Embed Size (px)
Citation preview

1
ANTICORPOS MONOCLONAIS CONTRA AS PRINCIPAIS TOXINAS DO VENENO DE
BOTHROPS ATROX: UMA PERSPECTIVA PARA O USO TERAPÊUTICO.
THIAGO SILVA FRAUCHES
CAMPOS DOS GOYTACAZES
AGOSTO 2007

2
Anticorpos Monoclonais contra as Principais Toxinas do
Veneno de Bothrops atrox: uma perspectiva para o uso
terapêutico.
Thiago Silva Frauches
”Tese apresentada ao Centro de Biociências
e Biotecnologia. Da Universidade Estadual
do Norte Fluminense Darcy Ribeiro, como
Parte das exigências para a obtenção do
título de Mestre em Biociências e
Biotecnologia.”
Orientador: Milton Masahiko Kanashiro
Campos dos Goytacazes – RJ
2007

3
Anticorpos Monoclonais contra as Principais Toxinas do
Veneno de Bothrops atrox: uma perspectiva para o uso
terapêutico.
Thiago Silva Frauches
”Tese apresentada ao Centro de Biociências
e Biotecnologia. Da Universidade Estadual
do Norte Fluminense Darcy Ribeiro, como
Parte das exigências para a obtenção do
título de Mestre em Biociências e
Biotecnologia.”
Aprovada em 31 de agosto de 2007
Comissão examinadora:
----------------------------------------------------------
Dr. André L. Fuly (UFF)
----------------------------------------------------------
Dr. Wilmar Dias da Silva (UENF)
----------------------------------------------------------
Dra Olga Lima T. Machado (UENF)
----------------------------------------------------------
Orientador: Dr. Milton M. Kanashiro (UENF)

4
Este trabalho foi desenvolvido no Laboratório de Biologia do Reconhecer do
Centro de Biociências e biotecnologia, da Universidade Estadual do Norte
Fluminense Darcy Ribeiro, sob a orientação do Prof. Dr. Milton M. Kanashiro.
Apoio Financeiro:
Fundo de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ)
Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq)
Parque de Alta Tecnologia do Norte Fluminense (TECNORTE)
Universidade Estadual do Norte Fluminense (UENF)

5
Dedico esta tese àqueles que acreditaram em mim,
que me apoiaram e me ajudaram a crescer profiossionalmente.
Dedico também à minha família,
aos meus amigos e ao meu amor.

6
AGRADECIMENTOS
Agradeço primeiramente aos meus pais, pelo esforço e muitas vezes pelo
sacrifício que realizaram e ainda os realizam para que hoje seus filhos tenham
condições de enfrentar o mundo com todas as suas dificuldades.
Agradeço principalmente ao Prof. Milton Kanashiro, pela orientação, pela
confiança e por todo conhecimento passado pela sua orientação, sendo fundamental
na minha formação acadêmica.
À Profa. Thereza Kipnis e ao Prof. Wilmar pelos esforços realizados para nos
proporcionar as condições necessárias para o desenvolvimento dos nossos projetos.
Ao Prof. Eulógio Q. Carvalho, pela realização das análises histológicas.
À Maria Aparecida A. Bohler, pela disponibilização dos camundongos, bem
como toda a infra-estrutura para a realização dos ensaios de soroproteção.
À Aline G. Cozendey por ter cedido com a maior boa vontade a sua máquina
fotográfica para a captura das fotos dos cortes histológicos.
Ao Prof. Jorge pelos toques nas metodologias, pelo apoio e pela
descontração.
Ao Fernando, à Juju, ao Sr. Jorge, à Rozângela, à Patrícia e à Rita pois sem
vocês no laboratório não teríamos condições de desenvolver nossos trabalhos.
Aos colegas (amigos) de laboratório, por terem feito parte do meu cotidiano e
por terem me ajudado de alguma forma na realização deste trabalho.
À Fernanda, o meu grande amor, por fazer parte da minha vida.
E a todos aqueles que, de alguma forma, me ajudaram na minha formação
acadêmica e na realização deste trabalho, o meu muito obrigado!

7
Índice
Lista de Figuras,Tabelas e Pranchas................................................................... IX Lista de Abreviaturas............................................................................................ XI Resumo................................................................................................................ XII Abstract................................................................................................................ XIII Introdução............................................................................................................ 2 Epidemiologia............................................................................................ 3 Gênero Bothrops....................................................................................... 4 O veneno................................................................................................... 4 Componentes do veneno.......................................................................... 5 Toxinas Hemorrágicas.................................................................... 6 Trombina-simile............................................................................... 9 Fosfolipase A2................................................................................. 10 O Envenenamento..................................................................................... 14 A Soroterapia............................................................................................. 17 Soro Antiofídico.......................................................................................... 19 Anticorpos Monoclonais Murinos Antitoxinas............................................. 22 Objetivo................................................................................................................ 24 Objetivos específicos................................................................................. 24 Justificativa........................................................................................................... 25 Materiais e Métodos............................................................................................. 27 Veneno e antiveneno................................................................................. 27 Anticorpos Monoclonais............................................................................. 27 Animais...................................................................................................... 27 Neutralização da Atividade Hemorrágica pelo mAb 59/2-E4..................... 28 Determinação da Atividade de Fosfolipase A2........................................... 28 Dosagem de creatina quinase em soro de camundongos inoculados com frações purificadas de PLA2 do veneno de B. atrox............................................. 29 Neutralização da atividade catalítica da trombina-simile pelo anticorpo monoclonal 6AD2-G5........................................................................................... 29 Neutralização da atividade coagulante do veneno de B. atrox pelo anticorpo monoclonal 6AD2-G5 in vitro................................................................................ 30 Produção de líquido ascítico in vivo........................................................... 30 Purificação dos anticorpos monoclonais.................................................... 30
Dosagem de Proteínas............................................................................... 31 Eletroforese em Gel de Poliacrilamida contendo Dodecil Sulfato de Sódio (SDS-PAGE)......................................................................................................... 31 Immunoblotting........................................................................................... 32 Reatividade dos anticorpos monoclonais................................................... 32 Determinação da DL50 do veneno de B. atrox............................................ 33 Neutralização da ação letal do veneno de B. atrox.................................... 33 Neutralização da atividade anticoagulante do veneno de B. atrox............ 34 Análises histológicas.................................................................................. 34 Resultados Avaliação das atividades neutralizantes dos anticorpos monoclonais ...... 37 Neutralização da atividade hemorrágica pelo mAb 59/2-E4...................... 37 Neutralização da atividade fosfolipase A2 pelo mAb A85/9-4.................... 39

8
Neutralização da atividade coagulante da trombina-simile pelo anticorpo monolonal 6AD2-G5............................................................................................. 42 Produção de líqüido ascítico...................................................................... 42 Purificação dos anticorpos monoclonais.................................................... 44 Avaliação da reatividade dos mAbs purificados......................................... 46 Ensaio de neutralização da atividade letal do veneno total in vivo pelo pool de mAbs............................................................................................................... 49 Determinação da DL50 do veneno de B. atrox........................................... 50 Neutralização da atividade anticoagulante do veneno de B. atrox............ 53 Análises Histológicas................................................................................. 56 Discussão............................................................................................................. 64 Conclusões........................................................................................................... 71 Referências Bibliográficas.................................................................................... 73

9
Lista de Figuras, Tabelas e Pranchas Figura 1: Atividade hemorrágica do veneno total de B. atrox....................... 38 Figura 2: Atividade proteolítica da hemorragina de baixo peso molecular pelo mAb 59/2-E4................................................................................ 38 Figura 3: Cromatografia de alta eficiência em coluna de fase reversa C4 da fração enriquecida em PLA2.................................................................................... 39 Figura 4: Análise por SDS-PAGE das frações obtidas do HPLC......................... 40 Figura 5: Dosagem de CK no soro de camundongos inoculados com as frações de PLA2 enriquecidas do veneno de B. atrox............................................................ 41 Figura 6: Atividade catalítica da trombina-simile pelo anticorpo monolonal 6AD2-G5 sobre o substrato TAME......................................................................... 43 Figura 7: Atividade coagulante do veneno de B. atrox in vitro..................... 44 Figura 8: Perfil eletroforético dos mAbs A85/9-4, 59/2-E4 e 6AD2-G5 purificados............................................................................................................ 45 Figura 9: Análise do percentual de pureza dos mAbs purificados........................ 47 Figura 10: Análise pelo teste de ELISA da atividade dos anticorpos purificados. 48 Figura 11: Neutralização da atividade anticoagulante do veneno de B. atrox...... 54 Figura 12: Neutralização da atividade anticoagulante do veneno de B. atrox pelo mAb 6AD2-G5....................................................................................................... 55 Tabela 1: Resumo das purificações dos mAbs..........................................................45
Tabela 2: Ensaio de soroproteção in vivo de camundongos desafiados com o pool
de mAbs e veneno................................................................................................ 49
Tabela 3: Ensaio de soroproteção in vivo de camundongos desafiados com
misturas de mAbs e veneno................................................................................. 50
Tabela 4: Cálculo da DL50 do veneno de B. atrox................................................ 51
Tabela 5: Ensaio de soroproteção in vivo de camundongos desafiados com
misturas de pool de mAbs e 5 DL50 de veneno.................................................... 52

10
Tabela 6: Ensaio de soroproteção in vivo de camundongos desafiados com
misturas de pool de mAbs e 4 DL50 de veneno.................................................... 52
Tabela 7: Ensaio de soroproteção in vivo de camundongos desafiados com
misturas de antiveneno e veneno de B. atrox....................................................... 53
Prancha 1: Análise histológica do peritônio-muscular do assoalho do abdominal 58
Prancha 2: Análise histológica do músculo do diafragma..................................... 59
Prancha 3: Análises histológicas do fígado, músculo do diafragma e baço......... 62

11
Lista de Abreviaturas
mAb Anticorpo Monoclonal
A85/9-4 Anticorpo Monoclonal anti fosfolipase A2
59/2-E4 Anticorpo Monoclonal anti metaloproteinase
6AD2-G5 Anticorpo Monoclonal anti trombina-simile
PLA2 Fosfolipase A2
SVMP Metaloproteinase de veneno de serpente
SDS-PAGE Eletroforese em Gel de Poliacrilamida com Dodecil Sulfato de Sódio
H&E Hematoxilina e Eosina
DL50 Dose Letal
kDa Quilo Dalton
TAME tert-Amyl Methyl Ether
NBD-PC 1-Palmitoyl-2-[6-[(7-nitro-2-1,3benzoxadiazol-4-yl)amino]hexanoyl]-sn- Glycero-3-Phosphocholine
BSA Albumina Sérica Bovina
BCA Ácido Biciconínico
CK Creatina quinase
D.O. Densidade Óptica
i.p. Intraperitoneal
s.c. Subcutânea
i.m. Intramuscular
min. Minutos
IgG Imunoglobulina G
ELISA Ensaio de Imunoadsorção Ligado á Enzima
Cont. Controle
PBS Tampão Salina Fosfato

12
TEMED Tetrametiletilenodiamina
APS Persulfato de Amônio

13
RESUMO
Este trabalho teve como objetivo avaliar a ação de anticorpos monoclonais
neutralizantes na proteção de camundongos desafiados com o veneno de B. atrox,
visando o desenvolvimento de um antiveneno ofídico baseado em um pool de
anticorpos monoclonais produzidos contra as principais toxinas do veneno desta
serpente. Os mAbs contra fosfolipase A2 (clone A85/9-4), metaloprotease (clone
59/2-E) e trombina-simile (clone 6AD2-G5) foram produzidos em grandes
quantidades e purificados. A purificação dos mAbs foi analisada por SDS-PAGE e a
capacidade de neutralização das respectivas toxinas também foram testadas.
Ensaios de soroproteção foram feitos em camundongos Suíços albinos (18 a 20g)
injetados com 13,5 mg do pool de mAbs pela via i.p., desafiados com 350µg de
veneno bruto pela via s.c. e a sobrevivência dos animais avaliada por 48 horas.
Demonstramos que no grupo tratado com mAbs todos os camundongos
sobreviveram, enquanto que no grupo não tratado somente um animal sobreviveu.
Experimentos utilizando mAbs e soro poliespecífico antibotrópico pré-incubados
com veneno de B. atrox e injetados pela via ip em camundongos também foram
realizados. Tecidos e órgãos dos animais submetidos ao teste de soroproteção
também foram analisados no nível histológico. Nos experimentos onde foram
avaliados tempo de sangramento dos animais tratados com o pool de mAbs, com o
mAb anti trombina-simile e soro antibothrópico pré-incubados com o veneno de B.
atrox, foi verificado que o tempo de sangramento dos animais tratado com os
anticorpos foi similar aos animais controles que não receberam o veneno.
Palavras-chaves: veneno, Bothrops atrox, antiveneno, anticorpos monoclonais

14
ABSTRACT
The main ojective of this work was the evaluation of monoclonals antibodies in
protecting mice challenged with Bothrops atrox snake venom, aiming the
development of an ophidian antivenom based on a pool of monoclonals antibodies
against the main toxic components of this snake venom. These mAbs against
phospholipase A2 (PLA2), hemorrhagic metalloprotease and thrombin-like enzymes
were produced in large quantity and purified. Purified mAbs were analyzed by SDS-
PAGE and tested for the capacity to neutralize the respective toxin. Serumprotection
assays were constituted by Swiss mice (18 to 20g) inoculated with 13,5mg of pool of
mAbs by via i.p., challenged with 350µg of crude venom by via s.c. and survival rate
scored during 48 hours. These results showed that groups treated with mAbs all mice
survived, while in the non treated group, only one animal survived. Experiments
employing mAbs and antibotropic polispecific antivenom pre-incubated with B. atrox
snake venom and injected via i.p. in mice were also realized. Organs and tissues of
animals submitted to serumprotection tests were collected for the histological
analysis. Bleeding time assays were realized in animals treated with pool of mAbs,
mAb anti thrombin-like and antibotropic serum pre-incubated with venom. These
results showed that the bleeding time of animals that received antibodies were
similar to control animals that not received venom.
Key-words: snake venom, Bothrop atrox, antivenom, monoclonals antibodies

15
INTRODUÇÃO

16
É possível que a forma alongada do corpo, sem patas e dotado de
movimentos ondulatórios ágeis, o olhar fixo, as cores vivas e, definitivamente, a
capacidade de algumas espécies de produzir e injetar um veneno mortal tenha feito
das serpentes uns dos animais mais temidos e rejeitados da humanidade
(MELGAREJO, 2003).
As serpentes são vertebrados, tetrápodos e amniotas. Entende-se por
vertebrados o grupo de animal cujos representantes possuem vértebras e, por
tetrápodos, aqueles vertebrados que possuem quatro membros locomotores. As
serpentes e alguns lagartos não apresentam membros locomotores, mas
descendem de animais com quatro patas, portanto são tetrápodos também.
Denominamos como aminiotas os tetrápodos que nascem de ovo amniótico, ou seja,
ovos que possuem as membranas córion, âmnion e alantóide, responsáveis pela
manutenção do ambiente aquático dentro do ovo e trocas gasosas (POUGH et. al,
1998).
As serpentes apresentam como características diagnósticas o corpo
extremamente alongado, sem apêndices locomotores e cintura escapular; perda da
sínfise mandibular (perda da sutura óssea das hemimandíbulas no mento,
substituída por um ligamento elástico); fechamento lateral da caixa craniana e perda
das pálpebras móveis, entre outras caracteríticas (FRANCO, 2003). Esses animais
alimentam-se de presas inteiras, e caçam utilizando estruturas quimiossensíveis.
São exclusivamente carnívoras, predando tanto vertebrados quanto invertebrados
(GREENE, 1997). A perda da sínfise mandibular e da cintura, entre outras
modificações cranianas, permite às várias serpentes abrirem a boca e engolirem
presas grandes, algumas com até 3,5 vezes o seu diâmetro. Existem serpentes
muito pequenas, como os escolecofídios (cobras-cegas), dentre os quais algumas
serpentes alcançam a maturidade sexual com pouco mais de 10 centímetros, até
serpentes gigantes, com mais de 10 metros de comprimento, caso de boídeos e
pitonídeos como a sucuri (Eunectes murinus) e a pitão-reticulada (Python reticulatus)
(FRANCO, 2003).

17
Esses animais distribuem-se por quase todos os ambientes do globo, com
exceção das calotas polares, onde o clima demasiadamente frio impossibilita a vida
de vertebrados ectotérmicos (CADLE, 1997). Há serpentes aquáticas e terrestres.
Dentre as aquáticas, temos as de água doce e as marinhas. No ambiente terrestre,
ocupam os habitats fossoriais, terrestre e arborícola. Vivem em matas, savanas ou
desertos (GREENE, 1997).
Há, atualmente, cerca de 2.900 espécies de serpentes catalogadas no
mundo, distribuídas em 465 gêneros e 20 famílias. A fauna de serpentes brasileiras
é muito diversa, sendo encontradas várias espécies por todo o território nacional. No
Brasil, temos representantes de 9 famílias, 75 gêneros e 321 espécies, ou seja,
cerca de 10% do total de espécies (FRANCO, 2003). Em nosso país, as serpentes
venenosas distribuem-se em quatro gêneros: Bothrops, Crotalus, Lachesis e
Micrurus. Os três primeiros pertencem à família Viperidae, subfamília Crotalinae e a
última Elapidae, subfamília Elapinae (MELGAREJO et. al, 2003).
EPIDEMIOLOGIA
A apresentação dos dados sobre acidentes ofídicos no Brasil é baseada na
notificação à Coordenação Nacional de Controle de Zoonoses e Animais
Peçonhentos (CNCZAP), que no período de janeiro de 1990 a dezembro de 1993,
foi de 81.611 acidentes, o que representou uma média de 20.000 casos/ano para o
país. A maioria dos acidentes foi registrada nas Regiões Sudeste e Sul, as mais
populosas e que contam com melhor organização de serviços de saúde e sistema
de informação. Os coeficientes anuais de incidência para o Brasil estiveram em
torno de 13 acidentes/10.000 habitantes, com distribuições desiguais nas diferentes
macrorregiões (ARAÚJO et. al, 2003). Nas diferentes regiões do país, o maior índice
foi no Centro-Oeste, ainda que apresente um alto coeficiente, é possível que ocorra
subnotificação na região Norte, tendo em vista as dificuldades de acesso aos
serviços de saúde, o mesmo ocorrendo para o Nordeste (MINISTÉRIO DA SAÚDE,
2001). A ocorrência do acidente ofídico está, em geral, relacionada a fatores
climáticos e aumento da atividade humana no campo. Com isso, nas regiões Sul,
Sudeste e Centro-Oeste, observa-se incremento do número de acidentes no período

18
de setembro a março. Na região Nordeste, os acidentes aumentam de janeiro a
maio, enquanto que, na região Norte, não se observa sazonalidade marcante,
ocorrendo uniformemente durante todo o ano (MINISTÉRIO DA SAÚDE, 2001). O
pé e a perna foram atingidos em 70,8% dos acidentes notificados e em 13,4% a mão
e o antebraço. A utilização de equipamentos individuais de proteção como sapatos,
botas, luvas de couro e outros poderiam reduzir em grande parte esses acidentes.
Em 52,3% das notificações, a idade dos acidentados variou de 15 a 49 anos, que
corresponde ao grupo etário onde se concentra a força de trabalho. O sexo
masculino foi acometido em 70% dos acidentes, o feminino em 20% e, em 10%, o
sexo não foi informado. Dos 81.611 casos notificados, houve registro de 359 óbitos.
Excluindo-se os 2.361 casos informados como “não peçonhentos”, a letalidade geral
para o Brasil foi de 0,45%. O maior índice foi observado nos acidentes por Crotalus,
onde em 5.072 acidentes ocorreram 95 óbitos (1,87%) (MINISTÉRIO DA SAÚDE,
2001).
GÊNERO BOTHROPS
Este gênero possui algumas das espécies mais importantes do ponto de vista
médico, já que produzem cerca de 90% dos 20.000 acidentes ofídicos anuais que o
Brasil registra (MINISTÉRIO DA SAÚDE, 2001). Das 20 espécies de Bothrops
reconhecidas no país, quatro delas são comumente responsáveis pelos acidentes
humanos: B. moojeni, no Centro Oeste; B. erythromelas, no Nordeste; B. jararaca
nas regiões Sul e Sudeste; e B. atrox na região Norte do país. Trata-se de uma
serpente ágil e ativa, que pode superar 1,5m de comprimento, de colorido variável,
que freqüenta bastante as beiras de rios, córregos e igarapés (Figura 1). È o
viperídeo mais freqüente no vale amazônico, sendo o principal causador de
acidentes da região Norte (MELGAREJO, 2003).
O VENENO
As peçonhas de serpentes são, provavelmente, os mais complexos de todos
os venenos. Contém vinte ou mais componentes diferentes, sendo que mais de 90%

19
do peso seco do veneno é constituído por proteínas, compreendendo grande
variedade de enzimas, toxinas não-enzimáticas e proteínas não-tóxicas. As frações
não-protéicas são representadas por carboidratos, lipídios, metais (freqüentemente
na forma de glicoproteínas e enzimas metaloprotéicas), aminas biogênicas,
nucleotídeos e aminoácidos livres. A função de cada um desses componentes, bem
como suas interações no envenenamento humano, ainda não foram totalmente
esclarecidas (MELGAREJO et. al, 2003). A maioria desses componentes é
produzida por duas glândulas de secreção exócrina situada uma de cada lado na
base da mandíbula, cujos ductos excretores principais se comunicam com as
correspondentes presas. No ato da picada as glândulas são comprimidas e lançam
suas secreções nos ductos que levam aos tecidos da vítima (FRANÇA et. al, 2003).
Além disso, os venenos de serpentes, que são ricos em componentes
farmacologicamente ativos, podem servir como ferramenta de estudo em várias
patologias, como doenças inflamatórias, nas quais as fosfolipases estão envolvidas
(VALENTIN e LAMBEAU, 2000), ou em metástases tumorais e angiogênses. Os
venenos também podem servir de protótipo para o estudo e desenvolvimento de
novas drogas, como foi o caso, por exemplo, da descoberta da bradicinina a partir
do veneno de B. jararaca (ROCHA e SILVA et al., 1949).
COMPONENTES DOS VENENOS
A variedade e complexidade dos elementos que compõem os venenos
ofídicos são responsáveis pelas dificuldades dos estudos das serpentes. Cerca de
90% do peso seco dos venenos é composto de proteínas. Carboidratos, lipídeos,
nucleotídeos, riboflavinas, cátions e ânions também compõem os venenos e,
embora nem todos eles sejam tóxicos por si, em conjunto podem ser responsáveis
pelos efeitos secundários de toxicidade (LOMONTE, 1994). Dentre os componentes
tóxicos dos venenos, três deles são os principais causadores de danos locais aos
tecidos: toxinas hemorrágicas, as trombinas-símiles e as fosfolipases A2.

20
TOXINAS HEMORRÁGICAS
Ohsaka (1979), estudando componentes do veneno de uma espécie de
serpente semelhante à Bothrops, mostrou que alguns deles induzem hemorragia,
promovendo abertura de junções intercelulares das células endoteliais dos capilares.
Entretanto, Ownby et al. (1990), estudando toxinas hemorrágicas de Crotalus atrox e
Crotalus horridus, propuseram um mecanismo diferente para atividade hemorrágica
destas toxinas: estas induziriam hemorragia rompendo a integridade das células
endoteliais e promovendo o escape de eritrócitos através dos espaços formados
entre elas.
Metaloproteinases de venenos de serpentes (SVMPs) estão presentes em
altas concentrações em venenos de serpentes pertencentes à família Viperidae e
mais recentemente, elas têm sido reportadas, embora em menores proporções,
também em venenos de serpentes da família Elapidae (MATSUI et al., 2000).
SVMPs são classificadas em classes PI a PIV de acordo com a organização de seus
multidomínios (BJARNASON and FOX, 1994; FOX and SERRANO, 2005). Estas
enzimas são sintetizadas in vivo como proteínas multimodulares que compreendem
um peptídeo sinal, um pró-domínio e um domínio metaloproteinase (classe PI).
Algumas SVMPs também exibem domínios C-terminal subseqüentes, denominados
desintegrinas (classe PII) ou domínio desintegrina-simile (classe PIII) e domínio
lectina-simile tipo-C (classe PIV) (HITE et al., 1994; BJARNASON and FOX, 1994).
Recentemente, Fox e Serrano (2005) atualizaram a proposta de classificação das
SVMPs, incluindo os seguintes precursores: precursor PIIa-PII cuja forma madura
não libera o domínio desintegrina; precursor PIIb-PII cuja forma madura compreende
SVMPs dimérica; precurusor D-I que não contém o domínio metaloproteinase;
precursor PIIIa-PIII que libera seus domínios ricos em cisteína; e precursos PIIIb-PIII
que forma SVMPs dimérica.
Todas as toxinas hemorrágicas isoladas até recentemente de crotalídeos e
viperídeos possuem atividades proteolíticas do tipo caseína ou dimetil caseína. As
caracterizações bioquímicas destas enzimas mostraram serem todas Zn-
metaloproteases (BJARNASON & FOX, 1988). Poucas destas toxinas foram
seqüenciadas. No entanto, a comparação das seqüências mostrou que apesar de

21
possuírem diferentes pesos moleculares (25-30 kDa a 60 kDa e 60-90 kDa) todas
estão relacionadas a um gene ancestral comum e não possuem similaridade a
nenhuma outra proteína conhecida, exceto uma significativa homologia com o sítio
de ligação do Zn em outras proteínas que o possuem (HITE , 1992 e PAINE , 1992).
A relação entre atividade proteolítica e mecanismos hemorrágicos está
relacionada com a degradação de componentes da membrana baso-lateral do
endotélio vascular. Usando substratos específicos como laminina, fibronectina,
colágeno tipo IV, os fatores hemorrágicos purificados mostraram-se altamente
proteolíticos. Maruyama e colaboradores (1992) e Bjarnason & Fox (1988)
propuseram um mecanismo de ação baseado na digestão enzimática da lâmina
basal da microvasculatura. Mais recentemente, usando-se células endoteliais em
cultura, foi demonstrada a participação de hemorraginas na organização de
componentes da matriz extracelular através de técnicas de imunohistoquímica e
microscopia eletrônica (TIMPL, 1989).
As toxinas hemorrágicas de alto peso molecular possuem um domínio do tipo
desintegrina (HITE, 1992 e PAINE, 1992). Desintegrinas são peptídeos ricos em
resíduos de cisteína, encontrados nos venenos de várias espécies de serpentes,
todos eles contendo a seqüência Arg-Gly-Asp (RGD). Esta seqüência conservada
permite-lhes inibir a agregação plaquetária e também interagir com as integrinas
celulares (GOULD et al.,1990) e (SCARBOROUGH et al., 1993). Especula-se
também, quanto à possibilidade desse domínio desintegrina, nas metaloproteinases
hemorrágicas, funcionar como unidade de direcionamento da atividade catalítica
lesando o endotélio em sítios específicos, aumentando assim a eficiência destas
enzimas. Um destes possíveis sítios de atuação seria as integrinas, localizadas na
superfície das células endoteliais, que estão envolvidas em processos de adesão à
matriz extracelular (RUOSLAHTI & PIERSCHBACHER ,1987) e (TASHIRO et
al.,1989). A proteólise destes receptores e/ou das proteínas da matriz extracelular a
eles associados causariam a desagregação das células do endotélio vascular. Um
outro possível sítio seria as metaloproteinases endógenas de matriz (MMPs), as
quais estão relacionadas com a degradação da membrana baso-lateral do endotélio
vascular em eventos como angiogênese e metástase tumoral (MURPHY et al.,1993)
e (BENELLI et al., 1994).

