Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DE PERNAMBUCO
CENTRO DE CIÊNCIAS DA SAÚDE
PROGRAMA DE PÓS-GRADUAÇÃO EM NUTRIÇÃO
MESTRADO EM NUTRIÇÃO
ÁREA DE CONCENTRAÇÃO: BASES EXPERIMENTAIS DA NUTRIÇÃO
CAMILA LIMA CHAGAS
Ação comportamental e eletrofisiológica cerebral da cafeína:
análise em ratos albinos em desenvolvimento
Recife
2017

CAMILA LIMA CHAGAS
Ação comportamental e eletrofisiológica cerebral da cafeína:
análise em ratos albinos em desenvolvimento
Dissertação apresentada ao Programa de Pós-Graduação em Nutrição do Centro de Ciências da Saúde da Universidade Federal de Pernambuco, para obtenção do título de Mestre em Nutrição.
Orientador: Dr. Rubem Carlos Araújo Guedes Co-orientador: Dr. Ricardo Abadie Guedes
Recife
2017


CAMILA LIMA CHAGAS
AÇÃO COMPORTAMENTAL E ELETROFISIOLÓGICA CEREBRAL DA CAFEÍNA: ANÁLISE
EM RATOS ALBINOS EM DESENVOLVIMENTO
Dissertação aprovada em 22/02/2017
_________________________________________________________________
Profª Dra. Raquel da Silva Aragão/ CAV-UFPE
_________________________________________________________________
Profº Dr. Marcelo Moraes Valença/ UFPE
_________________________________________________________________
Profª Dra. Rhowena Jane Barbosa de Matos/ CAV-UFPE
RECIFE
2017

AGRADECIMENTOS
À Deus, por estar presente em todos os momentos da minha vida e por não ter me
desamparado em nenhum momento.
Aos meus pais, Neide e Claudemir, por todo apoio, carinho, dedicação e educação.
Obrigada por estarem presente em todos os momentos da minha vida, pelas orações
e pelo exemplo de vida, sinto muito orgulho de vocês.
Ao meu irmão, Thiago, por estar ao meu lado sempre, pela amizade, confiança e por
tudo que representa em minha vida.
À minha tia Ana, minha prima Aline e minha cunhada Luciana por estarem presentes
em todas as minhas conquistas.
Ao meu namorado, Rômulo, principalmente pela dedicação, carinho e paciência
durante esse período do mestrado.
Aos amigos Paulo Neves e Caroline, meus sobrinhos, Sofia e Mateus, pelo carinho e
amor, pela presença constante em minha vida. Agradeço-os eternamente pelo apoio.
Ao professor Rubem Guedes, meu orientador, pelo acolhimento no laboratório, por
seu profissionalismo e competência. Admiro muito sua dedicação e amor ao trabalho.
Obrigada pelo apoio em todos os momentos do mestrado, pelos ensinamentos e por
ser essa pessoa iluminada. Tê-lo como orientador foi um presente de Deus.
Ao professor Ricardo Abadie Guedes, meu co-orientador, por todo o apoio desde o
início, por sempre acreditar em mim e fazer meu sonho se tornar realidade. Obrigada
por todas as orientações e ensinamentos que com certeza contribuíram para minha
formação profissional, pela amizade, conselhos e por ser essa pessoa especial em
minha vida. Jamais esquecerei tudo o que fez por mim.
A minha grande colaboradora e amiga, Regina de Deus. Não tenho palavras para
agradecer por todo o carinho, ensinamentos e os grandes momentos vividos durante
esses dois anos. Agradeço a Deus por ter encontrado uma pessoa tão querida nessa
caminhada, tenho certeza que é apenas o começo de uma grande amizade e que
Deus reservou um futuro brilhante para você.
À minha estagiária Cynthia, por toda colaboração durante a pesquisa.

Ao médico-veterinário Edeones França pela competência e cuidado com os animais.
A todos os amigos do LAFFINT pela ajuda em diversos momentos.
Às amigas do mestrado Maryane, Débora, Carolina, Daniela, Larissa e Jacqueline por
todo carinho e ajuda.
A todos que contribuíram direta ou indiretamente para a realização desse trabalho.

“Há homens que lutam um dia e são bons, há outros que lutam um ano e são melhores,
há aqueles que lutam muitos anos e são muito bons. Porém há os que lutam toda a
vida, estes são imprescindíveis. ”
Bertolt Brecht

RESUMO
A cafeína (1,3,7-trimetilxantina) é um psico-estimulante que parece agir através dos receptores da adenosina A1 e A2 e dos receptores de dopamina D2 e supõe-se que a sua administração possa influenciar na excitabilidade cerebral. Neste estudo avaliou-se a ação da cafeína sobre o comportamento semelhante ao de ansiedade e sobre a depressão alastrante cortical (DAC) em três grupos de ratos que receberam cafeína por gavagem nas doses de 15mg/Kg/dia, 30mg/Kg/dia e 45mg/Kg/dia, do 7º ao 27º dia de vida pós-natal (grupos Caf15, Caf30 e Caf45). Dois grupos controle receberam gavagem com o veículo (V) ou não receberam qualquer tratamento (ingênuo). Os seus pesos corporais foram registrados nos dias 7, 14, 21, 28 e no dia do registro da DAC. Aos 28 dias de vida, os animais foram submetidos ao teste comportamental do labirinto em cruz elevado e aos 30 dias ao teste do campo aberto. Aos 35 dias foram submetidos ao registro da DAC. Outro grupo de animais jovens (31-40 dias de vida) foi submetido à aplicação tópica cortical de uma solução de cafeína (15mg/ml) durante a sessão de registro da DAC. Os testes comportamentais sugeriram um efeito ansiolítico da cafeína: o tempo por cada entrada aumentou de forma dose-dependente nos animais tratados com cafeína no labirinto em cruz elevado e campo aberto, com valores significativamente maiores no grupo Caf45 em comparação ao grupo ingênuo. Aos 28 dias de vida os grupos Caf30 e Caf45 apresentaram pesos corporais menores (respectivamente 64,6±4,0g e 58,9±3,8g) em comparação aos grupos ingênuo (73,6±5,5g), veículo (72±3,7g) e Caf15 (70,5±4,5g). No LCE o número do comportamento de levantar-se foi significativamente menor no grupo Caf45 em relação aos grupos controles, o número desse comportamento nos grupos ingênuo, veículo, CAF15, CAF30 e CAF45 foram respectivamente, 10,6±1,4; 10,8±1,2; 9,5±1,3; 8,8±0,9; 6,9±1,4 (p<0,05). No campo aberto o número de comportamento de levantar-se foi significativamente menor no grupo Caf45 em relação aos grupos controles e Caf15. O número do comportamento de levantar-se dos grupos ingênuo, veículo, CAF15, CAF30 e CAF45 foram respectivamente, 9,5± 1,4; 9,8±1,4; 7,5±1,2; 6,6±1,4; 4,8±0,9 (p<0,05). Aos 35 dias de vida os grupos Caf30 e Caf45 apresentaram pesos corporais menores (respectivamente 103,3 ± 7,8 g e 99,4 ±6,4 g), quando comparados aos grupos V (124,2±7,1 g), ingênuo (120,9±9,5 g) e Caf15 (122,4 ± 12,9 g) (p < 0,05). Na DAC, os grupos tratados com cafeína tiveram velocidades (em mm/min) maiores (Caf15 = 4,06±0,10; Caf30 = 4,18 ± 0,18 e Caf45 = 4,74±0,18) do que os controles (ingênuo = 3,93±0,10; V = 3,92±0,16; p<0,05). A aplicação tópica de cafeína aumentou a velocidade e a amplitude da DAC e reduziu a sua duração. Os resultados indicam que a administração de cafeína durante o período crítico de desenvolvimento tem impacto sobre o comportamento de ansiedade, reduzindo-o, mas aumenta a velocidade de propagação da DAC, sugerindo, portanto, importante papel da cafeína na excitabilidade cerebral.
Palavras-chave: Cafeína. Ansiedade. Depressão cortical alastrante. Desenvolvimento cerebral.

ABSTRACT
Caffeine (1,3,7-trimethylxanthine) is a psycho-stimulant that acts through adenosine A1 and A2 receptors and dopamine D2 receptors. Evidence supports the assumption that it may influence brain excitability. This study evaluated in Wistar rats the action of caffeine on anxiety-like behavior and on the electrophysiological features of the excitability-related phenomenon known as cortical spreading depression (CSD). From postnatal day (PND) 7 to 27, animals received, via gavage, either caffeine (15, 30, or 45 mg/kg/day; groups Caf15, Caf30 and Caf45, respectively), or vehicle (group V) or no gavage (group naïve). Body weights were recorded at PND 7, 14, 21, 28 and P35. At PND28 the animals were behaviorally tested in the elevated plus-maze. At PND30 they were tested in the open field. At PND35, they were submitted to CSD recording. Another group of young animals (PND31-40) were subjected to topical cortical application of caffeine solution (15mg/ml) during the CSD recording session. Behavioral tests suggested an anxiolytic effect of caffeine: the time per entry increased in a dose-dependent manner in caffeine-treated animals in elevated plus-maze and open field, with significantly higher values in the Caf45 group compared to the naive group. At 28 days of age, the Caf30 and Caf45 groups had smaller body weights (64.6 ± 4.0g and 58.9 ± 3.8g, respectively) than the naive groups (73.6 ± 5.5g), vehicle (72 ± 3.7g) and Caf15 (70.5 ± 4.5g). In the elevated plus-maze the number of the rearing behavior was significantly lower in the Caf45 group than in the control groups, the number of this behavior in the naive, vehicle, CAF15, CAF30 and CAF45 groups were respectively 10.6 ± 1.4; 10.8 ± 1.2; 9.5 ± 1.3; 8.8 ± 0.9; 6.9 ± 1.4 (p<0.05). In the open field the number of rearing behavior was significantly lower in the Caf45 group than in the control and Caf15 groups. The number of the behavior of rearing from the naive, vehicle, Caf15, Caf30 and CAF45 groups were, respectively, 9.5 ± 1.4; 9.8 ± 1.4; 7.5 ± 1.2; 6.6 ± 1.4; 4.8 ± 0.9 (p <0.05). At 35 days of life the groups Caf30 and Caf45 had lower body weights (103.3 ± 7.8 g and 99.4 ± 6.4 g, respectively), compared to vehicle group (124.2 ± 7.1 g), naive group (120.9 ± 9.5 g) and Caf15 group (122.4 ± 12.9 g) (p <0.05). Caffeine-treated groups had higher CSD rates (in mm / min) (Caf15 = 4.06 ± 0.10; Caf30 = 4.18 ± 0.18 (p <0.05) and Caf45 = 4.74 ± 0.18) than controls (naive = 3.93 ± 0.10; V = 3.92 ± 0.16, p <0.05). Topical application of caffeine reversibly increased CSD velocity and amplitude, and reduced CSD duration. The results indicate that the administration of caffeine during the critical period of development reduced anxiety-like behavior, but increased the CSD propagation velocity, which suggests, therefore, an important role for caffeine on cerebral excitability.
Key words: Caffeine. Anxiety. Cortical spreading depression. Brain development.

LISTA DE ILUSTRAÇÕES
Figura 1 – Papel da cafeína na termogênese e ingestão energética................... 18
Figura 2 – Sequência de eventos da depressão alastrante cortical..................... 22
Figura 3 – Representação esquemática dos grupos experimentais ..................... 27
Figura 4 – Pesos corporais................................................................................... 32
Figura 5 – Avaliação comportamental do teste do labirinto em cruz elevado...... 33
Figura 6 – Avaliação comportamental do teste do campo aberto......................... 34
Figura 7 – Comportamento de rearing e stretched............................................... 35
Figura 8 - Registros eletrofisiológicos.................................................................. 36
Figura 9 – Velocidade de propagação da depressão alastrante cortical.............. 37
Figura 10 – Velocidade, amplitude e duração de aplicação tópica.......................39
LISTA DE TABELAS
Tabela 1 - Valores de cafeína encontrados em alguns produtos...........................17
Tabela 2 – Amplitude e duração das ondas de Depressão Alastrante Cortical.....38

LISTA DE ABREVIATURAS
ACTH Hormônio adenocorticotrópico
ANOVA Análise de Variância
CAF Cafeína
CT Cafeína tópica
DAC Depressão Alastrante Cortical
ECOG Eletrocorticograma
GABA Ácido gama-aminobutírico
HPA Eixo Hipotálamo-Pituitária-Adrenal
I Ingênuo
KCl Cloreto de Potássio
LAFFINT Laboratório de Fisiologia da Nutrição Naíde Teodósio
LCE Labirinto em Cruz Elevado
SNC Sistama Nervoso Central
UFPE Universidade Federal de Pernambuco
V Veículo
VLV Variação Lenta de Voltagem

SUMÁRIO
1 INTRODUÇÃO............................................................................................13
2 REVISÃO BIBLIOGRÁFICA.......................................................................15
2.1 NUTRIÇÃO, CAFEÍNA E SISTEMA NERVOSO.........................................15
2.2 CAFEÍNA E METABOLISMO......................................................................18
2.3 CAFEÍNA E ANSIEDADE............................................................................19
2.4 DEPRESSÃO ALASTRANTE CORTICAL.................................................. 20
3 HIPÓTESES............................................................................................... 24
4 OBETIVOS................................................................................................. 25
4.1 OBJETIVO GERAL..................................................................................... 25
4.2 OBJETIVOS ESPECÍFICOS...................................................................... 25
5 MÉTODOS.................................................................................................. 26
5.1 ANIMAIS..................................................................................................... 26
5.2 TRATAMENTO SISTÊMICO COM CAFEÍNA............................................ 26
5.3 TRATAMENTO TÓPICO COM CAFEÍNA.................................................. 27
5.4 DETERMINAÇÕES DO PESO CORPORAL.............................................. 27
5.5 AVALIAÇÃO COMPORTAMENTAL........................................................... 28
5.5.1 Avaliação comportamental no labirinto em cruz elevado (LCE)..................28
5.5.2 Avaliação comportamental no teste de campo aberto................................ 29
5.6 REGISTRO DA DEPRESSÃO ALASTRANTE CORTICAL........................ 29
5.7 APLICAÇÃO TÓPICA DE CAFEÍNA ..................................................... 30
5.8 EUTANÁSIA................................................................................................ 31
5.9 ANÁLISES ESTATÍSTICAS........................................................................ 31
6 RESULTADOS........................................................................................... 32
7 DISCUSSÃO............................................................................................... 40
8 CONSIDERAÇÕES FINAIS........................................................................ 46
REFERÊNCIAS.......................................................................................... 48
ANEXO 1 - Confirmação da submissão do artigo ao periódico........... 62
ANEXO 2 - Parecer do Comitê de Ética em Pesquisa da UFPE............ 63

13
1. INTRODUÇÃO
Ao final da década de 60, foi demonstrado que durante o desenvolvimento o
organismo, em geral, e o sistema nervoso, em particular, são vulneráveis a agressões
produzidas por fatores ambientais, nutricionais e farmacológicos; por isso a
denominação de período crítico do desenvolvimento. (MORGANE et al., 1993;
SAWAYA et al., 2012; GUEDES, 2011; et al., 2012). Nesse período ocorre um pico de
eventos como a neurogênese, gliogênese e migração celular. O conceito de
desenvolvimento crítico pode estar relacionado com a observação de que os
processos de desenvolvimento e organização são alterados com maior facilidade
durante a fase de desenvolvimento embrionário de maior atividade. (SAWAYA et al.,
2012). Quando ocorre algum dano que não seja durante o período de
desenvolvimento, o organismo pode ter prejuízos fisiológicos menores. (SAWAYA et
al., 2012).
Durante o desenvolvimento dos mamíferos, os nutrientes fornecidos pelos
alimentos são de extrema importância para a produção de energia, sínteses orgânicas
e desenvolvimento dos diversos sistemas do organismo. No sistema nervoso central,
uma alimentação inadequada pode ocasionar várias alterações permanentes na
formação de estruturas celulares e modificar negativamente os parâmetros
eletrofisiológicos do cérebro. (MORGANE et al., 1978, 1993; ALMEIDA et al., 2002;
GUEDES et al., 2013). Os desequilíbrios de alguns nutrientes da dieta, bem como o
elevado consumo de alimentos e bebidas com substâncias psico-estimulantes, como
a cafeína, podem influenciar o funcionamento neural. (SOUZA & SICHIERI, 2005;
AGUIAR et al., 2011).
A cafeína (1,3,7-trimetilxantina) é um psico-estimulante que parece agir através
dos receptores da adenosina dos tipos A1 e A2 e dos receptores de dopamina do tipo
D2 (AGUIAR et al., 2011). A cafeína é um alcaloide que pertence à estrutura de várias
espécies de plantas e sua estrutura molecular faz parte de um grupo de xantinas
metiladas como a treobromina e teofilina (DALY, 2007; SAWYNOK, 2011, TAVARES
& SACATA, 2012). Quando ingerida por via oral, atinge concentração plasmática
estável após cerca de 30 minutos a 2 horas, porém, esse período pode-se prolongar
com o consumo de alimentos (SAWYNOK,1993).

