
UNIVERSIDADE DE LISBOA
FACULDADE DE CIÊNCIAS
DEPARTAMENTO DE BIOLOGIA ANIMAL
Evolução das comunidades de macroinvertebrados bentónicos do estuário do Tejo
nas últimas duas décadas: efeitos das alterações nas redes de saneamento básico
e/ou resultado das alterações climáticas ?
Pedro Miguel Rodrigues de Sousa
Mestrado em Ecologia Marinha
Dissertação orientada por:
Doutor José Lino Costa
Doutora Paula Chainho
2016

i
Agradecimentos
Aos Doutores José Lino Costa e Paula Chainho pela orientação, conselhos e apoio prestados
ao longo deste ano de trabalho.
Ao Instituto MARE-ULisboa, nomeadamente todos os elementos e voluntários que estiveram
ligados aos projetos realizados em diversas áreas do estuário, e sem os quais também não teria sido
possível obter o material biológico e dados essenciais a este trabalho.
À Gilda Silva e Maria João por terem sido fundamentais na minha aprendizagem de todo o
procedimento de recolha de amostras e respetiva triagem.
Aos meus colegas de licenciatura e mestrado que me acompanharam ao longo desta jornada
árdua.
Aos meus professores que me inspiraram e deram-me as bases que me possibilitaram chegar
aqui.
Aos meus amigos que estiveram sempre lá para me apoiar.
À minha família, sobretudo à minha mãe, pelo amor incondicional, por tudo o que me deram e
ensinaram, e por me permitirem chegar até aqui.
À minha namorada, Verónica Cotrim, por ser a minha companheira, o meu porto de abrigo, a
minha melhor amiga, e a minha motivação para dar o meu melhor todos os dias.

ii
Resumo
Os estuários apresentam-se como um dos ecossistemas aquáticos mais importantes devido ao seu valor
ecológico e elevada produtividade, em comparação com outros ecossistemas. São ambientes com uma
variabilidade e dinâmica única, sendo essenciais para as comunidades de peixes, macroinvertebrados,
aves e plantas. Os estuários oferecem diversos serviços ambientais, algo que influencia a população
humana a se fixar nestas áreas e tem como resultado uma elevada pressão humana sobre os ecossistemas
estuarinos. Para além do fator Homem, também as alterações climáticas, das quais resultam mais
fenómenos meteorológicos extremos e um aumento genérico da temperatura, têm um papel na alteração
destes ecossistemas. De entre os vários organismos presentes nos estuários, podem-se destacar os
macroinvertebrados bentónicos, pelas suas características intrínsecas, que permitem determinar a
qualidade ecológica da água desses sistemas. Trata-se de um grupo com baixa mobilidade e que vive
em estreita associação com o substrato, apresentando espécies com diferentes ciclos de vida e diferentes
níveis de tolerância a contaminações ou outras situações de stress, o que permite usá-los com indicadores
ambientais. Este estudo incidiu na área do estuário do Tejo e teve como objetivo analisar a estrutura e
dinâmica das comunidades de marcoinvertebrados para perceber se estas têm vindo a mudar nos últimos
anos, e se tal se confirmasse, tentar perceber se estas alterações se deviam a alterações da qualidade da
água ou a mudanças de temperatura devido a alterações climáticas. Para tal, recorreu-se a dados
existentes em bases de dados do MARE - Faculdade de Ciências da Universidade de Lisboa, recolhidos
no âmbito de vários projetos levados a cabo em diversas áreas do estuário, desde 1996. Apesar de nestes
projetos terem sido recolhidas amostras em várias estações do ano, ao longo deste trabalho foram apenas
utilizadas as amostras recolhidas no verão, para garantir uma maior homogeneidade temporal. Para fazer
a caracterização das comunidades de macroinvertebrados bentónicos e perceber as possíveis variações
na qualidade da água, foi utilizado o Índice Biótico Marinho (AMBI). Já para detetar um eventual
impacto das alterações climáticas nestas comunidades, foi analisada a distribuição global de cada
espécie, tendo em conta a sua afinidade por climas mais temperados ou subtropicais. Os resultados do
presente trabalho mostraram a ocorrência de alterações nas comunidades de invertebrados bentónicos
no estuário do Tejo, nos últimos anos, tanto na área intermédia, como nas baías e área inferior do
estuário. Uma análise da relação dessas alterações com a qualidade ecológica e a distribuição geográfica
das espécies identificadas permitiu perceber que se verificou simultaneamente uma melhoria da
qualidade da água do estuário e uma alteração da composição das comunidades relativamente às suas
afinidades climáticas, com um aumento da representatividade das espécies com afinidades mais
meridionais. No entanto, essas variações foram mais notórias em áreas mais a montante e menos
profundas do estuário, onde a influência dos eventuais constrangimentos térmicos será sempre maior.
Palavas-chave: Estuário do Tejo, Macroinverterbrados bentónicos, Qualidade ecológica - AMBI,
Alterações climáticas, Distribuição geográfica das espécies.

iii
Abstract
Estuaries are considered as one of the most important aquatic ecosystems because of their ecological
value and high productivity, compared to other ecosystems. They are environments with a unique
variability and dynamics, and are essential for fish communities, macroinvertebrates, birds and plants.
Estuaries provide various environmental services, which influence the human population to settle in
these areas and results in a high human pressure on estuarine ecosystems. In addition to the human
factor, climate change, from which results more extreme weather events and a general rise in
temperature, also has a role in changing these ecosystems. Benthic macroinvertebrates have inherent
characteristics, which make them suitable to assess the ecological status of these systems. This
taxonomic group has low mobility, living in close association with the substrate, featuring species with
different life cycles and different levels of tolerance to contamination or other stress situations, providing
the suitable characteristics to be used as environmental indicators. This study focused on the Tagus
estuary area and aimed to analyze the structure and dynamics of macroinvertebrate communities, to
understand if there have been significant changes in these communities in recent years. On a second
stage, if these changes have been confirmed, trying to figure out if they were due to changes in water
quality or temperature changes due to climate change. Benthic macroinvertebrates databases available
at MARE - Faculty of Science, University of Lisbon were used for this assessment. This data was
collected by several different projects carried out in various areas of the estuary since 1996. Although
in these projects samples have been collected in various seasons, only samples collected during summer
were used in this study to ensure greater temporal uniformity. The AZTI Marine Biotic Index (AMBI)
was used to assess the ecological status based on benthic ivertebrate communities and identify possible
variations in water quality. The overall distribution of each species was examined to detect any impact
of climate change in these communities, ased on their affinity for more temperate or subtropical
climates. The results of this study showed the occurrence of changes in benthic invertebrate communities
in the Tagus estuary in recent years, both in the intermediate area, as in the bays and downstream area
of the estuary. An analysis of the relationship of these changes to the ecological quality and the
geographical distribution of species identified indicated a simultaneous improvement of the estuary
water quality and a change in the composition of communities in relation to their climate affinities, with
an increased representation of species with more southern affinities. However, these changes were more
noticeable in areas farther upstream and shallower estuary, where any influence of the thermal
constraints will always be greater.
Keywords: Tagus estuary; Benthic macroinvertebrates; Ecological quality - AMBI; Climate change;
Geographical distribution of species

iv
ÍNDICE
1. INTRODUÇÃO ..............................................................................................................................1
2. MATERIAL E MÉTODOS ...........................................................................................................5
2.1. Amostragem ........................................................................................................................... 5
2.1.1. Séries temporais na área inferior do estuário ................................................................. 5
2.1.2. Séries temporais na área intermédia do estuário ........................................................... 6
2.1.3. Série espacial do gradiente estuarino com um intervalo temporal longo ..................... 7
2.2 Processamento Laboratorial ................................................................................................ 8
2.3. Análise de dados .................................................................................................................... 9
3. RESULTADOS ............................................................................................................................ 12
3.1. Séries temporais na área inferior do estuário ................................................................... 12
3.1.1. Caracterização geral ....................................................................................................... 12
3.1.2 Evolução temporal .......................................................................................................... 13
3.2. Séries temporais na zona intermédia do estuário ............................................................. 22
3.2.1. Caracterização geral ....................................................................................................... 22
3.2.2. Evolução temporal .......................................................................................................... 23
3.3. Séries ao longo do gradiente estuarino com um intervalo temporal longo .................... 31
3.3.1. Caracterização geral ...................................................................................................... 31
3.3.2. Evolução temporal .......................................................................................................... 34
4. DISCUSSÃO ................................................................................................................................ 40
5. CONSIDERAÇÕES FINAIS ...................................................................................................... 44
6. REFERÊNCIAS BIBLIOGRÁFICAS ...................................................................................... 45

1
1. INTRODUÇÃO
Os estuários apresentam-se como um dos ecossistemas aquáticos mais importantes do Planeta
devido ao seu valor biológico e elevada produtividade, em comparação com outros ecossistemas
(Costanza et al., 1997). Isso deve-se à grande disponibilidade de nutrientes e de matéria orgânica e à
correspondente ação dos produtores primários que aí ocorrem. (McLuscky & Elliott, 2005). Por esse
motivo, as zonas costeiras, nas quais se incluem os estuários, proporcionam diversos e relevantes
serviços ambientais, o que contribuiu ao longo dos tempos para uma maior fixação da população humana
nestas áreas (Andrade et al., 2009). No entanto, durante este processo, o Homem acabou por modificar
significativamente esses sistemas salobros, tornando os estuários os meios aquáticos mais modificados
e ameaçados pela ação humana (Blaber et al., 2000).
Os estuários constituem meios aquáticos onde ocorre a transição entre ambientes fluviais e
marinhos, ou seja, são ambientes onde ocorre mistura de água doce e salgada. Estas condições fazem
com que tais ambientes apresentem uma variabilidade e uma dinâmica únicas (Wolanski, 2007). Para
além das variações constantes de salinidade e do nível das águas relacionadas com as marés, existem
outras variáveis ambientais que influenciam toda a dinâmica dos processos estuarinos, como flutuações
espaço-temporais na temperatura, turbidez, oxigénio dissolvido e tipo de substrato. (Vernberg, 1983).
Apesar desta variabilidade dos estuários, relacionada, sobretudo, com o fator salinidade, gerar
constrangimentos aos organismos vivos sob ponto de vista fisiológico, estes ecossistemas também
apresentam vantagens que beneficiam aquelas espécies de plantas e animais que conseguem colonizar
estes ambientes, tais como algumas aves, peixes e invertebrados, e no caso dos peixes e de alguns
invertebrados, sobretudo as suas formas juvenis, uma vez que a competição é aqui mais reduzida devido
à dificuldade inerente à sobrevivência neste tipo de sistemas (Vasconcelos et al., 2007). Isso acontece
porque os estuários oferecem abrigo à forte agitação marítima, devido à sua natureza abrigada, proteção
contra predadores devido à sua baixa profundidade e salinidade atenuada, grande disponibilidade de
alimento e grande diversidade de condições ambientais (Wolff, 1983; Haedrich, 1983; Miller et al.,
1985; Lenanton and Potter, 1987; Beck et al., 2001).
A presença humana junto aos estuários e áreas circundantes deixa estes sistemas vulneráveis às
atividades antropogénicas. Dentro destas, podem-se destacar como mais relevantes todas as alterações
físicas do estuário, as descargas de efluentes domésticos e industriais, a poluição de origem agrícola, a
navegação e atividades conexas, as dragagens para os mais diversos fins, a pesca e a aquacultura, a
introdução de espécies exóticas, entre outras. Estas ações afetam os estuários e resultam na degradação
geral do habitat e diminuição da qualidade da água, em particular. Consequentemente, estas alterações
nas condições do ambiente estuarino resultarão, muito frequentemente, em modificações na estrutura
das comunidades biológicas estuarinas, quer em termos espaciais, quer em termos temporais (Pearson
& Rosenberg, 1978).
No entanto, as alterações nos ecossistemas não se devem exclusivamente ao impacto direto das
atividades antropogénicas, pois atualmente estes encontram-se em constante mudança devido às
alterações climáticas, tratando-se normalmente de uma mudança mais lenta e gradual do que as
resultantes das atividades antropogénicas diretas, embora podendo provocar impactos pontuais
altamente significativos devido à ocorrência de eventos extremos cada vez mais frequentes. As
alterações climáticas são, por isso, eventos que alteram os ecossistemas à escala global, algo que afeta
naturalmente os seres vivos que aí ocorrem, obrigando-os a adaptarem-se para conseguirem sobreviver.

2
O presente estudo foi conduzido no estuário do Tejo, que é, não só o maior sistema estuarino de
Portugal, mas também o maior estuário da Europa Ocidental e um dos mais importantes da Europa.
Localizando-se perto da capital de Portugal, Lisboa (38º 44’ N, 9º 08’ W), apresenta uma área de cerca
de 325 km². Este estuário exibe também um comprimento bastante considerável, tendo uma extensão de
80 km desde o seu limite a montante, perto de Muge, até à foz, bem como uma largura máxima de 15
km (Costa, 1999).
A porção terminal do estuário está incluída na Área Metropolitana de Lisboa, região com maior
concentração populacional do país, com cerca de 2,5 milhões de habitantes. Pode-se, assim, concluir
que o estuário do Tejo é um sistema que se encontra bastante exposto à pressão humana, sobretudo
devido à presença do porto de Lisboa, diversos complexos industriais e várias outras infraestruturas
existentes à volta do estuário. A poluição resultante desta pressão humana e a redução dos caudais
dulciaquícolas, fruto da construção de barragens ao longo do rio, levou a alterações significativas neste
sistema salobro (Bettencourt et al., 2003).
O estuário do Tejo destaca-se pela sua elevada diversidade hidromorfológica, apresentando
grandes variações ao longo do seu percurso, nomeadamente em características como o tipo de
sedimento, profundidade e salinidade (ICNF, 2016). A área mais interior do estuário apresenta uma
morfologia deltaica, com baixa profundidade, extensas áreas de espraiado de maré e de sapal, e uma
reduzida intrusão salina. A área intermédia é mais profunda que a área interior, encontrando-se já
exposta a alguma intrusão salina. Por fim, a área terminal do estuário é aquela onde se faz sentir maior
influência das águas marinhas e do efeito da maré. Podendo atingir profundidades superiores a 40 m
(Oliveira, 1967), esta área do estuário apresenta normalmente uma salinidade muito semelhante à da
água do mar.
Esta elevada variabilidade das condições ambientais no estuário do Tejo está associada à
presença de diferentes comunidades de organismos em áreas diferentes do estuário. Portugal é uma área
de transição, biogeográfica, cuja fronteira ocorre sensivelmente entre o Cabo Carvoeiro, a norte, e o
estuário do Tejo, a sul (Hayden et al., 1984). De facto, nesta região ocorre uma mistura de floras e faunas
com maior influência setentrional, típicas de um clima temperado Atlântico, com outras de maior
influência meridional, típicas das regiões subtropicais do Mediterrâneo e do Atlântico (OSPAR
Commission, 2000; INAG, 2001). Estas características fazem do estuário do Tejo um ecossistema ideal
para estudar as potenciais alterações nas comunidades biológicas como resultado das alterações
climáticas.
De entre os vários grupos de organismos presentes nos estuários, podem-se destacar os
marcoinvertebrados como um grupo com características únicas. Estes desempenham um papel
importante na produtividade destes sistemas, sendo, não só bastante importantes para o funcionamento
das respetivas cadeias tróficas, mas também na dinâmica e reciclagem de nutrientes, na transformação
e decomposição de matéria orgânica e no fluxo de energia destes ecossistemas (Rhoads & Young, 1970;
Day et al., 1989). São ainda ferramentas muito úteis para avaliar o estado ecológico de um determinado
ecossistema e o impacto das atividades antropogénicas.
Os marcoinvertebrados apresentam uma mobilidade reduzida e habitam junto ao sedimento,
sendo que é nestes sedimentos que se acumulam as maiores concentrações de contaminantes, como por
exemplo partículas orgânicas ou inorgânicas e metais (Hartley & Dicks, 1987). A acumulação de
contaminantes e a baixa mobilidade faz com que a respetiva comunidade de macroinvertebrados
bentónicos acabe por refletir de forma relativamente fidedigna as condições ambientais deste meio, já
que estes organismos se encontram mais vulneráveis a perturbações locais. Para além disto, os
macroinvertebrados respondem rapidamente a stresses de origem antropogénica (Pearson and

3
Rosenberg, 1978), por terem ciclos de vida razoavelmente curtos, sendo por isso cada vez mais
utilizados para determinar a qualidade dos sistemas aquáticos (Attrill & Depledge, 1997).
Os macroinvertebrados integram diferentes taxa, logo, espécies com diferentes durações de
ciclo de vida e diferentes níveis de tolerância a contaminações ou outras situações de stress. Estas
características permitem usar os marcoinvertebrados como indicadores de alterações ambientais e do
impacto de contaminantes (Lyra et al., 2006). Os diferentes níveis de tolerância e adaptabilidade dos
macroinvertrebrados permitem, assim, obter uma resposta relativamente à extensão e magnitude dos
impactos criados pela ação do Homem, ao considerarem-se as diferentes características das várias
espécies em termos de adaptabilidade e resiliência, utilizando-se, deste modo, estes organismos como
indicadores de qualidade (Caeiro et al., 2005).
No entanto, a variabilidade espacial e temporal destas comunidades pode dificultar a distinção
clara entre o que são efeitos dessas diferenças espaciais e sazonais e efeitos da pressão antropogénica
(Boesch, 1973; Holland et al., 1987; Chainho et al., 2006).
A temperatura é um fator bastante relevante para a distribuição de cada espécie, tendo assim um
papel relevante na estrutura das comunidades e ecossistemas (Glynn, 1988). A temperatura afeta
parâmetros individuais dos organismos, tais como taxas de crescimento, longevidade e metabolismo,
bem como parâmetros populacionais, como a natalidade, a mortalidade ou o recrutamento (Pörtner et
al., 2008).
As alterações climáticas e os seus efeitos nos vários ecossistemas do Globo têm vindo a ser
analisadas com preocupação crescente, uma vez que se esperam alterações de temperatura e precipitação
significativas, associadas à ocorrência cada vez mais frequente de eventos meteorológicos extremos.
Em Portugal prevê-se que estas alterações se manifestem de forma mais acelerada em comparação com
a maioria das regiões do Globo (Intergovernmental Panel on Climate Change, 2001). Estes eventos
extremos, em particular a ocorrência de cheias e secas, afetam as características hidrológicas dos habitats
estuarinos, nomeadamente a temperatura, salinidade e regime de correntes.
Por esse motivo, os organismos aquáticos também são afetados por estes acontecimentos e
estima-se que não se encontram tão preparados para a adaptação a mudanças de temperatura, como a
maioria dos organismos terrestres. Isto deve-se ao facto da maioria dos organismos aquáticos serem
organismos poiquilotérmicos, ou seja, animais que não têm um mecanismo interno que regule a
temperatura do seu corpo.
As espécies marinhas só conseguem sobreviver dentro de certos limites térmicos, e visto que
não possuem um mecanismo interno que regule a temperatura corporal, esta acaba por influenciar o
habitat colonizado pelos organismos. Mesmo sem mecanismo regulador de temperatura, os animais
poiquilotérmicos podem apresentar comportamentos que influenciam a temperatura do corpo,
nomeadamente deslocarem-se para diferentes profundidades da coluna de água ou mesmo selecionarem
o habitat colonizado. Pode-se dizer, então, que a temperatura da água afeta e estabelece a distribuição e
abundância destes organismos, atuando sobretudo na sua fase mais precoce do desenvolvimento (Rozin
and Mayer, 1961; Neill et al., 1972; Neill and Magnuson, 1974).
Como referido anteriormente, os macroinvertebrados são bons indicadores da qualidade
ecológica da água, tendo por isso sido desenvolvidas inúmeras ferramentas (e.g. índices) utilizando estes
organismos para medir as alterações decorrentes da ação humana. Tem sido igualmente estudado o efeito
do stress natural resultante, por exemplo, da ocorrência de eventos extremos no uso dos indicadores de
qualidade (e.g. Chainho et al., 2007; Kröncke & Reiss, 2010; Neto et al., 2010). Já no que toca a
mudanças relacionadas com as alterações climáticas a longo prazo, alguns trabalhos realizados em

4
ambientes marinhos (e.g. Hiscock et al., 2004) apontam para uma expansão das áreas de distribuição de
algumas espécies, relacionada com as características do ciclo de vida dessas espécies e a sua capacidade
de dispersão. Já em ambientes estuarinos e para as espécies bentónicas, em particular, serão expectáveis
alterações mais lentas, uma vez estes organismos apresentam uma baixa mobilidade e muitos deles estão
relativamente confinados aos ambientes estuarinos e zonas costeiras adjacentes. Não existem estudos
sobre influência das alterações climáticas nas comunidades de macroinvertebrados do estuário do Tejo,
tendo sido estudados outros grupos de espécies existentes neste local, nomeadamente os peixes. Cabral
et al. (2001) e Vinagre et al. (2009) demonstraram que existe uma correlação entre a temperatura e a
distribuição geográfica de algumas espécies de peixes, afetando as variações térmicas sobretudo dos
indivíduos que se encontram nos limites da sua distribuição geográfica.
O objetivo principal deste estudo foi analisar as comunidades de macroinvertebrados em vários
locais do estuário do Tejo e testar a hipótese de nos últimos anos terem ocorrido alterações na estrutura
destas comunidades. E caso estas alterações se verificassem, investigar se tais mudanças se deveriam
principalmente a alterações na qualidade da água (devido ao investimento feito nos últimos anos para
melhorar as infraestruturas de saneamento da bacia) ou a uma expansão e/ou retração na distribuição
das espécies presentes (por causa de um incremento da temperatura devido ao fenómeno das alterações
climáticas).

5
2. MATERIAL E MÉTODOS
2.1. Amostragem
Este estudo foi realizado com base em séries temporais de dados previamente recolhidos no
âmbito de projetos desenvolvidos pelo Centro de Ciências do Mar e do Ambiente (MARE) da Faculdade
de Ciências da Universidade de Lisboa. Alguns destes projetos ainda estão em curso, pelo que o presente
estudo incluiu a realização de amostragens e triagens integradas nas equipas desses projetos.
Apesar de alguns dos projetos englobarem amostragens em diferentes épocas do ano, no
presente estudo foram utilizados apenas os dados correspondentes aos meses de verão, uma vez que é a
época que as comunidades de invertebrados refletem melhor as diferenças em termos de qualidade
ambiental (Chainho, 2008), e a consideração em simultâneo que amostras obtidas noutras épocas do ano
aumentaria a variabilidade intra-anual dos dados, tornando mais difícil a deteção dos padrões que se
pretendiam observar no âmbito do presente estudo.
Estas series temporais permitiram caracterizar a estrutura das comunidades de
macroinvertebrados em cada local e perceber a sua evolução, em contextos espácio-temporais distintos:
2.1.1. Séries temporais na área inferior do estuário
As amostragens realizadas na área inferior do estuário do Tejo foram enquadradas no âmbito do
projeto de Monitorização das comunidades biológicas da frente ribeirinha do Concelho de Almada.
Neste projeto fez-se o seguimento de duas zonas distintas, Portinho da Costa e Porto do Buxo, antes e
depois da entrada em funcionamento da ETAR municipal do Portinho da Costa, em 2003, e da
eliminação do efluente não tratado até então vertido diretamente para o meio estuarino no Porto do
Buxo, constituindo assim uma situação de referência imprescindível para acompanhar a evolução das
comunidades biológicas em função da alteração das condições da qualidade da água.
O Porto do Buxo e Portinho da Costa localizam-se na área terminal do estuário (Figura 1), sendo
esta a área do sistema que apresenta maior profundidade e uma influência maioritariamente marinha. Os
sedimentos de ambas as zonas são maioritariamente arenosos, como uma maior fração cascalhenta no
Porto do Buxo.
Em ambos os casos, a amostragem foi realizada com uma draga Day com uma área de ataque
de 0,1 m2. No Porto do Buxo foram recolhidas amostras em 9 estações de amostragem, distribuídas por
3 radiais e 3 transeptos (Figura 1). No Portinho da Costa as amostras foram recolhidas em 15 estações
de amostragem, distribuídas por 5 transeptos e 3 radiais. No entanto, relativamente ao Portinho da Costa,
é importante notar que, devido à ausência de dados em alguns anos, foram excluídos três dos 15 pontos
de amostragem. No presente estudo, para ambos os casos, foram utilizados os dados recolhidos no
período de 2002 a 2010.

6
Figura 1 – Localização das estações de amostragem da área inferior do estuário do Tejo, nas zonas do Porto do Buxo e
Portinho da Costa, concelho de Almada.
2.1.2. Séries temporais na área intermédia do estuário
Estas amostragens foram realizadas no âmbito dos projetos de Monitorização Biológica do
Parque das Nações (Expo) e dos efeitos das ETARs do Concelho de Lisboa, que tiveram como objetivos,
respetivos, acompanhar a evolução da situação ambiental da zona ribeirinha adjacente ao atual Parque
das Nações, após as intervenções de reabilitação que foram implementadas no âmbito da realização da
EXPO’98 e os efeitos do funcionamento das ETARs do Concelho de Lisboa no meio estuarino
adjacente. Neste caso, foram recolhidas amostras nas zonas intertidal e subtidal do Parque das Nações,
de 1998 a 2013, com exceção do ano 1999 para ambas as zonas, e 2002 para a zona subtidal. As recolhas
foram realizadas com uma draga van Veen, modelo SR, com uma área de ataque de 0,05 m2, em 12
estações de amostragem estabelecidas ao longo de uma grelha com 3 transeptos transversais à margem
e 4 radiais paralelos à margem, das quais 9 se localizaram na zona intertidal e as restantes 3 na zona
subtidal (Figura 2).
Esta zona do estuário do Tejo é caracterizada por zonas intertidais extensas, constituídas por
sedimentos predominantemente vasosos, e salinidade variável, de acordo com as flutuações diárias e
sazonais associadas à maior ou menor influência marinha ou dulciaquícola.

7
Figura 2 – Localização das estações de amostragem da área intermédia do estuário do Tejo, na zona intertidal e subtidal do
Parque das Nações (Expo).
2.1.3. Série espacial do gradiente estuarino com um intervalo temporal longo
Os dados incluídos nesta série foram recolhidos nas mesmas estações de amostragem em 2003
e 2014, com um intervalo temporal de 11 anos. Os primeiros foram obtidos no âmbito de uma tese de
doutoramento (Chainho, 2008), enquanto os últimos resultaram de amostragens realizadas no âmbito do
projeto Amêijoa-japonesa - Estado atual da população do estuário do Tejo, impactos e gestão da apanha.
As estações de amostragem foram agrupadas em diferentes zonas, de acordo com a sua
localização em diferentes áreas o estuário, com sete pontos correspondentes à zona intermédia do
estuário, quatro pontos às zonas das baías da margem sul, e as restantes três à zona inferior do estuário
(Figura 3).
Naturalmente, as estações da zona intermédia têm menos influência salina que as restantes. Pelo
contrário, as baías da margem sul apresentam genericamente batimetrias inferiores. O tipo de sedimento
é bastante variável mesmo dentro de cada agrupamento.

