parte do texto.
À Isabel Benedetti Figueiredo e ao Luis Carrazza, que forneceram contatos das
organizações de extrativistas de várias regiões. À Sandra Afonso, por ter me colocado em
contato com o Ariosto Pinheiro Lopes. Ao Ariosto, Maria José, Arlete Alves, José Antônio
e Giovanda Brandão que abriram as portas de cada uma das regiões visitadas, sendo
fundamentais para que eu conseguisse encontrar as áreas de estudo e por promoverem o uso
sustentável da biodiversidade do Cerrado!
Ao Ariosto, Elton, Erlandson, João, Jussyklebson, Leonino, Márcio, Mundeira,
Neuzimar, Rodrigo e Wellington que me guiaram na procura aos brejos. Aos moradores das
comunidades do Jalapão (principalmente Mumbuca, Prata e Boa Esperança) por terem nos

vi
recebido em suas casas e por terem colaborado com este estudo desde o início. Aos
proprietários de brejos e extrativistas de buriti das regiões de estudo, que me acolheram,
colaboraram com as entrevistas e pelas conversas sobre os brejos, sobre a vida no Sertão, e
sobre os encantos e mistérios do buriti.
À ONG Pequi - Pesquisa e Conservação do Cerrado por ter cedido um veículo para
as viagens de campo e pela infraestrutura em Brasília. À equipe do Parque Estadual do
Jalapão (PEJ), do Instituto Natureza do Tocantins e da Secretaria do Meio Ambiente e
Desenvolvimento Sustentável do Tocantins (especialmente Cassiana Moreira, Beatriz
Gonçalves, Rejane Nunes e Daiane Meyer), pela autorização para a realização do estudo no
PEJ e por terem auxiliado na organização de um seminário para apresentação dos
resultados em Palmas-TO em julho de 2011. Aos brigadistas do Parque Estadual do Jalapão
que fizeram os aceiros em 2009.
À todas as pessoas que me ajudaram e me fizeram companhia nas viagens de
campo, principalmente Ísis, Marco, Bernardo, Gabriel, Rebeca, Isabel, Saulo, Manuel da
Mumbuca, Waldir, Antonino, seu Manuel, Alexandre, André, Guilherme, e Alessandra. Ao
Sigmaringa por ter transmitido vários recados às comunidades do Jalapão pela Rádio
Senado.
Aos meus colegas de disciplinas da UNICAMP, principalmente Airton, Arildo,
Bruno Rosado, Christiane, Cristina, Débora, Flávia, Júlia, Juliana, Juliano, Kátia,
Leonardo, Marcelo, Patrícia, Pedro Cavalin, Pedro Eisenlohr, Rafael, Rubens e Talita. À
Cristina Baldauf pela parceria bem sucedida nos nossos dois projetos conjuntos. À amizade
do moleque mais doido de Brasília, Alexandre Rezende Tofeti.

vii
Aos professores da UNICAMP, principalmente Cristiana Seixas, Fernando Martins,
George Shepherd, Kikyo Yamamoto, Rafael Oliveira e Thomas Lewinsohn, que me
proporcionaram muitos conhecimentos novos. À Tamara Ticktin (University of Hawai'i at
Manoa) e suas alunas (Lisa, Anita e Daniela) pelas ótimas sugestões. Ao Aldicir Scariot,
Daniel Vieira (Embrapa Recursos Genéticos e Biotecnologia) e seus alunos (principalmente
Victor, Xitão, Aelton e Dannyel) pelas ótimas sugestões. À Carol Horvitz (University of
Miami) pelo curso de Modelos Matriciais para Ecologia de Populações. À Cristiana Seixas
e Tamara Ticktin pela contribuição ao terceiro capítulo.
À CAPES, CNPq (140813/2009-8; 472127/2008-0) e FAPESP (2008/08737-4)
pelas bolsas e auxílios concedidos. Ao Programa de Pequenos Projetos PPP-
ECOS/ISPN/GEF/PNUD (projeto BRA/04/21) pelos recursos financeiros. À Idea Wild
pelos equipamentos doados. Aos funcionários da secretaria de pós-graduação da Biologia
Vegetal, principalmente Maria Roseli.
Ao Alexandre Bonesso Sampaio e à Isabel Schmidt por me mostrarem que é
possível levar a vida com seriedade, muito trabalho e ao mesmo tempo aproveitar de forma
criativa os momentos de lazer. Muito obrigado pela ótima estadia no Hawaii!
À minha amada Ísis Meri Medri, pelo companheirismo em todos os momentos, por
ter proporcionado os melhores dias da minha vida e por me fazer ser uma pessoa melhor a
cada dia. Além disso, me deu segurança e confiança nos momentos que eu mais precisei
durante o trabalho de campo, me ajudou com a digitação dos dados e com a revisão do
texto.

viii
À minha mãe (Fátima Berenice Ferreira Bonesso), meu pai (Antônio Carlos Reis
Sampaio) e meus irmãos (Alexandre Bonesso Sampaio e Caroline Bonesso Sampaio) que
mesmo à distância fazem parte da minha vida todos os dias, a cada minuto. Muito obrigado
por todo o apoio e por torcerem por mim!

9
ÍNDICE
AGRADECIMENTOS .................................................................................................................. v
ÍNDICE ......................................................................................................................................... 9
RESUMO .................................................................................................................................... 11
ABSTRACT ................................................................................................................................ 13
INTRODUÇÃO GERAL ............................................................................................................ 15
Uso dos produtos da biodiversidade do Cerrado ..................................................................... 15
O buriti - descrição e usos ....................................................................................................... 16
Os brejos do Brasil Central ..................................................................................................... 22
REFERÊNCIAS BIBLIOGRÁFICAS .................................................................................... 25
CAPÍTULO1: Ontogenia, alometria e reprodução do buriti (Mauritia flexuosa; Arecaceae) .... 34
Resumo .................................................................................................................................... 34
Introdução ............................................................................................................................... 35
Material e métodos .................................................................................................................. 38
Resultados ............................................................................................................................... 43
Discussão................................................................................................................................. 52
Conclusões .............................................................................................................................. 58
REFERÊNCIAS BIBLIOGRÁFICAS .................................................................................... 58
CAPÍTULO 2: Impactos do extrativismo de frutos e do fogo na ecologia populacional do buriti (Mauritia flexuosa; Arecaceae) ................................................................................................... 64
Resumo .................................................................................................................................... 64
Introdução ............................................................................................................................... 65
Material e métodos .................................................................................................................. 69
Resultados ............................................................................................................................... 78
Discussão............................................................................................................................... 101

10
Conclusões ............................................................................................................................ 109
REFERÊNCIAS BIBLIOGRÁFICAS .................................................................................. 110
CAPÍTULO 3: Effects of socioeconomic conditions on the multiple uses of swamp forests ... 124
Abstract ................................................................................................................................. 125
Introduction ........................................................................................................................... 126
Material and methods ............................................................................................................ 130
Results ................................................................................................................................... 135
Discussion ............................................................................................................................. 137
References ............................................................................................................................. 144
CONSIDERAÇÕES FINAIS .................................................................................................... 160

11
RESUMO
Florestas geram subsistência e renda para comunidades rurais, por meio do extrativismo de
produtos florestais não madeireiros (PFNM), agricultura de coivara, criação de gado e
porcos, etc. Estas atividades podem ser alternativas econômicas de menor impacto do que o
corte seletivo de madeira e a conversão das florestas em monoculturas. Porém, a
sustentabilidade em longo prazo depende da intensidade de uso, que é influenciada por
fatores socioeconômicos dos usuários. É possível avaliar tanto os impactos ecológicos,
quanto os fatores socioeconômicos que influenciam o uso das florestas. Modelos matriciais
têm sido utilizados para avaliar os efeitos de perturbações na dinâmica populacional das
plantas. Nestes modelos são usadas matrizes de transição, construídas a partir de dados
demográficos de uma população, cujos indivíduos são classificados em estádios
ontogenéticos, classes de tamanho ou idade. Os estádios ontogenéticos são definidos por
características morfológicas dos indivíduos que se modificam durante o ciclo de vida. Os
parâmetros demográficos podem ser influenciados por perturbações naturais ou antrópicas.
As taxas de crescimento populacionais são calculadas e comparadas entre populações
sujeitas a diferentes intensidades de perturbações, como o extrativismo de PFNM. Outras
fontes de perturbação, como o fogo, podem interferir na avaliação dos impactos do
extrativismo de PFNM e precisam ser incluídas nos modelos matriciais para aumentar o seu
realismo. O buriti (Mauritia flexuosa) é uma palmeira sujeita à intenso extrativismo de
frutos nos brejos do Cerrado. Os frutos são utilizados para a produção de doces e óleo.
Além do extrativismo de frutos de buriti, os brejos são utilizados para a criação de gado e
porcos, para o estabelecimento de roças de coivara e são sujeitos a queimadas frequentes.

12
Os objetivos deste estudo foram: (i) avaliar se as alterações ao longo do ciclo de vida nas
características dos indivíduos de buriti estão relacionadas às variações nos parâmetros
demográficos; (ii) estimar a intensidade sustentável de extrativismo de frutos de buriti e a
frequência máxima de queimadas tolerada pelas populações de buriti; e (iii) avaliar a
influência das condições socioeconômicas das comunidades do entorno no uso dos brejos.
Identificamos quatro estádios ontogenéticos para o buriti utilizando duas características
macro-morfológicas (presença/ausência do estipe e atividade reprodutiva) e uma medida
quantitativa (altura). Cada estádio identificado tem contribuição particular para a dinâmica
das populações de buriti. O extrativismo de frutos de buriti não deve reduzir mais que 70%
da fecundidade das populações para ser sustentável em longo prazo. As queimadas bienais,
frequentes no Brasil Central, podem causar a mortalidade de indivíduos reprodutivos e
reduzir o tamanho das populações em longo prazo. Agricultura de coivara é praticada em
72% dos 75 brejos avaliados e mais da metade dos brejos são utilizados para a criação de
gado. Os recursos dos brejos são igualmente importantes para todas as classes sociais
avaliadas, mesmo os fazendeiros mais ricos. A alta intensidade de uso dos brejos verificada
sugere que para a sua conservação, as práticas de manejo atuais terão que ser modificadas,
principalmente em relação ao uso do fogo.

13
ABSTRACT
Forests are sources of income and subsistence to rural communities around the world by
means of activities such as harvesting of non-timber forest products (NTFPs), slash-and-
burn agriculture, cattle and pigs raising. These economic activities may be alternatives of
lower impact to selective logging or large monocultures. However, the long term
sustainability of these alternatives depends on the use intensity. Socioeconomic conditions
of users may influence the forest use intensity. It is possible to assess both the ecological
impacts of NTFPs harvesting and the socioeconomic conditions. Matrix models have been
used to evaluate the harvesting impacts on plant population dynamics. Lefkovitch matrix is
frequently required to apply these models. To construct this matrix is required demographic
data from a population, whose individuals are classified into ontogenetic stages or size
classes. Ontogenetic stages are defined by macro-morphological characteristics of
individuals that are correlated to the variation in demographic parameters along the life
cycle. Population growth rates are calculated and compared by matrix models among
populations with different intensities of disturbance, such as NTFP harvesting. Other
human disturbance sources, such as fire, may influence the evaluation of harvesting
impacts. Thus, these disturbances should be considered in the matrix models to improve
their realism. Buriti (Mauritia flexuosa) is a palm species subjected to high intensity fruit
harvesting in the swamp forests of Cerrado. Fruits are processed to candies and oil
production. In addition to fruit harvesting, swamp forests are used to cattle and pig raising,
to slash-and-burn agricultural practices and are frequently subjected to fire. This study
aimed to: (i) to assess the macro-morphological characteristics of individuals to define the

14
ontogenetic stages of buriti (Mauritia flexuosa; (ii) to assess the sustainable buriti fruit
harvesting intensity and the highest fire frequency tolerated by buriti populations; and (iii)
to assess the influence of buriti harvesters socioeconomic conditions on the multiple
resources use. We identified four ontogenetic stages to buriti using only two macro-
morphologic characteristics (presence/absence of aerial stem and reproductive activity) and
one quantitative variable (height). Each identified stage has a single contribution to buriti
population dynamics. Fruit harvesting may be sustainable only if it causes a reduction of up
to 70% in the population fecundity. Biennial burning of swamp forests, common in the
Central Brazil, may increase the mortality of reproductive individuals and decrease the
population size at long term. Slash-and-burn agriculture occurs in 72% of the 75 evaluated
swamp forests and more than a half of these forests are used to cattle raising. Swamp
forests resources are equally important to all socioeconomic classes, even richer farmers.
The high intensity of swamp forest uses suggests that their conservation will require change
to current management practices, mainly the fire frequency.

15
INTRODUÇÃO GERAL
Uso dos produtos da biodiversidade do Cerrado
Produtos florestais não-madeireiros (PFNMs) são todos os materiais biológicos,
com exceção da madeira, coletados nos ecossistemas naturais e utilizados pelos seres
humanos (De Beer & Mcdermott 1989). No domínio do Cerrado, a coleta de PFNMs ocorre
há pelo menos nove mil anos, desde a chegada dos primeiros povos coletores e caçadores
nômades à região (Klink & Moreira 2002).
Além de contribuir para a subsistência de muitas famílias, a comercialização de
PFNMs tem gerado renda para as comunidades rurais nas últimas décadas (Carvalho 2007;
Schmidt et al. 2007; Afonso & Ângelo 2009). A coleta de PFNMs é uma atividade
econômica que tende a causar menor impacto ao ecossistema do que outras formas de uso
da terra como a implantação de pastagens e a extração de madeira (Nepstad &
Schwartzman 1992; Sunderland et al. 2011). Por isso, o extrativismo tem sido considerado
uma importante ferramenta para a conservação do Cerrado (Klink & Machado 2005).
Diversas iniciativas governamentais, não-governamentais e privadas têm contribuído para
promover o uso sustentável da biodiversidade do Cerrado (Klink & Machado 2005;
Carrazza & Figueiredo 2010).
A intensidade de coleta dos PFNMs pode variar em função das condições
socioeconômicas dos extrativistas. Enquanto os mais pobres tendem a ser mais dependentes
dos PFNM para a sua subsistência, pois não possuem fontes alternativas de renda, os
usuários mais ricos tendem a se especializar no extrativismo dos produtos com maior valor
agregado, ou em outras fontes de renda, como a criação de gado (Godoy et al. 1995;

16
Barham et al. 1999). Além disso, a distância do domicílio do usuário ao recurso e o acesso
ao mercado consumidor são outras variáveis que podem influenciar a intensidade do
extrativismo (Hegde et al. 1996; Shanley et al. 2002; Marshall et al. 2003).
Há uma enorme riqueza de PFNMs no domínio do Cerrado, que são utilizados
como alimentos, remédios, ferramentas, materiais de construção, utensílios domésticos,
resinas, látex, óleos, ceras, fibras, aromáticos, artesanatos, entre outros. Estes produtos são
colhidos de inúmeras espécies vegetais (Botrel et al. 2006; Souza & Felfili 2006; Aquino et
al. 2007; Lima 2008). Entre os PFNMs do Cerrado com maior volume comercializado
estão as folhas e os frutos do buriti (Mauritia flexuosa L.f.; Arecaceae). De acordo com
estimativa conservadora do Instituto Brasileiro de Geografia e Estatística, em 2010, foram
comercializadas 465 toneladas de fibras retiradas das folhas de buriti, que renderam R$
1.693.000,00 (IBGE 2010). Não há estimativas confiáveis do volume comercializado de
frutos e de seus subprodutos para o Brasil.
O buriti - descrição e usos
Também conhecido como miriti, muriti, carangucha (Colômbia), moriche
(Venezuela) ou aguaje (Peru), o buriti é uma palmeira de vida longa, dióica, arborescente,
com estipe solitário, cilíndrico, com diâmetro na altura do solo entre 13 e 54 cm. Buritis
adultos podem alcançar até 40 m de altura e possuem de 8 a 20 folhas (Figura 1a;
Henderson et al. 1995).
O buriti tem sua área de distribuição na Bolívia, Peru, Equador, Colômbia,
Venezuela, Trinidad e Tobago, Guiana, Suriname e Guiana Francesa. No Brasil, ocorre nos
domínios do Cerrado, Caatinga, Pantanal e Amazônia. O limite sul da distribuição é o norte

17
do estado de São Paulo e a Cordilheira dos Andes a oeste (Figura 2; Henderson et al. 1995;
Oliveira Filho & Ratter 2000).
Figura 1. a) buriti (Mauritia flexuosa) no brejo Porcos (10°08''S; 46°30'''W), Jalapão - TO;
b) inflorescência feminina; c) inflorescência masculina; d) cacho com frutos maduros; e)
óleo e doces feitos com buriti; f) Orthopsittaca manilata (arara-de-ventre-vermelho),
psitacídeo especialista em frutos do buriti (Foto: Nick Athanas); g) fruto maduro com as
escamas; h) mesocarpo do fruto maduro; i) endocarpo; j) semente; k) extrativista colhendo
a folha jovem de um buriti não reprodutivo de 9 m de altura.

18
Figura 2. Distribuição de ocorrências do buriti (Mauritia flexuosa), adaptado de Oliveira
Filho & Ratter (2000).
O buriti possui múltiplos usos não só pelas comunidades rurais do Cerrado, mas em
todos os locais de ocorrência (Nascimento et al. 1999; Macía 2004; Sampaio et al. 2008;
Santos & Coelho-Ferreira 2012). O limbo foliar é utilizado para a cobertura de casas, para a
confecção de artesanatos e utensílios domésticos. A fibra retirada da face adaxial das pinas
das folhas jovens (Figura 1k) é utilizada para a confecção de cordas, tecidos, redes (Saraiva
2009) e para a costura do artesanato de capim-dourado - Syngonanthus nitens (Bong.)
Ruhland (Schmidt et al. 2007; Eichemberg & Scatena 2011). O pecíolo das folhas é
utilizado para a confecção de móveis, utensílios domésticos, artesanatos e brinquedos
(Santos & Coelho-Ferreira 2011). O palmito é comestível. Os frutos são consumidos in
natura, ou utilizados para a confecção de doces, sorvetes e para a extração do óleo (Figura

19
1e). O tronco é utilizado para construção, para fazer pontes e também para a corrida de
toras, que é um ritual de povos indígenas (Nascimento et al. 1999). O interior do caule é
esponjoso, de cor avermelhada e contém alta concentração de amido (Kahn 1988). A seiva
é doce e saborosa e é conhecida como vinho de buriti. O estipe, depois de apodrecido no
chão da floresta, pode apresentar até 500 larvas comestíveis do coleóptero suri,
Rhynchophorus palmarum (Linnaeus, 1758) (Curculionidae), que são utilizadas como
alimento por povos indígenas, principalmente na Amazônia (Gragson 1995). As raízes do
buriti são utilizadas como medicinal. Por todos esses usos e também por ser uma planta
muito majestosa, o buriti tem um grande valor histórico e sociocultural para as
comunidades rurais do Cerrado (Fernandes 2011).
Os frutos são os produtos do buriti com maior potencial econômico (Castro 1993).
Na polpa do fruto de buriti há entre 77,7 e 304 mg/100g de β-caroteno, precursor da
vitamina-A (Mariath et al. 1989; França et al. 1999). Além disso, os frutos são muito ricos
em óleos, principalmente na polpa (49,1 g/100g de massa fresca) e na casca (26,7 g/100g),
em proteínas e em vitaminas C e E (Castro 1993; Darnet et al. 2011). O óleo possui
atividade bactericida (Silveira et al. 2005) e é muito rico em antioxidantes, o que o torna
muito atrativo para a indústria de cosméticos, principalmente para a fabricação de protetor
solar (Zanatta et al. 2010). Além disso, o óleo pode ser utilizado para a produção de
plásticos fotoluminescentes (Durães et al. 2006).
A floração do buriti é sincrônica entre indivíduos pistilados (Figura 1b) e
estaminados (Figura 1c) e a espécie é xenogâmica, não ocorrendo apomixia. Abelhas do
gênero Trigona Jurine, 1807, coleópteros e dípteros são os principais polinizadores e não
ocorre anemofilia. O período de desenvolvimento dos frutos (Figura 1d) é longo, com

20
duração de mais de um ano desde a emissão das espatas até o completo amadurecimento e
dispersão dos frutos (Storti 1993; Abreu 2001). Existem variações regionais na época de
floração e frutificação e também nos polinizadores do buriti (Storti 1993; Peres 1994;
Abreu 2001; Fernandes 2002; Ponce Calderón 2002; Cabrera & Wallace 2007; Silva 2009).
No Brasil Central a floração se estende de novembro a abril. Abelhas do gênero Trigona
são as principais visitantes das flores pistiladas e estaminadas. Os primeiros frutos
começam a se desenvolver em fevereiro e permanecem em crescimento por pelo menos
oito meses. Em outubro começa a dispersão dos frutos, que só termina em julho do ano
seguinte (Abreu 2001). A produção de frutos é plurianual (Abreu 2001; Fernandes 2002). A
predação do pólen por microlepidópteros em anos de menor precipitação pode contribuir
para a diferença de produtividade de frutos entre anos (Abreu 2001). O sucesso reprodutivo
é naturalmente baixo, pois apenas 9,5% a 14% das flores produzem frutos, principalmente
devido à ausência de polinizadores (Storti 1993; Abreu 2001). Em áreas perturbadas, o
sucesso reprodutivo é menor e a frequência de sementes partenocárpicas é maior do que em
áreas conservadas, onde os polinizadores são mais abundantes (Abreu 2001).
Os frutos são envoltos por um pericarpo (ou casca), composto por escamas
triangulares castanho-avermelhadas (Figura 1g). O mesocarpo (ou polpa) é fino, amarelado
ou alaranjado (Figura 1h), carnoso, oleoso, e tem em média 11,4 g de massa. O endocarpo é
formado por um tecido esponjoso, delgado e branco (Figura 1i), que possui baixa
densidade, e possibilita ao fruto boiar quando submerso em água. O endosperma (ou
semente) é muito duro (Figura 1j), ovóide, possui 2,4 cm de longitude, ocupando a maior
parte do volume do fruto, com 4,86 g de massa e 57% de umidade. O embrião é pequeno,
em média 0,6 mm de comprimento, possui disposição lateral-basal, sendo totalmente

21
envolvido pelo endosperma. Geralmente há uma semente por fruto, mas podem ser
encontrados em baixa frequência frutos dispérmicos ou partenocárpicos (Ponce et al. 1999;
Fernandes 2002; Ponce Calderón 2002).
Os frutos são recursos importantes para várias espécies de animais. Entre as aves:
Ara manilata (Boddaert, 1783); Ara ararauna (Linnaeus, 1758); Anodorhynchus
hyacinthinus (Latham, 1790); Amazona aestiva (Linnaeus, 1758); Thraupis palmarum
(Wied, 1821); Schistochlamys melanopis (Latham, 1790); Cyanocorax cristatellus
(Temminck, 1823); Gnorimopsar chopi (Vieillot, 1819); Orthopsittaca manilata (Boddaert,
1783); Caracara plancus (Miller, 1777) e Porzana albicollis (Vieillot, 1819). Outras
espécies que também utilizam os frutos do buriti são os pequenos mamíferos: Nectomys
squamipes (Brants, 1827); Oxymycterus roberti Thomas, 1901; Oligoryzomys Bangs, 1900;
Sigmodon alstoni (Thomas, 1881); Didelphis marsupialis Linnaeus, 1758; Heteromys
anomalus (Thompson, 1815); Sciurus granatensis Humboldt, 1811 e Dasyprocta leporina
(Linnaeus, 1758). Entre os grandes mamíferos destacam-se: Mazama americana (Erxleben,
1777) – veado; Pecari tajacu (Linnaeus, 1758) – cateto; Tayassu pecari (Link, 1795) –
queixada; Tapirus terrestris (Linnaeus, 1758) – anta; Chrysocyon brachyurus (Illiger,
1815) – lobo-guará; Cuniculus paca (Linnaeus, 1766) – paca; Bos taurus Linnaeus, 1758 –
gado; Sus scrofa Linnaeus, 1758 – porcos domésticos e macacos (Bodmer 1991; Villalobos
1994; Fernandes 2002; Ponce Calderón 2002).
As aves e macacos são considerados dispersores primários, pois se alimentam dos
frutos no cacho e derrubam as sementes (Fernandes 2002). Indivíduos de Sigmodon alstoni,
Heteromys anomalus e Sciurus granatensis removem a polpa do fruto, carregam as
sementes para longe da planta mãe e as enterram, atuando como dispersores. Didelphis

22
marsupialis consome a polpa no mesmo local onde encontrou o fruto, sem carregar ou
enterrar as sementes e, portanto, não é considerado um dispersor. Porcos e catetos podem
quebrar as sementes com os dentes e são considerados predadores (Ponce Calderón 2002).
Algumas aves predam, no cacho, frutos em início de desenvolvimento. Veados e antas são
dispersores, pois as sementes são muito duras para serem predadas por esses animais
(Bodmer 1991; Gragson 1995). Além dos animais, a água é dispersora das sementes. O
endocarpo muitas vezes se mantém intacto após a retirada do mesocarpo pelos animais,
possibilitando a dispersão hidrocórica das sementes após a dispersão primária ou
secundária por animais (Fernandes 2002).
O buriti é considerado uma espécie chave para nidificação de psitacídeos. No
sudeste do Peru, pelo menos sete espécies fazem ninhos nos troncos de buritis adultos
mortos (Brightsmith 2005). Além disso, Orthopsittaca manilata (Figura 1f) é uma ave
especialista em frutos maduros de buriti. Como os frutos são disponíveis em épocas
distintas em diferentes regiões, a ave desloca-se a grandes distâncias para garantir a sua
alimentação durante o ano inteiro (Silva 2009). O buriti também é importante para a
manutenção da diversidade de artrópodes (Gurgel-Gonçalves et al. 2006).
Os sistemas de polinização zoocórica e a dispersão de sementes por animais e pela
água podem contribuir para o fluxo gênico a longas distâncias, que causa menor
variabilidade genética entre do que dentre populações de buriti (Gomes et al. 2011).
Os brejos do Brasil Central

23
O buriti é a espécie que caracteriza os brejos do Brasil Central. Os brejos são
compridos e estreitos (entre 10 e 100 m de largura), geralmente com estrutura florestal, que
se localizam ao redor de pequenos cursos d'água nas áreas de nascente, onde o relevo é
suave (Figura 3). O solo é hidromórfico e permanentemente inundado, com elevada acidez
e teor de alumínio, rico em matéria orgânica e pobre em cálcio e fósforo (Ivanauskas et al.
1997; Toniato et al. 1998). Em alguns locais, o nível da água está acima de 1 m do solo,
mesmo durante a estação seca. Os brejos são frequentemente margeados por campos
limpos, que não possuem o componente arbustivo-arbóreo e podem ser permanentemente
úmidos ou secos, dependendo da posição no relevo.
Figura 3. Brejo Atoladeira, Jalapão - TO (10°10'S; 46°28'W).

24
Os brejos com composição florística similar à atual surgiram no Brasil Central por
volta de 7.220 a 3.500 anos AP, com o aumento da umidade e condições climáticas
semelhantes às atuais (Ferraz-Vicentini & Salgado-Labouriau 1996; Barberi et al. 2000). O
estrato herbáceo dos brejos é ralo e dominado por pteridófitas e alguns indivíduos de
Poaceae e Cyperaceae nas porções mais secas (Ivanauskas et al. 1997). Apesar da alta
heterogeneidade florística entre brejos, as espécies mais frequentes deste tipo de vegetação
no sudeste do Brasil são Calophyllum brasiliense Cambess. (Calophillaceae), Protium
almecega L. Marchand (Burceraceae), Pera obovata (Klotzsch) Baill. (Euphorbiaceae),
Copaifera langsdorffii Desf. (Leguminosae) e Tapirira guianensis Aubl. (Anacardiaceae)
(Ivanauskas et al. 1997; Toniato et al. 1998; Marques et al. 2003). No Brasil Central, duas
outras espécies emergentes são muito frequentes, Mauritia flexuosa (Arecaceae) e Xylopia
emarginata Mart. (Annonaceae) (M. B. Sampaio; observação pessoal).
Além do extrativismo de frutos e folhas de buriti, os brejos do Brasil Central são
muito utilizados para agricultura de coivara, criação de gado e de porcos. Estes outros usos
ocorrem principalmente quando a paisagem é dominada por solos arenosos bem drenados,
inapropriados para as práticas de agricultura e pecuária. Nestas regiões, os brejos são a
principal fonte de recursos hídricos e solo fértil para as atividades de subsistência das
comunidades rurais. Queimadas frequentes ocorrem no interior desses brejos, causadas
principalmente pelo uso do fogo para renovação das pastagens nativas nos campos limpos
que margeiam os brejos.
O principal objetivo desta tese foi avaliar a sustentabilidade do extrativismo de
frutos de buriti da forma que é atualmente praticado no Brasil Central. O estudo foi
dividido em três capítulos com os seguintes objetivos específicos: (i) avaliar se as

25
alterações nas características macro-morfológicas ao longo do ciclo de vida dos indivíduos
de buriti estão relacionadas às variações nos parâmetros demográficos (Capítulo 1); (ii)
estimar a intensidade sustentável do extrativismo de frutos de buriti e a frequência máxima
de queimadas tolerada pelas populações desta espécie (Capítulo 2); e (iii) avaliar a
influência das condições socioeconômicas dos extrativistas na intensidade de uso múltiplo
dos brejos (Capítulo 3).
REFERÊNCIAS BIBLIOGRÁFICAS
Abreu, S. A. B. 2001. Biologia reprodutiva de Mauritia flexuosa L.f. (Arecaceae) em
vereda no município de Uberlândia - MG. Dissertação de Mestrado, Departamento de
Ecologia - UFU, Uberlândia - MG.
Afonso, S. R. & Ângelo, H. 2009. Mercado dos produtos florestais não-madeireiros do
Cerrado Brasileiro. Ciência Florestal 19(3)315-326.
Aquino, F. G.; Walter, B. M. T. & Ribeiro, J. F. 2007. Espécies vegetais de uso múltiplo
em reservas legais de Cerrado - Balsas, MA. Revista Brasileira de Biociências
5(1):147-149.
Barberi, M.; Salgado-Laboriau, M. L. & Suguio, K. 2000. Paleovegetation and
paleoclimate of "Vereda de Águas Emendadas", Central Brazil. Journal of South
American Earth Sciences 13:241-254.

26
Barham, B. L.; Coomes, O. T. & Takasaki, Y. 1999. Rain forest livelihoods: income
generation, household wealth and forest use. Unasylva 198:71.
Bodmer, R. E. 1991. Strategies of seed dispersal and seed predation in Amazonian
ungulates. Biotropica 23(3):255-261.
Botrel, R. T.; Rodrigues, L. A.; Gomes, L. J.; Carvalho, D. A. & Fontes, M. A. L. 2006.
Uso da vegetação nativa pela população local no município de Ingaí, MG, Brasil.
Acta Botânica Brasílica 20(1):143-156.
Brightsmith, D. J. 2005. Parrot nesting in Southeastern Peru: seasonal patterns and
keystone trees. Wilson Bulletin 117(3):296-305.
Cabrera, W. H. & Wallace, R. 2007. Patrones fenológicos de ocho especies de palmeras en
un bosque Amazónico de Bolivia. Revista Boliviana de Ecología y Conservación
Ambiental 21:1-18.
Carrazza, L. & Figueiredo, I. 2010. Cerrado que te quero vivo! Produtos e meios de vida
sustentáveis apoiados pelo Programa de Pequenos Projetos Ecossociais (PPP-ECOS).
Instituto Sociedade, População e Natureza, Brasília - DF.
Carvalho, I. S. H. 2007. Potenciais e limitações do uso sustentável da biodiversidade do
Cerrado: um estudo de caso da Cooperativa Grande Sertão no Norte de Minas.
Dissertação de Mestrado, CDS-UnB, Brasília - DF.
Castro; A. 1993. Buriti. In: Clay, J. W. & Clement, C. H. Selected species and strategies to
enhance income generation from Amazonian forests. FAO, Roma, Itália.

