Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DE SERGIPE
PRÓ-REITORIA DE PÓS-GRADUAÇÃO E PESQUISA
MESTRADO EM CIÊNCIAS FISIOLÓGICAS
GRACE KELLY MELO DE ALMEIDA
EFEITO ANTIOXIDANTE DA DIOSMINA EM MIOCÁRDIO
DE RATO SUBMETIDO À LESÃO DE REPERFUSÃO
SÃO CRISTÓVÃO
2014

i
GRACE KELLY MELO DE ALMEIDA
EFEITO ANTIOXIDANTE DA DIOSMINA EM
MIOCÁRDIO DE RATO SUBMETIDO À LESÃO DE
REPERFUSÃO
Dissertação apresentada ao Programa de Pós-
Graduação em Ciências Fisiológicas da
Universidade Federal de Sergipe como requisito à
obtenção do grau de Mestre em Ciências
Fisiológicas.
Orientadora:
Dra. Sandra Lauton Santos
SÃO CRISTÓVÃO
2014

ii
FICHA CATALOGRÁFICA ELABORADA PELA BIBLIOTECA CENTRAL
UNIVERSIDADE FEDERAL DE SERGIPE
Almeida, Grace Kelly Melo de
A447e
Efeito antioxidante da diosmina em miocárdio de rato submetido à lesão de reperfusão / Grace Kelly Melo de Almeida; orientadora Sandra Lauton Santos – São
Cristóvão, 2014. 54 f.: il.
Dissertação (mestrado em Ciências Fisiológicas) - Universidade Federal de Sergipe,
2014.
O 1. Reperfusão cardíaca. 2. Extresse oxidativo. 3. Morte celular. 4. Diosmina. I.
Santos, Sandra Lauton, orient. II. Título.
CDU: 612.17

iii
GRACE KELLY MELO DE ALMEIDA
EFEITO ANTIOXIDANTE DA DIOSMINA EM
MIOCÁRDIO DE RATO SUBMETIDO À LESÃO DE
REPERFUSÃO
Dissertação apresentada ao Programa de Pós-
Graduação em Ciências Fisiológicas da
Universidade Federal de Sergipe como requisito à
obtenção do grau de Mestre em Ciências
Fisiológicas.
Orientadora: Dra. Sandra Lauton Santos
______________________________________________________
1º Examinador: Dra. Cristiane Banni Corrêia
______________________________________________________
2º Examinador: Dr. Enilton Aparecido Camargo
______________________________________________________
1º Examinador Suplente: Dra. Virgínia Mara Pereira
SÃO CRISTOVÃO
2014

iv
Dedicatória
Minha Mãe, detentora da minha admiração.
Ao meu Pai, ausência de corpo e presença de
espírito diária.
Meu namorado, pelo amor e companheirismo.

v
AGRADECIMENTOS
A Deus por iluminar os meus caminhos e me dar forças para seguir sempre em frente.
A toda minha família e amigos, que mesmo não sabendo ao certo o que eu fazia, me
apoiavam.
À Dra. Sandra Lauton Santos, por ter me ajudado a conquistar mais uma etapa da
minha vida. Com você aprendi o compromisso com ensino e pesquisa, e, além disso, que as
dificuldades que nos são impostas precisam ser vencidas e jamais desistir dos nossos
objetivos. Enfim, agradeço pela confiança em meu trabalho e pela amizade que criamos
durante esses dois anos.
Ao Laboratório de Biofísica do Coração e seus membros, Dr. Eduardo Antônio Conde
Garcia, Dra. Carla Maria de Lins Vasconcelos e a Dra. Evaleide Diniz de Oliveira pelo
conhecimento passado a cada dia.
Aos alunos de mestrado, Jucilene Freitas, Péligris Henrique, José Marden, Rodrigo
Miguel, e aos alunos de doutorado Raquel Moreira e Thássio Mesquita pelo apoio e pela
nossa grande amizade, uma das melhores coisas que ganhei durante o mestrado. O recente
mestre, Evaldo, grande amigo e que fez meus dias no laboratório se tornarem mais divertidos.
Agradeço também a Itamar Couto, foi mais que um amigo, uma das pessoas que mais me
ajudou para o desenvolvimento desse trabalho. A recente mestra, Maraiza Bezerra, por suas
palavras de conforto durante os momentos de estresse.
Aos animais que doaram suas vidas inconscientemente em prol da evolução da
ciência.
À UFS e ao Programa de Pós-Graduação em Ciências Fisiológicas - PROCFIS e seus
membros pela contribuição da minha formação acadêmica.
A CAPES - FAPITEC, pelo suporte durante o período de mestrado.

vi
RESUMO
Efeito Antioxidante da Diosmina em Miocárdio de Rato Submetido á lesão de
Reperfusão. Grace Kelly Melo de Almeida, Mestrado em Ciências Fisiológicas, UFS, São
Cristóvão, 2014. A lesão de reperfusão é responsável por 50% do tamanho do infarto, sendo
a principal responsável pelas alterações cardíacas ocasionadas pela sobrecarga de cálcio e
estresse oxidativo que ocorrem no processo de isquemia-reperfusão. A prevenção ou
limitação desta área torna-se um alvo para a proteção cardíaca. Neste contexto, a diosmina por
ser um flavonoide que apresenta ampla atividade biológica, principalmente cardioprotetora,
apresenta-se como uma substância a ser utilizada para a prevenção dessas lesões. O objetivo
deste estudo foi avaliar o efeito antioxidante da diosmina na lesão de reperfusão. Utilizou-se o
sistema de perfusão aórtico do tipo Langendorff pressão constante para a indução do modelo
de isquemia global cardíaca. Foram utilizados ratos Wistar machos (250-300 g) e os
procedimentos foram aprovados pelo Comitê de Ética em Pesquisa com Animais da UFS
(04/2013). Os animais utilizados foram divididos em 4 grupos experimentais: Grupo 01 -
Sham: 20 minutos de estabilização, 100 minutos de perfusão com solução veículo (solução de
Krebs-Ringer acrescida de dimetilsulfóxido - DMSO 0,02 %); Grupo 02 - I-R + Veículo: 20
minutos de estabilização, 10 minutos de perfusão com solução veículo, seguido por 30
minutos de isquemia e, posteriormente, mais 60 minutos de reperfusão com a mesma solução;
Grupo 03 - I-R + Diosmina: 20 minutos de estabilização, perfusão por 10 minutos com
solução veículo, seguido por 30 minutos de isquemia e, posteriormente, mais 60 minutos de
reperfusão com solução de diosmina (0,1 µmol/L) e Grupo 04 - I-R + NAC: 20 minutos de
estabilização, perfusão por 10 minutos com solução veículo, seguido por 30 minutos de
isquemia e posteriormente, 60 minutos de reperfusão com o controle positivo NAC (N-
acetilcisteína - 24 µmol/L). Foi avaliado o efeito da diosmina sobre a contratilidade cardíaca,
através da mensuração da pressão intraventricular esquerda (PVE) e do índice de severidade
de arritmia (ASI). Além disso, foi analisada à área de lesão fazendo-se a marcação do infarto,
e mensuração da atividade das enzimas creatina quinase (CK) e lactato desidrogenase (LDH).
Também, foi analisado o efeito da diosmina na peroxidação lipídica (TBARS, hidroperóxidos
totais) e das enzimas antioxidantes superóxido dismutase (SOD), catalase (CAT), glutationa
peroxidase (GPx), glutationa redutase (GR) nos corações estudados. E por fim, foi observada
a expressão da proteína caspase-3 pela técnica de western blot. Como resultados obtivemos o
reestabelecimento da PVE quando comparada ao grupo Sham, assim como verificamos
redução do ASI (p< 0,01) em relação aos corações que foram reperfundidos com o veículo.
Observamos, ainda, diminuição (p< 0,01) da área de infarto e da atividade das enzimas CK
total e LDH. A peroxidação lipídica e a concentração de hidroperóxidos mostraram-se
reduzidas (p< 0,01) em relação aos corações reperfundidos com o veículo. Além disso, a
atividade das enzimas antioxidantes SOD, CAT, GPx e GR, também encontravam-se
reduzidas (p< 0,01) em relação aos corações reperfundidos com o veículo. Demonstramos
também redução (p< 0,01) da expressão da proteína caspase-3 quando comparado ao grupo
em que os corações foram reperfundidos com o veículo. Estes resultados em conjunto
evidenciam que a diosmina reduz as alterações decorrentes da isquemia-reperfusão cardíaca
por sua ação antioxidante.
Descritores: Reperfusão Cardíaca; Diosmina; Estresse Oxidativo; Morte Celular.

vii
ABSTRACT
Antioxidant Effect of Diosmin in Rat Myocardium Subjected to Reperfusion Injury.
Grace Kelly Melo de Almeida, MA in Physiological Sciences, UFS, São Cristóvão, 2014. The reperfusion injury is responsible for 50% of infarct size, being the main cause of cardiac
changes caused by calcium overload and oxidative stress occurring in the ischemia-
reperfusion process. Prevention or limitation of this area becomes a target for heart protection.
In this context, the diosmin to be a flavonoid that has wide biological activity, especially
cardioprotective, presents itself as a substance to be used for the prevention of these injuries.
The aim of this study was to evaluate the antioxidant effect of diosmin in reperfusion injury.
Was used aortic perfusion system of the type Langendorff constant pressure to induce cardiac
global ischemia model. Male Wistar rats (250-300 g) were used and the procedures were
approved by the Ethics Committee on Animal Research of the UFS (04/2013). The animals
were divided into 4 experimental groups: 01 - Sham: 20 minutes of stabilization, 100 minutes
of perfusion with vehicle solution (Krebs-Ringer solution plus dimethylsulfoxide - DMSO
0.02%); Group 02 - I-R + Vehicle: 20 minute of stabilization, 10 minute perfusion with
vehicle solution, followed by 30 minutes of ischemia and then reperfusion 60 minutes more
with the same solution, 10 minutes of perfusion with vehicle solution, followed by 30 minutes
of ischemia and then more 60 minutes of reperfusion with the same solution. Group 03 - I-R +
Diosmin: 20 minutes of stabilization, perfusion for 10 minutes with vehicle solution, followed
by 30 minutes of ischemia and subsequently a further 60 minutes of reperfusion with diosmin
solution (0.1 mol / L) and Group 04 - I-R + NAC: 20 minutes of stabilization, perfusion for 10
minutes with vehicle solution, followed by 30 min ischemia and after 60 minutes of
reperfusion with the positive control NAC (N-acetylcysteine - 24 mmol / L). Was evaluated
the effect of diosmin on cardiac contractility, by measuring the left intraventricular pressure
(PVE) and arrhythmia severity index (ASI). Furthermore, was analyzed the area of injury
making a mark on the infarct location, and measurement of the enzymes creatine kinase (CK)
and lactate dehydrogenase (LDH). Also, it was analyzed the effect of diosmin in lipid
peroxidation (TBARS, total hydroperoxides) and antioxidant enzymes superoxide dismutase
(SOD), catalase (CAT), glutathione peroxidase (GPx), glutathione reductase (GR) in the
hearts studied. Finally, we observed the expression of caspase-3 protein by western blot. As a
result we obtained the restoration of PVE when compared to the sham group, as well as
verified reduction of ASI (p< 0.01) compared to hearts were reperfused with the vehicle. We
also observed a decrease (p< 0.01) of the infarcted area and the overall activity of the
enzymes CK and LDH. Lipid peroxidation and the concentration of hydroperoxides shown to
be reduced (p< 0.01) compared to reperfused hearts with the vehicle. In addition, the activity
of antioxidant enzymes SOD, CAT, GPx and GR also were reduced (p< 0.01) compared to
reperfused hearts with the vehicle. Also demonstrated reduction (p< 0.01) the expression of
caspase-3 protein compared to the group in which the hearts were reperfused with the vehicle.
These results together show that the diosmin reduces the changes arising from cardiac
ischemia-reperfusion for their antioxidant activity.
Keywords: Cardiac Reperfusion; Diosmin; Oxidative Stress; Cell Death.

viii
LISTA DE FIGURAS
Figura 1. Eventos ocorridos durante a isquemia cardíaca...........................................................5
Figura 2. Eventos ocorridos durante a reperfusão cardíaca........................................................6
Figura 3. Ilustração da contratura cardíaca por sobrecarga de cálcio.........................................7
Figura 4. Esquema representativo da reação de Fenton (A) e Haber Weiss (B).........................9
Figura 5. Esquema do sistema antioxidante enzimático.............................................................9
Figura 6. Ilustração molecular da estrutura base dos flavonoides............................................12
Figura 7. Ilustração da estrutura molecular da diosmina..........................................................14
Figura 8. Representação esquemática do protocolo de perfusão com diosmina em
concentrações crescentes...........................................................................................................17
Figura 9. Esquema representativo dos grupos experimentais...................................................18
Figura 10. Ilustração do sistema de perfusão do tipo Langendorff pressão constante..............19
Figura 11. Efeito da diosmina na pressão intraventricular esquerda (PVE) em corações
isolados de ratos........................................................................................................................25
Figura 12. Efeito da diosmina na pressão intraventricular esquerda (PVE) em corações
isolados de ratos submetidos à isquemia-reperfusão................................................................26
Figura 13. Efeito da diosmina nas arritmias de reperfusão em corações isolados de ratos
submetidos à isquemia-reperfusão............................................................................................27

ix
Figura 14. Efeito da diosmina na limitação da área de infarto em corações isolados
submetidos à isquemia-reperfusão............................................................................................28
Figura 15. Efeito da diosmina na atividade da creatina quinase total (CK) em corações
isolados de ratos submetidos à isquemia-reperfusão................................................................29
Figura 16. Efeito da diosmina na atividade da lactato desidrogenase (LDH) em corações
isolados de ratos submetidos à isquemia-reperfusão................................................................29
Figura 17. Efeito da diosmina no grau de peroxidação lipídica (TBARS) em corações isolados
de ratos submetidos à isquemia-reperfusão..............................................................................31
Figura 18. Efeito da diosmina na concentração de hidroperóxidos totais em corações isolados
de ratos submetidos à isquemia-reperfusão..............................................................................32
Figura 19. Efeito da diosmina na atividade da superóxido dismutase (SOD) em corações
isolados de ratos submetidos à isquemia-reperfusão................................................................33
Figura 20. Efeito da diosmina na atividade da catalase (CAT) em corações isolados de ratos
submetidos à isquemia-reperfusão............................................................................................34
Figura 21. Efeito da diosmina na atividade da glutationa peroxidase (GPx) em corações
isolados de ratos submetidos à isquemia-reperfusão...............................................................35
Figura 22. Efeito da diosmina na atividade da glutationa redutase (GR) em corações isolados
de ratos submetidos à isquemia-reperfusão..............................................................................36
Figura 23. Efeito da diosmina na expressão proteica da caspase-3 em corações de ratos
submetidos à isquemia-reperfusão.........................................................................................38

x
LISTA DE SIGLAS
ANOVA Análise de Variância
ASI Índice de Severidade da Arritmia
ATP Adenosina Trifosfato
BHT Butil-hidroxitoluol
cAMP AMP Cíclico (Adenosina Monofosfato Cíclico)
CAT Catalase
CEPA Comitê de Ética em Pesquisa Animal
CK Creatina Quinase
CONCEA Conselho Nacional de Controle de Experimentação Animal
DMSO Dimetilsulfóxido
DNA Ácido Desoxiribonucleico
E.P.M. Erro Padrão da Média
EDTA Ácido Etilenediaminotetraacético
ELISA Do Inglês “Enzyme-linked Immunosorbent Assay”
FOX Oxidação Ferrosa do Xilenol Orange
GPx Glutationa Peroxidase
GR Glutationa Redutase
GSH Glutationa Reduzida
GSSH Glutationa Oxidada
IAM Infarto Agudo do Miocárdio
LDH Lactato Desidrogenase
MDA Malonaldeído
MPTP Poro de Transição de Permeabilidade Mitocondrial
MTT Sal de Tetrazolium
NAC N-acetilcisteína
NADP Nicotinamida Adenina Difosfato
NADPH Nicotinamida Adenina Difosfato Reduzida

xi
NADPH oxidase Nicotinamida Adenina Difosfato Oxidase
NCX Trocador Sódio-Cálcio
NHE Trocador Sódio-Hidrogênio
PBS Solução Tamponada de Fosfato
pH Potencial Hidrogeniônico
PMSF Fluoreto de Fenilmetilsulfonilo
PVE Pressão Intraventricular Esquerda
RNS Espécie Reativa de Nitrogênio
ROS Espécie Reativa de Oxigênio
RS Retículo Sarcoplasmático
RyR Receptor de Rianodina
SDS-PAGE Do Inglês “Sodium Dodecyl Sulfate Polyacrylamide Gel
Electrophoresis”
SERCA Cálcio-ATPase do Retículo Sarcoplasmático
SOD Superóxido Dismutase
TBA Ácido Tiobarbitúrico
TBARS Substâncias Reativas ao Ácido Tiobarbitúrico
TCA Ácido Tricloroacético
TTC Cloreto de Trifeniltetrazólio
u.a. Unidades Arbitrárias
UFS Universidade Federal de Sergipe
XO Xantina Oxidase
ΔE Variação de Absorbância
Δψ m Potencial Mitocondrial

