
UNIVERSIDADE FEDERAL DE MATO GROSSO
FACULDADE DE AGRONOMIA, MEDICINA VETERINÁRIA E
ZOOTECNIA
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS VETERINÁRIAS
SOROPREVALÊNCIA E FATORES DE RISCO PARA
Ehrlichia canis E Rickettsia spp. EM CÃES DA REGIÃO DO
PANTANAL DE MATO GROSSO, BRASIL
Andréia Lima Tomé Melo
CUIABÁ – MT
2011

UNIVERSIDADE FEDERAL DE MATO GROSSO
FACULDADE DE AGRONOMIA, MEDICINA VETERINÁRIA E
ZOOTECNIA
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS VETERINÁRIAS
SOROPREVALÊNCIA E FATORES DE RISCO PARA
Ehrlichia canis E Rickettsia spp. EM CÃES DA REGIÃO DO
PANTANAL DE MATO GROSSO, BRASIL
Autor (a): Andréia Lima Tomé Melo
Orientador: Prof. Dr. Daniel Moura de Aguiar
Co-Orientador: Prof. Dr. Luciano Nakazato
CUIABÁ – MT
2011
Dissertação apresentada ao Programa de Pós-graduação em Ciências Veterinárias, área de concentração: Medicina Veterinária, da Faculdade de Agronomia, Medicina Veterinária e Zootecnia, da Universidade Federal de Mato Grosso para a obtenção do título de Mestre em Ciências Veterinárias.

FICHA CATALOGRÁFICA
M528s Melo, Andréia Lima Tomé
Soroprevalência e fatores de risco para Ehrlichia canis e
Rickettsia spp. em cães da região do Pantanal de Mato Grosso,
Brasil / Andréia Lima Tomé Melo. – 2011.
95 f. : il. ; color. ; 30 cm.
Orientador: Prof. Dr. Daniel Moura de Aguiar.
Co-orientador: Prof. Dr. Luciano Nakazato.
Dissertação (mestrado) – Universidade Federal de Mato
Grosso, Faculdade de Agronomia, Medicina Veterinária e
Zootecnia, Pós-graduação em Ciências Veterinárias, Área de
Concentração: Medicina Veterinária, 2011.
Bibliografia: f. 80-94.
1. Cães – Doenças. 2. Carrapato – Cães. 3. Parasitologia
veterinária. 4. Cães – Parasitos - Soroprevalência. I. Título.
CDU – 619:616.993:636.7(043.3)
Ficha elaborada por: Rosângela Aparecida Vicente Söhn – CRB-1/931


FOLHA DE APROVAÇÃO
AUTOR (A): MELO, Andréia Lima Tomé
TÍTULO: SOROPREVALÊNCIA E FATORES DE RISCO PARA Ehrlichia canis E
Rickettsia spp. EM CÃES DA REGIÃO DO PANTANAL DE MATO GROSSO,
BRASIL
Aprovada em 25 de Fevereiro de 2011.
BANCA EXAMINADORA:
______________________________________________
Prof. Dr. Daniel Moura de Aguiar
(Departamento de Clínica Médica Veterinária/FAMEV/UFMT)
_____________________________________________
Prof. Drª. Valéria Régia Franco Sousa
(Departamento de Clínica Médica Veterinária/FAMEV/UFMT)
_____________________________________________
Prof. Dr. Marcelo Bahia Labruna
(Universidade de São Paulo – USP)
Dissertação apresentada ao Programa de Pós-graduação em Ciências Veterinárias, da Universidade Federal de Mato Grosso para a obtenção do título de Mestre em Ciências Veterinárias.

DEDICATÓRIA
Dedico este trabalho aos meus pais João e
Cristina, que me deram a vida e amor
incondicional, buscando sempre me
proporcionar uma educação exemplar,
constituída de princípios morais e éticos, que
moldaram o meu caráter e transformaram-me
no que sou hoje. Vocês me proporcionaram
todas as oportunidades possíveis para que eu
pudesse galgar novos ideais e realizar todos os
meus sonhos. Perdoem-me pelas ausências,
mas saibam que vocês, estão sempre junto a
mim, dentro do meu coração.
Dedico também à minha irmã Verônica, pelo
carinho e amor fraternal.

AGRADECIMENTO ESPECIAL
Ao meu orientador Daniel Moura de Aguiar, por toda paciência, atenção, incentivo e
amizade. Muito obrigada por sempre acreditar no sucesso desse experimento e por
despertar em mim a instigante vontade de trabalhar na área da pesquisa científica,
pela qual desenvolvi grande afeição. Obrigada também pela transmissão de tantos
ensinamentos, que pela maneira clara e didática com que foram passados,
contribuíram grandemente para a minha formação na pós-graduação.

AGRADECIMENTOS
Agradeço a Deus, por sempre me iluminar, concedendo a mim perseverança
e saúde para desenvolver esse experimento.
Nesses dois anos de Mestrado, pude conviver com pessoas maravilhosas,
que direta ou indiretamente contribuíram para a realização desse projeto. Foram
momentos inesquecíveis, de muita dedicação, trabalho e disciplina, para conseguir
concluir com êxito esse experimento. Portanto, dedico esses agradecimentos a
todos vocês:
- Professora Valéria Régia, por ceder o microscópio para leitura das lâminas de
imunofluorescência;
- Aos amigos do Laboratório de Microbiologia, em especial aos professores Luciano
Nakazato e Valéria Dutra, por auxiliarem em tudo que foi necessário;
- À professora Adriane e a todos do Laboratório de Patologia Clínica;
- Ao Laboratório de Parasitologia, na pessoa do Professor Richard (Tiozão), por toda
ajuda na identificação dos ectoparasitos e por ceder espaço para a realização das
análises sorológicas;
- Aos amigos do Laboratório de Parasitologia/Setor de Moléstia Infecciosas (MI):
Izabela, Rute, Thábata, Luana;
- Aos funcionários do HOVET: Taislayne, Roseane, Ravi, Ana Rita, Juliano,
colaboradores da limpeza, às enfermeiras Edna, Regina e Marlene, aos residentes;
- Ao funcionário da pós-graduação Jean;
- Aos colegas de pós-graduação (Turma 2009) em Ciências Veterinárias, em
especial ao casal Alison e Jociane, pelo auxílio na confecção dos mapas;
- Aos amigos e funcionários do VPS e dos Laboratórios de Doenças Parasitárias I e
II (Carrapatos) da USP: Jonas, Thiaguinho, João (Gaúcho), Herbert, Iara, Aline,
Aliny, Aline (LEB), Arley, Fernanda, Hilda, Renatinho, Danival, pela hospitalidade,
por todo auxílio e pelos agradáveis momentos de descontração compartilhados. Em
especial, ao professor Marcelo Bahia Labruna, por permitir que freqüentasse o VPS,
por todo apoio no decorrer desse projeto, pelos ensinamentos e também por
acreditar no sucesso do mesmo;
- Aos amigos de Poconé, funcionários da Associação Brasileira de Criadores de
Cavalo Pantaneiro, em especial à Alice (e família), pelo despreendimento em

sempre ajudar no que fosse preciso. Aos cães (urbanos e rurais) do município de
Poconé, bem como seus proprietários, por aceitarem partipar desse experimento;
- Ao CNPq e à CAPES, pelo fomento financeiro e pela concessão da bolsa de pós-
graduação, respectivamente;
Saibam que todos vocês também fazem parte dessa conquista, por isso,
muito obrigada.

RESUMO
SOROPREVALÊNCIA E FATORES DE RISCO PARA Ehrlichia canis E Rickettsia
spp. EM CÃES DA REGIÃO DO PANTANAL DE MATO GROSSO, BRASIL
Durante o ano de 2009, 320 cães das áreas urbana e rural do Pantanal do município
de Poconé/MT foram avaliados para a presença de anticorpos anti-Ehrlichia canis e
anti-Rickettsia spp. e para a presença de ectoparasitos. Fatores de risco para
infecção por E. canis ou Rickettsia spp. também foram determinados. Foi utilizado
como teste sorológico a Reação de Imunofluorescencia Indireta (RIFI). Para
Ehrlichia canis foi utilizado como antígeno a cepa São Paulo e para Rickettsia spp.
foram utilizados seis isolados brasileiros: R. rickettsii, R. parkeri, R. amblyommii, R.
rhipicephali, R. felis e R. bellii. Reações positivas para E. canis foram detectadas em
227 (70,9%) cães, sendo 119 (74,3%) para a área urbana e 108 (67,5%) para a área
rural (P>0,05), com títulos variando entre 40 a 327.680. Para Rickettsia spp., 152
(47,5%) cães foram positivos, sendo 31 (19,3%) da área urbana e 121 (75,6%) da
área rural (P<0.05), com títulos variando entre 64 a 32.768. Um total de 930
carrapatos ixodideos foram coletados dos cães, sendo 708 Rhipicephalus
sanguineus, 209 Amblyomma cajenennse e 13 Amblyomma ovale. Quatrocentos e
cinquenta e oito pulgas Ctenocephalides felis felis e nove piolhos Heterodoxus
spiniger também foram identificados. R. sanguineus foram detectados em 102
(63,7%) cães da área urbana e em 31 (19,3%) cães da área rural (P<0,001). Na área
urbana, ninfas de A. cajennense foram detectadas em apenas um cão (0,6%) sendo
que na área rural, foram identificadas ninfas e adultos dessa espécie de carrapato
em 59 (36,8%) cães. Carrapatos A. ovale foram detectados em cinco (3,1%) cães da
área rural. C. felis felis foram detectadas em 77 (48,1%) cães da area urbana e em
57 (35,6%) da área rural (P<0,05). O piolho H. spiniger foi identificado em seis cães
da cidade. O maior titulo de anticorpo anti-Rickettsia spp. foi observado para R.
amblyommii, sugerindo que cães da área rural têm sido infectados por esse agente.
A infecção por Rickettsia spp. foi significantemente maior em cães da área rural, em
cães com hábito de caça e/ou infestados por A. cajenennse. Implicações na saúde
pública foram discutidas.

Palavras-chaves: Ehrlichia canis, Rickettsia, soroprevalência, carrapato, cão,
Pantanal.

ABSTRACT
SEROPREVALENCE AND RISK FACTORS TO Ehrlichia canis AND Rickettsia
spp. IN DOGS FROM THE PANTANAL REGION OF MATO GROSSO STATE,
BRAZIL
During 2009, 320 dogs from urban and rural areas of a Pantanal region of Mato
Grosso State, Brazil, were evaluate for the prevalence of anti-Ehrlichia canis and
anti-Rickettsia spp. antibodies and for the presence of ectoparasites. Risk factors for
E. canis or Rickttsia spp. infection were also evaluated. Seropositive dogs were
determinate by immunofluorescence assay (IFA). For Ehrlichia canis test São Paulo
strain antigens were used, and for Rickettsia spp. antigens of six Brazilian species
were used: R. rickettsii, R. parkeri, R. amblyommii, R. rhipicephali, R. felis, and R.
bellii. Positive reaction against E. canis were detected in 227 (70.9%) dogs, being
119 (74.3%) from urban area and 108 (67.5%) from rural area (P>0.05), with
endpoint titers ranging from 40 to 327,680. For Rickettsia spp., 152 (47.5%) dogs
were positive, being 31 (19.3%) from urban and 121 (75.6%) from rural area
(P<0.05), with endpoint titers ranging from 64 to 32,768. A total of 930 ixodid ticks
were collected on dogs, being 708 Rhipicephalus sanguineus, 209 Amblyomma
cajennense and 13 Amblyomma ovale. Four hundred and fifty eight Ctenocephalides
felis felis fleas and nine Heterodoxus spiniger lices were also collected. R.
sanguineus ticks were found in 102 (63.7%) dogs in urban area and in 31 (19.3%)
dogs in rural area (P<0.001). While nymphs of the tick A. cajennense were found in a
single dog (0.6%) from the urban area, nymphs and adults of A. cajennense were
found in 59 dogs (36.8%) from rural area (P<0.05). Adults of A. ovale ticks were
detected in 5 (3.1%) dogs from rural area. Fleas C. felis felis, were found in 77
(48.1%) dogs from urban area and in 57 (35.6%) dogs from rural area (P<0.05). The
louse H. spiniger was identified in 6 urban dogs. The highest anti-Rickettsia spp.
endpoint titers were observed for R. amblyommii, suggesting that the rural dogs had
been infected by this agent. Infection Rickettsial infection was significantly higher in
rural dogs, in dogs with hunting practice, or in A. cajennense-infested dogs.
Implications to public health are discussed.

Keywords: Ehrlichia canis, Rickettsia, seroprevalence, ticks, dog, Pantanal.

LISTA DE TABELAS
Tabela 1: Prevalência de anticorpos anti-E. canis em cães provenientes de diversas
regiões do Brasil ......................................................................................... 23
Tabela 2: Número e freqüência de cães parasitados por ectoparasitos no município
de Poconé .................................................................................................. 40
Tabela 3: Ectoparasitos encontrados nos cães da área urbana e rural do município
de Poconé .................................................................................................. 42
Tabela 4: Distribuição dos títulos de anticorpos anti-Ehrlichia canis em cães
reagentes a Reação de Imunofluorescência Indireta (≥40) de área urbana e
rural do município de Poconé, MT – Cuiabá - 2011 ................................... 50
Tabela 5: Freqüência de cães reagentes a Reação de Imunofluorescência Indireta (≥
40) contra antígenos de Ehrlichia canis segundo a faixa etária e ambiente
urbano e rural de Poconé, MT .................................................................... 51
Tabela 6: Análise univariada e multivariada para detectar fator de risco associado à
soroprevalência de anticorpos anti-Ehrlichia canis no município de Poconé,
MT .............................................................................................................. 52
Tabela 7: Distribuição de cães soropositivos para Ehrlichia canis e espécies de
carrapatos identificados nos cães por localidade no município de Poconé,
MT .............................................................................................................. 53
Tabela 8: Distribuição dos títulos de anticorpos frente aos 6 antígenos de Rickettsia
spp. utilizados na RIFI (≥64) em cães das áreas urbana e rural do
município de Poconé, MT ........................................................................... 54
Tabela 9: Distribuição dos cães sororeagentes à Rickettsia spp. ............................. 56
Tabela 10: Distribuição dos cães sororeagentes à Rickettsia spp. e parasitismo de
carrapatos em diferentes localidades de Poconé, MT ................................ 64
Tabela 11: Freqüência de cães reagentes a Reação de Imunofluorescência Indireta
(≥ 64) contra antígenos de Rickettsia spp. segundo a faixa etária e
ambiente ..................................................................................................... 65
Tabela 12: Análise univariada e multivariada para detectar fator de risco associado à
soroprevalência de anticorpos anti-Rickettsia spp. no município de Poconé,
MT .............................................................................................................. 66

LISTA DE FIGURAS
Figura 1: Localização do município de Poconé, MT .................................................. 34
Figura 2: Cidade de Poconé, composta de 12 bairros, diferenciados por diferentes
tons de cores ............................................................................................. 35
Figura 3: Mapa da cidade de Poconé e localização dos 12 bairros .......................... 39
Figura 4: Propriedades e comunidades rurais participantes deste estudo no
município de Poconé ................................................................................. 40
Figura 5: Prevalência de ectoparasitos na cidade de Poconé ................................... 43
Figura 6: Prevalência de Rhipicephalus sanguineus na área rural de Poconé ......... 44
Figura 7: Prevalência de Amblyomma cajennense na área rural de Poconé ............ 45
Figura 8: Prevalência de Amblyomma ovale na área rural de Poconé ...................... 46
Figura 9: Prevalência de Ctenocephalides felis felis área rural de Poconé ............... 47
Figura 10: Distribuição dos cães sororeagentes para Ehlichia canis na cidade de
Poconé, MT ................................................................................................ 48
Figura 11: Distribuição dos cães sororeagentes para Ehlichia canis na área rural do
município de Poconé, MT ........................................................................... 49
Figura 12: Distribuição dos cães sororeagentes para Rickettsia spp. na cidade de
Poconé, MT ................................................................................................ 57
Figura 13: Prevalência das espécies do gênero Rickettsia spp. na área rural de
Poconé, MT ................................................................................................ 58
Figura 14: Prevalência de Rickettsia amblyommii na área rural de Poconé, MT....... 59
Figura 15: Prevalência de Rickettsia bellii na área rural de Poconé, MT .................. 60
Figura 16: Prevalência de Rickettsia parkeri na área rural de Poconé, MT ............... 61
Figura 17: Prevalência de Rickettsia rickettsii na área rural de Poconé, MT ............. 62
Figura 18: Prevalência de Rickettsia rhipicephali na área rural de Poconé, MT........ 63

SUMÁRIO
RESUMO
ABSTRACT
LISTA DE TABELAS
LISTA DE FIGURAS
1. INTRODUÇÃO ...................................................................................................... 16
2. REVISÃO DE LITERATURA ................................................................................ 20
2.1 Ehrlichia canis ............................................................................................. 20
2. Rickettsia spp. ............................................................................................... 26
2. MATERIAL E MÉTODOS ..................................................................................... 33
3.1 Local de estudo ........................................................................................... 33
3.2 Amostragem e questionário epidemiológico ............................................ 34
3.3 Colheita e identificação de ectoparasitos ................................................. 35
3.4 Reação de Imunofluorescência Indireta .................................................... 36
3.4.1 Ehrlichia canis ...................................................................................... 36
3.4.2 Rickettsia spp. ...................................................................................... 37
3.5 Análise estatística ....................................................................................... 38
4. RESULTADOS ...................................................................................................... 38
4.1 Ectoparasitos ............................................................................................... 41
4.2 Detecção de anticorpos anti-Ehrlichia canis ............................................ 48
4.3 Detecção de anticorpos anti-Rickettsia spp ............................................. 54
5. DISCUSSÃO ......................................................................................................... 67
6. CONCLUSÃO ....................................................................................................... 79
REFERÊNCIAS BIBLIOGRÁFICAS ......................................................................... 80
APÊNDICE A...............................................................................................................95

16
1 Introdução
Dentro do filo artrópode, os carrapatos são considerados vetores de um
grande número de agentes infecciosos (HOOGSTRAAL, 1967). Das
aproximadamente 825 espécies de carrapatos descritas no mundo, apenas 10%
assumem uma maior importância direta em saúde pública, devido às possibilidades
desses carrapatos parasitarem humanos (OLIVER, 1989). Obviamente, várias outras
espécies que nunca foram descritas parasitando humanos, assumem papel
importante indireto na saúde pública, pois contribuem para a manutenção enzoótica
de agentes infecciosos na natureza (HOOGSTRAAL, 1967).
As riquetsioses são enfermidades infecciosas transmitidas por artrópodes,
causadas por bactérias gram-negativas, parasitas intracelulares obrigatórios. Essas
doenças têm sido associadas a carrapatos desde o inicio do século 20, quando
foram confirmados como hospedeiros e transmissores das riquétsias aos seres
humanos (SANTOS, 2007). Desde então, as pesquisas sobre os carrapatos, como
fonte de patógenos para seres humanos, aumentaram em diversas partes do
mundo.
As riquétsias pertencem à ordem Rickettsiales, que estão divididas em duas
grandes famílias: Rickettsiaceae e Anaplasmataceae. As espécies da família
Rickettsiaceae patogênicas pertencem ao Grupo da Febre Maculosa (GFM), as
quais têm sido isoladas e/ou detectadas por ensaios moleculares e sorológicos em
artrópodes e animais, respectivamente, em várias regiões do Brasil. Aqui, a doença
causada por riquétsias do GFM é denominada Febre Maculosa Brasileira (FMB),
tendo sido notificada em todos os estados da região Sudeste do país (DIAS e
MARTINS, 1939; SEXTON et al., 1993; LABRUNA et al., 2009a). A espécie
envolvida na FMB é a Rickettsia rickettsii (RICKETTS, 1909), e sua transmissão tem
sido associada aos carrapatos Amblyomma cajenennse, A. aureolatum e
Rhipicephalus sanguineus (DIAS e MARTINS, 1939; PINTER e LABRUNA, 2006;
PACHECO et al., 2010; PIRANDA et al., 2011).
Contudo, com o avanço das pesquisas, principalmente no âmbito da biologia
molecular, outras riquétsias foram caracterizadas nos últimos anos na região
Sudeste, como R. bellii e R. parkeri em carrapatos A. dubitatum (LABRUNA et al.,
2004a); R. bellii em A. aureolatum (PINTER e LABRUNA, 2006), R. parkeri em A.

17
triste (SILVEIRA et al., 2007); e R. felis, em pulgas Ctenocephalides felis felis
(HORTA et al., 2005) e em carrapatos Amblyomma spp. (CARDOSO et al., 2006).
Na região Norte do país, R. bellii e R. amblyommii foram caracterizadas em
carrapatos do gênero Amblyomma (LABRUNA et al., 2004b) e R. rhipicephali em
carrapatos Haemaphysalis juxtakochi (LABRUNA et al., 2005a). Mais tarde, R.
amblyommii foi isolada em A. longirostre provenientes de aves silvestres da Mata
Atlântica do Estado de São Paulo (OGRZEWALSKA et al., 2008). Também já foi
relatada detecção molecular desta espécie em A. longirostre e A. geayi coletados de
aves silvestres da Amazônia (OGRZEWALSKA et al., 2010). Em humanos, a
espécie R. rickettsii já foi isolada e caracterizada, oriunda de pacientes de diferentes
localidades do Estado de São Paulo (NASCIMENTO et al., 2005; GEHRKE et al.,
2006). Recentemente, foi relatado um caso clínico de riquetsiose no interior de São
Paulo. A análise molecular do agente etiológico detectado pertence ao GFM,
apresentando-se intimamente relacionado com R. parkeri, R. africae e R. sibirica,
sendo denominado por cepa Mata Atlântica (SPOLIDORIO et al., 2010).
Além dos agentes correspondentes à família Rickettsiaceae, a família
Anaplasmataceae agrupa também importantes patógenos de importância em
medicina veterinária e saúde pública. No Brasil, a E. canis é a espécie responsável
pela Erliquiose Monocítica Canina (EMC), isolada in vitro a partir de cães
experimentalmente e naturalmente infectados (TORRES et al., 2002; AGUIAR et al.,
2008). A EMC é uma doença infecciosa severa, transmitida pelo carrapato R.
sanguineus, e responsável nos cães por transtornos hematológicos como anemia e
trombocitopenia. Os cães, em determinado estágio de infecção, se comportam como
portadores assintomáticos, pois são capazes de manter viáveis a E. canis por longo
período, sem apresentarem sinais clínicos (COHN, 2003). No carrapato é observado
apenas transmissão transestadial, fazendo do cão o principal reservatório (GROVES
et al., 1975).
Como o R. sanguineus encontra-se distribuído por quase todo território
urbano brasileiro (LABRUNA e PEREIRA, 2001), é provável que a EMC esteja
presente em todas as regiões do Brasil. Estudos epidemiológicos têm demonstrado
que as freqüências de infecção têm variado de 19% a 65%, em cães amostrados de
ambientes urbanos, rurais ou atendidos em clínicas e hospitais veterinários em
diversas regiões do Brasil, seja por testes sorológicos ou moleculares (DAGNONE et

18
al., 2003; LABARTHE et al., 2003; BULLA et al., 2004; TRAPP et al., 2006; AGUIAR
et al., 2007a; COSTA et al., 2007; SAITO et al., 2008).
Apesar da infecção por espécies dos gêneros Rickettsia spp. e Ehrlichia spp.
em animais serem frequentemente estudadas em diferentes regiões do Brasil,
observamos que na região do Pantanal matogrossense, essa informação é
desconhecida. Dados a respeito das doenças transmitidas por carrapatos, que
acometem os animais da região pantaneira são escassos; por outro lado, tem-se
relatado a presença de diferentes espécies de carrapatos nesta região (ITO et al.,
1998; BECHARA et al., 2000; MARTINS et al., 2004).
Espécies do gênero Amblyomma spp. têm sido freqüentemente observadas
em animais silvestres do Pantanal. Na região de Corumbá, MS, Ito et al. (1998)
identificaram A. cajennense e Rhipicephalus (Boophilus) microplus em queixadas
(Tayassu pecari) e capivaras (Hydrochoerus hydrochaeris). Martins et al. (2004)
registraram a ocorrência de carrapatos em Tamanduás-bandeira (Myrmecophaga
tridactyla) e Tamanduás-mirim (Tamandua tetradactyla). As espécies identificadas
foram A. cajennense, A. parvum e A. nodosum.
Bechara et al. (2000), na sub-região da Nhecolândia, identificaram as
espécies R. (Boophilus) microplus, A. cajennense, A. parvum, A. pseudoconcolor, A.
scalpturatum, A. nodosum, A. ovale e A. tigrinum. Segundo os autores, os
carrapatos parasitavam principalmente quatis (Nasua nasua), veados (Mazama
gouazoubira) e tamanduás (M. tridactyla).
De acordo com os trabalhos supracitados, o envolvimento de diferentes
espécies de carrapatos, inclusive R. (Boophilus) microplus, além de evidenciar uma
possível influência da intensa atividade pecuária no Pantanal, sugere que os animais
silvestres podem desempenhar importante papel na epidemiologia das doenças
transmitidas por carrapatos na região.
Localizado no interior da América do Sul, o Pantanal matogrossense é a
maior extensão úmida contínua do planeta. Hidrograficamente, todo o Pantanal faz
parte da bacia do rio Paraguai constituindo-se em uma imensa planície de áreas
alagáveis. O Pantanal é composto por três regiões distintas: amazônica, cerrado e
chaco (paraguaio e argentino). Durante a seca, que coincide com o inverno, a
temperatura pode chegar a 0°C, influenciada pelos ventos que chegam do sul do
continente. A vegetação do Pantanal não é homogênea e há um padrão diferente de
flora de acordo com a altitude. Nas partes mais baixas, predominam as gramíneas,

19
que são áreas de pastagens naturais para o gado. A vegetação de cerrado, com
árvores de porte médio entremeadas de arbustos e plantas rasteiras, aparece nas
alturas intermediárias. A poucos metros acima das áreas inundáveis, ficam os
capões de mato ou cordilheiras. Nas altitudes maiores, o clima árido e seco torna a
paisagem parecida com a da caatinga (ALHO et al., 1987).
Dentro desta imensa diversidade, mais de 80 espécies de mamíferos como
felídeos: onça-pintada (Panthera onca) e suçuarana (Puma concolor); canídeos:
cachorro do mato (Cerdocyon thous) e lobo guará (Crysocyon brachyurus);
cervídeos, veado campeiro (Ozotoceros bezoarticus), veado catingueiro (M.
gouazoupira), cervo do pantanal (Blastocerus dichotomus); suídeos: porco do mato
ou caititu (T. tajacu) e queixada (T. pecari); capivaras (H. hydrochaeris); tamanduás
(M. tridactyla e T. tetradactyla), antas (Tapirus terrestris) entre outras espécies
(ALHO et al., 1987), dividem o mesmo habitat, contribuindo para a manutenção de
diferentes espécies de ixodídeos bem como agentes infecciosos.
Como em toda região do cerrado do Brasil central, o meio ambiente vem
sendo modificado em nome do progresso; as matas nativas derrubadas, queimadas
indiscriminada, cursos d‘águas barrados, desviados ou aterrados, caça e/ou
espécies nativas trocadas pelas domésticas e tudo isso sem qualquer estudo prévio
do impacto que tais ações possam causar sobre as espécies que ali habitavam em
pleno equilíbrio.
Por outro lado, a beleza proporcionada pela paisagem pantaneira fascina
pessoas de todo o mundo, fazendo com que o turismo se desenvolva em vários
municípios da região, ocasionando desenvolvimento de um pensamento
ambientalista e social para o Pantanal matogrossense, o que tem levado vários
pesquisadores a discutirem o impacto da ocupação humana neste ecossistema.
Nesse sentido, o afluxo de turistas para a região pantaneira, aliado a ocupação
humana local susceptível, justifica a realização de pesquisas que enfocam e buscam
dados sobre saúde pública humana e animal.
Diante da estimativa que 75% das doenças infecciosas emergentes são de
ordem zoonótica (TAYLOR et al. 2001), estudos envolvendo animais domésticos
podem complementar ou elucidar estudos epidemiológicos em saúde pública. Como
no Brasil os cães vivem em contato direto com seres humanos, dados obtidos sobre
as doenças transmitidas por carrapatos nesses animais podem fornecer informações

20
a respeito de prevalência, fatores de risco para a exposição e infecção, antes da
ocorrência da doença em humanos.
Face a estes dados, o presente estudo objetiva investigar a prevalência da
infecção por espécies de Rickettsia spp. e E. canis em cães do município de
Poconé, situado no Pantanal do Estado de Mato Grosso, procurando associar
fatores de risco para a infecção, identificando inclusive espécies de ectoparasitos
que ocorrem nos animais estudados.
2 Revisão de Literatura
2.1 Ehrlichia spp.
As riquétsias fazem parte da ordem Rickettsiales, que está dividida em duas
grandes famílias: Rickettsiaceae e Anaplasmataceae. Essa nomenclatura foi
proposta recentemente por Dumler et al. (2001), que, ao utilizarem as descobertas
no campo de biologia molecular à cerca dos genes 16S rRNA e groESL, bem como
as características biológicas e antigênicas existentes nestes agentes, propuseram
uma reclassificação taxonômica dessa ordem. Sendo assim, a família Rickettsiaceae
passou a ter os gêneros Rickettsia e Orientia, enquanto que a família
Anaplasmataceae agrupa os gêneros Ehrlichia, Anaplasma, Wolbachia e
Neorickettsia. As mudanças também envolveram os gêneros Anaplasma, o qual
agora é composto pelas espécies Anaplasma phagocytophilum (originada da
reclassificação da E. equi, E. phagocytophila e do agente da erliquiose granulocitica
humana), A. platys (outrora E. platys) e A. bovis (antes E. bovis). O gênero
Neorickettsia, que agora contempla as espécies Neorickettsia risticii e N. sennetsu,
anteriormente nominadas de E. risticii e E. sennetsu, respectivamente. Além disso, a
espécie outrora denominada por Cowdria ruminantium passou a ser designada por
E. ruminantium.
Desse modo, o gênero Ehrlichia atualmente é composto por seis espécies: E.
canis, E. chaffeensis, E. ewingii, E. muris, E. ruminantium (DUMLER et al., 2001) e
Ehrlichia IOE (Ixodes ovatus Ehrlichia) (SHIBATA et al., 2000). São parasitas

21
intracelulares obrigatórios de diversas células hematopoiéticas animais, tais como
monócitos, macrófagos, neutrófilos e células endoteliais (DUMLER et al., 2001) e de
células do aparelho digestório, da glândula salivar e hemolinfa de carrapatos
(GROVES et al., 1975).
Em se tratando de saúde pública referente à infecção por bactérias do gênero
Ehrlichia, a Erliquiose Monocítica Humana (EMH), doença causada pela E.
chaffeensis tem sido relatada na América do Norte, sendo considerada doença
emergente, transmitida principalmente pelo carrapato A. americanum (OLANO e
WALKER, 2002). Representando também considerável potencial zoonótico, está a
espécie E. canis, que foi isolada pela primeira vez por Perez et al. (1996) a partir de
sangue humano na Venezuela, vindo a ser diagnosticada em 2006, a infecção pela
PCR em seis pacientes com doença clínica compatível com Erliquiose Monocítica
Humana (EMH) (PEREZ et al., 2006). Adicionalmente, no Brasil, dentre os relatos de
detecção molecular do agente, Diniz et al. (2007), demonstraram haver completa
similaridade entre a seqüência de nucleotídeos do gene 16S rRNA (1.434 pb) da E.
canis responsável pela EMH venezuelana e de uma amostra de erliquia proveniente
de sangue canino oriundo do Estado de São Paulo. Nesse sentido, tudo indica que o
mesmo agente causador de infecção humana na Venezuela, vem infectando cães
do Brasil.
Das espécies supracitadas, a única isolada até o momento acometendo cães
no Brasil é a E. canis, responsável por causar a doença denominada de Erliquiose
Monocítica Canina (EMC) (AGUIAR et al., 2008). Seu principal vetor é o carrapato R.
sanguineus, comumente chamado de ―carrapato marrom do cão‖ (KEEFE et al.,
1982; LABRUNA e PEREIRA, 2001). Outras duas espécies, E. chaffeensis,
causadora da Erliquiose Monocítica Humana (EMH) e E. ewingii, agente etiológico
da Erliquiose Granulocítica Humana (EGH) e Canina (EGC); também já foram
relatadas no país através de detecção molecular em cervos-do-pantanal e cães,
respectivamente (MACHADO et al., 2006; OLIVEIRA et al., 2009). Recentemente, foi
descrita a presença de anticorpos anti-E. canis em onças-pintadas do Pantanal de
Mato Grosso do Sul, bem como a detecção molecular de uma possível nova espécie
de Ehrlichia spp. em carrapatos A. cajennense e A. triste que parasitavam estes
carnívoros e, cujo seqüenciamento apontou 98% de similaridade com E.
ruminantium (WIDMER, 2009).

