Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DE SERGIPE
PRÓ-REITORIA DE PÓS-GRADUAÇÃO E PESQUISA
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS
FISIOLÓGICAS
FABÍULA FRANCISCA DE ABREU
AÇÕES FARMACOLÓGICAS DA RUTINA NA PANCREATITE AGUDA EM CAMUNDONGOS
SÃO CRISTÓVÃO
2014

i
FABÍULA FRANCISCA DE ABREU
AÇÕES FARMACOLÓGICAS DA RUTINA NA PANCREATITE AGUDA EM CAMUNDONGOS
Dissertação apresentada ao Programa de Pós-Graduação em Ciências Fisiológicas da Universidade Federal de Sergipe como requisito à obtenção do grau de Mestre em Ciências Fisiológicas. Orientador: Prof. Dr. Enilton Aparecido Camargo
SÃO CRISTÓVÃO
2014

ii
FICHA CATALOGRÁFICA ELABORADA PELA BIBLIOTECA CENTRAL UNIVERSIDADE FEDERAL DE SERGIPE
A162a
Abreu, Fabíula Francisca de Ações farmacológicas da rutina na pancreatite aguda em camundongos / Fabíula Francisca de Abreu ; orientador Enilton Aparecido Camargo. – São Cristóvão, 2014.
89 f. : il.
Dissertação (Mestrado em Ciências Fisiológicas) - Universidade Federal de Sergipe, 2014.
1. Farmacologia clínica. 2. Pancreatite. 3. Vitaminas – uso terapêutico. I. Camargo, Enilton Aparecido, orient. II. Título.
CDU 615.03:616.37

iii
FABÍULA FRANCISCA DE ABREU
AÇÕES FARMACOLÓGICAS DA RUTINA NA PANCREATITE AGUDA EM CAMUNDONGOS
Dissertação apresentada ao Programa de Pós-Graduação em Ciências Fisiológicas da Universidade Federal de Sergipe como requisito à obtenção do grau de Mestre em Ciências Fisiológicas.
______________________________________________________________
Orientador: Prof. Dr. Enilton Aparecido Camargo
_______________________________________________________________
1° Examinador: Profa. Dra. Flávia Teixeira Silva
_______________________________________________________________
2° Examinador: Profa. Dra. Soraia Kátia Pereira Costa
SÃO CRISTÓVÃO
2014

iv
AGRADECIMENTOS
Ao final de mais uma etapa em minha trajetória, gostaria de agradecer a todas as pessoas que contribuíram direta e indiretamente para que ela fosse concluída com êxito.
Ao prof. Enilton Camargo, meu orientador, por ter me aceitado como sua aluna, pelos ensinamentos, pela confiança e paciência, por não me deixar desanimar nos momentos de dificuldade e por não ter medido esforços para que este trabalho fosse executado com êxito. O senhor é um mestre exemplar e um modelo a ser seguido, quando eu “crescer” quero ser que nem você!
À minha família, em especial à minha mãe, pelo apoio incondicional, por estar sempre presente, por ser meu alicerce e pelo amor sempre implícito em todas as palavras e atitudes.
Aos colegas de LAFAPI, pelo companheirismo, pela colaboração e pela paciência nos surtos de chatice. Agradeço em especial, á grande amiga que ganhei logo ao entrar no laboratório, Ana Carla, por estar presente em todos os momentos, pela colaboração nos experimentos, pelas ricas e “discordantes” discussões, pelos inúmeros almoços maravilhosos, por tornar divertidas todas as situações (inclusive as mais tensas) e pela amizade. À Dani, por ter me guiado na condução dos primeiros experimentos e pelo suporte dado sempre que necessitei. Aos meus “curicos”, alunos de Iniciação Científica, Marcela, Allan, Janaíne, Daiane e Janaína, por suas contribuições específicas nas diversas fases dos “n” experimentos (que sempre eram os últimos...rsrs) e pelos finais de semana (e madrugadas, né Dai?) dedicados.
Aos professores Soraia Pereira Costa e Marcelo Nicolás Muscará, do Laboratório de Bioquímica dos Radicais Livres e Farmacorregulação da Inflamação Neurogênica do Instituto de Ciências Biomédicas da USP, por terem aberto as portas de seus laboratórios para mim, pelas ricas sugestões e por terem viabilizado a execução da segunda etapa deste trabalho.
Aos técnicos do ICB pelo carinho e empenho durante a realização dos experimentos e análises realizados em São Paulo. A Antônio, pela carinhosa recepção, pela competência ao realizar comigo o experimento às cegas para hiperalgesia e pelo feriado no escuro (e sem café!!!) dedicado á finalização deste. À Irene, pelas conversas, por ser sempre ser tão solícita em ajudar no que fosse necessário e por me “alimentar” com as inúmeras caixinhas de ponteiras preenchidas. À Simone, por todos os ensinamentos, pela confiança, amizade e principalmente pela determinação para que conseguíssemos cumprir todas as análises propostas a tempo... Não tenho palavras suficientes pra demonstrar como você foi importante!
Aos demais professores que contribuíram para realização deste trabalho: Sara Thomazzi, Waldeci de Lucca, Rosilene Calazans e Sandra Lauton.

v
Às minhas “curicas” tão especiais Ana Carla, Patrícia, Cibelle e Edênia pela amizade, pelas noites de estudos, pelo apoio, pela ajuda nos experimentos, por me inspirarem em tantos aspectos, pelos momentos de alegria e sufoco compartilhados, por serem minhas “orientadoras” particulares, principalmente durante a elaboração da dissertação; enfim, por tudo de bom que cada uma de vocês adicionou nesse processo.
Aos colegas de Mestrado e de Departamento, por tornarem esse período do mestrado enriquecedor, mais leve e divertido. À Rosana, Camilinha, Camilla, Valéria, Priscila, Luana, Rangel e Larissa, pelas conversas, discussões, almoços, saídas e momentos alegres. Aos lindos, “Nas manhas”, Patrícia, Ana Carla, Cibelle, Edênia, Lívia, Katty, Renan, Rafa, Matheus, Diogo, Priscila e Larissa pelos momentos de descontração, conversas científicas (ou não!) ...O Mestrado já valeu a pena só por ter conhecido todos vocês.
Aos ex-colegas de LEOPAR e grandes amigos, pelos ensinamentos e por serem minha inspiração de sucesso. Agradeço em especial aos meninos, Adriano, Lucas e Vanderson por me receberem generosamente em suas moradias no período do curso de Inverno e da realização dos Experimentos no ICB. À Isabella por estar sempre presente, me ouvindo e compartilhando momentos importantes.
A Daniel, pela proteção e companhia nas noites de sábado e domingo em que precisei ir tratar os animais.
Às minhas vizinhas: Monik, Sara e Fabi pela companhia, carinho e auxílio em momentos de crise.
À minha grande amiga Carla Mirelle, que apesar de estar longe, sempre mantém contato e a todo tempo tenta me situar no planeta terra, me dando sábios conselhos. Obrigada pela amizade e por estar sempre presente, compartilhando alegrias e tristezas.
Às amigas Karol, Lary, Edilaine, Paula, Morgana, Inaê, Priscila, Isabela, Mônica, Jack, Carol e demais amigos e amigas que não foram aqui citados.
À minha professora de Iniciação Científica Júnior, Maria Silene, por ter me apresentado o mundo da ciência e ter sido minha inspiração pra chegar até aqui.
Ao PROCFIS, pela oportunidade e viabilização desta pesquisa.
À Fundação de Apoio à Pesquisa e à Inovação Tecnológica do Estado de Sergipe (FAPITEC/SE), pelo apoio financeiro.
Aos camundongos, que ao terem suas vidas interrompidas, mesmo que involuntariamente, contribuíram para que no futuro, quem sabe, milhares de vidas humanas sejam poupadas.

vi
Só depende de nós...
"Hoje levantei cedo pensando no que tenho a fazer antes que o relógio marque meia noite. É minha função escolher que tipo de dia vou ter hoje.
Posso reclamar porque está chovendo ou agradecer às águas por lavarem a poluição. Posso ficar triste por não ter dinheiro ou me sentir encorajado para
administrar minhas finanças, evitando o desperdício.
Posso reclamar sobre minha saúde ou dar graças por estar vivo.
Posso me queixar dos meus pais por não terem me dado tudo o que eu queria ou
posso ser grato por ter nascido. Posso reclamar por ter que ir trabalhar ou agradecer por ter trabalho. Posso sentir tédio com o trabalho doméstico ou
agradecer a Deus por ter um teto para morar.
Posso lamentar decepções com amigos ou me entusiasmar com a possibilidade de fazer novas amizades. Se as coisas não saíram como planejei
posso ficar feliz por ter hoje para recomeçar.
O dia está na minha frente esperando para ser o que eu quiser. E aqui estou eu, o escultor que pode dar forma.
Tudo depende só de mim.”
(Charles Chaplin)

vii
RESUMO Ações Farmacológicas da Rutina na Pancreatite Aguda em Camundongos. Fabíula Francisca de Abreu, Mestrado em Ciências Fisiológicas, Universidade Federal de Sergipe, São Cristóvão/SE, 2014. A pancreatite aguda (PA) é uma doença grave em cerca de 20% dos casos, que causa hospitalização e óbito dos pacientes devido às complicações sistêmicas associadas. Seu tratamento clínico tem se mostrado insuficiente para controlar o processo inflamatório intrínseco da doença e é focado no manejo dos sintomas e complicações. Dentre os diversos fatores envolvidos na PA, destacam-se a resposta inflamatória e o estresse oxidativo. Neste contexto, a rutina, um flavonoide conhecido por suas atividades antioxidante e anti-inflamatória, apresenta-se como uma substância natural em potencial a ser utilizada no tratamento da PA. Assim, o objetivo do presente estudo foi investigar os efeitos farmacológicos da rutina em modelo de pancreatite aguda experimental induzida por L-arginina em camundongos. Foram utilizados camundongos Swiss machos adultos (25-30 g) e os procedimentos foram aprovados pelo Comitê de Ética em Pesquisa com Animais da UFS (43/2012). Para a indução da pancreatite os animais receberam duas injeções de L-arginina (8%, 4 g/kg, i.p., com intervalo de 1 h entre elas). O grupo controle recebeu duas injeções de salina (0,9%, i.p.). Os animais submetidos à indução da pancreatite foram tratados com rutina (37,5, 75 ou 150 mg/kg, v.o.) ou salina (veículo), após 24, 36, 48 e 60 h da 1° injeção de L-arginina. O grupo controle foi tratado com veículo nos mesmos tempos. A eutanásia dos animais foi realizada 72 h após a indução, com subsequente coleta de sangue e de órgãos (pâncreas, pulmão, fígado e rim). Foram avaliados parâmetros que permitiram inferir sobre o quadro inflamatório pancreático e sistêmico e avaliar as concentrações séricas de enzimas pancreáticas, a hiperalgesia abdominal e o estresse oxidativo. Os animais que receberam as injeções de L-arginina apresentaram aumento significativo dos parâmetros inflamatórios e bioquímicos preditivos de pancreatite, quando comparados aos animais que receberam salina. O tratamento com rutina reduziu a atividade de mieloperoxidase no pâncreas (p<0,001 para 37,5, 75 ou 150 mg/kg), mas não no pulmão, reduziu o índice de edema pancreático (p<0,001 para 37,5 mg/kg e p<0,05 para 75 e 150 mg/kg) e a concentração sérica de amilase (p<0,001 para 75 e 150 mg/kg). A partir destes resultados a dose de 75 mg/kg foi escolhida para ser utilizada nos experimentos posteriores. O tratamento com esta dose de rutina também diminuiu as concentrações séricas de lipase (p<0,001), proteína C reativa (p<0,001) e de interleucina 6 (p<0,001), bem como reduziu a hiperalgesia abdominal (p<0,05), após 72 h da injeção de L-arginina, quando comparados ao grupo L-arginina + Veículo. O tratamento com rutina (75 mg/kg) também reduziu a peroxidação lipídica induzida pela L-arginina em pâncreas, fígado e rim (p<0,001) e aumentou a atividade de catalase pancreática (p<0,001),de glutationa peroxidase (p<0,05) de superóxido dismutase no pâncreas (p<0,01) e no fígado (p<0,05), além de diminuir a expressão de 3-nitrotirosina no pâncreas (p<0,05). Estes resultados evidenciam que a rutina exerce efeito anti-inflamatório, antinociceptivo e antioxidante durante a PA induzida por L-arginina, os quais sugerem que este flavonoide seja de interesse para abordagens ou estudos futuros objetivando novas alternativas no tratamento da PA em humanos. Descritores: Pancreatite; Rutina; Inflamação; Dor; Estresse Oxidativo.

viii
ABSTRACT
Pharmacological Actions of Rutin on Acute Pancreatitis in Mice. Fabíula Francisca de Abreu, Mestrado em Ciências Fisiológicas, Universidade Federal de Sergipe, São Cristóvão/SE, 2014. Acute pancreatitis (AP) is a severe disease in about 20% of patients, causing hospitalization and death, mainly due to associated systemic complications. The treatment of this condition is still insufficient to control the intrinsic inflammatory process and is focused on managing the complications and symptoms of patients. Among the many factors involved in AP, the inflammatory response and oxidative stress can be highlighted. In this context, rutin is presented as a natural substance with important potential to treat AP, by considering its anti-inflammatory and antioxidant activities. The aim of this study is to investigate the pharmacological effects of rutin on experimental AP induced by L-arginine administration in mice. Adult male Swiss mice (25-30 g) were used in this study and all experiments were approved by this institution´s Ethics Committee in Animal Research (43/2012). For the induction of AP, mice received 2 injections of L-arginine (8%, 4 g/kg, i.p., with an interval of 1 h). The control group received the same volume of saline (0.9%) instead of L-arginine. Mice submitted to AP induction were treated with rutin (37.5, 75 or 150 mg/kg, p.o.) or saline (vehicle) after 24, 36, 48 and 60 h of the first injection of L-arginine. The control group was treated with vehicle at the same time points. The euthanasia occurred after 72 h of induction and was accompanied by blood and organ (pancreas, lung, liver and kidney) collection. We investigated parameters that permitted us to infer about pancreatic and systemic inflammation and evaluate serum concentrations of pancreatic enzymes, abdominal hyperalgesia and oxidative stress. In animals injected with L-arginine, it was detected the increase of inflammatory and biochemical parameters that confirmed the induction of AP, when compared with saline-injected animals. The treatment with rutin reduced the myeloperoxidase activity in pancreas (p<0.001 for 37.5, 75 or 150 mg/kg), but not in lung, reduced the pancreatic edema index (p<0.001 for 37.5 mg/kg and p<0.05 for 75 and 150 mg/kg) and the serum concentration of amylase (p<0.001 for 75 and 150 mg/kg). From these experiments, we chose the dose of 75 mg/kg for the next steps. In this way, treatment with this dose of rutin also reduced the serum lipase (p<0.001), C reactive protein (p<0.001) and interleukin-6 (p<0.001) concentrations, as well as decreased the abdominal hyperalgesia (p<0.05), when compared with Vehicle + L-arginine group after 72 h of L-arginine injection. The administration of rutin also diminished lipid peroxidation induced by L-arginine in pancreas, liver and kidney (p<0.001) and increased both the activity of catalase in pancreas (p<0.001), glutathione peroxidase in pancreas (p<0.05) and superoxide dismutase in pancreas (p<0.01) and liver (p<0.05). Besides, it decreased the expression of 3-nitrotyrosine in pancreas (p<0.05). Altogether, these results demonstrate that rutin exert anti-inflammatory, antinociceptive and antioxidant actions during AP induced by L-arginine, which are suggestive that this flavonoid is of interest for developing future studies or approaches focused on new alternatives to treat AP in humans. Keywords: Acute Panceatitis; Rutin; Inflammation; Pain; Oxidative Stress.

ix
LISTA DE FIGURAS
Figura 1 - Estrutura do Pâncreas ................................................................................ 4
Figura 2 – Estruturas químicas da quercetina e da rutina. ........................................ 18
Figura 3 - Linha temporal indicando a indução da pancreatite com L-arginina e o
esquema terapêutico de tratamento com a rutina. .................................................... 23
Figura 4 - Reações realizadas para mensuração da atividade de Catalase.. ........... 29
Figura 5 - Reação xantina oxidase/XTT .................................................................... 30
Figura 6 - Efeito da rutina sobre a concentração sérica de amilase em camundongos
com pancreatite aguda induzida por L-arginina.. ....................................................... 35
Figura 7 - Efeito da rutina sobre a atividade de mieloperoxidase (MPO) no pâncreas
(A) e no pulmão (B) em camundongos com pancreatite aguda induzida por L-
arginina. .................................................................................................................... 36
Figura 8 - Efeito da rutina sobre o índice de edema pancreático em camundongos
com pancreatite aguda induzida por L-arginina......................................................... 37
Figura 9 - Efeito da rutina sobre as concentrações séricas de lipase (A) e proteína C
reativa (PCR) (B) em camundongos com pancreatite aguda induzida por L-arginina.
.................................................................................................................................. 38
Figura 10 - Efeito da rutina sobre a concentração sérica de interleucina 6 (IL-6) de
camundongos com pancreatite aguda induzida por L-arginina ................................. 39
Figura 11 - Efeito da rutina na hiperalgesia abdominal durante a pancreatite aguda
induzida por L-arginina em camundongos. ............................................................... 40
Figura 12 - Efeito da rutina sobre as concentrações de malonaldeído (MDA) em
pâncreas (A), pulmão (B), fígado (C) e rim (D) de camundongos com pancreatite
aguda induzida por L-arginina. .................................................................................. 42
Figura 13 - Efeito da rutina sobre o conteúdo de 3-Nitrotirosina (3-NT) em pâncreas
de camundongos com pancreatite aguda induzida por L-arginina. ........................... 43
Figura 14 - Efeito da rutina sobre as atividades das enzimas antioxidantes catalase
(CAT), superóxido dismutase (SOD) e glutationa peroxidase (GSH-Px) no pâncreas
de camundongos com pancreatite aguda induzida por L-arginina ............................ 44

x
LISTA DE TABELAS
Tabela 1 - Relação dos grupos experimentais da Etapa 1. ....................................... 24
Tabela 2 - Relação dos grupos experimentais da Etapa 2. ....................................... 24
Tabela 3 - Efeito da rutina sobre as atividades das enzimas antioxidantes catalase
(CAT), superóxido dismutase (SOD) e glutationa peroxidase (GSH-Px) no pulmão,
fígado e rim de camundongos com pancreatite aguda induzida por L-arginina. ....... 45

xi
LISTA DE SIGLAS
BHT
BSA
Butil-hidroxitolueno
Albumina Bovina Sérica
CAT
CEPA
Catalase
Comitê de Ética em Pesquisa Animal
CGRP Peptídeo Relacionado ao Gene da Calcitonina
cNOS
COX2
DNA
Óxido Nítrico Sintase Constitutiva
Ciclo-oxigenase 2
Ácido Desoxirribonucleico
eNOS Óxido Nítrico Sintase Endotelial
EPM Erro Padrão da Média
GPx Glutationa Peroxidase
GSH Glutationa Reduzida
GSSG Glutationa Oxidada
HTAB
HRP
Hexadeciltrimetilamônio
“Horseradish” peroxidase
i.p. Intraperitoneal
IL Interleucina
IL-6R Receptor de Interleucina 6
iNOS Enzima Óxido Nítrico Sintase Induzível
LDL Lipoproteína de Baixa Densidade
LPS Lipopolissacarídeo
MDA Malonodialdeído
MPO
NADP+
Mieloperoxidase
Fosfato de Dinucleotídio de Nicotinamida e Adenina Oxidada
NADPH Fosfato de Dinucleotídio de Nicotinamida e Adenina Reduzida
NF-B
3-NT
Fator de Transcrição Nuclear kappa B
3-Nitrotirosina
NK-1 Neurocinina 1
NO
O.D
Óxido Nítrico
o-Dianisidina
PA Pancreatite Aguda

xii
PBS Tampão Fosfato Salina
PCR
PGE2
PL
PLA2
Proteína C Reativa
Prostaglandina E2
Peroxidação Lipídica
Fosfolipase A2
RNS Espécies Reativas de Nitrogênio
ROS Espécies Reativas de Oxigênio
SIRS Síndrome da Resposta Inflamatória Aguda
SOD Superóxido Dismutase
TBA
TBST
Ácido Tiobarbitúrico
Tampão Tris Fosfato Salina
TBARS Substâncias Reativas ao Ácido Tiobarbitúrico
TCA
TGO
TGP
TMB
Ácido Tricloroacético
Transaminase Glutâmica Oxalacética
Transaminase Glutâmica Pirúvica
3.3’5.5’-tetrametilbenzidina
TNF-α Fator de Necrose Tumoral Alfa
TRPA1 Receptor de Potencial Transitório Anquirina 1
TRPV1 Receptor de Potencial Transitório Vanilóide do Tipo 1
UFS Universidade Federal de Sergipe
v.o. Via Oral
XTT 3’-1- [(fenilamino)-carbonil]-3,4-tetrazólio-bis(4-metóxi-6-nitro)
ácido benzenosulfónico hidratado

xiii
SUMÁRIO
1 INTRODUÇÃO ..................................................................................................... 1
2 REVISÃO DE LITERATURA ................................................................................ 4
2.1 Pancreatite Aguda .......................................................................................... 4
2.2 Modelos de Pancreatite Aguda Experimental ................................................ 9
2.3 Radicais livres e Estresse Oxidativo na Pancreatite Aguda ......................... 12
2.4 Flavonoides .................................................................................................. 15
2.5 Rutina ........................................................................................................... 17
3 OBJETIVOS ....................................................................................................... 21
3.1 Geral ............................................................................................................ 21
3.2 Específicos ................................................................................................... 21
4 MATERIAL E MÉTODOS ................................................................................... 22
4.1 Animais ........................................................................................................ 22
4.2 Indução da Pancreatite Aguda por L-arginina .............................................. 22
4.3 Delineamento Experimental ......................................................................... 23
4.4 Avaliação de Parâmetros Inflamatórios ........................................................ 25
4.4.1 Determinação da atividade de Mieloperoxidase (MPO) ......................... 25
4.4.2 Índice de Edema .................................................................................... 25
4.5 Avaliação de Parâmetros Bioquímicos no Soro ........................................... 26
4.5.1 Determinação de Amilase e Lipase Séricas .......................................... 26
4.5.2 Concentração de proteína C reativa (PCR) ........................................... 26
4.5.3 Dosagem de interleucina 6 (IL-6) ........................................................... 26
4.6 Avaliação Comportamental .......................................................................... 27
4.6.1 Medida da Hiperalgesia Abdominal ....................................................... 27
4.7 Avaliação de Parâmetros Oxidantes e Antioxidantes ................................... 28
4.7.1 Determinação do grau de peroxidação lipídica ...................................... 28
4.7.2 Determinação da atividade de Catalase (CAT) ..................................... 28
4.7.3 Determinação da atividade de superóxido dismutase (SOD) ................ 30
4.7.4 Atividade de Glutationa Peroxidase (GSH-Px) ...................................... 31
4.7.5 Dosagem de proteínas .......................................................................... 32
4.7.6 Quantificação de resíduos proteicos de 3-nitrotirosina (3-NT) ............... 32
4.8 Análise Estatística ........................................................................................ 33

xiv
5 RESULTADOS ................................................................................................... 34
5.1 Efeito da Rutina sobre parâmetros inflamatórios e bioquímicos da
pancreatite aguda induzida por L-arginina ............................................................. 34
5.2 Efeito da Rutina sobre a Hiperalgesia Abdominal ........................................ 39
5.3 Efeito da Rutina sobre o Estresse Oxidativo na Pancreatite Aguda induzida
por L-arginina ......................................................................................................... 40
5.3.1 Peroxidação Lipídica ............................................................................. 40
5.3.2 Resíduos de 3-Nitrotirosina (3-NT) ........................................................ 42
5.3.3 Atividade das enzimas antioxidantes ..................................................... 43
6 DISCUSSÃO ...................................................................................................... 46
7 CONCLUSÃO .................................................................................................... 59
8 REFERÊNCIAS .................................................................................................. 60
ANEXO 1 ................................................................................................................... 75

1
1 INTRODUÇÃO
A pancreatite aguda é definida como um processo inflamatório agudo do
pâncreas que envolve variavelmente tecidos regionais ou sistemas de órgãos à
distância, e apresenta como principais características clínicas dor abdominal crônica
e aumento nas concentrações séricas das enzimas pancreáticas lipase e/ou amilase
(BOLLEN et al., 2008; YADAV; LOWENFELS, 2013). As principais causas
apontadas são o abuso de álcool e cálculos biliares (GRANGER; REMICK, 2005).
Independentemente do fator etiológico para o dano primário à célula acinar,
ocorre uma ativação prematura das enzimas digestivas pancreáticas que levam a
um processo de autodigestão responsável por lesão celular e tecidual,
desencadeando as respostas inflamatórias locais e sistêmicas da doença (BHATIA
et al., 2000). As principais enzimas envolvidas neste processo são a tripsina, a
amilase e a lipase pancreáticas (BHATIA, 2005).
Em cerca de 80% dos casos, ocorre resolução própria da doença, sem que
ocorra uma evolução do quadro inflamatório inicial (LUND et al., 2006). Nestes casos
a pancreatite é considerada autolimitada e de gravidade leve. Porém, nos outros
20% dos casos, a resposta inflamatória não é controlada e adquire gravidade
sistêmica, levando à morte celular e generalização da resposta inflamatória
(FROSSARD, 2009). Essa resposta pode levar à hospitalização ou mesmo ao óbito
do paciente, devido às complicações sistêmicas da doença, que envolvem
principalmente o pulmão, o fígado e os rins (GRANGER; REMICK, 2005),
determinando o curso e a gravidade da pancreatite aguda (SCHÜTTE;
MALFERTHEINER, 2008).
Essa amplificação da inflamação ocorre devido à ação de radicais livres, à
estimulação de citocinas inflamatórias (como o fator de necrose tumoral-α [TNF-α] e
a interleucina-6 [IL-6]) e a ação de células do sistema imune (BHATIA, 2005). Os
radicais livres são representados principalmente pelas espécies reativas de oxigênio
(ROS) e de nitrogênio (RNS), que são gerados no metabolismo oxidativo fisiológico,
mas que são produzidos excessivamente em diversas condições patológicas, a

2
exemplo da pancreatite (DASURI; ZHANG; KELLER, 2013). Eles são responsáveis
por acentuar e contribuir diretamente para a lesão celular e aumentar a resposta
inflamatória, além de ativar a transcrição do fator nuclear B (NF-B), que é um
evento crucial para estimular a expressão de diversos mediadores inflamatórios
envolvidos na resposta inflamatória aguda, como o TNF-α (ARJUMAND; SETH;
SULTANA, 2011;CHVANOV; PETERSEN; TEPIKIN, 2005).
Estudos realizados em modelos de pancreatite experimental demonstraram
que compostos que atuam bloqueando ou inibindo a atividade de mediadores
inflamatórios, como o TNF-α e a IL-6, podem atuar como potentes agentes
terapêuticos no seu tratamento (MAEDA et al., 2003; GLOOR et al., 1998; ZHANG et
al., 2013; GRANGER; REMICK, 2005). Compostos com propriedades antioxidantes
vêm se mostrando eficazes em reduzir a resposta inflamatória na pancreatite aguda,
inibindo alguns estágios que levam ao processo inflamatório ou bloqueando
diretamente a geração de RNS e ROS, responsáveis pelo dano celular na
pancreatite (CARVALHO et al., 2010; COMERT et al., 2007; CARRASCO et al.,
2013).
Apesar dos recentes avanços no diagnóstico e tratamento, a pancreatite
aguda continua sendo um desafio para os cirurgiões, clínicos e intensivistas, por
falta de um tratamento específico e eficaz no controle da doença, contribuindo para
índices de mortalidade entre 10% a 25% dos casos graves (TONSI, 2009).
Um grupo de substâncias que vêm se destacando, principalmente por seus
efeitos antioxidantes, são os flavonoides, uma classe de polifenois encontrados em
frutas, folhas e sementes que incluem seis grupos de substâncias principais:
antocianinas, proantocianinas, isoflavonas, flavononas, flavonóis e flavonas
(CERQUEIRA; MEDEIROS; AUGUSTO, 2007).
O primeiro flavonoide isolado que se tem conhecimento é a rutina, uma
flavona, obtida através da extração de laranjas que passou a ser amplamente
explorada por seus efeitos terapêuticos (MIYASHIRO, 2010), dentre os quais
destacam-se as propriedades antioxidante, anti-inflamatória e lipolítica (ARJUMAND;
SETH; SULTANA, 2011; OLIVEIRA et al., 2002).