8
Figura 3 – Localização das estações de amostragem da série espacial do gradiente do estuário do Tejo, localizadas nas zonas
intermédia, inferior e baías.
2.2. Processamento Laboratorial
Após serem recolhidas, as amostras foram preservadas em formaldeído a 4%. No laboratório,
procedeu-se à lavagem das amostras de modo a retirar o fixador e o máximo de sedimento possível,
tendo este processo sido efetuado com recurso a um crivo com malha de 0,5 mm e sob fluxo de água.
Após o processo de lavagem, a fração da amostra que ficou retida no crivo foi colocada em
álcool a 70%, ao qual foi adicionado o corante Rosa de Bengala. Este corante fixa-se aos organismos,
dando a estes um tom rosa, o que facilita assim o processo da sua recolha e identificação durante o
processo da triagem.
As amostras foram então triadas, seguindo-se a contagem e identificação dos organismos ao
menor nível taxonómico possível.

9
2.3. Análise de dados
A caracterização da comunidade de macroinvertebrados bentónicos foi efetuada com base na
determinação do seu estado ecológico e da distribuição biogeográfica das espécies presentes nos
diversos locais de amostragem.
Tendo em conta que eventuais modificações nas comunidades de macroinvertebrados
bentónicos ao longo do período em estudo podem ter estado relacionadas com alterações em termos das
pressões de origem humana, começou por se analisar o estado ecológico destas comunidades ao longo
dos anos. Assim, para determinar o estado ecológico das diferentes estações de amostragem foi utilizado
o Índice Biótico Marinho - AMBI (AZTI’s Marine Biotic Index) (Borja et al., 2000). O AMBI foi
desenvolvido com o objetivo de avaliar o estado ecológico das águas estuarinas e costeiras, tendo como
base modelos ecológicos, como o desenvolvido por Glémarec & Hily (1981). A aplicabilidade deste
índice para medir as respostas das comunidades de macroinvertebrados a diversos tipos de pressão
antropogénica e em diferentes tipos de habitat tem vindo a ser testada em todo o Mundo (e.g. Muniz et
al., 2005; Muxika et al., 2005; Teixeira et al., 2012). É também um dos índices que integra grande parte
das abordagens multivariadas utilizadas atualmente na avaliação do estado ecológico pelos diversos
Estados Membros da UE, no âmbito da implementação da Diretiva-Quadro da Água (Borja et al., 2009).
O AMBI tem como base a classificação da macrofauna bentónica em cinco grupos, de acordo
com a sua sensibilidade e/ou tolerâncias às pressões antropogénicas. Foi inicialmente desenvolvido com
base na tolerância ao enriquecimento orgânico. Os cinco grupos resumem-se da seguinte forma: espécies
muito sensíveis a enriquecimento orgânico e presentes em condições de baixa poluição, espécies
indiferentes a enriquecimento orgânico e presentes em condições de baixa poluição, espécies tolerantes
a enriquecimento orgânico excessivo, espécies oportunistas de segunda ordem e espécies oportunistas
de primeira ordem.
Para além das espécies, os organismos que foram classificados apenas até ao género também
foram tidos em conta nesta análise, visto que o AMBI também oferece uma classificação a este nível
taxonómico. Para as espécies para as quais o AMBI não atribui uma classificação, por atualmente ainda
não existir informação suficiente para tal, foi atribuído a sigla NA (não atribuído).
Foi calculado um valor de AMBI para cada estação de amostragem e determinado um valor
médio e respetivo erro padrão para cada ano, em cada uma das áreas examinadas.
Tendo em conta que também foi considerada a hipótese de possíveis alterações das comunidades
de macroinvertebrados estudadas poderem estar relacionadas com alterações climáticas, foi identificada
a distribuição global das espécies recolhidas no estuário do Tejo, para perceber se a eco-região deste
sistema aquático faz parte da área de distribuição conhecida para as mesmas ou se estas espécies se
tratam de novas ocorrências nesta eco-região.
Para tal, recorreu-se à informação fornecida pelo World Register of Marine Species
(http://www.marinespecies.org/index.php), complementada com uma pesquisa aprofundada de outras
fontes de informação sobre a distribuição geográfica das espécies, incluindo artigos científicos,
dissertações académicas e relatórios de projetos, entre outros.
Para perceber os possíveis padrões de mudança das espécies foi considerada a escala geográfica
da eco-região, de acordo com Spalding et al. (2007). Segundo estes autores, o estuário do Tejo encontra-
se na Costa Atlântica do Sul da Europa, dentro da Província Lusitânia (Figura 4).

10
Figura 4 – As eco-regiões à escala global, de acordo com Spalding et al. (2007).
Foi atribuído o valor 1 às espécies cuja a distribuição conhecida inclui a Costa Atlântica do Sul
da Europa, sendo que espécies cuja distribuição é indicada exclusivamente a norte desta eco-região
foram classificados com o valor 0, e espécies com distribuição exclusivamente a sul, nas quais se
incluem espécies do Mar Mediterrâneo, foram classificadas com o valor 2. Este método de classificação
facilita uma interpretação integrada dos dados para o conjunto das espécies presentes, já que a atribuição
de valores médios às amostras é indicadora da maior representatividade de espécies de distribuição norte
quando o valor é inferior a 1, enquanto valores superiores a 1 indicam uma maior representatividade de
espécies de distribuição a sul da Costa Atlântica do Sul da Europa.
Para determinar os padrões temporais das comunidades bentónicas, assim como a relação desses
padrões com a qualidade ecológica e a distribuição geográfica das espécies presentes em cada zona,
dentro das áreas estudadas, efetuou-se uma Análise de Coordenadas Principais (PCO), usando o
software Primer 6 + PERMANOVA (Clarke & Gorley, 2006; Anderson et al., 2008). Para isso,
procedeu-se a uma transformação por raiz quadrada dos dados biológicos e construiu-se uma matriz de
semelhanças utilizando o coeficiente de Bray-Curtis.
Para identificar quais espécies que estavam associadas aos padrões identificados procedeu-se à
sobreposição de vetores sobre a ordenação, que representam as correlações de Pearson (p<0,05) das
espécies identificadas com os eixos da PCO. Foram ainda sobrepostos à ordenação vetores que
representam a correlação entre os valores da qualidade ecológica (AMBI: 0-7) e da distribuição
geográfica (0-2) com os eixos da matriz, para perceber a sua relação com os padrões identificados
Foram também realizadas Análises de Variância Permutacional (Permanova) (Anderson, 2001),
para testar se existiam diferenças temporais significativas para a composição taxonómica, AMBI e
distribuição geográfica usando o ano como o único fator fixo para as séries temporais, sendo que para
as séries espaciais teve-se como fatores o ano e as áreas. Os dados foram previamente normalizados e
foi construída uma matriz de semelhanças com base em distâncias Euclidianas. Nos casos em que foram
encontradas diferenças significativas entre os anos (p<0,05) foram realizados testes pareados a
posteriori para determinar quais os anos significativamente diferentes relativamente à composição
taxonómica, qualidade ecológica e distribuição geográfica das espécies.

11
Foram ainda efetuados testes G-de-independência (Sokal & Rohlf, 1995) para determinar se a
proporção de espécies e de indivíduos com maior afinidade setentrional e meridional era independente
do ano de colheita.

12
3. RESULTADOS
3.1. Séries temporais na área inferior do estuário
3.1.1. Caracterização geral
Nas amostragens realizadas no Porto do Buxo e Portinho da Costa, correspondentes às séries
temporais da área inferior do estuário, foram capturados um total de 340 taxa diferentes e um total de
44 904 indivíduos (Tabela 1).
Os Polychaeta foram o grupo taxonómico dominante, tanto em número de espécies (43,8%)
como em abundância (65,6%), seguindo-se os Bivalvia com 13,2% do total de taxa identificados e
13,1% de abundância, e Malacostraca com 16,1% do total de taxa identificados e 6,9% de abundância
(Figura 5). É importante ainda salientar que apesar de os Clitellata serem representados apenas por um
taxa (i.e. Oligochaeta), a sua abundância é considerável (7,7%). Os taxa dominantes foram Mediomastus
fragilis (26,9%), Pomatoceros sp. (17,5%), Oligochaeta (7,7%), Venerupis corrugata (5,6%) e
Nemertea (3,4%).
Tabela 1 – Número de taxa incluídos nas classes representadas e respectiva abundância, nos dados recolhidos durante as
amostragens realizadas entre 2002 e 2010 no Porto do Buxo e Portinho da Costa, área inferior do estuário do Tejo
Classe Número de taxa Número de indivíduos
Anthozoa 22 533
Bivalvia 45 5885
Clitellata 1 3476
Echinoidea 3 39
Gastropoda 42 1107
Malacostraca 55 3110
Maxillopoda 6 887
Ophiuroidea 4 224
Polychaeta 149 29496
Polyplacophora 2 16
Pycnogonida 7 72
Rhabditophora 4 59
Total 340 44904

13
Figura 5 – Composição relativa (número de taxa e abundância) das amostras recolhidas entre 2002 e 2010 no Porto do Buxo
e Portinho da Costa, área inferior do estuário do Tejo.
3.1.2. Evolução temporal
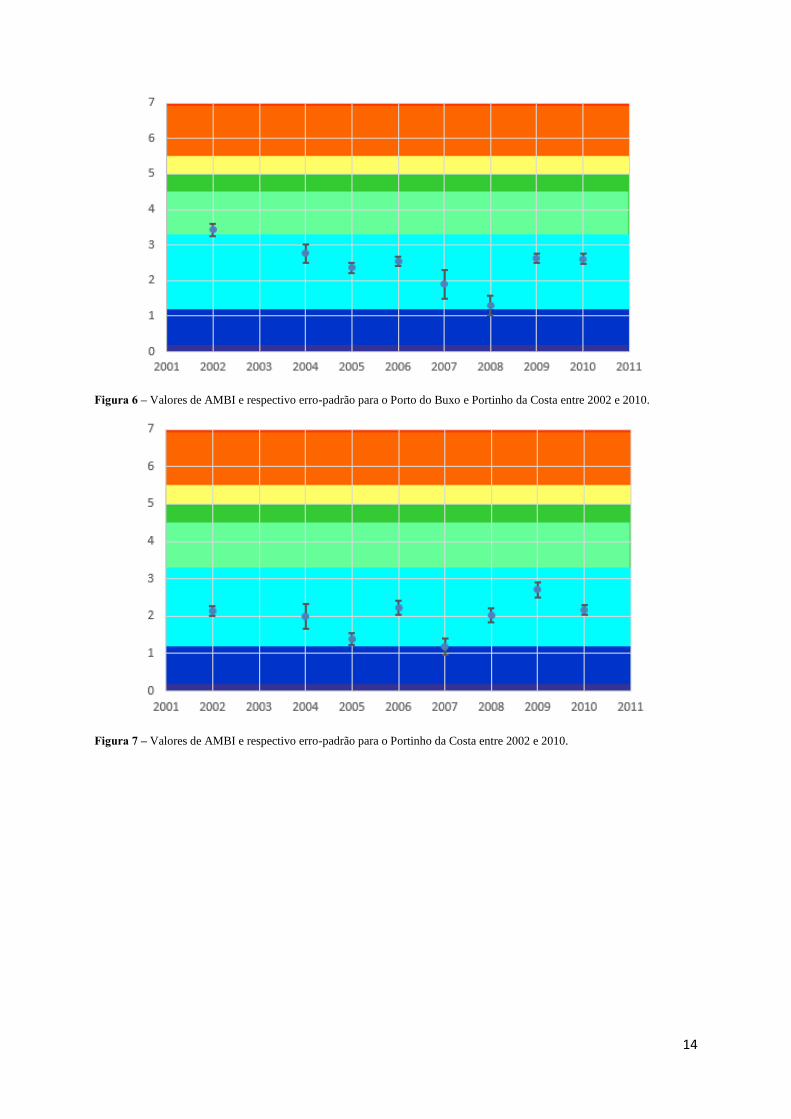
Os valores de AMBI obtidos para o Porto do Buxo e Portinho da Costa, as zonas onde foi
eliminada a descarga de efluentes e construída uma ETAR, respetivamente, mostram a ocorrência de
flutuações ao longo do tempo (Figuras 6 e 7). Os valores de AMBI oscilaram entre 1,30 e 3,43 no Porto
do Buxo e entre 1,19 e 2,71 no Portinho da Costa mas, apesar das variações temporais, esses valores
estiveram sempre dentro da gama de valores correspondentes à classificação do estado ecológico Bom.
Não obstante as flutuações de valores, no Porto do Buxo os valores do AMBI não voltam a atingir os
valores de 2002, havendo por isso uma melhoria relativamente ao estado inicial. O erro padrão mostra
que a variabilidade dentro das estações amostradas em cada local é relativamente baixa, sendo as
variações temporais muito mais acentuadas do que a variabilidade entre as estações de amostragem do
mesmo local.
14
Figura 6 – Valores de AMBI e respectivo erro-padrão para o Porto do Buxo e Portinho da Costa entre 2002 e 2010.
Figura 7 – Valores de AMBI e respectivo erro-padrão para o Portinho da Costa entre 2002 e 2010.

15
A distribuição geográfica com base no número de espécies para o Porto do Buxo não mostra
tendência evidente para aumento ou diminuição das espécies 0 e 2 (Figura 8). Em ambas as zonas
verificou-se uma maior percentagem de espécies 0 do que 2, sendo que no ano 2002 não se registou a
presença de espécies 2.
Figura 8 – Percentagem relativa do número de espécies do norte e espécies do sul no Porto do Buxo, concelho de Almada, no
período de 2002 e 2010: Espécies 0- Espécies com uma distribuição exclusivamente a norte da Costa Atlântica do Sul da
Europa; Espécies 2 – Espécies cuja a distribuição é exclusivamente a sul da Costa Atlântica do Sul da Europa.
As variações no número de indivíduos 0 e 2 são semelhantes ao que foi observado para o número
de espécies, com a ausência de uma tendência concreta (Figura 9). A abundância de espécies com diferentes
distribuições geográficas reforça ainda a maior abundância de espécies 0 do que 2, em particular nos anos
2005 e 2009.
Figura 9 – Percentagem relativa do número de indivíduos do norte e indivíduos do sul no Porto do Buxo, no Concelho de
Almada, entre 2002 e 2010: Indivíduos 0- Indivíduos com uma distribuição exclusivamente a norte da Costa Atlântica do Sul
da Europa; Indivíduos 2 – Indivíduos com uma distribuição exclusivamente a sul da Costa Atlântica do Sul da Europa.
16
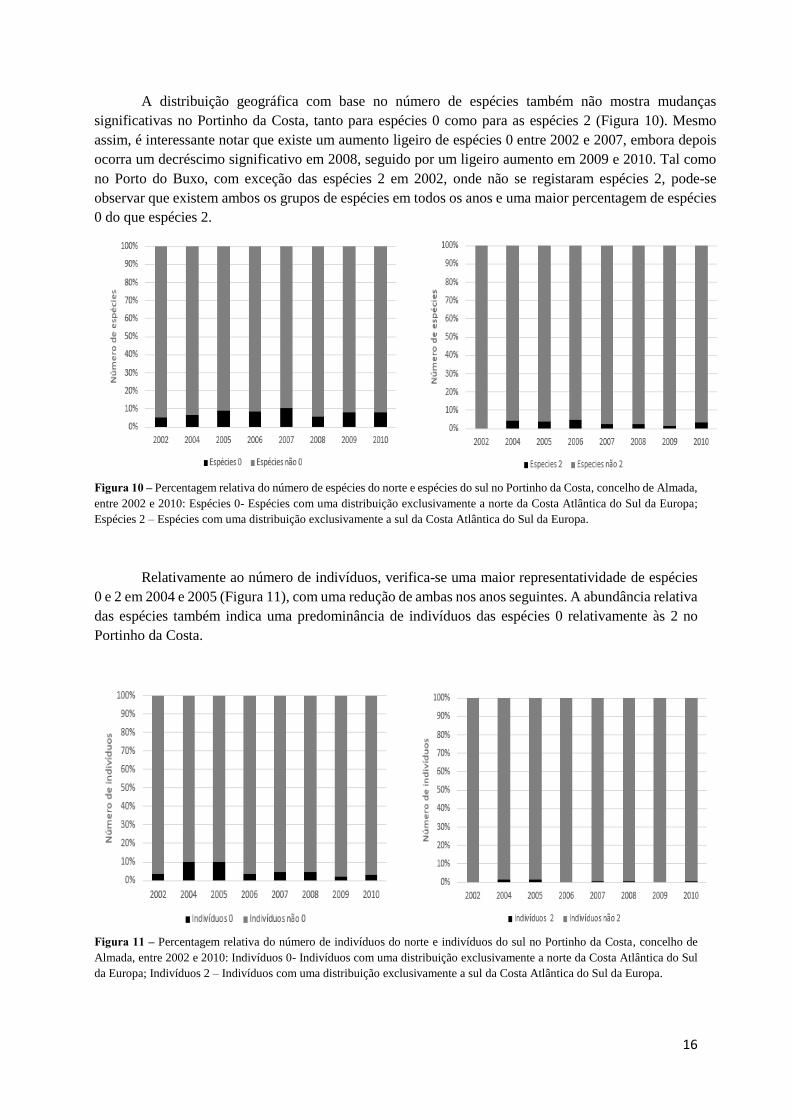
A distribuição geográfica com base no número de espécies também não mostra mudanças
significativas no Portinho da Costa, tanto para espécies 0 como para as espécies 2 (Figura 10). Mesmo
assim, é interessante notar que existe um aumento ligeiro de espécies 0 entre 2002 e 2007, embora depois
ocorra um decréscimo significativo em 2008, seguido por um ligeiro aumento em 2009 e 2010. Tal como
no Porto do Buxo, com exceção das espécies 2 em 2002, onde não se registaram espécies 2, pode-se
observar que existem ambos os grupos de espécies em todos os anos e uma maior percentagem de espécies
0 do que espécies 2.
Figura 10 – Percentagem relativa do número de espécies do norte e espécies do sul no Portinho da Costa, concelho de Almada,
entre 2002 e 2010: Espécies 0- Espécies com uma distribuição exclusivamente a norte da Costa Atlântica do Sul da Europa;
Espécies 2 – Espécies com uma distribuição exclusivamente a sul da Costa Atlântica do Sul da Europa.
Relativamente ao número de indivíduos, verifica-se uma maior representatividade de espécies
0 e 2 em 2004 e 2005 (Figura 11), com uma redução de ambas nos anos seguintes. A abundância relativa
das espécies também indica uma predominância de indivíduos das espécies 0 relativamente às 2 no
Portinho da Costa.
Figura 11 – Percentagem relativa do número de indivíduos do norte e indivíduos do sul no Portinho da Costa, concelho de
Almada, entre 2002 e 2010: Indivíduos 0- Indivíduos com uma distribuição exclusivamente a norte da Costa Atlântica do Sul
da Europa; Indivíduos 2 – Indivíduos com uma distribuição exclusivamente a sul da Costa Atlântica do Sul da Europa.
17
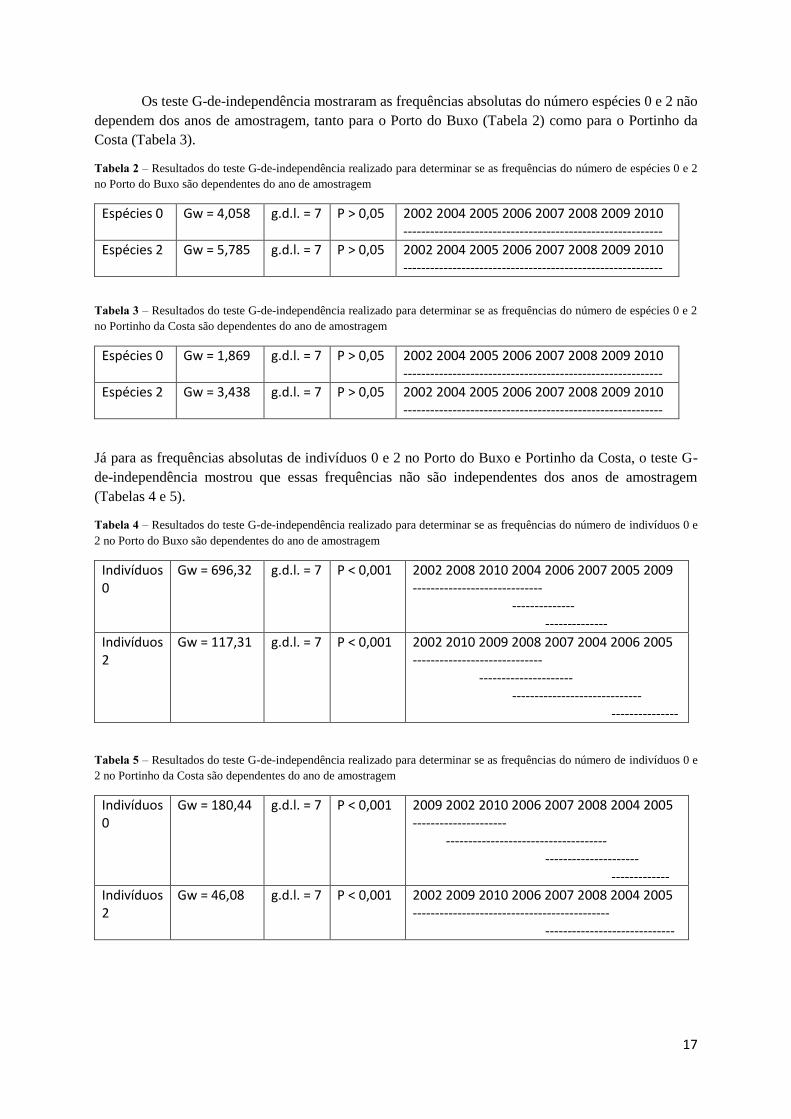
Os teste G-de-independência mostraram as frequências absolutas do número espécies 0 e 2 não
dependem dos anos de amostragem, tanto para o Porto do Buxo (Tabela 2) como para o Portinho da
Costa (Tabela 3).
Tabela 2 – Resultados do teste G-de-independência realizado para determinar se as frequências do número de espécies 0 e 2
no Porto do Buxo são dependentes do ano de amostragem
Espécies 0 Gw = 4,058 g.d.l. = 7 P > 0,05 2002 2004 2005 2006 2007 2008 2009 2010 ----------------------------------------------------------
Espécies 2 Gw = 5,785 g.d.l. = 7 P > 0,05 2002 2004 2005 2006 2007 2008 2009 2010 ----------------------------------------------------------
Tabela 3 – Resultados do teste G-de-independência realizado para determinar se as frequências do número de espécies 0 e 2
no Portinho da Costa são dependentes do ano de amostragem
Espécies 0 Gw = 1,869 g.d.l. = 7 P > 0,05 2002 2004 2005 2006 2007 2008 2009 2010 ----------------------------------------------------------
Espécies 2 Gw = 3,438 g.d.l. = 7 P > 0,05 2002 2004 2005 2006 2007 2008 2009 2010 ----------------------------------------------------------
Já para as frequências absolutas de indivíduos 0 e 2 no Porto do Buxo e Portinho da Costa, o teste G-
de-independência mostrou que essas frequências não são independentes dos anos de amostragem
(Tabelas 4 e 5).
Tabela 4 – Resultados do teste G-de-independência realizado para determinar se as frequências do número de indivíduos 0 e
2 no Porto do Buxo são dependentes do ano de amostragem
Indivíduos 0
Gw = 696,32 g.d.l. = 7 P < 0,001 2002 2008 2010 2004 2006 2007 2005 2009 ----------------------------- -------------- --------------
Indivíduos 2
Gw = 117,31 g.d.l. = 7 P < 0,001 2002 2010 2009 2008 2007 2004 2006 2005 ----------------------------- --------------------- ----------------------------- ---------------
Tabela 5 – Resultados do teste G-de-independência realizado para determinar se as frequências do número de indivíduos 0 e
2 no Portinho da Costa são dependentes do ano de amostragem
Indivíduos 0
Gw = 180,44 g.d.l. = 7 P < 0,001 2009 2002 2010 2006 2007 2008 2004 2005 --------------------- ------------------------------------ --------------------- -------------
Indivíduos 2
Gw = 46,08 g.d.l. = 7 P < 0,001 2002 2009 2010 2006 2007 2008 2004 2005 -------------------------------------------- -----------------------------
18
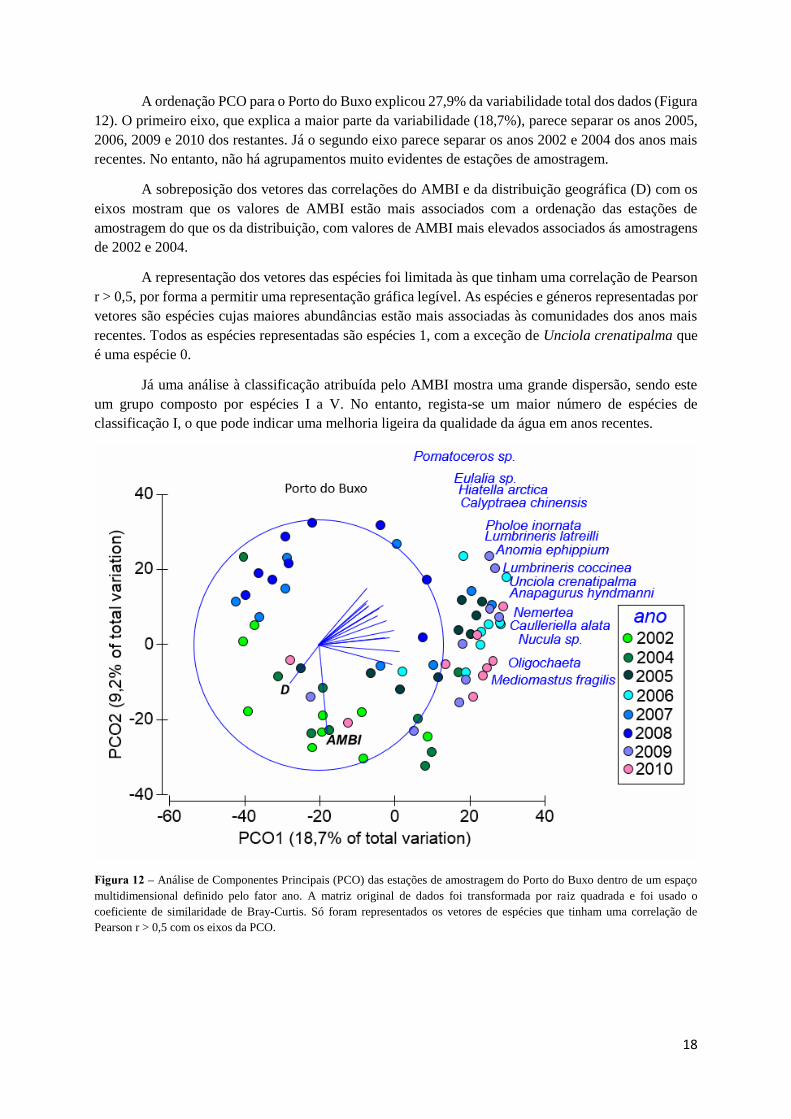
A ordenação PCO para o Porto do Buxo explicou 27,9% da variabilidade total dos dados (Figura
12). O primeiro eixo, que explica a maior parte da variabilidade (18,7%), parece separar os anos 2005,
2006, 2009 e 2010 dos restantes. Já o segundo eixo parece separar os anos 2002 e 2004 dos anos mais
recentes. No entanto, não há agrupamentos muito evidentes de estações de amostragem.
A sobreposição dos vetores das correlações do AMBI e da distribuição geográfica (D) com os
eixos mostram que os valores de AMBI estão mais associados com a ordenação das estações de
amostragem do que os da distribuição, com valores de AMBI mais elevados associados ás amostragens
de 2002 e 2004.
A representação dos vetores das espécies foi limitada às que tinham uma correlação de Pearson
r > 0,5, por forma a permitir uma representação gráfica legível. As espécies e géneros representadas por
vetores são espécies cujas maiores abundâncias estão mais associadas às comunidades dos anos mais
recentes. Todos as espécies representadas são espécies 1, com a exceção de Unciola crenatipalma que
é uma espécie 0.
Já uma análise à classificação atribuída pelo AMBI mostra uma grande dispersão, sendo este
um grupo composto por espécies I a V. No entanto, regista-se um maior número de espécies de
classificação I, o que pode indicar uma melhoria ligeira da qualidade da água em anos recentes.
Figura 12 – Análise de Componentes Principais (PCO) das estações de amostragem do Porto do Buxo dentro de um espaço
multidimensional definido pelo fator ano. A matriz original de dados foi transformada por raiz quadrada e foi usado o
coeficiente de similaridade de Bray-Curtis. Só foram representados os vetores de espécies que tinham uma correlação de
Pearson r > 0,5 com os eixos da PCO.
19
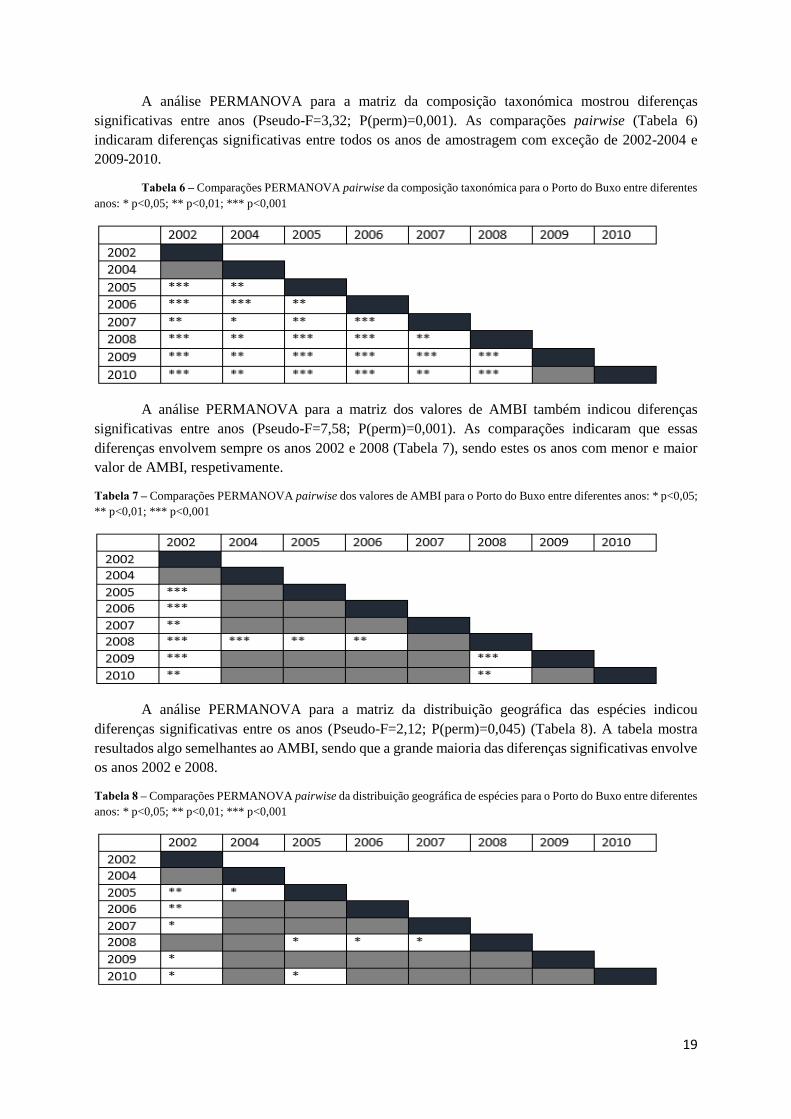
A análise PERMANOVA para a matriz da composição taxonómica mostrou diferenças
significativas entre anos (Pseudo-F=3,32; P(perm)=0,001). As comparações pairwise (Tabela 6)
indicaram diferenças significativas entre todos os anos de amostragem com exceção de 2002-2004 e
2009-2010.
Tabela 6 – Comparações PERMANOVA pairwise da composição taxonómica para o Porto do Buxo entre diferentes
anos: * p<0,05; ** p<0,01; *** p<0,001
A análise PERMANOVA para a matriz dos valores de AMBI também indicou diferenças
significativas entre anos (Pseudo-F=7,58; P(perm)=0,001). As comparações indicaram que essas
diferenças envolvem sempre os anos 2002 e 2008 (Tabela 7), sendo estes os anos com menor e maior
valor de AMBI, respetivamente.
Tabela 7 – Comparações PERMANOVA pairwise dos valores de AMBI para o Porto do Buxo entre diferentes anos: * p<0,05;
** p<0,01; *** p<0,001
A análise PERMANOVA para a matriz da distribuição geográfica das espécies indicou
diferenças significativas entre os anos (Pseudo-F=2,12; P(perm)=0,045) (Tabela 8). A tabela mostra
resultados algo semelhantes ao AMBI, sendo que a grande maioria das diferenças significativas envolve
os anos 2002 e 2008.
Tabela 8 – Comparações PERMANOVA pairwise da distribuição geográfica de espécies para o Porto do Buxo entre diferentes
anos: * p<0,05; ** p<0,01; *** p<0,001
20
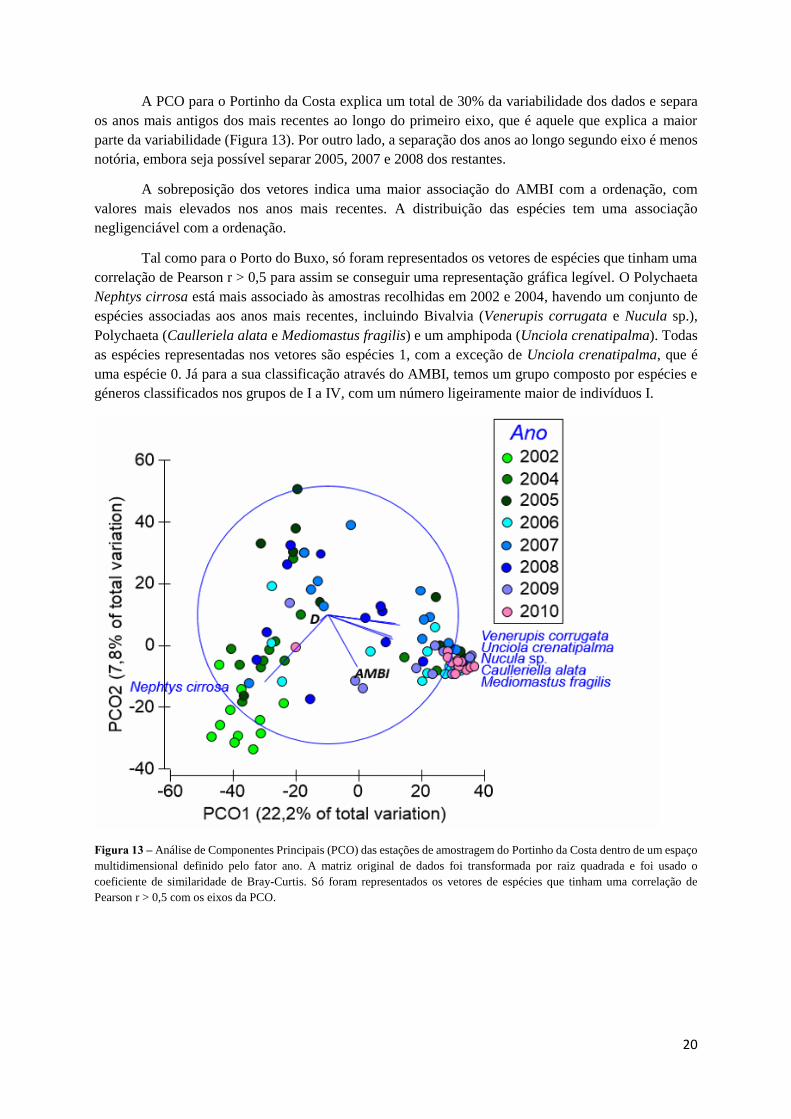
A PCO para o Portinho da Costa explica um total de 30% da variabilidade dos dados e separa
os anos mais antigos dos mais recentes ao longo do primeiro eixo, que é aquele que explica a maior
parte da variabilidade (Figura 13). Por outro lado, a separação dos anos ao longo segundo eixo é menos
notória, embora seja possível separar 2005, 2007 e 2008 dos restantes.
A sobreposição dos vetores indica uma maior associação do AMBI com a ordenação, com
valores mais elevados nos anos mais recentes. A distribuição das espécies tem uma associação
negligenciável com a ordenação.
Tal como para o Porto do Buxo, só foram representados os vetores de espécies que tinham uma
correlação de Pearson r > 0,5 para assim se conseguir uma representação gráfica legível. O Polychaeta
Nephtys cirrosa está mais associado às amostras recolhidas em 2002 e 2004, havendo um conjunto de
espécies associadas aos anos mais recentes, incluindo Bivalvia (Venerupis corrugata e Nucula sp.),
Polychaeta (Caulleriela alata e Mediomastus fragilis) e um amphipoda (Unciola crenatipalma). Todas
as espécies representadas nos vetores são espécies 1, com a exceção de Unciola crenatipalma, que é
uma espécie 0. Já para a sua classificação através do AMBI, temos um grupo composto por espécies e
géneros classificados nos grupos de I a IV, com um número ligeiramente maior de indivíduos I.
Figura 13 – Análise de Componentes Principais (PCO) das estações de amostragem do Portinho da Costa dentro de um espaço
multidimensional definido pelo fator ano. A matriz original de dados foi transformada por raiz quadrada e foi usado o
coeficiente de similaridade de Bray-Curtis. Só foram representados os vetores de espécies que tinham uma correlação de
Pearson r > 0,5 com os eixos da PCO.