14
Seu consumo tem sido associado à melhora da cognição e desempenho de
atletas, além de ser utilizada no tratamento da sonolência e redução da fadiga
(GIESBRECHT, 2010; BLOMS et al., 2010; LISKO, 2016). No entanto, quando
consumida em doses altas (acima de 40mg/Kg) pode provocar sintomas como insônia,
irritabilidade e/ou cefaleia e até alterações nutricionais (AGUIAR et al., 2011;
FREDHOLM et al., 1999). Outros efeitos negativos observados seriam aumento da
pressão sanguínea e ansiedade (WINSTON et al., 2005; LISKO, 2016). Em ratos,
observa-se que certas alterações comportamentais devidas ao consumo de cafeína
dependem da quantidade consumida (FREDHOLM et al.,1999; MCKIM, 1996;
AGUIAR et al., 2011).
A cafeína deve ser utilizada com cautela em indivíduos com algumas doenças,
como úlcera péptica, refluxo gastroesofágico, função renal ou hepática prejudicadas
e doença cardiovascular. (SAWYNOK, 2011). Outro caso que merece atenção é no
momento da gestação, pois estudos com animais mostraram relação entre alto
consumo materno de cafeína e alterações esqueléticas, retardo do crescimento
intrauterino e baixo peso ao nascer; algumas fontes limitam o consumo de cafeína por
gestantes a 5mg/kg/dia (DALY, 2007). A meia-vida da cafeína é prolongada durante a
gestação, especificamente durante o segundo e terceiro trimestre. (ZHANG, 2001).
No entanto, vale ressaltar que uma mesma dose de cafeína pode provocar reações
adversas em um indivíduo e em outro ter boa aceitação (JULIANO e GRIFFITHS,
2004; SAWYNOK, 2011)
Os efeitos de alterações nutricionais sobre as funções cerebrais podem ser
estudados através de modificações da atividade elétrica cortical cerebral, utilizando
como instrumento de investigação o fenômeno da depressão alastrante cortical
(DAC), descrito primeiramente por Leão (LEÃO, 1944). Esse fenômeno representa
uma resposta despolarizante do córtex cerebral provocada pela estimulação
mecânica, elétrica ou química de um ponto do tecido cortical. A DAC caracteriza-se
pela redução da atividade eletrocorticográfica espontânea, que se propaga pelo córtex
cerebral (MAIA et al., 2009, LIMA et al., 2009; AGUIAR et al., 2011).
Em um estudo preliminar no qual 30mg/Kg de cafeína foram administrados a
ratos adultos por apenas 04 dias, não se observou diferenças significativas na
propagação da DAC (AGUIAR et al., 2011). Portanto, tendo em vista a escassez de
estudos acerca de efeitos da cafeína sobre a DAC, principalmente durante o
desenvolvimento cerebral, justifica-se a relevância do presente estudo.

15
2. REVISÃO BIBLIOGRÁFICA
2.1 NUTRIÇÃO, CAFEÍNA E SISTEMA NERVOSO
Como mencionado na introdução, no sistema nervoso de mamíferos existe um
período crítico para o desenvolvimento. (MORGANE et al., 1993). Em humanos, esse
período começa no terceiro trimestre de gravidez e dura entre dois e quatro anos; no
rato, esse período envolve tanto a gestação quanto a lactação. (MORGANE et al.,
1993; BATISTA-OLIVEIRA et al., 2012). Esse é um período em que vários processos
fisiológicos ocorrem com velocidade máxima, ou seja, o cérebro está mais susceptível
a insultos ambientais e nutricionais nessa fase (MORGANE et al., 1993; BATISTA-
OLIVEIRA et al., 2012; SAWAYA et al., 2012).
A nutrição exerce um papel fundamental na maturação e desenvolvimento do
sistema nervoso central, para que se tenha um fornecimento de energia e nutrientes
essenciais para o desenvolvimento de estruturas celulares e sistemas metabólicos. O
desenvolvimento do cérebro e as suas consequentes reações fisiológicas dependem
tanto da genética como de fatores ambientais, nutricionais e culturais (MORGANE et
al., 1993).
No início da vida, o sistema nervoso está susceptível à ação de diversos fatores,
incluindo as deficiências nutricionais, estas, por sua vez, podem influenciar
negativamente os parâmetros estruturais, bioquímicos e eletrofisiológicos do cérebro.
(MORGANE et al., 1978, 1993; GUEDES et al., 2002; ALMEIDA et al., 2002)
Dependendo da intensidade e do tipo de desnutrição, os seus efeitos deletérios
podem ser permanentes; e estes efeitos têm sido muito estudados através do
fenômeno da DAC. (GUEDES et al., 2005; GUEDES et al., 2013). Vale ressaltar que
as alterações são mais graves, quando a desnutrição ocorre no período de
crescimento rápido do cérebro (DOBBING, 1968; GUEDES et al., 2013).
Há algumas centenas de anos, surgia na Europa o consumo de bebidas fontes
de cafeína, principalmente o café e, desde então, o consumo dessas bebidas se
tornou hábito em nossas culturas (FREDHOLM et al., 1999). O cultivo do café teve
início no Iêmen, próximo a Meca, no século XIX (GUERRA et al., 2000). A cafeína foi
isolada em 1820 e os seus efeitos não foram claramente reconhecidos até 1981,
quando o bloqueio de receptores da adenosina foi correlacionado às propriedades

16
estimulantes da cafeína e de seus análogos (MCKIM, 1996; DALY, 2007). Em grande
parte do mundo, as pessoas consomem cafeína em grande quantidade,
independentemente da idade e do status econômico (MCKIM).
O interesse em estudar a cafeína ganhou destaque quando estudos em animais
relacionaram o consumo de cafeína durante a gestação à diminuição no crescimento
intra-uterino fetal, redução do peso ao nascer, reabsorção fetal e teratogênese
(SOUZA; SICHIERI, 2005). Desde então, estudos sugeriram uma diminuição do
consumo de cafeína por mulheres grávidas. (SOUZA; SICHIERI, 2005). No entanto,
outros estudos indicam que o consumo moderado de cafeína durante a gestação não
está relacionado a tais efeitos negativos. (ZHANG, 2001; TAVARES e SAKATA,
2012), ou seja, a sua segurança do consumo de cafeína durante os períodos de
gestação e lactação ainda é controversa (TEMPLE et al., 2009; PORCIÚNCULA et al.,
2013).
Atualmente a cafeína é uma das substâncias mais consumidas no mundo e está
relacionada a vários processos fisiológicos, envolvendo resistência física, alteração
de humor e sono. (FREDHOLM et al., 1999). A cafeína é uma potente estimulador do
Sistema Nervoso Central e está presente em vários alimentos e bebidas,
principalmente em café, chá e refrigerantes tipo cola, além de ser utilizada em
medicamentos, seja como inibidor de apetite ou com finalidade estimulante (Tabela
1). Se ingerida em doses moderadas (10 a 40mg/kg/dia) pode levar à sonolência,
fadiga ou até mesmo à sensação de bem-estar, porém, em doses altas (>40mg/kg/dia)
pode produzir sintomas como nervosismo, insônia ou mesmo ataques de pânico.
(FREDHOLM et al., 1999; MCKIM, 1996). Em ratos e camundongos, as doses altas
podem levar a crises epilépticas. (FREDHOLM et al., 1999; SOUZA e SICHIERI, 2005;
GUEDES et al., 2012).
A cafeína é absorvida pelo trato gastrointestinal de forma rápida tanto em
humanos como em ratos (MAGKOS e KAVOURAS 2005). Em relação à sua ação
sobre o sistema nervoso central, a cafeína parece passar livremente do sangue para
o cérebro, exercendo um efeito protetor dose-dependente, principalmente em doenças
como Parkinson. (CHEN, 2010). Estudos demonstraram que doses moderadas de
cafeína atenuam o declínio cognitivo durante o envelhecimento (CONCANNON et al.,
1983), na doença de Alzheimer (ARENDASH et al., 2006; DALL’IGNA et al., 2007;
RITCHIE et al., 2007; ESPINOSA et al., 2013) e em distúrbios psiquiátricos
(PANDOLFO et al., 2013; KASTER et al.,2015). Por outro lado, os prejuízos causados

17
pela cafeína durante o período de desenvolvimento podem acarretar déficits a longo
prazo na função cerebral. (CONCANNON et al., 1983; PERUZZI et al., 1985).
(OFLUOGLU, 2009; CHEN et al., 2010; AGUIAR et al., 2011; TAVARES; SAKATA,
2012).
Tabela 1. Valores de cafeína encontrados em alguns produtos
Adaptado de GUEDES et al., 2012.
Os efeitos estimulantes da cafeína sobre o sistema nervoso central, estão
relacionados, principalmente ao seu papel no bloqueio de receptores de adenosina
A1 e A2A (CHEN et al., 2010), sendo a cafeína um antagonista não seletivo dos
receptores de adenosina A1 e A2A. A adenosina age em diversos sistemas
fisiológicos e os efeitos geralmente, são opostos aos da cafeína. (DAVIS e GREEN,
2009; CARRILLO e BENITEZ, 2000). Além disso, a cafeína influencia o metabolismo
normal de nucleotídeos cíclicos através da ação da enzima fosfodiesterase, pois
quando consumida em doses altas inibe essa enzima, bloqueando receptores do
ácido gama-aminobutírico tipo A (GABAA) e causando liberação de Ca2+ intracelular.
(POLESZAK et al., 2015). Em geral, as propriedades da cafeína ocorrem devido à
sua capacidade de interagir com neurotransmissores em diferentes regiões do
encéfalo podendo, dessa forma, influenciar suas propriedades eletrofisiológicas
(DALY, 2007; CARRILLO e BENITEZ, 2000; LORIST e TOPS, 2003; POLESZAK et
al., 2015).
Produto Quantidade de cafeína
Café 11 a 250 mg/100ml
Chá 2 a 540 mg/100ml
Chocolate 21 a 76 mg/100g
Refrigerante 10 a 20 mg/100ml
Bebidas energéticas 30 a 112 mg/ml
Medicamentos 30 a 220 mg/Comprimido

18
2.2 CAFEÍNA E METABOLISMO
Geralmente a obesidade é relacionada com o aumento de ingestão de alimentos
calóricos, com baixo valor nutricional e redução da atividade física, ou seja, um
desequilíbrio entre a ingestão e o gasto energético (DIEPVENS et al., 2007). O
sistema nervoso simpático exerce um papel importante na manutenção da
homeostase energética, através do controle neural e hormonal (HURSEL e
WESTERTERP-PLANTENGA, 2010). Sua ativação tem sido estudada no aumento da
saciedade, estímulo do gasto energético, em parte pelo aumento da oxidação de
gordura (ASTRUP et al., 1991).
O consumo moderado de cafeína por humanos tem sido associado a benefícios,
como, o aumento do gasto energético diário (BRACCO et al., 1995), diminuição da
ingestão energética (JESSEN et al., 2005) e diminuição do risco de desenvolvimento
de diabetes tipo 2 (SANTOS e LIMA, 2016). Alguns estudos demonstram que o
consumo de cafeína em pacientes diabéticos pode levar a perda de peso, através do
aumento da termogênese (GOLDSTEIN e JACOBS, 2010) e devido às propriedades
ergogênicas da cafeína (PALUSKA, 2003). A figura 1 mostra o papel da cafeína na
termogênese e ingestão energética.
Figura 1. Papel da cafeína na termogênese e na ingestão de energia (Adaptado de Harpaz et al., 2017).
SNS, Sistema Nervoso Simpático; cAMP, adenosina monofosfato cíclica; CA, Catecolaminas; GE,
gasto energético.
O consumo de café também tem sido associado à prevenção de doenças
cardiovasculares e câncer. (BIDEL et al. 2010; ZHANG et al., 2009). Esses efeitos

19
podem estar associados, pelo menos em parte, aos efeitos inibitórios da cafeína nos
receptores de adenosina (MULLER e JACOBSON, 2011) e também aos efeitos
antioxidantes dos ácidos fenólicos e clorogênicos, provenientes de compostos
gerados durante a torrefação (BOETTLER et al., 2011). No entanto, foram observados
tais efeitos em estudos que utilizaram café descafeinado, sugerindo que outros
compostos também podem estar envolvidos em tais benefícios (THOM, 2007;
VINSON et al., 2012). Os efeitos da cafeína na perda de peso ainda não estão
totalmente claros. Tais efeitos podem estar relacionados com restrição energética.
(FORTE E SCHNEIDER, 2012). Portanto, mais estudos são necessários para
esclarecer não só o real papel da cafeína na perda de peso como também a dosagem
necessária e a frequência do consumo (FORTE E SCHNEIDER, 2012).
2.3 CAFEÍNA E ANSIEDADE
Os primeiros modelos animais de ansiedade surgiram devido a ajustes
realizados através de procedimentos de aprendizagem consagrados do behaviorismo,
com o objetivo de verificar efeitos de drogas ansiolíticas. (HALL, 1934). Neste
contexto, testes comportamentais como o labirinto em cruz elevado (LCE) e o campo
aberto, são muito utilizados. (LORENZ, 1995, CRUZ e FERNANDEZ, 2012) O campo
aberto tem sido bastante utilizado para avaliar o estado emocional e atividade
locomotora de animais. O LCE explora o conflito, presente nos braços abertos do
labirinto, entre a tendência dos animais em explorar espaços novos e o seu medo de
altura e dos braços abertos (HALL, 1934, CRUZ e FERNANDEZ, 2012).
Alguns estudos demonstraram que o consumo agudo e crônico da cafeína em
ratos adultos leva a alterações comportamentais sugestivas de ansiogênese, tanto no
campo aberto (NOSCHANG et al., 2009) quanto no labirinto em cruz elevado (EL
YACOUBI et al., 2000). Estudos comportamentais em ratos, demonstraram ainda que
quando a cafeína é ingerida por via oral pode causar aversão, devido ao seu gosto
amargo, que por sua vez pode comprometer os resultados dos estudos, pois, estudos
em ratos tem demonstrado que os mesmos ingerem livremente apenas baixas doses
de cafeína. (HEPPNER et al., 1986). Entretanto, quando administrada em forma de
medicação pode originar sintomas como náuseas e irritação gástrica. (HEPPNER et
al., 1986; MCKIM, 1996; DRIFFITHS; MUMFORD, 1995; GUEDES et al., 2012). No

20
entanto, a literatura carece de estudos experimentais sobre efeitos comportamentais
da cafeína em animais em desenvolvimento
Um interesse renovado pelo estado de abstinência da cafeína surgiu a partir da
observação de que o seu consumo crônico seguido por um período de abstinência de
pelo menos 48 horas, aumenta o risco de ansiedade e provoca sintomas como dor de
cabeça e estresse, além de diminuição do estado de alerta (BHATTACHARYA et al.,
1997; SIMON et al., 2009; O’NEILL et al., 2016).
Evidencias apontam que a exposição à cafeína durante o desenvolvimento
cerebral leva a um comprometimento da memória seletiva, que parece ser maior em
ratas do que em ratos (SELL et al., 2000; CARROLL et al., 2004). Estudos clínicos
têm apontado para uma maior susceptibilidade de mulheres aos efeitos de outros
psicoestimulantes. No entanto, com relação à exposição à cafeína, as comparações
entre os gêneros ainda são controversas (TEMPLE e ZIEGLER, 2011; ARDAIS et al.,
2016).
Alguns estudos analisaram o efeito da administração da cafeína aguda no eixo
hipotálamo-pituitária-adrenal (HPA). Embora não se tenha comprovação de um papel
causal do HPA na ansiedade (ARMARIO et al, 2012), admite-se que ele pode
contribuir de forma importante para a patogênese da depressão e ansiedade.
(JEZOVA e HLAVACOVA, 2008;). Tanto em seres humanos (LIN et al., 1997), quanto
em ratos, a cafeína aumenta expressivamente o hormônio adenocorticotrópico
(ACTH). (PATZ et al., 2006; O’NEILL, 2016)
Estudos apontam que o antagonismo farmacológico seletivo dos receptores A1
e A2A, que estão envolvidos no mecanismo de ação da cafeína, está relacionado a
alterações comportamentais. (RANDALL et al., 2011) Em ratos, o aumento da
atividade locomotora e da frequência do comportamento de se levantar (“rearing”) teria
relação com o efeito estimulante da cafeína, mediado pela ativação dos receptores
A2A. Por outro lado, o efeito ansiogênico da cafeína estaria provavelmente relacionado
à sua ação sobre o receptor A1 (RANDALL et al., 2011; GUEDES et al., 2012)
2.4 DEPRESSÃO ALASTRANTE CORTICAL