22
A E. canis é classificada como α-proteobactéria e morfologicamente
caracteriza-se como pleomórfica, gram-negativa, com parede celular não protéica,
ausente de lipopolissacarídeo e peptídeoglicano e com capacidade de incorporação
de colesterol de membrana, o que poderia ser um facilitador no processo de
adaptação às células do carrapato vetor e do hospedeiro vertebrado (YU e
WALKER, 2006; RIKIHISA et al., 2006).
Apresenta tropismo pelos monócitos e macrófagos, na qual seu ciclo biológico
inicia com sua inoculação no cão pelo vetor R. sanguineus durante seu repasto
sanguíneo. Então, o agente entra nas células mononucleares por endocitose, em
forma de estruturas conhecidas como corpos elementares (0,2 a 0,6μm de
diâmetro), que realizam multiplicação nos fagolisossomos celulares. Cerca de três a
cinco dias após o início da infecção, os corpos elementares se agrupam, formando
grânulos subesféricos de coloração rósea a púrpura, designada por corpúsculos
iniciais (0,4 a 2,0μm). Na sequência, entre sete a doze dias, ocorre a replicação e
diferenciação dos corpúsculos iniciais em inclusões denominadas de mórulas (2,0 a
4,0 μm). Assim, os corpúsculos elementares podem deixar as células
mononucleares através de exocitose ou lise e com isso, infectar novas células,
disseminando-se pelo hospedeiro vertebrado, principalmente ao que concernem os
órgãos linfóides, nos quais predomina esse tipo celular (POPOV et al., 1998;
SANTARÉM, 2003).
A primeira descrição do agente foi em 1935, na Argélia, por Donatien e
Lestoquard, na qual um cão da raça Pastor Alemão encontrava-se infectado por
essa bactéria (MACHADO, 2004). Mais tarde, a doença recebeu notoriedade
mundial ao levar a óbito vários cães militares dos Estados Unidos que combatiam
durante a Guerra do Vietnã (HUXSOLL et al., 1970; KEEFE et al., 1982). Já seu
primeiro relato no Brasil ocorreu em Minas Gerais, onde foram observadas inclusões
citoplasmáticas compatíveis com mórulas de E. canis em linfócitos de cães (COSTA
et al., 1973).
Atualmente, a doença apresenta distribuição cosmopolita e o crescente
aumento de sua ocorrência nas diferentes regiões do Brasil está relacionado à
significativa presença do vetor R. sanguineus. Em função do seu hábito nidícola (do
latim nidi= ninho; cola= que permanece), esse artrópode encontra-se amplamente
disseminado e adaptado às condições de moradia dos cães domésticos do país
(LABRUNA e PEREIRA, 2001). Estudos demonstram que a presença do vetor

23
representa um fator de risco para a ocorrência da enfermidade (HARRUS et al.,
1997; DAGNONE et al., 2003). Vários estados têm relatado a erliquiose canina,
acometendo tanto animais atendidos em clínicas e hospitais veterinários quanto os
domiciliados nas áreas urbanas e rurais, conforme a Tabela 1 (TRAPP et al., 2002;
DAGNONE et al., 2003; LABARTHE et al., 2003; BULLA et al., 2004; AGUIAR et al.,
2007a; UENO et al., 2009; SOUZA et al., 2010; SILVA et al, 2010).
Tabela 1 – Prevalência de anticorpos anti-E. canis em cães provenientes de diversas
regiões do Brasil – Cuiabá - 2011
Local Ano Cães Referência
Testados Positivos (%)
Região Norte 2007 314 98 (31,2) Aguiar et al.,
2007ª
Região Sudeste 2007 226 101 (44,7) Costa et al., 2007
Região Sul 2008 389 19 (4,8) Saito et al., 2008
Região Centro-Oeste 2010 254 108 (42,5) Silva et al., 2010
Região Nordeste 2010 472 168 (35,6) Souza et al., 2010
A transmissão de E. canis no carrapato R. sanguineus é do tipo transestadial,
onde somente as fases de ninfa e adulto do vetor transmitem o agente. No que se
referem às larvas, elas não têm relevância neste processo porque a transmissão
transovariana não ocorre nos carrapatos. Todavia, novos estudos relatam a
capacidade de machos (adultos) infectados transmitirem (transmissão intraestadial)
a bactéria a diferentes cães de um mesmo lugar, sem que haja a presença da fêmea
(BREMER et al., 2005; STICH et al., 2008). Após infectar o carrapato, a bactéria se
multiplica no intestino e nas glândulas salivares, de onde será inoculada no cão no
momento de seu repasto sanguíneo (GROVES et al., 1975). A transmissão da
bactéria do carrapato para cães susceptíveis pode ser realizada em até 155 dias
após a infecção do carrapato. Outra forma de transmissão descrita é a transfusão
sanguínea (LEWIS et al., 1977).
A enfermidade caracteriza-se por ser multissistêmica, com período de
incubação de 8 a 20 dias, sintomatologia clínica complexa e que se encontra
distribuída entre as fases aguda, subclínica (ou assintomática) e crônica, na qual

24
somente em infecções experimentais é possível diferenciar estes estágios.
(HARRUS et al., 1997; SANTARÉM, 2003).
A patogenia verificada na fase aguda caracteriza-se por multiplicação do
agente em células mononucleares, principalmente dos órgãos do sistema fagocítico
mononuclear, como linfonodos, fígado, baço e medula óssea, causando
organomegalia e hiperplasia dessas células. Clinicamente, os cães nesta fase
apresentam febre, depressão, anorexia, letargia, esplenomegalia, linfoadenopatia e
tendências hemorrágicas. Verifica-se também leucopenia, anemia e
trombocitopenia, resultante da destruição periférica das plaquetas (HARRUS et al.,
1997; CASTRO et al., 2004).
Terminada esta etapa, os animais imunocompetentes podem evoluir para a
cura ou entrar numa fase assintomática (subclínica), que inicia entre 40 a 120 dias
após a infecção, podendo durar de 6 a 9 semanas ou persistir por alguns anos em
áreas consideradas enzoóticas (WANER et al., 1997; HARRUS e WANER, 2011). Já
na fase crônica da doença, é possível observar que a sintomatologia clínica
assemelha-se à observada na fase aguda, porém, com manifestações mais graves,
além de palidez de mucosas, perda de peso, fraqueza, edemas periféricos,
neuropatias, oftalmopatias e nefropatias (HARRUS et al., 1997; NEER, 1998). Nos
casos mais graves, pode haver um quadro hematológico de pancitopenia, associado
à infecções secundárias e hemorragias sistêmicas, podendo apresentar evolução
fatal (COHN, 2003; BOWMAN et al., 2009).
Segundo Harrus et al. (1997), a patogenia da enfermidade envolve
mecanismos imunológicos e inflamatórios, caracterizados muitas vezes por
hemaglutinação, hipergamaglobulinemia, infiltração leucocitária em órgãos
parenquimatosos, manguitos peri-vasculares em diversos locais, como rins, baço,
meninges, pulmões e olhos; além da presença de anticorpos anti-plaquetários em
cães experimentalmente infectados. A extinção da infecção pela E. canis depende
basicamente da resposta imune celular, muito embora a resposta imune humoral
também tenha alguma relevância (GANTA et al., 2004). Atualmente, é sabido que
esse patógeno tem como alvo o sistema imune, onde promove alguns desajustes na
resposta imunológica. Esse tipo de infecção causa redução significativa na
expressão de moléculas do complexo de histocompatibilidade principal de classe II
(MHCII), cuja função compreende a maturação de células T em linfócito T CD4+,
que possui importante atuação na elaboração e potencialização da resposta imune

25
celular e humoral. Neste tipo de infecção, há redução nas concentrações de linfócito
T CD4+, células sintetizadoras de IFN-γ, que atuam na estimulação da atividade
microbicida dos macrófagos, sendo, portanto, imprescindíveis para mecanismo de
proteção e eliminação das infecções causadas por essa bactéria (HASEGAWA,
2005).
O diagnóstico da erliquiose canina envolve vários métodos, tais como: história
clínica e anamnese, análises clínicas e laboratoriais, detecção direta (microscopia)
do agente nas células hematopoiéticas infectadas ou em tecidos, isolamento em
cultura celular, técnicas sorológicas como reação de imunofluorescência indireta
(RIFI), western immunoblot e ensaio de imunoabsorção por ligação enzimática
(ELISA), bem como a técnica molecular de Reação em Cadeia pela Polimerase
(PCR) (HARRUS e WANER, 2011).
A realização da citologia através do esfregaço de sangue periférico baseia-se
na identificação das mórulas nas células mononucleares infectadas. No entanto,
como a presença destas estruturas na corrente sanguínea é observada mais
frequentemente na fase aguda da doença e em pequena quantidade dos casos,
considera-se esse tipo de diagnóstico com baixa sensibilidade (HARRUS et al.,
1997). Outra alternativa para o diagnóstico seria o isolamento do agente em cultura
celular. A E. canis pode ser cultivada in vitro em células DH82 (dog histiocytosis),
provenientes de monócitos caninos a partir de um caso de histiocitoma (WELLMAN
et al., 1988).
Quanto ao uso do ELISA e do western immunoblot, ambos apresentam
elevada sensibilidade, sendo que este último tem sido utilizado para caracterizar e
distinguir os diferentes agentes no caso de reação cruzada entre os patógenos da
família Anaplasmataceae (HARRUS e WANER, 2011). Já a PCR vem se mostrando
um método sensível e específico para o diagnóstico da erliquiose canina,
principalmente nos casos de infecção aguda, haja vista que a detecção da bactéria
pode ser realizada antes da formação de mórulas ou da soroconversão (MCBRIDE
et al., 1996).
Apesar de inúmeros métodos de diagnóstico, a RIFI é considerada o teste
―padrão ouro‖ para identificação e quantificação de anticorpos da classe IgG anti-
E.canis, na qual títulos ≥ 40 indicam soropositividade (AGUIAR et al. 2007b;
HARRUS e WANER, 2011). Ao contrário da PCR, a RIFI é indicada para a detecção

26
de casos mais crônicos, uma vez que a formação de anticorpos inicia somente entre
1 a 3 semanas após a infecção (MCBRIDE et al., 1996).
Quanto ao tratamento, este se baseia na utilização de antimicrobianos, como
principalmente tetraciclinas e cloranfenicol, bem como terapia de suporte. O
prognóstico é considerado favorável quando o tratamento é iniciado o mais cedo
possível (WANER et al., 1997).
Ao que se refere à profilaxia, o controle do carrapato vetor é a medida mais
eficaz e capaz de evitar a infecção pelo agente (HARRUS et al., 1997). Levando em
conta a distribuição de uma população de carrapatos, considera-se que a menor
parte está em parasitose no cão, enquanto que a maioria está nas fases de vida livre
no ambiente, é imprescindível que a terapia carrapaticida atue tanto sobre o
hospedeiro como no ambiente em que ele está inserido (LABRUNA e PEREIRA,
2001).
2.2 Rickettsia spp.
As espécies da família Rickettsiaceae estão divididas em dois grupos,
baseados nos padrões antigênicos, moleculares e ecológicos: i grupo do Tifo (GT),
que inclui as espécies R. prowazekii, causadora do tifo epidêmico, e R. typhi, agente
do tifo endêmico ou tifo murino, ambas transmitidas por piolhos e pulgas
respectivamente (EREMEEVA e DASCH, 2000); ii grupo da Febre Maculosa (GFM),
constituído por mais de 30 espécies, dentre as quais pelo menos 15 causam
diferentes doenças ao homem (R. aeschlimannii, R. africae, R. akari, R. australis, R.
conorii, R. felis, R. helvetica, R. honei, R. japonica, R. marmionii, R.
mongolotimonae, R. parkeri, R. rickettsii, R. sibirica, R. slovaca). Com exceção de R.
akari e R. felis transmitidas por pequenos ácaros e pulgas, respectivamente, todas
as outras representantes desse grupo são veiculadas por carrapatos (BROUQUI et
al., 2004; RAOULT e ROUX, 1997). E sem pertencer a nenhum dos grupos citados,
estão as espécies R. bellii e R. canadensis, ambas de patogenicidade desconhecida
(YU e WALKER, 2006; LABRUNA, 2009a).
A doença causada pela R. rickettsii foi relatada pela primeira vez nos Estados
Unidos, onde foi denominada de Febre Maculosa das Montanhas Rochosas

27
(FMMR), na qual o pesquisador Howard Taylor Ricketts isolou a bactéria em 1909, e
estabeleceu a participação do carrapato da espécie Dermacentor andersoni no
processo de transmissão deste agente (RICKETTS, 1909). Nos Estados Unidos, R.
rickettsii é vetoriada pelo D. variabilis na costa leste, enquanto que na costa oeste o
carrrapato D. andersoni é o responsável por desempenhar essa função (MACDADE
e NEWHOUSE, 1986); entretanto, alguns estudos apontam o carrapato R.
sanguineus como um importante vetor desta riquétsia em algumas áreas (DEMMA et
al., 2005). Outros países, como Canadá, México, Costa Rica, Panamá, Colômbia e
Argentina também relataram a ocorrência deste patógeno, onde primariamente era
transmitida por carrapatos do gênero Amblyomma (MACDADE e NEWHOUSE,
1986; DUMLER e WALKER, 2005).
No Brasil, a doença causada por riquétsias do GFM é denominada Febre
Maculosa Brasileira (FMB), sendo causada principalmente pela bactéria R. rickettsii.
Esta enfermidade tem sido notificada em todos os estados da região Sudeste do
país (DIAS e MARTINS, 1939; SEXTON et al., 1993; LEMOS et al., 2001;
ROZENTAL et al., 2002; GALVÃO et al., 2003).
O carrapato A. cajennense é reconhecido como o principal vetor da R.
rickettsii na América Central e do Sul; e na região metropolitana da cidade de São
Paulo, é o A. aureolatum que desempenha esse papel (PINTER e LABRUNA, 2006;
PINTER et al., 2008; LABRUNA, 2009a). Recentemente, Pacheco et al. (2010)
realizaram o isolamento desta bactéria em R. sanguineus, o que reforça a idéia da
existência, aqui no país, de mais este vetor. Adicionalmente, Piranda et al. (2011)
conseguiram demonstrar, experimentalmente, que o cão pode atuar como
hospedeiro amplificador da R. rickettsiii para o carrapato R. sanguineus. Além disso,
esse estudo relata que ninfas e adultos de R. sanguineus transmitiram com sucesso
a bactéria às cobaias, confirmando a existência de uma competência vetorial, por
parte desse carrapato após aquisição da infecção a partir de cães riquetsêmicos.
Contudo, à medida que se intensificaram as pesquisas, principalmente no
âmbito da biologia molecular, outras riquétsias foram caracterizadas nos últimos
anos na região Sudeste, como R. bellii e R. parkeri em carrapatos A. dubitatum
(LABRUNA et al., 2004a); R. bellii em A. aureolatum (PINTER e LABRUNA, 2006);
R. parkeri em A. triste (SILVEIRA et al., 2007); e R. felis, em pulgas C. felis felis
(HORTA et al., 2005) e em carrapatos Amblyomma spp (CARDOSO et al., 2006). Na
região Norte do país, R. bellii e R. amblyommii foram isoladas em carrapatos do

28
gênero Amblyomma (LABRUNA et al., 2004b) e R. rhipicephali em carrapatos H.
juxtakochi (LABRUNA et al., 2005a). R. amblyommii foi isolada em A. longirostre
provenientes de aves silvestres da Mata Atlântica do estado de São Paulo
(OGRZEWALSKA et al., 2008). Também já foi relatada detecção molecular desta
espécie em A. longirostre e A. geayi coletados de aves silvestres da Amazônia
(OGRZEWALSKA et al., 2010). Em humanos, a espécie R. rickettsii já foi isolada e
caracterizada, oriunda de pacientes de diferentes localidades do Estado de São
Paulo (NASCIMENTO et al., 2005; GEHRKE et al., 2006). Recentemente, foi
relatado um caso clínico de riquetsiose no interior de São Paulo. A análise molecular
de amostras de pele do paciente, indicou que o agente etiológico pertence ao GFM,
apresentando-se intimamente relacionado com R. parkeri, R. africae e R. sibirica,
sendo denominado por cepa Mata Atlântica (SPOLIDORIO et al., 2010). E na
espécie canina, LABRUNA et al. (2009b) relataram pela primeira vez no Brasil
infecção natural nessa espécie animal.
A necessidade de reservatórios para a circulação do agente e a manutenção
do agente na natureza é fundamental. A capivara é um potencial amplificador da
Rickettsia spp. uma vez que foi demonstrada através de infecção experimental, a
sua capacidade de mantê-la circulante em seu organismo, sem apresentar sinais
clínicos da doença (TRAVASSOS e VALLEJO, 1942). Isolamentos do agente foram
realizados também em gambás (Didelphis marsupialis) no estado de Minas Gerais e
São Paulo (DIAS e MARTINS, 1939). Em outro estudo, foi demonstrado
experimentalmente que a R. rickettsii conseguiu infectar gambás, porém sem causar
doença clínica; além disso, observou-se o desenvolvimento riquetsemia capaz de
causar infecção em cobaias e carrapatos (HORTA et al., 2009). Souza et al. (2009)
verificaram que a R. rickettsii foi capaz de infectar capivaras, também sem causar
doença clínica, induzindo riquetsemia capaz de causar infecção em cobaias e
carrapatos. Esses resultados resultados indicam que as capivaras atuam como
hospedeiro amplificador desta bactéria em carrapatos A. cajennense no Brasil.
Os eqüinos assumem importante papel de sentinela da febre maculosa
(HORTA et al., 2004; LEMOS et al., 1996; SANGIONI et al., 2005). Em áreas
endêmicas para FMB nos Estados de Minas Gerais e São Paulo, estes animais se
apresentaram com altos títulos de anticorpos frente a R. rickettsii, possivelmente
correlacionados ao alto parasitismo pelo carrapato vetor A. cajennense (HORTA et
al., 2004; SANGIONI et al., 2005).

29
Os cães apresentam papel importante como hospedeiros de carrapatos e
pulgas, por isso, também podem ser considerados sentinelas da infecção (HORTA
et al., 2004; PINTER et al., 2008). Sangioni et al. (2005) analisando áreas
endêmicas e não endêmicas no estado de São Paulo, observaram presença de
anticorpos em cães oriundos da primeira área, indicando contato prévio com o
agente. Em outro estudo realizado em área endêmica do estado de São Paulo,
verificou-se uma significativa presença de anticorpos anti-R. rickettsii nos cães do
local analisado, os quais apresentavam parasitismos por carrapatos R. sanguineus e
Amblyomma spp. Além disso, recentemente houve também a detecção molecular
desse agente em R. sanguineus na região metropolitana de São Paulo (MORAES-
FILHO et al., 2009); e assim como os seres humanos, os cães também são
susceptíveis à infecção por R. rickettsii (LABRUNA et al., 2009b).
Em se tratando da espécie R. parkeri, atualmente ela é reconhecida por ser
patogênica ao ser humano (WHITMAN et al., 2007), sendo identificada nos Estados
Unidos infectando carrapatos A. maculatum. Na América do Sul, a mesma já foi
isolada em A. triste no Uruguai (VENZAL et al., 2004; PACHECO et al., 2006;
SAMMER et al., 2007). E no Brasil, a R. parkeri também já foi detectada por
isolamento e PCR em carrapatos A. triste (SILVEIRA et al., 2007) e por sorologia em
capivaras do estado de São Paulo (PACHECO et al., 2007) e cães do Rio Grande do
Sul (SAITO et al., 2008). Em A. dubitatum foi identificada uma Rickettsia spp., que é
filogeneticamente muito próxima à R. parkeri e que temporariamente foi denominada
de amostra COOPERI (= R. parkeri) (LABRUNA et al., 2004a). Como a doença
causada por R. parkeri é relativamente mais branda e associada a uma lesão
papular típica no local da picada do carrapato, haveria uma certa dificuldade no
desfecho do seu diagnóstico. Possivelmente as infecções humanas por este agente
na América do Sul não têm sido diagnosticadas ou talvez elas estejam sendo
diagnosticadas como sendo causadas por R. rickettsii. Ademais, há indícios de que
talvez os carrapatos sejam eficientes reservatórios dessa bactéria, já que a mesma
possui uma estreita relação com seus principais vetores, bem como elevadas taxas
de infecção nestes artrópodes (WHITMAN et al., 2007; LABRUNA, 2009a).
No que se refere à R. amblyommii, sabe-se que apresenta uma estreita
relação com carrapatos norte-americanos da espécie A. americanum (―lone star
tick‖), onde se processa transmissão ovariana e transestadial da bactéria e a partir
do qual ela foi inicialmente isolada nos Estados Unidos (BURGDORFER et al.,

30
1981). Na América do Sul, uma estirpe de Rickettsia amblyommii genotipicamente
semelhante já foi relatada na região norte do Brasil em carrapatos A. longirostre
(LABRUNA et al., 2004c). Posteriormente, foi detectada e isolada em carrapatos A.
coelebs e A. cajennense, respectivamente, do estado de Rondônia (LABRUNA et al.,
2004b). Recentemente, houve isolamento da mesma em A. neumanni na Argentina
(LABRUNA et al., 2007a). Apesar de ainda não ter sido isolada em seres humanos,
já foi verificado a maior presença de anticorpos para R. amblyommii do que à R.
rickettsii em pacientes febris oriundos de áreas endêmicas (APPERSON et al.,
2008).
Quanto à R. rhipicephali, seu primeiro relato foi em R. sanguineus, nos
Estados Unidos, onde, mais tarde, foi isolada também em três espécies de carrapato
pertencentes ao gênero Dermacentor, não havendo comprovação de sua
patogenicidade à humanos. No Brasil, a espécie já foi isolada, no estado de São
Paulo, a partir de H. juxtakochi (LABRUNA et al., 2007b)
Outra espécie que tem sido relatada em alguns países da América do Sul é a
R. felis, identificada em pulgas Ctenocephalides spp., considerado seu principal
hospedeiro (LABRUNA et al., 2007c; NAVA et al., 2008). Estudos demonstram que a
transmissão transovariana e transestadial dessa riquétsia nessas pulgas pode ser
possível (WEDINCAMP e FOIL, 2002). Além disso, um trabalho realizado com
pulgas C. felis felis coletadas de cães residentes na região metropolitana de São
Paulo detectou, pela PCR, a presença dessa riquétsia em 13 pulgas (nT= 24),
sugerindo a circulação deste agente nos locais avaliados (HORTA et al., 2005).
Entretanto, apesar do elevado parasitismo por essa espécie de pulgas nos
cães e também nos gatos, e do grande número de pulgas infectadas por essa
bactéria, talvez devesse haver mais casos de riquetsioses do que os relatados
atualmente. O mecanismo de transmissão deste patógeno pela pulga ainda não está
completamente elucidado, uma vez que se desconhece a presença de R. felis na
saliva destas pulgas, apesar de haver sido encontrada na glândula salivar das
mesmas, sugerindo outras possibilidades neste processo de vetoriação sugerindo
outras possibilidades neste processo de vetoriação. Uma hipótese seria o contato
direto de lesões cutâneas e/ou mucosas com fezes frescas contaminadas com este
agente infeccioso ou até mesmo a ingestão de pulgas infectadas (MACALUSO et al.,
2008; LABRUNA, 2009a).

