Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DA PARAÍBA
CENTRO DE CIÊNCIAS DA SAÚDE
PROGRAMA DE PÓS-GRADUAÇÃO EM PRODUTOS
NATURAIS E SINTÉTICOS BIOATIVOS
ANA CAROLINE DE LIMA SILVA
Investigação da atividade espasmolítica de uma série de derivados
do lapachol em íleo de cobaia: um estudo comparativo
João Pessoa-PB
2014

2
ANA CAROLINE DE LIMA SILVA
Investigação da atividade espasmolítica de uma série de derivados
do lapachol em íleo de cobaia: um estudo comparativo
Dissertação apresentada ao Programa de Pós-graduação em Produtos Naturais e Sintéticos Bioativos, do Centro de Ciências da Saúde, da Universidade Federal da Paraíba, como parte dos requisitos para obtenção do título de MESTRE EM PRODUTOS NATURAIS E SINTÉTICOS BIOATIVOS. Área de Concentração: FARMACOLOGIA
Orientadora: Profa. Dra. Bagnólia Araújo da Silva
Coorientadora: Profa. Dra. Fabiana de Andrade Cavalcante
João Pessoa-PB
2014

Catalogação na Publicação
Seção de Catalogação e Classificação
S586i Silva, Ana Caroline de Lima.
Investigação da atividade espasmolítica de uma série de
derivados do lapachol em íleo de cobaia: um estudo compara-
tivo / Ana Caroline de Lima Silva. - João Pessoa, 2014.
119 f. : il.
Orientadora: Bagnólia Araújo da Silva.
Coorientadora: Fabiana de Andrade Cavalcante.
Dissertação (Mestrado) - UFPB/CCS/ PgPNSB
1. Farmacologia. 2. Derivados do lapachol. 3. Nafto-
quinona. 4. Canais de potássio. 5. Ação espasmolítica.
I. Título.
UFPB/BC CDU - 615(043)


4
Dedicatória

5
Ao meu pai e ao meu avô, que hoje já não se
encontram comigo fisicamente, mas o amor de
vocês, esse levarei enquanto vida eu tiver. O
amor mais lindo e sincero.
Aos amores da minha vida, Eliene C. de Lima e
Leonízia Cordeiro, Mainha e Voinha, essa
vitória não é só minha, é também de vocês. Por
me amarem tanto e pela confiança depositada
desde sempre. Sou eternamente grata por
tanto amor e dedicação recebidos, muitas
vezes sem merecer. Sem vocês na minha vida,
nada disso faria sentido. Vocês são meus
exemplos, mulheres fortes, meu alicerce, minha
paz.
Aos que sempre estiveram comigo e acreditam
em mim.

6
Agradecimentos

7
Não conseguimos nada sozinhos, e precisamos sim da ajuda de várias
pessoas, seja com uma palavra de apoio em um momento oportuno, broncas
quando necessárias e um ombro que está sempre ali pronto a suportar as lágrimas.
Gratidão, essa é a palavra de ordem.
Ao meu Deus, por seu amor e fidelidade, fazendo-me perceber que são nos
momentos mais difíceis que os milagres acontecem, por dar-me forças pra seguir em
frente quando penso em desistir. Por colocar alguns anjos na minha vida, tornando
assim a caminhada mais leve e feliz.
À minha querida orientadora, Dra. Bagnólia Araújo da Silva, por me aceitar
como sua aluna desde a iniciação científica, um grande exemplo de pessoa,
educadora e profissional. Pela amizade, paciência, compreensão, risadas e
ensinamentos para a vida. Por sempre tentar extrair de nós o melhor que podemos
dar e por acreditar no meu potencial.
À Fabiana Cavalcante, coorientadora deste trabalho, por estar sempre
disposta a ajudar, tirando dúvidas, solucionando problemas. Sua ajuda foi muito
importante para a realização desse trabalho.
Ao Prof. Dr. Celso Camara pela gentileza em ceder os compostos testados
neste trabalho.
Aos Professores Caliandra Luna (UFPB) e Luis Fernando Marques (UFPB)
por aceitarem participar desta etapa tão importante na minha formação, pela
compreensão e pelas contribuições a este trabalho. Assim como as Professoras
Leônia Batista e Liana Clébia, por estarem dispostas à suplência e pelas
contribuições na escrita deste trabalho.
A todos os Professores que passaram na minha vida acadêmica, seja na
graduação de Farmácia ou no Programa de Pós-graduação em Produtos Naturais e
Sintéticos Bioativos (PPgPNSB) e que com certeza fizeram o diferencial na minha
formação, em especial a: Adalberto Coelho, Bagnólia Araújo, Demétrius Araújo, Inês
Freitas, Luis Fernando Marques e Tatjana Keesen, por me mostrar que ser
professor vai muito além de um título e sim de ter o poder de transformar pessoas,
extraindo do aluno aquilo que de melhor ele tem.
Ao coordenador do PPgPNSB, Prof. José Maria Barbosa Filho, e o
vice-coordenador, Prof. Josean Fechine Tavares, por conduzir o PgPNSB com
tamanha habilidade e competência.

8
À secretária do PPgPNSB Caroline Mangueira, pela paciência nas inúmeras
vezes que eu chegava lá aperreando, seja pra tirar dúvidas burocráticas ou pedir
papel. Por sua eficiência em conduzir seu trabalho.
A Luiz Henrique C. Vasconcelos, amigo querido que muito me ajudou na
realização desse trabalho, você foi fundamental. Serei sempre grata por tudo que
você fez para que essa dissertação saísse.
Ao meu aluno de iniciação científca, José Lucas, pela ajuda nos
experimentos, pela amizade, disposição e boa vontade em sempre me ajudar, seja
na bancada ou apenas em me ouvir.
À Rosimeire Santos pelos ensinamentos nos meus primeiros passos na vida
acadêmica, pela amizade e pelos sorrisos.
A todos os amigos e colegas que estão ou passaram pelo Laboratório de
Farmacologia Funcional Prof. George Thomas, entre eles, Alice Bezerra, Aline Brito,
Ana Carolina Correia, Anne Abreu, Cibério Macêdo, Fabiana Cavalcante, Fabio
Monteiro, Fernando Queiroga, Filipe Oliveira, Giuliana Amanda, Giulyane Moreno,
Hannah Olga, Iara Luna, Italo Martins, Joedna Cavalcante, Joelmir Silva, José Lucas
Galvãoo, Juliana Carreiro, Kimã Meira, Layanne Cabral, Luiz Henrique César, Luiz
Henrique Agra, Maria da Conceição, Milena Medeiros, Paula Benvindo, Rafael
Travassos, Renata Sampaio, Rosimeire Ferreira, Sarah Rebeca, Tamyris Freire pela
ajuda, risadas no cafezinho, discussões científicas, amizade e boa convivência.
Aos amigos que fiz na graduação em Farmácia-Bioquímica, em especial a
Ana Letícia, Anderson Moura, Filipe Rolim, Elton Falcão, Rômulo Sampaio, Fabio
Dias, Gislaine Oliveira, Waleska Viana e Lyndemberg Silveira.
A todos os amigos e colegas das turmas de Mestrado e Doutorado do
PgPNSB por dividirem as angústias, sonhos, lágrimas e muita felicidade, em
especial a Paula Benvindo, Élida Vieira, Sávio Benvindo, Tassiana Dantas, Danilo
Lemos, Milen Souza, Itamar Barbosa, Kivia Sales, Talissa Mozzini, Jacqueline Alves,
Leônidas Jr., Luciano Leite, Yuri Mangueira, Denise Leite.
A vida sempre nos mostra que não estamos nunca sozinhos, agradeço a
alguns que surgiram em minha vida para tornar a caminhada mais leve, feliz e
fazendo transbordar amor, verdadeiros encontros de alma.
À Vitória Mola, Natália Cândido, Misma Carolina e Suelen Alves amigas
queridas que sempre me apoiaram e me incentivaram. Vocês são muito importantes
nisso tudo, já pensamos em desistir, mas a força e a coragem é o que nos move.

9
A Rafael Travassos, meu anjo, amado amigo e irmão. Pela amizade,
companheirismo em dividir comigo lágrimas, sonhos e sorrisos. Por sempre acreditar
em mim, mesmo quando nem eu o fazia.
A Jailson Rocha, Ulrich Vasconcelos, Ian Porto, Sildivane Valcácia e Tatjana
Keesen, amigos amados, verdadeiros presentes que ganhei nesse último ano.
Obrigada por todo incentivo, pelos momentos deliciosos juntos, regados a cafés,
sorrisos e muita alegria.
A Italo Rossi e Paula Benvindo, amigos que muito me ajudaram na reta final
de elaboração desse trabalho, obrigada pelos bons momentos, risadas e pela
grande amizade de vocês.
Aos “legais sem futuro”; quantos momentos felizes, alguns aperriados
também mas tudo é para o nosso crescimento. Em especial a Sávio, Tassiana e
Danilo obrigada pela amizade de vocês e por tudo que passamos juntos.
Aos meus familiares, por todo o incentivo e por me apoiar em todos os
momentos, compreendendo minha ausência e vibrando com as conquistas.
Ao José Crispim Duarte, pela sua amizade, competência e auxílio técnico nos
mais variados problemas no Laboratório e no Biotério.
Ao Sr. Luís C. Silva e Adriano S. Cordeiro por trabalharem tão arduamente
para que possamos realizar nossos experimentos, por sua dedicação e cuidado no
trato dos animais no Biotério e no laboratório.
À Mônica e D. Evani pelo carinho, palavras de apoio, orações tendo sempre
uma palavra de incentivo nos momentos de estresse e serem sempre tão atenciosas
com todos.
À Sra. Luzinete, pelo trabalho realizado no laboratório, na limpeza de material
e equipamentos úteis à rotina do mesmo.
Ao Professor e Conselheiro Federal de Farmácia, Samuel Meira, pelo apoio
financeiro concedido para participação em congressos científicos.
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq),
pelo apoio financeiro.
À Coordenação de Aperfeiçoamento de Pessoal de Ensino Superior
(CAPES) pelo suporte técnico-científico.
À Direção do Centro de Biotecnologia (CBiotec), na pessoa do Prof. Dr. Valdir
de Andrade Braga, pelo apoio Institucional.

10
À Universidade Federal da Paraíba, instituição responsável pela minha
formação, responsável por me fazer enxergar o mundo de um novo jeito.
A todos que, de maneira direta ou indireta, contribuíram para a produção
dessa Dissertação de Mestrado.
Ana Caroline de Lima Silva

11
Resumo

12
RESUMO
As naftoquinonas são substâncias de grande interesse, pois apresentam uma variedade de atividades biológicas. Pertencente a esse grupo, destaca-se o lapachol, uma naftoquinona isolada de espécies de Tabebuia spp., conhecidas popularmente como “ipês”. Uma vez que muitas atividades farmacológicas, como a espasmolítica, são relacionadas ao lapachol e aos seus derivados naturais ou sintéticos, pesquisadores tem voltado sua atenção para esta molécula a fim de criar compostos com potencial terapêutico. Nessa perspectiva decidiu-se investigar um possível efeito espasmolítico de uma nova série de derivados do lapachol (UFRPE 15, UFRPE 17, UFRPE 31, UFRPE 114, UFRPE 115 e UFRPE 116) e do norlapachol (UFRPE 106, UFRPE 107, UFRPE 111, UFRPE 112, UFRPE 113 e UFRPE 117) em íleo isolado de cobaia, bem como investigar o mecanismo de ação do composto que apresentar uma maior eficácia. Na triagem farmacológica, os compostos derivados do lapachol apresentaram uma baixa eficácia farmacológica em inibir as contrações fásicas induzidas por carbacol e por histamina, apenas o composto UFRPE 115 inibiu as contrações fásicas induzidas por histamina de maneira dependente de concentração. Entretanto, os derivados do norlapachol mostraram-se mais promissores, uma vez que todos inibiram as contrações fásicas induzidas por carbacol e por histamina de maneira dependente de concentração, onde os compostos UFRPE 107, UFRPE 112 e UFRPE 117 apresentaram uma maior potência e eficácia relativas. O efeito relaxante destes foi avaliado no íleo pré-contraído com KCl, CCh ou histamina, e observou-se que estes 3 derivados relaxaram o íleo de cobaia de maneira dependente de concentração, sendo o derivado UFRPE 117 o que apresentou maior potência relativa, assim foi selecionado para o estudo do seu mecanismo de ação. Uma vez que UFRPE 117 inibiu as contrações induzidas por histamina, bem como relaxou o íleo pré-contraído pelo mesmo agonista, avaliou-se a participação dos receptores histaminérgicos nesse efeito. UFRPE 117 inibiu as curvas cumulativas à histamina, desviando-as para direita, de maneira não paralela e com redução do Emax, sugerindo um antagonismo do tipo não competitivo pseudo irreversível. Como a potência relaxante deste composto foi reduzida quando o íleo era pré-contraído com 40 mM de KCl, investigou-se a participação dos canais de K+ utilizando 5 mM de CsCl, um bloqueador não seletivo destes canais, onde a sua potência relaxante foi atenuada em cerca de 12 vezes na presença desse bloqueador, sugerindo a participação dos canais de K+ na ação espasmolítica de UFRPE 117. Para avaliar quais os subtipos de canais de K+ estariam envolvidos nesse efeito, utilizou-se 10-5 M de glibenclamida, um bloqueador dos KATP que não alterou a potência relaxante de UFRPE 117, descartando a participação desses canais. Entretanto na presença de 100 nM de apamina um bloqueador seletivos dos SKCa, e 1 mM de TEA+, um bloqueador seletivo dos BKCa a potência relaxante de UFRPE 117 foi atenuada em cerca de 4 e 5 vezes respectivamente, confirmando a participação destes subtipos de canais de K+. Assim o mecanismo de ação espasmolítica do derivado sintético do norlapachol, UFRPE 117 envolve a ativação/modulação positiva dos SKCa e BKCa, o que levaria a uma hiperpolarização de membrana, bloqueio indireto dos Cav, redução da [Ca2+]c e consequente relaxamento desse órgão.
Palavras-chave: derivados do lapachol, naftoquinona, canais de potássio, ação espasmolítica, íleo
de cobaia.
INVESTIGAÇÃO DA ATIVIDADE ESPASMOLÍTICA DE UMA SÉRIE DE DERIVADOS DO LAPACHOL EM ÍLEO DE COBAIA: UM ESTUDO COMPARATIVO
SILVA, A.C.L. Pós-Graduação em Produtos Naturais e Sintéticos Bioativos,
Dissertação de Mestrado, CCS/UFPB (2014).

13
Abstract

14
ABSTRACT The naphthoquinones are substances of great interest, since they have a variety of biological activities. Belonging to this group, there is lapachol, an isolated naphthoquinone species of Tabebuia spp., popularly known as "ipê". Many pharmacological activities, such as spasmolytic, are related to lapachol and its natural or synthetic derivatives. Researchers have turned their attention to this molecule in order to create compounds with therapeutic potential. From this perspective it was decided to investigate the spasmolytic effect of a new series of lapachol derivatives (UFRPE 15, UFRPE 17, UFRPE 31, UFRPE 114, UFRPE 115 e UFRPE 116) and norlapachol (UFRPE 106, UFRPE 107, UFRPE 111, UFRPE 112, UFRPE 113 e UFRPE 117) in the isolated guinea pig ileum, as well as investigate the mechanism of action of the compound which show a better efficacy. In the pharmacological screening, the lapachol derivatives showed low efficacy in inhibiting the phasic contractions induced by carbachol (CCh) and histamine. Only UFRPE 115 inhibited the phasic contractions induced by histamine in a concentration-dependent manner. However, derivatives from norlapachol were more promising, since all of them inhibited the phasic contractions induced by CCh and histamine in a concentration-dependent manner, where UFRPE 107, UFRPE 112 and UFRPE 117 compounds showed a better potency and efficacy. The spasmolytic effect of these has been reported in the ileum pre-contracted with KCl, CCh or histamine, and it was noted that these three derivatives relaxed guinea pig ileum in a concentration-dependent manner, and in special the UFRPE 117 showed the highest relative potency, thus, it was selected to study its mechanism of action. Once UFRPE 117 inhibited histamine-induced contractions and relaxed the ileum pre-contracted by the same agonist, we evaluated the involvement of histamine receptors in this effect. UFRPE 117 inhibited the cumulative curves to histamine, shifting them to right and in a nonparallel manner with Emax reduction, suggesting a noncompetitive antagonism pseudo irreversible type. Since the spasmolytic potency of this compound was reduced when the ileum was pre-contracted with 40 mM KCl, we investigated the involvement of K + channels using 5 mM CsCl, a non-selective blocker of these channels, where its relaxing power was attenuated by approximately 12 folds in the presence of this blocker, suggesting the involvement of K+ channels in spasmolytic action UFRPE 117. To assess which of K+ subtypes channels were involved in this effect, we used 10-5 M glibenclamide, a KATP blocker, whose did not alter the relaxing potency of UFRPE 117, discarding the involvement of these channels. However, in the presence of 100 nM apamin, a selective blocker of SKCa and 1 mM TEA+, a selective blocker of BKCa, The UFRPE 117 relaxant potency was attenuated around 4 to 5 folds, respectively, confirming the involvement of SKCa and BKCa. Thus, the mechanism of spasmolytic action of the synthetic derivative of norlapachol, UFRPE 117, involves activation/positive modulation of SKCa and BKCa, which would lead to a hyperpolarization of the membrane, indirect blocking Cav, reduction of [Ca2+]c and consequent relaxation of this organ.
Keywords: lapachol derivatives, naftoquinone, potassium channels, spasmolytic action, guinea pig
ileum
INVESTIGATION OF SPASMOLYTIC ACTIVITY OF A SERIES DERIVATIVE OF LAPACHOL IN GUINEA PIG ILEUM: A COMPARATIVE STUDY SILVA, A.C.L.
Pós-Graduação em Produtos Naturais e Sintéticos Bioativos, Dissertação de Mestrado, CCS/UFPB (2014).

15
LISTA DE FIGURAS
Figura 1 - Tabebuia avellanedae Lorentz e Griseb........................................... 28
Figura 2 –. Estruturas químicas do lapachol (1) e de seus derivados naturais, -
lapachona (2) e -lapachona (3)............................................................................30
Figura 3 – Reação de Hooker para obtenção do norlapachol (4) a partir do lapachol (1) .........................................................................................................................32
Figura 4 (A) – Estrutura química dos derivados do núcleo fenazina obtidos a partir do lapachol.............................................................................................................33
Figura 4 (B) – Estrutura química dos derivados do núcleo fenazina obtidos a partir do norlapachol........................................................................................................34
Figura 5 – Esquema do mecanismo fármaco-mecânico da contração no músculo liso pela ativação da via Gq/11-PLCβ1...............................................................................................................37
Figura 6 – Esquema do mecanismo de manutenção da contração no músculo liso pela ativação da via G12/13-ROCK...........................................................................39
Figura 7 – Estrutura dos canais de K+ sensíveis ao ATP (KATP)............................42
Figura 8 – Estrutura das subunidades α e β1 dos canais de K+ ativados por Ca2+ de grande condutância (BKCa) (A); associação das quatro subunidades dos BKCa
(B)............................................................................................................................43
Figura 9 – Estrutura dos canais de K+ ativados por Ca2+ de pequena condutância (SKCa) (A); mecanismo de ativação do canal pela ligação do Ca2+ à CaM
(B)............................................................................................................................44
Figura 10 – Esquema do mecanismo fármaco-mecânico do relaxamento no músculo liso pela ativação da via Gs -AC-PKA e NO-GCs-PKG............................................45
Figura 11 –.Registros típicos do efeito relaxante de UFRPE 112 sobre as contrações tônicas induzidas por 40 mM de KCl (A) e 10-5 M de carbacol (B) em íleo de cobaia......................................................................................................................72 Figura 12 –. Registros típicos do efeito relaxante de UFRPE 107 sobre as contrações tônicas induzidas por 40 mM de KCl (A) e 10-6 M de histamina (B) em íleo de cobaia....................................................................................................................74 Figura 13 –. Registros típicos do efeito relaxante de UFRPE 117 sobre as contrações tônicas induzidas por 40 mM de KCl (A) e 10-6 M de histamina (B) em íleo de cobaia..........................................................................................................76
Figura 14 –. Registros típicos do efeito relaxante de UFRPE 117 sobre as contrações tônicas induzidas por 10-6 M de histamina na ausência (A) e na presença (B) de 5 mM de CsCl em íleo de cobaia.................... ............................................79

16
Figura 15 –. Registros típicos do efeito relaxante de UFRPE 117 sobre as contrações tônicas induzidas por 10-6 M de histamina na ausência (A) e na presença (B) de 10-5 M de glibenclamida em íleo de cobaia................................................82 Figura 16 –. Registros típicos do efeito relaxante de UFRPE 117 sobre as contrações tônicas induzidas por 10-6 M de histamina na ausência (A) e na presença (B) de 100 nM de apamina em íleo de cobaia.......................................................84
Figura 17 –. Registros típicos do efeito relaxante de UFRPE 117 sobre as contrações tônicas induzidas por 10-6 M de histamina na ausência (A) e na presença (B) de 1 mM de TEA+ em íleo de cobaia..............................................................86
Figura 18 –. Proposta de mecanismo de ação espasmolítica para o derivado sintético do norlapachol UFRPE 117 em íleo isolado de cobaia...........................99