21
A análise PERMANOVA para a matriz da composição taxonómica mostrou diferenças
significativas entre anos (Pseudo-F=4,35; P(perm)=0,001). As comparações pairwise (Tabela 9)
indicaram diferenças significativas entre todos os anos de amostragem com exceção de 2005-2007 e
2007-2008.
Tabela 9 – Comparações PERMANOVA pairwise da composição taxonómica para o Portinho da Costa entre diferentes anos:
* p<0,05; ** p<0,01; *** p<0,001
A análise PERMANOVA para a matriz dos valores de AMBI também indicou diferenças
significativas entre anos (Pseudo-F=7,68; P(perm)=0,001). Pode-se observar que 2009 é
significativamente diferente de todos os outros anos (Tabela 10). Isto pode ser explicado pelo facto de
2009 se destacar como o ano com maior valor de AMBI. Já 2005 e 2007 serem diferentes de outros anos
está com certeza ligado ao facto de apresentarem valores de AMBI inferiores em comparação com os
restantes anos.
Tabela 10 – Comparações PERMANOVA pairwise dos valores de AMBI para o Portinho da Costa entre diferentes anos: *
p<0,05; ** p<0,01; *** p<0,001
Para os valores da distribuição geográfica das espécies a PERMANOVA mostrou que não há
diferenças significativas entre os vários anos (p < 0,05) no Portinho da Costa.
22
3.2. Séries temporais na zona intermédia do estuário
3.2.1. Caracterização geral
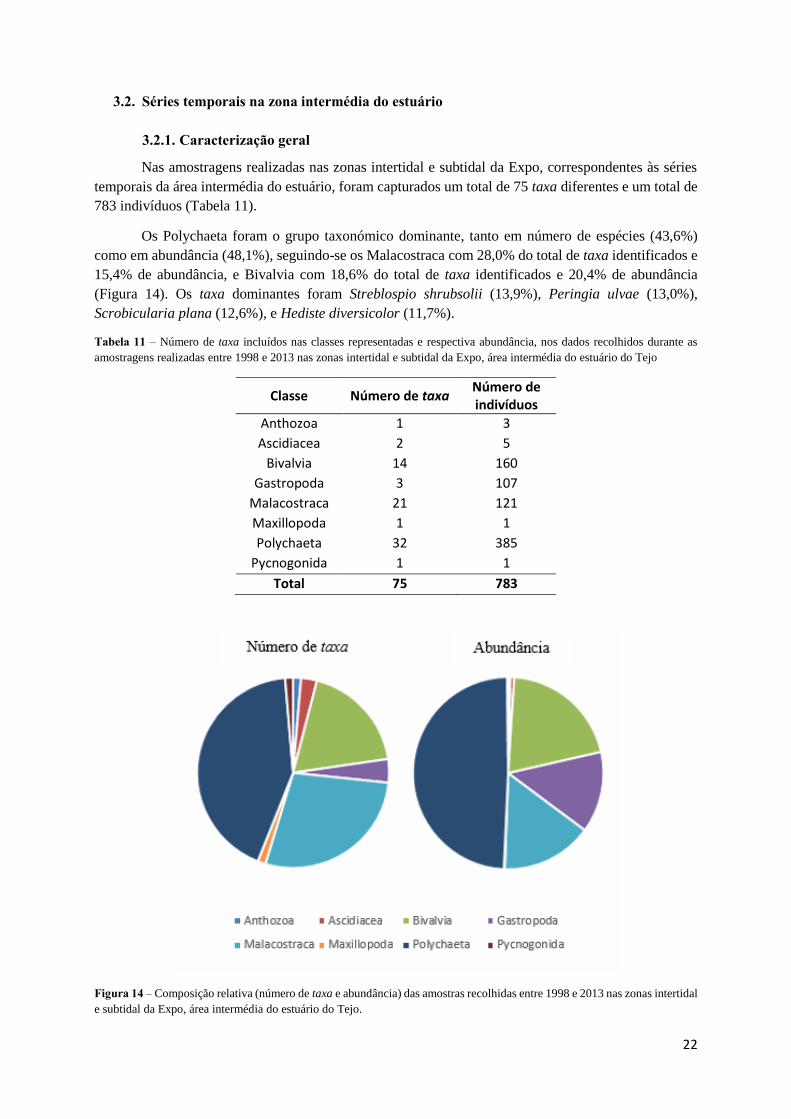
Nas amostragens realizadas nas zonas intertidal e subtidal da Expo, correspondentes às séries
temporais da área intermédia do estuário, foram capturados um total de 75 taxa diferentes e um total de
783 indivíduos (Tabela 11).
Os Polychaeta foram o grupo taxonómico dominante, tanto em número de espécies (43,6%)
como em abundância (48,1%), seguindo-se os Malacostraca com 28,0% do total de taxa identificados e
15,4% de abundância, e Bivalvia com 18,6% do total de taxa identificados e 20,4% de abundância
(Figura 14). Os taxa dominantes foram Streblospio shrubsolii (13,9%), Peringia ulvae (13,0%),
Scrobicularia plana (12,6%), e Hediste diversicolor (11,7%).
Tabela 11 – Número de taxa incluídos nas classes representadas e respectiva abundância, nos dados recolhidos durante as
amostragens realizadas entre 1998 e 2013 nas zonas intertidal e subtidal da Expo, área intermédia do estuário do Tejo
Classe Número de taxa Número de indivíduos
Anthozoa 1 3
Ascidiacea 2 5
Bivalvia 14 160
Gastropoda 3 107
Malacostraca 21 121
Maxillopoda 1 1
Polychaeta 32 385
Pycnogonida 1 1
Total 75 783
Figura 14 – Composição relativa (número de taxa e abundância) das amostras recolhidas entre 1998 e 2013 nas zonas intertidal
e subtidal da Expo, área intermédia do estuário do Tejo.
23
3.2.2. Evolução temporal
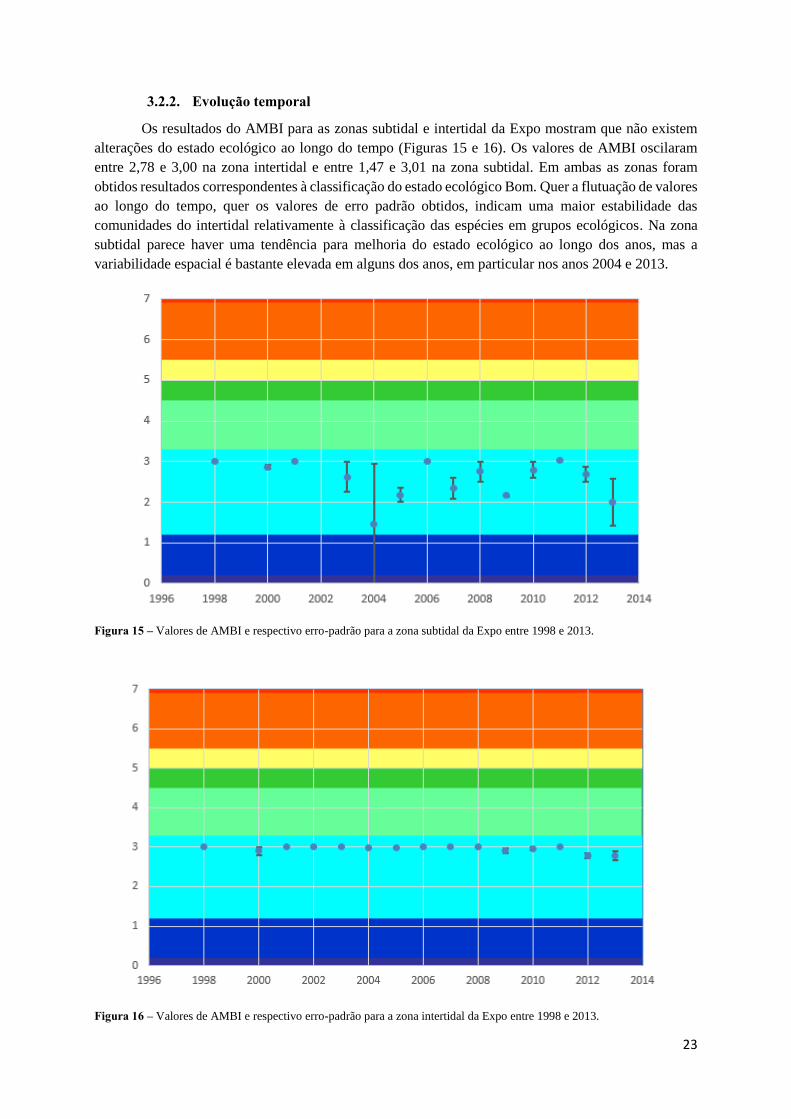
Os resultados do AMBI para as zonas subtidal e intertidal da Expo mostram que não existem
alterações do estado ecológico ao longo do tempo (Figuras 15 e 16). Os valores de AMBI oscilaram
entre 2,78 e 3,00 na zona intertidal e entre 1,47 e 3,01 na zona subtidal. Em ambas as zonas foram
obtidos resultados correspondentes à classificação do estado ecológico Bom. Quer a flutuação de valores
ao longo do tempo, quer os valores de erro padrão obtidos, indicam uma maior estabilidade das
comunidades do intertidal relativamente à classificação das espécies em grupos ecológicos. Na zona
subtidal parece haver uma tendência para melhoria do estado ecológico ao longo dos anos, mas a
variabilidade espacial é bastante elevada em alguns dos anos, em particular nos anos 2004 e 2013.
Figura 15 – Valores de AMBI e respectivo erro-padrão para a zona subtidal da Expo entre 1998 e 2013.
Figura 16 – Valores de AMBI e respectivo erro-padrão para a zona intertidal da Expo entre 1998 e 2013.

24
A distribuição geográfica com base no número de espécies e abundância na zona subtidal,
mostra a ocorrência pontual das espécies 0 e 2 a partir de 2007 e 2005, respetivamente (Figuras 17 e
18). Apesar de não se identificar uma tendência temporal clara, a presença de espécies 2 é consistente a
partir de 2011. De uma forma geral, a ocorrência de espécies 2 predomina em relação às espécies 0.
Figura 17 – Percentagem relativa do número de espécies do norte e espécies do sul na zona subtidal da Expo, Parque das
Nações, entre 1998 e 2013: Espécies 0- Espécies com uma distribuição exclusivamente a norte da Costa Atlântica do Sul da
Europa; Espécies 2 – Espécies com uma distribuição exclusivamente a sul da Costa Atlântica do Sul da Europa.
Os resultados obtidos relativamente ao número de indivíduos (Figura 18) complementam os
dados referentes à percentagem de espécies da zona subtidal da Expo. Os indivíduos 0 raramente são
registados neste local, e mesmo quando tal acontece o seu número é sempre muito baixo, sendo a única
exceção o ano 2007. Já para as espécies 2, estes dados obtidos também salientam a ideia que as espécies
2 tem aumentado neste local. Surgem em 2009, desaparecem em 2010, são de novo registadas em 2011,
sendo que depois ocorre um aumento nos anos seguintes.
Figura 18 – Percentagem relativa do número de indivíduos do norte e indivíduos do sul na zona subtidal da Expo, Parque das
Nações, entre 1998 e 2013: Indivíduos 0- Indivíduos com uma distribuição exclusivamente a norte da Costa Atlântica do Sul
da Europa; Indivíduos 2 – Indivíduos com uma distribuição exclusivamente a sul da Costa Atlântica do Sul da Europa.