21
A DAC foi descrita, pela primeira vez, em coelhos anestesiados como sendo uma
resposta propagável, de despolarização celular com redução (depressão) da atividade
elétrica do tecido cerebral cortical. (LEÃO, 1944). Essa resposta é consequência da
estimulação de um ponto do tecido, sendo totalmente reversível. A estimulação, por
sua vez, pode ser mecânica, química ou elétrica (LEÃO, 1944; GUEDES, 2005).
A atividade elétrica deprimida começa a se recuperar a partir do ponto onde se
originou e, ao final de cerca de 5 a 10 minutos, a região deprimida encontra-se
recuperada. Durante a DAC, simultaneamente à depressão da atividade elétrica
espontânea, ocorre uma variação lenta de voltagem (VLV) e o potencial do ponto
cortical começa a mudar, em relação a um ponto de potencial fixo, como o osso nasal,
e se torna progressivamente mais negativo. Essa mudança pode atingir valores que
variam de -5 a -30mV após 1 a 2 minutos de seu início, sendo revertido após alguns
minutos. (Figura 2) (LEÃO, 1944; GUEDES, 2011).
O termo depressão não está relacionado à doença psiquiátrica e sim a um
significado eletrofisiológico (redução da amplitude das ondas eletrocorticográficas).
No laboratório de Fisiologia da Nutrição Naíde Teodósio (LAFINNT), do Departamento
de Nutrição da UFPE, tem-se demonstrado que várias condições, de interesse para a
saúde humana, aumentam a suscetibilidade do cérebro para o fenômeno da DAC,
como é o caso da redução do cloreto extracelular (GUEDES e DO CARMO, 1980),
privação do sono (AMORIM et al., 1988), ingestão crônica de etanol (GUEDES e
FRADE, 1993; ABADIE-GUEDES et al., 2008; 2016), hipoglicemia (COSTA-CRUZ e
GUEDES, 2001), desnutrição (ROCHA-DE-MELO et al., 2006), hipertermia (FARIAS-
SANTOS et al., 2009),e tratamento com Arginina (FRAZÃO et al., 2008), com
glutamato mono-sódico (LIMA et al., 2014), com dexametasona (LOPES-DE-MORAIS
et al., 2014) e com Glutamina (LIMA et al., 2017). Por outro lado, outras condições
diminuem essa suscetibilidade, como por exemplo o tratamento dietético com lítio
(GUEDES et al., 1989), o hipotireoidismo (GUEDES e PEREIRA-DA-SILVA, 1993), o
envelhecimento (GUEDES et al., 1996), a estimulação ambiental (SANTOS–
MONTEIRO et al., 2000), a hiperglicemia (COSTA-CRUZ et al., 2006), o exercício
físico (BASTISTA-DE-OLIVEIRA, 2012).
Diferentes técnicas são usadas para determinar a atividade elétrica, entre elas o
eletroencefalograma (EEG), que é uma forma muito utilizada de se avaliar o registro
da atividade simultânea de milhares de neurônios localizados em uma região

22
específica cerebral. (GUEDES, 2005) É uma técnica empregada para diagnosticar
algumas doenças neurológicas em humanos e, em animais, pode ser realizada
através de eletrodos colocados sobre a dura-máter, via pequenos orifícios no crânio
com os animais sob efeito de anestesia (neste caso, chama-se eletrocorticograma;
EcoG). Assim, pode-se avaliar a atividade elétrica cortical (GUEDES, 2005).
Figura 2. Esquema da sequência de eventos da depressão alastrante cortical (DAC; adaptado
de Guedes, 2011). Em “a”, um córtex normal e nele o ponto estimulado (x), iniciando a DAC. Na
sequência, indicada pelas setas, “b” a “d”, a propagação concêntrica do fenômeno da DAC está
ilustrada. As áreas em branco representam porções do tecido cortical invadidas pelo fenômeno em
tempos sucessivos. As áreas quadriculadas, “c” a “f” indicam regiões que já sofreram a DAC e agora
estão se recuperando (áreas refratárias a uma nova estimulação). De “b” a “f”, observa-se que
propagação (área branca) e recuperação (área escura) dão-se de forma concêntrica, sendo o ponto
inicialmente estimulado o primeiro a se recuperar totalmente. Finalmente, todo o tecido se mostra
completamente recuperado, retornando à condição inicial em “a”. No centro da figura, um traçado de
registro demonstrando o eletrocorticograma (ECoG) e a variação lenta de voltagem (VLV), a qual
sempre aparece durante a DAC, quando o ECoG diminui sua amplitude. As letras “A” a “F”
correspondem a sequência das etapas representadas nos desenhos externos.

23
O fenômeno da DAC é utilizado há vários anos no Laboratório de Fisiologia da
Nutrição Naíde Teodósio (LAFINNT), como modelo experimental para avaliar a
excitabilidade cortical. Diversas situações clínicas também são estudadas,
principalmente as variáveis nutricionais e suas repercussões no sistema nervoso
(ROCHA-DE-MELO e GUEDES, 1997; BASTISTA-DE-OLIVEIRA, 2012). Vale
ressaltar que diversas funções cerebrais são consequência direta ou indireta da
atividade elétrica produzida pelas células do cérebro. O registro da atividade elétrica
cerebral espontânea, em humanos e animais de laboratório representa um meio
importante para o estudo do funcionamento cerebral. (GUEDES et al., 2012)
Os efeitos da cafeína são comumente investigados em diversos sistemas
fisiológicos, porém, no cérebro ainda não estão claramente definidos. (BLAHA et al.,
2007). Diante disto, no presente estudo, ratos machos da linhagem Wistar foram
submetidos, durante o período de desenvolvimento, a diferentes doses de cafeína
através de gavagem e os efeitos desse tratamento sobre funções neurais foram então
observados através de testes comportamentais e da DAC. Consideramos esta
proposta altamente pertinente, visto que irá estender e aprofundar o estudo inicial de
Aguiar e colaboradores (AGUIAR et al., 2011), analisando a interação da cafeína com
o estado nutricional e aspectos eletrofisiológicos em animais.

24
3. HIPÓTESES
A cafeína, administrada em diferentes doses ao longo do desenvolvimento,
alteraria o peso corporal dos animais.
O tratamento com cafeína resultaria em respostas comportamentais sugestivas
de ansiedade.
O tratamento sistêmico e tópico, aumentaria a susceptibilidade cortical ao
fenômeno eletrofisiológico da DAC.

25
4. OBJETIVOS
4.1 OBJETIVO GERAL
Investigar, no rato albino, a ação de diferentes doses de cafeína, aplicadas
durante o desenvolvimento do sistema nervoso, no que se refere aos seus efeitos
sobre o ganho de peso, o comportamento de ansiedade e a atividade elétrica cerebral.
4.2 OBJETIVOS ESPECÍFICOS
Analisar a evolução do peso corporal dos filhotes, como indicador do
desenvolvimento do organismo;
Avaliar o desempenho no labirinto em cruz elevado e no aparelho de campo
aberto, como indicador do comportamento de ansiedade e comportamento
exploratório, respectivamente.
Investigar a atividade elétrica cerebral através do fenômeno da DAC,
caracterizando seus parâmetros (velocidade de propagação, amplitude e
duração) em função das doses de cafeína empregadas.
Analisar em ratos jovens, a influência do tratamento tópico, no córtex cerebral,
com cafeína, sobre as características eletrofisiológicas da DAC, comparando
tais efeitos com aqueles do tratamento sistêmico.

26
5. MÉTODOS
O projeto foi desenvolvido no Laboratório de Fisiologia da Nutrição Naíde
Teodósio do Departamento de Nutrição da Universidade Federal de Pernambuco. Os
protocolos experimentais foram inicialmente submetidos à análise para aprovação da
Comissão de Ética no uso de animais da Universidade Federal de Pernambuco,
protocolo nº 23076.034601/2015-22 (Anexo B). As normas adotadas são sugeridas
pelo Colégio Brasileiro para Experimentação Animal e as normas estabelecidas pelo
National Institute of Health Guide for Care and Use of Laboratory Animals (Bethesda,
USA).
5.1 ANIMAIS
Foram utilizados ratos albinos Wistar machos (N=83) em ninhadas com 9 filhotes
por mãe, provenientes do Biotério do Departamento de Nutrição da Universidade
Federal de Pernambuco. Durante a gestação e lactação, as mães receberam dieta
padrão do biotério (“Presence”; Purina do Brasil Ltda., com 23% de proteína) e água,
ad libitum. O desmame foi realizado no 21º dia de vida, considerando o dia do
nascimento como 0 (zero; LIMA et al., 2009). Após o desmane, os filhotes também
foram alimentados com a dieta padrão do biotério. Os animais foram mantidos em sala
à temperatura de 22ºC±1ºC e em um ciclo claro-escuro de 12 horas (o escuro
iniciando-se as 18:00h).
5.2 TRATAMENTO SISTÊMICO COM CAFEÍNA
A cafeína foi obtida da Sigma Aldrich®. Do 7º ao 27º dia de vida pós-natal, três
grupos de filhotes de ratos Wistar (n=15 por grupo) receberam gavagem diária com
solução contendo respectivamente 15, 30 ou 45 mg de cafeína/Kg de peso
corporal/dia (grupos CAF15, CAF30 e CAF45, respectivamente). Esses três grupos
foram comparados a dois grupos em condições controle: uma dessas condições
consistiu na gavagem diária com água destilada, isto é, o veículo usado para diluir a
cafeína (grupo V; n= 15) e a outra condição foi aquela em que o animal não recebeu
qualquer gavagem (grupo ingênuo – I; n=15). O volume inicial administrado pela
gavagem foi de 0,5ml/dia na segunda e terceira semana de vida (correspondente a

27
primeira e segunda semana de tratamento) e 1.0ml/dia na quarta semana de vida
(terceira semana de tratamento), como já foi previamente descrito para outros
compostos (LIMA et al., 2009; FRANCISCO e GUEDES, 2015). O procedimento de
gavagem foi realizado sempre no mesmo horário, entre 12 e 14 horas, com auxílio de
uma cânula conectada a uma seringa de vidro, como descrito previamente (MAIA et
al, 2009).
5.3. TRATAMENTO TÓPICO COM CAFEÍNA
Um grupo de animais ingênuos (n=8), com idades entre 31-40 dias, foi tratado
durante o registro da DAC com aplicação tópica (T) no córtex cerebral, de cafeína
(concentração de 15mg/ml; grupo CT15). A aplicação de cafeína foi realizada após,
no mínimo, uma hora de registro basal da DAC, o qual era utilizado como controle,
como descrito previamente (AMANCIO-DOS-SANTOS et al., 2013)
5.4 DETERMINAÇÃO DO PESO CORPORAL
Os pesos corporais foram aferidos com auxílio de uma balança eletrônica da
marca Filizola (capacidade de 3,0 kg e escala em divisão de 0,5 g), aos 7, 14, 21, 28
dias de vida, e no dia do registro eletrofisiológico (35 dias).
Dias
07-27
28-30
35
Filhotes
Cafeína Controle
Divisão dos GruposDivisão dos Grupos
CAF 45CAF 30CAF 15
Gavagem
Testes
comportamentais
Registro da DAC
IV
Figura 3. Representação esquemática dos grupos experimentais e curso temporal dos
procedimentos a eles aplicados.

28
5.5 AVALIAÇÃO COMPORTAMENTAL
Os testes comportamentais foram filmados através de uma câmera de vídeo
instalada no teto, sobre os aparelhos comportamentais (labirinto em cruz elevado e o
campo aberto). Os vídeos foram analisados utilizando o software ANYmaze® (vesion
4.99m). Entre um teste e outro foi respeitado um período de 48 horas.
5.5.1 Avaliação comportamental no labirinto em cruz elevado (LCE)
O teste foi realizado colocando-se o animal em um labirinto em forma de cruz,
elevado do solo. O labirinto é formado por dois braços fechados e dois abertos (cada
um medindo 49cm de comprimento x 10cm de largura), com elevação de 55cm do
solo. Uma plataforma central (10x10cm de largura) conecta os braços abertos e
fechados. Os animais percorrem os dois tipos de braços, e são analisados a
frequência de entrada e o tempo gasto em cada tipo de braço, e outros tipos de
comportamento como deslocamento, levantar-se e esticar-se (FRANCISCO,
GUEDES, 2015). Segundo os estudiosos, existe uma relação inversa entre a
frequência de entrada e tempo gasto nos braços abertos, de um lado, e o grau de
ansiedade, do outro: quanto maiores os níveis de ansiedade, menor a porcentagem
de entrada nos braços abertos e de tempo gasto nos mesmos e vice-versa (HANDLEY
e MITHANI, 1984; PELLOW e FILE, 1986)
O procedimento foi realizado entre as 10:00 as 12:00h, utilizando a prole aos 28
dias de vida da prole. No dia do experimento, os ratos foram colocados na sala
experimental, individualmente, em gaiola apropriada e permaneceram 30 minutos
antes do início do teste, para adaptação ao novo ambiente. O animal foi colocado no
centro do aparelho cuidadosamente com o focinho voltado para um dos braços
abertos, sendo então permitida a livre exploração por 5 minutos. Após cada animal
ser testado, o labirinto foi limpo com uma mistura de álcool+água (70%+30%), para
eliminar pistas olfativas que pudessem interferir com o teste seguinte, respeitando-se
um intervalo de 1 minuto para que secasse completamente.
Posteriormente foram analisadas, nos vídeos gravados, as seguintes categorias
comportamentais:

29
a) Distância percorrida pelo animal no LCE;
b) Número de entradas nos braços abertos e fechados, considerando-se
uma entrada quando o animal entrava com a cabeça e o centro do corpo no
braço;
c) Tempo gasto em cada um dos braços;
d) Relação do tempo por entrada nos braços abertos.
e) Comportamento de levantar-se (“rearing”) e esticar-se (“stretched”)
5.5.2 Avaliação comportamental no teste de campo aberto
O teste do campo aberto consiste em se analisar, por vídeo, o comportamento
do animal numa arena circular (100 cm de diâmetro), envolvida com paredes de 52
cm de altura, de modo que o animal não possa fugir. O teste foi realizado entre as
10:00 as 12:00h, utilizando animais aos 30 dias de vida da prole. No dia do
experimento, os ratos foram colocados na sala experimental, individualmente, em
gaiola apropriada e permaneceram 5 minutos antes do início do teste, para adaptação
ao novo ambiente.
Após a adaptação, o animal foi colocado no centro do aparelho, onde também
foi permitida a livre exploração por 5 minutos. Após cada animal ter sido testado, o
aparelho foi limpo com mistura de álcool+água (70%+30%), respeitando um intervalo
de 1 minuto para que secasse completamente.
Nesse teste foram analisadas (posteriormente nos vídeos gravados) as seguintes
categorias comportamentais:
a) Distância percorrida pelo animal na arena;
b) Número de entradas na área central e na periferia;
c) Tempo gasto na área central e na periferia;
d) Relação do tempo por entrada na área central.
e) Comportamento de se levantar-se (“rearing”) e esticar-se (“stretched”)
5.6 REGISTRO DA DEPRESSÃO ALASTRANTE CORTICAL
O registro da DAC foi realizado quando os animais atingiram 35 dias de vida,
conforme descrito previamente (FRANCISCO e GUEDES, 2015). Inicialmente, sob

