
UNIVERSIDADE DE SÃO PAULO
ESCOLA DE ENGENHARIA DE LORENA
DÉBORA DUTRA MENEZES LEAL
Estudo da influência de espécies de Lactobacillus sobre a resposta humoral em
camundongos BALB/c
Lorena - SP
2007

DÉBORA DUTRA MENEZES LEAL
Estudo da influência de espécies de Lactobacillus sobre a resposta humoral em
camundongos BALB/c
Tese apresentada à Escola de Engenharia de Lorena
da Universidade de São Paulo para a obtenção do
título de Doutor em Biotecnologia Industrial.
Área de Concentração: Microbiologia Aplicada
Orientador: Prof. Dr. Ismael Maciel de Mancilha
Lorena – SP
2007

AUTORIZO A REPRODUÇÃO E DIVULGAÇÃO TOTAL OU PARCIAL DESTE TRABALHO, POR QUALQUER MEIO CONVENCIONAL OU ELETRÔNICO, PARA FINS DE ESTUDO E PESQUISA, DESDE QUE CITADA A FONTE.
Catalogação na Publicação Biblioteca Universitária
Escola de Engenharia de Lorena da Universidade de São Paulo
Leal, Débora Dutra Menezes
Estudo da influência de espécies de Lactobacillus sobre a resposta humoral em camundongos BALB/c / Débora Dutra Menezes Leal ; orientador Ismael Maciel de Mancilha. -- 2007
83 f. : fig.
Tese (Doutorado – Programa de Pós-Graduação em Biotecnologia Industrial. Área de Concentração: Microbiologia Aplicada) – Escola de Engenharia de Lorena da Universidade de São Paulo
1. Microbiologia Aplicada 2. Lactobacillus 3. Camundongos BALB/c 4. Resposta humoral. I. Título.
579.62 - CDU

DEDICATÓRIA
Aos meus pais, Amauri (in memorian) e Enancy, por terem sido os pais da minha filha
enquanto estive ausente, pelo incentivo constante me apoiando em todos os momentos com
infindável carinho e por terem sido um grande exemplo.

AGRADECIMENTOS
À Mariana e Henrique, pela compreensão, paciência e incentivo.
Aos meus irmãos Denise, Amauri e Alberto e cunhados, Inácio, Elisângela e Janaína,
pelas palavras certas nas horas difíceis.
Ao Prof. Dr. Ismael Maciel de Mancilha, pelo profissionalismo, oportunidade e
valiosa orientação durante a realização deste trabalho.
Aos professores Dr. Antonio José Piantino Ferreira e Drª. Claudete Serrano Astolfi
Ferreira do Departamento de Ornitopatologia da Faculdade de Medicina Veterinária e
Zootecnia da Universidade de São Paulo (FMVZ/USP), pelos ensinamentos, apoio técnico e
carinho com que me receberam.
Aos amigos e colegas de laboratório, Laura, André, Eliana, Amarílis, Marcelo, Sibele,
Jorge, Liliana, e em especial Larissa e Carina, pelo carinho e horas de trabalho compartilhadas.
Ao amigo Antonio Carlos Pedroso pelo auxílio durante a análise estatística.
Aos funcionários da FMVZ/USP, Alexandre (Laboratório de Virologia), Maria José
(Laboratório de Ornitopatologia), Claudia, Lílian (Laboratório de Técnicas Imunológicas),
Cláudio e Luciano (Laboratório de Histologia) e Claudia Mori (Biotério) pela colaboração.
Aos Professores da Escola de Engenharia de Lorena (EEL), Drª. Bernadete, Drª. Maria das
Graças, Dr. Arnaldo Márcio, Drª. Adriani e Dr. João Batista, pelos conhecimentos transmitidos.
Aos funcionários da EEL, Lilian Marton, Sandra Borges, Regina Horta e Maria Auxiliadora
Midões pela colaboração.
Aos amigos do Debiq da EEL, em especial, Carlos Sanches e Daniela Cortez, pelo
carinho e por compartilharem os momentos difíceis.
À Coordenadoria de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) pela
bolsa concedida.

RESUMO
LEAL, D.D.M. Estudo da influência de espécies de Lactobacillus sobre a resposta humoral em camundongos BALB/c. 2007. 83f. Tese de Doutorado (Biotecnologia Industrial), Escola de Engenharia de Lorena, Universidade de São Paulo.
No presente trabalho avaliou-se a influência de sete cepas de Lactobacillus pertencentes às espécies L. casei pseudoplantarum (30b e 30c), L. plantarum (11fb, 22c e 41b) e L. reuteri (18fa e 19fa) sobre a resposta humoral de camundongos BALB/c. Para tanto as cepas foram cultivadas individualmente e em co-cultura e caracterizadas quanto à susceptibilidade a antibióticos; resistência ao pH; produção de ácido láctico; proteínas secretadas e produção de IgA e IgG total sérica e na mucosa intestinal. Utilizou-se o método de difusão em disco para determinação do padrão de susceptibilidade das cepas de lactobacilos a 17 antibióticos de importância clínica em humanos e animais, observando-se que os lactobacilos cultivados em co-cultura (pool) apresentaram resistência a 76% dos antibióticos testados. A tolerância ao pH foi avaliada através do cultivo das cepas em caldo Man Rogosa e Sharpe (MRS) com valores de pH variando entre 4,0 e 6,0. Foi verificado crescimento de todas as cepas estudadas em todos os valores de pH testados. A avaliação da produção de ácido láctico foi realizada através do método de Cromatografia Líquida de Alta Eficiência, permitindo-se verificar que o pool de lactobacilos apresentou a maior concentração de ácido láctico (19,14 g/L). A análise do gel de poliacrilamida pelo método da eletroforese revelou poucas bandas protéicas no pool de lactobacilos em relação às cepas de L. casei pseudoplantarum (30b e 30c) e L. reuteri (18fa e 19fa), cultivadas individualmente. Para a determinação da resposta humoral utilizou-se os lactobacilos cultivados em co-cultura. Cento e vinte camundongos BALB/c foram submetidos a oito diferentes tratamentos: a) suspensão bacteriana contendo 109 UFC/mL (SB) via oral; b) sobrenadante sem tratamento térmico (SN1) via oral; c) SN1 via subcutânea (SC); d) sobrenadante tratado termicamente (SN2); e) SB via oral + 100 �g de ovalbumina (OVA) via SC; f) SN1 via oral + 100 �g de ovalbumina (OVA) via SC; g) SN1 via SC + 100 �g de ovalbumina (OVA) via SC; h) SN2 via SC + 100 �g de ovalbumina (OVA) via SC e determinada a produção de IgA e IgG sérica e da mucosa intestinal através do método de ELISA. Demonstrou-se que as espécies de lactobacilos utilizadas nesta pesquisa foram ineficazes na indução da resposta humoral em camundongos BALB/c.
Palavras-chave: Lactobacillus, camundongos BALB/c, resposta humoral

ABSTRACT
LEAL, D.D.M. Study of the influence of Lactobacillus species on the humoral immune response in BALB/c mice. 2007. 83f. Thesis (Doctoral in Industrial Biotechnology), Escola de Engenharia de Lorena, Universidade de São Paulo.
The present work evaluated the effect of seven strains of Lactobacillus: L. casei pseudoplantarum (30b and 30c), L. plantarum (11fb, 22c and 41b) and L. reuteri (18fa e 19fa) on the immune humoral response of BALB/c mice. For this purpose, the strains were cultivated individually and in co-culture. They were characterized in relation to the susceptibility to antibiotics; resistance to pH; lactic acid production; production of total IgA and IgG in serum and in the gut mucosal and proteins. The method of disc diffusion for determination of the susceptibility standard of Lactobacillus strains to 17 antibiotics of clinical importance in human beings and animals was used. The Lactobacillus cultivated in co-culture (pool) presented resistance to 76% of the tested antibiotics. The tolerance to pH was evaluated through the strains grown in MRS (Man Rogosa and Sharpe) broth pH range between 4.0 and 6.0. Growth of all studied strains was verified for all tested pH values. The evaluation of lactic acid production was carried out by High Efficiency Liquid Chromatography. The pool of Lactobacillus showed the highest concentration of lactic acid (19,14 g/L). The analysis by polyacrylamide protein gel electrophoresis revealed few proteinaceous bands in the pool of lactobacillus in comparison to the L. casei pseudoplantarum (30b and 30c) and L. reuteri (18fa and 19fa) strains cultivated individually. For the determination of the immune response, lactobacillus cultivated in co-culture were used. One hundred-twenty BALB/c mice were submitted to eight different treatments: a) the bacterial suspension with 109 UFC/mL (SB) via oral; b) supernatants without thermal treatment (SN1) via oral; c) SN1 via subcutaneous (SC); d) thermically treated supernatants (SN2); e) SB via oral +100 µg of ovalbumim (OVA) via SC; f) SN1 via oral +100 µg of ovalbumim (OVA) via SC; g) SN1 via SC +100 µg of ovalbumim (OVA) via SC; h) SN2 via SC +100 µg of ovalbumim (OVA) via SC. The production of IgA and IgG in serum and in the gut mucosal was determinated by ELISA method. The Lactobacillus species used in this research were found to be inefficacious in the induction of humoral immune response in BALB/c mice.
Key-words: Lactobacillus, BALB/c mice, humoral immune response

LISTA DE ILUSTRAÇÕES
Figura 1 – Colônias de espécies de Lactobacillus em agar MRS......................................... 43
Figura 2 – Perfil eletroforético das proteínas secretadas pelas cepas de Lactobacillus....... 55

LISTA DE TABELAS
Tabela 1 – Fermentação de carboidratos pelas cepas de Lactobacillus.................................. 44
Tabela 2 – Perfil de susceptibilidade das cepas de Lactobacillus a antibióticos.................... 45
Tabela 3 – Crescimento de Lactobacillus em caldo MRS a diferentes condições de pH...... 50
Tabela 4 – População de Lactobacillus cultivados em caldo MRS a 37ºC/18 h – 40 rpm..... 51
Tabela 5 – Produção de ácido láctico pelas cepas de Lactobacillus em caldo MRS a
37ºC/18 h – 40 rpm................................................................................................
52
Tabela 6 – Concentração média de IgA total no soro e mucosa intestinal de camundongos 57
Tabela 7 - Concentração média de IgG total no soro e mucosa intestinal de camundongos 58

SUMÁRIO
1 INTRODUÇÃO................................................................................................................ 11
2 REVISÃO DA LITERATURA....................................................................................... 14
2.1 PROBIÓTICOS – CONCEITO...................................................................................... 14
2.2 CONSTITUIÇÃO E UTILIZAÇÃO DE PROBIÓTICOS............................................. 15
2.3 MODO DE AÇÃO DOS PROBIÓTICOS..................................................................... 18
2.4 FATORES QUE INTERFEREM NA UTILIZAÇÃO DE PROBIÓTICOS.................. 20
2.5 AÇÃO IMUNOMODULADORA DOS PROBIÓTICOS............................................. 20
2.5.1 Sistema imunológico.................................................................................................. 20
2.5.2 Interferência no sistema imunológico...................................................................... 23
3 MATERIAIS E MÉTODOS........................................................................................... 30
3.1 MICRORGANISMOS.................................................................................................... 30
3.2 TESTES PARA CONFIRMAÇÃO DAS CARACTERÍSTICAS DAS ESPÉCIES
DE Lactobacillus...........................................................................................................
30
3.3 AVALIAÇÃO DA SUSCEPTIBILIDADE A ANTIBIÓTICOS................................... 32
3.4 ADAPTAÇÃO E AVALIAÇÃO DA TOLERÂNCIA AO pH DAS CEPAS DE
Lactobacillus..................................................................................................................
32
3.5 CARACTERIZAÇÃO QUANTO À PRODUÇÃO DE ÁCIDO LÁCTICO................. 33
3.6 FORMAÇÃO DO POOL DE Lactobacillus.................................................................. 33
3.7 OBTENÇÃO DO SOBRENADANTE DOS CULTIVOS DE Lactobacillus................ 34
3.8 TRATAMENTO DOS ANIMAIS.................................................................................. 34
3.9 OBTENÇÃO E DETERMINAÇÃO DE PROTEÍNAS DE Lactobacillus.................... 36
3.10 CARACTERIZAÇÃO DAS PROTEÍNAS EM GEL DE POLIACRILAMIDA
COM DODECIL SULFATO DE SÓDIO (SDS-PAGE)............................................
37

3.11 AVALIAÇÃO DA RESPOSTA IMUNOLÓGICA..................................................... 38
3.11.1 Obtenção do soro..................................................................................................... 39
3.12 OBTENÇÃO DO MUCO INTESTINAL DOS ANIMAIS......................................... 39
3.13 QUANTIFICAÇÃO DE IgA E IgG TOTAL SÉRICA E DE MUCOSA
INTESTINAL..............................................................................................................
40
3.14 ANÁLISE ESTATÍSTICA........................................................................................... 41
4 RESULTADOS E DISCUSSÃO..................................................................................... 42
4.1 CONFIRMAÇÃO DA IDENTIDADE DAS ESPÉCIES DE Lactobacillus.................. 42
4.2 SUSCEPTIBILIDADE A ANTIBIÓTICOS.................................................................. 44
4.3 TOLERÂNCIA AO pH.................................................................................................. 49
4.4 PRODUÇÃO DE ÁCIDO LÁCTICO............................................................................ 51
4.5 PERFIL ELETROFORÉTICO DAS PROTEÍNAS SECRETADAS PELAS CEPAS
DE Lactobacillus...........................................................................................................
54
4.6 AVALIAÇÃO DA RESPOSTA IMUNOLÓGICA....................................................... 56
4.6.1 Concentração de IgA e IgG total presente no soro e na mucosa intestinal.......... 56
5 CONCLUSÕES................................................................................................................ 63
REFERÊNCIAS.................................................................................................................. 64
APÊNDICES....................................................................................................................... 80

11
1 INTRODUÇÃO
As propriedades benéficas conferidas aos lactobacilos foram observadas por
Metchinikoff, no início do século passado, quando este verificou melhor qualidade de vida e
longevidade em indivíduos que consumiam diariamente iogurte contendo tais bactérias. Desde
então, vários estudos foram desenvolvidos buscando esclarecer os mecanismos responsáveis
pela ação das bactérias lácticas na promoção do bem-estar do hospedeiro.
Sabe-se, atualmente, que outras espécies microbianas como leveduras e fungos
também são capazes de estabelecer o equilíbrio da microbiota intestinal, e conseqüentemente,
o benefício à saúde do hospedeiro. Este grupo de organismos é denominado probiótico, termo
que se refere aos suplementos microbianos vivos, não patogênicos, que afetam beneficamente
o hospedeiro, por melhorar o balanço da microbiota do trato digestivo.
Vários efeitos têm sido atribuídos aos probióticos, dentre os quais destacam-se a
reposição da microbiota intestinal; efeito hipocolesterolêmico, aumento da absorção de alguns
minerais, efeito anticarcinogênico e modulação do sistema imune.
A contribuição dos probióticos sobre o sistema imune é de grande interesse na
medicina humana e animal, pois o sistema imune que apresenta uma homeostasia normal
contribui para que indivíduos sejam mais resistentes às doenças a que estão constantemente
expostos.
De acordo com Nicoli e Vieira (2000), investimentos expressivos têm sido envolvidos na
comercialização de probióticos para aplicações em seres humanos e animais, principalmente na
Europa, Estados Unidos e Japão, bem como em países em desenvolvimento como o Brasil. No
Brasil grandes empresas comercializam produtos lácteos fermentados ou suplementados, além de
produtos farmacêuticos com propriedades probióticas.

12
No setor animal, como conseqüência à restrição ao uso de antibióticos como
promotores de crescimento, houve uma intensificação em pesquisas envolvendo probióticos.
Na avicultura os probióticos foram muito utilizados no final dos anos 90, entretanto, os resultados
sobre os benefícios de seu uso em rações avícolas foram contraditórios (ROSTAGNO et al.,
2003).
Estudos realizados com ruminantes e suínos, visando a melhoria no desempenho
zootécnico, características de carcaça e controle de diarréias em animais jovens, também
demonstraram a necessidade de maiores esforços na investigação para o entendimento do
mecanismo de ação desses microrganismos. Estudos têm evidenciado a eficácia da utilização de
camundongos como modelo para predizer a imunogenicidade em bovinos (VALLE et al., 2001).
É importante considerar que o mercado consumidor tornou-se mais exigente,
preocupando-se com a qualidade dos alimentos que consomem, pois existe a consciência de que a
origem e qualidade dos alimentos estão diretamente relacionadas à qualidade de vida. O mercado
externo de carnes tem revelado a grande necessidade de investimento em pesquisas para que
possa garantir um produto saudável para o consumidor, e ao mesmo tempo contribuir com a
economia do país.
De acordo com o Ministério da Agricultura, Pecuária e Abastecimento, (ABEF, 2005),
no ano de 2005 o Brasil apresentou uma receita cambial de US$ 8,195 bilhões,
correspondendo a 7% do volume total das vendas externas do País. A carne de frango teve
uma participação de 42,8%, a bovina de 37,9%, a suína de 14,3%, a de peru 3,1% e de outras
a participação foi de 1,9%, evidenciando, portanto, um mercado promissor, ávido pela
aplicação de novas tecnologias para atender às exigências do mercado externo.
Visando contribuir para o entendimento do mecanismo de ação dos probióticos sobre o
sistema imunológico de camundongos, e promover a obtenção de informações que possam

13
colaborar com avanços tecnológicos nesta área de conhecimento, o presente trabalho teve
como objetivos caracterizar espécies de Lactobacillus quanto à susceptibilidade a antibióticos,
resistência ao pH e produção de ácido láctico. Determinou-se também o perfil de proteínas
secretadas pelas espécies de lactobacilos e as concentrações de IgA e IgG totais presentes no
soro e mucosa intestinal dos animais.