25
A distribuição geográfica com base no número de espécies mostra que na zona intertidal ao
longo deste longo período não foi registada uma única espécie 0 (Figura 19). Já para as espécies 2, os
resultados obtidos mostram um padrão algo semelhante ao da zona subtidal. Entre 1998 e 2010 não foi
registada nenhuma espécie 2, sendo que estas só surgiram em 2011, com o seu número a aumentar
ligeiramente nos anos seguintes.
Figura 19 – Percentagem relativa do número de espécies do norte e espécies do sul na zona intertidal da Expo, Parque das
Nações, entre 1998 e 2013: Espécies 0- Espécies com uma distribuição exclusivamente a norte da Costa Atlântica do Sul da
Europa; Espécies 2 – Espécies com uma distribuição exclusivamente a sul da Costa Atlântica do Sul da Europa.
Os dados para o número de indivíduos na zona intertidal são muito claros relativamente á sua
constituição. Não se observam nenhuns indivíduos 0, enquanto os indivíduos 2 existem em pouca
quantidade. Tendo em consideração os dados da figura 20, podemos dizer que ocorreu um aumento de
indivíduos 2 nos últimos anos, mas o seu número não é significativo o suficiente para dizer que este
grupo está em crescimento neste local.
Figura 20 – Percentagem relativa do número de indivíduos do norte e indivíduos do sul na zona intertidal da Expo, Parque das
Nações, entre 1998 e 2013: Indivíduos 0- Indivíduos com uma distribuição exclusivamente a norte da Costa Atlântica do Sul
da Europa; Indivíduos 2 – Indivíduos com uma distribuição exclusivamente a sul da Costa Atlântica do Sul da Europa.
26
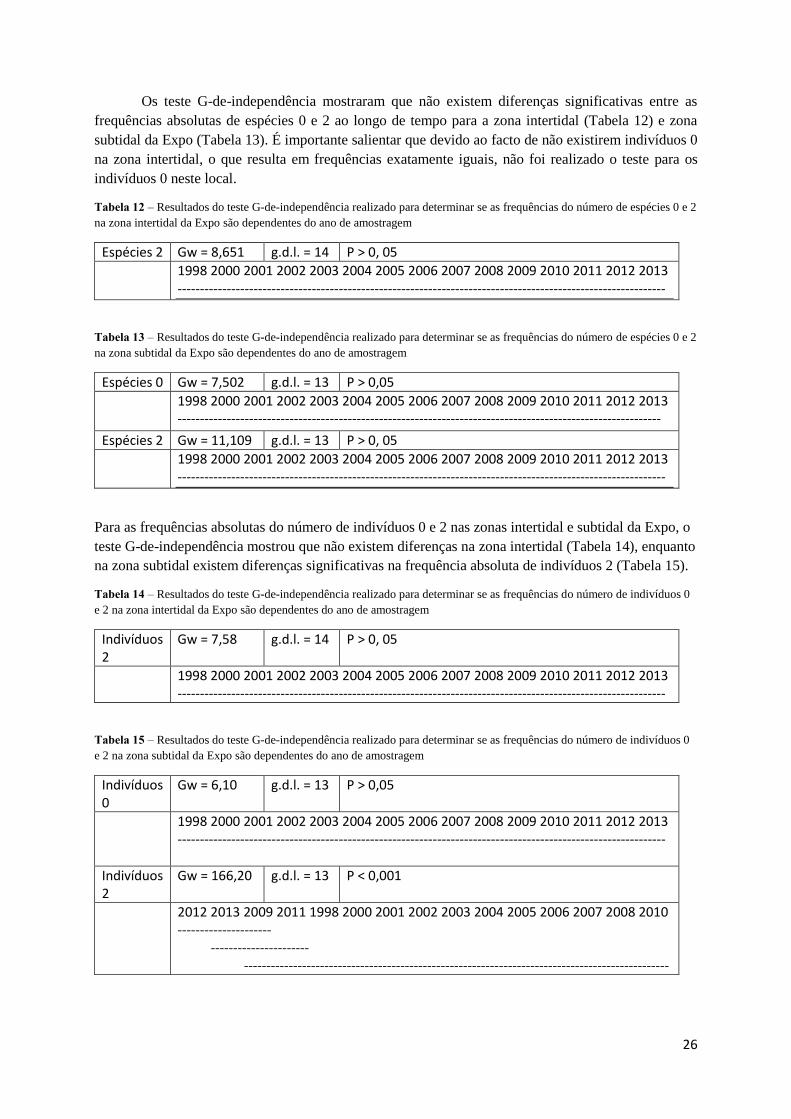
Os teste G-de-independência mostraram que não existem diferenças significativas entre as
frequências absolutas de espécies 0 e 2 ao longo de tempo para a zona intertidal (Tabela 12) e zona
subtidal da Expo (Tabela 13). É importante salientar que devido ao facto de não existirem indivíduos 0
na zona intertidal, o que resulta em frequências exatamente iguais, não foi realizado o teste para os
indivíduos 0 neste local.
Tabela 12 – Resultados do teste G-de-independência realizado para determinar se as frequências do número de espécies 0 e 2
na zona intertidal da Expo são dependentes do ano de amostragem
Espécies 2 Gw = 8,651 g.d.l. = 14 P > 0, 05
1998 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 2010 2011 2012 2013 -------------------------------------------------------------------------------------------------------------
Tabela 13 – Resultados do teste G-de-independência realizado para determinar se as frequências do número de espécies 0 e 2
na zona subtidal da Expo são dependentes do ano de amostragem
Espécies 0 Gw = 7,502 g.d.l. = 13 P > 0,05
1998 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 2010 2011 2012 2013 ------------------------------------------------------------------------------------------------------------
Espécies 2 Gw = 11,109 g.d.l. = 13 P > 0, 05
1998 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 2010 2011 2012 2013 -------------------------------------------------------------------------------------------------------------
Para as frequências absolutas do número de indivíduos 0 e 2 nas zonas intertidal e subtidal da Expo, o
teste G-de-independência mostrou que não existem diferenças na zona intertidal (Tabela 14), enquanto
na zona subtidal existem diferenças significativas na frequência absoluta de indivíduos 2 (Tabela 15).
Tabela 14 – Resultados do teste G-de-independência realizado para determinar se as frequências do número de indivíduos 0
e 2 na zona intertidal da Expo são dependentes do ano de amostragem
Indivíduos 2
Gw = 7,58 g.d.l. = 14 P > 0, 05
1998 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 2010 2011 2012 2013 -------------------------------------------------------------------------------------------------------------
Tabela 15 – Resultados do teste G-de-independência realizado para determinar se as frequências do número de indivíduos 0
e 2 na zona subtidal da Expo são dependentes do ano de amostragem
Indivíduos 0
Gw = 6,10 g.d.l. = 13 P > 0,05
1998 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 2010 2011 2012 2013 -------------------------------------------------------------------------------------------------------------
Indivíduos 2
Gw = 166,20 g.d.l. = 13 P < 0,001
2012 2013 2009 2011 1998 2000 2001 2002 2003 2004 2005 2006 2007 2008 2010 --------------------- ---------------------- -----------------------------------------------------------------------------------------------
27
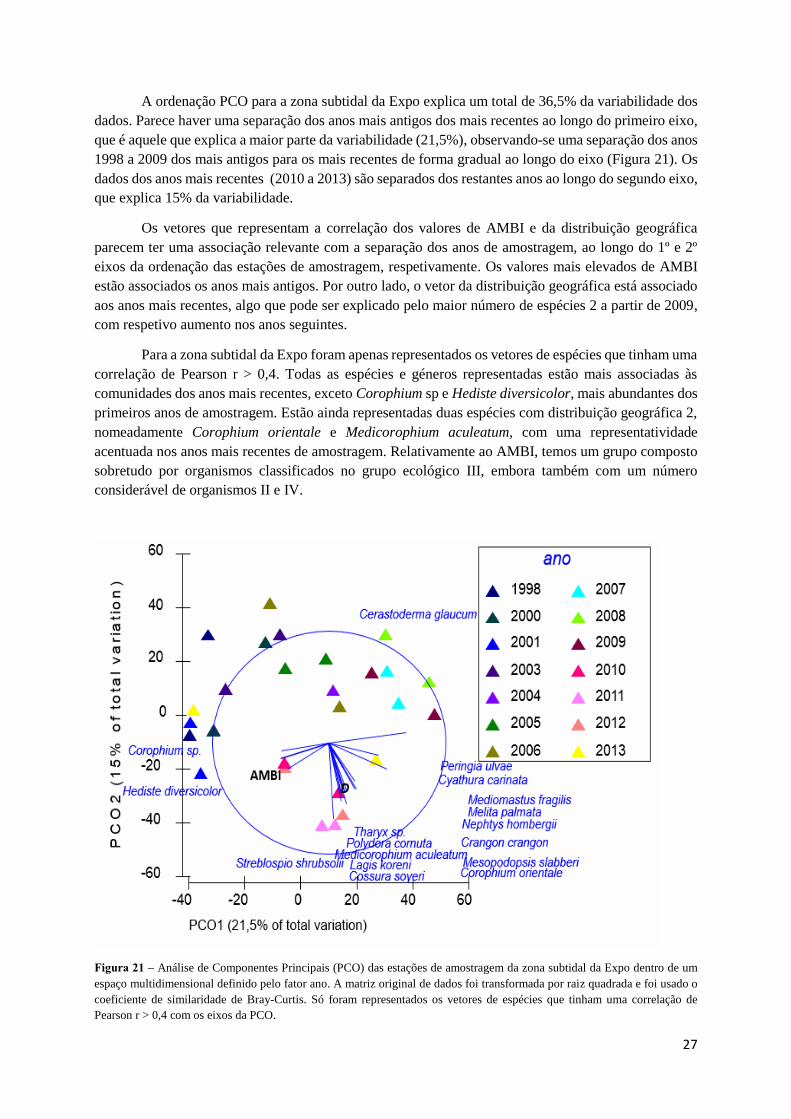
A ordenação PCO para a zona subtidal da Expo explica um total de 36,5% da variabilidade dos
dados. Parece haver uma separação dos anos mais antigos dos mais recentes ao longo do primeiro eixo,
que é aquele que explica a maior parte da variabilidade (21,5%), observando-se uma separação dos anos
1998 a 2009 dos mais antigos para os mais recentes de forma gradual ao longo do eixo (Figura 21). Os
dados dos anos mais recentes (2010 a 2013) são separados dos restantes anos ao longo do segundo eixo,
que explica 15% da variabilidade.
Os vetores que representam a correlação dos valores de AMBI e da distribuição geográfica
parecem ter uma associação relevante com a separação dos anos de amostragem, ao longo do 1º e 2º
eixos da ordenação das estações de amostragem, respetivamente. Os valores mais elevados de AMBI
estão associados os anos mais antigos. Por outro lado, o vetor da distribuição geográfica está associado
aos anos mais recentes, algo que pode ser explicado pelo maior número de espécies 2 a partir de 2009,
com respetivo aumento nos anos seguintes.
Para a zona subtidal da Expo foram apenas representados os vetores de espécies que tinham uma
correlação de Pearson r > 0,4. Todas as espécies e géneros representadas estão mais associadas às
comunidades dos anos mais recentes, exceto Corophium sp e Hediste diversicolor, mais abundantes dos
primeiros anos de amostragem. Estão ainda representadas duas espécies com distribuição geográfica 2,
nomeadamente Corophium orientale e Medicorophium aculeatum, com uma representatividade
acentuada nos anos mais recentes de amostragem. Relativamente ao AMBI, temos um grupo composto
sobretudo por organismos classificados no grupo ecológico III, embora também com um número
considerável de organismos II e IV.
Figura 21 – Análise de Componentes Principais (PCO) das estações de amostragem da zona subtidal da Expo dentro de um
espaço multidimensional definido pelo fator ano. A matriz original de dados foi transformada por raiz quadrada e foi usado o
coeficiente de similaridade de Bray-Curtis. Só foram representados os vetores de espécies que tinham uma correlação de
Pearson r > 0,4 com os eixos da PCO.

28
Os testes de Permanova mostram que há diferenças significativas na composição taxonómica
ao longo dos anos (Pseudo-F=1,66; P(perm)= 0,002). No entanto, os testes de pairwise mostraram que
não havia diferenças entre anos específicos (P(perm)<0,05), o que parece indicar que as alterações das
comunidades ao longo do tempo ocorreram de forma pouco acentuada, não se obtendo diferenças
estatisticamente significativas entre anos específicos.
Para o AMBI e distribuição geográfica os testes de Permanova mostram que não há diferenças
significativas (p < 0,05) ao longo do tempo.
29
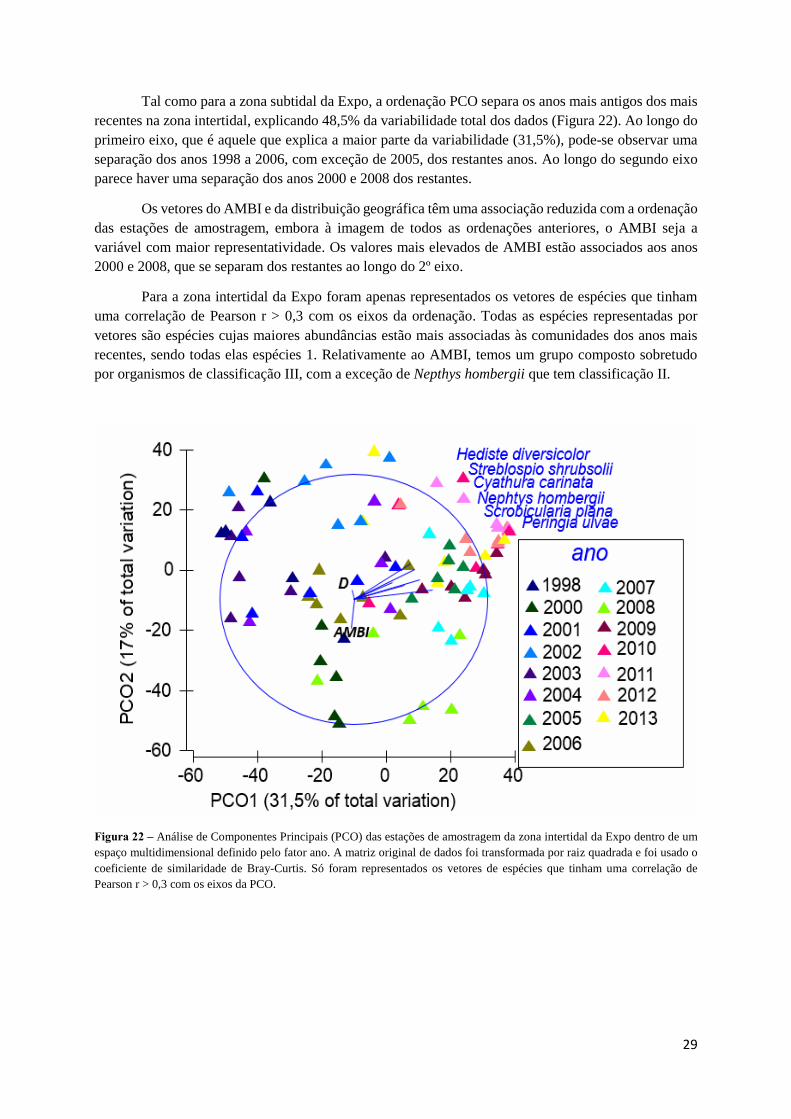
Tal como para a zona subtidal da Expo, a ordenação PCO separa os anos mais antigos dos mais
recentes na zona intertidal, explicando 48,5% da variabilidade total dos dados (Figura 22). Ao longo do
primeiro eixo, que é aquele que explica a maior parte da variabilidade (31,5%), pode-se observar uma
separação dos anos 1998 a 2006, com exceção de 2005, dos restantes anos. Ao longo do segundo eixo
parece haver uma separação dos anos 2000 e 2008 dos restantes.
Os vetores do AMBI e da distribuição geográfica têm uma associação reduzida com a ordenação
das estações de amostragem, embora à imagem de todos as ordenações anteriores, o AMBI seja a
variável com maior representatividade. Os valores mais elevados de AMBI estão associados aos anos
2000 e 2008, que se separam dos restantes ao longo do 2º eixo.
Para a zona intertidal da Expo foram apenas representados os vetores de espécies que tinham
uma correlação de Pearson r > 0,3 com os eixos da ordenação. Todas as espécies representadas por
vetores são espécies cujas maiores abundâncias estão mais associadas às comunidades dos anos mais
recentes, sendo todas elas espécies 1. Relativamente ao AMBI, temos um grupo composto sobretudo
por organismos de classificação III, com a exceção de Nepthys hombergii que tem classificação II.
Figura 22 – Análise de Componentes Principais (PCO) das estações de amostragem da zona intertidal da Expo dentro de um
espaço multidimensional definido pelo fator ano. A matriz original de dados foi transformada por raiz quadrada e foi usado o
coeficiente de similaridade de Bray-Curtis. Só foram representados os vetores de espécies que tinham uma correlação de
Pearson r > 0,3 com os eixos da PCO.
30
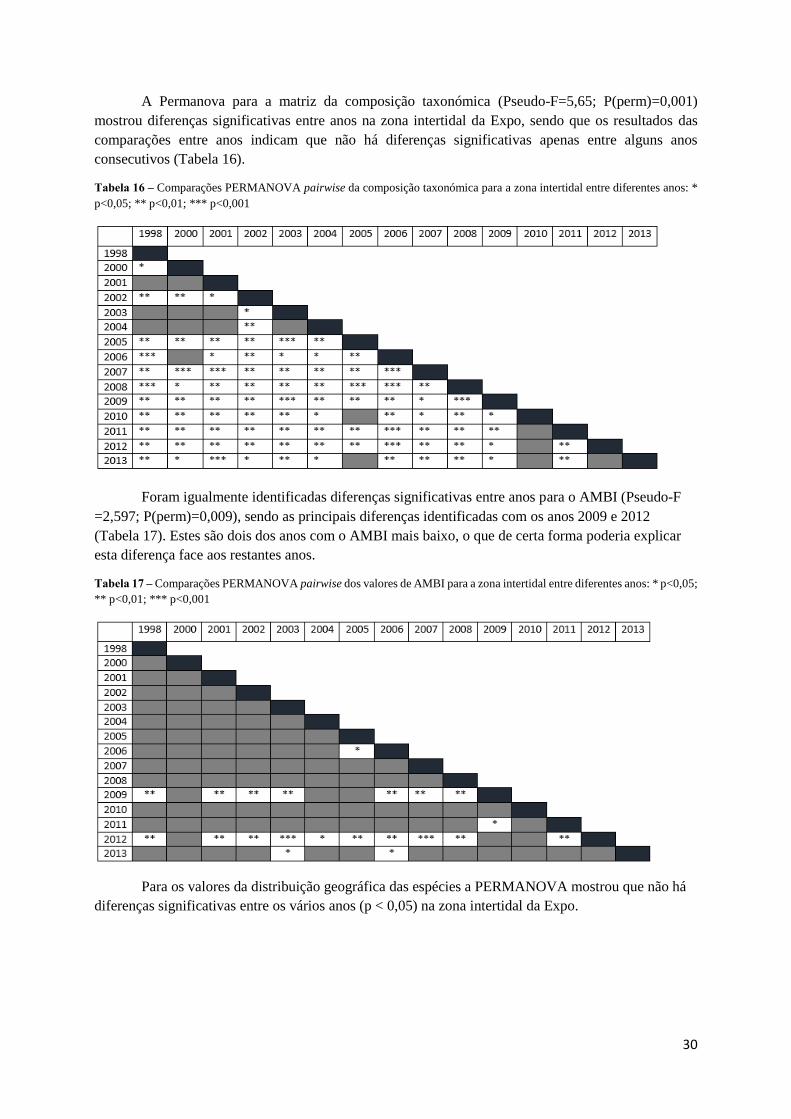
A Permanova para a matriz da composição taxonómica (Pseudo-F=5,65; P(perm)=0,001)
mostrou diferenças significativas entre anos na zona intertidal da Expo, sendo que os resultados das
comparações entre anos indicam que não há diferenças significativas apenas entre alguns anos
consecutivos (Tabela 16).
Tabela 16 – Comparações PERMANOVA pairwise da composição taxonómica para a zona intertidal entre diferentes anos: *
p<0,05; ** p<0,01; *** p<0,001
Foram igualmente identificadas diferenças significativas entre anos para o AMBI (Pseudo-F
=2,597; P(perm)=0,009), sendo as principais diferenças identificadas com os anos 2009 e 2012
(Tabela 17). Estes são dois dos anos com o AMBI mais baixo, o que de certa forma poderia explicar
esta diferença face aos restantes anos.
Tabela 17 – Comparações PERMANOVA pairwise dos valores de AMBI para a zona intertidal entre diferentes anos: * p<0,05;
** p<0,01; *** p<0,001
Para os valores da distribuição geográfica das espécies a PERMANOVA mostrou que não há
diferenças significativas entre os vários anos (p < 0,05) na zona intertidal da Expo.
31
3.3. Séries ao longo do gradiente estuarino com um intervalo temporal longo
3.3.1. Caracterização geral
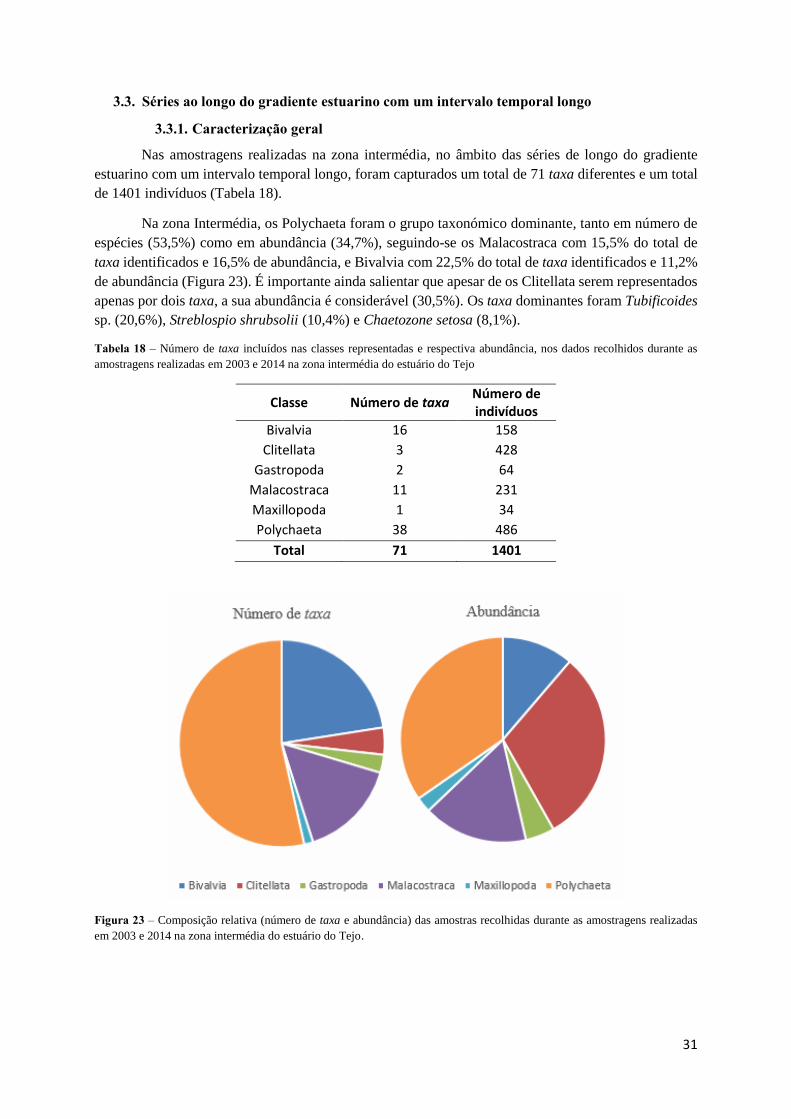
Nas amostragens realizadas na zona intermédia, no âmbito das séries de longo do gradiente
estuarino com um intervalo temporal longo, foram capturados um total de 71 taxa diferentes e um total
de 1401 indivíduos (Tabela 18).
Na zona Intermédia, os Polychaeta foram o grupo taxonómico dominante, tanto em número de
espécies (53,5%) como em abundância (34,7%), seguindo-se os Malacostraca com 15,5% do total de
taxa identificados e 16,5% de abundância, e Bivalvia com 22,5% do total de taxa identificados e 11,2%
de abundância (Figura 23). É importante ainda salientar que apesar de os Clitellata serem representados
apenas por dois taxa, a sua abundância é considerável (30,5%). Os taxa dominantes foram Tubificoides
sp. (20,6%), Streblospio shrubsolii (10,4%) e Chaetozone setosa (8,1%).
Tabela 18 – Número de taxa incluídos nas classes representadas e respectiva abundância, nos dados recolhidos durante as
amostragens realizadas em 2003 e 2014 na zona intermédia do estuário do Tejo
Classe Número de taxa Número de indivíduos
Bivalvia 16 158
Clitellata 3 428
Gastropoda 2 64
Malacostraca 11 231
Maxillopoda 1 34
Polychaeta 38 486
Total 71 1401
Figura 23 – Composição relativa (número de taxa e abundância) das amostras recolhidas durante as amostragens realizadas
em 2003 e 2014 na zona intermédia do estuário do Tejo.
32
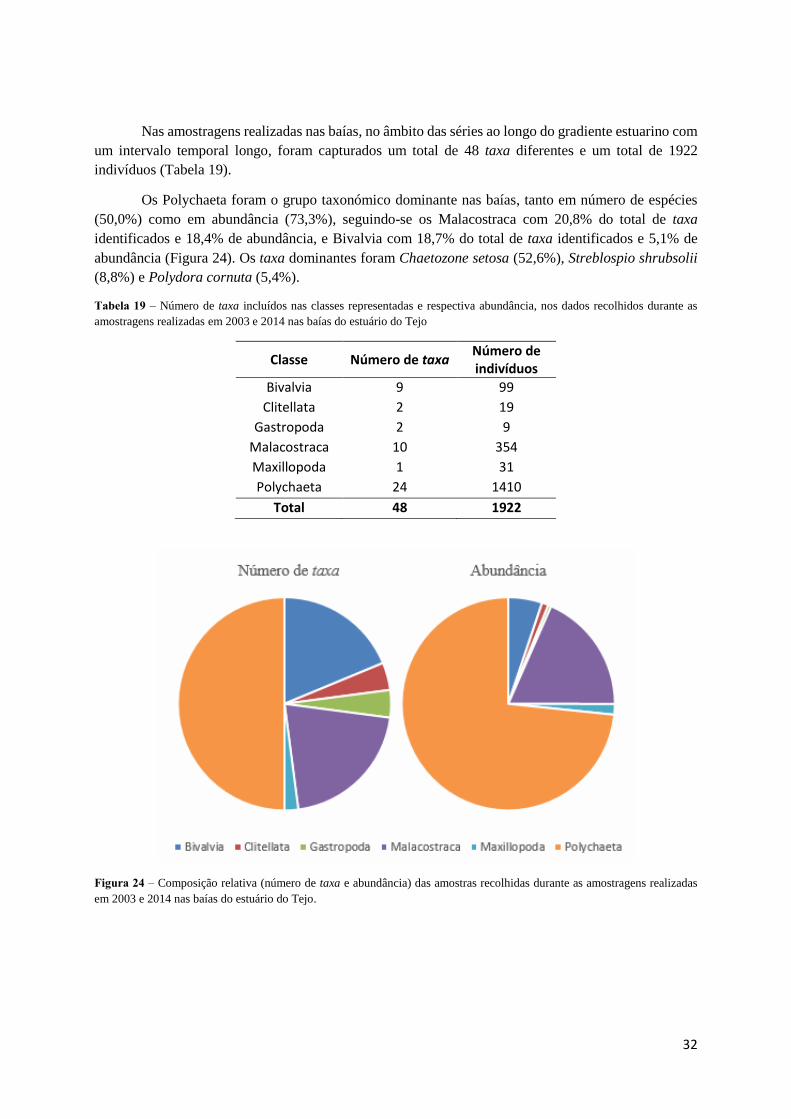
Nas amostragens realizadas nas baías, no âmbito das séries ao longo do gradiente estuarino com
um intervalo temporal longo, foram capturados um total de 48 taxa diferentes e um total de 1922
indivíduos (Tabela 19).
Os Polychaeta foram o grupo taxonómico dominante nas baías, tanto em número de espécies
(50,0%) como em abundância (73,3%), seguindo-se os Malacostraca com 20,8% do total de taxa
identificados e 18,4% de abundância, e Bivalvia com 18,7% do total de taxa identificados e 5,1% de
abundância (Figura 24). Os taxa dominantes foram Chaetozone setosa (52,6%), Streblospio shrubsolii
(8,8%) e Polydora cornuta (5,4%).
Tabela 19 – Número de taxa incluídos nas classes representadas e respectiva abundância, nos dados recolhidos durante as
amostragens realizadas em 2003 e 2014 nas baías do estuário do Tejo
Classe Número de taxa Número de indivíduos
Bivalvia 9 99
Clitellata 2 19
Gastropoda 2 9
Malacostraca 10 354
Maxillopoda 1 31
Polychaeta 24 1410
Total 48 1922
Figura 24 – Composição relativa (número de taxa e abundância) das amostras recolhidas durante as amostragens realizadas
em 2003 e 2014 nas baías do estuário do Tejo.