3
Estudo realizado com o extrato etanólico da entrecasca da Caesalpinia
pyramidalis Tul. (Fabaceae), demonstrou que este reduziu parâmetros inflamatórios,
oxidativos e nociceptivos, em modelo de pancreatite induzida por obstrução do ducto
biliopancreático em ratos (SANTANA et al., 2012). A análise cromatográfica indicou
a presença de rutina nesse extrato, sugerindo que este composto poderia estar
participando das ações farmacológicas verificadas.
Adicionalmente, a quercetina, flavonoide que se apresenta na forma aglicona,
e que consiste em um dos metabólitos ativos da rutina após passagem pelo trato
gastrointestinal (AFANAS’EV et al., 1989), atenua o quadro inflamatório pancreático
e pulmonar na pancreatite aguda induzida por ceruleína em camundongos
(CARVALHO et al., 2010).
Diante dos resultados prévios obtidos com o extrato etanólico da entrecasca
da Caesalpinia pyramidalis, do potencial antioxidante e anti-inflamatório da rutina e
das informações limitadas sobre o efeito deste composto em modelos de pancreatite
aguda, o presente estudo buscou investigar a hipótese de que a rutina possa
contribuir para o tratamento da pancreatite aguda experimental induzida por
L-arginina em camundongos.
4
2 REVISÃO DE LITERATURA
2.1 Pancreatite Aguda
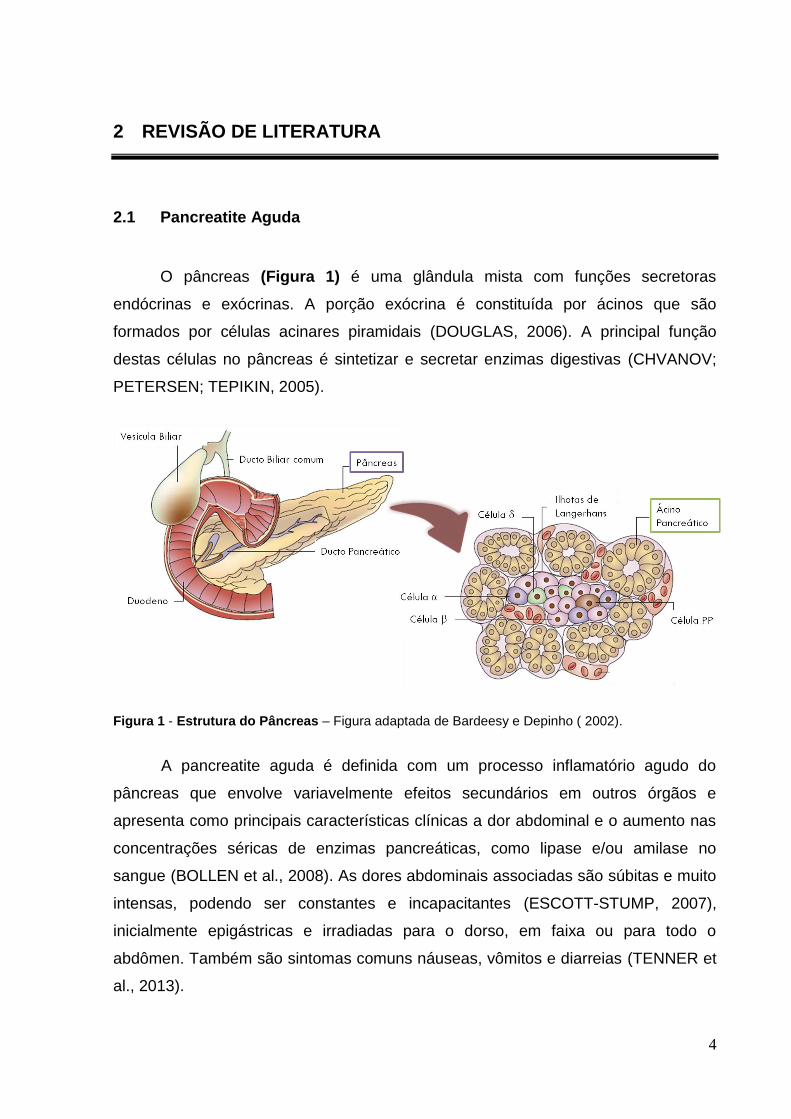
O pâncreas (Figura 1) é uma glândula mista com funções secretoras
endócrinas e exócrinas. A porção exócrina é constituída por ácinos que são
formados por células acinares piramidais (DOUGLAS, 2006). A principal função
destas células no pâncreas é sintetizar e secretar enzimas digestivas (CHVANOV;
PETERSEN; TEPIKIN, 2005).
Figura 1 - Estrutura do Pâncreas – Figura adaptada de Bardeesy e Depinho ( 2002).
A pancreatite aguda é definida com um processo inflamatório agudo do
pâncreas que envolve variavelmente efeitos secundários em outros órgãos e
apresenta como principais características clínicas a dor abdominal e o aumento nas
concentrações séricas de enzimas pancreáticas, como lipase e/ou amilase no
sangue (BOLLEN et al., 2008). As dores abdominais associadas são súbitas e muito
intensas, podendo ser constantes e incapacitantes (ESCOTT-STUMP, 2007),
inicialmente epigástricas e irradiadas para o dorso, em faixa ou para todo o
abdômen. Também são sintomas comuns náuseas, vômitos e diarreias (TENNER et
al., 2013).

5
A incidência da pancreatite aguda varia de acordo com região, atingindo uma
faixa de 13 a 45 pessoas para cada 100.000 habitantes (YADAV; LOWENFELS,
2013b), com taxa de mortalidade de 9% a 20%, sendo apontadas como causas mais
comuns o abuso de álcool e litíase, além de causas idiopáticas (GRANGER;
REMICK, 2005). A incidência no Brasil ainda não tem um consenso devido às
dificuldades no diagnóstico e subnotificações no levantamento epidemiológico,
porém números recentes do estado de São Paulo mostram que 20% dos casos de
pancreatite avaliados na Santa Casa de São Paulo são graves, sendo que metade
destes vão a óbito, e a prevalência é de 20 casos para cada 100 mil habitantes
(ALBUQUERQUE, 2013).
Uma das hipóteses propostas para a patogênese da pancreatite aguda é que
uma lesão primária às células acinares, ainda não elucidada, leva à ativação do
tripsinogênio intracelular e inibição da secreção enzimática, mecanismo que pode
ser potencializado por ação do álcool, drogas ou hipertrigliceridemia (FROSSARD;
STEER; PASTOR, 2008; GRANGER; REMICK, 2005). Essa ativação do
tripsinogênio converge para um processo de autodigestão tecidual mediado pela
ativação prematura de outras enzimas digestivas, como quimiotripsina,
carboxipolipeptidases, ribonucleases, desorribonucleases, esterases, fosfolipases,
lipase e amilase, com formação de mediadores biologicamente ativos e indução de
resposta inflamatória (GARDNER, 1979; DOUGLAS, 2006). A ativação destas
enzimas parece estar envolvida na lesão aos tecidos e às membranas celulares
causando edema, alterações vasculares, hemorragia e necrose (BHATIA, 2005).
Esse processo de lesão tecidual leva a ativação local do sistema imune,
incluindo células dendríticas, macrófagos, fibroblastos e células T (MEIRELLES JR.
et al., 2003). Na maioria dos casos, a pancreatite avança até este estágio e a
inflamação local é revertida per se (BOLLEN et al., 2008). Entretanto, em parte dos
casos, a doença evolui adquirindo gravidade sistêmica em decorrência do
descontrole da inflamação local que leva a uma resposta inflamatória generalizada
(BHATIA, 2005). Este quadro agrava o risco de ocorrer insuficiência múltipla dos
órgãos, hospitalização prolongada e até a morte (GRANGER; REMICK, 2005).
O recrutamento exacerbado de leucócitos para o pâncreas consiste em um
dos eventos críticos no desenvolvimento da pancreatite aguda (BHATIA, 2005) e

6
acredita-se que este seja regulado por moléculas de adesão e quimiocinas (MERZA
et al., 2013). É conhecido que os neutrófilos estão envolvidos no desenvolvimento
da necrose acinar em pancreatite aguda, por conta da ação de seus produtos
oxidantes (H2O2, O2-, e NO), os quais são potencialmente danosos às estruturas
celulares (SANDOVAL et al., 1996).
Assim, nos casos em que a resposta inflamatória progride ocorre um aumento
da produção de espécies reativas de oxigênio (ROS), que por sua vez atuam
exacerbando o dano celular e causando lesões às membranas celulares e
citoesqueleto, afetando a atividade de enzimas intracelulares, danificando o DNA,
depletando os níveis de antioxidantes e ativando o fator de transcrição NF-B
(RAKONCZAY, 2003). A ativação deste fator é responsável por induzir a expressão
do fator de necrose tumoral α (TNF-α) e outros mediadores inflamatórios, que
estimulam a invasão e ativação de células inflamatórias, as quais também estão
relacionados ao aumento da formação de ROS (ARJUMAND; SETH; SULTANA,
2011; ALTAVILLA et al., 2003).
O TNF-α é considerado a principal citocina inflamatória envolvida no
desencadeamento dos eventos iniciais das respostas inflamatórias, e conjuntamente
com a interleucina (IL-6) participa na patogênese da pancreatite aguda (CZAKÓ et
al., 2000a) e na geração de mediadores que levam à complicações de órgãos
distantes, a exemplo da indução da síntese hepática de proteínas de fase aguda
(ROMANO et al., 1997;SCHÜTTE; MALFERTHEINER, 2008;ZHANG et al., 2013).
Outro mediador importante envolvido nessa resposta inflamatória da
pancreatite aguda é o óxido nítrico (NO), o qual tem sido apontado por estar
envolvido no desenvolvimento das complicações sistêmicas e locais da pancreatite,
bem como no estresse oxidativo em vários órgãos, que é característico dos casos
mais graves (TAKÁCS et al., 2002). Porém, estudos também têm sugerido possíveis
efeitos benéficos do NO nos casos de pancreatite necrosante ou edematosa, por
ocasionar melhora da microcirculação, oxigenação pancreática e redução da adesão
(LEUNG; CHAN, 2009; SÁNCHEZ-BERNAL et al., 2004), de modo que, o papel
exato desse mediador na pancreatite ainda é incerto.

7
É importante ressaltar que em 80% dos casos de pancreatite aguda, ocorre
resolução própria da doença, sem que ocorra uma evolução do quadro inflamatório
inicial (LUND et al., 2006). Nestes casos a pancreatite é considerada autolimitada e
de gravidade leve (BOLLEN et al., 2008). Porém, nos outros 20%, a resposta
inflamatória não é controlada e adquire gravidade sistêmica, levando à morte celular
e generalização da resposta inflamatória (FROSSARD, 2009), o que pode levar o
paciente a hospitalização ou mesmo a óbito, devido às complicações sistêmicas da
doença, que envolvem principalmente o pulmão, o fígado e os rins (GRANGER;
REMICK, 2005). A disfunção múltipla dos órgãos e o desenvolvimento de
complicações sistêmicas ou locais determinam o curso e a gravidade da pancreatite
aguda (SCHÜTTE; MALFERTHEINER, 2008).
É sugerido que a dor oriunda da pancreatite estaria relacionada com a
inflamação, pois ocorre exposição das aferências pancreáticas à ação de
mediadores inflamatórios, neuropeptídeos endógenos, substâncias derivadas de
células do sistema imune e outras células, como as citocinas. Assim, além desses
mediadores levarem à lesão do tecido pancreático, eles podem eventualmente
causar danos as fibras aferentes sensoriais e hiperexcitabilidade (SCHWARTZ et al.,
2013). Estudos experimentais em modelos de pancreatite tem demonstrado o
envolvimento do neuropeptídeo CGRP (peptídeo relacionado ao gene da calcitonina)
liberado no corno dorsal da medula espinhal (WINSTON et al., 2003; WICK et al.,
2006), da expressão de receptores TRPV1 (WICK, 2005) e TRPA1 (SCHWARTZ et
al., 2011) e da ativação de receptores NK-1 (CAMARGO et al., 2011) nas aferências
pancreáticas. Por exemplo, no modelo de pancreatite induzida por fosfolipases A2
secretórias em ratos foi sugerido que a ação da substância P e da bradicinina na
sinalização do processo inflamatório decorrente da pancreatite (CAMARGO et al.,
2008) parecem contribuir para a indução da hiperalgesia abdominal tanto via
estimulação de receptores NK-1 (CAMARGO et al., 2011), quanto via produção de
NO (CAMARGO et al., 2013).
O reconhecimento de pacientes com risco de desenvolver as formas graves
da pancreatite numa fase ainda inicial da doença, é crucial para otimizar a terapia e
o monitoramento do quadro clínico (SCHÜTTE; MALFERTHEINER, 2008). Assim,
diversos marcadores vêm sendo identificados e correlacionados, com o objetivo de

8
facilitar esse reconhecimento e prevenir a progressão e as complicações da
pancreatite, que envolve disfunção múltipla dos órgãos e sepse (MCKAY; BUTER,
2003), as quais são as principais causas de morte no curso final dessa doença (AL-
BAHRANI; AMMORI, 2005; FROSSARD; HADENGUE; PASTOR, 2001; LEMPINEN;
PUOLAKKAINEN; KEMPPAINEN, 2005).
A proteína C reativa (PCR) é uma proteína de fase aguda, sintetizada
principalmente pelo fígado e sua liberação está relacionada com a ação de citocinas
circulantes (PUOLAKKAINEN et al., 1987). Ela é liberada em diversos processos
inflamatórios agudos, estando envolvida no reconhecimento de substâncias
autógenas potencialmente tóxicas liberadas no plasma por tecidos lesionados, com
o objetivo de detoxificar e/ou facilitar a eliminação destes (PUOLAKKAINEN et al.,
1987), sendo sua determinação um importante parâmetro para inferir sobre a
extensão da lesão tecidual. Em quadros de pancreatite aguda em humanos
apresenta-se como marcador bioquímico importante, pois sua elevação no soro é
considerada como um parâmetro altamente específico para a gravidade da
pancreatite, bem como para o diagnóstico, quando associado com alterações de
outros parâmetros séricos como amilase, lipase e IL-6 (KEIM et al., 1998;AL-
BAHRANI; AMMORI, 2005), indicando necrose pancreática (SCHÜTTE;
MALFERTHEINER, 2008) e disfunção múltipla dos órgãos (AKAY et al., 2008;
ANDERSSON, 2010).
Outro marcador importante para inferir sobre a ocorrência e a gravidade da
pancreatite é interleucina 6 (IL-6), a qual também é reportada por estar envolvida na
progressão da pancreatite e na disfunção múltipla dos órgãos associada (OSMAN et
al., 2002; FARKAS et al., 1998; MAEDA et al., 2003). É sabido que essa citocina
estimula a síntese e liberação de PCR hepática (GANAPATHI et al., 1988),
consistindo assim um importante parâmetro preditivo para pancreatite aguda grave
(BHATIA et al., 2000; ZHANG et al., 2013). Estudo recente evidenciou que pacientes
com pancreatite aguda e com disfunção respiratória, circulatória e renal
apresentaram valores mais altos dessa citocina juntamente com IL-8, em detrimento
de IL-18 e TNF-α (MALMSTRØM et al., 2012).
Ademais, a fisiopatologia e os mecanismos de ação dos mediadores
envolvidos na iniciação e progressão da pancreatite aguda ainda não estão bem

9
elucidados. Isso resulta na dificuldade de tratamento desta condição, principalmente
nos casos mais graves, nos quais há a necessidade de terapias anti-inflamatórias
clinicamente efetivas.
Segundo Tonsi et al. (2009), mesmo com os recentes avanços no diagnóstico
e tratamento, a pancreatite aguda ainda é um desafio para os cirurgiões, clínicos e
intensivistas, por não possuir um tratamento específico. Nenhuma terapia atual é
capaz de neutralizar o processo inflamatório da pancreatite, de modo que os
tratamentos utilizados buscam controlar basicamente os sintomas e as
complicações, em particular a dor, que inicialmente é controlada com a utilização de
analgésicos não-opíódes (GRIGSBY; RODRIGUEZ-RILO; KHAN, 2012). Porém, a
utilização destes na maioria das vezes falha, com consequente necessidade do uso
de narcóticos, que a longo prazo podem causar dependência.
Assim, evidencia-se a necessidade de desenvolvimento de novos e eficazes
agentes terapêuticos para o tratamento da pancreatite aguda.
2.2 Modelos de Pancreatite Aguda Experimental
Existem diversos modelos animais de pancreatite aguda descritos na
literatura, os quais variam de acordo com a gravidade, tipo de indução de edema,
progressão da lesão para múltiplos órgãos ou tipo de morte celular (GRANGER;
REMICK, 2005; LEUNG; CHAN, 2009; YILDIZ, 2010). Eles têm sido utilizados com o
objetivo de investigar os mecanismos fisiopatológicos da doença e para estudar os
efeitos de diferentes agentes terapêuticos. A seguir, estão descritos alguns modelos
que mimetizam as características da pancreatite aguda em humanos:
1. Administração de secretagogos: neste modelo ocorre estimulação da
secreção exócrina por meio da administração de um indutor de secreção. O
secretagogo mais utilizado é o análogo da colecistocinina, a ceruleína, o qual
geralmente é administrado por injeção intravenosa ou intraperitoneal. Causa
pancreatite edematosa por aumento da secreção pancreática (GRADY et al., 1996).
2. Obstrução do ducto biliopancreático: a ligação cirúrgica do ducto
biliopancreático resulta em aumento da pressão intraductal e vazamento de bile no

10
pâncreas, levando à subsequente inflamação, desencadeada pela ativação das
enzimas digestivas pancreáticas. Esse modelo causa pancreatite hemorrágica aguda
(LERCH et al., 1993).
3. Injeção intraductal: substâncias que induzem pancreatite podem ser
injetadas diretamente no ducto biliopancreático. São comumente utilizados sais
biliares, como o tauracolato de sódio (AHO et al., 1980) ou enzimas, como as
fosfolipases A2 (CAMARGO et al., 2005). Pode originar pancreatite edematosa ou
hemorrágica, a depender da substância injetada e apresenta boa reprodutibilidade e
relevância clínica.
4. Injeção de doses excessivas de L-arginina: a administração, por via
intraperitoneal, de altas doses deste aminoácido resulta em pancreatite necrosante,
apresentando como principais alterações edema intersticial, infiltração inflamatória e
degranulação acinar (TANI et al., 1990; CZAKÓ et al., 2000a; DAWRA et al., 2006).
A indução da pancreatite por L-arginina, é um modelo não invasivo, de fácil
indução e altamente reprodutível, que leva a uma forma de pancreatite aguda grave
necrosante que mimetiza a doença em humanos (CZAKÓ et al., 2000a; DAWRA;
SALUJA, 2012).
A administração de doses excessivas de L-arginina para indução de
pancreatite aguda em ratos foi caracterizado por (MIZUNUMA; KAWAMURA;
KISHINO, 1984), quando estes buscavam investigar os efeitos de altos níveis desse
aminoácido em diversos tecidos e observaram a presença de lesão,
especificamente, no pâncreas. Em seus resultados foram encontradas alterações
nas células acinares, como degeneração vacuolar, necrose celular e subsequente
fibrose, no tratamento com uma dose de 500 mg/kg de peso corporal de L-arginina.
A partir destes resultados diversos trabalhos começaram a ser conduzidos no intuito
de investigar o mecanismo de ação desta substância e a administração de altas
doses de L-arginina passou a ser utilizada como modelo de indução de pancreatite
aguda.
Czakó et al., (2000) demonstraram que altas doses de L-arginina provocam
alterações laboratoriais e morfológicas, após 24 horas da indução, como aumento da
concentração de amilase e necrose das células acinares, semelhantes às

11
encontradas na pancreatite aguda 24 horas após a indução. Eles também avaliaram
o papel do estresse oxidativo na patogênese da pancreatite nesse modelo e
verificaram um aumento nas concentrações de malondialdeído (MDA), como
marcador da peroxidação lipídica, e redução do sistema de defesa antioxidante
endógeno, com diminuição das atividades de catalase, superóxido dismutase (SOD)
e glutationa peroxidase.
Além disso, a evidência de que as óxido nítrico sintases (NOS) estão
presentes nas células acinares e o fato delas realizarem a metabolização da L-
arginina, as tornam alvo importante não só no modelo de pancreatite induzida por L-
arginina (DAWRA; SALUJA, 2012), mas também em outros modelos (CAMARGO et
al., 2011; CUZZOCREA et al., 2004; DIMAGNO, 2004). Outros estudos confirmam o
potencial do NO como mediador da iniciação e do desenvolvimento da pancreatite
aguda (DAWRA et al., 2007). Acredita-se que o NO produzido fisiologicamente
nessas células está implicado na sua homeostase (CHVANOV; PETERSEN;
TEPIKIN, 2005).
No contexto da pancreatite induzida por L-arginina, a expressão de isoformas
constitutiva (cNOS) e induzível (iNOS) foi aumentada 24 horas após indução de
pancreatite aguda em ratos e o tratamento com L-NAME, um inibidor da síntese de
NO, reverteu apenas a atividade cNOS, não sendo eficaz sobre iNOS,
provavelmente por causa da atividade exacerbada desta (TAKÁCS et al., 2002).
Essa exacerbação foi relacionada à atuação de citocinas inflamatórias, como TNF-α
e IL-6, que apresentaram concentrações séricas elevadas após indução de
pancreatite (TAKÁCS et al., 1996; SCHÜTTE; MALFERTHEINER, 2008)
Até pouco tempo, a maioria dos estudos realizados utilizando o modelo de
pancreatite aguda induzida por L-arginina foram com ratos, sendo poucos os
realizados em camundongos. Nesse contexto, (DAWRA et al., 2007) propuseram a
padronização de um protocolo de indução de pancreatite aguda por injeção de L-
arginina em camundongos, de forma que este apresentasse boa reprodutibilidade.
Até então, havia uma baixa reprodutibilidade deste modelo em camundongos, em
virtude da variabilidade das doses de L-arginina que eram testadas, as quais eram
similares às utilizadas em ratos. Deste modo, estes autores mostraram que a
administração de 2 doses de L-arginina de 4 g/kg em camundongos, com um

12
intervalo de 1 hora entre elas, levou a um aumento significativo de amilase no
plasma, de mieloperoxidase (MPO) pancreática, ativação de tripsina e alterações
histológicas, como vacuolização e necrose extensiva das células acinares,
semelhantemente à pancreatite aguda em humanos. Estas alterações se mostraram
mais pronunciadas no tempo de 72 h. O mesmo estudo também demonstrou que
houve alterações histológicas e infiltrado inflamatório no pulmão, característicos do
dano pulmonar encontrado em outros modelos de indução de pancreatite (DAWRA
et al., 2007).
Os mecanismos envolvidos nestes eventos são pouco compreendidos e
podem estar associados a vários fatores, como a produção de radicais livres. Uma
das hipóteses é o envolvimento de NO, estresse oxidativo e mediadores
inflamatórios como agentes da lesão acinar pancreática (DAWRA; SALUJA, 2012;
HEGYI et al., 2004). Outro mecanismo proposto é o envolvimento de metabólitos da
L-arginase, umas das enzimas que metabolizam a L-arginina, hidrolisando-a a
ornitina e ureia, os quais poderiam exercer algum efeito tóxico específico às células
acinares (BICZÓ et al., 2010; RAKONCZAY et al., 2008). Entretanto, o exato
mecanismo ainda não foi esclarecido.
2.3 Radicais livres e Estresse Oxidativo na Pancreatite Aguda
Os radicais livres são átomos, íons ou moléculas que apresentam um elétron
não pareado em sua órbita externa, caracterizados por grande instabilidade e
elevada reatividade, que podem interagir por meio de reações de óxido-redução com
quaisquer moléculas, inclusive biomoléculas como proteínas, fosfolipídios
insaturados e DNA, causando danos às células e tecidos (REIS et al., 2008). As
principais fontes destas espécies são a cadeia de transporte de elétrons
mitocondrial, o complexo NADPH oxidase, especialmente o encontrado em
neutrófilos e células fagocíticas, a xantina oxidase e as isoenzimas do citocromo 450
(CERQUEIRA; MEDEIROS; AUGUSTO, 2007; DASURI; ZHANG; KELLER, 2013).
Além da lesão direta, essas moléculas, representadas principalmente por
espécies reativas provenientes do metabolismo do oxigênio (ROS), funcionam como

