Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DA PARAÍBA
CENTRO DE CIÊNCIAS DA SAÚDE
PROGRAMA DE PÓS-GRADUAÇÃO EM PRODUTOS NATURAIS E
SINTÉTICOS BIOATIVOS
LUIZ HENRIQUE CÉSAR VASCONCELOS
Suplementação alimentar com óleo de coco virgem previne as
alterações na reatividade contrátil e relaxante em traqueia de
cobaia submetida à inflamação pulmonar alérgica crônica
João Pessoa-PB
2017

LUIZ HENRIQUE CÉSAR VASCONCELOS
Suplementação alimentar com óleo de coco virgem previne as
alterações na reatividade contrátil e relaxante em traqueia de
cobaia submetida à inflamação pulmonar alérgica crônica
Tese apresentada ao Programa de Pós-graduação em Produtos Naturais e Sintéticos Bioativos, do Centro de Ciências da Saúde, da Universidade Federal da Paraíba, como parte dos requisitos para a obtenção do título de DOUTOR EM PRODUTOS NATURAIS E SINTÉTICOS BIOATIVOS. Área de Concentração: FARMACOLOGIA
ORIENTADORA: Profa. Dra. Bagnólia Araújo da Silva
COORIENTADORA: Profa. Dra. Fabiana de Andrade Cavalcante
João Pessoa-PB
2017



Dedicatória

Às pessoas mais importantes da minha vida: meu pai, Osvaldo
César Galdino (in memoriam), maior exemplo de idoneidade
que tive; e minha mãe, Francisca das Chagas Vasconcelos, que
nunca mediu esforços para me prover de uma primorosa
criação. A vocês sou grato pela vida e por todos os valores
morais que possuo. Sei que onde imagino ser capaz de chegar
não se compara à certeza que vocês sempre tiveram.
Aos meus queridos irmãos: Domingos Sávio V. Galdino,
Maria César V. G. Silva e Rosenira V. Galdino. Meu amor
por vocês independe de consanguinidade.

Agradecimentos

Ninguém faz nada sozinho; sempre é necessário, pelo menos, mais um. Esse trabalho
foi fruto de muito esforço, confiança, ajuda e incentivo de muitas pessoas, sem as
quais ele não teria sido concretizado com tamanho primor. Por isso, quero agradecer
a todos que estiveram ao meu lado nesses quatro anos de caminhada.
A Deus, por permitir que tais pessoas fizessem parte da minha história.
À minha orientadora, Profa. Bagnólia A. da Silva, pelo maior exemplo de professor
que tive, em quem me espelho para seguir a docência. Pelas injeções de confiança
nos momentos de desânimo, por me mostrar que eu sempre poderia mais, pelas
muitas discussões científicas e por todo o suporte que me foi dado, especialmente
nos últimos dois anos, pelo qual sempre serei grato.
À minha coorientadora, Profa. Fabiana de A. Cavalcante, pela disponibilidade em
contribuir com o que fosse necessário e pela confiança em me entregar sua sala de
aula durante os estágios-docência em Fisiologia.
À Profa. Marta Suely Madruga e sua equipe de trabalho, pela disponibilidade para a
realização dos testes de controle de qualidade do óleo de coco virgem em seu
laboratório no Programa de Tecnologia e Ciência dos Alimentos no Centro de
Tecnologia da UFPB.
Ao Prof. Alexandre Sérgio Silva e sua equipe, especialmente as alunas Glêbia Alexa
Cardoso e Ana Paula Urbano Ferreira, que me acompanharam nos experimentos
bioquímicos de estresse oxidativo em seu laboratório no Departamento de Educação
Física do CCS/UFPB.
À Profa. Patricia Mirella da Silva Scardua, por ter praticamente feito de seu laboratório,
no Departamento de Biologia Molecular/CCEN/UFPB, uma extensão do nosso para
que pudéssemos processar as amostras para a análise histológica; e sua equipe de
trabalho, em especial seu aluno e meu grande amigo Dr. Fernando Ramos Queiroga,
que me acompanhou durante todo o processo de execução de preparo do tecido para
histologia.

À Profa. Giciane Carvalho Vieira, por ter prontamente aceito fazer a análise das
lâminas histológicas.
À Profa. Iolanda de Fátima Lopes Calvo Tibério, que me recebeu em seu laboratório,
na Universidade de São Paulo, com toda hospitalidade para aprender e executar a
técnica de imunohistoquímica; e sua equipe de trabalho, especialmente a Renato
Fraga Riguetti e a Flavia C. Ribas de Souza, que me acompanharam durante os
experimentos e me deram o suporte necessário durante o tempo de estágio no
laboratório.
Aos Professores Camille de Moura Balarini, Iolanda de Fátima Lopes Calvo Tibério,
Margareth de Fátima Formiga Melo Diniz e Tatjana Keesen de Souza Lima, titulares
da banca de defesa de Tese; Marcia Regina Piuvezam e Temilce Simões de Assis
Cantalice, suplentes da banca de defesa de Tese; Enéas Ricardo de Morais Gomes,
juntamente com Camille de Moura Balarini e Marcia Regina Piuvezam, titulares do
exame de qualificação; e Robson Cavalcante Veras, suplente do exame de
qualificação; pela disponibilidade em participar da avaliação desse trabalho, pelas
suas contribuições, assim como a participação nessa etapa da minha formação
acadêmica.
Aos Professores que me lecionaram durante a vida escolar, a graduação em Farmácia
e pelo Programa de Pós-graduação em Produtos Naturais e Sintéticos Bioativos
(PPgPNSB), especialmente a Adalberto Costa, Bagnólia Araújo da Silva, Demétrius
Antonio Machado de Araújo, Eduardo de Jesus Oliveira, Sandra Rodrigues
Mascarenhas e Tatjana Keesen de Souza Lima, pela visão de ciência e pela forma
única de lecionar que me despertou admiração.
Ao coordenador do PPgPNSB, Prof. Josean Fechine Tavares, pela competência com
que coordena esse Programa.
A Caroline Mangueira, pela sempre disponibilidade e pela eficiência com que conduz
seu trabalho na secretaria do PPgPNSB.

A Ana Carolina de Carvalho Correia e Cibério Landim Macêdo, meus primeiros
“mentores” de bancada no Laboratório de Farmacologia Funcional Prof. George
Thomas (LFF) durante a Iniciação Científica.
A todos que estão ou estiveram no LFF: Alana Costa, Aline Brito, Ana Caroline de
Lima, Anderson F. Diniz, Antônio Raphael Lima, Bárbara Cavalcanti, Elba dos Santos,
Filipe R. Oliveira, Giuliana Amanda, Giulyane Targino, Iana Oliveira, Iara Luna, Indyra
Figueiredo, Italo Martins, Joedna Cavalcante, José Lucas Galvão, Júnior Lacerda, Laiz
Aline, Layanne Cabral, Luan Diniz, Maria da Conceição Correia, Millena Medeiros,
Paula Benvindo, Pedro Moura, Rafael Lima, Rayane Fernandes, Renata Sampaio,
Sarah Rebeca, Thaynan Carvalho e Thiago Afonso. A todos pela ajuda, pelas
discussões, pela excelente convivência, pelos momentos de descontração e pelo
espírito de equipe que marca nosso grupo.
Aos queridos amigos Airlla Laana, Alana Costa, Aline Brito, André de Lima, Antônio
Raphael Lima, Bárbara Cavalcanti, Cínthia Dias, Daniele Ramos, Dayvid Farias,
Fernando Queiroga, Filipe Rodolfo Oliveira, Giuliana Amanda, Giulyanne Targino,
Heveline Medeiros, Indyra Figueiredo, Layanne Cabral, Luan Diniz, Luiz Henrique
Agra, Marcíllia Poncyanna, Sandro Filho, Sarah Rebeca, Tatyanna Kelvia, Thaynan
Carvalho, Thiago Melo e Vanessa da Nóbrega. Membros da “quinta série”, dos
“paueremba” ou sem grupo definido, agradeço a vocês por todo o amor, pelo apoio de
sempre, pelo companheirismo, pelas conversas, pelas “trollagens”, pelas farras, pelos
choros, pelas brigas... absolutamente tudo isso me impulsionam na busca daquilo que
pretendo alcançar. Sei que posso contar com cada um em todos os momentos, em
qualquer distância.
A Alana Costa, Giuliana Amanda, Iara Luna, Luan Diniz e Maria da Conceição Silva,
por terem sido parte fundamental na execução e obtenção dos dados aqui presentes,
seja na parte experimental, nas discussões de metodologias ou pontos de vista que
contribuíram para o amadurecimento desse trabalho, cada um conforme suas
possibilidades, mas dispostos a qualquer tempo.
A todos os colegas e amigos da turma do Doutorado outubro/2013 do PPgPNSB,
pelos bons momentos e pelo crescimento acadêmico e pessoal compartilhado.

A José Crispim Duarte e Roberta Nunes Parentoni, pela competência com que
conduzem o Biotério “Prof. Thomas George” e pelo auxílio técnico sempre que
necessário.
À Sra. Luzinete, pelo trabalho na limpeza de material e equipamentos úteis à rotina
do laboratório.
A Luís Cordeiro (Sr. Luís) e Adriano Cordeiro, pela colaboração no Biotério e no
laboratório, pela presteza e dedicação sempre.
A Mônica Rodrigues, nosso primeiro “bom dia” no laboratório, por toda sua alegria e
bom humor, por sempre nos receber carinhosamente e pelas palavras de conforto nas
horas difíceis no cotidiano do laboratório.
Ao Professor e Conselheiro Federal de Farmácia, João Samuel de Morais Meira, pelo
apoio financeiro concedido para participação em eventos científicos.
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) e à
Coordenação de Aperfeiçoamento de Pessoal de Ensino Superior (CAPES), pela
bolsa concedida e pelo suporte técnico-científico através do Portal Periódicos,
respectivamente.
À Universidade Federal da Paraíba, instituição responsável pela minha formação
profissional.
A todos que de maneira direta ou indireta contribuíram para a produção desta Tese
de Doutorado e pela minha formação profissional.
Obrigado!
Luiz Henrique César Vasconcelos

Resumo

A asma é uma doença inflamatória crônica das vias aéreas caracterizada pelo infiltrado de células imunes Th2, hiper-responsividade brônquica e declínio da função pulmonar, com muitos pacientes não respondendo apropriadamente à farmacoterapia. Nesse sentido, visando a busca por uma forma alternativa para a terapia antiasmática, avaliou-se os possíveis efeitos do óleo de coco virgem (OCV) em um modelo de inflamação pulmonar alérgica crônica (IPAC) induzida por ovalbumina (OVA). Os procedimentos experimentais foram aprovados pela Comissão de Ética no Uso de Animais da UFPB (certidão no 0410/13). Os animais foram divididos em grupos controle (GC), com IPAC (GASM), com IPAC tratados com dexametasona (2 mg/kg/dia, i.p.) (GASM/DEXA) ou suplementados com OCV 1 (GASM/OC1), 2 (GASM/OC2) ou 4 g/kg/dia (p.o.) (GASM/OC4). Foram analisados a morfologia do tecido pulmonar e aéreo, a reatividade contrátil e relaxante da traqueia, os níveis de malondialdeído (MDA) e a capacidade antioxidante total (CAT) no plasma e no pulmão, e a expressão proteica no pulmão por Western blot. Os cobaias com IPAC apresentaram infiltrado inflamatório peribrônquico, hiperplasia epitelial e espessamento da camada muscular lisa, que foram prevenidos pela dexametasona e pelo OCV nas três doses. O tempo para a resposta aguda à OVA também foi reduzido nesses animais, sendo prevenido apenas pelo OCV 4 g/kg. Nos animais com IPAC, a traqueia contraiu em resposta à OVA, sendo parcialmente prevenida pelo OCV 4 g/kg, indicando uma atenuação na liberação de mediadores contráteis. Os animais com IPAC apresentaram maior reatividade contrátil ao carbacol (CCh) e à histamina, mas não ao KCl, na presença de epitélio, sendo essa resposta prevenida pela dexametasona e pelo OCV 2 e 4 g/kg para o CCh, e 4 g/kg para a histamina. A potência relaxante da traqueia foi dificultada frente à isoprenalina, mas não ao nifedipino, na presença de epitélio, que foi prevenida pelo OCV 4 g/kg, indicando um comprometimento do acoplamento fármaco-, mas não do eletro-mecânico, de contração e relaxamento, dependente do epitélio. Evidenciou-se a participação do ânion superóxido e do peróxido de hidrogênio na hiper-contratilidade ao CCh, sendo o aumento, apenas do peróxido de hidrogênio, prevenido pelo OCV. Além disso, observou-se que a IPAC promoveu um aumento na liberação de produtos da 5-lipoxigenase (5-LO), enquanto no GASM/OC4 houve um provável equilíbrio entre os prostanoides relaxantes com os contráteis e os cisteinil-leucotrienos (Cys-LTs). Foi também evidenciado um possível aumento na atividade e/ou expressão da sintase de óxido nítrico induzida (iNOS), mas não da endotelial (eNOS), o qual foi prevenido pelo OCV 4 g/kg. Dados com o inibidor da Rho cinase (ROCK) sugerem um aumento na expressão ou na atividade da ROCK na IPAC, revertida pelo OCV. Já na investigação dos mecanismos envolvidos na reatividade relaxante, observou-se uma possível modulação negativa da via receptores β/BKCa pelo TGF-β na IPAC, e que o OCV previne apenas os efeitos do TGF-β sobre os receptores β, mas não a redução na atividade/expressão dos BKCa. Além disso, a IPAC parece aumentar a liberação de Cys-LTs, a qual é prevenida pelo OCV. Na análise do balanço estresse oxidativo/defesas antioxidantes, a CAT foi reduzida nos pulmões dos animais com IPAC, a qual foi prevenida pelo OCV 2 e 4 g/kg. Por fim, a expressão da PI3K foi aumentada pela IPAC, mas não a ERK1/2 e a SOD, e o tratamento com a dexametasona não preveniu o aumento da PI3K, mas elevou a ERK 1/2, e a suplementação com OCV também não preveniu o aumento da PI3K. Portanto, a IPAC gerou infiltrado inflamatório peribrônquico, hiperplasia epitelial, espessamento muscular liso aéreo e hiper-contratilidade muscular lisa via estresse oxidativo e suas interações com metabólitos do AA, as vias do NO, da RhoA/ROCK e do TGF-β, bem como um aumento na expressão da PI3K; e essas alterações foram prevenidas pelo óleo de coco virgem. Palavras-chave: 1. Cocus nucifera L.. 2. Asma. 3. Óxido nítrico. 4. Prostanoides. 5. Leucotrienos. 6. RhoA-ROCK. 7. TGF-β.
Suplementação alimentar com óleo de coco virgem previne as alterações na reatividade contrátil e relaxante em
traqueia de cobaia submetida à inflamação pulmonar alérgica crônica.
VASCONCELOS, L. H. C. Pós-graduação em Produtos Naturais e Sintéticos Bioativos,
Tese de Doutorado, CCS/UFPB (2017).

Abstract

Asthma is a chronic inflammatory disease of the airways characterized by Th2 immune cell infiltrates, bronchial hyperresponsiveness and declining lung function, with many patients failing to respond appropriately to pharmacotherapy. In this sense, aiming to search for an alternative form of antiasthmatic therapy, the possible effects of virgin coconut oil (VCO) on a chronic allergic lung inflammation (CALI) model induced by ovalbumin (OVA) were evaluated. The experimental procedures were approved by the Ethic Committee on Animal Use off UFPB (certificate 0410/13). The animals were divided into control (CG), CALI (ASTG), CALI treated with dexamethasone (2 mg/kg/ day, i.p) (AST/DEXAG) or supplemented with CVO 1 (AST/CO1G), 2 (AST/CO2G) or 4 g/kg/day (p.o) (AST/CO4G). Were analysed the morphology of lung and air tissue, the contractile and relaxing tracheal reactivity, the levels of malondialdehyde (MDA) and the total antioxidant capacity (TAC) in plasma and lung and protein expression in the lung by Western blot. Guinea pigs with CALI exhibited peribronchial inflammatory infiltrate, epithelial hyperplasia and smooth muscle thickening, which were prevented by dexamethasone and VCO. The time for acute response to OVA was also reduced in these animals, being prevented by VCO 4 g/kg. In animals with CALI, trachea contracted in response to OVA administration and was partially prevented by VCO 4 g/kg, indicating an attenuation in the release of contractile mediators. Animals with CALI showed a greater contractile response to carbachol (CCh) and histamine, but not to KCl, in the presence of epithelium, and this response was prevented by dexamethasone and VCO 2 and 4 g/kg for CCh, and 4 g/kg for histamine. The relaxing potency to isoprenaline was reduced, but not to nifedipine, in the presence of epithelium, which was prevented by VCO 4 g/kg, indicating that CALI interfered with the fármaco-, but not the electromechanical coupling of contraction and relaxation, in an epithelium-dependent manner. It was evidenced the participation of of superoxide anion and hydrogen peroxide in the hypercontractility, and the increase on peroxide hydrogen peroxide was prevented by VCO. Also, it was observed that IPAC promoted an increase in releasing 5-lipoxygenase (5-LO) products in CALI, while in AST/CO4G there was a balance between relaxing prostanoids with contractile and the cysteinyl leukotrienes (Cys-LTs). It was also observed a possible increase in activity and/or expression. of inducible NOS (iNOS), but not in endotelial NOS, that wwas prevented by VCO 4 g/kg. Data with the Rho kinase (ROCK) inhibitor suggest an increase in ROCK activity or expression in CALI, which was reverted by VCO. In the investigation of mechanism of relaxant reactivity, it was observed a possible negative modulation of β receptors/BKCa pathway by TGF-β, and that VCO prevents only the effects of TGF-β on β receptors, but not on the reduction of activity/expression of BKCa. Besides that, CALI appears to increase the Cys-LTs release, that is prevented by VCO. In the analysis of the oxidative stress and antioxidant defenses balance, TAC was reduced in the lungs of animals with CALI, which was reversed only by VCO (2 and 4 g/kg). Finally, the expression of PI3K was increased by CALI, but not ERK1/2 and SOD, and treatment with dexamethasone did not prevent the PI3K increase, but elevated ERK 1/2 levels, and VCO did not prevent the PI3K increase. Therefore, CALI generated peribronchial inflammatory infiltrate, epithelial hyperplasia, smooth muscle thickening and hypercontractility through oxidative stress and its interactions with AA metabolites, the NO, RhoA/ROCK and TGF-β pathways, and na increase in PI3K expression; and such changes were prevented by virgin coconut oil. Keywords: 1. Cocus nucifera L.. 2. Asthma. 3. Nitric oxide. 4. Prostanoids. 5. Leukotrienes. 6. RhoA-ROCK. 7. TGF-β.
Alimentary supplementation with virgin coconut oil prevents the alterations in contractile and relaxant reactivity in
trachea of guinea pigs submited to a chronic allergic lung inflammation.
VASCONCELOS, L. H. C. Pós-graduação em Produtos Naturais e Sintéticos Bioativos,
Tese de Doutorado, CCS/UFPB (2017).

LISTA DE FIGURAS
Figura 1 - Mapa da prevalência mundial da asma em crianças entre 13-14 anos .... 47
Figura 2 - Mapa da prevalência mundial da asma em adultos entre 18-45 anos ...... 47
Figura 3 - Mecanismo imunológico da asma alérgica ................................................. 57
Figura 4 - Morfologia das vias aéreas normais e asmáticas ....................................... 58
Figura 5 - Mecanismo do acoplamento fármaco-mecânico de contração muscular lisa
pela via da PLCβ1/2 ......................................................................................................... 76
Figura 6 - Mecanismos de transporte de Ca2+ nas células musculares lisas das vias
aéreas.............................................................................................................................. 78
Figura 7 - Mecanismo do acoplamento fármaco-mecânico de contração muscular lisa
pela via de sensibilização ao Ca2+................................................................................. 81
Figura 8 - Acoplamento eletromecânico de contração muscular lisa ......................... 83
Figura 9 - Mecanismo do acoplamento fármaco-mecânico de relaxamento do músculo
liso pelas vias da AC e da sGC ..................................................................................... 86
Figura 10 - Passos para o tratamento da asma, de acordo com sua apresentação
clínica .............................................................................................................................. 93
Figura 11 - Aparato utilizado para nebulização dos cobaias .................................... 102
Figura 12 - Protocolo de indução da inflamação pulmonar alérgica crônica em cobaias
....................................................................................................................................... 103
Figura 13 - Parte do sistema respiratório do cobaia dissecado, com vista da superfície
mediastinal pulmonar ................................................................................................... 106

Figura 14 - Anel de traqueia suspenso em cuba de banho para órgão isolado (A),
sistema de cubas de banho conectados a transdutores de força isométricos e
amplificador (B) e sistema de aquisição digital de dados (C) .................................... 107
Figura 15 - Fotomicrografias de pulmão de cobaias dos grupos GC (A), GASM (B),
GASM/DEXA (C), GASM/OC1 (D), GASM/OC2 (E) e GASM/OC4 (F) evidenciando o
infiltrado inflamatório .................................................................................................... 121
Figura 16 - Fotomicrografias de pulmão de cobaias dos grupos GC (A), GASM (B),
GASM/DEXA (C), GASM/OC1 (D), GASM/OC2 (E) e GASM/OC4 (F) evidenciando o
músculo liso dos brônquios intrapulmonares .............................................................. 122
Figura 17 - Fotomicrografias de brônquios extrapulmonares de cobaias dos grupos
GC (A), GASM (B), GASM/DEXA (C), GASM/OC1 (D), GASM/OC2 (E) e GASM/OC4
(F) evidenciando o epitélio e o músculo liso ............................................................... 123
Figura 18 - Registros originais representativos da resposta da traqueia de cobaia à
estimulação com 10 µg/mL de OVA em cobaias dos grupos GC (A), GASM (B),
GASM/DEXA (C), GASM/OC1 (D), GASM/OC2 (E) e GASM/OC4 (F) ..................... 126
Figura 19 - Registros originais representativos da reatividade contrátil ao CCh da
traqueia de cobaia dos grupos GC (A e B), GASM (C e D), na presença (A e C) e na
ausência (B e D) de epitélio funcional, GASM/DEXA (E), GASM/OC1 (F), GASM/OC2
(G) e GASM/OC4 (H), na presença de epitélio funcional. ......................................... 129
Figura 20 - Registros originais representativos da reatividade contrátil à histamina da
traqueia de cobaia dos grupos GC (A e B), GASM (C e D), na presença (A e C) e na
ausência (B e D) de epitélio funcional, GASM/DEXA (E), GASM/OC1 (F), GASM/OC2
(G) e GASM/OC4 (H), na presença de epitélio funcional. ......................................... 134
Figura 21 - Registros originais representativos da reatividade contrátil ao KCl da
traqueia de cobaia dos grupos GC (A e B) e GASM (C e D), na presença (A e C) e na
ausência (B e D) de epitélio funcional ......................................................................... 139

Figura 22 - Registros representativos do efeito da isoprenalina sobre o tônus basal da
traqueia de cobaia dos grupos GC (A e B) e GASM (C e D), na presença (A e C) e na
ausência (B e D) de epitélio funcional ......................................................................... 142
Figura 23 - Registros originais representativos da reatividade relaxante à isoprenalina
da traqueia de cobaia dos grupos GC (A e B), GASM (C e D), na presença (A e C) e
na ausência (B e D) de epitélio funcional, GASM/DEXA (E), GASM/OC1 (F),
GASM/OC2 (G) e GASM/OC4 (H), na presença de epitélio funcional ...................... 145
Figura 24 - Registros originais representativos da reatividade relaxante ao nifedipino
da traqueia de cobaia dos grupos GC (A e B) e GASM (C e D), na presença (A e C) e
na ausência (B e D) de epitélio funcional. .................................................................. 150
Figura 25 - Registros originais representativos da reatividade contrátil ao CCh em
traqueia de cobaia com epitélio funcional dos grupos GC (A, B, C), GASM (D, E, F) e
GASM/OC4 (G, H, I), na ausência e na presença da apocinina (A, D, G), tempol (B, E,
H) ou catalase (C, F, I) ................................................................................................. 154
Figura 26 - Registros originais representativos da reatividade contrátil ao CCh em
traqueia de cobaia com epitélio dos grupos GC (A, B), GASM (C, D) e GASM/OC4 (E,
F), na ausência e na presença da indometacina (A, C, E) ou da zileutona (B, D, F).
....................................................................................................................................... 160
Figura 27 - Registros originais representativos da reatividade contrátil ao CCh em
traqueia de cobaia com epitélio dos grupos GC (A), GASM (B) e GASM/OC4 (C), na
ausência e na presença do L-NAME ........................................................................... 165
Figura 28 - Registros originais representativos da reatividade contrátil ao CCh em
traqueia de cobaia com epitélio dos grupos GC (A, B, C), GASM (D, E, F) e GASM/OC4
(G, H, I) na ausência e na presença do Y-27632 ....................................................... 169
Figura 29 - Registros originais representativos da reatividade relaxante à isoprenalina
em traqueia de cobaia com epitélio dos grupos GC (A, B), GASM (C, D) e GASM/OC4
(E, F) na ausência e na presença do SB43154 (A, C, E) ou da IbTX (B, D, F) ....... 175

Figura 30 - Registros originais representativos da reatividade relaxante ao ácido
araquidônico em traqueia de cobaia com epitélio dos grupos GC (A, B, C), GASM (D,
E, F) e GASM/OC4 (G, H, I) na ausência (A, D, G) e na presença da indometacina (B,
E, H) ou da zileutona (C, F, I) ...................................................................................... 180
Figura 31 - Bandas proteicas de corrida em gel de eletroforese representativas da
expressão de PI3K, ERK 1/2, SOD e β-actina no pulmão de cobaias dos grupos GC,
GASM, GASM/DEXA e GASM/OC4 ............................................................................ 187

LISTA DE GRÁFICOS
Gráfico 1 - Tempo médio de contato dos cobaias com a OVA da primeira à quarta, na
quinta, na sexta e na sétima nebulizações ................................................................. 119
Gráfico 2 - Medida do infiltrado de células inflamatórias no pulmão (A), da hiperplasia
epitelial (B) e da espessura da camada de músculo liso das vias aéreas (C) de cobaias
dos grupos GC, GASM, GASM/DEXA, GASM/OC1, GASM/OC2 e GASM/OC4..... 124
Gráfico 3 - Efeito da estimulação da traqueia de cobaias com 10 µg/mL de OVA . 127
Gráfico 4 - Curvas concentrações-resposta cumulativas ao CCh em traqueia de
cobaia dos grupos GC e GASM, na presença e na ausência de epitélio funcional . 131
Gráfico 5 - Curvas concentrações-resposta cumulativas ao CCh em traqueia de
cobaia dos grupos GC, GASM, GASM/DEXA, GASM/OC1, GASM/OC2 e GASM/OC4,
na presença de epitélio funcional ................................................................................ 131
Gráfico 6 - Curvas concentrações-resposta cumulativas à histamina em traqueia de
cobaia dos grupos GC e GASM, na presença e na ausência de epitélio funcional . 136
Gráfico 7 - Curvas concentrações-resposta cumulativas à histamina em traqueia de
cobaia dos grupos GC, GASM GASM/DEXA, GASM/OC1, GASM/OC2 e GASM/OC4,
na presença de epitélio funcional ................................................................................ 136
Gráfico 8 - Curvas concentrações-resposta cumulativas ao KCl em traqueia de cobaia
dos grupos GC e GASM, na presença e na ausência de epitélio funcional ............. 140
Gráfico 9 - Curvas concentrações-resposta cumulativas à isoprenalina em traqueia de
cobaia dos grupos GC e GASM, na presença e na ausência de epitélio funcional . 143
Gráfico 10 - Curvas concentrações-resposta cumulativas à isoprenalina em traqueia
de cobaia pré-contraída com 10-6 M de CCh dos grupos GC e GASM, na presença e
na ausência de epitélio funcional................................................................................. 147

Gráfico 11 - Curvas concentrações-resposta cumulativas à isoprenalina em traqueia
de cobaia pré-contraída com 10-6 M de CCh, dos grupos GC, GASM, GASM/DEXA,
GASM/OC1, GASM/OC2 e GASM/OC4 na presença de epitélio funcional ............. 147
Gráfico 12 - Curvas concentrações-resposta cumulativas ao nifedipino em traqueia de
cobaia pré-contraída com 10-6 M de CCh, dos grupos GC e GASM, na presença e na
ausência de epitélio funcional ...................................................................................... 151
Gráfico 13 - Curvas concentrações-resposta cumulativas ao CCh em traqueia de
cobaia com epitélio funcional dos grupos GC (A), GASM (B) e GASM/OC4 (C), na
ausência e na presença da apocinina, do tempol ou da catalase ............................. 157
Gráfico 14 - Curvas concentrações-resposta cumulativas ao CCh em traqueia de
cobaia com epitélio dos grupos GC (A), GASM (B) e GASM/OC4 (C), na ausência e
na presença da indometacina ou da zileutona ........................................................... 162
Gráfico 15 - Curvas concentrações-resposta cumulativas ao CCh em traqueia de
cobaia com epitélio dos grupos GC (A), GASM (B) e GASM/OC4 (C), na ausência ()
e na presença () do L-NAME .................................................................................... 166
Gráfico 16 - Curvas concentrações-resposta cumulativas ao CCh em traqueia de
cobaia com epitélio dos grupos GC (A), GASM (B) e GASM/OC4 (C), na ausência e
na presença do Y-27632, e plot de Schild (D) para a curva de contração do CCh na
presença do Y-27632 ................................................................................................... 172
Gráfico 17 - Curvas concentrações-resposta cumulativas à isoprenalina em traqueia
de cobaia com epitélio dos grupos GC (A), GASM (B) e GASM/OC4 (C), na ausência
e na presença do SB431542 ou da IbTX .................................................................... 177
Gráfico 18 - Curvas concentrações-resposta cumulativas ao ácido araquidônico em
traqueia de cobaia com epitélio dos grupos GC, GASM e GASM/OC4, na ausência (A)
e na presença (B, C, D) da indometacina ou da zileutona ........................................ 183

Gráfico 19 - Valores dos níveis de MDA (μmol/L tecido) (A e B) e de inibição da
oxidação (%) (C e D) no plasma (A e C) e no homogenato pulmonar (B e D) de cobaias
dos grupos GC, GASM, GASM/DEXA, GASM/OC1, GASM/OC2 e GASM/OC4..... 186
Gráfico 20 - Expressão (u.a) das proteínas PI3K (A), ERK 1/2 (B), SOD (C) no pulmão
de cobaias dos grupos GC, GASM, GASM/DEXA e GASM/OC4. ............................ 188

LISTA DE TABELAS
Tabela 1 - Tempo médio de contato (s) dos cobaias com a ovalbumina da primeira à
quarta (1-4), na quinta (5), na sexta (6) e na sétima (7) nebulizações ..................... 119
Tabela 2 - Valores de Emax e pD2 do CCh em traqueia de cobaia dos grupos GC e
GASM, na presença e na ausência de epitélio funcional, e de GASM/DEXA,
GASM/OC1, GASM/OC2 e GASM/OC4, na presença de epitélio funcional ............ 132
Tabela 3 - Valores de Emax e pD2 da histamina em traqueia de cobaia dos grupos GC
e GASM, na presença e na ausência de epitélio funcional, e de GASM/DEXA,
GASM/OC1, GASM/OC2 e GASM/OC4, na presença de epitélio funcional ............ 137
Tabela 4 - Valores de Emax e pD2 do KCl em traqueia de cobaia dos grupos GC e
GASM, na presença e na ausência de epitélio funcional ........................................... 140
Tabela 5 - Valores de Emax e pD2 da isoprenalina em traqueia de cobaia dos grupos
GC e GASM, na presença e na ausência de epitélio ................................................. 143
Tabela 6 - Valores de Emax e pD2 da isoprenalina em traqueia de cobaia dos grupos
GC e GASM, na presença e na ausência de epitélio funcional, e de GASM/DEXA,
GASM/OC1, GASM/OC2 e GASM/OC4, na presença de epitélio funcional ............ 148
Tabela 7 - Valores de Emax e pD2 do nifedipino em traqueia de cobaia pré-contraída
com 10-6 M de CCh, dos grupos GC e GASM, na presença e na ausência de epitélio
funcional ........................................................................................................................ 151
Tabela 8 - Valores de Emax e pD2 do CCh em traqueia de cobaia com epitélio dos
grupos GC, GASM e GASM/OC4, na ausência e na presença de apocinina, tempol ou
catalase ......................................................................................................................... 158
Tabela 9 - Valores de Emax e pD2 do CCh em traqueia de cobaia com epitélio dos
grupos GC, GASM e GASM/OC4, na presença de epitélio funcional, na ausência e na
presença da indometacina ou da zileutona ................................................................ 163

Tabela 10 - Valores de Emax e pD2 do CCh em traqueia de cobaia com epitélio dos
grupos GC, GASM e GASM/OC4, na ausência e na presença do L-NAME ............ 167
Tabela 11 - Valores de Emax e pD2 do CCh em traqueia de cobaia dos grupos GC,
GASM e GASM/OC4, na presença de epitélio funcional, na ausência e na presença
do Y-27632 .................................................................................................................... 173
Tabela 12 - Valores de Emax e pD2 da isoprenalina em traqueia de cobaia dos grupos
GC, GASM e GASM/OC4, na presença de epitélio funcional, na ausência e na
presença do SB431542 ou da IbTX ............................................................................ 178
Tabela 13 - Valores de Emax e pD2 do ácido araquidônico em traqueia de cobaia dos
grupos GC, GASM e GASM/OC4, na presença de epitélio funcional, na ausência e na
presença da indometacina ou da zileutona ................................................................ 184

LISTA DE QUADROS
Quadro 1 - Expressão de receptores muscarínicos nas células das vias aéreas e seus
principais efeitos ............................................................................................................. 62
Quadro 2 - Sumário dos principais mediadores lipídicos na asma ............................. 67
Quadro 3 - Principais agonistas, receptores e acoplamentos envolvidos na contração
do músculo liso das vias aéreas .................................................................................... 73

LISTA DE ABREVIATURAS
[Ca2+]c concentração de cálcio citosólica
[cAMP]c concentração de cAMP citosólica
4Ca2+-CaM complexo 4 íons cálcio-calmodulina
5-LO 5-lipoxigenase
AA ácido araquidônico
AC ciclase de adenilil
ACh acetilcolina
AMP monofosfato de adenosina
ANOVA análise de variância
Anvisa Agência Nacional de Vigilância Sanitária
APC célula apresentadora de antígeno
ATP trifosfato de adenosina
BKCa canais de potássio sensíveis ao Ca2+ de grande condutância
CaM calmodulina
cAMP monosfofato cíclico de adenosina
CAT capacidade antioxidante total
CaV canais de cálcio dependentes de voltagem
CCh carbacol
CEUA Comissão de Ética no Uso de Animais
cGMP monofosfato cíclico de guanosina
CICR liberação de Ca2+ induzida por Ca2+
COX cicloxigenase
CPI-17 preptídio de 17 kDa inibidor de fosfatase ativada por PKC
Cys-LT cisteinil-leucotrieno
DAG diacilglicerol
DAMP padrão molecular associado ao dano
DATASUS Departamento de Informática do Sistema Único de Saúde
DC célula dendrítica
e.p.m. erro padrão da média
ECP proteína catiônica eosinofílica
EGF fator de crescimento epidérmico
Emax efeito máximo

EMTU unidade trófica epitelial mesenquimal
EPO peroxidase eosinofílica
ERK cinase regulada por sinal extracelular
FcεRI receptor Fcε tipo I
FGF fator de crescimento de fibroblastos
GATA-3 fator de transcrição de ação trans específico de células T tipo 3
GDI inibidor de dissociação do GDP
GDP difosfato de guanosina
GEF fator de troca de nucleotídio de guanina
Gi/o proteína Gi ou proteína Go
GINA Global Initiative of Asthma
GPCR receptor acoplado à proteína G
Gq/11 proteína Gq ou proteína G11
Gs proteína Gs
GTP trifosfato de guanosina
HDL lipoproteína de alta densidade
ICS corticosteroide inalatório
IgE imunoglobulina do tipo E
IL interleucina
ILC células linfoides inatas
IP3 1,4,5-trisfosfato de inositol
IP3R receptor de IP3
IPeFarM Instituto de Pesquisa em Fármacos e Medicamentos
ISAAC Estudo Internacional de Asma e Alergias na Infância
KATP canais de potássio sensíveis ao ATP
KV canais de potássio dependentes de voltagem
LABA agonista β de ação longa
LDL lipoproteína de baixa densidade
LT leucotrieno
MAPK proteína cinase ativada por mitógeno
MBP proteína básica principal
MDA malondialdeído
MHC complexo principal de histocompatibilidade
MLC cadeia leve da miosina

MLCK cinase da cadeia leve da miosina
MLCP fosfatase da cadeia leve da miosina
MMP metaloproteinase de matriz extracelular
NADPH nicotinamida adenina dinucleotídeo fosfato
NCX trocador Na+/Ca2+
NO óxido nítrico
NOS sintase de óxido nítrico
OCS corticosteroide oral
OCV Óleo de coco virgem
OMS Organização Mundial da Saúde
OVA ovalbumina
PAF fator de ativação plaquetária
PAMP padrão molecular associado ao patógeno
pD2 logaritmo negativo, na base 10, da concentração molar de uma
substância que produz 50% de seu efeito máximo
PDE fosfodiesterase
PGD2 prostaglandina D2
PGE2 prostaglandina E2
PGF2α prostaglandina F2α
PGI2 prostaglandina I2
PI3K fosfatidilinositol-3-cinase
PIP2 4,5-bisfosfato de fosfatidilinositol
PKA proteína cinase A
PKC proteína cinase C
PKG proteína cinase G
PLA2 fosfolipase A2
PLC-β1 fosfolipase Cβ1
PLC-β2 fosfolipase Cβ2
PMCA Ca2+-ATPase da membrana plasmática
PRR receptor de reconhecimento de padrão
RBD refino, branqueamento e desodorização
RDC Resolução de diretoria colegiada
ROC canal operado por receptor
ROCK proteína cinase associada à Rho

ROS espécie reativa de oxigênio
RS retículo sarcoplasmático
RTK receptor tirosina cinase
RyR receptor de rianodina
SABA agonista β de ação curta
SCF fator de células tronco
SERCA Ca2+-ATPase do retículo endo-sarcoplasmático
SOC canal de Ca2+ operado por estoque
SOD superóxido dismutase
SUS Sistema Único de Saúde
TCA ácido tricloroacético
TCR receptor de células T
TGF-β fator de crescimento transformador beta
Th17 linfócito T auxiliar tipo 17
Th2 linfócito T auxiliar tipo 2
Th9 linfócito T auxiliar tipo 9
TIMP inibidor tecidual de metaloproteinase
TLR receptor do tipo toll
TNF fator de necrose tumoral
Treg linfócito T regulatório
TSLP linfopoietina estromal tímica
TxA2 tromboxano A2
UFPB Universidade Federal da Paraíba
VLDL lipoproteína de muito baixa densidade
Vm potencial de membrana
VRGF fator de crescimento do endotélio vascular
OBS: as abreviaturas e os símbolos utilizados neste trabalho e que não constam nesta
relação, encontram-se descritas no texto ou são convenções adotadas
universalmente.

SUMÁRIO
1 INTRODUÇÃO ...................................................................................................... 35
2 FUNDAMENTAÇÃO TEÓRICA ............................................................................ 39
2.1 Os produtos naturais no contexto dos alimentos funcionais ............................... 40
2.2 Óleo de coco: potencial alimento funcional com propriedade de saúde ............. 43
2.3 Asma .................................................................................................................. 46
2.3.1 Epidemiologia da asma ................................................................................... 46
2.3.2 Características gerais da asma ....................................................................... 49
2.3.3 Fisiopatologia da asma ................................................................................... 51
2.3.4 Remodelamento das vias aéreas .................................................................... 57
2.3.5 Mediadores neurais e humorais envolvidos na asma ...................................... 61
2.3.5.1 Vias neurais colinérgicas .............................................................................. 61
2.3.5.2 Eicosanoides ................................................................................................ 62
2.3.5.3 Óxido nítrico ................................................................................................. 67
2.3.5.4 Estresse oxidativo ........................................................................................ 69
2.3.6 O papel do músculo liso no remodelamento e na hiper-responsividade das vias
aéreas na asma ....................................................................................................... 72
2.3.7 Modelos experimentais de asma ..................................................................... 88
2.3.8 Tratamento atual da asma e novas abordagens terapêuticas ......................... 90
3 OBJETIVOS ......................................................................................................... 95
3.1 Objetivo geral .................................................................................................... 96
3.2 Objetivos específicos ......................................................................................... 96
4 MATERIAL E MÉTODOS ..................................................................................... 97
4.1 MATERIAL ........................................................................................................ 98
4.1.1 Animais ........................................................................................................... 98
4.1.2 Substâncias e reagentes ................................................................................. 98
4.1.4 Soluções ......................................................................................................... 99
4.1.5 Aparelhos ...................................................................................................... 100
4.2 MÉTODOS ....................................................................................................... 101
4.2.1 Grupos experimentais ................................................................................... 101
4.2.2 Aquisição do óleo de coco virgem ................................................................. 102
4.2.3 Indução da inflamação pulmonar alérgica crônica em cobaias ...................... 102
4.2.4 Tratamento e suplementação dos animais .................................................... 103

4.2.5 Avaliação dos efeitos da inflamação pulmonar alérgica crônica e do tratamento
com a dexametasona ou da suplementação com o óleo de coco virgem sobre o tempo
de nebulização de cobaia ...................................................................................... 104
4.2.6 Avaliação dos efeitos da inflamação pulmonar alérgica crônica e do tratamento
com a dexametasona ou da suplementação com o óleo de coco virgem sobre a
morfologia pulmonar e brônquica de cobaia ........................................................... 104
4.2.7 Avaliação dos efeitos da inflamação pulmonar alérgica crônica e do tratamento
com a dexametasona ou da suplementação com o óleo de coco virgem sobre a
reatividade contrátil e relaxante da traqueia de cobaia........................................... 106
4.2.7.1 Obtenção dos anéis de traqueia ................................................................. 106
4.2.7.2 Avaliação da reatividade contrátil à ovalbumina em traqueia de cobaia, na
presença de epitélio funcional ................................................................................ 107
4.2.7.3 Avaliação da reatividade contrátil ao CCh, à histamina ou ao KCl em traqueia
de cobaia, na presença e na ausência de epitélio funcional ................................... 107
4.2.7.4 Avaliação do efeito da isoprenalina sobre o tônus basal em traqueia de cobaia,
na presença e na ausência de epitélio funcional .................................................... 108
4.2.7.5 Avaliação da reatividade relaxante à isoprenalina ou ao nifedipino em traqueia
de cobaia pré-contraída com 10-6 M de CCh, na presença e na ausência de epitélio
funcional ................................................................................................................ 108
4.2.8 Investigação do mecanismo de ação envolvido nas alterações induzidas pela
inflamação pulmonar alérgica crônica e pelo óleo de coco virgem sobre a reatividade
contrátil da traqueia de cobaia ............................................................................... 109
4.2.8.1 Avaliação do envolvimento das espécies reativas de oxigênio (ROS) ........ 109
4.2.8.1.1 Investigação da participação das enzimas nicotinamida adenina
dinucleotídeo fosfato (NADPH) oxidase, superóxido dismutase (SOD) e catalase . 109
4.2.8.1.1.1 Obtenção de curvas cumulativas concentrações-resposta ao CCh, na
ausência e na presença da apocinina, do tempol ou da catalase, em traqueia de cobaia
dos grupos GC, GASM e GASM/OC4 .................................................................... 109
4.2.8.2 Avaliação da participação dos produtos derivados do metabolismo do ácido
araquidônico pelas enzimas cicloxigenase (COX) e 5-lipoxigenase (5-LO) ............ 110
4.2.8.2.1 Obtenção de curvas cumulativas concentrações-resposta ao CCh, na
ausência e na presença da indometacina ou da zileutona, em cobaias dos grupos GC,
GASM e GASM/OC4 .............................................................................................. 110
4.2.8.3 Avaliação da participação da via do óxido nítrico (NO) ............................... 110

4.2.8.3.1 Obtenção de curvas cumulativas concentrações-resposta ao CCh, na
ausência e na presença do L-NAME, em cobaias dos grupos GC, GASM e GASM/OC4
............................................................................................................................... 110
4.2.8.4 Avaliação da participação da via da RhoA/Rho cinase (ROCK) ................. 110
4.2.8.4.1 Obtenção de curvas cumulativas concentrações-resposta ao CCh, na
ausência e na presença do Y-27632, em cobaias dos grupos GC, GASM e GASM/OC4
............................................................................................................................... 110
4.2.9 Investigação do mecanismo de ação envolvido nas alterações induzidas pela
inflamação pulmonar alérgica crônica e pelo óleo de coco virgem sobre a reatividade
relaxante da traqueia de cobaia ............................................................................. 111
4.2.9.1 Avaliação da participação do fator de crescimento transformador β (TGF-β) e
dos canais de potássio sensíveis ao Ca2+ de grande condutância (BKCa) .............. 111
4.2.9.1.1 Obtenção de curvas cumulativas concentrações-resposta à isoprenalina, na
ausência e na presença do SB431542 ou da iberiotoxina (IbTX), em traqueia de cobaia
pré-contraída com 10-6 M de CCh dos grupos GC, GASM e GASM/OC4 .............. 111
4.2.9.2 Avaliação da participação dos produtos derivados do metabolismo do ácido
araquidônico pelas enzimas cicloxigenase (COX) e 5-lipoxigenase (5-LO) ............ 112
4.2.9.2.1 Obtenção de curvas cumulativas concentrações-resposta ao ácido
araquidônico (AA), na ausência e na presença da indometacina ou da zileutona, em
traqueia de cobaia pré-contraída com 10-6 M de CCh dos grupos GC, GASM e
GASM/OC4 ............................................................................................................ 112
4.2.10 Avaliação dos efeitos da inflamação pulmonar alérgica crônica e do tratamento
com a dexametasona ou com o óleo de coco virgem sobre o balanço entre o estresse
oxidativo e as defesas antioxidantes de cobaia...................................................... 112
4.2.10.1 Obtenção do plasma ................................................................................ 112
4.2.10.2 Obtenção do homogenato do tecido pulmonar ......................................... 113
4.2.10.3 Análise dos níveis de malondialdeído (MDA) e da capacidade antioxidante
total (CAT) no plasma e no tecido pulmonar .......................................................... 113
4.2.11 Avaliação dos efeitos da inflamação pulmonar alérgica crônica e do tratamento
com a dexametasona ou da suplementação com o óleo de coco virgem sobre a
expressão de proteínas das vias de sinalização da proliferação muscular lisa e das
defesas antioxidantes ............................................................................................ 114

4.2.11.1 Análise dos níveis da fosfatidilinositol 3 cinase (PI3K), das proteínas cinases
reguladas por sinal extracelular 1/2 (ERK1/2) e da superóxido dismutase (SOD) no
tecido pulmonar ..................................................................................................... 114
4.2.11.2 Extração do pulmonar e corrida de eletroforese em gel de poliacrilamida 114
4.2.11.3 Western blot ............................................................................................. 115
4.2.12 Análise Estatística ....................................................................................... 115
5 RESULTADOS ................................................................................................... 116
5.1 Avaliação dos efeitos da inflamação pulmonar alérgica crônica e do tratamento
com a dexametasona ou da suplementação com o óleo de coco virgem sobre o tempo
de nebulização de cobaia ...................................................................................... 117
5.2 Avaliação dos efeitos da inflamação pulmonar alérgica crônica e do tratamento
com a dexametasona ou da suplementação com o óleo de coco virgem sobre a
morfologia pulmonar e brônquica de cobaia ........................................................... 120
5.3 Avaliação dos efeitos da inflamação pulmonar alérgica crônica e do tratamento
com a dexametasona ou da suplementação com o óleo de coco virgem sobre a
reatividade contrátil e relaxante da traqueia de cobaia........................................... 125
5.3.1 Avaliação da reatividade contrátil à ovalbumina em traqueia de cobaia, na
presença de epitélio funcional ................................................................................ 125
5.3.2 Avaliação da reatividade contrátil ao CCh em traqueia de cobaia, na presença e
na ausência de epitélio funcional ........................................................................... 128
5.3.3 Avaliação da reatividade contrátil à histamina em traqueia de cobaia, na
presença e na ausência de epitélio funcional ......................................................... 133
5.3.4 Avaliação da reatividade contrátil ao KCl em traqueia de cobaia, na presença e
na ausência de epitélio funcional ........................................................................... 138
5.3.5 Avaliação do efeito da isoprenalina sobre o tônus basal em traqueia de cobaia,
na presença e na ausência de epitélio funcional .................................................... 141
5.3.6 Avaliação da reatividade relaxante à isoprenalina em traqueia de cobaia
pré-contraída com 10-6 M de CCh, na presença e na ausência de epitélio funcional
............................................................................................................................... 144
5.3.7 Avaliação da reatividade relaxante ao nifedipino em traqueia de cobaia
pré-contraída com 10-6 M de CCh, na presença e na ausência de epitélio funcional
............................................................................................................................... 149

5.4 Investigação do mecanismo de ação envolvido nas alterações induzidas pela
inflamação pulmonar alérgica crônica e pelo óleo de coco virgem sobre a reatividade
contrátil da traqueia de cobaia ............................................................................... 152
5.4.1 Avaliação do envolvimento das espécies reativas de oxigênio (ROS) ........... 152
5.4.1.1 Investigação da participação das enzimas nicotinamida adenina dinucleotídeo
fosfato (NADPH) oxidase, superóxido dismutase (SOD) e catalase ....................... 152
5.4.1.1.1 Obtenção de curvas cumulativas concentrações-resposta ao CCh, na
ausência e na presença da apocinina, do tempol ou da catalase, em traqueia de cobaia
dos grupos GC, GASM e GASM/OC4 .................................................................... 152
5.4.2 Avaliação da participação dos produtos derivados do metabolismo do ácido
araquidônico pelas enzimas cicloxigenase (COX) e 5-lipoxigenase (5-LO) ............ 159
5.4.2.1 Obtenção de curvas cumulativas concentrações-resposta ao CCh, na ausência
e na presença da indometacina ou da zileutona, em cobaias dos grupos GC, GASM e
GASM/OC4 ............................................................................................................ 159
5.4.3 Avaliação da participação da via do óxido nítrico (NO).................................. 164
5.3.3.1 Obtenção de curvas cumulativas concentrações-resposta ao CCh, na ausência
e na presença do L-NAME, em cobaias dos grupos GC, GASM e GASM/OC4 ..... 164
5.4.4 Avaliação da participação da via da RhoA/Rho cinase (ROCK) .................... 168
5.4.4.1 Obtenção de curvas cumulativas concentrações-resposta ao CCh, na ausência
e na presença do Y-27632, em cobaias dos grupos GC, GASM e GASM/OC4 ..... 168
5.5 Investigação do mecanismo de ação envolvido nas alterações induzidas pela
inflamação pulmonar alérgica crônica e pelo óleo de coco virgem sobre a reatividade
relaxante da traqueia de cobaia ............................................................................. 174
5.5.1 Avaliação da participação do fator de crescimento transformador β (TGF-β) e
dos canais de potássio sensíveis ao Ca2+ de grande condutância (BKCa) .............. 174
5.5.1.1 Obtenção de curvas cumulativas concentrações-resposta à isoprenalina, na
ausência e na presença do SB431542 ou da iberiotoxina (IbTX), em traqueia de cobaia
pré-contraída com 10-6 M de CCh dos grupos GC, GASM e GASM/OC4 .............. 174
5.5.2 Avaliação da participação dos produtos derivados do metabolismo do ácido
araquidônico pelas enzimas cicloxigenase (COX) e 5-lipoxigenase (5-LO) ............ 179
5.5.2.1 Obtenção de curvas cumulativas concentrações-resposta ao ácido
araquidônico (AA), na ausência e na presença da indometacina ou da zileutona, em
traqueia de cobaia pré-contraída com 10-6 M de CCh dos grupos GC, GASM e
GASM/OC4 ............................................................................................................ 179

5.6 Avaliação dos efeitos da inflamação pulmonar alérgica crônica e do tratamento
com a dexametasona ou com o óleo de coco virgem sobre o balanço entre o estresse
oxidativo e as defesas antioxidantes de cobaia...................................................... 185
5.6.1 Análise dos níveis de malondialdeído (MDA) e da capacidade antioxidante total
(CAT) no plasma e no tecido pulmonar .................................................................. 185
5.7 Avaliação dos efeitos da inflamação pulmonar alérgica crônica e do tratamento
com a dexametasona ou da suplementação com o óleo de coco virgem sobre a
expressão de proteínas das vias de sinalização da proliferação muscular lisa e das
defesas antioxidantes ............................................................................................ 187
5.7.1 Análise dos níveis da fosfatidilinositol-3-cinase (PI3K), das proteínas cinases
reguladas por sinal extracelular 1/2 (ERK1/2) e da superóxido dismutase (SOD) no
tecido pulmonar ..................................................................................................... 187
6 DISCUSSÃO ....................................................................................................... 189
7 CONCLUSÕES ................................................................................................... 220
REFERÊNCIAS ..................................................................................................... 222
APÊNDICES .......................................................................................................... 273
ANEXOS ................................................................................................................ 277

1 Introdução

36 VASCONCELOS, L. H. C. Introdução
Durante muito tempo o homem se beneficiou dos produtos naturais na busca
por alívio e/ou cura de doenças (VIEGAS-JÚNIOR; BOLZANI; BARREIRO, 2006). A
natureza tem servido como fonte de medicamentos por milênios, sendo muitas drogas
desenvolvidas a partir de fontes vegetais (CRAGG; NEWMAN, 2013). Em virtude dos
seus efeitos terapêuticos, esses produtos eram comumente utilizados pela população
para fins medicinais (VIDO, 2009).
Diante da riqueza de produtos potencialmente terapêuticos presentes na
natureza e a fatores como a busca da população por um modelo de vida mais saudável
e natural, as pesquisas envolvendo plantas medicinais aumentaram mundialmente
(NEGRI, 2007).
No contexto dos produtos naturais, se inserem os alimentos funcionais, aqueles
que apresentam propriedades funcionais no organismo, com relação ao papel
metabólico e fisiológico, os quais podem ou não apresentar propriedade de saúde,
isto é, uma relação benéfica entre o alimento e uma determinada condição de saúde
(BRASIL, 1999c). Diversos alimentos são enquadrados dentro dessa classificação
pela legislação brasileira, destacando-se o óleo de coco, o qual é reconhecido pela
Agência Nacional de Vigilância Sanitária (Anvisa) como alimento próprio para
suplementação, com base em dados de segurança e eficácia (BRASIL, 2017a).
O óleo de coco (espécie: Cocos nucifera L., família: Arecaceae) é um produto
obtido da semente madura do coco ou da copra (polpa seca do coco), que é utilizada
principalmente para a obtenção do óleo, sendo constituído entre 65-75% dele (NMCE,
2007), largamente utilizado na alimentação e na indústria (DIA et al., 2005;
DEBMANDAL; MANDAL, 2011). O consumo de óleo de coco vem crescendo
mundialmente re razão de suas diversas propriedades já descritas, destacando-se a
melhora do perfil lipídico e a redução da gordura corporal (YOUSEFI; NATEGHI;
REZAEE, 2013).
Esse óleo é rico em ácidos graxos saturados de cadeia média, eficazes contra
o desenvolvimento de doenças cardiovasculares e inflamatórias
(CHANDRASHEKAR; LOKESH; KRISHNA, 2010), bem como compostos
antioxidantes (carotenoides e tocoferóis) e vitaminas (YOUSEFI; NATEGHI; REZAEE,
2013). Dentre as propriedades farmacológicas descritas para esse óleo, estão a
preventiva de doença coronariana (INAYAT; BAIG; BAQAI, 2013), antioxidante
(ARUNIMA; RAJAMOHAN, 2013), anti-inflamatória (ZAKARIA et al., 2011),
anti-hipertensiva (ALVES et al., 2015) e cardioprotetora (KAMISAH et al., 2015).

37 VASCONCELOS, L. H. C. Introdução
Muitos desses efeitos têm sido atribuídos, principalmente, ao constituinte majoritário
do óleo, o ácido láurico (KEYS; ANDERSON; GRANDE, 1965; FAO, 2014).
Assim, em função de suas ações sobre o processo inflamatório, o óleo de coco
é um potencial candidato na terapia adjuvante de diversas doenças inflamatórias
crônicas, como a asma alérgica.
A asma é uma doença inflamatória crônica das vias aéreas em que muitas
células inatas e adaptativas do sistema imune agem em conjunto com células
epiteliais para promover hiper reatividade brônquica, caracterizada como a tendência
das células musculares lisas em reagir de forma exacerbada a estímulos
inespecíficos, como ar frio e exercício físico; além de excesso de produção de muco,
remodelamento da parede das vias aéreas e estreitamento do lúmen dessas vias
condutoras. Em pacientes susceptíveis, ela leva a quadros de dispneia e períodos
repetidos de falta de ar, sibilos durante a respiração e opressão torácica
(LAMBRECHT; HAMMAD, 2015).
A asma é uma das condições crônicas mais comuns em crianças e adultos,
representando 7,7% da população mundial. Segundo dados da Organiação Mundial
de Saúde (OMS) e do Global Initiative of Asthma (GINA), a asma acomete entre 235
e 300 milhões de pessoas no mundo (GINA, 2017; OMS, 2017). Estima-se que, em
2025, existirão 100 milhões de novos casos de asma no mundo (GINA, 2010). Ainda
dentro dessa problemática, a asma acarreta curstos ao governo devido ao grande
número de internações, assim como o alto índice de mortalidade (GINA, 2017).
Atualmente, o tratamento da asma é baseado principalmente no uso de
broncodilatadores, como agonistas adrenérgicos β, antagonistas colinérgicos,
antileucotrienos e inibidores inespecíficos de fosfodiesterases ou ainda baseado na
supressão da inflamação, sendo os glicocorticoides os principais fármacos
disponíveis. Essas estratégias auxiliam no controle dos sintomas da asma, no entanto
apresentam diversos efeitos colaterais, como glaucoma, desequilíbrio metabólico e
imunosupressão (WALSH, 2005). Em muitos pacientes, a doença pode ser controlada
com a combinação de corticosteroides com agonistas adrenérgicos β2; todavia, em
5-10%, a doença é refratária ao uso de corticosteroides e, frequentemente, leva a
hospitalizações devido a infecções virais do sistema respiratório (LAMBRECHT;
HAMMAD, 2015).
Apesar da grande diversidade de fármacos para tal tratamento, esse ainda é
feito de forma paliativa e/ou preventiva; desse modo, fazem-se necessárias novas

38 VASCONCELOS, L. H. C. Introdução
abordagens terapêuticas que limitem ou, ao menos, tornem menos frequentes as
crises agudas de pacientes asmáticos, o que pode ser feito com o desenvolvimento
de novos fármacos com potente atividade moduladora sobre o processo
fisiopatológico da doença e menor incidência de efeitos colaterais, ou por meio de
formas alternativas que potencializem os efeitos dos fármacos atualmente disponíveis
para o tratamento da asma e que reduzam o desenvolvimento de crises agudas.
Diante do exposto, o óleo de coco virgem apresenta potencial como alimento
funcional com propriedade de saúde e, em caso de comprovação de sua atividade
sobre as vias aéreas, é possível sua utilização como terapia complementar para
prevenir ou diminuir as crises asmáticas. Além disso, apesar da grande quantidade de
estudos a respeito das atividades farmacológicas do óleo de coco, sua avaliação em
modelo animal de cobaia ainda é inexistente.
Com esse intuito, nesse trabaho, avaliou-se uma provável atividade
moduladora do óleo de coco virgem sobre os parâmetros de contratilidade do músculo
liso das vias aéreas, a inflamação pulmonar e o estresse oxidativo, a fim de
caracterizar seus efeitos sobre o processo fisiopatológico da inflamação pulmonar
alérgica crônica, contribuindo com evidências científicas complementares às já
existentes de modo a caracterizá-lo como um alimento funcional com propriedade de
saúde, especialmente por suas ações sobre o processo inflamatório das vias aéreas
que caracterizam a asma.

2 Fundamentação teórica

40 VASCONCELOS, L. H. C. Fundamentação Teórica
2.1 Os produtos naturais no contexto dos alimentos funcionais
Sabe-se que os produtos naturais têm sido uma fonte valiosa de agentes
terapêuticos por milênios e, até hoje, muitos medicamentos são produtos naturais ou
seus derivados (ATANASOV et al., 2015). As civilizações antigas dependiam de
plantas ou de outros recursos naturais para sustentar ou restaurar a saúde, e em
várias situações ainda representam alternativas terapêuticas interessantes para as
drogas sintéticas. Segundo a OMS, mais de 100 milhões de pessoas da Europa,
África, Ásia, Austrália e América do Norte são usuários de terapêuticas
complementares tradicionais. Especialmente na África e em alguns países em
desenvolvimento, a medicina tradicional é muitas vezes a principal fonte de cuidados
de saúde (OMS, 2013).
De acordo com esta visão, o número de estudos científicos na área de pesquisa
de produtos naturais está aumentando rapidamente (ATANASOV et al., 2015), de
modo que, nas últimas décadas, remédios à base de plantas e outros produtos
naturais têm atraído indubitavelmente muita atenção da pesquisa no contexto da
prevenção ou no tratamento de diversas doenças (HEBER, 2001; CEFALU; YE;
WANG, 2008; DONG; LU; ZHAO, 2012).
Inseridos nesse contexto estão os nutracêuticos e os alimentos funcionais. O
termo nutracêutico é um híbrido ou contração de “nutrição” e “farmacêutico”.
Atualmente, não existem definições universalmente aceitas para nutracêuticos e
alimentos funcionais, embora existam diferenças entre as definições oferecidas por
diferentes organizações profissionais orientadas para a saúde. Os alimentos
funcionais podem ser definidos como alimentos ou componentes dietéticos que, em
razão de seus componentes alimentares fisiologicamente ativos, fornecem benefícios
para a saúde além da nutrição básica, reduzindo o risco de desenvolvimento de
doenças crônicas (IFIC, 2006). Os alimentos funcionais incluem cereais enriquecidos,
pães, bebidas esportivas, lanches fortificados, alimentos para bebés, refeições
preparadas, dentre outros (WILDMAN, 2016).
O interesse nesses produtos continua a crescer, impulsionado por esforços
progressivos da pesquisa para identificar propriedades e aplicações potenciais de
substâncias nutracêuticas e combinados com o interesse público e a demanda do
consumidor. Os alimentos funcionais são considerados uma nova tendência do
mercado alimentício (RAUD-MATTEDI, 2008). Dentre as razões relacionadas ao
crescente mercado de alimentos funcionais, destacam-se: a busca por dietas

41 VASCONCELOS, L. H. C. Fundamentação Teórica
saudáveis valorizando componentes específicos do alimento em detrimento do
alimento como um todo, o reconhecimento pelas agências reguladoras dos benefícios
dos alimentos funcionais para a saúde e a possibilidade de redução de custos do
Estado no combate das doenças crônicas (MARINS; ARAUJO; JACOB, 2011).
Embora a genética desempenhe um papel importante no desenvolvimento de
diversas doenças, em geral, a maioria pode ser minimizada por iniciativas
relacionadas a uma dieta adequada e um estilo de vida mais saudável. Além disso, as
pessoas podem otimizar as capacidades de promoção da saúde de sua dieta por meio
de suplementação e consumo de alimentos que foram formulados ou fortificados para
incluir fatores que melhoram o estado de saúde (WILDMAN, 2016).
Essas definições para alimentos funcionais diferem dos nutracêuticos pois
estes referem-se aos constituintes ativos isolados de alimentos que apresentam
funções fisiológicas e de saúde no organismo – no contexto de prevenção de doenças
- e, ainda mais, diferem da definição de medicamento, pois este contempla os
produtos utilizados para fins de diagnóstico, cura, tratamento ou prevenção de
doenças (IFIC, 2006).
As alegações ou exigências para se enquadrar um alimento com funcional
variam entre países e são estabelecidas em normas específicas (STRINGHETA et al.,
2007). No Brasil, a Anvisa aprovou em 1999, por meio de Resoluções de Diretoria
Colegiada (RDC) e Portaria, os regulamentos que tratam das diretrizes básicas para
registro de alimentos e/ou novos ingredientes (RDC n°16/99) (BRASIL, 1999a); para
avaliação do risco e segurança dos alimentos (RDC n°17/99) (BRASIL, 1999b); para
análise e comprovação de alegação de propriedade funcional e/ou de saúde
descriminadas em rotulagem de alimentos (RDC nº 18/99 e Portaria 398/99)
(BRASIL,1999c); para registro de alimento com alegação de propriedades funcionais
e/ou de saúde (RDC n°19/99) (BRASIL, 1999d); e sobre substâncias bioativas e
probióticos isolados com alegação de propriedade funcional e/ou de saúde (RDC n°
02/2002) (BRASIL, 2002).
A legislação brasileira vigente não define o termo “alimentos funcionais”, mas
alegação de propriedade funcional, referindo como aquela relativa ao papel
metabólico ou fisiológico que o nutriente ou não nutriente tem no crescimento,
desenvolvimento, manutenção e outras funções normais do organismo humano”; bem
como alegação de propriedade de saúde, sendo “aquela que afirma, sugere ou implica
a existência da relação entre o alimento ou ingrediente com doença ou condição

42 VASCONCELOS, L. H. C. Fundamentação Teórica
relacionada à saúde” (BRASIL, 1999c). Uma vez avaliada pela Gerência Geral de
Alimentos (GGALI) da Anvisa e comprovado que seu uso é seguro e eficaz, o alimento
detentor da alegação pode ser disponibilizado para consumo. Tais alegações podem
então ser veiculadas em alimentos e ingredientes para consumo humano, em rótulos
e propagandas de produtos elaborados, embalados e prontos para a comercialização
e oferta ao consumidor (PEREIRA et al., 2014).
Segundo a RDC 18/99 e a Portaria 389/99 da Anvisa, para serem comprovadas
as alegações de alimento funcional e/ou propriedades de saúde, o produto deve
possuir dados de ensaios nutricionais, fisiológicos e toxicológicos em animais de
experimentação, ensaios bioquímicos, estudos epidemiológicos, seguidos de ensaios
clínicos, comprovação de uso tradicional, sem danos à saúde e evidências
abrangentes na literatura científica, nos organismos internacionais de saúde e
legislação internacionalmente reconhecida sobre as propriedades e características do
produto (BRASIL, 1999c).
Em 2013, diversos produtos foram aprovados pela Anvisa com alegações de
propriedade funcional; dentre esses, destacam-se os ácidos graxos Omega 3, com
alegação de manutenção de níveis saudáveis de triglicerídios; fibras alimentares
(como a dextrina resistente, goma guar parcialmente hidrolisada, lactulose e
polidextrose), para o auxílio no funcionamento do intestino; probióticos (como as
bactérias do gênero Lactobacillus, Bifidobacterium e Enterococcus), para o equilíbrio
da microbiota intestinal; e a proteína da soja (Glycine max), no auxílio da redução de
doenças crônicas e degenerativas (BRASIL, 2016).
No contexto dos alimentos funcionais, escontram-se os suplementos
alimentares, designados, em uma definição cunhada com base nas diretrizes do
Codex Alimentarius da Organização das Nações Unidas (ONU), e nas normas dos
Estados Unidos e da União Europeia, como produtos para ingestão oral, apresentados
em formas farmacêuticas, e destinado a suplementar a alimentação de indivíduos
saudáveis com nutrientes, substâncias bioativas, enzimas ou probióticos, isolados ou
combinados (CE, 2002; CAC, 2005; FDA, 2009).
Recentemente, a Anvisa publicou um documento base para a regulamentação
dos suplementos alimentares, dentre os quais se insere o óleo de coco. Os
constituintes autorizados para comercialização tiveram sua análise baseada em dados
de segurança e eficácia e a existência de especificações de referência, como as

43 VASCONCELOS, L. H. C. Fundamentação Teórica
Farmacopeias reconhecidas oficialmente, o Codex Alimentarius (ONU) e o Food
Chemical Codex (BRASIL, 2017a).
2.2 Óleo de coco: potencial alimento funcional com propriedade de saúde
Inserido nesse contexto dos alimentos funcionais, o óleo de coco vem
despertando muita atenção da população e da comunidade científica em razão das
diversas propriedades benéficas para a saúde que lhe vem sendo atribuídas,
especialmente nos últimos anos (MARINA; CHE MAN; AMIN, 2009).
Durante milhares de anos, os países tropicais usaram o coco da árvore Cocos
nucifera, Família Aracaceae (família das palmeiras), como parte integrante de sua
dieta e como meio de subsistência, uma vez que todas as partes da palha do coco
são úteis, com valor econômico significativo (BRUCE, 2004). O óleo de coco ou de
copra (polpa seca do coco) é um óleo comestível extraído da semente do coco maduro
da palmeira de coco (coqueiro) (KUMAR, 1997).
Vários métodos foram desenvolvidos para extrair o óleo de coco, seja através
de processamento seco ou úmido. O processamento a seco é a forma de extração
mais utilizada. A copra limpa, moída e cozida no vapor é pressionada por uma prensa
para obter óleo de coco, que passa pelos processos de refino, branqueamento e
desodorização (RBD). Durante o processo RBD, é aplicado aquecimento ao óleo,
especialmente durante o processo de desodorização, que é realizado à temperatura
entre 204 e 245 °C (O'BRIEN, 2004).
Atualmente, há uma tendência para a produção de óleo de coco que não
precisa passar pelo processo de RBD. Desse modo, ao invés de ser obtido por
processo a seco, este óleo é obtido por processamento úmido, que implica a extração
do creme do leite do coco fresco e, em seguida, quebra da emulsão de creme,
fornecendo o óleo. Após a obtenção da emulsão, outros processos, como fermentação
e separação por centrifugação, refrigeração e ação enzimática, permitem a separação
da fase oleosa da fase aquosa. Em alguns casos, a microextração é utilizada, isto é,
é realizada a fervura do óleo de coco fresco, seguido pela evaporação da água, ou é
feita a extração do óleo pela compressão direta da copra fresca (MARINA; CHE MAN;
NAZIMAH, 2009).
Este processo é mais desejável, pois nenhum tratamento químico ou térmico é
imposto no óleo, mantendo sua composição natural, sendo conhecido como óleo de
coco virgem (OCV). Dessa forma, com base na definição de óleo virgem, entende-se

44 VASCONCELOS, L. H. C. Fundamentação Teórica
que, enquanto o óleo não passar pelo processo de RBD e que não leve à alteração
da sua natureza química, o óleo pode ser considerado OCV (VILLARINO; LIZADA,
2007).
Diferentemente do óleo de coco refinado (extra virgem), que é produzido,
normalmente, para fins de cozimento, o OCV é comercializado ultimamente como um
óleo funcional, devido à sua rica composição química que é preservada durante o
processo de obtenção (MARINA; CHE MAN; AMIN, 2009).
O OCV consiste principalmente em ácidos graxos de cadeia média (MCFA),
que são resistentes à peroxidação. Eles diferem da gordura animal, que consiste em
ácidos graxos saturados de cadeia longa e é o principal fator de risco para doenças
cardiovasculares (BEZARD; BUGAUT; CLEMENT, 1971). Os MCFA diferem dos
ácidos graxos de cadeia longa na medida em que ajudam a proteger contra doenças
do sistema cardiovascular. Os MCFAs foram relatado para diminuir o risco de
aterosclerose e doença cardíaca em ratos, e esses ácidos graxos são os principais
responsáveis pelos efeitos benéficos do OCV (CONRADO, 2003).
A composição dos ácidos graxos no OCV, determinada por cromatografia
gasosa, inclui gorduras saturadas: ácido láurico (45-52%), ácido mirístico (16-21%),
ácido palmítico (7-10%), ácido caprílico (5-10%), ácido cáprico (4-8%), ácido esteárico
(2-4%), ácido capróico (0,5-1%) e ácido palmitoleico (traços); e gorduras insaturadas:
ácido oleico (5-8%), ácido linoleico (1-3%) e ácido linolênico (até 0,2%) (KAPPALLY;
SHIRWAIKAR; SHIRWAIKAR, 2015).
O óleo de coco também é rico em compostos fenólicos, o que lhe confere
propriedade antioxidante (SENEVIRATNE; DISSANAYAKE, 2008). Em comparação
ao óleo de coco refinado, o OCV contém maior conteúdo fenólico total, sugerindo-se
que o processo de RBD destrua consideravelmente os compostos fenólicos no óleo
de coco. Dentre os compostos fenólicos identificados no OCV, estão os ácidos
protocatecuico, vanílico, cafeico, siríngico, ferúlico e p-cumarico (MARINA; CHE MAN;
NAZIMAH, 2009; MARINA et al., 2009).
Já é relatada uma extensa literatura de efeitos farmacológicos do óleo de coco
virgem, seja em humanos ou em animais. Nesse contexto, Nevin; Rajamohan (2004)
relataram que o OCV reduz o colesterol total, os níveis de triglicerídios, fosfolípidios,
lipoproteína de baixa densidade (LDL) e de muito baixa densidade (VLDL) e aumenta
os níveis de lipoproteína de alta densidade (HDL) no soro e nos tecidos, em ratos.
Adicionalmente, estudo conduzido com mulheres com obesidade abdominal revelou

45 VASCONCELOS, L. H. C. Fundamentação Teórica
que a suplementação diária com OCV reduz o acúmulo de gordura nessa região
(ASSUNÇÃO et al., 2009).
Além disso, a administração de OCV a ratos aumenta as enzimas antioxidantes
e reduz o teor de peroxidação lipídica (NEVIN; RAJAMOHAN, 2006) e apresenta efeito
antitrombótico mais significativo que o óleo de coco extravirgem (NEVIN;
RAJAMOHAN, 2008). Também é descrito efeito hepatoprotetor para o OCV em ratos
submetidos a dano hepático por aumento das enzimas antioxidantes hepáticas,
melhora na função das enzimas transaminases e redução nos níveis de ácidos graxos
livres (ZAKARIA et al., 2011). Também foi descrita atividade antioxidante do OCV
associada aos seus polifenóis (MARINA et al., 2009).
Intahphuak; Khonsung; Panthong (2010) e Nevin; Rajamohan (2010)
descreveram atividade anti-inflamatória aguda, em modelo de inflamação induzida por
carragenina, e crônica, em artrite reumatoide, para o OCV em ratos, bem como efeito
cicatrizante em ratos com ferida dérmica.
É relatada também atividade hipotensora, observada em modelo induzido por
ingestão repetida de óleo de palma (conhecido popularmente como azeite de dendê)
em ratos. Foi observado efeito preventivo na elevação da pressão arterial e melhora
da função endotelial por aumentar a biodisponibilidade de NO (NURUL-IMAN et al.,
2013). Em adição a isso, ALVES et al. (2015) observou que a suplementação com
OCV associado ao exercício físico de natação normaliza a pressão arterial, melhora o
barorreflexo e reduz o estresse oxidativo em ratos espontaneamente hipertensos,
efeitos esses reproduzidos pelo ácido láurico (ALVES et al., 2017).
Desde a sua aparição no mercado, o OCV é bem aceito pelo consumidor como
óleo alimentar funcional e a sua demanda continua a aumentar (MARINA; CHE MAN;
AMIN, 2009). Dessa forma, como o óleo de coco já é reconhecido pelas suas diversas
atividades farmacológicas e seu potencial benéfico para a saúde, destacando-se,
ainda, sua atividade anti-inflamatória e antioxidante, é de se esperar os benefícios de
sua aplicação como suplemento alimentar, adjuvante ao tratamento convencional de
doenças inflamatórias, como a asma alérgica.

46 VASCONCELOS, L. H. C. Fundamentação Teórica
2.3 Asma
2.3.2 Epidemiologia da asma
A asma é um problema mundial que acomete uma grande parcela de indivíduos
em diferentes países; no entanto, por ser uma síndrome clínica, estudos de sua
epidemiologia têm sido difíceis, já que diversos pesquisadores usam diferentes
critérios de inclusão para identificar os casos da doença (SAPEY; WILSON, 2012).
Uma evidência disso é que, enquanto a Organização Mundial de Saúde (OMS)
estima, em valores absolutos, que 235 milhões de pessoas no mundo são afetadas
pela asma, sendo uma doença frequente e a mais comumente não notificada em
crianças (OMS, 2017), o GINA, em sua última atualização, estima que existam mais
de 300 milhões de casos de asma no mundo (GINA, 2017). Apesar disso, fica claro
que a asma é uma doença que afeta uma grande parcela da população mundial,
acarrentando uma sobrecarga de custos aos sistemas de saúde, às famílias e aos
pacientes (MASOLI et al., 2004).
Um aumento na prevalência de asma e outras alergias tem sido observado nas
últimas décadas (MASOLI et al., 2004; PAWANKAR et al., 2011). Esse aumento
rápido, com padrões sociais e demográficos claramente observáveis, sugere
mudanças no estilo de vida como uma possível explicação (VON HERTZEN;
HAAHTELA, 2005). A prevalência global da asma atualmente varia entre 1-16% da
população em diferentes países (MASOLI et al., 2004; LAI et al., 2009), e estima-se
que em 2025 existirão 100 milhões de novos casos de asma no mundo (GINA, 2010).
Os mapas de prevalência da asma costumam dividir esse parâmetro entre faixas
etárias que contemplam a fase infantil/adolescência (Figura 1) e a fase adulta
(Figura 2).

47 VASCONCELOS, L. H. C. Fundamentação Teórica
Figura 1 – Mapa da prevalência mundial da asma em crianças entre 13-14 anos.
Fonte: adaptado de GINA, 2017. Figura 2 – Mapa da prevalência mundial da asma em adultos entre 18-45 anos.
Fonte: adaptado de TO et al., 2012.
Um fator determinante da prevalência da asma é o sexo. Maior incidência e
prevalência de asma foram relatadas em homens na infância e em mulheres na
adolescência e na fase adulta (STRACHAN; BUTLAND; ANDERSON, 1996; VINK et
al., 2010). O Estudo Internacional de Asma e Alergias na Infância, fases I e II (ISAAC
I e II) concluiu que a prevalência de asma foi maior nos homens na faixa etária de 6-7
anos, enquanto que as mulheres apresentaram maior prevalência na faixa etária
13-14, com variações consideráveis entre países (ISAAC, 1998).
De acordo com diversos estudos, a América Latina se destaca por possuir,
junto com os países de língua inglesa, os maiores índices de prevalência da asma no
mundo (LAI et al., 2009; CHONG-NETO et al., 2012). Essas elevadas taxas de
prevalência foram associadas principalmente às diferenças na exposição a fatores

48 VASCONCELOS, L. H. C. Fundamentação Teórica
ambientais, mais do que a outros fatores, como características socioeconômicas
locais, latitude ou grau de higiene (CHONG-NETO et al., 2012).
O Brasil apresenta alta prevalência de asma, inclusive a grave (LAI et al., 2009;
KUSCHNIR et al., 2016). Com base em dados do Departamento de Informática do
Sistema Único de Saúde (DATASUS), a prevalência de asma diagnosticada no Brasil
é de 4,4%. Analisando-se adolescentes de 12-17 anos, faixa etária de elevado
acometimento por asma, a prevalência nacional de sintomas de asma é de 13,1%,
enquanto a de asma diagnosticada é de 8,7% (KUSCHNIR et al., 2016). Nesse
sentido, estima-se que, no Brasil, existam aproximadamente 20 milhões de asmáticos,
considerando-se uma prevalência global de 10% (SOLÉ et al., 2006; 2007).
A taxa de mortalidade por asma no mundo ainda é alta, apesar dos avanços
nos serviços de saúde público e a maior disponibilidade de medicamentos para o
tratamento da doença (GINA, 2017). De acordo com a última estimativa da OMS,
realizada em dezembro de 2016, 383 mil pessoas morreram por asma em 2015, sendo
esses óbitos maiores entre os adultos e em países em desenvolvimento (OMS, 2017).
No Brasil, a taxa de mortalidade por asma encontra-se estável, após sofrer
variação ao longo do tempo. Entre 1998 e 2007, a taxa média foi de 1,52/100.000
habitantes (variação de 0,85-1,72/100.000) (CRUZ et al., 2012), seguido por um
período de platô e uma tendência a queda, tendo atingido, em 2010, a marca de
0,44/100.000 habitantes (LOTUFO; BENSENOR, 2012). Nesse contexto, a asma
ainda é uma das 20 razões mais comuns para cuidados primários no Brasil,
representando um custo elevado para as famílias, comprometendo cerca de 25% da
renda familiar (CRUZ et al., 2012).
A taxa de hospitalizações pode servir como um indicador dos impactos da
doença sobre a população e, indiretamente, como indicador do nível de controle da
doença. Em uma pesquisa de hospitalizações anuais no Brasil entre 1998 e 2009, a
asma foi avaliada como a terceira causa mais freqüente de hospitalizações no SUS.
Todavia, de 2008 a 2013, houve uma redução no número de hospitalizações de
aproximadamente 205 mil para 130 mil, acarretando uma redução de quase 30% no
custo da doença ao SUS e que a colocou como a quarta causa de hospitalizações no
país. Além disso, foi observada uma redução de 10% na taxa de mortalidade por asma
no Brasil nesse período (CARDOSO et al., 2017). Estes dados refletem a influência
de programas que fornecem medicamentos gratuitos para asma no Brasil durante
esse período (SOUZA-MACHADO; SANTOS; CRUZ, 2010).

49 VASCONCELOS, L. H. C. Fundamentação Teórica
Como descrito, apesar da evolução na redução dos casos de asma no Brasil e no
mundo, essa doença ainda acomete uma grande parcela da população e é
responsável por muitos internamentos, gastos públicos e casos de mortalidade. Nesse
sentido, fazem-se necessários estudos que busquem uma maior compreensão dos
mecanismos patofisiológicos envolvidos na asma e a sua heterogeneidade, bem como
a busca por formas de tratamento e alternativas complementares, na tentativa de
impedir o ônus da doença àqueles que são acometidos.
2.3.2 Características gerais da asma
A asma é uma doença inflamatória crônica de natureza complexa, na qual
fatores genéticos e ambientais podem contribuir para o seu desenvolvimento (SILVA
et al., 2015). À medida que a fisiopatologia da asma vem se tornando mais clara e
novas descobertas vem sendo feitas, as definições da doença são alteradas a fim de
se incluir suas características patológicas (SAPEY; WILSON, 2012).
Essa doença caracteriza-se pelos seus diferentes fenótipos, os quais
apresentam mecanismos fisiopatológicos e manifestações clínicas distintos, tornando
o diagnóstico difícil (MOORE et al., 2010). Nesse sentido, baseada em considerações
acerca das características típicas da asma e que as distingue de outras condições
respiratórias, a Global Initiative for Asthma (GINA) a definiu como uma doença
caracterizada pela inflamação crônica das vias aéreas que resulta em sintomas
respiratórios, como sibilos, dispneia, opressão no peito ou tosse que variam ao longo
do tempo e em intensidade, juntamente com a limitação variável do fluxo de ar
expiratório (GINA, 2017).
Essas variações no fluxo de ar são muitas vezes desencadeadas por fatores
como exercício, exposição a alérgenos ou irritantes das vias aéreas, alterações
climáticas ou infecções virais das vias respiratórias. Esses sintomas e a limitação do
fluxo de ar podem se resolver espontaneamente ou em resposta à farmacoterapia,
podendo estar ausentes por semanas ou meses. Por outro lado, pacientes asmáticos
podem sofrer crises de episódios agudos (exacerbações) de asma que podem ser
fatais (GINA, 2017).
As reações obstrutivas da asma que limitam o fluxo respiratório são decorrentes
de um aumento na resistência das vias respiratórias (BUSSE et al., 2007). Esta, por
sua vez, está associada à hiper-responsividade aérea, uma das características
clínicas que diferenciam a asma de outras doenças pulmonares obstrutivas, que se

50 VASCONCELOS, L. H. C. Fundamentação Teórica
apresenta como um aumento na sensibilidade e na responsividade das vias aéreas a
estímulos broncoconstritores (LOTVALL; INMAN; O’BYRNE, 1998). Além disso, outra
característica da asma é a diminuição da broncodilatação em resposta à inspiração
profunda, comum em pacientes não asmáticos (KAPSALI et al., 2000).
O curso natural da asma depende de fatores tanto genéticos como ambientais.
Aspectos relacionados com a maturação imunológica, bem como o curso da
exposição a agentes locais, especialmente nos primeiros anos de vida, são
determinantes sobre o risco do aparecimento e a evolução da asma em indivíduos
geneticamente susceptíveis (BUSSE; LEMANSKE, 2001).
Normalmente, a doença se inicia em um estágio prodrômico, no qual o indivíduo
apresenta-se livre de manifestações sintomáticas da doença, mas possui
susceptibilidade para desenvolver a condição patológica, isto é, predisposição
genética. O que determina a manifestação da doença é a exposição aos fatores
ambientais, os quais atuam como gatilho; nesse caso, a doença progride sem
interrupção, até que medidas preventivas sejam adotadas, como evitar expor-se aos
agentes desencadeadores, ou a adoção de terapia medicamentosa (GUERRA;
MARTINEZ, 2009).
Uma vez que a asma é uma doença de forte influência genética, trata-se de
uma condição não curável por meio das estratégias terapêuticas atualmente
disponíveis. Assim, além da expansão das opções de tratamento, a fim de se
minimizar sua manifestação e exacerbação, existe também a necessidade de se
identificar fatores de susceptibilidade, permitindo que pacientes possam ser
diagnosticados ainda na fase prodrômica, prevenindo o aparecimento dos sintomas
(SAPEY; WILSON, 2012).
Além dos fatores genéticos, várias comorbidades e fatores de risco estão
relacionados com o desenvolvimento da asma, especialmente naqueles com asma
grave ou descontrolada. Dentre eles, destacam-se como fatores de risco a exposição
a antígenos ambientais, como o ácaro da poeira doméstica e de baratas, de pelos de
gato e cachorro e de fungos do gênero Aspergillus; o sexo, o fumo e a prática de
exercício físico; e como comorbidades, a obesidade, infecções respiratórias, doenças
psiquiátricas e estresse, rinossinusites e o uso de anti-inflamatórios não esteroidais.
Dessa forma, medidas para evitar tais agravantes são recomendadas, uma vez que
eles contribuem para o rápido aparecimento da doença, a exacerbação dos sintomas,

51 VASCONCELOS, L. H. C. Fundamentação Teórica
redução da qualidade de vida, além da necessidade de polifarmacoterapia e
consequente risco de interações medicamentosas (BOULET, 2009).
Embora a asma tenha sido considerada como uma doença única por anos,
estudos recentes se concentraram cada vez mais em sua heterogeneidade. A
caracterização desta heterogeneidade promoveu o conceito de que a asma consiste
em múltiplos fenótipos, isto é, grupos característicos específicos com variadas formas
de expressão (WENZEL, 2012). Muitos fenótipos da asma já foram identificados, e o
mais comum deles é a asma alérgica, que muitas vezes se inicia na infância e está
associada a história familiar de doenças alérgicas, como eczema, rinite alérgica ou
alergia a alimentos ou medicamentos (BEL, 2004). Outros fenótipos incluem a asma
não alérgica, a induzida por obesidade e por exercício físico (MOORE et al., 2010;
WENZEL, 2012).
Diante disso, apesar de a maioria dos estudos de asma focar apenas em um
processo altamente específico relacionado à inflamação alérgica das vias aéreas, o
que acaba por privilegiar o conceito clássico de doença alérgica do sistema
respiratório, os principais periódicos clínicos internacionais sugerem que o termo
asma está desatualizado, incentivando a evolução de definições mais detalhadas,
biologica e clinicamente, da doença (LANCET, 2006).
2.3.3 Fisiopatologia da asma
A asma é uma doença inflamatória crônica das vias aéreas em que muitas
células do sistema imune inato e adaptativo agem em conjunto com células epiteliais
para causar hiper-reatividade brônquica, superprodução de muco, remodelamento da
parede e estreitamento das vias aéreas (LAMBRECHT; HAMMAD, 2015).
Tradicionalmente, duas formas de asma foram definidas na clínica. A maioria
das crianças e cerca de 50% dos adultos apresentam asma alérgica (extrínseca), na
qual a doença coincide com a sensibilização alérgica, definida pela presença de
anticorpos IgE sérico e/ou um teste de sensibilização cutânea positivo para
(lipo)proteínas de alérgenos comumente inalados ou ingeridos, como ácaros
domésticos, esporos fúngicos, pólen de plantas ou amendoim. Em crianças, a doença
começa com a sensibilização alérgica, muitas vezes acompanhada de eczema no
primeiro ano de vida. Mais tarde elas desenvolvem rinite alérgica, podendo progredir
para a asma. Este aumento gradual dos sintomas foi denominado de "marcha atópica"
(SIMPSON et al., 2010).

