Embed Size (px)
Citation preview

Caracterização da resposta de defesa local e sistêmica, induzida
por lesão mecânica em plântulas de feijão-de-corda.
Márcio dos Santos Teixeira Pinto
Universidade Estadual do Norte Fluminense Darcy Ribeiro – UENF
Campos dos Goytacazes – RJ
Abril – 2008


Caracterização da resposta de defesa local e sistêmica, induzida
por lesão mecânica em plântulas de feijão-de-corda.
Márcio dos Santos Teixeira Pinto.
“Tese de Doutorado apresentada ao Centro de Biociências e Biotecnologia da
Universidade Estadual do Norte Fluminense Darcy Ribeiro, como parte das
exigências para a obtenção do título de Doutor em Biociências e
Biotecnologia.”
Campos dos Goytacazes – RJ
Abril - 2008

Caracterização da resposta de defesa local e sistêmica, induzida por lesão
mecânica em plântulas de feijão-de-corda.
Márcio dos Santos Teixeira Pinto.
“Tese de Doutorado apresentada ao Centro de Biociências e
Biotecnologia da Universidade Estadual do Norte Fluminense Darcy Ribeiro,
como parte das exigências para a obtenção do título de Doutor em Biociências
e Biotecnologia.”
Aprovada em 09 / 04 / 2008
Comissão Examinadora:
César Luis Siqueira Junior - CEFET/ RJ
Valdirene Moreira Gomes – LFBM; UENF / RJ
Olga Lima Tavares Machado – LQFPP; UENF / RJ
Kátia Valevski Sales Fernandes (Orientadora) LQFPP; UENF / RJ

À minha irmã Rose, por nunca ter desistido dos seus sonhos. E mesmo tendo a morte em seu futuro, lutou pela a alegria de sua vida.

Agradecimentos
Ofereço um agradecimento especial a minha orientadora, a professora Dra
Kátia. Por ter aceitado me orientar por tanto tempo, muitas vezes em assuntos que
vão além do âmbito acadêmico. Sendo agora, mais do que uma mestra, uma grande
amiga.
Agradeço à Nathália pela ajuda indireta, mas essencial, na conclusão da
minha tese, pela paciência em conviver com meus defeitos, e pela sua amizade
incondicional.
Agradeço ao Fabrício pela ajuda na conclusão deste trabalho. E lamento que
esta ação não o tenha ajudado tão bem como deveria.
Agradeço à Lucilene pela mútua ajuda e boa convivência no laboratório, e
pelos momentos de humor proporcionados pela sua presença.
Agradeço à Elaine, aluna da professora Elenir, pela boa convivência e ajuda
durante minha permanência.
Agradeço à Dra Elenir, pelo esforço de aceitar revisar esta tese, em tão pouco
tempo, e pela grande colaboração dada. Algo que não seria possível se não
houvesse um elo de amizade.
Agradeço a minha banca: Dr(s) Olga Machado, Valdirene Moreira e César
Siqueira, pela preciosa contribuição, e por aceitarem avaliar esta tese, em tempo tão
limitado.
Agradeço a minha irmã Rosana, pelo sacrifício em manter a integridade da
minha família durante momentos tão difíceis da minha vida. Coisa que não poderia
ter feito, eu mesmo.

Agradeço a minha mãe pelo imenso esforço em continuar existindo, e assim
poder me ajudar e me admirar naquilo que eu faço.
Agradeço a Deus por poder ajudar os meus amigos quando possível. E por
poder ter esperança em viver um futuro melhor.

I
INDICE
LISTA DE FIGURAS..................................................................................................IV
LISTA DE TABELAS..................................................................................................VI
LISTA DE ABREVIATURAS.................................................................................... VII
Resumo....................................................................................................................VIII
Abstract…………………………………………………………………………..…………IX
1 – INTRODUÇÃO.......................................................................................................1
1.1 - Estudo de respostas de plantas a ferimento...................................................1
1.1.2 – O papel do ácido jasmônico e seus derivados............................................9
1.1.3 – O estresse de ferimento mecânico como ativador de defesa e suas
limitações em comparação ao estímulo da herbivoria.........................................10
1.2 – Detecção da indução de proteínas de defesa como indicação de resposta
a estresses................................................................................................................11
1.2.1- Polifenol oxidases – PFOs............................................................................12
1.2.2- Inibidores de proteinases serínicas.............................................................14
1.2.3 - Inibidores de proteinases cisteínicas - cistatinas .....................................15
1.3 – O esxudado de floema como fonte de hormônios vegetais.......................16
1.4- A espécie Vigna unguiculata no estudo de defesa vegetal..........................18
2 – OBJETIVOS GERAIS..........................................................................................20
2.1 – Objetivos específicos......................................................................................20
3 – MATERIAIS..........................................................................................................21
3.1 – Material biológico............................................................................................21
3.1.1 – Sementes.......................................................................................................21
3.2 – Reagentes........................................................................................................21
3.3 – Equipamentos..................................................................................................21
4 – METODOLOGIA……………………………………………………...………………..23
4.1 – Detecção, isolamento e caracterização de polifenol oxidases induzidas
por ferimento............................................................................................................23
4.1.1 – Condições de cultivo de plantas.................................................................23
4.1.2 – Tratamento por ferimento e obtenção de extratos protéicos..................23
4.1.3 – Quantificação da atividade polifenol oxidásica.........................................24
4.1.4 – Quantificação protéica.................................................................................25
4.1.5 – Eletroforese em gel de poliacrilamida sob condições semi-
desnaturantes (SDS-PAGE).....................................................................................25

II
4.1.6 – Ensaio de atividade enzimática em gel......................................................26
4.1.7 – Isolamento de PFO induzida por ferimento...............................................26
4.1.8 – Parâmetros cinéticos da F- 0,2....................................................................27
4.1.8.1 - Ativação com SDS e estabilidade. ...........................................................27
4.1.8.2 – Medição de estabilidade térmica.............................................................27
4.1.8.3 - Determinação de pH ótimo de atividade .................................................28
4.1.8.4. – Especificidade a substratos e determinação de Km e Vmax .............28
4.1.8.5 – Ensaio de inibição de atividade polifenol oxidásica.............................28
4.2 – Detecção e caracterização de cistatinas induzidas por ferimento e
análises de parâmetros relacionadas.....................................................................29
4.2.1 – Condições de cultivo de plantas.................................................................29
4.2.2 – Tratamento por ferimento e obtenção de extrato protéico......................29
4.2.3 – Determinação da curva de atividade enzimática ......................................30
4.2.4 – Ensaio de inibição de atividade de proteinase cisteínica (dosagem de
cistatinas) .................................................................................................................30
4.2.5 – Eletroforese em gel de poliacrilamida sob condições desnaturantes e
redutoras (SDS-PAGE) ............................................................................................31
4.2.6 – Detecção de proteínas imunologicamente relacionadas a cistatinas por
Western Blotting.......................................................................................................31
4.2.7 – Ensaio enzimático para proteases cisteínicas totais................................32
4.2.8 - Eletroforese semi-desnaturante em gel de poliacrilamida com gelatina
(SDS-PAGE-Gelatina) ..............................................................................................32
4.2.9 – Ensaio de ativação de senescência............................................................33
4.3 – Análise do efeito eliciador de esxudado de floema. ...................................33
4.3.1 - Extração de esxudado de floema................................................................33
4.3.2 – Teste de atividade indutora.........................................................................34
4.3.3 – Análise de esxudado de floema por espectrometria de massas.............35
5 – RESULTADOS ....................................................................................................36
5.1 - Quantificação e caracterização da atividade polifenol oxidásica induzida
por ferimento. ..........................................................................................................36
5.2 – Visualização de proteínas totais e polifenol oxidases em unifoliatos de
feijão-de-corda..........................................................................................................36

III
5.3 – Purificação parcial de PFOs induzidas por ferimento. ...............................37
5.4 – Ensaios de cinética enzimática da fração F- 0,2..........................................43
5.4.1 - Ativação e estabilidade com SDS. ..............................................................43
5.4.2 – Medição de estabilidade térmica................................................................43
5.4.3 - Determinação de pH ótimo de atividade de F- 0,2.....................................43
5.4.4 - Especificidade a substratos e determinação de Km e Vmax....................47
5.4.5 - Ensaio de inibição de atividade...................................................................49
5.5 – Detecção e caracterização de cistatinas induzidas por ferimento.............49
5.5.1 – SDS-PAGE e Western blot de cistatinas induzidas por ferimento. ........50
5.5.2 – Atividade proteinásica do tipo cisteínica em unifoliatos de feijão-de-
corda submetidos ou não a lesão mecânica.........................................................50
5.5.3 – Proteinases cisteínicas em unifoliatos de feijão-de-corda submetidos ou
não a lesão mecânica...............................................................................................50
5.5.4- Ensaio de ativação de senescência............................................................50
5.6– Análise da atividade indutora e caracterização de esxudado de floema....55
5.6.1 - Teste de indução de PFOs por esxudado de floema ................................55
5.6.2 – Detecção da presença de AJ em esxudado de floema por
espectrometria de massas......................................................................................55
6- Discussão....................................................... .....................................................58
6.1 – Análise da indução e caracterização de PFOs ............................................58
6.2 – Análise da indução, caracterização e efeito de cistatinas em unifoliatos de
V. unguiculata ..........................................................................................................62
6.3 – Análise da composição e efeito eliciador de defesa de esxudado de
floema........................................................................................................................65
7 - CONCLUSÕES..................... ...............................................................................68
8- REFERÊNCIAS BIBLIOGRÁFICAS......................................................................70

IV
LISTA DE FIGURAS
Figura 1 – Indução sistêmica hipotética de genes de inibidores de proteinases do
tipo 2 (pin2) estimulada por um sinal elétrico de propagação (Adaptado de Roberts,
1992) AJ – Ácido Jasmônico, ABA - Ácido Abscísico..................................................4
Figura 2 – Modelo do efeito sistêmico da liberação e transporte de sistemina sobre a
ativação de genes de defesa (Ryan, 2000)……………………………………………… 5
Figura 3 – Teste do potencial do mutante Spr1 para resposta sistêmica a ferimento.
…………......………............................................................….......................................7
Figura 4 – Modelo de sinalização sistêmica de defesa vegetal, evidenciando a ação
do AJ como sinalizador sistêmico de defesa................................................................8
Figura 5 – Interferência de diversos hormônios vegetais e moléculas sinalizadoras
sobre a sinalização de defesa causada por ferimento.................................................9
Figura 6 - Análise eletroforética mostrando variação de proteínas obtidas de floema
provocada por diferentes métodos de extração.........................................................18
Figura 7 - Esquema de tratamento por lesão mecânica e coleta das regiões de
plantas de feijão-de-corda..........................................................................................24
Figura 8 - Esquema de coleta de esxudado de floema de plantas de feijão-de-
corda...........................................................................................................................34
Figura 9 – Procedimento para teste de atividade indutora de PFO dos exsudados de
floema de feijão-de-corda...........................................................................................35
Figure 10 – Atividade de polifenol oxidase de extratos protéicos obtidos de
unifoliatos feridos (barras brancas), e laterais aos feridos (barras cinzas) de plantas
de feijão-de-corda.......................................................................................................39
Figura 11 – SDS-PAGE (I) e gel de atividade para PFO na ausência (II) e na
presença de tropolona 1mM (III)...............................................................................40
Figura 12 – Cromatograma de troca iônica em DEAE-celulose da fração 30-80%
clarificada de unfoliatos ferido de feijão-de-corda......................................................41
Figura 13 - Efeito do SDS sobre ativação da atividade de F-0,2. o ensaio foi feito
com 5 mM de 4-metil-catecol.....................................................................................44
Figura 14 - Estabilidade térmica de F-0,2 extraída de V. unguiculata.......................44
Figura 15 - Efeito da variação de pH sobre a atividade de F-0,2..............................46
Figura 16 - Gráfico de Lineweaver-Burk, mostrando dois diferentes perfis de cinética
enzimática..................................................................................................................48

V
Figura 17 - Indução de cistatinas em unifoliatos de V.unguiculata............................51
Figura 18 – A, SDS-PAGE dos extratos protéicos clarificados obtidos de unifoliatos
de V.unguiculata............................. ...........................................................................52
Figura 19- Atividade total de proteases cisteínicas em unifoliatos de plantas de V.
unguiculata 24 horas após ferimento.........................................................................53
Figura 20 - Visualização eletroforética de proteases cisteínicas totais de uniofoliatos
de plantas de V. unguiculata, 24 horas após ferimento.............................................54
Figura 21 - Unifoliatos de plantas de feijão-de-corda, submetidos a estresse de
senescência. ..............................................................................................................54
Figura 22- Teste de indução de atividade de PFOs por exsudados de floema.........56
Figura 23 – Espectrograma obtido a partir da análise por espectrometria de massas
de amostras de esxudados de feijão-de-corda..........................................................57

VI
LISTA DE TABELAS
Tabela 1 - Perfil de atividade de PFOs de unifoliatos de feijão-de-
corda..................38
Tabela 2 - Etapas de purificação de F-0,2 de V.
unguiculata....................................42
Tabela 3 - Valores de atividade especifica de PFO de Vigna unguiculata, sobre 6
diferentes substratos comumente usados para caracterização de PFOs..................47
Tabela 4 - Valores de parâmetros de cinéticos medidos para F-0,2 de V.
unguiculata.................................................................................................................49
Tabela 5 - Inibição percentual de PFO parcialmente purificada de V. unguiculata..49

VII
LISTA DE ABREVIATURAS
AOS - Aleno Oxido Sintase
AOC - Aleno Óxido Ciclase
ABA - Ácido Abscísico
AJ - Ácido Jasmônico
EDTA - Ácido Etileno Diamino Tetracetico
IP - Inibidor de Proteinase.
IPs - Inibidores de Proteinases.
Ja1 - Mutante insensível a AJ – 1
MeJa - Metil-Jasmonato.
MAPK - Proteína Quinase Ativadora de Mitose
LOX - Lipoxigenase.
α-LeA - Ácido Linolenico.
OPDA - Ácido 12-oxofitodienoico.
OPR - OPDA redutase.
PFOs - Polifenol oxidases.
PM - Membrana Plasmática
PMSF - Fenilmetil-sulfonil fluoreto
PLA - Fosfolipase A
spr1 - Supressor de sistemina 1
spr2 - Supressor de sistemina 2
R - Receptor
V - Vacúolo

VIII
Resumo
A espécie Vigna unguiculata foi amplamente estudada a respeito de proteínas
de proteção constitutiva, porém a indução de proteínas de defesa nesta espécie vem
sendo estudada só recentemente. Neste trabalho, plantas de V. unguiculata, quando
feridas, aumentaram os níveis de polifenol oxidases (PFOs) e cistatinas, proteínas
classicamente relacionadas com defesa, tanto local como sistemicamente. PFOs
induzidas em unifoliatos de Vigna unguiculata foram purificadas por cromatografia de
troca iônica (DEAE-celulose) e vistas como sendo proteínas solúveis e inativas,
sendo otimamente ativadas por SDS 5 mM. A enzima é altamente termo-estável e
seu pH ótimo foi 6,0. Maiores atividades específicas foram direcionadas aos
substratos 4-4-metil-catecol e catecol, além de ser fortemente inibida pelo inibidor
específico de polifenol oxidases, tropolona. Os parâmetros cinéticos de constante de
afinidade e velocidade máxima contra 4-metil-catecol e catecol foram Km = 9,86 mM,
Vmax = 24,66 UE [�A min-1] e Km = 3,44 mM e Vmax = 6,64 EU [�A min-1],
respectivamente. Também foram constatados aumentos nos níveis de cistatinas
(cerca de 33 %), 24 horas após o ferimento, em relação a plantas controle e
cistatinas de 22 kDa e 29 kDa foram vistas como induzidas, por técnica de Western
blotting. Atividade total de proteinases cisteínicas, por sua vez, foi inibida em plantas
feridas, e uma forma minoritária de protease cisteínica de 50 kDa, observada em gel
semi-desnaturante de gelatina-PAGE, foi a principal atividade inibida em unifoliatos
feridos. Interessantemente, tais unifoliatos de plântulas de feijão-de-corda feridas
foram vistos como mais resistentes à senescência promovida por escuridão, que os
de plantas controle. Uma correlação positiva entre a indução de cistatinas, inibição
específica de protease cisteínica e resistência à senescência é inferida a partir de
tais dados. Solução de exsudado de floema de plantas feridas de feijão-de-corda
mostrou-se capaz de induzir o aumento da atividade de PFO em plantas não feridas
ao nível de unifoliatos e a análise dessa fração por espectrometria de massas
revelou a presença de AJ nesta mesma fração, sugerindo que esta molécula atue
como um sinal sistêmico de resposta à lesão mecânica em plantas de V.
unguiculata.

IX
Abstract
The species Vigna unguiculata (cowpea) has been widely studied in respect to
constitutive defense proteins, but the induction of defensive proteins has only
recently received some attention. At the present work, V. unguiculata seedlings,
when injured, have increased their levels of polyphenol oxidases (PPO) and
cystatins, classical defense-related proteins, both locally and systemically. Unifoliate
induced PPOs were purified by ion-exchange chromatography (DEAE-cellulose) and
were seen to be soluble inactive proteins better activated at 5 mM SDS. The enzyme
was highly thermostable and optimum pH was 6.0. Highest specific activities were
directed towards the substrates 4-methyl-catechol and catechol, besides being
strongly inhibited by the PPO-specific inhibitor, tropolone. The affinity constant and
maximum velocity kinetic parameters towards methyl-catechol and catechol were Km
= 9.86 mM, Vmax = 24.66 EU [�A min-1], and Km = 3.44 mM, Vmax = 6.64 EU [�A
min-1], respectively. We have also detected increases at the levels of cystatins (ca.
33 %), 24 hours after the injury, when compared to control plants, and 22 kDa and 29
kDa cystatins were seen as induced forms, by Western blotting technique. Total
cysteine proteinase activity, by its turn, was inhibited in wounded plants, and a less
predominant 50 kDa cysteine protease form, detected in semi-denaturing gelatin-
PAGE gels, was the major inhibited activity in wounded unifoliates. Interestingly, such
unifoliates of wounded cowpea plants were seen to be more resistant to darkness-
promoted senescence than those of control plants. A positive correlation between
cystatin induction, specific cysteine protease inhibition and higher senescence
resistance in wounded plants is inferred from these data. A solution of phloem
exudates from wounded cowpea plants was shown to be able of inducing increase of
PPO activity when injected in plants of the same species, not-wounded at unifoliates
and the analysis of this exudate by mass spectrometry revealed the presence of JA,
suggesting that this molecule may act as a systemic signal during wounding
response in V. unguiculata.