13
sinalizadores intracelulares que ativam vários processos fisiológicos e bioquímicos,
resultando em alterações celulares e teciduais (LEUNG; CHAN, 2009). As ROS
referem-se a todos os estados de excitação e oxidação de moléculas de oxigênio
resultantes de condições fisiológicas e incluem o radical ânion superóxido (O2•−),
radical hidroxila (•OH), oxigênio singlete, peróxido de hidrogênio (H2O2) e outros
peróxidos. Também atuam como radicais livres as espécies reativas de nitrogênio
(RNS) que são constituídas pelo óxido nítrico (NO) e seus intermediários de
oxidação ou produtos de redução, incluído nitrosil, óxido nitroso, peroxinitrito e
alguns adutos instáveis de NO com biomoléculas (VALKO et al., 2007). Estes são
igualmente participantes diretos da rede de sinalização intracelular e quando
produzidos em altas concentrações são altamente danosos (CERQUEIRA;
MEDEIROS; AUGUSTO, 2007; CHVANOV; PETERSEN; TEPIKIN, 2005).
O radical superóxido constitui uma das principais ROS, produzido em
processos celulares como respiração celular e sinalização via NADPH oxidase não
fagocítica (HALLIWELL; GUTTERIDGE, 1984). É capaz de reagir com outros
radicais, gerando espécies reativas secundárias igualmente tóxicas. Por exemplo, a
interação de NO e O2•−, pode gerar radicais peroxinitrito que são capazes de
modificar proteínas, alterando sua estrutura e função, levando diretamente ao dano
celular (SUGIURA; ICHINOSE, 2011; VALKO et al., 2007). Já a reação com o H2O2,
é capaz de gerar radicais hidroxila, que são altamente reativos e são reportados por
apresentar alta especificidade de reação com bases nitrogenadas de DNA, alterando
sua função e igualmente levando a dano celular (HALLIWELL, 2006).
Esses compostos também podem atuar sobre membranas celulares, de modo
a iniciar um processo denominado peroxidação lipídica. Nesse processo os ácidos
graxos das cadeias laterais dos lipídeos de membrana, especialmente aqueles que
contem duas ou mais ligações duplas, são oxidados a hidroperóxidos, os quais
podem interagir com outros radicais ou íons metálicos, gerando aldeídos citotóxicos
como o 4-hidroxi-2-nonenal (HNE) e o malonaldeído (MDA) (HALLIWELL;
GUTTERIDGE, 1984;VALKO et al., 2007).
Para equilibrar a resposta oxidativa, originada tanto em condições fisiológicas
quanto patológicas, o organismo desenvolveu uma série de mecanismos de defesa,
que inclui antioxidantes enzimáticos e não-enzimáticos, que exercem seus efeitos

14
através de múltiplas vias para tentar controlar as espécies reativas em um sistema
celular (DASURI; ZHANG; KELLER, 2013). Uma primeira linha de defesa atua como
detoxificadora do agente antes que ocorra alguma lesão, sendo representada pela
glutationa reduzida (GSH), tioredoxina e pela superóxido dismutase (SOD), enzima
que metaboliza o radical ânion superóxido produzindo peróxido de hidrogênio, que
por sua vez é convertido em água e oxigênio molecular pela catalase (CAT) e
peroxidases, como a glutationa peroxidase (GSH-Px), que atuam destruindo
peróxidos (HUBER; ALMEIDA; FÁTIMA, 2008; VALKO et al., 2007). Uma outra linha
de defesa importante tem como função reparar a lesão ocorrida, e é representada
pela glutationa redutase (GSH-Rd), vitamina C e também pela GSH-Px (FERREIRA;
MATSUBARA, 1997).
Recentemente, o aumento na expressão e/ou atividade de CAT e GSH-Px
são sugeridas por apresentar efeito neuroprotetor (DASURI; ZHANG; KELLER,
2013), bem como sua depleção parece aumentar a susceptibilidade a dano neuronal
(DASURI; ZHANG; KELLER, 2013;MAO et al., 2012).
Embora estejam envolvidos na regulação de processos fisiológicos, o excesso
na produção de ROS e RNS está relacionado à estimulação de algumas vias
intracelulares associadas ao desenvolvimento de diversas doenças (DASURI;
ZHANG; KELLER, 2013). Em condições normais ocorre um balanço entre a
produção de radicais livres e os mecanismos de defesa antioxidante, que mantêm o
sistema redox controlado. Porém, se ocorre uma exacerbação na produção de ROS
e RNS e os mecanismos de defesa antioxidantes não são suficientes para supri-la, é
estabelecido um quadro denominado estresse oxidativo, o qual está relacionado a
uma série de doenças degenerativas e inflamatórias (CERQUEIRA; MEDEIROS;
AUGUSTO, 2007).
Mutações no DNA decorrentes de dano oxidativo foram encontradas em
diversos tipos de tumores, sugerindo forte participação dessas espécies na etiologia
do câncer (MENA; ORTEGA; ESTRELA, 2009) e modificações oxidativas em
proteínas que participam de processos importantes de regulação da função
homeostática estão implicadas no desenvolvimento de doenças como Alzheimer’s,
Diabetes e Aterosclerose (DASURI; ZHANG; KELLER, 2013; VALKO et al., 2007). A
ROS e RNS também são reportadas por contribuir para a formação de fatores

15
quimiotáxicos que acentuam a atração de neutrófilos para o sítio inflamatório (“Metal
chelation therapy, oxygen radicals, and human disease”, 1985).
Investigações realizadas nos diversos modelos de pancreatite aguda
experimental têm demonstrado que ocorre uma diminuição das enzimas
antioxidantes SOD e CAT e aumento da peroxidação lipídica, representado por altas
concentrações teciduais de MDA (CZAKÓ et al., 2000b; LONG et al., 2005);
(SOWJANYA; SANDHYA; VEERESH, 2012), confirmando que as ROS e RNS
desempenham papel importante na patogênese da pancreatite aguda, sendo
relatados por participarem na iniciação e progressão desta doença (CZAKÓ et al.,
1998; GUICE et al., 1986). O infiltrado inflamatório tecidual decorrente da resposta
inflamatória inicial é em parte responsável pela grande produção destas espécies, as
quais juntamente com as citocinas levam ao dano tecidual pancreático (LONG et al.,
2005).
Trabalhos investigando o efeito de compostos com atividade antioxidante no
tratamento da pancreatite aguda vêm demonstrando que o controle do estresse
oxidativo mediado por estes, consiste em uma importante estratégia para controlar o
quadro inflamatório e a lesão tecidual desta doença (CARVALHO et al., 2010;
SOWJANYA; SANDHYA; VEERESH, 2012; SZABOLCS et al., 2006b) os quais
constituem o principal desafio encontrado pelo tratamento convencional utilizado.
Assim, as ROS e RNS estão envolvidas em vários quadros patológicos, a
exemplo da pancreatite aguda (QUE et al., 2010), e substâncias que possam auxiliar
a combater o estresse oxidativo que acompanha tais patologias podem ser de
importante uso terapêutico, como é o caso de flavonoides.
2.4 Flavonoides
Uma qualidade de vida mais saudável e a prevenção de doenças estão cada
vez mais relacionados a dietas ricas em frutas e vegetais. Dentre os componentes
destas dietas, evidências experimentais apontam que carotenoides e polifenois
poderiam estar exercendo um importante papel nos mecanismos de proteção e
diminuição do risco de incidência de doenças como câncer, diabetes e hipertensão,

16
atribuídos a esse tipo de alimentação (STAHL; SIES, 1996). Nesse contexto,
substâncias pertencentes a essas classes de compostos bioativos vêm sendo
evidenciadas por suas atividades biológicas protetoras, apresentando-se como
agentes terapêuticos em potencial para o tratamento de inúmeras patologias
(BIRADAR; VEERESH, 2012).
Os flavonoides consistem na classe de polifenois mais bem estudada por
seus efeitos antioxidantes e são encontrados naturalmente em frutas, folhas e
sementes na forma de glicosídeos ou agliconas (ESCOTT-STUMP, 2007).
Apresentam em comum a estrutura C6-C3-C6, constituída de dois anéis aromáticos
ligados por heterocíclico oxigenado e uma ou mais hidroxilas que lhe conferem
características antioxidantes, por serem capazes de reduzir radicais livres
(CERQUEIRA; MEDEIROS; AUGUSTO, 2007). Incluem seis grupos de substâncias
principais: antocianinas, proantocianinas, isoflavonas, flavononas, flavonóis e
flavonas (ESCOTT-STUMP, 2007), que variam estruturalmente de acordo com o
nível de hidrogenação, hidroxilação, metilação e sulfonação das moléculas
(BEHLING et al., 2008).
Os efeitos biológicos dos flavonoides tem sido extensivamente estudados e
elucidados, revelando importantes propriedades de interesse farmacológico, como
por exemplo, inibição do sistema NADPH-oxidase de neutrófilos (TAUBER; FAY;
MARLETTA, 1984), inibição de fosfolipase A2 (LINDAHL; TAGESSON, 1997),
inibição de processos geradores de radicais livres, mediada pela capacidade de
reagir com íons ferro, ânion superóxido e radicais peroxil (AFANAS’EV et al., 1989),
inibição da mieloperoxidase (MEOTTI et al., 2008), inibição da função plaquetária
(WRIGHT et al., 2013) e modulação de citocinas participantes da hiperalgesia, como
IL-6 e IL-10 (BORGHI et al., 2013).
Ensaios biológicos usando compostos isolados revelaram que os flavonoides
exibem uma larga faixa de efeitos nos sistemas biológicos (ROTELLI et al., 2003).
Destacam-se a capacidade antioxidante (mecanismo mais bem elucidado), atividade
anti-inflamatória e efeito vasodilatador, ação antialérgica, atividade hepatoprotetora e
anti-ulcerogênica, dentre outras (BEHLING et al., 2008; FASINU et al., 2013;
FONTANA PEREIRA et al., 2011).

17
A capacidade dos flavonoides de sequestrar ROS é resultante de sua ação
redutora sobre estar, que leva a formação de moléculas menos reativas e mais
estáveis (NIJVELDT et al., 2001). Além do sequestro de radicais livres, eles atuam
diminuindo a mobilização leucocitária e a adesão de células inflamatórias no
endotélio, inibindo sistemas enzimáticos, sendo assim considerados potenciais
agentes anti-inflamatórios (TAUBER; FAY; MARLETTA, 1984; RASO et al., 2001).
Estudo recente demonstrou que a quercetina nas doses de 25, 50 e 100
mg/kg foi capaz de reduzir todos os marcadores de dano pancreático em modelo de
pancreatite induzida por ceruleína, incluindo redução dos níveis séricos de TNF-α,
IL-1β e IL-6 e da marcação imunohistoquímica de TNF-α em pâncreas (CARVALHO
et al., 2010). Diante deste dado é possível que a rutina, um flavonoide
estruturalmente relacionado à quercetina (CRESPY et al., 2001), possa igualmente
exercer efeitos protetores sobre a pancreatite aguda experimental.
Outros flavonoides também foram reportados por atuarem atenuando a
resposta inflamatória e controlando a progressão da pancreatite aguda em modelos
experimentais em ratos e camundongos, a exemplo da baicaleina (ZHANG et al.,
2005) e da apigenina (LAMPROPOULOS et al., 2013)
Devido à amplitude dos estudos mostrando os efeitos benéficos dos polifenois
encontrados nas plantas e na dieta humana frente a patologias inflamatórias, é
sugerido que a investigação do efeito de flavonoides na pancreatite aguda pode
permitir o desenvolvimento de novas substâncias com potencial terapêutico.
2.5 Rutina
A rutina, quercetina-3-O-ramnosilglicosídeo, é um flavonoide da classe das
flavonas encontrado abundantemente em alimentos usados diariamente como alho,
soja e frutas, vinho, tomate e chocolate (ARJUMAND; SETH; SULTANA, 2011; LIMA
et al., 2003). Sua estrutura consiste na quercetina aglicona com uma porção
rutinosídeo na posição 3 do anel C (Figura 2). Ambas as formas, quercetina aglicona
e rutina, são descritas por exercerem diversas atividades farmacológicas, porém
estas atividades em muitos casos se mostraram divergentes em modelos in vivo e in

18
vitro, o que pode ser atribuído à presença do grupo açúcar na estrutura da rutina
(DOMITROVIĆ et al., 2012; KODA; KURODA; IMAI, 2009; SHEN et al., 2002).
Figura 2 – Estruturas químicas da quercetina e da rutina.
A quercetina consiste em um dos metabólitos ativos da rutina após passagem
pelo trato gastrointestinal (AFANAS’EV et al., 1989), sendo que as formas
glicosiladas da quercetina (a exemplo da rutina) são mais facilmente absorvidas e
apresentam melhor biodisponibilidade que a forma aglicona, devido à sua maior
afinidade pela membrana das células epiteliais (OLTHOF et al., 2000; BECHO;
MACHADO; DE OLIVEIRA GUERRA, 2010); (DE ARAÚJO et al., 2013) e à ação de
β-glUcosidases (HOLLMAN et al., 1995)
A rutina e a quercetina atuaram protegendo o fígado em modelo de lesão
hepática induzida por etanol e tetracloreto de carbono (CCL4), por meio de seus
efeitos antioxidantes e sequestradores de radicais livres, porém a rutina se mostrou
mais efetiva que a quercetina em restaurar as atividades das enzimas antioxidantes
CAT, SOD, GSH-Px, GR e GST, diminuídas nos grupos controles não tratados (LEE
et al., 2013).
Suas possíveis propriedades tóxicas foram investigadas por (LIMA et al.,
2003) em coelhos hiperlipidêmicos, que testaram doses de 600 mg/kg administradas
via oral por 28 dias, e não foram encontrados efeitos deletérios quando avaliados
marcadores de dano hepático e renal como creatina, uréia, transaminase glutâmico
oxalacética (TGO) e transaminase glutâmico pirúvica (TGP).

19
A rutina é descrita por sequestrar radicais superóxido e quelar íons metálicos
como ferro, que está envolvido na reação de Fenton, formando radicais hidroxila
altamente reativos (NIJVELDT et al., 2001). Também é reportada por seu efeito
inibidor da atividade da xantina oxidase, uma das principais vias de produção de
ROS (NIJVELDT et al., 2001). É relacionada à inibição da fosfolipase A2 humana
(PLA) do fluido sinovial (LINDHAL & TAGESSON, 1997 apud (SELLOUM et al.,
2003), redução dos níveis de colesterol e triglicérides em ratos hiperlipidêmicos
(OLIVEIRA et al., 2002), à proteção do dano gástrico induzido por etanol, por meio
da prevenção da peroxidação lipídica e do aumento na atividade das enzimas
antioxidantes (LA CASA et al., 2000).
Em modelo de inflamação aguda induzida por carragenina, a rutina reduziu a
formação de edema na pata, quando administrada na dose única de 100 mg/kg
antes da indução (SELLOUM et al., 2003). Na dose de 75 mg/kg reduziu o processo
inflamatório crônico da artrite experimental, induzida por carragenina (ROTELLI et
al., 2003). Controlou o estresse oxidativo e a redução da expressão de NF-B e
TNF-α em tratamento profilático da resposta inflamatória aguda renal, em dose única
de 75 ou 150 mg/kg (ARJUMAND; SETH; SULTANA, 2011). Também foi reportada
por reduzir a nocicepção em modelos de dor neuropática e inflamatória em
camundongos (AZEVEDO et al., 2013; LAPA et al., 2009)
Esses resultados mostram que a rutina possui potencial para tratar a
inflamação, nocicepção e/ou estresse oxidativo, inclusive em condições clínicas mais
complexas, nas quais o tratamento é dificultado, dada a sua complexidade. Este é o
caso da pancreatite aguda, doença que é objeto de estudo nesse projeto e cujo
possível tratamento com flavonoides ainda é pouco explorado nos modelos
experimentais.
Associado a estas evidências, destaca-se que estudo realizado por
(SANTANA et al., 2012) demonstrou que o tratamento prévio de ratos com o extrato
etanólico da entrecasca de uma planta endêmica na Caatinga, a Caesalpinia
pyramidalis Tul. (Fabaceae) reduziu os componentes inflamatórios (edema e MPO
pancreáticos), oxidativo (lipoperoxidação pancreática) e nociceptivo (hiperalgesia
abdominal) da pancreatite induzida por obstrução do ducto biliopancreático em ratos.
Além disso, análise cromatográfica indicou a presença da rutina no extrato desta

20
planta, sugerindo que este composto poderia ser o responsável por parte das ações
farmacológicas.
Recentemente foi demonstrado que a rutina apresentou atividade anti-
inflamatória em modelo de pancreatite crônica experimental induzida por álcool e
dieta hiperlipídica em ratos, onde reduziu o dano acinar, a inflamação e a
hemorragia pancreática (ARUNA; GEETHA; SUGUNA, 2013a).
Diante do potencial antioxidante e anti-inflamatório da rutina, da limitação de
informações na literatura sobre o efeito deste composto em modelos de pancreatite
aguda e dos resultados prévios obtidos com a quercetina e o extrato da Caesalpinia
pyramidalis, o presente estudo buscou avaliar a hipótese de que a rutina possa
contribuir para o tratamento da pancreatite aguda experimental induzida por L-
arginina em camundongos.

21
3 OBJETIVOS
3.1 Geral
Investigar as ações farmacológicas da rutina na pancreatite aguda induzida
por L-arginina em camundongos.
3.2 Específicos
Investigar o efeito da rutina sobre a infiltração de neutrófilos e a
formação de edema pancreático na pancreatite aguda induzida pela
L-arginina;
Verificar o efeito da rutina sobre as concentrações séricas de enzimas
pancreáticas, proteína C reativa e interleucina-6 em camundongos com
pancreatite aguda induzida por L-arginina;
Avaliar o efeito da rutina sobre a hiperalgesia abdominal na pancreatite
induzida por L-arginina;
Avaliar a atividade antioxidante da rutina na pancreatite aguda induzida
por L-arginina.

22
4 MATERIAL E MÉTODOS
4.1 Animais
Foram utilizados camundongos Swiss, adultos, machos, pesando entre 30-35
g, provenientes do Biotério Central da Universidade Federal de Sergipe (UFS) ou do
Biotério de criação de camundongos do Departamento de Farmacologia/Instituto de
Ciências Biomédicas da Universidade de São Paulo (USP), São Paulo. Os animais
foram mantidos em caixas plásticas, contendo 5 camundongos cada uma, em
ambiente com temperatura controlada (22-24°C), em ciclos de claro-escuro de 12/12
h e com água e ração ad libitum em todas as etapas experimentais. Os protocolos
experimentais foram aprovados pelo CEPA (Comitê de Ética na Pesquisa Animal)
desta Universidade (Protocolo nº 43/12 - ANEXO 1), que é regido pelo Conselho
Nacional de Controle de Experimentação Animal (CONCEA).
4.2 Indução da Pancreatite Aguda por L-arginina
Os camundongos (n=6-7 por grupo) receberam duas injeções de L-arginina
(8%, 4 g/kg, pH 7,0, Sigma-Aldrich, Saint Louis, MO) por via intraperitoneal (i.p.),
com intervalo de uma hora entre elas. Os controles receberam o mesmo volume de
solução salina 0,9% administradas pela mesma via (DAWRA et al., 2007).
Após 72 horas da primeira injeção os camundongos foram eutanasiados com
dose excessiva de isoflurano 5% por via inalatória, seguida por exsanguinação.
Após coleta de sangue da veia abdominal, os animais foram submetidos à perfusão
transcardíaca com solução salina (0,9%) heparinizada (5 UI/L). Em seguida foram
coletadas amostras de pâncreas, pulmão, fígado e rim, as quais foram devidamente
armazenadas em freezer à -80°C. O sangue foi centrifugado a 4000 rpm por 10 min
4°C, para a separação do soro, o qual foi estocado à -20°C.

23
4.3 Delineamento Experimental
Os camundongos foram tratados oralmente com veículo (salina 0,9%) ou
rutina (Sigma-Aldrich, Fluka, Brasil, SP) nos tempos de 24, 36, 48 e 60 h após a
indução da pancreatite, conforme delineamento apresentado na Figura 3 .
Os experimentos foram realizados em duas etapas, sendo que na primeira
foram conduzidos experimentos com três doses de rutina para determinar o possível
efeito desta sobre a pancreatite aguda induzida por L-arginina. Na segunda etapa
foram realizados experimentos para melhor investigar ações da rutina na pancreatite
aguda, inclusive em relação ao provável envolvimento do estresse oxidativo.
Na primeira etapa foram testadas terapeuticamente três doses de rutina,
sendo elas 37,5, 75 e 150 mg/kg (equivalentes a 55, 110 e 165 µmol/kg deste
flavonoide), escolhidas de acordo com dados da literatura que mostram efeitos
farmacológicos protetores desta em condições patológicas diversas (CERQUEIRA;
MEDEIROS; AUGUSTO, 2007; ARJUMAND; SETH; SULTANA, 2011; ROTELLI et
al., 2003; LA CASA et al., 2000).
Figura 3 - Linha temporal indicando a indução da pancreatite com L-arginina e o esquema terapêutico de tratamento com a rutina.
Nesta etapa animais foram divididos em 5 grupos experimentais com n=6-7
animais, conforme apresentado na Tabela 1.

24
Tabela 1 - Relação dos grupos experimentais da Etapa 1.
GRUPOS TRATAMENTO
Salina + Veículo Salina (0,9%, i.p., 2 administrações) + Veículo (v.o.)
L-arginina + Veículo L-arginina (4 g/kg, i.p., 2 administrações)+ Veículo (v.o.)
L-arginina + Rutina L-arginina (4 g/kg, i.p., 2 administrações) + Rutina 37,5 mg/kg (v.o.)
L-arginina (4 g/kg, i.p., 2 administrações) + Rutina 75 mg/kg (v.o.)
L-arginina (4 g/kg, i.p., 2 administrações) + Rutina 150 mg/kg (v.o.)
Os animais receberam administração da droga teste por via oral, utilizando-se
a técnica de gavagem, com volume administrado de 10 mL/kg, em quatro tempos
conforme mostrado na linha do tempo (Figura 3). Após 72 horas os animais foram
eutanasiados conforme descrito anteriormente. Para estes grupos, foram avaliados
parâmetros inflamatórios no pâncreas e no pulmão, além de amilase sérica.
Na segunda etapa, foi realizado o mesmo esquema terapêutico, utilizando-se
apenas a dose de rutina escolhida de acordo com os resultados da primeira etapa.
Deste modo, os animais foram divididos em 3 grupos experimentais de 6 animais
cada, de acordo com grupos apresentados na tabela 2.
Tabela 2 - Relação dos grupos experimentais da Etapa 2.
GRUPOS TRATAMENTO
Salina + Veículo Salina (0,9%, i.p., 2 administrações) + Veículo (v.o.)
L-arginina + Veículo L-arginina (4 g/kg, i.p., 2 administrações)+ Veículo (v.o.)
L-arginina + Rutina L-arginina (4 g/kg, i.p., 2 administrações) + Rutina 75 mg/kg (v.o.)
Para estes grupos foram avaliados adicionalmente a concentração sérica de
lipase, proteína C reativa (PCR) e de interleucina 6 (IL-6) séricas, a hiperalgesia
abdominal e o estresse oxidativo.

25
4.4 Avaliação de Parâmetros Inflamatórios
4.4.1 Determinação da atividade de Mieloperoxidase (MPO)
Nesse trabalho, a atividade de MPO foi mensurada utilizando-se a técnica
descrita por (BRADLEY et al., 1982) modificada por (CAMARGO et al., 2008), que
mede a velocidade de oxidação do substrato о-dianisidina pelo ânion superóxido,
resultante da degradação do H2O2, que gera um composto de cor marrom, e a
variação da densidade óptica deste entre as amostras em função do tempo é
detectada na absorbância de 460 nm.
Amostras de pâncreas e pulmão (~100 mg) dos animais foram coletadas,
pesadas e cortadas em pedaços pequenos e mantidas em tubos teste na presença
de tampão fosfato (50 mM, pH 6,0 contendo 0,5% de brometo de
hexadeciltrimetilamônio). Cada amostra foi dispersa em homogeneizador de tecidos
Ultra Turrax durante 15 s, e alíquotas de 1 mL do homogenato foram centrifugadas
(2 min, 14000 rpm, 4ºC), para obtenção do sobrenadante, o qual foi usado para a
determinação da atividade da enzima. Em uma placa de 96 poços, foram pipetados
de 10 a 20 µL do sobrenadante (a depender do tecido) e a estas foi adicionado 200
µL de uma solução contendo dihidrocloreto de ο-dianisidina (0,167 mg/mL,
preparada em tampão fosfato de potássio 50 mM contendo 0,005% de H2O2). As
alterações nos valores de absorbância (460 nm) foram registradas com leitor de
microplaca (Synergy MX®, Biotek, USA), com intervalos de 15 segundos por 5-10
min. Os resultados foram expressos como unidades de MPO por mg de tecido
(UMPO/mg de tecido). Uma UMPO foi considerada como a quantidade de enzima
que degrada 1 μmol de O2/min.
4.4.2 Índice de Edema
O índice de edema foi calculado pela razão entre o peso úmido e o peso seco
das amostras de tecido pancreático. As amostras de pâncreas foram submetidas à
secagem em estufa à temperatura de 90°C por 12 h e a massa dos tecidos,
mensurada antes e após a secagem (SANTANA et al., 2012).

