Embed Size (px)
DESCRIPTION
melhoramento de plantas
Citation preview
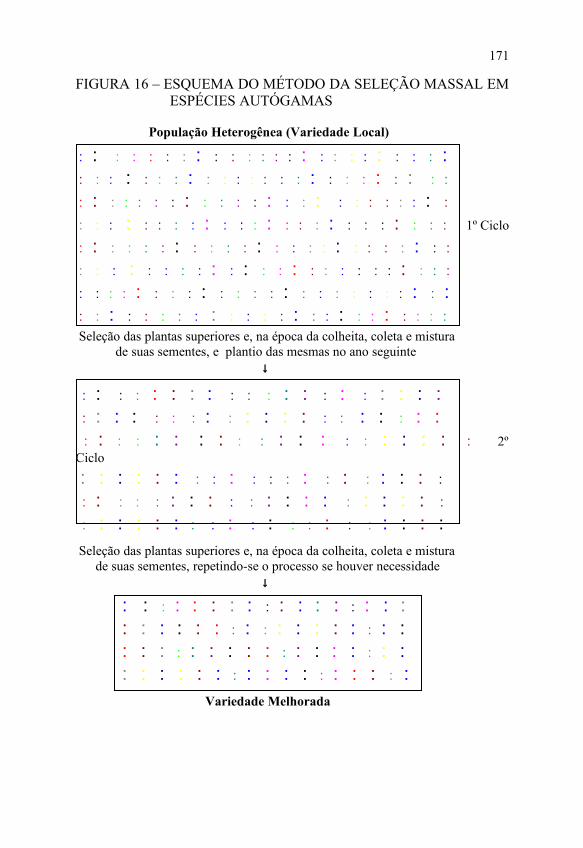
FIGURA 16 – ESQUEMA DO MÉTODO DA SELEÇÃO MASSAL EM ESPÉCIES AUTÓGAMAS
População Heterogênea (Variedade Local)
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 1º Ciclo
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ Seleção das plantas superiores e, na época da colheita, coleta e mistura de suas sementes, e plantio das mesmas no ano seguinte ↓
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 2º Ciclo ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ Seleção das plantas superiores e, na época da colheita, coleta e mistura de suas sementes, repetindo-se o processo se houver necessidade ↓
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ Variedade Melhorada
171

3.1.2 Método da seleção massal em espécies alógamas
Nas espécies alógamas, o método da seleção massal é bastante semelhante ao executado nas espécies autógamas, existindo, porém, poucas diferenças em função do seu modo de reprodução, o que, com certeza, irá reduzir a sua eficiência. São elas:
a) nas espécies alógamas, apesar de terem uma maior variância genética total, a variância genética aditiva é menor pela existência da dominância decorrente da heterozigose;
b) nas espécies alógamas, a seleção é baseada unicamente no genitor feminino, e não há controle da polinização, o que não é problema nas espécies autógamas, pois a autofecundação impede que plantas geneticamente inferiores polinizem as plantas selecionadas.
Em função disso, a intensidade de seleção utilizada nas espécies alógamas não deve ser forte, pois elas não toleram populações pequenas por causa do efeito da endogamia.
O referido método tem sido efetivo nas espécies alógamas para aumentar as freqüências gênicas de caracteres que são facilmente observados ou medidos. Por exemplo, na cultura do milho, foi possível desenvolver variedades diferindo na cor dos grãos, altura da planta, tamanho da espiga, localização da espiga no colmo, ciclo de maturação, resistência a várias doenças e porcentagem de óleo e de proteína.
Portanto, o método da seleção massal tem sido útil no desenvolvimento de variedades de espécies alógamas para propósitos especiais e na mudança da adaptação de variedades para torná-las adequadas ao cultivo em novas áreas.
No entanto, o método da seleção massal não tem sido efetivo na modificação de caracteres quantitativos, tais como a produção de grãos, os quais são governados por muitos genes
172

e não podem ser precisamente avaliados com base no aspecto de plantas individuais.
Dessa forma, o referido método tem-se mostrado ineficiente na melhoria desses caracteres em variedades adaptadas em função de três causas principais:
a) dificuldade de se identificarem genótipos superiores apenas pelo aspecto fenotípico de plantas individuais;
b) polinização não controlada completamente, de tal modo que as plantas selecionadas recebem grãos de pólen tanto de plantas superiores como inferiores;
c) seleção muito rigorosa, levando a uma população reduzida em tamanho, o que, por sua vez, conduz à perda de vigor e à redução da capacidade produtiva, por causa do efeito depressivo da endogamia.
O método também consiste em selecionar, numa população heterogênea, grande número de plantas de fenótipos semelhantes e, na colheita, misturar as sementes, efetuar, no ano seguinte, o plantio de tais sementes e proceder à nova seleção para aqueles caracteres pré-estabelecidos; e, por fim, repetir esse procedimento tantas vezes quanto necessário até que a população se torne bem homogênea para tais caracteres; as sementes, então, são multiplicadas e distribuídas aos agricultores.
Também, nesse caso, cada ciclo de seleção envolve apenas uma etapa, ou seja, plantio da população original, seleção de um grande número de plantas, colheita e mistura das sementes (não há teste de progênie), repetindo o processo, conforme apresentado na FIGURA 16.
Essa etapa inicia-se com o plantio de uma população alógama, em que a densidade de plantio e os tratos culturais são os mesmos do cultivo comercial da espécie em consideração.
Geralmente o tamanho da população inicial fica em torno de 2.000 a 10.000 plantas, havendo influência de vários
173

fatores. Entre eles, destacam-se os meios materiais de que dispõe o fitomelhorista e a heterogeneidade da população original.
Na época da colheita, selecionam-se em torno de 200 a 2.000 plantas (i varia de 10 a 20%) para os caracteres pré-estabelecidos; as sementes são colhidas, misturadas e guardadas na câmara de sementes para plantio no ano seguinte, repetindo-se o processo.
Vale ressaltar que, em função de o referido método ter-se mostrado ineficiente na melhoria de caracteres quantitativos em variedades adaptadas de espécies alógamas, procurou-se estudar as suas causas para torná-lo mais eficiente. Em função disso, algumas modificações surgiram no método original (também denominado de método de seleção massal simples) visando, principalmente, controlar a heterogeneidade do solo, visto que o método original, além de representar o sistema em que é mais reduzido o controle parental, apresenta também reduzido controle do meio ambiente.
As principais modificações ocorreram na cultura do milho, destacando-se a seleção massal estratificada, idealizada por GARDNER (1961). No procedimento foram incorporados quatro pontos em relação ao método original:
a) no local de plantio, o terreno deve ser o mais uniforme possível e estar isolado de outros cultivos de milho;
b) na época da colheita, a área é dividida em parcelas ou estratos, geralmente de 10 m2; a seleção de plantas é feita para cada estrato separadamente;
c) na seleção, apenas as plantas competitivas, que não apresentam falhas vizinhas, são selecionadas;
d) após a colheita das espigas, retira-se o mesmo número de sementes de cada espiga para garantir-se uma amostragem adequada.
A FIGURA 17 apresenta o esquema do método da seleção massal estratificada na cultura do milho.
174
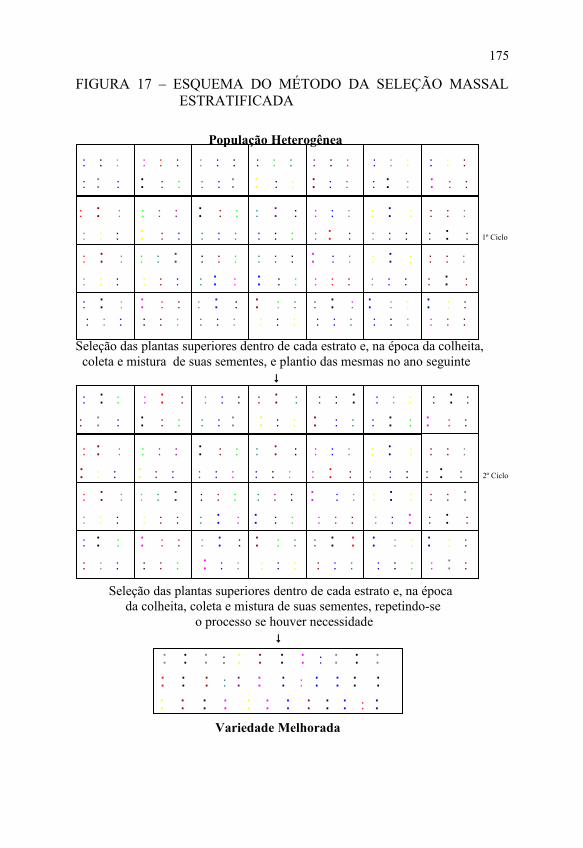
FIGURA 17 – ESQUEMA DO MÉTODO DA SELEÇÃO MASSAL ESTRATIFICADA
População Heterogênea
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 1º Ciclo
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
Seleção das plantas superiores dentro de cada estrato e, na época da colheita, coleta e mistura de suas sementes, e plantio das mesmas no ano seguinte ↓
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 2º Ciclo
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
Seleção das plantas superiores dentro de cada estrato e, na época da colheita, coleta e mistura de suas sementes, repetindo-se o processo se houver necessidade ↓
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ Variedade Melhorada
175