17
LISTA DE GRÁFICOS
Gráfico 1 – Efeito do derivado UFRPE 31 frente às contrações fásicas induzidas por 10-6 M de carbacol (A) e de histamina (B) em íleo de cobaia....................................59
Gráfico 2 – Efeito do derivado UFRPE 15 frente às contrações fásicas induzidas por 10-6 M de carbacol (A) e de histamina (B) em íleo de cobaia....................................60
Gráfico 3 – Efeito do derivado UFRPE 17 frente às contrações fásicas induzidas por 10-6 M de carbacol (A) e de histamina (B) em íleo de cobaia....................................61
Gráfico 4 – Efeito do derivado UFRPE 114 frente às contrações fásicas induzidas por 10-6 M de carbacol (A) e de histamina (B) em íleo de cobaia..............................62
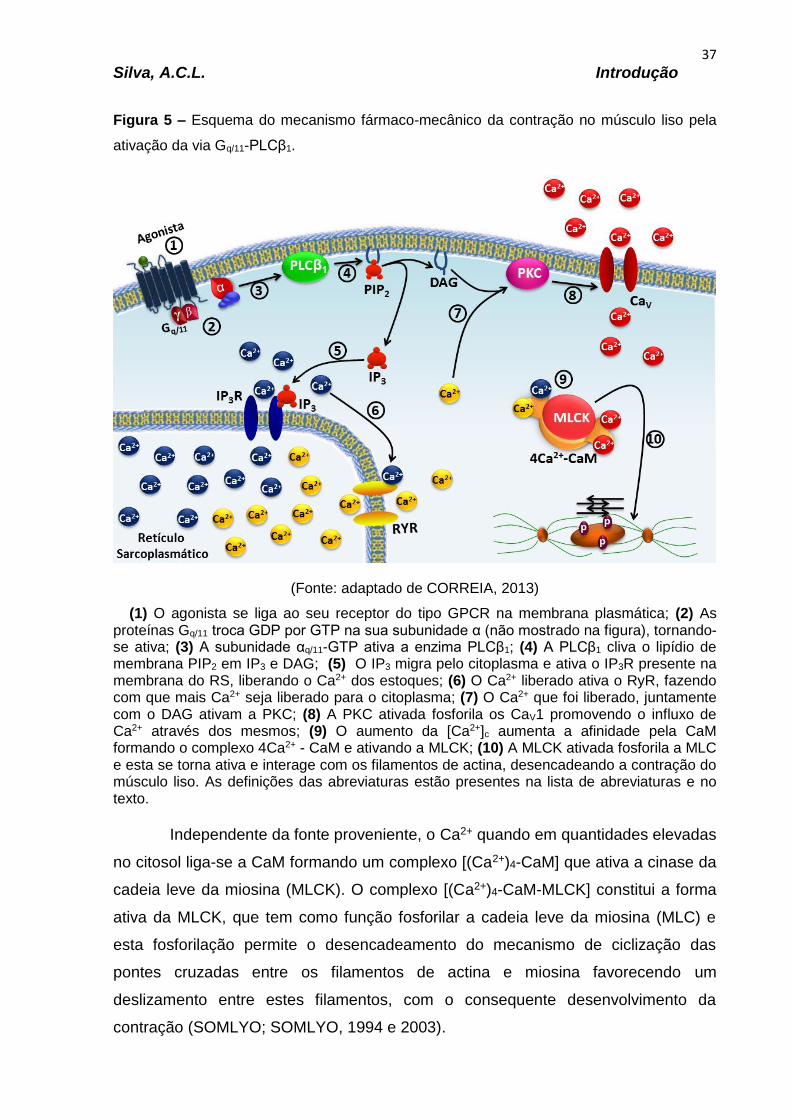
Gráfico 5 – Efeito do derivado UFRPE 115 frente às contrações fásicas induzidas por 10-6 M de carbacol (A) e de histamina (B) em íleo de cobaia..............................63
Gráfico 6 – Efeito do derivado UFRPE 116 frente às contrações fásicas induzidas por 10-6 M de carbacol (A) e de histamina (B) em íleo de cobaia..............................64
Gráfico 7 – Efeito do derivado UFRPE 106 frente às contrações fásicas induzidas por 10-6 M de carbacol (A) e de histamina (B) em íleo de cobaia..............................65
Gráfico 8 – Efeito do derivado UFRPE 107 frente às contrações fásicas induzidas por 10-6 M de carbacol (A) e de histamina (B) em íleo de cobaia..............................66
Gráfico 9 – Efeito do derivado UFRPE 111 frente às contrações fásicas induzidas por 10-6 M de carbacol (A) e de histamina (B) em íleo de cobaia..............................67
Gráfico 10 – Efeito do derivado UFRPE 112 frente às contrações fásicas induzidas por 10-6 M de carbacol (A) e de histamina (B) em íleo de cobaia..............................68
Gráfico 11 – Efeito do derivado UFRPE 113 frente às contrações fásicas induzidas por 10-6 M de carbacol (A) e de histamina (B) em íleo de cobaia..............................69
Gráfico 12 – Efeito do derivado UFRPE 117 frente às contrações fásicas induzidas por 10-6 M de carbacol (A) e de histamina (B) em íleo de cobaia..............................70
Gráfico 13 – Efeito de UFRPE 112 sobre as contrações tônicas induzidas por 40 mM de KCl (■) e 10-5 M de CCh (□) em íleo de cobaia..............................................73
Gráfico 14 – Efeito de UFRPE 107 sobre as contrações tônicas induzidas por 40 mM de KCl () e 10-6 M de histamina (○) em íleo de cobaia.....................................75
Gráfico 15 – Efeito de UFRPE 117 sobre as contrações tônicas induzidas por 40 mM de KCl () e 10-6 M de histamina () em íleo de cobaia...................................77
Gráfico 16 – Curvas concentrações-resposta cumulativas à histamina na ausência (controle) () e na presença de UFRPE 117 nas concentrações de 3 x 10-6(□); 10-5
(); 3 x 10-5 (○); 10-4 () e 3 x 10-4 M () em íleo de cobaia...................................78

18
Gráfico 17 – Efeito de UFRPE 117 sobre as contrações tônicas induzidas por 10-6 M de histamina na ausência () e na presença () de 5 mM de CsCl, em íleo de cobaia....................................................................................................................80
Gráfico 18 – Efeito de UFRPE 117 sobre as contrações tônicas induzidas por 10-6 M de histamina na ausência () e na presença () de 10-5 M de glibenclamida em íleo de cobaia.......................................................................................................83
Gráfico 19 – Efeito de UFRPE 117 sobre as contrações tônicas induzidas por 10-6 M de histamina na ausência () e na presença () de 100 nM apamina em íleo de cobaia..............................................................................................................................85
Gráfico 20 – Efeito de UFRPE 117 sobre as contrações tônicas induzidas por 10-6 M de histamina na ausência () e na presença () de 1 mM de TEA+ em íleo de cobaia....................................................................................................................87
Gráfico 21 – Valores de CE50 (M) de UFRPE 117 na ausência (controle) e na presença de bloqueadores de canais de K+ em íleo de cobaia.................................88

19
LISTA DE TABELAS
Tabela 1 - Composição da solução de Krebs modificado por Sun e Benish (1994) ...................................................................................................................................50
Tabela 2 - Valores de Emax (%) e de CI50 (M) dos derivados do lapachol frente às
contrações fásicas induzidas por carbacol ou por histamina em íleo de
cobaia.........................................................................................................................71
Tabela 3 - Valores de Emax (%) e de CI50 (M) dos derivados do norlapachol frente às
contrações fásicas induzidas por carbacol ou por histamina em íleo de
cobaia.........................................................................................................................71

20
LISTA DE ABREVIATURAS
[(Ca2+)4-CaM] complexo cálcio-calmodulina
[Ca2+] concentração de Ca2+
[Ca2+]c concentração de Ca2+ citosólico
[K+]e concentração extracelular de potássio
4-AP 4-aminopiridina
AC ciclase de adenilil
ACh acetilcolina
AMPc monosfofato cíclico de adenosina
ANOVA análise de variância
ATP trifosfato de adenosina
BKCa canais de potássio ativados por cálcio de grande condutância
CaM calmodulina
CaV canais de cálcio dependentes de voltagem
CaV1 canais de cálcio dependentes de voltagem do tipo 1
CaVL canais de cálcio dependentes de voltagem do tipo L
CCh carbacol
CE50 concentração de uma substância necessária para produzir 50%
de seu efeito máximo
CPI-17 inibidor da fosfatase da miosina potencializado pela PKC
CsCl cloreto de césio
DAG diacilglicerol
e.p.m. erro padrão da média
Emax efeito máximo
Gq/11 proteína Gq ou proteína G11

21
Gq subunidade α da proteína Gq
G12/13 proteína G12 ou proteína G13
GCs ciclase de guanilil solúvel
GMPc monofosfato cíclico de guanosina
GPCR receptor acoplado à proteína G
GTP trifosfato de guanosina
IP3 1,4,5-trisfosfato de inositol
IP3 R receptor de IP3
KATP canais de potássio sensíveis ao ATP
KCa canais de potássio ativados por cálcio
MLC cadeia leve da miosina
MLCK cinase da cadeia leve da miosina
MLCP fosfatase da cadeia leve da miosina
MYPT1 subunidade 1 condutora da fosfatase da miosina
PA ácido fosfatídico
PC fosfatidilcolina
PIP2 4,5-bisfosfato de fosfatidilinositol
PKA proteína cinase dependente de AMPc
PKG proteína cinase dependente de GMPc
PLC fosfolipase C
PLD fosfolipase D
PMCA Ca2+-ATPase da membrana plasmática
RhoA pequena proteína G ligante de GTP
ROCK cinase dependente de RhoA
RyR receptor de rianodina
RS retículo sarcoplasmático

22
SERCA Ca2+-ATPase do retículo endo-sarcoplasmático
SKCa canais de potássio ativados por cálcio de pequena condutância
SUR receptor para sulfonilureia
TEA+ íon tetraetilamônio
Vm potencial de membrana
ZIPK proteína cinase de interação zíper

23
SUMÁRIO
1 INTRODUÇÃO....................................................................................................26
2 OBJETIVOS........................................................................................................47
2.1 Geral.................................................................................................................47
2.2 Específicos.......................................................................................................47
3 MATERIAL E MÉTODOS...................................................................................49
3.1 MATERIAL.......................................................................................................49
3.1.1 Produto-teste.................................................................................................49
3.1.2 Animais..........................................................................................................49
3.1.3 Drogas e Reagentes......................................................................................49
3.1.4 Solução Nutritiva............................................................................................50
3.1.5 Aparelhos......................................................................................................51
3.2 MÉTODOS.......................................................................................................51
3.2.1 Preparação da solução-estoque....................................................................51
3.2.2 Triagem farmacológica .................................................................................52
3.2.2.1 Efeito dos derivados sintéticos frente às contrações fásicas induzidas por carbacol
ou por histamina em íleo de cobaia........................................................................52
3.2.2.2 Efeito de UFRPE 112, UFRPE 107 e UFRPE 117 sobre as contrações tônicas
induzidas por KCl, CCh ou histamina.....................................................................53
3.2.3 Investigação do mecanismo de ação relaxante de UFRPE 117 em íleo de
cobaia.....................................................................................................................53
3.2.3.1 Caracterização do bloqueio da contração induzida por
histamina................................................................................................................53

24
3.2.3.2 Avaliação do envolvimento dos canais de potássio no efeito relaxante de UFRPE
117........................................................................................................................54
3.2.3.2.1 Efeito de UFRPE 117 sobre as contrações tônicas induzidas por histamina na
presença de cloreto de césio (CsCl).....................................................................54
3.2.3.2.2 Efeito de UFRPE 117 sobre as contrações tônicas induzidas por histamina na
presença de glibenclamida, apamina ou tetraetilamônio......................................54
3.3 ANÁLISE ESTATÍSTICA................................................................................. 55
4 RESULTADOS....................................................................................................56
4.1 Triagem farmacológica.....................................................................................57
4.1.1 Efeito dos derivados sintéticos frente às contrações fásicas induzidas por CCh ou
por histamina em íleo de cobaia.............................................................................57
4.1.2 Efeito dos derivados sintéticos, UFRPE 112, UFRPE 107 e UFRPE 117, sobre as
contrações tônicas induzidas por KCl, por carbacol ou por
histamina................................................................................................................71
4.2 Investigação do mecanismo de ação relaxante de UFRPE 117 em íleo de
cobaia.....................................................................................................................77
4.2.1 Caracterização do bloqueio da contração induzida por
histamina................................................................................................................77
4.2.2 Avaliação do envolvimento dos canais de potássio no efeito relaxante de UFRPE
117 em íleo de cobaia............................................................................................78
4.2.2.1 Efeito de UFRPE 117 sobre as contrações tônicas induzidas por histamina na
presença de cloreto de césio..................................................................................78
4.2.2.2 Efeito de UFRPE 117 sobre as contrações tônicas induzidas por histamina na
presença de glibenclamida, apamina ou tetraetilamônio........................................80
5DISCUSSÃO......................................................................90
6CONCLUSÃO..................................................................101
REFERÊNCIAS.................................................................103
ANEXO................................................................................................................119

1 Introdução

26
Silva, A.C.L. Introdução
1.1 Produtos naturais: fonte para a descoberta de substâncias bioativas
Durante a maior parte da história, a natureza, em especial as plantas, tem
sido fonte de compostos potencialmente ativos usados para o tratamento de um
amplo espectro de doenças (CRAGG; GROTHAUS; NEWMAN, 2014). Os produtos
de origem natural desempenham um papel importante nos processos de descoberta
e desenvolvimento de medicamentos, seja como produtos diretos ou a partir de
substâncias fornecidas por eles, desempenhando um papel fundamental na
pesquisa farmacológica e na proteção à saúde (NEWMAN e CRAGG, 2012). Além
de serem usados pela população como fonte de remédios, os produtos naturais têm
servido como fonte e inspiração para uma grande fração da atual farmacopeia
(KINGSTON, 2011), fornecendo um vasto arsenal terapêutico contra inúmeras
enfermidades compreendendo, assim, uma fonte de inestimável valor às
necessidades medicinais da humanidade.
Diversas razões são apresentadas para explicar o sucesso dos produtos
naturais na descoberta de drogas: alta diversidade química, criação de moléculas
biologicamente ativas através da evolução e a semelhança estrutural com alvos
proteicos em muitas espécies (FEHER; SCHMIDT, 2003), além disso, eles são
tradicionalmente empregados na identificação, classificação de receptores e
investigação de funções fisiopatológicas e de sítios de ação de fármacos, atuando,
assim, como ferramentas farmacológicas. Exemplos clássicos incluem a nicotina,
muscarina, pilocarpina e atropina utilizadas no estudo de receptores nicotínicos e
muscarínicos (SIMMONDS, 2003).
Os produtos naturais e seus derivados representam mais de 50% de todas as
drogas em utilização clínica no mundo, com as plantas contribuindo com 25% desse
total (GURIB-FAKIM, 2006). Em um estudo publicado por Newman e Cragg (2012),
são descritos diversas novas moléculas originárias de produtos naturais durante um
período de 30 anos (1981-2010), e utilizando uma definição bastante ampla do que
constitui uma “droga derivada de produto natural” indicou que quase 50% das novas
drogas introduzidas neste período eram originadas de produtos naturais com os
agentes antineoplásicos, antibacterianos, antivirais, anti-hipertensivos e anti-
inflamatórios correspondendo a, aproximadamente, 45% dessas novas substâncias.
Adicionalmente, 13 drogas relacionadas aos produtos naturais foram aprovadas

27
Silva, A.C.L. Introdução
entre 2005 e 2007, das quais 05 destas são mencionadas como sendo o primeiro
representante (protótipo) de novas classes de drogas (HARVEY, 2008).
Dessa forma, a natureza é uma fonte de compostos com potencial para servir
de base e inspiração para gerar novos fármacos, seja diretamente, por semissíntese
ou síntese total e que ainda mostra-se uma alternativa bastante interessante e viável
uma vez que muitos produtos naturais, e derivados de produtos naturais
sinteticamente modificados, tem sido desenvolvidos com sucesso para uso clínico e
para o tratamento de inúmeras doenças humanas.
1.2 A família Bignoniaceae, a espécie Tabebuia avelanedae e as naftoquinonas
Muitas famílias botânicas são fornecedoras de produtos naturais para
obtenção de novas drogas, nesse contexto pode-se destacar a família Bignoniaceae
onde esta possui cerca de 120 gêneros e 800 espécies arbóreas, arbustivas e
trepadeiras com uma distribuição pantropical, e pronunciada ocorrência nos
neotrópicos, o Brasil apresenta alguns táxons endêmicos o também é o país onde a
maior diversidade desta família é encontra da, apresentando cerca de 32 gêneros e
350 espécies ocorrendo em vários tipos de ambiente, desde o cerrado até florestas
úmidas perenifólias (GENTRY, 1980; SILVA; QUEIROZ, 2003; SOUZA; LORENZI,
2008). As plantas pertencentes a essa família apresentam diferentes classes de
metabólitos secundários, incluindo saponinas, taninos, flavonoides, quinonas, ,
alcaloides, terpenos e etc (CHOUDHURY et al., 2011); várias espécies da família
Bignoniaceae são utilizadas na medicina popular, por exemplo, Jacaranda
cuspidifolia Mart. como um agente antibacteriano (ARRUDA et al., 2011) Tecoma
stans (L.) Juss. ex Kunth como hipoglicemiante(AGUILAR-SANTAMARIA et al.,
2009) e Arrabidaea brachypoda (DC.) por apresentar atividade anti-inflamatória e
antinociceptiva, entre outras (DA ROCHA et al., 2011).
Inserida na família Bignoniaceae, podemos destacar a espécie Tabebuia
avellanedae Lorentz ex Griseb (Figura 1) conhecida na medicina popular como “ipê-
roxo”, “pau-d'arco”, “lapacho”, entre outros nomes. Durante várias décadas,
preparações obtidas a partir dessa planta foram utilizadas na América do Sul e do
Norte como antineoplásico, antifúngico, antiviral, antimicrobiano, antiparasitário e
anti-inflamatório(SANTANA et al., 1968; UEDA et al., 1994;; MIRANDA et al., 2001;
RIFFEL et al., 2002; MACHADO et al., 2003; PEREIRA et al., 2006). A sua
entrecasca é utilizada como analgésico, antineoplásico, anti-inflamatória e diurética

28
Silva, A.C.L. Introdução
pela população em alguns lugares do norte do Brasil e as atividades farmacológicas
relacionadas a essa espécie se deve a presença de flavonoides, saponinas,
cumarinas e quinonas, onde em estudos para avaliar o perfil químico nas atividades
relatadas, as quinonas são as principais subtâncias ativas encontradas. (MIRANDA
et al., 2001; MACHADO et al., 2003 HUSSAIN et al., 2007). Portanto, Tabebuia
avellanedae é digna de atenção devido aos seus efeitos terapêuticos altamente
promissores e deve ser extensamente investigada como um importante recurso
medicamentoso (YAMASHITA et al., 2007).
Figura 1: Tabebuia avellanedae Lorentz e Griseb.
Fonte: www.bibvirt.futuro.usp.br/imagens/enciclopedia_de_plantas_flores/ipe_roxo.
As quinonas representam uma variada classe de metabólitos secundários que
possuem participação em alguns processos bioquímicos vitais, além de
apresentarem cada vez mais um maior destaque em estudos farmacológicos O
interesse nessas substâncias tem aumentado com o passar dos anos principalmente
pelo seu envolvimento em etapas importantes do ciclo de vida dos seres vivos,

29
Silva, A.C.L. Introdução
principalmente a nível de cadeia respiratória, fotossíntese e coagulação sanguínea
(MAHLER et al 1971., GOODWIN et al., 1972; SILVA et al., 2003).
Com base na sua estrutura molecular as quinonas são divididas em diferentes
grupos, utilizando-se como critério o tipo de sistema aromático que sustenta o anel
quinonoídico: (1) benzoquinonas – um anel benzênico; 2) naftoquinonas– um anel
naftalênico e 3) antraquinonas – um anel antracênico linear ou angular. (SILVA et al.,
2003)
As naftoquinonas representam uma grande classe de compostos naturais e
são encontradas em uma ampla variedade de famílias botânicas, fungos e bactérias
exemplos notáveis de naftoquinonas de origem natural incluem a juglona, a
plumbagina e a vitamina K, que desempenha um importante papel no processo de
coagulação sanguínea ,o lapachol (1), a -lapachona (2) e a -lapachona (3) (Figura
2), sendo a -lapachona e a -lapachona análogos naturais do lapachol. Esses três
isômeros constitucionais estão presentes nas plantas da família Bignoniaceae,
principalmente as pertencentes ao gênero Tabebuia, sendo o lapachol a
naftoquinona mais abundante entre as três. O lapachol também já foi relatado em
muitas espécies de outras famílias, como por exemplo, Verbenaceae, Proteaceae,
Leguminosae, Sapotaceae, entre outras (EPIFANO et al., 2014; FONSECA; BRAGA;
SANTANA, 2003).
Derivados de naftoquinonas têm apresentado efeitos farmacológicos valiosos,
como citotóxico, antibacteriano, antiviral, antifúngico, anti-inflamatório e anti-
protozoário (GROLIG AND WAGNER, 2005) e o mecanismo de ação desses efeitos
observados envolve a inibição das topoisomerases que desencadeia a apoptose
celular e a capacidade de formar radicais semiquinonas e espécies reativas de
oxigênio dentro da célula (SILVA, 2003), nessa perspectiva de importância
farmacológica das naftoquinonas ou seus derivados, um medicamento contendo um
derivado do lapachol, Atovaquone® , foi aprovado para ser utilizado no tratamento
da pneumocistose, toxoplasmose e malária (EYONG et al. 2008).

30
Silva, A.C.L. Introdução
Figura 2 – Estruturas químicas do lapachol (1) e de seus derivados naturais, -lapachona
(2) e -lapachona (3) (CAVALCANTE, 2008).
Neste contexto, o lapachol [2-hidroxi-3-prenil-1,4-naftoquinona] é um dos
melhores exemplos de substância com potencial para estudos fitoquímicos e
farmacológicos, pois é uma molécula biologicamente ativa versátil e que pode ser
encontrada em várias espécies de diferentes famílias (HUSSAIN, et al, 2007). Foi
isolado pela primeira vez da árvore Tabebuia avellanedae Lorentz ex Griseb.
(Bignoniaceae) pelo químico italiano Emanuele Paternò em 1882, primeiramente
recebeu o nome de ácido lapáchico devido as suas propriedades ácidas e o termo
“lapachol” foi atribuído por Hooker (HOOKER, 1936).
Espécies que contêm o lapachol e várias naftoquinonas são extensamente
usadas na medicina popular para o tratamento de câncer, lupus, infecções,
ferimentos e muitas outras doenças (DUKE, 1985), o lapachol já foi comercializado
no Brasil como coadjuvante no tratamento de diversos tipos de tumores (RAO;
McBRIDE; OLESON, 1968; SANTANA et al., 1968), não sendo mais disponível no
mercado (SILVA; FERREIRA; SOUZA, 2003) devido a restrições no seu uso clínico
uma vez que foram observados diversos efeitos colaterais, como problemas
gastrintestinais, anemia e aumento no tempo de coagulação (OLIVEIRA et al.,
1990).Dessa maneira, alterações químicas na estrutura da molécula do lapachol
podem ser realizadas visando a obtenção de novas substâncias, onde estas
modificações podem minimizar possíveis efeitos colaterais preservando a atividade
farmacológica, levando ao surgimento de novoss efeitos biológicos e também a
diminuição de uma atividade.
As naftoquinonas têm sido alvo de muitos estudos, uma vez que diversas
atividades biológicas são relacionadas a este grupo de substâncias e muitas dessas
são atribuídas ao lapachol e aos seus derivados naturais ou sintéticos, tais como
anti-inflamatória (MOON et al., 2007; SITÔNIO et al., 2013; TSENG et al, 2013)
(2)
O
O
O
6
7
5
8
1
4
2
3 2 ' 1 '
3 '
4 '
5 ' O
O
O
5' 4'
3'
1'
2'
3
2
4
1
8
5
7
6
(3)
OH
O
O
4'
5'
3'1'
2'3
2
4
1
8
5
7
6
(1)

31
Silva, A.C.L. Introdução
atividade antibacteriana e antifúngica ( MACHADO et al., 2003; PARK et al., 2005;
ANTUNES et al., 2006 ; PEREIRA et al., 2006; PINTO e CASTRO, 2009, SOUZA et
al., 2013).; antimalárica (FONSECA, BRAGA E SANTANA 2003; HUSSAIN et al.,
2007).); leishmanicida (TEIXEIRA et al., 2001 SOUZA-SILVA et al., 2014);
moluscicida (CAMARA et al., 2008) antineoplásica (RAO, 1974; HUSSAIN, et al.,
1994; LI et al., 1999; SILVA-JÚNIOR et al., 2007; EYONG et al., 2008;
SALUSTIANO et al., 2010; COSTA et al., 2011); inibição das topoisomerases I e II (
NEDER et al., 1998; KRISHNAN; BASTOW, 2000; COELHO-CERQUEIRA et al.,
2014) e citotoxicidade em células cancerosas (BALASSIANO et al., 2005;
SUNASSEE et al., 2013; FIORITO et al, 2014).
Dentre as diversas atividades biológicas apresentadas pelas naftoquinonas,
várias apresentaram atividade espasmolítica, como exemplos de naftoquinonas,
tem-se a juglona em artérias coronárias de coelho, útero de rata e intestino de rato
(AUYONG; WESTFALL; RUSSELL, 1963) as vitaminas K1 (2-metil-3-fitil-1,4-
naftoquinona) e K3 (3-metil-1,4-naftoquinona) em vasos coronários (LIDER;
SHISHKIN; FEOFILAKTOVA, 1987); o lapachol, -lapachona e -lapachona em íleo
de cobaia (CAVALCANTE et al., 2008), -lapachona em corpos cavernosos (BAE et
al, 2011). Outras quinonas apresentaram efeito espasmolítico em íleo isolado de
cobaia como, por exemplo p-benzoquinona, 2-metilbenzoquinona, e 2-
etilbenzoquinona (MOUSSATCHE et al., 1967) e os derivados sintéticos do lapachol:
norlapachol, -norlapachona, -norlapachona e hidro-hidroxi-norlapachol
(CAVALCANTE et al., 2010).
Uma vez que muitas atividades farmacológicas são relacionadas ao lapachol
e aos seus derivados naturais ou sintéticos, pesquisadores tem voltado sua atenção
para esta molécula a fim de criar compostos com potencial farmacológico. Tendo a
molécula do lapachol e do norlapachol que é um derivado sintético do lapachol como
ponto de partida, pesquisadores utilizaram diferentes aminas primárias para reagir
com o lapachol e o norlapachol a temperatura ambiente e sem solvente, e essa
reação forneceu novos compostos identificados como derivados do núcleo fenazina
(Santos et al., 2011) (Figura 4). Ainda não há relatos na literatura de atividades
farmacológicas relacionadas a estes novos compostos derivados do lapachol e do
norlapachol. A reação forneceu 6 compostos derivados do lapachol (A) codificados
como UFRPE 15, UFRPE 17, UFRPE 31, UFRPE 114, UFRPE 115 e UFRPE 116 e.

