
vii
UNIVERSIDADE ESTADUAL DO CENTRO-OESTE, UNICENTRO – PR
PROGRAMA DE PÓS-GRADUAÇÃO EM AGRONOMIA – PPGA
MESTRADO
MANEJO DA COBERTURA DO SOLO EM POMAR DE
PEREIRAS CV. HOSUI: EFEITOS NO SOLO,
NUTRIÇÃO E CRESCIMENTO DAS PLANTAS E
OCORRÊNCIA DE PLANTAS DANINHAS
DISSERTAÇÃO DE MESTRADO
EDINA COSTA DELONZEK
GUARAPUAVA, PR
2017

viii
EDINA COSTA DELONZEK
MANEJO DA COBERTURA DO SOLO EM POMAR DE PEREIRAS CV. HOSUI:
EFEITOS NO SOLO, NUTRIÇÃO E CRESCIMENTO DAS PLANTAS E
OCORRÊNCIA DE PLANTAS DANINHAS
Dissertação apresentada à Universidade
Estadual do Centro-Oeste, como parte das
exigências do Programa de Pós-Graduação em
Agronomia, área de concentração em
Produção Vegetal, para obtenção do título de
Mestre.
Prof. Dr. Marcelo Marques Lopes Muller
Orientador
Prof. Dr. Renato Vasconcelos Botelho
Co-orientador
GUARAPUAVA, PR
2017

ix
Catalogação na Fonte
Biblioteca da UNICENTRO

x

xi
AGRADECIMENTOS
Agradeço a Deus por tudo que tem me concedido principalmente, persistência para
cumprir essa meta.
Ao professor Dr. Renato Vasconcelos Botelho, pela ajuda, pelo incentivo e
principalmente por acreditar no meu trabalho.
Ao meu orientador, Dr. Marcelo Marques Lopes Muller, pelas orientações.
Ao Prof. Dr. Cleber Daniel de Goes Maciel, pela orientação, ajuda e incentivo.
À minha família, pelo amor, pela compreensão e pelo incentivo.
À Universidade Estadual do Centro-Oeste – UNICENTRO, pela oportunidade de
ingresso e conclusão do curso de Pós-Graduação em Agronomia.
A Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – CAPES, pela
concessão da bolsa de mestrado.
Aos grupos de Fruticultura e de Solos da UNICENTRO – Universidade Estadual do
Centro-Oeste, por toda ajuda nos trabalhos laboratoriais.
A todas as pessoas que, de forma direta ou indiretamente contribuíram para a
realização desse trabalho.
Obrigado!

xii
SUMÁRIO
LISTA DE FIGURAS............................................................................................................viii
LISTA DE TABELAS............................................................................................................ ix
RESUMO................................................................................................................................. xi
ABSTRACT............................................................................................................................ xii
1. INTRODUÇÃO.................................................................................................................... 1
2. OBJETIVOS......................................................................................................................... 3
2.1 Geral.................................................................................................................................... 3
2.2 Específicos........................................................................................................................... 3
3. REEFERENCIAL TEÓRICO............................................................................................ 4
3.1. Importância econômica da pereira.................................................................................. 4
3.2. Classificação botânica de pereira.................................................................................... 4
3.2.1. Cultivar Hosui................................................................................................................ 5
3.3. Plantas daninhas............................................................................................................... 5
3.4. Diferentes formas de manejo do solo............................................................................... 7
3.5. Efeitos da cobertura do solo na população de plantas daninhas.................................. 8
3.6.Caracterização de materiais que podem ser utilizados na cobertura de
solo........................................................................................................................................... 10
3.7. Fauna do Solo................................................................................................................. 12
3.7.1 Caracterização da fauna do solo.................................................................................. 13
3.8. Exigências nutricionais em plantas frutíferas.............................................................. 14
3.8.1 Macronutrientes............................................................................................................ 14
3.8.2 Micronutrientes............................................................................................................. 17
4. MATERIAL E MÉTODOS............................................................................................... 19
4.1. Local do experimento...................................................................................................... 19
4.2. Material experimental.................................................................................................... 19
4.3. Avaliações........................................................................................................................ 21
4.3.1. Coleta, quantificação e identificação de plantas daninhas....................................... 21
4.3.2. Altura das plantas de pereira...................................................................................... 22
4.3.3. Diâmetro do tronco..................................................................................................... 22
4.3.4. Diâmetro e numero de ramos...................................................................................... 23
4.3.5. Teor de clorofila........................................................................................................... 23
4.3.6. Temperatura do solo.................................................................................................... 23

xiii
4.3.7. Macrofauna do solo...................................................................................................... 23
4.3.8. Análises químicas do solo e nutricionais das plantas................................................ 24
4.3.9. Análise estatística......................................................................................................... 24
5. RESULTADOS................................................................................................................... 26
5.1.Plantas daninhas em função de diferentes tipos de coberturas de
solo........................................................................................................................................... 26
5.2.Crescimento vegetativo de pereiras cv. Hosui em diferentes coberturas de
solo........................................................................................................................................... 35
5.3. Temperatura do solo em resposta à cobertura vegetal................................................ 38
5.4. Efeito das diferentes coberturas sobre a macrofauna edáfica.................................... 39
5.5. Características químicas do solo.................................................................................... 40
5.6. Análise do conteúdo mineral das folhas........................................................................ 43
6. DISCUSSÃO....................................................................................................................... 45
6.1. Plantas daninhas em função de diferentes tipos de coberturas do solo..................... 45
6.2. Crescimento vegetativo de pereiras cv. Hosui em diferentes coberturas de solo..... 46
6.3 Temperatura do solo em resposta à cobertura vegetal................................................. 47
6.4. Efeito das diferentes coberturas sobre a macrofauna edáfica.................................... 47
6. 5. Características químicas do solo .................................................................................. 48
6.6. Análise do teor mineral das folhas................................................................................. 49
7. CONCLUSÃO.................................................................................................................... 52
8. REFERÊNCIAS BIBLIOGRÁFICAS............................................................................. 53

viii
LISTA DE FIGURAS
Figura 1. Tipos de coberturas sobre o solo, instaladas logo após a implantação do experimento,
constituídas por palhada de azevém+milheto (T2), serragem de pinus (T3), tela de
sombreamento 70% (T4), casca de arroz (T5), fibra de curauá (T6) e capina
(T7)................................................................................................................................
20
Figura 2. Valores de frequência relativa (Frr), densidade relativa (Der), abundância relativa (Abr) e
Índice de valor de importância (Ivi) das espécies de plantas daninhas em duas épocas de
manejo, quinze dias após implantação das coberturas realizado com sete tratamentos,
observados em levantamento fitossociológico na cultura da Pera. Cruz Machado-PR,
2015/2016..............................................................................................................................................
29
Figura 3. Valores de frequência relativa (Frr), densidade relativa (Der), abundância relativa (Abr) e
Índice de valor de importância (Ivi) das espécies de plantas daninhas em duas épocas de
manejo, quarenta e cinco dias após implantação das coberturas realizado com sete tratamentos,
observados em levantamento fitossociológico na cultura da Pera. Cruz Machado-PR,
2015/2016............................................................................................................................
30
Figura 4. Valores de frequência relativa (Frr), densidade relativa (Der), abundância relativa (Abr) e
Índice de valor de importância (Ivi) das espécies de plantas daninhas em duas épocas de
manejo, setenta e cinco dias após implantação das coberturas realizado com sete tratamentos,
observados em levantamento fitossociológico na cultura da Pera. Cruz Machado-PR,
2015/2016........................................................................................................................................
30
Figura 5. Valores de Valores de frequência relativa (Frr), densidade relativa (Der), abundância relativa
(Abr) e Índice de valor de importância (Ivi) das espécies de plantas daninhas em duas épocas
de manejo, quinze dias após implantação das coberturas realizado com sete tratamentos,
observados em levantamento fitossociológico na cultura da Pera. Cruz Machado-PR,
2016/2017...............................................................................................................................................
32
Figura 6. Valores de frequência relativa (Frr), densidade relativa (Der), abundância relativa (Abr) e
Índice de valor de importância (Ivi) das espécies de plantas daninhas em duas épocas de
manejo, setenta e cinco dias após implantação das coberturas realizado com sete tratamentos,
observados em levantamento fitossociológico na cultura da Pera. Cruz Machado-PR,
2015/2016........................................................................................................................................
33
Figura 7. Valores de frequência relativa (Frr), densidade relativa (Der), abundância relativa (Abr) e
Índice de valor de importância (Ivi) das espécies de plantas daninhas em duas épocas de
manejo, setenta e cinco dias após implantação das coberturas realizado com sete tratamentos,

ix
observados em levantamento fitossociológico na cultura da Pera. Cruz Machado-PR,
2015/2016........................................................................................................................................
34
Figura 8. Teor de clorofila (índice Spad) em folhas de pereiras das cv. Hosui sob diferentes
coberturas de solo no primeiro ciclo 2015/2016 (Figura A), segundo ciclo
(2016/2017) (Figura B). Letras distintas para uma mesma época de avaliação indicam
diferença significativa pelo teste Student Newman-Keuls ao nível de 5% de
probabilidade; ns = não significativo ao nível de 5% de probabilidade.........................
37
Figura 9.
Média mensal da temperatura do solo (ºC) a 5,0 cm de profundidade considerando-se
a cobertura do solo e o horário da determinação 7: 30 (Figura A), 12:30 (Figura B) e
17:30 (Figura C), no período de outubro de 2016 a março de 2017. Cruz Machado-
PR, 2017...........................................................................................................................
39
Figura 10.
Distribuição dos grupos taxonômicos por tipo de cobertura de solo em pereiras da cv.
Hosui, segundo ciclo (2016/2017). Cruz Machado – PR, 2017.............................................
39

x
LISTA DE TABELAS
Tabela 1. Análise química do solo, nas profundidades de 0-20 cm antes da implantação
das coberturas (setembro de 2015), e do pomar de pereiras cv. Hosui Cruz
Machado-PR, 2017............................................................................................................
19
Tabela 2. Espécies de plantas daninhas pertencentes às suas respectivas famílias e
classes identificadas em levantamento fitossociológico na cultura da Pera nos
períodos de 2015/16 e 2017/17, em diferentes coberturas de solo. Cruz
Machado-PR, 2017....................................................................................................................
26
Tabela 3. Médias de produção de massa seca de plantas daninhas (g/0,5 m2) em função
da cobertura na avaliação realizada aos 90 dias após a colocação das
coberturas, no pomar de pereiras cv. Hosui nos ciclos 2015/16 e 2016/17. Cruz
Machado-PR, 2017...............................................................................................
35
Tabela 4. Altura e diâmetro do caule de pereiras cv. Hosui nos ciclos 2015/16 e 2016/17,
número de ramos e diâmetro dos ramos no ciclo 2016/17, sob diferentes
coberturas de solo. Cruz Machado-PR, 2017..........................................................
36
Tabela 5. Distribuição do numero de indivíduos por unidade de área, riqueza total e
índices de Shannon e Pielou sob diferentes coberturas de solo, nas linhas de
plantio de pereira cv. Hosui. Cruz Machado-PR, 2017...........................................
40
Tabela 6. Valores médios de matéria orgânica, pH e alumínio em diferentes coberturas de
solo, nas profundidades de 0-10 cm e 10-20 cm, nas linhas de plantio de pereiras
cv. Hosui Cruz Machado-PR, 2017.........................................................................
41
Tabela 7. Valores médios de fósforo, potássio, cálcio e magnésio em diferentes coberturas
de solo, nas profundidades de 0-10 cm e 10-20 cm, nas linhas de plantio de
pereiras cv. Hosui. Cruz Machado-PR, 2017..........................................................
41
Tabela 8. Valores médios de ferro, cobre, manganês e zinco em diferentes coberturas de
solo, nas profundidades de 0-10 cm e 10-20 cm, nas linhas de plantio de pereiras
cv. Hosui. Cruz Machado-PR, 2017........................................................................
43
Tabela 9. Teores de nitrogênio, fósforo, potássio, cálcio, magnésio e enxofre (g kg-1
) em
folhas de pereira cv. Hosui sob diferentes coberturas de solo. Cruz Machado –
PR, 2017..................................................................................................................
43
Tabela 10. Teores de cobre, ferro, manganês e zinco (mg kg-1
) em folhas de pereira cv.
Hosui sob diferentes coberturas de solo. Cruz Machado – PR, 2017.....................
44

xi
RESUMO
DELONZEK. E. C. Manejo da cobertura do solo em pomar de pereiras cv. Hosui: efeitos
no solo, nutrição e crescimento das plantas e ocorrência de plantas daninhas
O objetivo foi avaliar o efeito de diferentes tipos de cobertura de solo no controle de
plantas daninhas e no desenvolvimento da pereira cv. Hosui. O experimento foi conduzido a
campo no município de Cruz Machado - PR, durante as safras de 2015/2016 e 2016/2017. O
pomar foi implantado em outubro 2015, com a cv. Hosui enxertadas sobre o porta-enxerto
Pyrus calleriana. O delineamento experimental foi em blocos casualisados com sete
tratamentos, quatro repetições e parcela experimental constituídas por quatro plantas. Os
tratamentos foram: 1) Cobertura natural; 2) palhada de azevém (Lolium multiflorum) e
milheto (Pennisetum glaucum); 3) serragem de pinus; 4) tela de sombreamento (70%); 5)
cobertura do solo com casca de arroz; 6) fibra de curauá (Ananas erectifolius) e 7) capina
(todo o ciclo). Foram realizadas as seguintes avaliações: levantamento florístico das plantas
daninhas, crescimento vegetativo das pereiras, temperatura e a composição faunística do solo,
características químicas do solo e estado nutricional das pereiras. Os resultados indicaram que
as coberturas com a fibra de curauá ou com palha de azevém+milheto tiveram os maiores
efeitos de supressão sobre as plantas daninhas e teores mais elevados no solo de alguns
elementos minerais como fósforo e zinco. Também houve interferência das coberturas com
serragem de pinus e a cobertura natural apresentando menores alturas e diâmetros das
pereiras. As coberturas também influenciaram a temperatura do solo, sendo que no tratamento
com capina manteve o solo com temperatura mais elevada nas medições realizadas nos
horários das 7:30 e 12:30 h. As maiores densidades de organismos da fauna edáfica foram
encontrados nas coberturas com casca de arroz e palhada de azevém+milheto. A análise foliar
demonstrou teores de Mg e Mn em níveis deficientes para todos os tratamentos.
Palavras-Chave: Pyrus spp., plantas espontâneas, sistemas de manejo, fertilidade do solo.

0
ABSTRACT
DELONZEK, E. C. Development of the Hosui pear trees and their different mulchs.
The present research aims to evaluate the effect of different kinds of mulch in the
control of spontaneous plants and in the development of the „Hosui‟ pear tree. The experiment
was conducted in an area in Cruz Machado/ PR, during the harvest of 2016 and 2017. The
orchard was introduced in October 2015 with a Hosui‟ pear tree grafted onto a rootstock
Pyrus calleriana. The experimental design was in randomized blocks, with seven treatments
and four repetitions and experimental parts constituted by four plants. The treatments were: 1)
witness (without cover), 2-) ryegrass (Lolium multiflorum) 3-)wooden dust, and millet
(Pennisetum glaucum), 4-)shading material (70%), 5-) coverages of the soil with mulch of
rice husk 6-) fiber of „curauá‟ (ananás erectifolius) and 7-) weeding (always clean). There
were accomplished the following evaluations: Floristic data of the weed, pear tree vegetative
growth, soil temperature and faunistic composition, soil chemical characteristics and
nutritional status of pear trees. The results indicated that the coverings with curauá fiber or
ryegrass straw + millet had higher suppression effects on the weeds and higher soil content of
some mineral elements such as phosphorus and zinc. There was also interference of the
coverings with pine sawdust and the natural cover showing lower heights and diameter of the
pear trees. The coverings also influenced the soil temperature, on the other hand in the
treatment with weeding, the soil was maintained with higher temperature in the measurements
performed in the hours of 7:30 a.m and 12:30 p.m. The highest densities of organisms of the
edaphic fauna were found in the coverings with rice husk and straw of ryegrass + millet. The
foliar analysis showed levels of Mg and Mn at deficient levels for all the treatments.
Key words: Pyrus spp., Spontaneous plants, management systems, soil fertility.
xii

1
1. INTRODUÇÃO
No Brasil a cultura da pereira apresentou crescimento nos anos 2010, atingindo em
2015 uma produção em torno de 22.000 toneladas anuais de peras das espécies Pyrus
communis L. e P. pyrina, em uma área de 1.680 hectares (FAO, 2016). No entanto, a
produção é ainda incipiente para atender a demanda do País, que ocupa a terceira posição
entre os maiores importadores de pera no mundo, ficando atrás apenas da Rússia e da União
Europeia (FAO, 2016).
O aumento do cultivo em território nacional tem encontrado entraves devido a vários
fatores, tais como: baixa tecnologia de produção, indefinição ou inexistência de cultivares e
porta-enxertos adaptados às diferentes condições edafoclimáticas e ainda, falta de
recomendação de adubação (MACHADO et al., 2012). A cultura da pereira está sujeita à ação
de vários fatores abióticos e bióticos que afetam direta ou indiretamente sua rentabilidade
agrícola, dentre esses fatores, tem grande importância a interferência das plantas daninhas.
As plantas daninhas podem interferir indiretamente no crescimento e na produção das
culturas perenes (PITELLI, 1987), por competirem por recursos essenciais ao crescimento e
liberarem substâncias alelopáticas (BLANCO e OLIVEIRA, 1978; SINGH e
ACHHIREDDY, 1987) e, indiretamente, por atuarem como hospedeiras intermediárias de
pragas e patógenos (LEITE JÚNIOR e MOHAN, 1990).
O uso de coberturas no solo pode contribuir no manejo de plantas espontâneas em
sistemas de produção sustentável. Nesse caso, o solo é manejado com espécies vegetais de
rápido crescimento e boa produção de biomassa, como também, podem ser utilizados
materiais como plástico preto, tela de sombreamento, casca de arroz, serragem de pinus e
fibras vegetais. O manejo cultural pode assim, reduzir a dependência por herbicidas, o que
seria benéfico em termos ecológicos, com possível diminuição nos custos e melhoramento
dos recursos naturais (CARVALHO e VARGAS, 2004).
As práticas convencionais intensivas de manejo na fruticultura, como a utilização de
fertilizantes sintéticos e agroquímicos para controle de pragas, doenças e plantas daninhas,
tem provocado preocupações em relação ao meio ambiente e à saúde pública, levando a busca
por práticas agrícolas mais sustentáveis (SAMPAIO et al., 2008). Tais práticas podem ter
influência sobre os organismos do solo, uma vez que alteram as características de seu habitat
e a composição da cadeia trófica (RUIZ-CAMACHO et al., 2009).
A cobertura do solo é uma técnica que traz benefícios aos sistemas de produção,
especialmente no que diz respeito ao manejo do solo e das plantas, podendo envolver o uso de
diferentes resíduos orgânicos (OLIVEIRA et al., 2008; SEDIYAMA et al., 2010). O manejo

2
adequado da cobertura do solo pode proporcionar melhoria de atributos físicos, químicos e
biológicos do solo (PELIZZA e FERRARI, 2009), o que auxilia na produção das espécies
frutíferas (RAGOZO et al., 2006), merecendo especial atenção em sistemas de produção
orgânica (WALSH et al., 1996; GOH et al., 2000). Pouco se conhece em relação aos efeitos
do manejo das coberturas de solo em termos de ocorrência de plantas daninhas e sobre a
competição com a cultura da pereira nas condições do Sul do Brasil. Dessa forma, pode variar
a eficiência dessa prática no manejo das plantas daninhas e sua resposta no crescimento da
pereira.
No entanto, poucos trabalhos de pesquisa têm sido desenvolvidos no Brasil,
procurando avaliar os efeitos das coberturas do solo sobre a ocorrência ou controle das plantas
daninhas. Além disso, devido à dificuldade operacional e aos custos envolvidos na obtenção e
distribuição no pomar, a aplicação de cobertura de solo ainda é uma técnica pouco utilizada
pelos produtores (DIXON et al., 2005). As hipóteses do trabalho são de que (i) as diferentes
coberturas de solo como forma de manejo podem ser usadas no controle de plantas daninhas,
e inclusive (ii) gerar uma melhoria na fertilidade do solo, no estado nutricional e crescimento
da pereira.

3
2. OBJETIVOS
2.1 Geral
Avaliar diferentes manejos da cobertura do solo na infestação de plantas daninhas, nos
atributos do solo no crescimento e nutrição da pereira cv. Hosui.
2.2 Específicos
Verificar os efeitos das diferentes coberturas no crescimento e desenvolvimento das
pereiras cv. Hosui;
Analisar os efeitos das diferentes coberturas, sobre as características químicas do solo
camadas 0-10 e 10-20 cm de profundidade do solo;
Analisar teores foliares de macro e micronutrientes;
Avaliar a influência das coberturas sobre a temperatura do solo;
Verificar a variação da clorofila foliar (índice SPAD) sobre diferentes coberturas do
solo;
Verificar o efeito das coberturas na composição faunística do solo;

4
3. REFERENCIAL TEÓRICO
3.1. Importância econômica da cultura da pereira
A produção mundial de peras é de aproximadamente 22,64 milhões de toneladas,
sendo a China o maior produtor, com participação de 67,3% da produção global; seguida pela
Itália, que responde por apenas 3,2% da produção mundial; os Estados Unidos, apresenta-se
com um percentual bem próximo ao da Itália sendo o terceiro maior produtor (FAO, 2016). O
Brasil ocupa a 49º lugar com uma produção de aproximadamente 22.000 toneladas em uma
área plantada de aproximadamente 1.870 hectares (IBGE, 2015).
No Brasil, o Estado do Rio Grande do Sul é o maior produtor da fruta com
aproximadamente 10.000 toneladas, o que corresponde a 48,8% da produção nacional,
seguido do Estado de Santa Catarina, com 6.500 toneladas (29,7%) e do Paraná, com 4.000
toneladas (18,17%) (IBGE, 2015).
Em relação ao mercado interno, destacam se as cultivares William‟s, Max Red
Bartlett, Packham‟s Triumph, Abate Fetel, Rocha, Hosui, Nijisseiki e Kousui que são
plantadas em pequenas áreas nas regiões de clima frio que compreende o sul do Brasil
(FAORO e ORTH, 2010). No Estado do Paraná as cultivares chinesa Yali, Kieffer, os clones
de Pera d‟Água e tipos de Pera Pedra são as variedades mais plantadas (AYUB e GIOPPO,
2009).
Apesar dos dados das últimas safras demonstrarem que a produção nacional cresceu, a
pera é a fruta fresca mais importada pelo País, uma vez que a produção não atende à demanda
de consumo interno, sendo necessário adquirir cerca de 200.000 toneladas de peras ao ano de
outros países, o que torna o Brasil o quarto maior importador da fruta no mundo (IBRAF,
2014).
3.2. Classificação botânica da pereira
A pereira é uma planta frutífera que faz parte da divisão Angiospermae, classe
Dicotyledoneae e pertence à família Rosácea subfamília Pomoideae e gênero Pyrus. O gênero
Pyrus diferenciou-se no período terciário em um território montanhoso da atual China
ocidental, dispersando-se a leste e a oeste e adaptando-se as mais diversas condições
edafoclimáticas, diferenciando-se até as espécies atualmente conhecidas (FIDEGHELI, 2007).
No processo para formação das espécies domesticadas de P. communis, houve
ocorrência de hibridizações interespecíficas (KUMAR e NEGI, 2010). No Ocidente, P.