1
1 - INTRODUÇÃO
1.1 - Estudo de respostas de plantas a ferimento.
Plantas, quando feridas, possuem a capacidade de mudarem a sua
constituição protéica, como um mecanismo de resposta a esse tipo de estresse.
Muitas dessas alterações podem estar diretamente relacionadas com defesa e
proteção. Entretanto outras modificações estão relacionadas com aspectos
diferentes do metabolismo vegetal, que deve adaptar-se a tal condição de estresse
(Ryan, 2000).
O trabalho que deu inicio aos estudos da resposta de defesa induzida em
plantas foi desenvolvido por Green & Ryan, quando em 1972, detectaram uma forte
indução, local e sistêmica, de um inibidor de quimiotripsina em folhas de plantas de
tomate, quando predadas pelo besouro da batata do Colorado (Leptinotarsa
decemlineata) ou quando mecanicamente injuriadas. A partir deste estudo pioneiro,
diversos trabalhos foram direcionados para o estudo de proteínas de defesa,
expressas, em plantas, frente não somente ao estresse de ferimento, como também
a resposta a pragas e patógenos. Porém havia uma importante questão a ser
respondida desde 1972: Qual seria o agente de sinalização responsável pela
transmissão do sinal, iniciado com o ferimento, para o restante da planta, uma vez
que a indução de defesa não se limitava ao sítio da lesão? Tal agente, antes mesmo
de ser descoberto, já era nomeado com a sigla PIIF, que quando traduzida significa
“Fator Indutor de Inibidores de Proteinases” (Ryan, 1974). Nesta mesma época,
também se constatou que plantas de tomate não eram as únicas que induziam a
expressão de inibidores de proteinases (IPs), quando estimuladas com extrato
vegetal contendo PIIF. Extratos foliares de mais 37 espécies de 20 famílias vegetais
também apresentaram esta mesma propriedade indutora de defesa (McFarland &
Ryan, 1974). Na época os autores atribuíram estes dados a possível presença de
substâncias similares a PIIF (PIIF-like) nos extratos protéicos testados, uma vez que
ainda não se conhecia nada a respeito da natureza dos eliciadores de indução
possivelmente presentes nestes extratos brutos, apesar de sugeridos vários
candidatos tais como o ácido jasmônico (AJ) e seu éster, metil jasmonato (MeJa).
Por algum tempo vários estudos concentraram-se na tentativa de identificação de tal
fator. Alguns trabalhos sugeriram que agentes de natureza não química, tais como

2
diferenças de potencial elétrico de membranas (Wildon et al. 1992; Volkov & Haack,
1995) e de potenciais hidráulicos (Tyree, 1970) fossem responsáveis pela
sinalização e aumento da expressão de proteínas de defesa. No entanto, a
descoberta de um componente protéico funcionando como ativador e sinalizador de
resposta sistêmica de defesa em plantas foi uma extraordinária conquista de Dr.
Clarence Ryan e seu grupo de pesquisa, da Washington State University (Pullman-
EUA). A descoberta de uma molécula capaz de aumentar intensamente a síntese de
inibidores de proteinases, denominada sistemina, um dos primeiros hormônios
peptídicos vegetais descobertos, foi publicada por Pearce et al. (1991). Roberts
apresentou, em sua revisão (figura 1), um modelo de ativação de defesa no qual o
estimulo de ferimento promoveria um sinal elétrico, que atingiria em áreas vizinhas,
interferindo na ação dos hormônios de defesa ABA, AJ e sistemina. Além disso, essa
via também é inibida por ácido salicílico (aspirina) e auxina. Posteriormente foi visto
que sistemina poderia ser conduzida pelo floema após a lesão (Ryan et al., 1995),
induzindo assim a síntese de inibidores de proteinases em células de tecidos
distantes do ponto lesionado, mediando respostas de defesa sistêmica em plantas
(Ryan, 2000). Confirmavam-se, pois, as suspeitas dos pesquisadores sobre a
existência de um fator molecular responsável pela transmissão de um sinal, iniciado
por injúria mecânica, anteriormente descrito como PIIF.
No início dos anos 90, o PIIF foi identificado e caracterizado como sendo um
hormônio peptídico, encontrado por Pearce et al. (1991) em folhas de tomate, e
assim denominado sistemina. A sistemina foi o primeiro sinalizador peptídico
descrito em plantas, e estudos posteriores revelaram que este peptídeo origina-se
de um precursor de 200 aminoácidos denominado pró-sistemina (Ryan et al., 1995),
que é clivada quando o tecido vegetal é ferido, liberando o peptídeo de 18
aminoácidos (sistemina), via tecido condutor (floema) para todo o organismo vegetal.
Os dados obtidos, desde então, mostram que a indução de proteínas de
defesa é um processo complexo que segue um determinado cronograma. Estudos
feitos com tomate mostram que o ferimento inicial induz a liberação de sistemina
logo nos primeiros minutos (Ryan, 2000). Este hormônio migra via floema, até ligar-
se a seu respectivo receptor em células vegetais de várias regiões da planta,
induzindo a síntese de metil-jasmonato que, por sua vez, induz a produção de
proteínas relacionadas com defesa, tais como inibidores de proteinases (Farmer &
Ryan, 1992). Uma longa cascata de sinalização iniciada pela sistemina torna este

3
hormônio muito potente, bastando lembrar que foram necessários 30 kg de folha
para o isolamento de 1g do material, mas que apenas essa quantidade deste
peptídeo foi capaz de induzir a produção máxima de inibidores de proteinases
serínicas em 4 x 104 plantas de tomate (Pearce et al., 1991).
Após a descoberta da sistemina não houve grandes avanços no sentido de
estudar outras formas protéicas relacionadas com indução de proteínas de defesa.
Em 2000, Pearce conseguiu isolar, de plantas de tabaco, dois peptídeos capazes de
induzir a expressão de inibidores de proteinases serínicas nesta espécie vegetal.
Curiosamente, assim como a sistemina não é capaz de induzir expressão de IPs em
plantas de tabaco, os hormônios “sistemina-like” de tabaco, denominados
tobsistemina 1 e tobsistemina 2, não são capazes de exercer qualquer atividade
indutora em plantas de tomate, mesmo sendo estas duas espécies vegetais
pertencentes à mesma família (Solanaceae). Além desse fato, as seqüências
peptídicas destes hormônios não apresentam qualquer homologia com a sistemina
de tomate, o que evidencia que a sistemina de tomate não é um hormônio universal
de indução de defesa, e que diferentes espécies podem ter suas defesas induzidas
por diferentes agentes eliciadores (Pearce et al., 2000). Tal sugestão foi confirmada
com a descoberta e caracterização de hormônios peptídicos, em diferentes espécies
da família das solanáceas, tais como batata, petúnia e beladona, apresentando as
mesmas diferenças estruturais e semelhanças de atividade biológica mencionadas
entre sisteminas de tomate e tabaco. Com base nestes dados foi criada uma nova
classificação para estes hormônios denominada família das sisteminas que é
constituída por peptídeos de baixa massa molecular, entre 15 e 20 resíduos de
aminoácidos, e um elevado efeito de indução de resposta a ferimento, podendo
atuar em concentrações abaixo da ordem de nanomolar (Pearce et al., 2006).
Este trabalho, bem como outros do mesmo grupo, nos quais experimentos
indicativos também haviam revelado a participação do fitohormônio ácido jasmônico,
e seu éster volátil, metil-jasmonato (MeJa), reproduziram-se de forma similar em
várias espécies vegetais (Ryan, 2000), sendo um modelo para os estudos deste tipo.
Neste novo modelo de indução de defesa, o ferimento causado em uma área da
planta causaria a liberação de sistemina, que migraria e se ligaria ao seu respectivo
receptor ativando uma cascata de sinalização, iniciada por despolarização de
membrana, e cujos eventos subseqüentes acreditam-se sejam: efluxo de cálcio do
4
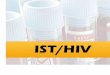
vacúolo (V); ativação de enzima MAPK (Proteína quinase ativadora de mitose);
ativação de uma fosfolipase A2 (PLA2) pela MAPK; liberação de ácido linoléico da
membrana plasmática por ação da PLA2; aumento da síntese de ácido jasmônico,
que atuaria diretamente na ativação de genes de defesa (figura 2).
Figura 1 – Indução sistêmica hipotética de genes de inibidores de proteinases do
tipo 2 (pin2) estimulada por um sinal elétrico de propagação (Adaptado de Roberts,
1992) AJ – Ácido Jasmônico, ABA - Ácido Abscísico.
Em 2003, foi publicado um trabalho (Lee & Howe, 2003) no qual um elegante
experimento foi feito com enxertos entre plantas do tipo selvagem e o mutante spr1 -
recessivo ao gene que codifica o receptor para sistemina SR160. Na figura 3
mostram-se os seguintes detalhamentos experimentais: (a) Um porta-enxerto do tipo
selvagem gera um sinal, quando ferido, que é percebido por um enxerto do mesmo
tipo inserido, resultando em um alto nível de transcrição de IPs. (b) A transcrição de
IPs em porta-enxertos spr1 é forte, porém drasticamente reduzida na parte
enxertada do mesmo tipo. (c) No híbrido de enxerto em que o tipo selvagem é
inserido sobre um porta-enxerto do tipo spr1, o porta-enxerto responde ao ferimento
como em (b). Entretanto, níveis de transcrição de IPs são fortemente reduzidos na
inserção do tipo selvagem. Porém como visto em (d), em uma condição recíproca
(spr1 - tipo selvagem), IPs acumulam-se em ambas as partes, indicando que
Ferim ento
Fus icoc ina
AspirinaC aule
Auxina [D ]
p in2
[S is tem ina]AJ
[A B A]
C otilédone P rim eira folhaverdadeira
S ina l e létrico propagado
Ferim ento
Fus icoc ina
AspirinaC aule
A ux ina
p in2
S is tem inaAJ
AB A
C otilédone P rim eira folhaverdadeira
S ina l e létrico propagado

5
incisões de enxertos spr1 podem perceber um sinal sistêmico iniciado por ferimento
em um porta-enxerto do tipo selvagem (adaptado de Stratmann, 2003).
Figura 2 – Modelo do efeito sistêmico da liberação e transporte de sistemina sobre a
ativação de genes de defesa (Adaptado de Ryan, 2000). O aumento de Ca++
citossólico e a ativação de MAPK são considerados como intermediários do
processo. MAPK - Proteína quinase ativadora de mitose; PLA2 - Fosfolipase A – 2;
PDA – Ácido fitodienóico; R – Receptor; V – Vacúolo.
O resultado do trabalho descrito acima revelou que a indução de proteínas de
defesa (IPs) ocorre mesmo em mutantes de plantas incapazes de reconhecer
sistemina (spr1). Por outro lado, em mutantes spr2 (recessivos para síntese de ácido
jasmônico) e Jai1 (recessivo para o reconhecimento do sinal AJ) a defesa sistêmica
foi totalmente extinta (figura 4). Tais dados mostraram que, diferentemente do que
Prosistemina
Liberação
Sistemina
Transporte
Etileno
Acido Jasmônico
Acido Linoleico
Ativação de Genes de Defesa
Pró-sistemina
Liberação
Sistemina
Transporte
Etileno
Ácido Jasmônico
Ácido Linolênico
Ativação de Genes de Defesa

6
se cogitava, o ácido jasmônico, e não a sistemina, teria um papel de indutor
sistêmico da resposta de defesa vegetal (Lee & Howe, 2003; Stratmann, 2003).
Neste novo paradigma, novamente estímulos elétricos e hidráulicos liberados
durante o ferimento poderiam estar afetando a produção de AJ, mas de uma forma
ainda desconhecida, deixando margem a novas pesquisas sobre o assunto
(Stratmann, 2003). Neste mesmo trabalho constata-se que a sistemina tem um papel
central na sinalização sistêmica, onde agiria como um estimulador local da produção
de AJ, controlando sua produção por um sistema de “feedback” positivo, e atuando,
assim, como um hormônio parácrino na defesa vegetal. Porém a sua ação mais
importante estaria em estimular o envio de AJ para outras partes não feridas. A falta
de resposta ao hormônio sistemina em mutantes spr1 não afeta a indução local de
inibidores de proteinases (Stratmann, 2003) (figura 3). Isto porque a produção local
de AJ pode também estar sendo aumentada por outros estímulos oriundos do
ferimento, em um processo ainda não bem caracterizado.
Consta na literatura que uma diversidade de hormônios agem juntamente
com a sistemina no estímulo das enzimas da via de síntese de ácido jasmônico em
plantas de tomate, espécie vegetal que representa o modelo inicial deste tipo de
estudo (Wasternack et al., 2006) (figura 4). Sabe-se hoje que enzimas responsáveis
pela via de síntese do ácido jasmônico tais como aleno óxido sintase (AOS), aleno
óxido ciclase (AOC) e lipoxigenase (LOX) têm suas expressões e atividades
intrinsecamente ligadas à ativação da via de defesa, representada pela liberação de
sistemina e aumento de vários hormônios, que antes estavam apenas relacionados
com outras vias (figura 4). Ocorreria assim o aumento da síntese de ácido jasmônico
e de seus precursores, quase que exclusivamente em elementos de tubo crivado e
células companheiras dos tecidos condutores do floema (Hause et al., 2003).
No final da primeira década do século XXI ainda restam muitas questões
acerca da ocorrência mútua e interferência entre diferentes vias de sinalização
reguladas por diferentes hormônios. Porém já é estabelecido que o processo de
sinalização de defesa acionado por ferimento não é algo linear, mas sim ocorrendo
em uma complexa rede hormonal (Wasternack et al., 2006) (figura 5). Outra questão
importante é reconsiderada a partir dos dados recém obtidos: AJ é considerado o
candidato atualmente mais evidente para sinalizador sistêmico de defesa, mas seria
este o único? Embora estudos feitos com mutantes acx1 (recessivos para enzima
Acil–CoA oxidase, responsável pela oxidação de ácidos graxos relacionados com a