52 VASCONCELOS, L. H. C. Fundamentação Teórica
Já a asma não alérgica (intrínseca) geralmente se desenvolve mais tardiamente
na vida e, por sua definição, não possui IgE reativos aos alérgenos no soro nem
qualquer envolvimento de células T auxiliares tipo 2 (células Th2). Esta forma da
doença é mais comum nas mulheres, muitas vezes é associada a rinossinusite crônica
e pólipos nasais, bem como à obesidade, e é difícil de tratar, requerendo tratamento
a longo prazo com corticosteroides sistêmicos (LAMBRECHT; HAMMAD, 2015).
Desde o início do conceito de que a imunidade pode ser dividida em processos
do tipo T helper (auxiliares) 1 e 2 (Th1 e Th2), a asma foi considerada um processo
patológico de perfil Th2 que está ligada fortemente a reações de hipersensibilidade do
tipo I (alergia). De fato, os dados de diversos estudos sugerem que a maioria dos
casos de asma se encaixa nesta visão tradicional (MOORE et al., 2010; SIROUX et
al., 2011). Apesar disso, embora o paradigma clássico Th1/Th2 permaneça como uma
estrutura útil para a compreensão da heterogeneidade das células T, esse conceito
foi ampliado pela descoberta de vários outros subconjuntos de células T, como células
Th17, Th9 e as células T regulatórias (TReg). Dados sugerem que as células T não se
tornam totalmente comprometidas com um subconjunto específico após a exposição
ao antígeno, mas podem apresentar plasticidade e serem influenciadas pelo
microambiente que as envolve (LLOYD; HESSEL, 2010).
Diante disso, até o presente, diferentes mecanismos imunológicos da asma já
foram descritos, como a asma eosinofílica de perfil Th2, a asma causada por células
linfoides inatas do tipo 2 (ILC2), a neutrofílica de perfil Th17 e a de perfil Th9
(LAMBRECHT; HAMMAD, 2015). Todavia, existe apenas um endótipo bem definido
proposto para a asma, o “Th2-high”, uma vez que o perfil “Th2-low” engloba diferentes
mecanismos imunológicos (FAHY, 2015).
De modo geral, pacientes com asma “Th2-high” têm eosinofilia e outros sinais
de inflamação alérgica das vias aéreas (DOUGHERTY et al., 2010). As células
específicas do perfil Th2 estão presentes nos pulmões de quase todos os pacientes
com asma, particularmente em pacientes com asma alérgica (ROBINSON et al.,
1992). Essas células produzem citocinas que induzem a síntese específica de IgE,
regulada pela IL-4, o recrutamento de eosinófilos (IL-5), de mastócitos (IL-9) e a
hiper-responsividade das vias aéreas, uma característica cardinal da asma, pela IL-13
(HOLGATE; POLOSA, 2008).
A indução da imunidade adaptativa requer células apresentadoras de antígeno
(APCs), e as células dendríticas (DCs) são o principal tipo de APC envolvido na

53 VASCONCELOS, L. H. C. Fundamentação Teórica
indução da resposta Th2 a alérgenos na asma, por meio da apresentação via molécula
do complexo principal de histocompatibilidade do tipo II (MHC II). No pulmão, as DCs
podem ser encontradas em todas as vias aéreas, no interstício pulmonar, na
vasculatura, na pleura e nos linfonodos brônquicos. Além disso, as DCs do pulmão
podem responder diretamente ao peptídio alergênico por expressarem receptores de
reconhecimento de padrão (PRRs), como o do tipo Toll (TLR), semelhantes a Nod e
de lectina de tipo C, que reconhecem padões moleculares associados ao patógeno
ou ao dano (PAMPs e DAMPs), e regulam a expressão de várias moléculas
coestimulatórias (como CD80 e CD86), citocinas e quimiocinas (como CCL17 e
CCL22), que atraem células T, eosinófilos e basófilos para os pulmões (LAMBRECHT;
HAMMAD, 2009).
Todas essas células formam uma rede interconectada de apresentação e
processamento de antígenos com os macrófagos no epitélio e na submucosa
brônquica, desempenhando um papel crítico no processo de sensibilização inicial por
apresentarem o peptídio antigênico, via MHC II, ao receptor de células T (TCR) dos
linfócitos T auxiliares. Em pacientes alérgicos, cuja resposta imune é direcionada ao
perfil Th2, dependente da ativação do fator de transcrição de ação trans específico de
células T do tipo 3 (GATA-3), ocorre a síntese e a secreção de citocinas
características, que incluem IL-4, IL-5 e IL-13. As IL-4 e IL-13 induzem a mudança de
isotipo de imunoglobulina G (IgG) para IgE pelos plasmócitos (ROMAGNANI, 2000),
além do remodelamento estrutural observado na asma crônica. Estas mudanças
estruturais incluem a metaplasia das células epiteliais em células caliciformes e
hipersecreção de muco, fibrose subepitelial, diferenciação de fibroblastos em
miofibroblastos e proliferação do músculo liso (DOUCET et al., 1998).
Os linfócitos T ativados também recrutam e ativam eosinófilos, promovendo sua
migração para as vias aéreas. A eosinofilia no tecido pulmonar é conduzida pela IL-5,
que apoia o desenvolvimento de eosinófilos na medula óssea e o seu recrutamento
para a mucosa pulmonar e o interstício através da produção de quimiocinas, como as
eotaxinas 1, 2 e 3 (CCL11, CCL24 e CCL26, respectivamente), que se ligam ao
receptor de quimiocinas CCR3, que é altamente expresso nos eosinófilos. Além disso,
as eotaxinas também estimulam a expressão de α4- e β1-integrinas nos eosinófilos,
permitindo sua ligação ao endotélio vascular e transmigração para o sistema
respiratório (PONATH et al., 1996).

54 VASCONCELOS, L. H. C. Fundamentação Teórica
Após serem estimulados, os eosinófilos desempenham um papel
pró-inflamatório importante, produzindo cisteinil-leucotrienos, bem como mais
citocinas Th2 (AKUTHOTA et al., 2008). Os produtos derivados de eosinófilos, como
a peroxidase eosinofílica (EPO), a proteína básica principal (MBP) e a e a proteína
catiônica eosinofílica (ECP), além da IL-13, causam dano tecidual e
hiper-responsividade brônquica diretamente, além de ativarem a imunidade
adaptativa através de efeitos nas DCs (COYLE et al., 1994; 1995; CHU et al., 2014).
Outras células importantes na asma alérgica são os mastócitos, que expressam
os receptores Fcε do tipo I (FcεRI) e residem próximo à superfície mucosa e em vasos
sanguíneos. Essas células sofrem maturação por influência de fatores locais,
especialmente o fator de células tronco (SCF, também conhecida como Kit) e a IL-3
(GURISH; AUSTEN, 2012). Essas células podem iniciar reações de hipersensibilidade
imediata, durante os desafios subsequentes com o alérgeno, degranulando em
resposta a sinais imunes tanto adaptativos, como através da ligação cruzada do
antígeno às IgEs ligadas aos FcεRIs (PRUSSIN; METCALFE, 2006), quanto inatos,
em resposta a agonistas dos TLRs, ou a citocinas, como a IL-33 (SILVER et al., 2010).
Após ativadas, essas células liberam histamina, cisteinil-leucotrienos,
prostaglandina D2, fator de ativação plaquetária (PAF), além de citocinas e
quimiocinas (METCALFE; BARAM; MEKORI, 1997; GALLI; NAKAE; TSAI , 2005;
BARRETT; AUSTEN, 2009). Os basófilos, granulócitos circulantes que também
expressam o receptor de IgE de alta afinidade FcεRI, amplificam as respostas
imediatas de hipersensibilidade ao liberar grânulos contendo histamina e produzindo
grandes quantidades de IL-4 (YOSHIMOTO et al., 2009).
Os grânulos de mastócitos contêm uma grande quantidade de constituintes
pré-formados. Dentre eles, a histamina é provavelmente a mais conhecida, mas
aminas adicionais, como a serotonina, também são encontradas nos grânulos. A
histamina tem papel fundamental na resposta aguda da asma por promover
broncoconstrição, vasodilatação e edema pulmonar, além de agir sobre terminações
nervosas sensoriais (RILEY, 1953; SJOERDSMA; WAALKES; WEISSBACH, 1957).
Além desses mediadores, os grânulos dos mastócitos também contêm um
grande número de hidrolases lisossomais (ARVAN; CASTLE, 1998; BLOTT;
GRIFFITHS, 2002) e a capacidade única de armazenar certas citocinas pré-formadas,
como IL-4 (REED; ALBINO; MCNUTT, 1995), IL-5 e IL-6 (BRADDING et al., 1993), e
fatores de crescimento, como o fator de necrose tumoral (TNF) (GORDON; GALLI,

55 VASCONCELOS, L. H. C. Fundamentação Teórica
1990), o fator de crescimento do endotélio vascular (VEGF) (BOESIGER et al., 1998;
GRUTZKAU et al., 1998) e o fator de crescimento transformador (TGF-β)
(LINDSTEDT et al., 2001). Várias proteases específicas de mastócitos, incluindo
triptases, quimase e carboxipeptidase A3 (CPA3) são também constituintes dos
grânulos maduros de mastócitos (PEJLER et al., 2007).
Diversas citocinas estão envolvidas nas respostas celulares da asma para
desencadear ou amplificar o processo fisiopatológico, além das clássicas citocinas
Th2. A linfopoietina estromal tímica (TSLP) é liberada por células epiteliais em
resposta a produtos microbianos, lesões físicas, citocinas inflamatórias (como IL-1β e
TNF) (ALLAKHVERDI et al., 2007), alérgenos proteolíticos (como o Der p 1, que age
em receptores PAR2) (PICHAVANT et al., 2005; ADAM et al., 2006), partículas de
diesel e de cigarro (KOUZAKI et al., 2009; BLECK et al., 2010).
A IL-25 é expressa no epitélio das vias aéreas após exposição a alérgenos,
partículas e infecção por helmintos (HURST et al., 2002; ANGKASEKWINAI et al.,
2007; HAMMAD et al., 2009), também é produzida por eosinófilos ativados, mastócitos
e basófilos e encontra-se elevada na submucosa brônquica infiltrada por eosinófilos
de pacientes asmáticos (DOLGACHEV et al., 2009).
A IL-33 é liberada do epitélio brônquico após desafio alergênico (CHUSTZ et
al., 2011; RAMADAS et al., 2011). Essa citocina atua em sinergismo com o SCF e o
receptor de IgE para ativar mastócitos e basófilos (SILVER et al., 2010). A IL-33
também aumenta a sobrevivência e a degranulação dos eosinófilos (CHERRY et al.,
2008).
Essas citocinas derivadas do epitélio (apesar de também serem liberadas por
células inflamatórias) podem ativar células imunes inatas, como basófilos, eosinófilos
e mastócitos, além de ILC2 (FORT et al., 2001; SCHNEIDER et al., 2009; NEILL et
al., 2010; SAENZ et al., 2010). Desse modo, uma vez iniciadas, as respostas epiteliais
aos alérgenos e poluentes são mantidas através da ação de citocinas efetoras Th2.
Além disso, as células epiteliais das vias aéreas respondem às citocinas IL-4 e IL-13
produzindo GM-CSF e as quimiocinas IL-8, CCL11 e CCL17, que atraem neutrófilos,
eosinófilos e mais células Th2, respectivamente, fornecendo um ciclo de feedback
importante que pode perpetuar a doença (MATSUKURA et al., 2001; LORDAN et al.,
2002).
Além das células Th2, outro grupo de células também participa no
desenvolvimento e, especialmente, na amplificação da resposta alérgica da asma: as

56 VASCONCELOS, L. H. C. Fundamentação Teórica
células linfoides inatas (ILCs), as quais foram inicialmente classificadas como células
efetoras não-T, não-B em muitos modelos de doenças de perfil Th2. As células ILC2
se assemelham às células Th2 e, apesar de não apresentarem receptores específicos
de antígeno (TCR), elas reagem às citocinas derivadas do epitélio IL-25, IL-33 e TSLP
(FORT et al., 2001; FALLON et al., 2006; KANG et al., 2012) e produzem citocinas
Th2 (MORO et al., 2010).
As ILC2s também contribuem para o desenvolvimento de hiper-responsividade
brônquica em resposta a vírus respiratórios, como o vírus da gripe e o rinovírus, em
camundongos adultos e neonatais, respectivamente (CHANG et al., 2011; HONG et
al., 2014). No contexto da infecção pelo vírus da gripe, as células ILC2, via liberação
de IL-5, contribuem para o acúmulo de eosinófilos nos pulmões. A participação dessas
células em infecções virais ajuda a explicar como a exposição viral pode causar ou
exacerbar sintomas de asma em algumas pessoas (GORSKI; HAHN; BRACIALE,
2013) (Figura 3).

57 VASCONCELOS, L. H. C. Fundamentação Teórica
Figura 3 – Mecanismo imunológico da asma alérgica.
Na presença de alérgeno, há a liberação de citocinas do epitélio, particularmente IL-25, IL-33 e linfopoietina estromal tímica (TSLP), a qual induz a expressão do ligante (OX40L) da molécula co-estimulatória de linfócitos T (OX40) em células dendríticas (DCs), que atuam para promover a sobrevivência e a expansão de células T. As DCs são mobilizadas para os linfonodos locais, onde apresentam o peptídeo antigênico às células T CD4+ virgens, ativando e convertendo-as em um estado competente produtoras de IL-4. Essas células T migram para zonas de células B onde se diferenciam em células T auxiliares foliculares (TFH) e se deslocam para a circulação para completar a sua maturação como células T auxiliares 2 (Th2). As células TFH secretoras de IL-4 em áreas de células B medeiam a troca de classe de IgE em linfócitos B, os quais se fixam na superfície de mastócitos, basófilos e eosinófilos (não mostrado na figura), mediando a degranulação dessas células. As células TH2 migram para o epitélio das vias aéreas e para a camada submucoca, onde secretam IL-5 e IL-13 para mediar a resposta inflamatória, com o acúmulo de mastócitos, basófilos e eosinófilos, e o remodelamento das vias aéreas. As células linfoides inatas do grupo 2 (ILC2) também participam desse processo, por serem ativadas pelas citocinas liberadas pelo epitélio (IL-25, IL-33 e TSLP), mediando as mesmas respostas inflamatórias e de remodelamento, mas por uma via não TH2. Fonte: adaptado de FAHY, 2015.
2.3.4 Remodelamento das vias aéreas
A inflamação das vias aéreas, a lesão tecidual e subsequente processo
anormal de reparo levam a mudanças estruturais nas paredes das vias respiratórias
de pacientes asmáticos, designados coletivamente como remodelamento das vias
aéreas. Essas alterações ocorrem em toda a árvore brônquica (SAETTA et al., 1991),
mas de maneira mais pronunciada nas pequenas vias aéreas (ELIAS et al., 1999).

58 VASCONCELOS, L. H. C. Fundamentação Teórica
Uma vez que a asma é caracterizada pela inflamação crônica das vias aéreas,
esse processo é considerado o evento inicial para o remodelamento (PASCUAL;
PETERS, 2005). Existe uma forte relação entre as citocinas Th2 e as alterações das
vias aéreas patognomônicas do remodelamento característico da asma. Nesse
contexto, a presença de IL-5 e IL-13 resulta em fibrose subepitelial, metaplasia das
células mucosas e infiltrado inflamatório rico em eosinófilos e macrófagos (LEE et al.,
1997; ZHU; HOMER; WANG, 1999). Em contrapartida, enquanto a IL-4 promove
eosinofilia e metaplasia mucosa, em modelos animais de superexpressão de IL-4 não
foi observada fibrose subepitelial, indicando que essa citocina não apresenta papel
importante nesse processo (RANKIN; PICARELLA; GEBA, 1996; JAIN-VORA et al.,
1997) (Figura 4).
Figura 4 – Morfologia das vias aéreas normais e asmáticas.
As vias aéreas na asma sofrem um importante remodelamento estrutural. As vias aéreas médias de um indivíduo normal e de um paciente asmático grave seccionadas e coradas com pentacromo de Movat. O epitélio na asma mostra hiperplasia e hipersecreção mucosa (azul) e espessamento significativo da membrana basal. O volume do músculo liso também encontra-se aumentado na asma. Fonte: adaptado de WADSWORTH; SIN; DORSCHEID, 2011.
Acredita-se cada vez mais que a predisposição à asma está relacionada com
um epitélio estrutural e funcionalmente defeituoso que conecta o ambiente externo às
estruturas subjacentes das vias aéreas. Este fenômeno é explicado pelo modelo da
unidade trófica epitelial mesenquimal (EMTU) (HOLGATE et al., 2000; 2004;

59 VASCONCELOS, L. H. C. Fundamentação Teórica
HOLGATE, 2007). Neste modelo, tanto a inflamação das vias aéreas como o
remodelamento são consequências das lesões ambientais repetitivas sobre o epitélio
das vias aéreas provocadas por vírus, poluentes atmosféricos ou tabaco (HOLGATE,
2010). Essas lesões levam à interação entre o epitélio disfuncional e o mesênquima
que resulta em amplificação das respostas inflamatórias e o remodelamento nas
camadas subjacentes da parede das vias aéreas com posterior reparo defeituoso
(DAVIES, 2009).
Em pacientes asmáticos, o epitélio aéreo é frágil e, em algumas áreas, ausente
em células ciliadas (LACKIE et al., 1997). A integridade da barreira epitelial depende
das junções aderentes que mantêm as células epiteliais brônquicas unidas (XIAO,
2011). As alterações epiteliais das vias aéreas envolvem a descamação, a perda de
células ciliadas e a hiperplasia de células caliciformes (AIKAWA et al., 1992;
CARROLL et al., 1993). Em razão do reparo anormal, a descamação epitelial leva ao
aumento da sua proliferação, contribuindo para o espessamento do epitélio e um
aumento da lâmina reticular (ou fibrose subepitelial) (COHEN et al., 2007),
acompanhado da hiperplasia das células de Goblet e um aumento na área das
glândulas mucosas subepiteliais (ORDOÑEZ et al., 2001; JENKINS et al., 2003).
A lâmina reticular, formada principalmente por fibras colágenas do tipo I, III e V
(HOMER; ELIAS, 2005), bem como outras proteínas de matriz extracelular, como
elastina, fibronectina, tenascina-C, lumicana, além de proteoglicanos (ROCHE et al.,
1989; LAITINEN et al., 1997; HUANG et al., 1999), possui uma espessura de 4-5 μm
em indivíduos não asmáticos, mas pode apresentar-se com 7-23 μm na asma
(HOMER; ELIAS, 2000). Os miofibroblastos, células especializadas com
características fenotípicas de fibroblastos e miócitos, são os principais efetores da
fibrose subepitelial; eles expressam a α-actina do músculo liso, produzem mediadores
inflamatórios e são grandes produtores de proteínas da matriz extracelular
necessárias para o reparo e remodelação dos tecidos (HUANG et al., 1999).
Evidências mostram que a ação dos miofibroblastos depende da participação
do TGF-β, liberado em resposta à ação da IL-13 sobre o epitélio (LEE et al., 2001). O
TGF-β é uma citocina produzida por diversas células pulmonares, incluindo as células
epiteliais, macrófagos, fibroblastos, linfócitos e eosinófilos (TAGAYA; TAMAOKI,
2007). Ele induz os fibroblastos a expressarem α-actina e assumir um fenótipo de
miofibroblasto (BATRA et al., 2004). Como parte do reparo anormal, TGF-β induz a

60 VASCONCELOS, L. H. C. Fundamentação Teórica
expressão e a secreção de múltiplas proteínas da matriz extracelular, enquanto
também inibe sua degradação (BRANTON; KOPP, 1999).
As metaloproteinases, endopeptidases dependentes de zinco capazes de
degradar moléculas de matriz extracelular, são mediadores da fibrose subepitelial. O
equilíbrio dinâmico entre as MMP e seus inibidores (inibidores teciduais de
metaloproteinases, TIMP) é um determinante crítico do remodelamento, de modo que
na asma, existe um equilíbrio profibrótico entre os dois (VISSE; NAGASE, 2003). A
metaloproteinase mais importante no remodelamento tecidual das vias aéreas é a
MMP-9, e o seu inibidor, o TIMP-1 (HOMER; ELIAS, 2005), apesar de outras
isoformas, como MMP-1, -2, -3 e -12 já terem sido descritas (LEE et al., 2001; SUZUKI
et al., 2001; BELLEGUIC et al., 2002).
Outro evento do processo de remodelamento é a metaplasia das células de
Goblet. Esse termo refere-se não à proliferação dessas células, mas à
transdiferenciação de células ciliadas e células de Clara em células de Goblet,
resultando na maior produção de muco (LE CRAS et al., 2011). Essas células
mucosas produzem a glicoproteína mucina (MUC), sendo a forma predominante nas
vias aéreas a MUC5AC; e as glândulas submucosas produzem, majoritariamente, a
MUC5B, sendo a MUC5B a predominante sob condições normais e a MUC5AC
super-regulada na asma (KIRKHAM et al., 2002; ROSE; VOYNOW, 2006; VOYNOW;
GENDLER; ROSE, 2006; EVANS et al., 2009). Esse processo está relacionado à ação
de citocinas, como a IL-13 (SONG et al., 2003) e IL-17A (TAKEYAMA et al., 2001;
SHAO; NAKANAGA; NADEL, 2004).
Biópsias brônquicas de pacientes asmáticos demonstram um aumento no
número e na área transversal dos vasos sanguíneos, predominantemente capilares e
vênulas, especialmente na lâmina própria, evento definido como neovascularização
brônquica, mediado principalmente pelo fator de crescimento do endotélio vascular
(VEGF) (HOSHINO; TAKAHASHI; AOIKE, 2001; ORSIDA et al., 2001). Acredita-se
que a neovascularização agrava a hiper-responsividade brônquica através do
aumento da congestão vascular, edema e inflamação e espessamento global da
parede das vias aéreas (HOSHINO; TAKAHASHI; AOIKE, 2001; HOMER; ELIAS,
2005).
Dentre as alterações decorrentes do remodelamento, a característica mais
proeminente é o aumento da massa de músculo liso (CARROLL et al., 1993),
decorrente da sua hipertrofia e hiperplasia, além da diferenciação e migração de

61 VASCONCELOS, L. H. C. Fundamentação Teórica
células mesenquimais para a camada de células musculares lisas (SCHMIDT et al.,
2003; HIRST et al., 2004; BERGERON; AL-RAMLI; HAMID, 2009). Essa hipertrofia
muscular lisa é responsável pelo aumento na contratilidade característica da asma,
principal fator causal da hiper-responsividade brônquica (MARTIN; DUGUET;
EIDELMAN, 2000).
2.3.5 Mediadores neurais e humorais envolvidos na asma
2.3.5.1 Vias neurais colinérgicas
O sistema nervoso parassimpático é a principal via neural broncoconstritora nas
vias aéreas (BARNES, 1992), sendo responsável pela manutenção do seu tônus basal
(WIDDICOMBE, 1963). As fibras colinérgicas atravessam o nervo vago nos gânglios
parassimpáticos e penetram na parede das vias aéreas. A acetilcolina (ACh), principal
neurotransmissor liberado pelas terminações parassimpáticas, age nos receptores
muscarínicos M3, acoplados às proteínas Gq/11, na junção neuromuscular,
responsáveis pela contração muscular lisa; mas também nos receptores
pré-sinápticos M2, acoplados às proteínas Gi/o, responsáveis pelo controle negativo da
liberação de ACh (ROUX et al., 1998).
Devido ao papel da ACh em induz a contração do músculo liso nas vias aéreas,
os fármacos anticolinérgicos são usados como broncodilatadores em doenças
obstrutivas das vias aéreas, como a asma (TASHKIN et al., 2008). No entanto, o papel
fisiopatológico da ACh excede a contração muscular lisa, tendo em vista que células
residentes e inflamatórias que controlam a inflamação e o remodelamento produzem
esse mediador e expressam receptores muscarínicos (WESSLER; KIRKPATRICK,
2008).
O quadro 1 sumariza os receptores muscarínicos expressos nas células das
vias aéreas e os principais efeitos decorrentes de sua ativação.
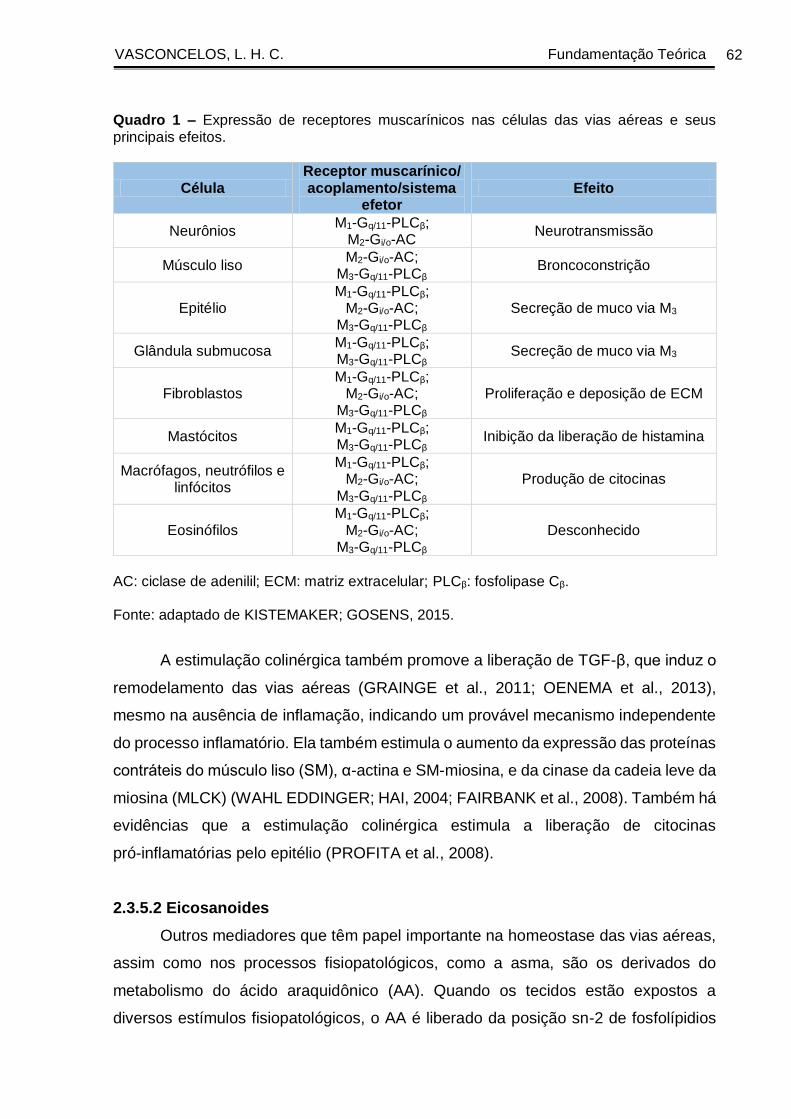
62 VASCONCELOS, L. H. C. Fundamentação Teórica
Quadro 1 – Expressão de receptores muscarínicos nas células das vias aéreas e seus principais efeitos.
Célula Receptor muscarínico/ acoplamento/sistema
efetor Efeito
Neurônios M1-Gq/11-PLCβ;
M2-Gi/o-AC Neurotransmissão
Músculo liso M2-Gi/o-AC;
M3-Gq/11-PLCβ Broncoconstrição
Epitélio M1-Gq/11-PLCβ;
M2-Gi/o-AC; M3-Gq/11-PLCβ
Secreção de muco via M3
Glândula submucosa M1-Gq/11-PLCβ; M3-Gq/11-PLCβ
Secreção de muco via M3
Fibroblastos M1-Gq/11-PLCβ;
M2-Gi/o-AC; M3-Gq/11-PLCβ
Proliferação e deposição de ECM
Mastócitos M1-Gq/11-PLCβ; M3-Gq/11-PLCβ
Inibição da liberação de histamina
Macrófagos, neutrófilos e linfócitos
M1-Gq/11-PLCβ; M2-Gi/o-AC;
M3-Gq/11-PLCβ Produção de citocinas
Eosinófilos M1-Gq/11-PLCβ;
M2-Gi/o-AC; M3-Gq/11-PLCβ
Desconhecido
AC: ciclase de adenilil; ECM: matriz extracelular; PLCβ: fosfolipase Cβ. Fonte: adaptado de KISTEMAKER; GOSENS, 2015.
A estimulação colinérgica também promove a liberação de TGF-β, que induz o
remodelamento das vias aéreas (GRAINGE et al., 2011; OENEMA et al., 2013),
mesmo na ausência de inflamação, indicando um provável mecanismo independente
do processo inflamatório. Ela também estimula o aumento da expressão das proteínas
contráteis do músculo liso (SM), α-actina e SM-miosina, e da cinase da cadeia leve da
miosina (MLCK) (WAHL EDDINGER; HAI, 2004; FAIRBANK et al., 2008). Também há
evidências que a estimulação colinérgica estimula a liberação de citocinas
pró-inflamatórias pelo epitélio (PROFITA et al., 2008).
2.3.5.2 Eicosanoides
Outros mediadores que têm papel importante na homeostase das vias aéreas,
assim como nos processos fisiopatológicos, como a asma, são os derivados do
metabolismo do ácido araquidônico (AA). Quando os tecidos estão expostos a
diversos estímulos fisiopatológicos, o AA é liberado da posição sn-2 de fosfolípidios

63 VASCONCELOS, L. H. C. Fundamentação Teórica
de membrana pela ação da fosfolipase A2 (DENNIS et al., 2011). O AA liberado pela
ação da PLA2 é então convertido em mediadores lipídicos, como os prostanoides, os
leucotrienos (LTs) e os ácidos hidroxi-eicosatetraenoicos (HETEs), através das suas
sintases específicas (NARUMIYA; SUGIMOTO; USHIKUBI, 1999).
Os prostanoides são biossintetizados a partir da enzima ciclo-oxigenase (COX),
a qual possui duas isoformas: a COX-1 e a COX-2. A expressão de COX-1 é
constitutiva na maioria dos tecidos, onde tem um papel na homeostase; já a COX-2
tem sua expressão de forma induzível tanto em células hematopoiéticas como não
hematopoiéticas em resposta a uma variedade de estímulos e está associada a
processos inflamatórios (SMITH; LANGENBACH, 2001; WARNER; MITCHELL, 2004;
KANG et al., 2007).
As COX-1 e 2 catalisam duas reações sequenciais para iniciar a via COX.
Primeiro, duas moléculas de oxigênio são inseridas no AA, ciclizando-o para produzir
PGG2. Em seguida, uma reação de endoperoxidase reduz PGG2 a PGH2, um
endoperóxido cíclico altamente instável e rapidamente convertido em 1 a 5
prostanoides bioativos pelas sintases específicas. A prostaglangina D2 (PGD2) é
formada pela sua sintase (PGDS) hematopoiética (H-PGDS) ou pela sintase do tipo
lipocalina (L-PGDS); a PGE2 por sua sintase (PGES) microsomal (mPGES) ou
citosólica (cPGES); a PGF2α pela sintase de PGF (PGFS); a PGI2 pela sintase de PGI
(PGIS) e o tromboxano A2 (TxA2) pela sintase de tromboxano (TXS) (HIRATA;
NARUMIYA, 2012).
Os prostanoides são liberados das células imediatamente após a formação.
Uma vez que são química e metabolicamente instáveis, geralmente funcionam apenas
localmente através de receptores de membrana em células-alvo (NARUMIYA;
SUGIMOTO; USHIKUBI, 1999).
A PGD2 é o principal prostanoide liberado pelos mastócitos em resposta à
ativação mediada por IgE (ROBERTS et al., 1990), sendo liberado em crises agudas
de asma (ROBINSON; HARDY; HOLGATE, 1985). Outros leucócitos, incluindo
células Th2 (TANAKA et al., 2000) e células dendríticas (URADE et al., 1989)
produzem PGD2, porém em níveis mais baixos. A PGD2 age em receptores DP1 e DP2
(também conhecido como CRTH2 ou molécula homóloga quimioatratora expressa em
células Th2) (TOWNLEY; AGRAWAL, 2012), sendo DP1 acoplado a Gs e DP2 a Gi/o
(ALEXANDER et al., 2013). A PGD2 é uma forte agente broncoconstritora,
apresentando papel importante na hiper-responsividade brônquica da asma.

64 VASCONCELOS, L. H. C. Fundamentação Teórica
Interessantemente, essa prostaglandina age preferencialmente sobre os receptores
de TxA2 (TP) do que sobre os receptores DP1/DP2 para promover contração muscular
lisa das vias aéreas (JOHNSTON et al., 1995).
O TxA2 é produzido por plaquetas ativadas, granulócitos e mastócitos e é um
potente indutor da agregação plaquetária (WHITTLE; MONCADA, 1983), além de ser
um forte agente broncoconstritor, por se ligar aos receptores TP e DP, acoplados às
proteínas Gq/11 (ALEXANDER et al., 2013).
A enzima PGFS, que catalisa a síntese da PGF2α, é expressa em linfócitos
pulmonares e do sangue periférico, sugerindo um possível papel em doenças
alérgicas, como a asma (SUZUKI-YAMAMOTO et al., 1999); no entanto, a distribuição
tecidual do mRNA do receptor FP é baixa nos pulmões, diferentemente de outros
órgãos, como os ovários e os rins (BREYER; BREYER, 2001). Dessa forma, em
contraste com as outras prostaglandinas, a sinalização PGF2α-receptor FP, acoplado
às proteínas Gq/11 (ALEXANDER et al., 2013), não parece ter um papel importante nos
processos inflamatórios e imunológicos; todavia, por ser um receptor de baixa
seletividade, já que outras prostaglandinas, como PGD2 e PGE2, se ligam a ele, sua
sinalização pode mediar respostas pulmonares independentemente da PGF2α (HATA;
BREYER, 2004).
Em contrapartida, a PGE2 possui um papel majoritariamente anti-inflamatório e
broncodilatador nas vias aéreas. Ela atua em quatro GPCRs distintos (receptores EP
1-4), resultando em efeitos complexos, uma vez que esses subtipos de receptores são
expressos em várias combinações por essencialmente todos os tipos celulares
(BREYER et al., 2001). Os receptores EP1 são acoplados às proteínas Gq/11, os EP3
acoplam a Gi/o, enquanto os receptores EP2 e EP4 acoplam à proteína Gs
(ALEXANDER et al., 2013). O efeito broncodilatador, em humanos, da PGE2 se dá
pela sua ação principalmente sobre os receptores EP4, enquanto em cobaias, se dá
pelo receptor EP2, o que pode explicar a ineficácia de agonistas EP2 em promover
broncodilatação em estudos clínicos (BUCKLEY et al., 2011).
A prostaciclina (PGI2) tem envolvimento na homeostase brônquica e pulmonar
(FANNING; BOYCE, 2013). Ela sinaliza através do seu receptor IP, acoplado às
proteinas Gs (ALEXANDER et al., 2013) e, semelhantemente à PGE2, possui papel
anti-inflamatório na asma (TAKAHASHI et al., 2002). Em contrapartida, apesar de seu
potente efeito relaxante sobre o músculo liso vascular, sendo seus análogos utilizados

65 VASCONCELOS, L. H. C. Fundamentação Teórica
no tratamento da hipertensão pulmonar; no músculo liso das vias aéreas, a PGI2 não
possui efeito pronunciado (BREYER et al., 2001).
Os leucotrienos são gerados a partir da metabolização do AA pela 5-LO,
enzima expressa principalmente em granulócitos, macrófagos e mastócitos
(SAMUELSSON et al., 1987). O AA se liga à proteína ativadora da 5-LO (FLAP),
ficando disponível para a ação da 5-LO. Esta, então, atua sobre o AA ligado ao FLAP,
juntamente com o oxigênio molecular, resultando na formação do ácido
5-hidroperoxieicosatetraenoico (5-HPETE), com subsequente formação do
intermediário instável leucotrieno A4 (LTA4). O LTA4 é rapidamente convertido a LTB4
pela enzima citosólica LTA4 hidrolase (MILLER et al., 1990; FUNK, 2001;
RINALDO-MATTHIS; HAEGGSTROM, 2010). As células inflamatórias, como
eosinófilos, basófilos, mastócitos e macrófagos alveolares, algumas das quais não
possuem a 5-LO, podem sintetizar LTC4 por uma via alternativa, por possuírem a LTC4
sintase na membrana, sendo então capazes de formar leucotrienos
independentemente da enzima 5-LO. A LTC4 sintase conjugada com a glutationa
reduzida na posição C6 do LTA4 para formar LTC4. LTB4 e LTC4 são exportados para
o espaço extracelular por proteínas transportadoras, onde o LTC4 é rapidamente
clivado pela transpeptidase para produzir LTD4 e, finalmente, o LTD4 é convertido em
LTE4 pela remoção do aminoácido glicina pela dipeptidase (LEWIS; ROBIN, 1985;
DRAZEN; AUSTEN, 1987; LEE; WOSZCZEK; FAROOQUE, 2009).
Os LTs são divididos em duas classes: o fator quimioatrativo LTB4, que contém
apenas uma porção hidroxila; e os CysLTs, que são aqueles formados pelo
metabolismo do LTB4 a LTC4, LTD4 e LTE4, que carregam porções de aminoácidos
(LUSTER; TAGER, 2004; YOKOMIZO; IZUMI; SHIMIZU, 2001). Os LTs estão
envolvidos em várias doenças inflamatórias, incluindo asma, rinite alérgica, doença
pulmonar obstrutiva crônica, além de doenças pulmonares intersticiais (OHNISHI et
al., 2008) e exercem os seus efeitos biológicos ligando-se aos seus receptores
CysLT1-2, acoplados às proteínas Gq/11 (SAMUELSSON, 1983; ALEXANDER et al.,
2013).
A função primária do LTB4 é a quimiotaxia leucocitária, atuando como potente
fator quimioatrativo para eosinófilos e neutrófilos (TAGER; LUSTER, 2003; LUSTER;
TAGER, 2004), como também para monócitos e células dendríticas (HUANG et al.,
1998; SERHAN; PRESCOTT, 2000), todas essas células expressando receptor BLT1
(SERHAN; PRESCOTT, 2000).

66 VASCONCELOS, L. H. C. Fundamentação Teórica
Os CysLTs são potentes agentes broncoconstritores e um dos componentes
inflamatórios mais importantes da asma. Eles são biossintetizados por eosinófilos,
basófilos, mastócitos e macrófagos alveolares, sendo produzidos principalmente
pelos eosinófilos e pelos mastócitos no início rápido da asma alérgica (ARM, 2004).
A contração do músculo liso dos brônquios foi a primeira propriedade biológica
reconhecida para os CysLTs, mesmo antes de sua estrutura ser elucidada. LTC4 e
LTD4 se mostraram potentes indutores de broncoconstrição em cobaia, in vitro e in
vivo, e em humanos (BACK et al., 2000).
O Quadro 2 sumariza as ações dos prostanoides e dos leucotrienos no sistema
respiratório.
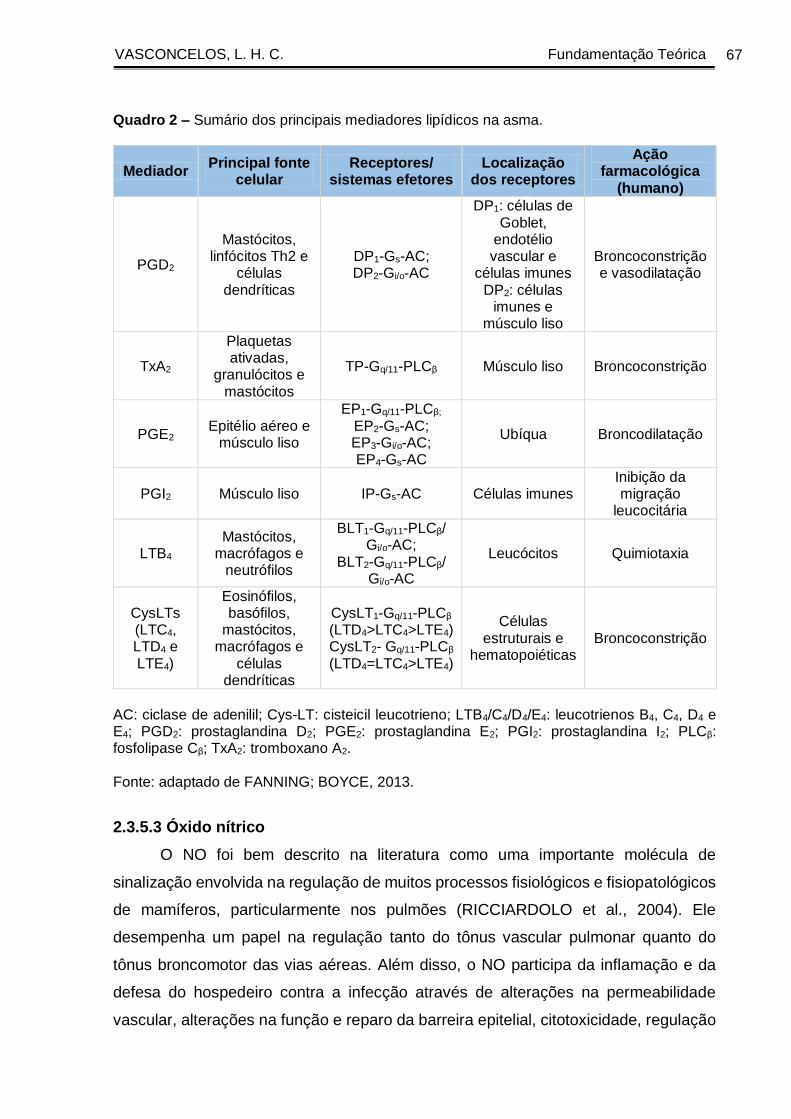
67 VASCONCELOS, L. H. C. Fundamentação Teórica
Quadro 2 – Sumário dos principais mediadores lipídicos na asma.
Mediador Principal fonte
celular Receptores/
sistemas efetores Localização
dos receptores
Ação farmacológica
(humano)
PGD2
Mastócitos, linfócitos Th2 e
células dendríticas
DP1-Gs-AC; DP2-Gi/o-AC
DP1: células de Goblet,
endotélio vascular e
células imunes DP2: células
imunes e músculo liso
Broncoconstrição e vasodilatação
TxA2
Plaquetas ativadas,
granulócitos e mastócitos
TP-Gq/11-PLCβ Músculo liso Broncoconstrição
PGE2 Epitélio aéreo e
músculo liso
EP1-Gq/11-PLCβ; EP2-Gs-AC; EP3-Gi/o-AC; EP4-Gs-AC
Ubíqua Broncodilatação
PGI2 Músculo liso IP-Gs-AC Células imunes Inibição da migração
leucocitária
LTB4 Mastócitos,
macrófagos e neutrófilos
BLT1-Gq/11-PLCβ/ Gi/o-AC;
BLT2-Gq/11-PLCβ/ Gi/o-AC
Leucócitos Quimiotaxia
CysLTs (LTC4, LTD4 e LTE4)
Eosinófilos, basófilos,
mastócitos, macrófagos e
células dendríticas
CysLT1-Gq/11-PLCβ (LTD4>LTC4>LTE4) CysLT2- Gq/11-PLCβ (LTD4=LTC4>LTE4)
Células estruturais e
hematopoiéticas Broncoconstrição
AC: ciclase de adenilil; Cys-LT: cisteicil leucotrieno; LTB4/C4/D4/E4: leucotrienos B4, C4, D4 e E4; PGD2: prostaglandina D2; PGE2: prostaglandina E2; PGI2: prostaglandina I2; PLCβ: fosfolipase Cβ; TxA2: tromboxano A2. Fonte: adaptado de FANNING; BOYCE, 2013.
2.3.5.3 Óxido nítrico
O NO foi bem descrito na literatura como uma importante molécula de
sinalização envolvida na regulação de muitos processos fisiológicos e fisiopatológicos
de mamíferos, particularmente nos pulmões (RICCIARDOLO et al., 2004). Ele
desempenha um papel na regulação tanto do tônus vascular pulmonar quanto do
tônus broncomotor das vias aéreas. Além disso, o NO participa da inflamação e da
defesa do hospedeiro contra a infecção através de alterações na permeabilidade
vascular, alterações na função e reparo da barreira epitelial, citotoxicidade, regulação

68 VASCONCELOS, L. H. C. Fundamentação Teórica
da motilidade ciliar, secreção de muco e infiltração de células inflamatórias
(REDINGTON, 2006). Essas múltiplas funções de NO foram implicadas na
patogênese de doenças inflamatórias crônicas das vias aéreas, como a asma
(BENSON; HARDY; MORRIS, 2011).
O NO é produzido por uma família de sintases de óxido nítrico (NOS), que
metabolizam a L-arginina através do intermediário N-hidroxi-L-arginina (NOHA) para
formar NO e L-citrulina usando oxigênio molecular e NADPH como cossubstratos.
Três isoenzimas de NOS foram identificadas em mamíferos, com distribuições
variáveis. As NOSs neuronal (nNOS ou NOS I) e endotelial (eNOS ou NOS III) são as
formas constitutivamente expressas (cNOS) no epitélio das vias aéreas, neurônios
não colinérgicos não adrenérgicos inibitórios (iNANC) e em células endoteliais dos
vasos das vias aéreas.
Sua atividade é regulada pelo cálcio intracelular, com início rápido da atividade
e produção de pequenas quantidades de NO na ordem de picomolar. Já a NOS
induzível (iNOS ou NOS II) é regulada transcricionalmente por estímulos
pró-inflamatórios, com a capacidade de produzir grandes quantidades (concentrações
nanomolares) de NO por longo período de tempo (REDINGTON, 2006; RACKE;
WARNKEN, 2010). Acerca da iNOS, sua regulação positiva, seja na expressão ou na
sua atividade, é observada nos pulmões asmáticos, e os níveis aumentados de NO
exalado são bem descritos em pacientes com asma (HAMID et al., 1993).
O substrato essencial que determina a formação desse mediador, a L-arginina,
deve apresentar biodisponibilidade suficiente para sua conversão a NO (RACKE;
WARNKEN, 2010). Esse processo se inicia a partir da absorção intestinal de
L-glutamina e L-prolina, os quais são convertidos a L-ornitina e esta a L-citrulina nos
hepatócitos. Em seguida, a L-citrulina é convertida a L-arginina no citosol das células
hepáticas, sendo então liberada para ficar biodisponível para a conversão a NO. A
L-citrulina é captada pelas células que expressam as isoformas de NOS pelo
transportador de aminoácido catiônico (CAT) (HAMMERMANN et al., 2001). Após ser
convertida, L-citrulina é novamente produzida, podendo ser reciclada a L-arginina,
fornecendo uma fonte prolongada para a iNOS (RACKE; WARNKEN, 2010).
Diante do papel dual do NO nas vias aéreas, ora protetor ora danoso, a
depender da sua fonte, tempo de formação e níveis de concentração atingidos,
mecanismos são propostos para explicar essa diferença de efeitos. Na fase precoce
da resposta asmática, o NO tem papel importante na redução da hiper-responsividade

69 VASCONCELOS, L. H. C. Fundamentação Teórica
brônquica, sendo a resposta hiper-reativa relacionada à deficiência de NO, seja por
maior atividade da arginase ou pela menor atividade das isoformas constitutivas da
NOS. Nesse sentido, diminuição da expressão da eNOS ou da nNOS já foi observada
em cobaias expostas a desafio alergênico repetido e em pacientes asmáticos,
respectivamente (RICCIARDOLO et al., 2001; SAMB et al., 2001). Em contrapartida,
na fase tardia da resposta asmática, prevalece a atividade aumentada da iNOS, que
produz grandes quantidades de NO, respondendo pela hiper-responsividade
brônquica (KHARITONOV et al., 1995; YAN et al., 1995).
Essa atividade aumentada (ou o aumento da sua expressão) da iNOS ocorre
devido à ação de citocinas inflamatórias (HAMID et al., 1993; RICCIARDOLO et al.,
2004). Corroborando essas proposições, foi observado em modelos de asma aguda
em cobaias que, na fase rápida da resposta asmática, os níveis de NO exalados
diminuem, assim como a expressão de iNOS não é alterada; contrariamente à fase
tardia, em que os níveis de NO exalados são elevados, assim como a expressão da
iNOS (PERSSON; GUSTAFSSON , 1993; YAN et al., 1995; DE BOER et al., 1996).
A inflamação e a hiper-responsividade das vias aéreas na asma não é resultado
do aumento da produção de NO em si, mas sim devido à formação do radical livre,
forte agente oxidante, peroxinitrito (ONOO-), resultante da reação do NO com o ânion
superóxido (O2-) nas vias aéreas (SADEGHI-HASHJIN et al., 1998; SALEH et al.,
1998). O peroxinitrito ativa os eosinófilos, aumenta a expressão de MUC5AC,
aumenta a permeabilidade microvascular, induz o dano epitelial e aumenta a
contração do músculo liso nas vias aéreas (SADEGHI-HASHJIN et al., 1996;
SUGIURA et al., 1999; DE BOER et al., 2001; FISCHER;VOYNOW, 2002). Nesse
contexto, estudos mostram que as células epiteliais das vias aéreas e as células
inflamatórias de biópsias brônquicas de pacientes asmáticos, bem como de cobaias
sensibilizadas, apresentam aumento da marcação de nitrotirosina (um marcador da
nitrosilação do resíduo de tirosina nas proteínas), que também está correlacionado
com o aumento do NO exalado, expressão de iNOS, AHR e inflamação eosinofílica,
tendo como intermediário a ação de radicais livres e o estresse oxidativo (SALEH et
al., 1998; SUGIURA et al., 1999; HANAZAWA; KHARITONOV; BARNES, 2000).
2.3.5.4 Estresse oxidativo
O sistema respiratório é um ambiente que apresenta risco aumentado de sofrer
por danos provocados pelo estresse oxidativo devido à grande susceptibilidade de

70 VASCONCELOS, L. H. C. Fundamentação Teórica
interações com fatores ambientais. Os pulmões são expostos a espécies reativas de
oxigênio (ROS) de origem exógena, como poluentes e fumaça de cigarro, e endógena,
produzidas principalmente por células inflamatórias, como eosinófilos, macrófagos e
neutrófilos (BISHOPP et al., 2017). Nesse sentido, pacientes asmáticos são
deficientes em defesas antioxidantes, com a resultante carga de estresse oxidativo
levando à disfunção pulmonar (MORWOOD et al., 2005; BISHOPP et al., 2017).
Os locais de produção de ROS no sistema pulmonar incluem sítios celulares,
como mitocôndrias, microssomos e enzimas (como a xantina oxidase,
mono-oxigenase P450, COX, LOX, indolamina dioxigenase e monoamina oxidase)
(FRIDOVICH, 1984; VALLYATHAN; SHI, 1997). Destas, as mitocôndrias são os
maiores contribuintes para a formação das ROS. Estima-se que 1-3% do fluxo de
elétrons nessa organela formem O2- (EVANS; LYRAS; HALLIWELL, 1999).
No ambiente pulmonar, células inflamatórias e epiteliais produzem diversas
ROS que englobam uma variedade de radicais livres de oxigênio, como o O2- e
radicais hidroxila (OH-), bem como derivados de oxigênio que não contém elétrons
desemparelhados, tais como peróxido de hidrogênio (H2O2), ácido hipocloroso
(HClO), ONOO- e ozônio (O3) (HENRICKS; NIJKAMP, 2001; LOXHAM; DAVIES;
BLUME, 2014).
O passo inicial para a formação desses radicais livres é ativação do complexo
enzimático da nicotinamida adenina dinucleotídio fosfato reduzida (NADPH) oxidase
presente na membrana celular e a geração do O2- (BABIOR, 1999). Esse composto
pode ser espontanea ou enzimaticamente dismutado a H2O2. Tanto o O2- como o H2O2
podem reagir, na presença de ferro ou outros metais, para formar o mais potente
radical OH- (BRUNORI; ROTILIO, 1984).
Além disso, granulócitos contém peroxidases (como mieloperoxidase e
peroxidase eosinofílica), que catalisam a reação do H2O2 com haletos, levando à
formação de hipo-haletos, como o HClO (BABIOR, 1978). Adicionalmente, o O2- pode
reagir com o NO para formar o ânion ONOO- (SADEGHI-HASHJIN et al., 1998).
Estudos recentes têm descrito uma nova fonte de O2-, a eNOS desacoplada, um
processo que ocorre quando a eNOS não está associada a cofator ou substrato (LUO
et al., 2014). Esse processo pode ocorrer por ação oxidativa do ONOO- sobre o cofator
tetrahidrobiopterina (FÖRSTERMANN; SESSA, 2012) ou por uma redução na
disponibilidade da L-arginina, comum na asma devido ao aumento na expressão da
enzima L-arginase (BENSON; HARDY; MORRIS, 2011).

71 VASCONCELOS, L. H. C. Fundamentação Teórica
As ROS promovem danos aos lipídios de membrana e às proteínas celulares,
levando à liberação de diversos mediadores inflamatórios e marcadores do dano
oxidativo. Dentre esses marcadores nos pulmões estão os produtos aldeídicos
malondialdeído (MDA), hexanal, heptanal, nonanal, acroleína, 4-hidroxihexanal
(4-HHE) e 4-hidroxinonenal (4-HNE); e o 8-isoprostano, este último já descrito
anteriormente. Dentre esses compostos, o MDA é o mais frequentemente estudado
em diversas doenças pulmonares, assim como um marcador sistêmico, sendo um
composto formado pela decomposição oxidativa de ácidos graxos poliinsaturados da
membrana (DEL RIO; STEWART; PELLEGRINI, 2005).
Para contrabalançar a formação dessas ROS, o organismo como um todo,
assim como o sistema respiratório, dispõe de sistemas antioxidantes enzimáticos e
nao enzimáticos, de modo a manter níveis normais desses radicais livres, necessários
para o perfeito funcionamento celular. Os maiores sistemas enzimáticos presentes
nas vias aéreas são o complexo da superóxido dismutase (SOD), que converte o O2-
em H2O2; a catalase, a qual converte o H2O2 em água e oxigênio molecular; e as
glutationas peroxidase e redutase, que inativam o H2O2 e outros hidroperóxidos
(KINNULA et al., 1994; REPINE; BAST; LANKHORST, 1997; PIETARINEN-RUNTTI
et al., 2000; LOXHAM; DAVIES; BLUME, 2014). Dentre esses sistemas antioxidantes,
a SOD apresenta expressão elevada nos pulmões, em torno das vias aéreas e do
músculo liso, sendo abundante no epitélio (SU et al., 1997), e desempenha papel
importante em combater o estresse oxidativo na asma.
Já é sabido que as células inflamatórias são as principais fontes de ROS na
asma. Desafios antigênicos em pacientes asmáticos aumentam a formação de ROS
pelos eosinófilos (SANDERS et al., 1995). Além disso, os leucócitos sanguíneos
aumentam a produção de O2-, indicando que tanto as células inflamatórias pulmonares
como intravasculares contribuem para o estresse oxidativo na asma (VACHIER et al.,
1992; NADEEM et al., 2003). Nesse caso, o epitélio das vias aéreas de asmáticos
produz poucos antioxidantes, o que explica a perda da eficácia do epitélio na
manutenção da homeostase das vias aéreas (XIAO, 2011)
Os alvos das ROS incluem receptores catalíticos, fosfatases, fosfolipídios, além
de proteínas da via das MAPKs (BOWLER, 2004). As ROS compreendem um tipo de
DAMP, que pode ativar as células dendríticas, estimulando o NF-κB (CKLESS et al.,
2011). Além disso, a produção de ROS pela NADPH oxidase está relacionada com a

72 VASCONCELOS, L. H. C. Fundamentação Teórica
proliferação do músculo liso das vias aéreas em resposta a fatores de crescimento,
seguida da ativação do NF-κB (BRAR et al., 2002).
2.3.6 O papel do músculo liso no remodelamento e na hiper-responsividade das
vias aéreas na asma
Apesar do grande número de novas funções descobertas para as células
musculares lisas, sua função primária é regular o tônus e o estreitamento das vias
respiratórias (MITCHELL, 2009). Elas estão localizadas na parede das vias aéreas, e
sua contração desempenha um importante papel fisiológico, na medida em que induz
uma redução no lúmen e, portanto, uma maior resistência ao fluxo de ar
(OUEDRAOGO; ROUX, 2014), controlando a ventilação intrapulmonar e,
consequentemente, a relação ventilação-perfusão (OTIS, 1983), confere estabilidade
mecânica às vias aéreas não cartilaginosas e evita que agentes tóxicos inalados
alcançem os espaços alveolares (PELAIA et al., 2008).
O músculo liso está localizado na parede da árvore traqueobrônquica,
extendendo-se desde a traqueia até os bronquíolos terminais. Na traqueia e nos
brônquios extralobares, a faixa muscular lisa conecta as duas extremidades do anel
de cartilagem aberto em forma de ferradura; nos bronquios intralobares, a organização
da cartilagem e do músculo liso muda, uma vez que o músculo forma uma camada
contínua na parede brônquica, enquanto que a cartilagem não constitui uma estrutura
contínua e está ausente nos brônquios periféricos (OUEDRAOGO; ROUX, 2014).
O estado contrátil do músculo liso está sob o controle de muitos mensageiros
extracelulares atuando em receptores específicos de membrana. Os principais são os
neurotransmissores do sistema nervoso autônomo, mediadores epiteliais e
mediadores liberados pelas células inflamatórias (ROUX, 2013).
O tônus do músculo liso das vias aéreas é mediado de forma sinificativa pela
atividade nervosa ganglionar e pós-ganglionar da inervação parassimpática. As
populações de receptores no músculo liso das vias aéreas são uma mistura de
receptores M2 e M3, predominando o subtipo M2, acoplado à proteínas Gi/o,
particularmente nas vias aéreas maiores. A contração, no entanto, é mediada
principalmente por receptores M3, acoplados às proteínas Gq/11 (ROFFEL; MEURS;
ZAAGSMA, 2001). Nesse sentido, apesar de serem menos expressos, os receptores
M3 são o principal subtipo muscarínico responsável pela contração do músculo liso
traqueobrônquico (ROFFEL et al, 1988).

73 VASCONCELOS, L. H. C. Fundamentação Teórica
A liberação de ACh das terminações nervosas parassimpáticas é regulada por
uma variedade de receptores pré-juncionais que podem inibir ou facilitar a liberação
do neurotransmissor. Os receptores M2 autoinibitórios, ativados pela própria ACh,
representam um importante feedback negativo, limitando a liberação adicional,
particularmente nas frequências mais altas. Nesse contexto, a disfunção desses
receptores contribui para a liberação exagerada de acetilcolina pelo nervo vago na
asma, aumentando a atividade reflexa colinérgica em resposta a estímulos inalatórios
(SANTING et al., 1992). A maioria das disfunções do autorreceptor M2 é causada por
eosinófilos ativados que migram para nervos colinérgicos e liberam MBP, que atua
como um antagonista alostérico dos receptores M2 (JACOBY; GLEICH; FRYER, 1993;
COSTELLO et al., 1997; ADAMKO et al., 1999) (Quadro 3).
Quadro 3 – Principais agonistas, receptores e acoplamentos envolvidos na contração do músculo liso das vias aéreas.
Agonista Receptor Sistemas efetores
ACh M3 Gq/11-PLCβ
Adenosina A1 Gi/o-AC
Bradicinina B1 Gq/11-PLCβ
CysLTs CysLT1R Gq/11-PLCβ
Endotelina-1 ETA/ETB ETA: Gq/11-PLCβ; ETB: Gq/11-PLCβ e Gi/o-AC
Histamina H1 Gq/11-PLCβ
Neurocininas NK1/NK2 Gq/11-PLCβ
PGD2 DP2 Gi/o-AC
PGF2α FP Gq/11-PLCβ
Serotonina 5-HT2A/5-HT2C Gq/11-PLCβ
TxA2 TP Gq/11-PLCβ
AC: ciclase de adenilil; PLCβ: fosfolipase Cβ. Fonte: adaptado de PELAIA et al., 2008 e de ALEXANDER et al., 2013.
Componentes i-NANC do sistema nervoso autônomo também regulam o tônus
das vias aéreas liberando outros agentes contráteis, como substância P, trifosfato de
adenosina (ATP) e neurocinina A, que podem agir em seus receptores específicos
diretamente no músculo liso, além de facilitar a neurotransmissão colinérgica, como
já descrito (BURNSTOCK, 2013). Por fim, diversas células localizadas nas paredes
das vias aéreas, como as células epiteliais, as células inflamatórias e os próprios
miócitos, podem liberar uma diversidade de mediadores, que também possuem papel
contrátil direto, como a histamina, metabólitos do AA, (como os prostanoides, o

74 VASCONCELOS, L. H. C. Fundamentação Teórica
CysLTC4 e o CysLTD4) (BARNES; PIPER; COSTELLO, 1984; LIU et al., 1990;
CHAMBERS et al., 2003), a endotelina, liberada pelas células epiteliais, dentre outros
(PEREZ-ZOGHBI; SANDERSON, 2007) (Quadro 3).
Todos esses agonistas compartilham uma característica, que é a necessidade
de se ligar a receptores presentes na membrana plasmática das células musculares
lisas. Após ativar seus receptores, desencadeiam uma cascata de sinalização
intracelular que culmina com a contração muscular. Desse modo, diz-se que esses
agonistas agem por um mecanismo fármaco-mecânico de contração (WEBB, 2003);
diferentemente, a contração pode ocorrer por mecanismos que independem da
ligação a receptores, mas que, por sua vez, promovem a contração por induzirem uma
despolarização de membrana, levando à abertura dos canais de Ca2+ dependentes de
voltagem (CaV), seguido do influxo de Ca2+ por esses canais, resultando na contração
muscular lisa (REMBOLD, 1996).
De todo modo, tanto no acoplamento fármaco-mecânico como no
eletromecânico, o evento chave para desencadear a contração muscular é o influxo
de Ca2+, de forma que esse íon é essencial para desencadear não apenas processos
celulares como secreção glandular e transmmissão sináptica, mas também a
contração muscular (CATTERALL, 2011). Somado a esse influxo de Ca2+, outro
evento responsável por iniciar a contração é a liberação desse íon dos seus estoques
intracelulares pelos receptores de rianodina (RyR). O retículo sarcoplasmático (RS)
representa a fonte intracelular mais importante de Ca2+; a liberação desse íon se dá
principalmente em decorrência da ativação dos receptores de 1,4,5-trisfosfato de
inositol (IP3) (IP3R), formado a partir do 4,5-bisfosfato de fosfatidilinositol (PIP2) da
membrana, seguida da abertura dos receptores de rianodina (RyR) pelo próprio Ca2+,
processo denominado de liberação de Ca2+ induzida por Ca2+ (CICR) (DELLIS et al.,
2006).
O mecanismo fármaco-mecânico de contração ocorre pela ligação do agonista
ao seu receptor acoplado à proteína G, mudança conformacional do receptor e
interação de sua terceira alça intracelular à subunidade α dessa proteína; essa
interação induz a troca do difosfato de guanosina (GDP) pelo trifosfato de guanosina
(GTP) pela subunidade Gα, a qual gera um impedimento estérico e sua consequente
dissociação do dímero Gβγ; a subunidade Gα na sua forma ativa estimula a enzima
fosfolipase Cβ1, que hidrolisa o fosfolipídio de membrana PIP2 a IP3 e diacilglicerol
(DAG). O IP3 induz a liberação de Ca2+ do RS por ativar os IP3R, e o Ca2+ ativa os

75 VASCONCELOS, L. H. C. Fundamentação Teórica
RyR, levando à liberação adicional de Ca2+; por sua vez, o DAG ativa a proteína cinase
C (PKC), translocada para a membrana pelo Ca2+, e ativa por fosforilação os CaV na
membrana, levando ao aumento na concentração citosólica de íons Ca2+ ([Ca2+]c) ou,
indiretamente, por inibição dos canais de K+ presentes na membrana plasmática,
levando à despolarização da membrana e consequente abertura dos CaV (FUKATA;
AMANO; KAIBUCHI, 2001) O DAG também ativa diretamente canais catiôicos não
seletivos, permeáveis ao Na+ e Ca2+, geralmente referidos como canais operados por
receptor (ROCs), formados por TRPC6 (SNETKOV et al., 2001; CORTELING et al.,
2004; PEREZ-ZOGHBI et al., 2009) (Figura 5).

76 VASCONCELOS, L. H. C. Fundamentação Teórica
Figura 5 – Mecanismo do acoplamento fármaco-mecânico de contração muscular lisa pela via da PLCβ1/2
1. O agonista se liga ao seu receptor do tipo GPCR na membrana plasmática, que pode estar acoplado às proteínas Gq/11 (1a) ou Gi/o (1b); 2. as subunidades α das proteínas G trocam GDP por GTP na sua subunidade (processo não mostrado na figura), tornando-se ativas (2a e 2b); 3. a subunidade αq/11-GTP ativa a enzima PLCβ1 (3a) enquanto o dímero βγi/o ativa a PLCβ2 (3b); 4. as PLCβ1/2 clivam o fosfolipídio de membrana PIP2 em IP3 e DAG (4a e 4b); O 5. o IP3 migra pelo citoplasma e ativa os IP3R presentes na membrana do RS, liberando o Ca2+ do RS; 6. o Ca2+ liberado ativa o RyR, aumentando a liberação de Ca2+ para o citosol; 7. O DAG formado pode ativar diretamente os ROCs, promovendo o influxo de Ca2+; 8. o Ca2+ que foi liberado, juntamente com o DAG ativam a PKC; 9. a PKC ativada fosforila os CaV, promovendo o influxo de Ca2+; 10. A elevação da [Ca2+]C aumenta a afinidade pela CaM, formando o complexo 4Ca2+-CaM, e ativando a MLCK; 11. a MLCK ativada fosforila a rMLC e esta se torna ativa, levando à interação dos filamentos de miosina com os de actina, desencadeando a contração do músculo liso. DAG: diacilglicerol; IP3R: receptor de IP3; RyR: receptor de rianodina; GDP: difosfato de guanina; GTP: trifosfato de guaninia; RS: retículo sarcoplasmático; PKC: proteína cinase C; MLCK: cinase da cadeia leve da miosina; CaM: calmodulina; ROC: canais operados por receptor; CaV: canais de Ca2+ dependentes de voltagem; IP3: 1,4,5-trisfosfato de inositol; PLCβ1/2: fosfolipase Cβ 1 e 2; PIP2: 4,5-bisfosfato de fosfatidilinositol. Fonte: Vasconcelos, 2017.
O ponto chave para a contração muscular é atingido com o aumento na [Ca2+]c,
que favorece a ligação desse íon à proteína calmodulina (CaM), na proporção de
quatro íons Ca2+ para uma CaM, formando o complexo 4Ca2+-CaM. Este complexo
ativa a cinase da cadeia leve da miosina (MLCK), que fosforila a cadeia leve
regulatória da miosina (rMLC) no resíduo Ser-19, promovendo a a formação das
pontes cruzadas entre os filamentos de miosina e os de actina, desencadeando assim
a contração muscular lisa. A MLCK é superexpressa em indivíduos asmáticos,
explicando a maior tensão desenvolvida pelo músculo liso brônquico (MA et al., 2002).

77 VASCONCELOS, L. H. C. Fundamentação Teórica
Esse acoplamento também é nomeado de acoplamento misto, já que pode determinar
a despolarização de membrana seja promover a abertura de canais catiônicos não
seletivos ou pela inibição dos canais de K+ pela PKC (WEBB, 2003) (Figura 5).
Após a exposição aos agonistas contráteis, o pico inicial e transitório na
concentração intracelular de Ca2+ é seguido rapidamente por um declínio para um
padrão estacionário caracterizado por um leve aumento prolongado do Ca2+ livre
citosólico acima dos níveis de repouso. Diferentemente da primeira fase da resposta,
devido à liberação de Ca2+ do RS, o estado estável (tônico) depende do influxo de
Ca2+ (MURRAY; FLEISCHMANN; KOTLIKOFF, 1993).
A depleção do Ca2+ do RS pelos processos já descritos estimula a entrada de
Ca2+ induzida por estoque via ação da STIM1 presente na membrana do RS que,
associada à proteína Orai, na membrana plasmática, promove a formação dos canais
de Ca2+ operados por estoque (SOCs) (HEWAVITHARANA et al., 2007; PEEL; LIU;
HALL, 2008). Além disso, o Ca2+ livre no citosol é bombeado de volta pela bomba de
Ca2+ do RS (SERCA) e para fora da célula pela bomba de Ca2+ da membrana
plasmática (PMCA), assim como também é transferido para o exterior da célula pelo
trocador Na+/Ca2+ (NCX), que transporta três íons Na+ para o interior da célula em
troca de um íon Ca2+ para fora (BERRIDGE, 2008).
Os mecanismos de entrada e liberação, e de extrusão e recaptação de Ca2+
nas células musculares lisas das vias aéreas encontram-se esquematiados na
Figura 6.

78 VASCONCELOS, L. H. C. Fundamentação Teórica
Figura 6 – Mecanismos de transporte de Ca2+ nas células musculares lisas das vias aéreas.
Os quadros pontilhados azuis representam os mecanismos de entrada e liberação de Ca2+, e as linhas pontilhadas vermelhas representam os mecanismos de extrusão e recaptação de Ca2+. Os níveis citosólicos de Ca2+ ([Ca2+]c) podem aumentar na célula muscular lisa das vias aéreas por meio do influxo através de Canais de Ca2+ operados por receptor (ROCs) ou por voltagem (CaV), que podem ser ativados, respectivamente, pelo diacilglicerol (DAG) ou pela proteína cinase C (PKC). A entrada também pode ocorrer pelo trocador Na+/Ca2+ reverso (NCX), que traansporta 1 Ca2+ para dentro da célula em troca de 3 Na+ para fora; ou pelos canais operados por estoque (SOCs), mediante ativação pela proteína de interação estromal tipo 1 (STIM1). O aumento do Ca2+ também se dá pela liberação a partir dos estoques no retículo sarcoplasmático (RS), pelos receptores de IP3 (IP3R) e de rianodina (RyR). Já a redução da [Ca2+]c ocorre pelo transporte de Ca2+ pelas Ca2+-ATPase da membrana plasmática (PMCA), para fora da célula, ou do RS (SERCA), para ser reestocadado; além da ação do trocador Na+/Ca2+, que transporta 1 íon Ca2+ para fora da célula em troca de 3 íons Na+ para o interior da célula muscular lisa. Fonte: Vasconcelos, 2017.
Apesar desses mecanismos de recaptação e efluxo de Ca2+, de forma
semelhante a outros músculos lisos, a contração muscular lisa das vias aéreas
persiste mesmo duranto estado estacionário do Ca2+; esse comportamento funcional
implica uma sensibilidade aumentada ao Ca2+ do aparato contrátil (SOMLYO;
SOMLYO, 1994) que permite que o músculo permaneça contraído, mesmo quando os
níveis intracelulares de Ca2+ são apenas ligeiramente aumentados acima das
concentrações iniciais. Os principais mecanismos de sinalização responsáveis por
essa sensibilização ao Ca2+ incluem a via RhoA/Rho cinase (ROCK) e o sistema efetor
PKC/CPI-17 (preptídio inibidor de fosfatase ativada por PKC, de 17 kDa) (SOMLYO;
SOMLYO, 2003).

79 VASCONCELOS, L. H. C. Fundamentação Teórica
A RhoA é uma pequena proteína monomérica que, no repouso, encontra-se
ligada ao GDP (por isso é denominada de “pequena proteína G”) e complexada no
citosol à proteína inibidora da dissociação do GDP (GDI). A interação de agonistas
contráteis com seus receptores também ativa as proteínas G12/13, cuja subunidade Gα
ativa diretamente o fator trocador de nucleotídio de guanina da RhoA (RhoGEF), que
dissocia a RhoA da GDI, permitindo a troca do GDP pelo GTP e sua translocação para
a face interna da membrana (SYLVESTER, 2004), onde ela ativa a sua cinase ROCK;
essa interação entre a RhoA e a ROCK induz uma mudança conformacional na porção
catalítica da ROCK, causando uma autofosforilação e ativação da enzima (SOMLYO;
SOMLYO, 2003), que então fosforila a subunidade de ligação à miosina (MYPT1) da
MLCP, tornando-a inativa. Dessa forma, a cadeia leve regulatória da miosina (rMLC)
não pode ser desfosforilada, mantendo o estado de “trava” das pontes cruzadas (ITO,
2001).
Diversos estudos mostram que a expressão de RhoA é aumentada na asma
(CHIBA et al., 2005) e que essa proteína também é superexpressa por citocinas, como
TNF-α, potencializando a contração induzida por ACh (SAKAI et al., 2004).
A atividade da MLCP também é modulada negativamente pela CPI-17, um
peptídio que serve como substrato tanto para a ROCK como para a PKC. A propósito,
a expressão desse peptídio, bem como sua fosforilação pela PKC induzida por ACh,
são aumentadas em ratos hiper-responsivos (SAKAI et al., 2004). Quando fosforilada
no resídio Thr38, a CPI-17 se liga à subunidade catalítica da proteína fosfatase 1
(PP1c) da MLCP para inibir a sua atividade enzimática. Logo, assim como ocorre na
via da ROCK, a rMLC se mantém em no estado fosforilado, permitindo a manutenção
da contração muscular lisa (KITAZAWA et al., 2000).
A RhoA-GTP também estimula a fosfolipase D (PLD), enzima associada a
membranas intracelulares e à membrana plasmática, específica para fosfatidilcolina
(PC) que, quando hidrolisada, libera o ácido fosfatídico (PA) que, através da ação da
enzima fosfo-hidrolase, é desfosforilado a DAG, que promove a ativação sustentada
da PKC (BERRIDGE, 2009). A ativação da PKC pela via da PLCβ1 sinergiza com a
via da PLD, de modo que a PKC ativada de maneira sustentada pode fosforilar o
resíduo Thr38 da CPI-17, aumentando assim sua potência inibitória sobre a PP1c por
mais de 1000 vezes, inibindo a ação da MLCP e mantendo a contração (SOMLYO;
SOMLYO, 2003) (Figura 7).

80 VASCONCELOS, L. H. C. Fundamentação Teórica
A maioria dos GPCRs conhecidos preferencialmente se acopla aos membros
da família Gi/o, sendo a proteína G mais abundantemente expressa na maioria dos
tipos celulares. A ativação de Gαi é tipicamente associada à inibição da atividade da
ciclase de adenilil (AC), que é estimulada por Gαs e, portanto, reduzindo a geração de
cAMP, cuja consequência funcional é a contração do músculo liso ou dificultar o seu
relaxamento. A subunidade Gβγ também pode ativar a PLCβ2, resultando em geração
de IP3 e DAG, com consequente mobilização de Ca2+ dos estoques e ativação da
PKC, respectivamente (BILLINGTON; PENN, 2003). Além disso, a proteína Gi parece
ser capaz de ativar a RhoA pela ativação do RhoGEF, mediando tanto a polimerização
de actina como, possivelmente, a sensibilização ao Ca2+; no entanto, ainda não está
claro se a ativação da RhoA é mediada pela subunidade α ou βγ das proteínas Gi/o
(HIRSHMAN; EMALA, 1999; CHIBA; SAKAI; MISAWA, 2001).

81 VASCONCELOS, L. H. C. Fundamentação Teórica
Figura 7 – Mecanismo do acoplamento fármaco-mecânico de contração muscular lisa pela via de sensibilização ao Ca2+.
1. O agonista se liga ao seu receptor do tipo GPCR na membrana plasmática, que pode estar acoplado às proteínas G12/13 (1a) ou Gi/o (1b); 2. as subunidades α das proteínas G trocam GDP por GTP na sua subunidade (processo não mostrado na figura), tornando-se ativas (2a e 2b); 3. a subunidade α12/13-GTP (3a) ou via ativação de GPCR acoplado a Gi/o (3b), a RhoGEF é ativada; 4. A RhoA se dissocia da GDI e troca o GDP pelo GTP, sendo translocada à membrana plasmática; 5. a RhoA ativa sua cinase associada, a RhoK; 6. A RhoK fosforila a MLCP na subunidade MYPT1, tornando-a inativa; ou 7. fosforila diretamente a cadeia leve da miosina; 8. a RhoA-GTP ativa diretamente a PLD que 9. cliva a PC a PA, que é então 10. convertido a DAG para, juntamente com o DAG proveniente da via da PLCβ1, 11. ativar a PKC que, por sua vez, 12. fosforila e ativa a CPI-17, que 13. interage com a subunidade PP1c da MLCP, inativando-a, contribuindo, então, para a manutenção do estado fosforilado da rMLC e das pontes cruzadas de actina e miosina. DAG: diacilglicerol; GDI: proteína inibidora da dissociação do GDP; GPCR: receptor acoplado à proteína G; IP3: 1,4,5-trisfosfato de inositol; MLCP: fosfatase da cadeia leve da miosina; MYPT1: subunidade de ligação à miosina; PA: ácido fosfatídico; PC: fosfatidilcolina; PIP2: 4,5-bisfosfato de fosfatidilinositol; PLD: fosfolipase D; PP1c: região catalítica da proteína fosfatase 1; RhoGEF: fator de troca de nucleotídeo guanina da RhoA; RhoK: Rho cinase. Fonte: Vasconcelos, 2017.
Apesar de poucas vezes lembrada, o papel da via das proteínas Gi/o na
contração muscular lisa, especialmente das vias aéreas, se torna evidente quando se
leva em conta que um dos mediadores contráteis mais potentes da asma, a PGD2,
promove forte broncoconstrição por se ligar a receptores DP2, os quais são acoplados
às proteínas Gi/o, além dos receptores TP, dependente da via da PLCβ1, dessa forma,
promovendo um efeito sinérgico entre as duas vias de sinaliação (JOHNSTON et al.,
1995).

82 VASCONCELOS, L. H. C. Fundamentação Teórica
O músculo liso aéreo apresenta características peculiares que o diferenciam
dos músculos esqueléticos e cardíacos, bem como dos músculos lisos vasculares.
Nesses tecidos, o acoplamento excitação-contração resulta em grande parte da
despolarização da membrana, levando ao influxo de Ca2+ através dos CaV, enquanto
nas vias aéreas, essa contribuição é pequena. De fato, bloqueadores de CaV exercem
apenas efeitos discretos na broncoconstrição induzida experimentalmente (MARSICO
et al., 1990; SANDERSON et al., 2008).
O potencial de membrana (Vm) dos miócitos das vias aéreas, de cerca
de -60 mV, varia espontaneamente, dando origem às chamadas ondas lentas, cuja
amplitude é de 8 a 12 mV, em uma frequência de 20 a 50 eventos/min, (TOMITA,
1989; THIRSTRUP, 2000). Esta atividade miogênica é provavelmente devido a uma
produção constitutiva de metabólitos do AA (HIROTA; HELLI; JANSSEN, 2007). No
entanto, essas ondas geralmente não são convertidas em potenciais de ação por
causa de um forte contrabalanço promovido pelo efluxo de íons K+. Essas correntes
retificadoras são mediadas pela abertura de canais de K+ ativados por Ca2+ de grande
condutância (BKCa), bem como de canais de K+ retificadores atrasados dependentes
de voltagem (KV), e de canais de K+ sensíveis ao ATP (KATP) responsáveis pelos fluxos
repolarizantes de íons K+ (HIROTA; HELLI; JANSSEN, 2007; ROUX et al., 2006).
O Vm das células musculares lisas das vias aéreas é ligeiramente maior que o
potencial de equilíbrio do K+ (EK), indicando a importância desses canais na
manutenção do Vm dessas células (KIRKPATRICK, 1981), e a despolariação da
membrana do músculo liso pode ser desencadeada pela inibição desses canais
(MOUNKAÏLA; MARTHAN; ROUX, 2005; ROUX et al., 2006). Outrossim, outros
canais também estão envolvidos na regulação/alteração do Vm do músculo liso aéreo,
como os canais de Cl- ativados por Ca2+ (CaCCs) e, em alguns casos, a entrada de
íons Na+ por vários canais catiônicos não seletivos (MOUNKAÏLA; MARTHAN; ROUX,
2005; ROUX et al., 2006).
Desse modo, o acoplamento eletromecânico da contração muscular lisa
depende da despolarização da membrana plasmática, que pode ocorrer pelo bloqueio
dos canais de K+ ou pelo aumento da concentração desse íon no meio extracelular;
nesse último caso, a ferramenta padrão utilizada é a estimulação com KCl, que
promove uma redução no gradiente eletroquímico do K+, reduzindo seu efluxo e
consequente acúmulo no interior celular, gerando a despolarização da membrana e
consequente abertura dos CaV, influxo de íons Ca2+ e consequente ativação da

83 VASCONCELOS, L. H. C. Fundamentação Teórica
maquinaria contrátil muscular lisa, como já descrito anteriormente (BOLTON, 1979)
(Figura 8).
Figura 8 – Acoplamento eletromecânico de contração muscular lisa.
No repouso, as células musculares lisas apresentam um efluxo constante de íons K+ devido ao gradiente eletroquímico desse íon, gerando uma carga residual negativa na região perimembranar interna, sendo o principal determinante do potencial de repouso de cerca de -60 mV (A). Um aumento na concentração de K+ no meio extracelular (pela adição de KCl, por exemplo), gera uma redução no gradiente eletroquímico desse íon, reduzindo seu efluxo e promovendo o seu acúmulo no interior da célula. Essa concentração de cargas positivas no espaço perimembranar interno determina uma despolarização de membrana, que leva à abertura dos CaV, influxo de Ca2+ e consequente ativação da maquinaria contrátil muscular lisa. CaV: canais de Ca2+ dependentes de voltagem. Fonte: Vasconcelos, 2017.
Se opondo à estimulação colinérgica, a estimulação adrenérgica estimula o
relaxamento do músculo liso aéreo. Apesar da inervação adrenérgica ser escassa, os
receptores β2, acoplados à proteína Gs (ALEXANDER et al., 2013), são altamente
expressos nas células musculares lisas (PACK; RICHARDSON, 1984; IND, 1994).
Além disso, a inervação adrenérgica, apesar de ser fracamente direcionada ao
músculo liso, age sobre os gânglios parassimpáticos, promovendo o controle da
estimulação colinérgica (DANIEL; KANNAN; DAVIS, 1986; RHODEN; MELDRUM;
BARNES, 1988). Além disso, a inervação i-NANC libera substâncias relaxantes, como
o peptídio intestinal vasoativo (VIP) e o óxido nítrico (NO), que se somam aos efeitos
da estimulação adrenérgica (JARTTI, 2001).O relaxamento no músculo liso ocorre
como resultado da remoção do estímulo contrátil ou pela ação de substâncias que

84 VASCONCELOS, L. H. C. Fundamentação Teórica
estimula a inibição do mecanismo contrátil, cujos principais mediadores foram
descritos (MORGAN, 1990).
O relaxamento muscular liso das vias aéreas ocorre devido à hiperpolarização
da membrana pela ação do NO, liberado pelo epitélio das vias aéreas ou pelas
terminações dos neurônios i-NANC, que ativa diretamente a ciclase de guanilil solúvel
(sGC); ou pale ação de agonistas relaxantes, como a adrenalina e a PGE2, sobre seus
receptores (já descritos), acoplados à proteína Gs; uma vez ligada ao GTP, a Gαs ativa
a enima ciclase de adenilil (AC). A sGC converte o GTP em monofosfato cíclico de
guanina (cGMP), que, por sua vez ativa a proteína cinase G (PKG), enquanto que a
AC converte o ATP em cAMP, que ativa a proteína cinase A (PKA). Ambas as
proteínas cinases ativam os canais de K+, com consequente efluxo desse íon,
hiperpolarização de membrana e fechamento dos CaV (REMBOLD, 1996). Além disso,
a hiperpolarização da membrana das células musculares lisas pode ser produzida por
substâncias que abrem diretamente os canais de K+, como por exemplo, a
cromacalina, a levocromacalina e o nicorandil e, consequentemente, aumentam o
efluxo de K+ (GURNEY, 1994). Dessa maneira, a hiperpolarização diminui o influxo de
Ca2+ através dos CaV, diminuindo, portanto, a [Ca2+]c, inativando o aparato contrátil e,
como consequência, levando ao relaxamento do músculo liso (REMBOLD, 1996).
Além desses alvos, a PKA ou PKG fosforilam outros substratos, promovendo:
o aumento na atividade da SERCA e da Ca2+-ATPase da membrana plasmática
(PMCA), aumentando assim, o sequestro e a extrusão de Ca2+, respectivamente; a
inativação dos IP3R, pela PKG, reduzindo a liberação de Ca2+ a partir do RS; a inibição
dos CaV, causando uma redução do influxo de Ca2+; a ativação dos canais de K+ que,
por repolarização ou por hiperpolarização, causam o bloqueio dos CaV e, por fim, a
inibição da MLCK, reduzindo sua afinidade pelo complexo 4Ca2+-CaM, diminuindo a
fosforilação da rMLC e, dessa forma, do processo contrátil (WOODRUM; BROPHY,
2001; DUTTA et al., 2002; DANILA; HAMILTON, 2004) (Figura 9).
Dentre os canais de K+ importantes na manutenção do tônus das células
musculares lisas das vias aéreas, destacam-se os BKCa, que apresentam uma grande
expressão na membrana dessas células e conduzem uma corrente de K+ de elevada
amplitude, uma vez que apresentam grande condutância a esse íon (KUME et al.,
1990). Esses canais são ativados por fosforilação por PKA (KUME et al., 1994; KUME,
2013), via agentes como agonistas adrenérgicos β2, e inativados por agonistas

85 VASCONCELOS, L. H. C. Fundamentação Teórica
muscarínicos M2, via Gαi/o, por inibição da AC e, consequentemente, da PKA (KUME;
KOTLIKOFF, 1991; KUME; GRAZIANO; KOTLIKOFF, 1992).
Além disso, o dímero βγ das proteínas Gi/o podem ativar a PLCβ2, como já foi
descrito, e, subsequentemente, a PKC pode fosforilar e inibir os BKCa, contribuindo
para dificultar o relaxamento (ZHOU et al., 2008). Nesse contexto, existem evidências
que, na asma, os receptores adrenérgicos β2 sofrem dessensibilização relacionada ao
processo inflamatório, como a ação da citocina IL-1β (KOTO et al., 1996) e do TGF-β
(ISHIKAWA et al., 2003), de modo que esse processo está intimamente relacionado
com a menor responsividade relaxante devido à inativação da via Gs/BKCa, importante
determinante do relaxamento muscular liso das vias aéreas (KUME, 2015).

