Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DO PARÁ
INSTITUTO DE ESTUDOS COSTEIROS
FACULDADE DE CIÊNCIAS BIOLÓGICAS
SUZANE SERRANO CERVEIRA
ANÁLISE MORFOMÉTRICA DA POPULAÇÃO DE MORCEGO HEMATÓFAGO
Desmodus rotundus (CHIROPTERA, PHYLLOSTOMIDAE), PROVENIENTES DOS
ESTADOS DO PARÁ E SÃO PAULO
BRAGANÇA
2011

SUZANE SERRANO CERVEIRA
ANÁLISE MORFOMÉTRICA DA POPULAÇÃO DE MORCEGO HEMATÓFAGO
Desmodus rotundus (CHIROPTERA, PHYLLOSTOMIDAE), PROVENIENTES DOS
ESTADOS DO PARÁ E SÃO PAULO
Trabalho de Conclusão de Curso
apresentado à Faculdade de Ciências
Biológicas, Instituto de Estudos Costeiros,
Campus Universitário de Bragança,
Universidade Federal do Pará, como
requisito parcial para obtenção do grau de
Licenciado Pleno em Ciências Biológicas.
Orientador: Prof. Dr. Marcus Emanuel
Barroncas Fernandes
BRAGANÇA – PARÁ
2011

SUZANE SERRANO CERVEIRA
ANÁLISE MORFOMÉTRICA DA POPULAÇÃO DE MORCEGO HEMATÓFAGO
Desmodus rotundus (CHIROPTERA, PHYLLOSTOMIDAE), PROVENIENTES DOS
ESTADOS DO PARÁ E SÃO PAULO
Banca Examinadora
__________________________________ Prof. Dr. Marcus Emanuel Barroncas Fernandes (Orientador)
Universidade Federal do Pará, Campus de Bragança - UFPA
__________________________________ Profa. Dra. Claudia Nunes-Santos
Universidade Federal do Pará, Campus de Bragança - UFPA
___________________________________ Profa. Dra. Janice Muriel Cunha
Universidade Federal do Pará, Campus de Bragança - UFPA
BRAGANÇA
2011

i
DEDICATÓRIA
Dedico este trabalho aos meus pais Laércio Cerveira Filho e Darcy Serrano Cerveira,
pois sem eles não estaria onde estou, à minha irmã Fabiane Serrano Cerveira, com quem moro
em Bragança e ao meu querido sobrinho que está quase nascendo Davi Cerveira.
Aos meus grandes amigos da graduação Edna Lima que sempre esteve ao meu lado
nos quatro anos de curso, ao Paulo Rodolfo pelas belas risadas e apoio sempre, Mayara Costa
uma surpresa agradável no ultimo ano do curso. Ao Bruno Andrade e Pedro Henrique que me
proporcionaram grandes e belos momentos e Allan Rodrigo com sua companhia agradável
sempre.
A ítala Maria, Karen Dayanne, Luciane e Larissa. As meninas super alegres Ádila,
Marília e Anete, ao meu querido Jackson que juntos viajamos. Ao seu Rubens e Dona Ângela
que me acolheram e sempre me trataram como uma filha, enfim dedico a todos da Biologia
2008. Aos amigos do LAMA que me fizeram companhia durante esses meses de estágio,
principalmente aos queridos Cleidson e Lanna.
Não esquecendo os meus grandes amigos Fernando (papito), Danielle, Adriane Maria,
Carol, Thayanne, Carla Solange, wenderson, Muller e Tadeu que sempre me deram força para
continuar nessa batalha.

ii
AGRADECIMENTOS
Agradeço ao CNPq e a Universidade Federal do Pará pelo apoio financeiro através da
bolsa de iniciação cientifica.
Ao meu orientador Prof. Dr. Marcus Emanuel Barroncas Fernandes, cujo empenho,
apoio e auxilio foram de inestimável valor durante a realização deste trabalho.
A Profa. Dra. Fernanda Atanaena Gonçalves de Andrade pelos conselhos e
ensinamentos, profissionais e pessoais, que cada dia me fortaleceu mais.
Ao Prof. Dr. Wilson Uieda por fornecer a possibilidade de fazer o treinamento de
morcegos e pela doação dos morcegos oriundos de São Paulo para a realização deste trabalho.
Aos meus grandes amigos que ajudaram na mensuração dos morcegos Danielle
Nascimento de Castro e Fernando Augusto Bessa Campos.

iii
“Alcançou o sucesso aquele que viveu bem, riu com freqüência e amou muito”
Autor Desconhecido

iv
RESUMO
Desmodus rotundus, o vampiro comum da América, é o morcego cujas análises
morfométricas têm levado a diversas reavaliações, que defendem a existência de subespécies.
Sendo que, para alguns autores, essas espécies não existiriam. Tais variações são,
provavelmente, por conseqüência da grande variação individual e análises com combinação
sexual, bem como de certa variação geográfica, e talvez até mesmo da existência de ecótipos.
Assim, o presente estudo investigou 39 marcadores, quanto à sua influencia em algum dos
eventos acima destacados. Com esse propósito foram obtidas medidas de caracteres externos
(n=16) e cranianos (n=22) em 243 espécimes de D. rotundus divididos em três grupos:
sudeste do Pará, outras mesorregiões do Estado (Nordeste, Sudoeste, metropolitana e Marajó)
e duas localidades do estado de São Paulo. Do total de amostras foram removidos os outliers,
resultando em um menor número de indivíduos machos (n=111), e fêmeas (n=59). A análise
de ordenação fatorial resultou em um total de 11 caracteres com cargas representativas acima
de 0,70, dos quais somente um foi comum para ambos os sexos. No geral a análise
discriminante entre as variáveis foi significativa tanto para os machos [λ=0,75, F
(28,19)=17.961, p<0,001], quanto para as fêmeas [λ=1,38, F (22,90)=6,89, p<0,001]. As
funções discriminantes revelaram boa precisão preditiva geral (razão de sucesso) para machos
(91%) e fêmeas (88%), separando os três grupos pré-estabelecidos na análise, onde São Paulo
separa facilmente das regiões do Pará. Os resultados do presente estudo mostram que há um
grande viés em utilizar no mesmo grupo indivíduos de sexos diferentes no intuito de
discriminar populações diferentes, como é o caso das duas regiões do Pará e São Paulo, além
de corroborar com estudos anteriores, cujos resultados já apontavam uma grande variabilidade
intraespecíficas nessa espécie de morcego vampiro.
Palavras-chave: Morcegos, Morfometria, Pará, São Paulo.