30
anestesia (uretana+cloralose; 1 g/kg + 40 mg/kg, respectivamente, injetados por via
intraperitoneal), foram trepanados três orifícios (2-4 mm de diâmetro) no crânio, ao
nível do hemisfério cerebral direito. Esses orifícios (um na região frontal, para deflagrar
a DAC e os outros dois na região parietal, para registrar a sua propagação) foram
alinhados no sentido ântero-posterior e paralelos à linha média. A DAC foi deflagrada
a intervalos de 20 min aplicando-se, por 1 min, uma bolinha de algodão (1-2 mm de
diâmetro) embebida em solução de KCl a 2% em um ponto do córtex frontal.
A variação lenta de voltagem (VLV) que acompanha a DAC foi registrada
continuamente por um período de 4 horas. Os dois eletrodos utilizados para o registro
e o eletrodo de referência comum (localizado no osso nasal) foram do tipo “prata-
cloreto de prata” de 3 cm de comprimento e 1 mm de diâmetro. Duas pipetas de
plástico (5cm de comprimento, diâmetro inferior de 1mm e superior de 5mm) já fixadas
entre si e com distância entre as suas pontas conhecida (variando entre 4 e 6 mm),
foram preenchidas com solução de Agar-Ringer a 0,5%, na qual os fios cloretados
foram imersos. Esse par de eletrodos foi fixado ao segurador de eletrodo do aparelho
estereotáxico. Dessa maneira, pelo giro de um parafuso, os eletrodos puderam ser
baixados e postos em contato com a superfície cortical de forma suave, evitando-se
pressão excessiva sobre a mesma.
A velocidade de propagação foi avaliada com base no tempo gasto pela DAC
para percorrer a distância entre os dois pontos de registro. Durante o registro, a
temperatura retal foi monitorada e mantida em 37 ± 1oC por meio de um aquecedor
elétrico regulável. Todas as informações sobre o animal foram anotadas em protocolo
individual.
5.7 APLICAÇÃO TÓPICA DE CAFEÍNA
Os animais desse grupo não receberam qualquer tratamento prévio; os mesmos
foram submetidos a aplicação tópica de cafeína durante o registro da DAC.
Previamente a esta aplicação, foi realizado, por um período de, no mínimo, uma hora,
registro eletrofisiológico da DAC (registro basal), com estimulação com KCl a cada 20
minutos. Durante esse período basal, a velocidade da DAC foi medida. Cerca de 10
minutos após a última medida do registro basal, a solução de cafeína (15mg/ml) foi
aplicada com o auxílio de uma seringa, sobre a superfície de um dos orifícios parietais,
usados para o registro eletrofisiológico. A cafeína permaneceu em contato com a

31
superfície cortical durante aproximadamente 10 minutos. Após esse tempo, o córtex
foi enxugado com um pedaço de algodão, e então, era realizada uma nova
estimulação com KCl para se obter outra medida de velocidade de propagação da
DAC. Essa aplicação tópica de cafeína foi repetida por três vezes a intervalos de 20
minutos. Em seguida, a região cortical tratada topicamente foi lavada com solução de
Ringer e registraram-se mais três episódios de DAC para evidenciar a recuperação
dos efeitos da cafeína sobre a DAC. Essa sequência ‘aplicação de cafeína por três
vezes mais recuperação por mais três episódios de DAC’, realizada inicialmente no
ponto de registro 1, foi em seguida repetida no ponto 2.
5.8 EUTANÁSIA
Ao final do registro eletrofisiológico, os animais, ainda anestesiados, foram
submetidos à eutanásia através da overdose de anestésico com subsequente
verificação da parada cárdio-respiratória.
5.9 ANÁLISES ESTATÍSTICAS
A análise estatística foi feita com o auxílio do programa ‘Sigmastat’(R) (versão
3.1). Para comparação intergrupos, os dados foram analisados utilizando-se
ANOVA, seguida, quando necessário, pelo teste de Holm-Sidak. Nas comparações
intragrupos (experimentos de aplicação tópica), utilizou-se o teste t para medidas
pareadas. Foram consideradas significativas as diferenças em que p<0,05.
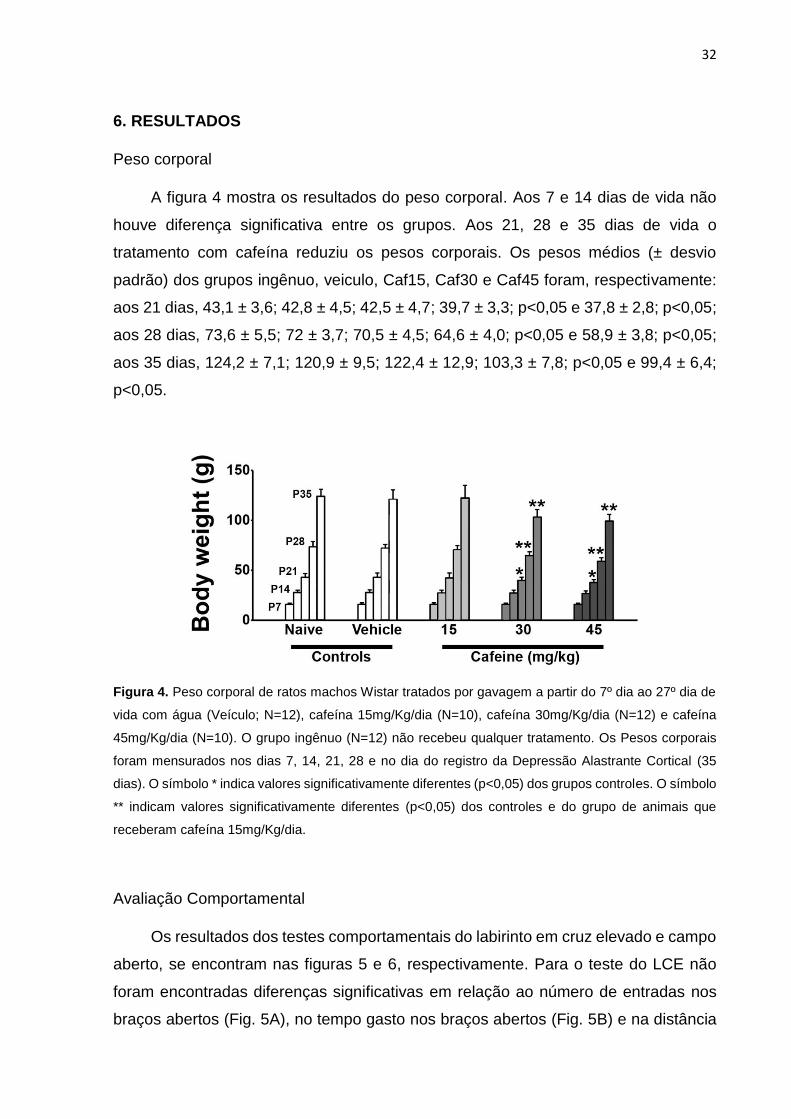
32
6. RESULTADOS
Peso corporal
A figura 4 mostra os resultados do peso corporal. Aos 7 e 14 dias de vida não
houve diferença significativa entre os grupos. Aos 21, 28 e 35 dias de vida o
tratamento com cafeína reduziu os pesos corporais. Os pesos médios (± desvio
padrão) dos grupos ingênuo, veiculo, Caf15, Caf30 e Caf45 foram, respectivamente:
aos 21 dias, 43,1 ± 3,6; 42,8 ± 4,5; 42,5 ± 4,7; 39,7 ± 3,3; p<0,05 e 37,8 ± 2,8; p<0,05;
aos 28 dias, 73,6 ± 5,5; 72 ± 3,7; 70,5 ± 4,5; 64,6 ± 4,0; p<0,05 e 58,9 ± 3,8; p<0,05;
aos 35 dias, 124,2 ± 7,1; 120,9 ± 9,5; 122,4 ± 12,9; 103,3 ± 7,8; p<0,05 e 99,4 ± 6,4;
p<0,05.
Figura 4. Peso corporal de ratos machos Wistar tratados por gavagem a partir do 7º dia ao 27º dia de
vida com água (Veículo; N=12), cafeína 15mg/Kg/dia (N=10), cafeína 30mg/Kg/dia (N=12) e cafeína
45mg/Kg/dia (N=10). O grupo ingênuo (N=12) não recebeu qualquer tratamento. Os Pesos corporais
foram mensurados nos dias 7, 14, 21, 28 e no dia do registro da Depressão Alastrante Cortical (35
dias). O símbolo * indica valores significativamente diferentes (p<0,05) dos grupos controles. O símbolo
** indicam valores significativamente diferentes (p<0,05) dos controles e do grupo de animais que
receberam cafeína 15mg/Kg/dia.
Avaliação Comportamental
Os resultados dos testes comportamentais do labirinto em cruz elevado e campo
aberto, se encontram nas figuras 5 e 6, respectivamente. Para o teste do LCE não
foram encontradas diferenças significativas em relação ao número de entradas nos
braços abertos (Fig. 5A), no tempo gasto nos braços abertos (Fig. 5B) e na distância

33
percorrida (Fig. 5D) entre os grupos tratados com cafeína e os grupos controles
ingênuo e veículo. As distâncias percorridas (em metros) dos grupos ingênuo, veículo,
Caf15, Caf30 e Caf45 foram, respectivamente, de: 10,98 ± 1,96; 10,47 ± 2,10; 12,61
± 2,35; 11,72 ± 2,44; e 11,41 ± 2,69. O número de entradas nos braços abertos, na
mesma sequência dos grupos acima, foi de 6,60 ± 2,46; 6,00 ± 2,45; 6,25 ± 1,75; 8,00
± 2,61 e 6,17 ± 3,06, respectivamente. O tempo que os grupos ingênuo, veículo,
Caf15, Caf30 e Caf45 permaneceram nos braços abertos foi de 30,4 ± 8,2; 29,2 ±
12,8; 36,5 ± 17,0; 41,0 ± 13,3 e 46,2 ± 18,4, respectivamente. A relação do tempo (em
segundos) por cada entrada (Fig 5C) aumentou de forma dose-dependente nos
animais tratados com cafeína com valores significativamente maiores no grupo CAF45
em comparação com o grupo ingênuo. Os valores dos grupos ingênuo, veículo, Caf15,
Caf30 e Caf45, foram de 4,7 ± 1,2; 5,0 ± 1,2; 6,2 ± 2,9; 6,5 ± 1,17 e 8,6 ± 2,59 (p<0,05),
respectivamente.
Figura 5. Atividade comportamental do teste de labirinto em cruz elevado dos animais com 28 dias de
vida. Os grupos foram submetidos ao tratamento por gavagem com água destilada (veículo), cafeína
(15mg/Kg/dia, 30mg/Kg/dia ou 45mg/Kg/dia) e grupo ingênuo (sem qualquer tratamento). As barras
representam os valores de média ± desvio padrão para 5 a 10 ratos por grupo. O símbolo * indica
valores significativamente diferentes (p<0,05).

34
No teste do campo aberto (Fig. 6) não foram encontradas diferenças
significativas no número de entradas no centro (Fig. 6A), no percentual de tempo no
centro (Fig. 6B) e na distância percorrida (Fig. 6D). Os valores do número de entrada
dos grupos ingênuo, veículo, CAF15, CAF30 e CAF45 4 foram de: 5,57 ± 2,23; 6,14 ±
2,19; 5,33 ± 2,58; 6,67± 2,94 e 4,43 ± 1,40, respectivamente. O tempo na mesma
sequência dos grupos acima, foi de: 13,73 ± 4,50; 19,76 ± 7;60; 15,30 ± 4,50; 19,20 ±
8,0 e 23,10 ± 12,80, respectivamente. As distâncias percorridas no campo aberto (em
metros) pelos grupos ingênuo, veículo, CAF15, CAF30 e CAF45 foram,
respectivamente, de: 23,61 ± 5,11; 20,10 ± 6,03; 22,92 ± 4,64; 26,36 ± 5,84; e 21,65
± 2,86. A relação do tempo (em segundos) por cada entrada (Fig. 6C) aumentou nos
animais tratados com cafeína e o grupo CAF45 apresentou valores significativamente
maiores em comparação com o grupo ingênuo. Os valores dos grupos ingênuo,
veículo, Caf15, Caf30 e Caf45, foram de 2,41 ± 0,48; 3,06 ± 0,34; 2,80 ± 1,20; 2,90 ±
0,50 e 5,50 ± 2,10 (p<0,05), respectivamente.
Figura 6. Atividade comportamental no campo aberto dos animais com 30 dias de vida. Os grupos
foram submetidos ao tratamento por gavagem a partir do 7 ao 27 dia de vida pós-natal, com água
destilada (veículo), cafeína (15mg/Kg/dia, 30mg/Kg/dia ou 45mg/Kg/dia) e grupo ingênuo (sem
qualquer tratamento). As barras representam os valores de média ± desvio padrão para 5 a 10 ratos
por grupo. O símbolo * indica valores significativamente diferentes (p<0,05).

35
No teste do LCE, os animais do grupo CAF45 diminuíram significativamente o
número de respostas do comportamento de levantar-se (“rearing) em comparação
com os grupos controles. No LCE o número do comportamento de levantar-se dos
grupos ingênuo, veículo, CAF15, CAF30 e CAF45 foram respectivamente, 10,6 ± 1,4;
10,8 ± 1,2; 9,5 ± 1,3; 8,8 ± 0,9; 6,9 ± 1,4. Em relação ao número do comportamento
de esticar-se (“stretched”) não houve diferenças significativas entre os grupos. O
número do comportamento de esticar-se (Figura 7) dos grupos ingênuo, veículo,
CAF15, CAF30 E CAF45 foram respectivamente, 1,9 ± 1,0; 1,8 ± 0,8; 1,7 ± 0,8; 1,4 ±
1,0; 1,2 ± 0,7 (p<0,05).
No campo aberto, os animais do grupo CAF45 diminuíram significativamente o
número de respostas do comportamento de levantar-se em comparação com os
grupos controles e CAF15. O número do comportamento de levantar-se dos grupos
ingênuo, veículo, CAF15, CAF30 e CAF45 foram respectivamente, 9,5 ± 1,4; 9,8 ± 1,4;
7,5 ± 1,2; 6,6 ± 1,4; 4,8 ± 0,9 (p<0,05). Em relação ao número do comportamento de
esticar-se (“stretched”) não houve diferenças significativas entre os grupos. O número
do comportamento de esticar-se (Figura 7) dos grupos ingênuo, veículo, CAF15,
CAF30 E CAF45 foram respectivamente, 2,1 ± 0,7; 1,9 ± 0,7; 2,0 ± 1,0; 2,0 ± 0,9; 1,6
± 0,7.
Figura 7. Números de respostas do comportamento de levantar-se (rearing) e esticar-se (stretched)
nos grupos com água destilada (veículo), cafeína (15mg/Kg/dia, 30mg/Kg/dia ou 45mg/Kg/dia) e grupo
ingênuo (sem qualquer tratamento) com 28 dias de idade (teste do labirinto em cruz elevado) e 30 dias
de idade (teste do campo aberto). As barras representam os valores de média ± desvio padrão para 5
a 10 ratos por grupo. O símbolo * indica valores significativamente diferentes (p<0,05).