7
via do AJ) mostrem uma forte evidência de que AJ ou compostos relacionados
tenham tal papel, uma dúvida permanece: Como um sinal produzido no floema pode
ativar a expressão de IPs que ocorrem exclusivamente em células do mesófilo? Por
outro lado, ainda não é conhecido o mecanismo pelo qual a sistemina, presente no
mesófilo, atua na ativação da síntese de AJ no floema (Wasternack et al., 2006).
Outro dado marcante do modelo atual é que a concentração de AJ em folhas não
feridas é muito inferior à encontrada em folhas feridas de tomate (Laudert & Weiler,
1998; Bowles, 1998; Rojo et al. 1999), a ponto de somente este hormônio não
exercer todo efeito indutor predito, o que leva a hipótese da existência de elementos
auxiliares ainda desconhecidos nesta rota de sinalização. Peróxido de hidrogênio
tem sido considerado como uma importante molécula sinalizadora; é um bom
candidato, pois apresenta alta mobilidade entre tecidos migrando inclusive via
apoplasto, e tem forte potencial de ativação sobre a via de formação de AJ (Orozco-
Cárdena et al. 2001). No entanto, são necessários estudos adicionais a respeito do
envio deste sinal em processo de resposta sistêmica a ferimento.
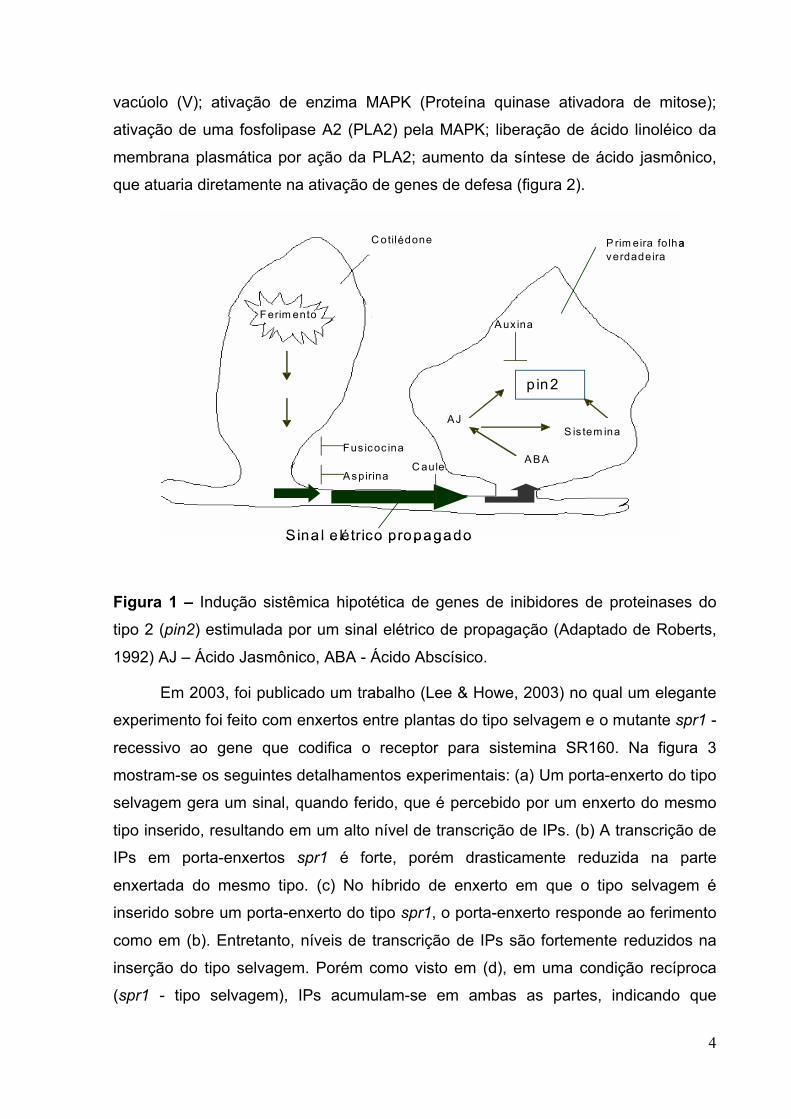
Figura 3 – Teste do potencial do mutante Spr1 para resposta sistêmica a ferimento.
Plantas foram enxertadas em quatro combinações distintas. Linhas horizontais
paralelas representam o ferimento e retângulos no caule representam a junção do
Tipo selvagem
Tipo selvagem
Tipo selvagem
Tipo selvagem
Porta-enxerto
enxerto
8
enxerto. Folhas verde claro e verde escuro indicam níveis baixos e altos de acúmulo
de transcrição de Ips, respectivamente (adaptado de Stratmann, 2003).
Figura 4 – Modelo de sinalização sistêmica de defesa vegetal, evidenciando a ação
do AJ como sinalizador sistêmico de defesa. Dados de trabalhos com mutantes de
plantas insensíveis à ação de sistemina (spr1 e spr2), e ao efeito do AJ (jai1) foram
usados (adaptado de Stratmann, 2003).
9
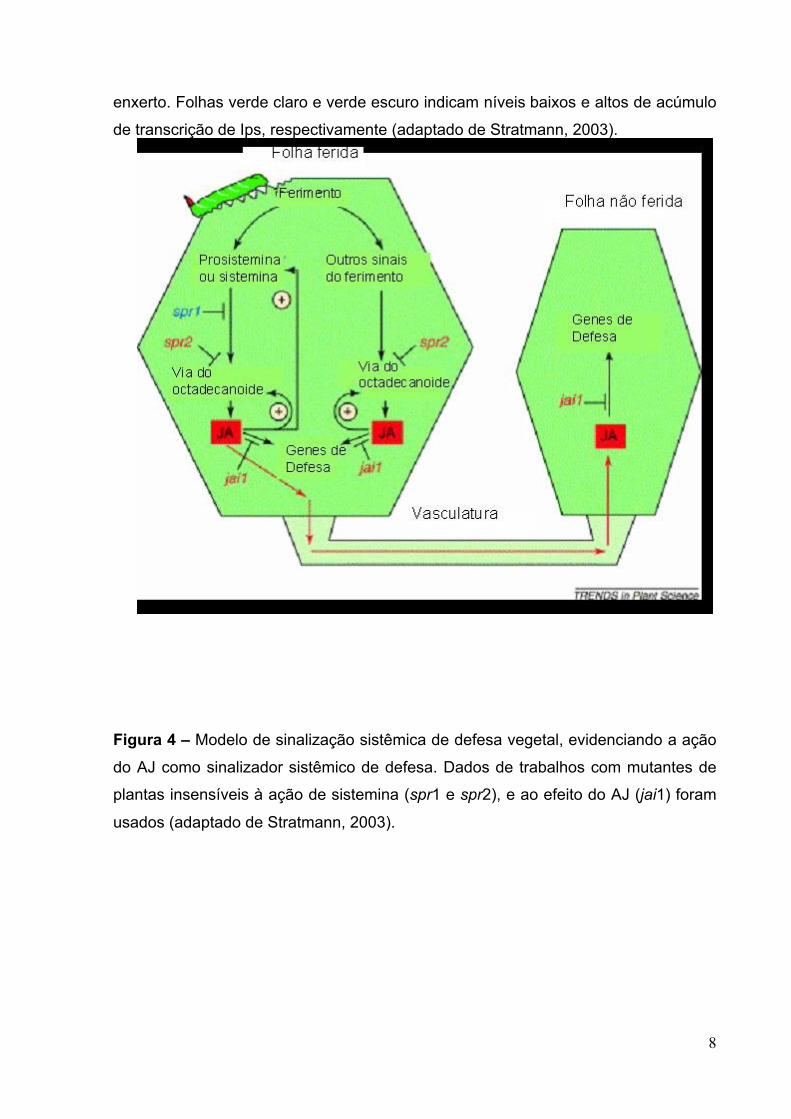
Figura 5 – Interferência de diversos hormônios vegetais e moléculas sinalizadoras
(Ácido salicílico - SA; Ácido abscísico – ABA; Etileno; Peróxido de hidrogênio –
H2O2; Óxido nítrico - NO; Oligogalacturonídeos – OGAs; e F Sintase AOC - Aleno
Óxido Ciclase; α-LeA - Ácido Linolênico, OPR3 - Ácido 12-Oxofitodienóico Redutase
– 3 (Wasternack et al., 2006).
1.1.2 – O papel do ácido jasmônico e seus derivados
O primeiro trabalho demonstrando o efeito do ácido jasmônico como um
hormônio de ação vegetal foi publicado em 1971. Neste, o ácido jasmônico isolado
de culturas do fungo Lasiodiplodia theobromae mostrou efeito inibitório sobre o
desenvolvimento de calos de plantas (Adridge et al., 1971). Este hormônio vegetal é
sintetizado a partir do ácido linoléico, um ácido graxo tri-insaturado, através de uma
rota metabólica, conhecida como rota octadecanóide. Seu derivado mais conhecido
é o metil-jasmonato, um éster volátil capaz de induzir defesa em outras plantas
próximas a de origem (Farmer & Ryan, 1990). Foi descrito como um componente de
M esófilo
S istêm ica
Prosistem ina
S istem ina
VA
S
CULATURA
IP2
G enes de defesa
IP2
G enes de defesa
M esófilo
S istêm ica
Prosistem ina
S istem ina
VA
S
CULATURA
IP2
G enes de defesa
IP2
G enes de defesa

10
óleos essenciais de Jasminum grandiflorum (jasmim), o que deu origem ao seu
nome. A primeira evidência mostrando a participação do AJ como hormônio de
defesa foi descrita em 1990, quando seu éster foi capaz de induzir a expressão de
inibidores de proteinases em espécies de fabáceas e solanáceas, em uma câmara
fechada (Farmer & Ryan, 1990). Embora tenha sido mostrado que metil-jasmonato
era capaz de induzir a expressão de IP em plantas de tomate dentro de câmaras
fechadas, em ambiente aberto, este efeito somente ocorre quando plantas de tomate
estão próximas a plantas da espécie Artemisia tridentata, que produz grande
quantidade de MeJa. A produção e liberação desse hormônio em plantas de tomate
feridas não ocorre em quantidade suficiente para estimular a indução de IPs em
plantas vizinhas (De Bruxelles & Roberts, 2001). Por outro lado, a importância do AJ
como molécula sinalizadora foi vista desde 1992 quando o seu acúmulo foi capaz de
modular genes relacionados com defesa estimulados por ferimento (Greelman et al.,
1992). Os níveis de AJ atingem o pico de formação logo nas primeiras horas após o
ferimento (Ryan, 2000). Muitos destinos metabólicos foram identificados para o AJ
após esse momento, além da formação de MeJa; AJ pode sofrer sulfatação,
glicosilação, metilação, além de reação com etileno (Wasternack et al., 2006). Tais
destinos refletem a importância da degradação deste, como um controle da
sinalização, evitando-se um estímulo excessivamente prolongado. Porém,
conjugados de jasmonatos com os aminoácidos isoleucina e leucina apresentaram
um poder de ativação de genes de defesa em plantas de Phaseolus lunatus cinco
vezes maior que ácido jasmônico não conjugado (Krumm et al., 1995). Tais dados
reforçam a importância da busca por novas moléculas sinalizadoras de defesa.
1.1.3 – O estresse de ferimento mecânico como ativador de defesa e suas
limitações em comparação ao estímulo da herbivoria
A técnica de exercer um ferimento mecânico em plantas como uma mímica à
ação de insetos mastigadores é estratégia que muito contribuiu para o estudo do
processo de sinalização de defesa em plantas (Ryan, 2000). No entanto, é evidente
que há distintas diferenças entre estes dois estresses de indução; primeiramente, o
ferimento mecânico não consegue imitar a liberação de FACs (aminoácidos
conjugados a ácidos graxos), comumente liberados pela saliva de insetos
mastigadores e que são importantes indutores da via octadecanóide (Turlings et al.,

11
1995). Além disso, um dos dados mais conflitantes entre estes dois estresses vem
de experimentos com plantas de Arabidopsis thaliana (Reymond et al., 2000), as
quais foram tratadas com três diferentes estresses: hídrico, ferimento e mastigação
por inseto. Neste trabalho a expressão de genes de defesa foi analisada por técnica
de “microarrays” e foram constatadas as ativações de mais de 150 genes
relacionados com ferimento mecânico e mastigação de insetos, sendo mais da
metade destes genes também dependentes da via do AJ. Porém, neste mesmo
trabalho foi visto que apenas um gene, dos 150 examinados, foi induzido por ação
de insetos e não por ferimento mecânico. Um outro intrigante trabalho feito com
plantas de tomate revelou que a expressão de uma RNase era aumentada frente ao
tratamento por ferimento mecânico, mas não sob o efeito da aplicação de sistemina
ou AJ. Esta enzima teve a sua expressão rapidamente aumentada nas primeiras dez
horas, somente a nível local do ferimento. Os estudos feitos a respeito de sua função
revelaram que ela estava envolvida no processo de senescência, sugerindo-se que
uma nova via de sinalização provocada por ferimento, independente de sistemina e
AJ, poderia estar atuando (Groβ et al., 2004). Tais informações são muito úteis, por
validarem o estimulo ferimento mecânico como tratamento para o estudo da defesa
vegetal em comparação à ação de insetos. Ao mesmo tempo, evidencia a clara
limitação de um tratamento, quando comparado aos demais mencionados acima.
1.2 – Detecção da indução de proteínas de defesa como indicação de resposta
a estresses
Plantas submetidas a estresses, como durante o ato de herbivoria,
desenvolveram estratégias de proteção específicas. Dentre estas se destacam:
formação de inibidores de proteinases; aumento da produção de compostos tóxicos
do metabolismo secundário, tais como nicotina ou outros fenóis ou alcalóides; e
emissão de compostos voláteis que atraem predadores dos insetos herbívoros
(Wasternack et al., 2006). Dentre muitas proteínas de defesa sistemicamente
induzidas em resposta a herbivoria, foram tomados, como parâmetros de análise, o
aumento de inibidores de proteases e o aumento da expressão de polifenol
oxidases, ambas selecionadas como proteínas indicadoras de resposta de defesa
induzida, no presente trabalho.

12
1.2.1- Polifenol oxidases – PFOs
Polifenol oxidases são enzimas que catalisam a oxidação dependente de
oxigênio de monofenóis (ex. L-tirosinases ou cresolases) ou orto-difenóis (ex.
catecolases, lacases) a ο-diquinonas (Steffens et al., 1994). Em plantas elas estão
fracamente aderidas à face lumenal da membrana interna ou no lúmen de tilacóides
(Steffens et al. 1994) e são encontradas em múltiplas isoformas. Atuam produzindo
ο-diquinonas, que são compostos altamente reativos que se ligam a aminoácidos
livres ou em resíduos de aminoácidos de moléculas protéicas, tornando-as
insolúveis e com baixo potencial nutritivo (Matheis & Whitaker, 1984; Garcia-
Carmona et al., 1988). Muitas polifenol oxidases de plantas apresentam latência,
identificada por um aumento de atividade quando em pH 6,0, ou pela ativação por
tratamento com proteases (tripsina ou protease K) ou concentrações milimolares de
SDS (Sellés-Marchart et al., 2006).
Vários genes de PFOs de plantas codificam proteínas maduras entre 52-65
kDa, e um peptídeo de endereçamento de 8 a 12 kDa, que são responsáveis pelo
transporte da enzima para dentro do lúmen dos tilacóides de cloroplastos (Demeke &
Morris, 2002). Embora PFOs já tenham sido encontradas em plantas há mais de um
século (Bertrand, 1896), um conhecimento completo sobre seu papel nesses
organismos ainda não foi alcançado. Funções distintas foram atribuídas a PFOs, tais
como: pigmentação e enegrecimento do tecido vegetal (Boonsiri et al., 2007;
Valentines et al., 2005); ação na reação de Mehler, uma vez que atua transferindo
elétrons de um substrato fenólico para o oxigênio, em uma cadeia transportadora de
elétrons, aumentando o consumo deste durante a foto-respiração (Thipyapong et al.,
2004); e proteção de plantas contra pragas e patógenos (Constabel & Ryan, 1998).
Este amplo espectro de idéias sobre os papéis de PFOs reflete o parcial
entendimento das funções de PFOs em plantas (Mayer, 2006). A habilidade natural
de PFOs de oxidarem fenóis ou polifenóis reflete seu mecanismo de ação como
proteína de defesa, já que as �-diquinonas originadas têm maior capacidade de
ligação e precipitação de proteínas e são mais tóxicas aos predadores de plantas do
que os fenóis originais (Felton et al., 1992). Já foi relatada a presença de PFOs
latentes em diversas espécies de plantas (Sellés-Marchart et al., 2007; Laveda et al.,
2002; Chazarra et al., 1997; Moore & Flurkey, 1988). Tal estado latente de algumas
PFOs reforça mais ainda a sugestão de um papel de defesa atuando posteriormente

13
ao ato de herbivoria em plantas, ao contrário do que é visto em formas ativas de
PFOs presentes em frutos maduros ou tecidos vegetais pigmentados. Por esse
raciocínio, PFOs latentes poderiam ser alvo de processos acionados pela injúria de
tecidos, através dos quais seriam ativadas por proteólise, conduzindo a ação da
enzima sobre os seus substratos, e evidenciando uma função protetora somente
durante o ataque de predadores (Mazzafera & Robinson, 2000). Dado que reforça
esta idéia é o fato de que PFOs se encontram em tilacóides, enquanto seus
substratos fenólicos estão presentes no vacúolo, de forma que estas moléculas
entram em contato somente após o ato de ferimento mecânico ou mastigação dos
tecidos por um herbívoro (Whitaker & Lee, 1995).
PFO é a proteína de defesa mais intensamente induzida em plantas de
tomate submetidas a ferimento ou a tratamento com vapor de metil-jasmonato. De
modo ainda mais geral, o aumento dos níveis de PFOs em plantas, causado por
ferimento ou por borrifações de MeJa, parece ser uma estratégia de defesa induzida
muito difundida entre diversas espécies vegetais, tal como foi visto em plantas de
tomate, tabaco, soja, ervilha dentre outras (Constabel & Ryan, 1998). No entanto a
resposta de indução desta enzima frente a estas duas formas de ativação é, na
maioria das vezes, diferente (Constabel et al., 1995; Constabel & Ryan, 1998).
A própria habilidade de PFOs de oxidarem fenóis ou polifenóis reflete seu
mecanismo de ação como uma proteína de defesa, já que as quinonas originadas
ligam-se e precipitam proteínas, sendo mais tóxicas ao predador que os fenóis
inicialmente encontrados nos tecidos da planta. Estas moléculas derivadas de fenóis
conferem características antinutricionais aos tecidos vegetais, inibindo, por exemplo,
a digestão de insetos roedores (Doan et al., 2004; Felton et al., 1992). O aumento da
resistência da parede celular é um outro aspecto da ação defensiva de PFOs, uma
vez que estas enzimas colaboram com a formação de lignina (Yuan et al., 2002;
Aquino-bolaño & Mercado-Silva, 2004). PFOs também têm um marcante papel de
defesa contra patógenos, sendo a ação destas enzimas uma das principais
respostas de defesa à infecção fúngica (Mohammadi & Kazemi, 2002), através de
sua participação no processo de síntese de melanina, a qual forma complexos
insolúveis com os fenóis oxidados pelas próprias PFOs, gerando uma barreira
resistente a entrada e espalhamento de patógenos. Inclusive a própria resposta de
hipersensibilidade e formação de área pigmentada durante o processo de resistência
sistêmica adquirida (SAR), é decorrente da indução e ação de PFOs. Aliando-se às

14
informações mencionadas acima, é certo destacar que a hipótese do papel
defensivo desta classe de proteínas foi marcantemente reforçada pela confirmação
da regulação da sua expressão, através da via octadecanóide, estando vinculada à
síntese de AJ e do sinal de defesa a ferimento, a sistemina (Pearce et al. 1991;
Constabel et al., 1995; Constabel & Ryan., 1998; Ryan, 2000). Além destes dados, o
seu efetivo efeito de proteção contra a lagarta militar (Spodoptera litura) em plantas
de tomate geneticamente modificadas para superexpressão de PFO, reforça essa
idéia (Mahanil et al., 2008).
1.2.2 - Inibidores de proteinases serínicas
Os inibidores de proteinases são potentes inativadores de enzimas
proteolíticas, mas também podem ativar mecanismos de “feedback” causando
superexpressão de proteases digestivas e perda de apetite em insetos herbívoros
(Ryan, 1990). No reino vegetal, a maioria dos inibidores de proteinases estudados é
das famílias Leguminosae, Solanaceae e Gramineae (Richardson, 1991).
Inibidores de proteinases serínicas são proteínas de defesa há muito tempo
estudadas. Este grupo representa o mais estudado entre todos os outros inibidores
de proteases conhecidos e se divide em dois grandes subgrupos: inibidores do tipo
Kunitz e do tipo Bowman–Birk. O marco deste estudo foi o trabalho de Mickel &
Standish, de 1947, no qual foi visto que larvas de alguns insetos não eram capazes
de se desenvolver em produtos derivados de soja. Posteriormente, essas proteínas
de defesa também representaram uma referência inicial nos estudos de indução de
defesa vegetal por ferimentos (Green & Ryan, 1972), onde foi vista a indução local e
sistêmica destes inibidores em plantas de tomate quando predadas pelo besouro
colorado da batata. Nesta mesma ocasião também foi constatado que o ferimento
mecânico nas folhas do tomateiro causava um efeito similar de indução. O primeiro
trabalho de transformação de plantas com inibidores de proteinases serínicas foi
desenvolvido em 1987 (Hilder et al., 1987), onde o gene codificante destes
inibidores, originário de sementes de Vigna unguiculata, foi transferido para plantas
de tabaco, conferindo, a estas, resistência contra diversas pragas, incluindo
lepidópteros dos gêneros Heliothis e Spodoptera, coleópteros dos gêneros
Diabrotica e Anthonomus e ortópteros do gênero Locust (Lawrence & Koundal,
2002).