v
LISTA DE FIGURAS
Figura 1 - Espécies de morcegos hematófagos. (A) Diphylla ecaudata, (B) Diaemus youngi e
(C) Desmodus rotundus .............................................................................................................1
Figura 2 – Distribuição geográfica do morcego Desmodus rotundus.......................................2
Figura 3 – Medidas cranianas realizadas em espécimes de Desmodus rotundus provenientes
do Pará e São Paulo................................................................................................................... 6
Figura 4 - Medidas externas realizadas em espécimes de Desmodus rotundus provenientes do
Pará e São Paulo ........................................................................................................................7
Figura 5 – Escores da função discriminante em fêmeas (A) e machos (B) de Desmodus
rotundus....................................................................................................................................12

vi
LISTA DE TABELAS
Tabela 1- Número de espécimes em ambos os sexos detinados as análises...............................8
Tabela 2- Matriz de análise fatorial com conjunto reduzido de variáveis dos componentes da
última rotação com VARIMAX normalizado............................................................................9
Tabela 3: Sumário da análise de função discriminante de 19 variáveis craniométricas e
externas de fêmeas e machos de Desmodus rotundus...............................................................10
Tabela 4: Coeficiente padronizado para variáveis canônicas dos caracteres avaliados que mais
contribuíram para a função discriminante em fêmeas e machos...............................................11
Tabela 5: Média das variáveis canônicas em machos e fêmeas de Desmodus rotundus,
representando as segregações entre grupos...............................................................................11
Tabela 6: Matriz de distâncias de Mahalanobis para morcego vampiro comum, sem
combinação das categorias sexuais...........................................................................................13

1
1.INTRODUÇÃO
Na ordem Chiroptera, a família Phyllostomidae, endêmica no continente
americano, apresenta o segundo maior numero espécies de morcegos no mundo,
enquanto que na região neotropical é a primeira em diversidade, com cerca de 160
espécies reconhecidas em 57 gêneros (REISet al., 2011). Os filostomídeos perfazem
uma variedade de hábitos alimentares que incluem como itens: frutos, néctar, polén,
folhas, insetos, vertebrados e sangue (REIS et al., 2007). Dessa maneira, os morcegos
influenciam diretamente na estruturae manutenção dos ecossistemas em que habitam
(WHITTAKER&JONES, 1994).
Ahematofagia em morcegos gerou uma simbologia de animal sugador, que
contribuiu para que todos os táxons fossem vistos como ofensivos. Mas no geral,
existem apenas três gêneros e três espécies de hematófagos (Figura 1), que estão
reunidas em uma subfamília, Desmodontinae, sendo essas:
Desmodusrotundus(Geoffroy, 1810), Diphyllaecaudataspix, 1823 e
Diaemusyoungi(Jentink, 1893). Dentre essas apenas D. rotundus, conhecido como
“vampiro comum”, alimenta-se, preferencialmente, de sangue de mamíferos de grande
porte, e eventualmente de aves. Já a demais espécie tem como fonte alimentar
preferencial sangue de aves, enquanto, mamíferos, seria uma fonte eventual (BRASS,
1994).
Figura 1 – Espécies de morcegos hematófagos. (A) Diphyllaecaudata, (B)
Diaemusyoungi e (C) Desmodusrotundus. Fotos: W. Uieda.
A espécie D. rotunduspossui uma ampla distribuição geográfica no continente
americano (Figura 2), abrangendo desde o norte do México até o norte da Argentina
(KOOPMAN, 1988; EMMONS, 1990). Morfologicamente, os adultos apresentam
C B A

2
tamanho mediano, com cabeça atingindo entre 69 a 90 mm, o antebraço pode ter
medidas entre 50 a 63 mm, e o peso pode alcançar de 25 a 40 g. A pelagem geralmente
apresenta coloração cinza-brilhante, mas já foram capturados indivíduos com variações
na cor entre o avermelhado, dourado e até mesmo alaranjado (BERNARD, 2005).
Outros caracteres incluem apresença de folha nasal reduzida, focinho curto, membrana
interfemural pequena, ausência de cauda e calcâneo, e principalmente, presença de
polegares compridos com três calosidades (GREENHALL et al., 1983; NOWAK,
1994). A fórmula dentária é I 1/2, C 1/1, PM 1/2 e M 1/1 (REISet al., 2011).
Figura 2. Distribuição geográfica do
morcego hematófago Desmodusrotundus.
Fonte:http://www.iucnredlist.org/apps/redli
st/search
Quanto ao seu hábitat, é preferencialmente em áreas silvestres, D. rotundus é
uma espécie rara, que pode habitar ocos de árvores, covas, cavernas, dentre outras
possibilidades capazes de suportar em média 30 indivíduos (UIEDAet al., 1996).
Contudo, já foram registradas em cavernas, colônias com mais 300 indivíduos
(SAZIMA, 1978; TADDEI et al., 1991 ; BREDT et al., 1998), sendo que abrigos com
mais de 100 espécimes são considerados atípicos por UIEDAet al. (1996). Para alguns
autores, a redução dos abrigos naturais ao vampiro comum parece ter favorecido sua
adaptação a novos refúgios artificiais, bem como o acesso a exposição de presas mais
passivas (GREENHALL & SCHMIDH, 1998).
Em regiões como no sul do Brasil, onde os rebanhos de gado são fartos, D.
rotundus tem se tornado bastante comum (TADDEI et al., 1991; UIEDA, 1996),

3
provavelmente, também em virtude da elevação de suas taxas de sobrevivência, devido
a fácil acessibilidade ao alimento e à disponibilidade de novos abrigos para a formação
de colônias.
Na Amazônia no ano de 1998, os morcegos ocupavam o segundo lugar como
transmissores da raiva em humanos, sendo superados apenas pelos cães. Em 2004, o
Ministério da Saúde notificou o registro de dois focos de raiva humana transmitidas por
morcegos na região (MINISTÉRIO DA SAÚDE 2005). Neste período foram totalizados
mais de 30 casos de raiva causados pelo morcegos, proporcionando uma mudança na
situação dos transmissores da raiva humana no Brasil, onde o principal agente passa a
ser o morcego. Portanto,atualmente D. rotundus é assunto de constantes discussões
referentes à vigilância em saúde e economia local, uma vez que também tem sido
relacionado a grandes prejuízos causados a pecuária (MÁLAGA- ALBA, 1954; ACHA
& MÁLAGA-ALBA, 1988; BRASS, 1994; GREENHAL & SCHMIDT, 1998).
Não somente os fatos observados no Brasil, mas também em outras áreas das
Américas, tem culminado em várias investigações sobre D. rotundus (BRASS, 1994;
FLORES-CRESPO & ARELLANO-SOTA, 1988; UIEDA, 1996; SCHNEIDER et al.,
2009). Contudo alguns estudos, ainda apresentam certas divergências.
Taxonomicamente são conhecidas mais três sinonímias para o gênero Desmodus e oito
para a espécie, (GREENHALL et al., 1988; WILSON & REEDER, 2005).
Entretanto, segundo KOOPMAN (1988), desde 1938 todos os nomes propostos
passaram a ser tratados como sinônimos de D. rotundus, com exceção de dois: D.
murinus (México) e D. dorbignyi (Chile). Para OSGOOD (1943),murinusseria na
verdade uma subespécie de menor tamanho, enquanto dorbignyiseria maior e de
coloração conspícua. KOOPMAN (1988) também inferiu que na verdade não existiriam
subespécies para D. rotundus, sobretudo, devido à ausência de bases científicas
significativas que validassem o acontecimento. Para o autor, muitas validações em nível
de subespécies seriam conseqüências da grande variação individual, bem como de certa
variação geográfica. MANN & AULAGNIER (1993), também citaram como indutores
de registro de subespécies para D. rotundus, a ampla variação individual (mas diante da
baixa amostragem), bem como as análises que desconsiderem o dimorfismo sexual.
Recentemente, MARTINS (2008), retomou a discussão sobre as subespécies
inferindo a respeito de prováveis padrões morfológicos de segregação filogeográfica.

