Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DE SÃO JOÃO DEL-REI – UFSJ
CAMPUS CENTRO-OESTE DONA LINDU – CCO
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS DA SAÚDE
ALINE DE FREITAS LOPES
AVALIAÇÃO DE CITOTOXICIDADE DE ANÁLOGOS DE
ALCALÓIDES MARINHOS DO TIPO 3-
ALQUILPIRIDÍNICOS EM LINHAGEM DE LEUCEMIA
MIELÓIDE AGUDA
DIVINÓPOLIS

UNIVERSIDADE FEDERAL DE SÃO JOÃO DEL-REI – UFSJ
CAMPUS CENTRO-OESTE DONA LINDU – CCO
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS DA SAÚDE
ALINE DE FREITAS LOPES
Dissetação apresentada ao Programa de Pós
Graduação em Ciências da Saúde, da
Universidade Federal de São João Del Rei, como
requisito para obtenção do título de Mestre.
Orientador: Prof.Dr.Adriano de Paula Sabino
DIVINÓPOLIS
DEZEMBRO/2016

AGRADECIMENTOS
Sinto-me verdadeiramente grata a todo apoio e ajuda que recebi de inúmeras pessoas
durante minha vida e que me deram força para cumprir mais esse trabalho.
Ao Gustavo, meu marido, pelo amor, companheirismo e paciência durantes esses anos
de estudos. Obrigada por estar sempre ao lado, me incentivando e me animando nas
horas difícieis.
Aos meus pais, pelo cuidado, carinho e incentivo dedicados à mim.
Aos meus familiares, em especial meu irmão, tios e primos, também pelo apoio e pelas
horas felizes que desfrutamos sempre que nos encontramos.
Ao meu orientador Prof. Adriano de Paula Sabino, pelos ensinamentos, contribuição e
ajuda na elaboração desse trabalho.
Ao Prof. Fernando de Pilla Varotti pela amizade, ensinamentos, apoio e ajuda, sem os
quais esse trabalho também não seria possível de ser realizado.
Aos amigos que fiz nos laboratórios por onde passei, em especial a Kecya,
contemporânea minha no programa, mas também à Fernanda, Silmara e Deise, pela
mentoria e auxílio da condução dos experimentos.

iii
RESUMO
A Leucemia Mielóide Aguda (LMA) se caracteriza pela proliferação anormal de
células progenitoras imaturas da linhagem mielóide (mieloblastos), que perderam a
habilidade de se diferenciar e a responder adequadamente a fatores de crescimento.
Essas células leucêmicas extravasam para o sangue e levam a falência medular, com
evolução rápida, levando ao óbito se não tratada. Nos últimos 20 anos não houve
grandes mudanças no esquema de tratamento da LMA, que envolve poliquimioterapia
citotóxica, porém as taxas de remissão completa e sobrevida livre de doença são baixas,
principalmente em pacientes acima de 65 anos que de forma geral toleram pouco
quimioterapia mais intensa. Baseado nesses dados, a busca e otimização de novos
compostos com potencial uso anti-leucêmico torna-se necessária. O objetivo deste
estudo foi avaliar o efeito de análogos de alcaloides marinhos do tipo 3-alquilpiridínicos
em uma linhagem humana de leucemia mielóide aguda (THP-1). Os resultados dos
experimentos demonstraram que o composto THP-002 foi citotóxico, apresentando
baixo IC50 (determinado através do ensaio in vitro de 3-(4,5-dimetil-2-tiazolil)-2,5-
difenil-tetrazólio (MTT)). Ele também foi capaz de causar dano ao DNA, provovando a
formação de micronúcleos e induzindo apoptose através da ativação de Caspase 3. A
análise de expressão gênica de TP53, p21 e BAK após exposição à diferentes
concentrações desse composto mostrou aumento significativa da expressão de BAK mas
não de p21. A expressão de TP53 foi demonstrada de forma estatisticamente
significativa apenas após 48h de exposição à concentração do composto-teste em 2x do
IC50, sugerindo que o composto induz apoptose mas por via diferente da p53.
Palavras - chave: Leucemia mielóide aguda, citotoxicidade, alcaloide marinho, 3-
alquilpiridínicos e THP-1

iv
ABSTRACT
Acute myeloid leucemia (AML) is characterized by abnormal proliferation of
immature progenitor cells of the myeloid lineage (myeloblasts), which have lost the
ability to differentiate and respond appropriately to growth factors. These leukemic cells
spill into the blood and lead to bone marrow failure with rapid evolution, leading to
death if not treated quickly. Over the past 20 years there have been no major changes in
AML treatment regimen. Current AML treatment protocols include cytotoxic multidrug
therapy, but complete remission rates and progression-free survival are low, especially
in patients over 65 years old who generally don´t tolerate intense chemotherapy . Based
on these information, the search and optimization of new compounds with potential
anti-leukemic use becomes necessary. Herein we presente an initial study of the
cytotoxicity effect of analogs of marine alkaloids type 3-alquilpiridínicos in AML
(THP-1 human cell line). The results of the experiments showed that the compound
THP-002 was cytotoxic, with low IC50 value (determined by vitro assay 3- (4,5-
dimethyl-2-thiazolyl) -2,5-diphenyl tetrazolium bromide (MTT)). It was also able to
induce DNA damage through micronucleus formation inducing apoptosis through
caspase 3 activation. Gene expression analysis of TP53, p21 and BAK after exposure to
different concentrations of this compound showed significant increased expression of
BAK but not in p21. The TP53 expression was demonstrated statistically significantly
only after 48 hours of exposure to the concentration of assayed compound at 2x the
IC50, suggesting that the compound induces apoptosis but different pathwayof p53.
Key-words: Acute myeloid leukemia, cytotoxicity, marine alkaloid, pyridine 3-alkyl
and THP-1

v
Sumário
RESUMO....................................................................................................................... III
LISTA DE FIGURAS................................................................................................... VI
LISTA DE SIGLAS E ABREVIATURAS ................................................................. IX
1. INTRODUÇÃO ......................................................................................................11
1.1 Hematopoese ............................................................................................................... 11
1.2 Leucemias Agudas ...................................................................................................... 13
1.2.1 Leucemia Linfoblástica Aguda ............................................................................... 13
1.2.2 Leucemia Mielóide Aguda ...................................................................................... 13
1.3 Apoptose ..................................................................................................................... 17
1.4 Terapia na Leucemia Mielóide Aguda ........................................................................ 18
1.5 Novas drogas: Desenvolvimento ................................................................................. 21
1.5.1 ALCALOIDES MARINHOS.............................................................................. 21
2 JUSTIFICATIVA ...................................................................................................22
3 OBJETIVOS ...........................................................................................................22
3.1 Objetivo geral .............................................................................................................. 22
3.2 Objetivos específicos................................................................................................... 23
4 MATERIAIS E MÉTODOS..................................................................................23
4.1 Obtenção dos compostos ............................................................................................. 23
4.2 Solubilização dos compostos ...................................................................................... 24
4.3 Cultivo de linhagens celulares ..................................................................................... 25
4.4 Ensaio de viabilidade celular ....................................................................................... 26
4.5 Ensaio de Genotoxicidade e Mutagênese .................................................................... 27
4.5.1 Ensaio de micronúcleo com bloqueio de citocinese (CBMN) ............................ 27
4.6 Detecção de Caspase 3 ................................................................................................ 28
4.7 Análise de expressão gênica ........................................................................................ 29
4.7.1 Extração de RNA ................................................................................................ 29
4.7.2 Confecção de cDNA ............................................................................................ 30
4.7.3 PCR em tempo real ............................................................................................. 30
4.8 Análises estatísticas ..................................................................................................... 32

vi
5 RESULTADOS E DISCUSSÃO ...........................................................................33
5.1 Ensaio de citotoxicidade e viabilidade celular ............................................................ 33
5.2 Ensaio de Genotoxicidade e Mutagênese .................................................................... 34
5.3 Detecção de caspase 3 ................................................................................................. 37
5.4 Análise da expressão gênica ........................................................................................ 38
6 CONCLUSÕES ......................................................................................................40
LISTA DE FIGURAS
Figura 1: Hematopoese . ............................................................................................................ 12

vii
Figura 2:- Frequência média de micronúcleos observados por 1000 células binucleadas de
TPH-1, após o tratamento com diferentes concentrações do composto THP-002. ..................... 35
Figura 3:- Índice de Divisão Nuclear das células THP-1, após o tratamento com diferentes
concentrações do composto THP-002.. ....................................................................................... 36
Figura 4: Quantificação da atividade da Caspase 3 por ensaio colorimétrico na linhagem THP-1
..................................................................................................................................................... 37
Figura 5: Gráfico da quantificação relativa do gene TP53 após tratamento com THP-002 na
linhagem THP-1 em diferentes concentrações ............................................................................ 38
Figura 6: Gráfico da quantificação relativa do gene p21 após tratamento com THP-002 na
linhagem THP-1 em diferentes concentrações ............................................................................ 39
Figura 7: Gráfico da quantificação relativa do gene BAK após tratamento com THP-002 na
linhagem THP-1 em diferentes concentrações ............................................................................ 39

viii
LISTA DE TABELAS
Tabela 1– Classificação das Leucemias Mielóides Agudas segundo o grupo Franco-Americano-
Britânico (FAB). ......................................................................................................................... 14
Tabela 2 – Classificação das Leucemias Mielóides Agudase neoplasias relacionadas, segundo a
Organização Mundial de Saúde (2008) ....................................................................................... 15
Tabela 3 - Estrutura dos 5 compostos análogos de alcaloides marinhos do tipo 3-
alquilpiridínicos ........................................................................................................................... 23
Tabela 4 - Sequência dos oligonucleotídeos iniciadores diretos (F) e reversos (R) utilizados na
qRT-PCR. .................................................................................................................................... 33
Tabela 5 - Valores de IC50 obtidos em ensaios in vitrocom linhagens celulares de THP-1 e WI-
26VA-4.. ...................................................................................................................................... 31

ix
LISTA DE SIGLAS E ABREVIATURAS
FLT-3 - FMS-like Tirosina Kinase-3
LA – Leucemia aguda
FAB - grupo cooperativo Franco-Americano-Britânico
LMA – Leucemia mielóide aguda
LLA – Leucemia linfocítica aguda
OMS – Organização mundial de saúde
FNT – Fator de necrose tumoral
Apaf1 - apoptic protease activating fatot-1
Bcl-2 – B-cell lymphoma 2
FISH - Hibridização in situ por Imunofluorescência
DNA - Ácido desoxirribonucléico(DesoxirribonucleicAcid)
PCR - Reação em cadeia da polimerase
RNA - Ácido ribonucléico
qRT-PCR - Reação em cadeia da polimerase em tempo real
HIDAC - Citarabina em alta dose
TMO – Transplante de medula óssea
RC – Remissão completa
ATRA - Ácido transretinóico
FLT3-DIT - Duplicação interna em tandem no gene FLT3
NPM – Nucleofosmina
EMEA - European Medicine Agency
3-APA - 3-alquilpiridínicos
DMSO – Dimetilsulfóxido
SFB – Soro fetal bovino
ATCC - American Type Culture Collection
CO2 – Dióxido de Carbono
THP-1 –Linhagem humana de Leucemia Mielóide Aguda (Leucemia Monocítica
Aguda) ATCC TIB-202
oC – Graus Celsius

x
h - Hora
MTT - 3-(4,5-dimetil-2-tiazolil)-2,5-difenil-tetrazólio
PBS - Tampão fosfato-salino (phosphatebuffered saline)
RPM – Rotações por minuto
RPMI - Meio de cultura (desenvolvido no Roswell Park Memorial Institute –RPMI)
WI-26-VA4 – Fibroblasto de pulmão humano
IC50 - Concentração que reduz a viabilidade celular em 50%
IS – Índice de Seletividade
MMS - metanossulfonato de metila
DAPI - 4 ', 6-diamidino-2-fenilindol
BNC - células binucleadas
MN – Micronúcleo
NDI - Índice de divisão nuclear
RNAm - RNA mensageiro
GAPDH - gliceraldeído-3-fosfato-desidrogenase