27
Darnet, S. H.; Silva, L. H. M.; Rodrigues, A. M. C. & Lins, R. T. 2011. Nutritional
composition, fatty acid and tocopherol contents of buriti (Mauritia flexuosa) and
patawa (Oenocarpus bataua) fruit pulp from the Amazon region. Ciência e
Tecnologia de Alimentos 31(2):488-491.
De Beer, J. H. & Mcdermott, M. 1989. The economic value of non-timber forest products
in south East Asia. Amsterdam, The Netherlands Committee for IUCN.
Durães, J. A.; Drummond, A. L.; Pimentel, T. A. P. F.; Murta, M. M.; Bicalho, F. S.;
Moreira, S. G. C. & Sales, M. J. A. 2006. Absorption and photoluminescence of
Buriti oil/polystyrene and Burti oil/poly(methyl methacrylate) blends. European
Polymer Journal 42:3324-3332.
Eichemberg, M. T. & Scatena, V. L. 2011. Handicrafts from Jalapão (TO), Brazil, and their
relationship to plant anatomy. Journal of the Torrey Botanical Society 138(1):34-40.
Fernandes, M. R. 2011. "A tree with much authority": the place of the buriti palm (Mauritia
flexuosa L.f.) in the sertaneja culture of Terra Ronca, Goiás state, Central Brazil.
Bioremediation, Biodiversity and Bioavailability 5:80-91.
Fernandes, N. M. P. 2002. Estratégias de produção de sementes e estabelecimento de
plântulas de Mauritia flexuosa L. f. (Arecaceae) no Vale do Acre/Brasil. Tese de
doutorado. UA/INPA, Manaus - AM.
Ferraz-Vicentini, K. R. & Salgado-Laboriau, M. L. 1996. Palynological analysis of a palm
swamp in Central Brazil. Journal of South American Earth Sciences 9:207-219.

28
França, L. F.; Reber, G.; Meireles, M. A. A.; Machado, N. T. & Brunner, G. 1999.
Supercritical extraction of carotenoids and lipids from buriti (Mauritia flexuosa), a
fruit from the Amazon region. Journal of Supercritical Fluids 14:247-256.
Godoy, R.; Brokaw, N. & Wilkie, D. 1995. The effect of income on the extraction of non-
timber tropical forest products: model, hypotheses, and preliminary findings from the
Sumu Indians of Nicaragua. Human Ecology 23:29-52.
Gomes, L. R. P.; Lopes, M. T. G.; Bentes, J. L. S.; Barros, W. S.; Neto, P. Q. C. & Contim,
L. A. S. 2011. Genetic diversity in natural populations of Buriti (Mauritia flexuosa L.
f.). Crop Breeding and Applied Biotechnology 11: 216-223.
Gragson, T. L. 1995. Pumé exploitation of Mauritia flexuosa (Palmae) in the Llanos of
Venezuela. Journal of Ethnobiology 15(2):177-188.
Gurgel-Gonçalves, R.; Palma, A. R. T.; Motta, P. C.; Bar, M. E. & Cuba, C. A. C. 2006.
Arthropods associated with the crown of Mauritia flexuosa (Arecaceae) palm trees in
three different environments from Brazilian Cerrado. Neotropical Entomology
35(3)302-312.
Hegde, R.; Suryaprakash, S.; Achoth, L. & Bawa, K. S. 1996. Extraction of non-timber
forest products in the forests of Biligiri Rangan Hills, India .1. Contribution to rural
income. Economic Botany 50:243-251.
Henderson, A.; Galeano, G. & Bernal, R. 1995. Field guide to the palms of the Americas.
Princeton University Press, Princeton, New Jersey.

29
IBGE. 2010. Produção da extração vegetal e da silvicultura. Volume 25. Rio de Janeiro -
RJ.
Ivanauskas, N. M.; Rodrigues, R. R. & Nave, A. G. 1997. Aspectos ecológicos de um
trecho de floresta de brejo em Itatinga, SP: florística, fitossociologia e seletividade de
espécies. Revista Brasileira de Botânica 20(2):139-153.
Kahn, F. 1988. Ecology of economically important palms in Peruvian Amazonia. Advances
in Economic Botany 6:42-49.
Klink, C. A. & Machado, R. B. 2005. A conservação do Cerrado brasileiro.
Megadiversidade 1(1):147-155.
Klink, C. A. & Moreira, A. G. 2002. Past and current human occupation, and land use. In:
Oliveira, P. S. & Marquis, R. J. (ed.) The Cerrados of Brazil: ecology and natural
history of a Neotropical Savanna. Columbia University Press, New York.
Lima, I. L. P. 2008. Etnobotânica quantitativa de plantas do Cerrado e extrativismo de
Mangaba (Hancornia speciosa Gomes) no Norte de Minas Gerais: implicações para o
manejo sustentável. Dissertação de Mestrado, Departamento de Ecologia – UnB,
Brasília - DF.
Macía, M. J. 2004. Multiplicity in palm uses by the Huaorani of Amazonian Ecuador.
Botanical Journal of the Linnean Society 144:149-159.

30
Mariath, J. G. R.; Lima, M. C. C. & Santos, L. M. P. 1989. Vitamin A activity of buriti
(Mauritia vinifera Mart) and its effectiveness in the treatment and prevention of
xerophthalmia. The American Journal of Clinical Nutrition 49:849-853.
Marques, M. C. M.; Silva, S. M. & Salino, A. 2003. Florística e estrutura do componente
arbustivo-arbóreo de uma floresta higrófila da bacia do rio Jacaré-Pepira, SP, Brasil.
Acta Botanica Brasilica 17(4):495-506.
Marshall, E.; Newton, A. C. & Schreckenberg, K. 2003. Commercialisation of non-timber
forest products: first steps in analyzing the factors influencing success. International
Forestry Review 5:128-137.
Nascimento, A. R. T.; Santos, A. A.; Martins, R. C.; & Dias, T. A. B. 2009. Palm
communities in the Kraho indigenous territory, Tocantins, Brazil: biodiversity and
ethnobotanical features. Interciencia 34:182-188.
Nepstad, D. C. & Schwartzman, S. 1992. Non-timber products from tropical forests:
evaluation of a conservation and development strategy. New York Botanical Garden,
New York.
Oliveira Filho, A. T. & Ratter, J. A. 2000. Padrões florísticos das matas ciliares da região
do Cerrado e a evolução das paisagens do Brasil central durante o quaternário tardio.
In: Rodrigues, R. R. & Leitão Filho, H. F. (ed.) Matas ciliares: conservação e
recuperação. Editora da USP, São Paulo - SP.
Peres, C. A. 1994. Composition, density, and fruiting phenology of arborescent palms in an
Amazonian Terra Firme Forest. Biotropica 26(3):285-294.

31
Ponce Calderón, M. E. 2002. Patrones de caída de frutos en Mauritia flexuosa L.f. y fauna
involucrada en los procesos de remoción de semillas. Acta Botanica Venezuelica
25(2):119-142.
Ponce, M. E.; Brandin, J.; Ponce, M. A. & Gonzales, V. 1999. Germinacion y
establecimiento de plantulas de Mauritia fluxuosa L.f. (Arecaceae) en los Llanos Sur-
Orientales del estado Guarico, Venezuela. Acta Botanica Venezuelica 22(1):167-183.
Sampaio, M. B.; Schmidt, I. B. & Figueiredo, I. B. 2008. Harvesting effects and population
ecology of the buriti palm (Mauritia flexuosa L. f., Arecaceae) in the Jalapão region,
Central Brazil. Economic Botany 62:171-181.
Santos, R. S. & Coelho-Ferreira, M. 2011. Artefatos de miriti (Mauritia flexuosa L. f.) em
Abaetetuba, Pará: da produção à comercialização. Boletim do Museu Paraense Emílio
Goeldi. Ciências Humanas 6(3):559-571.
Santos, R. S. & Coelho-Ferreira, M. 2012. Estudo etnobotânico de Mauritia flexuosa L. f.
(Arecaceae) em comunidades ribeirinhas do Município de Abaetetuba, Pará, Brasil.
Acta Amazônica 24(1):1-10.
Saraiva, N. A. 2009. Manejo sustentável e potencial econômico da extração do buriti nos
Lençois Maranhenses, Brasil. Dissertação de Mestrado, CDS-UnB, Brasília - DF.
Schmidt, I. B.; Figueiredo, I. B. & Scariot, A. 2007. Ethnobotany and effects of harvesting
on the population ecology of Syngonanthus nitens (Bong.) Ruhland (Eriocaulaceae), a
NTFP from Jalapão region, Central Brazil. Economic Botany 61(1):73-85.

32
Shanley, P.; Luz, L. & Swingland, I. R. 2002. The faint promise of a distant market: a
survey of Belem's trade in non-timber forest products. Biodiversity and Conservation
11:615-636.
Silva, P. A. 2009. Orthopsittaca manilata (Boddaert, 1783) (Aves: Psittacidae): abundância
e atividade alimentar em relação à frutificação de Mauritia flexuosa L. f. (Arecaceae)
numa vereda no Triângulo Mineiro. Dissertação de Mestrado, Ecologia e
Conservação de Recursos Naturais, UFU, Uberlândia - MG.
Silveira, C. S.; Pessanha, C. M.; Lourenço, M. C. S.; Neves Junior, I.; Menezes, F. S. &
Kaplan, M. A. C. 2005. Atividade antimicrobiana dos frutos de Syagrus oleracea e
Mauritia vinifera. Revista Brasileira de Farmacognosia 15(2):143-148.
Souza, C. D. & Felfili, J. M. 2006. Uso de plantas medicinais na região de Alto Paraíso de
Goiás, GO, Brasil. Acta Botanica Brasilica 20(1):135-142.
Storti, E. F. 1993. Biologia floral de Mauritia flexuosa Lin. Fil, na região de Manaus, AM,
Brasil. Acta Amazônica 23:371-381.
Sunderland, T. C. H; Ndoye, O. & Harrison-Sanchez, S. 2011. Non-timber forest products
and conservation: what prospects? In: Shackleton, S; Shackleton, C. & Shanley, P.
(ed.) Non-timber forest products in the global context. Springer-Verlag, Berlin.
Toniato, M. T. Z.; Leitão-Filho, H. F. & Rodrigues, R. R. 1998. Fitossociologia de um
remanescente de floresta higrófila (mata de brejo) em Campinas, SP. Revista
Brasileira de Botânica 21(2)197-210.

33
Villalobos, M. P. 1994. Guilda de frugivoros associada com o buriti (Mauritia flexuosa:
Palmae) numa vereda no Brasil Central. Dissertação de Mestrado, Departamento de
Ecologia – UnB, Brasília - DF.
Zanatta, C. F.; Mitjans, M.; Urgatondo, V.; Rocha-Filho, P. A. & Vinardell, M. P. 2010.
Photoprotective potential of emulsions formulated with Buriti oil (Mauritia flexuosa)
against UV irradiation on keratinocytes and fibroblasts cell lines. Food and Chemical
Toxicology 48:70-75.

34
CAPÍTULO1: Ontogenia, alometria e reprodução do buriti ( Mauritia flexuosa; Arecaceae)
Maurício Bonesso Sampaio
Flavio Antonio Maës dos Santos
Resumo
Estádios ontogenéticos de uma espécie podem ser descritos através das características
macro-morfológicas e variáveis quantitativas que os indivíduos apresentam durante o ciclo
de vida. O objetivo deste estudo foi avaliar a influência das variações macro-morfológicas
dos indivíduos durante o ciclo de vida na sobrevivência, alometria, produção de folhas e
reprodução do buriti (Mauritia flexuosa; Arecaceae). Observamos características
morfológicas e fizemos medidas biométricas em indivíduos de buriti de 16 brejos no Brasil
Central. Identificamos quatro estádios para o buriti utilizando apenas duas características
macro-morfológicas (presença/ausência do estipe e atividade reprodutiva) e uma medida
quantitativa (altura). Ao contrário de várias espécies de palmeiras, a taxa de produção de
folhas, a morfologia foliar e a atividade reprodutiva não se alteram com o crescimento dos
buritis. Porém, a taxa de sobrevivência e as relações alométricas variam durante o ciclo de
vida. Desta forma, cada estádio ontogenético pode ter uma contribuição particular para a
dinâmica das populações de buriti.
Palavras-chave: palmeira, ciclo de vida, biometria, crescimento, vereda

35
Introdução
As plantas de uma população são sujeitas a diferenças nas condições ambientais e
recursos disponíveis no micro-hábitat conforme avançam no ciclo de vida. As plantas
podem possuir adaptações evolutivas, como mudanças morfológicas, anatômicas,
fisiológicas e bioquímicas ao longo do ciclo de vida, como resposta a essas diferenças no
micro-hábitat (Begon et al. 2007). Essas mudanças intrínsecas nos indivíduos podem
ocasionar a aquisição ou perda de estruturas externas macro-morfológicas (Gatsuk et al.
1980). Assim, um estádio ontogenético pode ser caracterizado pelo conjunto dessas
estruturas que os indivíduos possuem em uma mesma fase do ciclo de vida.
As diferenças no micro-hábitat também geram alterações nas taxas de
sobrevivência, crescimento e reprodução durante o ciclo de vida de um indivíduo. Por
exemplo, as plântulas são dependentes das reservas da semente e são sujeitas a altas taxas
de mortalidade (Frost & Rydin 1997). Conforme a planta avança entre estádios, podem
ocorrer modificações na morfologia da folha ou na filotaxia, que possibilitam maior acesso
à luz, contribuindo para aumentar a taxa de crescimento e sobrevivência (Carvalho et al.
1999; Souza et al. 2003). Em uma planta imatura, o surgimento de raízes caulinígenas
possibilita maior sustentação e absorção de nutrientes, contribuindo para aumentar ainda
mais a sobrevivência e o crescimento do indivíduo (Bernacci et al. 2008). Quando atinge a
maturidade sexual, a planta passa a produzir sementes, com possibilidades de contribuir
para as gerações futuras. O investimento em atividade reprodutiva e a probabilidade de
sobreviver podem ser reduzidos quando o indivíduo adulto atinge a senilidade (Souza et al.

36
2003). Desta forma, as plantas de cada estádio ontogenético podem ter diferentes
contribuições para a dinâmica de uma população.
A idade cronológica pode ser uma medida direta da fase de desenvolvimento que
um indivíduo está no ciclo de vida, pois o tempo de duração de cada fase é programado
geneticamente (Gatsuk et al. 1980). Entretanto, o ambiente que um indivíduo está sujeito
pode sofrer alterações ao longo do tempo, modificando a duração das fases de
desenvolvimento das plantas (Barthélémy & Caraglio 2007). Por exemplo, a abertura de
uma clareira na floresta pode acelerar o crescimento de plântulas, que rapidamente tornam-
se reprodutivas. Ao passo que plântulas sob o dossel fechado, sujeitas a alta restrição
luminosa, demoram mais para atingirem a maturidade sexual. Portanto, neste caso a idade é
pouco relacionada às características ontogenéticas. Um conjunto de características macro-
morfológicas (como forma e filotaxia das folhas, presença/ausência de estipe,
presença/ausência de estruturas reprodutivas), que sejam facilmente visualizadas, somadas
a medidas quantitativas (como tamanho do indivíduo, número de folhas e tamanho das
folhas), podem ser mais bem relacionadas do que a idade à variação na probabilidade de
uma planta sobreviver, crescer e reproduzir durante o ciclo de vida (Gatsuk et al. 1980).
Desta forma, a classificação dos indivíduos em estádios ontogenéticos pode ser uma
ferramenta útil para descrever a estrutura de uma população e, também, para determinar a
influência que cada período do ciclo de vida possui para a dinâmica da população.
O crescimento das palmeiras ocorre principalmente pela produção e acumulação de
folhas durante o ciclo de vida (Carvalho et al. 1999). O aumento no número de folhas é
geralmente acompanhado pelo aumento no tamanho das folhas, na taxa de produção de
folhas e no crescimento do estipe em diâmetro e em comprimento (Souza et al. 2003).

37
Porém, essas relações alométricas tendem a variar durante a ontogenia, pois o ritmo de
crescimento pode mudar entre estádios. Além disso, as relações são diferentes entre
espécies. Por exemplo, os indivíduos dos estádios iniciais de várias palmeiras tendem a
apresentar crescimento rápido no diâmetro e crescimento lento na altura, enquanto que o
inverso ocorre no final do ciclo de vida (Kimura & Simbolon 2002). Já outras espécies,
principalmente do gênero Euterpe, podem apresentar taxa de crescimento diamétrico
semelhante durante todo o ciclo de vida (Alves et al. 2004; Avalos & Sylvester 2010).
A atividade reprodutiva das palmeiras pode ser influenciada pelo tamanho do
indivíduo de três formas: (i) a reprodução aumenta linearmente com o crescimento do
indivíduo durante o ciclo de vida, pois quanto maior o tamanho do indivíduo, maior a
disponibilidade de recursos para investir em reprodução (Cunningham 1997); (ii) a
atividade reprodutiva pode aumentar até alcançar um pico nos indivíduos de tamanhos
intermediários, e reduzir no final do ciclo de vida, quando os indivíduos tornam-se senis
(Barot & Gignoux 1999; Souza et al. 2003); ou (iii) não há relação entre a reprodução e o
tamanho do indivíduo (Sampaio & Scariot 2008; Rojas-Robles & Stiles 2009).
A descrição das alterações macro-morfológicas e das taxas demográficas dos
indivíduos de uma espécie durante a ontogenia pode gerar informações fundamentais para
os estudos de dinâmica de populações, e também, contribuir para elucidar os fatores que
influenciam a variação da sobrevivência, crescimento e reprodução durante o ciclo de vida.
Os objetivos deste estudo foram: (i) identificar e caracterizar estádios ontogenéticos do
buriti (Mauritia flexuosa L.f; Arecaceae) através da descrição das variações nas estruturas
macro-morfológicas dos indivíduos ao longo do ciclo de vida; (ii) descrever características
dos indivíduos que permitam identificar quando as plântulas passam a ser independentes

38
das reservas da semente; (iii) avaliar a influência de variações ontogenéticas dos indivíduos
na alometria e produção de folhas; e (iv) avaliar se o tamanho do indivíduo influencia a
reprodução. Para alcançar o quarto objetivo, nós testamos a hipótese de que não há
correlação entre a reprodução dos buritis e o tamanho dos indivíduos, pois a produção de
inflorescências dos buritis é irregular entre anos (Storti 1993; Abreu 2001), similar ao que
ocorre para a palmeira Oenocarpus bataua Mart. (Rojas-Robles & Stiles 2009). Assim, os
buritis reprodutivos podem acumular reservas durante os períodos de baixa produção de
inflorescências, independentemente do seu tamanho, para investimento nos eventos futuros
de reprodução.
Material e métodos
Espécie estudada
No início do ciclo de vida do buriti o estipe é subterrâneo e com o passar do tempo
torna-se aéreo. Cicatrizes foliares podem permanecer conspícuas no estipe. Indivíduos
adultos possuem até 20 folhas em leque, do tipo costapalmada (Henderson et al. 1995). A
lâmina foliar é dividida em duas partes por uma raque de 7 a 100 cm de comprimento. Cada
folha composta possui até 250 segmentos, com ápice muito agudo e comprimento de até 2,4
m. O pecíolo pode ter até 7 m de comprimento em indivíduos com estipe subterrâneo, é
triangular, com até 6,7 cm de largura. As folhas são persistentes, podendo permanecer
presas ao indivíduo durante meses por uma bainha aberta, mesmo depois de mortas. Os
frutos são elipsóides e possuem 5 cm de diâmetro e 7 cm de comprimento (Henderson et al.
1995). As folhas do buriti são colhidas pelos moradores locais e utilizadas como materiais

39
de construção, utensílios domésticos, brinquedos e artesanatos. O pecíolo é utilizado na
fabricação de móveis. Além disso, os frutos são utilizados como alimento (Sampaio et al.
2008).
Coleta de dados
Nós fizemos observações das características macro-morfológicas e medições de
variáveis quantitativas em indivíduos de 16 brejos no leste e nordeste do Tocantins e sul do
Piauí, Brasil (Tabela 1).
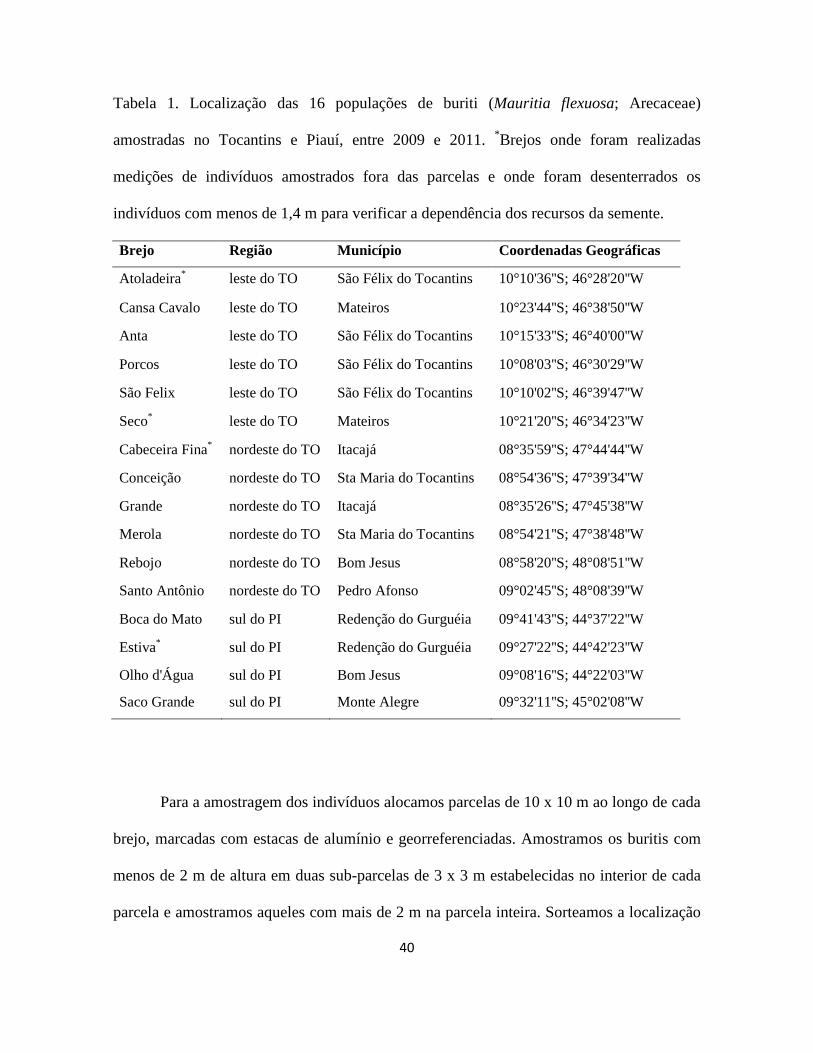
40
Tabela 1. Localização das 16 populações de buriti (Mauritia flexuosa; Arecaceae)
amostradas no Tocantins e Piauí, entre 2009 e 2011. *Brejos onde foram realizadas
medições de indivíduos amostrados fora das parcelas e onde foram desenterrados os
indivíduos com menos de 1,4 m para verificar a dependência dos recursos da semente.
Brejo Região Município Coordenadas Geográficas
Atoladeira* leste do TO São Félix do Tocantins 10°10'36''S; 46°28'20''W
Cansa Cavalo leste do TO Mateiros 10°23'44''S; 46°38'50''W
Anta leste do TO São Félix do Tocantins 10°15'33''S; 46°40'00''W
Porcos leste do TO São Félix do Tocantins 10°08'03''S; 46°30'29''W
São Felix leste do TO São Félix do Tocantins 10°10'02''S; 46°39'47''W
Seco* leste do TO Mateiros 10°21'20''S; 46°34'23''W
Cabeceira Fina* nordeste do TO Itacajá 08°35'59''S; 47°44'44''W
Conceição nordeste do TO Sta Maria do Tocantins 08°54'36''S; 47°39'34''W
Grande nordeste do TO Itacajá 08°35'26''S; 47°45'38''W
Merola nordeste do TO Sta Maria do Tocantins 08°54'21''S; 47°38'48''W
Rebojo nordeste do TO Bom Jesus 08°58'20''S; 48°08'51''W
Santo Antônio nordeste do TO Pedro Afonso 09°02'45''S; 48°08'39''W
Boca do Mato sul do PI Redenção do Gurguéia 09°41'43''S; 44°37'22''W
Estiva* sul do PI Redenção do Gurguéia 09°27'22''S; 44°42'23''W
Olho d'Água sul do PI Bom Jesus 09°08'16''S; 44°22'03''W
Saco Grande sul do PI Monte Alegre 09°32'11''S; 45°02'08''W
Para a amostragem dos indivíduos alocamos parcelas de 10 x 10 m ao longo de cada
brejo, marcadas com estacas de alumínio e georreferenciadas. Amostramos os buritis com
menos de 2 m de altura em duas sub-parcelas de 3 x 3 m estabelecidas no interior de cada
parcela e amostramos aqueles com mais de 2 m na parcela inteira. Sorteamos a localização

41
das parcelas ao longo do comprimento do brejo, respeitando uma distância mínima de 5 m
entre as parcelas. Não alocamos parcelas nos locais onde o brejo tinha menos do que 10 m
de largura e onde não havia formação de dossel contínuo. Alocamos de 13 a 20 parcelas em
cada brejo, sendo que a quantidade foi dependente do comprimento do brejo propício para a
amostragem.
Amostramos os indivíduos de cada parcela de janeiro a abril em 2009, 2010 e 2011.
Em cada amostragem medimos a altura dos indivíduos com menos de 2 m do solo até o
ápice da folha mais alta, usando uma trena. Para indivíduos com estipe aéreo nós medimos
apenas a circunferência à altura do solo (CAS), que não foi medida em 2011.
Marcamos a folha mais nova de cada indivíduo, quando possível, para contar o
número de folhas produzidas entre as amostragens. Para o cálculo da taxa anual de
produção de folhas excluímos os indivíduos que tiveram folhas colhidas por extrativistas ou
folhas predadas por animais. Além disso, excluímos da análise o brejo Porcos entre 2009 e
2010, pois houve uma queimada acidental neste brejo em setembro de 2008. Entre 2010 e
2011, além do brejo Porcos, excluímos os indivíduos dos brejos Santo Antônio, Rebojo,
Cabeceira Fina e Cansa Cavalo, que queimaram na seca de 2010. Não calculamos a taxa de
produção de folhas para os indivíduos reprodutivos, pois não foi possível marcar as folhas
devido à altura destes.
Além dos buritis amostrados dentro das parcelas, selecionamos 113 indivíduos com
altura entre 0,11 m e cerca de 10 m em quatro dos brejos amostrados (Tabela 1). Para cada
indivíduo contamos o número de folhas e coletamos a folha mais nova totalmente
expandida. Desta folha coletada medimos o comprimento do pecíolo, a maior largura do
pecíolo na porção central, contamos o número de segmentos foliares e medimos o

42
comprimento do limbo e da raque. Não amostramos indivíduos com mais de 10 m de altura
fora das parcelas, pois não foi possível coletar as folhas.
Desenterramos cuidadosamente 46 buritis com altura entre 0,11 m e 1,40 m (média
= 0,44 m; DP = 0,28 m) fora das parcelas em quatro brejos (Tabela 1) para verificar a
presença ou a ausência de ligação da plântula à semente. Nós verificamos a dependência da
plântula às reservas da semente utilizando a metodologia proposta por Fernandes (2002).
Este método consiste em pressionar a semente com a mão, quando ligada à plântula, para
avaliar a espessura do tegumento e do endosperma restante. Quando a semente estava
ausente, ou quando a semente quebrou-se sob leve compressão, nós consideramos o
indivíduo independente das reservas da semente.
Análises estatísticas
Para descrever as relações alométricas nós testamos análises de regressão
exponenciais, lineares, potenciais, logarítmicas e quadráticas, mas apresentamos somente o
resultado do modelo com melhor ajuste, avaliado através do coeficiente de determinação
(r2). Transformamos as variáveis por log10, quando necessário, para corrigir desvios da
normalidade tanto nos valores brutos das variáveis, quanto nos resíduos das regressões.
Apesar dos indivíduos terem sido amostrados em 16 populações, os dados foram
analisados conjuntamente para descrever a ontogenia da espécie. Estudos realizados com as
palmeiras Lytocaryum hoehnei (Burret) Toledo e Attalea humilis Mart. demonstraram que a
caracterização dos estádios não varia entre populações (Souza et al. 2000; Ferraz 2004).
Além disso, algumas relações alométricas foram comparadas entre populações de buriti e
não foram detectadas diferenças significativas (resultados não mostrados).

43
Resultados
Estádios ontogenéticos
Nós observamos duas modificações qualitativas nas características macro-
morfológicas dos indivíduos durante o ciclo de vida do buriti (Mauritia flexuosa): (i)
inicialmente o estipe é subterrâneo e com o passar do tempo passa a ser aéreo; (ii) a partir
de um determinado momento do ciclo de vida os indivíduos passam a produzir estruturas
reprodutivas.
Inicialmente nós identificamos três estádios ontogenéticos para o buriti utilizando
estas características qualitativas. O estádio 1 inclui indivíduos sem estipe acima do solo,
que possuem meristema apical subterrâneo, os pecíolos das folhas são parcialmente
enterrados e não apresentam atividade reprodutiva. Indivíduos do estádio 2 apresentam
estipe acima do solo, geralmente encoberto pelas bainhas das folhas, meristema apical e
pecíolos totalmente aéreos e não possuem atividade reprodutiva. No estádio 3, os
indivíduos possuem estipe aéreo e apresentam atividade reprodutiva. Os indivíduos
reprodutivos podem ser reconhecidos mesmo que não estejam reproduzindo no momento
da amostragem, pois as inflorescências antigas permanecem presas ao indivíduo por vários
meses após estarem secas. Depois de cair, a estrutura ainda pode permanecer no solo, sob o
indivíduo, por anos antes de se decompor completamente. O sexo (macho ou fêmea) do
buriti pode ser determinado facilmente, pois as inflorescências são bastante distintas
(Henderson et al. 1995).

44
Não observamos nenhuma característica qualitativa não-destrutiva que nos
permitisse identificar quais indivíduos estavam, ou não, ligados à semente. Entre os 46
indivíduos desenterrados, 52% ainda estavam ligados à semente, e tiveram em média 36 cm
(DP = 22 cm) de altura. As sementes estavam presas aos indivíduos, em média 12 cm
abaixo do nível do solo (DP = 7 cm; mínimo = 0 cm; máximo = 28 cm; n = 23). Entre os
indivíduos ligados à semente, 88% tiveram apenas uma folha e 12% tiveram duas folhas. O
maior indivíduo desenterrado que ainda mantinha conexão com a semente teve 90 cm de
altura.
Houve relação positiva significativa entre altura e sobrevivência tanto no período
entre 2009 e 2010 (Regressão Logística; χ2 = 263,1; n = 2.271; p < 0,001), quanto entre
2010 e 2011 (Regressão Logística; χ2 = 118,5; n = 787; p < 0,001). Os indivíduos com
menos de 100 cm tiveram taxa de sobrevivência crescente com a altura e geralmente menor
do que 65% (Figura 1). A taxa de sobrevivência dos indivíduos com altura entre 100 cm e
190 cm se manteve entre 80 e 100% e foi significativamente mais alta do que para os
indivíduos com menos de 100 cm de altura (χ2 = 360,3; gl = 1; p < 0,001). Essa relação foi
semelhante entre os dois períodos avaliados (Kolmogorov-Smirnov; p > 0,10; Figura 1).