Entre as vantagens do método da seleção massal estratificada, PATERNIANI e MIRANDA FILHO (1980) apresentam as seguintes:
a) apresenta relativa facilidade de condução; b) tamanho efetivo da população é grande; c) pode ser aplicada forte intensidade de seleção,
podendo chegar a 1%; d) o material é avaliado todos os anos, o que reduz os
problemas de interação por anos; e) obtém-se um ciclo por ano.Os mesmos autores apresentam as seguintes
limitações: a) não há controle da descendência, ou seja, plantas
fenotipicamente boas devido a condições de ambiente ou a interações gênicas especiais, poderão dar descendentes inferiores;
b) o material é avaliado num só local; por isso, com o tempo, a tendência é aumentar a adaptação da população para áreas específicas;
c) devido à forte competição entre as plantas, há a tendência das plantas mais altas serem as mais produtivas; assim, plantas baixas, potencialmente boas, ficam prejudicadas e não expressam o seu potencial. Depois dos ganhos genéticos obtidos com o método da seleção massal estratificada, esse método passou a interessar muitos fitomelhoristas; várias modificações foram sugeridas com o propósito de aumentar a sua eficiência, algumas das quais experimentadas em pequena escala, como foi o caso da seleção massal estratificada geneticamente, citado por PATERNIANI e MIRANDA FILHO (1980). O método visa controlar a heterogeneidade do solo através do plantio intercalado de plantas de um genótipo constante (híbrido simples), conforme apresentado na FIGURA 18.
176
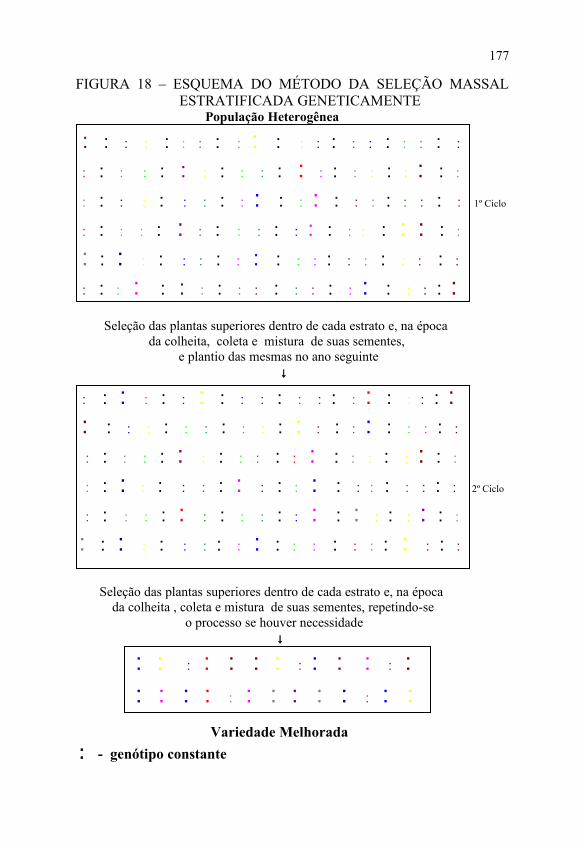
FIGURA 18 – ESQUEMA DO MÉTODO DA SELEÇÃO MASSAL ESTRATIFICADA GENETICAMENTE
População Heterogênea
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 1º Ciclo
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣♣ ♣ ♣
♣♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣♣ Seleção das plantas superiores dentro de cada estrato e, na época da colheita, coleta e mistura de suas sementes, e plantio das mesmas no ano seguinte ↓
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣♣ ♣
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 2º Ciclo
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣♣ ♣
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
Seleção das plantas superiores dentro de cada estrato e, na época da colheita , coleta e mistura de suas sementes, repetindo-se o processo se houver necessidade ↓
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ Variedade Melhorada ♣ - genótipo constante
177

Nesse caso, cada estrato é formado por duas plantas da população original adjacentes ao genótipo constante, e a seleção será feita em cada estrato em função das produções relativas das plantas em relação ao genótipo constante. Assim, se num determinado estrato a planta do genótipo constante produziu 200 gramas de grãos, e as plantas adjacentes produziram 220 e 190 gramas de grãos, respectivamente, isso indica que essas plantas produziram 10% a mais e 5% a menos que o genótipo constante. Num outro estrato, o genótipo constante produziu 150 gramas e as plantas adjacentes produziram 170 e 160 gramas de grãos, respectivamente, indicando que essas plantas produziram 13 e 7% a mais que o genótipo constante. Dessa forma, a seleção será feita em função desses valores relativos.
A fim de evitar os cruzamentos do genótipo constante com as plantas da população a serem selecionadas, duas alternativas podem ser usadas:
a) na primeira alternativa, a população a ser selecionada deve ser homozigótica para um gene recessivo com expressão no endosperma (grãos brancos ou milho opaco) e que, portanto, exibem o efeito de xênia (quando ocorre segregação na mesma geração de fertilização); nesse caso, os grãos resultantes do cruzamento são identificados e eliminados;
b) na segunda alternativa, pode-se utilizar um genótipo constante macho estéril.
3.2 Método do retrocruzamento
Esse método é especialmente adequado para se transferirem genes específicos para variedades que são muito boas, com relação a um grande número de atributos, sendo, porém, deficientes em uma ou algumas características. Inicialmente, o método foi usado praticamente no
178

melhoramento visando à resistência às doenças e aos insetos-praga, em função do efeito dramático que os microorganismos fitopatogênicos e os insetos-praga podem ter sobre a produção e a qualidade.
Em alguns casos, constitui-se na principal e/ou única medida de controle viável. Posteriormente, o método passou a ter uma utilização mais abrangente, pois, além da resistência às doenças e aos insetos-praga, outros caracteres foram incorporados no melhoramento, tais como caracteres morfológicos, características de cor e caracteres quantitativos de herança relativamente simples (precocidade, altura da planta, tamanho e forma da semente, etc.).
No referido método, tanto nas espécies autógamas como nas espécies alógamas, procura-se a transferência de um ou dois genes de um progenitor para outro que carece das qualidades condicionadas por esses genes. Em função disso, uma série de retrocruzamentos são feitos para o progenitor mais desejável ou recorrente, ao mesmo tempo em que é feita seleção para os caracteres que estão sendo transferidos do progenitor doador. À medida que a série de retrocruzamentos avança, a população híbrida vai-se tornando cada vez mais semelhante ao progenitor recorrente, conforme apresentado na TABELA 1.TABELA 1 – PORCENTAGENS DE GENES DOS PROGENITORES
INCORPORADOS À POPULAÇÃO HÍBRIDA EM CADA RETROCRUZAMENTO SUCESSIVO
_________________________________________________________________________________
Geração de Retrocruzamento Progenitor Doador Progenitor Recorrente____________________________________________________________________________________________________________
F1 50,000 50,000 1º Retrocruzamento 25,000 75,000 2º Retrocruzamento 12,500 87,500 3º Retrocruzamento 6,250 93,750 4º Retrocruzamento 3,125 96,875 5º Retrocruzamento 1,563 98,437 6º Retrocruzamento 0,781 99,219 7º Retrocruzamento 0,391 99,609 8º Retrocruzamento 0,195 99,805 9º Retrocruzamento 0,098 99,90210º Retrocruzamento 0,049 99,951____________________________________________________________________________________________________________
179

No método do retrocruzamento, a homozigose é atingida na mesma proporção da autofecundação, conforme a seguinte fórmula:
Proporção de homozigose = (2m−12m )
n
onde:m – número de gerações de autofecundação ou de
retrocruzamento;n – número de genes.
Entretanto, existe uma diferença básica, visto que, no
retrocruzamento, espera-se que metade da descendência seja homozigota para cada gene particular do progenitor recorrente, enquanto na autofecundação, embora metade da descendência seja homozigota para cada gene em consideração, somente metade desses homozigotos tem o mesmo genótipo do progenitor recorrente.
Se, por exemplo, os progenitores diferirem entre si, em quatro pares de alelos, e nenhuma seleção é praticada, seis retrocruzamentos produzirão uma população na qual 93,89% dos indivíduos serão homozigotos e idênticos ao progenitor recorrente para todos os quatro loci.
Quanto à proporção dos genes do progenitor doador que são eliminados durante a série de retrocruzamentos, o fenômeno da ligação gênica tem uma influência bastante significativa. Por exemplo, num determinado programa de retrocruzamento, que tem por objetivo transferir o gene dominante A do progenitor doador para o progenitor recorrente, o gene indesejável b está ligado ao gene A. O genótipo do híbrido F1 será Ab/aB, e a seleção para A, nas primeiras gerações de retrocruzamento, tenderá a levar junto o gene b, tornando difícil obter a recombinação desejada AB.
180

Mas, como o gene B é reintroduzido novamente em cada retrocruzamento, haverá muitas oportunidades para a ocorrência de permuta genética e, em conseqüência, o surgimento da recombinação desejada AB, cujas possibilidades são melhores que nas autofecundações.
Considerando-se que nenhuma seleção é praticada, exceto para o gene A que está sendo transferido, pode-se mostrar que a probabilidade de eliminar o gene indesejável b [P (b)] é calculada pela seguinte fórmula:
P (b) = 1 - (1 - p)m + 1
onde:p – fração de recombinação;m – número de retrocruzamentos.
Se, por exemplo, o gene b estiver localizado a 20 unidades de permuta genética de A, com seis retrocruzamentos a probabilidade de se eliminar o gene indesejável b será de 79,03%.
Naturalmente que, quanto mais ligados estiverem os genes A e b, menores são as probabilidades de eliminação de b nos retrocruzamentos, como, por exemplo, se o referido gene estivesse localizado a 5 unidades de permuta genética de A. Nesse caso, com seis retrocruzamentos, a probabilidade de eliminá-lo seria de 30,17%. Se os genes A e b estiverem próximos demais, de modo a não ocorrer a permuta genética entre eles, então b será transferido para o progenitor recorrente juntamente com o gene A, e a nova variedade poderá ser menos desejável que o progenitor recorrente.
No caso das espécies autógamas, o método do retrocruzamento difere dos outros métodos de melhoramento que envolve hibridação, porque não visa à obtenção de novos genótipos que reúnam as características desejáveis dos progenitores, mas ao melhoramento de variedades superiores
181

já bastante melhoradas, pela transferência de um ou dois genes do progenitor doador para essas variedades superiores.
No caso das espécies alógamas, o referido método tem o mesmo objetivo do melhoramento das espécies autógamas. Contudo, o procedimento usado para essas espécies é ligeiramente diferente quando se trata do melhoramento de populações, visto que é necessário um número muito maior de plantas do progenitor recorrente, em torno de 150 a 200 plantas, em cada retrocruzamento, de modo que represente as freqüências gênicas características desse progenitor.
Mas, quando se trata da introdução de genes específicos em linhagens autofecundadas (melhoramento de linhagens), as quais se destinam, primariamente, à produção de híbridos, o procedimento é essencialmente igual àquele aplicado às espécies autógamas.
Vale ressaltar que o sucesso do método do retrocruzamento depende das seguintes condições:
a) Deve-se ter um progenitor recorrente satisfatório – O progenitor recorrente deve ser uma “variedade de elite”, ou seja, uma variedade de grande aceitação comercial, produtiva, bem adaptada e com outras características de valor agronômico, cujo comportamento é comprovado em várias regiões, apresentando, porém, algumas deficiências que podem ser corrigidas. São materiais que já passaram pelo crivo do melhoramento e que são difíceis de serem substituídos. A cenoura Tropical, o feijão Carioca, o milho Centralmex, a soja Tropical e o tomate Santa Cruz são exemplos de progenitores recorrente satisfatórios.
b) Deve-se ter um progenitor doador adequado – O progenitor doador deve ser aquele que possui os genes que governam as características que faltam no progenitor recorrente, as quais devem ser facilmente identificadas por mera inspeção visual ou por testes simples. Contudo, quando esses genes são encontrados tanto em espécies cultivadas
182

como em espécies selvagens, é preferível optar pelo primeiro caso, pois reduz-se o número de retrocruzamentos e, conseqüentemente, o tempo e os gastos financeiros, em função da presença de muitas características semelhantes às do progenitor recorrente (menor diversidade genética), fazendo com que, no cruzamento entre os dois progenitores, o F1 tenha mais de 50% de genes do progenitor recorrente no início do programa de melhoramento, o que não acontece com as espécies selvagens.
c) Deve-se manter, com boa intensidade, o caráter em transferência por meio de vários retrocruzamentos – Independentemente do número de genes que governam o caráter a ser transferido, é preciso que uma boa intensidade do caráter seja mantida por meio de vários retrocruzamentos, sob pena de não se alcançarem, em toda a sua plenitude, os objetivos do programa de melhoramento. Entretanto, é necessário que o caráter a ser transferido tenha uma alta herdabilidade, para que se tenha facilidade e certeza na identificação do mesmo nas populações segregantes.
d) Deve-se utilizar um número suficiente de retrocruzamentos para reconstituir, num alto grau, o progenitor recorrente – O emprego de seis retrocruzamentos tem provado serem suficientes para reconstituir, num alto grau, o progenitor recorrente quando acompanhados de rígida seleção nas primeiras gerações de retrocruzamentos. É claro que, primariamente, isso é uma função do número de retrocruzamentos. No entanto, a seleção para o tipo do progenitor recorrente, nas primeiras gerações de retrocruzamento (até o terceiro retrocruzamento), tem-se mostrado eficiente mudar a população para as características da planta desse progenitor. Também, deve-se utilizar suficiente número de plantas do progenitor recorrente para manter toda sua variabilidade genética, de modo que o progenitor recorrente seja reconstituído em toda sua plenitude.
183