11
1. INTRODUÇÃO
As leucemias agudas são um grupo de neoplasias malignas que derivam da transformação
de células progenitoras hematopoéticas indiferenciadas ou parcialmente diferenciadas (Greer
et al. 2003). Durante a evolução de uma célula progenitora, ou seja, durante a hematopoese, é
possível que ocorra uma anormalidade genômica, como uma mutação somática, além de
outros fatores celulares e/ou moleculares que favoreçam a expansão desse clone defeituoso.
Na maioria dos casos, essas células leucêmicas extravasam para o sangue, onde podem ser
vistas em grande número, além de também poder infiltrar o fígado, baço, linfonodos e outros
tecidos (Pilger et al. 2006).
O entendimento do processo de hematopoese é necessário para a discussão da
fisiopatogênese da doença e seus desdobramentos.
1.1 Hematopoese
O tecido sanguíneo dos mamíferos é constituído por um conjunto de diferentes tipos de
células, incluindo eritrócitos, células mielóides (granulócitos, monócitos/macrófagos,
megacariócitos e plaquetas), células linfoides (linfócitos B e T e células Natural Killers),
células dendríticas e mastócitos, sendo todos originados de um único tipo de célula, chamada
célula tronco hematopoética. Ela possui a capacidade de se dividir, se auto-regenerando e
também de gerar uma célula precursora comprometida com uma linhagem celular específica
(Rossi et al. 2012; Zago et al. 2004). Os processos envolvidos na produção dessas células são
denominados coletivamente de hematopoese (Greer et al. 2003).
A hematopoese se inicia na embriogênese, nas ilhotas sanguíneas do saco vitelino, já nas
primeiras semanas de gestação. Nessa fase, é denominada hematopoese primitiva e é restrita à
produção de eritrócitos. Por volta da oitava semana e até aproximadamente o quinto mês de
gestação, o fígado fetal passa a ser o principal sítio de hematopoese. Os eritrócitos produzidos
nessa etapa já estão anucleados, diferente dos produzidos durante a hematopoese primitiva. A
partir da metade da gestação, a medula óssea toma lugar como o principal órgão
hematopoético, sendo responsável pela produção de todas as linhagens hematopoéticas. Após
o nascimento e por toda a vida, sob condições fisiológicas, o único sítio de hematopoese é a
medula óssea (Zago et al. 2004).

12
A célula tronco hematopoética origina um número de células progenitoras multipotentes,
que ainda retêm o potencial de gerar linhagens completas, mas com capacidade de auto-
renovação mais limitada. Progenitores multipotentes, por sua vez, levam a formação de
progenitores oligopotentes que possuem um potencial de desenvolvimento mais restrito. Nessa
fase, o progenitor linfocítico comum tem o potencial de gerar células efetoras linfóides
maduras, como o linfócito B, T, células dendríticas e Natural Killers. As células progenitoras
comprometidas com a linhagem mielóide, por sua vez, dão origem a células como os
granulócitos e monócitos (Gekas et al. 2006; Rossi et al. 2012) (Figura 1).
Legenda: PMC: Progenitor Mielóide Comum; PGML: Progenitor Granulócito/Macrófago/Linfócito; PEM:
Progenitor Megacariócito/Eritrócito; PGM: Progenitor Granulócito/Macrófago; PLC: Progenitor Linfócito
Comum.
O desenvolvimento das células hematopoéticas normais é um processo ordenado em várias
fases, bem regulamentado por uma complexa rede de fatores instrísecos e direcionamento do
microambiente que controlam os destinos das células no interior da medula óssea (Enciso et
al. 2015). São fatores importantes que regulam esses processos: fatores de transcrição,
reguladores do ciclo celular, moléculas sinalizadoras, receptores de superfície e citocinas.
Figura 1: Hematopoese*.
* Adaptado de Lara Rossi e colaboradores (Rossi et al. 2012)

13
Perturbações dessa fina homeostase podem induzir uma expansão celular sem controle,
levando a prejuízo da hematopoese normal e instalação de processo leucêmico. Os receptores
de Tirosina Kinase tipo III, como o FMS-like Tirosina Kinase-3 (FLT-3), c-KIT, cFMS e
PDGFR, por exemplo, desempenham um papel importante tanto na hematopoese normal
quanto na maligna, estando altamente expressos nas células progenitoras hematopoéticas mas
também na superfície de blastos leucêmicos (Enciso et al. 2015; Cells et al. 2014).
1.2 Leucemias Agudas
As Leucemias Agudas (LA) são um grupo significativamente heterogêneo de neoplasias
malignas incorporando subgrupos com comportamento biológico, fisiopatologia e abordagem
terapêuticas distintas. O primeiro sistema de classificação das LA aceito internacionalmente
foi proposto porum grupo cooperativo Franco-Americano-Britânico (FAB) em 1976. Essa
classificação sofreu várias revisões ao longo dos anos, mas ainda considera apenas as
características morfológicas e citoquímicas dos blastos como critério de classificação, sendo
que dois importantes grupos foram individualizados: Leucemia Mielóide Aguda (LMA) e
Leucemia Linfocítica Aguda (LLA) (Bennett et al. 1976; Bennett et al. 1985; Bennett et al.
1991; Bennett 1985).
1.2.1 Leucemia Linfoblástica Aguda
A LLA se origina da transformação maligna de uma célula progenitora linfóide durante
seu amadurecimento. Os sinais e sintomas da LLA são bem parecidos com os da LMA, pois
esse tipo de leucemia também se caracteriza pela proliferação de células blásticas, só que da
linhagem linfóide. As células doentes se acumulam na medula óssea, competindo com as
células normais, ocasionando anemia, trombicitopenia e neutropenia. O clone maligno alcança
a corrente sanguínea infiltrando outros órgãos como baço, linfonodos e sistema nervoso
central. Assim, o aparecimento de febre, fraqueza, sangramentos, perda de apetite, manchas
roxas e dor abdominal são característicos dessa doença (Greer et al. 2003; Lichtman 2006).
1.2.2 Leucemia Mielóide Aguda
A LMA se caracteriza pela proliferação anormal de células progenitoras imaturas da
linhagem mielóide (mieloblastos), que perderam a habilidade de se diferenciar e a responder
adequadamente a fatores de crescimento. Essas perdas levam a produção insuficiente de
células sangüíneas maduras, com conseqüente substituição do tecido normal. Em associação,
ocorre a perda da capacidade de desenvolvimento dos precursores normais da medula óssea.

14
Dessa forma, a infiltração neoplásica da medula óssea é acompanhada de sua falência
funcional. Essa é a principal causa de óbito dentre os portadores de LMA, já que essa
condição predispõe principalmente a sangramentos e infecções fatais. Também é possível
ocorrer infiltração de outros órgãos pelas células leucêmicas (Hall & Guyton 2011; Estey &
Döhner 2006).
Por definição, para o diagnostico de LMA é necessário a presença de 20% de mieloblastos
ou mais na medula óssea ou sangue períférico, segundo critério da Organização Mundial de
Saúde (OMS) (Vardiman et al. 2009). Vários sistemas de subclassificação da LMA foram
propostos ao longo dos últimos 40 anos. A classificação FAB, considerando critérios
morfológicos e citoquímicos, reconhece oito subtipos diferentes, em função da linhagem
mielóide afetada e do grau de diferenciação encontrado (Tabela 1) (Bennett et al. 1976;
Bennett et al. 1985; Bennett 1985; Zago et al. 2004).
Tabela 1– Classificação das Leucemias Mielóides Agudas segundo o grupo Franco-
Americano-Britânicom(FAB).
Subtipo FAB Sinonímia
M0 LMA sem diferenciação morfológica
M1 LMA com mínima diferenciação morfológica
M2 LMA com diferenciação
M3 Leucemia Aguda promielocítica
M4 Leucemia Mielomonocítica aguda
M5 Leucemia Monocítica Aguda
M6 Eritroleucemia
M7 LMA megacarioblástica
Na década de 90, a Organização Mundial de Saúde propôs uma outra forma de
classificação, sendo a mesma atualizada em 2002 e 2008. Foram incorporados nesse sistema
de classificação, além dos aspectos citomorfológicos e citoquímicos, também características
de imunofenotipagem e aspectos genético-moleculares (Tabela2).

15
A imunofenotipagem permite identificar antígenos de superfície, intracitoplasmáticos e
nucleares com o uso de anticorpos monoclonais marcados, sendo empregado principalmente
com o citômetro de fluxo e permitem a caracterização das células leucêmicas quanto à sua
origem e o grau de diferenciação, sendo atualmente técnica indispensável para o diagnóstico
de vários subtipos de LMA e também para a caracterização da Leucemia Aguda de Linhagem
Ambígua.
A citogenética e os estudos moleculares detectam anormalidades dentro do clone
leucêmico. A citogenética é avaliada através do cariótipo ou, em casos em que o cariótipo não
fornece informação adequada, pode-se lançar mão da Hibridização in situ por
Imunofluorescência (FISH) ou outros testes. A análise de genética molecular pode ser
realizada através da análise do ácido desoxirribonucléico (DNA) ou do ácido ribonucléico
(RNA) por Southern blot ou reação em cadeia da polimerase (PCR) e também por PCR em
tempo real (qRT-PCR) (Pilger et al. 2006).
Essa estratificação permite avaliar prognóstico, auxiliam no monitoramento após o
tratamento e pode direcionar a escolha da terapia mais adequada (Vardiman et al. 2002;
Vardiman et al. 2009; Pilger et al. 2006).
Tabela 2– Classificação das Leucemias Mielóides Agudas e neoplasias relacionadas, segundo
a Organização Mundial de Saúde (2008)
Leucemia Mielóide Aguda com anormalidades genéticas recorrentes
• LMA com t (8:21) (q22;q22);RUNX1-RUNX1T1
• LMA com inv (16) (p13.1q22) ou com t(16;16) (p13.1;q22);CBFB-MYH11
• LPA com t (15:17) (q22;q12);PML-RARA
• LMA com t (9;11) (p22;q23); MLLT3-MLL
• LMA com t (6:9) (p23;q34);DEK-NUP214
• LMA com inv (3) (q21q26.2) ou com t (3;3) (q21;q26.2);RPN1-EVI1
• LMA (megacarioblástica) t (1;22) (p13;q13);RBM15-MKL1
• LMA com mutação NPM1
• LMA com mutação CEBPA
Leucemia Mielóide Aguda com alterações relacionadas à mielodisplasia
Neoplasias Mielóides relacionadas à terapias

16
Leucemias Mielóides não especificadas
• LMA com diferenciação mínima
• LMA sem maturação
• LMA com maturação
• Leucemia mielomonocítica aguda
• Leucemia monoblástica/monocítica aguda
• Leucemia eritroide aguda
• Leucemia megacarioblástica aguda
• Leucemia basofílica aguda
• Panmielose aguda com mielofibrose
Sarcoma Mielóide
Proliferação Mielóide associada à Síndrome de Down
Neoplasia blástica plasmocitóide de células dentríticas
A LMA possui uma taxa de incidência global de 4 por 100.000 pessoas, mas com aumento
progressivo da incidência ao avançar da idade chegando a cerca de 20 casos por 100.000 entre
os adultos com 65 anos ou mais. A média de idade à apresentação da doença é de cerca de 70
anos, com relação de três homens para cada 2 mulheres afetadas. Em crianças com menos de
15 anos, é responsável por 20% a 25% das leucemias. Além da idade e do sexo, também são
fatores de risco conhecidos para a LMA: exposição à radiação ionizante e a substâncias
químicas como o benzeno (sendo o tabagismo a forma mais comum de exposição ao benzeno)
e agentes alquilantes. Tratamento quimioterápico prévio também é um importante fator de
risco (Estey & Döhner 2006; Queliane et al. 2011; Schürch et al. 2013; Pilger et al. 2006).
Cerca de 10 a 15% dos pacientes portadores de LMA, a adquiriram após tratamento
quimioterápico com drogas citotóxicas (agentes alquilantes ou com ação sobre a DNA
Topoisomerase II) previamente, geralmente para tratamento de tumor sólido (Estey & Döhner
2006). Entretanto, na maioria dos casos, a LMA surge sem motivo aparente, o que leva à
suposição de que fatores individuais e genéticos possam também estar relacionados com a
predisposição para a manifestação da doença (Pilger et al. 2006).
Os eventos moleculares precisos e responsáveis pela transformação leucêmica ainda não
são totalmente conhecidos, no entanto, a ativação de protoncogenes e mutações em genes