45
Figura 1. Taxa de sobrevivência dos indivíduos de buriti (Mauritia flexuosa) nos períodos
entre 2009 e 2010 e entre 2010 e 2011. Os indivíduos foram agrupados em classes de altura
com intervalos de 10 cm.
O estádio 1 foi dividido em duas classes de altura (Tabela 2): 1.1 - indivíduos com
menos de 100 cm de altura e; 1.2 - indivíduos com 100 cm ou mais de altura. Nós fizemos
essa divisão, pois os indivíduos com mais de 100 cm de altura já haviam esgotado as
reservas da semente e possuíam maior taxa de sobrevivência do que indivíduos com menos
de 100 cm de altura.

46
Tabela 2. Caracterização dos estádios ontogenéticos do buriti (Mauritia flexuosa) em
relação à altura, tipo de estipe e presença/ausência de estruturas reprodutivas.
Período ontogenético Estádios Altura Estipe Inflorescências
Pré-reprodutivo 1.1 < 1 m subterrâneo ausentes
1.2 ≥ 1 m subterrâneo ausentes
2 - aéreo ausentes
Reprodutivo 3 - aéreo presentes
Alometria
As relações alométricas foram diferentes entre os estádios 1 e 2. Para os buritis sem
estipe aéreo (estádio 1), conforme há crescimento no comprimento do pecíolo, há aumento
na largura do pecíolo, no número de segmentos foliares, no comprimento do limbo e no
comprimento da raque (Figura 2; Tabela 3). O número de folhas aumentou com a altura no
estádio 1 (Tabela 3).
Os indivíduos do estádio 2 (com estipe aéreo) não tiveram relação significativa
entre comprimento do pecíolo e largura do pecíolo, comprimento do limbo foliar e
comprimento da raque (Figura 2; Tabela 3), mas o número de segmentos foliares foi
negativamente relacionado ao comprimento do pecíolo (Figura 2b).
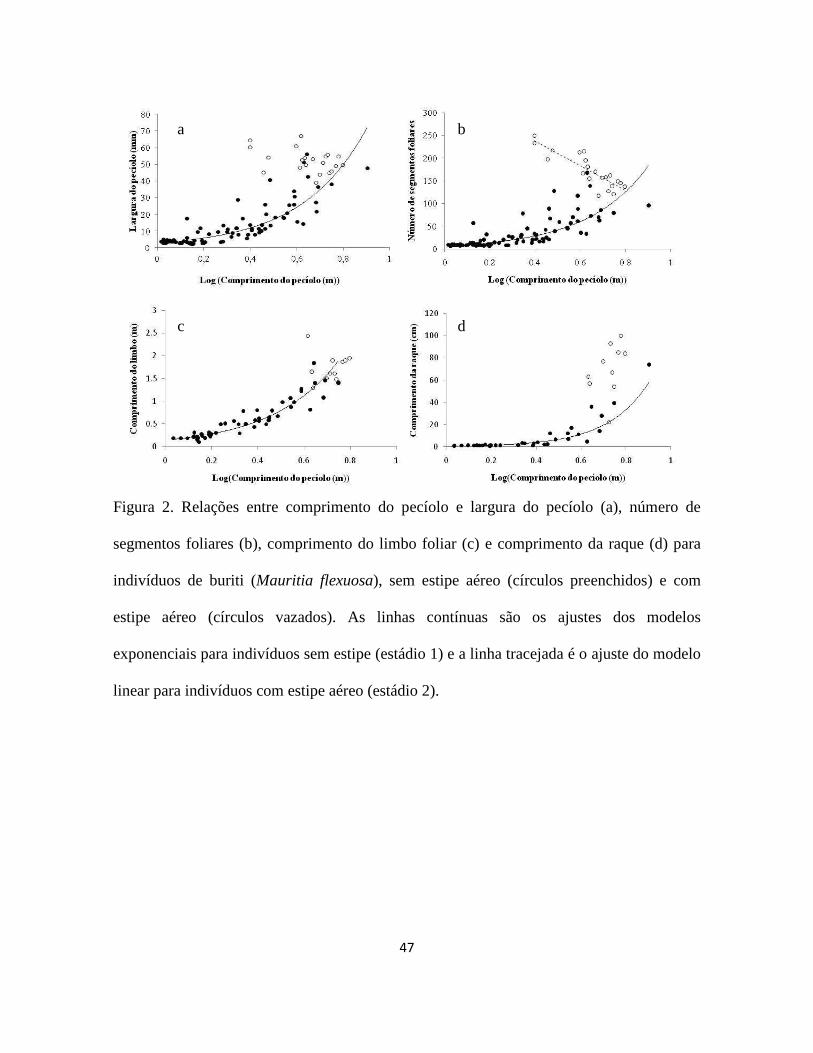
47
Figura 2. Relações entre comprimento do pecíolo e largura do pecíolo (a), número de
segmentos foliares (b), comprimento do limbo foliar (c) e comprimento da raque (d) para
indivíduos de buriti (Mauritia flexuosa), sem estipe aéreo (círculos preenchidos) e com
estipe aéreo (círculos vazados). As linhas contínuas são os ajustes dos modelos
exponenciais para indivíduos sem estipe (estádio 1) e a linha tracejada é o ajuste do modelo
linear para indivíduos com estipe aéreo (estádio 2).
a b
c d

48
Tabela 3. Equações das relações alométricas entre altura (m), comprimento do pecíolo (m),
largura do pecíolo (cm), número de segmentos foliares, comprimento do limbo (m),
comprimento da raque (cm) e número de folhas dos buritis (Mauritia flexuosa) sem (estádio
1) e com estipe aéreo (estádio 2). Foram utilizadas regressões lineares, exponenciais e
logarítmicas para o ajuste dos dados, onde: (*) p < 0,05; (**) p < 0,001; (ns) p > 0,05.
Var. ind. Var. dep. a b c Modelo r2 N
Est
ádio
1
compr. pec. larg. pec. 4,2 3,0 - y = a*e(bx) 0,69** 88
compr. pec. segm. foliar 11,6 2,8 - y = a*e(bx) 0,56** 92
compr. pec. compr. limbo 0,2 2,7 - y = a*e(bx) 0,87** 48
compr. pec. compr. raque 0,5 5,5 - y = a*ln(x)+b 0,90** 35
altura num. folha 1,0 3,0 - y = a*ln(x)+b 0,45** 50
Est
ádio
2 compr. pec. larg. pec. -24,3 68,1 - y = ax+b 0,18ns 21
compr. pec. segm. foliar -7,4 345,8 - y = ax+b 0,74** 21
compr. pec. compr. limbo -0,1 -0,2 - y = ax+b 0,01ns 12
compr. pec. compr. raque 170,1 -53,3 - y = ax+b 0,10 ns 10
A CAS médio diferiu significativamente entre os estádios 2 e 3 nos dois períodos
avaliados (Tabela 4). No estádio 2 o estipe teve em média 4 cm a menos de CAS do que no
estádio 3.

49
Tabela 4. Comparação da CAS (m) entre indivíduos dos estádios 2 e 3 e entre machos e
fêmeas (estádio 3), em 2009 e 2010. O número de indivíduos amostrados está entre
parênteses.
Ano Estádio 2 Estádio 3 t p
2009 1,1 (173) 1,1 (605) -3,1 0,002
2010 1,1 (164) 1,2 (630) -2,7 0,007
Produção de folhas
A mediana da taxa anual de produção de folhas foi igual a dois, independentemente
do estádio e do período avaliado. A taxa de produção de folhas novas teve apenas 1% da
sua variância explicada pela altura (r2 < 0,01; p < 0,001). A taxa de produção de folhas
novas também teve fraca relação com o número de folhas do indivíduo (Tabela 5; Figura
3), sendo que o melhor ajuste das regressões foi obtido utilizando-se o modelo quadrático.
De acordo com este modelo, os indivíduos do estádio 1.1 que inicialmente possuem apenas
uma folha, tendem a apresentar taxa de produção de folhas maior do que aqueles que
possuem de duas a seis folhas. Os indivíduos do estádio 2 com mais de seis folhas são os
que apresentam maior taxa de produção de folhas, podendo produzir até quatro folhas
novas por ano (Figura 3).

50
Tabela 5: Ajuste de modelos quadráticos entre a taxa de produção de folhas novas e o
número de folhas nos buritis (Mauritia flexuosa). Para todos os modelos ajustados p <
0,001.
Estádio Ano n r2 F
1.1 2009 – 2010 632 0,03 921
1.2 2009 – 2010 1.457 0,04 3.149
2 2009 – 2010 54 0,18 133
1.1 2010 – 2011 306 0,02 485
1.2 2010 – 2011 1.126 0,01 3.088
2 2010 – 2011 43 0,14 157
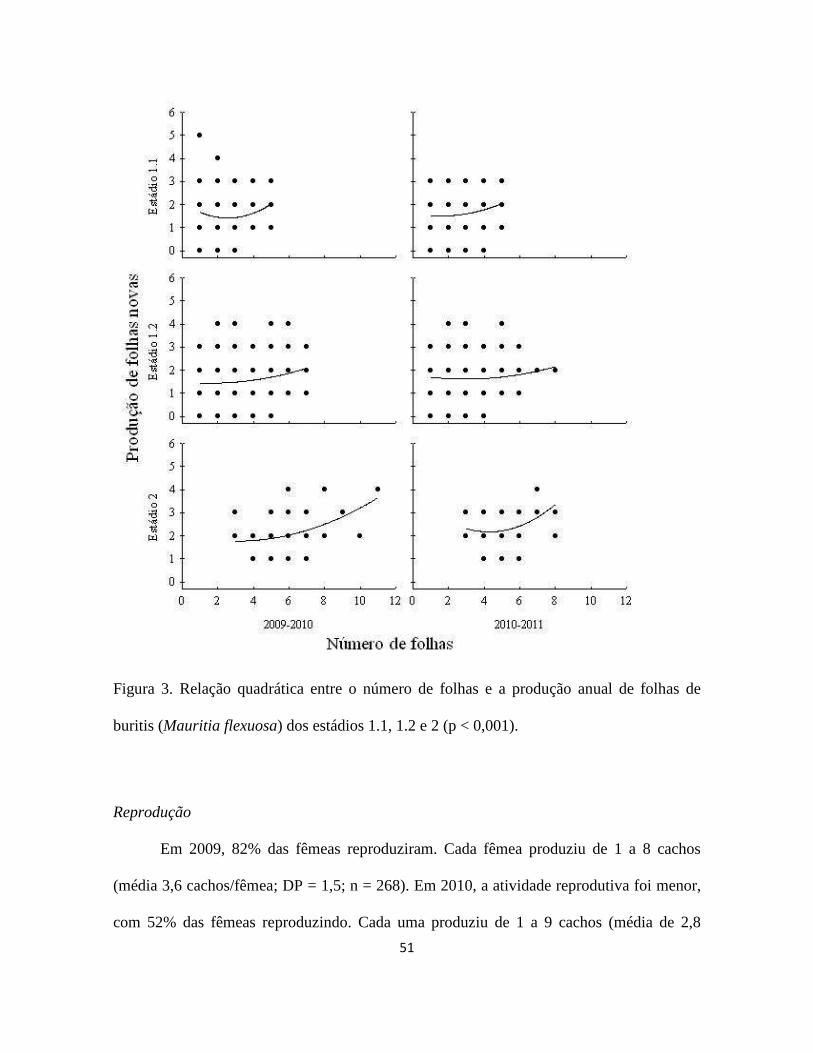
51
Figura 3. Relação quadrática entre o número de folhas e a produção anual de folhas de
buritis (Mauritia flexuosa) dos estádios 1.1, 1.2 e 2 (p < 0,001).
Reprodução
Em 2009, 82% das fêmeas reproduziram. Cada fêmea produziu de 1 a 8 cachos
(média 3,6 cachos/fêmea; DP = 1,5; n = 268). Em 2010, a atividade reprodutiva foi menor,
com 52% das fêmeas reproduzindo. Cada uma produziu de 1 a 9 cachos (média de 2,8

52
cachos/fêmea; DP = 1,7; n = 339). Em 2011, a atividade reprodutiva foi a mais fraca do
período de estudo, sendo que apenas 10% das fêmeas reproduziram. Cada fêmea em
atividade reprodutiva produziu de 1 a 7 cachos (média 2,4 cachos/fêmea; DP = 1,5; n =
341). O número de cachos produzidos por fêmea não teve relação com a CAS dos
indivíduos nos dois períodos de estudo (2009: r2 = 0,05; p < 0,001; n = 270; 2010: r2 = 0,01;
p = 0,69; n = 171).
Discussão
Estádios ontogenéticos
O buriti possui quatro estádios ontogenéticos durante o ciclo de vida. Este número
de estádios é o mínimo encontrado para palmeiras (Tabela 6). O surgimento do estipe aéreo
e a produção de estruturas reprodutivas são as únicas alterações consistentes em
características macro-morfológicas durante o ciclo de vida do buriti. Além dessas duas
alterações, a maioria das espécies de palmeiras apresenta variações conspícuas na
morfologia das folhas, que podem contribuir para reduzir o auto-sombreamento da copa de
um indivíduo, aumentando a eficiência de captação de luz (Kimura & Simbolon 2002). Por
exemplo, em palmeiras que ocorrem no subosque de florestas tropicais úmidas, como
Geonoma brevispatha Barb. Rodr. e G. schottiana Mart., as folhas são bífidas no início do
ciclo de vida, parcialmente pinadas em estádios intermediários e completamente pinadas
nos estádios reprodutivos (Souza et al. 2003; Sampaio & Scariot 2010). Ao contrário destas
duas espécies, e da maioria das palmeiras que ocorrem no subosque dessas florestas, as
folhas do buriti permanecem palmadas durante todo o ciclo de vida. Uma possível

53
explicação para isso seria a de que os buritis ocorrem em ambientes como os brejos do
Brasil Central e as florestas de várzea da Amazônia, que possuem cobertura do dossel
irregular, com maior intensidade de radiação no subosque do que em outras florestas
tropicais de dossel contínuo (Maurenza et al. 2012). Portanto, o auto-sombreamento das
folhas do buriti pode ter menor importância para a captação de luz do que para outras
espécies que apresentam alterações na morfologia foliar ao longo da ontogenia.
Tabela 6. Número de estádios ontogenéticos identificados, uso de variáveis quantitativas e
uso da morfologia das folhas para diferenciar os estádios ontogenéticos de várias espécies
de palmeiras.
Espécies Estádios Variável
quantitativa
Morf.
folha
Referências
Astrocaryum aculeatissimum 5 não sim Portela & Santos 2011
Attalea humilis 4 não sim Souza et al. 2000
Borassus aethiophum 4 não sim Barot & Gignoux 1999
Euterpe edulis 7 sim não Silva Matos et al. 1999
Euterpe precatoria 4 sim não Rocha 2004
Geonoma brevispatha 4 não sim Souza et al. 2003
Geonoma macrostachys 11 sim não Svenning & Macía 2002
Geonoma orbygniana 11 sim sim Rodríguez-Butiricá et al. 2005
Geonoma schottiana 5 não sim Sampaio & Scariot 2010
Lytocaryum hoehnei 4 não sim Ferraz 2004
Mauritia flexuosa 4 sim não Presente estudo
Phytelephas seemannii 5 sim sim Bernal 1998
Syagrus romanzoffiana 5 sim sim Bernacci et al. 2008
Apesar da morfologia da folha da maioria das palmeiras variar ao longo da
ontogenia, nem sempre esta característica é adequada para classificar os indivíduo em
estádios (Tabela 6). Nestes casos, geralmente a morfologia das folhas é substituída por

54
variáveis quantitativas, como altura, CAS, número de folhas e comprimento da raque. Para
o buriti, a altura pode ser um critério de separação de estádios, pois os indivíduos a partir de
1 m já estão estabelecidos, não dependendo das reservas da semente.
Na fase inicial da ontogenia os nutrientes do endosperma podem ser transferidos
para a plântula jovem por uma espécie de haustório formado pela parte distal do embrião
que permanece dentro da semente do buriti (Fernandes 2002). Em casa de vegetação, o
estabelecimento das plântulas – momento a partir do qual os recursos da semente estão
exauridos – coincide com o momento em que a primeira folha lançada alcança sua extensão
completa, com 35 cm de comprimento, aos 105 dias (Fernandes 2002). Porém, neste estudo
nós registramos em campo 11 de 26 indivíduos amostrados com altura superior a 35 cm,
que ainda dependiam das reservas da semente. Entre esses indivíduos, três tiveram duas
folhas e um deles teve 90 cm de altura.
Alometria
As variações nas características macro-morfológicas das palmeiras ao longo da
ontogenia frequentemente estão relacionadas às variações alométricas (Souza et al. 2000;
Kimura & Simbolon 2002; Souza et al. 2003; Watanabe & Suzuki 2007). Para o buriti
ocorrem várias alterações nas relações alométricas entre os estádios 1 e 2. No estádio 1, os
indivíduos tendem a apresentar relações entre as características biométricas das folhas e o
tamanho dos indivíduos (representado pela variável comprimento do pecíolo neste estudo).
Essas relações passam a ser inexistentes, ou são completamente modificadas quando os
indivíduos ingressam no estádio 2.

55
As relações não-lineares encontradas no estádio 1 (Figura 2) provavelmente são
causadas por um rápido crescimento inicial no comprimento do pecíolo acompanhadas por
pequenas alterações nas demais características biométricas avaliadas nas folhas. Quando o
comprimento do pecíolo alcança um determinado limite, é iniciado o aumento da largura do
pecíolo, do número de segmentos foliares, do comprimento do limbo e do comprimento da
raque.
Além de haver um rápido crescimento no comprimento do pecíolo no início do ciclo
de vida (estádio 1), ainda ocorre uma rápida acumulação de folhas conforme o buriti cresce.
Um indício desse vigoroso aumento no número de folhas é o fato de que encontramos três
indivíduos que já haviam produzido duas folhas quando ainda dependiam das reservas da
semente. Este fato também torna a variável número de folhas inadequada como critério de
classificação dos indivíduos nos estádios 1.1 e 1.2. A acumulação de folhas no estádio 1
provavelmente ocorre devido à maior longevidade das folhas, já que a taxa de produção de
folhas tem fraca correlação com a altura do indivíduo.
O tamanho das folhas (medido indiretamente pelas variáveis número de segmentos
foliares, comprimento da raque e comprimento do limbo) também aumenta conforme os
indivíduos do estádio 1 crescem. Porém, no estádio 2 o tamanho da folha não tem relação
com o tamanho do indivíduo, exceto quanto ao número de segmentos foliares.
Provavelmente, a folha alcança o seu tamanho ótimo em indivíduos no final do estádio 1.2.
Entre os estádios 2 e 3, o estipe cresce em média apenas 4 cm de circunferência.
Assim como em Geonoma brevispatha (Souza et al. 2003), o diâmetro do estipe do buriti
deve crescer rapidamente no início do ciclo de vida, quando há um lento crescimento do
estipe em comprimento. Quando atinge os estádios 2 e 3, o comprimento do estipe pode

56
aumentar rapidamente, enquanto o diâmetro se mantém constante. Assim, o diâmetro
alcançado no início do ciclo de vida pode ser suficiente para suportar o crescimento em
comprimento do estipe durante toda a ontogenia, sem que haja um aumento proporcional no
diâmetro (Alves et al. 2004). Além disso, apesar das palmeiras não possuírem crescimento
secundário, podem apresentar aumento na densidade dos tecidos na porção basal do tronco,
que contribui para manter a estabilidade mecânica (Kimura & Simbolon 2002; Alves et al.
2004).
Produção de folhas
As folhas das palmeiras são produzidas uma por vez no centro da copa (Carvalho et
al. 1999). A taxa de produção de folhas do buriti é pouco relacionada com o tamanho do
indivíduo e variou pouco entre os estádios iniciais (1.1, 1.2 e 2), assim como o encontrado
para Euterpe edulis Mart. (Carvalho et al. 1999) e Geonoma congesta H. Wendl. ex Spruce
(Chazdon 1992). Para outras espécies, a taxa de produção de folhas aumenta durante a
ontogenia, como Geonoma brevispatha (Souza et al. 2003) e G. schottiana (Sampaio &
Scariot 2008).
O buriti geralmente precisa acumular entre 4 e 5 folhas e ter no mínimo 4 m de
altura (do solo até o ápice da folha mais alta) antes de iniciar o crescimento do estipe em
comprimento. Com estipe aéreo, o número de folhas aumenta pouco até atingir o máximo
de nove folhas no estádio 2. Apesar do número de folhas ter sido contado somente até o
estádio 2 neste estudo, os buritis provavelmente permanecem acumulando folhas até o final
do ciclo de vida, pois no estádio 3 podem apresentar até 20 folhas (Henderson et al. 1995).
A área foliar total do buriti aumenta linearmente com o tamanho do indivíduo até o final do

57
ciclo de vida (Renninger & Philips 2010). Como o tamanho da folha estabiliza em estádios
intermediários, provavelmente é a acumulação de folhas que possibilita o aumento contínuo
na área foliar total. Para que haja a acumulação de folhas, a longevidade das folhas deve
aumentar conforme o indivíduo cresce, já que a taxa de produção de folhas se mantém
constante, assim como em Euterpe edulis (Carvalho et al. 1999).
Reprodução
O evento de reprodução por um indivíduo de buriti requer alto investimento em
recursos. Cada cacho tem em média 19,4 kg de frutos (Barbosa et al. 2010) e alguns buritis
podem produzir até 10 cachos com frutos durante um período reprodutivo. Apesar do alto
investimento, há fraca relação entre o tamanho do buriti (representado pela CAS) e o
número de infrutescências produzidas, assim como o encontrado em Geonoma schottiana
(Sampaio & Scariot 2008) e Oenocarpus bataua (Rojas-Robles & Stiles 2009). Esse
resultado difere da maioria das espécies de palmeiras, que apresentam relação positiva
linear (Oyama 1990; Scariot et al. 1995; Cunningham 1997; Svenning 2000) ou não-linear
(Barot & Gignoux 1999; Souza et al. 2003) entre o tamanho do indivíduo e a atividade
reprodutiva. Esta ausência de relação pode ocorrer, pois os buritis produzem inflorescências
irregularmente entre anos, com anos de alto investimento seguidos por anos de baixo ou
nenhum investimento, resultado similar ao encontrado por Storti (1993) e Abreu (2001).
Este padrão de reprodução é semelhante também com o que ocorre para Oenocarpus
bataua (Rojas-Robles & Stiles 2009). Durante o período inativo, o indivíduo pode
acumular as reservas necessárias para investimento nos eventos futuros de produção de
frutos, independentemente do seu tamanho.

58
Conclusões
O estádio 1.1 pode ser caracterizado pela dependência do indivíduo às reservas da
semente. Nessa fase os indivíduos geralmente estão sujeitos a altas taxas de mortalidade,
tendem a acumular folhas e podem apresentar rápido crescimento no comprimento do
pecíolo. Os indivíduos do estádio 1.2 podem ter sobrevivência acima de 80%. Conforme
crescem em tamanho, tendem a ter mais folhas, cada vez maiores. Quando um indivíduo
acumula entre 4 e 5 folhas pode produzir estipe aéreo e passar para o estádio 2. Nesta fase,
o comprimento do pecíolo pode diminuir conforme o estipe cresce e o tamanho das folhas
tende a ser independente do tamanho do indivíduo. Quando o buriti atinge um determinado
tamanho, pode tornar-se reprodutivo (estádio 3). O número de folhas tende a aumentar
conforme o indivíduo reprodutivo cresce, mas a atividade reprodutiva independe do
tamanho. As mudanças nas características macro-morfológicas dos indivíduos durante o
ciclo de vida do buriti podem estar relacionadas às alterações na sobrevivência e na
alometria dos indivíduos. Cada estádio ontogenético pode ter contribuição diferente para a
dinâmica das populações de buriti.
REFERÊNCIAS BIBLIOGRÁFICAS
Abreu, S. A. B. 2001. Biologia reprodutiva de Mauritia flexuosa L.f (Arecaceae) em vereda
no município de Uberlândia - MG. Dissertação de Mestrado, Departamento de
Ecologia - UFU, Uberlândia - MG.

59
Alves, L. F.; Martins, F. R. & Santos, F. A. M. 2004. Allometry of a neotropical palm,
Euterpe edulis Mart. Acta Botânica Brasílica 18(2):369-374.
Avalos, G. & Sylvester, O. 2010. Allometric estimation of total leaf area in the neotropical
palm Euterpe oleracea at La Selva, Costa Rica. Trees 24:969-974.
Barbosa, R. I.; Lima, A. D. & Junior, M. M. 2010. Biometria de frutos do buriti (Mauritia
flexuosa L.f. - Arecaceae): produção de polpa e óleo em uma área de savana em
Roraima. Amazônia: Ciência & Desenvolvimento 5(10):71-85.
Barot, S. & Gignoux, J. 1999. Population structure and life cycle of Borassus aethiopum
Mart.: evidence of early senescence in a palm tree. Biotropica 31:439-448.
Barthélémy, D. & Caraglio, Y. 2007. Plant architecture: a dynamic, multilevel and
comprehensive approach to plant form, structure and ontogeny. Annals of Botany
99:375-407.
Begon, M.; Townsend, C. R. & Harper, J. L. 2007. Ecologia de indivíduos a ecossistemas.
4° Edição. Artmed, Porto Alegre - RS. 752p.
Bernacci, L. C.; Martins, F. R. & Santos, F. A. M. 2008. Estrutura de estádios
ontogenéticos em população nativa da palmeira Syagrus romanzoffiana (Cham.)
Glassman (Arecaceae). Acta Botânica Brasílica 22(1):119-130.
Bernal, R. 1998. Demography of the vegetable ivory palm Phytelephas seemannii in
Colombia, and the impact of seed harvesting. Journal of Applied Ecology 35(1):64-
74.
Carvalho, R. M.; Martins, F. R. & Santos, F. A. M. 1999. Leaf ecology of pre-reproductive
ontogenetic stages of the palm tree Euterpe edulis Mart. (Arecaceae). Annals of
Botany 83:225-233.

60
Chazdon, R. L. 1992. Patterns of growth and reproduction of Geonoma congesta, a
clustered understory palm. Biotropica 24:43-51.
Cunningham, S. A. 1997. The effect of light environment, leaf area, and stored
carbohydrates on inflorescence production by a rain forest understory palm.
Oecologia 111:36-44.
Fernandes, N. M. P. 2002. Estratégias de produção de sementes e estabelecimento de
plântulas de Mauritia flexuosa L. f. (Arecaceae) no Vale do Acre/Brasil. Tese de
doutorado. UA/INPA, Manaus - AM.
Ferraz, D. K. 2004. Ecologia de populações de Lytocaryum hoehnei (BURRET) TOLEDO
(Arecaceae) na Reserva Florestal do Morro Grande, Cotia, SP. Tese de Doutorado.
Instituto de Biologia-UNICAMP, Campinas - SP.
Frost, I. & Rydin, H. 1997. Effects of competition, grazing and cotyledon nutrient supply
on growth of Quercus robur seedlings. Oykos 79:53-58.
Gatsuk, L. E.; Smirnova, O. V.; Vorontzova, L. I.; Zaugolnova, L. B. & Zhukova, L. A.
1980. Age states of plants of various growth forms - a review. Journal of Ecology
68:675-696.
Henderson, A.; Galeano, G. & Bernal, R. 1995. Field guide to the palms of the Americas.
New Jersey, Princeton University Press. 351p.
Kimura, M. & Simbolon, H. 2002. Allometry and life history of a forest understory palm
Pinanga coronata (Arecaceae) on Mount Halimun, West Java. Ecological Research
17:323-338.

61
Maurenza, D.; Marenco, R. A.; Parolin, P. & Piedade, M. T. F. 2012. Physiological
responses to flooding and light in two tree species native to the Amazonian
floodplains. Aquatic Botany 96:7-13.
Oyama, K. 1990. Variation in growth and reproduction in the neotropical dioecius palm
Chamaedorea tepejilote. Journal of Ecology 78:648-663.
Portela, R. C. Q. & Santos, F. A. M. 2011. Caracterização dos estádios ontogenéticos de
três espécies de palmeiras: uma proposta de padronização para estudos de dinâmica
populacional. Revista Brasileira de Botânica 34(4):523-535.
Renninger, H. J. & Phillips, N. 2010. Intrinsic and extrinsic hydraulic factors in varying
sizes of two Amazonian palm species (Iriartea deltoidea and Mauritia flexuosa)
differing in development and growing environment. American Journal of Botany
97(12):1926-1936.
Rocha, E. 2004. Potencial ecológico para o manejo de frutos de açaizeiro (Euterpe
precatoria Mart.) em áreas extrativistas no Acre, Brasil. Acta Amazônica 34(2):237-
255.
Rodríguez-Buriticá, S.; Orjuela, M. A. & Galeano, G. 2005. Demography and life history
of Geonoma orbignyana: an understory palm used as foliage in Colombia. Forest
Ecology and Management 211:329-340.
Rojas-Robles, R. & Stiles, F. G. 2009. Analysis of a supra-annual cycle: reproductive
phenology of the palm Oenocarpus bataua in a forest of the Colombian Andes.
Journal of Tropical Ecology 25:41-51.

62
Sampaio, M. B. & Scariot, A. 2008. Growth and reproduction of the understory palm
Geonoma schottiana Mart. in the gallery forest in Central Brazil. Revista Brasileira
de Botânica 31(3):433-442.
Sampaio, M. B. & Scariot, A. 2010. Effects of stochastic herbivory events on population
maintenance of an understorey palm species (Geonoma schottiana) in riparian
tropical forest. Journal of Tropical Ecology 26:151-161.
Sampaio, M. B.; Schmidt, I. B. & Figueiredo, I. B. 2008. Harvesting effects and population
ecology of the buriti palm (Mauritia flexuosa L.f., Arecaceae) in the Jalapão region,
Central Brasil. Economic Botany 62(2):171-181.
Scariot, A.; Lleras, E. & Hay, J. D. 1995. Flowering and fruiting phenologies of the palm
Acrocomia aculeata: patterns and consequences. Biotropica 27:168-173.
Silva Matos, D. M.; Freckleton, R. P. & Watkinson, A. R. 1999. The role of density
dependence in the population dynamics of a tropical palm. Ecology 80: 2635-2650.
Souza, A. F.; Martins, F. R. & Bernacci, L. C. 2003. Clonal growth and reproductive
strategies of the understory tropical palm Geonoma brevispatha: an ontogenetic
approach. Canadian Journal of Botany 81:101-112.
Souza, A. F.; Martins, F. R. & Matos, D. M. S. 2000. Detecting ontogenetics stages of the
palm Attalea humilis in fragments of the Brazilian Atlantic forest. Canadian Journal
of Botany 78:1227-1237.
Storti, E. F. 1993. Biologia floral de Mauritia flexuosa LIN. FIL, na região de Manaus,
AM, Brasil. Acta Amazônica 23(4):371-381.
Svenning, J. C. 2000. Growth strategies of clonal palms (Arecaceae) in a neotropical
rainforest, Yasuni, Ecuador. Australian Journal of Botany 48:167-178.

63
Svenning, J. C. & Macía, M. J. 2002. Harvesting of Geonoma macrostachys Mart. leaves
for thatch: an exploration of sustainability. Forest Ecology and Management 167:251-
262.
Watanabe, N. M. & Suzuki, E. 2007. Ontogenetic development in architecture and biomass
allocation of 13 rattan species in Indonesia. Journal of Plant Research 120(4):551-
561.