15
Embora inibidores de proteinases serínicas tenham grande potencial para
inibir proteinases serínicas, o besouro da batata do Colorado é capaz de induzir a
expressão de enzimas digestivas do tipo cisteínicas, quando alimentado com altos
níveis desses inibidores (Michaud et al., 1995). Fenômenos de adaptação similares
a esse põem em cheque a utilização destas proteínas e de seu potencial inseticida
em programas de melhoramento vegetal via engenharia genética.
1.2.3 - Inibidores de proteinases cisteínicas - cistatinas
O termo cistatina foi atribuído pela primeira vez por Barrett & Kirschke (1981)
a uma proteína de clara de ovo de galinha, dando origem ao nome dessa classe de
inibidores (“cisto = ovo”). Tais inibidores apresentam a propriedade de inibir
proteinases cisteínicas. Outras proteínas que apresentavam propriedades biológicas
e estruturais semelhantes a esta cistatina foram agrupadas (Barrett et al., 1987) e
desta forma, a superfamília das cistatinas foi dividida em três famílias constituintes:
família 1, com cistatinas A e B; família 2, com cistatinas C, D, E/M, F, S, SN e AS; e
família 3, com os cininogênios de alto e baixo peso molecular. Embora tenham sido
inicialmente encontradas em animais, também foram encontradas em plantas, não
muito tempo depois (Abe et al., 1987). As cistatinas de todos os grupos são
caracterizadas por apresentarem em comum, uma resíduo de glicina na porção N-
terminal, um “motif” constituído da seqüência QxVxG, que interage diretamente com
o sitio catalítico da enzima, e um resíduo de triptofano na porção carboxi-terminal
(Machleidt et al., 1989). Porém, todas as fitocistatinas diferem das demais por uma
seqüência específica de cistatinas de plantas (LVI]-[AGT]-[RKE]-[FY]-[AS]-[VI]-x-
[EDQV]-[HYFQ]-N), presente na porção amino-terminal e com estrutura em α-hélice
prevista (Margis et al., 1998).
Muitos coleópteros possuem proteinases cisteínicas como principais enzimas
proteolíticas em seu trato digestivo. Outros, como o besouro da batata do Colorado,
podem induzir proteinases cisteínicas em resposta a uma alimentação rica em
inibidores de proteinases serínicas (Jongsma et al., 1995; Cloutier et al., 2000; Zhu-
Salzman et al., 2003). A produção de plantas transgênicas com genes de cistatinas
vem sendo uma boa alternativa para proteção contra insetos que utilizam
proteinases cisteínicas em sua digestão protéica (Michaud et al., 1993). Entretanto, o
fenômeno de adaptação, anteriormente citado para o caso de inibidores de

16
proteinases serínicas, põe em cheque a utilização destas proteínas e de seu
potencial inseticida em programas de melhoramento vegetal via engenharia
genética. Por outro lado, não há evidências de que cistatinas atuem contra
mamíferos, sendo este um ponto positivo para sua utilização (Lawrence & Koundal,
2002). Além disso, trabalhos feitos com plantas transformadas com essas proteínas,
evidenciaram significativa proteção contra nematóides e patógenos (Haq et al.,
2004). Isto porque proteinases cisteínicas estão presentes em grande quantidade
nestes organismos. Também é constatado, na literatura, o aumento da resistência
aos vírus TEV e TVY de plantas de tabaco e batata, respectivamente, transformadas
com genes exógenos de orizacistatinas (Gutierrez-Campos et al., 1999). No entanto
fatos como os casos de adaptação acima destacados preocupam os cientistas
envolvidos em pesquisas direcionadas a estratégia de transformação de plantas
(Fernandes et al., 1993; Flores et al., 2001).
Além desta função direta de defesa contra pragas e patógenos, há trabalhos
recentes apontando para o papel desta proteína como atuante no processo de morte
celular programada em plantas (Belenghi et al., 2003). Segundo estes, cistatinas de
massa molecular em torno de 10 kDa estariam, juntamente com proteinases
cisteínicas, fazendo parte de uma cascata de sinalização ainda não bem elucidada,
na qual as cistatinas estariam reprimindo o desencadeamento de morte celular
programada em células vegetais, via liberação de óxido nítrico (NO). Estes trabalhos
citados apontam uma retomada ao estudo de transformações com estas proteínas,
porém com um enfoque não tão limitado, quando comparado ao de uma década
atrás, onde cistatinas eram vistas somente como biopesticidas.
1.3 – O esxudado de floema como fonte de hormônios vegetais
Em 1991, Pearce et al. idealizaram um experimento com o intuito de
investigar a natureza do processo de indução sistêmica em plantas de tomates,
bem como identificar a molécula sinalizadora responsável pelas respostas
sistêmicas destas plantas. Submergiram folhas de tomate destacadas da planta
íntegra em solução contendo diferentes frações protéicas obtidas de plantas de
tomate, e observaram a indução (ou não) de inibidores de proteinases serínicas

17
nos extratos protéicos obtidos destas folhas. A indução destes inibidores serviu
como constatação para a presença de um agente eliciador de defesa, na fração
testada como positiva, e que mais tarde seria caracterizada como sendo o
hormônio peptídico sistemina, que atualmente tem sua via de transporte muito
bem elucidada, sendo um hormônio conduzido pelo floema.
A via do floema é conhecida há muito tempo como a principal via de
transporte de nutrientes elaborados (fotoassimilatos) pelo organismo vegetal.
Recentes trabalhos mostram que o floema não está limitado apenas ao transporte
de sacarose; aminoácidos estão entre seus principais componentes sólidos,
dentre outros nutrientes. Existe uma verdadeira rede de comunicação molecular
fluindo dentro do floema, ativando e desativando diversos processos metabólicos
inclusive aqueles relacionados com defesa (Ryan, 2000). Sistemina, por exemplo,
flui do ponto de lesão para todo o organismo vegetal pela via do floema. A
principal vantagem do fluxo de moléculas sinalizadoras por esta via é a alta
velocidade de transporte, podendo chegar a cem centímetros por hora. Outra
característica importante desse fluxo é que este ocorre nos dois sentidos pelo
caule, o que permite uma melhor comunicação entre folhas (Buchanan et al.,
2000), diferentemente do fluxo do xilema que só ocorre no sentido raiz folha.
Apesar da sistemina e outros hormônios peptídicos serem transferidos via floema,
o conteúdo deste é muito pobre em componentes de constituição protéica e
peptídica, podendo chegar a valores de 0,2 a 2 mg / L, na maioria das espécies
vegetais, exceto em cucurbitáceas, onde os valores atingem de 40 a 100 mg / L
(Buchanan et al., 2000). Uma característica marcante da composição do floema é
a sua alta variação de componentes protéicos, frente à ação de um ferimento,
como visto na figura 5, onde é possível perceber que o perfil protéico do
esxudado coletado diretamente de ferimento impingido ao tecido vegetal é muito
diferente daquele coletado via extração por estilete de um inseto sugador. Tal
diferença é atribuída ao fato que insetos sugadores, afídios de um modo geral,
não disparam nenhuma cascata de sinalização de defesa uma vez que seu
aparelho de sucção não atravessa nenhuma célula do mesófilo, apenas
perfurando e sugando células do floema (Buchanan et al., 2000). Apesar das
baixas concentrações mencionadas, o esxudado de floema pode ser um
excelente meio para o isolamento de hormônios vegetais por apresentar uma
baixa variedade de compostos, de toda natureza, quando comparados a extratos

18
vegetais brutos, cuja diversidade protéica ultrapassa facilmente a faixa de
dezenas de milhares de proteínas diferentes. Em floema estas se restringem a
poucas centenas. Lembrando-se que a principal via de formação e condução de
AJ são elementos de tubo crivado do floema, a análise deste material traria uma
grande vantagem de se obter um meio enriquecido com as moléculas hormonais
eliciadoras de defesa sistêmica, facilitando assim o isolamento e, principalmente,
a quantificação destas.
Figura 6 - Análise eletroforética mostrando variação de proteínas, obtidas de
floema, provocada por diferentes métodos de extração: “wound exudate” -
obtenção por corte com lâmina no caule antes da coleta. “Stylet exudate” -
esxudado produzido pela drenagem por um estilete de um afídio (adaptado de
Buchanan et al., 2000). B – a composição da seiva do floema de mamona
(Ricinus communis), coletado como esxudado de cortes no floema (Taiz & Zeiger,
2004).
B Componentes Concentração mg/mL
Açucares 80,0 - 106Aminoácidos 5,2
Ácidos Orgânicos 2,0 – 3,2Proteínas 1,45 – 2,2Potássio 2,3 –4,4Cloreto 0,355 - 0,675Fosfato 0,350 - 0,550
Magnésio 0,109 –0,122
A

19
1.4- A espécie Vigna unguiculata no estudo de defesa vegetal
O presente projeto visa o estudo da ação do ferimento como indutor de defesa
vegetal em plantas de feijão-de-corda, investigando tanto a presença e
características de proteínas induzidas por tal estímulo, como a presença de um
agente eliciador de resposta sistêmica de defesa, fazendo uso da estratégia
descrita por Pearce et al. (1991). Tendo em vista que PFOs são enzimas
expressas em resposta a ferimento por diversas espécies de plantas (Constabel &
Ryan, 1998), esta enzima foi selecionada como marcadora de indução por
ferimento em plantas de feijão-de-corda, a exemplo dos inibidores de proteinases
serínicas induzidos em tomates.
A planta modelo do nosso estudo, o feijão-de-corda (Vigna unguiculata) é uma
leguminosa originária da África, e consiste em uma espécie cultivada de grande
importância nutricional para muitos povos africanos, asiáticos e para o nordeste
brasileiro (May et al., 1982). Esta espécie de fabácea já foi extensivamente estudada
no que se refere às suas formas constitutivas de defesa. Dentre estas estão os
inibidores de proteinases serínicas em frutos em formação (Carasco & Xavier-Filho,
1981), cistatinas (Fernandes et al., 1993), quitinases e ,1-3-glucanases (Gomes et
al., 1995; Ye et al., 2000), em sementes. Recentemente foi detectada em sementes
a presença constitutiva de proteínas de defesa pouco estudadas, tais como
defensinas e LTPs (Carvalho et al., 2001), bem como a forma mais particular e
característica de defesa constitutiva dessa espécie - formas variantes de vicilinas.
Vicilinas são a principal forma de proteínas de reserva de sementes de feijão-de-
corda (Xavier-Filho et al., 1989) e uma série de trabalhos do grupo (Gomes et al.,
1998; Sales et al.,1992; Firmino et al., 1996; Macedo et al.,1995; Fernandes &
Xavier-Filho, 1993), demonstram que tais proteínas possuem função dual, de
estocagem de aminoácidos e defesa contra predadores, assim como o são os
inibidores de proteases serínicas (Haq et al., 2004).
A participação de proteínas induzidas nos processos de defesa em V.
unguiculata, no entanto, é ainda muito pouco relatada. Simões-Araújo et al. (2002),
trabalhando com uma biblioteca de cDNAs de nódulos de V. unguiculata
termicamente estressados, encontraram transcritos homólogos a uma proteína
induzida por ferimento em Medicago sativa. Porém nenhum papel de defesa foi

20
associado a este produto de expressão gênica. Outro trabalho recente mostrou
indução de cistatinas, proteínas que são classicamente relacionadas com proteção
vegetal, frente à ação de estresse hídrico (Diop et al., 2004).
2 – OBJETIVOS GERAIS
- Investigar a indução de proteínas de defesa, em níveis local e sistêmico, em
plantas de feijão-de-corda, quando submetidas a ferimento.
- Investigar a variação da composição do esxudado do floema de plantas de
feijão-de-corda após o tratamento por lesão mecânica.
2.1 – Objetivos específicos
- Detectar e caracterizar polifenol oxidases induzidas, local e sistemicamente,
em plantas jovens de feijão-de-corda (V. unguiculata) mecanicamente injuriadas ao
nível de unifoliatos;
- Detectar e caracterizar a indução de cistatinas, local e sistemicamente, em
plantas jovens de feijão-de-corda (V. unguiculata) mecanicamente injuriadas ao nível
de unifoliatos;
- Investigar lesão mecânica, na prevenção de senescência decorrente de morte
celular programada;
- Comparar o potencial indutor de exsudatos de floema de plantas de feijão-de-
corda feridas e não feridas ao nível de unifoliato sobre a indução de polifenol
oxidases.
- Analisar por espectrometria de massas a presença de AJ no esxudado do
floema de plantas submetidas ao estresse de ferimento.

21
3 – MATERIAIS
3.1 – Material biológico
3.1.1 – Sementes
As sementes de feijão-de-corda (Vigna unguiculata L. Walp.), variedade
EPACE-10, foram fornecidas pelo Departamento de Fitotecnia do Centro de Ciências
Agrárias, Fortaleza – Ceará e posteriormente propagadas na cidade de Campos dos
Goytacazes.
3.2 – Reagentes
Ácido clorogênico (Sigma) L-Dopa (Sigma)
Azida sódica (Vetec) 4-metil-catecol (Sigma)
Azocaseína (Sigma) Papaína (Merck)
Acrilamida (Sigma) PMSF (Sigma)
Bis-acrilamida (Sigma) PVPP (Sigma)
-mercaptoetanol – (Sigma) Persulfato de amônio (Merck)
Catecol (Sigma) Pirogalol (Sigma)
Catequina (Sigma) Resorcinol (Vetec)
Coomassie Brilhante Blue G (Amersham) TEMED (Sigma)
Coomassie Brilhante Blue R (Sigma) L-Tirosina (Sigma)
DAB (Amersham) Triton X 100 – (Sigma)
DTT (sigma) Tropolona (Fluka)
EDTA (Vetec) Uréia (Vetec)
Gelatina (Sigma) L-Dopa (Sigma)
Outros reagentes utilizados foram de grau analítico e obtidos comercialmente.
3.3 - Equipamentos
Balança analítica AX 200 (Shimadzu)
Centrífuga refrigerada 5417 R (Eppendorf)

22
Espectrofotômetro UVMini 1240 (Shumadzu)
Ultracentrífuga CP 85 (Hitachi)
Mini-sistema de eletroforese vertical (Bio-Rad)
Espectrômetro de massas Q-TOF (Micromass)
Sistema de eletrotransferência CE (Amersham)

23
4 – METODOLOGIA
4.1 – Detecção, isolamento e caracterização de polifenol oxidases (PFOs)
induzidas por ferimento
4.1.1 – Condições de cultivo de plantas
Substrato orgânico para o cultivo de plantas foi tratado com aquecimento a
100 OC por 3 minutos para fazer assepsia, e usado no cultivo das plantas de V.
unguiculata. O solo foi posto em potes plásticos de 120 mL previamente limpos com
solução alcoólica 70 %. As sementes de Vigna unguiculata usadas eram novas, não
ultrapassando os 6 meses de idade após a colheita, e livres de contaminações
aparentes por bruquídeos ou patógenos. Estas tiveram suas superfícies previamente
desinfetadas com solução de hipoclorito de sódio 0,2%, por 1 minuto. Depois da
assepsia, as sementes foram plantadas, em número de 6 por pote contendo
substrato orgânico, a uma profundidade de 1 cm, devidamente adubado com
fertilizante líquido completo “Ouro verde” (6 gotas por pote), para supressão de
carências de macro e microelementos. Os potes foram levados para estufa onde as
plantas desenvolveram-se em condições de iluminação de 100 mM de fótons / cm2 /
s, seguindo um fotoperíodo de 18 horas de luz e 6 horas de escuro, ocorrendo uma
variação de temperatura de 25 a 30 ºC. A hidratação do substrato foi feita no início
do plantio, e regas diárias foram feitas mantendo-o úmido. Plantas foram submetidas
ao ensaio de indução por ferimento ao atingirem a idade de 7 dias após emergência
do substrato, estágio em que podiam ser visualizados o primeiro par de unifoliatos.
4.1.2 – Tratamento por ferimento e obtenção de extratos protéicos
O tratamento por ferimento foi feito usando-se um hemostato para fazer
injúrias, circulando somente um dos unifoliatos (figura 7). Após o tratamento, plantas
foram submetidas à luz com intensidade igual a 100 mM de fótons / cm2 / s e
unifoliatos foram coletados nos tempos de 0, 3, 8, 24, 48 e 72 horas após o
ferimento. Além de unifoliatos feridos (F), foram também coletados, nestes tempos,
unifoliatos laterais aos feridos (LF) e de plantas não feridas, considerados unifoliatos
controle (C). Os unifoliatos foram macerados em gral resfriado em gelo (0°C), com

24
tampão fosfato de sódio 10 mM, EDTA 5 mM, pH 6, na proporção de 1:2, obtendo-se
um extrato bruto que foi centrifugado a 20.000 x g, a 4oC, por 20 minutos, seguindo
um protocolo adaptado de Shin et al. (1997), obtendo-se assim a primeira fração
com proteínas solúveis. O sedimento da primeira centrifugação foi ressuspenso e
homogeneizado em tampão com a mesma composição descrita na primeira
extração, diferindo somente na presença de 1 % de Triton X 100. O material foi
centrifugado (20.000 x g, a 4oC, por 20 minutos) e o sobrenadante definido como
fração insolúvel protéica.
Figura 7 - Esquema de tratamento por lesão mecânica e coleta das regiões de
plantas de feijão-de-corda. F – unifoliato ferido; LF – unifoliato lateral ao ferido. Nas
plantas controles a região LF é considerada igual à região F, já que nenhum
ferimento foi provocado à planta.
4.1.3 – Quantificação da atividade polifenol oxidásica
As frações solúveis e insolúveis obtidas no item 4.1.2 foram dosadas em
relação à presença de polifenol oxidases (PFO), seguindo-se um protocolo adaptado
de Shin et al. (1997), sendo tomada por ensaio alíquotas de 300 �l para plantas
controles, com o tempo de 0 horas após ferimento, e 50 �l para os extratos das
demais plantas. O teste de atividade foi feito usando-se como substrato 4-metil-
catecol, em meio de ensaio apresentando a seguinte composição: tampão fosfato de
sódio 100 mM pH 6,0, catecol 5 mM, em um volume total de ensaio de 1000 �l. A

25
leitura espectrofotométrica foi feita, medindo-se a variação de absorbância em um
comprimento de onda de 410 nm, durante os primeiros minutos de reação. Todas as
medidas foram feitas em temperatura ambiente, sendo estabelecido como 1 UE o
aumento de absorbância em leitura de atividade correspondente a 0,01 por minuto.
4.1.4 – Quantificação protéica
Os ensaios de quantificação protéica dos extratos vegetais obtidos foram
feitos segundo metodologia descrita por Bradford (1976), usando-se albumina
bovina como padrão, fazendo-se leitura espectrofotométrica a 595 nm.
4.1.5 – Eletroforese em gel de poliacrilamida sob condições semi-
desnaturantes (SDS-PAGE)
As amostras de proteínas extraídas conforme descrito no item 4.1 foram
analisadas por eletroforese em gel de poliacrilamida, segundo método de Laemmli
(1970) com diversas modificações, a fim de proporcionar uma condição semi-
desnaturante, na qual uréia e -mercaptoetanol não foram usados, além do fato das
amostras não serem fervidas. Extratos protéicos de plantas controle e feridas, 48
horas após a lesão do unifoliato, foram equilibrados na proporção de 1:2 em tampão
de amostra, submetidos a uma corrida de duas horas e meia em um sistema
descontínuo de gel de poliacrilamida, aplicando-se 40 �g de proteína por poço, sob
uma voltagem constante de 200 V a uma temperatura de 8 oC. Após o processo, o
gel foi corado com Coomassie Brilliant Blue R e então descorado com uma solução
de metanol:ácido acético:água (40:10:50).