14
2 REVISÃO BIBLIOGRÁFICA
2.1 PROBIÓTICOS – CONCEITO
No início do século 20, foi proposto que bactérias específicas exerciam benefícios à
saúde do hospedeiro. Elie Metchnikoff, prêmio Nobel por sua descoberta da fagocitose e
diretor do Instituto Pasteur, lançou a idéia de que as bactérias lácticas presentes no iogurte
exerciam efeito benéfico ao organismo receptor, após observar que camponeses búlgaros, cuja
dieta incluía a ingestão diária de iogurte, apresentavam uma vida mais prolongada e saudável
em relação às populações que não consumiam iogurte. A partir de então, vários estudos foram
desenvolvidos para esclarecer a relação destas bactérias na promoção da saúde do hospedeiro
(SHORTT, 1999).
A palavra probiótico foi citada pela primeira vez por Lilley e Stillwell em 1965,
caracterizando substâncias secretadas por um microrganismo, as quais estimulavam o
crescimento de outro (VANBELLE; TELLER; FOCANT, 1990; FULLER, 1992).
Vários conceitos foram atribuídos à palavra probiótico desde então, a fim de ajustá-los
às descobertas científicas. Em 1971, Sperti, definiu probiótico como extrato de tecidos que
estimulavam o crescimento microbiano. Posteriormente, Parker em 1974 redefiniu o conceito
como sendo organismos e substâncias que contribuíam para o equilíbrio microbiano intestinal.
Em 1995, na Alemanha, em um seminário sobre probióticos, surgiram dois novos
conceitos, compreendendo, “uma preparação de microrganismos vivos ou estimulantes
microbianos que afetavam a microbiota indígena de um animal, planta ou alimento receptor
de uma forma benéfica” e “uma preparação microbiana que contém bactérias vivas ou
inativadas, incluindo seus componentes e produtos, que administrada por via oral ou por outra

15
superfície (mucosa) melhorava o balanço microbiano ou enzimático nessas superfícies ou
estimulava mecanismos imunes específicos ou não” (JANSEN; VAN DER WAAIJ, 1995).
Entretanto, a definição atual aceita internacionalmente descreve que se trata de
microrganismos vivos, não patogênicos, que administrados em quantidades adequadas,
conferem benefícios à saúde do hospedeiro (FOOD AND AGRICULTURE
ORGANIZATION OF UNITED NATIONS, WORLD HEALTH ORGANIZATION, 2001;
SANDERS, 2003).
Qualquer fator que leve ao desequilíbrio da microbiota intestinal, como o uso indevido
de antimicrobianos e estresse de qualquer natureza do hospedeiro, poderá permitir a instalação
e a multiplicação de microrganismos patogênicos. Dessa forma, o equilíbrio da microbiota
intestinal terá reflexo direto sobre o estado de saúde do hospedeiro (MILES, 1993).
Segundo Nicoli et al. (2003), os probióticos apresentam possibilidades de aplicação
para compensar uma perturbação prevista (prevenção) ou instalada (tratamento) das funções
da microbiota digestiva.
2.2 CONSTITUIÇÃO E UTILIZAÇÃO DE PROBIÓTICOS
A colonização do trato intestinal ocorre logo após o nascimento, sendo que os
primeiros microrganismos a colonizarem o intestino, são as bactérias aeróbicas e anaeróbicas,
como Escherichia coli, Clostridium spp., Streptococcus spp., Lactobacillus spp., Bacterióides
spp. e Bifidobacterium spp. Todos os componentes da microbiota do trato gastrintestinal
(TGI) são necessários para o intestino desenvolver suas funções específicas (SERVIN, 2004;
ISOLAURI; SALMINEN; OUWEHAND, 2004).

16
Nas primeiras investigações com bactérias probióticas, avaliou-se Lactobacillus
acidophilus porque pensava-se ser esta espécie dominante no intestino, e era preciso assegurar
que o organismo sobrevivesse nesse ambiente. Estudos posteriores demonstraram a existência
de outras espécies de lactobacilos no intestino contrariando a idéia inicial (FULLER, 1992).
As bactérias lácticas são os principais constituintes de um produto probiótico. Essas
bactérias apresentam algumas características comuns, são Gram positivas, normalmente
catalase negativa, asporogênicas e produtoras de ácido láctico como resultado de seu
metabolismo primário (FERREIRA, 2003).
No mercado, os probióticos disponibilizados são aqueles cujas bactérias pertencem ao
grupo das bactérias lácticas, embora algumas leveduras (Saccharomyces boulardii) e fungos
(Aspergillus sp.) possam ser usados. O termo “bactéria láctica” inclui atualmente 15 gêneros:
Aerococcus, Atopobium, Bifidobacterium, Brochothrix, Carnobacterium, Enterococcus,
Lactobacillus, Lactococcus, Leuconostoc, Oenococcus, Pediococcus, Streptococcus,
Tetrogenococcus, Vagococcus e Weissella (FERREIRA, 2003).
Os suplementos probióticos para alimentos destinados aos humanos são constiuídos de
bactérias pertencentes aos gêneros Lactobacillus e Bifidobacterium e, em menor escala,
Enterococcus faecium, uma vez que elas têm sido isoladas de todas as porções do trato
gastrintestinal de humano saudável. O íleo terminal e o cólon parecem ser, respectivamente, o
local de preferência para a colonização intestinal dos lactobacilos e bifidobactérias
(CHARTERIS et al., 1998; BIELECKA; BIEDRZYCKA; MAJKOWSKA, 2002). Entretanto,
deve ser salientado que o efeito de uma bactéria é específico para cada cepa, não podendo ser
extrapolado, inclusive para outras cepas da mesma espécie (GUARNER; MALAGELADA,
2003).

17
Os probióticos, geralmente, são usados em terapia oral e profilaxia de desordens
gastrintestinais, promovendo o equilíbrio da microbiota intestinal. Uma microbiota desejável
equilibrada influencia a transformação de moléculas produzidas pelo hospedeiro e o estado
fisiológico do seu trato intestinal, por meio da modulação da renovação de células epiteliais e do
estado imunológico do hospedeiro. Distúrbios nesse equilíbrio, resultante da dieta, consumo de
drogas, situações de estresse, tratamentos quimioterápicos ou tratamentos com antibióticos podem
provocar diferentes problemas gastrintestinais (FERREIRA, 2001a; FORESTIER et al., 2001;
MAIA et al., 2001).
Para que o probiótico seja efetivo, além da seleção cuidadosa de cepas, as bactérias
deverão ser capazes de passar pelo trato gástrico sem sofrer perda de sua função e atingir a
mucosa intestinal, onde desempenharão papel benéfico pela promoção de respostas
imunológicas, e/ou combatendo organismos patogênicos no trato intestinal do hospedeiro
(KOZASA, 1989).
Frente a isso, pode-se dizer que a eficácia de produtos probióticos se dará pelas
seguintes características: capacidade de ser preparado como produto viável em escala
industrial; permanecer estável e viável por longos períodos em condições de estocagem e a
campo; capacidade de sobreviver, não necessariamente crescer, no intestino e produzir efeito
benéfico no hospedeiro (FULLER, 1992; APOSTOLOU et al., 2001).
Estas são as condições desejáveis de um probiótico, entretanto, outros cuidados,
muitas vezes determinantes para o sucesso de um produto, também devem ser observados,
como a origem dos microrganismos probióticos. Há necessidade de que as bactérias sejam
não patogênicas, ou seja, possuam status GRAS (generally recognised as safe). No caso de
uso em humanos, o gênero ao qual pertence a bactéria deverá ser de origem humana, não estar

18
associada a doenças como endocardite, além da ausência de genes de resistência aos
antibióticos (COLLINS; THORNTON; SULLIVAN, 1998; SAARELA et al., 2000).
2.3 MODO DE AÇÃO DOS PROBIÓTICOS
O conhecimento da microbiota intestinal e suas interações levaram ao desenvolvimento
de estratégias alimentares, objetivando a manutenção e o estímulo de bactérias normais ali
presentes (GIBSON; FULLER, 2000). Segundo Crittenden (1999), é possível aumentar o
número de microrganismos promotores da saúde no trato gastrintestinal (TGI), através da
introdução de probióticos pela alimentação ou com o consumo de suplemento alimentar
prebiótico, que irá modificar seletivamente a composição da microbiota, fornecendo ao
probiótico vantagem competitiva sobre outras bactérias do ecossistema.
Estas vantagens competitivas já haviam sido observadas por Fuller (1989), que
enumerou três possíveis mecanismos de atuação dos probióticos. O primeiro mecanismo é a
supressão do número de células viáveis mediante a produção de compostos com atividade
antimcrobiana, a competição por nutrientes e por sítios de adesão. O segundo é a alteração do
metabolismo microbiano pelo aumento ou diminuição de atividade enzimática, e o terceiro
mecanismo é o estímulo da imunidade do hospedeiro, por meio do aumento dos níveis de
anticorpos e o aumento da atividade dos macrófagos.
A partir dos mecanismos enumerados por Fuller, diversos autores propuseram os
seguintes mecanismos de ação aos probióticos (DE SIMONE et al., 1986; VANBELLE;
TELLER; FOCANT, 1990; HAVENAAR; HUIS INT VELD., 1992; LINK-AMSTER et al.,
1994; ISOLAURI; SALMINEN; OUWEHAND, 1998; OUWEHAND, 1998; NAIDU;
BIDLACK; CLEMENS, 1999; NICOLI et al., 2003):

19
• Inibição da proliferação de bactérias patogênicas pela produção de ácidos
orgânicos e bacteriocinas, ou pela redução do pH;
• Produção de H2O2 prevenindo a adesão das bactérias patogênicas;
• Produção de metabólitos capazes de neutralizar toxinas bacterianas in situ ou
pela inibição de sua produção;
• Produção de enzimas que melhorariam a digestão e utilização dos alimentos, ou
promoveriam a detoxificação resultante da presença de metabólitos produzidos
pela microbiota intestinal;
• Estímulo do sistema imunológico;
• Produção de vitaminas e aumento da atividade celular no tocante à síntese de
lactase, sacarase e maltase;
• Proliferação no trato digestivo do hospedeiro e competição com as bactérias
patogênicas;
• Redução do catabolismo microbiano, promovendo um melhor equilíbrio da
microbiota, tendo como conseqüência a redução na produção e adsorção de
substâncias como amônia e aminas;
• Alívio dos sintomas de intolerância à lactose;
• Melhora da motilidade intestinal;
• Efeito anti-hipertensivo, hipocolesterolêmico; anti-diabético e anticarcinogênico.
O mecanismo pelo qual bactérias probióticas previnem a instalação e multiplicação de
microrganismos patogênicos na mucosa intestinal é conhecido como exclusão competitiva, ou
seja, bactérias probióticas irão competir por sítios de ligação e nutrientes, impedindo a ação de
bactérias patogênicas (HAVENAAR; HUIS INT VELD, 1992; OUWEHAND et al., 1999).

20
2.4 FATORES QUE INTERFEREM NA UTILIZAÇÃO DE PROBIÓTICOS
De acordo com O’Sullivan et al. (1992), existem poucos dados consistentes
cientificamente para evidenciar os efeitos benéficos dos probióticos incorporados em produtos
comerciais. Isso está relacionado a desenhos experimentais insatisfatórios, análise estatística
inadequada dos resultados, escolha inadequada da cepa probiótica e controle de qualidade
insatisfatório da cultura e do produto. O ceticismo existente em relação ao uso de probióticos
pode ser explicado em parte às pretensões exageradas e insubstanciadas usadas na propaganda
e venda destes produtos.
2.5 AÇÃO IMUNOMODULADORA DOS PROBIÓTICOS
2.5.1 Sistema imunológico
O sistema imunológico compreende células e moléculas responsáveis pela imunidade
de um indivíduo, ou seja, conferem proteção ao indivíduo através da ativação da resposta
imunológica. O contato do indivíduo com substâncias irritantes, toxinas, agentes infecciosos e
células neoplásicas ativam mecanismos imunológicos apropriados, incluindo a inflamação,
fagocitose, síntese de anticorpos ou ativação de linfócitos T para eliminar esses desafios
(CARLOS et al., 2003). O sistema imune apresenta diferentes funções como conseqüência de
interações que ocorrem nos órgãos linfóides (PHILLIPS; MURRAY; CROCKER, 1995).
Este sistema apresenta células específicas para o antígeno, compreendendo os linfócitos
timo-dependentes (linfócitos T) e linfócitos timo-independentes (linfócitos B). Funcionalmente
associadas a estes linfócitos encontram-se as células apresentadoras de antígenos, tais como

21
macrófagos, células dendríticas e as células epiteliais dos órgãos linfóides primários (ABBAS;
LICHTMAN, 2005).
Os linfócitos B constituem 5-10% do total de linfócitos do sangue; estes quando ativados
pelo antígeno diferenciam-se em plasmócitos que são células formadoras de anticorpos ou
imunoglobulinas. Os anticorpos são moléculas glicoprotéicas, apresentam dois pares de cadeias
polipeptídicas, denominadas cadeias leves e cadeias pesadas. Classificam-se em diferentes
isótipos e subtipos com base nas diferenças de amoinoácidos que compõem a região constante das
cadeias pesadas, estas classes e subclassses possuem propriedades funcionais diferentes. As
classes ou isótipos de anticorpos são: IgG, IgA, IgM, IgD e IgE (ROITT; DELVES, 2004;
ABBAS, LICHTMAN, 2005).
Os linfócitos T diferenciam-se no timo e compreendem os linfócitos T auxiliares (Th),
linfócitos citotóxicos (Tc) e linfócitos T reguladores, constituem 65-75% dos linfócitos do sangue
(JUNQUEIRA; CARNEIRO, 2001; ROITT; DELVES, 2004; ABBAS, LICHTMAN, 2005; LA
CAVA et al., 2006).
As células B são diferenciadas na medula óssea e consistem de duas populações, a primeira
constitui 15% dos linfócitos B, identificadas por um marcador de superfície celular denominado
CD5+, encontrado na cavidade peritoneal. A outra população constitui 85% das células B, sendo
encontrada no sangue e em todos os órgãos do sistema imune. Existe também a população de
células nulas, ou seja, não são consideradas células B nem células T, e sua diferenciação não está
estabelecida. Estas células, chamadas células natural killer (NK), apresentam importantes funções
na defesa do hospedeiro contra tumores (UYTDE HAAG; VAN DER HEIJDEN; OSTERHAUS,
1991, PERDIGÓN, 1995).
A entrada de antígenos pelas vias naturais de infecção (oral, respiratória, genital) garantem
a sua exposição a grandes regiões de tecidos linfóides, promovendo a ativação imunológica,

22
mediada pela secreção de IgA. A IgA é a principal classe de anticorpo que pode ser
eficientemente secretada através dos epitélios, desempenhando papel crítico na defesa contra
patógenos, neutralizando estes agentes (MCGHEE et al., 1992; SALMINEN; VON WRIGHT,
1998). Além da IgA sérica, a IgA secretora (sIgA) está presente na forma dimérica no intestino. A
sIgA é resistente à proteólise, não participa da resposta inflamatória, sua principal função é mediar
a exclusão imune de antígenos invasores, impedindo sua ligação às células epiteliais e penetração
de microrganismos (ERICKSON; HUBBARD, 2000).
Dentre os mecanismos de defesa inespecífico do hospedeiro inclui-se a fagocitose, que
é efetuada por monócitos, macrófagos, leucócitos polimorfonucleares e histiócitos, que atuam
removendo antígenos estranhos. O estado de ativação de macrófagos é uma medida da
resposta imune inespecífica, e pode ser quantificada pelo cálculo da atividade de enzimas
liberadas como uma conseqüência da ativação celular, ou por um teste funcional
(PERDIGÓN; ALVARES, 1992; PERDIGÓN et al., 1995).
As células fagocíticas dos mamíferos pertencem a dois sistemas complementares,
denominados mielóide e mononuclear. O mielóide consiste principalmente dos neutrófilos,
que agem e morrem rapidamente, enquanto que as células do sistema mononuclear agem mais
lentamente, mas são capazes de uma fagocitose repetida (TIZARD, 1998).
Os macrófagos recebem nomes diferentes conforme sua localização. Àqueles imaturos
encontrados no sangue são chamados de monócitos; no tecido conjuntivo são chamados histiócitos;
células de Kupffer no fígado; micróglia no cérebro; macrófagos alveolares nos alvéolos pulmonares
e macrófagos intravasculares pulmonares quando presentes nos capilares pulmonares (TIZARD,
1998; ROITT & DELVES, 2004). Estas células apresentam várias funções, como a endocitose,
citotoxidade (NORTH, 1978; ADAMS; HAMILTON, 1992), geração de óxido nítrico, liberação de
peróxido de hidrogênio, secreção de muitas proteínas diferentes como lisozima, componentes do

23
complemento, proteases lisossomais, colagenases, elastases e ativadores do plasminogênio; na
presença de antígeno secretam citocinas (interleucinas e fator de necrose tumoral-alfa) que regulam
a imunidade (TIZARD, 1998; CARLOS et al., 2003).
As mudanças no delicado equilíbrio do sistema imune podem ser refletidas por
mudanças nos níveis de citocinas detectadas no plasma ou soro. Todas as citocinas
desempenham várias atividades, e todas as células imunes apresentam receptores de membrana
específicos para determinadas citocinas, permitindo uma interação precisa e íntima entre os
vários tipos de células. As citocinas atuam de maneira similar aos hormônios, no entanto,
regulam o sistema imune (MIYAJIMA, et al., 1992; MATSUURA; FIOCCHI1, 1993 apud
PERDIGÓN et al., 1995, p.1598; CARLOS et al., 2003).
Os antígenos que atravessam o trato intestinal, representado pela microbiota natural que
funciona como barreira no processo de defesa do organismo, são absorvidos através da camada
epitelial, podendo sofrer redução da imunogenicidade ou serem transportados e induzirem uma
resposta imune. A integridade desta barreira é necessária para prevenir o transporte de antígenos
inadequados (PERDIGÓN; FULLER; RAYA, 2001).
2.5.2 Interferência no sistema imunológico
Em condições normais, inúmeras espécies de bactérias estão presentes no intestino, a
maioria delas anaeróbias estritas. Essa composição torna o intestino capaz de responder a
possíveis variações anatômicas e físico-químicas (LEE et al., 1999).
De acordo com Fuller (1992), a base científica para o isolamento de microrganismos
1MATSUURA, T.; FIOCCHI, C. Cytokine production in the gastrointestinal tract during inflammation. In: WALKER, W.; HAEMATZ, P.; WESHILL, B. Immunophysiology of the gut. NewYork: Acad. Press, 1993. p.145.