xii
SUMÁRIO
1. INTRODUÇÃO......................................................................................................................1
2. REVISÃO DA LITERATURA..............................................................................................3
2.1 Isquemia-Reperfusão Cardíaca.................................................................................3
2.2 Consequências da Lesão de Isquemia-Reperfusão Cardíaca....................................4
2.3 Radicais Livres e Estresse Oxidativo na Lesão de Reperfusão Cardíaca.................8
2.4 Flavonoides.............................................................................................................11
2.5 Diosmina.................................................................................................................14
3. OBJETIVOS.........................................................................................................................16
3.1 Geral........................................................................................................................16
3.2 Específicos..............................................................................................................16
4. MATERIAL E MÉTODOS..................................................................................................17
4.1 Animais...................................................................................................................17
4.2 Grupos Experimentais.............................................................................................17
4.3 Modelo de Isquemia-Reperfusão Cardíaca ............................................................18
4.4 Avaliação da Pressão Intraventricular Esquerda (PVE) .........................................19
4.5 Avaliação do Indice de Severidade da Arritmia (ASI) ..........................................19
4.6 Análises da Lesão Tecidual.....................................................................................20
4.6.1 Marcação da Área de Infarte...................................................................20
4.6.2 Determinação da Atividade da Enzima Creatina Quinase Total
(CK)...................................................................................................................20
4.6.3 Determinação da Atividade da Enzima Lactato Desidrogenase
(LDH)................................................................................................................20
4.7Avaliações dos Parâmetros Antioxidantes...............................................................21

xiii
4.7.1Determinação do Grau de Peroxidação Lipídica (TBARS) .....................21
4.7.2 Mensuração de Hidroperóxidos Totais.....................................................21
4.7.3Determinação da Atividade da Enzima Superóxido Dismutase
(SOD)................................................................................................................22
4.7.4 Determinação da Atividade da Enzima Catalase (CAT) .........................22
4.7.5 Avaliação da Atividade da Enzima Glutationa Peroxidase (GPx)...........22
4.7.6 Avaliação da Atividade da Enzima Glutationa Redutase (GR)................23
4.8 Determinação da Expressão Proteica de Caspase-3 por Western Blot....................23
4.9 Determinação da Concentração Total de Proteínas................................................24
4.10 Análises Estatísticas..............................................................................................24
5. RESULTADOS ....................................................................................................................24
6. DISCUSSÃO........................................................................................................................38
7. CONCLUSÃO......................................................................................................................46
8. REFERENCIAL BIBLIOGRÁFICO....................................................................................47
9. ANEXO.................................................................................................................................54

1
1. INTRODUÇÃO
O número de pessoas portadoras de patologias cardíacas, a exemplo da Doença
Arterial Coronariana e, consequentemente, o Infarto Agudo do Miocárdio (IAM), vem
crescendo nos últimos anos em todo o mundo. No Brasil, segundo o órgão brasileiro
DATASUS (2012) somente no mês de julho deste mesmo ano, 896 pessoas morreram em
hospitais vítimas do IAM, sendo a região nordeste a segunda maior contribuidora, 22.271.
Considerando que as doenças cardiovasculares constituem a principal causa de morbidade,
incapacidade e morte no mundo e no Brasil, o Ministério da Saúde através da Portaria nº
2.994/2011 aprovou a Linha de Cuidado do Infarto Agudo do Miocárdio e o Protocolo de
Síndromes Coronarianas Agudas (BRASIL, 2011).
Como meio de intervenção, a reperfusão é utilizada nas síndromes coronarianas
agudas. No entanto, a reperfusão tem o potencial de exacerbar a lesão tecidual, um processo
designado por "lesão de reperfusão”, sendo responsável por 50% do tamanho do infarto. A
lesão de reperfusão é representada por alterações tais como arritmias, disfunção mecânica ou
"atordoamento do miocárdio", lesão microvascular, respostas inflamatórias e apoptose
(ARTIGOU et al., 1993; BRASILEIRO, 1997; PERRELLI et al., 2011).
Os mecanismos envolvidos na fisiopatologia da lesão da reperfusão são complexos. As
teorias mais aceitas são conhecidas como “O Paradoxo do Cálcio” e o “Paradoxo do
Oxigênio” ou “Hipótese Oxiradical”. A primeira teoria tenta explicar que as alterações
decorrentes da lesão de reperfusão ocorrem devido ao grande influxo de cálcio (Ca2+
) para o
interior da célula. Já a segunda, tenta explanar que essas modificações são resultado do
“estresse oxidativo” (BRASILEIRO, 1997; PERRELLI et al., 2011).
Um dos principais focos em pesquisa cardiovascular tem sido desenvolver abordagens
para minimizar os danos no músculo cardíaco que são gerados pós-isquemia, através de
mecanismos que reduzam o tamanho do infarto assim como, todas as manifestações da lesão,
com consequente melhora no prognóstico. Assim, recentemente voltaram-se as atenções para
a pesquisa de drogas cardioprotetoras, utilizadas isoladamente ou em associação, para reduzir
ou prevenir o dano sobre a célula cardíaca (PARANG; SINGH; ARORA, 2005; WOLFF &
BUDKER, 2005).

2
Drogas derivadas de produtos naturais podem apresentar-se como substâncias com
efeitos terapêuticos nas lesões de reperfusão. Um grupo de substâncias que vêm se destacando
são os flavonoides, uma classe de polifenois encontrados em frutas, vegetais, grãos, cascas,
raízes, caules, flores, chá e vinho. Essas substâncias possuem uma série de propriedades
farmacológicas, que os fazem atuarem sobre sistemas biológicos de forma benéfica para a
saúde humana (NIJVELDT et al., 2011).
Diversas funções são atribuídas aos flavonoides, entre elas pode-se citar a capacidade
antioxidante, esta constitui a atividade mais elucidada pelos estudos até agora desenvolvidos;
atividade anti-inflamatória; vasodilatodora; analgésica; anticancerígena; atuação
antiplaquetária; bem como ações antimicrobianas e antivirais (LIN et al., 1997; VOLP et al.,
2008).
Há evidências que indicam os efeitos benéficos dos flavonoides em corações
submetidos à isquemia-reperfusão in vitro, o que pode ser de uso em situações agudas de
isquemia-reperfusão tais como, cirurgias cardíacas e transplantes (BHANDARY et al., 2012;
LEBEAU; NEVIERE; COTELLE, 2001). Além disso, estudos experimentais evidenciaram
que a administração oral de flavonoides pode fornecer proteção contra a isquemia-reperfusão
no miocárdio, o que seria um benefício para pessoas com doenças crônicas como doença
isquêmica do coração (SUZUKI et al., 2007; TOUFEKTSIAN et al., 2008).
A diosmina (3',5,7-tri-hidroxi-4'-metoxiflavona-7-ramnoglucosídeo) é um flavonoide
encontrado em frutas cítricas que possui atividades biológicas, tais como atividade anti-
inflamatória, antioxidante, anti-hipertensiva, entre outras (SRINIVASAN & PARI, 2012;
SILAMBARASAN & RAJA, 2012; TAHIR et al., 2013).
Devido à diversidade de propriedades apresentadas pela diosmina, assim como há
necessidade do surgimento de drogas cardioprotetoras contra o surgimento das lesões de
reperfusão, decidimos avaliar no presente estudo a hipótese de que a diosmina possa
contribuir para a prevenção das lesões decorrentes da reperfusão cardíaca.

3
2. REVISÃO DE LITERATURA
2.1 Isquemia-Reperfusão Cardíaca
O quadro clínico e diagnóstico do IAM foi descrito há cerca de cem anos quase
simultaneamente por dois autores russos, Obraztsov e Strazhesko, em 1910, numa revista
científica Alemã e, nos Estados Unidos, por Herrick num artigo publicado no JAMA em 1912
(MULLER, 1977).
Normalmente, a isquemia cardíaca ocorre quando o suprimento sanguíneo coronariano
para o miocárdio é interrompido devido à obstrução da vasculatura. Como o sangue através
das hemácias leva o oxigênio às células, a isquemia dá origem à hipóxia e contribui para a
fisiopatologia das lesões provocadas pelo IAM, insuficiência vascular periférica e choque
hipovolêmico (CENTURION et al., 2002).
A reperfusão compreende a restauração do fluxo sanguíneo com o consequente
restabelecimento das condições normóxicas após um período de hipóxia ou isquemia. Essa
restauração do fluxo sanguíneo produz aceleração das mudanças estruturais que se associam à
morte celular (RUIXING; WENWU; AL-GHAZALI, 2007). Existem diferentes hipóteses em
relação aos danos condicionados pela reperfusão, sendo as principais o aumento brusco na
oferta de O2 e na concentração intracelular de Ca2+
(FRÖHLICH et al., 2013; PIPER;
GARCÑA-DORADO; OVIZE, 1998b;).
Em 1972, Shen e Jennings demonstraram pela primeira vez que a lesão de reperfusão
do miocárdio resulta em sobrecarga de Ca2+
no citosol das células cardíacas. Além da
sobrecarga desse íon, a lesão de reperfusão é também ocasionada pelo estresse oxidativo
gerado, onde ambos levam a hipercontratura dos cardiomiócitos e abertura do poro de
transição de permeabilidade mitocondrial (MPTP) (FRÖHLICH et al., 2013).
As observações a respeito das mudanças decorrentes da reperfusão foram direcionadas
ao campo da cirurgia cardíaca com circulação extracorpórea, onde foi observado que na
cirurgia de revascularização miocárdica há presença de disfunção contrátil do coração, que
ocorre após o restabelecimento da circulação coronariana (LERCH, 1993; MAXWELL &
LIP, 1997).
As alterações decorrentes da reperfusão são representadas de várias maneiras: (1)
Arritmias, com a reoxigenação miocárdica, onde frequentemente se tem a presença de
arritmias graves como taquicardia e fibrilação ventricular; (2) Atordoamento do miocárdio ou

4
lesão funcional, fenômeno caracterizado por uma disfunção do miócito que irá se recuperar
em alguns dias ou horas; (3) Lesão letal, consiste da morte celular como consequência da
reperfusão (BRASILEIRO, 1997).
2.2 Consequências da Lesão de Isquemia-Reperfusão Cardíaca
Durante a isquemia do miocárdio, o metabolismo mitocondrial é deprimido e a
produção de ATP é diminuída. A ausência de O2 e substratos oxidáveis provoca a diminuição
na atividade da cadeia respiratória, sendo este o mecanismo pelo qual a síntese de ATP é
reduzida. Dessa forma, para aumentar as fontes energéticas, a célula ativa o metabolismo
anaeróbio, porém, essa estratégia se mostra insuficiente para suprir as necessidades
energéticas da célula miocárdica, o que resulta em grande aumento na produção de piruvato e
lactato. Assim, a acidificação do meio intracelular durante a isquemia, combinada com a
depleção energética, determina a inibição de quase todos os mecanismos enzimáticos da
célula, produzindo uma importante alteração iônica (LOPASCHUK et al., 2000).
Em suma, essas alterações levam ao acúmulo de prótons no citoplasma da célula e a
posterior ativação do trocador sódio-hidrogênio (NHE) para tentar normalizar a acidez
celular. A ativação do trocador NHE, por sua vez, aumenta à concentração de sódio (Na+) no
citosol, resultando na inversão do trocador sódio-cálcio (NCX), com consequente sobrecarga
de Ca2+
citosólico (WALTERS; PORTER; BROOKES, 2014). Devido a redução de ATP,
essas alterações são agravadas ainda mais pela redução da atividade da cálcio-ATPase da
membrana celular e da cálcio-ATPase do retículo sarcoplasmático (SERCA), que são
responsáveis pela remoção destes íons, contribuindo ainda mais com a sobrecarga de Ca2+
no
citosol (PIPER; KASSECKERT; ABDALLAH, 2006c). Além das alterações no transporte
iônico celular, ocorre ainda uma redução no influxo de Ca2+
para o interior das mitocôndrias
devido à redução do potencial mitocondrial (Δψm). Todavia, apesar de todas essas alterações,
o MPTP permanece no seu estado fisiológico, ou seja, no estado fechado (Figura 1)
(WALTERS; PORTER; BROOKES, 2014).

5
Figura 1 – Eventos ocorridos durante à isquemia cardíaca, adaptado de Walters et al., 2014. NHE: trocador
sódio-hidrogênio; NCX: trocador sódio-cálcio; Ca2+
-ATPase: bomba de cálcio-ATPase; Poro MPTP: poro de
transição de permeabilidade mitocondrial; ROS: espécies reativas de oxigênio; Fos - Ox: cadeia de transporte de
elétrons; Δψ m: potencial de membrana mitocondrial; NADH: nicotinamida adenina dinucleotídeo hidreto; CL:
cardiolipina; Ca2+
: íons cálcio; ATP: adenosina trifosfato; ANT: adenina nucleotídeo translocase; RYR: receptor
de rianodina.
Com a reperfusão e a consequente reintrodução de O2 a atividade do NCX é
restaurada, levando à saída de Ca2+
para o meio extracelular em troca de Na+
para o meio
intracelular (WALTERS; PORTER; BROOKES, 2014). Além disso, ocorre a restauração da
produção ATP e a posterior ativação da SERCA e da Ca2+
-ATPase da membrana plasmática,
que promovem a redução na concentração de Ca2+
no citosol que foi acumulado durante o
período de isquemia (PIPER; KASSECKERT; ABDALLAH, 2006c) (Figura 2).
Todavia, com a volta da atividade das mitocôndrias e a consequente reativação da
cadeia de transporte de elétrons, ocorre uma grande produção de espécies reativas de oxigênio
(ROS) e de espécies reativas de nitrogênio (RNS) (WALTERS; PORTER; BROOKES,
2014). Essas espécies reativas, por sua vez, promovem um ataque à cardiolipina, fosfolipídio
localizado na membrana mitocondrial interna, alterando a fluidez da membrana, a
permeabilidade iônica, a estrutura e a função dos componentes da cadeia transportadora de
elétrons mitocondrial, resultando em redução da eficiência da fosforilação oxidativa
mitocondrial (PARADIES et al., 2009). Além disso, as ROS e RNS ocasionam a ativação dos
receptores de rianodina (RyR), localizados no retículo sarcoplasmático (RS), o que culmina
em uma grande liberação de Ca2+
e contribui ainda mais para a sobrecarga deste íon no citosol
(KÖHLER; SAG; MAIER, 2014; PIPER; KASSECKERT; ABDALLAH, 2006c).

6
Os efeitos combinados da sobrecarga de Ca2+
no citosol e produção de ROS e RNS na
matriz mitocondrial favorecem a abertura do MPTP (WALTERS; PORTER; BROOKES,
2014). Com a abertura do MPTP há alteração nas membranas mitocondriais levando a uma
liberação de citocromo c. Este, por sua vez, induz a cascata de caspases que promovem a
morte celular por apoptose (MONASSIER, 2008). Tanto a apoptose quanto a necrose estão
presentes em corações isquêmicos. Na maioria dos casos, a morte celular apoptótica é mais
prevalente em lesão de isquemia-reperfusão, especificamente é iniciada durante a fase inicial
da isquemia e concluída na reperfusão (IBANEZ et al., 2011). Assim, todos esses fatores
somados são responsáveis pelo surgimento das lesões durante o período de reperfusão.
Figura 2 – Eventos ocorridos durante à reperfusão cardíaca, adaptado de Walters et al., 2014. NHE: trocador
sódio-hidrogênio; NCX: trocador sódio-cálcio; Ca2+
-ATPase: bomba de cálcio-ATPase; Poro PT: poro
mitocondrial; ROS: espécies reativas de oxigênio; Fos-Ox : cadeia de transporte de elétrons; Δψ m: potencial de
membrana mitocondrial; NADH: nicotinamida adenina dinucleotídeo hidreto; CL: cardiolipina; Ca2+
: íons
cálcio; ATP: adenosina trifosfato; ANT: adenina nucleotídeo translocase; RYR: receptor de rianodina.
Cirurgiões cardíacos observaram que a reperfusão após uma isquemia prolongada,
ocasionalmente provoca o fenômeno denominado de "coração de pedra”, ou seja, um coração
duro e pálido resultante da contratura muscular e da perda concomitante do teor de proteína
celular, representado histologicamente por cardiomiócitos hipercontraídos e membranas
celulares rompidas (GANOTE, 1983).
A contratura cardíaca está relacionada à redução sustentada e enrijecimento do
miocárdio decorrente de várias causas. A patogênese de contratura induzida por reperfusão