32
Silva, A.C.L. Introdução
6 compostos derivados do norlapachol (B) codificados como UFRPE 106, UFRPE
107, UFRPE 111, UFRPE 112, UFRPE 113 e UFRPE 117
Figura 3 – Reação de Hooker para obtenção do norlapachol (4) a partir do lapachol (1)
(CAVALCANTE, 2008).
OH
O
O
4'
5'
3'1'
2'3
2
4
1
8
5
7
6
(1)
KMnO4
NaOH6
7
5
8
1
4
2
3 2'
1'
3'
4'OH
O
O
(4)
(35 %)

33
Silva, A.C.L. Introdução
Figura 4 .(A) – Estrutura química dos derivados do núcleo fenazina obtidos a partir do
lapachol
N
N
O
OH
OH
N
N
O
OH
OH
UFRPE 15 UFRPE 17
N
N
O
OH
N
N
O
OH
OH
UFRPE 31 UFRPE 114
N
N
O
OH
OH
N
N
O
OH
OH
UFRPE 115 UFRPE 116

34
Silva, A.C.L. Introdução
Figura 4 .(B) – Estrutura química dos derivados do núcleo fenazina obtidos a partir
do norlapachol
N
N
O
OH
OH
N
N
O
OH
OH
UFRPE 106 UFRPE 107
N
N
O
OH
N
N
O
OH
OH
UFRPE 111 UFRPE 112
N
N
O
OH
OH
N
N
O
OH
OH
UFRPE 113 UFRPE 117

35
Silva, A.C.L. Introdução
1.3 Regulação da contração e do relaxamento do músculo liso
A regulação da contratilidade do músculo liso é um mecanismo biológico
crucial para entender funções fisiológicas como a manutenção da pressão arterial,
responsividade aérea e a motilidade gastrointestinal (STOTT, JEPPS,
GREENWOOD, 2014). Além disso, a desregulação da contração desse músculo
acarreta em várias desordens clínicas e doenças como hipertensão, asma,
dispepsia, diarreia, cólicas intestinais e uterinas, incontinência urinária, trabalho de
parto prematuro e disfunção erétil (KIM et al., 2008; STOTT, JEPPS,
GREENWOOD, 2014). O interesse em investigar substâncias com atividade sobre a
musculatura lisa reside no fato de que substâncias espasmolíticas têm uma
importante aplicação nesses processos fisiopatológicos. Dessa forma, os ensaios
farmacológicos utilizando como modelo experimental a musculatura lisa contribuem
para o desenvolvimento de fármacos candidatos a ser utilizados no tratamento das
diversas patologias que acometem esse músculo (CORREIA, 2013).
Bioquimicamente, a entrada de íons Ca2+ nas células é responsável por iniciar
numerosos eventos intracelulares, incluindo contração, secreção, transmissão
sináptica, regulação enzimática, fosforilação e desfosforilação de proteínas e
transcrição gênica (CATTERALL, 2011) e na busca por substâncias que interfiram
na sinalização intracelular do Ca2+, o músculo liso se tornou uma alternativa para o
desenvolvimento de técnicas que possibilitem essa investigação (KARAKI et al.,
1997).
Para que ocorra a contração no músculo liso, é necessário o aumento na
concentração citosólica de Ca2+ e esse aumento na [Ca2+]c é o fator primordial para
a produção da contração. Esta elevação no conteúdo citosólico de Ca2+ também
está envolvido na proliferação celular do músculo liso (HILL-EUBANKS et al., 2011).
A regulação funcional da [Ca2+]c, para dar início a uma resposta contrátil no músculo
liso depende de dois estímulos que levam a dois tipos de acoplamentos: o
acoplamento eletromecânico, que está envolvido com a mudança do potencial de
membrana (Vm) e o acoplamento fármaco-mecânico, que acontece quando a
contração promovida por um agonista é maior que a observada só com a mudança
do Vm (REMBOLD, 1996).

36
Silva, A.C.L. Introdução
Os mecanismos que podem levar a contração a partir do acoplamento
eletromecânico é resultante de uma despolarização de membrana diretamente
associada ao aumento da concentração extracelular de potássio ([K+]e) ou
indiretamente pela ligação dos agonistas aos seus receptores levando à abertura de
canais de cálcio dependentes de voltagem (Cav), na membrana plasmática,
causando aumento de Ca2+ no citosol e, consequentemente, a contração
(REMBOLD, 1996).
Os mecanismos fármacomecânicos da contração do músculo liso (Figura 5)
envolvem a ligação de um agonista como carbacol (CCh) ou histamina a seus
receptores acoplados à proteína G (GPCRs) e ativam a cascata do inositol, através
da proteína Gq ou G11, cujas suas subunidades ativam a fosfolipase C β1 (PLCβ1)
e, consequentemente, promovem hidrólise do 4,5-bisfosfato de fosfatidilinositol
(PIP2), produzindo o 1,4,5-trisfosfato de inositol (IP3) e o diacilglicerol (DAG)
(BERRIDGE, 2008; BILLINGTON; PENN, 2003). O retículo sarcoplasmático (RS)
representa a principal fonte de Ca2+ intracelular. O IP3 se liga aos receptores (IP3R),
e libera Ca2+ do RS. Além disso, no RS há os receptores de rianodina (RyR), que
são canais de Ca2+ ativados pelo próprio Ca2+ e são sensíveis à cafeína, eles são
ativados pelo Ca2+ previamente liberado via IP3R num processo denominado de
liberação de Ca2+ induzida pelo Ca2+ (CICR). Ambos os receptores permitem a
liberação de Ca2+ do RS para o citosol aumentando a [Ca2+]c (MCHALE et al., 2006;
DELLIS et al., 2006). A liberação de Ca2+ do RS induz a translocação da proteína
cinase C (PKC) para a membrana plasmática, onde é ativada quando entra em
contato com o DAG. Várias isoformas de PKC já foram encontradas no músculo liso,
onde a ativação dessa cinase leva a fosforilação de proteínas-alvo específicas,
como os CaV presentes na membrana plasmática ativando-os, promovendo o influxo
de Ca2+ (BERRIDGE, 2009a).
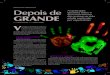
37
Silva, A.C.L. Introdução
Figura 5 – Esquema do mecanismo fármaco-mecânico da contração no músculo liso pela
ativação da via Gq/11-PLCβ1.
(Fonte: adaptado de CORREIA, 2013)
(1) O agonista se liga ao seu receptor do tipo GPCR na membrana plasmática; (2) As proteínas Gq/11 troca GDP por GTP na sua subunidade α (não mostrado na figura), tornando-se ativa; (3) A subunidade αq/11-GTP ativa a enzima PLCβ1; (4) A PLCβ1 cliva o lipídio de membrana PIP2 em IP3 e DAG; (5) O IP3 migra pelo citoplasma e ativa o IP3R presente na membrana do RS, liberando o Ca2+ dos estoques; (6) O Ca2+ liberado ativa o RyR, fazendo com que mais Ca2+ seja liberado para o citoplasma; (7) O Ca2+ que foi liberado, juntamente com o DAG ativam a PKC; (8) A PKC ativada fosforila os CaV1 promovendo o influxo de Ca2+ através dos mesmos; (9) O aumento da [Ca2+]c aumenta a afinidade pela CaM formando o complexo 4Ca2+ - CaM e ativando a MLCK; (10) A MLCK ativada fosforila a MLC e esta se torna ativa e interage com os filamentos de actina, desencadeando a contração do músculo liso. As definições das abreviaturas estão presentes na lista de abreviaturas e no texto.
Independente da fonte proveniente, o Ca2+ quando em quantidades elevadas
no citosol liga-se a CaM formando um complexo [(Ca2+)4-CaM] que ativa a cinase da
cadeia leve da miosina (MLCK). O complexo [(Ca2+)4-CaM-MLCK] constitui a forma
ativa da MLCK, que tem como função fosforilar a cadeia leve da miosina (MLC) e
esta fosforilação permite o desencadeamento do mecanismo de ciclização das
pontes cruzadas entre os filamentos de actina e miosina favorecendo um
deslizamento entre estes filamentos, com o consequente desenvolvimento da
contração (SOMLYO; SOMLYO, 1994 e 2003).

38
Silva, A.C.L. Introdução
A redução da concentração de Ca2+ citoplasmáticos para os níveis basais
desativa a MLCK. O relaxamento do músculo liso ocorre através da ação da fosfatase
da cadeia leve da miosina (MLCP), que desfosforila a MLC. Esta reação é
relativamente lenta, por isso as contrações do músculo liso são tipicamente mais
sustentadas e se dissipam mais lentamente que as do músculo estriado (GARRET;
GRISHAM, 1995).
A diferença fundamental entre a contração do músculo liso inicial e a
manutenção dessa contração é o equilíbrio entre MLCK e a atividade da MLCP,
esse balanço age como um mecanismo regulador reversível de
fosforilação/desfosforilação que integra a contração no músculo liso (WATTERSON;
RATZ; SPIEGEL, 2005). A MLCP consiste em 3 subunidades: uma catalítica, PP1c;
uma regulatória, MYPT1 e outra de 20 kDa de função ainda desconhecida
(SOMLYO; SOMLYO, 2003). A ROCK, uma cinase de serina/treonina, fosforila a
subunidade de ligação da miosina da MLCP, inibindo a sua atividade e, assim,
promovendo o estado fosforilado da MLC. Inibidores farmacológicos da ROCK,
como o Y-27632, bloqueiam a sua atividade por competir com o sítio de ligação do
nucleotídio trifosfato de guanosina (GTP) na enzima (CHITALEY; WEBER; WEBB,
2001; UEHATA et al., 1997).
Os mecanismos que levam a inibição da MLCP (Figura 6) são iniciados pela
ativação da pequena proteína G ligante de GTP (RhoA), dependente das proteínas
G heterotriméricas G12/13 e Gq/11, via uma pequena proteína G associada a um fator
de troca de nucleotídios de guanina (RhoGEFs). A RhoGEFs faz com que a RhoA
troque nucleotídios difosfato de guanosina (GDP) por GTP, sendo translocada para
a membrana, e uma vez RhoA-GTP ligado estimula a sua cinase associada (ROCK)
(EXTON, 1997; MURTHY, 2006). Embora a ROCK fosforile diretamente a MLC
(TOTSUKAWA et al., 2000), sua ação principal na sensibilização ao cálcio parece
ser a inibição da MLCP, esta ação se dá pela fosforilação direta dos resíduos de
Thr696 e Thr853 da MYPT1, causando a sua dissociação e inibição da PP1c da MLCP
(MURTHY, 2006; SOMLYO; SOMLYO, 2003). A ROCK também pode ativar uma
proteína cinase independente de Ca2+, mais conhecida como proteína cinase de
interação zíper (ZIPK). A ZIPK pode fosforilar diretamente a MLC, no entanto seu
alvo principal é o resíduo de Thr696 da MYPT1 o qual é fosforilado inibindo a ação da
MLCP (MURTHY, 2006).

39
Silva, A.C.L. Introdução
Figura 6 – Esquema do mecanismo de manutenção da contração no músculo liso pela ativação da via G12/13-ROCK.
(Fonte: adaptado de CORREIA, 2013)
.
(1) O agonista se liga ao seu receptor do tipo GPCR na membrana plasmática; (2) As proteínas G12/13 troca GDP por GTP na sua subunidade α (não mostrado da figura), tornando-se ativa; (3) A subunidade α12/13-GTP ativa o RhoGEF; (4) O RhoGEF faz com que a RhoA troque GDP por GTP, sendo deslocada para a membrana; (5) A RhoA-GTP ligado estimula a sua cinase ROCK; A ROCK possui vários alvos: (6) Inibe a MLCP; (7) Fosforila a MLC; (8) Ativa a ZIPK; (9) A ZIPK fosforila diretamente a MLC; (10) A ZIPK fosforila a MLCP, inibindo-a; (11) A RhoA-GTP ligado estimula a PLD; (12) A PLD cliva o lípídio de membrana PC em PA; (13) O PA é desfosforilado a DAG pela ação da enzima fosfo-hidrolase; (14) O DAG ativa a PKC, de maneira sustentada; (15) A PKC fosforila a CPI-17, ativando-a; (16) A CPI-17 inibe a ação da MLCP. Todos esses mecanismos favorecem o estado fosforilado a MLC e sua interação com os filamentos de actina, mantendo o músculo liso contraído. As definições das abreviaturas estão presentes na lista de abreviaturas e no texto.
A RhoA-GTP também estimula a fosfolipase D (PLD), essa enzima está
principalmente associada a membranas intracelulares, mas também é encontrada na
membrana plasmática e é específica para fosfatidilcolina (PC), liberando ácido
fosfatídico (PA) que através da ação da enzima fosfo-hidrolase é desfosforilado a
DAG levando a ativação sustentada da PKC (BERRIDGE, 2009a). A ativação da
PKC também pode ser dependente de Gq/11, da PLC que forma DAG a partir da
hidrólise do PIP2. A PKC pode fosforilar o resíduo de Thr38 da proteína inibitória

40
Silva, A.C.L. Introdução
endógena da MLCP (CPI-17), aumentando assim sua potência inibitória sobre a
PP1c por mais de 1000 vezes, inibindo assim a ação da MLCP (MURTHY, 2006;
SOMLYO; SOMLYO, 2003).
Sendo o aumento na [Ca2+]c o sinal que leva à contração,
consequentemente o relaxamento ocorre por diminuição dos níveis deste íon no
citosol do músculo (SOMLYO; SOMLYO, 1994). Esta diminuição na [Ca2+]c pode
ocorrer por um mecanismo eletromecânico, caracterizado pela repolarização (ou
hiperpolarização) da membrana, ou pelo mecanismo fármaco-mecânico, que se dá
pela ativação de receptores de membrana e inibição das vias bioquímicas que levam
a contração (WOODRUM; BROPHY, 2001).
O acoplamento eletromecânico de relaxamento também envolve a abertura
de canais de K+, que desempenham um papel chave na regulação do potencial de
repouso da membrana e na excitabilidade celular, sendo que a contração no
músculo liso depende do balanço entre o aumento da condutância ao K+, levando a
uma repolarização/hiperpolarização, enquanto a diminuição de sua condutância
levaria a uma despolarização (KNOT; BRAYDEN; NELSON, 1996). Os canais de K+
conduzem íons K+ através da membrana plasmática a favor do seu gradiente
eletroquímico, que normalmente constitui uma corrente de saída, no sentido do meio
intracelular para o meio extracelular (MACKINNON, 2003). Estas correntes são
produzidas porque a concentração de K+ no interior da célula (140 mM) é maior do
que no meio extracelular (2-5 mM), havendo o efluxo desse íon pelos canais
seletivos ao K+. Portanto, mantendo o potencial de membrana das células no estado
de repouso para valores mais negativos, aproximando-se do potencial de equilíbrio
de K+ (cerca de -90 mV) (VOGALIS, 2000).
Dessa forma, a abertura dos canais de K+ está associada com a
manutenção do potencial de repouso e de inibição da atividade contrátil (VOGALIS,
2000). Além de vários outros processos celulares incluindo controle do volume
celular, secreção hormonal e controle da formação e propagação de sinais elétricos
em células excitáveis (MACKINNON, 2003). Sua atividade pode ser regulada por
voltagem, Ca2+ ou por neurotransmissores e, consequentemente, as vias de
sinalização que eles estimulam (ALEXANDER; MATHIE; PETERS, 2011).
Os canais de cálcio e os de potássio estão intimamente relacionados com a
regulação do tônus do músculo liso (WATTERSON; RATZ; SPIEGEL, 2005). O

41
Silva, A.C.L. Introdução
movimento dos íons K+ através de canais localizados na membrana celular regula o
influxo de Ca2+ através dos CaV (THORNELOE; NELSON, 2005).
Os canais de potássio são um dos maiores e mais diversos grupos de canais
iônicos, com mais de 75 genes que codificam subunidades de canais de K+
identificados em humanos, uma família de proteínas de membrana diversa e
extensamente distribuída em células excitáveis e não excitáveis (STOTT, JEPPS,
GREENWOOD, 2014). Membros dessa família de canais exercem papel crítico em
processos intracelulares, regulando a secreção de hormônios e de
neurotransmissores, frequência cardíaca, secreção de insulina, excitabilidade
neuronal, transporte epitelial de eletrólitos, regulação do volume celular e contração
muscular lisa (SHIEH, et al., 2000) e sua atividade pode ser regulada por voltagem,
por Ca2+ ou por neurotransmissores e, consequentemente, as vias de sinalização
que eles estimulam (ALEXANDER; MATHIE; PETERS, 2011).
O músculo liso gastrointestinal pode ser regulado e expressa vários subtipos
de canais de K+, entre eles: os canais de K+ sensíveis ao ATP (KATP); os canais de
K+ ativados por Ca2+ (KCa) e os canais de K+ dependentes de voltagem (KV);
(VOGALIS, 2000; SENDERS, 2012).
Os KATP (Figura 7) são complexos existentes na forma de hetero-octâmeros
contendo dois tipos distintos de subunidades proteicas. Os canais consistem de
quatro subunidades dos canais de K+ retificadores de entrada (Kir), chamadas de Kir
6.1 ou Kir 6.2, cada uma contendo dois domínios (TM1 e TM2), ligados a uma região
de formação do poro extracelular (H5). Cada subunidade de Kir está associada com
uma porção regulatória que é um receptor para sulfonilureia (SUR)
(BABENKO; AGUILAR-BRYAN; BRYAN, 1998, QUAN, et al., 2011). Cada
subunidade SUR tem 17 regiões transmembranares agrupados em três domínios:
TMD0 (TM1-5), TMD1 (TM 6-11), e DTM2 (TM 12-17). Cada subunidade SUR
também tem dois domínios de ligação ao ATP (NBD) (QUAN, et al., 2011). A
diversidade molecular dos KATP nos diversos tecidos e espécies é decorrente da
presença de múltiplas isoformas do SUR (SUR1, SUR2A, SUR2B) (BRAYDEN,
2002).

42
Silva, A.C.L. Introdução
Figura 7 – Estrutura dos canais de K+ sensíveis ao ATP (KATP).
Fonte: adaptado de QUAN, et al., 2011.
Uma característica primária deste canal é ser inativado pelo aumento de
ATP intracelular, por sulfonilureias como a glibenclamida e a tolbutamida, e por uma
baixa concentração de Ba2+ extracelular (FLAGG et al., 2010; SANBORN, 2000;
STANDEN et al., 1989). Porém hoje se sabe que existem subtipos de KATP que são
relativamente insensíveis a níveis basais de ATP, mas são bloqueados
seletivamente pela glibenclamida do mesmo modo que os demais (FLAGG et al.,
2010).
Outra família de canais de K+ existente é a dos KCa, a qual é amplamente
expressa em tecidos neuronais e não neuronais, como epitélios, células sensoriais e
músculos lisos onde estes podem responder a estímulos vindos de alterações do
potencial de membrana, e também por alterações da [Ca2+]c, contribuindo para a
repolarização do potencial de ação (BERKEFELD; FAKLER; SCHULTE, 2010;
PETERSEN; MARUYAMA, 1984; VERGARA et al., 1998).
Esta família de canais de K+ é dividida em três grandes grupos com base em
sua condutância, cinética e farmacologia (SAH; FABER, 2002): 1) canais de K+
ativados por Ca2+ de pequena condutância (SKCa), bloqueados pela apamina; 2)
canais de K+ ativados por Ca2+ de condutância intermediária (IKCa), bloqueados pelo
clotrimazol e 3) canais de K+ ativados por Ca2+ de grande condutância (BKCa) que
são bloqueados por toxinas, como caribdotoxina e iberiotoxina, ou por baixas
concentrações do íon tetraetilamônio (TEA+). Os canais do tipo SKCa e IKCa são
insensíveis à voltagem e ativados por baixa concentração de Ca2+ citoplasmático

43
Silva, A.C.L. Introdução
(< 1,0 M), em contraste aos BKCa que são ativados tanto por voltagem como por
Ca2+ (WEI et al., 2005).
Os canais KCa foram subdivididos da seguinte forma: no grupo dos BKCa são
chamados de KCa1.1, enquanto que no grupo dos SKCa estão incluídos os KCa2.1,
2.2 e 2.3, e os IKCa passaram a ser chamados de KCa3.1. Três outros membros deste
grupo, KCa4.1, 4.2 e 5.2, foram incluídos no grupo dos KCa, uma vez que todos
apresentam uma semelhança estrutural, apesar de não serem ativados por Ca2+. Os
canais KCa4.1 e KCa4.2 são ativados por Na+ e Cl- intracelular, enquanto o KCa5.1 é
sensível ao pH (ALDRICH et al., 2009).
Em relação a sua estrutura, os BKCa (Figura 8) são compostos por quatro
subunidades formadoras de poro, onde cada uma dessas é formada por 7
segmentos transmembranares (S0-S6) e uma grande região citoplasmática
carboxiterminal (SALKOFF et al., 2006). S1-S4 formam o domínio sensor de
voltagem primário e S5 e S6 juntos irão formar o principal domínio de permeação
para os íons (JIANG et al., 2002). A subunidade β é composta por dois domínios
transmembranares com uma grande alça extracelular, e as caudas NH2 e COOH
terminais localizadas no citoplasma (BOYLE; TOMASIC; KOTLIKOFF, 1992;
FLEISCHMANN; WASHABAU; KOTLIKOFF 1993; LEE; CUI, 2010). As propriedades
fisiológicas dos BKCa, incluindo a sensibilidade ao Ca2+, são moduladas pelas
subunidades acessórias. As estimativas da sua densidade média, em músculo liso
variam entre 1 a 4 canais/m2 (BENHAM; BOLTON, 1986; SINGER; WALSH, 1987).
Figura 8 – Estrutura das subunidades α e β1 dos canais de K+ ativados por Ca2+ de grande condutância (BKCa) (A); associação das quatro subunidades dos BKCa (B).
Fonte: adaptado de LEDOUX et al., 2006.

44
Silva, A.C.L. Introdução
Os SKCa (Figura 9 ) são formados por quatro subunidades, cada uma delas
constituídas por seis domínios transmembranares (S1-S6), com caudas NH2 e
COOH terminais intracelulares. A região do poro (S5-S6) é a única região homóloga
das outras famílias de canais de K+. A CaM está presa a região COOH terminal de
cada subunidade do canal e a sensibilidade ao Ca2+ citoplasmático depende da
CaM. A ligação do Ca2+ à CaM induz a alteração conformacional do complexo
proteico levando à abertura do poro do canal (LEDOUX, et al., 2006).
Figura 9 – Estrutura dos canais de K+ ativados por Ca2+ de pequena condutância (SKCa) (A); mecanismo de ativação do canal pela ligação do Ca2+ à CaM (B).
Fonte: adaptado de LEDOUX et al., 2006.
Os mecanismos de relaxamento do músculo liso que estão envolvidos no
acoplamento fármaco-mecânico (Figura 9) incluem a fosforilação, via PKA ou PKG,
de vários substratos, ocasionando: (1) o aumento na atividade da Ca2+-ATPase tanto
do RS (SERCA) como da membrana plasmática (PMCA), aumentando, assim, o
sequestro e a extrusão de Ca2+, respectivamente, diminuindo a [Ca2+]c; (2) a
inativação IP3R, pela PKG, reduzindo sua capacidade de liberar o Ca2+ do RS; (3) a
diminuição da formação do IP3, levando à diminuição da liberação de Ca2+ dos RS;
(4) a inibição da MLCK, reduzindo sua afinidade pelo complexo 4Ca2+-CaM,
causando uma redução nos níveis de rMLC fosforilada e, dessa forma, do processo
contrátil; (5) a inibição direta dos CaV, causando uma redução da [Ca2+]c por diminuir
o influxo de Ca2+ e (6) a ativação de canais de K+ que, por repolarização ou por
hiperpolarização, causam o bloqueio dos CaV (WOODRUM; BROPHY, 2001;
DANILA; HAMILTON, 2004).