24
probióticos parte do conhecimento de que a microbiota intestinal esteja envolvida na proteção do
hospedeiro contra a colonização do trato intestinal por microrganismos não comensais. Segundo
este autor, três tipos de estudos experimentais fornecem este embasamento: animais livres de
germes são mais susceptíveis às infecções intestinais do que os animais com a microbiota
estabelecida; a administração de antibióticos aos animais os tornam mais susceptíveis a doenças
intestinais; o efeito protetor da microbiota intestinal pode ser restaurado pela administração ao
animal de uma suspensão preparada a partir das fezes de animais adultos da mesma espécie.
O trato gastrintestinal é um complexo ecossistema constituído de três principais
componentes que estão permanentemente em contato e interagem entre si, células do
hospedeiro, nutrientes e microbiota. A ecologia intestinal resultante de um equilíbrio
dinâmico, remodelado pela dieta ingerida, entre os habitantes do lúmen intestinal, abriga mais
de 400 espécies bacterianas diferentes (DOGI; PERDIGÓN, 2006).
A microbiota intestinal exerce influência considerável sobre uma série de reações
bioquímicas do hospedeiro, em contrapartida, quando em desequilíbrio pode favorecer a
proliferação de patógenos com conseqüente infecção bacteriana (ZIEMER; GIBSON, 1998).
De acordo com Isolauri et al. (1995), a barreira intestinal representada pela microbiota
comensal, quando perturbada por antígenos, patógenos, substância química ou radioativa,
sofre alterações na sua permeabilidade e no processo inflamatório da mucosa.
Acredita-se que as células de defesa sejam ativadas no intestino e transportadas para
outros locais como as mucosas do trato respiratório e urogenital (HAVENAAR; HUIS INT
VELD, 1992). O tecido linfóide associado ao intestino (GALT), o qual conecta-se a outros
sistemas de mucosa e imunidade sistêmica, é um dos primeiros contatos dos componentes da
dieta, antígeno dos alimentos, bactérias patogênicas e benéficas, além de outros componentes

25
estranhos. No trato intestinal, a bactéria probiótica aderente entrará em contato com esse
tecido (ISOLAURI; SALMINEN; SALMINEN, 1998).
A mucosa intestinal só pode exercer seu papel na assimilação de antígenos porque
possui mecanismos de transporte especializados presentes nas vilosidades do epitélio e
particularmente nas placas de Peyer (aglomerados de nódulos linfóides localizados na parede
do intestino delgado), essenciais para estimular a resposta imune específica (ABBAS;
LICHTMAN, 2005). Perdigón et al. (1999) demonstraram que Lactobacillus casei e L.
plantarum interagiam com as células das placas de Peyer, além de promoverem aumento nas
concentrações de IgA, células CD4+ e anticorpos específicos para a cepa estimulada.
A parede intestinal também contém macrófagos e linfócitos (células T e B) em íntimo
contato com os alimentos no processo de absorção. Os linfócitos na cavidade peritonial fornecem
uma proteção imediata contra microrganismos invasores que podem penetrar na área abdominal
devido à agressão intestinal ou lesão (PERDIGÓN et al., 1995; ELIAS; SOUZA, 2001).
Várias pesquisas têm ressaltado o caráter imunomodulador às bactérias probióticas.
Bloksma et al. (1979), observaram que a administração de Lactobacillus plantarum (células
viáveis e não viáveis) a camundongos induzia o aumento dos volumes do fígado e baço sugerindo
um aumento de atividade imunoadjuvante pelo sistema reticuloendotelial. Em estudo conduzido
em humanos acometidos de diarréia aguda, tratados com L. acidophilus LB demonstrou-se que a
duração da diarréia era reduzida significativamente comparada com os indivíduos reidratados
oralmente (MICHIELUTTI et al., 1996). Link-Amster et al. (1994), observaram ação
imunoadjuvante de L. acidophilus (La1) e bifidobacteria fornecidos no leite fermentado para
humanos num período de três semanas.
Perdigón et al. (1986a, b, 1987) constataram que bactérias lácticas administradas oral ou
intraperitonealmente ativam macrófagos intraperitoniais de camundongos, e que a dose ótima

26
para a via oral era de 6,0 x 109 células, e 2,4 x 109 células para a via intraperitonial. Estes
autores não observaram diferenças na ativação de macrófagos quando utilizaram células viáveis
e não viáveis nos animais tratados com L. delbrueckii bulgaricus e S. sallivarius thermophilus,
embora no caso de camundongos tratados com L. casei e L. acidophilus a resposta tenha sido
um pouco maior quando bactérias viáveis foram usadas. Em estudo posterior, Perdigón et al.
(1991), verificaram que a mistura de L. casei e L. acidophilus potencializava a resposta imune, e
também, que L. casei mostrou-se eficiente como imunoadjuvante na prevenção de infecções
entéricas pela secreção de IgA no lúmen intestinal, sendo este efeito de L. casei observado
também para animais desnutridos (PERDIGÓN et al., 1995).
Plant e Conway (2002), observaram que as linhagens de Lactobacillus fermentum
KLD e L. casei 393, interagiram com o trato gastrintestinal de camundongos e o colonizaram
por um período extenso comparado com outras linhagens. Considerando-se o fato que L.
acidophilus e L. casei são capazes de sobreviver no trato intestinal, o efeito sobre a resposta
imune específica e inespecífica da mistura desses microrganismos em leite fermentado e não
fermentado, também foi estudado. Os resultados mostraram que a administração oral de L.
casei, L. acidophilus e L. delbrueckii bulgaricus aumentaram as respostas imunes celular e
humoral (PERDIGÓN et al., 1986c, d, 1988a, b).
Em experimento conduzido com camundongos, Iannello et al. (1984) observaram que
a microbiota associada à mucosa pode influenciar o nível de ativação de macrófagos
peritoniais. Gauffin, Agüero e Perdigón (2002), demonstraram que a administração de L.
casei em animais desnutridos, promoveu a recuperação da função imune da mucosa e também
da barreira intestinal que se encontrava debilitada.

27
No entanto, Vitiñi et al. (2000) verificaram que o uso de bactérias lácticas como
imunoadjuvantes não deve ser generalizado para todos os gêneros e espécies, pois apresentam
diferentes comportamentos, em especial, sobre a resposta imune na mucosa intestinal.
Estudos em camundongos com L. casei Shirota e L. casei administrados via oral e intravenosa,
respectivamente, demonstraram efeito imunopotencializador (SAITO et al., 1983; HORI et al., 2002).
Em humanos também foi observado aumento nos mecanismos de defesa do hospedeiro, em
voluntários que consumiram leite fermentado com as cepas La1 (L. acidophilus) e Bb12
(Bifidobacterium bifidum) por um período de três semanas (SCHIFFRIN et al., 1995).
O polissacarídeo peptideoglicano de várias espécies bacterianas, incluindo o da microbiota
normal e patogênica, pode induzir respostas imunológicas no hospedeiro (STEWART-TULL, 1980;
STIMPSON; SCHWAB, 1989; SCHWAB, 1993). Em estudo com porquinhos da índia Namba et
al. (1981), chegaram a resultados que sugeriam que as bactérias intestinais deviam ser solubilizadas
por enzimas bacteriolíticas administradas oralmente, e que o N-acetilmuramil-L-alanil-D-
isoglutamina (MDP) presente na parede celular dessas bactérias seria o responsável pelo aumento da
resposta imune. Sabe-se que a lisozima age no peptideoglicano da parede celular e que a
mieloperoxidase dependente de peróxido de hidrogênio, na presença de íons cloreto, remove os
peptídeos da parede celular (STEWART-TULL, 1980). Em Candida albicans, a estrutura mínima
necessária para que atue como adjuvante é o componente insolúvel presente na parede celular, β-
glucan (CASSONE et al., 1981). Estruturas como o ácido teicóico e lectina, presentes nas cepas de
Propionibacterium acidopropionici CRL 1188 e L. casei CRL 431, podem participar do processo
de adesão às células intestinais, conferindo caráter imunoadjuvante (AMBROSINI et al., 1998).
O efeito anticancerígeno da fração celular de L. acidophilus 606 inativado por aquecimento
foi observado em estudo conduzido por Choi et al. (2006). Os pesquisadores observaram que os

28
polissacarídeos solúveis derivados da cepa exerceram atividade antioxidante significativa, assim
como, atividade anticarcinogênica sobre uma variedade de linhagens de células cancerígenas.
LeBlanc et al. (2006) estudaram a ação anticarcinogênica de kefir (bebida láctea fermentada
que contém uma mistura de microrganismos benéficos) e sua fração protéica em camundongos
Balb/c, e verificaram que ambos aumentaram os níveis séricos de IL-10 e diminuíram os níveis de
IL-6 (envolvidos na síntese de estrogênio) em glândulas mamárias, concluíram que os tratamentos
foram capazes de retardar o crescimento do tumor devido à sua capacidade imunoreguladora.
Segundo Carlos et al. (2003), ratos alimentados com produto de soja fermentado
(Enterococcus faecium e Lactobacillus helveticus), estimulados com lipopolissacarídeos
(LPS), apresentaram maior capacidade de indução de liberação das citocinas IL-1, IL-6, IL-
12, INF-γ, TNF-α e da indução da liberação de peróxido de hidrogênio e de ácido nítrico.
Constatou-se também que os componentes da soja participariam da liberação de citocinas.
Shu e Gill (2002) estudaram a proteção imunológica mediada por Lactobacillus
rhamnosus HN001 (DR20�) frente à infecção por Escherichia coli 0157:H7 em
camundongos, e observaram menor severidade da infecção sugerindo que este efeito poderia
estar relacionado com o aumento das respostas imune celular e humoral.
A via de degradação de carboidratos utilizados pelas bactérias lácticas reduz a
quantidade de carboidratos disponíveis, e conseqüentemente, acumula moléculas orgânicas
que exibem atividade antimicrobiana, sendo as mais comuns os ácidos láctico, acético e
propiônico (BLOM; MÖRTVEDT, 1991). Entretanto, outros componentes antimicrobianos,
como o peróxido de hidrogênio, dióxido de carbono, diacetil, bacteriocinas e óxido nítrico,
podem ser formados ou estimulados pela presença de diferentes bactérias lácticas
(OUWEHAND, 1998). De acordo com Freese; Shen e Galliers (1973), os ácidos láctico,

29
acético e propiônico interagiriam com as células da membrana neutralizando o potencial
eletroquímico.
Segundo Mercenier, Muller-Alouf e Grangette (2000), existe um considerável interesse
em comparar as respostas imunes induzidas pelas linhagens recombinantes de bactérias lácticas
produtoras de antígenos em três localizações celulares diferentes: citoplasma, meio extracelular e
próxima à parede celular, para, então analisar qual forma seria mais imunogênica e ainda observar
a duração da resposta induzida por essas linhagens quando administradas nas mucosas.
Nos últimos anos a contribuição do sistema imune das mucosas na defesa contra uma
grande variedade de microrganismos patogênicos aliado ao conhecimento da microbiota intestinal
levou ao desenvolvimento de estratégias alimentares, objetivando a manutenção e o estímulo das
bactérias normais ali presentes (PERDIGÓN et al., 1990; GIBSON; FULLER, 2000).
Erickson e Hubbard (2000), revisaram a função dos probióticos na imunomodulação.
Segundo os autores, os probióticos que possuem influências favoráveis nos processos
fisiológicos e patológicos no hospedeiro por seu efeito na microbiota intestinal, poderiam agir
na promoção da saúde humana. Embora sua aplicação tenha demonstrado tendências em
relação à alteração de alguns aspectos relativos à resposta imune, os mecanismos pelos quais
isto ocorre não estão esclarecidos.

30
3 MATERIAIS E MÉTODOS
O presente trabalho foi realizado no Laboratório de Ornitopatologia da Faculdade de
Medicina Veterinária e Zootecnia da Universidade de São Paulo – São Paulo-SP.
3.1 MICRORGANISMOS
Foram avaliadas no presente trabalho sete cepas de Lactobacillus caracterizadas
parcialmente como probióticas, pertencentes às espécies L. casei pseudoplantarum (30b e
30c), L. plantarum (11fb, 22c e 41b) e L. reuteri (18fa e 19fa), da coleção de culturas do
Laboratório de Ornitopalogia da FMVZ – USP, São Paulo, isoladas de cama de frango por
Paço et al. (2003). Estas cepas foram mantidas em Leite Desnatado Reconstituído (LDR) 10%
contendo 5% de glutamato de sódio. Para ativação as culturas foram repicadas três vezes
consecutivas em caldo Man Rogosa e Sharpe (MRS) por 18 horas a 37°C sob agitação de 40
rpm.
3.2 TESTES PARA CONFIRMAÇÃO DAS CARACTERÍSTICAS DAS ESPÉCIES DE
Lactobacillus
As cepas foram ativadas em agar MRS constituído por 10 g/L de peptona
(BACTOTM), 8 g/L de extrato de carne (BBLTM), 4 g/L de extrato de levedura (OXOID), 20
g/L de glicose (SYNTH), 2 g/L de fosfato dipotássico (RIEDEL-DE HAËN), 5 g/L de acetato
de sódio.H2O (SYNTH), 2 g/L de citrato triamoniacal (SYNTH), 0,2 g/L de sulfato de
magnésio.7H2O (RIEDEL-DE HAËN), 0,05 g/L de sulfato de manganês.4H2O (ECIBRA), 1
mL/L de Tween 80 (SYNTH) e ágar 15 g/L, a pH 6,2. Em seguida as cepas foram

31
caracterizadas quanto ao tamanho da colônia (2 - 5 mm), formato convexo, superfície lisa,
lustrosa e sem pigmentos (KANDLER; WEISS, 1986), e foram submetidas às provas
bioquímicas incluindo verificação da produção de catalase, coloração de Gram (MADIGAN
et al., 2000) e fermentação dos principais açúcares.
Para avaliação das propriedades de diferentes açúcares empregou-se meio
constituído de 10 g/L de citrato de amônio, 5 g/L de extrato de levedura, 5 g/L de fosfato
dipotássico , 2 g/L de citrato de amônio, 5 g/L de acetato de sódio, 0,5 g/L sulfato de
magnésio.7H2O, 0,2 g/L de sulfato de manganês.4H2O, 1 ml/L de Tween 80, 0,1% de
vermelho de fenol (MERCK) e 10 g/L do açúcar teste (arabinose, celebiose, frutose,
galactose, glicose, lactose, maltose, manitol, manose, melezitose, melibiose, rafinose,
rhamnose, salicina, sorbitol, sacarose, trealose e xilose) a partir de soluções a 20%
esterilizadas por filtração em membranas de 0,2 �m (AC020DB3, Sartorius, Minisrt High-
Flow). Um volume de 1,9 mL deste meio foi transferido para tubos de ensaio esterilizados
que receberam em seguida 0,1 mL da solução do açúcar teste e duas alçadas da suspensão
dos microrganismos previamente ativados. Em seguida foram incubadas a 37oC por um
período de sete dias para observação da mudança de coloração, em função da redução do
vermelho de fenol pelo ácido presente no meio em que o açúcar foi metabolizado
(KANDLER; WEISS, 1986).
A atividade da catalase foi determinada pelo gotejamento de peróxido de hidrogênio
(H2O2) 30% sobre as colônias frescas dos lactobacilos e observação do desprendimento de
oxigênio (MAC FADDIN, 1980).

32
3.3 AVALIAÇÃO DA SUSCEPTIBILIDADE A ANTIBIÓTICOS
Para a determinação do padrão de susceptibilidade a antibióticos das cepas avaliadas
empregou-se o método de difusão em disco (Dispens-o-Disc-Susceptibility Test System –
DIFCO).
Dezessete antibióticos foram testados, a saber: aztreonam (30 �g), canamicina (30 �g),
ceftazidima (30 �g), cefalotina (30 �g), cefoxitina (30 �g), ciprofloxacina (5 �g),
cloranfenicol (30 �g), enrofloxacina (5 �g), eritromicina (15 �g), fosfomicina (200 �g),
gentamicina (10 �g), imipenem (10 �g), neomicina (30 �g), nitrofurantoína (300 �g),
teicoplanina (30 �g), tetraciclina (30 �g) e trimetropim (5 �g).
As cepas foram cultivadas em agar MRS e uma colônia foi transferida para 2 mL de
caldo MRS por 24 horas a 37ºC, sob agitação de 40 rpm. Uma suspensão bacteriana com
densidade 1 na escala de McFarland foi obtida diluindo-se as cepas em PBS 0,1M, pH 7,4.
Após a semeadura de 0,1 mL desta suspensão, com o auxílio de alça de Drigalsky, em placa de
Petri contendo 20 ml de agar MRS, acrescentou-se os discos com os respectivos antibióticos. A
placa foi incubada a 37ºC por 24h e a susceptibilidade a antibiótico foi verificada por meio da
formação de halo de inibição (DANIELSEN; WIND, 2003), medidos e interpretados conforme
tabela fornecida pelo fabricante dos respectivos antimicrobianos (DIFCO).
3.4 ADAPTAÇÃO E AVALIAÇÃO DA TOLERÂNCIA AO PH DAS CEPAS DE
Lactobacillus
Neste experimento, as células de lactobacilos foram cultivadas em tubos de ensaio
contendo 5 mL de caldo MRS pH 6,5 e incubados sob agitação de 40 rpm por 18h a 37ºC. Em

33
seguida foram repicadas para tubos contendo 5 mL de caldo MRS pH 6,0 (corrigido com
ácido clorídrico 10%) nas mesmas condições e assim sucessivamente numa escala decrescente
de 0,5 até pH 4,0. Nesta condição as células foram repicadas três vezes consecutivas para se
adaptarem a este meio.
3.5 CARACTERIZAÇÃO QUANTO À PRODUÇÃO DE ÁCIDO LÁCTICO
As suspensões de células foram centrifugadas a 10.000 x g por 10 min, 4ºC cepas e os
sobrenadantes avaliados quanto à produção de ácido láctico que foi determinado por
Cromatografia Líquida de Alta Eficiência (Waters 786), nas seguintes condições: coluna BIO
RAD Aminex HPX-87H (300 x 7,8 mm); temperatura da coluna 45ºC; detector de índice de
refração WATERS 410, eluente H2SO4 0,01M, fluxo de 0,6 mL/min; volume de amostra
injetada, 20 µL. As amostras devidamente diluídas foram filtradas em filtro Sep Pak C18
(Millipore) e o eluente filtrado a vácuo em membrana HAVP 0,45 µm de poro, 47 mm de
diâmetro (Millipore) e simultaneamente desgaseificado por ultra-som (Thorton) por 25
minutos.
3.6 FORMAÇÃO DO POOL DE Lactobacillus
Para obtenção do pool das respectivas cepas de Lactobacillus cada cultura foi
individualmente ativada três vezes consecutivas em caldo MRS. Na primeira ativação, cada
isolado foi inoculado separadamente em tubos contendo 5 mL de caldo MRS, sendo que após
18 horas de incubação a 37ºC sob agitação (40 rpm), alçadas das respectivas suspensões
celulares foram transferidas, individualmente, para tubos contendo 10 mL de caldo MRS.

