Embed Size (px)
Citation preview

UNIVERSIDADE ESTADUAL DE CAMPINAS
INSTITUTO DE BIOLOGIA
MARCOS MACIEL JUNIOR
TERAPIA FARMACOLÓGICA DA DISTROFIA MUSCULAR: ASSOCIAÇÃO DO
ÔMEGA-3 E DEFLAZACORTE NA DISTROFINOPATIA DO CAMUNDONGO MDX
Campinas
2017

MARCOS MACIEL JUNIOR
TERAPIA FARMACOLÓGICA DA DISTROFIA MUSCULAR: ASSOCIAÇÃO DO
ÔMEGA-3 E DEFLAZACORTE NA DISTROFINOPATIA DO CAMUNDONGO MDX
Dissertação apresentada ao Instituto de Biologia da
Universidade Estadual de Campinas como parte dos
requisitos exigidos para obtenção do Título de Mestre
em Biologia Celular e Estrutural, na área de anatomia.
Orientadora: Profª. Drª. Maria Julia Marques
Campinas
2017
ESTE ARQUIVO DIGITAL CORRESPONDE À
VERSÃO FINAL DA DISSERTAÇÃO DEFENDIDA
PELO ALUNO MARCOS MACIEL JUNIOR E
ORIENTADA PELA PROFª. DRª. MARIA JULIA
MARQUES.

Agência(s) de fomento e nº(s) de processo(s): CAPES
Informações para Biblioteca Digital
Título em outro idioma: Pharmacological therapy of muscular dystrophy : association of
omega-3 and deflazacort in dystrophinopathy of mdx mouse
Palavras-chave em inglês: Muscular dystrophy, Duchenne
Mice, Inbred mdx
Fibrosis
Deflazacort
Omega-3 fatty acids
Área de concentração: Anatomia
Titulação: Mestre em Biologia Celular e Estrutural
Banca examinadora:
Maria Julia Marques [Orientador]
Luiz Fernando Takase
Ricardo Noboro Isayama
Data de defesa: 17-02-2017
Programa de Pós-Graduação: Biologia Celular e Estrutural

Campinas,17 de fevereiro de 2017.
COMISSÃO EXAMINADORA
Profª. Drª. Maria Julia Marques
Prof. Dr. Luiz Fernando Takase
Prof. Dr. Ricardo Noboro Isayama
Os membros da Comissão Examinadora acima assinaram a Ata de Defesa, que se encontra
no processo de vida acadêmica do aluno.

Dedico
Aos meninos com DMD, seus pais e familiares,
enfermeiros, fisioterapeutas, terapeutas ocupacionais,
psicólogos, médicos e pesquisadores que direta ou
indiretamente sofrem, cuidam e lutam na busca pela cura e
qualidade de vida desses meninos.

Agradecimento Especial
À Professora Dra. Maria Julia Marques, pela orientação, acolhimento, confiança, atenção,
paciência e pelo aprimoramento didático-científico compartilhado.
“Se deres um peixe a um homem faminto, vais alimentá-lo por um dia. Se o ensinares a
pescar, vais alimentá-lo toda a vida.”
(Anne Isabella Thackeray Ritchie)
Obrigado pela paciência que tem comigo nas nossas pescarias.

AGRADECIMENTOS
À Energia Superior, Deus, Cosmos, ou o nome que cada um queira dar àquele que me deu
belas oportunidades e colocou pessoas incríveis para que eu conseguisse aproveitar cada uma
delas, todas listadas abaixo.
À Marlene Ramos Rosa Bertolli, minha Mãe, meu pai, minha companheira, melhor amiga,
minha fada madrinha, aquela que sempre deu força e apoiou nas minhas escolhas, aquela que
vive meus sonhos comigo e faz meus pesadelos virarem paraíso.
Ao Marcos Maciel, meu pai, que confia cegamente nas minhas escolhas e nunca hesitou em
me ajudar.
Ao Robson William Bertolli, meu padrasto, que é companheiro e cuida da minha família em
São Carlos, viabilizando minha tranquilidade durante o mestrado.
Ao José Paulo Maciel Neto, meu irmão, criança que me faz acreditar na pureza das atitudes
dos seres humanos.
Aos meus avós Hermínia Ramos Ângulo, José Rosa, Benedicta Maria dos Santos Maciel
e José Paulo Maciel (in memoriam), que mesmo sem entender minha vinda pra Campinas,
confiam na minha escolha e me ajudam com suas orações para que tudo fique bem.
À Cilene Aparecida Ramos Rosa, minha tia, que deixa minha vida agridoce.
Aos meus tios e primos, em especial ao meu tio Mauro de Jesus Maciel, pelos encontros
breves e carregados de maturidade e sabedoria.
À todos os meus professores, fontes de profunda inspiração e sabedoria, representados aqui
pela “tia” Ana Sígari, minha primeira e eterna professora.
Aos professores do Laboratório de Anatomia da Universidade Federal de São Carlos
(UFSCar), Maria José Salete Viotto, Maíra Aparecida Stefanini, Luiz Fernando Takase e
Marcelo Martinez, minhas primeiras inspirações anatômicas, que tanto tiveram paciência e
me direcionaram para que eu pudesse realizar este sonho.

À professora Maria José Salete Viotto, pelo acolhimento no Laboratório de Anatomia da
UFSCar, orientação e paciência durante o início da minha jornada científica.
À professora Maíra Aparecida Stefanini, pelo acolhimento no Laboratório de Anatomia da
UFSCar e orientação durante minhas atividades como monitor de anatomia.
Aos amigos, colegas, copeiras e técnicos com quem compartilhei momentos de aprendizado e
alegria enquanto monitor no Laboratório de Anatomia da UFSCar, em especial à Marina
Augusto Silveira, Edvaldo Costa e Tábata Mariz Bohlen.
Às amigas enfermeiras que a graduação me deu e que estão sempre presentes na minha vida,
Aline Natalia Domingues, Isabella Nascimbeni Matos, Bianca Ruiz Oliveira, Camila
Félix Francisco, Beatriz de Oliveira Deniz, Suzana Maria Nery Carraschi, Daniela
Falcão, Ana Carolina Belmonte Assalin, Mariana Ramos da Silva, Camila Casagrande
Mela e Bruna Marski.
À amiga, Aline Natalia Domingues, pela amizade e companheirismo desde sempre.
À amiga e irmã, Marcia Keiko Nosse Araújo, pela amizade, confiança, respeito,
companheirismo, e por ter cedido seu espaço para que eu pudesse estudar.
Aos amigos que a UFSCar me deu, Fernanda Duarte Rosa, Luciane Sacramento, Maria
Augusta Sabadine, Havner Scherrer Cruz e Willians Indalécio, que continuam presentes
apesar da distância.
Aos amigos que a vida me deu, com quem muito aprendi, José Renato Gonçalves, José
Carlos Tavares da Motta Filho e Antônio Carlos Tadiello, vocês são muito importantes!
Ao Programa de Pós-graduação em Biologia Celular e Estrutural pela contribuição no
desenvolvimento profissional.
Ao Prof. Humberto Santo Neto pela colaboração, conhecimento e convívio durante a
execução deste trabalho.
Aos Profs. Valéria Helena Alves Cagnon Quitete, André Luis Bombeiro e Adriana
Fogagnolo Maurício pelas excelentes contribuições dadas durante o exame de qualificação.

Aos docentes do Departamento de Anatomia, Maria Julia Marques, Humberto Santo Neto,
Valéria Helena Alves Cagnon Quitete, Elaine Minatel, José Ângelo Camilli e Luciana
Bolsoni Lourenço Morandini, pela contribuição dada à minha formação e pelo
conhecimento compartilhado nas disciplinas cursadas durante toda essa jornada.
À Sra. Liliam Alves Senne Panagio pela atenção, auxilio e demasiada paciência antes e
durante o mestrado.
Aos funcionários do Departamento de Anatomia, à Sra. Rosalina de Fátima Telles, Sr.
Norivaldo Celestino, Sr. Marco Aurélio Ribeiro de Paula, Sr. Paulo Francisco dos
Santos, Sr. Walter Ferreira pelo auxílio técnico durante os estudos, amizade, e pelos
momentos de alegria e descontração.
Aos amigos do Laboratório de Plasticidade Muscular, Lab. Biologia de Reprodução,
Lab. Regeneração Nervosa, Lab. Plasticidade e Regeneração Óssea, Lab.
Eletromiografia e Controle Motor e Lab. Carcinogênese Urogenital e Imunoterapia pela
amizade, companheirismo e por tornarem o mestrado mais leve.
Aos amigos do Laboratório de Biologia Estrutural da Junção Neuromuscular, Adriana
Fogagnolo Maurício, Juliano Alves Pereira, Samara Camaçari de Carvalho, Jéssica
Silva Ferreira, Camila de Jesus Saqueli, Renato Rissi, Stephanie Furlan, Renata Acunha
e Paula Andrea Saenz, pelos bons momentos compartilhados.
Aos amigos Adriana Fogagnolo Maurício, Juliano Alves Pereira e Samara Camaçari de
Carvalho pela amizade, disponibilidade e extrema paciência que tiveram para me ensinar
desde a segurar um camundongo até o amedrontador Western Blot.
Aos amigos do Laboratório de Plasticidade Muscular, Daniela Sayuri Mizobuti, Túlio
Hermes, Rafael Mancio, Aline Macedo, pela amizade e pelo auxílio durante os
experimentos.
Aos amigos proporcionados pelas disciplinas da pós-graduação, Catharina Nucci Martins,
Luciana Ramalho, Luciana Sobral Moreira, Rodrigo Scarpari Carraro, Raphael
Casseb, Flávia Jacob, Julio Cesar Santini e Patrícia Ribeiro, cada um de vocês contribuiu
de maneira única para que o aprendizado durante as disciplinas ultrapassasse o que eu
aprenderia apenas nos livros e artigos.

Aos amigos com quem dividi teto em Campinas, Fernanda Duarte Rosa, Luciana Regina e
seu filhinho Alexandre, Renato Rissi, George Azevedo Lemos, Lucas de Souza, Leonardo
Staffa, Igor Franceschini, Carpegiani Borges e Ramon Rodrigues, obrigado por ajudarem
a tornar Campinas um bom lugar para se viver, morar e dormir.
Aos amigos que a UNICAMP me deu, Isadora Ribeiro, Jéssica Mendes, Luã Marcelo
Muriana e todos os colegas da musculação da Faculdade de Educação Física (FEF-
UNICAMP).
À Jéssica Silva Ferreira, pela amizade, cumplicidade, por me incentivar a sair da zona de
conforto do aprendizado, da pesquisa e da vida. Pelas madrugadas de estudo e reflexão para
tentar entender o blot. Por ter sido minha companheira nesses dois anos, muito obrigado!
À Catharina Nucci Martins, pela amizade, cumplicidade, pelas risadas, pela descontração e
pela irreverência. Também por todo aprendizado que me proporcionou, na neuroanatomia, na
pesquisa e na vida. Você é a pós-graduanda mais pesquisadora que eu conheço!
À Camila de Jesus Saqueli, pela amizade, companheirismo, pelas risadas e por cuidar de
mim. Por me acompanhar nas técnicas, por tirar os ependorfs do banho seco, por ser minha
memória operacional e me lembrar de guardar anticorpos, equipamentos, reuniões, obrigado
também por me lembrar que eu preciso beber água. Não sei o que seria de mim sem você!
Ao René Juliano Martins, pela amizade, pelos momentos de descontração e cultura, e por
me ajudar na edição deste trabalho!
Aos camundongos, por terem sido instrumentos fundamentais desta pesquisa.
Aos senhores Georges Bizet, Antonio Vivaldi, Wolfgang Amadeus Mozart, Frédéric
Chopin, Johann Sebastian Bach, Ludwig van Beethoven, pelas belas composições que
tanto me inspiraram durante a jornada resumida nas páginas do presente trabalho.
À CAPES, pela concessão da bolsa de mestrado.
À FAPESP, CNPq, CAPES, FAEPEX, CAPES/PROEX e CAPES/DS pelo apoio
financeiro indispensável.
Muito Obrigado!

“Por vezes sentimos que aquilo que fazemos não
é senão uma gota de água no mar. Mas o mar
seria menor se lhe faltasse uma gota.”
(Santa Madre Teresa de Calcutá)

RESUMO
A Distrofia Muscular de Duchenne (DMD) é uma doença genética muscular caracterizada
pela ausência da proteína distrofina. Na DMD e no seu modelo experimental, o camundongo
mdx, a falta da distrofina leva a degeneração muscular progressiva e insuficiência
cardiorrespiratória. Corticóides, tal como o deflazacorte, são a droga de escolha para
tratamento da DMD. Contudo, os efeitos adversos decorrentes do uso contínuo de corticóides,
motivam a busca por fármacos alternativos para o tratamento da DMD. O ômega-3 emerge
como uma terapia complementar para a distrofia, por seus efeitos anti-inflamatórios.
Entretanto, uma vez que os corticóides são a droga de escolha, torna-se importante verificar
os efeitos da associação de um corticóide com o ômega-3 em estágios tardios da doença (13
meses de idade). Estudando músculos afetados em diferentes intensidades pela doença
(diafragma e quadríceps), verificamos que os maiores efeitos da terapia combinada
(deflazacorte + ômega-3) foram observados no diafragma, provavelmente por ser o músculo
mais afetado. A terapia combinada foi mais efetiva que a monoterapia (deflazacorte apenas)
em reduzir os níveis de CK, em promover ganho de força muscular e em diminuir os níveis do
stress oxidativo, demonstrado por diminuição do marcador de peroxidação lipídica 4-HNE.
Moléculas pró-inflamatórias (TNF-α, NF-kB) e pró-fibróticas (TGF-β e fibronectina), foram
igualmente reduzidas por ambas terapias (combinada e monoterapia). No quadríceps, o efeito
mais evidente da terapia combinada em relação à monoterapia foi um aumento do TGF-β, o
que pode indicar aumento de fibrose em estágios posteriores ao do presente estudo, devendo
ser investigado. Este trabalho sugere que o ômega-3, combinado ao deflazacorte, oferece um
adicional terapêutico em relação à monoterapia, ao modular a peroxidação lipídica e estresse
oxidativo, sendo assim uma opção viável para futuros estudos clínicos em que o paciente já
esteja sob ação da terapia com corticóides.
Palavras-chave: Distrofia Muscular de Duchenne; mdx; fibrose; deflazacorte; ômega 3.

ABSTRACT
Duchenne muscular dystrophy (DMD) is a genetic muscle disease characterized by the lack of
dystrophin. In DMD and in the mdx mice model of DMD, lack of dystrophin leads to
progressive skeletal muscle degeneration and cardiorespiratory failure. Corticoids are the only
medication so far available to retard the progression of DMD. However, the secondary
adverse effects of corticoids stimulate the search for new drugs. Omega-3 emerges as a
potential therapy for DMD, due to its anti-inflammatory effects. Given that corticoids, such as
deflazacort, are the golden standard therapy for DMD, it is important to investigate the effects
of a combined therapy of deflazacort and omega-3 on dystrophy progression, at later stages of
the disease (13 months of age). By analyzing muscles that are differently affected by the
absence of dystrophin (diaphragm and quadriceps), we observed that the main effects of the
combined therapy were seen in the diaphragm, possibly for being the most affected muscle at
this period. The combined therapy was more effective than the monotherapy (deflazacort
alone) in reducing the serum levels of CK, in improving anterior limb muscle force and in
decreasing the levels of 4-HNE, a marker of membrane lipid peroxidation and oxidative
stress. Pro inflammatory (TNF-α and NF-kB) and pro fibrotic (TGF-β and fibronectin)
markers were equally reduced by both therapies (combined and monotherapy). In the
quadriceps muscle, the main effect of the combined therapy was to increase the levels of
TGF-β, suggesting a potential increase in fibrosis in the long-run, deserving further
investigation. The present study suggests that omega-3, combined to deflazacort, may offer an
additional therapeutic effect represented by a modulation of the oxidative stress, therefore
being a possible option for future clinical trials in patients that are already under corticoid
therapy.
Keywords: Duchenne muscular dystrophy; mdx; fibrosis; deflazacort; omega-3.

LISTA DE FIGURAS
Figura 1 - Complexo Distrofina-Glicoproteínas. ...................................................................... 19
Figura 2 - Via do ácido araquidônico. ...................................................................................... 22
Figura 3 - Equipamento para obtenção da medida de força dos membros anteriores. ............. 32
Figura 4 - Médias das massas corporais anterior (pré-tratamento) e posteriormente (pós-
tratamento) aos tratamentos. ..................................................................................................... 37
Figura 5 - Medidas de força realizadas anterior (pré-tratamento) e posteriormente (pós-
tratamento) aos tratamentos. ..................................................................................................... 38
Figura 6 - Médias dos níveis de CK obtidos após os tratamentos. ........................................... 39
Figura 7 - Cortes transversais de músculo diafragma corados com Hematoxilina e Eosina. ... 40
Figura 8: Cortes transversais de músculo diafragma corados com Tricrômico de Masson. .... 40
Figura 9 - Porcentagem da área ocupada por fibras com núcleo periférico (NP) no músculo
diafragma. ................................................................................................................................. 41
Figura 10 - Porcentagem da área ocupada por fibrose no músculo diafragma......................... 42
Figura 11 - Porcentagem da área ocupada por inflamação no músculo diafragma. ................. 42
Figura 12 - Porcentagem da área ocupada por fibras com núcleo central (NC) no músculo
diafragma. ................................................................................................................................. 43
Figura 13 - Cortes transversais de músculo quadríceps corados com Hematoxilina e Eosina. 44
Figura 14 - Cortes transversais de músculo quadríceps corados com Tricrômico de Masson. 44
Figura 15 - Porcentagem da área ocupada por fibras com núcleo central (NC) no músculo
quadríceps. ................................................................................................................................ 45
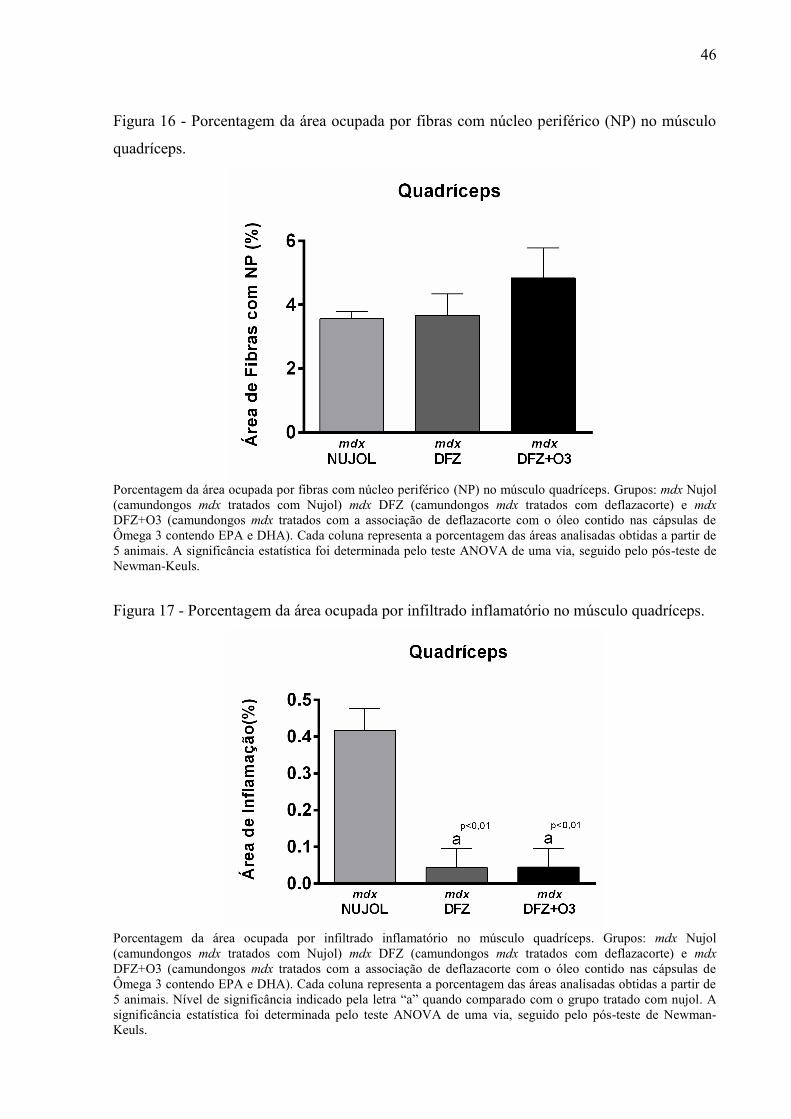
Figura 16 - Porcentagem da área ocupada por fibras com núcleo periférico (NP) no músculo
quadríceps. ................................................................................................................................ 46
Figura 17 - Porcentagem da área ocupada por infiltrado inflamatório no músculo quadríceps.
.................................................................................................................................................. 46
Figura 18 - Porcentagem da área ocupada por fibrose. ............................................................ 47
Figura 19 - Western Blot do músculo diafragma. .................................................................... 48
Figura 20 - Western Blot do músculo quadríceps. ................................................................... 50