26
4.1.6 – Ensaio de atividade enzimática em gel
O ensaio de atividade em gel foi feito usando-se um protocolo adaptado de
Shin et al. (1997), onde foi adicionado tampão de amostra (Tris-HCl 0,5 M, pH 6,8,
SDS 2 % e azul de bromofenol 1 %) na proporção de 1:2 (v/v) aos extratos obtidos
no item 5.1.2. Foi feita então uma SDS-PAGE conforme descrito no item 4.1.5 e o
gel obtido foi submetido à revelação para PFO, por submersão em tampão fosfato
100 mM, catecol 10 mM, até o aparecimento de bandas de atividade, o que ocorreu
no período de 3 a 5 minutos de incubação. O efeito inibitório de tropolona, um
inibidor especifico para PFOs, também foi testado. Para isso uma incubação prévia
com tropolona a 1 mM foi feita por 10 minutos, antes da visualização da atividade de
PFO sobre catecol.
4.1.7 – Isolamento de PFO induzida por ferimento
O extrato bruto protéico da fração solúvel, obtida de unifoliatos feridos após
48 horas desde a lesão, conforme o item 4.1.2, foi submetido à precipitação com
sulfato de amônio até uma concentração de saturação de 30 %. O material foi então
guardado a 4 oC por uma noite a fim de otimizar a precipitação. Após essa etapa o
material foi centrifugado a 10.000 g por 30 minutos a 4 0C. O sedimento foi
descartado e o sobrenadante foi re-precipitado com sulfato de amônio até uma
saturação de 80%, seguindo um protocolo estabelecido por Shi et al. (2002). A
fração 30-80 % obtida foi dessalinisada através de sua suspensão em solução
gelada de etanol 30% (0 oC) por 30 minutos, e centrifugada a 20.000 g por 10
minutos a 4 oC. O sedimento obtido foi seco a 8 oC por 24 horas e ressuspendido em
tampão fosfato 100 mM pH 7,0, Triton X-100 0,1 % , também empregado como
tampão de equilíbrio durante a fase de purificação por cromatografia de troca iônica.
O extrato clarificado foi estão submetido à separação cromatográfica em
coluna de troca iônica em DEAE-celulose. A cromatografia de troca iônica foi
efetuada usando-se concentrações crescentes de NaCl (0,0; 0,1; 0,2 e 0,5) no
tampão de eluição (tampão fosfato 100 mM pH 7,0 Triton X-100 0,1 %), sendo o
volume de 50 mL usado em cada concentração de sal no tampão de eluição. A
atividade das frações foi identificada por ensaio enzimático conforme descrito em

27
4.1.6. O pico de maior atividade obtido foi dialisado exaustivamente contra água,
liofilizado e guardado a -20 oC. Tal material foi definido como fração F-0,2, por ser
eluído na concentração de 0,2 M de NaCl, tendo elevado nível de atividade polifenol
oxidásica e tendo sido, portanto, usado posteriormente nas etapas de caracterização
enzimática.
4.1.8 – Parâmetros cinéticos da fração F-0,2
Todos os parâmetros cinéticos foram determinados usando-se o volume de
20 �l da fração F-0,2, que corresponde a valor de atividade de 15 UE, sendo
estabelecido como 1 UE o aumento de absorbância em leitura de atividade
correspondente a 0,01 por minuto.
4.1.8.1 - Ativação com SDS e estabilidade
O efeito ativador de SDS sobre F-0,2 foi testado usando-se 5 mM de 4-metil-
catecol como substrato em tampão fosfato de sódio 100 mM, pH 6,0 e uma variação
de concentrações de SDS de 0 a 140 mM, sendo o volume total do ensaio de 1000
�l. As demais condições para medição de atividade foram executados conforme o
protocolo descrito o item 4.1.3.
4.1.8.2 – Medição de estabilidade térmica
A energia de ativação para desnaturação de F-0,2 foi determinada por
medição da atividade enzimática, em banho circular, sobre um intervalo de
tratamentos térmicos variando de 30 a 100 oC, com um intervalo de 10 oC de
aumento em cada tratamento. O tempo de incubação de todos os tratamentos foi de
5 minutos. Após incubação, o material foi rapidamente resfriado em gelo. As
incubações foram efetuadas na presença e ausência de SDS 10 mM. As
concentrações de substrato e SDS usadas no ensaio foram as mesmas descritas no
item 4.1.3.

28
4.1.8.3 - Determinação de pH ótimo de atividade
O pH ótimo de atividade para F-0,2 foi determinado usando tampão citrato-
fosfato 100mM para um intervalo de medições de pH de 3 a 6,5 e tampão fosfato
100 mM para o intervalo de pH de 7 a 9. As demais condições de ensaio usadas
foram as mesmas descritas no item 4.1.3.
4.1.8.4. – Especificidade a substratos e determinação de Km e Vmax
A determinação da especificidade da fração F-0,2 a diferentes substratos foi
feita usando-se diferentes substratos comumente empregados para PFOs. Sendo
estes: 4-metil-catecol (M-catecol), pirogalol, catecol, ácido clorogênico e L-
dioxifenilalanina (L-DOPA). Todas as determinações foram efetuadas usando 100
mM de tampão fosfato pH 6,0, SDS 5 mM, sendo cada substrato ensaiado na
concentração de 5 mM. Foram tiradas medições no intervalo de tempo de cada
minuto até 5 minutos de reação.
Os valores de Km e Vmax foram calculados a partir do gráfico duplo recíproco
de Lineweaver & Burke, com os dados obtidos dos ensaios feitos com os substratos
catecol e 4-M-catecol usando-se concentrações crescentes de 1, 2, 4, 6, 8, 12 e 16
mM.
4.1.8.5 – Ensaio de inibição de atividade polifenol oxidásica
Quatro inibidores foram usados para determinação do perfil de inibição
enzimática da fração F-0,2: EDTA, resorcinol, azida sódica e tropolona. As
concentrações de inibidores usadas em ensaio foram 0,1; 1 e 10 mM, exceto para
tropolona (0, 01; 0,1 e 1 mM). As concentrações de substrato e SDS usadas no
ensaio, bem como outros parâmetros, foram os mesmos descritos no item 4.1.3

29
4.2 – Detecção e caracterização de cistatinas induzidas por ferimento e
análises de parâmetros relacionadas
4.2.1 – Condições de cultivo de plantas
As mesmas empregadas no item 4.1.1, com algumas modificações; as plantas
desenvolveram-se em condições de iluminação de valores iguais ou superiores a
300 mM de fótons / cm2 / s, seguindo um fotoperíodo de 12 horas de luz e 12 horas
de escuro.
4.2.2 – Tratamento por ferimento e obtenção de extrato protéico
O tratamento por ferimento foi feito usando-se um hemostato para fazer
injúrias em somente um dos unifoliatos das plantas teste plantas controle não foram
feridas. Após o tratamento plantas foram submetidas à luz com intensidade superior
ou igual a 300 mM de fótons / cm2 / s e unifoliatos foram coletados no tempo de 24
horas após ferimento. Logo após a coleta os unifoliatos foram macerados em
nitrogênio líquido e extraídos com tampão Tris-HCl 100 mM pH 6,8; PMSF 2 mM,
PVPP 5% na proporção de 1:1. O material foi centrifugado a 20.000 g por 20
minutos a 4 oC, o sobrenadante (extrato total) foi então fervido a 100 oC por 3
minutos. Após fervura o material foi centrifugado nas mesmas condições descritas
acima. O sobrenadante foi precipitado, adicionando-se um volume de solução de
TCA 50 % suficiente para se obter uma concentração final de 10 %. O material
permaneceu em gelo por 2 horas e foi centrifugado nas mesmas condições descritas
acima. Após centrifugação, o sobrenadante foi descartado e o sedimento re-
suspenso em metanol 100 %, para remoção do excesso de TCA. Após uma nova
centrifugação a 20.000g por 10 minutos e 4 oC, a amostra permaneceu secando por
16 horas a 8 oC. Após ter sido seco, o material foi re-extraído em tampão citrato
fosfato 100 mM pH 5,0, DTT 2 mM, e Triton X-100 0,1 %, o mesmo usado em ensaio
enzimático, na proporção de 60 μl por amostra, considerando-se uma quantidade
inicial de 1 g de tecido vegetal fresco. Após re-extração com vigorosa agitação do
material este foi centrifugado a 20000 por 30 minutos a 4 oC. O sobrenadante final foi
usado em medições de concentração protéica, segundo o item 4.1.4 e análise da
presença de cistatinas segundo o item 4.2.4.

30
4.2.3 – Determinação da curva de atividade enzimática
O ensaio de inibição de atividade cisteínica foi feito segundo Michaud et al.
(1987). Inicialmente foi estabelecida uma curva de atividade proteolítica com a
enzima modelo papaína, com o intuito de se estabelecer uma concentração ótima da
enzima no ensaio de inibição. A curva de atividade foi feita usando-se concentrações
crescentes de papaína na ordem de 1, 2, 4, 8 e 16 μg de papaína a partir de uma
solução estoque de 1 mg / mL, diluída em tampão citrato/fosfato de sódio
monobásico 100 mM pH 5,0, DTT 1,5 mM, e Triton X-100 0,1 %, mais a adição de
80 �l de azocaseína 1% até um volume total de incubação de 120 μl, o material foi
então incubado em temperatura ótima de reação para papaína (37 oC) por uma hora,
e a reação foi interrompida com a adição de 300 μl de TCA 10 %, aguardando por
meia hora para otimizar precipitação protéica. O material foi então centrifugado a
15000 g por 5 minutos à temperatura ambiente (25 oC). Posteriormente uma alíquota
de 350 μl do sobrenadante foi transferida para outro tubo e adicionados 350 μl de
solução NaOH 1 M. Paralelamente ao procedimento de medição de atividade, foram
feitos brancos de leitura nos quais foram repetidos todos os procedimentos, diferindo
apenas na ordem de aplicação da azocaseína 1%, sendo esta adicionada somente
após a adição do TCA 10 %. O produto de reação de hidrólise formado foi
mensurado por espectrofotometria a um comprimento de onda de 440 nm.
4.2.4 – Ensaio de inibição de atividade de proteinase cisteínica (dosagem de
cistatinas)
Uma concentração de papaína de 8 g por ensaio, foi escolhida uma vez que a
medição de seus produtos de hidrólise indicou valores de absorbância superiores a
0,45, O ensaio de inibição foi feito usando-se 10 μl de papaína mais 20 μl de
amostras tratadas para cistatinas, completando o volume com tampão de atividade
(citrato fosfato de sódio monobásico 100 mM pH 5,0, DTT 1,5 mM, e Triton X-100
0,1 %) até o valor de 40 μl. Todos os procedimentos posteriores foram feitos como
no item 4.2.3 (Michaud et al., 1987).

31
4.2.5 – Eletroforese em gel de poliacrilamida sob condições desnaturantes e
redutoras (SDS-PAGE)
As amostras proteínas extraídas conforme item 4.2.4 foram analisadas por
eletroforese em gel de poliacrilamida, sob condições desnaturantes, e redutoras
usando-se uréia na 8 M e -mercaptoetanol 4% no tampão de amostra, com
aquecimento a 100 oC, adaptado do método de Laemmli (1970). Foram aplicados 50
�g de proteína em cada poço, e a amostras foram submetidos a uma corrida de
duas horas e meia em um sistema descontínuo sob uma voltagem constante de 200
V a uma temperatura de 8 oC. Após o processo, o gel foi corado e descorado
conforme o item 4.1.5.
4.2.6 – Detecção de proteínas imunologicamente relacionadas a cistatinas por
Western Blotting
A metodologia de Western Blotting utilizada baseou-se em Towbin et al.
(1979). Os extratos protéicos obtidos segundo o item 4.2.2, foram submetidos a
eletroforese segundo item 4.2.5, as amostras foram corridas em duplicatas, sendo
uma gel corado para proteínas segundo item 4.2.5, e o outro transferido para
membrana de nitrocelulose com um tamanho de poro de 0,45 m. A transferência foi
feita num sistema semi-seco. O sanduíche consistia de 3 papéis de filtro embebidos
em tampão de transferência (Tris-HCl 25 mM, glicina 192 mM, metanol 20 % pH
8,3), a membrana de nitrocelulose, o gel contendo as proteínas a serem transferidas
e mais 6 papéis de filtro embebidos no mesmo tampão. A membrana e o gel foram
também previamente embebidos na mesma solução de transferência por 20
minutos. Quanto às condições de transferência foi empregada uma corrente elétrica
de 0,8 mA/cm2 de membrana durante aproximadamente 1 hora. Após transferência
foi feito coramento reversível com corante Ponceau para conferir a eficiência da
transferência.
Posteriormente, a membrana foi imersa em tampão bloqueador (gelatina 1%
em PBS [tampão fosfato 0,1 M NaCl 0,5 M pH 7,6, Tween 20 0,2 %]) por 1 hora. As
alíquotas dos anticorpos primários anti-cistatina de sementes de Vigna unguiculata
foi diluído 1:2000 no tampão bloqueador e a membrana foi imersa (16 horas) neste

32
tampão. Após este tempo lavou-se a membrana por 6 vezes com PBS, durante 10
minutos a cada lavagem, sendo a ultima feita com PBS sem a adição de Tween 20.
A membrana foi incubada por 3 horas com anticorpo anti-IgG de coelho, conjugado a
peroxidase, diluído 1:2000 em tampão bloqueador. Uma nova série de 6 lavagens de
10 minutos cada, com PBS, foi feita. A revelação para peroxidase foi feita
dissolvendo-se 5 mg de DAB (Diaminobenzidina) em uma mistura de 100 μl de
tampão Tris-HCl, 2 M pH 7,5, 300 l de imidazol 0,1 M e 4,9 mL de água destilada.
Após solubilização do DAB, 5 l de peróxido de hidrogênio 20 % foram adicionados
à mistura, sendo esta vertida sobre a membrana, cobrindo-a completamente. Após
rápida e leve agitação, o material permaneceu no escuro por 15 minutos, até
completar revelação que foi interrompida lavando-se a membrana com água
destilada em abundancia. A membrana foi seca e guardada em papel de filtro no
escuro.
4.2.7 – Ensaio enzimático para proteases cisteínicas totais
A medição da atividade total de proteinases cisteínicas foi feita a partir de
extratos brutos, obtidos de unifoliatos de plantas controle e feridas. Plantas foram
crescidas conforme o item 4.2.1, e plantas testes foram feridas em um dos
unifoliatos, e submetidas a 24 horas de luz constante; plantas controles foram
submetidas às mesmas condições, diferindo somente na ausência do tratamento por
ferimento. A extração protéica foi feita com tampão Tris-HCl 0,1 M, pH 6,8, PVPP
5%, EDTA 1 mM, na proporção de 1:1 (m / v), em gelo (0 oC), e procedeu-se por 2
horas. A suspensão foi centrifugada a 20000 x g por 10 min e o sobrenadante foi
usado para detecção de cistatinas. A análise de atividade foi feita usando-se 20 l
de extrato total de unifoliatos de plantas controle e testes, seguindo a mesma
metodologia descrita no item 4.2.3. Foram feitas 3 repetições para cada medição.
4.2.8 - Eletroforese semi-desnaturante em gel de poliacrilamida com gelatina
(SDS-PAGE-Gelatina)
Este gel foi preparado aplicando-se 1ml de gelatina (1%) ao volume da água,
deixando-o em uma concentração final de gelatina de 0,1% (para 10 ml de gel 12%).