17
supressores que regulam o ciclo celular, parecem estar envolvidas na patogênese das
leucemias, uma vez que levam à perda dos mecanismos normais controladores da proliferação
(divisão celular), diferenciação-maturação e/ou da morte celular programada (apoptose)
(Pilger et al. 2006).
1.3 Apoptose
A morte celular programada, que ocorre predominantemente via apoptose, exerce um
papel crucial no desenvolvimento da vida animal e vegetal, uma vez que trata-se de
mecanismo de defesa do organismo contra células danificadas, estressadas ou estimuladas por
quaisquer agentes (Alberts 2008). A indução de apoptose por ocasião de ocorrência de dano ao
DNA remove essas células prejudiciais, evitando um crescimento tumoral. Dessa forma,
falhas no mecanismo de apoptose está associado a proliferação celular inadequada,
desenvolvimento e progressão de tumores e sua resistência a quimioterápicos (Adan et al.
2016; Pistritto et al. 2016). Além disso, a morte celular por apoptose tem a vantagem de não
induzir processo inflamatório (Alberts 2008). Assim, a compreensão dos mecanismos e das
falhas nas vias apoptóticas e sua correlação com a ocorrência do câncer são importantes para o
desenvolvimento de novas terapias e métodos de prevenção.
A maquinaria envolvida na sinalização e efetivação da apoptose é complexa e envolve
muitas vias, sendo iniciada por duas vias principais, a via intrínseca ou a via extrínseca. A via
extrínseca é disparada após ativação de receptores específicos na membrana celular
(receptores de morte celular). Esses receptores pertencem à família do Fator de Necrose
Tumoral (que inclue o receptor para o TNF e os receptores Fas e TRAIL) (Pistritto et al.
2016). Já a via intrínseca é dependente da mitocôndria que libera Citocromo C para o citosol.
Esse se liga a uma proteína adaptadora chamada Apaf1, iniciando a ativação da cascata de
proteases chamadas caspases. Algumas vezes a via extrínseca recruta a via intrínseca para
amplificar o sinal, através da ativação um membro da família Bcl2. Por fim, ambas as vias
convergem para um final em comum, que envolve a ativação das caspases, que clivam
moléculas regulatórias e estruturais, culminando na morte da célula (Adan et al. 2016).
Nesse contexto em que a falha na execução da maquinaria para indução de apoptose
está ligada ao desenvolvimento de neoplasias malignas, o gene supressor tumoral TP53 (que
codifica a proteína p53) tem um papel crucial, pois regula uma extensa rede que controla a
integridade do genoma frente a danos celulares como alterações cromossômicas, hipóxia,
ativação de oncogenes celulares, oncoproteínas virais e depleção de metabólitos (Menendez et

18
al. 2007; Maximov & Maximov 2008). Mais de 50% de todos os tumores malignos são
causados por mutações no gene TP53 e a inativação de vias reguladas pela p53
dramaticamente aumenta a suscetibilidade ao desenvolvimento de câncer (Kim 2016). Muitos
quimioterápicos induzem apoptose por mecanismo dependente de p53 e a falta de p53 faz o
câncer menos sensível a eles.
A proteína p53 tanto está relacionada ao bloqueio do ciclo celular, em caso de dano ao
DNA quanto também, como já citado, desencadeia mecanismos para efetuar o reparo ou
induzir a apoptose. O bloqueio do ciclo se dá através de sua ação regulando e estimulando a
expressão do gene p21, cujo produto atua inibindo a atividade de cinases dependentes de
ciclinas necessárias para a transição entre as fases do ciclo celular G1 e S (Karimian et al.
2016).
A proteína p53 também regula a expressão de mediadores anti e pró apoptóticos, como
os membros da família Bcl-2, sendo os alvos mais importantes as proteínas pró-apoptóticas:
BAX, Noxa e PUMA (Norbury & Zhivotovsky 2004). A apoptose regulada pela p53, portanto,
quando ativada pela via intrínseca, é dependente da ativação de Bcl-2, que controla a liberação
de Citocromo C da mitocôndria, desencadeando a morte celular programada (Rossi et al.
2015).
A proteína BAK também pertence à família Bcl-2, subfamília BH123 e tem ação pró-
apoptótica. De forma semelhante à BAX, após receber o gatilho apoptótico pela via intrínseca,
ativa e forma oligômeros na membrana externa da mitocrôdria induzindo a liberação de
citocromo C e outras proteínas para o citosol (Alberts 2008).
1.4 Terapia na Leucemia Mielóide Aguda
A LMA tem início clínico abrupto e evolução rápida para óbito, caso o paciente não seja
tratado, sendo que o tratamento visa eliminar ou controlar a proliferação de clones leucêmicos,
e baseia-se no uso de poliquimioterapia (Bittencourt et al. 2003; Queliane et al. 2011). A
terapia é realizada em fases, sendo a primeira denominada indução, com o objetivo de induzir
a remissão completa da doença (definida como a presença de < de 5% de blastos na medula
óssea e contagem de neutrófilos e plaquetas em sangue periférico, maiores de 1.000 e
100.000/dL de sangue, respectivamente). Atingindo-se a remissão completa, seguem-se as

19
fases de pós remissão e consolidação, com o objetivo prolongar a remissão completa (Estey &
Döhner 2006).
O tratamento padrão para pacientes menores de 60 anos na fase de indução, inclui o uso de
Citarabina e um antracíclico, através da combinação 7+3, que envolve a infusão contínua por
7 dias de 100 a 200 mg/m2 de superfície corporal por dia do D1 ao D7 e 3 dias de um
antracíclico (Daunorrubicina 60mg/m2 por dia,Idarrubicina 12 mg/m2 por dia ou Mitoxantrona
10-12mg/m2 por dia), do D1 ao D3. Esquemas alternativos como associação da combinação
7+3 com Cladribina ou Fludarabina ou até Citarabina em alta dose (high-dosecytarabine -
HIDAC) associado a Idarrubicina ou Daunorrubicina também podem ser usados (Burnett et al.
2013; Holowiecki et al. 2012). Na fase pós indução, havendo resposta completa, pode-se
repetir mais um ciclo de 7+3 ou Citarabina em alta dose.
Na fase de consolidação, o esquema a ser utilizado envolve o conhecimento da
estratificação da LMA em baixo, intermediário e alto risco. Para baixo risco, HIDAC por 3 a 4
ciclos. Casos classificados de alto risco são elegíveis para o transplante de medula ossea
alogênico (TMO) aparentado ou não. A consolidação para pacientes com risco intermediário
pode envolver HIDAC ou TMO (Mayer 1994; Putten et al. 2011; Willemze et al. 2014; Stein
& Tallman 2012; Schlenk 2014).
Esse regime de indução induz a remissão completa (RC) em cerca de 60 a 80% dos
pacientes mais jovens (≤ 55-60 anos). No entanto, apenas 30% a 40% desses pacientes estão
vivos e livre de doença em 5 anos.
Pacientes acima de 60 anos podem não ser elegíveis para receber quimioterapia intensiva
com Antracíclico e Citarabina. Essa decisão envolve a idade do paciente, a presença de
comorbidades e status funcional. Caso o paciente não seja elegível para receber o esquema de
7+3, pode-se lançar mão de agentes hipometilantes (Azacitidina ou Decitabina) ou Citarabina
em baixas doses até a progressão.
Entre os idosos (> 55–60 anos), a RC é conseguida em 40% a 55% dos casos, com poucos
sobreviventes a longo prazo (10-15%) (Estey & Döhner 2006; Gong et al. 2015; Döhner et al.
2010).
Nos últimos 20 anos não houve grandes mudanças no esquema de tratamento da LMA,
consistindo de forma geral, de quimioterapia citotóxica na indução da remissão, sendo a
Citarabina associada a um antracíclico, como a Daunorrubicina, ou a Idarrubicina, o padrão-

20
ouro para a maioria dos pacientes. Os pacientes mais jovens se beneficiam de várias sessões
de quimioterapia intensiva de consolidação, em sequência. Já aqueles pacientes com fatores
citogenéticos de alto risco ou risco intermediário podem ser direcionados para o Transplante
de Medula Óssea (TMO). A LMA M3, é abordada de forma um pouco diferente dos demais
subtipos sendo associado um agente de diferenciação, o ácido transretinóico (ATRA), em
combinação com a quimioterapia padrão (Schürch et al. 2013; Tallman 2005).
Estratégias foram elaboradas para melhorar a taxa de RC como, por exemplo, o uso doses
mais altas e/ou alternativas de antracíclicos, doses maiores de Citarabina na indução ou na fase
de consolidação, em sequencia de uma indução convencional, a associação de agentes
citotóxicos convencionais, como o Etoposídeo e a associação de novos agentes com diferentes
mecanismos de ação, como a Fludarabina, um análogo de purina ou o Topotecano, ou o uso de
fatores de crescimentos. Para a maioria dos pacientes, nenhuma dessas alternativas se mostrou
mais eficiente do que o regime padrão de duas drogas, embora alguns subgrupos tenham
apresentado um aumento na sobrevida livre de doença (Bradstock et al. 2005; Estey & Döhner
2006; Estey 2001; Tallman 2005).
Os recentes avanços no estudo da biologia e das particularidades genético-moleculares da
Leucemia Mielóide Aguda levam a um subconjunto de LMAs distintas, que podem conduzir à
individualização da terapia, adaptando-a a doença de cada paciente. A estratificação da LMA
em alto, médio e baixo risco, por exemplo, pode auxiliar a determinar qual tratamento pós
remissão poderia ser mais apropriado para cada caso e identificar subgrupos com melhor
resposta a um determinado esquema quimioterápico.
No que se refere à análise citogenética, os pacientes portadores das anormalidades t(8;21),
inv(16) ou t(16;16), coletivamente chamadas do tipo core binding factor, possuem geralmente
menor risco de recaída quando tratados com três ou quatro ciclos de Citarabina em alta dose
após a indução, e geralmente não se beneficiam do TMO alogênico na primeira remissão
completa. No entanto, alterações como deleção do cromossomo 5 ou 7, del(5q), anormalidades
do 3q ou cariótipos com alterações complexas estão associados a alto risco de recaída e, nesse
caso, o TMO alogênico deve ser considerado, uma vez que aumenta a taxa de sobrevida. O
paciente com cariótipo normal, que corresponde a cerca de 45% dos casos é considerado como
risco intermediário (Copelan et al. 2015).
Atualmente já existem identificadas diversas mutações genéticas e expressões gênicas
desrreguladas em portadores de LMA, algumas das quais também relacionadas com o