22
A análise da seqüência de ácidos nucléicos que codifica para SVMPs indica
que elas são sintetizadas in vivo como zimógenos inativos (BJARNASON e FOX,
1994; SELISTRE-DE-ARAUJO e OWNBY, 1995; KINI, 1996). A ativação in vitro
destas enzimas pode ser executada tanto por serino-proteases extracelulares
(calicreína, plasmina, tripsina e elastase) quanto por metaloproteinases. Uma
exceção aparente é a pró-MMP-2 (gelatinase A), que não contém os sítios
necessários de clivagem para serino-proteases em seu domínio pró-proteína
(SPRINGMAN et al., 1990) dependendo da ação de uma metaloproteinase
associada à membrana (Membrane Type Matrix Metalloproteinase 1, MT-MMP1)
para a sua ativação. Grams et al. (1993) sugeriram que ativação in vivo ocorra pela
troca de cisteínas ou mecanismo do tipo velcro. De acordo com esse mecanismo, o
motivo conservado “PKMCGVT” encontrado no pró-domínio, interage com sítio ativo.
O grupo tiol do resíduo conservado da cisteína presente neste motivo atua como
quarto ligante do zinco catalítico no sítio ativo, portanto bloqueando a formação do
complexo enzima substrato. Esta idéia foi reforçada pela inibição da atividade
adamalsina II pela L-cisteína (IC50=117µM) e também pelo peptídeo correspondente
ao motivo presente no pró-domínio (IC50=3,2µM) (GRAMS et al., 1993).
A ativação enzimática pode ser alcançada tanto pela atividade autocatalítica
residual quanto por outras proteases do veneno (SHIMOKAWA et al., 1996). O
remodelamento da forma purificada do zimógeno de (Pro-ACLF) produzido em
bactéria produced gerou valiosa evidência para suportar a hipótese de ativação
autocatalítica (RAMOS et al., 2003). Durante o processo de remodelamento, a
ativação de intermediários com sucessiva perda de peso molecular foi gerada. A
seqüência N-terminal de três intermediários ativos revelou que duas delas, com alta
massa molecular, tiveram sucessivas perdas de segmentos N-terminais, mas
mantendo o motivo pró-domínio relacionado ao mecanismo de troca de cisteínas, e
embora, os três intermediários não apresentem este motivo e correspondem ao N-
terminal geralmente encontrado em espécies ativas. Em todos intermediários ativos,
a hidrólise ocorreu na região N-Terminal de resíduos hidrofóbicos (RAMOS et al.,
2003).

23
TROMBINA - SIMILE
A ação coagulante, também conhecida como síndrome da desfibrinação, é
resultante da propriedade que o veneno das serpentes do gênero Bothrops, Crotalus
e Lachesis têm de transformar diretamente o fibrinogênio em fibrina (KAMIGUTI E
CARDOSO, 1989). Esta ação é conhecida como “ação coagulante do tipo trombina”.
Além disso, a maioria dos venenos botrópicos tem capacidade de ativar o fator X
(atividade pró-coagulante) e a pró-trombina da cascata da coagulação, cujo
resultado final é o consumo de fibrinogênio e conseqüentemente a incoagulabilidade
sangüínea (SANO-MARTINS et al., 1995). Componentes do veneno das serpentes
que possuem atividade biológica semelhantes à trombina, denominados trombina-
simile, têm sido encontrados no veneno de serpentes dos gêneros Bothrops,
Crotalus e Lachesis. A maioria desses componentes age, apenas, sobre a
extremidade N-terminal da cadeia Aa do fibrinogênio originando o fibrinogênio A.
Como, geralmente, não agem sobre a cadeia Bb do fibrinogênio, os monômeros
formados carecem de um sítio de polimerização na extremidade N-terminal (SANO-
MARTINS et al., 1995). Sendo serino-proteinases, tais enzimas são inibidas com
maior ou menor eficiência por PMSF (Phenyl Methyl Sulfonyl
Fluoride), porém mostram-se insensíveis à heparina, hirudina, SBTI (Soybean
Trypsin Inhibitor), aprotinina (STOCKER e MEYER, 1989), além disso, essas
enzimas possuem outras características como ativação da proteína C (KISIEL et
al.,1987), clivagem do componente C3 do sistema complemento (TAMBOURGI et
al., 1994) e atividade girotóxica (ROSING e TASS, 1992). Trombina-simile de B.
jararaca, B. asper e B. atrox (CAVINATO,et al., 1998; PETRETSKI et al. 2000) entre
outras, já foram descritas. Embora possuindo algumas características particulares no
que se refere à massa molecular (22.000 kDa a 62.000kDa), ao conteúdo de
carboidratos, à especificidade do sítio de clivagem, às atividades biológicas e à
conversão de fibrinogênio, todas as trombinas-simile compartilham algumas
características como: a presença de um resíduo de serina no sítio ativo; uma baixa
capacidade de hidrolisar substratos como a caseína, hemoglobina, insulina ou
glucagon e uma alta atividade esterásica sobre substratos sintéticos como TAME
(tert -Amyl Methyl Ether) e BapNa (Na-Benzoil-DL-Arginine-p-Nitroaniline) (SANO-
MARTINS et al., 1995). Além disso, recentemente, Pérez et al. (2007) demonstraram

24
atividade girotóxica atribuída à trombina-símile BjussuSP-I, isolada do veneno de B.
jararacussu. Essa peculiaridade também já foi demonstrada em trombinas-símile
presente em venenos de Crotalus (MARTINS et al., 2002) e Lachesis (MAGALHÃES
et al., 1993 e SANCHES et al., 2000).
FOSFOLIPASE A2 (PLA2)
O papel biológico das PLA2s de mamíferos ainda não está claramente
definido, mas a sua distribuição específica em vários tecidos e em diferentes
estágios de desenvolvimento sugere que elas estejam envolvidas em vários
processos biológicos, como a clivagem de fosfolipídeos gerando intermediários que
atuam em processos inflamatórios, sinalização celular e sistema complemento. Mais
recentemente, vários estudos têm demonstrado o envolvimento de PLA2 de
mamíferos nos mecanismos de defesa, tais como: inflamação, proliferação, adesão
e contração celular. Com respeito às PLA2 de venenos animais já foram descritas as
atividades farmacológicas que incluem neurotoxicidade, miotoxicidade, hemorrágica,
hemolítica, edematogênica, hipotensiva, cardiotóxica, agregadora de plaquetas,
convulsivante, proliferação celular, antibacteriana e anti-HIV, o que sugere que PLA2
de veneno possa interferir em vários processos fisiológicos ou patológicos de
mamíferos, principalmente quando se leva em consideração a presença de
receptores de PLA2 em células de mamíferos (VALENTIN & LAMBEAU, 2000).
A superfamilia das fosfolipases A2 (PLA2; fosfatidilcolina-2-acilhidrolase, EC
3.1.1.4), consiste em uma grande variedade de enzimas definidas pela sua
habilidade de catalisar, especificamente, a hidrólise da ligação acil-éster na posição
sn-2 de fosfolipídeos liberando ácidos graxos livres e lisofosfolipídeos (DENNIS,
1994). Estes ácidos graxos livres, araquidônico (AA) e oléico (AO), podem funcionar
como reserva de energia, mas o AA pode também funcionar, principalmente, como
segundo mensageiro e como precursor de eicosanóides que são potentes
mediadores inflamatórios e de transdução de sinais (HANASAKI e ARITA, 1999). O
outro produto resultante da ação hidrolítica da PLA2, lisofosfolipídeo, desempenha
um papel importante na sinalização celular, remodelagem de fosfolipídeos e
pertubação da membrana celular (SIX E DENNIS, 2000).

25
As PLA2s, atualmente consideradas uma superfamília de enzimas
classificadas principalmente em três grupos: PLA2 citosólica (cPLA2), PLA2
secretórias Ca2+ dependentes (sPLA2) e PLA2 intracelular Ca2+ independentes
(CHAKRABORTI, 2003). Essas enzimas, ainda são classificadas com base em suas
seqüências de aminoácidos (BALSINDE et al., 1999), que resultou na divisão das
PLA2 em 10 grupos. E associado a novos critérios, Six e Dennis (2000)
reclassificaram as PLA2 dos venenos de serpentes, insetos e moluscos nos grupos I,
II, III e IX sendo os venenos de viperídeos e crotalídeos inseridos no grupo IIA. As
PLA2s de veneno botrópico, são ainda, subdivididas em dois grupos, as Asp49 e
Lys49. A presença do ácido aspártico na posição 49 é bastante conservada e está
intimamente envolvida na ligação do co-fator Ca++, essencial para a atividade
catalítica da enzima (SCOTT, et al., 1992) e faz com que estas PLA2s tenham alta
atividade sobre substratos sintéticos; por outro lado, a presença da lisina na posição
49, faz com que as atividades catalíticas dessas enzimas sobre substratos
sintéticos, sejam desprezíveis ou mesmo ausentes. Apesar disso, em geral, as PLA2
são extremamente ativas na indução de mionecrose (OWNBY, et al. 1999;
MARAGANORE et al. 1984). A presença de um grande número de resíduos básicos,
principalmente lisina (em torno de 15% do total de aminoácidos), é bastante
característica das PLA2s Lys49, que inclui também uma região rica em lisina na
porção C-terminal da molécula e dois sítios consenso de ligação à heparina, que
estão associados com a indução da mionecrose (OWNBY et al. 1999; SELISTRE-
DE-ARAUJO et al. 1996a e b; DUA e CHO, 1994).
A proposta de que a atividade miotóxica da PLA2 se expresse pela hidrólise
de lipídeos da membrana alvo é respaldada por algumas evidências que
correlacionam a miotoxicidade com a atividade catalítica da enzima (TAKASAKI et
al. 1990; DIAZ et al. 1991). Entretanto, a grande maioria dos dados experimentais,
obtidos principalmente na inibição química das PLA2s ou seu bloqueio com
anticorpos monoclonais ou policlonais, demonstram claramente que a atividade
farmacológica (miotoxicidade, edematogênica) de várias PLA2s independe ou
depende parcialmente da atividade catalítica da enzima (WARD et al. 1998a).
Os mecanismos pelos quais as PLA2s exercem toxicidade ainda não foram
bem esclarecidas. Kini e Evans (1989) propuseram um modelo, no qual sugerem

26
que a PLA2 se liga preferencialmente à célula ou no tecido alvo por afinidade para
alguma proteína de superfície ao invés do domínio lipídico. Após a ligação inicial, as
PLA2s podem induzir vários efeitos farmacológicos que são dependentes ou
independentes da sua atividade enzimática. Ward et al. (1998b) identificaram,
através de análise por “SequenceSpace”, nove aminoácidos bastante conservados
na sub-família das PLA2s Lys49. Estes aminoácidos eram agrupados três a três e
localizados no sítio ativo, no canal hidrofóbico de ligação com o substrato e na
interface de formação de homodímeros, sugerindo que estes três grupos de
aminoácidos estejam envolvidos na atividade farmacológica Ca++ independente.
O sítio de interação da miotoxina II de B. asper com a heparina foi mapeado
em uma região que compreende aos resíduos 115-129 próximo da região C-terminal
da molécula. A injeção intramuscular em camundongo do peptídeo recombinante
correspondente a esta seqüência não induziu mionecrose (CALDERÓN e
LOMONTE, 1998). Este peptídeo também demonstrou ter atividade bactericida
(PÁRAMO et al. 1998). Posteriormente, Calderón e Lomonte (1999) demonstraram
que camundongos imunizados com o peptídeo 115-129 da miotoxina II e desafiados
com a miotoxina II nativa apresentaram necrose tecidual reduzida em comparação
com os controles. Considerando que a ação farmacológica do peptídeo 115-129 da
miotoxina II é dependente da sua característica anfifílica, dada pela combinação de
resíduos catiônicos e hidrofóbicos, Lomonte et al. (1999b), substituiram as três
moléculas de tirosina por triptofano no peptídeo recombinante 115-129 da miotoxina
II. Os estudos farmacológicos demonstraram que esta substituição foi capaz de
restaurar a atividade miotóxica e edematogênica observada na miotoxina II.
A atividade edematogênica já foi observada em várias miotoxinas purificadas
do veneno de serpentes do gênero Bothrops. O edema geralmente se inicia entre
10-15 min após a inoculação e se estende até 6-8 h após a injeção (YAMAGUCHI et
al. 2001; OWNBY et al. 1999; LIU et al. 1991). Através de estudos com
videomicroscopia, Lomonte et al. (1994a) observaram que o extravasamento de
plasma de vênulas e capilares inicia-se a partir de 2min e é intenso após 4 –5min
após o tratamento. O edema inicial (até 30min após a injeção) é inibido pelo
tratamento com ciproheptadina, indicando o envolvimento da histamina e/ou
serotonina. O edema tardio (5h após a injeção) é inibido pelo pré-tratamento com

27
aspirina, sugerindo a participação da prostaglandina neste tipo de edema
(GUTIÉRREZ et al. 1986a).
Embora alguns autores tenham associado o edema à capacidade das PLA2s
de hidrolisar fosfolipídeos (LLORET e MORENO, 1993; VISHWANATH et al. 1988),
vários outros autores demonstraram que a atividade edematogênica independe da
atividade catalítica das PLA2s (DIAZ-OREIRO e GUTIÉRREZ, 1997; DANIELE et al.
1997; GUTIÉRREZ e LOMONTE, 1995; LIU et al. 1991). O fato de que a miotoxina
II, PLA2 Lys-49 cataliticamente inativa, de B. asper induz a formação de edema,
sugere que esta atividade farmacológica seja causada por mecanismo independente
da catálise enzimática (LOMONTE et al. 1993). Inclusive, Ambrosio et al. (2005)
propuseram um mecanismo baseado na estabilização de uma estrutura canônica
dentro da região C-Terminal da molécula que é favorecida pela presença de um
ligante de cadeia longa dentro do sítio ativo/canal hidrofóbico. Esta estrutura
provavelmente pode ser um ácido graxo ou fosfolipídeo in vivo. A estrutura canônica
leva à formação e exposição de extensões hidrofóbicas que se inserem na
membrana através de interações eletrostáticas envolvendo as lisinas da porção C-
Terminal, isso poderia levar a uma perturbação da membrana. Além disso, a
possibilidade de que o edema possa ser decorrente da necrose muscular causada
pelas miotoxinas parece ser infundada, uma vez que venenos com alta atividade
miotóxica (Ex. Micrurus spp) não induzem edema significativo em camundongos
(GUTIÉRREZ et al. 1983).
Modificações químicas da bothropstoxin-I (Lys49) e bothropstoxin-II (Asp49)
realizadas por Andrião-Escarso et al. (2000), demonstraram que para a
bothropstoxin-I os aminoácidos histidina, tirosina e triptofano desempenham um
papel importante na miotoxicidade, mas não interferem na indução de edema e
ruptura de lipossomos. Enquanto que para a bothropstoxin-II, esses resíduos
parecem ser relevantes nas atividades anticoagulante, miotóxica, citotóxica e
indutora de edema, e sem efeito na ruptura de lipossomos. Diaz et al. (1993)
mostraram que a alquilação da histidina 48 da miotxina II de B. asper não provocou
alterações na carga nem nas propriedades imunológicas, porém reduziu a sua
atividade miotóxica.

28
Os anti-soros convencionais não são capazes de neutralizar adequadamente
a atividade edematogênica do veneno, mesmo quando estes anti-soros são
previamente incubados com o veneno e inoculados em animais (GUTIERRÉZ et al.
1986b). Além do edema, uma grande quantidade de células inflamatórias é
observada no local da lesão (FARSKY et al., 1999 e 1997; ZULIANIA et al., 2005).
O ENVENENAMENTO
Os sintomas do envenenamento e lesões variam conforme a espécie de
serpente e/ou quantidade de veneno injetada. Venenos de serpentes dos gêneros
Crotalus (AZEVEDO-MARQUES et. al, 2003) e Micrurus (DA SILVA JR. et. al, 2003),
que são ricos em neurotoxinas, praticamente não lesam os tecidos; porém, às
vezes, induzem graves sintomas neurológicos irreversíveis causados pela ação
desses componentes nas sinapses nervosas. Venenos de serpentes dos gêneros
Bothrops e Lachesis, pobres em neurotoxinas, porém ricos em enzimas de largo
espectro de especificidade, produzem intensas lesões teciduais no local da picada,
geralmente, seguidas por sintomas sistêmicos decorrentes de complexas alterações
dos sistemas de coagulação, das calicreínas-cininas, do complemento e das
paredes vasculares (FRANÇA et. al, 2003 e MÁLAQUE et. al, 2003).
Os efeitos locais do veneno botrópico aparecem logo após sua injeção e são
caracterizados por hemorragia, edema, dermonecrose, mionecrose, (GUTIÉRREZ,
1990). Euchbaum (1947), Amorim et al. (1951) demonstraram que no
envenenamento por B. jararaca a hemorragia e o edema instalam-se poucos
minutos após a inoculação do veneno, seguindo-se então, necrose coagulativa da
musculatura e dos vasos sangüíneos, acompanhada de uma intensa reação
inflamatória aguda. Os venenos botrópicos causam também liberação de
substâncias farmacologicamente ativas que agem ou interagem com receptores
sobre as membranas celulares.
O edema induzido por venenos de serpentes do gênero Bothrops
provavelmente seja devido à ação combinada de componentes farmacologicamente
ativos e bioquimicamente heterogêneos (GUTIÉRREZ et al., 1989). Dentre eles

29
encontram-se: (a) toxinas hemorrágicas que rompem a microvasculatura induzindo
extravasamento de fluidos (OHSAKA, 1979; OWNBY, 1982); (b) toxinas que atuam
diretamente nas células endoteliais das vênulas e capilares, aumentando a
permeabilidade destes (OHSAKA, 1979; OWNBY, 1982); (c) componentes do
veneno, fosfolipases ou citotoxinas, que induzem a liberação de histamina de
mastócitos; (d) PLA2s que liberam ácido araquidônico e ácido graxo através da
hidrólise de fosfolipídeos de membranas celulares, iniciando a via que leva à síntese
de prostaglandinas (ROBBINS et al., 1987); (e) proteases que atuam sobre
cininogênios, liberando cininas, por exemplo, bradicinina (ROCHA e SILVA et al.,
1949). Calicreínas podem ser ativadas pelo fator XII da cascata de coagulação, uma
vez que este fator é ativado depois que a vasculatura tenha sofrido danos e (f)
componentes da cascata do complemento, particularmente C3a e C5a, que
participam da reação inflamatória (DIAS DA SILVA et al., 1967), um problema que
pode ser acentuado pela capacidade das metaloproteinases (BaP-1) em ativar o
sistema complemento (FARSKY et al., 2000). O edema pode contribuir para outros
efeitos do veneno, como compressão do tecido e subseqüente isquemia. A
mionecrose, causada principalmente pelo componente PLA2 do veneno, pode
resultar em perda tecidual permanente, perda da habilidade motora e amputação do
membro acometido (GUTIERREZ e LOMONTE, 1995).
A injeção subcutânea de veneno ou da miotoxina II de B. asper em
camundongos, induziu um aumento dos níveis séricos de IL-6 entre 3 e 6 horas após
a injeção, enquanto que IL-1a e TNF-a não foram detectados (LOMONTE et al.,
1993). Barros et al. (1998) demonstraram que em camundongos BALB/c inoculados
com veneno de B. atrox por via intraperitoneal apresentaram níveis séricos elevados
de IL-6, IL-10, INF-g, TNF-a e oxido nítrico (NO), e in vitro, células esplênicas
desafiadas com veneno produziram somente IL-10, enquanto que as células
peritoneais aderentes produziram IL-6, IL-10 e INF-g, sugerindo que estas citocinas e
NO podem estar envolvidos na lesão tecidual. Mais recentemente, Petricevich et al.
(2000) utilizando camundongos BALB/c inoculados com venenos de B. asper ou B.
jararaca, também detectaram altos níveis das citocinas TNF-a, IL-1, IL-6, IL-10 e
INF-g no soro destes animais. A concentração máxima de IL-6 no soro foi atingida
entre 4 e 6 horas, sugerindo que esta citocina poderia estar modulando a secreção

30
de TNF-a, IL-1 e a síntese de proteínas de fase aguda. Além disso, estes autores
sugerem também que o TNF-a, IL-1 e NO podem desempenhar um papel relevante
nas alterações patofisiologicas sistêmicas causadas por estes dois venenos. Zuliania
et al. (2005) observaram distinção na produção de citocinas quando frações
purificadas de Asp e Lys49 do veneno de B. asper foram injetadas em camundongos
suíços, houve a produção de IL-1 e IL-6 para ambas isoformas, porém só houve
produção de TNF-a quando injetados com Asp49.
Em pacientes humanos picados por serpentes do gênero Bothrops ou
Crotalus durissus terrificus, Barraviera (1994) detectou a IL-1 somente em 37,5%
dos pacientes acidentados com C. d. terrrificus, a IL-6 foi detectada em 75% e 100%
dos pacientes acometidos por Bothrops e C. d. terrificus, respectivamente e a IL-8 foi
observada em 12,5% e 62,5% dos pacientes picados por Bothrops e C.d. terrificus,
respectivamente. Enquanto que o TNF-a não foi detectado no soro de nenhum
paciente, sugerindo que a IL-6 e a IL-8 podem estar envolvidos no aumento dos
níveis séricos de proteínas de fase aguda e na inibição da produção de albuminas.
Dosagens de citocinas séricas em pacientes humanos que sofreram
acidentes escorpiônicos realizadas por Meki e El-Dean, (1998), detectaram níveis
elevados de IL-1b, IL-6, NO e a1-antitripsina, que declinavam após 24 horas de
internação, enquanto que nos casos fatais a IL-1b e IL-6 se mantinham elevados
após 24 horas, sugerindo que estas citocinas estão envolvidas na patogênese do
envenenamento e se correlacionam com a severidade do envenenamento.
Barravieira (1997), comparando o perfil bioquímico sérico observado durante o
envenamento ofídico e escorpiônico, sugeriu que este quadro se assemelha ao
trauma agudo, induzindo uma típica resposta de fase aguda.
A importância do papel das citocinas e quimiocinas no desenvolvimento e
regulação das reações inflamatórias, observadas em várias condições patológicas,
tem sido extensivamente estudada nestes últimos anos, porém o envolvimento de
citocinas e quimiocinas na patofisiologia do envenenamento por serpentes, bem
como os mecanismos de sinalização intracelular induzida por componentes do
veneno em células alvo, tais como, endotélio vascular e musculatura estriada
esquelética, ainda não foram esclarecidos. Um melhor entendimento destes

31
mecanismos moleculares envolvidos na patogênese do envenenamento botrópico
pode abrir novas abordagens de interesse na terapêutica do envenenamento ofídico.
A SOROTERAPIA
A preocupação com o envenenamento ofídico e seu tratamento é bastante
antiga. No Brasil, já durante o período de colonização, o ofidismo era considerado
responsável por um número significativo de óbitos, sendo catalogado como uma das
grandes pragas existentes até então. Com relação ao tratamento, a literatura
produzida na época referia-se basicamente a ervas, rituais e manipulações utilizados
pela população para neutralizar os efeitos do veneno (SANTOS FILHO, 1991).
Esse tipo de abordagem sofreu modificação substancial na transição entre os
séculos XIX e XX, com o desenvolvimento da Microbiologia e Imunologia. As
grandes descobertas científicas vieram privilegiar a observação e a experiência
sistemáticas que deixavam de ser uma prática circunstancial (WEN, 2003).
Uma das grandes descobertas da imunologia foi à evidência de que um alto
grau de resistência a agentes biológicos de certas doenças poderia ser conseguido
pela injeção dos próprios organismos ou de extratos produzidos a partir deles. Era o
princípio da imunização ativa artificialmente induzida. Descobriu-se, também, que o
sangue dos animais imunizados continha os chamados anticorpos, que não só
destruíam os organismos invasores como apresentavam poder terapêutico e
profilático quando injetados no indivíduo doente. Era o princípio da imunização
passiva artificialmente adquirida, obtida por meio de soros hiperimunes, inicialmente
para difteria, tétano, botulismo e envenenamento ofídico (WEN, 2003). Foi assim
que Sewall, em 1887, pôde demonstrar que, em pombas, a inoculação repetida de
veneno de serpente do gênero Sistrurus, um tipo de cascavel norte-americana, em
doses bastante diluídas, produzia resistência contínua e gradativa contra os efeitos
da peçonha, sem aparente influência sobre o estado dos animais.