5
communis var. pyrastere e P. communis var. caucasica foram provavelmente os ancestrais da
pereira europeia, e assim descendendo a grande maioria das cultivares comerciais no mundo
(GIAYETTO e VILLARREAL, 2010). Em termos de classificação, as cultivares de pereiras
mais consumidas mundialmente podem ser classificadas em dois grupos: Europeias (Pyrus
communis) e Asiáticas, que são subdivididas em pera japonesa (Pyrus pyrifolia) e pera
chinesa (Pyrus ussuriensis e Pyrus bretschneideri) (FAORO, 2009).
No Brasil, a Pyrus communis, popularmente conhecida como pera europeia é a espécie
mais difundida. A frutificação ocorre em cerca de três ou mais anos, a fruta é um pomo, de
formato arredondado ou piriforme, de textura carnuda, suculenta e doce (NAKASU e
FAORO, 2003).
Cada cultivar de pera deve ser escolhida conforme suas características fenológicas,
pois são exigentes em frio, o que é um fator limitante para produção. Além disso, há outros
fatores como os edafoclimáticos e a comercialização para cada região produtora (BETTIOL
NETO et al., 2014).
3.2.1. Cultivar Hosui
De procedência japonesa, esta cultivar produz frutas de tamanho grande, com
coloração marrom-dourada quando maduros. Sua polpa é branca, crocante, macia, suculenta,
doce e de boa qualidade. É uma planta vigorosa, de hábito ereto e a sua maturação ocorre do
final de janeiro ao início de fevereiro, pode ser conservada de dois a três meses em câmara
fria. Apresenta suscetibilidade as doenças entomosporiose e sarna (FAORO e ORTH, 2010).
A cultivar Hosui necessita de um período de frio (721 horas) para a quebra de
dormência, assim como apresenta intensidade mediana de florescimento e reduzido número
de flores por gemas. No entanto, a produtividade média está em torno de 36 toneladas ha-1
(OLIVEIRA et al., 2008). As plantas são vigorosas, com frutos de massa em torno de 285 g,
os quais tem boa aceitação no mercado (FAORO, 2009).
3.3. Plantas daninhas
Entre os grupos de plantas existentes no meio ambiente, as plantas espontâneas,
também conhecidas como plantas daninhas, que nascem espontaneamente no meio, sendo
adaptáveis a qualquer tipo de clima e solo, ocorrem naturalmente e têm crescimento
espontâneo em áreas de cultivo, tem elevada importância, pois prejudicam as culturas
comerciais (ALTIERI et al., 2003). Alguns estudos de ecologia de plantas daninhas enfatizam

6
as características e ajustes de crescimento, como o que ocorre nos sistemas convencionais de
cultivo, que permitem a essas espécies explorar os nichos ecológicos deixados em aberto nas
terras cultivadas e os mecanismos de adaptação que lhes possibilitam sobreviver mesmo sob
condições de perturbações máximas do solo (RIZZARDI et al., 2001).
Segundo Gomes e Christoffoleti (2008) as características que permitem as infestantes
colonizarem com sucesso os agroecossistemas incluem: necessidades para germinação
atendidas em muitos ambientes, uma vez que, o cultivo melhora a germinação das sementes
porque aumenta o número de pontos com microclima favorável; longevidade das sementes
por existir espécies cujas sementes podem permanecer viáveis até por mais de 80 anos;
dormência variável das sementes, rápido crescimento, considerando que a produção de
sementes geralmente começa após um curto período de crescimento vegetativo; alta produção
de sementes sob condições favoráveis, autocompatíveis, mas não completamente autógamas
ou apomíticas, muitas infestantes anuais podem produzir sementes sem polinização cruzada;
adaptação para dispersão a curta e longas distâncias, reprodução vegetativa vigorosa ou
regeneração de fragmentos das ervas perenes, capacidade de competir interespecificamente de
maneira especializada, superbrotamento, crescimento de choque, substâncias alelopáticas;
capacidade de se adaptar e tolerar ambientes diversos.
O resultado da competição com as plantas daninhas e as culturas é a redução na
produção ou na qualidade do produto. As plantas daninhas podem causar perdas na
produtividade, variando de 10 a mais de 80% em função da espécie competidora, do grau de
infestação, do período de convivência, bem como do estádio de desenvolvimento da cultura e
das condições climáticas durante a convivência (SILVA e SILVA, 2007).
Segundo Kuva et al. (2007), o grau de interferência das plantas daninhas nas culturas
agrícolas pode ser definido como a redução percentual da produção econômica provocada
pela convivência com a comunidade infestante. No entanto, o grau de interferência depende
de fatores ligados à própria cultura (espécie ou variedade, idade, espaçamento e densidade de
plantio), à comunidade infestante (composição específica, densidade e distribuição) e à época
e extensão do período de convivência, podendo, ser influenciado pelas condições edáficas,
climáticas e pelos tratos culturais, tornando-se fundamental a determinação das espécies
presentes através de índices fitossociológicos para se entender a dinâmica da flora infestante
(PITELLI, 1985).
Avaliando períodos de convivência de plantas daninhas com a cultura dos citros, no
Estado de São Paulo, em diferentes épocas do ano, Carvalho et al. (1993) demonstraram que o
período de convivência mais prejudicial à produtividade ocorre entre os meses de agosto e
janeiro quando a redução de produtividade é de 25% em relação às plantas livres da

7
interferência durante todo o ano, período de convivência que abrange os meses de maio a
julho, não interferiu significativamente com a produtividade da cultura.
Pelizza e Ferrari (2009), avaliando o crescimento inicial de plantas de macieiras
implantadas em um pomar, verificaram que a presença de plantas daninhas na linha de plantio
afeta o crescimento da cultura, com redução na altura e no diâmetro de plantas, evidenciando
interferência no período de crescimento das macieiras.
3.4. Diferentes formas de manejo do solo
Práticas que favoreçam a manutenção de água, com redução da evaporação são
componentes importantes de sustentabilidade agrícola. Isso se dá principalmente pelo fato de
que a evaporação é, em geral, mais facilmente manejada em agroecossistemas do que a
transpiração de uma planta cultivada (GLIESSMAN, 2005). O uso de coberturas é uma
estratégia de manejo do solo que pode influenciar no desenvolvimento de plantas de espécies
frutíferas, podendo ser utilizadas espécies vegetais de crescimento rápido para produção de
fitomassa, ou outros materiais, tais como plástico preto, tela de sombreamento, serragem de
pinus, casca de arroz entre outros. No passado, as intervenções culturais eram direcionadas à
maximização da produção, com o passar do tempo, os objetivos mudaram e visam a melhoria
da qualidade do produto e maior integração entre o cultivo e o ambiente (MARANGONI et
al., 1995). Através do uso das coberturas mortas se propõe uma alternativa ecológica,
econômica e viável no manejo do solo e na adoção de práticas conservacionistas, pois
propicia redução progressiva no consumo energético (RUFATO, 2004).
No manejo de infestantes em sistema orgânico, recomenda-se o uso de práticas que
evitem a ressemeadura de invasoras, tais como o uso de plantas com grande capacidade de
abafamento das plantas daninhas, para formação da cobertura morta, de quantidade de palha
suficiente para recobrir o solo, o uso de plantas de coberturas com efeito alelopático e o
plantio em época propícia (SKORA NETO, 1998).
Os métodos manuais combinados com mecânicos, como é o caso do uso de roçadeiras
são os que substituem o químico na maioria das vezes (VARGAS e BERNARDI, 2003).
Aspecto a ser considerado no plantio direto orgânico é a substituição dos herbicidas
dessecantes e dos herbicidas durante o ciclo da cultura. Para esta substituição, no sistema
orgânico são empregadas plantas de grande capacidade de abafamento das infestantes para a
formação da cobertura morta no final do período chuvoso e que são roladas ou roçadas na fase
de formação de grãos (DAROLT e SKORA NETO, 2002).

8
A cobertura do solo é uma prática agrícola que consiste em manter protegida a
superfície do solo, nas linhas de plantio, com uma camada de material orgânico, resultante de
sobras de culturas como as palhas ou cascas (PEREIRA et al., 2012). A cobertura do solo com
capim, palha, casca, plástico, entre outros materiais, promove melhorias e benefícios para as
plantas cultivadas com a redução da insolação direta ao solo, evitando a alta evaporação e
aumentando a flora e fauna (VARGAS e OLIVEIRA, 2005).
Além disso, a cobertura do solo pode ser bastante efetiva no controle da erosão hídrica
(AMADO et al., 1989; SCHICK et al., 1999), na redução da infestação por plantas
espontâneas (DERKSEN et al., 2002; BALBINOT et al., 2003; BALBINOT et al., 2005) e
algumas espécies podem apresentar efeito sobre a população de nematóides (MOJTAHEDI et
al., 1993). A cobertura do solo também pode influenciar a temperatura e consequentemente,
nos processos físicos, químicos e biológicos que nele ocorrem (BRADY, 1989).
Alguns trabalhos confirmam os efeitos positivos da aplicação de cobertura morta sobre
o aumento da produção das culturas agrícolas (MOORE-GORDON et al., 1997). Isto pode
estar relacionado a vários fatores, como na melhoria da densidade do sistema radicular de
plantas frutíferas (DIXON et al., 2007). Espíndola (2007) afirma que o sistema radicular da
planta deve ser protegido das altas temperaturas e das variações diárias de umidade e
temperatura, para promover um bom desenvolvimento vegetativo e alta produção de frutas.
Em um estudo realizado por Moura et al. (2014), verificou-se que a cobertura do solo
proporciona um bom crescimento da planta de mirtilo cv. bluegem, influenciando na
qualidade dos frutos no segundo ano após sua aplicação da cobertura vegetal na superfície do
solo. No entanto, para a cobertura das linhas de plantio, pode ser utilizado material sintético
como plásticos e/ou de resíduos orgânicos, disponíveis na região e de baixo custo
(CAPISTRANO, 2008). A preferência é por resíduos vegetais com decomposição mais lenta,
para mantê-lo protegendo o solo por maior período de tempo. Os materiais mais usados para
cobertura do solo são acículas de pinus, cascas, serragens e filme plástico (ESPÍNDOLA,
2007). Em um estudo realizado durante 5 anos, com cultivares de mirtilo Highbush em solo
com e sem cobertura, Clark e Moore (1991) observaram que a produção de frutos e o
crescimento das plantas foram superiores em solos com cobertura vegetal concluindo que
plantas frutíferas respondem positivamente a cobertura do solo.
3.5. Efeitos da cobertura do solo na população de plantas daninhas
A cobertura colocada na superfície do solo serve como um elemento isolante, podendo
filtrar os feixes de luz de ondas longas inibindo o processo de germinação das plantas

9
daninhas (ADEGAS, 1997). O processo de germinação das plantas daninhas, estando ligados
a esses fatores, reduz substancialmente devido à presença de grande quantidade de cobertura
do solo. Os efeitos da cobertura sobre as plantas daninhas devem ser analisados sob três
aspectos: físico, biológico e químico (PITELLI e PITELLI, 2004).
Os efeitos físicos causam sombreamento e, consequente redução da amplitude térmica
e hídrica do solo e à filtragem da luz que atinge a palha, afetando a dormência e,
consequentemente, a germinação das plantas daninhas (SEVERINO e CHRISTOFFOLETI,
2001). Também a supressão da emergência de plantas daninhas pela cobertura pode ocorrer
devido à produção de metabólitos secundários, denominados aleloquímicos, os quais são
liberados no ambiente em função da decomposição dos resíduos vegetais, podendo interferir
na germinação, pela inativação dos mecanismos de dormência, e também no crescimento
inicial de plantas daninhas (GOMES JR. e CHRISTOFFOLETI, 2008; MONQUERO et al.,
2009).
No efeito biológico, a presença da cobertura vai criar condições para a instalação de
uma densa e diversificada microbiocenose na camada superficial do solo. Na composição
desta microbiocenose, há uma grande quantidade de organismos que podem utilizar sementes
de plantas daninhas como fonte de energia, para sua multiplicação. De maneira geral, os
microrganismos exercem importantes funções de deterioração e perda da viabilidade dos
diversos tipos de propágulos no solo (PITELLI e DURIGAN, 2001). Além disso, a cobertura
do solo cria um abrigo seguro para alguns inimigos naturais, como roedores, insetos e outros
pequenos animais que são predadores de sementes e plântulas das espécies de plantas
daninhas (ALVES e PITELLI, 2001).
Em relação aos efeitos químicos, a cobertura morta pode ter influência alelopática
sobre as plantas daninhas. A alelopatia consiste na liberação de substâncias químicas no
ambiente por um dado organismo, as quais irão interagir com outros, inibindo ou estimulando
o seu crescimento e desenvolvimento (RICE, 1984). Essa influência pode ocorrer entre
microrganismos, entre microrganismos e plantas, entre plantas cultivadas, entre plantas
daninhas, e entre plantas daninhas e plantas cultivadas.
A palhada que compõe a cobertura morta têm importância alelopática sobre as
infestantes, em função da liberação lenta dos compostos aleloquímicos se comparado ao
material incorporado ao solo, propiciando efeitos mais pronunciados e prolongados
(ALMEIDA, 1988). Os aleloquímicos sintetizados pelas plantas mantêm-se nos tecidos
mesmo depois da morte delas (ALMEIDA, 1991). Por ação da chuva e do orvalho, vão sendo
lixiviados para o solo, onde podem afetar a germinação de sementes ou o desenvolvimento de
plântulas. Assim, a decomposição do material vegetal é um dos fatores que influenciam nesse

10
aspecto, sendo que a maioria das poaceas tem decomposição lenta e, consequentemente, ação
alelopática longa, enquanto as leguminosas têm decomposição rápida e ação alelopática alta
no início, porém, de curta duração.
A liberação desses produtos ocorre à medida que os tecidos do material vegetal vão
sendo degradados pelos micro e macroinvertebrados do solo. Assim, a taxa de decomposição
da cobertura morta é, até certo ponto, indicativo de até quando as toxinas estão sendo
liberadas no terreno (ALMEIDA, 1988). O estudo do efeito alelopático de resíduos vegetais
utilizados para cobertura morta sobre plantas infestantes e sobre as espécies cultivadas é
realizado, principalmente, em bioensaios que utilizam extratos aquosos ou alcoólicos em
testes em câmaras de germinação, avaliando a percentagem de germinação das sementes e o
alongamento de radículas (CARVALHO et al., 2012).
De acordo com Gravena et al. (2004) é possível considerar que a composição da flora
infestante e a eficiência do seu controle pela cobertura do solo são influenciadas pela
quantidade, pela composição, pela periodicidade de produção e pelo tempo de permanência da
cobertura morta em uma determinada área.
Em termos de manejo da vegetação espontânea, a forma mais usual de controle é com
uso de herbicidas. No entanto, outras formas de controle podem ser utilizadas, especialmente
nas áreas de pomares. Neste caso, métodos mecânicos e culturais, como uso da roçadeira
manual ou a enxada, roçadeira acoplada ao trator representam uma forma alternativa de
manejo das plantas daninhas na linha e na entrelinha das fruteiras (CARVALHO e VARGAS,
2004).
3.6. Caracterização de materiais que podem ser utilizados na cobertura de solo
As constantes buscas por materiais alternativos de baixo custo e com elevada
eficiência para a cobertura do solo é um fator importante tendo em vista que, o
aproveitamento de resíduos que possam ser utilizados pode ser uma saída economicamente
viável. Com relação as palhadas oriundas de resíduos vegetais na utilização como coberturas
dá-se preferência por aquelas que tem uma lenta decomposição.
O azevém anual (Lolium multiflorum) é uma gramínea originária do Mediterrâneo, de
inverno, com ciclo anual, utilizada principalmente como forrageira e para fornecimento de
palhadas (MORAES, 1995). Essa espécie se adapta bem a solos de baixa e média fertilidade,
com boa resposta à adubação, de fácil dispersão e, por isso, está presente e se caracteriza
como planta daninha em praticamente todas as lavouras de inverno, em pomares e em
vinhedos da região Sul do Brasil (VARGAS et al., 2007). Além disso, pode ser utilizada

11
isoladamente ou em consórcios e representa excelente alternativa para compor sistemas de
integração lavoura-pecuária (FLORES et al., 2008).
O milheto caracteriza-se pela alta resistência à seca, com crescimento inicial rápido e
boa capacidade de perfilhamento (CHAVES e CALEGARI, 2001). A facilidade de semeadura
e de se obter sementes fazem do milheto uma das espécies de maior uso como planta de
cobertura do solo e formadora de palhada, embora seus resíduos tenham apresentado
decomposição mais acelerada (ALVARENGA et al., 2001).
O milheto contribui para o controle de plantas infestantes, principalmente pela
competição por água, luz e nutrientes, tendo rápido crescimento inicial e cobrindo
rapidamente o solo (CARVALHO e AMABILE, 2006). Quando semeado na primavera, seu
uso é mais específico para cobertura do solo, preenchendo um espaço em que o solo,
geralmente, fica ocioso e descoberto, exposto à degradação (PITOL, 1993).
A casca de arroz descartada em moinhos de beneficiamento pode ser uma alternativa
no controle das espontâneas. Constituída de quatro camadas fibrosas, esponjosas ou celulares,
são altamente silicosas. Acredita-se que a sílica da casca de arroz exista na forma opalina (a
opala é uma fase hidro-amorfa da sílica). Aparentemente, a sílica é transportada a partir do
solo pela planta como ácido monossílico, o qual concentra-se na casca e no caule da planta
por evaporação da água e, finalmente, se polimeriza para formar a membrana sílico-celulósica
(HOUSTON, 1972).
De acordo com Levy Neto e Pardini (2006), as fibras vegetais possuem baixa massa
específica, maciez e abrasividade reduzida, são recicláveis, não tóxica e biodegradáveis
apresentando baixo custo e baixo consumo de energia na produção. O curauá (Ananas
erectifolius) é uma planta da mesma família do abacaxi (bromeliácea) nativa da região
amazônica. Existem duas variedades distintas do curauá, uma de folha roxo-avermelhada
(curauá-roxo) e outra de folha verde-claro (curauá-branco) (SANTOS, 2013).
O curauá encontra-se nas regiões dos rios Xingu, Tocantins, Tapajós, Marcuru,
Trombetas, Paru, Acará e Guamá, nas partes altas da Ilha do Marajó e no Amapá. É também
certa sua existência nos Estados de Goiás, Mato Grosso e no Acre. Ocorre em lugares onde a
precipitação pluviométrica ultrapassa a 2.000 mm anuais. Encontram-se pés isolados, nas
margens dos rios e nas clareiras dos matos, sombreados ou não, formando agrupamento mais
ou menos pequenos e ralos. Suas folhas são rígidas, eretas e planas, sua medida chega a 1 m
ou mais de comprimento, medem cerca de 3,5 cm de largura podendo ser até mais larga
(MEDINA, 1959). A fibra desta planta está sendo utilizada para reforçar matrizes de
polipropileno em compósitos utilizados na parte interna de veículos do mercado nacional

12
(ROSA et al., 2012). E também seu efeito está sendo testado no desempenho de compósitos
cimentícios utilizados pela construção civil (TEIXEIRA, 2015).
A serragem, resíduo de pinus produzido pelas madeireiras são em grande parte
descartados e apenas parcialmente reciclados, são na maioria das vezes geralmente queimadas
ou descartadas no ambiente, provocando poluição do solo, ar e água. A parte aproveitada dos
resíduos é formada por costaneiras (tábua de qualidade inferior) e cavacos maiores
(WANDER, 2001).
Tais problemas tendem a se agravar pelo aumento projetado da produção de madeira
serrada, já que atualmente diversas empresas florestais estão envolvidas no manejo de pinus e
eucalipto, com vista ao seu uso múltiplo. Podendo ser sua utilização como cobertura morta o
que minimizaria a poluição.
Em um estudo realizado por Kozinski (2006), avaliando diferentes coberturas de solo
na cultura do mirtileiro cv. Bluecrop, comprovou-se que a serragem teve influência positiva
no rendimento de frutos. Rufato et al. (2004) verificaram que as coberturas vegetais de ervilha
forrageira não tiveram efeito sobre o diâmetro do tronco das plantas de pessegueiro cv.
Maciel, porém interferiram no volume de copa, na produção e no índice de fertilidade das
plantas de pessegueiro.
3.7. Fauna do Solo
As relações entre a fauna edáfica e os atributos do solo têm despertado um interesse
cada vez maior em virtude da sua eficiência como indicador das alterações promovidas pelos
sistemas de manejo. Diversas pesquisas desenvolvidas têm demonstrado que esses
organismos são bastante sensíveis às alterações que ocorrem em seu habitat, sejam elas de
ordem natural ou antrópicas (OLIVEIRA et al., 2014).
O acompanhamento das comunidades que compõem a fauna do solo, em diferentes
sistemas agrícolas, é um importante instrumento que pode ser integrado às práticas de
monitoramento da qualidade do solo, fornecendo informações sobre o manejo aplicado, o
desenvolvimento e funcionamento dos agroecossistemas (SILVEIRA, 2016). Os processos de
fragmentação da serrapilheira e decomposição da matéria orgânica pelos microrganismos,
com a ciclagem de nutrientes no ambiente do solo, são grandemente influenciados pela ação
dos invertebrados da fauna edáfica, como resultado direto ou indireto de suas atividades
alimentares (WINK et al., 2005).
Neste sentido o conhecimento da composição, abundância e distribuição dos
organismos edáficos, têm um papel relevante na manutenção da capacidade produtiva do solo,