21
prognóstico. Esses marcadores são particulamente importantes no entendimento do
comportamento heterogêneo dentre pacientes com cariótipo normal, por exemplo.
A FMS-related tirosina kinase 3 (FLT3) é um receptor tirosino-kinase transmembrana que,
quando ativado pelo seu ligante, estimula a proliferação celular. A Duplicação Interna em
Tandem no gene FLT3 (FLT3-DIT) provoca ativação sustentada desse receptor. Portadores de
LMA, com cariótipo normal, mas portando FLT3-DIT tem resposta inferior ao tratamento do
que aqueles que não possuem essa mutação, sendo essa mutação a segunda mais frequente
encontrada em pacientes com LMA.
Já a NPM1 é uma proteína nucleolar codificada pelo gene responsável pela síntese da
nucleofosmina (NPM). É uma proteína multifuncional, agindo durante o ciclo celular.
Mutação isolada da NPM1, na ausência de mutação FLT3-DIT, confere prognóstico favorável
a esses pacientes.
Assim, os desafios atuais impostos pelas alternativas terapêuticas limitadas e muitos
efeitos adversos da quimioterapia da LMA, bem como a heterogeneidade de subgrupos de
pacientes portadores de LMA, considerando a diversidade citogenética e molecular, justificam
o desenvolvimento urgente de compostos baseados em novas classes de moléculas para o
tratamento dessa doença. Analisando-se a situação sob esta perspectiva pode-se constatar a
necessidade imediata de se obter novos protótipos de medicamentos que possam ser incluídos
futuramente no arsenal terapêutico em uso (Döhner et al. 2010; Copelan et al. 2015; Licínio &
Silva 2010).
1.5 Novas drogas: Desenvolvimento
1.5.1 ALCALOIDES MARINHOS
Os alcalóides marinhos são uma das principais classes de metabólitos secundários isolados
de diferentes organismos marinhos tais como cianobactérias, tunicados, ascídios, anêmonas e
esponjas (Mehbub et al. 2014). Esses compostos possuem diferentes atividades biológicas já
descritas na literatura tais como: anti-inflamatória, antimalárica e antiviral. No entanto, os
alcalóides marinhos bem como os seus análogos sintéticos destacam-se pela sua marcante
citotoxicidade, o que os torna atraentes para o desenvolvimento de novas drogas para o
tratamento do câncer. Recentemente foi relatado que cerca de 36% dos compostos compostos

22
isolados de esponjas marinhas capazes de induzir apoptose pertencem à classe dos alcalóides
(Essack et al. 2011).
A Trabectedina, por exemplo, é um alcaloide tetrahidroisoquinolínico isolado do tunicato
caribenho Ecteinascidiaturbinata, utilizado como agente antineoplásico. Desenvolvido pela
PharmaMar em 1996, a Trabectedina (comercializada sob o nome de Yondelis®), foi o
primeiro fármaco a ser aprovado para o uso clínico no tratamento de sarcomas de tecidos
moles, autorizado pela European Medicine Agency (EMEA) para a comercialização em 2007
(Costa-Lotufo et al. 2009). Atualmente, está em fase de testes pré-clínicos para o tratamento
de câncer de ovário, próstata, mama e tumores sólidos pediátricos (Oliveira et al. 2012). A
Trabectedina teve seu uso aprovado em 57 países aumentando as vendas em 17 milhões de
euros no primeiro trimestre de 2010 (Felício et al. 2012).
Esponjas marinhas da ordem Haplosclerida em especial (gênero Haliclona, Xestopongia e
Amphimedon) são fontes ricas de diversos alcalóides estruturalmente complexos. Muitos dos
alcalóides da classe dos 3-alquilpiridínicos (3-APA) foram isolados desses organismos. O que
caracteriza os alcalóides do tipo 3-APA é sua maior simplicidade estrutural. Eles possuem
uma cadeia alifática longa diversamente funcionalizada ligada a posição 3 do anel piridínico.
A teoneladina C é um exemplo de alcalóide do tipo 3-APA de baixa complexidade estrutural
para o qual já foi descrita atividade antimicrobiana e citotóxica (Gonçalves et al. 2014).
Análogos sintéticos de alcalóides do tipo 3-APA já apresentaram atividade pró-apoptótica
contra linhagens tumorais de carcinoma de colon humano (RKO-AS-45-1) (Gonçalves et al.
2014; Pereira et al. 2012).
2 JUSTIFICATIVA
A busca de novos compostos com potencial atividade antitumoral que possam
substituir ou melhorar os atuais tem sido uma necessidade médica e prioridade em pesquisa.
Diante da limitação terapêutica para o tratamento da LMA, associada a efeitos adversos
significativos, principalmente em pacientes maiores de 65 anos, é importante o estudo e
investigação de novos compostos que poderão ser incorporados no arsenal terapêutico.
3 OBJETIVOS
3.1 Objetivo geral

23
Testar análogos de alcaloides marinhos do tipo 3-alquilpiridínicos, em linhagem de LMA
tendo como perspectiva a obtenção de novos agentes terapêuticos.
3.2 Objetivos específicos
• Realizar os testes de avaliação da citotoxicidade, através de ensaio de viabilidade
celular, com os seguintes análogos de alcaloides: TEOC-5B, THP-002, OH-002, SCN-
002, SCN-003;
• Comparar os resultados obtidos nos testes de análogos de alcaloides marinhos com os
resultados de testes com quimioterápico padrão;
• Realizar ensaio de Genotoxicidade e Mutagênese com os compostos estudados para
identificar os diferentes riscos genéticos que podem estar associados à exposição
destes ;
• Realizar ensaios adicionais por metodologia molecular para confirmar apoptose e
tentar rastrear pistas do mecanismo de ação.
4 MATERIAIS E MÉTODOS
4.1 Obtenção dos compostos
Os compostos utilizados neste estudo foram cedidos pelo Prof. Dr. Gustavo
Henrique Ribeiro Viana do Laboratório de Química Orgânica da Universidade Federal
de São João Del Rei (USFJ). No total, foram testados 05 compostos análogos de
alcalóides marinhos do tipo 3-alquilpiridínicos. As estruturas dos compostos estão
apresentadas na Tabela 3.
Tabela 3 - Estrutura dos 5 compostos análogos de alcaloides marinhos do tipo 3-
alquilpiridínicos
Código da Amostra Estrutura e M.M.
TEOC-5b

24
THEOC(ANALOG)
THP-002
THEOC(ANALOG)
OH-002
THEOC(ANALOG)
SCN-002
THEOC(ANALOG)
SCN-003
4.2 Solubilização dos compostos
Os compostos foram solubilizados para a realização dos testes biológicos, sendo
utilizado o solvente dimetilsulfóxido (DMSO) (Sigma-Aldrich, St. Louis, MO, EUA)
resultando numa concentração de 10 mg/mL. Essa solução foi mantida a uma
temperatura próxima de 4 ºC. O composto antitumoral utilizado como controle positivo,
foi o Etoposídeo (Sigma/Aldrich), o qual passou por uma diluição equivalente dos
compostos sintéticos utilizando o mesmo solvente, DMSO e obtendo a mesma
concentração de 10 mg/mL. Para o controle negativo foram utilizadas células sem o
tratamento.
No momento dos testes de citotoxicidade, foram feitas cinco diluições seriadas
utilizando-se o meio de cultura RPMI-1640 (Sigma/Aldrich) com 1% de Soro Fetal
Bovino, (SFB) (Gibco, EUA) nas seguintes concentrações a partir da solução estoque:
1000 μg/mL, 100 μg/mL, 10 μg/mL, 1 μg/mL e 0,1 μg/mL. No volume final a
concentração de DMSO foi de 0,01%. Os meios de cultura possuem em sua
composição além do SFB, 1% de antibióticos (penicilina e estreptomicina 100 U/mL).

25
4.3 Cultivo de linhagens celulares
A linhagem celular humana utilizada para análise de toxicidade foi a THP-1
(Leucemia Monocítica Aguda ATCC TIB-202). Esta linhagem foi cedida pelo
Laboratório de Hematologia Clínica e Molecular da Faculdade de Farmácia – UFMG.
A linhagem foi mantida congelada em criotubos com 5% de DMSO e meio de
cultura RPMI-1640 com 10% de SFB. Os criotubos foram mantidos em nitrogênio
líquido no banco de criopreservação (CryoPlus 7405 / ThermoScientific, EUA). Para
começar o cultivo, os conteúdos dos criotubos foram descongelados e transferidos para
um tubo de centrifuga de 15 mL contendo 2 mL de meio RPMI-1640 com 10% Soro
fetal bovino (SFB), e então submetidos à centrifugação de 988g durante 5 minutos
(mod. CS- 6R, Beckman, EUA). Em seguida descartou-se o sobrenadante. A massa
celular presente no fundo do tubo foi ressuspendida em 5 mL de meio de cultura. A
suspensão celular foi transferida para uma garrafa plástica de cultivo celular T75 (75
cm3, volume de 250 mL) (CorningCostar Inc., EUA) e mantida em estufa
(Thermoelectronco. EUA) a 37 ºC com atmosfera úmida de 5% de CO2.
Em seguida, realizou-se a manutenção da linhagem, no qual o meio foi trocado a
cada 48-72 horas para garantir a renovação dos nutrientes. Com o auxílio de um
microscópio invertido (mod. Olympus, CKX 41) a morfologia celular foi observada
diariamente como forma de acompanhamento da cultura. Para a contagem de células, o
conteúdo da garrafa plástica de cultivo celular era transferido para tubo de centrifuga de
15ml, centrifugadas por 5 minutos a 988g e o sobrenadante descartado. A massa celular
foi ressuspendida em 5 ml de meio RPMI-1640 com 10% de SFB. Desta suspensão de
células foi retirado 50 µL para misturar com 50 µL de azul de tripan para contagem
manual de células viáveis em câmara de neubauer. O azul de tripan cora o citoplasma de
células mortas possibilitando assim a contagem de células viáveis. Posteriormente à
contagem, a suspensão celular foi distribuída em placas de 96 poços (1 x 105
células/100 µL por poço) (CorningCostar Inc., EUA) para realização do ensaio de
viabilidade celular.
O grupo de pesquisa do Laboratório de Bioquímica Medicinal da UFSJ também
utilizou previamente a linhagem não tumoral de células de pulmão humano WI-26VA-
4, cujos dados foram citados nesse trabalho para fins de comparação com os dados
obtidos.

26
4.4 Ensaio de viabilidade celular
As moléculas sintéticas de análogos de alcalóides marinhos tiveram suas
citotoxicidade avaliadas através do ensaio de 3-(4,5-dimetil-2-tiazolil)-2,5-difenil-
tetrazólio (MTT). Este método é baseado na atividade da enzima mitocondrial
succinatodesidrogenase que se encontra ativa apenas em células viáveis
metabolicamente funcionais. Essa enzima reduz o sal de tetrazólio solúvel de cor
amarela em um produto insolúvel de cor violeta, cristais de formazan cuja quantidade
pode ser determinada por espectrofotometria, onde a intensidade da cor resultante da
dissolução dos cristais é proporcional à atividade da enzima e, por conseguinte ao
número de células viáveis (Carmichael et al. 1987; Park et al. 1987).
Para a avaliação da atividade citotóxica foi feita um plaqueamento de 3,095 x
106 µL de células em placas de 96 poços em meio RPMI-1640 acrescido de 10% de
SFB. Após o plaqueamento celular, as placas foram incubadas em estufa por 24 horas a
37 ºC em atmosfera de 5% de CO2 em ambiente úmido, para conseguinte adição das
amostras a serem testadas.
Após o período de incubação, as células foram lavadas com solução salina
tamponada (PBS). Os testes de citotoxicidade foram realizados em cinco diluições
seriadas na escala decimal a partir da solução estoque (compostos e controle positivo),
utilizando RPMI-1640 com 1% de suplementação de SFB. Cada concentração foi
testada em triplicata e cada ensaio foi também repetido em triplicata. Após uma
incubação de 48 horas, o meio em cada poço foi recolhido e as células foram lavadas
com PBS novamente. O efeito citotóxico foi medido com o ensaio colorimétrico MTT.
Para determinar a viabilidade das células, 100 µL do sal tetrazólico MTT (5 mg/mL) foi
adicionado a cada poço e as células foram cultivadas em adicional incubação durante 3
horas. Retirou-se o sobrenadante e aplicou-se 50 µL de DMSO em cada poço, para
solubilizar o produto insolúvel de formazan. A leitura foi realizada em um leitor de
microplacas através da espectrofotometria no comprimento de onda de 550 nm. A
concentração que reduz a viabilidade celular em 50% (IC50) na presença dos compostos
testes e do controle positivo, foi determinada em comparação com células cultivadas
sem a presença de compostos (consideradas 100% de viabilidade).