LISTA DE ABREVIATURAS
AA – Ácido araquidônico
CDG – Complexo distrofina-glicoproteína
CK – Enzima Creatina Quinase
COX – Cicloxigenase
DFZ - Deflazacorte
DHA – Ácido Docosahexaenóico
DIA – Diafragma
DMD – Distrofia Muscular de Duchenne
EPA – Ácido Eicosapentanóico
ERMO - Espécies reativas do metabolismo do oxigênio
FLA - Fosfolipases A2
GAPDH – gliceraldeído 3-fosfato dehidrogenase
HeE – Hematoxilina e Eosina
mdx – X chromossome-linked muscular dystrophy
LT - Leucotrienos
MEC – Matriz extracelular
NC – Núcleo central
NF-kB - Fator Nuclear-kappa B
NP – Núcleo periférico
O3 – Ômega 3
PG - Prostaglandinas
QUA – Quadríceps
RL – Radical Livre
TGF-β1 - Fator de crescimento transformador beta-1
TM – Tricrômico de Masson
TNF-α _ Fator de necrose tumoral-alfa
TX - Tromboxanos
U/L – Unidade Internacional
4HNE- 4-hidroxinonenal

SUMÁRIO
1 INTRODUÇÃO ...................................................................................................... 18
1.1 DISTROFIA MUSCULAR DE DUCHENNE...................................................... 18
1.2 CAMUNDONGO MDX ....................................................................................... 20
1.3 FISIOPATOLOGIA .............................................................................................. 21
1.3.1 Inflamação ............................................................................................................ 21
1.3.2 Estresse Oxidativo ............................................................................................... 23
1.3.3 Fibrose .................................................................................................................. 24
1.4 DEFLAZACORTE (DFZ) .................................................................................... 25
1.5 ÔMEGA 3 (O3) ..................................................................................................... 26
2 OBJETIVOS .......................................................................................................... 29
2.1 OBJETIVO GERAL.............................................................................................. 29
2.2 OBJETIVOS ESPECÍFICOS ................................................................................ 29
3 MATERIAIS E MÉTODOS ................................................................................. 30
3.1 ANIMAIS .............................................................................................................. 30
3.2 PROTOCOLO EXPERIMENTAL ....................................................................... 30
3.2.1 Grupos tratados com Nujol ................................................................................ 30
3.2.2 Grupo tratado com DFZ ..................................................................................... 30
3.2.3 Grupo tratado com associação de DFZ e O3 .................................................... 31
3.3 MASSA CORPÓREA ........................................................................................... 31
3.4 MEDIDA DA FORÇA .......................................................................................... 31
3.5 ANÁLISE DE CREATINA QUINASE (CK) NO PLASMA SANGUÍNEO ...... 32
3.6 OBTENÇÃO DE MÚSCULOS PARA MORFOMETRIA .................................. 32
3.7 PREPARO DOS MÚSCULOS PARA A TÉCNICA DE HEMATOXILINA E
EOSINA (HE) ....................................................................................................... 33
3.8 PREPARO DOS MÚSCULOS PARA A TÉCNICA DE TRICRÔMICO DE
MASSON (TM) ..................................................................................................... 33
3.9 ANÁLISE MORFOMÉTRICA ............................................................................. 34
3.10 QUANTIFICAÇÃO PROTEICA PELA TÉCNICA DE WESTERN BLOT....... 34
3.10.1 Preparo do extrato total ...................................................................................... 34
3.10.2 Quantificação das proteínas ............................................................................... 34
3.10.3 Anticorpos Primários .......................................................................................... 35
3.10.4 Anticorpos Secundários ...................................................................................... 35
3.11 ANÁLISE ESTATÍSTICA.................................................................................... 36
4 RESULTADOS ...................................................................................................... 37
4.1 MASSA CORPORAL ........................................................................................... 37

4.2 ANÁLISE FUNCIONAL: MEDIDA DE FORÇA ............................................... 37
4.3 ANÁLISE BIOQUÍMICA: NÍVEIS SÉRICOS DE CREATINA QUINASE
(CK) ....................................................................................................................... 38
4.4 ANÁLISE MORFOLÓGICA................................................................................ 39
4.4.1 Músculo Diafragma ............................................................................................. 39
4.4.1.1 Análise Qualitativa ................................................................................................ 39
4.4.1.2 Análise Quantitativa .............................................................................................. 41
4.4.2 Músculo Quadríceps............................................................................................ 43
4.4.2.1 Análise Qualitativa ................................................................................................ 43
4.4.2.2 Análise Quantitativa .............................................................................................. 45
4.5 WESTERN BLOT ................................................................................................. 47
4.5.1 Músculo Diafragma ............................................................................................. 47
4.5.2 Músculo Quadríceps............................................................................................ 49
5 DISCUSSÃO .......................................................................................................... 51
5.1 QUANTO AOS EFEITOS DAS TERAPIAS NA MASSA CORPORAL E
MEDIDA DE FORÇA. ......................................................................................... 51
5.2 QUANTO AOS EFEITOS DAS TERAPIAS NOS NÍVEIS DE CREATINA
QUINASE (CK). ................................................................................................... 52
5.3 QUANTO AOS EFEITOS DAS TERAPIAS NO MÚSCULO DIAFRAGMA .. 53
5.4 QUANTO AOS EFEITOS DAS TERAPIAS NO MÚSCULO QUADRÍCEPS . 59
6 CONCLUSÃO ........................................................................................................ 64
REFERÊNCIAS BIBLIOGRÁFICAS ................................................................. 65
ANEXO A ............................................................................................................... 74
ANEXO B ............................................................................................................... 75

18
1 INTRODUÇÃO
1.1 DISTROFIA MUSCULAR DE DUCHENNE
As distrofias musculares designam um grupo heterogêneo de doenças hereditárias e
progressivas que se manifestam principalmente pela degeneração dos músculos e consequente
fraqueza. Dependendo do tipo de distrofia, os grupos musculares afetados e o prognóstico da
doença são variáveis. Dentre as distrofias, a Distrofia Muscular de Duchenne (DMD) destaca-
se por ser a mais comum e grave, afetando uma em cada 3300 crianças do sexo masculino
(ENGEL et al., 2004).
A DMD foi inicialmente estudada pelo médico francês Guillaume Benjamin Amand
Duchenne em 1868. Duchenne a descreveu como uma paralisia muscular pseudo-hipertrófica.
Para seu diagnóstico, os seguintes sinais deveriam ser observados: enfraquecimento muscular
que se inicia nos membros inferiores, seguido por pseudo-hipertrofia dos mesmos;
comprometimento da deambulação e postura e enfraquecimento muscular conforme a
progressão da doença. William Gowers, neurologista britânico, notou o caráter hereditário da
DMD e descreveu que crianças afetadas pela doença sustentavam-se pelos membros
superiores e escalavam o próprio corpo para assumirem posição ortostática (ENGEL et al.,
2004).
A DMD é uma doença causada pela mutação do gene que codifica a distrofina, uma
proteína de 427 kDa, compreendendo 79 exons, localizado no braço curto do cromossomo X
(locus Xp21) (KOENIG et al., 1987). A distrofina é expressa principalmente nos músculos
esquelético e cardíaco e, em menor proporção, no músculo liso e no tecido nervoso
(HOFMANN et al., 1987). A ocorrência de deleções, duplicações e mutações pontuais pode
resultar em completa perda da expressão de distrofina (RAHIMOV e KUNKEL, 2013). A
distrofina desempenha papel essencial dentro de um complexo oligomérico composto por
outras glicoproteínas, como as sarcoglicanas (α, β, ɤ e δ), distroglicanas (α e β), distrobrevina
e sintrofinas (α e β), formando o complexo distrofina-glicoproteínas (CDG) (figura 1). O
CDG conecta o citoesqueleto de actina da fibra muscular à matriz extracelular circundante
através do sarcolema (MRAK, 1985; HOFFMAN et al., 1987; ENGEL et al., 2004;
MARQUES, 2004). Na ausência de distrofina a formação do CDG fica prejudicada, tornando
o sarcolema instável e susceptível a lesões, levando ao influxo exacerbado de cálcio,
mionecrose, instalação de processo inflamatório com ciclos de degeneração e regeneração
muscular e fibrose (MARIOL e SÉGALAT, 2001).

19
Figura 1 - Complexo Distrofina-Glicoproteínas.
Esquema do complexo distrofina-glicoproteínas representando a α e β distroglicanas; α, β, ɤ, e δ sarcoglicanas; α
e β1 sintrofinas; distrobrevina e distrofina. A distrofina exerce papel importante na estabilização do sarcolema
conectando actina citoesquelética à cadeia da laminina-2 localizada na lâmina basal via complexo de
glicoproteínas (Laboratório de Junção Neuromuscular – IB - UNICAMP).
Os sinais iniciais de comprometimento e/ou atraso motor se manifestam nos primeiros
cinco anos de vida, sendo que as crianças apresentam dificuldade para correr ou subir degraus
e quedas frequentes. A rápida progressão da doença somada ao comprometimento precoce da
musculatura proximal dos membros, perda da deambulação, contraturas articulares e cifo-
escoliose resultam na dependência de cadeira de rodas aproximadamente na primeira década
de vida (ENGEL et al., 2004). Entre a primeira e a segunda década de vida os pacientes
desenvolvem cardiomiopatia (ENGEL et al., 2004; MACHADO, 2011, SPURNEY, 2011).
Embora a utilização de órteses e de ventilação mecânica aumentem a sobrevida do doente, a
insuficiência respiratória e a cardiomiopatia são as principais causas do óbito dos pacientes,
ocorrendo em torno da segunda ou terceira décadas da vida (BOGDANOVICH et al., 2004;
MOXLEY et al., 2005). Não obstante, o uso de anti-inflamatórios esteroidais prolonguem a
sobrevida dos pacientes, ainda não existe cura para a DMD.
Biópsias musculares de pessoas acometidas pela DMD apresentam importantes
alterações morfológicas. Fibras musculares em degeneração são comumente observadas em
músculos distróficos. Observa-se variação do diâmetro das fibras musculares nos estágios
iniciais da doença. O músculo distrófico é progressivamente substituído por tecidos
conjuntivo e adiposo, o que aumenta o seu volume, dando a falsa impressão que o músculo

20
está hipertrofiado, justificando a pseudo-hipertrofia inicialmente proposta por Duchenne
(ENGEL et al., 2004).
1.2 CAMUNDONGO MDX
Após a identificação da alteração genética causadora da DMD foi observado que
outros animais, como aves, cães, peixes, gatos e camundongos também apresentavam
mutações genéticas que resultavam na não expressão de distrofina. Enquanto que em cães a
doença se manifesta de forma mais severa, em que a degeneração muscular se sobrepõe a
regeneração, em camundongos e gatos a fibrose do tecido muscular é mais amena, com maior
processo de regeneração. Embora a progressiva degeneração muscular seja característica em
todos esses animais, alguns aspectos como o quadro clínico, início do processo e capacidade
de regeneração muscular podem ser divergentes (MRAK, 1985; SEIXAS et al., 1997).
Os camundongos mdx (x chromossome-linked muscular dystrophy) foram inicialmente
identificados na colônia C57BL10/ScSn por apresentarem níveis elevados de creatina quinase
(CK), enzima muscular que atua como catalisadora para obtenção de energia a partir de
creatinafosfoquinase e que é liberada no sangue mediante mionecrose (BULFIELD et al.,
1984). Estudos posteriores revelaram que o camundongo mdx apresenta mutação específica
do gene da distrofina (HOFFMAN et al., 1987).
A ausência de distrofina provoca respostas distintas em diferentes músculos. Os
músculos de contração rápida são mais susceptíveis à distrofia e ao envelhecimento em todas
as espécies (LYNCH et al., 2001). No camundongo mdx há um início de necrose a partir do
21º dia pós-natal, aproximadamente, em músculos do membro posterior, como os músculos
tibial anterior (RADLEY e GROUNDS, 2006; SHAVLAKADZE et al., 2004) e sóleo
(PASSAQUIN et al., 2002). Durante a fase inicial as principais alterações histológicas
encontradas no tecido muscular são a necrose e áreas com inflamação. Ao redor da quinta
semana é possível observar que a necrose é seguida por fase de regeneração (GROUNDS,
2008; PASTORET e SEBILLE, 1995; SEIXAS et al., 1997; WHITEHEAD et al., 2006b).
Neste período, diferentemente dos músculos dos membros posteriores, o músculo diafragma
não apresenta necrose. Miofibras são evidentes a partir de 15 dias após o nascimento e o dano
é menor em comparação com os músculos dos membros por pelo menos até 30 dias
(SHAVLAKADZE et al., 2004). Com 12 semanas de idade ainda é possível observar ciclos
de regeneração em resposta à necrose (SHAVLAKADZE et al., 2004). Ao longo do tempo, o
diafragma apresenta mionecrose mais grave do que a musculatura dos membros, com

21
distribuição extensiva de tecido conjuntivo fibroso, de forma semelhante à encontrada na
DMD (LYNCH et al., 1997, STEDMAN et al., 1991)
Por outro lado, alguns músculos da região da cabeça e pescoço, como os músculos
extraoculares (FISHER et al., 2005), masseter (MULLER et al., 2001) e músculos laríngeos
(MARQUES et al., 2007), apresentam quadro patológico leve e são relativamente poupados
da mionecrose. As razões para essa distrofia branda ainda não estão muito esclarecidas,
embora seja observado que esses músculos podem ter uma capacidade melhorada para regular
a homeostase do cálcio (FERRETTI et al., 2009; KHURANA et al., 1995) e propriedades
mecânicas que oferecem resistência aos danos (WIESEN et al., 2007).
Ainda que o camundongo mdx apresente infiltrado inflamatório nas áreas de
mionecrose, difere da DMD por não conter intensa fibrose com depósito de tecido adiposo na
maioria dos músculos esqueléticos. Todavia o diafragma exibe degeneração e fibrose
semelhantes às encontradas em alguns músculos de humanos distróficos. Camundongos mdx
com oito meses de idade, que estão na fase tardia da doença, também apresentam deposição
de tecido adiposo, diminuição da sobrevida, além de cardiomiopatia (MOREIRA et al., 2013;
LEFAUCHEUR et al., 1995;PASTORET e SEBILLE, 1995; SEIXAS et al., 1997).
Embora existam diferenças entre a fisiopatologia e quadro clínico da distrofia entre
humanos e camundongos, o camundongo mdx é considerado excelente modelo para a
investigação dos mecanismos patogênicos da mionecrose e regeneração na DMD. Os
camundongos encontram-se em maior disponibilidade, baixo custo e fácil manuseio, o que
favorece o estudo da doença e desenvolvimento de novas estratégias terapêuticas para uso
potencial na DMD (GROUNDS, 2008; HATHOUT et al., 2016; MARQUES, 2004; SEIXAS
et al., 1997).
1.3 FISIOPATOLOGIA
1.3.1 Inflamação
Considerando o papel estrutural da distrofina, sua deficiência ou ausência torna o
sarcolema instável, susceptível à pequenas lesões que permitirão o aumento do influxo de
cálcio na fibra muscular. O excesso de cálcio altera a homeostase da fibra, levando à
mionecrose (ENGEL, 2004; MARIOL e SÉGALAT, 2001). Consecutivamente, células de
defesa como macrófagos e neutrófilos são recrutadas para o local da injúria, instalando-se um
processo inflamatório crônico (SPENCER e TIDBALL, 2001; TIDBALL, 2004).