31
R. bellii já foi relatada em diversas espécies de carrapatos na América do
Norte (Dermacentor, Haemaphysalis, Argas e Ornithodoros) (PHILLIP et al., 1983),
no Brasil (A. aureolatum, A. dubitatum, A. humerale, A. rotundatum, A.
oblongoguttatum, A. scalpturatum, A. ovale, Ixodes loricatus e H. juxtakochi)
(LABRUNA et al., 2004a; LABRUNA et al., 2004b; HORTA et al., 2006; PINTER e
LABRUNA, 2006) e na Argentina (A. neumanni) (LABRUNA et al., 2007a). Sua
patogenicidade ao ser humano é desconhecida. Pelo fato deste agente ter sido
relatado em várias espécies de carrapatos, ele pode ser considerado como a
riquétsia com maior número de relatos em diferentes carrapatos pertencentes às
famílias Ixodidae e Argasidae (LABRUNA et al., 2007b).
A transmissão da riquétsia causadora de Febre Maculosa ocorre através da
picada por carrapatos, que devem permanecer fixados à pele do hospedeiro por um
período varíavel de cinco a vinte horas, tempo necessário para uma possível
reativação da bactéria na glândula salivar do carrapato. Dessa forma, a partir da
picada do carrapato infectado, a riquétsia se dissemina pelo organismo através dos
vasos linfáticos e pequenos vasos sanguíneos, atingindo pele, cérebro, pulmões,
coração, fígado, baço, pâncreas e trato gastrointestinal (LABRUNA e PEREIRA,
1998; DANTAS-TORRES, 2007). O período de incubação da bactéria dura em
média 7 dias, seguido por aparecimento de manifestações clínicas de caráter
inespecífico. Os principais sintomas incluem febre alta, mialgias, insuficiência renal
aguda, prejuízos ao sistema nervoso central, como cefaléia, déficit neurológico,
meningite/meningoencefalite, icterícia, exantema, manifestações hemorrágicas, dor
abdominal e problemas respiratórios. As lesões cutâneas caracterizam-se pelo
aparecimento de máculas, além de necrose e gangrena da pele (ANGERAMI et al.,
2006; BRASIL, 2005). Nos cães, a infecção pela bactéria R. rickettsii é caracterizada
por febre, letargia, emese, anorexia, diarréia e comprometimento do sistema nervoso
central (síndrome vestibular e ataxia) (LABRUNA et al., 2009b). Num estudo com
infecção experimental de cães por R. rickettsii, além desses sinais supracitados, foi
observado, lesões oculares, anemia e trombocitopenia (PIRANDA et al., 2008).
Quanto ao diagnóstico da enfermidade, atualmente, a sorologia, é o método
considerado ―padrão ouro‖, e, apesar da elevada sensibilidade, este teste não
consegue distinguir entre uma infecção causada por R. rickettsii de outra causada
por outra espécie do GFM. Por isso, neste teste deve ser utilizado antígenos locais e
considera-se o provável antígeno responsável pela infecção (PARI), como aquele

32
que obteve um título quatro vezes superior ao observado para qualquer outra
espécie de Rickettsia spp. testada. A detecção molecular é outra alternativa para o
diagnóstico, porém, sua realização esbarra no fato de haver baixa quantidade de
riquétsia circulante na corrente sanguínea. A técnica de imunohistoquímica
geralmente é usada em pacientes com lesões cutâneas através de biópsia da pele.
Também é possível realizar o isolamento da bactéria através do cultivo in vitro, que
requer uma boa estrutura laboratorial, haja vista que esse agente é classificado
como nível 3 com relação à biosseguridade (BRASIL, 2005; BROUQUI, et al., 2004).
Quanto ao tratamento, antibióticos como as tetraciclinas e o cloranfenicol são
considerados os mais eficazes. Em relação à profilaxia, há várias medidas que
incluem combate aos carrapatos vetores através da utilização de carrapaticidas
comerciais, capacitação dos profissionais de saúde, alertando para a importância do
diagnóstico precoce e diferencial com outras doenças e esclarecimento da
população sobre a circulação sazonal do carrapato, como forma educativa e
preventiva (respeitando o ano biológico do carrapato e, conseqüentemente, a
ocorrência dos casos). Além disso, é importante também orientar a população
quanto ao uso de barreiras físicas quando houver a exposição principalmente em
áreas consideradas endêmicas para FMB e com possibilidade de existir carrapatos;
e isso consiste em: usar roupas claras e com mangas compridas, para facilitar a
visualização de carrapatos; utilizar calças compridas, inserindo a parte inferior por
dentro de botas, preferencialmente de cano longo e vedadas com fita adesiva de
dupla face, examinar o próprio corpo em intervalos de duas a três horas, a fim de
verificar a presença de carrapatos, pois quanto mais rápido os mesmos forem
retirados, menor a chance de infecção, haja vista que a transmissão é verificada
quando os carrapatos permanecem fixados à pele do hospedeiro por um período
variável de cinco a vinte horas, tempo necessário para uma possível reativação do
agente na glândula salivar do carrapato. A retirada dos carrapatos (caso sejam
encontrados no corpo) deve ser feita, preferencialmente, com o auxílio de uma
pinça. Nos locais públicos conhecidamente infestados por carrapatos devem ser
obrigados (pela vigilância sanitária) a informar seus freqüentadores, por meio de
placas, da presença de carrapatos e das formas de proteção (LABRUNA e
PEREIRA, 1998; BRASIL, 2005).
Para o controle em áreas de ocorrência de foco, a equipe de zoonoses deve
ser acionada para orientar as medidas específicas a serem implementadas, tais

33
como informações quanto ao ciclo do carrapato, transmissão da doença e atividades
que devem ser realizadas. (BRASIL; 2005).
3 MATERIAL E MÉTODOS
3.1 Local de estudo
O município de Poconé (16º15‘24‖ Sul; 56º37‘22‖ Oeste) está localizado no
nordeste da região do Pantanal, 100 Km a sudoeste da cidade de Cuiabá, capital do
estado de Mato Grosso, tendo como vizinhos os municípios de Cáceres (oeste),
Nossa Senhora do Livramento (norte), Barão de Melgaço (leste) e estado de Mato
Grosso do Sul na borda sul (Figura 1).
Poconé tem uma população estimada de 32.162 habitantes e 3.111 cães,
dispostos dentro de uma área total de 17.261 Km2, na qual 81% (~14.000 Km2) ficam
inundados, anualmente, durante a estação chuvosa. (IBGE; 2007). Esta área
apresenta altitude de 142 m, com clima quente e chuvoso nos meses de primavera e
verão, quando a temperatura média varia em torno de 32°C, enquanto que no
outono e no inverno predominam o frio e a seca, com temperatura média de 21°C. A
média da precipitação anual varia entre 1.000 e 1.400 mm, com o período chuvoso
concentrado entre os meses de outubro a maio. O ambiente do Pantanal consiste de
três regiões distintas: amazônia, cerrado e chaco, com uma vegetação heterogênia,
variando de acordo com a altitude: em áreas mais baixas as gramíneas são
predominantes, em altitude intermediária o cerrado é predominante, e em altitudes
mais elevadas, a vegetação torna-se árida e seca, assemelhando-se ao bioma
caatinga (FERNANDES et al., 2010).

34
3.2 Amostragem e questionário epidemiológico
As colheitas de soro dos cães foram realizadas durante os meses de julho a
setembro de 2009. Utilizando o programa EpiInfo versão 6.0 e considerando uma
prevalência estimada de 50%, erro padrão de 10% e intervalo de confiança de 99%,
determinou-se que uma amostra representativa da população canina do local seria
de 160 cães, com idade superior a 2 meses. Para melhor efeito de comparação,
foram amostrados 160 cães da área urbana e 160 do ambiente rural. Na área
urbana, foram colhidas amostras de cães de todos os bairros (Figura 2), localizados
tanto na região central quanto na região peri-urbana; enquanto que na área rural, as
colheitas de amostras sanguíneas contemplaram 25 fazendas e três comunidades
rurais. Durante as visitas no ambiente rural, as coordenadas geográficas de cada
localidade foram obtidas através do eTREX Vista HCx GPS (Garmin®, Kansas,
USA).
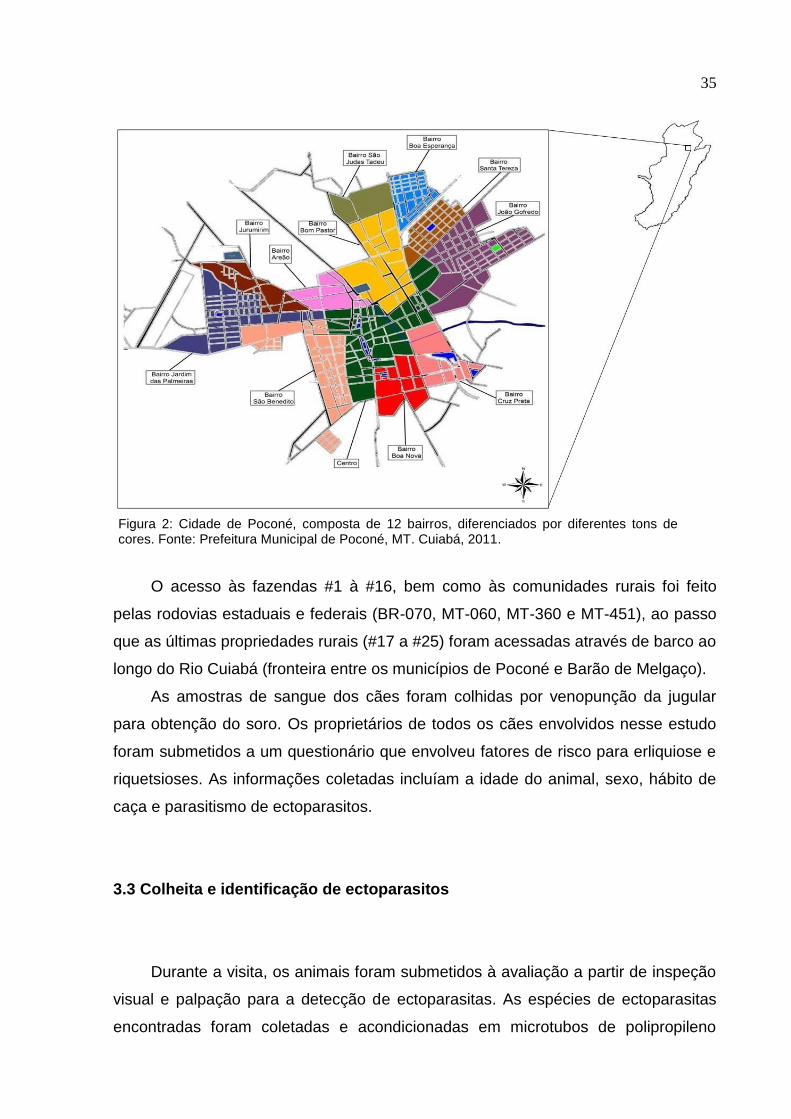
Figura 1: Localização do município de Poconé, MT. Cuiabá, 2011.
35
O acesso às fazendas #1 à #16, bem como às comunidades rurais foi feito
pelas rodovias estaduais e federais (BR-070, MT-060, MT-360 e MT-451), ao passo
que as últimas propriedades rurais (#17 a #25) foram acessadas através de barco ao
longo do Rio Cuiabá (fronteira entre os municípios de Poconé e Barão de Melgaço).
As amostras de sangue dos cães foram colhidas por venopunção da jugular
para obtenção do soro. Os proprietários de todos os cães envolvidos nesse estudo
foram submetidos a um questionário que envolveu fatores de risco para erliquiose e
riquetsioses. As informações coletadas incluíam a idade do animal, sexo, hábito de
caça e parasitismo de ectoparasitos.
3.3 Colheita e identificação de ectoparasitos
Durante a visita, os animais foram submetidos à avaliação a partir de inspeção
visual e palpação para a detecção de ectoparasitas. As espécies de ectoparasitas
encontradas foram coletadas e acondicionadas em microtubos de polipropileno
Figura 2: Cidade de Poconé, composta de 12 bairros, diferenciados por diferentes tons de cores. Fonte: Prefeitura Municipal de Poconé, MT. Cuiabá, 2011.

36
contendo álcool isopropílico absoluto, entretanto, algumas ninfas vivas ingurgitadas
de carrapatos foram acondicionadas vivas em tubos de polietileno e transportadas
ao Laboratório a fim de procederem a ecdise para o estágio adulto em incubadora a
25 ºC e umidade relativa (UR) igual a 90%. As ninfas que morreram no laboratório
antes de atingirem a fase adulta foram preservadas em álcool isopropílico absoluto.
Os carrapatos (adultos e ninfas), piolhos e pulgas foram identificadas de acordo
com as chaves taxonômicas de Onofrio et al. (2006), Martins et al. (2010),
Guimarães et al. (2001) e Linardi e Guimarães (2000), respectivamente.
3.4 Reação de Imunofluorescência Indireta
3.4.1 Ehrlichia canis
A Reação de Imunofluorescência Indireta (RIFI) foi realizada a partir da cepa
São Paulo de E. canis. As amostras sorológicas foram utilizadas seguindo uma
diluição inicial de 1:40 (AGUIAR et al. 2007b) em PBS (pH 7,2), com 1% de
soroalbumina bovina, e aplicadas em lâminas contendo antígeno previamente
fixado. Em cada lâmina foram incluídos soros controles não reativo (controle
negativo) e soro reativo (controle positivo). Após aplicação das amostras, as lâminas
foram incubadas por 30 minutos a 37 ºC em câmara úmida, seguida por lavagem em
solução salina tamponada - PBS (pH 7,2). Depois da secagem em temperatura
ambiente, foi adicionado conjugado de coelho anti-IgG de cão (Sigma® Diagnostics,
St. Louis, Mo) com diluição de 1:1000. As lâminas foram novamente incubadas a 37º
C por 30 minutos em câmara úmida, lavadas em PBS (pH 7,2) por 10 minutos e
submetidas a secagem. Posteriormente, adicionou-se glicerina (pH 8,5) nas lâminas
para então, serem examinadas em microscópio (objetiva de 40x) de
epifluorescência. As amostras consideradas positivas passaram por sucessivas
diluições na razão dois com objetivo de obter o título final.

37
3.4.2 Rickettsia spp.
Anticorpos contra Rickettsia spp. foram investigados usando seis antígenos
derivados de isolados de Rickettsia spp. do Brasil: R. bellii cepa Mogi, R.
amblyommii cepa Ac37, R. rhipicephali cepa HJ5, R. rickettsii cepa Taiaçu, R. felis
cepa Pedreira, e R. parkeri cepa At24, como descrito por Labruna et al. (2007d). Em
cada lâmina foram incluídos soros controles não reativo (controle negativo) e soro
reativo (controle positivo). Inicialmente as lâminas contendo antígeno fixado foram
imersas em cuba contendo PBS (pH 7,4), seguidas de secagem em temperatura
ambiente. Os soros dos cães foram diluídos a 1:64 e adicionados às lâminas, que
foram incubadas a 37 ºC por 30 minutos em câmara úmida. Após incubação, as
lâminas foram lavadas duas vezes em ―washing buffer‖ (PBS com Triton a 1%),
seguida por nova secagem. Depois, foi adicionado a elas conjugado de coelho anti-
IgG de cão (Sigma® Diagnostics, St. Louis, Mo) na diluição de 1:1000, sendo
novamente incubadas a 37 ºC por 30 minutos em câmara úmida e lavadas conforme
descrito acima. Após a secagem, aplicou-se glicerina nas lâminas para serem
observadas em microscópio (objetiva de 40x) de epifluorescência.
As amostras consideradas positivas passaram por sucessivas diluições na
razão dois com objetivo de obter o título final. Para determinar o provável antígeno
responsável pela infecção (PARI), considerou-se o resultado cujo título foi quatro
vezes maior do que o observado para qualquer outra espécie de Rickettsia spp.
testada, sendo então o anticorpo considerado homólogo à espécie ou a um genótipo
estreitamente relacionado de maior titulação (SAITO et al., 2008).

38
3.5 Análise estatística
A prevalência de anticorpos anti-E. canis e anti-Rickettsia spp. foi estimada
com intervalo de confiança de 95% (THRUSFIELD, 1995), utilizando o programa
EpiInfo, versão 6.0. As diferenças observadas entre os ectoparasitas encontrados
nas áreas urbana e rural foram avaliadas pelo teste do Qui-Quadrado (2). A
associação entre de anticorpos caninos anti-E. canis e anti-Rickettsia spp. e as
variáveis foi realizada pela regressão logística em duas etapas: análise univariada e
multivariada. Na análise univariada, foi utilizado o teste do Qui-Quadrado (2) ou o
teste exato de Fischer. Na análise multivariada, realizou-se avaliação pela técnica de
regressão logística, com apenas as variáveis que apresentaram valor de p menor
que 20% (P<0,20) na primeira etapa. No modelo final de regressão logística, foram
classificadas como fator de risco as variáveis que apresentaram p<0,05, com
respectivos valores de odds ratio (OR) e intervalo de confiança (IC) de 95%. O 2
(―goodness-of-fit‖) foi utilizado conforme estatística de Hosmer e Lemeshow (1989),
considerado significativo quando P ≤ 0,05. O programa estatístico SPSS, versão 16,
foi utilizado para estas análises. O teste do 2 por tendência foi utilizado para testar a
associação entre a soroprevalência e a faixa etária dos cães com auxilio do
programa EpiInfo 6.04.
4. RESULTADOS
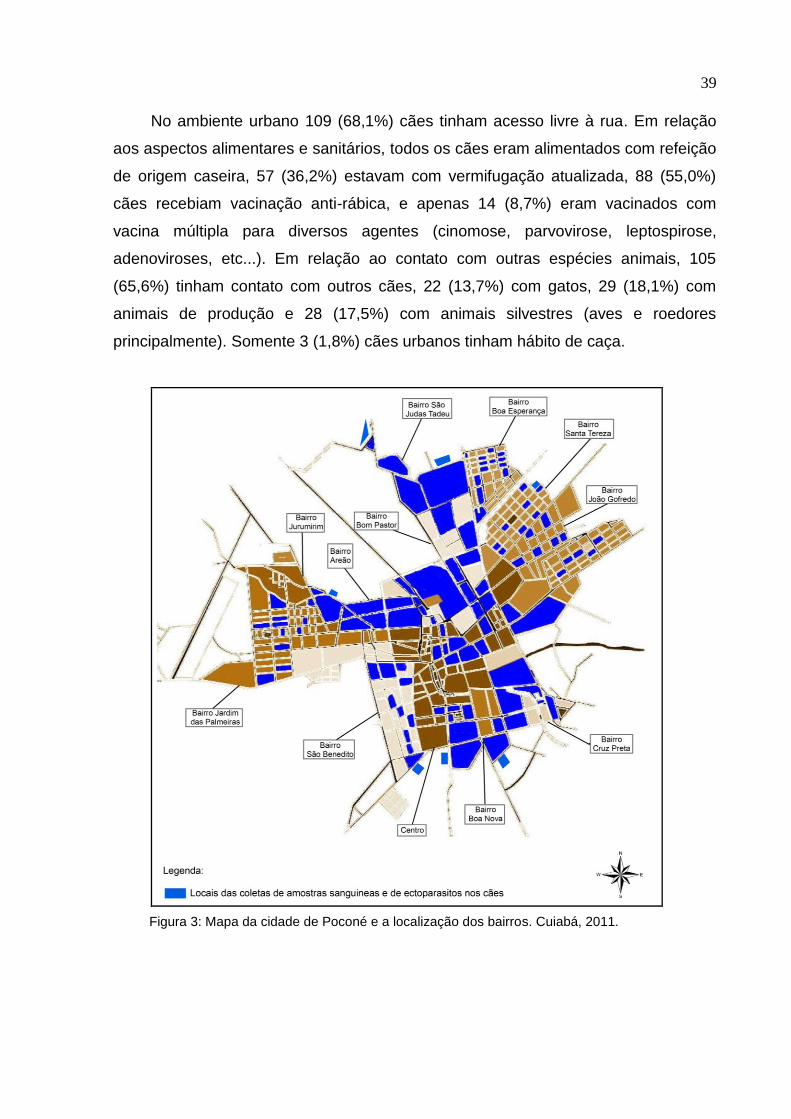
Um total de 320 cães (160 da área urbana e 160 da área rural) foram avaliados
neste estudo. Na área urbana, foram visitados 12 bairros (~13 cães por bairro)
(Figura 3) e no ambiente rural, foram 118 cães de 25 fazendas (~4,7 cães por
fazenda) e 42 de três comunidades rurais (~14 cães por comunidade) (Figura 4). Os
animais amostrados estavam aparentemente saudáveis, com idade variando entre 1
e 72 meses, e abrangendo diferentes raças — embora em sua maioria eram
mestiços, 180 (56,2%) cães eram machos e 140 (43,8%) eram fêmeas.
39
No ambiente urbano 109 (68,1%) cães tinham acesso livre à rua. Em relação
aos aspectos alimentares e sanitários, todos os cães eram alimentados com refeição
de origem caseira, 57 (36,2%) estavam com vermifugação atualizada, 88 (55,0%)
cães recebiam vacinação anti-rábica, e apenas 14 (8,7%) eram vacinados com
vacina múltipla para diversos agentes (cinomose, parvovirose, leptospirose,
adenoviroses, etc...). Em relação ao contato com outras espécies animais, 105
(65,6%) tinham contato com outros cães, 22 (13,7%) com gatos, 29 (18,1%) com
animais de produção e 28 (17,5%) com animais silvestres (aves e roedores
principalmente). Somente 3 (1,8%) cães urbanos tinham hábito de caça.
Figura 3: Mapa da cidade de Poconé e a localização dos bairros. Cuiabá, 2011.

40
Quanto aos cães do ambiente rural, 18 (11,2%) cães tinham acesso à cidade
de Poconé. Todos os cães também tinham a comida caseira como base alimentar.
Noventa e sete (60,6%) cães eram utilizados diretamente na prática da caça no
campo ou na floresta. Do ponto de vista sanitário, 74 (46,2%) estavam com
vermifugação atualizada, 135 (84,3%) cães recebiam vacinação anti-rábica, e 44
(27,5%) eram vacinados com vacina múltipla para diversos agentes. Em relação ao
contato com outras espécies animais, 155 (96,8%) tinham contato com outros cães,
123 (76,8%) com gatos, 156 (97,5%) com animais de produção e todos os cães
(100%) tinham contato com animais silvestres.
Na área rural, quatro localidades estavam localizadas fora do município de
Poconé. Uma grande área das fazendas # 7 e # 8 pertence ao município de Poconé,
estando inclusive registrados no escritório rural do município, porém as respectivas
sedes situam-se no município de Nossa Senhora do Livramento. A fazenda # 21
encontra-se nas margens do rio Cuiabá e apresenta coordenadas geográficas
registradas dentro dos limites do município de Barão de Melgaço; já as áreas de uso
doméstico da comunidade # 3 estão entre Poconé e Nossa Senhora do Livramento
(Figura 4).
Figura 4: Propriedades e comunidades rurais participantes deste estudo no município de
Poconé, MT. Cuiabá, 2011.

41
4.1 Ectoparasitos
Ao que se refere aos ectoparasitos, foram identificados carrapatos da espécie
R. sanguineus em 102 (63,7%) cães de todos os bairros da área urbana (P<0,001).
A. cajennense (7 ninfas) foram detectadas em 1 (0,6%) cão do ambiente urbano.
Além do parasitismo por carrapatos também verificou-se a presença de pulgas C.
felis felis, encontradas em 77 (48,1%) cães da área urbana (P<0,05). E em 6 (3,7%)
cães urbanos foram identificados piolhos da espécie Heterodoxus spiniger (Tabela
2).
Tabela 2 – Número e frequência de cães parasitados por ectoparasitos no
município de Poconé – Cuiabá - 2011
* P<0,01; ** P<0,05
Ao todo, 930 carrapatos foram coletados, sendo 708 (76,1%) R. sanguineus
(100 ninfas e 608 adultos), 209 (22,4%) A. cajennense (200 ninfas e 9 adultos) e 13
(1,3%) adultos de A. ovale. Também foram identificadas 458 pulgas C. felis felis e 9
piolhos H. spiniger (Tabela 3).
Ectoparasitas
Ambiente urbano Ambiente rural
Nº de cães infestados
% Nº de cães infestados
%
R. sanguineus 102 63,7* 31 19,3*
A. cajenennse (ninfas) 1 0,6* 54 33,7
A. cajenennse (adultos) 0 0 08 5.0
A. ovale (adultos) 0 0 05 3,1
C. felis felis 77 48,1** 57 35,6**
H. spiniger 06 3,7 0 0