13
graças as diferentes funções que estes desempenham. Vários critérios são utilizados para
classificar os organismos que compõem a fauna do solo, e procuram descrever suas funções
neste habitat.
3.7.1 Caracterização da comunidade faunística do solo
A decomposição dos resíduos orgânicos e a ciclagem de nutrientes são evidenciadas
como parte da atividade de microrganismos e um conjunto composto por diversos animais
que influencia de maneira decisiva no funcionamento da flora decompositora, como resultado
direto e indireto de sua atividade de alimentação (LOPES ASSAD, 1997).
O solo é um dos ambientes mais complexo e diverso do planeta, este fato é decorrente
da adaptação de diversos organismos à vida no solo. Diversas espécies de organismos podem
viver em um solo agrícola, desde os microscópicos até a macrofauna facilmente visível
(BROW e FRAGOSO, 2003).
A fauna edáfica é composta por animais invertebrados que podem passar uma parte do
ciclo vital ou todo o ciclo no solo. O aumento do número de indivíduos, a diversidade e a
uniformidade de espécies da fauna do solo ocorre pela disponibilidade de condições
ambientais, que favorecem a reprodução dos invertebrados como a melhoria das deposições
de resíduos vegetais (NUNES, 2010).
A fauna do solo é constituída pelos vários grupos de organismos que vivem no solo,
que variam de tamanho a partir de poucos micrômetros (Protozoa) até vários centímetros ou
mais (grandes minhocas ou diversas espécies de Diplopoda próprias de ambientes tropicais)
os quais exercem as mais variadas funções e são responsáveis por alterações nas propriedades
edáficas (CORREIA et al., 1997).
Segundo Swift et al. (1979), a fauna do solo é classificada de acordo com seu
diâmetro, em microfauna (0,004 a 0,01 mm), mesofauna (0,01 a 2 mm) e macrofauna (2 a 20
mm), considerando aspectos fisiológicos tais como o regime alimentar, em saprófagos,
fitófagos e predadores (PINHEIRO, 1996).
A influência da cobertura morta na biologia do solo tem sido avaliada por seus efeitos
globais em processos biológicos de liberação de CO2 (respirometria), mineralização do
nitrogênio orgânico, população e biomassa microbiana (SILVA et al., 2007). Poucos trabalhos
referem-se a grupos fisiológicos mais específicos ou parâmetros qualitativos da biologia do
solo e não são relatados os efeitos seletivos do tipo de cobertura morta (como p.ex. serragem
de pínus, gramíneas, cascas, etc) na macro e microbiota. No entanto, sabe-se que a

14
decomposição de um dado tipo de cobertura morta induz, inicialmente, a atividade de alguns
organismos para os quais serve como fonte de energia e nutrientes (BOTELHO et al., 2007).
Os produtos residuais dessa decomposição inicial realizada pelos organismos edáficos
tornam-se então úteis a outros tipos de vida, estabelecendo-se relações sintróficas e
antagônicas que mantêm o equilíbrio da comunidade biológica como um todo que são
essenciais para a conservação da fertilidade e produtividade do ecossistema (CORREIA e
OLIVEIRA, 2005). Esses fatores sucessivos de ciclos biológicos constitui o solo vivo, que é a
chave da fertilidade, fornecendo os nutrientes necessários à nutrição das plantas e evitando
perdas por lixiviação e erosão, ou seja, promove a potencialização dos nutrientes que passam
para a forma orgânica (imobilização) e, à morte do organismo, retomam ao solo por
mineralização.
3.8. Exigências nutricionais em plantas frutíferas
A fim de garantir a capacidade de qualidade na produção de frutíferas, é recomendado
buscar o equilíbrio entre o desenvolvimento vegetativo e reprodutivo, sendo que o manejo da
adubação é um dos melhores meios (TAGLIAVINI e MARANGONI, 2000). A nutrição
mineral é uma condição importante no aumento da produção de frutíferas, deste modo, os
minerais são cumpridor de várias funções nas plantas como processos energéticos, ativação
enzimática e regulação osmótica das membranas (MARSCHNER, 1995; FAUST, 1989).
De acordo com Kangueehi (2008), a exigência nutricional para produção de frutas é
fundamental especialmente para plantios de alta densidade. Um método eficaz para
determinar a necessidade nutricional é baseado na análise mineral de toda a planta
(WEINBAUM et al., 2001). Sendo assim, vários estudos têm sido realizados em culturas
como a pereira (STASSEN e NORTH, 2005), macieira (NEILSEN e NEILSEN, 2003;
HAYNES e GOH, 1980), pessegueiro (STASSEN, 1987), entre outros.
3.8.1. Macronutrientes
Dentre os macronutrientes o nitrogênio (N) é o elemento que as plantas exigem em
maior quantidade em relação aos demais minerais. Sendo este um componente da célula
vegetal que faz parte de moléculas de aminoácidos, proteínas, ácidos nucléicos e outros
compostos orgânicos e, portanto, desempenha um papel importante em processos metabólicos
das plantas (SOUZA e FERNANDES, 2006; TAIZ e ZEIGER, 2009).

15
Acessível no solo o N situa-se sobre várias formas, envolvendo amônio, nitrato,
aminoácidos, peptídios, mas sua forma de agir ainda é desconhecida. De modo geral, os
vegetais diferem na sua preferência por fontes de N, mas o absorvem sobretudo sob formas
inorgânicas, como nitrato ou amônio (SOUZA e FERNANDES, 2006).
Na assimilação do N pelas raízes, os aminoácidos são transportados para as folhas por
fluxo transpiratório, via xilema (MARSCHNER, 1995). No entanto, também pode ser
transportado através da membrana plasmática de certas células, em outras formas, como
peptídios menores, bases purinas, pirimidinas e seu derivados (GILLISSEN et al., 2000).
A deficiência de N nas plantas é caracterizada pela redução do crescimento e pela
coloração verde-clara nas folhas, os sintomas são visíveis em toda a planta. As folhas têm a
senescência prematura. A partir da senescência das folhas o N é transportado via floema para
órgãos perenes das árvores antes da abscisão folias (CASTAGNOLI et al, 1990).
Em um estudo realizado por Stassen e North (2005), em pereiras cv. Forelle, os
maiores teores de N foram encontrados nas folhas. Portanto, a aplicação de N deve ser durante
o outono por resultar no seu acúmulo em partes permanentes da árvore como reservas e
redistribuídos no início da temporada para o novo fluxo de crescimento (STASSEN et al.,
1981). Nas outras culturas, como no pessegueiro, todo o N utilizado para um novo
crescimento durante os primeiros 25 a 30 dias a partir da brotação provem da remobilização
das partes permanentes (RUFAT e DEJONG, 2001).
Os teores foliares de fósforo (P) na maioria das plantas ocorrem, em concentrações
entre 0,1 e 0,4%, sendo mais baixos que os teores normalmente encontrados para N e K
(TISDALE et al., 1993). A função do P é a constituição de macromoléculas estruturais e na
maioria dos ácidos nucléicos participa da transferência de energia (ARAÚJO e MACHADO,
2006; MARSCHNER, 1995).
Os solos brasileiros são geralmente pobres em P, sendo que este elemento é pouco
móvel, pois sua disponibilidade é afetado pelo pH, pela capacidade de absorção e de fixação
no solo. As formas de P absorvidas pelas plantas são HPO4-2 e H2PO4-, variando a
disponibilidade conforme o pH do solo (BASSO et al., 2003).
A insuficiência de P é marcada pelo atraso no crescimento das plantas e de raízes,
atraso no florescimento, inibição da brotação de gemas laterais e diminuição do número de
frutos. A toxidez de P pode ser acentuada com a deficiência de Zn (ARAÚJO e MACHADO,
2006).
O Potássio (K) é o cátion mais abundante no citoplasma das células e desempenha a
ativação de enzimas, na síntese de proteínas, no movimento dos estômatos, na fotossíntese e
na extensão celular. É também o principal cátion envolvido no estabelecimento do turgor

16
celular e na manutenção da eletroneutralidade celular (TAIZ e ZEIGER, 2009;
MARSCHNER, 1995).
O K é móvel no floema, resultando em uma boa distribuição para os frutos carnosos.
Estudos indicam que as árvores frutíferas têm maior absorção de K por unidade de massa seca
da raiz do que as árvores não frutíferas (KANGUEEHI, 2008). No solo, a disponibilidade de
K é interferida pela capacidade de troca de cátions (CTC), da matéria orgânica e das
adubações. Altas concentrações de K no solo inibem a absorção de Ca, causando “bitter pit”
em maçãs e em peras (BASSO et al., 2003). Em estudos realizados com peras cv. Forelle,
Stassen e North (2005) verificaram maiores quantidades de K, Ca e Mg nas raízes, quando
enxertadas sobre marmeleiro “A”, em comparação àquelas enxertadas sobre o híbrido de
pereira “BP1”.
Os resultados dos efeitos de K são similares em peras e maçãs, aumentando os
atributos de qualidade tais como o tamanho dos frutos, firmeza da polpa, teor de sólidos
solúveis, açúcares e ácidos orgânicos, rendimento em suco, sabor, enquanto o excesso de K
estimula uma série de distúrbios fisiológicos, tais como “bitter pit” e pingo de mel (HUDINA
e STAMPAR, 2002).
O Enxofre (S) é elementar sendo incorporado em aminoácidos, proteínas e co-enzimas
e também é necessário para a síntese de outros compostos, incluindo a coenzima A e a
vitamina B (NEILSEN e NEILSEN, 2003; TAIZ e ZEIGER, 2009).
O S é absorvido pelas raízes das plantas quase exclusivamente na forma de íon sulfato
(SO42-), porém, em pequenas quantidades o sulfato pode ser absorvido pelas folhas das
plantas, destacando que em altas concentrações este elemento pode ser tóxico (TISDALE et
al., 1993). O processo de armazenamento e mobilização deste elemento exige mudanças
sazonais na regulação da absorção de sulfato e no seu transporte pelo xilema, que são
parcialmente independentes do estado do enxofre armazenado (HERSCBACH et al., 2000).
A deficiência de S provoca atraso no crescimento e clorose nas plantas. Maioria das
vezes, a deficiência de S é confundida com deficiência de N. No entanto, embora semelhantes
os sintomas de deficiência de S ocorrem primeiro nas folhas mais novas das plantas
(TISDALE et al., 1993; MARSCHNER, 1995), enquanto que o N é translocados para as parte
novas.
O Cálcio (Ca) é o elemento mineral mais importante na conservação pós-colheita em
frutos, principalmente em maçãs e peras que são armazenadas por longos períodos. Também é
importante em outros tipos de frutos que precisam de elevado níveis de Ca para garantir um
amadurecimento lento e maior vida útil de prateleira (KANGUEEHI, 2008).

17
Este elemento mineral é absorvido pelas plantas na forma de íon Ca+2, sendo
encontrado nas folhas em concentrações entre 0,2 a 1,0%. Tem uma função importante na
estrutura e permeabilidade das membranas celulares, na divisão celular. Na presença de Ca
ocorre também a absorção regulada de outros cátions, devido a sua carga iônica (TAIZ e
ZEIGER, 2009).
O Ca é pouco móvel na planta. O suprimento adequado de N favorece a absorção de
Ca. Baixo pH e alta disponibilidade de K e Mg induzem a falta de Ca na planta. No entanto, a
disponibilidade de B auxilia na translocação de Ca na planta e para os frutos. A utilização do
gesso agrícola é prática recomendável como fonte complementar de cálcio, para melhorar a
relação de Ca e Mg em solos desequilibrados (BASSO et al., 2003).
A deficiência de Ca é manifestada pelo crescimento retardado e necrose de brotos e
ápice das raízes. Baixas concentrações de Ca no interior dos frutos podem ocasionar
distúrbios fisiológicos tais como mancha de cortiça, pingo de mel e degenerescência
senescente em frutos de peras e maçãs (KANGUEEHI, 2008; MARSCHNER, 1995).
O Magnésio (Mg) é um elemento mineral exigido e absorvido pelas plantas frutíferas
em quantidades inferiores ao Ca+2, sendo que a absorção de Mg é reduzida pela concorrência
com os cátions como o K+, NH4+, Ca+2 e Mn+2 bem como o H+ (MARSCHNER, 1995).
Sendo o principal constituinte da molécula de clorofila e este íon tem a importante função na
ativação enzimática da ribulose-difosfato carboxilase/oxigenase (rubisco) (TAIZ e ZEIGER,
2009; MALAVOLTA, 1980).
A concentração de Mg nas folhas aumenta durante as últimas seis semanas antes da
queda das folhas, a partir da translocação das partes permanentes da planta, quando é perdido
na queda das folhas. De acordo com Kangueehi (2008), a perda de aproximadamente 44% do
Mg presente na árvore está ligada à queda das folhas.
3.8.2 Micronutrientes
Os micronutrientes são considerados elementos essenciais para o crescimento das
plantas e se caracterizam por serem absorvidos em quantidades pequenas (da ordem de
miligramas por quilograma de matéria seca da planta). Sendo que pulverizações foliares ou
aplicações no solo podem garantir a necessidade da planta. A maioria dos micronutrientes é
imóvel no floema e os sintomas aparecem principalmente nas folhas novas (NEILSEN e
NEILSEN, 2003).
O Cobre (Cu) localizado nos cloroplastos das plantas onde participa das reações
fotossintéticas. Também é encontrado em enzimas envolvidas com proteínas e metabolismo

18
de carboidratos. O cobre é absorvido na forma iônica Cu2+
e se move lentamente na planta. A
deficiência de Cu causa inibição da lignificação do xilema, que pode levar à murcha e
enrolamento das folhas novas, com flacidez nos pecíolos e talos, e clorose (GIL SALAYA,
2000; KANGUEEHI, 2008).
As plantas raramente apresentam deficiência de Cu, sendo a sua disponibilidade
adequada na maioria dos solos. No entanto, a deficiência de Cu pode ocorrer em plantas
cultivadas em solos com baixo teor total de Cu ou em solos com altos teores de matéria
orgânica. Dentre os micronutrientes, a deficiência de Cu é a mais difícil de diagnosticar,
devido à interferência de outros elementos, como: P, Fe, Mo, Zn e S. Em citros e outras
frutíferas, aplicações em excesso de fertilizantes fosfatados podem provocar a deficiência de
Cu (DECHEN e NACHTIGALL, 2006).
O Ferro (Fe) está associado com os cloroplastos onde desempenha papel na síntese de
clorofila, cadeia de transporte de elétrons (citocromo, ferrodoxina), fixação de N2 e
respiração. Sua deficiência diminui a clorofila e fotossíntese, afetando a atividade das enzimas
como rubisco e outras, na respiração, e síntese de proteínas, reduz o descarregamento do
tecido celular no floema. Nos frutos reduz fortemente seu tamanho e qualidade (GIL
SALAYA, 2000; TAIZ e ZEIGER, 2009).
As plantas exudam substâncias que formam complexos com ferro para a absorção,
como o ácido málico e outras, que mobilizam o ferro do solo para as raízes. O Fe3+
deve ser
reduzido a Fe2+
nas membranas das células absorventes (TAIZ e ZEIGER, 2009).
O manganês (Mn) tem participação em vários processos na planta, tais como:
oxirredução (fotólise da água nos cloroplastos), redução do nitrato e ativação de várias
enzimas, entre elas a síntese da clorofila (TAIZ e ZEIGER, 2009). Sendo considerado
geralmente um elemento imóvel na planta, é fornecido preferencialmente para o tecido jovem
ou em crescimento, manifestando assim sua deficiência nas folhas mais velhas na forma de
manchas irregulares nas margens entre as nervuras (KANGUEEHI, 2008).
A quantidade de Zinco (Zn) é pequena nas plantas, porém já foram identificadas cerca
de 60 enzimas com participação do Zn e, portanto, desempenham funções importantes na
planta. Tem grande importância no papel de co-fator de enzimas na produção do hormônio
vegetal ácido indolacético (IAA) (MENGEL e KIRKBY, 1982). A deficiência de Zn resulta
em internódios curtos e as folhas apresentam tamanho menor e clorose internerval. A brotação
também é deficiente ao longo dos ramos. A disponibilidade varia de acordo com o tipo de
solo, o teor de matéria orgânica e o pH (BASSO et al., 2003).

19
4. MATERIAL E MÉTODOS
4.1. Local do experimento
O experimento foi conduzido em pomar comercial na região do Palmital do Meio,
pertencente ao Município de Cruz Machado, Paraná, Brasil, (25º 59' 47" S. 51º 08' 17" O) e a
1094 metros de altitude (MAACK, 2002).
O solo da área experimental é classificado como Latossolo Bruno distrófico típico, de
textura argilosa (EMBRAPA, 2013), constituído por teores de argila, silte e areia de 500, 300
e 200 g kg-1
, respectivamente.
O clima da região, segundo a classificação de Köppen, é Cfb, apresentando
temperatura moderada com chuva bem distribuída e verão brando, o mês mais frio apresenta
temperatura média inferior a 18ºC, no mês mais quente a temperatura média é inferior a 22ºC,
sem estação seca definida, e a precipitação média anual é de 1400 mm (IAPAR, 2013).
4.2. Material experimental
O pomar foi implantado em outubro de 2015, com a cultivar de pereira Hosui
enxertada sobre porta-enxerto Pyrus calleriana. A polinizadora utilizada foi a cv. Packham`s
Triumph na proporção de 8:1, com linhas espaçadas em 3,0 m e espaçamento entre plantas de
2,5 m, equivalente à densidade de plantio de 1.333 plantas ha-¹.
O delineamento experimental utilizado foi em blocos casualizados, com sete tratamentos e
quatro repetições. As unidades experimentais foram constituídas por parcelas de 10 m2 (1,0 x
10 m) com 4 plantas de pereira, onde utilizou-se como área útil 5 m2, nas duas plantas
centrais. A adubação básica de plantio foi realizada de acordo com a análise do solo (Tabela
1) e com as recomendações para o cultivo da pereira em sistema orgânico. Foram aplicados na
cova 5,0 kg de esterco de galinha curtido e 500 g de fosfato natural (SOUZA e RESENDE,
2003).
Tabela 1. Análise química do solo, na profundidade de 0-20 cm antes da implantação das
coberturas (setembro de 2015) e do pomar de pereiras cv. Hosui (Cruz Machado-
PR).
Prof. K Ca Mg Al P MO pH
cm cmolc/dm3
mg/dm3 g/dm
3 CaCl
0-20 0,10 3,60 2,00 0,20 2,70 41,10 4,70

20
A calagem foi definida após a interpretação da análise de solo, seguindo as
recomendações para a cultura das frutíferas (EMBRAPA, 2007).
Os tratamentos foram representados pelos seguintes sistemas de produção como
coberturas do solo: 1) Cobertura natural (espécies vegetais nativas encontradas em área de
cultivo de frutíferas na região); 2) Cobertura de Azevém + milheto; 3) Serragem de pinus; 4)
Tela de sombreamento (70 %); 5) Casca de arroz; 6) fibra de curauá e 7) capina manual
(manutenção da parcela sem nenhuma cobertura, durante todo o ciclo) (Figura 1).
Figura 1. Tipos de coberturas sobre o solo, instaladas logo após a implantação do
experimento, constituídas por palhada de azevém+milheto (T2), serragem de
pinus (T3), tela de sombreamento 70% (T4), casca de arroz (T5), fibra de curauá
(T6) e com capina manual (T7).
Logo após o plantio das mudas foram realizadas podas, com o objetivo de padronizar
o tamanho das pereiras (com desponte a 1m do solo). O manejo da cobertura vegetal na
entrelinha da cultura constituiu-se de roçadas realizadas com periodicidade mensal.
Para as coberturas do solo de origem vegetal foram utilizados os seguintes materiais:
casca de arroz (proveniente do beneficiamento do arroz em um moinho da região, equivalente
a massa de 30.000 kg por ha-¹); serragem de pinus (resíduos de uma madeireira local,
equivalente a 40.000 kg por ha-¹); palha de azevém (Lolium multiflorum) associado á palha de