22
Mediadores químicos da inflamação são produzidos e colaboram para a progressão do
processo inflamatório. Dentre estes mediadores, destacam-se as prostaglandinas e os
tromboxanos, cuja síntese se dá a partir dos ácidos graxos da membrana, tal como
descreveremos a seguir. O ácido linoleico é um ácido graxo essencial pertencente à família do
ômega-6, encontrado nos fosfolipídios presentes na composição da membrana celular
(FOGAGNOLO MAURÍCIO et al., 2013). Com a alteração da homeostase na fibra muscular
proveniente do excesso de cálcio, as fosfolipases A2 (FLA2), uma família de proteínas
capazes de hidrolisar os fosfolipídios da membrana, são liberadas convertendo o ácido
linoleico em ácido araquidônico (AA). A cascata do ácido araquidônico (AA) é uma
importante via metabólica que utiliza este ácido para síntese de uma gama de mediadores
lipídicos chamados genericamente de eicosanoides (RANG et al., 2007). A presente cascata
pode ser dividida em duas vias principais, uma dependente de ciclo-oxigenase (COX) e outra
dependente de lipo-oxigenase (LOX). A via COX oxidará o AA em prostaglandinas (PG) e
tromboxanos (TX), mediadores da inflamação, enquanto que a via LOX terá como produtos
as lipoxinas e leucotrienos (LT) da série par, que são pró-inflamatórios. (BABCOCK,
HELTON e ESPAT, 2000; CALDER, 2011; FABIAN, KIMLER e HURSTING, 2015;
RANG et al., 2007; SCHELLACK, 2006). (figura 2)
Figura 2 - Via do ácido araquidônico.
Vias de síntese de eicosanoides a partir do AA. COX, ciclo-oxigenases; LOX, lipo-oxigenases; HETE, ácido
hidroxieicosatetraenóico; HPETE ácido hidroperoxieicosatetraenóico; PG, prostaglandina; TX, tromboxano; LT,
leucotrieno. (Adaptado de CALDER et al., 2008)
A produção destes mediadores químicos estimula a chegada de mais células
inflamatórias nos locais com mionecrose, exacerbando a degeneração muscular e aumentando

23
a produção de radicais livres (FOGAGNOLO MAURÍCIO et al., 2013; MACHADO et al.,
2011; SPENCER e TIDBALL, 2001), culminando na ativação da via do fator nuclear-kappa
B (NF-kB), resultando no aumento da expressão de citocinas pró-inflamatórias, tais como o
fator de necrose tumoral alfa (TNF-α) e o fator de transformação do crescimento beta (TGF-β)
(SPENCER e TIDBALL, 2001). O aumento dessas citocinas contribui para a lesão de outras
fibras musculares, que é intensificada pelo aumento do estresse oxidativo (SPENCER e
TIDBALL, 2001; WHITEHEAD et al., 2006).
1.3.2 Estresse Oxidativo
Radical livre (RL) é um termo genérico que designa átomo ou molécula que apresenta
número impar de elétrons na sua última camada eletrônica. Este não emparelhamento de
elétrons confere alta reatividade a esses átomos ou moléculas, possibilitando a captação
(oxidação) ou a doação (redução) de elétrons para outras moléculas (FERREIRA e
MATSUBARA, 1997). A geração de RLs constitui um processo fisiológico uma vez que
esses radicais desempenham papel como mediadores para transferência de elétrons em várias
reações bioquímicas (FERREIRA e MATSUBARA, 1997). Contudo, a sua produção
excessiva desencadeia danos oxidativos (BARBOSA et al., 2010; FERREIRA e
MATSUBARA, 1997; SHAMI e MOREIRA, 2004). A literatura sugere que o termo “radical
livre” não seja o ideal para nomear tais agentes reativos que são patogênicos, uma vez que,
em sua maioria, são derivados do metabolismo do oxigênio (O2) e que nem sempre
apresentam elétrons desemparelhados na sua última camada eletrônica, podendo então ser
nomeados de “espécies reativas do metabolismo do oxigênio” (ERMO) (FERREIRA e
MATSUBARA, 1997).
A produção desequilibrada de ERMOs durante os processos metabólicos resultou no
desenvolvimento de mecanismos de defesa antioxidantes, como forma de controlar a
ocorrência dos danos oxidativos (BIANCHI e ANTUNES, 1999; FERREIRA e
MATSUBARA, 1997). Quando há desequilíbrio entre os compostos oxidantes e
antioxidantes, seja pela demasiada produção de ERMOs ou pela diminuição da remoção dos
mesmos, instala-se um processo denominado estresse oxidativo. Tal processo resulta na
oxidação de biomoléculas e consequente desequilíbrio da homeostase, culminando no dano de
células e tecidos. Quando ocorre a oxidação dos fosfolipídios presentes na membrana celular,
tal processo é denominado de peroxidação lipídica (BARBOSA et al., 2010; HALLIWELL e
WHITEMAN, 2004), processo que resulta na formação de um produto final específico, o 4-

24
hidroxinonenal (4-HNE) (ZHONG e YIN, 2015). Alguns estudos demonstraram que o
camundongo mdx apresenta níveis elevados de 4-HNE quando comparados ao camundongo
controle (FOGAGNOLO-MAURÍCIO et al., 2013; RAPUCCI MORAES et al., 2015). Desta
forma, o 4-HNE se revela um importante marcador do estresse oxidativo resultante da
peroxidação lipídica da membrana celular (FOGAGNOLO-MAURÍCIO et al., 2013;
RAPUCCI MORAES et al., 2015; UCHIDA, 2003; ZHONG e YIN, 2015).
1.3.3 Fibrose
Com o fenômeno da mionecrose, na tentativa de promover a regeneração muscular,
ocorre ativação das células satélites (CS), que são células tronco musculares capazes de se
diferenciar em outras CS ou em mioblastos (MAURO, 1961). Com a progressão da doença
observa-se declínio da capacidade regenerativa dessas células, com consequente deposição de
tecido fibro-adiposo (CONTRERAS et al., 2016).
Existem diversas teorias que tentam justificar a diminuição da capacidade regenerativa
do músculo distrófico. A que parece ser mais aceita sugere que há exaustão de CS devido aos
ciclos repetitivos de degeneração e regeneração (BLAU et al., 1983; SACCO et al., 2010). Tal
exaustão acarreta a perda da habilidade de regeneração do músculo distrófico e após
sucessivos ciclos de degeneração muscular, as fibras musculares são gradualmente
substituídas por componentes de matriz extracelular (MEC) como os tecidos conjuntivos
denso (fibroso) e adiposo, que são produzidos por fibroblastos após ativação de progenitores
fibro-adiposos, como o fator de transformação do crescimento beta (TGF-β1) (CONTRERAS
et al., 2016). A fibrose excessiva prejudica a contratilidade muscular, a vascularização e a
regeneração tecidual (SERRANO et al., 2011). Após a inflamação crônica, os fibroblastos
ativados continuam produzindo proteínas da MEC em excesso, levando à progressiva fibrose
muscular em músculos distróficos (MA et al., 2016).
O TGF-β1 é um mediador pró-fibrótico presente no paciente com DMD e no
camundongo mdx (CHEN et al., 2005). Estudos in vivo com uso de antagonistas ao TGF-β1
confirmaram o papel deste mediador na fibrose muscular e o apontou como um potencial alvo
terapêutico (ACUNA et al., 2014; COHN et al., 2007; MOREIRA et al., 2013).
Outro conhecido mediador pró-fibrótico é a fibronectina. A fibronectina é uma família
de glicoproteínas de elevado peso molecular que está presente na MEC, atuando na adesão
celular à matriz. Estudos recentes demonstraram que os níveis de fibronectina estavam
aumentados no camundongo distrófico quando comparados ao controle (C57BL10),

25
revelando-se um potencial marcador de fibrose (CONTRERAS et al., 2016; SUN et al.,
2016).
1.4 DEFLAZACORTE (DFZ)
Em 1974, Drachman e colaboradores propuseram o uso de esteróides como forma de
tratamento temporário e paliativo para pacientes com DMD, ressaltando os efeitos colaterais e
as variações encontradas no prognóstico da doença mediante seu uso (DRACHMAN et al.,
1974). Desde então, foram realizados estudos randomizados em diversos países na tentativa
de descobrir qual o esteróide e a dosagem mais eficazes para tratamento da DMD
(ANGELINI e PETERLE, 2012).
Há aproximadamente duas décadas, um grupo italiano foi o primeiro a testar o
deflazacorte (DFZ), uma oxazolidina derivada da prednisona. Utilizaram 0,9 mg/kg/dia de
DFZ que seria a dose equivalente a 0,75 mg/kg/dia de prednisona, droga até então conhecida e
utilizada como terapia da DMD (ANGELINI et al., 1994; ANGELINI e PETERLE, 2012).
Foi demonstrado que o deflazacorte produzia efeitos semelhantes ao da prednisona,
porém com menor risco de ganho de peso, fator que contribui para o aumento do período de
deambulação (BONIFATI et al., 2000). Outro estudo demonstrou que meninos tratados com
DFZ apresentaram preservação da função respiratória e possível proteção contra
desenvolvimento de cardiomiopatia em comparação com os meninos do grupo controle, além
de menor perda de massa óssea e redução da escoliose (BIGGAR et al., 2001).Vários ensaios
clínicos randomizados evidenciaram que o uso de deflazacorte promoveu melhora da força
muscular e prolongamento da deambulação (ANGELINI et al., 1994; BIGGAR et al., 2001;
BONIFATI et al., 2000; HOUDE et al., 2008; MCADAM et al., 2012). Estudos em
camundongos mdx revelaram que o uso de DFZ em longo prazo também retarda a progressão
de cardiomiopatia (ACHER et al., 2006; MARQUES et al., 2009; SKRABEK e
ANDERSON, 2001).
Não se sabe exatamente de que forma os corticóides atuam na DMD. Algumas
hipóteses apontam que os esteroides reduzem a mionecrose e inflamação ao interagirem com
receptores intracelulares para facilitar a síntese de lipocortina, que inibe diretamente a FLA2,
impedindo a liberação do ácido araquidônico e consequentemente a formação de todos os
endoperóxidos cíclicos de prostanóides (SCHELLACK, 2006; SPULER e ENGEL, 1998).
Outra hipótese é a de que os esteróides podem atuar na proliferação de células-tronco
precursoras miogênicas, aumentando a regeneração muscular devido a um efeito anabólico

26
(PASQUINI et al., 1995; SKLAR e BROWN et al., 1991). Outro estudo concluiu que o efeito
benéfico dos esteróides na DMD é associado ao aumento da massa muscular mediada pela
inibição da proteólise (RIFAI et al., 1995). Os esteroides também podem atuar aumentando
moléculas sinérgicas, tais como glicoproteínas musculares que complementam a ação da
distrofina. Foi demonstrado que a metilprednisona aumenta a expressão de utrofina, uma
proteína embrionária que apresenta função semelhante à da distrofina (SKLAR e BROWN et
al., 1991).
A eficácia clínica do uso de corticosteroides, como o DFZ, para tratamento da DMD já
está estabelecida. Entretanto, devido ao uso contínuo desta droga, vários efeitos colaterais são
observados, tais como aumento de peso, alterações na puberdade, osteoporose, dentre outros
(ANGELINI e PETERLE, 2012), que afetam a qualidade de vida do paciente. Assim, a busca
por fármacos alternativos é de grande relevância para a terapia farmacológica da DMD
(BEYTÍA et al., 2012; MANZUR et al., 2008).
1.5 ÔMEGA 3 (O3)
Os ácidos graxos são formados por cadeias de átomos de carbono que se ligam a
átomos de hidrogênio com um radical ácido em uma de suas extremidades e podem ser
classificados em três tipos: saturado, monoinsaturado e poli-insaturado de acordo com o tipo
de ligação entre as cadeias (CALDER, 2008). O ácido eicosapentaenóico (EPA)(C20H30O2) e
o ácido docosaexaenóico (DHA)(C22H32O2) são ácidos graxos poli-insaturados em que a
dupla ligação localiza-se no terceiro carbono a partir da extremidade oposta à carboxila.
Portanto, fazem parte da família do ômega 3 (CALDER, 2008). Não são produzidos pelo
organismo humano, devendo assim ser obtido através da dieta. O ômega 3 pode ser
encontrado em castanhas ou na linhaça, sendo o óleo de peixe sua principal fonte (JAMES et.
al., 2000; LEWIS et al., 2016).
Vários ensaios clínicos relataram benefícios com o emprego do ômega-3 em muitas
enfermidades, incluindo doenças cardiovasculares (HARPER et al., 2006), epilepsia
(SCHLANGER et al., 2002), dislipidemias (DAVIDSON e BENES, 2016), doença
inflamatória do intestino (CALDER 2008) e caquexia associada ao câncer (BABCOCK et al.,
2000). O EPA, isoladamente, também se mostrou eficaz na inibição do fator de transcrição
fator nuclear kappa B (NF-kB) (BABCOCK et al., 2000), redução na produção de TNF-α por
macrófagos (BABCOCK et al., 2002) e para evitar os efeitos prejudiciais do TNF-α durante a
diferenciação do músculo esquelético in vitro (MAGEE et al., 2008).

27
No organismo, o EPA atua gerando eicosanoides da série impar, que são anti-
inflamatórios, e na diminuição de citocinas pró-inflamatórias, como o TNF-α (CALDER et
al., 2008). O mecanismo do EPA na diminuição dos níveis desta citocina não está muito
esclarecido. Uma hipótese é que ele diminui a transcrição do gene do TNF-α através da
redução da translocação de NF-kB, elemento que regula o processo inflamatório (CALDER,
2008). Os efeitos de proteção do EPA podem estar relacionados com a sua habilidade de
modular as respostas inflamatórias e imunológicas. O EPA inibe a dissociação entre IkB
(proteína inibitória) e NF-kB, impedindo que este último seja translocado para o núcleo,
inibindo a transcrição do gene do TNF-α (BABCOCK et. al., 2000; CALDER, 2008;
MACHADO et al., 2011). Outra hipótese é que o EPA e o DHA também podem inibir a
produção de eicosanoides da série par (pró-inflamatórios) gerados a partir do AA, uma vez
que são capazes de atuar como substrato nas vias COX e LOX, dando origem a eicosanóides
da série impar, que tem ação anti-inflamatória (CALDER, 2008).
O DHA é o ácido graxo mais insaturado presente em membranas. Esse ácido graxo
influencia as propriedades biofísicas de membranas, tais como fluidez e capacidade de
deformação (JUDÈ et al., 2006; TUAZON e HENDERSON, 2012). Estudos em nosso
laboratório (MACHADO et al., 2011; FOGAGNOLO MAURÍCIO et al., 2012)
demonstraram que o EPA purificado ou a combinação de EPA com DHA em camundongos
mdx na fase inicial da doença, melhorou a distrofinopatia ao promover a estabilidade do
sarcolema, talvez como resultado da incorporação do DHA na membrana. Tal incorporação
também pode proteger a fibra distrófica contra o aumento do influxo de cálcio, naturalmente
observado nessas fibras, possivelmente por efeitos diretos do DHA nos canais de cálcio da
membrana (FOGAGNOLO MAURÍCIO et al., 2012; MACHADO et al., 2011). Observou-se
ainda que o ômega 3 reduziu os níveis de TNF-α e 4-HNE, além de diminuir a mionecrose e
as áreas com infiltrado inflamatório (FOGAGNOLO MAURÍCIO et al., 2012).
A DMD é extensamente estudada, tanto sob o ponto de vista molecular quanto
genético. Terapias genéticas encontram-se em franco desenvolvimento (BEYTÍA et al.,
2012), envolvendo exon skipping (KOLE e KRIEG, 2015) e novas abordagens terapêuticas
em humanos, com uso do Ataluren (MUNTONI et al., 2016) e Eteplirsen (KINANE et al.,
2016).
Entretanto, estas novas terapias estão disponíveis somente para casos de mutações
específicas, sendo de uso bastante restrito. Desta forma, abordagens terapêuticas com
fármacos já em uso para outras doenças humanas, com efeitos colaterais e doses bem
estabelecidas para humanos, são de grande interesse para a DMD. Uma vez que os corticóides

28
ainda são a droga de escolha para a maioria dos pacientes com DMD, torna-se de grande
relevância estudar os efeitos de outras drogas combinadas ao deflazacorte.
Considerando as propriedades anti-inflamatórias do ácido eicosapentaenóico e seus
efeitos positivos na distrofia do camundongo mdx (FOGAGNOLO MAURÍCIO et al., 2012;
MACHADO et al., 2011), levantamos a hipótese de que a associação do DFZ com o ômega-3
poderia ser mais eficaz do que a terapia com o DFZ isolado, para retardar a progressão da
doença.

29
2 OBJETIVOS
2.1 OBJETIVO GERAL
O objetivo do presente trabalho é verificar se a terapia com ômega 3 (EPA + DHA) em
associação ao deflazacorte (DFZ) tem melhores efeitos que a terapia com DFZ, na proteção
dos músculos estriados esqueléticos quadríceps e diafragma em camundongos mdx na fase
tardia da doença (13 meses).
2.2 OBJETIVOS ESPECÍFICOS
Verificar se a terapia com ômega 3 (EPA + DHA) em associação ao deflazacorte (DFZ)
tem melhores efeitos que a monoterapia com DFZ contra a mionecrose em
camundongos mdx na fase tardia da doença.
Observar se a terapia combinada proporciona melhor efeito anti-inflamatório que a
terapia com DFZ, a ser demonstrado pelos níveis de mediadores inflamatórios TNF-α e
NF-kB e pela área de inflamação dos músculos quadríceps e diafragma.
Investigar se a terapia combinada assegura maior diminuição da fibrose quando
comparada com a monoterapia, na avaliação dos níveis de TGF-β1 e de fibronectina, e
da área de fibrose dos músculos quadríceps e diafragma.
Verificar se a terapia combinada possibilita maior diminuição do estresse oxidativo
quando comparada com a terapia com DFZ na avaliação dos níveis de 4-HNE.

30
3 MATERIAIS E MÉTODOS
3.1 ANIMAIS
No presente trabalho foram utilizados camundongos com oito meses de idade, de
ambos os sexos, da linhagem C57BL/10-Dmdmdx/PasUnib (mdx) e da linhagem
C57BL/10ScCr/PasUnib (controle). Os animais foram adquiridos no Biotério Central da
UNICAMP e mantidos no Biotério do Laboratório de Anatomia do Instituto de Biologia da
UNICAMP em caixas padrão (28,5cm de comprimento, 18cm de largura e 13cm de altura)
em ambiente controlado (23º C, 12 horas de ciclo claro/escuro) com ração Nuvilab CR-1
(Nuvital, Paraná, Brasil) e água ad libitum. O uso de animais para o presente trabalho está de
acordo com os Princípios Éticos na Experimentação Animal, sob protocolo nº 3806-1,
aprovado pelo Comitê de Ética no Uso de Animais da Universidade Estadual de Campinas
(CEUA-UNICAMP) (ANEXO A).
Os animais foram divididos em quatro grupos experimentais:
Camundongos C57BL/10 tratados com nujol (n=10)
Camundongos mdx tratados com nujol (n=15);
Camundongos mdx tratados com DFZ (n=15);
Camundongos mdx tratados com associação de DFZ e O3(n=15);
3.2 PROTOCOLO EXPERIMENTAL
3.2.1 Grupos tratados com Nujol
Os camundongos da linhagem C57BL/10 e mdx foram tratados com óleo mineral
(Nujol, Mantecorp, São Paulo, Brasil) via gavagem. Tal tratamento não apresenta efeito
farmacológico algum sobre a fisiologia do animal, sendo realizado apenas para equiparar o
estresse do procedimento de gavagem entre os grupos (FOGAGNOLO et al., 2012). O
tratamento teve início quando os animais completaram oito meses de idade e foram tratados
três vezes por semana durante cinco meses.
3.2.2 Grupo tratado com DFZ

31
Os camundongos distróficos foram tratados com 1,2 mg/kg de DFZ (Libbs, Brasil) em
meio aquoso via gavagem, três vezes por semana durante cinco meses (CARVALHO et al.,
2013). O tratamento teve início quando os animais completaram oito meses de idade.
3.2.3 Grupo tratado com associação de DFZ e O3
Os camundongos distróficos foram tratados com óleo obtido das cápsulas de Omega-3
EPA 1000mg (Fabricado por FDC vitamins, Inc., Miami, Flórida, EUA. Importado e
distribuído exclusivamente por Fedco Ind. Ltda.), sendo que receberam 300mg/kg de ômega 3
contendo 0,4g de EPA, 0,2g de DHA, 2mg de vitamina E, 0,9g de proteínas, 2,0g de gorduras
totais, 0,4g de gordura saturada, 0,0g de gordura trans, 0,0g de gordura monossaturada e 1,0g
de gordura poli-insaturada (FOGAGNOLO MAURÍCIO et al., 2012). Concomitantemente,
foi administrado 1,2 mg/kg de DFZ (Libbs, Brasil) em meio aquoso (CARVALHO et al.,
2013). O tratamento iniciou-se quando os animais completaram oito meses de idade e ambas
as drogas foram administradas via gavagem três vezes por semana durante cinco meses.
3.3 MASSA CORPÓREA
Para todos os grupos analisados, foi realizada a medida das massas corpóreas dos
camundongos antes do início do tratamento para ajuste de dosagem da terapia farmacológica,
e após o término dos tratamentos propostos para análise da massa antes e após o tratamento.
Para tal foi utilizada uma balança modelo AS2000C (Marte balanças e equipamentos®).
3.4 MEDIDA DA FORÇA
Os camundongos de todos os grupos analisados tiveram as medidas de força obtidas
antes do início e após o término dos tratamentos sugeridos. A força foi quantificada no
aparelho de medida de força horizontal (Gripstrength; Newprimer®). Durante a avaliação os
animais foram estabilizados pela cauda e com as patas dianteiras seguraram uma tela de nylon
ligada a um transdutor (figura 3), que transmitiu a força de contração dos músculos (em
quilogramas) para o aparelho. O pico de força de cinco tentativas mostradas no visor do
aparelho foram colhidas e utilizadas para composição da média de cada animal, sendo o
resultado final expresso em força/massa (g/g) (MESSINA et al., 2009; TANIGUTI et al.,
2011).