26
4.5 Avaliação de Parâmetros Bioquímicos no Soro
4.5.1 Determinação de Amilase e Lipase Séricas
A determinação da amilase e lipase sérica foram realizadas por método
colorimétrico utilizando-se kits comerciais (kit cinético para determinação de amilase
[Labtest®] e kit colorimétrico para determinação de lipase [Human do Brasil®]), de
acordo com as instruções do fabricante.
4.5.2 Concentração de proteína C reativa (PCR)
A concentração de PCR foi determinada no soro pela técnica
imunoturbidimétrica, utilizando-se kit comercial (PCRTEST ULTRASENSÍVEL,
Doles), contendo uma suspensão de partículas de látex sensibilizada com anticorpos
anti-PCR, que ao reagir com PCR da amostra aglutinam e geram uma reação
túrbida, cuja intensidade é detectada em espectrofotômetro (540 nm) e é
proporcional a concentração de PCR na amostra.
4.5.3 Dosagem de interleucina 6 (IL-6)
A concentração sérica de IL-6 foram mensuradas por ensaio de ELISA,
conforme instruções do fabricante (BioLegend, San Diego, USA).
Para isto, uma placa de 96 poços foi sensibilizada com 100 μL de anticorpo
monoclonal anti-IL-6 diluído 1:200 em tampão carbonato de sódio (0,1 M; pH 9,5), e,
a seguir, foi incubada a 4ºC por 18 h. Para o bloqueio do anticorpo primário, a placa
foi submetida a quatro lavagens com o tampão fosfato salina contendo 0,05% de
Tween 20 (PBST) e depois preenchidas com 200 μL/poço de solução de bloqueio
(BSA 1% em PBS), incubada a 37ºC por 1 h, sob agitação a 200 rpm. Em seguida
foi realizado novo ciclo de 4 lavagens. Ao término do período de bloqueio, seguiu-se
a adição de 100 μL/poço das amostras e da curva padrão de concentração de IL-6
recombinante fornecidos no kit, com incubação de 18 h a 4ºC.
Após novo ciclo de lavagens, o volume de 100 μL do anticorpo de detecção
anti-IL-6 biotinilado foi acrescido aos poços da placa que foi mantida em temperatura
ambiente por 1 h. Em seguida, a placa foi submetida a novo ciclo de lavagem e o

27
volume de 100 μL de solução com conjugado HRP-avidina (1:1000) foi adicionado
em cada poço e a placa foi mantida em temperatura ambiente por 30 min sob
agitação. Após novos ciclos de lavagens da placa, a reação antígeno-anticorpo foi
revelada seguida a adição de 100 μL por poço da solução de 3.3’5.5’-
tetrametilbenzidina (TMB). A reação foi interrompida após 30 min pela adição de 100
μL de ácido sulfúrico (2 N) em cada poço.
A leitura foi realizada em espectrofotômetro Espectra Max plus 384
(Sunnyvale, CA, EUA) em comprimento de onda de 450 nm com correção para 570
nm. As concentrações das amostras foram calculadas a partir das curvas-padrão
obtidas com as citocinas recombinantes utilizando-se o software Softmax® Pro
(Version 5; Molecular Devices). Os resultados foram expressos em pg/mL.
4.6 Avaliação Comportamental
4.6.1 Medida da Hiperalgesia Abdominal
Para a realização deste teste, utilizou-se o modelo eletrônico de von Frey,
(EFF31, Insight®, Ribeirão Preto-SP, VIVANCOS et al., 2004). Os animais foram
transferidos para gaiolas individuais e ambientados por 3 dias seguidos (1 vez ao dia
por 30 min). Os animais receberam estímulos mecânicos aplicados no abdômen, em
triplicata, nos diferentes grupos experimentais, antes e após a injeção da L-arginina
nos tempos -1 (basal) e 72 h. Para cada animal e em cada um dos tempos de
avaliação após a indução da pancreatite registrou-se o limiar da resposta de retirada
do abdômen (em g) resultante da média de três medidas consecutivas com
intervalos de pelo menos 1 min entre elas (CAMARGO et al., 2011).
O teste foi realizado de maneira encoberta, de modo que o experimentador
que realizou as medidas não tinha conhecimento sobre os grupos experimentais. Os
valores foram expressos como a diferença entre os valores de intensidade do
estímulo inicial e final ( em g).

28
4.7 Avaliação de Parâmetros Oxidantes e Antioxidantes
4.7.1 Determinação do grau de peroxidação lipídica
O complexo formado por substâncias reativas ao ácido tiobarbitúrico (TBARS)
é amplamente usado como biomarcador de peroxidação lipídica em sistemas
biológicos, pois a determinação destes produtos envolve a quantificação dos
aldeídos formados durante a peroxidação lipídica, resultantes da degradação de
hidroperóxidos, que reagem com o ácido tiobarbitúrico, incluindo o malonaldeído
(MDA) (BOSE; SUTHERLAND; PINSKY, 1989).
Amostras de pâncreas, pulmão, fígado e rim foram pesadas e
homogeneizadas em 10 volumes de solução de fosfato de potássio (50 mM, pH 7,4)
contendo butil-hidroxitoluol (BHT; 12,6 mM) utilizado para prevenir a oxidação
durante o preparo. A seguir, alíquotas contendo 200 µL do homogenato (em
duplicata) foram incubadas a 90°C durante 45 min com 400 µL de solução contendo
TBA a 0,37%, em meio ácido (ácido tricloroacético (TCA) a 15% e ácido clorídrico a
0,25 N). No final da incubação, as amostras foram centrifugadas por 5 min à 14000
rpm, e 300 µL do sobrenadante foram retirados e submetidos à extração do produto
de reação com igual volume de n-butanol e 30 µL solução saturada de NaCl,
seguida pela agitação em vórtex por 30 s e centrifugação durante 2 min a 14000
rpm. Os sobrenadantes (fase orgânica) foram pipetados em placa de 96 poços para
leitura dos valores de absorbância em leitor de microplaca Espectra Max plus 384
(Sunnyvale, CA, EUA) a 535 nm (corrigidos pelos valores de absorbância a 572 nm).
Os resultados foram expressos como pmol de MDA/mg de tecido (BOSE,
SUTHERLAND, PINSKY, 1989), utilizando-se, para o cálculo, o coeficiente de
extinção molar do MDA de 1,55 x 105 M-1 cm-1.
4.7.2 Determinação da atividade de Catalase (CAT)
A determinação da atividade da CAT ocorreu em duas etapas, consistindo de
duas reações (Figura 4). Na primeira reação (R1), a catalase contida na amostra
dismuta o H2O2 (de concentração conhecida) em água e oxigênio molecular. Em
seguida, na segunda reação (R2), o H2O2 remanescente é determinado pela
oxidação do reagente o-dianisidina (O.D) em uma reação catalisada pela enzima

29
peroxidase (HRP), conforme método modificado de (FOSSATI; PRENCIPE; BERTI,
1980).
Figura 4 - Reações realizadas para mensuração da atividade de Catalase. Reação 1: dismutação do peróxido de hidrogênio pela Catalase. Reação 2: formação do cromóforo A por pela reação do peróxido restante com a o-dianisidina catalisada pela HRP.
Amostras de pâncreas, pulmão, fígado e rins foram pesadas e
homogeneizadas em tampão fosfato (50 mM, pH 7,0; 1:10, p/v).
Em seguida, os homogenatos foram centrifugados a 10.000 rpm por 10 min a
4ºC e o sobrenadante foi separado para o ensaio. Metade do volume do
sobrenadante foi incubado a 60ºC por 2 h, com objetivo de inativar a catalase e
assim ser utilizado como branco da reação.
Uma curva padrão de H2O2 foi preparada em tampão fosfato 50 mM, pH 7,0
nas concentrações: 8820, 4410, 2205, 1125, 882, 441, 221, 113, 88, 44, 22 e 11 µM.
Reação 1: foi realizada incubando-se 30 µL de sobrenadante das amostras
(brutos e inativados) com 500 µL de H2O2 (10 mM) em temperatura ambiente (de 2 a
5 min a depender do tecido). A reação foi interrompida com 500 µL de NaN3 1 mM.
Em paralelo, 30 µL de tampão fosfato (50 mM, pH 7,0) foram incubados com 500 µL
de H2O2 (nas diferentes concentrações da curva) com os respectivos tempos de
incubação.
Reação 2: foi realizada em placa de 96 poços incubando-se 20 µL do produto
da primeira reação com 200 µL de reagente contendo 0,167 mg/mL de o-dianisidina
e 0,095 µg/mL de HRP em tampão fosfato 5 mM, pH 6,0.
A monitorização da velocidade de formação do produto de oxidação da o-
dianisidina foi realizada registrando-se o aumento da absorbância da mistura a 460
nm, por meio de leituras coletadas em intervalos de 10 s durante 10 min. O valor da

30
atividade da catalase foi calculado a partir da velocidade máxima por minuto de cada
reação e extrapolado na curva de H2O2. O resultado foi expresso em U Catalase/mg
de proteína. Uma unidade de catalase foi definida como a quantidade de H2O2 (em
µmol) degradada por minuto.
4.7.3 Determinação da atividade de superóxido dismutase (SOD)
A atividade enzimática específica total das isoenzimas da SOD (CuZnSOD e
MnSOD) teciduais foi baseada no método da xantina oxidase/XTT {3’-1-
[(fenilamino)-carbonil]-3,4-tetrazólio-bis(4-metóxi-6-nitro) ácido benzenosulfónico
hidratado} descrito por (UKEDA et al., 1997). Neste método ocorre a formação do
produto XTT-formazan que apresenta cor vermelha detectada à 470 nm, conforme
reação mostrada na Figura 5.
Figura 5 - Reação xantina oxidase/XTT. A enzima xantina oxidase (XO) gera radicais livres ao reagir com a xantina, incluindo ânions superóxido. Estes por sua vez podem reduzir o XTT tetrazólium em formazan. Na presença de SOD o ânion superóxido é sequestrado, diminuindo assim a formação do produto XTT-formazan.
Deste modo, a atividade da superóxido dismutase é determinada através do
grau de inibição desta reação.
Amostras de pâncreas, pulmão, fígado e rins foram pesadas e
homogeneizadas em tampão fosfato 50 mM, pH 7,0 (1:10, p/v). Em seguida, os
homogenatos foram centrifugados a 10.000 rpm por 10 min a 4ºC e o sobrenadante
foi separado para o ensaio. Deste, foram retirados 250 µL e adicionados 400 µL de
etanol:clorofórmio (62,5:37,5 - v/v), para remoção de compostos interferentes. A
mistura foi agitada por 30 seg e centrifugada a 4000 rpm por 10 min a 4ºC.

31
A reação foi realizada em duplicata, pipetando-se em placa de 96 poços: 10
µL do sobrenadante obtido (diluído quando necessário), 280 µL do meio de reação
(tampão carbonato, xantina 3 mM e XTT 0,75 mM) e 10 µL de XO (56,1 mU/mL). As
leituras foram realizadas durante 20 min à 470 nm. Os resultados foram expressos
como mUSOD/mg de proteína. Uma unidade de SOD foi definida como a quantidade
de SOD capaz de transformar 1 µmol de O2/min.
4.7.4 Atividade de Glutationa Peroxidase (GSH-Px)
A atividade da enzima glutationa peroxidase (GSH-Px) foi determinada de
acordo com o método descrito por (PAGLIA; VALENTINE, 1967), no qual a molécula
oxidada de glutationa (GSSG), produzida na reação primária com a GSH-Px, é
reduzida por intermédio de uma segunda enzima, a glutationa redutase (GR), com o
auxílio do cofator NADPH, como mostram as reações abaixo:
GSH-Px
R – O – O – H + 2GSH R – O – H + GSSH + H2O
GR
GSSG + NADPH + H+ 2GSH + NADP+
Esse ensaio é uma medida indireta da atividade da glutationa peroxidase
(GSH-Px). A glutationa oxidada (GSSG), produzida pela redução de um peróxido
orgânico (terc-butil - hidroperóxido estável que não reage com a catalase) por ação
da GSH-Px, é reciclada para sua forma reduzida pela (GR). Na presença de GSH-Px
ocorre oxidação de NADPH para NADP+ acompanhada pelo decréscimo da
absorbância a 340 nm, o qual é utilizado como forma de monitorar indiretamente a
atividade desta enzima, calculada a partir o coeficiente de extinção molar do NADPH
(ε = 6,22 mM-1.cm-1).
As amostras de pâncreas, pulmão, fígado e rins foram pesadas e
homogeneizadas em tampão fosfato 50 mM, pH 7,0 (1:10, p/v) contendo EDTA 0,1
M. Em seguida, os homogenatos foram centrifugados a 10.000 rpm por 10 min a 4ºC
e o sobrenadante foi separado para o ensaio.

32
A reação foi realizada em duplicata, consistindo de duas etapas. Na primeira
etapa incubou-se em placa de 96 poços 30 µL da amostra (diluída quando
necessário) com 155 µL do meio de reação (Tampão fosfato 50 mM, pH 7,0
contendo EDTA 0,1 M, GSH 0,08 M e GR 9,6 U/mL) por 5 min a 37ºC. Na segunda
etapa, pipetou-se 5 µL de terc-butil hidroperóxido e em seguida 10 µL de NADPH 1,2
mM.
A absorbância foi monitorada em 340 nm à 37°C por 10 min. Os resultados
foram expressos em µmol GSH/min/mg de proteína.
4.7.5 Dosagem de proteínas
Em todos os experimentos de medida de atividade enzimática (CAT, SOD, e
GSH-Px), a concentração total de proteínas nos homogenatos foi medida pelo
método de (BRADFORD, 1976), utilizando-se o reagente Bio-Rad Protein Assay.
Para traçar a curva de calibração, foram utilizadas concentrações de 10, 50, 100,
200, 300, 400 e 500 μg/mL de albumina do soro bovino (BSA - Bovine Serum
Albumin), diluídas a partir de uma solução mãe inicial de 1000 μg/mL.
4.7.6 Quantificação de resíduos proteicos de 3-nitrotirosina (3-NT)
A avaliação dos resíduos de 3-nitrotirosina como índice de dano oxidativo
e/ou produção de peroxinitrito foi realizada em homogenatos de pâncreas por slot
blot conforme método descrito por (SULTANA; BUTTERFIELD, 2008).
As amostras foram pesadas e homogeneizadas em 5 volumes de tampão
Tris-HCl 50 mM (Tris-HCl 50 mM, pH 7,4 contendo 1 mM de PMSF, leupeptina 10
µg/ml e inibidor de tripsina 10 µg/ml) e centrifugadas durante 5 minutos a 2.500 rpm
à 4°C. Os sobrenadantes foram separados e a concentração de proteínas dos
mesmos, determinadas pelo método de Bradford conforme descrito anteriormente.
Subsequentemente, as amostras (0,25 μg de proteina) foram transferidas
para uma membrana de nitrocelulose através do sistema slot blot utilizando-se
vácuo. As membranas foram então coradas com solução de vermelho de Ponceau S
(0,1% em ácido acético a 5%), e fotografadas a fim de verificar-se a eficiência da
transferência, sendo em seguida descoradas com água.

33
O bloqueio de sítios inespecíficos presentes nas membranas de nitrocelulose
foi realizado através da incubação da membrana com uma solução de caseína 0,2%
em TBST (20 mM de Tris-HCl, 137 mM de NaCl, pH 7,4, contendo 0,1% de Tween-
20) por 1 hora, sob agitação contínua. Após esse período, as membranas foram
incubadas com o anticorpo primário anti-3-nitrotirosina para camundongo (Upstate,
Biotecnology, NY, USA), na diluição de 500 ng/mL em TBST, durante 16 horas sob
agitação contínua a 18°C.
Em seguida, as membranas foram lavadas com TBST (6 x10min) e incubadas
com o anticorpo secundário de cabra IgG anti-camundongo conjugado com
peroxidase (Signalway, College Park, MD, USA) na diluição de 1:3000 em TBST por
2 horas. Posteriormente, a membrana foi lavada com TBST e submetida à revelação
por quimioluminescência. A captação dos sinais de quimioluminescência
(SuperSignal West Pico; Thermo Scientific, Rockford, IL, USA) e suas intensidades
foram estimadas por densitometria utilizando-se o software Softmax® Pro (Version
5; Molecular Devices). A nitração de proteínas foi quantificada pela comparação com
um padrão de BSA nitrada com peroxinitrito (NTBSA) e os resultados normalizados
pelos valores de intensidade das bandas obtidos pela coloração com Ponceau S. Os
resultados foram expressos como variação percentual da razão entre a
densitometria da nitrotirosina e a quantidade de proteínas coradas com Ponceau S
em relação ao grupo controle.
4.8 Análise Estatística
Os resultados experimentais foram expressos como média ± erro padrão da
média (E.P.M.). O teste de Kolmogorov-Smirnov foi utilizado para avaliar a
distribuição normal. Em todos os testes, os dados apresentaram distribuição normal
e foram analisados por ANOVA de uma via. O nível de significância entre os grupos
foi determinado pelo pós-teste de Bonferroni, calculado com auxílio do programa
GraphPad Prism (versão 5.0). Valores de p<0,05 foram considerados
estatisticamente significativos.