4
Estes padrões de morfometria craniana e dos pêlos, analisados com funções
discriminantes e variáveis canônicas, propõem que se reconheçam duas linhagens
distintas para D. rotundus. Neste caso, uma estaria a leste na Mata Atlântica, com o
restabelecimento D. r.rotundus, e outra a oeste do Brasil (incluindo a Amazônia),
identificada como D. r. murinus. Ainda para o autor, esta última subespécie teria mais
viabilidade do que D. r.rotundus, para a caracterização em termos de morfometria
craniana, devido a apresentar maior estabilidade fenotípica. Até o momento, o trabalho
de MARTINS (2008) foi o único no Brasil a inferir sobre tais variações diferentes
biomas. No entanto, o autor cita que ainda é preciso uma amostragem mais cuidadosa
das regiões incluindo a Amazônia, cuja quantidade de espécimes foi baixa (n=22), além
do fato de que as análises realizadas foram com os sexos combinadas. Visto isso, o
presente trabalho pretende realizar uma caracterização morfológica, do tipo craniano e
corpóreo, para testar a viabilidade quanto à segregação entre e dentre os indivíduos dos
grupos da espécie D. rotundusde áreas do Estado do Pará e de São Paulo.
2. OBJETIVOS
2.1 Objetivo geral
Investigar as variações dos caracteres cranianos e corpóreos de espécimes de D.
rotundus oriundos dos estados do Pará e São Paulo.
2.2 Objetivos específicos
Verificar se os indivíduos oriundos da mesorregião Sudeste formam um grupo
separado das outras mesorregiões do Estado do Pará.
No caso da separação dos espécimes do Estado do Pará em dois grupos, verificar
se estes também são coletivamente distintos morfologicamente dos indivíduos
provenientes do Estado de São Paulo.

5
3.Material e Métodos
Foram examinados 243 indivíduos de D. rotundus, 158 machos e 85 fêmeas,
oriundas de 14 localidades do Pará e 2 de São Paulo.ANDRADE (2011) realizou o
trabalho com morcegos hematófagos de cinco mesorregiões do estado do Pará e
observou uma pequena diferença entre os morcegos da mesorregião sudeste com outras
mesorregiões deste estado. Este trabalho usou como referencia Andrade para separar os
grupos do Pará. Do total das amostras, cerca de 50% foram cedidas pela Coleção
Científica do Departamento de Zoologia, UNESP- Botucatu, 15% são provenientes do
Laboratório de Diagnóstico de Raiva, da Seção de Arbovirologia e Febres Hemorrágicas
(SAARB) do Instituto Evandro Chagas (IEC) em Belém- Pará. As demais amostras
foram cedidas pela Coleção Científica do Departamento de Zoologia, UNESP-
Botucatu, coletados entre 1999 e 2000 e outras espécimes desse morcego foram
adquiridos através de coletas de campo pela Fernanda Andrade.
Para cada exemplar, foi verificado o município, data da coleta, sexo e classe
etária (adulto). Todos os espécimes incluídos na análise apresentam características
diagnósticas de adultos, tais como epífises dos antebraços completamente ossificadas e
articulações metacarpo- falangeanas proeminentes. A retirada do crânio foi realizada
através da abertura bucal com o rebatimento da pele e limpeza através do uso de larvas e
adultos de insetos do gênero Dermestes (Coleoptera, Dermestidae), no criadouro de
dermestáriodo Laboratório de Mastozoologia (UFPA/ Bragança). Depois de limpos os
crânios foram clareados com água oxigenada Volume dez, registrados e acondicionados
em frascos identificados.
Foram obtidas 16 medidas corpóreas e 22 craniométricas de todos os exemplares
(figuras 3 e 4), sendo estas adaptadas de VIZZOTO & TADDEI (1973) e MARTINS
(2008). Comprimento total do crânio (Ct), Comprimento do crânio até os incisivos
(CInc), Comprimento da série de dentes superiores (CM2/3S), Comprimento côndilo-
canino (Cc),Largura zigomática (Lz), Largura externa dos molares (Lm), Largura
externa dos cingula-caninos (Lc), Largura palatal (Lpt), Largura bula timpânica na
porção mais larga (LBTil), Largura bula timpânica na porção mais próxima (LBTpp),
Largura posorbitária (Lp), Largura interorbital (Li), Largura mastóidea (Lmt),
Comprimento do canino ao processo mastóideo oposto (CaPMO), Comprimento do
nasal ao forâmen (CNaFo), Comprimento palatal (Cpt), Comprimento basal (B), Altura

6
da caixa craniana (Acx), Altura do occipital (Ao), Comprimento da série de dentes
inferiores(CM2/3I), Comprimento da mandíbula (Cm), Altura da base do crânio
a crista sagital (BcCs), Antebraço esquerdo (AtE),Antebraço direito (AtD), Polegar
(Po), III metacarpo (IIIMe), 1ª falange do terceiro metacarpo (1F-III), 2ª falange do
terceiro metacarpo (2F-III), IV metacarpo (IVMe) 1ª falange do quarto metacarpo (1F-
IV), 2ª falange do quarto metacarpo (2FIV), V metacarpo (VMe), 1ª falange do quinto
metacarpo (1FV), 2ª falange do quinto metacarpo (2F-V), Tíbia (Ti), Uropatágio altura
(Ua), Uropatágio comprimento (Uc) e Pé (P).
Figura 3: Medidas cranianas realizadas em espécimes de D. rotundus provenientes do Pará e São
Paulo, Brasil.

7
3.1Análise dos dados
Tendo em vista a presença de dimorfismo sexual, as análises foram efetuadas com
sexos separados, o que causou a diminuição do número amostral de cada grupo geográfico.
Para cada medida nos indivíduos de ambos os sexos, foram calculadas a média, desvio padrão
e erro padrão. Após tais cálculos, medidas com valores fora do limite padrão foram
removidas, por apresentarem outliers e indivíduos de medidas discrepantes.
Inicialmente foram realizadas análises multivariadas para uma matriz com dados
originais das três populações de D. rotundus. Em seguida, uma analise fatorial de ordenação
exploratória foi aplicada no intuito de investigar a existência de padrões entre as medidas, já
que a análise fatorial é uma técnica de interdependência que fornece as ferramentas para
analisar as estruturas das correlações em um grande número de variáveis definindo conjunto
de variáveis que são fortemente inter-relacionadas. Neste caso é empregado o método de
rotação ortogonal Varimax normalizado, pois maximiza a soma de variâncias de cargas
exigidas na matriz fatorial, simplificando as colunas da mesma.
Logo, depois de identificadas as variáveis que fazem a diferença para machos e
fêmeas de D. rotundus, estas foram avaliadas quanto à hipótese da existência de diferenças
significativas entre os valores amostrais. Para tanto foi realizada Análise Canônica
Discriminante, baseada na Matriz de DistânciaMahalanobis (MD).
Figura 4: Medidas externas realizadas em espécimes de D.rotundus provenientes dos estados do Pará
e São Paulo, Brasil.