32
Figura 3 - Equipamento para obtenção da medida de força dos membros anteriores.
O animal segura a argola tracionando-a posteriormente, exercendo força muscular que é registrada no visor
digital, em quilogramas. (Laboratório de Junção Neuromuscular – IB – UNICAMP).
3.5 ANÁLISE DE CREATINA QUINASE (CK) NO PLASMA SANGUÍNEO
A quantificação de CK no plasma sanguíneo é um indicador do processo de
mionecrose. Para coleta de sangue os animais foram sacrificados com dose letal de anestésico
composto por uma mistura de cloridrato de cetamina (130mg/kg Francotar, Virbac, São Paulo,
Brasil) e cloridrato de xilazina (6,8 mg/kg, 2%, Virbaxyl, Virbac), por via intraperitoneal.
Após anestesia foi realizada toracotomia com exposição do coração, e no ventrículo direito foi
realizada punção utilizando-se uma agulha com seringa contendo anticoagulante. Um volume
de aproximadamente 1 ml de sangue foi aspirado e transferido para um ependorf. Em seguida,
o sangue foi centrifugado (centrifuga refrigerada Sigma® 3-18k) nas seguintes condições:
936g, 4°C por 15 minutos, separando-se o plasma dos elementos figurados. O plasma foi
utilizado para análise. Para quantificação da CK foi utilizado o Kit CK da Bioclin®. As
absorbâncias das amostras foram lidas a 25°C utilizando-se espectrofotômetro U.V. (Thermo
Electron Corporation® Spectrophotometer Genesys 20) com comprimento de onda de 340 nm
e cubetas de quartzo de 1 cm de caminho óptico. Os valores foram expressos em U/L
(FOGAGNOLO MAURÍCIO et al., 2012).
3.6 OBTENÇÃO DE MÚSCULOS PARA MORFOMETRIA
Após a coleta de sangue para quantificação de CK plasmático foi realizada perfusão
vascular com injeção de 10ml de solução tampão fosfato (PBS) em ventrículo esquerdo,
obedecendo-se o trajeto da circulação sistêmica. Uma pequena incisão foi realizada no átrio
direito para ejeção do sangue remanescente na circulação. Em seguida, os músculos
diafragma e quadríceps (terço médio) foram dissecados, retirados e fixados em suportes de

33
madeira com uma cola resultante da mistura de Tissue-Tek com cola Tragacanth e imersos em
isopentano à -80ºC por 40 segundos e imediatamente colocados em nitrogênio líquido à -
159ºC. Os suportes com os músculos foram retirados do nitrogênio e armazenados à -80ºC em
Biofreezer (BioFreezer Jouan, VX380). Para obtenção dos cortes histológicos, os músculos
foram mantidos por aproximadamente 30 minutos no criostato até atingirem a temperatura de
-26ºC e seccionados transversalmente na espessura de 8μm utilizando criostato (Leica
CM1860).
3.7 PREPARO DOS MÚSCULOS PARA A TÉCNICA DE HEMATOXILINA E EOSINA
(HE)
Os cortes congelados foram inicialmente fixados com solução de Bouin por três horas
e em seguida, imersos em etanol durante duas horas. Após esta etapa, as lâminas foram
lavadas em água corrente por dez minutos e os cortes corados com hematoxilina de Harris e
em seguida, com eosina. Posteriormente, os cortes foram desidratados em séries de etanol e
diafanizados em xilol e as lâminas foram montadas com uso de Entellan e lamínulas para
posterior observação em microscopia de luz. Para esta coloração foram quantificadas as áreas
com presença de infiltrado inflamatório, evidenciada pela presença de células
polimorfonucleares, assim como a área de fibras regeneradas, indicada pelas fibras com
núcleo central. (BRIGUET et al., 2004)
3.8 PREPARO DOS MÚSCULOS PARA A TÉCNICA DE TRICRÔMICO DE MASSON
(TM)
Os cortes foram fixados conforme protocolo de HE e em seguida foram lavados em
água corrente por dez minutos e corados com hematoxilina de Harris por oito minutos. Após
este período, as lâminas foram lavadas novamente em água corrente e imersas em solução de
Masson por 15 minutos. As lâminas foram então banhadas em solução de ácido acético a
0,2% e rapidamente mergulhadas em solução de azofloxina (AFO) por cinco minutos e em
seguida, em solução de verde luz (light green) por 30 segundos. Após esta etapa, os cortes
foram banhados em ácido acético a 0,2% para serem submetidos à desidratação em série de
etanol e à diafanização com xilol. As lâminas foram montadas com Entellan e os cortes
observados em microscópio de luz e submetidos à morfometria. Para esta coloração foi
quantificada área ocupada por tecido conjuntivo, indicativo de fibrose.

34
3.9 ANÁLISE MORFOMÉTRICA
A captação de imagens foi realizada com fotomicroscópio (Nikon Eclipse E-400) com
objetiva de 10X acoplado a um computador e vídeo câmera (Nikon Express Series,
Shinagawa, Tokyo, Japão) para documentação fotográfica. As análises morfométricas
propostas foram realizadas com software Image-Pro Plus® Version 6.0.0.260.
3.10 QUANTIFICAÇÃO PROTEICA PELA TÉCNICA DE WESTERN BLOT
Foi verificado se o tratamento proposto alterou a quantidade de proteínas relacionadas
à inflamação (TNF-α, NF-kB), fibrose (TGF-β1, fibronectina) e estresse oxidativo (4-HNE).
Os valores foram normalizados com a proteína gliceraldeído 3-fosfato dehidrogenase
(GAPDH). Para isso, foram utilizados nove animais de cada grupo.
3.10.1 Preparo do extrato total
Após o término dos tratamentos, os animais foram sacrificados e foi realizada perfusão
vascular com PBS conforme apresentado nos itens 3.4 e 3.5, e os músculos diafragma e
quadríceps foram retirados. Tais músculos foram seccionados em pequenos fragmentos e
homogeneizados por 30 segundos em velocidade máxima (homogeneizador Polytron PTA
20S modelo PT10/35; Brinkmann Instruments, Westbury, NY, EUA) com 2 ml de solução
tampão a 4ºC (Triton X-100 1%, tris-HCl 100mM [pH 7,4], pirofosfato de sódio 100mM,
fluoreto de sódio 100mM, ETDA 10mM, ortovanadato de sódio 10 mM, PMSF 2 mM e
0,1mg/ml de aprotinina). Os extratos foram então centrifugados a 11000 rpm, à 4ºC por 30
minutos, sendo o sobrenadante utilizado para análise do extrato total. A quantificação proteica
total foi realizada pelo método de Bradford (1976).
3.10.2 Quantificação das proteínas
Às amostras do extrato proteico foi adicionado tampão Laemmli (azul de bromofenol
0,04 mg/ml eTris-HCl 0,12 M, glicerol 20%, SDS 2% e ß-mercaptoetanol 0,28 M) e
aquecidas por 5 minutos a 100ºC. Em seguida, 60µg de proteína foram aplicados em gel SDS-
poliacrilamida 8-15% em aparelho para eletroforese (mini-Protean, Bio-Rad Laboratories,
Richmond, CA, EUA). A eletrotransferência do gel para a membrana de nitrocelulose foi
realizada em 90 minutos a 120 V em aparelho de transferência (mini-Protean, Bio-Rad

35
Laboratories, Richmond, CA, EUA). As membranas foram incubadas com solução basal
(Trisma base 10 mM, cloreto de sódio 150 mM e Tween-20 0,02%) com os respectivos
anticorpos primários a 4ºC, durante 12 horas. Após este período as membranas foram lavadas
por 30 minutos com solução basal, para depois serem incubadas com anticorpos secundários,
diluídos em 10 ml de solução basal por duas horas, em temperatura ambiente. Posteriormente,
as membranas foram lavadas por 30 minutos com solução basal. As diluições dos anticorpos
primários e secundários foram definidas de acordo com a marcação obtida.
Para detectar as bandas imunorreativas, as membranas foram expostas a solução de
quimiluminescência (Super Signal West Pico Chemiluminescente, Pierce Biotechnology,
Rockford, Illinois, USA) por 5 minutos, seguida de exposição no G-Box A detecção das
bandas imunofluorescentes e a quantificação foram realizadas no aparelho G-Box Chemi pelo
software de aquisição de imagem GeneSnap (Syngene, Maryland-EUA). As densidades das
bandas foram quantificadas pelo software de análise GeneTools (Syngene, Maryland-EUA).
3.10.3 Anticorpos Primários
TNF-α (Rabbit anti-mouse tumor necrosis factor-alpha polyclonal antibody–TNF-α-
Millipore);
NF-kB (rabbit/goat polyclonal Santa Cruz Biotechnology. Santa Cruz, California,
USA);
TGF-β1 (mouse monoclonal; Sigma-Aldrich, St Louis, Missouri, USA);
4-HNE (goat polyclonal; Santa Cruz Biotechnology, Santa Cruz, California, USA);
Fibronectina (Abcam discover more, Cambridge)
GAPDH Glyceraldehyde 3-phosphate dehydrogenase- (Santa Cruz Biotechnology,
Santa Cruz, California, USA).
3.10.4 Anticorpos Secundários
IgG rabbit, mouse e goat conjugados à peroxidase, correspondente do anticorpo
primário (KPL, Gaithersburg, Maryland, USA).

36
3.11 ANÁLISE ESTATÍSTICA
Para verificação da normalidade dos dados foi utilizado o teste Kormogorov-Smirnov.
Todos os resultados estão apresentados como média ± desvio padrão. Quando necessária
comparação direta entre as médias de dois grupos experimentais foi realizado o teste t de
Student. A comparação entre três grupos foi realizada através de análise de variância
(ANOVA) de uma via, seguido pelo pós-teste de Newman-Keuls. Em todas as análises, os
valores de p foram considerados estatisticamente significativos apenas quando inferiores a
0,05 (p<0,05). Para todos os cálculos estatísticos foi utilizado o software GraphPad Prism
versão 6.01 (GraphPad Software, Inc., San Diego, CA).

37
4 RESULTADOS
4.1 MASSA CORPORAL
Para analisar se os tratamentos com DFZ e DFZ+O3 afetaram o crescimento somático
dos animais, foi realizada comparação de suas massas corporais antes do início e após o
término do tratamento (Figura 4). Não foi observada diferença significativa na média da
massa corporal entre o início e o final do tratamento de todos os grupos analisados.
Figura 4 - Médias das massas corporais anterior (pré-tratamento) e posteriormente (pós-
tratamento) aos tratamentos.
Médias das massas corporais anterior (pré-tratamento) e posteriormente (pós-tratamento) aos tratamentos
propostos em cada grupo. Grupos: C57BL10 (camundongos C57BL10 tratados com Nujol), mdx Nujol
(camundongos mdx tratados com nujol), mdx DFZ (camundongos mdx tratados com deflazacorte) e mdx
DFZ+O3 (camundongos mdx tratados com a associação de deflazacorte com o óleo contido nas cápsulas de
Ômega 3 contendo EPA e DHA). Cada coluna representa a média das massas corporais mensuradas a partir de
12 animais. A significância estatística foi determinada pelo teste t de Student.
4.2 ANÁLISE FUNCIONAL: MEDIDA DE FORÇA
Comparando as medidas de força obtidas antes do início do tratamento com as obtidas
após o término do tratamento de cada grupo estudado, nota-se que o animal normal
(C57BL/10) apresenta discreto aumento (9,8% de aumento; p<0,05 em relação ao início) da
força muscular dos membros anteriores (Figura 5). Este aumento não foi observado nos
animais distróficos tratados com nujol e com deflazacorte. Os animais mdx tratados com a
combinação apresentaram aumento significativo (14,6%) da força dos membros anteriores ao
final da terapia (p<0,05).

38
Figura 5 - Medidas de força realizadas anterior (pré-tratamento) e posteriormente (pós-
tratamento) aos tratamentos.
Medidas de força realizadas anterior (pré-tratamento) e posteriormente (pós-tratamento) aos tratamentos
propostos em cada grupo. Grupos: C57BL10 (camundongos C57BL10 tratados com Nujol), mdx Nujol
(camundongos mdx tratados com nujol), mdx DFZ (camundongos mdx tratados com deflazacorte) e mdx
DFZ+O3 (camundongos mdx tratados com a associação de deflazacorte com o óleo contido nas cápsulas de
Ômega 3 contendo EPA e DHA). Cada coluna representa a média das medidas de força obtidas a partir de 10-11
animais. Nível de significância indicado pela letra “a” quando comparado com o início do tratamento (P<0,05).
A significância estatística foi determinada pelo teste t de Student.
4.3 ANÁLISE BIOQUÍMICA: NÍVEIS SÉRICOS DE CREATINA QUINASE (CK)
Como esperado, os camundongos mdx apresentaram elevação significativa (8,6 vezes
de aumento) da CK em relação ao animal normal (C57BL/10). Os camundongos mdx tratados
com DFZ apresentaram diminuição significativa de 68% da CK quando comparados aos
camundongos mdx tratados com nujol. Já os níveis de CK dos camundongos mdx tratados
com a associação de DFZ+O3 apresentaram diminuição de 84% quando comparados ao grupo
tratado com nujol e de 57% quando comparados ao grupo tratado com DFZ, sendo essa
diminuição significativa em ambas as comparações. Não foi observada diferença significativa
dos níveis de CK entre os grupos CT e DFZ+O3 (figura 6).

39
Figura 6 - Médias dos níveis de CK obtidos após os tratamentos.
Médias dos níveis de CK obtidos após os tratamentos propostos em cada grupo. Grupos: C57BL10
(camundongos C57BL10 tratados com Nujol), mdx Nujol (camundongos mdx tratados com nujol), mdx DFZ
(camundongos mdx tratados com deflazacorte) e mdx DFZ+O3 (camundongos mdx tratados com a associação de
deflazacorte com o óleo contido nas cápsulas de Ômega 3 contendo EPA e DHA). Cada coluna representa a
média dos níveis de CK obtidas a partir de 9-15 animais. Nível de significância indicado pela letra “a” quando
comparado com o grupo C57BL10, “b” quando comparado com o grupo mdx tratado com Nujol, “c” quando
comparado com o grupo mdx tratado com DFZ. A significância estatística foi determinada pelo teste ANOVA de
uma via, seguido pelo pós-teste de Newman-Keuls.
4.4 ANÁLISE MORFOLÓGICA
4.4.1 Músculo Diafragma
4.4.1.1 Análise Qualitativa
No músculo diafragma do grupo C57BL10, foram observadas majoritariamente fibras
com núcleo periférico, característica do músculo normal, enquanto que no diafragma dos
animais distróficos foi possível visualizar fibras com núcleo periférico e com núcleo central,
característica de fibras normais e regeneradas, respectivamente, além de pequenas áreas com
celularidade aumentada, representando áreas com presença de inflamação (figura 7). A
presença de fibrose foi evidenciada em todos os grupos dos animais mdx, através do tecido
conjuntivo corado por verde-luz pela coloração de Tricrômico de Masson (figura 8).

40
Figura 7 - Cortes transversais de músculo diafragma corados com Hematoxilina e Eosina.
Cortes transversais de músculo diafragma corados com Hematoxilina e Eosina. Em A, músculo de camundongo
C57BL10. Em B, músculo de camundongo mdx tratado com nujol. Em C, músculo de camundongo mdx tratado
com DFZ. Em D, músculo de camundongo mdx tratado com DFZ+O3. Indicados: área de inflamação (asterisco),
fibra com núcleo periférico (seta) e fibra com núcleo central (cabeça de seta). Escala 100µm.
Figura 8: Cortes transversais de músculo diafragma corados com Tricrômico de Masson.
Cortes transversais de músculo diafragma corados com Tricrômico de Masson. Em A, músculo de camundongo
C57BL10. Em B, músculo de camundongo mdx tratado com nujol. Em C, músculo de camundongo mdx tratado
com DFZ. Em D, músculo de camundongo mdx tratado com DFZ+O3. Área de fibrose representada pela cor
verde-luz. Escala 100µm.

41
4.4.1.2 Análise Quantitativa
Ambos os tratamentos protegeram o músculo diafragma da distrofinopatia no estágio
tardio da doença, promovendo aumento significativo das fibras com núcleo periférico (50%
de aumento no grupo tratado com DFZ e 40% no grupo tratado com DFZ+O3 quando
comparados ao grupo tratado com nujol) (figura 9). Foi observada redução da área de fibrose
dos animais tratados com DFZ+O3 (40,6±5,1%) e dos animais tratados com DFZ
(36,1±4,4%), quando comparadas ao grupo tratado com Nujol (57,9±5,9%) (figura 10). Não
foi observada diferença significativa nas áreas com inflamação (figura 11) e fibras
regeneradas (figura 12), indicadas por centralização nuclear.
Figura 9 - Porcentagem da área ocupada por fibras com núcleo periférico (NP) no músculo
diafragma.
Porcentagem da área ocupada por fibras com núcleo periférico (NP) no músculo diafragma. Grupos: mdx Nujol
(camundongos mdx tratados com Nujol), mdx DFZ (camundongos mdx tratados com deflazacorte) e mdx
DFZ+O3 (camundongos mdx tratados com a associação de deflazacorte com o óleo contido nas cápsulas de
Ômega 3 contendo EPA e DHA). Cada coluna representa a porcentagem das áreas analisadas obtidas a partir de
5 animais. Nível de significância indicado pela letra “a” quando comparado com o grupo tratado com nujol. A
significância estatística foi determinada pelo teste ANOVA de uma via, seguido pelo pós-teste de Newman-
Keuls.