34
5 RESULTADOS
5.1 Efeito da Rutina sobre parâmetros inflamatórios e bioquímicos da
pancreatite aguda induzida por L-arginina
Inicialmente foram avaliados os efeitos da rutina sobre a hiperamilasemia e a
inflamação em pâncreas e pulmão de animais com pancreatite aguda induzida por L-
arginina. Para isso, a rutina, nas doses de 37,5, 75 ou 150 mg/kg, foi administrada
por via oral (v.o.), nos tempos 24, 36, 48 e 60 horas, e seus efeitos foram avaliados
72 h após a indução.
A concentração sérica de amilase foi aumentada (p<0,001) na pancreatite
aguda induzida por L-arginina em relação ao grupo Salina + Veículo (Figura 6). O
tratamento com rutina, nas doses de 75 ou 150 mg/kg, reverteu (p<0,001) o aumento
da concentração de amilase (Figura 6), quando comparado ao tratamento com L-
arginina + Veículo. Não foram verificadas diferenças significativas entre estes grupos
e o grupo Salina + Veículo.
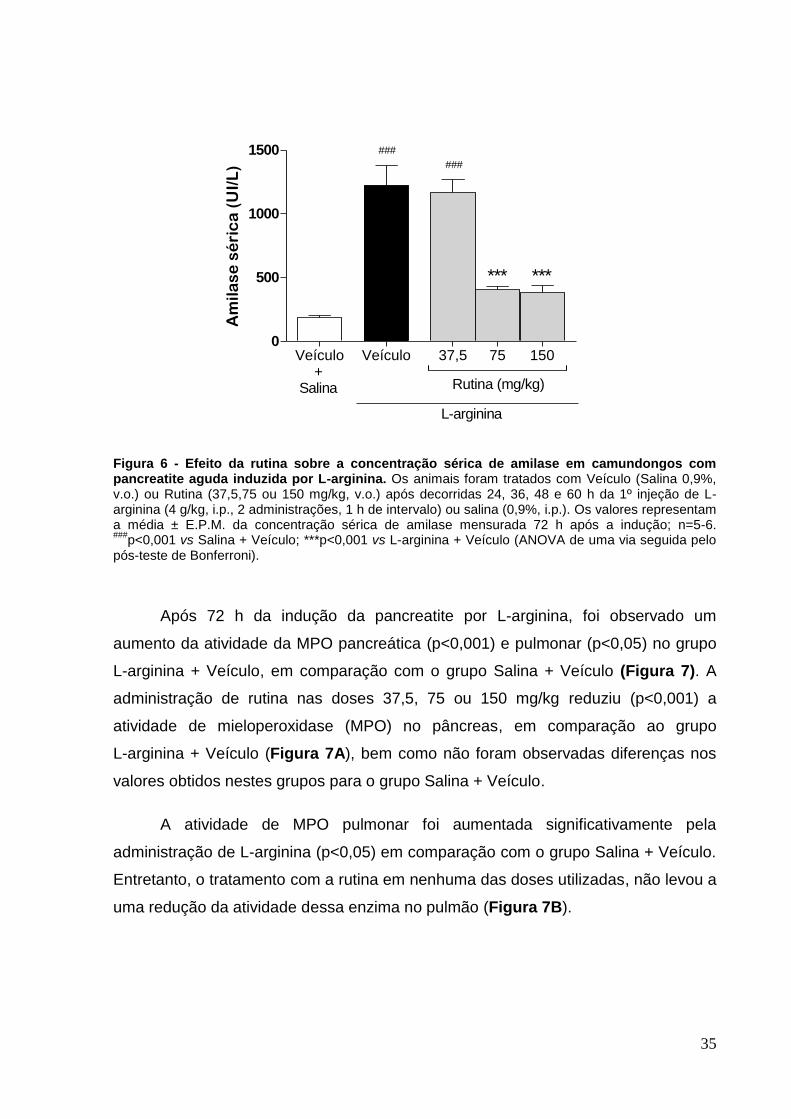
35
Veículo Veículo 37,5 75 1500
500
1000
1500
Rutina (mg/kg)
L-arginina
###
***
###
***
+Salina
Am
ila
se
sé
ric
a (
UI/
L)
Figura 6 - Efeito da rutina sobre a concentração sérica de amilase em camundongos com pancreatite aguda induzida por L-arginina. Os animais foram tratados com Veículo (Salina 0,9%, v.o.) ou Rutina (37,5,75 ou 150 mg/kg, v.o.) após decorridas 24, 36, 48 e 60 h da 1º injeção de L-arginina (4 g/kg, i.p., 2 administrações, 1 h de intervalo) ou salina (0,9%, i.p.). Os valores representam a média ± E.P.M. da concentração sérica de amilase mensurada 72 h após a indução; n=5-6. ###
p<0,001 vs Salina + Veículo; ***p<0,001 vs L-arginina + Veículo (ANOVA de uma via seguida pelo pós-teste de Bonferroni).
Após 72 h da indução da pancreatite por L-arginina, foi observado um
aumento da atividade da MPO pancreática (p<0,001) e pulmonar (p<0,05) no grupo
L-arginina + Veículo, em comparação com o grupo Salina + Veículo (Figura 7). A
administração de rutina nas doses 37,5, 75 ou 150 mg/kg reduziu (p<0,001) a
atividade de mieloperoxidase (MPO) no pâncreas, em comparação ao grupo
L-arginina + Veículo (Figura 7A), bem como não foram observadas diferenças nos
valores obtidos nestes grupos para o grupo Salina + Veículo.
A atividade de MPO pulmonar foi aumentada significativamente pela
administração de L-arginina (p<0,05) em comparação com o grupo Salina + Veículo.
Entretanto, o tratamento com a rutina em nenhuma das doses utilizadas, não levou a
uma redução da atividade dessa enzima no pulmão (Figura 7B).
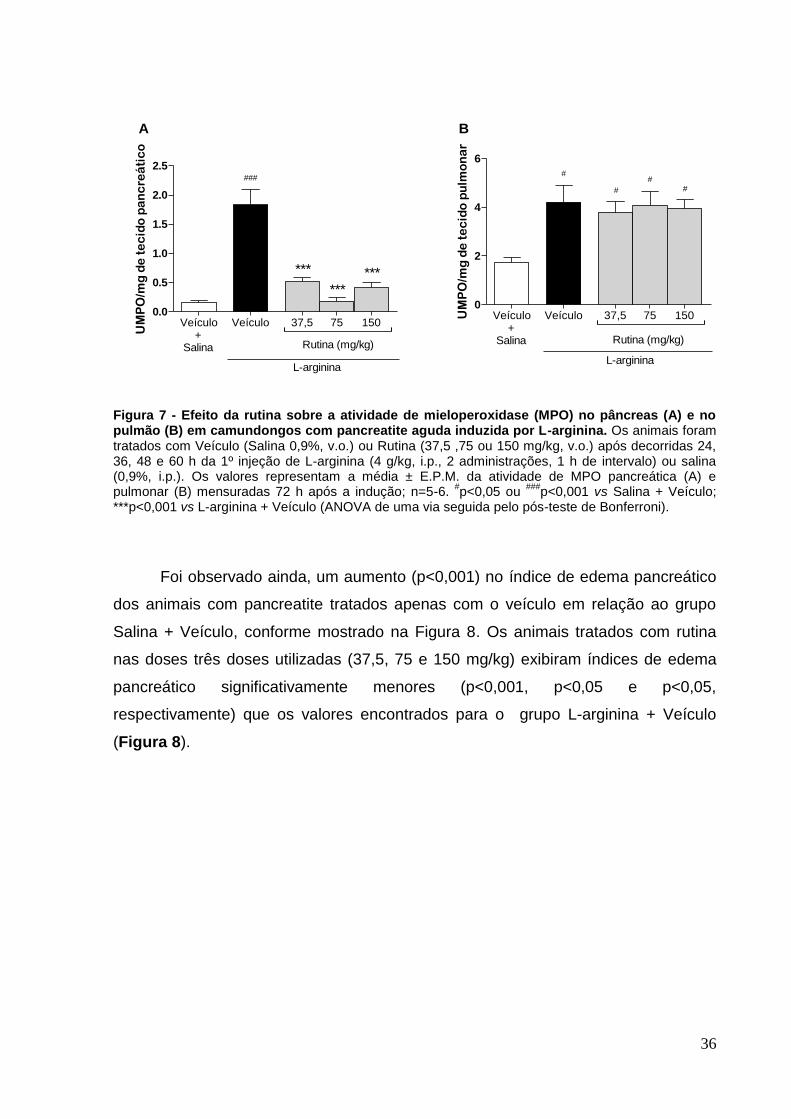
36
Veículo Veículo 37,5 75 1500
2
4
6
Rutina (mg/kg)
L-arginina
#
# #
#
+Salina
UM
PO
/mg
de
te
cid
o p
ulm
on
ar
Veículo Veículo 37,5 75 1500.0
0.5
1.0
1.5
2.0
2.5
Rutina (mg/kg)
L-arginina
###
***
***
***
+Salina
UM
PO
/mg
de
te
cid
o p
an
cre
áti
co
A B
Figura 7 - Efeito da rutina sobre a atividade de mieloperoxidase (MPO) no pâncreas (A) e no pulmão (B) em camundongos com pancreatite aguda induzida por L-arginina. Os animais foram tratados com Veículo (Salina 0,9%, v.o.) ou Rutina (37,5 ,75 ou 150 mg/kg, v.o.) após decorridas 24, 36, 48 e 60 h da 1º injeção de L-arginina (4 g/kg, i.p., 2 administrações, 1 h de intervalo) ou salina (0,9%, i.p.). Os valores representam a média ± E.P.M. da atividade de MPO pancreática (A) e pulmonar (B) mensuradas 72 h após a indução; n=5-6.
#p<0,05 ou
###p<0,001 vs Salina + Veículo;
***p<0,001 vs L-arginina + Veículo (ANOVA de uma via seguida pelo pós-teste de Bonferroni).
Foi observado ainda, um aumento (p<0,001) no índice de edema pancreático
dos animais com pancreatite tratados apenas com o veículo em relação ao grupo
Salina + Veículo, conforme mostrado na Figura 8. Os animais tratados com rutina
nas doses três doses utilizadas (37,5, 75 e 150 mg/kg) exibiram índices de edema
pancreático significativamente menores (p<0,001, p<0,05 e p<0,05,
respectivamente) que os valores encontrados para o grupo L-arginina + Veículo
(Figura 8).
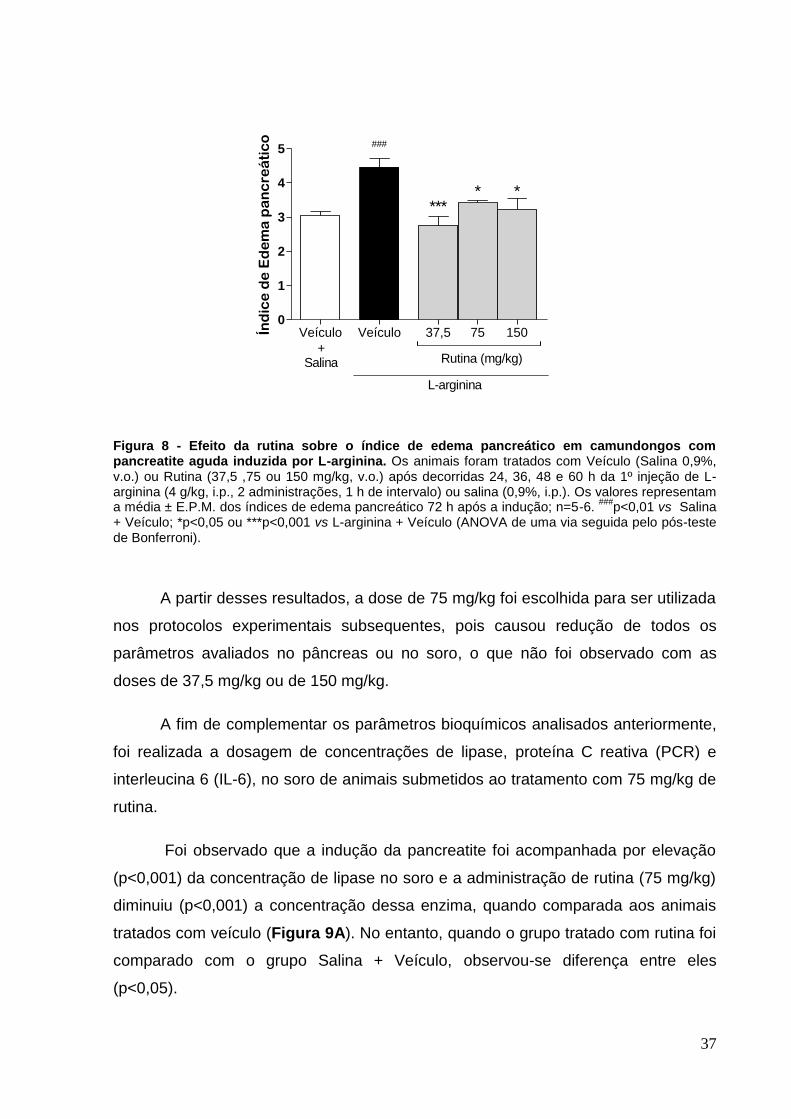
37
Veículo Veículo 37,5 75 1500
1
2
3
4
5
Rutina (mg/kg)
L-arginina
###
****
+Salina
*
Índ
ice
de
Ed
em
a p
an
cre
áti
co
Figura 8 - Efeito da rutina sobre o índice de edema pancreático em camundongos com pancreatite aguda induzida por L-arginina. Os animais foram tratados com Veículo (Salina 0,9%, v.o.) ou Rutina (37,5 ,75 ou 150 mg/kg, v.o.) após decorridas 24, 36, 48 e 60 h da 1º injeção de L-arginina (4 g/kg, i.p., 2 administrações, 1 h de intervalo) ou salina (0,9%, i.p.). Os valores representam a média ± E.P.M. dos índices de edema pancreático 72 h após a indução; n=5-6.
###p<0,01 vs Salina
+ Veículo; *p<0,05 ou ***p<0,001 vs L-arginina + Veículo (ANOVA de uma via seguida pelo pós-teste de Bonferroni).
A partir desses resultados, a dose de 75 mg/kg foi escolhida para ser utilizada
nos protocolos experimentais subsequentes, pois causou redução de todos os
parâmetros avaliados no pâncreas ou no soro, o que não foi observado com as
doses de 37,5 mg/kg ou de 150 mg/kg.
A fim de complementar os parâmetros bioquímicos analisados anteriormente,
foi realizada a dosagem de concentrações de lipase, proteína C reativa (PCR) e
interleucina 6 (IL-6), no soro de animais submetidos ao tratamento com 75 mg/kg de
rutina.
Foi observado que a indução da pancreatite foi acompanhada por elevação
(p<0,001) da concentração de lipase no soro e a administração de rutina (75 mg/kg)
diminuiu (p<0,001) a concentração dessa enzima, quando comparada aos animais
tratados com veículo (Figura 9A). No entanto, quando o grupo tratado com rutina foi
comparado com o grupo Salina + Veículo, observou-se diferença entre eles
(p<0,05).
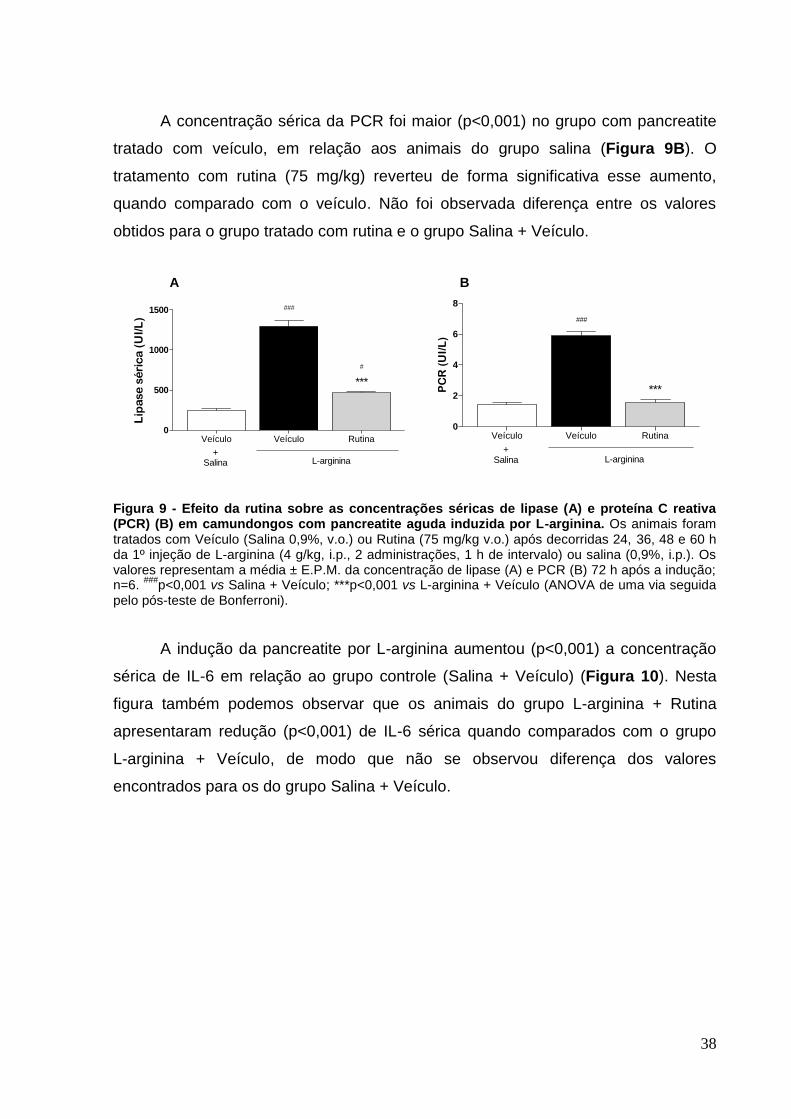
38
A concentração sérica da PCR foi maior (p<0,001) no grupo com pancreatite
tratado com veículo, em relação aos animais do grupo salina (Figura 9B). O
tratamento com rutina (75 mg/kg) reverteu de forma significativa esse aumento,
quando comparado com o veículo. Não foi observada diferença entre os valores
obtidos para o grupo tratado com rutina e o grupo Salina + Veículo.
Veículo Veículo Rutina0
500
1000
1500
L-arginina
***
###
+Salina
Lip
as
e s
éri
ca
(U
I/L
)
#
Veículo Veículo Rutina0
2
4
6
8
L-arginina
***
###
+Salina
PC
R (
UI/
L)
BA
Figura 9 - Efeito da rutina sobre as concentrações séricas de lipase (A) e proteína C reativa (PCR) (B) em camundongos com pancreatite aguda induzida por L-arginina. Os animais foram tratados com Veículo (Salina 0,9%, v.o.) ou Rutina (75 mg/kg v.o.) após decorridas 24, 36, 48 e 60 h da 1º injeção de L-arginina (4 g/kg, i.p., 2 administrações, 1 h de intervalo) ou salina (0,9%, i.p.). Os valores representam a média ± E.P.M. da concentração de lipase (A) e PCR (B) 72 h após a indução; n=6.
###p<0,001 vs Salina + Veículo; ***p<0,001 vs L-arginina + Veículo (ANOVA de uma via seguida
pelo pós-teste de Bonferroni).
A indução da pancreatite por L-arginina aumentou (p<0,001) a concentração
sérica de IL-6 em relação ao grupo controle (Salina + Veículo) (Figura 10). Nesta
figura também podemos observar que os animais do grupo L-arginina + Rutina
apresentaram redução (p<0,001) de IL-6 sérica quando comparados com o grupo
L-arginina + Veículo, de modo que não se observou diferença dos valores
encontrados para os do grupo Salina + Veículo.
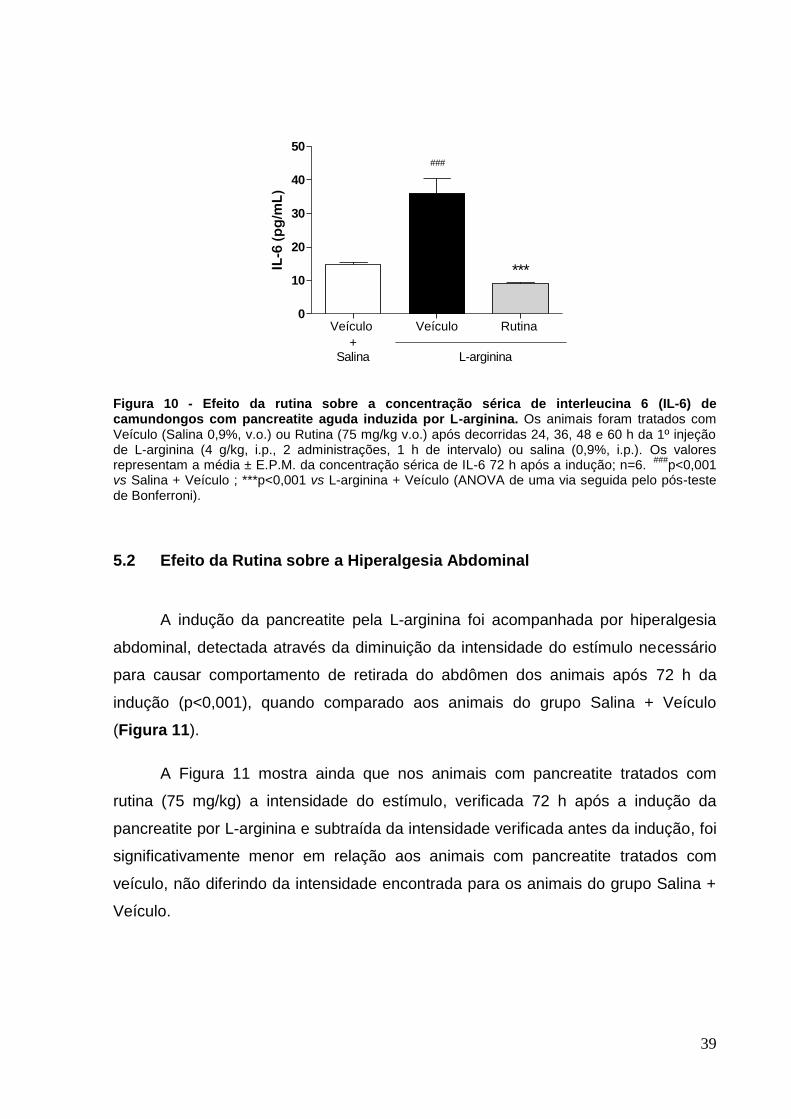
39
Veículo Veículo Rutina 0
10
20
30
40
50
L-arginina
###
***
+Salina
IL-6
(p
g/m
L)
Figura 10 - Efeito da rutina sobre a concentração sérica de interleucina 6 (IL-6) de camundongos com pancreatite aguda induzida por L-arginina. Os animais foram tratados com Veículo (Salina 0,9%, v.o.) ou Rutina (75 mg/kg v.o.) após decorridas 24, 36, 48 e 60 h da 1º injeção de L-arginina (4 g/kg, i.p., 2 administrações, 1 h de intervalo) ou salina (0,9%, i.p.). Os valores representam a média ± E.P.M. da concentração sérica de IL-6 72 h após a indução; n=6.
###p<0,001
vs Salina + Veículo ; ***p<0,001 vs L-arginina + Veículo (ANOVA de uma via seguida pelo pós-teste de Bonferroni).
5.2 Efeito da Rutina sobre a Hiperalgesia Abdominal
A indução da pancreatite pela L-arginina foi acompanhada por hiperalgesia
abdominal, detectada através da diminuição da intensidade do estímulo necessário
para causar comportamento de retirada do abdômen dos animais após 72 h da
indução (p<0,001), quando comparado aos animais do grupo Salina + Veículo
(Figura 11).
A Figura 11 mostra ainda que nos animais com pancreatite tratados com
rutina (75 mg/kg) a intensidade do estímulo, verificada 72 h após a indução da
pancreatite por L-arginina e subtraída da intensidade verificada antes da indução, foi
significativamente menor em relação aos animais com pancreatite tratados com
veículo, não diferindo da intensidade encontrada para os animais do grupo Salina +
Veículo.
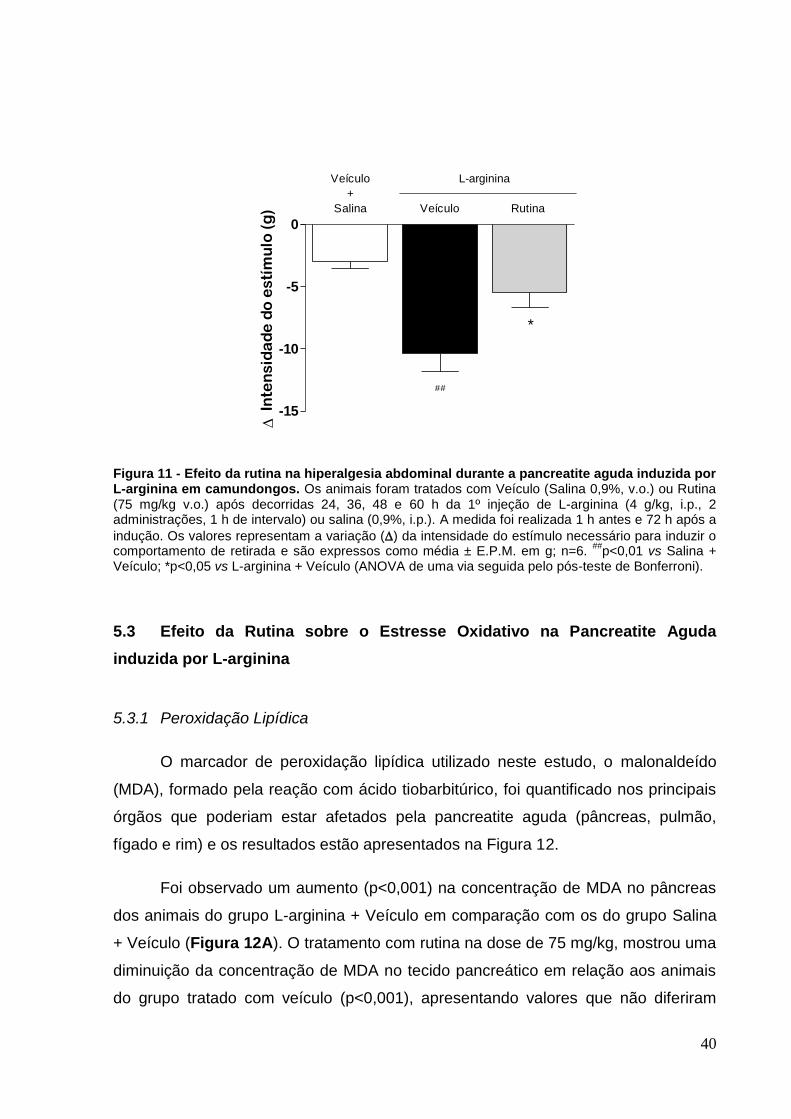
40
-15
-10
-5
0
##
*
L-argininaVeículo
+
Salina Veículo Rutina
I
nte
ns
ida
de
do
es
tím
ulo
(g
)
Figura 11 - Efeito da rutina na hiperalgesia abdominal durante a pancreatite aguda induzida por L-arginina em camundongos. Os animais foram tratados com Veículo (Salina 0,9%, v.o.) ou Rutina (75 mg/kg v.o.) após decorridas 24, 36, 48 e 60 h da 1º injeção de L-arginina (4 g/kg, i.p., 2 administrações, 1 h de intervalo) ou salina (0,9%, i.p.). A medida foi realizada 1 h antes e 72 h após a
indução. Os valores representam a variação () da intensidade do estímulo necessário para induzir o comportamento de retirada e são expressos como média ± E.P.M. em g; n=6.
##p<0,01 vs Salina +
Veículo; *p<0,05 vs L-arginina + Veículo (ANOVA de uma via seguida pelo pós-teste de Bonferroni).
5.3 Efeito da Rutina sobre o Estresse Oxidativo na Pancreatite Aguda
induzida por L-arginina
5.3.1 Peroxidação Lipídica
O marcador de peroxidação lipídica utilizado neste estudo, o malonaldeído
(MDA), formado pela reação com ácido tiobarbitúrico, foi quantificado nos principais
órgãos que poderiam estar afetados pela pancreatite aguda (pâncreas, pulmão,
fígado e rim) e os resultados estão apresentados na Figura 12.
Foi observado um aumento (p<0,001) na concentração de MDA no pâncreas
dos animais do grupo L-arginina + Veículo em comparação com os do grupo Salina
+ Veículo (Figura 12A). O tratamento com rutina na dose de 75 mg/kg, mostrou uma
diminuição da concentração de MDA no tecido pancreático em relação aos animais
do grupo tratado com veículo (p<0,001), apresentando valores que não diferiram

41
significativamente dos encontrados para os animais do grupo Salina + Veículo, como
observado na Figura 12A.
Como se observa na Figura 12B, a indução da pancreatite por L-arginina não
modificou as concentração de MDA detectada no tecido pulmonar, bem como não
houve alteração nos valores deste marcador encontrados nos grupos tratados com
rutina quando comparados com os animais dos outros grupos. Em contrapartida,
houve aumento nas concentrações de MDA nos tecidos hepático e renal (p<0,001)
dos animais do grupo L-arginina + Veículo quando comparados ao grupo Salina +
Veículo (Figura 12C e 12D). Tanto no fígado quanto no rim, as concentrações de
MDA foram diminuídas (p<0,001) pelo tratamento com rutina na dose de 75 mg/kg,
conforme mostrado na Figura 12, painéis C e D, não apresentando diferenças em
relação aos animais do grupo Salina + Veículo.
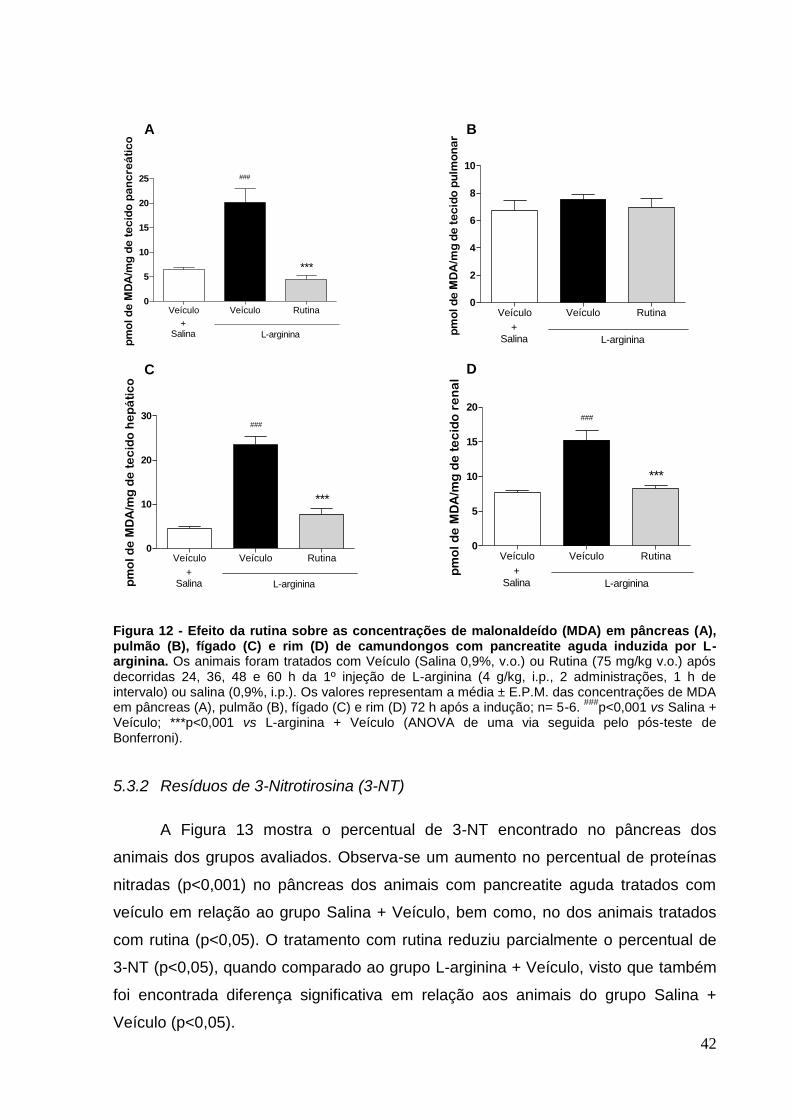
42
Veículo Veículo Rutina0
5
10
15
20
25
L-arginina
###
***
+Salina
pm
ol
de
MD
A/m
g d
e t
ec
ido
pa
nc
reá
tic
o
Veículo Veículo Rutina0
2
4
6
8
10
L-arginina
+Salina
pm
ol
de
MD
A/m
g d
e t
ec
ido
pu
lmo
na
r
Veículo Veículo Rutina0
10
20
30
L-arginina
###
***
+Salinap
mo
l d
e M
DA
/mg
de
te
cid
o h
ep
áti
co
Veículo Veículo Rutina0
5
10
15
20
L-arginina
###
***
+Salina
pm
ol
de
MD
A/m
g d
e t
ec
ido
re
na
l
A B
C D
Figura 12 - Efeito da rutina sobre as concentrações de malonaldeído (MDA) em pâncreas (A), pulmão (B), fígado (C) e rim (D) de camundongos com pancreatite aguda induzida por L-arginina. Os animais foram tratados com Veículo (Salina 0,9%, v.o.) ou Rutina (75 mg/kg v.o.) após decorridas 24, 36, 48 e 60 h da 1º injeção de L-arginina (4 g/kg, i.p., 2 administrações, 1 h de intervalo) ou salina (0,9%, i.p.). Os valores representam a média ± E.P.M. das concentrações de MDA em pâncreas (A), pulmão (B), fígado (C) e rim (D) 72 h após a indução; n= 5-6.
###p<0,001 vs Salina +
Veículo; ***p<0,001 vs L-arginina + Veículo (ANOVA de uma via seguida pelo pós-teste de Bonferroni).
5.3.2 Resíduos de 3-Nitrotirosina (3-NT)
A Figura 13 mostra o percentual de 3-NT encontrado no pâncreas dos
animais dos grupos avaliados. Observa-se um aumento no percentual de proteínas
nitradas (p<0,001) no pâncreas dos animais com pancreatite aguda tratados com
veículo em relação ao grupo Salina + Veículo, bem como, no dos animais tratados
com rutina (p<0,05). O tratamento com rutina reduziu parcialmente o percentual de
3-NT (p<0,05), quando comparado ao grupo L-arginina + Veículo, visto que também
foi encontrada diferença significativa em relação aos animais do grupo Salina +
Veículo (p<0,05).
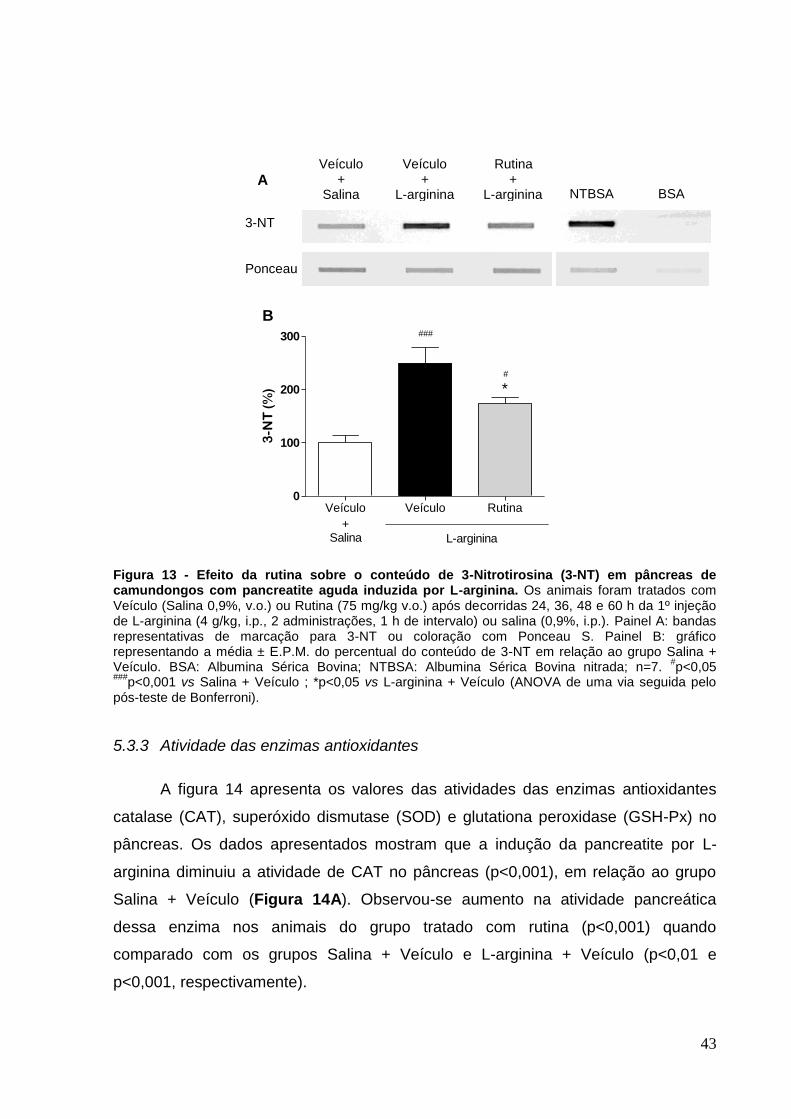
43
Veículo Veículo Rutina0
100
200
300
L-arginina
###
*#
+Salina
3-N
T (
%)
B
Figura 13 - Efeito da rutina sobre o conteúdo de 3-Nitrotirosina (3-NT) em pâncreas de camundongos com pancreatite aguda induzida por L-arginina. Os animais foram tratados com Veículo (Salina 0,9%, v.o.) ou Rutina (75 mg/kg v.o.) após decorridas 24, 36, 48 e 60 h da 1º injeção de L-arginina (4 g/kg, i.p., 2 administrações, 1 h de intervalo) ou salina (0,9%, i.p.). Painel A: bandas representativas de marcação para 3-NT ou coloração com Ponceau S. Painel B: gráfico representando a média ± E.P.M. do percentual do conteúdo de 3-NT em relação ao grupo Salina + Veículo. BSA: Albumina Sérica Bovina; NTBSA: Albumina Sérica Bovina nitrada; n=7.
#p<0,05
###p<0,001 vs Salina + Veículo ; *p<0,05 vs L-arginina + Veículo (ANOVA de uma via seguida pelo
pós-teste de Bonferroni).
5.3.3 Atividade das enzimas antioxidantes
A figura 14 apresenta os valores das atividades das enzimas antioxidantes
catalase (CAT), superóxido dismutase (SOD) e glutationa peroxidase (GSH-Px) no
pâncreas. Os dados apresentados mostram que a indução da pancreatite por L-
arginina diminuiu a atividade de CAT no pâncreas (p<0,001), em relação ao grupo
Salina + Veículo (Figura 14A). Observou-se aumento na atividade pancreática
dessa enzima nos animais do grupo tratado com rutina (p<0,001) quando
comparado com os grupos Salina + Veículo e L-arginina + Veículo (p<0,01 e
p<0,001, respectivamente).
3-NT Ponceau
NTBSA BSA
A
Veículo + Salina
Veículo + L-arginina
Rutina + L-arginina