33
Nas amostragens realizadas na zona inferior do estuário, no âmbito das séries ao longo do
gradiente estuarino com um intervalo temporal longo, foram capturados um total de 46 taxa diferentes
e um total de 591 indivíduos (Tabela 20).
Os Polychaeta foram o grupo taxonómico dominante na zona inferior, tanto em número de
espécies (50,0%) como em abundância (77,1%), seguindo-se os Bivalvia com 21,7% do total de taxa
identificados e 6,5% de abundância (Figura 25). Os taxa dominantes foram Chaetozone setosa (48,0%),
Streblospio shrubsolii (8,6%) e Melita Palmata (7,9%).
Tabela 20 – Número de taxa incluídos nas classes representadas e respectiva abundância, nos dados recolhidos durante as
amostragens realizadas em 2003 e 2014 na zona inferior do estuário do Tejo
Classe Número de taxa Número de indivíduos
Bivalvia 10 39
Clitellata 2 11
Gastropoda 3 21
Malacostraca 8 64
Polychaeta 23 456
Total 46 591
Figura 25 – Composição relativa (número de taxa e abundância) das amostras recolhidas durante as amostragens realizadas
em 2003 e 2014 na zona inferior do estuário do Tejo.
34
3.3.2. Evolução temporal
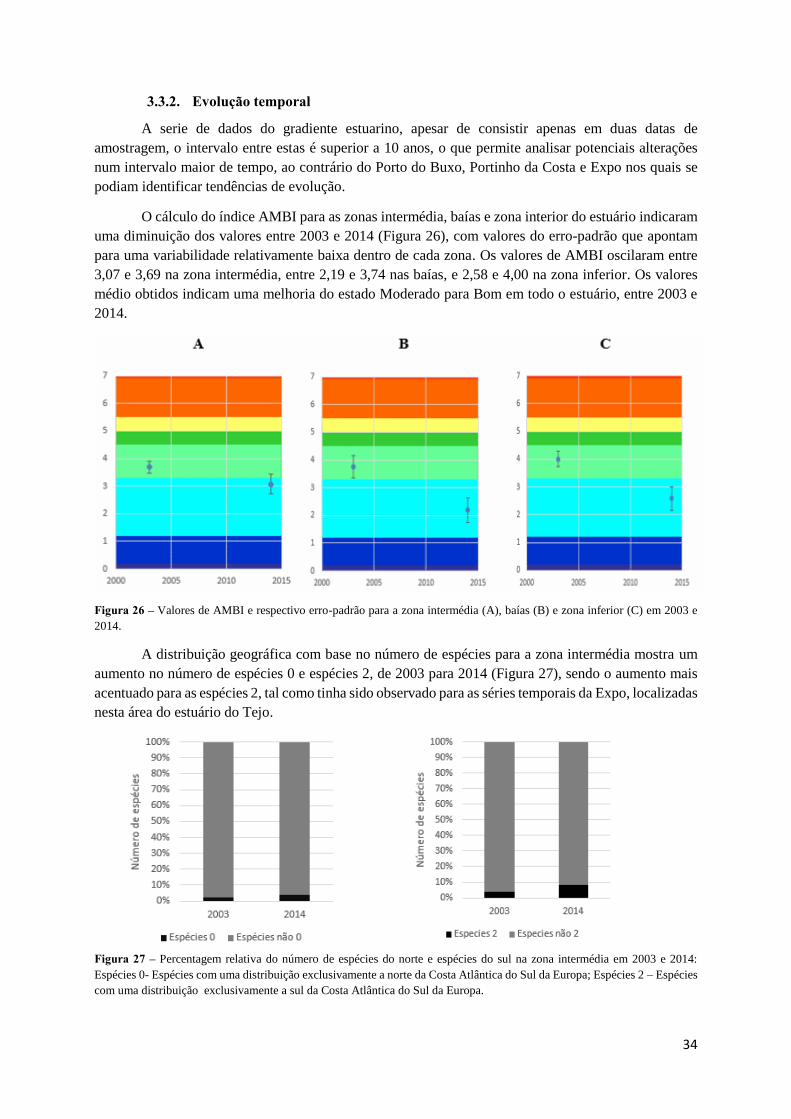
A serie de dados do gradiente estuarino, apesar de consistir apenas em duas datas de
amostragem, o intervalo entre estas é superior a 10 anos, o que permite analisar potenciais alterações
num intervalo maior de tempo, ao contrário do Porto do Buxo, Portinho da Costa e Expo nos quais se
podiam identificar tendências de evolução.
O cálculo do índice AMBI para as zonas intermédia, baías e zona interior do estuário indicaram
uma diminuição dos valores entre 2003 e 2014 (Figura 26), com valores do erro-padrão que apontam
para uma variabilidade relativamente baixa dentro de cada zona. Os valores de AMBI oscilaram entre
3,07 e 3,69 na zona intermédia, entre 2,19 e 3,74 nas baías, e 2,58 e 4,00 na zona inferior. Os valores
médio obtidos indicam uma melhoria do estado Moderado para Bom em todo o estuário, entre 2003 e
2014.
Figura 26 – Valores de AMBI e respectivo erro-padrão para a zona intermédia (A), baías (B) e zona inferior (C) em 2003 e
2014.
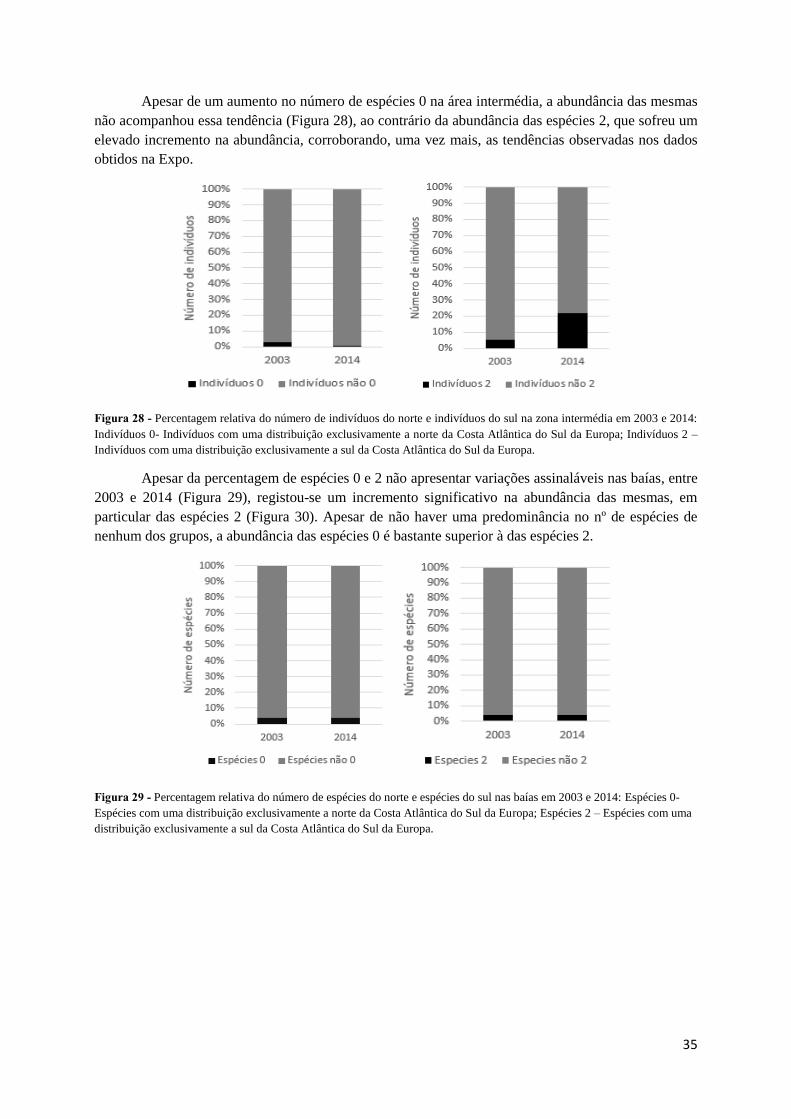
A distribuição geográfica com base no número de espécies para a zona intermédia mostra um
aumento no número de espécies 0 e espécies 2, de 2003 para 2014 (Figura 27), sendo o aumento mais
acentuado para as espécies 2, tal como tinha sido observado para as séries temporais da Expo, localizadas
nesta área do estuário do Tejo.
Figura 27 – Percentagem relativa do número de espécies do norte e espécies do sul na zona intermédia em 2003 e 2014:
Espécies 0- Espécies com uma distribuição exclusivamente a norte da Costa Atlântica do Sul da Europa; Espécies 2 – Espécies
com uma distribuição exclusivamente a sul da Costa Atlântica do Sul da Europa.
35
Apesar de um aumento no número de espécies 0 na área intermédia, a abundância das mesmas
não acompanhou essa tendência (Figura 28), ao contrário da abundância das espécies 2, que sofreu um
elevado incremento na abundância, corroborando, uma vez mais, as tendências observadas nos dados
obtidos na Expo.
Figura 28 - Percentagem relativa do número de indivíduos do norte e indivíduos do sul na zona intermédia em 2003 e 2014:
Indivíduos 0- Indivíduos com uma distribuição exclusivamente a norte da Costa Atlântica do Sul da Europa; Indivíduos 2 –
Indivíduos com uma distribuição exclusivamente a sul da Costa Atlântica do Sul da Europa.
Apesar da percentagem de espécies 0 e 2 não apresentar variações assinaláveis nas baías, entre
2003 e 2014 (Figura 29), registou-se um incremento significativo na abundância das mesmas, em
particular das espécies 2 (Figura 30). Apesar de não haver uma predominância no nº de espécies de
nenhum dos grupos, a abundância das espécies 0 é bastante superior à das espécies 2.
Figura 29 - Percentagem relativa do número de espécies do norte e espécies do sul nas baías em 2003 e 2014: Espécies 0-
Espécies com uma distribuição exclusivamente a norte da Costa Atlântica do Sul da Europa; Espécies 2 – Espécies com uma
distribuição exclusivamente a sul da Costa Atlântica do Sul da Europa.
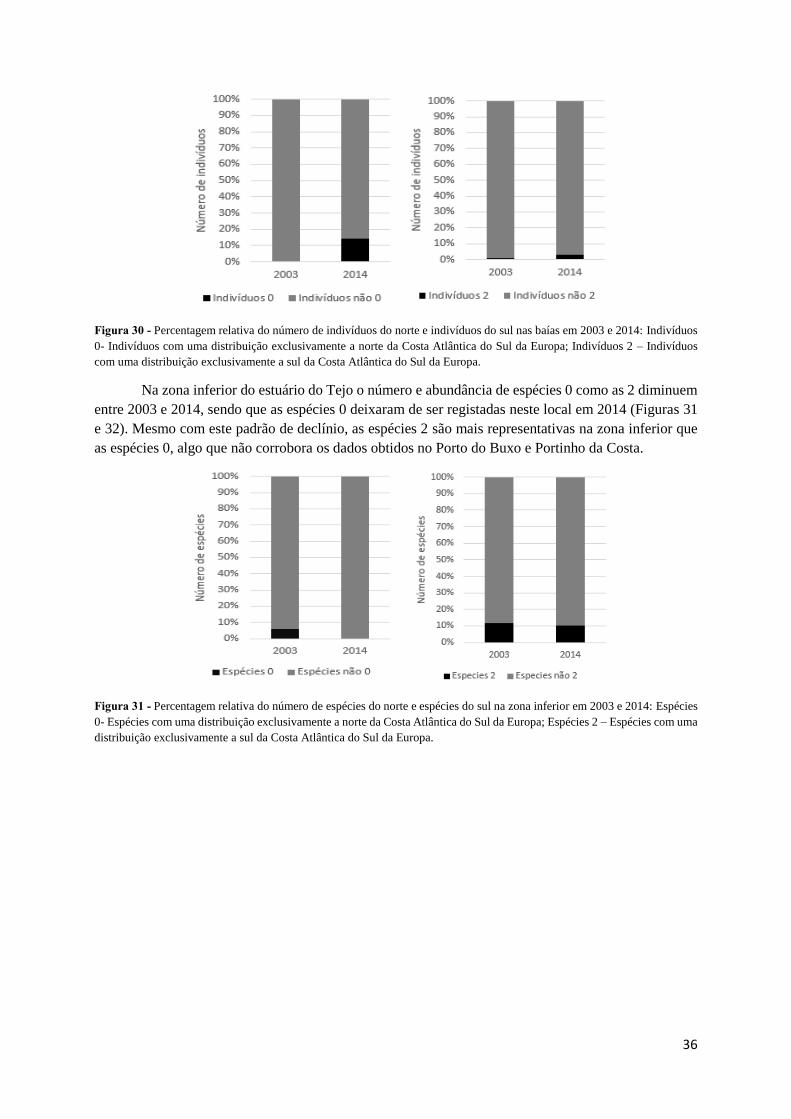
36
Figura 30 - Percentagem relativa do número de indivíduos do norte e indivíduos do sul nas baías em 2003 e 2014: Indivíduos
0- Indivíduos com uma distribuição exclusivamente a norte da Costa Atlântica do Sul da Europa; Indivíduos 2 – Indivíduos
com uma distribuição exclusivamente a sul da Costa Atlântica do Sul da Europa.
Na zona inferior do estuário do Tejo o número e abundância de espécies 0 como as 2 diminuem
entre 2003 e 2014, sendo que as espécies 0 deixaram de ser registadas neste local em 2014 (Figuras 31
e 32). Mesmo com este padrão de declínio, as espécies 2 são mais representativas na zona inferior que
as espécies 0, algo que não corrobora os dados obtidos no Porto do Buxo e Portinho da Costa.
Figura 31 - Percentagem relativa do número de espécies do norte e espécies do sul na zona inferior em 2003 e 2014: Espécies
0- Espécies com uma distribuição exclusivamente a norte da Costa Atlântica do Sul da Europa; Espécies 2 – Espécies com uma
distribuição exclusivamente a sul da Costa Atlântica do Sul da Europa.
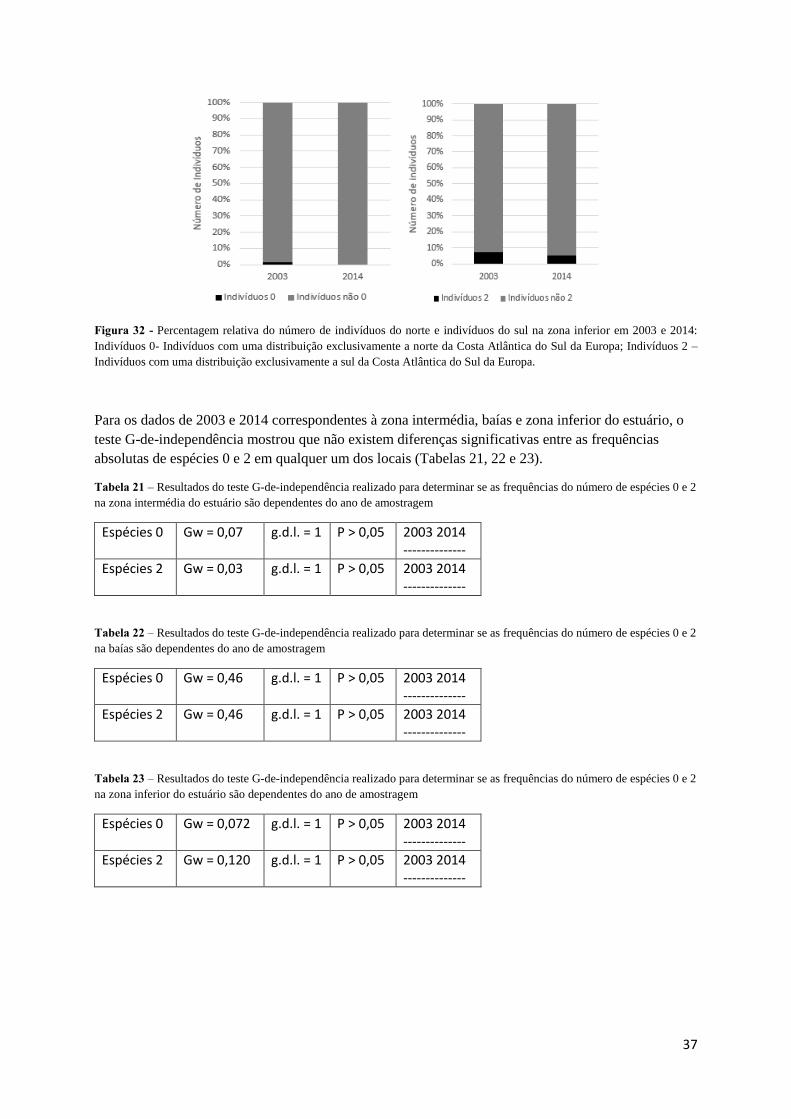
37
Figura 32 - Percentagem relativa do número de indivíduos do norte e indivíduos do sul na zona inferior em 2003 e 2014:
Indivíduos 0- Indivíduos com uma distribuição exclusivamente a norte da Costa Atlântica do Sul da Europa; Indivíduos 2 –
Indivíduos com uma distribuição exclusivamente a sul da Costa Atlântica do Sul da Europa.
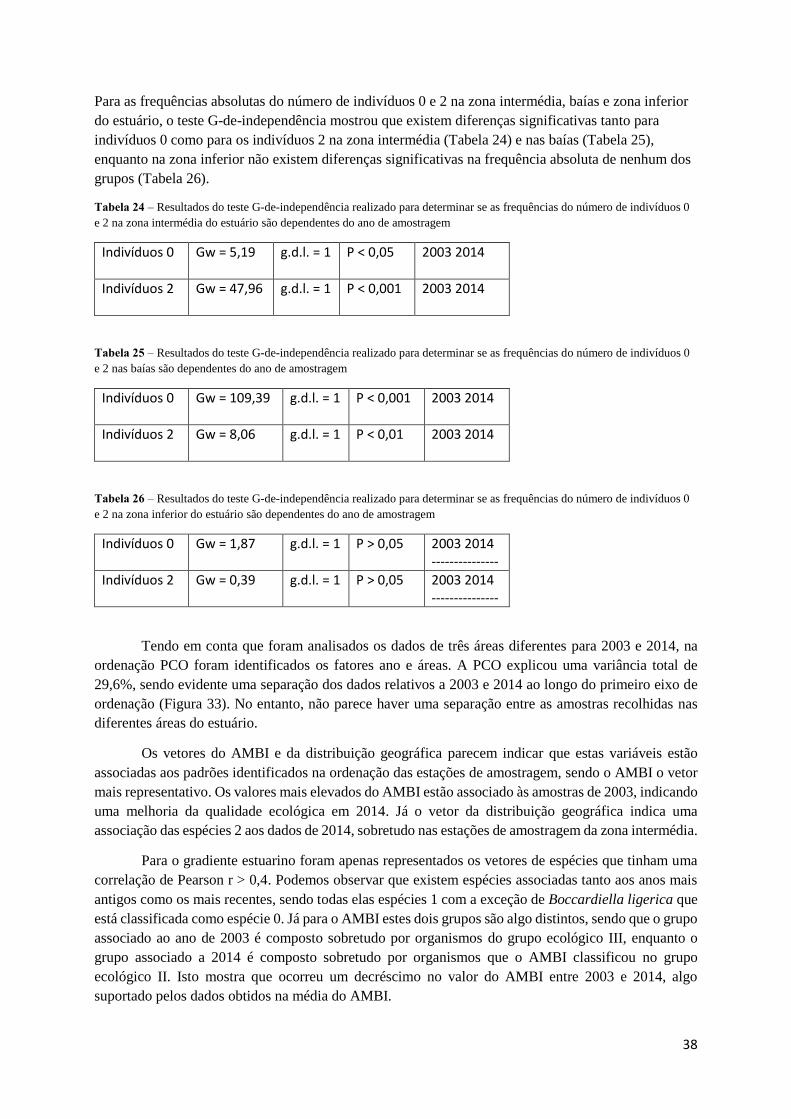
Para os dados de 2003 e 2014 correspondentes à zona intermédia, baías e zona inferior do estuário, o
teste G-de-independência mostrou que não existem diferenças significativas entre as frequências
absolutas de espécies 0 e 2 em qualquer um dos locais (Tabelas 21, 22 e 23).
Tabela 21 – Resultados do teste G-de-independência realizado para determinar se as frequências do número de espécies 0 e 2
na zona intermédia do estuário são dependentes do ano de amostragem
Espécies 0 Gw = 0,07 g.d.l. = 1 P > 0,05 2003 2014 --------------
Espécies 2 Gw = 0,03 g.d.l. = 1 P > 0,05 2003 2014 --------------
Tabela 22 – Resultados do teste G-de-independência realizado para determinar se as frequências do número de espécies 0 e 2
na baías são dependentes do ano de amostragem
Espécies 0 Gw = 0,46 g.d.l. = 1 P > 0,05 2003 2014 --------------
Espécies 2 Gw = 0,46 g.d.l. = 1 P > 0,05 2003 2014 --------------
Tabela 23 – Resultados do teste G-de-independência realizado para determinar se as frequências do número de espécies 0 e 2
na zona inferior do estuário são dependentes do ano de amostragem
Espécies 0 Gw = 0,072 g.d.l. = 1 P > 0,05 2003 2014 --------------
Espécies 2 Gw = 0,120 g.d.l. = 1 P > 0,05 2003 2014 --------------
38
Para as frequências absolutas do número de indivíduos 0 e 2 na zona intermédia, baías e zona inferior
do estuário, o teste G-de-independência mostrou que existem diferenças significativas tanto para
indivíduos 0 como para os indivíduos 2 na zona intermédia (Tabela 24) e nas baías (Tabela 25),
enquanto na zona inferior não existem diferenças significativas na frequência absoluta de nenhum dos
grupos (Tabela 26).
Tabela 24 – Resultados do teste G-de-independência realizado para determinar se as frequências do número de indivíduos 0
e 2 na zona intermédia do estuário são dependentes do ano de amostragem
Indivíduos 0 Gw = 5,19 g.d.l. = 1 P < 0,05 2003 2014
Indivíduos 2 Gw = 47,96 g.d.l. = 1 P < 0,001 2003 2014
Tabela 25 – Resultados do teste G-de-independência realizado para determinar se as frequências do número de indivíduos 0
e 2 nas baías são dependentes do ano de amostragem
Indivíduos 0 Gw = 109,39 g.d.l. = 1 P < 0,001 2003 2014
Indivíduos 2 Gw = 8,06 g.d.l. = 1 P < 0,01 2003 2014
Tabela 26 – Resultados do teste G-de-independência realizado para determinar se as frequências do número de indivíduos 0
e 2 na zona inferior do estuário são dependentes do ano de amostragem
Indivíduos 0 Gw = 1,87 g.d.l. = 1 P > 0,05 2003 2014 ---------------
Indivíduos 2 Gw = 0,39 g.d.l. = 1 P > 0,05 2003 2014 ---------------
Tendo em conta que foram analisados os dados de três áreas diferentes para 2003 e 2014, na
ordenação PCO foram identificados os fatores ano e áreas. A PCO explicou uma variância total de
29,6%, sendo evidente uma separação dos dados relativos a 2003 e 2014 ao longo do primeiro eixo de
ordenação (Figura 33). No entanto, não parece haver uma separação entre as amostras recolhidas nas
diferentes áreas do estuário.
Os vetores do AMBI e da distribuição geográfica parecem indicar que estas variáveis estão
associadas aos padrões identificados na ordenação das estações de amostragem, sendo o AMBI o vetor
mais representativo. Os valores mais elevados do AMBI estão associado às amostras de 2003, indicando
uma melhoria da qualidade ecológica em 2014. Já o vetor da distribuição geográfica indica uma
associação das espécies 2 aos dados de 2014, sobretudo nas estações de amostragem da zona intermédia.
Para o gradiente estuarino foram apenas representados os vetores de espécies que tinham uma
correlação de Pearson r > 0,4. Podemos observar que existem espécies associadas tanto aos anos mais
antigos como os mais recentes, sendo todas elas espécies 1 com a exceção de Boccardiella ligerica que
está classificada como espécie 0. Já para o AMBI estes dois grupos são algo distintos, sendo que o grupo
associado ao ano de 2003 é composto sobretudo por organismos do grupo ecológico III, enquanto o
grupo associado a 2014 é composto sobretudo por organismos que o AMBI classificou no grupo
ecológico II. Isto mostra que ocorreu um decréscimo no valor do AMBI entre 2003 e 2014, algo
suportado pelos dados obtidos na média do AMBI.
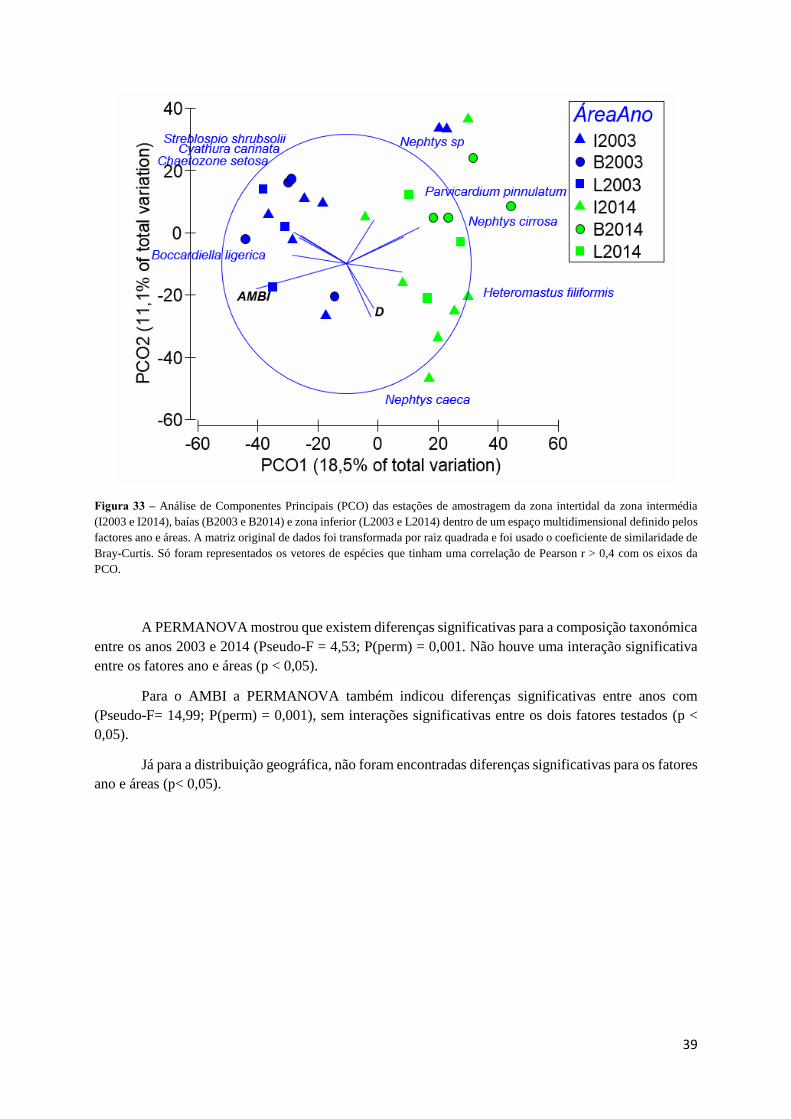
39
Figura 33 – Análise de Componentes Principais (PCO) das estações de amostragem da zona intertidal da zona intermédia
(I2003 e I2014), baías (B2003 e B2014) e zona inferior (L2003 e L2014) dentro de um espaço multidimensional definido pelos
factores ano e áreas. A matriz original de dados foi transformada por raiz quadrada e foi usado o coeficiente de similaridade de
Bray-Curtis. Só foram representados os vetores de espécies que tinham uma correlação de Pearson r > 0,4 com os eixos da
PCO.
A PERMANOVA mostrou que existem diferenças significativas para a composição taxonómica
entre os anos 2003 e 2014 (Pseudo-F = 4,53; P(perm) = 0,001. Não houve uma interação significativa
entre os fatores ano e áreas (p < 0,05).
Para o AMBI a PERMANOVA também indicou diferenças significativas entre anos com
(Pseudo-F= 14,99; P(perm) = 0,001), sem interações significativas entre os dois fatores testados (p <
0,05).
Já para a distribuição geográfica, não foram encontradas diferenças significativas para os fatores
ano e áreas (p< 0,05).