64
CAPÍTULO 2: Impactos do extrativismo de frutos e do fogo na ecologia populacional do buriti ( Mauritia flexuosa; Arecaceae)
Maurício Bonesso Sampaio
Flavio Antonio Maës dos Santos
Resumo
A coleta de produtos florestais não-madeireiros (PFNMs) em alta intensidade pode causar
impactos negativos na estrutura e na dinâmica da população explorada. Outras perturbações
antrópicas como as queimadas, podem aumentar ou diminuir a sustentabilidade ecológica
do extrativismo de PFNMs. Avaliamos se os impactos do extrativismo de frutos e de
queimadas praticados por comunidades rurais modificam a ecologia populacional do buriti
(Mauritia flexuosa; Arecaceae) no Brasil Central. Amostramos 16 populações de buriti,
sendo nove com histórico de coleta de frutos (extrativismo) e sete sem histórico de coleta
(controle). Em cinco populações, sendo três controle e duas sujeitas ao extrativismo,
ocorreram queimadas acidentais durante o estudo. O tamanho das populações de buriti pode
não reduzir em longo prazo caso o extrativismo de frutos cause no máximo 70% de redução
na fecundidade. Já as queimadas frequentes podem causar a mortalidade de buritis
reprodutivos e reduzir o recrutamento de plântulas, contribuindo para a diminuição no
tamanho das populações. Evitar a entrada do fogo no interior dos brejos é fundamental para
a conservação dos buritis e ao mesmo tempo, para manter uma importante fonte de renda
para as comunidades rurais do Brasil Central.
Palavras-chave: extrativismo sustentável, LTRE, reprodução plurianual, queimadas,
Cerrado

65
Introdução
Produtos florestais não madeireiros (PFNM) são utilizados por 1,4 a 1,6 bilhões de
pessoas no mundo. De 0,5 a 1 bilhão de pessoas são pequenos proprietários de terras que
manejam áreas naturais remanescentes para obter fontes de subsistência e renda
(Shackleton et al. 2011). Além de contribuir para amenizar a pobreza de comunidades
rurais (Babulo et al. 2009), o extrativismo de PFNM também pode ser uma alternativa
econômica (Byron & Arnold 1997) de menor impacto do que o corte seletivo de madeira e
a conversão das florestas em pastagens (Nepstad & Schwartzman 1992) ou monoculturas.
Porém, para ser uma alternativa viável em longo prazo, o extrativismo precisa ter
sustentabilidade econômica, ecológica e social (Arnold & Pérez 2001; Newton 2008;
Sunderland et al. 2011).
Em alguns sistemas de extrativismo de PFNM em áreas naturais pode ocorrer
sobrexploração, causando a redução no tamanho das populações manejadas e a perda de
sustentabilidade ecológica (Murali et al. 1996; Ganeshaiah et al. 1998; Bhattacharya &
Hayat 2004; Nakazono et al. 2004; Rist et al. 2010; Fedele et al. 2011). O impacto do
extrativismo em populações naturais depende da história de vida da planta, do tipo de
PFNM coletado, das práticas de manejo adotadas e de características da população como
abundância de indivíduos, atividade reprodutiva, taxa de crescimento populacional e
resiliência da população às perturbações naturais (Ticktin 2004; Ticktin & Schackleton
2011).
Para determinar a sustentabilidade do extrativismo, o primeiro passo é avaliar os
impactos da colheita do PFNM na estrutura e dinâmica das populações manejadas. Isso

66
pode ser feito experimentalmente comparando-se a dinâmica de populações sujeitas ao
extrativismo com populações controle, utilizando-se réplicas em cada tratamento, de forma
a captar a heterogeneidade ambiental (Gaoue & Ticktin 2010; Schmidt et al. 2011).
Também é importante considerar o histórico de variação das atividades de extrativismo ao
longo do tempo em uma população (Nantel et al. 1996; Ticktin et al. 2002; Gaoue et al.
2011). Nos sistemas reais de extrativismo, a intensidade de coleta pode variar
temporalmente, pois os extrativistas decidem quando, onde e quanto coletar, dependendo
das suas necessidades e do seu conhecimento a respeito do tempo de resposta da população
aos eventos de extrativismo anteriores (Gaoue et al. 2011). Efeitos compensatórios podem
minimizar o impacto do extrativismo em curto prazo, porém ao longo do tempo o efeito
acumulado da coleta de PFNM pode causar impacto na população manejada (Endress et al.
2006). Assim, ao contrário das simulações experimentais de extrativismo, resultados mais
realistas podem ser obtidos quando é feita a comparação da dinâmica de populações
controle com a dinâmica de populações sujeitas aos sistemas de extrativismo de PFNMs,
que possuam histórico de exploração.
As plantas manejadas por comunidades locais podem ser sujeitas a outras
perturbações, além do extrativismo, como fogo (Ratsirarson et al. 1996; Sinha & Brault,
2005; Schmidt et al. 2007), pastejo (Sullivan et al. 1995; Ratsirarson et al. 1996; Lopez-
Toledo et al. 2011), proliferação de parasitas (Rist et al. 2008), de lianas (Kainer et al.
2007) e a fragmentação dos hábitats (Portela et al. 2010). Estas perturbações também
podem influenciar a dinâmica das populações alvo do extrativismo e torná-las mais ou
menos tolerantes à coleta de PFNMs.

67
Os efeitos do fogo em uma população são muito dependentes da autoecologia da
espécie, da frequência, época e intensidade de fogo (Hoffmann 1999; Duchesne & Wetzel
2004). Em alguns casos a queimada aumenta a mortalidade de indivíduos de diversos
estádios do ciclo de vida, diminui o tamanho dos indivíduos e reduz a fecundidade, o que
pode causar um decréscimo na taxa de crescimento populacional (Barrow et al. 1993;
Hoffmann 1999; Liddle et al. 2006; Souza & Martins 2004). Em outros casos, a
produtividade do PFNM pode ser estimulada pelo fogo, como o que ocorre com os escapos
de capim-dourado, Syngonanthus nitens (Bong.) Ruhland, utilizados para a produção de
artesanatos no domínio do Cerrado (Schmidt et al. 2007). Para palmeiras adaptadas a
ambientes sujeitos a queimadas frequentes, como Serenoa repens (W. Bartram) Small e
Sabal etonia Swingle ex Nash na Flórida, o fogo não causa a mortalidade de indivíduos e
pode estimular a produção de folhas, a floração e a produção de frutos (Abrahamson 1999).
A palmeira buriti, Mauritia flexuosa L.f, é intensivamente utilizada pelas
comunidades rurais em todos os locais de ocorrência (Gragson 1992; Macía 2004;
Nascimento et al. 2009; Saraiva 2009; Horn et al. 2012). O buriti ocorre dos Andes, no
noroeste da América do Sul, até o litoral brasileiro no Pará e Maranhão. O limite sul da sua
distribuição é o norte do estado de São Paulo (Henderson et al. 1995). Os frutos e as folhas
desta palmeira estão entre os PFNMs mais importantes para a subsistência das comunidades
rurais e também para a cultura local (Fernandes 2011). As fibras obtidas das folhas novas
são utilizadas para costurar os escapos de capim-dourado na confecção de artesanatos nos
estados de Tocantins, Goiás e Bahia (Schmidt 2011). Os frutos são utilizados para a
produção de doces, polpa congelada e óleo, produzidos em unidades familiares ou em

68
pequenas agroindústrias em todo o domínio do Cerrado (Carrazza & Figueiredo 2010;
Sampaio 2011).
O extrativismo das folhas novas não causa impactos no crescimento dos indivíduos,
na estrutura e na dinâmica das populações de buriti (Sampaio et al. 2008; Saraiva 2009). O
sistema tradicional de extrativismo de frutos utilizado na Amazônia Peruana, que requer a
derrubada dos reprodutivos (Manzi & Coomes 2009; Horn et al. 2012), pode ser
sustentável se causar a mortalidade de no máximo 22,5% das fêmeas em intervalos de no
mínimo 20 anos (Holm et al. 2008). No Brasil, os frutos do buriti são colhidos
principalmente no chão, após a dispersão natural, ou os cachos com frutos maduros são
coletados diretamente do indivíduo reprodutivo, sem causar a sua mortalidade. A
sustentabilidade deste sistema de extrativismo de frutos pode ser maior do que o praticado
no Peru, porém, isso ainda não foi avaliado.
De forma geral, a coleta de frutos de espécies arbóreas de vida longa tende a causar
baixo impacto negativo nas populações manejadas, pois os indivíduos geralmente
produzem muitos frutos durante um longo período de tempo (Ticktin 2004), mas ainda há
pouca informação disponível, principalmente para palmeiras. De acordo com resultados já
encontrados para palmeiras, o extrativismo pode ser sustentável em uma ampla faixa de
porcentagens de frutos colhidos anualmente, que vai de 25% a 95% dos frutos produzidos
em uma população (Ratsirarson et al. 1996; Bernal 1998; Rocha 2004; Lima 2011). Para
outras espécies, como Lodoicea maldivica Pers., que possui baixa produtividade de frutos, a
intensidade da coleta praticada atualmente é maior do que a capacidade tolerada pelas
populações remanescentes nas Ilhas Seicheles, que são pequenas e sujeitas a perturbações
antrópicas (Rist et al 2010).

69
O buriti pode gerar renda e segurança alimentar para comunidades locais, e também,
é uma espécie importante para o funcionamento ecossistêmico dos brejos. Entender como o
extrativismo de frutos e o fogo influenciam a dinâmica populacional desta palmeira é
fundamental para subsidiar as ações de manejo e conservação. Portanto, este estudo teve
como objetivo avaliar o impacto do extrativismo de frutos e do fogo na ecologia
populacional do buriti. Nós testamos duas hipóteses: (i) a coleta de frutos de buriti não
influencia o recrutamento de plântulas, a estrutura e a dinâmica das populações, assim
como encontrado para algumas espécies de palmeiras (Ratsirarson et al. 1996; Bernal 1998;
Rocha 2004; Lima 2011); (ii) o fogo não causa impactos negativos na estrutura e na
dinâmica das populações de buriti, assim como encontrado para as palmeiras Serenoa
repens e Sabal etonia, que são tolerantes às queimadas (Abrahamson 1999).
Material e métodos
Espécie de estudo
O buriti (Mauritia flexuosa) é uma palmeira arborescente, com estipe solitário,
dióica, xenogâmica, cujos principais polinizadores são abelhas do gênero Trigona Jurine,
1807, coleópteros e dípteros, e não ocorre anemofilia. O desenvolvimento das
inflorescências até o amadurecimento dos frutos demora mais de um ano (Abreu 2001;
Storti 1993). O amadurecimento e a dispersão dos frutos no Brasil Central iniciam em
outubro e só terminam em julho do ano seguinte (Abreu 2001). A atividade reprodutiva é
plurianual, tanto em nível individual, quanto populacional, sendo que o número de

70
inflorescências é cerca de duas vezes maior em um ano de alta produção do que em anos de
baixa produção, mas há produção de frutos anualmente (Abreu 2001; Fernandes 2002).
Cada infrutescência produz de 479 a 1.889 frutos (Storti 1993; Silva 2009). No Brasil
Central, os frutos são ovais, de 4 a 5 cm de comprimento, com peso médio de 38,2 g (EP =
0,6 g; n=80) (M. B. Sampaio; dados não publicados). Uma infrutescência produz de 19,4 a
36 kg de frutos (Torres et al. 2006; Barbosa et al. 2010). O endosperma é muito duro e em
geral há apenas uma semente por fruto, mas pode ocorrer dispermia ou partenocarpia em
baixa frequência (Fernandes 2002). A dispersão das sementes é hidrocórica ou zoocórica
(Ponce et al. 1999; Fernandes 2002; Ponce Calderón 2002). Em campo, a germinabilidade
das sementes é de 77 a 86%. As sementes começam a germinar 65 dias após a dispersão e
permanecem viáveis por pelo menos 1 ano e 2 meses (Ponce et al. 1999).
O buriti é a espécie dominante nos brejos do Brasil Central. Os brejos são
compridos e estreitos, geralmente florestais, que se localizam ao redor de pequenos cursos
d'água nas áreas de nascente, onde o relevo é geralmente suave. O solo é hidromórfico e
permanentemente inundado (Ivanauskas et al. 1997; Toniato et al. 1998). Os brejos são
frequentemente margeados por campos limpos, que não possuem o componente arbóreo-
arbustivo e podem ter o solo permanentemente úmido ou seco dependendo da posição no
relevo.
Coleta dos dados
Nós amostramos as populações de buriti em 16 brejos (Tabela 1) de três regiões do
Brasil Central: leste de Tocantins, no Jalapão, (10°13'S; 46°34'W), nordeste do Tocantins

71
(08°51'S; 47°51'W) e sul do Piauí (09°12'S; 44°36'). Nestas três regiões, os brejos ocorrem
em abundância e existem associações ou cooperativas que coordenam as atividades de
extrativismo de PFNM. Em nove brejos havia histórico de coleta de frutos de buriti em alta
intensidade, ou seja, mais de 100 kg de polpa processada por safra e por brejo há pelo
menos três safras (ou seis anos) anteriores ao início do estudo. Nos demais sete brejos, que
foram considerados como pertencentes ao tratamento controle, não havia histórico de
intensa coleta de frutos, ou seja, não havia relatos de colheita, ou a produção era de menos
do que 100 kg de polpa por safra, e a colheita estava ocorrendo apenas para consumo da
família. Além disso, nós amostramos brejos que tinham a menor intensidade possível de
outras perturbações no seu interior, como queimadas, criações de porcos e/ou de gado.
Apesar do tempo decorrido desde a última queimada ter sido variável entre brejos, todas as
áreas estavam há pelo menos dois anos sem queimar antes do início do estudo (Tabela 1),
de acordo com as informações obtidas com os proprietários dos brejos e por observações de
campo.

72
Tabela 1. Localização dos 16 brejos amostrados, ano de ocorrência da última queimada e
uso do brejo para criação de gado, porcos e para o extrativismo de frutos de buriti
(quantidade estimada de polpa processada por safra). L = Leste, NE = Nordeste, S = Sul.
Brejo Região Última queimada Gadod Porcoe Tratamentof
Atoladeira L-Tocantins 2005a ausente ausente controle
Cansa Cavalo L-Tocantins 2006a, b ausente ausente controle
Anta L-Tocantins antes de 1998a ausente ausente controle
Porcos L-Tocantins 2000c pouco ausente coletag
São Félix L-Tocantins antes de 1998 ausente pouco coletag
Seco L-Tocantins antes de 1993 pouco ausente coletag
Cabeceira Fina NE-Tocantins 2006b ausente ausente controle
Conceição NE-Tocantins antes de 1989 pouco ausente coleta (≈400 kg)
Grande NE-Tocantins parte do brejo em 2005 pouco ausente coleta (≈200 kg)
Merola NE-Tocantins 1999 pouco ausente controle
Rebojo NE-Tocantins 1999b ausente pouco coletag
Santo Antônio NE-Tocantins 1999b ausente ausente controle
Boca do Mato S-Piauí parte do brejo em 2006 ausente pouco coleta (≈150 kg)
Estiva S-Piauí antes de 1989 ausente ausente controle
Olho d'Água S-Piauí antes de 1989 pouco pouco coleta (≈600 kg)
Saco Grande S-Piauí antes de 1989 pouco ausente coleta (≈1.000 kg) a Em junho de 2009 esses brejos foram aceirados para evitar a entrada de fogo. b Brejos que foram acidentalmente queimados na seca de 2010. c Brejo queimado em setembro de 2008 por incêndio acidental. d Pouco significa que menos de 10 cabeças de gado entram em alguns locais do brejo. e Pouco significa que os porcos são criados confinados, ao lado da área do brejo amostrada. Eventualmente alguns animais podem sair do cercado e utilizar a área amostrada. f Brejos controle foram os que não tinham histórico de coleta de frutos, ou a produção de polpa era menor do que 100 kg por safra e exclusivamente para consumo da família. g Não foi possível estimar a quantidade de polpa processada, pois os frutos são colhidos simultaneamente por várias famílias.
Amostramos as populações de buriti em parcelas de 10 x 10 m marcadas com
estacas de alumínio e georreferenciadas. Sorteamos a localização das parcelas ao longo do
comprimento e da largura do brejo de forma que a distância mínima entre parcelas fosse de

73
5 m. Não alocamos parcelas nos locais onde o brejo tinha menos que 10 m de largura e
onde não havia formação de dossel contínuo. Marcamos de 13 a 20 parcelas em cada brejo,
sendo o número de parcelas dependente do comprimento do brejo que poderia ser utilizado
para a amostragem. Medimos os indivíduos com menos de 2 m de altura em duas sub-
parcelas de 3 x 3 m estabelecidas sistematicamente nos cantos de cada parcela, sendo que
os demais indivíduos amostramos na parcela inteira. Cada indivíduo amostrado foi marcado
com uma placa de alumínio numerada. Medimos a altura do solo até o ápice do pecíolo da
folha mais alta e quando o buriti estava no estádio reprodutivo nós determinamos o sexo e
contamos o número de cachos.
Monitoramos as populações da região do Jalapão semestralmente durante três anos,
em julho de 2008, janeiro de 2009, junho de 2009, janeiro de 2010, junho de 2010, janeiro
de 2011 e julho de 2011. Nas demais regiões, Nordeste do Tocantins e Sul do Piauí,
realizamos apenas três amostragens durante dois anos, a primeira entre fevereiro e abril de
2009, a segunda entre fevereiro e março de 2010 e a última em março de 2011.
Análise dos dados
Para avaliar a dinâmica das populações, calculamos as taxas vitais anuais para cada
um dos 16 brejos e construímos matrizes de transição (A) estruturadas por estádio (Figura
1) de acordo com Morris e Doak (2002). Classificamos os buritis amostrados em quatro
estádios ontogenéticos (Capítulo 1): 1.1 ─ indivíduos sem estipe acima do solo, não-
reprodutivos e com altura inferior a 1 m; 1.2 ─ sem estipe acima do solo, não-reprodutivos

74
e com 1 m ou mais de altura; 2 ─ com estipe acima do solo e não-reprodutivos; 3 ─ com
estipe acima do solo e reprodutivos.
A=
�����s1(1-g21) 0 0 s4f4
s1g21 s2(1-g32-r12) 0 00 s2g32 s3(1-g43) 00 0 s3g43 s4 �
����
Figura 1. Ciclo de vida e matriz de transição de Lefkovitch para as populações de buriti
(Mauritia flexuosa), cujas taxas vitais dos estádios ontogenéticos (1.1, 1.2, 2 e 3) são
representadas pelas setas com linhas contínuas e a fecundidade é representada pela seta
tracejada. As taxas vitais são sobrevivência (si), crescimento (gij) e fecundidade (f4).
Calculamos as taxas vitais de sobrevivência (si), crescimento (gij) e fecundidade (f4)
de acordo com Morris e Doak (2002). A fecundidade foi calculada dividindo-se o número
de plântulas recrutadas no tempo t+1 pelo número de indivíduos reprodutivos (machos e
fêmeas) no tempo t. Calculamos a taxa de crescimento populacional assintótica (λ) para a
população amostrada em cada brejo e em cada ano utilizando-se o método power (Caswell
2001). Quando λ > 1 a população tem taxa de crescimento positiva, negativa para λ < 1 e
nula para λ = 1. Estimamos os intervalos de confiança do λ com viés corrigido, a 95% de
probabilidade, utilizando bootstrap com 10.000 aleatorizações para cada matriz A (Caswell
1.1 1.2 2 3
a11 a22 a33 a44
a21 a32 a43
a14

75
2001). Em cada aleatorização os indivíduos foram sorteados com reposição dentro de cada
estádio. Os λs foram considerados significativamente diferentes quando não houve
sobreposição dos intervalos de confiança. Em cada aleatorização nós calculamos os
elementos aij da matriz resultante (Ak) pela média das taxas vitais de indivíduos sorteados
dentro de um mesmo estádio com igual probabilidade e com reposição. As populações com
poucos buritis nos estádios 2 e 3 tiveram taxas de permanência dos indivíduos nestes
estádios iguais a 1 em muitas das aleatorizações, independentemente das demais taxas
vitais. Para minimizar este problema, sempre que ocorreu, nós substituímos as taxas vitais
dos estádios 2 e 3 destas populações pelas respectivas taxas médias do conjunto de
populações do mesmo ano independentemente do tratamento. Não houve diferenças
significativas nas taxas vitais dos indivíduos deste estádio entre os tratamentos extrativismo
e controle (resultados não mostrados).
Como a fecundidade do buriti varia entre anos, nós avaliamos o efeito da atividade
reprodutiva plurianual na dinâmica populacional do buriti. Este efeito foi incorporado nos
modelos matriciais estocásticos. Para isso, nós construímos matrizes de transição (A) para
as populações controle do Jalapão agregadas, calculamos as taxas demográficas e
montamos matrizes controle agregadas, como se todos os indivíduos dos brejos controle
pertencessem a uma única população, para cada um dos três períodos: (A1) junho de 2008 a
junho de 2009; (A2) junho de 2009 a junho de 2010; e (A3) junho de 2010 a julho de 2011.
Calculamos a elasticidade de λ às variações em cada taxa vital, sobrevivência, crescimento
e fecundidade (Caswell 2001). Para avaliar a contribuição de cada taxa vital para a
diferença do λ entre os três períodos utilizamos a análise life table response experiments
(LTRE) em desenho fixo (Caswell 2001). Com esta análise calculamos a contribuição de

76
cada taxa vital (cij) para a diferença entre as taxas de crescimento populacional de cada
período (t) utilizando a fórmula: cij = S*ij(vtij – vt+1ij); onde S* ij é a sensibilidade da matriz
média (M), calculada por: M = (At+At+1)/2, e vij são as taxas vitais de uma das três matrizes
de transição agregadas dos dois períodos de estudo, A1, A2 ou A3. As matrizes foram
comparadas duas a duas. Quando cij teve valor positivo, a taxa vital teve contribuição
positiva para a diferença no λ entre períodos, ou seja, a taxa demográfica foi maior para o
período t+1 do que t. O valor negativo de cij ocorreu quando a taxa vital foi maior para o
período t do que t+1.
Nós calculamos a taxa de crescimento populacional estocástica (rs) (Caswel 2001)
pela simulação da variação do tamanho populacional sob diferentes frequências de
ocorrência de anos bons (alta taxa de recrutamento de indivíduos do estádio 1.1) e anos
ruins (baixo recrutamento) em longo prazo para as populações controle do Jalapão. Esta
taxa foi calculada pela equação rs = log10(λs). Utilizamos 50.000 aleatorizações para
calcular cada valor de rs e calculamos o intervalo de confiança com base na distribuição dos
valores calculados. As matrizes A1 e A3 representaram anos bons, enquanto A2 representou
o ano ruim (veja os resultados).
Para avaliar o efeito do extrativismo de frutos na dinâmica populacional do buriti
em cada ano, nós comparamos o λ das populações controle agregadas com o λ das
populações sujeitas ao extrativismo de frutos agregadas. Excluímos desta análise as
populações que queimaram acidentalmente no ano da queimada e nos anos posteriores
(Tabela 1). Utilizamos a análise LTRE para calcular a contribuição de cada taxa vital para a
diferença das taxas de crescimento entre tratamentos para cada ano. Os valores de

77
contribuição (cij) positivos indicam que a taxa vital em questão foi mais alta para o controle
do que para o extrativismo e os negativos indicam o inverso.
Para estimar a taxa de crescimento populacional estocástica (rs) em diferentes
cenários de intensidade de extrativismo de frutos nós utilizamos 50.000 aleatorizações. Em
cada aleatorização nós alteramos os valores de fecundidade nas matrizes das populações
controle para simular diferentes intensidades de extrativismo de frutos. Usamos as matrizes
dos dois períodos de estudo (2009-2010 e 2010-2011) para incluir no modelo a variação
anual na atividade reprodutiva.
Para avaliar o efeito do fogo na dinâmica das populações nós comparamos a
dinâmica do brejo Porcos, que queimou acidentalmente em setembro de 2008, com a
dinâmica das populações controle do Jalapão agregadas. Também comparamos a dinâmica
das populações dos brejos que queimaram acidentalmente na estação seca (de agosto a
outubro) de 2010, Santo Antônio, Rebojo, Cabeceira Fina e Cansa Cavalo, com a dinâmica
das populações controle das três regiões agregadas. O brejo Porcos foi analisado
separadamente dos demais brejos queimados, pois os anos que ocorreram as queimadas
foram diferentes entre brejos.
Calculamos o efeito de diferentes intervalos entre queimadas (2, 3, 4, 5, 20 e 50
anos) e da ausência de queimadas (intervalo = ∞ anos) na taxa de crescimento populacional
estocástica (rs) utilizando 50.000 aleatorizações. Nesta análise nós utilizamos os dados
demográficos de cada uma das populações que queimaram para simular o ano de queimada
e as matrizes dos controles agregadas para simular os anos sem fogo. Usamos as matrizes
controle dos dois períodos de estudo separadas (2009-2010 e 2010-2011) para incluir no
modelo a variação anual na atividade reprodutiva.

78
Para simular o efeito da queimada e do extrativismo de frutos simultaneamente em
uma população nós calculamos a taxa de crescimento populacional estocástica utilizando
diferentes intervalos entre queimadas (2, 3, 4, 5, 20, 50, e ∞ anos) nas populações sujeitas
ao extrativismo. Nesta análise nós usamos os dados dos dois períodos de amostragem para
inserir no modelo a variação da fecundidade entre anos e utilizamos 50.000 aleatorizações.
Nós também simulamos o efeito da queimada em diferentes intervalos em populações
sujeitas à intensidade máxima de extrativismo sustentável encontrada, ou seja, quando há a
redução de 70% da fecundidade.
Resultados
Influência da atividade reprodutiva plurianual
A atividade reprodutiva variou durante o período de estudo e influenciou o
recrutamento de indivíduos do estádio 1.1 (Figuras 2, 3 e ANEXO 1). Em janeiro de 2009
houve alto recrutamento nas populações do Jalapão (Figura 2), que provavelmente foi
causado por um evento de produção de frutos maduros anterior ao início do estudo (entre
outubro de 2007 e fevereiro de 2008, de acordo com informações de extrativistas). Em
junho de 2008 foram encontrados poucos cachos na população. A quantidade de cachos
aumentou a partir de janeiro de 2009. Em janeiro e junho de 2010 foram encontrados
muitos cachos com frutos maduros (Figura 3). De janeiro de 2009 a junho de 2010 houve
baixo recrutamento, que coincidiu com o período de entressafra de frutos. Em janeiro de

79
2011 e julho de 2011 houve alto recrutamento novamente (Figura 2), resultado da safra de
frutos de outubro de 2009 a junho de 2010 (ANEXO 1).
Figura 2. Densidade (média ± EP) de recrutas de buriti (Mauritia flexuosa) do estádio 1.1
em seis brejos do Jalapão de janeiro de 2008 a julho de 2011.

80
Figura 3. Média (±EP) do número de cachos/100 m2, contendo frutos maduros, frutos
imaturos ou botões florais, contados semestralmente em buritis (Mauritia flexuosa) fêmeas
de seis populações do Jalapão, de junho de 2008 a junho de 2011.
A taxa de crescimento das populações controle foi significativamente maior entre
junho de 2008 e junho de 2009, quando houve alto recrutamento, do que entre junho 2009 e
junho de 2010, quando houve baixo recrutamento. Porém, não houve diferença significativa
no λ do período entre junho de 2010 e julho de 2011 (alto recrutamento) em relação aos
demais (Figura 4).
frutos maduros
botões
frutos imaturos
botões
botões
frutos maduros
frutos maduros

81
Figura 4. Taxa anual de crescimento populacional (± intervalo de confiança a 95% de
probabilidade) de populações de buriti (Mauritia flexuosa) controle, no Jalapão, para três
períodos: junho de 2008 a junho de 2009, junho de 2009 a junho de 2010 e de junho de
2010 a julho de 2011. A linha horizontal representa a taxa de crescimento na qual o
tamanho populacional se mantém constante.
A taxa vital que mais contribuiu para a diferença na dinâmica populacional entre os
três períodos foi a fecundidade (Figura 5). A contribuição da fecundidade entre o primeiro e
o segundo período foi negativa, pois esta taxa foi maior no primeiro período (f = 4 ind. do
estádio 1.1/ind. do estádio 3) do que no segundo (f = 0,34 ind. do estádio 1.1/ind. do estádio
3). A contribuição da fecundidade foi positiva para as outras duas comparações, pois houve
maior fecundidade no terceiro período (f = 11,32 ind. do estádio 1.1/ind. do estádio 3) do
que no primeiro e no segundo períodos.
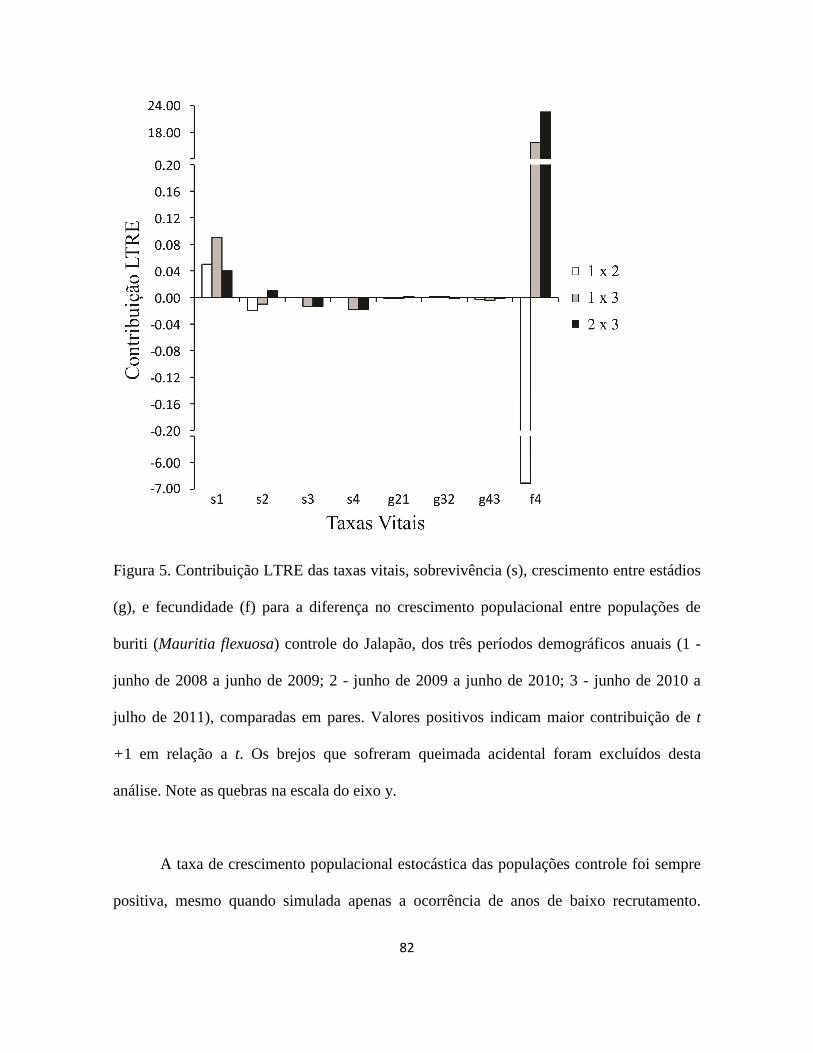
82
Figura 5. Contribuição LTRE das taxas vitais, sobrevivência (s), crescimento entre estádios
(g), e fecundidade (f) para a diferença no crescimento populacional entre populações de
buriti (Mauritia flexuosa) controle do Jalapão, dos três períodos demográficos anuais (1 -
junho de 2008 a junho de 2009; 2 - junho de 2009 a junho de 2010; 3 - junho de 2010 a
julho de 2011), comparadas em pares. Valores positivos indicam maior contribuição de t
+1 em relação a t. Os brejos que sofreram queimada acidental foram excluídos desta
análise. Note as quebras na escala do eixo y.
A taxa de crescimento populacional estocástica das populações controle foi sempre
positiva, mesmo quando simulada apenas a ocorrência de anos de baixo recrutamento.