44
A atividade de SOD pancreática não foi alterada pela administração de L-
arginina (Figura 14B) porém o tratamento com rutina aumentou a atividade de SOD,
tanto em relação ao grupo Salina + Veículo (p<0,001) quanto em relação ao grupo L-
arginina + Veículo (p<0,01). Igualmente, a atividade de GSH-Px no pâncreas não foi
alterada pela administração de L-arginina (Figura 14c) e o tratamento com rutina
aumentou a atividade de SOD em relação ao grupo Salina + Veículo (p<0,05), não
apresentando diferença significativa em relação aos animais do grupo Salina +
Veículo.
Salina Veículo Rutina 0
5
10
15
20
25
L-arginina
##
***
##
UC
AT
/mg
de
pro
teín
a
Salina Veículo Rutina 0
5
10
15
20
25
L-arginina
##
**
###
mU
SO
D/m
g d
e p
rote
ína
Salina Veículo Rutina 0
5
10
15
20
L-arginina
##
*
um
ol
GS
H/m
in/m
g p
rote
ína
A B
C
Figura 14 - Efeito da rutina sobre as atividades das enzimas antioxidantes catalase (CAT), superóxido dismutase (SOD) e glutationa peroxidase (GSH-Px) no pâncreas de camundongos com pancreatite aguda induzida por L-arginina. Os animais foram tratados com Veículo (Salina 0,9%, v.o.) ou Rutina (75 mg/kg v.o.) após decorridas 24, 36, 48 e 60 h da 1º injeção de L-arginina (4 g/kg, i.p., 2 administrações, 1 h de intervalo) ou salina (0,9%, i.p.). Os valores representam a média ± E.P.M. das atividades de CAT, SOD e GSH-Px no pâncreas 72 h após a indução; n=5-6.
##p<0,01 ou
###p<0,001 vs Salina + Veículo ; *p<0,05, **p<0,01 ou ***p<0,001 vs L-arginina + Veículo (ANOVA de
uma via seguida pelo pós-teste de Bonferroni).
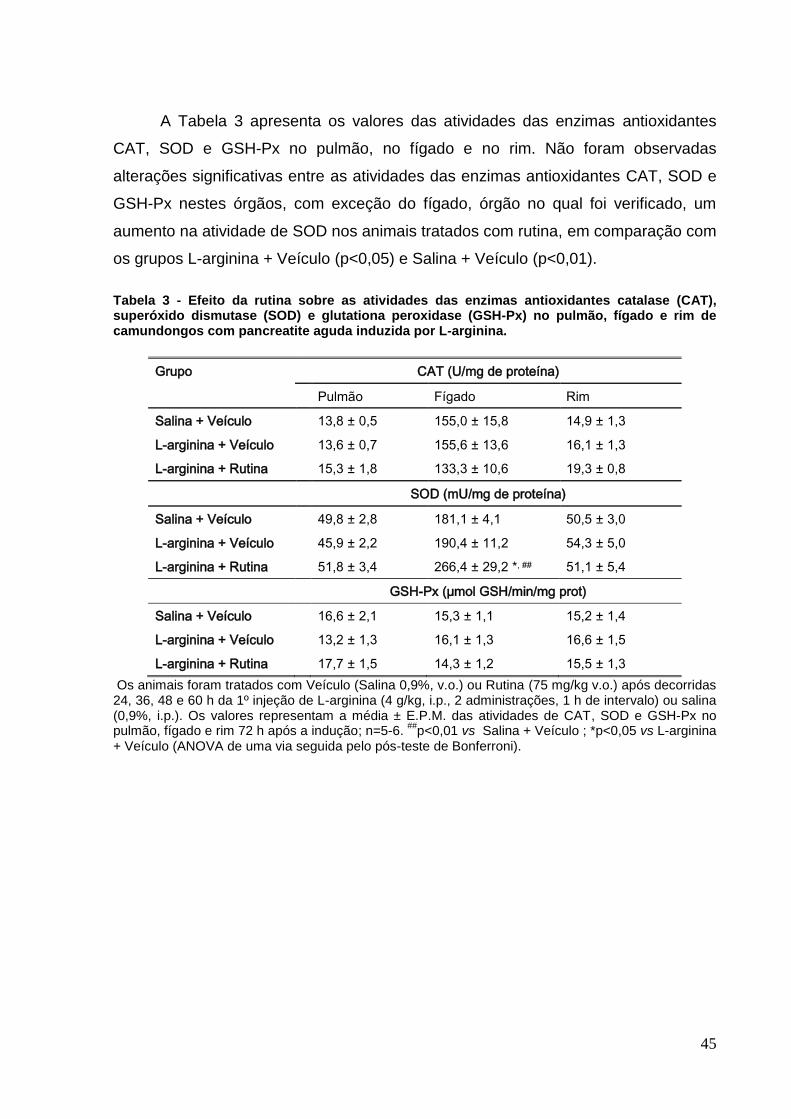
45
A Tabela 3 apresenta os valores das atividades das enzimas antioxidantes
CAT, SOD e GSH-Px no pulmão, no fígado e no rim. Não foram observadas
alterações significativas entre as atividades das enzimas antioxidantes CAT, SOD e
GSH-Px nestes órgãos, com exceção do fígado, órgão no qual foi verificado, um
aumento na atividade de SOD nos animais tratados com rutina, em comparação com
os grupos L-arginina + Veículo (p<0,05) e Salina + Veículo (p<0,01).
Tabela 3 - Efeito da rutina sobre as atividades das enzimas antioxidantes catalase (CAT), superóxido dismutase (SOD) e glutationa peroxidase (GSH-Px) no pulmão, fígado e rim de camundongos com pancreatite aguda induzida por L-arginina.
Grupo CAT (U/mg de proteína)
Pulmão Fígado Rim
Salina + Veículo 13,8 ± 0,5 155,0 ± 15,8 14,9 ± 1,3
L-arginina + Veículo 13,6 ± 0,7 155,6 ± 13,6 16,1 ± 1,3
L-arginina + Rutina 15,3 ± 1,8 133,3 ± 10,6 19,3 ± 0,8
SOD (mU/mg de proteína)
Salina + Veículo 49,8 ± 2,8 181,1 ± 4,1 50,5 ± 3,0
L-arginina + Veículo 45,9 ± 2,2 190,4 ± 11,2 54,3 ± 5,0
L-arginina + Rutina 51,8 ± 3,4 266,4 ± 29,2 *, ## 51,1 ± 5,4
GSH-Px (μmol GSH/min/mg prot)
Salina + Veículo 16,6 ± 2,1 15,3 ± 1,1 15,2 ± 1,4
L-arginina + Veículo 13,2 ± 1,3 16,1 ± 1,3 16,6 ± 1,5
L-arginina + Rutina 17,7 ± 1,5 14,3 ± 1,2 15,5 ± 1,3
Os animais foram tratados com Veículo (Salina 0,9%, v.o.) ou Rutina (75 mg/kg v.o.) após decorridas 24, 36, 48 e 60 h da 1º injeção de L-arginina (4 g/kg, i.p., 2 administrações, 1 h de intervalo) ou salina (0,9%, i.p.). Os valores representam a média ± E.P.M. das atividades de CAT, SOD e GSH-Px no pulmão, fígado e rim 72 h após a indução; n=5-6.
##p<0,01 vs Salina + Veículo ; *p<0,05 vs L-arginina
+ Veículo (ANOVA de uma via seguida pelo pós-teste de Bonferroni).

46
6 DISCUSSÃO
O tratamento atual da pancreatite aguda tem se mostrado ineficaz no controle
do processo inflamatório intrínseco da doença, justificando a necessidade da
realização de estudos que busquem descobrir e caracterizar substâncias com
potencial terapêutico no tratamento desta doença. No presente estudo são
mostrados resultados que apontam para os efeitos benéficos da rutina, relacionados
com a redução da resposta inflamatória e nociceptiva, bem como do estresse
oxidativo, em modelo de pancreatite aguda experimental induzida por L-arginina.
Inicialmente, é importante ressaltar que os resultados obtidos no presente
estudo comprovam a indução da pancreatite com administração de L-arginina em
camundongos. Foi verificado um aumento das concentrações séricas de lipase,
amilase, PCR e IL-6, da atividade de mieloperoxidase em pâncreas e pulmão, do
índice de edema pancreático, bem como, a ocorrência de peroxidação lipídica, no
pâncreas, fígado e rim, evidenciada por aumento nas concentrações de MDA.
Também foi demonstrada, pela primeira vez, a ocorrência de hiperalgesia
abdominal, em camundongos com pancreatite induzida por L-arginina. Os resultados
relacionados a hiperamilasemia e à atividade de MPO pancreática e pulmonar
corroboram o estudo de (DAWRA et al., 2007), o qual mostrou que a administração
de L-arginina, na mesma dose e via de administração utilizadas no presente estudo,
induziu pancreatite aguda em camundongos com maiores alterações inflamatórias
no tempo de 72 h pós-indução.
O aumento nas concentrações séricas de amilase e lipase são reportados
como marcadores preditivos no diagnóstico da pancreatite aguda em humanos
(KEIM et al., 1998). Em humanos, a amilase se eleva dentro de poucas horas após o
desenvolvimento da dor, permanecendo elevada por 3 a 5 dias (FORSMARK;
BAILLIE, 2007; FAN; ANDRÉN-SANDBERG, 2010). Observou-se que o tratamento
com rutina nas doses de 75 e 150 mg/kg preveniu a elevação de amilase no soro
dos animais com pancreatite aguda (Figura 6). O estudo realizado por Carvalho et
al. (2010) avaliou o pré-tratamento de camundongos com pancreatite aguda induzida
por ceruleína, com o flavonoide quercetina, nas doses de 25, 50 e 100 mg/kg, e

47
evidenciou a redução da amilase sérica, nas 3 doses utilizadas, 6 h após a indução,
corroborando o efeito supracitado para a rutina.
Na pancreatite aguda, a concentração sérica de lipase se eleva
posteriormente ao aumento da amilase, diminuindo em tempos mais tardios.
Associada com a hiperamilasemia, a hiperlipasemia é considerada como parâmetro
altamente específico para doenças pancreáticas (ABELA; CARTER, 2010; FAN;
ANDRÉN-SANDBERG, 2010). Foi observado que, no tempo de 72 horas, os animais
com pancreatite aguda exibiram concentrações de lipase marcadamente elevadas
em relação ao grupo controle e que o tratamento com rutina na dose de 75 mg/kg
impediu esta elevação (Figura 9A), em consonância com dados da literatura que
verificaram efeitos inibidores expressivos sobre a lipase pancreática para diversos
flavonoides (GATTO et al., 2002; BIRARI; BHUTANI, 2007), inclusive para rutina
(PRINCE; KANNAN, 2006). Esse achado, juntamente com o observado para os
níveis de amilase, apontam um importante efeito protetor da rutina no pâncreas,
diminuindo a gravidade da pancreatite aguda, visto que estas enzimas participam do
processo autodigestivo característico da lesão pancreática nessa doença (BHATIA,
2005).
Aliado à redução das hiperamilasemia e hiperlipasemia, neste estudo
verificou-se que nos animais com pancreatite tratados com rutina a atividade de
MPO estava diminuída no pâncreas, em relação aos animais do grupo L-arginina +
Veículo (Figura 7A). Ratos com pancreatite crônica experimental suplementados
com rutina na dose de 100 mg/kg por 31 dias, também tiveram a atividade de MPO
reduzida, quando avaliada no soro em comparação com os respectivos controles
não tratados (ARUNA; GEETHA; SUGUNA, 2013b). Estes resultados concordam
com os obtidos por outros autores, que avaliaram a atividade anti-inflamatória da
rutina em diversos modelos de inflamação e observaram diminuição da atividade de
MPO, em relação aos controles não tratados (CRUZ et al., 1998; ABDEL-RAHEEM,
2010; LIU et al., 2013). Vale destacar que a MPO é uma peroxidase secretada dos
grânulos azurófilos de neutrófilos, por exemplo, durante processos inflamatórios
agudos. A determinação da atividade desta enzima é amplamente utilizada para
inferir sobre o conteúdo tecidual de neutrófilos e a intensidade da inflamação, não só

48
no pâncreas, mas também em vários outros tecidos (BRADLEY et al., 1982; ZHANG,
2002; CAMARGO et al, 2005; DAWRA et al., 2007).
A rutina foi recentemente descrita por atuar na inibição da expressão de
moléculas de adesão, o que pode resultar na diminuição ou bloqueio da adesão e
infiltração de leucócitos (LEE; KU; BAE, 2012). Ela também pode estar impedindo a
quimioatração de leucócitos para o tecido pancreático, devido a sua ação inibitória
em macrófagos ativados e consequente inibição da liberação de mediadores pró-
inflamatórios por estes (KAUSS et al., 2008).
O acúmulo de neutrófilos ativos na microvasculatura pulmonar é um dos
eventos marcadamente importantes na resposta sistêmica da pancreatite aguda
grave, o qual foi comprovado em diversos modelos de pancreatite aguda, inclusive
no modelo de indução por L-arginina em camundongos (DAWRA et al., 2008;
MERZA et al., 2013), respaldando os resultados aqui encontrados, que confirmam
atividade de MPO aumentada no pulmão dos animais com pancreatite (Figura 7B).
Curiosamente, não foi observado efeito da rutina sobre a atividade de MPO
pulmonar em nenhuma das doses utilizadas, indicando que ela não afetou a
migração de leucócitos para o pulmão. Esse achado discorda dos resultados obtidos
por Carvalho et al. (2010) que verificaram que o pré-tratamento com quercetina
causou redução da atividade de MPO no pulmão de camundongos com pancreatite
aguda induzida por ceruleína. Estudo realizado por (JUNG et al., 2007) em cobaias
com asma, demonstrou que ambas, a quercetina e a rutina, foram capazes de inibir
o recrutamento de leucócitos na resposta asmática, porém a quercetina apresentou
efeito superior ao da rutina, sendo que o mecanismo para tal diferença não foi
esclarecido.
A explicação para essa ausência de efeito da rutina no pulmão, no modelo de
pancreatite utilizado não está clara. Sugere-se que essa ausência ocorra devido a
diferenças na microvasculatura pulmonar em detrimento da do pâncreas, o que pode
ser responsável por uma menos absorção do metabólitos ativos da rutina e
consequentemente pela ausência do efeito farmacológico verificado no pâncreas.
O acúmulo de leucócitos na microcirculação pancreática durante a lesão
inflamatória é acompanhado de alterações na permeabilidade vascular, resultando

49
em uma desordem microcirculatória caracterizada por vasodilatação e edema
(TAKÁCS et al., 2002). Em concordância, os resultados obtidos evidenciaram um
índice de edema pancreático elevado nos animais com pancreatite aguda, 72 h após
a indução, confirmando achados da literatura para o modelo de L-arginina em
camundongos (CHEN et al., 2012; AWLA et al., 2012). Os animais com pancreatite
aguda que foram tratados com rutina em todas as doses utilizadas apresentaram
menor índice de edema pancreático (Figura 8), corroborando o efeito protetor já bem
evidenciado da rutina sobre células endoteliais, cujo mecanismo, ainda pouco
esclarecido, parece estar relacionado à regulação da produção de óxido nítrico e de
H2O2 (HAN, 2009;GONG et al., 2010) que estão envolvidos tanto na vasodilatação
quanto na lesão direta às membranas destas células, alterando sua permeabilidade
(KUO; HEIN, 2013).
O presente estudo demonstrou, pela primeira vez, que a pancreatite aguda
induzida por L-arginina leva a uma elevação da PCR no soro, bem como evidenciou
que o tratamento com rutina na dose de 75 mg/kg reverteu tal aumento (Figura 9B).
Dessa forma a redução da concentração da PCR, correlacionada com o efeito sobre
as concentrações de lipase e amilase, reforça a hipótese de que a rutina se
apresenta como agente terapêutico em potencial para controlar a gravidade da
pancreatite aguda. Outro aspecto importante a se considerar no que se refere ao
aumento da concentração de PCR, é que sua liberação ocorre em consequência da
estimulação hepática por citocinas pró-inflamatórias (LE; VILCEK, 1989; SCHÜTTE;
MALFERTHEINER, 2008), principalmente pela IL-6 (GANAPATHI et al., 1988;
FROSSARD; HADENGUE; PASTOR, 2001).
A IL-6 é liberada no início da resposta inflamatória, principalmente por
macrófagos, em resposta à lesão tecidual (AL-BAHRANI; AMMORI, 2005b) e a
subsequente liberação de PCR por sua vez, estimula uma maior expressão de
receptores de IL-6 (IL-6R) em neutrófilos, facilitando a formação do complexo IL-
6/IL-6R e amplificando os efeitos inflamatórios da IL-6 (JONES et al., 1999;ZHANG
et al., 2013). Corroborando esta correlação, os dados obtidos no presente trabalho
mostram que os animais com pancreatite aguda, apresentaram índices elevados de
IL-6 circulante (Figura 10), de acordo com o aumento da PCR sérica, bem como com

50
o maior infiltrado neutrofílico, evidenciado pelo aumento da atividade de MPO no
pulmão e no pâncreas.
É válido acrescentar que, a elevação de IL-6 circulante acompanhada do
aumento da PCR, consiste em um marcador conjunto altamente preditivo para a
gravidade da pancreatite aguda, bem como para falência múltipla dos órgãos
associada à doença (LESER et al., 1991; SATHYANARAYAN et al., 2007). Deste
modo, os resultados encontrados no presente estudo comprovam a gravidade da
pancreatite induzida por L-arginina, bem como ressaltam a possibilidade de outros
órgãos, além do pulmão, estarem sendo afetados no modelo utilizado.
Igualmente ao observado para PCR, a rutina impediu a elevação de IL-6,
reforçando seu efeito atenuante da inflamação no modelo de pancreatite aguda
utilizado, corroborando achado encontrado por (ARUNA; GEETHA; SUGUNA,
2013b) no soro de ratos com pancreatite crônica suplementados com rutina. O
mecanismo pelo qual esta estaria inibindo a liberação de IL-6 e consequentemente
de PCR pode estar relacionado com a redução da migração de neutrófilos,
demonstrada pela diminuição da atividade de MPO, visto que estas células são
importantes liberadoras dessa citocina durante a resposta inflamatória
(YAMAMOTO; ROSE-JOHN, 2012). Outro mecanismo possível pode ser devido a
redução da formação das ROS e RNS direta ou indiretamente, visto que estas
também são descritas por estimularem a liberação de citocinas pró-inflamatórias
pelas células do sistema imune (KUO; HEIN, 2013).
Esse efeito inibidor da rutina sobre IL-6 também foi evidencia por (LEE et al.,
2013) em modelos de hepatotoxicidade induzida por etanol e CCL4 em
camundongos, quando administrada por 4 semanas consecutivas na dose de 11,5
mg/kg. Este efeito foi atribuído à capacidade da rutina de regular os níveis de
expressão de RNAm de citocinas pró-inflamatórias, demonstrada em estudos
anteriores. O tratamento com rutina 0,1% na dieta causou redução da expressão de
mRNA de IL-6 na mucosa colônica de camundongos submetidos a indução de colite
por sulfato sódico de dextrana na água de beber (KWON et al., 2005), efeito também
evidenciado em hipocampo de ratos com neurotoxicidade induzida por
trimetilestanho (TMT) tratados com rutina 0,75% na dieta (KODA; KURODA; IMAI,
2009).

51
A atividade anti-inflamatória demonstrada pela rutina no presente estudo,
sinalizou para o fato de que o tratamento com este flavonoide poderia interferir com
o possível quadro de hiperalgesia abdominal, que comumente está presente não só
em animais de experimentação, mas também em humanos que são acometidos pela
pancreatite. Este parâmetro vem sendo caracterizado em modelos de pancreatite
experimental em ratos e camundongos (WINSTON et al., 2003; NISHIMURA et al.,
2010; CAMARGO et al., 2011), e no presente estudo observou-se pela primeira vez,
a ocorrência de hiperalgesia abdominal em modelo de pancreatite induzida por L-
arginina em camundongos. Esta foi caracterizada pela diminuição significativa do
limiar de estímulo mecânico, aplicado na região abdominal, necessária para induzir
um comportamento de retirada nos animais. Esse reflexo de retirada após estímulo
mecânico na região abdominal é descrito na literatura por ser um comportamento
que indica uma maior sensibilização dessa região, mimetizando a dor visceral
exibida em humanos com pancreatite aguda (VERA-PORTOCARRERO; LU;
WESTLUND, 2003).
Também foi verificado no presente estudo que, o tratamento com rutina na
dose de 75 mg/kg reduziu a hiperalgesia abdominal nos animais com pancreatite
após 72 h da indução, quando comparado ao tratamento com veículo. Ressalte-se
que nesse tempo avaliado foi observada maior resposta inflamatória pancreática,
segundo descrito anteriormente (DAWRA et al., 2007).
O resultado observado demonstra que a rutina causou um efeito
antinociceptivo (Figura 11), possivelmente relacionado com a sua atividade anti-
inflamatória pancreática e concorda com estudo prévio que evidenciou que o
tratamento com o extrato etanólico da Caesalpinia pyramidalis, que acredita-se
conter rutina, reduziu a hiperalgesia abdominal em ratos (SANTANA et al., 2012).
Adicionalmente, o efeito antinociceptivo da rutina também foi observado em outros
estudos. (LAPA et al., 2009) mostraram que a rutina (3-100 mg/kg) isolada do
Polygala paniculata L. reduziu a nocicepção induzida por glutamato na pata de
camundongos, mensurada pelo tempo de lambida na pata. De maneira semelhante,
a rutina (10-20 mg/kg) isolada do extrato hidroalcoólico de Phyllanthus niruri reduziu
o comportamento nociceptivo e a inflamação vesical em camundongos com cistite
hemorrágica induzida por ciclofosfamida (BOEIRA et al., 2011).

52
Azevedo et al. (2013) também demonstraram efeito antinociceptivo da rutina
nas doses 25, 50 e 100 mg/kg em modelo de dor neuropática periférica crônica
induzida por oxaliplatina em camundongos. Esta ação foi atribuída à proteção contra
peroxidação lipídica e nitrosilação proteica e à modulação da expressão de iNOS
pela rutina, visto que nesse modelo de dor utilizado, o dano neural parece estar
associado ao dano oxidativo no corno da raiz dorsal. Entretanto, o mecanismo
evidenciado ainda não pode ser extrapolado para o efeito verificado para rutina no
presente estudo, visto que os eventos e mediadores envolvidos na origem da
estimulação das aferências pancreáticas e na consequente resposta
comportamental verificada, ainda não estão bem esclarecidos, dificultando a
identificação de alvos específicos para intervenção farmacológica.
É interessante observar que um estudo piloto tentou relacionar a dor
abdominal com o estresse oxidativo, pela avaliação de parâmetros oxidantes e
antioxidantes no sangue de pacientes com dor abdominal derivada de etiologias
diversas (CHI; SHIESH; LIN, 2002). Este estudo verificou uma correlação entre
capacidade antioxidante total, as concentrações elevadas de PCR e a gravidade das
doenças destes pacientes. Deste modo, mais estudos precisam ser realizados, para
caracterizar e identificar os mediadores envolvidos na hiperalgesia abdominal na
pancreatite aguda, não descartando a possibilidade de que ROS e RNS estejam
relacionados nessa resposta.
O efeito anti-hiperalgésico da rutina nos animais com pancreatite poderia ser
confundido por uma ação depressora central ou relaxante muscular causada por
este composto. Entretanto, esta ação é pouco provável de ter ocorrido, pois o estudo
prévio de (MACHADO et al., 2008) demonstrou que a rutina (0,3-3 mg/kg) não
produziu alteração da atividade locomotora de camundongos em campo aberto ou
no tempo de nado forçado e reduziu o tempo de imobilidade no teste de suspensão
da cauda. Além disso, um estudo direcionado para avaliar o efeito de flavonoides no
comprometimento cognitivo induzido por lipopolissacarideo de parede bacteriana
mostrou que animais jovens ou velhos submetidos ao tratamento crônico com a
quercetina (25-100 mg/kg) não tiveram alteração da atividade locomotora, sendo que
no teste do Rota-rod os animais mais velhos tiveram melhor performance quando
tratados com a quercetina (PATIL et al., 2003).