36
Velocidade de propagação da depressão alastrante cortical
Como descrito na sessão de métodos, nos experimentos com a aplicação
sistêmica de cafeína, a DAC foi registrada continuamente durante 4 horas. O
fenômeno foi deflagrado a cada 20 minutos, aplicando-se uma bolinha de algodão (1-
2 mm de diâmetro) embebida em solução de KCl a 2% (aproximadamente 270 mM)
em um ponto da superfície cortical frontal durante 1 minuto. A propagação da DAC foi
registrada nos 2 pontos de registro, na região parietal. O aparecimento da variação
lenta de voltagem, que é típica da DAC, confirmou a presença do fenômeno após a
estimulação com KCl (Figura 8).
Figura 8. Registros da variação lenta de potencial (P) durante a depressão alastrante
cortical (DAC) em dois pontos corticais (1 e 2) em ratos machos, aos 35 dias de idade. O
diagrama do crânio mostra as posições 1 e 2 onde foram obtidos os traçados marcados
com esses mesmos números. A posição do eletrodo de referência (R) nos ossos nasais e
o ponto de aplicação do estímulo com KCl, que desencadeia a DAC, também são
mostrados. O grupo ingênuo não recebeu qualquer tipo de tratamento. Os grupos veículo,
cafeína 15mg/Kg/dia, cafeína 30mg/Kg/dia e cafeína 45mg/Kg/dia foram tratados por
gavagem dos 07 aos 27 dias de vida. As barras horizontais e verticais indicam,
respectivamente, 1 minuto e -10mV. As latências para a DAC se propagar entre os pontos
1 e 2 são indicadas pelas linhas tracejadas.
Observou-se diferenças significativas entre os grupos analisados. Os grupos
tratados com cafeína apresentaram aumento, dose-dependente, da velocidade de
propagação da DAC, em comparação com os grupos controles ingênuo, veículo e
Caf15. As velocidades médias ± DP (em mm/min), para os grupos ingênuo, veículo,
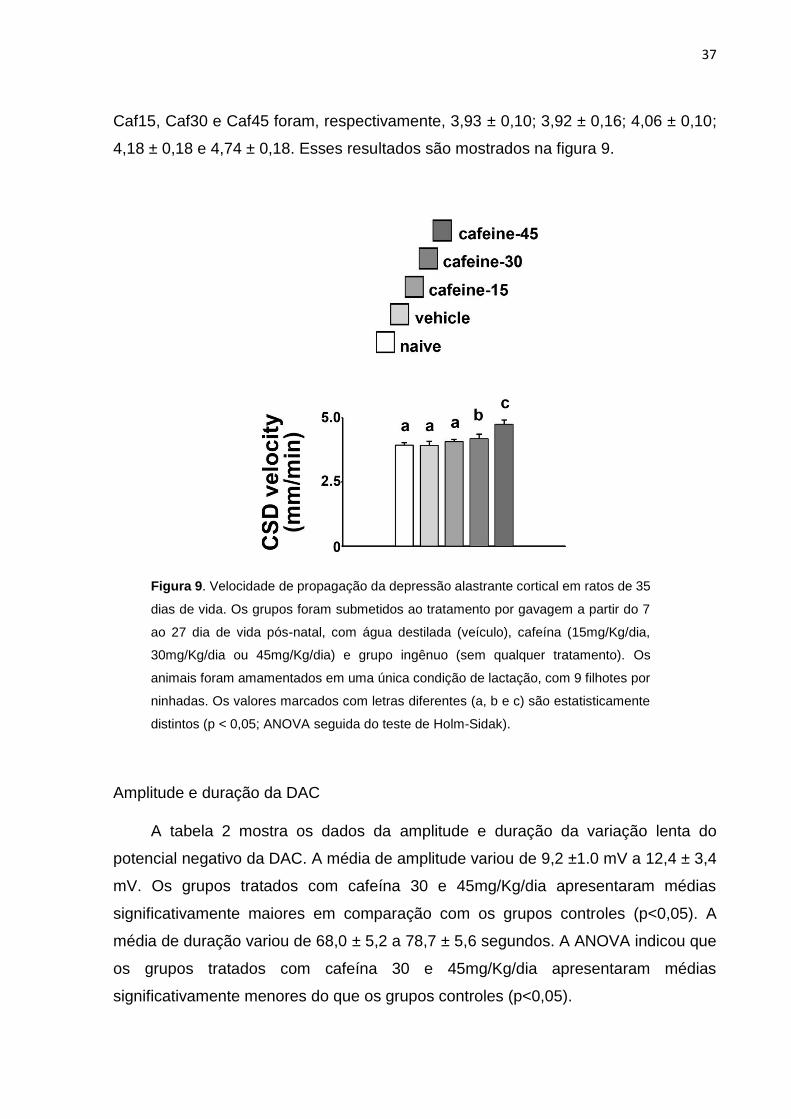
37
Caf15, Caf30 e Caf45 foram, respectivamente, 3,93 ± 0,10; 3,92 ± 0,16; 4,06 ± 0,10;
4,18 ± 0,18 e 4,74 ± 0,18. Esses resultados são mostrados na figura 9.
Figura 9. Velocidade de propagação da depressão alastrante cortical em ratos de 35
dias de vida. Os grupos foram submetidos ao tratamento por gavagem a partir do 7
ao 27 dia de vida pós-natal, com água destilada (veículo), cafeína (15mg/Kg/dia,
30mg/Kg/dia ou 45mg/Kg/dia) e grupo ingênuo (sem qualquer tratamento). Os
animais foram amamentados em uma única condição de lactação, com 9 filhotes por
ninhadas. Os valores marcados com letras diferentes (a, b e c) são estatisticamente
distintos (p < 0,05; ANOVA seguida do teste de Holm-Sidak).
Amplitude e duração da DAC
A tabela 2 mostra os dados da amplitude e duração da variação lenta do
potencial negativo da DAC. A média de amplitude variou de 9,2 ±1.0 mV a 12,4 ± 3,4
mV. Os grupos tratados com cafeína 30 e 45mg/Kg/dia apresentaram médias
significativamente maiores em comparação com os grupos controles (p<0,05). A
média de duração variou de 68,0 ± 5,2 a 78,7 ± 5,6 segundos. A ANOVA indicou que
os grupos tratados com cafeína 30 e 45mg/Kg/dia apresentaram médias
significativamente menores do que os grupos controles (p<0,05).

38
Tabela 2 – Amplitude e duração da variação lenta de potencial negativo da depressão alastrante
cortical (DAC) em ratos com 35 dias tratados dos 7-27 dia de vida com 15/mg/Kg/dia ou
30mg/Kg/dia ou 45mg/Kg/dia de cafeína (grupos Caf15, Caf30 3 Caf45, respectivamente). Esses
grupos foram comparados a dois grupos controles, tratados com veículo (água; grupo V) ou sem
qualquer tratamento (grupo ingênuo). Os dados estão expressos em média ± desvio padrão, com
o número de animais por grupo descrito entre parênteses. Os valores marcados com letras
diferentes são significativamente diferentes (p<0,05; ANOVA, seguido pelo teste de Holm-Sidak)
Group Amplitude (mV) Duration (s)
Naive 9.2 ± 1.0 (12)
a 77.5 ± 5.2 (11)
a
Vehicle 9.4 ± 1.0 (11)
a 78.7 ± 5.6 (11)
a
Caf15 9.3 ± 1.9 (14)
a 74.7 ± 5.4 (15)
a
Caf30 11.9 ± 2.6 (13)
b 69.7 ± 3.2 (13)
b
Caf45 12.4 ± 3.4 (12)
b 68.0 ± 5.2 (13)
b
Aplicação tópica de cafeína
A aplicação tópica de cafeína por três vezes, durante 10 minutos cada vez, a
intervalos de 20 minutos, na concentração de 15mg/ml, no córtex parietal (ponto de
registro 1), aumentou significativamente a velocidade de propagação da DAC para
4,71 ± 0,20 mm/min, quando comparada ao seu valor basal (antes da aplicação de
cafeína), que foi 4,26 ± 0,08 mm/min (p < 0,05). Em seguida, ao longo de uma hora,
a velocidade da DAC recuperou seu valor basal (media de 4,25 ± 0,06 mm/min). Uma
segunda aplicação de cafeína, agora no ponto de registro 2, com os mesmos
parâmetros descritos para o ponto 1, acelerou significativamente (p < 0,05) a DAC
(velocidade média de 4,70 ± 0,19 mm/min), a qual, após um período de recuperação
de uma hora, voltou a um valor médio de 4,25 ± 0,07 mm/min (Figura 10-A).
A aplicação tópica de cafeína no ponto de registro 1, conforme descrito no
parágrafo anterior, diminuiu significativamente a duração da variação lenta de

39
potencial negativo da DAC, que passou de um valor basal (em segundos) de 75,65 ±
4,42 para 65,90 ± 4,39 (p < 0,05). Em seguida, ao longo de uma hora de recuperação,
a duração aumentou para 73,23 ± 3,95. Uma segunda aplicação de cafeína, agora no
ponto de registro 2, reduziu a duração da DAC para 67,42 ± 4,08, a qual, após mais
uma hora de recuperação, aumentou para 73,23 ± 3,59 (p < 0,05; Figura 10-B).
A aplicação tópica de cafeína no ponto de registro 1 aumentou significativamente
a amplitude da variação lenta de potencial negativo da DAC, que passou de um valor
basal de 8,35 ± 1,56 mV para 10,18 ± 1,8 mV (p < 0,05). Em seguida, ao longo de
uma hora de recuperação, a amplitude decresceu para 8,53 ± 1,64 mV. Uma segunda
aplicação de cafeína, agora no ponto de registro 2, elevou a amplitude para 10,67 ±
1,74 mV, a qual, após mais uma hora de recuperação, diminuiu para 8,51 ± 1,30 mV
(p < 0,05; Figura 10-C).
Figura 10 – Velocidade (A), duração (B) e amplitude (C) das ondas de Depressão Alastrante Cortical,
em ratos jovens (31-40 dias) submetidos à aplicação tópica de cafeína (15mg/ml; n=8). A solução foi
aplicada topicamente durante 10 minutos imediatamente antes da deflagração da DAC com KCl na
superfície do córtex parietal. Basal, indica as ondas iniciais, antes da aplicação tópica com cafeína;
topic-1, aplicação tópica com cafeína no ponto de registro 1; recov-1, recuperação dos efeitos da
cafeína em 1; topic-2, aplicação tópica com cafeína no ponto de registro 2; recov-2, recuperação dos
efeitos da cafeína em 2. O símbolo * indica diferença significativa em relação ao valor basal (p<0,05;
teste t pareado).

40
7. DISCUSSÃO
Nossos dados demostraram que o tratamento crônico com cafeína, em suas
maiores doses, resultou em redução do ganho de peso corporal. O tratamento
sistêmico com cafeína na dose de 15mg/Kg/dia não alterou significantemente a
velocidade de propagação da DAC e as doses maiores de 30 e 45mg/Kg/dia
aumentaram significantemente essa velocidade. Nos animais tratados de forma tópica
com a concentração de 15mg/ml, foi detectado aumento da velocidade de propagação
do fenômeno. Sobre as avaliações comportamentais, houve efeito sugestivo de
redução da ansiedade nos grupos tratados com cafeína, somente os que receberam
a dose de 45mg/Kg/dia; estes apresentaram valores significativamente maiores em
relação ao grupo ingênuo. Nos testes do LCE e campo aberto, os animais do grupo
CAF45 diminuíram significativamente o número de respostas do comportamento de
levantar-se (“rearing) em comparação com os grupos controles, demonstrando um
possível efeito ansiolítico da cafeína. Embora represente um estresse para o animal,
o procedimento de gavagem não deve ser a causa das alterações ponderais e
eletrofisiológicas descritas neste estudo, visto que os animais do grupo veículo
também foram submetidos à gavagem durante o mesmo período e no entanto
apresentaram pesos e velocidades semelhantes ao grupo ingênuo, o qual não
recebeu qualquer tratamento.
Diferentemente dos nossos resultados, O´Neill et al. (2016) expuseram ratos
dos 28 aos 55 dias de vida ao consumo de cafeína em água (0,3g/L) e demonstraram
que não houve diferença significativa no peso corporal dos animais em relação ao
grupo controle. Ardais et al. (2014) também expuseram animais dos 28 aos 48 dias
de vida ao consumo de cafeína dissolvida em água, durante o período noturno
(período ativo dos animais); as doses recebidas pelos animais eram de 0,1mg/L,
0,3mg/L ou 1,0mg/L e não foram encontradas diferenças significativas no peso
corporal. Essa discrepância pode ser explicada pelo consumo de cafeína na água,
pois seu sabor amargo pode inibir o consumo dos animais (HEPPNER et al., 1986).
Outro fator que poderia explicar as diferenças entre nossos resultados e os dois
estudos é representado pelas diferentes fases do desenvolvimento em que os animais
se encontravam no período do tratamento com cafeína (HEPPNER et al., 1986).
Zhao et al. (2016) submeteram ratos de 4 semanas de idade ao consumo de
dietas contendo cafeína (0,03% e 0,06%) e catequinas (0,3% e 0,6%) isoladas e

41
combinadas, durante 24 semanas. Após esse período foi verificado diminuição do
peso corporal dos animais, além da redução dos níveis séricos de triglicerídeos e
colesterol em relação ao grupo controle. Ao comparar os grupos de tratamento isolado
e combinados, foi observado que tanto a cafeína como as catequinas apresentam
atividades anti-obesidade e podem ativar a enzima proteína-quinase ativada por AMP
(AMPK). Esta enzima está envolvida por sua vez em várias vias metabólicas, com um
importante papel na regulação da homeostase, além de estimular rapidamente o
metabolismo dos lipídeos (LEE et al., 2012; SANDERS et al., 2007).
A redução no ganho de peso corporal dos animais do nosso estudo pode ser
consequência do equilíbrio energético, pois a cafeína pode reduzir a ingestão
energética e aumentar o gasto energético (HARPAZ et al., 2017). A cafeína parece
ainda potencializar o efeito termogênico e aumentar a oxidação lipídica através da
enzima fosfodiesterase. (BELZA et al., 2008). No entanto, a relação entre a cafeína,
ingestão alimentar e perda de peso não está totalmente esclarecida (SWEENEY et
al., 2016). Pettenuzzo et al. (2008) demonstraram que o consumo crônico de cafeína
em ratos não altera o consumo de alimentos, ao mesmo tempo que outros estudos
demonstram que a cafeína diminui o consumo de alimentos e consequentemente leva
a uma redução do peso corporal. (GAVRIELI et al., 2013; RACOTTA et al., 1994;
SCHUBERT et al., 2014).
Dados da literatura, em humanos, demonstraram que a administração aguda
de cafeína pode exercer um efeito anoréxico (TREMBLAY et al., 1988), e a
reintrodução de cafeína após um período de abstinência pode reduzir a ingestão diária
de energia, diminuindo assim, o número de refeições (RACOTTA et al., 1994; COMER
et al., 1997; JESSEN et al., 2005). Por outro lado, estudos demonstram que a cafeína
pode aumentar a entrada sináptica excitatória para neurônios do peptídeo relacionado
a Agouti (AgRP) (YANG et al., 2011), que quando ativado pode aumentar o
comportamento alimentar e o apetite (APONTE et al., 2011; STERNSON e ATASOY,
2014). A cafeína também pode aumentar os níveis de dopamina e então ativar
neurônios dopaminérgicos no núcleo accumbens e esta região cerebral está
relacionada com o comportamento alimentar (O'CONNOR et al., 2015; RETZBACH et
al., 2014; SOLINAS et al., 2002), ou seja, a ingestão de cafeína poderia aumentar a
ingestão alimentar.

42
Vale ressaltar que alguns estudos não avaliam a ingestão dietética dos
indivíduos durante o período da pesquisa, além de acompanhar os indivíduos durante
um curto período de tempo, dificultando a análise sob a real relação de perda de peso
e consumo de cafeína (FORTE e SCHNEIDER, 2012). Essas discrepâncias podem
ser devidas às diferentes doses consumidas e distintas vias de administração
empregadas nos diversos estudos (SWEENEY et al, 2016). Além disso, o controle da
alimentação pode ser alterado por diferentes fatores, como nutrientes disponíveis e
estresse (SILVEIRA et al., 2004).
Estudos tem demonstrado que o consumo moderado de cafeína produz efeito
analgésico e sensação de bem-estar (GUNJA e BROWN, 2012; CANO-MARQUINA
et al., 2013; MI-JOO et al., 2016). Os benefícios da cafeína em adultos têm sido
associados ao antagonismo dos receptores de adenosina A2A (CUNHA e
AGOSTINHO, 2010) e mais recentemente com o controle da função astrocítica
(MATOS et al., 2015). Por outro lado, altas doses podem levar a taquicardia, insônia,
depressão e aumento do risco de ansiedade, provavelmente devido à ação da cafeína
sobre o receptor adenosinérgico tipo A1 (TEMPLE, 2011; RANDALL et al., 2011.
GUEDES et al., 2012; O’NEILL et al., 2016).
Em nosso estudo foram observadas diferenças significativas nos testes
comportamentais do campo aberto e labirinto em cruz elevado quando avaliado o
tempo por entrada no braço aberto, no grupo que recebeu cafeína 45mg/Kg/dia em
relação ao grupo ingênuo e no número do comportamento de levantar-se também do
grupo que recebeu a maior dose de cafeína. Por outro lado, a administração crônica
de cafeína tem sido relatada como ansiogênica (BHATTACHARYA et al, 1997),
ansiolítica (PECHLIVANOVA et al., 2012) ou sem efeitos no comportamento de
ansiedade (FILE et al., 1988).
O’Neill et al. (2016) observaram que animais que consumiram cafeína (0,3g/L)
na água, entre 28 e 55 dias de vida, tiveram um aumento da ansiedade em testes
comportamentais, incluindo labirinto em cruz elevado e campo aberto, resultados que
também foram encontrados em outros estudos, tanto no campo aberto (NOSCHANG
et al., 2009) quanto no labirinto em cruz elevado (EL YACOUBI et al., 2000). A
administração de cafeína pode exercer efeitos ansiolíticos ou ansiogênicos, porém
tais efeitos dependem de alguns fatores como o teste de ansiedade utilizado e do sexo
dos indivíduos (HUGHES e HANCOCK, 2014). Nesse contexto, Noschang et al.