34
Decorridas 18 horas nas mesmas condições, os conteúdos destes tubos foram transferidos
conjuntamente para frascos com tampa rosqueável contendo 400 mL de caldo MRS e
incubados novamente por 18 horas nas mesmas condições para a formação do pool de
lactobacilos. Posteriormente, a suspensão celular resultante deste pool foi centrifugada a
10.000 x g por 10 min a 4ºC, lavadas três vezes consecutivas em solução PBS 0,1M, pH 7,4,
ressuspensa em solução de sacarose a 20% para conferir uma concentração de 109 UFC/mL e
estocadas a -80ºC em microtubos de 1,5 ml (Eppendorf).
3.7 OBTENÇÃO DO SOBRENADANTE DOS CULTIVOS DE Lactobacillus
Ao sobrenadante resultante da centrifugação da suspensão celular referente ao pool de
lactobacilos, conforme descrito no item anterior, foram adicionados individualmente
inibidores de proteases, nas concentrações de 0,3 mg de ácido etilenodiamino tetra acético
(EDTA), 75 µg de fluoreto de fenilmetilsulfonil (PMSF), 2 µg de aprotinina e 0,7 µg de
pepstatina por mL de sobrenadante. Estas concentrações foram pré-determinadas para evitar a
degradação das proteínas produzidas pelas cepas de Lactobacillus (BOLLAG; ROZYCKI;
EDELSTEIN, 1996). Em seguida o sobrenadante foi filtrado em membrana 0,2 µm
(AC020DB3, Sartorius, Minisart High-Flow) e acondicionado em microtubos de 1,5 ml
(Eppendorf) a -80ºC.
3.8 TRATAMENTO DOS ANIMAIS
Cento e vinte camundongos Balb/c fêmeas, com idade entre 8 e 14 semanas, foram
divididos em 2 grupos (A e B) de 60 animais, sendo que cada grupo foi subdividido em 4

35
grupos com 15 animais cada e submetidos aos respectivos tratamentos. Os grupos controle,
totalizando 48 animais foram subdivididos em grupos de 6 animais. Os animais foram
mantidos em gaiolas plásticas em temperatura ambiente, receberam água e ração ad libitum.
Cada grupo constituiu uma unidade experimental.
Os experimentos, A e B, foram conduzidos paralelamente. Os animais tratados oralmente
receberam o pool de células ou sobrenadante por sonda intragástrica conforme proposto por Perdigón et
al., (1988a, 2002). O período de experimentação teve duração de 42 dias.
Os animais devidamente agrupados foram submetidos a oito diferentes tratamentos,
onde foram testados suspensão bacteriana (SB) contendo 109 UFC/mL, sobrenadante sem
tratamento (SN1), sobrenadante tratado a 100 oC / 10 minutos (SN2) e solução salina (0,85%
de NaCl) da seguinte forma:
Grupo A:
a) Tratamento 1: 0,1mL de SB via oral por 8 dias consecutivos.
Controle: 0,1 mL de solução salina via oral por 8 dias consecutivos.
b) Tratamento 2: 0,2 mL de SN1 via oral por 8 dias consecutivos.
Controle: 0,1 mL de solução salina via oral por 8 dias consecutivos.
c) Tratamento 3: 0,2 mL de SN1 via subcutânea (SC) por 3 dias consecutivos a intervalos
de 24 horas.
Controle: 0,2 mL de solução salina via SC por 3 dias consecutivos a intervalos de 24
horas.
d) Tratamento 4: 0,2 mL de SN2, via SC durante 3 dias consecutivos a intervalos de 24
horas.
Controle: 0,2 mL de solução salina via SC por 3 dias consecutivos com intervalos de 24
horas.

36
Grupo B:
Neste grupo os animais foram imunizados via SC com 100 �g de ovalbumina (OVA) no 1o
e 28o dias de tratamento conforme proposto por Lauterslager, Luuk e Hilgers (2003), e
submetidos aos seguintes tratamentos:
e) Tratamento 5: 0,1mL de SB via oral por 8 dias consecutivos + 0,1 mL de OVA via SC.
Controle: 0,1 mL de solução salina via oral por 8 dias consecutivos.
f) Tratamento 6: 0,2 mL de SN1 via oral + 0,1 mL OVA via SC .
Controle: 0,1 mL de solução salina via oral por 8 dias.
g) Tratamento 7: 0,2 mL de SN1 via SC por 3 dias consecutivos a intervalos de 24 horas +
0,1 mL OVA via SC.
Controle: 0,2 ml de solução salina via SC por 3 dias consecutivos a intervalos de 24
horas .
h) Tratamento 8: 0,2 ml de SN2, via SC durante 3 dias consecutivos a intervalso de 24
horas + 0,1 mL OVA via SC.
Controle: 0,2 ml de solução salina via SC por 3 dias consecutivos a intervalos de
24 horas.
3.9 OBTENÇÃO E DETERMINAÇÃO DE PROTEÍNAS DE Lactobacillus
As cepas de lactobacilos foram cultivadas em frascos contendo 100 mL de caldo MRS
a 37ºC/18h sob agitação de 40 rpm e centrifugadas a 10.000 x g por 10 minutos, 4oC. Em
seguida, foram adicionados, individualmente, ao sobrenadante os inibidores de proteases, nas
concentrações de 0,3 mg de ácido etilenodiamino tetra acético (EDTA), 75 µg de fluoreto de
fenilmetilsulfonil (PMSF), 2 µg de aprotinina e 0,7 µg de pepstatina por mL de sobrenadante

37
(BOLLAG; ROZYCKI; EDELSTEIN, 1996).
O sobrenadante obtido foi filtrado em membrana milipore 0,2 µm (AC020DB3,
Sartorius, Minisrt High-Flow), subdividido em alíquotas e mantido a –80°C. Para
confirmação da ausência de células no sobrenadante, o mesmo foi plaqueado em ágar MRS
(FERREIRA, 2001b).
A concentração de proteína no sobrenadante foi determinada por meio da reação com
o ácido bicinconínico empregando-se kit comercial Micro BCA Protein Assay Reagent Kit,
Pierce, conforme proposto por Ferreira (2001b).
3.10 CARACTERIZAÇÃO DAS PROTEÍNAS EM GEL DE POLIACRILAMIDA COM
DODECIL SULFATO DE SÓDIO (SDS-PAGE)
As proteínas presentes no sobrenadante foram devidamente caracterizadas por
eletroforese conforme metodologia proposta por Laemmli (1970). Para tanto, o gel de
poliacrilamida foi preparado com solução de acrilamida e bis-acrilamida (30% e 0,8%,
respectivamente) obtendo-se uma porosidade de 12,5% para o gel de separação e 5% para o gel
de empilhamento. A este gel foi aplicado um volume de sobrenadante correspondente a 80 µg
das proteínas produzidas pelas cepas de lactobacilos. A eletroforese foi conduzida com
aplicação de voltagem constante de 100 volts enquanto as cepas passavam pelo gel de
empilhamento, sendo aumentada para 200 volts quando as cepas alcançaram o gel de
separação. Utilizou-se como marcador de massa molecular a fosforilase b (94 KDa), albumina
sérica bovina (67 KDa), ovalbumina (43 KDa), anidrase carbônica (30 KDa), inibidor de
tripsina (20,1 KDa) e α-lactoalbumina (14,4 KDa) (Amersham Pharmacia Biotech. Sweden).

38
Após o término da eletroforese, o gel foi tratado em soluções específicas e impregnado com
nitrato de prata a fim de revelar as bandas protéicas, de acordo com metodologia descrita por Blum
et al. (1987).
3.11 AVALIAÇÃO DA RESPOSTA IMUNOLÓGICA
A resposta imunológica nos camundongos foi avaliada por meio da determinação da
concentração de IgA e IgG sérica e da mucosa intestinal, conforme demonstrado no Quadro 1.
Quadro 1 – Avaliação da preparação de probiótico como imunomodulador
Experimentos Fração
Probiótica
Vias de
Administração
Dose e
Freqüência
Resposta
Imune
Lactobacillus In natura Oral 1,0 x 109 UFC/8dias
Oral 0,2 mL/8dias SN1
Grupo A
Sobrenadante
SN2 SC 0,2 mL/3dias
Lactobacillus
In natura
+
OVA
Oral
1,0 x 109 UFC/8dias
+
100µg
Oral 0,2 mL/8dias + 100µg SN1 +
OVA
Grupo B
Sobrenadante SN2 +
OVA
SC 0,2 mL/3dias
+ 100µg
IgA/IgG sérica
IgA/IgG mucosa
intestinal
SC = subcutânea; SN1 = sobrenadante sem tratamento térmico; SN2 = sobrenadante tratado termicamente; OVA = ovalbumina

39
3.11.1 Obtenção do soro
Para obtenção do soro sangüíneo 5 camundongos foram sacrificados a intervalos de 14
dias de tratamento por deslocamento cervical, e sangrados por punção cardíaca. O sangue
coletado foi centrifugado em microtubos de 1,5 mL (Eppendorf) a 3.000 x g por 5 minutos a
4ºC. Os soros obtidos foram acondicionados em microtubos de 1,5 mL (Eppendorf) e
mantidos a –20ºC até a realização dos testes.
a) Soro controle positivo
O soro controle positivo para o teste de ELISA foi preparado a partir do sangue de
camundongos que receberam por via oral o pool de lactobacilos contendo 109 UFC/mL. Os
soros obtidos do sacrifício dos cinco animais foram juntados e armazenados a –20ºC até
realização das análises.
b) Soro controle negativo
O soro controle negativo foi obtido a partir do sangue de camundongos no primeiro dia do
experimento. Os respectivos soros foram juntados e armazenados a -20°C até serem analisados.
3.12 OBTENÇÃO DO MUCO INTESTINAL DOS ANIMAIS
Para a avaliação da concentração de anticorpos presentes na mucosa intestinal dos
animais, realizou-se a extração do órgão seguindo as seguintes etapas:
a) remoção do intestino desde o duodeno até o jejuno;

40
b) remoção de toda gordura lateral do intestino;
c) remoção do conteúdo intestinal injetando-se 1mL de tampão fosfato (PBS 0,1 M pH
7,4) e inibidores de proteases PMSF 0,01 M (0,3 �g/mL), Pepstatina A (0,7 �g/mL)
e EDTA (0,3 mg/mL), sendo recolhido em um tubo de boca larga para facilitar
manuseio, mantendo-o sempre em banho de gelo;
d) centrifugação do conteúdo intestinal a 10.000 x g por 10 minutos a 4°C;
e) armazenamento do sobrenadante em microtubo de 1 mL a –20°C até a realização
das análises.
3.13 QUANTIFICAÇÃO DE IgA E IgG TOTAL SÉRICA E DE MUCOSA INTESTINAL
Os soros obtidos de cada grupo foram misturados em igual volume, para a realização
do teste de ELISA. A mistura resultante, cujo volume totalizou 100 µL, foi diluída de acordo
com a recomendação do fabricante do kit para a detecção de IgG ou IgA total (ELISA Starter
Accessory Package, Bethyl Laboratories, Inc), e os soros correspondentes aos controles
positivo e negativo foram distribuídos nos respectivos poços da placa de ELISA.
Após a realização das cinco etapas descritas no protocolo do kit para a detecção dos
anticorpos produzidos, procedeu-se a leitura de absorbância a 450 nm em leitor de ELISA
(Elx800, Universia Microplate Reader, BIO-TECK Instruments) e os resultados foram
calculados de acordo com a curva padrão fornecida pelo kit e expressos em mg de IgA ou IgG
por mL de soro.
3.14 ANÁLISE ESTATÍSTICA

41
Os dados obtidos foram analisados por meio do software Graphpad InStat
(GRAPHPAD, 1998). Utilizou-se o teste de Kolmogorov-Smirnov para verificar se os dados
encontravam-se dentro da distribuição normal e o teste “t” de Student (p<0,05) para
demonstrar a diferença entre os grupos.

42
4 RESULTADOS E DISCUSSÃO
4.1 CONFIRMAÇÃO DA IDENTIDADE DAS ESPÉCIES DE lactobacillus
As colônias de lactobacilos cultivadas em agar MRS apresentaram características
morfológicas descritas por Kandler e Weiss (1986), como coloração esbranquiçada, lisas,
convexas lustrosas, bem delimitadas e diâmetros variando entre 2 a 4 mm (Figura 1). Dentre
as espécies estudadas no presente trabalho a cepa de L. plantarum (22c) apresentou colônia
com diâmetro de 4 mm, o maior observado, e as cepas de L. reuteri (18fa e 19fa)
apresentaram colônias com o menor diâmetro, 2 mm. Foi verificado também que as cepas
pertencentes à mesma espécie apresentaram diâmetros de colônias semelhantes. Por meio do
teste de coloração de Gram foi possível confirmar que as espécies estudadas eram Gram positivas e
apresentavam células em forma de bacilo.

43
11fb 22c
18 fa 19 fa
30b 30c
41b Pool
Figura 1 – Colônias de espécies de Lactobacillus em agar MRS
Na Tabela 1 encontram-se apresentados os resultados referentes à fermentação de
açúcares e atividade da catalase das cepas de lactobacilos. Observa-se que todas as cepas
foram catalase negativas e apresentaram perfil de fermentação de açúcares conforme descrito
por Kandley e Weiss (1986) confirmando, portanto, a identidade das respectivas espécies.

44
Tabela1 - Fermentação de carboidratos pelas cepas de Lactobacillus
Lactobacillus plantarum Lactobacillus reuteri Lactobacillus casei pseudoplantarum Carboidratos
11fb 22c 41b 18fa 19fa 30b 30c Amygdalin + + + - - + +
Arabinose - + - - - - -
Celobiose + + + - - + +
Frutose + + + + + + +
Galactose + + + + + + +
Glicose + + + + + + +
Lactose + + + + + + +
Maltose + + + + + + +
Manitol + + + - - + +
Manose + + + - - + +
Melezitose - + + - - + +
Melibiose + + + + + - -
Rafnose + + + + + - -
Ramose - - - - - - -
Ribose + + + + + + +
Salicina + + + - - + +
Sacarose + + + + + + +
Sorbitol + + + - - + +
Trealose + + + - - + +
Xilose + - - - - - -
Catalase - - - - - - -
4.2 SUSCEPTIBILIDADE A ANTIBIÓTICOS
Várias pesquisas (TEUBER; MEILE; SCHWARZ, 1999; ROJO-BEZARES et al., 2006;
HUMMEL et al, 2007) têm demonstrado que a resistência a antibióticos pode ser intrínseca e não
transmissível, no entanto, genes de resistência presentes no DNA bacteriano de algumas cepas podem
ser transmitidos às bactérias comensais e estas podem transferir horizontalmente às bactérias
patogênicas presentes na microbiota intestinal do hospedeiro.
Desta forma, a susceptibilidade das cepas de lactobacilos em estudo, isolados e na forma de

45
pool (cultura mista), a antibióticos mais relevantes na terapia humana e animal, foi analisada e os
resultados encontram-se apresentados na Tabela 2.
Tabela 2 - Perfil de susceptibilidade das cepas de Lactobacillus a antibióticos
Lactobacillus plantarum
Lactobacillus reuteri
L. casei pseudoplantarum Pool Antibióticos
11fb 22c 41b 18fa 19fa 30b 30c P Beta lactâmicos Cefalosporina de 1a geração Cefalotina S R R S S R MS MS
Cefalosporina de 2ª geração Cefoxitina R R R S R R R R
Cefalosporina de 3a geração Ceftazidima R R R S S R R R
Imipenem Imipenem S S S S S S R S Fluorquinolonas Ciprofloxacina R R R R R R R R Enrofloxacina R R R R R R R R Aminoglicosídeos Gentamicina R R R R R R R R Canamicina R R R R R R R R Neomicina R R R R R R R R Vancomicina Teicoplanina R R R R R R R R Tetraciclina Tetraciclina R R R R S R S R Cloranfenicol Cloranfenicol S R S S S R S S Monobactâmicos Aztreonam R R R R R R R R Macrolídeo Eritromicina R R R MS MS R MS S Fosfomicina Fosfomicina R R R R R R R R Derivados Nitrofurânico Nitrofurantoína MS R R R S R R R
Inibidor de redutase Trimetropim R R R R R R R R R= resistente; S= sensível; MS= moderadamente sensível.
Nota-se que 100% das cepas de L. plantarum (11fb, 22c e 41b) apresentaram
sensibilidade ao imipenem, e 67% (11fb e 41b) ao cloranfenicol. As cepas de L. reuteri (18fa
e 19fa) apresentaram perfil de susceptibilidade diferenciado para cefoxitina, tetraciclina e
nitrofurantoína, enquanto que L. casei pseudoplantarum (30b e 30c) revelou diferenças na

46
susceptibilidade por cefalotina, imipenem, tetraciclina, cloranfenicol e eritromicina.
O pool formado pela co-cultura das cepas de lactobacilos apresentou sensibilidade a
imipenem, cloranfenicol e eritromicina, e moderadamente sensível a cefalotina. Verifica-se
também que a susceptibilidade aos antimicrobianos cefoxitina, tetraciclina, cloranfenicol,
eritromicina e nitrofurantoína, apresentou-se diferenciada para cepas da mesma espécie,
sugerindo, portanto, que a resistência seja específica da cepa.
Os antimicrobianos imipenem e cloranfenicol são compostos que têm ação sobre a
síntese da parede celular e síntese de proteína, respectivamente, apresentando atividade
bactericida e bacteriostática. Sabe-se que o imipenem apresenta maior atividade contra
bactérias Gram positivas, justificando a sensibilidade do lactobacilo a este agente
(GÓRNIACK, 2002).
Em geral as cefalosporinas são os antibióticos de preferência para o tratamento de
infecções bacterianas mistas, entretanto, muitas bactérias têm se apresentado resistentes, daí a
necessidade da administração do imipenem como alternativa viável para o controle de
infecções graves, demonstrando a importância de se conhecer a susceptibilidade bacteriana a
este composto.
No que se refere ao cloranfenicol, a susceptibilidade das cepas de L. plantarum (11fb e
41b), L. reuteri (18fa e 19fa), L. casei pseudoplantarum (30c) e do pool de lactobacilos foi
condizente com seu modo de ação, uma vez que se trata de antibiótico que apresenta amplo
espectro de atuação, entretanto, as cepas 22c e 30b exibiram resistência ao antimicrobiano,
conforme proposto por Delgado, Flórez e Mayo (2005) e Vay et al. (2007).
Segundo Spinosa (2002) a resistência ao cloranfenicol em geral é mediada por
plasmídeos, os quais são capacitados a produzir a cloranfenicol-acetiltransferase que inativa o
antibiótico. Neste sentido, as cepas 22c e 30b não seriam recomendadas para compor um