8
As análises univariadas e multivariadas, utilizadas no presente trabalho, foram
executadas utilizando-se os seguintes programas: BioEstat 5.0 e Statistic 5.0.
4.RESULTADOS
Um total de 170 indivíduos de D. rotundus foi analisado, sendo 111 Machos e 59
fêmeas(Tabela 1). Dos três grupos a serem testados, as fêmeas apresentaram as médias mais
altas em 36 de 38 mensurações cranianas e externas. O mesmo pode ser afirmado quando se
compara os grupo pois os espécimes de São Paulo apresentaram as maiores médias do que as
amostras do estado do Pará , observadas em ambos os sexos. Quanto às avaliações dentro dos
grupos, estas foram realizadas apenas para os exemplares do Pará( incluindo o Sudoeste),
únicos a possuir representatividade geográfica (ANDRADE 2011). Neste caso, os indivíduos
que apresentaram os menores valores medidos pertenciam à mesorregião Sudeste, sendo estes
seguidos pelos dados das demais amostras de hematófagos do Pará.
Tabela 1: Número de espécimes em ambos os sexos destinados as análises
Grupos
Machos Fêmeas Total
Sudeste doPará 16 18 34
Pará
44 12 56
São Paulo
51 29 80
Total
111 59 170
No presente trabalho, a análise fatorialmostrou que para as fêmeas 11 medidas
explicaram cerca de 80% de toda a variação encontrada, sendo as variáveis CT, CB, B, CPT,
CC+Inc, CC+PMO e CNaFo foram as mais importantes (FATOR-1) (Tabela 2). Para os
machos foi observado que 14 medidas representam também cerca de 80% da variação
encontrada nos indivíduos analisados (Tabela 2). As variáveis que se destacaram nessa análise
foram utilizadas na análise discriminante, no intuito de verificar os três grupos de D.
rotundus. Dentre estas variáveis nos machos e fêmeas 5 delas são comuns para ambos os
sexos CT, CB,B, CC+Inc e CNaFo.