53
O dano à célula acinar na fase inicial e durante o curso da pancreatite
também está relacionado à ação de ROS e de nitrogênio RNS que se apresentam
em altas concentrações devido ao “burst” oxidativo do processo inflamatório iniciado
(BABU et al., 2009), o qual vem sendo demonstrado nos diversos modelos de
pancreatite experimental (LEUNG; CHAN, 2009). Para verificar o possível papel da
rutina sobre esse processo, foram avaliados parâmetros comumente utilizados como
forte indicadores da presença de dano oxidativo e nitrosativo, como a peroxidação
lipídica (PL) e a nitração proteica.
A formação excessiva de produtos de PL, provenientes do ataque de ROS e
RNS às membranas celulares, consiste num importante mecanismo de iniciação ao
dano celular e tecidual em diversas doenças (NEGRE-SALVAYRE et al., 2010),
inclusive na pancreatite aguda. Aumento nas concentrações teciduais de MDA, um
dos produtos da PL, foram encontrados em modelos de indução por ceruleína,
taurocolato, obstrução do ducto biliopancreático e L-arginina (LEUNG; CHAN,
2009;CZAKÓ et al., 2000b). Em consonância, os resultados encontrados nesse
estudo evidenciaram que os animais com pancreatite aguda induzida por L-arginina
exibiram concentrações aumentadas de MDA no pâncreas, fígado e rim (Figura 12),
confirmando que o desenvolvimento da pancreatite aguda está associada com a
geração de radicais livres não só no pâncreas, mas também em outros órgãos.
Quando comparado ao grupo com pancreatite tratado com veículo, o
tratamento com rutina na dose de 75 mg/kg restaurou as concentrações de MDA no
pâncreas, fígado e rim (Figura 12A, C e D), demonstrando o efeito protetor da rutina
contra a lesão tecidual. Esse efeito da rutina sobre a PL é bem caracterizado na
literatura para diversas condições experimentais em múltiplos órgãos, a destacar: a
isquemia cerebral em ratos por oclusão da artéria cerebral média (KHAN et al.,
2009), a lesão intestinal induzida por isquemia e reperfusão em ratos (ITAGAKI et
al., 2010), a inflamação renal induzida por cisplatina em camundongos (ARJUMAND;
SETH; SULTANA, 2011), a hepatotoxicidade induzida por tetracloreto de carbono
em ratos (KHAN; KHAN; SAHREEN, 2012) e a lesão de mucosa gástrica induzida
por isquemia reperfusão gástrica em ratos (LIU et al., 2013).
No caso do tecido pancreático, este resultado também concorda com os
efeitos da rutina sobre o edema e a atividade de MPO descritos anteriormente nesse

54
trabalho, de modo que esta enzima é descrita por reagir com espécies radicalares e
mediar o processo de iniciação e propagação da PL, gerando produtos capazes de
lesar fosfolipídeos e lipoproteínas de membranas e alterar a função destas
(PODREZ et al., 1999; ZHANG, 2002; KOSTYUK et al., 2003). Esses produtos,
incluindo o MDA, podem causar lesão nas células endoteliais dos capilares,
resultando em aumento da permeabilidade microvascular e edema (ABDIN et al.,
2010).
Estudo in vitro, realizado por (KOSTYUK et al., 2003), com moléculas de LDL
isoladas de sangue de voluntários humanos, demonstrou que diversos flavonoides,
inclusive a rutina, podem co-interagir com a MPO durante a PL mediada por esta,
inibindo ou reduzindo a fase de propagação responsável pela geração excessiva de
hidroperóxidos tóxicos. Interessantemente, estes autores observaram que de vários
flavonoides analisados (quercetina, rutina, taxifolina, luteolina e (-)-epicatequina), a
oxidação da rutina por essa enzima foi dependente da presença de nitrito na reação.
Este achado sugere que a rutina, por sua atividade sequestradora de nitrito,
apresenta forte capacidade protetora contra a peroxidação lipídica por atuar sobre o
sistema MPO/nitrito.
Outros mecanismos propostos para esse efeito protetor da rutina sobre a PL
incluem seu potencial de reagir diretamente com ânions superóxido (O2)
(AFANAS’EV et al., 1989), sua capacidade de quelar íons metálicos, impedindo a
geração de ROS pela reação de Fenton (AFANAS’ EV et al., 1995) e redução
indireta da formação dessas espécies reativas, por impedir a infiltração de
neutrófilos, que também são importantes liberadores de ROS e de RNS (POCH et
al., 1999).
Estas evidências também podem explicar por outro ângulo a baixa atividade
de MPO encontrada nos animais com pancreatite tratados com rutina, sugerindo
que, talvez esse resultado represente o bloqueio das reações catalisadas pela MPO.
Isto seria resultante da interação da rutina com a enzima ou pelo sequestro dos
substratos utilizados, alternativamente ao impedimento da migração de leucócitos
sugerida anteriormente, ou mesmo pela associação dos dois mecanismos. Assim, se
fazem necessárias investigações adicionais para entender o mecanismo de ação da

55
rutina, analisando, por exemplo, a expressão de moléculas de adesão e a liberação
de outros fatores quimiotáxicos.
Interessantemente, não foram encontradas alterações nas concentrações de
MDA no pulmão dos animais com pancreatite aguda induzida por L-arginina, a
despeito do aumento da atividade MPO neste tecido. Não ficou esclarecido, o
porquê dessa ausência de efeito da rutina no tecido pulmonar. É possível que algum
fator protetor característico do tecido pulmonar possa estar atuando de modo a
protegê-lo da peroxidação lipídica mediada pela MPO, salientando que também não
foi verificada a presença de edema pulmonar nesses animais (dados não
mostrados).
Análise temporal realizada por (ELDER et al., 2011) em ratos com pancreatite
aguda induzida por L-arginina, demonstrou que o pico da resposta inflamatória no
pâncreas destes animais ocorre no tempo de 36 horas, enquanto que a atividade de
MPO pulmonar aumentou 12 horas após a indução, porém só houve formação de
edema pulmonar e aumento na contagem de células inflamatórias no lavado bronco-
alveolar após 48 h, indicando uma lesão pulmonar tardia. Como no modelo em
camundongos, já foi descrito que o pico da resposta inflamatória pancreática ocorre
no tempo de 72 h (DAWRA et al., 2007), é possível que a lesão pulmonar venha a
ser instalada em tempo posterior, justificando a ausência de edema e PL nos
animais com pancreatite aguda avaliados no presente estudo.
O mecanismo envolvido na migração de leucócitos para o pulmão parece ser
independente do espécies reativas, ressaltando que a lesão pulmonar secundária a
pancreatite é complexa, envolvendo múltiplos mediadores e mecanismos ainda
pouco elucidados (CLOSA et al., 1999; ELDER et al., 2011).
A lesão pulmonar resultante da pancreatite foi descrita por estar relacionada
com a passagem pelo fígado de substâncias liberadas pelo pâncreas na circulação,
sugerindo que mediadores solúveis liberados pelo fígado possam ser responsáveis
pela resposta inflamatória no pulmão (CLOSA et al., 1996; CLOSA et al., 1999).
Seguindo essa linha, recentemente foi sugerido que a IL-6 parece exercer seus
efeitos principalmente na pancreatite aguda na forma grave, desempenhando um
papel importante no desenvolvimento da lesão pulmonar associada (HUANG et al.,

56
2012). Ela estaria atuando por meio de um mecanismo de trans-sinalização em
conjunto com IL-6R, o qual ativa uma cascata de sinalização capaz de regular
processos inflamatórios no sítio principal da inflamação, amplificando a resposta
para o pulmão e outros órgãos, por meio do recrutamento de leucócitos (ZHANG et
al., 2013).
Estas evidências reforçam a participação desta interleucina na fisiopatologia
da doença. Entretanto, mais estudos devam ser realizados, para melhor esclarecer
esses achados, visto que a síndrome da dificuldade respiratória aguda contribui
significativamente para as altas taxas de morbidade e mortalidade dos casos de
pancreatite grave (ZHANG et al., 2013).
A formação de resíduos de nitrotirosina foi descrita por estar relacionada com
a formação endógena de peroxinitrito, originado da reação entre NO e o radical O2,
que induz uma série de modificações oxidativas em proteínas, a exemplo da nitração
de resíduos de tirosina (GOW et al., 1996). Adicionalmente também foi demonstrado
que a reação do nitrito com o ácido hipocloroso e da MPO com o H2O2 e/ou com
espécies derivadas também levam a nitração de tirosina em condições inflamatórias
agudas e crônicas (HAZEN et al., 1999; GAUT et al., 2002; VLASOVA; SOKOLOV;
ARNHOLD, 2012).
Os camundongos com pancreatite aguda induzida por ceruleína exibiram uma
maior expressão de resíduos de nitrotirosina no tecido pancreático, em relação aos
controles e o tratamento com extrato de chá verde, rico em flavonoides, reverteu
esse aumento (BABU et al., 2009). Similarmente, o presente estudo verificou o
aumento do conteúdo pancreático de resíduos de nitrotirosina durante a pancreatite
induzida por L-arginina, o que não havia sido mostrado por estudo prévio. O
tratamento com rutina, na dose de 75 mg/kg, reduziu este conteúdo elevado de 3-
nitrotirosina (Figura 13), o que sustenta a hipótese de que o efeito protetor da rutina
na pancreatite aguda, é decorrente de sua ação sequestradora de espécies
radicalares e/ou de sua capacidade de atuar junto a MPO minimizando os efeitos
resultantes da reação desta com nitritos e seus produtos.
Em relação à atividade das enzimas antioxidantes, os resultados observados
(Figura 14 e Tabela 3) sugerem que a L-arginina leva a depleção da atividade de

57
CAT no pâncreas, os quais estão de acordo com estudos prévios utilizando esse
modelo em ratos e camundongos (CZAKÓ et al., 2000;SOWJANYA; SANDHYA;
VEERESH, 2012; PANDURANGAN, 2013). Essa depleção é referida como
resultante do aumento de ROS e RNS durante a progressão da doença (CZAKÓ et
al., 1998), as quais podem atacar o sítio ativo das enzimas antioxidantes, levando a
uma perda de função (SZABOLCS et al., 2006a). Porém, as atividades de SOD e
GSH-Px pancreáticas não foram alteradas pela indução da pancreatite, contrastando
os dados da literatura, já citados para catalase.
O tratamento com rutina na dose de 75 mg/kg levou a um aumento da
atividade de CAT, provavelmente por inibir esse “auto ataque’’, reagindo diretamente
com as espécies reativas. Curiosamente, o tratamento com a rutina na dose de 75
mg/kg também aumentou as atividades de SOD e GSH-Px, a despeito destas não
terem sido diminuídas nos animais que receberam L-arginina, sugerindo a
estimulação de uma resposta protetora pela rutina no pâncreas dos animais com
pancreatite aguda.
De maneira geral, o efeito inibitório da rutina na LP, associado a diminuição
do conteúdo de 3-NT pancreático e ao aumento da atividade da catalase, SOD e
GSH-Px neste tecido, indicam que o estresse oxidativo tem um papel importante na
pancreatite induzida pela L-arginina e pode ser modulado pela administração
terapêutica de rutina.
O pâncreas é relatado na literatura por ser mais sensível aos efeitos tóxicos
destas moléculas, em virtude de apresentar uma menor atividade e/ou expressão
das enzimas antioxidantes que o fígado e o rim, de modo que nos pâncreas estas
enzimas acabam sendo depletadas mais rapidamente (CZAKÓ et al., 2000b). Os
dados obtidos nesse estudo demonstram que a atividade de CAT, SOD ou GSH-Px
não foi alterada no fígado e no rim dos animais com pancreatite aguda em
comparação com os do grupo controle, enquanto que no pâncreas houve um
diminuição significativa da atividade de CAT.
Os resultados apresentados no presente estudo mostram que a rutina tem
potencial para o tratamento da pancreatite e este fato está atrelado as suas ações
anti-inflamatórias e nociceptiva, que de várias formas parecem estar relacionadas

58
com a redução do estresse oxidativo. Como este é um estudo inicial, outros
precisam ser conduzidos para melhor caracterizar estas e outras atividades
potenciais desse flavonoide, que podem direcionar, ou mesmo dirimir, o uso deste
composto natural no tratamento da pancreatite.

59
7 CONCLUSÃO
Esse estudo mostra que a rutina apresenta efeitos anti-inflamatórios, anti-
nociceptivo e antioxidante no modelo de pancreatite aguda induzida por L-arginina
em camundongos. Esses efeitos sugerem que a rutina representa uma alternativa
terapêutica em potencial para ser utilizada no tratamento da pancreatite aguda em
humanos. É importante ressaltar que investigações adicionais precisam ser
realizadas para comprovar seu potencial clínico.

60
8 REFERÊNCIAS
ABDEL-RAHEEM, I. T. Gastroprotective Effect of Rutin against Indomethacin-Induced Ulcers in Rats: GASTROPROTECTIVE EFFECT OF RUTIN IN RATS. Basic & Clinical Pharmacology & Toxicology, v. 107, n. 3, p. 742–750, set. 2010.
ABDIN, A. A. et al. Effect of pentoxifylline and/or alpha lipoic acid on experimentally induced acute pancreatitis. European Journal of Pharmacology, v. 643, n. 2-3, p. 289–296, set. 2010.
ABELA, J. E.; CARTER, C. R. Acute pancreatitis – a review. Surgery (Oxford), v. 28, n. 5, p. 205–211, maio 2010.
AFANAS’ EV, I. B. et al. Different antioxid ant activities of bioflavonoid rutin in normal and ironoverloading rats. Biochemical pharmacology, v. 50, n. 5, p. 627–635, 1995.
AFANAS’EV, I. B. et al. Chelating and free radical scavenging mechanisms of inhibitory action of rutin and quercetin in lipid peroxidation. Biochemical Pharmacology, v. 38, n. 11, p. 1763–1769, jun. 1989.
AHO, H. J. et al. Experimental pancreatitis in the rat. The role of phospholipase A in sodium taurocholate-induced acute haemorrhagic pancreatitis. Scandinavian journal of gastroenterology, v. 15, n. 8, p. 1027–1031, 1980.
AKAY, S. et al. Use of activated protein C has no avail in the early phase of acute pancreatitis. HPB, v. 10, n. 6, p. 459–463, 2008.
AL-BAHRANI, A. Z.; AMMORI, B. J. Clinical laboratory assessment of acute pancreatitis. Clinica chimica acta; international journal of clinical chemistry, v. 362, n. 1-2, p. 26–48, dez. 2005.
ALBUQUERQUE, F. Agência Brasil. São Paulo tem 20 casos de pancreatite aguda para cada 100 mil habitantes, http://www.ebc.com.br/noticias/saude/2013/06/sao-paulo-tem-20-casos-de-pancreatite-aguda-para-cada-100-mil-habitantes. Acesso em: 20/01/2013.
ALTAVILLA, D. et al. Attenuated Cerulein-Induced Pancreatitis in Nuclear Factor–κB–Deficient Mice. Laboratory Investigation, v. 83, n. 12, p. 1723–1732, dez. 2003.
ANDERSSON, E. Tissue factor in predicted severe acute pancreatitis. World Journal of Gastroenterology, v. 16, n. 48, p. 6128, 2010.
ARJUMAND, W.; SETH, A.; SULTANA, S. Rutin attenuates cisplatin induced renal inflammation and apoptosis by reducing NFκB, TNF-α and caspase-3 expression in wistar rats. Food and Chemical Toxicology, v. 49, n. 9, p. 2013–2021, set. 2011.

61
ARUNA, R.; GEETHA, A.; SUGUNA, P. Expression of caspase activation recruitment and pyrin domain levels of apoptosis-associated speck-like protein complex in the pancreas of rats subjected to experimental pancreatitis: Influence of rutin administration. Human & experimental toxicology, 28 nov. 2013a.
ARUNA, R.; GEETHA, A.; SUGUNA, P. Expression of caspase activation recruitment and pyrin domain levels of apoptosis-associated speck-like protein complex in the pancreas of rats subjected to experimental pancreatitis: Influence of rutin administration. Human & Experimental Toxicology, 28 nov. 2013b.
AWLA, D. et al. NFATc3 Regulates Trypsinogen Activation, Neutrophil Recruitment, and Tissue Damage in Acute Pancreatitis in Mice. Gastroenterology, v. 143, n. 5, p. 1352–1360.e7, nov. 2012.
AZEVEDO, M. et al. The antioxidant effects of the flavonoids rutin and quercetin inhibit oxaliplatin-induced chronic painful peripheral neuropathy. Molecular Pain, v. 9, n. 1, p. 53, 2013.
BABU, B. I. et al. Green Tea Polyphenols Ameliorate Pancreatic Injury in Cerulein-Induced Murine Acute Pancreatitis: Pancreas, v. 38, n. 8, p. 954–967, nov. 2009.
BARDEESY, N.; DEPINHO, R. A. Pancreatic cancer biology and genetics. Nature Reviews Cancer, v. 2, n. 12, p. 897–909, dez. 2002.
BECHO, J. R. M.; MACHADO, H.; DE OLIVEIRA GUERRA, M. Rutina–Estrutura, Metabolismo e Potencial Farmacológico. Revista Interdisciplinar de Estudos Experimentais-Animais e Humanos Interdisciplinary Journal of Experimental Studies, v. 1, n. 1, 2010.
BEHLING, E. et al. Flavonóide quercetina: aspectos gerais e a\ccões biológicas. Alimentos e Nutri\ccão Araraquara, v. 15, n. 3, p. 285–292, 2008.
BHATIA, M. et al. Inflammatory mediators in acute pancreatitis. The Journal of pathology, v. 190, n. 2, p. 117–125, fev. 2000.
BHATIA, M. Inflammatory response on the pancreatic acinar cell injury. Scandinavian journal of surgery: SJS: official organ for the Finnish Surgical Society and the Scandinavian Surgical Society, v. 94, n. 2, p. 97–102, 2005.
BICZÓ, G. et al. Inhibition of arginase activity ameliorates L-arginine-induced acute pancreatitis in rats. Pancreas, v. 39, n. 6, p. 868–874, ago. 2010.
BIRADAR, S.; VEERESH, B. International Journal of Drug Development & Research. Screening of Natural Antioxidants by using L-Arginine induced acute Pancreatitis Model, v. 4, n. 4, 2012.
BIRARI, R. B.; BHUTANI, K. K. Pancreatic lipase inhibitors from natural sources: unexplored potential. Drug Discovery Today, v. 12, n. 19-20, p. 879–889, out. 2007.

62
BOEIRA, V. T. et al. Effects of the hydroalcoholic extract of Phyllanthus niruri and its isolated compounds on cyclophosphamide-induced hemorrhagic cystitis in mouse. Naunyn-Schmiedeberg’s Archives of Pharmacology, v. 384, n. 3, p. 265–275, 23 jul. 2011.
BOLLEN, T. L. et al. The Atlanta Classification of acute pancreatitis revisited. The British journal of surgery, v. 95, n. 1, p. 6–21, jan. 2008.
BORGHI, S. M. et al. Vitexin inhibits inflammatory pain in mice by targeting TRPV1, oxidative stress, and cytokines. Journal of natural products, v. 76, n. 6, p. 1141–1149, 28 jun. 2013.
BOSE, R.; SUTHERLAND, G. R.; PINSKY, C. Biological and methodological implications of prostaglandin involvement in mouse brain lipid peroxidation measurements. Neurochemical research, v. 14, n. 3, p. 217–220, mar. 1989.
BRADFORD, M. M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical biochemistry, v. 72, p. 248–254, 7 maio 1976.
BRADLEY, P. P. et al. Measurement of Cutaneous Inflammation: Estimation of Neutrophil Content with an Enzyme Marker. Journal of Investigative Dermatology, v. 78, n. 3, p. 206–209, mar. 1982.
CAMARGO, E. A. et al. Characterization of the acute pancreatitis induced by secretory phospholipases A2 in rats. Toxicon: official journal of the International Society on Toxinology, v. 46, n. 8, p. 921–926, 15 dez. 2005.
CAMARGO, E. A. et al. Role of substance P and bradykinin in acute pancreatitis induced by secretory phospholipase A2. Pancreas, v. 37, n. 1, p. 50–55, jul. 2008.
CAMARGO, E. A. et al. Abdominal hyperalgesia in secretory phospholipase A2-induced rat pancreatitis: distinct roles of NK1 receptors. European journal of pain (London, England), v. 15, n. 9, p. 900–906, out. 2011.
CAMARGO, E. A. et al. Inhibition of inducible nitric oxide synthase-derived nitric oxide as a therapeutical target for acute pancreatitis induced by secretory phospholipase A2. European journal of pain (London, England), 24 out. 2013.
CARRASCO, C. et al. Chemopreventive effects of resveratrol in a rat model of cerulein-induced acute pancreatitis. Molecular and Cellular Biochemistry, 15 nov. 2013.
CARVALHO, K. M. M. B. et al. The natural flavonoid quercetin ameliorates cerulein-induced acute pancreatitis in mice. Biological & pharmaceutical bulletin, v. 33, n. 9, p. 1534–1539, 2010.

63
CERQUEIRA, F. M.; MEDEIROS, M. H. G. DE; AUGUSTO, O. Antioxidantes dietéticos: controvérsias e perspectivas. Química Nova, v. 30, n. 2, p. 441–449, abr. 2007.
CHEN, J. et al. Netrin-1 Protects against L-Arginine-Induced Acute Pancreatitis in Mice. PLoS ONE, v. 7, n. 9, p. e46201, 27 set. 2012.
CHI, C.-H.; SHIESH, S.-C.; LIN, X.-Z. Total antioxidant capacity and malondialdehyde in acute abdominal pain. The American journal of emergency medicine, v. 20, n. 2, p. 79–82, mar. 2002.
CHVANOV, M.; PETERSEN, O. H.; TEPIKIN, A. Free radicals and the pancreatic acinar cells: role in physiology and pathology. Philosophical Transactions of the Royal Society B: Biological Sciences, v. 360, n. 1464, p. 2273–2284, 29 dez. 2005.
CLOSA, D. et al. Hepatic involvement in pancreatitis-induced lung damage. The American journal of physiology, v. 270, n. 1 Pt 1, p. G6–13, jan. 1996.
CLOSA, D. et al. Activation of alveolar macrophages in lung injury associated with experimental acute pancreatitis is mediated by the liver. Annals of surgery, v. 229, n. 2, p. 230–236, fev. 1999.
COMERT, B. et al. Combination of allopurinol and hyperbaric oxygen therapy: A new treatment in experimental acute necrotizing pancreatitis? World Journal of Gastroenterology, v. 13, n. 46, p. 6203, 2007.
CRESPY, V. et al. Comparison of the intestinal absorption of quercetin, phloretin and their glucosides in rats. The Journal of nutrition, v. 131, n. 8, p. 2109–2114, 2001.
CRUZ, T. et al. Oral administration of rutoside can ameliorate inflammatory bowel disease in rats. Life sciences, v. 62, n. 7, p. 687–695, 1998.
CUZZOCREA, S. et al. REDUCTION IN THE DEVELOPMENT OF CERULEIN-INDUCED ACUTE PANCREATITIS BY TREATMENT WITH M40401, A NEW SELECTIVE SUPEROXIDE DISMUTASE MIMETIC: Shock, v. 22, n. 3, p. 254–261, set. 2004.
CZAKÓ, L. et al. Involvement of oxygen-derived free radicals in L-arginine-induced acute pancreatitis. Digestive diseases and sciences, v. 43, n. 8, p. 1770–1777, ago. 1998.
CZAKÓ, L. et al. The pathogenesis of L-arginine-induced acute necrotizing pancreatitis: inflammatory mediators and endogenous cholecystokinin. Journal of Physiology-Paris, v. 94, n. 1, p. 43–50, 2000a.
CZAKÓ, L. et al. Oxidative stress in distant organs and the effects of allopurinol during experimental acute pancreatitis. International journal of pancreatology:

64
official journal of the International Association of Pancreatology, v. 27, n. 3, p. 209–216, jun. 2000b.
DASURI, K.; ZHANG, L.; KELLER, J. N. Oxidative stress, neurodegeneration, and the balance of protein degradation and protein synthesis. Free Radical Biology and Medicine, v. 62, p. 170–185, set. 2013.
DAWRA, R. et al. Development of a new mouse model of acute pancreatitis induced by administration of L-arginine. AJP: Gastrointestinal and Liver Physiology, v. 292, n. 4, p. G1009–G1018, 7 dez. 2007.
DAWRA, R. et al. An improved method for extracting myeloperoxidase and determining its activity in the pancreas and lungs during pancreatitis. Pancreas, v. 37, n. 1, p. 62–68, 2008.
DAWRA, R.; SALUJA, A. K. L-arginine-induced experimental acute pancreatitis. The Pancreapedia: Exocrine Pancreas Knowledge Base, 2012.
DE ARAÚJO, M. E. M. B. et al. Enzymatic de-glycosylation of rutin improves its antioxidant and antiproliferative activities. Food Chemistry, v. 141, n. 1, p. 266–273, nov. 2013.
DIMAGNO, M. J. Endothelial nitric oxide synthase is protective in the initiation of caerulein-induced acute pancreatitis in mice. AJP: Gastrointestinal and Liver Physiology, v. 287, n. 1, p. G80–G87, 25 mar. 2004.
DOMITROVIĆ, R. et al. Differential hepatoprotective mechanisms of rutin and quercetin in CCl(4)-intoxicated BALB/cN mice. Acta pharmacologica Sinica, v. 33, n. 10, p. 1260–1270, out. 2012.
DOUGLAS, C. R. Fisiologia aplicada á nutrição. 2. ed. Guanabara: Koogan, 2006.
ELDER, A. S. F. et al. L-Arginine-induced acute pancreatitis results in mild lung inflammation without altered respiratory mechanics. Experimental lung research, v. 37, n. 1, p. 1–9, fev. 2011.
ESCOTT-STUMP, S. Nutrição relacionada ao diagnóstico e tratamento. 5. ed. Barueri, SP: Manole, 2007.
FAN, B.-G.; ANDRÉN-SANDBERG, A. Acute pancreatitis. North American journal of medical sciences, v. 2, n. 5, p. 211–214, maio 2010.
FARKAS, G. et al. Experimental acute pancreatitis results in increased blood–brain barrier permeability in the rat: a potential role for tumor necrosis factor and interleukin 6. Neuroscience letters, v. 242, n. 3, p. 147–150, 1998.
FASINU, P. et al. Flavonoids and polymer derivatives as CYP3A4 inhibitors for improved oral drug bioavailability. Journal of Pharmaceutical Sciences, v. 102, n. 2, p. 541–555, fev. 2013.