40
4. DISCUSSÃO
Os estudos sobre as variações temporais das comunidades de invertebrados bentónicos a
longo prazo são bastante limitados, quer pela morosidade do processamento de amostras, quer
pelos custos envolvidos. Por esse motivo, são muito raros os trabalhos disponíveis com séries
temporais longas, que permitam perceber as tendências de evolução das comunidades bentónicas
a longo prazo (e.g. Dauvin, 2000; Frid et al., 2009). Alguns dos trabalhos a longo prazo realizados
indicam que as tendências nos dados das comunidades bentónicas são consistentes entre épocas
do ano (e.g. Alden et al., 1997), no entanto, outros estudos sobre estas comunidades demonstram
que a avaliação das tendências do estado ecológico pode ser influenciada pela variabilidade
sazonal (e.g. Reiss and Kröncke 2005), sendo necessário selecionar as épocas em que as
comunidades refletem mais os efeitos das pressões antropogénicas do que das variações naturais
(Chainho et al,, 2007). As diferenças nas comunidades bentónicas ao longo do gradiente estuarino
(variabilidade espacial), são outro fator que dificulta a identificação de tendências temporais
(Chainho et al., 2008). Por esse motivo, o presente estudo teve como objetivo a identificação de
possíveis alterações temporais nas comunidades de invertebrados bentónicos, considerando
diferentes escalas temporais e espaciais no estuário do Tejo. Esta abordagem permitiu identificar
variações temporais significativas, quer no âmbito de séries temporais continuadas e localizadas
em áreas específicas do estuário, quer quando foram analisados dados de um gradiente estuarino
mais lato, com um intervalo temporal longo.
As ordenações das várias séries de dados analisadas mostram, de uma forma geral, uma
evolução temporal que tende a separar os dados mais antigos dos mais recentes, com maior
destaque para as diferenças entre os dados amostrados com um intervalo temporal longo (2003-
2014).
Qualidade ecológica
Tendo em conta que nos últimos 20 anos ocorreram diversas intervenções que
promoveram a recuperação do estuário do Tejo no que respeita à carga orgânica, nomeadamente
a construção de várias Estações de Tratamento de Águas Residuais (ETAR), um dos objetivos
deste estudo foi determinar se as alterações observadas nas comunidades bentónicas estariam
associadas a alterações da qualidade da água. Para isso foi identificada a qualidade ecológica para
os dados das várias séries temporais, com base no cálculo do AMBI, tendo-se verificado algumas
diferenças entre as séries estudadas. Na série temporal da zona inferior do estuário, apesar de se
terem verificados flutuações nos valores obtidos, esses estiveram sempre na amplitude da
classificação em Bom estado, exceto no local onde foi cessada a descarga de efluentes não
tratados (Porto do Buxo), onde se observou uma melhoria do estado a sequência da remoção dessa
fonte de contaminação orgânica. Embora não se observem mudanças muito significativas entre
2002 e 2010, observou-se uma melhoria entre 2002 e 2005, que parece estar associada á remoção
do esgoto, algo que foi registado em outros trabalhos da mesma natureza e que usaram o índice
AMBI (Borja, 2003; Borja, 2006). As pequenas oscilações observadas no Portinho da Costa ao
longo do tempo parecem enquadrar-se no conceito de “distúrbios de pulso” (Bender et al., 1984),
que são pequenas perturbações que aumentam ou diminuem certas populações temporariamente.
Estes pulsos são de curta duração e tem origens imprevisíveis e intensidades variadas, podendo
ser de origem natural ou antropógenica. Embora estas flutuações possam ser assosicadas a eventos
como variações sazonais ou eventos extremos que ocorreram nesta zona no estuário, a presença
de uma ETAR no Portinho da Costa poderá explicar estas variações temporais, uma vez que é
possível que a ETAR não tenha apresentado sempre a mesma eficácia de tratamento neste espaço

41
de tempo, algo que pode ter-se refletido pontualmente nas descargas e qualidade da água junto a
este local.
Para a série temporal da zona intermédia do estuário (Expo), não se observaram alterações
significativas nos valores de AMBI entre 1998 e 2013, tanto em ambientes intertidais, como
subtidais. Na zona intertidal a amplitude de variação dos valores de AMBI foi diminuta e na zona
subtidal as variações ao longo do tempo foram semelhantes às observadas no Portinho da Costa.
Os resultados obtidos parecem estar relacionados com clara dominância de espécies do
grupo ecológico III, as quais são tolerantes ao enriquecimento orgânico (Grall and Glémarec,
1997). A dominância de espécies pertencentes a este grupo é característica de ecossistemas como
os estuários (Borja et al., 2000), algo que está de acordo com as amostras recolhidas na área da
Expo. A utilização de índices para avaliar o estado ecológico nestas áreas tem revelado inúmeros
constrangimentos, que requerem a adaptação das ferramentas existentes (Chainho, 2008) e a
utilização de indicadores que integrem vários tipos de métricas (Borja et al., 2009; Quintino et
al., 2006; Dauvin, 2007; Pinto et al., 2009; Borja et al., 2011). A resposta das espécies às
condições variáveis e imprevisíveis dos sistemas de transição também é apontada como um
problema porque torna-se difícil separar stresses de origem natural daqueles de origem
antropogénica (Elliott and Quintino, 2007).
Para além disto, a robustez do AMBI é reduzida quando temos um número reduzido de
taxa e/ou indivíduos por amostra (Borja et al., 2004), algo que se verificou nas amostras
recolhidas na Expo, em certos momentos, sobretudo para as zonas intertidais.
De uma forma geral, a comparação dos resultados dos anos mais antigos com os mais
recentes, indica uma diminuição dos valores do AMBI, o que corresponde a uma melhoria da
qualidade da água. O conjunto de dados usados foi limitado pelo facto de se terem selecionado as
estações de amostragens que permitiram ter a série temporal mais longa possível, mas trabalhos
anteriores que usaram um conjunto maior de estações de amostragem mostram uma tendência
significativa para a melhoria do estado ecológico nesta área, considerando vários atributos da
comunidade bentónica, incluindo os valores de AMBI (Chainho et al., 2010). Esta melhoria
parece estar associadas à requalificação da antiga área industrial, mas também ao funcionamento
da ETAR de Beirolas, situada nas imediações, que foi ampliada e melhorada em 2000, sendo
precisamente a partir desta altura que se começaram a observar maiores variações nos valores de
AMBI.
Os dados de 2003-2014, correspondentes a várias zonas do estuário, apontam para uma
melhoria geral na qualidade da água, com os valores de AMBI a diminuírem entre 2003 e 2014
para a zona intermédia, baías e zona inferior. Para o Porto do Buxo, Portinho da Costa e Expo
observou-se uma evolução gradual da qualidade da água com amostras realizadas em vários anos,
enquanto estes dados correspondem a apenas dois anos e com um intervalo de 10 anos entre si, o
que permite observar variações de longo prazo. De facto, se compararmos o primeiro e ultimo
ano de amostragem para cada local, pode-se observar que o primeiro ano tem sempre valores de
AMBI superiores ao do último, sendo a exceção o caso do Portinho da Costa, onde os valores são
semelhantes.
Alterações climáticas
As alterações climáticas têm levado ao aumento da temperatura dos sistemas aquáticos a
nível global. As espécies podem responder às alterações climáticas ao mudar a sua abundância e
distribuição de acordo com as mudanças que ocorrem nos ecossistemas (Holt, 1990). Entre 1930

42
e 1950, ocorreu uma diminuição na abundância relativa no número de espécies intertidais
pertencentes á classe Cirripedia, na costa do Reino Unido, sendo este fenómeno uma resposta
destas espécies de águas frias a um aumento de temperatura na ordem dos 0,5oC (Southward and
Crisp, 1954). Segundo Austin et al. (2010), o número de espécies invasoras em estuários
aumentou nos últimos 20 anos devido às alterações climáticas. Isto levanta a hipótese que
pequenas variações de temperatura são suficientes para afetar a abundância relativa de espécies,
sobretudo aquelas que se encontram nos seus limites de distribuição.
No caso do estuário do Tejo, a temperatura da água subiu mais de um grau entre 1980 e
meados dos anos 90, um período de aproximadamente 15 anos (Cabral et al., 2001). Este aumento
de temperatura seria de aproximadamente dois graus em 2008, em comparação com 1980.
Adicionando o facto de o estuário do Tejo ser considerado um limite biogeográfico (Hayden et
al., 1984), seria de esperar que caso se registassem alterações nas comunidades devido às
mudanças de temperatura, estas se refletissem com um aumento de abundância e alargamento da
distribuição das espécies com maiores afinidades meridionais, e um decréscimo na abundância e
recuo na distribuição das espécies com maiores afinidades setentrionais.
Este cenário não foi registado na série temporal da área inferior do estuário do Tejo, uma
vez que não se verificaram tendências significativas na representatividade das espécies com
distribuição mais meridional e setentrional, sendo estas últimas dominantes comparativamente às
primeiras. Já na área intermédia do estuário, observou-se um aumento da representatividade das
espécies com maior afinidade meridional, tanto na zona intertidal, como na subtidal, em particular
a partir de 2009. Os dados de 2003 e 2014 também corroboram os resultados das séries temporais,
indicando um claro crescimento no número de espécies e indivíduos com distribuição mais
subtropical na área intermédia do estuário. Nas baías observou-se um ligeiro aumento do número
de indivíduos com afinidades meridionais, mas na área inferior observou-se uma ligeira
diminuição, tanto do número de espécies, como de indivíduos com características subtropicais,
embora continuem a ser dominantes relativamente às espécies setentrionais.
Estes resultados parecem indicar que a dispersão geográfica das espécies em meio
estuarino está condicionada pelas condições ambientais específicas das diferentes áreas do
gradiente estuarino. Segundo Hiscook et al. (2004), as alterações nas comunidades devido a
mudanças de temperatura serão mais notáveis em sistemas de água parada ou pouco
movimentada. No caso de algumas espécies com afinidade meridional, as temperaturas mais
elevadas que ocorrem, nas épocas mais quentes, em águas superficiais de sistemas como lagoas e
estuários são importantes e benéficas para a reprodução e possível aumento das populações destas
espécies.
A área inferior do estuário encontra-se na zona terminal deste sistema, com características
muito semelhantes ou idênticas às da água do mar, incluindo parâmetros determinantes para as
comunidades bentóncias, como a temperatura e salinidade. Por sua vez, a área intermédia e as
baías são mais vulneráveis a alterações globais, uma vez que a massa de água é bastante menos
profunda, tendo sido nessas áreas que ocorreram as maiores alterações na representatividade das
espécies com afinidade meridional, com uma aumento das mesmas, tanto no âmbito da série
temporal, como dos dados recolhidos em 2003 e 2014, ao longo do gradiente estuarino. A série
temporal da área intermédia do estuário permitiu ainda perceber que as alterações no balanço de
espécies meridionais versus setentrionais são diferenciais entre as zonas intertidais e subtidais.
No intertidal, não foi registado um número considerável de espécies meridionais e verificou-se a
ausência de espécies setentrionais, o que parece estar relacionado com o facto dos ambientes
intertidais serem mais vulneráveis a mudanças de temperatura, podendo ser letais para muitos

43
indivíduos devido a temperaturas extremamente baixas no inverno (e.g. Crisp, 1964; Todd and
Lewis, 1984) e altas no verão (e.g. Hawkins and Hartnoll, 1985).
Ao contrário da área terminal do estuário, onde a temperatura é mais estável e há maior
agitação marítima, a zona interior do estuário apresenta menor movimento das massas de água, o
que, associado às elevadas temperaturas registadas durante o verão, pode levar a um aumento da
temperatura das águas superficiais nestes locais. Estas temperaturas ligeiramente mais elevadas
parece constituir um fator favorável ao estabelecimento de espécies com características mais
subtropicais nestes ecossistemas, permitindo assim a sua sobrevivência e reprodução (Hiscook et
al., 2004).
A dispersão de espécies meridionais para norte e o recuo das espécies setentrionais
também para norte será progressivo, uma vez que, para além da temperatura, os organismos são
ainda condicionados pelas características das massas de água, qualidade da água, mecanismos de
dispersão, ciclo de vida, interações bióticas e presença de barreiras geográficas (Clark and Frid,
2001; Hiscock et al., 2004; Thuiller, 2004). Isto é ainda salientado pelos testes G-de-
independência que mostram que embora ocorram diferenças significativas no número de
indivíduos 0 e 2 entre anos em alguns pontos do estuário, o mesmo nunca se registou para o
número de espécies.
As variações de temperatura devido às alterações climáticas decorrem de forma lenta e
gradual, algo que dificilmente se consegue refletir em alterações significativas das comunidades,
de forma óbvia, em 10-15 anos de amostragem, pelo que foram pouco significativas do ponto de
vista estatístico as alterações verificadas para a distribuição geográfica das espécies analisadas no
estuário do Tejo. Barry et al. (1995) referem um aumento no número de espécies intertidais de
sul e um decréscimo na abundância de espécies do norte em Monterey, California. Estas
alterações nas comunidades ocorreram no período entre 1932 e 1993, sendo que durante este
intervalo de tempo a temperatura das águas costeiras aumentou cerca de 0,75 ºC (Sagarin et al.,
1999). Os resultados do presente estudo, apesar de ter sido realizado num intervalo de tempo
relativamente curto, já providenciam algumas evidências de possíveis alterações relacionadas
com alterações climáticas em curso na região biogeográfica onde se localiza o estuário do Tejo.
Alguns estudos anteriores, realizados com peixes, já tinham demonstrado que existe uma
correlação entre a temperatura e a distribuição geográfica de algumas espécies, afetando
sobretudo os indivíduos que se encontram nos limites de distribuição da sua espécie (Cabral et
al., 2001; Vinagre et al., 2009).

44
5. CONSIDERAÇÕES FINAIS
Os resultados do presente trabalho mostraram a ocorrência de alterações nas comunidades
de invertebrados bentónicos no estuário do Tejo, nos últimos anos, tanto na área intermédia, como
nas baías e área inferior do estuário, sendo essas alterações igualmente identificadas no âmbito da
análise de séries temporais contínuas, como de dados recolhidos com um intervalo longo. Uma
análise da relação dessas alterações com a qualidade ecológica e a distribuição geográfica das
espécies identificadas permitiu perceber que se verificou simultaneamente uma melhoria da
qualidade da água do estuário e uma alteração da composição das comunidades relativamente às
suas afinidades climáticas, com um aumento da representatividade das espécies com
características meridionais.
Esta tendência para a melhoria da qualidade reforça a importância de implementar
medidas mitigadores, como construção e melhoria das ETARs e remoção de efluentes não
tratados, como aconteceu nos locais dos casos de estudo do Portinho da Costa, Porto do Buxo e
Expo.
Apesar da sua baixa mobilidade, os macroinvertebrados bentónicos também forneceram
dados interessantes relativamente às alterações climáticas e a sua influência sobre a distribuição
geográfica das espécies aquáticas. Apesar de se terem verificado diferenças entre locais de
amostragem, os resultados obtidos apontam para um maior aumento no número de espécies
meridionais do que setentrionais. Para além disso, o número de espécies e indivíduos com
afinidades temperadas e subtropicais tem vindo a aumentar nos últimos anos nas zonas
intermédias do estuário, algo que poderá estar associado ao facto de serem águas mais paradas e
menos profundas e que assim aquecem mais durante a estação do verão.
Estes resultados reforçam a importância da realização de estudos de longo termo, uma
vez que as grandes alterações na composição das comunidades ocorrem naturalmente em períodos
que podem variar entre meses e décadas. Muitas destas alterações resultam de processos
estocásticos, outras refletem as interações entre as espécies ou resultados de pressões humanas e
outras podem ainda estar relacionadas com alterações de longo prazo, como é o caso das
alterações climáticas (Gray and Christie, 1983).
Para futuros trabalhos que abordem as temáticas da qualidade da água e efeito das alterações
globais na distribuição das espécies seria também importante analisar a evolução das
comunidades para as restantes estações do ano. Neste trabalho abordou-se apenas o verão por
ser a estação mais “regular e estável” e ser aquela onde não é comum ocorrer fenómenos
meteorológicos extremos. Tendo em consideração os resultados obtidos na componente da
distribuição geográfica, ter dados para as restantes estações do ano podia ter fornecido
informação adicional que pudesse explicar não só o aumento de espécies do sul nos últimos
anos, bem como o maior domínio de espécies do norte na zona exterior do estuário.

45
6. REFERÊNCIAS BIBLIOGRÁFICAS
Abd-Elnaby, F.A. (2009). New records of Polychaetes from the south part of Suez Canal, Egypt.
World J. Fish Mar. Sci. 1(1): 7-19
Adema, J.P.H.M. (1991). De krabben van Nederland en Belgie (Crustacea, Decapoda, Brachyura)
[The crabs of the Netherlands and Belgium (Crustacea, Decapoda, Brachyura)]. Nationaal
Natuurhistorisch Museum: Leiden, The Netherlands. ISBN 90-73239-02-8. 244 pp
Aloui-Bejaoui, N.; Afli, A. (2012). Functional diversity of the macro-invertebrate community in
the port area of Kerkennah Islands (Tunisia). Mediteranean Marine Science. 13 (1): 93-102.
Anderson, M. J., R. N. Gorley & K. R. Clarke, 2008. PERMANOVA+ for PRIMER: Guide to
Software and Statistical Methods. PRIMER-E, Plymouth: 213.
Andrade, F., Cabral, H., e Borges, M.F., 2009. Ambientes Costeiros. In Pereira, H.; Domingos,
T.; Vicente L., e Proença, V. (editores), Ecossistemas e Bem-Estar Humano. Avaliação para
Portugal do Millenium Ecosystem Assessment. Lisboa, Portugal: Escolar Editora, pp. 413-435
Antit, M.; Gofas, S.; Salas, C.; Azzouna, A. (2011). One hundred years after Pinctada: an update
on alien Mollusca in Tunisia. Mediterranean Marine Science 12(1): 53-73.
Attrill, M.J. & Depledge, M.H. (1997). Community and population indicators of ecosystem
health: targeting links between levels of biological organisation. Aquatic Toxicology 38, 183-
197.
Austin GE, Rehfisch HA, Viles HA, Berry PM. 2001. Impacts on coastal environments. In
Climate Change and Nature Conservation in Britain and Ireland: Modelling Natural Resource
Responses to Climate Change (the MONARCH project). Harrison, PA, Berry PM, Dawson, TE
(eds). UKCIP Technical Report: Oxford, 177-228.
Ayari, R. 2004. Les peuplements macro-zoobenthiques du petit golfe de Tunis: Structure,
Organisation et Etat sanitaire. MSc Thesis. University of Carthage, Faculte des Sciences de
Bizerte
Bachelet, G.; Dauvin, J.-C.; Sorbe, J.C. (2003). An updated checklist of marine and brackish water
Amphipoda (Crustacea: Peracarida) of the southern Bay of Biscay (NE Atlantic). Cah. Biol. Mar.
44(2): 121-151
Backeljau, T. (1986). Lijst van de recente mariene mollusken van België [List of the recent marine
molluscs of Belgium]. Koninklijk Belgisch Instituut voor Natuurwetenschappen: Brussels,
Belgium. 106 pp.
Barnard, K.H. (1925). A Revision of the Family Anthuridae (Crustacea Isopoda), with Remarks
on certain Morphological Peculiarities. Journ. Limn. Soc (London) (Zool.) 36: 109-160.
Barry JP, Baxter CH, Sagarin RD, Gilman SE. 1995. Climate-related, long-term faunal changes
in a California rocky intertidal community. Science 267: 672-675.

46
Beck MW, Heck KL, Able KW, Childers DL, Eggleston DB, Gillanders BM, et al. The
identification, conservation, and management of estuarine and marine nurseries for fish and
invertebrates. 2001 Bioscience 51;633 – 641.
Bellan, G. (2001). Polychaeta, in: Costello, M.J. et al. (Ed.) (2001). European register of marine
species: a check-list of the marine species in Europe and a bibliography of guides to their
identification. Collection Patrimoines Naturels, 50: pp. 214-231
Beneden. (1867a). Recherches sur la faune littorale de Belgique : polypes / par P.-J. van Beneden.
Bettencourt, A., Gomes, V., Dias, A., Ferreira, G., Silva, M.C., Costa, L., 2003. Estuários
Portugueses. Direcção dos Serviços de Planeamento, Instituto da Água, Ministério das Cidades,
Ordenamento do Território e Ambiente, Lisboa.
Beyst, B. (2001). Epi- en hyperbenthische gemeenschappen van Belgische zandstranden [Epi-
and hyperbenthic communities of Belgian sandy beaches]. PhD Thesis. Universiteit Gent.
Instituut voor Dierkunde. Vakgroep morfologie, systematiek en ecologie: Gent, Belgium. 351 pp.
Bird, G.J. (2001). Tanaidacea. In: Costello, M.J., C.S. Emblow, & R. White (eds.), European
Register of Marine Species. A Check-List of the Marine Species in Europe and a Bibliography of
Guides to Their Identification. Patrimoines Naturels. 50: 310-315
Blaber SJM, Cyrus DP, Albaret JJ, Ching CV, Day JW, Elliott M, et al. Effects of fishing on the
structure and functioning of estuarine and nearshore ecosystems. ICES J Mar 2000;57:590-602.
Boesch, D.F. (1973). Classifictaion and community structure of macrobenthos in the Hampton
Roads area, Virginia. Marine Biology 21, 226-244.
Bogdanos, C.; Fredj, G. (1983). Sur la presence de Cossura costa Kitamori, (Polychaeta,
Cossuridae) dans les eaux cotieres Grecques (Mer Egee, Mer Ionienne). Thalassographica. 6: 5-
15
Borges, P.A.V., Costa, A., Cunha, R., Gabriel, R., Gonçalves, V., Martins, A.F., Melo, I., Parente,
M., Raposeiro, P., Rodrigues, P., Santos, R.S., Silva, L., Vieira, P. & Vieira, V. (Eds.) (2010). A
list of the terrestrial and marine biota from the Azores. Princípia, Oeiras, 432 pp.
Borja, A ., Franco, J., Muxika, I., 2004b. The biotic indices and the Water Framework Directive:
the required consensus in the new benthic monitoring tools (correspondence). Mar. Poll. Bull. 48,
405 -408.
Borja, A., Franco, J., Pérez, V., 2000. A marine biotic index to establish the ecological quality of
soft-bottom benthos within European estuarine and coastal environments. Marine Pollution
Bulletin 40, 1100e1114.
Borja, A., Miles, A., Occhipinti-Ambrogi, A., Berg, T., 2009. Current status of macroinvertebrate
methods used for assessing the quality of European marine waters: implementing the Water
Framework Directive. Hydrobiologia 633, 181–196
Bouvier, E.-L. (1923). Pycnogonides [Pycnogonida]. Fauna of France, 7. Paul Lechevalier: Paris,
France. 69 pp.
Byrne, P. e O’Halloran, J. (2001). The role of bivalve molluscs as tools in estuarine sediment
toxicity testing: a review. Hydrobiologia, 465: 209-217.

47
Cabral, H.N., Costa, M.J., Salgado, J.P., 2001. Does the Tagus estuary fish community reflect
environmental changes? Climate Research 18, 119–126.
Caeiro, S., Costa, M., Goovaerts, P. e Martins, F. (2005). Benthic biotope index for classifying
habitats in the sado estuary: Portugal. Marine Environmental Research, 60: 570-593.
Cardigos, F.; Tempera, F.; Ávila, S.; Gonçalves, J.; Colaço, A.; Santos, R. S. (2006). Non-
indigenous marine species of the Azores. Helgoland Marine Research. 60(2), 160-169.
Carpenter, G.H. (1905). The marine fauna of the coast of Ireland: 6. Pycnogonida. Fisheries,
Ireland, Sci. Invest. 1904(IV, [1905]): 1-8, plates I-III
Cattrijsse, A.; Vincx, M. (2001). Biodiversity of the benthos and the avifauna of the Belgian
coastal waters: summary of data collected between 1970 and 1998. Sustainable Management of
the North Sea. Federal Office for Scientific, Technical and Cultural Affairs: Brussel, Belgium. 48
pp.
Chainho, P., Costa, J.L., Chaves, M.L., Dauer, D.M., Costa, M.J., 2007. Influence of seasonal
variability in benthic invertebrate community structure on the use of biotic indices to assess the
ecological status of a Portuguese estuary. Marine Pollution Bulletin 54, 1586-1597.
Chainho, P., 2008. Contribution to the development of biotic integrity assessmenttools for
Portuguese estuaries based on benthic communities. Ph.D. Thesis. University of Lisbon.
Christie, H.; Jørgensen, N.M.; Norderhaug, K.M.; Waage-Nielsen, E. (2003). Species distribution
and habitat exploitation of fauna associated with kelp (Laminaria hyperborea) along the
Norwegian Coast. J. Mar. Biol. Ass. U.K. 83(4): 687-699
Clark, R.A., Frid, C.L.J., 2001. Long-term changes in the North Sea ecosystem. Environmental
Review 9, 131e187.
Clarke, K.R. & R.N. Gorley, 2006. PRIMER v6: User Manual/Tutorial. PRIMER-E, Plymouth.
Costa, M. J. 1999. O Estuário do Tejo. Edições Cotovia Lda. Lisboa.
Costanza, R., dArge, R., deGroot, R., Fraber, S., Grasso, M., Hannon B., et al. (1997). The value
of the world’s ecosystem services and natural capital. Nature 387, 253-60.
Crisp D J (ed.). 1964. The effects of the severe winter of 1962/63 on marine life in Britain. Journal
of Animal Ecology 33: 165-210.
Crisp, D.J., Southward, A.J. & Southward, E.C., 1981. On the distribution of the intertidal
barnacles Chthamalus stellatus, Chthamalus montagui and Euraphia depressa. Journal of the
Marine Biological Association of the United Kingdom, 61, 359-380.
Daglı, E.; Cınar, M. E. (2008). Invasion of polluted soft substratum of Izmir Bay (Aegean Sea,
eastern Mediterranean) by the spionid polychaete worm, Pseudopolydora paucibranchiata
(Polychaeta: Spionidae). Cahiers de Biologie Marine. 49(1): 87-96.
Darmoul, B., Hadj Ali, S., Vitello, P. 1980. Effets des rejets industriels de la région de Gabès
(Tunisie) sur le milieu marin recepteur.Bull. Inst. Nat. Sc. Oceanogr. Pêche, Salammbô, 7: 5-61.
Davoult, D.; Dewarumez, J.-M.; Luczak, C.; Migné, A. (1999). Nouvelles signalisations d'espèces
benthiques sur les côtes françaises de la Manche orientale et de la Mer du Nord [New reports of

48
benthic species from the french coasts of the eastern English Channel and the North Sea]. Cah.
Biol. Mar. 40(2): 121-127
Davoult, D. (2002). Faune et flore du littoral du Pas-de-Calais et de la Manche orientale: mise à
jour de la liste des espèces d'Echinodermes [Fauna and flora of the Pas-de-Calais coast and the
eastern English Channel: review of the echinoderm species list]. Rev. Trav. Stat. Mar. Wimereux
2001(24): 1-3
Dauer, D. M. (1993) Biological criteria, environmental health and estuarine macrobenthic
community structure. Marine Pollution Bulletin 26 (5), 249±257.
Dauvin, J.-C. (1999). Mise a jour de la liste des espèces d'Amphipodes (Crustacea: Peracarida)
présents en Manche [An up-to-date list of the amphipods (Crustacea: Peracarida) from the English
Channel. Cah. Biol. Mar. 40(2): 165-183
Dauvin, J.C. 2000. The muddy fine sand Abra alba–Melinna palmate community of the Bay of
Morlaix twenty years after the Amoco Cadiz oil spill. Marine Pollution Bulletin 40: 528–536.
Dauvin, J.-C.; Dewarumez, J.-M.; Gentil, F. (2003). Liste actualisée des espèces d’Annélides
Polychètes présentes en Manche [An up to date list of polychaetous annelids from the English
Channel]. Cah. Biol. Mar. 44(1): 67-95
Day, J.W.Jr., Hall, C.A.S., Kemp, W. M. & Yañez-Arancibia, A. (1989). Estuarine Ecology. John
Wiley, New York, 558 pp.
de Bruyne, R.H. (1991). Schelpen van de Nederlandse kust [Shells of the Dutch coast].
Jeugdbondsuitgeverij/KNNV Uitgeverij: Utrecht, The Netherlands. ISBN 90-5107-017-9. III,
165 pp.
De Maersschalck, V. (2004). An inventory of the macrofauna of the Sluice Dock at Oostende:
contribution to an integrated management [Een inventarisatie van de macrofauna van de Spuikom
te Oostende: bijdrage tot een geïntegreerd beheer]. MSc Thesis. Universiteit Gent, Vakgroep
Biologie: Gent, Belgium. 71, tables, figures pp
Dore I. & Frimodt C. (1987). An Illustrated Guide to Shrimp of the World. Van Nostrand
Reinhold Company,, New York, 1-229.
Dumoulin, E. (1989). Micromolluscs in sand suppletions from the Goote Bank [Micromollusken
uit opgespoten zand afkomstig van de Goote Bank]. De Strandvlo 9(1): 21-31
Dyer, K. R. 1996 The definition of the Severn estuary. Proceedings of the Bristol Naturalists’
Society 56, 53–66.
Dyntaxa. (2013). Swedish Taxonomic Database.
d'Udekem d'Acoz, C. (1986). Report on a walk between De Haan and Blankenberge (Belgium)
[Rapport sur une promenade entre De Haan et Blankenberge]. De Strandvlo 6(3): 68-69
d'Udekem d'Acoz, C. (1990). Notes on some organisms collected between Wenduine and De
Haan on 3 March 1990 [Notes sur quelques organismes recueillis entre Wenduine et De Haan le
3 mars 1990]. De Strandvlo 10(3): 74-78
d'Udekem d'Acoz, C. (1999). Inventory and distribution of the decapod crustaceans from the
northeastern Atlantic, the Mediterranean and the adjacent continental waters north of 25°N.