21
milheto (Pennisetum glaucum), cortado antes de produzir sementes sendo (provenientes de
uma área utilizada para a formação de palhada, 45 dias com palha de azevém (35.500 kg de
palha por ha-¹) e após esse período foi adicionada a palha de milheto (40.000 kg por ha
-¹).
Antes da inserção dos materiais de cobertura, realizou-se uma capina e a retirada dos resíduos
vegetais que se encontravam em toda área experimental.
As coberturas de solo foram inicialmente instaladas em 02/10/2015, com uma camada
de 10 cm por 1 m de largura. Para os tratamentos em que se utilizou como cobertura a tela de
sombreamento 70% (Sol Pack Ltda, Osasco, SP, Brasil) e a fibra de curauá (Pematec Triangel
do Brasil Ltda, Santarém, PA, Brasil), os materiais foram recortados no tamanho da parcela,
colocado sobre o solo e, em seguida, presos por uma camada de solo nas laterais. Com
exceção da fibra de curauá, as demais coberturas com material vegetal, foram repostas
noventa dias após a instalação após nova capina. No segundo ciclo, realizou-se novamente a
instalação de todas as coberturas em 01/10/16 e reinstalação no dia 03/01/2017, setenta e
cinco dias após a infestação (exceto a fibra de curauá) após nova capina.
A indução de brotação das gemas de pereira foi realizada com base na metodologia de
Botelho et al., (2006), em 23/09/2016 utilizando-se óleo mineral (Iharol®
, Iharabrás, S.A.) e
óleo vegetal (Aureo®, Bayer, S.A.) em uma aplicação, nas concentrações de 4% e 4%,
respectivamente.
4.3. Avaliações
As avaliações foram realizadas nos ciclos de 2015/2016 e 2016/2017, primeiro e
segundo ano após a implantação do pomar de pereira, respectivamente.
4.3.1 Coleta, quantificação e identificação das plantas daninhas
As amostragens foram realizadas próximo às pereiras, lançando-se aleatoriamente dois
quadrados vazado com área interna de 0,25 m² em todas as parcelas a cada 30 dias, durante os
períodos de outubro/2015 a março de 2016 e outubro/2016 a março de 2017, conforme
metodologia descrita por Maciel et al. (2010). Para cada tratamento, a área amostrada nos dois
períodos de seis meses com o quadrado vazado correspondeu a 24 m², e no total das
avaliações a 168 m².
Para a classificação e quantificação das espécies das plantas daninhas por família,
nome científico e nome popular, foi utilizada literatura específica (KISSMAN e GROTH,
1997, 1999 e 2000; LORENZI, 2008; LORENZI, 2014), tomando como base o sistema

22
Angiosperm Phylogeny Group II (2003), com auxílio nas delimitações das famílias e
ordenamento de alguns gêneros, conforme Souza e Lorenzi (2005).
Aos 75 dias da implantação das coberturas de solo, mais de 50% da infestação das
plantas daninhas apresentavam-se em estádio reprodutivo, quando foi efetuada a coleta da
parte aérea das plantas daninhas em 0,5 m² da área útil das parcelas. Esse procedimento foi
realizado colhendo-se separadamente todas as plantas daninhas por espécie, as quais foram
acondicionadas em sacos de papel e levadas para laboratório, para secagem em estufa de
circulação de ar forçada por 72 horas em temperatura de 65 °C, e posterior determinação da
massa seca das plantas.
Com os dados obtidos foram determinados os seguintes parâmetros fitossociológicos:
número de indivíduos por espécie; número de parcelas em que a espécie estava presente;
frequência (índice da ocorrência das espécies em cada quadrado); densidade (índice da
quantidade de indivíduos de uma mesma espécie em cada quadrado); abundância relativa
(concentração das espécies nos diferentes pontos da área total - m²); índice de valor de
importância (IVI), conforme Müeller-Dombois e Ellenberg (1974).
Para o cálculo dos parâmetros estudados foram utilizadas as seguintes fórmulas:
Freqüência (Fre) = [Nº de parcelas que contém a espécie/ Nº total de parcelas utilizadas]
Densidade (Den) = [Nº total de indivíduos por espécie/ Área total coletada]
Abundância (Abu) = [Nº total indivíduos por espécie/ Nº total parcelas contendo a espécie]
Freqüência Relativa (Frr) = [Frequência da espécie x 100/ Frequência total das espécies]
Densidade Relativa (Der) = [Densidade da espécie x 100/ Densidade total das espécies]
Abundância Relativa (Abr) = [Abundância da espécie x 100/ Abundância total das espécies]
Índice de Valor de Importância (IVI) = [Frr + Der + Abr]
4.3.2. Altura das plantas de pereira
As medições de altura de cada pereira foram realizadas mensalmente de janeiro de
2016 a fevereiro de 2017(exceto, no período de maio a setembro). Para estas medições foi
utilizada uma fita métrica presa a uma régua de madeira com 3 m de comprimento.
4.3.3. Diâmetro do tronco
Procedeu-se a mensuração do diâmetro do tronco mensalmente de janeiro de 2016 a
fevereiro de 2017 (exceto, no período de maio a setembro). Para isto, todas as plantas de cada

23
parcela foram medidas 10 cm acima do ponto de enxertia. Esta mensuração foi efetuada com
o uso de um paquímetro digital, sendo os resultados expressos em mm.
4.3.4. Diâmetro e número de ramos
Essas avaliações foram realizadas de novembro de 2016 a fevereiro 2017, quando
foram selecionados casualmente 5 ramos por planta para a medição do diâmetro dos ramos
(DR), com o uso de um paquímetro digital a 10 cm da inserção do ramo com o tronco. Para o
número de ramos (NR) foi realizada contagem total dos ramos por planta (BOTELHO et al.,
2010).
4.3.5. Teor de clorofila
O teor relativo de clorofila (SPAD) foi determinada mediante leituras com
clorofilômetro, modelo SPAD-502 (Minolta, Osaka, Japão). Foram realizadas quatro
medições por folha, com dois pontos de cada lado da nervura central, na face adaxial em
quatro folhas por planta de duas plantas centrais de cada parcela. Ao longo do ciclo foram
feitas medições mensais do índice SPAD, sendo a primeira determinação realizada aos 30 dias
após plena brotação utilizando folhas do terço médio da planta. As medições ocorreram até a
planta começar a apresentar a queda natural das folhas (mês de abril), sempre após as 16 horas
período com temperaturas mais amenas.
4.3.6. Temperatura do solo
As medições de temperatura do solo foram efetuadas diariamente às 7:30, 12:30 e
17:30 horas, de outubro de 2016 a março de 2017, na profundidade de 5 cm, com um
Termohigrômetro digital (Incoterm, Porto Alegre, RS, Brasil). O aparelho foi protegido por
um abrigo meteorológico constituído de garrafa PET, com livre passagem de ar, pendurado à
altura de 1,5 metros acima da superfície do solo (AZEVEDO e GALVANI, 2003).
4.3.7. Macrofauna do solo
A macrofauna edáfica foi avaliado mediante a caracterização da diversidade e riqueza
de espécies de organismos subterrâneos presentes em áreas de cultivo de pereira.
A coleta dos organismos da macrofauna foi realizada a partir da escavação de

24
monólitos de solo (25 cm × 25 cm de largura, 20 cm de profundidade) de acordo com o
protocolo padrão do Tropical Soil Biology and Fertility (TSBF) Programme (ANDERSON e
INGRAM, 1993), a cerca de 20 cm de distância da planta. Cada monólito coletado,
correspondendo à profundidade de 0-20 cm, com o uso de uma pá reta sendo o material
armazenado em sacos plásticos e identificados conforme o tratamento e número da amostra.
Estas amostras de solo foram levadas ao laboratório e inspecionadas, utilizando-se
bandejas plásticas e espátulas para a triagem dos organismos, os quais foram armazenados em
frascos contendo álcool 70% para posterior contagem e identificação. Após a contagem, foi
determinada a densidade total (número de indivíduos por metro quadrado), os índices de
riqueza de Margalef, equitabilidade de Pielou e diversidade de Shannon (ODUM, 1988).
4.3.8. Análises químicas do solo e nutricionais das plantas
As amostras de solo foram coletadas em fevereiro de 2017. Foram realizadas
amostragens em duas profundidades diferentes: 0-10 e 10-20 cm. Para avaliar as propriedades
químicas, foram realizadas oito amostragens por parcela com trado calador, para formar a
amostra composta. Foram determinadas as seguintes propriedades químicas das amostras de
solos: pH, matéria orgânica, alumínio e teor de nutrientes, usando métodos descritos pela
EMBRAPA (1997).
Em fevereiro de 2017, foram coletadas 10 folhas por parcela experimental, retiradas
do terço médio das plantas na porção mediana de ramos do ano, de acordo com metodologia
descrita por Basso et al. (2003). Posteriormente, o material foi imerso em solução com
detergente neutro a 1%, enxaguado em água destilada e seco a 65°C (até peso constante) para
posterior moagem, utilizando-se moinho tipo Willey e peneira com malha de 1 mm (BASSO
et al., 2003). O nitrogênio (N) foi determinado pelo método Kjeldahl após digestão sulfúrica.
Os nutrientes fósforo (P), potássio (K), cálcio (Ca), magnésio (Mg), enxofre (S), ferro (Fe),
cobre (Cu), manganês (Mn) e zinco (Zn) foram extraídos por digestão nítrico-perclórica. A
determinação dos elementos Ca, Mg, Fe, Cu, Mn e Zn foi realizada por espectrometria de
absorção atômica em chama (SpectrAA 220, Varian, Austrália), o K por fotometria de chama
(B462, Micronal, São Paulo, SP, Brasil) e o S e P por espectrofotometria UV-VIS (UV-1800,
Shimadzu, Kioto, Japão) (EMBRAPA, 2009). Todas as análises químicas foram realizadas no
Laboratório de Solo e Nutrição de Plantas da UNICENTRO.
4.3.9. Análises estatísticas

25
Os dados foram submetidos à análise de variância e, em caso de significância
(p<0,05), as médias foram comparadas pelo teste de Student Newman-Keuls, ao nível de 5%
de probabilidade de erro, utilizando-se o programa Sisvar 5.0 (FERREIRA, 2011).

26
5. RESULTADOS
5.1. Plantas daninhas em função de diferentes tipos de coberturas do solo
Foram identificadas 12 espécies de plantas daninhas, em sua maioria dicotiledôneas
(08), além de 03 espécies de folha estreita (poaceae) e uma da família cyperaceae (Cyperus
rotundus) dentre as monocotiledôneas infestando a área do pomar distribuídas em 7 famílias
no primeiro (2015/2016) e segundo ciclo (2016/2017) (Tabela 2). As famílias mais
representativas dos levantamentos fitossociológicos, ou seja, com maior número de espécies,
foram Asteraceae e Poaceae no primeiro ciclo e segundo ciclo, com um total de 4 e 3 espécies
cada uma, respectivamente. As famílias Cyperaceae, Amaranthaceae, Malvaceae,
Plantaginaceae e Solanaceae apareceram em ambos os ciclos com as respectivas coberturas de
solo: cobertura natural (T1), palhada de azevém+milheto (T2), serragem de pinos (T3), tela de
sombreamento 70% (T4) e casca de arroz (T5).
Tabela 2. Espécies de plantas daninhas pertencentes às suas respectivas famílias e classes
identificadas em levantamento fitossociológico na cultura da pereira nos períodos de
2015/16 e 2017/17. Cruz Machado - PR, 2017.
Família Espécie Nome popular Classe
Asteraceae Eupatorium maximilianii Mata pasto Dicotiledônea
Asteraceae Sonchus oleraceus Serralha Dicotiledônea
Asteraceae Galinsoga parviflora Picão branco Dicotiledônea
Asteraceae Bidens pilosa Picão preto Dicotiledônea
Cyperaceae Cyperus meyenianus Tiririca Monocotiledônea
Poaceae Setaria geniculata Capim-rabo-de-raposa Monocotiledônea
Poaceae Digitaria horizontalis Capim colchão Monocotiledônea
Poaceae Urochloa plantaginea Capim marmelada Monocotiledônea
Plantaginaceae Plantago tomentosa Tanchagem Dicotiledônea
Solanaceae Solanum americanum Maria pretinha Dicotiledônea
Malvaceae Malvastrum arvensis Guanxuma Dicotiledônea
Amaranthaceae Amaranthus spinosus Caruru Dicotiledônea
As principais espécies encontradas, representando os maiores índices de toda a
comunidade de infestantes (Galisonga parviflora, Bidens pilosa, Cyperus meyenianus
Digitaria horizontalis e Urochloa plantaginea), e os parâmetros fitossociológicos calculados
para cada tratamento nos quatro períodos de avaliação estão apresentados nas figuras 3 a 8.
De modo geral, as espécies de plantas daninhas que apresentaram as maiores
porcentagens de frequências, densidades e abundâncias relativas, independentemente do tipo
cobertura do solo e época de avaliação, foram G. parviflora (Asteraceae) e B. pilosa
(Asteraceae), constituindo assim os maiores índices de valor de importância.
Na avaliação do tratamento cobertura natural do solo, as espécies com os maiores

27
índices de valor de importância (IVI) foram G. parviflora e B. pilosa, sendo que registraram
as maiores frequências, densidades e abundâncias relativas, independentemente da época de
avaliação. Entre as demais espécies que se destacaram em um segundo grupo foram a
Digitaria horizontalis e Urochloa plantaginea, ambas pertencentes a família Poaceae. Para a
condição da cobertura natural do solo, em relação aos demais tratamentos, observou-se o
maior número de espécies e famílias de plantas daninhas em todas as épocas e períodos de
avaliação.
Para a cobertura do solo com palhada de azevém+milheto ocorreu a inibição inicial de
infestação, uma vez que não foram encontradas plantas daninhas aos 15 dias após a primeira
implantação da cobertura nos períodos de 2015/16 e 2016/17, assim como aos 15 dias após a
reinstalação das coberturas em 2016/17.
Foram C. meyenianus, G. parviflora e B. pilosa foram as espécies que constituíram os
maiores (IVI‟s) nas avaliações realizadas no ciclo 2015/16. Embora também tenha ocorrido a
presença de C. meyenianus, as espécies que se destacaram principalmente pelas maiores
densidade e abundâncias relativas, e consequentemente IVI's, foram em ordem decrescente:
G. parviflora, U. plantaginea, D. horizontalis, S. genicula e B. pilosa, no período de 2016/17.
A palhada de azevém+milheto proporcionou redução do número de espécies por família em
relação à condição de ausência de cobertura do solo, mantido com capina no inicio da
instalação das coberturas.
Na cobertura do solo com serragem de pinus, observou a inibição total do surgimento
inicial de infestação apenas aos 15 dias após a primeira implantação em 2016/17 (Figura 6).
De modo geral, quanto ao no número de espécies por família, houve pequeno aumento no
número de espécies por família e quantidade de famílias nas avaliações de 2015/16 e redução
expressiva do período de 2016/17 (Figura 6). As espécies G. parviflora e B. pilosa foram as
que mais se destacaram pelos maiores IVI's nas avaliações de 2015/16, assim como as
espécies G. parviflora, B. pilosa, D. horizontalis S. genicula e C. meyenianus nas avaliações
de 2016/17 (Figura 6).
As epécies G. parviflora e B. pilosa, D. horizontalis S. genicula e U. plantaginea se
destacaram com os maiores frequências, densidades e abundâncias relativas em condição da
cobertura do solo com tela de sombreamento 70% , independentemente da época de avaliação
(Figura 7), mesmo havendo redução plena do surgimento de infestantes aos 15 dias após a
segunda reinstalação da tela 70% em 2016/17. Para a cobertura do solo com casca de arroz,
ocorreu aumento do número de espécies por família e quantidade de famílias nas avaliações
de 2015/16 e 2016/17 (Figura 8). As espécies G. parviflora e B. pilosa foram as que mais se
destacaram pelos maiores IVI's nas avaliações de 2015/16 e 2016/17 (Figura 8). É importante

28
ressaltar que para a cobertura do solo com fibra de curauá não foram identificadas plantas
daninhas em nenhuma das avaliações, nos dois períodos estudados, constituindo entre os
tratamentos estudados, uma ferramenta promissora e ideal para o manejo de plantas daninhas
em sistema de produção de culturas perenes. Além disso, a cobertura de solo proveniente da
combinação de palhada de azevém+milheto também apresentou excelente performance na
supressão do surgimento de plantas daninhas, com índices gerais de redução da população
infestante, considerando todos os meses de avaliação, em torno de 93% e 74%,
respectivamente para os períodos de período de 2015/16 e 2016/17 (Figura 2).
Com exceção da fibra do curauá, de forma geral os demais tratamentos pouco
influenciaram em relação a diversidade da infestação, com relação ao número de famílias e
espécies por famílias (Figuras 2, 3, 4, 5, 6 e 7), sendo constatado apenas menor número de
famílias de plantas daninhas e espécies por família para a cobertura do solo com tela de
sombreamento 70% (Figura 6).

29
Figura 2. Valores de frequência relativa (Frr), densidade relativa (Der), abundância relativa (Abr) e
Índice de valor de importância (Ivi) das espécies de plantas daninhas em duas épocas de
manejo, quinze dias após implantação das coberturas realizado com sete tratamentos,
observados em levantamento fitossociológico na cultura da Pera. Cruz Machado-PR.
2015/2016.
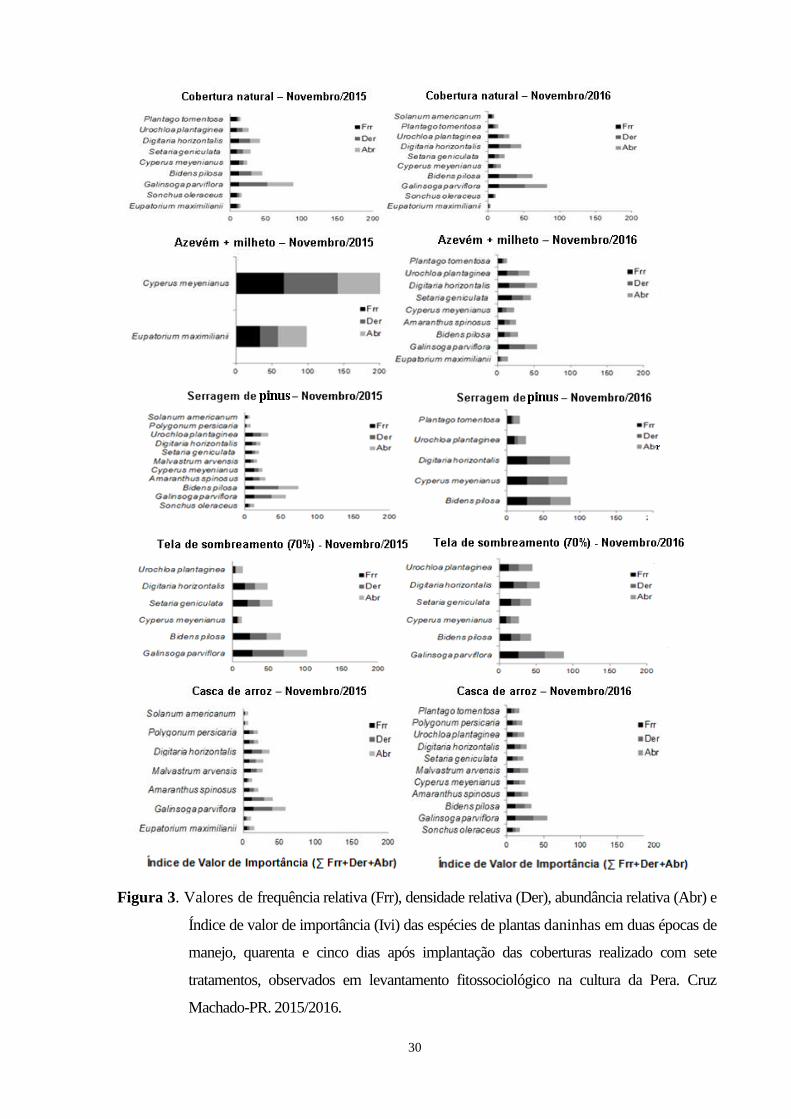
30
Figura 3. Valores de frequência relativa (Frr), densidade relativa (Der), abundância relativa (Abr) e
Índice de valor de importância (Ivi) das espécies de plantas daninhas em duas épocas de
manejo, quarenta e cinco dias após implantação das coberturas realizado com sete
tratamentos, observados em levantamento fitossociológico na cultura da Pera. Cruz
Machado-PR. 2015/2016.

31
Figura 4. Valores de frequência relativa (Frr), densidade relativa (Der), abundância relativa (Abr) e
Índice de valor de importância (Ivi) das espécies de plantas daninhas em duas épocas de
manejo, setenta e cinco dias após implantação das coberturas realizado com sete
tratamentos, observados em levantamento fitossociológico na cultura da Pera. Cruz
Machado-PR. 2015/2016.
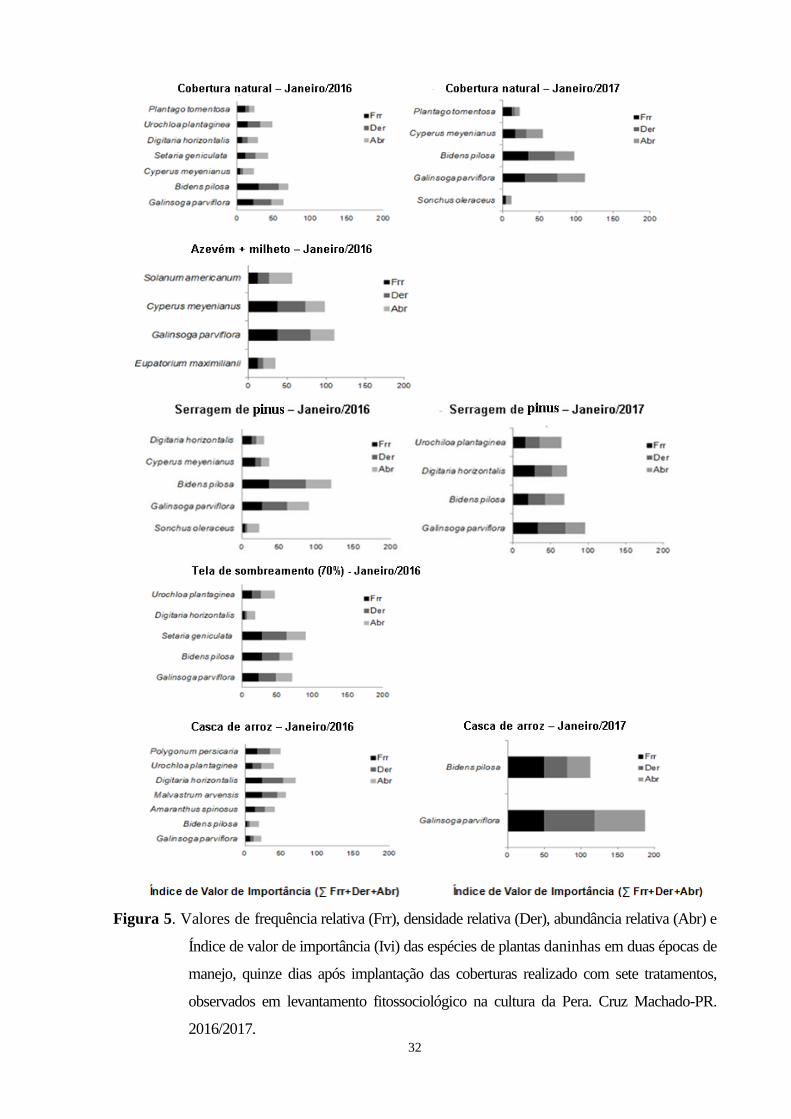
32
Figura 5. Valores de frequência relativa (Frr), densidade relativa (Der), abundância relativa (Abr) e
Índice de valor de importância (Ivi) das espécies de plantas daninhas em duas épocas de
manejo, quinze dias após implantação das coberturas realizado com sete tratamentos,
observados em levantamento fitossociológico na cultura da Pera. Cruz Machado-PR.
2016/2017.