27
4.5 Ensaio de Genotoxicidade e Mutagênese
Os ensaios de genotoxicidade e mutagênese foram realizados no Laboratório de
Biologia Celular e Mutagênese - UFSJ do prof. Dr. Fábio Vieira dos Santos.
4.5.1 Ensaio de micronúcleo com bloqueio de citocinese (CBMN)
O ensaio de micronúcleo foi baseado na metodologia de Fenech (Fenech 2000),
com o objetivo de analisar o potencial mutagênico do composto que apresentou melhor
resultado no ensaio de citotoxicidade nas células da linhagem THP-1. As células foram
distribuídas em placas de 24 poços (2,5 × 105 células/poço) e as placas foram incubadas
a 37 °C numa atmosfera úmida de 5% de CO2.
Após 24 horas, o meio de cultura foi substituído e as células foram tratadas com
o composto-teste em três concentrações diferentes e diluídos em meio de cultura. Em
seguida permaneceram incubadas por 3 h. O grupo referente ao controle negativo foi
tratado com PBS, e o grupo de controle positivo recebeu o tratamento com
metanossulfonato de metila (MMS-400 uM).
Ao término do tempo de tratamento descrito, as células foram lavadas duas
vezes com PBS e centrifugadas durante 5 min a 1200 rpm. O pellet foi suspenso em
meio de cultura completo contendo citocalasina-B a uma concentração final de 3 µg/mL
e essa suspensão permaneceu na incubadora sob as mesmas condições descritas
anteriormente.
Passadas 24 horas do tratamento com citocalasina-B e consequente bloqueio da
citocinese, esse material foi centrifugado por 5 minutos a 1.200 rpm e o sedimento foi
ressuspendido em solução hipotônica gelada (1% de citrato de sódio), juntamente com
uma gota de formaldeído a 1% e homogeneizada cuidadosamente com uma pipeta de
Pasteur. Foi realizado nova centrifugação durante 5 min a 1200 rpm com o pellet
resultante ressuspendido em 5 mL de solução fixadora (metanol/ácido acético 3:1 (v/v)).
Em seguida, os tubos foram novamente centrifugados durante 5 min, e o sobrenadante
foi descartado. As lâminas utilizadas para disposição do material celular estavam
limpas, geladas e com um filme de água destilada em sua superfície, onde colocou-se
três gotas da suspensão celular.

28
Foi confeccionado um total de 10 lâminas em cada experimento, sendo o mesmo
realizado em três repetições independentes. As lâminas foram fixadas em metanol
gelado durante 5 minutos, secas ao ar e armazenadas até à coloração.
No momento da análise citogenética, as lâminas foram coradas com DAPI (4 ',
6-diamidino-2-fenilindol) diluído em tampão fosfato (0,06 M de Na2HPO4 e 0,06 M de
KH2PO4, pH 6,8) durante 2 min, e lavadas com água destilada. Foram analisadas sob
um microscópio de fluorescência (Zeiss, Axioscope) com um filtro de excitação de 365
nm e um filtro barreira de 445/450 nm. Mil células binucleadas (BNC) com citoplasma
bem preservado foram analisadas para cada concentração da droga teste, com o objetivo
de estabelecer a frequência de células micronucleadas, em um teste cego. As células que
continham 1-3 micronúcleos foram consideradas para análise.
A influência dos compostos sobre a divisão celular foi avaliada calculando-se o
Índice de Divisão Nuclear (NDI) nas células THP-1. As mesmas lâminas analisadas
para o teste do micronúcleo foram utilizadas para essa determinação. Foram analisadas
500 células íntegras, contadas utilizando microscopia de fluorescência tal como descrito
acima, verificando-se a presença de 1ou mais núcleos em cada uma delas.
O índice de divisão nuclear (NDI) foi calculado para concentração, de acordo
com a fórmula:
NDI = (Número de células mononucleares + 2 x Número de células binucleadas
+ 3 x No. de células trinucleados + 4 x No. de células quadrinuclear) / Nº total de
células.
4.6 Detecção de Caspase 3
Foi selecionado para esse experimento, o composto que apresentou melhor
resultado no ensaio de citotoxicidade. Para a investigação da ativação da Caspase 3 após
o tratamento com o composto-teste foi utilizado o Kit colorimétrico da R&D Systems
(Wiesbaden-Nordenstadt, Alemanha) de acordo com as instruções do próprio fabricante.
Neste ensaio, 3x106 células foram semeadas em garrafas de cultura 25 cm2 e
incubadas por período de 48 horas na ausência ou presença do composto-teste. Para o
controle positivo, as células foram tratadas com Etoposideo. Após incubação as células
foram inicialmente centrifugadas e o pellet de células formado foi incubado com 75µl
tampão de lise em banho de gelo por 10 min. Os homogeneizados foram centrifugados a
20.000 g durante 01 min e o sobrenadante foi incubado com o substrato para caspase-3

29
(DEVD-pNA) a 37°C por 2h. A absorbância de cada amostra foi registrada a 405nm e o
nível da atividade da caspase-3 foi diretamente proporcional à reação de cor.
4.7 Análise de expressão gênica
4.7.1 Extração de RNA
A extração do RNA total foi realizada com o reagente Trizol LS (Introgen,
EUA) seguindo-se rigorosamente as recomendações do fabricante. A extração foi obtida
com o composto selecionado no ensaio de viabilidade celular.
Essa extração foi realizada 24 e 48 horas após o tratamento com o composto-
teste nas concentrações de IC50, 2 x IC50 e IC50/2. Para o controle foi realizado a extração
de RNA das células não tratadas. As preparações das placas de 96 poços para a extração
foram equivalentes às preparadas para o teste de citotoxicidade, com uma quantidade de
plaqueamento de 3x106 células por poço.
O meio de cultura presente na placa foi removido para a adição direta de 10 µL
de reagente Trizol LS, para que houvesse a lise das células. Homogeneizou-se e
incubou-se por 5 minutos em temperatura ambiente para permitir a completa
dissociação do complexo nucleoproteína sendo o conteúdo dos poços transferidos para
um tubo de 1,5 mL. Depois foram adicionados 3 µL de clorofórmio (Merck), para
separação das fases orgânica e aquosa.
As amostras foram agitadas vigorosamente por 15 segundos e incubadas por 10
minutos à temperatura ambiente. A seguir as amostras foram centrifugadas a 12000 g
por 15 minutos a uma temperatura de 4 ºC. A fase aquosa (camada superior) foi
transferida para outro tubo estéril e o restante foi descartado. Na fase aquosa, contendo
o RNA total, foram adicionados 7μL de isopropanol 100% (Merck) para precipitação
do RNA. Seguiu-se uma nova incubação por 10 minutos a temperatura ambiente. As
amostras foram novamente centrifugadas a 12000 xg por 10 minutos (4ºC) e o
sobrenadante descartado. O sedimento formado foi lavado com 14 µL de etanol (Merck)
75% (diluído em água DEPC 0,1% estéril). As amostras foram homogeneizadas e
centrifugadas a 7500 xg por 5 minutos (4 ºC) e logo após o sobrenadante descartado
novamente. O pellet de RNA formado foi ressuspendido com 40 µL de água livre de
RNAse. Em seguida foram incubados por 15 minutos a uma temperatura de 60 ºC para

30
completa dissolução do RNA. Por fim o RNA total foi usado para síntese de cDNA ou
armazenado a -80 ºC até sua utilização.
Para quantificar o RNA total obtido após o processo de extração, as amostras
foram analisadas por espectrofotometria (Nanovue, GE Healthcare Life Sciences, UK),
através da leitura simultânea da densidade ótica nos comprimentos de onda 260 e 280
nm. Para a verificação da pureza do RNA extraído foi calculado a razão entre a
absorbância medida a 260 nm e a 280 nm. Valor entre 1,5 e 2,0 indicou grau de pureza
satisfatória.
4.7.2 Confecção de cDNA
Os RNAs extraídos foram submetidos à reação de transcriptase reversa para
obtenção de cDNA, seguindo-se rigorosamente as instruções do fabricante ( High
CapacitycDNA Reverse Transcription kit® (AppliedBiosystems, USA). Ao término
dessa etapa foram obtidos 20 µL de cDNA, que foram armazenados a -20 ºC para
serem posteriormente utilizados para a análise de expressão gênica. Para verificar a
integridade dos RNAs extraídos e a qualidade dos cDNAs sintetizados realizou-se a
amplificação através da reação em cadeia de polimerase (PCR) do fragmento de cDNA
correspondente ao RNA mensageiro (RNAm) da gliceraldeído-3-fosfato-desidrogenase
(GAPDH) e visualizou-se os resultados através do gel de poliacrilamida a 8%.
4.7.3 PCR em tempo real
Para que os experimentos de qRT - PCR, que utilizam o fluoróforo SYBR®
Green, ocorram com alta eficiência, foi necessário realizar uma série de padronizações,
e a primeira delas foi a busca por uma temperatura ideal de anelamento para cada
sequência de oligonucleotídeos (Tabela 4) a serem analisados. Essa padronização foi
realizada através de PCR convencional, onde as reações foram processadas em um
termociclador (AppliedBiosystems, Foster, CA, USA). As reações de amplificação
compreendem uma desnaturação inicial a 94°C por 3 minutos, seguida de 35 ciclos de
desnaturação a 94°C por 0,5 minutos, anelamento no qual a temperatura vai ser
determinada por 1 minuto e extensão a 72°C por 2 minutos, com extensão final a 72 °C
por 10 minutos.

31
A segunda padronização foi a titulação dos oligonucleotídeos iniciadores, ou
seja, realizou-se uma série de reações em que se variou a concentração dos iniciadores,
o que possibilitou a escolha da concentração mais adequada. Para isso, foram feitas
diluições dos iniciadores, que variaram entre as concentrações de 100 a 600 nanomolar
(nM). Todas as reações foram feitas em triplicatas, com controles negativos para cada
combinação de iniciadores. As condições encontradas foram posteriormente testadas na
construção de uma curva padrão, construída a partir de diluições seriadas do pool de
cDNA. As curvas padrão serviram como teste da eficiência da reação, onde uma curva
padrão ótima apresenta valor de eficiência de 1 (100% de amplificação por ciclo),
situação na qual as concentrações da curva devem corresponder com as diluições
realizadas.
Tabela 4 -Sequência dos oligonucleotídeos iniciadores diretos (F) e reversos (R)
utilizados na qRT-PCR.
Primer Sequências Tamanho do
Amplicon (pb)
TP53 - F 5′- TGCAGCTGTGGGTTGATTCC -3′ 396
TP53 - R 5′- AAACACGCACCTCAAAGCTGTTC -3
P21 - F 5′- TGAGCCGCGACTGTGATG -3′ 82
P21 – R
BAK – F
BAK – V
5- GTCTCGGTGACAAAGTCGAAGTT -3′
5′-TCTGGCCCTACAC -3′
5′-ACAAACTGGCCCAACAGAAC -3′
201
Após o término da padronização foram realizadas reações de PCR em tempo real
para o cálculo da expressão gênica relativa, utilizando-se o método do ΔΔCT
comparativo. Todas as reações de qRT - PCR foram realizadas em equipamento
StepOne Real-Time PCR (AppliedBiosystems, Foster, CA, USA), utilizando kit SYBR
Green Master Mix (AppliedBiosystems, Foster, CA, USA). Este kit contém todos os
componentes (exceto oligonucleotídeos iniciadores, amostras e água), necessários para
as reações de PCR: tampão 2X, dNTPs, MgCl2, SYBR Green I Dye, AmpliTaq Gold®
DNA polimerase e ROX como referência passiva. Os passos do ciclo de amplificação
foram: 50 °C por 2 minutos, 95 °C por 10 minutos, 60 °C por 1 minuto, em um total de
40 ciclos.