42
Figura 10 - Porcentagem da área ocupada por fibrose no músculo diafragma.
Porcentagem da área ocupada por fibrose no músculo diafragma. Grupos: mdx Nujol (camundongos mdx
tratados com Nujol), mdx DFZ (camundongos mdx tratados com deflazacorte) e mdx DFZ+O3 (camundongos
mdx tratados com a associação de deflazacorte com o óleo contido nas cápsulas de Ômega 3 contendo EPA e
DHA). Cada coluna representa a porcentagem das áreas analisadas obtidas a partir de 5 animais. Nível de
significância indicado pela letra “a” quando comparado com o grupo tratado com nujol. A significância
estatística foi determinada pelo teste ANOVA de uma via, seguido pelo pós-teste de Newman-Keuls.
Figura 11 - Porcentagem da área ocupada por inflamação no músculo diafragma.
Porcentagem da área ocupada por inflamação no músculo diafragma. Grupos: mdx Nujol (camundongos mdx
tratados com Nujol), mdx DFZ (camundongos mdx tratados com deflazacorte) e mdx DFZ+O3 (camundongos
mdx tratados com a associação de deflazacorte com o óleo contido nas cápsulas de Ômega 3 contendo EPA e
DHA). Cada coluna representa a porcentagem das áreas analisadas obtidas a partir de 5 animais.. A significância
estatística foi determinada pelo teste ANOVA de uma via, seguido pelo pós-teste de Newman-Keuls.

43
Figura 12 - Porcentagem da área ocupada por fibras com núcleo central (NC) no músculo
diafragma.
Porcentagem da área ocupada por fibras com núcleo central (NC) no músculo diafragma. Grupos: mdx Nujol
(camundongos mdx tratados com Nujol), mdx DFZ (camundongos mdx tratados com deflazacorte) e mdx
DFZ+O3 (camundongos mdx tratados com a associação de deflazacorte com o óleo contido nas cápsulas de
Ômega 3 contendo EPA e DHA). Cada coluna representa a porcentagem das áreas analisadas obtidas a partir de
5 animais. A significância estatística foi determinada pelo teste ANOVA de uma via, seguido pelo pós-teste de
Newman-Keuls.
4.4.2 Músculo Quadríceps
4.4.2.1 Análise Qualitativa
O músculo quadríceps do grupo C57BL10 apresentou fibras com núcleo periférico,
característica histológica do músculo normal, enquanto que nos quadríceps dos animais
distróficos foi notada predominância de fibras com núcleo central, de diâmetros variados,
característica de fibras regeneradas (figura 13). Aparentemente, nenhuma área de inflamação
ou fibrose foram observadas no músculo quadríceps. (figura 14)

44
Figura 13 - Cortes transversais de músculo quadríceps corados com Hematoxilina e Eosina.
Cortes transversais de músculo quadríceps corados com Hematoxilina e Eosina. Em A, músculo de camundongo
C57BL10. Em B, músculo de camundongo mdx tratado com nujol. Em C, músculo de camundongo mdx tratado
com DFZ. Em D, músculo de camundongo mdx tratado com DFZ+O3. Indicados: fibra com núcleo periférico
(seta) e fibra com núcleo central (cabeça de seta). Escala 100µm.
Figura 14 - Cortes transversais de músculo quadríceps corados com Tricrômico de Masson.
Cortes transversais de músculo quadríceps corados com Tricrômico de Masson. Em A, músculo de camundongo
C57BL10. Em B, músculo de camundongo mdx tratado com nujol. Em C, músculo de camundongo mdx tratado
com DFZ. Em D, músculo de camundongo mdx tratado com DFZ+O3. Área de fibrose representada pela cor
verde-luz. Escala 100µm.

45
4.4.2.2 Análise Quantitativa
Todos os grupos distróficos apresentaram predomínio de área regenerada, representada
por fibras com centralização nuclear (95,1± 0,6%, mdx Nujol; 95,7± 0,2%, mdx DFZ; 95,3±
1,3% mdx DFZ+O3), porém não houve diferença significativa entre os grupos (figura 15). A
área ocupada por fibras com o núcleo periférico representou aproximadamente 4% da área
total do músculo quadríceps, enquanto que o restante esteve ocupado por diminuta área de
inflamação e fibrose, que estão quantificadas nas figuras 16, 17 e 18, respectivamente.
Figura 15 - Porcentagem da área ocupada por fibras com núcleo central (NC) no músculo
quadríceps.
Porcentagem da área ocupada por fibras com núcleo central (NC) no músculo quadríceps. Grupos: mdx Nujol
(camundongos mdx tratados com Nujol), mdx DFZ (camundongos mdx tratados com deflazacorte) e mdx
DFZ+O3 (camundongos mdx tratados com a associação de deflazacorte com o óleo contido nas cápsulas de
Ômega 3 contendo EPA e DHA). Cada coluna representa a porcentagem das áreas analisadas obtidas a partir de
5 animais. A significância estatística foi determinada pelo teste ANOVA de uma via, seguido pelo pós-teste de
Newman-Keuls.
46
Figura 16 - Porcentagem da área ocupada por fibras com núcleo periférico (NP) no músculo
quadríceps.
Porcentagem da área ocupada por fibras com núcleo periférico (NP) no músculo quadríceps. Grupos: mdx Nujol
(camundongos mdx tratados com Nujol) mdx DFZ (camundongos mdx tratados com deflazacorte) e mdx
DFZ+O3 (camundongos mdx tratados com a associação de deflazacorte com o óleo contido nas cápsulas de
Ômega 3 contendo EPA e DHA). Cada coluna representa a porcentagem das áreas analisadas obtidas a partir de
5 animais. A significância estatística foi determinada pelo teste ANOVA de uma via, seguido pelo pós-teste de
Newman-Keuls.
Figura 17 - Porcentagem da área ocupada por infiltrado inflamatório no músculo quadríceps.
Porcentagem da área ocupada por infiltrado inflamatório no músculo quadríceps. Grupos: mdx Nujol
(camundongos mdx tratados com Nujol) mdx DFZ (camundongos mdx tratados com deflazacorte) e mdx
DFZ+O3 (camundongos mdx tratados com a associação de deflazacorte com o óleo contido nas cápsulas de
Ômega 3 contendo EPA e DHA). Cada coluna representa a porcentagem das áreas analisadas obtidas a partir de
5 animais. Nível de significância indicado pela letra “a” quando comparado com o grupo tratado com nujol. A
significância estatística foi determinada pelo teste ANOVA de uma via, seguido pelo pós-teste de Newman-
Keuls.

47
Figura 18 - Porcentagem da área ocupada por fibrose.
Porcentagem da área ocupada por fibrose. Grupos: mdx Nujol (camundongos mdx tratados com Nujol) mdx DFZ
(camundongos mdx tratados com deflazacorte) e mdx DFZ+O3 (camundongos mdx tratados com a associação de
deflazacorte com o óleo contido nas cápsulas de Ômega 3 contendo EPA e DHA). Cada coluna representa a
porcentagem das áreas analisadas obtidas a partir de 5 animais. Nível de significância indicado pela letra “a”
quando comparado com o grupo tratado com nujol. A significância estatística foi determinada pelo teste
ANOVA de uma via, seguido pelo pós-teste de Newman-Keuls.
4.5 WESTERN BLOT
4.5.1 Músculo Diafragma
No músculo diafragma, observou-se aumento dos níveis de TNF-α, NF-kB, TGF-β1,
Fibronectina e 4-HNE nos animais mdx tratados com Nujol, em comparação aos músculos de
animais do grupo controle, C57BL/10. Ambos tratamentos reduziram significativamente os
níveis de NF-kB e de Fibronectina. A terapia combinada revelou diminuição dos níveis de
TNF-α quando comparado ao grupo tratado com Nujol e dos níveis de 4-HNE quando
comparado aos grupos tratados com Nujol e DFZ (figura 19).

48
Figura 19 - Western Blot do músculo diafragma.
Western Blot do músculo diafragma. Grupos: C57BL10 (camundongos C57BL10 tratados com Nujol), mdx
Nujol (camundongos mdx tratados com Nujol) mdx DFZ (camundongos mdx tratados com deflazacorte) e mdx
DFZ+O3 (camundongos mdx tratados com a associação de deflazacorte com o óleo contido nas cápsulas de
Ômega 3 contendo EPA e DHA). Gráfico representando os níveis de TNF-α, NF-kB, TGF-β1, Fibronectina e 4-
HNE normalizados pelo GAPDH. Nível de significância indicado pela letra “a” quando comparado com o grupo
C57BL10, “b” quando comparado com o grupo mdx tratado com Nujol, “c” quando comparado com o grupo
mdx tratado com DFZ. A significância estatística foi determinada pelo teste ANOVA de uma via, seguido pelo
pós-teste de Newman-Keuls.

49
4.5.2 Músculo Quadríceps
No músculo quadríceps, foi observado aumento dos níveis de NF-kB, TGF-β1 e
Fibronectina nos animais mdx tratados com Nujol, em comparação aos músculos de animais
do grupo controle, C57BL/10. Ambos os tratamentos reduziram significativamente os níveis
de Fibronectina e a terapia com DFZ diminuiu os níveis de NF-kB quando comparado ao
grupo distrófico tratado com Nujol. Os níveis de TGF- β1 aumentaram significativamente no
grupo tratado com DFZ quando comparado com o grupo mdx tratado com Nujol e no grupo
tratado com DFZ+O3 quando comparado com os grupos mdx tratado com Nujol e com DFZ
(figura 20).

50
Figura 20 - Western Blot do músculo quadríceps.
Western Blot do músculo quadríceps. Grupos: C57BL10 (camundongos C57BL10 tratados com Nujol), mdx
Nujol (camundongos mdx tratados com Nujol) mdx DFZ (camundongos mdx tratados com deflazacorte) e mdx
DFZ+O3 (camundongos mdx tratados com a associação de deflazacorte com o óleo contido nas cápsulas de
Ômega 3 contendo EPA e DHA). Gráfico representando os níveis de TNF-α, NF-kB, TGF-β1, Fibronectina e 4-
HNE normalizados pelo GAPDH. Nível de significância indicado pela letra “a” quando comparado com o grupo
C57BL10, “b” quando comparado com o grupo mdx tratado com Nujol, “c” quando comparado com o grupo
mdx tratado com DFZ. A significância estatística foi determinada pelo teste ANOVA de uma via, seguido pelo
pós-teste de Newman-Keuls.

51
5 DISCUSSÃO
5.1 QUANTO AOS EFEITOS DAS TERAPIAS NA MASSA CORPORAL E MEDIDA
DE FORÇA.
Atualmente, sabe-se da eficácia do uso de anti-inflamatórios esteróides, como o
deflazacorte, para a terapia da DMD (BIGGAR et al., 2001; MESA et al. 1991). Entretanto,
devido ao uso contínuo, inúmeros efeitos colaterais interferem na qualidade de vida dos
pacientes (ANGELINI e PETERLE, 2012), fazendo-se necessária a busca por fármacos
alternativos para terapia da DMD (MOXLEY et al., 2005; MANZUR et al., 2008; BEYTÍA et
al., 2012).
Estudos prévios em nosso laboratório demonstraram que o EPA purificado
(MACHADO et al., 2011) ou cápsulas de ômega-3 contendo a combinação de EPA com DHA
(FOGAGNOLO MAURÍCIO et al., 2013), atenuaram a mionecrose em músculos esqueléticos
de camundongos mdx durante os estágios iniciais da doença, possivelmente por promover a
estabilidade do sarcolema, uma vez que já foi demonstrado que o DHA reduz o influxo de
cálcio induzido pelo estresse oxidativo, mecanismo que está associado à distribuição de
“transient receptor potential channel 1” (TRPC-1), proteína transmembrana relacionada com
o controle de influxo e efluxo de cálcio, presente também no sarcolema (YE et al., 2010).
Esses autores observaram ainda que o ômega 3 reduziu os níveis de citocinas que modulam a
inflamação. No presente trabalho verificamos se a associação do deflazacorte com o ômega-3
apresenta melhores efeitos do que a monoterapia com deflazacorte no estágio tardio da
doença.
A dosagem de ômega-3 recomendada para humanos é de 3 a 5 g/dia, sendo que acima
deste valor começa a apresentar toxicidade (TISDALE e DHESI et al., 1990). Convertendo a
dose utilizada em humanos para camundongos de acordo com a normalização proposta por
Reagn-Shaw e colaboradores (2008), teríamos uma dose de 300mg/kg de ômega 3 que
corresponde a dose utilizada neste trabalho. O mesmo é válido para a dose de deflazacorte
(1,2 mg/kg) (REAGAN-SHAW, NIHAL e AHMAD, 2008; FOGAGNOLO MAURÍCIO et
al., 2012; CARVALHO et al., 2013). Embora tenha-se que ter cuidado ao se transpor doses de
animais para humanos, a dose utilizada no presente trabalho está dentro de limites
compatíveis com doses para humanos. De fato, durante todo o período de tratamento não foi
observada diferença entre os grupos estudados no que tange à mortalidade e alterações da
massa corporal (figura 4). Isto sugere que as dosagens utilizadas nos tratamentos realizados

52
não provocaram nenhum efeito colateral perceptível nos camundongos mdx no estágio tardio
da distrofinopatia.
Foi observado discreto aumento de força nos membros anteriores ao final do
tratamento em todos os grupos estudados. Todavia, o ganho da força foi significativo somente
no grupo tratado com a associação de Deflazacorte com Ômega 3 (14%) (figura 5). Já foi
relatado que a suplementação com óleo de peixe contendo EPA melhora a força muscular e
capacidade funcional em humanos, mas não foi encontrado relato de melhora da força em
camundongos (RODACKI et al., 2012). Têm-se conhecimento do efeito anti-inflamatório do
Ômega 3 com consequente diminuição da mionecrose no camundongo mdx, observada no
presente trabalho pela diminuição da CK. Sugere-se que a diminuição da inflamação e
mionecrose, esteja associada ou ao efeito anti-inflamatório do DFZ e do O3, ou pela proteção
do sarcolema proveniente do DHA presente no ômega 3 (CARVALHO et al., 2013;
FOGAGNOLO MAURÍCIO et al., 2012; MACHADO et al., 2011). Desta forma, com a
diminuição da mionecrose, pode ter havido maior preservação de fibras musculares,
justificando o aumento da força de tração observado no grupo tratado com a associação das
drogas.
5.2 QUANTO AOS EFEITOS DAS TERAPIAS NOS NÍVEIS DE CREATINA QUINASE
(CK).
Sabe-se que tanto pacientes com DMD como camundongos mdx, apresentam aumento
nos níveis séricos de creatina quinase, enzima liberada na corrente sanguínea após a
mionecrose, indicativo de degeneração muscular (BULFIELD et al., 1984; ENGEL et al.,
2004; HUSSEIN et al., 2014). Embora não seja um biomarcador específico de distrofias
musculares, é amplamente utilizada desde a descoberta da doença durante a triagem realizada
para a confirmação do diagnóstico da DMD (ENGEL et al., 2004).
No presente trabalho observamos que os níveis séricos da CK estavam bastante
aumentados no grupo tratado com nujol. Uma vez que este tratamento não apresenta efeito
farmacológico algum sobre a saúde do animal, este aumento indica a presença de degeneração
muscular, resultado esperado na distrofinopatia do camundongo mdx (BULFIELD et al.,
1984; ENGEL et al., 2004; GROUNDS, 2008; PASTORET e SEBILLE, 1995; SEIXAS et
al., 1997). O grupo tratado com DFZ apresentou diminuição de 64% dos níveis de CK quando
comparados com o grupo tratado com nujol, resultado que concorda com um estudo realizado
em humanos, que verificou diminuição dos níveis de CK após tratamento com prednisona,

53
corticóide de ação semelhante a do DFZ (HUSSEIN et al., 2014). Já o grupo tratado com a
associação de DFZ+O3 apresentou diminuição de 84% quando comparado ao grupo tratado
com nujol (figura 6). O efeito do EPA purificado (MACHADO et al., 2011) ou do óleo
contido nas cápsulas de ômega 3, contendo EPA e DHA (CARVALHO et al., 2013;
FOGAGNOLO MAURÍCIO et al., 2012) na redução dos níveis de CK no camundongo mdx
já é bem conhecido. Este dado sugere que a combinação teve melhor efeito em diminuir a
mionecrose, do que a monoterapia (DFZ), possivelmente por serem fármacos com efeito anti-
inflamatório, que podem ter apresentado efeito somatório na diminuição da inflamação,
corroborando de forma importante para a redução da mionecrose (SPENCER e TIDBALL,
2001).
A diminuição da CK pode estar relacionada com a proteção contra a mionecrose, que
foi evidenciada no presente trabalho pelo aumento da área ocupada por fibras com núcleo
periférico presente no músculo diafragma, indicativo de fibras que não sofreram degeneração.
Embora não tenha sido observada diferença significativa entre as terapias no aumento de área
ocupada por NP, a associação de DFZ+O3 foi mais eficaz na redução de CK do que a terapia
com DFZ.
Uma hipótese para explicar o mecanismo de proteção contra mionecrose decorrente do
tratamento com ômega 3 seria a de que o DHA, ao se incorporar na membrana, pode
estabilizar o sarcolema (MACHADO et al., 2011). Whitehead e colaboradores (2006)
sugerem que o aumento da concentração de cálcio no músculo distrófico ativa proteases que
desencadeiam a mionecrose (WHITEHEAD et al., 2006). Com a incorporação do DHA no
sarcolema, poderia haver aumento da sua estabilidade, diminuição do influxo de cálcio (JUDÈ
et al., 2006; POUND et al., 2001), e consequentemente redução da mionecrose (MACHADO
et al., 2011; MARIOL e SÉGALAT, 2001).
5.3 QUANTO AOS EFEITOS DAS TERAPIAS NO MÚSCULO DIAFRAGMA
A ausência da distrofina acarreta danos à fibra muscular gerando inflamação crônica e
mionecrose (ENGEL, 2004). Posterior ao processo inflamatório ocorre a ativação de células
satélites as quais se fundem para formar miotubos. A diferenciação de miotubos em miofibras
e a deposição de tecido conjuntivo ocorrem como etapa final da regeneração (TIDBALL,
2005). Sabe-se que concomitantemente à resposta inflamatória, quando não há eficiente
regeneração, ocorre a formação de fibrose (BERNASCONI et al., 1995; CONTRERAS et al.,
2016). A fibrose muscular é uma característica morfológica encontrada tanto em indivíduos