32
No Brasil, os trabalhos de Wücherer (1866) permitiram que o envenenamento
ofídico transpusesse os limites da cultura popular e da farmacopéia de botica para
integrar-se às pesquisas acadêmicas. Acreditando que a imunidade poderia advir do
próprio envenenamento, o autor achava que era possível obter uma “profilaxia”
semelhante à da varíola. Essa idéia não pôde ser comprovada, mas deu impulso a
uma série questionamentos e pesquisas na área (WEN, 2003).
Para a descoberta de um imunizante realmente eficaz contra o
envenenamento ofídico, foram de grande importância as pesquisas de Behring e
Kitasato (1890) observando que animais tratados com culturas de bacilos de tétano
e difteria produziam substâncias no seu próprio sangue capazes de neutralizar as
toxinas produzidas pelos microrganismos. A constatação de que um animal poderia
ser protegido contra o envenenamento se recebesse o soro de outros animais que
haviam sido previamente imunizados com o veneno foi feita, na mesma época, por
Phisalix e Bertrand (1894) e Calmette (1894). Este último foi o primeiro a utilizar o
antiveneno no tratamento de pacientes picados por serpentes. Tendo verificado que
a substância tinha ação preventiva e curativa contra a peçonha, deu início à
fabricação do primeiro soro antiofídico em Instituto Pasteur em Lille, na França.
No Brasil, o desenvolvimento da maioria das pesquisas em saúde pública
esteve fortemente associado ao combate de doenças transmissíveis, que
constituíam grave problema no final do século XIX (WEN, 2003). Surgiram, assim, os
institutos de pesquisa, entre os quais o Instituto Bacteriológico (1892), Instituto
Oswaldo Cruz (1900) e o Instituto Serumtherapico do Estado de São Paulo (1901),
atual Instituto Butantan (FONSECA, 1954). Foi neste último que Vital Brazil (1901)
demonstrou, pela primeira vez, que os envenenamentos causados por cascavel e
jararaca apresentavam manisfetações clínicas distintas, dando início às experiências
de imunização em cães, cabritos, bois e cavalos. Graças ao pesquisador brasileiro,
ficou comprovado que a especificidade dos soros estava relacionada ao gênero da
serpente (BRAZIL, 1903), contrariamente ao que supunha o pesquisador francês
Calmette, que difundiu o uso do antiveneno, acreditando que o soro produzido com o
veneno de algumas espécies protegeria o indivíduo contra o envenenamento por
qualquer serpente.

33
Desde então, vários estudos têm demonstrado o valor indiscutível dos
antivenenos na terapêutica dos acidentes por animais peçonhentos. Em 1901, Vital
Brazil iniciou a produção dos soros antiofídicos. Naquela ocasião, estimava a
letalidade dos acidentes ofídicos em 25%, proporção essa que mostrava redução de
50% dos casos de morte atribuída ao uso do soro. Passadas quatro décadas desde
o início da produção dos soros antiofídicos no Brasil, estatísticas do Instituto Vital
Brazil indicavam uma letalidade variável de 2,6 a 4,6% em acidentes humanos
(BARROSO, 1943).
SORO ANTIOFÍDICO
A utilização da soroterapia para o tratamento de acidentes por animais
peçonhentos data das últimas décadas do século XIX e a produção de soros
antitoxinas animais ainda é baseada nos métodos originalmente descritos por Vital
Brazil que imunizou cavalos com os venenos das espécies Bothrops jararaca e
Crotalus durissus terrificus e mostrou que a utilização do soro desses animais podia
neutralizar a ação dos venenos dessas serpentes (BRAZIL, 1903 e1905).
A produção de soros antitoxinas animais ainda é baseada nos métodos
originalmente descritos. Animais de grande porte são imunizados com venenos de
uma ou mais espécies de animais peçonhentos de importância médica. O soro
desses animais contém os anticorpos com capacidade de neutralizar as toxinas dos
venenos e é classificado como mono-específico ou poli-específico (monovalente ou
polivalente, respectivamente), segundo o número de venenos empregados na
imunização (CARDOSO et al., 2003)
Dessa forma, a escolha dos antígenos utilizados na imunização dos animais é
um fator primordial para a obtenção dos produtos ativos. A composição dos venenos
animais é bastante variável. Assim, vários soros devem ser produzidos, com a
finalidade específica de utilização em certo tipo de acidente. De modo geral, os
principais grupos de animais peçonhentos com importância médica causam
manifestações clínicas de envenenamento distintas, com exceção das serpentes do

34
gênero Bothrops e Lachesis. Nesses casos, usa-se o recurso da administração do
soro poli-específico, que tem as propriedades de neutralizar os efeitos induzidos
pelos diferentes venenos e, embora a utilização desses soros seja uma decisão
correta para essas circunstâncias, esse procedimento leva a uma série de
desvantagem (CARDOSO et al., 2003).
Os soros poliespecífico podem ser obtidos pela imunização dos animais com
uma mistura de diferentes venenos ou pela mistura de soros monoespecíficos
(obtido pela imunização independente com venenos de uma única espécie ou
gênero). As desvantagens desse processo consistem principalmente na baixa
eficiência do soro resultante. Dadas às características estruturais de cada toxina,
algumas delas são muito mais eficientes em induzir resposta imune. A imunização
de animais com misturas complexas de antígenos pode então levar a produção de
anticorpos predominantemente contra as toxinas presentes no veneno de uma das
espécies, deixando sem proteção os efeitos causados por toxinas menos
imunogênicas. Por outro lado, a mistura de soros mono-específicos leva,
necessariamente, a uma diluição do conteúdo total de anticorpos contra o veneno de
cada espécie. Dessa forma, a eficiência dos soros poli-específicos é sempre muito
reduzida quando comparado aos mono-específicos (CARDOSO et al., 2003). Outro
fenômeno que também pode ocorrer é a redução da resposta imune, e
conseqüentemente, redução da produção de anticorpos, devido à presença de
proteínas com atividade imunossupressoras, como as descritas no veneno de
Crotalus durissus terríficus (CARDOSO et al., 1997).
Outro aspecto a ser considerado na escolha dos antígenos é a variabilidade
que existe nos venenos de animais mesmo dentro do mesmo gênero, ou ainda
dentro da mesma espécie. Essa variabilidade intra-específica está geralmente
relacionada com a distribuição geográfica do animal, sua idade, ou época do ano em
que o animal foi capturado. A mistura de imunização deve incluir venenos de
indivíduos de diferentes idades, coletados em diferentes épocas do ano, nas regiões
onde o soro vai ser utilizado (WARRELL, 1997).

35
Além disso, recentemente no Encontro Nacional dos Laboratórios Produtores
de Soro e do Programa de Controle de Acidentes por Animais Peçonhentos em
setembro de 2003”, realizado no Instituto Butantan, foram analisadas as vantagens e
as desvantagens do uso de anticorpos heterólogos na soroterapia dos
envenenamentos causados por animais peçonhentos. Nos Dados do Ministério da
Saúde do Brasil constam que a presteza na administração do soro específico
(primeiras horas após o acidente), na dose correta e única (por exemplo, 10 ampolas
de 10mL nos acidentes considerados graves) e pela via correta (via intravenosa),
como indicado Manual de Diagnóstico e Tratamento dos Acidentes por Animais
Peçonhentos (Brasília: Ministério da Saúde. Fundação Nacional de Saúde, 1998, p.
13), reduziu o número de óbitos decorrentes de acidentes ofídicos. Apesar dessa
aparente melhora obtida com a soroterapia, os inconvenientes da utilização de
anticorpos heterólogos induzem reações adversas imediatas (RAIs). A incidência de
RAIs oscila: 14% em Belo Horizonte; 24% em Ribeirão Preto; 46% em Campinas e
25-80% em São Paulo. As RAIs representam a expressão clínica dos mecanismos
de hipersensibilidade às proteínas heterólogas do antiveneno. São classificadas em
reações precoces e tardias. As reações precoces se manifestam na pele (urticária e
angiodema), nos tratos gastrointestinal (náuseas, vômitos, cólicas abdominais e
diarréia), respiratório superior (obstrução por edema da laringe) e inferior
(broncoespasmo) e cardiovascular (hipotensão e choque). As reações tardias, bem
menos freqüentes, são reações de hipersensibilidade do tipo III produzidas por
imunocomplexos formados por anticorpos humanos antianticorpos eqüinos (doença
do soro a mais freqüente) (WEN, 2003). Glomerulopatias podem ocorrer
principalmente em crianças cujo limite de tolerância a proteínas heterólogas
circulantes seria da ordem de 60mg (MAZZUCCONI et al., 1989). A maioria dos
pacientes submetidos à soroterapia heteróloga fica quase definitivamente
impossibilitada de receber anticorpos da mesma origem devido ao risco de
desenvolver doença do soro e/ou choque anafilático. A esses grandes
inconvenientes acrescem-se a administração endovenosa de grandes volumes de
soro principalmente para crianças, as dificuldades de transporte e armazenamento
nos postos de saúde de grandes quantidades de soro em temperaturas de ordem de
4ºC, o custo de produção desses soros que exigem renovação periódica dos

36
estoques armazenados nos postos de saúde. Além disso, nos soros heterólogos
apenas 10-20% dos anticorpos são específicos para toxina importantes nos
envenenamentos por animais peçonhentos (WEN, 2003).
ANTICORPOS MONOCLONAIS MURINOS ANTITOXINAS
A partir dos anos 70, Kohler e Milstein (1975) desenvolveram um método para
imortalizar e clonar células produtoras de anticorpos específicos para um
determinante antigênico particular, revolucionando a imunologia e abrindo inúmeras
perspectivas para a área clínica. O método desenvolvido se baseia no fato de uma
célula B produzir anticorpos de uma única especificidade e, portanto, um tumor
monoclonal derivado da fusão entre um linfócito B e uma célula mielomatosa, produz
indefinidamente um único tipo de anticorpo.
Alguns aspectos fazem do anticorpo monoclonal uma das mais importantes
ferramentas de pesquisa: é o melhor método conhecido para preservar
indefinidamente uma célula produtora de anticorpos dirigidos contra um
determinante antigênico conhecido. A possibilidade de se ter uma célula produzindo
sempre o mesmo anticorpo é extremamente útil dada à homogeneidade do produto
obtido. Os anticorpos policlonais, resultantes da imunização de animais variam, nas
diferentes partidas, dadas a variabilidade dos animais e aos protocolos de
imunização. Além disso, o anticorpo monoclonal se constitui em um método valioso
para a identificação de antígenos desconhecidos, contidos dentro de uma mistura de
antígenos, visto que cada hibridoma é específico para um único determinante
antigênico. Em razão de sua alta especificidade, os anticorpos monoclonais são
reagentes padronizados que podem descriminar com precisão as diferenças
existentes na mesma ou em diferentes moléculas (por exemplo, a substituição de
único aminoácido), se constituindo em importantes ferramentas de pesquisa nas
áreas de pesquisa básica, imunodiagnóstico e clínica (CARDOSO et al., 2003).

37
Anticorpos monoclonais murinos específicos para toxinas de venenos animais
também já foram descritos, como, por exemplo, anticorpos monoclonais dirigidos
contra a-neurotoxina do veneno de Naja nigricollis (BOULAIN et al., 1982), Mojave
toxina (RAEL et al., 1986), b-bunga-rotoxina do veneno de Bungarus multicinctus
(DANSE et al., 1989), fatores hemorrágicos presentes no veneno de Crotalus atrox
(PEREZ et al., 1984 e MARTINEZ et al., 1989) ou no veneno de C. viridis viridis (LI
et al., 1993), ou ainda anticorpos monoclonais anti-PLA2 do veneno de C. durissus
terrificus (CHOUMET et al., 1992). Embora ainda não estejam em uso terapêutico,
tais anticorpos monoclonais têm permitido o isolamento de antígenos purificados, a
caracterização de epítopos presentes nas toxinas, os quais estão relacionados com
sua atividade patológica e/ou letal, possibilitando o desenho de drogas mais
potentes. O entendimento do modo de ação das toxinas, bem como a elucidação
dos mecanismos pelos quais os antivenenos promovem a neutralização destas
(CARDOSO et al., 2003).
As desvantagens apresentadas pelos monoclonais, que dificultam sua
utilização para fins terapêuticos, referem-se principalmente a sua alta
imunogenicidade. Pacientes tratados por longos períodos com anticorpos
monoclonais murinos, como nos casos de tratamento de tumores, acabam por
desenvolver uma resposta imune humoral contra anticorpos administrados (HAMA –
human antimouse antibody) (CARDOSO et al., 2003).
A terapia baseada na administração de anticorpos monoclonais murinos ou de
monoclonais recombinantes (anticorpos monoclonais humanizados) poderia,
teoricamente, tornar-se muito mais potente que a soroterapia atual, dada à presença
majoritária de anticorpos específicos e da conseqüente eliminação de proteínas
desnecessárias ou até indutoras de reações adversas. O grande desafio a ser
vencido refere-se ao estabelecimento de estudos mais conclusivos sobre as
vantagens em substituir a soroterapia atual.

38
OBJETIVO
Este projeto tem como objetivo avaliar a ação de anticorpos monoclonais
neutralizantes na proteção de camundongos desafiados com o veneno de B. atrox,
visando o desenvolvimento de um antiveneno ofídico baseado em um pool de
anticorpos monoclonais produzidos contra as principais toxinas do veneno desta
serpente.
OBJETIVOS ESPECÍFICOS
· Purificar os principais componentes tóxicos do veneno de Bothrops atrox,
hemorraginas, trombina-simile e PLA2, através de métodos conjugados de
cromatografia de exclusão molecular com a cromatografia líquida de alta
eficiência (HPLC);
· Analisar a pureza bem como dosar a atividade enzimática de cada uma das
frações purificadas;
· Produzir e purificar grandes quantidades de anticorpos monoclonais.
· Testar a capacidade dos anticorpos monoclonais em bloquear os efeitos
tóxicos dos respectivos componentes em testes in vitro e in vivo;
· Elaborar misturas de anticorpos monoclonais contendo diferentes
concentrações de cada um dos componentes;
· Avaliar a capacidade das misturas de anticorpos monoclonais em neutralizar
a letalidade do veneno bruto in vivo;
· Avaliar nos animais tratados com misturas de anticorpos monoclonais e
desafiados com veneno de B. atrox os seguintes parâmetros: tempo de
coagulação sangüínea e histopatologia do fígado, rins, baço, pulmões,
coração e músculos do diafragma e do assoalho do peritônio.

39
JUSTIFICATIVA
A grande quantidade de informações sobre a natureza dos venenos animais,
obtidas pelas mais diferentes abordagens, ainda não foi capaz de adicionar
alternativas na terapia dos envenenamentos por toxinas animais. A utilização de um
soro sintético baseado em anticorpos monoclonais poderá tornar a soroterapia
antiofídica mais eficaz, reduzindo-se a concentração de proteínas injetadas no
paciente e dessa forma minimizar os efeitos colaterais. O sucesso dessa proposta
servirá de base para a elaboração de um soro antiofídico com anticorpos
humanizados. Além disso, este trabalho talvez possa criar uma alternativa à
produção de soro antiofídico utilizando animais.

40
MATERIAIS E
MÉTODOS

41
VENENO E ANTIVENENO
O veneno da serpente Bothrops atrox utilizado neste trabalho foi fornecido
pelo Laboratório de Hepertologia do Instituto Butantan, São Paulo, SP, Brasil, sob
forma liofilizada e consistia de um pool do veneno de serpentes provenientes de
Tucuruí, PA. Para o uso, o veneno foi pesado e diluído em água destilada a uma
concentração de 10mg/mL e estocado em freezer a –20ºC, em alguns experimentos,
o veneno foi diluído em solução salina (NaCl a 0,9%). O antiveneno botrópico (lote
0512219/B, 0611203-A/B/C e 3584), com potência para neutralizar 5mg de veneno
para cada 1mL de soro, foi fornecido pelo Instituto Butantan, São Paulo, SP, Brasil e
acondicionado a 4oC.
ANTICORPOS MONOCLONAIS
Os anticorpos monoclonais utilizados nesta dissertação foram produzidos em
trabalhos anteriores no Laboratório de Biologia do Reconhecer. O mAb 59/2-E4
(antiZn-metaloproteinase) desenvolvido por Barros et al. (1998); o mAb 6AD2-G5
(antitrombina-simile); produzido por Petretski et al. (2000a) e o mAb A85/9-4
(antifosfolipase A2) gerado por Kanashiro et al. (2002).
ANIMAIS
Foram utilizados camundongos suíços albinos de ambos os sexos com cerca
de dois meses de vida, pesando entre 18 e 22g. Foram também utilizados
camundongos isogênicos BALB/c de ambos os sexos com aproximadamente 6
meses de idade. Os animais foram fornecidos pelo Biotério de Camundongos
Isogênicos de Centro de Biociências e Biotecnologia da Universidade Estadual do
Norte Fluminense Darcy Ribeiro e pelo Biotério do Serviço de Animais de
Laboratório (SAL) do Instituto Nacional de Controle de Qualidade em Saúde
(INCQS). Todos os animais foram mantidos em ambiente com renovação de ar,
alimentação e água ad libitum, obedecendo às normas éticas para uso de animais
de experimentação.

42
NEUTRALIZAÇÃO DA ATIVIDADE HEMORRÁGICA PELO MAB 59/2-E4
O ensaio de neutralização da atividade hemorrágica foi realizado incubando-
se diferentes concentrações (100, 50 e 25mg) de mAb 59/2-E4 com 2mg de veneno
de B. atrox durante 30min. à temperatura ambiente. Em seguida, as misturas foram
inoculadas por via intradérmica nos dorsos de camundongos Suíços albinos
previamente depilados. Após trinta minutos, os animais foram sacrificados, a pele da
região dorsal foi rebatida e a área hemorrágica na região subcutânea dos animais foi
avaliada.
O anticorpo monoclonal 59/2-E4 também foi testado quanto a sua capacidade
de inibir a atividade catalítica da Zn-metaloproteinase de baixo peso molecular
purificada do veneno de B. atrox, segundo metodologia descrita por Petretski et al,
2000b. Trinta microgramas da enzima purificada foram pré-incubada com 300µg do
mAb 59/2-E4 ou a mesma concentração de anticorpo policlonal de coelho durante
30min. à temperatura ambiente. Após a incubação, a atividade catalítica
remanescente da enzima foi avaliada utilizando-se como substrato a azocaseína
(Sigma, USA) de acordo com metodologia descrita por Petretski et al. (2000b). O
controle positivo foi constituído de enzima incubada com IgG não relacionada. A
reação foi lida em espectrofotômetro com comprimento de onda a 440nm.
DETERMINAÇÃO DA ATIVIDADE DE FOSFOLIPASE A2
A atividade fosfolipásica foi determinada utilizando-se o substrato sintético
fluorogênico (1-palmitoil-2-[6-(7-nitro-2-1,3-benzoxadiazol-4-yl)amino]caproyl]-sn-
Glycero-3-phosphocholine) (Avanti Polar-Lipids Inc., USA) (NBDPC). A reação foi
realizada em 2ml de tampão NBD (50mM Tris-HCl pH 8,0; 3mM CaCl2; 100mM
NaCl) acrescido de 5µl do substrato NBDPC (1mg/ml, diluído em tampão NBD) e 5µl
de amostra. A reação foi monitorada em fluorímetro (Hitachi - F4500, Japão), onde o
fluoróforo foi excitado a 460nm e o incremento de fluorescência mensurado a
534nm.

43
DOSAGEM DE CREATINA CINASE (CK) EM SORO DE CAMUNDONGOS TRATADOS
COM FRAÇÕES ENRIQUECIDAS EM PLA2 DO VENENO DE B. ATROX
Neste ensaio, empregamos metodologia descrita por Kanashiro et al., 2002.
Brevemente, camundongos Suíços albinos adultos foram injetados com as 40µg
frações de PLA2 por via intramuscular (i.m.) no músculo gastrocnêmio. Uma hora
após a injeção, os animais foram sacrificados com CO2 e o sangue colhido por
punção cardíaca. O soro foi separado e a enzima CK sérica foi dosada utilizando o
kit CK (NAC) Granutest 2,5 (Diagnostica Merk, Germany). Para avaliar a atividade
neutralizante do mAb A85/9-4 (antiPLA2), 40 µg da enzima PLA2 foram previamente
incubada com 140 µg do mAb durante 30 minutos a temperatura ambiente e, em
seguida, inoculada pela via i.m. em camundongos. Uma hora após a inoculação os
animais foram sacrificados, o soro colhido e a enzima CK sérica foi dosada. Cada
experimento foi realizado com pelo menos três animais em cada grupo.
NEUTRALIZAÇÃO DA ATIVIDADE CATALÍTICA DA TROMBINA-SIMILE PELO
ANTICORPO MONOLONAL 6AD2-G5
A inibição da atividade catalítica da trombina-simile de B. atrox pelo mAb
6AD2-G5 foi avaliada pré-incubando-se 40mg da fração enriquecida de trombina-
simile (1,4 mg/mL) com concentrações variadas (5, 10, 20 e 50mg) do anticorpo
6AD2-G5 (5,4mg/mL) durante 30min. à temperatura ambiente. Logo a seguir, as
amostras foram incubadas com 2,9mL de solução composta de substrato TAME
0,2M (Sigma Chemical Company – USA) diluído em tampão (0,2M Tris pH 8,1;
0,01M CaCl2 · 2H2O) por uma hora a temperatura ambiente. Em seguida, a
atividade remanescente da enzima foi medida em espectrofotômetro sob
comprimento de onda de 247nm.

44
NEUTRALIZAÇÃO DA ATIVIDADE COAGULANTE DE VENENO DE B. ATROX PELO
ANTICORPO MONOCLONAL 6AD2-G5 IN VITRO
A inibição da atividade coagulante foi realizada incubando concentrações
variadas (0.1, 0.5, 1.0, 3.0, 5.0, 10mg respectivamente) do mAb 6AD2-G5
(4,62mg/mL) com 20mg de veneno B. atrox durante 5min. à temperatura ambiente.
Em seguida, a essas misturas foram adicionados 500mL de solução composta de
fibribogênio bovino (Sigma, EUA) a 2,7mg/mL, diluído em tampão (50mM Tris-HCl
pH 8,0; 100mM NaCl2; 5mM CaCl2 ). Logo após a adição do fibrinogênio, por análise
visual, o tempo de formação do coágulo foi cronometrado.
PRODUÇÃO DE LÍQÜIDO ASCÍTICO IN VIVO
Células de hibridomas produtoras de anticorpos monoclonais anti-
serinoprotease (trombina-simile) 6AD2-G5 (PETRETSKI et al, 2000a), antifosfolipase
A2 A85/9-4 (KANASHIRO et al, 2002) e anti-hemorragina (Zn-metaloproteinase)
59/2-E4 (BARROS et al, 1998) do veneno de B. atrox, foram descongeladas e
cultivadas em garrafas de cultura com meio DMEM-F12 suplementado com 10% de
SFB e 10µg/mL de gentamicina. Uma vez amplificada a cultura, 1 x 106 células
foram inoculadas por via intraperitoneal (i.p.) em camundongos BALB/c previamente
injetados via i.p. com 0,4 mL de óleo mineral com 7 dias de antecedência. Após a
inoculação os animais foram observados diariamente e com aproximadamente 10
dias após a inoculação, o líquido ascítico foi colhido por punção abdominal
PURIFICAÇÃO DOS ANTICORPOS MONOCLONAIS
A purificação foi realizada segundo o método descrito por Steinbuch e Audran
(1969) através de precipitação com ácido caprílico. O líqüido ascítico ou o soro de
camundongos normais foi diluído 1:3 em tampão acetato de sódio 60 mM, pH 4.0,
em seguida, foi acrescentado, lentamente, 0.4mL de ácido caprílico para cada 10mL
de líqüido ascítico sob agitação, a temperatura ambiente. A agitação foi mantida por
30 min e, a seguir, o material foi centrifugado a 5000g durante 1h a 4ºC. Após a

45
centrifugação, o sobrenadante foi coletado, o pH ajustado para 7.0 com NaOH 3M e
a suspensão, novamente centrifugada nas mesma condições acima. Ao
sobrenandante resultante foi adicionado sulfato de amônio a 45% p/v,
vagarosamente, e a precipitação foi realizada sob agitação a 4ºC, overnight. Por
último, a suspensão foi centrifugada a 5000g durante 30 min. e o precipitado
resultante foi suspenso em solução salina e dialisado contra solução salina.
DOSAGEM DE PROTEÍNAS
Para determinação da concentração de proteínas nas amostras purificadas,
foi utilizada a metodologia descrita por Bradford (1976). Resumidamente, amostras
de mAb purificado foram incubadas com 1mL de solução de Bradford por 10min. em
seguida, as densidades ópticas (D.O.) das amostras foram determinadas em
espectrofotômetro (Shimadzu, Japão) sob comprimento de onde de 492ηm. Como
curva padrão foi usada albumina sérica bovina (BSA). Também foi aplicada, por ser
mais sensível a metodologia do ácido bicinconínico (BCA). Brevemente, amostras de
mAb foram incubados com 200µL de solução BCA (CuSO4 4% diluído 1:49 em
BCA), por 30min. a 37ºC. A seguir, as D.O. das amostras foram determinadas em
espectrofotômetro com comprimento de onda de 592ηm. Utilizou-se novamente BSA
como curva padrão.
ELETROFORESE EM GEL DE POLIACRILAMIDA CONTENDO DODECIL SULFATO
DE SÓDIO (SDS PAGE)
Os géis de SDS-PAGE foram feitos em sistema descontínuo (Bio Rad Mini
Protean II, USA), constituído de gel separador com 15% de acrilamida (1,5 mL de
Tris-HCl 3M pH 8.8; 2,4 mL de Acrilamida/bisacrilamida 30%; 20µL de SDS a 10%;
45 µL de APS a 10%; 5 µL de TEMED; 2 mL de água destilada) e o gel de
empilhamento com 4% de acrilamida (1,5mL de Tris-HCl pH 6.8; 400µL de
Acrilamida/Bisacrilamida; 30µL SDS a 10%; 30µL de APS a 10%; 6µL de TEMED;
1,049mL de água destilada) em tampão de corrida Tris-Glicina (Tris-base 1,5%;