42
Tabela 3 – Ectoparasitos encontrados nos cães da área urbana e rural do município
de Poconé – Cuiabá - 2011
Ectoparasitos Área urbana Área rural Total
Rhipicephalus sanguineus 97 N; 212 F; 237 M 03 N; 85 F; 74 M 708
Amblyomma cajennense (ninfas) 07 N 193 N 200
Amblyomma cajennense (adultos) - 03 F; 06 M 09
Amblyomma ovale - 09 F; 04 M 13
Ctenocephalides felis felis 185 F; 78 M 139 F; 56 M 458
Heterodoxus spiniger 03 F; 05 M - 09*
F: fêmea; M: macho; N: ninfa. * O sexo de um espécime não foi determinado devido à morfologia deformada
43
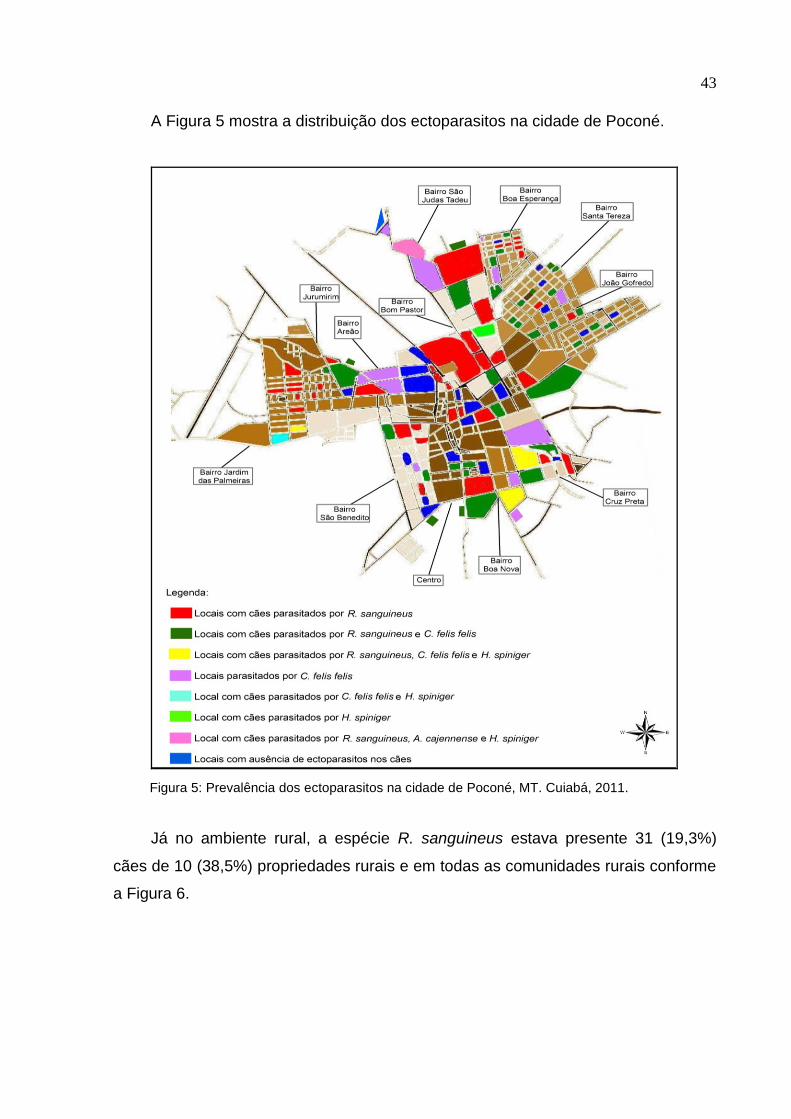
A Figura 5 mostra a distribuição dos ectoparasitos na cidade de Poconé.
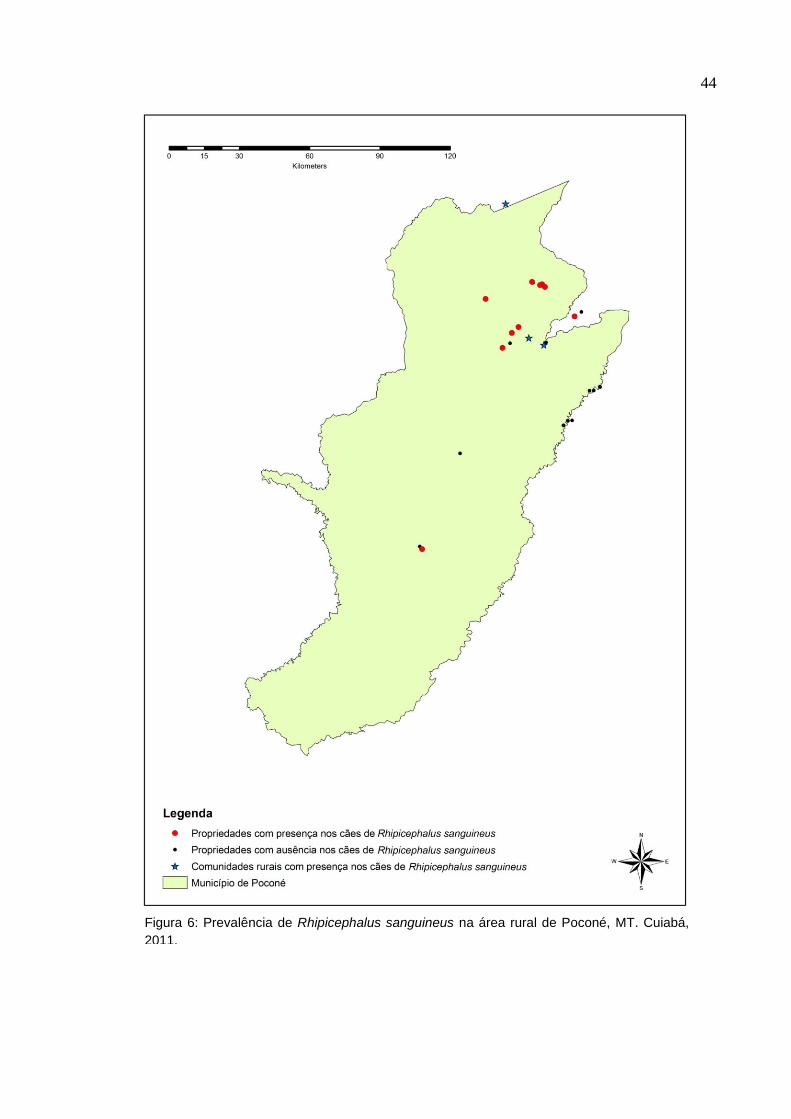
Já no ambiente rural, a espécie R. sanguineus estava presente 31 (19,3%)
cães de 10 (38,5%) propriedades rurais e em todas as comunidades rurais conforme
a Figura 6.
Figura 5: Prevalência dos ectoparasitos na cidade de Poconé, MT. Cuiabá, 2011.
44
Figura 6: Prevalência de Rhipicephalus sanguineus na área rural de Poconé, MT. Cuiabá,
2011.
45
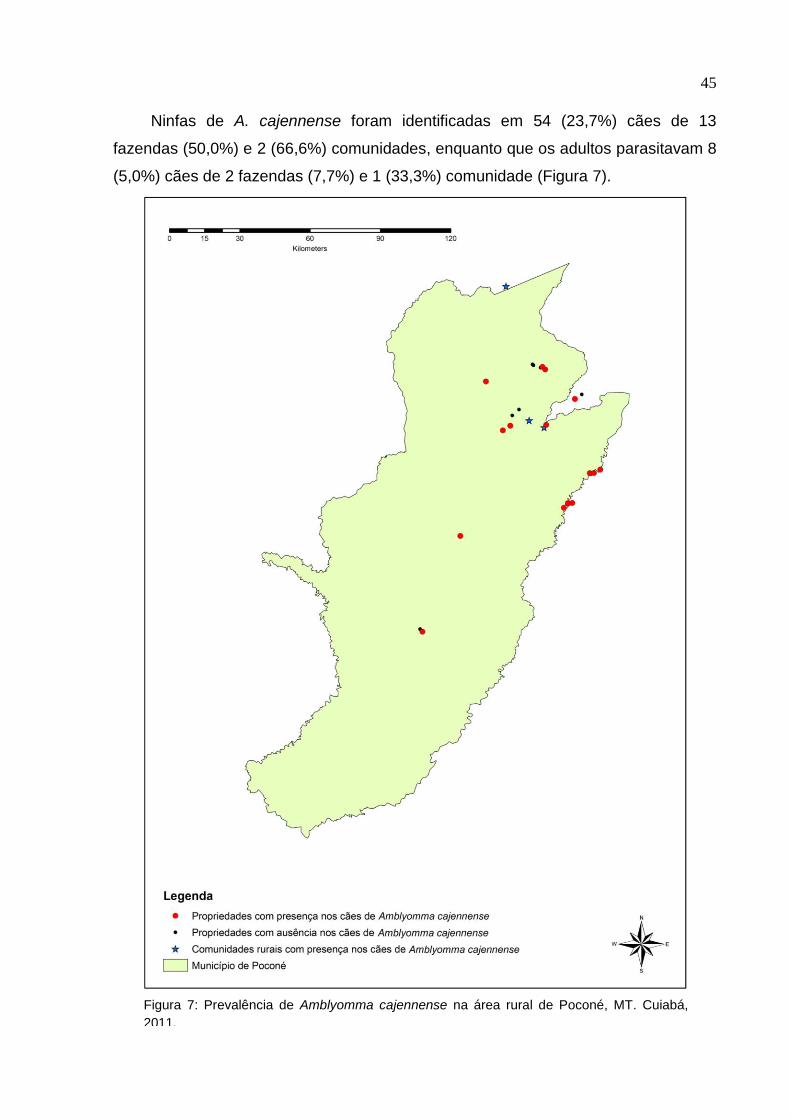
Ninfas de A. cajennense foram identificadas em 54 (23,7%) cães de 13
fazendas (50,0%) e 2 (66,6%) comunidades, enquanto que os adultos parasitavam 8
(5,0%) cães de 2 fazendas (7,7%) e 1 (33,3%) comunidade (Figura 7).
Figura 7: Prevalência de Amblyomma cajennense na área rural de Poconé, MT. Cuiabá,
2011.
46
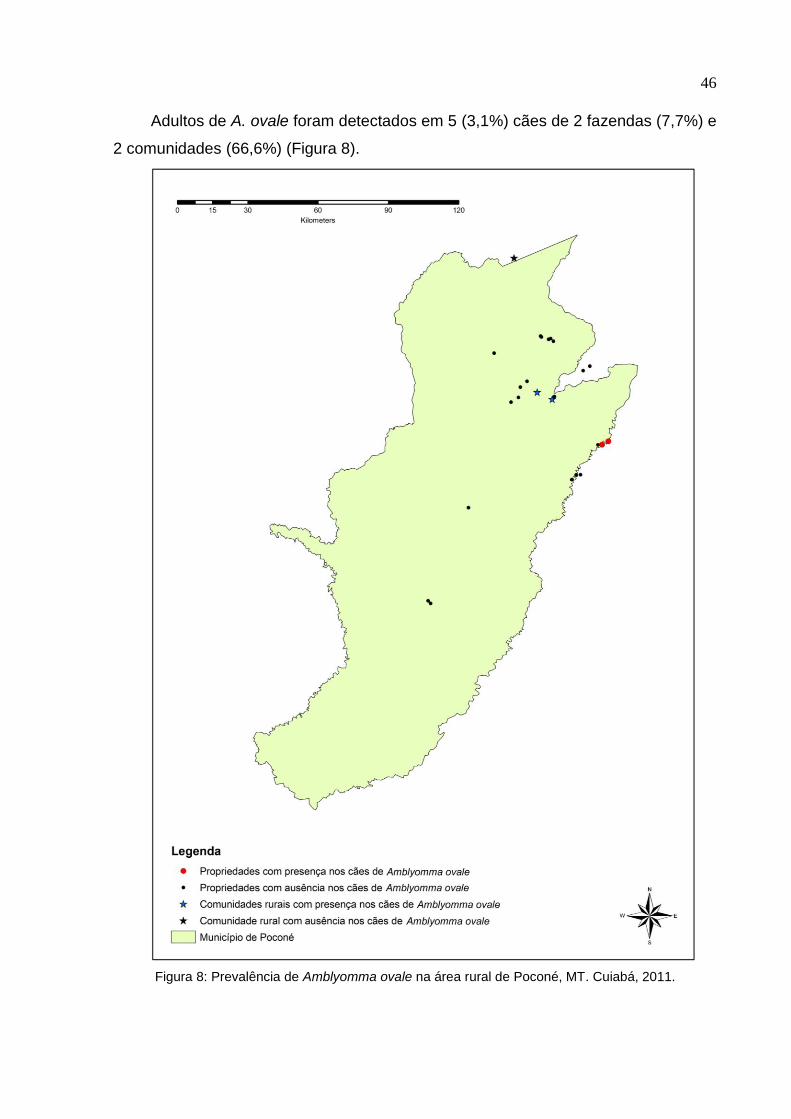
Adultos de A. ovale foram detectados em 5 (3,1%) cães de 2 fazendas (7,7%) e
2 comunidades (66,6%) (Figura 8).
Figura 8: Prevalência de Amblyomma ovale na área rural de Poconé, MT. Cuiabá, 2011.
47
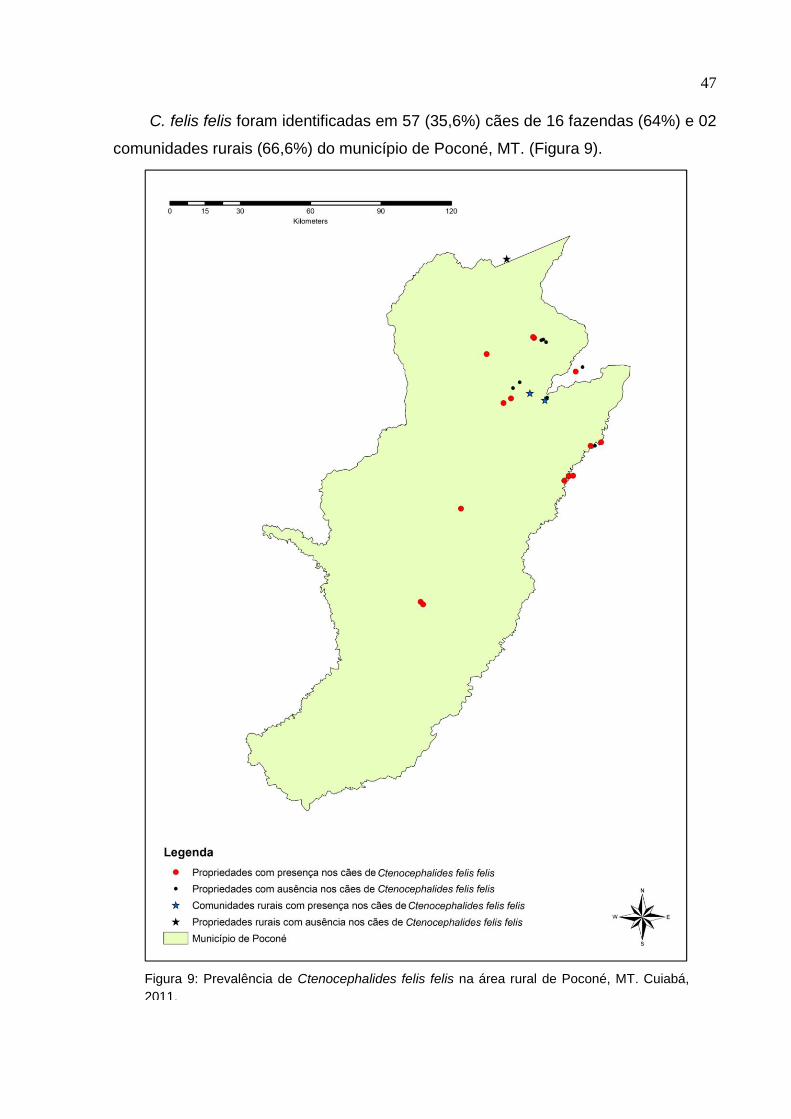
C. felis felis foram identificadas em 57 (35,6%) cães de 16 fazendas (64%) e 02
comunidades rurais (66,6%) do município de Poconé, MT. (Figura 9).
Figura 9: Prevalência de Ctenocephalides felis felis na área rural de Poconé, MT. Cuiabá,
2011.

48
4.2 Detecção de anticorpos anti-Ehrlichia canis
Anticorpos contra E. canis foram detectados em 227 (70,9%; IC 95%: 65,7 – 75,7%)
cães, dos quais 119 (74,3%; IC 95%: 67,1 – 80,6%) cães positivos eram da área urbana e
108 (67,5%; IC 95%: 59,9 – 74,4%) da área rural (P>0,05). Na cidade de Poconé, todos os
bairros (100%) apresentaram cães soropositivos (Figura 10).
Figura 10: Distribuição de cães sororeagentes para Ehrlichia canis na cidade de
Poconé, MT. Cuiabá, 2011.

49
Na área rural, os cães sororeagentes estavam presentes em 21 (84%)
fazendas e em todas (100%) as comunidades rurais (Figura 11).
Figura 11: Distribuição dos cães sororeagentes para Ehrlichia canis na área rural do
município de Poconé, MT. Cuiabá, 2011.
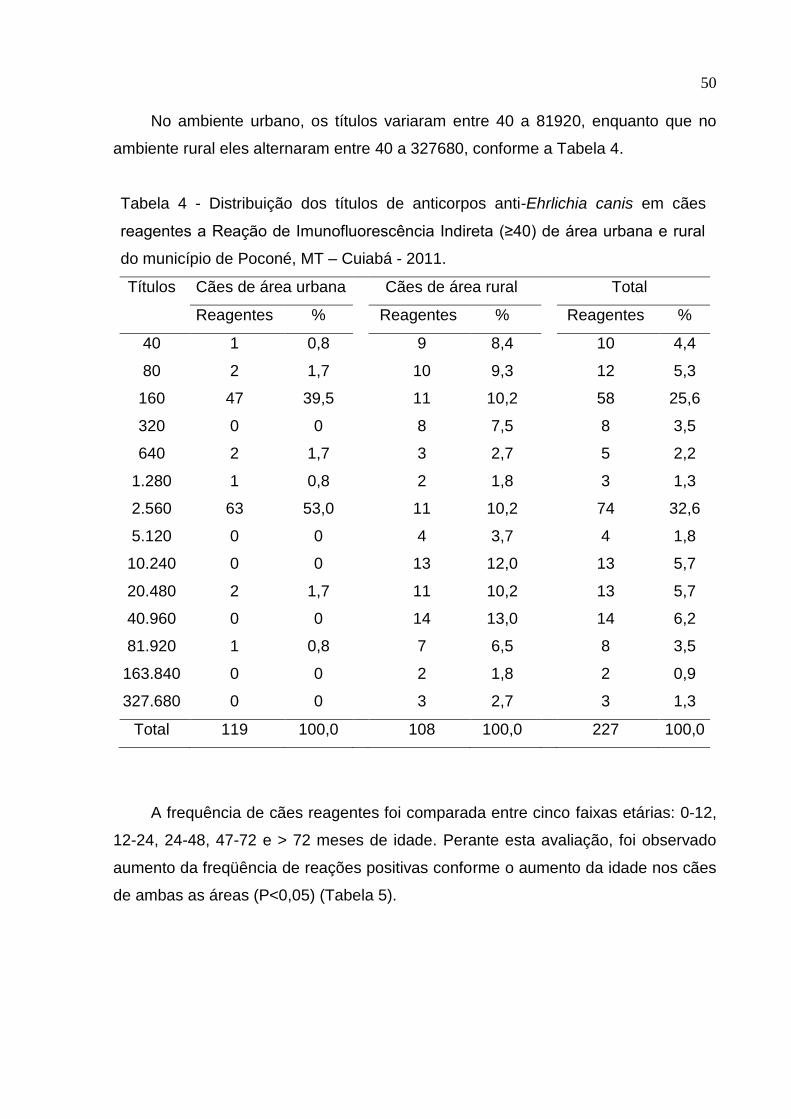
50
No ambiente urbano, os títulos variaram entre 40 a 81920, enquanto que no
ambiente rural eles alternaram entre 40 a 327680, conforme a Tabela 4.
Tabela 4 - Distribuição dos títulos de anticorpos anti-Ehrlichia canis em cães
reagentes a Reação de Imunofluorescência Indireta (≥40) de área urbana e rural
do município de Poconé, MT – Cuiabá - 2011.
Títulos Cães de área urbana Cães de área rural Total
Reagentes % Reagentes % Reagentes %
40 1 0,8 9 8,4 10 4,4
80 2 1,7 10 9,3 12 5,3
160 47 39,5 11 10,2 58 25,6
320 0 0 8 7,5 8 3,5
640 2 1,7 3 2,7 5 2,2
1.280 1 0,8 2 1,8 3 1,3
2.560 63 53,0 11 10,2 74 32,6
5.120 0 0 4 3,7 4 1,8
10.240 0 0 13 12,0 13 5,7
20.480 2 1,7 11 10,2 13 5,7
40.960 0 0 14 13,0 14 6,2
81.920 1 0,8 7 6,5 8 3,5
163.840 0 0 2 1,8 2 0,9
327.680 0 0 3 2,7 3 1,3
Total 119 100,0 108 100,0 227 100,0
A frequência de cães reagentes foi comparada entre cinco faixas etárias: 0-12,
12-24, 24-48, 47-72 e > 72 meses de idade. Perante esta avaliação, foi observado
aumento da freqüência de reações positivas conforme o aumento da idade nos cães
de ambas as áreas (P<0,05) (Tabela 5).
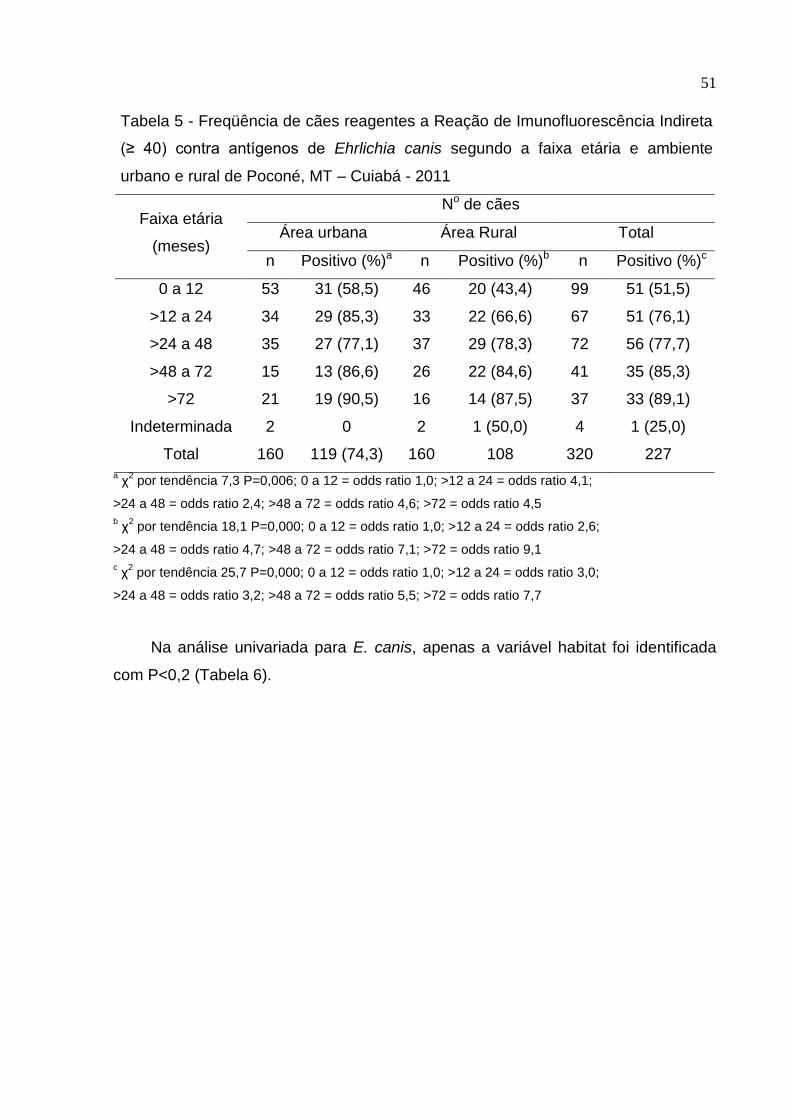
51
Tabela 5 - Freqüência de cães reagentes a Reação de Imunofluorescência Indireta
(≥ 40) contra antígenos de Ehrlichia canis segundo a faixa etária e ambiente
urbano e rural de Poconé, MT – Cuiabá - 2011
Faixa etária
(meses)
No de cães
Área urbana Área Rural Total
n Positivo (%)a n Positivo (%)b n Positivo (%)c
0 a 12 53 31 (58,5) 46 20 (43,4) 99 51 (51,5)
>12 a 24 34 29 (85,3) 33 22 (66,6) 67 51 (76,1)
>24 a 48 35 27 (77,1) 37 29 (78,3) 72 56 (77,7)
>48 a 72 15 13 (86,6) 26 22 (84,6) 41 35 (85,3)
>72 21 19 (90,5) 16 14 (87,5) 37 33 (89,1)
Indeterminada 2 0 2 1 (50,0) 4 1 (25,0)
Total 160 119 (74,3) 160 108 320 227
a χ
2 por tendência 7,3 P=0,006; 0 a 12 = odds ratio 1,0; >12 a 24 = odds ratio 4,1;
>24 a 48 = odds ratio 2,4; >48 a 72 = odds ratio 4,6; >72 = odds ratio 4,5
b χ
2 por tendência 18,1 P=0,000; 0 a 12 = odds ratio 1,0; >12 a 24 = odds ratio 2,6;
>24 a 48 = odds ratio 4,7; >48 a 72 = odds ratio 7,1; >72 = odds ratio 9,1
c χ
2 por tendência 25,7 P=0,000; 0 a 12 = odds ratio 1,0; >12 a 24 = odds ratio 3,0;
>24 a 48 = odds ratio 3,2; >48 a 72 = odds ratio 5,5; >72 = odds ratio 7,7
Na análise univariada para E. canis, apenas a variável habitat foi identificada
com P<0,2 (Tabela 6).
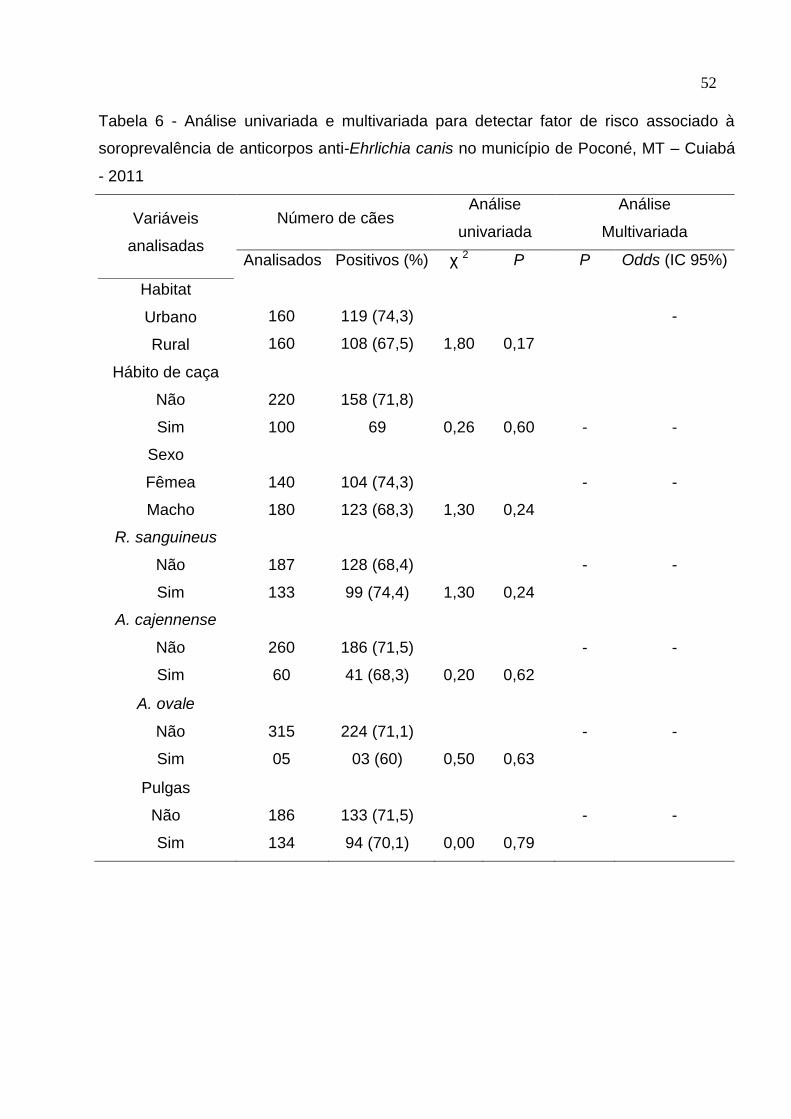
52
Tabela 6 - Análise univariada e multivariada para detectar fator de risco associado à
soroprevalência de anticorpos anti-Ehrlichia canis no município de Poconé, MT – Cuiabá
- 2011
Variáveis
analisadas
Número de cães Análise
univariada
Análise
Multivariada
Analisados Positivos (%) χ 2 P P Odds (IC 95%)
Habitat
Urbano
Rural
160
160
119 (74,3)
108 (67,5)
1,80
0,17
-
Hábito de caça
Não
Sim
220
100
158 (71,8)
69
0,26
0,60
-
-
Sexo
Fêmea
Macho
140
180
104 (74,3)
123 (68,3)
1,30
0,24
- -
R. sanguineus
Não
Sim
187
133
128 (68,4)
99 (74,4)
1,30
0,24
- -
A. cajennense
Não
Sim
260
60
186 (71,5)
41 (68,3)
0,20
0,62
- -
A. ovale
Não
Sim
315
05
224 (71,1)
03 (60)
0,50
0,63
- -
Pulgas
Não
Sim
186
134
133 (71,5)
94 (70,1)
0,00
0,79
- -
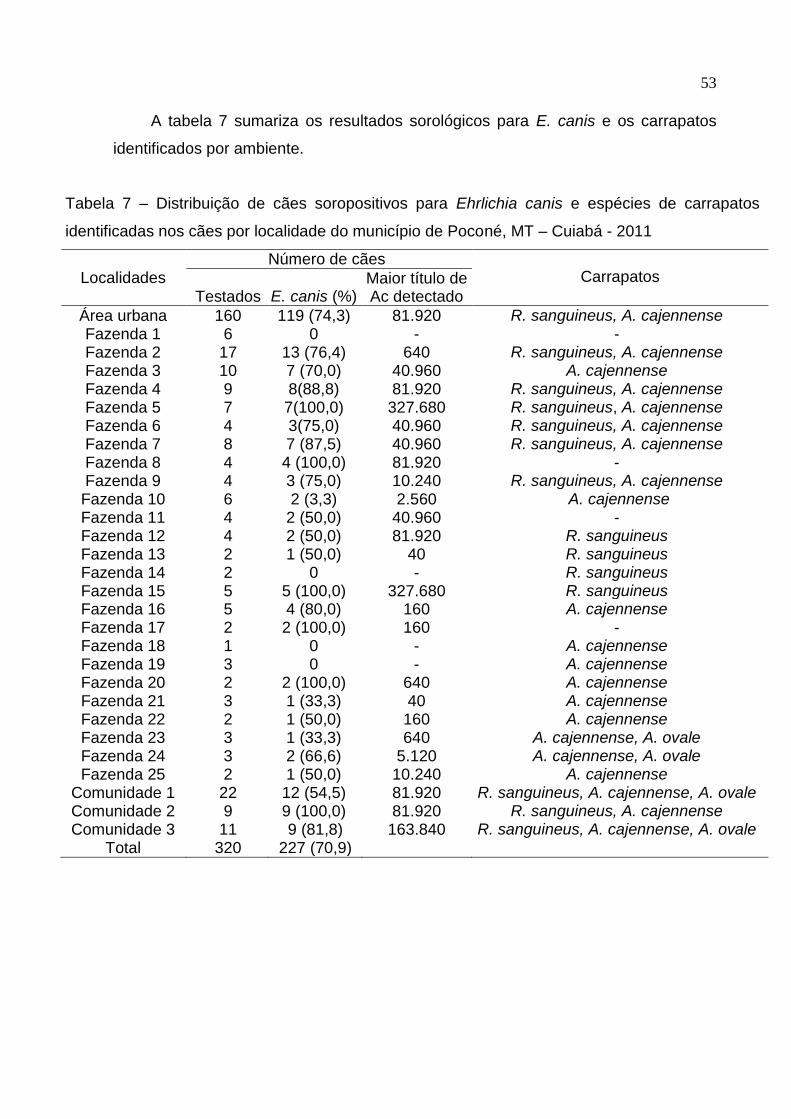
53
A tabela 7 sumariza os resultados sorológicos para E. canis e os carrapatos
identificados por ambiente.
Tabela 7 – Distribuição de cães soropositivos para Ehrlichia canis e espécies de carrapatos
identificadas nos cães por localidade do município de Poconé, MT – Cuiabá - 2011
Localidades Número de cães
Carrapatos
Testados E. canis (%) Maior título de Ac detectado
Área urbana 160 119 (74,3) 81.920 R. sanguineus, A. cajennense Fazenda 1 6 0 - - Fazenda 2 17 13 (76,4) 640 R. sanguineus, A. cajennense Fazenda 3 10 7 (70,0) 40.960 A. cajennense Fazenda 4 9 8(88,8) 81.920 R. sanguineus, A. cajennense Fazenda 5 7 7(100,0) 327.680 R. sanguineus, A. cajennense Fazenda 6 4 3(75,0) 40.960 R. sanguineus, A. cajennense Fazenda 7 8 7 (87,5) 40.960 R. sanguineus, A. cajennense Fazenda 8 4 4 (100,0) 81.920 - Fazenda 9 4 3 (75,0) 10.240 R. sanguineus, A. cajennense Fazenda 10 6 2 (3,3) 2.560 A. cajennense Fazenda 11 4 2 (50,0) 40.960 - Fazenda 12 4 2 (50,0) 81.920 R. sanguineus Fazenda 13 2 1 (50,0) 40 R. sanguineus Fazenda 14 2 0 - R. sanguineus Fazenda 15 5 5 (100,0) 327.680 R. sanguineus Fazenda 16 5 4 (80,0) 160 A. cajennense Fazenda 17 2 2 (100,0) 160 - Fazenda 18 1 0 - A. cajennense Fazenda 19 3 0 - A. cajennense Fazenda 20 2 2 (100,0) 640 A. cajennense Fazenda 21 3 1 (33,3) 40 A. cajennense Fazenda 22 2 1 (50,0) 160 A. cajennense Fazenda 23 3 1 (33,3) 640 A. cajennense, A. ovale Fazenda 24 3 2 (66,6) 5.120 A. cajennense, A. ovale Fazenda 25 2 1 (50,0) 10.240 A. cajennense
Comunidade 1 22 12 (54,5) 81.920 R. sanguineus, A. cajennense, A. ovale Comunidade 2 9 9 (100,0) 81.920 R. sanguineus, A. cajennense Comunidade 3 11 9 (81,8) 163.840 R. sanguineus, A. cajennense, A. ovale
Total 320 227 (70,9)