43
(2009) observaram que o consumo crônico de cafeína na água, aumentou a
ansiedade de ratos machos, porém, tal efeito não foi observado nas fêmeas. Em
contrapartida, efeitos benéficos do consumo crônico de cafeína sobre a memória têm
sido observados em mulheres, mas não em homens (JOHNSON-KOZLOW et al.,
2002; RITCHIE et al., 2007), da mesma maneira esses resultados também são
encontrados em animais (FISHER e GUILLET, 1997).
Alguns estudos demonstram que os efeitos da cafeína durante o
desenvolvimento do cérebro podem levar a déficits duradouros na função cerebral
(CONCANNON et al., 1983). Nesse contexto, animais que receberam cafeína durante
o período de gestação e lactação através de suas mães, apresentaram atividade
locomotora reduzida (CONCANNON et al., 1983; PERUZZI et al., 1985), perfil
ansiogênico (HUGHES et al, 2014) e comprometimento do desempenho em tarefas
de aprendizagem e memória (SILVA et al., 2013; SINTON et al., 1981; SOELLNER et
al., 2009.) Uma interpretação alternativa do efeito ansiolítico da ingestão de cafeína
poderia envolver a ação da cafeína sobre os receptores de adenosina e dopamina
(Cunha et al., 2008). Isto também foi sugerido para o comportamento em ratos
(Randall et al., 2011). Apesar de todas as evidências, uma investigação adicional é
necessária para esclarecer os mecanismos subjacentes à ação ansiolítica da cafeína.
Parte substancial do presente estudo foi dirigido especialmente à avaliação das
alterações eletrofisiológicas após o tratamento sistêmico e tópico com cafeína durante
o período crítico de desenvolvimento no sistema nervoso. Nossos resultados
mostraram que a cafeína facilita a propagação da DAC, tanto em relação ao
tratamento sistêmico quanto tópico. Duas possibilidades podem explicar o efeito da
administração crônica de cafeína na Depressão Alastrante Cortical observada no
presente estudo. A primeira possibilidade diz respeito à ação direta da cafeína sobre
o tecido cortical. A segunda envolve à ação indireta, através da participação de
metabolitos de cafeína ou um ou mais sistemas de neurotransmissores sensíveis à
cafeína, como por exemplo o sistema de óxido nítrico (FERRÉ et al., 2013).
Independente da ação da cafeína na DAC, seja ela direta ou indireta, vários
mecanismos podem estar envolvidos, como por exemplo a modulação do equilíbrio
redox (BARCELOS et al., 2014a, DI GIROLAMO et al., 2016). Mesmo que não
tenhamos medido o estado redox do cérebro, a cafeína pode ter influenciado o
equilíbrio redox cerebral, como foi sugerido para o fígado de ratos (BARCELOS et al.,

44
2014a) e plasma humano (DI GIROLAMO et al., 2016). No entanto, no momento não
podemos descartar a participação de outros mecanismos como a ação da cafeína em
marcadores de inflamação (BARCELOS et al., 2014b) e em aminoácidos (JORDÁ et
al., 1989).
Considerando que a cafeína influencia a atividade elétrica cerebral
(TCHEKALAROVA et al., 2010), alguns estudos avaliaram tal efeito, porém,
diferentemente dos nossos achados não encontraram diferenças significativas. Por
exemplo, Aguiar et al., (2011), administraram 30mg/Kg de cafeína de forma aguda
(por apenas 4 dias), via intraperitoneal durante o registro da DAC, em ratos Wistar
com 75-85 dias de vida. Nenhuma diferença foi encontrada na velocidade de
propagação da DAC, antes e depois da administração da cafeína. É importante
ressaltar que os efeitos da cafeína dependem da dose, com a eficácia variando entre
10 a 40mg/Kg. Nosso estudo utilizou várias doses e uma delas superior aos estudos
acima, assim como a via de administração diferente e um tempo maior de
administração da droga (21 dias). Essas discrepâncias metodológicas podem explicar
as diferenças nos resultados dos diversos autores.
Um mecanismo que provavelmente está envolvido nos efeitos cerebrais
provocados pela cafeína é a interação entre essa droga e os receptores de Adenosina
A1 e dopamina D2, pois a mesma aumenta a liberação de vários neurotransmissores
como noradrenalina e ácido γ-amino-butírico (GABA) (GOLDBERG et al., 1985; DALY,
2007; GUEDES et al., 2012). Os efeitos associados a doses baixas de cafeína estão
relacionados a um efeito semelhante ao de um antagonista seletivo de receptor A2A
(SVENNINGSSON e FREDHOLM, 1997; SVENNINGSSON et al.,1997).
Vale ressaltar que a modulação da depressão alastrante cortical por fatores que
também modulam o comportamento (Lima et al., 2017), sugere uma relação entre a
atividade comportamental e o fenômeno eletrofisiológico. Além disso, o fenômeno da
DAC parece ser relevante para doenças neurológicas importantes (SCHRAEGLE e
TITUS, 2017), onde podem ocorrer alterações comportamentais (GORJI, 2001;
TORRENTE et al., 2014, KRAMER et al., 2016). Futuras experiências são necessárias
para esclarecer ainda mais os mecanismos subjacentes da ação comportamental e
eletrofisiológica da cafeína no cérebro.

45
Nossos resultados sugerem que o tratamento com cafeína, em sua maior dose,
durante o período de desenvolvimento cerebral, causa redução do ganho do peso
corporal, acelera a velocidade de propagação da DAC e reduz o efeito de ansiedade.
Por fim deve-se levar em consideração que extrapolar para seres humanos os dados
encontrados nesse estudo é algo que requer cuidado, assim como a dose de cafeína
consumida, visto que, em excesso, essa substância pode ter efeitos não desejáveis.

46
8. CONSIDERAÇÕES FINAIS
Com base nos resultados dessa dissertação, podemos concluir que:
O tratamento sistêmico com cafeína, reduziu o ganho de peso corporal dos
animais, de forma proporcional às doses empregadas e aumentou a velocidade
de propagação da DAC. Tais efeitos não devem ser atribuídos ao procedimento
de gavagem, visto que, os animais do grupo veículo também foram submetidos
ao procedimento de gavagem e apresentam médias semelhantes ao grupo
ingênuo.
O tratamento tópico com cafeína, também alterou os parâmetros da DAC
(aumentou velocidade de propagação e amplitude, e reduziu sua duração),
refletindo um resultado novo para a ação eletrofisiológica da cafeína.
Nossos resultados sugerem um efeito ansiolítico da cafeína, o qual precisa ser
mais estudado para confirmação.
Este estudo é considerado pioneiro no que diz respeito à ação eletrofisiológica
da cafeína, e portanto mais estudos são necessários para entender melhor a
relação entre cafeína e excitabilidade cerebral.
Nossos dados permitem que sugiramos cautela no consumo prolongado de
cafeína por parte de gestantes, lactantes e crianças nos primeiros anos de vida,
pois tal consumo pode levar a efeitos neurais não desejáveis.
Os resultados obtidos nesse trabalho demonstram a influência da cafeína no
fenômeno da DAC. No entanto, as complexas interações entre cafeína, peso corporal,
ansiedade e sistema nervoso necessitam de mais investigações. Então, visando dar
continuidade nesse trabalho, sugere-se como perspectivas:

47
Investigar os efeitos do tratamento crônico com cafeína durante o período
crítico de desenvolvimento sobre a memória (por meio de testes
comportamentais específicos, como o labirinto aquático de Morris);
Comparar os efeitos do tratamento com cafeína em diferentes períodos de
desenvolvimento e gêneros;
Analisar parâmetros bioquímicos dos animais tratados com cafeína a longo
prazo;
Comparar a ação do tratamento agudo e crônico da cafeína sobre efeitos
eletrofisiológicos;
Em modelos animais de consumo crônico de cafeína, estudar o impacto da sua
abstinência sobre a depressão alastrante cortical.
Investigar os efeitos da cafeína em condições muito favoráveis, ou
desfavoráveis de lactação, representadas respectivamente pelo aleitamento
em ninhadas com número reduzido, ou excessivo de filhotes.

48
REFERÊNCIAS ABADIE-GUEDES R.; SANTOS S.D.; CAHÚ T.B.; GUEDES R.C.A.; BEZERRA R.S.
Dose-dependent effects of astaxanthin on cortical spreading depression in chronically ethanol-treated adult rats. Alcoholism: Clinical and Experimental Research, v.32, n.8, p.1417-1421, 2008.
ABADIE-GUEDES, R.; BEZERRA, R.S.; GUEDES, R.C.A. Alpha-tocopherol
counteracts spreading depression in rats of various ages, with and without ethanol abstinence. Alcoholism: Clinical and Experimental Research, v. 40, p. 728-733, 2016.
AGUIAR, M.J.L.; AGUIAR, C.R.A.; GUEDES, R.C.A. Caffeine/Nutrition interaction in
the rat brain: Influence on latent inhibition and cortical spreading depression. European Journal of Pharmacology, v. 650, p. 268-274, 2011.
ALMEIDA, S.S.; DUNTAS, L.H.; DYE, L.; NUNES, M.L.; PRASAD, C.; ROCHA,
J.B.T.; WAINWRIGHT, P.; ZAIA, C.T.B.V.; GUEDES, R.C.A. Nutrition and Brain Function: A Multidisciplinary Virtual Symposium. Nutritional Neuroscience, v. 5 (5), p. 311-320, 2002.
AMORIM, L.F.; GUEDES, R.C.A.; MEDEIROS, M.C.; SILVA, A.T.; CABRAL-FILHO,
J.E. Apomorphine does not mimic the effects of REM-sleep deprivation on cortical spreading depression. Brazilian Journal of Medical and Biological Research, v. 21, p. 611-614, 1988.
APONTE, Y.; ATASOY, D.; STERNSON, S. M. AGRP. Neurons are sufficient to
orchestrate feeding behavior rapidly and without training. Nature Neuroscience, v. 14, p. 351- 355, 2011.
ARDAIS A.P.; ROCHA A.S..; BORGES M.F.; FIOREZE G.T.; SALLABERRY C.;
MIORANZZA S.; NUNES F.; PAGNUSSAT N.; BOTTON P.H.S.; CUNHA R.A.; LISIANE DE OLIVEIRA PORCIÚNCULA L.O. Caffeine exposure during rat brain development causes memory impairment in a sex selective manner that is offset by caffeine consumption throughout life. Behavioural Brain Research, v. 303, p. 76-84, 2016.
ARDAIS, A.P.; BORGES, M.F.; ROCHA, A.S.; SALLABERRY, C.; CUNHA R.A.;
PORCIÚNCULA, L.O. Caffeine triggers behavioral and neurochemical alterations in adolescent rats. Neuroscience, v. 270, p. 27–39, 2014.
ARENDASH, G.W.; SCHLEIF, W.; REZAI-ZADEH, K.; JACKSON, E.K.; ZACHARIA,
L.C.; CRACCHIOLO, J.R.; SHIPPY, D.; TAN, J. Caffeine protects Alzheimer’s mice against cognitive impairment and reduces brain beta-amyloid production. Neuroscience, v. 142, p. 941–952, 2006.
ARMARIO, A.; DAVIU, N.; MUNOZ-ABELLAN, C.; RABASA, C.; FUENTES, S.;
BELDA, X.; GAGLIANO, H.; NADAL, R. What can we know from pituitary-adrenal

49
hormones about the nature and consequences of exposure to emotional stressors?. Cellular Molecular Neurobiology, v. 32, p. 749-758, 2012.
ASTRUP, A.; TOUBRO, S.; CANNON, S.; HEIN, P.; MADSEN, J. Thermogenic
synergism between ephedrine and caffeine in healthy volunteers: a double-blind, placebo-controlled study. Metabolism Clinical and Experimental, v. 40, p. 323-329, 1991.
BARCELOS, R.P.; SOUZA, M.A.; AMARAL, G.P.; STEFANELLO, S.T.; BRESCIANI,
G.; FIGHERA, M.R.; SOARES, F.A.A.; BARBOSA, N.V. Caffeine supplementation modulates oxidative stress markers in the liver of trained rats. Life Sciences, v. 96, p. 40–45, 2014a.
BARCELOS, R.P.; SOUZA, M.A.; AMARAL, G.P.; STEFANELLO, S.T.; BRESCIANI,
G.; FIGHERA, M.R.; SOARES, F.A.A.; BARBOSA, N.V. Caffeine intake may modulate inflammation markers in trained rats. Nutrients, v. 6, p. 1678-1690, 2014b.
BATISTA-DE-OLIVEIRA, M.; LOPES, A.A.C.; MENDES-DA-SILVA, R.F.; GUEDES,
R.C.A. Aging-dependent brain electrophysiological effects in rats after distinct lactation conditions, and treadmill exercise: A spreading depression analysis. Experimental Gerontology, v. 47, p.452-457, 2012.
BELZA, A.; FRANDSEN, E.; KONDRUP, J. Body fat loss achieved by stimulation of
thermogenesis by a combination of bioactive food ingredients: a placebo-controlled, double-blind 8-week intervention in obese subjects. International Journal Obesity, v. 31, p. 121-130, 2008.
BHATTACHARYA, S.K.; SATYAN, K.S.; CHAKRABARTI, A. Anxiogenic action of
caffeine: an experimental study in rats. Journal Psychopharmacology, v. 11, p. 219–224, 1997.
BIDEL, S.; HU, G.; JOUSILAHTI, P.; ANTIKAINEN, R.; PUKKALA, E.; HAKULINEN,
T.; TUOMILEHTO, J. Coffee consumption and risk of colorectal cancer. European Journal of Clinical Nutrition, v. 64, p. 917–923, 2010.
BLAHA, M.; BENES V.; DOUVILLE C.M.; NEWELL D.W. The effect of caffeine on
dilated cerebral circulation and on diagnostic CO2 reactivity testing. Journal of Clinical Neuroscience, v. 14, p. 464-467, 2007.
BLOMS, L.P. FITZGERALD JS, SHORT MW, WHITEHEAD JR. The effects of
caffeine on vertical jump height an execution in collegiate athletes. Journal Strenght Conditions Research, v. 30, p. 1855-1861, 2016.
BOETTLER, U.; SOMMERFELD, K.; VOLZ, N.; PAHLKE, G.; TELLER, N.;
SOMOZA, V.; LANG, R.; HOFMANN, T.; MARKO, D. Coffee constituents as modulators of Nrf2 nuclear translocation and ARE (EpRE)-dependent gene expression. Journal of Nutritional Biochemistry, v. 22, p. 426–440, 2011.

50
BRACCO, D.; FERRARRA, J.M.; ARNAUD, M.J.; JEQUIER, E.; SCHUTZ, Y. Effects of caffeine on energy metabolism, heart rate, and methylxanthine metabolism in lean and obese women. American Journal of Physiology, v. 269, p. 671–8, 1995.
CANO-MARQUINA, A.; TARÍN, J.J.; CANO, A. The impact of coffee on health.
Maturitas, v. 75, p. 7-21, 2013. ÇAKIR, O.K.; ELLEK, N.; SALEHIN, N.; HAMAMCI, R.; KELEŞ, H.; KAYALI, D.G.;
AKAKIN, D.; YÜKSEL, M.; ÖZBEYLI, D. Protective effect of low dose caffeine on psychological stress and cognitive function. Physiology and Behavior, v. 168, p. 1–10, 2017.
CARRILLO, J.A.; BENITEZ, J. Clinically significant pharmacokinetic interactions
between dietary caffeine and medications. Clinical Pharmacokinetics, v. 39, p. 127-153, 2000.
CARROLL, M.E.; LYNCH, W.J.; ROTH, M.E.; MORGAN, A.D.; COSGROVE, K.P.
Sex and estrogen influence drug abuse. Trends in Pharmacological Science, v. 25, p. 273–279, 2004.
CHEN, J. F.; YU L.; SHEN H.Y.; HE J.C.; WANG X.; ZHENG R. What knock-out
animals tell us about the effects of caffeine. Journal of Alzheimer’s Diseases, v. 20, p. S17-S24, 2010.
COELHO, C.F.; CORRENTE, J.E.; MICHELIN, E.; RAVAGNANI, F. C. P.; PEREIRA,
A.F.; BURINI, R.C. Impacto de um programa de intervenção para mudança do estilo de vida sobre indicadores de aptidão física, obesidade e ingestão alimentar de indivíduos adultos. Revista Brasileira de Atividade Física e Saúde, v. 15, p. 21-27, 2010.
COMER, S.D.; HANEY, M.; FOLTIN, R.W.; FISCHMAN. Effects of caffeine
withdrawal on humans living in a residential laboratory. Experimental Clinical Psychopharmacology, v. 5, p. 399-403, 1997.
CONCANNON, J.; BRAUGHLER, J.M.; SCHECHTER, M.D. Pre- and postnatal
effects of caffeine on brain biogenic amines, cyclic nucleotides and behavior in developing rats, Journal Pharmacology Experimental Therapeutics, n. 226, p. 673–679, 1983.
COSTA-CRUZ, R.R.G.; AMÂNCIO-DOS-SANTOS, A.; GUEDES, R.C.A.
Characterization of cortical spreading depression in adult well-nourished and malnourished rats submitted to the association of pilocarpine-induced epilepsy plus streptozotocin-induced hyperglycemia. Neuroscience Letters, v. 401, p. 271-275, 2006.
COSTA-CRUZ, R.R.G.; GUEDES, R.C. A. Cortical spreading depression during
streptozotocin-induced hyperglycemia in nutritionally normal and early-malnourished rats. Neuroscience Letters, v. 303, p. 177-180, 2001.