83
Porém, a redução da frequência de anos de baixo recrutamento aumentou o valor do rs
(Figura 6).
Figura 6. Simulação da taxa de crescimento populacional estocástica (± intervalo de
confiança) influenciada por diferentes frequências de ocorrência de anos de baixo
recrutamento de indivíduos do estádio 1.1. Os dados foram coletados em populações de
buriti (Mauritia flexuosa) controle amostradas no Jalapão em três períodos, junho de 2008 a
junho de 2009 (alto recrutamento), junho de 2009 a junho de 2010 (baixo recrutamento) e
junho de 2010 a julho de 2011 (alto recrutamento). Os brejos que sofreram queimada
acidental foram excluídos desta análise no ano da queimada e posteriormente.

84
Influência da razão sexual e da produção de cachos
A estrutura variou entre populações no período de estudo (Figura 7) desde a
ausência de indivíduos no estádio 1.1, com predominância de indivíduos no estádio 1.2 no
brejo Seco, até mais de 90% dos indivíduos amostrados pertencentes ao estádio 1.1 no brejo
Grande em 2009 (Figura 7).

85
Figura 7. Porcentagem dos indivíduos nas classes 1.1, 1.2, 2 e 3 para 16 populações de
buriti (Mauritia flexuosa) em três regiões do Brasil Central em 2009, 2010 e 2011. (*)
populações controle.
2009
2010
2011

86
A razão sexual dos indivíduos reprodutivos variou significativamente de 0,38 (brejo
Grande) a 3,33 machos/fêmeas (Atoladeira) entre populações (χ2 = 90,8; p < 0,001; gl = 15;
Figura 8), mas não variou entre anos (χ2 = 2,0; p > 0,05; gl = 30).
Figura 8. Média da razão sexual dos buritis reprodutivos (estádio 3) no período de 2009 a
2011, em 16 populações de buriti (Mauritia flexuosa) de três regiões do Brasil Central. (*)
populações controle. A linha vertical indica a razão sexual 1:1.
A porcentagem de indivíduos no estádio 1.1 em janeiro de 2011 não foi
significativamente correlacionada à porcentagem de fêmeas em janeiro de 2010 (r = 0,12; p
= 0,65; n = 16) e nem à densidade de cachos maduros nas populações em janeiro de 2010 (r
= 0,40; p = 0,13; n = 16; Figuras 7 e 9).

87
Figura 9. Densidade média de cachos maduros por 100 m2 contados em janeiro de 2010 em
16 populações de buriti (Mauritia flexuosa) de três regiões do Brasil Central. (*)
populações controle.
Efeitos do extrativismo de frutos
Entre os oito brejos amostrados com menor proporção de indivíduos no estádio 1.1
e maior proporção de indivíduos nos estádios 2 e 3, seis eram alvo de extrativismo de frutos
em 2009 e 2010 (Figura 7). Em 2011, os quatro brejos com menor proporção de indivíduos
no estádio 1.1 foram sujeitos à intensa coleta de frutos (Figura 7). De forma geral, a
densidade de indivíduos no estádio 1.1 foi significativamente menor em brejos sujeitos ao
extrativismo do que nos brejos controle nos três períodos avaliados (2009: χ2 = 86,94; p <
0,001; 2010: χ2 = 144,76; p < 0,001; 2011: χ2 = 38,69; p < 0,001; Figura 10).
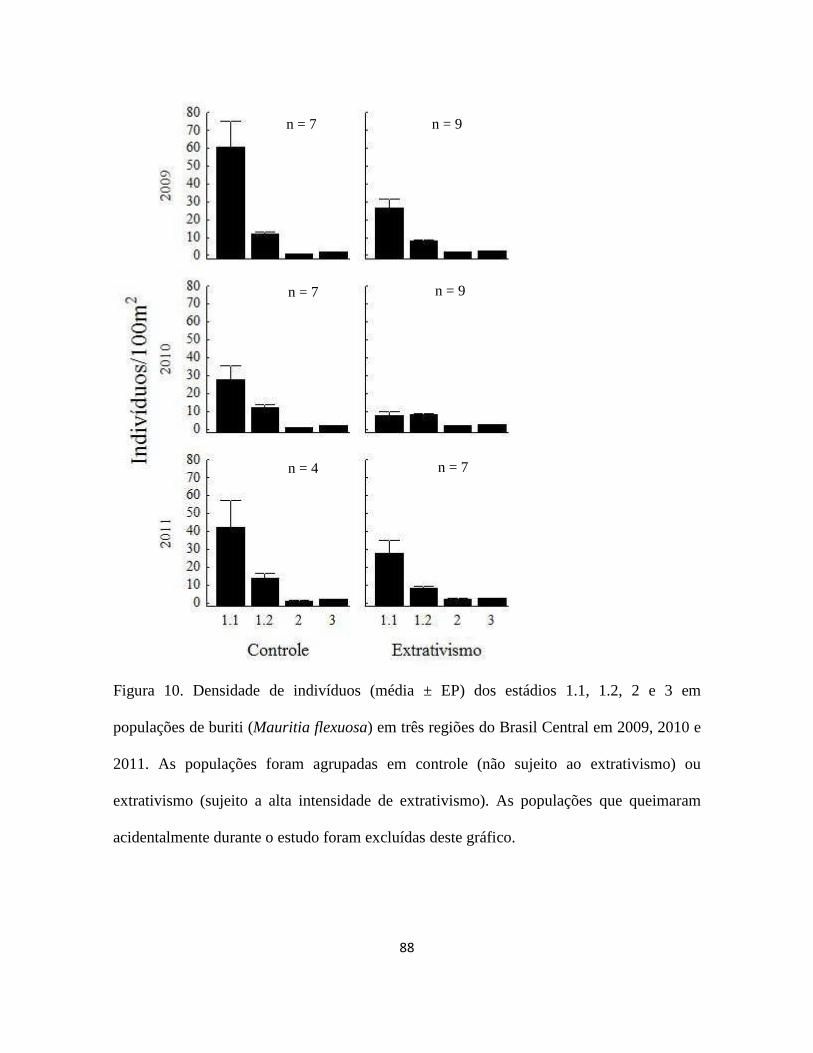
88
Figura 10. Densidade de indivíduos (média ± EP) dos estádios 1.1, 1.2, 2 e 3 em
populações de buriti (Mauritia flexuosa) em três regiões do Brasil Central em 2009, 2010 e
2011. As populações foram agrupadas em controle (não sujeito ao extrativismo) ou
extrativismo (sujeito a alta intensidade de extrativismo). As populações que queimaram
acidentalmente durante o estudo foram excluídas deste gráfico.
n = 7
n = 7
n = 4
n = 9
n = 9
n = 7
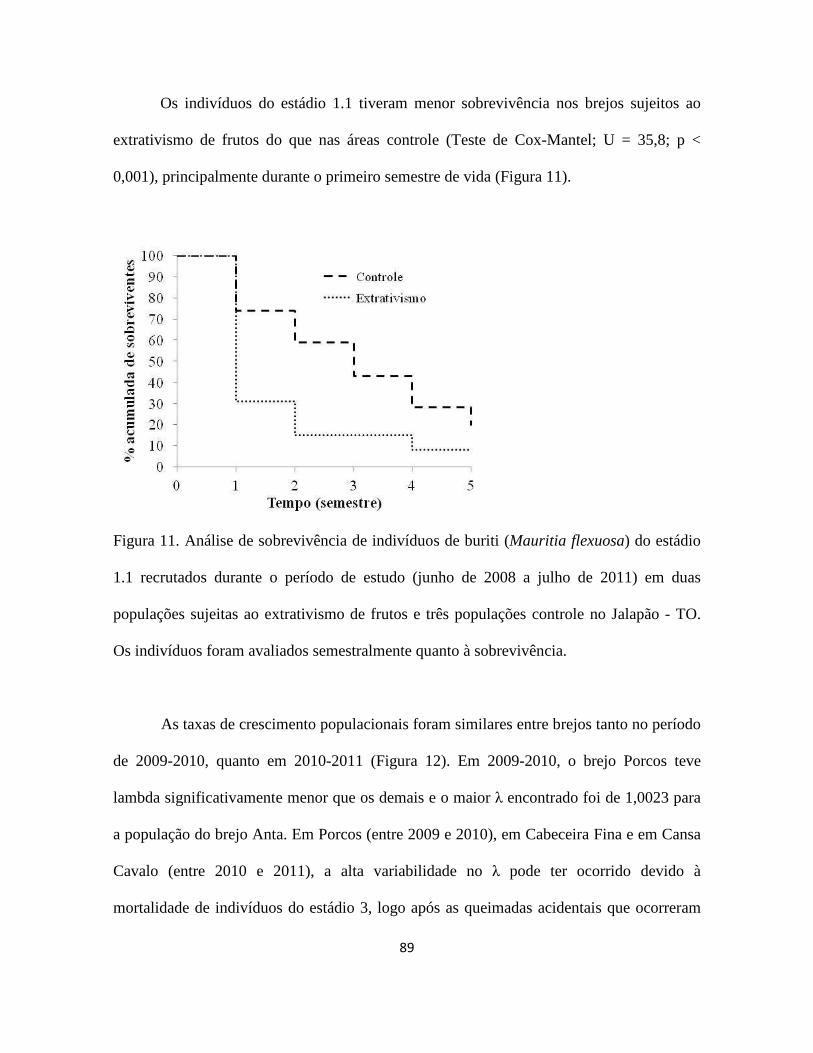
89
Os indivíduos do estádio 1.1 tiveram menor sobrevivência nos brejos sujeitos ao
extrativismo de frutos do que nas áreas controle (Teste de Cox-Mantel; U = 35,8; p <
0,001), principalmente durante o primeiro semestre de vida (Figura 11).
Figura 11. Análise de sobrevivência de indivíduos de buriti (Mauritia flexuosa) do estádio
1.1 recrutados durante o período de estudo (junho de 2008 a julho de 2011) em duas
populações sujeitas ao extrativismo de frutos e três populações controle no Jalapão - TO.
Os indivíduos foram avaliados semestralmente quanto à sobrevivência.
As taxas de crescimento populacionais foram similares entre brejos tanto no período
de 2009-2010, quanto em 2010-2011 (Figura 12). Em 2009-2010, o brejo Porcos teve
lambda significativamente menor que os demais e o maior λ encontrado foi de 1,0023 para
a população do brejo Anta. Em Porcos (entre 2009 e 2010), em Cabeceira Fina e em Cansa
Cavalo (entre 2010 e 2011), a alta variabilidade no λ pode ter ocorrido devido à
mortalidade de indivíduos do estádio 3, logo após as queimadas acidentais que ocorreram

90
nestes brejos (veja as matrizes de transição de cada brejo em cada período de amostragem
no ANEXO 2). Em São Félix, a alta variabilidade no λ entre 2010 e 2011 pode ter ocorrido
devido ao maior recrutamento de indivíduos do estádio 2 para 3 do que nas demais
populações. Nos brejos Anta e Grande, houve alta taxa de fecundidade, que pode ter
contribuído para a alta variabilidade no λ destes brejos.
Entre 2010 e 2011, o λ teve maior variabilidade entre populações (Figura 12),
porém não houve diferença significativa entre λs. O valor de λ mais baixo encontrado foi de
0,9755 para as populações dos brejos Cabeceira Fina e Cansa Cavalo, que sofreram
queimada acidental na seca de 2010. Cabeceira Fina foi a única população que teve λ
significativamente menor do que 1 no período entre 2010 e 2011. O maior λ deste período
foi de 1,0211 para o brejo São Félix, que não diferiu de 1.
O λ dos brejos agregados por tratamento diferiu significativamente entre
extrativismo e controle no período entre 2009 e 2010, mas não entre 2010 e 2011 (Figura
12).

91
Figura 12. Taxa anual de crescimento populacional (± intervalo de confiança) de nove
populações de buriti (Mauritia flexuosa) sujeitas ao extrativismo, de sete populações
controle e das populações agregadas nos tratamentos controle e extrativismo, em três
regiões do Brasil Central entre 2009 e 2011. As setas indicam as populações que sofreram
queimadas.

92
A elasticidade de λ foi maior para a sobrevivência dos indivíduos no estádio 3 (s4)
do que para as outras taxas demográficas, tanto para os brejos controle, quanto para o
extrativismo nos dois períodos de estudo (Figura 13). O λ teve baixa elasticidade à
fecundidade.
Figura 13. Elasticidade de λ a pequenas variações nas taxas vitais, sobrevivência (s),
crescimento entre estádios (g), e fecundidade (f) para os estádios 1.1 (1), 1.2 (2), 2 (3) e 3
(4) das populações de buriti (Mauritia flexuosa) controle e extrativismo em dois períodos
2009-2010 e 2010-2011. As populações que queimaram foram excluídas desta análise.
A fecundidade dos brejos sujeitos ao extrativismo (média = 0,20; EP = 0,06) foi
54% menor que nos brejos controle (média = 0,44; EP = 0,14) em 2009-2010. Em 2010-
2011, a fecundidade foi 36% menor para o extrativismo (média = 1,75; EP = 0,81) do que
para o controle (média = 2,73; EP = 1,40). O maior λ no controle do que no extrativismo no
período entre 2009 e 2010 pode ter ocorrido devido a maior sobrevivência dos indivíduos
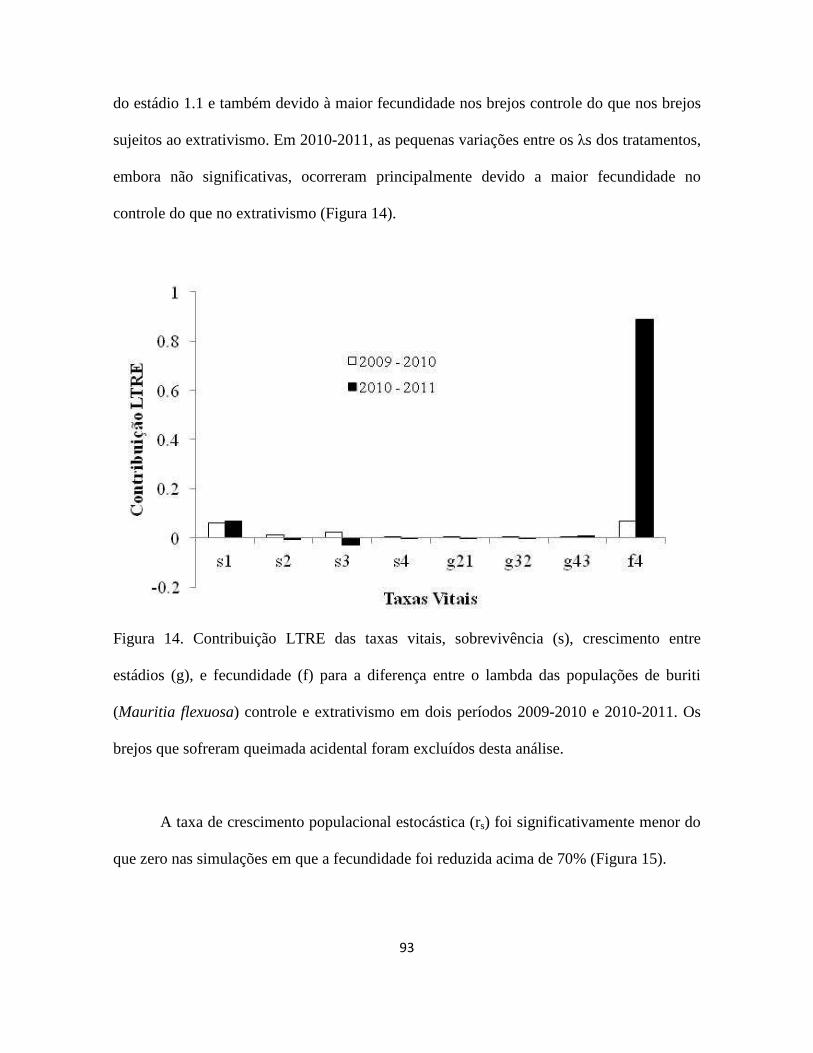
93
do estádio 1.1 e também devido à maior fecundidade nos brejos controle do que nos brejos
sujeitos ao extrativismo. Em 2010-2011, as pequenas variações entre os λs dos tratamentos,
embora não significativas, ocorreram principalmente devido a maior fecundidade no
controle do que no extrativismo (Figura 14).
Figura 14. Contribuição LTRE das taxas vitais, sobrevivência (s), crescimento entre
estádios (g), e fecundidade (f) para a diferença entre o lambda das populações de buriti
(Mauritia flexuosa) controle e extrativismo em dois períodos 2009-2010 e 2010-2011. Os
brejos que sofreram queimada acidental foram excluídos desta análise.
A taxa de crescimento populacional estocástica (rs) foi significativamente menor do
que zero nas simulações em que a fecundidade foi reduzida acima de 70% (Figura 15).
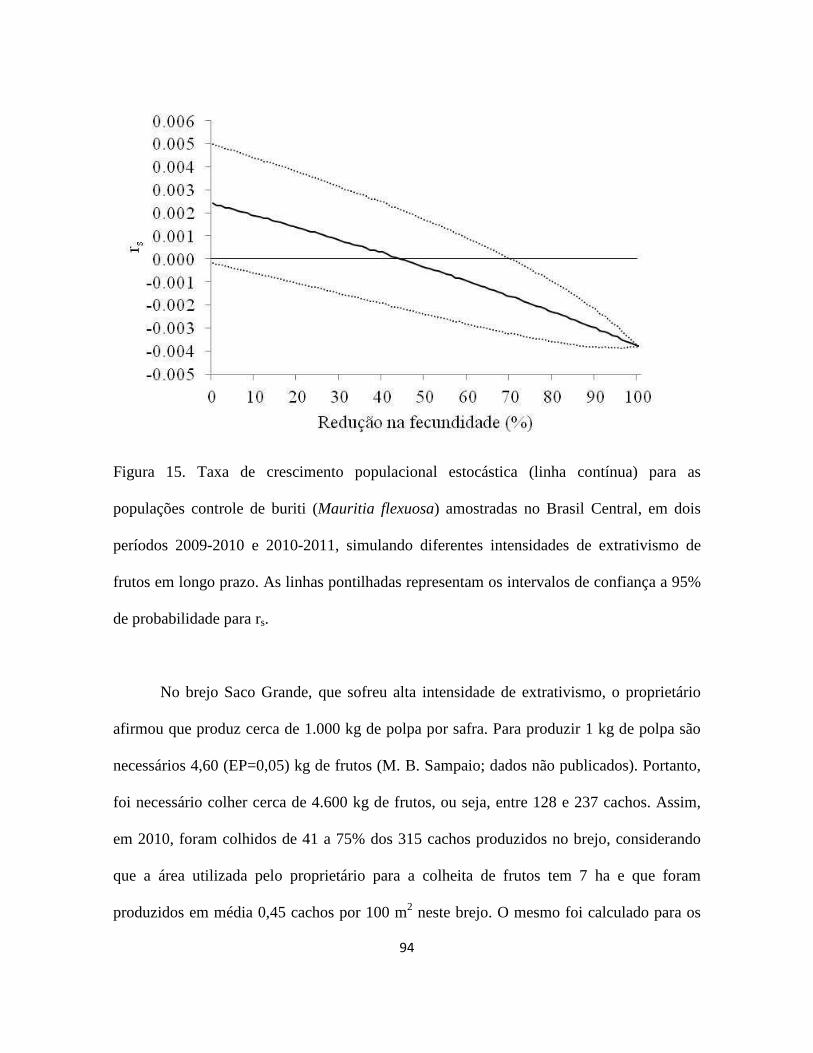
94
Figura 15. Taxa de crescimento populacional estocástica (linha contínua) para as
populações controle de buriti (Mauritia flexuosa) amostradas no Brasil Central, em dois
períodos 2009-2010 e 2010-2011, simulando diferentes intensidades de extrativismo de
frutos em longo prazo. As linhas pontilhadas representam os intervalos de confiança a 95%
de probabilidade para rs.
No brejo Saco Grande, que sofreu alta intensidade de extrativismo, o proprietário
afirmou que produz cerca de 1.000 kg de polpa por safra. Para produzir 1 kg de polpa são
necessários 4,60 (EP=0,05) kg de frutos (M. B. Sampaio; dados não publicados). Portanto,
foi necessário colher cerca de 4.600 kg de frutos, ou seja, entre 128 e 237 cachos. Assim,
em 2010, foram colhidos de 41 a 75% dos 315 cachos produzidos no brejo, considerando
que a área utilizada pelo proprietário para a colheita de frutos tem 7 ha e que foram
produzidos em média 0,45 cachos por 100 m2 neste brejo. O mesmo foi calculado para os

95
outros brejos que havia informações sobre a intensidade de coleta (Tabela 3). A coleta de
cachos variou de 0,8 a 75% dos cachos produzidos nos brejos (Tabela 3).
Tabela 3. Quantidade de polpa de buriti produzida para comercialização, porcentagem dos
cachos produzidos que foram colhidos, e produção máxima sustentável de polpa,
considerando a redução de até 70% na fecundidade para a safra de 2010 em brejos sujeitos
à colheita de frutos de buriti (Mauritia flexuosa) no Brasil Central, utilizando duas
estimativas de produção de frutos por cacho (A) 19,4 kg (Barbosa et al. 2010) e (B) 36 kg
(Torres et al. 2006).
Brejo Polpa (kg) Cachos colhidos (%) Produção máxima (kg)
A B A B
Boca do Mato 150 1,4 0,8 7.439 13.805
Conceição 400 57,7 31,1 486 901
Grande 200 2,5 1,3 5.619 10.426
Olho d'Água 600 68,7 37,0 611 1.134
Saco Grande 1.000 75,3 40,6 930 1.726
Efeito do fogo
A queimada no brejo Porcos (em setembro de 2008) causou a mortalidade de 8%
dos indivíduos do estádio 3 (n = 37) e 7% dos indivíduos do estádio 2 (n = 58). Além disso,
64% dos indivíduos do estádio 1.2 (n = 203) tiveram a folha mais alta queimada e sofreram

96
redução na altura. Entretanto, após a queimada, a taxa de produção de folhas foi
significativamente maior no brejo Porcos do que nas populações controle (Kolmogorov-
Smirnov; p < 0,001). Nas amostragens de janeiro e junho de 2011, houve intenso
recrutamento de indivíduos do estádio 1.1 no brejo Porcos. Entretanto, não houve diferença
significativa entre os λs da população de Porcos durante o período de estudo (Figura 16).
Nos quatro brejos que queimaram na seca de 2010 (Santo Antônio, Cabeceira Fina,
Cansa Cavalo e Rebojo), a queimada causou a morte de 3% dos indivíduos do estádio 3 (n
= 162) e 7% dos indivíduos do estádio 2 (n = 54). Houve duas diferentes respostas ao fogo
entre as quatro populações que queimaram na seca de 2010. Cabeceira Fina (único brejo
que teve λ significativamente menor que 1) e Cansa Cavalo sofreram maior impacto da
queimada do que os brejos Rebojo e Santo Antônio (Figura 12). Porém, a taxa de
crescimento populacional dos quatro brejos agregados que queimaram (λ = 0,9691; IC =
0,9444 - 1,0000) não diferiu significativamente do controle (λ = 0,9924; IC = 0,9849 -
1,0000), no período que houve a queimada.

97
Figura 16. Taxas de crescimento de populações (± intervalo de confiança a 95% de
probabilidade) de buriti (Mauritia flexuosa) estimadas entre junho de 2008 e junho de 2011
para os brejos controle (n = 3) e para o brejo Porcos, que queimou acidentalmente em
setembro de 2008. O brejo Cansa Cavalo queimou no final da seca de 2010 e foi excluído
do controle no período entre junho de 2010 e junho de 2011.
Entre 2008 e 2009, a fecundidade foi menor para a população de Porcos do que para
o controle, que pode ter contribuído para a diferença entre as taxas de crescimento das
populações (Figura 17). Entre 2009 e 2010, houve maior sobrevivência de indivíduos no
estádio 1 em Porcos do que no controle, o que pode ter amenizado os efeitos do fogo. Entre
2010 e 2011, houve intensa fecundidade no brejo Porcos, que contribuiu muito para
amenizar os efeitos do fogo. Para as quatro populações que queimaram em 2010, a
fecundidade, que foi menor nos brejos queimados do que no controle, foi a taxa
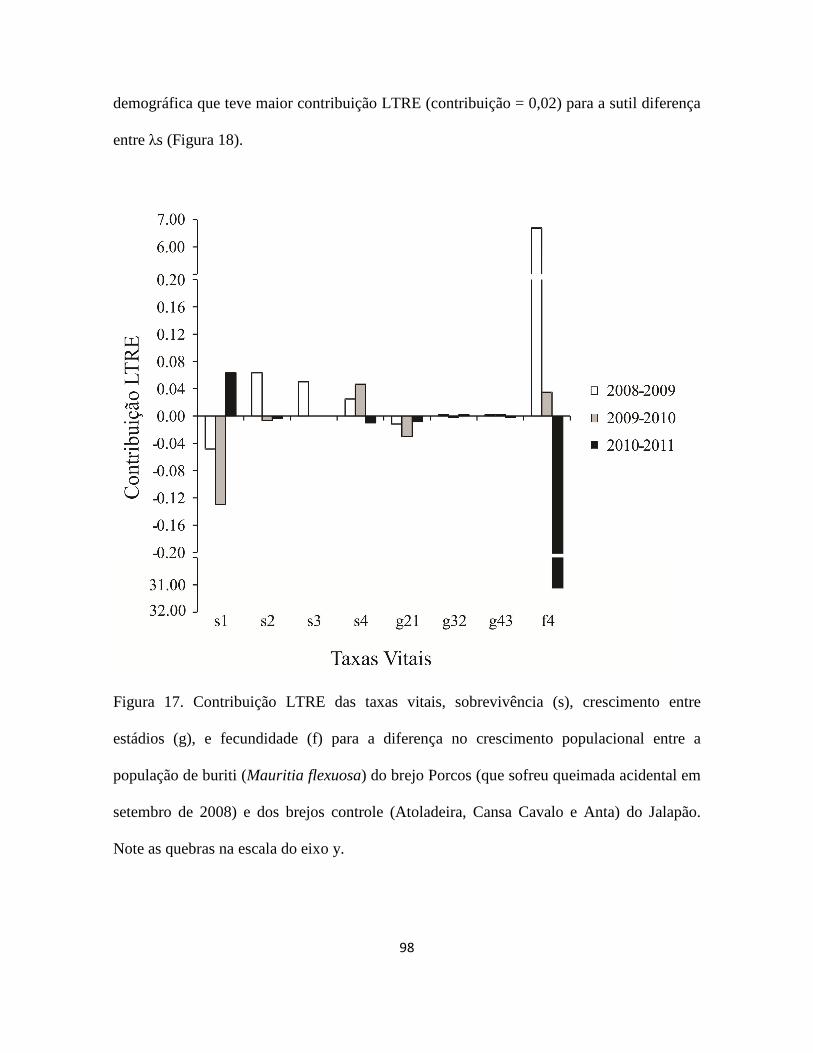
98
demográfica que teve maior contribuição LTRE (contribuição = 0,02) para a sutil diferença
entre λs (Figura 18).
Figura 17. Contribuição LTRE das taxas vitais, sobrevivência (s), crescimento entre
estádios (g), e fecundidade (f) para a diferença no crescimento populacional entre a
população de buriti (Mauritia flexuosa) do brejo Porcos (que sofreu queimada acidental em
setembro de 2008) e dos brejos controle (Atoladeira, Cansa Cavalo e Anta) do Jalapão.
Note as quebras na escala do eixo y.

99
Figura 18. Contribuição LTRE das taxas vitais, sobrevivência (s), crescimento entre
estádios (g), e fecundidade (f) para a diferença no crescimento populacional entre as
populações de buriti (Mauritia flexuosa) controle e as que queimaram na seca de 2010
(Rebojo, Santo Antônio, Cansa Cavalo e Cabeceira Fina) no período entre 2010 e 2011.
A resposta à simulação estocástica considerando diferentes probabilidades de
ocorrência do fogo foi diferente entre populações (Figura 19). Em Porcos, Santo Antônio e
Rebojo pode ocorrer estabilidade no tamanho populacional em longo prazo mesmo quando
as queimadas ocorrem a cada 3 ou 4 anos. As populações de Cansa Cavalo e Cabeceira
Fina foram as mais afetadas pelas queimadas. A população de Cabeceira Fina teve rs
significativamente menor do que zero para frequências de queimadas maiores do que uma
vez a cada 10 anos (Figura 19). Porém, em todos os brejos avaliados poderá ocorrer
redução significativa no tamanho da população caso a queimada ocorra a cada dois anos.

100
Figura 19. Taxa de crescimento populacional estocástica (± intervalo de confiança a 95%
de probabilidade) estimada a diferentes intervalos entre queimadas para cinco populações
de buriti (Mauritia flexuosa) sujeitas a queimadas durante o período de estudo.
Efeitos simultâneos do fogo e do extrativismo de frutos
O crescimento populacional não diferiu significativamente entre populações
queimadas do tratamento controle e nas populações queimadas do tratamento extrativismo,
na intensidade de coleta de frutos observada (Figura 20). Nestas duas simulações, o modelo
prediz que haverá redução significativa no tamanho populacional em longo prazo, caso
queimadas ocorram em intervalos menores do que 10 anos. Porém, na simulação de 70% de

101
redução na fecundidade, houve taxa de crescimento populacional negativa,
independentemente da frequência de queimadas.
Figura 20. Taxa de crescimento populacional estocástica (± intervalo de confiança a 95%
de probabilidade) estimada em diferentes intervalos entre queimadas para a média das
populações de buriti (Mauritia flexuosa) sem extrativismo, para populações sujeitas à
intensidade de extrativismo observada e para populações sujeitas a 70% de redução da
fecundidade.
Discussão
Atividade reprodutiva plurianual e extrativismo de frutos

102
A estrutura das populações de buriti é muito heterogênea entre brejos,
principalmente em relação à proporção de indivíduos no estádio 1.1. Parte desta variação
ocorre devido às diferenças entre populações na razão sexual dos reprodutivos e na
produtividade de cachos. Porém, as diferentes intensidades de extrativismo de frutos
praticadas nos brejos contribuem muito para a heterogeneidade da estrutura das populações,
de forma consistente entre anos. Populações sujeitas à intensa coleta tendem a apresentar
menor densidade de indivíduos dos estádios iniciais do que as não coletadas, da mesma
forma que para outras espécies sujeitas ao extrativismo de frutos (Runk 1998; Peres et al.
2003; Souza 2007; Avocèvou-Ayisso et al. 2009; Rist et al. 2010).
Para o buriti, há menor densidade de indivíduos do estádio 1.1 na maioria das áreas
sujeitas ao extrativismo do que nas áreas controle. A exceção ocorre em alguns brejos,
como em Atoladeira, que não é sujeito ao extrativismo, mas que possui baixa proporção de
indivíduos no estádio 1.1. Isso pode ter ocorrido em função da baixa proporção relativa de
fêmeas (Figura 8). Em Atoladeira ocorreu também uma forte enxurrada em janeiro de 2009,
alguns dias antes da amostragem, que pode ter causado a mortalidade de indivíduos do
estádio 1.1. Além disso, neste brejo há baixa produção de cachos com frutos (Figura 9). No
brejo Grande, onde a intensidade de coleta de frutos é alta, há uma alta proporção de
indivíduos no estádio 1.1 (Figura 7). Isso porque neste brejo a alta densidade de cachos
maduros (Figura 9) pode compensar a intensa colheita de frutos pelos extrativistas.
Além da redução no recrutamento, há também menor sobrevivência dos indivíduos
do estádio 1.1 nas populações sujeitas ao extrativismo do que no controle. A redução na
sobrevivência dos indivíduos do estádio 1.1 pode ter sido causada pelo pisoteio das plantas
durante as atividades de coleta e também, por atividades de limpeza do chão ao redor dos

103
reprodutivos, realizadas pelos extrativistas para facilitar a localização dos frutos durante a
coleta e para a abertura de trilhas de coleta no interior do brejo.
Além de variar entre populações, a estrutura também varia temporalmente,
influenciada principalmente pela produção de frutos. A frequência bienal observada para a
safra de buriti concorda com a percepção dos extrativistas e também com os resultados de
um estudo fenológico realizado no Cerrado (Abreu 2001). Os períodos de produção e
amadurecimento dos frutos na população coincidem com os eventos de recrutamento de
indivíduos do estádio 1.1, com atraso de alguns meses entre o amadurecimento e a
germinação das sementes em campo, assim como o encontrado por Ponce et al. (1999).
Apesar de influenciar bastante a estrutura das populações, a variação temporal na
atividade reprodutiva do buriti influencia a dinâmica populacional em longo prazo,
entretanto, mesmo que os anos de baixo recrutamento ocorram frequentemente, o tamanho
das populações tenderá a crescer, ainda que mais lentamente do que na frequência bienal de
eventos de recrutamento. Porém, nos anos de alto recrutamento parece haver maior
variabilidade nas taxas de crescimento das populações do que nos anos de baixo
recrutamento (Figuras 4 e 12).
A menor densidade de recrutas em algumas populações não implica
necessariamente em impactos negativos na dinâmica populacional (Virillo et al. 2011). Os
sistemas de extrativismo de frutos de buriti podem ter pouca influencia para a dinâmica das
populações, mesmo nas áreas com longo histórico de intensa colheita, onde há baixa
densidade de indivíduos do estádio 1.1.
Dois fatores contribuem para que o extrativismo de buriti tenha alto potencial de
sustentabilidade ecológica. Primeiro, há alta produtividade de cachos de buriti nos brejos.