45
Silva, A.C.L. Introdução
Figura 10 – Acoplamento fármaco-mecânico do relaxamento muscular liso.
(Fonte: adaptado de CORREIA, 2013)
(1) O agonista se liga ao seu receptor do tipo GPCR na membrana plasmática; (2) a
proteína Gs troca GDP por GTP na sua subunidade α (não mostrado na figura), tornando-se
ativa; (3) a subunidade αs-GTP ativa a AC; (4) a AC converte o ATP em cAMP; (5) o NO
gerado tanto dos nervos como das células musculares lisas estimula a atividade da sGC;
a sGC converte o GTP em cGMP; (6) os nucleotídios cíclicos, (7) cAMP e cGMP, ativam
suas respectivas proteínas cinases, PKA e PKG. Ambas as proteínas cinases fosforilam
vários substratos: (8) ativam os canais de K+; (9) inibem os CaV; (10) aumentam a atividade
da SERCA e da PMCA; (11) ativam do trocador Na+/Ca2+. Todos esses mecanismos
diminuem a [Ca2+]c; (12) inibem a MLCK, reduzindo sua afinidade pelo complexo 4Ca2+-
CaM. Todos esses mecanismos impedem a ativação da rMLC e, consequentemente, a
interação dos filamentos de miosina com os de actina, promovendo o relaxamento do
músculo liso. As definições das abreviaturas estão presentes na lista de abreviaturas e no
texto.

2 Objetivos

47
Silva, A.C.L. Objetivos
2.1 Gerais
Contribuir com o estudo farmacológico das naftoquinonas em nível
funcional, especificamente de uma série de novos compostos derivados do
lapachol e do norlapachol em íleo de cobaia.
2.2 Específicos
Investigar a atividade espasmolítica desses compostos em íleo isolado
de cobaia.
Determinar e comparar a potência farmacológica dos compostos
testados com base nos valores de CI50 (concentração de uma substância que
inibe 50% do efeito máximo de um agonista) ou de CE50 (concentração de uma
substância que produz 50% de seu efeito máximo).
Determinar e comparar a eficácia farmacológica dos compostos com
base nos valores de Emax (efeito máximo).
Elucidar o mecanismo de ação espasmolítica em íleo isolado de cobaia
avaliando:
A característica do antagonismo da contração induzida por
histamina;
A modulação dos canais de potássio pelo composto;
Determinar quais os subtipos desses canais estão envolvidos;

3 Material e Métodos

49
Silva, A.C.L. Material e Métodos
3.1 MATERIAL
3.1.1 Produto-teste
A série de compostos sintéticos derivados do lapachol e do norlapachol
codificados como UFRPE 15, UFRPE 17 UFRPE 31, UFRPE 106, UFRPE 107,
UFRPE 111. UFRPE 112, UFRPE 113, UFRPE 114, UFRPE 115, UFRPE 116,
UFRPE 117 foram gentilmente cedidos pelo Prof. Dr. Celso de Amorim Camara
do Departamento de Ciências Moleculares da Universidade Federal Rural de
Pernambuco.
3.1.2 Animais
Eram utilizados cobaias (Caviaporcellus) de ambos os sexos, pesando
entre 300 e 500 g, provenientes do Biotério Prof. Thomas George do Centro de
Biotecnologia (CBiotec) da UFPB.
Antes dos experimentos, os animais eram mantidos sob rigoroso
controle alimentar com uma dieta a base de ração tipo pellets (Labina®) com
livre acesso a água, ventilação e temperatura (21 1 C) controladas e
constantes, submetidos diariamente a um ciclo claro-escuro de 12 h, sendo o
período claro das 6 h às 18 h. Os experimentos eram realizados no período das
8 às 20 h. Todos os procedimentos experimentais foram realizados seguindo
os princípios de cuidados com animais, submetidos e aprovados pela
Comissão de Ética no Uso de Animais do CBiotec (CEUA/CBiotec) sob Nº de
certidão 0805/13.
3.1.3 Drogas e reagentes
O cloreto de cálcio di-hidratado (CaCl2.2H2O), e o sulfato de magnésio
hepta-hidratado (MgSO4.7H2O) foram adquiridos da Vetec (Brasil). O
bicarbonato de sódio (NaHCO3), o cloreto de sódio (NaCl), o cloreto de
potássio (KCl) e o álcool etílico absoluto foram adquiridos da Fmaia (Brasil). A
glicose, o fosfato de sódio monobásico (NaH2PO4), o fosfato de sódio dibásico
(Na2HPO4), o hidróxido de sódio (NaOH) e o ácido clorídrico (HCl) foram
adquiridos da Nuclear (Brasil). Estas substâncias, exceto a glicose, o
bicarbonato de sódio e o cloreto de sódio, eram dissolvidas e diluídas em água

50
Silva, A.C.L. Material e Métodos
destilada para obtenção de cada solução-estoque, que era mantida sob
refrigeração.
O cloridrato de carbamilcolina (CCh) e o diidrocloridrato de histamina
foram adquiridos da Merck (Brasil). Estas substâncias foram dissolvidas e
diluídas em água destilada para obtenção de suas soluções-estoque (10-2 M),
que eram mantidas a 0 °C em refrigerador. Tais soluções eram diluídas em
água destilada para obtenção de concentrações apropriadas a cada protocolo
experimental.
O cloreto de césio (CsCl), o tetraetilamônio (TEA+), a glibenclamida, a
apamina, a 4-aminopiridina (4-AP) e o óleo de castor (Cremophor), foram
obtidos da Sigma-Aldrich (Brasil). Estas substâncias eram mantidas em um
“freezer” à temperatura de -20° C, dissolvidas e diluídas em água destilada,
exceto a glibenclamida, que era dissolvida em álcool etílico absoluto.
A mistura carbogênica (95% de O2 e 5% de CO2)foi adquirida da White
Martins (Brasil).
3.1.4 Soluções nutritivas
Era utilizada a solução nutritiva de Krebs modificado, sendo a mesma
ajustada ao pH 7,4 com NaOH ou HCl(1 N), aerada com carbogênio e mantida
a 37° C, cuja composição está discriminada a seguir:
Composição Concentração (mM)
NaCl 117,0
KCl 4,7
MgSO4 1,3
NaH2PO4 1,2
CaCl2 2,5
NaHCO3 25,0
Glicose 11,0
Tabela 1 - Composição da solução de Krebs modificado por Sun e Benish (1994).

51
Silva, A.C.L. Material e Métodos
3.1.5 Aparelhos
Para o registro das contrações isotônicas os órgãos eram suspensos
em cubas de 5 mL e conectados a uma alavanca isotônica de inscrição frontal
em cilindros esfumaçados de um quimógrafo (DTF, Brasil). As contrações
isométricas eram registradas utilizando transdutores de força isométricos
FORT-10 conectados a um amplificador modelo TMB4M, ambos da World
PrecisionInstruments, Sarasota, FL, EUA), os quais estavam conectados a uma
placa conversora analógico/digital instalada em um microcomputador contendo
o programa BioMed© versão Rv2 (BioData, João Pessoa, PB, Brasil). A
temperatura das cubas era controlada com uma bomba termostática modelo
(POLYSTAT 12002 - Cole-Palmer, EUA).
Os valores de pH eram aferidos através de um pHmetro digital modelo
PG2000 (GEHAKA, São Paulo, SP, Brasil).
3.2 MÉTODOS
3.2.1 Preparação da solução-estoque
Os compostos eram solubilizados em Cremophor (3%) e diluídos em
água destilada para obtenção da solução estoque a 10-2 M. Essa solução era
estocada a 0° C. A concentração final de Cremophor nas cubas nunca excedia
0,01% (v/v); nessa concentração, o Cremophor não apresenta efeito relaxante
ou contrátil significante em íleo de cobaia, de acordo com dados obtidos em
experimentos anteriores realizados em nosso laboratório.
No momento da realização dos experimentos, esta solução era diluída
de acordo com a exigência do protocolo. As concentrações das substâncias
para a realização dos experimentos foram escolhidas de acordo com o padrão
estabelecido no Laboratório de Farmacologia Funcional, onde estas são as
concentrações de 3 x 10-5 e 10-4 M consideradas concentrações submáxima e
máxima respectivamente para os testes in vitro. Se houver atividade
espasmolítica nessas concentrações numa potência superior a 50% de efeito
os experimentos serão realizados com diferentes concentrações das
substâncias, de modo a achar a concentração inicial que promove 0% de

52
Silva, A.C.L. Material e Métodos
relaxamento e a concentração final que promove o efeito máximo de
relaxamento, sendo adicionadas as cubas em incrementos de 3.
3.2.2 Triagem farmacológica
3.2.2.1 Efeito dos derivados sintéticos do lapachol e norlapachol frente às
contrações fásicas induzidas por carbacol ou por histamina em íleo de
cobaia
O íleo de cobaia era preparado de acordo com Daniel et al. (2001). Os
cobaias eram mantidos em jejum por um período de 18 horas (dando-lhe
somente água nesse período). Após este período os animais eram
eutanasiados por deslocamento cervical seguido de secção dos vasos
cervicais. O abdômen era aberto e um segmento do íleo de aproximadamente
15 cm de comprimento era retirado e colocado em uma placa de Petri contendo
solução nutritiva de Krebs modificado a 37 C sob aeração com carbogênio.
Após a dissecação, o segmento do íleo era seccionado em fragmentos de 2-3
cm de comprimento, suspensos individualmente em cubas de vidro (5 mL) e
deixados em repouso por 30 minutos, durante este período a solução nutritiva
era trocada a cada 15 minutos. Após o período de estabilização, era induzida
uma contração isotônica com KCl 40 mM para verificar a funcionalidade do
órgão. A preparação era lavada e após 15 minutos, eram induzidas duas
contrações isotônicas de magnitude similares com 10-6 M de CCh ou de
histamina. Os derivados sintéticos eram incubados por 15 minutos em
preparações diferentes e em diferentes concentrações, e na presença dos
mesmos era induzida uma terceira contração utilizando um dos agonistas. A
inibição da resposta submáxima de CCh e de histamina era avaliada por
comparação das respostas antes (controle) e após a adição dos derivados à
cuba. Os valores da CI50 eram obtidos por regressão não linear.

53
Silva, A.C.L. Material e Métodos
3.2.2.2 Efeito de UFRPE 112, UFRPE 107 e UFRPE 117 sobre as
contrações tônicas induzidas por KCl, CCh ou histamina
O íleo era montado como descrito no item 3.2.2.1. Após o período de
estabilização, uma contração submáxima, registrada utilizando transdutores
isométricos acoplados a um sistema de aquisição digital era obtida com 40 mM
de KCl, 10-5 M de CCh ou 10-6 M de histamina e durante a fase tônica
sustentada os compostos eram adicionados de maneira cumulativa à cuba e
em preparações diferentes. O relaxamento produzido pelos derivados sintéticos
que apresentaram maior potência frente às contrações fásicas foi expresso
como a porcentagem reversa da contração inicial produzida pelos agentes
contráteis. Os valores de CE50 foram expressos como a média erro padrão da
média (e.p.m.) dos valores individuais calculados pelo uso da regressão não
linear.
3.2.3 Investigação do mecanismo de ação relaxante de UFRPE 117 em íleo
de cobaia
3.2.3.1 Caracterização do bloqueio da contração induzida por histamina
Após o período de estabilização, era induzida uma contração isotônica
com KCl 40 mM para verificar a funcionalidade do órgão. A preparação era
lavada e, após 15 minutos, eram induzidas duas curvas concentrações-
resposta cumulativas similares à histamina (10-9 - 10-4 M). Em seguida, na
ausência da histamina, UFRPE 117 era incubado por 15 minutos em diferentes
concentrações e em experimentos independentes. Após este período, na
presença do composto, uma nova curva concentração-resposta cumulativa à
histamina era obtida. Os resultados foram avaliados comparando-se a
percentagem da resposta contrátil na presença do composto com aquela obtida
pela média das amplitudes máximas obtidas das curvas controle (100%), na
ausência do composto. O tipo de antagonismo exercido por UFRPE 117 foi
avaliado com base na análise dos valores da CE50 e dos valores do efeito
máximo (Emax) da histamina calculados a partir das curvas concentrações-
resposta, na ausência (controle) e na presença de UFRPE 117.

54
Silva, A.C.L. Material e Métodos
3.2.3.2 Avaliação do envolvimento dos canais de potássio no efeito
relaxante de UFRPE 117
3.2.3.2.1 Efeito de UFRPE 117 sobre as contrações tônicas induzidas por
histamina na presença de cloreto de césio (CsCl)
O íleo era montado como descrito no item 3.2.2.1. Após o período de
estabilização do órgão,era adicionado à cuba 5 mM de CsCl, um bloqueador
não seletivo dos canais de K+ (CECCHI et al., 1987), após 20 minutos na
presença do bloqueador era induzida uma contração com 10-6 M de histamina
e sob o componente tônico dessa contração era adicionado o composto
UFRPE 117 de forma cumulativa. O relaxamento produzido por UFRPE 117 foi
expresso como a percentagem reversa da contração inicial induzida pelo
agonista. Os valores de CE50 de UFRPE 117 foram calculados a partir das
curvas concentrações-resposta na presença do bloqueador e comparados com
os valores de CE50 de UFRPE 117 na ausência do bloqueador.
3.2.3.2.2 Efeito de UFRPE 117 sobre as contrações tônicas induzidas por
histamina na presença de glibenclamida, apamina ou tetraetilamônio
(TEA+)
O íleo era montado como descrito no item 3.2.2.1. Após o período de
estabilização, era adicionado à cuba 10-5 M de glibenclamida, um bloqueador
seletivo dos canais de K+ sensíveis ao ATP (KATP) (SUN; BENISHN, 1994),
100 nM de apamina, um bloqueador seletivo de canais de K+ ativados pelo
Ca2+ de pequena condutância (SKCa) (ISHII; MAYLIE; ADELMAN, 1997; VAN
DER STAAY et al., 1999) ou 1 mM de TEA+, que nessa concentração é um
bloqueador seletivo dos canais de K+ ativados pelo Ca2+ de grande condutância
(BKCa) (KNOT, BRAYDEN, NELSON, 1996), após 20 minutos na presença dos
bloqueadores, em preparações diferentes, uma contração com 10-6 M de
histamina era obtida. Em seguida o derivado UFRPE 117 era adicionado
cumulativamente sob o componente tônico da contração. O relaxamento
produzido por UFRPE 117 e seus valores de CE50 foram calculados a partir das

55
Silva, A.C.L. Material e Métodos
curvas concentrações-resposta na presença dos bloqueadores e comparados
com os valores de CE50 de UFRPE 117 na ausência do bloqueador.
3.3 Análise estatística
Todos os resultados obtidos foram expressos como média ± e.p.m. e
analisados estatisticamente empregando-se o teste t (não-pareado) ou análise
de variância (ANOVA) one-way seguido do pós-teste de Bonferroni e as
diferenças entre as médias foram consideradas significantes quando os valores
de p < 0,05. Como parâmetro de eficácia foi utilizado o efeito máximo (Emax) e
como parâmetro de potência foram utilizados as CE50 ou CI50. Os valores de
CE50 e CI50 foram calculados por regressão não linear para todos os
experimentos realizados(NEUBIG et al., 2003). Todos os resultados foram
analisados com auxílio do programa GraphPadPrism® versão 5.01 (GraphPad
Software Inc., San Diego CA, EUA).

4 Resultados

57
Silva, A.C.L. Resultados
4.1 Triagem farmacológica
4.1.1 Efeito dos derivados sintéticos do lapachol e norlapachol frente às
contrações fásicas induzidas por CCh ou por histamina em íleo de cobaia
O derivado UFRPE 31 (Gráfico 1A) não inibiu as contrações fásicas induzidas
por 10-6 M de CCh nas concentrações utilizadas (3x10-5 M e 10-4 M), já os compostos
UFRPE 15(Gráfico 2A), UFRPE 17 (Gráfico 3A), UFRPE 114 (Gráfico 4A), UFRPE
115 (Gráfico 5A), e UFRPE 116 (Gráfico 6A), inibiram as contrações fásicas
induzidas por 10-6 M de CCh apenas na concentração de 10-4 M, apresentando
Emax de 25,4 ± 5,3; 30,7 ± 6,7; 39,6 ± 4,5; 25,9 ± 2,7 e 30,7 ± 2,9% respectivamente
(n = 3);resultado semelhante foi observado para os derivados UFRPE 31(Gráfico
1B), UFRPE 15 (Gráfico 2B), UFRPE 114(Gráfico 4B), e UFRPE 116 (Gráfico 6B),
nas contrações fásicas induzidas por 10-6 M de histamina com Emax de 35,6 ± 6,0,
37,6 ± 6,6; 39,7 ± 1,9 e 37,8 ± 2,9%, respectivamente (n = 3).
O derivado UFRPE 17 na concentração de 3x10-5 M, inibiu as contrações
fásicas induzidas por 10-6 M de histamina com Emax= 38,8 ± 6,2%, (n = 3) (Gráfico
3B). Já o derivado UFRPE 115 (Gráfico 5B), inibiu as contrações fásicas induzidas
por 10-6 M de histamina de maneira significante e dependente de concentração (10-5
M – 3x10-4 M) com CI50 = 8,3 ± 0,8 10-5 M e Emax= 93,4 ± 1,3 % (n = 5). Todos os
valores de CI50 e Emax dos derivados do lapachol são expressos na Tabela 2
Por sua vez, os compostos UFRPE 106 (Gráfico 7A) e UFRPE 107 apenas na
concentração de 10-4 M, inibiram as contrações fásicas induzidas por 10-6 M de CCh
apresentando Emax de 36,4 ±4,4 e 29,8 ± 5,2 %, respectivamente (Gráfico 8A). O
composto UFRPE 113 (Gráfico 9A), inibiu as contrações fásicas induzidas por 10-6 M
de CCh nas concentrações de 3x10-5 M e 10-4 M apresentando Emax= 28,1 ± 2,8%
(n=3). Já os derivados UFRPE 111 (Gráfico 10A), UFRPE 112 (Gráfico 11A) e
UFRPE 117(Gráfico 12A), inibiram as contrações fásicas induzidas por 10-6 M de
CCh de maneira dependente de concentração (3x10-6 M – 10-4 M) apresentando CI50
de 9,5 ± 0,08 x 10-5 ; 2,7 ± 0,4 x 10-5 e 4,2 ± 0,6 x 10-5 M e Emax de 89,9 ± 4,3; 97,2 ±
1,3 e 99,6 ± 0,4% respectivamente (n = 5).
O composto UFRPE 106 (Gráfico 7B), inibiu as contrações fásicas induzidas
por 10-6 M de histamina apenas na concentração de 10-4 M apresentando
Emax de 19,9 ± 2,2% (n = 3). Já os derivados UFRPE 107 (Gráfico 8B), UFRPE 111

58
Silva, A.C.L. Resultados
(Gráfico 10B), UFRPE 112 (Gráfico 11B), UFRPE 113 (Gráfico 9B) e UFRPE 117
(Gráfico 12B) inibiram as contrações fásicas induzidas por 10-6 M de histamina de
maneira significante e dependente de concentração (10-5 - 3 x 10-4 M) apresentando
CI50 de 4,4 ± 0,6 x 10-5; 1,0 ± 0,02 x 10-4; 8,8 ± 0,02 x 10-5;8,1 ± 0,6 x 10-5 e 4,6 ± 0,6
x 10-5 M (n = 5) respectivamente. Todos os valores de CI50 e Emax dos derivados do
norlapachol são expressos na Tabela 3.

59
Silva, A.C.L. Resultados
Gráfico 1- Efeito de UFRPE 31 frente às contrações fásicas induzidas por 10-6 M de carbacol (A) e de histamina(B) em íleo de cobaia.
As colunas e as barras verticais representam a média e o e.p.m., respectivamente, (n = 3). ANOVA one-way seguido do pós-teste de Bonferroni. *p < 0,05 (controle vs.UFRPE 31).
Contr
ole -5
3 x
10-4
10
0
25
50
75
100
UFRPE [31] M
A
Co
ntr
ação
(%
)
Contr
ole -5
3 x
10-4
10
0
25
50
75
100
*
[UFRPE 31] M
B
Co
ntr
ação
(%
)

60
Silva, A.C.L. Resultados
Gráfico 2- Efeito de UFRPE 15 frente às contrações fásicas induzidas por 10-6 M de carbacol (A) e de histamina(B) em íleo de cobaia.
As colunas e as barras verticais representam a média e o e.p.m., respectivamente, (n = 3). ANOVA one-way seguido do pós-teste de Bonferroni. *p < 0,05 e **p < 0,01 (controle vs. UFRPE 15).
Contr
ole -5
3 x
10-4
10
0
25
50
75
100
*
[UFRPE 15] M
A
Co
ntr
ação
(%
)
Contr
ole -5
3 x
10-4
10
0
25
50
75
100
**
[UFRPE 15] M
B
Co
ntr
ação
(%
)

61
Silva, A.C.L. Resultados
Gráfico 3- Efeito de UFRPE 17 frente às contrações fásicas induzidas por 10-6 M de
carbacol (A) e de histamina(B) em íleo de cobaia.
As colunas e as barras verticais representam a média e o e.p.m., respectivamente, (n = 3). ANOVA one-way seguido do pós-teste de Bonferroni. *p < 0,05; **p < 0,01 e ***p < 0,001(controle vs. UFRPE 17).
Contr
ole-5
3 x
10-4
10
0
25
50
75
100
**
[UFRPE 17] M
A
Co
ntr
ação
(%
)
Contr
ole -5
3 x
10-4
10
0
25
50
75
100
***
*
[UFRPE 17] M
B
Co
ntr
ação
(%
)