7
pode ser de dois tipos: contratura por sobrecarga de Ca2+
e a contratura rigor (PIPER;
ABDALLAH; SCHÄFER, 2004a).
A contratura rigor se desenvolve na isquemia-reperfusão no momento em que há
baixas concentrações citosólicas de ATP, durante o período de isquemia, com consequente
alteração das ligações entre os miofilamentos que resulta em encurtamento muscular. Com a
reperfusão, a diminuição de energia é restaurada e a contratura rigor geralmente é reversível.
A contratura desenvolvida por este mecanismo na verdade não causa grandes danos
estruturais, mas leva a defeitos no citoesqueleto tornando os cardiomiócitos mais frágeis e,
portanto, susceptíveis à danos mecânicos (PIPER; MEUTER; SCHÄFER, 2003d).
A contratura por sobrecarga de Ca2+
surge em virtude das altas concentrações
citosólicas de Ca2+
, resultante do transporte iônico do NCX em modo inverso durante a
isquemia assim como, da saída destes íons do RS durante a reperfusão (PIPER;
KASSECKERT; ABDALLAH, 2006c; WALTERS; POTER; BROOKES, 2014). Essa
sobrecarga provoca uma ativação miofibrilar descontrolada, que também é alimentada pelo
reabastecimento de ATP (Figura 3). Estes mecanismos celulares contribuem para
hipercontratura dos cardiomiócitos, que é uma das causas de lesão celular letal que ocorre
durante a reperfusão (PIPER; ABDALLAH; SCHÄFER, 2004a).
Figura 3 – Ilustração da contratura cardíaca por sobrecarca de Ca2+
, adaptada de Piper et al. (2004a). NCE:
trocador sódio-cálcio; SR: retículo sarcoplasmático; ATP: adenosina trifosfato; Ca2+
: íons cálcio.
Foi evidenciado em experimento in situ que corações que sofreram contratura
apresentaram uma grande região necrótica e que as mesmas correlacionavam-se com o

8
aumento de enzimas antioxidantes durante os primeiros minutos de reperfusão (BARRABÉS
et al., 1996). Estas observações indicam que o processo de morte celular e desenvolvimento
de hipercontraturas pós-isquêmicas são fenômenos relacionados, e são fatores responsáveis
para o desenvolvimento de lesões de reperfusão.
2.3 Radicais Livres e Estresse Oxidativo na Lesão de Reperfusão Cardíaca
Radical livre é um grupo de átomos ou moléculas em um estado particular que contém
um ou mais elétrons não pareados no seu orbital. São caracterizados por apresentar uma
grande instabilidade e reatividade, pode interagir por meio de reações de óxido-redução com
qualquer molécula, tais como proteínas, fosfolipídios insaturados e ácidos
desoxirribonucleicos (DNA), causando danos às células e tecidos. Outra característica
importante dos radicais livres é a tendência em gerar reação de cadeia na produção de radicais
livres que, em última análise, amplifica seu efeito destrutivo (FERREIRA & MATSUBARA,
1997b; PHAM-HUY & PHAM-HUY, 2008).
Os radicais livres incluem hidroxila (OH•), ânion superóxido (O2
•-), o óxido nítrico
(NO•), dióxido de nitrogênio (NO2
•), peroxil (ROO
•) e peroxil lipídico (LOO
•). Além disso, o
peróxido de hidrogênio (H2O2), o ozônio (O3), o oxigênio singleto (1O2), o ácido hipocloroso
(HOCl), o ácido nitroso (HNO2), o peroxinitrito (ONOO-), o trióxido de diozônio (N2O3), o
peróxido lipídico (LOOH) não são radicais livres, no entanto, podem induzir reações
radicalares no organismo, sendo por isso também considerados como espécies reativas
(PHAM-HUY & PHAM-HUY, 2008).
As ROS e as RNS, em cardiomiócitos e outras células, são geradas por diversas fontes,
dentre as quais, as mais importantes são: as mitocôndrias, a nicotinamida adenina
dinucleotídeo fosfato-hidrogênio oxidase da família Nox (NADPH oxidase), xantina oxidase
(XO) e as óxido nítrico sintases (NOS) (DUNCAN et al., 2005; TZIOMALOS & HARE,
2009).
As espécies reativas também podem ser geradas sem o auxílio de enzimas como, por
exemplo, nas reações de Fenton e Haber Weiss (Figura 4). Na reação de Fenton, ocorre à
formação de duas OH•, um radical livre altamente reativo, pela reação de íons ferroso (Fe
2+)
ou cobre (Cu+) com o H2O2. Na reação de Haber Weiss ocorre à interação do H2O2 com o O2
•-

9
e também são formados dois radicais OH• (HALLIWELL & CROSS, 1994; TZIOMALOS &
HARE, 2009).
Figura 4 - A) Reação de Fenton - Formação de dois radicais hidroxila (OH•) pela reação de íos ferroso (Fe
2+) ou
cobre (Cu+) com o peróxido de hidrogênio (H2O2). B) Reação de Haber Weiss – Interação do peroxido de
hidrogênio (H2O2) com o ânion superóxido (O2•-) com a formação de dois radicais hidroxilas (OH
•).
Para proteger-se dessas espécies reativas, a célula possui um sistema antioxidante, que
compreendem as enzimas antioxidantes (1) superóxido dismutase (SOD), enzima que age
dismutando o O2•- em H2O2; (2) catalase (CAT), enzima que promove a dismutação do H2O2
em água (H2O) e O2, impedindo a formação do radical OH•; (3) glutationa peroxidase (GPx),
atua em conjunto com a CAT impedindo a formação do radical OH• (BRASILEIRO, 1997;
DUNCAN et al., 2005) (Figura 5).
Figura 5 - Esquema do sistema antioxidante enzimático, adaptado de Griendling e Fitzgerald (2003). Conversão
do oxigênio (O2) em ânion superóxido (O2•-) pelas enzimas NADPH oxidase. O O2
•- é dismutado pela superóxido
dismutase (SOD), formando peróxido de hidrogênio (H2O2), que pode ser convertido em uma molécula de água
(H2O) e oxigênio (O2) pela catalase (Cat) e/ou glutationa peroxidase (GPx). O H2O2 pode formar radical
hidroxila (OH•) após reação reagir com o ferro (Fe
+2) na reação de Fenton. A glutationa redutase (GR), mantêm o
equilíbrio entre a glutationa reduzida (GSH) e a glutationa oxidada (GSSG). O óxido nítrico (NO) é formado por
ação da enzima óxido nítrico sintase (NOS), a partir do aminoácido L-arginina que produz NO e L-citrulina. O
O2•- também pode reagir com o NO para formar peroxinitrito (OONO
-)
Além dos antioxidantes enzimáticos, existem os antioxidantes não enzimáticos que são
divididos em endógenos e exógenos. Os antioxidantes endógenos são produzidos pelo

10
metabolismo no corpo, tais como o ácido lipoide, glutationa, L-arginina, coenzima Q10, a
melatonina, ácido úrico, bilirrubina, proteínas de quelante de metal, transferrina, etc. Os
antioxidantes exógenos são os compostos que não podem ser produzidos no corpo e devem
ser fornecidos através de alimentos ou suplementos, tais como a vitamina E, vitamina C,
carotenoides, metais vestigiais (selênio, magnésio, zinco), flavonoides, ácido graxo ômega-3 e
ômega-6, etc. (DROGE, 2002).
A produção de ROS e de RNS desempenha um papel vital em muitos processos
biológicos, incluindo o crescimento celular, a sinalização celular, o relaxamento do músculo
liso, as respostas imunes, a síntese de moléculas biológicas, e modulação da pressão
sanguínea (DASURI; ZHANG; KELLER, 2013). Entretanto, quando produzido em excesso,
os radicais livres geram um fenômeno chamado de estresse oxidativo. Este é ocasionado
quando há o desequilíbrio entre os sistemas de defesa antioxidante com predomínio dos
oxidantes, culminando no surgimento de danos celulares (PHAM-HUY & PHAM-HUY,
2008).
A ação desses compostos nas membranas celulares estimula o processo de
peroxidação lipídica, onde os ácidos graxos das cadeias laterais dos lipídeos, especialmente
aqueles que contêm duas ou mais ligações duplas, são oxidados a hidroperóxidos. Estes
compostos, por sua vez, podem interagir com outros radicais ou íons metálicos gerando
aldeídos citotóxicos como o malonaldeído (MDA) (VALKO et al., 2007).
Os danos oxidativos induzidos nas células e tecidos decorrentes do estresse oxidativo,
têm sido implicados em vários processos patológicos tais como na isquemia-reperfusão
(obstrução arterial aguda, infarto do miocárdio, clampeamento vascular, cardioplegia na
circulação extracorpórea), pancreatites, insuficiência renal aguda, insuficiência hepática,
colites ulcerativas e outras doenças inflamatórias. Além disso, as espécies reativas também
estão implicadas na carcinogênese devido a danos ao DNA (VALKO et al., 2007; YOSHIDA,
1996).
Na isquemia-reperfusão cardíaca diversas alterações metabólicas são produzidas e
condicionam o aumento na produção de radicais livres. Essas alterações compreendem: (1) a
redução dos componentes da cadeia respiratória como o citocromo 1; (2) o aumento de cAMP
e de seus metabólitos (adenosina, inosina e hipoxantina) que promove a ativação da XO; (3)
acumulação de NADH, NADPH, flavoproteínas e lactato, que reagem com o O2 induzindo a

11
formação de radicais livres; (4) alterações do metabolismo dos lipídeos, em especial, a
ativação de fosfolipases pela acumulação de Ca2+
citosólico, as quais induzem a liberação de
ácido araquidônico que estimula à formação de radicais livres pela cicloxigenase (MAUPOIL
et al., 1993).
Investigações realizadas em modelo experimental de isquemia - reperfusão cardíaca
têm demonstrado que o dano celular gerado nessa patogênese é acompanhado por alteração na
atividade das enzimas SOD, CAT, GPx, bem como, na peroxidação lipídica (GE et al., 2014;
QIAO; MA; LIU, 2011) . Estes indicativos confirmam que a ROS e RNS desempenham um
papel importante na lesão de reperfusão cardíaca.
Atualmente, vários trabalhos vêm investigando o efeito de compostos com atividade
antioxidante no modelo de isquemia - reperfusão cardíaca, com a finalidade de reduzir os
danos que são gerados (HE et al., 2012; JIN, 2012; ZHANG et al., 2013). Tem-se observado
que tais substâncias, tem a capacidade de reduzir o dano celular que é ocasionado, podendo
estas ser de importante uso terapêutico.
2.4 Flavonoides
Alguns compostos fenólicos são originados do metabolismo secundário de plantas,
derivados dos aminoácidos aromáticos fenilalanina e também da tirosina. Essas substâncias
possuem anéis aromáticos com um ou mais grupos de hidroxila, incluindo seus derivados.
Existe uma grande variedade de compostos fenólicos, incluindo os fenóis simples, derivados
do ácido benzoico, ligninas, flavonoides, entre outros (SHAHIDI & NACZK, 2003).
Os flavonoides foram descobertos em 1530 pelo Szent-György, conquistando, assim, o
prêmio Nobel. Este pesquisador extraiu a citrina da casca do limão, substância essa,
apresentando capacidade de regulação da permeabilidade dos capilares sanguíneos. Desta
forma, essa classe de produtos naturais foi denominada como vitamina P (de permeabilidade)
e também por vitamina C2, visto que alguns flavonoides apresentavam propriedades
semelhantes às da vitamina C. Porém, devido a não confirmação destas substâncias como
pertencentes à classe das vitaminas, essa classificação foi abandonada em 1950 (MARTÍNEZ-
FLÓREZ et al., 2002).
O termo flavonoides deriva do latim “flavus” que significa amarelo, e define uma
classe de pigmentos responsáveis pela tonalidade das flores e folhas nas cores amarelo,

12
laranja e vermelho (NANDAVE; OJHA; ARYA, 2005). Estão presentes em várias partes das
plantas, desde a raiz até as flores e frutos, sendo encontrados nos vacúolos das células (YAO,
2004).
Os flavonoides ocorrem na forma livre (aglicona) ou ligada a açucares (glicosídeos).
São componentes de baixo peso molecular com estrutura base C6-C3-C6 (dois anéis fenil – A
e B – ligados através de um anel pirano – C) (Figura 6). Dependendo da substituição e do
grau de oxidação no anel C3, os flavonoides podem ser classificados em 10 classes de
compostos: antocianinas, leucoantocianidinas, flavonóis, flavonas, glicoflavonas, biflavonilas,
chalconas, aronas, flavononas e isoflavonas (YAO, 2004).
Figura 6- Ilustração molecular da estrutura plana base dos flavonoides (YOKOZAWA et al., 1997).
Esses compostos são consumidos em grandes proporções dentro de uma dieta humana
regular, e são encontrados em vegetais, legumes, frutas, chás de ervas, mel, entre outros
produtos de consumo cotidiano. Também são encontrados em várias plantas medicinais, e
vêm sendo utilizados pela medicina popular em todo o mundo, principalmente em
substituição a remédios no combate e prevenção de doenças tais como no câncer, doenças
cardiovasculares, doenças neurodegenerativas, processos inflamatórios, entre outros (LOPES
et al., 2000; YOKOZAWA et al., 1997).
Em relação aos interesses farmacêuticos, os flavonoides ocupam lugar de destaque
devido suas diversas interferências nos sistemas biológicos, tais como inibindo os sistemas
NADPH oxidase de neutrófilos, fosfolipase A2, mieloperoxidase, entre outras (AKHLAGHI
& BANDY, 2009).
A propriedade mais bem descrita desse grupo de substâncias é a capacidade de atuar
como antioxidante. As flavonas e catequinas parecem ser os mais poderosos dessa classe, para
a proteção do organismo contra as ROS. Essa proteção é conferida devido à capacidade desses

13
compostos em inativar os radicais livres, devido à presença do grupo hidroxila em sua
composição que sequestram tais espécies reativas. Além disso, alguns dos efeitos
antioxidantes dos flavonoides estão relacionados à sua capacidade em quelar íons metálicos,
tais como ferro e cobre, que são cofatores críticos da reação de Fenton na geração de radicais
livres (NIJVELDT et al., 2001).
Pesquisas sobre os flavonoides receberam impulso adicional com a descoberta do
“Paradoxo Francês”, que faz um link entre um maior consumo de vinho tinto com a redução
da incidência de doenças cardíacas. Neste estudo, indivíduos que apresentavam elevada
concentração de colesterol sanguíneo, semelhantes aos dos americanos e ingleses, e que
consumiam vinho tinto diariamente, a incidência de mortalidade por doenças coronárias
isquêmicas eram tão baixas quanto a dos japoneses e chineses. Esse efeito foi atribuído aos
componentes do vinho tinto, que diminuíam a oxidação do colesterol LDL, agregação
plaquetária e a formação de trombose na população francesa (RENAUD & LORGERIL,
1992; VOLP et al., 2008).
Recentemente, investigações epidemiológicas sugerem um papel protetor dos
flavonoides na dieta contra a doença arterial coronariana. Em um estudo, por exemplo, foi
analisada a relação entre o consumo de chá e café com a incidência de infarto do miocárdio
(SESSO et al., 1999). Os indivíduos que ingeriam mais de uma xícara de chá por dia
apresentaram menor risco de desenvolver a doença, enquanto o consumo de café não foi
significantemente associado com a redução do risco cardiovascular.
Trabalhos em modelo de isquemia-reperfusão cardíaca experimental, também tem
demonstrado os benefícios dos flavonoides na atenuação das lesões de reperfusão que são
geradas. Sun et al. (2012) estudando os efeitos da luteolina em corações de ratos diabéticos
submetidos à isquemia-reperfusão observaram que esse polifenol foi capaz de reduzir o
tamanho do infarto e melhorar a hemodinâmica do coração.
Em outro ensaio, foi investigado o efeito do pré-tratamento com quercetina em
corações submetidos à isquemia cardíaca in vivo, e pode-se evidenciar uma diminuição dos
indicadores de estresse oxidativo e inflamatórios relacionados com a lesão de isquemia do
miocárdio (LIU et al., 2014).

14
Devido aos extensos efeitos benéficos dos flavonoides frente a diversas patologias,
especificamente no sistema cardiovascular em modelos experimentais de isquemia -
reperfusão cardíaca, em que há excesso de radicais livres e sobrecarga de Ca2+
, substâncias
que atuem nestas desordens são de grande relevância, pois podem ser potencialmente
utilizadas terapeuticamente na prevenção das lesões de reperfusão cardíaca.
2.5 Diosmina
A diosmina (3',5,7-tri-hidroxi-4'-metoxiflavona-7-ramnoglucosídeo) (Figura 7) é um
flavonoide encontrado em frutas cítricas. Foi isolada pela primeira vez em 1925, a partir da
Scrophularia nodosa. Em 1969, foi utilizada pela primeira vez como agente terapêutico
(COVA, 1992).
Figura 7 – Ilustração da estrutura molecular da diosmina (SILAMBARASAN & RAJA ,2012).
Investigações farmacocinéticas têm demonstrado que a diosmina é rapidamente
transformada pela flora intestinal em sua forma aglicona, diosmetin. Diosmetin é absorvida e
rapidamente distribuída por todo o corpo, com tempo de meia-vida de 26-43 horas. Diosmetin
é degradada em ácidos fenólicos ou em derivados de glicina conjugado, que são eliminados
através da urina (LYSENG-WILLIAMSON & PERRY, 2003).
A diosmina é utilizada, por ser um agente protetor vascular, para o tratamento de
insuficiência venosa crônica, linfodema e varizes (JANTET, 2002; PECKING et al., 1997).
Como um flavonoide, a diosmina também exibe propriedades anti-inflamatórias, antioxidante,
entre outras (KILICOGLU et al., 2012; TONG et al., 2012) .
Silambarasan e Raja (2012) avaliaram o efeito anti - hipertensivo da diosmina e
constataram que esse flavonoide foi capaz de reduzir a pressão arterial sistólica e diastólica, e
o grau de peroxidação lipídica no plasma e tecidos (fígado, rim, coração e aorta).