33
Esta eletroforese correu utilizando-se uma voltagem constante de 150 V por 2 horas,
8 oC, aplicando-se 50 �g de proteína por poço, com as amostras de extrato total
obtidas conforme o item 4.2.2. Após a corrida, o gel foi lavado com Triton X-100
2,5% por 2 vezes, durante 30 minutos cada vez, e incubado em um banho-maria a
30°C em tampão citrato fosfato de sódio monobásico 100 mM pH 5,0, DTT 1,5 mM,
e Triton X-100 0,1 % durante 16 horas, com o intuído de detectar bandas de
atividade de proteinases cisteínicas. Após o processo, o gel foi corado e descorado
segundo item 4.2.5. A revelação das bandas de atividade consiste no aparecimento
de bandas brancas em um fundo azul.
4.2.9 – Ensaio de ativação de senescência
Plantas foram crescidas como em 4.2.1, até 5 dias de idade. Foram feridas
como em 4.3.7 e submetidas à iluminação constante, por 24 h. Visando induzir
senescência foliar, plantas foram deixadas no escuro por 5 dias a 28 oC, segundo
metodologia de Giridhar & Thimann (1984), com modificações.
4.3 – Análise do efeito eliciador de esxudado de floema
4.3.1 - Extração de esxudado de floema
A extração do esxudado de floema foi feita segundo Narváez-Vasquéz et al.
(1995). As plantas foram previamente feridas nos unifoliatos, e cortadas abaixo do
par de unifoliatos, no caule, com uma lâmina virgem bem afiada; foram colocadas
em microtubos com água destilada por 10 min, para eliminação de restos celulares e
foram transferidas para outros microtubos contendo 0,5 mL de uma solução tampão
de EDTA 1mM, Hepes 5 mM pH 7,0 (figura 8). Os experimentos foram conduzidos
no escuro, em câmara úmida (100% umidade relativa), a 25o C. Após 24 h de
esxudação, as plantas foram removidas e o esxudado de floema foi obtido. Também
foram extraídos esxudados de plantas não feridas no unifoliato para obtenção de um
controle. Os esxudados foram liofilizados, sendo então concentrados dez vezes.

34
4.3.2 – Teste de atividade indutora
Os esxudados de floema (4.3.1) foram submetidos a teste de sua capacidade
de induzirem a síntese de PFOs, segundo metodologia adaptada de Ryan (1974). O
teste consiste na inoculação de solução contendo as amostras testes através de
uma seringa hipodérmica de aplicação de insulina, no pecíolo de unifoliatos de
plantas de feijão-de-corda (figura 9). Após inoculação as plantas foram mantidos sob
luminosidade de 90 mM fótons/cm2/s por 24 horas. Posteriormente os unifoliatos
foram submetidos à extração protéica e determinação dos níveis de PFO conforme
os itens 4.1.2 e 4.1.3, respectivamente. O esquema abaixo (figura 9) resume o
procedimento para detecção do efeito eliciador de esxudado de floema.
Figura 8 - Esquema de coleta de esxudado de floema de plantas de feijão-de-corda.
A – Ferimento no unifoliato. B - caule cortado. C – Solução contendo tampão de
extração de esxudado (EDTA 1mM, Hepes 5 mM pH 7,0).
C B

35
Figura 9 – Procedimento para teste de atividade indutora de PFO dos exsudados de
floema de feijão-de-corda.
4.3.3 – Análise de esxudado de floema por espectrometria de massas
Uma alíquota de 10 μl de esxudado de floema concentrada por liofilização foi
transferida para um microtubo, sendo diluída em 100 μl de solução de acetonitrila
50 % / ácido fórmico 0,1%. A solução resultante foi então submetida a análise por
espectrometria de massas em um espectrômetro do tipo Q-Tof (“Electrospray”) da
marca Micromass. A solução de acetonitrila / ácido fórmico, pura, foi usada como
branco na análise do aparelho.
Extração protéica
Verificação da indução de PFO por ensaio de atividade enzimática.
Amostra sendo testada ou tampão no controle
24 h

36
5 – RESULTADOS
5.1 - Quantificação e caracterização da atividade polifenol oxidásica induzida
por ferimento
A análise da atividade polifenol oxidásica dos extratos protéicos obtidos
conforme item 5.1.2 foram feitas na presença e na ausência de SDS (tabela 1).
Frações solúveis de PFOs apresentaram grande diferença de atividade entre
extratos de plantas controles e plantas teste (unifoliatos feridos e laterais aos
feridos), somente quando ativadas com SDS. Nenhuma diferença é evidenciada
entre frações insolúveis de PFO, incubadas ou não com SDS 5 mM. As atividades
de todos os extratos PFOs solúveis representaram somente 5 % da atividade
polifenol oxidásica, total quando medida na ausência de SDS. Porém este
percentual atinge os 50 % somente em extratos de plantas feridas (unifoliato ferido e
lateral ao ferido) em incubação com SDS 5mM.
Os ensaios enzimáticos revelaram um significativo aumento da atividade de
PFOs solúveis no tempo de 24 horas após ferimento, em ambos unifoliatos feridos e
laterais aos feridos de plantas testes, quando comparados aos controles de 0 horas.
A indução máxima de atividade foi atingida 48 horas após o tratamento em ambos
unifoliatos feridos (13 vezes os valores de 0 horas) e laterais aos feridos (15 vezes).
No tempo de 72 horas ambos unifoliatos feridos e laterais ainda mantinham altos
níveis quando comparados aos unifoliatos de plantas controles de 0 horas (figura 8).
5.2 – Visualização de proteínas totais e polifenol oxidases em unifoliatos de
feijão-de-corda
A análise de extratos protéicos por SDS-PAGE semi-desnaturante foi feita
usando-se plantas controle e feridas no estágio de maior diferença dos níveis de
expressão de PFOs (48 horas, figura 10). Não foi observada nenhuma diferença
visível entre os perfis das amostras de unifoliatos controles, feridos e laterais aos
feridos (figura 11 I). Entretanto, quando o gel foi incubado com catecol para
detecção de atividade de PFO, foi facilmente observada a indução de bandas de

37
atividade, com massas de 58, 73 e superior a 200 kDa respectivamente. Estas
estavam presentes em amostras de unifoliatos feridos e laterais aos feridos de
plantas testes (figura 11 II - setas). Incubação do gel com tropolona (1mM), um
inibidor de PFOs, seguida de revelação com catecol conforme descrito acima,
mostrou inibição das bandas de atividade anteriormente observadas (figura 11 III).
5.3 – Purificação parcial de PFOs induzidas por ferimento
A purificação parcial e caracterização da PFOs induzidas por ferimento foi
processada a partir de unifoliatos feridos de 48 horas, tempo relativo à sua indução
máxima. O extrato clarificado (EC) obtido após precipitação com sulfato de amônio e
clarificação com etanol foi separado através de cromatografia de troca iônica em
DEAE-celulose. A figura 12 mostra o perfil de separação do EC onde se observa que
o maior pico de atividade (frações 34, 35 e 36) ficaram retidos na matriz. Embora
tenham sido detectados quatro picos de proteína, a maior atividade de PFO foi
associada ao terceiro, eluído com 0,2 M de NaCl. Devido ao elevado nível de
enriquecimento deste pico, este foi usado para caracterização de parâmetros
cinéticos e identificado como PFO de feijão-de-corda induzida por ferimento (F-0,2).
Uma atividade residual foi detectada em frações eluídas com 0,1 M de NaCl, as
quais não foram usadas neste tipo de análise.
O processo de purificação de PFOs de feijão-de-corda teve um rendimento de
purificação máximo de 11,18, como mostrado na tabela 2. A etapa de cromatografia
por DEAE-celulose foi a responsável pelo maior nível de purificação alcançado.
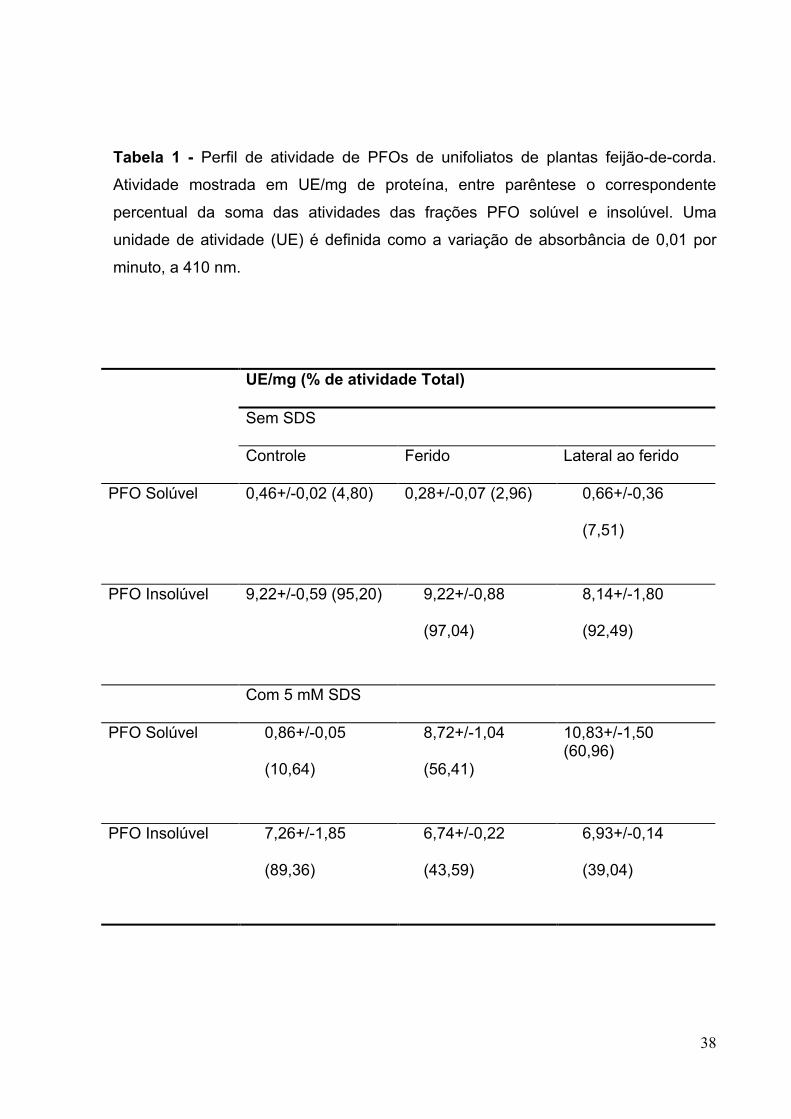
38
Tabela 1 - Perfil de atividade de PFOs de unifoliatos de plantas feijão-de-corda.
Atividade mostrada em UE/mg de proteína, entre parêntese o correspondente
percentual da soma das atividades das frações PFO solúvel e insolúvel. Uma
unidade de atividade (UE) é definida como a variação de absorbância de 0,01 por
minuto, a 410 nm.
UE/mg (% de atividade Total)
Sem SDS
Controle Ferido Lateral ao ferido
PFO Solúvel 0,46+/-0,02 (4,80) 0,28+/-0,07 (2,96) 0,66+/-0,36
(7,51)
PFO Insolúvel 9,22+/-0,59 (95,20) 9,22+/-0,88
(97,04)
8,14+/-1,80
(92,49)
Com 5 mM SDS
PFO Solúvel 0,86+/-0,05
(10,64)
8,72+/-1,04
(56,41)
10,83+/-1,50 (60,96)
PFO Insolúvel 7,26+/-1,85
(89,36)
6,74+/-0,22
(43,59)
6,93+/-0,14
(39,04)
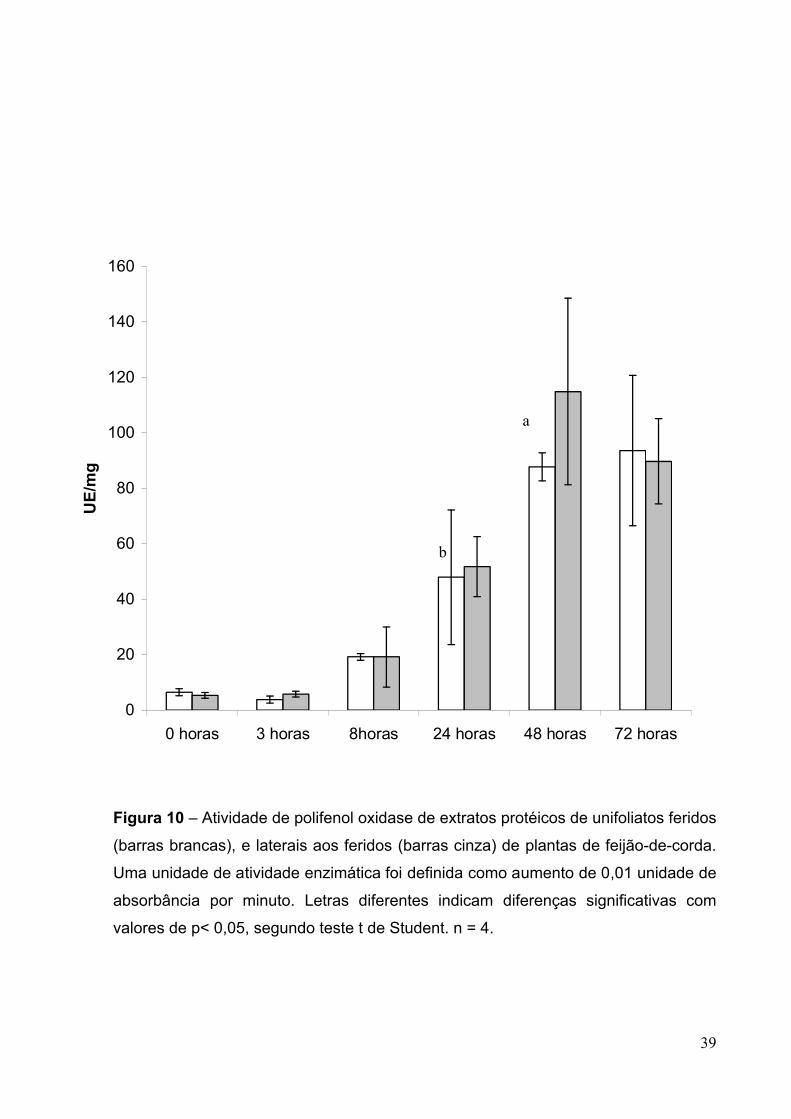
39
Figura 10 – Atividade de polifenol oxidase de extratos protéicos de unifoliatos feridos
(barras brancas), e laterais aos feridos (barras cinza) de plantas de feijão-de-corda.
Uma unidade de atividade enzimática foi definida como aumento de 0,01 unidade de
absorbância por minuto. Letras diferentes indicam diferenças significativas com
valores de p< 0,05, segundo teste t de Student. n = 4.
0
20
40
60
80
100
120
140
160
0 horas 3 horas 8horas 24 horas 48 horas 72 horas
UE
/mg
b
a
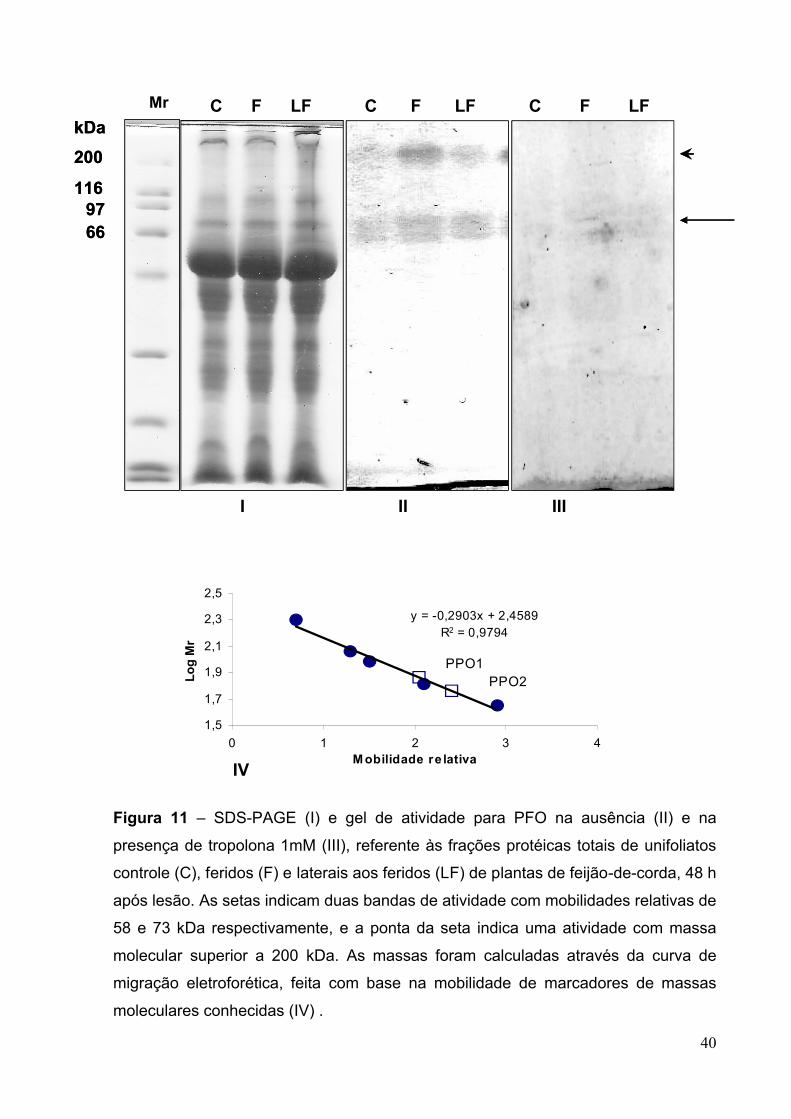
40
Figura 11 – SDS-PAGE (I) e gel de atividade para PFO na ausência (II) e na
presença de tropolona 1mM (III), referente às frações protéicas totais de unifoliatos
controle (C), feridos (F) e laterais aos feridos (LF) de plantas de feijão-de-corda, 48 h
após lesão. As setas indicam duas bandas de atividade com mobilidades relativas de
58 e 73 kDa respectivamente, e a ponta da seta indica uma atividade com massa
molecular superior a 200 kDa. As massas foram calculadas através da curva de
migração eletroforética, feita com base na mobilidade de marcadores de massas
moleculares conhecidas (IV) .
y = -0,2903x + 2,4589R2 = 0,9794
1,5
1,7
1,9
2,1
2,3
2,5
0 1 2 3 4M obilidade re lativa
Lo
g M
r
PPO2PPO1
Mr
kDa
200
1169766
m
C F LF C F LF C F LFkDa
200
1169766
IV
I II III
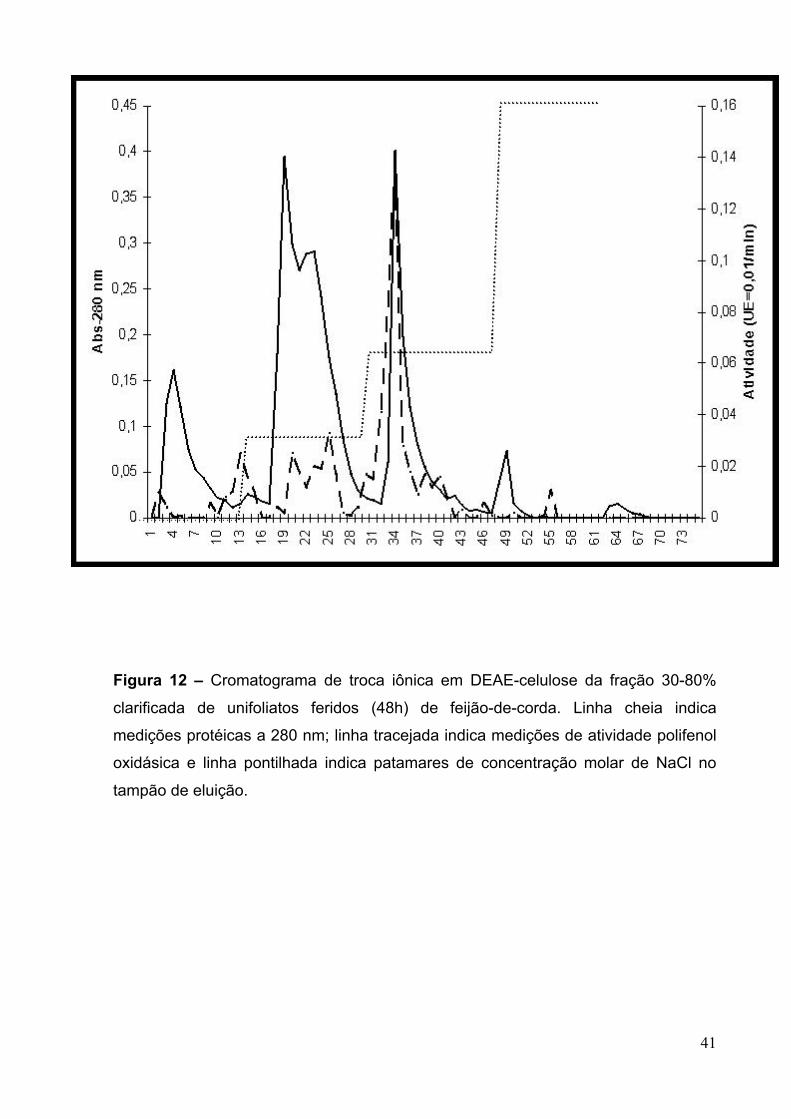
41
Figura 12 – Cromatograma de troca iônica em DEAE-celulose da fração 30-80%
clarificada de unifoliatos feridos (48h) de feijão-de-corda. Linha cheia indica
medições protéicas a 280 nm; linha tracejada indica medições de atividade polifenol
oxidásica e linha pontilhada indica patamares de concentração molar de NaCl no
tampão de eluição.
0,5 M
0,1 M
0,2 M