51
CRUZ, A.P.M.; LANDEIRA-FERNANDEZ, J. Modelos animais de ansiedade e o estudo experimental de Drogas serotonérgicas. In: LANDEIRA-FERNANDEZ, J.; FUKUSIMA, S. (org). Métodos em Neurociência. São Paulo: Manole, p. 192-217, 2012.
CUNHA, R.A.; AGOSTINHO, P.M. Chronic caffeine consumption prevents memory
disturbance in different animal models of memory decline, Journal Alzheimers Diseases, v. 20, p. 95–116, 2010.
DALL’IGNA, O.P.; FETT, P.; GOMES, M.W.; SOUZA, D.O.; CUNHA, R.A.; LARA,
D.R. Caffeine and adenosine A(2a) receptor antagonists prevent beta-amyloid (25–35)-induced cognitive deficits in mice, Experimental Neurology, n. 203, p. 241–245, 2007.
DALY, J.M. Caffeine analogs: biomedical impact. Cellular and Molecular Life
Science, v. 62, p. 2153-2169, 2007. DAVIS, J.K.; GREEN, J.M. Caffeine and anaerobic performance: ergogenic value
and mechanisms of action. Sports Medicine, v. 39, p. 813-832, 2009. DI GIROLAMO, F.G.; MAZZUCCO, S.; SITULIN, R.; MOHORKO, N.; JENKO-
PRAZNIKAR, Z.; PETELIN, A.; TENCE, M.; PISOT, R.; NAVARINI, L.; BIOLO, G. Roasting intensity of naturally low-caffeine Laurina coffee modulates glucose metabolism and redox balance in humans. Nutrition, v. 32, p. 928–936, 2016.
DIEPVENS, K.; WESTERTERP, K.R.; WESTERTERP-PLANTENGA, M.S. Obesity
and thermogenesis related to the consumption of caffeine, ephedrine, capsaicin, and green tea. American Journal Physiology Regulatory Integrative Comparative Physiology, v. 292, p. 77–85, 2007.
DOBBING, J. Vulnerable periods in developing brain. In: Davison, A.N., Dobbing, J.
(Eds.), Applied Neurochemistry. Blackwell, Oxford, p. 287–316, 1968. EL YACOUBI, M. et al. The anxiogenic-like effect of caffeine in two experimental
procedures measuring anxiety in the mouse is not shared by selective A(2A) adenosine receptor antagonists. Psychopharmacology, v. 148, p. 153-163, 2000.
ESPINOSA, J.; ROCHA, A.; NUNES, F.; COSTA, M.S.; SCHEIN, V.; KAZLAUCKAS,
V.; KALININE, E.; SOUZA, D.O.; CUNHA, R.A.; PORCIÚNCULA, R.O. Caffeine consumption prevents memory impairment, neuronal damage, and Adenosine A2A receptors upregulation in the hippocampus of a rat model of sporadic dementia, Journal Alzheimers Disease, v. 34, p. 509–518, 2013.
FARIAS-SANTOS, R.C.; LIRA, M.C.A.; PEREIRA, D.E.S.; SÁ, I.R.; PIMENTEL,
M.R.F.; ARAÚJO, L.L.; GUEDES, R.C.A. Exposure of developing well-nourished and malnourished rats to environmental heating facilitates cortical spreading depression propagation at adulthood. Neuroscience Letters. v. 454, p. 218-222, 2009.

52
FILE, S.E.; BALDWIN, H.A.; JOHNSTON, A.L.; WILKS, L.J. Behavioral effects of acute and chronic administration of caffeine in the rat. Pharmacology Biochemistry and Behavior, v. 30, p. 809–815, 1988.
FISHER, S.; GUILLET, R. Neonatal caffeine alters passive avoidance retention in
rats in an age- and gender-related manner. Developmental Brain Research, v. 98, p. 145–149, 1997.
FORTE, G.C.; SCHENEIDER, A.P. Modulação do gasto energético pela ingestão de
capsaicina, cafeína e catequinas: uma revisão sistemática. Revista brasileira de obesidade, nutrição e emagrecimento, v. 6, p. 207-215, 2012.
FRANCISCO, E.S.; GUEDES, R.C.A. Neonatal taurine and alanine modulate
anxiety‑like behavior and decelerate cortical spreading depression in rats previously suckled under different litter sizes. Amino Acids, v. 47, p. 2437-2445, 2015.
FRAZÃO, M.F.; MAIA, L.M.S.S.; GUEDES, R.C.A. Early malnutrition, but not age,
modulates in the rat the L-arginine facilitating effect on cortical spreading depression. Neuroscience Letters, v.447, p. 26-30, 2008.
FREDHOLM, B.B. et al. Actions of Caffeine in the Brain with Special Reference to
Factors that Contribute to Its Widespread Use. Pharmacological Reviews, v. 51, p. 83-133, 1999.
GAVRIELI, A.; KARFOPOULOU, E.; KARDATOU, E.; SPYRELI, E.;
FRAGOPOULOU, E.; MANTZOROS, C.S.; YANNAKOULIA, M. Effect of Different Amounts of Coffee on Dietary Intake and Appetite of Normal-Weight and Overweight/Obese Individuals. Obesity, v. 21, p. 1127-1132, 2013.
GIESBRECHT, T.; RYCROFT JA, ROWSON MJ, DE BRUIN EA. The combination of
L-theanine and caffeine improves cognitive performance and increases subjective alertness. Nutritional Neurosciences, v. 13, p. 283-290 2010.
GOLDBERG SR, PRADA JA, KATZ JL. Stereoselective behavioral effects of N6-
phenylisopropyl- adenosine and antagonism by caffeine. Psychopharmacology, v. 87, p. 272–277, 1985.
GOLDSTEIN, E.; JACOBS, P.L.; WHITEHURST, M.; PENHOLLOW, T.; ANTONIO,
J. Caffeine enhances upper body strength in resistance-trained women. Journal International Society Sports Nutrition, p. 7-8, 2010.
GORJI, A. Spreading depression: a review of the clinical relevance. Brain Research, p.33–60, 2001. GREENBERG, J. A.; BOOZER, C. N.; GELIEBTER, A. Coffee, diabetes, and weight
control. American Society for Clinical Nutrition, v. 84, p. 682–693, 2006.

53
GRIFFITHS, R. R.; MUMFORD, G. K. Caffeine-a drug of abuse? In: Bloom, F. E. and Kupfer, D. J. (ed.). Psychopharmacology: The Fourth Generation of Progress. Raven Press, New York, p. 1699–1713, 1995.
GUEDES, R.C.A.; AMORIM, L.F.; MEDEIROS, M.C.; SILVA, A.T.; TEODÓSIO, N.R.
Effect of dietary lithium on cortical spreading depression. Brazilian Journal of Medical and Biological Research, v. 22, p. 923–925, 1989.
GUEDES R.C.A.; FRADE S.F. Effect of ethanol on cortical spreading depression.
Journal Medical Biological Research, v. 26, p. 1241-4, 1993. GUEDES, R.C.A Cortical Spreading Depression: A Model for Studying Brain
Consequences of Malnutrition. In: Victor R. Preedy, Ronald R Watson and Colin R Martin (eds.), Handbook of Behavior, Food and Nutrition, Springer, London, p. 2343-2355, 2011.
GUEDES, R.C.A Electrophysiological Methods: Aplication in Nutritional
neuroscience. In: Liebermann, H.; Kanarek, R.; Prasad, C. (eds), Nutritional Neurosciences: Overview of emerging field, CRC Press, New York, p. 39-54, 2005.
GUEDES, R.C.A., ROCHA-DE-MELO, A.P., BORBA, J.M.C., ALVES, C.R.R.,
SANTOS, A.A., TEODÓSIO, N.R. Nutrição e fisiologia do sistema nervoso. In: De Angelis, R.; Tirapegui, J. (eds.), Fisiologia da Nutrição Humana: Aspectos básicos, aplicados e funcionais: Editora Atheneu, São Paulo, 24, p. 417-430, 2007.
GUEDES, R.C.A.; AGUIAR, M.J.L.; ALVES-DE-AGUIAR, C.P.P. Caffeine and
Nutrition: an Overview. In: Preedy VR (editor) Caffeine: Chemistry, Analysis, Function and Effects, Food and Nutritional Components, The Royal Society of Chemistry Press, London, UK, n. 2, p. 4-21, 2012.
GUEDES, R.C.A.; AMORIM, L.F.; TEODÓSIO, N.R. Effect of aging on cortical
spreading depression. Brazilian Journal of Medical and Biological Research, v. 29, p. 1407-1412, 1996.
GUEDES, R.C.A.; DO CARMO, R.J. Influence of ionic alterations produced by gastric
washing on cortical spreading depression. Experimental Brain Research, v. 39, p. 341-349, 1980.
GUEDES, R.C.A.; PEREIRA-DA-SILVA, M.S. Effect of pre and postnatal
propythiouracil administration on propagation cortical spreading depression of adults rats. . Brazilian Journal of Medical and Biological Research, v. 26, p. 1123-1128, 1993.
GUEDES, R.C.A.; ROCHA-DE-MELO, A.P.; LIMA, K.R.; ALBUQUERQUE, J.M.;
FRANCISCO, E.S. Early malnutrition attenuates the impairing action of naloxone on spreading depression in Young rats. Nutritional Neuroscience, v. 16, p. 1340-1352, 2013.

54
GUEDES, R.C.A.; VASCONCELOS, C.A.C. Sleep deprivation enhances in adult rats the antagonistic effects of pilocarpine on cortical spreading depression: a dose-response study. Neurocience Letters, v. 442, p.118-122, 2008.
GUNJA, N.; BROWN, J.A. Energy drinks: health risks and toxicity. Medical Journal
Australia v. 196, p. 46–49, 2012. HALL, C.S.; Emotional behavior in the rat. I. Defecation and urination as measures of
individual differences in emotionality. Journal comparative psychology, v. 18, p. 385-403, 1934.
HANDLEY, S.L.; MITHANI, S. Effects of alpha-adrenoreceptor agonist in a maze-
exploration model of fear motivated behavior. Naunyn Schmiedebergs Archives Pharmacology, v. 327, p. 1-5, 1984.
HARPAZ, E.; TAMIR, S.; WEINSTEIN, A.; WEINSTEIN, Y. The effect of caffeine on
energy balance. Journal of Basic and Clinical and Physiology and Pharmacology, v. 28, p.1–10, 2017.
HEPPNER, C.C.; KEMBLE, E.D.; COX, W.N. Effects of blood deprivation on caffeine
consumption in male and female rats. Pharmacology Biochemistry and Behavior, v. 24, p. 1555-1559, 1986.
HUANG, Z. L.; URADE, Y.; HAYAISHI, O. The role of adenosine in the regulation of
sleep. Current Topics in Medicinal Chemistry, v. 11, p. 1047 - 1057, 2011. HUGHES, N.J.; HANCOCK, N.J.; HENWOOD, G.A.; RAPLEY, S.A. Evidence for
anxiolytic effects of acute caffeine on anxiety-related behavior in male and female rats tested with and without bright light. Behavioral Brain Research, v. 271, p. 7–15, 2014.
HURSEL R.; WESTERTERP-PLANTENGA, M.S. Thermogenic ingredients and body
weight regulation. International Journal Obesity, v. 34, p. 659–69, 2010. JESSEN, A.; BUEMANN, B.; TOUBRO, S.; SKOVGAARD, I.M.; ASTRUP, A. The
appetite-suppressant effect of nicotine is enhanced by caffeine. Diabetes Obesity and Metabolism, v.7, p. 327–33, 2005.
JEZOVA, D.; HLAVACOVA, N. Endocrine factors in stress and psychiatric disorders:
focus on anxiety and salivary steroids. Academic Science, v. 1148, p. 495-503, 2008.
JOHNSON-KOZLOW, M.; KRITZ-SILVERSTEIN, D.; BARRETT-CONNOR, E.;
MORTON, D. Coffee consumption and cognitive function among older adults. American Journal of Epidemiology, v. 156, p. 842–850, 2002.
JORDA, A.; PORTOLÉS, M.; GUASCH, R.; BERNAL, D.; SAEZ, G.T. Effect of
caffeine on urea biosynthesis and some related processes, ketone bodies, ATP and liver amino acids. Biochemical Pharmacology, v. 38, p. 2727-2732, 1989.

55
JULIANO, L.M.; GRIFFITHS, R.R. A critical review of caffeine withdrawal: empirical Validation of symptoms and signs, incidence severity and associated features. Psychopharmacology, v. 176, p. 1-29, 2004.
KASTER, M.P.; MACHADO, N.J.; SILVA, H.B.; NUNES, A.; ARDAIS, A.P.;
SANTANA, M.; BAQI, Y.; MÜLLER, C.E.; RODRIGUES, A.L.; PORCIÚNCULA, L.O.; CHEN, J.F.; TOMÉ, A.R.; AGOSTINHO, P.; CANAS, P.M.; CUNHA, R.A. Caffeine acts through neuronal adenosine A2A receptors to prevent mood and memory dysfunction triggered by chronic stress, Proceed National Academy Sciences, v. 112, p. 7833–7838, 2015.
KRAMER, D.R.; FUJII, T; OHIORHENUAN, I., LIU C.Y. Cortical spreading
depolarization: pathophysiology, implications, and future directions. Journal of Clinical Neuroscience, p. 22–27, 2016.
LEÃO, A.A.P. Spreading depression of activity in the cerebral cortex. Journal of Neurophysiology, v. 7, p.359–390, 1944a.
LEÃO, A.A.P. Pial circulation and spreading depression of activity in the cerebral
cortex. Journal of Neurophysiology, v. 7, p.391-396, 1944b.
LEE, C.E.; HJ, H.; HWANG, J.T.; SUNG, M.J.; YANG, H.J.; KIM, H.J.; PARK, J.H.;
KWON, D.Y.; KIM, M.S. Long-term consumption of platycodi radix ameliorates
obesity and insulin resistance via the activation of AMPK pathways. Evidence-
Based Complementary and Alternative Medicine, v. 2012, 2012 DOI:
10.1155/2012/759143.
LIMA, D.S.C.; FRANCISCO, E.S.; LIMA, C.B.; GUEDES. R.C.A. Neonatal L-
glutamine modulates anxiety-like behavior, cortical spreading depression and
microglial immunoreactiviy: analysis in developing rats suckled on normal size-
and large size litters, Amino Acids 49:337–346, 2017. Doi: 10.1007/s00726-
016-2365-2 LIMA, D.S.C.; Maia, L.M.S.S.; Barboza, E.A.; Duarte, R.A.; Sousa, L.S.; GUEDES,
R.C.A. L-glutamine supplementation during the lactation period facilitates cortical spreading depression in well-nourished and early-malnourished rats. Life Sciences, v. 85, p. 241-247, 2009.
LIMA, C.B.; SOARES, G.S.; VITOR, S.M.; ANDRADE-DA-COSTA, B.L.;
CASTELLANO, B.; GUEDES, R.C.A. Spreading depression features and Iba 1 immunoreactivity in the cerebral cortex of developing rats submitted to treadmill exercise after treatment with monosodium glutamate. International Journal of Developmental Neuroscience, v. 33, p. 98-105, 2014.
LIN, A.S.; UHDE, T.W.; SLATE, S.O.; MCCANN, U.D. Effects of intravenous
caffeineadministered to healthy males during sleep. Depression Anxiety, v. 5, p. 21–28 1997.