32
A avaliação da expressão do RNAm do gene de interesse foi realizada por PCR em
tempo real com o uso de ensaioTaqMan (AppliedBiosystems, EUA) seguindo-se
rigorosamente as recomendações do fabricante. As reações continham um volume total
de mistura de 20 µL, contendo 1X TaqMan Universal Master Mix II sem UNG 1X
Assay, Mix 20x (AppliedBiosystems) e cDNA (100 ng ). A PCR foi realizada no
equipamento AppliedBiosystems StepOne - Real Time PCR System a 50 °C durante 2
minutos, 95 °C durante 10 minutos, seguido de 40 ciclos a 95 °C durante 15 segundos e
60 °C durante 1 minuto. A quantificação de cada amostra foi realizada em triplicata e o
método utilizado foi o de comparação de ΔΔCT. Todas as reações foram submetidas às
mesmas condições de análise e normalizadas pelo sinal do corante de referência passiva
ROX para correção de flutuações na leitura decorrentes a variações de volume e
evaporação ao longo da reação.
O resultado, expresso em valor de CT (cyclethreshold), se refere ao número de ciclos
de PCR necessários para que o sinal fluorescente atinja o limiar de detecção e é
inversamente proporcional ao número de cópias da sequência alvo presente na amostra.
Após finalização dos ciclos, um threshold de 0,1 foi estabelecido e os valores CT foram
gerados através do software do equipamento AppliedBiosystems. Todos os transcritos
foram normalizados com o controle endógeno GAPDH. Este atua como controle interno
tendo a finalidade de remover ou reduzir as diferenças da amostragem que muitas vezes
estão relacionadas à quantidade e qualidade do RNA extraído.
Para validar a amplificação de cada gene alvo, foram realizadas diluições seriadas
em escala decimal de cDNA, que foram submetidas à amplificação nas mesmas
condições anteriormente citados, para a montagem da curva padrão. O gráfico gerado
pela concentração de cDNA e os respectivos valores de CT foram empregados para o
cálculo da equação de regressão e para a determinação do coeficiente de relação (R2) e
da eficiência do ensaio.
Os resultados obtidos foram analisados pelo software StepOne v 2.3
(AppliedBiosystems, EUA) e os resultados individuais expressos em valores de
quantificação relativa foram transferidos para planilhas e agrupados de acordo com a
linhagem celular, concentrações e o tempo de tratamento para realização da análise
estatística.
4.8 Análises estatísticas

33
Os valores de IC50 foram calculados usando-se o programa OriginPro 8.0
(OriginLab Corporation, Northampton, MA, USA). Para a análise dos dados com
distribuição normal, foi usado análise de variância (ANOVA) seguido pelo do ensaio de
comparação múltipla de Tukey. Para aqueles sem distribuição normal, utilizamos
ANOVA para comparação de mais grupos: o teste não paramétrico de Kruskal-Wallis
seguido pelo teste de comparação múltipla Holm-Sidak. Foi considerado significativo
valor de p<0,05.
5 RESULTADOS E DISCUSSÃO
5.1 Ensaio de citotoxicidade e viabilidade celular
A fim de se avaliar a atividade citotóxica dos compostos, foram realizados ensaios
de MTT na linhagem celular humana THP-1. Em trabalho prévio realizado pelo grupo
de pesquisa do Laboratório de Bioquímica Medicinal da UFSJ, esses compostos
também foram testados previamente em uma linhagem não tumoral: WI-26VA-4 e
utilizando o etoposídeo como composto padrão (Gonçalves et al. 2014). O
quimioterápico Citarabina também foi usado na linhagem THP-1 para cálculo do IC50,
para efeito comparativo. Na Tabela 5 estão apresentados os dados dos testes de
citotoxidade e viabilidade celular.
Tabela 5 -Valores de IC50 obtidos em ensaios in vitro com linhagens celulares de THP-
1 e WI-26VA-4.
Código da Amostra Estrutura e M.M.
IC50 (µM) ± DP
THP-1 WI-26-
VA4
TEOC-5b
40.31±12.93 45.52±2.71
THEOC(ANALOG)THP-
002
4,05±0,1 6,4±0,7

34
THEOC(ANALOG)OH-
002
71.12±5.79 99,1±11,2
THEOC(ANALOG)SCN-
002
41.93±2.02 13,28±4,24
THEOC(ANALOG)SCN-
003
69.92±3.47 14,64±2,39
Etoposídeo ___ 5.8±0.63 4,3 ± 1,34
Citarabina ----- 39,7±5,8 ----
Observou-se que para a linhagem THP-1, com exceção do composto de código
THEOC(ANALOG)THP-002, todos os outros quatro compostos apresentaram valores de
IC50 acima do obtido pela Citarabina e pelo Etoposídeo, sendo esse último já usado como
quimioterápico (Sinkule 1984), se mostrando inefetivos. No entanto o composto
THEOC(ANALOG)THP-002 apresentou um valor de IC50 particulamente baixo, sugerindo
ser altamente citotóxico para essa linhagem.
Para a linhagem WI-26-VA4, os valores de IC50 variaram de 4,3 µM
(Etoposideo) até 99 µM. Não foi realizado experimento com Citarabina nessa linhagem.
Análogos sintéticos de 3APA vem sendo testados contra linhagens celulares com
resultados promissores e em muitos estudos, sua ação citotóxica vem decorrente de
indução de apoptose (Essack et al. 2011; Pereira et al. 2012; Gonçalves et al. 2014). De
fato, a morte celular induzida por apoptose é extremamente vantajosa em um contexto
de desenvolvimento de novos fármacos, uma vez que ocorre sem liberação de conteúdo
citosólico no meio extracelular e evita o início de cascata inflamatória.
5.2 Ensaio de Genotoxicidade e Mutagênese
Após avaliação e análise prévia da atividade citotóxica dos análogos sintéticos de
alcalóides do tipo 3-APA e com os resultados demonstrados na Tabela 4, foi visualizado
que o composto THP-002, apresentou uma alta atividade citotóxica, sendo utilizado na
realização do teste de micronúcleo, testado em 3 diferentes concentrações.
35
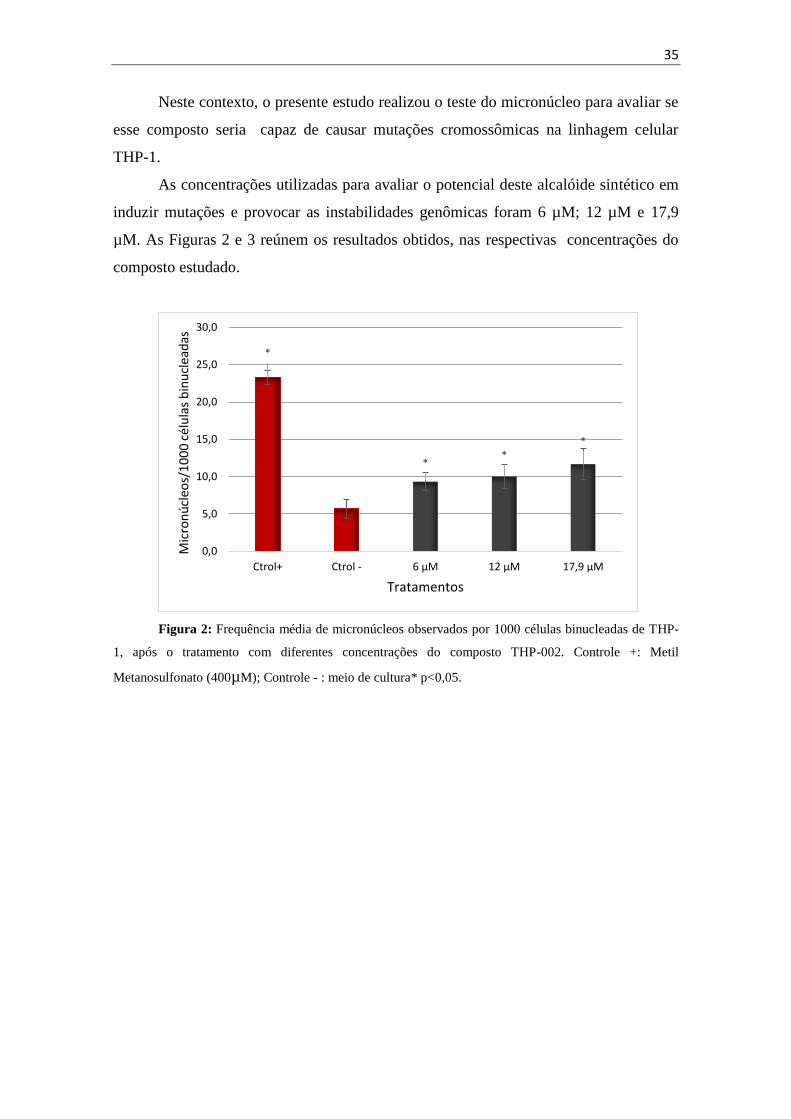
Neste contexto, o presente estudo realizou o teste do micronúcleo para avaliar se
esse composto seria capaz de causar mutações cromossômicas na linhagem celular
THP-1.
As concentrações utilizadas para avaliar o potencial deste alcalóide sintético em
induzir mutações e provocar as instabilidades genômicas foram 6 µM; 12 µM e 17,9
µM. As Figuras 2 e 3 reúnem os resultados obtidos, nas respectivas concentrações do
composto estudado.
Figura 2: Frequência média de micronúcleos observados por 1000 células binucleadas de THP-
1, após o tratamento com diferentes concentrações do composto THP-002. Controle +: Metil
Metanosulfonato (400µM); Controle - : meio de cultura* p<0,05.
*
**
*
0,0
5,0
10,0
15,0
20,0
25,0
30,0
Ctrol+ Ctrol - 6 µM 12 µM 17,9 µM
Mic
ron
úcl
eos/
10
00
célu
las
bin
ucl
ead
as
Tratamentos

36
Figura 3: Índice de Divisão Nuclear das células THP-1, após o tratamento com diferentes
concentrações do composto THP-002. Controle +: Metil Metanosulfonato (400µM); Controle - : meio de
cultura * p<0,05.
Como pôde ser observado na figura 2, o tratamento com composto THP-002
resultou em um aumento significativo da frequência de células binucleadas com
micronúcleos em comparação ao controle negativo nas três concentrações estudadas,
ocorrendo mais acentuadamente na maior concentração testada, de 17,9 µM. Além
disso, foi observado que o tratamento com esse composto, nas três concentrações
estudadas, levou a uma diminuição do Índice de Divisão Nuclear, com o relação ao
observado no controle negativo (figura 3).
Esses resultados indicam que o composto THP-002 induziu mutação
cromossômica nas células da linhagem THP-1, provavelmente levando à parada do ciclo
celular, reduzindo a taxa de proliferação das mesmas.
A ocorrência dos micronúcleos representa uma resposta integrada da
instabilidade de cromossomos, fenótipos e alterações celulares causadas por defeitos
genéticos e ou exposição exógena a agentes genotóxicos. Frente a isto, os estudos de
mutagenicidade ajudam na avaliação da segurança e eficácia dos produtos estudados
(Fenech 2000; Fenech et al. 1999).
Além disso, considerando que a atividade anticâncer de quimioterápicos e da
radioterapia é em grande extensão diretamente relacionada com sua propriedade de
induzir dano no DNA, a identificação de um composto capaz de induzir a formação de
MN permite inferir que esse composto tem o potencial de ativar a morte celular por
*
* * *
1,15
1,20
1,25
1,30
1,35
1,40
1,45
1,50
1,55
1,60
Ctrol+ Ctrol - 6 µM 12 µM 17,9 µM
Índ
ice
de
Div
isão
Nu
clea
r
Tratamento