47
produto probiótico, porém, analisando-se o pool de lactobacilos verifica-se que este exibiu
susceptibilidade ao cloranfenicol, demonstrando que o cultivo das cepas de lactobacilos em
co-cultura apresentou o mesmo comportamento de 71% das cepas cultivadas individualmente.
Este comportamento frente ao cloranfenicol provavelmente ocorreu porque as cepas de
lactobacilos estudadas não apresentaram genes de resistência, à semelhança do estudo
conduzido por Hummel et al. (2007) que verificaram que as espécies L. pentosus BFE 7437 e
L. plantarum BFE 7440, embora apresentassem resistência ao agente não foi constatado a
presença do gene de resistência.
A ação da cefoxitina sobre as cepas 18fa e 19 fa pode ser explicada levando-se em
consideração o seu modo de atuação que se dá principalmente sobre as bactérias Gram
negativas, embora possa atuar sobre Gram positivas como demonstrado na Tabela 2, à
semelhança do proposto na literatura (CEBECI; GÜRAKAN, 2003; DANIELSEN; WIND,
2003; VAY et al., 2007).
No tocante a resistência à tetraciclina e nitrofurantoína pela cepa 18fa e 30b também
foram verificadas por outros autores (TEUBER; MEILE; SCHWARZ, 1999; CEBECI;
GÜRAKAN, 2003; DANIELSEN; WIND, 2003; ROJO-BEZARES et al., 2006). Em
contrapartida, Hamilton-Miller e Shah (1998) constataram em cepas de lactobacilos mais de
90% de sensibilidade à nitrofurantoína e tetraciclina. Scott et al. (2000) relacionaram a
resistência à tetraciclina pelas bactérias intestinais ao aumento do uso destes antimicrobianos.
Teuber, Meile e Schwarz. (1999), publicaram uma revisão sobre este tema onde citam
que a resistência à tetraciclina por L. plantarum pode ser observada em queijos fabricados
com leite in natura. A resistência à tetraciclina também foi demonstrada em estudo conduzido
com diferentes cepas de L. plantarum, realizado por Cebeci e Gürakan (2003). Estes autores
verificaram que 100% das cepas avaliadas apresentaram resistência a cefoxitina, aztreonam e

48
nitrofurantoína, enquanto que para eritromicina o perfil apresentado foi de sensível a
moderadamente sensível.
Rojo-Bezares et al. (2006) fazem uma consideração importante com relação à detecção
de genes de resistência em lactobacilos, pois estes podem ocorrer com outras funções
metabólicas, além daquela de proteção da bactéria frente aos antimicrobianos, e desta forma,
chamam a atenção para a necessidade da realização de estudos visando o entendimento das
funcionalidades das proteínas codificadas pelos genes de resistência a antibióticos.
Segundo Teuber, Meile e Schwarz (1999) a resistência a eritromicina está ligada à
presença de plasmídeos, conforme verificado por Hummel et al. (2007), que detectaram a
presença de gene de resistência a eritromicina em L. salivarius BFE 7441.
É interessante notar que o pool de lactobacilos apresentou sensibilidade moderada a
cefalotina e sensibilidade a eritromicina, enquanto que 43% e 57% das cepas cultivadas
individualmente foram resistentes aos respectivos antibióticos, demonstrando que a
resistência a estes antimicrobianos apresentada pelas cepas individuais provavelmente não
está ligada a gene de resistência.
O pool de lactobacilos apresentou resistência a 76% dos antimicrobianos testados. É
importante salientar que dois fatores podem ter influenciado nestes resultados, sendo que o
primeiro diz respeito a ocorrência de resistência intrínseca, como é o caso da teicoplanina e
aminoglicosídeos, e o segundo diz respeito à origem das cepas avaliadas no presente trabalho,
que foram isoladas de cama de frango. É sabido que na avicultura os antimicrobianos estão
presentes nas dietas das aves para controle e tratamento de doenças infecciosas e também
como promotores de crescimento (MOREIRA; MORAES, 2002; DIBNER RICHARDS,
2005; SMIRNOV et al., 2005)
Embora os antibióticos utilizados como promotores de crescimento tenham tido seu

49
uso proibido em muitos países, problemas decorrentes da aplicação indiscriminada dessas
substâncias continuam causando prejuízos à saúde pública e no controle de infecções
bacterianas, devido à resistência as várias classes desses compostos (ENDTZ et al., 1991;
REINA; ROSS; SERRA, 1994; SÁNCHEZ et al.1994; VELÁZQUEZ et al., 1995;
AARESTRUP et al., 1997; MOREIRA; MORAES, 2002; MONET et al., 2000).
Uma vez que a resistência aos antibióticos seja uma característica passível de ser
transmitida para bactérias patogênicas, este é um parâmetro importante na seleção de bactérias
probióticas. Embora não se tenha estudado a presença de plasmídeos nos lactobacilos, o perfil
de resistência encontrado para o pool das cepas (Tabela 2) aos antimicrobianos cefoxitina,
ceftazidima, ciprofloxacina, enrofloxacina, gentamicina, canamicina, neomicina, teicoplanina,
tetraciclina, aztreonam, fosfomicina, nitrofurantoína e trimetropim, também é reportado na
literatura (TEUBER; MEILE; SCHWARZ, 1999; HUMMEL et al, 2007), e na maioria das
vezes, sem ligação à presença de plasmídeos.
4.3 TOLERÂNCIA AO pH
As bactérias probióticas durante crescimento no trato intestinal competem com outras
bactérias pela disponibilidade de substrato, assim como liberando produtos metabólicos,
acetato e lactato, que promovem a redução do pH e conseqüente inibição de espécies
patogênicas (NAIDU; BIDLACK; CLEMENS, 1999). A habilidade de resistir ao stress ácido
é um importante fator para colonização e estabelecimento dos organismos comensais no trato
gastrintestinal do hospedeiro (LORCA; VALDEZ, 2001). Desta forma, analisou-se no
presente trabalho o crescimento das cepas de Lactobacillus em caldo MRS a diferentes pH,
cujos resultados encontram-se apresentados na Tabela 3.
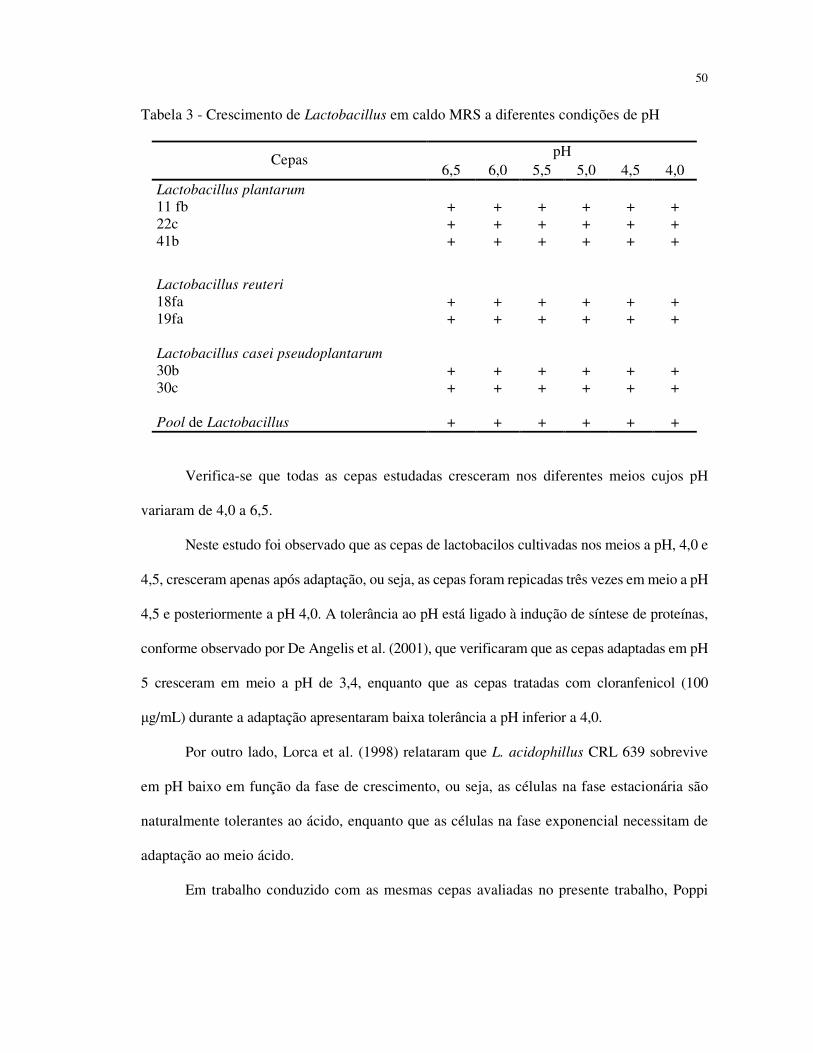
50
Tabela 3 - Crescimento de Lactobacillus em caldo MRS a diferentes condições de pH
pH Cepas 6,5 6,0 5,5 5,0 4,5 4,0
Lactobacillus plantarum 11 fb + + + + + + 22c + + + + + + 41b + + + + + +
Lactobacillus reuteri 18fa + + + + + + 19fa + + + + + + Lactobacillus casei pseudoplantarum 30b + + + + + + 30c + + + + + + Pool de Lactobacillus + + + + + +
Verifica-se que todas as cepas estudadas cresceram nos diferentes meios cujos pH
variaram de 4,0 a 6,5.
Neste estudo foi observado que as cepas de lactobacilos cultivadas nos meios a pH, 4,0 e
4,5, cresceram apenas após adaptação, ou seja, as cepas foram repicadas três vezes em meio a pH
4,5 e posteriormente a pH 4,0. A tolerância ao pH está ligado à indução de síntese de proteínas,
conforme observado por De Angelis et al. (2001), que verificaram que as cepas adaptadas em pH
5 cresceram em meio a pH de 3,4, enquanto que as cepas tratadas com cloranfenicol (100
�g/mL) durante a adaptação apresentaram baixa tolerância a pH inferior a 4,0.
Por outro lado, Lorca et al. (1998) relataram que L. acidophillus CRL 639 sobrevive
em pH baixo em função da fase de crescimento, ou seja, as células na fase estacionária são
naturalmente tolerantes ao ácido, enquanto que as células na fase exponencial necessitam de
adaptação ao meio ácido.
Em trabalho conduzido com as mesmas cepas avaliadas no presente trabalho, Poppi

51
(2005) determinou a população individual e em co-cultura (pool após 18 horas de cultivo) em
tubos de ensaio contendo 5 mL de caldo MRS e pH final variando entre 3,83 e 4,29, e
observou que a população bacteriana ao final da fermentação (Tabela 4) apresentou uma
variação na ordem de 107 a 109 UFC/mL à semelhança dos resultados obtidos por Avonts et
al. (2004), embora estes autores tenham conduzido a fermentação a pH constante 6,5.
Tabela 4: População de Lactobacillus cultivados em caldo MRS a 37ºC/18 h - 40 rpm
Cepas de Lactobacillus População pH
L. plantarum – 11fb 6,2 x 108 UFC/mL 4,10
L. plantarum – 22c 8,0 x 108 UFC/mL 3,83
L. plantarum – 41b 3,9 x 108 UFC/mL 3,88
L. reuteri– 18fa 2,2 x 108 UFC/mL 4,29
L. reuteri – 19fa 1,0 x 107 UFC/mL 3,96
L.casei pseudoplantarum –30b 3,3 x 109 UFC/mL, 3,95
L. casei. pseudoplantarum – 30c 7,9 x 109 UFC/mL 3,98
Pool (co-cultura): 8,3 x 108 UFC/mL 3,90
Fonte: POPPI (2005)
Em estudo conduzido por Lorca e Valdez (1999) foi constatado maior tolerância das células
de L. acidophilus CRL 639 ao pH 3,0 em cultivo a 25ºC, tanto na fase exponencial quanto na fase
estacionária de crescimento quando comparada ao cultivo a 37ºC. Estes autores concluíram que a
temperatura e idade da cultura de Lactobacillus interferiram na tolerância da cepa ao ambiente
ácido.
4.4 PRODUÇÃO DE ÁCIDO LÁCTICO
A fermentação envolvendo as bactérias lácticas resulta no acúmulo de ácidos

52
orgânicos (acético, fórmico e láctico), destacando-se o ácido láctico como principal
metabólito gerado a partir do piruvato pela lactato desidrogenase (NAIDU; BIDLACK;
CLEMENS, 1999).
A ação inibitória dos ácidos orgânicos na forma não dissociada é de 100 a 600 vezes
maior do que a forma dissociada. O ácido láctico não dissociado pode permear a membrana
celular por difusão e liberar prótons na célula. O influxo de prótons parece induzir a
acidificação do citoplasma e dissipar o potencial de prótons na membrana (EKLUND, 1983;
GOBBETTI et al., 2005).
A produção de ácido láctico pelas cepas de Lactobacillus estudadas encontra-se
apresentada na Tabela 5.
Tabela 5 – Produção de ácido láctico pelas cepas de Lactobacillus em caldo MRS a 37ºC/18 h - 40 rpm
Cepas Ácido Láctico (g/L)
Lactobacillus plantarum -11 fb 15,00
Lactobacillus plantarum – 22c 18,39
Lactobacillus plantarum – 41b 18,36
Lactobacillus reuteri – 18 fa 16,12
Lactobacillus reuteri – 19 fa 17,79
Lactobacillus casei pseudoplantarum – 30b 8,71
Lactobacillus casei pseudoplantarum – 30c 16,72
Pool de Lactobacillus 19,14
As concentrações de ácido láctico produzido pelas cepas estudadas variaram entre 15 e
19,14 g/L, com exceção da cepa L. casei pseudoplantarum (30b) que apresentou produção de
8,71 g/L de ácido láctico. O pool constituído por sete cepas de lactobacilos apresentou a maior
produção de lactato.
Sabe-se que o gênero Lactobacillus apresenta três grupos distintos quanto à forma de

53
degradação de hexoses, homofermentativos obrigatórios que degradam as hexoses e
apresentam conversão quase que exclusiva de ácido láctico; heterofermentativos obrigatórios,
os quais convertem as hexoses em quantidades equimolares de ácido láctico, etanol e/ou ácido
acético e gás carbônico, e por fim os heterfermentativos facultativos que fermentam as
hexoses de maneira semelhante ao grupo homofermentativo, contudo, sob condições de
limitação de glicose, a bactéria pode converter as hexoses em ácido láctico, ácido acético e/ou
etanol (KANDLER; WEISS, 1986).
Desta forma pode-se inferir que a maior concentração de ácido láctico obtida pelas
cepas quando foram cultivadas em co-cultura (19,4 g/L), provavelmente se deve ao
metabolismo heterofermentativo facultativo encontrado em 71,43% das cepas estudadas, L.
plantarum (11fb, 22c e 41b) e L. casei pseudoplantarum (30b e 30c), os quais seguiram o
metabolismo homofermentativo, uma vez que a glicose não foi fator limitante.
No presente trabalho, a maior produção de ácido láctico foi observada pelo pool de
lactobacilos, o que também foi verificado por Lee (2005) que constatou uma produção de 110
g/L de ácido láctico em cultivo contendo cinco cepas de Lactobacillus após 70 horas de
fermentação contra 90 g/L no mesmo tempo para L. casei (NRRL-B1445). Segundo o autor,
este resultado se deve a um provável sinergismo entre as cepas avaliadas.
As concentrações de ácido láctico obtidas pelo cultivo individual das cepas de L.
plantarum 11fb, 22c e 41b, estão de acordo com os resultados encontrados por Ogunbanwo et
al. (2004) que estudaram a produção de compostos antimicrobianos produzidos por
lactobacilos isolados de mandioca e verificaram que a cepa de L. plantarum F1 produziu 16,4
g/L de ácido láctico em meio MRS após 48 horas de fermentação e pH final de 3,1.

54
Zhao et al. (2007) estudaram as propriedades funcionais de duas cepas de L.
acidophilus, La-1 e La-2, e verificaram uma produção de ácido láctico de 12,73 g/L e 13,33
g/L, respectivamente, em leite desnatado a 37ºC por 36 horas. Foi observado também que a
conversão da lactose a ácido láctico foi superior a 90%, à semelhança dos resultados
encontrados para as cepas 22c, 41b e pool de lactobacilos estudados no presente trabalho.
Devido à ação antagônica do ácido láctico sobre bactérias patogênicas, Poppi (2005),
analisou este efeito sobre Escherichia coli O157:H7 e Listeria monocytogens, utilizando as
mesmas cepas de lactobacilos avaliadas neste trabalho, e verificou que o ácido láctico não
influenciou no efeito bacteriostático ou bactericida sobre as bactérias patogênicas.
4.5 PERFIL ELETROFORÉTICO DAS PROTEÍNAS SECRETADAS PELAS CEPAS DE
Lactobacillus
Por meio da eletroforese em gel de poliacrilamida (SDS-PAGE), foi possível estudar o
perfil de proteínas produzidas pelas cepas cultivadas individualmente e em co-cultura (pool).
Dessa forma, pode-se observar uma grande concentração de proteínas com massa molecular
entre 43-94 KDa nas cepas de L. reuteri, L. casei pseudoplantarum e no pool de
Lactobacillus.

55
Nota-se que existe uma banda com massa molecular próximo a 67 KDa em todas as
cepas estudadas, sugerindo que esta seja uma proteína normalmente presente neste gênero de
bactérias. As cepas de mesma espécie, L. reuteri (18fa e 19fa) ou L. casei pseudoplantarum
(30b e 30c) apresentaram um perfil protéico muito semelhante. Kanashiro (2002) determinou
as massas moleculares de proteínas de Lactobacillus sp. (amostras 55-1 e 129) e verificou
maior concentração de proteínas entre 25,74 – 93,02 KDa, semelhante, portanto, ao obtido
neste estudo.
O pool de Lactobacillus expressou poucas proteínas em relação às cepas individuais, a
cepa 18 fa apresentou a maior diversidade de proteínas, enquanto as amostras de L. plantarum
11fb, 22c e 41b expressaram a menor concentração de proteínas. Estes resultados encontram-
se na figura 2.
11fb 18fa 19fa 22c 41b 30b 30c Pool
FIGURA 2 - Perfil eletroforético das proteínas secretadas pelas cepas de Lactobacillus
94KDa 67KDa 43KDa 30KDa 20,1KDa 14,4KDa

56
A caracterização protéica representou as proteínas que podem atuar como antígenos
estimuladores da resposta imune detectada, ou seja, de anticorpos, IgA e IgG, sendo, portanto,
fundamental para as análises de quantificação de imunoglobulinas nos camundongos.
4.6 AVALIAÇÃO DA RESPOSTA IMUNOLÓGICA
O efeito dos probióticos sobre a resposta imune foi bastante estudado (DE SIMONE, et al,
1986; MAASEN et al., 1998; PERDIGÓN et al., 1999; PERDIGÓN et al., 2002; PAVAN;
DESREUMAUX; MERCENIER, 2003). Existem evidências em sistemas in vitro e em modelos
animais e humanos que sugerem que os probióticos podem estimular tanto a resposta imune
específica quanto a inespecífica. Acredita-se que esses efeitos sejam mediados por uma ativação de
macrófagos, aumento nos níveis de citocinas e aumento da atividade de células natural killer e/ou
dos níveis de imunoglobulinas (SAAD, 2006).
4.6.1 Concentração de IgA e IgG total presente no soro e mucosa intestinal
Embora se tenha observado pequenas diferenças na concentração de imunoglobulinas
sérica e de mucosa intestinal com relação aos controles, a análise estatística revelou que não
houve diferenças significativas (p < 0,05), portanto, as proteínas produzidas pelo pool de
lactobacilos não estimularam a resposta imune nos camundongos. Não houve diferenças
significativas (p< 0,05) entre os grupos A e B, demonstrando que as proteínas de
Lactobacillus também foram ineficazes como adjuvantes da resposta humoral.
Os resultados da concentração de IgA e IgG presentes no soro e mucosa intestinal
estão apresentados nas tabelas 6 e 7, respectivamente.