15
Em outro estudo, foram avaliados os efeitos anti-hiperlipidêmicos em miocárdio de
ratos infartados induzidos com isoproterenol. O pré-tratamento com diosmina minimizou as
alterações eletrocardiográficas e diminuiu o grau de peroxidação lipídica (QUEENTHY &
JOHN, 2013).
Um recente trabalho demonstrou o efeito benéfico no perfil oxidativo do coração de
animais tratados com diosmina e submetidos à isquemia e reperfusão tecidual
(SENTHAMIZHSELVAN et al., 2014), prevenindo, desta forma, os danos cardíacos. Esses
resultados aqui citados evidenciam que a diosmina possui potencial para tratar patologias em
que há alterações cardíacas e estresse oxidativo. Alterações estas, que ocorrem no infarto
agudo do miocárdio, objeto do presente estudo.
Apesar de Senthamizhselvan et al. (2014) demonstrarem o papel preventivo da
diosmina no coração submetido à isquemia e reperfusão, não é conhecido se essa substância é
capaz de melhorar as lesões que são geradas no coração após a isquemia. Sabe-se que os
primeiros minutos de reperfusão são cruciais para o desenvolvimento das lesões, assim,
propusemos verificar a hipótese de que a diosmina possa contribuir para a prevenção do
surgimento das lesões de reperfusão em coração de ratos pós-isquêmicos.

16
3. OBJETIVOS
3.1 Geral
Avaliar os efeitos da diosmina na prevenção das lesões de reperfusão cardíaca em
ratos Wistar.
3.2 Específicos
Verificar o efeito antioxidante da diosmina em tecido cardíaco pós-isquêmico.
Avaliar o possível efeito antiarrítmico da diosmina em coração pós-isquêmico.
Analisar se a diosmina atenua a morte celular em coração pós-isquêmico.

17
4. MATERIAL E MÉTODOS
4.1 Animais
Foram utilizados ratos Wistar, pesando entre 250 e 300 g, provenientes do Biotério
Central da Universidade Federal de Sergipe (UFS) e do Biotério do Laboratório de Biofísica
do Coração da UFS. Os animais tiveram livre acesso à alimentação e água, foram submetidos
a ciclos claro/escuro de 12/12 horas, em temperatura controlada (23 ± 3ºC). Durante a
execução dos experimentos foram obedecidas as normas de manipulação dos animais
propostas pelo Conselho Nacional de Controle de Experimentação Animal (CONCEA).
Esse projeto foi aprovado pelo Comitê de Ética em Pesquisa Animal (CEPA) com o
número de protocolo 04/2013 (ANEXO 1).
4.2 Grupos Experimentais
Para determinar a dose da diosmina a ser utilizada no modelo experimental de
isquemia-reperfusão cardíaca, foi analisado o efeito contrátil desse flavonoide. Para isso,
foram utilizados corações isolados de ratos (n = 4). Esses, após a canulação e estabilização no
sistema de perfusão do tipo Langendorff pressão constante, foram perfundidos com solução
de Krebs-Ringer em concentrações crescentes de diosmina (0,1 µmol/L – 10 µmol/L) por um
período de 15 minutos (Figura 8).
Figura 8 - Representação esquemática do protocolo de perfusão com diosmina em concentrações
crescentes.
Para os experimentos de isquemia-reperfusão, os animais utilizados foram divididos
em 4 grupos experimentais (Figura 9):

18
Grupo 01 - Sham: Após um período de estabilização (20 minutos), os corações foram
perfundidos por mais 100 minutos com solução veículo (solução de Krebs-Ringer acrescida
de dimetilsulfóxido - DMSO 0,02%).
Grupo 02 - I-R + Veículo: Após um período de estabilização os corações foram
perfundidos por 10 minutos com solução veículo, seguido por um período de 30 minutos de
isquemia e, posteriormente, mais 60 minutos de reperfusão com a mesma solução.
Grupo 03 - I-R + Diosmina: Após um período de estabilização, os corações foram
perfundidos por 10 minutos com solução veículo, seguido por um período de 30 minutos de
isquemia e, posteriormente, mais 60 minutos de reperfusão com solução de diosmina (Krebs-
Ringer + diosmina 0,1 µmol/L diluída em DMSO 0,02%).
Grupo 04 - I-R + NAC: Após um período de estabilização, os corações foram
perfundidos por um período de 10 minutos com solução veículo, seguido por um período de
30 minutos de isquemia e, posteriormente, mais 60 minutos de reperfusão com o controle
positivo NAC (N- acetilcisteína 24 µmol/L em solução de Krebs- Ringer).
Figura 9 – Esquema representativo das etapas realizadas nos grupos experimentais.
4.3 Modelo de Isquemia-Reperfusão Cardíaca
Os animais foram decapitados após 10-15 minutos da injeção intraperitoneal de 400UI
de heparina. O tórax foi aberto e o coração cuidadosamente removido e perfundido no sistema
de perfusão do tipo Langendorff de pressão constante (Figura 10). O fluido de perfusão
utilizado foi à solução de Krebs-Ringer contendo (em mmol/L): 118,4 NaCl, 4,7 KCl, 1,2 de
KH2PO4, 1,2 MgSO4. 7H2O, 2,5 CaCl2. 2H2O, 11,7 glicose e 26,5 NaHCO3. Esta solução foi
mantida a 37°C, com pressão de 65 mmHg, pH 7,4 e oxigenação (5% de O2 e 95% de CO2)
constantes. Foi aguardado um período de estabilização de 20 minutos com solução Krebs-

19
Ringer, seguido de um período de perfusão de 10 minutos com solução veículo. Foi realizada
isquemia global por 30 minutos pela interrupção da disponibilização da solução nutritiva e da
oxigenação. Após este período, o fluxo foi restabelecido e mantido por um período de 60
minutos (BELL; MOCONU; YELLON, 2011).
Figura 10 - Ilustração do sistema de perfusão do tipo Langendorff pressão, adaptada de Skrzypiec-Spring et al.
(2007). VL: ventrículo esquerdo; O2: oxigênio; CO2: gás carbônico.
4.4 Avaliação da Pressão Intraventricular Esquerda
A avaliação da pressão intraventricular esquerda (PVE) foi determinada em coração
isolado de rato, por meio de um balonete insuflado com água até uma pressão de 15 cmHg. O
balonete foi introduzido no ventrículo esquerdo do coração através da abertura oriunda da
remoção do átrio esquerdo. Este sistema estava acoplado a um transdutor de pressão (HP
1290A), cujos sinais foram amplificados (HP 7754A), digitalizados (DATAQ DI 710,
WINDAQ PRO Acquision) e gravados em computador. Todo o sistema hidráulico usado para
determinar a pressão intraventricular foi preenchido com água destilada e calibrado com uma
coluna de mercúrio
4.5 Avaliação do Índice de Severidade da Arritmia
As arritmias cardíacas foram avaliadas como a presença de taquicardia e/ou fibrilação
ventricular nos primeiros 30 minutos de reperfusão. Para obter uma medida quantitativa, as
arritmias foram classificadas arbitrariamente por sua duração. A ocorrência de arritmias

20
cardíacas de até 3 minutos foi classificada com o fator 2; 3 - 6 minutos foi classificada com
fator 4; 6 - 10 minutos foi classificada com fator 6; 10-15 minutos foi classificada com o fator
8; 15 - 20 minutos foi classificada com o fator 10; 20 - 25 minutos foi classificada com o fator
11; e de 25 - 30 minutos classificada com o fator 12. Entretanto, se maior ou igual há 30
minutos foi considerada uma arritmia irreversível (FERREIRA; SANTOS; ALMEIDA,
2001a).
4.6 Análises da Lesão Tecidual
4.6.1 Marcação da Área de Infarte
Para avaliação macroscópica do tamanho do infarto, os corações foram tratados com
cloreto de trifeniltetrazólio (TTC). Para isso, os corações foram seccionados (± 2 mm) em
criostato e os cortes obtidos foram incubados em solução de TTC 1% (diluído em solução de
Krebs-Ringer) durante 10 minutos. Após esse período, os cortes foram mergulhados em
solução de formalina tamponada por 30 minutos, para a remoção do excesso da coloração. Em
seguida, os tecidos foram digitalizados em um scanner (EPSON TX 105) e as imagens obtidas
foram analisadas com o software ImageJ 1.38x (NIH) (BELL; MOCANU; YELLON, 2011) .
4.6.2 Determinação da Atividade da Enzima Creatina Quinase Total
A atividade da creatina quinase total (CK) foi determinada no perfusato, obtido nos
períodos da estabilização do coração e reperfusão, utilizando-se o kit comercial (VIDA
biotecnologia). Foi utilizado o método cinético, de acordo com as instruções do fabricante.
4.6.3 Avaliação da Atividade da Enzima Lactato Desidrogenase
A determinação da atividade da lactato desidrogenase (LDH) foi realizada em
perfusato, obtido nos períodos da estabilização do coração e reperfusão. Foi utilizado o
método cinético, de acordo com as instruções do fabricante (Labteste).

21
4.7 Avaliações dos Parâmetros Antioxidantes
4.7.1 Determinação do Grau de Peroxidação Lipídica
As mensurações dos produtos de peroxidação lipídica foram feitas de acordo com
Bose et al.(1989). Resumidamente, as amostras foram pesadas e homogeneizadas em 10 vezes
o volume de solução de tampão fosfato (50 mmol/L, pH 7,4), contendo butil-hidroxitoluol
(BHT; 12,6 mmol/L). Em seguida, 200 µL do homogenato foram incubados a 90ºC por 45
minutos com solução contendo ácido tiobarbitúrico (TBA 0,37%), em meio ácido (15% de
ácido tricloroacético- TCA e 0,25 N de ácido clorídrico). As amostras, em seguida, foram
centrifugadas por 5 minutos a 14.000 rpm. Ao sobrenadante, acrescentou-se n-butanol e
solução saturada de NaCl. A mistura foi agitada em vórtex por 30 segundos e novamente
centrifugada a 14.000 rpm (Heal Force, Neofuge 15R) por 2 minutos. Alíquotas do
sobrenadante foram pipetadas em placas de 96 poços para a leitura de absorbância em leitor
de microplaca (Biotek, ELx800 Absorbance Microplate Reader) a 535 nm, corrigindo pelos
valores de absorbância a 572 nm. A quantidade de MDA produzida foi expressa em
nanomoles por gramas de tecido, e foi interpretada como marcador de peroxidação lipídica
formado pela reação com o ácido tiobarbitúrico (TBARS).
4.7.2 Mensuração de Hidroperóxidos Totais
A quantificação dos hidroperóxidos totais foi realizada de acordo com Jiang et al.
(1992). O método se baseia basicamente na oxidação de íons ferroso (Fe2+
) a íons férricos
(Fe3+
) em condições ácidas, pelos hidroperóxidos lipídicos. O indicador utilizado é o xilenol
orange que reage com os íons Fe 3+
produzindo um cromóforo azul-arroxeado, o qual pode ser
medido espectrofotometricamente a 560 nm. O ensaio foi realizado em reagente de FOX, o
qual é composto por xilenol orange (0,25 mmol/L), sulfato ferroso amoniacal (Fe
(NH4)2(SO4)2.6H2O, 0,25 mmol/L), hidroxitolueno butilado (BHT, 4,4 mmol/L), metanol
(CH4O) e ácido sulfúrico (H2SO4, 97%). Em um microtubo foi adicionado reagente de FOX
ao homogenato. Após 30 minutos, a amostra foi centrifugada e o sobrenadante colocado em
microplaca. A quantificação dos hidroperóxidos foi expressa em mol/L, foi utilizado o
coeficiente de extinção molar 4,3 x 10-4
M-1
cm-1.

22
4.7.3 Determinação da Atividade da Enzima Superóxido Dismutase
A atividade da superóxido dismutase (SOD) foi mensurada através da formação de
O2•-
pela auto-oxidação do pirogalol e a inibição da redução do sal de tetrazolium (MTT)
segundo Madesh e Balasubramanian (1998). Resumidamente, o tecido foi homogeneizado em
tampão fosfato (PBS 50 mmol/L, pH 7,4), centrifugado a 12000 rpm (Heal Force, Neofuge
15R) por 30 minutos. A reação foi realizada pipetando em placa de Elisa: o sobrenadante
obtido, PBS, MTT (1,25 mmol/L) e pirogalol (100 µmol/L). A microplaca foi agitada por 5
minutos e logo após acrescentou-se DMSO. A leitura foi realizada em leitor de microplaca
(Biotek, ELx800 Absorbance Microplate Reader) a 570 nm. A atividade da SOD foi expressa
em unidade de SOD por miligrama de proteína.
4.7.4 Determinação da Atividade da Enzima Catalase
O ensaio da atividade da catalase (CAT) foi realizado conforme o método descrito por
Nelson e Kiesow (1972) e adaptado por Gioda et al. (2010). Resumidamente, as amostras
foram homogeneizadas em PBS e em seguida os homogenatos foram centrifugados (Heal
Force, Neofuge 15R) a 12000 rpm por 30 minutos a 4 °C. Em cubeta de quartzo foi pipetado
tampão fosfato (50 mmol/L, pH 7,0) e o homogeneizado. A reação foi iniciada com a adição
de H2O2 (0,3 mol/L). As medidas foram realizadas em espectrofotômetro (Hitachi, Japão), em
intervalos de 15 segundos, a 25°C, no comprimento de onde de 240 nm. A atividade da
enzima foi expressa pela diferença da variação das absorbâncias (ΔE) /minuto/miligrama de
proteínas.
4.7.5 Avaliação da Atividade da Enzima Glutationa Peroxidase
A atividade da enzima glutationa peroxidase (GPx) foi determinada de acordo com o
método descrito por Paglia e Valentine (1967). Em suma, as amostras foram pesadas e
homogeneizadas em tampão fosfato de sódio (50 mmol/L), acrescido de KCl (140 mmol/L),
pH 7,4 (1:10, p/v). Em seguida, os homogenatos foram centrifugados (Heal Force, Neofuge
15R) a 12000 rpm por 30 minutos a 4°C e o sobrenadante separado para o ensaio. Na
microplaca foi colocado tampão fosfato (100 mmol/L, pH 7,0), NADPH (8,4 µmol/L), GR
(10 U/mg de proteína/mL), azida sódica (NaN3, 1,125 mol/L), GSH (0,15 mmol/L), H2O2 (2,2
mmol/L) e a amostra. O monitoramento foi feito a 340 nm, 25ºC, por 8 minutos. A atividade

23
da Gpx foi avaliada pela oxidação do NADPH. Os resultados foram expressos em
nanomol/NADPH/minuto/miligrama de proteínas.
4.7.6 Avaliação da Atividade da Enzima Glutationa Redutase
A atividade da enzima glutationa redutase (GR) foi determinada de acordo com o
método de Mannervik e Carlberg (1985), no qual a atividade da redutase é proporcional ao
consumo de NADPH monitorado a 340 nm. Em síntese, a amostra foi homogeneizada em
tampão fosfato (0,2 mol/L, pH 7,5) contendo ácido etilenodiamino tetra-acético (EDTA, 6,3
mmol/L), acrescido de Leupeptina (5 mg/mL) e fluoreto de fenilmetilsulfonilo (PMSF, 100
mmol/L). O homogenato foi centrifugado por 30 minutos a 12.000 rpm (Heal Force, Neofuge
15R), a 4ºC. Para o ensaio, ao homogenato foram adicionados solução de albumina (0,5
mg/mL de tampão) e GSSH (10 mmol/L). A reação foi iniciada com a adição de NADPH (1,2
mg/mL). O monitoramento foi feito a 340 nm, 37ºC, por 8 minutos. O resultado foi expresso
em miliunidade de GR/minuto/ miligrama de proteínas.
4.8 Determinação da Expressão Proteica da Caspase - 3 por Western Blot
As amostras foram diluídas em tampão da amostra (Tris HCl/SDS pH = 6,8, 3%
Glycerol, 1% SDS, 0,6% β-mercaptoetanol e 0,1% Azul de Bromofenol) e aquecidas à 98°C
por 5 minutos. Para separação, foram aplicados 60 µg de proteínas em gel de SDS-PAGE (do
inglês “sodium dodecyl sulfate polyacrylamide gel electrophoresis”) a 7,5% ou 10%.
Após serem separadas no gel de poliacrilamida, as proteínas foi transferida para uma
membrana de nitrocelulose (Millipore®, USA) com poros de 0,44 μm. A qualidade da
transferência foi monitorada através da coloração da membrana com solução de Ponceau
0,3%. A membrana foi então lavada em solução salina tamponada com Tris-Base acrescido
com 0,05% de Tween 20% (TBS-T) e colocada por 1 a 2 horas em solução de bloqueio (4%
de albumina em TBS-T). Após o bloqueio, a membrana foi incubada em temperatura de 6-
8ºC, com o anticorpo primário específico diluído em 1% de albumina em TBS-T, overnight.
Os seguintes anticorpos primários foram utilizados: anti-caspase-3 (1:5000; policlonal
feito em coelho; Santa Cruz Biotechnology Inc) e anti-GAPDH (1:10000; policlonal feito em
camundongo; Sigma). Em seguida, a membrana foi lavada três vezes com TBS-T durante 5
minutos e incubada por 2 horas com anticorpos secundários específicos conjugados à
peroxidase (1:2000 - Sigma). Após o período de incubação, a membrana foi novamente