Uma particularidade muito interessante do método do retrocruzamento é que pode ser conduzido em condições de meio ambiente bem diferentes daquelas para as quais se destinará a nova variedade, desde que permita a expressão do caráter em transferência. Nesse caso, por exemplo, pode-se desenvolver uma variedade de uma determinada espécie vegetal para o Estado de Alagoas, conduzindo o programa de melhoramento no Estado de São Paulo ou até em outro país, desde que o caráter a ser transferido do progenitor doador para o progenitor recorrente possa expressar-se.
Uma outra particularidade também interessante do referido método é que o mesmo dispensa o uso de ensaios comparativos, desde que se tenha certeza de que o progenitor recorrente foi plenamente reconstituído.
O método consiste em cruzar dois progenitores, sendo um recorrente e o outro doador, para obter o híbrido, cruzar o híbrido com o progenitor recorrente e selecionar as plantas que carregam os genes do progenitor doador, repetindo-se o processo por várias vezes até que seja reconstituído o progenitor recorrente, e, por fim, autofecundar as plantas para obter homozigose para o caráter transferido e selecionar as plantas homozigotas, que resultam na nova variedade.
O método do retrocruzamento envolve três etapas distintas:
a) cruzamento do progenitor recorrente com o progenitor doador, a fim de obter o híbrido como ponto de partida para o início da série de retrocruzamentos;
b) retrocruzamento do híbrido com o progenitor recorrente e seleção das plantas que carregam os genes do progenitor doador, repetindo-se o processo, por várias vezes, até reconstituir-se o progenitor recorrente;
c) autofecundação das plantas, no final do último retrocruzamento, para obter homozigose para o caráter transferido e seleção das plantas homozigotas, que resultam na nova variedade.
184

As FIGURAS 19 e 20 apresentam os esquemas do referido método na transferência de um gene dominante e de um gene recessivo, respectivamente, podendo-se visualizar as três etapas distintas.
A primeira etapa inicia-se com a escolha dos progenitores recorrente e doador, levando-se em conta as condições apresentadas anteriormente, plantio de sementes dos mesmos e posterior cruzamento entre eles, a fim de obter-se o híbrido como ponto de partida para o início da série de retrocruzamentos.
A segunda etapa, que é a mais demorada, inicia-se com o plantio de sementes do híbrido e do progenitor recorrente e posterior cruzamento entre eles (1º retrocruzamento). Em seguida, faz-se o plantio de sementes do 1º retrocruzamento e do progenitor recorrente. Posteriormente, dentre as plantas do 1º retrocruzamento, é feita a seleção de plantas que carregam os genes de interesse do progenitor doador, as quais são cruzadas com o progenitor recorrente (2º retrocruzamento). Esse processo é repetido até o último retrocruzamento, que geralmente corresponde ao 6º retrocruzamento.
Vale ressaltar que esse procedimento é válido para a transferência de um gene dominante. No caso da transferência de um gene recessivo, há necessidade de autofecundação das plantas do híbrido, nos retrocruzamentos de ordem ímpar, antes da seleção, para que se identifiquem as plantas que carregam os genes de interesse do progenitor doador.
A terceira etapa inicia-se com o plantio de sementes do último retrocruzamento; depois é feita a seleção de plantas que carregam os genes de interesse do progenitor doador, seguida da autofecundação dessas plantas para obter homozigose para o caráter transferido; e seleção das progênies homozigotas, mistura de suas sementes, resultando na nova variedade.
185
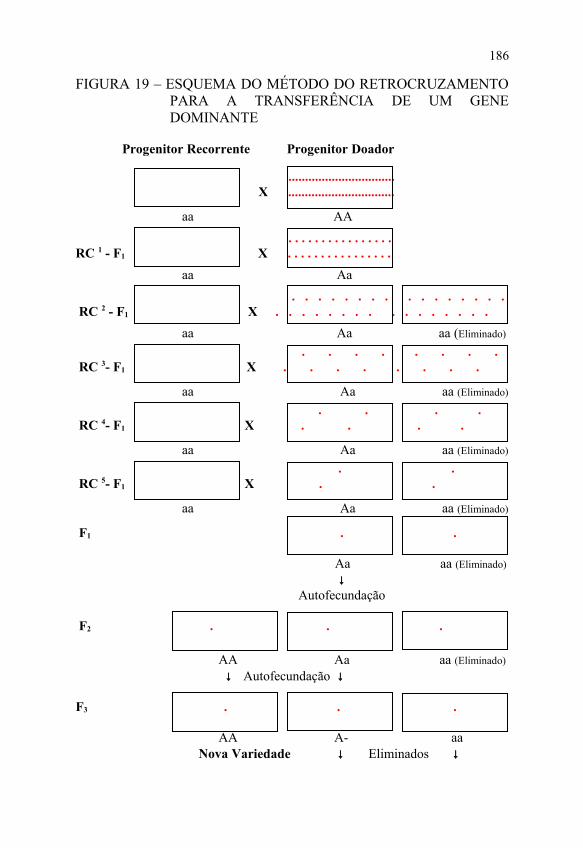
FIGURA 19 – ESQUEMA DO MÉTODO DO RETROCRUZAMENTO PARA A TRANSFERÊNCIA DE UM GENE DOMINANTE
Progenitor Recorrente Progenitor Doador
................................ X ................................
aa AA . . . . . . . . . . . . . . . .RC 1 - F1 X . . . . . . . . . . . . . . . . aa Aa . . . . . . . . . . . . . . . . RC 2 - F1 X . . . . . . . . . . . . . . . . aa Aa aa (Eliminado)
. . . . . . . . RC 3- F1 X . . . . . . . .
aa Aa aa (Eliminado)
. . . . RC 4- F1 X . . . .
aa Aa aa (Eliminado)
. . RC 5- F1 X . .
aa Aa aa (Eliminado)
F1 . .
Aa aa (Eliminado) ↓ Autofecundação
F2 . . .
AA Aa aa (Eliminado) ↓ Autofecundação ↓ F3 . . . AA A- aa Nova Variedade ↓ Eliminados ↓
186
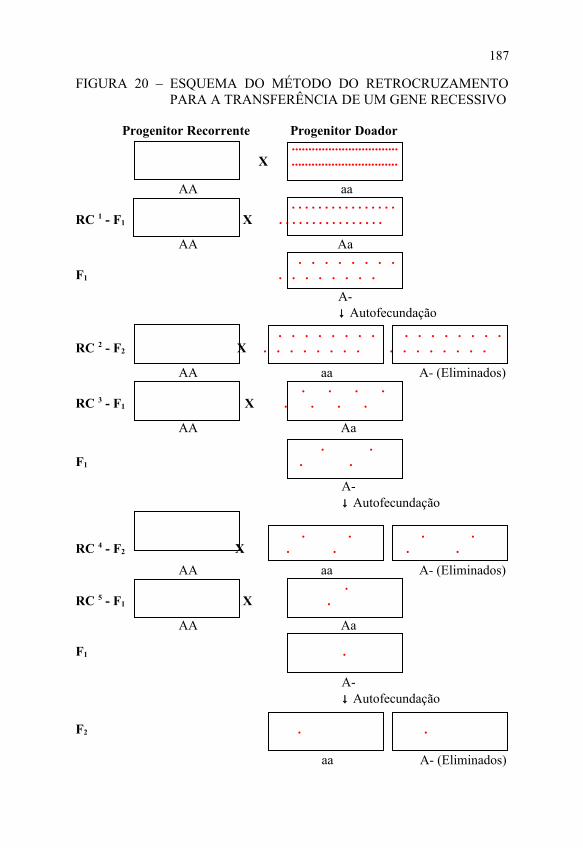
FIGURA 20 – ESQUEMA DO MÉTODO DO RETROCRUZAMENTO PARA A TRANSFERÊNCIA DE UM GENE RECESSIVO
Progenitor Recorrente Progenitor Doador ................................ X ................................
AA aa . . . . . . . . . . . . . . . .RC 1 - F1 X . . . . . . . . . . . . . . . .
AA Aa . . . . . . . .F1 . . . . . . . . A- ↓ Autofecundação
. . . . . . . . . . . . . . . .RC 2 - F2 X . . . . . . . . . . . . . . . .
AA aa A- (Eliminados) . . . .RC 3 - F1 X . . . .
AA Aa
. .F1 . .
A- ↓ Autofecundação
. . . .RC 4 - F2 X . . . . AA aa A- (Eliminados) .RC 5 - F1 X .
AA Aa
F1 .
A- ↓ Autofecundação F2 . . aa A- (Eliminados)
187

Nova VariedadeOs esquemas do método do retrocruzamento
apresentados anteriormente se referem ao melhoramento de apenas um caráter. Quando, porém, o melhoramento envolve dois ou mais caracteres, há três alternativas:
a) melhoramento por etapas; b) transferência paralela; c) transferência simultânea.No melhoramento por etapas, o progenitor recorrente
é inicialmente melhorado para um caráter; depois de melhorado, vai ser utilizado como progenitor recorrente para melhorar o outro caráter, e assim por diante. É uma alternativa muito demorada, porém é mais precisa e menos trabalhosa que as outras, sendo, portanto, a mais utilizada .
Na transferência paralela, os programas de retrocruzamentos são conduzidos ao mesmo tempo; os produtos finais são cruzados, obtendo-se a geração F1 e, em seguida, a geração F2, em que são selecionadas as plantas que carregam os genes desejáveis dos progenitores doadores. É uma alternativa rápida, porém é menos precisa e mais trabalhosa que o melhoramento por etapas.
Na transferência simultânea, um único programa de retrocruzamento é utilizado na transferência dos genes desejáveis do progenitor doador. É uma alternativa tão rápida quanto a transferência paralela. Também é tão trabalhosa quanto a anterior, pois obriga o fitomelhorista a trabalhar com populações maiores. Quanto à precisão, tem a mesma da anterior, se ambas forem conduzidas na mesma área experimental.
Quando, porém, na transferência paralela, os programas são conduzidos em ambientes propícios para a expressão de cada caráter, a transferência simultânea torna-se menos precisa, porque, nem sempre, o ambiente propicia as condições necessárias para a expressão dos vários caracteres ao mesmo tempo.
188