57
Tabela 6 – Concentração média de IgA total no soro e mucosa intestinal de camundongos
Concentração de IgA (mg/mL)
Soro Mucosa Intestinal
Tratamentos
Grupo A 14 dias 28 dias 42 dias 14 dias 28 dias 42 dias
1 0,3906 0,5923 0,2879 2,5138 1,6454 2,9141
Controle 0,3748 0,5304 0,2326 2,3274 1,6189 2,4378
2 0,3562 0,4792 0,1346 2,8117 1,7970 2,2037
Controle 0,3310 0,4364 0,1306 2,2605 1,5135 1,9373
3 0,4435 0,3713 0,4565 1,5391 1,5271 2,4685
Controle 0,4322 0,3475 0,4312 1,4416 1,4782 2,1290
4 0,3809 0,4498 0,2249 1,7504 1,4136 2,3320
Controle 0,3529 0,4038 0,2222 1,5985 1,2210 2,4145
Grupo B 14 dias 35 dias 42 dias 14 dias 35 dias 42 dias
5 0,3788 0,3465 0,1777 2,4732 1,4648 2,7383
Controle 0,3555 0,3029 0,1975 2,3274 1,2780 2,3410
6 0,1931 0,2187 0,1878 2,5770 2,3229 2,7905
Controle 0,1691 0,1937 0,2151 2,1650 1,8500 2,2849
7 0,3749 0,3996 0,1750 2,5186 1,6101 2,6315
Controle 0,3535 0,3856 0,1883 2,4285 1,4515 2,3342
8 0,2256 0,3996 0,1663 2,9361 1,2378 2,5549
Controle 0,2182 0,3896 0,1826 2,6016 1,2042 2,3274
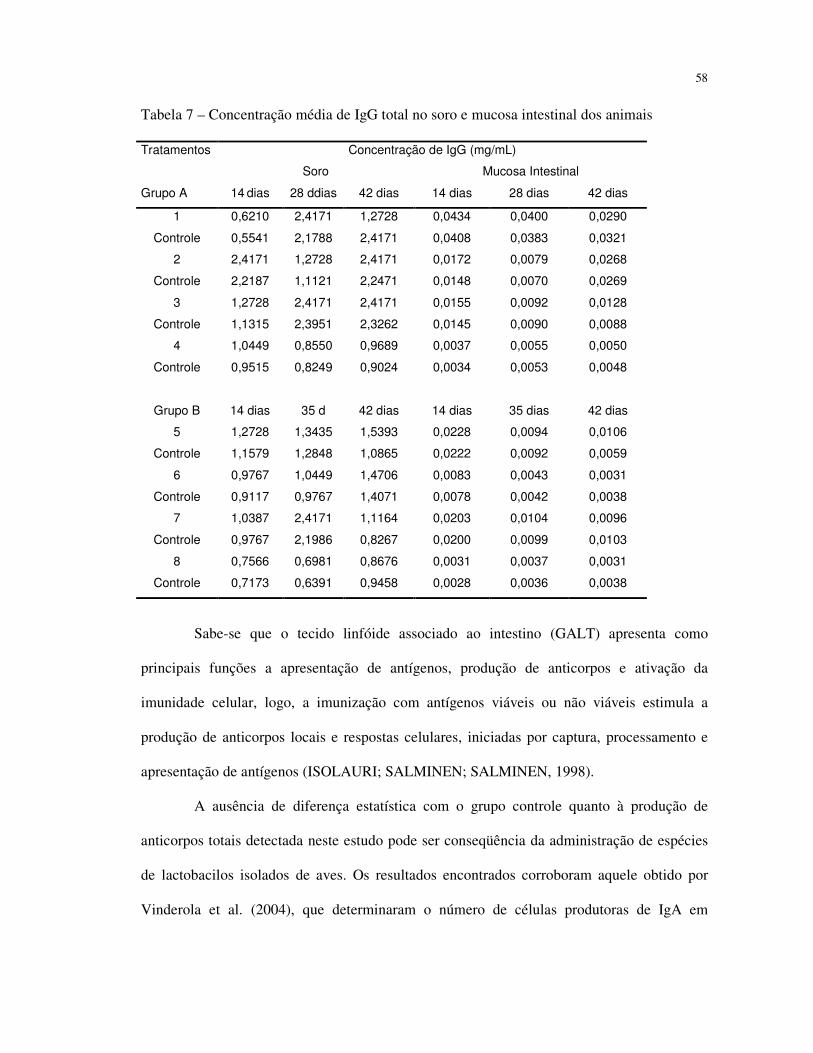
58
Tabela 7 – Concentração média de IgG total no soro e mucosa intestinal dos animais
Concentração de IgG (mg/mL)
Soro Mucosa Intestinal
Tratamentos
Grupo A 14 dias 28 ddias 42 dias 14 dias 28 dias 42 dias
1 0,6210 2,4171 1,2728 0,0434 0,0400 0,0290
Controle 0,5541 2,1788 2,4171 0,0408 0,0383 0,0321
2 2,4171 1,2728 2,4171 0,0172 0,0079 0,0268
Controle 2,2187 1,1121 2,2471 0,0148 0,0070 0,0269
3 1,2728 2,4171 2,4171 0,0155 0,0092 0,0128
Controle 1,1315 2,3951 2,3262 0,0145 0,0090 0,0088
4 1,0449 0,8550 0,9689 0,0037 0,0055 0,0050
Controle 0,9515 0,8249 0,9024 0,0034 0,0053 0,0048
Grupo B 14 dias 35 d 42 dias 14 dias 35 dias 42 dias
5 1,2728 1,3435 1,5393 0,0228 0,0094 0,0106
Controle 1,1579 1,2848 1,0865 0,0222 0,0092 0,0059
6 0,9767 1,0449 1,4706 0,0083 0,0043 0,0031
Controle 0,9117 0,9767 1,4071 0,0078 0,0042 0,0038
7 1,0387 2,4171 1,1164 0,0203 0,0104 0,0096
Controle 0,9767 2,1986 0,8267 0,0200 0,0099 0,0103
8 0,7566 0,6981 0,8676 0,0031 0,0037 0,0031
Controle 0,7173 0,6391 0,9458 0,0028 0,0036 0,0038
Sabe-se que o tecido linfóide associado ao intestino (GALT) apresenta como
principais funções a apresentação de antígenos, produção de anticorpos e ativação da
imunidade celular, logo, a imunização com antígenos viáveis ou não viáveis estimula a
produção de anticorpos locais e respostas celulares, iniciadas por captura, processamento e
apresentação de antígenos (ISOLAURI; SALMINEN; SALMINEN, 1998).
A ausência de diferença estatística com o grupo controle quanto à produção de
anticorpos totais detectada neste estudo pode ser conseqüência da administração de espécies
de lactobacilos isolados de aves. Os resultados encontrados corroboram aquele obtido por
Vinderola et al. (2004), que determinaram o número de células produtoras de IgA em

59
camundongos tratados oralmente com cepas de lactobacilos comensais e não comensais, e
observaram que as bactérias exógenas administradas em uma concentração de 104 células/dia
por 5 dias tiveram valores similares aos encontrados para o grupo controle, demonstrando que
não houve efeito deste tratamento.
Revolledo Pizarro (2005) estudou a resposta imune em frangos desafiados com
Salmonella enterica sorotipo Typhimurium Nalr e tratados com probiótico composto por
Bifidobacterium bifidus, quatro espécies de Enterococcus e doze espécies de Lactobacillus
isolados de aves, dentre as quais, duas cepas de L. reuteri, duas cepas de L. plantarum e duas
de L. casei pseudoplantarum, foram as mesmas cepas utilizadas no presente trabalho, e
verificou que os níveis de IgG total no conteúdo intestinal mantiveram-se estáveis durante o
tratamento, enquanto que a produção de IgA total apresentou-se significativa.
Outra possibilidade da ineficácia de indução da resposta imune poderia ser a falta de
adesão bacteriana ao epitélio intestinal, assim como o tempo de passagem pelo trato
gastrintestinal insuficiente para que pudesse exercer ação imunomoduladora (NAIDU;
BIDLACK; CLEMENS, 1999). Vários componentes da superfície celular bacteriana
participam da aderência das cepas às células epiteliais intestinais. Fernández, Boris e Barbés
(2003) estudaram as propriedades probióticas de L. gasseri e L. acidophilus e observaram que
a aderência dependia da presença de proteínas, carboidratos e cátions divalentes,
provavelmente o Ca2+. Outros pesquisadores (CHAUVIÉRE et al., 1992; COCONIER et al.,
1992; BERNET, et al., 1993) reportaram que Lactobacilli aderem às células intestinais
humana através de mecanismos que envolvem diferentes combinações de carboidrato e
proteína na superfície da parede celular.
Muitos organismos procarióticos, incluindo-se algumas espécies de lactobacilos,
secretam um material viscoso denominado glicocálix que é depositado na superfície celular. O

60
glicocálix apresenta em sua constituição o ácido N-acetilglucosamina, que também está
presente na parede celular das bactérias Gram positivas. A composição monossacáride do
hidrolisado celular tem sido um critério para a diferenciação química deste grupo de bactérias.
Estudos envolvendo a análise de carboidrato em Lactobacilli têm mostrado que
monossacarídeos como N-acetilglucosamina e N-acetilgalactosamina estão presentes no
glicocálix em quantidades significativas. Além disso, a presença destes e outros
monossacarídeos em glicopolímeros sobre o glicocálix de Lactobacilli foram associados às
propriedades adesivas das cepas (POMAZANOV et al., 1998 apud ANNUK et al., 2001).
Alguns pesquisadores concordam que além da propriedade de adesão à mucosa
intestinal, uma cepa probiótica também precise ser comensal, pois nesta situação, é esperado
que apresente maior eficácia no ambiente em que foi isolada (APOSTOLOU et al, 2001;
SAARELA et al., 2000). Em contrapartida, Dogi, Perdigón (2006) demonstraram que tanto
cepas comensais como não comensais foram capazes de induzir respostas imunes no intestino
sem produzir efeitos colaterais como uma resposta imune inflamatória.
O efeito probiótico de bactérias lácticas está relacionado às propriedades da
superfície celular, que determina sua capacidade em aderir às células entéricas, aumentando a
capacidade bacteriana de seqüestrar compostos tóxicos ou enviar sinais de ativação para a
produção de citocinas (GÓMEZ-ZAVAGLIA et al., 2002), prevenindo-se sua eliminação pelo
peristaltismo (GRANATO et al., 1999).
A adesão de microrganismos, ingeridos oralmente, ao intestino é um fenômeno
complexo que pode ser um processo constituído de várias etapas, envolvendo mecanismos
específicos e inespecíficos. O efeito sobre o hospedeiro dependerá da internalização de
POMAZANOV, V.V.; OXIPOV, G.A; SHABANOVA, E.A; DEMINA, A,M.; MOROZOV, OV. Potential use of the monosaccharide composition of bactéria for their identification. Mikrobiologia, v.57, p.107-113, 1998.

61
alimentos ou espécies bacterianas para interagir com as células linfóides associadas aos sítios
indutores de uma resposta imune no intestino (PERDIGÓN et al., 2000).
Por outro lado, a falta de uma produção aumentada de anticorpos no grupo tratado
poderia ser devido à constituição do pool bacteriano usado, pois existiam duas cepas da
espécie L. reuteri e de acordo com Christensen et al. (2002), esta espécie inibe a síntese de IL-
6, IL-12 e TNF-� por células dendríticas, ou seja, inibe a síntese de IL-6 que está associada à
produção de anticorpos. Maasen et al. (2000) administraram a camundongos, por via oral, L.
reuteri e verificaram que não houve indução de citocinas IL-4 e IL-10, a não ser quando os
animais foram imunizados parenteralmente com antígenos. A administração de L. plantarum
também não influenciou os níveis de IL-4 no cólon e íleo de camundongos (PAVAN et al.,
2003). As citocinas IL-6 e IL-4 são responsáveis pela multiplicação de linfócitos B e
diferenciação em plasmócitos que produzem anticorpos. Dessa forma, a presença destas
espécies bacterianas no pool administrado pode ter interferido na ação destas citocinas, que
em conseqüência inibiu a síntese de anticorpos. Entretanto, Perdigón et al. (2002) observaram
que o tratamento de camundongos com L. casei, L. delbrueckii bulgaricus e L. acidophilus
afetou a resposta humoral sistêmica e foram detectados altos níveis de IL-4 em animais
tratados com L. delbrueckii bulgaricus e L. casei.
Kanashiro (2000) observou que frangos tratados com cepas de Lactobacillus sp
isolados de aves livres de patógenos específicos (SPF) apresentaram pouco estímulo de
produção de IgG específica para as proteínas bacterianas e relacionou este resultado à
capacidade de colonização da espécie e produção diferenciada de proteínas in vitro e in vivo.
O grupo B (Tabela 6 e Tabela 7) demonstra que o pool de lactobacilos não agiu
como adjuvante, pois os níveis de IgA e IgG em relação ao grupo A não se apresentaram
aumentados. He et al. (2005) verificaram que a administração oral de Lactobacillus GG

62
inativado termicamente aumentava a resposta imune sistêmica e de mucosa favorecendo a
produção de IgA secretora.
A administração oral de um antígeno protéico freqüentemente induz acentuada
supressão das respostas imunes sistêmicas celulares e humorais à imunização pelo mesmo
antígeno, fenômeno este conhecido como tolerância (ABBAS; LICHTMAN, 2005). A
tolerância está ligada a microbiota natural do hospedeiro, uma vez que camundongos normais
apresentaram tolerância a antígenos da dieta por um período de aproximadamente três meses,
enquanto que em camundongos Germ-Free a tolerância observada foi de 20 dias (MOREAU;
CORTHIER, 1988).
É importante salientar que os animais utilizados no experimento foram criados sem
isoladores, podendo justificar as diferenças nas concentrações de IgA e IgG observadas entre
os grupos A e B.

63
5 CONCLUSÕES
Os resultados obtidos no presente trabalho permitem concluir que:
• O pool de lactobacilos apresentou resistência a treze antimicrobianos, representando
76,47% dos antibióticos testados;
• Todas as cepas de lactobacilos cultivadas invidualmente e em co-cultura apresentaram
crescimento nos diferentes meios com valores de pH variando entre 4,0 e 6,5;
• O cultivo em co-cultura de Lactobacillus (pool) apresentou a maior concentração de
ácido láctico quando comparado com as cepas individuais;
• O perfil protéico do pool de Lactobacillus expressou poucas proteínas em relação às
cepas individuais de L. reuteri (18fa e 19fa) e L. casei pseudoplantarum (30b e 30c);
• O pool de Lactobacillus foi ineficaz tanto para induzir a resposta humoral pela
produção de IgA e IgG, quanto para atuar como adjuvante.

64
REFERÊNCIAS
AARESTRUP, F.M.; NIELSEN; E.M.; MADSEN, M.; ENGBERG, J. Antimicrobial susceptibility patterns of thermophilic Campylobacter spp. from humans pigs cattle and broilers in Denmark. Antimicrobial Agents and Chemotherapy, v.41, n.10, p.2244-2250, oct.1997.
ABBAS, A.K.; LICHTMAN, A.H. Imunologia Celular e Molecular. 5. ed. Rio de Janeiro: Elsevier Editora Ltda., 2005
ABEF. Relatório Anual 2005. Disponível em: <http://www.abef.com.br>. Acesso em: 04 abr. 2007
ADAMS, D.O.; HAMILTON, T.A. Macrophages as destructive cells in host defense. In: GALLIN, J.I.; GOLDSTEIN, R.S.; SNYDERMAN, R. Inflamation: Basic Principles and Clinical Correlates. New York: Raven, 1992. p.637-632.
AMBROSINI, M.V.; GONZALEZ, S.; RUIZ HOLGADO, A.P.; OLIVER, G. Study of the morphology of the cell walls of some strains of lactic acid bacteria and related species. Journal of Food Protection, v.61, n.5, p.557-562, may1998.
ANNUK, H.; HYNES, S.O.; HIRMO, S.; MIKELSAAR, M.; WADSTRÖM, T. Characterisation and differentiation of lactobacilli by lectin typing. Journal of Medical Microbiology, v.50, p.1069-1074, 2001.
APOSTOLOU, E.; KIRJAVAINEN, P.V.; SAXELIN, M.; RAUTELIN, H.; VALTONEN, V.; SALIMINEN, S.J.; OUWEHAND, A.C. Good adhesion properties of probiotics: a potential risk for bacteremia? FEMS Immunology and Medical Microbiology, v.31, n.1, p.35-39, jul.2001.
AVONTS, L.; VAN UYTVEN, E.; DE VUYST, L. Cell growth and bacteriocin production of probiotic Lactobacillus strains in different media. International Dairy Journal, v.14, n.11, p.947-955, nov.2004.
BIELECKA, M.; BIEDRZYCKA, E.; MAJKOWSKA, A. Selection of probiotics and prebiotics for synbiotics and confirmation of their in vivo effectiveness. Food Research International, v.35, n.2/3, p.125-131, 2002.