104
Em 2010 houve em média 167 cachos/ha (EP = 36). Segundo, um sistema de extrativismo
de PNFM tem potencial de sustentabilidade quando as taxas vitais com alta elasticidade
possuem baixa contribuição LTRE para a diferença no λ entre controle e extrativismo
(Zuidema et al. 2007). Para o buriti, a elasticidade do λ é alta para a sobrevivência de
reprodutivos e baixa para a fecundidade. Portanto, apesar das altas contribuições LTRE
encontradas para a redução da fecundidade devido ao extrativismo de frutos, pode haver
pouca diferença nas taxas de crescimento populacionais entre controle e extrativismo, assim
como o esperado para espécies de vida longa (Ticktin 2004), como o buriti.
Apesar do alto potencial de sustentabilidade, se o extrativismo de frutos provocar
uma redução maior que 70% na fecundidade, o tamanho das populações de buriti poderá
diminuir em longo prazo. Algumas famílias de extrativistas conseguem produzir e
comercializar até duas toneladas de polpa de buriti colhidos de um brejo (Capítulo 3), o que
nas populações pequenas e menos produtivas, como em Saco Grande, seria suficiente para
reduzir a fecundidade em mais do que 70% (Tabela 3). Em outros brejos, para que
houvesse a redução de 70% ou mais na fecundidade, a intensidade de extrativismo de frutos
teria que ser muito maior do que a atual capacidade de processamento e comercialização da
polpa pelos produtores. Os extrativistas dos brejos Boca do Mato e Grande dificilmente
conseguirão produzir e comercializar entre 9 e 13 toneladas de polpa para que haja redução
de 70% na fecundidade destas populações de buriti (Tabela 3). Mesmo nos brejos que se
localizam próximos às comunidades rurais e que são utilizados simultaneamente por várias
famílias, como em Seco, Porcos, São Félix e Rebojo, o extrativismo parece não estar
afetando negativamente a taxa de crescimento das populações.

105
A limitação da produtividade dos extrativistas geralmente não é a disponibilidade de
frutos, mas sim a dificuldade de acesso ao mercado (Capítulo 3), pouca organização entre
produtores, falta de infraestrutura para a produção, baixa tecnologia de processamento,
baixa capacidade de armazenamento da produção, alta competição por mercados no auge
da safra e a falta de capital de giro (M. B. Sampaio; observação pessoal).
Ainda que não haja limitação ao processamento da polpa dos frutos, as populações
de buriti podem tolerar maior intensidade de extrativismo, caso as sementes sejam levadas
de volta para os brejos onde foram colhidas, após a retirada da polpa. Esta ação de manejo
pode ser bem sucedida por quatro fatores, (i) o processamento dos frutos não danifica as
sementes; (ii) a taxa de germinação das sementes de buriti em campo é acima de 77%
(Ponce et al. 1999); (iii) muitas vezes são encontrados amontoados de sementes de buriti
germinando nos quintais das casas dos extrativistas em locais inadequados ao crescimento e
estabelecimento das mudas (Sampaio 2011); e (iv) geralmente as áreas de colheita ficam
próximas às casas dos extrativistas (Sampaio et al. 2008) e portanto, não seria dispendioso
aos extrativistas carregar as sementes até o brejo. Porém nenhum dos extrativistas visitados
durante o estudo adotava esta prática de manejo.
Apesar do baixo impacto nas populações de buriti, o extrativismo de frutos não deve
afetar negativamente os processos ecossistêmicos para que esta atividade seja sustentável
(Ticktin 2004; Ticktin & Shackleton 2011). Podem ocorrer variações na composição de
espécies de aves e redução na abundância de aves e de mamíferos frugívoros em áreas
sujeitas ao extrativismo de frutos de palmeiras (Moegenburg & Levey, 2003). Portanto, o
próximo passo é avaliar os impactos da colheita de frutos para a assembleia de frugívoros
que dependem dos frutos de buriti. Bodmer (1991), Fernandes (2002), Ponce Calderón

106
(2002), Villalobos (1994) e Zona & Henderson (2005), encontraram diversas espécies de
animais que se alimentam dos frutos do buriti, incluindo aves, roedores, mamíferos de
médio porte e répteis, incluindo espécies ameaçadas de extinção, que podem ser afetadas
negativamente pela coleta de frutos, como anta - Tapirus terrestris (Linnaeus, 1758),
veado-mateiro - Mazama americana (Erxleben, 1777), veado-catingueiro - M. gouazoubira
(Fisher, 1814), cateto - Pecari tajacu (Linnaeus, 1758), queixada - Tayassu pecari (Link,
1795), lobo-guará - Chrysocyon brachyurus (Illiger, 1815), jabutis - Geochelone
carbonaria (Spix, 1824) e G. denticulata (Linnaeus, 1766) e arara-azul - Anodorhynchus
hyacinthinus (Latham, 1790). Além dessas espécies, o psitacídeo Orthopsittaca manilata
(Boddaert, 1783) alimenta-se somente dos frutos maduros de buriti ainda no cacho (Silva
2009).
A coleta dos frutos do chão pode ter maior potencial de sustentabilidade do que o
corte dos cachos com frutos maduros, pois os animais que usam as sementes antes da
dispersão (como as aves e os macacos) não são afetados, enquanto que os animais que
comem os frutos no chão podem utilizar parte dos frutos caídos antes de serem colhidos
pelos extrativistas. A manutenção de populações viáveis destes frugívoros é importante
para a dinâmica, estrutura espacial e variabilidade genética das populações de buriti, pois
vários deles atuam como dispersores das sementes (Bodmer 1991; Villalobos 1994;
Fernandes 2002; Ponce Calderón 2002).
Efeitos das queimadas

107
Os efeitos causados pelo extrativismo de frutos na dinâmica das populações de
buriti podem ser muito menores do que o efeito das queimadas. O grau de perturbação
causado pelo fogo varia principalmente em função da frequência de ocorrência e da
intensidade da queimada (Duchesne & Wetzel 2004; Sinha & Brault 2005). A intensidade
do fogo é diretamente relacionada à altura que a chama alcança (Alexander 1982). Assim,
quando o fogo no interior dos brejos é intenso, as labaredas podem alcançar as folhas secas
que permanecem durante muito tempo penduradas no estipe dos buritis dos estádios 2 e 3.
Conforme estas folhas queimam, o fogo sobe nos buritis e queima as estruturas
reprodutivas, as folhas verdes e o meristema apical, causando a morte do indivíduo. Em
outros casos, o fogo pode ser mais brando, com chamas rente ao solo, que tende a causar
menor mortalidade dos indivíduos reprodutivos (M. B. Sampaio; observação pessoal).
Portanto, as queimadas intensas podem causar maior influência no λ das populações de
buriti do que as brandas.
As populações de buriti queimadas podem permanecer por três anos tendo sua
dinâmica influenciada pelo fogo, como o que ocorreu para a população do brejo Porcos. As
variações na fecundidade, que provavelmente ocorrem devido às queimadas, podem ser os
principais fatores que influenciam o crescimento populacional após o fogo, enquanto que a
mortalidade de reprodutivos pode ter pouca contribuição. Em geral, o fogo pode causar
pouca mortalidade de buritis sem estipe aéreo, pois o meristema apical destes geralmente
está protegido do fogo debaixo da água do brejo. Porém, a queimada pode reduzir o
tamanho destes indivíduos, atrasando o seu crescimento. A baixa mortalidade de indivíduos
dos estádios mais jovens, devido ao meristema subterrâneo e o aumento na taxa de
produção de folhas após a queimada, são fatores que contribuem para a resistência das

108
populações de buriti ao fogo. As mesmas estratégias ocorrem nas palmeiras Attalea humilis
Mart. (Souza & Martins 2004), Sabal palmmeto (Walter) Lodd. ex Schult. & Schult. f.
(McPherson & Williams 1998), Serenoa repens e Sabal etonia (Abrahamson 1999). As
duas últimas espécies ainda apresentam aumento da floração após a queimada
(Abrahamson 1999), o que ameniza os efeitos do fogo nas populações.
A resistência do buriti ao fogo tem sido considerada como um fator importante para
a dominância desta espécie nos brejos da América do Sul. Esta dominância iniciou-se há
2.000 anos, quando as queimadas passaram a ser mais frequentes no Holoceno (Montoya et
al. 2011). Apesar da resistência do buriti ao fogo, o tamanho das populações pode reduzir
em longo prazo caso ocorram queimadas em alta intensidade e em intervalos menores do
que 10 anos. Este intervalo entre queimadas é semelhante ao tolerado (7 a 9 anos) por
populações de Phyllanthus emblica L. (Phyllanthaceae; Sinha & Brault 2005). Mesmo as
queimadas de baixa intensidade podem causar a redução no tamanho das populações de
buriti se ocorrerem a cada dois ou três anos. Nas três regiões de estudo, as comunidades
locais utilizam o fogo principalmente para renovar as pastagens que ficam ao redor dos
brejos (Capítulo 3) e muitas vezes, a queimada não é controlada adequadamente e acaba
atingindo os brejos. Na região do Jalapão, queimadas em larga escala tendem a ser mais
frequentes do que nas outras duas áreas, pois além de ser usado para renovar as pastagens, o
fogo é utilizado para estimular a floração do capim-dourado. Além disso, no Jalapão há
baixa densidade populacional e grandes áreas contínuas de cerrado conservado, o que
contribui para a propagação das queimadas. Por isso, nesta região, é comum o fogo entrar
nos brejos a cada dois anos (Schmidt et al. 2007; Schmidt 2011). Caso as queimadas
ocorram na frequência bienal em longo prazo, as populações de buriti poderão sofrer rápida

109
redução no seu tamanho. O modelo utilizado nesta análise não considerou o efeito
acumulado de queimadas sucessivas. Portanto, o impacto das queimadas pode não ter sido
adequadamente avaliado, principalmente nas frequências mais altas de fogo. A capacitação
de pessoas das comunidades rurais e o fornecimento de equipamentos adequados ao
combate a incêndios podem ajudar a reduzir a frequência de queimadas nos brejos e
melhorar o controle do uso do fogo para a renovação das pastagens.
Conclusões
A sustentabilidade dos sistemas de extrativismo deve ser avaliada levando-se em
conta outros fatores que possam causar perturbações às populações sujeitas à coleta de
PFNM (Ratsirarson et al. 1996; Sinha & Brault 2005; Schmidt et al. 2007). Para as
populações de buriti, o fogo tem potencial de causar mais impactos negativos do que o
extrativismo de frutos. Isto porque as queimadas podem aumentar a mortalidade de buritis
reprodutivos e reduzir o tamanho dos indivíduos com estipe subterrâneo. As populações de
buriti podem não ser viáveis se ocorrerem queimadas a cada dois anos, que é a frequência
comumente empregada pelas comunidades rurais no Brasil Central. Portanto, esta fonte de
perturbação antrópica precisa ser considerada na análise da sustentabilidade de um sistema
de extrativismo de frutos, sujeito ao fogo, para melhorar a precisão das estimativas.
A intensidade de coleta de frutos de buriti atualmente praticada no Brasil Central
pode ser sustentável, desde que o fogo ocorra em intervalos de mais do que 10 anos. Já o

110
histórico das atividades de coleta de frutos de buriti pode ter pouca influência na
sustentabilidade da atividade, ao contrário do encontrado em outros sistemas de
extrativismo de PFNMs (Nantel et al. 1996; Ticktin et al. 2002; Gaoue et al. 2011). O
potencial de sustentabilidade da coleta de frutos de buriti será ainda maior caso a simples
ação de retornar as sementes para o brejo após a remoção da polpa dos frutos seja adotada
pelos extrativistas. Assim, a coleta de frutos de buriti poderá continuar a ser uma
importante fonte de renda para inúmeras famílias de comunidades rurais, ao mesmo tempo
em que promove a conservação dos brejos no Brasil Central.
REFERÊNCIAS BIBLIOGRÁFICAS
Abreu, S. A. B. 2001. Biologia reprodutiva de Mauritia flexuosa L.f. (Arecaceae) em
vereda no município de Uberlândia - MG. Dissertação de Mestrado, Departamento de
Ecologia - UFU, Uberlândia - MG.
Abrahamson, W. G. 1999. Episodic reproduction in two fire-prone palms, Serenoa repens
and Sabal etonia (Palmae). Ecology 80(1):100-115.
Alexander, M. E. 1982. Calculating and interpreting forest fire intensities. Canadian Journal
of Botany 60:349-357.
Arnold, J. E. M. & Pérez, M. R. 2001. Can non-timber forest products match tropical forest
conservation and development objectives? Ecological Economics 39:437-447.
Avocèvou-Ayisso, C.; Sinsin, B.; Adégbidi, A; Dossou, G. & Van Damme, P. 2009.
Sustainable use of non-timber forest products: impact of fruit harvesting on

111
Pentadesma butyracea regeneration and financial analysis of its products trade in
Benin. Forest Ecology and Management 257:1930-1938.
Babulo, B.; Muys, B.; Nega, F.; Tollens, E.; Nyssen, J.; Deckers, J. & Mathijs, E. 2009.
The economic contribution of forest resource use to rural livelihoods in Tigray,
Northern Ethiopia. Forest Policy and Economics 11(2):109-117.
Barbosa, R. I.; Lima, A. D. & Junior, M. M. 2010. Biometria de frutos do buriti (Mauritia
flexuosa L.f. - Arecaceae): produção de polpa e óleo em uma área de savana em
Roraima. Amazônia: Ciência & Desenvolvimento 5(10):71-85.
Barrow, P.; Duff, G.; Liddle, D. & Russell-Smith, J. 1993. Threats to monsoon rainforest
habitat in northern Australia: the case of Ptychosperma bleeseri Burret (Arecaceae).
Australian Journal of Ecology 18:463-471.
Bernal, R. 1998. Demography of the vegetable ivory palm Phytelephas seemannii in
Colombia, and the impact of seed harvesting. Journal of Applied Ecology 35(1): 64-
74.
Bhattacharya, P. & Hayat, S. F. 2004. Sustainable NTFP management for rural
development: a case from Madhya Pradesh, India. International Forestry Review
6(2):161-168.
Bodmer, R. E. 1991. Strategies of seed dispersal and seed predation in Amazonian
ungulates. Biotropica 23(3):255-261.
Byron, N. & Arnold, M. 1997. What futures for the people of the tropical forests? Working
Paper, num. 19, CIFOR, Bongor, Indonesia.

112
Carrazza, L. & Figueiredo, I. 2010. Cerrado que te quero vivo! - Produtos e meios de vida
sustentáveis apoiados pelo Programa de Pequenos Projetos Ecossociais (PPP-ECOS).
Instituto Sociedade População e Natureza, Brasília - DF.
Caswell, H. 2001. Matrix population models: construction, analysis and interpretation.
Sunderland, MA, Sinauer Associates, Inc. 722p.
Duchesne, L. C. & Wetzel, S. 2004. Effect of fire intensity and depth of burn on Lowbush
Blueberry, Vaccinium angustifolium, and Velvet Leaf Blueberry, Vaccinium
myrtilloides, production in Eastern Ontario. The Canadian Field-Naturalist 118:195-
200.
Endress, B. A.; Gorchov, D. L. & Berry, E. J. 2006. Sustainability of a non-timber forest
product: effects of alternative leaf harvest practices over six years on yield and
demography of the palm Chamaedorea radicalis. Forest Ecology and Management
234: 181-191.
Fedele, G.; Urech, Z. L.; Rehnus, M. & Sorg, J. P. 2011. Impact of women's harvest
practices on Pandanus guillaumetii in Madagascar's Lowland Rainforests. Economic
Botany 65(2):158-168.
Fernandes, N. M. P. 2002. Estratégias de produção de sementes e estabelecimento de
plântulas de Mauritia flexuosa L. f. (Arecaceae) no Vale do Acre/Brasil. Tese de
doutorado. UA/INPA, Manaus - AM.
Fernandes, M. R. 2011. "A tree with much authority": the place of the buriti palm (Mauritia
flexuosa L.f.) in the sertaneja culture of Terra Ronca, Goiás state, Central Brazil.
Bioremediation, Biodiversity and Bioavailability 5:80-91.

113
Ganeshaiah, K. N.; Shaanker, R. U.; Murali, K. S.; Shankar, U. & Bawa, K. S. 1998.
Extraction of non-timber forest products in the forest of Bilingiri Rangan Hills, India.
5. Influence of dispersal mode on species response to anthropogenic pressures.
Economic Botany 52:316-319.
Gaoue, O. G.; Horvitz, C. C. & Ticktin, T. 2011. Non-timber forest product harvest in
variable environments: modeling the effect of harvesting as a stochastic sequence.
Ecological Applications 21(5):1604-1616.
Gaoue, O. G. & Ticktin, T. 2010. Effects of harvest of nontimber forest products and
ecological differences between sites on the demography of African Mahogany.
Conservation Biology 24(2):605-614.
Gragson, T. L. 1992. The use of palms by the Pume indians of Southwestern Venezuela.
Principes 36(3):133-142.
Henderson, A.; Galeano, G. & Bernal, R. 1995. Field guide to the palms of the Americas.
New Jersey, Princeton University Press. 351p.
Hoffmann, W. A. 1999. Fire and population dynamics of woody plants in a Neotropical
Savanna: matrix model projections. Ecology 80(4):1354-1369.
Holm, J. A.; Miller, C. J. & Cropper, W. P. 2008. Population dynamics of the dioecious
Amazonian palm Mauritia flexuosa: simulation analysis of sustainable harvesting.
Biotropica 40:550-558.
Horn, C. M.; Gilmore, M. P. & Endress, B. A. (2012). Ecological and socio-economic
factors influencing aguaje (Mauritia flexuosa) resource management in two
indigenous communities in the Peruvian Amazon. Forest Ecology and Management
267:93-103.

114
Ivanauskas, N. M.; Rodrigues, R. R. & Nave, A. G. 1997. Aspectos ecológicos de um
trecho de floresta de brejo em Itatinga, SP: florística, fitossociologia e seletividade de
espécies. Revista Brasileira de Botânica 20(2):139-153.
Kainer, K. A.; Wadt, L. H. O. & Staudhammer, C. L. 2007. Explaining variation in Brazil
nut fruit production. Forest Ecology and Management 250:244-255.
Liddle, D. T.; Brook, B. W.; Matthews,J.; Taylor, S. M. & Caley, P. 2006. Threat and
response: a decade of decline in a regionally endangered rainforest palm affected by
fire and introduced animals. Biological Conservation 132:326-375.
Lima, V. V. F. 2011. Estrutura e dinâmica de populações de coquinho-azedo (Butia
capitata (Mart.) Beccari; Arecaceae) em áreas de extrativismo no norte de Minas
Gerais, Brasil. Dissertação de Mestrado, Departamento de Engenharia Florestal, UnB,
Brasília - DF.
Lopez-Toledo, L.; Horn, C. & Endress, B. A. 2011. Distribution and population patterns of
the threatened palm Brahea aculeata in a tropical dry forest in Sonora, Mexico.
Forest Ecology and Management 261(11):1901-1910.
Macía, M. J. 2004. Multiplicity in palm uses by the Huaorani of Amazonian Ecuador.
Botanical Journal of the Linnean Society 144:149-159.
Manzi, M. & Coomes, O. T. 2009. Managing Amazonian palms for community use: a case
of aguaje palm (Mauritia flexuosa) in Peru. Forest Ecology and Management
257:510-517.
McPherson, K. & Williams, K. 1998. The role of carbohydrate reserves in the growth,
resilience, and persistence of cabbage palm seedlings (Sabal palmetto). Oecologia
117:460-468.

115
Moegenburg, S. M. & Levey, D. J. 2003. Do frugivores respond to fruit harvest? An
experimental study of short-term responses. Ecology 84(10):2600-2612.
Montoya, E.; Rull, V.; Stansell, N. D.; Abbott, C. M. B.; Nogué, S.; Bird, B. W. & Díaz,
W. A. 2011. Forest-savanna-morichal dynamics in relation to fire and human
occupation in the southern Gran Sabana (SE Venezuela) during the last millennia.
Quaternary Research 76:335-344.
Morris, W. F. & Doak, D. F. 2002. Quantitative conservation biology: theory and practice
of population viability analysis. Sinauer Associates, Inc., Sunderland, Massachussets,
USA.
Murali, K. S.; Shankar, U.; Shaanker, R.; Ganeshaiah, K. N. & Bawa, K. S. 1996.
Extraction of non-timber forest products in the forests of Biligiri Rangan Hills, India.
2. Impact of NTFP extraction on regeneration, population structure, and species
composition. Economic Botany 50:252-269.
Nakazono, E. M.; Bruna, E. M. & Mesquita, R. C. G. 2004. Experimental harvesting of the
non-timber forest product Ischnosiphon polyphyllus in central Amazonia. Forest
Ecology and Management 190:219-225.
Nantel, P.; Gagnon, D. & Nault, A. 1996. Population viability analysis of American
Ginseng and Wild Leek harvested in stochastic environments. Conservation Biology
10(2):608-621.
Nascimento, A. R. T.; Santos, A. A.; Martins, R. C.; & Dias, T. A. B. 2009. Palm
communities in the Kraho indigenous territory, Tocantins, Brazil: biodiversity and
ethnobotanical features. Interciencia 34:182-188.

116
Nepstad, D. C. & Schwartzman, S. 1992. Non-timber products from tropical forests:
evaluation of a conservation and development strategy. New York Botanical Garden,
New York.
Newton, A. C. 2008. Conservation of tree species through sustainable use: how can it be
achieved in practice? Oryx 42(2):195-205.
Peres, C. A.; Baider, C; Zuidema, P. A.; Wadt, L. H. O.; Kainer, K. A.; Gomes-Silva, D. A.
P.; Salomão, R. P.; Simões, L. L.; Franciosi, E. R. N.; Valverde, F. C.; Gribel, R.;
Shepard Jr., G. H.; Kanashiro, M.; Coventry, P.; Yu, D. W.; Watkinson, A. R. &
Freckleton, R. P. 2003. Demographic threats to the sustainability of Brazil nut
exploitation. Science 302:2112-2114.
Ponce, M. E.; Brandin, J.; Ponce, M. A. & Gonzales, V. 1999. Germinacion y
establecimiento de plantulas de Mauritia fluxuosa L.f. (Arecaceae) en los Llanos Sur-
Orientales del estado Guarico, Venezuela. Acta Botanica Venezuelica 22(1):167-183.
Ponce Calderón, M. E. 2002. Patrones de caída de frutos en Mauritia flexuosa L.f. y fauna
involucrada en los procesos de remoción de semillas. Acta Botanica Venezuelica
25(2):119-142.
Portela, R. C. Q.; Bruna, E. M. & Santos, F. A. M. 2010. Demography of palm species in
Brazil's Atlantic forest: a comparison of harvested and unharvested species using
matrix models. Biodiversity and Conservation 19:2389-2403.
Ratsirarson, J.; Silander, J. A. & Richard, A. F. 1996. Conservation and management of a
threatened Madagascar palm species, Neodypsis decaryi, Jumelle. Conservation
Biology 10(1):40-52.

117
Rist, L.; Kaiser-Bunbury, C. N.; Fleischer-Dogleyd, F.; Edwards, P.; Bunbury, N. &
Ghazoul, J. 2010. Sustainable harvesting of coco de mer, Lodoicea maldivica, in the
Vallée de Mai, Seychelles. Forest Ecology and Management 260:2224-2231.
Rist, L.; Uma Shaanker, R.; Milner-Gulland, E. J. & Ghazoul, J. 2008. Managing
mistletoes: the value of local practices for a non-timber forest resource. Forest
Ecology and Management 255:1684-1691.
Rocha, E. 2004. Potencial ecológico para o manejo de frutos de açaizeiro (Euterpe
precatoria Mart.) em áreas extrativistas no Acre, Brasil. Acta Amazônica 34(2): 237-
255.
Runk, J. V. 1998. Productivity and sustainability of a vegetable ivory palm (Phytelephas
aequatorialis, Arecaceae) under three management regimes in Northwestern Ecuador.
Economic Botany 52(2):168-182.
Sampaio, M. B. 2011. Boas práticas de manejo para o extrativismo sustentável do buriti.
Instituto Sociedade População e Natureza, Brasília - DF, 80 p.
Sampaio, M. B.; Schmidt, I. B. & Figueiredo, I. B. 2008. Harvesting effects and population
ecology of the buriti palm (Mauritia flexuosa L. f., Arecaceae) in the Jalapão region,
Central Brazil. Economic Botany 62:171-181.
Saraiva, N. A. 2009. Manejo sustentável e potencial econômico da extração do buriti nos
Lençois Maranhenses, Brasil. Dissertação de Mestrado, CDS-UnB, Brasília - DF.
Schmidt, I. B. 2011. Effects of local ecological knowledge, harvesting and fire on golden-
grass (Syngonanthus nitens, Eriocaulaceae), a non-timber forest product (NTFP)
species from the Brazilian savanna. PhD Thesis, Botany department, University of
Hawai`i at Manoa, Honolulu.

118
Schmidt, I. B.; Figueiredo, I. B. & Scariot, A. 2007. Ethnobotany and effects of harvesting
on the population ecology of Syngonanthus nitens (Bong.) Ruhland (Eriocaulaceae), a
NTFP from Jalapão region, Central Brazil. Economic Botany 61(1):73-85.
Schmidt, I. B.; Mandle, L.; Ticktin, T. & Gaoue, O. G. 2011. What do matrix population
models reveal about the sustainability of non-timber forest product harvest? Journal
of Applied Ecology 48:815-826.
Shackleton, S.; Delang, C. O. & Angelsen, A. 2011 From subsistence to safety nets and
cash income: exploring the diverse values of non-timber forest products for
livelihoods and poverty alleviation. In: Shackleton, S; Shackleton, C.; Shanley, P.
(ed.) Non-timber forest products in the global context. Springer-Verlag, Berlin.
Silva, P. A. 2009. Orthopsittaca manilata (Boddaert, 1783) (Aves: Psittacidae): abundância
e atividade alimentar em relação à frutificação de Mauritia flexuosa L.f (Arecaceae)
numa vereda no Triângulo Mineiro. Dissertação de Mestrado, Universidade Federal
de Uberlândia, Uberlândia - MG.
Sinha, A. & Brault, S. 2005. Assessing sustainability of nontimber forest product
extractions: how fire affects sustainability. Biodiversity and Conservation 14:3537-
3563.
Souza, A. F. 2007. Ecological interpretation of multiple population size structures in trees:
the case of Araucaria angustifolia in South America. Austral Ecology 32:524-533.
Souza, A. F. & Martins, F. R. 2004. Population structure and dynamics of a neotropical
palm in fire-impacted fragments of the Brazilian Atlantic Forest. Biodiversity and
Conservation 13:1611-1632.

119
Storti, E. F. 1993. Biologia floral de Mauritia flexuosa Lin. Fil, na região de Manaus, AM,
Brasil. Acta Amazônica 23:371-381.
Sullivan, S.; Konstant, T. L. & Cunningham, A. B. 1995. The impact of utilization of palm
products on the population structure of the vegetable ivory palm (Hyphaene
petersiana, Arecaceae) in North-Central Namibia. Economic Botany 49(4):357-370.
Sunderland, T. C. H; Ndoye, O. & Harrison-Sanchez, S. 2011. Non-timber forest products
and conservation: what prospects? In: Shackleton, S; Shackleton, C.; Shanley, P. (ed.)
Non-timber forest products in the global context. Springer-Verlag, Berlin.
Ticktin, T. 2004. The ecological implications of harvesting non-timber forest products.
Journal of Applied Ecology 41:11-21.
Ticktin, T.; Nantel, P.; Ramirez, F. & Johns, T. 2002. Effects of variation on harvest limits
for nontimber forest species in Mexico. Conservation Biology 16(3):691-705.
Ticktin, T. & Shackleton, C. 2011. Harvesting non-timber forest products sustainably:
opportunities and challenges. In: Shackleton, S; Shackleton, C.; Shanley, P. (ed.)
Non-timber forest products in the global context. Springer-Verlag, Berlin.
Toniato, M. T. Z.; Leitão-Filho, H. F. & Rodrigues, R. R. 1998. Fitossociologia de um
remanescente de floresta higrófila (mata de brejo) em Campinas, SP. Revista
Brasileira de Botânica 21(2):197-210.
Torres, D. C.; Acevedo, E. O. & Alvarado, L. F. 2006. Aguaje: la maravillosa palmera de la
Amazonía. Instituto de Investigaciones de la Amazonía Peruana.
Villalobos, M. P. 1994. Guilda de frugivoros associada com o buriti (Mauritia flexuosa:
Palmae) numa vereda no Brasil Central. Dissertação de Mestrado, Departamento de
Ecologia - UnB, Brasília - DF.

120
Virillo, C. B.; Martins, F. R.; Tamashiro, J. Y. & Santos, F. A. M. 2011. Is size structure a
good measure of future trends of plant populations? An empirical approach using five
woody species from the Cerrado (Brazilian savanna). Acta Botanica Brasilica
25(3):593-600.
Zona, S. & Henderson, A. 2005. Additions to "A review of animal-mediated seed dispersal
of palms". http://www.virtualherbarium.org/palms/psdispersal.html
Data de acesso: dezembro de 2011.
Zuidema, P. A.; De Kroon, H. & Werger, M. J. A. 2007. Testing sustainability by
prospective and retrospective demographic analyses: evaluation for palm leaf harvest.
Ecological Applications 17(1):118-128.