46
Glicina 7,2%; SDS 0.5%: pH 8,3) segundo a metodologia descrita por Laemmli
(1970). A corrente elétrica utilizada foi de 80v até as amostras cruzarem o gel de
empilhamento, em seguida, ajustada para 120v até o fim da corrida. Os géis foram
corados com azul de Coomassie R250 (Gibco BRL, USA) a 0,1% em solução
metanol-acético por overnigth e descorados em solução descorante (30% de
metanol e 10% de ácido acético) até a visualização das bandas.
IMMUNOBLOTTING
Primeiramente, realizou-se uma SDS-PAGE conforme anteriormente descrito.
O gel foi carregado com 15µg de cada mAb, A85/9-4, 59/2-E4 e 6AD2-G5
purificados. Em seguida, as proteínas contidas no gel foram transferidas para uma
membrana de nitrocelulose (Bio Rad, USA) utilizando o sistema Bio Rad Mini
Transblot II sob voltagem constante de 10v por um período de 2h em tampão fosfato
(Na2HPO4 1,5%; NaH2PO4 1,2%). Após a transferência, a membrana foi corada com
Ponceau S (Sigma Chemical Co, EUA) (Ponseau S 1g; ác. acético 1mL; H2O
destilada 100mL), lavada com água destilada e bloqueada com 15mL de leite
desnatado (leite Molico a 5% em PBST) (por overnight a 4ºC. Após o bloqueio, a
membrana sofreu duas lavagens com PBST durante 5min. cada. Em seguida, a
membrana foi incubada com anticorpo anti-IgG de camundongo marcado com
peroxidase (Southern Biotecnologies, EUA), diluído 1:2000, durante 2h a
temperatura ambiente. A membrana foi , novamente, lavada 3 vezes com PBST
(NaCl 0,8%; Na2HPO4 1,15%; KCl 0,2%; KH2PO4 0,2%; 0,5% tween) por 5min. Após
as lavagens, a membrana foi revelada com solução 3,3-Diaminobenzidina (DAB)
(4,9mL H20; 0,1mL Tris 2M pH 7,5; 0,3mL Imidazol 0,1mL, 5mg DAB, 0,05mL H202),
a reação foi interrompida com adição de água destilada, logo após a visualização
das bandas.
REATIVIDADE DOS ANTICORPOS MONOCLONAIS
A reatividade dos mAbs foi estudada pelo teste de ELISA. Os antígenos,
10mg/mL de uma fração enriquecida de trombina-simile e 30mg/mL do veneno bruto

47
de Bothrops atrox diluídos em tampão carbonato/bicarbonato 0,05M, pH 9.6
(50mL/poço) foram incubados overnight a 4ºC. Após a adsorção do antígeno, as
placas foram lavadas duas vezes com PBST e bloqueados com gelatina 1%
(100mL/poço) diluída em PBS por uma hora, a temperatura ambiente. Após o
bloqueio e uma nova lavagem com PBST, foram acrescentados os anticorpos
monoclonais (75mL/poço) diluídos em PBST, começando com uma concentração
inicial de 1mg/mL seguindo de diluições (fator 5) até o limite de 0,0128 x 10-3mg/mL.
Como controle, foi utilizado IgG inespecífica de camundongos normais. A incubação
da placa foi feita a 37ºC por 45 min. Após três lavagens com PBST, foi adicionado o
anticorpo conjugado anti-IgG de camundongo (Southern Biotechnologies, EUA)
marcado com peroxidase diluído 1:2000 (50mL/poço) em PBST e nova incubação foi
realizada a 37ºC por 45min. Esta reação foi revelada pela adição do substrato H2O2
e ortofenildiamina – OPD (Sigma Chemical Co, EUA) diluídos em tampão (5,75mL
H20 dest.; 3,25mL ác. cítrico 0,1M; 3,5mL fosfato de sódio 0,2M; 5mg OPD; 0,005
H202). A reação foi interrompida adicionando-se H2SO4 3N (50mL/poço) e lida em
comprimento de onda de 492 nm em espectrofotômetro de placas (Tittertek
Multiskan).
DETERMINAÇÃO DA DL50 DO VENENO DE B. ATROX
A determinação da DL50 do veneno de B. atrox foi relizada conforme Finney,
(1971). Brevemente, grupos de dez camundongos suíços albinos com peso entre 18
e 22g, foram inoculados com cada uma das diferentes concentrações de veneno de
B. atrox por via i.p. A sobrevivência dos animais foi avaliada num período de 48h. O
cálculo da DL50 foi realizado, utilizando-se o programa Probit versão 97.1 (F.R.
Marsman, Holanda).
NEUTRALIZAÇÃO DA AÇÃO LETAL DO VENENO DE B. ATROX
Diferentes concentrações do pool de mAbs neutralizantes foram incubados
com 2-3DL50 do veneno de B. atrox, a seguir, esta mistura foi inoculada em

48
camundongos Suíços albinos de 18 a 20g por via i.p. Alternativamente, o pool de
mAbs foi inoculado por via i.p. e em seguida, os camundongos foram desafiados
com o veneno pela via subcutânea (s.c.). Este ensaio foi também realizado segundo
metodologia preconizada pela Farmacopéia Brasileira. Diluições seriadas (fator 1,3)
do pool de mAbs foi incubado com 4 ou 5DL50 de veneno de B. atrox a 37ºC por 30
min. Em seguida, a mistura foi inoculada por via i.p nos animais. A sobrevivência dos
camundongos foi avaliada por até 48 horas. Neste ensaio, foi também testado um
soro antiofídico comercial, fornecido pelo Instituto Butantan, pré-incubado com
veneno de B. atrox. No grupo controle os animais foram inoculados somente com
veneno.
NEUTRALIZAÇÃO DA ATIVIDADE ANTICOAGULANTE DO VENENO DE B. ATROX
A neutralização da atividade anticoagulante foi realizada conforme adaptação
de Otero et al. (2000). Para determinar a cinética da atividade anticoagulante do
veneno de B. atrox, camundongos Suíços albinos de 18 a 22g (n=5) foram
previamente anestesiados com 250µg de cloridrato de quetamina (Vetbrand, Brasil)
via s.c. e inoculados com 200µg de veneno bruto por via i.p. Em diferentes intervalos
de tempo, as pontas das caudas dos animais foram cortadas e imersas em 10mL de
água destilada até o fim do sangramento. Para a determinação do tempo normal de
coagulação, foram utilizados animais não inoculados com veneno. A D.O. da água
foi determinada utilizando espectrofotômetro com comprimento de onda a 410nm.
Para o ensaio de neutralização, camundongos suíços albinos (n=8) anestesiados,
foram inoculados por via i.p. com 200µg de veneno bruto previamente incubado com
13,5mg do pool de mAbs durante 30 min a temperatura ambiente. Experimentos
adicionais, também foram realizados, utilizando-se dois grupos de animais (n=4)
inoculados com veneno de B. atrox previamente incubado com 3,5mg do mAb
6AD2-G5 (antitrombina-simile) ou com 500µL de soro antibotrópico durante 30min. à
temperatura ambiente. Após 1 hora, as pontas das caudas dos animais foram
cortadas e imersas em água destilada até cessar a hemorragia. A D.O. da água foi
determinada utilizando espectrofotômetro com comprimento de onda a 410nm.

49
ANÁLISES HISTOLÓGICAS
Grupos de camundongos submetidos ao ensaio de neutralização, em tempos
determinados (2h, 24h e 48h), foram sacrificados e amostras de coração, pulmões,
fígado, rins, baço e músculos do abdômen e diafragma foram colhidas e
imediatamente embebidas em solução de formol 10% tamponado para fixação. Após
a fixação, as amostras foram desidratadas em concentrações seriadas de álcool (70,
90, 100%), imersas em Xilol e inclusas em parafina de acordo com os protocolos de
rotina para microscopia óptica. Os cortes de 5mm feitos em micrótomo semi-
automático (Leica, Brasil) e coradas com hematoxilina e eosina (H&E). As análises
das lâminas foram feitas em microscópio óptico Axioplan Zeiss (Alemanha). Nas
análises, as lesões foram classificadas como: discreta, moderada e grave.

50
RESULTADOS

51
AVALIAÇÃO DAS ATIVIDADES NEUTRALIZANTES DOS ANTICORPOS
MONOCLONAIS
Os anticorpos monoclonais neutralizantes contras as principais toxinas do
veneno de B. atrox hemorragina, fosfolipase A2 e trombina-simile, foram produzidos
e purificados a partir de líquido-ascítico produzidos em camundongos BALB/c. As
atividades neutralizantes contra as respectivas toxinas foram avaliadas.
NEUTRALIZAÇÃO DA ATIVIDADE HEMORRÁGICA PELO MAB 59/2-E4
Na Figura 1 podemos observar que as três concentrações de anticorpos
utilizados foram capazes de inibir em diferentes graus a hemorragia induzida pelo
veneno, sendo que a concentração de 100µg do mAb 59/2-E4 foi mais eficiente na
inibição da atividade hemorrágica.
Esta mesma preparação de anticorpo monoclonal 59/2-E4 também foi testada
quanto a sua capacidade de inibir a atividade catalítica da Zn-metaloproteinase de
baixo peso molecular purificada do veneno de B. atrox (PETRETSKI et al, 2000b).
Podemos observar na Figura 2 que o anticorpo monoclonal 59/2-E4 ou anticorpos
policlonais produzidos em coelho quando pré-incubados com a hemorragina de
baixo peso do veneno, não foram capazes de inibir a atividade proteásica desta
enzima. A atividade catalítica da enzima incubada com os anticorpos foi similar à
atividade da amostra controle positiva que foi incubada com um anticorpo não
relacionado. Cinéticas utilizando soros hiperimunes produzidos em camundongos,
coelhos e galinhas, também foram realizadas, sendo que nenhum dos soros foi
capaz de inibir a atividade catalítica das metoloproteinases do veneno de B. atrox
(dado não mostrado).

52
Figura 1: Atividade hemorrágica do veneno total de B. atrox. Concentrações de
100, 50 e 25 mg de mAb 59/2-E4 purificado e foram pré-incubados com 2 mg de
veneno durante 30 minutos. A seguir, esta mistura foi inoculada intradermicamente
no dorso de camundongos Suíços albinos e trinta minutos após, os animais foram
sacrificados e a área hemorrágica no subcutâneo dos animais foi avaliada. 1-
Controle (100 mg de mAb IgG1 não relacionado); 2- mAb 59/2-E4 25 mg; 3- mAb
59/2-E4 50 mg; 4- mAb 59/2-E4 100 mg.
Figura 2: Atividade proteolítica da hemorragina de baixo peso molecular pelo
mAb 59/2-E4. Trinta microgramas da enzima purificada foram pré-incubada com
300µg do mAb 59/2-E4 ou a mesma concentração de anticorpo policlonal de coelho
durante 30min. a temperatura ambiente. Após a incubação a atividade catalítica
remanescente da enzima foi avaliada utilizando-se como substrato a azocaseína. O
controle positivo foi constituído de enzima incubado com IgG inespecífica. A reação
foi lida em espectrofotômetro com comprimento de onda a 440 nm.
0
0,20,4
0,60,8
1
Cont (-) Cont (+) Soro 59/2-E4
DO
440
nm
0
0,20,4
0,60,8
1
Cont (-) Cont (+) Soro 59/2-E4
DO
440
nm
1 2
4 3
2 1
4 3
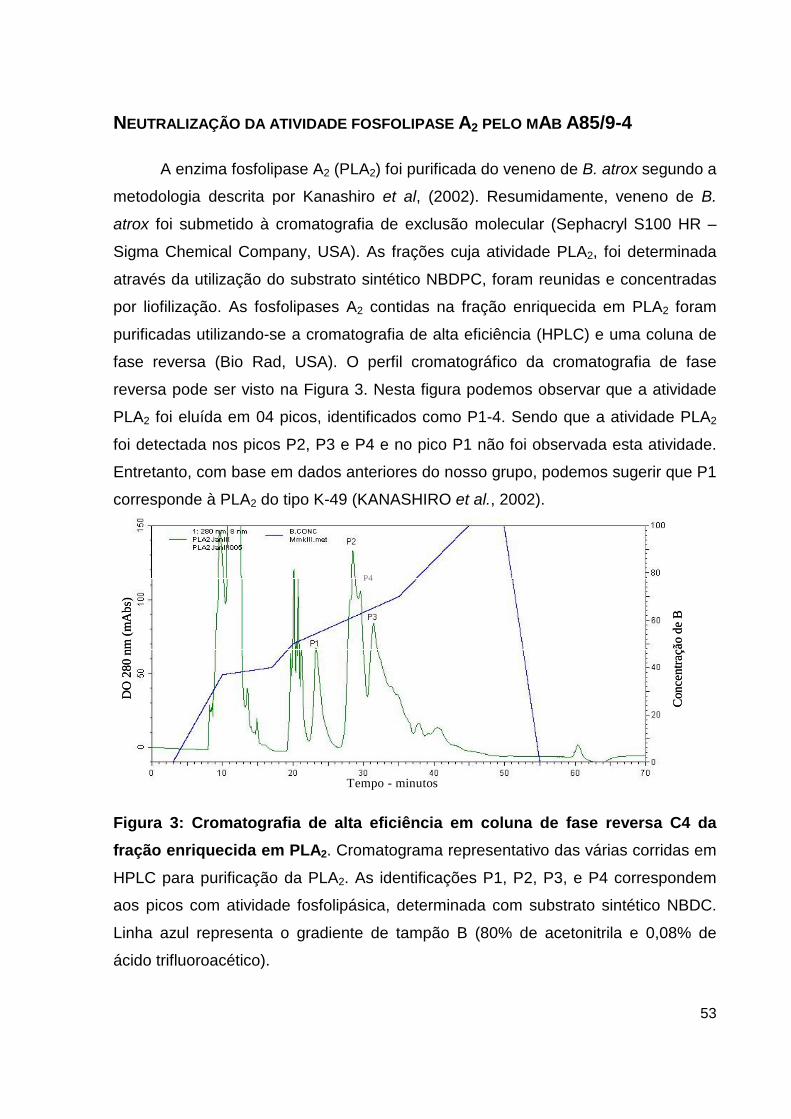
53
NEUTRALIZAÇÃO DA ATIVIDADE FOSFOLIPASE A2 PELO MAB A85/9-4
A enzima fosfolipase A2 (PLA2) foi purificada do veneno de B. atrox segundo a
metodologia descrita por Kanashiro et al, (2002). Resumidamente, veneno de B.
atrox foi submetido à cromatografia de exclusão molecular (Sephacryl S100 HR –
Sigma Chemical Company, USA). As frações cuja atividade PLA2, foi determinada
através da utilização do substrato sintético NBDPC, foram reunidas e concentradas
por liofilização. As fosfolipases A2 contidas na fração enriquecida em PLA2 foram
purificadas utilizando-se a cromatografia de alta eficiência (HPLC) e uma coluna de
fase reversa (Bio Rad, USA). O perfil cromatográfico da cromatografia de fase
reversa pode ser visto na Figura 3. Nesta figura podemos observar que a atividade
PLA2 foi eluída em 04 picos, identificados como P1-4. Sendo que a atividade PLA2
foi detectada nos picos P2, P3 e P4 e no pico P1 não foi observada esta atividade.
Entretanto, com base em dados anteriores do nosso grupo, podemos sugerir que P1
corresponde à PLA2 do tipo K-49 (KANASHIRO et al., 2002).
Figura 3: Cromatografia de alta eficiência em coluna de fase reversa C4 da
fração enriquecida em PLA2. Cromatograma representativo das várias corridas em
HPLC para purificação da PLA2. As identificações P1, P2, P3, e P4 correspondem
aos picos com atividade fosfolipásica, determinada com substrato sintético NBDC.
Linha azul representa o gradiente de tampão B (80% de acetonitrila e 0,08% de
ácido trifluoroacético).
P4
DO
280
nm
(m
Abs
)
Con
cent
raçã
o de
B
Tempo - minutos
P4
DO
280
nm
(m
Abs
)
Con
cent
raçã
o de
B
Tempo - minutos
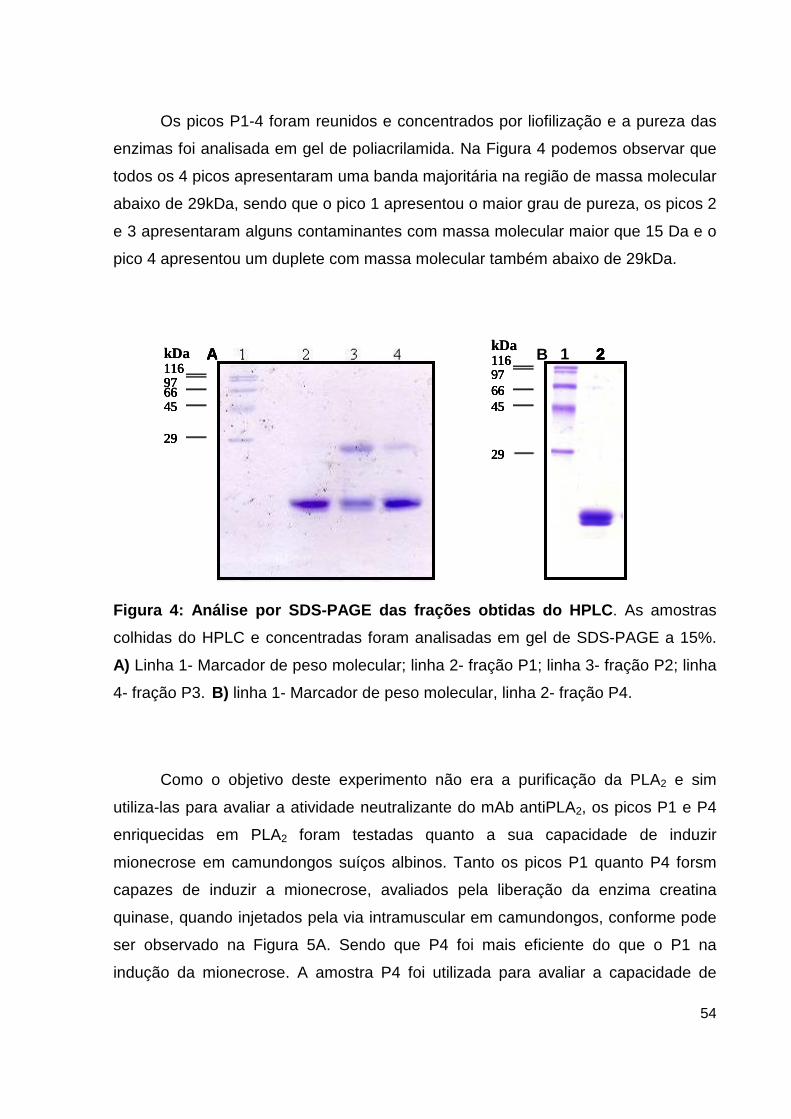
54
Os picos P1-4 foram reunidos e concentrados por liofilização e a pureza das
enzimas foi analisada em gel de poliacrilamida. Na Figura 4 podemos observar que
todos os 4 picos apresentaram uma banda majoritária na região de massa molecular
abaixo de 29kDa, sendo que o pico 1 apresentou o maior grau de pureza, os picos 2
e 3 apresentaram alguns contaminantes com massa molecular maior que 15 Da e o
pico 4 apresentou um duplete com massa molecular também abaixo de 29kDa.
Figura 4: Análise por SDS-PAGE das frações obtidas do HPLC. As amostras
colhidas do HPLC e concentradas foram analisadas em gel de SDS-PAGE a 15%.
A) Linha 1- Marcador de peso molecular; linha 2- fração P1; linha 3- fração P2; linha
4- fração P3. B) linha 1- Marcador de peso molecular, linha 2- fração P4.
Como o objetivo deste experimento não era a purificação da PLA2 e sim
utiliza-las para avaliar a atividade neutralizante do mAb antiPLA2, os picos P1 e P4
enriquecidas em PLA2 foram testadas quanto a sua capacidade de induzir
mionecrose em camundongos suíços albinos. Tanto os picos P1 quanto P4 forsm
capazes de induzir a mionecrose, avaliados pela liberação da enzima creatina
quinase, quando injetados pela via intramuscular em camundongos, conforme pode
ser observado na Figura 5A. Sendo que P4 foi mais eficiente do que o P1 na
indução da mionecrose. A amostra P4 foi utilizada para avaliar a capacidade de
A116976645
29
B 1 2116976645
29
kDa kDaA
116976645
29
AAA116976645
29
116976645
29
B 1 2116976645
29
B 1 2B 1 2B 1 2B 1 2116976645
29
116976645
29
kDa kDa
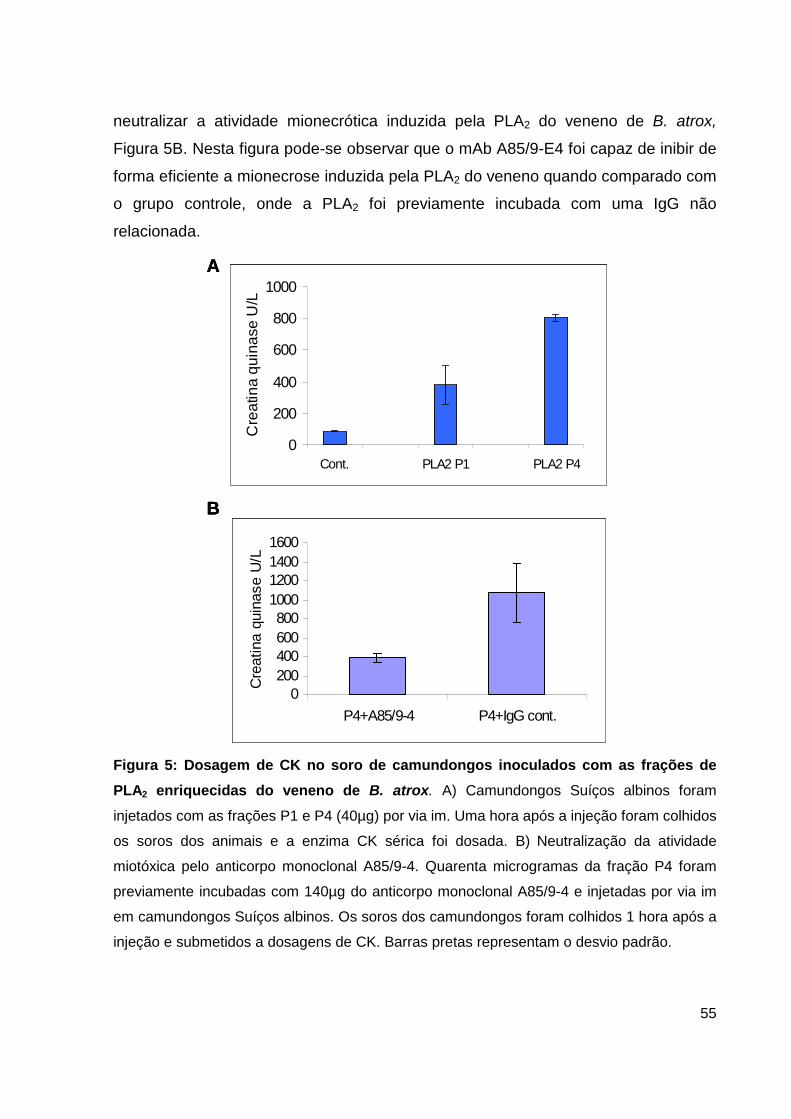
55
neutralizar a atividade mionecrótica induzida pela PLA2 do veneno de B. atrox,
Figura 5B. Nesta figura pode-se observar que o mAb A85/9-E4 foi capaz de inibir de
forma eficiente a mionecrose induzida pela PLA2 do veneno quando comparado com
o grupo controle, onde a PLA2 foi previamente incubada com uma IgG não
relacionada.
Figura 5: Dosagem de CK no soro de camundongos inoculados com as frações de
PLA2 enriquecidas do veneno de B. atrox. A) Camundongos Suíços albinos foram
injetados com as frações P1 e P4 (40µg) por via im. Uma hora após a injeção foram colhidos
os soros dos animais e a enzima CK sérica foi dosada. B) Neutralização da atividade
miotóxica pelo anticorpo monoclonal A85/9-4. Quarenta microgramas da fração P4 foram
previamente incubadas com 140µg do anticorpo monoclonal A85/9-4 e injetadas por via im
em camundongos Suíços albinos. Os soros dos camundongos foram colhidos 1 hora após a
injeção e submetidos a dosagens de CK. Barras pretas representam o desvio padrão.
0
200
400
600
800
1000
Cont. PLA2 P1 PLA2 P4
Cre
atin
a qu
inas
e U
/LA
0200400600800
1000120014001600
P4+A85/9-4 P4+IgG cont.
Cre
atin
a qu
inas
e U
/L
B
0
200
400
600
800
1000
Cont. PLA2 P1 PLA2 P4
Cre
atin
a qu
inas
e U
/LA
0
200
400
600
800
1000
Cont. PLA2 P1 PLA2 P4
Cre
atin
a qu
inas
e U
/LA
0200400600800
1000120014001600
P4+A85/9-4 P4+IgG cont.
Cre
atin
a qu
inas
e U
/L
B
0200400600800
1000120014001600
P4+A85/9-4 P4+IgG cont.
Cre
atin
a qu
inas
e U
/L
B