54
afetados pela DMD (ENGEL, 2004), quanto em camundongos mdx na fase tardia da doença,
como observado no presente estudo.
O diafragma é um dos músculos mais afetados pela fibrose na DMD (ENGEL, 2004) e
no camundongo mdx (BARBIN et al., 2016; CARVALHO et al., 2013; FOGAGNOLO
MAURÍCIO et al., 2012; STEDMAN et al., 1991; PEREIRA et al., 2015). Seu acometimento
corrobora de forma importante para o prejuízo da função respiratória, resultando na
insuficiência respiratória (ALLEN, 2010), uma das principais complicações da DMD, levando
ao óbito (BOGDANOVICH et al., 2004; MOXLEY et al., 2005).
Com base nos nossos resultados, observamos que ambas as terapias aumentaram a
área ocupada por fibras com núcleo periférico, um indicativo de fibras que não sofreram
degeneração, representando a ação protetora das drogas contra a mionecrose, já demonstrada,
no caso do deflazacorte, em humanos (MESA et al., 1991) e animais distróficos (PEREIRA et
al., 2015), e no caso do ômega 3, apenas em animais distróficos (CARVALHO et al., 2013;
FOGAGNOLO MAURÍCIO et al., 2012; MACHADO et al., 2011).
Também foi observado que tanto a monoterapia com DFZ quanto a associação de DFZ
+ O3 diminuíram a área de fibrose no músculo diafragma, provavelmente pelo efeito anti-
inflamatório das terapias, uma vez que a inflamação é um fenômeno precursor da fibrose (MA
et al., 2016). O efeito do ômega 3 na redução da fibrose do músculo diafragma já foi
demonstrado no camundongo mdx (FOGAGNOLO MAURÍCIO et al., 2016). Porém, parece
não haver um consenso sobre a ação dos corticóides na fibrose presente na distrofinopatia do
camundongo mdx. Divergindo dos nossos resultados, um trabalho que utilizou camundongos
mdx jovens (21 dias de vida), tratados com prednisolona durante três semanas, revelou que o
tratamento com corticóide não alterou a fibrose presente no músculo diafragma (WELLER et
al., 2012). Também parece não haver um acordo sobre a ação dos corticóides em outros
tecidos. Um estudo demonstrou que a terapia crônica com DFZ reduziu a fibrose no músculo
cardíaco de camundongos distróficos (MARQUES et al., 2009), enquanto que outro estudo
demonstrou que a prednisona e a prednisolona, corticóides de ação semelhante ao DFZ,
aumentaram a fibrose no miocárdio (SALI et al., 2012).
A literatura sugere que os mecanismos pelos quais o ômega-3 protege contra a
mionecrose sejam decorrentes da ação anti-inflamatória do EPA (CALDER et al., 2008;
FOGAGNOLO MAURÍCIO et al., 2012) e da incorporação do DHA na membrana,
promovendo a estabilidade do sarcolema (FOGAGNOLO-MAURÍCIO et al., 2013).
No que se refere a inflamação, sabe-se que o EPA atua na geração de eicosanoides da
série impar, que são anti-inflamatórios, e na redução de citocinas pró-inflamatórias, como o

55
TNF-α (CALDER et al., 2008). Foi possível observar uma redução da área de inflamação na
análise histológica (figura 11), que ainda que não tenha sido significativa para nenhuma das
terapias, está de acordo com os níveis dos marcadores relacionados à inflamação encontrados
no presente trabalho. Houve redução significativa do nível de TNF-α apenas no grupo tratado
com a associação de DFZ+O3 quando comparada ao grupo mdx tratado com Nujol (figura
19).
Uma hipótese sobre o mecanismo do EPA na diminuição dos níveis de TNF-α está
relacionada com a sua habilidade de modular as respostas inflamatórias e imunológicas. O
EPA inibe a dissociação entre IkB (proteína inibitória) e NF-kB, impedindo a translocação do
NF-kB para o núcleo, diminuindo a transcrição do gene do TFN-α (BABCOCK et. al., 2000;
CALDER, 2008; MACHADO et al., 2011). Além disso, o EPA e o DHA inibem a produção
de eicosanoides da série par (pró-inflamatórios) gerados a partir do AA, pois são capazes de
atuar como substrato nas vias COX e LOX, originando eicosanóides da série impar, que tem
ação anti-inflamatória (CALDER, 2008).
Também observamos que ambas terapias foram eficazes na redução dos níveis de NF-
kB no músculo diafragma. O NF-kB é um importante fator de transcrição que, dentre outras
funções, está diretamente relacionado a regulação do processo inflamatório, uma vez que,
quando ativado, aumenta a expressão de citocinas pró-inflamatórias, tais como o fator de
necrose tumoral alfa (TNF-α) (MESSINA et al., 2006; SPENCER e TIDBALL, 2001). Um
estudo com emprego de tratamento crônico com EPA e DHA em camundongos mdx revelou
diminuição dos níveis de NF-kB no músculo diafragma (APOLINÁRIO et al., 2015). Outro
estudo demonstrou que o DFZ foi eficaz na redução dos níveis de NF-kB no músculo
diafragma de camundongos distróficos (PEREIRA et al., 2015), concordando com nossos
resultados. Alguns autores sugerem que os esteroides reduzem a inflamação através da
interação com receptores intracelulares que sintetizam lipocortina, que inibe diretamente a
FLA2, impedindo a liberação do ácido araquidônico e consequentemente a formação de todos
os endoperóxidos cíclicos de prostanóides (SCHELLACK, 2006; SPULER e ENGEL, 1998),
desta maneira, impedem a formação de mediadores inflamatórios. Sugerimos que a redução
dos níveis de NF-kB no grupo tratado com a associação de DFZ+O3 pode ser justificada pela
ação anti-inflamatória de ambos os fármacos, que embora tenham ação em diferentes
momentos da cascata do AA, podem ter apresentado efeito somatório na redução dos
mediadores inflamatórios estudados no presente trabalho.
Um outro fenômeno importante na fisiopatologia da DMD é a formação da fibrose.
Admite-se que, com a progressão da doença e vários ciclos de degeneração-regeneração,

56
perde-se a capacidade regenerativa por exaustão das células satélites (BLAU et al., 1983;
SACCO et al., 2010). Nesse momento, as fibras musculares passam a ser substituídas por
fibrose, o que caracteriza os estágios mais avançados da doença (CONTRERAS et al., 2016;
MA et al., 2016). O TGF-β1 é um importante mediador da fibrose, atuando em receptores
específicos nos fibroblastos e promovendo a formação e deposição de colágeno na matriz
extracelular (CONTRERAS et al., 2016). O estudo do TGF-β1 fornece dados valiosos sobre a
fibrose nos músculos distróficos (ACUNA et al., 2014; BERNASCONI et al., 1995; CHEN et
al., 2005). Terapias com drogas anti-inflamatórias e antifibróticas diminuem a expressão de
TGF-β1 e consequentemente a formação de fibrose no músculo distrófico (HARTEL et al.,
2001; TANIGUTI et al., 2011; MOREIRA et al., 2013). Observamos redução significativa da
área de fibrose no músculo diafragma dos animais que receberam DFZ e a associação de
DFZ+O3 quando comparados com os animais mdx que receberam nujol (figura 10).
Entretanto, não foi observada alteração significativa dos níveis de TGF-β1 entre esses grupos.
Uma explicação para este achado pode residir no fato de que, em condições mais estáveis, em
que a fibrose já esteja depositada, os níveis do TGF-β1 não estão necessariamente elevados,
ou seja, a elevação do TGF-β1 pode preceder a deposição de grande quantidade de fibrose, tal
como observada na análise histopatológica, nos estágios mais tardios da doença. De fato,
Contreras e colaboradores (2016) observaram que os níveis de TGF- β1 estão mais elevados
durante a deposição do tecido fibro-adiposo na MEC do músculo esquelético, não
apresentando diferenças quando o tecido fibroso já está instalado (CONTRERAS et al., 2016)
Durante o fenômeno inflamatório na DMD, temos a produção de células inflamatórias
como os macrófagos e de fatores pró-fibróticos, como o TGF-β1 (MOYER e WAGNER ,
2011). Observações recentes demonstram que, sob dano muscular, o TNF-α secretado por
macrófagos durante o processo inflamatório induz a apoptose de moléculas progenitoras de
tecido fibro-adiposos. Foi demonstrado ainda que em condições de dano crônico, os
macrófagos expressam elevados níveis de TGF-β1, evitando a apoptose dessas moléculas,
induzindo a formação de MEC (LEMOS et al., 2015). Recentemente (CONTRERAS et al.,
2016), encontraram níveis elevados de mRNA de TGF-β1 e TNF-α no diafragma de animais
distróficos, quando comparados com os animais controle (C57BL10), ambos com oito meses
de idade. Curiosamente, os mesmo autores detectaram níveis de TGF-β1 mais elevados
quando comparados com os níveis de TNF-α no músculo gastrocnêmio duas semanas após
desnervação, onde uma quantidade pronunciada de fibrose foi observada (CONTRERAS et
al., 2016). Esses achados sugerem que os níveis de TGF- β1 estão mais elevados durante a
deposição do tecido fibro-adiposo na MEC do músculo esquelético, não apresentando

57
diferenças quando o tecido fibroso já está instalado, o que está de acordo com os nossos
achados no presente trabalho.
Uma outra molécula que serve como indicativo de fibrose é a fibronectina. A
fibronectina compreende uma família de glicoproteínas de elevado peso molecular presente na
MEC, e é produzida por fibroblastos durante a instalação da fibrose no músculo distrófico
(SUN et al., 2016). A fibronectina atua na adesão celular à MEC e desempenha papel
importante no desenvolvimento da fibrose no músculo distrófico, regulando a organização e
deposição de alguns componentes da MEC como o colágeno do tipo I (BERNASCONI et al.,
1995).
Estudos recentes demonstraram elevação dos níveis de fibronectina em camundongos
mdx na fase tardia da doença (6- 8 meses) quando comparados com os animais controle
(CONTRERAS et al., 2016; SUN et al., 2016), dados que concordam com o aumento dos
níveis de fibronectina observado no grupo mdx tratado com Nujol quando comparado com o
grupo C57BL10, conforme demonstrado no presente estudo (figura 19). Ambas terapias
reduziram significativamente os níveis de fibronectina no músculo diafragma, o que está de
acordo com a redução significativa da área ocupada por fibrose observada nos grupos tratados
com DFZ e DFZ+O3 (figura 10). Esses dados sugerem que a fibronectina pode agravar o
fenótipo distrófico, conforme sugerido anteriormente (CONTRERAS et al., 2016; SUN et al.,
2016). Sugerimos que a diminuição deste marcador possa estar relacionada com a ação das
drogas em receptores que podem regular a expressão da fibronectina. Uma vez que ambas as
terapias não alteraram significativamente os níveis de TGF-β1 no diafragma distrófico, é
possível que não haja uma relação direta entre o TGF-β1 e a redução da expressão da
fibronectina, necessitando de mais estudos que possam elucidar o mecanismo de ação das
drogas na diminuição da fibrose.
Vimos anteriormente que além da inflamação e da deposição de tecido fibroso no
músculo distrófico, outro fenômeno diretamente relacionado com a progressão da DMD e da
distrofinopatia do camundongo mdx é o estresse oxidativo (FOGAGNOLO-MAURÍCIO et
al., 2013; KOZAKOWSKA et al., 2015). Quando este fenômeno resulta na oxidação dos
fosfolipídios presentes na membrana celular, tal processo é denominado de peroxidação
lipídica (BARBOSA et al., 2010; HALLIWELL e WHITEMAN, 2004), que tem como
resultado a formação de alguns produtos finais, como o 4-hidroxinonenal (4-HNE) (ZHONG
e YIN, 2015). Alguns estudos demonstraram que o camundongo mdx apresenta níveis
elevados de 4-HNE quando comparados ao camundongo controle (FOGAGNOLO-
MAURÍCIO et al., 2013; RAPUCCI MORAES et al., 2015), conforme observamos no

58
presente estudo (figura 19). Observamos que a associação de DFZ+O3 diminuiu
significativamente os níveis de 4-HNE quando comparado aos grupos distróficos tratados
com Nujol e com DFZ. O DFZ sozinho não apresentou esse efeito.
Este dado sugere que a associação de DFZ+O3 além de diminuir a mionecrose e a
inflamação no músculo diafragma, conforme visto até então, também diminuiu o estresse
oxidativo, evidenciado pela diminuição dos níveis de 4-HNE. Um estudo anterior realizado
em nosso laboratório já havia demonstrado a ação da terapia com EPA+DHA na redução do
4-HNE em camundongos mdx jovens (FOGAGNOLO-MAURÍCIO et al., 2013). Além disso,
foi demonstrado que o consumo de ômega-3 promove a diminuição de ERMOs, contribuindo
para redução do dano tecidual (KAJIKAWA et al., 2010; XIE et al., 2011), que se reflete na
diminuição da inflamação e fibrose observadas no presente trabalho. Desta forma, a
associação de DFZ+O3 foi mais eficiente que terapia com DFZ na diminuição do estresse
oxidativo.
Em síntese, observamos que o músculo diafragma distrófico, na idade estudada (13
meses), apresenta-se com grande quantidade de fibrose. Dos cinco fenômenos avaliados
(função, mionecrose, inflamação, fibrose e estresse oxidativo) a associação de DFZ+O3
apresentou melhores efeitos do que a terapia com DFZ em três fenômenos: função,
representada pela medida de força; redução da mionecrose, através da diminuição dos níveis
de CK e redução da peroxidação lipídica, exibida pela diminuição dos níveis de 4-HNE. Desta
forma, o emprego do ômega 3 como suplementação do tratamento com DFZ, pode se
apresentar como uma potencial terapia para DMD. Entretanto, os estudos clínicos devem estar
atentos para possíveis efeitos adversos desta combinação, uma vez que podem ocorrer
especialmente em pacientes nos estágios mais tardios, que já estejam sob uso do corticóide há
mais tempo.

59
5.4 QUANTO AOS EFEITOS DAS TERAPIAS NO MÚSCULO QUADRÍCEPS
A mionecrose presente no músculo distrófico leva à ativação das células satélites, na
tentativa de promover a regeneração muscular (MAURO, 1961). As CS são células tronco
musculares localizadas entre a lâmina basal e a fibra muscular, capazes de se diferenciar em
outras CS ou em mioblastos (MAURO, 1961). Com a progressão da doença observam-se
ciclos de degeneração e regeneração, presentes na DMD (MARIOL e SÉGALAT, 2001) e no
camundongo mdx (GROUNDS, 2008; SHAVLAKADZE et al., 2004). A degeneração-
regeneração contínua leva ao declínio da capacidade regenerativa das CSs, culminando na
deposição de tecido fibro-adiposo (CONTRERAS et al., 2016). Diversas teorias procuram
justificar a diminuição da capacidade regenerativa do músculo distrófico; a que parece ser
mais aceita sugere que há exaustão de CS devido aos ciclos repetitivos de degeneração e
regeneração (BLAU et al., 1983; SACCO et al., 2010).
No músculo quadríceps dos animais distróficos, foi observada uma área
majoritariamente (aproximadamente 95%) ocupada por fibras regeneradas (figura 15),
representadas pela centralização nuclear (figura 13), não havendo diferença significativa desta
área entre os grupos. Um estudo prévio realizado em nosso laboratório demonstrou que o
músculo quadríceps de camundongos mdx apresentou elevada regeneração tanto em
camundongos na fase inicial (um mês de idade) como na fase tardia (oito meses de idade) da
doença (APOLINÁRIO et al., 2015). Foi observado ainda, que no músculo quadríceps de
camundongos jovens havia grande área ocupada por fibras, aparentemente de menor diâmetro
e com núcleo periférico, envolvidas por infiltrado inflamatório, classificando essa área como
“área em regeneração”, enquanto que em camundongos na fase tardia da doença foi observada
extensa área ocupada por fibras com núcleo central, porém, aparentemente com maior
diâmetro e sem inflamação, classificada como “área regenerada” (APOLINÁRIO et al.,
2015). No presente estudo foi observado predomínio de área regenerada (figura 15) e
classificamos toda a área ocupada por fibras com centralização nuclear, independente do
diâmetro das fibras como “área regenerada”, ou seja, em um estágio mais avançado do
processo regenerativo.
A capacidade regenerativa do músculo esquelético se deve à atividade miogênica das
CS, que originam células progenitoras necessárias para a reparação muscular e para a auto-
renovação das próprias CS (YIN et al., 2013). A auto-renovação é possível através da divisão
celular simétrica (gerando duas células-tronco filhas idênticas) ou através da divisão celular
assimétrica (gerando uma célula-tronco e uma célula progenitora) (CHANG e RUDNICKI,

60
2014). A adequada polarização dos centríolos e a orientação do fuso mitótico parece ser o
ponto crucial para a divisão celular assimétrica (SCHOBER et al., 1999; SILLER et al.,
2006), porém, a deficiência de distrofina nas CSs distróficas interfere na expressão de
proteínas relacionadas à orientação do fuso mitótico, resultando na redução das divisões
celulares assimétricas e, consequentemente, na perda gradual de células miogênicas
progenitoras, refletindo em deficiente regeneração muscular (DUMONT et al., 2015). Por
alguma razão desconhecida, diferentemente da maioria dos músculos estudados em
camundongos mdx, o músculo quadríceps apresenta ciclos de degeneração e regeneração em
animais distróficos jovens (APOLINÁRIO et al., 2015), porém em camundongos na fase
tardia da doença, ele se revela quase que completamente regenerado, apresentando mais de
80% de área ocupada por fibras com núcleo central aos oito meses de idade (APOLINÁRIO
et al.; 2015) e 95% de área regenerada aos 13 meses, conforme observamos no presente
estudo.
As vias de sinalização Notch e Wnt são importantes para a especificação da divisão
celular das CS (JIANG et al., 2014). Uma vez que a sinalização destas vias ocorre juntamente
com as proteínas do complexo PAR (nome oriundo de “partition-defective”- partição
defeituosa), que estão relacionadas à orientação do fuso mitótico (OSSIPOVA et al., 2009),
especula-se que a desregulação dessas vias seja uma consequência direta da perda da
polaridade celular devido a ausência de distrofina. Curiosamente, um recente estudo descobriu
que Jagged1, um gene regulador da via Notch, foi expresso em dois cães distróficos da raça
Golden Retriver (GRMD). Estes cães, diferentemente do habitual, não apresentavam o
fenótipo distrófico esperado (VIEIRA et al., 2015). Os mioblastos derivados dos músculos
destes cães apresentaram maior capacidade proliferativa, concordando com a descoberta de
Collesi e colaboradores (2008), que demonstraram que a expressão de Jagged1 estimulou a
proliferação de cardiomiócitos de camundongos. Interessantemente, a expressão ectópica de
Jagged1 no peixe-zebra (outro modelo experimental da DMD) foi suficiente para recuperar o
fenótipo distrófico (VIEIRA et al., 2015). Ainda não se sabe o mecanismo exato de como o
Jagged1 atua na melhora da distrofia e como a sua expressão afeta a função das CS. Uma vez
que a sinalização Notch está diretamente relacionada com a divisão celular assimétrica, parece
provável que os efeitos melhoradores de Jagged1 são um resultado da restauração da divisão
celular assimétrica em CS distróficas (CHANG e RUDNICKI, 2014). Desta forma, parece
interessante que as vias Notch e Wnt, bem como o gene Jagged1 sejam estudados na tentativa
de elucidar o mecanismo de regeneração do músculo quadríceps distrófico.