121
ANEXO 1. Número de parcelas amostradas (Parc.) e número de indivíduos amostrados de
cada estádio (1.1, 1.2, 2 e 3) em 16 populações de buriti (Mauritia flexuosa) em 2009, 2010
e 2011. (*) Populações controle.
2009 2010 2011
Brejo Parc. 1.1 1.2 2 3 1.1 1.2 2 3 1.1 1.2 2 3
Seco 15 0 160 45 26 3 160 41 31 10 169 43 31
Atoladeira* 13 24 174 17 26 6 173 22 26 48 177 25 26
Saco Grande 20 50 307 53 41 37 312 56 38 429 339 58 38
Boca do Mato 20 18 79 3 53 2 77 2 53 1 77 1 53
São Felix 17 45 195 99 44 32 198 98 44 119 191 105 44
Olho D'Água 20 53 238 66 19 10 234 66 21 27 229 69 23
Merola* 16 49 107 30 66 8 104 31 66 0 100 34 66
Rebojo 20 65 79 15 49 34 79 15 49 38 75 12 48
Cabeceira Fina* 20 51 76 11 28 15 65 9 30 33 47 12 30
Santo Antônio* 20 94 137 11 56 43 134 13 57 119 121 17 53
Porcos 18 118 81 23 35 17 82 17 39 52 71 14 39
Cansa Cavalo* 17 292 151 16 34 148 165 14 36 162 163 14 35
Estiva* 20 222 115 26 27 27 114 20 33 60 106 20 33
D'Anta* 20 595 225 13 23 369 248 15 24 386 259 18 24
Conceição 20 147 32 8 51 27 32 5 51 142 31 5 51
Grande 20 339 62 13 50 106 61 11 52 360 55 14 51
Total 296 2162 2218 449 628 884 2238 435 650 1986 2210 461 645
122
ANEXO 2. Matrizes de transição para 16 populações de buriti (Mauritia flexuosa) em dois
períodos, 2009 a 2010 e 2010 a 2011.
Brejo 2009 - 2010 2010 - 2011 Atoladeira 0,25 0,00 0,00 0,00 0,17 0,00 0,00 1,81 0,04 0,96 0,00 0,00 0,00 0,98 0,00 0,00 0,00 0,03 0,92 0,00 0,00 0,02 0,98 0,00 0,00 0,00 0,07 1,00 0,00 0,00 0,01 1,00 Boca do Mato 0,31 0,00 0,00 0,41 0,31 0,00 0,00 2,48 0,00 0,96 0,00 0,00 0,00 0,96 0,00 0,00 0,00 0,00 0,99 0,00 0,00 0,04 0,98 0,00 0,00 0,00 0,00 1,00 0,00 0,00 0,01 1,00 Cabeceira Fina 0,16 0,00 0,00 0,50 0,07 0,00 0,00 2,04 0,00 0,94 0,00 0,00 0,00 0,90 0,00 0,00 0,00 0,03 0,82 0,00 0,00 0,03 0,98 0,00 0,00 0,00 0,09 1,00 0,00 0,00 0,01 0,93 Cansa Cavalo 0,41 0,00 0,00 0,85 0,22 0,00 0,00 2,94 0,01 0,99 0,00 0,00 0,01 0,93 0,00 0,00 0,00 0,00 0,88 0,00 0,00 0,00 0,98 0,00 0,00 0,00 0,13 1,00 0,00 0,00 0,01 0,97 Conceição 0,06 0,00 0,00 0,35 0,30 0,00 0,00 2,63 0,01 0,97 0,00 0,00 0,00 0,97 0,00 0,00 0,00 0,00 0,63 0,00 0,00 0,00 0,98 0,00 0,00 0,00 0,00 1,00 0,00 0,00 0,01 1,00 Anta 0,58 0,00 0,00 0,91 0,60 0,00 0,00 6,83 0,02 0,97 0,00 0,00 0,01 0,98 0,00 0,00 0,00 0,01 0,92 0,00 0,00 0,01 0,98 0,00 0,00 0,00 0,08 1,00 0,00 0,00 0,01 1,00 Estiva 0,07 0,00 0,00 0,44 0,11 0,00 0,00 1,73 0,00 0,98 0,00 0,00 0,00 0,92 0,00 0,00 0,00 0,00 0,77 0,00 0,00 0,00 0,98 0,00 0,00 0,00 0,23 1,00 0,00 0,00 0,01 1,00 Grande 0,25 0,00 0,00 0,40 0,51 0,00 0,00 5,88 0,00 0,97 0,00 0,00 0,01 0,87 0,00 0,00 0,00 0,00 0,85 0,00 0,00 0,05 0,98 0,00 0,00 0,00 0,15 1,00 0,00 0,00 0,01 1,00

123
Brejo 2009 - 2010 2010 - 2011 Merola 0,29 0,00 0,00 0,31 0,29 0,00 0,00 0,57 0,05 0,94 0,00 0,00 0,06 0,91 0,00 0,00 0,00 0,00 0,92 0,00 0,00 0,00 0,80 0,00 0,00 0,00 0,07 1,00 0,00 0,00 0,00 1,00 Olho D'Água 0,06 0,00 0,00 0,08 0,00 0,00 0,00 0,00 0,02 0,96 0,00 0,00 0,00 0,95 0,00 0,00 0,00 0,01 0,92 0,00 0,00 0,03 0,98 0,00 0,00 0,00 0,07 1,00 0,00 0,00 0,01 1,00 Porcos 0,50 0,00 0,00 0,29 0,35 0,00 0,00 10,92 0,20 0,97 0,00 0,00 0,14 0,98 0,00 0,00 0,00 0,02 0,96 0,00 0,00 0,01 0,98 0,00 0,00 0,00 0,00 0,93 0,00 0,00 0,00 1,00 Rebojo 0,24 0,00 0,00 0,11 0,33 0,00 0,00 0,93 0,00 0,86 0,00 0,00 0,00 0,72 0,00 0,00 0,00 0,00 0,82 0,00 0,00 0,06 0,89 0,00 0,00 0,00 0,18 1,00 0,00 0,00 0,00 1,00 Saco Grande 0,06 0,00 0,00 0,02 0,00 0,00 0,00 0,02 0,00 0,94 0,00 0,00 0,00 0,99 0,00 0,00 0,00 0,00 0,67 0,00 0,00 0,00 0,50 0,00 0,00 0,00 0,33 1,00 0,00 0,00 0,00 1,00 Santo Antônio 0,14 0,00 0,00 0,03 0,35 0,00 0,00 1,18 0,04 0,95 0,00 0,00 0,06 0,85 0,00 0,00 0,00 0,00 0,74 0,00 0,00 0,00 0,82 0,00 0,00 0,00 0,17 1,00 0,00 0,00 0,00 1,00 São Felix 0,15 0,00 0,00 0,11 0,30 0,00 0,00 1,14 0,02 0,95 0,00 0,00 0,10 0,95 0,00 0,00 0,00 0,01 0,97 0,00 0,00 0,03 0,95 0,00 0,00 0,00 0,03 1,00 0,00 0,00 0,03 1,00 Seco 0,19 0,00 0,00 0,12 0,00 0,00 0,00 0,13 0,00 0,96 0,00 0,00 0,00 0,97 0,00 0,00 0,00 0,01 0,89 0,00 0,00 0,02 0,98 0,00 0,00 0,00 0,11 1,00 0,00 0,00 0,00 1,00

124
CAPÍTULO 3: Effects of socioeconomic conditions on the multiple uses of swamp forests
Maurício Bonesso Sampaio (corresponding author)
Programa de Pós Graduação em Biologia Vegetal, Instituto de Biologia, CP6109,
Universidade Estadual de Campinas – UNICAMP, 13083-970, Campinas, SP, Brasil
and
Pequi - Pesquisa e Conservação do Cerrado (www.pequi.org.br), SCLN 408, Bloco E, Sala
201, 70856-570, Brasília, DF, Brasil.
Tamara Ticktin
University of Hawai`i at Manoa, Botany Department, 3190 Maile Way, Honolulu, HI
96822, EUA
Cristiana Simão Seixas
Núcleo de Estudos e Pesquisas Ambientais – NEPAM, Universidade Estadual de Campinas
– UNICAMP, 13083-867, Campinas, SP, Brasil
Flavio Antonio Maës dos Santos
Departamento de Biologia Vegetal, Instituto de Biologia, CP6109, Universidade Estadual
de Campinas – UNICAMP, 13083-970, Campinas, SP, Brasil

125
Abstract
Local communities in central Brazil use swamp forests to harvest buriti palm (Mauritia
flexuosa) fruit as well as for agriculture, cattle raising and pig farming. This study aimed to
assess the intensity of forest use by buriti fruit harvesters and identify how their
socioeconomic conditions influence resource use. We visited 75 swamp forests where buriti
fruits are harvested and interviewed the head of the nearest household. Agriculture was
practiced in 72% of forests and cattle raising in 52%. Almost a half (48%) of households
had agriculture and buriti fruit harvest as the main income source. Forests resources were
equally important to all socioeconomic classes, even richer farmers. The intensity of fruit
harvest did not differ between collective and private use regimes of forests. Market access
was a limitation to fruit harvest intensity. The high intensity of swamp forest use suggests
that their conservation will require change to current management practices.
Keywords: NTFP, local people, livelihood, Cerrado, forest conservation

126
Introduction
Worldwide, people living in and around forests often depend on non-timber forest products
(NTFPs) for their livelihoods (Narendran et al. 2001; Marshall and Newton 2003;
Paumgarten and Shackleton 2009). NTFPs are biological materials other than timber which
are harvested from wild species from natural ecosystems for local human communities use
(De Beer and McDermott 1989; Shackleton et al. 2011). As pointed out by Davidson-Hunt
et al. (2001) the concept of NTFP also refers to a consideration of the interests, values and
activities of people who have largely been excluded from forest research, planning and
management. Local people collect NTFPs mainly for food, fuelwood, medicine,
construction materials and domestic utensils. In addition to NTFP harvesting, local people
also carry out other commercial and subsistence activities inside forests, for example, slash-
and-burn agriculture, cattle raising and pig farming.
The harvest of forest resources such as NTFP has the potential to increase income
(Hegde et al. 1996; Davidar et al. 2008; Babulo et al. 2009) and decrease rural poverty and
income inequality in local communities (Pyhälä et al. 2006; Babulo et al. 2009). Moreover,
NTFP marketing is part of a local economy, whose commercialization is generally done by
small farmers and harvesters. In this way, NTFP may strengthen community organization
and improve social justice (Marshall et al. 2003).
Socioeconomic conditions can influence the type and intensity of forest use. For
example, poorer households often have fewer livelihood options than richer households,
and thus have higher NTFP home consumption and participation in NTFP

127
commercialization (Shackleton and Shackleton 2006). On the other hand, if given a choice,
many forest users would choose not to depend on wild resources for their survival (Hegde
and Enters 2000; Fisher et al. 2005). Therefore, richer households can become specialized
in the most valuable wild forest resources or replace harvesting activities with other income
sources, as cattle raising (Godoy et al. 1995; Barham et al. 1999).
In addition to user income source, other factors that may influence the intensity of
forest resource use by local people include distance from the household to the resource,
access to the market, and forest tenurial regime. The distance from the household to the
resource is directly related to the opportunity cost of NTFP harvesting (Hegde et al. 1996).
This tends to be an important factor when NTFP are used for household consumption only,
because at greater distances, people may buy the available products at local markets instead
of harvesting them from the forest. However, distance to resource may not be an important
limitation when NTFP are harvested for commercial purposes (Davidar et al. 2008).
Access to the market is often the main obstacle for NTFP harvesters, mainly due to
the high transportation costs associated with long-distances to the final consumers and poor
transportation infrastructure (Marshall et al. 2003). Additionally, some products are highly
perishable and require quick means of transportation (Shanley et al. 2002).
NTFPs are often common-pool resources, and therefore may be subject to
degradation as a result of overexploitation by multiple users and for which exclusion of
users is costly (Ostrom 1990, Dietz et al. 2002). The way in which harvesters extract and
manage NTFP may vary according to the forest tenurial regime. According to the
Commons Theory, common pool resources such as NTFP may be managed under the
following tenurial regimes or a combination of them: private-property, state-property,

128
common-property or they may be exploited under no regime (i.e., open access system)
(Ostrom 1990, Feeny et al. 1990).
In open-access systems, resources are exploited by several users; access exclusion is
very difficult, and resource use involves subtractability, i.e., resource exploitation by one
user decrease resource availability for others (Ostrom et al. 1999). In open-access systems,
competition among resource users may be so intensive that it leads to resource depletion in
short time (Hardin 1968; Pandit and Thapa 2003). Additionally, the access to NTFPs may
be unequal between poorer and richer harvesters, with the later usually having greater
means for harvesting (Adhikari et al. 2004).
In other NTFP extraction systems, communities have developed management
regulations for accessing and using resources; these are known as common property
regimes (Ostrom et al. 1999). Under such regulations, the harvesting intensity of NTFP
resources may be sustainable, i.e. it may avoid resource overexploitation (Yang et al.
2009).
When forests are privately owned, resource use is regulated by landowner.
Restricting access may lead to greater economic returns and be more sustainable than open
access system, but often the landowner is the sole beneficiary of the resources (Yang et al.
2009). In some cases though, the resources in private property may be shared among other
users with the landowner’s permission. This is the case of mangaba (Hancornia speciosa
Gomes; Apocynaceae) fruit harvesting in Northeastern Brazil (Schmitz et al. 2009) as well
as of buriti palm (Mauritia flexuosa L. f.; Arecaceae) fruit harvest in the central Brazil. In
both cases, de jure private property may function either as open access systems or common
property depending on the establishment of access and management rules among users.

129
Understanding the socioeconomic factors that influence the type and intensity of
forest resource utilization is key to designing management plans to forest conservation
(Gavin and Anderson 2007). Most of studies that have assessed the effects of
socioeconomic variables on patterns of NTFP harvest have focused on only one or few
variables, and on only one or few NTFP resources (e.g. Godoy et al. 1995; Bista and Webb
2006). However, to identify bottlenecks in sustainable resource management it is important
to simultaneously evaluate the factors that influence NTFP harvest as well as other kinds of
forest use and management. In addition, most studies to date have been carried out on a
very small scale (among one or a few communities), and thus the results have limited
representativeness of general patterns (Basurto and Ostrom 2009).
This study focuses on the multiple uses of swamp forests in five regions of central
Brazil. Swamp forests are important for regulation of groundwater (Lima and Zakia 2000),
to supply drinking water for human beings and wildlife, and to provide nesting areas and
ecological corridors to animals (Marinho-Filho and Gastal 2000). Swamp forests are also
important to local people for harvesting buriti fruit, practicing slash-and-burn agriculture,
cattle raising and pig farming, but little is known about the extent and intensity of use these
activities.
Our objective was to describe the type and intensity of swamp forest uses over a
large geographic area, and identify if socioeconomic conditions of NTFP harvesters
influence these uses. We tested three hypotheses: 1 – The type and intensity of swamp
forest use differs between poorer and richer households. We expected that households more
dependent on NTFP harvesting and subsistence agriculture (poorer households) practice
less cattle rising and pig farming than those involved in mechanized agriculture or wage

130
labor (richer households); 2 – The level of buriti fruit harvesting depends on the distance to
resource, marketing access and main income source of the household; 3 – The intensity of
NTFP harvest is higher in forests exploited by multiple users (owners and outsiders) than in
those exploited privately (by owners e/or their employees only). We discuss the
implications of our findings for the sustainable management of swamp forests.
Material and methods
Study area
This study was carried out in five regions of the central Brazil (Figure 1). The regions were
selected based on two conditions, a high abundance of swamp forests, and the presence of
associations or cooperatives that coordinate NTFP harvesting activities. The inhabitants of
the study regions are mainly descendants of settlers who arrived in the region in the past
two centuries. The fruits of buriti palm (Mauritia flexuosa) are harvested for consumption
by people of most households, but are commercialized by a few of them. The ripe fruits are
often collected on the ground of the swamp forests after they are naturally dispersed. The
fruits are usually harvested from natural buriti populations, and only a few buriti plantations
can be found. The main goal of this cultivation is mostly to control erosion or improve
water quality, and not fruit production.
Buriti fruit sold as fresh pulp, candies or oil are among the most important NTFPs
of central Brazil. Several other uses of buriti by local communities, such as medicine,

131
domestic utensils, handicrafts and construction materials, have been described in central
Brazil (Sampaio et al. 2008; Nascimento et al. 2009) and the Amazon (Balick 1979; Peters
et al. 1989; Kahn 1991; Gragson 1995; Manzi and Coomes 2009; Santos and Coelho-
Ferreira 2012). Fruits are produced annually from October to February, but high fruit
production occurs every other year (Storti 1993).
Buriti palm is one of the most frequent and abundant emergent species of swamp
forests, with an average of more than 300 adult individuals per ha (Sampaio et al. 2008).
Swamp forests occur in lowlands associated with the headwaters of small streams. The soils
of swamp forests are frequently hydromorphic with high acidity and aluminum content,
rich in organic matter and poor in calcium and phosphorous (Toniato et al. 1998). In some
places the water level is more than 1 m above the soil surface. Swamp forests are
surrounded by humid grasslands and scrublands, which occur in the elevated portions of the
landscape. Sandy soils are predominant in the landscape of all five study regions.
In addition to buriti fruit harvesting, swamp forests are also used by people of most
households of the five study regions for other commercial and subsistence activities
including slash and burn agriculture, cattle raising and pig farming. Cattle raising occurs
mainly in the humid grasslands surrounding the swamp forests, but the animals frequently
use the interior of swamp forests to drink water and for shade. In some swamp forests the
water table is low and allows for livestock grazing and the soil humidity is useful for
raising pigs. Local people grow rice, corn, beans or sugarcane in swamp forests for
subsistence and to sell the surplus crop in the local markets. Fire is used to promote
clearing of vegetation and mineralization of the organic matter. Local people also use fire

132
to renew pastures and to supply palatable forage to cattle in the surroundings of the swamp
forests and fire may escape and reach the interior of the swamps.
Data collection
We selected 75 swamp forests in the five regions we visited (Figure 1): South of Piauí state
(n = 35); Northeast of Tocantins state (n = 27); North of Minas Gerais state (n = 8); East of
Tocantins state (n = 4); and West of Bahia state (n = 1). The swamp forests were smaller
than 100 ha and were selected if buriti fruit harvesting occurred there, and if it was located
close to a road with easy access. We visited the swamp forests from January to April 2009,
accompanied by a person familiar with the local communities. We located the closest
household to each swamp forest and asked permission to the head of the household or the
person responsible for the land to conduct our research. We carried out a semi-structured
interview with the head of the closest household about the uses of the swamp forest to
obtain information about the present and past agricultural activities inside of the forest; the
number of cattle head; pig farming (absent, intensive or extensive); amount of buriti pulp
commercialized (sum of all users of the swamp forest); and the frequency of swamp forest
burning. After the interview, we visited the swamp forest accompanied by the household
head to cross-check the information he provided. We use this data to classify the swamp
forests into categories of use intensity, according to Table 1. We also asked the interviewee
about the type of market access for buriti products; the main income source of the
household; and the type of users of the swamp forest, according to the categories of the
Table 1. We grouped the data for each variable into categories to reduce possible

133
imprecision of the responses we received. The collection of more precise data would
require intensive data collection over a long period for each swamp forest, and this was not
possible in our case due to the large amount of swamp forests we sampled.
Local people frequently practice selective timber logging and game hunting for
subsistence and income generation in swamp forests, but we did not assess these activities
in this study. It was not possible to obtain reliable answers from local people about these
activities using a short term survey because both activities are illegal according to Brazilian
Federal laws. All management activities in swamp forests are illegal, even NTFP
harvesting, according to the Brazilian Forest Code (law number 4771, September 15, 1965).
However, law enforcement is much less stringent on activities that cause lower
environmental impacts, such as NTFP harvesting, than on hunting and logging. We
strongly believe that the illegality of swamp forest use did not bias our data, because of the
low law enforcement. In fact, among the activities we assessed, users were only concerned
about enforcement regarding the slash-and-burn agriculture plots and uncontrolled fire on
swamp forest.
Data analysis
To assess the relationships among all variables (socioeconomic conditions and type and
intensity of use) at the same time, we did a Correspondence Analysis (CA). We then carried
out a Canonical Correspondence Analysis (CCA) and a Monte Carlo test (with 10,000
matrix permutations) to assess the correlation between the socioeconomic conditions matrix
(access to fruit marketing, distance to swamp forest, type of user and main income source)

134
and forest uses matrix (agriculture practices, cattle raising, pig farming and fruits
harvesting), according to Legendre and Legendre (1998).
To test for differences in type and intensity of swamp forest use between richer
versus poorer households we used the income source as a proxy for relative income level.
Thus, we classified the households in two groups: those whose income comes from in-farm
or out-farm employment or from cattle raising or mechanized agriculture (richer
households); and those whose the main income source are slash-and-burn agriculture or
NTFP harvesting (poorer households). We performed a Discriminant Function Analysis
(DFA) to test for differences between the two groups based on type and intensity of swamp
forest use (Tabachnick and Fidell 2001).
We carried out an Frequency Table Analysis to identify differences in distance to
resource, marketing access and income source among households who carry out buriti fruit
harvest at different levels of intensity: low intensity of fruit harvest and absence of
commercialization (low-intensity harvest); low buriti pulp commercialization (moderate-
intensity harvest); and high buriti pulp commercialization (high-intensity harvest).
Buriti fruits are often shared among neighbors, when a swamp forest owner allows
other people to harvest the fruits. We carried out a Frequency Table Analysis to test the
effect of resource use regime (collective or private) on buriti fruit harvest intensity. We did
not test the difference between the two resource use regimes on the intensity of swamp
forest use for agriculture practices, cattle raising, pig farming and fire, because in general
swamp forest owners do not allow other people to practice these activities on their lands.
CA and CCA analysis were performed using Fitopac 1.6.4 (Shepherd 2006), DFA
and Frequency Table Analysis were performed using STATISTICA 7.0 (StatSoft 2004).

135
Results
Buriti fruit was harvested for commercial purposes in the majority (63%) of swamp forests
we sampled. Cattle was raised in 72% and slash-and-burn agriculture was carried out in
52% of them (Figure 2). A total of 33% of the swamp forests were used simultaneously for
fruit harvesting, cattle raising and agricultural practices (Figure 3). One quarter of the
swamp forests were used for pig farming (Figure 2) and in 4% of them fruit harvesting,
cattle raising, agriculture and pig farming were carried out simultaneously (Figure 3). Only
6% of the swamp forests were used exclusively for buriti fruit harvesting. The fire return
interval was every two or three years in 11% of the swamp forests (Figure 2).
For almost a half (48%) of the households, slash-and-burn agriculture or NTFP
harvest was the main income source (Figure 4). The highest distance between a household
and a swamp forest in our sampling was about 3,000 m, but 48% of households were closer
than 500 m to the swamp forest (Figure 4).
The use intensity of swamp forests was negatively correlated among agriculture, pig
farming and fire frequency, according to the Correspondence Analysis (CA) (Figure 5).
Thus, swamp forests with a large proportion of the area used for agricultural practices
generally were not used for high intensity pig farming, nor are they frequently burned. Fruit
harvest intensity was not correlated with any other variable in the first two CA axes (Figure
5) and in the other axes (data not showed). Fire frequency had a low positive correlation
with cattle raising (Figure 5). The type and intensity of use and the socioeconomic

136
conditions of the household were not more similar within a region than among regions
(Figure 5).
There were no significant correlations (Monte Carlo test; p = 0.15 for the first
canonical axis) between the socioeconomic conditions of users (access to fruit market,
distance to swamp forest, type of users and income source) and the type and intensity of
swamp forest uses (agriculture practices, cattle raising, pig farming and fruits harvesting),
according to the Canonical Correspondence Analysis (CCA). Additionally, the first three
canonical axis of CCA had only 9.6% of accumulated inertia. However, according to CA
results there was a low positive correlation between pig raising and income source (Figure
5).
There was no significant difference (Wilk’s λ = 0.92; p = 0.35) in the use intensity
of swamp forests between the households whose the income comes from employment or
from managing small to large productive farms and those whose the income comes from
subsistence activities as slash-and-burn agriculture or NTFP harvesting. Both household
groups practiced similar intensities of fire, agriculture, cattle raising, pigs raising and fruits
harvesting in the swamp forests (Figure 6).
Market access increased significantly from low-intensity harvesting to high-
intensity harvesting (χ2 = 59.99; d.f. = 6; p < 0.001; Figure 7). However, the harvesting
intensity level was not dependent of the distance from the household to the swamp forest
(χ2 = 2.37; d.f. = 4; p = 0.67) or the main income source (χ2 = 3.32; d.f. = 6; p = 0.77).
Buriti fruit harvest by multiple users takes place in 31% of sampled swamp forests,
despite being de jure private property; while in 69% of sampled swamp forests the
harvesting was restricted only to the interviewed household. In 48% of the swamp forests

137
exploited by multiple users the landowner was not a fruit harvester himself. In 81% of the
privately used swamp forests the landowner was the fruit harvester, while in the remaining
19% of swamps the fruit was only harvested by employees. High intensity fruit harvesting
(> 100 kg of pulp sold per year) occurred in 47% of swamp forests exploited by multiple
users and in 27% of those only exploited by the interviewed household. However, the
intensity of fruit harvest did not differ significantly (χ2 = 2.40; d.f. = 2; p = 0.30) between
the two use regimes.
Discussion
Use and importance of swamp forest resources
Buriti fruits are an important resource to people living close to swamp forests in central
Brazil, especially during periods of high fruit production. Households may produce up to
2000 kg of buriti fresh pulp in a harvesting season earning up to U$ 4,800 in four or five
months of work (M. B. Sampaio and F. A. M. Santos; unpublished data). This represents an
earning about four times the Brazilian minimum wage in each month of the production
season. Buriti fruits are also an important NTFP for people from other regions where this
species occurs. In the Peruvian Amazon close to Iquitos, up to 40% of households
commercialize buriti fruits; this is the third largest source of income, after agriculture and
fishing (Pyhälä et al. 2006), and the production value is U$ 1,525 per ha.year-1 (Peters et al.
1989). In the central Brazil, buriti fruit harvesting is not the main income source for most

138
households, essentially because: (i) buriti fruit production has a biannual peak and occurs
seasonally (only between October and February); and (ii), as our results illustrate, swamp
forests are suitable for other activities that can offer more reliable income sources for
households such as cattle raising, agriculture and pig farming.
We found that the use of swamp forests in central Brazil is very intense. More than
half of the swamp forests are used for agriculture and about three quarters for cattle raising.
In 28% of swamp forests, agricultural activities cover most of the forest. The vegetation of
some of the swamp forests, mainly in the South of Piauí state, was almost totally cleared
for sugar-cane and corn production, and only a few buriti individuals of the original forest
remained. In 13% of sampled swamp forests, cattle were allowed to use all of the forest.
The understory of these intensively used forests may be cleared and the soil compacted,
which may prevent the buriti seedling growth and recruitment (M. B. Sampaio; personal
observation).
Pig farming is less frequent in the swamp forests than agriculture and cattle raising,
but it is very important to the poorest people in rural communities of Brazil, mainly as a
source of low-cost protein (Silva-Filha et al. 2008). Pigs raised in swamp forests can forage
buriti fruits on the ground, reducing fruit availability to harvesters. Cattle can also forage
buriti fruits, but at a lower intensity than pigs. Even so, roaming pigs were raised in many
swamp forests that had high intensity of fruit harvesting. While detrimental for buriti
harvesting, raising pigs during the buriti fruiting season is probably advantageous for
households, since pigs fatten quicker during this season.
Influence of income source on type and intensity of swamp forest uses

139
Our hypothesis that the type and intensity of swamp forest use differs between poorer and
richer households was not confirmed. The intensity of swamp forest used for cattle raising,
pig farming, fruit harvesting, agriculture and fire frequency was very similar between
households whose main income comes from employment or from cattle raising and/or
mechanized agriculture (richer households), and those whose income are based on
subsistence activities as slash-and-burning agriculture or NTFP harvesting (poorer
households). This lack of difference may be a result of two factors. First, the soils of
swamp forests have higher fertility, organic matter and humidity than the most common
soils in the landscape, which are generally sandy and dry. Thus, independently of the user
income source, the swamp forests soils have better conditions for cattle raising, pig farming
and agriculture practices in the landscape. Even for the large farmers, the swamp forests are
important for agricultural activities and cattle raising due to water and shade availability.
Second, as in the Peruvian Amazon (Coomes et al. 2004), buriti fruits are a resource so
abundant and valuable that may be harvested intensively both by richer and poorer
households. However, when richer farmers do not harvest the buriti fruits, or when the
harvesting is of low intensity, they allow the access of poorer harvesters to the swamp
forest. Thus, the fruit harvesting intensity in the swamp forest is independent on the
households income source.
Buriti harvest intensity and other swamp forest uses

140
Our hypothesis that the level of buriti fruit harvesting depends on the distance to resource,
marketing access and income source of the household was partially confirmed. Households
intensively harvesting buriti in the nearby swamp forest had higher market access than
those harvesting only for their own consumption. However, the level of buriti fruit
harvesting is independent of the distance to resource and the household income source.
Marketing of buriti fruits in the studied regions is promoted mainly by local
harvester associations or cooperatives. In the South of Piauí, the pulp is collected at
harvesters’ homes by a truck and carried to the industry. The harvesters engaged in this
process were the ones involved in the highest intensity of fruit collection in our study.
Those not engaged in these associations have to transport their products to the local markets
by themselves. When harvesters are located close to the market or have means to transport
their product, they can commercialize large amounts of fruit. When they live far from the
market and have no easy and cheap means of transportation, they often harvest fruits only
for consumption. Similar to the NTFP fruit markets in the Amazon (Shanley et al. 2002),
market access, mainly transportation infrastructure, is a key factor influencing the intensity
of buriti fruit harvesting and marketing in the central Brazil. Poor market access is also a
limiting factor to buriti fruit harvesting in the Peruvian Amazon (Horn et al. 2012).
Collective vs. private use of swamp forests
The swamp forests investigated in this study are all located inside de jure private property,
although some could be either de facto open-access or common property systems. The
methodology we used did not aim to differentiate property regimes, nevertheless we were

141
able to identify swamp forests used by only their owners and/or employees (private
property), and those used by multiple users (either open access or common property).
Theory predicts that in open access systems, competition among users can be so intensive
that resources are depleted in short time (Hardin 1968; Pandit and Thapa 2003).
In our study, buriti fruit harvest intensity did not differ between collective (multiple
users) and private use systems. This indicates that some management rules are likely to
occur among buriti harvesters to avoid high intensity of fruit harvest in swamp forests with
collective use. That is, swamp forests used collectively are likely under a common-property
arrangement than in an open-access system. Collective use of swamp forests by several
fruit harvesters may not be a constraint to forest conservation or to maintain a sustainable
buriti fruit harvest. In fact, locally-devised rules have been described for sustainable
managing several common-pool resources worldwide (Ostrom et al. 1999; Schmitz et al.
2009; Yang et al. 2009; Baird 2010). Investigating use and access management rules
among buriti harvesters is an important subject for future research.
Implications for swamp forest conservation
We found that swamp forests are intensively used across central Brazil for NTFP
harvesting and agricultural activities, in both collective and private use systems, and by
households with different socioeconomic conditions. Although our study was not designed
to assess the environmental impacts of swamp forest resources use, our quantification of the
intensity of different kinds of uses provides an indirect measure of these impacts; and the
heavy use we recorded suggests that conservation actions for swamp forests will require

142
changes to current practices. We observed a negative correlation between use intensity and
environmental impacts in some swamp forests intensively used for agricultural activities,
cattle raising and/or pig farming, as these had highly compacted soil. The latter affects
vegetation dynamics, forest structure, forest regeneration and the water quality, which was
turbid. Conservation actions should limit these activities in some portions of the swamp
forests. Improving agricultural practices to increase soil productivity may reduce the
necessity of continually clearing old-growth swamp forest areas for agriculture (Kato et al.
1999; Aguiar et al. 2011). Fencing swamp forests may reduce the impacts of cattle and pigs
on forest integrity and water quality. However, animal husbandry is important to
households' food security. Thus, animals access to water and shade should be allowed in
some parts of the swamp forest, since in the study regions the dominant vegetation consists
of low stature plants on dry soils, which do not provide enough shade and water for pigs
and cattle.
We found a correlation between cattle raising intensity and fire frequency. This may
occur because sometimes cattle farmers lose control of fire used for pasture renewal and it
extends into the swamp forests. A large area of the grassland surrounding the swamp
forests generally needs to be burned biannually for pasture renewal, mainly in the Jalapão
region (Schmidt et al. 2007; Schmidt 2011). The fire spreads fast in grasslands and is
difficult to control, especially when it is lit in the hottest periods of the driest days, and
where the grassland has not been burnt for more than two years (Govender et al. 2006). To
protect swamp forests from undesirable fires, local people should employ firebreaks when
burning their pastures.