62
Silva, A.C.L. Resultados
Gráfico 4- Efeito de UFRPE 114 frente às contrações fásicas induzidas por 10-6 M de carbacol (A) e de histamina(B) em íleo de cobaia
As colunas e as barras verticais representam a média e o e.p.m., respectivamente, (n = 3). ANOVA one-way seguido do pós-teste de Bonferroni. ***p < 0,001 (controle vs. UFRPE 114).
Contr
ole -5
3 x
10-4
10
0
25
50
75
100
***
[UFRPE 114] M
A
Co
ntr
ação
(%
)
Contr
ole -5
3 x
10-4
10
0
25
50
75
100
***
***
[UFRPE 114] M
B
Co
ntr
ação
(%
)
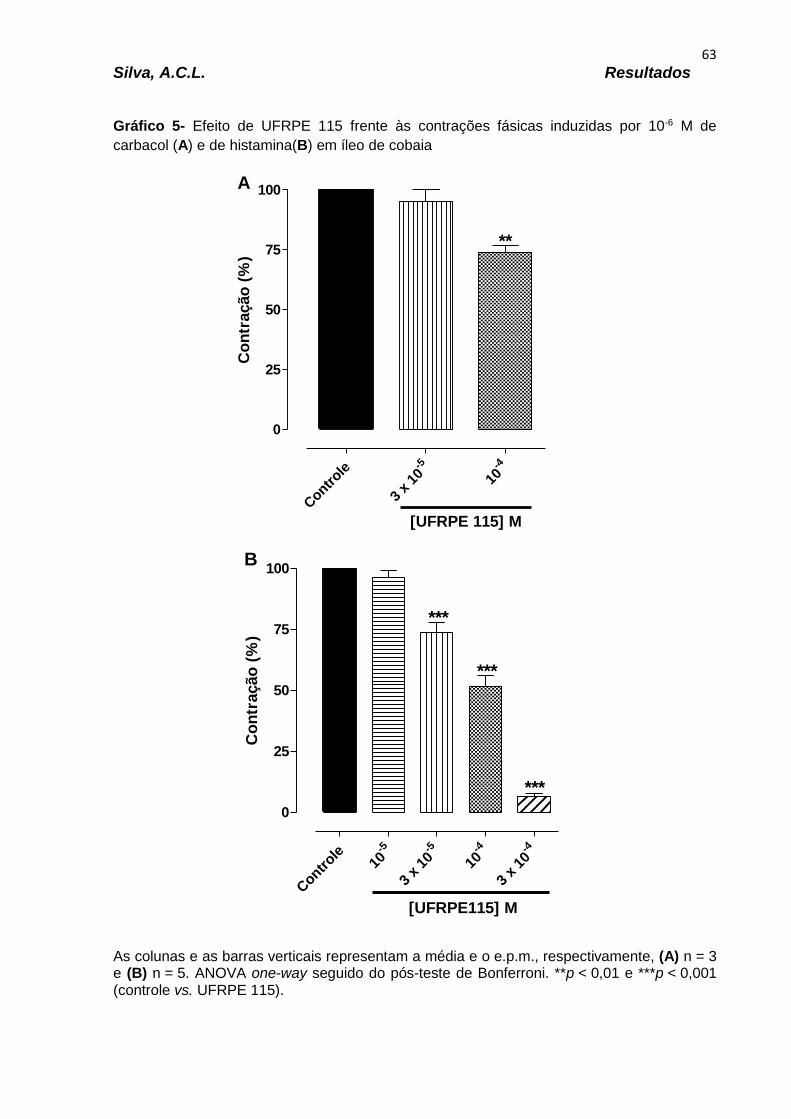
63
Silva, A.C.L. Resultados
Gráfico 5- Efeito de UFRPE 115 frente às contrações fásicas induzidas por 10-6 M de
carbacol (A) e de histamina(B) em íleo de cobaia
As colunas e as barras verticais representam a média e o e.p.m., respectivamente, (A) n = 3 e (B) n = 5. ANOVA one-way seguido do pós-teste de Bonferroni. **p < 0,01 e ***p < 0,001 (controle vs. UFRPE 115).
Contr
ole -5
3 x
10-4
10
0
25
50
75
100
**
[UFRPE 115] M
A
Co
ntr
ação
(%
)
Contr
ole -5
10-5
3 x
10-4
10-4
3 x
10
0
25
50
75
100
[UFRPE115] M
***
***
***
B
Co
ntr
ação
(%
)

64
Silva, A.C.L. Resultados
Gráfico 6- Efeito de UFRPE 116 frente às contrações fásicas induzidas por 10-6 M de
carbacol (A) e de histamina(B) em íleo de cobaia.
As colunas e as barras verticais representam a média e o e.p.m., respectivamente, (n = 3). ANOVA one-way seguido do pós-teste de Bonferroni. ***p < 0,001 (controle vs. UFRPE 116).
Contr
ole -5
3 x
10-4
10
0
25
50
75
100
***
[UFRPE 116] M
A
Co
ntr
ação
(%
)
Contr
ole -5
3 x
10-4
10
0
25
50
75
100
***
***
[UFRPE 116] M
B
Co
ntr
ação
(%
)

65
Silva, A.C.L. Resultados
Gráfico 7- Efeito de UFRPE 106 frente às contrações fásicas induzidas por 10-6 M de
carbacol (A) e de histamina(B) em íleo de cobaia.
As colunas e as barras verticais representam a média e o e.p.m., respectivamente, (n = 3). ANOVA one-way seguido do pós-teste de Bonferroni. ***p < 0,001 (controle vs. UFRPE 106).
Contr
ole -5
3 x
10-4
10
0
25
50
75
100
***
[UFRPE 106] M
A
Co
ntr
ação
(%
)
Contr
ole -5
3 x
10-4
10
0
25
50
75
100
***
[UFRPE 106] M
B
Co
ntr
ação
(%
)

66
Silva, A.C.L. Resultados
Gráfico 8- Efeito de UFRPE 107 frente às contrações fásicas induzidas por 10-6 M de
carbacol (A) e de histamina(B) em íleo de cobaia.
As colunas e as barras verticais representam a média e o e.p.m., respectivamente, (A) n = 3 e (B) n = 5. ANOVA one-way seguido do pós-teste de Bonferroni. **p < 0,01 e ***p < 0,001 (controle vs. UFRPE 107).
Contr
ole -5
3 x
10-4
10
0
25
50
75
100
[UFRPE 107] M
**
A
Co
ntr
ação
(%
)
Contr
ole -5
10-5
3 x
10-4
10-4
3 x
10
0
25
50
75
100
***
***
[UFRPE 107] M
***
B
Co
ntr
ação
(%
)

67
Silva, A.C.L. Resultados
Gráfico 9- Efeito de UFRPE 113 frente às contrações fásicas induzidas por 10-6 M de
carbacol (A) e de histamina(B) em íleo de cobaia.
As colunas e as barras verticais representam a média e o e.p.m., respectivamente, (A) n = 3 e (B) n = 5.ANOVA one-way seguido do pós-teste de Bonferroni. *p < 0,05; **p < 0,01 e ***p < 0,001 (controle vs. UFRPE 113).
Contr
ole -5
3 x
10-4
10
0
25
50
75
100
**
***
[UFRPE 113] M
A
Co
ntr
ação
(%
)
Contr
ole -5
10-5
3 x
10-4
10-4
3 x
10
0
25
50
75
100
[UFRPE 113] M
*
***
***
B
Co
ntr
ação
(%
)

68
Silva, A.C.L. Resultados
Gráfico 10- Efeito de UFRPE 111 frente às contrações fásicas induzidas por 10-6 M de carbacol (A) e de histamina(B) em íleo de cobaia.
As colunas e as barras verticais representam a média e o e.p.m., respectivamente, (n = 5). ANOVA one-way seguido do pós-teste de Bonferroni. ***p < 0,001 (controle vs. UFRPE 111).
Contr
ole -5
3 x
10-4
10-4
3 x
10
0
25
50
75
100
[UFRPE 111] M
***
***
A
Co
ntr
ação
(%
)
Contr
ole -5
3 x
10-4
10-4
3 x
10
0
25
50
75
100
***
[UFRPE111] M
***
B
Co
ntr
ação
(%
)
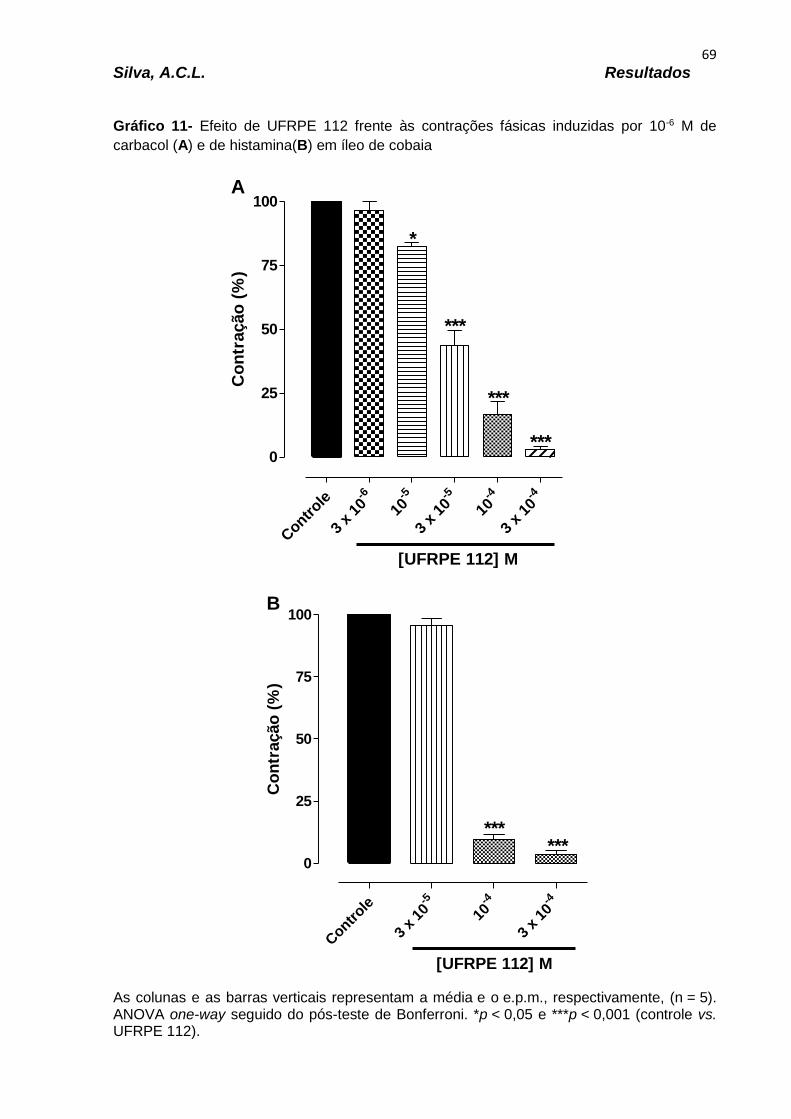
69
Silva, A.C.L. Resultados
Gráfico 11- Efeito de UFRPE 112 frente às contrações fásicas induzidas por 10-6 M de
carbacol (A) e de histamina(B) em íleo de cobaia
As colunas e as barras verticais representam a média e o e.p.m., respectivamente, (n = 5). ANOVA one-way seguido do pós-teste de Bonferroni. *p < 0,05 e ***p < 0,001 (controle vs. UFRPE 112).
Contr
ole -6
3 x
10-5
10-5
3 x
10-4
10-4
3 x
10
0
25
50
75
100
*
***
***
***
[UFRPE 112] M
A
Co
ntr
ação
(%
)
Contr
ole -5
3 x
10-4
10-4
3 x
10
0
25
50
75
100
[UFRPE 112] M
******
B
Co
ntr
ação
(%
)

70
Silva, A.C.L. Resultados
Gráfico 12- Efeito de UFRPE 117 frente às contrações fásicas induzidas por 10-6 M de
carbacol (A) e de histamina(B) em íleo de cobaia
As colunas e as barras verticais representam a média e o e.p.m., respectivamente, (n = 5). ANOVA one-way seguido do pós-teste de Bonferroni. **p < 0,01 e ***p < 0,001 (controle vs. UFRPE 117).
Contr
ole -5
10-5
3 x
10-4
10-4
3 x
10
0
25
50
75
100
***
***
***
[UFRPE117] M
A
Co
ntr
ação
(%
)
Contr
ole -5
10-5
3 x
10-4
10-4
3 x
10
0
25
50
75
100
***
***
***
[UFRPE 117] M
B
Co
ntr
ação
(%
)

71
Silva, A.C.L. Resultados
Tabela 2- Valores de Emax (%) e de CI50 (M) dos derivados do lapachol frente às contrações
fásicas induzidas por carbacol ou por histamina em íleo de cobaia.
Composto Carbacol Histamina
CI50 (M) Emax (%) CI50 (M) Emax (%)
UFRPE 15 Nd 25,5 ± 5,3 Nd 37,6 ± 6,6
UFRPE 17 Nd 30,7 ± 6,7 Nd 38,8 ± 6,2
UFRPE 31 Nd 0,0 Nd 35,6 ± 6,0
UFRPE 114 Nd 39,6 ± 4,5 Nd 39,7 ± 1,9
UFRPE 115 Nd 25,9 ± 2,7 8,3 ± 0,8 x 10-5 93,4 ± 1,3
UFRPE 116 Nd 30,7 ± 2,9 Nd 37,8 ± 2,9
Teste t, **p< 0,01 e ***p< 0,001 (CCh vs. histamina). Nd: não determinado. Fonte: Autor, 2014.
Tabela 3- Valores de Emax (%) e de CI50 (M) dos derivados do norlapachol frente às
contrações fásicas induzidas por carbacol ou por histamina em íleo de cobaia.
Composto Carbacol Histamina
CI50 (M) Emax (%) CI50 (M) Emax (%)
UFRPE 106 Nd 36,4 ± 4,4 Nd 19,9 ± 2,2
UFRPE 107 Nd 29,8 ± 5,2 4,4 ± 0,6 x10-5 95,8 ± 1,6
UFRPE 111 9,5 ± 0,08 10-5 89,9 ± 4,3 1,0 ± 0,02 x 10-4** 92,7 ± 4,4
UFRPE 112 2,7 ± 0,4 x 10-5 97,2 ± 1,3 8,8 ± 0,02 x 10-5 *** 96,3 ± 1,4
UFRPE 113 Nd 28,1 ± 2,8 8,1 ± 0,6 x10-5 97,5 ± 1,2
UFRPE 117 4,2 ± 0,6 x 10-5 99,6 ± 0,4 4,6 ± 0,6 x10-5 99,6 ± 0,4
Teste t, **p< 0,01 e ***p< 0,001 (CCh vs. histamina). Nd: não determinado. Fonte: Autor, 2014.
4.1.2 Efeito dos derivados sintéticos, UFRPE 112, UFRPE 107 e UFRPE 117,
sobre as contrações tônicas induzidas por KCl, por carbacol ou por histamina
O composto UFRPE 112 (10-7- 3x10-4 M, n = 5) relaxou de maneira
dependente de concentração e significante o íleo de cobaia (Figura 11)
pré-contraído com 40 mM de KCl (CE50= 3,4 ± 0,2 x 10-5 M e Emax = 90,5 ± 1,2%) e
com 10–5 M de CCh (CE50 = 4,7 ± 1,1 x 10-6 M e Emax = 100%)(Gráfico 13). De
maneira semelhante, o composto UFRPE 107 (10-7 - 3 x 10-4 M, n = 5) relaxou de
maneira dependente de concentração e significante o íleo de cobaia (Figura 12)
pré-contraído com 40 mM de KCl (CE50= 3,9 ± 0,5 x 10-5 M e Emax= 93,8 ± 2,0%) e
por 10–6 M de histamina (CE50 = 1,5 ± 0,5 x 10-5 M e Emax= 99,6 ± 0,4%) (Gráfico 14).
De maneira semelhante, o composto UFRPE 117 (10-8- 10-4 M, n = 5)
relaxou de maneira significante e dependente de concentração o íleo de cobaia
(Figura 13) pré-contraído com 40 mM de KCl (CE50= 3,3 ± 0,8 x 10-5 M e

72
Silva, A.C.L. Resultados
Emax = 91,6± 3,7%) e por 10–6 M de histamina (CE50= 4,2 ± 0,9 x 10-6 M e
Emax = 100%) (Gráfico 15).
Figura 11 - Registros típicos do efeito relaxante de UFRPE 112 sobre as contrações tônicas induzidas por 40 mM de KCl (A) e 10-5 M de carbacol (B) em íleo de cobaia.
CCh = carbacol e as setas para baixo representam as concentrações cumulativas de
UFRPE 112, respectivamente.
Fonte: Autor, 2014.
A
B
B

73
Silva, A.C.L. Resultados
Gráfico 13 – Efeito de UFRPE 112 sobre as contrações tônicas induzidas por 40 mM de KCl (■) e 10-5 M de CCh (□)em íleo de cobaia.
Os símbolos e as barras verticais representam a média e o e.p.m., respectivamente, n = 5.
-7 -6 -5 -4 -3
0
25
50
75
100
log [UFRPE 112] M
Rela
xam
en
to (
%)

74
Silva, A.C.L. Resultados
Figura 12 - Registros típicos do efeito relaxante de UFRPE 107 sobre as contrações tônicas induzidas por 40 mM de KCl (A) e 10-6 M de histamina (B) em íleo de cobaia.
hist = histamina e as setas para baixo representam as concentrações cumulativas de UFRPE 107, respectivamente. Fonte: Autor, 2014.
A
B

75
Silva, A.C.L. Resultados
Gráfico 14 – Efeito de UFRPE 107 sobre as contrações tônicas induzidas por 40 mM de KCl () e 10-6 M de histamina (○)em íleo de cobaia.
Os símbolos e as barras verticais representam a média e o e.p.m., respectivamente(n = 5).
-7 -6 -5 -4 -3
0
25
50
75
100
log [UFRPE 107] M
Rela
xam
en
to (
%)

76
Silva, A.C.L. Resultados
Figura 13 - Registros típicos do efeito relaxante de UFRPE 117 sobre as contrações tônicas induzidas por 40 mM de KCl (A) e 10-6M de histamina (B) em íleo de cobaia.
Hiist = histamina e as setas para baixo representam as concentrações cumulativas de UFRPE 117, respectivamente. Fonte: Autor, 2014.
A
A
B
B

77
Silva, A.C.L. Resultados
Gráfico 15 – Efeito de UFRPE 117 sobre as contrações tônicas induzidas por 40 mM de KCl () e 10-6 M de histamina () em íleo de cobaia.
Os símbolos e as barras verticais representam a média e o e.p.m., respectivamente (n = 5.)
4.2 Investigação do mecanismo de ação relaxante de UFRPE 117 em íleo de
cobaia
4.2.1 Caracterização do bloqueio da contração induzida por histamina
O composto UFRPE 117 (10-5,3 x 10-5, 10-4 e 3 x 10-4 M, n = 5) inibiu de
maneira significante e dependente de concentração as contrações induzidas por
histamina (Gráfico 16). As curvas cumulativas à histamina foram desviadas para a
direita de forma não paralela e com redução do Emax de 100% (controle) para70,0 ±
6,8; 52,4 ± 10,2; 26,9 ± 6,3 e 5,5 ± 2,2% respectivamente. Os valores de CE50 da
histamina passaram de 2,9 ± 0,5 x 10-7 M (controle) para 5,8 ± 1,1,8,2 ± 1,3 e 3,9 ±
0,8 x 10-7M, nas concentrações de 10-5,3 x 10-5e 10-4 M respectivamente.
-8 -7 -6 -5 -4 -3
0
25
50
75
100
log [UFRPE 117] M
Rela
xam
en
to (
%)

78
Silva, A.C.L. Resultados
Gráfico 16 –Curvas concentrações-resposta cumulativas à histamina na ausência (controle)
() e na presença de UFRPE 117 nas concentrações de 3 x 10-6(□);10-5();3 x 10-5(○);
10-4() e 3 x 10-4 M () em íleo de cobaia.
4.2.2 Avaliação do envolvimento dos canais de potássio no efeito relaxante de
UFRPE 117 em íleo de cobaia
4.2.2.1 Efeito de UFRPE 117 sobre as contrações tônicas induzidas por
histamina na presença de cloreto de césio
O composto UFRPE 117 relaxou o íleo de cobaia pré-contraído por 10-6 M de
histamina na ausência (10-8- 10-4 M, n = 5) e na presença (10-8- 3 x 10-4 M, n = 5) de
5 mM de CsCl, um bloqueador não seletivo dos canais de potássio (Figura 14). A
potência relaxante de UFRPE 117 (CE50 = 5,1 ± 0,7 x 10-5 M ) na presença de CsCl
foi reduzida em 12 vezes quando comparada ao relaxamento produzido por UFRPE
117 na ausência de CsCl (CE50 = 4,2 ± 0,9 x 10-6 M), bem como sua eficácia foi
atenuada na presença de CsCl (Emax= 90,9 ± 2,5%) (Gráfico 17).
-9 -8 -7 -6 -5 -4
0
25
50
75
100
Log [histamina] M
Co
ntr
ação
(%
)

79
Silva, A.C.L. Resultados
Figura 14 – Registros típicos do efeito relaxante de UFRPE 117 sobre as contrações tônicas induzidas por 10-6 M de histamina na ausência (A) e na presença (B) de 5 mM de CsCl em íleo de cobaia.
‘
Hist = Histamina e as setas para baixo representam a adição cumulativa de UFRPE 117. Fonte: Autor, 2014.
A
B

80
Silva, A.C.L. Resultados
Gráfico 17 – Efeito de UFRPE 117 sobre as contrações tônicas induzidas por 10-6 M de histamina na ausência () e na presença () de 5 mM de CsCl, em íleo de cobaia.
Os símbolos e as barras verticais representam a média e o e.p.m., respectivamente, (n = 5).
4.2.2.2 Efeito de UFRPE 117 sobre as contrações tônicas induzidas por
histamina na presença de glibenclamida, apamina ou tetraetilamônio (TEA+)
O composto UFRPE 117 (10-8- 3 x 10-4 M) relaxou o íleo de cobaia pré-
contraído por 10-6 M de histamina de maneira significante e dependente de
concentração tanto na ausência (controle) como na presença (10-8 - 3 x 10-4 M) de
diferentes bloqueadores seletivos de canais de potássio.
Na presença de 10-5 M de glibenclamida (bloqueador seletivo dos KATP) a
curva de relaxamento não foi atenuada quando comparada a curva controle (Figura
15 e Gráfico 18). Diferentemente na presença de 100 nM de apamina (bloqueador
seletivo dos SKCa) e de 1 mM de TEA+ (nessa concentração, bloqueador seletivo dos
BKCa) as curvas controle do UFRPE 117 foram desviadas para a direita de maneira
significante (Figuras 17 e 18, Gráficos 19 e 20, respectivamente).
-8 -7 -6 -5 -4 -3
0
25
50
75
100
log [UFRPE 117] M
Rela
xam
en
to (
%)

81
Silva, A.C.L. Resultados
Observa-se que a potência relaxante de UFRPE 117
(CE50 = 4,2 ± 0,9 x 10-6 M, n = 5) não foi reduzida na presença de 10-5 M de
glibenclamida, bloqueador seletivo dos KATP (CE50 = 4,6 ± 0,8 x 10-6 M) , enquanto
que, na presença de 100 nM de apamina, bloqueador seletivo dos SKCa,
(CE50 = 1,7 ± 0,06 x 10-5 M, n = 4) ou 1 mM de TEA+, (CE50 = 2,0 ± 0,4 x 10-5 M
n = 4), a potência relaxante reduzida em cerca de 4 vezes na presença de apamina
e cerca de 5 vezes na presença de 1 mM de TEA+, bem como sua eficácia
farmacológica também foi reduzida na presença desses bloqueadores(Gráfico 21).