24
lavada por mais três vezes em TBS-T durante 5 minutos. As bandas proteicas foram
detectadas por uma reação de quimioluminescência (Luminata strong™ - Western HRP
substrate, Merck-Millipore, Darmstad, Germany) e a intensidade das mesmas foi avaliada por
análise densitométrica através do software ImageJ 1.38x (NIH).
Para estes experimentos foram utilizados o sistema Mini Protean III Tetracell e Mini
Trans-Blot Electrophoretic Transfer Cell (BIORAD®).
4.9 Determinação da Concentração Total de Proteínas
A concentração total de proteínas foi determinada pelo método de Lowry et al. (1951).
Resumidamente, às amostras foi adicionado NaOH 0,5 mol/L, e após 15 minutos
acrescentado 2% Na2CO3, 1% CuSO4 e 2% KNaC4H4O6.4H2O (100:1:1). Ao final foi
adicionado o reagente de Folin e após 30 minutos foi realizada a leitura a 630 nm em
espectrofotômetro de placa (ELx800, BIOTEK Instruments®). Uma curva padrão de
albumina bovina (Sigma-Aldrich®) foi construída para comparação.
4.10 Análises Estatísticas
Para tratamento estatístico, os resultados obtidos foram analisados em software Prism
5.1 (GraphPad). Os dados experimentais foram expressos com média ± erro padrão da média
(E.P.M.). Para a comparação múltipla dos dados paramétricos foi utilizado análise de
variância (ANOVA) de uma via e o grau de significância entre os grupos foi determinado pelo
pós-teste de Bonferroni. Para efeito estatístico, foram considerados significativos os valores
que apresentaram p < 0,01.
5. RESULTADOS
5.1 Efeito da Diosmina na Pressão Intraventricular Esquerda
Inicialmente avaliamos os efeitos da diosmina sobre a PVE (Figura 11). Para isso,
foram testadas as concentrações de 0,1 µmol/L, 1 µmol/L e 10 µmol/L dessa substância em
miocárdio de ratos.
A diosmina, nas concentrações de 1 µmol/L e 10 µmol/L, desencadeou elevação da
PVE (3,2 ± 0,3 cmHg, n = 4; 2,4 ± 0,2 cmHg, n = 4) ( p< 0,01) em relação ao período de

25
estabilização (1,20 ± 0,09 cmHg, n = 4). Em contrapartida, a concentração de 0,1 µmol/L
(Figura 14B) não apresentou tal resposta (1,3 ± 0,1 cmHg, n = 4).
Para avaliar o papel antioxidante da diosmina nos corações pós-isquêmicos, objetivo
principal do presente trabalho, buscamos analisar o efeito de uma concentração desta droga
que não interferisse na PVE. Então, a concentração de 0,1 µmol/L, por não ter promovido esta
alteração, foi escolhida para darmos continuidade ao presente estudo.
Figura 11 - Efeito da diosmina na pressão intraventricular esquerda (PVE) em corações isolados de ratos.
PVE desenvolvida nos corações nos períodos de estabilização (n= 4) e perfusão com diosmina nas concentrações
de 0,1, 1 e 10 µmol/L (n= 4). Os valores foram representados como média ± E.P.M. e expressos em centímetros
de mercúrio (cmHg) *** p< 0,01 ou *p< 0,01 vs Estabilização; ### p< 0,01 ou #p< 0,01 vs Diosmina 0,1
µmol/L (ANOVA de uma via seguida do pós-teste de Bonferroni).
Assim, posteriormente avaliamos o efeito da diosmina na concentração de 0,1 µmol/L
na PVE em miocárdios que sofreram isquemia-reperfusão (Figura 12). Foi observado que
houve uma redução (p< 0,01) da PVE dos corações do grupo I-R + Veículo (0,19 ± 0,04
cmHg, n= 4) em relação aos corações do grupo Sham ( 0,97 ± 0,18 cmHg, n= 4).
Nos corações reperfundidos com a diosmina, houve um aumento desta medida (1,06 ±
0,03 cmHg, n= 3, p< 0,01) em relação aos corações do grupo I-R + Veículo (0,19 ± 0,04
cmHg, n= 4), sendo que este aumento se aproximou aos valores basais de PVE dos corações
do grupo Sham (0,97 ± 0,18 cmHg, n= 4). Assim como ocorreu com o grupo I-R + Diosmina,
quando os corações foram reperfundido com o controle positivo houve um aumento da PVE
(1,00 ± 0,03 cmHg, n= 4, p< 0,01) em relação aos corações que foram reperfundidos com o
veículo (0,19 ± 0,04 cmHg, n= 4). Porém, não houve diferença significativa entre os corações
do grupo I-R + Diosmina e o grupo I-R + NAC.

26
Figura 12 - Efeito da diosmina na pressão intraventricular esquerda (PVE) em corações isolados de ratos
submetidos á isquemia-reperfusão. PVE desenvolvida nos corações dos grupos Sham (n= 4), I-R + Veículo
(n= 4), I-R + Diosmina (n= 3), I-R + NAC (n= 3). Os valores foram representados como média ± E.P. M e
expressos em centímetros de mercúrio (cmHg). ***p< 0,01 vs Sham; ### p< 0,01 ou #p< 0,01 vs I-R + Veículo
(ANOVA de uma via seguida do pós-teste de Bonferroni).
5.2 Efeito da Diosmina nas Arritmias de Reperfusão
A fim de analisarmos também as alterações contráteis existentes durante á reperfusão,
mensuramos á presença de arritmias, através do ASI, durante os primeiros 30 minutos de
reperfusão. Foi evidenciado que o grupo I-R + Veículo apresentou um índice de 9,6 ± 0,9
(n= 6). O grupo I-R + Diosmina teve a medida do ASI reduzida (3,3 ± 0,9, n= 6, p< 0,01) em
relação ao grupo I-R + Veículo. Assim como o grupo I-R + Diosmina, o grupo I-R + NAC
também apresentou baixos índices de ASI (3,7 ± 0,6, n= 7, p< 0,01) quando comparado com o
grupo I-R + Veículo. Porém, não houve diferença estatística entre os grupos I-R + Diosmina e
I-R + NAC (Figura 13).

27
Figura 13 - Efeito da diosmina nas arritmias em corações isolados de ratos induzidas pela isquemia-
reperfusão. As arritmias de reperfusão foram quantificadas durante os primeiros 30’ de reperfusão, através do
índice de severidade de arritmia (ASI) nos grupos I-R + Veículo (n= 6), I-R + Diosmina (n= 6) e I-R + NAC
(n= 7). Os valores foram representados como média ± E.P. M e expressos em unidade arbitrária (u.a). ###p<
0,01 vs I-R + Veículo (ANOVA de uma via seguida pelo pós-teste de Bonferroni).
5.3 Efeito da Diosmina na Limitação da Área de Infarto
Na figura 14A podemos observar a área de lesão do infarto nos corações dos grupos
estudados. Estas regiões lesadas foram quantificadas e apresentadas na figura 14B. Nos
resultados obtidos podemos verificar que o grupo Sham não apresentou área isquêmica
(0,0 ± 0,0, n= 4).
Entretanto, no grupo I-R + Veículo os corações apresentaram uma área isquêmica
(21,5 ± 3,6, n= 4, p< 0,01). Porém, ao reperfundir os corações isquêmicos com diosmina ou
NAC a área lesionada foi reduzida (6,9 ± 0,8 % e 6,3 ± 1,1% respectivamente, n= 4; n= 5,
p< 0,01). A redução da área isquêmica observada no grupo I-R + Diosmina não foi diferente
do grupo I-R + NAC.

28
A
B
Figura 14 - Efeito da diosmina na limitação da área de infarto em corações isolados submetidos à
isquemia-reperfusão. Painel A: Imagem representativa da área de infarto nos grupos experimentais estudados.
Sham (n= 4), I-R + Veículo (n= 4), I-R + Diosmina (n= 4) e I-R + NAC (n= 5). Painel B: Percentual da área de
infarto dos grupos estudados representado como média ± E.P.M. ***p< 0,01 vs Sham; ###p< 0,01 vs I-R +
Veículo (ANOVA de uma via seguida pelo pós-teste de Bonferroni).

29
5.4 Efeito da Diosmina na Atividade da Enzima Creatina Quinase Total
Avaliamos, no perfusato, a atividade da enzima CK Total. Esta enzima consiste em
uma macromolécula intracelular encontrada no miocárdio, que extravasa das células
miocárdicas devido à lesão da membrana sarcolemal (LOZOVOY; PRIESNITZ; SILVA,
2008).
Nos corações não submetidos à isquemia-reperfusão, a atividade desta enzima foi de
55,9 ± 6,4 U/L (n= 12). Já os corações submetidos à isquemia e reperfundidos com o veículo,
tiveram a atividade da CK elevada (141,5 ± 10,1 U/L, n= 9, p< 0,01) em relação aos corações
não isquêmicos.
Em contrapartida, nos corações do grupo I-R + Diosmina observamos que a atividade
desta enzima estava reduzida (77,9 ± 5,2 U/L, n= 9, p< 0,01) quando comparada aos corações
do grupo I-R + Veículo. Os corações reperfundidos com NAC também tiveram a atividade da
CK total (56,1 ± 6,6 U/L, n= 8, p< 0,01) reduzida em relação aos reperfundidos com veículo.
Contudo, essa redução não foi significativa em relação aos corações reperfundidos com
diosmina (Figura 15).
Figura 15 - Efeito da diosmina na atividade da creatina quinase (CK) em corações isolados de ratos
submetidos á isquemia-reperfusão. A atividade da CK foi observada nos grupos Sham (n= 12), I-R + Veículo
(n= 9), I-R + Diosmina (n= 9) e I-R + NAC (n= 8). Os valores foram representados como média ± E.P.M e
expressos em unidade por litro (U/L). ***p< 0,01 vs Sham; ###p< 0,01 vs I-R + Veículo (ANOVA de uma via
seguida do pós-teste de Bonferroni).

30
5.5 Efeito da Diosmina na Atividade da Enzima Lactato Desidrogenase
Também avaliamos a atividade da enzima LDH no perfusato. Assim como a CK, esta
enzima consiste em um dos marcadores que definem a possibilidade de IAM.
A atividade da LDH mostrou-se elevada (p< 0,01) nos corações reperfundidos com
veículo (29,7 ± 2,0 U/L, n= 11) em comparação ao grupo Sham (10,5 ± 1,2 U/L, n= 10)
evidenciando, desta forma, que os corações sofreram lesão tecidual. No grupo I-R + Diosmina
a atividade da LDH reduziu (14,4 ± 2,6 U/L, n= 7, p< 0,01) em relação ao grupo I-R +
Veículo (29,7 ± 2,0 U/L, n= 11). Além disso, nos corações reperfundidos com NAC a
atividade desta enzima, também, se mostrou reduzida (10,9 ± 3,2 U/L, n= 7, p< 0,01).
Contudo, a redução da atividade da LDH no grupo controle positivo não foi diferente do
grupo diosmina (Figura 16).
Figura 16 - Efeito da diosmina na atividade da lactato desidrogenase (LDH) em corações isolados de ratos
submetidos á isquemia-reperfusão. A atividade da LDH foi analisada nos grupos Sham (n= 10), I-R + Veículo
(n= 11), I-R + Diosmina (n= 7) e I-R + NAC (n= 7). Os valores foram representados como média ± E.P.M e
expressos em unidade por litro (U/L). ***p< 0,01 vs Sham; ###p< 0,01 vs I-R + Veículo (ANOVA de uma via
seguida pelo pós-teste de Bonferroni).
5.6 Efeito da Diosmina no Grau de Peroxidação Lipídica
Na perspectiva de analisarmos o efeito da diosmina sobre o dano causado nas
membranas celulares pela alta concentração de ROS e RNS, investigamos a formação do
produto indireto da peroxidação lipídica, o MDA, nos grupos experimentais estudados. Foi
observado que no grupo I-R + Veículo houve aumento (9,1 ± 0,8 nmol de MDA/mg de tecido,

31
n= 13, p< 0,01) da medida de TBARS quando comparado ao grupo Sham (3,0 ± 0,5 nmol de
MDA/mg de tecido, n= 10). No grupo I-R + Diosmina verificamos uma redução na
concentração de MDA (5,4 ± 0,5 nmol de MDA/mg de tecido, n= 11, p< 0,01) em relação ao
grupo I-R + Veículo.
O grupo I-R + NAC apresentou baixo grau de peroxidação lipídica (2,8 ± 0,5 nmol de
MDA/mg de tecido, n= 7, p< 0,01) quando comparado ao grupo I-R + Veículo. Todavia, não
houve diferença significativa na concentração de TBARS entre os grupos I-R + Diosmina e
I-R + NAC (Figura 17).
Figura 17 - Efeito da diosmina no grau de peroxidação lipídica em corações isolados de ratos submetidos à
isquemia-reperfusão. Foram mensuradas as concentrações de malondealdeído (MDA) nos grupos Sham
(n= 10), I-R + Veículo (n= 13), I-R + Diosmina (n= 11) e I-R + NAC (n= 7). Os valores foram representados
como média ± E.P.M. e expressos em nanomoles de MDA por grama de tecido (nmol de MDA/ g de tecido). ***
p< 0,01 vs Sham; ##p< 0,01 ou ###p< 0,01 vs I-R + Veículo (ANOVA de uma via seguida pelo pós-teste de
Bonferroni).
5.7 Efeito da Diosmina na Concentração de Hidroperóxidos Totais
Ainda avaliando os danos ocasionados nas membranas celulares durante a isquemia-
reperfusão cardíaca, mensuramos os hidroperóxidos totais nos quatro grupos estudados. A
medida de hidroperóxidos totais no grupo Sham foi de 4,90 x 10-6
±0, 93 x 10-6
mol/L (n= 13,
p< 0,05). O grupo I-R + Veículo apresentou um aumento nesta medida (9,5 x10-6
± 2,5 x 10-7
mol/L, n = 7, p< 0,01) em relação ao grupo Sham. O grupo I-R + Diosmina apresentou
quantidades reduzidas de hidroperóxidos (3,40 x 10-6
± 0, 69 x 10-6
mol/L, n= 11, p< 0,01) em
relação ao grupo I-R + Veículo. Quanto ao grupo I-R + NAC ocorreu, também, redução
destes compostos (1,90 x 10-6
± 0,38 x10-6
mol/ L, n= 8, p< 0,01) em relação ao grupo I-R +

32
Veículo. Em contrapartida, não houve diferença estatística entre os grupos I-R + NAC e I-R +
Diosmina (Figura 18).
Figura 18 - Efeito da diosmina na concentração de hidroperóxidos em corações isolados de ratos
submetidos à isquemia-reperfusão. Os hidroperóxidos totais foram mensurados nos grupos Sham (n= 13),
I-R+ Veículo (n= 7), I-R + Diosmina (n= 11) e I-R + NAC (n= 8). Os valores foram representados como média ±
E.P. M e expressos em moles por litro (mol/L). **
p< 0,01 vs Sham; ###p < 0,01 vs I-R + Veículo (ANOVA de
uma via seguida do pós-teste de Bonferroni).
5.8 Efeito da Diosmina na Atividade da Enzima Superóxido Dismutase
Com o intuito de avaliarmos a atividade das enzimas antioxidantes no modelo de
isquemia-reperfusão cardíaca foi observada a atividade da SOD nos corações estudados.
Ocorreu aumento da atividade desta enzima no grupo I-R+ Veículo (2,0 ± 0,2 U/mg de
proteína, n= 6, p< 0,01) em relação ao grupo Sham (0,3 ± 0,1 U/ mg de proteína, n= 9).
Quando reperfundidos com diosmina a atividade desta enzima foi reduzida (0,2 ± 0,1 U/ mg
de proteína, n= 8, p< 0,01). Além disso, o grupo reperfundido com NAC também apresentou
uma diminuição da atividade da SOD (0,10 ± 0,03 U/ mg de proteína, n= 9, p< 0,01) em
relação ao grupo I-R + Veículo . Mas, não houve diferença entre os corações reperfundidos
com a diosmina e os reperfundidos com o NAC (Figura 19).