33
Figura 6. Valores de frequência relativa (Frr), densidade relativa (Der), abundância relativa (Abr) e
Índice de valor de importância (Ivi) das espécies de plantas daninhas em duas épocas de
manejo, quarenta e cinco dias após implantação das coberturas realizado com sete
tratamentos, observados em levantamento fitossociológico na cultura da Pera. Cruz
Machado-PR. 2016/2017.
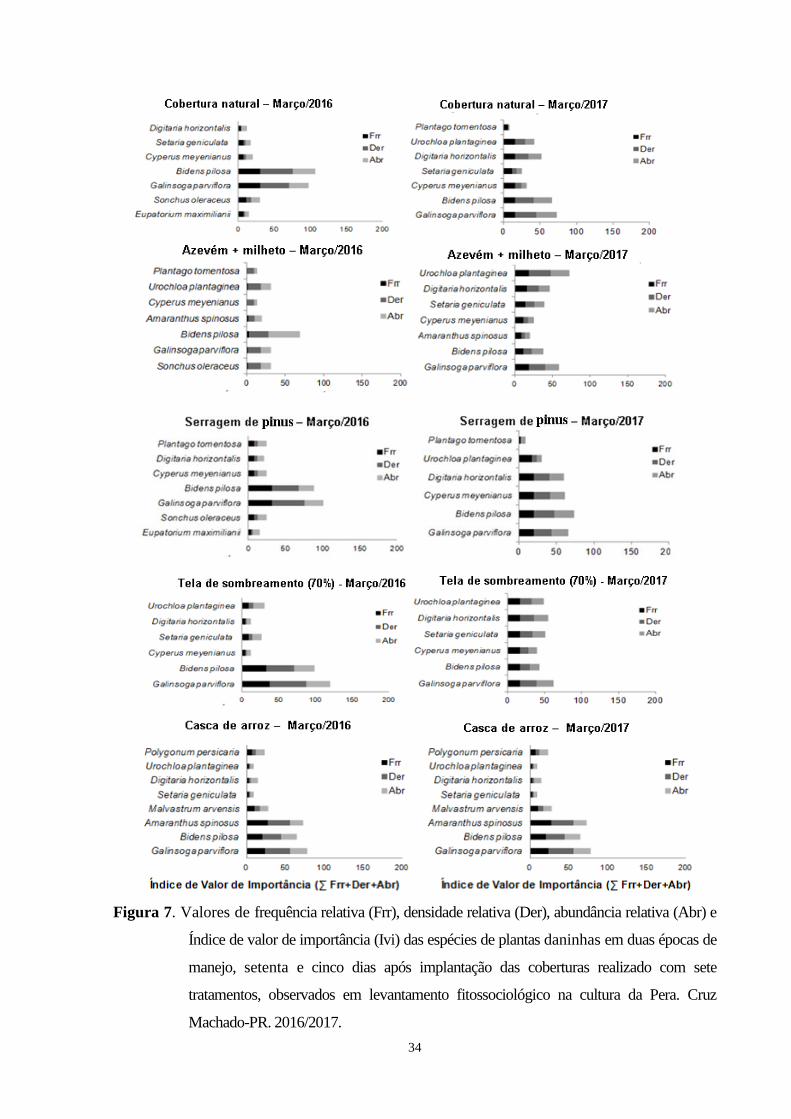
34
Figura 7. Valores de frequência relativa (Frr), densidade relativa (Der), abundância relativa (Abr) e
Índice de valor de importância (Ivi) das espécies de plantas daninhas em duas épocas de
manejo, setenta e cinco dias após implantação das coberturas realizado com sete
tratamentos, observados em levantamento fitossociológico na cultura da Pera. Cruz
Machado-PR. 2016/2017.

35
Na Tabela 3 estão apresentadas as médias de produção de massa seca de plantas
daninhas (0,5 m2) aos 75 dias após a aplicação das coberturas mortas no solo. Pode-se
perceber que a cobertura de azevém+milheto (2015/16) interferiu na produção da massa seca
das plantas daninhas quando comparadas com os tratamentos cobertura natural, serragem, tela
de sombreamento e casca de arroz. Para a fibra de curauá, não foi detectado nenhum tipo de
infestação em ambos os períodos. Dentre as coberturas, no segundo ciclo (2016/17) a
cobertura de casca de arroz apresentou a maior quantidade de massa seca de plantas daninhas,
sendo similar à cobertura natural. Com exceção da fibra de curauá que não apresentou
nenhuma incidência de plantas daninhas, as demais coberturas de solo tiveram redução em
relação à casca de arroz, com significância em 2017/17, mas não se diferiram da cobertura
natural, em exceção a azevém+milheto (2015/16).
Tabela 3. Produção de massa seca de plantas daninhas (g/0,5 m2) em função da cobertura do
solo, na avaliação aos 75 dias após a colocação das coberturas, em pomar de
pereiras cv. Hosui nos ciclos 2015/16 e 2016/17. Cruz Machado-PR, 2017.
Massa seca das plantas daninhas
Coberturas 2015/16 2016/17
Cobertura natural 131,8 a 107,4 ab
Azevém+milheto 62,3 b 64,6 b
Serragem de pinus 137,2 a 70,7 b
Tela de sombreamento (70%) 111,0 a 52,8 b
Casca de arroz 156,0 a 141,9 a
Fibra de curauá 0 c 0 c
Capina (sem cobertura) 0 c 0 c Letras distintas na coluna indicam diferença significativa pelo teste de Student
Newman-Keuls ao nível de 5% de probabilidade.
5.2. Crescimento vegetativo de pereiras cv. Hosui em diferentes coberturas de
solo
A resposta da pereira à cobertura do solo demonstrou diferenças entre os tratamentos
nos dois ciclos (Tabela 4). A altura de planta no primeiro ciclo (2015/16) após o plantio teve
as maiores alturas observadas nos tratamentos com coberturas de fibra de curauá, capina,
azevém+milheto e tela de sombreamento se diferiram significativamente da cobertura com
serragem de pinus. A altura de planta avaliada no segundo ciclo (2016/17) após o plantio
evidenciou efeito dos tratamentos, passando de 1,98 cm no sistema com cobertura natural para
2,48 cm no manejo com fibra de curauá diferindo-se estatisticamente. Os tratamentos com
tela de sombreamento, capina, azevém+milheto e casca de arroz foram semelhante neste

36
período e os menores valores correspondeu a cobertura com serragem de pinus e a cobertura
natural os quais diferiram-se dos demais tratamentos.
Para o diâmetro do caule, houve diferenças entre os tratamentos para os dois ciclos
vegetativos, sendo que para o ciclo (2015/16) o tratamento com fibra de curauá diferenciou-se
estatisticamente dos demais tratamentos. Para as avaliações do segundo ciclo (2016/17) as
maiores dimensões foram verificadas para o tratamento com a fibra de curauá, capina, tela de
sombreamento, palhada de azevém + milheto e casca de arroz e o menor diâmetro para a
cobertura de serragem de pinus em ambos os períodos (Tabela 4).
Em relação ao número de ramos, não houve diferenças significativas entre as
coberturas para o período de avaliação. Para o diâmetro de ramos, houve efeito significativo
dos tipos de coberturas do solo, sendo que os maiores valores foram verificados para os
tratamentos com tela de sombreamento, capina, fibra de curauá e casca de arroz. E o menor
foi observado em solo que recebeu a cobertura de serragem de pinus durante o período
avaliado (Tabela 4).
Tabela 4. Altura e diâmetro do caule de pereiras cv. Hosui nos ciclos 2015/16 e 2016/17,
número de ramos e diâmetro dos ramos no ciclo 2016/17, sob diferentes
coberturas de solo. Cruz Machado-PR, 2017.
Alturas das
plantas (m)
Diâm. do
caule (mm)
Nº de
ramos
Diâm.dos
ramos (mm)
Tratamentos 2015/2016 2016/2017 2015/2016 2016/2017 2016/2017 2016/2017
Cobertura natural 1,17 bc 1,98 c 17,4 bc 25,4 ab 7,87 ns
8,94 ab
Azevém+milheto 1,22 b 2,24 ab 17,4 bc 27,1 ab 8,50 8,81 ab
Serragem de pinus 1,13 c 1,82 c 16,5 c 24,0 b 8,12 8,45 b
Tela de sombreamento (70%) 1,21 ab 2,32 b 17,6 bc 28,3 ab 8,75 9,53 a
Casca de arroz 1,19 bc 2,20 ab 17,5 bc 26,6 ab 7,37 9,33 a
Fibra de curauá 1,26 a 2,48 a 20,5 a 30,9 a 8,37 9,41 a
Capina 1,25 a 2,28 ab 18,9 b 28,6 ab 8,12 9,51 a
CV (%) 21,53 11,08 18,72 14,53 21,71 14,75
Letras distintas na coluna indicam diferença significativa pelo teste de Student Newman-Keuls ao nível de
5% de probabilidade; ns = não significativo ao nível de 5% de probabilidade.
Considerando as três épocas de avaliação do teor de clorofila (índice Spad) das folhas
de pereira cv. Hosui no primeiro ciclo vegetativo houve diferenças significativas entre os
tratamentos na primeira e na terceira época (Figura 8 A). No mês de fevereiro os tratamentos
com as coberturas de fibra de curauá e palhada de azevém+milheto apresentaram os maiores
índices Spad e foram significativamente superiores aos demais. A cobertura com tela de
sombreamento apresentou menor valor de índice Spad, não se diferindo do tratamento com
cobertura natural. Os demais tratamentos tiveram valores intermediários.
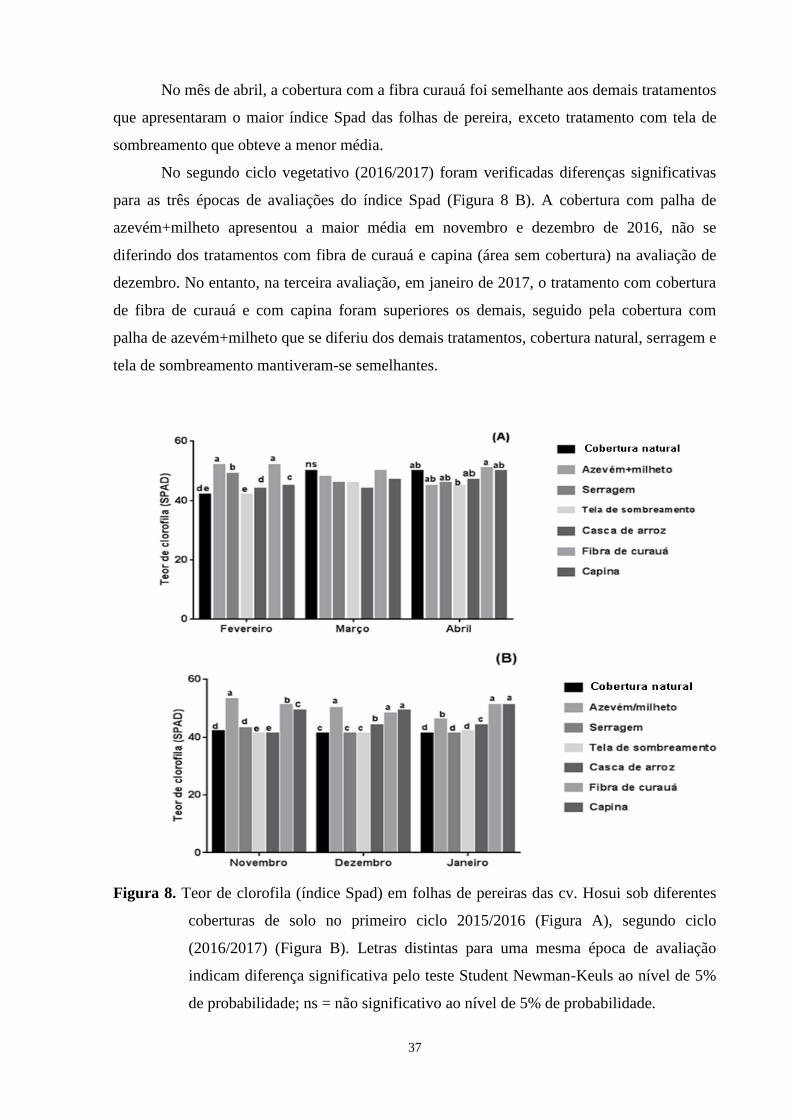
37
No mês de abril, a cobertura com a fibra curauá foi semelhante aos demais tratamentos
que apresentaram o maior índice Spad das folhas de pereira, exceto tratamento com tela de
sombreamento que obteve a menor média.
No segundo ciclo vegetativo (2016/2017) foram verificadas diferenças significativas
para as três épocas de avaliações do índice Spad (Figura 8 B). A cobertura com palha de
azevém+milheto apresentou a maior média em novembro e dezembro de 2016, não se
diferindo dos tratamentos com fibra de curauá e capina (área sem cobertura) na avaliação de
dezembro. No entanto, na terceira avaliação, em janeiro de 2017, o tratamento com cobertura
de fibra de curauá e com capina foram superiores os demais, seguido pela cobertura com
palha de azevém+milheto que se diferiu dos demais tratamentos, cobertura natural, serragem e
tela de sombreamento mantiveram-se semelhantes.
Figura 8. Teor de clorofila (índice Spad) em folhas de pereiras das cv. Hosui sob diferentes
coberturas de solo no primeiro ciclo 2015/2016 (Figura A), segundo ciclo
(2016/2017) (Figura B). Letras distintas para uma mesma época de avaliação
indicam diferença significativa pelo teste Student Newman-Keuls ao nível de 5%
de probabilidade; ns = não significativo ao nível de 5% de probabilidade.
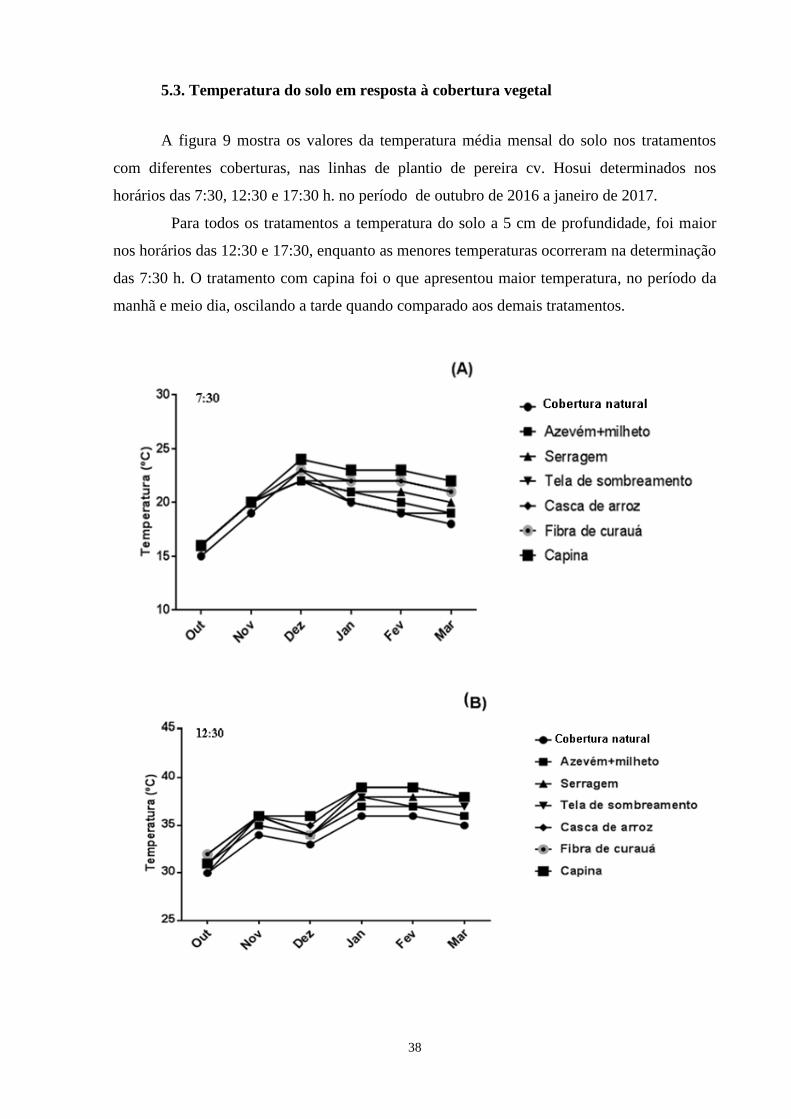
38
5.3. Temperatura do solo em resposta à cobertura vegetal
A figura 9 mostra os valores da temperatura média mensal do solo nos tratamentos
com diferentes coberturas, nas linhas de plantio de pereira cv. Hosui determinados nos
horários das 7:30, 12:30 e 17:30 h. no período de outubro de 2016 a janeiro de 2017.
Para todos os tratamentos a temperatura do solo a 5 cm de profundidade, foi maior
nos horários das 12:30 e 17:30, enquanto as menores temperaturas ocorreram na determinação
das 7:30 h. O tratamento com capina foi o que apresentou maior temperatura, no período da
manhã e meio dia, oscilando a tarde quando comparado aos demais tratamentos.

39
Figura 9. Média mensal da temperatura do solo (ºC) a 5,0 cm de profundidade considerando-
se a cobertura do solo e o horário da determinação 7: 30 (Figura A), 12:30 (Figura
B) e 17:30 (Figura C), no período de outubro de 2016 a março de 2017. Cruz
Machado-PR, 2017.
5.4. Efeito das diferentes coberturas sobre a macrofauna edáfica
Os organismos edáficos que mais se destacaram foram o grupo Isoptera presente em
todos os tipos de coberturas e em segundo lugar aparece o grupo Formicidae. Quanto ao
grupo Coleóptera, este foi mais representativo para a cobertura de palhada de
azévém+milheto. O grupo taxonômico Diplopoda somente foi encontrado na cobertura de
serragem de pinus e do grupo Aranae nas coberturas de azevém+milheto, serragem e tela de
sombreamento (Figura 10).
Figura 10. Distribuição dos grupos taxonômicos por tipo de cobertura de solo em pereiras da
cv. Hosui, segundo ciclo (2016/2017). Cruz Machado - PR.

40
A cobertura de solo que apresentou maior densidade total de indivíduos por metro
quadrado foi a cobertura com a casca de arroz (280 indiv. m²), seguido da palhada de
azevém+milheto (212 indiv. m²). Entre os sistemas de manejo houve diferenciação para a
densidade total de organismos edáficos em função do tipo de cobertura (Tabela 5).
Dentre os grupos taxonômicos a riqueza total não apresentou diferenças para as
coberturas de azevém+milheto, serragem e tela de sombreamento. Os valores do índice de
Shannon indicaram que dentre as coberturas, a palhada de azevém+milheto foi a que
apresentou maior diversidade. O índice de Pielou mostrou que a cobertura com a casca de
arroz possui a maior uniformidade de espécies e a tela de sombreamento a menor (Tabela 5).
Tabela 5. Distribuição do numero de indivíduos por unidade de área, riqueza total e índices
de Shannon e Pielou sob diferentes coberturas de solo, nas linhas de plantio de
pereira cv. Hosui. Cruz Machado-PR, 2017.
COBERTURAS
Nº de
Indivíduos (m-2)
Riqueza
total
Índice de
Shannon
Índice
de Pielou
Cobertura natural 3,7 c 3 b 0,34 d 0,21 c
Azevém+milheto 13,2 b 4 a 0,77 a 0,38 b
Serragem de pinus 4,5 c 4 a 0,40 d 0,20 c
Tela de sombreamento 4,0 c 4 a 0,37 d 0,19 c
Casca de arroz 17,5 a 4 a 0,68 b 0,68 a
Fibra de curauá 7,7 bc 3 b 0,57 c 0,36 b
Capina 6,7 bc 3 b 0,50 c 0,32 b
Letras distintas na linha indicam diferença significativa pelo teste Student Newman-Keuls ao nível
de 5% de probabilidade.
5.5. Características químicas do solo
Para a matéria orgânica, os maiores teores foram determinados no solo com as
coberturas de casca de arroz, fibra de curauá e capina na camada superficial (0-10 cm) bem
como na camada 10-20 diferindo-se significativamente das demais coberturas de solos e nas
duas profundidades. Em relação ao pH do solo na linha do plantio, foram observadas
diferenças entre os tratamentos de palhada de azevém+milheto (5,67) e tela de sombreamento
(5,18) na profundidade de 0-10 cm. Em geral, os tratamentos sem cobertura (testemunha),
serragem de pínus, tela de sombreamento e fibra de curauá apresentaram pH abaixo de 5,5
para a profundidade de 0-10 cm com exceção do tratamento com palhada de azevém+milheto
e capina. Na camada de 10-20 cm, não houve diferenças significativas entre os tratamentos.
Não houve diferenças significativas para o teor Al3+
em nenhuma das profundidades (Tabela
6).