86 VASCONCELOS, L. H. C. Fundamentação Teórica
Figura 9 – Mecanismo do acoplamento fármaco-mecânico de relaxamento do músculo liso pelas vias da AC e da sGC.
1. O agonista se liga ao seu receptor do tipo GPCR na membrana plasmática; 2. a proteína Gs troca GDP por GTP na sua subunidade α (processo não mostrado na figura), tornando-se ativa; 3. a subunidade Gαs-GTP ativa a AC; 4. a AC converte o ATP em cAMP; 5. o NO gerado tanto dos nervos como das células epiteliais estimula a atividade da sGC; 6. a sGC converte o GTP em cGMP; 7. os nucleotídios cíclicos, cAMP e cGMP, ativam suas respectivas proteínas cinases, PKA e PKG. Ambas as proteínas cinases fosforilam vários substratos: 8. ativam os canais de K+; 9. inibem os CaV; 10. aumentam a atividade da SERCA e da PMCA; 11. ativam o NCX; 12. A PKG inibe os IP3R. Todos esses mecanismos diminuem a [Ca2+]C; 13. inibem a MLCK, reduzindo sua afinidade pelo complexo 4Ca2+-CaM. Todos esses mecanismos impedem a fosforilação da rMLC e, consequentemente, a interação dos filamentos de miosina com os de actina, promovendo o relaxamento do músculo liso. AC: ciclase de adenilil; ATP: trifosfato de adenosina; cAMP: monofosfato cíclico de adenosina; CaV: canais de cálcio dependentes de voltagem; cGMP: monofosfato cíclico de guanosina; GPCR: receptor acoplado à proteína G; IP3R: receptor de IP3; MLCK: cinase da cadeia leve da miosina; NCX: trocador Na+/Ca2+; NO: óxido nítrico; PKA: proteína cinase A; PKG: proteína cinase G; PMCA: Ca2+-ATPase da membrana plasmática; SERCA: Ca2+-ATPase do retículo sarcoplasmático; sGC: ciclase de guanilil solúvel. Fonte: Vasconcelos, 2017.
As células musculares lisas têm um papel fundamental no remodelamento das
vias aéreas, pois o aumento na sua massa é um dos responsáveis pelo maior
estreitamento do lúmen bronquiolar (MARTIN; RAMOS-BARBON, 2003;
MCPARLAND; MACKLEM; PARE, 2003). Esse aumento na massa de músculo liso é
explicado, principalmente, pela hiperplasia celular (PANETTIERI et al., 1998a,b;
SALMON et al., 1999), mas também ocorre pela hipertrofia (EBINA et al., 1993). Além
disso, essas células também alteram seu fenótipo para um estado hiper-contrátil, que
contrai mais rapidamente, resultando em um estreitamento exagerado das vias aéreas

87 VASCONCELOS, L. H. C. Fundamentação Teórica
(HALAYKO; STEPHENS, 1994; MA et al., 1998; HALAYKO et al., 1999). Assim, esse
estado é resultante de um aumento nas proteínas da maquinaria contrátil, actina e
miosina (HALAYKO et al., 1996), bem como de proteínas regulatórias, como a MLCK
e a calponina, e de receptores, como o receptor M3 (HALAYKO; STEPHENS, 1994;
HALAYKO et al., 1999).
Adicionalmente ao fenótipo hiper-contrátil, as células musculares lisas
adquirem um fenótipo secretório, produzindo citocinas, quimiocinas e proteínas da
matriz extracelular, contribuindo e promovendo um ciclo de inflamação e
remodelamento que contribui para a progressão da asma (JOHNSON; KNOX, 1997;
McKAY; SHARMA, 2001; BLACK; BURGESS; JOHNSON, 2003; HIRST, 2003;
PANETTIERI, 2003).
Nesse contexto, GPCRs e receptores tirosina-cinase (RTKs) para fatores de
crescimento são os principais reguladores desses processos de mudança fenotípica.
Os GPCRs induzem a proliferação via ativação das proteínas Gi/o ou Gq/11, e agonistas,
como a trombina, são mais eficazes que outros, como a acetilcolina, a histamina e os
Cys-LTs, em promover o crescimento celular (SHIMIZU et al., 2000; TRAN;
STEWART, 2003). Dentre os fatores de crescimento, destacam-se o fator de
crescimento epidérmico (EGF), o fator de crescimento do endotélio vascular (VEGF),
o fator de crescimento de fibroblastos (FGF) e o TGF-β (RUOCCO et al., 1996;
REDINGTON et al., 1997; AMISHIMA et al., 1998; HOSHINO; TAKAHASHI; AOIKE,
2001).
Os fatores de crescimento agem para estimular a proliferação muscular lisa
pela ativação da PI3K, que fosforila o PIP2, produzindo o 3,4,5-trisfosfato de
fosfatidilinositol (PIP3), o qual ativa diretamente a proteína cinase B (PKB ou Akt) ou
indiretamente pela ativação da proteína cinase dependente de PIP3 (PDK), que
fosforila a PKB (ALESSI et al., 1997; FRANKE et al., 1997; TOKER; CANTLEY, 1997).
A PKB inibe a cinase da glicogênio sintase tipo 3 (GSK3) ou ativa a cinase S6, que
ativa a p70. Ambas atividades levam à proliferação (BURGERING; COFFER, 1995;
FRAME; COHEN, 2001). A PI3K também ativa a proteína cinase Src, a qual ativa
outras vias de sinalização ativadas por RTKs, como a via das MAPK (HU et al., 1995).
Gi/o e Gq/11 também podem ativar a PI3K ou a MAPK p42/p44 (ERK 1/2). O
dímero βγ da Gi/o ativa a PI3K diretamente, e esta, por sua vez, pode ativar a ERK
1/2 via Src (LOPEZ-ILASACA et al., 1997). Via Gq, a PKC também pode levar à
transativação de RTKs via proteína tirosina-cinase dependente de Ca2+ (Pyk2), ou

88 VASCONCELOS, L. H. C. Fundamentação Teórica
pode ativar diretamente a Raf-1, que ativa a MEK1 e esta a ERK1/2 (KOLCH et al.,
1993; HAWES; LEV et al., 1995; DELLA-ROCCA et al., 1997; MACK et al., 2001).
Uma possível explicação para o fato dos agonistas de GPCRs não serem fortes
mitógenos é que a ativação da PI3K ou da ERK 1/2 por essa via é fraca e lenta para
induzir proliferação (KELLEHER et al., 1995; ORSINI et al., 1999). Todavia, em
combinação, agonistas GPCRs e fatores de crescimento apresentam forte atividade
mitogênica (PANETTIERI et al., 1998a,b; KRYMSKAYA et al., 2000).
Por fim, além do papel majoritário e independente da PI3K e da ERK1/2,
GPCRs podem induzir proliferação pela ativação da proteína RhoA, que causa a
ativação e translocação nuclear do fator de resposta ao soro (SRF), o qual atua como
fator de transcrição para genes específicos do músculo liso (CAMORETTI-MERCADO
et al., 2000; FUKATA; AMANO; KAIBUCHI, 2001; MACK et al., 2001; LIU et al., 2003).
2.3.7 Modelos experimentais de asma
Diversos modelos animais de asma foram estabelecidos para se pesquisar os
seus mecanismos patológicos e potenciais terapias (NIALS; UDDIN, 2008). Esses
modelos variam com relação à espécie animal, o antígeno, as vias de administração
e o tempo de exposição a esse antígeno. Todavia, a maioria desses modelos
concentra-se na inflamação aguda baseada na exposição a alérgeno por curto período
de tempo; devem-se, no entanto, considerar as diferenças entre a inflamação
pulmonar aguda e crônica, uma vez que a asma é considerada uma doença
inflamatória crônica (McMILLAN; LLOYD, 2004).
As reações precoces e tardias na asma aguda envolvem alterações no controle
neurogênico e não neurogênico da função do músculo liso das vias aéreas, bem como
danos epiteliais, espessamento da mucosa devido à vasodilatação local e ao edema
e secreções mucosas no lúmen das vias aéreas, todos esses envolvidos no
desenvolvimento da hiper-reatividade das vias aéreas (BOUSQUET, et al., 2000).
Além disso, a resposta inflamatória aguda é limitada às vias aéreas proximais e o perfil
de citocinas dos pulmões também varia com o número de desafios alergênicos (DI
VALENTIN et al., 2009). Já na asma crônica, a hiper-responsividade persistente das
vias aéreas, assim como o declínio da função pulmonar, estão relacionados com o
remodelamento tecidual devido a alterações estruturais irreversíveis de suas paredes
(BOUSQUET et al., 2000; MEURS; MAARSINGH; ZAAGSMA, 2003).

89 VASCONCELOS, L. H. C. Fundamentação Teórica
A OVA é tradicionalmente utilizada para mimetizar um modelo animal
sensibilizado; apesar desse agente não ser um antígeno adequado para a patogênese
da asma em humanos, a exposição a esse antígeno exógeno provoca inflamação das
vias respiratórias característica da asma humana de maneira semelhante em animais.
Além disso, apesar de muitos modelos animais induzirem a sensibilização por via
intraperitoneal, essa via não é apropriadamente representativa da história espontânea
de início da asma humana (WALSH et al., 2008).
Uma vez estabelecida a doença pulmonar alérgica dependente de OVA, o
fenótipo da doença não pode ser mantido por períodos prolongados, presumivelmente
por causa do desenvolvimento de mecanismos de tolerância que podem ser
exclusivos do pulmão. Nesse sentido, durante o protocolo de indução da asma,
atenção se faz necessária na tentativa de se evitar o desenvolvimento de tais
mecanismos; isto é, um estado em que o sistema respiratório do animal não mais
progride na direção de uma resposta imune suficiente para promover o
remodelamento tecidual (LOCVVKE et al., 2007; POSSA et al., 2014). Nesse sentido,
em um modelo de asma crônica, o aumento gradual nas concentrações da OVA é
necessário para evitar que os animais desenvolvam tolerância e as alterações
teciduais decorrentes do processo fisiopatológico não sejam desencadeados
(TIBÉRIO et al., 1997; PIGATI et al., 2015; IKEDA, 2016).
Outro aspecto importante no delineamento do protocolo de indução de asma é
com relação à espécie animal. Vários estudos em camundongos têm usado
exposições repetidas ao alérgeno para induzir a inflamação crônica, na qual são
observadas alterações estruturais irreversíveis nas vias aéreas (KUMAR et al., 2003;
HOYMANN et al., 2009; RIESENFELD, 2010). Todavia, apesar da importância dos
roedores (camundongos e ratos) na elucidação dos mecanismos da inflamação
alérgica do sistema respiratório, os mediadores que regulam o tônus do músculo liso
e determinam a responsividade brônquica nesses animais diferem consideravelmente
daqueles encontrados em humanos e cobaias (CANNING, 2003).
Nesse sentido, as desvantagens no uso de camundongos residem na
dificuldade em se medir a hiper-responsividade brônquica devido a características
anatômicas e fisiológicas. Sua estrutura pulmonar é caracterizada por vias aéreas
largas e escassas em tecido muscular liso, assim como em glândulas mucosas
(ZOSKY; SLY, 2007; IRVIN; BATES, 2003). Além disso, o músculo liso aéreo desses
animais é pouco responsivo a diversos estímulos broncoconstritores implicados na

90 VASCONCELOS, L. H. C. Fundamentação Teórica
fisiopatologia da asma, como histamina, cisteinil leucotrienos, neurocininas,
bradicinina e prostanoides (CANNING, 2003).
Semelhantemente, os ratos exibem fraca resposta broncoconstritora,
comparado a humanos, seus mastócitos liberam principalmente serotonina em
resposta ao estímulo antigênico, e a inervação iNANC é ausente nessa espécie.
Dessa forma, a asma em roedores apresenta-se mais útil no estudo da imunologia
que da reatividade contrátii da musculatura lisa das vias aéreas. (CANNING, 2003;
KUCHAREWICZ; BODZENTA-ŁUKASZYK; BUCZKO, 2007).
Sendo assim, do ponto de vista fisiológico e farmacológico, cobaias
sensibilizadas são preferíveis como modelo animal para investigar os mecanismos
das reações asmáticas rápidas e tardias (MEURS; GOSENS; ZAAGSMA, 2008).
Similaridade com humanos tem sido observadas nas reações asmáticas nesses
animais, incluindo inflamação eosinofílica das vias aéreas e hiper-responsividade
brônquica, tanto quantitativa como qualitativamente (MEURS et al., 2006).
Comparado com roedores, cobaias também apresentam maior proximidade
com humanos com relação à resposta muscular lisa e ao reflexo autonômico. Eles
liberam, majoritariamente, histamina e leucotrienos em resposta à estimulação
mastocitária, assim como o perfil de receptores farmacológicos aproxima-se dos
humanos, o que explica a grande similaridade de respostas após exposição a drogas
(CANNING, 2003; RESSMEYER et al., 2006).
Diante disso, o emprego de cobaias como modelo animal de asma é preferível
para se investigar os mecanismos da hiper-responsividade das vias aéreas na asma
comparado a ratos e camundongos (MEURS et al., 2006).
2.3.8 Tratamento atual da asma e novas abordagens terapêuticas
A farmacoterapia atual da asma se baseia em duas frentes: a
broncodilatação/broncoproteção, por inibição direta da contração ou pelo relaxamento
do músculo liso brônquico; e controle da inflamação de modo a limitar a geração de
estímulos que causam a broncoconstrição (PERA; PENN, 2016).
O controle da inflamação é feito pelo uso de anti-inflamatórios esteroidais, os
corticosteroides, por via oral (OCS) ou inalatória (ICS), que agem a longo prazo
inibindo o processo inflamatório, a exemplo da fluticasona e da budesonida (PERA;
PENN, 2016). Desde o seu surgimento e introdução no tratamento da asma, os
corticoides têm apresentado problemas relacionados, especialmente, aos seus efeitos

91 VASCONCELOS, L. H. C. Fundamentação Teórica
colaterais, de modo que novos fármacos da classe foram e são desenvolvidos com o
intuito de melhorar sua potência e, principalmente, seus efeitos sistêmicos (COLICE,
2006).
Os efeitos colaterais mais comuns dos corticosteroides são a candidíase oral,
catarata, glaucoma, hematomas faciais da pele e osteoporose. O risco dessas
complicações é muito menor com os glicocorticoides inalatórios em comparação com
os glucocorticoides orais (GINA, 2017).
Outras drogas anti-inflamatórias são os cromoglicatos que agem estabilizando
os mastócitos, como a cromolina, impedindo a degranulação mastocitária. Tanto os
corticosteroides como os cromoglicatos são empregados para tratar a asma leve
persistente (LANE; LEE, 1996). Outra classe emergente no tratamento da asma grave
são os anticorpos anti-IgE, como o omalizumab, especialmente para pacientes
resistentes aos corticosteroides (HUMBERT et al., 2014).
Outras classes que exibem, além do efeito anti-inflamatório, atividade
broncodilatadora direta são os antagonistas de CysLT1R, que podem complementar
os efeitos dos corticosteroides (CURRIE; LIPWORTH, 2002; DAVIS et al., 2009;
RAMSAY et al., 2009).
Os agentes broncodilatadores agem, principalmente, por ativação de GPCR-Gs
ou antagonismo de GPCR-Gq/11. Esses agentes são formados pelos agonistas
adrenérgicos β, como salbutamol, salmeterol e formoterol; os antagonistas dos
receptores muscarínicos M3, como ipratrópio e tiotrópio; e dos receptores de CysLTs,
como montelucaste e zafirlucaste. Além desses, outra classe empregada no
tratamento da asma, mas que não age diretamente sobre os GPCSs, são os inibidores
de fosfodiesterases, enzimas que degradam os nucleotídios cíclicos, a exemplo da
teofilina (PRADO et al., 2014).
Os agonistas β de ação curta (SABA), a exemplo do salbutamol e do formoterol,
são os principais fármacos utilizados para o controle rápido dos sintomas de asma. A
grande maioria dos pacientes com a forma mais leve de asma ou o tipo intermitente
utiliza esses fármacos por ocasião de crises agudas de asma. São, por isso, usados
para o alívio dos sintomas da asma ou preventivamente em situações de atividades
que são conhecidas por provocar esses sintomas. Já os pacientes com asma
persistente necessitam fazer o uso diário de medicamentos, conhecidos como
"controladores de longo prazo". Nesse contexto, os corticosteroides inalatórios agem
para diminuir a inflamação alérgica pulmonar crônica. Além disso, a adição de um

92 VASCONCELOS, L. H. C. Fundamentação Teórica
agonista β de ação longa (LABA), como o salmeterol ou o formoterol, é muitas vezes
necessária em combinação com um corticosteroide inalatório para adultos com asma
persistente (GINA, 2017).
Os modificadores de leucotrienos, como montelucaste, zafirlucaste e zileutona,
são uma alternativa aos glicocorticoides inalatórios. Eles têm um efeito
broncodilatador, diminuindo a inflamação e a produção e secreção de muco (BERGER
et al., 2009). Esses fármacos são usados para evitar a asma induzida por exercício.
No entanto, para o tratamento convencional da asma, os modificadores de
leucotrienos são geralmente menos efetivos do que os corticosteroides inalatórios
(GINA, 2017).
Já o grupo de pacientes com asma grave, que engloba cerca de 10% dos
indivíduos asmáticos, requer doses elevadas de glicocorticoide inalatório ou
tratamento contínuo ou quase contínuo por via oral para manter seu controle de asma
(PELAIA; VATRELLA; MASELLI, 2012). Para estes pacientes, o tratamento é baseado
em glicocorticoides orais, altas doses de corticoide inalatório e a adição de um terceiro
agente controlador, como um antileucotrieno, teofilina ou mesmo terapia anti-IgE
(BOUSQUET et al., 2004; KERSTJENS et al., 2012). Os agonistas β de ação
prolongada devem ser adicionados e o tiotrópio deve ser reservado para pacientes
com sintomas de asma em curso e limitação do fluxo aéreo mesmo com a combinação
de terapia com glicocorticoide inalado/LABA (BATEMAN et al., 2011).
Diante disso, os medicamentos primários para o tratamento da asma variam de
acordo com a idade e a gravidade da avaliação clínica e funcional. Esses fatores
devem ser revisados e ajustados para o nível requerido de controle de sintomas
(PRADO et al., 2014). A terapia é aumentada de forma gradual até o controle da asma
(com base nos sintomas, necessidade de medicação de resgate e exacerbações) e a
quantidade de classes terapêuticas requeridas define a gravidade da asma
(Figura 10).

93 VASCONCELOS, L. H. C. Fundamentação Teórica
Figura 10 – Passos para o tratamento da asma, de acordo com sua apresentação clínica.
ICS: costicosteroide inalatório; LABA: agonista β de ação longa; OCS: corticosteróide oral; SABA: agonista β de ação curta. Fonte: redesenhado de GINA, 2017.
Novos fármacos estudados até o presente para o tratamento da asma incluem
anticorpos anti-IL-13, que vem mostrando resultados promissores em estudos clínicos
(HODSMAN et al., 2013; NOONAN et al., 2013; DE BOEVER et al., 2014) e anti-IL-5
(POWELL et al., 2015). No Brasil, foi aprovado em 2017 o uso do mepolizumabe,
anticorpo anti-IL-5, para o tratamento da asma grave (BRASIL, 2017b). Além disso,
dentro das classes já existentes, incluem formulações de inaladores triplos, que
contemplam ICS, LABA e antagonista muscarínico de ação longa (LAMA), para o
tratamento da asma grave; inibidores de cinases, como a MAPK p38, PI3K, JAK;
antagonistas de receptores de quimiocinas; além de compostos antioxidantes
(GROSS; BARNES, 2017).
Até o momento, nenhuma terapia contra a asma foi utilizada para direcionar
principalmente contra o remodelamento das vias aéreas. Os fármacos
anti-inflamatórios continuam a ser a categoria de drogas mais importante utilizada na
asma e, apesar dos avanços recentes em novas terapias biológicas altamente
direcionadas no desenvolvimento da fase tardia, ainda há uma necessidade de novas
terapias efetivas de asma (BERAIR; BRIGHTLING, 2014).
Incluídas nesse contexto, novas formas de abordagem terapêutica, pouco
estudadas até então, têm sido desenvolvidas, a exemplo dos alimentos funcionais
que, apesar de não poderem ser utilizados no tratamento efetivo de doenças, podem
servir de adjuvante às terapias convencionais para melhorar as funções fisiológicas,

94 VASCONCELOS, L. H. C. Fundamentação Teórica
permitindo ao organismo responder adequadamente aos estímulos patológicos que
lhe acometem (IFIC, 2006).
Nesse sentido, diante das diversas atividades farmacológicas descritas e seus
efeitos benéficos sobre diversos parâmetros bioquímicos e fisiológicos, bem como sua
atividade anti-inflamatória e seus efeitos sobre doenças crônicas, esse trabalho foi
desenvolvido com o intuito de se avaliar um possível efeito da suplementação de
cobaias com o óleo de coco virgem em um modelo de inflamação pulmonar alérgica
crônica, à luz das suas prováveis ações benéficas sobre a asma e seu potencial como
alimento funcional.

3 Objetivos

96 VASCONCELOS, L. H. C. Objetivos
3.1 Objetivo geral
Avaliar os efeitos da suplementação de cobaias com inflamação pulmonar
alérgica crônica com óleo de coco virgem sobre a reatividade contrátil e relaxante do
músculo liso da traqueia, a morfologia pulmonar e brônquica, o balanço estresse
oxidativo/defesas antioxidantes e o crescimento do músculo liso aéreo.
3.2 Objetivos específicos
Padronizar e implantar um modelo de inflamação pulmonar alérgica crônica em
cobaias no Laboratório de Farmacologia Funcional Prof. George Thomas;
Avaliar os efeitos da inflamação pulmonar alérgica crônica em cobaias e da
suplementação com óleo de coco virgem sobre a(o):
o infiltrado de células inflamatórias no interstício pulmonar, a hiperplasia
epitelial e o crescimento muscular liso das vias aéreas;
o reatividade contrátil da traqueia, in vitro, frente à ovalbumina, aos agonistas
carbacol e histamina, e ao KCl;
o reatividade relaxante da traqueia, in vitro, ao agonista isoprenalina e ao
nifedipino;
o envolvimento do estresse oxidativo, dos produtos da COX e da 5-LO, da via
do NO e da via da RhoA/ROCK na reatividade contrátil da traqueia ao CCh;
o participação do TGF-β, dos BKCa e dos produtos da COX e da 5-LO na
reatividade relaxante da traqueia;
o balanço estresse oxidativo/defesas antioxidantes;
o expressão das proteínas relacionadas à hipertrofia/hiperplasia muscular lisa
PI3K e ERK1/2, e da enzima antioxidante SOD.

4 Material e Métodos

98 VASCONCELOS, L. H. C. Material e Métodos
4.1 MATERIAL
4.1.1 Animais
Cobaias (Cavia porcellus) (n = 123) da linhagem Dunkan-Hartley de ambos os
sexos, com 3 meses de vida, pesando inicialmente entre 300 e 350 g, provenientes
do Biotério Prof. Thomas George do Instituto de Pesquisa em Fármacos e
Medicamentos do Campus I da Universidade Federal da Paraíba (IPeFarM/UFPB)
foram utilizados nesse estudo.
Antes dos experimentos, os animais eram mantidos sob rigoroso controle
alimentar com uma dieta balanceada à base de ração tipo pellets (Privilege®)
acrescida de capim e cenoura para reposição do conteúdo de ácido ascórbico, com
acesso à água ad libitum, com ventilação e temperatura (21 ± 1 °C) controladas e
constantes, submetidos diariamente a um ciclo claro-escuro de 12 h, sendo o período
claro das 6 às 18 h. Os experimentos eram realizados no período das 8 às 22 h. Todos
os procedimentos experimentais foram realizados seguindo os princípios de cuidados
com animais, de acordo com o “Guidelines for the ethical use of animals in applied
ethology studies” (SHERWIN et al., 2003) e o Guia Brasileiro de Produção,
Manutenção ou Utilização de Animais em Atividades de Ensino ou Pesquisa Científica,
do Conselho Nacional de Controle de Experimentação Animal (CONCEA) (BRASIL,
2016) e aprovados pela Comissão de Ética no Uso de Animais da UFPB
(CEUA n° 0410/13) (Anexo 1).
4.1.2 Substâncias e reagentes
O ácido acético (C2H4O2), o cloreto de cálcio di-hidratado (CaCl2.2H2O), o
clorofórmio (CHCl3), o hidróxido de potássio e o sulfato de magnésio hepta-hidratado
(MgSO4.7H2O) foram adquiridos da Vetec (Brasil); o ácido clorídrico (HCl), o
bicarbonato de sódio (NaHCO3), o cloreto de potássio (KCl) e o cloreto de sódio (NaCl)
foram adquiridos da Fmaia (Brasil); o fosfato de potássio monobásico (KH2PO4), o
fosfato de sódio dibásico (Na2HPO4), a glicose e o hidróxido de sódio (NaOH) foram
adquiridos da Nuclear (Brasil); o ácido araquidônico (AA), o ácido etilenodiamino
tetracético (EDTA), o ácido perclórico, o ácido tiobarbitúrico (TBA), a albumina, a
apocinina, a aprotinina, o cloridrato de carbamilcolina (carbacol, CCh), o cloridrato de
Nω-Nitro-L-arginina metil éster (L-NAME), o di-hidrocloridrato de histamina, o
ditiotreitol (DTT), a eosina, o fenil-metil-sulfonil fluoreto (PMSF), a fenolftaleína, o

99 VASCONCELOS, L. H. C. Material e Métodos
formaldeído, a hematoxilina, a indometacina, a isoprenalina, o malondialdeido (MDA),
o metano hidroximetilamina (tris), o nifedipino, a ovalbumina (OVA) (grade V), o
tempol, o tween 20, o xileno e o 1,1-difenil-2-picril hidrazil (DPPH) foram adquiridos
da Sigma-Aldrich (Brasil); a catalase, o di-hidrocloreto de
(R)-(+)-trans-4-(1-aminoetil)-N-(4-piridil) ciclohexanocarboxamida (Y-27632), o hidrato
de 4-(5-benzol[1,3]dioxol-5-il-4-piridina-2-il-1H-imidazol-2-il)-benzamida (SB431542),
a iberiotoxina (IbTX) e a zileutona foram adquiridos da Cayman Chemical (Brasil); a
dexametasona foi adquirida da Aché Laboratórios Farmacêuticos (Brasil); e o soro
fisiológico (NaCl 0,9%) foi adquirido da Fresenius Kabi LTDA (Brasil).
Estas substâncias, exceto o bicarbonato de sódio, o cloreto de sódio e a glicose,
eram dissolvidas e diluídas em água destilada; o ácido araquidônico, a apocinina, a
indometacina e o Y-27632 eram dissolvidos em álcool absoluto (96 °GL) para
obtenção de cada solução-estoque, as quais eram mantidas sob refrigeração a 4
ou -20 °C. A OVA era dissolvida em soro fisiológico (NaCl 0,9%).
A mistura carbogênica (95% de O2 e 5% de CO2) foi adquirida da White Martins
(Brasil).
Todos os reagentes utilizados na eletroforese em gel de poliacrilamida
contendo sódio dodecil sulfato (SDS-PAGE), o reagente de Bradford, o tampão de
Laemmli, o triton X-100, o pirofosfato de sódio, o fluoreto de sódio e o ortovanadato
de sódio foram adquiridos da Bio-Rad (EUA). A membrana de nitrocelulose e os kits
para detecção de quimioluminescência foram adquiridos da Amersham (Reino Unido).
Os anticorpos anti-ERK1/2, -SOD e -β-actina foram adquiridos da Santa Cruz
Biotechnology (EUA), o anticorpo anti-PI3K foi adquirido da Abcam plc. (Reino Unido)
e o anticorpo secundário biotinilado de cabra anticoelho foi adquirido da Vector
Laboratories (EUA).
4.1.3 Soluções
Para a manutenção dos anéis de traqueia, era utilizada a solução de Krebs
(ajustada ao pH 7,4 com NaOH ou HCl 1N), gaseificada com carbogênio e mantida à
temperatura de 37 °C, contendo a seguinte composição (mM): NaCl (118,0); KCl (4,6);
MgSO4 (5,7); KH2PO4 (1,1); CaCl2 (2,5); glicose (11,0); NaHCO3 (25,0).
Para conservação das amostras para procedimentos histológicos, era utilizada
solução de formaldeído tamponado 10% com a seguinte composição: formaldeído
(1:10 v/v), NaH2PO4 (0,04:10 m/v), Na2HPO4 (0,005:10 m/v) e H2O (9:10 v/v).

100 VASCONCELOS, L. H. C. Material e Métodos
As amostras para separação em eletroforese eram extraídas em tampão
contendo a seguinte composição: triton X-100 1%, tris 100 mM (pH 7,4), pirofosfato
de sódio 100 mM, fluoreto de sódio 100 mM, EDTA 10 mM, ortovanadato de sódio 10
mM, PMSF 2 mM e aprotinina 0,01 mg/mL.
Para o bloqueio de ligações inespecíficas de proteínas à membrana de
nitrocelulose era utilizada uma solução de bloqueio contendo leite desnatado Molico®,
tris 10 mM, NaCl 150 mM e tween 20 0,02%.
4.1.4 Aparelhos
Para o registro das contrações isométricas, a traqueia era suspensa em cubas
de banho (6 mL) para órgãos isolados, modelo BOI-04, e conectados a transdutores
de força isométricos modelo TIM 05, acoplados a um amplificador modelo AECAD04F.
Este, por sua vez, era conectado a um sistema de aquisição digital de dados com o
software AQCAD versão 2.1.6, para aquisição dos dados, e ANCAD, para análise. O
sistema continha uma bomba termostática modelo BT-60 que controlava a
temperatura das cubas. Todos os aparelhos foram adquiridos junto a AVS Projetos
(São Paulo, SP, Brasil).
Para indução da inflamação pulmonar alérgica crônica, era utilizado um
nebulizador ultrassônico (Soniclear®, São Paulo/SP), com um fluxo variável entre 0,7
a 1,25 mL/min e tamanho médio da partícula de 4 μm, conectado a um recipiente
fechado de poliacrílico (20x13x15 cm) contendo um orifício para a renovação do ar.
Para a realização dos cortes histológicos, as amostras eram submetidas ao
processador de tecidos modelo PT05 (LUPTEC, Brasil), seccionadas em micrótomo
rotativo modelo RM 2125 RT (Leica, Alemanha), fotografadas em microscópio ótico
com câmera acoplada modelo BX41 (Olympus, Formosa) e posteriormente analisadas
por programa analisador de imagem (Leica Qwin 3.1).
O pH das soluções era aferido através de um pHmetro digital, modelo PG2000
(GEHAKA, São Paulo, SP, Brasil).
As substâncias eram pesadas em balança analítica modelo AG 200 e os
animais em balança semianalítica (ambas da GEHAKA, São Paulo, SP, Brasil).
Para a centrifugação das amostras, eram utilizadas centrífuga refrigerada,
modelo VS-15000 CFN II (VISION, Coreia do Sul) e centrífuga de bancada, modelo
NI 1811-A (NOVA instruments, Piracicaba, SP, Brasil).

101 VASCONCELOS, L. H. C. Material e Métodos
As medidas de absorbância eram feitas utilizando um espectrofotômetro
ultravioleta modelo SP 22 (Bioespectro, Salvador, BA, Brasil).
As amostras eram agitadas em vórtex Maxi Mix II, Thermolyne, type 37600
Mixer (Dubuque, Iowa, EUA). Para aquecer as amostras, eram utilizados
banhos-maria modelo BM-02 (Kacil, Recife, PE, Brasil) e modelo BM-20 (Megapor,
Zaragoza, Espanha).
O processamento das amostras para extração do conteúdo proteico era
realizado com o auxílio do processador de tecidos do tipo Polytron PTA 20S
(Brinkmann Instruments, USA). Além disso, a eletroforese em gel de poliacrilamida
(SDS-PAGE) e transferência de proteínas para a membrana de nitrocelulose foi
realizada em equipamentos da Bio-Rad (Richmond, CA). Adicionalmente, a detecção
da quimiluminescência foi realizada no leitor MF-CHEMIBIS (Bio-Imaging Systems
Ltda., Jerusalem, Israel) e a densitometria óptica foi quantificada no programa ImageJ
1.43u (National Institute of Health, USA).
4.2 MÉTODOS
4.2.1 Grupos experimentais
Os animais foram divididos em 6 grupos (n = 5-8, cada):
GC: grupo controle, submetido a nebulização com soro fisiológico;
GASM: grupo com inflamação pulmonar alérgica crônica, submetido a
nebulização com OVA;
GASM/DEXA: grupo com inflamação pulmonar alérgica crônica tratado com
dexametasona (2 mg/kg/dia);
GASM/OC1: grupo com inflamação pulmonar alérgica crônica suplementado
com óleo de coco virgem (1 g/kg/dia);
GASM/OC2: grupo com inflamação pulmonar alérgica crônica suplementado
com óleo de coco virgem (2 g/kg/dia);
GASM/OC4: grupo com inflamação pulmonar alérgica crônica suplementado
com óleo de coco virgem (4 g/kg/dia);

102 VASCONCELOS, L. H. C. Material e Métodos
4.2.2 Aquisição do óleo de coco virgem
O óleo de coco virgem foi adquirido de loja de produtos naturais do município
de João Pessoa/PB, Brasil, marca Dr. Orgânico®, lote n° V15 245, extraído por
prensagem a frio, não refinado.
Os testes de determinação do perfil lipídico (Apêndices 1 e 2) e de controle de
qualidade (índice de acidez, de peróxidos e de saponificação) (Apêndice 3) foram
conduzidos no Laboratório de Análises Químicas de Alimentos (LAQA), do Centro de
Tecnologia (CT) da UFPB, sob a supervisão da Profa. Dra. Marta Suely Madruga.
4.2.3 Indução da inflamação pulmonar alérgica crônica em cobaias
Para a nebulização, os cobaias eram colocados individualmente em uma caixa
poliacrílica fechada acoplada a um nebulizador ultrassônico (Figura 11). Logo após,
eram nebulizados com solução de OVA em soro fisiológico por um tempo máximo de
15 minutos ou até o aparecimento de desconforto respiratório, definido pela presença
de espirros, coriza, tosse e/ou tiragem da parede torácica; o grupo controle recebia
apenas soro fisiológico. O tempo que os cobaias permaneciam em nebulização foi
definido como “tempo de inalação”.
Figura 11 – Aparato utilizado para nebulização dos cobaias.
Fonte: Vasconcelos, 2017.
O protocolo consistia de sete inalações realizadas durante um período de
quatro semanas, em intervalos de 96 h entre cada inalação, com a concentração de

103 VASCONCELOS, L. H. C. Material e Métodos
OVA sendo aumentada (1-5 mg/mL) a fim de se evitar tolerância. Nas quatro primeiras
inalações, os cobaias eram submetidos à OVA na concentração de 1 mg/mL; na quinta
e na sexta inalações, os animais recebiam solução de OVA 2,5 mg/mL; e na sétima
inalação, a concentração de OVA era aumentada para 5 mg/mL. Após 72 h da última
inalação, os animais eram eutanasiados para experimentação (Figura 12). Os cobaias
do grupo controle eram submetidos ao mesmo procedimento de inalação, porém
recebendo apenas soro fisiológico (Adaptado de TIBÉRIO et al., 1997; ANGELI et al.,
2008; PIGATI et al., 2015).
Figura 12 - Protocolo de indução da inflamação pulmonar alérgica crônica em cobaias.
DEXA: dexametasona; OCV: óleo de coco virgem; OVA: ovalbumina. Fonte: Vasconcelos, 2017.
A análise, in vitro, da indução do quadro de inflamação pulmonar alérgica
crônica era feita pela Reação de Schultz-Dale, por meio da qual as preparações de
traqueia eram estimuladas com solução de OVA 10 µg/mL, que induz a liberação de
mediadores contráteis pelas células do microambiente, e, na presença de reatividade
contrátil, o animal era considerado sensibilizado (SCHULTZ, 1910; DALE, 1913;
CHAND; EYRE, 1978; JONES; CHARETTE; DENIS, 1988). Adicionalmente, foi feita
a análise histológica dos tecidos pulmonar e brônquico, para comprovar o
desenvolvimento da inflamação pulmonar pelos animais, conforme descrito adiante.

104 VASCONCELOS, L. H. C. Material e Métodos
4.2.4 Tratamento e suplementação dos animais
Os grupos suplementados com óleo de coco virgem recebiam, por via oral, por
meio de gavagem, doses diárias de 1, 2 ou 4 g/kg (INTAHPHUAK; KHONSUNG;
PANTHONG, 2010; NANDAKUMARAN et al., 2011; ALVES et al., 2015;
VASCONCELOS, 2015) durante todo o período de indução da asma, finalizando no
dia anterior ao da eutanásia do animal. Os animais do grupo que recebeu a droga
padrão (dexametasona) foram tratados com o fármaco na dose de 2 mg/kg/dia, por
via intraperitoneal, iniciando-se 24 horas após a quarta nebulização, para garantir que
os animais estavam sensibilizados. Durante esse período, o tratamento com
dexametasona era feito 5 horas antes da exposição ao antígeno
(LEICK-MALDONADO et al., 2004).
4.2.5 Avaliação dos efeitos da inflamação pulmonar alérgica crônica e do
tratamento com a dexametasona ou da suplementação com o óleo de coco
virgem sobre o tempo de nebulização de cobaia
O tempo decorrido para que os cobaias começassem a apresentar sinais de
desconforto respiratório, durante o tempo de inalação, que durava até o máximo de
15 minutos, era avaliado, assim como o desenvolvimento de restrições respiratórias,
tais como dispneia ou apneia. As comparações eram feitas entre os grupos GC,
GASM, GASM/DEXA, GASM/OC1, GASM/OC2 e GASM/OC4.
4.2.6 Avaliação dos efeitos da inflamação pulmonar alérgica crônica e do
tratamento com a dexametasona ou da suplementação com o óleo de coco
virgem sobre a morfologia pulmonar e brônquica de cobaia
Os cobaias eram eutanasiados por deslocamento cervical seguido de secção
dos vasos cervicais, procedimento que foi aprovado pela CEUA/UFPB, uma vez que
o uso de guilhotina promove dano à traqueia. Além disso, o exsanguinamento garante
a total retirada do sangue do leito vascular, que poderia interferir nos experimentos
pela presença de substâncias que modulam a contratilidade muscular lisa quando em
contato com o lúmen da traqueia. Por fim, o uso de anestésicos poderia interferir com
o potencial de membrana da célula muscular lisa, alterando a atividade de canais
iônicos. Tais medidas são amparadas pela Resolução nº 1000/2012 do Conselho
Federal de Medicina Veterinária, capítulo III, Art. 14, § 2º (BRASIL, 2012).

105 VASCONCELOS, L. H. C. Material e Métodos
Após a eutanásia dos cobaias, o pulmão e o brônquio extrapulmonar direito
eram isolados e fixados em solução de formaldeído tamponado 10% durante três dias
e, em seguida, transferidos para etanol 70%. As amostras eram então submetidas a
procedimento padrão de inclusão em parafina para procedimentos histológicos. Este
procedimento incluía os seguintes passos: 1) desidratação do tecido em álcool 70%
por 24 h, em seguida a 80, 96 e 100% durante 1 h cada; 2) diafanização, na qual o
tecido era imerso em solução de álcool absoluto e xileno (1:1) durante 1 h, seguida de
duas imersões em xileno puro por 1 h cada; 3) impregnação em parafina, em que a
amostra era imersa em dois banhos de parafina líquida (aquecida a 50 °C) durante 1
h cada.
Para o preparo das lâminas, os blocos obtidos eram cortados transversalmente
em segmentos de 5 µm de espessura utilizando um micrótomo rotativo. As
preparações eram submetidas a procedimento de coloração com hematoxilina e
eosina de Mayer. As lâminas eram analisadas em um microscópio óptico com uma
câmara anexa. Para uma análise panorâmica do corte histológico, as lâminas eram
analisadas no aumento total (A.T.) x40 (1500 µm) e x100 (600µm).
Os aspectos avaliados foram a estrutura da parede dos brônquios, a
integridade do tecido e a migração celular. Para cada grupo experimental, eram
analisadas 5 lâminas de 5 animais diferentes. As fotomicrografias das lâminas foram
capturadas através de câmera acoplada a um microscópio óptico. As imagens foram
calibradas através do programa Motic Plus de acordo com as objetivas utilizadas: 4x
e 10x. A análise histológica das lâminas foi efetuada por um operador capacitado que
analisou qualitativamente os parâmetros histológicos e depois quantificou
estatisticamente através de score, sendo as lâminas codificadas para que o
analisados não tivesse conhecimento do grupo analisado.
As alterações no infiltrado inflamatório, na hiperplasia epitelial e na espessura
do músculo liso foram comparadas entre os grupos controle e asmático e com os
tratamentos com a dexametasona ou com o óleo de coco virgem nas doses de 1, 2 e
4 g/kg. Essas alterações foram pontuadas de acordo com os níveis (scores) descritos
abaixo:
1 – ausência de alterações histológicas;
2 – grau leve: aumento de menos de 25% em relação ao controle;
3 – grau moderado: aumento de 25 a 49% em relação ao controle;
4 – grau acentuado: aumento de 50 a 75% em relação ao controle;
106 VASCONCELOS, L. H. C. Material e Métodos
5 – grau muito acentuado: aumento de mais de 75% em relação ao controle.
4.2.7 Avaliação dos efeitos da inflamação pulmonar alérgica crônica e do
tratamento com a dexametasona ou da suplementação com o óleo de coco
virgem sobre a reatividade contrátil e relaxante da traqueia de cobaia
4.2.7.1 Obtenção dos anéis de traqueia
Após a eutanásia do cobaia, a cavidade torácica era aberta e era feita a
dissecação de parte do seu sistema respiratório (desde a laringe até os pulmões)
(Figura 13). A traqueia era isolada, limpa do tecido adjacente e segmentada em
fragmentos contendo 3 ou 4 anéis cartilaginosos. Esses segmentos eram suspensos
individualmente por hastes de aço inoxidável em cubas de banho para órgão isolado
(6 mL) contendo solução nutritiva de Krebs, à temperatura de 37 °C, gaseificados com
carbogênio e mantidos sob tensão de repouso de 1 g. Os órgãos permaneciam em
repouso por um período de 60 minutos, sendo a solução trocada a cada 15 minutos
para evitar a influência de metabólitos liberados pelo órgão no meio (TSCHIRHART et
al., 1987) (Figura 14).
Figura 13 – Parte do sistema respiratório do cobaia dissecado, com vista da superfície mediastinal pulmonar.
Fonte: Vasconcelos, 2017

107 VASCONCELOS, L. H. C. Material e Métodos
Figura 14 – Anel de traqueia suspenso em cuba de banho para órgão isolado (A), sistema de cubas de banho conectados a transdutores de força isométricos e amplificador (B) e sistema de aquisição digital de dados (C).
Fonte: Vasconcelos, 2017.
Após o período de estabilização, a integridade do epitélio da traqueia era
verificada pela adição de 10-4 M de AA à cuba durante a fase tônica da primeira
contração induzida por 10-6 M de CCh. Os anéis cujo relaxamento fosse igual ou
superior a 50% (em relação à força de contração inicial) eram considerados com
epitélio funcional; já aqueles com relaxamento igual ou inferior a 10% eram
considerados sem epitélio funcional. A remoção do epitélio era feita através do atrito
do lúmen do órgão com uma haste envolta em algodão e embebida com solução de
Krebs (TSCHIRHART et al., 1987).
4.2.7.2 Avaliação da reatividade contrátil à ovalbumina em traqueia de cobaia,
na presença de epitélio funcional
A traqueia era montada como descrito no item anterior. Após o período de
estabilização, quando a linha de base se mantinha constante, os anéis de traqueia
eram estimulados com OVA 10 µg/mL (SCHULTZ, 1910; DALE, 1913; CHAND; EYRE,
1978; JONES; CHARETTE; DENIS, 1988), e a amplitude da contração era comparada
entre os grupos GC, GASM, GASM/DEXA, GASM/OC1, GASM/OC2 e GASM/OC4.
4.2.7.3 Avaliação da reatividade contrátil ao CCh, à histamina ou ao KCl em
traqueia de cobaia, na presença e na ausência de epitélio funcional
A traqueia era montada como descrito no item 4.2.7.1. Após a verificação da
integridade do epitélio, a preparação era lavada e, após 30 minutos, era induzida uma
curva concentrações-resposta cumulativa ao CCh, agonista dos receptores
muscarínicos (OUEDRAOGO; ROUX, 2014), à histamina, agonista dos receptores

108 VASCONCELOS, L. H. C. Material e Métodos
histaminérgicos (OUEDRAOGO; ROUX, 2014) ou ao KCl, agente contrátil de
acoplamento eletromecânico (DU et al., 2006).
A reatividade contrátil era calculada com base na amplitude média da resposta
da traqueia de cobaia do grupo controle. As comparações eram feitas entre os grupos
GC, GASM, GASM/DEXA, GASM/OC1, GASM/OC2 e GASM/OC4, na presença e na
ausência de epitélio funcional, com base nos valores de efeito máximo (Emax) e de pD2
[logaritmo negativo na base 10 da concentração de uma substância que produz 50%
do seu Emax (CE50)] dos agentes contráteis, calculados a partir das curvas cumulativas
concentrações-resposta obtidas.
4.2.7.4 Avaliação do efeito da isoprenalina sobre o tônus basal em traqueia de
cobaia, na presença e na ausência de epitélio funcional
A traqueia era montada como descrito no item 4.2.7.1. Após o período de
estabilização, quando a linha de base se mantinha constante, a isoprenalina, agonista
dos receptores adrenérgicos β (NALINE et al., 1994), era adicionada à cuba em
concentrações cumulativas até o seu Emax ser atingido.
A reatividade relaxante era calculada com base na amplitude média da resposta
da traqueia de cobaia do grupo controle. As comparações eram feitas entre os grupos
GC, GASM, GASM/DEXA, GASM/OC1, GASM/OC2, e GASM/OC4, na presença e na
ausência de epitélio funcional, com base nos valores de Emax e de pD2 da isoprenalina,
calculados a partir das curvas cumulativas concentrações-resposta obtidas.
4.2.7.5 Avaliação da reatividade relaxante à isoprenalina ou ao nifedipino em
traqueia de cobaia pré-contraída com 10-6 M de CCh, na presença e na ausência
de epitélio funcional
A traqueia era montada como descrito no item 4.2.7.1. Após a verificação da
integridade do epitélio, a preparação era lavada e, após 30 minutos, era induzida uma
contração com 10-6 M de CCh. Após a formação do componente tônico da contração,
eram adicionados à cuba a isoprenalina, agonista dos receptores β (NALINE et al.,
1994), ou o nifedipino, bloqueador dos CaV (AHMED; FOSTER; SMALL, 1985), de
maneira cumulativa.
A reatividade relaxante era expressa como a percentagem reversa da
contração inicial produzida pelo CCh. As comparações eram feitas entre os grupos
GC, GASM, GASM/DEXA, GASM/OC1, GASM/OC2, GASM/OC4, na presença e na

109 VASCONCELOS, L. H. C. Material e Métodos
ausência de epitélio funcional, com base nos valores de Emax e de pD2 dos agentes
relaxantes, calculados a partir das curvas cumulativas concentrações-resposta
obtidas.
4.2.8 Investigação do mecanismo de ação envolvido nas alterações induzidas
pela inflamação pulmonar alérgica crônica e pelo óleo de coco virgem sobre a
reatividade contrátil da traqueia de cobaia
4.2.8.1 Avaliação do envolvimento das espécies reativas de oxigênio (ROS)
4.2.8.1.1 Investigação da participação das enzimas nicotinamida adenina
dinucleotídeo fosfato (NADPH) oxidase, superóxido dismutase (SOD) e catalase
4.2.8.1.1.1 Obtenção de curvas cumulativas concentrações-resposta ao CCh, na
ausência e na presença da apocinina, do tempol ou da catalase em traqueia de
cobaia dos grupos GC, GASM e GASM/OC4
A traqueia era montada como descrito no item 4.2.7.1. Após a verificação da
presença de epitélio, a preparação era lavada e, após 30 minutos, era induzida uma
curva concentrações-resposta cumulativa ao CCh, considerada como controle. A
preparação era lavada e, após 30 minutos, a apocinina 10-5 M, um inibidor da NADPH
oxidase (SUTCLIFFE et al., 2012) ou o tempol, um mimético da SOD (DE BOER et
al., 2001; adaptado de SCHNACKENBERG; WILCOX, 2001), eram incubados por 30
minutos; ou a catalase 100 UI/mL (RHODEN; BARNES, 1989) era incubada por 10
min. Após esse tempo, era induzida uma nova curva cumulativa ao CCh.
A reatividade contrátil da traqueia na presença do inibidor era calculada com
base na amplitude máxima da primeira curva controle obtida. A reatividade contrátil
era avaliada tendo como base os valores de Emax e pD2 do CCh, na ausência e na
presença da apocinina, do tempol ou da catalase, e o efeito do inibidor sobre a curva
cumulativa concentrações-resposta ao CCh era comparada entre os grupos GC,
GASM e GASM/OC4.

110 VASCONCELOS, L. H. C. Material e Métodos
4.2.8.2 Avaliação da participação dos produtos derivados do metabolismo do
ácido araquidônico pelas enzimas cicloxigenase (COX) e 5-lipoxigenase (5-LO)
4.2.8.2.1 Obtenção de curvas cumulativas concentrações-resposta ao CCh, na
ausência e na presença da indometacina ou da zileutona, em cobaias dos
grupos GC, GASM e GASM/OC4
A traqueia era montada como descrito no item 4.2.7.1. Em seguida, após a
obtenção da curva controle ao CCh, como descrito no item anterior, a indometacina
10-5 M, um inibidor da COX (OREHEK; DOUGLAS; BOUHUYS, 1975), ou a zileutona
10-5 M, um inibidor da 5-LO (MALO et al., 1994), eram incubadas por 30 minutos. Após
esse tempo, era induzida uma nova curva cumulativa ao CCh.
A reatividade contrátil da traqueia na presença do inibidor era calculada e
avaliada como descrito no item anterior.
4.2.8.3 Avaliação da participação da via do óxido nítrico (NO)
4.2.8.3.1 Obtenção de curvas cumulativas concentrações-resposta ao CCh, na
ausência e na presença do L-NAME, em cobaias dos grupos GC, GASM e
GASM/OC4
A traqueia era montada como descrito no item 4.2.7.1. Em seguida, após a
obtenção da curva controle ao CCh, como descrito no item anterior, L-NAME 10-4 M,
um inibidor da NOS (DE BOER et al., 2001) era incubado por 45 minutos. Após esse
tempo, era induzida uma nova curva cumulativa ao CCh.
A reatividade contrátil da traqueia na presença do L-NAME era calculada e
avaliada como descrito no item 4.2.8.1.1.1.
4.2.8.4 Avaliação da participação da via da RhoA/Rho cinase (ROCK)
4.2.8.4.1 Obtenção de curvas cumulativas concentrações-resposta ao CCh, na
ausência e na presença do Y-27632, em cobaias dos grupos GC, GASM e
GASM/OC4
A traqueia era montada como descrito no item 4.2.7.1. Em seguida, após a
obtenção da curva controle ao CCh, como descrito no item anterior, o Y-27632
(10-7-10-4 M), um inibidor da ROCK (SCHAAFSMA et al., 2004) era incubado por 30
minutos, em diferentes preparações. Após esse tempo, era induzida uma nova curva
cumulativa ao CCh.

111 VASCONCELOS, L. H. C. Material e Métodos
A reatividade contrátil da traqueia na presença do Y-27632 era calculada e
avaliada como descrito no item 4.2.8.1.1.1, e o efeito do inibidor sobre a curva
cumulativa concentrações-resposta ao CCh era comparada entre os grupos GC,
GASM e GASM/OC4, com base no valor do logaritmo negativo na base 10 da
concentração molar de um antagonista necessária para duplicar a concentração do
agonista que promove a resposta submáxima original obtida na ausência de
antagonista (pA2) (SCHILD, 1947, 1949).
A medida empírica da interação droga-receptor (CCh-receptores M3) foi
calculada como log (DR-1). A inclinação da reta do plot do Schild (log DR-1 vs. log
[Y-27632]), que determina o quanto a reatividade contrátil do CCh foi alterada pelo
Y-27632, foi calculada por regressão linear (ARUNLAKSHANA; SCHILD, 1959), e a
correlação entre as concentrações do Y-27632 e a resposta (medida pelo log DR-1)
foi determinada por meio do valor do r2, que mede a qualidade do ajuste da reta obtida
pela regressão linear.
4.2.9 Investigação do mecanismo de ação envolvido nas alterações induzidas
pela inflamação pulmonar alérgica crônica e pelo óleo de coco virgem sobre a
reatividade relaxante da traqueia de cobaia
4.2.9.1 Avaliação da participação do fator de crescimento transformador β
(TGF-β) e dos canais de potássio sensíveis ao Ca2+ de grande condutância
(BKCa)
4.2.9.1.1 Obtenção de curvas cumulativas concentrações-resposta à
isoprenalina, na ausência e na presença do SB431542 ou da iberiotoxina (IbTX),
em traqueia de cobaia pré-contraída com 10-6 M de CCh dos grupos GC, GASM
e GASM/OC4
A traqueia era montada como descrito no item 4.2.7.1. Após a verificação da
presença de epitélio, a preparação era lavada e, após 30 minutos, o SB431542 10-5 M,
um antagonista dos receptores do tipo I de TGF-β (INMAN et al., 2002; adaptado de
KIMURA et al., 2014), era incubado por 60 minutos; ou a IbTX 10-7 M, um bloqueador
seletivo dos BKCa (YAGI; KUWAHARA; TSUBONE, 2002), era incubado por 20
minutos, em diferentes preparações. Após esse tempo, uma nova curva cumulativa à
isoprenalina era induzida.

112 VASCONCELOS, L. H. C. Material e Métodos
A reatividade relaxante da traqueia à isoprenalina na presença do SB431542
ou da IbTX era calculada com base na amplitude máxima da contração, e a resposta
era avaliada tendo de acordo com os valores de Emax e pD2 da isoprenalina, na
ausência e na presença dos inibidores, e comparada entre os grupos GC, GASM e
GASM/OC4.
4.2.9.2 Avaliação da participação dos produtos derivados do metabolismo do
ácido araquidônico pelas enzimas cicloxigenase (COX) e 5-lipoxigenase (5-LO)
4.2.9.2.1 Obtenção de curvas cumulativas concentrações-resposta ao ácido
araquidônico (AA), na ausência e na presença da indometacina ou da zileutona,
em traqueia de cobaia pré-contraída com 10-6 M de CCh dos grupos GC, GASM
e GASM/OC4
A traqueia era montada como descrito no item 4.2.7.1. Após a verificação da
presença de epitélio, a preparação era lavada e, após 30 minutos, uma contração com
CCh 10-6 M era induzida e, sobre o componente tônico dessa contração, o AA,
substrato para as enzimas COX e 5-LO, era adicionado de maneira cumulativa
(TSCHIRHART et al., 1987). Em seguida, a preparação era lavada e, após 30 minutos,
a indometacina 10-5 M, um inibidor da COX (OREHEK; DOUGLAS; BOUHUYS, 1975),
ou a zileutona 10-5 M, um inibidor da 5-LO (MALO et al., 1994), eram incubados por
30 minutos, em diferentes preparações. Após esse tempo, uma nova curva cumulativa
à isoprenalina era induzida.
A reatividade relaxante da traqueia ao AA na ausência e na presença da
indometacina ou da zileutona era calculada e avaliada como descrito no item anterior.
4.2.10 Avaliação dos efeitos da inflamação pulmonar alérgica crônica e do
tratamento com a dexametasona ou com o óleo de coco virgem sobre o balanço
entre o estresse oxidativo e as defesas antioxidantes de cobaia
4.2.10.1 Obtenção do plasma
Após a eutanásia dos animais, eram coletados 10 mL de sangue por meio de
secção dos vasos cervicais, colocados imediatamente em tubos de ensaio contendo
anticoagulante (EDTA) e levados à centrifugação a 1.198 x g durante 10 minutos. O
sobrenadante era então transferido para tubos Eppendorf® e refrigerado a -20 °C até
a análise (OKAFOR et al., 2011; SILVA et al., 2012).

113 VASCONCELOS, L. H. C. Material e Métodos
4.2.10.2 Obtenção do homogenato do tecido pulmonar
Após a eutanásia dos animais, os pulmões eram isolados e congelados a -20 °C
até o preparo do homogenato. Para isso, o tecido era pesado, macerado e
homogeneizado com KCl 10% na proporção de 1:1. Em seguida, as amostras eram
centrifugadas (1.198 x g/10 min) e o sobrenadante era separado para análise
posterior.
4.2.10.3 Análise dos níveis de malondialdeído (MDA) e da capacidade
antioxidante total (CAT) no plasma e no tecido pulmonar
A medida da produção de MDA era feita por meio do método de quantificação
das espécies reativas ao ácido tiobarbitúrico (TBARS) (OHKAWA; OHISHI; YAGI,
1979), em que duas moléculas do TBA se condensam com uma molécula de MDA
através de uma reação colorimétrica, cujo produto pode ser facilmente detectado por
espectrofotometria (GIERA; LINGEMAN; NIESSEN, 2012). Após a obtenção do
plasma e do homogenato pulmonar, alíquotas de 250 µL eram incubadas a 37 °C em
banho-maria por 60 minutos. Depois, as amostras eram precipitadas com 400 µL de
ácido perclórico 35% e centrifugadas a 16.851 x g por 20 minutos a 4 °C. O
sobrenadante era transferido para tubos Eppendorf®, 400 µL de ácido tiobarbitúrico
(TBA) 0,6% eram adicionados às amostras e incubadas à temperatura de 95-100 °C
por 1 h. Em seguida, após o resfriamento, as amostras eram lidas em
espectrofotômetro a 532 nm. As amostras turvas após o banho-maria eram
centrifugadas novamente a 1.198 x g por 10 minutos antes da leitura. A determinação
dos níveis de MDA (μmol/L de amostra) nas amostras era feita substituindo os valores
de absorbância na curva-padrão de MDA obtida para a solução padrão (1 µL de
1,1,3,3-tetrametoxipropano em 70 mL de água destilada, diluída em séries de 250,
500, 750, 1000, 1250, 1500, 1750, 2000, 2250, 2500, 2750 e 3000 µL de água
destilada).
A medida da capacidade antioxidante total (CAT) era feita utilizando o método
colorimétrico da redução do 1,1-difenil-2-picril hidrazil (DPPH) (BRAND-WILLIAMS;
CUVELIER; BERSET, 1995), que baseia-se na habilidade da amostra em reduzir o
radical DPPH, que apresenta coloração roxa, a 1,1-difenil-2-picril hidrazina, de cor
transparente, a qual é detectada por espectrofotometria (FLOEGEL et al., 2011). Em
tubos apropriados para centrífuga, eram adicionados 50 µL do plasma ou do
homogenato pulmonar e 2 mL da solução de DPPH dissolvido em etanol absoluto

114 VASCONCELOS, L. H. C. Material e Métodos
(0,012 g/L) e protegido da luz; em seguida, os tubos eram agitados em vórtex por 10
segundos e, então, mantidos em repouso por 30 minutos. A seguir, as amostras eram
centrifugadas a 7.489 x g por 15 minutos a 20 °C. O sobrenadante era lido em
espectrofotômetro a 515 nm. Os resultados eram expressos como percentual de
inibição da oxidação, através da seguinte equação: CAT = 100-([DPPH·R] x
T)/([DPPH·R] x B) x 100; sendo [DPPH·R] x T e [DPPH·R] x B correspondentes à
concentração de DPPH· remanescente após 30 minutos, avaliados na amostra (T) e
no branco (B) preparado com água destilada.
As análises eram feitas comparando-se os níveis de MDA (μmol/L de amostra)
ou da CAT (%) entre os grupos GC, GASM, GASM/DEXA, GASM/OC1, GASM/OC2
e GASM/OC4.
4.2.11 Avaliação dos efeitos da inflamação pulmonar alérgica crônica e do
tratamento com a dexametasona ou da suplementação com o óleo de coco
virgem sobre a expressão de proteínas das vias de sinalização da proliferação
muscular lisa e das defesas antioxidantes
4.2.11.1 Análise dos níveis da fosfatidilinositol-3-cinase (PI3K), das proteínas
cinases reguladas por sinal extracelular 1/2 (ERK1/2) e da superóxido dismutase
(SOD) no tecido pulmonar
4.2.11.2 Extração do pulmonar e corrida de eletroforese em gel de poliacrilamida
Após a eutanásia dos cobaias, o pulmão era isolado da caixa torácica e
rapidamente congelado em N2 líquido. Os órgão eram mantidos sob congelamento a
-80 °C até as análises. Os pulmões eram homogeneizados em tampão de extração,
cuja composição está descrita no item 3.1.3, utilizando um processador de tecidos do
tipo Polytron PTA 20S (Brinkmann Instruments, EUA). Em seguida, os extratos
teciduais eram centrifugados a 17.530 g a 4 °C por 20 minutos. Após esse processo,
o conteúdo proteico das amostras era quantificado no sobrenadante utilizando o
reagente de Bradford (Bio-Rad, Richmond, CA), e as amostras eram conservadas pela
adição do tampão de Laemmli (Bio-Rad, Richmond, CA) (LAEMMLI, 1970), acrescido
de DTT 200 mM (4:1 v/v). Posteriormente, 50 a 100 μg de proteínas totais eram
submetidas a eletroforese em gel de poliacrilamida contendo SDS-PAGE 15% em
aparelho de minigel (Bio-Rad, Richmond, CA). Em cada gel era adicionado um
marcador com peso molecular com valores conhecidos (Bio-Rad, Richmond, CA).

115 VASCONCELOS, L. H. C. Material e Métodos
4.2.11.3 Western blot
Após a separação, as proteínas eram transferidas para uma membrana de
nitrocelulose por estimulação elétrica a 120 V por 2 h (TOWBIN; STAEHELIN;
GORDON, 1992). Em seguida, a ligação inespecífica de proteínas à membrana de
nitrocelulose era reduzida pela incubação com uma solução de bloqueio, cuja
composição está descrita no item 3.1.3, em temperatura ambiente durante 2 h sob
leve agitação.
Subsequentemente, as membranas de nitrocelulose eram incubadas com os
anticorpos anti-PI3K, -ERK1/2 e -SOD, preparados em solução bloqueadora
(substituindo o leite pela albumina na sua composição), pernoite à temperatura de
4 °C, sob leve agitação. Após esse período, as membranas eram lavadas com esta
solução decrescida de leite ou albumina por 30 minutos e então o anticorpo secundário
de coelho conjugado com peroxidase era incubado por 1 h à temperatura ambiente.
Para a revelação das membranas, era adicionada uma solução para detecção
por quimioluminescência conforme descrito no kit comercial. Nesse protocolo, a
emissão da luz era detectada pelo sistema de detecção MF-CHEMIBIS (Bio-Imaging
Systems Ltda., Jerusalem, Israel) e a determinação da intensidade das bandas
reveladas era realizada pela leitura da densitometria óptica das imagens analisadas
no programa ImageJ 1.43u (National Institute of Health, EUA).
4.2.12 Análise Estatística
Os dados foram expressos como média ± erro padrão da média (e.p.m.), a
normalidade da variância foi verificada pelo teste de Shapiro-Wilk, e foram analisados
estatisticamente empregando-se o teste t, para comparações entre dois grupos
experimentais, ou a análise de variância (ANOVA) one-way, seguido do pós-teste de
Tukey, para comparações múltiplas entre os grupos experimentais. As diferenças
entre as médias foram consideradas significantes (hipótese nula descartada) quando
p < 0,05.
Todos os dados foram analisados utilizando o programa GraphPad Prism®
versão 5.01 (GraphPad Software Inc., San Diego CA, U.S.A.).

5 Resultados

117 VASCONCELOS, L. H. C. Resultados
5.1 Avaliação dos efeitos da inflamação pulmonar alérgica crônica e do
tratamento com a dexametasona ou da suplementação com o óleo de coco
virgem sobre o tempo de nebulização de cobaia
Padronizou-se para o os cobaias do grupo GC o tempo de inalação de 900 s
em todas as nebulizações (Gráfico 1, n = 8).
Nas quatro primeiras nebulizações, os animais dos grupos GASM e
GASM/DEXA e GASM/OC4 apresentaram tempos de 900 s, e os dos grupos
GASM/OC1 e GASM/OC2 apresentaram tempos de inalação de 857,3 ± 25,6 e
842,7 ± 24,1 s, não havendo diferença estatística entre todos eles (Gráfico 1, Tabela
1, n = 8).
A inflamação pulmonar reduziu o tempo da quinta nebulização (456,4 ± 96,7 s),
em relação ao grupo GC. O tratamento com dexametasona não alterou esse tempo
(627,9 ± 108,2 s), seja em relação ao grupo GC ou ao GASM. A suplementação dos
cobaias com o óleo de coco virgem nas doses de 1 ou 2 g/kg não preveniu a redução
desse tempo (416,3 ± 126,3 e 424,8 ± 74,7 s, respectivamente), que foram menores
que os do grupo GC, sem diferir do grupo GASM; já a suplementação na dose de
4 g/kg preveniu a redução do tempo de nebulização (815,3 ± 42,4 s), que foi
semelhante ao grupo GC, diferindo do GASM (Gráfico 1, Tabela 1, n = 8).
Na sexta nebulização, a inflamação pulmonar reduziu o tempo de contato dos
cobaias com a OVA (400,8 ± 51,0 s), sendo menor que o do grupo GC. O tratamento
com a dexametasona não alterou esse tempo (707,8 ± 74,6 s), valor que não diferiu
nem do GC nem do GASM. A suplementação com o óleo de coco virgem nas doses
de 1 ou 2 g/kg não alterou esse tempo, em relação a GASM (511,8 ± 120,2 e
682,5 ± 106,3 s, respectivamente), enquanto a suplementação com a dose de 4 g/kg
preveniu a redução desse tempo (861,9 ± 38,1 s), diferindo do GASM (Gráfico 1,
Tabela 1, n = 8).
No modelo de inflamação pulmonar, o tempo da sétima nebulização foi
reduzido (273,3 ± 26,6 s), sendo menor que o do grupo GC. O tratamento com
dexametasona não preveniu a redução desse tempo (567,5 ± 98,5 s), valor que não
diferiu do grupo GC e do GASM. A suplementação com óleo de coco virgem nas doses
de 1 ou 2 g/kg não preveniu a redução desse tempo (453,9 ± 106,8 e 389,0 ± 58,8 s,
respectivamente), que foram semelhantes ao grupo GASM, enquanto a
suplementação na dose de 4 g/kg preveniu a redução do tempo de contato dos

118 VASCONCELOS, L. H. C. Resultados
cobaias com a OVA (715,0 ± 48,1 s), o qual não diferiu do grupo GC (Gráfico 1, Tabela
1, n = 8).
Nenhum cobaia do grupo GC apresentou sinais de desconforto respiratório;
diferentemente, os cobaias do GASM apresentaram tosse, espirros e, especialmente
na sétima nebulização, quadros de dispneia e de tiragem torácica. O mesmo foi
observado nos cobaias dos grupos GASM/DEXA, GASM/OC1, GASM/OC2 e
GASM/OC4, porém com menor intensidade nos grupos GASM/DEXA e GASM/OC4.

119 VASCONCELOS, L. H. C. Resultados
Gráfico 1 – Tempo médio de contato dos cobaias com a OVA da primeira à quarta, na quinta, na sexta e na sétima nebulizações.
1-4 5 6 7
1-4 5 6 7
1-4 5 6 7
1-4 5 6 7
1-4 5 6 7
1-4 5 6 7
0
150
300
450
600
750
900 GC
GASM
GASM/DEXA
GASM/OC1
GASM/OC2
GASM/OC4*
*
*
*
**
*
*
#
#
#
#
##
#
#
Ordem da nebulização
Tem
po
de n
eb
ulização
(s)
As colunas e as barras verticais representam a média e o e.p.m., respectivamente (n = 8). ANOVA one-way seguido do pós-teste de Bonferroni: *p < 0,05 (1-4 vs. 5/6/7 intragrupos); ANOVA one-way seguido do pós-teste de Tukey: #p < 0,05 (GC vs. demais grupos); γp < 0,05 (GASM vs. demais grupos). Fonte: Vasconcelos, 2017.
Tabela 1 - Tempo médio de contato (s) dos cobaias com a ovalbumina da primeira à quarta (1-4), na quinta (5), na sexta (6) e na sétima (7) nebulizações.
Grupo 1-4 5 6 7
GC 900,0 900,0 900,0 900,0
GASM 900,0 456,4 ± 96,7 * # 400,8 ± 51,0 * # 273,3 ± 26,6 * #
GASM/DEXA 900,0 627,9 ± 108,2 707,8 ± 74,6 567,5 ± 98,5 *
GASM/OC1 857,3 ± 25,6 416,3 ± 126,3 * 511,8 ± 120,2 * 453,9 ± 106,8 * #
GASM/OC2 842,7 ± 24,1 424,8 ± 74,7 * # 682,5 ± 106,3 389,0 ± 58,8 * #
GASM/OC4 900,0 815,3 ± 42,4 γ 861,9 ± 38,1 γ 715,0 ± 48,1 * γ
As colunas e as barras verticais representam a média e o e.p.m., respectivamente (n = 8). ANOVA one-way seguido do pós-teste de Bonferroni: *p < 0,05 (1-4 vs. 5/6/7 intragrupos); ANOVA one-way seguido do pós-teste de Tukey: #p < 0,05 (GC vs. demais grupos); γp < 0,05 (GASM vs. demais grupos). Fonte: Vasconcelos, 2017.

120 VASCONCELOS, L. H. C. Resultados
5.2 Avaliação dos efeitos da inflamação pulmonar alérgica crônica e do
tratamento com a dexametasona ou da suplementação com o óleo de coco
virgem sobre a morfologia pulmonar e brônquica de cobaia
A análise histológica permitiu uma avaliação da estrutura morfológica dos vasos
sanguíneos, brônquios intrapulmonares, bronquíolos e alvéolos pulmonares (Figuras
15 e 16) e dos brônquios extrapulmonares (Figura 17).
Comparando-se os cortes histológicos dos pulmões no A.T. x40, observou-se
que o grupo GC apresentou aspecto histológico normal e histoarquitetura pulmonar
preservada (Figura 15A), enquanto os pulmões dos cobaias do grupo GASM
apresentaram grande quantidade de infiltrado de células inflamatórias nas regiões
peribronquiolar, perivascular e ausência de infiltrado nos alvéolos pulmonares
(Figura 15B, Gráfico 2A). Com o tratamento com a dexametasona (Figura 15C,
Gráfico 2A) ou com a suplementação com o óleo de coco virgem nas doses de 1, 2 e
4 g/kg (Figura 15D-F, Gráfico 2A), foi possível observar uma diminuição da migração
de células para o pulmão, sendo a dose de 4 g/kg a mais eficaz em alterar esse
parâmetro.
O A.T. x100 confirmou com maior detalhe estrutural as mudanças observadas
no A.T. x40. Comparado ao grupo GC (Figura 16A, Gráfico 2B), foi observada
hiperplasia epitelial nos animais do grupo GASM (Figura 16B, Gráfico 2B), e o
tratamento com a dexametasona (Figura 16C, Gráfico 3) ou a suplementação com o
óleo de coco virgem nas doses de 1, 2 e 4 g/kg (Figura 16D-F, Gráfico 2B) reduziu a
hiperplasia epitelial.
Semelhantemente, conforme observado no A.T. x100, comparado ao grupo GC
(Figura 16A, Gráfico 2C), os animais do grupo GASM apresentaram maior espessura
do músculo liso brônquico intrapulmonar por hipertrofia e/ou hiperplasia (Figura 16B,
Gráfico 2C). O tratamento com a dexametasona (Figura 16C, Gráfico 2C) ou com o
óleo de coco virgem nas doses de 1, 2 e 4 g/kg (Figura 16D-F, Gráfico 2C) reduziu
esse espessamento, sendo a dose de 1 g/kg a mais eficaz.
Quando analisadas os cortes histológicos de brônquio extrapulmonar no A.T.
x40, foi possível observar que, comparando aos cortes histológicos do grupo GC
(Figura 17A, Gráfico 2C), os cobaias do grupo GASM apresentaram desenvolvimento
do músculo liso brônquico por hipertrofia e/ou hiperplasia (Figura 17B, Gráfico 2C), e
os tratamentos com a dexametasona (Figura 17C, Gráfico 2C) ou a suplementação
com o óleo de coco virgem nas doses de 1, 2 e 4 g/kg (Figura 17D-F, Gráfico 2C)
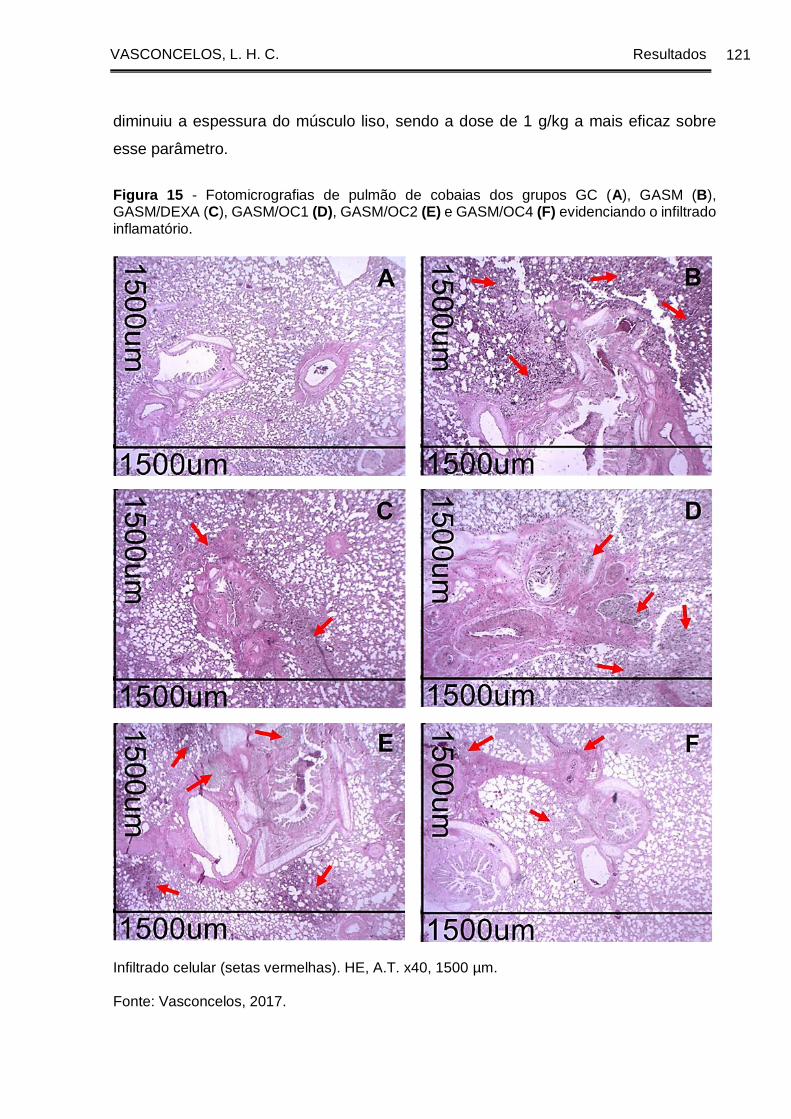
121 VASCONCELOS, L. H. C. Resultados
diminuiu a espessura do músculo liso, sendo a dose de 1 g/kg a mais eficaz sobre
esse parâmetro.
Figura 15 - Fotomicrografias de pulmão de cobaias dos grupos GC (A), GASM (B), GASM/DEXA (C), GASM/OC1 (D), GASM/OC2 (E) e GASM/OC4 (F) evidenciando o infiltrado inflamatório.
Infiltrado celular (setas vermelhas). HE, A.T. x40, 1500 µm. Fonte: Vasconcelos, 2017.