33
Figura 19 - Efeito da diosmina na atividade da enzima superóxido dismutase (SOD) em corações isolados
de ratos submetidos à isquemia-reperfusão. A atividade da SOD foi mensurada nos grupos Sham (n= 9), I-R
+ Veículo (n= 6), I-R + Diosmina (n= 8) e I-R + NAC (n= 9). Os valores foram representados média ± E.P. M e
expressos em unidade de SOD por miligrama de proteína (U/mg de proteína). ***p< 0,01 vs Sham; ###p< 0,01
vs I-R + Veículo (ANOVA de uma via seguida pelo pós-teste de Bonferroni).
5.9 Efeito da Diosmina na Atividade da Enzima Catalase
Analisamos, também, atividade da CAT nos grupos experimentais estudados e
observamos um aumento (p< 0,01) na atividade desta enzima no grupo I-R + Veículo (0,20 ±
0,01 ΔE/min/mg de proteína, n= 8) em relação ao grupo Sham (0,020 ± 0,005 ΔE/min/mg de
proteína, n= 10). Entretanto, o grupo I-R+ Diosmina e o grupo I-R + NAC apresentaram a
atividade da CAT reduzida (0,020 ± 0,004 ΔE/min/mg de proteína, n= 9 e 0,04 ± 0,01
ΔE/min/mg de proteína, n= 7, p< 0,01, respectivamente) em relação ao grupo I-R + Veículo.
O grupo I-R + Diosmina não apresentou diferença significativa em relação ao grupo I-R +
NAC (Figura 20).

34
Figura 20 - Efeito da diosmina na atividade da catalase (CAT) em corações isolados de ratos submetidos à
isquemia-reperfusão. A atividade da CAT foi mensurada nos grupos Sham (n= 10), I-R + Veículo (n= 8), I-R +
Diosmina (n= 9) e I-R + NAC (n= 7). Os valores foram representados como média ± E.P. M e expressos pela
diferença de absorbância por minuto, por miligrama de proteína (ΔE/min/mg de proteína). ***p< 0,01 vs Sham;
###p< 0,01 vs I-R + Veículo (ANOVA de uma via seguida pelo pós-teste de Bonferroni).
5.10 Efeito da Diosmina na Atividade da Enzima Glutationa Peroxidase
A enzima antioxidante GPx foi avaliada e pudemos verificar um aumento (p< 0,01) da
atividade desta enzima no grupo I-R + Veículo (0,20 ± 0,02 nmol de NADPH/min/mg de
proteína, n= 5) quando comparada com a do grupo Sham (0,05 ± 0,01 nmol de
NADPH/min/mg de proteína, n= 5). Porém, ao avaliarmos a atividade enzimática da GPx no
grupo I-R + Diosmina foi evidenciada uma redução (0,020 ± 0,009 nmol de NADPH/ min/mg
de proteína, n= 6, p< 0,01) quando comparada com a atividade desta enzima no grupo I-R +
Veículo. No grupo controle positivo foi observado, também, que a atividade da GPx estava
reduzida (0,030 ± 0,008 nmol de NADPH/min/mg de proteína, n= 7, p< 0,01) em relação ao
grupo I-R + Veículo. Entretanto, a atividade da GPx no grupo NAC não apresentou diferença
em relação a do grupo diosmina (Figura 21).

35
Figura 21 - Efeito da diosmina na atividade da glutationa peroxidase (GPx) em corações isolados de ratos
submetidos à isquemia-reperfusão. A atividade da GPx foi avaliada nos corações dos grupos Sham (n= 5), I-R
+ Veículo (n= 5), I-R + Diosmina (n= 6) e I-R + NAC (n= 7). Os valores foram representados média ± E.P. M e
expressos em nanomoles de NADPH por minuto, por miligrama de proteína (nmol NADPH/min/mg proteína).
***p< 0,01 vs Sham; ###p< 0,01 vs I-R + Veículo (ANOVA de uma via seguida do pós-teste de Bonferroni).
5.11 Efeito da Diosmina na Atividade da Enzima Glutationa Redutase
No grupo Sham, a atividade da GR foi de 0,030 ± 0,007 nmol de NADPH/min/mg de
proteína (n= 9). Os corações reperfundidos com o veículo apresentaram a atividade elevada
desta enzima (0,20 ± 0,02 nmol de NADPH/min/mg de proteína, n= 9, p< 0,01) em relação ao
grupo Sham. Porém, nos corações reperfundios com a diosmina a atividade desta enzima
reduziu (0,040 ± 0,007 nmol de NADPH/min/mg de proteína, n= 9, p< 0,01) em relação ao
grupo I-R + Veículo. A atividade da GR mensurada nos corações reperfundidos com o NAC,
também se mostrou reduzida (0,010 ± 0,005 nmol de NADPH/min/mg de proteína, n= 7,
p< 0,01). Todavia, esta enzima manteve sua atividade semelhante nos corações reperfundidos
tanto com a diosmina quanto com o NAC (Figura 22).

36
Figura 22 - Efeito da diosmina na atividade da glutationa redutase (GR) em corações isolados de ratos
submetidos à isquemia-reperfusão. A atividade da GR foi mensurada nos grupos Sham (n= 9), I-R +Veículo
(n= 7), I-R + Diosmina (n= 9) e I-R + NAC (n= 7). Os valores foram representados como média ± E.P.M e
expressos em miliunidade de GR por minuto ,por miligramas de proteína ( mU/min/mg proteína). ***p< 0,01 vs
Sham; ###p< 0,01 vs I-R + Veículo (ANOVA de uma via seguida pelo pós-teste de Bonferroni).
5.12 Efeito da Diosmina na Expressão Proteica de Caspase-3
Na figura 23A podemos observar a expressão proteica de caspase-3 nos corações dos
grupos experimentais avaliados. Observamos um aumento na expressão proteica de caspase-3
nos corações reperfundidos com veículo (3,5 ± 0,6, n= 4). Porém, essa expressão foi reduzida
(p< 0,01) nos corações reperfundidos com diosmina (1,7 ± 0,2, n= 4). O mesmo foi observado
nos corações reperfundidos com NAC (1,1 ± 0,1, n= 5). Todavia, a redução da expressão de
caspase-3 não se mostrou diferente entre os grupos I-R + Diosmina e I-R + NAC (Figura
23B).
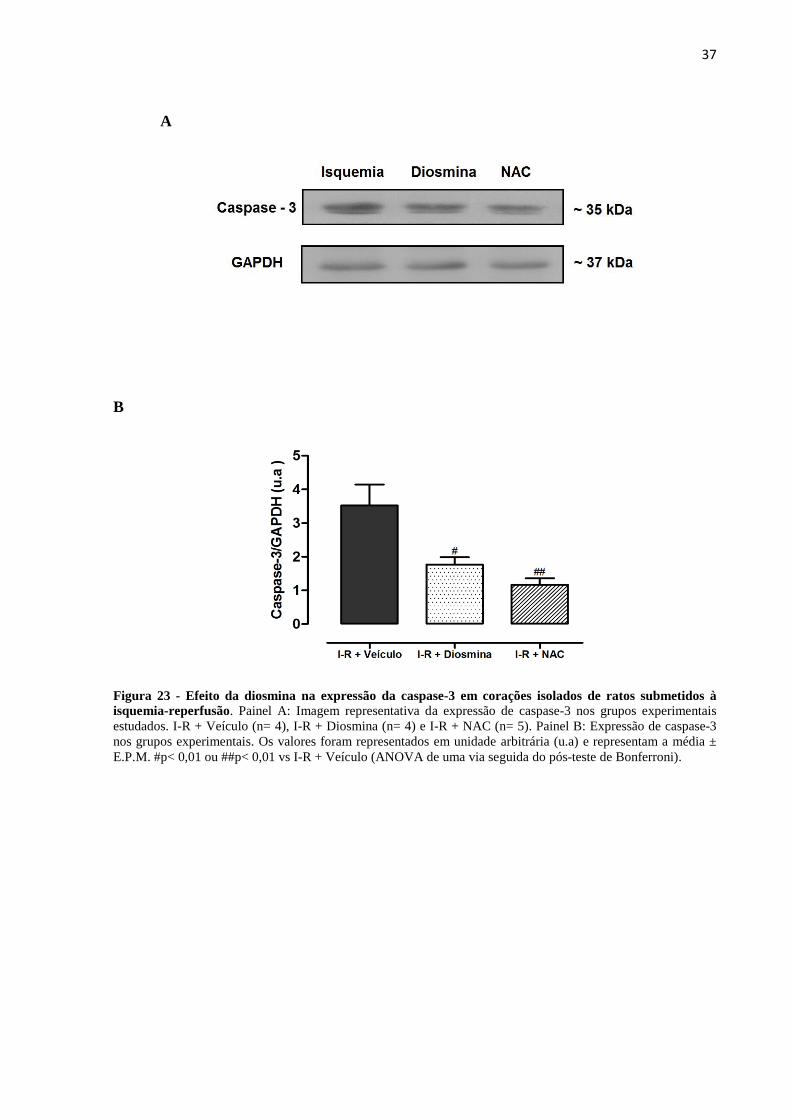
37
A
B
Figura 23 - Efeito da diosmina na expressão da caspase-3 em corações isolados de ratos submetidos à
isquemia-reperfusão. Painel A: Imagem representativa da expressão de caspase-3 nos grupos experimentais
estudados. I-R + Veículo (n= 4), I-R + Diosmina (n= 4) e I-R + NAC (n= 5). Painel B: Expressão de caspase-3
nos grupos experimentais. Os valores foram representados em unidade arbitrária (u.a) e representam a média ±
E.P.M. #p< 0,01 ou ##p< 0,01 vs I-R + Veículo (ANOVA de uma via seguida do pós-teste de Bonferroni).

38
6 DISCUSSÃO
A doença cardíaca isquêmica secundária ou IAM é um dos problemas de saúde mais
prevalentes no mundo, e uma das principais causas de morbidade e mortalidade. A
reoxigenação miocárdica desencadeia as lesões de reperfusão que são responsáveis pelo
surgimento de alterações tanto contráteis quanto oxidantes, as quais são representadas por
arritmias ventriculares e até mesmo por morte celular (BRASILEIRO, 1997; ZWEIER;
TALUKDER, 2006).
Essa lesão, inicia-se durante a isquemia por insuficiência do metabolismo anaeróbico
em manter o aporte energético do cardiomiócito que ativa a glicólise, resultando na redução
do pH e consequente elevação de Na+ e Ca
2+ intracelular. Com a reperfusão, ocorrem
distúrbios no transporte iônico acarretando sobrecarga de Ca2+
no citoplasma e há formação
exacerbada de ROS e RNS pelas mitocôndrias, levando a mudanças estruturais e funcionais
dos constituintes celulares e ativação de vias sinalizadoras de morte celular (AKAHLAGHI;
BANDY, 2009).
A elevação da concentração de ROS e RNS associada ao acúmulo de Ca2+
no
citoplasma causa lesões de isquemia-reperfusão (WALTERS; POTER; BROOKES, 2014).
Essa lesão é estabelecida nos primeiros minutos da restauração do fluxo sanguíneo, tornando
esse tempo importante para modulação de ações que previnam o seu surgimento (PIPER;
ABDALLAH; SCHÄFER, 2004a).
Em humanos foi demonstrado que durante a isquemia e reperfusão do miocárdio, para
a defesa há uma depleção de antioxidantes endógenos não enzimáticos tais como, ácido
ascórbico e vitamina E contra o estresse oxidativo. Este estudo indica a suplementação com
antioxidantes como terapia alternativa ao tratamento convencional da isquemia cardíaca
(AKAHLAGHI & BANDY, 2009).
Muitos antioxidantes encontrados em produtos naturais podem intervir e ajudar na
prevenção de lesões de reperfusão (SUZUKI et al., 2007; TOUFEKTSIAN, 2008), além
disso, apresentam baixo custo e diversas vantagens em relação aos compostos sintéticos.
Abordagens da eficácia de agentes farmacológicos para atenuar as consequências da
lesão de isquemia-reperfusão mostram limitações experimentais ou não conseguem traduzir-

39
se em tratamentos clínicos úteis (FERDINANDY; SCHULZ; BAXTER, 2007). Desta forma,
necessita-se de alternativas que possam ser utilizadas para essa finalidade.
No presente trabalho, utilizamos um modelo de isquemia-reperfusão global, no qual
foi estudado o efeito da diosmina, flavonoide com característica antioxidante, na prevenção da
lesão de reperfusão.
Evidenciamos que os corações do grupo I-R + Veículo, tiveram uma redução
acentuada da função contrátil cardíaca (Figura 12). Já é conhecido que a depressão da
contratilidade cardíaca é uma consequência precoce da isquemia miocárdica, e pode resultar
em insuficiência cardíaca aguda. Além disso, a taxa e/ou tempo de recuperação da função
cardíaca pós-isquêmica é amplamente utilizada como índice de lesão de isquemia e reperfusão
experimental (FERDIANDY; SCHULZ; BAXTER, 2007).
Todavia, nos corações isquêmicos, a reperfusão com diosmina (Figura 12) ofereceu
uma proteção cardíaca por uma melhor recuperação pós-isquêmica do ventrículo esquerdo.
Resultado semelhante ao nosso foi observado por Senthamizhselvan et al. (2014), que
utilizaram como pré-tratamento a diosmina nas concentrações de 50 e 100 mg/Kg,
identificando que esta substância promoveu recuperação da força contrátil ventricular,
aumentando-a em relação ao controle. Assim como a diosmina, em outro estudo, evidenciou-
se o papel protetor da diogenina contra a lesão de isquemia-reperfusão. Foi avaliado o efeito
desta substância (1nM) na função contrátil cardíaca, e pode-se observar que a mesma
melhorou durante a reperfusão via recuperação da PVE (BADALZADEH, R.; YOUSEFI, B.;
MAJIDINIA, M.; EBRAHIMI, H, 2014).
Durante a isquemia-reperfusão, arritmias ventriculares graves como taquicardia
ventricular e fibrilação ventricular, podem se desenvolver contribuindo para a morte súbita
cardíaca após a oclusão coronária (JANSE & WIT, 1989).
Em nosso estudo, observamos no grupo I-R + Veículo que ocorreu um aumento do
ASI (Figura 13). Esta evidência pode ser explicada pela redução da concentração de ATP
celular durante o período de isquemia, onde muitas ATPases reduzem sua atividade
resultando numa elevada sobrecarga de Ca2+
durante a diástole, que culmina em pós-
despolarizações ocasionando arritmias ventriculares (DEL MONTE et al., 2004). Além disso,
durante a isquemia ocorre elevação da concentração de prótons no espaço intracelular,
ocasionando o influxo de Na+ pelo trocador Na
+/H
+. Tal elevação da concentração intracelular

40
deste íon, por sua vez, aumenta a concentração citosólica de Ca2+
por meio da inversão do
trocador Na+/Ca
2+, contribuindo também para o surgimento de arritmias (BERS, 2002).
Quando os corações pós-isquêmicos foram reperfundidos com diosmina, essa
substância apresentou um efeito antiarritmogênico (Figura 13), o qual foi representado pela
redução do ASI. Corroborando com nosso resultado, Liao et al. (2010) investigaram o efeito
da luteolina na arritmia ventricular em corações submetidos a isquemia e reperfusão. Nesse
estudo, o pré-tratamento com a luteolina (1 a 10 μg/kg - via intravenosa) reduziu a incidência
e a duração da taquicardia ventricular, bem como a fibrilação ventricular durante a isquemia
do miocárdio e reduziu arritmias ventriculares.
Em nosso estudo, realizamos a marcação do infarto dos corações dos grupos em
estudo com TTC (Figura 14). Este corante tem por finalidade marcar o tecido viável onde há
desidrogenases ativas que convertem o composto solúvel em precipitado insolúvel, formazan
vermelho. Como as desidrogenases estão localizadas principalmente nas mitocôndrias, a
intensidade de coloração está correlacionada com o número de mitocôndrias funcionais e
pode facilmente diferenciar o tecido viável do tecido não viável, este representado pela
coloração esbranquiçada (FERRERA et al., 2009). Observamos que os corações pós-
isquêmicos reperfundidos com o veículo apresentaram a área de infarto maior, verificada por
um aumento acentuado da área esbranquiçada. Todavia, os corações que foram reperfundidos
com a diosmina tiveram a área lesionada reduzida, evidenciado que esta substância tem a
capacidade de prevenir o surgimento das lesões de reperfusão.
Sun et al. (2012) tiveram como um dos objetivos no seu estudo, investigar os efeitos
biológicos da luteolina sobre o tamanho do infarto do miocárdio em ratos diabéticos
submetidos a isquemia-reperfusão. Foi observado que o pré-tratamento com a luteolina foi
capaz de reduzir a área de infarto de maneira significativa em relação ao controle. Liao et al.
(2010) também observaram o efeito protetor do pré-tratamento com luteolina em corações de
ratos submetidos a isquemia-reperfusão, e evidenciaram que esta substância foi capaz de
reduzir a área de infarto dos mesmos, quando comparada ao grupo controle.
Em nossos experimentos, ao analisamos a atividade das enzimas CK e LDH nos
corações do grupo isquêmico, observamos que havia uma elevação da atividade destas
enzimas (Figuras 15 e 16). Na isquemia, com a cessação da glicólise aeróbica e
consequentemente estabelecimento da glicólise anaeróbica, ocorre uma produção ineficiente

41
de fosfatos de alta energia e ao acúmulo de ácido láctico, o que culmina na diminuição do pH
celular e alterações metabólicas importantes. Sem energia para a manutenção da sua atividade
metabólica normal e integridade da membrana celular, a célula morre liberando suas
macromoléculas na circulação, tais como CK e LDH (LOZOVOY; PRIESNITZ; SILVA,
2008).
Os corações que foram reperfundidos com a diosmina tiveram a atividade da CK e
LDH reduzidas, sugerindo que este flavonoide possui uma propriedade protetora contra os
danos celulares ocasionados pela isquemia-reperfusão. Resultados semelhantes aos nossos
foram identificados no estudo de Senthamizhselvan et al. (2014), os quais observaram que os
corações pré-tratados com diosmina apresentaram uma redução da atividade das enzimas CK
e LDH no perfusato coronário. Badalzadeh et al. (2014), avaliaram o efeito diogenina em
corações submetidos à isquemia global, e verificaram que este flavonoide reduziu a
concentração de LDH no perfusato coronário. Liu et al. (2014) avaliaram o efeito protetor da
quercetina no dano celular miocárdico pós-isquêmico, e observaram que este flavonoide
reduziu as concentrações séricas de CK e LDH.
Com a volta da circulação de sangue nos corações submetidos à isquemia ocorre o
estresse oxidativo devido à formação exacerbada de radicais livres, sendo considerado como
um dos principais fatores que contribuem para a lesão de reperfusão. Moléculas reativas de
oxigênio e nitrogênio estão envolvidas em alguns efeitos deletérios à célula, tais como a
peroxidação lipídica das membranas celulares. Este fenômeno é considerado como o principal
mecanismo de lesão em uma variedade de doenças, principalmente na isquemia-reperfusão
(CECONI et al., 1992). Em consonância, os nossos resultados revelaram que os corações
submetidos à isquemia e reperfusão, na situação controle, exibiram concentrações aumentadas
de MDA e hidroperóxidos totais, indicando claramente a elevação do estresse oxidativo
(Figuras 17 e 18).
Ao investigamos a ação da diosmina no grau da peroxidação lipídica, constatamos que
esta substância atua reduzindo a formação de MDA (Figura 17). Este resultado está em
concordância com os efeitos da diosmina observados na produção de hidroperóxidos totais
(Figura 18), os quais são produtos primários da peroxidação lipídica (LIMA & ABDALLA,
2001). Assim, a diosmina apresenta um efeito protetor contra a lesão tecidual pós-isquêmica.