41
Tabela 6. Valores médios de matéria orgânica, pH e alumínio em diferentes coberturas de
solo, nas profundidades de 0-10 cm e 10-20 cm, nas linhas de plantio de pereiras
cv. Hosui. Cruz Machado - PR, 2017.
M.O pH Al
---------g/dm3
------- ------CaCl2 ---- ---cmolc/dm3 ---
Profundidade (cm)
Coberturas 0 -10 10-20 0-10 10-20 0-10 10-20
Cobertura natural 33,97 b 30,16 b 5,43 ab 5,25ns
0,08 ns
0,10 ns
Azevém+milheto 34,93 b 31,31 b 5,67 a 5,4 0,06 0,08
Serragem 30,31 b 27,41 b 5,45 ab 5,32 0,09 0,06
Tela de sombreamento 34,47 b 29,80 b 5,18 c 5,18 0,16 0,22
Casca de arroz 42,06 a 33,34 ab 5,60 a 5,33 0,04 0,11
Fibra de curauá 39,91 ab 36,51 ab 5,40 ab 5,33 0,20 0,09
Capina 39,79 ab 38,18 a 5,51 b 5,43 0,07 0,06
CV (%) 11,41 9,22 10,42 13,52 12,51 29,78 Letras distintas na linha indicam diferença significativa pelo teste Student Newman-Keuls
ao nível de 5% de probabilidade; ns = não significativo ao nível de 5% de probabilidade.
Para o fósforo todos os tratamentos apresentaram teores maiores, na camada
superficial (0-10 cm), sendo que o tratamento com a cobertura de palhada de azevém+milheto
incrementou significativamente o teor deste elemento. Para camada de 10-20 cm, o tratamento
de cobertura com fibra de curauá conferiu o maior teor de P no solo, não se diferindo,
contudo, dos tratamentos com a cobertura de palha de azevém+milheto ou com capina (área
sem cobertura) (Tabela 7).
Os teores de potássio foram maiores na profundidade de 10-20 em relação à camada
superficial. Na camada de 0-10 cm o maior teor de potássio foi verificado para o tratamento
com cobertura de palha de azevém+milheto, que se diferiu significativamente dos demais.
Para a profundidade 10-20 cm, os maiores teores de potássio foram observados para os
tratamentos com coberturas de casca de arroz, serragem de pinus e fibra de curauá, que se
diferiram significativamente dos tratamentos cobertura natural e tela de sombreamento
(Tabela 7).
Em relação ao cálcio, na profundidade de 0-10 cm, os valores encontrados foram
acima de 4,0 cmolc/dm3, sendo que o tratamento cobertura natural apresentou o menor teor, se
diferindo significativamente da maioria dos tratamentos, com exceção da cobertura com palha
de azevém+milheto. Na camada de 10-20 cm o tratamento cobertura natural, também
apresentou o menor teor de Ca2+
, diferindo-se estatisticamente dos tratamentos com cobertura
de fibra de curauá e da capina que tiveram os maiores teores. Os teores de magnésio
apresentaram redução no solo com aumento da profundidade para todos os tratamentos. Na
camada mais superficial de 0-10 cm, os tratamentos com cobertura de fibra de curauá e a

42
capina tiveram teores significativamente superiores deste elemento. Na camada de 10-20 cm,
os maiores teores foram verificados para o tratamento com cobertura de casca de arroz
(Tabela 7).
Tabela 7. Valores médios de fósforo, potássio, cálcio e magnésio em diferentes coberturas de
solo, nas profundidades de 0-10 cm e 10-20 cm, nas linhas de plantio de pereiras
cv. Hosui. Cruz Machado - PR, 2017.
P K Ca Mg
mg/dm3
cmolc/dm3
Profundidade(cm)
Coberturas 0 -10 10-20 0-10 10-20 0-10 10-20 0-10 10-20
Cobertura natural 2,19 b 0,37 c 0,69 c 0,51 c 4,85 c 7,55 b 3,12 b 1,02 c
Azevém+milheto 5,18 a 3,10 a 1,29 a 1,29 a 8,95 b 9,72 ab 3,80ab 1,49 ab
Serragem 2,50 b 0,70 c 0,92 b 0,52 c 10,0 a 7,92 b 4,09 a 1,28 ab
Tela de sombreamento 1,31 c 0,69 c 0,86 b 0,62 c 11,6 a 8,03 b 4,17 a 1,49 ab
Casca de arroz 2,57 b 1,00 bc 0,98 b 0,66 c 11,3 a 10,1 ab 3,37 b 1,75 a
Fibra de curauá 3,57 ab 2,52 b 1,11 b 0,85 b 12,8 a 12,2 a 1,72 c 1,82 a
Capina 2,31 b 1,11 b 0,87 b 0,67 c 11,0 a 12,0 a 1,96 c 1,58 b
CV (%) 10,21 24,92 23,97 29,60 27,97 19,15 21,62 26,17 Letras distintas na linha indicam diferença significativa pelo teste Student Newman-Keuls ao nível de 5% de
probabilidade.
Na profundidade de 0-10 cm, não houve diferenças para os teores de Fe e Mn,
enquanto que na camada de 10-20 cm não houve diferenças para os tores de Cu e Zn. Na
camada de 0-10 cm, o teor de Cu foi significativamente superior no tratamento com cobertura
de fibra de curauá e, o teor de zinco foi estatisticamente maior para os tratamentos com
cobertura de palha de azevém+milheto ou fibra de curauá.
Tabela 8. Valores médios de ferro, cobre, manganês e zinco em diferentes coberturas de solo,
nas profundidades de 0-10 cm e 10-20 cm, nas linhas de plantio de pereiras cv.
Hosui. Cruz Machado-PR, 2017.
Fe Cu Mn Zn
mg/dm
3
Profundidade (cm)
Coberturas 0 -10 10-20 0-10 10-20 0-10 10-20 0-10 10-20
Cobertura natural 21,8 ns
17,69 a 1,57 b 1,52 ns
27,27 ns
21,46 ab 0,10 c 0,14 ns
Azevém+milheto 18,17 16,75 a 1,50 b 1,57 36,35 27,04 ab 0,58 a 0,36
Serragem 20,74 10,18 b 1,68 b 1,35 27,92 20,03 b 0,06 d 0,08
Tela de sombreamento 22,62 17,26 a 1,45 b 1,46 23,58 21,08 ab 0,17 c 0,06
Casca de arroz 18,14 11,54 ab 1,41 b 1,32 34,24 32,56 ab 0,10 c 0,22
Fibra de curauá 16,85 7,06 c 1,86 a 1,60 35,72 35,06 a 0,42 b 0,32
43
Capina 16,51 14,42 ab 1,47 b 1,44 35,84 35,85 a 0,15 c 0,19
CV (%) 18,85 26,53 18,28 16,27 25,00 24,41 18,40 22,50 Letras distintas na linha indicam diferença significativa pelo teste Student Newman-Keuls ao nível de 5% de
probabilidade; ns = não significativo ao nível de 5% de probabilidade.
5.6. Análise do teor mineral das folhas
Os teores médios de nutrientes nas folhas de pereiras cv. Hosui são apresentados nas
Tabelas 9 e 10. Não houve efeito significativo dos tratamentos para a maioria dos
macronutrientes, com exceção dos elementos S e P (Tabela 9). Para o teor de S os tratamentos
com cobertura natural e com capina foram significativamente superiores, enquanto que nas
pereiras sob coberturas com serragem e tela de sombreamento apresentaram os menores
teores. O teor de P foi maior nas pereiras sob cobertura de fibra de curauá, que se diferiu
estatisticamente do tratamento com cobertura de tela.
Tabela 9. Teores de nitrogênio, fósforo, potássio, cálcio, magnésio e enxofre (g kg-1
) em
folhas de pereira cv. Hosui sob diferentes coberturas de solo. Cruz Machado – PR,
2017.
Tratamentos N P Ca K Mg S
--------------------------------- g kg-1
-------------------------------
Cobertura natural 26,03 ns
3,14 ab 14,61 ns
19,01 ns
1,41 ns
4,19 bc
Azevém+milheto 30,51 2,91 ab 14,61 19,54 1,73 3,55 c
Serragem 28,74 3,55 ab 13,13 19,01 1,63 2,88 d
Tela de sombreamento 29,66 2,83 b 15,13 17,95 1,84 2,98 d
Casca de arroz 30,23 3,49 ab 12,9 18,48 1,63 3,88 bc
Fibra de curauá 30,49 4,03 a 16,09 17,95 1,74 5,27 a
Capina 30,51 3,37 ab 12,1 17,15 1,60 4,81b
CV (%) 14,21 15,25 19,61 10,96 12,96 14,80 Letras distintas na coluna indicam diferença significativa pelo teste de Student Newman-Keuls ao
nível de5% de probabilidade; ns = não significativo ao nível de 5% de probabilidade.
Para todos os micronutrientes analisados foram verificadas diferenças significativas
entre os tratamentos (Tabela 10). O maior teor de Cu foi verificado para o tratamento com
capina (área limpa) que foi significativamente superior ao tratamento cobertura natural. Para
os elementos Fe e Mn, os maiores teores foram verificados para o tratamento com cobertura
de fibra de curauá. A cobertura com casca de arroz apresentou o maior teor de Zn, que se
diferiu estatisticamente do tratamento de capina (área limpa).

44
Tabela 10. Teores de cobre, ferro, manganês e zinco (mg kg-1
) em folhas de pereira cv. Hosui
sob diferentes coberturas de solo. Cruz Machado – PR, 2017.
Tratamentos Cu Fe Mn Zn
-------------------------- mg kg-1
----------------------
Cobertura natural 2,14 c 36,70 b 11,76 ab 26,59 ab
Azevém+milheto 6,18 b 39,60 ab 7,37 ab 50,18 a
Serragem 4,74 ab 51,37 ab 7,30 ab 39,71 ab
Tela de sombreamento 4,92 ab 48,15 ab 19,85 b 44,13 b
Casca de arroz 3,49 ab 44,77 ab 6,50 ab 54,50 a
Fibra de curauá 5,02 ab 70,35 a 25,50 a 19,37 ab
Capina 7,33 a 66,47 ab 2,94 c 7,00 c
CV (%) 20,85 27,34 25,05 24,47 Letras distintas na coluna indicam diferença significativa pelo teste de Student Newman-Keuls
ao nível de 5% de probabilidade.

45
6. DISCUSSÃO
6.1. Plantas daninhas em função de diferentes tipos de coberturas do solo
Em função dos resultados obtidos no levantamento fitossociológico para as diferentes
coberturas de solo na cultura da pereira, observou-se que nos dois anos de avaliação ocorreu a
incidência das mesmas infestantes para ambos os períodos estudados, variando apenas o
numero de daninhas em função do manejo da cobertura.
De acordo com Roush e Radosevich, (1985), normalmente, as plantas que apresentam
maior porte e crescimento mais rápido são as que causam maior interferência competitiva. Em
um estudo Correia, Durigan e Klink (2006), observaram que a composição específica e as
densidades populacionais das comunidades infestantes foram influenciadas pelos sistemas de
produção de cobertura morta.
A cobertura do solo com fibra de curauá foi altamente eficiente no controle das plantas
daninhas, constituindo a melhor estratégia para o manejo de infestantes em sistema de
produção de pereira entre os tratamentos estudados. Esse resultado certamente está
diretamente relacionado com o sombreamento total da superfície do solo durante todo o
período de avaliação, o que pode ter influenciado a dinâmica do banco de sementes das
plantas daninhas (VOLL et al., 2005), por reduzir a quantidade da luz, a amplitude térmica e
hídrica do solo, assim por existir possivelmente efeito supressor no solo. Entretanto, não
existem informação dessas propriedades da fibra de curauá em relação ao desenvolvimento de
plantas daninhas.
Além disso, a cobertura de solo constituída por palhada de azevém+milheto também
apresentou supressão das plantas daninhas pela utilização de culturas de cobertura no solo,
sendo essas informações relacionadas à dinâmica do banco de sementes de plantas daninhas,
relacionadas principalmente em relação a quantidade da luz sobre espécies fotoblásticas
positivas e efeito físicos e alelopáticos, além de outros (ALMEIDA, 1988).
No entanto, houve o predomínio da produção de maior quantidade de matéria seca da
parte aérea das plantas para a cobertura de casca de arroz seguida da área com cobertura
natural do solo. Este efeito pode estar relacionado com a quantidade de concentrações
alelopáticas, baixas favorecem a germinação, intermediárias inibem e altas concentrações
favorecem o potencial germinativo (SPIASSI et al., 2011). Com relação aos períodos,
verificou-se um decréscimo gradativo para a massa seca das daninhas com o decorrer do
tempo. Resultado esperado, uma vez que as coberturas sufocam as daninhas, restringindo o
seu desenvolvimento, devido ao preenchimento das partículas, proporcionando um

46
sombreamento no solo, impossibilitando a sobrevivência das plantas daninhas e até mesmo
seu surgimento.
6.2. Crescimento vegetativo de pereiras cv. Hosui em diferentes coberturas de
solo
As avaliações realizadas nos dois ciclos após o plantio evidenciaram que tanto para
altura da planta quanto para diâmetro do caule e de ramos houve interferência das plantas
daninhas no desenvolvimento das plantas de pereiras cv. Hosui. A interferência negativa das
plantas daninhas sobre as fruteiras foi observado por Favaro (2015) que avaliaram diferentes
métodos de controle de plantas daninhas em citros, e observou redução no desenvolvimento e
redução significativa nos teores foliares de clorofila das plantas que sofreram interferência.
Desta forma, verificou-se que o manejo adequado da cobertura vegetal é imprescindível para
garantir um bom desenvolvimento das plantas frutíferas.
De forma semelhante, Pelizza et al. (2009) verificaram que a presença de vegetação
espontânea na linha de plantio afetou o crescimento de macieiras cultivar Galaxy, com
redução na altura e no diâmetro de plantas, evidenciando competição da vegetação de
cobertura do solo no período de crescimento das macieiras. De acordo com Matheis et al.
(2006) dentre as práticas de manejo mais utilizada nas áreas frutícolas destaca-se a roçada,
que controla o desenvolvimento das plantas daninhas por deixar uma camada de biomassa
protegendo a superfície do solo.
De acordo com os resultados desta pesquisa, uma alternativa interessante para impedir
o crescimento das plantas daninhas na linha de plantio de pereiras seria a colocação de
coberturas mortas. Dentre os materiais testados, a fibra de curauá e a capina mostraram-se ser
o método mais eficiente para a cultura da pereira, restringindo o desenvolvimento de todas as
espécies infestantes, seguido pela palha de azevém+milheto que trouxe também resultados
positivos em termos de redução do número de indivíduos de plantas daninhas. Embora, a
capina manual tenha proporcionado um bom controle das plantas daninhas, esta prática
demanda bastante mão-de-obra o que pode ser economicamente inviável.
A eficácia do uso de coberturas no manejo de plantas daninhas verificado neste
trabalho estão de acordo com os resultados relatados por Shribbs e Skroch (1986), em
experimento conduzido em pomar de macieiras, no qual observaram que houve maior
crescimento do diâmetro do tronco das plantas frutíferas e comprimento de suas brotações
com o uso de cobertura morta. Em outro trabalho, Dal Bó e Becker (1994), verificaram que o
uso de plantas de cobertura de inverno em parreirais com a cultivar Isabel com 16 anos de

47
idade, aumentou a produtividade de uvas em relação ao solo capinado, possivelmente por
melhorias nas características físico-químicas do solo.
Em relação aos resultados das análises de clorofila, verificou-se para a cv. Hosui uma
redução dos teores em função dos períodos avaliados , podendo este fato estar relacionado à
senescência das folhas. Além disso, as coberturas também afetaram o índice Spad, sendo que
os maiores valores foram encontrados nos tratamentos com coberturas de fibra de curauá,
azevém+milheto e a área capinada, demonstrando que as pereiras que foram submetidas a
uma menor competição com plantas daninhas, mantiveram-se fotossinteticamente mais ativas,
o que se refletiu diretamente no crescimento vegetativo.
6.3. Temperatura do solo em resposta à cobertura vegetal
De maneira geral, a temperatura do solo mostrou variação, porém com oscilações mais
atenuadas com o uso dos diferentes tipos de coberturas mortas no solo. O uso de cobertura
morta de solo proporciona temperaturas mais amenas e teores de umidade mais elevados do
que o solo nu (COSTA et al., 1997), o que possivelmente deve ter ocorrido no tratamento com
capina. Resultados semelhantes foram observados por Faria (1996), que relataram maiores
valores de temperatura do solo no tratamento sem cobertura e menores para as coberturas
testadas com cobertura natural.
Segundo Amado et al. (1990), os principais fatores ambientais que afetam a
temperatura do solo são a radiação solar direta e difusa, a duração diária do brilho solar e a
cobertura do solo, enquanto os fatores do solo são: o albedo, a condutância térmica, a
atividade biótica, a estrutura e a umidade do solo, dentre outros. Oliveira et al. (2005) também
verificaram oscilações mais expressivas quando o solo se encontrava sem cobertura.
Os resultados mostraram que a cobertura do solo e o nível de sombreamento
influenciam diretamente nas flutuações de temperatura do solo, quando terreno se encontra
sem cobertura facilita a exposição à insolação.
6.4. Efeito das diferentes coberturas sobre a macrofauna edáfica
A área que apresentou maior densidade de indivíduos da macrofauna do solo por
metro quadrado (indiv. m²) foi a cobertura com casca de arroz seguida da palhada de
azevém+milheto, número superior às demais coberturas estudadas, que de acordo com Silva
et al. (2006) pode estar relacionado ao ambiente mais favorável em termos de variedade de
micro-habitats e oferta de recursos explica. Entre os demais sistemas não houve diferenciação

48
para a densidade total de organismos edáficos. Ainda, pode-se observar que a cobertura com a
palhada de azevém+milheto teve a presença do grupo taxonômico coleóptera que são
consideradas indesejáveis na agricultura, porém podem ser benéficas para a fertilidade e a
física do solo (CORREIA e OLIVEIRA, 2005).
Em geral, observou-se a presença de grupos taxonômicos Hymenoptera (Formicidae) e
Isoptera (cupins), em todos os tipos de coberturas. Estes organismos são considerados
“engenheiros do ecossistema”, pelos diferentes efeitos nos processos que condicionam a
fertilidade do solo, pela regulação das populações microbianas responsáveis pela humificação
e mineralização e pela formação de agregados, que podem proteger parte da matéria orgânica
do solo de uma mineralização rápida, por meio de sua ação mecânica (SANTOS et al., 2008).
Os grupos Isoptera e Formicidae foram os grupos mais representativos nas coberturas
da palhada de azevém+milheto e casca de arroz. A presença de cobertura no solo contribui para
o acréscimo da disponibilidade de energia e requer a criação de novos habitats favoráveis à
colonização de organismos edáficos, o que pode beneficiar a sustentabilidade ecológica dos
sistemas de produção agroecológicos (SILVA et al., 2007).
6.5. Características químicas do solo
As diferentes coberturas são determinantes da quantidade e qualidade dos resíduos
orgânicos, influenciando diretamente na dinâmica da matéria orgânica (M.O) do solo. Como
as alterações nos estoques da M.O são lentas, os curtos períodos experimentais dificultam a
visualização do seu comportamento a médio e longo prazo (BAYER et al., 2000). O aumento
significativo da matéria orgânica do solo para as coberturas com casca de arroz ou fibra de
curauá em relação ás outras coberturas, pode estar associado, entre outros fatores, aos níveis
de resíduos depositados ao solo e para o tratamento com capina apresentar alto teor de MO,
pode estar relacionado à incorporação da cobertura natural pelo revolvimento do solo. O teor de
matéria orgânica do solo é bastante variável em função das condições de manejo adotado nos
sistemas agrícolas, porém, tende a um valor de equilíbrio de acordo com as características
peculiares de cada agroecossistema. Sendo considerada uma fonte de nutrientes às plantas,
influencia a infiltração, a retenção de água, a estruturação e a susceptibilidade do solo à
erosão, atuando também sobre outros atributos, tais como: capacidade de troca de cátions,
ciclagem de nutrientes, complexação de elementos tóxicos do solo e estimulação da biota do
solo (CONCEIÇÃO et al., 2005; UNGERA et al., 1991).
Na camada superficial, as concentrações médias de cálcio no solo foram superiores
nas coberturas com serragem de pinus, tela de sombreamento, casca de arroz, fibra de curauá

49
e capina. Com o aumento da profundidade também houve diferenças, sendo que a casca de
arroz e a fibra de curauá mantiveram-se com os maiores teores e o tratamento com capina
permaneceu com a quantidade menor, ou seja, com o mesmo padrão da camada superficial.
Esse fato indicou que o Ca2+
foi deslocado dos sítios de troca e movimentou-se ao longo do
perfil. Os valores médios dos teores de Ca2+
na camada superficial do solo decresceram em
alguns tipos de coberturas. No solo sobre diferentes coberturas, as variações existentes na
solução de solo podem ser decorrentes de variações do teor da matéria orgânica, lixiviação,
atividade microbiana e sazonalidade na absorção de nutrientes pela vegetação (MIRANDA et
al., 2006).
As concentrações de magnésio no solo se mantiveram semelhantes para todos os
tratamentos, exceto, para a cobertura com fibra de curauá que apresentou valor mais elevado.
Houve elevação do teor de magnésio do solo, principalmente na camada superficial reduzindo
com o aumento da camada. Este fato pode ser atribuído provavelmente, à absorção pelas
plantas e aos restos de material orgânico em vários estádios de decomposição, os quais,
possivelmente, retardaram a solubilização desse nutriente.
O potássio do solo apresentou diferenças significativas, com maior concentração para
a cobertura com azevém+milheto, em relação às outras formas de cobertura. No entanto na
camada de 0-10 cm, foi possível observar maiores teores desse nutriente. Provavelmente esse
fato este relacionado com a maior eficiência na reciclagem de nutrientes, já que com a
mineralização dos resíduos vegetais, os cátions são liberados na solução do solo nas formas
inorgânicas, podendo ser absorvidos novamente pelas raízes ou adsorvidos às cargas
negativas do solo dependentes de pH (MALUF et al., 2015). A menor lixiviação de NO3- com
íons acompanhantes causa maior acúmulo de K+ na camada superior do solo (CABRERA,
2000). Além disso, acredita-se que a diminuição nos teores de K+ no solo se deve ao maior
acúmulo de K nas plantas, como foi observado por Teixeira et al. (2002).
O zinco do solo foi significativamente mais elevado para as coberturas de
azevém+milheto e fibra de curauá na camada superficial não diferindo entre os tratamentos na
camada mais profunda. O zinco é um dos micronutrientes, cuja deficiência é relativamente
comum, principalmente por ser baixa a sua quantidade no solo (SENGIK, 2003). O alumínio,
ferro e manganês no solo foram os três elementos que não mostraram diferença significativa
entre os tratamentos aplicados (p ≤ 0,05). Em função das coberturas aplicadas era de se
esperar diferenças no teor no solo, principalmente para o ferro. Uma hipótese para explicar o
ocorrido é que a planta tenha absorvido o ferro de forma diferente entre os tratamentos.
6.6. Análise do teor mineral das folhas