9
Tabela 2: Matriz de análise fatorial com conjunto reduzido de variáveis dos componentes da última rotação
com VARIMAX normalizado.
Caractere Fêmeas Machos
Fator 1 Fator 2 Fator 3 h² Fator 1 Fator 2 Fator 3 h²
CT 0,89 - - 0,81 0,88 - - 0,84
CB 0,88 - - 0,82 0,88 - - 0,81
CC - - - - 0,88 - - 0,79
B 0,91 - - 0,88 0,88 - - 0,86
CPT 0,89 - - 0,81 - - - -
C-M2/3S - - 0,86 0,35 - - - -
CM - - - - 0,87 - - 0,83
LC - - 0,84 0,37 - - - -
Acx - - - - - - 0,85 0,24
Ao - - - - - - 0,77 0,38
CC+Inc 0,92 - - 0,84 0,88 - - 0,90
Cca+ PMO 0,90 - - 0,85 - - - -
CNaFo 0,88 - - 0,77 0,88 - - 0,85
1ºFal. III - - - - - 0,84 - 0,52
1º Fal. IV - 0,86 - 0,27 - 0,83 - 0,51
V Met - - - - 0,73 - - 0,78
1º Fal. V - 0,83 - 0,29 - - - -
Ti - - - - - 0,74 - 0,60
Ur.Al - - - - - - 0,77 0,13
Eigenvalue 6,15 1,70 1,05 7,93 1,45 1,27
% Total Variance 55,91 15,46 9,56 56,97 13,70 11,32
Cumulative % 55,91 71,37 80,94 56,94 69,64 80,05
4.1Análise discriminante
A presente análise revelou pelo método stepwiseas médias das variáveis que foram
significativamente discriminantes entre si, tanto para as fêmeas (λ de Wilks=1,38; F
(22,90)=6,89, p<0,001) quanto para os machos (λ de Wilks=0,75; F (28,190)=17,96;
p<0,001). Na Tabela 3 estão expostos os valores de λ parcial referentes à contribuição
específica de cada variável para a discriminação entre os grupos, onde as variáveis de menor
valor são aquelas com maior poder discriminante. Sendo assim, os caracteres que
demonstraram valores de λ parcial, cujas distribuições foram razoavelmente mais expressivas
para a distinção entre grupos, foram os seguintes: comprimento Basal (CB), Comprimento
côndilo-canino (CC), Altura da caixa craniana (Acx), Comprimento do nasal ao forâmen
(CNaFo), V metacarpo (VMet) e Uropatágio altura (UrAl) para os machos, já para as fêmeas
o caráter de maior poder discriminante é o Comprimento Basal (CB).
10
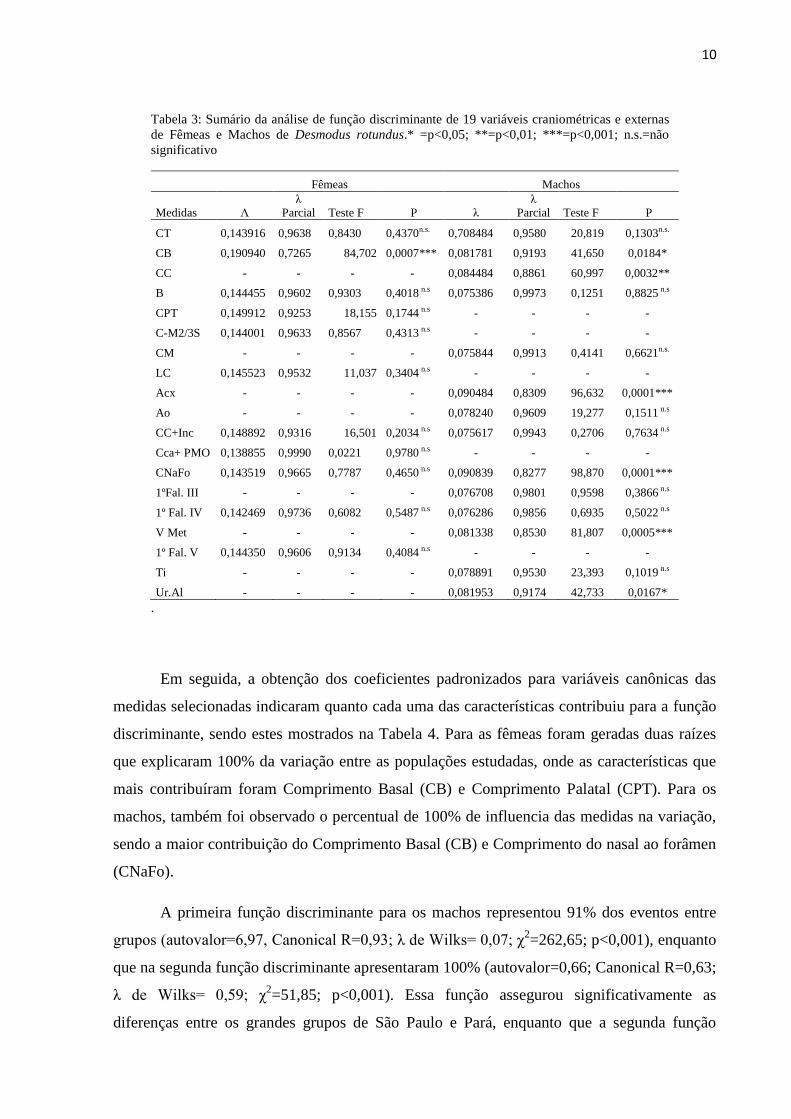
Tabela 3: Sumário da análise de função discriminante de 19 variáveis craniométricas e externas
de Fêmeas e Machos de Desmodus rotundus.* =p<0,05; **=p<0,01; ***=p<0,001; n.s.=não
significativo
Fêmeas Machos
Medidas Λ
λ
Parcial Teste F P λ
λ
Parcial Teste F P
CT 0,143916 0,9638 0,8430 0,4370n.s. 0,708484 0,9580 20,819 0,1303n.s.
CB 0,190940 0,7265 84,702 0,0007*** 0,081781 0,9193 41,650 0,0184*
CC - - - - 0,084484 0,8861 60,997 0,0032**
B 0,144455 0,9602 0,9303 0,4018 n.s 0,075386 0,9973 0,1251 0,8825 n.s
CPT 0,149912 0,9253 18,155 0,1744 n.s - - - -
C-M2/3S 0,144001 0,9633 0,8567 0,4313 n.s - - - -
CM - - - - 0,075844 0,9913 0,4141 0,6621n.s.
LC 0,145523 0,9532 11,037 0,3404 n.s - - - -
Acx - - - - 0,090484 0,8309 96,632 0,0001***
Ao - - - - 0,078240 0,9609 19,277 0,1511 n.s
CC+Inc 0,148892 0,9316 16,501 0,2034 n.s 0,075617 0,9943 0,2706 0,7634 n.s
Cca+ PMO 0,138855 0,9990 0,0221 0,9780 n.s - - - -
CNaFo 0,143519 0,9665 0,7787 0,4650 n.s 0,090839 0,8277 98,870 0,0001***
1ºFal. III - - - - 0,076708 0,9801 0,9598 0,3866 n.s
1º Fal. IV 0,142469 0,9736 0,6082 0,5487 n.s 0,076286 0,9856 0,6935 0,5022 n.s
V Met - - - - 0,081338 0,8530 81,807 0,0005***
1º Fal. V 0,144350 0,9606 0,9134 0,4084 n.s - - - -
Ti - - - - 0,078891 0,9530 23,393 0,1019 n.s
Ur.Al - - - - 0,081953 0,9174 42,733 0,0167*
.
Em seguida, a obtenção dos coeficientes padronizados para variáveis canônicas das
medidas selecionadas indicaram quanto cada uma das características contribuiu para a função
discriminante, sendo estes mostrados na Tabela 4. Para as fêmeas foram geradas duas raízes
que explicaram 100% da variação entre as populações estudadas, onde as características que
mais contribuíram foram Comprimento Basal (CB) e Comprimento Palatal (CPT). Para os
machos, também foi observado o percentual de 100% de influencia das medidas na variação,
sendo a maior contribuição do Comprimento Basal (CB) e Comprimento do nasal ao forâmen
(CNaFo).
A primeira função discriminante para os machos representou 91% dos eventos entre
grupos (autovalor=6,97, Canonical R=0,93; λ de Wilks= 0,07; χ2=262,65; p<0,001), enquanto
que na segunda função discriminante apresentaram 100% (autovalor=0,66; Canonical R=0,63;
λ de Wilks= 0,59; χ2=51,85; p<0,001). Essa função assegurou significativamente as
diferenças entre os grandes grupos de São Paulo e Pará, enquanto que a segunda função
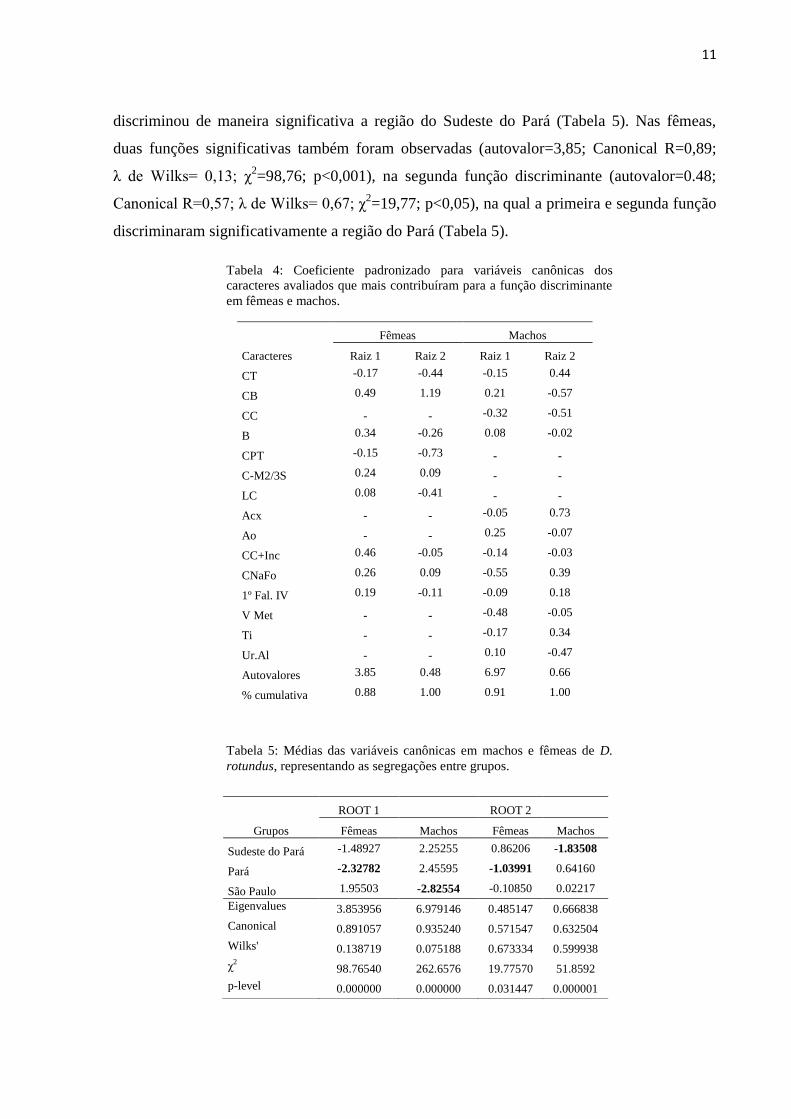
11
discriminou de maneira significativa a região do Sudeste do Pará (Tabela 5). Nas fêmeas,
duas funções significativas também foram observadas (autovalor=3,85; Canonical R=0,89;
λ de Wilks= 0,13; χ2=98,76; p<0,001), na segunda função discriminante (autovalor=0.48;
Canonical R=0,57; λ de Wilks= 0,67; χ2=19,77; p<0,05), na qual a primeira e segunda função
discriminaram significativamente a região do Pará (Tabela 5).
Tabela 4: Coeficiente padronizado para variáveis canônicas dos
caracteres avaliados que mais contribuíram para a função discriminante
em fêmeas e machos.
Fêmeas Machos
Caracteres Raiz 1 Raiz 2 Raiz 1 Raiz 2
CT -0.17 -0.44 -0.15 0.44
CB 0.49 1.19 0.21 -0.57
CC - - -0.32 -0.51
B 0.34 -0.26 0.08 -0.02
CPT -0.15 -0.73 - -
C-M2/3S 0.24 0.09 - -
LC 0.08 -0.41 - -
Acx - - -0.05 0.73
Ao - - 0.25 -0.07
CC+Inc 0.46 -0.05 -0.14 -0.03
CNaFo 0.26 0.09 -0.55 0.39
1º Fal. IV 0.19 -0.11 -0.09 0.18
V Met - - -0.48 -0.05
Ti - - -0.17 0.34
Ur.Al - - 0.10 -0.47
Autovalores 3.85 0.48 6.97 0.66
% cumulativa 0.88 1.00 0.91 1.00
Tabela 5: Médias das variáveis canônicas em machos e fêmeas de D.
rotundus, representando as segregações entre grupos.
ROOT 1
ROOT 2
Grupos Fêmeas Machos Fêmeas Machos
Sudeste do Pará -1.48927 2.25255 0.86206 -1.83508
Pará -2.32782 2.45595 -1.03991 0.64160
São Paulo 1.95503 -2.82554 -0.10850 0.02217
Eigenvalues 3.853956 6.979146 0.485147 0.666838
Canonical 0.891057 0.935240 0.571547 0.632504
Wilks' 0.138719 0.075188 0.673334 0.599938
χ2 98.76540 262.6576 19.77570 51.8592
p-level 0.000000 0.000000 0.031447 0.000001
12
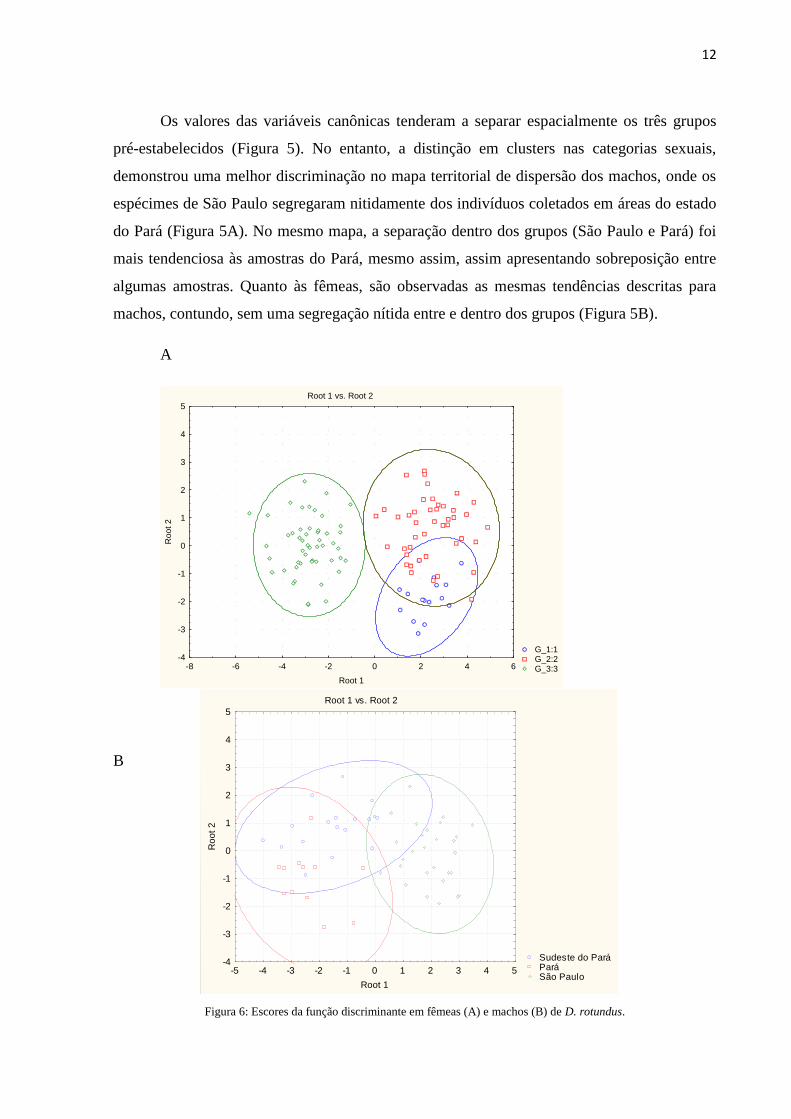
Os valores das variáveis canônicas tenderam a separar espacialmente os três grupos
pré-estabelecidos (Figura 5). No entanto, a distinção em clusters nas categorias sexuais,
demonstrou uma melhor discriminação no mapa territorial de dispersão dos machos, onde os
espécimes de São Paulo segregaram nitidamente dos indivíduos coletados em áreas do estado
do Pará (Figura 5A). No mesmo mapa, a separação dentro dos grupos (São Paulo e Pará) foi
mais tendenciosa às amostras do Pará, mesmo assim, assim apresentando sobreposição entre
algumas amostras. Quanto às fêmeas, são observadas as mesmas tendências descritas para
machos, contundo, sem uma segregação nítida entre e dentro dos grupos (Figura 5B).
A
Root 1 vs. Root 2
G_1:1 G_2:2 G_3:3-8 -6 -4 -2 0 2 4 6
Root 1
-4
-3
-2
-1
0
1
2
3
4
5
Roo
t 2
B
Figura 6: Escores da função discriminante em fêmeas (A) e machos (B) de D. rotundus.
Root 1 vs. Root 2
Sudeste do Pará Pará São Paulo
-5 -4 -3 -2 -1 0 1 2 3 4 5
Root 1
-4
-3
-2
-1
0
1
2
3
4
5
Ro
ot
2