122 VASCONCELOS, L. H. C. Resultados
Figura 16 - Fotomicrografias de pulmão de cobaias dos grupos GC (A), GASM (B), GASM/DEXA (C), GASM/OC1 (D), GASM/OC2 (E) e GASM/OC4 (F) evidenciando o músculo
liso dos brônquios intrapulmonares.
Músculo liso brônquico intrapulmonar (setas vermelhas), infiltrado inflamatório (setas verdes). HE, A.T. x100, 600 µm. Fonte: Vasconcelos, 2017.
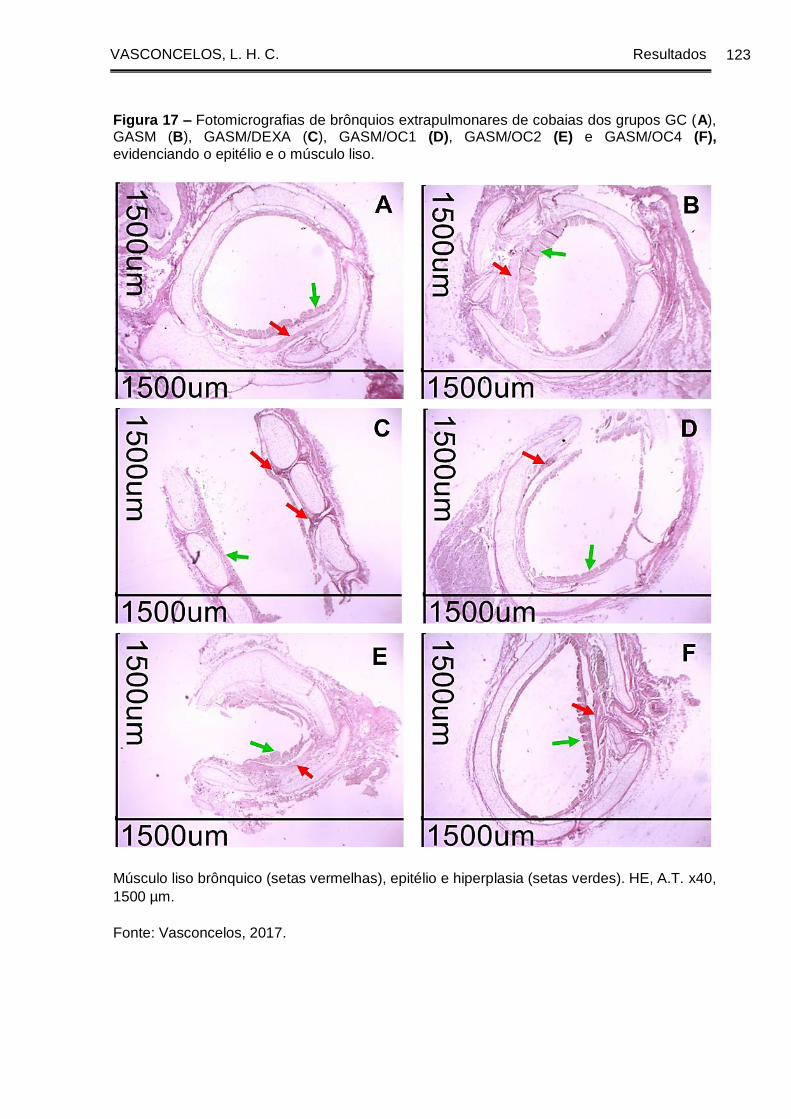
123 VASCONCELOS, L. H. C. Resultados
Figura 17 – Fotomicrografias de brônquios extrapulmonares de cobaias dos grupos GC (A), GASM (B), GASM/DEXA (C), GASM/OC1 (D), GASM/OC2 (E) e GASM/OC4 (F),
evidenciando o epitélio e o músculo liso.
Músculo liso brônquico (setas vermelhas), epitélio e hiperplasia (setas verdes). HE, A.T. x40,
1500 µm.
Fonte: Vasconcelos, 2017.
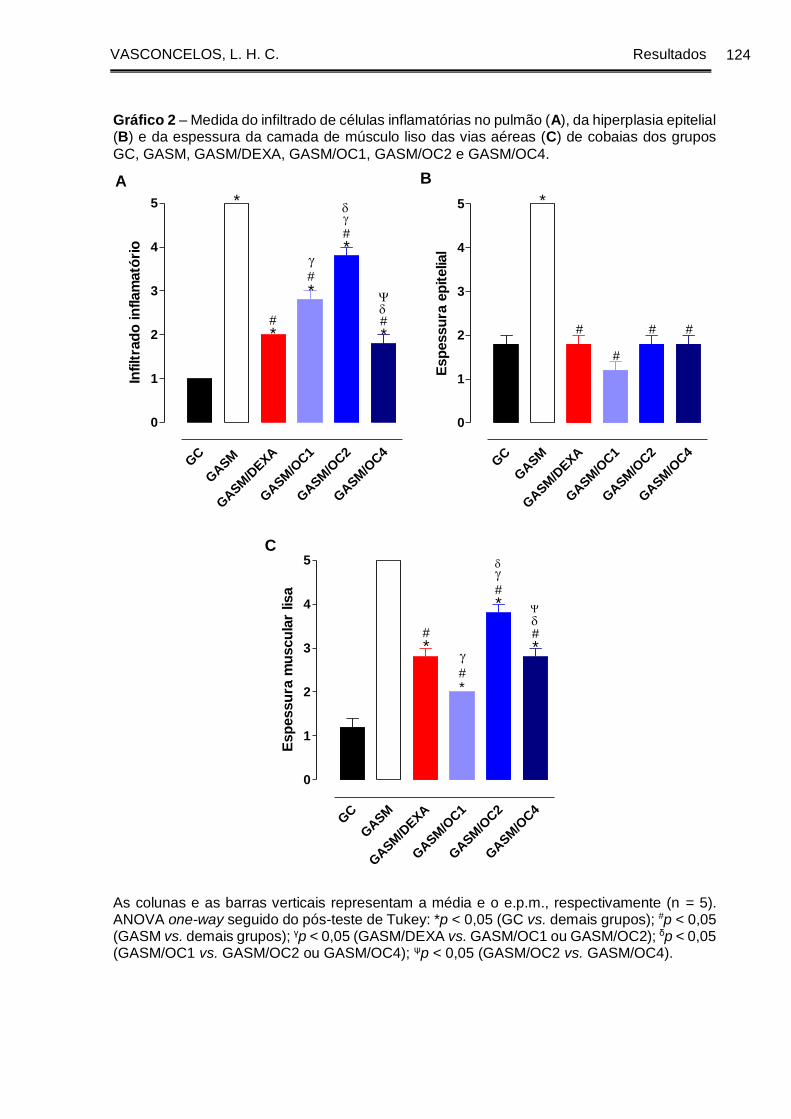
124 VASCONCELOS, L. H. C. Resultados
Gráfico 2 – Medida do infiltrado de células inflamatórias no pulmão (A), da hiperplasia epitelial (B) e da espessura da camada de músculo liso das vias aéreas (C) de cobaias dos grupos
GC, GASM, GASM/DEXA, GASM/OC1, GASM/OC2 e GASM/OC4.
GC
GASM
GASM
/DEXA
GASM
/OC1
GASM
/OC2
GASM
/OC4
0
1
2
3
4
5
#*
*
*
*
#
#
#
*
Infilt
rad
o in
flam
ató
rio
GC
GASM
GASM
/DEXA
GASM
/OC1
GASM
/OC2
GASM
/OC4
0
1
2
3
4
5
#
#
# #
*
Esp
essu
ra e
pit
elia
l
GC
GASM
GASM
/DEXA
GASM
/OC1
GASM
/OC2
GASM
/OC4
0
1
2
3
4
5
#
*
*
*
*#
#
#
Esp
essu
ra m
uscu
lar
lisa
As colunas e as barras verticais representam a média e o e.p.m., respectivamente (n = 5). ANOVA one-way seguido do pós-teste de Tukey: *p < 0,05 (GC vs. demais grupos); #p < 0,05 (GASM vs. demais grupos); γp < 0,05 (GASM/DEXA vs. GASM/OC1 ou GASM/OC2); δp < 0,05 (GASM/OC1 vs. GASM/OC2 ou GASM/OC4); ψp < 0,05 (GASM/OC2 vs. GASM/OC4).
A B
C

125 VASCONCELOS, L. H. C. Resultados
5.3 Avaliação dos efeitos da inflamação pulmonar alérgica crônica e do
tratamento com a dexametasona ou da suplementação com o óleo de coco
virgem sobre a reatividade contrátil e relaxante da traqueia de cobaia
5.3.1 Avaliação da reatividade contrátil à OVA em traqueia de cobaia, na
presença de epitélio funcional
A traqueia de cobaia dos animais do grupo GC não apresentaram reatividade
contrátil à estimulação com OVA (Emax = 1,8 ± 0,8%) (Figura 18A, Gráfico 3);
diferentemente, a inflamação pulmonar promoveu reatividade contrátil significante
(Emax = 100%) (Figura 18B, Gráfico 5).
O tratamento com a dexametasona não alterou a reatividade contrátil da
traqueia (Emax = 89,1 ± 10,5%), em comparação com o grupo GASM (Figura 18C,
Gráfico 3, n = 5)
Do mesmo modo, a suplementação com o óleo de coco virgem nas doses de 1
(Emax = 65,4 ± 8,3%) e de 2 g/kg (Emax = 81,6 ± 13,3%) não reduziu a reatividade
contrátil da traqueia, não diferindo do grupo GASM (Figuras 18D e E, Gráfico 3, n =
5). Diferentemente, a suplementação com o óleo na dose de 4 g/kg reduziu a
amplitude da contração da traqueia (Emax = 40,3 ± 2,1%) em relação ao grupo GASM,
não diferindo do grupo GC (Figura 18F, Gráfico 3, n = 5).
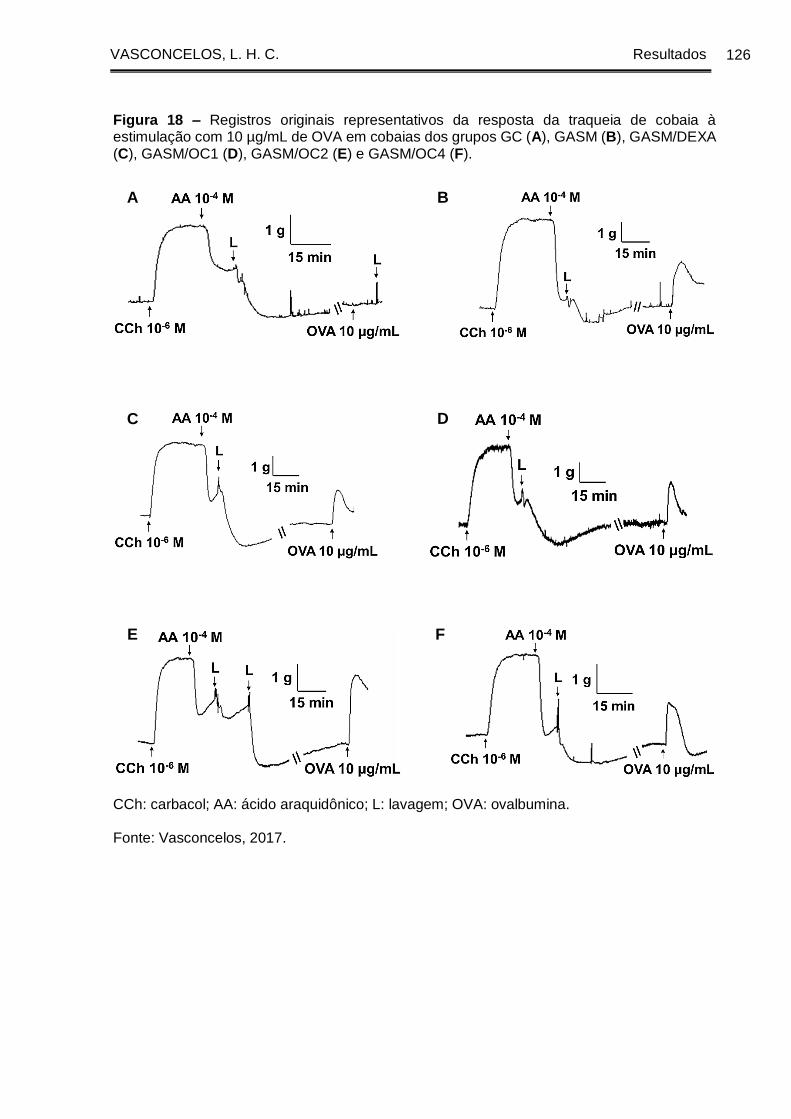
126 VASCONCELOS, L. H. C. Resultados
Figura 18 – Registros originais representativos da resposta da traqueia de cobaia à estimulação com 10 µg/mL de OVA em cobaias dos grupos GC (A), GASM (B), GASM/DEXA (C), GASM/OC1 (D), GASM/OC2 (E) e GASM/OC4 (F).
CCh: carbacol; AA: ácido araquidônico; L: lavagem; OVA: ovalbumina. Fonte: Vasconcelos, 2017.
A B
C D
E F
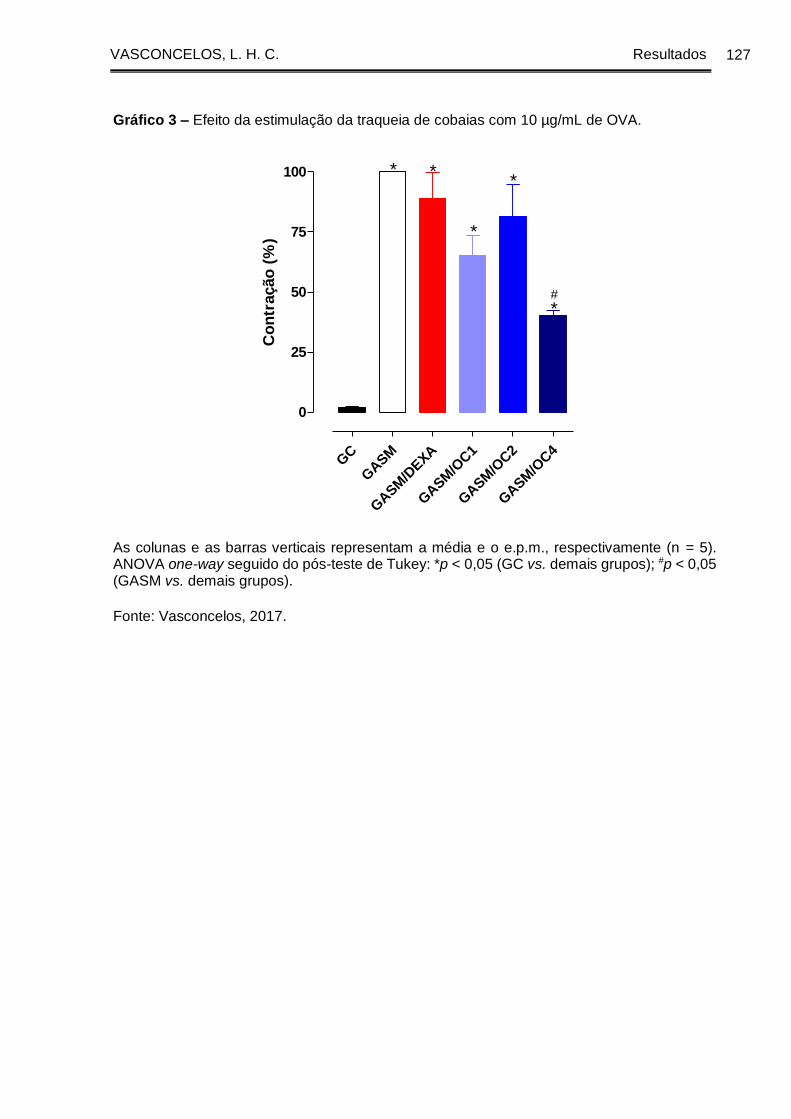
127 VASCONCELOS, L. H. C. Resultados
Gráfico 3 – Efeito da estimulação da traqueia de cobaias com 10 µg/mL de OVA.
GC
GASM
GASM
/DEXA
GASM
/OC1
GASM
/OC2
GASM
/OC4
0
25
50
75
100 * *
*
*
#
*
Co
ntr
ação
(%
)
As colunas e as barras verticais representam a média e o e.p.m., respectivamente (n = 5). ANOVA one-way seguido do pós-teste de Tukey: *p < 0,05 (GC vs. demais grupos); #p < 0,05 (GASM vs. demais grupos).
Fonte: Vasconcelos, 2017.

128 VASCONCELOS, L. H. C. Resultados
5.3.2 Avaliação da reatividade contrátil ao CCh em traqueia de cobaia, na
presença e na ausência de epitélio funcional
A traqueia de cobaia do grupo GC contraiu em resposta à adição de
concentrações cumulativas de CCh (10-9-10-4 M) tanto na presença (Emax = 100%,
pD2 = 6,63 ± 0,10) com na ausência (Emax = 100%, pD2 = 6,85 ± 0,09) de epitélio
funcional. A inflamação pulmonar aumentou a eficácia contrátil do CCh
(Emax = 185,3 ± 16,1%), mas não alterou sua potência (pD2 = 6,74 ± 0,03) na presença
de epitélio funcional; enquanto na ausência de epitélio, não houve diferença na
eficácia (Emax = 89,2 ± 9,8%) e na potência do CCh (pD2 = 6,62 ± 0,07), comparados
ao grupo GC. Não houve diferença na potência contrátil do CCh entre os anéis de
traqueia com e sem epitélio em ambos os grupos (Figura 19A-D, Gráfico 4, Tabela 2,
n = 5).
O tratamento com a dexametasona preveniu parcialmente o aumento da
eficácia contrátil do CCh (Emax = 133,4 ± 4,9%), mas não alterou a sua potência
(pD2 = 6,59 ± 0,11), em relação aos grupos GC e GASM, em traqueia de cobaia com
inflamação pulmonar na presença de epitélio (Figura 19E, Gráfico 5, Tabela 2, n = 5).
A suplementação com o óleo de coco virgem na dose de 1 g/kg não preveniu o
aumento da eficácia (Emax = 153,2 ± 9,3%) e não alterou a potência contrátil do CCh
(pD2 = 6,61 ± 0,08), comparadas aos grupos GC e GASM, em traqueia de cobaia com
inflamação pulmonar na presença de epitélio (Figura 19F, Gráfico 5, Tabela 2, n = 5).
Diferentemente, com a suplementação com o óleo nas doses de 2 e 4 g/kg, o
aumento da eficácia contrátil do CCh foi completamente prevenido
(Emax = 108,6 ± 13,7 e 98,3 ± 18,5%, respectivamente), sem alteração na sua potência
(pD2 = 6,60 ± 0,07 e 6,49 ± 0,09, respectivamente), em relação ao GC e ao GASM,
em traqueia de cobaia com inflamação pulmonar na presença de epitélio (Figuras 19G
e H, Gráfico 5, Tabela 2, n = 5).
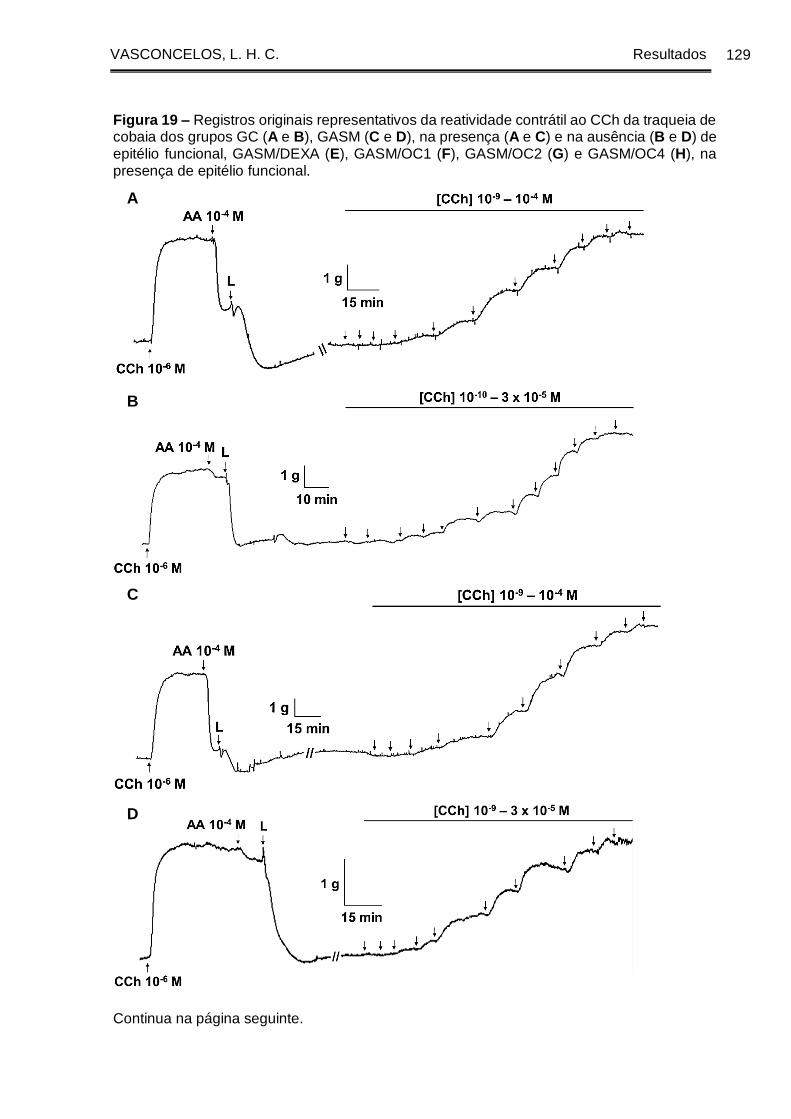
129 VASCONCELOS, L. H. C. Resultados
Figura 19 – Registros originais representativos da reatividade contrátil ao CCh da traqueia de cobaia dos grupos GC (A e B), GASM (C e D), na presença (A e C) e na ausência (B e D) de epitélio funcional, GASM/DEXA (E), GASM/OC1 (F), GASM/OC2 (G) e GASM/OC4 (H), na presença de epitélio funcional.
Continua na página seguinte.
A
B
C
D
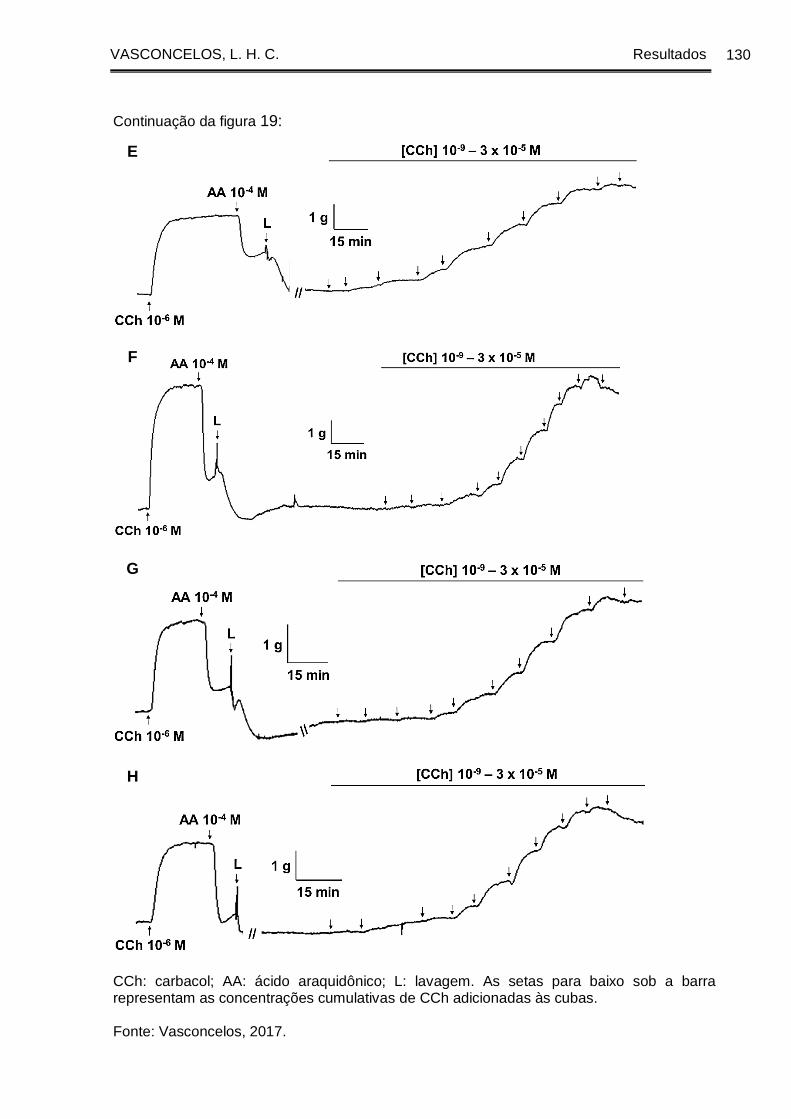
130 VASCONCELOS, L. H. C. Resultados
Continuação da figura 19:
CCh: carbacol; AA: ácido araquidônico; L: lavagem. As setas para baixo sob a barra representam as concentrações cumulativas de CCh adicionadas às cubas. Fonte: Vasconcelos, 2017.
E
F
G
H
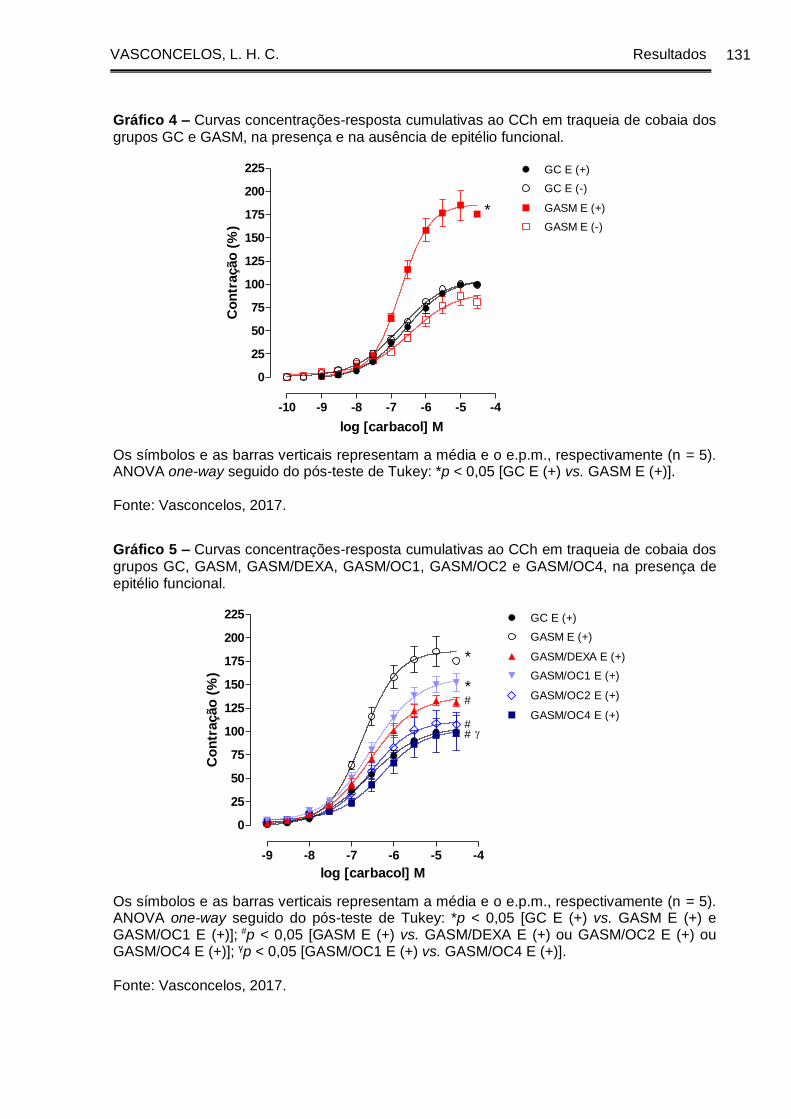
131 VASCONCELOS, L. H. C. Resultados
Gráfico 4 – Curvas concentrações-resposta cumulativas ao CCh em traqueia de cobaia dos grupos GC e GASM, na presença e na ausência de epitélio funcional.
-10 -9 -8 -7 -6 -5 -4
0
25
50
75
100
125
150
175
200
225
*GASM E (-)
GASM E (+)
GC E (-)
GC E (+)
log [carbacol] M
Co
ntr
ação
(%
)
Os símbolos e as barras verticais representam a média e o e.p.m., respectivamente (n = 5). ANOVA one-way seguido do pós-teste de Tukey: *p < 0,05 [GC E (+) vs. GASM E (+)]. Fonte: Vasconcelos, 2017.
Gráfico 5 – Curvas concentrações-resposta cumulativas ao CCh em traqueia de cobaia dos grupos GC, GASM, GASM/DEXA, GASM/OC1, GASM/OC2 e GASM/OC4, na presença de epitélio funcional.
-9 -8 -7 -6 -5 -4
0
25
50
75
100
125
150
175
200
225
GASM/OC2 E (+)
GC E (+)
GASM E (+)
GASM/OC4 E (+)
GASM/OC1 E (+)
GASM/DEXA E (+)
*#
*
##
log [carbacol] M
Co
ntr
ação
(%
)
Os símbolos e as barras verticais representam a média e o e.p.m., respectivamente (n = 5). ANOVA one-way seguido do pós-teste de Tukey: *p < 0,05 [GC E (+) vs. GASM E (+) e GASM/OC1 E (+)]; #p < 0,05 [GASM E (+) vs. GASM/DEXA E (+) ou GASM/OC2 E (+) ou GASM/OC4 E (+)]; γp < 0,05 [GASM/OC1 E (+) vs. GASM/OC4 E (+)]. Fonte: Vasconcelos, 2017.
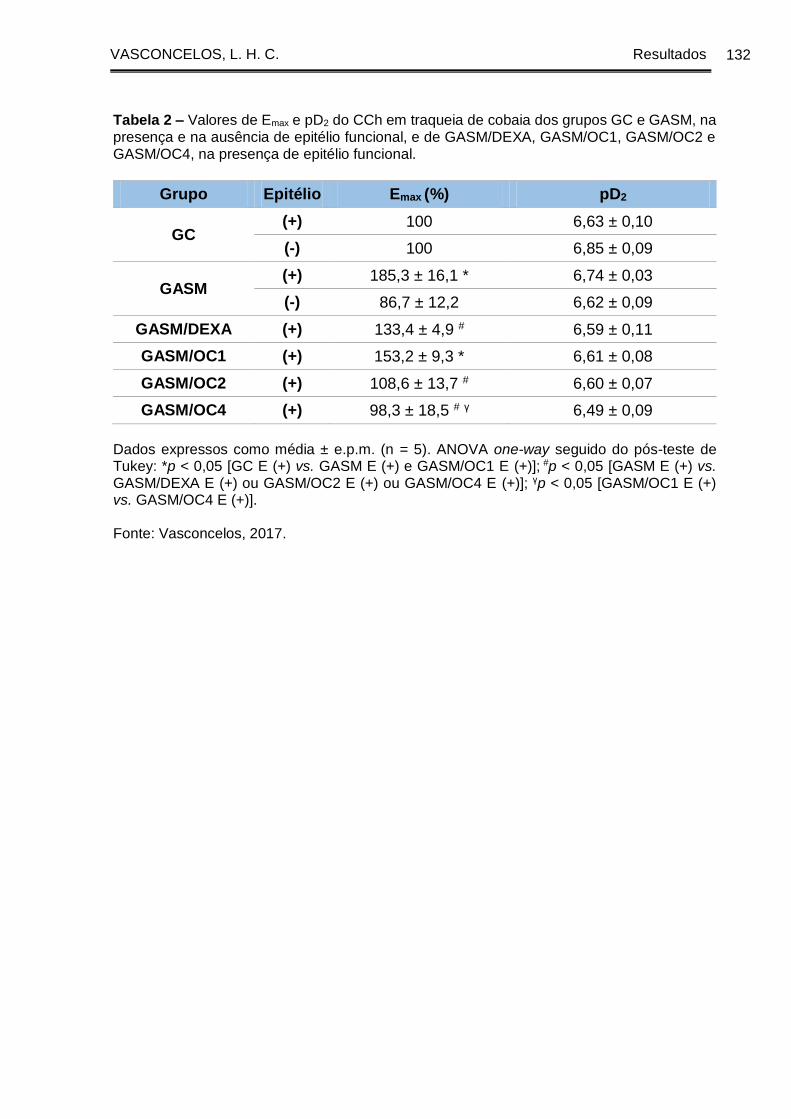
132 VASCONCELOS, L. H. C. Resultados
Tabela 2 – Valores de Emax e pD2 do CCh em traqueia de cobaia dos grupos GC e GASM, na presença e na ausência de epitélio funcional, e de GASM/DEXA, GASM/OC1, GASM/OC2 e GASM/OC4, na presença de epitélio funcional.
Grupo Epitélio Emax (%) pD2
GC (+) 100 6,63 ± 0,10
(-) 100 6,85 ± 0,09
GASM (+) 185,3 ± 16,1 * 6,74 ± 0,03
(-) 86,7 ± 12,2 6,62 ± 0,09
GASM/DEXA (+) 133,4 ± 4,9 # 6,59 ± 0,11
GASM/OC1 (+) 153,2 ± 9,3 * 6,61 ± 0,08
GASM/OC2 (+) 108,6 ± 13,7 # 6,60 ± 0,07
GASM/OC4 (+) 98,3 ± 18,5 # γ 6,49 ± 0,09
Dados expressos como média ± e.p.m. (n = 5). ANOVA one-way seguido do pós-teste de Tukey: *p < 0,05 [GC E (+) vs. GASM E (+) e GASM/OC1 E (+)]; #p < 0,05 [GASM E (+) vs. GASM/DEXA E (+) ou GASM/OC2 E (+) ou GASM/OC4 E (+)]; γp < 0,05 [GASM/OC1 E (+) vs. GASM/OC4 E (+)]. Fonte: Vasconcelos, 2017.

133 VASCONCELOS, L. H. C. Resultados
5.3.3 Avaliação da reatividade contrátil à histamina em traqueia de cobaia, na
presença e na ausência de epitélio funcional
A traqueia de cobaias do grupo GC contraiu em resposta à adição de
concentrações cumulativas de histamina (10-8-3 x 10-3 M) tanto na presença
(Emax = 100%, pD2 = 5,05 ± 0,04) com na ausência (Emax = 100%, pD2 = 5,80 ± 0,15)
de epitélio funcional. A inflamação pulmonar aumentou a eficácia contrátil da
histamina na presença de epitélio (Emax = 132,6 ± 5,9%), mas não alterou sua potência
(pD2 = 5,11 ± 0,12). Já na ausência de epitélio, não houve diferença na eficácia
(Emax = 102,2 ± 11,4%) e na potência contrátil da histamina (pD2 = 5,80 ± 0,15) entre
os grupos GASM e GC. Tanto no GC quanto no GASM, houve diferença na potência
contrátil da histamina entre os anéis de traqueia com e sem epitélio, sendo o agonista
mais potente 6,4 e 4,2 vezes na ausência de epitélio, nos grupos GC e GASM,
respectivamente (Figura 20A-D, Gráfico 6, Tabela 3, n = 5).
O tratamento com dexametasona preveniu completamente o aumento da
eficácia contrátil da histamina (Emax = 89,5 ± 9,2%), mas não alterou sua potência
(pD2 = 5,21 ± 0,15), em comparação aos grupos GC e GASM, em traqueia de cobaia
com inflamação pulmonar na presença de epitélio (Figura 20E, Gráfico 7, Tabela 3,
n = 5).
A suplementação com o óleo de coco virgem na dose de 1 g/kg não preveniu o
aumento da eficácia contrátil da histamina (Emax = 158,8 ± 16,4%) e não alterou sua
potência (pD2 = 5,23 ± 0,05), comparadas aos grupos GC e GASM, em traqueia de
cobaia com inflamação pulmonar na presença de epitélio (Figura 20F, Gráfico 7,
Tabela 3, n = 5).
De maneira semelhante, a suplementação com o óleo na dose de 2 g/kg não
alterou a eficácia (Emax = 101,5 ± 5,8%) e a potência contrátil da histamina
(pD2 = 4,99 ± 0,13), em comparação aos grupos GC e GASM, em traqueia de cobaia
com inflamação pulmonar na presença de epitélio (Figura 20G, Gráfico 7, Tabela 3,
n = 5).
Contrariamente, com a suplementação com o óleo na dose de 4 g/kg, o
aumento da eficácia contrátil da histamina foi completamente prevenido
(Emax = 87,6 ± 8,0%), mas não houve alteração na sua potência (pD2 = 4,99 ± 0,16),
em relação aos grupos GC e GASM, em traqueia de cobaia com inflamação pulmonar
na presença de epitélio (Figura 20H, Gráfico 7, Tabela 3, n = 5).
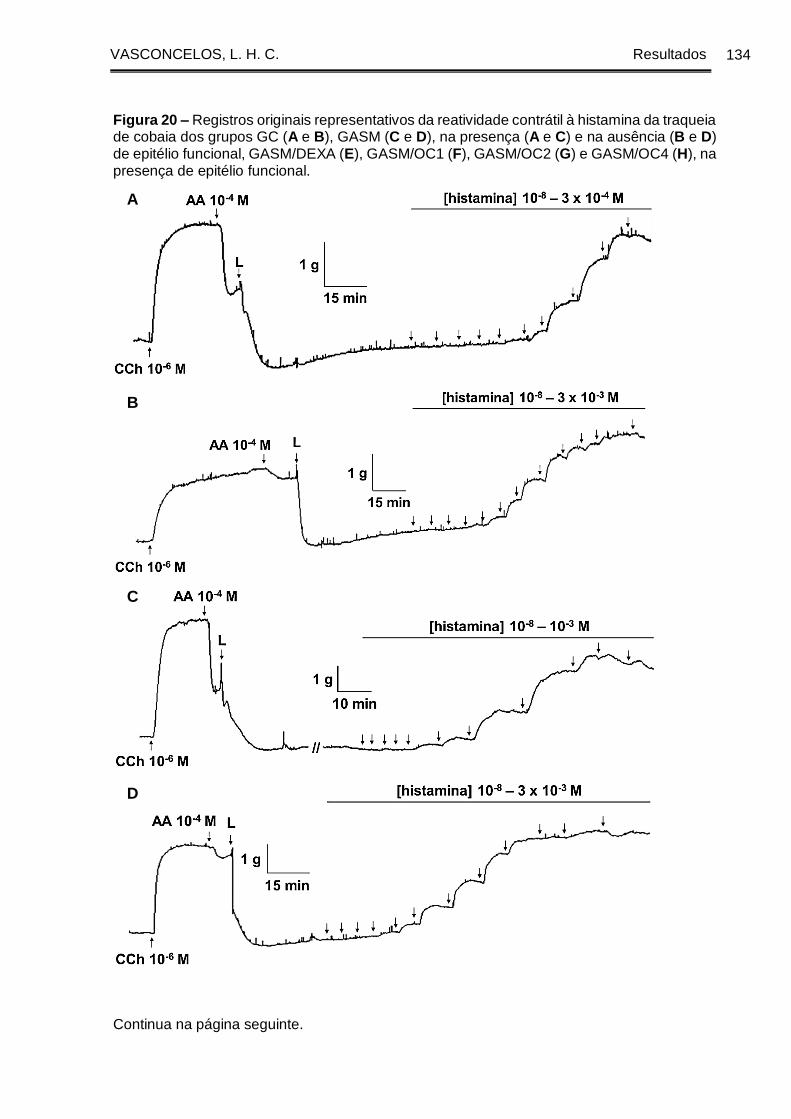
134 VASCONCELOS, L. H. C. Resultados
Figura 20 – Registros originais representativos da reatividade contrátil à histamina da traqueia de cobaia dos grupos GC (A e B), GASM (C e D), na presença (A e C) e na ausência (B e D) de epitélio funcional, GASM/DEXA (E), GASM/OC1 (F), GASM/OC2 (G) e GASM/OC4 (H), na presença de epitélio funcional.
Continua na página seguinte.
A
B
C
D
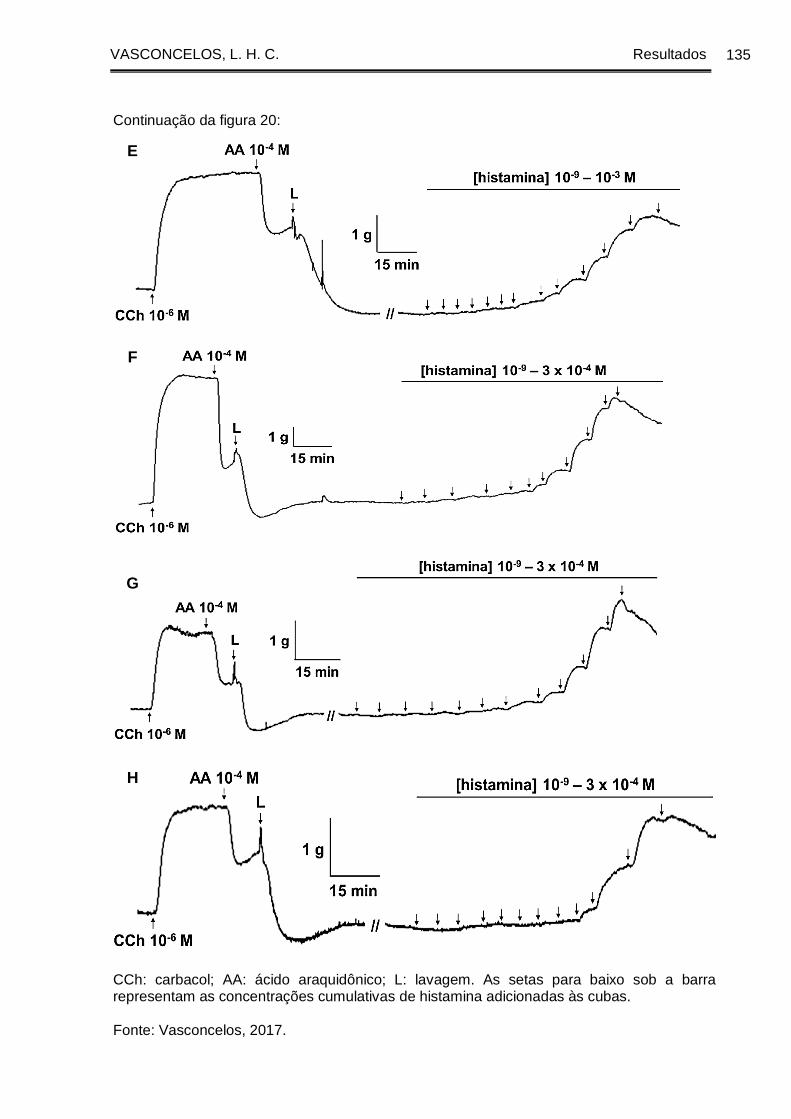
135 VASCONCELOS, L. H. C. Resultados
Continuação da figura 20:
CCh: carbacol; AA: ácido araquidônico; L: lavagem. As setas para baixo sob a barra representam as concentrações cumulativas de histamina adicionadas às cubas. Fonte: Vasconcelos, 2017.
E
F
G
H
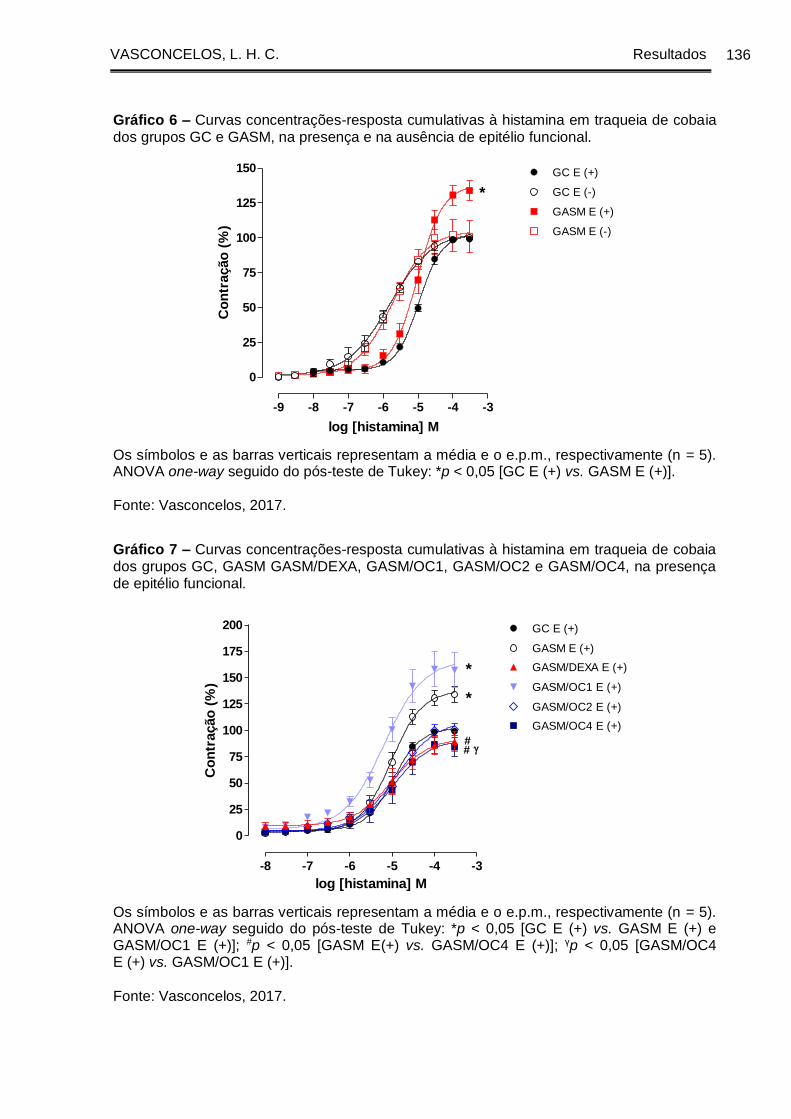
136 VASCONCELOS, L. H. C. Resultados
Gráfico 6 – Curvas concentrações-resposta cumulativas à histamina em traqueia de cobaia dos grupos GC e GASM, na presença e na ausência de epitélio funcional.
-9 -8 -7 -6 -5 -4 -3
0
25
50
75
100
125
150 GC E (+)
GC E (-)
GASM E (+)
GASM E (-)
*
log [histamina] M
Co
ntr
ação
(%
)
Os símbolos e as barras verticais representam a média e o e.p.m., respectivamente (n = 5). ANOVA one-way seguido do pós-teste de Tukey: *p < 0,05 [GC E (+) vs. GASM E (+)]. Fonte: Vasconcelos, 2017.
Gráfico 7 – Curvas concentrações-resposta cumulativas à histamina em traqueia de cobaia dos grupos GC, GASM GASM/DEXA, GASM/OC1, GASM/OC2 e GASM/OC4, na presença de epitélio funcional.
-8 -7 -6 -5 -4 -3
0
25
50
75
100
125
150
175
200
GASM/DEXA E (+)
GASM/OC2 E (+)
GC E (+)
GASM E (+)
GASM/OC4 E (+)
GASM/OC1 E (+)
*
*
##
log [histamina] M
Co
ntr
ação
(%
)
Os símbolos e as barras verticais representam a média e o e.p.m., respectivamente (n = 5). ANOVA one-way seguido do pós-teste de Tukey: *p < 0,05 [GC E (+) vs. GASM E (+) e GASM/OC1 E (+)]; #p < 0,05 [GASM E(+) vs. GASM/OC4 E (+)]; γp < 0,05 [GASM/OC4 E (+) vs. GASM/OC1 E (+)]. Fonte: Vasconcelos, 2017.
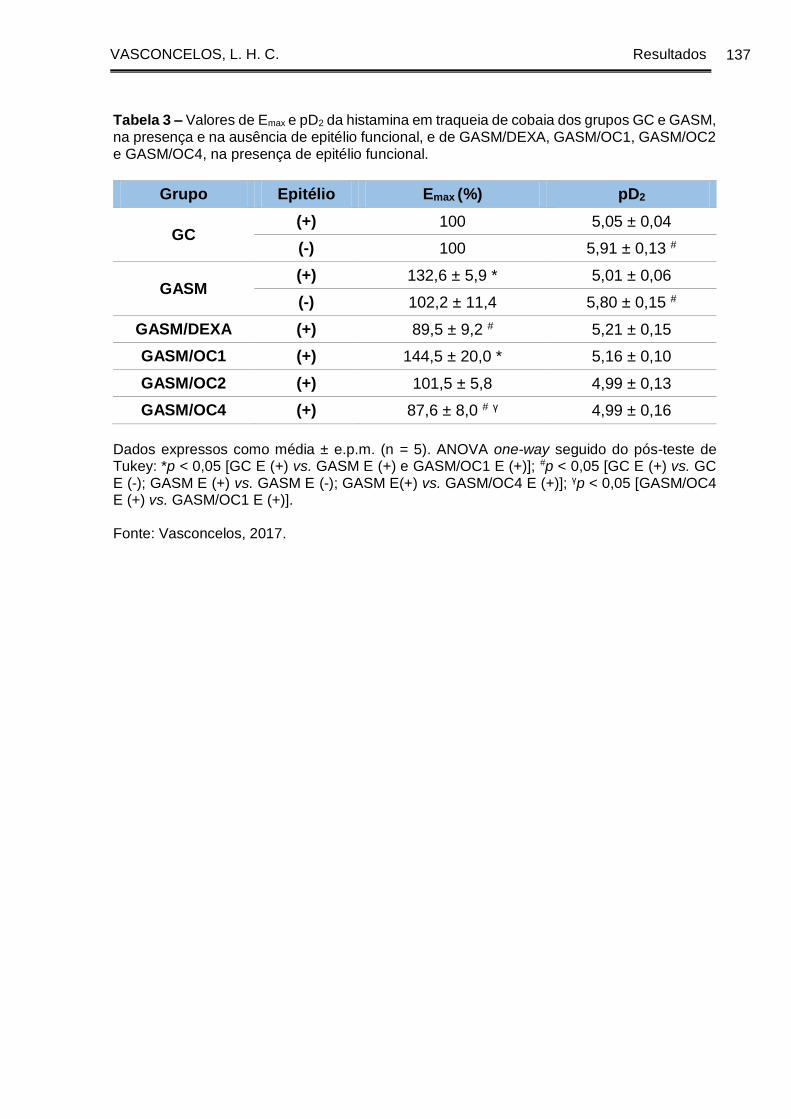
137 VASCONCELOS, L. H. C. Resultados
Tabela 3 – Valores de Emax e pD2 da histamina em traqueia de cobaia dos grupos GC e GASM, na presença e na ausência de epitélio funcional, e de GASM/DEXA, GASM/OC1, GASM/OC2 e GASM/OC4, na presença de epitélio funcional.
Grupo Epitélio Emax (%) pD2
GC (+) 100 5,05 ± 0,04
(-) 100 5,91 ± 0,13 #
GASM (+) 132,6 ± 5,9 * 5,01 ± 0,06
(-) 102,2 ± 11,4 5,80 ± 0,15 #
GASM/DEXA (+) 89,5 ± 9,2 # 5,21 ± 0,15
GASM/OC1 (+) 144,5 ± 20,0 * 5,16 ± 0,10
GASM/OC2 (+) 101,5 ± 5,8 4,99 ± 0,13
GASM/OC4 (+) 87,6 ± 8,0 # γ 4,99 ± 0,16
Dados expressos como média ± e.p.m. (n = 5). ANOVA one-way seguido do pós-teste de Tukey: *p < 0,05 [GC E (+) vs. GASM E (+) e GASM/OC1 E (+)]; #p < 0,05 [GC E (+) vs. GC E (-); GASM E (+) vs. GASM E (-); GASM E(+) vs. GASM/OC4 E (+)]; γp < 0,05 [GASM/OC4 E (+) vs. GASM/OC1 E (+)]. Fonte: Vasconcelos, 2017.

138 VASCONCELOS, L. H. C. Resultados
5.3.4 Avaliação da reatividade contrátil ao KCl em traqueia de cobaia, na
presença e na ausência de epitélio funcional
A traqueia de cobaia do grupo GC contraiu em resposta à adição de
concentrações cumulativas de KCl (10-3 - 3 x 10-1 M) tanto na presença (Emax = 100%,
pD2 = 1,60 ± 0,01) como na ausência de epitélio funcional (Emax = 100%,
pD2 = 1,89 ± 0,08). A inflamação pulmonar não alterou a eficácia e a potência contrátil
do KCl, tanto na presença (Emax = 119,0 ± 14,2%, pD2 = 1,56 ± 0,03) como na ausência
de epitélio funcional (Emax = 107,6 ± 11,5%, pD2 = 1,73 ± 0,07). Entretanto, no grupo
GC, a potência do KCl foi 1,8 vezes maior na ausência de epitélio, não diferindo no
grupo GASM (Figura 21A-D, Gráfico 8, Tabela 4, n = 5).
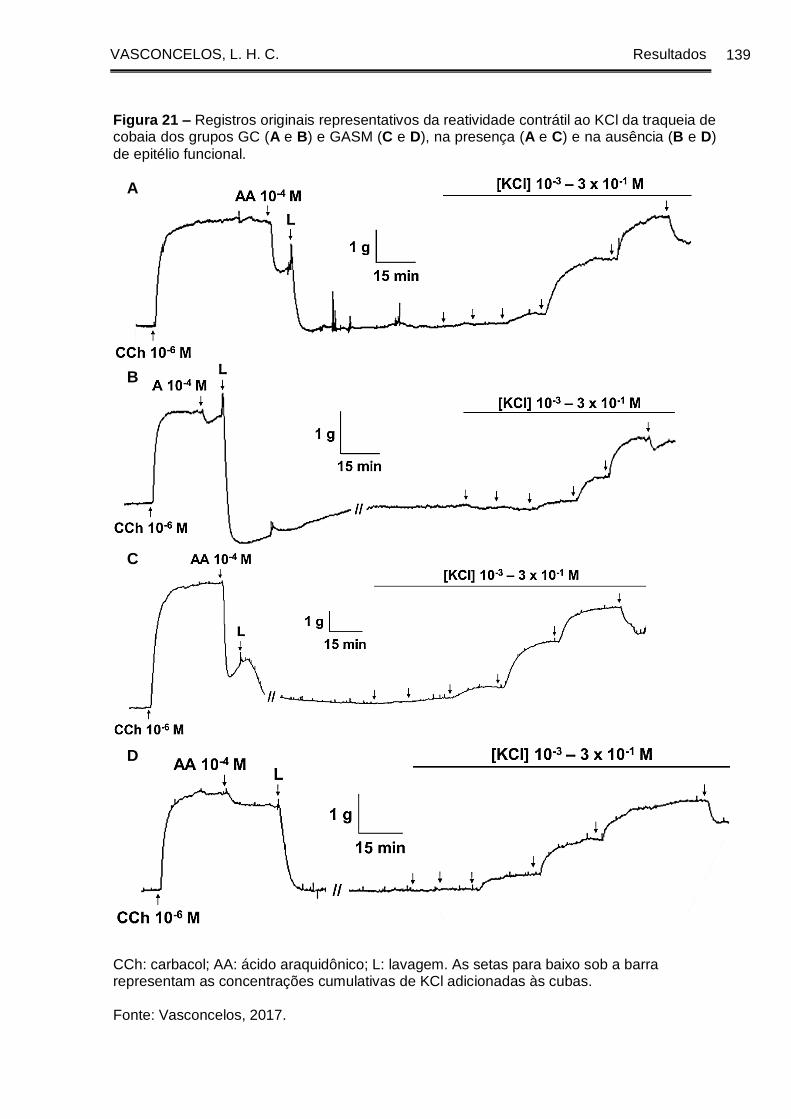
139 VASCONCELOS, L. H. C. Resultados
Figura 21 – Registros originais representativos da reatividade contrátil ao KCl da traqueia de cobaia dos grupos GC (A e B) e GASM (C e D), na presença (A e C) e na ausência (B e D)
de epitélio funcional.
CCh: carbacol; AA: ácido araquidônico; L: lavagem. As setas para baixo sob a barra representam as concentrações cumulativas de KCl adicionadas às cubas. Fonte: Vasconcelos, 2017.
A
B
C
D
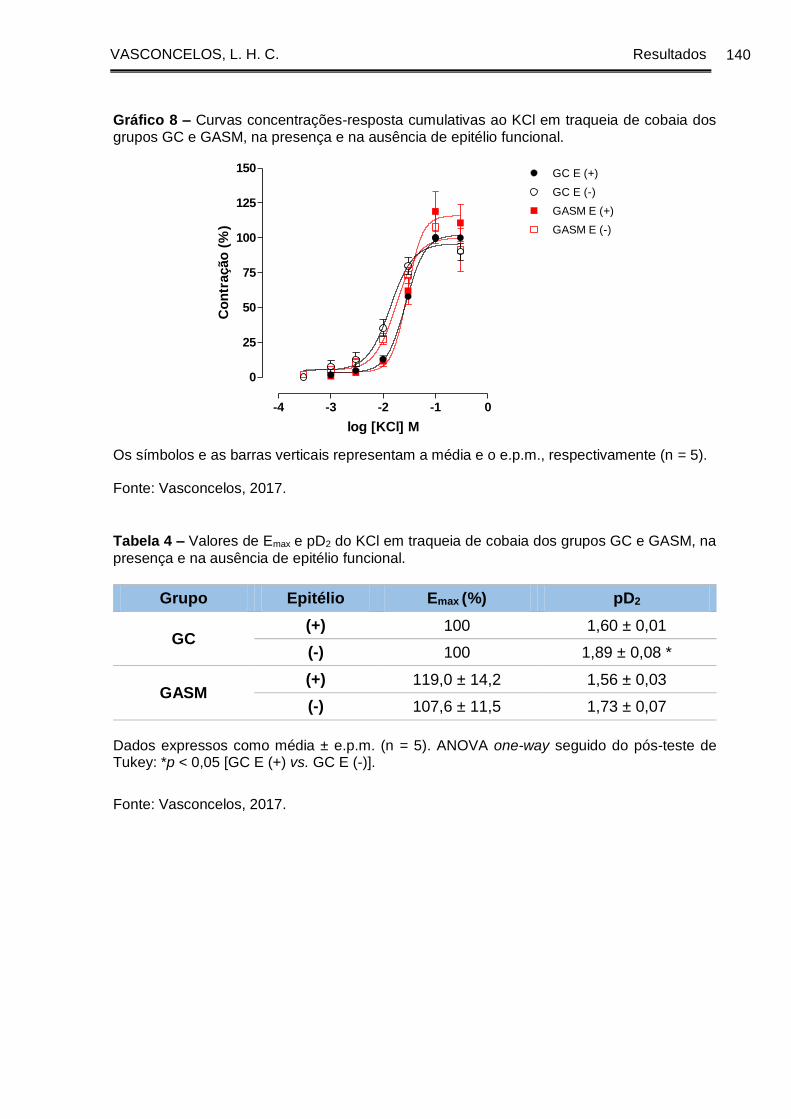
140 VASCONCELOS, L. H. C. Resultados
Gráfico 8 – Curvas concentrações-resposta cumulativas ao KCl em traqueia de cobaia dos grupos GC e GASM, na presença e na ausência de epitélio funcional.
-4 -3 -2 -1 0
0
25
50
75
100
125
150 GC E (+)
GC E (-)
GASM E (+)
GASM E (-)
log [KCl] M
Co
ntr
ação
(%
)
Os símbolos e as barras verticais representam a média e o e.p.m., respectivamente (n = 5). Fonte: Vasconcelos, 2017.
Tabela 4 – Valores de Emax e pD2 do KCl em traqueia de cobaia dos grupos GC e GASM, na
presença e na ausência de epitélio funcional.
Grupo Epitélio Emax (%) pD2
GC (+) 100 1,60 ± 0,01
(-) 100 1,89 ± 0,08 *
GASM (+) 119,0 ± 14,2 1,56 ± 0,03
(-) 107,6 ± 11,5 1,73 ± 0,07
Dados expressos como média ± e.p.m. (n = 5). ANOVA one-way seguido do pós-teste de Tukey: *p < 0,05 [GC E (+) vs. GC E (-)].
Fonte: Vasconcelos, 2017.

141 VASCONCELOS, L. H. C. Resultados
5.3.5 Avaliação do efeito da isoprenalina sobre o tônus basal da traqueia de
cobaia, na presença e na ausência de epitélio funcional
A isoprenalina reduziu o tônus basal da traqueia de cobaias do grupo GC tanto
na presença (Emax = 100%, pD2 = 8,10 ± 0,09) como na ausência (Emax = 100%,
pD2 = 8,35 ± 0,08) de epitélio funcional. A inflamação pulmonar não alterou a resposta
da traqueia à isoprenalina sobre seu tônus basal tanto na presença
(Emax = 111,8 ± 7,1%, pD2 = 8,24 ± 0,12) quanto na ausência (Emax = 92,7 ± 14,4%,
pD2 = 8,62 ± 0,14) de epitélio funcional. Não houve diferença nem na eficácia nem na
potência da isoprenalina entre as preparações com e sem epitélio, tanto no grupo GC
quanto no GASM (Figura 22A-D, Gráfico 9, Tabela 5, n = 5).

142 VASCONCELOS, L. H. C. Resultados
Figura 22 – Registros representativos do efeito da isoprenalina sobre o tônus basal da traqueia de cobaia dos grupos GC (A e B) e GASM (C e D), na presença (A e C) e na ausência (B e D) de epitélio funcional.
CCh: carbacol; AA: ácido araquidônico; L: lavagem. As setas para baixo sob a barra representam as concentrações cumulativas de isoprenalina adicionadas às cubas. Fonte: Vasconcelos, 2017.
A
B
D
C
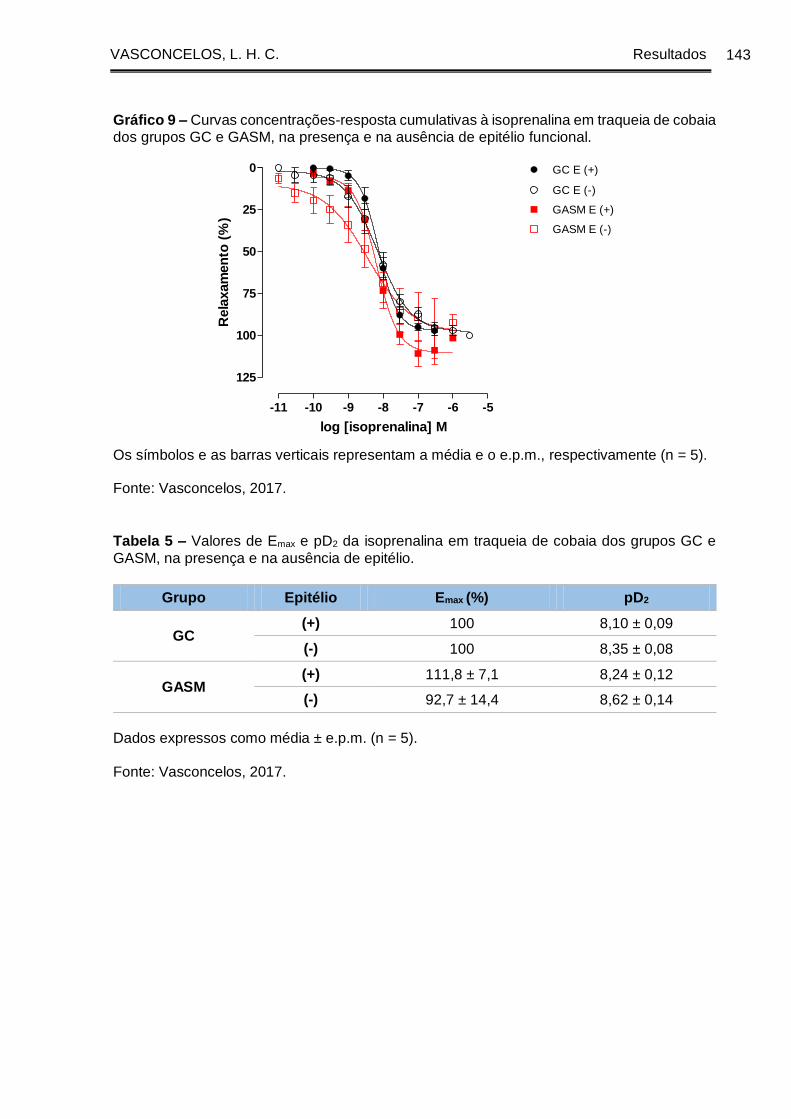
143 VASCONCELOS, L. H. C. Resultados
Gráfico 9 – Curvas concentrações-resposta cumulativas à isoprenalina em traqueia de cobaia dos grupos GC e GASM, na presença e na ausência de epitélio funcional.
-11 -10 -9 -8 -7 -6 -5
0
25
50
75
100
125
GC E (+)
GC E (-)
GASM E (+)
GASM E (-)
log [isoprenalina] M
Rela
xam
en
to (
%)
Os símbolos e as barras verticais representam a média e o e.p.m., respectivamente (n = 5). Fonte: Vasconcelos, 2017.
Tabela 5 – Valores de Emax e pD2 da isoprenalina em traqueia de cobaia dos grupos GC e
GASM, na presença e na ausência de epitélio.
Grupo Epitélio Emax (%) pD2
GC (+) 100 8,10 ± 0,09
(-) 100 8,35 ± 0,08
GASM (+) 111,8 ± 7,1 8,24 ± 0,12
(-) 92,7 ± 14,4 8,62 ± 0,14
Dados expressos como média ± e.p.m. (n = 5). Fonte: Vasconcelos, 2017.

144 VASCONCELOS, L. H. C. Resultados
5.3.6 Avaliação da reatividade relaxante à isoprenalina em traqueia de cobaia,
na presença e na ausência de epitélio funcional
A isoprenalina relaxou a traqueia de cobaia do grupo GC pré-contraída com
10-6 M de CCh, tanto na presença (Emax = 135,7 ± 9,1%, pD2 = 7,79 ± 0,10) como na
ausência (Emax = 115,9 ± 8,7%, pD2 = 7,81 ± 0,09) de epitélio funcional. A inflamação
pulmonar não alterou a eficácia relaxante da isoprenalina tanto na presença
(Emax = 124,4 ± 6,6%) como na ausência (Emax = 118,2 ± 7,3%) de epitélio funcional.
No entanto, a potência relaxante do agonista foi reduzida cerca de 3,5 vezes no grupo
GASM (pD2 = 7,24 ± 0,09) em relação ao grupo GC na presença, mas não na ausência
de epitélio (pD2 = 7,89 ± 0,12) (Figura 23A-D, Gráfico 10, Tabela 6, n = 5).
O tratamento com dexametasona não alterou a eficácia da isoprenalina
(Emax = 122,3 ± 6,3%) nem preveniu a redução da sua potência (pD2 = 7,20 ± 0,06),
em relação aos grupos GC e GASM, em traqueia de cobaia com inflamação
pulmoanar na presença de epitélio (Figura 23E, Gráfico 11, Tabela 6, n = 5).
A suplementação com o óleo de coco virgem na dose de 1 g/kg não alterou a
eficácia (Emax = 130,4 ± 5,9%) e não preveniu a redução da potência relaxante da
isoprenalina (pD2 = 7,14 ± 0,13), em relação aos grupos GC e ao GASM, em traqueia
de cobaia com inflamação pulmonar na presença de epitélio (Figura 23F, Gráfico 11,
Tabela 6, n = 5).
A suplementação com o óleo de coco virgem na dose de 2 g/kg, a eficácia
relaxante da isoprenalina não foi modificada (Emax = 134,2 ± 11,6%), assim como a
redução da sua potência não foi prevenida (pD2 = 7,37 ± 0,08), em relação aos grupos
GC e GASM, em traqueia de cobaia com inflamação pulmonar na presença de epitélio
(Figura 23G, Gráfico 11, Tabela 6, n = 5).
Já a suplementação com o óleo de coco virgem na dose de 4 g/kg também não
alterou a eficácia relaxante da isoprenalina (Emax = 117,8 ± 12,0%), mas preveniu a
redução da sua potência (pD2 = 7,93 ± 0,14), em relação aos grupos GC e GASM, em
traqueia de cobaia com inflamação pulmonar na presença de epitélio (Figura 23H,
Gráfico 11, Tabela 6, n = 5).
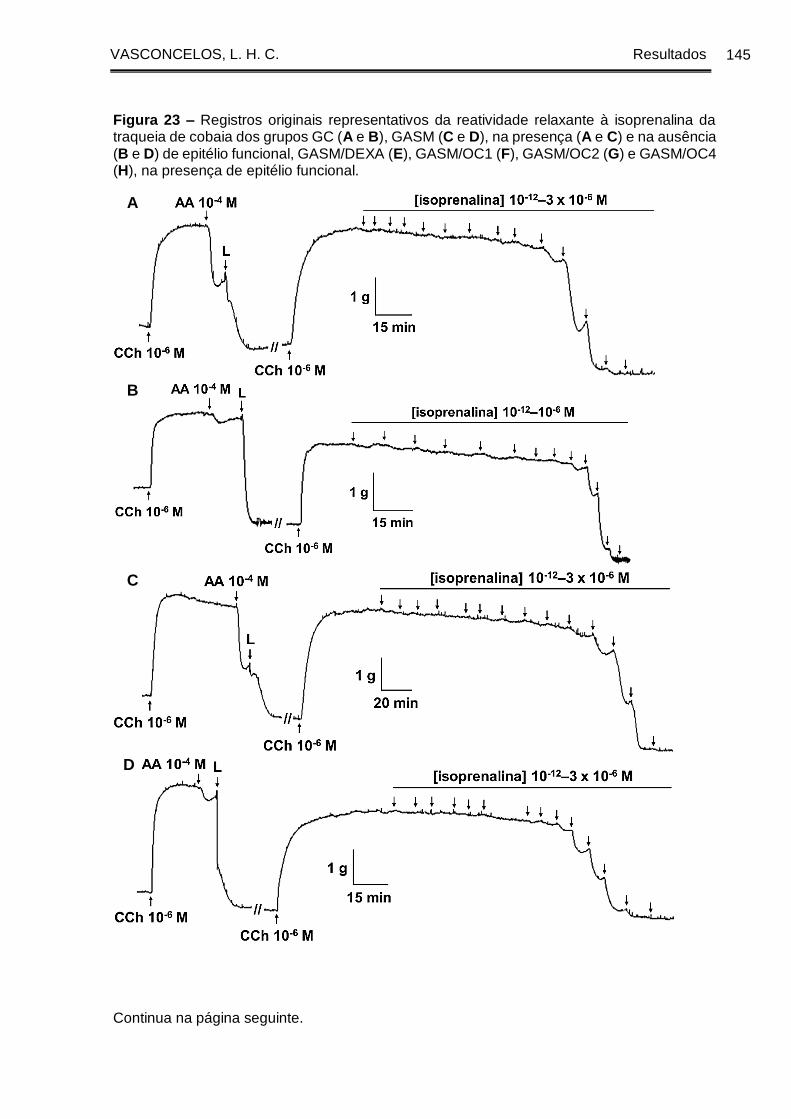
145 VASCONCELOS, L. H. C. Resultados
Figura 23 – Registros originais representativos da reatividade relaxante à isoprenalina da traqueia de cobaia dos grupos GC (A e B), GASM (C e D), na presença (A e C) e na ausência (B e D) de epitélio funcional, GASM/DEXA (E), GASM/OC1 (F), GASM/OC2 (G) e GASM/OC4 (H), na presença de epitélio funcional.
Continua na página seguinte.
D
A
B
C
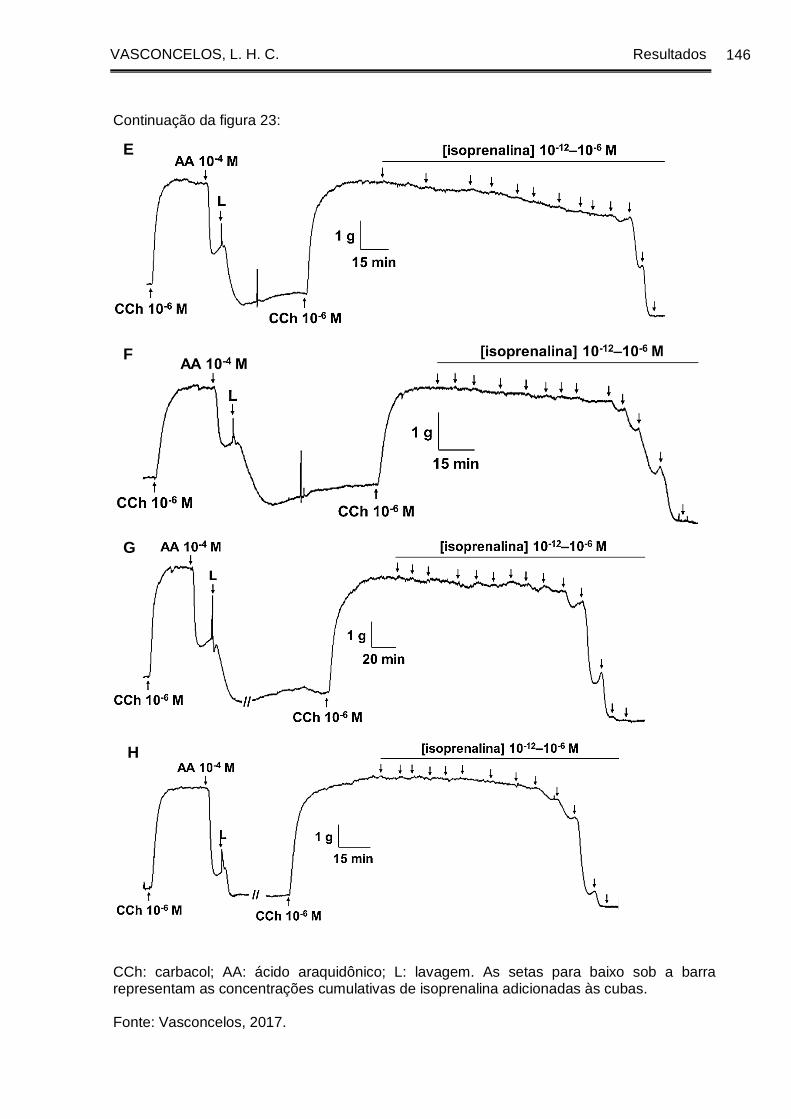
146 VASCONCELOS, L. H. C. Resultados
Continuação da figura 23:
CCh: carbacol; AA: ácido araquidônico; L: lavagem. As setas para baixo sob a barra representam as concentrações cumulativas de isoprenalina adicionadas às cubas. Fonte: Vasconcelos, 2017.
E
F
G
H
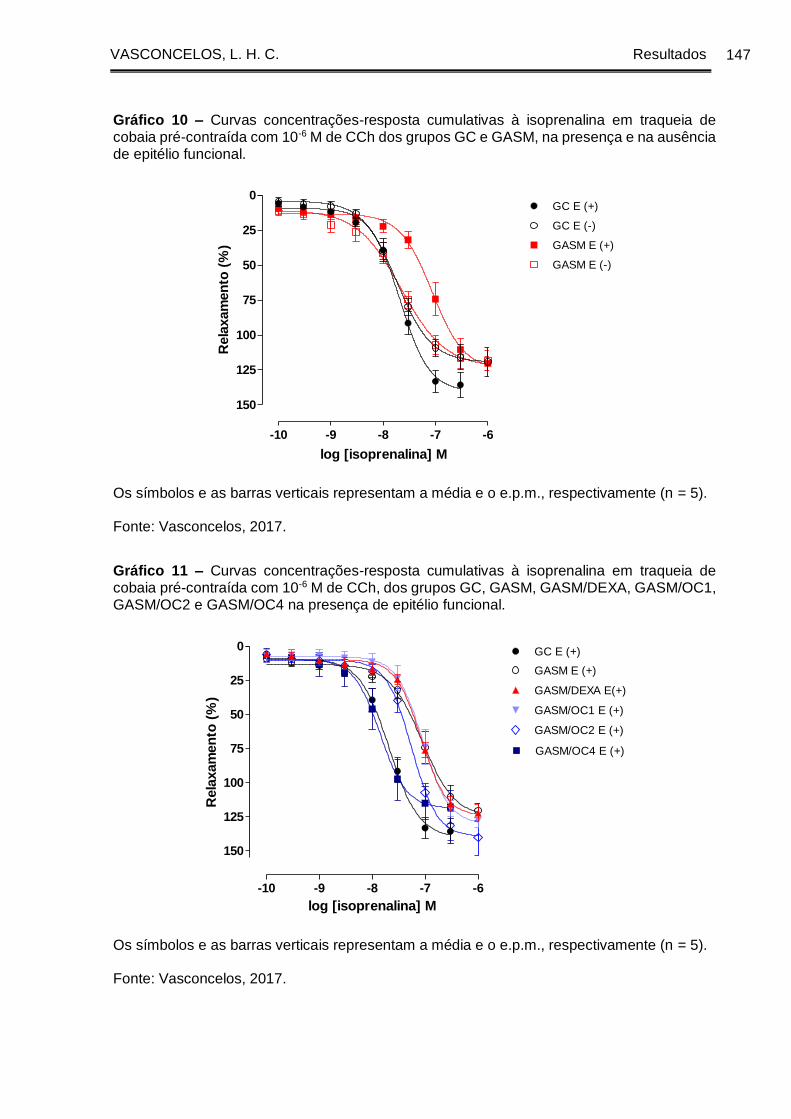
147 VASCONCELOS, L. H. C. Resultados
Gráfico 10 – Curvas concentrações-resposta cumulativas à isoprenalina em traqueia de cobaia pré-contraída com 10-6 M de CCh dos grupos GC e GASM, na presença e na ausência de epitélio funcional.
-10 -9 -8 -7 -6
0
25
50
75
100
125
150
GC E (+)
GC E (-)
GASM E (+)
GASM E (-)
log [isoprenalina] M
Rela
xam
en
to (
%)
Os símbolos e as barras verticais representam a média e o e.p.m., respectivamente (n = 5). Fonte: Vasconcelos, 2017.
Gráfico 11 – Curvas concentrações-resposta cumulativas à isoprenalina em traqueia de cobaia pré-contraída com 10-6 M de CCh, dos grupos GC, GASM, GASM/DEXA, GASM/OC1, GASM/OC2 e GASM/OC4 na presença de epitélio funcional.
-10 -9 -8 -7 -6
0
25
50
75
100
125
150
GASM/OC2 E (+)
GC E (+)
GASM E (+)
GASM/OC4 E (+)
GASM/OC1 E (+)
GASM/DEXA E(+)
log [isoprenalina] M
Rela
xam
en
to (
%)
Os símbolos e as barras verticais representam a média e o e.p.m., respectivamente (n = 5). Fonte: Vasconcelos, 2017.
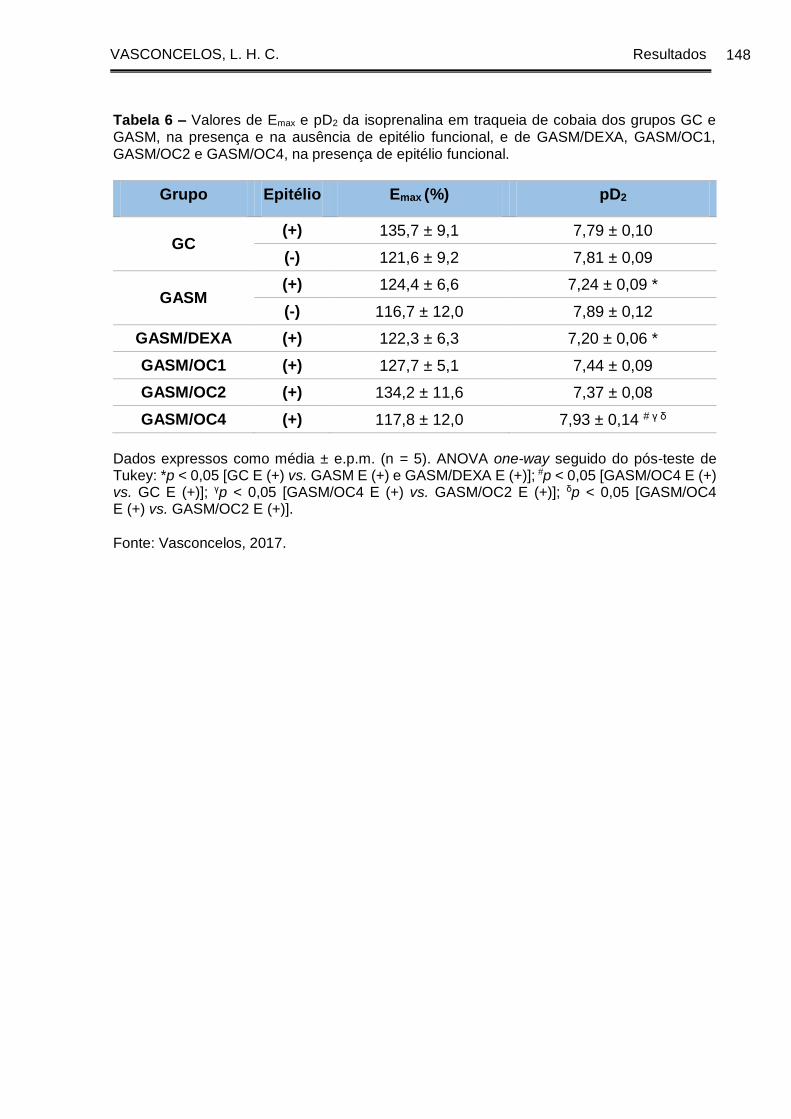
148 VASCONCELOS, L. H. C. Resultados
Tabela 6 – Valores de Emax e pD2 da isoprenalina em traqueia de cobaia dos grupos GC e GASM, na presença e na ausência de epitélio funcional, e de GASM/DEXA, GASM/OC1, GASM/OC2 e GASM/OC4, na presença de epitélio funcional.
Grupo Epitélio Emax (%) pD2
GC (+) 135,7 ± 9,1 7,79 ± 0,10
(-) 121,6 ± 9,2 7,81 ± 0,09
GASM (+) 124,4 ± 6,6 7,24 ± 0,09 *
(-) 116,7 ± 12,0 7,89 ± 0,12
GASM/DEXA (+) 122,3 ± 6,3 7,20 ± 0,06 *
GASM/OC1 (+) 127,7 ± 5,1 7,44 ± 0,09
GASM/OC2 (+) 134,2 ± 11,6 7,37 ± 0,08
GASM/OC4 (+) 117,8 ± 12,0 7,93 ± 0,14 # γ δ
Dados expressos como média ± e.p.m. (n = 5). ANOVA one-way seguido do pós-teste de Tukey: *p < 0,05 [GC E (+) vs. GASM E (+) e GASM/DEXA E (+)]; #p < 0,05 [GASM/OC4 E (+) vs. GC E (+)]; γp < 0,05 [GASM/OC4 E (+) vs. GASM/OC2 E (+)]; δp < 0,05 [GASM/OC4 E (+) vs. GASM/OC2 E (+)].
Fonte: Vasconcelos, 2017.

149 VASCONCELOS, L. H. C. Resultados
5.3.7 Avaliação da reatividade relaxante ao nifedipino em traqueia de cobaia, na
presença e na ausência de epitélio funcional
O nifedipino relaxou a traqueia de cobaia pré-contraída com 10-6 M de CCh do
grupo GC tanto na presença (Emax = 138,2 ± 8,3%, pD2 = 4,54 ± 0,15) como na
ausência (Emax = 132,9 ± 7,2, pD2 = 4,73 ± 0,10) de epitélio funcional. A inflamação
pulmonar não alterou a eficácia e a potência relaxante do nifedipino, tanto na presença
(Emax = 122,8 ± 8,8%, pD2 = 4,54 ± 0,15) como na ausência (Emax = 123,9 ± 10,4%,
pD2 = 4,71 ± 0,23) de epitélio funcional. Não houve diferença na eficácia e na potência
relaxante do nifedipino entre as preparações com e sem epitélio nos grupos GC e
GASM (Figura 24A-D, Gráfico 12, Tabela 7, n = 5).
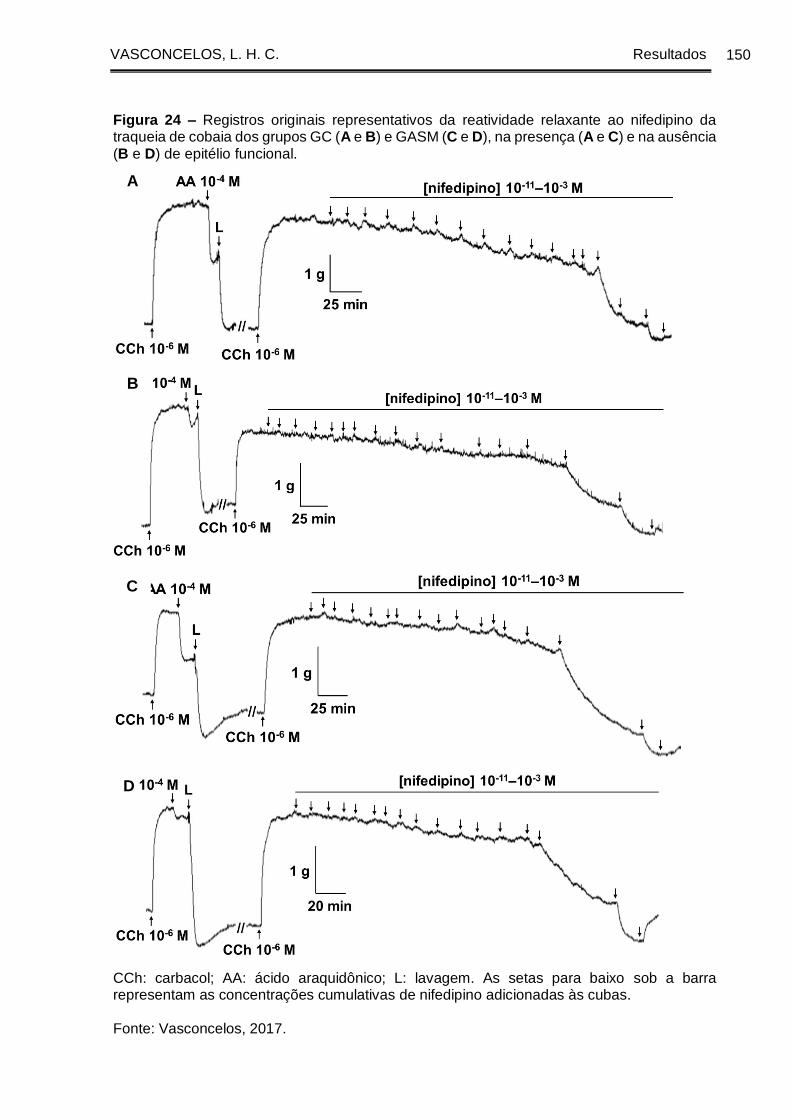
150 VASCONCELOS, L. H. C. Resultados
Figura 24 – Registros originais representativos da reatividade relaxante ao nifedipino da traqueia de cobaia dos grupos GC (A e B) e GASM (C e D), na presença (A e C) e na ausência (B e D) de epitélio funcional.
CCh: carbacol; AA: ácido araquidônico; L: lavagem. As setas para baixo sob a barra representam as concentrações cumulativas de nifedipino adicionadas às cubas. Fonte: Vasconcelos, 2017.
A
B
C
D

151 VASCONCELOS, L. H. C. Resultados
Gráfico 12 – Curvas concentrações-resposta cumulativas ao nifedipino em traqueia de cobaia pré-contraída com 10-6 M de CCh, dos grupos GC e GASM, na presença e na ausência de epitélio funcional.
-11 -10 -9 -8 -7 -6 -5 -4 -3
0
25
50
75
100
125
150
GC E (+)
GC E (-)
GASM E (+)
GASM E (-)
log [nifedipino] M
Rela
xam
en
to (
%)
Os símbolos e as barras verticais representam a média e o e.p.m., respectivamente (n = 5). Fonte: Vasconcelos, 2017.
Tabela 7 – Valores de Emax e pD2 do nifedipino em traqueia de cobaia pré-contraída com 10-6 M de CCh, dos grupos GC e GASM, na presença e na ausência de epitélio funcional.
Grupo Epitélio Emax (%) pD2
GC (+) 138,2 ± 8,3 4,54 ± 0,15
(-) 132,9 ± 7,2 4,73 ± 0,10
GASM (+) 122,8 ± 8,8 4,30 ± 0,02
(-) 123,9 ± 10,4 4,71 ± 0,23
Dados expressos como média ± e.p.m. (n = 5).
Fonte: Vasconcelos, 2017.

152 VASCONCELOS, L. H. C. Resultados
5.4 Investigação do mecanismo de ação envolvido nas alterações induzidas pela
inflamação pulmonar alérgica crônica e pelo óleo de coco virgem sobre a
reatividade contrátil da traqueia de cobaia
5.4.1 Avaliação do envolvimento das espécies reativas de oxigênio (ROS)
5.4.1.1 Investigação da participação das enzimas nicotinamida adenina
dinucleotídeo fosfato (NADPH) oxidase, superóxido dismutase (SOD) e catalase
5.4.1.1.1 Obtenção de curvas cumulativas concentrações-resposta ao CCh, na
ausência e na presença da apocinina, do tempol ou da catalase, em traqueia de
cobaia dos grupos GC, GASM e GASM/OC4
A curva concentrações-resposta cumulativa ao CCh (10-10-3 x 10-5 M) do grupo
GC (Emax = 100%; pD2 = 6,82 ± 0,06) não foi alterada na presença da apocinina,
inibidora da NADPH oxidase, nem com relação à eficácia (Emax = 115,6 ± 9,4%) nem
à potência (pD2 = 6,52 ± 0,13), em traqueia de cobaia na presença de epitélio (Figura
25A, Gráfico 13A, Tabela 8, n = 5). Diferentemente, na presença do tempol, mimético
da SOD, a curva de contração do CCh (10-10-3 x 10-5 M) foi deslocada para a direita
3,2 vezes (pD2 = 6,30 ± 0,07), sem alteração da eficácia contrátil do agonista
(Emax = 99,4 ± 12,3%) (Figura 25B, Gráfico 13A, Tabela 8, n = 5). Semelhantemente,
na presença da catalase, enzima que converte H2O2 em H2O e O2, a potência contrátil
do CCh (10-10-10-4 M) foi reduzida em 4 vezes (pD2 = 6,23 ± 0,09), sem alteração da
sua eficácia (Emax = 116,6 ± 11,9%) (Figura 25C, Gráfico 13C, Tabela 8, n = 5).
Nos cobaias com inflamação pulmonar, a curva cumulativa de contração ao
CCh (10-10-10-4 M) (Emax = 100%; pD2 = 6,35 ± 0,11) não foi modificada pela apocinina,
tanto com relação à eficácia (Emax = 105,6 ± 11,7%) como à potência
(pD2 = 6,54 ± 0,11), em traqueia de cobaia na presença de epitélio (Figura 25D,
Gráfico 13B, Tabela 8, n = 5). Semelhantemente, na presença do tempol, a curva de
contração do CCh não foi modificada (Emax = 101,0 ± 11,7%; pD2 = 6,62 ± 0,27) (Figura
25E, Gráfico 13B, Tabela 8, n = 5), assim como na presença da catalase
(Emax = 102,4 ± 9,1%; pD2 = 5,99 ± 0,06) (Figura 25F, Gráfico 13C, Tabela 8, n = 5).
Com a suplementação com o óleo de coco virgem na dose de 4 g/kg, foi
observado o mesmo dos grupos GC e GASM, na presença da apocinina, em que a
curva cumulativa de contração ao CCh (10-10-3 x 10-5 M) (Emax = 100%;
pD2 = 6,88 ± 0,07) não foi alterada, com relação aos parâmetros de eficácia
(Emax = 102,3 ± 4,3%) e de potência (pD2 = 6,81 ± 0,14), em traqueia de cobaia

153 VASCONCELOS, L. H. C. Resultados
asmático na presença de epitélio (Figura 25G, Gráfico 13A, Tabela 8, n = 5). Na
presença do tempol, observou-se o mesmo do grupo GASM, em que a potência
(pD2 = 6,44 ± 0,12) e a eficácia contrátil do CCh (Emax = 91,9 ± 13,0%) não foram
alteradas (Figura 25H, Gráfico 13B, Tabela 8, n = 5). Diferentemente, na presença da
catalase, a potência contrátil do CCh foi reduzida 6,6 vezes (pD2 = 6,07 ± 0,08), sem
que a eficácia contrátil do agonista fosse alterada (Emax = 111,4 ± 8,8%) (Figura 25I,
Gráfico 13C, Tabela 8, n = 5).
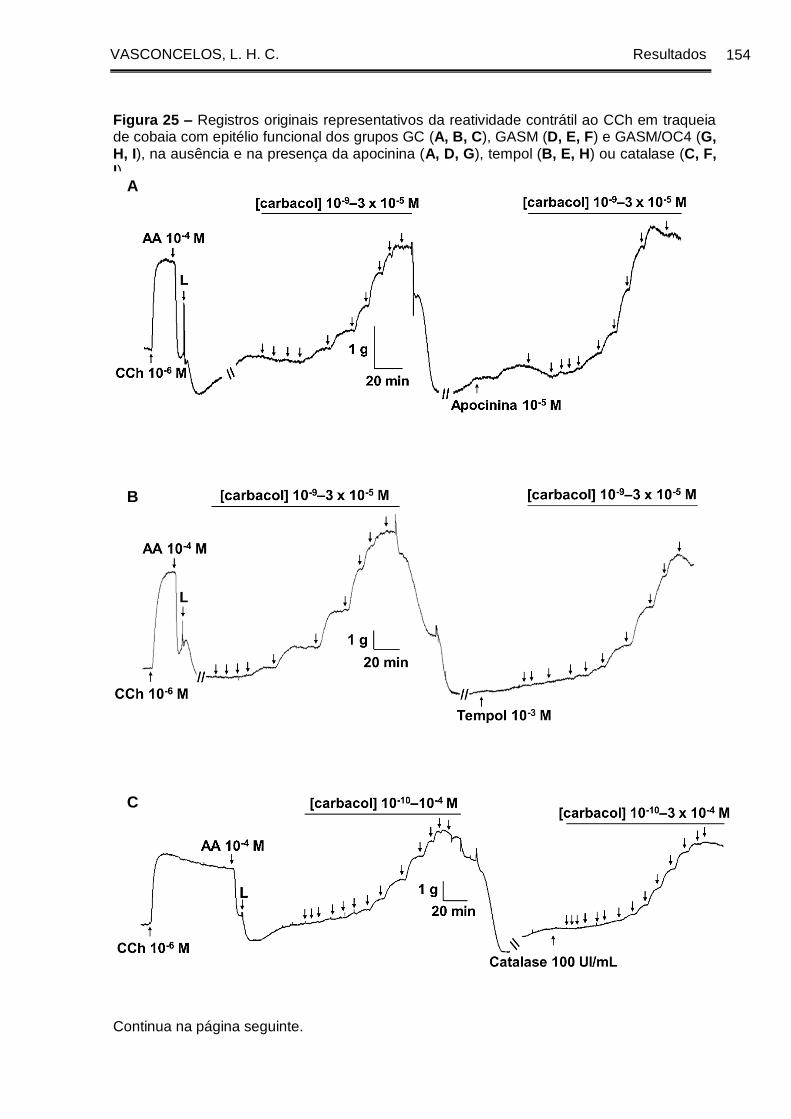
154 VASCONCELOS, L. H. C. Resultados
Figura 25 – Registros originais representativos da reatividade contrátil ao CCh em traqueia de cobaia com epitélio funcional dos grupos GC (A, B, C), GASM (D, E, F) e GASM/OC4 (G, H, I), na ausência e na presença da apocinina (A, D, G), tempol (B, E, H) ou catalase (C, F, I).
Continua na página seguinte.
A
B
C

155 VASCONCELOS, L. H. C. Resultados
Continuação da Figura 25:
Continua na página seguinte.
E
F
D

156 VASCONCELOS, L. H. C. Resultados
Continuação da Figura 25:
CCh: carbacol; AA: ácido araquidônico; L: lavagem. As setas para baixo sob a barra representam as concentrações cumulativas de carbacol adicionadas às cubas. Fonte: Vasconcelos, 2017.
G
H
I

157 VASCONCELOS, L. H. C. Resultados
Gráfico 13 – Curvas concentrações-resposta cumulativas ao CCh em traqueia de cobaia com epitélio funcional dos grupos GC (A), GASM (B) e GASM/OC4 (C), na ausência e na presença
da apocinina, do tempol ou da catalase.
-10 -9 -8 -7 -6 -5 -4
0
25
50
75
100
125
150 GC
GC + apocinina
GC + tempol
GC + catalase
log [carbacol] M
Co
ntr
ação
(%
)
-10 -9 -8 -7 -6 -5 -4
0
25
50
75
100
125
150 GASM
GASM + apocinina
GASM + tempol
GASM + catalase
log [carbacol] M
Co
ntr
ação
(%
)
-10 -9 -8 -7 -6 -5 -4
0
25
50
75
100
125
150 GASM/OC4
GASM/OC4 + apocinina
GASM/OC4 + tempol
GASM/OC4 + catalase
log [carbacol] M
Co
ntr
ação
(%
)
Os símbolos e as barras verticais representam a média e o e.p.m., respectivamente (n = 5).
Fonte: Vasconcelos, 2017.
A B
C

158 VASCONCELOS, L. H. C. Resultados
Tabela 8 – Valores de Emax e pD2 do CCh em traqueia de cobaia com epitélio dos grupos GC, GASM e GASM/OC4, na ausência e na presença de apocinina, tempol ou catalase.
Grupo Bloqueador/
ativador Emax (%) pD2
GC
Controle 100,0 6,82 ± 0,06
Apocinina 115,6 ± 9,4 6,52 ± 0,13
Tempol 99,4 ± 12,3% 6,30 ± 0,07 *
Catalase 116,6 ± 11,9% 6,23 ± 0,09 *
GASM
Controle 100,0 6,35 ± 0,11
Apocinina 105,6 ± 11,7 6,54 ± 0,11
Tempol 101,0 ± 11,7% 6,62 ± 0,27
Catalase 91,9 ± 13,0% 5,99 ± 0,06 #
GASM/OC4
Controle 100 6,88 ± 0,07
Apocinina 102,3 ± 4,3 6,81 ± 0,14
Tempol 99,8 ± 11,4 6,54 ± 0,13
Catalase 111,4 ± 8,8% 6,07 ± 0,08 γ
Dados expressos como média ± e.p.m. (n = 5). ANOVA one-way seguido do pós-teste de Tukey: *p < 0,05 (GC controle vs. GC + tempol/catalase); #p < 0,05 (GASM controle vs. GASM
+ catalase); γp < 0,05 (GASM/OC4 controle vs. GASM/OC4 + catalase). Fonte: Vasconcelos, 2017.