37
apoptose, de forma que passa a ser um potencial candidato a quimioterápico (Utani et al.
2010; Abbotts, Rachel; Thompson, Nicola; Madhusudan 2014).
5.3 Detecção de caspase 3
Conforme já citado, o tratamento do câncer convencional envolve agentes que
promovem dano ao DNA. No entanto, podem potencialmente atingir células normais
além das tumorais, que se dividem indiscriminadamente. Dentre tais agentes, cita-se a
radiação ionizante, agentes alquilantes, inibidores da DNA topoisomerase, etc (Essack
et al. 2011).
Os antracíclicos, dentre os quais a Daunorrubicina e a Idarrubicina que fazem
parte do esquema de tratamento standart para LMA, e parecem atuar como inibidores
da DNA topoisomerase. Já a Citarabina, sendo um análogo do nucleosídeo pirimidina,
age como um anti-metabólito inibindo a síntese do DNA (Martin et al. 2009; Cros et al.
2004).
Com o objetivo de elucidar o mecanismo de ação pelo qual o composto THP-002
leva a morte celular, após o tratamento das células THP-1 com ele, foi avaliada a
presença de atividade de caspase 3. A Figura 4 reúne os resultados obtidos nesse
experimento.
Figura 4: Quantificação da atividade da Caspase 3 por ensaio colorimétrico na linhagem THP-
1(*p<0.05), observados com o Controle – (ausência do composto-teste), com o composto-teste THP-002
em concentração de 4,05±0,1µM/mL e com o Controle + (Etoposídeo).
0
0,05
0,1
0,15
0,2
0,25
Control - 4.05±0.1 Control+
Ati
vid
ad
e d
e C
asp
ase
-3
38
Esses dados sugerem que a atividade enzimática da Capase-3 aumentou nas
células que receberam tratamento com o composto THP-002, na concentração de 4,0
µM/mL. Dessa forma, infere-se que a citotoxicidade observada com esse composto
envolve dano ao DNA e indução de morte por apoptose.
5.4 Análise da expressão gênica
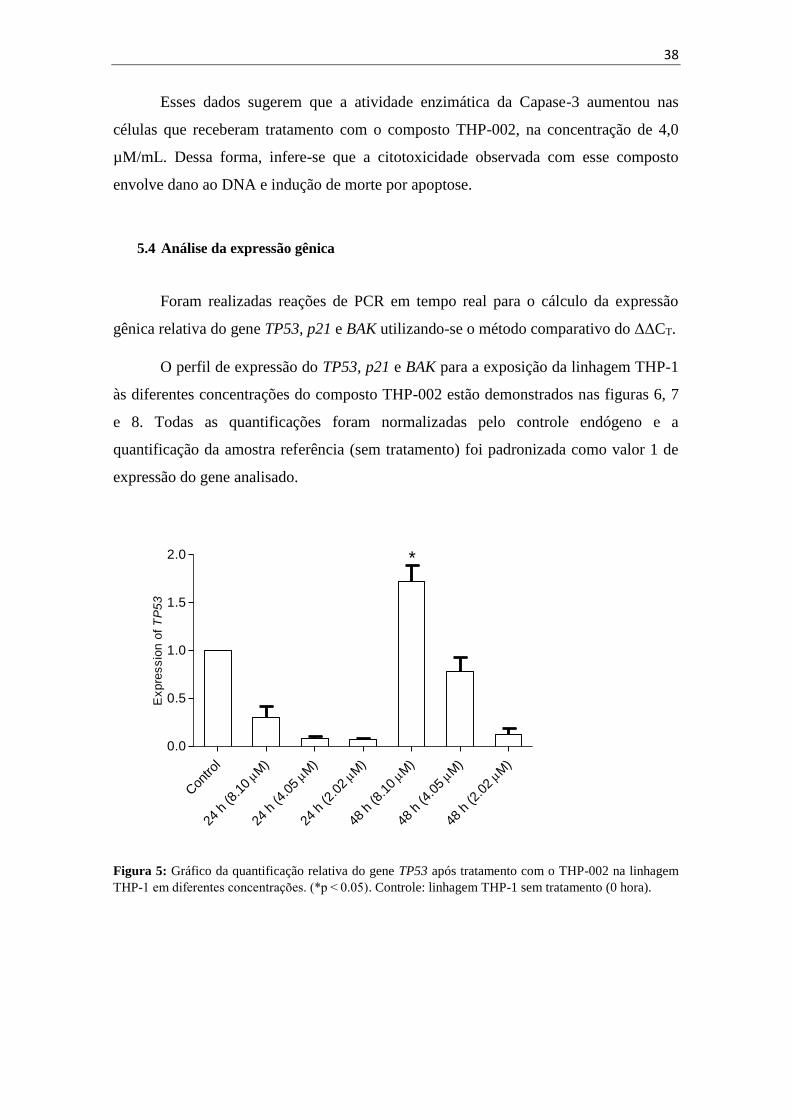
Foram realizadas reações de PCR em tempo real para o cálculo da expressão
gênica relativa do gene TP53, p21 e BAK utilizando-se o método comparativo do ΔΔCT.
O perfil de expressão do TP53, p21 e BAK para a exposição da linhagem THP-1
às diferentes concentrações do composto THP-002 estão demonstrados nas figuras 6, 7
e 8. Todas as quantificações foram normalizadas pelo controle endógeno e a
quantificação da amostra referência (sem tratamento) foi padronizada como valor 1 de
expressão do gene analisado.
Con
trol
M)
24 h
(8.1
0 M
)
24 h
(4.0
5 M
)
24 h
(2.0
2 M
)
48 h
(8.1
0 M
)
48 h
(4.0
5 M
)
48 h
(2.0
2
0.0
0.5
1.0
1.5
2.0 *
Ex
pre
ss
ion
of
TP
53
Figura 5: Gráfico da quantificação relativa do gene TP53 após tratamento com o THP-002 na linhagem
THP-1 em diferentes concentrações. (*p < 0.05). Controle: linhagem THP-1 sem tratamento (0 hora).

39
Con
trol
M)
24 h
(8.1
0 M
)
24 h
(4.0
5 M
)
24 h
(2.0
2 M
)
48 h
(8.1
0 M
)
48 h
(4.0
5 M
)
48 h
(2.0
2
0.0
0.5
1.0
1.5
* * *
*
Ex
pre
ss
ion
of
P2
1
Figura 6: Gráfico da quantificação relativa do gene p21 após tratamento com THP-002 na linhagem
THP-1 em diferentes concentrações. (*p < 0.05). Controle: linhagem THP-1 sem tratamento (0 hora).
Con
trol
M)
24 h
(8.1
0 M
)
24 h
(4.0
5 M
)
24 h
(2.0
2 M
)
48 h
(8.1
0 M
)
48 h
(4.0
5 M
)
48 h
(2.0
2
0
2
4
6
8*
*
* * *
Ex
pre
ss
ion
of
BA
K
Figura 7: Gráfico da quantificação relativa do gene BAK após tratamento com THP-002 na linhagem
THP-1 em diferentes concentrações. (*p < 0.05). Controle: linhagem THP-1 sem tratamento (0 hora).
Ao avaliar o perfil de expressão do gene TP53 (figura 5) e do gene p21 (figura
6), observou-se que o nível de transcritos não aumentou após exposição de 24 h nem de
48 h com a concentração do composto de 4,0 µM. Já o resultado do experimento de
expressão de BAK (figura 7) desmonstra claramente um aumento em sua expressão, em
comparação ao controle após o tratamento por 24 h e 48 h com o composto THP-002.

40
Conforme já citado, o gene TP53, atravé da codificação da fosfoproteína nuclear
p53, tem papel ativo na parada do ciclo celular e indução da expressão de genes de
reparo de DNA danificado, bem como indução de apoptose atuando como um supressor
tumoral. No entanto, as células da linhagem THP-1 apresentam deficiência na expressão
de TP53 (Reisman 2002), sendo que, em modelos de LMA, a detecção de mutações e
delações cromossômicas nesse locus está associado a resistência ao tratamento (Wattel
et al. 1994; Yin et al. 2006). Dessa forma, o tratamento da LMA deve ser focado em
vias independentes de p53 (Ng 2011).
A proteína BAK é uma reguladora chave na sinalização pró-apoptótica. E de
acordo com os resultados apresentados nesse estudo, parece estar envolvida no
mecanismo de apoptose ativando caspase 3 por via independente de p53, fato esse já
descrito também em estudo in vitro com cultura de células de neoplasia gástrica (Tong
et al. 2004; Degenhardt et al. 2002).
Dessa forma, infere-se que o composto THP-002 é um agente potencial de uso
contra a LMA e estudos adicionais são necessários para elucidação mais detalhada dessa
via independente de p53 pela qual é induzida a morte celular.
6 CONCLUSÕES
Dos 5 compostos análogos de alcalóides marinhos do tipo 3-alquilpiridínicos
testados, um deles: THP-002 se mostrou ativo contra a linhagem THP-1.
O ensaio de genotoxicidade evidenciou que o tratamento com composto THP-
002 levou aumento significativo da frequência de células binucleadas com micronúcleos
bem como levou a uma diminuição do Índice de Divisão Nuclear, com o relação ao
observado no controle negativo, concluindo-se que esse composto tem o potencial de
gerar mutação cromossômica.
Houve aumento na atividade da Caspase 3 após tratamento com o composto THP-
002, sugerindo que a morte celular se dá por indução de apoptose.
A análise de expressão gênica evidenciou aumento na expressão de BAK, após
exposição da linhagem THP-1 ao composto THP-002, reforça a hipótese de indução de
apoptose por ativação da via intrínseca, no entanto não houve aumento da expressão de
TP53 nem p21 após 24h de exposição, de forma de se supõe que a ativação da apoptose

41
seja por via independente da p53.
7 Referências
Abbotts, Rachel; Thompson, Nicola; Madhusudan, S., 2014. DNA repair in cancer :
emerging targets for personalized therapy. Cancer Management and Research,
pp.77–92.
Adan, A., Yandim, M.K. & Baran, Y., 2016. Major apoptotic mechanisms and genes
involved in apoptosis. Tumor Biol.
Alberts, B., 2008. Molecular Biology of The Cell 5th ed.,
Bennett, J.M., 1985. Criteria for the Diagnosis of Acute Leukemia of Megakaryocyte
Lineage (M7). Annals of Internal Medicine, 103(3), p.460. Available at:
http://annals.org/article.aspx?articleid=699926 [Accessed January 16, 2016].
Bennett, J.M. et al., 1991. Proposal for the recognition of minimally differentiated acute
myeloid leukaemia (AML-MO). British Journal of Haematology, 78(3), pp.325–
329. Available at: http://doi.wiley.com/10.1111/j.1365-2141.1991.tb04444.x
[Accessed January 14, 2016].
Bennett, J.M. et al., 1976. Proposals for the Classification of the Acute Leukaemias
French-American-British (FAB) Co-operative Group. British Journal of
Haematology, 33(4), pp.451–458. Available at:
http://doi.wiley.com/10.1111/j.1365-2141.1976.tb03563.x [Accessed October 12,
2015].
Bennett, J.M. et al., 1985. Proposed revised criteria for the classification of acute
myeloid leukemia. A report of the French-American-British Cooperative Group.
Annals of internal medicine, 103(4), pp.620–5. Available at:
http://www.ncbi.nlm.nih.gov/pubmed/3862359 [Accessed November 25, 2015].
Bittencourt, R. et al., 2003. Leucemia Mielóide Agura: perfil de duas décadas do
Serviço de Hematologia do Hospital das Clínicas de Porto Alegre - RS. Revista
Brasileira de Hematologia e Hemoterapia, 25(1), pp.17–24. Available at:
http://www.scielo.br/scielo.php?script=sci_arttext&pid=S1516-

42
84842003000100004&lng=en&nrm=iso&tlng=pt [Accessed May 9, 2014].
Bradstock, K.F. et al., 2005. A randomized trial of high- versus conventional-dose
cytarabine in consolidation chemotherapy for adult de novo acute myeloid
leukemia in first remission after induction therapy containing high-dose cytarabine.
Blood, 105(2), pp.481–488.
Burnett, A.K. et al., 2013. Optimization o Chemotherapy for Younger Patients With
Acute Myeloid Leukemia: Results of the Medical Research Council AML 15 Trial.
Journal of Clinical Oncology, 31(27).
Carmichael, J. et al., 1987. Evaluation of a Tetrazolium-based of Chemosensitivity
Testing1 Semiautomated Colorimetrie Assay : Assessment. Cancer Research, 47,
pp.936–942.
Cells, P. et al., 2014. Stem Cell Reports. Stem Cell Reports, 2(6), pp.866–880. Available
at: http://dx.doi.org/10.1016/j.stemcr.2014.03.014.
Copelan, E.A. et al., 2015. Use of molecular markers to determine postremission
treatment in acute myeloid leukemia with normal cytogenetics. Hematology/
Oncology and Stem Cell Therapy, 8(4), pp.143–149. Available at:
http://dx.doi.org/10.1016/j.hemonc.2015.09.003.
Costa-Lotufo, L.V. et al., 2009. Organismos marinhos como fonte de novos fármacos:
histórico & perspectivas. Química Nova, 32(3), pp.703–716. Available at:
http://www.scielo.br/scielo.php?script=sci_arttext&pid=S0100-
40422009000300014&lng=en&nrm=iso&tlng=pt [Accessed May 31, 2016].
Cros, E. et al., 2004. Problems Related to Resistance to Cytarabine in Acute Myeloid
Leukemia. , 45(June), pp.1123–1132.
Degenhardt, K. et al., 2002. BAX and BAK mediate p53-independent suppression of
tumorigenesis. Cancer Cell, 2, pp.193–203.
Döhner, H. et al., 2010. Diagnosis and management of acute myeloid leukemia in
adults: recommendations from an international expert panel, on behalf of the
European LeukemiaNet. Blood, 115(3), pp.453–474. Available at:
http://bloodjournal.hematologylibrary.org/content/115/3/453.short%5Cnhttp://ww
w.bloodjournal.org/content/115/3/453.abstract.