54
4.3 Detecção de anticorpos anti-Rickettsia spp.
Ao todo, 152 (47,5%; IC 95%: 42,0 – 52,9%) cães foram sororeagentes a pelo
menos uma espécie de Rickettsia spp., com títulos variando entre 64 a 32.768
(Tabela 8).
Tabela 8 - Distribuição dos títulos de anticorpos frente aos 6 antígenos de Rickettsia
spp. utilizados na RIFI (≥64) em cães das áreas urbana e rural do município de
Poconé, MT – Cuiabá - 2011
Espécies
Ambientes
Urbano Rural
Títulos detectados (n*) Títulos detectados (n*)
R. amblyommii 128 (3), 512 (2), 1.024 (1),
8.192 (1)
512 (3), 1.024 (5), 2.048 (7),
4.096 (10), 8.192 (13), 16.384
(12), 32.768 (6)
R. parkeri 128 (1), 256 (1) 128 (1), 512 (1)
R. rickettsii 512 (1), 8.192 (1) 512 (1)
R. rhipicephali 256 (1), 512 (1) 256 (1)
R. belli 0 128 (1), 256 (1), 512 (1), 1.024
(1)
R. felis 0 0
*n: número de cães
Enquanto 31 (19,3%; 95% IC: 13,8 – 26,0%) cães da área urbana foram
sororeagentes, na área rural 121 (75,6%; 95% IC: 68,5 – 81,8%) cães reagiram
(P<0,05). Ao todo, 127 (39,6%), 108 (33,7%), 108 (33,7%), 62 (19,3%), 43 (13,4%),
e 42 (13,16%) cães foram reagentes a R. amblyommii, R. rhipicephali, R. parkeri, R.
rickettsii, R. bellii e R. felis, respectivamente.
Um total de 63 (41,4%) cães, de 23 localidades apresentaram título final para
R. amblyommii pelo menos quatro vezes maior do que os outros cinco antígenos. Os
títulos de anticorpos observados nestes cães foram considerados como sendo
estimulados pela R. amblyommii ou uma espécie próxima. Com base nesses

55
critérios, somente 3 a 4 soros foram considerados estimulados por R. parkeri, R.
bellii, R. rickettsii ou R. rhipicephali. Para os outros 75 cães sororeagentes, não foi
possível determinar o agente infeccioso, porque eles apresentaram títulos
semelhantes (diferença inferior a 4 vezes) para duas ou mais espécies de Rickettsia
spp. ou tiveram um único título de 64 para somente uma espécie Rickettsia spp.
(Tabela 9).
.
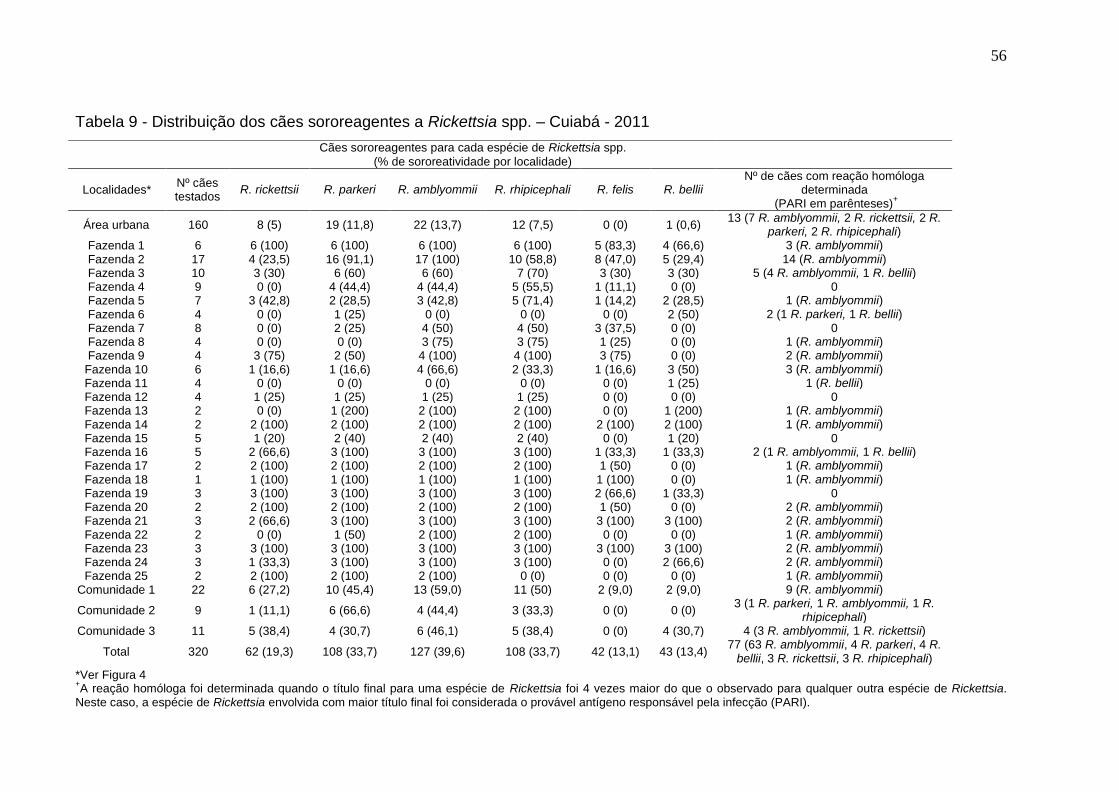
56
Tabela 9 - Distribuição dos cães sororeagentes a Rickettsia spp. – Cuiabá - 2011
*Ver Figura 4 +A reação homóloga foi determinada quando o título final para uma espécie de Rickettsia foi 4 vezes maior do que o observado para qualquer outra espécie de Rickettsia.
Neste caso, a espécie de Rickettsia envolvida com maior título final foi considerada o provável antígeno responsável pela infecção (PARI).
Cães sororeagentes para cada espécie de Rickettsia spp.
(% de sororeatividade por localidade)
Localidades* Nº cães testados
R. rickettsii R. parkeri R. amblyommii R. rhipicephali R. felis R. bellii Nº de cães com reação homóloga
determinada (PARI em parênteses)
+
Área urbana 160 8 (5) 19 (11,8) 22 (13,7) 12 (7,5) 0 (0) 1 (0,6) 13 (7 R. amblyommii, 2 R. rickettsii, 2 R.
parkeri, 2 R. rhipicephali) Fazenda 1 6 6 (100) 6 (100) 6 (100) 6 (100) 5 (83,3) 4 (66,6) 3 (R. amblyommii)
Fazenda 2 17 4 (23,5) 16 (91,1) 17 (100) 10 (58,8) 8 (47,0) 5 (29,4) 14 (R. amblyommii) Fazenda 3 10 3 (30) 6 (60) 6 (60) 7 (70) 3 (30) 3 (30) 5 (4 R. amblyommii, 1 R. bellii) Fazenda 4 9 0 (0) 4 (44,4) 4 (44,4) 5 (55,5) 1 (11,1) 0 (0) 0 Fazenda 5 7 3 (42,8) 2 (28,5) 3 (42,8) 5 (71,4) 1 (14,2) 2 (28,5) 1 (R. amblyommii)
Fazenda 6 4 0 (0) 1 (25) 0 (0) 0 (0) 0 (0) 2 (50) 2 (1 R. parkeri, 1 R. bellii) Fazenda 7 8 0 (0) 2 (25) 4 (50) 4 (50) 3 (37,5) 0 (0) 0 Fazenda 8 4 0 (0) 0 (0) 3 (75) 3 (75) 1 (25) 0 (0) 1 (R. amblyommii) Fazenda 9 4 3 (75) 2 (50) 4 (100) 4 (100) 3 (75) 0 (0) 2 (R. amblyommii)
Fazenda 10 6 1 (16,6) 1 (16,6) 4 (66,6) 2 (33,3) 1 (16,6) 3 (50) 3 (R. amblyommii) Fazenda 11 4 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 1 (25) 1 (R. bellii) Fazenda 12 4 1 (25) 1 (25) 1 (25) 1 (25) 0 (0) 0 (0) 0 Fazenda 13 2 0 (0) 1 (200) 2 (100) 2 (100) 0 (0) 1 (200) 1 (R. amblyommii)
Fazenda 14 2 2 (100) 2 (100) 2 (100) 2 (100) 2 (100) 2 (100) 1 (R. amblyommii) Fazenda 15 5 1 (20) 2 (40) 2 (40) 2 (40) 0 (0) 1 (20) 0 Fazenda 16 5 2 (66,6) 3 (100) 3 (100) 3 (100) 1 (33,3) 1 (33,3) 2 (1 R. amblyommii, 1 R. bellii) Fazenda 17 2 2 (100) 2 (100) 2 (100) 2 (100) 1 (50) 0 (0) 1 (R. amblyommii)
Fazenda 18 1 1 (100) 1 (100) 1 (100) 1 (100) 1 (100) 0 (0) 1 (R. amblyommii) Fazenda 19 3 3 (100) 3 (100) 3 (100) 3 (100) 2 (66,6) 1 (33,3) 0 Fazenda 20 2 2 (100) 2 (100) 2 (100) 2 (100) 1 (50) 0 (0) 2 (R. amblyommii) Fazenda 21 3 2 (66,6) 3 (100) 3 (100) 3 (100) 3 (100) 3 (100) 2 (R. amblyommii)
Fazenda 22 2 0 (0) 1 (50) 2 (100) 2 (100) 0 (0) 0 (0) 1 (R. amblyommii) Fazenda 23 3 3 (100) 3 (100) 3 (100) 3 (100) 3 (100) 3 (100) 2 (R. amblyommii) Fazenda 24 3 1 (33,3) 3 (100) 3 (100) 3 (100) 0 (0) 2 (66,6) 2 (R. amblyommii) Fazenda 25 2 2 (100) 2 (100) 2 (100) 0 (0) 0 (0) 0 (0) 1 (R. amblyommii)
Comunidade 1 22 6 (27,2) 10 (45,4) 13 (59,0) 11 (50) 2 (9,0) 2 (9,0) 9 (R. amblyommii)
Comunidade 2 9 1 (11,1) 6 (66,6) 4 (44,4) 3 (33,3) 0 (0) 0 (0) 3 (1 R. parkeri, 1 R. amblyommii, 1 R.
rhipicephali) Comunidade 3 11 5 (38,4) 4 (30,7) 6 (46,1) 5 (38,4) 0 (0) 4 (30,7) 4 (3 R. amblyommii, 1 R. rickettsii)
Total 320 62 (19,3) 108 (33,7) 127 (39,6) 108 (33,7) 42 (13,1) 43 (13,4) 77 (63 R. amblyommii, 4 R. parkeri, 4 R.
bellii, 3 R. rickettsii, 3 R. rhipicephali)
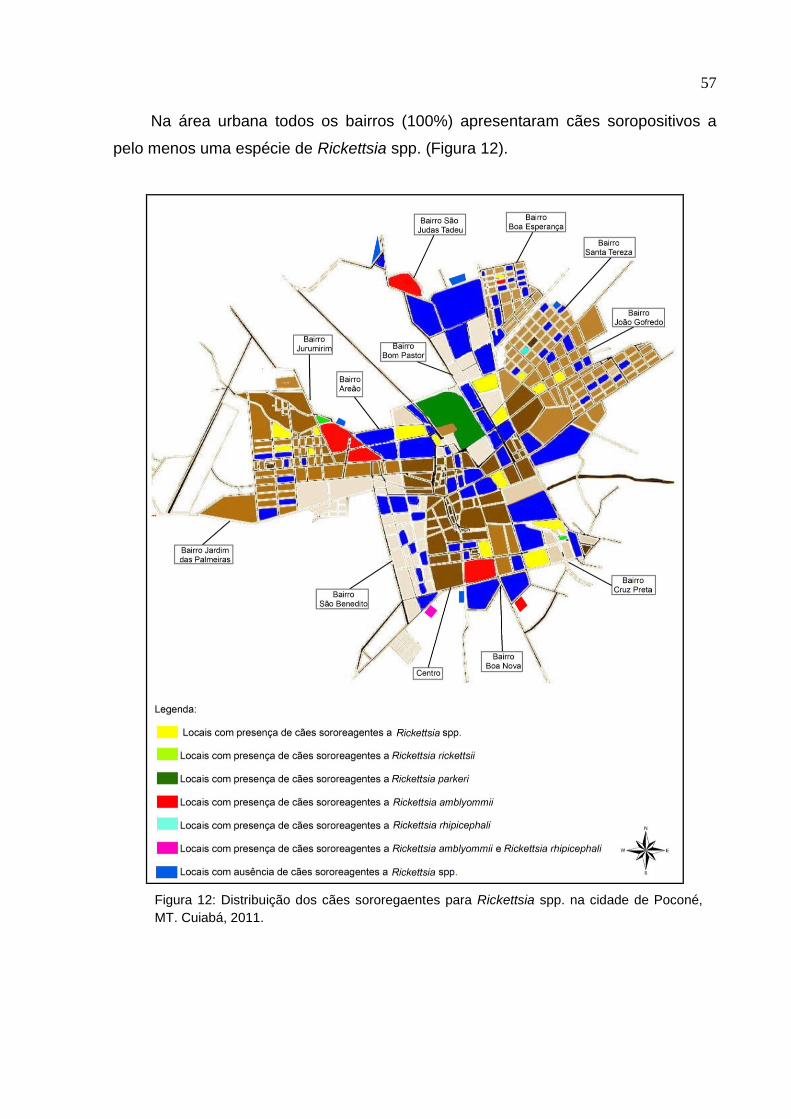
57
Na área urbana todos os bairros (100%) apresentaram cães soropositivos a
pelo menos uma espécie de Rickettsia spp. (Figura 12).
Figura 12: Distribuição dos cães sororegaentes para Rickettsia spp. na cidade de Poconé,
MT. Cuiabá, 2011.

58
Na área rural, todas as fazendas e todas as comunidades rurais apresentaram
cães reagentes para Rickettsia spp. (Figura 13).
Figura 13: Prevalência das espécies do gênero Rickettsia na área rural do municcípio de
Poconé, MT. Cuiabá, 2011.
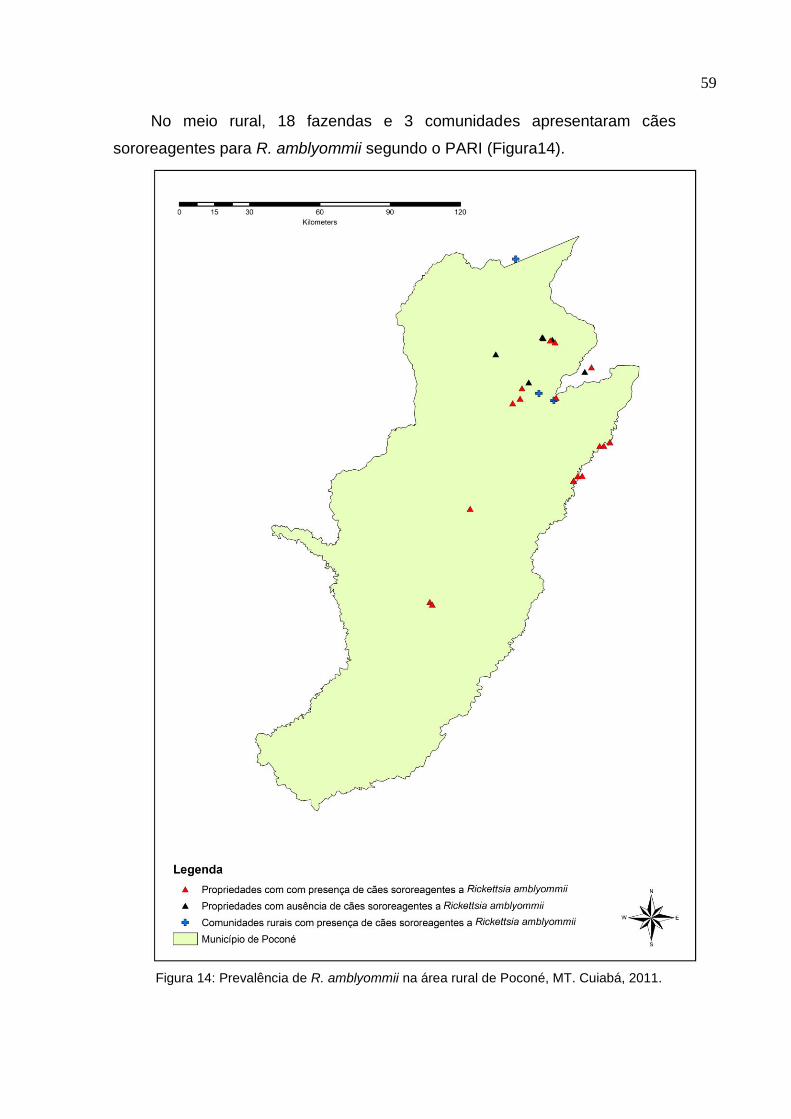
59
No meio rural, 18 fazendas e 3 comunidades apresentaram cães
sororeagentes para R. amblyommii segundo o PARI (Figura14).
Figura 14: Prevalência de R. amblyommii na área rural de Poconé, MT. Cuiabá, 2011.

60
Quatro fazendas apresentaram cães com anticorpos contra R. bellii segundo
o PARI (Figura 15).
Figura 15: Prevalência de R. bellii na área rural de Poconé, MT. Cuiabá, 2011.

61
Contra R. parkeri, 1 fazenda e em 1 comunidade rural apresentaram cães
sororeagentes segundo o PARI (Figura 16).
Figura 16: Prevalência de R. parkeri na área rural de Poconé, MT. Cuiabá, 2011.
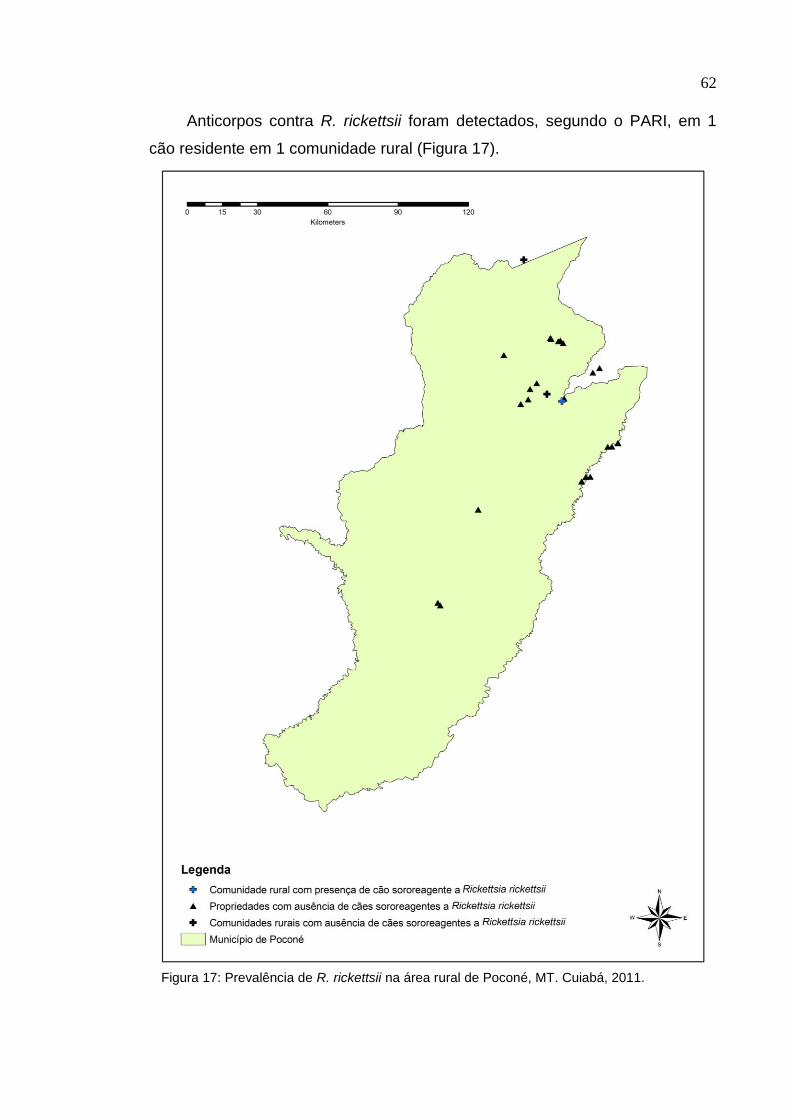
62
Anticorpos contra R. rickettsii foram detectados, segundo o PARI, em 1
cão residente em 1 comunidade rural (Figura 17).
Figura 17: Prevalência de R. rickettsii na área rural de Poconé, MT. Cuiabá, 2011.
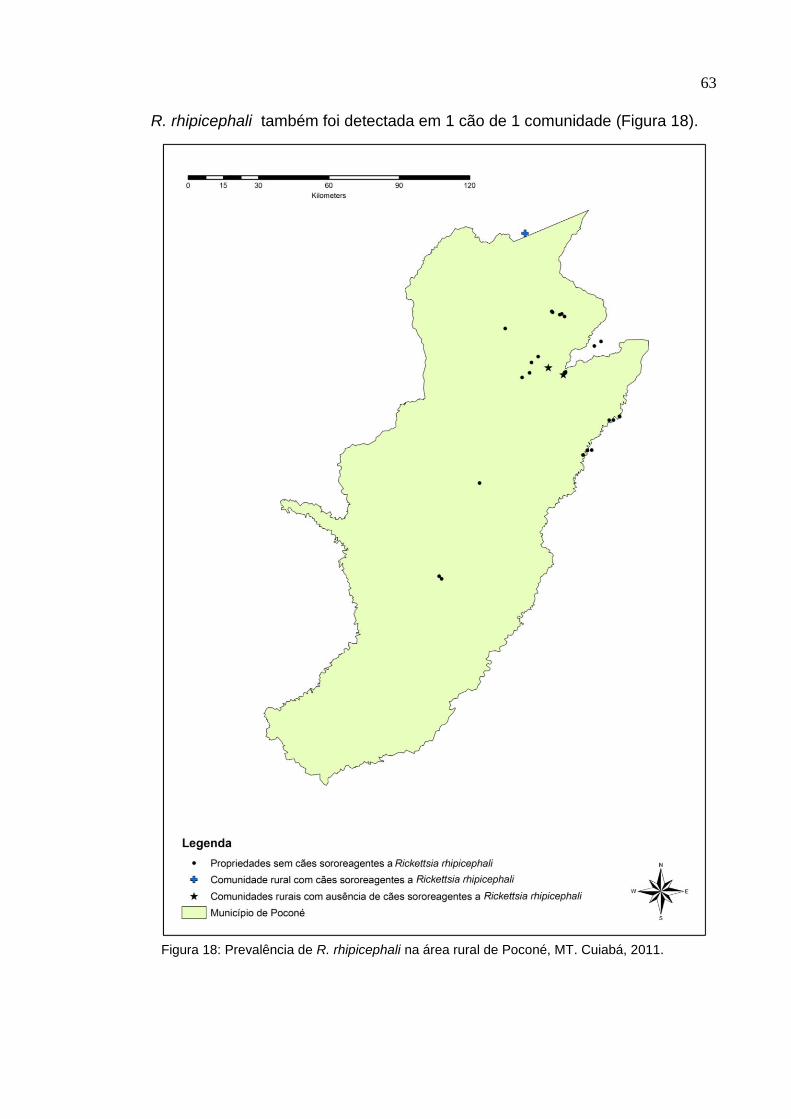
63
R. rhipicephali também foi detectada em 1 cão de 1 comunidade (Figura 18).
Figura 18: Prevalência de R. rhipicephali na área rural de Poconé, MT. Cuiabá, 2011.