159 VASCONCELOS, L. H. C. Resultados
5.4.2 Avaliação da participação dos produtos derivados do metabolismo do
ácido araquidônico pelas enzimas cicloxigenase (COX) e 5-lipoxigenase (5-LO)
5.4.2.1 Obtenção de curvas cumulativas concentrações-resposta ao CCh, na
ausência e na presença da indometacina ou da zileutona, em cobaias dos
grupos GC, GASM e GASM/OC4
A curva concentrações-resposta cumulativa ao CCh (10-10-3 x 10-5 M) do grupo
GC (Emax = 100%; pD2 = 6,82 ± 0,06) teve sua resposta máxima cerca de 1,5 vezes
aumentada pela indometacina, inibidora da COX (Emax = 148,5 ± 6,6%), e 1,4 vezes
pela zileutona, inibidora da 5-LO (Emax = 138,7 ± 12,9%), mas sem alterar a potência
(pD2 = 6,62 ± 0,09 e 6,55 ± 0,07, respectivamente), em traqueia de cobaia na presença
de epitélio (Figura 26A-B, Gráfico 14A, Tabela 9, n = 5).
Nos cobaias com inflamação pulmonar, a eficácia contrátil do CCh (10-10-10-4 M)
(Emax = 100%) foi aumentada cerca de 1,6 vezes pela indometacina
(Emax = 160,2 ± 11,9%), mas não foi alterada pela zileutona (Emax = 99,6 ± 4,4%). Já a
sua potência (pD2 = 6,35 ± 0,11) foi aumentada 2,2 vezes pela indometacina
(pD2 = 6,77 ± 0,09), mas não foi alterada pela zileutona (pD2 = 5,98 ± 0,11), em
traqueia de cobaia na presença de epitélio (Figura 26C-D, Gráfico 14B, Tabela 9,
n = 5).
Nos animais suplementados com o óleo de coco virgem na dose de 4 g/kg, a
eficácia contrátil do CCh (10-10-3 x 10-5 M) (Emax = 100%) não foi alterada pela
indometacina (Emax = 129,5 ± 13,7%) ou pela zileutona (Emax = 102,7 ± 6,3%). Já a
potência do CCh (pD2 = 6,88 ± 0,07) não foi alterada pela indometacina
(pD2 = 6,78 ± 0,07), mas foi reduzida 5,3 vezes pela zileutona (pD2 = 6,16 ± 0,08), em
traqueia de cobaia asmático na presença de epitélio (Figura 26E-F, Gráfico 14C,
Tabela 9, n = 5).
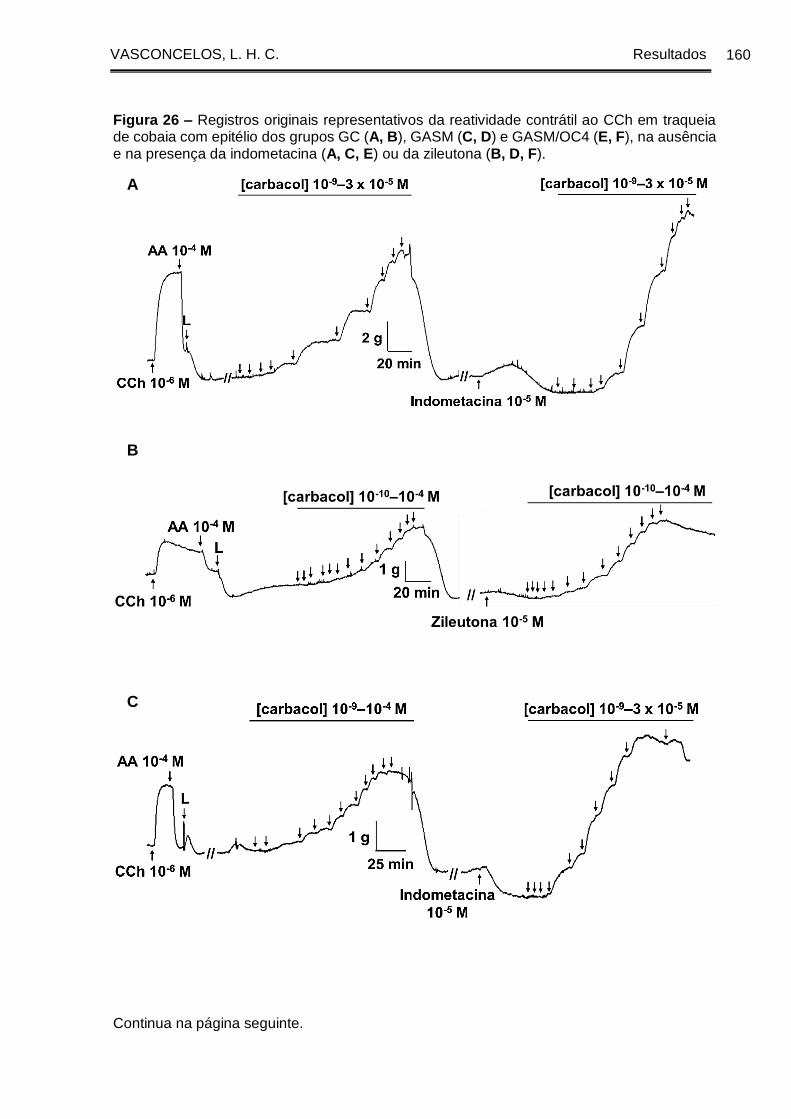
160 VASCONCELOS, L. H. C. Resultados
Figura 26 – Registros originais representativos da reatividade contrátil ao CCh em traqueia de cobaia com epitélio dos grupos GC (A, B), GASM (C, D) e GASM/OC4 (E, F), na ausência e na presença da indometacina (A, C, E) ou da zileutona (B, D, F).
Continua na página seguinte.
A
B
C
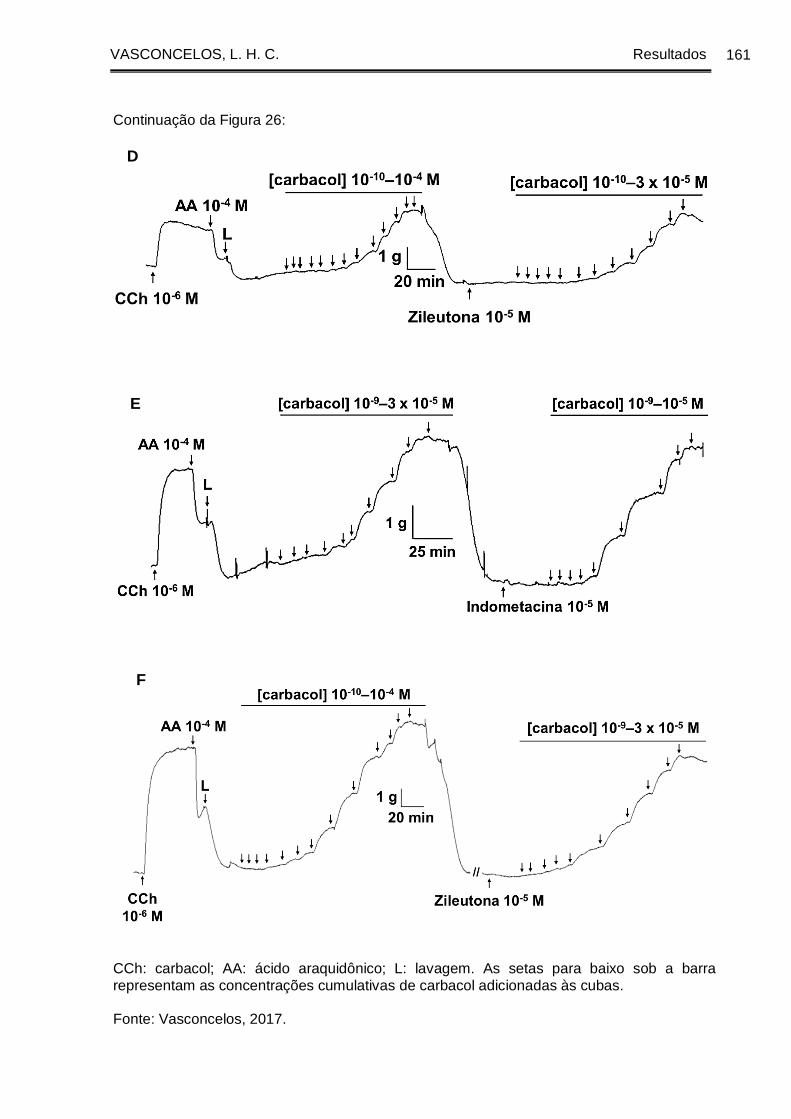
161 VASCONCELOS, L. H. C. Resultados
Continuação da Figura 26:
CCh: carbacol; AA: ácido araquidônico; L: lavagem. As setas para baixo sob a barra representam as concentrações cumulativas de carbacol adicionadas às cubas. Fonte: Vasconcelos, 2017.
D
E
F
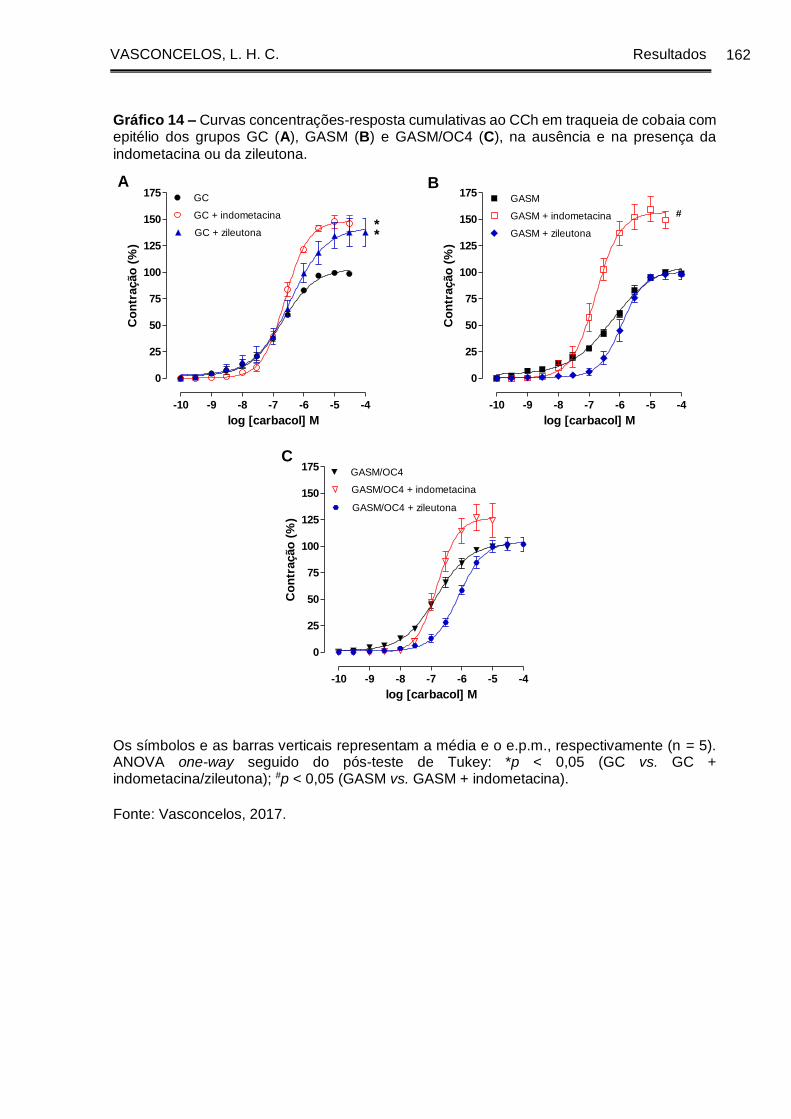
162 VASCONCELOS, L. H. C. Resultados
Gráfico 14 – Curvas concentrações-resposta cumulativas ao CCh em traqueia de cobaia com epitélio dos grupos GC (A), GASM (B) e GASM/OC4 (C), na ausência e na presença da
indometacina ou da zileutona.
-10 -9 -8 -7 -6 -5 -4
0
25
50
75
100
125
150
175GC
GC + indometacina
GC + zileutona**
log [carbacol] M
Co
ntr
ação
(%
)
-10 -9 -8 -7 -6 -5 -4
0
25
50
75
100
125
150
175GASM
GASM + indometacina
GASM + zileutona
#
log [carbacol] M
Co
ntr
ação
(%
)
-10 -9 -8 -7 -6 -5 -4
0
25
50
75
100
125
150
175GASM/OC4
GASM/OC4 + indometacina
GASM/OC4 + zileutona
log [carbacol] M
Co
ntr
ação
(%
)
Os símbolos e as barras verticais representam a média e o e.p.m., respectivamente (n = 5). ANOVA one-way seguido do pós-teste de Tukey: *p < 0,05 (GC vs. GC + indometacina/zileutona); #p < 0,05 (GASM vs. GASM + indometacina).
Fonte: Vasconcelos, 2017.
A
C
B

163 VASCONCELOS, L. H. C. Resultados
Tabela 9 – Valores de Emax e pD2 do CCh em traqueia de cobaia com epitélio dos grupos GC, GASM e GASM/OC4, na presença de epitélio funcional, na ausência e na presença da indometacina ou da zileutona.
Grupo Inibidor Emax (%) pD2
GC
Controle 100,0 6,82 ± 0,06
Indometacina 148,5 ± 6,6% * 6,62 ± 0,09
Zileutona 138,7 ± 12,9% * 6,55 ± 0,07
GASM
Controle 100,0 6,35 ± 0,11
Indometacina 160,2 ± 11,9% # 6,77 ± 0,09 #
Zileutona 99,6 ± 4,4% 5,98 ± 0,11
GASM/OC4
Controle 100 6,88 ± 0,07
Indometacina 129,5 ± 13,7 6,78 ± 0,07
Zileutona 129,5 ± 13,7% 6,16 ± 0,08 γ
Dados expressos como média ± e.p.m. (n = 5). ANOVA one-way seguido do pós-teste de Tukey: *p < 0,05 (GC controle vs. GC + indometacina/zileutona); #p < 0,05 (GASM +
indometacina vs. GASM controle); γp < 0,05 (GASM/OC4 + zileutona vs. GASM/OC4 controle). Fonte: Vasconcelos, 2017.

164 VASCONCELOS, L. H. C. Resultados
5.4.3 Investigação da participação da via do óxido nítrico (NO)
5.4.3.1 Obtenção de curvas cumulativas concentrações-resposta ao CCh, na
ausência e na presença do L-NAME, em cobaias dos grupos GC, GASM e
GASM/OC4
A curva concentrações-resposta cumulativa ao CCh (10-10-3 x 10-5 M) do grupo
GC (Emax = 100%; pD2 = 6,82 ± 0,06) não foi alterada pelo L-NAME, inibidor da NOS,
tanto com relação à eficácia (Emax = 109,5 ± 10,4%) como à potência contrátil
(pD2 = 6,54 ± 0,11), em traqueia de cobaia na presença de epitélio (Figura 27A, Gráfico
15A, Tabela 10, n = 5).
Nos cobaias com inflamação pulmonar, tanto a eficácia (Emax = 100%) como a
potência contrátil (pD2 = 6,35 ± 0,11) do CCh (10-10-10-4 M) foi aumentada pelo
L-NAME (Emax = 144,4 ± 16,0%; pD2 = 6,74 ± 0,03), cerca de 1,4 e 2,2 vezes,
respectivamente, em traqueia de cobaia na presença de epitélio (Figura 27B, Gráfico
15B, Tabela 10, n = 5).
A suplementação com o óleo de coco virgem na dose de 4 g/kg promoveu uma
resposta semelhante ao observado no grupo GC, não sendo a curva de contração ao
CCh (10-10-3 x 10-5 M) (Emax = 100%; pD2 = 6,88 ± 0,07) alterada pelo L-NAME com
relação aos parâmetros de eficácia (Emax = 87,4 ± 7,9%) e potência (pD2 = 6,98 ±
0,19), em traqueia de cobaia asmático na presença de epitélio (Figura 27C, Gráfico
15C, Tabela 10, n = 5).
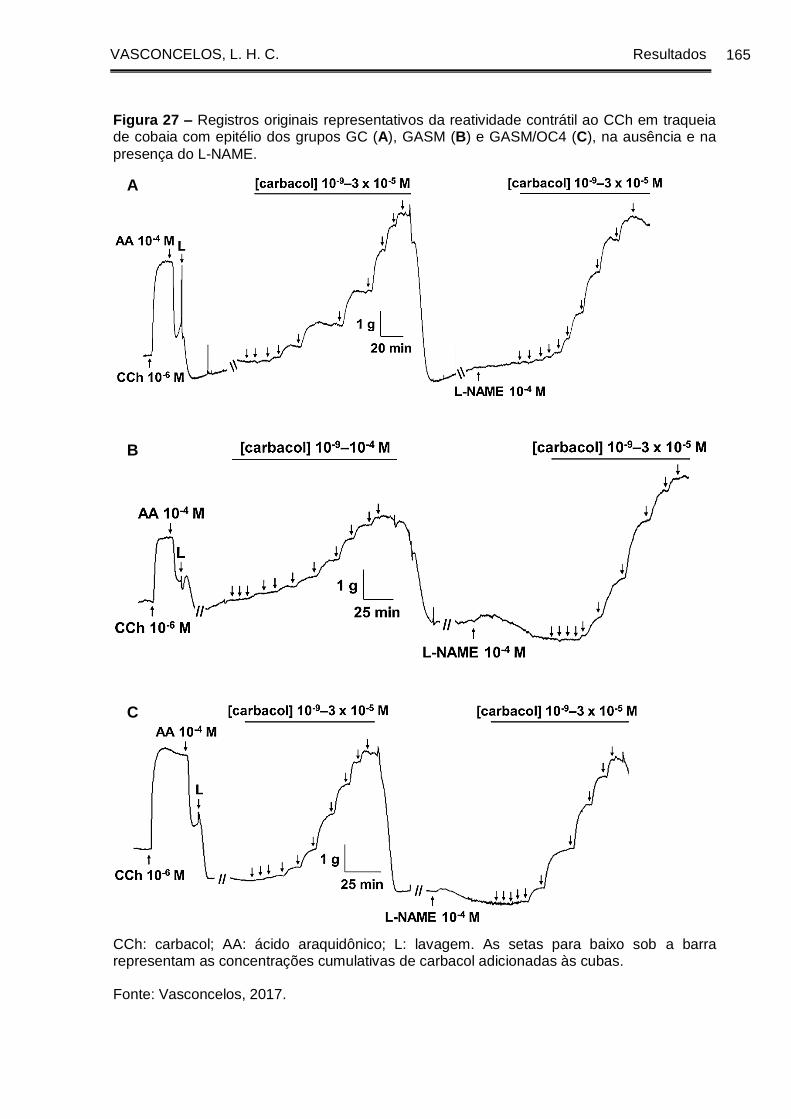
165 VASCONCELOS, L. H. C. Resultados
Figura 27 – Registros originais representativos da reatividade contrátil ao CCh em traqueia de cobaia com epitélio dos grupos GC (A), GASM (B) e GASM/OC4 (C), na ausência e na
presença do L-NAME.
CCh: carbacol; AA: ácido araquidônico; L: lavagem. As setas para baixo sob a barra representam as concentrações cumulativas de carbacol adicionadas às cubas. Fonte: Vasconcelos, 2017.
A
B
C
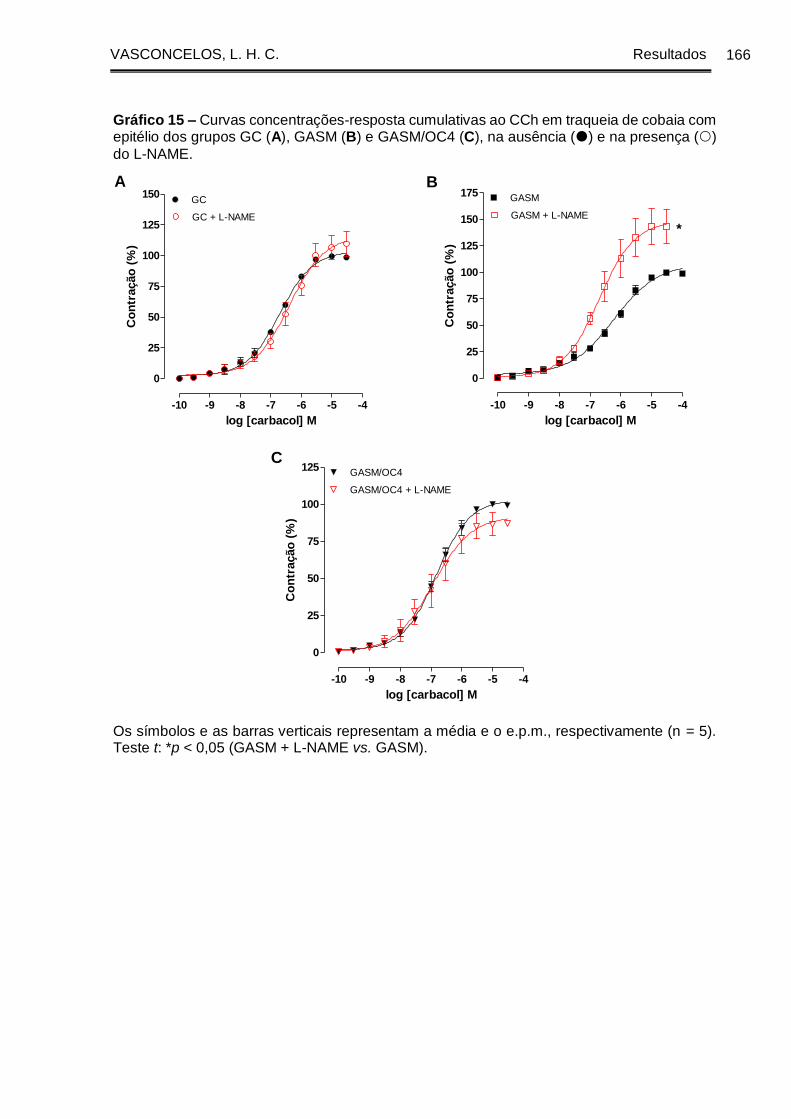
166 VASCONCELOS, L. H. C. Resultados
Gráfico 15 – Curvas concentrações-resposta cumulativas ao CCh em traqueia de cobaia com epitélio dos grupos GC (A), GASM (B) e GASM/OC4 (C), na ausência () e na presença ()
do L-NAME.
-10 -9 -8 -7 -6 -5 -4
0
25
50
75
100
125
150GC
GC + L-NAME
log [carbacol] M
Co
ntr
ação
(%
)
-10 -9 -8 -7 -6 -5 -4
0
25
50
75
100
125
150
175GASM
GASM + L-NAME
*
log [carbacol] M
Co
ntr
ação
(%
)
-10 -9 -8 -7 -6 -5 -4
0
25
50
75
100
125GASM/OC4
GASM/OC4 + L-NAME
log [carbacol] M
Co
ntr
ação
(%
)
Os símbolos e as barras verticais representam a média e o e.p.m., respectivamente (n = 5). Teste t: *p < 0,05 (GASM + L-NAME vs. GASM).
A B
C

167 VASCONCELOS, L. H. C. Resultados
Tabela 10 – Valores de Emax e pD2 do CCh em traqueia de cobaia com epitélio dos grupos GC, GASM e GASM/OC4, na ausência e na presença do L-NAME.
Grupo L-NAME Emax (%) pD2
GC - 100,0 6,82 ± 0,06
+ 109,5 ± 10,4% 6,54 ± 0,11
GASM - 100,0 6,35 ± 0,11
+ 144,4 ± 16,0% * 6,74 ± 0,03 *
GASM/OC4 - 100 6,88 ± 0,07
+ 87,4 ± 7,9 6,98 ± 0,19
Dados expressos como média ± e.p.m. (n = 5). Teste t: *p < 0,05 [GASM L-NAME (+) vs. GASM L-NAME (-)]. Fonte: Vasconcelos, 2017.

168 VASCONCELOS, L. H. C. Resultados
5.4.4 Investigação da participação da via da RhoA/Rho cinase (ROCK)
5.4.4.1 Obtenção de curvas cumulativas concentrações-resposta ao CCh, na
ausência e na presença do Y-27632, em cobaias dos grupos GC, GASM e
GASM/OC4
A curva concentrações-resposta cumulativa ao CCh (10-10-3 x 10-5 M) do grupo
GC (Emax = 100%; pD2 = 6,82 ± 0,06) teve sua eficácia inalterada pelo Y-27632 (10-7,
10-6 e 10-5 M) (Emax = 121,6 ± 7,5; 91,3 ± 9,9 e 104,3 ± 7,5%, respectivamente); Na
presença da inibidor da concentração de 10-7 M, a potência contráil do CCh não foi
alterada (pD2 = 6,66 ± 0,12); entretanto, a curva foi deslocada para a direita, com a
redução da sua potência contrátil, cerca de 5 vezes na presença do Y-27632 10-6 M,
inibidor da ROCK, (pD2 = 6,13 ± 0,05) e 14 vezes na presença de 10-5 M (pD2 = 5,73
± 0,13), em traqueia de cobaia na presença de epitélio (Figura 28A, Gráfico 16A,
Tabela 11, n = 5).
Nos animais com inflamação pulmonar, tanto a eficácia (Emax = 100%) como a
potência contrátil (pD2 = 6,35 ± 0,11) do CCh (10-10-10-3 M) não foi alterada pelo
Y-27632 10-6 M (Emax = 104,4 ± 6,9%; pD2 = 6,39 ± 0,09); entretanto, na concentração
de 10-5 M, a eficácia do CCh foi aumentada (Emax = 121,8 ± 5,4%), mas sua potência
contrátil foi reduzida cerca de 7 vezes (pD2 = 5,64 ± 0,11); e na presença do inibidor
na concentração de 10-4 M, a eficácia do CCh permaneceu inalterada, em relação
àquela na ausência do inibidor (Emax = 86,7 ± 5,4%), mas sua potência contrátil foi
reduzida cerca de 60 vezes (pD2 = 4,73 ± 0,13) em traqueia de cobaia na presença
de epitélio (Figura 28B, Gráfico 16B, Tabela 11, n = 5).
Nos animais suplementados com o óleo de coco virgem na dose de 4 g/kg, a
curva de contração ao CCh (10-10-10-4 M) (Emax = 100%; pD2 = 6,88 ± 0,07) não foi
alterada com relação ao parâmetro de eficácia pelo Y-27632 nas concentrações de
10-7, 10-6 ou 10-5 M (Emax = 104,3 ± 10,4; 89,7 ± 9,1 e 104,5 ± 10,7%, respectivamente).
Todavia, a potência contrátil do CCh foi cerca de 11, 6 e 22 vezes reduzida pelo
Y-27632 nas concentrações de 10-7 (pD2 = 5,88 ± 0,12), 10-6 (pD2 = 6,14 ± 0,12) e
10-5 M (pD2 = 5,52 ± 0,06), respectivamente, em traqueia de cobaia asmático na
presença de epitélio (Figura 28C, Gráfico 16C, Tabela 11, n = 5).
A inclinação do plot de Schild para foi 1,06 ± 0,10 para o GC, 1,20 ± 0,24 para
o GASM e 0,24 ± 0,04 para o GASM/OC4; o valor de r2 foi 0,92 ± 0,04 para o GC,
0,94 ± 0,02 para o GASM e 0,24 ± 0,11 para o GASM/OC4; e o valor do pA2 foi
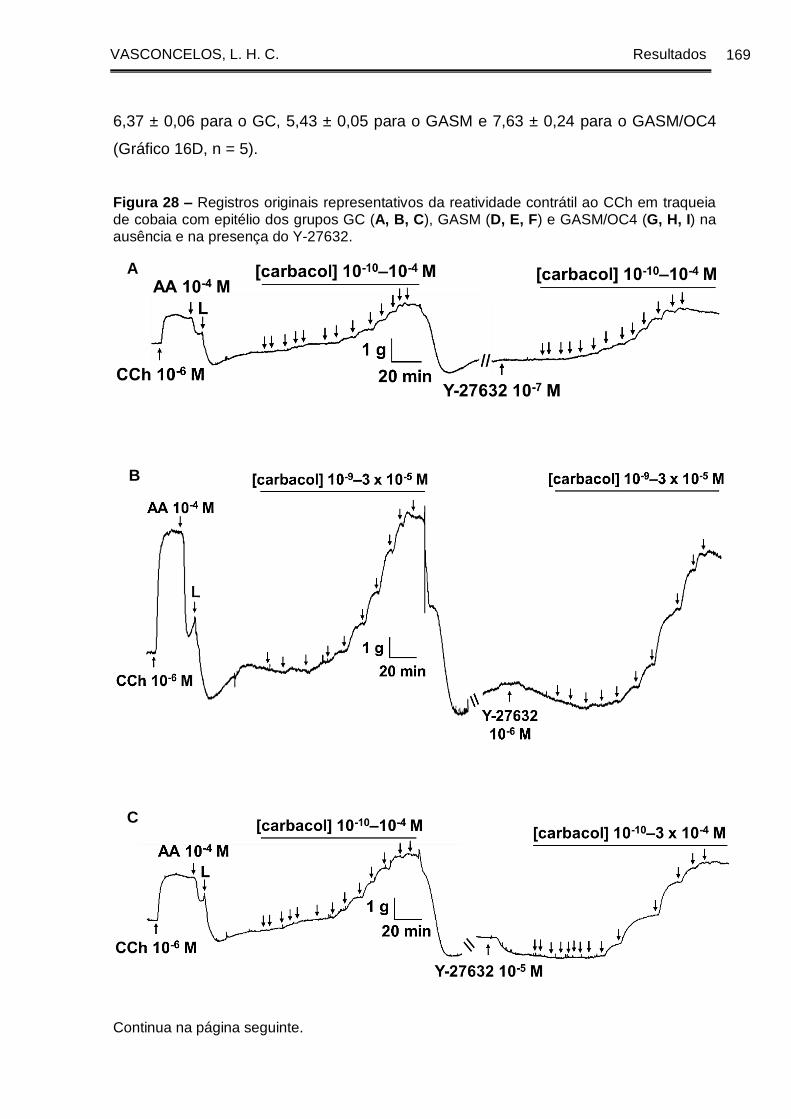
169 VASCONCELOS, L. H. C. Resultados
6,37 ± 0,06 para o GC, 5,43 ± 0,05 para o GASM e 7,63 ± 0,24 para o GASM/OC4
(Gráfico 16D, n = 5).
Figura 28 – Registros originais representativos da reatividade contrátil ao CCh em traqueia de cobaia com epitélio dos grupos GC (A, B, C), GASM (D, E, F) e GASM/OC4 (G, H, I) na ausência e na presença do Y-27632.
Continua na página seguinte.
A
B
C

170 VASCONCELOS, L. H. C. Resultados
Continuação da Figura 28:
Continua na página seguinte.
D
E
F
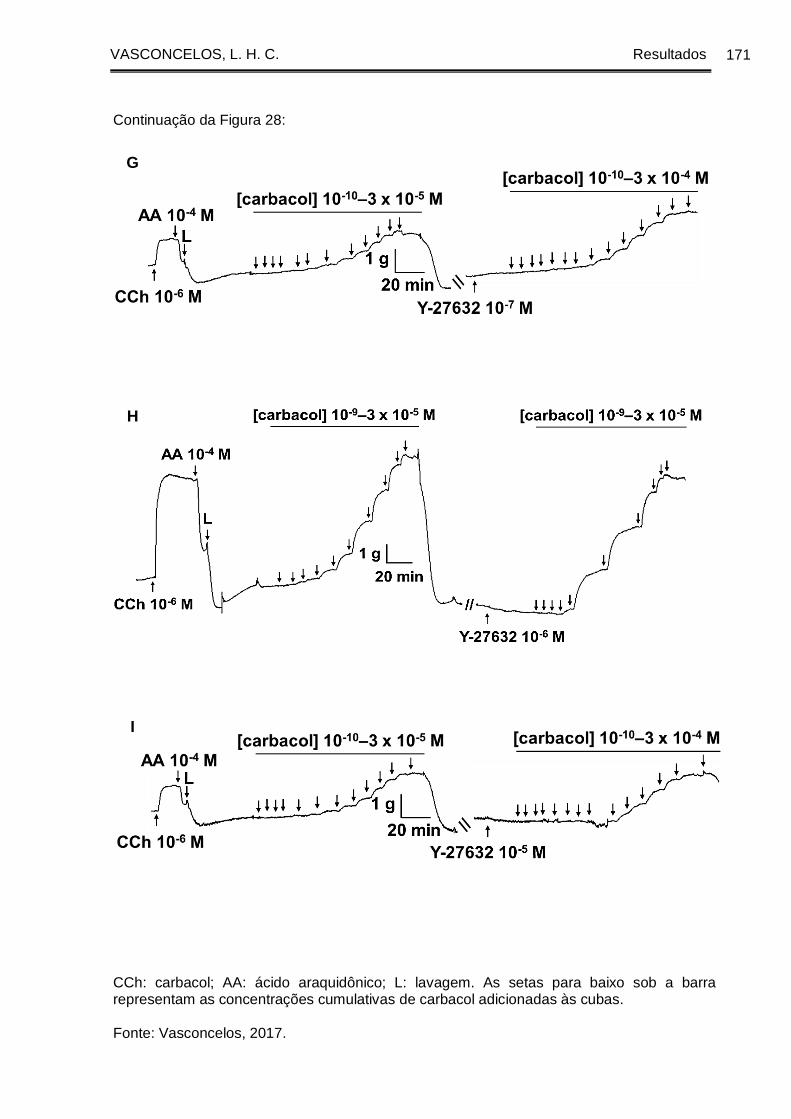
171 VASCONCELOS, L. H. C. Resultados
Continuação da Figura 28:
CCh: carbacol; AA: ácido araquidônico; L: lavagem. As setas para baixo sob a barra representam as concentrações cumulativas de carbacol adicionadas às cubas. Fonte: Vasconcelos, 2017.
H
G
I

172 VASCONCELOS, L. H. C. Resultados
Gráfico 16 – Curvas concentrações-resposta cumulativas ao CCh em traqueia de cobaia com epitélio dos grupos GC (A), GASM (B) e GASM/OC4 (C), na ausência e na presença do Y-27632, e plot de Schild (D) para a curva de contração do CCh na presença do Y-27632.
-10 -9 -8 -7 -6 -5 -4
0
25
50
75
100
125
150 GC
GC + Y-27632 10-6
M
GC + Y-27632 10-7
M
GC + Y-27632 10-5
M
log [carbacol] M
Co
ntr
ação
(%
)
-10 -9 -8 -7 -6 -5 -4 -3
0
25
50
75
100
125
150 GASM
GASM + Y-27632 10-6
M
GASM + Y-27632 10-5
M
GASM + Y-27632 10-4
M
*
log [carbacol] M
Co
ntr
ação
(%
)
-10 -9 -8 -7 -6 -5 -4
0
25
50
75
100
125 GASM/OC4
GASM/OC4 + Y-27632 10-6 M
GASM/OC4 + Y-27632 10-7 M
GASM/OC4 + Y-27632 10-5 M
log [carbacol] M
Co
ntr
ação
(%
)
Os símbolos e as barras verticais representam a média e o e.p.m., respectivamente (n = 5). ANOVA one-way seguido do pós-teste de Tukey: *p < 0,05 (GASM + Y-27632 10-5 M vs. GASM).
Fonte: Vasconcelos, 2017.
-7.0 -6.5 -6.0 -5.5 -5.0
-2
-1
0
1
GC
GASM
GASM/OC4
log [Y-27632] M
log
(D
R-1
)
A
C
B
D

173 VASCONCELOS, L. H. C. Resultados
Tabela 11 – Valores de Emax e pD2 do CCh em traqueia de cobaia dos grupos GC, GASM e GASM/OC4, na presença de epitélio funcional, na ausência e na presença do Y-27632.
Grupo Y-27632 (M) Emax (%) pD2
GC
Controle 100,0 6,82 ± 0,06
10-7 121,6 ± 7,5% 6,66 ± 0,12
10-6 91,3 ± 9,9% 6,13 ± 0,05 *
10-5 104,3 ± 7,5% 5,73 ± 0,13 * #
GASM
Controle 100,0 6,35 ± 0,11
10-6 104,4 ± 6,9% 6,39 ± 0,09
10-5 121,8 ± 5,4% * 5,64 ± 0,11 *
10-4 86,7 ± 5,4% 4,73 ± 0,13 * #
GASM/OC4
Controle 100 6,88 ± 0,07
10-7 104,3 ± 10,4 5,88 ± 0,12 *
10-6 89,7 ± 9,1 6,14 ± 0,12 *
10-5 104,5 ± 10,7% 5,52 ± 0,06 * #
Dados expressos como média ± e.p.m. (n = 5). ANOVA one-way seguido do pós-teste de Tukey: *p < 0,05 (GC Controle vs. GC + Y-27632 10-6/10-5 M ou GASM Controle vs. GASM + Y-27632 10-5/10-4 ou GASM/OC4 Controle vs. GASM/OC4 + Y-27632 10-7/10-6/10-5 M); #p < 0,05 (GC + Y-27632 10-5 M vs. GC + Y-27632 10-4 M ou GASM + Y-27632 10-4 M vs. GASM + Y-27632 10-5 ou GASM/OC4 + Y-27632 10-5 vs. GASM/OC4 + Y-27632 10-6/10-7 M). Fonte: Vasconcelos, 2017.

174 VASCONCELOS, L. H. C. Resultados
5.5 Investigação do mecanismo de ação envolvido nas alterações induzidas pela
inflamação pulmonar alérgica crônica e pelo óleo de coco virgem sobre a
reatividade relaxante da traqueia de cobaia
5.5.1 Avaliação da participação do fator de crescimento transformador β (TGF-β)
e dos canais de potássio sensíveis ao Ca2+ de grande condutância (BKCa)
5.5.1.1 Obtenção de curvas cumulativas concentrações-resposta à isoprenalina,
na ausência e na presença do SB431542 ou da iberiotoxina (IbTX), em traqueia
de cobaia pré-contraída com 10-6 M de CCh dos grupos GC, GASM e GASM/OC4
No grupo controle, a curva de relaxamento da isoprenalina (10-10-10-6 M)
(Emax = 135,7 ± 9,1%, pD2 = 7,79 ± 0,10) não foi alterada pelo SB431542, antagonista
dos receptores do tipo I de TGF-β, tanto com relação à eficácia (Emax = 132,4 ± 7,8%)
quanto à potência relaxante (pD2 = 7,96 ± 0,08). Diferentemente, na presença da IbTX,
bloqueador dos BKCa, a potência relaxante da isoprenalina foi reduzida 2,3 vezes
(pD2 = 7,39 ± 0,06), mas sua eficácia foi aumentada (Emax = 175,7 ± 11,0%) (Figura
29A-B, Gráfico 17A, Tabela 12, n = 5).
Nos animais com inflamação pulmonar, a curva de relaxamento da isoprenalina
(10-10-10-6 M) (Emax = 120,7 ± 5,2%; pD2 = 7,24 ± 0,09) foi potencializada cerca de 7
vezes pelo SB431542 (pD2 = 8,38 ± 0,08), sem alteração da sua eficácia relaxante
(Emax = 108,9 ± 5,6%). Já na presença da IbTX, tanto a eficácia (Emax = 137,5 ± 3,9%)
quanto a potência (pD2 = 7,44 ± 0,12) da isoprenalina não foi alterada (Figura 29C-D,
Gráfico 17B, Tabela 12, n = 5).
Nos animais suplementados com o óleo de coco virgem, a reatividade relaxante
à isoprenalina (10-10-10-6 M) (Emax = 117,8 ± 12,0%; pD2 = 7,94 ± 0,14) não foi alterada
pelo SB421542 ou pela IbTX, seja com relação à eficácia (Emax = 114,5 ± 5,5;
136,5 ± 7,6%, respectivamente) ou à potência relaxante (pD2 = 8,08 ± 0,10;
7,58 ± 0,03, respectivamente) (Figura 29E-F, Gráfico 17C, Tabela 12, n = 5).
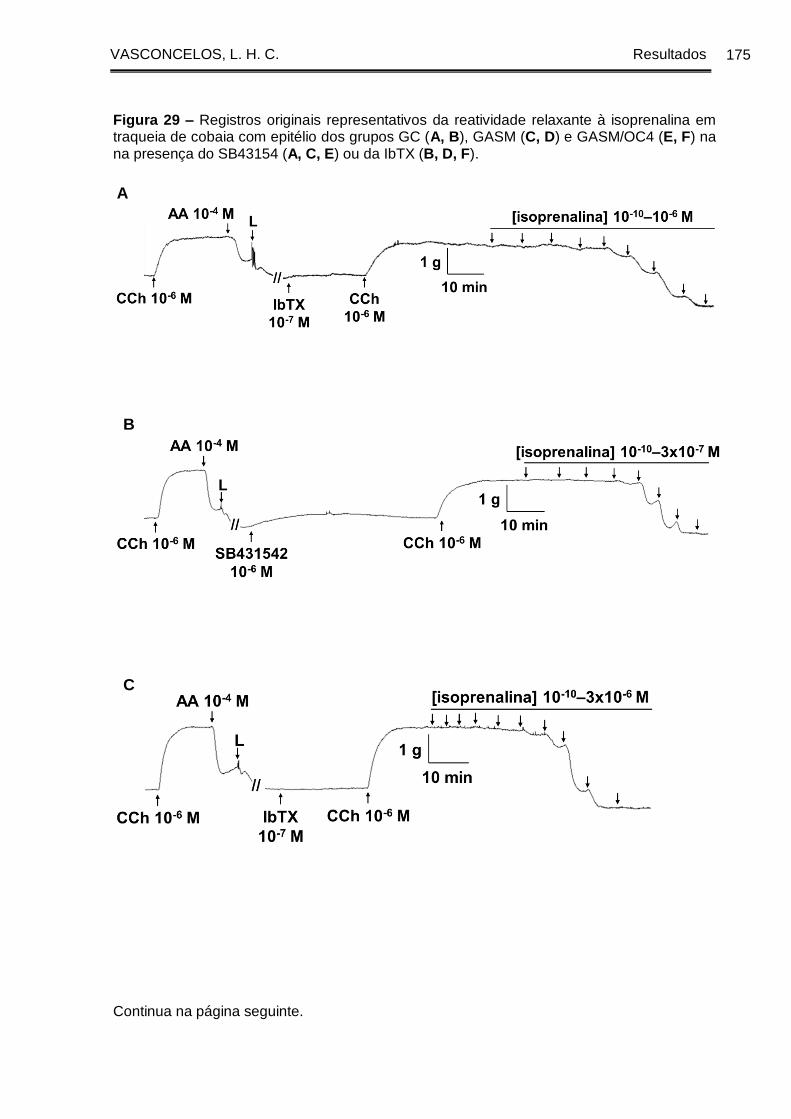
175 VASCONCELOS, L. H. C. Resultados
Figura 29 – Registros originais representativos da reatividade relaxante à isoprenalina em traqueia de cobaia com epitélio dos grupos GC (A, B), GASM (C, D) e GASM/OC4 (E, F) na na presença do SB43154 (A, C, E) ou da IbTX (B, D, F).
Continua na página seguinte.
A
B
C

176 VASCONCELOS, L. H. C. Resultados
Continuação da Figura 29:
CCh: carbacol; AA: ácido araquidônico; IbTX: iberiotoxina; L: lavagem. As setas para baixo sob a barra representam as concentrações cumulativas de isoprenalina adicionadas às cubas. Fonte: Vasconcelos, 2017.
D
E
F
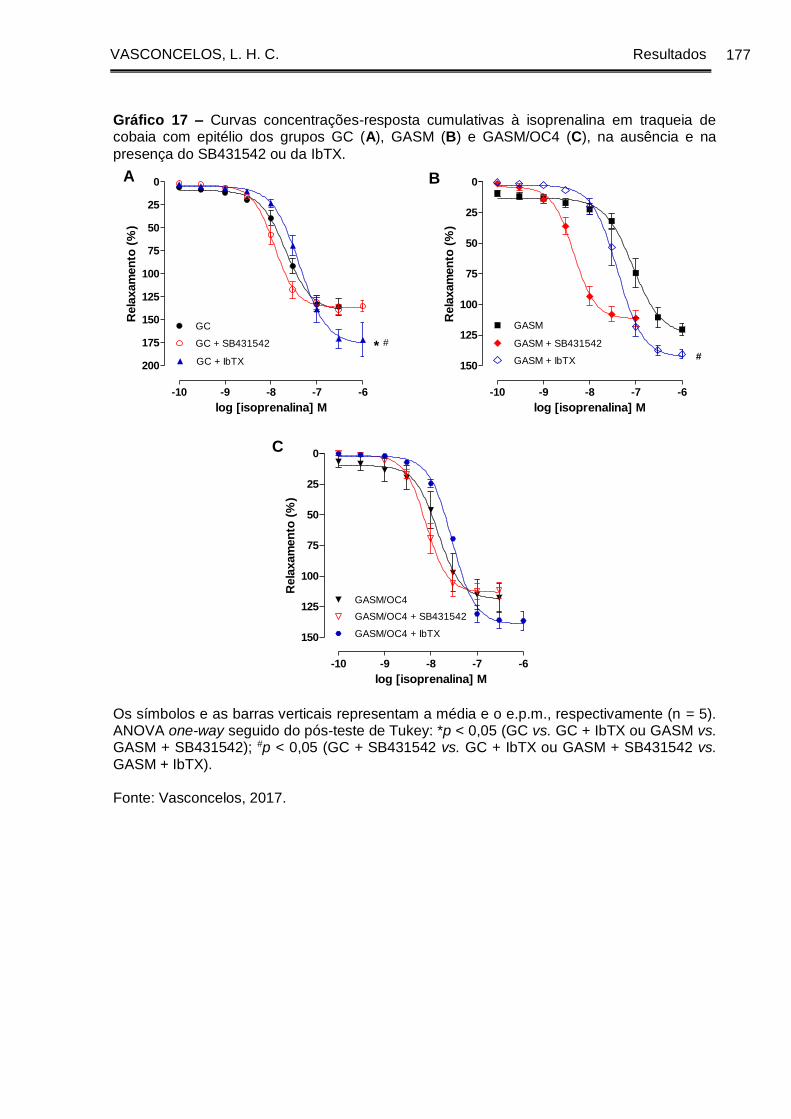
177 VASCONCELOS, L. H. C. Resultados
Gráfico 17 – Curvas concentrações-resposta cumulativas à isoprenalina em traqueia de cobaia com epitélio dos grupos GC (A), GASM (B) e GASM/OC4 (C), na ausência e na
presença do SB431542 ou da IbTX.
-10 -9 -8 -7 -6
0
25
50
75
100
125
150
175
200
GC
GC + SB431542
GC + IbTX
* #
log [isoprenalina] M
Rela
xam
en
to (
%)
-10 -9 -8 -7 -6
0
25
50
75
100
125
150
GASM + SB431542
GASM + IbTX #
GASM
log [isoprenalina] M
Rela
xam
en
to (
%)
-10 -9 -8 -7 -6
0
25
50
75
100
125
150
GASM/OC4
GASM/OC4 + SB431542
GASM/OC4 + IbTX
log [isoprenalina] M
Rela
xam
en
to (
%)
Os símbolos e as barras verticais representam a média e o e.p.m., respectivamente (n = 5). ANOVA one-way seguido do pós-teste de Tukey: *p < 0,05 (GC vs. GC + IbTX ou GASM vs. GASM + SB431542); #p < 0,05 (GC + SB431542 vs. GC + IbTX ou GASM + SB431542 vs. GASM + IbTX). Fonte: Vasconcelos, 2017.
A B
C

178 VASCONCELOS, L. H. C. Resultados
Tabela 12 – Valores de Emax e pD2 da isoprenalina em traqueia de cobaia dos grupos GC, GASM e GASM/OC4, na presença de epitélio funcional, na ausência e na presença do SB431542 ou da IbTX.
Grupo Inibidor/
bloqueador Emax (%) pD2
GC
Controle 135,7 ± 9,1% 7,79 ± 0,10
SB431542 132,4 ± 7,8% 7,96 ± 0,08
IbTX 175,7 ± 11,0% * # 7,39 ± 0,06 * #
GASM
Controle 120,7 ± 5,2% 7,24 ± 0,09
SB431542 108,9 ± 5,6% 8,38 ± 0,08 *
IbTX 137,5 ± 3,9% # 7,44 ± 0,12 #
GASM/OC4
Controle 117,8 ± 12,0% 7,94 ± 0,14
SB431542 114,5 ± 5,5% 8,08 ± 0,10
IbTX 136,5 ± 7,6% 7,58 ± 0,03 #
Dados expressos como média ± e.p.m. (n = 5). ANOVA one-way seguido do pós-teste de Tukey: *p < 0,05 (GC Controle vs. GC + IbTX ou GASM Controle vs. GASM + SB431542); #p < 0,05 (GC + SB431542 vs. GC + IbTX ou GASM + SB431542 vs. GASM + IbTX ou GASM/OC4 + SB431542 vs. GASM/OC4 + IbTX).
Fonte: Vasconcelos, 2017.

179 VASCONCELOS, L. H. C. Resultados
5.5.2 Avaliação da participação dos produtos derivados do metabolismo do
ácido araquidônico pelas enzimas cicloxigenase (COX) e 5-lipoxigenase (5-LO)
5.5.2.1 Obtenção de curvas cumulativas concentrações-resposta ao ácido
araquidônico (AA), na ausência e na presença da indometacina ou da zileutona,
em traqueia de cobaia pré-contraída com 10-6 M de CCh dos grupos GC, GASM
e GASM/OC4
A traqueia dos animais dos grupos GC, GASM e GASM/OC4 relaxou em
resposta à adição de concentrações cumulativas de AA (10-7-10-3 M) sobre o órgão
pré-contraído com 10-6 M de CCh. Comparando-se os valores de Emax, é possível
observar que não houve diferença na eficácia relaxante do AA entre os três grupos
avaliados (Emax = 85,4 ± 12,0; 92,9 ± 6,6 e 89,5 ± 6,3%, respectivamente). Entretanto,
comparado com o grupo GC (pD2 = 4,91 ± 0,09), a inflamação pulmonar promoveu
redução de cerca de 4 vezes na potência relaxante do AA (pD2 = 4,27 ± 0,06), e a
suplementação com o óleo de coco virgem na dose de 4 g/kg preveniu a redução da
potência (pD2 = 4,79 ± 0,09), não diferindo do GC, e não alterou a eficácia relaxante
do AA (Emax = 94,0 ± 5,7%) (Figura 30A, D, G, Gráfico 18A, Tabela 13, n = 3-5).
Na presença da indometacina ou da zileutona, a curva de relaxamento do AA
do grupo GC não foi alterada, tanto com relação à eficácia (Emax = 70,3 ± 13,4 e
86,3 ± 16,0%, respectivamente) quanto à potência (pD2 = 4,65 ± 0,11 e 4,67 ± 0,13,
respectivamente) (Figura 30B-C, Gráfico 18B, Tabela 13, n = 4). Diferentemente, no
grupo GASM, a eficácia do AA foi diminuída (Emax = 50,9 ± 4,5%), assim como a
potência foi reduzida 2,2 vezes (pD2 = 3,88 ± 0,06) pela indometacina. No entanto,
na presença da zileutona, nenhum desses dois parâmetros foi alterado
(Emax = 83,5 ± 10,5%; pD2 = 4,45 ± 0,28) (Figura 30E-F, Gráfico 18C, Tabela 13, n = 5).
Já nos animais suplementados com o óleo de coco virgem na dose de 4 g/kg, a
indometacina reduziu a eficácia (Emax = 60,2 ± 4,9%) e a potência relaxante do AA
(pD2 = 4,05 ± 0,15), cerca de 8 vezes; todavia, diferentemente, a zileutona
potencializou o relaxamento induzido pelo AA cerca de 2 vezes (pD2 = 5,26 ± 0,16),
sem alterar sua eficácia (Emax = 77,6 ± 8,2%) (Figura 30H-I, Gráfico 18D, Tabela 13,
n = 4).
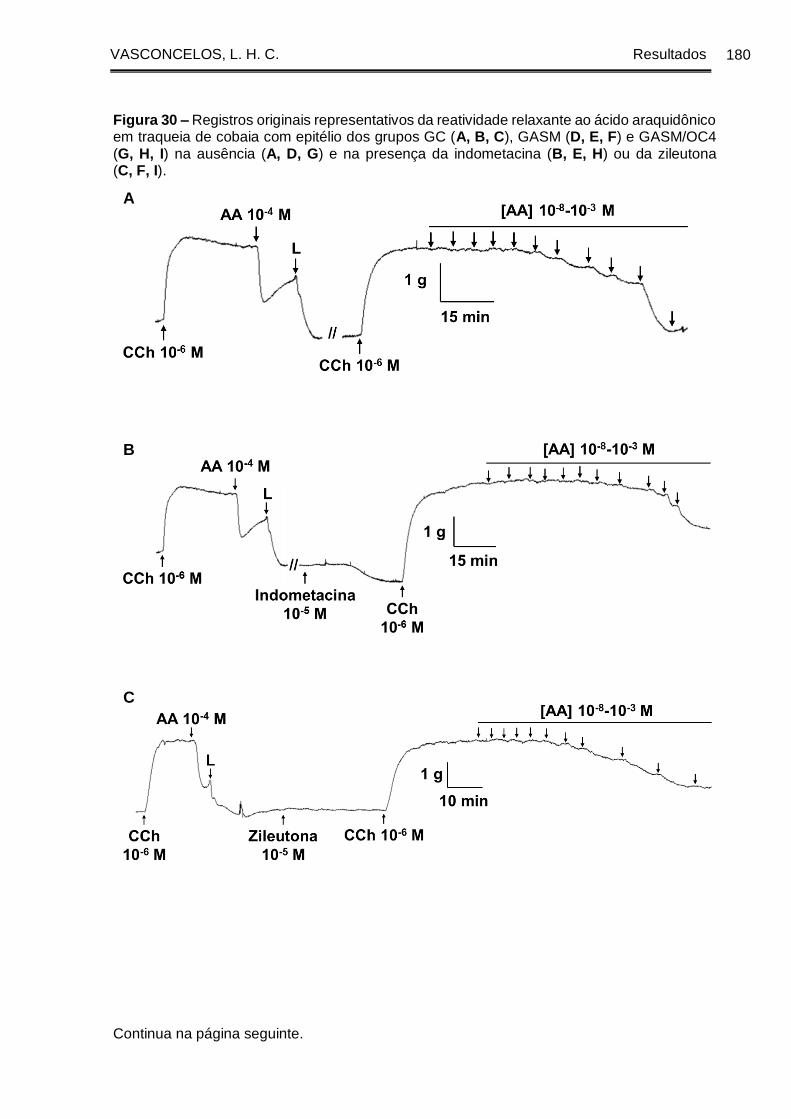
180 VASCONCELOS, L. H. C. Resultados
Figura 30 – Registros originais representativos da reatividade relaxante ao ácido araquidônico em traqueia de cobaia com epitélio dos grupos GC (A, B, C), GASM (D, E, F) e GASM/OC4 (G, H, I) na ausência (A, D, G) e na presença da indometacina (B, E, H) ou da zileutona (C, F, I).
Continua na página seguinte.
A
B
C
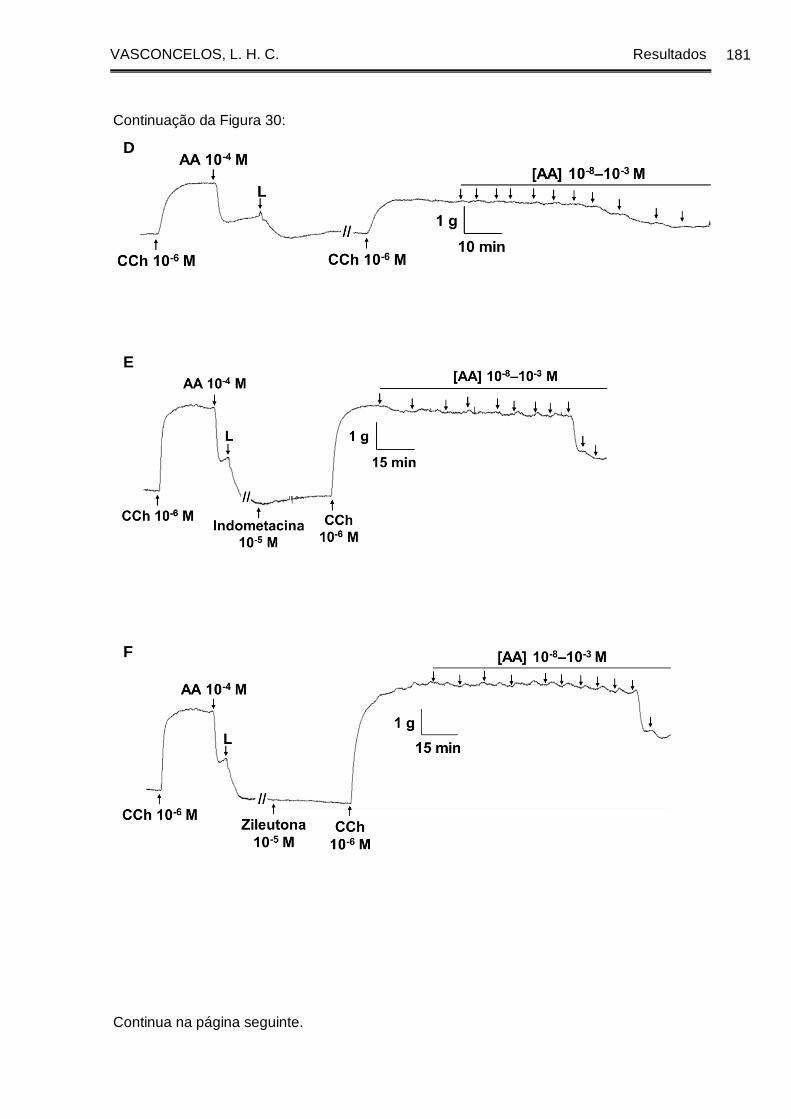
181 VASCONCELOS, L. H. C. Resultados
Continuação da Figura 30:
Continua na página seguinte.
D
E
F
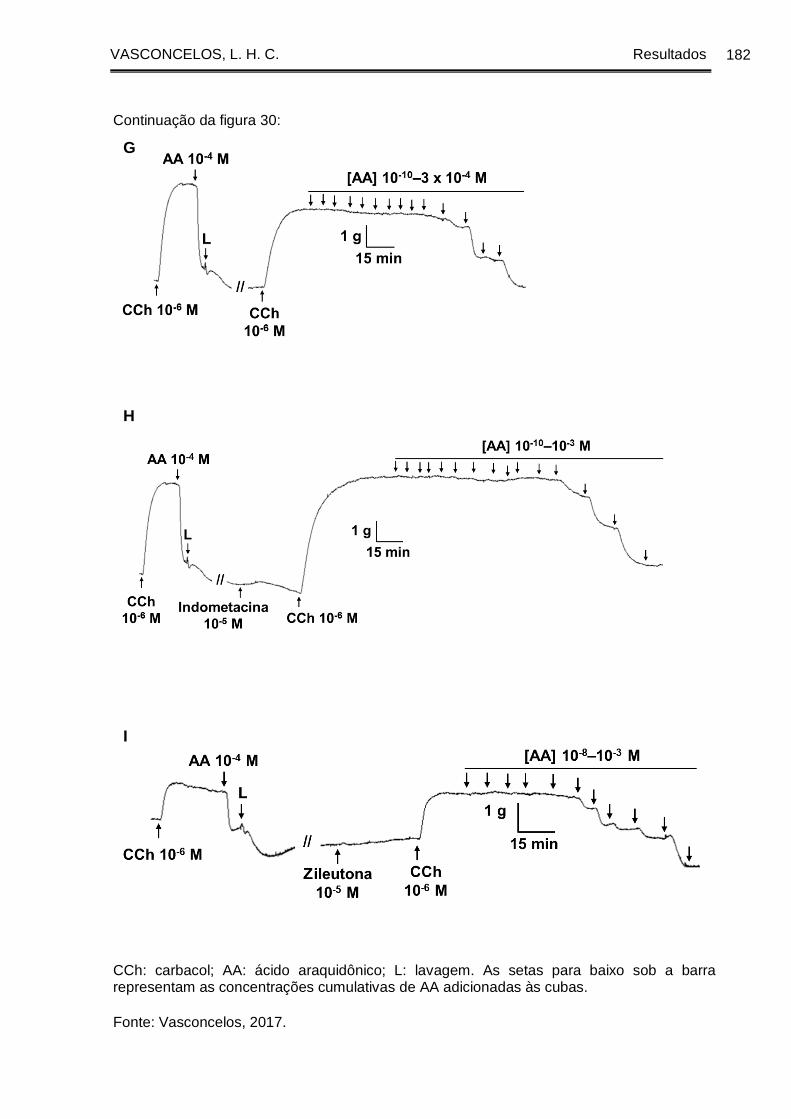
182 VASCONCELOS, L. H. C. Resultados
Continuação da figura 30:
CCh: carbacol; AA: ácido araquidônico; L: lavagem. As setas para baixo sob a barra representam as concentrações cumulativas de AA adicionadas às cubas. Fonte: Vasconcelos, 2017.
G
H
I
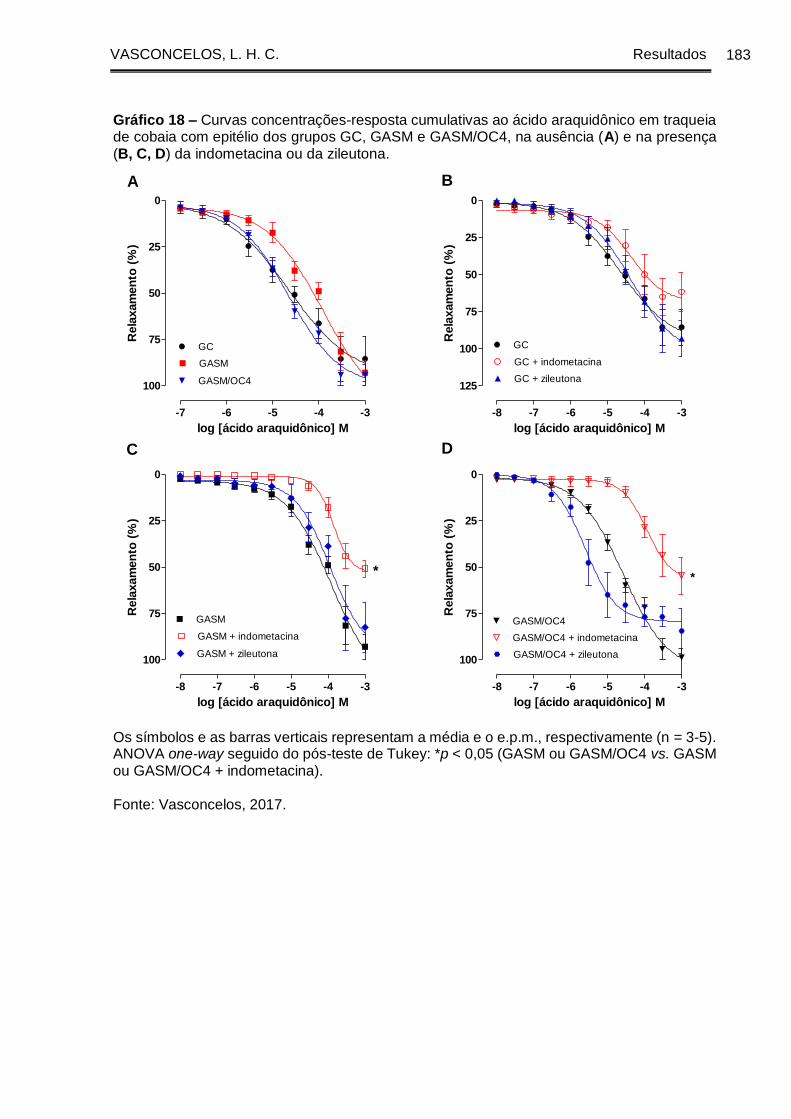
183 VASCONCELOS, L. H. C. Resultados
Gráfico 18 – Curvas concentrações-resposta cumulativas ao ácido araquidônico em traqueia de cobaia com epitélio dos grupos GC, GASM e GASM/OC4, na ausência (A) e na presença (B, C, D) da indometacina ou da zileutona.
-7 -6 -5 -4 -3
0
25
50
75
100
GASM
GC
GASM/OC4
log [ácido araquidônico] M
Rela
xam
en
to (
%)
-8 -7 -6 -5 -4 -3
0
25
50
75
100
125
GC
GC + zileutona
GC + indometacina
log [ácido araquidônico] M
Rela
xam
en
to (
%)
-8 -7 -6 -5 -4 -3
0
25
50
75
100
GASM
GASM + indometacina
GASM + zileutona
*
log [ácido araquidônico] M
Rela
xam
en
to (
%)
-8 -7 -6 -5 -4 -3
0
25
50
75
100
GASM/OC4
GASM/OC4 + indometacina
GASM/OC4 + zileutona
*
log [ácido araquidônico] M
Rela
xam
en
to (
%)
Os símbolos e as barras verticais representam a média e o e.p.m., respectivamente (n = 3-5). ANOVA one-way seguido do pós-teste de Tukey: *p < 0,05 (GASM ou GASM/OC4 vs. GASM ou GASM/OC4 + indometacina). Fonte: Vasconcelos, 2017.
A B
C D

184 VASCONCELOS, L. H. C. Resultados
Tabela 13 – Valores de Emax e pD2 do ácido araquidônico em traqueia de cobaia dos grupos GC, GASM e GASM/OC4, na presença de epitélio funcional, na ausência e na presença da indometacina ou da zileutona.
Grupo Inibidor Emax (%) pD2
GC
Controle 85,4 ± 12,0% 4,91 ± 0,09
Indometacina 70,3 ± 13,4% 4,65 ± 0,11
Zileutona 86,3 ± 16,0% 4,67 ± 0,13
GASM
Controle 92,9 ± 6,6% * 4,27 ± 0,06
Indometacina 50,9 ± 4,5% # 3,88 ± 0,06 #
Zileutona 83,5 ± 10,5% 4,45 ± 0,28
GASM/OC4
Controle 94,0 ± 5,7% δ 4,79 ± 0,09
Indometacina 60,2 ± 4,9% # 4,05 ± 0,15 #
Zileutona 77,6 ± 8,2% 5,26 ± 0,16 # γ
Dados expressos como média ± e.p.m. (n = 3-5). ANOVA one-way seguido do pós-teste de Tukey: *p < 0,05 (GC controle vs. GASM controle); #p < 0,05 (GASM ou GASM/OC4 vs. GASM
ou GASM/OC4 + indometacina ou GASM/OC4 + zileutona); γp < 0,05 (GASM/OC4 +
indometacina vs. GASM/OC4 + zileutona) δp < 0,05 (GASM controle vs. GASM/OC4 controle).
Fonte: Vasconcelos, 2017.

185 VASCONCELOS, L. H. C. Resultados
5.6 Avaliação dos efeitos da inflamação pulmonar alérgica crônica e do
tratamento com a dexametasona ou com o óleo de coco virgem sobre o balanço
entre o estresse oxidativo e as defesas antioxidantes de cobaia
5.6.1 Medida dos níveis de malondialdeído (MDA) e da capacidade antioxidante
total (CAT) no plasma e no tecido pulmonar
Os animais do grupo GC apresentaram níveis plasmáticos de MDA de
2,1 ± 0,3 μmol/L, e a inflamação pulmonar não alterou esses níveis (2,0 ± 0,3 μmol/L).
Semelhantemente, os animais do grupo GASM/DEXA apresentaram níveis de MDA
no plasma de 2,2 ± 0,2 μmol/L, não diferindo de GC e GASM, assim como os animais
suplementados com as três doses do óleo de coco virgem, cujos valores de MDA
foram de 2,0 ± 0,2; 2,0 ± 0,2 e 1,5 ± 0,3 μmol/L, respectivamente, não diferindo de
nenhum dos demais grupos (Gráfico 19A).
No tecido pulmonar, os animais dos grupos GC e GASM apresentaram níveis
de MDA de 3,0 ± 0,3 e 2,8 ± 0,1 μmol/L, respectivamente, que não diferiram entre si.
Tanto o tratamento com a dexametasona como a suplementação com o óleo de coco
virgem nas três doses não alterou os níveis de MDA no tecido pulmonar (2,6 ± 0,2;
2,8 ± 0,1; 3,0 ± 0,1 e 2,9 ± 0,2 μmol/L, respectivamente), em relação aos grupos GC
e GASM (Gráfico 19B)
Com relação à capacidade antioxidante total, no plasma, os animais do grupo
GC apresentaram valor de inibição da oxidação de 23,6 ± 3,6%, que não diferiu do
grupo GASM, que apresentou valor de inibição de 19,8 ± 3,2%. O tratamento com
dexametasona ou a suplementaç ão com o óleo de coco virgem nas três doses não
alterou a capacidade antioxidante plasmática dos cobaias (16,8 ± 3,0; 21,2 ± 1,5%;
16,2 ± 3,4%; 17,4 ± 4,5, respectivamente (Gráfico 19C).
Já nos pulmões, os animais do GC apresentaram valor de inibição da oxidação
de 83,0 ± 4,2%, enquanto a inflamação pulmonar reduziu a capacidade antioxidante
pulmonar para 59,8 ± 6,6%. Enquanto o tratamento com dexametasona não alterou
a capacidade antioxidante pulmonar dos cobaias com inflamação pulmonar
(74,6 ± 5,0%), assim como a suplementação com o óleo de coco virgem na dose de
1 g/kg (77,2 ± 2,8%); as doses de 2 e 4 g/kg aumentaram a capacidade antioxidante
pulmonar para 82,6 ± 5,0 e 87,8 ± 1,4%, respectivamente, em relação aos grupos GC
e GASM (Gráfico 19D).

186 VASCONCELOS, L. H. C. Resultados
Gráfico 19 – Valores dos níveis de MDA (μmol/L tecido) (A e B) e de inibição da oxidação (%) (C e D) no plasma (A e C) e no homogenato pulmonar (B e D) de cobaias dos grupos GC,
GASM, GASM/DEXA, GASM/OC1, GASM/OC2 e GASM/OC4.
GC
GASM
GASM
/DEXA
GASM
/OC1
GASM
/OC2
GASM
/OC4
0
1
2
3
4
MD
A (
mo
l/L
tecid
o)
GC
GASM
GASM
/DEXA
GASM
/OC1
GASM
/OC2
GASM
/OC4
0
1
2
3
4
MD
A (
mo
l/L
tecid
o)
GC
GASM
GASM
/DEXA
GASM
/OC1
GASM
/OC2
GASM
/OC4
0
25
50
75
100
Inib
ição
da o
xid
ação
(%
)
GC
GASM
GASM
/DEXA
GASM
/OC1
GASM
/OC2
GASM
/OC4
0
25
50
75
100
*
# #
Inib
ição
da o
xid
ação
(%
)
As colunas e as barras verticais representam a média e o e.p.m., respectivamente (n = 5). ANOVA one-way seguido do pós-teste de Tukey: *p < 0,05 (GC vs. GASM); #p < 0,05 (GASM vs. demais grupos).
Fonte: Vasconcelos, 2017.
A B
C D

187 VASCONCELOS, L. H. C. Resultados
5.7 Avaliação dos efeitos da inflamação pulmonar alérgica crônica e do
tratamento com a dexametasona ou da suplementação com o óleo de coco
virgem sobre a expressão de proteínas das vias de sinalização da proliferação
muscular lisa e das defesas antioxidantes
5.7.1 Análise dos níveis da fosfatidilinositol-3-cinase (PI3K), das proteínas
cinases reguladas por sinal extracelular 1/2 (ERK1/2) e da superóxido dismutase
(SOD) no tecido pulmonar
A expressão da PI3K (0,64 ± 0,11) foi aumentada nos animais com inflamação
pulmonar (1,34 ± 0,18), mas o tratamento com a dexametasona (1,42 ± 0,07) ou a
suplementação com OCV (1,22 ± 0,10) não preveniram esse aumento. Já as ERK 1/2
e a SOD não apresentaram diferença entre os animais dos grupos GC (2,85 ± 0,89 e
1,61 ± 0,62, respectivamente) e GASM (2,34 ± 051 e 0,65 ± 0,11, respectivamente); o
tratamento com a dexametasona ou a suplementação com OCV não alterou a
expressão das ERK 1/2 (3,25 ± 0,67 e 2,55 ± 0,37, respectivamente) ou da SOD
(0,89 ± 0,15 e 1,11 ± 0,57, respectivamente) (Figura 31, Gráfico 20A-C).
Figura 31 – Bandas proteicas de corrida em gel de eletroforese representativas da expressão
de PI3K, ERK 1/2, SOD e β-actina no pulmão de cobaias dos grupos GC, GASM,
GASM/DEXA e GASM/OC4.
Revelação das bandas realizada por quimioluminescência (n = 3). Fonte: Vasconcelos, 2017.
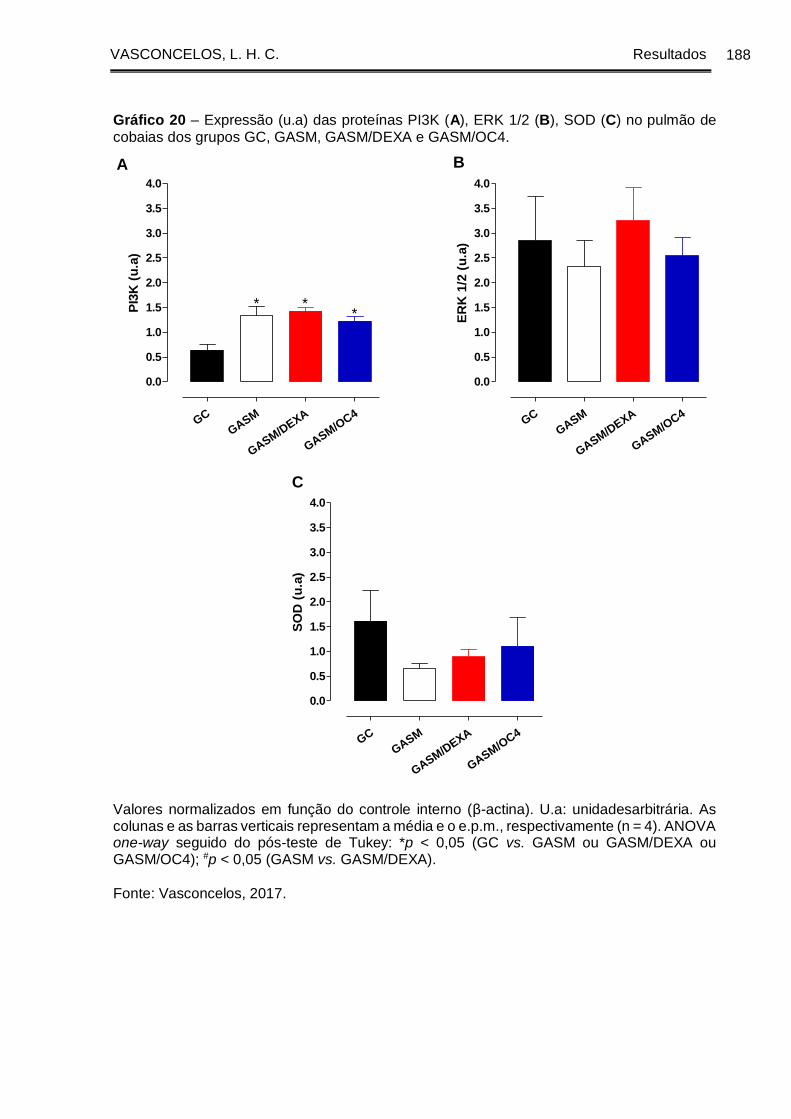
188 VASCONCELOS, L. H. C. Resultados
Gráfico 20 – Expressão (u.a) das proteínas PI3K (A), ERK 1/2 (B), SOD (C) no pulmão de cobaias dos grupos GC, GASM, GASM/DEXA e GASM/OC4.
GC
GASM
GASM/D
EXA
GASM/O
C4
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
* **P
I3K
(u
.a)
GC
GASM
GASM/D
EXA
GASM/O
C4
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
ER
K 1
/2 (
u.a
)
GC
GASM
GASM/D
EXA
GASM/O
C4
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
SO
D (
u.a
)
Valores normalizados em função do controle interno (β-actina). U.a: unidadesarbitrária. As colunas e as barras verticais representam a média e o e.p.m., respectivamente (n = 4). ANOVA one-way seguido do pós-teste de Tukey: *p < 0,05 (GC vs. GASM ou GASM/DEXA ou GASM/OC4); #p < 0,05 (GASM vs. GASM/DEXA).
Fonte: Vasconcelos, 2017.
A B
C

6 Discussão

190 VASCONCELOS, L. H. C. Discussão
Nesse trabalho, foi padronizado e implantado um modelo de inflamação
pulmonar alérgica crônica baseado na exposição repetida e prolongada das vias
respiratórias de cobaias à ovalbumina, e foram avaliados, primeira vez nesse modelo
animal, os efeitos da suplementação de cobaias com óleo de coco virgem. Foi
observado que a inflamação pulmonar promoveu as alterações características da
asma em humanos, como a infiltração de células inflamatórias na região
peribrônquica, hiperplasia epitelial e desenvolvimento muscular liso aéreo;
hiper-contratilidade muscular lisa por mecanismos relacionados com a promoção de
estresse oxidativo e suas interações com metabólitos do ácido araquidônico, a via do
NO e da RhoA/ROCK e do TGF-β, bem como um desbalanço entre ROS e defesas
antioxidantes nos pulmões e aumento da expressão da proteína PI3K; e que essas
alterações foram revertidas pela suplementação dos cobaias com inflamação
pulmonar com o óleo de coco virgem, mas sem alteração na expressão das proteínas
PI3K, ERK1/2 e SOD.
Devido às diferenças entre as características funcionais e teciduais da asma
aguda e crônica, nesse trabalho, buscou-se um modelo de inflamação pulmonar que
reproduzisse as características da asma alérgica crônica em humanos.
Experimentalmente, tem sido demonstrado que o remodelamento tecidual decorrente
dessa doença ocorre após exposições repetidas prolongadas do animal a soluções
antigênicas por via inalatória (HUTSON et al., 1988; SANTING et al., 1992;
WESTERHOF et al., 2001). Tais eventos fisiopatológicos envolvendo a resposta
pulmonar a exposições múltiplas a antígenos estão provavelmente mais relacionados
à asma clínica do que aqueles da resposta aguda a um único desafio em animais
sensibilizados (TIBÉRIO at al., 1997).
Nesse sentido, foi utilizado o modelo de inflamação pulmonar alérgica crônica
descrito por Tibério et al. (1997), baseado na exposição prolongada de cobaias à OVA
em concentrações crescentes por via inalatória, durante quatro semanas, em
intervalos de tempo curtos, a fim de se avaliar os efeitos desse modelo sobre
parâmetros ainda não investigados relativos à contratilidade muscular lisa das vias
aéreas e os processos subjacentes que interferem sobre essa função. Posteriormente,
buscou-se avaliar um possível efeito modulador do OCV nos mecanismos
fisiopatológicos do modelo de inflamação pulmonar, tanto à luz do processo
inflamatório crônico, como da hiper-reatividade brônquica.