3.3 Método de variedades híbridas
Esse método consiste em explorar o vigor de híbrido ou heterose na geração F1, a qual é obtida pelo cruzamento entre variedades de polinização livre, linhagens endógamas ou outras populações que sejam geneticamente divergentes. No entanto, na prática, explora-se mais a heterose a partir de linhagens endógamas. Mas, quando o uso de variedades híbridas é viável, o referido método permite aproveitar melhor a heterose do que qualquer outro método de melhoramento.
De acordo com PATERNIANI (1974), o método apresenta as seguintes vantagens:
a) permite a associação, no mesmo indivíduo, de caracteres que estão separados em progenitores distintos, sendo que, de outro modo, essa associação pode ser muito difícil ou mesmo impraticável;
b) permite a obtenção, em prazo relativamente curto, de genótipos superiores que podem ser utilizados na prática;
c) permite a obtenção de um melhoramento genético substancial, possivelmente em parte pela utilização de interações gênicas que só podem ser aproveitadas na geração híbrida;
d) permite obter produtos uniformes e bem padronizados;
e) a geração F1 geralmente apresenta menor interação por ambiente do que ocorre com genótipos homozigotos, decorrendo disso uma adaptabilidade mais ampla do híbrido, cujas produções oscilam menos entre locais e anos distintos, propriedade essa denominada homeostase;
f) possibilita e traz grande estímulo ao desenvolvimento de uma indústria de sementes, com reflexos em geral benéficos para a região, pois, indiretamente, contribui também para melhores práticas e condições de cultivo.
189

No entanto, o mesmo autor menciona as seguintes limitações, quando se considera apenas o uso da heterose:
a) dificilmente serão obtidos os melhores genótipos possíveis, pois, sem o concurso de métodos que aumentam a freqüência de genes favoráveis nas populações básicas, a heterose só utilizará uma parte dos genes úteis existentes na espécie;
b) a heterose é explorada de modo aleatório, atingindo um teto difícil de ser ultrapassado;
c) a heterose só pode ser empregada, na prática, para as espécies nas quais a semente híbrida é facilmente obtida a baixo custo, ou nas quais o produto comercial é muito valorizado;
d) seu uso só é viável em regiões onde existem facilidades para produção, processamento e distribuição de semente híbrida de boa qualidade.
Atualmente, o método de variedades híbridas é muito usado em hortaliças, tanto em espécies autógamas (beringela, pimenta, pimentão e tomate) como em espécies alógamas (beterraba, cebola, cebolinha, cenoura, brássicas, melancia, melão, pepino, etc.). Porém, o referido método teve o maior desenvolvimento na cultura do milho, sendo o grande responsável pelo intenso aumento da sua produtividade.
Neste momento, é oportuno fazer um breve comentário sobre a descoberta do método. O fenômeno do vigor de híbrido foi descoberto, pela primeira vez, na cultura do milho, mais ou menos por acaso, por volta de 1905 a 1908, pelos americanos Shull e East. Como se sabe, foi em 1900 que foram redescobertas as Leis de Mendel. Após isto, muitos geneticistas procuraram verificar se as leis mendelianas eram aplicadas a outras espécies.
Shull, que trabalhava com milho, procurou estudar a herança do número de fileiras de grãos na espiga. Ele autofecundou duas variedades por cinco gerações sucessivas, sendo uma com oito fileiras de grãos na espiga e a outra com
190

muitas fileiras de grãos na espiga. Em seguida, cruzou as duas linhagens S5 e verificou que o F1 era muito vigoroso, inclusive superior aos pais.
East descobriu a mesma coisa, mas ele estava mais interessado em caracteres quantitativos. Ele queria verificar a perda de vigor devido à autofecundação em altura de planta e produção de grãos. Nos dois casos, eles cruzaram linhagens de variedades diferentes e observaram aumento de vigor.
O próprio Darwin havia observado este aumento de vigor, só que não tinha quantificado o fenômeno. Na época dos trabalhos de Shull e de East, os fitomelhoristas tinham problemas em aumentar a produção de milho nos Estados Unidos pelos métodos de seleção massal e de seleção espiga por fileira. Pensaram, então, em utilizar o milho híbrido. Isso era um problema, porque as linhagens produziam poucas sementes.
Para se ter uma idéia, numa variedade que produzisse 2.000 kg/ha, as linhagens S5 produziriam em torno de 400 kg/ha. Se for plantado 1ha de terra, ter-se-ão 7.500 m2 de fêmeas e 2.500 m2 de machos, e colher-se-ão 300 kg/ha de sementes do híbrido simples. Assim, a quantidade de semente híbrida colhida é muito pequena e, com isso, o processo é antieconômico.
Em 1918, Jones, um discípulo de East, teve a idéia de produzir híbridos duplos, a qual foi muito eficiente e útil, visto que, em 1921, já existia plantio comercial de milho híbrido. Porém, só depois de um certo tempo foi que o milho híbrido se tornou um fator significativo na agricultura, já que, em 1933, representava menos de 1% da área plantada. Por volta de 1940, mais da metade da área destinada ao milho, nos Estados Unidos, era plantada com híbridos e, em 1944, os híbridos representavam mais de 80% da área. Na faixa do milho, a aceitação dos híbridos foi tão completa que virtualmente desapareceram as variedades de polinização livre. Foi assim que surgiu o método de variedades híbridas.
191

Vários tipos de híbridos podem ser produzidos, a saber:
a) Top-cross – híbrido resultante do cruzamento entre uma linhagem endogâmica e uma variedade de polinização livre (L x V). Esse tipo de híbrido não tem sido considerado de valor comercial, mas é amplamente utilizado nos programas de avaliação de linhagens para utilização em híbridos.
b) Híbrido simples – híbrido resultante do cruzamento entre duas linhagens endogâmicas (L1 x L2). Em geral é mais produtivo e mais uniforme do que os demais tipos de híbridos, porém apresenta menor estabilidade de produção e maior custo de produção de sementes, porque o progenitor feminino é uma linhagem endogâmica, a qual exibe produção muito baixa (geralmente em torno de 20 a 25% da população de origem). Também sua obtenção é mais rápida do que os demais híbridos de linhagens.
c) Híbrido simples modificado – híbrido resultante do cruzamento entre um híbrido originário de duas progênies afins da mesma linhagem endogâmica (progenitor feminino) e uma linhagem endogâmica ou um híbrido entre progênies afins (progenitor masculino), ou seja, (L1 x L1
’) x L2 ou (L1
x L1 ’) x (L2 x L2
’). Em qualquer caso, o custo de produção de sementes é menor do que o do híbrido simples, visto que o progenitor feminino apresenta um certo vigor que se manifesta em maior produção de sementes. Apresenta praticamente a mesma uniformidade do híbrido simples. Em termos de produtividade, só é inferior ao híbrido simples. O tempo para sua obtenção é igual ao do híbrido triplo e triplo modificado.
d) Híbrido triplo – híbrido resultante do cruzamento entre um híbrido simples (progenitor feminino) e uma linhagem endogâmica (progenitor masculino) , ou seja, (L1 x L2) x L3. Em geral é mais produtivo que o híbrido triplo modificado, duplo, múltiplo e intervarietal, enquanto o custo
192

de produção de sementes é menor que o do híbrido simples modificado. Apresenta boa uniformidade e maior estabilidade de produção do que os híbridos simples e simples modificado; porém, leva-se mais tempo para sua obtenção do que na do híbrido simples.
e) Híbrido triplo modificado – híbrido resultante do cruzamento entre um híbrido simples (progenitor feminino) e um híbrido oriundo de duas progênies afins da mesma linhagem endogâmica (progenitor masculino), ou seja, (L1 x L2) x (L3 x L3
’). Em geral, é mais produtivo que o híbrido duplo, múltiplo e intervarietal, enquanto o custo de produção de sementes é menor que o do híbrido triplo, visto que o progenitor masculino apresenta um certo vigor que se manifesta em maior produção de grãos de pólen e, em conseqüência, uma maior produção de sementes. Também apresenta boa uniformidade e maior estabilidade de produção do que os híbridos simples e simples modificado; porém, leva-se mais tempo para sua obtenção do que na do híbrido simples.
f) Híbrido duplo – híbrido mais largamente utilizado, sendo resultante do cruzamento entre dois híbridos simples (L1 x L2) x (L3 x L4). Em geral, é mais produtivo que o híbrido múltiplo e intervarietal, enquanto o custo de produção de sementes é menor que o híbrido triplo modificado. Apresenta menor uniformidade do que os híbridos triplo e triplo modificado e maior estabilidade de produção do que os híbridos simples e simples modificado; porém, leva-se o mesmo tempo dos híbridos triplo e triplo modificado para sua obtenção.
g) Híbrido múltiplo – híbrido muito pouco usado comercialmente, sendo resultante do cruzamento de um híbrido simples com um híbrido duplo (L1 x L2) x [(L3 x L4) x (L5 x L6)], de um híbrido triplo com um duplo [(L1 x L2) x L3] x [(L4 x L5) x (L6 x L7)], de dois híbridos duplos [(L1 x L2) x (L3 x L4)] x [(L5 x L6) x (L7 x L8)], etc. Em geral é mais
193

produtivo que um híbrido intervarietal; apresenta maior estabilidade do que qualquer híbrido de linhagens, mas é menos uniforme que tais híbridos. Para sua obtenção, leva-se mais tempo do que com os demais híbridos de linhagens. Suas gerações avançadas podem ser utilizadas como fonte de novas linhagens.
h) Híbrido Intervarietal – híbrido resultante do cruzamento entre duas variedades de polinização livre (V1 x V2). Esse tipo de híbrido pode ser utilizado comercialmente, pois permite a utilização da heterose sem a necessidade de obtenção de linhagens, o que torna o custo de obtenção bastante baixo, além de levar menos tempo para a sua síntese. É mais estável que qualquer híbrido de linhagens; porém, é menos uniforme que tais híbridos. Suas gerações avançadas podem ser utilizadas como população-base para o melhoramento intrapopulacional.
Como, na prática, o método de variedades híbridas explora mais a heterose a partir de linhagens endógamas, de agora em diante toda a discussão será feita nessa direção, usando a cultura do milho como padrão, visto que o referido método teve o maior desenvolvimento nessa espécie, sendo o grande responsável pelo intenso aumento da sua produtividade.
O método consiste em obter linhagens endógamas, avaliá-las quanto à capacidade de combinação, selecionando as mais adequadas, efetuar os cruzamentos em todas as combinações possíveis entre as linhagens selecionadas, e, por fim, avaliar os cruzamentos em ensaio de campo, selecionando-se os cruzamentos mais heteróticos, os quais irão constituir as novas variedades híbridas.
O método de variedades híbridas envolve três etapas distintas:
a) Obtenção das linhagens endogâmicas – Na primeira etapa, diversos métodos podem ser utilizados na obtenção das linhagens endogâmicas: método padrão, método
194