50
Nos teores médios de nutrientes nas folhas não houve efeito significativo dos
tratamentos para a maioria dos macronutrientes, exceto para Mg, cujo teor foi inferior em
todos os tipos de coberturas. O que pode ser justificado pelo fato que o Mg é exigido e
absorvido pelas plantas frutíferas em quantidades inferiores ao Ca, sendo que a absorção de
Mg é reduzida pela concorrência com os cátions como o K, NH4+, Ca e Mn bem como o H
(MARSCHNER, 1995). Independente disso, os teores foliares de Mg ficaram todos abaixo da
faixa de valores tidas como adequada para a pereira, que por exemplo para cv. Rocha em
Portugal está entre 1,8 e 2,0 g kg-1
(LQARS, 2006). De forma semelhante, Stassen e North
(2005) verificaram que a pereira cv. Forelle, quando enxertada sobre o porta-enxerto anão
marmeleiro „A‟ (Cydonia oblonga), apresentou menores teores foliares Mg. A cultivar Hosui
apresentou teores de N nas folhas elevados, estando ligeiramente acima do normal (20 a 25,0g
kg-1
), de acordo com recomendação da CQFS-RS/SC (2004) para a cultura da pereira no
Brasil.
Em relação ao P, os tratamentos apresentaram teores foliares na faixa considerada
normal (1,5 a 3,0 g kg-1
), isso se deve ao teor de P no solo em nível adequado (CQFS-RS/SC,
2004). Os teores de K se apresentaram na faixa normal (12,00 a 15,00 g kg-1
) para todos os
tratamentos. Estes resultados se devem aos teores médios de K no solo (CQFS-RS/SC, 2004).
Botelho et al. (2010) ao estudarem os teores foliares de nutrientes em pereiras
verificaram que as cultivares Cascatense e Tenra apresentaram os teores de K nas folhas de
9,8 e 10,5 g kg-1
respectivamente, teores estes situados abaixo do normal (8,00 a 11,90 g kg-
1), mesmo realizando as adubações recomendadas de plantio e de formação utilizando-se
como fonte de K o cloreto potássio e com altos teores de K no solo. Este fato pode estar
relacionado ao porta-enxerto diferente, as plantas deste experimento são enxertadas em Pyrus
calleryana, mais vigoroso que o marmelereiro CP.
Os teores de Ca ficaram acima da faixa suficiente em todos os tratamentos, ou seja,
acima de 11,0 a 17,0 g kg-1
(CQFS-RS/SC, 2004). Quanto aos teores de S nas folhas, também
apresentaram valores acima da faixa considerada normal (1,8 a 2,0 g kg-1
).
Para os micronutrientes foram verificadas diferenças significativas entre tratamentos.
Os teores de Cu foram abaixo da faixa suficiente para os tratamentos testemunha (sem
cobertura) e com cobertura de serragem, tela de sombreamento ou casca de arroz, sendo
suficientes apenas no tratamento com capina e nas coberturas de azevém+milheto e fibra de
curauá. Já para os teores de Fe, os tratamentos com serragem, fibra de curauá e capina
apresentaram suficiência na faixa considerada normal (0 a 250 g kg-1
). Todos os tratamentos
apresentaram teores foliares de Mn abaixo do normal (20 a 100 mg kg-1
). No entanto para o

51
Zn, os tratamentos com a fibra de curauá e capina apresentaram teores abaixo do normal,
enquanto que os tratamentos sem cobertura (testemunha) ou com cobertura de
azevém+milheto, serragem, tela de sombreamento e casca de arroz apresentaram teores
normais, situando-se entre 20 a 100 mg kg-1
(BASSO et al., 2003; CQFS-RS/SC, 2004) No
estudo realizado por Botelho et al. (2010), sobre o estado nutricional de diferentes cultivares
de pereira nas condições de Guarapuava-PR, a cv. Cascatense apresentou os seguintes teores
foliares: 90,3 mg kg-1
de Fe, 92,0 mg kg-1
de Mn, 28,3 mg kg-1
de Zn, 10,0 mg kg-1
de Cu e
19,3 mg kg-1
de B.
Singh et al. (2005) também verificaram diferenças significativas entre 13 cultivares de
pereiras asiáticas, com teores variando para os seguintes micronutrientes Mn (112 a 135 mg
kg-1
), Fe (148 a 209 mg kg-1
), Zn (24 a 50 mg kg-1
) e Cu (9 a 18 mg kg-1
).
Efeitos na melhoria dos atributos do solo também foram observados com o uso das
coberturas mortas, com aumento da matéria orgânica nas coberturas com casca de arroz, fibra
de curauá e capina e teores de nutrientes como P, K, Zn, Ca, Mg e Cu quando comparados a
cobertura natural.

52
7. CONCLUSÕES
Os tratamentos mais eficientes na supressão das plantas daninhas na linha de plantio da
pereira, num pomar na fase de implantação, foram capina, fibra de curauá e palhada de
azevém+milheto.
As coberturas com vegetação natural e serragem de pinus na linha de plantio afetou o
crescimento da pereira, com redução na altura e no diâmetro. Fibra de curauá, casca de arroz,
tela de sombreamento, capina manual e palhada de azevém+milheto também contribuíram
para o maior desenvolvimento vegetativo das pereiras cv. Hosui e maiores teores de clorofila
nas folhas.

53
8. REFERÊNCIAS BIBLIOGRÁFICAS
ADEGAS, F.S. Manejo integrado de plantas daninhas. In: CONFERÊNCIA ANUAL DE
PLANTIO DIRETO, 2. Pato Branco. Resumo de palestras. Passo Fundo: Ed. Aldeia Norte,
p. 17-26, 1997.
ALMEIDA, F. S. Efeitos alelopáticos de resíduos vegetais. Pesquisa Agropecuária
Brasileira, Brasília, v.26, n.2, p. 221-236, 1991.
ALMEIDA, F.S. A alelopatia e as plantas. Londrina: IAPAR, 68 p., 1988.
ALVARENGA, R. C.; CABEZAS, W. A. L.; CRUZ, J. C.; SANTANA, D. P. Plantas de
cobertura de solo para sistema plantio direto. Informe Agropecuário, Belo Horizonte, v.22,
n.208, p.25-36, 2001.
ALVES, P. L. C. A.; PITELLI, R. A. Manejo ecológico de plantas daninhas. Informe
Agropecuário, Belo Horizonte, v.22, n.212, p.29-39, 2001.
AMADO, T. J. C.; COGO, N. P.; LEVIEN, R. Eficácia relativa do manejo do resíduo cultural
de soja na redução das perdas do solo por erosão hídrica. Revista Brasileira de Ciência do
Solo, Campinas, v. 13, n. 2, p. 251-257, 1989.
ANDERSON, J. M.; INGRAM, J. S. I. Tropical Soil Biological and Fertility: a handbook
of methods. 2.ed. Wallingford: CAB International, 221p., 1993.
Angiosperm Phylogeny Group - APG II. An update of the Angiosperm Phylogeny Group
classification for the orders and families of flowering plants: APG II. Botanical Journal of
the Linnean Society, n. 141, p. 399-436, 2003.
AYUB, R. A.; GIOPPO, M. A Cultura da pereira. In: ENCONTRO DE FRUTICULTURA
DOS CAMPOS GERAIS, 2. Ponta Grossa. Anais... Ponta Grossa: Universidade Estadual de
Ponta Grossa, 2009. v.1, p.25-33, 2009.
AZEVEDO, T. R., GALVANI, E. Ajuste do ciclo médio mensal horário da temperatura do
solo em função da temperatura do ar. Revista Brasileira de Agrometeorologia, Santa Maria,
n. 11, p. 123-130, 2003.
BALBINOT JR, A. A.; FLECK, N. G.; BARBOSA NETO, J. F.; RIZZARDI, M. A.
Características de plantas de arroz e a habilidade competitiva com plantas daninhas. Planta
daninha, Viçosa, v.21 (2), p.165-174, 2003.
BALBINOT JR, A.; BACKES, R.; ALVES, A.; OGLIARI, J.; FONSECA, J. Contribuição de
componentes de rendimento na produtividade de grãos em variedades de polinização aberta
de milho. Current Agricultural Science and Technology, v.11, n. 2, p.131-148, 2005.
BASSO, C.; FREIRE, C.J.S.; SUZUKI, A. Solos, adubação e nutrição. In: QUEZADA, A.C.;
NAKASU, B.H.; HERTER, F.G. Pêra: Produção. Brasília: Embrapa, p.55-67, 2003.
BAYER, C.; MIELNICZUK, J.; MARTIN-NETO, L. Efeito de sistemas de preparo e de
cultura na dinâmica da matéria orgânica e na mitigação das emissões de CO2. Revista
Brasileira Ciência do Solo, Viçosa, v. 24, p. 599-607, 2000.

54
BETTIOL NETO, J. E.; CHAGAS, E. A.; SANCHES, J.; PIO, R.; ANTONIALI, S.; CIA, P.
Production and postharvest quality of pear tree cultivars in subtropical conditions at eastern of
São Paulo state, Brazil. Ciência Rural, v. 44 n.10, p. 1740-1746, 2014.
BLANCO, H.G.; OLIVEIRA, D.A. Estudos dos efeitos da época de controle do mato sobre a
produção de citrus e a decomposição da flora daninha. Arquivos do Instituto Biológico, São
Paulo, v. 45, p.25-36, 1978.
BOTELHO, R.V.; AYUB, R.A.; MULLER, M.M.L. Somatória de horas de frio e de unidades
de frio em diferentes regiões do estado do Paraná. Scientia Agraria, Curitiba, v.7, n.1-2,
p.89-96, 2006.
BOTELHO, S. A.; RAVA, C. A.; LEANDRO, W. M.; DA SILVA COSTA, J. L.
SUPRESSIVIDADE INDUZIDA A Rhizoctonia solani Kühn PELA ADIÇÃO DE
DIFERENTES RESÍDUOS VEGETAIS AO SOLO. Pesquisa Agropecuária Tropical
(Agricultural Research in the Tropics), v. 31 n.1, 43-46, 2007.
BOTELHO, V. R.; MÜLLER, M. M. L.; BASSO, C.; SUZUKI, A. Estado nutricional de
diferentes cultivares de pereira nas condições edafoclimáticas de Guarapuava-PR. Revista
Brasileira de Fruticultura, Jaboticabal, v.32, n.3, p.884-891, 2010.
BRADY, N. C. Natureza e propriedades dos solos. 7. ed. Rio de Janeiro: Freitas
Bastos, 878 p., 1989.
BROWN, G. G.; FRAGOSO, C. El uso de la macrofauna edáfica en la agricultura del siglo
XXI: problemas y perspectivas. In: G. G. Brown, C.; Fragoso.; L. J. Oliveira (Eds.). O uso da
macrofauna edáfica na agricultura do século XXI: a importância dos engenheiros do solo.
Embrapa Soja Série Documentos, Londrina, n.224, p.11-19, 2003.
CAPISTRANO, U. Implantação da cultura do mirtilo. In: SEMINÁRIO NACIONAL SOBRE
FRUTICULTURA DE CLIMA TEMPERADO, 8 (Ed.) NUNES, E. C. Resumos...São
Joaquim, Epagri, p. 47-52, 2008.
CARVALHO, A. M.; AMABILE, R. F. Plantas condicionadoras de solo: interações
edafoclimáticas, uso e manejo. In: CARVALHO, A. M.; AMABILE, R. F. (Ed.). Cerrado:
adubação verde. Planaltina: Embrapa Cerrados, p. 143-163, 2006.
CARVALHO, J. E. B. de.; VARGAS, L. Manejo e Controle de Plantas Daninhas em
Frutíferas. In: VARGAS, L.; ROMAN, E. S. Manual de Manejo e Controle de Plantas
Daninhas. 1. ed. Bento Gonçalves: EMBRAPA/CNPUV, 652 p., 2004.
CARVALHO, J. E. B.; CALDAS, R. C.; CARDOSO, S. S.; COSTA NETO, A. O. Influência
das épocas de controle das plantas daninhas na produção de laranja “Pera”. Planta Daninha,
Viçosa, v. 11, n. 1/2, p. 49-54, 1993.
CARVALHO, L. B.; ALVES, P. L. C. A.; GONZÁLEZ-TORRALVA, F.; CRUZHIPOLITO,
H. E.; ROJANO-DELGADO, A. M.; DE PRADO, R.; GIL-HUMANES, J.; BARRO, F.;
LUQUE DE CASTRO, M. D. Pool of resistance mechanisms to glyphosate in Digitaria
insularis. Journal of Agricultural and Food Chemistry, Washington, v. 60, n. 2, p. 615-
622, 2012.
CHAVES, J. C. D.; CALEGARI, A. Adubação verde e rotação de culturas. Informe
Agropecuário, Belo Horizonte: EPAMIG, v.3, n.25, p.53-60. 2001.

55
CLARK, J. R.; MOORE, J. N. Southern Higbush blueberry response to mulch. American
Society for Horticultural Science, Alexandria, v. 1, n.1, p.52-54, 1991.
CONCEIÇÃO, P.C.; AMADO, T.J.C.; MIELNICZUK, J.; SPAGNOLLO, E. Qualidade do
solo em sistemas de manejo avaliada pela dinâmica da matéria orgânica e atributos
relacionados. Revista Brasileira de Ciência do Solo, Viçosa, v.29, p.777-788, 2005.
CORREIA, M. E. F.; ANDRADE, A.G.; FARIA, S. M. Sucessão das comunidades de
macroartrópodos edáficos em plantações de três leguminosas arbóreas, In: CONGRESSO
BRASILEIRO DE CIÊNCIAS DO SOLO, 26. 1997, Rio de Janeiro, RJ. Resumos..., Rio de
Janeiro, RJ, 1997.
CORREIA, M. E. F.; OLIVEIRA, L. C. M. Importância da fauna para a ciclagem de
nutrientes. In: AQUINO, A. M; ASSIS, R. L. (Eds.). Processos biológicos no sistema solo-
planta: ferramentas para a agricultura sustentável. Brasília: Embrapa Informação
Tecnológica, p. 18-29, 2005.
COSTA, J. T. A.; SILVA, L. A. da; MELO, F. I. O. Efeitos do turno de rega e cobertura
morta na cultura do alho na serra da Ibiapaba, Ceará: I. umidade e temperatura do solo.
Ciência Agronômica, Fortaleza, v. 28, n. 1/2, p. 70-84, 1997.
CQFS-COMISSÃO DE QUÍMICA E FERTILIDADE DO SOLO-RS/SC. Manual de
adubação e de calagem para os estados do Rio Grande do Sul e de Santa Catarina. 10.
ed. Porto Alegre: SBCS-Núcleo Regional Sul, 400 p, 2004.
DAL BÓ, M.A.; BECKER, M. Avaliação de sistemas de manejo do solo para a cultura da
uva. Pesquisa Agropecuária Brasileira, Brasília, v.29, n.2, p.263- 266, 1994.
DAROLT, M. R.; SKORA NETO, F. Sistema de plantio direto em agricultura
orgânica. Revista Plantio Direto, v. 70, p. 28-31, 2002.
DERKSEN, D. A.; ANDERSON, R. L.; BLACKSHAW, R. E.; MAXWELL, B. Weed
dynamics and management strategies for cropping systems in the northern Great
Plains. Agronomy Journal, v. 94 n. 2, p.174-185, 2002.
DIXON, J.; ELMSLY, T.; FIELDS, F.; DEREK, S.; MANDEMAKER, A.J.; GREENWOOD,
A.C.; PAK, H.; CUTTING, J. Mulching – is it worth it? In: NEW ZEALAND AND
AUSTRALIA AVOCADO GROWER‟S CONFERENCE, 5., Tauranga. Proceedings…
Tauranga, 2005. p. 1-7, 2005.
DIXON, J.; ELMSLY, T.; FIELDS, F.; SMITH, D.B.; MANDEMAKER, A.J.;
GREENWOOD, A.C; PAK, H. What, when, where and how much mulch should be applied
to „Hass‟ avocado trees in the Western Bay of Plenty. New Zealand Avocado Grower’s
Association Annual Research Report, Tauranga, v. 7, p. 49-72, 2007.
EMBRAPA. EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA. A cultura da
pêra. Brasília: Embrapa Clima Temperado. Informação Tecnológica. 58 p. 2007.
EMBRAPA. EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA. Centro
Nacional de Pesquisa de Solos. Manual de métodos de análise de solo. 2. ed. Rio de Janeiro:
Embrapa Solos. 212 p. 1997.

56
EMBRAPA. EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA. Manual de
análises químicas de solos, plantas e fertilizantes. 2.ed. Brasília, Informação Tecnológica.
628 p. 2009.
ERASMO, E.A.L.; PINHEIRO, L.L.A.; COSTA, N.V. Levantamento fitossociológico das
comunidades de plantas infestantes em áreas de produção de arroz irrigado cultivado sob
diferentes sistemas de manejo. Planta Daninha, Viçosa-MG, v.22, n.2, p.195-201, 2004.
ESPÍNDOLA, L. Producción de arándanos na California. Revista Fruticola, v. 28, n. 3, p.
104-114, 2007.
FACHINELLO, J. C.; PASA, M. S. Porta enxertos na cultura da Pereira. In: III Reunião
técnica da cultura da pereira. Lages – SC, p.70-77, 2010.
FAO. FOOD AGRICULTURE ORGANIZATION OF THE UNITED NATIONS. Faostat
Database Prodstat. Disponível em: <http://faostat.fao.org/>. Acesso em: 11 dezembro 2016.
FAO. FOOD AND AGRICULTURE ORGANIZATION OF THE UNITED NATIONS -
FAO - CA Adoption Worldwide. Disponível em: < http://www.fao.org/ag/ca/6c.html>
Acesso em 23 setembro 2016.
FAORO, I. D. Biologia reprodutiva da pereira japonesa (pyrus pyrifolia var. culta) sob o
efeito do genótipo e do ambiente. Tese (Doutorado em Ciências) – Universidade Federal de
Santa Catarina, 2009.
FAORO, I. D.; ORTH, A. I. A cultura da pereira no Brasil. Revista Brasileira de
Fruticultura, Jaboticabal, v. 32, n. 1 p. 001-342, 2010.
FAORO, I. D.; ORTH, A. I. Qualidade de frutos da pereira-japonesa colhidos em duas regiões
de Santa Catarina, Brasil. Revista Brasileira de Fruticultura, Jaboticabal, v. 32, n. 1, p. 308-
315, 2010.
FARIA, J.C. Dinâmica de água, comportamento térmico e selamento de um Podzólico
Vermelho-Amarelo, em relação ao controle de plantas invasoras. Dissertação (Mestrado
em Fitotecnia) – Universidade Federal de Viçosa, Programa de Pós-Graduação em Fitotecnia,
1996.
FAVARO, C. G. influência da área de controle de plantas daninhas na laranjeira ‘Pera
Rio’. Dissertação (Mestrado em Ciências Agrárias e Veterinárias) – Unesp, Câmpus de
Jaboticabal, 2016.
FERREIRA, D. F. Sisvar: a computer statistical analysis system. Ciência Agrotecnologia.
Lavras, v.35, p.1039-1042, 2011.
FIDEGHELI, C. Il pero, In.: Botânica, Origine ed Evoluzione. ART Servizi Editoriali, p. 3-
16. 2007.
FLORES, R. A.; DALL‟AGNOL, M.; NABINGER, C.; MONTARDO, D. P. Produção de
forragem de populações de azevém anual no Rio Grande do Sul. Revista Brasileira de
Zootecnia, Viçosa, v. 37, n. 7, p. 1168-1175, 2008.
GIAYETTO, A.; VILLARREAL, P. Manual para el productor y el empacador. In.:

57
Enfermedades y su manejo en el cultivo. p.96-103. 2010.
GLIESSMAN, S. R. Agroecologia: processos ecológicos em agricultura sustentável. 3ª.
Ed. Porto Alegre: Editora da UFRGS, 653p. 2005.
GOH, K.M.; PEARSON, D.R.; DALY, M.J. Effects of apple orchard production systems on
some important soil physical, chemical and biological quality parameters ture, Bicester, v.18,
n.3, p.269-292, 2000.
GOMES JR, F. G.; CHRISTOFFOLETI, P. J. Biologia e manejo de plantas daninhas em áreas
de plantio direto. Planta Daninha, Viçosa-MG, v. 26, n. 4, p. 789-798, 2008.
GRAVENA, R.; RODRIGUES, J. P. R. G.; SPINDOLA, W.; PITELLI, R. A.; ALVES, P. L.
C. A. Controle de plantas daninhas através da palha de cana-de-açúcar associada à mistura
dos herbicidas trifloxysulfuron sodium + ametrina. Planta Daninha, Viçosa-MG, v. 22, n. 3,
p. 419-427, 2004.
HOUSTON, D.F. Rice, chemistry and technology. American Association of Cereal Chemical.
St. Paul. Minnesota, USA, p. 301-351, 1972.
IAPAR. Instituto Agronômico do Paraná. Disponível em: <http://www.iapar.br. > Acesso em
outubro de 2010.
IBRAF. Instituto Brasileiro de Frutas. Frutas frescas – Importação. Disponível
em:<http://www.ibraf.org.br/estatisticas/Exportação/Comparativo_das_Exportações_Brasileir
as_de_Frutas_frescas_2012-2013.pdf>. Acesso em: 10 dez. 2014.
INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA - IBGE. Produção
Agrícola Municipal. Disponível em: <http://www.sidra.ibge.gov.br>. Acesso em: 11 out.
2015.
KISSMANN K.G.; GROTH, D. Plantas infestantes e nocivas: Tomo I. 2 ed., São Paulo.
824p., 1997.
KISSMANN K.G.; GROTH, D. Plantas infestantes e Nocivas: Tomo III. 2. ed., São Paulo.
722p., 2000.
KISSMANN K.G.; GROTH, D.Plantas infestantes e Nocivas: Tomo II. 2. ed., São Paulo.
978p., 1999.
KOZINSKI, B. Influence of Mulching and Nitrogen Fertilization Rate on Growth and Yield
of Highbush Blueberry. In: Proceedings on Vaccinium Culture, 8 (Ed) FONSECA, L. L. da et
al. Acta Horticulturae, Wageningen, v. 715, p. 231-235, 2006.
KUMAR,K., NEGI, R.K. The pear In.: Origin and taxonomy, 1ed. Índia, IBDC publisher,
p.39-45. 2010.
KUVA, M. A.; PITELLI, R. A.; SALGADO, T. P.; ALVES, P. L. C. A. Fitossociologia de
comunidades de plantas daninhas em agroecossistema cana-crua. Planta Daninha, Viçosa-
MG, p. 501-511, 2007.