49
Collection Patrimoines Naturels, 40. Muséum national d'Histoire naturelle. Paris. ISBN 2-86515-
114-10. X, 383 pp.
D'Udekem d'Acoz C. & Wirtz P., (2000).Observations on some interesting coastal Crustacea
Decapoda from the Azores, with a key to the genus Eualus Thallwitz, 1892 in the Northeastern
Atlantic and the Mediterranean. Arquipélago, Life and Marine Sciences. Bull. Univ. of the
Azores, 19A, 67-84.
El Lakhrach, H., Hattour, A. 2010. Répartition des Crustacés Décapodes dans le Golfe de Gabès.
Rapp. Comm. int. Mer Médit., 39
Elliott, M., Quintino, V., 2007. The estuarine quality paradox. Environmental homeostasis and
the difficulty of detecting anthropogenic stress in naturally stressed areas. Marine Pollution
Bulletin 54, 640e645.
Emmerson W.D. A comparison between decapod Species common to both Mediterranean and
Southern African Waters.
Eneman, E. (1984). Uit het Natuurhistorisch Archief [From the Natural History Archive]. De
Strandvlo 4(1): 4-17
Enzenross, L.; Enzenross, R. (2001). Untersuchungen über das Vorkommen mariner Mollusken
in tunesichen Gewässern. Shriften für Malakozoologie. 17, 45-62.
Faasse, M.; Van Moorsel, G. (2000). Nieuwe en minder bekende vlokreeftjes van sublitorale
harde bodems in het Deltagebied (Crustacea: Amphipoda: Gammaridea) [New and lesser-known
amphipods of hard substrates in the Delta area of the Netherlands (Crustacea: Amphipoda:
Gammaridea)]. Ned. Faunist. Meded. 11: 19-44
Faubel, A.; Noreña, C. (2001). Turbellaria, in: Costello, M.J. et al. (Ed.) (2001). European register
of marine species: a check-list of the marine species in Europe and a bibliography of guides to
their identification. Collection Patrimoines Naturels, 50: pp. 123-136
Fauchald, K. (2007). World Register of Polychaeta.
Faulwetter, Sarah (2010). Check-list of marine Polychaeta from Greece. Aristotle University of
Thessaloniki. Assembled in the framework of the EU FP7 PESI project
Faune et flore du littoral du Nord, du Pas-de-Calais et de la Belgique: inventaire. Commission
Régionale de Biologie Région Nord Pas-de-Calais: France. 307 pp.
Fiege, D.; Licher, F.; Mackie, A.S.Y. (2000). A partial review of the European Magelonidae
(Annelida: Polychaeta): Magelona mirabilis redefined and M. johnstoni sp. nov. distinguished. J.
Mar. Biol. Ass. U.K. 80: 215-234
Fofonoff, P.W.; Ruiz, G.M.; Steves, B.; Carlton, J.T. (2014). National Exotic Marine and
Estuarine Species Information System (NEMESIS)
Fowler, G.H. 1897. Contributions to our knowledge of the plankton of the Faroe Channel. III.
The later development of Arachnactis albida (M. Sars) with notes on Arachnactis bournei (sp.n.).
Proceedings of the Zoological Society of London. 1897: 803-809
Frid, C.L.J., P.R. Garwood, and L.A. Robinson. 2009. Observing change in a North Sea benthic
system: a 33 year time series. Journal of Marine Systems 77: 227–236.

50
Giltay, L. (1929). Quelques Pycnogonides des environs de Banyuls (France) [On some
pycnogonids from Banyuls (France)]. Ann. Bull. Soc. Entomol. Belg. 69: 172-176
Glémarec, M. and Hily, C. (1981) Perturbations apportées á la macrofaune benthique de la baie
de Concarneau par les éuents urbains et portuaires. Acta Oecologica Oecologia Applicata 2,
139±150.
Gofas, S.; Le Renard, J.; Bouchet, P. (2001). Mollusca. in: Costello, M.J. et al. (eds), European
Register of Marine Species: a check-list of the marine species in Europe and a bibliography of
guides to their identification. Patrimoines Naturels. 50: 180-213.
Gollasch, S.; Riemann-Zurneck (1996) Transoceanic dispersal of benthic
macrofauna:Haliplanella luciae (Verrill, 1898) (Anthozoa, Actinaria) found on a ship's hull in a
shipyard dock in Hamburg, Germany, Helgoländer Meeresuntersuchungen 50: 253-258
Guiry, M.D. & Guiry, G.M. (2011). Species.ie version 1.0 World-wide electronic publication,
National University of Ireland, Galway (version of 15 March 2010).
Haedrich RL. Estuarine fishes. In: Ketchum B, editor. Ecosystems of the world. Estuarine and
Enclosed SeasAmsterdam: Elsevier; 1983. p. 183–207.
Hartmann-Schröder, G. (1996). Annelida, Borstenwürmer, Polychaeta [Annelida, bristleworms,
Polychaeta]. 2nd revised ed. The fauna of Germany and adjacent seas with their characteristics
and ecology, 58. Gustav Fischer: Jena, Germany. ISBN 3-437-35038-2. 648 pp.
Hartley, J.P., Dicks, B. 1987. Macrofauna of subtidal sediments using remote sampling. Pp. 106-
130. In: Baker, J.M., Wolff, W. (eds.). Biological surveys of estuaries and coasts. Cambridge
University Press, Cambridge, United Kingdom.
Hayden, B.P., G.C. Ray. & R. Dolan. 1984. Classification of coastal and marine environments.
Environmental Conservation, 11: 199-207.
Hayward, P.J.; Ryland, J.S. (Ed.). (1990). The marine fauna of the British Isles and North-West
Europe: 1. Introduction and protozoans to arthropods. Clarendon Press: Oxford, UK. ISBN 0-19-
857356-1. 627 pp.
Hansson, H.G. (Comp.) 1998. NEAT (North East Atlantic Taxa): Scandinavian Echinodermata
Hawkins SJ, Hartnoll RG.1985. Factors determining the upper limits of intertidal canopy-forming
algae. Marine Ecology Progress Series 20: 265-271.
Hiscock, K., Southward, A.J., Tittley, I. & Hawkins, S.J. ( 2004). Effect of changing temperature
on benthic marine life in Britain and Ireland . Aquatic Conservation 14 : 333-362.
Hoek, P.P.C. (1881). Nouvelles études sur les Pycnogonides [New studies on the Pycnogonida].
Arch. de Zool. Exp. et Gén. 9: 445-542, plates XXIII-XXX
Holland, A.F., Shaughnessy, A.T. & Hiegel, M.H. (1987). Long-term variation in mesohaline
Chesapeake Bay macrobenthos: spatial and temporal patterns. Estuaries 10, 227-245.
Holly, M. (1938). Polychaeten aus der Nordsee [Polychaetes from the North Sea]. Bull. Mus.
royal d'Hist. Nat. Belg./Med. Kon. Natuurhist. Mus. Belg. 14(12): 1-24

51
Holt, E.W.L., & W.I. Beaumont. 1900 (1902). Survey of fishing grounds, west coast of Ireland,
1890-1891. X.- Report on the Crustacea Schizopoda of Ireland.-- The Scientific Transactions of
the Royal Dublin Society, Ser. 2, 7 (7): 221-252 (1 plate).
Holt, R.D., 1990. The microevolutionary consequences of climate change. Trends in Ecology and
Evolution 5, 311e315.
Holtmann, S.E.; Groenewold, A.; Schrader, K.H.M.; Asjes, J.; Craeymeersch, J.A.; Duineveld,
G.C.A.; van Bostelen, A.J.; van der Meer, J. (1996). Atlas of the zoobenthos of the Dutch
continental shelf. Ministry of Transport, Public Works and Water Management: Rijswijk, The
Netherlands. ISBN 90-369-4301-9. 243 pp.
Holthuis, L.B. (1950). Isopodes et Tanaidacés marins de la Belgique; remarques sur quelques
espèces de la zone méridionale de la Mer du Nord [Marine Isopoda and Tanaidacea from Belgium:
remarks on some species from the southern North Sea]. Med. K. Belg. Inst. Nat. Wet. 26(53): 1-
19
Huwae, P.; Rappé, G. (2003). Waterpissebedden: een determineertabel voor de zoet-, brak- en
zoutwaterpissebedden van Nederland en België. Wetenschappelijke Mededelingen van de
Koninklijke Nederlandse Natuurhistorische Vereniging, 226. KNNV Uitgeverij: Utrecht, The
Netherlands. ISBN 90-5011-171-8. 55 pp.
INAG. 2001. Plano Nacional da Água. Instituto da Água, Lisboa, Portugal.
Ingle, R., 1993. Hermit crabs of the northeastern Atlantic Ocean and the Mediterranean Sea.
Intergovernmental Panel on Climate Change, 2001. Third assessment report of the working group
I. In: Houghton, J.T., et al. (Eds.), The Science of Climate Change. Cambridge University Press,
Cambridge, 578 pp.
ILVO macrobenthos in soft sediments: macrobenthos monitoring in the Belgian Part of the North
Sea since 1979.
Jones, A.M.; Baxter, J.M. (1987). Molluscs: Caudofoveata, Solenogastres, Polyplacophora and
Scaphopoda: keys and notes for the identification of the species. Synopses of the British fauna
(new series), 37. E.J. Brill; W. Backhuys: London, UK. ISBN 90-04-08197-6. vi, 154 pp.
Kato, T.; Pleijel, F. (2003). A revision of Paranaitis Southern, 1914 (Polychaeta: Phyllodocidae).
Zoological Journal of the Linnean Society. 138(4): 379-429.
Kerckhof, F., Haelters, J.; Gollasch, S. (2007). Alien species in the marine and brackish
ecosystem: the situation in Belgian waters. Aquatic Invasions. 2(3): 243-257
Koehler, R. (1921). Echinodermes. Faune de France, 1. Librairie de la Faculte des Sciences, Paris.
216 pp.
Koukouras, A. (2010). Check-list of marine species from Greece. Aristotle University of
Thessaloniki. Assembled in the framework of the EU FP7 PESI project.
Labrune, C., Amouroux, J.M., Sarda, R., Dutrieux, E., Thorin, S., Rosenberg, R., Gre´mare, A.,
2006. Characterization of the ecological quality of the coastal Gulf of Lions (NW Mediterranean).
A comparative approach based on three biotic indices. Mar. Pollut. Bull. 52, 34–47.

52
Legakis, A. (2001). Chelicerata, in: Costello, M.J. et al. (Ed.) (2001). European register of marine
species: a check-list of the marine species in Europe and a bibliography of guides to their
identification. Collection Patrimoines Naturels, 50: pp. 237
Leewis, R. (2002). Flora en fauna van de zee [Marine flora and fauna]. Veldgids, 16. KNNV
Uitgeverij: Utrecht, The Netherlands. ISBN 90-5011-153-X. 320 pp.
Legakis, A. (2001). Chelicerata, in: Costello, M.J. et al. (Ed.) (2001). European register of marine
species: a check-list of the marine species in Europe and a bibliography of guides to their
identification. Collection Patrimoines Naturels, 50: pp. 237
Leloup, E. (1952). Coelentérés [Coelenterata]. ---. Institut royal des Sciences naturelles de
Belgique: Brussels, Belgium. 283 pp.
Lenanton RCJ, Potter IC. Contribution of estuaries to commercial fisheries in temperate Western
Australia and the concept of estuarine dependence. Estuaries 1987;10:28–35.
Little, C.; Morritt, D.; Seaward, D.R.; Williams, G.A. (1989). Distribution of intertidal molluscs
in lagoonal shingle (The Fleet, Dorset, U.K.). J. Conch., Lond. 33: 225-232
Lyra, F., Carvalho, P., Antunes, C., Vasconcelos, T. e Basto, M. (2006). Resultados preliminares
sobre a contaminação de sedimentos do rio Minho por metais vestigiais e pesticidas
organoclorados. Em: Actas do III Simpósio sobre a bacia hidrográfica do rio Minho, Vila Nova
de Cerveira, 1 e 2 de Junho de 2006, Câmara Municipal de Vila Nova de Cerveira, Vila Nova de
Cerveira.
L. B. Holthuis & E. Gottlies(1958) An annotated list of Decapod Crustacea of the Mediterranean
Coast of Israel, with an appendix listing the Decapoda of the Eastern Mediterranean. Bulletin of
the Research Council of Israel. Haifa, Israel. 1-126pp
Massin, Cl.; De Ridder, Ch. (1989). Les échinodermes de Belgique [The Echinodermata from
Belgium], in: Wouters, K.; Baert, L. (Ed.) (1989). Proceedings of the Symposium "Invertebrates
of Belgium". pp. 395-402
Mauchline, J.; Murano, M. (1977). World list of the Mysidacea, Crustacea. J. Tokyo Univ. Fish.
64 (1): 39-88.
McLusky, D.S. 1971. Ecology of Estuaries. Heinemann, London, U.K.
McLusky, D.S. & Elliott, M. (2005). The Estuarine Ecosystem: Ecology, Threats and
Management (3rd ed.). Oxford University Press, 222 pp.
Mees, J. (1994). Het hyperbenthos van ondiepe kustgebieden en estuaria: gemeenschapsstruktuur
en biologie van de dominante soorten [The hyperbenthos of shallow coastal waters and estuaries:
community structure and biology of the dominant species]. PhD Thesis. Universiteit Gent.
Mariene Biologie. Instituut voor Dierkunde. Vakgroep Morfologie, Systematiek en Ecologie:
Gent, Belgium. 212 pp.
Molnar, J. L.; Gamboa, R. L.; Revenga, C.; Spalding, M. D. (2008). Assessing the global threat
of invasive species to marine biodiversity. Frontiers in Ecology and the Environment. 6(9): 485-
492.
Molodtsova T.N., 2001. On the systematic position of Cerianthus septentrionalis van Beneden,
1923. Zoosystematica Rossica, 10: 9-10.

53
Mora, C., Ospina, A., 2001. Tolerance to high temperatures and potential impact of sea warming
on reef fishes of Gorgona Island (tropical eastern Pacific). Marine Biology 139, 765–769.
Muller, Y. (2004). Faune et flore du littoral du Nord, du Pas-de-Calais et de la Belgique:
inventaire. [Coastal fauna and flora of the Nord, Pas-de-Calais and Belgium: inventory].
Commission Régionale de Biologie Région Nord Pas-de-Calais: France. 307 pp.
Muniz, P., Venturini, N., Pires-Vanin, A.M.S., Tommasi, L.R., Borja, Á ., 2005. Testing the
applicability of a Marine Biotic Index (AMBI) to assessing the ecological quality of soft-bottom
benthic communities, in the South America Atlantic region. Mar. Pollut. Bull. 50, 624–637.
Musco, L.; Giangrande, A. (2005). Mediterranean Syllidae (Annelida: Polychaeta) revisited:
biogeography, diversity and species fidelity to environmental features. Mar. Ecol. Progr. Ser. 304:
143-153
Muxika, I., Borja, A., Bonne, W., 2005. The suitability of the marine biotic index (AMBI) to new
impact sources along European coasts. Ecological Indicators 5, 19–31.
Neill, W.H., Magnuson, J.J., 1974. Distributional ecology and behavioral thermoregulation of
fishes in relation to heated effluent from a power plant at Lake Monona, Wisconsin. Transactions
of the American Fisheries Society 103, 663–710.
Neill, W.H., Magnuson, J.J., Chipman, G., 1972. Behavioral thermoregulation by fishes: a new
experimental approach. Science 176, 1443–1445.
O’ Connor, T. P. (1992) Recent trends in coastal environmental quality: results from the Ærst
Æve years of the NOAA Mussel Watch project. US Department of Commerce, NOAA, Rockville,
45 pp.
Oliveira, R., 1967. Contribuição para o estudo do estuário do Tejo. Sedimentologia. Memórias do
Laboratório Nacional de Engenharia Civil, 296, 1-61.
OSPAR Commission. 2000. Quality status report 2000. OSPAR Commission, London, U.K
Partridge, J.K. (1977). Littoral and benthic investigations on the west coast of Ireland - VI
(Section A Faunistic and ecological studies). Annotated bibliographies of the genus Tapes
(Bivalvia: veneridae): Part II Tapes semidecussatus. Proceedings of the Royal Irish Academy.
77B: 1-64.
Passos, F. e Lyra. C. (2007). Caracterização da Comunidade de Macroinvertebrados Bentónicos
do Estuário do Rio Minho – Sua Relação com a Distribuição de Poluentes no Sedimento.
Faculdade de Ciências da Universidade do Porto. Pp. 21-40.
Pearson, T.H., & Rosenberg, R. (1978). Macrobenthic succession in relation to organic
enrichment and pollution of marine environment. Oceanographic Marine Biological Annual
Review, 16: 229-311.
Petersen, M.E. (1998). Pholoe (Polychaeta: Pholoidae) from northern Europe: a key and notes on
the nearshore species. J. Mar. Biol. Ass. U.K. 78(4): 1373-1376
Pinto, R., Patricio, J., Baeta, A., Fath, B.D., Neto, J.M., Marques, J.C., 2009. Review and
evaluation of estuarine biotic indices to assess benthic condition. Ecological Indicators 9, 1e25.

54
Polk, Ph. (1976). Inventarisatie plankton: fauna en flora [Plankton inventory : fauna and flora],
in: Nihoul, J.C.J.; De Coninck, L. (Ed.) (1976). Project Sea final report: 7. Inventory of fauna and
flora. Project Sea final report, 7: pp. 233-311
Poore, G. C. B.; Bruce, N. L. (2012). Global Diversity of Marine Isopods (Except Asellota and
Crustacean Symbionts). PLoS ONE. 7(8): e43529.
Pörtner, H.O., Bock, C., Knust, R., Lannig, G., Lucassen, M., Mark, F.C., Sartoris, F.J., 2008.
Cod and climate in a latitudinal cline: physiological analyses of climate effects in marine fishes.
Climate Research 37, 253–270.
Pritchard, D.W. (1967). What is an estuary: physical viewpoint, in: Lauff, G.H. (Ed.) (1967).
Estuaries. American Association for the Advancement of Science Publication, 83: pp. 3-5
Quintino, V., Elliott, M., Rodrigues, A.M., 2006. The derivation, performance and role of
univariate and multivariate indicators of benthic change: case studies at differing spatial scales.
Journal of Experimental Marine Biology and Ecology 330, 368e382.
Ramos, M. (2010). IBERFAUNA. The Iberian Fauna Databank.
Rappé, G. (1984). Verslag excursie naar Wenduine op 19 februari 1984 [Report on the excursion
to Wenduine (Belgium), 19 February 1984]. De Strandvlo 4(1): 3
Renier, Stefano Andrea. 1804. Tavola alfabetica delle Conchiglie Adriatiche. Nominate dictro il
Sistemo di Linneo, Edizione di Gmelin. v-xiii.
Rhoads, D. C. & D.K. Young (1970). The influence of deposit-feeding organisms on sediment
stability and community trophic structure. Journal of Marine Research 28, 150-178.
Risso, J.C. 1978. Faune malacologique de la plateforme tunisienne. Etude de quelques dragages
et carottages effectués à l'intérieur et au large du golfe de Gabès. Bull. Inst. Nat. Sc. Oceanogr.
Pêche, Salammbô, 5 (1-4): 17-41.
Rolán Mosquera E. (1983). Moluscos de la ria de Vigo 1. Gasteropodes. Thalassas 1 (1) Anexo
1: 1-383
Rolán E., 2005. Malacological Fauna From The Cape Verde Archipelago. Part 1, Polyplacophora
and Gastropoda.
Rozin, P.N., Mayer, J., 1961. Thermal reinforcement and thermoregulatory behavior in the
goldfish, Carassius auratus. Science 134, 942–943.
Sagarin RD, Barry JP, Gilman SE, Baxter CH. 1999. Climate-related change in an intertidal
community over short and long time scales. Ecological Monographs 69: 465-490.
San Martín, G. and López, E. 2002. New species of Autolytus Grube, 1850, Paraprocerastea San
Martín & Alós, 1989, and Sphaerosyllis Claparède, 1863 (Syllidae, Polychaeta) from the Iberian
Peninsula. Sarsia, 87(2): 135-143
Schockaert, E.R.; Jouk, P.E.H.; Martens, P.M. (1989). Free-living Plathelminthes from the
Belgian coast and adjacent areas, in: Wouters, K.; Baert, L. (Ed.) (1989). Proceedings of the
Symposium "Invertebrates of Belgium". pp. 19-25

55
Simboura N, Zenetos A, 2002. Benthic indicators to use in ecological quality classification of
Mediterranean soft bottom marine ecosystems, including a new biotic index. Mediterranean
Marine Science 3: 77-111.
Simboura, N.; Sigala, K.; Voutsinas, E.; Kalkan, E. (2008). First Occurrence Of The Invasive
Alien Species Polydora cornuta Bosc, 1802 (Polychaeta: Spionidae) On The Coast Of Greece
(Elefsis Bay; Aegean Sea). Mediterranean Marine Science. 9(2): 119-124.
Simons, E.; Simons, G.; Corstanje, H. (1988). Report on the beach excursion at Oostduinkerke
(Belgium) on 2 November 1988 [Verslag van de strandexcursie te Oostduinkerke op 2 november
1988]. De Strandvlo 8(4): 206-209
Sokal, R. R., & Rohlf, F. J. (1995). Biometry (3rd ed.). New York: Freeman.
Southward AJ, Crisp DJ. 1954a. Recent changes in the distribution of the intertidal barnacles
Chthamalus stellatus Poli and Balanus balanoides L. in the British Isles. Journal of Animal
Ecology 23: 163-177.
Southward, A.J. (2001). Cirripedia - non-parasitic Thoracica, in: Costello, M.J. et al. (Ed.) (2001).
European register of marine species: a check-list of the marine species in Europe and a
bibliography of guides to their identification. Collection Patrimoines Naturels, 50: pp. 280-283
Spalding MD, Fox HE, Allen GR, Davidson NFerdana ZA et al., 2007.Marine ecoregions
of the world: a bioregionalization of coastal and shelf areas. BioScience 57 :573–583
Streftaris, N.; Zenetos, A.; Papathanassiou, E. (2005). Globalisation in marine ecosystems: the
story of non-indigenous marine species across European seas. Oceanogr. Mar. Biol. Ann. Rev.
43: 419-453.
Tattersall, W.M. & Tattersall, O.S. (1951). The British Mysidacea. Ray Society: London. VIII,
460 pp
Teixeira H, Weisberg SB, Borja A, Ranasinghe JA, Cadien DB, Velarde RG, Lovell LL, Pasko
D, Phillips CA, Montagne DE, Salas F, Marques JC, 2012. Calibration and Validation of the
AZTI's Marine Biotic Index (AMBI) for Southern California Marine Bays. Ecological Indicators
12, 84-95.
Terlizzi, A.; Scuderi, D.; Fraschetti, S.; Anderson, M.J. (2005). Quantifying effects of pollution
on biodiversity: a case study of highly diverse molluscan assemblages in the Mediterranean. Mar.
Biol. 148(2): 293-305
Thuiller, W., 2004. Patterns and uncertainties of species’ range shifts under climate change.
Global Change Biology 10, 2220e2227.
Todd CD, Lewis JR. 1984. Effects of low air temperature on Laminaria digitata in south-western
Scotland. Marine Ecology Progress Series 16: 199-201.
Trott, T. J. (2004). Cobscook Bay inventory: a historical checklist of marine invertebrates
spanning 162 years. Northeastern Naturalist. 11, 261-324.
Van Belle, R.A. (1988). De Europese keverslakken (Polyplacophora) [The European
Polyplacophora]. Vita Marina Jan.-Mrt: 9-132

56
van der Land, J.; den Hartog, J.H. (2001). Actiniaria, in: Costello, M.J. et al. (Ed.) (2001).
European register of marine species: a check-list of the marine species in Europe and a
bibliography of guides to their identification. Collection Patrimoines Naturels, 50: pp. 106-109
van Ofwegen, L.; Grasshoff, M.; van der Land, J. (2001). Octocorallia (excl. Pennatulacea), in:
Costello, M.J. et al. (Ed.) (2001). European register of marine species: a check-list of the marine
species in Europe and a bibliography of guides to their identification. Collection Patrimoines
Naturels, 50: pp. 104-105
Vanderperren, J.-P. (1989). The Natural History Archive 1988 [Het Natuurhistorisch Archief
1988]. De Strandvlo 9(4): 89-90
Vanhaelen, M.-Th. (1989). Some observations on the beach between Koksijde and Oostduinkerke
during and after the warm summer of 1989 [Enkele waarnemingen van het strand tussen Koksijde
en Oostduinkerke tijdens en na de warme zomer van 1989]. De Strandvlo 9(4): 117-120
Vanhaelen, M.-Th. (2003). Stranding van levende Aequipecten opercularis (L., 1758) en vondsten
van levende Pecten maximus (L., 1758) na zware oktoberstorm 2002 [Strandings of living
Aequipecten opercularis (L., 1758) and findings of living Pecten maximus (L., 1758) after severe
October storm 2002]. De Strandvlo 23(2): 42-46
Vanhaelen, M.-Th. (2005). Nog enkele zeldzame bivalve-vondsten op de Belgische Westkust in
de winter van 2004-2005 [More rare finds of bivalves at the Belgian west coast in the winter of
2004-2005]. De Strandvlo 25(2): 55-57
Vasconcelos, R.P., Reis-Santos, P., Fonseca, V., Maia, A., Ruano, M., França, S., Vinagre, C.,
Costa, M.J., Cabral, H. (2007). Assessing anthropogenic pressures on estuarine fish nurseries
along Portuguese coast: A multi-metric índex and conceptual approach. Science of the Total
Environment 374, 199-215.
Venice System 1958 Symposium on the classification of brackishwaters, Venice April 8–14,
1958. Archives Oceanography andLimnology 11, suppl, 1–248.
Vernberg, W.B. (1983). Responses to estuarine stress. In Estuaries and Enclosed Seas, pp 43-63.
Elsevier Science Publishing Company, Amesterdam, The Netherlands.
Vinagre, C., Santos, F., Cabral, H., Costa, M. (2009). Impact of climate and hydrology on juvenile
fish recruitment towards estuarine nursery grounds in the context of climate change.
Watling, L. (2001). Cumacea, in: Costello, M.J. et al. (Ed.) (2001). European register of marine
species: a check-list of the marine species in Europe and a bibliography of guides to their
identification. Collection Patrimoines Naturels, 50: pp. 309.
Wittmann, K.J. (1992). Morphogeographic variations in the genus Mesopodopsis Czerniavsky
with descriptions of three new species (Crustacea, Mysidacea). Hydrobiologia. 241: 71-89.
Wolff, W.J. (1983). Estuarine benthos, in: Ketchum, B.H. (Ed.) (1983). Estuaries and enclosed
seas. Ecosystems of the World, 26: pp. 151-182. Elsevier Science Publishing Company,
Amesterdam, The Netherlands.
Zamouri-Langar, N.; Chouba, L.; Ajjabi Chebil, L.; Mrabet, R.; El Abed, A. (2011). Les
coquillages bivalves des côtes tunisiennes. Institut National des Sciences et Technologies de la
Mer: Salammbô. ISBN 978-9938-9512-0-2. 128 pp.