da cova única, método genealógico, método do híbrido críptico, método da seleção zigótica, método de monoplóides duplicados, etc.
a.1) Método padrão – O método padrão, o mais usado na obtenção de linhagens, consiste em autofecundar centenas de plantas selecionadas dentro de uma variedade de polinização livre ou híbrido promissor, avaliar suas progênies em fileiras, fazendo-se a seleção entre e dentro delas, repetindo-se o processo durante, no mínimo, cinco gerações sucessivas de autofecundação. As plantas são selecionadas pelos seus caracteres fenotípicos, tais como vigor, resistência às doenças e aos insetos-praga, resistência ao acamamento, altura da planta, rendimento, etc.
a.2) Método da cova única – O método da cova única é muito semelhante ao método padrão. A única diferença é que cada progênie é representada por uma única cova com três plantas ao invés de uma fileira com várias plantas (geralmente em torno de 25 - 30 plantas). Em função disso, reduz-se a área plantada e diminui-se o custo de obtenção de linhagens endogâmicas. Além disso, faculta ao fitomelhorista trabalhar com um número muito maior de progênies (dez vezes mais em relação ao método padrão, que é de 1.000 linhagens), aumentando-se, assim, a possibilidade de seleção entre progênies. Mesmo assim, esse método não tem sido muito usado na prática.
a.3) Método genealógico – O método genealógico consiste no cruzamento entre duas linhagens que se combinam bem, a fim de obter novas linhagens de segundo ciclo do híbrido resultante por endogamia.
a.4) Método do híbrido críptico – O método do híbrido críptico corresponde, essencialmente, a um teste precoce de combinação através de cruzamentos entre plantas individuais. O método se baseia, portanto, na avaliação de famílias de irmãos germanos obtidas pelo cruzamento de plantas individuais S0 (não endógamas) e prolíficas. Ao
195

mesmo tempo, as plantas envolvidas no cruzamento são autofecundadas, utilizando-se a segunda espiga de cada planta.
As progênies de irmãos germanos (S0 x S0), assim obtidas, são avaliadas em ensaio de produção com repetições, para a identificação dos melhores cruzamentos. As progênies S1 correspondentes aos melhores cruzamentos são plantadas aos pares na geração seguinte, para a obtenção de novas progênies de irmãos germanos (S1 x S1), por cruzamentos entre plantas individuais de cada par de progênies S1. Esse processo continua até que se consiga o grau de endogamia desejado nas linhagens que serão utilizadas para a obtenção de híbridos simples (Sn x Sn).
O método foi delineado para a obtenção de pares de linhagens com boa capacidade de combinação, tais que, quando cruzadas, produzam híbridos superiores. Porém, as linhagens superiores podem, eventualmente, produzir bons híbridos quando em cruzamento com outras linhagens que não os seus pares específicos.
a.5) Método da seleção zigótica – O método da seleção zigótica, também chamado de método do híbrido meio críptico, é utilizado quando se dispõe de uma linhagem comercial, e o objetivo é desenvolver uma nova linhagem, a partir de uma população heterogênea e prolífica, para a obtenção de um híbrido superior.
As plantas da população são cruzadas com a linhagem padrão (L) e, ao mesmo tempo, autofecundadas. As progênies de irmãos germanos (S0 x L) são avaliadas em ensaio de produção com repetições, para a identificação dos melhores cruzamentos. As progênies S1 correspondentes aos melhores cruzamentos são cruzadas novamente com a linhagem padrão (S1 x L) e autofecundadas. O processo continua até que se consiga o grau de endogamia desejado na nova linhagem, para obtenção do híbrido (Sn x L).
196

Este método pode ser utilizado para a obtenção de um híbrido triplo, utilizando-se um híbrido simples comercial, em vez de linhagem, como progenitor constante.
a.6) Método de monoplóides duplicados – O método de monoplóides duplicados tem por objetivo a obtenção de linhagens 100% homozigóticas, a partir de monoplóides duplicados.
O procedimento do método é o seguinte: inicialmente é feito o cruzamento das plantas de uma população heterogênea com o marcador genético Rnf (embrião roxo), o qual funcionará como progenitor masculino, e seleção das sementes com embrião branco (monoplóides), ou seja, aquelas sementes que se originaram da não-fertilização na oosfera.
Em seguida, tais sementes são imersas em solução de colchicina por um determinado período de tempo, para promover-se a duplicação do número de cromossomos, e, posteriormente, serem plantadas. Como as plantas resultantes dessas sementes são menos vigorosas, apresentando folhas estreitas, etc., produzem em torno de 5 a 6 sementes, as quais serão plantadas para multiplicação, já como plantas diplóides (linhagens homozigóticas). Posteriormente, tais linhagens serão cruzadas com um testador (linhagem superior ou híbrido simples comercial), a fim de identificar as linhagens superiores.
Vale ressaltar que a freqüência de monoplóides é de 1/1.000, de que 1/10 é duplicado; desses 1/100 serve para milho híbrido, ou seja, por esse método de obtenção de linhagens, o fitomelhorista obtém uma linhagem 100% homozigótica para cada milhão de plantas da população original.
Trata-se de um método que, com possibilidade de se adotar na prática, permitiria economizar de um a três anos no processo de obtenção de linhagens, quando comparado com o método padrão.
197

b) Avaliação das linhagens endogâmicas – Na segunda etapa, a avaliação das linhagens endogâmicas é feita de duas maneiras: cruzamentos ou topcross.
b.1) Cruzamentos – A avaliação das linhagens por meio de cruzamentos é um processo muito trabalhoso e dispendioso, visto que as linhagens devem ser cruzadas entre si em todas as combinações possíveis e, com um número relativamente pequeno de linhagens, é grande o número de híbridos simples possíveis de serem sintetizados; tais híbridos deverão ser avaliados em ensaios de campo, os quais devem ser repetidos em mais de um ano agrícola e em mais de um local, para que se identifiquem as linhagens superiores. Por exemplo, com 15 linhagens, é possível sintetizar 105 híbridos simples, valor esse obtido pela seguinte fórmula (já apresentada no Capítulo 1 do Volume 3 – Estimação de Parâmetros Genéticos):
NHS = n (n - 1)/2
onde:n – número de linhagens.
Em função disso, esse método de avaliação de linhagens quase que não é utilizado na prática.
b.2) Topcross – O topcross é o método de avaliação de linhagens mais usado, visto que o mesmo não apresenta a limitação do método anterior. Consiste em cruzar todas as linhagens com um testador de ampla base genética, sendo, geralmente, uma variedade de polinização livre, em que se avalia a capacidade geral de combinação. Em seguida, os híbridos simples são avaliados em ensaios de campo, os quais devem ser repetidos em mais de um ano agrícola e em mais de um local, para se identificarem as linhagens superiores, ou seja, aquelas linhagens que dão maior vigor de híbrido com o testador.
198

Por exemplo, com 50 linhagens obtêm-se 50 híbridos simples para avaliar por esse método, número muito pequeno quando comparado com o método anterior (1.225 híbridos simples para avaliar).
c) Obtenção da variedade híbrida – A terceira etapa consiste na obtenção da variedade híbrida. Inicialmente, as linhagens superiores, que foram selecionadas na etapa anterior, são cruzadas entre si em todas as combinações possíveis para a produção dos híbridos simples. Em seguida, são sintetizados os híbridos duplos e/ou os híbridos triplos possíveis.
Entretanto, mesmo utilizando o topcross para a avaliação preliminar das linhagens (etapa anterior), o número final de linhagens geralmente ainda é muito grande para permitir uma avaliação dos híbridos duplos e/ou triplos possíveis. Se, por exemplo, tem-se 10 linhagens, é possível sintetizar 630 híbridos duplos e/ou 360 híbridos triplos, valores esses obtidos, respectivamente, pelas seguintes fórmulas (já apresentadas no Capítulo 1 do Volume 3 – Estimação de Parâmetros Genéticos):
NHD = n (n - 1)(n - 2)(n - 3)/8
e
NHT = n (n - 1)(n - 2)/2
onde:n – número de linhagens.
Vê-se, logo, que é uma tarefa muito difícil para o fitomelhorista sintetizar e avaliar os híbridos duplos e/ou híbridos triplos possíveis. Em função disso, pesquisas foram realizadas no sentido de encontrar um meio de prever o comportamento dos híbridos duplos e/ou híbridos triplos a
199

partir do comportamento dos híbridos simples, o que iria facilitar bastante o trabalho do fitomelhorista, além de reduzir bastante o custo de obtenção da variedade híbrida. Assim, chegou-se a um método de prever o comportamento dos híbridos duplos e/ou híbridos triplos a partir do comportamento dos híbridos simples não-parentais, por meio da seguinte expressão (já apresentada no Capítulo 1 do Volume 3 – Estimação de Parâmetros Genéticos):
M̄ =( X )(Y )
onde:M̄ – média da população descendente do cruzamento;X e Y – proporção de alelos de cada um dos genótipos que
serão cruzados para produzir a população de média M̄ .
Dessa forma, estima-se o comportamento de tais híbridos. Com base nas predições, o fitomelhorista irá sintetizar e avaliar somente aqueles híbridos que, de antemão, mostraram ser agronomicamente superiores, dos quais surgirão as novas variedades híbridas.
As FIGURAS 21, 22 e 23 apresentam os esquemas das etapas para a obtenção de híbridos simples, triplos e duplos na cultura do milho, respectivamente.
200