191 VASCONCELOS, L. H. C. Discussão
A hipersensibilidade do tipo I medeia muitas das reações asmáticas, incluindo
a hiper-responsividade das vias aéreas desencadeada após estimulação antigênica.
Esse mecanismo está relacionado à captura de IgE por células expressando
receptores de alta afinidade (FCεRI) na sua superfície, especialmente mastócitos,
células dendríticas e eosinófilos, que liberam o conteúdo de seus grânulos
citoplasmáticos após exposição ao antígeno, incluindo os agentes contráteis
histamina, cisteinil leucotrienos (LTC4, LTD4 e LTE4) e prostanoides (PGF2α, PGD2 e
TxA2) (OZIER et al., 2011), além de outras substâncias e citocinas pró-inflamatórias,
mediando a obstrução dos brônquios (ISHIZAKA; ISHIZAKA; TOMIOKA, 1977;
TURNBULL et al., 1977; FIREMAN, 2003).
Após a exposição ao alérgeno, a resposta asmática em humanos é dividida em
duas fases: uma rápida, que ocorre nos primeiros 20 minutos de exposição; e uma
tardia, que se inicia após 2-6 horas de estimulação antigênica (FIREMAN, 2003). Em
cobaias, a resposta asmática é dividida em uma fase imediada, que ocorre até 5 horas
após a exposição ao alérgeno, e uma tardia, que se inicia 8 horas após a exposição e
pode se extender até 23 horas (TEN BERGE et al., 1995). Sendo assim, a estimulação
de animais sensibilizados com antígeno por via inalatória é um método eficaz de se
avaliar a hiper-responsividade das vias aéreas, a fim de se confirmar a efetividade da
indução do quadro de inflamação pulmonar.
Os animais com inflamação pulmonar apresentaram menor tempo em contato
com o antígeno durante as quinta, sexta e sétima nebulizações, uma vez que esse
tempo foi suficiente para desencadear os sinais de estresse respiratório, resultante da
resposta aguda após reexposição antigênica. Além disso, observou-se uma queda
gradual no tempo de nebulização dos animais desse grupo, indicando uma
intensificação do quadro inflamatório pelas exposições às concentrações crescentes
de OVA (Gráfico 1, Tabela 1).
Esses dados são corroborados por estudos anteriores, que observaram a
presença de hiper-reatividade das vias aéreas em animais submetidos ao mesmo
protocolo de inflamação pulmonar (TIBÉRIO et al., 1997; LEICK-MALDONADO et al.,
2004; PIGATI et al., 2015). Analisando-se no contexto do modelo animal, esse dado
suporta a utilização de cobaias como modelo para investigar a hiper-responsividade
das vias aérea na inflamação pulmonar alérgica crônica, dada a intensidade das
respostas apresentadas pelos animais durante os desafios com OVA as quais

192 VASCONCELOS, L. H. C. Discussão
incluiam tosse, espirro, retração torácica com encurtamento do tempo de respiração
e, em alguns casos, óbito devido a obstrução completa das vias aéreas.
Através da análise morfológica (Figuras 15-17, Gráficos 2A-C), observou-se
infiltrado peribrônquico de células inflamatórias, além de algumas características do
remodelamento tecidual, a saber a hiperplasia das células epiteliais das vias aéreas e
o espessamento da camada de músculo liso brônquico por processo de hiperplasia
ou hipertrofia. Esses achados estão de acordo com o que ocorre na asma alérgica em
humanos, em que o processo de remodelamento promove as alterações observadas
nos pulmões dos cobaias (AIKAWA et al., 1992; CARROLL et al., 1993; COHEN et
al., 2007; BERGERON; AL-RAMLI; HAMID, 2009).
Por fim, a efetividade da indução da inflamação pulmonar foi confirmada, in
vitro, por meio da Reação de Schultz-Dale, em que o músculo liso de um animal
sensibilizado antigenicamente contrai-se após a reexposição a este antígeno
mediante a liberação de mediadores contráteis pelas células do microambiente
(SCHULTZ, 1910; DALE, 1913). Inclusive, muitos processos fundamentais,
mediadores e reguladores da patogênese da asma foram demonstrados
primeiramente em cobaias através desse método de hipersensibilidade imediata
(CANNING; CHOU, 2010). O fato dos cobaias com inflamação pulmonar
apresentarem uma significativa resposta contrátil à OVA, diferentemente dos animais
do grupo GC, confirmou o processo de sensibilização (Figura 18A-B, Gráfico 3). Esse
dado é respaldado por estudos anteriores, que mostraram que a traqueia de cobaias
asmáticos contrai em resposta à estimulação antigênica in vitro (SOTMRADA;
DICKEY, 1976; SHAUKAT et al., 2015; MARTINS, 2016).
Apesar do grande número de modelos animais que reproduzem as
características da asma alérgica, esses estudos não têm levado ao surgimento de
novas terapias para o crescente número de pacientes asmáticos (LLOYD; HESSEL,
2010). Diante desse cenário, fazem-se necessárias novas abordagens terapêuticas
que limitem ou, ao menos, tornem menos frequentes as crises agudas de pacientes
asmáticos, o que pode ser feito com o desenvolvimento de novas drogas que modulem
o processo fisiopatológico da doença associado a uma menor incidência de efeitos
colaterais, ou por meio de formas alternativas que potencializem os efeitos dos
medicamentos atualmente disponíveis para o tratamento da asma.
Nesse contexto, insere-se o óleo de coco virgem (OCV), produto que vem
sendo largamente utilizado na alimentação e na indústria e cujo consumo vem

193 VASCONCELOS, L. H. C. Discussão
crescendo principalmente devido às suas várias atividades benéficas sobre o perfil
lipídico (NEVIN; RAJAMOHAN, 2004; FERANIL et al., 2011) e à redução da gordura
corporal (KASAI et al., 2003; ASSUNÇÃO et al., 2009). O OCV é rico em ácidos graxos
saturados de cadeia média que, além de serem ricas fontes de energia pelo seu rápido
metabolismo hepático, apresentam atividade preventiva sobre o desenvolvimento de
doenças cardiovasculares (NURUL-IMAN et al., 2013; ALVES et al., 2015). Diversos
estudos relatam os efeitos pré-clínicos desse óleo, especialmente sobre doenças
crônicas, destacando-se as atividades antitrombótica (NEVIN; RAJAMOHAN, 2008),
hipotensora (ALVES et al., 2015; KAMISAH et al., 2015), anti-inflamatória
(INTAHPHUAK; KHONSUNG; PANTHONG, 2010; ZAKARIA et al., 2011; VYSAKH et
al., 2014) e no tratamento da obesidade (ASSUNÇÃO et al., 2009), sendo esses
efeitos atribuídos ao ácido láurico, bem como aos compostos fenólicos presentes no
óleo.
Não são descritos na literatura efeitos do óleo de coco em parâmetros
fisiológicos de cobaias, sendo esse trabalho pioneiro nessa investigação. Nesse
sentido, decidiu-se avaliar os potenciais efeitos benéficos da suplementação com
OCV sobre a inflamação pulmonar alérgica crônica em cobaias, diante de um possível
efeito modulador desse óleo sobre os mecanismos fisiopatológicos da doença.
Até o presente, não há dados na literatura a respeito da toxicidade aguda ou
crônica do óleo de coco virgem; todavia, a partir dos dados observacionais desse
trabalho, é possível indicar uma baixa toxicidade desse óleo em cobaias, uma vez
que, mesmo na dose mais elevada (4 g/kg), não foram observados sinais de toxicidade
ou letalidade nos animais.
Posto que a avaliação do tempo de nebulização com a OVA se mostra um
protocolo eficaz na determinação do tempo necessário para que a resposta aguda da
asma seja desencadeada, e que os cobaias com inflamação pulmonar desenvolveram
essa resposta em tempo mais rápido, avaliou-se se a suplementação com o óleo de
coco virgem e com o fármaco padrão, a dexametasona, retardaria o tempo para que
essa resposta fosse desencadeada.
O aumento do tempo de contato com a OVA induzido pela dexametasona,
apenas na sétima nebulização (Gráfico 1, Tabela 1), está em conformidade com dados
da literatura (LEICK-MALDONADO et al., 2004; PIGATI et al., 2015). A suplementação
com o OCV na maior dose (4 g/kg) preveniu completamente essa resposta aguda na
quinta, sexta e sétima nebulizações (Gráfico 1, Tabela 1), indicando um efeito

194 VASCONCELOS, L. H. C. Discussão
atenuante do OCV sobre a resposta rápida broncoconstritora da crise asmática. Por
se tratar de uma avaliação in vivo, em que há a influência de diversos fatores, não é
possível inferir por qual mecanismo específico o OCV está agindo para promover essa
resposta; todavia, é provável que ele reduza a liberação de mediadores contráteis
pelas células inflamatórias, a própria contratilidade do músculo liso das vias aéreas
e/ou a produção de muco pelas células caliciformes.
Diante dessas hipóteses, foi feita a análise histológica do tecido pulmonar e
brônquico, a fim de se evidenciar um possível efeito da dexametasona e do óleo de
coco virgem sobre esses parâmetros. Assim, observou-se que o tratamento com a
dexametasona, bem como a suplementação com o OCV (especialmente na dose de
4 g/kg), reduziram a migração de células inflamatórias para o interstício pulmonar e
para a região peribrônquica. Além disso, houve redução da hiperplasia epitelial, na
mesma proporção tanto para a dexametasona como para as três doses do OCV, e
uma prevenção do espessamento muscular liso; sendo, para esse parâmetro, a dose
de 1 g/kg a mais efetiva (Figuras 15-17, Gráfico 2). Associando-se esses resultados
aos dados já descritos, sugere-se que o efeito do OCV observado até aqui se dê,
principalmente, por uma atenuação do processo inflamatório, mais que sua própria
ação sobre a musculatura lisa brônquica, e da hiperplasia epitelial, os quais agem
através da liberação de citocinas, fatores de crescimento e mediadores contráteis para
promover a hiper-contratilidade muscular lisa característica da crise asmática
(METCALFE; BARAM; MEKORI, 1997; GALLI; NAKAE; TSAI , 2005; BARRETT;
AUSTEN, 2009); além de serem corroborados por estudos que demonstraram um
efeito anti-inflamatório do OCV (INTAHPHUAK; KHONSUNG; PANTHONG, 2010;
NEVIN; RAJAMOHAN, 2010).
As interações indivíduo-meio ambiente que levam à hiper-responsividade
brônquica são múltiplas e a explicação para esse fenômeno é, provavelmente,
multifatorial. Nesse contexto, o tempo de nebulização, per se, não permite identificar
o componente responsável pelo efeito broncoconstritor; todavia, deve-se considerar a
previsibilidade da participação central do músculo liso nesse problema, uma vez que
ele é o principal componente envolvido no estreitamento do lúmen das vias aéreas
(LAUZON; MARTIN, 2016).
Assim, avaliou-se a resposta contrátil da traqueia de cobaia, in vitro, à
estimulação com OVA, de modo a averiguar se essa hiper-responsividade observada

195 VASCONCELOS, L. H. C. Discussão
in vivo ocorria por um aumento da contratilidade muscular lisa, pela Reação de
Schultz-Dale (SCHULTZ, 1910; DALE, 1913).
Como já descrito, os cobaias com inflamação pulmonar apresentaram uma
significativa resposta contrátil à estimulação com OVA, enquanto os cobaias não
sensibilizados não responderam ao estímulo, indicando a hiper-contratilidade das vias
aéreas (Figura 18A-B, Gráfico 3). Observou-se ainda que os cobaias tratados com a
dexametasona responderam de maneira semelhante aos com inflamação pulmonar à
estimulação com a OVA (Figura 18C, Gráfico 3). Apesar de controverso, dados na
literatura mostram que os corticosteroides tem pepel importante em conter o processo
inflamatório, enquanto seu papel sobre musculatura lisa ainda é incerto (PRADO et
al., 2012). Desse modo, pode-se inferir que o efeito observado in vivo para a
dexametasona sobre o tempo de nebulização se dê por uma redução na inflamação,
e não por reduzir a contratilidade muscular lisa, dado esse de acordo com o observado
na análise histológica (Figuras 15-17, Gráfico 2).
Nos animais suplementados com OCV, observou-se que, nas doses de 1 e
2 g/kg, não houve alteração na resposta contrátil da traqueia (Figura 18D e E,
Gráfico 3), enquanto a dose de 4 g/kg (Figura 18F, Gráfico 3) reduziu em cerca de
60% a contração em relação aos animais com inflamação pulmonar. Esses dados
indicam que o OCV pode inibir, parcialmente, a liberação de fatores contráteis pelas
células imunes estimuladas com OVA ou modular negativamente a contratilidade
muscular lisa para reduzir a broncoconstrição observada na asma. Esse efeito estaria
relacionado à inibição do recrutamento de células inflamatórias e da atividade
pró-contrátil do epitélio sobre o músculo liso, conforme observado pela análise
histológica do tecido pulmonar e brônquico (Figura 15-17, Gráfico 2).
A habilidade em prontamente modificar o comprimento muscular liso em
experimentos com órgãos isolados se mostra útil na demonstração de características
importantes da função contrátil do músculo liso aéreo. Dessa maneira, é possível
determinar se a hiper-responsividade observada em pacientes asmáticos está
associada com alterações nas propriedades contráteis do músculo liso isolado.
Resultados contraditórios são observados quando são sumarizados estudos
que avaliaram a resposta contrátil do músculo liso a diferentes agentes contráteis em
distintos modelos animais de asma. Nesse contexto, hiper-contratilidade foi observada
para a histamina (SCHELLENBERG; FOSTER, 1984; JONGSTE et al., 1987;
CERRINA et al., 1989; BAI, 1991), estimulação colinérgica (JONGSTE et al., 1987;

196 VASCONCELOS, L. H. C. Discussão
BAI, 1991; BRAMLEY et al., 1994; CHIBBA et al., 2000; CHESNÉ et al., 2015;
MATUSOVSKY et al., 2016), LTC4 (JONGSTE et al., 1987), adenosina (BJORCK;
GUSTAFSSON; DAHLÉN, 1992) e KCl (CHESNÉ et al., 2015). No entanto,
hipocontratilidade também foi observada em outros modelos animais de asma para a
histamina (WHICKER; ARMOUR; BLACK, 1988) e para estimulação colinérgica
(GOLDIE et al., 1986; WHICKER; ARMOUR; BLACK, 1988).
Ainda nesse contexto, dados da literatura mostram que a liberação de citocinas
pró-inflamatórias aumenta a contratilidade muscular lisa. IL-5, IL-13, IL-33, GM-CSF,
IFN-γ e IL-1β foram relacionados com o aumento da amplitude máxima de contração,
bem como uma maior sensibilidade a agentes contráteis in vitro (HAKONARSON et
al., 1999; BARLOW et al., 2013; DELMOTTE et al., 2014); o TNF-α ao aumento da
amplitude de contração por estimulação elétrica (ANTICEVICH et al., 1995;
DELMOTTE et al., 2014); a IL-17A ao aumento da contratilidade ao KCl e à metacolina
(KUDO et al., 2012), além de efeito sinérgico dessa citocina com o TNF-α sobre a
contração muscular lisa a esses agentes contráteis (MASUDA et al., 2013). Além
disso, maior contratilidade à bradicinina foi observada em preparações in vitro de
músculo liso brônquico após estimulação de TLR4 (DI CANDIA et al., 2017).
Assim, apesar de muitos estudos suportarem a hiper-contratilidade muscular
lisa na asma devido a hiper-responsividade a diferentes agentes contráteis, há dados
que sugerem que não há diferença nessa resposta entre não asmáticos e asmáticos.
É importante observar, no entanto, que essa variabilidade pode ser explicada pelas
diferenças fenotípicas da asma, o modelo experimental adotado, bem como a
preparação do tecido empregada (WRIGHT, 2013).
À vista do já conhecido papel primário do músculo liso na hiper-responsividade
brônquica, nesse trabalho foi investigada a reatividade da traqueia de cobaias em
resposta a estímulos contráteis e relaxantes. Foi observado um aumento na
reatividade contrátil da traqueia de cobaias com inflamação pulmonar, em relaçao aos
animais não sensibilizados, frente aos agonistas carbacol (Figura 19A-D, Gráfico 4,
Tabela 2) e à histamina (Figura 20A-D, Gráfico 6, Tabela 3), mas não ao KCl (Figura
21A-D, Gráfico 8, Tabela 4), apenas na presença de epitélio funcional, indicando uma
maior participação do componente fármaco-mecanico na hipercontratilidade da
traqueia de cobaia na inflamação pulmonar, sem alteração no componente
eletromecânico, e que esse efeito depende da funcionalidade epitelial.

197 VASCONCELOS, L. H. C. Discussão
Já com relação à potência dos agentes contráteis, não houve diferença entre
os animais com e sem inflamação pulmonar. Estudos anteriores mostraram que
citocinas, como IL-5 e IL-13, aumentadas na asma, induzem hiper-responsividade in
vitro à acetilcolina em músculo liso isolado de coelho (HAKONARSON et al., 1999;
LAPORTE et al., 2001). Adicionalmente, a triptase liberada por mastócitos induz
hipercontratilidade à histamina em brônquio humano sensibilizado possivelmente
devido à sua atividade enzimática sobre receptores ativados por protease (JOHNSON
et al., 1997).
Outras hipóteses podem ser postuladas para explicar essa resposta, como a
maior sensibilidade do aparato contrátil ao Ca2+ (SAVINEAU; MARTHAN, 1994) ou
maior liberação de Ca2+ dos estoques intracelulares, uma vez que já foi descrito que
citocinas pró-inflamatórias, como IL-1β e IL-13, aumentam a expressão da proteína
CD38 (ciclase de ADP-ribosil), formadora de ADP-ribose cíclica que, por sua vez, ativa
os receptores de rianodina (DESHPANDE et al., 2003; 2004; 2005). De maneira
similar, a IL-13 promove um aumento na expressão de proteínas da via RhoA/ROCK2,
aumenta o co-alinhamento das fibras de actina dentro do tecido e induz
hiper-reatividade à acetilcolina, semelhante aos resultados observados tanto em
estudos clínicos como em modelos animais (NESMITH et al., 2014).
Além disso, também já foi demonstrado que a maior geração de força
isométrica está relacionada com o aumento da massa de tecido muscular liso em
preparações de aneis de brônquio humano (ARMOUR; DIMENT; BLACK, 1988) e que
a redução concomitante da hiper-responsividade das vias aéreas e da massa de
músculo liso estão correlacionadas, sugerindo que essa mudança estrutural poderia
explicar a mudança funcional na responsividade contrátil muscular lisa (DANEK et al.,
2004). Diante disso, fica evidente que a massa excessiva desse tecido e o
espessamento da parede das vias aéreas estão associados à hiper-reatividade
brônquica (NIIMI et al., 2000; LITTLE et al., 2002; WOODRUFF et al., 2004).
Esses dados até aqui fornecem subsídios à hipótese de que essa
hipercontratilidade seja decorrente de um aumento na massa de tecido muscular liso
decorrente da inflamação pulmonar crônica. Ademais, uma vez que a sensibilidade a
um fármaco está relacionada tanto à afinidade do receptor como ao número deste em
si na membrana da célula (NEUBIG et al., 2003), não é descartada a possibilidade de
um aumento na expressão dos receptores M3 e H1 no músculo liso aéreo, assim como
das proteínas downstream de sinalização desses receptores, uma vez que a

198 VASCONCELOS, L. H. C. Discussão
hiper-eficácia contrátil só foi observada para os agentes de acoplamento
farmaco-mecânico, mesmo sem alteração na potência contrátil. Os mecanismos
funcionais relacionados com esse efeito foram investigados, sendo abordados
subsequentemente.
Ainda no contexto da reatividade contrátil da traqueia de cobaia,
comparando-se a potência entre os dois agonistas, é importante observar que a
sensibilidade ao carbacol foi maior que à histamina, nos cobaias com e sem
inflamação pulmonar, fato esse explicado pela reconhecida heterogeneidade regional
intrínseca do músculo liso das vias aéreas periféricas e centrais. Enquanto as vias
aéreas periféricas são mais sensíveis à histamina, a estimulação colinérgica é mais
pronunciada nas vias aéreas centrais, como a traqueia (DANIEL et al., 1986; LEFF et
al., 1988).
Além disso, comparando-se a potência dos três agentes contráteis entre as
preparações na presença e na ausência de epitélio funcional, observa-se que, para a
histamina e o KCl, ambos foram mais potentes na ausência de epitélio funcional;
enquanto, de maneira controversa, o carbacol foi equipotente. Esses dados estão de
acordo, em parte, com Holroyde (1986), que observou que a retirada do epitélio de
preparações de traqueia de cobaia aumentava a sensibilidade aos três agentes
contráteis; essa diferença foi atribuída não à eliminação de possíveis fatores
relaxantes, mas à remoção da barreira física de permeabilidade imposta pelo epitélio
traqueal.
Estudos têm relatado que o epitélio das vias aéreas desempenha papel
fundamental na promoção da resposta alérgica, bem como no remodelamento tecidual
e na alteração da contratilidade muscular lisa. Postula-se que o dano epitelial
decorrente da inflamação alérgica resulta na geração de sinais que agem no tecido
mesenquimal para propagar e amplificar a inflamação e o remodelamento. Por
exemplo, esse estímulo inflamatório pode induzir a produção de mediadores pelo
epitélio, como o TGF-β, liberado em resposta à estimulação por citocinas, como a
IL-13 (DAVIES, 2009) que estimula a diferenciação de fibroblastos da matriz
extracelular em miofibroblastos, os quais medeiam a hipertrofia muscular lisa na
asma, além de promoverem contração mais rápida e sustentada (BOXALL;
HOLGATE; DAVIES, 2006).
As células epiteliais são também uma fonte de fatores contráteis que
contribuem para a broncoconstrição, como leucotrienos; prostanoides, como PGF2α,

199 VASCONCELOS, L. H. C. Discussão
PGD2 e TxA2; assim como endotelina (BARNES, PIPER, COSTELLO, 1984; LIU et
al., 1990; HOWARTH, 1995; PEREZ-ZOGHBI, SANDERSON, 2007). Diante disso, é
razoável afirmar que o aumento da resposta contrátil observada se dê como
consequência dos efeitos do epitélio como liberador de mediadores hipertróficos, a
longo prazo, e pró-contráteis, a curto prazo, para a musculatura lisa das vias aéreas.
Tendo em vista que a asma levou a um aumento na contratilidade do músculo
liso da traqueia de cobaia frente aos agonistas CCh e histamina, mas não ao KCl, na
presença de epitélio, decidiu-se investigar se a dexametasona e o OCV preveniriam a
hiper-contratilidade desse músculo em preparações com epitélio íntegro, e foi
observado que a dexametasona reverteu a resposta contrátil da traqueia dos cobaias
asmáticos tanto ao CCh (Figura 19E, Gráfico 5, Tabela 2) quanto à histamina,
conforme observado pela redução da eficácia desses dois agonistas, mas sem alterar
suas potências contráteis (Figura 20E, Gráfico 7, Tabela 3).
Os efeitos dos glicocorticoides sobre a contratilidade da musculatura lisa ainda
são controversos. Nabishah et al. (1990) observaram que o tratamento com
dexametasona por 7 dias, na dose de 0,12 mg/kg/dia, reduziu a sensibilidade do
brônquio de ratos à estimulação com acetilcolina, fato esse justificado pela redução
na expressão no número de receptores muscarínicos. Pavlonic et al. (1998), no
entanto, avaliando a resposta contrátil da traqueia de ratos ao carbacol, verificaram
que o tratamento em dose única (1,2 mg/kg) com triancinolona reduziu a tensão
máxima do agente contrátil, sem, entretanto, alterar sua potência. Lefcoe (1956) havia
observado que a hidrocortisona antagonizava as contrações induzidas tanto por
histamina quanto por carbacol em preparações de anéis de traqueia de cobaia.
Além disso, Scherrer et al. (1997) observaram que o tratamento com
metilprednisona, na dose de 10 mg/kg, reduzia a densidade de receptores
muscarínicos após 3 h da administração. Todavia, essa densidade era reestabelecida
após 24 h da administração da droga. Adicionalmente, de maneira controversa, os
autores verificaram que a resposta contrátil do parênquima pulmonar dos cobaias não
foi afetada pelo tratamento com o glicocorticoide após 96 h de tratamento. Diante
disso, os dados se mostram de difícil comparação e conclusão, não sendo possível
dizer, de maneira precisa, o real efeito do tratamento com glicocorticoide sobre a
reatividade muscular lisa das vias aéreas, uma vez que este pode variar conforme o
regime de tratamento, o modelo animal adotado, bem com a preparação de órgão
utilizada.

200 VASCONCELOS, L. H. C. Discussão
A suplementação com o OCV na dose de 1 g/kg não preveniu a
hiper-reatividade contrátil da traqueia de cobaia com inflamação pulmonar frente ao
CCh (Figura 19F, Gráfico 5, Tabela 2) ou à histamina (Figura 20F, Gráfico 7, Tabela 3),
mas preveniu ao CCh, nas doses de 2 e 4 g/kg (Figura 19G e H, Gráfico 5, Tabela 2),
e à histamina, apenas na maior dose testada (Figura 20H, Gráfico 6, Tabela 3). Esses
resultados se correlacionam com o que foi observado para a estimulação com OVA,
cuja amplitude da contração foi reduzida pelo OCV na dose de 4 g/kg. Tais dados
apontam para um possível efeito modulador negativo do OCV sobre os mecanismos
contráteis que foram alterados pela inflamação pulmonar, em especial, a expressão
de receptores muscarínicos e histaminérgicos, bem como as proteínas das suas vias
downstream de sinalização intracelular. Também pode ser apontado como alvo de
modulação do OCV o próprio processo inflamatório crônico, responsável pelas
alterações, a longo prazo, no tecido muscular liso das vias aéreas na asma, além do
efeito do óleo em reduzir a hipertrofia muscular lisa, porém em menor extensão, já que
esses dados são respaldados pelo observado na analise histológica pulmonar e
brônquica.
Até aqui, observou-se que o modelo de inflamação pulmonar alérgica crônica
em cobaias induz hiper-reatividade contrátil do músculo liso traqueal à estimulação
antigênica e com os agonistas carbacol e histamina; e que a suplementação com o
OCV, especialmente na dose de 4 g/kg, reverte esses parâmetros, relacionando-se
tais efeitos com a atividade anti-inflamatória do óleo, além da modulação de proteínas
das vias de sinalização de contração e da maquinaria contrátil, bem como de
mecanismos intracelulares hipertróficos do músculo liso.
Por meio de uma conversão direta, essa dose de 4 g/kg de OCV corresponderia
a um consumo humano, considerando-se um adulto de 70 kg, de 280 g de óleo.
Entretanto, a Food and Drug Administration (FDA) sugere que a extrapolação da dose
animal para humano é corretamente calculada por meio da normalização em relação
à área de superfície corporal, representada em mg/kg2 (REAGAN-SHAW; NIHAL;
AHMAD, 2007). Nesse sentido, a maior dose de OCV corresponde a uma dose de
0,86 g/kg. Considerando-se ainda que o modelo animal que mimetiza a inflamação
pulmonar é limitada ao período de indução até a eutanásia, é de se convir que, em
humanos, cuja suplementação com o óleo duraria tempo mais prolongado, essa dose
poderia ser ajustada sem que os efeitos benéficos sejam comprometidos.

201 VASCONCELOS, L. H. C. Discussão
Complementar aos mecanismos contráteis que medeiam a
hiper-responsividade brônquica, esse fenômeno também está relacionado com uma
menor liberação de agentes relaxantes para o músculo liso, sejam eles hormônios,
neurotransmissores ou mediadores inflamatórios, como adrenalina (BARNES, 1986),
peptídeo intestinal vasoativo (BEN-JEBRIA et al., 1993) e prostanoides, como PGE2
e PGI2 (WALTERS et al., 1982). A dessensibilização de receptores adrenérgicos β2
também está associada ao aumento da responsividade muscular lisa a estímulos
contráteis na asma (BARNES, 1995).
Avaliando-se a responsividade do músculo liso das vias aéreas em modelos
animais de asma, tem sido observado hipo-responsividade a agentes relaxantes,
como a teofilina e agonistas β2 (CERRINA et al., 1986; GOLDIE et al., 1986;
WHICKER; ARMOUR; BLACK, 1988; BAI, 1991; NESMITH et al., 2014).
Tomando esses dados como premissa, decidiu-se investigar a reatividade
relaxante da traqueia de cobaias, a fim de determinar se a inflamação pulmonar
promove alterações na responsividade relaxante das vias aéreas, além de obter
indícios de quais mecanismos estariam por trás desse efeito, e se a suplementação
com o OCV reverteria essa alteração.
Não foram observadas diferenças tanto na eficácia quanto na potência
relaxante da isoprenalina sobre o tônus basal da traqueia de cobaia com inflamação
pulmonar, tanto na presença como na ausência de epitélio, comparados ao grupo GC
(Figura 22A-D, Gráfico 9, Tabela 5). Entretanto, contrariamente a esse dado, quando
o órgão era pré-contraído com carbacol, foi observado um deslocamento da curva de
relaxamento da isoprenalina para a direita, isto é, uma redução, em cerca de 3,5
vezes, na sua potência relaxante em traqueia de cobaia com inflamação pulmonar na
presença (Figura 23A e C, Gráfico 10, Tabela 6), mas não na ausência de epitélio
(Figura 23B e D, Gráfico 10, Tabela 6), sem alteração na sua eficácia relaxante,
fortalecendo os indícios da participação fundamental do epitélio na hiper-reatividade
do modelo de inflamação pulmonar.
Apesar da aparente contradição desses dados, uma vez que a asma leva a
aumento do tônus basal com consequente estreitamento do lúmen das vias aéreas,
estudos anteriores em animais mostram a existência de um antagonismo funcional
entre a contração mediada pela estimulação colinérgica e o relaxamento induzido por
agonistas adrenérgicos β (FERNANDES; FRYER; HIRSHMAN, 1992; WATSON;
EGLEN, 1994). Receptores M3 influenciam de modo crucial a função desses

202 VASCONCELOS, L. H. C. Discussão
receptores por mecanismo que envolve a ativação da PKC e fosforilação direta dos
receptores β2, bem como da proteína Gs, promovendo desacoplamento do receptor
com sua proteína G, bem como dessensibilização (MEURS et al., 2001); além da
fosforilação da cinase dos adrenoceptores β (BARK), amplificando a
dessensibilização desses receptores (DE BLASI; PARRUTI; SALLESE, 1995;
BOTERMAN et al., 2006). Tais processos explicam a bem conhecida atenuação da
eficácia de agonistas β durante episódios de broncoconstrição grave (MEURS;
GOSENS; ZAAGSMA, 2008).
Conhecida essa relação, estudo foi conduzido para determinar se uma das
razões para a menor responsividade aos agonistas β sobre a contração por ACh é a
exposição a citocinas específicas, como a IL-13, produzida, dentre outras células, pelo
epitélio das vias aéreas na inflamação alérgica pulmonar, tendo sido demonstrado
uma redução do relaxamento por esse mecanismo (GRUNSTEIN et al., 2002). Além
disso, Nesmith et al. (2014) observaram que as vias aéreas de pacientes não
asmáticos respondiam de forma semelhante àqueles com perfil fenotípico asmático,
induzido por IL-13, à estimulação com isoprenalina quando esta era adicionada às
preparações na ausência de estimulação contrátil prévia. Todavia, quando o órgão
era pré-contraído com acetilcolina, a resposta relaxante era atenuada, quando
comparado com as preparações de indivíduos não asmáticos. Esses dados confirmam
estudo anterior que mostrou que a IL-13 reduz a responsividade dos receptores
adrenéricos β do músculo liso das vias aéreas (LAPORTE et al., 2001).
Além da IL-13, outros mediadores inflamatórios já foram relacionados com a
hiporresponsividade à estimulação adrenérgica nas vias aéreas e à dessensibilização
dos receptores β2 (GUPTA et al., 2016). A IL-1β, uma citocina produzida por diversas
células do pulmão, reduz a responsividade adrenérgica β de células musculares lisas
humanas (LAPORTE et al., 2000). Além disso, fatores de crescimento, como o
TGF-β1, produzido também pelo epitélio brônquico, reduz o número de
adrenoceptores β em células musculares lisas de traqueia humana, além de reduzir
os níveis de cAMP aumentados por agonistas β2 (NOGAMI et al., 1994).
Uma vez que foi observado uma menor sensibilidade da traqueia de cobaia à
isoprenalina, apenas na presença de epitélio, decidiu-se avaliar se a suplementação
com OCV reverteria essa resposta em preparações com epitélio íntegro. Inicialmente,
foi visto que o fármaco padrão, dexametasona, não alterou a resposta a esse agonista,
uma vez que a curva de relaxamento se manteve sobreposta com a dos animais com

203 VASCONCELOS, L. H. C. Discussão
inflamação pulmonar (Figura 23E, Gráfico 11, Tabela 6). Esse dado se contrapõe com
dados da literatura, que mostram que glicocorticoides melhoram a responsividade do
músculo liso das vias aéreas a agonistas adrenérgicos β por aumentar a transcrição
gênica e expressão desses receptores (LEFCOE, 1956; SCHERRER et al., 1997;
SCHRAMM, 2000). Todavia, é importante observar que em nenhum desses trabalhos
foi utilizado um modelo animal de asma alérgica, o que torna difícil sua extrapolação
para os obtidos no nesse trabalho. Ademais, é provável que a upregulation promovida
pelos glicocorticoides em animais ou cultura de células musculares lisas não
asmáticos não seja suficiente para reverter a dessensibilização desencadeada pela
inflamação pulmonar observada no presente estudo.
Diferentemente, apesar da suplementação com o OCV nas doses de 1 e 2 g/kg
não ter alterado a curva de relaxamento à isoprenalina em traqueia de cobaia com
inflamação pulmonar (Figura 23F e G, Gráfico 11, Tabela 6), foi observado que na
dose de 4 g/kg, a curva foi deslocada para a esquerda, se sobreponto ao controle
(Figura 23H, Gráfico 11, Tabela 6), indicando assim que a suplementação com o óleo
na maior dose testada previne a hipo-responsividade da traqueia de cobaia a esse
agonista adrenérgico β. Tal efeito pode ter diversas explicações, destacando-se uma
possível inibição da dessensibilização dos receptores β, provável responsável pela
menor sensibilidade da traqueia de cobaia asmático à isoprenalina, por mecanismos
dependentes da participação do epitélio, como a inibição da liberação de citocinas e
fatores de crescimento, como IL-13 e TGF-β, os quais já foram relacionados com a
redução da expressão desses receptores e consequente menor responsividade
relaxante do músculo liso das vias aéreas a agonistas adrenérgicos β (NOGAMI et al.,
1994).
Tendo em vista que foi avaliada a reatividade contrátil da traqueia de cobaia ao
KCl, substância que induz contração por mecanismo eletromecânico que, por sua vez,
leva à despolarização de membrana e consequente abertura dos CaV (DU et al.,
2006), cuja amplitude da contração não diferiu entre os animais com e sem inflamação
pulmonar, decidiu-se comprovar esse resultado avaliando-se a resposta relaxante
desse órgão ao nifedipino, um bloqueador dos CaV (AHMED; FOSTER; SMALL,
1985).
A reatividade relaxante da traqueia de cobaias dos grupos GC e GASM não
diferiu, tanto em termos de potência como de eficácia, na presença e na ausência de
epitélio (Figura 24A-D, Gráfico 12, Tabela 7), indicando que a inflamação pulmonar

204 VASCONCELOS, L. H. C. Discussão
não altera o influxo de Ca2+ pelos CaV, seja pelo limiar de voltagem necessário para
levar à abertura desses canais, a probabilidade de abertura, a condutância ou a sua
expressão na membrana das células musculares lisas. Além disso, esses dados
confirmam o que foi observado para a reatividade contrátil da traqueia ao KCl, que
também não foi alterada pela inflamação pulmonar.
Diante desses dados, conclui-se parcialmente que o modelo de inflamação
pulmonar alérgica crônica promove alterações teciduais no sistema respiratório de
cobaias, levando a uma hiper-responsividade muscular lisa traqueal, caracterizada por
uma maior reatividade a agonistas contráteis e menor responsividade a agonista
relaxante, por mecanismos dependentes da participação do epitélio. Além disso, o
OCV previne a hiper-reatividade muscular lisa, provavelmente por modular as vias
desencadeadas a partir do epitélio das vias aéreas, além de impedir a progressão do
remodelamento tecidual do sistema respiratório. Por fim, tomadas as avaliações em
conjunto, a dose de 4 g/kg se mostra a mais efetiva em reverter todas essas
alterações. Nesse sentido, prosseguiu-se com a investigação dos mecanismos
funcionais relacionados com as alterações desencadeadas pela inflamação pulmonar,
bem como promovidos pelo óleo de coco virgem na dose de 4 g/kg para prevenir essas
alterações no músculo liso da traqueia de cobaia.
A asma é caracterizada por uma sobrecarga de espécies reativas de oxigênio,
que ocasionam estresse oxidativo e alterações nas funções de diversos componentes
do sistema respiratório (MORWOOD et al., 2005; BISHOPP et al., 2017). As principais
ROS envolvidas com a função desse sistema são o ânion superóxido, os radicais
hidroxila e o peróxido de hidrogênio, os quais levam à formação de outra variedade
de ROS (HENRICKS; NIJKAMP, 2001; LOXHAM; DAVIES; BLUME, 2014).
Diversos trabalhos relataram os efeitos das ROS no funcionamento do músculo
liso das vias aéreas. Peróxido de hidrogênio e níveis aumentados de oxigênio induzem
contração da traqueia de cobaia (RHODEN; BARNES, 1989). Além dos seus efeitos
diretos, estudos mostram que as ROS também influenciam a reatividade das vias
aéreas a agonistas contráteis e relaxantes; dentre eles, destacam-se a resposta
contrátil aumentada à acetilcolina e à metacolina (HENRICKS; NIJKAMP, 2001), à
histamina (SADEGHI-HASHJIN et al., 1996), à serotonina (SZAREK; SCHMIDT,
1990), e a diminuição no número e na função de receptores adrenérgicos β2
(NIJKAMP; HENRICKS, 1990; IKUTA et al., 1992).

205 VASCONCELOS, L. H. C. Discussão
Tendo em vista essas premissas, investigou-se a contribuição das ROS na
contração mediada pelo CCh em traqueia de cobaias com e sem inflamação
pulmonar. Observou-se que a apocinina, inibidor da NADPH oxidase, não alterou a
curva cumulativa de contração ao CCh, com relação à eficácia e à potência do
agonista, nas preparações de traqueia de cobaia dos grupos GC (Figura 25A, Gráfico
13A, Tabela 8), GASM (Figura 25D, Gráfico 13B, Tabela 8) e GASM/OC4 (Figura 25G,
Gráfico 13C, Tabela 8). Esse dado indica que o ânion superóxido proveniente da
NADPH oxidase não exerce papel importante na modulação do tônus contrátil da
traqueia de cobaia, independente se há ou não um quadro de inflamação pulmonar
alérgica, e que o OCV também não altera essa via de formação de ROS.
Diferentemente, o tempol, mimético da SOD, reduziu a potência contrátil do
CCh nos animais sem inflamação pulmonar (Figura 25B, Gráfico 13A, Tabela 8), mas
não nos animais com pulmões inflamados GASM (Figura 25E, Gráfico 13B, Tabela 8).
Semelhantemente ao GASM, no GASM/OC4 a potência do CCh também não foi
reduzida pelo tempol (Figura 25H, Gráfico 13C, Tabela 8), indicando, assim, que as
ROS, especialmente o ânion superóxido, estão aumentados nos animais asmáticos,
uma vez que o tempol foi incapaz de impedir a ação desse ânion no músculo liso
traqueal, mas que esse aumento se daria por vias independentes da NADPH oxidase,
como a xantina oxidase e a eNOS desacoplada, vias igualmente responsáveis pela
formação do ânion superóxido.
Corroborando essa participação do ânion superóxido na regulação do tônus
das vias aéreas, De Boer et al. (1998) evidenciaram que a contração mediada por
metacolina era atenuada na presença da SOD, indicando a influência dessa ROS na
contratilidade muscular lisa das vias aéreas. Além disso, o resultado semelhante ao
do grupo GASM observado nos animais com inflamação pulmonar suplementados
com OCV indicam que o óleo não modula essa via de formação das ROS; porém,
diante das evidências relatadas na literatura acerca da atividade antioxidante do OCV,
postula-se que ele aja nas etapas subsequentes da formação das ROS, como a
geração do peróxido de hidrogênio a partir do ânion superóxido.
Mátyás; Pucovský; Bauer (2002) avaliaram qual das ROS apresentaria maior
importância no contexto da contratilidade da traqueia de cobaia e foi observado que
tanto o ânion superóxido quanto os radicais hidroxila e o peróxido de hidrogênio
contraem o músculo liso traqueal, sendo este último a ROS responsável pelo maior
aumento no tônus muscular. Além disso, os efeitos desses radicais livres são contidos

206 VASCONCELOS, L. H. C. Discussão
pelas defesas antioxidantes presentes no epitélio íntegro e o dano epitelial
potencializa o efeito contrátil dessas ROS.
Nesse sentido, uma vez que a asma induz dano epitelial, é razoável inferir sobre
uma possível redução da capacidade do epitélio traqueal em capturar esses radicais
livres, levando assim a um desbalanço entre ROS/defesas antioxidantes, favorecendo
o dano muscular liso aéreo. Corroborando essa hipótese, De Raeve et al. (1997)
observaram que células epiteliais isoladas de asmáticos possuem menor atividade da
SOD em comparação ao epitélio de não asmáticos. Além disso, uma vez que o
peróxido de hidrogênio é a ROS de maior importância na contração muscular lisa das
vias aéreas, decidiu-se investigar a participação desse radical livre no efeito contrátil
ao CCh.
Enquanto a catalase, enzima que converte o peróxido de hidrogênio em água
e oxigênio molecular (RHODEN; BARNES, 1989), reduziu a potência contrátil do
CChh cerca de 4 vezes (Figura 25C, Gráfico 13A, Tabela 8), nos animais com
inflamação pulmonar, a potência do CCh não foi alterada (Figura 25F, Gráfico 13B,
Tabela 8), semelhante ao observado na presença do tempol; indicando um aumento
na produção do H2O2 desencadeada pelo processo inflamatório, refletindo a
inabilidade da catalase em reduzir os níveis dessa ROS. Interessantemente, nos
animais com inflamação pulmonar suplementados com OCV, houve uma forte
prevenção da formação desse radical livre, uma vez que a potência contrátil do CCh
foi reduzida em 6,6 vezes na presença da catalase (Figura 25I, Gráfico 13C, Tabela 8).
Assim, sugere-se que o OCV promove seu efeito antioxidante nos pulmões de cobaias
com inflamação pulmonar não por reduzir a formação ou a depuração do ânion
superóxido, mas os efeitos danosos do peróxido de hidrogênio sobre a contratilidade
muscular lisa.
O envolvimento dos prostanoides na patogênese da asma é bem reconhecido,
sendo descrito que, no fluido broncoalveolar de pacientes asmáticos, os níveis de
PGD2 e PGF2α são, respectivamente, 12 e 22 vezes maiores que em indivíduos não
asmáticos (LIU et al., 1990). É descrito, no entanto, que o uso de AINES, tais como o
AAS e a indometacina, induz broncoconstrição grave (VALLY; TAYLOR; THOMPSON,
2002; MASCIA et al., 2005), assim como camundongos deficientes em COX-1 exibem
inflamação aumentada das vias aéreas em modelo de asma (CAREY et al., 2003;
NAKATA et al., 2005). Além disso, foi evidenciado que a super-expressão da COX-1

207 VASCONCELOS, L. H. C. Discussão
no epitélio de camundongos reduz a hiper-responsividade das vias aéreas (CARD et
al., 2006).
Tais efeitos são explicados pelo desvio do metabolismo do AA da via da COX
para a via da 5-LO e consequente elevação dos níveis de leucotrienos (DAHLÉN et
al., 1998; DAHLÉN et al., 2002; HONDA; KABASHIMA, 2014), como os cisteinil
leucotrienos, responsáveis pela forte broncoconstrição e eosinofilia (DAVIDSON et al.,
1987; LAIDLAW et al., 2012), uma vez que estudos com camundongos deficientes em
5-LO mostraram haver uma redução na hiper-responsividade induzida por antígeno
(IRVIN et al., 1997). Semelhantemente, foi demonstrado que a zileutona, um inibidor
da 5-LO, assim como um inibidor da FLAP, reduzem a hiper-responsividade das vias
aéreas em camundongos (HENDERSON et al., 1996; VARGAFTIG; SINGER, 2003).
Os efeitos da inibição da COX sobre a contração do músculo liso das vias
aéreas já haviam sido descritos por Orehek; Douglas; Bouhuys (1975), que
observaram que a incubação de anéis de traqueia de cobaia com indometacina
promovia efeitos semelhantes aos observados nesse trabalho para o grupo GC:
redução do tônus basal, baixa resposta contrátil a pequenas concentrações de CCh e
aumento na resposta contrátil em altas concentrações desse agonista (Figura 26A,
Gráfico 14A, Tabela 9). Semelhantemente, a zileutona, inibidora da 5-LO, aumentou
a eficácia contrátil do CCh nos animais do grupo GC, na mesma proporção da
indometacina (Figura 26B, Gráfico 14A, Tabela 9), indicando um equilíbrio entre as
ações da COX e da 5-LO na ausência de inflamação pulmonar, responsáveis pela
manutenção da homeostase respiratória, de modo que a inibição de uma via promove
o desvio igualitário para a via colateral de metabolismo do AA.
Diferentemente, os animais com inflamação pulmonar apresentaram tanto a
resposta contrátil máxima quanto a potência do CCh aumentada pela indometacina
(Figura 24B, Gráfico 14B, Tabela 9), mas não pela zileutona (Figura 26D, Gráfico 14B,
Tabela 9), indicando que na inflamação pulmonar a liberação de prostaglandinas
relaxantes esteja deficiente, sendo tal hipótese respaldada por pesquisas que
demonstraram em modelos de asma uma hipo-responsividade muscular lisa das vias
aéreas à PGE2 e à PGI2 (LUNDEQUIST et al., 2010; MORI et al., 2011; JAIN et al.,
2014). Além disso, sugere-se que os produtos da 5-LO, os Cys-LTs, estejam sendo
formados em uma proporção significantemente maior que nos animais do grupo GC,
ou ainda que o tecido muscular encontra-se sensibilizado a esses mediadores, sendo
a zileutona ineficaz em impedir suas ações, corroborando dados da literatura

208 VASCONCELOS, L. H. C. Discussão
(DAVIDSON et al., 1987; HENDERSON et al., 1996; IRVIN et al., 1997; VARGAFTIG;
SINGER, 2003; LAIDLAW et al., 2012).
Por outro lado, nos animais do GASM/OC4, observou-se resposta diferente de
GC e GASM: a indometacina não alterou a eficácia e a potência do CCh (Figura 24C,
Gráfico 14C, Tabela 9); e, interessantemente a zileutona reduziu a potência contrátil
do CCh (Figura 24C, Gráfico 14C, Tabela 9). Esses dados levam a sugerir que a
suplementação com o OCV alterou o padrão de síntese e liberação dos mediadores
lipídicos provenientes da cascata de sinalização do AA, com um provável aumento da
formação de prostanoides relaxantes sobre os contráteis, que se equilibram com a
formação de Cys-LTs. Desse modo, o bloqueio da COX manteria uma produção ainda
suficiente de prostanoides relaxantes que se equilibra com os produtos da 5-LO, e a
inibição desta faz prevalecer a ação dos prostanoides relaxantes para reduzir a
capacidade contrátil do CCh. Apesar desses dados nortearem para um possível efeito
benéfico da suplementação com o OCV, do ponto de vista da asma como uma doença
inflamatória cujo agravamento é fortemente influenciado por tais mediadores, deve-se
ter cautela sobre até que ponto esse efeito pode ser benéfico ou danoso para a
homeostase respiratória.
Foi evidenciada a participação do ânion superóxido na hiper-contratilidade
traqueal de animais asmáticos, a qual não foi revertida pelo OCV. Interessantemente,
estudos demonstram a participação de prostanoides formados independentemente da
COX, via peroxidação do ácido araquidônico da membrana, promovida por radicais
livres (MORROW; ROBERTS, 1996; JANSSEN, 2000; 2001).
Tais compostos, os isoprostanos, como a 8-iso-PGF2α (HAZBUN et al., 1993),
são considerados marcadores do estresse oxidativo em pacientes asmáticos e podem
induzir contração do músculo liso das vias aéreas (JANSSEN, 2000; 2001). No modelo
de inflamação pulmonar, foi evidenciado em estudos anteriores a participação da
8-iso-PGF2α no aumento da resistência das vias aéreas associado ao estresse
oxidativo em cobaias (POSSA et al., 2012; PIGATI et al., 2015). Além disso, estudo
conduzido por Mátyás; Pucovský; Bauer (2002) demonstrou que a contratilidade da
traqueia de cobaia induzida por peróxido de hidrogênio, radicais hidroxila ou ânion
superóxido estava associada à ativação da 5-LO, preferencialmente, bem como da
COX, indicando o papel fundamental dos leucotrienos na reatividade contrátil do
músculo liso aéreo dependente das ROS.

209 VASCONCELOS, L. H. C. Discussão
Diante disso, pode-se sugerir também que, com a inibição da COX, o excedente
de ácido araquidônico ficou disponível para sofrer ação de radicais livres e formar
isoprostanos contráteis, explicando o aumento da sensibilidade contrátil ao CCh
observado no GASM, e que o provável efeito antioxidante do OCV foi um dos fatores
que contribuíram para a reversão desse efeito. Essa hipótese é suportada por dados
da literatura, que mostram que o OCV, e/ou seus polifenois, apresentam atividade
anti-inflamatória por inibição da formação de edema de pata induzida por ácido
araquidônico (INTAHPHUAK; KHONSUNG; PANTHONG, 2010) ou por redução da
expressão da COX-2 em ratos (VYSAKH et al., 2014); além de atividade antioxidante,
seja por inibir a peroxidação lipídica ou por aumentar a expressão de enzimas
antioxidantes (NEVIN; RAJAMOHAN, 2006; MARINA et al., 2009; ABUJAZIA et al.,
2012; ARUNIMA; RAJAMOHAN, 2013; VYSAKH et al., 2014; ALVES et al., 2015).
Além disso, estudo realizado por Henry et al. (2002) evidenciaram que ácidos
graxos saturados da dieta, especialmente aqueles entre C-10 e C-12, apresentam
capacidade antioxidante e inibitória das COXs I e II. Outrossim, recentemente foi
descrito que o ácido láurico reduz o estresse oxidativo em ratos hipertensos (ALVES
et al., 2017). Desse modo, baseado nos estudos já descritos, provavelmete os efeitos
observados para o OCV podem estar relacionados tanto com seus compostos
fenólicos como com seu ácido graxo majoritário, o ácido láurico.
Outro importante mediador da reatividade contrátil e relaxante do músculo liso,
de modo geral, é o NO, também produzido pelo epitélio traqueal e brônquico, além de
outras fontes, como neurônios nitrérgicos, células inflamatórias e o próprio músculo
liso (RICCIARDOLO, 2003).
O sistema respiratório apresenta as isoformas constitutivas de NOS tanto nos
neurônios iNANC (nNOS) como no endotélio dos vasos pulmonares e no epitélio
traqueobrônquico (eNOS), envolvidas na regulação do tônus das vias aéreas
(RICCIARDOLO et al., 2004). Apesar do NO produzido a partir da eNOS estar
associada com a inibição da inflamação por meio da supressão da ativação do NF-kB
e consequente redução da expressão da iNOS, bem como de citocinas inflamatórias
(CIRINO; FIORUCCI; SESSA, 2003; COOK et al., 2003; TEN BROEKE et al., 2006),
nas vias aéreas inflamadas de asmáticos, há uma elevada expressão da iNOS,
especialmente nas células epiteliais e inflamatórias, incluindo macrófagos, neutrófilos
e eosinófilos (RICCIARDOLO et al., 2004; HAMID et al., 1993; ASANO et al., 1994).
Altas concentrações de NO produzido a partir da iNOS tem sido considerado danoso

210 VASCONCELOS, L. H. C. Discussão
para as vias aéreas (RICCIARDOLO et al., 2004), uma vez que seus efeitos são
associados à formação do radical livre peroxinitrito (SADEGHI-HASHJIN et al., 1998),
havendo uma forte correlação entre os níveis de NO, a eosinofilia e a
hiper-responsividade das vias aéreas (DUPONT et al., 1998; JATAKANON et al.,
1998).
Diante dessa premissa, decidiu-se investigar se a hiper-reatividade da traqueia
de cobaias com inflamação pulmonar ao CCh estava associada com a via do NO, seja
por uma menor eficácia na sua produção a partir da eNOS ou por uma super atividade
da iNOS.
Evidenciou-se que, enquando o pré-tratamento com o inibidor de NOS
(L-NAME) não alterou a curva de contração do CCh nos animais do grupo GC (Figura
27A, Gráfico 15A, Tabela 10, no GASM, tanto a eficácia quanto a potência contrátil do
CCh foram aumentadas (Figura 27B, Gráfico 15B, Tabela 10). Baseado na premissa
de que o NO causa broncodilatação, seria esperado que a inibição da síntese de NO
pelo L-NAME aumentasse ou potencializasse o efeito contrátil do CCh, o que não foi
observado no presente trabalho para GC. Diante disso, assume-se que, nos animais
não asmáticos, não há ação tônica do NO sobre a contratilidade das vias aéreas,
diferentemente do que foi observado nos animais com inflamação pulmonar. Tal efeito
pode estar associado a uma expressão diferencial da eNOS e da iNOS entre os
animais com e sem inflamação pulmonar, assim como o efeito inibitório do L-NAME
sobre essas duas isoformas. Presume-se que o L-NAME bloqueia a eNOS, mas não
a iNOS, através da qual o NO está sendo predominantemente formado nos animais
do GASM, e nos quais a eNOS encontra-se deficiente, aumentando a efetividade da
inibição promovida pelo L-NAME.
Esses dados são corroborados por Antosova e Strapkova (2008), que
observaram efeitos semelhantes do L-NAME em cobaias aos deste trabalho. No
entanto, de modo controverso, Antosova et al. (2015) evidenciaram que o
pré-tratamento da traqueia de cobaia com L-NAME aumenta a resposta contrátil desse
órgão tanto em animais não asmáticos quanto asmáticos. Diante disso, é possível que
o efeito do L-NAME sobre a atividade das diferentes isoformas de NOS dependa do
modelo de inflamação pulmonar alérgica adotado e a habilidade deste em alterar a
expressão dessas isoformas nas vias aéreas.
De fato, para para o presente modelo de inflamação pulmonar, demonstrou-se
um aumento na expressão da iNOS no tecido pulmonar (PRADO et al., 2005; POSSA

211 VASCONCELOS, L. H. C. Discussão
et al., 2012) e que, enquanto o tratamento crônico de cobaias asmáticos com L-NAME
leva ao aumento da resistência das vias aéreas, o tratamento com o 1400W, um
inibidor da iNOS, atenua esse parâmetro (PRADO et al., 2005). Além disso, foi visto
por Angeli et al. (2008) que o efeito contrátil do NO produzido pela iNOS envolve a
liberação de 8-iso-PGF2α a partir da peroxidação do AA da membrana pelo radical
livre peroxinitrito e que o bloqueio da iNOS reduz os níveis desse isoprostano em
animais sensibilizados com ovalbumina (STARLING et al., 2009).
Interessantemente, nos animais do GASM/OC4, o L-NAME não alterou a curva
de contração do CCh (Figura 27C, Gráfico 15C, Tabela 10), de modo semelhante ao
observado no GC, indicando uma reversibilidade na sinalização do NO promovida pelo
OCV. Uma vez que o óleo também apresentou efeito benéfico sobre o estresse
oxidativo e a síntese e liberação dos prostanoides, é possível sugerir um efeito em
cascata promovido pela suplementação com o OCV: redução na formação de ROS e
de NO, via iNOS, com consequente diminuição da peroxidação lipídica e menor
formação de isoprostanos. Diante desses dados e as suposições levantadas a partir
deles, faz-se necessário investigar seus possiveis efeitos diretos sobre a expressão
de proteínas chave envolvidas nesse mecanismo, isto é, a COX, a iNOS e as enzimas
do sistema antioxidante pulmonar.
Ainda com relação à interface ROS-NO-isoprostanos, é relatado que esses
mediadores lipídicos induzem contração muscular lisa das vias aéreas agindo em
receptores tirosina cinase, bem como sobre a via de sinaliação RhoA/ROCK,
culminando com a diminuição da atividade da MLCP. Desse modo, a resposta final
está associada com um aumento no nível de fosforilação da cadeia leve da miosina e
hipercontratilidade das vias aéreas (JANSSEN, 2001).
A via RhoA/ROCK é apontada como uma das responsáveis por contribuir com
a hiper-contratilidade das vias aéreas na asma (CHIBA; MATSUSUE; MISAWA,
2010). Estudos mostram que a reatividade muscular brônquica aumentada a
agonistas muscarínicos, tanto em modelos animais como em humanos, se dá sem
alteração na expressão de seus receptores (LEE et al., 1994; CHIBA; MISAWA, 1995;
CHIBA et al., 2000) ou nos níveis de Ca2+ intracelulares (CHIBA et al., 1999a), mas
por uma maior sensibilização ao Ca2+ mediada pela RhoA e aumento da expressão
dessa proteína, tanto em modelo animal de asma em ratos como em camundongos
(CHIBA et al., 1999b; CHIBA; MISAWA, 2004). Nesse sentido, Kudo et al. (2012)

212 VASCONCELOS, L. H. C. Discussão
demonstraram que a hipercontratilidade muscular das vias aéreas induzida por IL-17A
está associada à sinalização intracelular mediada pela via NF-κB/RhoA/ROCK2.
Diante dessa premissa, decidiu-se investigar a participação da ROCK na
hiper-contratilidade observada na traqueia dos animais com inflamação pulmonar ao
CCh. Nos animais sem inflamação pulmonar, observou-se que a curva de contração
do CCh foi deslocada para a direita, sem redução da eficácia contrátil (Figura 28A-C,
Gráfico 16A, Tabela 11), indicando que o inibidor de ROCK, o Y-27632, reduziu a
potência contrátil do agonista. A curva do CCh no GASM também foi deslocada para
a direita, porém, esse descolamento só ocorreu em uma concentração 10 vezes maior
do inibidor da ROCK. Além disso, a eficácia contrátil do CCh foi aumentada pelo
inibidor, indicando um provável efeito compensatório mediado em resposta à inibição
da ROCK (Figura 28D-F, Gráfico 16B, Tabela 11). Além disso, comparando-se os
valores de pA2 do Y-27632, observa-se que a concentração do inibidor necessária
para reduzir pela metade a potência do CCh foi quase 10 vezes maior nos animais
com inflamação pulmonar. Esses dados suportam um provável aumento na expressão
das proteínas da via RhoA/ROCK induzido pela inflamação alérgica.
Analisando outros modelos animais de inflamação pulmonar alérgica, é
possível associar uma forte relação entre a hiper-responsividade muscular lisa das
vias aéreas com a super expressão da RhoA. Chiba et al. (1999b) evidenciaram níveis
aumentados de RhoA no músculo liso brônquico de ratos hiper-responsivos e Chiba
et al. (2003) observou níveis aumentados de RNAm dessa proteína nesse modelo
animal de asma brônquica, bem como em camundongos (CHIBA et al., 2005;
2009a,b,c). Em cobaias, também é conhecido o papel da via da ROCK na
contratilidade brônquica, conforme observado na hiper-responsividade das vias
aéreas utilizando esse modelo animal (HASHIMOTO et al., 2002; SCHAAFSMA et al.,
2004).
Com relação ao modelo de inflamação pulmonar desse trabalho, também foi
evidenciada a participação da via RhoA/ROCK na hiper-responsividade das vias
aéreas. Foi demonstrado recentemente que o tratamento de cobaias asmáticas com
o Y-27632 atenuava a sinalização mediada pela ROCK, melhorando a mecânica
pulmonar, o remodelamento e o estresse oxidativo (POSSA et al., 2012; PIGATI et al.,
2015). Além disso, foi observado que a inibição dessa via leva a uma redução da
expressão dos marcadores do estresse oxidativo iNOS e 8-iso-PGF2α (PIGATI et al.,
2015), sugerindo assim um crosstalk entre essas vias de sinalização.

213 VASCONCELOS, L. H. C. Discussão
Tendo em vista que a hiper-contratilidade da traqueia de cobaias com
inflamação pulmonar é dependente da liberação de mediadores contráteis e
inflamatórios pelo epitélio, a participação da ROCK nesse efeito é respaldada por
dados que mostram que, em animais asmáticos, essa via é estimulada por citocinas
liberadas pelo epitélio traqueobrônquico, como a IL-13 e o TNF-α, agindo em nível
transcricional, elevando os níveis do mRNA e da proteína no citosol tanto das células
inflamatórias residentes das vias aéreas como diretamente nas células musculares
lisas (CHIBA et al., 2009a, b; GOTO et al., 2010a, b), por meio da ativação das vias
do NF-kB (GOTO; CHIBA; MISAWA, 2009; GOTO et al., 2010a) e da STAT6 (CHIBA
et al., 2009a,b; GOTO et al., 2010a).
Curiosamente, quando foi utilizado o inibidor de ROCK nos animais do
GASM/OC4, a curva cumulativa do CCh também foi deslocada para a direita na
mesma faixa de concentrações do inibidor observado no grupo GC (Figura 28G-I,
Gráfico 16C, Tabela 11), sendo o pA2 do Y-27632 cerca de 22 vezes maior que o do
GASM, indicando não apenas uma provável reversão na expressão das proteínas
RhoA/ROCK – ou na sua atividade – aos níveis basais, mas para abaixo do basal
promovida pelo OCV. Esse efeito pode estar relacionado à ação do óleo sobre os
mediadores que ativam essa via de sinalização, especialmente aqueles provenientes
das células inflamatórias ou do epitélio das vias aéreas. Essa hipótese é suportada
por dados de Wallace et al. (2001) que demonstraram que camundongos C57BL/6
alimentados com dieta rica em óleo de coco apresentavam uma alteração no perfil de
citocinas produzidas por linfócitos T tímicos. Nesse estudo foi observado que o óleo
de coco induzia uma maior liberação de IL-2 e IFN-γ, além de aumentar a relação
IFN-γ/IL-4, direcionando assim para uma predominância do perfil Th1 de resposta
imune, o qual é reconhecido por inibir o perfil Th2, característico de doenças alérgicas,
como a asma.
Diante dos dados descritos até aqui, fica claro que o modelo de inflamação
pulmonar alérgica crônica em cobaias promoveu alterações na função pulmonar, com
resultante hiper-contratilidade muscular das vias aéreas, e que essas alterações foram
revertidas pelo OCV na maior dose utilizada. Tais modificações estão associadas com
a sinalização mediada pelo cruzamento das vias das ROS, prostanoides/isoprostanos,
NO e ROCK.
Como foi demonstrado, a reatividade relaxante da traqueia de cobaias com
inflamação pulmonar foi prejudicada (Figura 23A e C, Gráfico 10, Tabela 6), e esse

214 VASCONCELOS, L. H. C. Discussão
efeito foi prevenido pelo OCV (Figura 23H, Gráfico 11, Tabela 6). Nesse sentido,
decidiu-se avaliar os mecanismos envolvidos nesse efeito. Foi observado que a
redução na responsividade à isoprenalina se deu apenas nas preparações
pré-contraídas; e é relatado que a estimulação colinérgica promove a liberação de
TGF-β, que induz o remodelamento das vias aéreas (GRAINGE et al., 2011; OENEMA
et al., 2013). Além disso, na asma os receptores adrenérgicos β2 sofrem
dessensibilização devido à ação do TGF-β (ISHIKAWA et al., 2003), de modo que
esse processo está intimamente relacionado com a menor responsividade relaxante
devido à inativação da via Gs/BKCa, importante determinante do relaxamento muscular
liso das vias aéreas, tanto pela dessensibilização β adrenérgica, via TGF-β ou
receptores M3-PKC, como pela modulação negativa desses canais pela via
desencadeada pela estimulação colinérgica- Gβγi/o-BKCa (KUME, 2015).
Dessa forma, investigou-se um provável crosstalk entre TGF-β-receptores
β-BKCa. Nos animais sem inflamação pulmonar, observou-se que a curva de
relaxamento da isoprenalina não foi alterada pelo SB431542, antagonista dos
receptores do tipo I de TGF-β (Figura 29A, Gráfico 17A, Tabela 15), mas houve
redução da potência relaxante na presença da iberiotoxina, bloqueador dos BKCa
(Figura 29B, Gráfico 17A, Tabela 12), como esperado, uma vez que, nas vias aéreas
de não asmáticos, o relaxamento muscular liso via receptores β é mediado também
via abertura dos BKCa (KUME, 2013). Todavia, houve um aumento da eficácia
relaxante da isoprenalina na presença do bloqueador, efeito esse atribuído a um
provável mecanismo de compensação em resposta ao bloqueio desses canais. Além
disso, uma vez que nas vias aéreas não asmáticas a liberação de TGF-β é limitada e
não possui papel importante na regulação da expressão dos receptores β, conforme
o resultado obtido, espera-se que o antagonista não promova efeito na ausência do
agonista.
Diferentemente, nos cobaias com inflamação pulmonar, o relaxamento induzido
pela isoprenalina foi potencializado cerca de 7 vezes pelo antagonista dos receptores
de TGF-β do tipo I (Figura 29C, Gráfico 17B, Tabela 12), indicando que, de fato, a
sinalização mediada via TGF-β-RI leva à dessensibilização dos receptores β. Tal
resultado era esperado, uma vez que, no presente modelo de inflamação pulmonar,
foi demonstrado uma maior liberação de TGF-β nos pulmões (POSSA et al., 2012).
Adicionalmente, observou-se que o bloqueador dos BKCa não foi capaz de alterar o
relaxamento (Figura 29D, Gráfico 17B, Tabela 12), indicando que essa inabilidade se

215 VASCONCELOS, L. H. C. Discussão
dê pelo fato desses canais já se encontrarem pouco ativados por ocasião da
inflamação pulmonar, contribuindo para a hipo-responsividade relaxante observada.
Já nos animais com inflamação pulmonar suplementados com OCV, foi
observado que o relaxamento não foi alterado pelo antagonista dos TGF-β-RI (Figura
29E, Gráfico 17C, Tabela 12), semelhante ao observado nos animais sem inflamação
pulmonar, indicando uma possível redução na produção de TGF-β induzida pela
inflamação. No entanto, a iberiotoxina também não alterou a curva de relaxamento da
isoprenalina (Figura 29F, Gráfico 17C, Tabela 12), postulando a hipótese de que os
BKCa permaneceriam inativados – ou em nível transcricional, com sua expressão
reduzida - mesmo com a suplementação com o OCV.
Sabendo-se que, além da via de sinalização mediada pelos receptores β, o
tônus muscular liso aéreo é fortemente influenciado tanto pelos prostanoides
contráteis e relaxantes como pelos Cys-LTs (BACK et al., 2000) e que, como descrito,
os produtos do metabolismo do AA pela COX e pela 5-LO estão envolvidos na
hiper-reatividade contrátil traqueal, avaliou-se se esses metabólitos estariam
influenciando a responsividade relaxante da traqueia de cobaia.
Comparado aos animais do grupo GC, no GASM o relaxamento induzido pelo
AA foi prejudicado, sendo a potência reduzida, semelhante ao observado para a
isoprenalina, e essa redução da potência foi prevenida pela suplementação com o
OCV (Figura 30A, D, G, Gráfico 18A, Tabela 13). Esse efeito pode ter ocorrido por
uma menor formação de prostanoides relaxantes ou por uma maior formação de
Cys-LTs. Essas hipóteses foram avaliadas e observou-se que o relaxamento induzido
pelo AA nos animais sem inflamação pulmonar não foi alterado pelo inibidor da COX
ou da 5-LO (Figura 30B-C, Gráfico 18B, Tabela 13), mas sua potência e eficácia
relaxante foram reduzidas pela indometacina nos animais com inflamação pulmonar
(Figura 30E-F, Gráfico 18C, Tabela 13) indicando que o processo inflamatório alérgico
possivelmente altera o equilíbrio existente entre a produção de prostanoides
relaxantes e contráteis e Cys-LTs, favorecendo uma maior formação dos produtos
pró-contráteis da via da 5-LO. De fato, na asma crônica, os brônquios humanos
possuem um tônus muscular liso aumentado devido à liberação constitutiva de
CysLTs (BACK et al., 2000; CARAMORI, 2003). Esses dados se assemelham ao
observado para a curva de contração ao CCh, que sugeriu uma maior influência dos
Cys-LTs na contratilidade aumentada pela inflamação pulmonar.

216 VASCONCELOS, L. H. C. Discussão
Interessantemente, nos animais suplementados com OCV, a curva de
relaxamente foi alterada de maneira oposta: inibindo a COX, a potência relaxante foi
reduzida, enquanto que inibindo a 5-LO, a potência relaxante foi aumentada (Figura
30H-I, Gráfico 18D, Tabela 13). Assim como observado para a curva de contração ao
CCh, esses dados sugerem que o OCV altera o metabolismo do AA, seja favorecendo
a formação de prostanoides relaxantes ou a diminuição dos Cys-LTs. Todavia,
deve-se analisar com cautela esses dados, pois, apesar de aparentemente benéfico,
não se sabe como esse desbalanço, aparentemente a favor do relaxamento muscular
liso, pode interferir com a homeostase respiratória.
Em síntese, foi observado que, assim como a inflamação pulmonar aumentou
a contratilidade da traqueia de cobaia, também reduziu a reatividade relaxante desse
órgão, e que o OCV preveniu essa alteração, cujos mecanismos estão relacionados a
dois caminhos distintos: liberação de TGF-β, com consequente
inativação/dessensibilização de receptores β e BKCa; e redução da liberação de
prostanoides relaxante/aumento da liberação de Cys-LTs.
O aumento na produção de ROS aumenta a peroxidação lipídica, bem como o
dano às proteínas, agravando a inflamação das vias aéreas por múltiplos
mecanismos, incluindo a liberação de mediadores inflamatórios e efeitos sobre a
produção de muco e sobre o músculo liso (HOLGUIN, 2013). Por outro lado, os
mecanismos de defesa antioxidantes prejudicados contribuem para o dano oxidativo
na asma (GALLI et al., 2012). Dentre os marcadores do estresse oxidativo, estão
aqueles específicos dos pulmões (LOUHELAINEN et al., 2008; KOUTSOKERA et al.,
2013) e os sistêmicos (SHOKI et al., 2013; AGUSTI; SIN, 2014). Nesse contexto, é
importante se avaliar esses marcadores tanto localmente como sistemicamente uma
vez que, enquanto níveis aumentados desses marcadores no plasma indicam um
dano oxidativo sistêmico, o qual pode ou não ter sido originado do sistema respiratório,
a medida desses marcadores em amostras pulmonares reflete mais fielmente o
estresse oxidativo que ocorre no sistema respiratório (ANTUS, 2016).
Sabendo-se que as ROS podem estar relacionadas, como discutido
anteriormente, com a via da COX e do NO e, dessa forma, diante do provável papel
do estresse oxidativo como gatilho das alterações desencadeadas pela inflamação
pulmonar, resolveu-se avaliar o balanço estresse oxidativo/defesa antioxidante no
tecido pulmonar e no plasma dos cobaias, a fim de corroborar os dados funcionais
observados.

217 VASCONCELOS, L. H. C. Discussão
A inflamação pulmonar não promoveu estresse oxidativo sistêmico.
Semelhantemente, o tratamento com dexametasona ou a suplementação com OCV
nas três doses não alterou esse parâmetro (Gráfico 19A). Do mesmo modo, a
inflamação não alterou os níveis de MDA pulmonares, assim como o tratamento com
dexametasona ou a suplementação com OCV (Gráfico 19B).
Esses dados diferem da literatura acerca do efeito de modelos animais de asma
sobre os níveis desse marcador do estresse oxidativo. Huang; Fang; Liou (2017) e
Nesi et al. (2017) observaram um aumento nos níveis de MDA em modelo de asma
alérgica crônica em camundongos. Pinkerton et al. (2016) mediram os níveis dos
marcadores 3-nitrotirosina e 8-isoprostano em modelo de asma alérgica resistente a
esteroides e observaram níveis aumentados desses compostos nos animais. Além
disso, o aumento nos níveis de MDA também foi relatado em outros trabalhos como
característica do dano oxidativo promovido pela asma, tanto em camundongo como
em rato (BAO et al., 2016; SHAKERI; SOUKHTANLOO; BOSKABADY, 2017;
CHEKCHAKI et al., 2017).
Uma explicação para o resultado obtido é com relação ao modelo de inflamação
pulmonar e, principalmente, o tempo desde o último desafio antigênico até a obtenção
da amostra. Yalcin et al. (2012) observaram que pacientes asmáticos não apresentam
níveis aumentados de MDA em relação aos pacientes não asmáticos, e que os níveis
de MDA mostram-se elevados em períodos de exacerbação da asma, mas não em
períodos estáveis, além de sua capacidade antioxidante estar reduzida especialmente
durante esses períodos de crise aguda. Adicionalmente, em estudo clínico, Fatani
(2014) observou que, apesar dos níveis de MDA estarem elevados em pacientes
asmáticos, esse aumento se dava, especialmente, na fase aguda da resposta
asmática, decaindo progressivamente na sua fase tardia. Desse modo, é
preponderante afirmar que os níveis normais de MDA observados ocorram por uma
possível depuração tecidual e plasmática desse composto durante o intervalo (72 h)
até o isolamento da amostra e sua subsequente análise. Além disso, no caso do
plasma, pode-se inferir também a ausência de dano oxidativo sistêmico, o que é
menos provável para o tecido pulmonar.
O estresse oxidativo é definido como um desbalanço entre a formação de ROS
e os mecanismos de defesa antioxidante, de modo que tanto a elevada produção de
radicais livres como a atenuação da capacidade antioxidante podem induzir o estresse
oxidativo (ANTUS, 2016). Dentre as enzimas antioxidantes do sistema pulmonar, a

218 VASCONCELOS, L. H. C. Discussão
SOD, a catalase e a glutationa apresentam papel principal na defesa contra uma
grande variedade de radicais livres produzidos endogenamente nos pulmões ou
provenientes do meio, de modo que na asma essas defesas encontram-se
prejudicadas, contribuindo para o dano oxidativo na doença (DOZOR, 2010; NADEEM
et al., 2014).
Enquanto a capacidade antioxidante plasmática se manteve inalterada nos
animais com inflamação pulmonar, assim como o tratamento com a dexametasona ou
a suplementação com OCV não alterou esse parâmetro (Gráfico 19C), no tecido
pulmonar a capacidade antioxidante foi reduzida pela inflamação, e a suplementação
com o OCV nas doses de 2 e 4, mas não com a com a dose de 1 g/kg ou o tratamento
com dexametasona, não preveniu essa redução (Gráfico 19D).
A redução na capacidade antioxidante é corroborada por trabalhos que
mostram resultado semelhante com outros modelos animais de asma alérgica (BAO
et al., 2016; CHEKCHAKI et al., 2017; NESI et al., 2017). Todavia, apesar de
controverso o efeito observado para o grupo tratado com dexametasona, Pinkerton et
al. (2016) observou que o aumento no estresse oxidativo promovido pela asma não
era revertido pelo tratamento com a dexametasona.
Ademais, o efeito observado para o OCV está de acordo com dados da
literatura, que apontam o seu efeito antioxidante como um dos benefícios promovidos
pela sua suplementação (NEVIN; RAJAMOHAN, 2006; MARINA et al., 2009;
ABUJAZIA et al., 2012; ARUNIMA; RAJAMOHAN, 2013; VYSAKH et al., 2014; ALVES
et al., 2015).
Conforme foi demonstrado, a inflamação pulmonar promoveu espessamento
da camada de músculo liso, que pode ter ocorrido por processos de hipertrofia e/ou
hiperplasia. Esses processos de crescimento celular são dependentes de duas vias
de sinalização independentes que regulam o crescimento das células musculares lisas
das vias aéreas, a PI3K e as ERK 1/2 (AMMIT; PANETTIERI JR, 2001;
GERTHOFFER; SINGER, 2003; GOSENS et al., 2008).
Uma vez que a capacidade antioxidante foi prejudicada pela inflamação
pulmonar, sendo a SOD uma enzima-chave nesse processo, e que a PI3K, assim
como as MAPKs ERK1/2, ambos ativados por vias de sinalização mediadas pela
ativação de GPCRs ou de receptores tirosina cinase, estão envolvidas na proliferação
muscular lisa, investigou-se a expressão dessas proteínas nos pulmões de cobaia.

219 VASCONCELOS, L. H. C. Discussão
A expressão da PI3K foi aumentada nos pulmões de cobaias com inflamação
pulmonar (Figura 31, Gráfico 20A), conforme esperado, uma vez que é descrito que
essa proteína está relacionada com a patogênese da asma e está envolvido com o
processo de remodelamento tecidual e a hiperresponsividade brônquica (TAKEDA et
al., 2010), além de coordenar a ativação e a proliferação de linfócitos T na doença
(ZHANG et al., 2013). Por esse motivo, se intensifica a busca por drogas que inibam
essa proteína para o tratamento da asma (DUAN et al., 2005; DOUKAS et al., 2008;
PARK; MIN; LEE, 2008). Todavia, o tratamento com a dexametasona ou a
suplementação com OCV (Figura 31, Gráfico 20A) não preveniram esse aumento,
indicando que devem agir em outros alvos para atenuar o quadro asmático.
Já a expressão da SOD e das ERK 1/2 não foi alterada pela inflamação
pulmonar, descartando seus envolvimentos na patogênese da doença (Figura 31,
Gráfico 20B-C). O tratamento com a dexametasona ou a suplementação com OCV
(Figura 31, Gráfico 20B-C) não alterou a expressão das ERK 1/2 e da SOD, o que
pode ser explicado pelo fato de cobaias apresentarem resistência parcial aos
glicocorticoides devido à baixa afinidade de seus receptores (cerca de 20 vezes menor
que humanos) (KEIGHTLEY et al., 1998).
Como nos dados funcionais foi observado uma forte participação da ROCK na
hiper-contratilidade da traqueia de cobaia, e que essa proteína também apresenta
papel-chave na regulação da proliferação muscular lisa (CAMORETTI-MERCADO et
al., 2000; FUKATA et al., 2001; MACK et al., 2001; LIU et al., 2003), associado ao fato
de que a via RhoA/ROCK está envolvida na proliferação muscular lisa da aorta de rato
induzida por trombina (SEASHOLTZ et al., 1999; IWAMOTO et al., 2000), sugere-se
que essa enzima também esteja relacionada com o espessamento do músculo liso
brônquico. Porém, faz-se necessário medir a expressão dessa proteína para confirmar
essa hipótese.
Tomados em conjunto, demonstra-se pela primeira vez, com os dados obtidos
neste estudo, que o OCV desponta como uma droga promissora, à luz do seu
potencial papel como alimento funcional, na terapia adjuvante da inflamação pulmonar
alérgica crônica. No entanto, estudos adicionais, tanto em ensaios pré-clínicos como
de fase clínica, são indispensáveis para confirmar a reprodutibilidade dos dados e a
confiabilidade acerca de seus possíveis efeitos benéficos na asma alérgica.

7 Conclusões

221 VASCONCELOS, L. H. C. Conclusões
Na padronização e implantação de modelo de inflamação pulmonar alérgica
crônica em cobaias e a avaliação dos efeitos da sua suplementação com óleo de coco
virgem, pode-se concluir que:
a. a inflamação pulmonar alérgica crônica promove hiper-contratilidade
muscular lisa a agentes contráteis de acoplamento fármaco-mecânico por
mecanismos dependentes da participação do epitélio traqueal e pela
interface entre o estresse oxidativo, a produção de mediadores contráteis
derivados do metabolismo do ácido araquidônico, o óxido nítrico e a via da
RhoA/ROCK;
b. a hiporresponsividade relaxante da traqueia é mediada pelo TGF-β e pela
inibição dos BKCa;
c. a inflamação pulmonar reduz apenas a capacidade antioxidante pulmonar;
d. O OCV não apresenta efeito tóxico, até a dose de 4 g/kg, em cobaias;
e. o OCV, nas doses de 2 e 4 g/kg/dia, reverte a hiper-contratilidade muscular
lisa, in vitro, à estimulação com os agonistas CCh, histamina e antigênica
com ovalbumina, mas apenas a dose de 4 g/kg/dia reverte a responsividade
relaxante;
f. o efeito atenuante do OCV sobre a hiper-contratilidade muscular lisa ocorre
pela modulação negativa das vias ativadas pelo estresse oxidativo, os
prostanoides, o óxido nítrico e da RhoA/ROCK;
g. a suplementação com OCV melhora a capacidade antioxidante total
pulmonar de cobaias com inflamação pulmonar;
h. a inflamação pulmonar aumenta a expressão da PI3K, mas não das ERK
1/2 e da SOD; o tratamento com dexametasona ou a suplementação com
OCV não altera a expressão de nenhuma dessas proteínas.

Referências

223 VASCONCELOS, L. H. C. Referências
ABUJAZIA, M. A.; MUHAMMAD, N.; SHUID, A. N.; SOELAIMAN, I. N. The effects of virgin coconut oil on bone oxidative status in ovariectomised rat. Evidence-Based Complementary and Alternative Medicine, v. 2012, p. 1-6, 2012.
ADAM, E.; HANSEN, K. K.; ASTUDILLO, O. F.; COULON, L.; BEX, F.; DUHANT, X.; JAUMOTTE, E.; HOLLENBERG, M. D.; JACQUET, A. The house dust mite allergen Der p 1, unlike Der p 3, stimulates the expression of interleukin-8 in human airway epithelial cells via a proteinase-activated receptor-2–independent mechanism. Journal of Biological Chemistry, v. 281, p. 6910–6923, 2006.
ADAMKO, D. J.; YOST, B. L.; GLEICH, G. J.; FRYER, A. D.; JACOBY, D. B. Ovalbumin sensitization changes the inflammatory response to subsequent parainfluenza infection. Eosinophils mediate airway hyperresponsiveness, M2 muscarinic receptor dysfunction, and antiviral effects. Journal of Experimental Medicine, v. 190, n. 10, p. 1465-1478, 1999.
AGUSTI, A.; SIN, D. D. Biomarkers in COPD. Clinics in Chest Medicine, v. 35, n. 1, p. 131-141, 2014.
AHMED, F.; FOSTER, R. W.; SMALL, R. C. Some effects of nifedipine in guinea‐pig isolated trachealis. British Journal of Pharmacology, v. 84, n. 4, p. 861-869, 1985.
AIKAWA, T.; SHIMURA, S.; SASAKI, H.; EBINA, M.; TAKISHIMA, T. Marked goblet cell hyperplasia with mucus accumulation in the airways of patients who died of severe acute asthma attack. Chest, v. 101, n. 4, p. 916–921, 1992.
AKUTHOTA, P.; WANG, H. B.; SPENCER, L. A.; WELLER, P. F. Immunoregulatory roles of eosinophils: a new look at a familiar cell. Clinical & Experimental Allergy, v. 38, p. 1254-1263, 2008.
ALESSI, D. R.; JAMES, S. R.; DOWNES, C. P.; HOLMES, A. B.; GAFFNEY, P. R. J.; REESE, C. B.; COHEN, P. Characterization of a 3-phosphoinositide-dependent protein kinase which phosphorylates and activates protein kinase B alpha. Current Biology, v. 7, p. 261-269, 1997.
ALEXANDER, S. P.; BENSON, H. E.; FACCENDA, E.; PAWSON, A. J.; SHARMAN, J. L.; SPEDDING, M.; PETERS, J. A.; HARMAR, A. J. The Concise Guide to PHARMACOLOGY 2013/14: G protein‐coupled receptors. British journal of pharmacology, v. 170, n. 8, p. 1459-1581, 2013.
ALLAKHVERDI, Z.; COMEAU, M. R.; JESSUP, H. K.; YOON, B. R. P.; BREWER, A.; CHARTIER, S.; PAQUETTE, N.; ZIEGLER, S. F.; SARFATI, M.; DELESPESSE, G. Thymic stromal lymphopoietin is released by human epithelial cells in response to microbes, trauma, or inflammation and potently activates mast cells. Journal of Experimental Medicine, v. 204, p. 253-258, 2007.
ALVES, N. F. B.; PORPINO, S. K.; MONTEIRO, M. M.; GOMES, E. R.; BRAGA, V. A. Coconut oil supplementation and physical exercise improves baroreflex sensitivity and oxidative stress in hypertensive rats. Applied Physiology, Nutrition, and Metabolism, v. 40, n. 4, p. 393-400, 2015.

224 VASCONCELOS, L. H. C. Referências
ALVES, N. F. B.; QUEIROZ, T. M.; TRAVASSOS, R. A.; MAGNANI, M., BRAGA, V. A. Acute Treatment with Lauric Acid Reduces Blood Pressure and Oxidative Stress in Spontaneously Hypertensive Rats. Basic & Clinical Pharmacology & Toxicology, v. 120, n. 4, p. 348-353, 2017.
AMISHIMA, M.; MUNAKATA, M.; NASUHARA, Y.; SATO, A.; TAKAHASHI, T.; HOMMA, Y.; KAWAKAMI, Y. Expression of epidermal growth factor and epidermal growth factor receptor immunoreactivity in the asthmatic human airway. American Journal of Respiratory and Critical Care Medicine, v. 157, p. 1907-1912, 1998.
AMMIT, A. J.; PANETTIERI, R. A. Invited review: the circle of life: cell cycle regulation in airway smooth muscle. Journal of Applied Physiology, v. 91, p. 1431-1437, 2001.
ANGELI, P.; PRADO, C. M.; XISTO, D. G.; SILVA, P. L.; PÁSSARO, C. P.; NAKAZATO, H. D.; LEICK-MALDONADO, E. A.; MARTINS, M. A.; ROCVVCO, P. R.; TIBÉRIO, I. F. L. C. Effects of chronic L-NAME treatment lung tissue mechanics, eosinophilic and extracellular matrix responses induced by chronic pulmonary inflammation. The American Journal of Physiology-Lung Cellular and Molecular Physiology, v. 294, p. 1197–205, 2008.
ANGKASEKWINAI, P.; PARK, H.; WANG, Y. H.; WANG, Y. H.; CHANG, S. H.; CORRY, D. B.; LIU. Y. J.; ZHU, Z.; DONG, C. Interleukin 25 promotes the initiation of proallergic type 2 responses. Journal of Experimental Medicine, v. 204, n. 7, p. 1509-1517, 2007.
ANTICEVICH, S. Z.; HUGHES, J. M.; BLACK, J. L.; ARMOUR, C. L. Induction of human airway hyperresponsiveness by tumour necrosis factor-alpha. European Journal of Pharmacology,v. 284, p. 221-225, 1995.
ANTOSOVA, M.; STRAPKOVA, A. Dual effect of L-NAME on airway hyperreactivity. Acta Medica Martiniana, v. 8, p. 8-16, 2008.
ANTOSOVA, M.; STRAPKOVA, A.; MIKOLKA, P.; MOKRY´, J.; MEDVEDˇOVA´, I.; MOKRA, D. The Influence of L-NAME on iNOS Expression and Markers of Oxidative Stress in Allergen-Induced Airway Hyperreactivity. Advances in Experimental Medicine and Biology-Neuroscience and Respiration, v. 7, p. 1-10, 2015.
ANTUS, B. Oxidative stress markers in sputum. Oxidative Medicine and Cellular Longevity, v. 2016, p. 1-12, 2016.
ARM, J. Leukotriene generation and clinical implications. Allergy and Asthma Proceedings, v. 25, No. 1, p. 37-42, 2004.
ARMOUR, C. L.; DIMENT, L. M.; BLACK, J. L. Relationship between smooth muscle volume and contractile response in airway tissue. Isometric versus isotonic measurement. Journal of Pharmacology and Experimental Therapeutics, v. 245, n. 2, p. 687-691, 1988.
ARUNIMA, S.; RAJAMOHAN, T. Effect of virgin coconut oil enriched diet on the antioxidant status and paraoxonase 1 activity in ameliorating the oxidative stress in rats-a comparative study. Food & Function, v. 4, n. 9, p. 1402-1409, 2013.