61
Os efeitos do ômega-3 e dos corticóides sobre a miogênese já foram investigados em
estudos anteriores. No que se refere aos efeitos miogênicos dos corticóides, um estudo
realizado in vitro revelou que a dexametasona, um conhecido corticóide, reduziu a miogênese
e inibiu a ativação de CSs através da diminuição dos níveis de MyoD e miogenina, fatores de
transcrição relacionados à diferenciação celular e miogênese (BRUSCOLI et al., 2010). Em
nosso estudo, não observamos redução da área regenerada no grupo distrófico tratado com
DFZ e no grupo tratado com associação de DFZ+O3. Sugerimos que na idade estudada, o
músculo quadríceps de camundongos mdx não apresenta mais ciclos de degeneração e
regeneração, apresentando-se regenerado.
No que se refere ao ômega-3, nosso laboratório demonstrou que a terapia com
EPA+DHA aumentou a regeneração nos músculos diafragma e quadríceps de camundongos
mdx com 1 mês de idade. Este efeito, entretanto, não foi observado em animais mais velhos
com oito meses de idade (APOLINÁRIO et al., 2015) ou com 13 meses de idade, tal como
observado no presente estudo, em que não houve diferenças significativas da área regenerada
entre os grupos estudados, em ambos músculos (DIA e QUA). Em conjunto, estes dados
sugerem que os efeitos pró-regenerativos do ômega 3 são observados preferencialmente nas
fases iniciais da doença, provavelmente pelo fato das CS ainda estarem hábeis a regeneração
nesta etapa inicial. Possivelmente, o ômega-3 também se incorpore em suas membranas e
restaure, pelo menos em parte, sua capacidade regenerativa através da estabilização de suas
membranas (das CS) e da cascata de sinalização que envolve Notch e Wnt. Já nos estágios
mais tardios da doença, provavelmente as CS já estejam exauridas, com a população reduzida,
não havendo assim células suficientes para responder ao ômega-3.
O ômega-3 tambem parece atuar na cascata de sinalização MAPK (“Mitogen
Activated Protein Kinases”- Proteína-quinases ativadas por mitógenos), que possui
componentes inibitórios e excitatórios da miogênese (GUTTRIDGE et al., 2000; SHI et al.,
2010) e que pode ser modulada pelo NF-kB e TNF-α. Nos estágios iniciais da doença, o
ômega-3 diminuiu os níveis de NF-kB e TNF-α, o que também poderia contribuir para a
regeneração (MOON et al., 2007; ZGHEEL et al., 2014). No presente estudo não observamos
efeitos da terapia combinada no NF-kB e TNF-α, que pudessem ser correlacionados a melhora
da regeneração, provavelmente pelo músculo QUA já se encontrar completamente
regenerado.
Embora a área ocupada por infiltrado inflamatório observada seja ínfima, observamos
que houve redução significativa desta área nos grupos distróficos tratados com DFZ e
DFZ+O3 (aproximadamente 0,1% da área total), quando comparados com o grupo distrófico

62
tratado com nujol (0,4%) (figura 17). Entretanto, para os mediadores inflamatórios estudados
no presente estudo, só foi observada redução significativa nos níveis de NF-kB para o grupo
tratado com DFZ. Não foi observada diferença significativa nos níveis de TNF-α entre os
grupos (figura 20). Também não foi observada diferença significativa nos níveis de 4-HNE
entre os grupos. Esses dados confirmam a hipótese de que o músculo quadríceps distrófico, na
idade estudada, sofre pouca ou nenhuma influência dos fármacos, uma vez que assume
fenótipo regenerado, não ocorrendo ciclos de degeneração e regeneração, com consequentes
episódios de inflamação e peroxidação lipídica, como ocorre no início da doença
(APOLINÁRIO et al., 2015).
Além da pequena área de inflamação observada no músculo quadríceps, também
observamos diminuta área ocupada por tecido fibroso (figura 14) que corresponde
aproximadamente a menos de 1% da área total do músculo no grupo mdx tratado com nujol e
menos de 0,6 % e 0,1% nos grupos tratados com DFZ e DFZ+O3, respectivamente (figura
18). Houve redução significativa da fibrose nos grupos tratados com DFZ e DFZ+O3 em
comparação com o grupo distrófico tratado com nujol, o que está de acordo com os níveis de
fibronectina observados, que reduziram de forma proporcional à redução da área de fibrose.
Conforme vimos anteriormente, a fibronectina é produzida por fibroblastos e atua na adesão
celular à MEC e está relacionada com o desenvolvimento da fibrose no músculo distrófico
(BERNASCONI et al., 1995; SUN et al., 2016)
Na figura 14, também é possível observar que no quadríceps dos animais tratados com
DFZ+O3, a MEC encontra-se abundante, porém não consideramos tal característica como
área ocupada por tecido fibroso. Sugerimos que tal conformação possa ser ou um
espessamento do perimísio, que futuramente poderá progredir para fibrose, ou as fibras do
quadríceps estão em fase de regeneração e ainda não atingiram seu diâmetro final. Caso a
primeira proposição seja verdadeira, o elevado nível de TGF-β1 observado no grupo tratado
com a associação de DFZ+O3, poderia ser justificado pela deposição de tecido fibro-adiposo
na MEC, não excluindo a possibilidade de um possível início de fibrose (CONTRERAS et al.,
2016).
Em suma, observamos que o músculo quadríceps distrófico, na idade estudada,
apresenta-se quase que completamente regenerado, sofrendo pouca ou nenhuma ação dos
fármacos estudados no presente estudo. Embora, na análise histológica, tenha sido possível
observar diferenças significativas dos tratamentos nas áreas do músculo ocupadas por
infiltrado inflamatório e fibrose, tais áreas correspondiam a valores inferiores a 0,5% e 1% da
área total do músculo, respectivamente, o que provavelmente não deve afetar a função do

63
músculo no camundongo. Portanto, a associação de DFZ+O3 não apresentou diferenças com
relação à terapia com DFZ no músculo quadríceps de camundongos mdx, na idade estudada.
Em tempo, sugerimos que mais estudos sejam realizados no quadríceps, a fim de esclarecer
seu mecanismo de regeneração, na esperança de encontrar a chave para a cura da DMD.

64
6 CONCLUSÃO
Concluímos, que dos cinco fenômenos avaliados (função muscular, mionecrose,
inflamação, fibrose e estresse oxidativo) a associação de DFZ+O3 apresentou melhores
efeitos do que a terapia com DFZ em três fenômenos: função muscular (melhorou a força
muscular); redução da mionecrose (diminuição da CK) e redução da peroxidação lipídica
(redução do 4-HNE no músculo diafragma). Estes efeitos podem resultar da ação anti-
inflamatória de ambas drogas, somada à ação inibitória do estresse oxidativo exercida pelo
O3. Desta forma, o emprego do ômega 3 como suplementação do tratamento com DFZ, pode
ser considerado como uma potencial terapia para a DMD, sendo uma opção viável para
futuros estudos clínicos em que o paciente já esteja sob ação da terapia com corticóides.

65
REFERÊNCIAS BIBLIOGRÁFICAS
ACUNA, M.J.; PESSINA, P.; OLGUIN, H. et al. Restoration of muscle strength in dystrophic
muscle by angiotensin-1-7 through inhibition of TGF beta signaling. Human Molecular
Genetics, v. 23, p. 1237- 1249, 2014.
ALLEN, J. Pulmonary complications of neuromuscular disease: A Respiratory mechanics
perspective. Pediatric respiratory reviews, v. 11, ed. 1, p. 18-23, 2010.
ANGELINI, C.; PEGORARO, E.; TURELLA, E.; INTINO, M.T.; PINI, A.; COSTA, C.
Deflazacort in Duchenne dystrophy: study of long-term effect. Muscle e Nerve, v. 17, n. 4, p.
386-391, 1994.
ANGELINI, C.; PETERLE, E. Old and new therapeutic developments in steroid treatment in
Duchenne muscular dystrophy. Acta Myologica, v.31, n. 1, p. 9-15, 2012.
APOLINÁRIO, L.M.; CARVALHO, S.C.; SANTO-NETO, H.; MARQUES, M.J. Long-
Term Therapy With Omega-3 Ameliorates Myonecrosis and Benefits Skeletal Muscle
Regeneration in Mdx Mice. The Anatomical Record, v. 298, p. 1589–1596, 2015.
ARCHER, J.D.; VARGAS, C.C.; ANDERSON, J.E. Persistent and improved functional gain
in mdx dystrophic mice after treatment with L-arginine and deflazacort. FASEB Journal, v.
20, n. 6, p. 738-740, 2006.
BABCOCK T.A.; HELTON W. S; HONG D. AND JOSEPH ESPAT N. Omega-3 Fatty Acid
Lipid Emulsion Reduces LPS-Stimulated Macrophage TNF-alpha Production. Mary Amm
Liebert, Inc, v. 3, n. 2, 2002.
BABCOCK, T.; HELTON, W. S.; JOSEPH, E. N. Eicosapentaenoic Acid (EPA): An
Antiinflammatory Omega-3 Fat With Potential Clical Applications. Nutrition, v.16, p.1116-
1118, 2000.
BARBOSA, KIRIAQUE BARRA FERREIRA et al . Estresse oxidativo: conceito,
implicações e fatores modulatórios. Revista de Nutrição, Campinas, v. 23, n. 4, p. 629-643,
Aug. 2010.
BERNASCONI, P.; TORCHIANA, E.; CONFALONIERI, P.; BRUGNONI, R.; BARRESI,
R.; MORA, M.; CORNELIO, F.; MORANDI, L.; MANTEGAZZA, R. Expression of
transforming growth factor-β1 in dystrophic patient muscles correlates with fibrosis. The
Journal of Clinical Investigation, v. 96, p. 1137-1144, 1995.
BEYTÍA, M. L.; VRY, J.; KIRSCHNER, J. Drug treatment of Duchenne muscular dystrophy:
available evidence and perspectives. Acta Myologica, v. 31, n. 1, p.4-8, 2012.
BIANCHI, M.L.P.; ANTUNES, L.M.G. Radicais livres e os principais antioxidantes da dieta.
Revista de Nutrição, v. 12, n. 12, p. 123-130, 1999.

66
BIGGAR, W.D.; GINGRAS, M.; FEHLINGS, D.L.; HARRIS, V.A.; STEELE, C.A.
Deflazacort treatment of Duchenne muscular dystrophy. Journal of Pediatrics, v. 38, p. 45–
50, 2001.
BLAU, H.M.; WEBSTER, C.; PAVLATH, G.K. Defective myoblasts identified in Duchenne
muscular dystrophy. Proceedings of the National Academy of Sciences, n. 80, p. 4856–
4860, 1983.
BOGDANOVICH, S.; PERKINS, K.J.; KRAG, T.O.; KHURANA, T.S. Therapeutics for
Duchenne muscular dystrophy: current approaches and future directions. Journal of
Molecular Medicine, v. 82, n. 2, p.102-115, 2004.
BONIFATI, D.M.; RUZZA, G. BONOMETTO, P. A multicenter double-blind randomized
trial of deflazacort versus prednisone in Duchenne muscular dystrophy. Muscle e Nerve, v.
23, p. 1344-1347, 2000.
BRADFORD, M. M.A rapid and sensitive method for quantitation of microgram quantities of
protein utilizing the principle of protein-dye-binding. Analls of Biochemistry, v.7, n.72,
p.248-54, 1976
BRIGUET, A.; COURDIER-FRUH, I.; FOSTER, M.; MEIER, T.; MAGYAR, J. P.
Histological parameters for the quantitative assessment of muscular dystrophy in the mdx-
mouse. Neuromuscular Disorders, v. 14, p. 675–682, 2004.
BRUSCOLI, S.; DONATO, V.; VELARDI, E.; DI SANTE, M.; MIGLIORATI, G.;
DONATO, R.; RICCARDI, C. Glucocorticoid-induced Leucine Zipper (GILZ) and Long
GILZ Inhibit Myogenic Differentiation and Mediate Anti-myogenic Effects of
Glucocorticoids. The journal of biological chemistry, v. 285, n. 14, p. 10385–10396, 2010.
BULFIELD, G.; SILLER, W.G.; WIGHT, P.A.L.; MOORE, K. X-chromosome-linked
muscular dystrophy (mdx) in the mouse. Processings of the National Academy of Sciences
of United States of America, v. 81, n. 4, p. 1189-1192, 1984
CALDER, P.C. Polyunsaturated fatty acids, inflammatory processes and inflammatory bowel
diseases. Molecular Nutrition e Food Research, v. 52, p. 885–897, 2008.
CARVALHO, S.C.; APOLINÁRIO, L.M.; MATHEUS, S.M.; SANTO NETO, H.;
MARQUES, M.J. EPA protects against muscle damage in the mdx mouse model of Duchenne
muscular dystrophy by promoting a shift from the M1 to M2 macrophage phenotype. Journal
of Neuroimmunology, n.15, v. 264, p. 41-47. 2013.
CHANG, N.C. e RUDNICKI, M.A. Satellite cells: the architects of skeletal muscle. Current
Topics in Developmental Biology, v. 107, p. 161–181, 2014.
CHEN, Y.W.; NAGARAJU, K.; BAKAY, M.; MCINTYRE, O.; RAWAT, R.; SHI, R.;
HOFFMAN, E.P. Early onset of inflammation and later involvement of TGFbeta in Duchenne
muscular dystrophy. Neurology, v. 65, p. 826-834, 2005.

67
COHN, R.D.; VAN, E.R.P.; C.; HABASHI, J.P. et al. Angiotensin II type 1 receptor blockade
attenuates TGFbeta- induced failure of muscle regeneration in multiple myopathic states.
Nature Medicine, v. 13, p. 204-210, 2007
COLLESI, C.; ZENTILIN, L.; SINAGRA, G. et al. Notch1 signaling stimulates proliferation
of immature cardiomyocytes. The Journal of Cell Biology, v. 183, p. 117–128, 2008.
CONTRERAS, O.; REBOLLEDO, D.L.; OYARZÚN, J.E.; OLGUÍN, H.C.; BRANDAN, E.
Connective tissue cells expressing fibro/adipogenic progenitor markers increase under chronic
damage: relevance in fibroblast-myofibroblast differentiation and skeletal muscle fibrosis.
Cell and Tissue Research. v. 364, p.647–660, 2016.
DAVIDSON, M.H.; BENES, L. B. The future of n-3 polyunsaturated fatty acid therapy.
Current opinion in lipidology. v. 27, n. 6, 2016.
DRACHMAN, D.B.; TOYKA, K.V.; MYER, E. Prednisone in Duchenne muscular
dystrophy. The Lancet, v. 14, n. 2, p. 1409-1412, 1974.
DUMONT, N.A.; WANG, Y.X.; VON MALTZAHN, J. et al. Dystrophin expression in
muscle stem cells regulates their polarity and asymmetric division. Nature Medicine, n. 21,
p. 1455–1463, 2015.
ENGEL, A. G. Muscular Dystrophies. In: Engel A. G.; Franzini-Armstrong, C. Myology:
basic and clinical. Ed. 3, v.2, p.961-1012, 2004.
FERREIRA, A.L.A.; MATSUBARA, L.S. Radicais livres: conceitos, doenças relacionadas,
sistema de defesa e estresse oxidativo. Revista da Associação Médica Brasileira, v. 43, n. 1,
p. 61-68, 1997.
FERRETTI, R.; MARQUES, M. J. PERTILLE, A.; SANTO-NETO, H. Sarcoplasmic-
endoplasmic-reticulum ca(2+)-atpase and calsequestrin are overexpressed in spared intrinsic
laryngeal muscles of dystrophin-deficient mdx mice. Muscle eNerve, v. 39 p. 609 -615, 2009.
FOGAGNOLO MAURÍCIO, A.; MINATEL, E.; SANTO NETO, H.; MARQUES, M.J.
Effects of fish oil containing eicosapentaenoic acid and docosahexaenoic acid on dystrophic
mdx mice. Clinical Nutrition, v. 32, n. 636-642, 2013.
GROUNDS M.; RADLEY H.; LYNCH G.; NAGARAJU K.; LUCA A. Towards developing
standard operating procedures for pre-clinical testing in the mdx mouse model of Duchenne
muscular dystrophy. Neurobiology of Disease, v. 19, 2008.
GUTTRIDGE, D.C.; MAYO, M.W.; MADRID, L.V.; WANG, C.Y.; BALDWIN, A.S.J. NF-
kappaB- induced loss of MyoD messenger RNA: possible role in muscle decay and cachexia.
Science, v. 29, p. 2363–2366, 2000.
HALLIWELL, B.; WHITEMAN, M. Measuring reactive species and oxidative damage in
vivo and in cell culture: how should you do it and what do the results mean? British Journal
of Pharmacology, v. 142, n. 2, p. 231-255, 2004

68
HARPER, C.R., EDWARDS, M.J., DEFILIPIS, A.P., JACOBSON, T.A. Flaxseed oil
increases the plasma concentrations of cardioprotective (n-3) fatty acids in humans. Journal
of Nutrition, v. 136, p. 83–87, 2006.
HARTEL, J.V.; GRANCHELLI, J.A.; HUDECKI, M.S.; POLLINA, C.M.; GOSSELIN, L.E.
Impact of prednisone on TGF-β1 and collagen in diaphragm muscle from mdx mice. Muscle
e Nerve, v. 24, p. 428-432, 2001.
HATHOUT, Y.; SEOL,H.; HAN, M.H.J.; ZHANG, A.; BROWN, K.J.; HOFFMAN, E.P.
Clinical utility of serum biomarkers in Duchenne muscular dystrophy. Clinical Proteomics,
v. 13, n. 9, 2016.
HOFFMAN E.P.; BROWN R.H.; KUNKEL L.M. Dystrophin: The protein product of
Duchenne muscular dystrophy locus. Cell, v. 51, 1987.
HOUDE, S.; FILIATRAULT, M.; FOURNIER, A., DUBÉ, J., D'ARCY, S.; BÉRUBÉ, D.,
BROUSSEAU, Y.; LAPIERRE, G.; VANASSE, M. Deflazacort use in Duchenne muscular
dystrophy: an 8-year follow-up. Pediatric Neurology, v.38, n. 3, p. 200-206, 2008.
HUSSEIN, G.; MANSOUR, L.; GHAFAR, H.A.; MOSTAFA, F.A.; FAWAZ, L. Short-term
Effects of Corticosteroid Therapy on Cardiac and Skeletal Muscles in Muscular Dystrophies.
Journal Of Investigative Medicine, v. 62, ed. 6, p. 875-879, 2014.
JAMES, M.J., GIBSON, R.A., CLELAND, L.G. Dietary polyunsaturated fatty acids and
inflammatory mediator production. American Journal of Clinical Nutrition, v. 71, p. 343–
348, 2000.
JIANG, C.; WEN, Y.; KURODA, K. et al. Notch signaling deficiency underlies age
dependent depletion of satellite cells in muscular dystrophy. Disease Models e Mechanisms,
v.7, p. 997–1004, 2014.
JUDÈ, S.; ROGER, S.; MARTEL, E.; BESSON, P.; RICHARD, S.; BOUGNOUX, P. Dietary
long-chain omega-3 fatty acids of marine origin: a comparison of their protective effects on
coronary heart disease and breast cancers. Progress in Biophysics e Molecular Biology, v.
90, p. 299-325, 2006.
KINANE, B.; MAYER, O.; LOWES, L.; KHAN, N.; GALLITANO, K.; DUDA, P.;
MENDELL, J. Respiratory function in eteplirsen-treated Duchenne muscular dystrophy
(DMD) patients compared to natural history. Abstracts 2016 / Neuromuscular Disorders, n.
26 S88–S212, 2016.
KOLE, R.; KRIEG, A.M. Exon skipping therapy for Duchenne muscular dystrophy.
Advanced Drug Delivery Reviews, v. 87, n. 104–107, 2015.
KOZAKOWSKA, M.; PIETRASZEK-GREMPLEWICZ, K.; JOZKOWICZ, A.; DULAK, J.
The role of oxidative stress in skeletal muscle injury and regeneration: focus on antioxidant
enzymes. Journal of muscle research and cell motility, v. 36, ed. 6, p. 377-393, 2015.
LEFAUCHER J.P.; PASTORET C.; SEBILLE A. Phenotype of distrophinophathy in old mdx
mice. The Anatomical Record, v.242, p. 70 –76, 1995.