FIGURA 21 – ESQUEMA DAS ETAPAS PARA OBTENÇÃO DE HÍBRIDOS SIMPLES NA CULTURA DO MILHO
Variedade de Polinização Livre ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
utofecundação das plantas mais vigorosas para obtenção das progênies S1, dando início a síntese de linhagens endogâmicas ↓
Avaliação das progênies autofecundadas até a 5ª geração, em fileiras ↓ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣S1 ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 1 2 3 4 5 6 7 8 9 10 ↓ ↓ ↓ ↓ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣S2 ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 1.2 3.2 3.4 6.1 6.4 8.3 9.4 10.2 ↓ ↓ ↓ ♣ ♣ ♣ ♣ ♣ ♣ ♣
S3 ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
1.2.3 3.2.1 3.4.1 3.4.4 8.3.3 8.3.4 10.2.2 ↓ ↓ ↓ ↓ ♣ ♣ ♣ ♣ ♣ ♣ ♣
S4 ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 1.2.3.2 1.2.3.4 3.4.1.3 3.4.4.2 8.3.3.2 8.3.3.3 10.2.2.4 ↓ ↓ ↓ ↓ ↓ ↓ ♣ ♣ ♣ ♣ ♣ ♣ ♣
S5 ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 1.2.3.2.3 1.2.3.4.1 3.4.1.3.1 3.4.4.2.4 8.3.3.2.2 10.2.2.4.1 10.2.2.4.3 (1) (2) (3) (4) (5) (6) (7) ↓ Lote de Despendoamento ↓ ♠ ♣ ♣ ♠ ♣ ♣ ♠ ♣ ♣ ♠ ♣ ♠ ♠ ♣ ♣ ♠ ♣ ♣ ♠ ♣ ♣ ♠ ♣ ♠ ♠ ♣ ♣ ♠ ♣ ♣ ♠ ♣ ♣ ♠ ♣ ♠ ♠ ♣ ♣ ♠ ♣ ♣ ♠ ♣ ♣ ♠ ♣ ♠ T 1 2 T 3 4 T 5 6 T 7 T
Plantio das linhagens endogâmicas e cruzamento de todas elas com o testador de ampla base genética ↓
Avaliação dos híbridos simples em ensaios de campo H7 H5 H2 H4 H1 H3 H6
H5 H7 H6 H1 H4 H2 H3
H1 H2 H3 H6 H7 H5 H4
Seleção das linhagens endogâmicas superiores ↓ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 1 3 4 6 7 Plantio das linhagens endogâmicas selecionadas e cruzamento entre si em todas as combinações possíveis ↓ Avaliação dos híbridos simples em ensaios de campo HS13 HS14 HS16 HS17 HS34 HS36 HS37 HS46 HS47 HS67
HS17 HS37 HS67 HS13 HS47 HS14 HS46 HS34 HS16 HS36
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ HS34 HS47
Variedades Híbridas
201

FIGURA 22 – ESQUEMA DAS ETAPAS PARA OBTENÇÃO DE HÍBRIDOS TRIPLOS NA CULTURA DO MILHO
Variedade de Polinização Livre ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
utofecundação das plantas mais vigorosas para obtenção das progênies S1, dando início a síntese de linhagens endogâmicas ↓
Avaliação das progênies autofecundadas até a 5ª geração, em fileiras ↓ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣S1 ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 1 2 3 4 5 6 7 8 9 10 ↓ ↓ ↓ ↓ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣S2 ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 1.2 3.2 3.4 6.1 6.4 8.3 9.4 10.2 ↓ ↓ ↓ ♣ ♣ ♣ ♣ ♣ ♣ ♣
S3 ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
1.2.3 3.2.1 3.4.1 3.4.4 8.3.3 8.3.4 10.2.2 ↓ ↓ ↓ ↓ ♣ ♣ ♣ ♣ ♣ ♣ ♣
S4 ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 1.2.3.2 1.2.3.4 3.4.1.3 3.4.4.2 8.3.3.2 8.3.3.3 10.2.2.4 ↓ ↓ ↓ ↓ ↓ ↓ ♣ ♣ ♣ ♣ ♣ ♣ ♣
S5 ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 1.2.3.2.3 1.2.3.4.1 3.4.1.3.1 3.4.4.2.4 8.3.3.2.2 10.2.2.4.1 10.2.2.4.3 (1) (2) (3) (4) (5) (6) (7) ↓ Lote de Despendoamento ↓ ♠ ♣ ♣ ♠ ♣ ♣ ♠ ♣ ♣ ♠ ♣ ♠ ♠ ♣ ♣ ♠ ♣ ♣ ♠ ♣ ♣ ♠ ♣ ♠ ♠ ♣ ♣ ♠ ♣ ♣ ♠ ♣ ♣ ♠ ♣ ♠ ♠ ♣ ♣ ♠ ♣ ♣ ♠ ♣ ♣ ♠ ♣ ♠ T 1 2 T 3 4 T 5 6 T 7 T
lantio das linhagens endogâmicas e cruzamento de todas elas com o testador de ampla base genética ↓
Avaliação dos híbridos simples em ensaios de campo H7 H5 H2 H4 H1 H3 H6
H5 H7 H6 H1 H4 H2 H3
H1 H2 H3 H6 H7 H5 H4
Seleção das linhagens endogâmicas superiores ↓ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 1 3 4 6 7 Plantio das linhagens endogâmicas selecionadas e cruzamento entre si em todas as combinações possíveis ↓ Cruzamento dos híbridos simples com as linhagens endogâmicas para obtenção dos híbridos triplos ↓
Avaliação dos híbridos triplos em ensaios de campo HT5 HT12 HT9 HT11 HT2 HT10 HT7 HT1 HT8 HT6 HT4 HT3 HT11 HT10 HT8 HT3 HT1 HT4 HT7 HT5 HT2 HT9 HT6 HT12
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ HT(13) 7 HT(34) 7
Variedades Híbridas
202

FIGURA 23 – ESQUEMA DAS ETAPAS PARA OBTENÇÃO DE HÍBRIDOS DUPLOS NA CULTURA DO MILHO
Variedade de Polinização Livre ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
utofecundação das plantas mais vigorosas para obtenção das progênies S1, dando início a síntese de linhagens endogâmicas ↓
Avaliação das progênies autofecundadas até a 5ª geração, em fileiras ↓ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣S1 ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 1 2 3 4 5 6 7 8 9 10 ↓ ↓ ↓ ↓ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣S2 ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 1.2 3.2 3.4 6.1 6.4 8.3 9.4 10.2 ↓ ↓ ↓ ♣ ♣ ♣ ♣ ♣ ♣ ♣
S3 ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
1.2.3 3.2.1 3.4.1 3.4.4 8.3.3 8.3.4 10.2.2 ↓ ↓ ↓ ↓ ♣ ♣ ♣ ♣ ♣ ♣ ♣
S4 ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 1.2.3.2 1.2.3.4 3.4.1.3 3.4.4.2 8.3.3.2 8.3.3.3 10.2.2.4 ↓ ↓ ↓ ↓ ↓ ↓ ♣ ♣ ♣ ♣ ♣ ♣ ♣
S5 ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 1.2.3.2.3 1.2.3.4.1 3.4.1.3.1 3.4.4.2.4 8.3.3.2.2 10.2.2.4.1 10.2.2.4.3 (1) (2) (3) (4) (5) (6) (7) ↓ Lote de Despendoamento ↓ ♠ ♣ ♣ ♠ ♣ ♣ ♠ ♣ ♣ ♠ ♣ ♠ ♠ ♣ ♣ ♠ ♣ ♣ ♠ ♣ ♣ ♠ ♣ ♠ ♠ ♣ ♣ ♠ ♣ ♣ ♠ ♣ ♣ ♠ ♣ ♠ ♠ ♣ ♣ ♠ ♣ ♣ ♠ ♣ ♣ ♠ ♣ ♠ T 1 2 T 3 4 T 5 6 T 7 T
Plantio das linhagens endogâmicas e cruzamento de todas elas com o testador de ampla base genética ↓
Avaliação dos híbridos simples em ensaios de campo H7 H5 H2 H4 H1 H3 H6
H5 H7 H6 H1 H4 H2 H3
H1 H2 H3 H6 H7 H5 H4
Seleção das linhagens endogâmicas superiores ↓ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 1 3 4 6 7 Plantio das linhagens endogâmicas selecionadas e cruzamento entre si em todas as combinações possíveis ↓
Cruzamento dos híbridos simples para obtenção dos híbridos duplos ↓
Avaliação dos híbridos duplos em ensaios de campo HD(13)(46) HD(13)(47) HD(13)(67) HD(14)(36) HD(14)(37) HD(14)(67) HD(16)(34) HD(16)(37) HD(16)(47) HD(17)(34) HD(17)(36) HD(17)(46) HD(34)(67) HD(36)(47) HD(37)(46)
HD(13)(47) HD(13)(46) HD(34)(67) HD(17)(36) HD(14)(67) HD(13)(67) HD(16)(47) HD(16)(34) HD(36)(47) HD(17)(46) HD(17)(34) HD(37)(46) HD(16)(37) HD(14)(36) HD(14)(37)
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ HD(13)(4 7) HD(34)(6 7)
Variedades Híbridas
203

4 Métodos de Melhoramento de Espécies Propagadas Assexuadamente
No Grupo 4, incluem-se os seguintes métodos de melhoramento: seleção clonal; autofecundação; hibridação; e policruzamento.
4.1 Método da seleção clonal
Esse método deve ser usado para se obterem novas variedades, a partir de populações heterogêneas, formadas por diversos clones, as quais são mantidas pelos agricultores por muitos anos. O sucesso do método depende da existência de genótipos superiores na população original. Esse método pode ser conduzido de forma massal ou individual, ou seja, seleção clonal massal ou seleção clonal individual.
4.1.1 Método da seleção clonal massal
Esse método consiste em selecionar, numa população heterogênea, as melhores plantas de fenótipos semelhantes, retirar das mesmas os órgãos vegetativos usados na multiplicação da espécie, os quais podem ser, dependendo da espécie, rizomas, tubérculos, colmo, estolho, ramo, bubilho, etc., e misturá-los, dando origem à nova variedade.
A variedade obtida por esse método é constituída por uma mistura de clones, conforme apresentado na FIGURA 24.
O referido método inicia-se com o plantio de uma população heterogênea, em que a densidade de plantio e os tratos culturais são os mesmos do cultivo comercial da espécie em consideração.
Geralmente, o tamanho da população inicial fica em torno de 2.000 a 3.000 plantas, o qual dependerá de vários fatores, entre eles, os meios materiais de que dispõe o fitomelhorista e a heterogeneidade da população original.
204
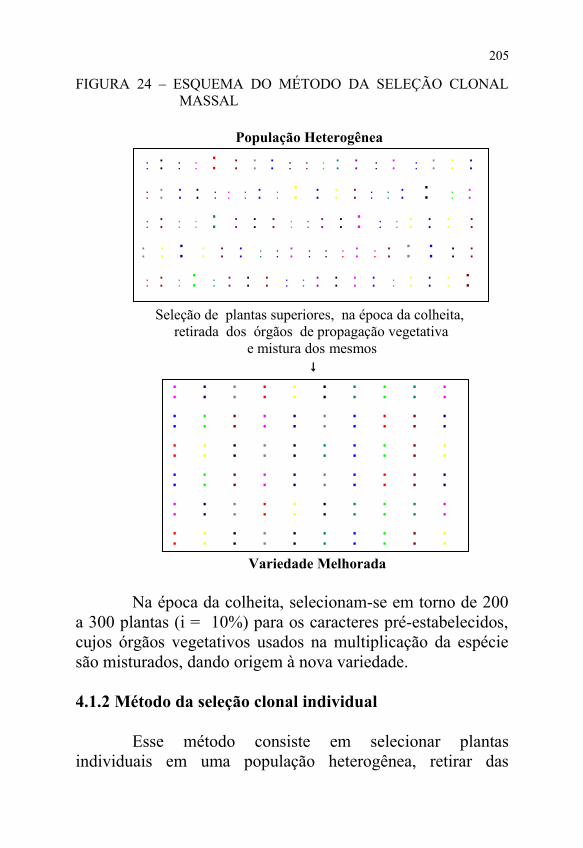
FIGURA 24 – ESQUEMA DO MÉTODO DA SELEÇÃO CLONAL MASSAL
População Heterogênea
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ Seleção de plantas superiores, na época da colheita, retirada dos órgãos de propagação vegetativa e mistura dos mesmos ↓
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
Variedade Melhorada
Na época da colheita, selecionam-se em torno de 200 a 300 plantas (i = 10%) para os caracteres pré-estabelecidos, cujos órgãos vegetativos usados na multiplicação da espécie são misturados, dando origem à nova variedade.
4.1.2 Método da seleção clonal individual
Esse método consiste em selecionar plantas individuais em uma população heterogênea, retirar das
205