42
Corroborando nosso estudo, Senthamizhselvan et al. (2014) analisaram o efeito do
pré-tratamento com diosmina em relação ao grau de hidroperóxidos totais e de MDA no
coração de ratos submetidos a isquemia-reperfusão, identificando que a substância apresenta
um potencial de proteção contra a peroxidação lipídica. Liu et al. (2014) investigaram o efeito
protetor da quercetina sobre lesões de isquemia-reperfusão, in vivo, em corações de ratos. Os
autores demonstraram que este flavonoide reduziu o grau de MDA em relação ao grupo
controle, sugerindo uma característica em comum dos flavonoides, a forte capacidade
protetora contra a peroxidação lipídica.
Esse efeito da diosmina sobre a peroxidação lipídica é bem caracterizado na literatura
para diversas condições experimentais, a destacar: neuropatia diabética experimental em ratos
(JAIN et al., 2014), hepatocarcinogenese em modelo murino (THAIR et al., 2013a), lesão
pancreática induzida por isquemia-reperfusão hepática de rato (KILICOGLU et al., 2012) e
isquemia-reperfusão de retina de ratos (TONG et al., 2012). Os mecanismos propostos para
estes efeitos incluem seu caráter antioxidante.
A produção de ROS e RNS durante a isquemia é geralmente baixa e seu significado
patológico é incerto. Tem sido proposto que estas espécies geradas durante a isquemia podem
desempenhar um papel importante na sinalização, e que elas podem provocar um pré-
condicionamento. Entretanto, a geração de radicais livres é produzida em larga escala durante
os primeiros minutos de reperfusão pós-isquêmica, contribuindo para a patogênese de
diversas doenças, especialmente quando os mecanismos de defesa normais estão prejudicados
(PENNA et al., 2009).
Em nosso modelo de isquemia e reperfusão, na ausência de diosmina, houve uma
elevação da atividade das enzimas antioxidantes SOD, Cat, Gpx e GR (Figuras 19 a 22),
provavelmente devido ao aumento da geração de ROS e RNS. Esses dados quando
comparados a literatura se mostram distintos. Os estudos desenvolvidos por Liu et al. (2014) e
Senthamizhselvan et al. (2014) apontaram que os danos ocasionados pelas espécies reativas
são responsáveis pela redução da atividade dessas enzimas antioxidantes. A divergência entre
nossos dados possivelmente é em decorrência das distinções entre os parâmetros
metodológicos adotados por esses autores, que evidenciaram o efeito das substâncias luteolina
e diosmina, respectivamente, em pré-tratamentos de ratos.
A geração de ROS e RNS determina mudanças dependentes do tempo na
disponibilidade de antioxidantes no miocárdio após isquemia-reperfusão. Esse desbalanço

43
pode ser ocasionado pelos diferentes modelos experimentais empregados por distintos grupos
de pesquisadores. A maioria dos estudos parece encontrar esgotamento ou elevação de
antioxidantes enzimáticos e isso é atribuído a diferentes espécies animais, grau de fluxo
sanguíneo colateral, tempo de isquemia, horário da administração de drogas e via de
administração de substâncias (MARCZIN et al., 2003).
Em relação ao tempo, 30 a 35 minutos de isquemia são o ideal para ocasionar a lesão
tecidual. Para a progressão das lesões, a duração da reperfusão deve refletir o curso de tempo
da fisiopatologia da lesão miocárdica, a qual parece variar de espécie para espécie. Para o
coração de rato, muitos grupos relatam que 30 minutos de reperfusão é suficiente. Além disso,
há evidências que não existe nenhuma diferença no tamanho do infarto nos tempos de 60 a
120 minutos de reperfusão (BELL; MOCONU; YELLON, 2011; FERRERA et al., 2009).
Assim, utilizamos em nosso modelo experimental, o tempo de 30 minutos para isquemia e 60
minutos para a reperfusão.
A SOD é uma enzima responsável pela remoção do O2•-, primeiro radical livre
produzido durante o estresse oxidativo. Após a formação do H2O2, dois mecanismos de
defesas endógenos são ativados, a CAT e a GPx, sendo esta última, um dos agentes mais
importantes dos sistemas de defesas antioxidantes (DHALLA et al., 2000). Na presença de
baixas quantidades de H2O2, os peróxidos orgânicos são eliminados preferencialmente por
esta última. Já em condições onde há altas concentrações de H2O2 predomina a ação da Cat
(CHUNG-MAN et al., 2001).
Após a exposição da glutationa reduzida (GSH) ao agente oxidante, ocorre sua
oxidação à glutationa oxidada (GSSH) via enzima GPx, que a utiliza como substrato para
catalisar a reação para remoção dos peróxidos, gerando uma molécula de água. A recuperação
da GSH é então feita pela enzima GR, uma flavoproteína dependente de NADPH, sendo esta,
uma etapa essencial para manter íntegro o sistema de proteção celular (DHALLA et al.,
2000).
Evidenciamos, que nos corações isquêmicos que foram reperfundidos com diosmina
as enzimas SOD, Cat, GPx e GR tiveram suas atividades reduzidas. Acreditamos que isso
tenha ocorrido devido à capacidade sequestradora de radicais livres da diosmina. Essa
proteção é característica dos flavonoides, devido à presença de grupo hidroxila em sua
estrutura que inativa os radicais livres (NIJVELDT et al., 2001). Senthamizhselvan et al.

44
(2014) observaram o efeito quelante da diosmina. Este flavonoide foi capaz de eliminar o O2•-
e OH•, de maneira dependente, nas concentrações de 20, 40, 60, 80 e 100 µmol/L. Assim, os
radicais livres tais como O2•- e OH
•, produzidos durante a isquemia-reperfusão, foram
eliminados pela diosmina. Queenthy e John (2013) também observaram a capacidade
sequestradora dos radicais livres por essa substância. A eliminação de O2•- e OH
• foi realizada
de maneira dependente e uma pequena quantidade desta substância também pode produzir
impacto positivo.
Na isquemia-reperfusão cardíaca o aumento acentuado da produção de ROS e RNS,
associado à sobrecarga de Ca2+
são as causas mais importantes para a abertura do MPTP. Esse
poro é um canal não específico, formado no local do contato entre as membranas interna e
externa mitocondriais. A sua composição molecular exata permanece controversa e este
parece possuir um regulador importante, a ciclofilina D. Como resultado da abertura do
MPTP, há rupturas da membrana externa que facilitam a liberação do citocromo c, o que
culmina em morte celular (MOZAFFARI et al., 2013; WALTERS; POTTER; BROOKES,
2014).
Observamos em nosso estudo, que os corações do grupo isquêmico apresentaram um
aumento na expressão da proteína caspase-3 (Figura 23). A apoptose de cardiomiócitos é um
dos principais contribuintes para o desenvolvimento de infarto do miocárdio, e está
relacionada à insuficiência cardíaca (BIALIK et al., 1997; GOTTLIEB et al., 1994). Na
cardiomiopatia humana, a apoptose de células do músculo cardíaco é associada com a
liberação de citocromo c e ativação de caspase-3. Esta ativação ocorre durante a fase final da
via de sinalização pró-apoptótica e representa um potencial alvo de intervenção para a
supressão da apoptose em cardiomiócitos (NARULA et al., 1999).
Nossos dados evidenciam que nos corações reperfundidos com diosmina a expressão
da caspase-3 apresentou-se reduzida (Figura 23). A isquemia-reperfusão induz apoptose no
miocárdio, enquanto que a inibição da atividade de caspase atenua a apoptose em
cardiomiócitos e, consequentemente, a lesão decorrente da isquemia-reperfusão
(BORUTAITE et al., 2003; GUSTAFSSON et al., 2002 ). Assim, acreditamos que a diosmina
é capaz de impedir o desencadeamento de morte celular nos cardiomiócitos pós-isquêmicos.

45
A maioria dos estudos sugerem que a família Bcl-2 é reguladora chave da apoptose
fisiológica e patológica. A família é composta por dois promotores de morte celular, tais
como Bax, Bad e inibidores da morte celular, que incluem a proteína Bcl-2, Bcl-X, etc
(CHAE et al. 2004; JIAN WEN et al. 2003). Entretanto, interessantemente no estudo de
Senthamizhselvan et al. (2014), ao investigarem o papel protetor da diosmina em corações
isquêmicos no processo de morte celular não foi observada um aumento da expressão da
proteína Bcl-2, o que não corrobora com a literatura.
Dong et al. (2011) investigaram o papel da vitexin em coração de ratos isolados
submetidos á isquemia global na atenuação da apoptose. Estes autores observaram que a
perfusão com essa substância antes da isquemia, promoveu uma redução da expressão da
proteína Bax e aumento da expressão da proteína Bcl-2, evidenciando o papel anti-apoptótico
desse flavonoide nas lesões de isquemia-reperfusão.
Em conjunto, os resultados apresentados no presente estudo sugerem que a diosmina
reduziu as lesões decorrentes da reperfusão e este fato possivelmente está atrelado às suas
ações antioxidantes, e que a mesma não apresenta divergências de efeitos em relação a outras
substâncias pertencentes à mesma classe (QI et al., 2011; LIU et al., 2014).
É importante destacar que mais estudos sobre os mecanismos de ação protetor deste
flavonoide necessitam ser desenvolvidos para que o mesmo venha ser utilizado em
tratamentos clínicos para atenuar as consequências da lesão de reperfusão.

46
7 CONCLUSÃO
Esse estudo indica que a diosmina apresenta efeito protetor quanto ao surgimento das
lesões de reperfusão, sugerindo que a mesma representa uma alternativa terapêutica em
potencial a ser utilizada no tratamento ou como coadjuvante pós-infarto agudo do miocárdio
em humanos. Vale ressaltar que investigações adicionais precisam ser realizadas para
comprovar seu potencial clínico.

47
8 REFERENCIAL BIBLIOGRÁFICO
AKHLAGHI, M.; BANDY, B. Mechanisms of flavonoid protection against myocardial
ischemia–reperfusion injury.Journal of molecular and cellular cardiology, v. 46, n. 3, p.
309-317, 2009.
ARTIGOU, J. Y. et al. Letraitement antioxidant dans lìschemie reperfusion myocardique.
Arch Mal Couer, v. 86, n. 105-109, 1993.
BADALZADEH, R.; YOUSEFI, B.; MAJIDINIA, M.; EBRAHIMI, H. Anti-arrhythmic
effect of diosgenin in reperfusion-induced myocardial injury in a rat model: activation of
nitric oxide system and mitochondrial KATP channel. The Journal of Physiological
Sciences, p. 1-8, 2014. (2014).
BARRABÉS, J. A. et al. Myocardial segment shrinkage during coronary reperfusion in situ.
Pflügers Archiv, v. 431, n. 4, p. 519-526, 1996.
BELL, R. M.; MOCANU, M. M.; YELLON, D. M. Retrograde heart perfusion: the
Langendorff technique of isolated heart perfusion. Journal of molecular and cellular
cardiology, v. 50, n. 6, p. 940-950, 2011.
BERS, D. M. Calcium and cardiac rhythms physiological and pathophysiological.
Circulation research, v. 90, n. 1, p. 14-17, 2002.
BHANDARY, B. et al. The protective effect of rutin against ischemia/reperfusion-associated
hemodynamic alteration through antioxidant activity. Arch Pharm Res, v. 35, n. 6, p. 1091-
1097, 2012.
BIALIK, S. et al. Myocyte apoptosis during acute myocardial infarction in the mouse
localizes to hypoxic regions but occurs independently of p53. J. Clin. Invest, v.100, p.1363–
1372, 1997.
BORUTAITE, V. et al. Inhibition of mitochondrial permeability transition prevents
mitochondrial dysfunction, cytochromec release and apoptosis induced by heart ischemia. J.
Mol. Cell. Cardiol., v. 35, p. 357–366, 2003.
BOSE, R.; SUTHERLAND, G. R.; PINSKY, C. Biological and methodological implications
of prostaglandin involvement in mouse brain lipid peroxidation measurements.
Neurochemical research, v. 14, n. 3, p. 217-220, 1989.
BRASIL. Portaria nº 2.994 do Ministério da Saúde, de 13 de dezembro de 2011.
BRASILEIRO, A. L. S. A injúria de reperfusão miocárdica. Rev SOCERJ, v. 10, p. 79-88,
1997.
CECONI, C. et al. Lipid peroxidation during myocardial reperfusion. Molecular and cellular
biochemistry, v. 111, n. 1-2, p. 49-54, 1992.
CENTURION, S. et al. Aspectos básicos da lesão de isquemia e reperfusão e do pré-
condicionamento isquêmico. Acta Cirúrgica Brasileira, v. 17, n. Suplemento 3, 2002.

48
CHAE, I. et al. Nitric oxide-induced apoptosis is mediated by Bax/Bcl-2 gene expression,
transition of cytochrome c, and activation of caspase-3 in rat vascular smooth muscle cells.
Clinica chimica acta, v. 341, n. 1, p. 83-91, 2004.
CHUNG-MAN, H. J. et al. Differential expression of manganese superoxide dismutase and
catalase in lung cancer. Cancer Res, v. 61, p. 8578-8585, 2001.
COVA, D. et al. Pharmacokinetics and metabolism of oral diosmin in healthy volunteers. Int
J Clin Pharmacol Ther Toxicol, v. 30, p. 29-33, 1992.
DASURI, K.; ZHANG, L; KELLER, J. N. Oxidative stress, neurodegeneration, and the
balance of protein degradation and protein synthesis. Free Radical Biology and Medicine, v.
62, p. 170-185, 2013.
DATASUS Disponível em: http://tabnet.datasus.gov.br/cgi/tabcgi.exe?sim/cnv/obt10uf.def.
Acessado em 01 de setembro de 2014.
DEL MONTE, F. et al. Abrogation of ventricular arrhythmias in a model of ischemia and
reperfusion by targeting myocardial calcium cycling. Proceedings of the National Academy
of Sciences of the United States of America, v. 101, n. 15, p. 5622-5627, 2004.
DHALLA, N. S. et al. Status of myocardial antioxidants in ischemia–reperfusion injury.
Cardiovascular Research, v. 47, n. 3, p. 446-456, 2000.
DONG, L. et al. Vitexin protects against myocardial ischemia/reperfusion injury in
Langendorff-perfused rat hearts by attenuating inflammatory response and apoptosis. Food
and Chemical Toxicology, v. 49, n. 12, p. 3211-3216, 2011.
DROGE, W. Free radicals in the physiological control of cell function. Review. Physiol Rev.,
v. 82, p. 47-95, 2002.
DUNCAN, J. G. et al. Chronic xanthine oxidase inhibition prevents myofibrillar protein
oxidation and preserves cardiac function in a transgenic mouse model of cardiomyopathy.
American Journal of Physiology - Heart and Circulatory Physiology, v. 289, p. 1512–
1518, 2005.
FERDINANDY, P.; SCHULZ, R.; BAXTER, G. F. Interaction of cardiovascular risk factors
with myocardial ischemia/reperfusion injury, preconditioning, and
postconditioning. Pharmacological reviews, v. 59, n. 4, p. 418-458, 2007.
FERREIRA, A. J.; SANTOS, R. A. S; ALMEIDA, A. P. Angiotensin-(1-7): cardioprotective
effect in myocardial ischemia/reperfusion. Hypertension, v. 38, n. 3, p. 665-668, 2001a.
FERREIRA, A. L. A.; MATSUBARA, L. S. Radicais livres: conceitos, doenças relacionadas,
sistema de defesa e estresse oxidativo. Rev Ass Med Brasil, v. 43, p. 61-68, 1997b.
FRÖHLICH, G. M. et al. Myocardial reperfusion injury: looking beyond primary PCI.
European heart journal, v. 34, n. 23, p. 1714-1722, 2013.
GANOTE, C. E. Contraction band necrosis and irreversible myocardial injury. J Mol Cell
Cardiol, v. 15, p. 67-73, 1983.
GE, G., et al. Protective effect of Salvia miltiorrhiza aqueous extract on myocardium
oxidative injury in ischemic–reperfusion rats. Gene, v. 546, p. 97-103, 2014.