82
Silva, A.C.L. Resultados
Figura 15 – Registros típicos do efeito relaxante de UFRPE 117 sobre as contrações tônicas induzidas por 10-6 M de histamina na ausência (A) e na presença (B) de 10-5 M de glibenclamida em íleo de cobaia
.
Hist = Histamina e Glib = glibenclamida, as setas para baixo representam a adição
cumulativa de UFRPE 117
Fonte: Autor, 2014.
A
B

83
Silva, A.C.L. Resultados
Gráfico 18 – Efeito de UFRPE 117 sobre as contrações tônicas induzidas por 10-6 M de histamina na ausência () e na presença () de 10-5 M de glibenclamida em íleo de cobaia.
Os símbolos e as barras verticais representam a média e o e.p.m., respectivamente, (n = 5).
-8 -7 -6 -5 -4 -3
0
25
50
75
100
log [UFRPE 117] M
Rela
xam
en
to (
%)

84
Silva, A.C.L. Resultados
Figura 16 – Registros típicos do efeito relaxante de UFRPE 117 sobre as contrações tônicas induzidas por 10-6 M de histamina na ausência (A) e na presença (B) de 100 nM de apamina em íleo de cobaia.
Hist. = Histamina e as setas para baixo representam a adição cumulativa de UFRPE 117.
Fonte: Autor, 2014.
A
B

85
Silva, A.C.L. Resultados
Gráfico 19- Efeito de UFRPE 117 sobre as contrações tônicas induzidas por
10-6 M de histamina na ausência () e na presença () de 100 nM apamina em íleo de
cobaia.
Os símbolos e as barras verticais representam a média e o e.p.m., respectivamente.
Controle (n = 5) e apamina (n = 3).
-8 -7 -6 -5 -4 -3
0
25
50
75
100
log [UFRPE 117] M
Rela
xam
en
to (
%)

86
Silva, A.C.L. Resultados
Figura 17 – Registros típicos do efeito relaxante de UFRPE 117 sobre as contrações tônicas induzidas por 10-6 M de histamina na ausência (A) e na presença (B) de 1 mM de TEA+ em íleo de cobaia.
Hist. = Histamina e as setas para baixo representam a adição cumulativa de UFRPE 117
Fonte: Autor, 2014.
A
B

87
Silva, A.C.L. Resultados
Gráfico 20 – Efeito de UFRPE 117 sobre as contrações tônicas induzidas por 10-6 M de histamina na ausência () e na presença () de 1 mM de TEA+ em íleo de cobaia.
Os símbolos e as barras verticais representam a média e o e.p.m., respectivamente.
Controle.(n = 5) e TEA+ (n = 4).
-8 -7 -6 -5 -4 -3
0
25
50
75
100
log [UFRPE 117] M
Rela
xam
en
to (
%)

88
Silva, A.C.L. Resultados
Gráfico 21 – Valores de CE50 (M) de UFRPE117 na ausência (controle) e na presença de bloqueadores de canais de K+ em íleo de cobaia.
CsCl = cloreto de césio, TEA+ = íon tetraetilamônio. Teste t, **p < 0,01; ***p < 0,001 (controle
vs. presença de bloqueadores de canais de potássio)
hista
min
a
CsC
l
gliben
clam
ida
apam
ina
1 m
M
+
TEA
0
1.010-5
2.010-5
3.010-5
4.010-5
5.010-5
6.010-5
Histamina
***
***
**
CE
50
(M)

5 Discussão

90
Silva, A.C.L. Discussão
O modelo do músculo liso intestinal é um meio importante para se investigar o
efeito e os mecanismos de ação de substâncias que possam ser utilizadas em
processos fisiopatológicos, como diarréias e cólicas intestinais. Neste trabalho foi
descrito pela primeira vez o efeito espasmolítico de uma nova série de derivados
sintéticos tendo como moléculas precursoras o lapachol e e o norlapachol.
Tanto o lapachol, quanto os seus derivados naturais e sintéticos, tem sido
alvo de constante interesse farmacológico devido à variedade de atividades
biológicas que essas substâncias apresentam. Uma vez que já é relatado na
literatura que o lapachol e alguns derivados tanto naturais quanto sintéticos
apresentam efeito espasmolítico em diferentes modelos de músculo liso (BAE et al.,
2011) inclusive em íleo de cobaia, fomos investigar neste trabalho se esta nova série
derivados do lapachol e do norlapachol chamados de derivados do núcleo fenazina,
apresentariam efeito espasmolítico em íleo de cobaia, uma vez que em estudos
realizados anteriormente em nosso laboratório neste modelo de músculo liso
(CAVALCANTE et al., 2008; CAVALCANTE et al., 2010), um de seus precursores, o
lapachol, apresentou efeito espasmolítico e o norlapachol que é um derivado
sintético do lapachol não apresentou este efeito.
Em íleo de cobaia, vários agentes contráteis, como os agonistas
muscarínicos, a histamina (TRIGGLE et al., 1989) e o KCl (TRIGGLE; TRIGGLE,
1976; HURWITZ et al., 1980), causam uma resposta contrátil bifásica que consiste
em uma contração inicial transiente, chamada componente fásico onde este dura
menos que 30s, seguido de uma contração sustentada denominada componente
tônico, onde este se desenvolve lentamente ao longo de 15 minutos e é mantido
durante o período de exposição ao agente contrátil (TRIGGLE; TRIGGLE, 1976;
BOLTON, 1979; VAN BREEMEN; AARONSON; LOUTZENHISER, 1979).
Esta resposta bifásica observada é devido à fonte dual de Ca2+ no músculo
liso. É sugerido que a contração fásica é causada por liberação de Ca2+ dos
estoques intracelulares mediados por IP3 (ABDELLATIF, 1989; KOBAYASHI et al.,
1989), enquanto que a contração tônica é atribuída ao influxo de Ca2+ através dos
canais para cálcio dependentes de voltagem (CaV), uma vez que a contração tônica
é suprimida pelo bloqueador de CaV, verapamil (JIM et al., 1981).
Diante dessas premissas, primeiramente realizou-se uma triagem
farmacológica, onde foi investigado se os derivados sintéticos do lapachol (UFRPE

91
Silva, A.C.L. Discussão
15, UFRPE 17, UFRPE 31, UFRPE 114, UFRPE 115 e UFRPE 116) e do
norlapachol (UFRPE 106, UFRPE 107, UFRPE 111, UFRPE 112, UFRPE 113 e
UFRPE 117) inibiriam as contrações fásicas induzidas por CCh ou por histamina no
íleo de cobaia.
Em estudos realizados anteriormente (CAVALCANTE et al., 2008) observou
que o lapachol apresentou atividade espasmolítica em íleo de cobaia, onde esse
composto foi capaz de inibir tanto as contrações fásicas induzidas por CCh (CI50 =
1,5 ± 0,2 x 10-4 ) quanto por histamina (CI50 = 3,6 ± 0,5 x 10-5 ). Na triagem
farmacológica realizada, os resultados obtidos mostraram que quando as contrações
eram induzidas por carbacol, todos os novos derivados sintéticos do lapachol
apresentaram efeito inibitório com baixa eficácia espasmolítica, resultados
semelhantes foram observados para os compostos derivados do lapachol UFRPE
31, UFRPE 15, UFRPE 17, UFRPE 114 e UFRPE 116 frente às contrações fásicas
induzidas por histamina, onde os mesmos apresentaram efeito espasmolítico
significante em inibir estas contrações, no entanto não ultrapassaram 40% de
inibição mesmo nas maiores concentrações utilizadas (Tabela 2), mostrando
novamente que as modificações estruturais realizadas na molécula do lapachol
levaram a uma diminuição do efeito espasmolítico do lapachol em inibir as
contrações fásicas induzidas por histamina.
Interessantemente, o composto UFRPE 115 inibiu as contrações induzidas
por histamina (CI50= 8,3 ± 0,8 x 10-5 M) porém com uma potência farmacológica
cerca de 2 vezes menor, comparado ao efeito do lapachol em inibir as mesmas
contrações (CI50= 3,6±0,5 x 10-5 M; CAVALCANTE, 2008). Sugerindo assim que as
modificações estruturais realizadas na molécula do lapachol para formar novos
compostos nitrogenados, levaram a uma diminuição do efeito espasmolítico
apresentado pelo lapachol anteriormente, e isso pode ser atribuído a presença de
dois grupos prenila que estão posicionados próximos a cadeia lateral alifática que
está ligada ao núcleo fenazina. A presença desses dois grupos prenila que são
volumosos, pode resultar em um impedimento estérico, levando assim, a uma
possível diminuição da interação da cadeia lateral alifática com o alvo receptor e
consequente diminuição da atividade farmacológica. .
Em se tratando dos novos compostos derivados do norlapachol, que é um
derivado sintético não nitrogenado do lapachol, os derivados nitrogenados UFRPE

92
Silva, A.C.L. Discussão
106, UFRPE 107 e UFRPE 113 apresentaram efeito espasmolítico significante em
inibir as contrações fásicas induzidas por 10-6 M de CCh, porém com baixa eficácia
farmacológica, uma vez que seus efeitos não ultrapassaram 40% de inibição mesmo
em altas concentrações, de maneira interessante foi observado que apenas os
derivados UFRPE 111, UFRPE 112 e UFRPE 117 inibiram as contrações fásicas
induzidas por 10-6 M de CCh de maneira dependente de concentração com Emax
superior a 90% de inibição, e ao analisar os valores de CI50 para estes 3 compostos
(Tabela 3) pode-se observar que os compostos UFRPE 112 e UFRPE 117 foram os
mais potentes em inibir as contrações frente ao agonista carbacol, comparado aos
outros dois compostos.
Quando as contrações eram induzidas por 10-6 M histamina, o derivado
UFRPE 106 apresentou uma baixa eficácia farmacológica, diferentemente, os
demais compostos (UFRPE 107, UFRPE 111, UFRPE 112, UFRPE 113 e UFRPE
117) inibiram as contrações fásicas induzidas por 10-6 M de histamina de maneira
dependente de concentração e com Emax superior a 90% de inibição, ao analisar os
valores de CI50 para estes 5 compostos (Tabela 3) observa-se que os compostos
UFRPE 107 e UFRPE 117 foram os mais potentes em inibir essas contrações,
mostrando que as modificações estruturais no norlapachol foram essenciais para o
aumento na potência relativa destes derivados, e este é um achado muito
importante, uma vez que em estudos anteriores realizados por CAVALCANTE et al.,
2010, o norlapachol inibiu fásicas induzidas tanto por CCh quanto por histamina em
íleo de cobaia com uma eficácia farmacológica baixa, inferior a 35%.
Ao analisarmos as estruturas dos derivados do norlapachol, podemos notar
que a perda de um carbono dos grupamentos prenila torna esses substituintes
menos volumosos, fazendo com que a cadeia lateral alifática ligada ao núcleo
fenazina , ficasse mais exposta. A ausência de um possível impedimento estérico
levou a uma possibilidade maior de interação dessa cadeia com o alvo receptor,
resultando em uma maior potência farmacológica desses derivados em relação ao
seu precursor, o norlapachol.
Dessa forma, ao final da triagem farmacológica, analisando os valores de CI50
pode-se observar que os compostos UFRPE 107, UFRPE 112 e UFRPE 117
apresentaram-se como os de maior potência farmacológica em inibir as contrações
fásicas induzidas por CCh e/ou histamina em íleo isolado de cobaia em relação a

93
Silva, A.C.L. Discussão
série testada (Tabela 3) e ainda que o UFRPE 107 apresentou uma maior
seletividade e potência em inibir as contrações induzidas por histamina, que o
UFRPE 112 inibiu as contrações fásicas induzidas tanto por CCh quanto por
histamina, sendo o mais potente em inibir as contrações induzidas por CCh e que o
UFRPE 117 demonstrou ser equipotente em inibir as contrações induzidas pelos
dois agonistas. Ao analisar a estrutura química dos três compostos, observou-se que
os 3 possuem a cadeia lateral alifática mais extensa que os outros compostos, além
da presença da hidroxila, sugerindo assim que esse grupamento é importante para o
efeito observado. Diante desses resultados prosseguimos com a avaliação do efeito
relaxante desses 3 compostos no íleo de cobaia pré-contraído por agentes
diferentes agentes contráteis.
No músculo liso de íleo de cobaia, tanto a contração fásica como a tônica são
dependes do Ca2+ extracelular, uma vez que ambas as contrações foram inibidas na
ausência de Ca2+ extracelular (HONDA; TAKANO; KAMIYA, 1996). A remoção de
íons Ca2+ do meio externo bloqueia totalmente a resposta contrátil tanto a agentes
despolarizantes como a agonistas em poucos segundos, sugerindo que as fontes
intracelulares de Ca2+ não contribuem significativamente para o nível de tensão
atingido (NOUAILHETAS et al., 1985). No entanto, a influência do Ca2+ extracelular é
relativamente maior na resposta contrátil tônica comparado com a fásica (TRIGGLE
et al., 1979).
O íleo é um órgão totalmente dependente de variação do potencial de
membrana (NOUAILHETAS et al., 1985) e como o componente tônico da contração
induzida por agonistas de acoplamento misto (UNNO; KOMORI; OHASHI, 1995;
BOLTON et al., 1981) ou por um agente despolarizante (KCl) é mantido quase que
exclusivamente por influxo de cálcio através dos CaV (BOLTON, 1979; REMBOLD,
1996; BOLTON et al., 2006), nos questionamos se estes compostos poderiam estar
agindo por bloqueio do influxo de Ca2+ através dos CaV, ou se estes poderiam estar
levando a uma ativação de canais de K+ promovendo hiperpolarização resultando
em um bloqueio indireto dos Cav.
Como os mecanismos envolvidos na manutenção da fase tônica da contração
diferem daqueles da fase fásica em íleo isolado (ABDELLATIF, 1989; KOBAYASHI
et al., 1989; HONDA; TAKANO; KAMIYA, 1996), resolveu-se verificar se os

94
Silva, A.C.L. Discussão
compostos UFRPE 107, UFRPE 112 e UFRPE 117 além de promover efeito
espasmolítico em inibir as contrações fásicas, ainda promoveriam o relaxamento do
íleo isolado de cobaia pré-contraído com KCl, um agente despolarizante que age
levando o músculo liso a contração por um mecanismo eletromecânico, por carbacol
ou por histamina, ambos agindo por um acoplamento do tipo misto (BOLTON et al.,
1981).
Observou-se que os três derivados, UFRPE 112 (Gráfico 13), UFRPE 107
(Gráfico 14) e UFRPE 117 (Gráfico 15) relaxaram o íleo de cobaia pré-contraído com
KCl, com CCh ou com histamina. Entretanto, ao analisar os valores de CE50
observou-se que os três compostos foram menos potentes em relaxar o órgão pré-
contraído com KCl comparados com os agonistas (CCh ou histamina).
Tendo em vista que um dos objetivos desse estudo foi comparar as eficácias
e potências dos compostos testados, evidenciou-se que o derivado UFRPE 117
apresentou os melhores parâmetros farmacológicos o qual além de inibir as
contrações fásicas induzidas por histamina (CI50= 4,6 ± 0,6 x10-5 M), relaxou o órgão
pré-contraído com o mesmo agonista com uma maior potência (CE50= 4,2 ± 0,9x10-6
M) e apresentou sua potência relaxante diminuída quando o íleo era pré-contraído
com 40mM de KCl (CE50 = 3,3 ± 0,8 x 10-5 M), assim decidiu-se prosseguir com o
estudo utilizando esse composto para se elucidar seu possível mecanismo de ação
envolvido na atividade espasmolítica, com a finalidade de descobrir drogas com um
maior potencial terapêutico ou que venham a servir como ferramenta farmacológica
para o melhor entendimento dos mecanismos envolvidos nos processos
fisiopatológicos envolvendo a musculatura lisa intestinal. ,
Como UFRPE 117 apresentou uma maior potência em relaxar o órgão pré-
contraído por histamina e o mesmo também inibiu as contrações induzidas por esse
agonista, levantou-se a hipótese de que o mesmo estaria agindo nos receptores
histaminérgicos (H1) para promover sua ação espasmolítica. Para verificar essa
hipótese, decidiu-se caracterizar o tipo de antagonismo exercido pelo composto
frente às curvas concentrações-respostas cumulativas induzidas por histamina.
Nesse protocolo experimental, observou-se um desvio para direita das curvas
controle cumulativas à histamina, de forma não paralela e com redução do Emax
(Gráfico 16), caracterizando dessa forma, um antagonismo do tipo não competitivo.

95
Silva, A.C.L. Discussão
O antagonismo do tipo não competitivo pode ocorrer quando existe a ligação
do antagonista ao mesmo sítio de ligação do agonista (antagonismo
pseudo-irreversível) ou em um sítio distinto (antagonismo alostérico)
(BLUMENTHAL, GARRISON, 2011). O perfil do Gráfico 16 representa um
antagonismo pseudo-irreversível, uma vez que a inibição da resposta contrátil da
histamina na presença do UFRPE 117 não atingiu um valor limitante, característica
observada no antagonismo alostérico.
Os canais de K+ são reguladores chave do tônus do músculo liso e da
atividade , e o aumento na sua atividade resulta em hiperpolarização da membrana
da célula, além disso onde o fluxo dos íons por canais presentes na membrana
plasmática regulam o influxo de Ca2+ através dos CaV e consequentemente a
contração do músculo liso (THORNELOE; NELSON, 2005).
Substâncias espasmolíticas podem promover seus efeitos por mecanismos
dependentes da abertura de canais para K+, onde essa abertura causa uma
hiperpolarização da membrana e leva à inibição dos canais de cálcio dependentes
de voltagem (Cav), diminuindo o influxo de cálcio e reduzindo a [Ca2+]c levando o
músculo ao relaxamento (WESTON; EDWARDS, 1992). É relatado na literatura que
substâncias ativadoras de canais para K+ são menos potentes em relaxar o músculo
liso quando o músculo se encontra estimulado por altas [K+]e, enquanto que as
substâncias bloqueadoras de canais para Ca2+ são mais potentes quando este
músculo está estimulado por altas [K+]e (GURNEY, 1994) e que ativadores de
canais de K+ são substâncias que incluem um grupo de diversas moléculas com um
largo potencial terapêutico (QUEST, 1992; EMPFIELD; RUSSELL; TRAINOR, 1995).
Diante dessas premissas e associado ao fato de que UFRPE 117 foi menos
potente em relaxar o íleo pré-contraído por 40 mM de KCl, hipotetizou-se que este
composto estaria modulando positivamente os canais de K+ para exercer seu efeito
relaxante. Para comprovar essa possibilidade, utilizou-se como ferramenta
farmacológica um bloqueador não seletivo desses canais, o CsCl (KNOT;
BRAYDEN; NELSON, 1996). Ao analisarmos o resultado, observou-se que o efeito
relaxante do composto UFRPE 117 foi atenuado na presença de 5 mM de CsCl,
tendo a sua potência relaxante diminuída em cerca de 12 vezes (Gráfico 17).
Sugerindo assim, a participação desses canais na atividade espasmolítica do
composto UFRPE 117 em íleo de cobaia.

96
Silva, A.C.L. Discussão
Uma vez que o tônus do músculo liso gastrintestinal pode ser regulado por
vários subtipos de canais de K+, entre eles: os KCa, os KATP, e os KV (VOGALIS,
2000), decidiu-se investigar quais subtipo(s) de canais de K+ que estariam sendo
modulados por esse derivado do norlapachol.
Os KATP, canais de K+ ativados por uma diminuição na concentração
intercelular de ATP, foram inicialmente identificados em miócitos ventriculares
(NOMA, 1983). Posteriormente, os KATP com características semelhantes foram
demonstrados em outros tecidos, tais como, células β-pancreáticas, músculo
esquelético, neurônios e músculos lisos (TERAMOTO, 2006). Esse canal tanto pode
ser inibido por ATP intracelular como por sulfonilureias como, por exemplo, a
glibenclamida e a tolbutamida, e por baixa concentração de Ba2+ extracelular
(STANDEN et al., 1989; SANBORN, 2000). Sun e Benishin (1994) mostraram que os
KATP estão presentes em músculo longitudinal de íleo de cobaia, pois a cromacalina,
nicorandil e pinacidil, ativadores desses canais, foram capazes de inibir as
contrações tônicas induzidas por histamina (0,5 M) ou por KCl (30 mM).
Para verificar a hipótese de que o composto UFRPE 117 poderia estar
ativando os canais KATP para exercer seu efeito relaxante em íleo de cobaia, avaliou-
se a atividade espasmolítica do mesmo na ausência e na presença de
glibenclamida, um bloqueador seletivo desses canais (SUN; BENISHIN, 1994).
Como o efeito relaxante de UFRPE 117 não foi alterado na presença de
glibenclamida, (Gráfico 18), essa hipótese foi descartada, demonstrando a não
participação dos KATP neste efeito.
Outra família de canais de K+ existente é a dos canais para K+ ativados por
Ca2+ (KCa). Estes contribuem para a repolarização do potencial de ação e são
divididos em três grandes famílias com base em suas condutância, cinética e
farmacologia: 1) canais de K+ ativados por Ca2+ de pequena condutância (SKCa), são
bloqueados por apamina; 2) canais de K+ ativados por Ca2+ de condutância
intermediária (IKCa) bloqueados pelo clotrimazole, e 3) canais de K+ ativados por
Ca2+ de larga condutância (BKCa) que são bloqueados por toxinas, como
caribdotoxina e iberiotoxina, ou por baixas concentrações de TEA+(SAH; FABER,
2002).
Os KCa estão presentes no músculo liso gastrintestinal e uma vez ativados,
pelo aumento da [Ca2+]c, dão início ao efluxo dos íons K+ que via

97
Silva, A.C.L. Discussão
repolarização/hiperpolarização do potencial de membrana restabelece as [Ca2+]c
através da limitação do influxo do íon Ca2+ seja pela desativação dos CaV ou pelo
aumento da atividade do trocador Na+/Ca2+ (FAKLER; ADELMAN, 2008).
Os canais KCa1.1 ou BKCa são encontrados em cérebro, cóclea, músculo
esquelético e músculo liso. Já os KCa2.1; 2.2 e 2.3 ou SKCa são largamente
encontrados em cérebro, coração e alguns tipos de músculo liso. Os canais de K+
ativados por Ca2+ de pequena condutância (SKCa) estão constitutivamente
associados a CaM, que medeia a abertura do canal através da ligação com o
Ca2+(BERKEFELD; FAKLER; SCHULTE, 2010). Os SKCa geram uma corrente
hiperpolarizante em células excitáveis após a geração do potencial de ação, e assim
levam ao relaxamento do músculo liso (BERKEFELD; FAKLER; SCHULTE, 2010;
BRAINARD; KOROVKINA; ENGLAND, 2007).
Assim, com o objetivo de investigar se UFRPE 117 estaria modulando
positivamente os SKCa para relaxar o íleo de cobaia, utilizou-se a apamina, um
bloqueador seletivo desses subtipos de canais (ISHII; MAYLIE; ADELMAN, 1997;
VAN DER STAAY et al., 1999), como ferramenta farmacológica. A curva de
relaxamento de UFRPE 117 (CE50 = 4,2 ± 0,9 x 10-6 M) foi deslocada para a direita
na presença de 100nM de apamina (CE50 = 1,7 ± 0,06 x 10-5 M) com redução da
potência relaxante em cerca de 4 vezes (Gráfico 18). Confirmando assim, a
participação dos canais SKCa na atividade espasmolítica do composto UFRPE 117
(Gráfico 19).
Em processos fisiológicos, a ativação dos BKCa provê um mecanismo de
feedback que limita a contração por causar hiperpolarização e redução da duração
do potencial de ação no músculo liso, limitando a entrada de Ca2+ através dos CaV
(PONTE et al., 2012). Romero et al. (1998), utilizando a técnica de patch-clamp,
caracterizaram uma população de BKCa que são indiretamente ativados pela
angiotensina II, um agonista contrátil que induz dessensibilização do componente
tônico da contração em íleo de cobaia. Posteriormente, Silva, Nouailhetas e
Aboulafia (1999) demonstraram que a dessensibilização do componente tônico
induzida pela angiotensina II era devido à abertura dos BKCa, levando a uma
repolarização e, consequentemente, fechando os CaV responsáveis pelo influxo de
Ca2+ que sustenta o componente tônico da contração em íleo de cobaia.

98
Silva, A.C.L. Discussão
Então diante dessas premissas, passamos a investigar se o composto
UFRPE 117 estaria agindo sobre os canais BKCa, ativando-os e consequentemente
levando a uma repolarização de membrana o que levaria a um bloqueio indireto dos
CaV. Em presença de 1 mM de TEA+, uma concentração específica para bloquear
apenas os BKCa (LATORRE et al., 1989; KNOT; BRAYDEN; NELSON, 1996), a
curva de relaxamento de UFRPE 117 (CE50 = 4,2 ± 0,9 x 10-6 M) foi deslocada para a
direita na presença de 1 mM de TEA+(CE50 = 2,0 ± 0,4 x 10-5 M), e sua a potência
relaxante foi reduzida cerca de 5 vezes na presença do bloqueador, indicando assim
a participação dos BKCa, no efeito relaxante de UFRPE 117 em íleo de cobaia
(Gráfico 20).
Outros subtipos de canais de K+ não investigados neste trabalho, como os
Kv, podem estar envolvidos nesse efeito relaxante, bem como outras vias de
sinalização relacionadas ao relaxamento do músculo liso, com a dos nucleotídeos
cíclicos ou RhoA/RhoK, que podem estar envolvidas nesse mecanismo de ação
espasmolítica do composto.
Diante dos resultados expostos, neste trabalho foi descrito pela primeira vez
o efeito espasmolítico de uma nova série de derivados sintéticos do lapachol e do
norlapachol. Um dos resultados mais relevantes desse estudo, mostrou que as
modificações estruturais nas moléculas do lapachol e norlapachol foi fundamental
para a diminuição ou aumento dessa atividade espasmolítica. Além disso, foi
possível comparar as potências e eficácias farmacológicas dos 12 compostos,
mostrando quais os mais potentes, permitindo inferir pela análise das estruturas
químicas das moléculas, os possíveis grupamentos responsáveis por esse efeito.
Evidenciou-se que o mecanismo de ação espasmolítica de UFRPE 117, um
novo derivado sintético do norlapachol, em íleo de cobaia se dá através de um
antagonismo não-competitivo dos receptores de histamina e da modulação positiva
dos canais de K+, em especial dos subtipos SKCa e BKCa, o que ocasiona uma
hiperpolarização de membrana com consequente fechamento dos CaV e provável
redução da [Ca2+]c que leva ao relaxamento da musculatura lisa intestinal(Figura 18),
mostrando que os derivados do norlapachol são moléculas promissoras para serem
utilizadas em doenças associadas a alterações na motilidade intestinal, sendo
necessários mais estudos para confirmar o seu uso.