mesmas os órgãos vegetativos usados na multiplicação da espécie, avaliar os clones em ensaios de campo e selecionar os mais satisfatórios, que vão constituir as novas variedades.
Cada variedade obtida por esse método é constituída por um só clone.
A seleção clonal individual envolve duas etapas distintas:
a) seleção de plantas individuais dentro da população heterogênea e plantio dos clones em linhas;
b) realização dos ensaios de produção, por dois a três anos, no mínimo, para serem eleitos os clones que vão constituir as novas variedades, conforme apresentado na FIGURA 25.
A primeira etapa inicia-se com o plantio da população heterogênea que vai sofrer seleção, espaçando-se bem as plantas para permitir um melhor desenvolvimento e facilitar o exame das mesmas.
O tamanho da população inicial deve ser o mesmo do método anterior , o qual dependerá de vários fatores, entre eles, os meios materiais de que dispõe o fitomelhorista e a heterogeneidade da população original.
Durante o desenvolvimento das plantas, cada uma é cuidadosamente examinada na busca das características desejáveis. Quando as plantas completarem o seu ciclo, ou seja, na época da colheita, serão selecionadas aquelas que apresentam as características desejáveis, cujo número é o mesmo do método anterior.
Essa é a fase mais crítica dessa etapa, pois cada planta selecionada corresponde a um clone e, se os mesmos não representarem os genótipos superiores da população inicial, todo o trabalho subseqüente não será capaz de compensar aquelas ausências iniciais.
Das plantas selecionadas, são retirados os órgãos vegetativos usados na multiplicação da espécie para plantio dos clones em linhas.
206
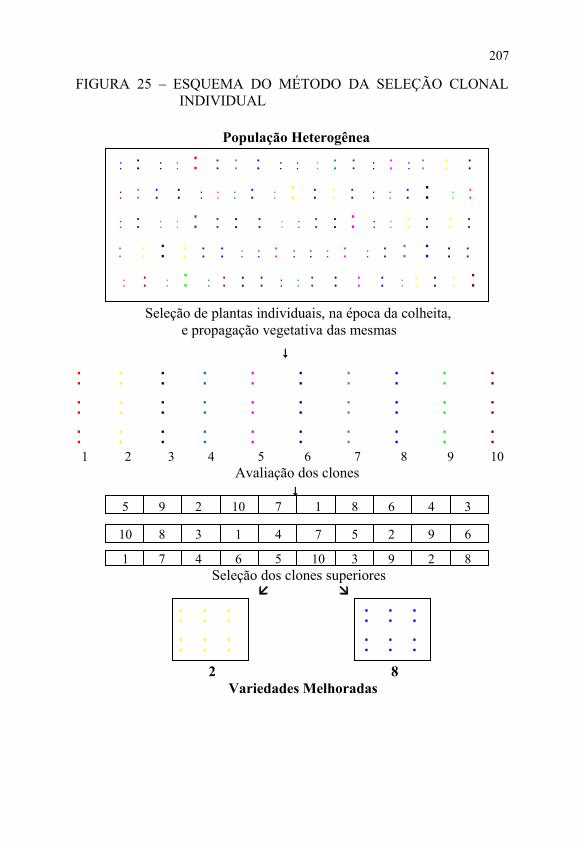
FIGURA 25 – ESQUEMA DO MÉTODO DA SELEÇÃO CLONAL INDIVIDUAL
População Heterogênea
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ Seleção de plantas individuais, na época da colheita, e propagação vegetativa das mesmas
↓
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 1 2 3 4 5 6 7 8 9 10 Avaliação dos clones ↓ 5 9 2 10 7 1 8 6 4 3
10 8 3 1 4 7 5 2 9 6
1 7 4 6 5 10 3 9 2 8 Seleção dos clones superiores
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
2 8 Variedades Melhoradas
207

A segunda etapa corresponde à realização dos ensaios de produção, por dois a três anos, no mínimo, a fim de serem comparados, entre si, e, também, com algumas das melhores variedades comerciais em cultivo na região, os clones quanto à capacidade produtiva e às outras características de valor agronômico. Esses ensaios são instalados em diversos locais da região para a qual se destinam as novas variedades.
Para esses ensaios, utiliza-se, geralmente, o delineamento em blocos casualizados, com quatro a seis repetições. Quando, porém, o número de clones é elevado, FERREIRA (2000) recomenda o uso do delineamento em blocos casualizados com alguns tratamentos comuns em vez dos reticulados, com duas a três repetições.
Os clones que tenham dado os melhores resultados nos ensaios de produção são multiplicados e distribuídos com os agricultores da região.
4.2 Método da autofecundação
Esse método consiste em se autofecundar plantas de uma população clonal, plantar as sementes botânicas e selecionar plantas individuais, retirar das mesmas os órgãos vegetativos usados na multiplicação da espécie, avaliar os clones em ensaios de campo e selecionar os mais satisfatórios, que vão constituir as novas variedades.
O referido método se baseia no fato de que a autofecundação, praticada em plantas propagadas assexuadamente, já libera a variabilidade genética presente na população na geração seguinte, uma vez que tais plantas são altamente heterozigóticas, dando imediata oportunidade à seleção.
O método da autofecundação envolve três etapas distintas:
a) autofecundação de plantas dentro da população clonal;
208

b) plantio das sementes botânicas, seleção de plantas individuais na população oriunda da autofecundação e plantio dos clones em linhas;
c) realização dos ensaios de produção, por dois a três anos, no mínimo, sendo eleitos os clones que irão constituir as novas variedades, conforme apresentado na FIGURA 26.
A primeira etapa inicia-se com o plantio da população clonal que vai sofrer autofecundação, espaçando-se bem as plantas para permitir um melhor desenvolvimento das mesmas.
O tamanho da população clonal deve ser tal que permita uma quantidade suficiente de sementes para produzir a geração seguinte no tamanho adequado.
Na época do florescimento, as plantas mais vigorosas serão autofecundadas; suas sementes serão colhidas, misturadas e guardadas para plantio no ano seguinte.
A segunda etapa inicia-se com o plantio das sementes botânicas, cuja população resultante vai sofrer seleção, num espaçamento mais amplo para permitir um melhor desenvolvimento das plantas e facilitar o exame das mesmas.
O tamanho dessa população deve ser o mesmo do método de seleção clonal, o qual dependerá dos fatores citados no método anterior.
Durante o desenvolvimento das plantas, cada uma é cuidadosamente examinada na busca das características desejáveis. Quando as plantas completarem o seu ciclo, ou seja, na época da colheita, serão selecionadas aquelas que apresentam as características desejáveis, cujo número é o mesmo do método de seleção clonal.
Essa é a fase mais crítica dessa etapa, pois cada planta selecionada corresponde a um clone e, se os mesmos não representarem os genótipos superiores da população resultante da autofecundação, todo o trabalho subseqüente não será capaz de compensar aquelas ausências iniciais.
209

FIGURA 26 - ESQUEMA DO MÉTODO DA AUTOFECUNDAÇÃO População Clonal ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
Autofecundação das plantas superiores, na época da colheita, coleta e mistura das sementes botânicas, e posterior plantio das mesmas ↓
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ Seleção de plantas individuais, na época da colheita, e propagação vegetativa das mesmas ↓
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 1 2 3 4 5 6 7 8 9 10 Avaliação dos clones ↓ 5 9 2 10 7 1 8 6 4 3
10 8 3 1 4 7 5 2 9 6
1 7 4 6 5 10 3 9 2 8 Seleção dos clones superiores
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 1 8
210

Variedades Melhoradas
Das plantas selecionadas, são retirados os órgãos vegetativos usados na multiplicação da espécie para plantio dos clones em linhas.
A terceira etapa corresponde à realização dos ensaios de produção, por dois a três anos, no mínimo, a fim de serem compados entre si, e, também, com algumas das melhores variedades comerciais em cultivo na região, os clones quanto à capacidade produtiva e às outras características de valor agronômico.
Esses ensaios são planejados e conduzidos de maneira idêntica ao método da seleção clonal individual.
Os clones que tenham dado os melhores resultados nos ensaios de produção são multiplicados e distribuídos com os agricultores da região.
4.3 Método da hibridação
Esse método consiste no cruzamento entre duas populações clonais, plantio das sementes botânicas e seleção de plantas individuais, sendo retirados das mesmas os órgãos vegetativos usados na multiplicação da espécie, avaliação dos clones em ensaios de campo e seleção dos mais satisfatórios, os quais vão constituir as novas variedades.
O referido método se baseia no fato de que o cruzamento praticado em plantas propagadas assexuadamente já libera a variabilidade genética presente na população na geração seguinte, uma vez que tais plantas são altamente heterozigóticas, dando imediata oportunidade à seleção.
O método da hibridação envolve três etapas distintas:a) cruzamento entre duas populações clonais; b) plantio das sementes botânicas, seleção de plantas
individuais na população oriunda do cruzamento e plantio dos clones em linhas;
211