65
FERREIRA, A. L. A.; MATSUBARA, L. S. Radicais livres: conceitos, doenças relacionadas, sistema de defesa e estresse oxidativo. Revista da Associação Médica Brasileira, v. 43, n. 1, mar. 1997.
FONTANA PEREIRA, D. et al. Effects of flavonoids on α-glucosidase activity: Potential targets for glucose homeostasis. Nutrition, v. 27, n. 11-12, p. 1161–1167, nov. 2011.
FORSMARK, C. E.; BAILLIE, J. AGA Institute Technical Review on Acute Pancreatitis. Gastroenterology, v. 132, n. 5, p. 2022–2044, maio 2007.
FOSSATI, P.; PRENCIPE, L.; BERTI, G. Use of 3,5-dichloro-2-hydroxybenzenesulfonic acid/4-aminophenazone chromogenic system in direct enzymic assay of uric acid in serum and urine. Clinical chemistry, v. 26, n. 2, p. 227–231, fev. 1980.
FROSSARD, J.-L. Experimental evidence of obesity as a risk factor for severe acute pancreatitis. World Journal of Gastroenterology, v. 15, n. 42, p. 5260, 2009.
FROSSARD, J.-L.; HADENGUE, A.; PASTOR, C. M. New serum markers for the detection of severe acute pancreatitis in humans. American journal of respiratory and critical care medicine, v. 164, n. 1, p. 162–170, 2001.
FROSSARD, J.-L.; STEER, M. L.; PASTOR, C. M. Acute pancreatitis. Lancet, v. 371, n. 9607, p. 143–152, 12 jan. 2008.
GANAPATHI, M. K. et al. Role of interleukin-6 in regulating synthesis of C-reactive protein and serum amyloid A in human hepatoma cell lines. Biochemical and biophysical research communications, v. 157, n. 1, p. 271–277, 30 nov. 1988.
GARDNER, J. D. Regulation of pancreatic exocrine function in vitro: initial steps in the actions of secretagogues. Annual review of physiology, v. 41, p. 55–66, 1979.
GATTO, M. T. et al. Antimicrobial and anti-lipase activity of quercetin and its C2-C16 3-O-acyl-esters. Bioorganic & medicinal chemistry, v. 10, n. 2, p. 269–272, fev. 2002.
GAUT, J. P. et al. Myeloperoxidase produces nitrating oxidants in vivo. Journal of Clinical Investigation, v. 109, n. 10, p. 1311–1319, 15 maio 2002.
GLOOR, B. et al. Mechanism of increased lung injury after acute pancreatitis in IL-10 knockout mice. Journal of Surgical Research, v. 80, n. 1, p. 110–114, 1998.
GONG, G. et al. Rutin inhibits hydrogen peroxide-induced apoptosis through regulating reactive oxygen species mediated mitochondrial dysfunction pathway in human umbilical vein endothelial cells. European Journal of Pharmacology, v. 628, n. 1-3, p. 27–35, fev. 2010.

66
GOW, A. J. et al. Effects of peroxynitrite-induced protein modifications on tyrosine phosphorylation and degradation. FEBS letters, v. 385, n. 1-2, p. 63–66, 29 abr. 1996.
GRADY, T. et al. Edema and intrapancreatic trypsinogen activation precede glutathione depletion during caerulein pancreatitis. The American journal of physiology, v. 271, n. 1 Pt 1, p. G20–26, jul. 1996.
GRANGER, J.; REMICK, D. Acute pancreatitis: models, markers, and mediators. Shock, v. 24, p. 45–51, 2005.
GRIGSBY, B.; RODRIGUEZ-RILO, H.; KHAN, K. Antioxidants and Chronic Pancreatitis: Theory of Oxidative Stress and Trials of Antioxidant Therapy. Digestive Diseases and Sciences, v. 57, n. 4, p. 835–841, 3 fev. 2012.
GUICE, K. S. et al. Superoxide dismutase and catalase: a possible role in established pancreatitis. American journal of surgery, v. 151, n. 1, p. 163–169, jan. 1986.
HALLIWELL, B. Oxidative stress and neurodegeneration: where are we now? Journal of Neurochemistry, v. 97, n. 6, p. 1634–1658, jun. 2006.
HALLIWELL, B.; GUTTERIDGE, J. M. Lipid peroxidation, oxygen radicals, cell damage, and antioxidant therapy. Lancet, v. 1, n. 8391, p. 1396–1397, 23 jun. 1984.
HAN, Y. Rutin has therapeutic effect on septic arthritis caused by Candida albicans. International Immunopharmacology, v. 9, n. 2, p. 207–211, fev. 2009.
HARDMAN, J. et al. Intravenous selenium modulates L-arginine-induced experimental acute pancreatitis. JOP, v. 6, n. 5, p. 431–437, 2005.
HAZEN, S. L. et al. Formation of Nitric Oxide-Derived Oxidants by Myeloperoxidase in Monocytes : Pathways for Monocyte-Mediated Protein Nitration and Lipid Peroxidation In Vivo. Circulation Research, v. 85, n. 10, p. 950–958, 12 nov. 1999.
HEGYI, P. et al. L-arginine-induced experimental pancreatitis. World Journal of Gastroenterology, v. 10, n. 14, p. 2003–2009, 2004.
HOLLMAN, P. C. et al. Absorption of dietary quercetin glycosides and quercetin in healthy ileostomy volunteers. The American journal of clinical nutrition, v. 62, n. 6, p. 1276–1282, dez. 1995.
HUANG, L. et al. Effects of Chai-Qin-Cheng-Qi decoction (柴芩承气汤) on acute
pancreatitis-associated lung injury in mice with acute necrotizing pancreatitis. Chinese Journal of Integrative Medicine, 21 set. 2012.

67
HUBER, P. C.; ALMEIDA, W. P.; FÁTIMA, Â. DE. Glutationa e enzimas relacionadas: papel biológico e importância em processos patológicos. Química Nova, v. 31, n. 5, p. 1170–1179, 2008.
ITAGAKI, S. et al. Protective effects of quercetin-3-rhamnoglucoside (rutin) on ischemia-reperfusion injury in rat small intestine. Food Chemistry, v. 118, n. 2, p. 426–429, jan. 2010.
JONES, S. A. et al. C-reactive Protein: A Physiological Activator of Interleukin 6 Receptor Shedding. Journal of Experimental Medicine, v. 189, n. 3, p. 599–604, 1 fev. 1999.
JUNG, C. H. et al. Anti-asthmatic action of quercetin and rutin in conscious guinea-pigs challenged with aerosolized ovalbumin. Archives of pharmacal research, v. 30, n. 12, p. 1599–1607, dez. 2007.
KAUSS, T. et al. Rutoside decreases human macrophage-derived inflammatory mediators and improves clinical signs in adjuvant-induced arthritis. Arthritis Research & Therapy, v. 10, n. 1, p. R19, 2008.
KEIM, V. et al. A comparison of lipase and amylase in the diagnosis of acute pancreatitis in patients with abdominal pain. Pancreas, v. 16, n. 1, p. 45–49, jan. 1998.
KHAN, M. M. et al. Rutin protects the neural damage induced by transient focal ischemia in rats. Brain Research, v. 1292, p. 123–135, set. 2009.
KHAN, R. A.; KHAN, M. R.; SAHREEN, S. CCl4-induced hepatotoxicity: protective effect of rutin on p53, CYP2E1 and the antioxidative status in rat. BMC Complementary and Alternative Medicine, v. 12, n. 1, p. 178, 2012.
KODA, T.; KURODA, Y.; IMAI, H. Rutin Supplementation in the Diet has Protective Effects Against Toxicant-Induced Hippocampal Injury by Suppression of Microglial Activation and Pro-Inflammatory Cytokines: Protective Effect of Rutin Against Toxicant-Induced Hippocampal Injury. Cellular and Molecular Neurobiology, v. 29, n. 4, p. 523–531, 21 jan. 2009.
KOSTYUK, V. A. et al. Myeloperoxidase/nitrite-mediated lipid peroxidation of low-density lipoprotein as modulated by flavonoids. FEBS Letters, v. 537, n. 1-3, p. 146–150, fev. 2003.
KUO, L.; HEIN, T. W. Vasomotor Regulation of Coronary Microcirculation by Oxidative Stress: Role of Arginase. Frontiers in Immunology, v. 4, 2013.
KWON, K. H. et al. Dietary rutin, but not its aglycone quercetin, ameliorates dextran sulfate sodium-induced experimental colitis in mice: attenuation of pro-inflammatory gene expression. Biochemical pharmacology, v. 69, n. 3, p. 395–406, 1 fev. 2005.

68
LA CASA, C. et al. Evidence for protective and antioxidant properties of rutin, a natural flavone, against ethanol induced gastric lesions. Journal of ethnopharmacology, v. 71, n. 1-2, p. 45–53, jul. 2000.
LAMPROPOULOS, P. et al. The role of apigenin in an experimental model of acute pancreatitis. The Journal of surgical research, v. 183, n. 1, p. 129–137, jul. 2013.
LAPA, F. DA R. et al. Antinociceptive properties of the hydroalcoholic extract and the flavonoid rutin obtained from Polygala paniculata L. in mice. Basic & clinical pharmacology & toxicology, v. 104, n. 4, p. 306–315, abr. 2009.
LE, J. M.; VILCEK, J. Interleukin 6: a multifunctional cytokine regulating immune reactions and the acute phase protein response. Laboratory investigation; a journal of technical methods and pathology, v. 61, n. 6, p. 588–602, dez. 1989.
LEE, C.-C. et al. Rutin and quercetin, bioactive compounds from tartary buckwheat, prevent liver inflammatory injury. Food & function, v. 4, n. 5, p. 794–802, 30 abr. 2013.
LEE, W.; KU, S.-K.; BAE, J.-S. Barrier protective effects of rutin in LPS-induced inflammation in vitro and in vivo. Food and Chemical Toxicology, v. 50, n. 9, p. 3048–3055, set. 2012.
LEMPINEN, M.; PUOLAKKAINEN, P.; KEMPPAINEN, E. Clinical value of severity markers in acute pancreatitis. Scandinavian journal of surgery, v. 94, n. 2, p. 118, 2005.
LERCH, M. M. et al. Pancreatic duct obstruction triggers acute necrotizing pancreatitis in the opossum. Gastroenterology, v. 104, n. 3, p. 853–861, mar. 1993.
LESER, H. G. et al. Elevation of serum interleukin-6 concentration precedes acute-phase response and reflects severity in acute pancreatitis. Gastroenterology, v. 101, n. 3, p. 782–785, set. 1991.
LEUNG, P. S.; CHAN, Y. C. Role of Oxidative Stress in Pancreatic Inflammation. Antioxidants & Redox Signaling, v. 11, n. 1, p. 135–166, jan. 2009.
LIMA, L. R. P. et al. Acta Farm. Bonaerense. Toxidade Aguda de Rutina e Bixina de Bixa orellana, v. 22, n. 1, p. 21–16, 2003.
LINDAHL, M.; TAGESSON, C. Flavonoids as phospholipase A2 inhibitors: importance of their structure for selective inhibition of group II phospholipase A2. Inflammation, v. 21, n. 3, p. 347–356, jun. 1997.
LIU, Y. et al. Protective effect of rutin against acute gastric mucosal lesions induced by ischemia-reperfusion. Pharmaceutical Biology, v. 51, n. 7, p. 914–919, jul. 2013.

69
LONG, J. et al. Nuclear factor-kappaB activation on the reactive oxygen species in acute necrotizing pancreatitic rats. World Journal of Gastroenterology, v. 11, n. 27, p. 4277, 2005.
LUND, H. et al. Long-term recurrence and death rates after acute pancreatitis. Scandinavian journal of gastroenterology, v. 41, n. 2, p. 234–238, fev. 2006.
MACHADO, D. G. et al. Antidepressant-like effect of rutin isolated from the ethanolic extract from Schinus molle L. in mice: Evidence for the involvement of the serotonergic and noradrenergic systems. European Journal of Pharmacology, v. 587, n. 1-3, p. 163–168, jun. 2008.
MAEDA, K. et al. The effect of TNF-α converting enzyme inhibitors on cytokine response in acute pancreatitis. International Congress Series, v. 1255, p. 173–175, ago. 2003.
MALMSTRØM, M. L. et al. Cytokines and Organ Failure in Acute Pancreatitis: Inflammatory Response in Acute Pancreatitis. Pancreas, v. 41, n. 2, p. 271–277, mar. 2012.
MAO, P. et al. Mitochondria-targeted catalase reduces abnormal APP processing, amyloid production and BACE1 in a mouse model of Alzheimer’s disease: implications for neuroprotection and lifespan extension. Human Molecular Genetics, v. 21, n. 13, p. 2973–2990, 5 abr. 2012.
MCKAY, C. J.; BUTER, A. Natural history of organ failure in acute pancreatitis. Pancreatology: official journal of the International Association of Pancreatology (IAP) ... [et al.], v. 3, n. 2, p. 111–114, 2003.
MEIRELLES JR., R. F. et al. Reactive oxygen species inactivation improves pancreatic capillary blood flow in caerulein-induced pancreatitis in rats. Acta Cirurgica Brasileira, v. 18, p. 29–33, 2003.
MENA, S.; ORTEGA, A.; ESTRELA, J. M. Oxidative stress in environmental-induced carcinogenesis. Mutation Research/Genetic Toxicology and Environmental Mutagenesis, v. 674, n. 1-2, p. 36–44, mar. 2009.
MEOTTI, F. C. et al. Myricitrin as a substrate and inhibitor of myeloperoxidase: Implications for the pharmacological effects of flavonoids. Free Radical Biology and Medicine, v. 44, n. 1, p. 109–120, jan. 2008.
MERZA, M. et al. Inhibition of geranylgeranyltransferase attenuates neutrophil accumulation and tissue injury in severe acute pancreatitis. Journal of leukocyte biology, v. 94, n. 3, p. 493–502, set. 2013.
Metal chelation therapy, oxygen radicals, and human disease. Lancet, v. 1, n. 8421, p. 143–145, 19 jan. 1985.

70
MIYASHIRO, C. A. H. Avaliação da atividade antioxidante e antiinflamatória do flavonóide rutina e derivados contendo metal de transição. Dissertação—São Paulo: Universidade de São Paulo, 2010.
MIZUNUMA, T.; KAWAMURA, S.; KISHINO, Y. Effects of injecting excess arginine on rat pancreas. The Journal of nutrition, v. 114, n. 3, p. 467–471, mar. 1984.
MOHSENI SALEHI MONFARED, S.-S. Antioxidant therapy in the management of acute, chronic and post-ERCP pancreatitis: A systematic review. World Journal of Gastroenterology, v. 15, n. 36, p. 4481, 2009.
NEGRE-SALVAYRE, A. et al. Pathological aspects of lipid peroxidation. Free Radical Research, v. 44, n. 10, p. 1125–1171, out. 2010.
NIJVELDT, R. J. et al. Flavonoids: a review of probable mechanisms of action and potential applications. The American journal of clinical nutrition, v. 74, n. 4, p. 418–425, out. 2001.
NISHIMURA, S. et al. The proteinase/proteinase-activated receptor-2/transient receptor potential vanilloid-1 cascade impacts pancreatic pain in mice. Life Sciences, v. 87, n. 19-22, p. 643–650, nov. 2010.
OLIVEIRA, T. T. DE et al. Efeito de diferentes doses de flavonóides em ratos hiperlipidêmicos. Revista de Nutrição, v. 15, n. 1, p. 45–51, jan. 2002.
OLTHOF, M. R. et al. Bioavailabilities of quercetin-3-glucoside and quercetin-4′-glucoside do not differ in humans. The Journal of nutrition, v. 130, n. 5, p. 1200–1203, 2000.
OSMAN, M. O. et al. PROFILES OF PRO-INFLAMMATORY CYTOKINES IN THE SERUM OF RABBITS AFTER EXPERIMENTALLY INDUCED ACUTE PANCREATITIS. Cytokine, v. 17, n. 1, p. 53–59, jan. 2002.
PAGLIA, D. E.; VALENTINE, W. N. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. The Journal of laboratory and clinical medicine, v. 70, n. 1, p. 158–169, jul. 1967.
PANDURANGAN, A. K. Modulatory influence of Green tea polyphenols on lipid peroxidation during L-Arginine induced acute pancreatitis in Wistar rats. IOSR Journal of Environmental Science, Toxicology and Food Technology, v. 3, n. 2, p. 77–82, 2013.
PATIL, C. S. et al. Protective effect of flavonoids against aging- and lipopolysaccharide-induced cognitive impairment in mice. Pharmacology, v. 69, n. 2, p. 59–67, out. 2003.
POCH, B. et al. The role of polymorphonuclear leukocytes and oxygen-derived free radicals in experimental acute pancreatitis: mediators of local destruction and activators of inflammation. FEBS letters, v. 461, n. 3, p. 268–272, 19 nov. 1999.

71
PODREZ, E. A. et al. Myeloperoxidase-generated reactive nitrogen species convert LDL into an atherogenic form in vitro. Journal of Clinical Investigation, v. 103, n. 11, p. 1547–1560, 1 jun. 1999.
PRINCE, P. S. M.; KANNAN, N. K. Protective effect of rutin on lipids, lipoproteins, lipid metabolizing enzymes and glycoproteins in streptozotocin-induced diabetic rats. Journal of Pharmacy and Pharmacology, v. 58, n. 10, p. 1373–1383, out. 2006.
PUOLAKKAINEN, P. et al. C-reactive protein (CRP) and serum phospholipase A2 in the assessment of the severity of acute pancreatitis. Gut, v. 28, n. 6, p. 764–771, jun. 1987.
QUE, R. et al. Correlation of nitric oxide and other free radicals with the severity of acute pancreatitis and complicated systemic inflammatory response syndrome. Pancreas, v. 39, n. 4, p. 536–540, maio 2010.
RAKONCZAY, Z. NF-κB activation is detrimental in arginine-induced acute pancreatitis. Free Radical Biology and Medicine, v. 34, n. 6, p. 696–709, 15 mar. 2003.
RAKONCZAY, Z. et al. A new severe acute necrotizing pancreatitis model induced by l-ornithine in rats: Critical Care Medicine, v. 36, n. 7, p. 2117–2127, jul. 2008.
RASO, G. M. et al. Inhibition of inducible nitric oxide synthase and cyclooxygenase-2 expression by flavonoids in macrophage J774A.1. Life sciences, v. 68, n. 8, p. 921–931, 12 jan. 2001.
REIS, J. S. et al. Estresse oxidativo: revisão da sinalização metabólica no diabetes tipo 1. Arquivos Brasileiros de Endocrinologia & Metabologia, v. 52, n. 7, out. 2008.
ROMANO, M. et al. Role of IL-6 and its soluble receptor in induction of chemokines and leukocyte recruitment. Immunity, v. 6, n. 3, p. 315–325, mar. 1997.
ROTELLI, A. E. et al. Comparative study of flavonoids in experimental models of inflammation. Pharmacological research: the official journal of the Italian Pharmacological Society, v. 48, n. 6, p. 601–606, dez. 2003.
SÁNCHEZ-BERNAL, C. et al. Nitric oxide protects against pancreatic subcellular damage in acute pancreatitis. Pancreas, v. 28, n. 1, p. e9–e15, 2004.
SANDOVAL, D. et al. The role of neutrophils and platelet-activating factor in mediating experimental pancreatitis. Gastroenterology, v. 111, n. 4, p. 1081–1091, out. 1996.
SANTANA, D. G. et al. Beneficial effects of the ethanol extract of Caesalpinia pyramidalis on the inflammatory response and abdominal hyperalgesia in rats with acute pancreatitis. Journal of ethnopharmacology, v. 142, n. 2, p. 445–455, 13 jul. 2012.

72
SATHYANARAYAN, G. et al. Elevated level of interleukin-6 predicts organ failure and severe disease in patients with acute pancreatitis. Journal of Gastroenterology and Hepatology, v. 22, n. 4, p. 550–554, abr. 2007.
SCHÜTTE, K.; MALFERTHEINER, P. Markers for predicting severity and progression of acute pancreatitis. Best Practice & Research Clinical Gastroenterology, v. 22, n. 1, p. 75–90, fev. 2008.
SCHWARTZ, E. S. et al. Synergistic role of TRPV1 and TRPA1 in pancreatic pain and inflammation. Gastroenterology, v. 140, n. 4, p. 1283–1291.e1–2, abr. 2011.
SCHWARTZ, E. S. et al. TRPV1 and TRPA1 Antagonists Prevent the Transition of Acute to Chronic Inflammation and Pain in Chronic Pancreatitis. Journal of Neuroscience, v. 33, n. 13, p. 5603–5611, 27 mar. 2013.
SELLOUM, L. et al. Anti-inflammatory effect of rutin on rat paw oedema, and on neutrophils chemotaxis and degranulation. Experimental and toxicologic pathology: official journal of the Gesellschaft für Toxikologische Pathologie, v. 54, n. 4, p. 313–318, mar. 2003.
SHEN, S.-C. et al. In vitro and in vivo inhibitory activities of rutin, wogonin, and quercetin on lipopolysaccharide-induced nitric oxide and prostaglandin E2 production. European Journal of Pharmacology, v. 446, n. 1-3, p. 187–194, jun. 2002.
SOWJANYA, J.; SANDHYA, T.; VEERESH, B. Ameliorating Effect of Eugenol on L-arginine Induced Acute Pancreatitis and Associated Pulmonary Complications in Rats. Pharmacologia, v. 3, n. 12, p. 657–664, 1 dez. 2012.
STAHL, W.; SIES, H. Lycopene: a biologically important carotenoid for humans? Archives of biochemistry and biophysics, v. 336, n. 1, p. 1–9, 1 dez. 1996.
SUGIURA, H.; ICHINOSE, M. Nitrative stress in inflammatory lung diseases. Nitric Oxide, v. 25, n. 2, p. 138–144, ago. 2011.
SULTANA, R.; BUTTERFIELD, D. A. Slot-blot analysis of 3-nitrotyrosine-modified brain proteins. Methods in enzymology, v. 440, p. 309–316, 2008.
SZABOLCS, A. et al. Effect of melatonin on the severity of L-arginine-induced experimental acute pancreatitis in rats. World Journal of Gastroenterology, v. 12, n. 2, p. 251, 2006a.
SZABOLCS, A. et al. Beneficial effect of resveratrol on cholecystokinin-induced experimental pancreatitis. European Journal of Pharmacology, v. 532, n. 1-2, p. 187–193, fev. 2006b.
TAKÁCS, T. et al. Cytokine level changes in L-arginine-induced acute pancreatitis in rat. Acta physiologica Hungarica, v. 84, n. 2, p. 147–156, 1996.

73
TAKÁCS, T. et al. The role of nitric oxide in edema formation in L-arginine-induced acute pancreatitis. Pancreas, v. 25, n. 3, p. 277–282, 2002.
TANI, S. et al. New model of acute necrotizing pancreatitis induced by excessive doses of arginine in rats. Digestive diseases and sciences, v. 35, n. 3, p. 367–374, 1990.
TAUBER, A. I.; FAY, J. R.; MARLETTA, M. A. Flavonoid inhibition of the human neutrophil NADPH-oxidase. Biochemical Pharmacology, v. 33, n. 8, p. 1367–1369, abr. 1984.
TENNER, S. et al. American College of Gastroenterology guideline: management of acute pancreatitis. The American journal of gastroenterology, v. 108, n. 9, p. 1400–1415; 1416, set. 2013.
TONSI, A.-F. Acute pancreatitis at the beginning of the 21st century: The state of the art. World Journal of Gastroenterology, v. 15, n. 24, p. 2945, 2009.
UKEDA, H. et al. Spectrophotometric assay for superoxide dismutase based on tetrazolium salt 3’--1--(phenylamino)-carbonyl--3, 4-tetrazolium]-bis(4-methoxy-6-nitro)benzenesulfonic acid hydrate reduction by xanthine-xanthine oxidase. Analytical biochemistry, v. 251, n. 2, p. 206–209, 5 set. 1997.
VALKO, M. et al. Free radicals and antioxidants in normal physiological functions and human disease. The International Journal of Biochemistry & Cell Biology, v. 39, n. 1, p. 44–84, jan. 2007.
VERA-PORTOCARRERO, L. P.; LU, Y.; WESTLUND, K. N. Nociception in persistent pancreatitis in rats: effects of morphine and neuropeptide alterations. Anesthesiology, v. 98, n. 2, p. 474–484, fev. 2003.
VLASOVA, I. I.; SOKOLOV, A. V.; ARNHOLD, J. The free amino acid tyrosine enhances the chlorinating activity of human myeloperoxidase. Journal of Inorganic Biochemistry, v. 106, n. 1, p. 76–83, jan. 2012.
WICK, E. C. Transient receptor potential vanilloid 1, calcitonin gene-related peptide, and substance P mediate nociception in acute pancreatitis. AJP: Gastrointestinal and Liver Physiology, v. 290, n. 5, p. G959–G969, 8 dez. 2005.
WICK, E. C. et al. Calcitonin gene-related peptide partially mediates nociception in acute experimental pancreatitis. Surgery, v. 139, n. 2, p. 197–201, fev. 2006.
WINSTON, J. H. et al. Acute pancreatitis results in referred mechanical hypersensitivity and neuropeptide up-regulation that can be suppressed by the protein kinase inhibitor k252a. The Journal of Pain, v. 4, n. 6, p. 329–337, ago. 2003.
WRIGHT, B. et al. Flavonoid inhibitory pharmacodynamics on platelet function in physiological environments. Food & Function, v. 4, n. 12, p. 1803, 2013.

74
YADAV, D.; LOWENFELS, A. B. The Epidemiology of Pancreatitis and Pancreatic Cancer. Gastroenterology, v. 144, n. 6, p. 1252–1261, maio 2013.
YAMAMOTO, K.; ROSE-JOHN, S. Therapeutic blockade of interleukin-6 in chronic inflammatory disease. Clinical pharmacology and therapeutics, v. 91, n. 4, p. 574–576, abr. 2012.
YILDIZ, B. D. Basic Experimental Pancreatitis Models for Beginners. Surgical Science, v. 01, n. 02, p. 31–39, 2010.
ZHANG, H. et al. IL-6 trans-signaling promotes pancreatitis-associated lung injury and lethality. Journal of Clinical Investigation, v. 123, n. 3, p. 1019–1031, 15 fev. 2013.
ZHANG, R. Myeloperoxidase Functions as a Major Enzymatic Catalyst for Initiation of Lipid Peroxidation at Sites of Inflammation. Journal of Biological Chemistry, v. 277, n. 48, p. 46116–46122, 30 set. 2002.
ZHANG, X.-P. et al. Effects of emodin and baicalein on rats with severe acute pancreatitis. World journal of gastroenterology: WJG, v. 11, n. 14, p. 2095–2100, 14 abr. 2005.

75
ANEXO 1





