42
Tabela 2 – Etapas de purificação de F-0,2 de V. unguiculata. Uma unidade de
atividade (UE) é definida como a variação de absorbância de 0,01 por minuto, a 410
nm.
Etapas de
purificação
Volume
(mL)
Concentra
ção protéica
(mg/mL)
Proteína
total
(mg)
Atividade
total (UE)
Atividade
específica
(UE/mg)
Rendi
mento
Purific
a
ção
Extrato
bruto
110 4,9 539,22 56100 1,04 100 1
35-80%
(NH4)2SO4
7,5 1,57 11,80 4050 3,43 7,22 3,3
F-0,2 4 0,62 2,48 2880 11,63 5,4 11,18

43
5.4 – Ensaios de cinética enzimática da fração F-0,2
5.4.1 - Ativação e estabilidade com SDS.
A fração F-0,2 foi usada para a investigação da influência do SDS como
ativador de PFOs de feijão-de-corda. SDS é um importante agente ativador para
varias PFOs. Tal habilidade está relacionada com a interação deste detergente com
o centro de reação da enzima, sendo intrinsecamente dependente do pH (Jiménez &
García-Carmona, 1996). A polifenol oxidase aqui estudada é uma enzima latente é
foi otimamente ativada em uma concentração de SDS de 10 mM. Aproximadamente
60 % da atividade foi mantida mesmo em concentrações de SDS tão elevadas
quanto 140 mM (figura 13). Esta característica é responsável pela manutenção de
sua atividade em eletroforese (SDS-PAGE) sob condições semi-desnaturantes.
5.4.2 – Medição de estabilidade térmica
A determinação de estabilidade térmica, da fração F-0,2, foi feita na presença
e ausência de SDS (figura 14). A atividade na ausência de SDS foi altamente
conservada, mantendo-se em 84 % do inicial, mesmo após 5 minutos a 80 oC,
reduzindo-se drasticamente a 13,4 % da atividade inicial, em 90 oC de incubação. Na
presença de SDS 10 mM, a estabilidade térmica foi diminuída; a redução da
atividade enzimática foi de 31 %, quando observada a 70 oC, e a total perda de
atividade foi detectada a 80 oC.
5.4.3 - Determinação de pH ótimo de atividade de F- 0,2
Para determinação do pH ótimo de atividade. A atividade de F-0,2 foi medida
na presença de 10 mM de SDS e 5 mM de 4-metil-catecol, e foram usados dois tipos
de tampões: tampão citrato-fosfato 100 mM para variação de pH de 3,0 a 6,0 e
tampão fosfato para a variação de pH entre 7,0 e 9,0. A fração F-0,2 apresentou
nenhuma atividade em pH 3,0 ocorrendo uma significativo aumento de atividade até
os valores de pH 5,5 e tendo o valor 6,0 como o valor ótimo de pH para atividade

44
desta enzima. Entre os valores de 7,0 a 9,0, a atividade reduziu e se manteve em
torno de 60% do valor máximo (figura 15).
0
20
40
60
80
100
120
0 10 20 30 40 50 60 70 80 90 100 110 120 130 140 150
SDS (mM)
Ati
vid
ad
e r
ela
tiv
a (
%)
Figura 13 - Efeito do SDS sobre ativação da atividade de F-0,2. Ensaio foi feito com
4-metil-catecol 5 mM.
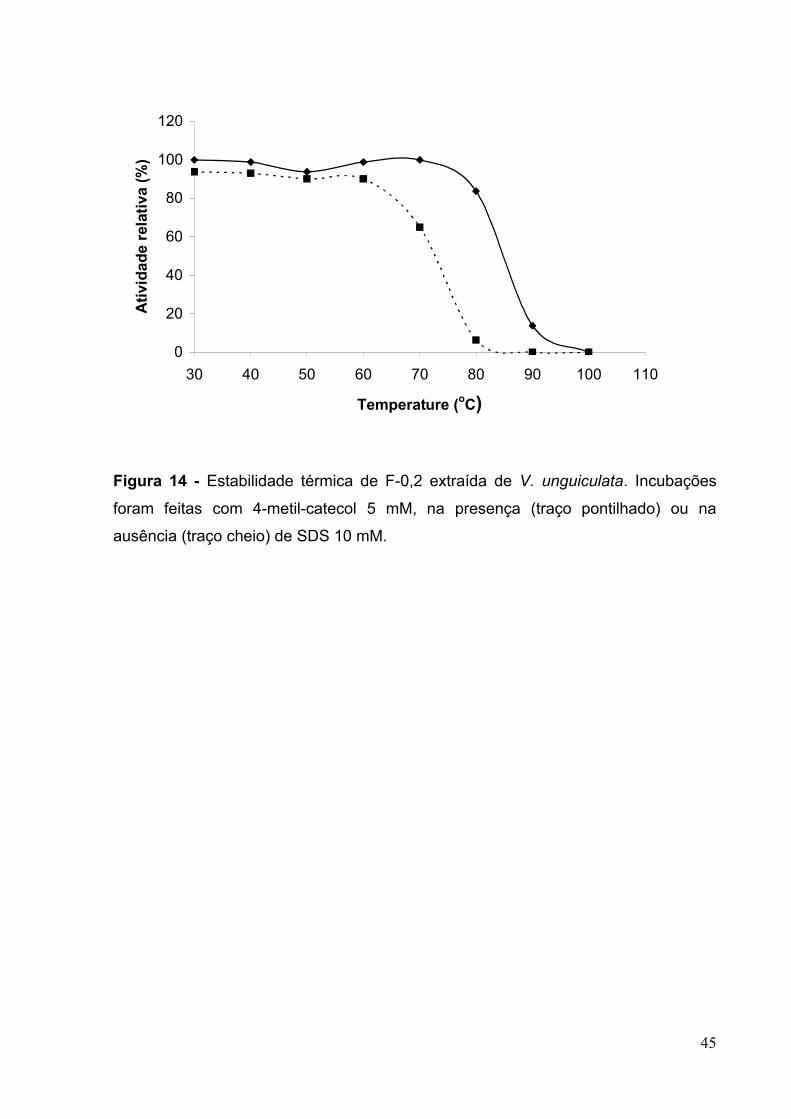
45
0
20
40
60
80
100
120
30 40 50 60 70 80 90 100 110
Temperature (oC)
Ati
vid
ade
rela
tiva
(%
)
Figura 14 - Estabilidade térmica de F-0,2 extraída de V. unguiculata. Incubações
foram feitas com 4-metil-catecol 5 mM, na presença (traço pontilhado) ou na
ausência (traço cheio) de SDS 10 mM.
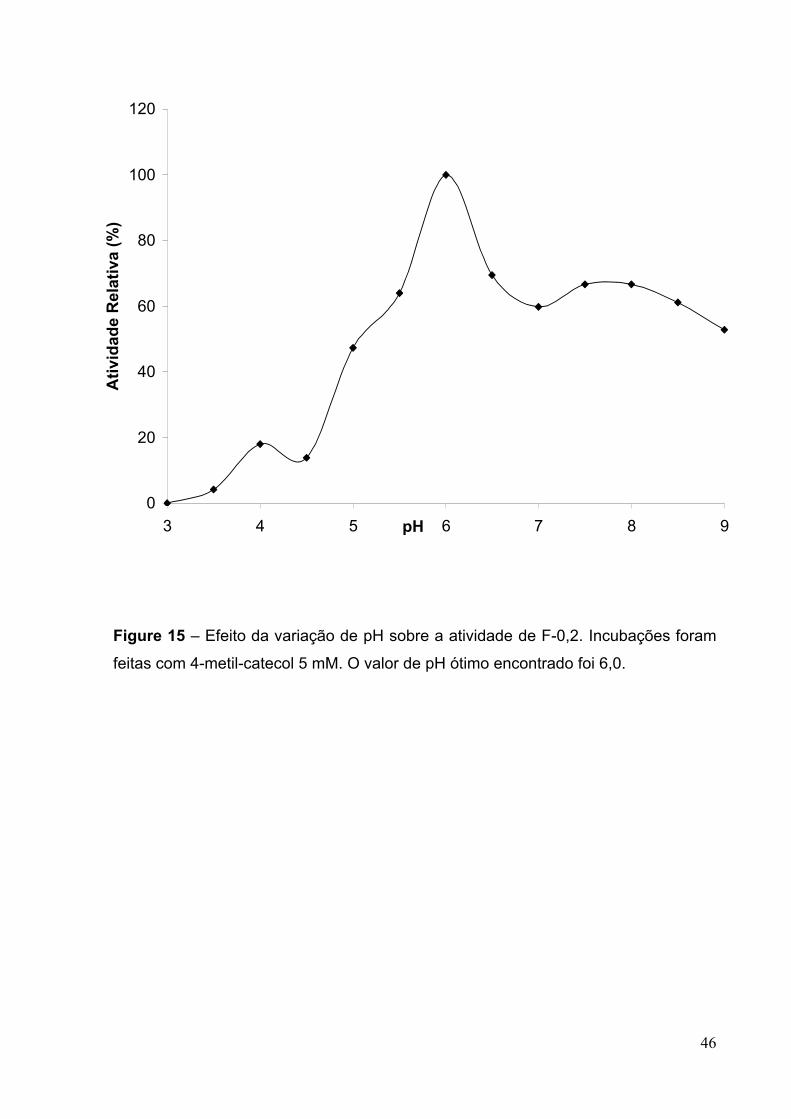
46
Figure 15 – Efeito da variação de pH sobre a atividade de F-0,2. Incubações foram
feitas com 4-metil-catecol 5 mM. O valor de pH ótimo encontrado foi 6,0.
0
20
40
60
80
100
120
3 4 5 6 7 8 9pH
Ati
vid
ade
Rel
ativ
a (%
)
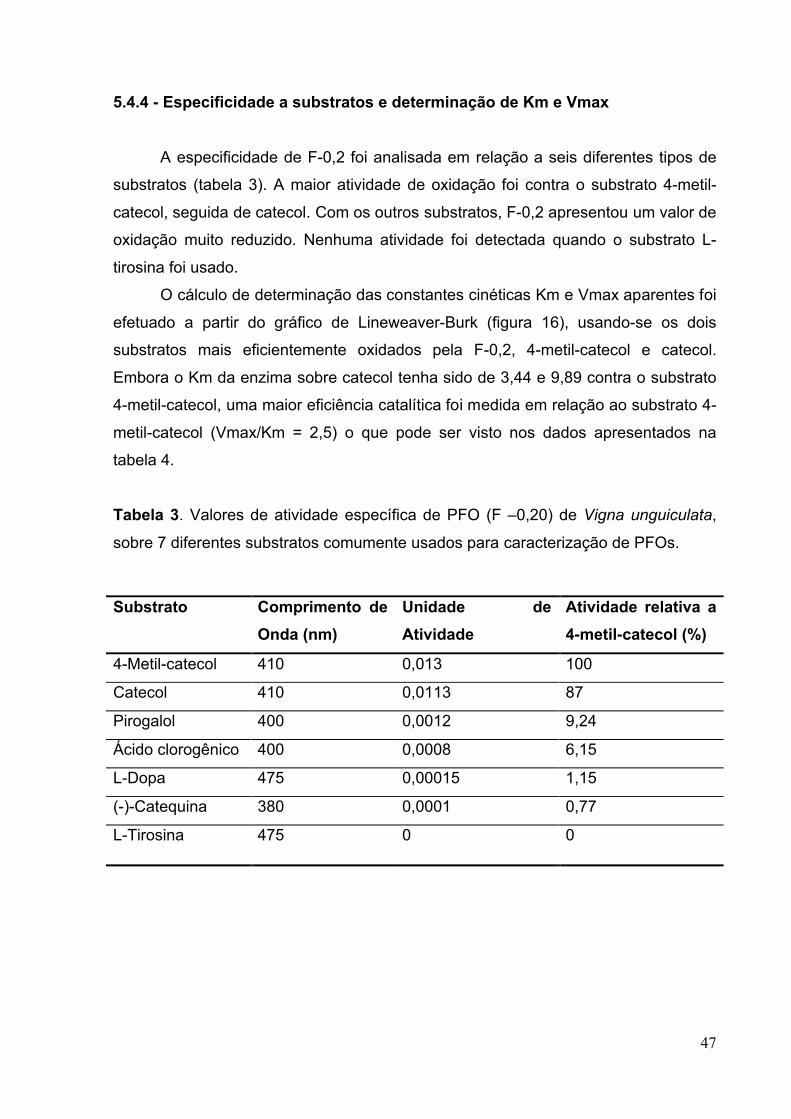
47
5.4.4 - Especificidade a substratos e determinação de Km e Vmax
A especificidade de F-0,2 foi analisada em relação a seis diferentes tipos de
substratos (tabela 3). A maior atividade de oxidação foi contra o substrato 4-metil-
catecol, seguida de catecol. Com os outros substratos, F-0,2 apresentou um valor de
oxidação muito reduzido. Nenhuma atividade foi detectada quando o substrato L-
tirosina foi usado.
O cálculo de determinação das constantes cinéticas Km e Vmax aparentes foi
efetuado a partir do gráfico de Lineweaver-Burk (figura 16), usando-se os dois
substratos mais eficientemente oxidados pela F-0,2, 4-metil-catecol e catecol.
Embora o Km da enzima sobre catecol tenha sido de 3,44 e 9,89 contra o substrato
4-metil-catecol, uma maior eficiência catalítica foi medida em relação ao substrato 4-
metil-catecol (Vmax/Km = 2,5) o que pode ser visto nos dados apresentados na
tabela 4.
Tabela 3. Valores de atividade específica de PFO (F –0,20) de Vigna unguiculata,
sobre 7 diferentes substratos comumente usados para caracterização de PFOs.
Substrato Comprimento de
Onda (nm)
Unidade de
Atividade
Atividade relativa a
4-metil-catecol (%)
4-Metil-catecol 410 0,013 100
Catecol 410 0,0113 87
Pirogalol 400 0,0012 9,24
Ácido clorogênico 400 0,0008 6,15
L-Dopa 475 0,00015 1,15
(-)-Catequina 380 0,0001 0,77
L-Tirosina 475 0 0
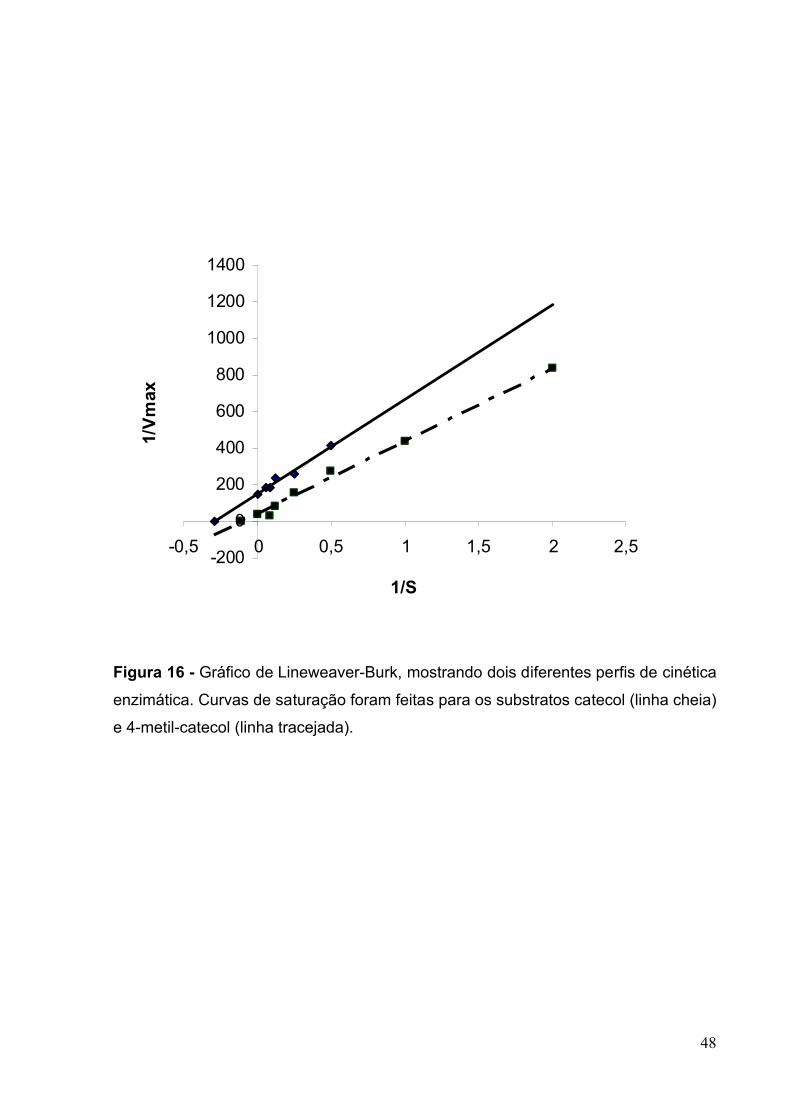
48
-200
0
200
400
600
800
1000
1200
1400
-0,5 0 0,5 1 1,5 2 2,5
1/S
1/V
max
Figura 16 - Gráfico de Lineweaver-Burk, mostrando dois diferentes perfis de cinética
enzimática. Curvas de saturação foram feitas para os substratos catecol (linha cheia)
e 4-metil-catecol (linha tracejada).