65
BLOKSMA, N.; HEER, E.; VAN DIJK, H.; WILLERS, J.M. Adjuvanticity of lactobacilli – I. Differential effects of viable and killed bacteria. Clinical and Experimental Immunology, v.37, n.2, p.367-375, 1979.
BLOM, H.; MÖRTVEDT, C. Anti-microbial substances produced by food associated micro-organisms. Biochemical Society Transactions, v.19, n.3, p.694-698, aug.1991.
BLUM, H.; BIER, H.; GROSS, H.J. Improved silver staining of plant protein, RNA and DNA in polyacrilamide gel. Electrophoresis, v.8, n.2, p.93-99, 1987.
BOLLAG, D.; ROZYCKI, M.D.; EDELSTEIN, S.J. Protein Methods. 2. ed.. New York: Wiley-Liss. 415p., 1996.
CARLOS, I.Z.; VENDRAMINI, A.P.; VENDRAMINI, R.C.; DÂMASO, E.R.; ROSSI, E.A. Influência de Nutrientes no Sistema Imune: Papel das Citocinas, Peróxido de Hidrogênio e Óxido Nítrico. In: FERREIRA, C.L.F. Prebióticos e Probióticos: Atualização e Prospecção. Viçosa: Universidade Federal de Viçosa, 2003. p.135-154.
CASSONE, A.; MARCONI, P.; BISTONI, F.; MATTIA, E.; SBARAGLIA, G.; GARACI, E.; BONMASSAR, E. Immunoadjuvant effects of Candida albicans and its cell wall fractions in a mouse lymphoma model. Cancer Immunology Immunotherapy, v.10, n.4, p.181-190, apr.1981.
CEBECI, A.; GÜRAKAN, C. Properties of potential probiotic Lactobacillus plantarum strains. Food Microbiology, v.20, n.5, p.511-518, oct. 2003.
CHARTERIS, W.P.; KELLY, P.M.; MORELLI, L.; COLLINS, J.K. Ingredient selection criteria for probiotic microorganisms in functional dairy foods. International Journal of Dairy Technology, v.51, n.4, p.123-136, nov.1998.
CHOI, S.S.; KIM, Y.; HAN, K.S.; YOU, S.; OH, S.; KIM, S.H. Effects of Lactobacillus strains on cancer cell proliferation and oxidative stress in vitro. Letters in Applied Microbiology, v.42, n.5, p.452-458, may 2006.
CHRISTENSEN, H.R.; FROKIAER, H.; PESTKA, J.J. Lactobacilli differentially modulate expression of cytokines and maturation surface markers in murine dendritic cells. The Journal of Immununology, v.168, n.1, p.171-178, jan. 2002.

66
COLLINS, J.K.; THORNTON, G.; SULLIVAN, G.O. Selection of probiotic strains for human applications. International Dairy Journal, v.8, n.5-6, p.487-490, may1998.
CRITTENDEN, R.G. Prebiotics. In: TANNOCK, G.W. Probiotics: a critical review. Wymondham: Horizon Scientific, 1999. p.141-156.
DANIELSEN, M.; WIND, A. Susceptibility of Lactobacillus spp. to antimicrobial agents. International Journal of Food Microbiology, v.82, n.1, p.1-11, apr.2003.
DE ANGELIS, M.; BINI, L.; PALLINI, V.; COCCONCELLI, P.S.; GOBBETTI, M. The acid-stress response in Lactobacillus sanfranciscensis CB1. Microbiology, v.147, n.7, p.1863-1873, jul. 2001.
DELGADO, S.; FLÓREZ, A.B.; MAYO, B. Antibiotic susceptibility of Lactobacillus and Bifidobacterium species from the human gastrointestinal tract. Current Microbiology, v.50, n.4, p.202-207, apr. 2005.
DE SIMONE, C.; BIANCHI SALVADORI, B.; NEGRI, R.; FERRAZZI, M.; BALDINELLI, L.; VESELY, R. The adjuvant effect of yogurt on productio of gama-interferon by com A-stimulated human peripheral blood lymphocytes. Nutrition Reports International, v.33, n.3, p.425-432, marc.1986.
DIBNER, J.J.; RICHARDS, J.D. Antibiotic growth promoters in agriculture: history and mode of action. Poultry Science, v.84, n.4, p.634-643, apr. 2005.
DOGI, C.A.; PERDIGÓN, G. Importance of the host specificity in the selection of probiotic bacteria. Journal of Dairy Research, v.73, n.3, p.357-366, aug. 2006.
EKLUND, T. The antimicrobial effect of dissociated and undissociated sorbic acid at different pH levels. Journal of Applied Bacteriology, v.54, n.3, p.383-389, jun. 1983.
ELIAS, C.C.; SOUZA, H.S.P. Imunologia da mucosa intestinal – Da bancada ao leito. Rio de Janeiro: Ateneu, 2001. 187p.

67
ENDTZ; H.P. ; RUIJS, G.J.; VAN KLINGEREN, B.; JANSEN, W.H.; VAN DERREYDEN, T.; MOUTON, R.P. Quinolone resistance in campylobacter isolated from man and poultry following the introduction fluorquinolones in veterinary medicine. The Journal of Antimicrobial Chemotherapy, v.27, n.2, p.199-208, feb. 1991.
ERICKSON, K.L.; HUBBARD, N.E. Probiotic immunomodulation in health and disease. The Journal of Nutrition, v.130, n.2, p.403S-409S, feb. 2000.
FERNÁNDES, M.F.; BORIS, S.; BARBÉS, C. Probiotic properties of human lactobacilli strains to be used in the gastrointestinal tract. Journal of Applied Microbiology, v.94, n.3, p.449-455, marc. 2003.
FERREIRA, C.L.L.F. Tecnologia para produtos lácteos funcionais: Probióticos. In: PORTUGAL, J.A.B.; CASTRO, M.C.D.; SILVA,J.H.F.; SAVINO, A.C.; NEVES, B.S.; ARCURI, E.F. O agronegócio do leite e os alimentos funcionais. Juiz de Fora: Epamig. v.181, 203p., 2001a.
FERREIRA, C.L.L.F. Grupo de Bactérias Lácticas – Caracterização e aplicação tecnológica de bactérias probióticas. In: FERREIRA, C.L.F. Prebióticos e Probióticos: Atualização e Prospecção, Viçosa: UFV. p.7-34, 2003.
FERREIRA, C.S.A. Detecção de imunoglobulinas séricas a proteínas de Salmonella enterica sorotipo Enteritidis em aves infectadas experimentalmente. 2001. 85f. Tese (Doutorado em Patologia Experimental e Comparada), Universidade de São Paulo, São Paulo, 2001b.
FOOD AND AGRICULTURE ORGANIZATION OF THE UNITED NATIONS, WORLD HEALTH ORGANIZATION. Evaluation of health and nutritional properties of probiotics in food including powder milk with live lactic acid bacteria. Córdoba, 2001. 34p. Disponível em: <ftp://ftp.fao.org/es/esn/food/probioreport_ en.pdf>. Acesso em: 03 fev. 2005.
FORESTIER, C.; DE CHAMPS, C.; VATOUX, C.; JOLY, B. Probiotic activities of Lactobacillus casei rhamnosus: in vitro adherence of intestinal cells and antimicrobial properties. Research in Microbiology, v.152, n.2, p.167-173, marc. 2001.
FREESE, A.C.; SHEN, C.W.; GALLIERS,E. Function of lipophilic acids as antimicrobial food additives. Nature, v.241, n.5388, p.321-325, feb. 1973.

68
FULLER, R. Probiotics in man and animals. Journal of Applied Bacteriology, v.66, n.5, p.365-378, may 1989.
FULLER, R. The effect of Probiotics on the gut micro-ecology of farm animals. In: WOOD, B.J.B. The Lactic acid bacteria – The lactic acid bacteria in health and disease. Vol I. London: Chapman & Hall. p.171-192. 1992.
GAUFFIN, C.P.; AGÜERO, G.; PERDIGÓN, G. Adjuvant effects of Lactobacillus casei added to a renutrition diet in a malnourished mouse model. Biocell, v.26, n.1, p.35-48, apr. 2002.
GIBSON, G.R.; FULLER, R. Aspects of in vitro and in vivo research approaches directed toward identifying probiotics and prebiotics for human use. The Journal of Nutrition, v.130, 2S Suppl., p.391S-394S, feb. 2000.
GOBBETTI, M; DE ANGELIS, M.; CORSETTI, A.; DI CAGNO, R. Biochemistry and physiology of sourdough lactic acid bacteria. Trends in Food Science & Technology, v.16, n.1-3, p.57-69, jan.-mar. 2005.
GÓMEZ-ZAVAGLIA, A.; KOCIUBINSKY, G.; PÉREZ, P.; DISALVO, E.; DE ANTONI, G.L. Effect of bile on the lipid composition and surface properties of bifidobacteria. Journal of Applied Microbiology, v.93, n.5, p.794-799, may 2002.
GÓRNIAK, S.L. Quimioterápicos. In: SPINOSA, H.S.; GÓRNIAK, S.L.; BERNARDI, M.M. Farmacologia Aplicada à Medicina Veterinária. 3. ed. Rio de Janeiro: Guanabara Koogan, 2002. p.398-408.
GRANATO, D., PEROTTI, F., MASSEREY, I., ROUVET, M., GOLLIARD, M., SERVIN, A., BRASSART, D. Cell surface-associated lipoteichoic acid acts as an adhesion factor for attachment of Lactobacillus johnsonii La1 to human enterocyte-like Caco-2 cells. Applied and Environmental Microbiology, v.65, n.3, p.1071–1077, mar. 1999.
GRAPHPAD INSTAT SOFTWARE. Statisitcal analysis systems for personal computers. San Diego: GraphPad Software, 1998.
GUARNER, F.; MALAGELADA, J.R. Gut flora in health and disease. Lancet, v.361, n.9356, p.512-519, feb. 2003.

69
HAMILTON-MILLER, J.M.T.; SHAH, S. Vancomycin susceptibility as an aid to the identification of lactobacilli. Letters in Applied Microbiology, v.26, n.3, p.153-154, feb. 1998.
HAVENAAR, R.; HUIS INT VELD, J.H.J. Probiotics: A General View. In: WOOD, B.J.B. The Lactic Acid Bacteria – The Lactic Acid Bacteria in Health and Disease. London: Chapman & Hall, 1992. v.1. p.151-170.
HE, F.; MORITA, H.; KUBOTA, A.; OUWEHAND, A.C.; HOSODA, M.; HIRAMATSU, M.; KURISAKI, J.C. Effect of orally administered non-viable Lactobacillus cells on murine humoral immune responses. Microbiology and Immunology, v. 49, n.11, p.993-997, nov. 2005.
HORI, T.; KIYOSHIMA, J.; SHIDA, K.; YASUI, H. Augmentation of cellular immunity and reduction of influenza virus titer in aged mice fed Lactobacillus casei strain shirota. Clinical and Diagnostic Laboratory Immunology, v.9, n.1, p.105-108, jan. 2002.
HUMMEL, A.S.; HERTE, C.; HOLZAPFEL, W.H.; FRANZ, C.M.A.P. Antibiotic resistances of starter and probiotic strains of lactic acid bacteria. Applied and Environmental Microbiology, v.73, n.3, p.730-739, feb. 2007.
IANELLO, D.; BONINA,L., DELFINO,D.; BERLINGHIERI, M.C.; GISMONDO, M.R.; MASTROENI, P. Effect of oral administration of a variety of bacteria on depressed macrophage functions in tumour-bearing rats. Annales d’Immunology, v.135C, n.3, p.345-352, may-jun. 1984.
ISOLAURI, E.; SALMINEN, S.; OUWEHAND, A.C. Probiotics. Best Practice & Clinical Gastroenterology, v.18, n.2, p.299-313, apr. 2004.
ISOLAURI, E., JOENSUU, J., SUOMALAINEN, H., LUOMALA, M., VESIKARI, T. Improved immunogenicity of oral D x RRV reassortant rotavirus vaccine by Lactobacillus casei GG. Vaccine, v.13, n.3, p.310-312, 1995.
ISOLAURI, E.; SALMINEN, E.; SALMINEN,S. Lactic Acid Bacteria and Immune Modulation In: SALMINEN,S.; VON WRIGHT, A. Lactic Acid Bacteria – Microbiology and Functional Aspects. New York: Marcel Dekker, 1998. p.255-68.

70
JANSEN & VAN DER WAAIJ, 1995. Disponível em: <http:www.microbiologia2000.hpg.ig.com.br/ciência_e_educacao/7/index_int_8.html>. Acesso em: 10 de fevereiro de 2003.
JOHNANSEN, F.E.; PEDNA, M.; NORDERHAUG, I.N.; HANEBERG, B.; HIETALA, M.A.; KRAJCI, P. Absence of epithelial immunoglobulin A transport, with increased mucosal leakiness, in polymeric immunoglobulin receptor/secretory component-deficient mice. The Journal of Experimental Medicine, v.190, n.7, p.915-921, oct. 1999.
JUNQUEIRA, L.C.; CARNEIRO, J. Histologia Básica. 8. ed. Rio de Janeiro: Guanabara & Koogan, 2001. p.220-240.
KANASHIRO, A.M.I. Influência da administração contínua de probiótico a frangos de corte (Gallus gallus, LINNAEUS, 1758) sobre a imunidade humoral específica, atividades, enzimáticas séricas e concentração de colesterol plasmático. 2000. 78f. Tese (Doutorado em Clínica Cirúrgica Veterinária). Universidade de São Paulo, São Paulo, 2000.
KANDLER, O.; WEISS, N. Bergey’s Manual Systematic Bacteriology. Baltimore: Williams, 1986. p.1208-1234.
KATLA, A.K.; KRUSE, H.; JOHNSEN, G.; HERIKSTAD, H. Antimicrobial susceptibility of starter culture bacteria used in Norwegian dairy products. International Journal of Food Microbiology, v.67, n.2-3, p.147-152, jan-feb. 2001.
KONEMAN, E.W.; ALLEM, S.D.; JANDA, W.M.; SCHRECKENBERGER, P.C.; WINN, P.C. Other miscellaneous fastidious gram-negative bacteria. Diagnostic microbiology. Philadelphia: J.B. Lippincott Company, p.433-486. 1997.
KOZASA, M. Probiotics for animal use in Japan. Ver. Sci. Tech. Off. Int. Epiz., v.8, n.2, p.517-531, 1989.
LA CAVA, A.; VAN KAER, L.; SHI, F.D. CD4+ CD25+ Tregs and NKT cells: regulators regulating regulators. TRENS in Immunology, v.27, n.7, p.322-327, jul.2006.
LAEMMLI, U.K. Cleavage of structural protein during the assembly of the head of bacteriophage T4. Nature, v.227, n.5259, p.680-685, aug. 1970.

71
LAUTERSLAGER, T.G.M; WIL STOK, L.A.T.H. Improvement of the systemic prime/oral boost strategy for systemic and local responses. Vaccine, v.21, n.13-14, p.1391-1399, mar. 2003.
LEBLANC, A.M; MATAR,C; FARNWORTH, E.; PERDIGÓN, G. Study of cytokines involved in the prevention of a murine experimental breast cancer by kefir. Cytokine, v.34, n.1-2, p.1-8, may 2006.
LEE, K. Comparision of fermentative capacities of Lactobacili in single and mixed culture in industrial media. Process Biochemistry, v.40, n.5, p.1559-1564, apr. 2005.
LEE, Y.K.; NOMOTO, K.; SALMINEN, S.; GORBACH, S.L. Handbook of probiotics. New York: Wiley, 1999. 211p.
LINK-AMSTER, H.; ROCHAT, F.; SAUDAN, K.Y.; MIGNOT, O.; AESCHLIMANN, J.M. Modulation of a specific humoral immune response and changes in intestinal flora mediated through fermented milk intake. FEMS Immunology and Medical Microbiology, v.10, n.5, p.55-63, nov. 1994.
LORCA, G.L.; RAYA, R.R.; TARANTO, M.P.; VALDEZ, G.F. Adaptative acid tolerance response in Lactobacillus acidophilus. Biotechnology Letters, v.20, n.3, p.239-241, marc. 1998.
LORCA, G.L.; VALDEZ, G.F. The effect of suboptimal growth temperature and growth phase on resistance of Lactobacillus acidophilus to environmental stress. Cryobiology, v.39, n.2, p.144-149, sep. 1999.
LORCA, G.L.; VALDEZ, G.F. A low-pH-inducible, stationary-phase acid tolerance response in Lactobacillus acidophilus CRL 639. Current Microbiology, v. 42, n.1, p.21-25, jan. 2001.
MAASEN, C.B.M.; VAN HOLTEN-NEELEN, C.; BALK, F.; DEN BAK-GLASHOUWE, M.J.; LEER, R.J.; LAMAN, J.D.; BOERSMA, W.J.A.; CLAASSEN, E. Strain-dependent induction of cytokine profiles in the gut by orally administered Lactobacillus strains. Vaccine, v.18, n.23, p.2613-2623, may 2000.
MAC FADDIN, J.F. Pruebas Bioquimicas para la Identificacion de Bacterias de Importancia Clinica. Buenos Aires: Panamericana, 1980, 301p.

72
MAIA O.B.; DUARTE, R.; SILVA, A.M.; CARA, D.C.; NICOLI, J.R. Evaluation of the components of a commercial probiotic in gnotobiotic mice experimentally challenged with Salmonella enterica subsp. enterica ser. Typhimurium. Veterinary Microbiology, v.79, n.2, p.183-189, mar. 2001.
MADIGAN, M.T.; MARTINKO, J.M.; PARKER, J. Brock Biology of Microorganisms. 9ª Ed., London: Prentice-Hall, 2000, p.51-53.
MARAGKOUDAKIS, P.A.; ZOUMPOPOULOU, G.; MIARIS, C.; KALANTZOPOULOS, G.; POT, B.; TSAKALIDOU, E. Probiotic potential of Lactobacillus strains isolated from dairy products. International Dairy Journal, v.16, n.3, p.189-199, mar.2006.
MCGHEE, J.R.; MESTECKY, J.; DERTZBAUGH, M.T.; ELDRIDGE, J.H.; HIRASAWA, M.; KIVONO, H. The mucosal imunu system from fundamental concepts to vaccine development. Vaccine, v.10, n.2, p.75-88, 1992.
MERCENIER, A.; MULLER-ALOUF, H.; GRANGETTE, C. Lactic acid bacteria as live vaccines. Current Issues in Molecular Biology, v. 2, n.2, p.17-25, jan. 2000.
MICHIELUTTI, F.; BERTINI, M.; PRESCIUTTINI, B.; ANDREOTTI, G. Valutazione clinica di un nuovo batterioterapico orale in pazienti di età pediatrica com diarrea acuta. Minerva Medica, v.87, p.545-550, 1996.
MILES, R.D. Manipulation of the microflora of the gastrointestinal tract.: natural ways to prevent colonization by pathogens. In: ALTECH BIOTECHNOLOGY IN THE FEED INDUSTRY, Florida, 1993. Proceedings. p.133-150.
MIYAJIMA, A.; KITAMURE, T.; HARADE, N.; YOKOTA, T.; ARAI, K. Cytokine receptors and signal transduction. Annual Review Immunology, v.10, p.295-331, apr.1992.
MONET, D.L.; EMBORG, H.D.; ANDERSEN, S.R.; SCHÖLLER, C.; SORENSEN, T.L.; BAGER, F. Vigilância da resistência aos antibióticos na Dinamarca. Eurosurveillance Monthly, v.5, n.12, p.129-132, dec. 2000.
MOREAU, M.C.; CORTHIER, G. Effect of the gastrointestinal microflora on induction and maintenance of oral tolerance to ovalbumin in C3H/HeJ mice. Infection and Immunity, v.56, n.10, p.2766-2768, oct. 1988.