56
NEUTRALIZAÇÃO DA ATIVIDADE COAGULANTE DA TROMBINA-SIMILE PELO
ANTICORPO MONOLONAL 6AD2-G5
A inibição da atividade catalítica da trombina-simile de B. atrox pelo mAb
6AD2-G5 foi avaliada incubando-se uma fração enriquecida de trombina-simile com
o anticorpo e a atividade remanescente foi quantificada com a utilização do substrato
TAME. Na Figura 6B podemos ver que o mAb 6AD2-G5 foi capaz de inibir a
atividade catalítica da trombina-simile sobre o substrato TAME em todas as
concentrações testadas.
O mAb 6AD2-G5 também foi avaliado quanto à capacidade de neutralizar a
conversão de fibrinogênio em fibrina in vitro. Assim, concentrações variadas de mAb
6AD2-G5 foram incubadas com 20mg de veneno de B. atrox durante 5min a
temperatura ambiente. Logo a seguir, acrescentou-se solução de fibrinogênio bovino
e o tempo de formação do coágulo foi cronometrado. Na figura 7 podemos observar
que o mAb 6AD2-G5 também foi capaz de inibir a formação de coágulo de forma
dose dependente.
PRODUÇÃO DE LÍQÜIDO ASCÍTICO
Aproximadamente 10 dias após a inoculação dos hibridomas 6AD2-G5,
A85/9-4 e 59/2-E4 em camundongos BALB/c, foram coletados, por punção
abdominal, os líquidos ascíticos produzidos pelos animais. As ascites colhidas foram
centrifugadas a 800 g e o sobrenadante foi colhido e armazenado no freezer –20ºC
para posterior purificação dos anticorpos.
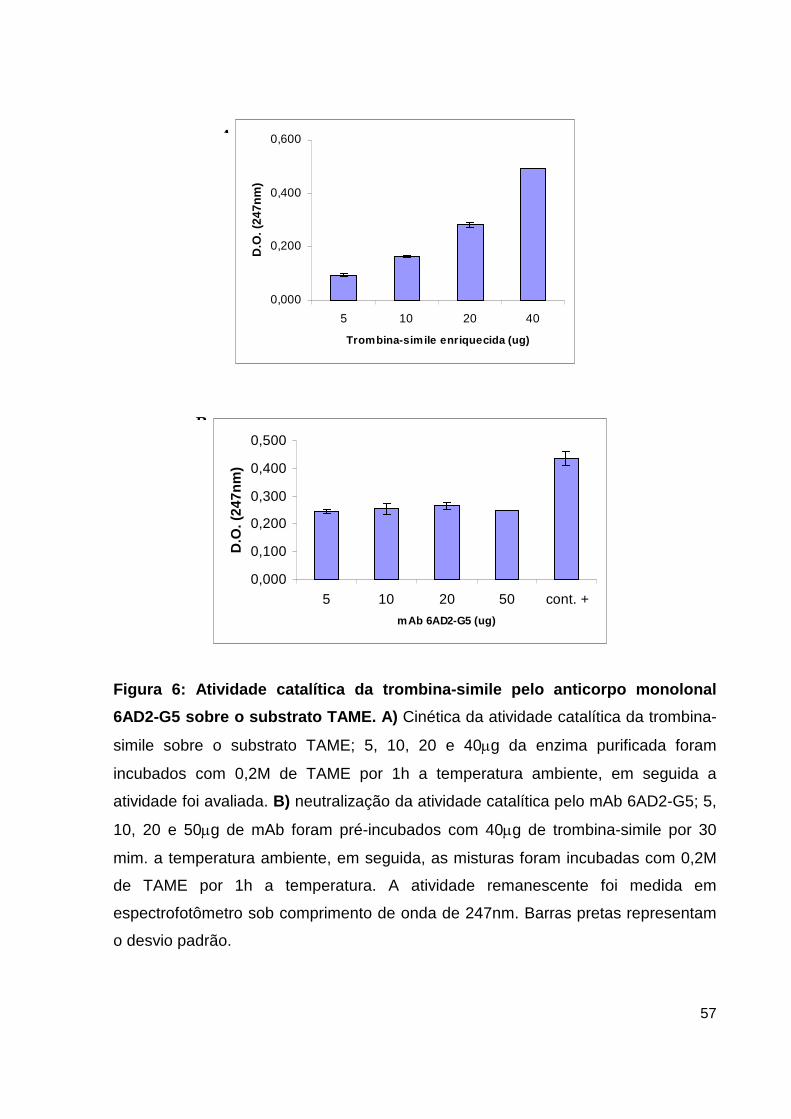
57
Figura 6: Atividade catalítica da trombina-simile pelo anticorpo monolonal
6AD2-G5 sobre o substrato TAME. A) Cinética da atividade catalítica da trombina-
simile sobre o substrato TAME; 5, 10, 20 e 40mg da enzima purificada foram
incubados com 0,2M de TAME por 1h a temperatura ambiente, em seguida a
atividade foi avaliada. B) neutralização da atividade catalítica pelo mAb 6AD2-G5; 5,
10, 20 e 50mg de mAb foram pré-incubados com 40mg de trombina-simile por 30
mim. a temperatura ambiente, em seguida, as misturas foram incubadas com 0,2M
de TAME por 1h a temperatura. A atividade remanescente foi medida em
espectrofotômetro sob comprimento de onda de 247nm. Barras pretas representam
o desvio padrão.
0,000
0,200
0,400
0,600
5 10 20 40
Trombina-simile enriquecida (ug)
D.O
. (24
7nm
)
A
0,000
0,100
0,200
0,300
0,400
0,500
5 10 20 50 cont. +
mAb 6AD2-G5 (ug)
D.O
. (24
7nm
)
B
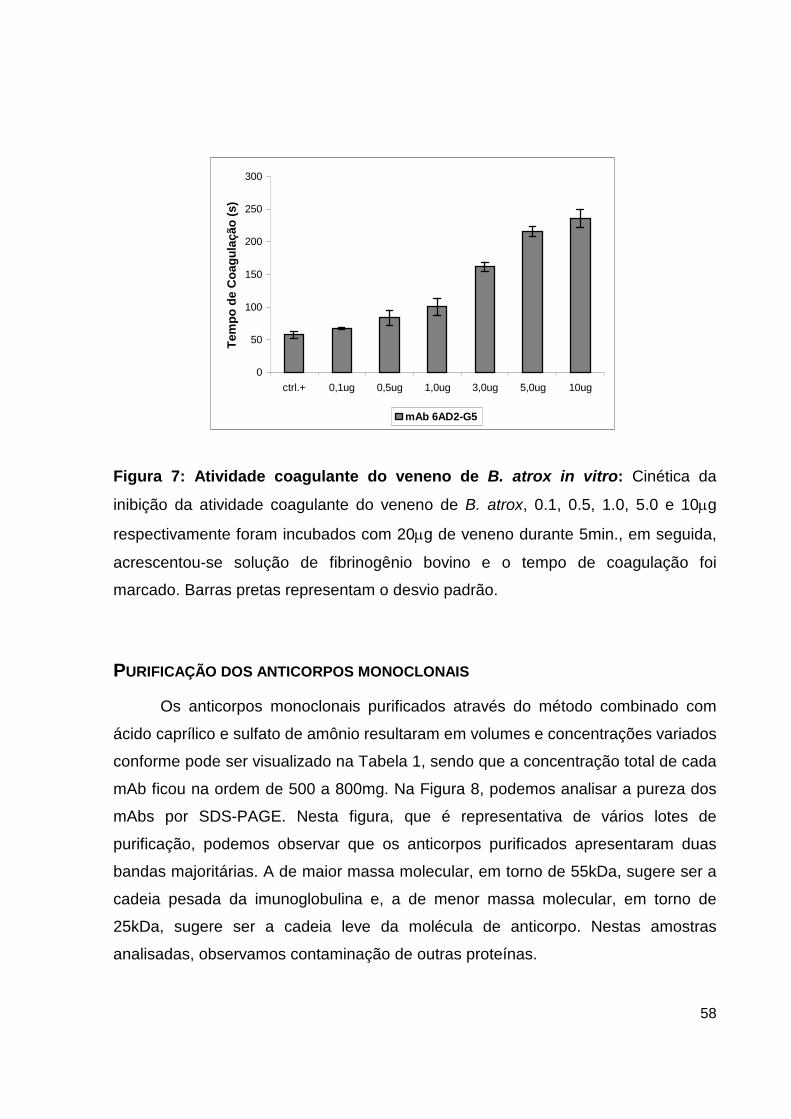
58
Figura 7: Atividade coagulante do veneno de B. atrox in vitro: Cinética da
inibição da atividade coagulante do veneno de B. atrox, 0.1, 0.5, 1.0, 5.0 e 10mg
respectivamente foram incubados com 20mg de veneno durante 5min., em seguida,
acrescentou-se solução de fibrinogênio bovino e o tempo de coagulação foi
marcado. Barras pretas representam o desvio padrão.
PURIFICAÇÃO DOS ANTICORPOS MONOCLONAIS
Os anticorpos monoclonais purificados através do método combinado com
ácido caprílico e sulfato de amônio resultaram em volumes e concentrações variados
conforme pode ser visualizado na Tabela 1, sendo que a concentração total de cada
mAb ficou na ordem de 500 a 800mg. Na Figura 8, podemos analisar a pureza dos
mAbs por SDS-PAGE. Nesta figura, que é representativa de vários lotes de
purificação, podemos observar que os anticorpos purificados apresentaram duas
bandas majoritárias. A de maior massa molecular, em torno de 55kDa, sugere ser a
cadeia pesada da imunoglobulina e, a de menor massa molecular, em torno de
25kDa, sugere ser a cadeia leve da molécula de anticorpo. Nestas amostras
analisadas, observamos contaminação de outras proteínas.
0
50
100
150
200
250
300
ctrl.+ 0,1ug 0,5ug 1,0ug 3,0ug 5,0ug 10ug
Tem
po
de
Co
agu
laçã
o (
s)
mAb 6AD2-G5
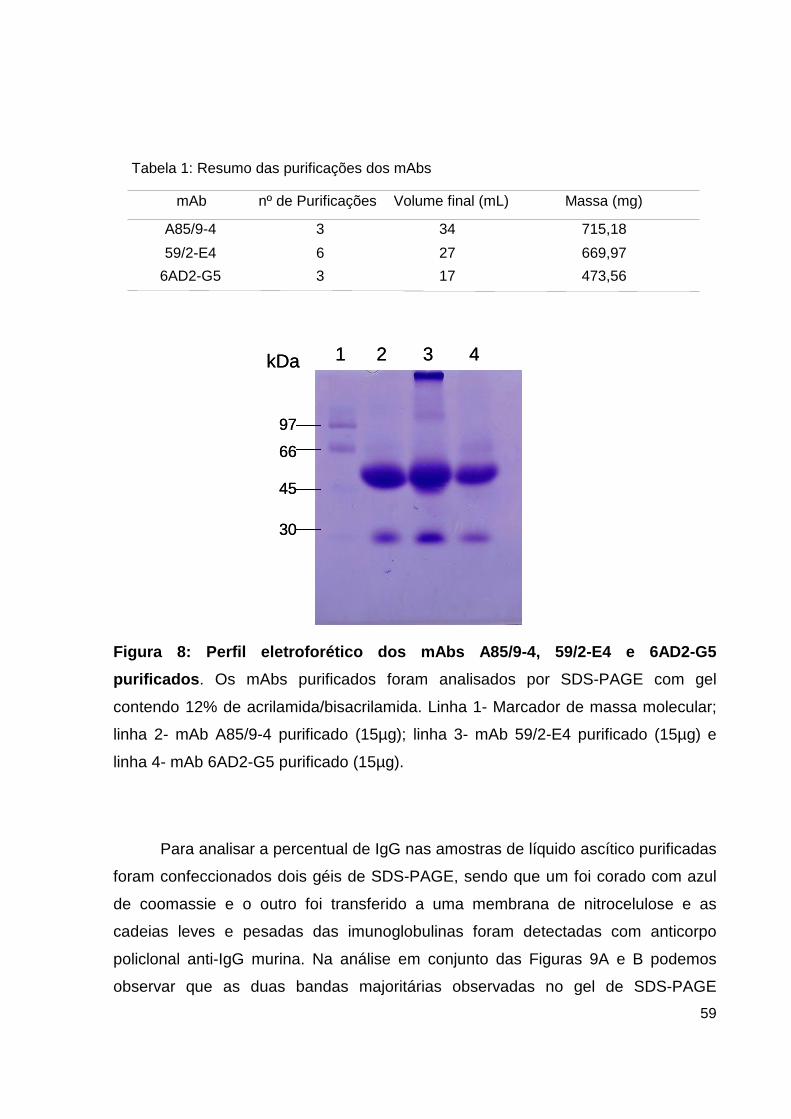
59
Figura 8: Perfil eletroforético dos mAbs A85/9-4, 59/2-E4 e 6AD2-G5
purificados. Os mAbs purificados foram analisados por SDS-PAGE com gel
contendo 12% de acrilamida/bisacrilamida. Linha 1- Marcador de massa molecular;
linha 2- mAb A85/9-4 purificado (15µg); linha 3- mAb 59/2-E4 purificado (15µg) e
linha 4- mAb 6AD2-G5 purificado (15µg).
Para analisar a percentual de IgG nas amostras de líquido ascítico purificadas
foram confeccionados dois géis de SDS-PAGE, sendo que um foi corado com azul
de coomassie e o outro foi transferido a uma membrana de nitrocelulose e as
cadeias leves e pesadas das imunoglobulinas foram detectadas com anticorpo
policlonal anti-IgG murina. Na análise em conjunto das Figuras 9A e B podemos
observar que as duas bandas majoritárias observadas no gel de SDS-PAGE
Tabela 1: Resumo das purificações dos mAbs
mAb nº de Purificações Volume final (mL) Massa (mg)
A85/9-4 3 34 715,18
59/2-E4 6 27 669,97
6AD2-G5 3 17 473,56
1 2 3 4kDa
97
66
45
30
1 2 3 4kDa
97
66
45
30

60
correspondem à cadeia pesada e cadeia leve dos anticorpos murinos.
Adicionalmente, podemos ver também que o mAb 6AD2-G5 apresentou o maior
percentual de contaminantes dentre os três mAbs analisados no gel. Na análise
densitométrica do gel de SDS-PAGE, Figura 9C, podemos ver que os mAbs A85/9-
4, 59/2-E4 e 6AD2-G5 apresentaram o percentual de 46, 45 e 37% de IgG,
respectivamente, obtida pela razão da somatória das densidades ópticas das
cadeias pesada e leve sobre a densidade óptica total e multiplicada por 100. Dessa
forma, foi possível estimar um percentual médio de 42% de IgG nas amostras
purificadas de mAbs.
AVALIAÇÃO DA REATIVIDADE DOS MABS PURIFICADOS
A capacidade dos mAbs purificados de identificar e se ligar aos respectivos
antígenos foi avaliada pelo teste de ELISA. Na Figura 10 pode ser visto o resultado
de um ELISA representativo dos vários lotes de anticorpos monoclonais submetidos
à purificação. Nas Figuras 10A e B podemos observar que os mAbs 59/2-E4 e
A85/9-4 purificados foram eficazes no reconhecimento dos respectivos antígenos,
mesmo em baixas concentrações. Enquanto que o mAb 6AD2-G5, Figura 10C, não
apresentou uma titulação similar aos outros dois anticorpos testados. Este fato se
justifica, provavelmente, porque o antígeno utilizado, fração enriquecida de trombina-
simile, não continha a concentração adequada de trombina-simile requerida para o
teste ou o antígeno apresentava-se degradado. O veneno bruto não foi utilizado
como antígeno neste caso porque a proporção de trombina-simile no veneno total é
baixa.
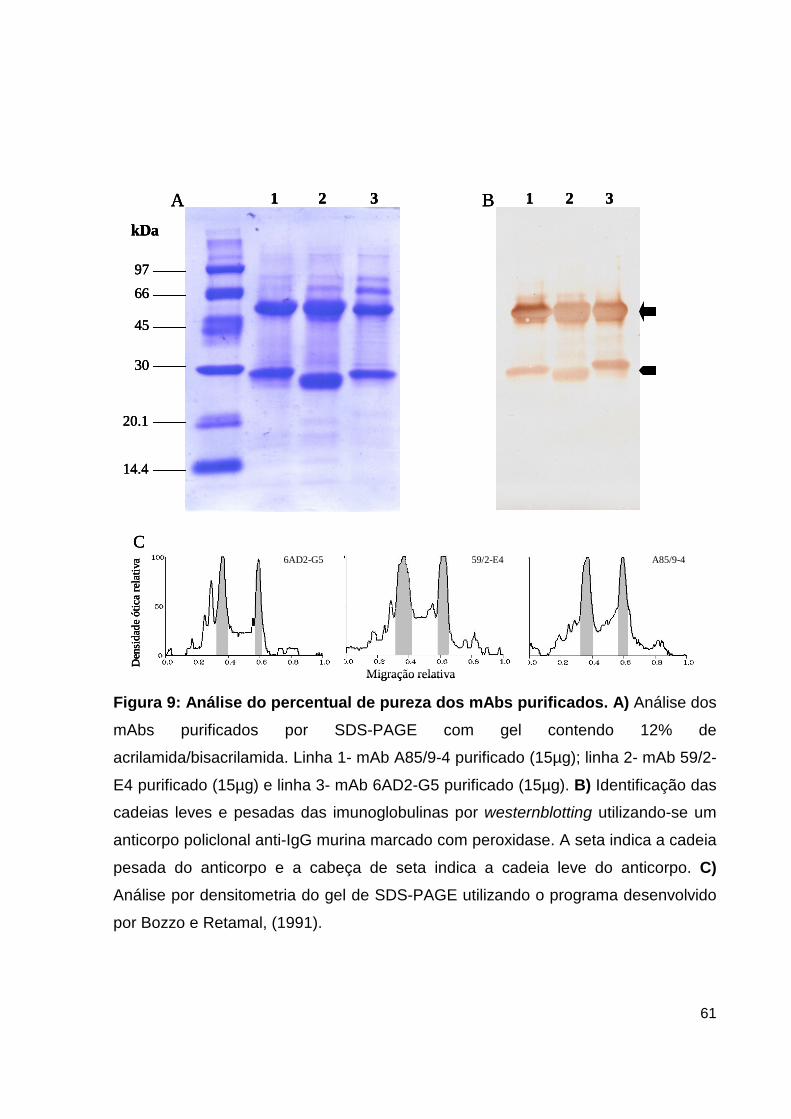
61
Figura 9: Análise do percentual de pureza dos mAbs purificados. A) Análise dos
mAbs purificados por SDS-PAGE com gel contendo 12% de
acrilamida/bisacrilamida. Linha 1- mAb A85/9-4 purificado (15µg); linha 2- mAb 59/2-
E4 purificado (15µg) e linha 3- mAb 6AD2-G5 purificado (15µg). B) Identificação das
cadeias leves e pesadas das imunoglobulinas por westernblotting utilizando-se um
anticorpo policlonal anti-IgG murina marcado com peroxidase. A seta indica a cadeia
pesada do anticorpo e a cabeça de seta indica a cadeia leve do anticorpo. C)
Análise por densitometria do gel de SDS-PAGE utilizando o programa desenvolvido
por Bozzo e Retamal, (1991).
Den
sida
de ó
tica
rela
tiva
Migração relativa
C6AD2-G5 A85/9-459/2-E4
Den
sida
de ó
tica
rela
tiva
Migração relativa
C
Den
sida
de ó
tica
rela
tiva
Migração relativa
C6AD2-G5 A85/9-459/2-E4
1 2 3B1 2 3A
14.4
97
66
30
20.1
45
kDa
1 2 3B 1 2 3B1 2 3A
14.4
97
66
30
20.1
45
kDa
1 2 3A
14.4
97
66
30
20.1
45
kDa
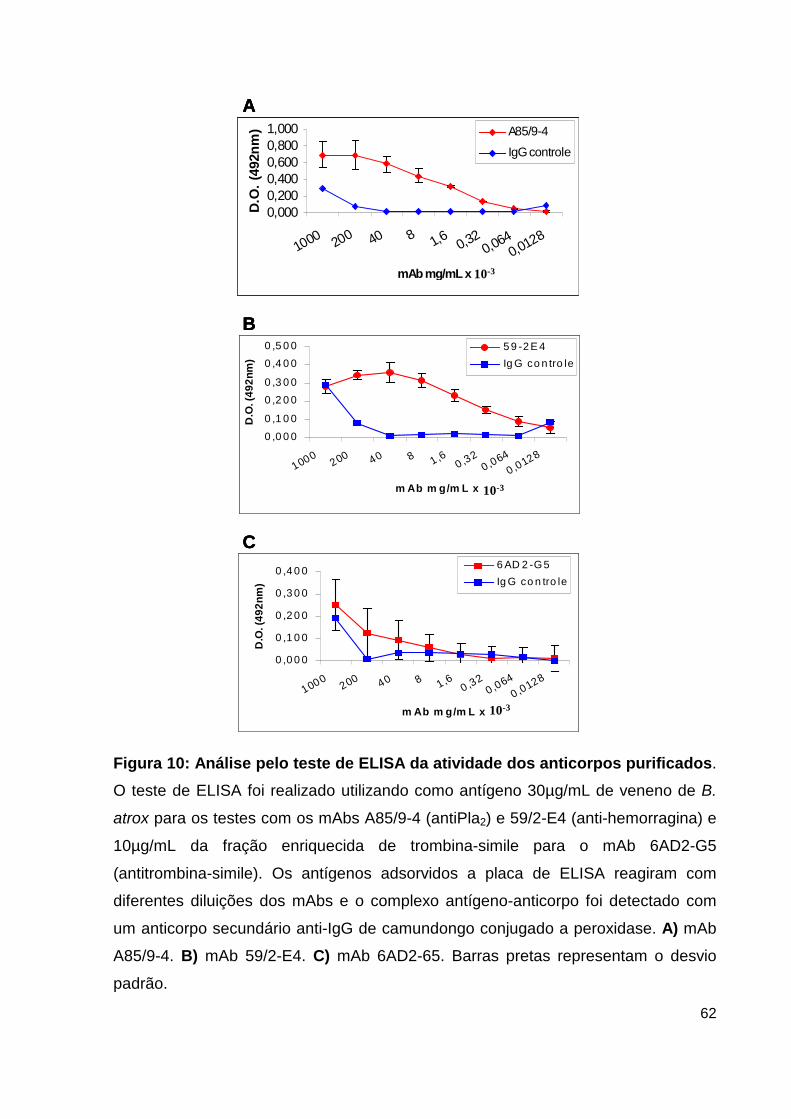
62
Figura 10: Análise pelo teste de ELISA da atividade dos anticorpos purificados.
O teste de ELISA foi realizado utilizando como antígeno 30µg/mL de veneno de B.
atrox para os testes com os mAbs A85/9-4 (antiPla2) e 59/2-E4 (anti-hemorragina) e
10µg/mL da fração enriquecida de trombina-simile para o mAb 6AD2-G5
(antitrombina-simile). Os antígenos adsorvidos a placa de ELISA reagiram com
diferentes diluições dos mAbs e o complexo antígeno-anticorpo foi detectado com
um anticorpo secundário anti-IgG de camundongo conjugado a peroxidase. A) mAb
A85/9-4. B) mAb 59/2-E4. C) mAb 6AD2-65. Barras pretas representam o desvio
padrão.
A
0,0000,2000,4000,6000,8001,000
1000 200 40 8 1,6 0,320,064
0,0128
mAb mg/mL x
D.O
. (4
92n
m) A85/9-4
IgG controle
10-3
B
0 ,0 0 0
0 ,1 0 0
0 ,2 0 0
0 ,3 0 0
0 ,4 0 0
0 ,5 0 0
1000200 40 8 1,6
0 ,320,064
0,0128
m Ab m g/m L x
D.O
. (4
92
nm
)
5 9 -2 E 4
Ig G co n tro le
10-3
C
0 ,0 0 0
0 ,1 0 0
0 ,2 0 0
0 ,3 0 0
0 ,4 0 0
1000200 40 8 1,6
0 ,320,064
0,0128
m Ab m g/m L x
D.O
. (4
92
nm
)
6 AD 2 -G5
Ig G co n tro le
10-3
A
0,0000,2000,4000,6000,8001,000
1000 200 40 8 1,6 0,320,064
0,0128
mAb mg/mL x
D.O
. (4
92n
m) A85/9-4
IgG controle
10-3
A
0,0000,2000,4000,6000,8001,000
1000 200 40 8 1,6 0,320,064
0,0128
mAb mg/mL x
D.O
. (4
92n
m) A85/9-4
IgG controle
10-3
0,0000,2000,4000,6000,8001,000
1000 200 40 8 1,6 0,320,064
0,0128
mAb mg/mL x
D.O
. (4
92n
m) A85/9-4
IgG controle
10-3
B
0 ,0 0 0
0 ,1 0 0
0 ,2 0 0
0 ,3 0 0
0 ,4 0 0
0 ,5 0 0
1000200 40 8 1,6
0 ,320,064
0,0128
m Ab m g/m L x
D.O
. (4
92
nm
)
5 9 -2 E 4
Ig G co n tro le
10-3
B
0 ,0 0 0
0 ,1 0 0
0 ,2 0 0
0 ,3 0 0
0 ,4 0 0
0 ,5 0 0
1000200 40 8 1,6
0 ,320,064
0,0128
m Ab m g/m L x
D.O
. (4
92
nm
)
5 9 -2 E 4
Ig G co n tro le
10-3
0 ,0 0 0
0 ,1 0 0
0 ,2 0 0
0 ,3 0 0
0 ,4 0 0
0 ,5 0 0
1000200 40 8 1,6
0 ,320,064
0,0128
m Ab m g/m L x
D.O
. (4
92
nm
)
5 9 -2 E 4
Ig G co n tro le
10-3
C
0 ,0 0 0
0 ,1 0 0
0 ,2 0 0
0 ,3 0 0
0 ,4 0 0
1000200 40 8 1,6
0 ,320,064
0,0128
m Ab m g/m L x
D.O
. (4
92
nm
)
6 AD 2 -G5
Ig G co n tro le
10-3
C
0 ,0 0 0
0 ,1 0 0
0 ,2 0 0
0 ,3 0 0
0 ,4 0 0
1000200 40 8 1,6
0 ,320,064
0,0128
m Ab m g/m L x
D.O
. (4
92
nm
)
6 AD 2 -G5
Ig G co n tro le
10-3
0 ,0 0 0
0 ,1 0 0
0 ,2 0 0
0 ,3 0 0
0 ,4 0 0
1000200 40 8 1,6
0 ,320,064
0,0128
m Ab m g/m L x
D.O
. (4
92
nm
)
6 AD 2 -G5
Ig G co n tro le
10-3

63
ENSAIO DE NEUTRALIZAÇÃO DA ATIVIDADE LETAL DO VENENO TOTAL IN VIVO
PELO POOL DE MABS
No primeiro ensaio preliminar o pool de mAbs foi constituído de 3,45mg de
cada mAb (A85/9-4, 59/2-E4 e 6AD2-G5) totalizando 10,35mg a ser administrado
para cada camundongo. O pool de mAbs foi incubado com 200 ou 300µg de veneno
de B. atrox por 30 minutos e inoculado via i.p. em camundongos de 18-20g. A
sobrevivência dos animais foi avaliada em 24 e 48 horas após a inoculação. Todos
os animais que receberam o pool de mAbs incubados com 300µg de veneno
morreram em até 48 horas, enquanto 3 de 5 animais inoculados que receberam o
pool de mAbs pré-incubados com 200µg de veneno sobreviveram, conforme pode
ser visto na Tabela 2.
Tabela 2: Ensaio de soroproteção in vivo de camundongos desafiados
com o pool de mAbs e veneno
* A mistura de mAbs utilizada corresponde a 3,45mg de cada anticorpo monoclonal
A85/9-4, 59/2-E4 e 6AD2-G5 totalizando 10,35mg/camundongo.
**A quantidade de veneno utilizada por camundongo
Grupos mAbs* Veneno** Vivos/Total
Grupo 1 10,35mg 200ug 3/5
Grupo 2 ---- 200ug 1/5
Grupo 3 10,35mg 300ug 0/5
Grupo 4 ---- 300ug 0/5