58
LEITE JÚNIOR, R.P.; MOHAN, S.K. Integrated management of citrus bacterial canker
disease caused by Xanthomonas campestris pv. citri in the State of Paraná, Brazil. Crop
protection, v.9, p.3-7, 1990.
LEVY NETO, F.; PARDINI, L. C. Compósitos estruturais: ciência e tecnologia. 1. ed. São
Paulo: Edgard Blücher. 2006.
LOPES ASSAD, M. L. Papel da macrofauna edáfica de invertebrados no comportamento de
solos tropicais. In CONGRESSO BRASILEIRO DE CIÊNCIA DO SOLO, 26, 1997, Rio de
Janeiro. Anais... Rio de Janeiro: EMBRAPA-CNPS, 1997.
LORENZI, H. Manual de identificação e controle de plantas daninhas: plantio direto e
convencional. 7.ed. Nova Odessa: Instituto Plantarum, 383p., 2014.
LORENZI, H. Plantas daninhas do Brasil: terrestres, aquáticas, parasitas e tóxicas. 4.ed.,
Nova Odessa, 640p., 2008.
LQARS, Manual de fertilização das culturas. INIAP/LQARS. Lisboa. 2006.
MARANGONI, B.; SCUDELLARI, D.; TACLIAVINI, M. Relazione tra nutrizione azotata e
metabolismo delle piante. Rivista di Frutticoltura e di Ortofloricoltura, v. 57, p. 7-8, 1995.
MACHADO, B.D. Aspectos vegetativos e produtivos de cultivares copa de pereira
europeia com combinações de porta enxertos. Dissertação (Mestrado em Produção
Vegetal) – Universidade do estado de Santa Catarina – UDESC. Lages, SC. 2011.
MACHADO, B. D.; RUFATO, A. De. R.; MARCONFILHO, J. L. Cultivares de pereiras
europeias. In: RUFATO, L.; KRETZSCHMAR, A. A.; BOGO, A. A cultura da Pereira.
Florianópolis: DIOESC, Série Fruticultura, p. 150 - 171, 2012.
MACIEL, C. D. G.; POLETINE, J. P.; OLIVEIRA NETO, A. M.; GUERRA, N.;
JUSTINIANO, W. Levantamento fitossociológico de plantas daninhas em cafezal orgânico.
Bragantia, Campinas, v. 69, p. 631-636, 2010.
MAACK, R. Geografia física do Estado do Paraná. 3º ed. Curitiba: Imprensa Oficial, 440
p. 2002.
MARSCHNER, H. Mineral nutrition of higher plants. San Diego: Academic Press, 889p.,
1995.
MATHEIS, H.A.S.M.; AZEVEDO, F. A.; VICTORIA FILHO, R. Adubação verde no manejo
de plantas daninhas na cultura de citros. Laranja, Cordeirópolis, v.27, n.1, p.101-110, 2006.
MEDINA, J. C. Plantas Fibrosas da Flora Mundial, Instituto Agronômico de Campinas, p.
787-792, 1959.
MALUF, H. J. G. M.; SOARES, E. M. B.; SILVA, I. R. D.; NEVES, J. C. L.; SILVA, M. F.
D. O. NUTRIENT AVAILABILITY AND RECOVERY FROM CROP RESIDUES IN SOIL
WITH DIFFERENT TEXTURES. Revista Brasileira de Ciência do Solo, v. 39, n. 6, p.
1690-1702, 2015.

59
MIRANDA, J.; COSTA, L. M. D.; RUIZ, H. A.; EINLOFT, R. Chemical composition of soil
solution under different land cover and soluble organic carbon in water from small
creeks. Revista Brasileira de Ciência do Solo, v. 30, n.4, p. 633-647, 2006.
MOJTAHEDI, H.; SANTO, G. S.; WILSON, J. H.; HANG, A. N. Managing Meloidogyne
chitwoodi on potato with rapeseed as green manure. Plant disease, v.77, n. 1, p.42-46, 1993.
MONQUERO, P. A.; AMARAL, L. R.; INÁCIO, E. M.; BRUNHARA, J. P.; BINHA, D. P.;
SILVA, P. V.; SILVA, A. C. Effect of green fertilizers on the suppression of different species
of weeds. Planta Daninha, Viçosa, v.27, n.1, p. 85-95, 2009.
MORAES, Y.J.B. Forrageiras: conceito, formação e manejo. Guaíba: Agropecuária,
215p., 1995.
MOORE-GORDON, C.; COWAN, A.K.; WOLSTENHOLME, B.N. Mulching of avocado
orchards to increase Hass yield and fruit size and boost financial rewards a three-year
summary of research findings. South African Avocado Growers' Association Yearbook,
Pietermaritzburg, v. 20, p. 46-49, 1997.
MUELLER-DOMBOIS, D.; H. ELLENBERG. Aims and Methods of Vegetation Ecology.
Wiley, New York. 547 p., 1974.
NAKASU, B.H.; LEITE, D.L. Indicação de porta enxerto e cultivares de pereira para o sul do
Brasil. Hortisul, Pelotas, v.1, n.2, p.20-24, 1990.
NUNES, J. S. Atributos Biológicos do Solo de Áreas em Diferentes Níveis de Degradação
no Sul do Piauí. Dissertação (Mestrado em Agronomia - Solos e Nutrição de Plantas) -
Universidade Federal do Piauí, Campus Professora Cinobelina Elvas, Bom Jesus - PI, 2010.
OLIVEIRA, C. A. P.; SOUZA, C.M. de. Influência da cobertura morta na umidade,
incidência de plantas daninhas e de broca-do-rizoma (Cosmopolites sordidus) em um pomar
de bananeiras (Musa ssp.). Revista Brasileira de Fruticultura, Jaboticabal, v. 25, n. 2, p.
345-347, 2003.
ODUM, E.P. Ecologia. Rio de Janeiro: Guanabara Koogan, 434 p. 1988.
OLIVEIRA, M. A. D.; GOMES, C. F. F.; PIRES, E. M.; MARINHO, C. G. S.; DELLA
LUCIA, T. M. C. Environmental bioindicators: insects as a tool for biodiversity
monitoring. Revista Ceres, v. 61, p. 800-807, 2014.
OLIVEIRA, M. L. D.; RUIZ, H. A.; COSTA, L. M. D.; SCHAEFER, C. E. Flutuações de
temperatura e umidade do solo em resposta a cobertura vegetal. Revista Brasileira de
Engenharia Agrícola e Ambiental, v. 9, n. 4, p. 535-539, 2005.
OLIVEIRA, O. R.; PERESSUTI, R. A.; SKALITZ, R.; ANTUNES, M. C.; LUIZ ANTONIO
BIASI, L. A.; ZANETTE, F. Quebra de dormência de pereira „hosui‟ com uso de óleo mineral
em dois tipos de condução. Revista Brasileira de Fruticultura, Jaboticabal, v.30, n.2, p.409-
413, 2008.
PELIZZA, L.; FERRARI, A. Anhedonia in schizophrenia and major depression: state or
trait. Ann Gen Psychiatry, v. 8, n. 1, p. 22, 2009.

60
PELIZZA, T. R.; MAFRA, A. L.; AMARANTE, C. V. T.; NOHATTO, M. A.; VARGAS, L.
Coberturas do solo e crescimento da macieira na implantação de um pomar em sistema
orgânico de produção. Revista Brasileira de Fruticultura, Jaboticabal, v. 31, n. 3, p. 739-
748, 2009.
PEREIRA, D. C.; GRUTZMACHER, P.; BERNARDI, F. H,.; MALLMANN, L. S.; COSTA,
L. A. M.; COSTA, M. S. S. M. Produção de mudas de almeirão e cultivo no campo, em
sistema agroecológico. Revista Brasileira de Engenharia Agrícola e Ambiental, v. 16, p.
1100-1106, 2012.
PINHEIRO, L. B. A. Estudo da macrofauna de solos cultivados com cana-de-açúcar, sob
diferentes manejos de colheita – crua e queimada. Dissertação (Mestrado em Agronomia –
Ciência do Solo) – Universidade Federal Rural do Rio de Janeiro, 1996.
PIO, R.; CHAGAS, E. A.; BARBOSA, W.;SIGNORINI, G.; ENTELMANN, F.A.;
FIORAVANÇO, J. C.; FACHINELLO, J. C.; BIANCHI, V.J. Desenvolvimento de 31
cultivares de marmeleiro enxertadas no porta-enxerto „Japonês‟. Revista Brasileira de
Fruticultura, Jaboticabal, v. 30, n.2, p. 466-470, 2008.
PITELLI, R. A. Interferência de plantas daninhas em culturas agrícolas. Informativo
Agropecuário, v. 11, n. 129, p. 16-27, 1985.
PITELLI, R. A.; DURIGAN, J. C. Ecologia das plantas daninhas no sistema plantio direto. In:
ROSSELLO, R. D. Siembra directa en el cono sur. Montevideo: PROCISUR, p. 203-210,
2001.
PITELLI, R. A.; PITELLI, R. L C. M. Biologia e ecofisiologia das plantas daninhas. In:
VARGAS, L.; ROMAM, E. S. (Eds.). Manual de manejo e controle de plantas daninhas.
Bento Gonçalves: Embrapa Uva e Vinho, p. 29-56, 2004.
PITELLI, R.A. Competição e controle de plantas daninhas em áreas agrícolas. IPEF, v.4,
n.12, p.25-35, 1987.
PITOL, C. Espécies vegetais para safrinha e inverno visando cobertura do solo. Maracaju:
Fundação MS, 6 p., 1993.
RAGOZO, C. R.; LEONEL, S.; CROCCI, A. J. Adubação verde em pomar cítrico. Revista
Brasileira de Fruticultura, Jaboticabal, v. 28, n. 1, p. 69-72, 2006.
RESENDE, F. V.; SOUZA, L. S. D.; OLIVEIRA, P. S. R. D.; GUALBERTO, R. Efficiency
of mulching on soil moisture and temperature, weed control and yield of carrot in summer
season. Ciência e Agrotecnologia, Lavras, v. 29, n. 1, p. 100-105, 2005.
RICE, E. L. Allelopathy. 2.ed. New York: Academic, 422 p., 1984.
RIZZARDI, M. A.; FLECK, N. G.; VIDAL, R. A.; MEROTTO, A.; AGOSTINETTO, D.
Competição por recursos do solo entre ervas daninhas e culturas. Ciência Rural, v.31, n.4,
p.707-714, 2001.
ROSA, C. C. DA.; DIAS, C. M. G.; K, B. Estudo das propriedades físicas da fibra de curauá.
In: XL Congresso Brasileiro de Educação em Engenharia, Belém do Pará, 2012.

61
ROUSH, M. L.; RADOSEVICH, S. R. Relationships between growth and competitiveness of
four annual weeds. Journal of Applied Ecology, v. 22, p. 895-905, 1985.
RUFATO, L.; MARCON FILHO, J. L.; MARODIN, G. A. B.; KRETZSCHMAR, A. A.;
MIQUELLUTI, D. J. Intensidade e épocas de poda verde em pereira'Abate Fetel'sobre dois
porta-enxertos. Revista Brasileira de Fruticultura, Jaboticabal, v. 34, n. 2, p. 475-481,
2012.
RUFATO, L.; DE ROSSI A.; GIACOBBO C. L. and FACHINELLO, J. C. Vegetative
propagation of seven quince cultivars for utilization as pear rootstocks in Brazil. Acta
Horticulturae, v. 658, p. 667- 671, 2004.
RUIZ-CAMACHO, V, N. E.; PANDO, A.; DUBS, T.D.F.; LAVELLE, P. Indicateurs
synthétiques de la qualité du sol. Etude et Gestion des Sols, v. 16, p. 323-338, 2009.
SANTOS, F. R. S. Desenvolvimento e aplicação de compósitos à base de matriz polimé-
rica reforçado com fibras de curauá (Ananás Erectifolius) e resíduos de madeiras
amazônicas. Dissertação (Mestrado) – Mestrado em Engenharia Civil, Universidade Federal
do Amazonas, 2013.
SAMPAIO, D. B.; ARAUJO, A. S. F.; SANTOS, V. B. Avaliação de indicadores biológicos
de qualidade do solo sob sistemas de cultivo convencional e orgânico de frutas. Ciência e
Agrotecnologia, v.32, n.2, p.353-359, 2008.
SANTOS, G. G.; SILVEIRA, P. M.; MARCHÃO, R. L.; BECQUER, T.; BALBINO, L. C.
Macrofauna edáfica associada a plantas de cobertura em plantio direto em um latossolo
vermelho do cerrado. Pesquisa Agropecuária Brasileira, v. 43, n. 1, p. 115-122, 2008.
SCHICK, J.; BERTOL, I.; BATISTELA, O. Erosão hídrica em Cambissolo Húmico
alumínico submetido a diferentes sistemas de preparo e cultivo do solo – I: Perdas de solo e
água. Revista Brasileira de Ciência do Solo, v.24, n.2, p.427-436, 1999.
SEDIYAMA, M. A.; SANTOS, M. R.; VIDIGAL, S. M.; SALGADO, L. T. Produtividade e
exportação de nutrientes em beterraba cultivada com cobertura morta e adubação
orgânica. Revista Brasileira de Engenharia Agrícola e Ambiental, v.15, n.9, p.883-890,
2010.
SENGIK, E. S. Os macronutrientes e os micronutrientes das plantas. 22 p., 2003.
Disponível em: <http://www.nupel.uem.br/nutrientes-2003.pdf> Acesso em maio 2017.
SHRIBBS, J.M.; SKROCH, W.A. Influence of 12 ground cover systems on young (cultivar)
Smoothee Golden Delicious apple (Malus domestica) trees: I. Growth. Journal of the
American Society for Horticultural Science, v.111, n.4, p.525-528, 1986.
SPIASSI, Ariane; FORTES, Andréa.M.T; PEREIRA, Dércio.C; SENEM, Jaqueline;
TOMAZONI, Dayana. Alelopatia de palhadas de coberturas de inverno sobre o crescimento
inicial de milho. Semina: Ciências Agrárias, Londrina, v.32, n.2, p.577-582, 2011.
SILVA, A.A.; SILVA, J.F. Tópicos em manejo de plantas daninhas. Editora UFV. 1ª ed.
367 p., 2007.
SILVA, M. B.; KLIEMANN, H. J.; DA SILVEIRA, P. M.; LANNA, A. C. Atributos

62
biológicos do solo sob influência da cobertura vegetal e do sistema de manejo. Pesquisa
Agropecuária Brasileira, v. 42, n.12, p.1755-1761, 2007.
SILVA, R.F.; AQUINO, A.M.; MERCANTE, F. M.; GUIMARÃES, M. F. Macrofauna
invertebrada do solo sob diferentes sistemas de produção em Latossolo da Região do Cerrado.
Pesquisa Agropecuária Brasileira, v. 41, p. 697-704, 2006.
SILVEIRA, E. R. DIVERSIDADE E PAPEL FUNCIONAL DA MACROFAUNA DO
SOLO NA INTEGRAÇÃO LAVOURA-PECUÁRIA. Revista Técnico-Científica, n. 4,
2016.
SINGH, M.; ACHHIREDDY, N.R. Influence of Lantana on growth of various citrus
rootstooks. Hortscience, v.22 n.3, p.385-386, 1987.
SINGH, T.; SANDHU, A.S.; SINGH, R.; DHILLON W.S. Vegetative and fruiting behavior
of semi-soft pear strains in relation to nutrient status. Acta Horticulturae, Wageningen,
n.696, p.289-293, 2005.
SOSNA, I.; KORTYLEWSKA, D. Estimation of Interstock and Intermediate Stock
Usefulness for Summer Pear Cvs. Budded on Two Rootstocks Journal of Horticultural
Research, v.21, n.1, p.79-82, 2013.
SOUZA V.C., LORENZI H. Botânica sistemática: guia ilustrado para identificação das
famílias de angiospermas da flora brasileira, baseado em APG II. Nova Odessa:
Plantarum, 2005.
SOUZA, J. L.; RESENDE, P. Manual de horticultura orgânica. Viçosa: Aprenda Fácil,
2003.
SKORA NETO, F. Manejo de plantas daninhas. Plantio direto: pequena propriedade
sustentável, 1998.
TAYLORSON, R. B.; BORTHWICK, H. A. Light filtration by foliar canopies: significance
for light-controlled weed seed germination. Weed Sciense., v. 17, n. 1, p. 48-51, 1969.
TEIXEIRA, L.A.J.; SPIRONELLO, A.; FURLANI, P.R.; SIGRIST, J.M.M. Parcelamento da
adubação NPK em abacaxizeiro. Revista Brasileira de Fruticultura, Jaboticabal, v.24, n.1,
p. 219-224, 2002.
TEIXEIRA, R. S. Efeito das fibras de curauá e de polipropileno no desempenho de
compósitos cimentícios produzidos por extrusão. Tese (Tese de Doutorado) - Universidade
de São Paulo, 2015.
TELLES, C. A. Compatibilidade e crescimento de mudas de pessegueiro interenxertadas
com ameixeiras, damasqueiro e cerejeira. Dissertação (Mestrado em Agronomia),
Faculdade de Ciências Agronômicas, Universidade Federal do Paraná, Curitiba, 2005.
UNGERA, P.W.; STEWARTA, B.A.; PARRB, J.F.; SINGHC, R.P. Crop residue
management and tillage methods for conserving soil and water in semi-arid regions. Soil
Tillage Research, v.20, p.219-240, 1991.

63
VARGAS, L.; BERNARDI, J. Manejo de plantas daninhas na produção orgânica de
frutas. Embrapa Uva e Vinho, 2003.
VARGAS, L.; MORAES, R. M. A.; BERTO, C. M. Herança da resistência de azevém
(Lolium multiflorum) ao glyphosate. Planta Daninha, v. 25, n. 3, p. 567-571, 2007.
VARGAS, L.; OLIVEIRA, O. L. P. Manejo da vegetação e cobertura. In: SISTEMA de
produção de uvas rústicas para processamento em regiões tropicais do Brasil. Bento
Gonçalves: Embrapa Uva e Vinho, 2005. Disponível em: <https://www.embrapa.br/uva-e-
vinho/>. Acesso em: outubro 2016.
VERÍSSIMO, V. Porta-enxertos para pereira (Pyrus sp.): implicações sobre a
dormência, biologia floral e conteúdo de carboidratos. Tese (Doutorado em Ciências)-
Universidade Federal de Pelotas, Programa de Pós-Graduação em Agronomia, 2008.
VOLL, E.; GAZZIERO, D. L. P.; BRIGHENTI, A. M.; ADEGAS, F. S.; GAUDÊNCIO, C.
D. A.; VOLL, C. E. A dinâmica das plantas daninhas e práticas de manejo. Embrapa
Soja. 2005.
ZENI, J. A.; SNYDER-MACKLER, L. Early postoperative measures predict 1-and 2-year
outcomes after unilateral total knee arthroplasty: importance of contralateral limb
strength. Physical therapy, v. 90, n. 1, p. 43-54, 2010.
SWIFT, M. J.; HEAL,O. W.; ANDERSON, J. M. Decomposition in terrestrial ecosystems.
Oxford: Blackwell, 372 p., 1979.
WALSH, B.D.; SALMINS, S.; BUSZARD, D.J.; MACKENZIE, A.F. Impact of soil
management systems on organic dwarf apple orchards and soil aggregate stability, bulk
density, temperature and water content. Canadian Journal of Soil Science, v.76, n.2, p.203–
209, 1996.
WANDER, P.R. Utilização de resíduo de madeira e lenha como alternativas de energia
renováveis para o desenvolvimento sustentável da região nordeste do Estado do Rio
Grande do Sul. Tese (Doutorado em Engenharia Mecânica) – Universidade Federal do Rio
Grande do Sul, 2001.
WERTHEIM, S. J. Pruning. In: Fundamentals of temperate zone tree fruit production. Leiden,
Holanda: Backhuys Publishers, p.176-189, 2005.
WINK, C.; GUEDES, J. V. C.; FAGUNDES, C. K.; ROVEDDER, A.P. Insetos edáficos
como indicadores da qualidade ambiental. Revista de Ciências Agroveterinárias, v. 4, p. 60-
71, 2005.
Recommended

