49
GIODA, C. R. et al. Cardiac oxidative stress is involved in heart failure induced by thiamine
deprivation in rats. Am J Physiol Heart Circ Physiol, v. 298, p. 2039-2045, 2010.
GOTTLIEB, R.A. et al. Reperfusion injury induces apoptosis in rabbit cardiomyocytes. J.
Clin. Invest., v. 94, p. 1621–1628, 1994.
GRIENDLING, K. K.; FITZGERALD, G. A. Oxidative stress and cardiovascular injury. Part
I: Basic mechanisms and in vivo monitoring of ROS. Circulation, v. 108, p.1912-1916, 2003.
GUSTAFSSON, A.B. et al. TAT protein transduction into isolated perfused hearts: TAT-
apoptosis repressor with caspase recruitment domain is cardioprotective. Circulation, v. 106,
p. 735–739, 2002.
HALLIWELL, B.; CROSS, C. E. Oxygen-derived species: Their relation to human disease
and environmental stress. Environmental Health Perspectives, v. 102, p. 5 1994.
HE, H. et al. Cardioprotective effects of saponins from Panax japonicus on acute myocardial
ischemia against oxidative stress-triggered damage and cardiac cell death in rats. Journal of
Ethnopharmacology, v. 140, p. 73-82, 2012.
IBANEZ, B. et al. Lethal myocardial reperfusion injury: A necessary evil? International
Journal of Cardiology, v. 151, p. 3–11, 2011.
JAIN, D. et al. Protective effect of diosmin against diabetic neuropathy in experimental rats.
Journal of integrative medicine, v. 12, n. 1, p. 35-41, 2014.
JANSE, Michiel J.; WIT, A. L. Electrophysiological mechanisms of ventricular arrhythmias
resulting from myocardial ischemia and infarction. Physiol Rev, v. 69, n. 4, p. 1049-1169,
1989.
JANTET, G. Chronic venous insufficiency: worldwide results of the relief study. Angiology,
v. 53, n. 3, p. 245-256, 2002.
JIAN WEN, D. et al. Intermittent hypoxia attenuates ischemia/reperfusion induced apoptosis
in cardiac myocytes via regulating Bcl-2/Bax expression. Cell research, v. 13, n. 5, p. 385-
391, 2003.
JIANG, Z. Y.; HUNT, J. V.; WOLFF, S. P. Ferrous ion oxidation in the presence of xylenol
orange for detection of lipid hydroperoxide in low density lipoprotein. Analytical
biochemistry, v. 202, n. 2, p. 384-389, 1992.
JIN, H. B. Protective roles of quercetin in acute myocardial ischemia and reperfusion injury in
rats. Mol Biol Rep, v. 39, p. 11005–11009, 2012.
KILICOGLU, S. S. et al. The effect of diosmin on pancreatic injury induced by hepatic
ischemia reperfusion in rats. Bratislavske lekarske listy, v. 114, n. 3, p. 119-124, 2012.
KÖHLER, A. C.; SAG,C.M.; MAIER, L. S. Reactive oxygen species and excitation–
contraction coupling in the context of cardiac pathology. Journal of Molecular and Cellular
Cardiology, v. 73, p. 92-102, 2014.
LEBEAU, J.; NEVIERE, R.; COTELLE, N. Os efeitos benéficos de diferentes flavonóides,
na recuperação funcional após isquemia e reperfusão em ratos isolado coração. Bioorganic &
letras química medicinal, v.11, n. 1, p. 23-27, 2001.

50
LERCH, R. Oxidative substrate metabolism during postischemic reperfusion. Basic Res
Cardiol, v. 88, p. 525-544, 1993.
LIAO, P. H. et al. Cardioprotective effects of luteolin during ischemia-reperfusion injury in
rats. Circulation journal: official journal of the Japanese Circulation Society, v. 75, n. 2,
p. 443-450, 2010.
LIMA, E. S.; ABDALLA, D. S. P. Peroxidação lipídica: mecanismos e avaliação em amostras
biológicas. Brazilian Journal of Pharmaceutical Sciences, v. 37, n. 3, p. 293-303, 2001.
LIN, M. et al. In vitro anti-HIV activity of bioflavonoids isolated from rhus succedanea and
Garciania multiflora. J Nat Prod, v. 60, p.884-888, 1997.
LIU, H. et al. Heart protective effects and mechanism of quercetin preconditioning on anti-
myocardial ischemia reperfusion (IR) injuries in rats. Gene, v. 545, p. 149–155, 2014.
LOPASCHUK, G. D. et al. The antianginal drug trimetazidine shifts cardiac energy
metabolism from fatty acid oxidation to glucose oxidation by inhibiting mitochondrial long-
chain 3-ketoacyl coenzyme A thiolase. Circ Reseach., v. 86, p. 580-588, 2000.
LOPES, R. M. et al. Flavonóides. Biotecnologia, Ciência & Desenvolvimento, v. 3, n. 14,
2000.
LOWRY, O. H. et al. Biol. Chem. v 193, p. 265-275, 1951.
LOZOVOY, M. A. B.; PRIESNITZ, J. C.; SILVA, S. A. Infarto agudo do miocárdio:
aspectos clínicos e laboratoriais. Revista Interbio, Dourados, v. 2, n. 1, p. 4-10, 2008.
LYSENG-WILLIAMSON, K. A.; PERRY, C. M. Micronised purified flavonoid fraction.
Drugs, v. 63, n. 1, p. 71-100, 2003.
MADESH, M.; BALASUBRAMANIAN, K. A. Microtiter plate assay for superoxide
dismutase using MTT reduction by superoxide. Indian J. Biochem. Biophys, v. 35, p. 184-
188, 1998.
MANNERVIK, B.; CARLBERG, I. Glutathione reductase. Methods in Enzymology, v. 113,
p. 484-490, 1985.
MARCZIN, N. et al. Antioxidants in myocardial ischemia–reperfusion injury: therapeutic
potential and basic mechanisms. Arch Biochem Biophys, v. 420, p. 222-236, 2003.
MARTÍNEZ-FLÓREZ, S. et al. Los flavonoides: propiedades y acciones antioxidantes.
Nutrición hospitalaria, v. 17, n. 6, 2002.
MAUPOIL, V. et al. Identification et evaluation des radicaux libres dans la sequence
ischémie-reperfusion cardiaque. Arch Mal Coeur., v. 86, p. 63-68, 1993.
MAXWELL, S. R. J; LIP, G. Y. H. Reperfusion injury: a review of the pathophysiology,
clinical manifestations and therapeutic options. International journal of cardiology, v. 58, n.
2, p. 95-117, 1997.
MONASSIER, J. P. Reperfusion injury in acute myocardial infarction: from bench to cath
lab. Part II: Clinical issues and therapeutic options. Archives of cardiovascular diseases, v.
101, n. 9, p. 565-575, 2008.

51
MOZAFFARI, M. S. et al. Mechanisms of load dependency of myocardial ischemia
reperfusion injury. American journal of cardiovascular disease, v. 3, n. 4, p. 180, 2013.
MULLER, J. E. Diagnosis of myocardial infarction: historical notes from the Soviet Union
and the United States. The American journal of cardiology, v. 40, n. 2, p. 269-271, 1977.
NANDAVE, M.; OJHA, S. K.; ARYA, D. S. Protective role of flavonoids in cardiovascular
diseases. Nat Prod Radiance, v. 4, p. 166-176, 2005.
NARULA, J. et al. Apoptosis in heart failure: release of cytochrome c from mitochondria and
activation of caspase-3 in human cardiomyopathy. Proc. Natl. Acad. Sci., v. 96, p. 8144–
8149, 1999.
NELSON, D. P.; KIESOW, L. A. Enthalpy of decomposition of hydrogen peroxide by
catalase at 25°C (with molar extinction coefficients of H2O2 solutions in the UV). Anal
Biochem, v. 49, p. 474–478, 1972.
NIJVELDT, R. J. et al. Flavonoids: a review of probable mechanisms of action and potential
applications. Am J Clin Nutr, v. 74, n. 4, p. 418-425, 2001.
PAGLIA, D. E. ; VALENTINE, W. N. J. Lab. Clin. Med. v. 70, p. 158-169, 1967.
PARADIES, G. et al. Role of cardiolipin peroxidation and Ca2+
in mitochondrial dysfunction
and disease. Cell calcium, v. 45, n. 6, p. 643-650, 2009.
PARANG, P.; SINGH, B.; ARORA, R. Metabolic modulators for chronic cardiac ischemia. J
Cardiovasc Pharmacol Ther, v. 10, p. 217-223, 2005.
PECKING, A. P. et al. Efficacy of Daflon 500 mg in the treatment of lymphedema (secondary
to conventional therapy of breast cancer). Angiology, v. 48, n. 1, p. 93-98, 1997.
PENNA, C. et al. Cardioprotection: a radical view: free radicals in pre and postconditioning.
Biochimica et Biophysica Acta (BBA)-Bioenergetics, v. 1787, n. 7, p. 781-793, 2009.
PERRELLI, M. G. et al. Ischemia/reperfusion injury and cardioprotective mechanisms: Role
of mitochondria and reactive oxygen species. World J Cardiol,v. 3, p. 186 -200, 2011.
PHAM-HUY, L. A.; HE, H.; PHAM-HUY, C. Free radicals, antioxidants in disease and
health. International journal of biomedical science, v. 4, n. 2, p. 89, 2008.
PIPER, H. M.; ABDALLAH, Y.; SCHÄFER, C. The first minutes of reperfusion: a window
of opportunity for cardioprotection. Cardiovascular research, v. 61, n. 3, p. 365-371, 2004a.
PIPER, H. M.; GARCÑA-DORADO, D.; OVIZE, M. A fresh look at reperfusion injury.
Cardiovascular research, v. 38, n. 2, p. 291-300, 1998b.
PIPER, H. M.; KASSECKERT, S.; ABDALLAH, Y. The sarcoplasmic reticulum as the
primary target of reperfusion protection. Cardiovascular research, v. 70, n. 2, p. 170-173,
2006c.
PIPER, H. M.; MEUTER, K.; SCHÄFER, C. Cellular mechanisms of ischemia-reperfusion
injury. The Annals of thoracic surgery, v. 75, n. 2, p. S644-S648, 2003d.

52
QI, L. et al. Luteolin improves contractile function and attenuates apoptosis following
ischemia–reperfusion in adult rat cardiomyocytes. European journal of pharmacology, v.
668, n. 1, p. 201-207, 2011.
QIAO, Z.; MA, J.; LIU, H. Evaluation of the antioxidant potential of Salvia miltiorrhiza
ethanol extract in a rat model of ischemia-reperfusion injury. Molecules, v. 16, n. 12, p.
10002-10012, 2011.
QUEENTHY, S. S.; JOHN, B. Diosmin exhibitsanti hyperlipidemiac effects in isoproterenol
induced myocardial infarcted rats..European Journal of Pharmacology, v. 718, p. 213-218,
2013.
RENAUD, S.; LORGERIL, M. Wine, alcohol, platelets, and the French paradox for coronary
heart disease. The Lancet, v. 339, p. 1523-1526, 1992.
RUIXING, Y.; WENWU, L.; AL-GHAZALI, R. Trimetazidine inhibits cardiomyocyte
apoptosis in a rabbit model of ischemia-reperfusion. Translational Research, v. 149, n. 3, p.
152-160, 2007.
SENTHAMIZHSELVAN, O. et al. Diosmin pretreatment improves cardiac function and
suppresses oxidative estress in rat heart after ischemia/reperfusion. European Journal of
Pharmacology, v. 736, p. 131-137, 2014.
SESSO, H. D. et al. Coffee and tea intake and the risk of myocardial infarction. American
journal of epidemiology, v. 149, n. 2, p. 162-167, 1999.
SHAHIDI, F.; NACZK, M. Phenolics in food and nutraceuticals. CRC press, 2003.
SHEN A. C.; JENNINGS, R. B. Myocardial calcium and magnesium in acute ischemic
injury. Am J Pathol, v. 67, p. 417–440, 1972.
SILAMBARASAN, T.; RAJA, B. Diosmin, a bioflavonoid reverses alterations in blood
pressure, nitric oxide, lipid peroxides and antioxidant status in DOCA-salt induced
hypertensive rats. European Journal of Pharmacology, v. 679, p. 81–89, 2012.
SKRZYPIEC-SPRING, M. et al. Isolated heart perfusion according to Langendorff-still
viable in the new millennium. Journal of pharmacological and toxicological methods, v.
55, n. 2, p. 113-126, 2007.
SRINIVASAN, S.; PARI, L. Ameliorative effect of diosmin, a citrus flavonoid against
streptozotocin- nicotinamide generated oxidative stress induced diabetic rats. Chemico -
Biological Interactions, v. 195, p. 43–51, 2012.
SUN, D. et al. Luteolin limits infarct size and improves cardiac function after myocardium
ischemia/reperfusion injury in diabetic rats. PLoS One, v. 7, n. 3, p. e33491, 2012.
SUZUKI, J. et al. Tea catechins attenuate chronic ventricular remodeling after myocardial
ischemia in rats. Journal of Molecular and Cellular Cardiology, v. 42, n. 2, p. 432–440,
2007.
TAHIR, M. et al. Diosmin abrogates chemically induced hepatocarcinogenesis via alleviation
of oxidative stress, hyperproliferative and inflammatory markers in murine model.
Toxicology letters, v. 220, n. 3, p. 205-218, 2013.

53
TONG, N. et al. Diosmin Protects Rat Retina from Ischemia/Reperfusion Injury. Journal of
ocular pharmacology and therapeutics, v. 28, p. 459-466, 2012.
TOUFEKTSIAN, M. C. et al. Chronic dietary intake of plant-derived anthocyanins protects
the rat heart against ischemia-reperfusion injury. Journal of nutrition, v. 138, n. 4, p. 747-
752, 2008.
TZIOMALOS, K.; HARE, J. M. Role of xanthine oxidoreductase in cardiac nitroso redox
imbalance. Frontiers in Bioscience, v. 14, p. 237–262, 2009.
VALKO, M. et al. Free radicals and antioxidants in normal physiological functions and
human disease. The International Journal of Biochemistry & Cell Biology, v. 39, p. 44-84,
2007.
VOLP, A. C. P. et al. Flavonóides antocianinas: características e propriedades na nutrição e
saúde. Rev Bras Nutr Clin, v. 23, p. 141-149, 2008.
WALTERS, A. M.; PORTER, G. A.; BROOKES, P. S. Mitochondria as a drug target in
ischemic heart disease and cardiomyopathy. Circulation research, v. 111, n. 9, p. 1222-1236,
2014.
WOLFF, J. A.; BUDKER, V. O mecanismo de absorção de DNA nu e expressão. Avanços
na genética, v. 54, p. 1-20, 2005.
YAO, L.H. et al. Flavonoids in food an their health benefitis. Plant. Foods for human
Nutrition, v.59.p.113-122, 2004.
YOKOZAWA, T. et al. Antioxidant activity of flavones and flavonols in vitro. Phytotherapy
Research, v. 11, p. 446-450, 1997.
YOSHIDA, W. Radicais livres na síndrome da isquemia e reperfusão. Circ Vasc Angiol, v.
12, p. 82-95, 1996.
ZHANG, R. et al. Clematichinenoside attenuates myocardial infarction in
ischemia/reperfusion injury both in vivo and in vitro. Planta Med, v. 79, n. 14, p. 1289-1297,
2013.
ZWEIER, J. L.; TALUKDER, M. A. H. The role of oxidants and free radicals in reperfusion
injury. Cardiovascular research, v. 70, n. 2, p. 181-190, 2006.

54
ANEXOI





























![(Fibras cardíacas I [Modo de Compatibilidade])](https://img.document.onl/doc/110x75/5571ff9b49795991699dab47/fibras-cardiacas-i-modo-de-compatibilidade.jpg)