64
Em outro modelo de análise, os camundongos foram inoculados pela via subcutânea
(s.c.) com o pool de mAbs e em seguida desafiados com veneno total de B. atrox pela
mesma via. Os três mAbs foram reunidos a uma concentração de 13,5 mg/camundongo
(5,0mg de A85/9-4 e de 59/2-E4; e 3,5mg de 6AD2-G5) em um volume de final de 0,7 mL e
inoculado via ip em camundongo Suíço albino com 18 a 20g. O grupo controle foi inoculado
com a mesma concentração de IgG inespecífica. Em seguida, os animais foram desafiados
com 350mg de veneno pela via subcutânea. A sobrevivência dos animais foi observada por
48 horas. Na Tabela 3 podemos observar que o grupo experimental constituído de
camundongos previamente tratados com os anticorpos específicos, todos os animais
sobreviveram. Enquanto que no grupo controle, que recebeu IgG inespecífica, somente um
camundongo sobreviveu.
DETERMINAÇÃO DA DL50 DO VENENO DE B. ATROX
A determinação da DL50 do veneno de B. atrox foi realizada inoculando-se
doses variadas do veneno em grupos de 10 camundongos Suíços albinos. Após a
inoculação a taxa de sobrevivência dos animais foi avaliada nos tempos de 24 e 48
horas. Na Tabela 4 podemos observar que no grupo 5 que foi inoculado com 62,2µg
de veneno todos os animais sobreviveram, enquanto que no grupo 1 que foi
inoculado com 129µg de veneno somente 4 animais sobreviveram. O resultado final
Grupos mAbs* Veneno** Vivos/Total
Grupo 1 13,5mg 350ug 5/5
Grupo 2 ----- 350ug 1/5
*A mistura de mAbs utilizada corresponde a 5,0mg dos mAbs A85/9-4 e 59/2-E4 e
3,5 mg de 6AD2-G5, por camundongo.
** A quantidade de veneno utilizada para cada camundongo.
Tabela 3: Ensaio de soroproteção in vivo de camundongos desafiados com misturas de mAbs e veneno

65
deste experimento foi analisado pelo método de Probit (Finney, 1971) que
determinou a concentração de 120,89µg a DL50 do veneno de B. atrox para
camundongos Suíços albinos de 18-22 g.
Uma vez demonstrado que os três mAbs em conjunto são capazes de inibir a
atividade letal do veneno de B. atrox, realizamos alguns ensaios de soroproteção de acordo
com a metodologia preconizada pela Farmacopéia Brasileira. No primeiro ensaio realizado
foram utilizados 4 grupos 8 de camundongos cada e com 18 a 22g de peso. Cada animal do
grupo 1 recebeu 13,5 mg do pool de mAbs previamente incubados com 5 DL50 de veneno de
B. atrox. Da mesma forma, cada animal do grupo 2 recebeu 10,5 mg do pool de mAbs, do
grupo 3 recebeu 7,9 mg do pool de mAbs e do grupo 4, cada animal recebeu 6,0 mg do pool
de mAbs. Na Tabela 5 estão sumarizados os resultados com 48 horas após a inoculação.
Nesta tabela podemos observar que somente no grupo 1 que recebeu 13,5 mg do pool de
mAbs apresentou dois animais sobreviventes. Nos grupos 2 e 4 houve somente um animal
Grupos Veneno(ug) Vivos/Total
Grupo 1 129 4/10
Grupo 2 120,7 7/10
Grupo 3 89,6 8/10
Grupo 4 74,6 9/10
Grupo 5 62,2 10/10
Tabela 4: Cálculo da DL50 do veneno de B. atrox
Grupo 6 51,8 10/10
Grupo 5 43,2 10/10

66
sobrevivente e no grupo 3 não houve camundongo sobrevivente. Todos os camundongos
sobreviventes foram sacrificados, necropsiados e amostras de coração, pulmões, fígado,
rins, baço e músculos do abdômen e do diafragma foram colhidas para análises
histológicas.
Grupos mAbs* Veneno** Vivos/Total
Grupo 1 13,5mg 5DL50 2/8
Tabela 5: Ensaio de soroproteção in vivo de camundongos desafiados com misturas de pool de mAbs e 5 DL50 de veneno
* A mistura de mAbs utilizada corresponde a 5,0mg dos mAbs A85/9-4 e 59/2-E4 e
3,5 mg de 6AD2-G5, por camundongo.
**A DL50 do veneno de B. atrox corresponde a 120,89µg.
Grupo 2 10,5mg 5DL50 1/8
Grupo 3 7,9mg 5DL50 0/8
Grupo 4 6,0mg 5DL50 1/8

67
Um ensaio similar ao apresentado na Tabela 6 foi repetido, sendo que foram
utilizados 4 DL50 do veneno de B. atrox e as concentrações do pool de mAbs por grupo
foram de 10,5mg , 7,9mg e 6,0mg por animal, respectivamente. Na Tabela 7 podemos
observar que no grupo 1 houve 3 animais que sobreviveram ao desafio com o veneno, no
grupo 2 houve apenas 1 animal e no grupo 3 não houve animal sobrevivente.
Com o intuito de fazer uma análise comparativa da potência do pool de mAbs com o
antiveneno comercial produzido pelo Instituto Butantan, foi realizado um ensaio de
soroproteção utilizando diferentes diluições do antiveneno antibotrópico e desafiado com 5
DL50 do veneno de B. atrox. Neste ensaio também não foi possível calcular a potencia do
antiveneno botrópico, pois houve somente dois animais sobreviventes nos grupos 1, 2 e 3 e,
no grupo 4 houve somente um animal vivo, conforme pode ser observado na Tabela 8.
Novos ensaios de soroproteção serão realizados para confirmar a potência do pool de mAbs
e a potência do antiveneno botrópico frente ao veneno de B. atrox.
Grupos mAbs* Veneno** Vivos/Total
Tabela 6: Ensaio de soroproteção in vivo de camundongos desafiados com
misturas de pool de mAbs e 4 DL50 de veneno
* A mistura de mAbs utilizada corresponde a 5,0mg dos mAbs A85/9-4 e 59/2-E4 e
3,5 mg de 6AD2-G5, por camundongo.
**A DL50 do veneno de B. atrox corresponde a 120,89µg.
Grupo 1 10,5mg 4DL50 3/8
Grupo 2 7,9mg 4DL50 1/8
Grupo 3 6,0mg 4DL50 0/8

68
NEUTRALIZAÇÃO DA ATIVIDADE ANTICOAGULANTE DO VENENO DE B. ATROX
Na Figura 11A podemos observar que o aumento do tempo de coagulação
em camundongos Suíços albinos inoculados com 200µg pela via ip do veneno de B.
atrox tem início aos 10 minutos após a inoculação do veneno e atinge o seu valor
máximo entre 30 e 60 minutos.
Para avaliar a capacidade do pool de mAbs neutralizar a atividade
anticoagulante do veneno de B. atrox, foi incubado 200µg do veneno com 13,5 mg
do pool de mAbs e inoculados via ip em camundongos Suíços albinos. Uma hora
após a inoculação a intensidade de sangramento dos animais foi avaliada. Na Figura
11B podemos observar que a intensidade de sangramento dos animais tratados com
o pool de mAbs foi similar à intensidade medida antes da inoculação do veneno pré-
incubado com o pool. Enquanto que os animais que receberam somente o veneno
apresentaram um sangramento muito mais intenso.
A ação isolada do mAb 6AD2-G5 antitrombina-simile foi também avaliada.
Camundongos Suíços albinos foram inoculados com 150µg de veneno previamente
incubado com 3,5mg do mAb 6AD2-G5 ou com 500µL de antiveneno antibotrópico.
A neutralização da atividade anticoagulante do veneno conferida pelo mAb 6AD2-G2
foi similar a atividade do antiveneno antibotrópico, conforme pode ser visto na Figura
12.
Grupos Antiveneno* Veneno** Vivos/Total
Grupo 1 0,750 mL 5DL50 2/8
Tabela 7: Ensaio de soroproteção in vivo de camundongos desafiados com
misturas de antiveneno e veneno de B. atrox.
* Antiveneno com potência para neutraliza 5mg de veneno por mL de soro.
** A DL50 do veneno de B. atrox corresponde a 120,89µg.
Grupo 2 0,570 mL 5DL50 2/8
Grupo 3 0,430 mL 5DL50 2/8
Grupo 4 0,330 mL 5DL50 1/8
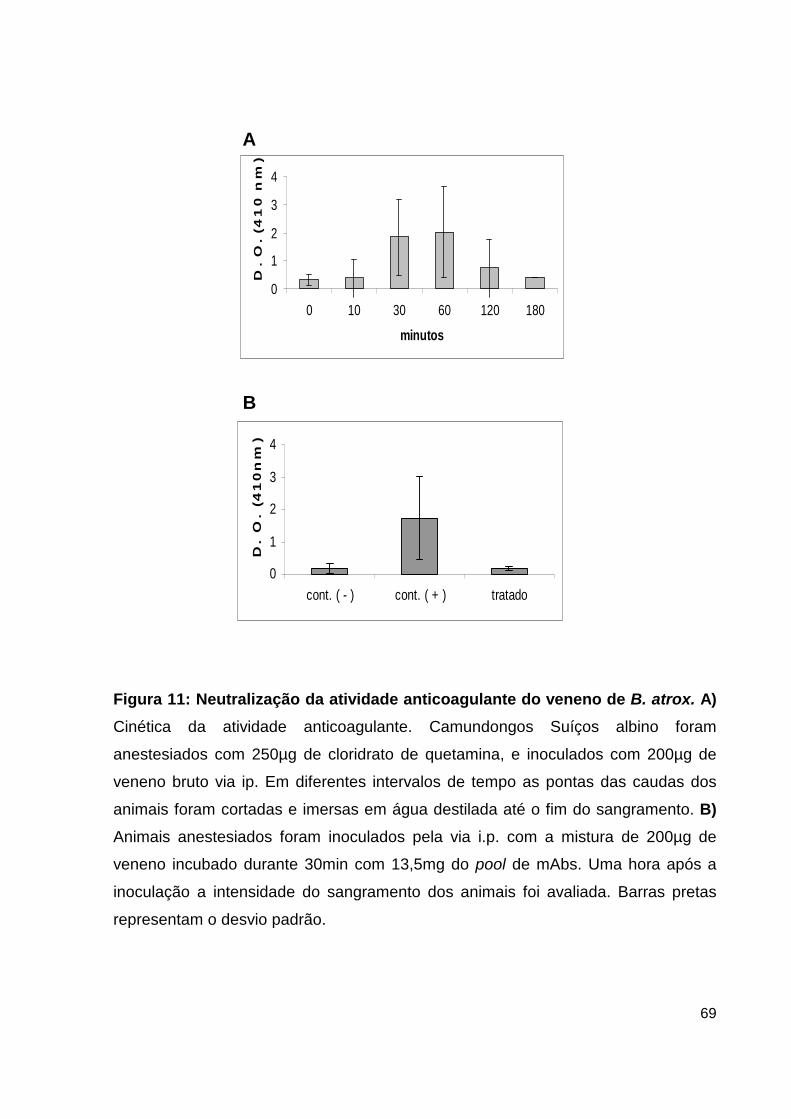
69
Figura 11: Neutralização da atividade anticoagulante do veneno de B. atrox. A)
Cinética da atividade anticoagulante. Camundongos Suíços albino foram
anestesiados com 250µg de cloridrato de quetamina, e inoculados com 200µg de
veneno bruto via ip. Em diferentes intervalos de tempo as pontas das caudas dos
animais foram cortadas e imersas em água destilada até o fim do sangramento. B)
Animais anestesiados foram inoculados pela via i.p. com a mistura de 200µg de
veneno incubado durante 30min com 13,5mg do pool de mAbs. Uma hora após a
inoculação a intensidade do sangramento dos animais foi avaliada. Barras pretas
representam o desvio padrão.
0
1
2
3
4
0 10 30 60 120 180
minutos
D.
O.
(41
0 n
m)
A
0
1
2
3
4
cont. ( - ) cont. ( + ) tratado
D.
O.
(41
0n
m)
B
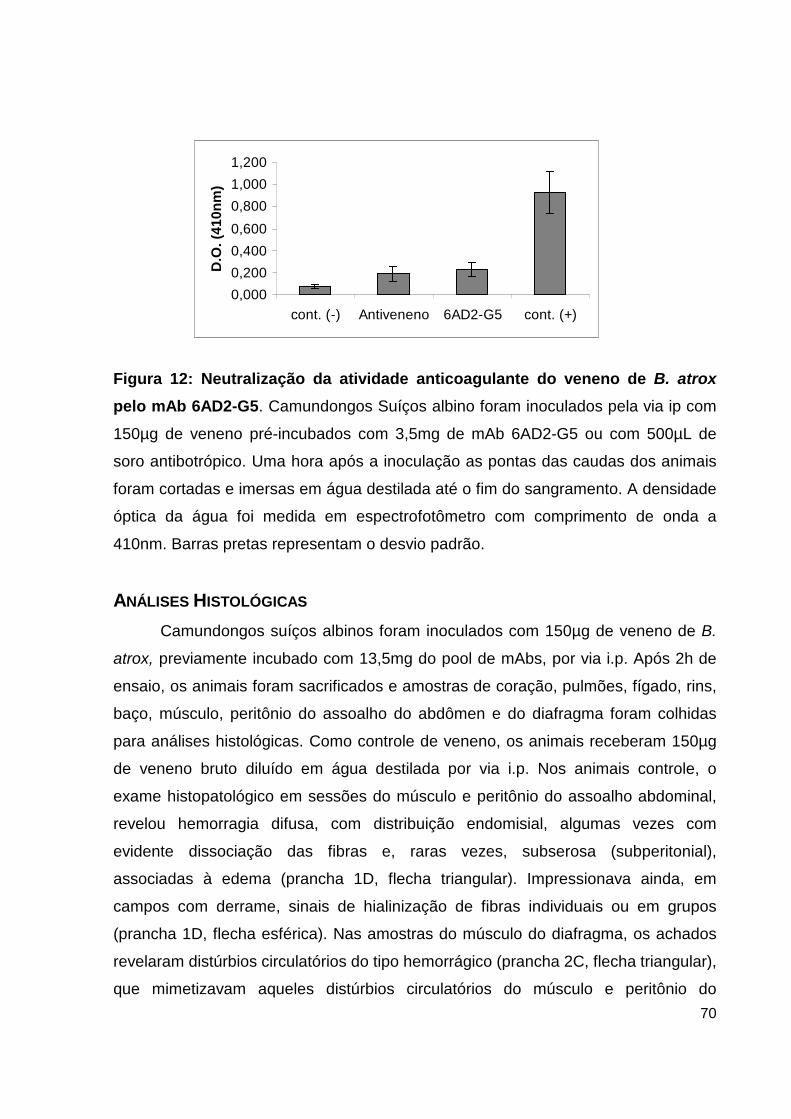
70
Figura 12: Neutralização da atividade anticoagulante do veneno de B. atrox
pelo mAb 6AD2-G5. Camundongos Suíços albino foram inoculados pela via ip com
150µg de veneno pré-incubados com 3,5mg de mAb 6AD2-G5 ou com 500µL de
soro antibotrópico. Uma hora após a inoculação as pontas das caudas dos animais
foram cortadas e imersas em água destilada até o fim do sangramento. A densidade
óptica da água foi medida em espectrofotômetro com comprimento de onda a
410nm. Barras pretas representam o desvio padrão.
ANÁLISES HISTOLÓGICAS
Camundongos suíços albinos foram inoculados com 150µg de veneno de B.
atrox, previamente incubado com 13,5mg do pool de mAbs, por via i.p. Após 2h de
ensaio, os animais foram sacrificados e amostras de coração, pulmões, fígado, rins,
baço, músculo, peritônio do assoalho do abdômen e do diafragma foram colhidas
para análises histológicas. Como controle de veneno, os animais receberam 150µg
de veneno bruto diluído em água destilada por via i.p. Nos animais controle, o
exame histopatológico em sessões do músculo e peritônio do assoalho abdominal,
revelou hemorragia difusa, com distribuição endomisial, algumas vezes com
evidente dissociação das fibras e, raras vezes, subserosa (subperitonial),
associadas à edema (prancha 1D, flecha triangular). Impressionava ainda, em
campos com derrame, sinais de hialinização de fibras individuais ou em grupos
(prancha 1D, flecha esférica). Nas amostras do músculo do diafragma, os achados
revelaram distúrbios circulatórios do tipo hemorrágico (prancha 2C, flecha triangular),
que mimetizavam aqueles distúrbios circulatórios do músculo e peritônio do
0,000
0,200
0,400
0,600
0,800
1,000
1,200
cont. (-) Antiveneno 6AD2-G5 cont. (+)
D.O
. (41
0nm
)

71
assoalho dorsal da cavidade abdominal. Nos animais tratados com o pool de mAbs,
as amostras de músculos do diafragma e do abdômen, o derrame se fazia presente,
notadamente na camada voltada para cavidade adominal (prancha 2B, flecha
triangular). Havia ainda associados ao músculo abdominal, sinais de leucocitose,
representados por numerosos polimorfos atulhando luzes dilatadas de capilares,
instalando um quadro de miosite sero-hemorrágica (prancha 1C, flecha triangular).
Nas demais amostras de órgãos colhidas, não houve alterações significativas
(resultado não mostrado).
Camundongos que sobreviveram as primeiras 24h do ensaio de soroproteção
com o pool de mAbs foram sacrificados, necropsiados e novamente amostras de
coração, pulmões, fígado, rins, baço e músculos do diafragma e do assoalho do
abdômen foram colhidas para análises histológicas. Nos animais controle positivo,
análises histopatológicas em sessões do fígado revelaram tumefação difusa discreta
(flecha triangular) com sinais discretos de leucositose (flecha esférica) nos vasos
hepáticos (prancha 3A) enquanto que nos animais tratados com o pool de mAbs
nenhuma alteração significativa foi observada (prancha 3B). Em sessões do músculo
do assoalho do peritôneo os achados revelaram hemorragia livre, algumas vezes
endomisial, a hemorragia discreta também se fazia presente no tecido adiposo
mesentérico.
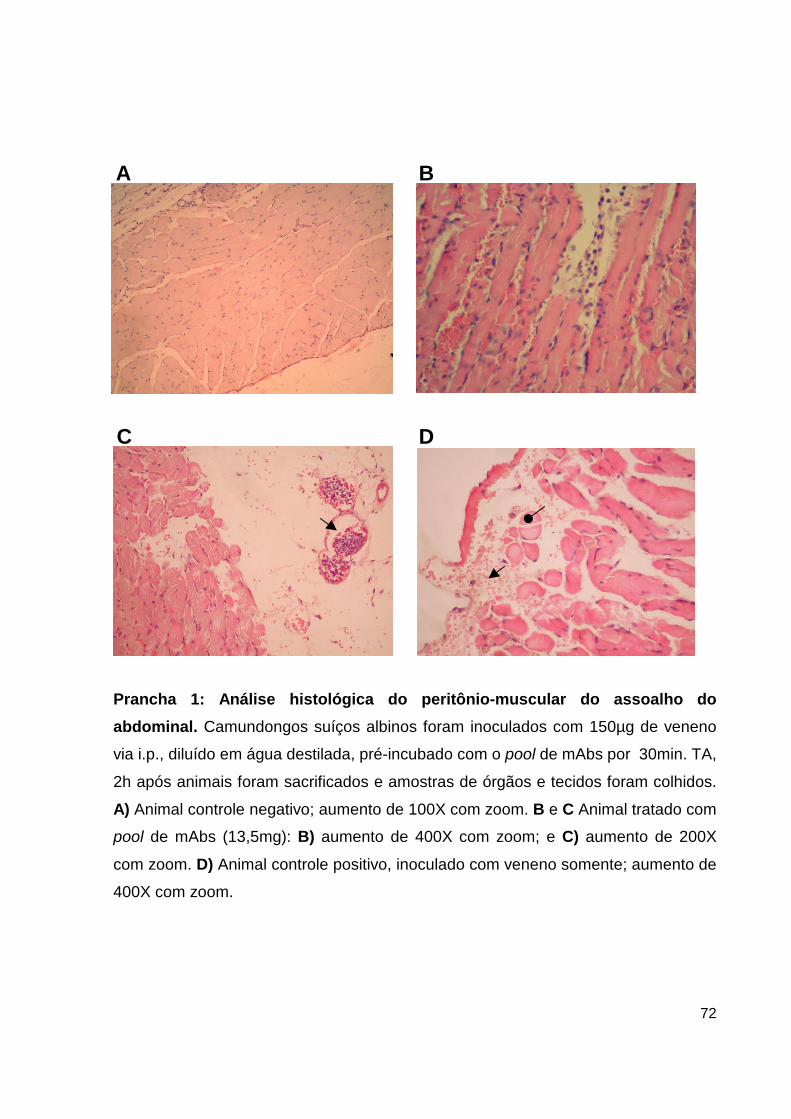
72
Prancha 1: Análise histológica do peritônio-muscular do assoalho do
abdominal. Camundongos suíços albinos foram inoculados com 150µg de veneno
via i.p., diluído em água destilada, pré-incubado com o pool de mAbs por 30min. TA,
2h após animais foram sacrificados e amostras de órgãos e tecidos foram colhidos.
A) Animal controle negativo; aumento de 100X com zoom. B e C Animal tratado com
pool de mAbs (13,5mg): B) aumento de 400X com zoom; e C) aumento de 200X
com zoom. D) Animal controle positivo, inoculado com veneno somente; aumento de
400X com zoom.
D C
A B
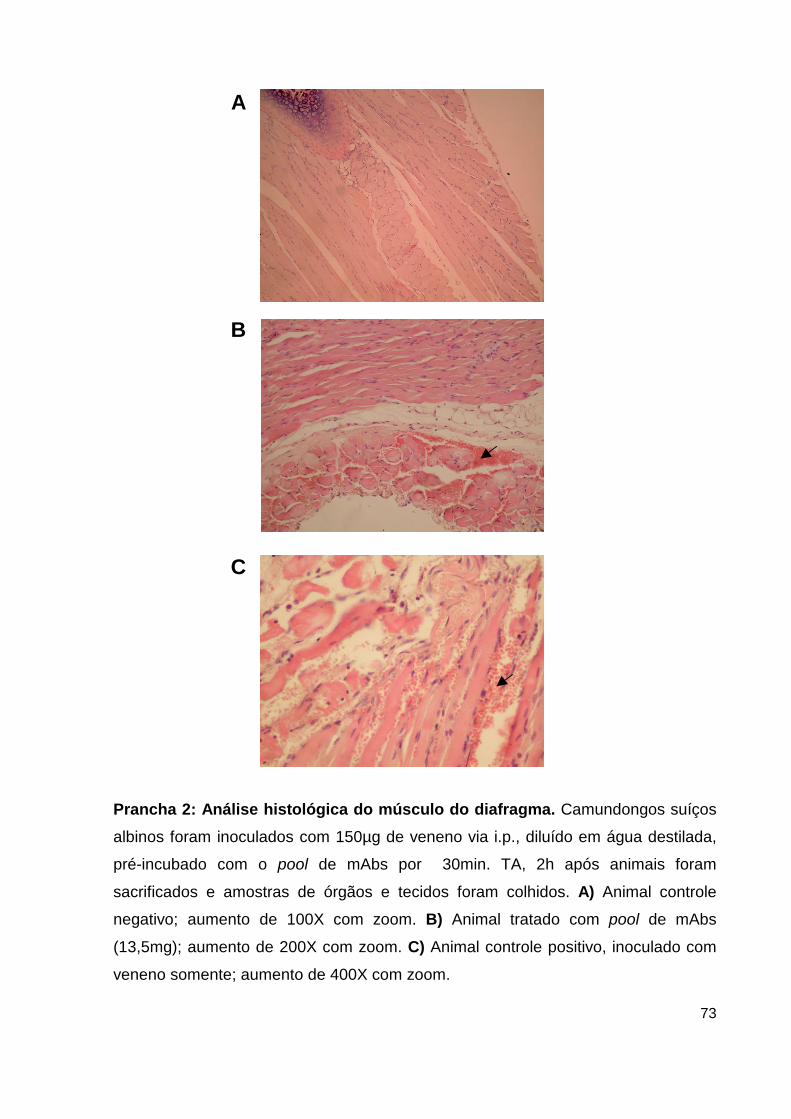
73
Prancha 2: Análise histológica do músculo do diafragma. Camundongos suíços
albinos foram inoculados com 150µg de veneno via i.p., diluído em água destilada,
pré-incubado com o pool de mAbs por 30min. TA, 2h após animais foram
sacrificados e amostras de órgãos e tecidos foram colhidos. A) Animal controle
negativo; aumento de 100X com zoom. B) Animal tratado com pool de mAbs
(13,5mg); aumento de 200X com zoom. C) Animal controle positivo, inoculado com
veneno somente; aumento de 400X com zoom.
B
A
C

74
No músculo do diafragma dos animais que receberam somente o veneno, a
análise revelou hemorragia moderada endomisial e perimisial, em alguns campos
havia sinais de edema discreto (prancha 3C, flecha triangular), sendo que lesões
similares, porém, em menor grau, foram observadas nos animais tratados com o
soro polivalente (prancha 3D, flecha triangular). Em sessões do baço, os achados
revelaram um comprometimento congestivo hemorrágico moderado (prancha 3E,
flecha triangular) e nos animais tratados com o pool de mAbs o mesmo padrão de
lesão foi observado, porem em menor grau. Nas demais amostras colhidas, não
foram encontradas alterações dignas de nota. Nos animais tratados com o pool de
mAbs, de uma forma geral, os exames histológicos revelaram alterações em menor
grau quando comparado com os animais controles que receberam somente veneno.
Animais submetidos ao ensaio de soroproteção com antiveneno comercial
também foram analisados. Após 24h de envenenamento, os animais sobreviventes
foram sacrificados e necropsiados. Sendo os tecidos e órgãos analisados
histologicamente. O exame histológico das sessões do músculo do diafragma
revelou hemorragia subperitonial e intramuscular discreta com sinais de processo
congestivo hemorrágico na região subpleural do diafragma (prancha 3D, felcha
esférica), o exame também revelou distúrbio circulatório associado a infiltrados
escassos de neutrófilos e hialinização de fibras individuais, em algumas vezes com
dissolução. O exame em sessões dos pulmões revelou derrame alveolar focal,
provavelmente devido ao método de sacrifício adotado (resultado não mostrado).
Nas amostras do assoalho abdominal os distúrbios eram semelhantes aos
encontrados nos músculo do diafragma com distúrbio congestivo hemorrágico
discreto e hialinização de fibras individuais. Em alguns animais, nos rins, os achados
revelaram congestão corticoglomerular com raros cilindros hialinos inclusive. No
fígado, a análise demonstrou tumefação turva discreta difusa.
Os animais que sobreviveram as 48h do ensaio de soroproteção com o pool
de mAbs foram sacrificados, necropsiados e mais uma vez amostras de coração,
pulmões, fígado, rins, baço e músculos do diafragma e do assoalho do abdômen
foram colhidas para análises histológicas. Nos animais controle, o exame
histopatológico no fígado revelou tumefação difusa discreta sem sinais de distúrbio
vascular, em alguns campos os achados revelaram degeneração hidrópica. Nas