225 VASCONCELOS, L. H. C. Referências
ARUNLAKSHANA, O. T.; SCHILD, H. O. Some quantitative uses of drug antagonists. British journal of pharmacology, v. 14, n. 1, p. 48-58, 1959.
ARVAN, P.; CASTLE, D. Sorting and storage during secretory granule biogenesis: looking backward and looking forward. Biochemical Journal, v. 332, n. 3, p. 593-610, 1998.
ASANO K, CHEE, C. B.; GASTON, B.; LILLY, C. M.; GERARD, C.; DRAZEN, J. M.; STAMLER, J. S. Constitutive and inducible nitric oxide synthase gene expression, regulation, and activity in human lung epithelial cells. Proceedings of the National Academy of Sciences, v. 91, p. 10089-10093, 1994.
ASSUNÇÃO, M. L.; FERREIRA, H. S.; SANTOS, A. F., CABRAL, C. R.; FLORÊNCIO, T. M. M. T. Effects of dietary coconut oil on the biochemical and anthropometric profiles of women presenting abdominal obesity. Lipids, v. 44, n. 7, p. 593-601, 2009.
ATANASOV, A.G.; WALTENBERGER, B.; PFERSCHY-WENZIG, E.M.; LINDER, T.; WAWROSCH, C.; UHRINE, P.; TEMMLF, V.; WANG, L.; SCHWAIGER, S.; HEISSA, E. H.; ROLLINGERA, J. M.; SCHUSTERF, D.; BREUSSE, J. M.; BOCHKOVG, V.; MIHOVILOVICD, M. D.; KOPPA, B.; BAUERC R.; DIRSCHA, V. M.; STUPPNERB, H. Discovery and resupply of pharmacologically active plant-derived natural products: A review. Biotechnology advances, v. 33, n. 8, p. 1582-1614, 2015.
BABIOR, B. M. NADPH oxidase: an update. Blood, v. 93, n. 5, p. 1464-1476, 1999.
BABIOR, B. M. Oxygen-dependent microbial killing by phagocytes. New England Journal of Medicine, v. 298, n. 12, p. 659-668, 1978.
BACK, M.; NOREL, X.; WALCH, L.; GASCARD, J.; DE MONTPREVILLE, V.; DAHLEN, S.; BRINK, C. Antagonist resistant contractions of the porcine pulmonary artery by cysteinyl-leukotrienes. European Journal of Pharmacology, v. 401, n. 3, p. 381-388, 2000.
BAI, T. R. Abnormalities in airway smooth muscle in fatal asthma. American Review of Respiratory Disease, v. 143, p. 441-443, 1991.
BAO, H. R.; LIU, X. J.; LI, Y. L.; MEN, X.; ZENG, X. L. Sinomenine attenuates airway inflammation and remodeling in a mouse model of asthma. Molecular Medicine Reports, v. 13, n. 3, p. 2415-2422, 2016.
BARLOW, J. L.; PEEL, S.; FOX, J.; PANOVA, V.; HARDMAN, C. S.; CAMELO, A.; BUCKS, C.; WU, X.; KANE, C. M.; NEILL, D. R.; FLYNN, R. J.; SAYERS, I.; HALL, I. P.; MCKENZIE, A. N. J. IL-33 is more potent than IL-25 in provoking IL-13–producing nuocytes (type 2 innate lymphoid cells) and airway contraction. Journal of Allergy and Clinical Immunology, v. 132, n. 4, p. 933-941, 2013.
BARNES, P. J. Adrenergic and non-adrenergic, non-cholinergic control of airways. Respiration, v. 50, n. 2, p. 9-16, 1986.
BARNES, P. J. Modulation of neurotransmission in airways. Physiol Ver, v. 72, p. 699-729, 1992.

226 VASCONCELOS, L. H. C. Referências
BARNES, P. J. β-adrenergic receptors and their regulation. American Journal of Respiratory and Critical Care Medicine, v. 152, n. 3, p. 838-860, 1995.
BARNES; N. C.; PIPER, P. J.; COSTELLO, J. F. Comparative Effects of Inhaled Leukotriene C4, Leukotriene D4, and Histamine in Normal Human Subjects. Thorax, v. 39, n. 7, p. 500-504, 1984.
BARRETT, N. A.; AUSTEN, K. F. Innate cells and T helper 2 cell immunity in airway inflammation. Immunity, v. 31, p. 425-437, 2009.
BATEMAN, E. D.; KORNMANN, O.; SCHMIDT, P.; PIVOVAROVA, A.; ENGEL, M.; FABBRI, L. M. Tiotropium is noninferior to salmeterol in maintaining improved lung function in B16-Arg/Arg patients with asthma. Journal of Allergy and Clinical Immunology, v. 128, n. 2, p. 315-322, 2011.
BATRA, V. ; MUSANI, A. I.; HASTIE, A. T.; KHURANA, S.; CARPENTER, K. A.; ZANGRILLI, J. G.; PETERS, S. P. Bronchoalveolar lavage fluid concentrations of transforming growth factor (TGF)-β1, TGF-β2, interleukin (IL) 4 and IL-13 after segmental allergen challenge and their effects on α smoothmuscle actin and collagen III synthesis by primary human lung fibroblasts. Clinical and Experimental Allergy, v. 34, n. 3, p. 437–444, 2004.
BEL, E. H. Clinical phenotypes of asthma. Current Opinion in Pulmonary Medicine, v. 10, p. 44-50, 2004.
BELLEGUIC, C.; CORBEL, M.; GERMAIN, N. Increased release of matrix metalloproteinase-9 in the plasma of acute severe asthmatic patients. Clin Exp Allergy, v. 31, p. 217–223, 2002.
BEN-JEBRIA, A.; MARTHAN, R.; ROSSETTI, M.; SAVINEAU, J. P. Effect of passive sensitization on the mechanical activity of human isolated bronchial smooth muscle induced by substance P, neurokinin A and VIP. British Journal of Pharmacology, v. 109, n. 1, p. 131-136, 1993.
BENSON, R. C.; HARDY, K. A.; MORRIS, C. R. Arginase and arginine dysregulation in asthma. Journal of Allergy, v. 2011, 2011.
BERAIR, R.; BRIGHTLING, C.E. Asthma therapy and its effect on airway remodelling. Drugs, v. 74, n. 12, p. 1345-1369, 2014.
BERGER, W. E.; KERWIN, E.; BERNSTEIN, D. I.; PEDINOFF, A.; BENSCH, G.; KARAFILIDIS, J. Efficacy and safety evaluation of ciclesonide in subjects with mild-to-moderate asthma not currently using inhaled corticosteroids. Allergy and asthma proceedings. OceanSide Publications, Inc, p. 304-314, 2009.
BERGERON, C.; AL-RAMLI, W.; HAMID, Q. Remodeling in asthma. Proceedings of the American Thoracic Society, v. 6, n. 3, p. 301-305, 2009.
BERRIDGE, M. J. Cell Signalling Pathways. In: Cell Signalling Biology. Portland Press Limited , p. 1-118, 2009.

227 VASCONCELOS, L. H. C. Referências
BERRIDGE, M. J. Smooth muscle cell calcium activation mechanisms. The Journal of physiology, v. 586, n. 21, p. 5047-5061, 2008.
BEZARD, J.; BUGAUT, M.; CLEMENT, G. Triglyceride composition of coconut oil. Journal of the American Oil Chemists' Society, v. 48, n. 3, p. 134-139, 1971.
BISHOPP, A.; SATHYAMURTHY, R.; MANNEY, S.; WEBBSTER, C.; KRISHNA, M.; MANSUR A. H. Biomarkers of oxidative stress and antioxidants in severe asthma: A Prospective Case-Control Study. Annals of Allergy, Asthma & Immunology, v. 118, n. 4, p. 445-451, 2017.
BJORCK, T.; GUSTAFSSON, L. E.; DAHLEN, S. E. Isolated bronchi from asthmatics are hyperresponsive to adenosine, which apparently acts indirectly by liberation of leukotrienes and histamine. American Review of Respiratory Disease, v. 145, p. 1087-1091, 1992.
BLACK, J. L.; BURGESS, J. K.; JOHNSON, P. R. A. Airway smooth muscle - its relationship to the extracellular matrix. Respiratory Physiology and Neurobiology, v. 137, p. 339-346, 2003.
BLECK, B., TSE, D.B., GORDON, T., AHSAN, M.R.; REIBMAN, J. Diesel exhaust particle-treated human bronchial epithelial cells upregulate Jagged-1 and OX40 ligand in myeloid dendritic cells via thymic stromal lymphopoietin. J. Immunol., v. 185, p. 6636–6645, 2010.
BLOTT, E. J.; GRIFFITHS, G. M. Secretory lysosomes. Nature Reviews Molecular Cell Biology, v. 3, n. 2, p. 122-131, 2002.
BOESIGER, J.; TSAI, M.; MAURER, M.; YAMAGUCHI, M.; BROWN, L. F.; CLAFFEY, K. P.; DVORAK, H. F.; GALLI, S. J. Mast cells can secrete vascular permeability factor/ vascular endothelial cell growth factor and exhibit enhanced release after immunoglobulin E‐dependent upregulation of Fcε receptor I expression. Journal of Experimental Medicine, v. 188, n. 6, p. 1135-1145, 1998.
BOLTON, T. B. Mechanisms of action of transmitters and other substances on smooth muscle. Physiological Reviews, v. 59, n. 3, p. 606-718, 1979.
BOTERMAN, M.; SMITS, S. R.; MEURS, H.; ZAAGSMA, J. Protein kinase C potentiates homologous desensitization of the β2-adrenoceptor in bovine tracheal smooth muscle. European Journal of Pharmacology, v. 529, n. 1, p. 151-156, 2006.
BOULET, L. P. Influence of comorbid conditions on asthma. European Respiratory Journal, v. 33, n. 4, p. 897-906, 2009.
BOUSQUET, J.; WENZEL, S.; HOLGATE, S.; LUMRY, W.; FREEMAN, P.; FOX, H. Predicting response to omalizumab, an anti-IgE antibody, in patients with allergic asthma. Chest, v. 125, n. 4, p. 1378-1386, 2004.
BOWLER, R. P. Oxidative stress in the pathogenesis of asthma. Current Allergy and Asthma Reports, v. 4, n. 2, p. 116-122, 2004.

228 VASCONCELOS, L. H. C. Referências
BOXALL, C.; HOLGATE, S. T.; DAVIES, D. E. The contribution of transforming growth factor-β and epidermal growth factor signalling to airway remodelling in chronic asthma. European Respiratory Journal, v. 27, n. 1, p. 208-229, 2006.
BRADDING, P.; FEATHER, I. H.; WILSON, S.; BARDIN, P. G.; HEUSSER, C. H.; HOLGATE, S. T.; HOWARTH, P. H. Immunolocalization of cytokines in the nasal
mucosa of normal and perennial rhinitic subjects. The mast cell as a source of IL‐4, IL‐5, and IL‐6 in human allergic mucosal inflammation. The Journal of Immunology, v. 151, n. 7, p. 3853-3865, 1993.
BRAMLEY, A. M.; THOMSON, R. J.; ROBERTS, C. R.; SCHELLENBERG, R. R. Hypothesis: excessive bronchoconstriction in asthma is due to decreased airway elastance. The European Respiratory Journal, v. 7, n. 2, p. 337-341, 1994.
BRAND-WILLIAMS, W.; CUVELIER, M. E.; BERSET, C. L. W. T. Use of a free radical method to evaluate antioxidant activity. LWT-Food Science and Technology, v. 28, n. 1, p. 25-30, 1995.
BRANTON, M. H.; KOPP, J. B. TGF-β and fibrosis. Microbes and Infection, v. 1, n. 15, p. 1349–1365, 1999.
BRAR, S. S.; KENNEDY, T. P.; STURROCK, A. B.; HUECKSTEADT, T. P.; QUINN, M. T.; MURPHY, T. M.; CHITANO, M.; HOIDAL, J. R. NADPH oxidase promotes NF-κB activation and proliferation in human airway smooth muscle. American Journal of Physiology-Lung Cellular and Molecular Physiology, v. 282, n. 4, p. L782-L795, 2002.
BRASIL. Conselho Federal de Medicina Veterinária. Resolução nº 1000, de 11 de maio de 2012. Dispõe sobre procedimentos e métodos de eutanásia em animais e dá outras providências. Brasília-DF, 2012.
BRASIL. Ministério da Ciência, Tecnologia e Inovação. Conselho Nacional de Controle de Experimentação Animal. Guia Brasileiro de Produção, Manutenção ou Utilização de Animais em Atividades de Ensino ou Pesquisa Científica: fascículo 1: introdução geral. Brasília-DF, 2016.
BRASIL. Ministério da Saúde. Agência Nacional de Vigilância Sanitária. 2016. Nutrientes e não nutrientes com as alegações padronizadas e os respectivos requisitos específicos. In: Alimentos com Alegações de Propriedades Funcionais e ou de Saúde. Disponível em: <http://portal.anvisa.gov.br/alimentos/alegacoes> Acesso em: 21 julho 2017.
BRASIL. Ministério da Saúde. Agência Nacional de Vigilância Sanitária. 2017b. Aprovado medicamento inédito para asma. Nucala (mepolizumabe) será indicado para o tratamento de asma grave em adultos. Disponível em: <http://portal.anvisa.gov.br/noticias/> Acesso em: 22 setembro 2017.
BRASIL. Ministério da Saúde. Agência Nacional de Vigilância Sanitária. Suplementos alimentares: Documento de base para discussão regulatória. Brasília-DF, 2017a.

229 VASCONCELOS, L. H. C. Referências
BRASIL. Ministério da Saúde. Agência Nacional de Vigilância Sanitária. Resolução nº 16, de 30 de abril de 1999. Aprova o Regulamento Técnico de Procedimentos para Registro de Alimentos e ou Novos Ingredientes. Brasília-DF, 1999a.
BRASIL. Ministério da Saúde. Agência Nacional de Vigilância Sanitária. Resolução nº 17, de 30 de abril de 1999. Aprova o Regulamento Técnico que Estabelece as Diretrizes Básicas para Avaliação de Risco e Segurança dos Alimentos. Brasília-DF, 1999b.
BRASIL. Ministério da Saúde. Agência Nacional de Vigilância Sanitária. Resolução nº 18, de 30 de abril de 1999. Aprova o Regulamento Técnico que estabelece as diretrizes básicas para análise e comprovação de propriedades funcionais e ou de saúde alegadas em rotulagem de alimentos, constante do anexo desta portaria. Diário Oficial da União, Brasília, DF, 03 mai. 1999c.
BRASIL. Ministério Da Saúde. Agência Nacional de Vigilância Sanitária. Resolução nº 19, de 30 de abril de 1999. Aprova o Regulamento Técnico de procedimentos para registro de alimento com alegação de propriedades funcionais e ou de saúde em sua rotulagem. Brasília-DF, 1999d.
BRASIL. Ministério da Saúde. Agência Nacional de Vigilância Sanitária. Resolução nº 02, de 07 de janeiro de 2002. Aprova o Regulamento Técnico de substâncias bioativas e probióticos isolados com alegação de propriedade funcional e/ou de saúde. Brasília-DF, 2002.
BREYER, M. D.; BREYER, R. M. G protein-coupled prostanoid receptors and the kidney. Annu Rev Physiol., v. 63, p. 579–605, 2001.
BREYER, R. M.; BAGDASSARIAN, C. K.; MYERS, S. A.; BREYER, M. D. Prostanoid receptors: subtypes and signaling. Annual Review of Pharmacology and Toxicology, v. 41, n. 1, p. 661-690, 2001.
BRUCE, F. The Coconut Oil Miracle, Avery, USA, 4th edn, 1-7, 2004.
BUCKLEY, J.; BIRRELL, M. A.; MAHER, S. A.; NIALS, A. T.; CLARKE, D. L.; BELVISI, M. G. EP4 receptor as a new target for bronchodilator therapy. Thorax, v. 66, p.1029-1035, 2011.
BURGERING, B. M. T.; COFFER, P. J. Protein-Kinase-B (C-Akt) in Phosphatidylinositol 3-OH kinase Signal-Transduction. Nature, v. 376, p. 599-602, 1995.
BURNSTOCK, G. Purinergic signalling: pathophysiology and therapeutic potential. Keio J Med, v. 62, p. 63-73, 2013.
BUSSE, W. W.; LEMANSKE, R. F. Asthma. The New England Journal of Medicine, v. 344, p. 350-362, 2001.
CAMORETTI-MERCADO, B.; LIU, H. W.; HALAYKO, A. J.; FORSYTHE, S. M.; KYLE, J. W.; LI, B.; FU, Y.; MCCONVILLE, J.; KOGUT, P.; VIEIRA, J. E.; PATEL, N. M.; HERSHENSON, M. B.; FUCHS, E.; SINHA, S.; MIANO, J. M.; PARMACEK, M. S.; BURKHARDT, J. K.; SOLWAY, J. Physiological control of smooth muscle-specific

230 VASCONCELOS, L. H. C. Referências
gene expression through regulated nuclear translocation of serum response factor. The Journal of Biological Chemistry, v. 275, p. 30387-30393, 2000.
CANNING, B. J. Modeling asthma and COPD in animals: a pointless exercise? Current Opinion in Pharmacology, v. 3, p. 244–250, 2003.
CANNING, B. J.; CHOU, Y. Using guinea pigs in studies relevant to asthma and COPD. Pulmonary Pharmacology and Therapeutics, v. 21, n. 5, p. 702-720, 2008.
CARAMORI, G.; ADCOCK, I. Pharmacology of airway inflammation in asthma and COPD. Pulmonary pharmacology & therapeutics, v. 16, n. 5, p. 247-277, 2003.
CARD, J. W.; CAREY, M. A.; BRADBURY, J. A.; GRAVES, J. P.; LIH, F. B.; MOORMAN, M. P.; MORGAN, D. L.; DEGRAFF, L. M.; ZHAO, Y.; FOLEY, J. F.; ZELDIN, D. C. Cyclooxygenase-1 overexpression decreases Basal airway responsiveness but not allergic inflammation. The Journal of Immunology, v. 177, n. 7, p. 4785-4793, 2006.
CARDOSO, T. A.; RONCADA, C.; SILVA, E. R.; PINTO, L. A.; JONES, M. H.; STEIN, R. T.; PITREZ, P. M. Impacto da asma no Brasil: análise longitudinal de dados extraídos de um banco de dados governamental brasileiro. Jornal Brasileiro de Pneumologia, v. 43, n. 3, p. 163-168, 2017.
CAREY, M. A.; GERMOLEC, D. R.; BRADBURY, J. A.; GOOCVVH, R. A.; MOORMAN, M. P.; FLAKE, G. P.; LANGENBACH, R.; ZELDIN, D. C. Accentuated T helper type 2 airway response after allergen challenge in cyclooxygenase-1−/− but not cyclooxygenase-2−/− mice. American Journal of Respiratory and Critical Care Medicine, v. 167, n. 11, p. 1509-1515, 2003.
CARROLL, N.; ELLIOT, J.; MORTON, A.; JAMES, A. The structure of large and small airways in nonfatal and fatal asthma. American Review of Respiratory Disease, v. 147, n. 2, p. 405–410, 1993.
CATTERALL, W. A. Voltage-Gated Calcium Channels. Cold Spring Harbor perspectives in biology, v. 3, n. 8, p. a003947, 2011.
CE – COMUNIDADE EUROPEIA. Diretiva 2002/46/CE - Relativa à aproximação das legislações dos Estados-Membros respeitantes aos suplementos alimentares. Jornal Oficial das Comunidades Europeias, v. 183, 2002, p. 51-57.
CEFALU, W. T.; YE, J.; WANG, Z. Q. Efficacy of dietary supplementation with botanicals on carbohydrate metabolism in humans. Endocrine, Metabolic & Immune Disorders-Drug Targets (Formerly Current Drug Targets-Immune, Endocrine & Metabolic Disorders), v. 8, n. 2, p. 78-81, 2008.
CERRINA, J.; LABAT, C.; HAYE-LEGRANDE, I.; RAFFESTIN, B.; BENVENISTE, J.; BRINK, C. H. Isolated bronchial muscle preparations from asthmatic patients: effects of indomethacin and contractile agonists. Prostaglandins, v. 37, p. 457-469, 1989.
CERRINA, J.; LADURIE, M. L. R.; LABAT, C.; RAFFESTIN, B.; BAYOL, A.; BRINK, C. Comparison of Human Bronchial Muscle Responses to Histamine in vivo with

231 VASCONCELOS, L. H. C. Referências
Histamine and Isoproterenol Agonists in vitro. American Review of Respiratory Disease, v. 134, n. 1, p. 57-61, 1986.
CHAMBERS, L. S.; BLACK, J. L.; GE, Q.; CARLIN, S. M.; AU, W. W.; PONIRIS, M.; THOMPSON, J.; JOHNSON, P. R.; BURGESS, J. K. PAR-2 Activation, PGE2, and COX-2 in Human Asthmatic and Nonasthmatic Airway Smooth Muscle Cells. American Journal of Physiology, v. 285, n. 3, p. L619-L627, 2003.
CHAND, N.; EYRE, P. The Schultz-Dale reaction. A review. Agents & Actions, v. 8, p. 171-184, 1978.
CHEKCHAKI, N.; KHALDI, T.; ROUIBAH, Z.; ROUAG, M.; SEKIOU, O.; MESSARAH, M.; BOUMENDJEL, A. Anti-inflammatory and antioxidant effects of two extracts from Pistacia lentiscus in liver and erythrocytes, in an experimental model of asthma. Methods, v. 18, p. 19, 2017.
CHERRY, W. B.; YOON, J.; BARTEMES, K. R.; IIJIMA, K.; KITA, H. A novel IL-1 family cytokine, IL-33, potently activates human eosinophils. Journal of Allergy and Clinical Immunology, v. 121, p. 1484-1490, 2008.
CHESNÉ, J.; BRAZA, F.; CHADEUF, G.; MAHAY, G.; CHEMINANT, M. A.; LOY, J.; BROUARD, S.; SAUZEAU, V.; LOIRAND, G.; MAGNAN, A. Prime role of IL-17A in neutrophilia and airway smooth muscle contraction in a house dust mite-induced allergic asthma model. Journal of Allergy and Clinical Immunology, v. 135, n. 6, p. 1643, 2015.
CHIBA, Y.; MATSUSUE, K.; MISAWA, M. RhoA, a possible target for treatment of airway hyperresponsiveness in bronchial asthma. Journal of Pharmacological Sciences, v. 114, n. 3, p. 239-247, 2010.
CHIBA, Y.; MISAWA, M. Characteristics of muscarinic cholinoceptors in airways of antigeninduced airway hyperresponsive rats. Comparative Biochemistry and Physiology, v. 111C, p. 351-357, 1995.
CHIBA, Y.; MISAWA, M. The role of RhoA-mediated Ca2+ sensitization of bronchial smooth muscle contraction in airway hyperresponsiveness. Journal of Smooth Muscle Research, v. 40, n. 4, 5, p. 155-167, 2004.
CHIBA, Y.; NAKAZAWA, S.; TODOROKI, M.; SHINOZAKI, K.; SAKAI, H.; MISAWA, M. Interleukin-13 augments bronchial smooth muscle contractility with an up-regulation of RhoA protein. American Journal of Respiratory Cell and Molecular Biology, v. 40, pp. 159-167, 2009a.
CHIBA, Y.; SAKAI, H.; MISAWA, M. Possible involvement of G(i3) protein in augmented contraction of bronchial smooth muscle from antigen-induced airway hyperresponsive rats. Biochemical Pharmacology, v. 61, n. 7, p. 921-924, 2001.
CHIBA, Y.; SAKAI, H.; SUENAGA, H.; KAMATA, K.; MISAWA, M. Enhanced Ca2+ sensitization of the bronchial smooth muscle contraction in antigen-induced airway hyperresponsive rats. Research Communications In Molecular Pathology And Pharmacology, v. 106, p. 77 85, 1999a.

232 VASCONCELOS, L. H. C. Referências
CHIBA, Y.; SAKAI, H.; WACHI, H.; SUGITANI, H.; SEYAMA, Y.; MISAWA, M. Upregulation of rhoA mRNA in bronchial smooth muscle of antigen-induced airway hyperresponsive rats. Journal of Smooth Muscle Research, v. 39, p. 221-228, 2003.
CHIBA, Y.; TAKADA, Y.; MIYAMOTO, S.; MITSUI-SAITO, M.; KARAKI, H.; MISAWA, M. Augmented acetylcholine-induced, Rho-mediated Ca2+ sensitization of bronchial smooth muscle contraction in antigen-induced airway hyperresponsive rats. British Journal of Pharmacology, v. 127, p. 597-600, 1999b.
CHIBA, Y.; TAKADA, Y.; SAKAI, H.; TAKEYAMA, H.; MISAWA, M. Acetylcholine-induced smooth muscle contraction of intrapulmonary small bronchi is augmented in antigen-induced airway hyperresponsive rats. The Japanese Journal of Pharmacology, v. 84, p. 221-224, 2000.
CHIBA, Y.; TANABE, M.; GOTO, K.; SAKAI, H.; MISAWA, M. Down-regulation of miR-133a contributes to up-regulation of RhoA in bronchial smooth muscle cells. American Journal of Respiratory Cell and Molecular Biology, v. 180, p. 713-719, 2009c.
CHIBA, Y.; TODOROKI, M.; NISHIDA, Y.; TANABE, M.; MISAWA, M. A novel STAT6 inhibitor AS1517499 ameliorates antigen-induced bronchial hypercontractility in mice. American Journal of Respiratory Cell and Molecular Biology, v. 41, p. 516-524, 2009b.
CHIBA, Y.; UENO, A.; SHINOZAKI, K.; TAKEYAMA, H.; NAKAZAWA, S.; SAKAI, H.; MISAWA, M. Involvement of RhoA-mediated Ca2+ sensitization in antigen-induced bronchial smooth muscle hyperresponsiveness in mice. Respiratory Research, v. 6, n. 1, p. 4, 2005.
CHONG-NETO, H. J.; ROSÁRIO, N. A.; SOLÉ, D.; LATIN AMERICAN ISAAC GROUP. Asthma and rhinitis in South America: how different they are from other parts of the world. Allergy, Asthma & Immunology Research, v. 4, n. 2, p. 62-67, 2012.
CHU, D. K.; JIMENEZ-SAIZ, R.; VERSCHOOR, C. P.; WALKER, T. D.; GONCHAROVA, S.; LLOP-GUEVARA, A.; SHEN, P.; GORDON, M. E.; BARRA, N. G.; BASSETT, J. D.; KONG, J.; FATTOUH, R.; MCCOY, K. D.; BOWDISH, D. M.; ERJEFÄLT, J. S.; PABST, O.; HUMBLES, A. A.; KOLBECK, R.; WASERMAN, S.; JORDANA, M. Indigenous enteric eosinophils control DCs to initiate a primary TH2 immune response in vivo. Journal of Experimental Medicine, v. 211, n. 8, p. 1657-1672, 2014.
CHUSTZ, R.T. Regulation and function of the IL-1 family cytokine IL-1F9 in human bronchial epithelial cells. American Journal of Respiratory Cell and Molecular Biology, v. 45, p. 145–153, 2011.
CIRINO, G.; FIORUCCI, S.; SESSA, W. C. Endothelial nitric oxide synthase: the Cinderella of inflammation? Trends in Pharmacological Sciences, v. 24, p. 91-95, 2003.
CKLESS, K.; HODGKINS, S. R.; ATHER, J. L.; MARTIN, R.; POYNTER, M. E. Epithelial, dendritic, and CD4+ T cell regulation of and by reactive oxygen and nitrogen species in allergic sensitization. Biochimica et Biophysica Acta (BBA)-General Subjects, v. 1810, n. 11, p. 1025-1034, 2011.

233 VASCONCELOS, L. H. C. Referências
CODEX ALIMENTARIUS COMMISSION. Guidelines for Vitamin and Mineral Food Supplements (CAC/GL 55 - 2005), Food and Agricultural Organization of the United Nations and the World Health Organization, 2005.
COHEN, L.; XUEPING, E.; TARSI, J.; RAMKUMAR, T.; HORIUCHI, T. K.; COCHRAN, R.; DEMARTINO, S.; SCHECHTMAN, K. B.; HUSSAIN, I.; HOLTZMAN, M. J.; CASTRO, M. Epithelial cell proliferation contributes to airway remodeling in severe asthma. American Journal of Respiratory and Critical Care Medicine, v. 176, n. 2, p. 138-145, 2007.
COLICE, G. L. New developments in inhaled corticosteroids. Allergy and asthma proceedings. OceanSide Publications, Inc, 2006. p. 332-340.
CONRADO, S. D. COCONUT OIL: Atherogenic or Not? (What therefore causes Atherosclerosis?). Philippines Journal of Cardiology, v. 31, n. 3, p. 97-104, 2003.
COOK, S.; VOLLENWEIDER, P.; MENARD, B.; EGLI, M.; NICOD, P.; SCHERRER, U. Increased eNO and pulmonary iNOS expression in eNOS null mice. European Respiratory Journal, v. 21, p. 770-773, 2003.
CORTELING, R. L.; LI, S.; GIDDINGS, J.; WESTWICK, J.; POLL, C.; HALL, I. P. Expression of transient receptor potential C6 and related transient receptor potential family members in human airway smooth muscle and lung tissue. American Journal of Respiratory Cell and Molecular Biology, v. 30, n. 2, p. 145-154, 2004.
COSTELLO, R. W.; SCHOFIELD, B. H.; KEPHART, G. M.; GLEICH, G. J.; JACOBY, D. B.; FRYER, A. D. Localization of eosinophils to airway nerves and effect on neuronal M2 muscarinic receptor function. American Journal of Physiology-Lung Cellular and Molecular Physiology, v. 273, n. 1, p. L93-L103, 1997.
COYLE, A. J.; ACKERMAN, S. J.; BURCH, R.; PROUD, D.; IRVIN, C. G. Human eosinophil-granule major basic protein and synthetic polycations induce airway hyperresponsiveness in vivo dependent on bradykinin generation. Journal of Clinical Investigation, v. 95, n. 4, p. 1735, 1995.
COYLE, A. J.; PERRETTI, F.; MANZINI, S.; IRVIN, C. G. Cationic protein-induced sensory nerve activation: role of substance P in airway hyperresponsiveness and plasma protein extravasation. Journal of Clinical Investigation, v. 94, n. 6, p. 2301, 1994.
CRAGG, G.M.; NEWMAN, D.J, Natural products: A continuing source of novel drug leads. Biochimica et Biophysica Acta, v. 1830, p. 3670-3695, 2013.
CRUZ, A. A.; FERNANDES, A. L. G.; PIZZICHINI, E.; FITERMAN, J.; PEREIRA, L. F. F.; PIZZICHINI, M. M. M.; JONES, M.; LIMA, M. A.; CAMARGOS, P. A. M.; STIRBULOV, R. Diretrizes da Sociedade Brasileira de Pneumologia e Tisiologia. Jornal Brasileiro de Pneumologia, v. 38, n. Suplemento 1, 2012.
CURRIE, G. P.; LIPWORTH, B. J. Bronchoprotective effects of leukotriene receptor antagonists in asthma: a meta-analysis. Chest, v. 122, n. 1, p. 146-150, 2002.

234 VASCONCELOS, L. H. C. Referências
DAHLÉN, B.; NIZANKOWSKA, E.; SZCZEKLIK, A.; ZETTERSTRÖM, O.; BOCVVHENEK, G.; KUMLIN, M.; MASTALERZ, L.; PINIS, G.; SWANSON, L. J.; BOODHOO, T. I.; WRIGHT, S.; DUBÉ, L. M.; DAHLÉN, S. E. Benefits from adding the 5-lipoxygenase inhibitor zileuton to conventional therapy in aspirin-intolerant asthmatics. American Journal of Respiratory and Critical Care Medicine, v. 157, n. 4, p. 1187-1194, 1998.
DAHLÉN, S. E.; MALMSTRÖM, K.; NIZANKOWSKA, E.; DAHLÉN, B.; KUNA, P.; KOWALSKI, M.; LUMRY, W. R.; PICADO, C.; STEVENSON, D. D.; BOUSQUET, J.; PAUWELS, R.; HOLGATE, S. T.; SHAHANE, A.; ZHANG, J.; REISS, T. F.; SZCZEKLIK, A. Improvement of aspirin-intolerant asthma by montelukast, a leukotriene antagonist: a randomized, double-blind, placebo-controlled trial. American Journal of Respiratory and Critical Care Medicine, v. 165, n. 1, p. 9-14, 2002.
DALE, H. H. The anaphylactic reaction of plain muscle in the guinea pig. Journal of Pharmacology and Experimental Therapeutics, v. 4, p. 167-223, 1913.
DANEK, C. J.; LOMBARD, C. M.; DUNGWORTH, D. L.; COX, P. G.; MILLER, J. D.; BIGGS, M. J.; KEAST, T. M.; LOOMAS, B. E.; WIZEMAN, W. J.; HOGG, J. C.; LEFF, A. R. Reduction in airway hyperresponsiveness to methacholine by the application of RF energy in dogs. Journal of Applied Physiology, v. 97, p, 1946-1953, 2004.
DANIEL, E. E.; KANNAN, M.; DAVIS, C.; POSEY-DANIEL, V. Ultrastructural studies on the neuromuscular control of human tracheal and bronchial muscle. Respiratory Physiology, v. 63, p. 109-128, 1986.
DANILA, C. I.; HAMILTON, S. L. Phosphorylation of ryanodine receptors. Biological Research, v. 37, n. 4, p. 521-525, 2004.
DAVIDSON, A. B.; LEE, T. H.; SCANLO, SOLWAY, J.; MCFADDEN, E. R. J.; INGRAM, R. H. J.; COREY, E. J.; AUSTEN, K. F.; DRAZEN, J. M. Bronchoconstrictor Effects of Leukotriene E4 in Normal and Asthmatic Subjects 1-4. American Review of Respiratory Disease, v. 135, n. 2, p. 333-337, 1987.
DAVIES, D. E. The role of the epithelium in airway remodeling in asthma. Proceedings of the American Thoracic Society, v. 6, n. 8, p. 678-682, 2009.
DAVIS, B. E.; ILLAMPERUMA, C.; GAUVREAU, G.M.; WATSON, R. M.; O'BYRNE, P. M.; DESCHESNES, F.; BOULET, L. P.; COCKCROFT, D.W. Single-dose desloratadine and montelukast and allergeninduced late airway responses. European Respiratory Journal, v. 33, n. 6, p. 1302-1308, 2009.
DE BLASI, A.; PARRUTI, G.; SALLESE, M. Regulation of G protein-coupled receptor kinase subtypes in activated T lymphocytes. Selective increase of beta-adrenergic receptor kinase 1 and 2. Journal of Clinical Investigation, v. 95, n. 1, p. 203, 1995.
DE BOER, J.; MEURS, H.; COERS, W.; KOOPAL, M.; BOTTONE, A. E.; VISSER, A. C.; TIMENS, W.; ZAAGSMA, J. Deficiency of nitric oxide in allergen-induced airway hyperreactivity to contractile agonists after the early asthmatic reaction: an ex vivo study. British Journal of Pharmacology, v. 119, n. 6, p. 1109-1116, 1996.

235 VASCONCELOS, L. H. C. Referências
DE BOER, J.; MEURS, H.; FLENDRIG, L.; KOOPAL, M.; ZAAGSMA, J. Role of nitric oxide and superoxide in allergen-induced airway hyperreactivity after the late asthmatic reaction in guinea-pigs. British Journal of Pharmacology, v. 133, n. 8, p. 1235-1242, 2001.
DE BOEVER, E. H.; ASHMAN, C.; CAHN, A. P.; LOCANTORE, N. W.; OVEREND, P.; POULIQUEN, I. J.; SERONE, A. P.; WRIGHT, T. J.; JENKINS, M. M.; PANESAR, I. S.; THIAGARAJAH, S. S. Efficacy and safety of an anti–IL-13 mAb in patients with severe asthma: a randomized trial. Journal of Allergy and Clinical Immunology, v. 133, n. 4, p. 989-996. e4, 2014.
DE RAEVE, H. R.; THUNNISSEN, F. B.; KANEKO, F. T.; GUO, F. H.; LEWIS, M.; KAVURU, M. S.; SECIC, M.; THOMASSEN, M. J.; ERZURUM, S. C. Decreased Cu, Zn-SOD activity in asthmatic airway epithelium: correction by inhaled corticosteroid in vivo. American Journal of Physiology-Lung Cellular and Molecular Physiology, v. 272, n. 1, p. L148-L154, 1997.
DEBMANDAL, M.; MANDAL, S. Coconut (Cocus nucifera L.: Arecaceae): in health promotion and disease prevention. Asian Pacific Journal of Tropical Medicine, v. 4, p. 241-247, 2011.
DEL RIO, D.; STEWART, A. J.; PELLEGRINI, N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutrition, Metabolism and Cardiovascular Diseases, v. 15, n. 4, p. 316-328, 2005.
DELLA-ROCCA, G. J.; VAN-BIESEN, T.; DAAKA, Y.; LUTTRELL, D. K.; LUTTRELL, L. M.; LEFKOWITZ, R. J. Ras-dependent mitogen-activated protein kinase activation by G protein coupled receptors: Convergence of G-i- and G-q-mediated pathways on calcium/calmodulin, Pyk2, and Src kinase. The Journal of Biological Chemistry, v. 272, p. 19125-19132, 1997.
DELLIS, O.; DEDOS, S. G.; TOVEY, S. C.; TAUFIQ-UR-RAHMAN, DUBEL, S. J.; TAYLOR, C. W. Ca2+ entry through plasma membrane IP3 receptors. Science, v. 313, n. 5784, p. 229-233, 2006.
DELMOTTE, P. F.; WALLERICH, S. F.; PRAKASH, Y. S.; SIECK, G. C. Inflammatory Cytokines Induced Endoplasmic Reticulum Stress In Airway Smooth Muscle. In: C109. LUNG BIOLOGY: AT A CROSSROAD BETWEEN INJURY AND REPAIR. American Thoracic Society, p. A5321-A5321, 2014.
DENNIS, E. A.; CAO, J.; HSU, Y. H.; MAGRIOTI, V.; KOKOTOS, G. Phospholipase A2 enzymes: physical structure, biological function, disease implication, chemical inhibition, and therapeutic intervention. Chem Rev., v. 111, n. 10, p. 6130–6185, 2011.
DESHPANDE, D. A.; DOGAN, S.; WALSETH, T. F.; MILLER, S. M.; AMRANI, Y.; PANETTIERI, R. A.; KANNAN, M. S. Modulation of calcium signaling by interleukin-13 in human airway smooth muscle: role of CD38/cyclic adenosine diphosphate ribose pathway. American Journal of Respiratory Cell andMolecular Biology, v. 31, n. 1, p. 36-42, 2004.

236 VASCONCELOS, L. H. C. Referências
DESHPANDE, D. A.; WALSETH, T. F.; PANETTIERI, R. A.; KANNAN, M. S. CD38/cyclic ADP-ribose-mediated Ca2+ signaling contributes to airway smooth muscle hyper-responsiveness. The Faseb Journal, v. 17, n. 3, p. 452-454, 2003.
DESHPANDE, D. A.; WHITE, T. A.; DOGAN, S.; WALSETH, T. F.; PANETTIERI, R. A.; KANNAN, M. S. CD38/cyclic ADP-ribose signaling: role in the regulation of calcium homeostasis in airway smooth muscle. American Journal of Physiology, v. 288, n. 5, p. L773-L788, 2005.
DI CANDIA, L.; GOMEZ, E.; VENEREAU, E.; CHACHI, L.; KAUR, D.; BIANCHI, M. E.; CHALLISS, R. A. J.; BRIGHTLING, C. E.; SAUNDERS, R. M. HMGB1 is upregulated in the airways in asthma and potentiates airway smooth muscle contraction via TLR4. The Journal of Allergy and Clinical Immunology, v. 140, n. 2, p. 584, 2017.
DI VALENTIN, E.; CRAHAY, C.; GARBACKI, N.; HENNUY, B.; GUEDERS, M.; NOEL, A.; FOIDART, J.; GROOTEN, J.; COLIGE, A.; PIETTE, J.; CATALDO, DIDIER. New asthma biomarkers: lessons from murine models of acute and chronic asthma. American Journal of Physiology Lung Cell and Molecular Physiology, v. 296, p. L185-197, 2009.
DIA, V. P.; GARCIA, V. V.; MABESA, R. C.; TECSON-MENDOZA, E. M. Comparative physiochemical characteristics of virgin coconut oil produced by different methods. Philippine Agricultural Scientist, v. 88, p. 462-475, 2005.
DOLGACHEV, V.; PETERSEN, B. C.; BUDELSKY, A. L.; BERLIN, A. A.; LUKACS, N. W. Pulmonary IL-17E (IL-25) production and IL-17RB+ myeloid cell-derived TH2 cytokine production are dependent upon stem cell factor-induced responses during chronic allergic pulmonary disease. The Journal of Immunology, v. 183, p. 5705-5715, 2009.
DONG, H.; LU, F. E.; ZHAO, L. Chinese herbal medicine in the treatment of nonalcoholic fatty liver disease. Chinese Journal of Integrative Medicine, v. 18, n. 2, p. 152-160, 2012.
DOUCET, C.; BROUTY-BOYÉ, D.; POTTIN-CLÉMENCEAU, C.; CANONICA, G. W.; JASMIN, C.; AZZARONE, B. Interleukin IL-4 and IL-13 act on human lung fibroblast implications on asthma. Journal of Clinical Investigation, v. 101, n. 10, p. 2129, 1998.
DOUGHERTY, R. H.; SIDHU, S. S.; RAMAN, K.; SOLON, M.; SOLBERG, O. D.; CAUGHEY, G. H.; WOODRUFF, P. G.; FAHY, J. V. Accumulation of intraepithelial mast cells with a unique protease phenotype in TH2-high asthma. Journal of Allergy and Clinical Immunology, v. 125, p. 1046-1053, 2010.
DOUKAS, J.; EIDE, L.; STEBBINS, K.; RACANELLI-LAYTON, A.; DELLAMARY, L.; MARTIN, M.; DNEPROVSKAIA, E.; NORONHA, G.; SOLL, R.; WRASIDLO, W.; ACEVEDO, L.; CHERESH, D. A. Aerosolized phosphoinositide 3-kinase γ/δ inhibitor TG100-115 [3-[2, 4-diamino-6-(3-hydroxyphenyl) pteridin-7-yl] phenol] as a therapeutic candidate for asthma and chronic obstructive pulmonary disease. Journal of Pharmacology and Experimental Therapeutics, v. 328, p. 758-765, 2009.

237 VASCONCELOS, L. H. C. Referências
DOZOR, A. J. The role of oxidative stress in the pathogenesis and treatment of asthma. Annals of the New York Academy of Sciences, v. 1203, p. 133-137, 2010.
DRAZEN, J.; AUSTEN, K. Leukotrienes and airway responses. The American Review of Respiratory Disease, v. 136, n. 4, p. 985-998, 1987.
DU, W.; MCMAHON, T. J.; ZHANG, Z. S.; STIBER, J. A.; MEISSNER, G.; EU, J. P. Excitation–contraction coupling in airway smooth muscle. The Journal of Biological Chemistry, v. 281, p. 30143-30151, 2006.
DUAN, W.; DATILES, A. M. A.; LEUNG, B. P.; VLAHOS, C. J.; WONG, W. F. An anti-inflammatory role for a phosphoinositide 3-kinase inhibitor LY294002 in a mouse asthma model. International Immunopharmacology, v. 5, p. 495-502, 2005.
DUPONT, L. J.; ROCVVHETTE, F.; DEMEDTS, M. G.; VERLEDEN, G. M. Exhaled nitric oxide correlates with airway hyperresponsiveness in steroid-naive patients with mild asthma. Americal Journal of Respiratory and Critical Care Medicine, v. 157, p. 894-898, 1998.
DUTTA, K.; CARMODY, M. W.; CALA, S. E.; DAVIDORFF, A. J. Depressed PKA activity contributes to impaired SERCA function and is linked to the pathogenesis of glucose-induced cardiomyophathy. Journal of Molecular and Cellular Cardiology, v. 34, n. 8, p. 985-996, 2002.
EBINA, M.; TAKAHASHI, T.; CHIBA, T.; MOTOMIYA, M. Cellular Hypertrophy and Hyperplasia of Airway Smooth Muscles Underlying Bronchial-Asthma - A 3-D Morphometric Study. The American Review of Respiratory Disease, v. 148, p. 720-726, 1993.
ELIAS, J. A.; ZHU, Z.; CHUPP, G.; HOMER, R. J. Airway remodeling in asthma. Journal of Clinical Investigation, v. 104, n. 8, p. 1001–1005, 1999.
EVANS, P.; LYRAS, L.; HALLIWELL, B. Measurement of protein carbonyls in human brain tissue. Methods in Enzymology, v. 300, p. 145-156, 1999.
FAHY, J. V. Type 2 inflammation in asthma - present in most, absent in many. Nature Reviews Immunology, v. 15, n. 1, p. 57-65, 2015.
FAIRBANK, N. J.; CONNOLLY, S. C.; MACKINNON, J. D.; WEHRY, K.; DENG, L.; MAKSYM, G. N. Airway smooth muscle cell tone amplifies contractile function in the presence of chronic cyclic strain. Am. J. Physiol. Lung Cell. Mol. Physiol., v. 295, p. 479–488, 2008.
FALLON, P. G.; BALLANTYNE, S. J.; MANGAN, N. E.; BARLOW, J. L.; DASVARMA, A.; HEWETT, D. R.; MCILGORM, A.; JOLIN, H. E.; MCKENZIE, A. N. J. Identification of an interleukin (IL)-25-dependent cell population that provides IL-4, IL-5, and IL-13 at the onset of helminth expulsion. Journal of Experimental Medicine, v. 203, n. 4, p. 1105-1116, 2006.
FANNING, L. B.; BOYCE, J. A. Basic Science for the Clinician: Lipid Mediators. Annals of allergy, asthma & immunology: official publication of the American College of Allergy, Asthma, & Immunology, v. 111, n. 3, p. 155, 2013.

238 VASCONCELOS, L. H. C. Referências
FAO - Food and Agriculture Organization of the United Nations. Disponível em: <http://faostat.fao.org/site/339/default.aspx>. Acesso em: 29 setembro 2017.
FATANI, S. H. Biomarkers of oxidative stress in acute and chronic bronchial asthma. Journal of Asthma, v. 51, n. 6, p. 578-584, 2014.
FERANIL, A. B.; DUAZO, P. L.; KUZAWA, C. W.; ADAIR, L. S. Coconut oil predicts a beneficial lipid profile in pre-menopausal women in the Philippines. Asia Pacific Journal of Clinical Nutrition, v. 20, n. 2, p. 190, 2011.
FERNANDES, L. B.; FRYER, A. D.; HIRSHMAN, C. A. M2 muscarinic receptors inhibit isoproterenol-induced relaxation of canine airway smooth muscle. Journal of Pharmacology and Experimental Therapeutics, v. 262, n. 1, p. 119-126, 1992.
FIREMAN, P. Understanding asthma pathophysiology. In: Allergy and Asthma Proceedings. OceanSide Publications, Inc, 2003. p. 79-83.
FISCHER, B. M.; VOYNOW, J. A. Neutrophil elastase inducesMUC5AC gene expression in airway epithelium via a pathway involving reactive oxygen species. American Journal of Respiratory Cell and Molecular Biology, v. 26, n. 4, p. 447-452, 2002.
FLOEGEL, A.; KIM, D. O.; CHUNG, S. J.; KOO, S. I.; CHUN, O. K. Comparison of ABTS/DPPH assays to measure antioxidant capacity in popular antioxidant-rich US foods. Journal of Food Composition and Analysis, v. 24, n. 7, p. 1043-1048, 2011.
FOOD AND DRUG ADMINISTRATION. FDA. Dietary Supplement Labeling Requirements and Recommendations under the Dietary Supplement and Nonprescription Drug Consumer Protection Act. Federal Register, v. 74, n. 35, 2009.
FÖRSTERMANN, U.; SESSA, W. C. Nitric oxide synthases: regulation and function. European Heart Journal, v. 33, n. 7, p. 829-837, 2012.
FORT, M. M.; CHEUNG, J.; YEN, D.; LI, J.; ZURAWSKI, S. M.; LO, S.; MENON, S.; CLIFFORD, T.; HUNTE, B.; LESLEY, R.; MUCHAMUEL, T.; HURST, S. D.; ZURAWSK, G.; LEACH, M. W.; GORMAN, D. M.; RENNICK, D. M. IL-25 induces IL-4, IL-5, and IL-13 and TH2-associated pathologies in vivo. Immunity, v. 15, n. 6, p. 985-995, 2001.
FRAME, S.; COHEN, P. GSK3 takes centre stage more than 20 years after its discovery. Biochemistry Journal, v. 359, p. 1-16, 2001.
FRANKE, T. F.; KAPLAN, D. R.; CANTLEY, L. C.; TOKER, A. Direct regulation of the Akt protooncogene product by phosphatidylinositol-3,4-bisphosphate. Science, v. 275, p. 665-668, 1997.
FRIDOVICH, I. Overview: Biological sources of O2. Methods in Enzymology v. 105, p. 59-61, 1984.
FUKATA, Y.; AMANO, M.; KAIBUCHI, K. Rho-Rho-kinase pathway in smooth muscle contraction and cytoskeletal reorganization of non-muscle cells. Trends in Pharmacological Sciences, v. 22, p. 32-39, 2001.

239 VASCONCELOS, L. H. C. Referências
FUNK, C. Prostaglandins and leukotrienes: advances in eicosanoid biology. Science, v. 294, n. 5548, p. 1871-1875, 2001.
GALLI, F.; BATTISTONI, A.; GAMBARI, R.; POMPELLA, A.; BRAGONZI, A.; PILOLLI, F.; IULIANO, L.; PIRODDI, M.; DECHECCHI, M. C.; CABRINI, G.; WORKING GROUP ON INFLAMMATION IN CYSTIC FIBROSIS. Oxidative stress and antioxidant therapy in cystic fibrosis. Biochimica et Biophysica Acta, v. 1822, n. 5, p. 690-713, 2012.
GALLI, S. J.; NAKAE, S.; TSAI, M. Mast cells in the development of adaptive immune responses. Nature immunology, v. 6, n. 2, p. 135, 2005.
GERTHOFFER, W. T.; SINGER, C. A. MAPK regulation of gene expression in airway smooth muscle. Respiratory Physiology and Neurobiology, v. 137, p. 237-250, 2003.
GIERA M.; LINGEMAN H.; NIESSEN W. M. A. Recent advancements in the LC- and GC-based analysis of malondialdehyde (MDA): a brief overview. Chromatographia, v. 75, n. 9-10, p. 433-440, 2012.
GINA (Global initiative for asthma). Global strategy for asthma management and prevention, 2010. Disponível em: <www.ginasthma.org>.
GLOBAL INITIATIVE FOR ASTHMA (GINA). Global strategy for asthma management and prevention. Updated 2017. Vancouver, USA, 2017.
GOLDIE, R. G.; SPINA, D.; HENRY, P. J.; LULICH, K. M.; PATERSON, J. W. In vitro responsiveness of human asthmatic bronchus to carbachol, histamine, beta-adrenoceptor agonists and theophylline. British Journal of Clinical Pharmacology, v. 22, p. 669-677, 1986.
GORDON, J. R.; GALLI, S. J. Mast cells as a source of both preformed and immunologically inducible TNFα/cachectin. Nature, v. 346, p. 274-276, 1990.
GORSKI, S. A.; HAHN, Y. S.; BRACIALE, T. J. Group 2 innate lymphoid cell production of IL-5 is regulated by NKT cells during influenza virus infection. PLoS Pathogens, v. 9, n. 9, p. e1003615, 2013.
GOSENS, R.; ROSCIONI, S. S.; DEKKERS, B. G.; PERA, T.; SCHMIDT, M.; SCHAAFSMA, D.; ZAAGSMA, J.; MEURS, H. Pharmacology of airway smooth muscle proliferation. European Journal of Pharmacology, v. 585, p. 385-397, 2008.
GOTO, K.; CHIBA, Y.; MATSUSUE, K.; HATTORI, Y.; MAITANI, Y.; SAKAI, H.; KIMURA, S.; MISAWA, M. The proximal STAT6 and NF-kappaB sites are responsible for IL-13- and TNF-alpha-induced RhoA transcriptions in human bronchial smooth muscle cells. Pharmacol Research, v. 61, p. 466-472, 2010a.
GOTO, K.; CHIBA, Y.; MISAWA, M. IL-13 induces translocation of NF-kappaB in cultured human bronchial smooth muscle cells. Cytokine, v. 46, p. 96-99, 2009.
GOTO, K.; CHIBA, Y.; SAKAI, H.; MISAWA, M. Mechanism of inhibitory effect of prednisolone on RhoA upregulation in human bronchial smooth muscle cells. Biological and Pharmaceutical Bulletin, v. 33, p. 710-713, 2010b.

240 VASCONCELOS, L. H. C. Referências
GRAINGE, C. L.; LAU, L. C. K.; WARD, J. A.; DULAY, V.; LAHIFF, G.; WILSON, S.; HOLGATE, S.; DAVIES, D. E.; HOWARTH, P. H. Effect of bronchoconstriction on airway remodeling in asthma. N. Engl. J. Med., v. 364, p. 2006–2015, 2011.
GROSS, N. J.; BARNES, P.J. New therapies for asthma and chronic obstructive pulmonary disease. American Journal of Respiratory and Critical Care Medicine, v. 195, n. 2, p. 159-166, 2017.
GRUNSTEIN, M. M.; HAKONARSON, H.; LEITER, J.; CHEN, M.; WHELAN, R.; GRUNSTEIN, J. S.; CHUANG, S. IL-13-dependent autocrine signaling mediates altered responsiveness of IgE-sensitized airway smooth muscle. American Journal of Physiology-Lung Cellular and Molecular Physiology, v. 282, n. 3, p. L520-L528, 2002.
GUERRA, S.; MARTINEZ, F. D. The natural history of asthma and COPD. In: BARNES, P.; DRAZEN, J. M.; RENNARD, S. I.; THOMSON, N. C. (Eds.). Asthma and COPD. Basic mechanisms and clinical management. Academic Press, 2009, p. 23-33.
GUPTA, M. K.; CAO, G.; PANETTIERI, R. A.; NAGA PRASAD, S. V. Inflammation leads to loss of β-adrenergic receptor function in asthma from human airway smooth muscle cells. In: A29. INFLAMMATION AND MECHANISMS OF AIRWAY SMOOTH MUSCLE CONTRACTION. American Thoracic Society, p. A1270-A1270, 2016.
GURISH, M. F.; AUSTEN, K. F. Developmental origin and functional specialization of mast cell subsets. Immunity, v. 37, p. 25-33, 2012.
GURNEY, A. M. Mechanisms of drug-induced vasodilation. Journal of Pharmacy and Pharmacology, v. 46, n. 4, p. 242-251, 1994.
HAKONARSON, H.; MASKERI, N.; CARTER, C.; GRUNSTEIN, M. M. Regulation of TH1- and TH2-type cytokine expression and action in atopic asthmatic sensitized airway smooth muscle, Journal of Clinical Investigation, v. 103, n. 7, p. 1077-1087, 1999.
HALAYKO, A. J.; CAMORETTI-MERCADO, B.; FORSYTHE, S. M.; VIEIRA, J. E.; MITCHELL, R. W.; WYLAM, M. E.; HERSHENSON, M. B.; SOLWAY, J. Divergent differentiation paths in airway smooth muscle culture: Induction of functionally contractile myocytes. American Journal of Physiology, v. 276, p. L197-L206, 1999.
HALAYKO, A. J.; SALARI, H.; MA, X.; STEPHENS, N. L. Markers of airway smooth muscle cell phenotype. American Journal of Physiology, v. 270, p. L1040-L1051, 1996.
HALAYKO, A. J.; STEPHENS, N. L. Potential role for phenotypic modulation of bronchial smooth muscle cells in chronic asthma. Canadian Journal of Physiology and Pharmacology, v. 72, p. 1448-1457, 1994.
HAMID, Q.; SPRINGALL, D. R.; RIVEROS-MORENO, V. CHANEZ, P. BOUSQUET, J.; HOWARTH, P.; REDINGTON, A.; BOUSQUET, J.; GODARD, P.; HOLGATE, S.; POLAK, J. M. Induction of nitric oxide synthase in asthma. Lancet, v. 342, n. 8886-8887, p. 1510-1513, 1993.

241 VASCONCELOS, L. H. C. Referências
HAMMAD, H.; CHIEPPA, M.; PERROS, F.; WILLART, M. A.; GERMAIN, R. N.; LAMBRECHT, B. N. House dust mite allergen induces asthma via Toll-like receptor 4 triggering of airway structural cells. Nature Medicine, v. 15, n. 4, p. 410-416, 2009.
HAMMERMANN, R.; STICHNOTE, C.; CLOSS, E. I.; NAWRATH, H.; RACK, K. Inhibition of nitric oxide synthase abrogates lipopolysaccharides-induced up-regulation of L-arginine uptake in rat alveolar macrophages. British Journal of Pharmacology, v. 133, n. 3, p. 379-386, 2001.
HANAZAWA, T.; KHARITONOV, S. A.; BARNES, P. J. Increased nitrotyrosine in exhaled breath condensate of patients with asthma. American Journal of Respiratory and Critical Care Medicine, v. 162, n. 4, p. 1273-1276, 2000.
HASHIMOTO, T.; NAKANO, Y.; YAMASHITA, M.; FANG, Y. I.; OHATA, H.; MOMOSE, K. Role of Rho-associated protein kinase and histamine in lysophosphatidic acid-induced airway hyperresponsiveness in guinea pigs. The Japanese Journal of Pharmacology, v. 88, n. 3, p. 256-261, 2002.
HATA, A. N.; BREYER, R. M. Pharmacology and signaling of prostaglandin receptors: multiple roles in inflammation and immune modulation. Pharmacol Ther., v. 103, n. 2, p. 147–166, 2004.
HAWES, B. E.; VAN BIESEN, T.; KOCH, W. J.; LUTTRELL, L. M.; LEFKOWITZ, R. J, Distinct pathways of Gi- and Gq-mediated mitogen-activated protein kinase activation. The Journal of Biological Chemistry, v. 270, p. 17148-17153, 1995.
HAZBUN, M. E.; HAMILTON, R.; HOLIAN, A.; ESCHENBACHER, W. L. Ozone Induced Increases in Substance P and 8-EP1-Prostaglandin F2α in the Airways of Human Subjects. American Journal of Respiratory Cell and Molecular Biology, v. 9, p. 568-568, 1993.
HEBER, D. Herbs and atherosclerosis. Current Atherosclerosis Reports, v. 3, n. 1, p. 93-96, 2001.
HENDERSON, W. R. J.; LEWIS, D. B.; ALBERT, R. K.; ZHANG, Y.; LAMM, W. J.; CHIANG, G. K.; JONES, F.; ERIKSEN, P.; TIEN, Y. T.; JONAS, M.; CHI, E. Y. The importance of leukotrienes in airway inflammation in a mouse model of asthma. Journal of Experimental Medicine, v. 184, n. 4, p. 1483-1494, 1996.
HENRICKS, P. A. J.; NIJKAMP, F. P. Reactive oxygen species as mediators in asthma. Pulmonary Pharmacology and Therapeutics, v. 14, n. 6, p. 409-421, 2001.
HENRY, G. E.; MOMIN, R. A.; NAIR, M. G.; DEWITT, D. L. Antioxidant and cyclooxygenase activities of fatty acids found in food. Journal of Agricultural and Food Chemistry, v. 50, n. 8, p. 2231-2234, 2002.
HEWAVITHARANA, T.; DENG, X.; SOBOLOFF, J.; GILL, D. L. Role of STIM and Orai proteins in the store-operated calcium signaling pathway. Cell calcium, v. 42, n. 2, p. 173-182, 2007.
HIRATA, T.; NARUMIYA, S. Prostanoids as regulators of innate and adaptive immunity. Adv Immunol., v. 116, p. 143–174, 2012.

242 VASCONCELOS, L. H. C. Referências
HIROTA, S.; HELLI, P.; JANSSEN, L. J. Ionic mechanisms and Ca2þ handling in airway smooth muscle. European Respiratory Journal, v. 30, n. 1, p. 114-133, 2007.
HIRSHMAN, C. A.; EMALA, C. W. Actin reorganization in airway smooth muscle cells involves Gq and Gi-2 activation of Rho. American Journal of Physiology-Lung Cellular and Molecular Physiology, v. 277, n. 3, p. L653-L661, 1999.
HIRST, S. J.; MARTIN, J. G.; BONACCI, J. V.; CHAN, V.; FIXMAN, E. D.; HAMID, Q. A.; HERSZBERG, B.; LAVOIE, J.; MCVICKER, C. G.; MOIR, L.; NGUYEN, T. T. B.; PENG, Q.; BARBÓN, D. R. Proliferative aspects of airway smooth muscle. Journal of Allergy and Clinical Immunology, v. 114, n. 2, p. S2-S17, 2004.
HIRST, S. J. Regulation of airway smooth muscle cell immunomodulatory function: role in asthma. Respiratory Physiology and Neurobiology, v. 137, p. 309-326, 2003.
HODSMAN, P.; ASHMAN, C.; CAHN, A.; DE BOEVER, E.; LOCANTORE, N.; SERONE, A.; POULIQUEN, I. A phase 1, randomized, placebo-controlled, dose-escalation study of an anti- IL-13 monoclonal antibody in healthy subjects and mild asthmatics. British Journal of Clinical Pharmacology, v. 75, n. 1, p. 118-128, 2013.
HOLGATE, S. T. Epithelium dysfunction in asthma. Journal of Allergy and Clinical Immunology, v. 120, n. 6, p. 1233–1244, 2007.
HOLGATE, S. T.; ARSHAD, H. S.; ROBERTS, G. C.; HOWARTH, P. H.; THURNER, P.; DAVIES, D. E. A new look at the pathogenesis of asthma. Clinical Science, v. 118, n. 7, p. 439-450, 2010.
HOLGATE, S. T.; DAVIES, D. E.; LACKIE, P. M.; WILSON, S. J.; PUDDICOMBE, S. M.; LORDAN, J. L. Epithelial-mesenchymal interactions in the pathogenesis of asthma. Journal of Allergy and Clinical Immunology. v. 105, n. 2, p. 193 204, 2000.
HOLGATE, S. T.; HOLLOWAY, V.; WILSON, S.; BUCCHIERI, F.; PUDDICOMBE, S.; DAVIES, D. E. Epithelial-mesenchymal communication in the pathogenesis of chronic asthma. Proceedings of the American Thoracic Society, v. 1, n. 2, p. 93-98, 2004.
HOLGATE, S. T.; POLOSA, R. Treatment strategies for allergy and asthma. Nature Reviews Immunology, v. 8, p. 218–230, 2008.
HOLGUIN, F. Oxidative stress in airway diseases. Annals of the American Thoracic Society, v. 10, p. S150-S157, 2013.
HOLROYDE, M. C. The influence of epithelium on the responsiveness of guinea‐pig isolated trachea. British Journal of Pharmacology, v. 87, n. 3, p. 501-507, 1986.
HOMER, R. J.; ELIAS, J. A. Airway remodeling in asthma: therapeutic implications of mechanisms. Physiology, v. 20, n. 1, p. 28–35, 2005.
HOMER, R. J.; ELIAS, J. A. Consequences of long-term inflammation: airway remodeling. Clinics in Chest Medicine, v. 21, n. 2, p. 331–343, 2000.

243 VASCONCELOS, L. H. C. Referências
HONDA, T.; KABASHIMA, K. Leukotrienes as key mediators and amplifiers in allergic inflammation: insights from the bench and clinic. Experimental Dermatology, v. 23, n. 2, p. 95-96, 2014.
HOSHINO, M.; TAKAHASHI, M.; AOIKE, N. Expression of vascular endothelial growth factor, basic fibroblast growth factor, and angiogenin immunoreactivity in asthmatic airways and its relationship to angiogenesis. Journal of Allergy and Clinical Immunology, v. 107, n. 2, p. 295–301, 2001.
HOWARTH, P. H.; REDINGTON, A. E.; SPRINGALL, D. R.; MARTIN, U.; BLOOM, S. R.; POLAK, J. M.; HOLGATE, S. T. Epithelially derived endothelin and nitric oxide in asthma. International Archives of Allergy and Immunology, v. 107, n. 1-3, p. 228-230, 1995.
HOYMANN, H. G.; WOLLIN, L.; MULLER, M.; KOROLEWITZ, R.; KRUG, N.; BRAUN, A.; BEUME, R. Effects of the phosphodiesterase type 4 inhibitor roflumilast on early and late allergic response and airway hyperresponsiveness in Aspergillus fumigatus sensitized mice. Pharmacology, v. 83, p. 188-195, 2009.
HU, Q. J.; KLIPPEL, A.; MUSLIN, A. J.; FANTL, W. J.; WILLIAMS, L. T. Ras-Dependent Induction of Cellular-Responses by Constitutively Active Phosphatidylinositol-3 Kinase. Science, v. 268, p. 100-102, 1995.
HUANG, J.; OLIVENSTEIN, R.; TAHA, R.; HAMID, Q.; LUDWIG, M. Enhanced proteoglycan deposition in the airway wall of atopic asthmatics. American Journal of Respiratory and Critical Care Medicine, v. 160, n. 2, p. 725–729, 1999.
HUANG, W. C.; FANG, L. W.; LIOU, C. J. Phloretin attenuates allergic airway inflammation and Oxidative stress in asthmatic Mice. Frontiers in Immunology, v. 8, p. 1-13, 2017.
HUANG, W. W.; GARCIA-ZEPEDA, E. A.; SAUTY, A.; OETTGEN, H. C.; ROTHENBERG, M. E.; LUSTER, A. D. Molecular and biological characterization of the murine leukotriene B4 receptor expressed on eosinophils. Journal of Experimental Medicine, v. 188, n. 6, p. 1063-1074, 1998.
HUMBERT,M.; BUSSE, W.; HANANIA, N. A.; LOWE, P. J.; CANVIN, J.; ERPENBECK, V. J.; HOLGATE, S. Omalizumab in asthma: an update on recent developments. Journal of Allergy and Clinical Immunology: In Practice, v. 2, p. 525-536, 2014.
HURST, S. D.; MUCHAMUEL, T.; GORMAN, D. M.; GILBERT, J. M.; CLIFFORD, T.; KWAN, S.; MENON, S.; SEYMOUR, B.; JACKSON, C.; KUNG, T. T.; BRIELAND, J. K.; ZURAWSKI, S. M.; CHAPMAN, R. W.; ZURAWSKI, G.; AND ROBERT L. COFFMAN. New IL-17 family members promote TH1 or TH2 responses in the lung: in vivo function of the novel cytokine IL-25. The Journal of Immunology, v. 169, n. 1, p. 443-453, 2002.
HUTSON, P. A.; CHURCH, M.;K.; CLAY, T. P.; MILLER, P.; HOLGATE, S. T. Early and late phase bronchoconstriction after allergen challenge of nonanesthetized guinea pigs: the association of disordered airway physiology to leukocyte infiltration. The American Review of Respirtory Diseases, v. 137, p. 548–557, 1988.

244 VASCONCELOS, L. H. C. Referências
IFIC. International Food Information Council Web site, <http://ific.org/nutrition/functional/index.cfm, 2006>. Acesso em 21 jul 2017.
IKEDA, M.; KATOH, S.; SHIMIZU, H.; OKA, M. Development of two types of mite-allergen induced murine model of chronic asthma with different severity. Kawasaki Medical Journal, v. 42, n. 1, p. e7, 2016.
IKUTA, N.; SUGIYAMA, S.; TAKAGI, K.; SATAKE, T.; OZAWA, T. Implication of oxygen radicals on airway hyperresponsiveness after ovalbumin challenge in guinea pigs. American Review of Respiratory Disease, v. 145, p. 561-565, 1992.
INAYAT, A.; BAIG, S. A.; BAQAI, T. Does coconut oil reduce the risk of coronary artery diseases? Journal of the Pakistan Medical Association, v. 63, n. 6, p. 797, 2013.
IND, P. W. Role of the sympathetic nervous system and endogenous catecholamines in the regulation of airways smooth muscle tone. In: RAEBURN, D.; GIEMBYCZ, M. (eds.). Airways smooth muscle: structure, innervation and neurotransmission. Edited by Basel: Birkhäuser Verlag, 1994.
INMAN, G. J.; NICOLAS, F. J.; CALLAHAN, J. F.; HARLING, J. D.; GASTER, L. M.; REITH, A. D.; LAPING, N. J.; HILL, C. S. SB-431542 is a potent and specific inhibitor of transforming growth factor β superfamily type I activin receptor-like kinase (ALK) receptors ALK4, ALK5, and ALK7. Molecular Pharmacology, v. 62, p. 65-74, 2002.
INTAHPHUAK, S.; KHONSUNG, P.; PANTHONG, A. Anti-inflammatory, analgesic, and antipyretic activities of virgin coconut oil. Pharmaceutical Biology, v. 48, n. 2, p. 151-157, 2010.
IRVIN, C. G.; BATES, J. H. T. Measuring the lung function in the mouse: the challenge of size. Respiratory Research, v. 4, n. 1, p. 1, 2003.
IRVIN, C. G.; TU, Y. P.; SHELLER, J. R.; FUNK, C. D. 5-Lipoxygenase products are necessary for ovalbumin-induced airway responsiveness in mice. American Journal of Physiology-Lung Cellular and Molecular Physiology, v. 272, n. 6, p. L1053-L1058, 1997.
ISAAC STEERING COMMITTEE. Worldwide variations in the prevalence of asthma symptoms: the International Study of Asthma and Allergies in Childhood (ISAAC). The European Respiratory Journal, v. 12, n. 2, p. 315-335, 1998.
ISHIKAWA, T.; KUME, H.; KONDO, M.; ITO, Y.; YAMAKI, K.; SHIMOKATA, K. Inhibitory effects of interferon-γ on the heterologous desensitization of β-adrenoceptors by transforming growth factor-β1 in tracheal smooth muscle. Clinical & Experimental Allergy, v. 33, n. 6, p. 808-815, 2003.
ISHIZAKA, T.; ISHIZAKA, K.; TOMIOKA, H. Release of histamine and slow reacting substance of anaphylaxis (SRS-A) by IgE-anti-IgE reactions on monkey mast cells. The Journal of Immunology, v. 108, n. 2, p. 513-520, 1972.
ITO, S.; KUME, H.; HONJO, H.; KATOH, H.; KODAMA, I.; YAMAKI, K.; HAYASHI, H. Possible involvement of Rho kinase in Ca2+ sensitization and mobilization by MCh

245 VASCONCELOS, L. H. C. Referências
in tracheal smooth muscle. American Journal of Physiology-Lung Cellular and Molecular Physiology, v. 280, n. 6, p. L1218-L1224, 2001.
IWAMOTO, H.; NAKAMUTA, M.; TADA, S.; SUGIMOTO, R.; ENJOJI, M.; NAWATA, H. A p160ROCK-specific inhibitor, Y-27632, attenuates rat hepatic stellate cell growth. Journal of Hepatology, v. 32, p. 762-770, 2000.
JACOBY, D. B.; GLEICH, G. J.; FRYER, A. D. Human eosinophil major basic protein is an endogenous allosteric antagonist at the inhibitory muscarinic M2 receptor. Journal of Clinical Investigation, v. 91, n. 4, p. 1314, 1993.
JAIN, P. P.; LEBER, R.; NAGARAJ, C.; LEITINGER, G.; LEHOFER, B.; OLSCHEWSKI, H.; OLSCHEWSKI, A.; PRASSL, R.; MARSH, L. M. Liposomal nanoparticles encapsulating iloprost exhibit enhanced vasodilation in pulmonary arteries. International Journal of Nanomedicine, v. 9, p. 3249, 2014.
JAIN-VORA, S.; WERT, S. E.; TEMANN, U. A.; RANKIN, J. A.; WHITSETT, J. A. Interleukin-4 alters epithelial cell differentiation and surfactant homeostasis in the postnatal mouse lung. American Journal of Respiratory Cell and Molecular Biology, v. 17, n. 5, p. 541–551, 1997.
JANSSEN, L. J. Isoprostanes: An Overview and Putative Roles in Pulmonary Pathophysiology. American Journal of Physiology and Lung Cellular Molecular Physiology, v. 280, n. 6, p. L1067-L1082, 2001.
JANSSEN, L. J. Isoprostanes: generation, pharmacology, and roles in free-radical-mediated effects in the lung. Pulmonary Pharmacology & Therapeutics, v. 13, n. 4, p. 149-155, 2000.
JARTTI, T. Asthma, asthma medication and autonomic nervous system dysfunction. Clin Physiol, v. 21, p. 260-269, 2001.
JATAKANON, A.; LIM, S.; KHARITONOV, S. A.; CHUNG, K. F.; BARNES, P. J. Correlation between exhaled nitric oxide, sputum eosinophils, and methacholine responsiveness in patients with mild asthma. Thorax, v. 53, p. 91-95, 1998.
JENKINS, H. A.; COOL, C.; SZEFLER, S. J.; COVAR, R.; BRUGMAN, S.; GELFAND, E. W.; SPAHN, J. D. Histopathology of severe childhood asthma: a case series. Chest, v. 124, n. 1, p. 32-41, 2003.
JOHNSON, P. R.; AMMIT, A. J.; CARLIN, S. M.; ARMOUR, C. L.; CAUGHEY, G. H.; BLACK, J. L. Mast cell tryptase potentiates histamine-induced contraction in human sensitized bronchus. European Respiratory Journal, v. 10, n. 1, p. 38-43, 1997.
JOHNSON, S. R.; KNOX, A. J. Synthetic functions of airway smooth muscle in asthma. Trends in Pharmacological Sciences, v. 18, p. 288-292, 1997.
JOHNSTON, S. L.; FREEZER, N. J.; RITTER, W.; O’TOOLE, S.; HOWARTH, P. H. Prostaglandin D2-induced bronchoconstriction is mediated only in part by the thromboxane prostanoid receptor. European Respiratory Journal, v. 8, n. 3, p. 411-415, 1995.

246 VASCONCELOS, L. H. C. Referências
JONES, T. R.; CHARETTE, L.; DENIS, D. Antigen-induced contraction of guinea-pig isolated trachea: studies with novel inhibitors and antagonists of arachidonic acid metabolites. British Journal of Pharmacology, v. 95, p. 309-321, 1988.
JONGSTE, J. C.; MONS, H.; BONTA, I. L.; KERREBIJN, K. F. In vitro responses of airways from an asthmatic patient. European Journal of Respiratory Diseases, v. 71, p. 23-29, 1987.
KAMISAH, Y.; PERIYAH, V.; LEE, K. T.; NOOR-IZWAN, N.; NURUL-HAMIZAH, A.; NURUL-IMAN, B. S.; SUBERMANIAM, K.; JAARIN, K.; AZMAN, A.; FAIZAH, O.; QODRIYAH, H. M. S. Cardioprotective effect of virgin coconut oil in heated palm oil diet-induced hypertensive rats. Pharmaceutical Biology, v. 53, n. 9, p. 1243-1249, 2015.
KANG, Y. J.; MBONYE, U. R.; DE LONG, C. J.; WADA, M.; SMITH, W. L. Regulation of intracellular cyclooxygenase levels by gene transcription and protein degradation. Progress in Lipid Research, v. 46, p. 108-125, 2007.
KANG, Z.; SWAIDANI, S.; YIN, W.; WANG, C.; BARLOW, J. L.; GULEN, M. F.; BULEK, K.; DO, J.; ARONICA, M.; MCKENZIE, A. N. J.; MIN, B.; LI, X. Epithelial cell-specific Act1 adaptor mediates interleukin- 25-dependent helminth expulsion through expansion of Lin-c-Kit+ innate cell population. Immunity, v. 36, n. 5, p. 821-833, 2012.
KAPPALLY, S.; SHIRWAIKAR, A.; SHIRWAIKAR, A. Coconut oil–a review of potential applications. Journal for Drugs and Medicines, v. 10, 2015.
KAPSALI, T.; PERMUTT, S.; LAUBE, B.; SCICHILONE, N.; TOGIAS, A. Potent bronchoprotective effect of deep inspiration and its absence in asthma. Journal of Applied Physiology,v. 89, p. 711-720, 2000.
KASAI, M.; NOSAKA, N.; MAKI, H.; NEGISHI, S.; AOYAMA, T.; NAKAMURA, M.; SUZUKI, Y.; TSUJI, H.; UTO, H.; OKAZAKI, M.; KONDO, K. Effect of dietary medium-and long-chain triacylglycerols (MLCT) on accumulation of body fat in healthy humans. Asia Pacific Journal of Clinical Nutrition, v. 12, n. 2, p. 151-160, 2003.
KEIGHTLEY, M. C.; CURTIS, A. J.; CHU, S.; FULLER, P. J. Structural determinants of cortisol resistance in the guinea pig glucocorticoid receptor. Endocrinology, v. 139, p. 2479-2485, 1998.
KELLEHER, M. D.; ABE, M. K.; CHAO, T. S. O.; JAIN, M.; GREEN, J. M.; SOLWAY, J.; ROSNER, M. R.; HERSHENSON, M. B. Role of MAP kinase activation in bovine tracheal smooth muscle mitogenesis. American Journal of Physiology, v. 268, p. L894-L901, 1995.
KERSTJENS, H. A.; ENGEL, M.; DAHL, R.; PAGGIARO, P.; BECK, E.; VANDEWALKER, M.; SIGMUND, R.; MATH., D.; SEIBOLD, W.; MORONI-ZENTGRAF, P.; BATEMAN, E. D. Tiotropium in asthma poorly controlled with standard combination therapy. New England Journal of Medicine, v. 367, n. 13, p. 1198-1207, 2012.
KEYS, A.; ANDERSON, J. T.; GRANDE, F. Serum cholesterol response to changes in the diet. 1. Iodine value of the fat versus 2S-P. Metabolism, v. 14, p.747–758, 1965.

247 VASCONCELOS, L. H. C. Referências
KHARITONOV, S. A.; O’CONNOR, B. J.; EVANS, D. J.; BARNES, P. J. Allergen-induced late asthmatic reactions are associated with elevation of exhaled nitric oxide. American Journal of Respiratory and Critical Care Medicine, v. 151, n. 6, p. 1894-1899, 1995.
KIMURA, C.; KONISHI, S.; HASEGAWA, M.; OIKE, M. Development of vascular smooth muscle contractility by endothelium-derived transforming growth factor β proteins. Pflügers Archiv-European Journal of Physiology, v. 466, n. 2, p. 369-380, 2014.
KINNULA, V. L.; YANKASKAS, J. R.; CHANG, L.; VIRTANEN, I.; LINNALA, A.; KANG, B. H.; CRAPO, J. D. Primary and immortalized (BEAS 2B) human bronchial epithelial cells have significant antioxidative capacity in vitro. American Journal of Respiratory Cell and Molecular Biology, v. 11, n. 5, p. 568-576, 1994.
KIRKHAM, S.; SHEEHAN, J. K.; KNIGHT, D.; RICHARDSON, P. S.; THORNTON, D. J. Heterogeneity of airways mucus: variations in the amounts and glycoforms of the major oligomeric mucins MUC5AC and MUC5B. Biochem J, v. 361, p. 537–546, 2002.
KIRKPATRICK, C. T. Tracheobronchial smooth muscle. Smooth Muscle: An Assessment of Current Knowledge. In: Bulbring, E.; Brading, A. F.; Jones, A. W.; Tomita, T. (eds.)., p. 385-395, 1981.
KISTEMAKER, L. E. M.; GOSENS, R. Acetylcholine beyond bronchoconstriction: roles in inflammation and remodeling. Trends in pharmacological sciences, v. 36, n. 3, p. 164-171, 2015.
KITAZAWA, T.; ETO, M.; WOODSOME, T. P.; BRAUNTINGAM, D. L. Agonists trigger G-protein mediated activation of the CPI-17 inhibitor phosphoprotein of myosin light chain phosphatase to enhance vascular smooth muscle contractility. Journal of Biological Chemistry, v. 275, p. 9897-9900, 2000.
KOLCH, W.; HEIDECKER, G.; KOCHS, G.; HUMMEL, R.; VAHIDI, H.; MISCHAK, H.; FINKENZELLER, G.; MARME, D.; RAPP, U. R. Protein kinase C alpha activates RAF-1 by direct phosphorylation. Nature, v. 364, p. 249-252, 1993.
KOTO, H.; MAK, J. C.; HADDAD, E. B.; XU, W. B.; SALMON, M.; BARNES, P. J.; CHUNG, K. F. Mechanisms of impaired β-adrenoceptor-induced airway relaxation by interleukin-1β in vivo in the rat. Journal of Clinical Investigation, v. 98, n. 8, p. 1780, 1996.
KOUTSOKERA, A.; KOSTIKAS, K.; NICOD, L. P.; FITTING, J. W. Pulmonary biomarkers in COPD exacerbations: a systematic review. Respiratory Research, v. 14, 2013.
KOUZAKI, H., O’GRADY, S.M., LAWRENCE, C.B.; KITA, H. Proteases induce production of thymic stromal lymphopoietin by airway epithelial cells through protease-activated receptor-2. J. Immunol., v. 183, p. 1427–1434, 2009.
KRYMSKAYA, V. P.; ORSINI, M. J.; ESZTERHAS, A. J.; BRODBECK, K. C.; BENOVIC, J. L.; PANETTIERI, R. A.; PENN, R. B. Mechanisms of proliferation synergy by receptor tyrosine kinase and G protein-coupled receptor activation in human airway

248 VASCONCELOS, L. H. C. Referências
smooth muscle. American Journal of Respiratory Cell and Molecular Biology, v. 23, p. 546-554, 2000.
KUCHAREWICZ, I.; BODZENTA-ŁUKASZYK, A.; BUCZKO, W. Experimental asthma in rats. Pharmacological Reports, v. 60, n. 6, p. 783-788, 2007.
KUDO, M.; MELTON, A. C.; CHEN, C.; ENGLER, M. B.; HUANG, K. E.; REN, X.; WANG, Y.; BERNSTEIN, X.; LI, J. T.; ATABAI, K.; HUANG, X; SHEPPARD, D. IL-17A produced by αβ T cells drives airway hyper-responsiveness in mice and enhances mouse and human airway smooth muscle contraction. Nature medicine, v. 18, n. 4, p. 547-554, 2012.
KUMAR, P. D. The role of coconut and coconut oil in coronary heart disease in Kerala, south India. Tropical Doctor, v. 27, p. 215-7,1997.
KUME, H. Ca2+ dynamics and Ca2+ sensitization in the regulation of airway smooth muscle tone. In: Muscle Cell and Tissue. InTech, 2015.
KUME, H. Large-conductance calcium-activated potassium channels. In: Wang, Y. X. (ed.). Calcium Signaling in Airway Smooth Muscle Cells. Springer, New York, p. 49-83, 2013.
KUME, H.; GRAZIANO, M. P.; KOTLIKOFF, M. I. Stimulatory and inhibitory regulation of calcium- activated potassium channels by guanine nucleotide-binding proteins. Proceedings of the National Academy of Sciences, v. 89, n. 22, p. 11051-11055, 1992.
KUME, H.; HALL, I. P.; WASHABAU, R. J.; TAKAGI, K.; KOTLIKOFF, M. I. β-adrenergic agonists regulate KCa channels in airway smooth muscle by cAMP-dependent and –independent mechanisms. Journal of Clinical Investigation, v. 93, p. 371-379,1994;.
KUME, H.; KOTLIKOFF, M. I. Muscarinic inhibition of single KCa channels in smooth muscle cells by a pertussis-sensitive G protein. American Journal Physiology-Cell Physiology, v. 261, p. C1204-9, 1991.
KUME, H.; TAKAGI, K.; SATAKE, T.; TOKUNO, H.; TOMITA, T. Effects of intracellular pH on calcium-activated potassium channels in rabbit tracheal smooth muscle. The Journal of physiology, v. 424, n. 1, p. 445-457, 1990.
KUSCHNIR, F. C.; GURGEL, R. Q.; SOLÉ, D.; COSTA, E.; FELIX, M. M.; OLIVEIRA, C. L.; VASCONCELLOS, M. T. L.; KUSCHNIR, M. C. C. ERICA: prevalence of asthma in Brazilian adolescents. Revista de Saude Publica, v. 50, Suppl 1, p. 1s-13s, 2016.
LACKIE, P.M., BAKER, J.E., GUNTHERT, U.; HOLGATE, S.T. Expression of CD44 isoforms is increased in the airway epithelium of asthmatic subjects. American Journal of Respiratory Cell and Molecular Biology, v. 16, p. 14-22, 1997.
LAEMMLI, U. K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature, v. 227, p. 680-685, 1970.

249 VASCONCELOS, L. H. C. Referências
LAI, C.; BEASLEY, R.; CRANE, J.; FOLIAKI, S.; SHAH, J.; WEILAND, S. Global variation in the prevalence and severity of asthma symptoms: phase three of the International Study of Asthma and Allergies in Childhood (ISAAC). Thorax, 2009.
LAIDLAW, T. M.; KIDDER, M. S.; BHATTACHARYYA, N.; XING, W.; SHEN, S.; MILNE, G. L.; CASTELLS, M. C.; CHHAY, H.; BOYCE, J. A. Cysteinyl leukotriene overproduction in aspirin-exacerbated respiratory disease is driven by platelet-adherent leukocytes. Blood, v. 119, n. 16, p. 3790-3798, 2012.
LAITINEN, A.; ALTRAJA, A.; K¨AMPE, M.; LINDEN, M.; VIRTANEN, I.; LAITINEN, L. A. Tenascin is increased in airway basement membrane of asthmatics and decreased by an inhaled steroid. American Journal of Respiratory and Critical Care Medicine, v. 156, n. 3, p. 951–958, 1997.
LAMBRECHT, B. N.; HAMMAD H. The immunology of asthma. Nature Immunology, v. 16, n. 1, p. 45-56, 2015.
LAMBRECHT, B. N.; HAMMAD, H. Biology of lung dendritic cells at the origin of asthma. Immunity, v. 31, p. 412-424, 2009.
LANCET. A plea to abandon asthma as a disease concept. The Lancet, 368, n. 9537, p. 705, 2006.
LANE, S. J.; LEE, T. H. Mast cell effector mechanisms. Journal of Allergy and Clinical Immunology, v. 98, n. 5, p. S67-S72, 1996.
LAPORTE, J. C.; MOORE, P. E.; BARALDO, S.; JOUVIN, M. H.; CHURCH, T. L.; SCHWARTZMAN, I. N.; PANETTIERI, R. J.; KINET, J. P.; SHORE, S. A. Direct effects of interleukin-13 on signaling pathways for physiological responses in cultured human airway smooth muscle cells. American Journal of Respiratory and Critical Care Medicine, v. 164, n. 1, p. 141-148, 2001.
LAPORTE, J. D.; MOORE, P. E.; LAHIRI, T.; SCHWARTZMAN, I. N.; PANETTIERI, R. A. J.; SHORE, S. A. p38 MAP kinase regulates IL-1β responses in cultured airway smooth muscle cells. American Journal of Physiology-Lung Cellular and Molecular Physiology, v. 279, n. 5, p. L932-L941, 2000.
LAUZON, A.; MARTIN, J. G. Airway hyperresponsiveness: smooth muscle as the principal actor. F1000Research, v. 5, 2016.
LE CRAS, T. D.; ACCIANI, T. H.; MUSHABEN, E. M.; KRAMER, E. L.; PASTURA, P. A.; HARDIE, W. D.; KORFHAGEN, T. R.; SIVAPRASAD, U.; ERICKSEN, M.; GIBSON, A. M.; HOLTZMAN, M. J.; WHITSETT, J. A.; HERSHEY, G. K. Epithelial EGF receptor signaling mediates airway hyperreactivity and remodeling in a mouse model of chronic asthma. Am. J. Physiol. Lung Cell. Mol. Physiol., v. 300, p. 414–421, 2011.
LEE, C. G.; HOMER, R. J.; ZHU, Z.; LANONE, S.; WANG, X.; KOTELIANSKY, V.; SHIPLEY, M. J.; GOTWALS, P.; NOBLE, P.; CHEN, Q.; SENIOR, R. M.; ELIAS, J. A. Interleukin-13 induces tissue fibrosis by selectively stimulating and activating transforming growth factor β1. Journal of Experimental Medicine, v. 194, n. 6, p. 809-821, 2001.