143
Buriti fruit harvesting has a much lower ecological impact than the other forest uses.
The effects of fruit harvesting on buriti population structure and dynamics may be
negligible over the long-term (Holm et al. 2008; Manzi and Coomes 2009), especially if
harvesters carry the seeds back to the interior of the swamp forest after removing the fruit
pulp (M. B. Sampaio and F. A. M. dos Santos; unpublished data). None of the harvesters
interviewed said they carry seeds back into the swamp forest, but most of them agreed that
this is easy to implement and may improve seedling recruitment. Buriti fruit harvesting
may be sustainable even when the swamp forests are shared among several users; as our
results indicate, harvest intensity does not vary between private use and collective use
arrangements. In sum, despite being an illegal activity, buriti fruit harvesting may
contribute to swamp forest conservation if it generates incomes that help to prevent the
conversion of swamp forests into other uses.
If our harvest recommendations above are followed, a potential way to increase
buriti harvesting, and therefore, household income, may be to improve harvesters’ access to
buriti fruit market, because the facility of market access was positively correlated to
harvesting intensity (Figure7). This is also the case in Peruvian Amazon (Horn et al. 2012)
and other NTFP harvesting systems (Ghate et al. 2009). Strengthening existing
cooperatives and organized groups of harvesters or creating new ones in the study regions
may help increase market access (Edwards 1996; Bista and Webb 2006) for buriti
harvesters. Moreover, promoting innovative ways to process and conserve buriti fruits
within each household may add value to their products and extend the storage period of the
buriti products, increasing the chances of accessing local and regional markets. Both
actions depend on building local capacity.

144
Although some actions may be taken to increase the income generated from fruit
harvesting, there will be still tradeoffs between swamp forest uses, particularly concerning
availability of animal protein for household consumption. These include raising pigs inside
of swamp forests and the inadequate fire management practiced mainly by cattle farmers.
These two management actions likely decrease the availability of buriti fruit to harvesters.
Natural resource management and conservation is not an easy task. Understanding
the multiple uses of swamp forests and the socioeconomic conditions that may influence
them is a first step towards identifying relevant conservation measures and practices.
Management actions should be balanced and negotiated with swamp forest users, so that
the use of forest resources can increase the income of local people and at the same time
better ensure swamp forest conservation.
References
Adhikari, B., Di Falco, S. and Lovett, J. C. (2004). Household characteristics and forest
dependency: evidence from common property forest management in Nepal.
Ecological Economics 48:245-257.
Aguiar, A. C. F., Freitas, I. C., Carvalho, C. S., Monroe, P. H. M. and Moura, E. G. (2011).
Efficiency of an agrosystem designed for family farming in the pre-Amazon region.
Renewable Agriculture and Food Systems 26(1):24-30.

145
Babulo, B., Muys, B., Nega, F. , Tollens, E., Nyssen, J., Deckers, J. and Mathijs, E. (2009).
The economic contribution of forest resource use to rural livelihoods in Tigray,
Northern Ethiopia. Forest Policy and Economics 11:109-117.
Baird, I. G. (2010). Private, small groups, or communal: Dipterocarpus wood resin tree
tenure and management in Teun Commune, Kon Mum district, Ratanakiri Province,
Northeastern Cambodia. Society & Natural Resources 23:1027-1042.
Balick, M. J. (1979). Economic Botany of the Guahibo. 1.Palmae. Economic Botany
33:361-376.
Barham, B. L., Coomes, O. T. and Takasaki, Y. (1999). Rain forest livelihoods: income
generation, household wealth and forest use. Unasylva 198:71.
Basurto, X. and Ostrom, E. (2009). The core challenges of moving beyond Garrett Hardin.
Journal of Natural Resources Policy Research 1:255-259.
Bista, S. and Webb, E. L. (2006). Collection and marketing of non-timber forest products in
the far western hills of Nepal. Environmental Conservation 33:244-255.
Coomes, O. T., Barham, B. L. and Takasaki, Y. (2004). Targeting conservation–
development initiatives in tropical forests: insights from analyses of rain forest use
and economic reliance among Amazonian peasants. Ecological Economics 51:47-
64.
Davidar, P., Arjunan, M. and Puyravaud, J. P. (2008). Why do local households harvest
forest products? A case study from the southern Western Ghats, India. Biological
Conservation 141:1876-1884.
Davidon-Hunt, I., Duchesne, L. C. and Zasada, J. C. (2001). Non-timber forest products:
local livelihoods and integrated forest management. In Davidon-Hunt, I., Duchesne,

146
L. C. and Zasada, J. C. Forest communities in the third Millennium: linking
research, business, and policy toward a sustainable non-timber forest product sector.
United States Department of Agriculture (USDA) General Technical Report NC-
217. St. Paul. 151 p.
De Beer, J. H. and McDermott, M. (1989). The economic value of non-timber forest
products in South East Asia. The Netherlands Committee for IUCN, Amsterdam.
Dietz, T., Dolsak, N., Ostrom, E., and Stern, P. C. (2002). The drama of the commons. In
Ostrom, E., Dietz, T., Dolsak, N., Stern, P. C., Stonich, S., and Weber, E. U. (eds.),
The dramas of the commons. Washington, D.C. National Academy Press.
Edwards, D. M. (1996). The trade in non-timber forest products from Nepal. Mountain
Research and Development 16:383-394.
Feeny, D., Berkes, F., McCay, B. J., and Acheson, J. M. (1990). The tragedy of the
commons: twenty-two years later. Human Ecology 18: 1–19.
Fisher, R., Maginnis, S., Jackson, W. J., Barrow, E. and Jeanrenaud, S. (2005). Poverty and
conservation: landscapes, people and power. IUCN, Gland, Switzerland and
Cambridge, UK.
Gavin, M. C., and Anderson, G. J. (2007). Socioeconomic predictors of forest use values in
the Peruvian Amazon: A potential tool for biodiversity conservation. Ecological
Economics 60:752-762.
Ghate, R., Mehra, D. and Nagendra, H. (2009). Local institutions as madiators of the
impact of markets on non-timber forest product extraction in central India.
Environmental Conservation 36(1):51-61.

147
Godoy, R., Brokaw, N. and Wilkie, D. (1995). The effect of income on the extraction of
non-timber tropical forest products: model, hypotheses, and preliminary findings
from the Sumu Indians of Nicaragua. Human Ecology 23:29-52.
Govender, N., Trollope, W. S. W. and Van Wilgen, B. W. (2006). The effect of fire season,
fire frequency, rainfall and management on fire intensity in savanna vegetation in
South Africa. Journal of Applied Ecology 43:748-758.
Gragson, T. L. (1995). Pumé exploitation of Mauritia flexuosa (Palmae) in the Llanos of
Venezuela. Journal of Ethnobiology 15:177-188.
Hardin, G. (1968). Tragedy of Commons. Science 162:1243-1248.
Hegde, R. and Enters, T. (2000). Forest products and household economy: a case study
from Mudumalai Wildlife Sanctuary, Southern India. Environmental Conservation
27:250-259.
Hegde, R., Suryaprakash, S., Achoth, L. and Bawa, K. S. (1996). Extraction of non-timber
forest products in the forests of Biligiri Rangan Hills, India. 1. Contribution to rural
income. Economic Botany 50:243-251.
Holm, J. A., Miller, C. J. and Cropper, W. P. (2008). Population dynamics of the dioecious
Amazonian palm Mauritia flexuosa: simulation analysis of sustainable harvesting.
Biotropica 40:550-558.
Horn, C. M.; Gilmore, M. P. and Endress, B. A. (2012). Ecological and socio-economic
factors influencing aguaje (Mauritia flexuosa) resource management in two
indigenous communities in the Peruvian Amazon. Forest Ecology and Management
267:93-103.

148
Kahn, F. (1991). Palms as key swamp forest resources in Amazonia. Forest Ecology and
Management 38:133-142.
Kato, M. S. A., Kato, O. R., Denich, M. and Vlek, P. L. G. (1999). Fire-free alternatives to
slash-and-burn for shifting cultivation in the eastern Amazon region: the role of
fertilizers. Field Crops Research 62:225-237.
Legendre, P. and Legendre, L. (1998). Numerical ecology. 2nd edition. Elsevier Science,
Amsterdan, The Netherlands.
Lima, W. P. and Zakia, M. J. B. (2000). Hidrologia de matas ciliares. In: Rodrigues, R. R.
and Leitão-Filho, H. F (ed.). Matas ciliares: conservação e recuperação. São Paulo:
Editora da Universidade de São Paulo: Fapesp, pp. 33-44.
Manzi, M. and Coomes, O. T. (2009). Managing Amazonian palms for community use: a
case of aguaje palm (Mauritia flexuosa) in Peru. Forest Ecology and Management
257:510-517.
Marinho-Filho, J. and Gastal, M. L. (2000). Mamíferos das matas ciliares dos Cerrados do
Brasil central. In: Rodrigues, R. R. and Leitão-Filho, H. F (ed.). Matas ciliares:
conservação e recuperação. São Paulo: Editora da Universidade de São Paulo:
Fapesp, pp. 209-221.
Marshall, E. and Newton, A. C. (2003). Non-timber forest products in the community of El
Terrero, Sierra de Manantlan Biosphere Reserve, Mexico: Is their use sustainable?
Economic Botany 57:262-278.
Marshall, E., Newton, A. C. and Schreckenberg, K. (2003). Commercialisation of non-
timber forest products: first steps in analysing the factors influencing success.
International Forestry Review 5:128-137.

149
Narendran, K., Murthy, I. K., Suresh, H. S., Dattaraja, H. S., Ravindranath, N. H. and
Sukumar, R. (2001). Nontimber forest product extraction, utilization and valuation:
a case study from the Nilgiri Biosphere Reserve, southern India. Economic Botany
55:528-538.
Nascimento, A. R. T., Santos, A. A., Martins, R. C. and Dias, T. A. B. (2009). Palm
communities in the Kraho indigenous territory, Tocantins, Brazil: biodiversity and
ethnobotanical features. Interciencia 34:182-188.
Ostrom, E. (1990). Governing the commons. The evolution of institutions for collective
action. Cambridge University Press, Cambridge.
Ostrom, E., Burger, J., Field, C. B., Norgaard, R. B. and Policansky, D. (1999). Revisiting
the commons: local lessons, global challenges. Science 284:278-282.
Pandit, B. H. and Thapa, G. B. (2003). A tragedy of non-timber forest resources in the
mountain commons of Nepal. Environmental Conservation 30:283-292.
Paumgarten, F. and Shackleton, C. M. (2009). Wealth differentiation in household use and
trade in non-timber forest products in South Africa. Ecological Economics 68:2950-
2959.
Peters, C. M., Balick, M. J., Kahn, F. and Anderson, A. B. (1989). Oligarchic forests of
economic plants in Amazonia: utilization and conservation of an important tropical
resource. Conservation Biology 3:341-349.
Pyhälä, A., Brown, K. and Adger, W. N. (2006). Implications of livelihood dependence on
non-timber products in Peruvian Amazonia. Ecosystems 9:1328-1341.

150
Sampaio, M. B., Schmidt, I. B. and Figueiredo, I. B. (2008). Harvesting effects and
population ecology of the buriti palm (Mauritia flexuosa L. f., Arecaceae) in the
Jalapão region, Central Brazil. Economic Botany 62:171-181.
Santos, R. S. and Coelho-Ferreira, M. (2012). Estudo etnobotânico de Mauritia flexuosa L.
f. (Arecaceae) em comunidades ribeirinhas do Município de Abaetetuba, Pará,
Brasil. Acta Amazônica 24(1):1-10.
Schmidt, I. B. (2011). Effects of local ecological knowledge, harvesting and fire on golden-
grass (Syngonanthus nitens, Eriocaulaceae), a non-timber forest product (NTFP)
species from the Brazilian savanna. PhD Thesis, Botany department, University of
Hawai`i at Manoa, Honolulu.
Schmidt, I. B., Figueiredo, I. B. and Scariot, A. (2007). Ethnobotany and effects of
harvesting on the population ecology of Syngonanthus nitens (Bong.) Ruhland
(Eriocaulaceae), a NTFP from Jalapão region, central Brazil. Economic Botany
61(1):73-85.
Schmitz, H., Mota, D. M. and Júnior, J. F. S. (2009). Gestão coletiva de bens comuns no
extrativismo da mangaba no nordeste do Brasil. Ambiente & Sociedade 12:273-292.
Shackleton. C., Delang, C. O., Shackleton. S. and Shanley, P. (2011). Non-timber forest
products: concept and definitions. In: Shackleton, S., Shackleton, C. and Shanley, P.
(Ed.). Non-timber forest products in the global context. Springer-Verlag, Berlin,
Heidelberg. pp. 3-21.
Shackleton. C. and Shackleton. S (2006). Household wealth status and natural resource use
in the Kat River valley, South Africa. Ecological Economics 57:306-317.

151
Shanley, P., Luz, L. and Swingland, I. R. (2002). The faint promise of a distant market: a
survey of Belem's trade in non-timber forest products. Biodiversity and
Conservation 11:615-636.
Shepherd, G. J. (2006). FitopacShell 1.6.4. Manual versão preliminar. Departamento de
Botânica, Universidade Estadual de Campinas, Campinas - SP.
Silva-Filha, O. L., Filho, E. C. P., Souza, J. F., Oliveira, A. S., Oliveira, R. J. F., Melo, M.,
Melo, L. M., Araújo, K. A. O. and Sereno, J. R. B. (2008). Caracterização do
sistema de produção de suínos locais na microrregião do Curimataú Paraibano.
Revista Brasileira de Saúde e Produção Animal 9:7-17.
StatSoft, Inc. (2004). STATISTICA (data analysis software system), version 7.
www.statsoft.com.
Storti, E. F. (1993). Biologia floral de Mauritia flexuosa Lin. Fil, na região de Manaus,
AM, Brasil. Acta Amazônica 23:371-381.
Tabachnick, B. G. and Fidell, L. S. (2001). Using multivariate statistics. 4 edition. Allyn &
Bacon, Boston, USA.
Toniato, M. T. Z., Leitão-Filho, H. D. F. and Rodrigues, R. R. (1998). Fitossociologia de
um remanescente de floresta higrófila (mata de brejo) em Campinas, SP. Revista
Brasileira de Botânica 21:197-210.
Yang, X. F., Wilkes, A., Yang, Y. P., Xu, J. C., Geslani, C. S., Yang, X. Q., Gao, F., Yang,
J. K. and Robinson, B. (2009). Common and privatized: conditions for wise
management of matsutake mushrooms in Northwest Yunnan Province, China.
Ecology and Society 14(2):30.

152
FIGURE 1
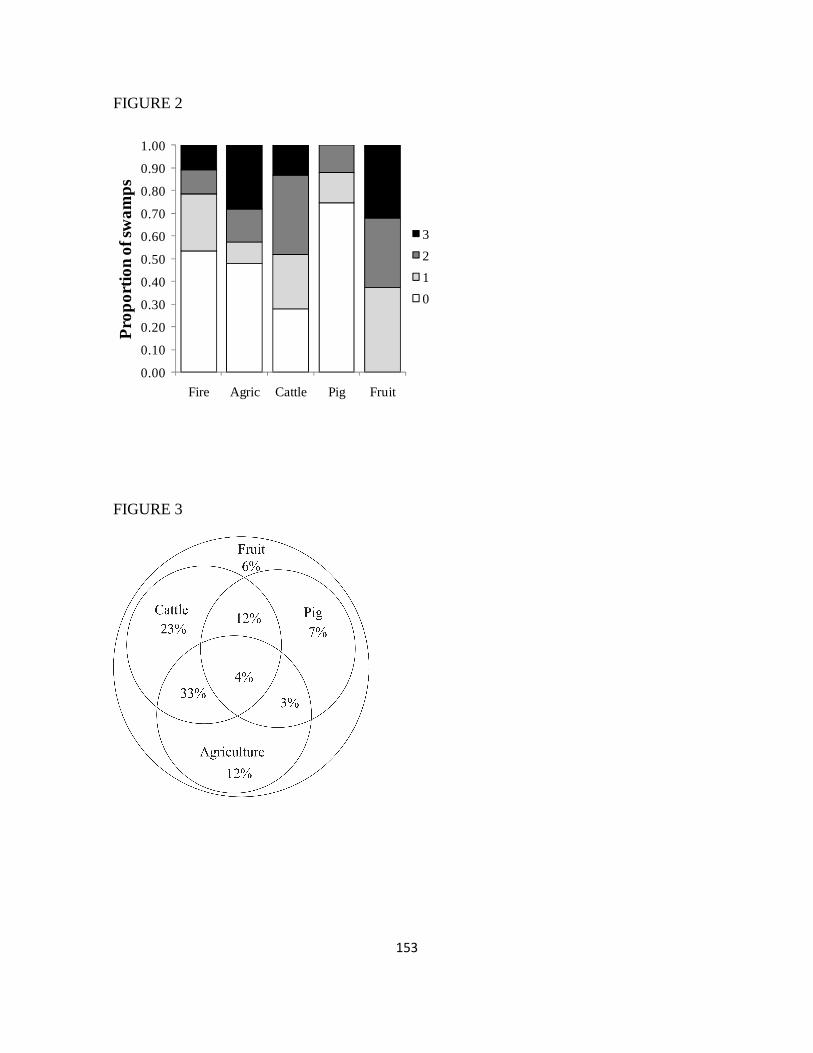
153
FIGURE 2
0.00
0.10
0.20
0.30
0.40
0.50
0.60
0.70
0.80
0.90
1.00
Fire Agric Cattle Pig Fruit
Pro
po
rtio
n o
f sw
am
ps
3
2
1
0
FIGURE 3

154
FIGURE 4
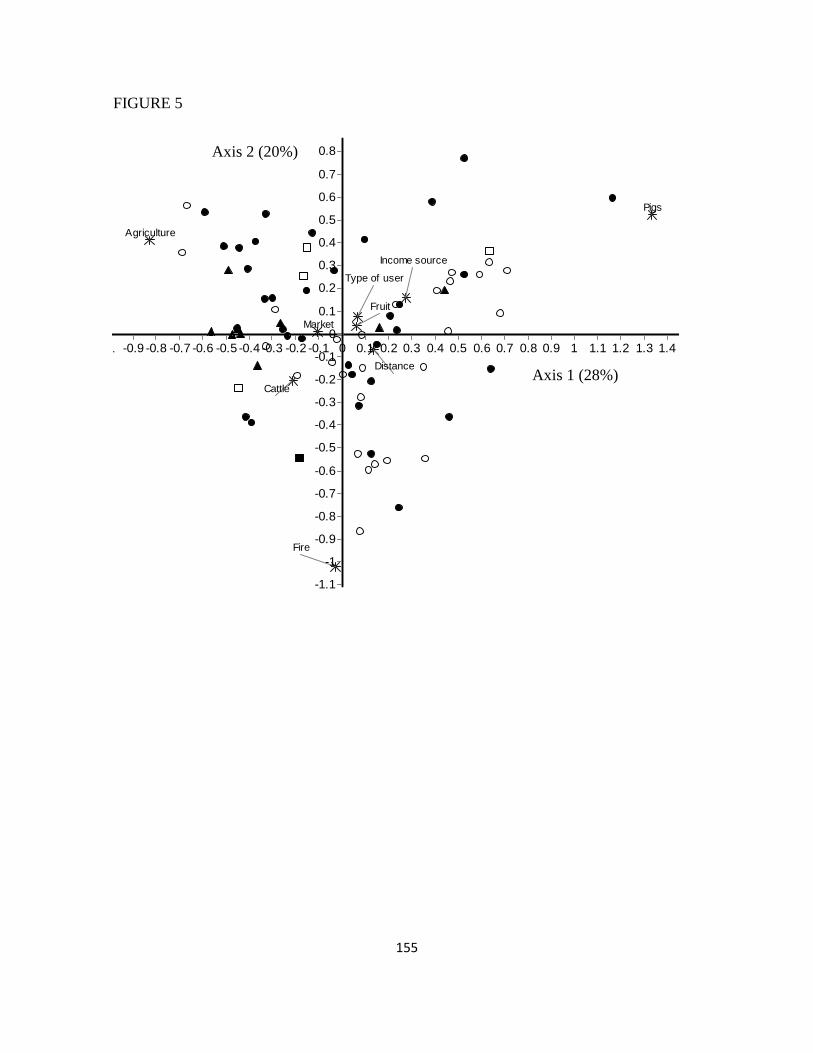
155
FIGURE 5
Fire
Agriculture
Cattle
Pigs
Fruit
Distance
Market
Type of user
Income source
1.41.31.21.110.90.80.70.60.50.40.30.20.10-0.1-0.2-0.3-0.4-0.5-0.6-0.7-0.8-0.9-1
0.8
0.7
0.6
0.5
0.4
0.3
0.2
0.1
0
-0.1
-0.2
-0.3
-0.4
-0.5
-0.6
-0.7
-0.8
-0.9
-1
-1.1
Axis 1 (28%)
Axis 2 (20%)
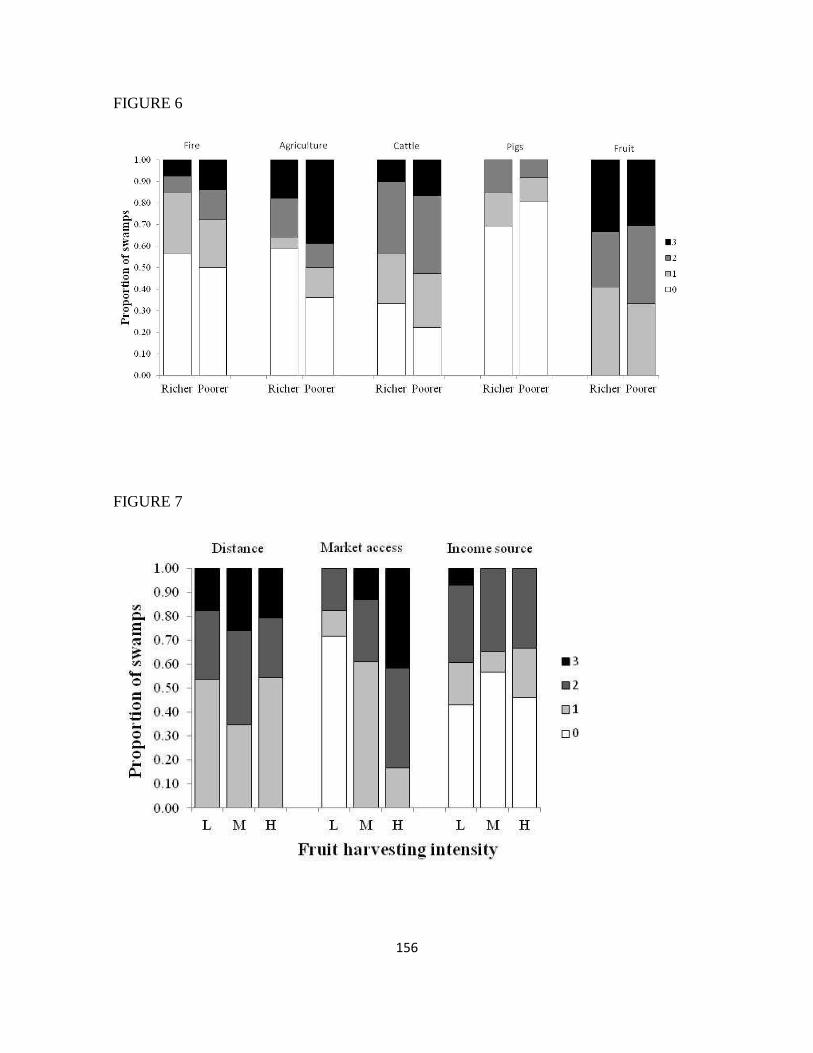
156
FIGURE 6
FIGURE 7

157
FIGURE LEGENDS:
Fig. 1 Location of 75 swamp forests sampled in five regions of the central Brazil.
Fig. 2 Frequency distribution of swamp forests (n = 75) subject to different types and
intensity of uses. Fire frequency from 0 (> 15 years without fire) to 3 (fire every 2 or 3
years); intensity of agriculture from 0 (absent) to 3 (occurs in a large swamp forest area);
cattle raising intensity from 0 (absent) to 3 (> 10 cattle use all the swamp forest); pig
raising from 0 (absent) to 2 (roaming pigs inside of the swamp forest) and; fruits harvesting
intensity from 1 (lowest) to 3 (highest), according to Table 1.
Fig. 3 Percentage of concurrent multiple uses of swamp forest (n = 75) by local people for
buriti fruits harvesting, cattle raising, slash-and-burn agriculture and pig farming.
Fig. 4 Frequency distribution of households (n = 75) in different categories of: distance to
swamp forests from 1 (nearest) to 3 (farthest); access to fruit marketing from 0 (no sale) to
3 (higher access); type of user, 1 (only users from the interviewed household) or 2 (multiple
users) and; income source from 0 (slash-and-burn and NTFP), to 3 (mechanized agriculture
on large farm), according to Table 1.
Fig. 5 First and second axis of the Correspondence Analysis (48% of accumulated inertia),
showing the swamp forests in five regions (black circles = South of Piauí; white circles =
Northeast of Tocantins; black square = West of Bahia; white squares = East of Tocantins;

158
triangles = North of Minas Gerais). The variables (asterisk) were the type and intensity of
swamp use for agriculture, market access, type of user, main income source, fruit
harvesting, distance from household to swamp forest, cattle raising, pigs farming and fire
frequency.
Fig. 6 Frequency distribution of swamp forests use by higher income households, where the
income comes from employment or from managing small to large productive farms
(richer); and by lower income households, where the income are based on subsistence
activities as slash-and-burning agriculture or NTFP harvesting (poorer). The fire frequency
categories ranged from 0 (> 15 years without fire) to 3 (fire every 2 or 3 years); intensity of
agriculture from 0 (absent) to 3 (occurs in a large swamp forest area); cattle raising
intensity from 0 (absent) to 3 (> 10 cattle use all the swamp forest); pig raising from 0
(absent) to 2 (roaming pigs inside of the swamp forest) and; fruits harvesting intensity from
1 (lowest) to 3 (highest), according to Table 1.
Fig. 7 Frequency distribution of swamp forests used by three groups of buriti fruits
harvesters: (L) low-intensity of harvesting (no commercialization); (M) moderate-intensity
of harvesting (commercialization of less than 100 kg of pulp per year); and (H) high
intensity of harvesting (commercialization of more than 100 kg of pulp per year). The
distance to swamp forests categories ranged from 1 (nearest) to 3 (farthest); access to fruit
market from 0 (no sale) to 3 (higher access) and; main income source from 0 (slash-and-
burn and NTFP) to 3 (large farmer), according to Table 1.

159
Table 1 Description of variables collected in 75 swamp forests in central Brazil.
Variable Intensity rating
Description
Typ
e an
d in
tens
ity o
f sw
amp
fore
st u
se Agriculture 0 never happened
1 was practiced in the past, but not currently 2 slash-and-burn in a small area of the swamp forest 3 occurs in most of the swamp forest area Cattle 0 absent raising 1 less than 10 cattle head use the swamp forest 2 more than 10 cattle head, but in a small area of the swamp 3 more than 10 cattle head use all the swamp Pig farming 0 absent 1 fenced pig farming in the swamp forest 2 roaming pigs in the swamp forest Buriti fruit 1 low harvesting intensity, no commercialization harvesting 2 low commercialization (<100 kg of pulp per year) 3 high commercialization (>100 kg of pulp per year) Fire 0 more than 15 years without fire 1 occasional (every 6 to 14 years) 2 every 4 or 5 years 3 every 2 or 3 years
Soc
ioec
onom
ic c
ondi
tions
Facility of 0 buriti products are not sold market 1 buriti products are carried to the market with great difficulty access 2 buriti products are carried to the market easily 3 purchasers come to buy at the household Main 1 slash-and-burn agriculture or NTFP harvesting income 2 salary (pensions, rural jobs, etc) source 3 cattle or mechanized agriculture in a small farm (<100 ha) 4 cattle or mechanized agriculture in a large farm (>100 ha) Distance 1 <500 m from swamp forest to the interviewed household 2 500 to 1,000 m from the interviewed household 3 >1,000 m from the interviewed household Type of 1 landowners and/or people from the interviewed household user 2 multiple users, including people from other farms/households

160
CONSIDERAÇÕES FINAIS
Este estudo traz uma avaliação de impactos de atividades de extrativismo de frutos
de buriti, uma importante palmeira da América do Sul. Além disso, análises de
sustentabilidade ecológica do extrativismo de PFNM similares a esta são escassas para
espécies do domínio do Cerrado, apesar do grande potencial de uso sustentável da sua
biodiversidade. As escalas espaciais e temporais utilizadas neste estudo são raras na
literatura especializada em ecologia de populações de plantas. Este trabalho também é
inovador ao avaliar simultaneamente vários fatores socioeconômicos que influenciam os
múltiplos usos da vegetação por comunidades rurais.
Três principais lacunas no conhecimento ainda precisam ser preenchidas para
compreender melhor os resultados aqui obtidos:
1 - Apesar de vários estudos de fenologia reprodutiva do buriti terem sido realizados, todos
os dados disponíveis foram coletados em curto prazo (um ou dois anos). Portanto, não há
informações suficientes para determinar com precisão a frequência temporal de ocorrência
dos eventos reprodutivos, tanto em indivíduos, quanto em populações. Um estudo
fenológico, em longo prazo e em larga escala, poderá contribuir também para entender
melhor os fatores (climáticos, biológicos ou intrínsecos) que causam a variação interanual e
a variação latitudinal na reprodução. Essas informações são importantes para melhorar a
precisão da simulação dos efeitos do extrativismo de frutos e para que os extrativistas
possam ter previsões mais acuradas da época de ocorrência das safras. Esta informação
poderá contribuir para o melhor planejamento e o controle das atividades de coleta.

161
2 - Investigar as regras de acesso e de uso dos brejos estabelecidas entre extrativistas e
proprietários das terras contribuirá para avaliar com maior confiança as implicações para a
sustentabilidade social, econômica e ecológica do atual sistema de coleta de frutos do
buriti;
3 - Avaliar os impactos do extrativismo de frutos para a assembleia de frugívoros que
utilizam este recurso será fundamental para estimar os impactos ecossistêmicos das
atividades de extrativismo. Além disso, esta informação será importante para determinar
com maior confiabilidade a sustentabilidade ecológica do extrativismo de frutos, já que a
coleta de frutos, em longo prazo, pode contribuir para a redução das populações de animais
dispersores das sementes de buriti;
4 - Avaliar os impactos ecossistêmicos das queimadas, das roças de coivara e das criações
de animais domésticos no interior dos brejos, para avaliar a sustentabilidade ecológica
destas atividades econômicas.
Apesar destas lacunas, os resultados deste trabalho demonstram claramente que é
possível a geração de renda por comunidades rurais por meio do extrativismo de frutos de
buriti, sem que haja a sobrexploração das plantas manejadas. A coleta de frutos de buriti
pode ser sustentável mesmo considerando o impacto de outras fontes de perturbação
provocadas pelos próprios extrativistas nas suas atividades econômicas primárias, como as
queimadas no interior dos brejos utilizadas para a renovação das pastagens.





