75
demais amostras, alterações significativas, não foram encontradas. Nos animais
tratados com o pool de mAbs, após 48h de ensaio, as análises histopatológicas
revelaram tumefação turva discreta no fígado. O músculo do diafragma encontrava-
se sem alterações. Em sessões do baço, foi encontrado um processo congestivo
hemorrágico discreto. Nas demais amostras, não foram encontradas alterações
dignas de nota.
Nos camundongos submetidos ao ensaio de soroproteção com antiveneno
comercial, após 48h de envenenamento, a análise histológica dos tecidos e órgãos
dos animais sobreviventes revelou lesões semelhantes às encontradas nos animais
tratados com o pool de mAbs. Em sessões do fígado, novamente tumefação discreta
se fazia presente. Nos músculos do diafragma e do peritônio, os achados revelaram
hemorragia discreta, em raros campos, no diafragma, havia hemorragia moderada
focal. No baço desses animais, processo congestivo hemorrágico também foi
encontrado. Nas demais amostras coletadas, nenhuma alteração significativa foi
encontrada.
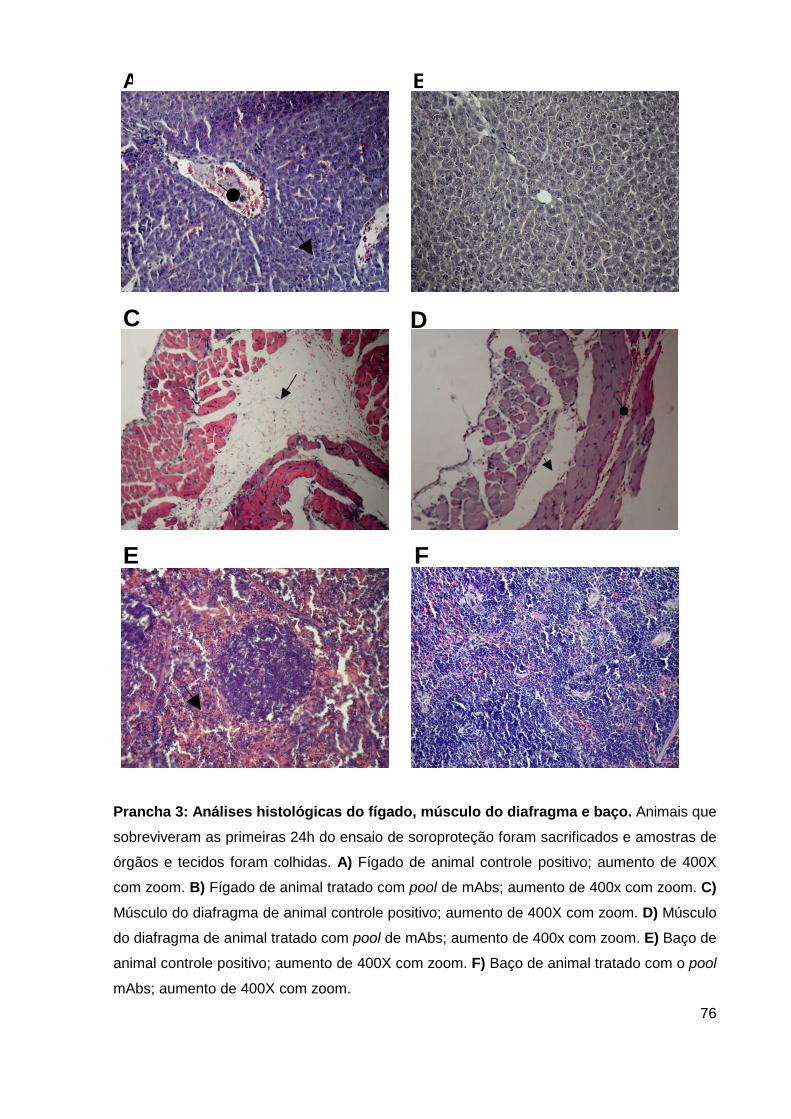
76
Prancha 3: Análises histológicas do fígado, músculo do diafragma e baço. Animais que
sobreviveram as primeiras 24h do ensaio de soroproteção foram sacrificados e amostras de
órgãos e tecidos foram colhidas. A) Fígado de animal controle positivo; aumento de 400X
com zoom. B) Fígado de animal tratado com pool de mAbs; aumento de 400x com zoom. C)
Músculo do diafragma de animal controle positivo; aumento de 400X com zoom. D) Músculo
do diafragma de animal tratado com pool de mAbs; aumento de 400x com zoom. E) Baço de
animal controle positivo; aumento de 400X com zoom. F) Baço de animal tratado com o pool
mAbs; aumento de 400X com zoom.
A B
C
FE
D

77
DISCUSSÃO

78
Acidentes ofídicos ocorrem freqüentemente no Brasil, com uma incidência
anual de 20.000 casos reportados, sendo que o acidente botrópico corresponde ao
acidente ofídico de maior importância epidemiológica no país, responsável por cerca
de 90% dos envenenamentos (MINISTÉRIO DA SAÚDE, 2001). Na região
amazônica, a espécie Bothrops atrox é responsável por 90% dos acidentes
(MELGAREJO, 2003). Venenos de serpentes contêm misturas complexas de
centenas de moléculas farmacologicamente ativas, incluindo componentes
orgânicos e minerais (histamina e outros alérgenos, poliaminas, alcalóides...),
pequenos peptídeos e proteínas (MARKLAND, 1998; FRY, 1999). Os efeitos
biológicos são complexos porque componentes diferentes possuem ações distintas
e devem, sinergicamente, agir com outras moléculas do veneno (CALVETE et al.,
2005).
Mesmo após mais de 100 anos da introdução da soroterapia por Calmette, a
administração de antivenenos, baseados em anticorpos heterólogos, permanece
como único tratamento contra picadas de serpentes (BON, 1996). A primeira
geração de antivenenos era constituída de soros de animais hiperimunizados com
os respectivos venenos. A próxima etapa surgiu com o desenvolvimento de métodos
bioquímicos capazes de enriquecer os preparados com anticorpos,
preferencialmente, separando a fração protetora de outros componentes séricos
conhecidos como causadores de reações de hipersensibilidades. Atualmente, os
antivenenos são constituídos de fragmentos de imunoglobulinas purificados e com
alta potencia neutralizante (STOCK et al., 2007). Ainda assim, administração de
soros heterólogos em pacientes induz reações alérgicas que podem ser inicias ou
tardias, sendo as EARs (early antivenom reactions) o principal problema associado à
soroterapia (OTERO-PATIÑO et al., 1998). Além disso, os soros antibotrópicos
poliespecíficos produzidos no Brasil não são eficientes na neutralização dos efeitos
do veneno de algumas espécies de Bothrops da floresta amazônica, principalmente
B. atrox (MUNIZ et al., 2000).
Neste trabalho tivemos como objetivo a produção de um antiveneno baseado
em anticorpos monoclonais (mAbs) contra os principais componentes tóxicos do
veneno de B. atrox. Para isso, anticorpos monoclonais contra fosfolipase A2 (PLA2),
trombina-simile e Zn-metaloproteinase foram produzidos em escala de miligramas,

79
purificados, agrupados em um pool e avaliados quanto à capacidade de
neutralização dos principais efeitos tóxicos do veneno.
A purificação de anticorpos monoclonais pelo método combinado do ácido
caprílico e precipitação com sulfato de amônio, utilizado neste trabalho, apresentou
um percentual relativamente baixo de pureza. Conforme pode ser observado na
Figura 9, o percentual médio de pureza nas diferentes preparações ficou em torno
de 42%. Em trabalhos anteriores, realizados em nosso grupo, utilizando esta mesma
metodologia, foi observado um grau de pureza maior do que o observado neste
trabalho. Possivelmente o grau de pureza dos mAbs pouco satisfatório pode ser
melhorado fazendo-se um acompanhamento mais adequado dos ajustes de pH
durante o processo de purificação. Além disso, estes resultados devem ser
considerados ao analisar os outros ensaios que utilizaram estes preparados.
Os mAbs anti-PLA2 (figura 10A) e Zn-metaloproteinase (figura10B) foram
eficazes no reconhecimento de seus respectivos antígenos, mesmo nas menores
diluições. Entretanto, o mAb anti-trombina-simile não apresentou o mesmo padrão
de reconhecimento daqueles mAbs (figura 10C). Esta baixa eficiência pode ter
ocorrido devido aos seguintes fatores: a) o antígeno utilizado no ensaio, trombina-
simile enriquecida, não se encontrava na concentração adequada para o teste; b)
sabe-se que o mAb 6AD2-G5 reconhece epítopos conformacionais (PETRETSKI et
al., 2000a), a adsorção do antígeno na placa de ELISA pode ter induzido a uma
mudança conformacional do antígeno, diminuindo, assim, a ligação do mAb 6AD2-
G5 ao seu epítopo; ou ainda, c) o antígeno poderia estar degradado.
A síndrome hemorrágica é uma das principais conseqüências graves do
envenenamento por serpentes pertencentes à família Viperidae (GUTIÉRREZ et al.,
2005). O principal componente responsável por este sangramento local, ou
sistêmico (em casos severos de envenenamento), é a metaloproteinase de serpente
(STOCKER et al., 1995). A neutralização da atividade hemorrágica induzida pelo
veneno de B. atrox foi analisada in vivo na Figura 1. O mAb antimetaloproteinase
59/2-E4 foi capaz de inibir a atividade hemorrágica de maneira dose dependente.
Porém, em ensaios in vitro, o mAb 59/2-E4 foi incapaz de inibir a atividade catalítica

80
da Zn-metaloproteinase de baixo peso molecular (Figura 2) purificada do veneno de
B. atrox (PETRETSKI et al., 2000b), sugerindo que o mAb 59/2-E4 não seja
direcionado contra o sítio catalítico da enzima, mas contra uma outra região da
molécula que seja responsável pelo acoplamento da metaloproteinase ao substrato,
o domínio desintegrina e o domínio enriquecido em cisteína. Conforme foi
demonstrado por Tanjoni et al., (2003) utilizando anticorpos monoclonais
desenvolvidos contra a hemorragina de Bothrops jararaca.
Mionecrose é outra complicação médica importante nos acidentes ofídicos
(LOMONTE et al., 2003). A miotoxidade descrita em envenenamentos por estas
serpentes é atribuída principalmente às PLA2 (TEIXEIRA et al., 2003). A purificação
da PLA2 segundo metodologia descrita por Kanashiro et al., (2002) foi compatível
com os resultados observados pelo autor (Figuras 4 e 5). A eficácia do mAb anti-
PLA2 na neutralização da atividade miotóxica provocada pela PLA2, foi avaliada
indiretamente pelos níveis séricos de creatina quinase. As análises mostraram que
o mAb A85/9-4 foi eficaz na neutralização da atividade miotóxica provocada pela
fração PLA2 -P4 em camundongos Suíços albinos (Figura 5B), conforme já
demonstrados em trabalhos anteriores do nosso grupo.
A ação coagulante, também conhecida como síndrome da desfibrinação, é
resultante da propriedade que o veneno das serpentes do gênero Bothrops, Crotalus
e Lachesis têm de transformar diretamente o fibrinogênio em fibrina (KAMIGUTI e
CARDOSO, 1989). A enzima responsável pela formação do coágulo na fase final da
coagulação sanguínea é a trombina-símile (ZAGANELLI et al. 1996). O mAb 6AD2-
G5 foi testado quanto à capacidade de inibição da atividade catalítica utilizando o
TAME como substrato. Na Figura 6B podemos observar que o mAb 6AD2-G5 foi
eficaz em neutralizar a atividade catalítica da trombina-simile em todas
concentrações testadas. O mAb 6AD2-G5 também foi testado quanto à capacidade
de neutralizar a conversão de fibrinogênio em fibrina. Na figura 7 podemos observar
que o mAb 6AD2-G5, foi capaz de inibir a formação do coágulo. Em algumas
amostras purificadas, o mAb foi capaz de inibir em até três vezes a atividade
coagulante do veneno (resultado não mostrado). Esses dados demonstram que,
apesar da baixa titulação observada no teste de ELISA (Figura 10C), o mAb 6AD2-

81
G5 foi capaz de neutralizar eficientemente a atividade catalítica da serinoproteinase
trombina-simile.
Após a comprovação da eficácia de cada um dos mAbs na neutralização de
suas respectivas toxinas, eles foram agrupados em pool e testados quanto à
capacidade de neutralização da ação letal do veneno de B. atrox. No primeiro ensaio
preliminar, onde o pool de mAbs foi pré-incubado com veneno bruto e inoculado por
via i.p. em camundongos Suíço albinos, o grupo de animais desafiado com 200mg de
veneno, a maioria dos camundongos sobreviveu, enquanto que no grupo controle,
que recebeu veneno somente, nenhum animal permaneceu vivo (Tabela 3). No
ensaio seguinte, onde o pool de mAbs foi inoculado nos animais pela via i.p.. e
desafiados com veneno bruto pela via s.c., foi observado que o grupo de
camundongos tratados e desafiados com 350mg de veneno, todos os camundongos
sobreviveram (Tabela 4). Enquanto que no grupo controle que não receberam o pool
de mAbs, somente um camundongo sobreviveu. Esses dados mostram que os três
mAbs em conjunto são capazes de inibir a ação letal do veneno de B. atrox.
Adicionalmente, poderíamos considerar que a concentração de 13,5mg de proteínas
administrada por animal seja um pouco elevada, entretanto, esta análise deve levar
em consideração de que somente 42% desta preparação correspondem a
anticorpos, reduzindo para aproximadamente 5,7mg de anticorpos administrados
para cada animal.
Ensaios de soroproteção realizados conforme metodologia preconizada pela
Farmacopéia Brasileira podem ser visualizada nas tabelas 6 e 7, onde foram
utilizados 4 e 5 DL50 do veneno de B. atrox por animal, respectivamente. Estas duas
tabelas demonstram que a concentração de anticorpos utilizada não foi capaz de
neutralizar a toxicidade letal da dose de veneno injetada. Foi realizado, também, um
ensaio de soroproteção utilizando um soro poliespecífico antibotrópico pré-incubado
com 5 DL50 de veneno de B. atrox (tabela 8), que também não foi capaz de
neutralizar a ação letal da dose de veneno injetada. Este resultado demonstra a
ineficácia dos antivenenos poliespecíficos nacionais na neutralização da ação letal
do veneno de B. atrox, conforme já demonstrado por Muniz et al. (2000). O baixo
nível de proteção conferido pelo pool mAbs, comparável ao nível de proteção

82
conferido pelo antiveneno poliespecífico, poderia estar associado à ação sinérgica
das outras toxinas presentes no veneno, além das três principais toxinas
neutralizadas pelos mAbs. Uma outra possível justificativa é de que a DL50 por nós
determinada (120,89µg) esteja acima da DL50 real, uma vez que a DL50 determinada
por Otero et al. (2000) foi de 66,2µg de veneno por animal. Entretanto, Barros et al.
(1998) utilizando várias linhagens de camundongos isogênicos, demonstraram
suscetibilidades diferenciadas entre os diversos padrões genéticos murinos
testados. Sendo a DL50 de 89,4µg de veneno de B. atrox para a linhagem A/J de
maior sensibilidade, e a DL50 de 137µg de veneno para linhagem mais resistente
DBA/2. Uma confirmação da DL50 para este lote de veneno será realizada e novos
ensaios de soroproteção serão realizados para confirmar as potências do pool de
mAbs e do antiveneno botrópico frente ao veneno de B. atrox.
O pool de mAbs também foi avaliado quanto à capacidade de neutralização
da atividade anticoagulante in vivo promovida pelo veneno de B. atrox. Lorìa et al.
(2003) demonstraram que a incoagulabilidade sangüínea perdura por três horas
após a inoculação do veneno de B. asper em camundongos. Similarmente, a
inoculação de veneno de B. atrox em camundongos causou o mesmo efeito (Figura
11A). O pool de mAbs neutralizou de forma eficiente a atividade anticoagulante,
conforme pode ser visto na Figura 11B a intensidade do sangramento permaneceu
similar à intensidade observada nos animais controles que não receberam o veneno.
O mAb 6AD2-G5 por si só também foi capaz de neutralizar a atividade
anticoagulante do veneno de B. atrox de forma eficiente, semelhante ao resultado
visto com o soro antibotrópico (Figura 12), confirmando os resultados observados
nas Figuras 6 e 7 nos ensaios in vitro e corroborando com os resultados
demonstrados por Petretski et al. (2000a) que o mAb antitrombina-simile é um
potente inibidor desta enzima.
As análises histológicas dos animais sobreviventes, de uma maneira geral,
demonstrou que tanto o pool de mAbs como o soro antiofídico foram capazes de
reduzir significativamente o nível de lesões causadas pelo veneno de B. atrox nos
tempos de 2hr, 24hr e 48hr, quando comparado com o grupo controle que foi

83
inoculado semente com o veneno. Os animais tratados com o pool de mAbs
apresentaram graus de lesões similares aos animais tratados com o soro antiofídico.
Nesta dissertação foi demonstrada a eficácia do pool de mAbs, composto por
anticorpos contra metaloproteinase, PLA2 e trombina-símile, na neutralização das
atividades letal e hemorrágica promovidas pelo veneno de B. atrox. Esses dados em
conjunto apontam para a possibilidade de desenvolvimento de um antiveneno
baseado em anticorpos monoclonais humanizados.

84
CONCLUSÕES

85
Os resultados aqui apresentados nos permitem concluir que:
· O mAb anti Zn-metaloproteinase foi capaz de inibir a atividade hemorrágica in
vivo de maneira dose dependente. Porém este mAb foi incapaz de inibir a
atividade catalítica da enzima in vitro;
· O mAb anti-PLA2 foi capaz de neutralizar a atividade miotóxica induzida pela
fração PLA2-P4 em camundongos Suíços albinos;
· O mAb anti trombina-simile neutralizou eficientemente a atividade catalítica da
enzima sobre o substrato TAME. O mAb também foi capaz de neutralizar a
conversão de fibrinogênio em fibrina, in vitro, induzida pela trombina-simile;
· O pool de mAbs neutralizou a ação letal do veneno de B. atrox quando pré-
incubado com veneno e inoculado via i.p. nos animais. Resultado similar foi
alcançado quando o pool foi inoculado via i.p. e, em seguida, os animais
foram desafiados com veneno por via s.c.;
· Ensaios de soroproteção conforme preconizado pela Farmacopéia Brasileira,
o pool de mAbs não foi capaz de conferir proteção aos animais. A proteção
também não foi atingida quando soros antibotrópicos poliespecíficos foram
empregados;
· O pool de mAbs neutralizou eficientemente a atividade anticoagulante
induzida pelo veneno de B. atrox. O mAb antitrombina-simile, por si só, foi
também capaz de inibir essa atividade.
· A análise histológica dos tecidos dos animais experimentais demonstrou que
o pool de mAbs e o soro antiofídico foram capazes de reduzir
significativamente as lesões teciduais induzidas pelo veneno.

86
REFERÊNCIAS
BIBLIOGRÁFICAS

87
AMBROSIO, ALB, Nonato, MC, Selistre-de-Araújo, HS, Arni, R, Ward, RJ, Ownby,
CL, de Souza, DHF, Garratt, RC. A Molecular Mechanism for Lys49-
Phospholipase A2 Activity Based on Ligand-induced Conformational Change. J.
Biol. Chemis. 280 (8) 25:7326–7335. 2005.
AMORIN, MF, de Melo, RF, Saliba, F. Envenenamento botrópico e crotálico.
Contribuição para o estudo experimental comparado das lesões. Mem. Instit.
Butantan, 23: 63-108. 1951.
ANDRIÃO-ESCARSO, SH, Soares, AM, Rodrigues, VM, Angulo, Y, Diaz, C
Lomonte, B, Gutiérres, JM, Giglio, JR. Myotoxic phospholipases A2 in Bothrops
snake venoms: effect of chemical modifications on the enzymatic and
pharmacological properties of bothropstoxins from Bothrops jararacussu.
Biochimie, 82: 755-763. 2000.
ARAÚJO, FAA, Santalúcia, M, Cabral, RF. Epidemiologia dos acidentes por animais
peçonhentos. In Animais Peçonhentos do Brasil. 1 ed. São Paulo:
Sarvier/FAPESP, 2003. p. 6
AZEVEDO-MARQUES, MM, Hering, SE, Cupo, P. Acidente Crotálico. In Animais
Peçonhentos do Brasil. 1 ed. São Paulo: Sarvier/FAPESP, 2003. p. 91
BALSINDE, J, Balboa, MA, Insel, PA, Dennis, EA. Regulation and inhibition of
phospholipase A2. Annu. Rev. Pharmacol. Toxicon, 39: 175-189. 1999.
BARRAVIERA, B. Systemic inflammatory response syndrome in envenoming.
Toxicon, 35(1):13-4. 1997.
BARRAVIERA , B. Acute-phase response in snakebite. Toxicon, 32(8):861-2. 1994.
BARROS, SF, Friedlanskaia, I, Petricevich, VL, Kipnis, TL. Local inflamation, lethality
and cytokines release in mice injected with Bothrops atrox venom. Mediators of
Inflamation, 7: 339. 1998.
BARROSO, RD. Ofidismo no Brasil. Considerações em torno de 2238 acidentes
ofídicos tratados com soro. Bol. Inst. Vital Brazil 1943/44;25-27:35-47.
BENELLI, R, Adati, R, Ensoli, B, Stetler-Stevenson, WG, Santi, L, Albini, A. Inhibition
of AIDS-Kaposi´s sarcoma cell induced endotelial cell invasión by TIMP-2 and a
synthetic peptide from the metalloproteinase propeptide: implications for an anti-
angiogenic therapy. Onco Res. 6(6):251-2577. 1994.
BEHRING, E, KITASATO, S. Über das Zustandekommen der Diptherie-Immunität bei

88
Tieren 1890. Dtsch med Wschr. 16:1113-4. 1890.
BJARNASON, JB, FOX, JW. Hemorrhagic toxins from snake venoms. Toxicol. Toxin
Rev 7: 121-209. 1988.
BJARNASON, JB, FOX, JW. Hemorrhagic metalloproteinases from snake venoms.
Pharmacol. Ther. 62, 325–372. 1994.
BRADFORD, M.A. A rapid and sensitive method for the quantization of microgram
amounts of protein utilizing the principle or protein-dye binding. Anal. Biochem.72,
248-254. 1976.
BON, C. The serum-therapie was discovered 100 years ago. Toxicon 34: 142–143.
1996.
BOULAIN, JC, Menez, A, Couderec, J, Faure G, Liacopoulos, P, Fromageot, P.
Neutralizing monoclonal antibody especific for Naja nigricollis toxin alpha:
preparation, characterization, and localization of the antigenic binding site.
Biochemistry. 21(12): 2910-5. 1982.
BOZZO, S, RETAMAL, C. Gel perfect. Geles unidimensionales, um nuevo método
densitométrico para computadores personales. Archivos de Biologia e Medicina
Experimental, 24: 181-184.
BRAZIL, V. Contribuição ao estudo do veneno ophidico. I. Tratamento das
mordeduras das cobras. Ver. Med. São Paulo 4. 4:375-80. 1901.
BRAZIL, V. Contribuição ao estudo do veneno ophidico. III. Tratamento das
mordeduras das cobras. Ver. Med. São Paulo 4. 6:265-78. 1903.
BRAZIL, V. Contribuition à l´étude d´origine ophidiene. Paris: A. Maloine, 1905.
CADLE, JE. Geographic distribution: problems in phylogeny and zoogeography. In
Snakes: Ecology and Evolutionary Biology. New York: Macmillian Publishing Co.,
1997. p 77-105.
CALDERÓN, L, LOMONTE, B. Immunochemical characterization and role in toxic
activities of region 115-129 of myotoxin II, a Lys49 phospholipase A2 from
Bothrops asper snake venom. Arch. Biochem. Biophys., 358: 343-350. 1998.
CALDERÓN, L, LOMONTE, B. Inhibition of the myotoxins acton of Bothrops asper
myotoxins II in mice by immunization with its synthetic peptide 115-129. Toxicon,
37: 683-687. 1999.
CALMETTE, A. Contribution a l´étude du venin des serpents. Immunisation des

89
animaux et traitement de l´envenimation. Ann. Inst. Pasteur. 8:275-91. 1894.
CALVETE, JJ, MArcinkiewicz, C, Monléon, D, Esteves, V, Celda, B Juárez, P, Sanz,
L. Snake venom disintegrins: evolution of structure and finction. Toxicon 45. 1063-
1084. 2005.
CAVINATO, RA, Remold, H, Kipnis, TL. Purifcation and variability in thrombine-like
activity of Bothrops atrox venom from different geographic regions. Toxicon, 36:
257-267.1998
CARDOSO, FD, Lafaye, P, Nato, F ET AL. Neutralizing human anti-crotoxin scFv
World Congress International Society Toxinology. Isla Margarita, O-24Tu. 1997.
CARDOSO, FD, Yamaguchi, IK, da Silva, AMM. Produção de Soros Antitoxinas e
Perspectivas de Modernização por Técnicas de Biologia Molecular. In Animais
Peçonhentos do Brasil. 1 ed. São Paulo: Sarvier/FAPESP, 2003. p. 367-379
CHAKRABORTI, S. Phospholipase A2 isoforms: a perspective. Cellular Signalling,
15: 637-665. 2003.
CHOUMET, V, Faure, G, Robbe-Vicent, A, Saliou, B, Mazie JC, Bon C.
Immunological analysis of a snake venom phosphilipase A2 neurotoxin, crotoxin,
with monoclonal antibodies. Mol. Immunol. 29(7-8):871-82. 1992.
DA SILVA JR., NJ, Bucaretchi, F. Mecanismos de Ação do Veneno Elapídico e
Aspectos Clínicos dos Acidentes. In Animais Peçonhentos do Brasil. 1 ed. São
Paulo: Sarvier/FAPESP, 2003. p. 99
DANIELE, JJ, Bianco, ID, Delgado, C, Carrillo, DB, Fidelio, GD. A new
phospholipase A2 isoform isolated from Bothrops neuwiedii (Yarara chica) venom
with novel kinetic and chromatographic properties. Toxicon 35 (8), 1205–1215.
1997.
DANSE, JM, KEMPF, J. Preparation and caracterization of monoclonal antibodies
against beta-bungarotoxin and its A- and B-chains. Toxicon. 27(9):1011-9. 1989.
DENNIS, EA. Diversity of groups types, regulation and function of phospholipase A2.
J. Biol. Chem., 269: 13057. 1994.
DIAS DA SILVA, W, Eisele, JW, Lepow, IH. Complement as madiator of
inlamation:III. Purification of the activity with anaphylatoxin properties generates
by interaction of the first four components end its identification as a cleavage
product of C3. J. Exp. Med., 126: 1027-1048. 1967.