250 VASCONCELOS, L. H. C. Referências
LEE, J. J.; MCGARRY, M. P.; FARMER, S. C. Interleukin-5 expression in the lung epithelium of transgenic mice leads to pulmonary changes pathognomonic of asthma. Journal of Experimental Medicine, v. 185, n. 12, p. 2143–2156, 1997.
LEE, J. Y.; UCHIDA, Y.; SAKAMOTO, T.; HIRATA, A.; HASEGAWA, S.; HIRATA, F. Alteration of G protein levels in antigen-challenged guinea pigs. Journal of Pharmacology and Experimental Therapeutics, v. 271, p. 1713–1720, 1994.
LEE, T. H.; WOSZCZEK, G.; FAROOQUE, S. P. Leukotriene E4: perspective on the forgotten mediator. Journal of Allergy and Clinical Immunology, v. 124, n. 3, p. 417-421, 2009.
LEE, Y.; LEE, B.; RHEE, Y.; SONG, C. The involvement of matrix metalloproteinase-9 in airway inflammation of patients with acute asthma. Clin Exp Allergy., v. 31, p. 623–1630, 2001.
LEFCOE, N. M. The effect of hydrocortisone hemisuccinate on tracheal smooth muscle of the guinea pig and cat. Journal of Allergy, v. 27, n. 4, p. 352-358, 1956.
LEFF, A. R. Endogenous Regulation of Bronchomotor Tone1-3. American Review of Respiratory Disease, v. 137, p. 1198-1216, 1988.
LEICK-MALDONADO, E. A.; KAY, F. U.; LEONHARDT, M. C.; KASAHARA, D. I.; PRADO, C. M.; FERNANDES, F. T.; MARTINS, M. A.; TIBÉRIO, I. F. L. C. Comparison of glucocorticoid and cysteinyl leukotriene receptor antagonist treatments in an experimental model of chronic airway inflammation in guinea-pigs. Clinical and Experimental Allergy, v. 34, p. 145–152, 2004.
LEWIS, R.; ROBIN, J. Arachidonic acid derivatives as mediators of asthma. Journal of Allergy and Clinical Immunology, v. 76, n. 2, p. 259-264, 1985.
LINDSTEDT, K. A.; WANG, Y.; SHIOTA, N.; SAARINEN, J.; HYYTIÄINEN, M.; KOKKONEN, J. O.; KOKKONEN, J.; KESKI-OJA, J.; KOVANEN, P. T. Activation of
paracrine TGF‐β1 signaling upon stimulation and degranulation of rat serosal mast cells: a novel function for chymase. The FASEB Journal, v. 15, n. 8, p. 1377-1388, 2001.
LITTLE, S. A.; SPROULE, M. W.; COWAN, M. D.; MACLEOD, K. J.; ROBERTSON, M.; LOVE, J. G.; CHALMERS, G. W.; MCSHARRY, C. P.; THOMSON, N. C. High resolution computed tomographic assessment of airway wall thickness in chronic asthma: reproducibility and relationship with lung function and severity. Thorax, v. 57, n. 3, p. 247-253, 2002.
LIU, H. W.; HALAYKO, A. J.; FERNANDES, D. J.; HARMON, G. S.; MCCAULEY, J. A.; KOCIENIEWSKI, P.; MCCONVILLE, J.; FU, Y.; FORSYTHE, S. M.; KOGUT, P.; BELLAM, S.; DOWELL, M.; CHURCHILL, J.; SOLWAY, J.; REMAINING, A. The RhoA/Rho kinase Pathway Regulates Nuclear Localization of Serum Response Factor. American Journal of Respiratory Cell and Molecular Biology, v. 29, p. 39-47, 2003.
LIU, M. C.; BLEECKER, E. R.; LICHTENSTEIN, L. M.; KAGEY-SOBOTKA, A.; NIV, Y.; MCLEMORE, T. L.; PERMUTT, S.; PROUD, D.; HUBBARD, W. C. Evidence for

251 VASCONCELOS, L. H. C. Referências
elevated levels of histamine, prostaglandin D2, and other bronchoconstricting prostaglandins in the airways of subjects with mild asthma. American Review of Respiratory Disease, v. 142, n. 1, p. 126-132, 1990.
LLOYD, C. M.; HESSEL, E. M. Functions of T cells in asthma: more than just TH2 cells. Nature Reviews Immunology, v. 10, n. 12, p. 838-848, 2010.
LOCVVKE, N. R.; ROYCE, S. G.; WAINEWRIGHT, J. S.; SAMUEL, C. S.; TANG, M. L. Comparison of airway remodeling in acute, subacute, and chronic models of allergic airways disease. American Journal of Respiratory Cell and Molecular Biology, v. 36, n. 5, p. 625-632, 2007.
LOPEZ-ILASACA, M.; CRESPO, P.; PELLICI, P. G.; GUTKIND, J. S.; WETZKER, R. Linkage of G protein-coupled receptors to the MAPK signaling pathway through PI 3-kinase gamma. Science, v. 275, p. 394-397, 1997.
LORDAN, J. L.; BUCCHIERI, F.; RICHTER, A.; KONSTANTINIDIS, A.; HOLLOWAY, J. W.; THORNBER, M.; PUDDICOMBE, S. M.; BUCHANAN, D.; WILSON, S. J.; DJUKANOVIĆ, R.; HOLGATE, S. T.; DAVIES, D. E. Cooperative effects of TH2 cytokines and allergen on normal and asthmatic bronchial epithelial cells. The Journal of Immunology, v. 169, p. 407–414, 2002.
LOTUFO, P. A.; BENSENOR, I. M. Temporal trends of asthma mortality rates in Brazil from 1980 to 2010. Journal of Asthma, v. 49, n. 8, p. 779-784, 2012.
LOTVALL, J.; INMAN, M.; O’BYRNE, P. Measurement of airway hyperresponsiveness: new considerations. Thorax, v. 53, p. 419-424, 1998.
LOUHELAINEN, N.; MYLLARNIEMI, M.; RAHMAN, I.; KINNULA V. L. Airway biomarkers of the oxidant burden in asthma and chronic obstructive pulmonary disease: current and future perspectives. International Journal of Chronic Obstructive Pulmonary Disease, v. 3, n. 4, p. 585-603, 2008.
LOXHAM, M.; DAVIES, D. E.; BLUME, C. Epithelial function and dysfunction in asthma. Clinical and Experimental Allergy, v. 44, n. 11, p. 1299-1313, 2014.
LUNDEQUIST, A.; NALLAMSHETTY, S. N.; XING, W.; FENG, C.; LAIDLAW, T. M.; UEMATSU, S.; AKIRA, S.; BOYCE, J. A. Prostaglandin E2 exerts homeostatic regulation of pulmonary vascular remodeling in allergic airway inflammation. The Journal of Immunology, v. 184, n. 1, p. 433-441, 2010.
LUO, S.; LEI, H.; QIN, H.; XIA, Y. Molecular mechanisms of endothelial NO synthase uncoupling. Current Pharmaceutical Design, v. 20, n. 22, p. 3548-3553, 2014.
LUSTER, A. D.; TAGER, A. M. T-cell trafficking in asthma: lipid mediators grease the way. Nature Reviews Immunology, v. 4, n. 9, p. 711, 2004.
MACK, C. P.; SOMLYO, A. V.; HAUTMANN, M.; SOMLYO, A. P.; OWENS, G. K. Smooth muscle differentiation marker gene expression is regulated by RhoA-mediated actin polymerization. The Journal of Biological Chemistry, v. 276. P. 341-347, 2001.

252 VASCONCELOS, L. H. C. Referências
MALO, P. E.; BELL, R. L.; SHAUGHNESSY, T. K.; SUMMERS, J. B.; BROOKS, D. W.; CARTER, G. W. The 5-lipoxygenase inhibitory activity of zileuton in in vitro and in vivo models of antigen-induced airway anaphylaxis. Pulmonary pharmacology, v. 7, n. 2, p. 73-79, 1994.
MARINA, A. M. CHE MAN, Y. B.; NAZIMAH, S. A. H.; AMIN, I. Antioxidant capacity and phenolic acids of virgin coconut oil. International Journal of Food Sciences and Nutrition, v. 60, n. sup2, p. 114-123, 2009.
MARINA, A. M.; CHE MAN, YB; AMIN, I. Virgin coconut oil: emerging functional food oil. Trends in Food Science & Technology. v. 20, n. 10, p. 481-487, 2009.
MARINS, R. M.; ARAUJO, I. S.; JACOB, S. C. A propaganda de alimentos: orientação, ou apenas estímulo de consumo? Ciência Saúde Coletiva, v.16, n.9, p. 3873-3882, 2011.
MARSICO, S. A.; PELAIA, G.; GREMBIALE, R. D.; PUGLIESE, C.; CELLAMARE, P. Calcium ions and airway smooth muscle contraction. In: Airway Obstruction and Inflammation. Karger Publishers, p. 26-34, 1990.
MARTIN, J. G.; DUGUET, A.; EIDELMAN, D. H. The contribution of airway smooth muscle to airway narrowing and airway hyperresponsiveness in disease. European Respiratory Journal, v. 16, n. 2, p. 349-354, 2000.
MARTIN, J. G.; RAMOS-BARBON, D. Airway smooth muscle growth from the perspective of animal models. Respiratory Physiology and Neurobiology v. 137, p. 251-261, 2003.
MARTINS, Italo Rossi Roseno. A ação relaxante de novos derivados N-sulfonilidrazônicos do LASSBio-448, inibidores de PDE4, em um modelo de asma alérgica em cobaias: caracterização funcional do mecanismo relaxante do LASSBio-1847. 220 f. Pós-graduação em Produtos Naturais e Sintéticos Bioativos, área de concentração Farmacologia (Tese) - Centro de Ciências da Saúde, Universidade Federal da Paraíba - UFPB, João Pessoa, 2016.
MASCIA, K.; HASELKORN, T.; DENIZ, Y. M.; MILLER, D. P.; BLEECKER, E. R.; BORISH, L; TENOR STUDY GROUP. Aspirin sensitivity and severity of asthma: evidence for irreversible airway obstruction in patients with severe or difficult-to-treat asthma. Journal of Allergy and Clinical Immunology, v. 116, n. 5, p. 970-975, 2005.
MASOLI, M.; FABIAN, D.; HOLT, S.; BEASLEY, R. The global burden of asthma: executive summary of the GINA Dissemination Committee report. Allergy, v. 59, n. 5, p. 469-478, 2004.
MASUDA, M.; KUDO, M.; MORIYAMA, Y.; YAMAMOTO, M.; KOBAYASHI, N.; TOMARU, K.; TANAKA, S.; ISHII, H.; KAKEGAWA, S.; SASAKI, M.; KANEKO, T.; SHEPPARD, D.; ISHIGATSUBO, Y. Interleukin-17A synergistically enhances mouse airway smooth muscle contraction with tumor necrosis factor-through RhoA/ROCK2 mRNA stability change. In: B33. ASTHMA MEDIATORS AND BIOMARKERS. American Thoracic Society, p. A2639-A2639, 2013.

253 VASCONCELOS, L. H. C. Referências
MATSUKURA, S.; STELLATO, C.; GEORAS, S. N.; CASOLARO, V.; PLITT, J. R.; MIURA, K.; KUROSAWA, S.; SCHINDLER, U.; SCHLEIMER, R. P. Interleukin-13 upregulates eotaxin expression in airway epithelial cells by a STAT6-dependent mechanism. American Journal of Respiratory Cell and Molecular Biology, v. 24, p. 755–761, 2001.
MATUSOVSKY, O. S.; KACHMAR, L.; IJPMA, G.; BATES, G.; ZITOUNI, N.; BENEDETTI, A.; LAVOIE, J. P.; LAUZON, A. M. Peripheral Airway Smooth Muscle, but Not the Trachealis, Is Hypercontractile in an Equine Model of Asthma. American Journal of Respiratory Cell and Molecular Biology, v. 54, n. 5, p. 718-727, 2016.
MÁTYÁS, Š.; PUCOVSKÝ, V.; BAUER, V. Effects of various reactive oxygen species on the guinea pig trachea and its epithelium. The Japanese Journal of Pharmacology, v. 88, n. 3, p. 270-278, 2002.
McKAY, S.; SHARMA, H. S. Autocrine regulation of asthmatic airway inflammation: role of airway smooth muscle. Respiratory Research, v. 3, p. 11, 2001.
McMILLAN, S. J.; LLOYD, C. M. Prolonged allergen challenge in mice leads to persistent airway remodeling. Clinical & Experimental Allergy, v. 34, p. 497-507, 2004.
MCPARLAND, B. E.; MACKLEM, P. T.; PARE, P. D. Airway wall remodeling: friend or foe? Journal of Applied Physiology, v. 95, p. 426-434, 2003.
METCALFE, D. D.; BARAM, D.; MEKORI, Y. A. Mast cells. Physiological Reviews, v. 77, n. 4, p. 1033-1079, 1997.
MEURS, H.; GOSENS, R.; ZAAGSMA, J. Airway hyperresponsiveness in asthma: lessons from in vitro model systems and animal models. European Respiratory Journal, v. 32, n. 2, p. 487-502, 2008.
MEURS, H.; MAARSINGH, H.; ZAAGSMA, J. Arginase and asthma: novel insights into nitric oxide homeostasis and airway hyperresponsiveness. Trends in Pharmacological Science, v. 24, p. 450-455, 2003.
MEURS, H.; ROFFEL, A. D. F.; ELZINGA, C. R. S.; ZAAGSMA, J. Muscarinic receptor-β-adrenoceptor cross-talk in airways smooth muscle. In: Muscarinic Receptors in Airways Diseases. Birkhäuser Basel, p. 121-157, 2001.
MEURS, H.; SANTING, R. E.; REMIE, R.; MARK, T. W. V. D.; WESTERHOF, F. J.; ZUIDHOF, A. B.; BOS, I. S. T.; Zaagsma, J. A guinea pig model of acute and chronic asthma using permanently instrumented and unrestrained animals. Nature Protocols, v. 1, n. 2, p. 840-847, 2006.
MILLER, D. K.; GILLARD, J. W.; VICKERS, P. J.; SADOWSKI, S.; LE´VEILLE´, C.; MANCINI, J. A.; CHARLESON, P.; DIXON, R. A. F.; FORD-HUTCHINSON, A. W.; FORTIN, R.; GAUTHIER, J. Y.; RODKEY, J.; ROSEN, R.; ROUZER, C.; SIGAL, I. S.; STRADER, C. D.; EVANS, J. F. Identification and isolation of a membrane protein necessary for leukotriene production. Nature, v. 343, n. 6255, p. 278, 1990.

254 VASCONCELOS, L. H. C. Referências
MITCHELL, H. W. Airway smooth muscle contraction–Perspectives on past, present and future. Pulmonary pharmacology & therapeutics, v. 22, n. 5, p. 363-369, 2009.
MOORE, W. C.; MEYERS, D. A.; WENZEL, S. E.; TEAGUE, W. G.; LI, H.; LI, X.; D’AGOSTINO JR., R.; CASTRO, M.; CURRAN-EVERETT, D.; FITZPATRICK, A. M.; GASTON, B.; JARJOUR, N. N.; SORKNESS, R.; CALHOUN, W. J.; CHUNG, K. F.; COMHAIR, S. A. A.; DWEIK, R. A.; ISRAEL, E.; PETERS, S. P.; BUSSE, W. W.; ERZURUM, S. C.; BLEECKER, E. R. Identification of asthma phenotypes using cluster analysis in the Severe Asthma Research Program. American Journal of Respiratory and Critical Care Medicine, v. 181, n. 4, p. 315-323, 2010.
MORGAN, K. The role of calcium in the control of vascular tone as assessed by the Ca2+ indicator aequorin. Cardiovascular drugs and therapy, v. 4, p.1355 1362, 1990.
MORI, A.; ITO, S.; MORIOKA, M.; ASO, H.; KONDO, M.; SOKABE, M.; HASEGAWA, Y. Effects of specific prostanoid EP receptor agonists on cell proliferation and intracellular Ca2+ concentrations in human airway smooth muscle cells. European Journal of Pharmacology, v. 659, n. 1, p. 72-78, 2011.
MORO, K.; YAMADA, T.; TANABE, M.; TAKEUCHI, T.; IKAWA, T.; KAWAMOTO, H.; FURUSAWA, J.; OHTANI, M.; FUJII, H.; KOYASU, S. Innate production of TH2 cytokines by adipose tissue-associated c-Kit+Sca-1+ lymphoid cells. The Journal of Immunology, v. 184, 2010.
MORROW, J. D.; ROBERTS, L. J. The isoprostanes: current knowledge and directions for future research. Biochemical Pharmacology, v. 51, n. 1, p. 1-9, 1996.
MORWOOD, K.; GILLIS, D.; SMITH, W.; KETTE, F. Aspirin‐sensitive asthma. Internal Medicine Journal, v. 35, n. 4, p. 240-246, 2005.
MOUNKAÏLA, B.; MARTHAN, R.; ROUX, E. Biphasic effect of extracellular ATP on human and rat airways is due to multiple P2 purinoceptor activation. Respiratory Research, v. 6, n. 1, p. 143, 2005.
MURRAY, R. K.; FLEISCHMANN, B. K.; KOTLIKOFF, M. I. Receptor-activated Ca influx in human airway smooth muscle: use of Ca imaging and perforated patch-clamp techniques. American Journal of Physiology-Cell Physiology, v. 264, n. 2, p. C485-C490, 1993.
NABISHAH, B. M.; MORAT, P. B.; KHALID, B. A.; KADIR, B. A. Effect of acetylcholine and morphine on bronchial smooth muscle contraction and its modulation by steroid hormones. Clinical and Experimental Pharmacology and Physiology, v. 17, n. 12, p. 841-847, 1990.
NADEEM, A.; CHHABRA, S. K.; MASOOD, A.; RAJ, H. G. Increased oxidative stress and altered levels of antioxidants in asthma. Journal of Allergy and Clinical Immunology, v. 111, n. 1, p. 72-78, 2003.
NADEEM, A.; SIDDIQUI, N.; ALHARBI, N. O.; ALHARBI, M. M. Airway and systemic oxidant-antioxidant dysregulation in asthma: a possible scenario of oxidants spill over from lung into blood. Pulmonary Pharmacology and Therapeutics, v. 29, n. 1, p. 31-40, 2014.

255 VASCONCELOS, L. H. C. Referências
NAKATA, J.; KONDO, M.; TAMAOKI, J.; TAKEMIYA, T.; NOHARA, M.; YAMAGATA, K.; NAGAI, A. Augmentation of allergic inflammation in the airways of cyclooxygenase-2- deficient mice. Respirology, v. 10, p. 149-156, 2005.
NALINE, E.; ZHANG, Y.; QIAN, Y.; MAIRON, N.; ANDERSON, G. P.; GRANDORDY, B.; ADVENIER, C. Relaxant effects and durations of action of formoterol and salmeterol on the isolated human bronchus. European Respiratory Journal, v. 7, n. 5, p. 914-920, 1994.
NANDAKUMARAN, M.; ANGELAKI, E.; AL-AZEMI, N., AL-SARRAF, H.; AL-SALEH, E. Influence of coconut oil administration on some hematologic and metabolic parameters in pregnant rats. The Journal of Maternal-Fetal and Neonatal Medicine, v. 24, p. 1254-1258, 2011.
NARUMIYA, S.; SUGIMOTO, Y.; USHIKUBI, F. Prostanoid receptors: structures, properties, and functions. Physiol Rev, v. 79, p. 1193e226, 1999.
NEGRI, M. L. S. Secagem das folhas de espinheira - santa - Maytenus ilicifolia Mart. ex Reiss. sob diferentes temperaturas e influência nos teores de polifenóis, na atividade antioxidante e nos aspectos microbiológicos. Dissertação (mestrado em Ciências Farmacêuticas). Universidade Federal do Paraná, 2007.
NEILL, D. R.; WONG, S. H.; BELLOSI, A.; FLYNN, R. J.; DALY, M.; LANGFORD, T. K. A.; BUCKS, C.; KANE, C. M.; FALLON, P. G.; PANNELL, R.; JOLIN, H. E.; MCKENZIE, A. N. J. Nuocytes represent a new innate effector leukocyte that mediates type-2 immunity. Nature, v. 464, p. 1367–1370, 2010.
NESI, R. T.; KENNEDY-FEITOSA, E.; LANZETTI, M.; ÁVILA, M. B.; MAGALHÃES, C. B.; ZIN, W. A.; FAFFE, D. S.; PORTO, L. C.; VALENÇA, S. S. Inflammatory and Oxidative Stress Markers in Experimental Allergic Asthma. Inflammation, p. 1-11, 2017.
NESMITH, A. P.; AGARWAL, A.; MCCAIN, M. L.; PARKER, K. K. Human airway musculature on a chip: an in vitro model of allergic asthmatic bronchoconstriction and bronchodilation. Lab on a Chip, v. 14, n. 20, p. 3925-3936, 2014.
NEUBIG, R. R.; SPEDDING, M.; KENAKIN, T.; CHRISTOPOULOS, A. International Union of Pharmacology Committee on Receptor Nomenclature and Drug Classification. XXXVIII. Update on terms and symbols in quantitative pharmacology. Pharmacological Reviews, v. 55, n. 4, p. 597-606, 2003.
NEVIN, K. G.; RAJAMOHAN, T. Beneficial effects of virgin coconut oil on lipid parameters and in vitro LDL oxidation. Clinical Biochemistry, v. 37, n. 9, p. 830-835, 2004.
NEVIN, K. G.; RAJAMOHAN, T. Effect of topical application of virgin coconut oil on skin components and antioxidant status during dermal wound healing in young rats. Skin Pharmacology and Physiology, v. 23, p. 290-7, 2010.
NEVIN, K. G.; RAJAMOHAN, T. Influence of virgin coconut oil on blood coagulation factors, lipid levels and LDL oxidation in cholesterol fed Sprague-Dawley rats. e-SPEN,

256 VASCONCELOS, L. H. C. Referências
the European e-Journal of Clinical Nutrition and Metabolism, v. 3, n. 1, p. e1-e8, 2008.
NEVIN, K. G.; RAJAMOHAN, T. Virgin coconut oil supplemented diet increases the antioxidant status in rats. Food Chemistry, v. 99, n. 2, p. 260-266, 2006.
NIIMI, A.; MATSUMOTO, H.; AMITANI, R.; NAKANO, Y.; MISHIMA, M.; MINAKUCHI, M.; NISHIMURA, K.; ITOH, H.; IZUMI, T. Airway wall thickness in asthma assessed by computed tomography. Relation to clinical indices. American Journal of Respiratory and Critical Care Medicine, v. 162, p. 1518-1523, 2000.
NIJKAMP, F. P.; HENRICKS, P. A. J. Beta-adrenoceptors in lung inflammation. American Review of Respiratory Disease, v. 141, p. S145-S150, 1990.
NMCE. Report on copra. National multi-commodity Exchange of India Limited, p. 1-14, 2007.
NOGAMI, M.; ROMBERGER, D. J.; RENNARD, S. I.; TOEWS, M. L. TGF-beta 1 modulates beta-adrenergic receptor number and function in cultured human tracheal smooth muscle cells. American Journal of Physiology-Lung Cellular and Molecular Physiology, v. 266, n. 2, p. L187-L191, 1994.
NOONAN, M.; KORENBLAT, P.; MOSESOVA, S.; SCHEERENS, H.; ARRON, J. R.; ZHENG, Y.; PUTNAM, W. S.; PARSEY, M.; BOHEN, S. P.; MATTHEWS, J. G. Dose-ranging study of lebrikizumab in asthmatic patients not receiving inhaled steroids. Journal of Allergy and Clinical Immunology, v. 132, n. 3, p. 567-574. e12, 2013.
NURUL-IMAN, B. S.; KAMISAH, Y.; JAARIN, K.; QODRIYAH, H. J. M. S. Virgin coconut oil prevents blood pressure elevation and improves endothelial functions in rats fed with repeatedly heated palm oil. Evidence-Based Complementary and Alternative Medicine, v. 2013, 2013.
O’BRIEN, R. D. Fats and oils: Formulating and processing for applications. New York: CRC Press, 2008.
OENEMA, T. A.; MAARSINGH, H.; SMIT, M.; GROOTHUIS, G. M.; MEURS, H.; GOSENS, R. Bronchoconstriction induces TGF-beta release and airway remodelling in guinea pig lung slices. PLoS ONE v. 8, p. 65580, 2013.
OHKAWA, H.; OHISHI, N.; YAGI, K. Assay for lipid peroxidation in animal tissues by thiobarbituric acid reaction. Analytical Biochemistry, v. 95, p. 351-358, 1979.
OHNISHI, H.; MIYAHARA, N.; DAKHAMA, A.; TAKEDA, K.; MATHIS, S.; HARIBABU, B.; GELFAND, E. W. Corticosteroids enhance CD8þ T cell-mediated airway hyperresponsiveness and allergic inflammation by upregulating leukotriene B4 receptor 1. Journal of Allergy and Clinical Immunology, v. 121, n. 4, p. 864-871. e4, 2008.
OKAFOR, O. Y.; ERUKAINURE, O. L.; AJIBOYE, J. A.; ADEJOBI, R. O.; OWOLABI, F. O.; KOSOKO, S. B. Modulatory effect of pineapple peel extract on lipid peroxidation, catalase activity and hepatic biomarker levels in blood plasma of alcohol–induced

257 VASCONCELOS, L. H. C. Referências
oxidative stressed rats. Asian Pacific Journal of Tropical Biomedicine, v. 1, n. 1, p. 12-14, 2011.
ORDOÑEZ, C. L.; KHASHAYAR, R.; WONG, H. H.; FERRANDO, R.; WU, R.; HYDE, D. M.; HOTCHKISS, J. A.; ZHANG, Y.; NOVIKOV, A.; DOLGANOV, G.; FAHY, J. V. Mild and moderate asthma is associated with airway goblet cell hyperplasia and abnormalities in mucin gene expression. American Journal of Respiratory and Critical Care Medicine, v. 163, n. 2, p. 517-523, 2001.
OREHEK, J. E. A. N.; DOUGLAS, J. S.; BOUHUYS, A. Contractile responses of the guinea-pig trachea in vitro: modification by prostaglandin synthesis-inhibiting drugs. Journal of Pharmacology and Experimental Therapeutics, v. 194, n. 3, p. 554-564, 1975.
ORSINI, M. J.; KRYMSKAYA, V. P.; ESZTERHAS, A. J.; BENOVIC, J. L.; PANETTIERI, R. A.; PENN, R. B. MAPK superfamily activation in human airway smooth muscle: mitogenesis requires prolonged p42/p44 activation. American Journal of Physiology, v. 277, n. 3, p. L479–L488, 1999.
OTIS, A. B. A perspective of respiratory mechanics. J Appl Physiol Respir Environ Exerc Physiol, v. 54, p. 1183-1187, 1983.
OUEDRAOGO, N.; ROUX, E. Physiology of Airway Smooth Muscle Contraction: An Overview. Journal of Pulmary and Respiratory Medicine, v. 4, n. 221, p. 2, 2014.
PACK, R. J.; RICHARDSON, P.S. The aminergic innervation of the human bronchus: a light and electron microscopic study. Journal of Anatomy, v. 138, p. 493-502, 1984.
PANETTIERI JR, R. A.; GOLDIE, R. G.; RIGBY, P. J.; ESZTERHAS, A. J.; HAY, D. W. P. Endothelin 1-induced potentiation of human airway smooth muscle proliferation: An ET-A receptormediated phenomeno. British Journal of Pharmacology, v. 118, p. 191-197, 1996.
PANETTIERI, R. A. Airway smooth muscle: immunomodulatory cells that modulate airway remodeling? Respiratory Physiology and Neurobiology, v. 137, p. 277-293, 2003.
PANETTIERI, R. A.; TAN, E. M. L.; CIOCCA, V.; LUTTMANN, M. A.; LEONARD, T. B.; HAY, D. W. P. Effects of LTD4 on human airway smooth muscle cell proliferation, matrix expression, and contraction in vitro: Differential sensitivity to cysteinyl leukotriene receptor antagonists. American Journal of Respiratory Cell and Molecular Biology, v. 19, p. 453-461, 1998a.
PANETTIERI, R. A.; MURRAY, R. K.; ESZTERHAS, A. J.; BILGEN, G.; MARTIN, J. G. Repeated allergen inhalations induce DNA synthesis in airway smooth muscle and epithelial cells in vivo. American Journal of Physiology, v. 274, p. L417-L424, 1998b.
PARK, S. J.; MIN, K. H.; LEE, Y. C. Phosphoinositide 3‐kinase δ inhibitor as a novel therapeutic agent in asthma. Respirology, v. 13, p. 764-771, 2008.

258 VASCONCELOS, L. H. C. Referências
PASCUAL, R. M.; PETERS, S. P. Airway remodeling contributes to the progressive loss of lung function in asthma: an overview. Journal of Allergy and Clinical Immunology, v. 116, n. 3, p. 477–486, 2005.
PAVLOVIC, D.; VIIRES, N.; ZEDDA, C.; FOURNIER, M.; AUBIER, M. Effects of corticosteroids on epithelial structure and smooth muscle function of rat trachea. European Respiratory Journal, v. 11, n. 3, p. 575-582, 1998.
PAWANKAR, R.; CANONICA, G. W.; HOLGATE, S. T.; LOCKEY, R. F. World Allergy Organization (WAO) white book on allergy, 2011. Wisconsin: World Allergy Organisation. Disponível em: <http://www. worldallergy. org/UserFiles/file/WAO-White-Book-on-Allergy web.pdf>.
PEEL, S. E.; LIU, B.; HALL, I. P. ORAI and store-operated calcium influx in human airway smooth muscle cells. American Journal of Respiratory Cell and Molecular Biology, v. 38, n. 6, p. 744-749, 2008.
PEJLER, G.; ÅBRINK, M.; RINGVALL, M.; WERNERSSON, S. Mast cell proteases. Advances in Immunology, v. 95, p. 167-255, 2007.
PELAIA, G.; RENDA, T.; GALLELLI, L.; VATRELLA, A.; BUSCETI, M. T.; AGATI, S.; CAPUTI, M.; CAZZOLA, M.; MASELLI, R.; MARSICO, S. A. Molecular mechanisms underlying airway smooth muscle contraction and proliferation: implications for asthma. Respiratory Medicine, v. 102, n. 8, p. 1173-1181, 2008.
PELAIA, G.; VATRELLA, A.; MASELLI, R. The potential of biologics for the treatment of asthma. Nature Reviews Drug Discovery, v. 11, n. 12, p. 958-972, 2012.
PERA, T.; PENN, R. B. Bronchoprotection and bronchorelaxation in asthma: New targets, and new ways to target the old ones. Pharmacology & Therapeutics, v. 164, p. 82-96, 2016.
PEREIRA, M. C. S.; AMARAL, M. P. H.; PINHATI, R. R.; PINTO, C. L. O.; MENDONÇA A. E.; FURTADO, M. M.; STRINGUETA, P. C.; PINTO, M. A. O. Proposta de guia simplificado para registro de alimento com alegações de propriedades funcionais. Vigilância Sanitária em Debate, v. 2, n. 2, p. 88-95, 2014.
PEREZ-ZOGHBI, J. F.; KARNER, C.; ITO, S.; SHEPHERD, M.; ALRASHDAN, Y.; SANDERSON, M. J. Ion channel regulation of intracellular calcium and airway smooth muscle function. Pulmonary Pharmacology & Therapeutics, v. 22, n. 5, p. 388-397, 2009.
PEREZ-ZOGHBI, J. F.; SANDERSON, M. J. Endothelin-induced contraction of bronchiole and pulmonary arteriole smooth muscle cells is regulated by intracellular Ca2+ oscillations and Ca2+ sensitization. American Journal of Physiology-Lung Cellular and Molecular Physiology, v. 293, n. 4, p. L1000-L1011, 2007.
PERSSON, M. G.; GUSTAFSSON, L. E. Allergen-induced airway obstruction in guinea-pigs is associatedwith changes in nitric oxide levels in exhaled air. Acta Physiologica Scandinavica, v. 149, n. 4, p. 461-466, 1993.

259 VASCONCELOS, L. H. C. Referências
PICHAVANT, M.; CHARBONNIER, A. S.; TARONT, S.; BRICHET, A.; WALLAERT, B.; PESTEL, J.; TONNEL, A. B.; GOSSET, P. Asthmatic bronchial epithelium activated by the proteolytic allergen Der p 1 increases selective dendritic cell recruitment. The Journal of Allergy and Clinical Immunology, v. 115, p. 771–778, 2005.
PIETARINEN-RUNTTI, P.; LAKARI, E.; RAIVIO, K. O.; KINNULA, V. L. Expression of antioxidant enzymes in human inflammatory cells. American Journal of Physiology-Cell Physiology, v. 278, n. 1, p. C118-C125, 2000.
PIGATI, P. A.; RIGHETTI, R. F.; POSSA, S. S.; ROMANHOLO, B. S.; RODRIGUES, A. P. D.; SANTOS, A. S. A.; XISTO, D. G.; ANTUNES, M. A.; PRADO, C. M.; LEICK, E. A.; MARTINS, M. A.; ROCVVCO, P. R. M.; TIBÉRIO, I. F. L. C. Y-27632 is associated with corticosteroid potentiated control of pulmonar remodeling and inflammation in guinea pigs with chronic allergic inflammation. BMC Pulmonary Medicine, v. 15, p. 85-100, 2015.
PINKERTON, J.; KIM, R.; ESSILFIE, A.; RAE, B.; MAYALL, J.; ALI, M. K.; STARKEY, M.; WOOD, L.; BISWAL, S.; HORVAT, J.; HANSBRO, P. Investigating antioxidant therapy for steroid-resistant asthma. European Respiratory Journal, 2016.
PONATH, P. D.; QIN, S.; POST, T.W.; WANG, J.; WU, L. Molecular cloning and characterization of a human eotaxin receptor expressed selectively on eosinophils. J. Exp. Med. v. 183, p. 2437-2448, 1996.
POSSA, S. S.; CHARAFEDDINE, H. T.; RIGHETTI, R. F.; SILVA, P. A.; ALMEIDA-REIS R.; SARAIVA-ROMANHOLO, B. M.; PERINI, A.; PRADO, C. M.; LEICK-MALDONADO, E. A.; MARTINS, M. A.; TIBÉRIO, I. F. L. C. Rho-kinase inhibition attenuates airway responsiveness, inflammation, matrix remodeling, and oxidative stress activation induced by chronic inflammation. American Journal of Physiology-Lung Cellular and Molecular Physiology, v. 303, n. 11, p. L939-L952, 2012.
POSSA, S. S.; RIGHETTI, R. F.; RUIZ-SCHÜTZ, V. C.; NAKASHIMA, A. S.; PRADO, C. M.; LEICK, E. A.; MARTINS, M. A.; TIBÉRIO, I. F. L. C. Influence of Oral Tolerance on Lung Cytokines Expression and Oxidative Stress Activation in Guinea Pigs with Chronic Inflammation. Journal of Allergy and Therapy, v. 5, p. 165, 2014.
POWELL, C.; MILAN, S. J.; DWAN, K.; BAX, L.; WALTERS, N. Mepolizumab versus placebo for asthma. Status and Date: New, published in, n. 7, 2015.
PRADO, C. M.; LEICK, E. A.; LOPES, F. D. T. Q. S.; MARTINS, M. A.; TIBÉRIO, I. F. L. C. Different modulators of airways and distal lung parenchyma contractile responses in the physiopathology of asthma. In: Haruo Sug, InTech-OpenAccess Publisher (Org.) (Eds.), Current Basic and Pathological Approaches to the Function of Muscle Cells and Tissues – From Molecules to Humans. , 1st ed. InTech – Open Access Publisher, Rijeka-Croatia, ISBN 978-953-51-0679-1, p. 1–30 (Chapter 13) 2012.
PRADO, C. M.; LEICK-MALDONADO, E. A.; KASAHARA, D. I.; CAPELOZZI, V. L.; MARTINS, M. A.; TIBÉRIO, I. F. Effects of acute and chronic nitric oxide inhibition in an experimental model of chronic pulmonary allergic inflammation in guinea pigs.

260 VASCONCELOS, L. H. C. Referências
American Journal of Physiology-Lung Cellular and Molecular Physiology, v. 289, n. 4, p. L677-L683, 2005.
PRADO, C. M.; RIGHETTI, R. F.; DA SILVA PIGATI, P. A.; POSSA, S. S.; DOS SANTOS, A. S. A. New Pharmacological Targets for Asthma Drug Development. Journal of Allergy and Therapy, v. 5, p. 170, 2014.
PROFITA, M.; BONANNO, A.; SIENA, L.; FERRARO, M.; MONTALBANO, A. M.; POMPEO, F.; RICCOBONO, L.; PIEPER, M. P.; GJOMARKAJ, M. Acetylcholine mediates the release of IL-8 in human bronchial epithelial cells by a NFkB/ERK-dependent mechanism. Eur. J. Pharmacol., v. 582, p. 145 153, 2008.
PRUSSIN, C.; METCALFE, D. D. 5. IgE, mast cells, basophils, and eosinophils. Journal of Allergy and Clinical Immunology, v. 117, p. S450-S456, 2006.
RACKE, K.; WARNKEN, M. L-arginine metabolic pathways. The Open Nitric Oxide Journal, v. 2, p. 9-19, 2010.
RAMADAS, R. A., EWART, S. L., MEDOFF, B. D.; LEVINE, A. M. Interleukin-1–family member 9 stimulates chemokine production and neutrophil influx in mouse lungs. American Journal of Respiratory Cell and Molecular Biology, v. 44, p. 134–145, 2011.
RAMSAY, C. F., SULLIVAN, P., GIZYCKI, M., WANG, D., SWERN, A. S., BARNES, N. C.; REISS, P. K.; JEFFERY, P. K. Montelukast and bronchial inflammation in asthma: a randomised, doubleblind placebo-controlled trial. Respiratory Medicine, v. 103, n. 7, p. 995-1003, 2009.
RANKIN, J. A.; PICARELLA, D. E.; GEBA, G. P. Phenotypic and physiologic characterization of transgenic mice expressing interleukin 4 in the lung: lymphocytic and eosinophilic inflammation without airway hyperreactivity. Proceedings of the National Academy of Sciences of the United States of America, v. 93, n. 15, p. 7821–7825, 1996.
RAUD-MATTEDI, C. Os alimentos funcionais: A nova fronteira da indústria alimentar. Revista Sociologia Política, v.16, n.31, p. 85-100, 2008.
REAGAN-SHAW, S.; NIHAL, M.; AHMAD, N. Dose translation from animal to human studies revisited. The FASEB Journal, v. 22, p. 659-661, 2007.
REDINGTON, A. E. Modulation of nitric oxide pathways: therapeutic potential in asthma and chronic obstructive pulmonary disease. European Journal of Pharmacology, v. 533, n. 1–3, p. 263–276, 2006.
REDINGTON, A. E.; MADDEN, J.; FREW, A. J.; DJUKANOVIC, R.; ROCHE, W. R.; HOLGATE, S. T.; HOWARTH, P. H. Transforming growth factor-beta 1 in asthma - Measurement in bronchoalveolar lavage fluid. American Journal of Respiratory and Critical Care Medicine, v. 156, p. 642-647, 1997.
REED, J. A.; ALBINO, A. P.; MCNUTT, N. S. Human cutaneous mast cells express basic fibroblast growth factor. Laboratory Investigation; a Journal of Technical Methods and Pathology, v. 72, n. 2, p. 215-222, 1995.

261 VASCONCELOS, L. H. C. Referências
REMBOLD, C. M. Electromechanical and pharmacomechanical coupling. In: BÁRÁNY, M. (ed.). Biochemistry of Smooth Contraction. San Diego, Academic Press, p. 227-239, 1996.
REPINE, J. E.; BAST, A. A. L. T.; LANKHORST, I. D. A. Oxidative stress in chronic obstructive pulmonary disease. American Journal of Respiratory and Critical Care Medicine, v. 156, n. 2, p. 341-357, 1997.
RESSMEYER, A. R.; LARSSON, A. K.; VOLLMER, E.; DAHLÉN, S. E.; UHLIG, S.; MARTIN, C. Characterisation of guinea pig precision-cut lung slices: comparison with human tissues. European Respiratory Journal, v. 28, n. 3, p. 603-611, 2006.
RHODEN, K. J.; BARNES, P. J. Effect of hydrogen peroxide on guinea‐pig tracheal smooth muscle in vitro: role of cyclo‐oxygenase and airway epithelium. British Journal of Pharmacology, v. 98, n. 1, p. 325-330, 1989.
RHODEN, K. J.; MELDRUM, L. A.; BARNES, P. J. Inhibition of cholinergic neurotransmission in human airways by beta 2-adrenoceptors. J Appl Physiol, v. 65, p. 700-705, 1988.
RICCIARDOLO, F. L. M.; STERK, P. J.; GASTON, B.; FOLKERTS, G. Nitric oxide in health and disease of the respiratory system. Physiological Reviews, v. 84, n. 3, p. 731-765, 2004.
RICCIARDOLO, F. L. Multiple roles of nitric oxide in the airways. Thorax, v. 58, p. 175-182, 2003.
RICCIARDOLO, F. L.; TIMMERS, M. C.; GEPPETTI, P.; VAN SCHADEWIJKD, A.; BRAHIM, J. J.; SONT, J. K.; GOUW, H. W. F. M.; HIEMSTRA, P. S.; VAN KRIEKEN, J. H. J. M.; STERK, P. J. Allergeninduced impairment of bronchoprotective nitric oxide synthesis in asthma. Journal of Allergy and Clinical Immunology, v. 108, n. 2, p. 198-204, 2001.
RIESENFELD, E. P.; SULLIVAN, M. J.; THOMPSON-FIGUEROA, J. A.; HAVERKAMP, H. C.; LUNDBLAD, L. K.; BATES, J. H.; IRVIN, C. G. Inhaled salmeterol and/or fluticasone alters structure/function in a murine model of allergic airways disease. Respiratory Research, v. 11, p. 22, 2010.
RILEY, J. F. Histamine in tissue mast cells. Science, v. 118, p. 332, 1953.
RINALDO-MATTHIS, A.; HAEGGSTROM, J. Z. Structures and mechanisms of enzymes in the leukotriene cascade. Biochimie, v. 92, n. 6, p. 676-681, 2010.
ROBINSON, C.; HARDY, C. C.; HOLGATE, S. T. Pulmonary synthesis, release, and metabolism of prostaglandins. J Allergy Clin Immunol, v. 76, p. 265-271, 1985.
ROBINSON, D. S.; HAMID, Q.; YING, S.; TSICOPOULOS, A.; BARKANS, J.; BENTLEY, A. M.; CORRIGAN, C.; DURHAM, S. R.; KAY, A. B. Predominant TH2-like bronchoalveolar T-lymphocyte population in atopic asthma. New England Journal of Medicine, v. 326, n. 5, p. 298-304, 1992.

262 VASCONCELOS, L. H. C. Referências
ROCHE, W. R.; WILLIAMS, J. H.; BEASLEY, R.; HOLGATE, S. T. Subepithelial fibrosis in the bronchi of asthmatics. Lancet, v. 1, n. 8637, p. 520–524, 1989.
ROFFEL, A. F.; ELZINGA, C. R.; VAN AMSTERDAM, R. G.; DE ZEEUW, R. A.; ZAAGSMA, J. Muscarinic M2 receptors in bovine tracheal smooth muscle: discrepancies between binding and function. European Journal of Pharmacology, v. 153, p. 73-82, 1988.
ROFFEL, A. F.; MEURS, H.; ZAAGSMA, J. Identification, localization and function of muscarinic receptor subtypes in the airways. In: ZAAGSMA, J.; MEURS, H.; ROFFEL, A. F. (eds.) Muscarinic Receptors in Airways Diseases. Basel, Birkhauser Verlag, p. 63-85, 2001.
ROMAGNANI, S. Role of lymphocytes in allergic disease. Journal of Allergy and Clinical Immunology, v. 105, p. 399–408, 2000.
ROSE, M. C.; VOYNOW, J. A. Respiratory tract mucin genes and mucin glycoproteins in health and disease. Physiol Ver, v. 86, p. 245–278, 2006.
ROUX, E. Temporal aspects of calcium signaling in airways smooth muscle cells. In Calcium Signaling In Airway Smooth Muscle Cells. Edited by Wang YX. New York: Springer, 2013.
ROUX, E.; MOLIMARD, M.; SAVINEAU, J. P.; MARTHAN, R. Muscarinic stimulation of airway smooth muscle cells. Gen Pharmacol, v. 31, p. 349-356, 1998.
ROUX, E.; NOBLE, P. J.; NOBLE, D.; MARHL, M. Modelling of calcium handling in airway myocytes. Progress in Biophysics and Molecular Biology, v. 90, n. 1, p. 64-87, 2006.
RUOCCO, S.; LALLEMAND, A.; TOURNIER, J. M.; GAILLARD, D. Expression and localization of epidermal growth factor, transforming growth factor-alpha, and localization of their common receptor in fetal human lung development. Pediatric Research, v. 39, p. 448-455, 1996.
SADEGHI-HASHJIN, G.; FOLKERTS, G.; HENRICKS, P. A. J.; MUIJSERS, R. B. R.; NIJKAMP, F. P. Peroxynitrite in airway diseases. Clinical and Experimental Allergy, v. 28, p. 1464-1473, 1998.
SADEGHI-HASHJIN, G.; FOLKERTS, G.; HENRICKS, P. A.; VERHEYEN, A. K.; VAN DER LINDE, H. J.; VAN ARK, I.; COENE, A.; NIJKAMP, F. P. Peroxynitrite induces airway hyperresponsiveness in guinea pigs in vitro and in vivo. American Journal of Respiratory and Critical Care Medicine, v. 153, n. 5, p. 1697-1701, 1996.
SAENZ, S. A.; SIRACUSA, M. C.; PERRIGOUE, J. G.; SPENCER, S. P.; URBAN JR, J. F.; TOCKER, J. E.; BUDELSKY, A. L.; KLEINSCHEK, M. A.; KASTELEIN, R. S.; KAMBAYASHI, T.; BHANDOOLA, A.; ARTIS, D. IL25 elicits a multipotent progenitor cell population that promotes TH2 cytokine responses. Nature, v. 464, p. 1362–1366, 2010.

263 VASCONCELOS, L. H. C. Referências
SAETTA, M.; DI STEFANO, A.; ROSINA, C.; THIENE, G.; FABBRI, L. M. Quantitative structural analysis of peripheral airways and arteries in sudden fatal asthma. American Review of Respiratory Disease, v. 143, n. 1, p. 138–143, 1991.
SAKAI, H.; OTOGOTO, S.; CHIBA, Y.; ABE, K.; MISAWA, M. Involvement of p42/44 MAPK and RhoA protein in augmentation of ACh-induced bronchial smooth muscle contraction by TNF-a in rats. Journal of Applied Physiology, v. 97, n. 6, p. 2154-2159, 2004.
SALEH, D.; ERNST, P.; LIM, S.; BARNES, P. J.; GIAID, A. Increased formation of the potent oxidant peroxynitrite in the airways of asthmatic patients is associated with induction of nitric oxide synthase: effect of inhaled glucocorticoid. FASEB Journal, v. 12, n. 11, p. 929-937, 1998.
SAMB, A.; PRETOLANI, M.; DINH-XUAN, A. T.; OUKSEL, H.; CALLEBERT, J.; LISDERO, C.; AUBIER, M.; BOCZKOWSKI, J. Decreased pulmonary and tracheal smooth muscle expression and activity of type 1 nitric oxide synthase (nNOS) after ovalbumin immunization and multiple aerosol challenge in guinea pigs. American Journal of Respiratory and Critical Care Medicine, v. 164, n. 1, p. 149-154, 2001.
SAMUELSSON, B. Leukotrienes: mediators of immediate hypersensitivity reactions and inflammation. Science, v. 220, p. 568-575, 1983.
SAMUELSSON, B.; DAHLEN, S. E.; LINDGREN, J. A.; ROUZER, C. A.; SERHAN, C. N. Leukotrienes and lipoxins: structures, biosynthesis, and biological effects. Science, v. 237, p. 1171-1176, 1987.
SANDERS, S. P.; ZWEIER, J. L.; HARRISON, S. J.; TRUSH, M. A.; REMBISH, S. J.; LIU, M. C. Spontaneous oxygen radical production at sites of antigen challenge in allergic subjects. American Journal of Respiratory and Critical Care Medicine, v. 151, n. 6, p. 1725-1733, 1995.
SANDERSON, M. J.; DELMOTTE, P.; BAI, Y.; PEREZ-ZOGBHI, J. F. Regulation of airway smooth muscle cell contractility by Ca2þ signaling and sensitivity. Proceedings of the American Thoracic Society, v. 5, n. 1, p. 23-31, 2008.
SANTING, R.E.; MEURS, H.; VAN DER MARK, T. W.; REMIE, R.; OOSTEROM, W. C.; BROUWER, F.; ZAAGSMA, J. A novel method to assess airway function parameters in chronically instrumented, unrestrained guinea pigs. Pulmonary Pharmacology, v. 5, p. 265–272, 1992.
SAPEY, E.; WILSON, D. The Natural History of Asthma. In: SAPEY, E. (Ed.). Bronchial Asthma – Emerging Therapeutic Strategies. INTECH Open Access Publisher, p. 3-18, 2012.
SAVINEAU, J. P.; MARTHAN, R. Activation properties of chemically skinned fibres from human isolated bronchial smooth muscle. Journal of Physiology, v. 474, n. 3, p. 433-438, 1994.
SCHAAFSMA, D.; GOSENS, R.; BOS, I. S. T.; MEURS, H.; ZAAGSMA, J.;
NELEMANS, S. A. Allergic sensitization enhances the contribution of Rho‐kinase to

264 VASCONCELOS, L. H. C. Referências
airway smooth muscle contraction. British Journal of Pharmacology, v. 143, n. 4, p. 477-484, 2004.
SCHELLENBERG, R. R.; FOSTER, A. In vitro responses of human asthmatic airway and pulmonary vascular smooth muscle. International Archives of Allergy and Applied Immunology, v. 75, p. 237-241, 1984.
SCHERRER, D.; LACH, E.; LANDRY, Y.; GIES, J. P. Glucocorticoid modulation of muscarinic and β‐adrenergic receptors in guinea pig lung. Fundamental and Clinical Pharmacology, v. 11, n. 2, p. 111-116, 1997.
SCHILD, H. O. pA, a new scale for the measurement of drug antagonism. British journal of pharmacology, v. 2, n. 3, p. 189-206, 1947.
SCHILD, H. O. pAx and competitive drug antagonism. British journal of pharmacology, v. 4, n. 3, p. 277-280, 1949.
SCHMIDT, M.; SUN, G.; STACEY, M. A.; MORI, L.; MATTOLI, S. Identification of circulating fibrocytes as precursors of bronchial myofibroblasts in asthma. Journal of Immunology, v. 171, n. 1, p. 380-389, 2003.
SCHNACKENBERG, C. G.; WILCOX, C. S. The SOD mimetic tempol restores vasodilation in afferent arterioles of experimental diabetes. Kidney International, v. 59, n. 5, p. 1859-1864, 2001.
SCHNEIDER, E.; PETIT-BERTRON, A. F.; BRICARD, R.; LEVASSEUR, M.; RAMADAN, A.; GIRARD, J. P.; HERBELIN, A.; DY, M. IL-33 activates unprimed murine basophils directly in vitro and induces their in vivo expansion indirectly by promoting hematopoietic growth factor production. The Journal of Immunology, v. 183, p. 3591–3597, 2009.
SCHRAMM, C. M. β-adrenergic relaxation of rabbit tracheal smooth muscle: a receptor deficit that improves with corticosteroid administration. Journal of Pharmacology and Experimental Therapeutics, v. 292, n. 1, p. 280-287, 2000.
SCHULTZ, W. H. Physiological studies in anaphylaxis. I. The reaction of smooth muscle of the guinea pig sensitized with horse serum. Journal of Pharmacology and Experimental Therapeutics, v. 1, p. 549-567, 1910.
SEASHOLTZ, T. M.; MAJUMDAR, M.; KAPLAN, D. D.; BROWN, J. H. Rho and Rho kinase mediate thrombin-stimulated vascular smooth muscle cell DNA synthesis and migration. Circulation Research, v. 84, p. 1186-1193, 1999.
SENEVIRATNE, K. N.; DISSANAYAKE, M. S. Variation of phenolic content in coconut oil extracted by two conventional methods. International Journal of Food Science & Technology, v. 43, n. 4, p. 597-602, 2008.
SERHAN, C. N.; PRESCOTT, S. M. The scent of a phagocyte: advances on leukotriene b(4) receptors. Journal of Experimental Medicine, v. 192, n. 3, p. F5-F8, 2000.

265 VASCONCELOS, L. H. C. Referências
SHAKERI, F.; SOUKHTANLOO, M.; BOSKABADY, M. H. The effect of hydro-ethanolic extract of Curcuma longa rhizome and curcumin on total and differential WBC and serum oxidant, antioxidant biomarkers in rat model of asthma. Iranian Journal of Basic Medical Sciences, v. 20, n. 2, p. 155, 2017.
SHAUKAT, A.; TARIQ, S.; SHARIF, M.; NAJMI, M. H. Effect of Nebivolol on tone of tracheal muscle of guinea pig. Journal of Ayub Medical College Abbottabad, v. 27, n. 3, p. 527-529, 2015.
SHERWIN, C. M.; CHRISTIANSEN, S. B.; DUNCAN, I. J.; ERHARD, H. W.; LAY, D. C.; MENCH, J. A.; O’CONNOR, C. E.; PETHERICK, J. C. Guidelines for the ethical use of animals in applied ethology studies. Applied Animal Behaviour Science, v. 81, n. 3, p. 291-305, 2003.
SHIMIZU, S.; GABAZZA, E. C.; HAYASHI, T.; IDO, M.; ADACHI, Y.; SUZUKI, K. Thrombin stimulates the expression of PDGF in lung epithelial cells. American Journal of Physiology, v. 279, p. L503-L510, 2000.
SHOKI, A. H.; MAYER-HAMBLETT, N.; WILCOX, P. G.; SIN, D. D.; QUON, B. S. Systematic review of blood biomarkers in cystic fibrosis pulmonary exacerbations. Chest Journal, v. 144, n. 5, p. 1659-1670, 2013.
SILVA, A. S.; PAIM, F. C.; SANTOS, R. C.; SANGOI, M. B.; MORESCO, R. N.; LOPES, S. T.; JAQUESE, J. A.; BALDISSARELLIF, J.; MORSCH, V. M.; MONTEIRO, S. G. Nitric oxide level, protein oxidation and antioxidant enzymes in rats infected by Trypanosoma evansi. Experimental Parasitology, v. 132, n. 2, p. 166-170, 2012.
SILVER, M. R.; MARGULIS, A.; WOOD, N.; GOLDMAN, S. J.; KASAIAN, M.; CHAUDHARY, D. IL-33 synergizes with IgE-dependent and IgE-independent agents to promote mast cell and basophil activation. Inflammation Research, v. 59, n. 3, p. 207-218, 2010.
SIMPSON, A.; TAN, V. Y.; WINN, J.; SVENSÉN, M.; BISHOP, C. M.; HECKERMAN, D. E.; BUCHAN, I.; CUSTOVIC, A. Beyond atopy: multiple patterns of sensitization in relation to asthma in a birth cohort study. American Journal of Respiratory and Critical Care Medicine, v. 181, n. 11, p. 1200-1206, 2010.
SIROUX, V.; BASAGAÑA, X.; BOUDIER, A.; PIN, I.; GARCIA-AYMERICH, J.; VESIN, A.; SLAMA, R.; JARVIS, D.; ANTO, J. M.; KAUFFMANN, F.; SUNYER, J. Identifying adult asthma phenotypes using a clustering approach. European Respiratory Journal, v. 38, p. 310-317, 2011.
SJOERDSMA, A.; WAALKES, T. P.; WEISSBACH, H. Serotonin and histamine in mast cells. Science, v. 125, p. 1202-1203, 1957.
SMITH, W. L.; LANGENBACH, R. Why there are two cyclooxygenase isozymes. J Clin Invest., v. 107, p. 1491–1495, 2001.
SNETKOV, V. A.; HAPGOOD, K. J.; MCVICKER, C. G.; LEE, T. H.; WARD, J. P. T. Mechanisms of leukotriene D4- induced constriction in human small bronchioles. British Journal of Pharmacology, v. 133, n. 2, p. 243-252, 2001.

266 VASCONCELOS, L. H. C. Referências
SOLÉ, D.; CAMELO-NUNES, I. C.; WANDALSEN, G. F.; PASTORINO, A. C.; JACOB, C. M.; GONZALEZ, C.; WANDALSEN, N. F.; ROSÁRIO FILHO, N. A.; FISCHER, G. B.; NASPITZ, C. K. Prevalence of symptoms of asthma, rhinitis, and atopic eczema in Brazilian adolescents related to exposure to gaseous air pollutants and socioeconomic status. Journal of Investigational Allergology and Clinical Immunology, v. 17, n. 1, p. 6-13, 2007.
SOLÉ, D.; WANDALSEN, G. F.; CAMELO-NUNES, I. C.; NASPITZ, C. K. ISAAC - BRAZILIAN GROUP. Prevalence of symptoms of asthma, rhinitis, and atopic eczema among Brazilian children and adolescents identified by the International Study of Asthma and Allergies in Childhood (ISAAC) - Phase 3. Jornal de Pediatria, v. 82, n. 5, 341-346, 2006.
SOMLYO, A. P.; SOMLYO, A. V. Ca2+ Sensitivity of Smooth Muscle and Nonmuscle Myosin II: Modulated by G Proteins, Kinases, and Myosin Phosphatase. Physiological Reviews, v. 83, n. 4, p. 1325-1358, 2003.
SOTMRADA, J. F.; DICKEY, D. W. Effect of Antigen Challenge on Sensitized Guinea-pig Trachea. Respiration Physiology, v. 27, p. 241-251, 1976.
SOUZA-MACHADO, A.; SANTOS, P. M.; CRUZ, A. A. Adherence to treatment in severe asthma: predicting factors in a program for asthma control in Brazil. World Allergy Organization Journal, v. 3, n. 3, p. 48-52, 2010.
STARLING, C. M.; PRADO, C. M.; LEICK-MALDONADO, E. A.; LANÇAS, T.; REIS, F. G.; ARISTÓTELES, L. R.; DOLHNIKOFF, M.; MARTINS, M. A.; TIBÉRIO, I. F. Inducible nitric oxide synthase inhibition attenuates lung tissue responsiveness and remodeling in a model of chronic pulmonary inflammation in guinea pigs. Respiratory Physiology and Neurobiology, v. 165, p. 185-194, 2009.
STRACHAN, D. P.; BUTLAND, B. K.; ANDERSON, H. R. Incidence and prognosis of asthma and wheezing illness from early childhood to age 33 in a national British cohort. BMJ, v. 312, n. 7040, p. 1195-1199, 1996.
STRINGHETA, P. C.; DE OLIVEIRA, T. T.; GOMES, R. C.; DO AMARAL, M. D. P. H.; DE CARVALHO, A. F.; VILELA, M. A. P. Políticas de saúde e alegações de propriedades funcionais e de saúde para alimentos no Brasil. Revista Brasileira de Ciências Farmacêuticas, v. 43, n. 2, p. 181-194, 2007.
SU, W. Y.; FOLZ, R.; CHEN, J. S.; CRAPO, J. D.; CHANG, L. Y. Extracellular superoxide dismutase mRNA expressions in the human lung by in situ hybridization. American Journal of Respiratory Cell and Molecular Biology, v. 16, n. 2, p. 162-170, 1997.
SUGIURA, H.; ICHINOSE, M.; OYAKE, T.; MASHITO, Y.; OHUCHI, Y.; ENDOH, N.; MIURA, M.; YAMAGATA, S.; KOARAI, A.; AKAIKE, T.; MAEDA, H.; SHIRATO, K. Role of peroxynitrite in airway microvascular hyperpermeability during late allergic phase in guinea pigs. American Journal of Respiratory and Critical Care Medicine, v. 160, n. 2, p. 663-671, 1999.
SUTCLIFFE, A.; HOLLINS, F.; GOMEZ, E.; SAUNDERS, R.; DOE, C.; COOKE, M.; CHALLISS, R. A. J.; BRIGHTLING, C. E. Increased nicotinamide adenine dinucleotide

267 VASCONCELOS, L. H. C. Referências
phosphate oxidase 4 expression mediates intrinsic airway smooth muscle hypercontractility in asthma. American Journal of Respiratory and Critical Care Medicine, v. 185, n. 3, p. 267-274, 2012.
SUZUKI, R.; KATO, T.; MIYAZAKI, Y. Matrix metalloproteinases and tissue inhibitors of matrix metalloproteinases in sputum from patients with bronchial asthma. Journal Asthma, v. 38, p. 477–484, 2001.
SUZUKI-YAMAMOTO, T.; NISHIZAWA, M.; FUKUI, M.; OKUDA-ASHITAKA, E.; NAKAJIMA, T.; ITO, S.; WATANABE, K. cDNA cloning, expression and characterization of human prostaglandin F synthase. FEBS Lett., v. 462, n. 3, p. 335-340, 1999.
SYLVESTER, J. T. The tone of pulmonary smooth muscle: ROK and Rho music? American Journal of Physiology-Lung Cellular and Molecular Physiology, v. 287, n. 4, p. L624-L630, 2004.
SZAREK, JOHN L.; SCHMIDT, NANCY L. Hydrogen peroxide-induced potentiation of contractile responses in isolated rat airways. American Journal of Physiology-Lung Cellular and Molecular Physiology, v. 258, n. 4, p. L232-L237, 1990.
TAGAYA, E.; TAMAOKI, J. Mechanisms of airway remodeling in asthma. Allergology International, v. 56, n. 4, p. 331-340, 2007.
TAGER, A. M.; LUSTER, A. D. BLT1 and BLT2: the leukotriene B(4) receptors. Prostaglandins Leukot Essent Fatty Acids, v. 69, p. 123-134, 2003.
TAKEDA, M.; ITO, W.; TANABE, M.; UEKI, S.; KIHARA, J.; KATO, H.; TANIGAI, T.; KAYABA, H.; SASAKI, T.; CHIHARA, J. The pathophysiological roles of PI3Ks and therapeutic potential of selective inhibitors in allergic inflammation. International Archives of Allergy and Immunology, v. 152, p. 90-95, 2010.
TANAKA, K.; OGAWA, K.; SUGAMURA, K.; NAKAMURA, M.; TAKANO, S.; NAGATA, K. Cutting edge: differential production of prostaglandin D2 by human helper T cell subsets. J Immunol., v. 164, p. 2277–2280, 2000.
TASHKIN, D. P.; CELLI, B.; SENN, S.; BURKHART, D.; KESTEN, S.; MENJOGE, S.; DECRAMER, M. A 4-year trial of tiotropium in chronic obstructive pulmonary disease. N. Engl. J. Med., v. 359, p. 1543–1554, 2008.
TEN BERGE, D. E.; SANTING, R. E.; HAMSTRA, J. J.; ROFFEL, A. F.; ZAAGSMA, J. Dysfunction of muscarinic M2 receptors after the early allergic reaction: possible contribution to bronchial hyperresponsiveness in allergic guinea‐pigs. British Journal of Pharmacology, v. 114, p. 881-887, 1995.
TEN BROEKE, R.; CROM, R.; HAPEREN, R. V.; VERWEIJ, V.; LEUSINK-MUIS, T.; ARK, I. V.; CLERCK, F.; NIJKAMP, F. P.; FOLKERTS, G. Overexpression of endothelial nitric oxide synthase suppresses features of allergic asthma in mice. Respiratory Research, v. 7, p. 58, 2006.
THIRSTRUP, S. Control of airway smooth muscle tone. I e electrophysiology and contractile mediators. Respiratory Medicine, v. 94, n. 4, p. 328-336, 2000.

268 VASCONCELOS, L. H. C. Referências
TIBÉRIO, I. F. L. C., TURCO, G. M. G., LEICK-MALDONADO, E. A., SAKAE, R. S., PAIVA, P. S. O., WARTH, M. P. T. N., SILVA, J. R. L., SALDIVA, P. H. N., MARTINS, M. A. Effects of neurokinin depletion on airway inflammation induced by chronic antigen exposure. American Journal of Respiratory and Critical Care Medicine, v. 155, n. 5, p. 1739-1747, 1997.
TO, T.; STANOJEVIC, S.; MOORES, G.; GERSHON, A. S.; BATEMAN, E. D.; CRUZ, A. A.; BOULET, L. P. Global asthma prevalence in adults: findings from the cross-sectional world health survey. BMC public health, v. 12, n. 1, p. 204, 2012.
TOKER, A.; CANTLEY, L. C. Signalling through the lipid products of phosphoinositide-3-OH kinase. Nature, v. 387, p. 673-676, 1997.
TOMITA, T. Electrical properties of airway smooth muscle. In: Airway smooth muscle in health and disease. Springer US, p. 151-167, 1989.
TOWBIN, H.; STAEHELIN, T.; GORDON, J. A method has been devised for the electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets. Biotechnology, v. 24, p. 145-149, 1992.
TOWNLEY, R. G.; AGRAWAL, S. CRTH2 antagonists in the treatment of allergic responses involving TH2 cells, basophils, and eosinophils. Annals of Allergy, Asthma & Immunology, v. 109, n. 6, p. 365-374, 2012.
TRAN, T.; STEWART, A. G. Protease-activated receptor (PAR)-independent growth and pro-inflammatory actions of thrombin on human cultured airway smooth muscle. British Journal of Pharmacology, v. 138, p. 865-875, 2003.
TSCHIRHART, E.; FROSSARD, N.; BERTRAND, C.; LANDRY, Y. Arachidonic acid metabolites and airway epithelium-dependent relaxant factor. Journal of Pharmacology and Experimental Therapeutics, v. 11, p. 310-316.
TURNBULL, L.; LEITCH, A. G.; TURNBULL, L.; CROFTON, J. W.; KAY, A. B. Mediators of immediate-type hypersensitivity in sputum from patients with chronic bronchitis and asthma. The Lancet, v. 310, n. 8037, p. 526-529, 1977.
URADE, Y.; UJIHARA, M.; HORIGUCHI, Y.; IKAI, K.; HAYAISHI, O. The major source of endogenous prostaglandin D2 production is likely antigen-presenting cells. Localization of glutathionerequiring prostaglandin D synthetase in histiocytes, dendritic, and Kupffer cells in various rat tissues. J Immunol., v. 143, p. 2982-2989, 1989.
VACHIER, I.; DAMON, M.; LE DOUCEN, C.; DE PAULET, A. C.; CHANEZ, P.; MICHEL, F. B.; GODARD, P. Increased oxygen species generation in blood monocytes of asthmatic patients. American Review of Respiratory Disease, v. 146, p. 1161-1161, 1992.
VALLY, H.; TAYLOR, M. L.; THOMPSON, P. J. The prevalence of aspirin intolerant asthma (AIA) in Australian asthmatic patients. Thorax, v. 57, n. 7, p. 569-574, 2002.

269 VASCONCELOS, L. H. C. Referências
VALLYATHAN, V.; SHI, X. The role of oxygen free radicals in occupational and environmental lung diseases. Environmental Health Perspectives, v. 105, n. 1, p. 165, 1997.
VARGAFTIG, B. B.; SINGER, M. Leukotrienes mediate part of Ova-induced lung effects in mice via EGFR. American Journal of Physiology-Lung Cellular and Molecular Physiology, v. 285, n. 4, p. L808-L818, 2003.
VASCONCELOS, Maria Helena Araujo. Avaliação da qualidade física e química de óleo de coco (Cocos nucifera L.) e seus efeitos sobre os parâmetros lipídicos e oxidativos em ratos dislipidêmicos sedentários e exercitados. 141 f. Pós-graduação em Ciência e Tecnologia de Alimentos (Dissertação) – Centro de Tecnologia, Universidade Federal da Paraíba – UFPB, 2015.
VIDO, D. L. R. Comparação da composição química e das atividades biológicas dos óleos essenciais de folhas de populações de Hedyosmum brasiliense Mart. ex Miq. provenientes da Serra do Mar e da Serra da Mantiqueira (Mata Atlântica). Dissertação (Mestrado em Biodiversidade Vegetal e Meio Ambiente). Instituto de Botânica da Secretaria do Meio Ambiente, São Paulo, 2009.
VIEGAS-JÚNIOR, C.; BOLZANI, V. S.; BARREIRO, E. J. Os produtos naturais e a química medicinal moderna. Química Nova, v. 29, n. 2, p. 326-337, 2006.
VILLARINO, B. J.; DY, L. M.; LIZADA, C. C. Descriptive sensory evaluation of virgin coconut oil and refined, bleached and deodorized coconut oil. LWT-Food Science and Technology, v. 40, n. 2, p. 193-199, 2007.
VINK, N. M.; POSTMA, D.S.; SCHOUTEN, J. P.; ROSMALEN, J. G. M.; BOEZEN, H. M. Gender differences in asthma development and remission during transition through puberty: The TRacking Adolescents' Individual Lives Survey (TRAILS) study. Journal of Allergy and Clinical Immunology, v. 126, n. 3, p. 498-504.e6, 2010.
VISSE, R.; NAGASE, H. Matrix metalloproteinases and tissue inhibitors of metalloproteinases: structure, function, and biochemistry. Circulation Research, v. 92, n. 8, p. 827–839, 2003.
VON HERTZEN, L.; HAAHTELA T. Signs of reversing trends in prevalence of asthma. Allergy, v. 60, p. 283-292, 2005.
VYSAKH, A.; RATHEESH, M.; RAJMOHANAN, T. P.; PRAMOD, C.; PREMLAL, S.; KUMAR, B. G.; SIBI, P. I. Polyphenolics isolated from virgin coconut oil inhibits adjuvant induced arthritis in rats through antioxidant and anti-inflammatory action. International Immunopharmacology, v. 20, n. 1, p. 124-130, 2014.
WADSWORTH, S. J.; SIN, D. D.; DORSCHEID, D. R. Clinical update on the use of biomarkers of airway inflammation in the management of asthma. Journal of Asthma and Allergy, v. 4, p. 77, 2011.
WAHL, M.; EDDINGER, T. J.; HAI, C. M. Sinusoidal length oscillation- and receptormediated mRNA expression of myosin isoforms and alpha-SM actin in airway smooth muscle. Am. J. Physiol. Cell. Physiol., v. 287, p. 1697–1708, 2004.

270 VASCONCELOS, L. H. C. Referências
WALLACE, F. A.; MILES, E. A.; EVANS, C.; STOCVVK, T. E.; YAQOOB, P.; CALDER, P. C. Dietary fatty acids influence the production of Th1-but not Th2-type cytokines. Journal of Leukocyte Biology, v. 69, n. 3, p. 449-457, 2001.
WALSH, E. R.; SAHU, N.; KEARLEY, J.; BENJAMIN, E.; KANG, B. H.; HUMBLES, A.; AUGUST, A. Strain-specific requirement for eosinophils in the recruitment of T cells to the lung during the development of allergic asthma. Journal of Experimental Medicine, v. 205, n. 6, p. 1285-1292, 2008.
WALSH, G.M. Novel therapies for asthma--advances and problems.Current Pharmaceutical Design, v. 11 p. 3027-3038, 2005.
WALTERS, E. H.; BEVAN, C.; PARRISH, R. W.; DAVIES, B. H.; SMITH, A P. Time-dependent effect of prostaglandin E2 inhalation on airway responses to bronchoconstrictor agents in normal subjects. Thorax, v. 37, n. 6, p. 438-442, 1982.
WARNER, T. D.; MITCHELL, J. A. Cyclooxygenases: new forms, new inhibitors, and lessons from the clinic. FASEB J., v. 18,p. 790–804, 2004.
WATSON, N.; EGLEN, R. M. Effects of muscarinic M2 and M3 receptor stimulation and antagonism on responses to isoprenaline of guinea‐pig trachea in vitro. British Journal of Pharmacology, v. 112, n. 1, p. 179-187, 1994.
WEBB, R. C. Smooth muscle contraction and relaxation. Advances in Physiology Education, v. 27, n. 4, p. 201-206, 2003.
WENZEL, S. E. Asthma phenotypes: the evolution from clinical to molecular approaches. Nature Medicine, v. 18, p. 716-725, 2012.
WESSLER, I.; KIRKPATRICK, C.J. Acetylcholine beyond neurons: the non neuronal cholinergic system in humans. Br. J. Pharmacol., v. 154, p. 1558–1571, 2008.
WESTERHOF, F. J.; WESTERHOF, F.; TIMENS, W.; OOSTEN, A. V.; ZUIDHOF, A. B.; NAUTA, N.; SCHUILING, M.; VOS, J. T. W. M.; ZAAGSMA, J.; MEURS, H.; COERS, W. Inflammatory cell distribution in guinea pig airways and its relationship to airway reactivity. Mediators of Inflammation, v. 10, p. 143–154, 2001.
WHICKER, S. D.; ARMOUR, C. L.; BLACK, J. L. Responsiveness of bronchial smooth muscle from asthmatic patients to relaxant and contractile agonists. Pulmonary Pharmacology, v. 1, n. 1, p. 25-31, 1988.
WHITTLE, B. J.; MONCADA, S. Pharmacological interactions between prostacyclin and thromboxanes. British Medical Bulletin, v. 39, p. 232-238, 1983.
OMS. Organização Mundial de Saúde. Asthma, 2017. Disponível em: <http://www.who.int/mediacentre/factsheets/fs307/en/>. Acesso em: 05 julho 2017.
WIDDICOMBE, J. G. Regulation of tracheobronchial smooth muscle. Physiol Rev, v. 43, p. 1-37, 1963.
WILDMAN, R. E. C. (Ed.). Handbook of nutraceuticals and functional foods. CRC press, 2016.

271 VASCONCELOS, L. H. C. Referências
WOODRUFF, P. G.; DOLGANOV, G. M.; FERRANDO, R. E.; DONNELLY, S.; HAYS, S. R.; SOLBERG, O. D.; CARTER, R.; WONG, H. H.; CADBURY, P. S.; FAHY, J. V. Hyperplasia of smooth muscle in mild to moderate asthma without changes in cell size or gene expression. American Journal of Respiratory and Critical Care Medicine, v. 169, p. 1001-1006, 2004.
WOODRUM, D. A.; BROPHY, C. M. The paradox of smooth muscle physiology. Molecular and Cellular Endocrinology, v. 177, n. 1-2, p.135-143, 2001.
OMS. Organização Mundial de Saúde. Traditional Medicine Strategy 2014-2023; WHO Geneva, Switzerland, 2013.
WRIGHT, D.; SHARMA, P.; RYU, M.; RISSÉ, P. A.; NGO, M.; MAARSINGH, H.; KOZIOL-WHITE, C.; JHA, A.; HALAYKO, A. J.; WEST, A. R. Models to study airway smooth muscle contraction in vivo, ex vivo and in vitro: implications in understanding asthma. Pulmonary Pharmacology & Therapeutics, v. 26, n. 1, p. 24-36, 2013.
XIAO, C. Defective epithelial barrier function in asthma. J. Allergy Clin. Immunol., v. 128, p. 549–556 e p. 1–12, 2011.
XIAO, C.; PUDDICOMBE, S. M.; FIELD, S.; HAYWOOD, J.; BROUGHTON-HEAD, V.; PUXEDDU, I.; HAITCHI, H. M.; VERNON-WILSON, E.; SAMMUT, D.; BEDKE, N.; CREMIN, C.; SONES, J.; DJUKANOVIĆ, R.; HOWARTH, PH.; COLLINS, J. E.; HOLGATE, S. T.; MONK, P.; DAVIES, D. E. Defective epithelial barrier function in asthma. The Journal of Allergy and Clinical Immunololy, v. 128, p. 549–556 e p. 1–12, 2011.
YAGI, Y.; KUWAHARA, M.; TSUBONE, H. Ca2+-activated K+ channel blockers induce PKC modulated oscillatory contractions in guinea pig trachea. Comparative Biochemistry and Physiology. Toxicology & Pharmacology: CBP, v. 131, n. 4, p. 511-519, 2002.
YALCIN, A.; GORCZYNSKI, R. M.; PARLAK, G. E.; KARGI, A.; BISGIN, A.; SAHIN, E.; KOSE, S.; GUMUSLU, S. Total antioxidant capacity, hydrogen peroxide, malondialdehyde and total nitric oxide concentrations in patients with severe persistent allergic asthma: its relation to omalizumab treatment. Clinical Laboratory, v. 58, n. 1-2, p. 89-96, 2012.
YAN, Z. Q.; HANSSON, G. K.; SKOOGH, B. E.; LOTVALL, J. O. Induction of nitric oxide synthase in a model of allergic occupational asthma. Allergy, v. 50, n. 9, p. 760-764, 1995.
YOSHIMOTO, T.; YASUDA, K.; TANAKA, H.; NAKAHIRA, M.; IMAI, Y.; FUJIMORI, Y.; NAKANISHI, K. Basophils contribute to TH2-IgE responses in vivo via IL-4 production and presentation of peptide-MHC class II complexes to CD4+ T cells. Nature Immunology, v. 10, n. 7, p. 706-712, 2009.
YOUSEFI, M.; NATEGHI, L.; REZAEE, K. Investigation of physiochemical properties, fatty acids profile and sterol content in Malaysian coconut and palm oil. Annals of Biological Research, v. 4, p. 214-219, 2013.

272 VASCONCELOS, L. H. C. Referências
ZAKARIA, Z. A.; Somchit, M. N.; Mat Jais, A. M.; Teh, L. K.; Salleh, M. Z.; Long, K. In vivo antinociceptive and anti-inflammatory activities of dried and fermented processed virgin coconut oil. Medical Principles and Practice, v. 20, n. 3, p. 231-236, 2011.
ZHANG, W.; NIE, Y.; CHONG, L.; CAI, X.; ZHANG, H.; LIN, B.; LIANG, Y.; LI, C. PI3K and Notch signal pathways coordinately regulate the activation and proliferation of T lymphocytes in asthma. Life sciences, v. 92, 890-895, 2013.
ZHOU, X. B.; WULFSEN, I.; LUTZ, S.; UTKU, E.; SAUSBIER, U.; RUTH, P.; WIELAND, T.; KORTH, M. M2 muscarinic receptors induce airway smooth muscle activation via a dual, Gβγ mediated inhibition of large conductance Ca2+-activated K+ channel activity. Journal of Biological Chemistry, v. 283, n. 30, p. 21036-21044, 2008.
ZHU, Z.; HOMER, R. J.; WANG, Z. Pulmonary expression of interleukin-13 causes inflammation, mucus hypersecretion, subepithelial fibrosis, physiologic abnormalities, and eotaxin production. Journal of Clinical Investigation, v. 103, n. 6, p. 779–788, 1999.
ZOSKY, G. R.; SLY, P. D. Animal models of asthma. Clinical & Experimental Allergy, v. 37, n. 7, p. 973-988, 2007.

Apêndices
274 VASCONCELOS, L. H. C. Apêndices
Apêndice 1 – Cromatograma do perfil de ácidos graxos do óleo de coco virgem, obtido por cromatografia gasosa.
RT: tempo de retenção. Análise realizada em duplicata.
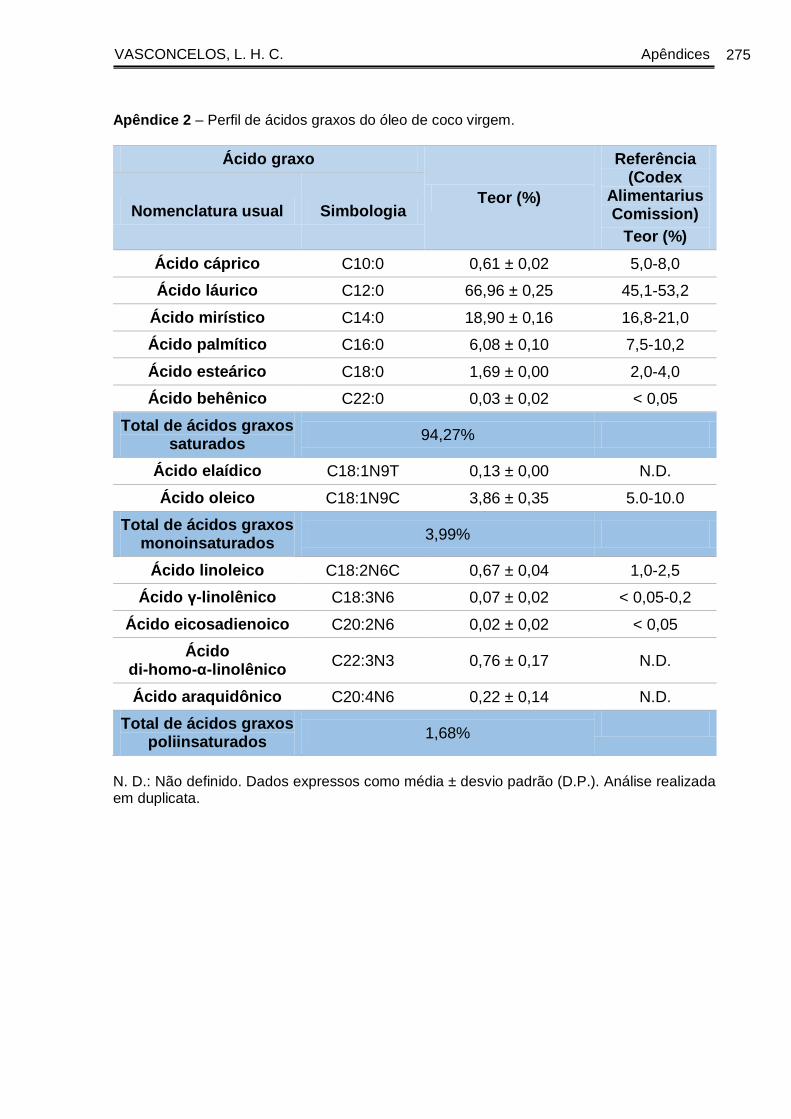
275 VASCONCELOS, L. H. C. Apêndices
Apêndice 2 – Perfil de ácidos graxos do óleo de coco virgem.
Ácido graxo
Teor (%)
Referência (Codex
Alimentarius Comission)
Teor (%)
Nomenclatura usual Simbologia
Ácido cáprico C10:0 0,61 ± 0,02 5,0-8,0
Ácido láurico C12:0 66,96 ± 0,25 45,1-53,2
Ácido mirístico C14:0 18,90 ± 0,16 16,8-21,0
Ácido palmítico C16:0 6,08 ± 0,10 7,5-10,2
Ácido esteárico C18:0 1,69 ± 0,00 2,0-4,0
Ácido behênico C22:0 0,03 ± 0,02 < 0,05
Total de ácidos graxos saturados
94,27%
Ácido elaídico C18:1N9T 0,13 ± 0,00 N.D.
Ácido oleico C18:1N9C 3,86 ± 0,35 5.0-10.0
Total de ácidos graxos monoinsaturados
3,99%
Ácido linoleico C18:2N6C 0,67 ± 0,04 1,0-2,5
Ácido γ-linolênico C18:3N6 0,07 ± 0,02 < 0,05-0,2
Ácido eicosadienoico C20:2N6 0,02 ± 0,02 < 0,05
Ácido di-homo-α-linolênico
C22:3N3 0,76 ± 0,17 N.D.
Ácido araquidônico C20:4N6 0,22 ± 0,14 N.D.
Total de ácidos graxos poliinsaturados
1,68%
N. D.: Não definido. Dados expressos como média ± desvio padrão (D.P.). Análise realizada em duplicata.

276 VASCONCELOS, L. H. C. Apêndices
Apêndice 3 – Perfil de ácidos graxos do óleo de coco virgem.
Parâmetro químico Valores
Referência
(Codex Alimentarius Comission e Anvisa)
Acidez (mg KOH/g óleo)
0,28 ± 0,00
< 4,0
(Codex Alimentarius Comission e Anvisa)
Peróxidos (meq/kg óleo)
2,28 ± 0,16
< 15,0
(Codex Alimentarius Comission e Anvisa)
Saponificação (mg KOH/g óleo)
230,57 ± 2,07
248,0-265,0
(Codex Alimentarius Comission)
Dados expressos como média ± D.P. Análises realizadas em triplicata.

Anexos

278 VASCONCELOS, L. H. C. Anexos
Anexo 1 – Certidão da Comissão de Ética no Uso de Animais da UFPB.

279 VASCONCELOS, L. H. C. Anexos
Anexo 2 – Artigo publicado na revista “Molecules”, FI: 2,861, em 2014.

280 VASCONCELOS, L. H. C. Anexos
Anexo 3 – Artigo publicado na revista “Planta Médica”, FI: 2,342, em 2015.

281 VASCONCELOS, L. H. C. Anexos
Anexo 4 – Artigo publicado na revista “BMC Complementary and Alternative Medicine”, FI: 2,288, em 2015.

282 VASCONCELOS, L. H. C. Anexos
Anexo 5 – Artigo publicado na revista “Zeitschrift fur Naturforschung. C, A Journal of Biosciences”, FI: 0,88, em 2015.

283 VASCONCELOS, L. H. C. Anexos
Anexo 6 – Artigo publicado na revista “Bioscience Reports”, FI: 2,906, em 2015.

284 VASCONCELOS, L. H. C. Anexos
Anexo 7 – Artigo publicado na revista “European Journal of Pharmacology”, FI: 2,896, em 2015.

285 VASCONCELOS, L. H. C. Anexos
Anexo 8 – Artigo publicado na revista “Frontiers in Pharmacology”, FI: 4,400, em 2016.

286 VASCONCELOS, L. H. C. Anexos
Anexo 9 – Artigo publicado na revista “Frontiers in Physiology”, FI: 4,134, em 2016.





