13
5.DISCUSSÃO
Segundo MANN & AULAGNIER (1993), para que haja alta representatividade dos
indivíduos estudados, a amostragem tem que ser numerosa. De fato, esta afirmação se reflete
nos resultados de MARTINS (2008) com relação à distribuição de D. rotundus na Amazônia,
pois segundo este autor, existe uma homogeneidade regional para esta espécie. Contudo, esta
conclusão é fruto de pouca amostragem na Amazônia(número baixo de indivíduos) ,
influenciando os resultados de forma tendenciosa. Ao contrário, os resultados do presente
estudo mostraram que existe uma distinção entre os grupos de D. rotundus analisados para a
Amazônia, sendo os indivíduos da mesorregião Sudeste diferentes daqueles do restante do
Estado do Pará. Adicionalmente, é importante ressaltar que ainda ocorrem nessa área
variações individuais suficientes nos caracteres de cada um dos sexos, ou seja, a análise dos
dados sem combinação sexual, promoveu a segregação em grupos, cujo resultado se
expressou melhor e mais e de maneira mais evidente entre os machos. O dimorfismo sexual
segundo HERSHKOVITZ (1949), quando desconsiderado, tende a mascarar as separações entre
os grupos. Portanto, é assegurada a diferença entre regiões e dentro das regiões, mas sob um
número muito baixo de variáveis que se distinguem entre os sexos, as quais são um pouco
mais expressivas quando combinados os sexos. Provavelmente, estas diferenças possam estar
relacionadas às variações individuais geográficas, principalmente, influenciadas pelo
comportamento sexual e por fatores ambientais, como disponibilidade de abrigo e alimento.
No entanto, tais bases morfológicas pouco contribuem para a formação de subespécies
distintas, como proposto por MARTINS (2008), bem como para o reconhecimento da
estabilidade morfológicas dos espécimes da Amazônia.
A regra de BERGMAN (1847) diz que a massa dos indivíduos tende a ser maior
conforme o aumento da latitude, presumivelmente por razões de termorregulação. ASHTON
et al. (2000) testaram tal afirmativa e confirmaram ter encontrado evidencias suficientes para
considerá-la como uma tendência geral para os mamíferos. Considerando este fato, o padrão
de variação geográfica em D. rotundus parece assumir essa tendência, uma vez que os
indivíduos de São Paulo apresentam maiores proporções corporais e cranianas do que os
outros dois grupos do Pará.
A hipótese para tal divergência diz respeito à variação do clima local e foi sugerido
pela Dra. Nancy Simmons- curadora do AMNH. As florestas tropicais úmidas e desertos do
México ocorrem separados por distâncias relativamente pequenas, comparadas com as