99
Silva, A.C.L. Discussão
Figura 18 – Proposta de mecanismo de ação espasmolítica para o derivado sintético
do norlapachol UFRPE 117 em íleo isolado de cobaia.
N
N
O
OH
OH
O mecanismo de ação espasmolítica de UFRPE 117em íleo de cobaia, envolve um antagonismo do tipo não competitivo dos receptores histaminérgicos (1) e uma modulação positiva dos canais de K+, particularmente os dos subtipos SKCa e BKCa (2) hiperpolarizando as células intestinais, resultando em um bloqueio indireto dos Cav1 (3), resultando na redução da [Ca2+]c (4) e, consequentemente, relaxamento desse órgão. As definições das abreviaturas estão presentes na lista de abreviaturas e no texto. Fonte: Autor, 2014.
UFRPE 117

6 Conclusão

101
Na investigação do efeito espasmolítico de novos derivados
sintéticos do lapachol e do norlapachol em íleo isolado de cobaia
concluímos que:
Os derivados do lapachol (UFRPE 15, UFRPE 17, UFRPE 31, UFRPE
114, UFRPE 115 e UFRPE 116) apresentaram efeito antiespasmódico,
porém com baixa eficácia farmacológica com relação ao seu precursor.
Os derivados do norlapachol (UFRPE 106, UFRPE 107, UFRPE 111,
UFRPE 112, UFRPE 113 e UFRPE 117) apresentaram efeito
antiespasmódico com uma maior potência e eficácia farmacológicas
quando comparados ao seu precursor
Os compostos UFRPE 107, UFRPE 112 e UFRPE 117 relaxaram o íleo
de cobaia de maneira dependente de concentração.
O composto UFRPE 117 mostrou ser o mais promissor dos derivados
testados, apresentando uma maior potência e eficácia farmacológica
No estudo do mecanismo de ação espasmolítica do composto
UFRPE 117 em íleo de cobaia, concluiu-se que:
Exerce um antagonismo do tipo não competitivo pseudo-
irreversível dos receptores histaminérgicos
Envolve a modulação positiva dos canais de K+
Havendo a participação dos subtipos de canais de potássio
envolvidos SKCa e BKCa

Referências

103
Silva, A.C.L. Referências
ABDEL-LATIF, A. A. Calcium mobilizing receptors, polyphospholinositides, generation of second messengers and contraction in mammalian smooth muscle: historical perspectives and current status. Life Science,v. 45, p. 757 – 786, 1989.
ABOULAFIA, J; SILVA, B. A.; NOUAILHETAS, V. L. A. Protein kinase C modulators enhance angiotensin II desensitization of guinea pig ileum via maxi-K+ channels. European Journal of Pharmacology, v. 442, p. 29-36, 2002. AGUILAR-SANTAMARÍA, L., RAMÍREZ, G., NICASIO, P., C. ALEGRÍA-REYES, C., HERRERA-ARELLANO, A. Antidiabetic activities of Tecoma stans (L.) Juss. ex Kunth. Journal of Ethnopharmacology, v.124, p. 284–288, 2009.
ALDRICH, R.; CHANDY, K. G.; GRISSMER, S.; GUTMAN, G. A.; WEI, A. D.; WULFF, H. Calcium-activated potassium channels, introductory chapter. Lastmodified on 15/10/2009. Acessoem 19/12/2013. IUPHAR database(IUPHAR-DB), http://www.iuphardb.org/DATABASE/FamilyIntroductionForward?familyId=69.
ALEXANDER, S. P. H.; MATHIE, A.; PETERS, J. A. Guide to Receptors and Channels (GRAC), 5rd edition. British Journal of Pharmacology, v. 164 Suppl 1, p. S142-143, 2011.
ANTUNES, R. M. P.; LIMA, E. O.; PEREIRA, M. S. V.; CAMARA, C. A.; ARRUDA, T. A.; CATÃO, R. M. R.; BARBOSA; T. P.; NUNES, X. P.; DIAS, C. S.; SILVA, T. M. S. Atividade antimicrobiana “in vitro” e determinação da concentração inibitória mínima (CIM) de fitoconstituintes e produtos sintéticos sobre bactérias e fungos leveduriformes.Rev. Bras. Farmacogn.,v. 16, n. 4, p. 517-524,2006.
ARRUDA, A.L.A., VIEIRA, C.J.B., SOUSA, D.G., OLIVEIRA, R.F., CASTILHO, R.O. Jacaranda cuspidifolia Mart. (Bignoniaceae) as an antibacterial agente. Journal of Medicinal Food, v. 12, p. 1604–1608, 2011.
ASHIDA,Y.; MATSUMOTO, T.; KURIKI, H.; SHIRAISHI, M.; KATO, K.; TERAO, S. A novel anti-asthmatic quinone derivative, AA-2414 with a potent antagonistic activity against a variety of spasmogenicprostanoids. Prostaglandins, v. 38, n. 1, p. 91-112, 1989.
AUYONG, T. K.; WESTFALL, B. A.; RUSSELL, R. L. Pharmacological aspects of juglone. Toxicon, v. 1, n. 4, p. 235-239, 1963.
BABENKO, A. P.; AGUILAR-BRYAN, L.; BRYAN, J. A view of sur/KIR6.X, KATP channels. Annual Review of Physiology, v. 60, p. 667-687, 1998.

104
Silva, A.C.L. Referências
BAE, J.H.; KIM, J.W.; KWEON, G.R.; JEONG, K.H.; KIM, J.J.;MOON, du G. Corpus cavernosal smooth muscle relaxation effect of a novel AMPK activator, beta-lapachone.The Journal of Sexual Medicine, v. 8, p. 2205–2214, 2011.
BALASSIANO, I.T., DE PAULO S.A., HENRIQUES-SILVA N. et al
Demonstration of the lapachol as a potential drug for reducing cancer
metastasis. Oncol Rep v. 13 p. 329–333, 2005.
BENHAM, C. D.; BOLTON, T. B. Spontaneous transient outward currents in single visceral and vascular smooth muscle cells of the rabbit. The Journal of Physiology, v. 381, p. 385-406, 1986.
BERKEFELD, H.; FAKLER, B.; SCHULTE, U. Ca2+-Activated K+ Channels: From Protein Complexes to Function. Physiological Reviews, v. 90, n. 4, p. 1437-1459, 2010.
BERRIDGE, M. J. Cell Signalling Pathways. In: Cell Signalling Biology. Portland Press Limited , p. 1-118, 2009a.
BERRIDGE, M. J. Smooth muscle cell calcium activation mechanisms. The Journal of Physiology, v. 586, n. 21, p. 5047-5061, 2008.
BILLINGTON, C. K.; PENN, R. B. Signaling and regulation of G protein-coupled receptors in airway smooth muscle. Respiratory Research, v. 4, n. 1, p. 2, 2003.
BLUMENTHAL, D. K.; GARRISON, J. C. Pharmacodynamics: molecular mechanisms of drug action. In: BRUNTON, L. L.; CHABNER, B. A.; KNOLLMANN, B. C. (Ed.) Goodman and Gilman's The Pharmacological Basis of Therapeutics. 12th ed. San Diego, CA: The McGraw-Hill Companies, Inc. 2011.
BOLTON, T. B. Calcium events in smooth muscles and their interstitial cells; physiological roles of sparks.The Journal of Physiology,v. 570, p. 5-11, 2006.
BOLTON, T. B. Mechanism of action of transmitters and other substances on smooth-muscle. Physiological Reviews, v. 59, n. 3, p. 606-718, 1979.
BOLTON, T. B.; CLARK, J. P.; KITAMURA, K.; LANG, R. J. Evidence that histamine and carbachol may open the same ion channels in longitudinal smooth muscle of guinea-pig ileum. The Journal of Physiology, v. 320, p.363-379, 1981.
BOYLE, J. P.; TOMASIC, M.; KOTLIKOFF, M. I. Delayed rectifier potassium channels in canine and porcine airway smooth muscle cells. The Journal of Physiology, v. 447, p. 329-350, 1992.
BRAINARD, A. M.; KOROVKINA, V. P.; ENGLAND, S. K. Potassium channels and uterine function. Seminars in cell & developmental biology, v. 18, n. 3, p. 332-339, 2007.

105
Silva, A.C.L. Referências
BRAYDEN, J. E. Functional Roles Of KATP Channels In Vascular Smooth Muscle. Clinical and Experimental Pharmacology and Physiology, v. 29, n. 4, p. 312-316, 2002.
CAMARA, C.A., SILVA, T.M.S. , DA-SILVA, T.G., MARTINS, R.M., BARBOSA,
T.P ., PINTO, A.C., VARGAS, M.D. Molluscicidal activity of 2-hydroxy-[1,4]
naphthoquinone and derivatives. Annals of the Brazilian Academy of
Sciences v. 80(2), p. 329-334, 2008.
CARVALHO, L. H. ; ROCHA, E. M.; RASLAN, D. S.; OLIVEIRA, A. B.; KRETTLI, A. U. In vitro activity of natural and synthetic naphthoquinones against erythrocytic stages of Plasmodium falciparum. Braz. J. Med. Biol. Res., v. 21, p. 485-487, 1988.
CATTERALL, W. A. Voltage-Gated Calcium Channels, Cold Spring Harbor Perspectives in Biology, v. 11, p. 1-24, 2011.
CAVALCANTE, F. A.; MONTEIRO, F. S.; MARTINS, I. R. R.; BARBOSA, T.P.; CAMARA, C. A.; PINTO, A. C.; VARGAS; M. D.; SILVA, B. A. Synthetic lapachol derivatives relax guinea-pig ileum by blockade of the voltage-gated calcium channels.Z. Naturforsch., v. 65c, p. 627-636, 2010.
CAVALCANTE, F. A.; SILVA, J. L. V.; CARVALHO, V. M. N.; CAMARA, C. A.; SILVA, T. M. S.; PINTO, A. C.; VARGAS; M. D.; SILVA, B. A.Spasmolytic
activity of lapachol and its derivatives, - and -lapachone, on guinea-pig ileum involves blockade of the voltage-operated calcium channel.Brazilian Journal of Pharmacognosy, v. 18b, p. 183-189, 2008.
CHITALEY, K., WEBER, D. & WEBB, R C. RhoA/Rho-kinase, vascular changes, and hypertension. Current Hypertension Reports, 3(2), p.139-144. 2001.
.CHOUDHURY S.; DATTA S.; DAS TALUKDAR A.; DUTTA, M. Phytochemistry of the Family Bignoniaceae- A review. Biological and Environmental Sciences, v. 7, Number I, p. 145-150, 2011.
COELHO-CERQUEIRA, E., NETZ, P.A., CANTO, V. P., PINTO, A.C.,
FOLLMER, C. Topoisomerase Inhibition: Antitumor 1,4-Naphthoquinones as
Potential Inhibitors of Human Monoamine Oxidase. Chem Biol Drug Des 2014;
v. 83, p. 401–410, 2014.
CORREIA, A. C. C. Estudo comparativo da atividade espasmolítica de óleos essenciais de espécies de Annonaceae: Rollinialeptopetala R. E. Fries, Xylopialangsdorfiana A. St.-Hil. &Tul. eXylopiafrutescensAubl. Tese (Doutorado). Programa de Pós-graduação em Produtos Naturais e Sintéticos Bioativos, da Universidade Federal da Paraíba, João Pessoa, Paraíba/PB, 2013.

106
Silva, A.C.L. Referências
COSTA, W.F., OLIVEIRA, A.B., NEPOMUCENO, J.C. Lapachol as na epithelial
tumor inhibitor agent in Drosophila melanogaster heterozygote for tumor
suppressor gene wts. Genetic Molecular Research. v. 10, p. 3236–3245,
2011.
CRAGG, G.M.; NEWMAN, D.J.; GROTHAUS, P.G. New Horizons for Old Drugs and Drug Leads. Journal of Natural Products, 70(3), p.461-477, 2014. DA ROCHA, C. Q., VILELA, F.C., CAVALCANTE, G. P., SANTA-CECÍLIA, F.V., LUCAS SANTOS-E-SILVA, L., SANTOS, M.H., GIUSTI-PAIVA, A. Anti-inflammatory and antinociceptive effects of Arrabidaea brachypoda (DC.) Bureau roots. Journal of Ethnopharmacology, v. 133 p. 396–401, 2011.
DANIEL, E. E.; KWAN, C.Y.; JANSSEN, L. Pharmacological techniques for the in vitro study of intestinal smooth muscle. Journal Pharmacological Toxicology, v. 45, p.141-159, 2001.
DELLIS, O.; DEDOS, S. G.; TOVEY, S. C.; TAUFIQ-UR-RAHMAN, DUBEL, S. J.; TAYLOR, C. W. Ca2+ entry through plasma membrane IP3 receptors. Science, v. 313, n. 5784, p. 229-233, 2006.
DONATO, R. Functional roles of S100 proteins, calcium-binding proteins of the EF-hand type. BiochimicaetBiophysicaActa, v. 1450, n. 3, p.191-231, 1999.
DONATO, R. S100: a multigenic family of calcium-modulated proteins of the EF-hand type with intracellular and extracellular functional roles. The International Journal of Biochemistry & Cell Biology, v. 33, n. 7, p. 637-668, 2001.
DUKE, J. A. CRC Hanbook of Medicinal Herbs; CRC: Boca Raton, p 470, 1985.
EPIFANO. F., GENOVESE, S., FIORITO, S. Lapachol and its congeners as anticancer agents: a review. Phytochemic Review, v. 13, p. 37–49, 2014.
EXTON, J. H. New developments in phospholipase D. The Journal of Biological Chemistry, v. 272, n. 25, p. 15579-15582, 1997.
EYONG, K.O., KUMAR, P.S., KUETE, V. Semisynthesis and antitumoral activity of 2-acetylfuranonaphthoquinone and other naphthoquinone derivatives from lapachol. Bioorg Med Chem Lett v.18, p. 5387–5390, 2008.
FAKLER, B.; ADELMAN, J. P. Control of K(Ca) channels by calcium nano/microdomains. Neuron, v. 59, n. 6, p. 873-881, 2008.
FEHER, M.; SCHMIDT, J. M. Property distributions: differences between drugs, natural products, and molecules from combinatorial chemistry. Journal of Chemical Information and Computer Sciences, v. 43, p. 218-227, 2003.

107
Silva, A.C.L. Referências
FELÍCIO, A. C.; CHANGA, C. V.; BRANDÃO, M. A.; PETERS, V. M.; GUERRA, M. O. Fetal growth in rats treated with lapachol. Contraception, v. 66, p. 289–293, 2002.
FERREIRA, V. F. Aprendendo sobre os conceitos de ácidos e bases: a extração do lapachol de serragem do ipê. Química Nova naEscola, v. 4, p. 35-36, 1996.
FIORITO, S.; EPIFANOA, F.; BRUYÈRE C.; MATHIEU, V.; KISS, R.; GENOVESE, S. Growth inhibitory activity for cancer cell lines of lapacholand its natural and semi-synthetic derivatives. Bioorganic & Medicinal Chemistry Letters, v. 24, p. 454-457, 2014.
FLAGG, T. P.; ENKVETCHAKUL, D.; KOSTER, J. C.; NICHOLS, C. G. Muscle KATP Channels: Recent Insights to Energy Sensing and Myoprotection. Physiological Reviews, v. 90, n. 3, p. 799-829, 2010.
FLEISCHMANN, B. K.; WASHABAU, R. J.; KOTLIKOFF, M. I. Control of resting membrane potential by delayed rectifier potassium currents in ferret airway smooth muscle cells. The JournalofPhysiology, v. 469, p. 625-638, 1993.
FOGLIO, M. A.; QUEIROGA, C. L.; SOUSA, I. M. O.; RODRIGUES, R. A. F. Plantas medicinais como fonte de recursos terapêuticos: um modelo multidisciplinar.MultiCiência: construindo a história dos produtos naturais, v. 7, 2006.
FONSECA, S. G. C.; BRAGA, R. M. C.; SANTANA, D. P. Lapachol – química, farmacologia e métodos de dosagem. Rev. Bras. Farm., v. 84, n. 1, p. 9-16, 2003.
GAFNER, S.; WOLFENDER, J.-L.; NIANGA, M.; STOECKLI-EVANS, H.; HOSTETTMANN, K. Antifungal and antibacterial naphthoquinones from newbouldialaevisroots. Phytochemistry, v. 42, p. 1315-1320, 1996.
GARRET, R. H.; GRISHAM, C. M. Muscle contraction. In: Molecular Aspects of Cell Biology. Phyladelphia, Saunders, p. 1156-1179, 1995.
GENTRY, A.H. Bignoniaceae. Part I. Tribes Crescentieae and Tourrentieae. Flora Neotropica. Monogr. 25. The New York Botanical Garden, New York. p.1-130 1980.
GIBBS, R. D. Chemotaxonomy of Flowering Plants, University Press: Montreal 1974.
GOEL, R. K.; PATHAK, N. K. R.; BISWAS, M.; PANDEY, V. B.; SANYAL, A. K. Effect of lapachol, a naphthoquinone isolated from Tectonagrandis, on

108
Silva, A.C.L. Referências
experimental peptic ulcer and gastric secretion J. Pharm. Pharmacol., v. 39, p. 138-140, 1987.
GUERRA, M. O.; MAZONI, A. S. B.; BRANDÃO, M. A. F.; PETERS, V. M. Interceptive effect of lapachol in rats. Contraception, v.60, p.305, 1999.
GUERRA, M. O.; MAZONI, A. S. B.; BRANDÃO, M. A. F.; PETERS, V. M. Toxicology of lapachol in rats: embryolethality. Rev. Brasil. Biol., n. 61, n. 1, p. 171-174, 2001.
GURIB-FAKIM, A. Medicinal plants: Traditions of yesterday and drugs of tomorrow. Molecular Aspects of Medicine, v. 27, p. 1-93, 2006.
GURNEY A. M. Mechanisms of drug-induced vasodilation. Journal of Pharmacy and Pharmacology, v. 46, p. 242 - 251, 1994. HARVEY, A.L.;. Natural products in drug discovery. Drug Discovery Today, 13(19-20), p.894-901, 2008.
HEIZMANN, C.W. (Ed.) Novel Calcium-Binding Proteins, Springer-Verlag, Berlin, Heidelberg, p. 65–99, 1991.
HONDA, K.; TAKANO, Y.; KAMIYA, H. Involvement of protein kinase C in muscarinic agonist-induced contractions of guinea pig ileal longitudinal muscle. General Pharmacology, v. 27, p. 957-961, 1996.
HOOKER, S. C. Condensation of aldehydes with -hydroxy--naphthoquinone. synthesis of hydrolapachol. J. Am. Chem. Soc., v. 58, p. 1163-1167, 1936.
HUSSAIN, H., KROHN, K., AHMAD, V.U., MIANA, G.A., GREEND, I.R. Lapachol: an overview. Special Issue Reviews and Accounts. v. (ii), p. 145-171, 2007. HUSSAIN, I.; MOHLER, J. L.; SEIGIER, H. F.; BESTERMAN. J. M. Elevation of topoisomerase I messenger RNA. protein, and catalytic activity in human tumors: demonstration of tumor-type specificity and implications for cancer chemotherapy. Cancer Res., v. 54, p. 539-546, 1994.
ISHII, T. M.; MAYLIE, J.; ADELMAN, J P. Determinants of apamin and d-tubocurarine block in SK potassium channels. The Journal of Biological Chemistry, v. 272, n. 37, p. 23195-23200, 1997.
JAN, L. Y.; JAN, Y. N. Structural elements involved in specific K+ channel functions. Annual Review of Physiology, v. 54, p. 537-555, 1992.
JIANG, Y.; LEE, A.; CHEN, J.; CADENE, M.; CHAIT, B. T.; MACKINNON, R. Crystal structure and mechanism of a calcium-gated potassium channel. Nature, v. 417, n. 6888, p. 515-522, 2002.

109
Silva, A.C.L. Referências
JIM, K.; HARRIS, A., ROSENBERGER L. B.; TRIGGLE, D. J. Stereoselective and non-sterenselective effects of D 600 (Methoxyverapamil) in smooth muscle. European Journal of Pharmacology,v. 76, p. 67-72, 1981.
JOHNSON, I. S.; ARMSTRONG, J. G.; GORMAN, M.; BURNETT, J. P., Jr. The Vinca Alkaloids: a New Class of Oncolytic Agents. Cancer Res., v. 23, n. 81, p. 1390-1427, 1963.
KARAKI H; MITSUI-SAITO, M.; AMANO, K.; HARADA, K.; MIYAMOTO, S.; NAKAZAWA, H.; WON, K.; SATO, K. Calcium movements, distribution, and functions in smooth muscle. Pharmacological Reviews, v. 49, n. 2, p.157-230, 1997.
KIM, H. R.; APPEL, S.; VETTERKIND, S.; GANGOPADHYAY, S. S.; MORGAN, K. G. Smooth muscle signalling pathways in health and disease. Journal of Cellular and Molecular Medicine, v. 12, n. 6a, p. 2165-2180, 2008.
KINGSTON, D. G. I. Modern Natural Products Drug Discovery and Its Relevance to Biodiversity Conservation. Journal of Natural Products, v. 74, n. 3, p. 496-511, 2011.
KLOTZ, L.; PATAK, P.; ALE-AGHA, N.; BUCHCZYK, D. P.; ABDELMOHSEN, K.; GERBER, P. A.; VON MONTFORT, C.; SIES, H. 2-Methyl-1,4-naphthoquinone, vitamin K3, decreases gap-junctional intercellular communication via activation of the epidermal growth factor receptor/extracellular signal-regulated kinase cascade. Cancer Res., v. 62, p. 4922–4928, 2002.
KNOT, H. T.; BRAYDEN, E. J.; NELSON, M. T. Calcium channels and potassium channels. In BÁRÁNY, M. Biochemistry of smooth muscle contraction.San Diego, Academic Press, 203-219, 1996.
KOBAYASHI, S.; KITAZAWA, T.; SOMLYO, A. V.; SOMLYO, A. P. Citosolic
heparin inhibits muscarinic and -adrenergic Ca2+-release in smooth muscle: physiological role of inositol 1,4,5-trisphosphate in pharmacomechanical coupling.Journal of Biological Chemistry, v. 264, p. 17997-18004, 1989.
KOYAMA, J.; MORITA, I.; TAGAHARA, K.; HIRAI, K. Ciclopentenedialdehydes from the Tabebuiaimpetiginosa. Phytochemistry, v. 53, p. 869-872, 2000.
KRISHNAN, P.; BASTOW, K. F. Novel Mechanisms of DNA Topoisomerase II
Inhibition by pyranonaphthoquinone derivatives – eleutherin, -lapachone, and
-lapachone. Biochem. Pharmacol., v. 60, p. 1367–1379, 2000.