64
Na Tabela 10 estão discriminados os prováveis antígenos do gênero Rickettsia
responsáveis pelas reações sorológicas e o parasitismo por diferentes espécies de
carrapatos.
Tabela 10 - Distribuição dos cães sororeagentes a Ricketttsia spp. e parasitismo de carrapatos
em diferentes localidades de Poconé, MT – Cuiabá - 2011
Localidades Número de cães
Carrapatos Testados Provável Rickettsia detectada (maior título por localidade)
Área urbana 160 7 R. amblyommii (8.192); 2 R. parkeri (256); 2 R. rickettsii (8.192),
2 R. rhipicephali (512); 18 Rickettsia spp. (1.024) R. sanguineus, A. cajennense
Fazenda 1 6 3 R. amblyommii (16.384); 3 Rickettsia spp. (8.192) -
Fazenda 2 17 14 R. amblyommii (16.384); 3 Rickettsia spp. (16.384) R. sanguineus, A. cajennense
Fazenda 3 10 4 R. amblyommii (16.384); 1 R. bellii (1.024); 3 Rickettsia spp.
(4.096) A. cajennense
Fazenda 4 9 5 Rickettsia spp. (4.096) R. sanguineus, A. cajennense
Fazenda 5 7 1 R. amblyommii (512); 5 Rickettsia spp. (2.048) R. sanguineus, A. cajennense
Fazenda 6 4 1 R. parkeri (512); 1 R. bellii (256); 1 Rickettsia spp. (64) R. sanguineus, A. cajennense
Fazenda 7 8 4 Rickettsia spp. (4.096) R. sanguineus, A. cajennense
Fazenda 8 4 1 R. amblyommii (2.048); 2 Rickettsia spp. (2.048) -
Fazenda 9 4 2 R. amblyommii (8.192); 2 Rickettsia spp. (16.384) R. sanguineus, A. cajennense
Fazenda 10 6 3 R. amblyommii (16.384); 1 Rickettsia spp. (512) A. cajennense
Fazenda 11 4 1 R.bellii (512) -
Fazenda 12 4 1 Rickettsia spp. (2.048) R. sanguineus
Fazenda 13 2 1 R. amblyommii (1.024); 1 Rickettsia spp. (1.024) R. sanguineus
Fazenda 14 2 1 R. amblyommii (32.768); 1 Rickettsia spp. (2.048) R. sanguineus
Fazenda 15 5 2 Rickettsia spp. (4.096) R. sanguineus
Fazenda 16 5 1 R. amblyommii (4.096); 1 R. bellii (128);
2 Rickettsia spp. (8,192) A. cajennense
Fazenda 17 2 1 R. amblyommii (4.096); 1 Rickettsia spp. (8.192) -
Fazenda 18 1 1 R. amblyommii (16.384) A. cajennense
Fazenda 19 3 3 Rickettsia spp. (16.384) A. cajennense
Fazenda 20 2 2 R. amblyommii (32.768) A. cajennense
Fazenda 21 3 2 R. amblyommii (16.384); 1 Rickettsia spp. (16.384) A. cajennense
Fazenda 22 2 1 R. amblyommii (16.384); 1 Rickettsia spp. (16.384) A. cajennense
Fazenda 23 3 2 R. amblyommii (32.768); 1 Rickettsia spp. (32.768) A. cajennense, A. ovale
Fazenda 24 3 2 R. amblyommii (32.768);1 Rickettsia spp. (16.384) A. cajennense, A. ovale
Fazenda 25 2 1 R. amblyommii (32.768); 1 Rickettsia spp. (8.192) A. cajennense
Comunidade 1 22 9 R. amblyommii (4.096); 4 Rickettsia spp. (1.024) R. sanguineus, A. cajennense,
A. ovale
Comunidade 2 9 1 R. amblyommii (512); 1 R. parkeri (128); 1 R. rhipicephali (256);
5 Rickettsia spp. (4.096) R. sanguineus, A. cajennense
Comunidade 3 11 1 R. rickettsii (512); 3 R. amblyommii (16.384);
3 Rickettsia spp. (8.192) R. sanguineus, A. cajennense,
A. ovale
Total 320 77 (63 R. amblyommii, 4 R. parkeri, 4 R. bellii, 3 R. rickettsii,
3 R. rhipicephali)
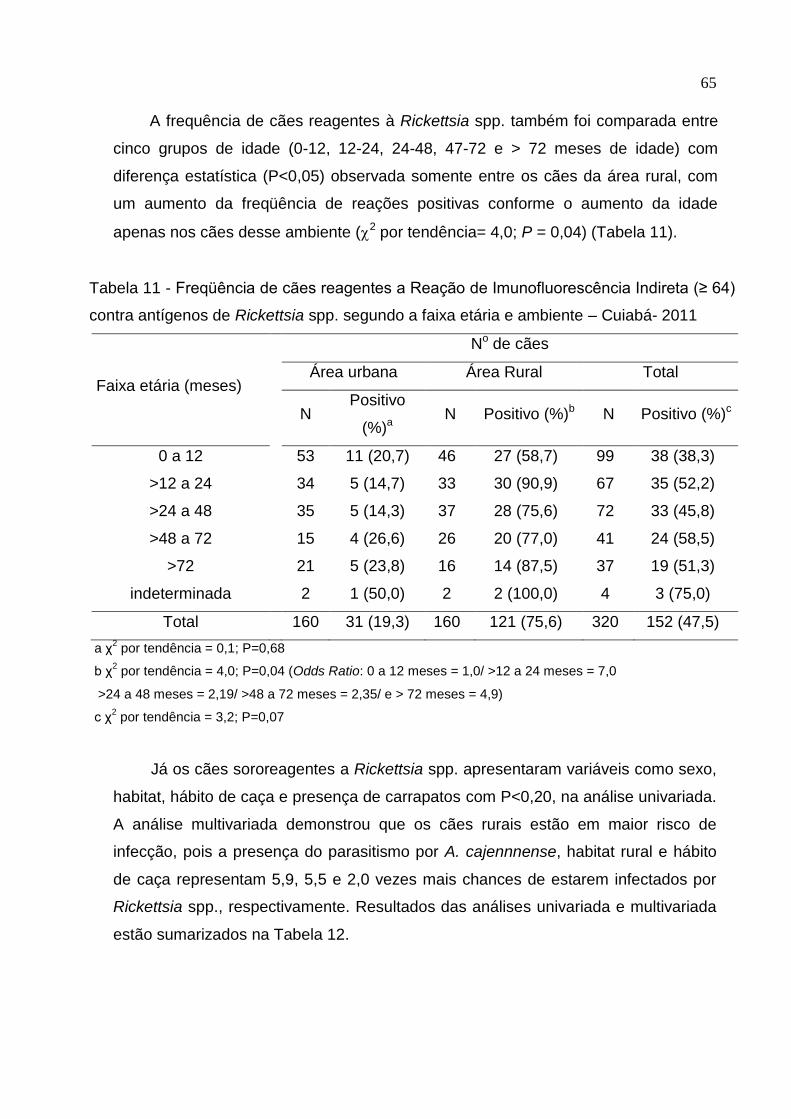
65
A frequência de cães reagentes à Rickettsia spp. também foi comparada entre
cinco grupos de idade (0-12, 12-24, 24-48, 47-72 e > 72 meses de idade) com
diferença estatística (P<0,05) observada somente entre os cães da área rural, com
um aumento da freqüência de reações positivas conforme o aumento da idade
apenas nos cães desse ambiente (2 por tendência= 4,0; P = 0,04) (Tabela 11).
Tabela 11 - Freqüência de cães reagentes a Reação de Imunofluorescência Indireta (≥ 64)
contra antígenos de Rickettsia spp. segundo a faixa etária e ambiente – Cuiabá- 2011
Faixa etária (meses)
No de cães
Área urbana Área Rural Total
N Positivo
(%)a N Positivo (%)b N Positivo (%)c
0 a 12 53 11 (20,7) 46 27 (58,7) 99 38 (38,3)
>12 a 24 34 5 (14,7) 33 30 (90,9) 67 35 (52,2)
>24 a 48 35 5 (14,3) 37 28 (75,6) 72 33 (45,8)
>48 a 72 15 4 (26,6) 26 20 (77,0) 41 24 (58,5)
>72 21 5 (23,8) 16 14 (87,5) 37 19 (51,3)
indeterminada 2 1 (50,0) 2 2 (100,0) 4 3 (75,0)
Total 160 31 (19,3) 160 121 (75,6) 320 152 (47,5)
a χ2 por tendência = 0,1; P=0,68
b χ2 por tendência = 4,0; P=0,04 (Odds Ratio: 0 a 12 meses = 1,0/ >12 a 24 meses = 7,0
>24 a 48 meses = 2,19/ >48 a 72 meses = 2,35/ e > 72 meses = 4,9)
c χ2 por tendência = 3,2; P=0,07
Já os cães sororeagentes a Rickettsia spp. apresentaram variáveis como sexo,
habitat, hábito de caça e presença de carrapatos com P<0,20, na análise univariada.
A análise multivariada demonstrou que os cães rurais estão em maior risco de
infecção, pois a presença do parasitismo por A. cajennnense, habitat rural e hábito
de caça representam 5,9, 5,5 e 2,0 vezes mais chances de estarem infectados por
Rickettsia spp., respectivamente. Resultados das análises univariada e multivariada
estão sumarizados na Tabela 12.
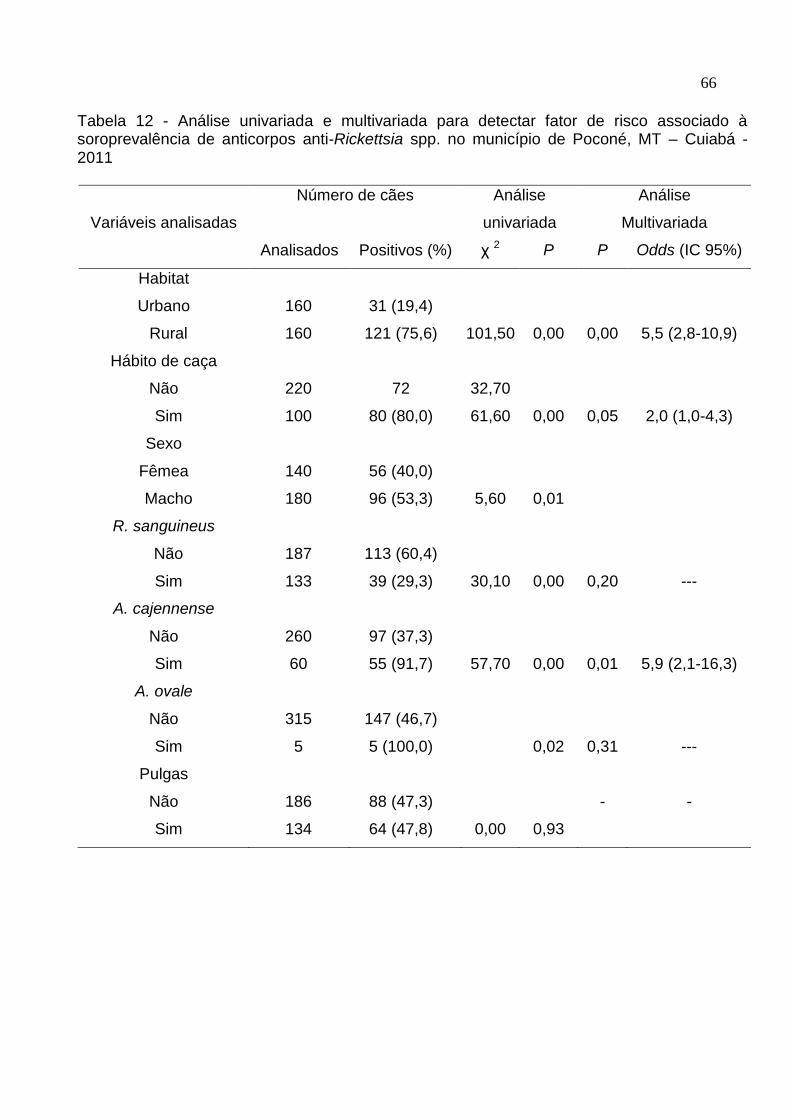
66
Tabela 12 - Análise univariada e multivariada para detectar fator de risco associado à soroprevalência de anticorpos anti-Rickettsia spp. no município de Poconé, MT – Cuiabá - 2011
Variáveis analisadas
Número de cães
Análise
univariada
Análise
Multivariada
Analisados Positivos (%) χ 2 P P Odds (IC 95%)
Habitat
Urbano
Rural
160
160
31 (19,4)
121 (75,6)
101,50
0,00
0,00
5,5 (2,8-10,9)
Hábito de caça
Não
Sim
220
100
72
80 (80,0)
32,70
61,60
0,00
0,05
2,0 (1,0-4,3)
Sexo
Fêmea
Macho
140
180
56 (40,0)
96 (53,3)
5,60
0,01
R. sanguineus
Não
Sim
187
133
113 (60,4)
39 (29,3)
30,10
0,00
0,20
---
A. cajennense
Não
Sim
260
60
97 (37,3)
55 (91,7)
57,70
0,00
0,01
5,9 (2,1-16,3)
A. ovale
Não
Sim
315
5
147 (46,7)
5 (100,0)
0,02
0,31
---
Pulgas
Não
Sim
186
134
88 (47,3)
64 (47,8)
0,00
0,93
- -

67
5 DISCUSSÃO
Este trabalho relata pela primeira vez a prevalência de anticorpos anti-E. canis
e Rickettsia spp., bem como a presença de ectoparasitos em cães da região norte
do Pantanal. De maneira geral, a prevalência (70,9%) de anticorpos anti-E. canis
encontrada neste trabalho é o maior índice detectado no Brasil. Estudos recentes no
país mostram que que a soroprevalência de E. canis nos cães varia conforme a
região estudada.
Em estudo realizado na região Norte do país, Aguiar et al. (2007a)
pesquisaram quanto à presença de anticorpos anti-E. canis em cães urbanos e
rurais do município de Monte Negro, RO, onde a soroprevalência encontrada foi de
31,2%., com diferença estatística significativa entre as prevalências das áreas
urbana (37,9%) e rural (24,8%). Ao passo que nesse estudo essa diferença não foi
observada.
Essa predominância de R. sanguineus no ambiente urbano, conforme
representado na Figura 5 também foi observada nos cães residentes em Cuiabá,
capital de Mato Grosso. O inquérito sorológico realizado com esses animais mostrou
que 42,5% eram reagentes à E. canis, e, igualmente ao nosso estudo, não foi
identificado associação entre a infestação de carrapatos e a presença de anticorpos
frente essa bactéria (SILVA et al., 2010). O resultado da análise sorológica indica
que o cão foi exposto ao agente infeccioso e por isso, desenvolveu anticorpos.
Entretando, o fato dele não apresentar parasitismo pelo carrapato no momento da
colheita da amostra sanguínea, não significa que ele não possa ter sido parasitado e
infectado outrora.
No Nordeste, Souza et al. (2010) detectaram 35,6% dos cães sororeagentes a
E. canis em dois distritos de Salvador, BA. Adicionalmente, foi realizado análise
molecular das amostras sanguíneas e dos carrapatos (todos da espécie R.
sanguineus) que parasitavam esses animais. Como resultado, 34,5% dos cães
foram PCR-positivos para este patógeno; enquanto que a infecção nos carrapatos
foi observada em 21, 9%. Apesar do presente estudo não ter por objetivo a detecção
molecular de E. canis, pode-se supor que existe a circulação dessa bactéria também
entre o carrapato vetor, uma vez que a mesma também foi relatada em R.

68
sanguineus de outras localidades do Brasil, com prevalência mínima de infecção
variando entre 2,3% a 3,7% (AGUIAR et al., 2007a).
Apesar de o ambiente rural apresentar maior infestação por espécies do
gênero Amblyomma spp. (A. cajennense: 38,7% de cães infestados e A. ovale: 3,1%
de cães parasitados) quando comparada ao R. sanguineus (19,3% de cães
infestados), nota-se que nesse local a prevalência de E. canis (67,5%) foi relevante,
superando a relatada em áreas rurais do estado de Minas Gerais, cujo valor ficou
em 44,7%, conforme apresentado por Costa et a. (2007). Com base neste achados,
este mesmo estudo aborda a hipótese do carrapato A. cajennense participar da
epidemiologia na transmissão de Ehrlichia spp., visto que assim como nós, essa foi
a espécie de carrapato mais prevalente nos cães rurais analisados naquela região.
Porém, é preciso levar em consideração que até o presente momento, o único vetor
incriminado na transmissão de E. canis é o R. sanguineus (DUMLER et al., 2001;
LABRUNA e PEREIRA, 2001).
Diferentemente da nossa prevalência, Saito et al. (2008) descreve que apenas
4,8% dos cães, provenientes em sua grande maioria de áreas rurais da região Sul
do Brasil, apresentam anticorpos anti-E. canis. Ao observarmos a distribuição
espacial da soroprevalência nos cães rurais do município de Poconé, fica evidente
no mapa (Figura 11) as 24 localidades (fazendas e comunidades rurais) com
presença de animais positivos, ao passo que somente 3 propriedades no Rio
Grande do Sul continham animais reagentes (SAITO et al., 2008).
A elevada taxa de anticorpos sugere a ocorrência da infecção por outra
possível espécie de Ehrlichia spp. na região, uma vez que os testes sorológicos não
podem distinguir a infecção por E. canis daquela causada por outras espécies deste
gênero, especialmente aquelas pertencentes ao mesmo genogrupo (HARRUS e
WANER, 2011). Analisando as características da região do Pantanal, como por
exemplo, o clima quente e úmido, é possível inferir que elas proporcionam condições
para a manutenção de agentes infecciosos como a Ehrlichia spp., haja vista que
essas características climáticas favorecem a instalação e disseminação de
carrapatos vetores.
Em se tratando da biologia do R. sanguineus, estudos apontam que ele pode
desenvolver-se em ampla faixa de temperatura e umidade relativa, podendo haver
maior rapidez na fixação em humanos e coelhos quando ele está exposto à altas
temperaturas, quando ele é considerado mais ativo, e portanto, apto a propagar a

69
infecção entre os hospedeiros vertebrados susceptíveis (HARRUS e WANER, 2011)
Desse modo, pode-se concluir que a região do Pantanal matogrossense propicia
condições ambientais favoráveis ao desenvolvimento deste carrapato, implicando
assim, na disseminação de patógenos.
Adicionalmente, a existência de uma grande variedade de espécies animais
também contribui para a sobrevivência destes artrópodes. A relevância da presença
desta grande variedade de espécies animais pode estar relacionada ao fato deles
servirem de hospedeiros capazes de manter espécies desconhecidas de Ehrlichia
spp.
Com base nos resultados da área rural, sugere-se que uma espécie
desconhecida de Ehrlichia poderia estar infectando os carrapatos e animais na
região. Nos locais onde o carrapato R. sanguineus está presente, os cães
apresentaram altos títulos (~20.480), que justifica uma reação homóloga à infecção
por E. canis. No entanto, em algumas fazendas, cães jovens (entre 3 e 12 meses)
parasitados somente por espécies de Amblyomma spp. (dados não apresentados)
apresentaram elevados títulos (~ 1.280) de anticorpos contra E. canis, sugerindo ser
conseqüência de um estímulo imunológico contra outras espécies de Ehrlichia,
tendo em vista a ocorrência de reação sorológica cruzada entre espécies do gênero
Ehrlichia (HARRUS E WANER, 2011).
Devido à adaptação da E. canis ao carrapato R. sanguineus, a prevalência de
anticorpos anti-E. canis nos cães urbanos era esperada (74,3%). Entretanto, não foi
observada diferença estatística significativa (P> 0,05) entre os resultados
sorológicos contra E. canis nos dois ambientes, uma vez que 67,5% dos cães rurais
foram soropositivos. Por outro lado, observou-se diferenças entre a prevalência de
R. sanguineus em cães urbanos (63,7%) e rurais (19,3%) (P <0,05). Apesar do R.
sanguineus preferir parasitar os cães em todas as três fases de alimentação, os
cães de áreas rurais do Brasil são parasitados principalmente por espécies do
gênero Amblyomma. É sabido que o gênero Amblyomma parasita naturalmente
várias espécies de mamíferos e aves provenientes de regiões de matas, sendo
considerados seus hospedeiros naturais. Nesse sentido, quando os cães domésticos
invadem esses ecossistemas eles comportam-se como hospedeiros acidentais
desses artrópodes, já que a presença do cão nestes ambientes não é condição
essencial para a manutenção de populações do gênero (LABRUNA e PEREIRA,

70
2001). O parasitismo dos cães por Amblyomma spp. pode estar possibilitando a
transmissão de uma espécie desconhecida de Ehrlichia spp.
Para reforçar, uma possível nova espécie de Ehrlichia spp., foi recentemente
identificada em A. cajennense e A. triste coletados de onças-pintadas (Panthera
onca) na região sul do Pantanal, que também apresentaram anticorpos contra E.
canis (WIDMER, 2009). Trata-se de um achado interessante, uma vez que o
sequenciamento das amostras PCR-positivas indicou 98% de similaridade com E.
ruminantium. Esses resultados instigam a realização de mais estudos a esse
respeito, pois a possibilidade de haver uma nova espécie do gênero Ehrlichia spp.
circulando entre os animais domésticos e silvestres na região pantaneira não pode
ser desconsiderada. Por conseguinte, como se pode observar nos mapas (Figuras 7
e 8), os cães rurais contemplados em nosso estudo estavam mais frequentemente
expostos à infestação por carrapatos Amblyomma spp., tanto aqueles provenientes
de localidades (propriedades e comunidades rurais) mais próximas ou mais
distantes da sede do município; confirmando que, embora o cão não seja
considerado hospedeiro primário desse artrópode, quando ele tem acesso às áreas
de matas, acaba sendo parasitado acidentalmente (LABRUNA e PEREIRA, 2001).
Além disso, é importante ressaltar que algumas espécies de Amblyomma spp. atuam
na epidemiologia de outras espécies de Ehrlichia spp., como, E. chaffeensis e E.
ewingii, ambas transmitidas pelo A. americanum na América do Norte, ou ainda E.
ruminantium, na África e Caribe por outros Amblyomma spp. (DUMLER et al., 2001).
Em estudo realizado na região Sul, o carrapato R. sanguineus foi considerado
um fator de risco para a erliquiose monocítica canina (DAGNONE et al., 2003). No
presente estudo, todos os bairros (100%) da área urbana e apenas 10 (38,5%)
fazendas e todas as comunidades rurais tinham cães parasitados por R. sanguineus;
porém, a existência dessa espécie de carrapato não foi associada à presença de
anticorpos anti-E. canis.
Por outro lado, encontramos associações entre idade e E. canis em ambas as
áreas, o que revela uma maior chance dos cães tornarem-se infestados por
carrapatos com o passar do tempo. Do mesmo modo, Aguiar et al. (2007a) e Silva et
al. (2010) também verificaram um aumento da freqüência de anticorpos conforme o
aumento da idade dos cães. A justificativa para esses resultados estaria no fato dos
cães mais velhos terem mais tempo de exposição ao vetor e consequetemente à E.
canis.

71
Anticorpos contra Rickettsia spp. foram encontrados em 152 (47,5%) cães de
ambas as áreas. Esse resultado retrata a ocorrência sorológica de pelo menos cinco
espécies do gênero Rickettsia no Pantanal de Mato Grosso. Diferença entre área
urbana e rural foram estatisticamente significativas e parece estar associada com
cães de áreas rurais. Segundo a totalidade de cães avaliados, o parasitismo por
Amblyomma spp., o hábito de caça e a origem rural apresentaram-se como fatores
de risco para a infecção por Rickettsia spp.
Esse resultado pode ser explicado pelas características biológicas das
espécies do gênero Amblyomma. É sabido que seus hospedeiros primários são o
eqüino, a anta e a capivara, sendo naturalmente encontrado em áreas de florestas.
Entretanto, o fato do ser humano e/ou o cão invadirem esse território, aliado ao
hábito de tocaia desses carrapatos, acabam tornando-se hospedeiros acidentais
(LABRUNA e PEREIRA, 2001). Nesse sentido, a origem rural e o hábito de caça dos
cães rurais do Pantanal matogrossense contribui, enfaticamente, para a infestação
por Amblyomma spp., o que representa um fator de risco para sororeação à
Rickettsia spp.
A associação de anticorpos contra Rickettsia spp. e a idade foi observada nas
áreas rurais, resultado este que pode ser atribuído à presença de Amblyomma spp.
nos cães, uma vez que essa infestação é considerada fator de risco para
soropositividade. Além disso, deve-se apreciar o fato de haver menor especificidade,
quanto os hospedeiros, pelos estágios imaturos da espécie A. cajennense, o que
também facilitaria maior parasitismo nos cães (ONOFRIO et al., 2006). Nosso estudo
reforça esse comportamento do carrapato, haja vista que de 209 A. cajennense
coletados dos cães, 200 eram ninfas e somente nove eram adultos
As análises sorológicas frente às espécies do gênero Rickettsia resultaram
numa predominância de anticorpos homólogos à R. amblyommii, ou uma espécie
estreitamente relacionada, cujos índices foram os maiores detectados em cães até
então, no país. A espécie foi responsável por 41,1% das reações de
imunofluorescência, na qual 4,3% (7/160) dos cães eram do ambiente urbano e 35%
(56/160) pertenciam à área rural. No Brasil, dois outros trabalhos, provenientes de
regiões geográficas distintas, relataram a presença de anticorpos contra R.
amblyommii em cães domésticos.
O primeiro foi realizado na região Norte, no estado de Rondônia, com cães
urbanos e rurais do município de Monte Negro (LABRUNA et al., 2007d). Ao todo,

72
317 cães foram investigados quanto à presença de anticorpos frente à espécies de
Rickettsia do GFM que ocorrem no Brasil. Destes, em apenas em dois cães do
ambiente rural, R. amblyommii foi considerado o provável antígeno responsável pela
infecção, demonstrando uma diferença considerável com os nossos resultados. Pela
Figura 14, observamos a ampla distribuição dessa espécie na área rural do
município de Poconé, que atingiu 35% dos cães das localidades (fazendas e
comunidades rurais) visitadas. Quanto aos cães urbanos, nosso estudo detectou
que sete cães tinham anticorpos homólogos a essa espécie, ao passo que no estudo
de Monte Negro isso não foi observado. Por outro lado, nessa mesma região
Amazônica, Labruna et al. (2004b) conseguiram isolarar R. amblyommii a partir de A.
cajennense, evidenciando a participação desse artrópode na epidemiologia do
patógeno.
Já em outro trabalho, realizado no estado do Rio Grande do Sul, Saito et al.
(2008), também pesquisaram anticorpos anti-Ricketttsia em cães urbanos e rurais,
onde somente um cão apresentou reação à R. amblyommii O fato de existir
prevalências tão diferentes entre os cães das regiões Norte e Sul, bem como entre
os cães avaliados neste estudo evidenciam que a distribuição desse agente possa
estar relacionada à presença do vetores já conhecidos ou supostamente, de
espécies de carrapatos que ainda não foram implicados no processo de transmissão
desses patógenos.
Aparentemente, R. amblyommii ocorre em todo o continente americano.
Recentemente, na província de Coclé, no Panamá, foi realizado um estudo
sorológico com cães, eqüinos e ectoparasitos coletados dos cães (R. sanguineus, A.
ovale e A. oblongoguttatum, e C. felis) e dos eqüinos (A. cajennense e Dermacentor
nitens). Após análise molecular, detectou-se o DNA de R. amblyommii nos
carrapatos A. cajennense, D. nitens, bem como, a presença de anticorpos nos cães
(6/20) e nos equinos (5/20). Este trabalho conclui afirmando que esses resultados
são indícios de que essa riquétsia possivelmente está circulando entre os carrapatos
e os animais domésticos da área estudada (BERMÚDEZ et al., 2011).
No Brasil, essa espécie já foi detectada em carrapatos A. cajennense e A.
coelebs, provenientes da Floresta Amazônica, em diferentes áreas do estado de
Rondônia (LABRUNA et al., 2004b). Também nesse estado, o agente foi encontrado
em A. longirostre (LABRUNA et al., 2004c). Já no estado de São Paulo, A.
longirostre coletados de aves silvestres da Mata Atlântica continham a bactéria

73
(OGRZEWALSKA et al., 2008). Mais recentemente, Ogrzewalska et al. (2010)
relatou novamente a infecção em A. longirostre e A. geayi que parasitavam aves
silvestres da Amazônia, enquanto que na Argentina, a mesma foi isolada a partir do
carrapato A. neumanni (LABRUNA et al., 2007a).
No centro e no sudeste dos EUA, R. amblyommii foi detectada em altas taxas
em carrapatos A. americanum (JIANG et al., 2010), na qual é suspeita de ser
patogênica para os seres humanos (APPERSON et al., 2008). Casos de pacientes
febris suspeitos de terem FMMR acabaram demonstrando maior reação sorológica
frente à R. amblyommii do que à R. rickettsii (APPERSON et al., 2008). Em nossa
região não há registros de doença em humanos transmitida por carrapatos nem
tampouco sobre alguma enfermidade febril desconhecida. Inegavelmente novos
estudos serão necessários para melhor compreensão a respeito das características
epidemiológicas da R. amblyommii no Pantanal.
Com relação à R. parkeri, ela vem sendo associada à doença em humanos nos
Estados Unidos. Trata-se de uma enfermidade com sintomatologia mais branda,
muitas vezes relacionada à linfoadenopatia e a uma lesão papular típica nos locais
de fixação do carrapato. A sua transmissão é realizada pelo A. maculatum (Estados
Unidos) e A. triste (Uruguai) (VENZAL et al., 2004; PACHECO et al., 2006), e foram
detectados no presente estudo em dois cães da área urbana e rural. No Brasil, a R.
parkeri também já foi detectada por isolamento e PCR em carrapatos A. triste
(SILVEIRA et al., 2007) e por sorologia em capivaras do estado de São Paulo
(PACHECO et al., 2007) e cães do Rio Grande do Sul (SAITO et al., 2008). Em A.
dubitatum foi identificada uma cepa de Rickettsia spp., filogeneticamente muito
próxima à R. parkeri e que temporariamente foi denominada de amostra COOPERI
(= R. parkeri) (LABRUNA et al., 2004a). Nossos resultados reforçam a afirmação de
que esta espécie poderia estar ocorrendo em áreas consideradas não endêmicas
para Febre Maculosa Brasileira (FMB) (HORTA et al., 2007).
No Brasil, a infecção em cães por R. parkeri foi detectada usando os mesmos
critérios do presente trabalho, onde soros de cães apresentaram resposta de
anticorpo predominante para R. parkeri em áreas endêmicas e não endêmicas dos
estados de São Paulo (HORTA et al., 2007). Em Rondônia, anticorpos anti-R.
parkeri foram detectados em cães tanto da área urbana quanto da área rural, assim
como observado em nosso estudo (LABRUNA et al., 2007d). Já no Rio Grande do
Sul, Saito et al. (2008) relataram a presença de cães sororeagentes a esse

74
patógeno, na qual, 39,6% (154/389) dos cães avaliados foram reagentes. Vale
ressaltar que nesse estudo, os cães que tinham acesso às áreas de pastagens ou
florestas tinham mais chances de serem sororeativos à Rickettsia spp..
Adicionalmente, a presença de populações de carrapatos A. triste e A. dubitatum no
Rio Grande do Sul poderia indicar que esses artrópodes seriam os responsáveis
pela transmissão dessa bactéria. Já em outro estudo, detectou-se que carrapatos A.
ovale estavam infectados por Rickettsia sp. cepa Mata Atlântica, uma nova cepa
capaz de provocar uma escara associada à febre maculosa no Estado de São
Paulo; sugerindo iclusive, que a espécie A. ovale possa ser o vetor da Rickettsia sp.
cepa Mata Atlântica no estado de São Paulo (SABATINI et al., 2010).
Anticorpos anti-R. bellii foram detectados em cães de quatro fazendas do
município de Poconé. Esta espécie tem sido relatada e isolada a partir de várias
espécies de carrapatos do gênero Amblyomma na América do Sul. Essa espécie já
foi isolada a partir de A. scalpturatum, A. ovale, A. rotundatum, A. humerale e A.
oblongoguttatum advindos da região Norte do Brasil (LABRUNA et al., 2004b). No
estado de São Paulo a bactéria foi isolada a partir de carrapatos da espécie A.
aureolatum que parasitavam cães provenientes de uma área endêmica para FMB
(PINTER e LABRUNA, 2006). Adicionalmente, a mesma foi identificada em
carrapatos H. juxtakochi provenientes do estado de São Paulo. Pelo fato de ter sido
identificada em tantas espécies de carrapatos, certamente essa espécie ser
considerada como a riquétsia que mais freqüentemente infecta esses artrópodes no
país, apesar de não haver nenhum registro de patogenicidade em animais ou em
seres humanos (LABRUNA et al., 2007b).
Ao que se refere à espécie R. rhipicephali, verificou-se que apenas três cães
foram sororeagentes, sendo 2 cães urbanos e apenas um da área rural. No Brasil,
R. rhipicephali foi detectada no carrapato H. juxtakochi de Rondônia e São Paulo
(LABRUNA et al., 2005a, 2007b), e foi encontrada infectando cães pertencentes ao
ambiente rural na Amazônia (LABRUNA et al., 2007d). Desde seu primeiro
isolamento em R. sanguineus nos Estados Unidos, esse agente nunca foi associado
à infecção humana (LABRUNA et al., 2007b).
R. rickettsii é o principal agente responsável pela FMB no Brasil, enfermidade
caracterizada pela gravidade clínica e elevada letalidade em casos registrados na
região sudeste (ANGERAMI et al., 2006). Em algumas áreas desta região
consideradas endêmicas, a soroprevalência de R. rickettsii nos cães pode atingir