57
Zimmer, C. (1933). Cumacea. The fauna of the North Sea and Baltic, 23(10.g4). Akademische
Verlagsgesellschaft: Leipzig, Germany. 70-120 pp.
Zintzen, V. (2007). Biodiversity of shipwrecks from the Southern Bight of the North Sea. PhD
Thesis, University of Louvain, Department of Biology, 343 p.
Zouari, S. 1985. Contribution à l'étude systématique des lamellibranches des côtes tunisiennes.
D.E.A. Faculté des Sciences de Tunis, 245pp.
Zühlke, R.; Alvsvåg, J.; De Boois, I.; Cotter, J.; Ehrich, S.; Ford, A.; Hinz, H.; Jarre-Teichmann,
A.; Jennings, S.; Kröncke, I.; Lancaster, J.; Piet, G.; Prince, P. (2001). Epibenthic diversity in the
North Sea. Senckenb. Marit. 31(2): 269-281

58
ANEXO
Distribuição AMBI
Abludomelita gladiosa (Bate, 1862) 1 III
Abludomelita obtusata (Montagu, 1813) 1 III
Abra alba (W. Wood, 1802) 1 III
Abra prismatica (Montagu, 1808) 1 I
Abra segmentum (Récluz, 1843) 2 III
Abra sp. - III
Achaeus cranchii Leach, 1817 1 I
Achelia echinata Hodge, 1864 1 I
Actinia equina (Linnaeus, 1758) 1 I
Actinothoe sphyrodeta (Gosse, 1858) 1 NA
Adamsia palliata (Müller, 1776) 1 II
Alkmaria romijni Horst, 1919 1 III
Alvania beanii (Hanley in Thorpe, 1844) 1 I
Amage adspersa (Grube, 1863) 1 I
Ammothella longipes (Hodge, 1864) 1 I
Ampelisca diadema (Costa, 1853) 1 II
Ampelisca sp. - I
Ampelisca spinipes Boeck, 1861 0 I
Ampharete acutifrons (Grube, 1860) 1 II
Amphibalanus improvisus (Darwin, 1854) 1 NA
Amphilochus spencebatei (Stebbing, 1876) 1 II
Amphipholis squamata (Delle Chiaje, 1828) 1 I
Amphipoda sp. - NA
Amphitritides gracilis (Grube, 1860) 1 II
Amphiura chiajei Forbes, 1843 1 II
Anapagurus hyndmanni (Bell, 1845) 1 I
Anapagurus laevis (Bell, 1845) 1 III
Anemonia viridis (Forskål, 1775) 1 NA
Anomia ephippium Linnaeus, 1758 1 I
Anoplodactylus petiolatus (Krøyer, 1844) 1 II
Anthopleura thallia (Gosse, 1854) 1 NA
Anthura gracilis (Montagu, 1808) 1 I
Aonides oxycephala (Sars, 1862) 1 III
Aphelochaeta sp. - IV
Apolochus neapolitanus (Della Valle, 1893) 1 II
Apolochus picadurus (J.L. Barnard, 1962) 2 NA
Apseudopsis latreillii (Milne-Edwards, 1828) 1 III
Arabella geniculata (Claparède, 1868) 1 I
Arabella iricolor (Montagu, 1804) 1 I
Arca tetragona Poli, 1795 1 I
Armandia cirrhosa Filippi, 1861 2 I
Armandia polyophthalma Kükenthal, 1887 1 I

59
Aulactinia verrucosa (Pennant, 1777) 1 NA
Austrominius modestus Darwin, 1854 1 II
Autolytus sp. - II
Balanus crenatus Bruguiére, 1789 0 NA
Balanus spongicola Brown, 1844 0 NA
Barnea candida (Linnaeus, 1758) 1 I
Bathyporeia nana Toulmond, 1966 0 I
Bela nebula (Montagu, 1803) 1 I
Bittium latreillii (Payraudeau, 1826) 2 I
Bittium reticulatum (da Costa, 1778) 1 I
Boccardiella ligerica (Ferronnière, 1898) 0 III
Bodotria scorpioides (Montagu, 1804) 1 I
Bolocera tuediae (Johnston, 1832) 1 II
Bosemprella incarnata (Linnaeus, 1758) 1 I
Branchiomma sp. - I
Buccinum sp. - II
Calliostoma zizyphinum (Linnaeus, 1758) 1 I
Callipallene brevirostris (Johnston, 1837) 1 I
Calyptraea chinensis (Linnaeus, 1758) 1 I
Capitella capitata (Fabricius, 1780) 1 V
Carcinus maenas (Linnaeus, 1758) 1 III
Cataphellia brodricii (Gosse, 1859) 0 NA
Caulleriella alata (Southern, 1914) 1 IV
Caulleriella bioculata (Keferstein, 1862) 1 IV
Cerastoderma edule (Linnaeus, 1758) 1 III
Cerastoderma glaucum (Bruguière, 1789) 1 III
Cereus pedunculatus (Pennant, 1777) 1 I
Cerianthus lloydii Gosse, 1859 0 I
Cerithiopsis minima (Brusina, 1865) 2 I
Chaetopleura angulata (Spengler, 1797) 1 NA
Chaetopterus variopedatus (Renier, 1804) 1 I
Chaetozone caputesocis (Saint-Joseph, 1894) 1 IV
Chaetozone gibber Woodham & Chambers, 1994 0 IV
Chaetozone setosa Malmgren, 1867 1 IV
Chaetozone zetlandica McIntosh, 1911 1 IV
Chamelea gallina (Linnaeus, 1758) 1 I
Chondrochelia savignyi (Kroyer, 1842) 1 III
Chone sp. - II
Chrysallida sp. - II
Chthamalus montagui Southward, 1976 1 I
Cirratulus sp. - IV
Cirriformia tentaculata (Montagu, 1808) 1 IV
Clausinella fasciata (da Costa, 1778) 1 I
Corbula gibba (Olivi, 1792) 1 IV
Corophium orientale Schellenberg, 1928
Corophium sp.
2
-
III
III

60
Cossura coasta Kitamori, 1960 2 IV
Cossura soyeri Laubier, 1964 1 IV
Crangon crangon (Linnaeus, 1758) 1 I
Crisilla semistriata (Montagu, 1808) 1 I
Cyathura carinata (Krøyer, 1847) 1 III
Cymodoce truncata Leach, 1814 1 I
Devonia perrieri (Malard, 1904) 1 I
Diadumene cincta Stephenson, 1925 0 NA
Diadumene lineata (Verrill, 1869) 1 NA
Dikoleps cutleriana (Clark, 1849) 1 I
Diogenes pugilator (Roux, 1829) 1 II
Diopatra micrura Pires, Paxton, Quintino & Rodrigues,
2010 1 NA
Diopatra neapolitana Delle Chiaje, 1841 1 I
Diplocirrus sp. - I
Dipolydora coeca (Örsted, 1843) 1 IV
Dipolydora flava (Claparède, 1870) 1 IV
Dipolydora socialis (Schmarda, 1861) 0 IV
Dodecaceria concharum Örsted, 1843 1 IV
Donax vittatus (da Costa, 1778) 1 I
Echinocyamus pusillus (O.F. Müller, 1776) 1 I
Edwardsia timida Quatrefages, 1842 0 NA
Epilepton clarkiae (Clark W., 1852) 1 I
Epitonium clathrus (Linnaeus, 1758) 1 I
Ericthonius punctatus (Bate, 1857) 1 I
Eulalia sp. - II
Eulimella cerullii (Cossmann, 1916) 1 NA
Eulimella sp. - I
Eumida sanguinea (Örsted, 1843) 1 II
Eunereis longissima (Johnston, 1840) 1 III
Eunice vittata (Delle Chiaje, 1828) 1 II
Eurydice spinigera Hansen, 1890 1 I
Euspira nitida (Donovan, 1804) 1 II
Eusyllis sp. - II
Exogone sp. - II
Fimbriosthenelais minor (Pruvot & Racovitza, 1895) 1 II
Flabelligera affinis M. Sars, 1829 1 II
Gammaropsis maculata (Johnston, 1828) 1 I
Gastrosaccus spinifer (Goës, 1864) 1 II
Gattyana cirrhosa (Pallas, 1766) 1 III
Gibbula cineraria (Linnaeus, 1758) 1 I
Gibbula umbilicalis (da Costa, 1778) 0 I
Gitana sarsi Boeck, 1871 1 II
Glycera alba (O.F. Müller, 1776) 1 IV
Glycera tesselata Grube, 1840 1 II
Glycera tridactyla Schmarda, 1861 1 II

61
Glycera unicornis Lamarck, 1818 1 II
Goniada maculata Ørsted, 1843 1 II
Gouldia minima (Montagu, 1803) 1 I
Gracilechinus acutus (Lamarck, 1816) 1 I
Gregariella semigranata (Reeve, 1858) 1 I
Gyptis propinqua Marion & Bobretzky, 1875 1 II
Hanleya hanleyi (Bean, 1844) 1 I
Harmothoe benthophila intermedia Støp-Bowitz, 1951 NA NA
Harmothoe extenuata (Grube, 1840) 1 II
Harmothoe fraserthomsoni McIntosh, 1897 1 II
Harmothoe imbricata (Linnaeus, 1767) 1 II
Harmothoe impar (Johnston, 1839) 1 II
Harmothoe longisetis (Grube, 1863) 1 II
Harmothoe serrata Day, 1963 2 NA
Harmothoe spinifera (Ehlers, 1864) 1 II
Harpinia crenulata (Boeck, 1871) 1 I
Harpinia pectinata Sars, 1891 1 I
Hediste diversicolor (O.F. Müller, 1776) 1 III
Hesione sp. - II
Heteromastus filiformis (Claparède, 1864) 1 IV
Heterotanais oerstedii (Krøyer, 1842) 1 III
Hiatella arctica (Linnaeus, 1767) 1 I
Hilbigneris gracilis (Ehlers, 1868) 1 II
Hormathia coronata (Gosse, 1858) 1 NA
Hyalinoecia brementi Fauvel, 1916 2 II
Hydroides sp. - III
Ianiropsis sp. - NA
Idotea emarginata (Fabricius, 1793) 0 II
Idunella picta (Norman, 1889) 0 I
Iphimedia gibbula Ruffo e Schiecke, 1979 NA NA
Iphimedia minuta G.O. Sars, 1882 1 I
Iphinoe tenella Sars, 1878 1 I
Jasmineira candela (Grube, 1863) 1 II
Jasmineira elegans Saint-Joseph, 1894 1 II
Kellia suborbicularis (Montagu, 1803)
Kirkegaardia heterochaeta (Laubier, 1961)
1
2
I
IV
Kurtiella bidentata (Montagu, 1803) 1 III
Labrorostratus de Saint-Joseph, 1888 NA NA
Lagis koreni Malmgren, 1866 1 IV
Lanice conchilega (Pallas, 1766) 1 II
Leitoscoloplos mammosus Mackie, 1987 0 III
Lepidochitona cinerea (Linnaeus, 1767) 1 II
Lepidonotus clava (Montagu, 1808) 1 II
Leptocheirus pilosus Zaddach, 1844 1 III
Leptoplana tremellaris (Müller OF, 1773) 1 II
Leucothoe incisa (Robertson, 1892) 1 I

62
Leucothoe procera Bate, 1857 1 I
Leucothoe spinicarpa (Abildgaard, 1789) 1 I
Lumbrineris coccinea (Renier, 1804) 1 II
Lumbrineris funchalensis (Kinberg, 1865) 0 II
Lumbrineris latreilli Audouin & Milne Edwards, 1834 1 II
Lygdamis muratus (Allen, 1904) 1 NA
Lysidice unicornis (Grube, 1840) 1 II
Macomangulus tenuis (da Costa, 1778) 1 I
Macrophthalmus sp. - NA
Macropodia rostrata (Linnaeus, 1761) 1 I
Maera grossimana (Montagu, 1808) 1 I
Magelona alleni Wilson, 1958 1 I
Magelona equilamellae Harmelin, 1964 2 I
Magelona filiformis Wilson, 1959 1 I
Magelona minuta Eliason, 1962 1 I
Magelona mirabilis (Johnston, 1865) 1 I
Magelona sp. - I
Malmgrenia andreapolis McIntosh, 1874 1 II
Malmgrenia polypapillata (Barnich & Fiege, 2001) 2 NA
Malmgreniella lunulata (Delle Chiaje, 1830) 1 II
Manayunkia aestuarina (Bourne, 1883) 0 III
Mangelia attenuata (Montagu, 1803) 1 II
Mangelia costata (Pennant, 1777) 1 II
Mangelia costulata Risso, 1826 1 I
Manzonia crassa (Kanmacher, 1798) 1 I
Marphysa sanguinea (Montagu, 1815) 1 II
Medicorophium aculeatum (Chevreux, 1908) 2 III
Medicorophium annulatum (Chevreux, 1908) 2 III
Medicorophium minimum (Schiecke, 1978) 2 III
Mediomastus fragilis Rasmussen, 1973 1 III
Megaluropus agilis Hoeck, 1889 1 I
Megaluropus massiliensis Ledoyer, 1976 2 I
Melinna palmata Grube, 1870 1 III
Melita palmata (Montagu, 1804) 1 I
Mesacmaea mitchellii (Gosse, 1853) 1 NA
Mesochaetopterus sp. - III
Mesopodopsis slabberi (van Beneden, 1861) 1 II
Metridium dianthus (Ellis, 1768) 0 NA
Microdeutopus algicola Della Valle, 1893 2 I
Microdeutopus anomalus (Rathke, 1843) 1 I
Microdeutopus armatus Chevreux, 1886 1 I
Microdeutopus obtusatus Myers, 1973 2 NA
Microphthalmus sp. - II
Microspio sp. - III
Mimachlamys varia (Linnaeus, 1758) 1 I
Modiolula phaseolina (Philippi, 1844) 1 I

63
Modiolus barbatus (Linnaeus, 1758) 1 I
Modiolus modiolus (Linnaeus, 1758) 1 II
Moerella donacina (Linnaeus, 1758) 1 I
Molgula occulta Kupffer, 1875 1 I
Molgula tubifera (Orstedt, 1844) 0 NA
Monocorophium acherusicum (Costa, 1857) 1 III
Monocorophium sextonae (Crawford, 1937) 1 III
Musculus costulatus (Risso, 1826) 1 I
Musculus discors (Linnaeus, 1767) 1 I
Musculus subpictus (Cantraine, 1835) 1 I
Myrianida quindecimdentata (Langerhans, 1884) 1 II
Myrianida sp. - II
Mysta picta (Quatrefagues, 1866) 1 III
Mytilus edulis Linnaeus, 1758 1 III
Mytilus galloprovincialis Lamarck, 1819 1 III
Nais sp. - IV
Nassarius sp.
Nemertea
-
-
II
III
Neomysis integer (Leach, 1814) 1 II
Nephtys caeca (Fabricius, 1780) 1 II
Nephtys cirrosa Ehlers, 1868 1 II
Nephtys hombergii Savigny in Lamarck, 1818 1 II
Nephtys hystricis McIntosh, 1900 1 II
Nephtys longosetosa Örsted, 1843 1 II
Nephtys sp. - II
Noemiamea dolioliformis (Jeffreys, 1848) 1 II
Notomastus latericeus Sars, 1851 1 III
Notomastus profondus (Eisig, 1887) 1 III
Nototropis guttatus Costa, 1853 1 I
Nucula hanleyi Winckworth, 1931 1 I
Nucula nucleus (Linnaeus, 1758) 1 I
Nucula sp. - I
Nucula sulcata Bronn, 1831 1 I
Ocenebra erinaceus (Linnaeus, 1758) 1 II
Odostomia lukisii Jeffreys, 1859 1 NA
Odostomia turrita Hanley, 1844 1 II
Odostomia unidentata (Montagu, 1803)
Oligochaeta
0
-
II
V
Ondina coarctata (G.O. Sars, 1878) 0 NA
Ophiothrix fragilis (Abildgaard, in O.F. Müller, 1789) 1 I
Ophiura ophiura (Linnaeus, 1758) 1 II
Ophryotrocha puerilis Claparède & Metschnikow, 1869 1 IV
Ostrea edulis Linnaeus, 1758 1 I
Othomaera othonis (Milne-Edwards, 1830) 0 I
Owenia fusiformis Delle Chiaje, 1844 1 II
Oxydromus flexuosus (Delle Chiaje, 1827) 1 II
Pagurus alatus Fabricius, 1775 1 II

64
Pagurus bernhardus (Linnaeus, 1758) 1 II
Pagurus cuanensis Bell, 1846 1 II
Paleanotus sp. - NA
Paradoneis lyra (Southern, 1914) 1 III
Paragnathia formica (Hesse, 1864) 1 III
Paranaitis kosteriensis (Malmgren, 1867) 1 II
Paraphoxus oculatus (G. O. Sars, 1879) 1 II
Parapionosyllis labronica Cognetti, 1965 2 II
Parapionosyllis sp. - II
Parthenina flexuosa (Monterosato, 1874) 2 NA
Parthenina juliae (de Folin, 1872) 2 NA
Parvicardium exiguum (Gmelin, 1791) 1 I
Parvicardium pinnulatum (Conrad, 1831) 1 I
Pecten maximus (Linnaeus, 1758) 1 I
Perforatus perforatus (Bruguière, 1789) 1 NA
Perinereis cultrifera (Grube, 1840) 1 III
Peringia ulvae (Pennant, 1777) 1 III
Perioculodes aequimanus (Korssman, 1880) 2 I
Perioculodes longimanus (Bate & Westwood, 1868) 1 II
Philine aperta (Linnaeus, 1767) 1 II
Philine catena (Montagu, 1803) 1 II
Pholoe inornata Johnston, 1839 1 IV
Pholoe minuta (Fabricius, 1780) 1 II
Phoxichilidium femoratum (Rathke, 1799) 0 I
Phtisica marina Slabber, 1769 1 I
Phyllodoce laminosa Savigny in Lamarck, 1818 1 II
Phyllodoce longipes Kinberg, 1866 0 II
Phyllodoce maculata (Linnaeus, 1767) 1 II
Phyllodoce mucosa Ørsted, 1843 1 III
Phyllodoce schmardaei Day, 1963 1 II
Phylo foetida (Claparède, 1869) 1 I
Pionosyllis sp. - II
Pisione remota (Southern, 1914) 1 I
Platynereis dumerilii (Audouin & Milne Edwards, 1833) 1 III
Pleioplana atomata (Müller OF, 1776) 1 NA
Poecilochaetus serpens Allen, 1904 1 I
Polititapes rhomboides (Pennant, 1777) 1 I
Polycirrus sp. - IV
Polydora ciliata (Johnston, 1838) 1 IV
Polydora cornuta Bosc, 1802
Pomatoceros sp.
1
-
IV
II
Pontocrates altamarinus (Bate & Westwood, 1862) 1 II
Pontocrates arenarius (Bate, 1858) 1 II
Potamilla torelli (Malmgren, 1866) 1 II
Prionospio cirrifera Wirén, 1883 1 IV
Prionospio multibranchiata Berkeley, 1927 1 III

65
Prionospio sp. - IV
Prionospio steenstrupi Malmgren, 1867 1 IV
Prosphaerosyllis sp. - II
Psamathe fusca Johnston, 1836 1 II
Psammechinus miliaris (P.L.S. Müller, 1771) 1 I
Pseudamussium peslutrae (Linnaeus, 1771) 1 II
Pseudopolydora antennata (Claparède, 1869) 1 IV
Pseudopolydora paucibranchiata (Okuda, 1937) 1 IV
Pseudopolydora pulchra (Carazzi, 1895) 1 IV
Pusillina inconspicua (Alder, 1844) 1 I
Pycnogonum litorale (Strom, 1762) 1 II
Pygospio elegans Claparède, 1863 1 III
Retusa truncatula (Bruguière, 1792) 1 II
Rocellaria dubia (Pennant, 1777) 1 I
Ruditapes philippinarum (Adams & Reeve, 1850) 1 III
Sabellaria alveolata (Linnaeus, 1767) 1 I
Sabellaria spinulosa (Leuckart, 1849) 1 I
Sagartia elegans (Dalyell, 1848) 1 NA
Sagartia troglodytes (Price in Johnston, 1847) 1 I
Sagartiogeton undatus (Müller, 1778) 1 NA
Scolelepis (Scolelepis) squamata (O.F. Muller, 1806) 1 III
Scoletoma impatiens (Claparède, 1868) 1 II
Scoloplos (Scoloplos) armiger (Müller, 1776) 1 III
Scrobicularia plana (da Costa, 1778) 1 III
Scyllarus arctus (Linnaeus, 1758) 1 I
Serpula lobiancoi Rioja, 1917 2 I
Setia pulcherrima (Jeffreys, 1848) 0 NA
Sigambra sp. - III
Solecurtus sp. - NA
Solen marginatus Pulteney, 1799 1 I
Sphaeroma serratum (Fabricius, 1787) 1 III
Sphaeroma sp. - III
Sphaerosyllis bulbosa Southern, 1914 1 II
Sphaerosyllis hystrix Claparède, 1863 1 II
Sphaerosyllis parabulbosa San Martín & López, 2002 1 NA
Sphaerosyllis pirifera Claparède, 1868 1 II
Sphaerosyllis sp. - II
Sphaerosyllis taylori Perkins, 1981 1 III
Spio decoratus Bobretzky, 1870 1 III
Spio filicornis (O.F. Müller, 1776) 1 III
Spiochaetopterus costarum (Claparède, 1869) 1 I
Spiophanes bombyx (Claparède, 1870) 1 III
Spiralinella spiralis (Montagu, 1803) 0 II
Spirobranchus lamarcki (Quatrefages, 1866) 1 II
Spisula elliptica (Brown, 1827) 0 I
Spisula solida (Linnaeus, 1758) 1 I

66
Spisula subtruncata (da Costa, 1778) 1 I
Stenothoe marina (Bate, 1856) 1 II
Sternaspis scutata Ranzani, 1817 1 III
Sthenelais boa (Johnston, 1833) 1 II
Stomphia coccinea (Müller, 1776) 0 NA
Streblospio shrubsolii (Buchanan, 1890) 1 III
Stylochoplana maculata (Quatrefage, 1845) 1 NA
Stylostomum ellipse (Dalyell, 1853) 1 NA
Syllides sp. - II
Syllidia armata Quatrefages, 1866 1 II
Syllis armillaris (O.F. Müller, 1776) 1 II
Syllis garciai (Campoy, 1982) 2 II
Syllis gracilis Grube, 1840 1 III
Syllis parapari San Martín & López, 2000 2 II
Tanais dulongii 1 II
Tanaissus lilljeborgi (Stebbing, 1891) 0 III
Tharyx sp. - IV
Timoclea ovata (Pennant, 1777) 1 I
Tritia incrassata (Strøm, 1768) 1 II
Tritia pygmaea (Lamarck, 1822) 1 II
Tritia reticulata (Linnaeus, 1758) 1 II
Trophonopsis muricata (Montagu, 1803) 1 I
Tubificoides sp. - V
Turbonilla acutissima Monterosato, 1884 2 I
Turbonilla lactea (Linnaeus, 1758) 1 I
Turbonilla magnifica Seguenza G., 1880 2 NA
Unciola crenatipalma (Bate, 1862) 0 I
Urothoe pulchella (Costa, 1853) 1 I
Urticina eques (Gosse, 1858) 0 NA
Urticina felina (Linnaeus, 1761) 0 I
Venerupis corrugata (Gmelin, 1791) 1 I
Venus casina Linnaeus, 1758 1 I
Venus verrucosa Linnaeus, 1758 1 I
Verruca stroemia O.F. Müller, 1776 1 I
Zeuxo sp. - NA
Recommended

