90
DIAZ, C, Gutiérrez, JM, Lomonte, B, Gené, JA. The effect of myotoxins isolated from
Bothrops snake venoms on multilamellar lipossomes: relationship to
phospholipase A2, aticoagulant and myotoxic activities. Biochim. Biophys. Acta,
1070: 455-460. 1991.
DIAZ, C, Gutiérrez, JM, Lomonte, B, Nuñez, J. p-Bromophenacyl bromide
modification of Bothrops asper myotoxin II, a lysine-49 phospholipase A2, affects
its pharmacological activities. Toxicon 31 (9), 1202–1206. 1993.
DUA, R, CHO, W. Inhibition of human secretory class II phospholipase A2 by
heparin. Eur. J. Biochem. 221: 481-490. 1994.
DIAZ-OREIRO, C, GUTIÉRREZ, JM. Chemical modification of histidine and lysine
residues of myotoxic phospholipases A2 isolated from Bothrops asper and
Bothrops godmani snake venoms: effects on enzymatic and pharmacological
properties.Toxicon 35 (2), 241–252. 1997.
EICHBAUM, F.W. Ação dermatóxica de venenos ofídicos e sua neutralização pelos
antivenenos. Mem. Instit. Butaatan, 20: 79-94. 1947.
FARSKY, SH, Walber, J, Costa-Cruz, M, Cury, Y, Teixeira, CF. Leukocyte response
induced by Bothrops jararaca crude venom: in vivo and in vitro studies. Toxicon.
35(2):185-93. 1997.
FARSKY, SH, Goncalves, LR, Cury, Y. Characterization of local tissue damage
evoked by Bothrops jararaca venom in the rat connective tissue microcirculation:
an intravital microscopic study. Toxicon, 37(7):1079-83.1999
FERREIRA, RN, Machado de Avila, RA, Sanchez, EF, Maria, WS, Molina, F,
Granier, C, Chávez-Olórtegui, C. Antibodies against synthetic epitopes inhibit the
enzymatic activity of mutalysin II, a metalloproteinase from bushmaster snake
venom. Toxicon 48, 1098-1103. 2006.
FINNEY, DJ. Statistical aspects of monitoring for dangers in drug therapy.
Methods Inf Med. 10(1):1-8. 1971.
FRANÇA, FOS, MÁLAQUE, CMS. Acidente Botrópico. In Animais Peçonhentos do
Brasil. 1 ed. São Paulo: Sarvier/FAPESP, 2003. p. 72
FRANCO, FL. Origem e diversidade das serpentes. In Animais Peçonhentos do
Brasil. 1 ed. São Paulo: Sarvier/FAPESP, 2003. p. 13 e 14.

91
FRY, B.G. Structure-function properties of venom components from Australian
elapids. Toxicon 37, 11–32. 1999.
FONSECA, F. Instituto Butantã: sua origem, desenvolvimento e contribuição ao
progresso de São Paulo. São Paulo: Comissão do IV Centenário da Cidade de
São Paulo, 1954.
FOX, JW, SERRANO, SM. Structural considerations of the snake venom
metalloproteinases, key members of the M12 reprolysin family of
metalloproteinases. Toxicon 45, 969–985. 2005.
GOULD, RJ, Polokoff, MA, Friedman, PA, Huang, TF, Holt, JC, Cook, JJ,
Niewiarawski, S. Desintegrins: a family of integrin inhibitory proteins from viper
venoms. Proc. Soc. Exp. Biol. Med., 195: 168-171. 1990
GREENE, HW. Snakes. The Evolution of Mystery in Nature. Berkeley: University of
California press, 1997. p 351.
GUTIÉRREZ, JM, Rucavado, A., Escalante, T, Dýaz, C. Hemorrhage induced by
snake venom metalloproteinases: biochemical and biophysical mechanisms
involved in microvessel damage. Toxicon 45, 997-1011. 2005.
GUTIÉRREZ, JM, Lomonte, B, Cerdas, L. Isolation and partial characterization of a
myotoxins from the venom of the snake Bothrops nunmifer. Toxicon, 24: 885-894.
1986a
GUTIÉRREZ, JM, Lomonte, B, Chaves, F, Moreno, E, Cerdas, L. Pharmacological
activities of a toxic phospholipase A isolated from venom of the snake Bothrops
asper. Comp. Biochem. Physiol., 84C: 159-164. 1986b.
GUTIÉRREZ, JM, Chaves, F, Gene, JA, Lomonte, B, Camacho, Z., Schosinsky, K.
Myonecrosis induced by a basic myotoxin isolated from the venom of the snake
Bothrops nunmifer (Jumping viper) from Costa Rica. Toxicon 27: 735-746. 1989
GUTIÉRREZ, JM. Local pathological effects induced by Bothrops snake venoms.
Mem. Instit. Butantan, 52 (supl.): 37-38. 1990
GUTIÉRREZ, JM, LOMONTE, B. PhosPholipases A2 myotoxins form Bothrops
venoms. Toxicon 33: 1405-1424.1995.
GUTIÉRREZ, JM, LOMONTE, B. Phospholipase A2 myotoxins from Bothrops snake
venoms. Toxicon, 33: 1405-1424. 1995.
GRAMS, F, Huber, R, Kress, LF, Moroder, L, Bode, W. Activation of snake venom

92
metalloproteinases by a cysteine switch-like mechanism. FEBS Lett. 335, 76–80.
1993.
GUTIERREZ JM, Lomonte B, Portilla E, Cerdas L, Rojas E. Local effects induced by
coral snake venoms: evidence of myonecrosis after experimental inoculations of
venoms from five species. Toxicon. 21(6):777-83. 1983.
HITE, LA, SHannon, JD, Bjarnason, JB, Fox, JW. Sequence of a cDNA clone
encoding the zinc metalloproteinase hemorrhagic toxin e from Crotalus atrox:
evidence for signal, zymogen, and disintegrin-like structures.
Biochemistry, 31(27):6203-6211. 1992
HITE, LA, Jia, LG, Bjarnason, JB, Fox, JW. cDNA sequences for four snake venom
metalloproteinases: structure, classification, and their relationship to mammalian
reproductive proteins. Arch. Biochem. Biophys. 308, 182–191. 1994.
HANASAKI, K, ARITA, H. Biological and pathological functions of phospholipase A(2)
receptor.Arch. Biochem. Biophys. 372: 215-223. 1999
KAMIGUTI, AS, CARDOSO, JL Haemostatic changes caused by the venoms of
South American snakes. Toxicon 27(9): 955-63. 1989.
KANASHIRO, MM, Escocard, RCM, Petretski, JH, Prates, MV, Alves, EW, Machado,
OLT, Dias da Silva, W, Kipnis, TL. Biochemical and biological properties of
phospholipase A2 isolated from Bothrops atrox snake venom. Biochem.
Pharmacol. 64, 1179-1186. 2002.
KINI, RM, EVANS, HJ. A model to explain the pharmacological effects of snake
venoms phospholipases A2. Toxicon, 37: 613-635.1989.
KINI, RM. Are C-type Lectin-related proteins derived by proteolysis of
metalloproteinase/disintegrin precursor proteins? Toxicon 34, 1287–1294. 1996.
KINI, RM. Excitement ahead: structure, function and mechanism of snake venom
phospholipase A2 enzymes. Toxicon, 15;42(8):827-40. 2003.
KISIEL, W, Kondo, S, Smith, KJ, Mcmullen, BA. Smith, L.F. Characterization of a
protein C activator from Agkistrodon contortrix contortrix venom.
J Biol Chem, 15;262(26):12607-13. 1987
KOHLER, G, MILSTEIN, C. Continous cultures of fused cells secreting antibody of
predefined specificity. Nature. 256(5517):495-7. 1975.
LAEMMLI, UK. Cleavage of structural proteins during the assembly of bacteriophage

93
T4. Nature, 227: 680-685. 1970.
LI, Q, Coldberg, TR, Ownby, CL. Purification and characterization of two high
molecular weight hemorrhagic toxins from Crotalus viridis viridis venom using
monoclonal antibodies. Toxicon. 31(6):711-22. 1993.
LIU, CS, Chen, JM, Chang, CH, Chen, SW, Teng, CM, Tsai, IH. The amino acid
sequence and properties of an edema-inducing Lys49 phospholipase A2 homolog
from the venom of Trimeresurus mucrosquamatus. Biochim. Biophys. Acta 1077:
362-370. 1991.
LLORET, S, MORENO, JJ. Oedema formation and degranulation of mast cells by
phospholipase A2 purified from porcine pancreas and snake venoms. Toxicon 31,
949–956. 1993.
LOMONTE, B, Tarkowski, A, Hanson, LA. Host response to Bothrops asper snake
venom: análisis of edema formation, inflammatory cells, and cytokine release in a
mouse model. Inflammation, 17: 93-105. 1993.
LOMONTE, B, Moreno, E, Tarkowski, A, Hanson, LA, Maccarana, M. Neutralizating
interaction between heparins and myotoxin II, a Lys49 phospholipase A2 from
Bothrops asper snake venom. Identification of a heparin-binding and cytolytic toxin
region by the use of synthetic peptides and molecular modeling. Biochem.
Pharmacol. 47, 1509–1518. 1994a.
LOMONTE, B, Pizarro-Cerdá, J, Ângulo, Y, Gorvel, JP, Moreno, E. Tyr – Trp-
substituted peptide 115-129 of a Lys49 phospholipase A2 expresses enhanced its
in vivo myotoxic effect. Biochim. Biophys. Acta, 1461, 19-16. 1999b.
LORÍA, GD, Rucavado, A, Kamiguti, AS, Theakston, RDG, Fox, JW, Alape, A,
Gutiérrez, JM. Characterization of “basparin A”, a prothrombin-activating
metalloproteinase, from the venom of the snake Bothrops asper that inhibits
platelet aggregation and induces defibrination and thrombosis. Arch Biochem
Biophys. 418(1): 13-24. 2003.
MAGALHÃES, A, da Fonseca, BCB, Diniz CR, Gilroy, J. Richardson M. The
complete amino acid sequence of a thrombin-like enzyme/gyroxin analogue from
venom of the bushmaster snake (Lachesis muta muta). FEBS, 39 (1,2): 116-120.
1993.
MÁLAQUE, CMS, França, FOS. Acidente Laquético. In Animais Peçonhentos do

94
Brasil. 1 ed. São Paulo: Sarvier/FAPESP, 2003. p. 87
MARKLAND, FS. Snake venoms and the hemostatic system. Toxicon 36, 1749–
1800. 1998.
MARAGANORE, JM, Merutka, G, Cho, W, Welches, W, Kézdy, FJ, Heinrikison, RL.
A new class of phospholipase A2 with Lys49 in place of aspartate 49. J. Biol.
Chem., 259: 13839-13843. 1984.
MARTINS, AMC, Toyama, MH, Havt, A, Novello, JM, Marangoni, S, Fonteles, MC,
Monteiro, HSA. Detemination of Crotalus durissus cascavella venom components
tha induce renal toxity in isolated rat kidneys. Toxicon 40: 1165-1171. 2002.
MARTINEZ, RA, Huang, SY, Perez, JC. Antigenic relationships of fractionated
western diamondback rattlesnake (Crotalus atrox) hemorrhagic toxins and other
rattlesnake venoms as indicated by monoclonal antibodies. Toxicon. 27(2):239-45.
1989.
MARUYAMA, M, Sugiki, M, Yoshida, E, Shimaya, K, Mihara, H. Broad substrate
specificity of snake venom fibrinolytic enzymes: possible role in haemorrhage.
Toxicon,30(11):1387-97.1992.
MATSUI, T, Fujimura, Y, Titani, K. Snake venom proteases affecting hemostasis and
thrombosis. Biochim. Biophys. Acta 1477, 146–156. 2000.
MAZZUCCONI, MG, Ferrari, A, Vignetti, M, Giona, F, Girolami, D, Martinelli, E,
Mandelli, F. High-dose intravenous gammaglobulin (HDIVG): therapy of refractory
ITP in adults and children. Haematologica. 74(1):63-6. 1989.
MEKI, AR, Mohey, El-Dean, ZM. Serum interleukin-1beta, interleukin-6, nitric oxide
and alpha1-antitrypsin in scorpion envenomed children. Toxicon, 36(12):1851-9.
1998.
MELGAREJO, AR. Serpentes Peçonhentas do Brasil. In Animais Peçonhentos do
Brasil. 1 ed. São Paulo: Sarvier/FAPESP, 2003. p. 43, 44 e 46.
MINISTÉRIO DA SAÚDE. Manual de diagnóstico e tratamento de acidentes por
animais peçonhentos. 2ª ed. – Brasília: Fundação Nacional de Saúde. 2001
MUNIZ, EG, Maria, WS, Estevão-Costa, MI, Buhrnheim, P, Chávez-Olórtegui, C.
Neutralizating potency of horses antibothropic Brazilian antivenom against
Bothrops snake venoms from Amazonian rain forest. Toxicon 38, 1859-1863.
2000.

95
MURPHY, AN, Unsworth, EJ, Stetler-Stevenson, WG. Tissue inhibitor of
metalloproteinases-2 inhibits bFGF-induced human microvascular endothelial cell
proliferation. J. Cell. Physiol, 157:351-358. 1993.
OHSAKA, A. Hemorrhagic, necrotizing and edema-inducing effects of snake venoms.
In Handbook of Experimental Pharmacology (edited by Lee, C.Y.), vol. 52: 481-
546. Springer, Berlin. 1979.
OTERO-PATIÑO, R, Cardoso, JLC, Higashi, HG, Nunez, V, Diaz A, Toro, MF,
Garcia, ME, Sierra, A, Garcia, LF, Moreno, AM, Medina, MC, Castañeda, N, Silva-
Diaz, JF, Murcia, M, Cardenas, SY, Dias da Silva, W, and the Regional Group on
Antivenom Therapy Research (REGATHER). A randomized, blinded, comparative
trial of one pepsin-digested and two whole IgG antivenoms for Bothrops snake
bites in Uraba, Colômbia. Am. J. Trop. Med. Hyg., 58(2), pp. 183-189. 1998.
OWNBY, CL, Gutierrez, JM, Colberg, TR, Odell, GV. Quantitation of myonecrosis
induced by myotoxin a from prairie rattlesnake (Crotalus viridis viridis) venom.
Toxicon, 20(5):877-85. 1982.
OWNBY, CL, Fletcher, JE, Coldberg, TR. Cardiotoxin 1 form cobra (Naja naja atra)
venom causes necrosis of skeletal muscle in vivo. Toxicon, 28: 837. 1990
OWNBY, CL, Selistre-de-Araujo, HS, Mhite, SP, Fletcher, JE. Lysine 49
phospholipase A2 proteins. Toxicon: 37: 411-445. 1999
OTERO, R, Núñez, V, Jiménez, SL, Fonnegra, R, Osorio, RG, García, ME, Díaz, A.
Snakebites and ethnobotany in the northwest region of Colombia Part II:
Neutralization of lethal and enzymatic effects of Bothrops atrox venom. J.
Ethnopharmacol. 71(3):505-11. 2000
PAINE, MJ, Desmond, HP, Theaskton, RD, Crampton, JM. Purification, cloning and
molecular characterization of a high molecular weight hemorrhagic
metalloprotease, jararhagin, from Bothrops jararacussu venom. J. Biol. Chem.
267:22869. 1992.
PÁRAMO, L, Lomonte, B, Pizarro-Cerdá, J, Bengoechea, JA, Gorvel, JP, Moreno, E.
Bactericidal activity of Lys49 and Asp49 myotoxic phospholipases A2 from
Bothrops asper snake venom: synthetic Lys49 myotoxin II-(115–129)-peptide
identifies its bactericidal region. Eur. J. Biochem. 253: 452–461. 1998.

96
PÉREZ, AV, Saraviac, P, Rucavado, A, Carolina D. Sant’ana, CD, Soares, AM,
Gutiérrez, JM. Local and systemic pathophysiological alterations induced by a
serine proteinase from the venom of the snake Bothrops jararacuss. Toxicon, 49:
1063-1069. 2007.
PEREZ, JC, Garcia, VE, Huang, SY. Production of a monoclonal antibody against
hemorragic activity of Crotalus atrox (western diamondback rattlesnake) venom.
Toxicon. 27(2):239-45. 1984.
PETRETSKI JH, Kanashiro M, Silva CP, Alves EW, Kipnis TL. Two related thrombin-
like enzymes present in Bothrops atrox venom. Braz J Med Biol Res. 33(11):1293-
300. 2000a.
PETRETSKI JH, Kanashiro MM, Rodrigues FR, Alves EW, Machado OL, Kipnis TL.
Edema induction by the disintegrin-like/cysteine-rich domains from a Bothrops
atrox hemorrhagin. Biochem Biophys Res Commun. 16; 276(1):29-34. 2000b.
PETRICEVICH, VL, Teixeira, CF, Tambourgi, DV, Gutierrez, JM. Increments in
serum cytokine and nitric oxide levels in mice injected with Bothrops asper and
Bothrops jararaca snake venoms. Toxicon 38 (9), 1253–1266. 2000.
PHISALIX, M, BERTRAND, G. Sur la proprieté antitoxique du sang des animaux
vaccinés contre le venin de vipère. C. R. Soc. Biol. 1:111-5. 1894.
POUGH, FH, Andrews, RM, Cadle, JE, Crump, ML, Savitsky, AH, Wells, KD.
Hepertology. Upper Saddle River: Prentice Hall, 1998. p 577.
RAEL, ED, Salo, RJ, Zepeda, H. Monoclonal antibodies to Mojave toxin and use for
isolation of cross-reacting proteins in Crotalus venoms. Toxicon. 24(7):661-8.
1986,
RAMOS, OHP, Carmona, AK, Selistre-de-Araujo, HS. Expression, refolding, and in
vitro activation of a recombinant snake venom prometalloprotease.Protein Expr.
Purif. 28, 34–41. 2003.
ROBBINS, SL, Cotran, RS, Kumar, V. Patolgia estrctural y functional. México, Nueva
Editorial Interamericana: 1434. 1987.
ROCHA E SILVA, M, Beraldo, WT, Rosenfeld, G. Bradykinin a hypotensive and
smooth muscular stimulating factor released from plasma globulin dy snake
venoms and trypisin. Am. J. Physiol. 156: 261-271. 1949.
ROSING, J, TANS, G. Structural and functional properties of snake venom

97
prothrombin activators. Toxicon. 30(12):1515-27.1992.
RUOSLAHTI, E, PIERSCHBACHE, RMD. New perspectives in cell adhesion: RGD
and integrins. Science, 238 (4826):491-497. 1987
SANCHEZ, EF, Santos, CI, Magalhães, A, Diniz, CR, Figueiredo, S, John G.
Richardson M. Isolation of a proteinase with plasminogen-activating activity from
Lachesis muta muta (Bushmster) snake venom. Arch. Biochem. Biophys. 378 (1):
131–141. 2000.
SANTOS FILHO, L. História geral da Medicina brasileira. São Paulo. Hucitec/Edusp.
1991.
SCARBOROUGH, RM, Rose, JW, Naughton, MA, Phillips, DR, Nannizzi, L, Arfsten,
A, Campbell, AM, Charo, IF. Characterization of the integrin specificities of
disintegrins isolated from American pit viper venoms. J. Biol.Chem., 268:1058-
1065. 1993.
SCOTT, DL, Achari, A, Vidal, JC, Sigler, PB. Crystallographic and biochemical
studies of the (inactive) Lys-49 phospholipase A2 from the venom of Agkistrodon
piscivorus piscivorus. J. Biol. Chem. 267, 22645–22657. 1992.
SELISTRE-DE-ARAÚJO, HS, White, SP, Ownby, CL. cDNA cloning and sequences
analisis of a lisina 49 phospholipase A2 myotoxin from Agkistrodon contortrix
laticinctus sanke venom. Arch. Biochem. Biophys. 326: 21. 1996a.
SELISTRE-DE-ARAÚJO, HS, White, SP, Ownby, CL. Sequence analysis of Lys49
phospholipase A2 myotoxins: a higly conserved class of proteins. Toxicon, 34:
1237. 1996b.
SELISTRE, HS, GIGLIO, JR. Isolation and characterization of a thrombin-like
enzyme from the venom of the snake Bothrops insularis (Jararaca ilhoa). Toxicon,
25(11): 1135–1144.1987.
Selistre-de-Araujo, HS, Ownby, CL. Molecular cloning and sequence analysis of
cDNAs for metalloproteinases from broad-banded copperhead Agkistrodon
contortrix laticinctus. Arch. Biochem. Biophys. 320, 141–148. 1995.
SEWALL, H. Experiments in the preventive inoculation of rattlesnake venom. J.
Physiol. (Lond). 21:241-3. 1887

98
SHIMOKAWA, K, JIA, LG,Wang, XM, Fox, JW. Expression, activation,and
processing of the recombinant snake venom metalloproteinase, proatrolysin E.
Arch. Biochem. Biophys. 335, 283–294. 1996
SIX, DA, DENNIS, EA. The expanding superfamily of phospholipase A2 enzymes:
classification and characterization. Biochim. Biophys. Acta 1488, 1–19. 2000.
SPRINGMAN, EB, Angleton, EL, Birkedal-Hansen, H, Van Wart, HE. Multiple modes
of activation of latent human fibroblast collagenase: evidence for the role of a
Cys73 active-site zinc complex in latency and a "cysteine switch" mechanism for
activation. Proc. Natl. Acad. Sci. USA, 87 (1):364-8. 1990.
STOCK RP, Massougbodjii A, Alagón A, Chippaux JP. Bringing antivenoms to Sub-
Saharan Africa. Nature Biotechonology, 25: 173-177. 2007
STOCKER, K, MEIER, J. Snake venom proteins in hemostasis: new results. Folia
Haematol Int Mag Klin Morphol Blutforsch. 116(6):935-53. 1989.
TAKASAKI, C, Sugama, A, Yanagita, A, Tamiya, N, Rowan, EG, Harvey, AL. Effects
of chemical modifications of Pa-11, a phospholipase A2 from the venom of
Australian king brown snake (Pseudechis australis), on its biological activities.
Toxicon, 28(1):107-17. 1990
TAMBOURGI, DV, Dos Santos, MC, Furtado, M de F, DE Freitas, MC, da Silva, WD,
Kipnis, TL. Pro-inflammatory activities in elapid snake venoms. Br J
Pharmacol.Jul;112(3):723-7.1994.
TANJONI I, Butera D, Bento L, Della-Casa MS, Marques-Porto R, Takehara HA,
Gutierrez JM, Fernandes I, Moura-da-Silva AM. Snake venom metalloproteinases:
structure/function relationships studies using monoclonal antibodies. Toxicon.
42(7): 801-808. 2003.
TASHIRO, K, Sephel, GC, Weeks, B, Sasaki, M, Martin, GR, Kleinman, HK. A
synthetic peptide containing the IKVAV sequence from the A chain of laminin
mediates cell attachment, migration, and neurite outgrowth. J. Biol. Chem., 264:
16174-16182. 1989.
TEIXEIRA, CFP, Landucci, ECT, Antunes, E, Chacur, M, Cury, Y. Inflammatory
effects of snake venom myotoxic phospholipases A2. Toxicon 42, 947–962. 2003.
TIMPL, R. Structure and biological activity of basement membrane proteins.
Eur.J.Biochem.,180 (3): 487-502. 1989.

99
VALENTIN, E, LAMBEAU, G. Increasing molecular diversity of secreted
phospholipase A2 and their receptors and binding proteins. Biochim. Biophys.
Acta, 1488 (1-2), 59-70. 2000.
VISHWANATH, BS, Kini, RM, Gowda, TV. Purification and partial biochemical
characterization of an edema inducing phospholipase A2 from Vipera russelli
(Russell's viper) snake venom. Toxicon, 26(8):713-20. 1988.
WARD, RJ, de Azevedo, WF Jr., Arni, RK. At the interface: crystal structure of
phospholipase A2. Toxicon 36: 1623, 1998a.
WARD, RJ, Rodrigues Alves, A, RUGGIERO Neto, J, Arni, RK, Casari, G. A
SequenceSpace analysis of Lys49 phospholipases A2: clues towards identification
of residues involved in a novel mechanism of membrane damage and in
myotoxicity. Protein Engng, 11: 285–294. 1998b.
WARRELL, DA. Geographical intraspecies variation in the clinical manifestation of
envenoming by snakes. In: Venomous snakes: ecology, evolution and snake bites.
Oxford: Claredon Press. 1997. 189-204.
WATANABE, L, Gava, LM, Angulo, Y, Lomonte, B, Arni, RK. Crystallization of the
Lys49 homologue, miotoxin II, from the venom of Atropoide nunmifer. Biophys.
Acta. 1703(1): 87-89. 2004.
WEN, FH. Soroterapia. In Animais Peçonhentos do Brasil. 1 ed. São Paulo:
Sarvier/FAPESP, 2003. p. 380-393.
WRAY, W, Boulikas, T, Wray, VP, Hancock, RN. Silver staining of proteins in
ployacrylamida gels. Analytical Biochemistry. 118: 197-203. 1981.
YAMAGUCHI Y, Shimohigashi Y, Chijiwa T, Nakai M, Ogawa T, Hattori S, Ohno M.
Characterization, amino acid sequence and evolution of edema-inducing, basic
phospholipase A2 from Trimeresurus flavoviridis venom.
Toxicon. 39 (7):1069-76. 2001.
ZAGANELLI, GL, Zaganelli, MGM, Magalhaes, A, Diniz CR, Lima, ME. Purification
and characterization of a fibrinogen-clotting enzyme from the venom of jararacuçu
(Bothrops jararacussu). Toxicon, 34(7): 807-819. 1996.
ZULIANIA, JP, Cristina, MF, Zamunera, SR, Gutiérrez, JM, Teixeira CFP.
Inflammatory events induced by Lys49 and Asp-49 phospholipases A2 isolated

100
from Bothrops asper snake venom: role of catalytic activity.Toxicon. 45, 335–346.
2005.





