75
cerca de 70% (MORAES-FILHO et al., 2009) ou ainda observa-se a predominância
de cães com elevados títulos de anticorpos nos testes sorológicos (HORTA et al.,
2007; PINTER et al., 2008). Em áreas não-endêmicas a soroprevalência é muito
menor e em geral, alguns cães reagem com títulos menores (SAITO et al., 2008).
Adicionalmente, R. rickettsii é capaz de causar infecção seguida de manifestação
clínica na espécie canina (LABRUNA et al., 2009b).
No presente estudo, a região é considerada não-endêmica, e os valores de
soroprevalência relatados têm similaridade com outros estudos realizados em áreas
onde a endemia não é observada (SANGIONI et al., 2005; SAITO et al., 2008).
Entretanto, os títulos detectados em um cão (8.192 para R. rickettsii, 2.048 para R.
rhipicephali, 1.024 para R. parkeri e 512 para R. amblyommii) foram semelhantes
aos cães positivos provenientes de áreas endêmicas, indicando reação homóloga
contra R. rickettsii ou uma espécie estreitamente relacionada. Esse resultado
desperta atenção pelo fato deste cão ser da área urbana, onde os carrapatos R.
sanguineus são predominantes. Até 2009, a transmissão da FMB estava associada
aos carrapatos A. cajennense (América do Sul) e A. aureolatum (região
metropolitana da cidade de São Paulo), no entanto, novos estudos apontam que a
espécie R. sanguineus participa da epidemiologia da R. rickettsii, onde o DNA da
bactéria foi identificado São Paulo (MORAES-FILHO et al., 2009).
Posteriormente, conseguiu-se isolar este agente a partir de R. sanguineus
provenientes de Minas Gerais (PACHECO et al., 2010). Piranda et al. (2011)
também demonstraram que em condições experimentais, os cães domésticos
podem atuar como hospedeiros amplificadores da R. rickettsii para os carrapatos R.
sanguineus. Nos Estados Unidos, essa espécie de carrapatos foi implicada na
transmissão de R. rickettsii em humanos com FMMR. Esses pacientes descreveram
a presença desse artrópode nos cães com os quais mantinham contato, e alguns
relataram histórico de picada de carrapato antes da ocorrência da enfermidade
(DEMMA et al., 2005). Apesar dos poucos relatos de parasitismo humano por R.
sanguineus na América do Sul (GUGLIELMONE et al., 2006), o fato desta espécie
de carrapato ter sido identificada nos cães urbanos e rurais deste estudo, denota a
ampla distribuição da espécie nos locais de moradia do cão, cuja proximidade do ser
humano apresenta grande notoriedade.
No caso de R. felis, apesar do considerável parasitismo de seu vetor - pulgas
C. felis felis - nenhum dos 320 cães apresentaram indícios de anticorpos homólogos

76
a este agente. Embora nosso objetivo não fosse identificar as bactérias encontradas
em pulgas, existem estudos que mostram, através de técnicas moleculares, a
presença de infecção nestes ectoparasitas. Da mesma forma, um estudo em
diferentes áreas do estado de São Paulo mostrou que não havia nenhuma evidência
sorológica de R. felis em cães, gatos e gambás que estavam parasitados por pulgas
infectadas por essa bactéria (HORTA et al., 2007).
No Brasil, as informações sobre a infecção por R. felis ou mecanismos de
transmissão para uma pulga ou a probabilidade de esse ectoparasita adquirir uma
nova infecção ainda não foi totalmente explicada. Estudos demonstram que a
transmissão transovariana e transestadial dessa riquétsia nos vetores pode ser
possível (WEDINCAMP e FOIL, 2002). Entretanto, apesar do elevado parasitismo
por essa espécie de pulgas nos cães e também nos gatos, e do grande número de
pulgas infectadas por essa bactéria, provenientes inclusive de áreas endêmicas para
FMB (HORTA et al., 2005), talvez devesse haver mais casos de riquetsioses do que
os relatados atualmente.
Diante do exposto, supõe-se que, por motivos desconhecidos, que somente
uma minoria de seres humanos e de animais são susceptíveis à essa infecção, uma
vez que o mecanismo de transmissão pelo vetor deste patógeno ainda não está
completamente elucidado, uma vez que não se conhece a presença de R. felis na
saliva destas pulgas, apesar de haver sido encontrada na glândula salivar das
mesmas, sugerindo outras possibilidades neste processo de vetorização. Uma
hipótese seria o contato direto de lesões cutâneas e/ou mucosas com fezes frescas
contaminadas com este agente infeccioso ou até mesmo a ingestão de pulgas
infectadas (MACALUSO et al., 2008; LABRUNA, 2009a).
Quanto aos carrapatos, a presença e predominância de R. sanguineus na área
urbana (63,7% de prevalência) já era esperada, uma vez que este carrapato
encontra-se disseminado nas áreas de todo o país, onde geralmente mantém ativa
sua capacidade parasitária durante todo ano, desde que a temperatura não se torne
muito baixa (LABRUNA e PEREIRA, 2001). Por outro lado, apenas 19,3% dos cães
rurais de 10 fazendas estavam parasitados por R. sanguineus. O parasitismo por
este carrapato nas áreas rurais geralmente é menor do que nas áreas urbanas, pois
o estabelecimento dele no meio rural está geralmente ligado aos cães que estão
confinados (mesmo que parcialmente) em pequenos locais, que de certo modo
simulam ambientes urbanos, onde se pode empreender um ciclo de vida nidícola,

77
típico dessa espécie de carrapato (LABRUNA e PEREIRA, 2001). Certamente, a
ampla disseminação do R. sanguineus verificada principalmente no ambiente urbano
corrobora com a elevada soroprevalência de anticorpos anti-E. canis, uma vez que
as características climáticas da região e as condições de moradia dos cães
analisados dão suporte ao parasitismo por este vetor, o que garantiria a circulação
do mesmo e quiçá, de outros patógenos entre os hospedeiros susceptíveis.
A baixa prevalência da espécie A. cajennense na área urbana (0,6%) contrasta
com o grande número de carrapatos encontrado na área rural, representado
principalmente por ninfas de A. cajennense e adultos de A. ovale. Um resultado
semelhante também foi descrito por Dantas-Torres (2009), que detectou A.
cajennense e A. ovale em cães de áreas rurais do Nordeste do Brasil. Os cães de
ambiente rural da região do Pantanal muitas vezes são utilizados no pastoreio do
gado ou na prática da caça; hábitos estes que permitem o acesso às pastagens do
gado e aos habitats da vida selvagem.
Como já foi comentado, a presença de cavalos, e de espécies selvagens, como
capivaras (H. hydrochaeris) e antas (T. terrestris) na área rural (dados não
apresentados) pode estar associado a presença de carrapatos do gênero
Amblyomma verificada neste estudo, pois estes animais são considerados os
principais hospedeiros desse artrópode (LABRUNA et al., 2001; SZABÓ et al.,
2001).
Além disso, A. ovale foi considerada uma das espécies de carrapatos mais
comuns parasitando carnívoros silvestres em cães rurais do Brasil, inclusive na
região do Pantanal (SZABÓ et al., 2001; LABRUNA et al., 2005b). Recentemente,
em estudo realizado em Minas gerais, Szabó et al. (2010) também verificou a
presença de A. cajennense e A. ovale em cães pertencentes ao ambiente rural,
reforçando que a distribuição de espécies do gênero Amblyomma possivelmente
esteja relacionada à presença dos seus hospedeiros primários .
C. felis felis foi a única espécie de pulga observada nos cães de ambas as
áreas, porém, a prevalência na área urbana foi maior que na área rural (P <0,05).
Esta espécie tem sido relatada em outras regiões do Brasil, incluindo dentre as
regiões Sul (BELLATTO et al., 2003), Nordeste (DANTAS-TORRES et al., 2004) e o
Sudeste (RODRIGUES et al., 2001; CARDOSO et al., 2006; OLIVEIRA et al., 2008).
H. spiniger foi observado em seis cães de área urbana. Estes piolhos são
comuns em países tropicais e subtropicais (URQUHART et al., 1996). No presente

78
estudo, os cães rurais não foram encontrados parasitados por qualquer espécie de
piolho, contrapondo aos resultados relatados por Dantas-Torres et al. (2009), que
verificou que 22% dos cães de ambiente rural de Pernambuco estavam parasitados
por essa espécie de piolho.
Nossos resultados reforçam a necessidade de novas pesquisas nesta região,
pois a presença de agentes da ordem Rickettsiales e de ectoparasitos nos cães
provenientes desta área de estudo, indubitavelmente, representam uma condição
primária para a presença de patógenos transmitidos por carrapatos. Nesse sentido,
o presente estudo destaca a possibilidade de diversas zoonoses transmitidas por
carrapatos estarem circulando na região norte do Pantanal. Além disso, se
considerarmos que as atividades de pecuária e turismo, ligado principalmente à
pesca e ao safari, estão entre as práticas econômicas mais importantes na área do
Pantanal, no qual, pesquisas sobre as zoonoses transmitidas por carrapatos nesta
região é de grande relevância para a saúde pública.

79
6 CONCLUSÃO
A partir dos resultados do presente estudo concluímos que:
Carrapatos R. sanguineus são mais freqüentes na área urbana do
município de Poconé, MT;
Carrapatos A. cajenennse e A. ovale são mais freqüentes na área rural do
município de Poconé, MT;
Estágios iniciais (ninfas) de Amblyomma cajennnense são mais
frequentemente encontrados nos cães rurais de Poconé, MT, do que as
formas adultas;
Pulgas C. felis felis são mais freqüentes na área urbana do município de
Poconé, MT.
A prevalência de anticorpos anti-Ehrlichia spp. em cães foi idêntica para
ambas áreas estudadas;
A infecção por Ehrlichia spp em cães do município de Poconé, MT não
está associada à infestação por R. sanguineus;
A ocorrência de anticorpos anti-Ehrlichia spp. em cães de ambiente rural
pode ser decorrente da infecção por outra espécie que não E. canis;
Cães mais velhos sofreram maior exposição a Ehrlichia spp;
A prevalência de anticorpos anti-Rickettsia spp. em cães é maior no
ambiente rural, quando comparado ao urbano;
Habitar áreas rurais, o hábito de caça e o parasitismo por A. cajenennse
são fatores de risco para infecção por Rickettsia spp. em cães do
município de Poconé, MT;
A espécie R. amblyommii ou uma mais próxima antigenicamente é a mais
prevalente nos cães do município de Poconé, MT;
O município de Poconé, MT pode ser considerado uma zona silenciosa
para FMB.

80
REFERÊNCIAS
AGUIAR, D. M.; CAVALCANTE, G. T.; PINTER, A.; GENNARI, S. M.; CAMARGO, L. M. A.; LABRUNA, M. B. Prevalence of Ehrlichia canis (Rickettsiales: Anaplasmataceae) in dogs and Rhipicephalus sanguineus (Acari: Ixodidae) ticks from Brazil. Journal of Medical Entomology, v. 44, p.126-132, 2007a. AGUIAR, D. M.; SAITO, T. B.; HAGIWARA, M. K.; MACHADO, R. Z.; LABRUNA, M. B. Diagnóstico sorológico de erliquiose canina com antígeno brasileiro de Ehrlichia canis. Ciência Rural, v.37, p.796-802, 2007b. AGUIAR, D. M. HAGIWARA, M. K.; LABRUNA, M. B. In vitro isolation and molecular characterization of an Ehrlichia canis strain from São Paulo, Brazil. Brazilian Journal of Microbiology, v. 39, p. 489-493, 2008. ANGERAMI, R. N.; RESENDE, M. R.; FELTRIN, A. F. C.; KATZ,.G.; NASCIMENTO, E. M.; STUCCHI, R. S. B.; SILVA, L. J. Brazilian spotted fever: a case series from an endemic area in Southeastern Brazil. Clinical aspects. Annals of the New York Academy of Sciences, v. 1078, p. 252–254, 2006. APPERSON, C. S.; ENGBER, B.; NICHOLSON, W. L.; MEAD, D. G.; MEAD, D. G.; ENGEL, J.; YABSLEY, M. J.; DAIL, K.; JOHNSON, J.; WATSON, D.W. Tick-borne diseases in North Carolina: is ―Rickettsia amblyommii‖ a possible cause of rickettsiosis reported as Rock Montain spotted fever? Vector-Borne and Zoonotic Diseases, v. 8, p. 597-606, 2008. ALHO, C. J. R.; JÚNIOR, T. E. L.; CAMPOS, Z. M. S.; GONÇALVES, H. C. Mamíferos da Fazenda Nhumirim, sub-região de Nhecolândia, Pantanal do Mato Grosso do Sul. I – Levantamento preliminar de espécies. Revista Brasileira de Zoologia, v. 4, p.151-164, 1987. BECHARA, G. H.; SZABÓ, M. P.; DUARTE, J. M.; MATUSHIMA, E. R.; PEREIRA, M. C.; RECHAV, Y.; KEIRANS, J. E.; FIELDEN, L. J. Ticks associated with wild animals in the Nhecolândia Pantanal, Brazil. Annals of the New York Academy of Sciences, v. 916, p.289-97, 2000. BELLATO, V.; SARTOR, A. A.; SOUZA, A. P.; RAMOS, B. C. Ectoparasitos em caninos do município de Lages, Santa Catarina, Brasil. Revista Brasileira de Parasitologia Veterinária, v. 12, p. 95-98, 2003.

81
BRASIL. Ministério da Saúde. Secretaria de Vigilância em Saúde. Guia de Vigilância Epidemiológica. 2005. Disponível em: <http://www.portal.saude.gov.brr> Acesso em: 10 jan. 2011. BREMER, W. G.; SCHAAEFER, J. J.; WAGNER, E. R.; EWING, S. A.; RIKIHISA, Y.; NEEDHAM, G. R.; JITTAPALAPONG, S.; MOORE, D. L.; STICH, R. W. Transstadial and intrastadial experimental transmition of Ehrlichia canis by male Rhipicephalus sanguineus. Veterinary Parasitology, v. 131, p. 95-105, 2005. BERMÚDEZ, C. S. E.; ZALDÍVAR, A. Y.; SPOLIDORIO, M. G.; MORAES-FILHO, J.; MIRANDA, R. J.; CABALLERO, C. M., MENDOZA, Y., LABRUNA, M. B. Rickettsial infection in domestic mammals and their ectoparasites in El Valle de Antón, Coclé, Panamá. Veterinary Parasitology, v. 177, p. 134-138, 2011 . BOWMAN, D.; LITTLE, S. E.; LORENTZEN, L.; SHIELDS, J.; SULLIVAN, M. P.; CARLIN, E. P. Prevalence and geographic distribuition of Dirofilaria immits, Borrelia burgdorferi, Ehrlichia canis, and Anaplasma phagocytophilum in dogs in the United States: Results of a national clinic-based serologic survey. Veterinary Parasitology, v. 160, p. 138-148, 2009. BROUQUI, P.; BACELLAR, F.; BARANTON, G.; BIRTLES, R. J.; BJOERSDORFF, A.; BLANCO, J. R.; CARUSO, G.; CINCO, M.; FOURNIER, P. E.; FRANCAVILLA E.; JENSENIUS, M.; KAZAR J.; LAFERL, H.; LAKOS, A.; LOTRIC-FURLAN, S.; MAURIN, M.; OTEO, J. A.; PAROLA, P.; PEREZ-EID, C.; PETER, O.; POSTIC, D.; RAOULT, D.; TELLEZ, A.; TSELENTIS, Y.; WILSKE, B. Guideline for the diagnosis of tick-borne bacterial diseases in Europe. Clinical Microbiology and Infection, v.10, p.1108-1132, 2004. BULLA, C.; TAKAHIRA, R. K.; ARAUJO JR., J. P.; TRINCA, L. A.; LOPES, R. S.; WEIDMEYER, C. E. The relationship between the degree of thrombocytopenia and infection with Ehrlichia canis in an endemic area. Veterinary Research, v.35, p.141-146, 2004. BURGDORFER, W.; HAYES, S.; THOMAS JR., L.; LANCASTER, J. L. A new spotted fever group rickettsia from the lone star tick, Amblyomma americanum. In: BURGDORFER, W.; ANACKER, R. L. (Ed). Rickettsiae and Rickettsial Diseases. New York: Academic Press, 1981, 595-602. CARDOSO, L. D.; FREITAS, R. N.; MAFRA, C. L.; NEVES, C. V. B.; FIGUEIRA, F. C. B.; LABRUNA, M. B.; GENNARI, S. M.; WALKER, D. H.; GALVÃO, M. A. M. Caracterização de Rickettsia spp circulante em foco silencioso de febre maculosa brasileira no Município de Caratinga, Minas Gerais, Brasil. Caderno de Saúde Pública, v.22, p.495-501, 2006.

82
CASTRO, M. B.; MACHADO, R. Z.; AQUINO, L. P. C. T.; ALESSI, A. C.; COSTA, M. T. Experimental, acute canine monocytic ehrlichiosis: clinicopathological and immunopathological findings. Veterinary Parasitology, v.119, p.73-86, 2004. COHN, L. A. Ehrlichiosis and related infections. Veterinary Clinic of Small Animal, v.33, p.863-884, 2003. COSTA, J. O.; SILVA, M.; BATISTA JR., J. A.; GUIMARÃES, M. P. Ehrlichia canis infection in dogs in Belo Horizonte – Brazil. Arquivos da Escola de Veterinária da Universidade de Minas Gerais, v. 25, p. 199-200, 1973. COSTA JR., L. M.; REMBECK, K.; RIBEIRO, M. F. BEELITZ, P.; PFISTER, K.; PASSOS, L. M. F. Sero-prevalence and risk indicators for canine ehrlichiosis in three rural areas of Brazil. The Veterinary Journal, v.174, p.673-6, 2007. DAGNONE, A. S.; AUTRAN, H. S. M.; VIDOTTO, M. C.; JOJIMA, F. S.; VIDOTTO, O. Ehrlichiosis in anemic, thrombocytopenic, or tick-infested dogs from a hospital population in south Brazil. Veterinary Parasitology, v.117, p.285-290, 2003. DANTAS-TORRES, F.; FIGUEREDO, L. A.; FAUSTINO, M. A. G. Ectoparasitos de caes provenientes de alguns municipios da região metropolitana do Recife, Pernambuco, Brasil. Revista Brasileira de Parasitologia Veterinária, v. 13, p. 151-154, 2004. DANTAS-TORRES, F. Rock mountain spotted fever. Lancet Infectious Diseases, v. 7; p. 724-732, 2007. DANTAS-TORRES, F. Ticks on domestic animals in Pernambuco, Northeastern Brazil. Revista Brasileira de Parasitologia Veterinária, v. 18, p. 22-28, 2009. DEMMA, L. J.; TRAEGER, M. S.; NICHOLSON, W. L.; PADDOCK, C. D.; BLAU, D. M.; EREMEEVA, M. E.; DASCH, G. A.; LEVIN, M. L.; SINGLETON, J. JR.; ZAKI, S. R.; CHEEK, J. E.; SWERDLOW, D. L.; MCQUISTON, J. H. Rocky Mountain spotted fever from an unexpected tick vector in Arizona. New England Journal of Medicine; v. 53, p. 587–594, 2005. DIAS, E.; MARTINS, A. V. Spotted Fever in Brazil. American Journal of Tropical Medicina, v.19, p.103-108, 1939.

83
DINIZ, P. P. V. P.; SCHWARTZ, D. S.; MORAIS, H. S. A.; BREITSCHWERDT, E. B. Surveillance for zoonotic vector-borne infections using sick dogs from southeastern Brazil. Vector Borne and Zoonotic Diseases, v.7, n.4, p. 689-97, 2007. DUMLER, J. S.; BARBET, A. F.; BEKKER, C. P. J.; DASCH, G. A.; PALMER, G. H.; RAY, S. C.; RIKIHISA, Y.; RURANGIRWA, F. R. Reorganization of genera in the families Rickettsiaceae and Anaplasmataceae in the order Rickettsiales: unification of some species of Ehrlichia with Anaplasma, Cowdria with Ehrlichia and Ehrlichia with Neorickettsia, descriptions of six new species combinations and designation of Ehrlichia equi and HGE agent as subjective synonyms of Ehrlichia phagocytophila. International Journal of Systematic and Evolutionary Microbiology, v. 51: p. 2145-2165. 2001. DUMLER, J. S.; WALKER, D. H. Rocky Mountain spotted fever-changing ecology and persisting persisting virulence. New England Journal of Medicine, v. 353, p. 551–553, 2005. EREMEEVA, M. E.; DASCH, G. A. Rickettsiae. In: Lederberg, J. Encyclopedia of Microbialogy, New York: Academic Press, 2000. v. 4, p. 140-180. FERNANDES, I. M.; SIGNOR, C. A.; PENHA, J. O Pantanal e o sistema de pesquisa. In: FERNANDES, I. M.; SIGNOR, C. A.; PENHA, J. (Ed). Biodiversidade no Pantanal de Poconé. Cuiabá: Attema; 2010: 13-23. GANTA, R. R.; CHENG, C.; WILKERSON, M. J.; CHAPES, S. K. Delayed clearance of Ehrlichia chaffeensis infection in CD4+ T-cell knockout mice. Infection and Immunity, v. 72, p.159-67, 2004.
GALVÃO, M. A. M., CALIC, S. B., CHAMONE, C. B., MAFRA, C. L., CESARINO- FILHO, C., OLANO, J. P., WALKER, D. H. Spotted fever rickettisiosis in Coronel Fabriciano, Minas Gerais State. Revista da Sociedade Brasileira de Medicina Tropical, v. 36, p. 479-81, 2003. GROVES, M. G., DENNIS, G. L., AMYX, H. L., HUXSOLL, D. L., 1975. Transmission of Ehrlichia canis to dogs by ticks (Rhipicephalus sanguineus). American Journal of Veterinary Research, v. 36, 937–940, 1975. GEHRKE, F.; NASCIMENTO, E. M. M.; SOUZA, E. R.; COLOMBO, S.; SILVA, L. J.; SCHUMAKER, T. T. S. Detection of Rickettsia rickettsii and Rickettsia sp in 24 patient blood clots from different municipalities of the State of São Paulo. Annals of the New York Academy of Sciences, v. 1078, p. 260-262, 2006.

84
GUIMARÃES, J. H., TUCCI, E. C., BARROS-BATTESTI, D. M. Phthiraptera. In: GUIMARÃES, J. H.; TUCCI, E. C.; BARROS-BATTESTI, D. M. (Ed) Ectoparasitos de Importância Veterinária. São Paulo: Plêiade; 2001, p. 175-193. GUGLIELMONE, A. A.; BEATI, L.; BARROS-BATTESTI., D. M.; LABRUNA, M. B.; NAVA, S.; VENZAL, J. M.; MANGOLD, A. J.; SZABÓ, M. P. J.; MARTINS, J. R.; GONZÁLEZ-ACUÑA, D.; ESTRADA- PEÑA, A. Ticks (Ixodidae) on humans in South America. Experimental and Applied Acarology, v. 40, 83-100, 2006. HARRUS, S.; WANER, T.; BARK, H. Canine monocytic ehrlichiosis: an update. Compendium on Continuing Education for the Practicing Veterinarian. v. 36, p. 431-447, 1997. HARRUS, S.; WANER, T. Diagnosis of canine monocytotropic ehrlichiosis (Ehrlichia canis): An overview. The Veterinary Journal, v. 187, p. 292-296, 2011. HASEGAWA, M. Y. Dinâmica da infecção experimental de cães por Ehrlichia canis: aspectos clínicos, laboratoriais e resposta imune humoral e celular. 2005. 134 f. Tese (Doutorado). Faculdade de Medicina veterinária e Zootecnia, Universidade de São Paulo, São Paulo, 2005. HOOGSTRAAL, H. Ticks in relation to humam diseases caused by rickettsia species. Annual Review Entomology, v. 12, p.377-420, 1967. HORTA, M. C.; SANGIONI, L. A.; VIANNA, M. C.; SCHUMAKER, T. T. S.; LABRUNA, M. B.; WALKER, D. H. Prevalence of antibodies to spotted fever group Rickettsiae in humans and domestic animals in a Brazilian spotted fever-endemic area in the State of São Paulo, Brazil: serologic evidence for infection by Rickettsia rickettsii and another spotted fever group Rickettsia. American Journal of Tropical Medicine and Hygiene, v.71, p. 93-97, 2004. HORTA, M. C.; PINTER, A.; CORTEZ, A.; SOARES, R. M.; GENNARI, S. M.; SCHUMAKER, T. T. S.; LABRUNA, M. B. Rickettsia felis (Rickettsiales: Rickettsiaceae) in Ctenocephalides felis felis (Siphonaptera: Pulicidae) in the State of São Paulo, Brazil. Arquivo Brasileiro de Medicina Veterinária e Zootecnia,v. 57, p. 321-325, 2005. HORTA, M. C., PINTER, A.; SCHUMAKER, T. T. S; LABRUNA, M. B. Natural infection, transovarial transmission, and transstadial survival of Rickettsia bellii in the tick Ixodes loricatus (Acari: Ixodidae) from Brazil. Annals of the New York Academy of Sciences. v. 1078, p. 285–290, 2006.

85
HORTA, M.C.; LABRUNA, M.B.; PINTER, A.; LINARDI, P.M.; SCHUMAKER, T.T.S. Rickettsia infection in five areas of the State of São Paulo, Brazil. Memórias do Instituto Oswaldo Cruz, v. 102, p. 793–801, 2007. HORTA, M. C.; MORAES-FILHO, J.; CASAGRANDE, R. A.; SAITO, T. B; ROSA, S. C.; OGRZEWALSKA, M.; MATUSHIMA, E. R.; LABRUNA, M. B.Experimental infection of opossums Didelphis aurita by Rickettsia rickettsii and evaluation of the transmission of the infection to ticks Amblyomma cajennense. Vector-Borne and Zoonotic Diseases, v. 9, p. 109-117, 2009. HOSMER, D. W.; LEMESHOW, S. Aplied logistic regression. Ney York: Wiley; 1989, 307p. HUXSOLL, D. L.; HILDEBRANT, P. K.; NIMS, R. M.; AMIX, H. L.; FERGUSON, J. A. Epizootilogy of tropical canine pancytopenia. Journal of Wildlife Diseases, v.6, p. 220-225, 1970. INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA. IBGE Cidades @. 2007. Disponível em: <http://www.ibge.gov.br> Acesso em: 20 nov. 2010. ITO, F. H.; VASCONCELLOS, S. A.; BERNARDI, F.; NASCIMENTO, A. A.; LABRUNA, M. B. Evidência sorológica de brucelose e leptospirose e parasitismo por ixodídeos em animais silvestres do pantanal Sul-matogrossense. Ars Veterinaria, v.14, p.302-310, 1998.
YU, X. J.; WALKER, D. H. The Order Rickettsiales. In: DWORKIN, M.; FALKOW, S.; ROSENBERG, E.; SCHLEIFER, K. H.; STACKEBRANDT, E. (Ed). The Prokaryotes: a handbook on the biology of bacteria. 5.ed. New York: Springer-Velag, 2006. Disponível em: <http://link.springer-ny.com/link/service/books/10125> Acesso em: 02 dez. 2010. JIANG, J.; YARINA, T.; MILLER, M. K.; STROMDAHL, E. Y.; RICHARDS, A. L. Molecular Detection of Rickettsia amblyommii in Amblyomma americanum Parasitizing Humans. Vector-Borne and Zoonotic Diseases, v. 10, p. 329-40, 2010. KEEFE, T. J.; HOLLAND, C. J.; SALIER, P. E.; RISTIC, M. Distribution of Ehrlichia canis among military working dogs in the world and selected civilian dogs in the United States. Journal of the American Veterinary Medical Association, v.181, p.236-238, 1982.