69
LEMOS, D.R.; BABAEIJANDAGHI, F.; LOW, M.; CHANG, C.K.; LEE, S.T.; FIORE, D.;
ZHANG, R.H.; NATARAJAN, A.; NEDOSPASOV, S.A.; ROSSI, F.M. Nilotinib reduces
muscle fibrosis in chronic muscle injury by promoting TNF-mediated apoptosis of
fibro/adipogenic progenitors. Nature Medicine, v. 21, p.786–794, 2015.
LEWIS, K.D.; HUANG, W.; ZHENG, X.; JIANG, Y.; FELDMAN, R.S.; FALK, M.C.
Toxicological evaluation of arachidonic acid (ARA)-rich oil and docosahexaenoic acid
(DHA)-rich oil. Food and Chemical Toxicology, v. 96, n. 133-144, 2016.
LYNCH, G.S.; HINKLE, R.T.; CHAMBERLAIN, J.S.; BROOKS, S.V.; FAULKNER, J.A.
Force and power output of fast and slow skeletal muscles from mdx mice 6–28 months old.
The Journal of Physiology, n. 535, p. 591–600, 2001.
MA, Z.Y.; ZHONG, Z.G.; QIU, M.Y.; ZHONG, Y.H.; ZHANG, W.X. TGF-β1 activates the
canonical NF-kB signaling to promote cell survival and proliferation in dystrophic muscle
fibroblasts in vitro. Biochemical and Biophysical Research Communications, v. 471, n.
576-581, 2016.
MACHADO, R.V.; MAURÍCIO, A.F.; TANIGUTI, A.P.; FERRETTI, R.; NETO, H.S.;
MARQUES, M.J. Eicosapentaenoic acid decreases TNF-a and protects dystrophic muscle of
mdx mice from regeneration. Journal of Neuroimmunology, v. 232, p. 145-150, 2011.
MAGEE P.; PEARSON S.: ALLEN J. The omega-3 fatty acid, eicosapentaenoic acid (EPA)
prevents the damaging effects of tumor necrosis factor (TNF)-alpha during murine skeletal
muscle cell differentiation. Lipids in health and disease, v. 7, n. 24, 2008.
MARIOL, M.C.; SÉGALAT, L. Muscular degeneration in the absence of dystrophin is a
calcium-dependent process. Current Biology, v. 30, n. 11, p. 1691-1694, 2001.
MARQUES M. J. Structural Biology of the distrophin – deficient muscle fiber. Brazilian
Journal of Morphological Science, v. 21, p. 145-152, 2004.
MARQUES, M.J.; OGGIAM, D.S.; BARBIN, I.C.; FERRETTI. R.; SANTO NETO, H.
Long-term therapy with deflazacort decreases myocardial fibrosis in mdx mice. Muscle e
Nerve, v.40, n. 3, p. 466-468, 2009.
MAURO, A. Satellite cell of skeletal muscle fibers. The Journal of Biophysical and
Biochemical Cytology, v. 9, p. 493–495, 1961.
MCADAM, L.C.; MAYO, A.L.; ALMAN, B.A.; BIGGAR, W.D. The Canadian experience
with long-term deflazacort treatment in Duchenne muscular dystrophy. Acta Myologica, v.
31, n. 1, p. 16-20, 2012.
MCGREEVY, J.W.; HAKIM, C.H.; MCINTOSH, M.A.; DUAN, D. Animal models of
Duchenne muscular dystrophy: from basic mechanisms to gene therapy. Disease Model
Mechanisms. v. 8, n. 3, p. 195-213, 2015.
MENDELL, J.R.; SHILLING, C.; LESLIE, N.D.; FLANIGAN, K.M.; AL-DAHHAK, R.;
GASTIER-FOSTER, J.; KNEILE, K.; DUNN, D.M.; DUVAL, B.; AOYAGI, A.; HAMIL,
C.; MAHMOUD, M.; ROUSH, K.; BIRD, L.; RANKIN, C.; LILLY, H.; STREET, N.;

70
CHANDRASEKAR, R.; WEISS, R.B. Evidence-based path to newborn screening for
Duchenne muscular dystrophy. Annals of Neurology. V. 71, n. 3, p.304-313, 2012.
MESA, L.E.; DUBROVSKY, A.L.; CORDERI, J.; MARCO, P.; FLORES, D. Steroids in
duchenne muscular dystrophy deflazacort trial. Neuromuscular Disorders, v. 1, n. 4. p. 261
266, 1991.
MESSINA, S.; BITTO, A.; AGUENNOUZ, M.; MAZZEO, A.; MIGLIORATO, A.;
POLIYOF.; IRRERA, N.; ALTAVILLA, D.; VATA, G.L.; RUSSO, M.; NARO, A.; DE
PASQUALEM, G.; RIZZUTO, E.; MUSARÒ, A.; SQUADRITO, F.; VITA, G. Flavocoxid
counteracts muscle necrosis and improves functional properties in mdx mice: a comparison
study with methylprednisolone. Experimental neurology, v. 220, n. 2, p. 349-58, 2009.
MESSINA, S.; BITTO, A.; AGUENNOUZ, M.; MINUTOLI, L.; MONICI, M.C.;
ALTAVILLA, D.; SQUADRITO, F.; VITA, G. Nuclear factor kappa-B blockade reduces
skeletal muscle degeneration and enhances muscle function in Mdx mice. Experimental
Neurology, n. 198, p. 234–241, 2006.
MOON, D.O.; KIM, K.C.; JIN, C.Y.; HAN, M.H.; PARK, C.; LEE, K.J.; PARK, Y.M.;
CHOI, Y.H.; KIM, G.Y. Inhibitory effects of eicosapentaenoic acid on lipopolysaccharide-
induced activation in bv2 microglia. International Immunopharmacology, v. 7, p. 222–229,
2007.
MOREIRA, D. O.; PEREIRA, J.A.; TANIGUTI, A.P.; MATSUMURA, C.Y.; RAMOS,
L.A.; AREAS, M.A.; NETO, H.S.; MARQUES, M.J. Suramin attenuates dystrophin-deficient
cardiomyopathy in the mdx mouse model of duchenne muscular dystrophy. Muscle e Nerve,
v. 48, n. 6, p. 911-91, 2013.
MOXLEY, R.T.; ASHWAL, S.; PANDYA, S.; CONNOLLY, A.; FLORENCE, J.;
MATHEWS, K.; BAUMBACH, L.; MCDONALD, C.; SUSSMAN, M.; WADE, C. Practice
parameter: corticosteroid treatment of Duchenne dystrophy: report of the Quality Standards
Subcommittee of the American Academy of Neurology and the Practice Committee of the
Child Neurology Society. Neurology, v. 64, n. 1, p.13-20, 2005.
MOYER, A.L.; WAGNER, K.R. Regeneration versus fibrosis in skeletal muscle. Current
Opinion Rheumatology, v. 23, p. 568–573, 2011.
MRAK, R.E. Muscle membranes in diseases of muscle. Boca Raton, FF:CRC Press, p. 24-52,
1985.
MUNTONI, F.; LUO, X.; ELFRING, G.; KROGER, H.; RIEBLING, P.; ONG, T.; SPIEGEL,
R.; PELTZ, S.; MCDONALD,C. Results of North Star ambulatory assessments in the phase 3
ataluren confirmatory trial in patients with nonsense mutation Duchenne muscular dystrophy.
(ACT DMD). Abstracts 2016 / Neuromuscular Disorders, n. 26, 2016.
OSSIPOVA, O. EZAN, J.; SOKOL, S.Y. PAR-1 phosphorylates Mind bomb to promote
vertebrate neurogenesis. Developmental Cell, v. 17, p. 222–233, 2009.
PASQUINI, F.; GUERIN, C.; BLAKE, D.; et al. The effects of glucocorticoids on the
accumulation of utrophin by cultured normal and dystrophic human skeletal muscle satellite
cells. Neuromuscular Disorders, v. 5, p. 105-114, 1995.

71
PASSAQUIN, A.C.; RENARD, M.; KAY, L.; CHALLET, C.; MOKHTARIAN, A.;
WALLIMANN, T.; RUEGG, U.T. Creatine supplementation reduces skeletal muscle
degeneration and enhances mitochondrial function in mdx mice. Neuromuscular Disorders,
v. 12, p. 174–182, 2002.
PASTORET, C.; SEBILLE, A. Mdx mice show weakness and muscle deterioration with age.
Journal of Neural Science, v. 129, p.97-105, 1995.
RADLEY, H.G.; GROUNDS, M.D. Cromolyn administration (to block mast cell
degranulation) reduces necrosis of dystrophic muscle in mdx mice. Neurobiology of Disease,
v. 23, p. 387–397, 2006.
RAPUCCI MORAES, L.H.; BOLLINELI, R.C.; MIZOBUTI, D.S.; SILVEIRA, L.R.;
MARQUES, M.J.; MINATEL, E. Effect of N acetylcysteine plus deferoxamine on oxidative
stress and inflammation in dystrophic muscle cells. Redox Report, v. 20 n. 3, 2015.
REAGAN-SHAW, S.; NIHAL, M.; AHMAD, N. Dose translation from animal to human
studies revisited. FASEB Journal, v. 22, n. 3, p. 659-661, 2008.
RIFAI, Z.; WELLE, S.; MOXLEY, R.T., et al. Effect of prednisone on protein metabolism in
Duchenne dystrophy. American Journal of Physiology, v. 268, p. 67-74, 1995.
RODACKI, C.L.; RODACKI, A.L.; PEREIRA, G.; NALIWAIKO, K.; COELHO, I.;
PEQUITO, D.; FERNANDES, L.C. Fish-oil supplementation enhances the effects of strength
training in elderly women. American Journal of Clinical Nutrition, v. 95, n. 2, p. 428-36,
2012.
SACCO, A.; MOURKIOTI, F.; TRAN, R. et al. Short telomeres and stem cell exhaustion
model Duchenne muscular dystrophy in mdx/mTR mice. Cell, v. 143, p. 1059–1071, 2010.
SCHLANGER, S., SHINITZKY, M., YAM, D. Diet enriched with omega-3 fatty acids
alleviates convulsion symptoms in epilepsy patients. Epilepsy, v. 43, p. 103–104, 2002.
SCHOBER, M.; SCHAEFER, M.; KNOBLICH, J.A. Bazooka recruits Inscuteable to oriente
asymmetric cell divisions in Drosophila neuroblasts. Nature, v. 402, p. 548–551, 1999.
SEIXAS, S.L.; LAGROTA-CÂNDIDO, J.; SAVINO, W.; QUIRICO-SANTOS, T. The
importance of mdx mouse in the physiopathology of Duchenne's muscular dystrophy. Arq
Neuropsiquiatry, p.610-617, 1997.
SERRANO, A.L.; MANN, C.J.; VIDAL, B.; ARDITE, E.; PERDIGUERO, E.; MUNOZ-
CANOVES. Cellular and molecular mechanisms regulating fibrosis in skeletal muscle repair
and disease. Current Topics in Developmental Biology, v. 96 , 2011.
SHAMI, N.J.I.E.; MOREIRA, E.A.M. Licopeno como agente antioxidante. Revista de
Nutrição, v.17, n. 2, p. 227-236, 2004.
SHAVLAKADZE, T.; WHITE, J.; HOH, J.F.; ROSENTHAL, N.; GROUNDS, M.D.
Targeted expression of insulin-like growth factor-1 reduces the necrosis of dystrophic mdx
muscle. Molecular Therapy, v. 10, p. 829–843, 2004.

72
SHI, H.; BOADU, E.; MERCAN, F.; LE, A.M.; FLACH, R.J.; ZHANG, L.; TYNER, K.J.;
OLWIN, B.B.; BENNETT, A.M. MAP kinase phosphatase-1 deficiency impairs skeletal
muscle regeneration and exacerbates muscular dystrophy. FASEB Journal, v. 24, p. 2985–
2997, 2010.
SILLER, K.H.; CABERNARD, C.; DOE, C.Q. The NuMA-related Mud protein binds Pins
and regulates spindle orientation in Drosophila neuroblasts. Nature Cell Biology, v. 8, p.
594–600, 2006.
SKLAR, R.M.; BROWN, R.H. Methylprednisolone increases dystrophin levels by inhibiting
myotube death during myogenesis of normal human muscle in vitro. Journal of the
Neurological Sciences, v. 101, p. 73-81, 1991.
SKRABEK, R.Q.; ANDERSON, J.E. Metabolic shifts and myocyte hypertrophy in
deflazacort treatment of mdx mouse cardiomyopathy. Muscle e Nerve, v. 24, n. 2, p. 192-
202, 2001.
SPENCER, M.J.; TIDBALL, J.G. Do immune cells promote the pathology of dystrophin
deficient myopathies? Neuromuscular Disorders, v. 11, p. 556- 564, 2001.
SPINAZZOLA, J.M.; KUNKEL, L.M. Pharmacological therapeutics targeting the secondary
defects and downstream pathology of Duchenne muscular dystrophy. Expert opinion on
orphan drugs, v. 4, ed. 11, p. 1179-1194, 2016.
SPULER, S.; ENGEL, A.G. Unexpected sarcolemmal complement membrane attack complex
deposits on nonnecrotic muscle fibers in muscular dystrophies. Neurology, v. 50, p. 41- 46,
1998.
SPURNEY CF. Cardiomyopathy of Duchenne muscular dystrophy: current understanding and
future directions. Muscle e Nerve, v. 44, p. 8-19, 2011.
STEDMAN, H.H.; SWEENY, H.L.; SHRAGER, J.B.; MAGUIRE, H.C.; PANETTIERI,
R.A.; PETROF, B.; NARUSAWA, M.; LEFEROVICH, J.M.; SLADKY, J.T.; KELLY, A.M.
The MDX mouse diaphragm reproduces the degenerative changes of Duchenne muscular
dystrophy. Nature, v. 352, p. 536–538, 1991.
SUN, C.; LI. S.; LI, D. Sulforaphane mitigates muscle fibrosis in mdx mice via Nrf2-
mediated inhibition of TGF-/Smad signaling. Journal of Applied Physiology, n. 120, p. 377–
390, 2016.
TANIGUTI, A.P.; PERTILLE, A.; MATSUMURA, C.Y.; SANTO-NETO, H.; MARQUES,
M.J. Prevention of muscle fibrosis and myonecrosis in mdx mice by suramin, a TGF-beta1
blocker. Muscle e Nerve, v. 43, n. 1, p. 82-87, 2011.
UCHIDA, K. 4-Hydroxy-2-nonenal: a product and mediator of oxidative stress. Progress in
Lipid Research, p. 1-26, 2003.
VIEIRA, N.M.; ELVERS, I.; ALEXANDER, M.S. et al. (2015) Jagged 1 rescues the
Duchenne muscular dystrophy phenotype. Cell, v. 163, p. 1204–1213, 2015.

73
WELLER, C.; ZSCHUNTZSCH, J.; MAKOSCH, G.; METSELAAR, J.M.; KLINKER, F.;
KLINGE, L.; LIEBETANZ, D.; SCHMIDT, J. Motor performance of young dystrophic mdx
mice treated with long-circulating prednisolone liposomes. Journal Of Neuroscience
Research, v. 90, ed. 5, p. 1067-1077, 2012.
WHITEHEAD N.P.; YEUNG E.W.; ALLEN D.G. Muscle damage in mdx (dystrophic) mice:
role of calcium and reactive oxygen species. Clinical and Experimental Pharmacology &
Physiology, v. 33, p. 657-662, 2006.
YE, S.; TAN, L.; MA, J.; SHI, Q.; LI, J. Polyunsaturated docosahexaenoic acid suppresses
oxidative stress induce endothelial cell calcium influx by altering lipid composition in
membrane caveolar rafts. Prostaglandins Leukot Essent Fatty Acids, v. 83, p. 37-43, 2010.
YIN, H.; PRICE, F.; RUDNICKI, M.A. Satellite cells and the muscle stem cell niche.
Physiological Reviews, v. 93, p. 23–67, 2013.
ZGHEEL, F.; ALHOSIN, M.; RASHID, S.; BURBAN, M.; AUGER, C.; SCHINI-KERTH,
V.B. Redox-sensitive induction of src/PI3-kinase/akt and MAPKs pathways activate eNOS in
response to EPA:DHA. PLoS One, v. 9, 2014.
ZHONG, H.; YIN, H. Role of lipid peroxidation derived 4-hydroxynonenal (4-HNE) in
cancer: Focus in go mitochondria. Redox Biology, v.4, p.193–199, 2015.

74
ANEXO A

75
ANEXO B












![MANUAL DE INSTRUÇÕES MINI HI-FI SYSTEM€¦ · 17 - Instalar usando o “Google Android Market "Rttlqj [qf~ ^ytwj-< 18 Função Rádio CK Zz{nsit jxyf °jx ij ] int CK Xjqmtwfsit](https://img.document.onl/doc/110x75/601cf848ecf4eb65944544c7/manual-de-instrues-mini-hi-fi-system-17-instalar-usando-o-aoegoogle-android.jpg)
