56
LISKO, J.G.; LEE, G.E.; KIMBRELL, J.B.; RYBAK, M.E.; BLASINI, L.V.; WATSON, C.H. Caffeine Concentrations in Coffee, Tea, Chocolate, and Energy Drink Flavored E-liquids. Nicotine & Tobacco Research, v. 00, p. 1-9, 2016.
LOPES-DE-MORAIS, A.A.C.; MENDES-DA-SILVA, R.F.; SANTOS, E.M.; GUEDES,
R.C.A. Neonatal dexamethasone accelerates spreading depression in the rat, and antioxidant vitamins counteract this effects. Brain Research, v. 1591, p. 93-101, 2014.
LORENZ, K.Z. Os fundamentos da etiologia. São Paulo: UNESP, 1995. LORIST, M.M.; TOPS, M. Caffeine fatigue, and cognition. Brain Cognition, v. 53, p.
82-94, 2003. MAGKOS, F.; KAVOURAS, S. A. Caffeine Use in Sports, Pharmacokinetics in Man,
and Cellular Mechanisms of Action. Critical Reviews in Food Science and Nutrition, v. 45, p. 535–562, 2005.
MAIA, L.M.S.S.; AMANCIO-DOS-SANTOS A.; DUDA-DE-OLIVEIRA D.; ANGELIM
M.K.C.; GERMANO P.C.P.; SANTOS S.F.; GUEDES R.C.A. L-Arginine administration during rat brain development facilitates spreading depression propagation: evidence for a dose- and nutrition-dependent effect. Nutritional Neuroscience, v.12, n.2, p. 73-80, 2009.
MATOS, M.; SHEN, H.Y.; AUGUSTO, E.; WANG, Y.; WEI, C.J.; WANG, Y.T.;
AGOSTINHO, P.; BOISON, D.; CUNHA, R.A.; CHEN, J.F. Deletion of adenosine A2A receptors from astrocytes disrupts glutamate homeostasis leading to psychomotor and cognitive impairment: relevance to schizophrenia, Biological Psychiatry, 2015.
MCKIM, W.A. Caffeine and the methylxanthines. In: Mckim, W.A. (ed). Drugs and
Behavior an introduction to behavioral pharmacology. Prentice Hall, p. 191-210, 1996.
MI-JOO, J.; CHANG-HO, Y.; HAE-JIN, K.; HYO-MIN, K.; A-SOL, K.; HA-NA, M.;
SEUNG-PIL, J. The Relationship of Caffeine Intake with Depression, Anxiety, Stress, and Sleep in Korean Adolescents. Korean Journal of Family Medicine, v. 37, p. 111-116, 2016.
MORGANE, P. J., MILLER, M., KEMPER, T., STERN, W., FORBES, W., HALL, R.,
BRONZINO J.; KISSANE J.; HAWRYLEWICZ E.; RESNICK O. The effects of protein
malnutrition on the developing central nervous system in the rat. Neuroscience and
Biobehavioral Reviews, v. 2, p. 137-230, 1978.
MORGANE, P.J.; AUSTIN-LAFRANCE R.; BRONZINO, J.D.; TONKISS, J.; DIAZ-
CINTRA, S.; CINTRA, L.; KEMPER, T.; GALLER J.R. Prenatal malnutrition and development of the brain. Neuroscience and Biobehavioral Reviews, v. 17, p. 91-128, 1993.

57
MULLER, C. E.; JACOBSON, K. A. Recent developments in adenosine receptor ligands and their potential as novel drugs. Biochimica et Biophysica Acta, v. 1808, p. 1290–1308, 2011.
NOSCHANG, C.G.; PETTENUZZO L.F.; VON POZZER TOIGO E.; ANDREAZZA,
A.C.; KROLOW, R.; FACHIN, A. ÁVILA, M.C.; ARCEGO, D. CREMA, L.M.; DIEHL, L.A.; GONÇALVES, C.A.; VENDITE, D.; DALMAZ, C. Sex-specific differences on caffeine consumption and chronic stress-induced anxiety-like behavior and DNA breaks in the hippocampus Pharmacology Biochemistry and Behavior, v. 94,p. 63–69, 2009.
O'CONNOR, E. C.; KREMER, Y.; LEFORT, S.; HARADA, M.; PASCOLI, V.;
ROHNER, C., Lüscher, C. Accumbal D1R neurons projecting to lateral
hypothalamus authorize feeding. Neuron, v. 88, p. 553-564, 2015.
OFLUOGLU, E.; PASAOGLU, H.; PASAOGLU, A. The effects of caffeine on L-
Arginine Metabolism in the brain of rats. Neurochemical Research, v. 34, p. 395-399, 2009.
O’NEILL, C.E., NEWSOM, R.J.; STAFFORD, J.; SCOTT, T.; ARCHULETA, S.;
LEVIS, S.C.; SPENCER, R.L.; CAMPEAU, S.; BACHTELL, R.K. Adolescent caffeine consumption increases adulthood anxiety-related behavior and modifies neuroendocrine signaling. Psyconeuroendocrinology, v. 67, p. 40-50, 2016.
PALUSKA, S.A. Caffeine and exercise. Current Sports Medicine Reports, v. 2, p.
213-219, 2003. PANDOLFO, P.; MACHADO, N.J.; KÖFALVI, A.; TAKAHASHI, R.N.; CUNHA, R.A.
Caffeine regulates frontocorticostriatal dopamine transporter density and improves attention and cognitive deficits in an animal model of attention deficit hyperactivity disorder, European Neuropsychopharmacol, v. 23, p. 317–328, 2013.
PATZ, M.D.; DAY, H.E.; BUROW. A.; CAMPEAU. S. Modulation of the hypothalamo-
pituitary-adrenocortical axis by caffeine. Psychoneuroendocrinology, v. 31, p. 493-500, 2006.
PECHLIVANOVA, D.M.; TCHEKALAROVA, J.D.; ALOVA, L.H.; PETKOV, V.V.;
NIKOLOV, R.P.; YAKI-MOVA, K.S. Effect of long-term caffeine administration on depressive-like behavior in rats exposed to chronic unpredictable stress. Behavioral Pharmacology, v. 23, p. 339–47, 2012.
PELLOW, S.; FILE, S.E. Anxiolytic and anxiogenic drug effects on exploratory activity
in an elevated plus-maze: a novel test of anxiety in the rat. Pharmacology Biochemistry and behavior, v. 24, p. 525-529, 1986.
PERUZZI, G.; LOMBARDELLI, G.; ABBRACCHIO, M.P.; COEN, M.; CATTABENI, F.
Perinatal caffeine treatment: behavioral and biochemical effects in rats before weaning, Neurobehavioral Toxicology and Teratology, v. 7, p. 453–460, 1985.

58
PETTENUZZO, L.F.; NOSCHANG, C.; TOIGO, E.V.P.; FACHIN, A.; VENDITE, D.;
DALMAZ, C. Effects of chronic administration of caffeine and stress on feeding behavior of rats. Physiology & Behavior, v. 95, p. 295–301, 2008.
POLESZAK et al., 2015. The influence of caffeine on the activity of moclobemide,
venlafaxine, bupropion and milnacipran in the forced swim test in mice. Life Science, v. 136, p. 13-18, 2015.
PORCIÚNCULA, L.O.; SALLABERRY, C.; MIORANZZA, S.; BOTTON, P.H.;
ROSEMBERG, D.B. The Janus face of caffeine, Neurochemistry, v. 63, p. 594–609, 2013.
RACOTTA, I.S.; LEBLANC, J.; RICHARD, D. The effect of caffeine on food intake in
rats: involvement of corticotropin-releasing factor and the sympatho-adrenal system. Pharmacology Biochemical Behavior, v. 48, p. 887-92, 1994.
RANDALL, P. A.; NUNES, E. J.; JANNIERE, S. L.; STOPPER, C. M.; FARRAR, A.
M.; SAGER, T. N.; BAQI, Y.; HOCKEMEYER, J.; MULLER, C. E.; SALAMONE, J. D. Stimulant effects of adenosine antagonists on operant behavior: differential actions of selective A2A and A1 antagonists. Psychopharmacology, V. 216, no. 2, pp 173–186. 2011.
RETZBACH, E. P.; DHOLAKIA, P. H.; DUNCAN-VAIDYA, E. The effect of daily
caffeine exposure on lever-pressing for sucrose and c-fos expression in the nucleus accumbens in the rat. Physiology & Behavior, v. 135, p. 1-6, 2014.
RITCHIE, K.; CARRIÈRE, I.; MENDONÇA, A.; PORTET, F.; DARTIGUES, J.F.;
ROUAUD, O.; BARBERGER-GATEAU, P.; ANCELIN, M.L. The neuroprotective effects of caffeine: a prospective population study (the Three City Study), Neurology, v. 69, p. 536–545, 2007.
ROCHA-DE-MELO, A.P.; CAVALCANTI, J.B; SANTOS BARROS, AM.; GUEDES,
R.C.A. Manipulation of rat litter size during suckling influences cortical spreading depression after weaning and at adulthood. Nutritional Neuroscience, v.9 (3/4), p. 155-160, 2006.
ROCHA-DE-MELO, A.P.; GUEDES, R.C.A. Spreading depression is facilitated in
adult rats previously submitted to short episodes of malnutrition during the lactation period. Brazilian Journal of Medical and Biological Research, v. 30: p. 663-669, 1997.
SANDERS, M.J.; GRONDIN, P.O.; HEGARTY, B.D.; SNOWDEN, M.A.; CARLING,
D. Investigating the mechanism for AMP activation of the AMP-activated protein kinase cascade. Biochemical Journal, v. 403, p. 139–148, 2007.
SANTOS, R.M.M.; LIMA, D.R.A. Coffee consumption, obesity and type 2 diabetes: a
mini‑review. European Journal Nutritional, v. 55, p. 1345–1358, 2016.

59
SANTOS-MONTEIRO, J.S.; TEODÓSIO, N.R.T.; GUEDES, R.C.A. Long-lasting effects of early environmental stimulation on cortical spreading depression in normal and early malnourished adult rats. Nutritional Nerusocience, v. 1, p. 29-40, 2000.
SAWAYA, A.L.; LEANDRO, C.G.; WAITZBERG, D.L. Fisiologia da Nutrição na
saúde e na doença. 1ªed. São Paulo: Editora Atheneu, 2012. SAWYNOK, J. Methylxanthines and pain. Handbook of Experimental
Pharmacology, v. 200, p. 311-329, 2011. SAWYNOK, J. YAKSH, T.L. Caffeine as an analgesic adjuvant: a review of
pharmacology and mechanisms of action. Pharmacological Reviews, v. 45, p. 43-45, 1993.
SCHRAEGLE, W.A.; TITUS, J.B. The relationship of seizure focus with depression,
anxiety, and health-related quality of life in children and adolescents with epilepsy. Epilepsy & Behavior, v.68, p. 115-122. 2017. doi:10.1016/j.yebeh.2016.12.009, 2017.
SCHUBERT, M.M.; HALL, S.; LEVERITT, M.; GRANT, G. SABAPATHY S,
DESBROW B. Caffeine consumption around an exercise bout: effects on energy expenditure, energy intake, and exercise enjoyment. Journal of Applied Physiology, v. 117, p. 745–54, 2014.
SELL, S.L.; SCALZITTI, J.M.; THOMAS, M.L.; CUNNINGHAM, K.A. Influence of
ovarian hormones and estrous cycle on the behavioral response to cocaine in female rats, Journal of Pharmacology and Experimental Therapeutics, v. 293, p. 879–886, 2000.
SIMON, S. C.; HERNING, R. I.; BETTER, W.; CADET, J. L.; GRIFFITHS, R. R.
Caffeine withdrawal, acute effects, tolerance, and absence of net beneficial effects of chronic administration: cerebral blood flow velocity, quantitative EEG, and subjective effects. Psychopharmacology, v. 204, p. 573–585, 2009.
SINTON, C.M.; VALATX, J.L.; JOUVET, M. Gestational caffeine modifies offspring
behaviour in mice. Psychopharmacology, v. 75, p. 69–74, 1981. SILVA, C.G.; MÉTIN, C.; FAZELI, W.; MACHADO, N.J.; DARMOPIL, S.; LAUNAY,
P.S.; GHESTEM, A.; NESA, M.P.; BASSOT, E.; SZABÓ, E.; BAQI, Y.; MÜLLER, C.E.; TOMÉ, A.R.; IVANOV, A.; ISBRANDT, D.; ZILBERTER, Y.; CUNHA, R.A.; ESCLAPEZ, M.; BERNARD, C. Adenosine receptor antagonists including caffeine alter fetal brain development in mice, Science Translational Medicine, v. 5, p. 104-197, 2013.
SOELLNER, D.E.; GRANDYS, T.; NU˜NEZ, J.L. Chronic prenatal caffeine exposure
impairs novel object recognition and radial arm maze behaviors in adult rats. Behavioral Brain Research, v. 205, p. 191–199, 2009.

60
SOLINAS, M.; FERRE, S.; YOU, Z. B.; KARCZ-KUBICHA, M.; POPOLI, P.; GOLDBERG, S. R. Caffeine induces dopamine and glutamate release in the shell of the nucleus accumbens. Journal of Neuroscience v. 22, pp. 6321- 6324, 2002.
SOUZA, R.A.G.; SICHIERI, R. Consumo de Cafeína e Prematuridade. Revista de
Nutrição, v. 8, n. 5, 2005. STERNSON, S. M.; ATASOY, D. Agouti-related protein neuron circuits that regulate
appetite. Neuroendrocrinology, v. 2, p. 95- 102, 2014. SVENNINGSSON, P.; FREDHOLM, B.B. Caffeine mimics the effect of a dopamine
D2/3 receptor agonist on the expression of immediate early genes in globus pallidus. Neuropharmacology, v. 36, p. 1309-1307, 1997.
SVENNINGSSON, P.; HALL, H.; SEDVALL, G.; FREDHOLM, B.B. Distribution of
adenosine receptors in the postmortem human brain: an extended autoradiographic study, Synapse, v. 27, P. 322–335, 1997.
SWEENEY, P.; LEVACK, R.; WATTERS, J.; XU, Z.; YANG, Y. Caffeine increases
food intake while reducing anxiety-related behavior. Appetite, v. 101, p. 171–7, 2016.
TAGLIABUE, A.; TERRACINA, D.; CENA, H.; TURCONI, G.; LANZOLA, E.;
MONTOMOLI, C. Coffee induced thermogenesis and skin temperature. International Journal of Obesity and Related Metabolic Disorders, v.18, p. 537–541, 1994.
TAVARES, C.; SAKATA, R. Cafeína para o tratamento de dor. Revista Brasileira de
Anestesiologia, v. 62, p. 387-401, 2012. TCHEKALAROVA, J.; KUBOVA, J.; MARES, P. Effects of early postnatal caffeine
exposure on seizure susceptibility of rats are age- and model-dependent. Epilepsy Research, v.88, p. 231-238, 2010.
TEMPLE, J.L. Caffeine use in children: what we know, what we have left to learn,
and why we should worry. Neuroscience & Biobehavioral Reviews, nº. 33, p. 793-806, 2009.
TEMPLE, J.L.; ZIEGLER, A.M. Gender differences in subjective and physiological
responses to caffeine and the role of steroid hormones, Journal of Caffeine Research, v. 1, p. 41–48, 2011.
THOM, E. The effect of chlorogenic acid enriched coffee on glucose absorption in
healthy volunteers and its effect on body mass when used long-term in overweight and obese people. The journal of International Medical Research, v. 35, p. 900–908, 2007.
TORRENTE D, CABEZAS R, AVILA MF, GARCÍA-SEGURA LM, BARRETO GE,
GUEDES RCA. Cortical spreading depression in traumatic brain injuries: Is there

61
a role for astrocytes? Neuroscience Letters, v. 565, p. 2-6, doi 10.1016/j.neulet.2013.12.058, 2014
TREMBLAY, A.; MASSON, E. LEDUC, S. HOUDE, A. DESPRES, J.P. Caffeine
reduces spontaneous energy intake in men but not in women. Nutrition Research, v. 8, p. 553-558, 1988.
VINSON, J. A.; BURNHAM, B. R.; NAGENDRAN, M. V. Randomized, double-blind,
placebo-controlled, linear dose, crossover study to evaluate the efficacy and safety of a green coffee bean extract in overweight subjects. Diabetes Metabolic Syndrome Obesity, v. 5, p. 21–27, 2012.
WINSTON, A.P. et al. Neurpsychiatric effects of caffeine. Advanced Psychiatric
treatment, v. 11, p. 432-439, 2005. ZHAO, Y.; YANG, L.; HUANG, Z.; LIN, L.; ZHENG, G. Synergistic effects of caffeine
and catechins on lipid metabolism in chronically fed mice via the AMP‑activated protein kinase signaling pathway. European Journal of Nutrition, 2016. DOI 10.1007/s00394-016-1271-4
YANG, Y.; ATASOY, D.; SU, H.; STERNSON, S. Hunger states switch a flip-flop
memory circuit via a synaptic AMPK-dependent positive feedback loop. Cell, v. 146, p. 992 – 1003, 2011.
ZHANG, W.Y. A benefit- risk assessment of caffeine as an analgesic adjuvant. Drug
Safety, v. 24, p. 1127-1142, 2001. ZHANG, W.; LOPEZ-GARCIA, E.; LI, T. Y.; HU, F. B.; VAN DAM, R. M. Coffee
consumption and risk of cardiovascular diseases and all-cause mortality among men with type 2 diabetes. Diabetes Care, v. 32, p. 1043–1045, 2009.

62
. ANEXOS
ANEXO 1 – Confirmação da submissão do artigo ao periódico

63
ANEXO 2 – Parecer do Comitê de Ética em Pesquisa da UFPE





