14
distâncias entre as amostras do Brasil (Pará e São Paulo). Morcegos habitando esses dois
ecossistemas podem apresentar modificações morfológicas relacionadas às diferentes
condições climáticas destas regiões (MARTINS, 2008). Desse modo, a comparação
morfológica entre os indivíduos analisados das duas diferentes formações vegetais poderia
demonstrar a existência de adaptações locais para essa espécie de morcego, considerando as
condições climáticas do Sudeste e Norte do Brasil. Além do mais, a variação no tamanho
pode ser um acontecimento de maior plasticidade, do que o observado na forma.
O tamanho craniano já foi demonstrado ter uma plásticidade fenotípica em outras
ordens da classe Mammalia, como no caso do roedor Thomonysbattae,onde a resposta foi a
qualidade nutricional (PATTON & BRYSKY, 1987). No estudo de LESSA et al. (2005)
também foi encontrada variação craniana estruturada geograficamente em roedores da espécie
Kerodonrupestris. Estes autores tiveram como base, as alterações ambientais ao longo de uma
escala geológica.
A variação geográfica em D. rotundus já foi constatada anteriormente, porém as
localidades analisadas eram muito próximas. ANDRADE (2011) estudou populações das
cinco mesorregiões do Estado do Pará e observou uma grande sobreposição de valores em
ambos os sexos. Sendo que a análise discriminante mostrou níveis aceitáveis de atribuições
geográficas para as fêmeas, ao contrário para os machos que não apresentaram caracterização
geográfica confiável. Em maior escala, MARTINS (2009) diz que existe uma clara separação
entre o sudeste e norte do país no período do pleistoceno, pois estudos com sedimentação
descrevem que estas regiões (Mata Atlântica e Amazônia) têm sido fragmentadas, com áreas
secas em aberto relacionadas com períodos de glaciação.
Martins (2009) também argumenta que os morcegos hemátofagos podem voar até 20
km por noite à procura de alimento e que habita todos os biomas da região Neotropical, então
as barreiras ecológicas severiam como uma possível explicação para tal acontecimento. No
entanto, os resultados apresentados no presente trabalho não corrobora com o de MARTINS
(2009), pois na região amazônica os grupos são formados, mas sua separação é sutil, já os
morcegos de São Paulo formam um único grupo. Esse autor também relata que a Mata
Atlântica é um grupo monofilético e a Amazônia é somente um grupo que pode estar
divergindo.
Por fim, é importante ressaltar que MARTINS (2009), em seu estudo, utilizou apenas
22 morcegos oriundos de várias localidades da Amazônia para realizar suas análises

15
estatísticas, porém , pequenas diferenças inter-populacionais podem ser resultado de diversos
fatores, e desta forma, a chance de se detectar tais variações é diretamente proporcional ao
número amostral (WILLIG, 1983), enquanto que este trabalho utilizou 90 individuos só para
o Estado do Pará. A pouca representatividade dos dados (n=22) pode ter enviesado os
resultados para a região amazônica, pois este é um dos maiores biomas da região neotropical e
a diferença entre ambientes é considerável. Assim, espera-se que novos estudos e mais
abrangentes ainda venham contribuir para o melhor esclarecimento da variação geográfica de
D. rotundus nos biomas da região Neotropical, bem como do nível de diferenciação entre as
populações neles existentes.

16
6. CONCLUSÕES
As variáveis cranianas(CT, CB, CC, B, CPT, C-M2/3S, CM, LC, Acx, Ao, CC+Inc,
Cca+ PMO e CNaFo)discriminaram melhor do que as corpóreas na separação entre os três
grupos previamente estabelecidos.
Existe uma diferença morfométrica entre indivíduos de D. rotundus de sexos
diferentes, que reflete no resultado quanto à separação de grupos de diversas regiões.
Morcegos hematófagos oriundos da mesorregião Sudeste formam um grupo separado
das outras localidades do Estado do Pará.
Os dois grupos formados para a região Amazônica também são separados
morfologicamente dos indivíduos provenientes do Estado de São Paulo.