110
Silva, A.C.L. Referências
LEDOUX, J.; WERNER, M. E.; BRAYDEN, J. E.; NELSON, M. T. Calcium-Activated Potassium Channels and the Regulation of Vascular Tone. Physiology,v. 21, p. 69-78, 2006.
LEE, U. S.; CUI, J. BK channel activation: structural and functional insights. Trends in Neurosciences, v. 33, n. 9, p. 415-423, 2010.
LIDER, V. A.;SHISHKIN, S. B.; FEOFILAKTOVA, S. N. Mechanism of action of naphthoquinones and hydroxycoumarins on the mechano-active properties of isolated coronary arteries.FarmakolToksikol, v. 50, n. 5, p. 31-34, 1987.
MACHADO TB, PINTO AV, PINTO MCFR, LEAL ICR, SILVA MG, AMARAL ACF, KUSTER RM, SANTOS KRN:In vitro activity of Brazilian medicinalplants, naturally occuringnaphthoquinones and their analogues,against methicillin-resistant Staphylococcus aureus.International Journal of Antimicrobial Agents,.p. 279-284, 2003.
MATHAN, V. I. Diarrhoeal diseases. British Medical Bulletin, v. 54, p. 402 – 419, 1998.
MACHADO, T.B.; PINTO, A.V.; PINTO, M.C.F.R.; LEAL, I.C.R.; SILVA, M.G.; AMARAL, A.C. F.; KUSTER, R.M. E NETTO-DOS SANTOS, K.R. In vitro medicinal plants, naturally occurring naphthoquinones and their analogues, against methicillin-resistant Staphylococcus aureus. International Journal of Antimicrobial Agents, v. 21, n.3, p. 279-284, 2003.
MACKINNON, R. Potassium channels. FEBS Letters, v. 555, n. 1, p. 62-65, 2003.
MCHALE, N.; HOLLYWOOD, M.; SERGEANT, G.; THORNBURY, K. Origin of spontaneous rhythmicity in smooth muscle. The Journal of Physiology, v. 570, n. 1, p. 23-28, 2006.
MIRANDA F. G. G., VILAR J. C., ALVES I. A. N., CAVALCANTI S. C. H., ANTONIOLLI A. R.: Antinociceptive and antiedematogenic properties and acutetoxicity of Tabebuiaavellanedae Lor. exGriseb. inner barkaqueous extract.BMC Pharmacology, v. 1, p. 6, 2001.
MOON, D. O.; CHOI, Y. H.; KIM, N. D.; PARK, Y. M.; KIM, G. Y. Anti-inflammatory effects of β-lapachone in lipopolysaccharide-stimulated BV2 microglia. Int. Immunopharmacol., v.7, p. 506–514, 2007.
MOURA, K. C. G.; EMERY, F. S.; NEVES-PINTO, C., PINTO, M. C. F. R.; DANTAS, A. P.; SALOMÃO, K.; CASTRO, S. L.; PINTO, A. V. Trypanocidal activity of isolated naphthoquinones from Tabebuiaand some heterocyclic derivatives: a review from an interdisciplinary study. J. Braz. Chem. Soc.,v. 12, n. 3, p. 325-338, 2001.

111
Silva, A.C.L. Referências
MOUSSATCHE, H.; CUADRA, J. L; RAMOS, P. R.; PERISSE, A. C. M. Pharmacological experiments with benzoquinones. Rev. Bras. Biol., v. 27, n. 4, p. 369-79, 1967.
MUÑOZ-MINGARRO D.; ACERO, N.; LLINARES, F.; POZUELO, J. M.; GALÁN DE MERA, A.; VICENTEN, J. A.; MORALES, L.; ALGUACIL, L. F.; PÉREZ, C. Biological activity of extracts from Catalpa bignonioides Walt. (Bignoniaceae). Journal of. Ethnopharmacology., v. 87, p. 163–167, 2003.
MURRAY, K. R.; GRANNER, D.K., MAYES, P.A.; RODWELL, V. W.Harper’s Illustrated Biochemistry. 26ª ed. Mcgraw-Hill. 2003.
MURTHY, K. S. Signaling for contraction and relaxation in smooth muscle of the gut. Annual Review of Physiology, v. 68, n. 1, p. 345-374, 2006.
NEDER, K.; MARTON, L. J.; LIU, L. F.; FRYDMAN, B. Reaction of -lapachone and related naphthoquinones with 2-mercaptoethanol: A biomimetic model of topoisomerase II poisoning by quinones. Cell Mol. Biol., v. 44, p. 465–474, 1998.
NELSON, D. L.; COX, M. M. Leningher Principles of Biochemistry. 4a ed. W. H. Freeman. 2004.
NEUBIG, R. R.; SPEDDING, M.; KENAKIN, T.; CHRISTOPOULOS, A. International Union of Pharmacology Committee on Receptor Nomenclature and Drug Classification. XXXVIII. Update on terms and symbols in quantitative pharmacology. Pharmacological Reviews, v. 55, n. 4, p. 597-606, 2003.
NEWMAN, D. J.; CRAGG, G. M. Natural products as sources of new drugs over the 30 years from 1981 to 2010. Journal of Natural Products, v. 75, p. 311−335, 2012.
NOMA, A. ATP-regulated K+ channels in cardiac muscle. Nature, v. 305, n. 5930, p. 147-148,1983.
NOUAILHETAS, V. L. A.; SHIMUTA, S. I.; PAIVA, A. C. M.; PAIVA, T. B. Calcium and sodium dependence of the biphasic response of the guinea-pig ileum agonists. EuropeanJournalofPharmacology, v. 116, p. 41-47, 1985.
OLIVEIRA, A. B.; RASLAN, D. S.; MIRAGLIA, M. C.; MESQUITA, A. A. L; ZANI, C. L; FERREIRA, D. T; MAIA, J. G. S. Estrutura química e atividade biológica de naftoquinonas de Bignoniáceas Brasileiras. Quimica. Nova, v. 13, n. 4, p. 302, 1990.
PARK, B. S.; KIM, J. R.; LEE, S. E.; KIM, K. S.; TAKEOKA, G. R.; AHN, Y. J.; KIM, J. H. Selective growth-inhibiting effects of compounds identified in

112
Silva, A.C.L. Referências
Tabebuiaimpetiginosainner bark on human intestinal bacteria. J. Agric. Food Chem., v. 53, p. 1152-1157, 2005.
PEREIRA, E.M., MACHADO, T.B., LEAL, I.C.R., JESUS, D.M., DAMAS, C.R.A.D., PINTO, A.V., MARVAL, M. G, KUSTER, R.M., SANTOS, K.R.N. Tabebuia avellanedae naphthoquinones: activity against methicillin-resistant staphylococcal strains, cytotoxic activity and in vivo dermal irritability analysis. Annals of Clinical Microbiology and Antimicrobials, 2006. PEREIRA, E.M., MACHADO, T.B., LEAL, I.C.R., JESUS, D.M., DAMAS, C.R.A.D., PINTO, A.V., MARVAL, M. G, KUSTER, R.M., SANTOS, K.R.N. Tabebuia avellanedae naphthoquinones: activity against methicillin-resistant staphylococcal strains, cytotoxic activity and in vivo dermal irritability analysis. Annals of Clinical Microbiology and Antimicrobials, 2006.
PETERSEN, O. H.; MARUYAMA, Y. Calcium-activated potassium channels and their role in secretion. Nature, v. 307, n. 5953, p. 693-696, 1984.
PINTO C. N.; DANTAS A. P., DE MOURA K. C., EMERY F. S., POLEQUEVITCH P. F., PINTO M. C., DE CASTRO S. L., PINTO A. V. Chemical reactivity studieswith naphthoquinones from Tabebuia with anti-trypanosomalefficacy Arzneimittelforschung, v. 50, n. 12, p. 1120-8, 2000.
PINTO, A. V.; PINTO, M. C.; LAGROTA, M. .H.; WIGG, M. D.; AGUIAR, A. N. Antiviral activityofnaphthoquinones. I. Lapachol derivatives against enteroviruses. Revista Latinoamericana de Microbiologia, v. 29, n. 1, p. 15-20, 1987.
PINTO, A.V. E CASTRO, S.L. The trypanocidal activity of naphthoquinones: a review. Molecules, v. 14, p. 4570- 4590, 2009. PLANCHON, S. M.; PINK, J. J.; TAGLIARINO, C.; BORNMANN, W. G.;
VARNES, M. E.; BOOTHMAN, D. A. -Lapachone-Induced Apoptosis in Human Prostate Cancer Cells: Involvement of NQO1/xip3. Exp. Cell Res., v. 267, p. 95–106, 2001. PLANCHON, S. M.; WUERZBERGER, S.; FRYDMAN, B.; WITIAK, D. T.; HUTSON, P.; CHURCH, D. R.; WILDING, G.; BOOTHMAN, D. A. Beta-lapachone-mediated apoptosis in human promyelocytic leukemia (HL-60) and human prostate cancer cells: A p53-independent response. Cancer Res., v. 55, p. 3706–3711, 1995.
PONTE, C. G.; MCMANUS, O. B.; SCHMALHOFER, W. A.; SHEN, D.-M.; DAI, G.; STEVENSON, A.; SUR, S.; SHAH, T.; KISS, L.; SHU, M.; DOHERTY, J. B.; NARGUND, R.; KACZOROWSKI, G. J.; SUAREZ-KURTZ, G.; GARCIA, M. L. Selective, direct activation of high-conductance, calcium-activated potassium channels causes smooth muscle reaxation. Molecular Pharmacology, v. 81, n. 4, p. 567-77, 2012.

113
Silva, A.C.L. Referências
QUAN, Y.; BARSZCZYK, A.; FENG, Z. P.; SUN, H. S. Current Understanding of KATP Channels in Neonatal Diseases: Focus on Insulin Secretion Disorders. Acta Pharmacologica Sinica, v. 32, p. 765–780, 2011.
QUEST, U.; COOK, N. S. Moving together: K+-channel openers and ATP-sensitive K+-channels. Trends in Pharmacological Science, v. 10, p. 431–435, 1989.
RAO, K. V.; McBRIDE, T. J.; OLESON, J. J. Recognition and evaluation of lapachol as an antitumor agent. Cancer Res., v. 28, p. 1952-1954, 1968.
RAO, K. V.; Quinone natural products: streptonigrin (nsc-45383) and lapachol (nsc-11905) structure-activity relationships. Cancer Chemother. Rep., v. 4, p. 11, 1974.
RAO, V. S. N. et al. Investigations on the gastroprotective and antidiarrhoeal properties of ternatin, a tetramethoxyflavone from Egletes viscose. PlantaMédica, v. 63, p. 146-149, 1997. REMBOLD, C. M. Electromechanical and pharmacomechanical coupling. In: Bárány; M. Biochemistry of smooth contraction. San Diego, Academic Press, p. 227-239, 1996.
SACAU, E. P.; ESTÉVEZ-BRAUN, A.; RAVELO, A. G.; FERRO, E. A.; TOKUDA, H.; MUKAINAKA, T.; NISHINO, H. Inhibitory effects of Lapachol derivatives on epstein-barr virus activation. Bioorganic. Medicinal. Chemistry., v. 11, n.4, p. 483-488, 2003.
SAH, P.; FABER, E. S. L. Channels underlying neuronal calcium-activated potassium currents. Progress in Neurobiology, v. 66, n. 5, p. 345-353, 2002.
SALKOFF, LAWRENCE; BUTLER, A.; FERREIRA, G.; SANTI, C.; WEI, AGUAN. High-conductance potassium channels of the SLO family. Nature Reviews Neuroscience, v. 7, n. 12, p. 921-931, 2006.
SANBORN, B. M. Relationship of ion channel activity to control of myometrial calcium. Journal of the Society for Gynecologic Investigation, v. 7, n. 1, p. 4-11, 2000.
SANDERS, K.M., KOH, S.D., RO, S., WARD, S.M Regulation of
gastrointestinal motility - insights from smooth muscle biology. Nat. Rev.
Gastroenterol. Hepatol. V. 9, p. 633–645 2012.
SANTANA, C. F.; LIMA, O.; D' ALBUQUERQUE, I. L.; LACERDA A. L.; MARTINS, D. G. Antitumoral and toxicological properties of extracts of bark and various wood components of Pau d'arco (Tabebuiaavellanedae). Rev. Inst. Antibiot. (Recife), v. 8, n. 1, p. 89-94, 1968.

114
Silva, A.C.L. Referências
SANTOS, M.D.F.; LITIVACK-JUNIOR,J.T.; ANTUNES,R.V.;SILVA, T.M.S.;CAMARA, C.A. New Adducts of Lapachol with Primary Amines. J. Braz. Chem. Soc., v. 22, p. 796-800, 2011.
SEOANE, S.; RODRÍGUEZ, P. D.; LAGO, J. S.; GALLEGO, R.; FERNÁNDEZ,
R. P.; LANDIN, M.Cytotoxicity of lapachol, -lapachone and related synthetic 1,4- naphthoquinones against oesophageal cancer cells. European Journal of Pharmaceutics and Biopharmaceutics, 2013.(Article in press)
SHIEH, C.; COGHLAN, M.; SULLIVAN, J. P.; GOPALAKRISHNAN, M. Potassium Channels: Molecular Defects, Diseases, and Therapeutic Opportunities. PharmacologicalReviews, v. 52, p. 557-593, 2000.
SILVA, M. F., LISBDA, P. L. B. AND LISBDA, R. C. L. In: Nomes vulgares de plantas amazônicas p. 163. Conselho National de Desenvolvimento Cientifico e Tecnológico. Instituto national de Pesquisas da Amazônia, Manaus. 1977.
SILVA, M. M. da; QUEIROZ, L. P. de. A família Bignoniaceae na região de Catolés, Chapada Diamantina, Bahia, Brasil. Sitientibus Série Ciências Biológicas, v. 3, n. 1/2, p. 3-21, 2003.
SILVA, M. N.; FERREIRA, V. F.; SOUZA, M. C. B. V. Um panorama atual da
química e da farmacologia de naftoquinonas, com ênfase na -lapachona e derivados. Química. Nova, v. 26, n. 3, p. 407-416, 2003.
SILVA-JÚNIOR, E. N.; SOUZA, M. C. B. V.; PINTO, A. V.; PINTO, M. C. F. R.; GOULART, M. O. F.; BARROS, F. W. A.; PESSOA, C.; COSTA-LOTUFO, L. F.; MONTENEGRO, R. C.; MORAES, M. O.; FERREIRA, V. F. Synthesisandpotentantitumoractivityof new arylamino derivatives of nor-β-lapachone and nor-α-lapachone. Bioorg. Med. Chem., v. 15, n. 22, p. 7035-7041, 2007
SINGER, J. J.; WALSH JR., J. V. Characterization of calcium-activated potassium channels in single smooth muscle cells using the patch-clamp technique. PflugersArchiv: European Journal of Physiology, v. 408, n. 2, p. 98-111, 1987.
SITÔNIO, M.M., CARVALHO-JÚNIOR, C.H.R., CAMPOS, I.A., SILVA, J.B.N.F., MARIA DO CARMO ALVES DE LIMA, M.C.A.L., GÓES A.J.S., MAIA, M.B.S., ROLIM NETO, P.J., SILVA, T.G. Anti-inflammatory and anti-arthritic activities of 3,4-dihydro-2,2-dimethyl-2H-naphthol[1,2-b]pyran-5,6-dione (b-lapachone). Inflammation Research v. 62, p. 107-113, 2013.
SOMLYO, A. P.; SOMLYO, A. V. Signal transduction and regulation in smooth muscle. Nature, v. 372, n. 6503, p. 231-236, 1994.

115
Silva, A.C.L. Referências
SOMLYO, A. P.; SOMLYO, A. V. Ca2+ Sensitivity of Smooth Muscle and Nonmuscle Myosin II: Modulated by G Proteins, Kinases, and Myosin Phosphatase. Physiological Reviews, v. 83, n. 4, p. 1325-1358, 2003.
SOMLYO, A. P.; WU, X.; LALKER, L. A.; SOMLYO, A. V. Pharmacomechanical coupling: the role of calcium, G-proteins, kinases and phosphatases. ReviewsofPhysiology, BiochemistryandPharmacology, v. 134, p. 201-234, 1999.
SOUZA, M. P.; MATOS, M. E. O.; MATOS, F. J. A.; MACHADO, M. I. L.; CRAVEIRO A. A. Constituintes químicos de plantas medicinais brasileiras, Fortaleza: UFC, 1991.
SOUZA, V. C.; LORENZI, H. Botânica Sistemática: guia ilustrado para identificação das famílias de Fanerógamas nativas e exóticas no Brasil, baseado em APG II. 2. ed. Nova Odessa: Instituto Plantarum, p 574, 2008. SOUZA-SILVA, F., NASCIMENTO, S.B., BOURGUIGNON, S.C., PEREIRA, B.A.S., CARNEIRO, P.F., SILVA, W.S., ALVES, C.R., PINHO, R.T. Evidences for leishmanicidal activity of the naphthoquinone derivative epoxy-α-lapachone Experimental Parasitology. v.147 p. 81–84, 2014.
STANDEN, N.; QUAYLE, J.; DAVIES, N.; BRAYDEN, J. E; HUANG, Y.; NELSON, M. T. Hyperpolarizing vasodilators activate ATP-sensitive K+ channels in arterial smooth muscle. Science, v. 245, n. 4914, p. 177-180, 1989.
STOTT, J.B., JEPPS, T.A., GREENWOOD, I.A. KV7 potassium channels: a new
therapeutic target in smooth muscle disorders. Drug Discovery Today. V. 19,
2014.
SUN, Y. -D. & BENISHIN, C. G. K+ channel openers relax longitudinal muscle of guinea-pig ileum. European Journal of Pharmacology, v. 271, p. 453-459, 1994.
SUNASSEE, S. N.; VEALE, C. G. L.; GOUDEN, N. S.; OSONIYI, O.; HENDRICKS, D. T.; CAIRA, M. R.; DE LA MARE; J. A.; EDKINS, A. L.; PINTO,
A.V.Cytotoxicity of lapachol, -lapachone and related synthetic 1,4- naphthoquinones against oesophageal cancer cells. European Journal of Medicinal Chemistry, v. 62, p. 98-110, 2013.
TANTRY, M. A. Plant natural products and drugs: a comprehensive study. Asian Journal of Traditional Medicines, v.4, p.241-249, 2009.
TAYLOR, L. Herbal secrets of the Rainflorest. Roaklin: Prima Health, 1998.
TEIXEIRA, M. J.; DE ALMEIDA, Y. M.; VIANA, J. R.; HOLANDA FILHA, J. G.; RODRIGUES, T. P.; PRATA, J. R. C. JR.; COELHO, I. C. B.; RAO, V. S.;

116
Silva, A.C.L. Referências
POMPEU, M. M. L. In vitro leishmanicidalactivityof 2-hydroxy-3-(3-methyl-2-butenyl)-1,4-naphthoquinone (lapachol). Phytoter. Res., v. 15, p. 44, 2001.
TERAMOTO, N. Physiological roles of ATP-sensitive K+ channels in smooth muscle. The Journal of Physiology, v. 572, n. Pt 3, p. 617-624, 2006.
THORNELOE, K. S.; NELSON, M. T. Ion channels in smooth muscle: regulators of intracellular calcium and contractility. Can. J. Physiol. Pharmacol., v. 83, p. 215–242, 2005.
TOTSUKAWA, G.; YAMAKITA, Y.; YAMASHIRO, S.; HARTSHORNE, D.J.; SASAKI, Y.; MATSUMURA, F. Distinct Roles of Rock (Rho-Kinase) and Mlck in Spatial Regulation of Mlc Phosphorylation for Assembly of Stress Fibers and Focal Adhesions in 3t3 Fibroblasts. The Journal of Cell Biology, v. 150, n. 4, p. 797-806, 2000.
TRIGGLE, C. R.; TRIGGLE, D. J. An analysis of the action of cations of the lanthanide series on the mechanical responses of guinea-pig ileal longitudinal muscle. The Journal of Physiology, v. 254, p. 39-54, 1976.
TRIGGLE, C. R.; SWAMY, V. C.; TRIGGLE, D. J. Calcium antagonists and contractile responses in rat vas deferens and guinea pig ileal smooth muscle. Canadian Journal of Physiology and Pharmacology, v. 57, p. 804-818, 1979.
TRIGGLE, D. J.; ZHENG, W.; HAWTHORN, M.; KWON, Y. W.; WEI, X. Y.; JOSLYN, A.; FERRANTE, J.; TRIGGLE, A. M. Calcium channels in smooth muscle. Annals of the New York Academy of Sciences, v. 560, p. 215-229, 1989.
TSENG, C. H; CHENG, C.M.; TZENG, C. C.; PENG, S. I.; Yang, C. L.; Chen, Y. L. Synthesis and anti-inflammatory evaluations of b-lapachone derivatives. Bioorganic & Medicinal Chemistry v. 21 (2013) 523–531, 2013.
UEHATA, M.; ISHIZAKI, T.; SATOH, H.; ONO, T.; KAWAHARA, T.; MORISHITA, T.; TAMAKAWA, H.; YAMAGAMI, K.; INUI, J.; MAEKAWA, M.; NARUMIYA, S. Calcium sensitization of smooth muscle mediated by a Rho-associated protein kinase in hypertension. Nature, v. 389, n. 6654, p. 990-994, 1997.
UNNO, T.; KOMORI, S.; OHASHI, H. Inhibitory effect of muscarinic receptor activation on Ca2+ channel current in smooth muscle cells of guinea-pig ileum. The Journal of Physiology, v. 484, n. 3, p. 567-581,1995.
VAN BREEMEN, C.; AARONSON, P.; LOUTZENHISER, R. Sodium-Calcium interaction in mammalian smooth muscle. Pharmacological Reviews, v. 30, p. 167-208, 1979.

117
Silva, A.C.L. Referências
VAN DER STAAY, F. J.; FANELLI, R. J.; BLOKLAND, A.; SCHMIDT, B. H. Behavioral effects of apamin, a selective inhibitor of the SKCa-channel, in mice and rats. Neuroscience and Biobehavioral Reviews. v. 23, n. 8, p. 1087-1110, 1999.
VERGARA, C.; LATORRE, R.; MARRION, N. V.; ADELMAN, J P. Calcium-activated potassium channels. Current Opinion in Neurobiology, v. 8, n. 3, p. 321-329, 1998.
VOGALIS, F. Potassium channels in gastrointestinal smooth muscle. Journal of Autonomic Pharmacology, v. 20, p. 207-219, 2000.
WARASHINA, T.; NAGATANI, Y.; NORO, T. Constituints from the bark of Tabebuiaimpetiginosa. Phytochemistry, v. 65, p. 2003-2011, 2004.
WATTERSON, K. R.; RATZ, P. H.; SPIEGEL, S. The role of sphingosine-1-phosphate in smooth muscle contraction. Cellular Signalling, v. 17, p. 289-298, 2005.
WEI, A. D.; GUTMAN, G. A.; ALDRICH, R.; CHANDY, K. G.; GRISSMER, S.; WULFF, H. International Union of Pharmacology. LII. Nomenclature and Molecular Relationships of Calcium-Activated Potassium Channels. Pharmacological Reviews, v. 57, n. 4, p. 463-472, 2005.
WESTON, A. H.; EDWARDS, G. Recent progress in potassium channel opener pharmacology. Biochemical Pharmacology, v. 43, n. 1, p. 47-54, 1992.
WOODRUM, D. A.; BROPHY, C. M. The paradox of smooth muscle physiology. Molecular and Cellular Endocrinology, v. 177, n. 1-2, p. 135-143, 2001.
YAMASHITA MITSUAKI; KANEKO MASAFUMI; IIDA AKIRA; TOKUDA HARUKUNI; NISHIMURA KATSUMI. Stereoselective synthesis and cytotoxicity of a cancer chemopreventivenaphthoquinone from Tabebuiaavellanedae. Bioorganic & Medicinal Chemistry Letters, V. 17, p. 6417-6420, 2007.

Anexo

119
Silva, A.C.L. Anexo