c) realização dos ensaios de produção, por dois a três anos, no mínimo, sendo eleitos os clones que vão constituir as novas variedades, conforme apresentado na FIGURA 27.
A primeira etapa inicia-se com o plantio das duas populações clonais que vão sofrer cruzamento, espaçando-se bem as plantas para permitir um melhor desenvolvimento das mesmas.
O tamanho das duas populações clonais deve ser tal que permita uma quantidade suficiente de sementes para produzir a geração seguinte no tamanho adequado.
Na época do florescimento, as plantas mais vigorosas serão cruzadas, e suas sementes serão colhidas, misturadas e guardadas para plantio no ano seguinte.
A segunda etapa inicia-se com o plantio das sementes botânicas, cuja população resultante vai sofrer seleção, num espaçamento mais amplo para permitir um melhor desenvolvimento das plantas e facilitar o exame das mesmas.
O tamanho dessa população deve ser o mesmo do método de seleção clonal, o qual dependerá dos fatores já citados no referido método.
Durante o desenvolvimento das plantas, cada uma é cuidadosamente examinada na busca das características desejáveis. Quando as plantas completarem o seu ciclo, ou seja, na época da colheita, serão selecionadas aquelas que apresentam as características desejáveis, cujo número é o mesmo do método de seleção clonal.
Essa é a fase mais crítica dessa etapa, pois cada planta selecionada corresponde a um clone e, se os mesmos não representarem os genótipos superiores da população resultante do cruzamento, todo o trabalho subseqüente não será capaz de compensar aquelas ausências iniciais.
Das plantas selecionadas, são retirados os órgãos vegetativos usados na multiplicação da espécie para plantio dos clones em linhas.
212
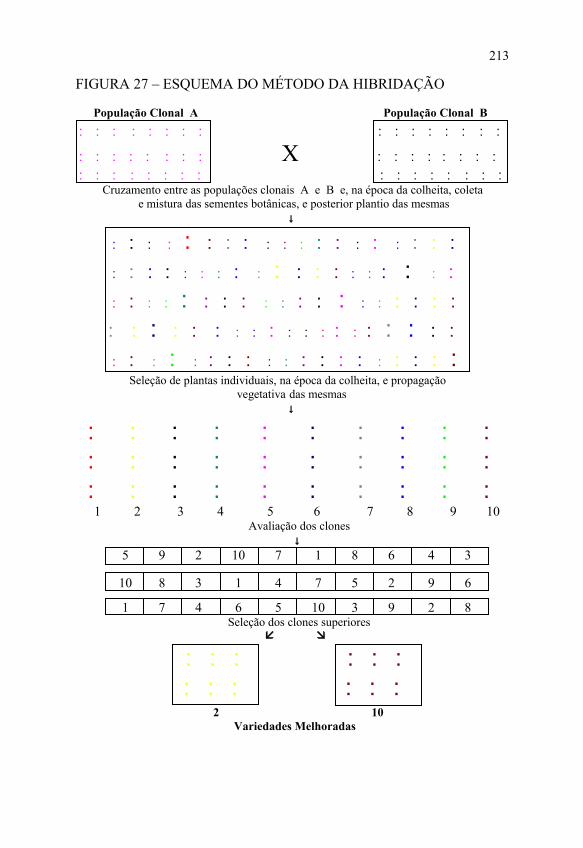
FIGURA 27 – ESQUEMA DO MÉTODO DA HIBRIDAÇÃO
População Clonal A População Clonal B ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ X ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ Cruzamento entre as populações clonais A e B e, na época da colheita, coleta e mistura das sementes botânicas, e posterior plantio das mesmas ↓
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ Seleção de plantas individuais, na época da colheita, e propagação vegetativa das mesmas ↓
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 1 2 3 4 5 6 7 8 9 10 Avaliação dos clones ↓ 5 9 2 10 7 1 8 6 4 3
10 8 3 1 4 7 5 2 9 6
1 7 4 6 5 10 3 9 2 8 Seleção dos clones superiores
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
2 10 Variedades Melhoradas
213

A terceira etapa corresponde à realização dos ensaios de produção, por dois a três anos, no mínimo, a fim de serem comparados entre si, e, também, com algumas das melhores variedades comerciais em cultivo na região, os clones quanto à capacidade produtiva e às outras características de valor agronômico.
Esses ensaios são planejados e conduzidos de maneira idêntica ao método da seleção clonal individual.
Os clones que tenham dado os melhores resultados nos ensaios de produção são multiplicados e distribuídos com os agricultores da região.
4.4 Método do policruzamento
Esse método consiste na seleção de clones numa população heterogênea, obtenção de progênies de meios-irmãos dos clones selecionados, avaliação dessas progênies em ensaios de campo, e recombinação dos melhores clones, que foram selecionados em função do comportamento de suas progênies, para formar a população melhorada, denominada de sintético, repetindo-se o processo.
Nesse método, cada ciclo envolve três etapas distintas:
a) obtenção das progênies de meios-irmãos dos clones selecionados;
b) avaliação das progênies de meios-irmãos em ensaio de produção;
c) recombinação dos melhores clones, conforme apresentado na FIGURA 28.
A primeira etapa inicia-se com o plantio de uma população heterogênea e posterior seleção de algumas plantas que vão ser propagadas vegetativamente. Dessa forma, de cada planta selecionada, são retirados os órgãos vegetativos usados na multiplicação da espécie.
214

FIGURA 28 – ESQUEMA DO MÉTODO DO POLICRUZAMENTO População Heterogênea
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
Seleção de plantas superiores e, na época da colheita, retirada dos órgãos de propagação vegetativa das mesmas, onde uma parte é usada para manter os clones em fileiras e a outra parte para formar o campo de policruzamento ↓ Clones em Fileira
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 1 2 3 4 5 6 7 8 9 10 ↓ Campo de Policruzamento
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ Na época da colheita, coleta das sementes botânicas de cada clone, individualmente ↓ Avaliação das progênies de meios-irmãos dos clones ↓
5 9 2 10 7 1 8 6 4 3
10 8 3 1 4 7 5 2 9 6
1 7 4 6 5 10 3 9 2 8 Seleção das progênies de meios-irmãos superiores e identificação dos melhores clones ↓ Recombinação dos melhores clones
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ 3 5 8 10 Na época da colheita, coleta das sementes botânicas de cada clone e mistura das mesmas, dando origem ao sintético ↓
♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣ ♣
Sintético (População Melhorada)
215

Uma parte desses órgãos vegetativos será plantada em fileira, para manter o clone, e a outra parte é usada para formar o campo de policruzamento.
No campo de policruzamento, serão colhidas as sementes botânicas de cada clone, individualmente, as quais vão representar as progênies de meios-irmãos.
A segunda etapa corresponde à realização do ensaio de produção, a fim de avaliar as progênies de meios-irmãos quanto à capacidade produtiva.
Nesse ensaio, utiliza-se, geralmente, o delineamento em blocos casualizados, com quatro a seis repetições. Quando, porém, o número de progênies é elevado, FERREIRA (2000) recomenda o uso do delineamento em blocos casualizados com alguns tratamentos comuns em vez dos reticulados, com duas a três repetições. Cada progênie de meios-irmãos, em geral, será representada por uma fileira de quatro a cinco metros de comprimento, em cada repetição.
Na época da colheita, cada fileira é colhida individualmente e avaliada a capacidade produtiva. A seguir, é feita a seleção das progênies que tenham dado os melhores resultados no ensaio de produção, a partir das quais serão identificados os melhores clones.
A terceira etapa corresponde à recombinação dos melhores clones. Órgãos vegetativos são retirados dos clones selecionados para plantio.
Na época do florescimento, esses clones são cruzados em todas as combinações possíveis, ou coletando-se os grãos de pólen de todas as plantas dos clones, misturando-os e polinizando manualmente todas as plantas dos referidos clones, ou deixando-se que eles se intercruzem naturalmente.
As sementes botânicas, resultantes da recombinação dos melhores clones, são colhidas e misturadas, dando origem ao sintético, que pode ser usado como nova população original para iniciar um novo ciclo de policruzamento.
216

REFERÊNCIAS
ALLARD, R. W. Princípios do melhoramento genético de plantas. Rio de Janeiro: USAID e Editora Edgard Blücher Ltda, 1971. 381p.
BRIM, C. A.; BURTON, J. W. Recurrent selection in soybeans. II. Selection for increased percent protein in seeds. Crop Science, v. 19, p.494-498, 1979.
BYRNE, I. ; RASMUSSON, D. C. Recurrente selection for mineral content in wheat and barley. Euphytica, v. 23, p.241-249, 1973.
CAVALERI, P. A. A semente. In: NEVES, O. S. et al. Cultura e adubação do algodoeiro. São Paulo: Instituto Brasileiro de Potassa, 1965. p.161-220.
DOGGETT, H. Recurrent selection in sorghum populations. Heredity, v.28, p.9-29, 1972.
ECKEBIL, J. P.Evaluation of sorghum random-mating populations trough S1 progeny testing. Lincoln: University of Nebraska, 1974. 160 p. Tese, Ph. D.
FERREIRA, P. V. Estatística experimental aplicada à agronomia. 3.ed. Maceió: Editora da Universidade Federal de Alagoas, 2000. 422p.
GARDNER, C. O. An evaluation of effects of mass selection and seed irradiation with thermal neutrons on yield of corn. Crop Science, v. 1, p.124-245, 1961.
HANSON, W. D.; PROBST, A. H.; CALDWELL, B. E. Evaluation of a population of soybeans genotypes with
217

implications for improving self-pollinated crops. Crop Science, v.7, p.99-103, 1967.
JAN-ORN, J. Estimates of genetic and environmental components of variance in some quantitative genetic traits from families derived from the NP3R random-mating sorghum population and their application in breeding systems. Lincoln: University of Nebraska, 1973. 163 p. Tese, Ph. D.
KENWORTHY, W. J.; BRIM, C. A. Recurrent selection in soybeans. I. Seed yield. Crop Science, v. 19, p.315-318, 1979.
KHADR, F. H.; FREY, K. J. Effectiveness of recurrent selection in oat breeding (Avena sativa L.). Crop Science, v. 5, p.349-354, 1965.
MATZINGER, D. F.; WERNSMAN, E. A.; COCKERHAM, C. C. Recurrent family selection and correlated response in Nicotiana tabacum L. I. “Dixie bright 244” X “Coker 139”. Crop Science, v. 12, p.40-43, 1972.
MEREDITH JUNIOR, W. R.; BRIDGE, R. R. Recurrent selection for lint percent within a cultivar of cotton (Gossypium hirsutum L.). Crop Science, v. 13, p.698-701, 1973.
MILLER, P. A.; RAWLINGS, J. O. Selection for increased lint yield and correlated responses in upland cotton (Gossypium hirsutum L.). Crop Science, v. 7, p.637-640, 1967.
PATERNIANI, E. Estudos recentes sobre heterose. Boletim da Fundação Cargill, n.1, p.1-36, 1974.
218

PATERNIANI, E.; IKUTA, H. Seleção recorrente recíproca em berinjela (Solanum melongena L.). Relatório Científico do Departamento e Instituto de Genética da ESALQ/USP, v. 7, p.168-173, 1973.
PATERNIANI, E.; MIRANDA FILHO, J. B. Melhoramento de populações. In: PATERNIANI, E. Melhoramento e produção de milho no Brasil. Piracicaba: Fundação Cargill, 1980. p.202-256.
TREVISAN, W. L. Avaliação de progênies de meios irmãos numa população de intercruzamento de sorgo (Sorghum bicolor (L.) Moench). Piracicaba: ESALQ/USP, 1976. 81 p. Dissertação, Mestrado.
VENCOVSKY, R. Herança quantitativa. In: PATERNIANI, E. Melhoramento e produção de milho no Brasil. Piracicaba: Fundação Cargill, 1980. p.122-201.
219

![Metodos y Modelos de Investigacion de Operaciones Vol I Modelos Deterministicos[1]](https://img.document.onl/doc/110x75/55cf96e3550346d0338e72d8/metodos-y-modelos-de-investigacion-de-operaciones-vol-i-modelos-deterministicos1.jpg)



