73
MOREIRA, M.A.S.; MORAES, C.A. Resistance to antibiotics in Gram-negative bactéria isolated from broiler carcasses. Arquivo Brasileiro de Medicina Veterinária, v.54, n.1, p.1-7, feb. 2002.
NAIDU, A.S.; BIDLACK, W.R.; CLEMENS, R.A. Probiotic spectra of lactic acid bacteria (LAB). Critical Reviews Food Science and Nutrition, v.38, n.1, p.13-126, jan. 1999.
NAMBA, Y.; HIDAKA, Y.; TAKI, K.; MORIMOTO, T. Effect of oral administration of lysozyme or digested bacterial cell walls on immunostimulation in guinea pigs. Infection and Immunity, v.31, n.2, p.580-583, feb. 1981.
NICOLI, J.R.; VIEIRA, L.Q. Probióticos, prebióticos e simbióticos – Moduladores do ecossistema digestivo. Ciência Hoje, v.28, n.163, p.34-38, ago. 2000.
NICOLI, J.R.; VIEIRA, E.C.; PENNA, F.J.; VIEIRA, L.Q.; RODRIGUES, A.C.P.; NEUMANN, E.; SILVA, A.M.; LIMA FILHO, J.V.M.; BAMBIRRA, E.A.; ARANTES, R.M.E.; MACHADO, D.C.C. Probióticos: Experiências com animais gnotobióticos. In: FERREIRA, C.L.L. Prebióticos e Probióticos: Atualização e Prospecção. Viçosa: UFV, 2003. p.123-133.
NORTH, R.J. The concept of activated macrophage. Journal of Immunology, v.121, n.3, p.806-809, sep. 1978.
O’SULLIVAN, M.G.; THORTON, G.; O’SULLIVAN, G.C.; COLLINS, J.K. Probiotic bacteria:myth or reality? Trends in Food Science & Technology, v.3, p.309-314, 1992.
OGUNBANWO, S.T.; SANNI; A.I.; ONILUDE, A.A. Effect of bacteriocinogenic Lactobacillus spp. on the shelf life of fufu, a tradiotional fermented cassava product. World Journal of Microbiology & Biotechnology, v.20, n.1, p.57-63, feb. 2004.
OUWEHAND, A.C. Antimicrobial components from lactic acid bacteria. In: SALMINEN,S.; VON WRIGHT, A. Lactic Acid Bacteria – Microbiology and Functional Aspects. New York: Marcel Dekker, 1998. p.139-159.
OUWEHAND, A.C.; KIRJAVAIEN, P.V.; SHORTT, C.; SALMINEN, S. Probiotics: mechanisms and established effects. International Dairy Journal, v.9, n.1, p.43-52, jan. 1999.

74
PAÇO, R.S.; LEME, I.L.; BOTTINO, J.A.; FERREIRA, A.J.P. Identification of Lactobacillus spp. from broiler litter in Brazil. Brazilian Journal of Microbiology, v.34, n.3, p.236-237, jul-sep. 2003.
PAVAN, S.; DESREUMAUX, P.; MERCENIER, A. Use of mouse models to evaluate the persistence, safety, and immune modulation capacities of lactic acid bacteria. Clinical and Diagnostic Laborotory Immunology, v.10, n.4, p.696-701, jul. 2003.
PERDIGÓN, G.; ALVARES, S. Probiotics and the immune state. In: FULLER, R. Probiotics – The cientific basis. London: Chapman & Hall.1992. p.145-180.
PERDIGÓN, G.; ALVAREZ, S.; NADER DE MACIAS, M.E.; MARGINI, R.A.; OLIVER, G.; PESCE DE RUIZ HOLGADO, A.A. Lactobacilli administered orally induce release of enzymes from peritoneal macrophages in mice. Milchwissenschaft, v.41, n.6, p.344, 1986a.
PERDIGÓN, G.; ALVAREZ, S.; NADER DE MACIAS, M.E.; PESCE DE RUIZ HOLGADO, A.A. Actividad adyuvante de bacterias bácticas: perspectivas para su uso en vacunas orales. Revista Argentina Microbiologia, v.20, p.141-146, 1988a.
PERDIGÓN, G.; ALVAREZ, S.; NADER DE MACIAS, M.E. et al. The oral administration of lactic acid bacteria increases the mucosal intestinal immunity in response to enteropathogens. Journal of Food Protection, v.53, v.5, p.404-410, may 1990.
PERDIGÓN, G.; ALVAREZ, S.; OLIVER, G. PESCE DE RUIZ HOLGADO, A.A. Immunoadjuvant activity of oral Lactobacillus casei: influence of dose on the secretory immune response and protective capacity in intestinal infections. The Journal of Dairy Research, v.58, n.4, p.485-496, nov. 1991.
PERDIGÓN,G.; ALVAREZ, S.; RACHID, M.; AGÜERO, G.; GOBBATO, N. Immune system stimulation by probiotics. Journal of Dairy Science, v. 78, n.7, p.1597-1606, jul. 1995.
PERDIGÓN, G.; FULLER, R.; RAYA, R. Lactic acid bacteria and their effect on the immune system. Current Issues in Intestinal Microbiology, v.2, n.1, p.27-42, 2001.
PERDIGÓN, G.; MALDONADO GALDEANO,C.; VALDEZ, J.C.; MEDICI, M. Interaction of lactic acid bacteria with the gut immune system. European Journal of Clinical Nutrition, v.56, Supl.4, p.S21-S26, 2002.

75
PERDIGÓN G.; NADER DE MACÍAS, M.E.; ALVAREZ, S.; MEDICI, M; OLIVER, G.; RUIZ HOLGADO, A Effect of a mixture of Lactobacillus casei and Lactobacillus acidophilus administered orally on the immune system in mice. Journal of Food Protection, v.49, n. 12, p.986-989, dec. 1986d.
PERDIGÓN, G.; NADER DE MACÍAS, M.E.; ALVAREZ, S. et al. Actividade imunopotenciadora de bacterias lácticas administradas por vía oral. Su efecto beneficioso en diarreas infantiles. Medicina, v.46, p.751-754, 1986c.
PERDIGÓN, G.; NADER DE MACÍAS, M.E.; ALVAREZ, S.; OLIVER, G.; PESCE DE RUIZ HOLGADO, A.A. Enhancement of immune response in mice fed with Streptococcus thermophilus and Lactobacillus acidophilus. Journal of Dairy Science, v. 70, n.5, p.919-926, may 1987.
PERDIGÓN, G.; NADER DE MACÍAS, M.E.; ALVAREZ, S. et al. Systemic augmentation of the immune response in mice by feeding fermented milks with Lactobacillus casei and Lactobacillus acidophilus. Immunology, v.63, n.1, p.17-23, jan. 1988b.
PERDIGÓN, G.; NADER DE MACIAS, M.E.; ALVAREZ, S.; OLIVER, G.; PESCE DE RUIZ HOLGADO, A.A. Effect of perorally administered lactobacilli on macrophage activation im mice. Infection and Immunity, v.53, n.2, p.404-410, feb. 1986b.
PERDIGÓN, G.; PESCE DE RUIZ HOLGADO, A. Mechanisms involved in the immunostimulation by lactic acid bacteria. In: Probiotics Immunomodulation by the gut microflora and probiotics. 3ed. Dordrecht: Kluwer Academic. 2000. p.213-233.
PERDIGÓN, G.; VINTIÑI, E.; ALVAREZ, S.; MEDINA, M; MEDICI, M. Study of the possible mechanisms involved in the mucosal immune system activation by lactic acid bacteria. Journal of Dairy Science, v.82, n.6, p. 1108-1114, jun. 1999.
PHILLIPS, J.; MURRAY, P.; CROCKER, J. The biology of disease. Cambridge: Blackwell Sci., 1995. 310p.
PLANT, L.J.; CONWAY, P.L. Adjuvant properties and colonization potential of adhering and non-adhering Lactobacillus spp. following oral administration to mice. FEMS Immunology and Medical Microbiology, v.34, n.2, p.105-111, jun. 2002.

76
POPPI, L.B. Avaliação do efeito antagônico de espécies de Lactobacillus sobre Escherichia coli O157:H7 e Listeria monocytogenes. 2005. 113f. Dissertação (Mestrado em Biotecnologia Industrial) – Departamento de Biotecnologia, Faculdade de Engenharia Química de Lorena, Lorena, 2005.
REINA,, J.; ROS, M.J.; SERRA, A. Susceptibilities to 10 antimicrobial agents of 1220 Campylobacter strains isolated from (1984) to (1987) from feces of pediatric patients. Antimicrobial Agents and Chemotherapy, v.38, n.12, p.2917-2920, dec. 1993.
REVOLLEDO PIZARRO, L. D.C. Estudo da resposta imune e da colonização e invasão por Salmonella enterica subsp enterica sorotipo Typhimurium Nalr, em frangos de corte, tratados com glucano, probióticos e produtos de exclusão competitiva. 2005. 116p. Tese (Doutorado em Patalogia Experimental e Comparada), FMVZ, Universidade de São Paulo, São Paulo, 2005.
ROITT, I.M.; DELVES, P.J. Fundamentos de Imonologia. 10. ed. Rio de Janeiro: Guanabara Koogan, 2004, 489p.
ROJO-BEZARES, B.; SÁENZ, Y.; POETA, P.; ZARAZAGA, M.; RUIZ-LARREA, F.; TORRES, C. Assessment of antibiotic susceptibility within lactic acid bacteria strains isolated from wine. International Journal of Food Microbiology, v.111, n.3, p.234-240, oct. 2006.
ROSTAGNO, H.S.; ALBINO, L.F.T.; FERES, F.A.; TOLEDO, R.S. Utilização de probióticos e prebióticos em aves: In: FERREIRA, C.L.F. Prebióticos e Probióticos: Atualização e Prospecção. Viçosa: UFV, 2003. p.181-202.
SAAD, S.M.I. Probióticos e prebióticos: o estado da arte. Revista Brasileira de Ciências Farmacêuticas, v.42, n.1, p.1-16, jan.-mar., 2006.
SAARELA, M.; MOGENSEN, G.; FONDÉN, R.; MÄTTÖ, J.; MATTILA-SANDHOLM, T. Probiotic bacteria: safety, functional and probiotic properties. Journal of Biotechnology, v.84, n.3, p.197-215, dec. 2000.
SAITO, H.; SATO, K., HORIKAWA, Y.; JIN, B.W.; TOMIOKA, H.; WATANABE, T. Enhanced humoral antibody production and delayed type hypersensitivity response in mice by Lactobacillus casei. Hiroshima Journal of Medical Sciences, v. 32, n.2, p.223-226, 1983.

77
SALMINEN, S.; VON WRIGHT, A. Lactic Acid Bacteria – Microbiology and Fuctional Aspects. New York: Marcel Dekka. 1998. 615p.
SÁNCHEZ, R.; FERNÁNDEZ-BACA, V.; DÍAZ, M.D.; MUNOZ, P.; RODRÍGUES-CRÉXEMS, M.; BOUZA, E. Evolution of susceptibilities of Campylobacter spp. to quinolones and macrolides. Antimicrobial Agents Chemotherapy, v.38, n.9, p.1879-1882, sep. 1994.
SANDERS, M.E. Overview of functional foods: emphasis on probiotic bacteria. International of Dairy Journal, v.8, p.341-347, 1998.
SANDERS, M.E. Probiotics: considerations for human health. Nutrition Reviews, v.61, n.3, p.91-99, marc. 2003.
SCHIFFRIN, E.J.; ROCHAT, F.; LINK-AMSTER, H.; AESCHLIMANN, J.M.; DONNET-HUGHES, D. Immunomodulation of human blood cells following the ingestion of lactic acid bacteria. Journal of Dairy Science, v.78, n.3, p.491-497, mar. 1995.
SCHWAB, J.H. Phlogistic properties of peptidoglycan-polysaccharide polymers from cell walls of pathogenic and normal-flora bacteria which colonize humans. Infection and Immunity, v.61, n.11, p.4535-4539, nov. 1993.
SCOTT, K.P.; MELVILLE, C.M.; BARBOSA, T.M.; FLINT, H.J. Occurrence of the new tetracycline resistance gene tet(W) in bacteria from the human gut. Antimicrobial Agents Chemotherapy, v.44, n.3, p.775-777, marc. 2000.
SERVIN, A.L. Antagonistic activities of lactobacilli and bifidobacteria against microbial pathogens. FEMS Microbiology Reviews, v.28, n.4, p.405-440, oct. 2004.
SHORTT, C. The probiotic century: historical and current perspectives. Trends in Food Science & Technology, v.10, n.12, p.411-417, dec. 1999.
SHU, Q.; GIL, H.S. Immune protection mediated by the probiotic Lactobacillus rhamnosus HN001 (DR20TM) against E. coli O157:H7 infection im mice. FEMS Immunology and Medical Microbiology, v.34, p.59-64, 2002.

78
SOZZI, T.; SMILEY, M.B. Antibiotic resistances of yogurt starter cultures Streptococcus thermophilus and Lactobacillus bulgaricus. Applied and Environmental Microbiology, v.40, n.5, p.862-865, nov. 1980.
SPINOSA, H.S. Antibióticos: Tetraciclinas, cloranfenicol e análogos. In: SPINOSA, H.S.; GÓRNIAK, S.L.; BERNARDI, M.M. Farmacologia Aplicada à Medicina Veterinária, 3. ed. Rio de Janeiro: Guanabara Koogan. 2002. p.420-424.
STAVRIC, S. Defined cultures and prospects. International Journal of Food Microbiology, v.15, n.3-4, p.245-263, mar-apr. 1992.
STEWART-TULL, D.E.S. The immunological activities of bacterial peptidoglycans. Ann. Ver. Microbiology, v.34, p.311-340, 1980.
STIMPSON, S.A.; SCHWAB, J.H. Chronic remitent erosive arthritis induced by bacterial peptidoglycan-polysaccharide strutures. In: CHANG, J.Y.; LEWIS, A.J. Pharmacological methods in the control of inflammation. New York: Alan R. Liss. p.381-394, 1989.
TEUBER, M.; MEILE, L.; SCHWARZ, F. Acquired antibiotic resistance in lactic acid bacteria from food. Antonie van Leeuwenhoek, v.76, p.115-137, 1999.
TIZARD, I.R. Imunologia Veterinária – Uma Introdução. 5. ed. São Paulo: Roca. 1998. 546p.
TOWBIN, H.; STAHELIN, T.; GORDON, J. Elctrophoretic transfer of proteins from plyacrilamide gels to nitrocellulose sheets> procedure and some applications. Procceedings National Academy Science USA, v.76, p.4350-4, 1979.
UYTDE HAAG, F.; VAN DER HEIJDEN, R.; OSTERHAUS, A. Maintenance of immunological memory a role for CD5+ Bcells? Immunology Today, v.12, p.439, 1991.
VALLE, M.R.; MONTERO, C.; MACHADO, H.; JOGLAR, M. The evaluation of yeast derivatives as adjuvants for the immune response to the Bm86 antigen in cattle. BMC Biotechnology, v.1, n.1-3., may 2001.
VANBELLE, M.; TELLER, E.; FOCANT, M. Probiotics in animal nutrition: a review. Arch. Anim. Ntri., v.40, n.7, p.543-567, 1990.

79
VAY, C.; CITTADINI, R.; BARBERIS, C.; RODRÍGUEZ, C.H.; MARTÍNEZ, H.P.; GENERO, F.; FAMIGLIETTI, A. Antimicrobial susceptibility of non- enterocociccal intrinsic glycopeptide-resistant Gram-positive organisms. Diagnostics Microbiology and Infectious Disease, v.57, n.2, p.183-188, feb.2007.
VELÁZQUEZ, J.B.; JIMÉNEZ, A.; CHOMÓN, B.; VILLA, T.G. Incidence and transmisión of antibiotic resístanse in Campylobacter jejuni and Campylobacter coli. Journal of Antimicrobial Chemotherapy, v.35, n.1, p.173-178, jan. 1995.
VINDEROLA, C.G.; MEDICI, M.; PERDIGÓN, G. Relationship between interactions sites in the gut, hydrophobicity, mucosal immunomodulating capacities and cell wall protein profiles in indigenous and exogenous bacteria. Journal of Applied Microbiology, v.96, p.230-243, 2004.
VITIÑI, E.; ALVAREZ, S.; MEDINA, M.; MEDICI, M.; DE BUDEGUER, M.V.; PERDIGÓN, G. Gut mucosal immunostimulation by lactic acid bacteria. Biocell, v.24, n.3, p.223-232, 2000.
WALKER, W.; HAEMATZ, P.; WESHILL, B. Immunophysiology of the gut. NewYork: Acad. Press, 1993. p.145.
ZHAO, R.; SUN, J.; MO, H.; ZHU, Y. Analysis of functional properties of Lactobacillus acidophilus. World Journal Microbiology Biotechnology, v.23, p.195-200, 2007.
ZIEMER, C.J.; GIBSON, G.R. An overview of probiotics, prebiotics and synbiotics in the functional food concept: perspectives and future strategies. International of Dairy Journal, v.8, p.473-479, 1998.

80
APÊNDICES

81
APÊNDICE A - Biografia
Débora Dutra Menezes Leal, filha de Amauri Menezes Leal e Enancy Dutra Menezes
Leal, nasceu em 30 de dezembro de 1971, na cidade de Lorena, São Paulo.
Graduou-se em Zootecnia pela Universidade Federal de Lavras em setembro de 1996.
Em setembro do mesmo ano iniciou o curso de pós graduação em nível de mestrado em
Ciência dos Alimentos na mesma Universidade, defendendo a dissertação intitulada
“Caracterização e Identificação de Listeria monocytogenes e Bactérias Gram Negativas em
Mastite Clínica e Subclínica” em maio de 1999.
Em maio de 2003 ingressou no curso de doutorado na Escola de Engenharia de Lorena
na área de Biotecnologia Industrial, defendendo a tese: “Estudo da influência de espécies de
Lactobacillus sobre a resposta humoral em camundongos BALB/c” em junho de 2007.

82
APÊNDICE B – Perfil eletroforético das proteínas secretadas pelas cepas de Lactobacillus
11fb 18fa 19fa 22c 30b 30c 41b Pool
Concentração de proteína aplicada no gel = 20 �g/mL
83
APÊNDICE C – Perfil eletroforético das proteínas secretadas pelas cepas de Lactobacillus sem
adição dos inibidores de proteases
Neste perfil eletroforético foi possível visualizar apenas o padrão de massa molecular, pois as
proteínas foram degradadas.
Recommended

