86
LABARTHE, N.; CAMPOS PEREIRA, M.; BALBARINI, O.; MCKEE, W.; COIMBRA, C. A.; HOSKINS, J. Serologic prevalence of Dirofilaria immintis, Ehrlichia canis and Borrelia burgdorferi infection in Brazil. Veterinary Therapeutics, v.4, p.67-75, 2003. LABRUNA, M. B.; PEREIRA, M. C. Febre Maculosa: aspectos clínico-epidemiológicos. Clínica Veterinária v.12, p.19-23, 1998. LABRUNA, M. B.; PEREIRA, M. C. Carrapatos em cães no Brasil. Clínica Veterinária v.30, p.24-32, 2001. LABRUNA, M. B; KERBER, C. E.; FACCINI, J. L. H.; DEWAAL, D. T.;.GENNARI, S. M. Risk factors to tick infestations and their occurrence on horses in the state of São Paulo. Brazilian Journal of Veterinary Parasitology; v. 97, p. 1–14, 2001. LABRUNA, M. B.; WHITWORTH, T.; HORTA, M. C.; BOUYER, D. H.; MACBRIDGE, J.; PINTER, A.; POPOV, V.; GENNARI, S. M.; WALKER, D. H. Rickettsia species infecting Amblyomma cooperi ticks from an endemic area for Brazilian spotted fever in the state of São Paulo, Brazil. Journal of Clinical Microbiology. v.42, p.90-98, 2004a. LABRUNA, M. B.; WHITWORTH, T.; BOUYER, D. H.; MCBRIDE, J.; CAMARGO, L. M. A.; CAMARGO, E. P.; POPOV, V.; WALKER, D. H. Rickettsia bellii and Rickettsia amblyommii in Amblyomma ticks from the state of Rondônia, western Amazon, Brazil. Journal of Medical Entomology, v.41, p.1073-1081, 2004b. LABRUNA, M. B.; MCBRIDE, J. W.; BOUYER, D. H.; CAMARGO, L. M.; WALKER, D. H. Molecular evidence for a spotted fever group Rickettsia species in the tick Amblyomma longirostre in Brazil. Journal of Medical Entomology; v.41, p. 533–537, 2004c. LABRUNA, M. B.; CAMARGO, L. M. A.; CAMARGO, E. P.; WALKER, D. H. Detection of a spotted fever group Rickettsia in the tick Haemaphysalis justakochi in Rondonia, Brazil. Veterinary Parasitology, v.127, p.169-174, 2005a. LABRUNA, M. B.; JORGE, R. S.; SANA, D. A.; JÁCOMO, A. T.; KASHIVAKURA, C. K.; FURTADO, M. M.; FERRO, C.; PEREZ, S. A.; SILVEIRA, L.; SANTOS, T. S. JR.; MARQUES, S. R.; MORATO, R. G.; NAVA, A.; ADANIA, C. H.; TEIXEIRA, R. H.; GOMES, A. A.; CONFORTI, V. A.; AZEVEDO, F. C.; PRADA, C. S.; SILVA, J. C.; BATISTA, A. F.; MARVULO, M. F.; MORATO, R. L.; ALHO, C. J.; PINTER, A.; FERREIRA, P. M.; FERREIRA, F.; BARROS-BATTESTI, D. M. Ticks (Acari: Ixodida) on wild carnivores in Brazil. Experimental and Applied Acarology, v.36, p.149-163, 2005b.

87
LABRUNA, M. B; PACHECO, R. C.; NAVA, S., BRANDÃO, P. E., RICHTZENHAIN, L. J.; GUGLIELMONE, A. A. Infection by Rickettsia bellii and Candidatus ‗‗Rickettsia amblyommii‘‘ in Amblyomma neumanni ticks from Argentina. Microbial Ecology, v. 54, p126–133, 2007a. LABRUNA, M. B. PACHECO, R. C; RICHTZENHAIN, L. J.; SZABÓ, M. P. J.. Isolation of Rickettsia rhipicephali and Rickettsia bellii from ticks Haemaphysalis juxtakochi in the state of Sao Paulo, Brazil. Applied and Environmental Microbiology, v.73, p.869–873, 2007b. LABRUNA, M. B.; OGRZEWALSKA, M.; MORAES-FILHO, J.; LEPE, P.; GALLEGOS, J. L.; LÓPEZ, J. Rickettsia felis in Chile. Emerging Infectious Diseases, v. 13, p.1794–1795, 2007c. LABRUNA, M. B.; HORTA, M. C.; AGUIAR, D. M.; CAVALCANTE, G. T.; PINTER, A.; GENNARI, S. M.; CAMARGO, L. M. A. Prevalence of Rickettsia Infection in Dogs from the Urban and Rural Areas of Monte Negro Municipality, Western Amazon, Brazil. Vector-Borne and Zoonotic Diseases, v.7, p.249-256, 2007d. LABRUNA, M. B. Ecology of rickettsia in South America. Annals of the New York Academy of Sciences, v. 1166, p. 156-66. 2009a. LABRUNA, M. B.; KAMAKURA, O.; MORAES-FILHO, J.; HORTA, M. C.; PACHECO, R. C. Rocky Mountain spotted fever in dogs, Brazil. Emerging Infectious Diseases, v. 15, p. 458-60. 2009b. LEWIS, G. E.; RICTIC, M.; SMITH, R. D.; LINCOLN, T; STEPHENSON, E. H. The brown dog tick Rhipicephalus sanguineus and the dog as experimental hosts of Ehrlichia canis. American Journal of Veterinariay Research, v. 38, p. 1953-1955, 1977. LEMOS, E. R. S.; MACHADO, R. D.; COURA, J. R.; GUIMARÃES, M. A. A. M.; CHAGAS, N. Epidemiological aspects of the Brazilian Spotted Fever: Serological survey of dogs and horses in an endemic area in the State of São Paulo, Brazil. Revista do Instituto de Medicina Tropical de São Paulo, v.36, p.427-430, 1996. LEMOS, E. R. S.; ALVARENGA, F. B. F.; CINTRA, M. L.; RAMOS, M. C.; PADDOCK, C. D.; FEREBEE, T. L.; ZAKI, S. R.; FERREIRA, F. C. C.; RAVAGNANI, R. C.; MACHADO, R. D.; GUIMARÃES, M. A. A. M.; COURA, J. R. Spotted fever in Brazil: a seroepidemiological study and description of clinical cases in na endemic área in the state of São Paulo. American Journal of Tropical Medicine and Hygiene, v. 65, p. 329-334, 2001.

88
LINARDI, P. M., GUIMARÃES, L. R. Sistemática dos sifonápteros brasileiros. In: LINARDI, P. M., GUIMARÃES, L. R. (Ed). Sifonápteros do Brasil. São Paulo: Museu de Zoologia USP/FAPESP; 2000, p. 37-48. MACHADO, R. Z. Erliquiose canina. Revista Brasileira de Parasitologia Veterinária, v. 13, suplemento 1, p. 53-57, 2004. MACHADO, R. Z.; DUARTE, J. M. B.; DAGNONE, A. S.; SZABÓ, M. P. J. Detection of Ehrlichia chaffeensis in Brazilia marsh deer (Blastocerurs dichotomus). Veterinary Parasitology, v. 139, p. 262-266, 2006. MACALUSO, K. R.; PORNWIROON, W.; POPOV, V. L., FOIL, L. D.Identification of Rickettsia felis in the salivary glands of cat fleas. Vector-Borne and Zoonotic Diseases,.v. 8, p. 391–396, 2008. MARTINS, J. R.; MEDRI, I. M.; OLIVEIRA, C. M.; GUGLIELMONE, A. Ocorrência de carrapatos em tamanduá-bandeira (Myrmecophaga tridactyla) e tamanduá-mirim (Tamandua tetradactyla) na região do Pantanal Sul Mato-Grossense, Brasil. Ciência Rural, v.34, p. 293-295, 2004. MARTINS, T. F.; ONOFRIO, V. C.; BARROS-BATTESTI, D. M.; LABRUNA, M. B. Nymphs of the genus Amblyomma (Acari: Ixodidae) of Brazil: descriptions, redescriptions, and identification key. Ticks and Tick Borne Disease, v. 1, p. 75–99, 2010. MCBRIDE, J.; CORSTVET, R.; GAUNT, S.; CHINSANGARAM, J.; AKITA, G.; OSBURN, B. PCR detection of acute Ehrlichia canis infection in dogs. Journal of Veterinary Diagnostic Investigation, v. 8, p. 441- 447, 1996. MCDADE, J. E.; NEWHOUSE, V. F. Natural history of Rickettsia rickettsii. Annual Review of Microbiology, v. 40, p. 287-309, 1986. MORAES-FILHO, J.; PINTER, A.; PACHECO, R. C.; GUTMANN, T. B.; BARBOSA, S. O.; GONZÁLES, M. A. R. M.; MURARO, M. A.; SILVIA, R. M. CECÍLIO, S. R. M.; LABRUNA, M. B. New Epidemiological Data on Brazilian Spotted Fever in an Endemic Area of the State of São Paulo, Brazil. Vector–Borne and Zoonotic Diseases, v. 9, p. 73-78, 2009 NASCIMENTO, E. M.; GEHRKE, F. S.; MALDONADO, R. A.; COLOMBO, S.; SILVA L. J.; SCHUMAKER, T. T. S. Detection of a Brazilian spotted fever infection by

89
polymerase chain reaction in a patient from the state of São Paulo. Memórias do Instituto Oswaldo Cruz, v.100, p.277-279, 2005. NAVA, S.; ÉREZ-MARTÍNEZ, L.; VENZAL, J. M.; PORTILLO, A.; ÁÑEZ, S.; OTEO, J. A. Rickettsia felis in Ctenocephalides felis from Argentina. Vector–Borne and Zoonotic Diseases. v. 8, p.465–466, 2008. NEER, T. M. Ehrlichiosis. Canine monocytic and granulocytic ehrlichiosis. In: GREENE, C. E. (Ed.), Infectious Diseases of the Dog and Cat. Saunders, Philadelphia; 1998: 139–154. OLANO, J. P.; WALKER, D. H. Human ehrlichioses. Medical Clinics of North America, v.86, p.375-92, 2002. OLIVER, J. H. Biology and systematics of ticks (Acari:Ixodida). Annual Review of Ecology, Evolution and Systematics, v. 20, p. 397-430, 1989. OLIVEIRA, K. A.; OLIVEIRA, L. S.; DIAS, C. C.; A. JR., SILVA, .; ALMEIDA, M. R; ALMADA, G.; BOUYER, D. H.; GALVÃO, M. A. M.; MAFRA, C. L. Molecular identification of Rickettsia felis in ticks and fleas from an endemic area for Brazilian Spotted Fever. Memórias do Instituto Oswaldo Cruz, v. 103, p.191-194, 2008. OLIVEIRA, L. S.; OLIVEIRA, K. A.; MOURÃO, L. C.; PESCATORE, A. M.; ALMEIDA, M. R.; CONCEIÇÃO, L. G.; GALVÃO, M. A .M.; MAFRA, C. First repor of Ehrlichia ewingii detected by molecular investigation in dogs from Brazil. Clinical Microbiology and Infection, Paris, v. 15, p. 55-56, 2009. OGRZEWALSKA, M.; PACHECO, R. C.; UEZU, A.; FERREIRA, F.; LABRUNA, M. B. Ticks (Acari: Ixodidae) infesting wild birds in a Atlantic Forest area in the state of São Paulo, Brazil, with isolation of Rickettsia from the tick Amblyomma longirostre. Journal of Medical Entomology,v. 45, p.770–774, 2008. OGRZEWALSKA, M; UEZU, A.; LABRUNA, M. B. Ticks (Acari: Ixodidae) infesting wild birds in the eastern Amazon, northern Brazil, with notes on rickettsial infection in ticks. Journal of Medical Entomology, v. 106, p. 809– 816, 2010. ONOFRIO, V. C., LABRUNA, M. B., PINTER, A.; GIACOMIN, F. G.; BARROS-BATTESTI, D. M. Comentários e chaves para as espécies do gênero Amblyomma. In: BARROS-BATTESTI, D. M.; ARZUA, M.; BECHARA, G. H. (Ed). Carrapatos de importância médico-veterinária da região neotropical: um guia ilustrado para identificação de espécies. São Paulo: Vox/ICTTD-3/Butantan; 2006: 53-113.

90
PACHECO, R. C.; VENZAL, J. M.; RICHTZENHAIN, L. J.; LABRUNA, M. B.; Rickettsia parkeri in Uruguay. Emerging Infectious Diseases, v. 12, p.1804-5, 2006. PACHECO, R. C.; HORTA, M. C.; MORAES-FILHO, J.; ATALIBA, A. C.; PINTER, A.; LABRUNA, M. B. Rickettsial infection in capybaras (Hydrochoerus hydrochaeris) from São Paulo, Brazil: serological evidence for infection by Rickettsia bellii and Rickettsia parkeri. Biomedica, v. 27, p. 364-371, 2007. PACHECO, R. C.; MORAES-FILHO, J.; GUEDES, E.; SILVEIRA, I.; RICHTZENHAIN, L.; LEITE, R.; LABRUNA, M. B. Rickettsial infections of dogs, horses and ticks in Juiz de Fora, southeastern Brazil, and isolation of Rickettsia rickettsii from Rhipicephalus sanguineus ticks. Medical and Veterinary Entomology, 2010 (No prelo). PEREZ, M.; RIKIHISA, Y.; WEN, B. Ehrlichia canis-like agent isolated from a man in Venezuela: antigenic and genetic characterization. Journal of Clinical Microbiology, v.34, p. 2133-2139, 1996. PEREZ, M.; BODOR, M.; ZHANG, C.; XIONG, Q.; RIKIHISA, Y. Human infection with Ehrlichia canis accompanied by clinical signs in Venezuela. Annals of the New York Academy of Sciences, v.1078, p.110-117, 2006. PHILIP, R. N.; CASPER, E. A.; ANACKER, R. L.; CORY, J.; HAYES, S. F.; BURGDORFER, W.; YUNKER, C. E. Rickettsia bellii sp. nov.: a Tick-Borne Rickettsia, widely distributed in the United States, that is distinct from the Spotted Fever and Typhus Biogroups. International Journal of Systematic Bacteriology, v. 33, p. 94-106, 1983. PINTER, A.; LABRUNA, M. B. Isolation of Rickettsia rickettsii and Rickettsia bellii in cell culture from the tick Amblyomma aureolatum in Brazil. Annals of the New York Academy of Sciences, v. 1078, p.523–529, 2006. PINTER, A., HORTA, M. C., PACHECO, R. C., MORAES-FILHO, J.; LABRUNA, M. B. Serosurvey of Rickettsia spp. in dogs and humans from an endemic area for Brazilian spotted fever in the State of São Paulo, Brazil. Caderno de Saúde Pública, v. 24, p. 247-252, 2008. PIRANDA, E. M; FACCINI, J. L. H.; PINTER, A.; SAITO, T. B.; PACHECO, R. C.; HAGIWARA, M. K.; LABRUNA, M. B. Experimental infection of dogs with a Brazilian strain of Rickettsia rickettsii: clinical and laboratory findings. Memórias do Instituto Oswaldo Cruz, v. 103, p. 696-701, 2008.

91
PIRANDA, E. M.; FACCINI, J. L. H.; PINTER, A.; PACHECO, R. C. Experimental infection of Rhipicephalus sanguineus ticks with the bacterium Rickettsia rickettsii, using experimentally infected dogs. Vector–Borne and Zoonotic Diseases, v. 11, p. 29-36, 2011. POPOV, V. L.; HAN, V. C.; CHEN, M.; DUMLER, J. S.; FENG, H. M.; ANDREADIS, T. G.; TESH, R. B.; WALKER, D. H. Ultrastructural diffentiation of the genogroups in the genus Ehrlichia. Journal of Medical Microbiology, v.47, p. 2235-2251, 1998. RAOULT, D.; ROUX, V. Rickettsioses as paradigms of new or emerging infectious diseases. Clinical Microbiology Review, v.10, p.694-719, 1997. RICKETTS, G. T. Some aspects of Rock Mountain Spotted Fever as shown by recent investigations. Medical Record. v. 36, p. 843-855, 1909. RIKIHISA, Y. Ehrlichia subversion of host innate responses. Current Opinion in Microbiology, v.9, p. 95-101, 2006. RODRIGUES, A. F. S. F; DAEMON, E.; D‗AGOSTO, M. Investigação sobre alguns ectoparasitos em cães de rua no Município de Juiz de Fora, Minas Gerais. Revista Brasileira de Parasitologia Veterinária, 2001; v. 10, p.13-19, 2001. ROZENTAL, T.; BUSTAMANTE, M. C.; AMORIM, M.; SERRA-FREIRE, N. M.; LEMOS, E. R. S. Evidence of spotted fever group rickettsiae in state of Rio de Janeiro, Brazil. Revista do Instituto de Medicina Tropical de São Paulo, v. 44, p.155-158, 2002. SABATINI, G. S.; PINTER, A.; NIERI-BASTOS, F. A.; MARCILI, A.; LABRUNA, M. B. Survey of ticks (Acari: Ixodidae) and their rickettsia in an Atlantic rain forest reserve in the State of São Paulo, Brazil. Journal of Medical Entomology, v. 47, p. 913-916, 2010. SAITO, T. B.; CUNHA-FILHO, N. A.; PACHECO, R. C.; FERREIRA, F.; PAPPEN, F. G.; FARIAS, N. A.; LARSSON, C. E.; LABRUNA, M. B. Canine Infection by Rickettsiae and Ehrlichiae in Southern Brazil. American Journal of Tropical Medicine and Hygiene, v.79, p.102-108, 2008. SANGIONI, L. A.; HORTA, M. C.; VIANNA, M. C. B.; GENNARI, S. M.; SOARES, R. M.; GALVÃO, M. A. M.; SCHUMAKER, T. T. S.; FERREIRA, F.; VIDOTTO, O.; LABRUNA, M. B. Rickettsial infection in animals and Brazilian Spotted Fever endemicity. Emerging Infectious Disisease, v.11, p.265-270, 2005.

92
SANTARÉM, V. A. Achados epidemiológicos, clínicos e hematológicos e comparação de técnicas para diagnóstico de Ehrlichia canis. 2003. 127 f. Tese (Doutorado) – Faculdade de Medicina Veterinária e Zootecnia. Universidade Estadual Paulista ―Júlio de Mesquita Filho‖, Botucatu, 2003. SANTOS, A. P. Aspectos ecológicos da febre maculosa brasileira em um foco endêmico no Estado de São Paulo. 2007, 86 f. Tese (Doutorado). Faculdade de Medicina Veterinária e Zootecnia, Universidade de São Paulo, São Paulo, 2007. SEXTON, D. J.; MUNIZ, M.; COREY, G. R.; BREITSCHWERDT, E. B.; HEGARTY, B. C.; DUMLER, S.; WALKER, D. H.; PECANHA, P. M.; DIETZE, R. Brazilian spotted fever in Espírito Santo, Brazil: description of a focus of infection in a new endemic region. American Journal of Tropical Medicine and Hygiene, v.49, p.222-226, 1993. SILVA, J. N.; ALMEIDA, A. B. P. F.; SORTE, E. C. B.; FREITAS, A. G.; SANTOS, L. G. F.; AGUIAR, D. M.; SOUSA, V. R. F.Soroprevalência de anticorpos anti-Ehrlichia canis em cães de Cuiabá, Mato Grosso. Revista Brasileira de Parasitologia Veterinária, v. 19, p. 34-37, 2010. SILVEIRA, I.; PACHECO, R. C.; SZABÓ, M. P.; RAMOS, H. G.; LABRUNA, M. B. Rickettsia parkeri in Brazil. Emerging Infectious Disiseases, v.13, p.1111-1113, 2007. SHIBATA, S. I.; KAWAHARA, M.; RIKIHISA, Y.; FUJITA, H.; WATANABE, Y.; SUTO, C.; ITO, T. New Ehrlichia species closely related to Ehrlichia chaffeensis isolated from Ixodes ovatus ticks in Japan. Journa of Clinical Microbiology, v.38, p.1331-1338, 2000. SOUZA, C. E.; MORAES-FILHO, J.; OGRZEWALSKA, M.; UCHOA, F. C.; HORTA, M. C.; SOUZA, S. S.; BORBA, R. C.; LABRUNA, M. B. Experimental infection of capybaras Hydrochoerus hydrochaeris by Rickettsia rickettsii and evaluation of the transmission of the infection to ticks Amblyomma cajennense. Veterinary Parasitology, v. 161, p. 116-121, 2009. SOUZA, B. M. P. S.; LEAL, D. C.; BARBOZA, D. C. P. M.; UZÊDA, R. S.; ALCÂNTARA, A. C.; FERREIRA, F.; LABRUNA, M. B.; GONDIM, L. F. P; FRANKE, C. R. Prevalence of ehrlichial infection among dogs and ticks in Northeastern Brazil. Revista Brasileira de Parasitologia Veterinária, v. 19, p.15-19, 2010.

93
SPOLIDORIO, M. G.; LABRUNA, M. B.; MANTOVANI, E.; BRANDAO, P. E.; RICHTZENHAIN, L. J.; YOSHINARI, N. H. Novel spotted fever group rickettsiosis, Brazil. Emerging Infectious Diseases, v.16, p. 521-523, 2010. STICH, R. W.; SCHAEFER, J. J.; BREMER, W. G.; NEEDHAM, G. R.; JITTAPALAPONG, S. Host surveys, ixodid tick biology and transmission scenarios as related to the tick-borne pathogen, Ehrlichia canis. Veterinary Parasitology, v. 158, p. 256-273, 2008. SUMNER, J. W.; DURDEN; L. A.; GODDARD, J.; STROMDAHL, E. Y.; CLARK, K. L.; REEVES, W. K.; PADDOCK, C. D. Gulf Coast ticks (Amblyomma maculatum) and Rickettsia parkeri, United States. Emerging Infectious Disiseases, v.13, p. 751–753, 2007. SZABÓ, M. P. J.; CUNHA, T. M; PINTER, A., VICENTINI, F. Ticks (Acari: Ixodidae) associated with domestic dogs in Franca region, São Paulo, Brazil. Experimental and Applied Acarology , v. 25, p. 909–916, 2001. SZABÓ, M. P. J.; SOUZA, L. G. A; OLEGÁRIO, M. M. M.; FERREIRA, F. A.; NETO, A. A. P. Ticks (Acari: Ixodidae) on dogs from Uberlândia, Minas Gerais, Brazil Transboundary and Emerging Diseases. v. 57, p. 72–74, 2010. TAYLOR, L. H.; LATHAM, S. M.; WOOLHOUSE, M. E. Risk factors for human disease emergence. Philosophical Transactions of the Royal Society Biological Sciences, v.356, p.983–989, 2001. TORRES, H. M.; MASSARD, C. L.; FIGUEIREDO, M. J.; FERREIRA, T.; ALMOSNY, N. R. P. Isolamento e propagação da Ehrlichia canis em células DH82 e obtenção de antígeno para a reação de imunofluorescência indireta. Revista Brasileira de Ciência. Veterinária, v.9, p.77-82, 2002. THRUSFIELD, M. SURVEYS. In: THRUSFIELD, M. (Ed). Veterinary Epidemiology. Oxford: Blackwell Science Ltd; 1995, p. 223-247. TRAPP, S. M.; DAGNONE, A. S.; VIDOTTO, O.; FREIRE, R. L.; AMUDE, A. M.; MORAIS, H. S. A. Seroepidemiology of canine babesiosis and ehrlichiosis in a Hospital population. Veterinary Parasitology, v.140, p.223-230, 2006. TRAVASSOS, J.; VALLEJO, A. Comportamento de alguns cavídeos (Cavia aperea e Hydrochoerus capybara) às inoculações experimentais do vírus da Febre Maculosa.

94
Possibilidade desses cavídeos representarem o papel de depositários transitórios do vírus na natureza. Memórias do Instituto Butantã, v.15, p.73-86, 1942. UENO; T. E. H.; AGUIAR, D. M.; PACHECO, R. C.; RICHTZENHAIN, L. J.; RIBEIRO, M. G.; PAES, A. C.; MEGID, J.; LABRUNA, M. B. Ehrlichia canis em cães, atendidos em hospital veterinário de Botucatu, Estado de São Paulo, Brasil. Revista Brasileira de Parasitologia Veterinária, v. 18, p. 57-61, 2009. URQUHART, G. M.; ARMOUR, J; DUNCAN, J. L.; DUNN, A. M.; JENNINGS, F. W. Order Phthiraptera. In: URQUHART, G. M.; ARMOUR, J.; DUNCAN, J. L.; DUNN, A. M.; JENNINGS, F. W. (Ed). Veterinary Parasitology. Oxford: Blackwell Science Ltd; 1996, p. 147-153. VENZAL, J. M.; PORTILLO, A.; ESTRADA-PEÑA, A., CASTRO, O., CABRERA, P. A.; OTEO, .J. A. Rickettsia parkeri in Amblyomma triste from Uruguay. Emerging Infectious Disiseases.v. 10, p.1493– 1495, 2004. WANER, T.; HARRUS, S.; BARK, H.; BOGIN, E.; AVIDAR, Y.; KEYSARY, A. Characterization of the subclinical phase of canine ehrlichiosis in experimentally infected beagle dogs. Veterinary Parasitology, v. 69, p. 307-317, 1997. WELLMAN, M.; KRAKOWKA, S.; JACOBS, R.; KOCIBA, G. A macrophage-monocyte cell line from a dog with malignant histiocytosis. In Vitro Cellular e Developmental Biology, v. 24, p. 223-229, 1988. WHITMAN, T. J.; RICHARDS, A. L., PADDOCK, C. D.; TAMMINGA, C. L., SNIEZEK, P. J.; JIANG, J.; BYERS, D. K.; SANDERS, J. W. Rickettsia parkeri infection after tick bite, Virginia. Emerging Infectious Disiseases, v. 13, p. 334–336, 2007. WEDINCAMP, J.; FOIL. L. D. Vertical transmission of Rickettsia felis in the cat flea (Ctenocephalides felis Bouché). Journal of Vector Ecology. v. 27, p. 96–101, 2002. WIDMER, C. E. Perfil sanitário de onças-pintadas (Phantera onça) de vida livre no Pantanal Sul do Mato Grosso do Sul – Brasil. 2009; 89f. Dissertação (Mestrado). Medicina Veterinária Preventiva e Saúde Animal, Universidade de São Paulo, São Paulo, 2009.

95
APÊNDICE A – Questionário epidemiológico Sangue n° _____________
DADOS DO PROPRIETÁRIO: Nome: Telefone: Rua: nº: Bairro: Complemento: CEP: Área: ( ) Urbana ( ) Rural DADOS DO ANIMAL: - Nome: Sexo: Idade: Raça: - Ambiente que o animal vive ou frequenta: ( ) Urbano ( ) Peri – urbano ( ) Rural - Tem acesso à rua? ( ) Sim ( ) Não ( ) Esporadicamente ( ) Frequentemente - Tem acesso à área rural? ( ) Sim ( ) Não ( ) Esporadicamente ( ) Frequentemente - Tem contato com animais silvestres? ( ) Sim ( ) Não ( ) Esporadicamente ( ) Frequentemente - Tem contato com animais de produção? ( ) Sim ( ) Não ( ) Esporadicamente ( ) Frequentemente - Tem hábitos de caça ou pastoreio? ( ) Sim ( ) Não ( ) Esporadicamente ( ) Frequentemente - Possui contactante (s): ( ) Sim ( ) Não. Se SIM, de qual espécie? - Vacinação: ( ) Sim, atualizada. ( ) Sim, desatualizada. ( ) Não. - Vermifugação: ( ) Sim, atualizada. ( ) Sim, desatualizada. ( ) Não. - Tipo de alimentação: ( ) Ração comercial ( ) Dieta caseira ( ) Mista - Infestação por ectoparasitos: Carrapatos: Momento da coleta: ( ) Sim ( ) Não Infestação recente (menos de 60 dias): ( ) Sim ( ) Não Pulgas: ( ) Sim ( ) Não - Condição fisiológica do cão (avaliação subjetiva): ( ) Boa ( ) Razoável ( ) Ruim - Animal está em tratamento para alguma doença intercorrente? ( ) Sim ( ) Não. Se SIM, para qual doença? - Já teve erliquiose? ( ) Sim ( ) Não. Se SIM, o animal se recuperou com o tratamento proposto? ( ) Sim (melhora) ( ) Não (recidiva) - Houve, ao final da visita domiciliar, alguma assistência veterinária? ( ) Sim ( ) Não. Se SIM, favor discriminar abaixo: ( ) Orientação sanitária (esquema vacinal e/ou vermifugação) ( ) Prescrição de medicamento (s). Qual(s):
Recommended

