43
Enciso, J., Mendoza, L. & Pelayo, R., 2015. Normal vs. Malignant hematopoiesis: the
complexity of acute leukemia through systems biology. Frontiers in Genetics,
6(September), pp.1–5. Available at:
http://journal.frontiersin.org/Article/10.3389/fgene.2015.00290/abstract.
Essack, M., Bajic, V.B. & Archer, J.A.C., 2011. Recently confirmed apoptosis-inducing
lead compounds isolated from marine sponge of potential relevance in cancer
treatment. Marine drugs, 9(9), pp.1580–606. Available at:
http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=3225937&tool=pmcent
rez&rendertype=abstract [Accessed May 31, 2016].
Estey, E. & Döhner, H., 2006. Acute myeloid leukaemia. Lancet, 368, pp.1894–907.
Available at: http://www.ncbi.nlm.nih.gov/pubmed/24749757.
Estey, E.H., 2001. Comparison of idarubicin + ara-C-, fludarabine + ara-C-, and
topotecan + ara-C-based regimens in treatment of newly diagnosed acute myeloid
leukemia, refractory anemia with excess blasts in transformation, or refractory
anemia with excess blasts. Blood, 98(13), pp.3575–3583. Available at:
http://www.bloodjournal.org/cgi/doi/10.1182/blood.V98.13.3575 [Accessed May
24, 2014].
Felício, R. de, Oliveira, A.L.L. de & Debonsi, H.M., 2012. Bioprospecção a partir dos
oceanos: conectando a descoberta de novos fármacos aos produtos naturais
marinhos. Ciência e Cultura, 64(3), pp.39–42. Available at:
http://cienciaecultura.bvs.br/scielo.php?script=sci_arttext&pid=S0009-
67252012000300013&lng=en&nrm=iso&tlng=pt [Accessed May 31, 2016].
Fenech, M. et al., 1999. Necrosis , apoptosis , cytostasis and DNA damage in human
lymphocytes measured simultaneously within the cytokinesis-block micronucleus
assay : description of the method and results for hydrogen peroxide. Mutagenesis,
14(6), pp.605–612.
Fenech, M., 2000. The in vitro micronucleus technique. Mutation Research, 455, pp.81–
95.
Gekas, C. et al., 2006. Hematopoietic Stem Cells. The American Journal of Pathology,
169(August), pp.338–346. Available at:
http://dx.doi.org/10.2353/ajpath.2006.060312.

44
Gonçalves, A.M.M.N. et al., 2014. Synthesis and Biological Evaluation of Novel 3-
Alkylpyridine Marine Alkaloid Analogs with Promising Anticancer Activity.
Marine drugs, 12(8), pp.4361–4378.
Gong, Q. et al., 2015. High doses of daunorubicin during induction therapy of newly
diagnosed acute myeloid leukemia: A systematic review and meta-analysis of
prospective clinical trials. PLoS ONE, 10(5), pp.1–14.
Greer, J.P., Foerster, J. & Lukens, J.N., 2003. Wintrobe â€TM s Clinical Hematology
11th ed.,
Hall, J.E. & Guyton, A.C., 2011. Tratado de Fisiología médica 12th ed.,
Holowiecki, J. et al., 2012. Cladribine , But Not Fludarabine , Added to Daunorubicin
and Cytarabine During Induction Prolongs Survival of Patients With Acute
Myeloid Leukemia : A Multicenter , Randomized Phase III Study. Journal of
Clinical Oncology, 30(20), pp.2441–2448.
Karimian, A., Ahmadi, Y. & Yousefi, B., 2016. Multiple functions of p21 in cell cycle ,
apoptosis and transcriptional regulation after DNA damage. DNA Repair, 42,
pp.63–71. Available at: http://dx.doi.org/10.1016/j.dnarep.2016.04.008.
Kim, S., 2016. Role of p53 isoforms and aggregations in cancer. Kin and An Medicine,
95(26), pp.1–6.
Lichtman, M., 2006. Williams Hematology 7th ed.,
Licínio, M. & Silva, M. da, 2010. Importância da detecção das mutações no gene FLT3
e no gene NPM1 na leucemia mieloide aguda–Classificação da Organização
Mundial de Saúde 2008. Rev Bras Hematol Hemoter, (55 48), pp.476–481.
Available at: http://www.scielo.br/pdf/rbhh/v32n6/12.pdf.
Martin, Š. et al., 2009. Anthracycline-induced cardiotoxicity : Overview of studies
examining the roles of oxidative stress and free cellular iron. Pharmacological
Reports, pp.154–171.
Maximov, G.K. & Maximov, K.G., 2008. The Role of p53 Tumor-Suppressor Protein in
Apoptosis and Cancerogenesis MB THE ROLE OF p53 TUMOR-SUPPRESSOR
PROTEIN IN APOPTOSIS AND. Biotechnol. & Biotechnol. EQ., 22.

45
Mayer, R.J.. D.T.B.. S.C.A.. B.D.T., 1994. Intensive Postremissions Chemotherapy in
Adults With Acute Myeloid Leukemia. The New England Journal of Medicine,
331(14), pp.896–903.
Mehbub, M.F. et al., 2014. Marine sponge derived natural products between 2001 and
2010: trends and opportunities for discovery of bioactives. Marine drugs, 12(8),
pp.4539–77. Available at:
http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=4145330&tool=pmcent
rez&rendertype=abstract [Accessed May 31, 2016].
Menendez, D. et al., 2007. Changing the p53 master regulatory network : ELEMENT
ary , my dear. Oncogene, 53, pp.2191–2201.
Ng, K.P., 2011. p53 independent epigenetic-differentiation treatment in xenotransplant
models of acute myeloid leukemia. Leukemia, 25(11), pp.1739–1750.
Norbury, C.J. & Zhivotovsky, B., 2004. DNA damage-induced apoptosis. Oncogene,
23, pp.2797–2808.
Oliveira, A.L.L. de, Felício, R. de & Debonsi, H.M., 2012. Marine natural products:
chemical and biological potential of seaweeds and their endophytic fungi. Revista
Brasileira de Farmacognosia, 22(4), pp.906–920. Available at:
http://www.scielo.br/scielo.php?script=sci_arttext&pid=S0102-
695X2012000400028&lng=en&nrm=iso&tlng=en [Accessed May 31, 2016].
Park, J. et al., 1987. Chemosensitivity Testing of Human Colorectal Carcinoma Cell
Lines Using a. Cancer Research, 47, pp.5875–5879.
Pereira, J.R.C.S. et al., 2012. Cytotoxicity evaluation of marine alkaloid analogues of
viscosaline and theonelladin C. Biomedicine & Preventive Nutrition, 2(2), pp.145–
148.
Pilger, D.A., Wagner, S.C. & Citogen, I., 2006. Diagnóstico laboratorial das leucemias
mielóides agudas. J Bras Patol Med Lab, 42(2), pp.77–84.
Pistritto, G. et al., 2016. Apoptosis as anticancer mechanism : function and dysfunction
of its modulators and targeted therapeutic strategies. Aging, 8(4), pp.603–619.
Putten, W. Van et al., 2011. Cytarabine Dose for Acute Myeloid Leukemia. The New
England Journal of Medicine, 364(11), pp.1027–1036.

46
Queliane et al., 2011. Leucemia mieloide aguda versus ocupação profissional : perfil
dos trabalhadores atendidos no Hospital de Hematologia de Recife *. Rev Esc
Enferm USP, 45(6), p.1446.
Reisman, D., 2002. Lack of p53 expression in human myeloid leukemias is not due to
mutations in transcriptional regulatory regions of the gene. Leukemia, 16, pp.2165–
2167.
Rossi, A. et al., 2015. DNA repair and aging : the impact of the p53 family. , 7(12),
pp.1050–1065.
Rossi, L. et al., 2012. Less Is More: Unveiling the Functional Core of Hematopoietic
Stem Cells through Knockout Mice. Cell Stem Cell, 11(3), pp.302–317. Available
at: http://linkinghub.elsevier.com/retrieve/pii/S1934590912004845.
Schlenk, R.F., 2014. Post-remission therapy for acute myeloid leukemia.
Haematologica, 99(11), pp.1663–1670.
Schürch, C.M., Riether, C. & Ochsenbein, A.F., 2013. Dendritic Cell-Based
Immunotherapy for Myeloid Leukemias. Frontiers in immunology, 4(December),
p.496. Available at:
http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=3876024&tool=pmcent
rez&rendertype=abstract [Accessed April 29, 2014].
Sinkule, J.A., 1984. Etoposide: a semisynthetic epipodophyllotoxin. Chemistry,
pharmacology, pharmacokinetics, adverse effects and use as an antineoplastic
agent. Pharmacotherapy, 4(2), pp.61–73. Available at:
http://www.ncbi.nlm.nih.gov/pubmed/6326063 [Accessed June 14, 2016].
Stein, E.M. & Tallman, M.S., 2012. Remission induction in acute myeloid leukemia. Int
J Hematol, 96, pp.164–170.
Tallman, M.S., 2005. New strategies for the treatment of acute myeloid leukemia
including antibodies and other novel agents. Hematology / the Education Program
of the American Society of Hematology. American Society of Hematology.
Education Program, 2002(312), pp.143–50. Available at:
http://www.ncbi.nlm.nih.gov/pubmed/16304372.
Tong, Q. et al., 2004. BAK overexpression mediates p53-independent apoptosis

47
inducing effects on human gastric cancer cells. BMC Cancer, 4(33), pp.1–8.
Utani, K. et al., 2010. Emergence of Micronuclei and Their Effects on the Fate of Cells
under Replication Stress. PLoS ONE, 5(4).
Vardiman, J.W. et al., 2009. The 2008 revision of the World Health Organization (
WHO ) classification of myeloid neoplasms and acute leukemia : rationale and
important changes. Blood, 114(5), pp.937–951.
Vardiman, J.W., Harris, N.L. & Brunning, R.D., 2002. Review article The World
Health Organization ( WHO ) classification of the myeloid neoplasms. Blood,
100(7), pp.2292–2302.
Wattel, B.E. et al., 1994. p53 Mutations Are Associated With Resistance to
Chemotherapy and Short Survival in Hematologic Malignancies. Blood, 84(9),
pp.3148–3157.
Willemze, R. et al., 2014. High-Dose Cytarabine in Induction Treatment Improves the
Outcome of Adult Patients Younger Than Age 46 Years With Acute Myeloid
Leukemia : Results of the. Journal of Clinical Oncology, 32(3), pp.219–229.
Yin, B. et al., 2006. Trp53 loss during in vitro selection contributes to acquired Ara-C
resistance in acute myeloid leukemia. Experimental Hematology, 34, pp.631–641.
Zago, M.A., Falcão, R.P. & Pasquini, R., 2004. Hematologia Fundamentos e Prática
1st ed.,