17
7. REFERENCIAS BIBLIOGRAFICAS
ACHA, P. N.; MALÁGA-ALBA, M. EconomiclossesduetoDesmodusrotundus. In: Natural
history of vampire bats. GREENHALL, A. M. &SCHIMDT, U. (Eds.). Florida: CRC Press, p.
207-214, 1988.
AGUIAR, L. M. S. Subfamília Desmodontinae. In: Morcegos do Brasil. REIS, N. R. dos;
PERACCHI, A. L.; PEDRO, W. A.; LIMA, I. P. (Org.). Londrina: UEL/SEMA, p. 37-44, 2007.
ALTENBACH, J. S. Locomotion. In: Natural history of vampire bats. GREENHALL, A. M.
&SCHIMDT, U. (Eds.). Florida: CRC Press, p. 71-83, 1988.
ANDRADE, F. A. G. Análise Morfométrica de espécie Desmodusrotundus
(CHIROPTERA, PHYLLOSTOMIDAE) coletados no estado do Pará, Norte do Brasil.
Tese (Doutorado). Programa de pós-graduação Emilio Goelgi. 152 f., 2011.
ARELLANO-SOTA, C. Biology, ecology, andcontrolofthe vampire bat. Rev. Infect. Dis, 10(4):
p. 615-619, 1988.
ASHTON, K.G.; M.C. TRACY& A. DE QUEIROZ. Is Bergmann´s Rule Valid for Mammals?
The American Naturalist.156(4), p. 390-415, 2000.
BERGMANN, C. UeberdieVerhaltnisse der Warmeokomie des Thierezuiher Grosse.
GottingerStudien. (3), p. 595-708, 1847.
BERNARD, E. Morcegos vampiros: sangue, raiva e preconceito. Ciência Hoje. v. 36, n. 214, p.
44-49, 2005.
BRASS, D. A. Rabies in bats: Natural History and Public Hearth Implications.
Connecticut: Livia Press, 342 f., 1994.
BREDT, A. Morcegos em áreas urbanas e rurais: Manual de Manejo e Controle. Brasília,
Ministério da Saúde, Fundação Nacional de Saúde, 117 f., 1998.
CABRERA A. Catalogo de losmamiferosdelsur. Rev. Museu Argentino Ciencia Natural
“BernadinoRivaldavia”, Zoologia (4): 93 f., 1958.
EMMONS, L. Neotropical rainforest mammals a Field guide. In: University of Chicago Press,
1990.

18
HERSHKOVITZP.. Mammals of northen Colombia. Preliminary report nº5: Bats (Chiroptera).
Proceedings of .S. Natural. Museum99:429-454, 1981
KOOPMAN, K F. Systematics and distribution,.In:Natural history of
vampirebats.GREENHALL, A M & U SCHMIDT (eds.).Boca Raton, CRC Press, 246 f., p. 7-17,
1988.
LESSA G, P R GONÇALVES& L M PESSOA. Variação geografica em caracteres cranianos
quantitativos de kerodonrupestris(WIED, 1820) (Rodentia, Cavidae). Arquivos do Museu
Nacional, Rio de Janeiro, 63(1): p.75-88, 2005
MALAGÁ-ALBA A. Vampire bat as a carrier of rabies. In:AmericanJournal of public health,
44: p. 909-918, 1954.
MANN, C.S. & S. AULAGNIER. Cranial and Brachial Biometry of
Desmodusrotundus(Chiroptera, Phyllostomidae) In:French-Guiana. Mammalia, (57): p.
589-599, 1993.
MARTINS, F M. Filogeografia intraespecífica do morcego hematófago Desmodusrotundus
(Chiroptera, Phyllostomidae). Tese (Doutorado) – Instituto de Biociências da Universidade
de São Paulo. Instituto de Biociências. Departamento de Biologia – Genética. 153 f., 2008.
MARTINS, F. M.; TEMPLETON, A. R.; PAVAN, A. C. O.; KOHLBACH, B. C.; MORGANTE, J. S.
Phylogeography of the common vampire bat (Desmodusrotundus): Marked population
structure, Neotropical Pleistocene vicariance and incongruence between nuclear and mtDNA
markers. BMC Evolutionary Biology. v. 9, 294 f., 2009.
OSGOOD, W H. The mammals of Chile.In:Field Mus. Nat. Hist. Zool., (30), pp. 61, 1943.
PATTON, J.L. &BRYLSKi, P.V. Pocket gophers in alfafa fields: causes and consequences of
habitat- related body size variation.In:The American Naturalist, Chicago, 130(4): p. 493-
505, 1987.
GREENHALL, A M, G JOERMANN& U SCHMIDT. Desmodusrotundus.In:Mammalian
Species,202: p. 1-6. 1983.
GREENHALL, A M & U SCHMIDT (eds.).Boca Raton, CRC Press, 246 f., 1988.

19
NOWAK, R M. Walker’s Mammals Of The World.Sixthy Edition. Baltimore: The Johns
Hopkins Univ. Press., 1996.
REIS, N. R.; SHIBATTA, O. A.; PERACCHI, A. L.; PEDRO, W. A.; LIMA, I. P. Sobre os morcegos
brasileiros. In: Morcegos do Brasil. REIS, N. R. dos; PERACCHI, A. L.; PEDRO, W. A.; LIMA,
I. P. (Org.). Londrina: UEL/SEMA, p. 17-25; 219-223, 2007.
REIS, N. R.Subfamília Desmodontinae. In: Mamíferos do Brasil. REIS, N. R.; PERACCHI,
A. L.; PEDRO, W. A.; LIMA, I. P. (Org.). Londrina: UEL/SEMA, 2ª Ed. p. 163-165, 2011.
SAZIMA, I. Aspectos do comportamento alimentar do morcego hematófago,
Desmodusrotundus. Boletim de Zoologia da Universidade de São Paulo. v. 3, p. 97-119,
1978.
SCHNEIDER, M. C.; ARON, J.; SANTOS-BURGOA, C.; UIEDA, W.; RUIZ-VELAZCO, S. Common
vampire bat attacks on humans in a village of the Amazon region of Brazil Cadernos de
SaúdePública. v. 17, n. 6, p. 1531-1536, 2009.
TADDEI, V. A.; GONÇALVES, C. A.; PEDRO, W. A. Distribuição do morcego vampiro
Desmodusrotundus(Chiroptera, Phyllostomidae) no Estado de São Paulo e a raiva dos
animais domésticos. Campinas: Secretaria de Agricultura e Abastecimento do Estado de São
Paulo, 107 f., 1991.
UIEDA, W.;HAYASHI, M. M; GOMES, L. M.; SILVA, M. M. S.. Espécies de quirópteros
diagnosticadas com raiva no Brasil. Boletim do Instituto Pasteur. v. 2, n. 1, p. 17-36, 1996.
VIZOTTO LD &V A TADDEI. Chave para determinação de quirópteros brasileiros. Revista da
faculdade de filosofia, ciências e Letras de São José do Rio Preto,Boletim de Ciências (1):
p.1-72, 1973.
WILLING, M.R. Composition, microgeographic variation, and sexual dimorphism in Caatingas
and Cerrado bat communities from northeastern Brazil. Bulletin of Carnegie Museum of
Natural Histoty(23): p.1-131, 1983.

20