49
Tabela 4. Valores de parâmetros de cinéticos medidos para F-0,2 de V. unguiculata.
Substrato Km Vmax Vmax/Km
4-metil-catecol 9,86 24,66 2,50
Catecol 3,44 6,64 1,93
5.4.5 - Ensaio de inibição de atividade
O perfil de inibição de F-0,2 foi caracterizado usando-se 4 diferentes
inibidores em 4 diferentes concentrações (0,01; 0,1; 1,0 e 10 mM). Sendo usado
como substrato 4-metil-catecol, numa concentração de 5 mM (tabela 5). Os dados
revelaram que tropolona foi o mais potente inibidor encontrado.
Tabela 5. Inibição percentual de PFO (F –0,20) parcialmente purificada de V.
unguiculata.
Percentual de inibição (%)
Inibidores/concentração 0,01 0,1mM 1 10
Resorcina nd 20 20 40
EDTA nd 15 15 15
Tropolona 50 70 77 nd
Azida sódica nd 20 20 100
5.5 – Detecção e caracterização de cistatinas induzidas por ferimento
O ferimento a nivel de unifoliato induziu um aumento do nível de cistatinas em
plantas de V. unguiculata, porém em patamares mais modestos. Na figura 17 nota-
se um aumento médio de 33 % nos níveis destes inibidores, em plantas feridas, 24 h
após lesão, comparando-se aos de plantas controle. Os dados indicam que, sob o
aspecto quantitativo, os níveis de cistatinas aumentam na mesma proporção, tanto a
nível local no unifoliato ferido, quanto a nível sistêmico, no lateral ao ferido,

50
evidenciando que a indução de cistatinas em feijão-de-corda ocorre a nível local e
sistêmico, frente ao estimulo de lesão mecânica.
5.5.1 – SDS-PAGE e Western blot de cistatinas induzidas por ferimento.
A análise por SDS-PAGE das frações protéicas de unifoliatos, 24 horas após
lesão mecânica (item 4.2.2), é vista na figura 18 A. A análise por Western blot dos
perfis protéicos foi feita usando-se anticorpos policlonais contra cistatina de
sementes de V. unguiculata, e mostrou um grande aumento de uma banda de 22
kDa em unifoliato ferido, e o aparecimento de outra de 29 kDa, em unifoliatos laterais
aos feridos, quando comparadas à amostra controle (figura 18 B)
5.5.2 – Atividade proteinásica do tipo cisteínica em unifoliatos de feijão-de-
corda submetidos ou não a lesão mecânica
Os dados da medição da atividade proteinásica do tipo cisteínica em extratos
de unifoliatos mostram uma ligeira redução de atividade em unifoliatos feridos
quando comparados à atividade de extratos obtidos de unifoliatos controle, redução
esta que corresponde a aproximadamente 20% enquanto que nos unifoliatos laterais
aos feridos unifoliatos não houve diferença significativa em relação ao controle
(figura 19).
5.5.3 – Proteinases cisteínicas em unifoliatos de feijão-de-corda submetidos ou
não a lesão mecânica
Análise de atividades de proteases cisteínicas por gel SDS-PAGE-gelatina
mostrou que uma banda de massa molecular de 50 kDa, que representa uma fração
minoritária da atividade total, foi fortemente inibida em unifoliatos ferido e lateral ao
ferido. Uma outra banda de atividade de 70 kDa foi parcialmente inibida em
unifoliatos feridos (figura 20).
5.5.4 - Ensaio de ativação de senescência
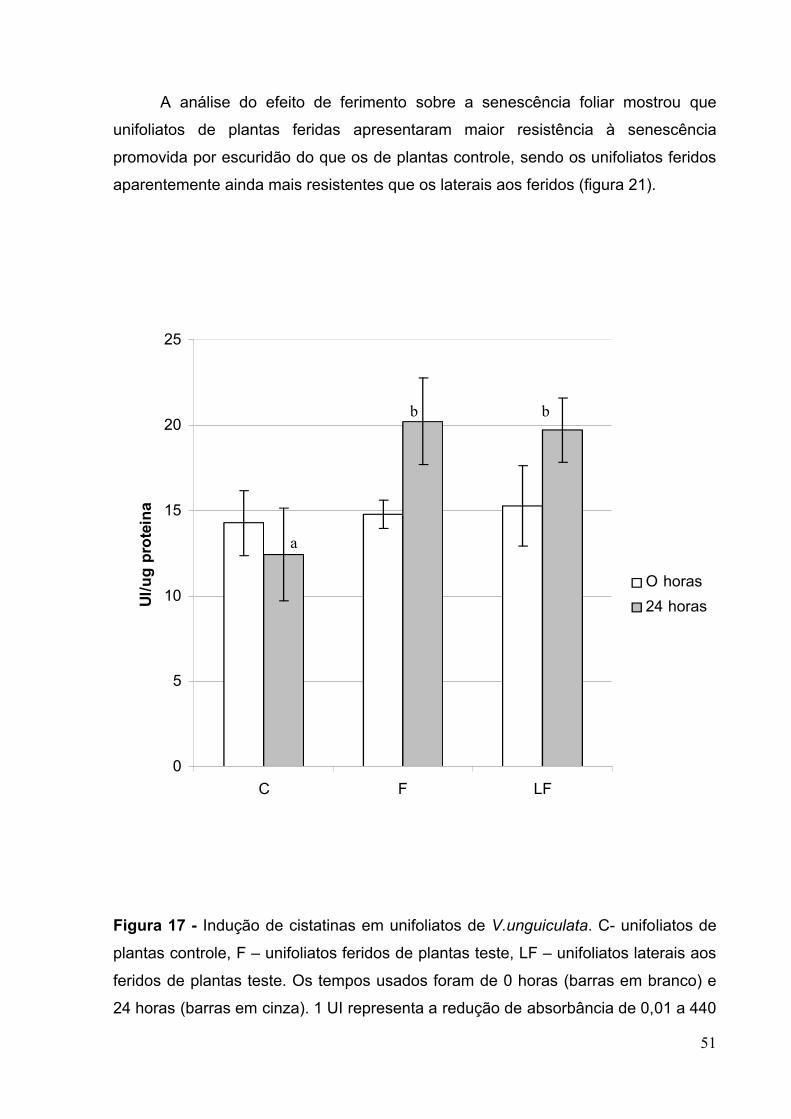
51
A análise do efeito de ferimento sobre a senescência foliar mostrou que
unifoliatos de plantas feridas apresentaram maior resistência à senescência
promovida por escuridão do que os de plantas controle, sendo os unifoliatos feridos
aparentemente ainda mais resistentes que os laterais aos feridos (figura 21).
0
5
10
15
20
25
C F LF
UI/
ug
pro
tein
a
O horas
24 horas
Figura 17 - Indução de cistatinas em unifoliatos de V.unguiculata. C- unifoliatos de
plantas controle, F – unifoliatos feridos de plantas teste, LF – unifoliatos laterais aos
feridos de plantas teste. Os tempos usados foram de 0 horas (barras em branco) e
24 horas (barras em cinza). 1 UI representa a redução de absorbância de 0,01 a 440
bb
a
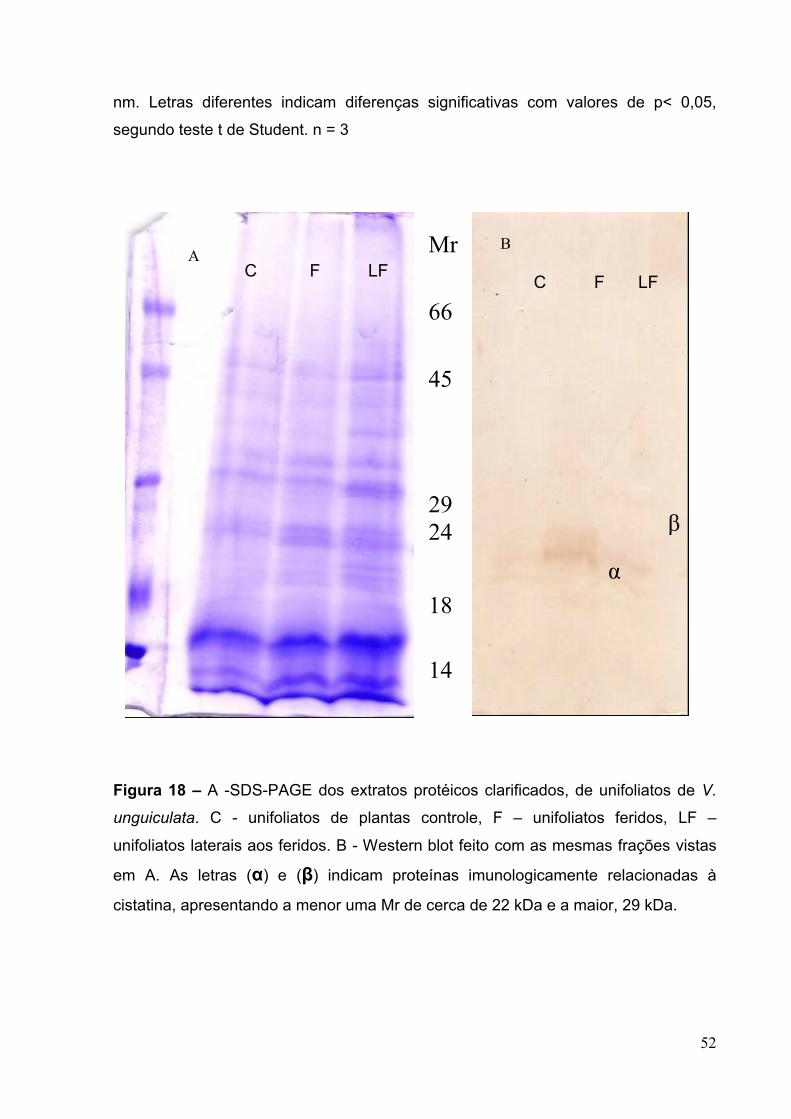
52
nm. Letras diferentes indicam diferenças significativas com valores de p< 0,05,
segundo teste t de Student. n = 3
Figura 18 – A -SDS-PAGE dos extratos protéicos clarificados, de unifoliatos de V.
unguiculata. C - unifoliatos de plantas controle, F – unifoliatos feridos, LF –
unifoliatos laterais aos feridos. B - Western blot feito com as mesmas frações vistas
em A. As letras (α) e (β) indicam proteínas imunologicamente relacionadas à
cistatina, apresentando a menor uma Mr de cerca de 22 kDa e a maior, 29 kDa.
A C F LF
Mr
66
45
2924
18
14
C F LF
B
α
β
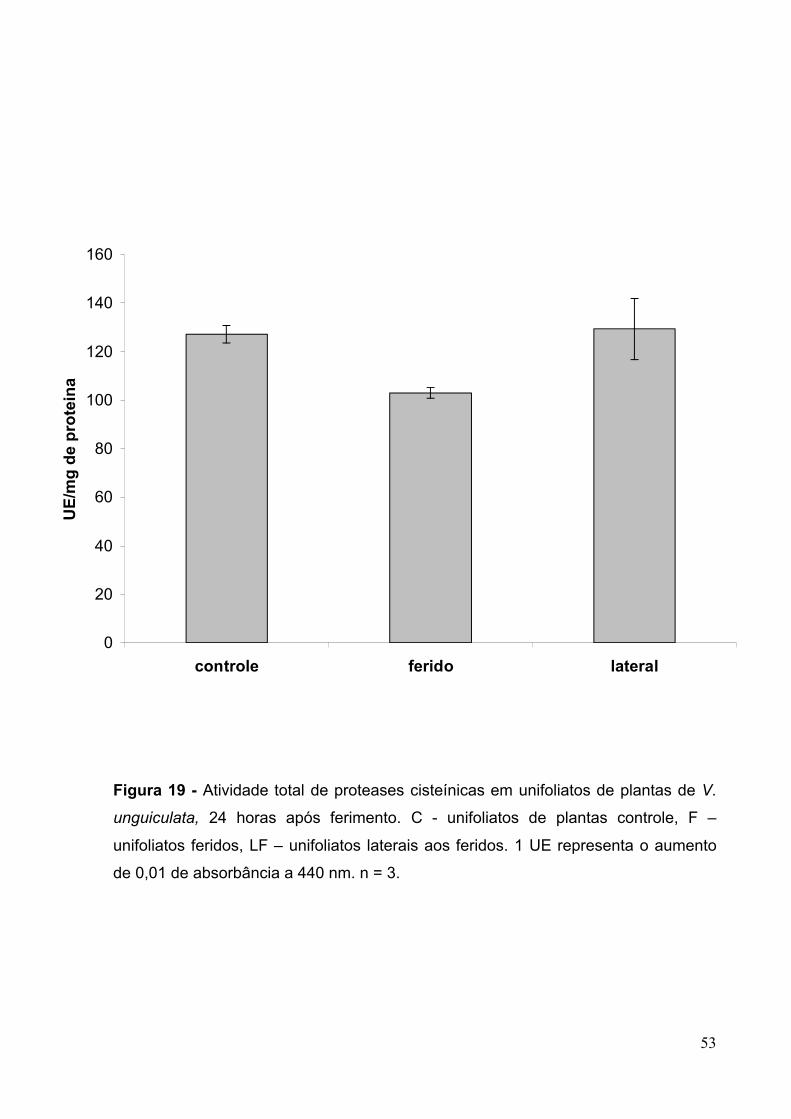
53
Figura 19 - Atividade total de proteases cisteínicas em unifoliatos de plantas de V.
unguiculata, 24 horas após ferimento. C - unifoliatos de plantas controle, F –
unifoliatos feridos, LF – unifoliatos laterais aos feridos. 1 UE representa o aumento
de 0,01 de absorbância a 440 nm. n = 3.
0
20
40
60
80
100
120
140
160
controle ferido lateral
UE
/mg
de
pro
tein
a

54
Figura 20 – Visualização eletroforética de proteases cisteínicas totais de uniofoliatos
de plantas de V. unguiculata, 24 horas após ferimento. A – SDS-PAGE. B - Atividade
gelatinolítica das amostras mostradas em A, em pH 5,0. � indica uma banda de
protease cisteínica de cerca de 70 kDa, reduzida parcialmente em unifoliatos feridos;
� mostra uma banda de 50 kDa, fortemente presente em unifoliatos controle, mas
quase ausente em unifoliatos feridos e laterais aos feridos.
Figura 21 - Unifoliatos de plantas de feijão-de-corda, submetidos a estresse de
senescência. C - unifoliatos de plantas controle, F – unifoliatos feridos, LF –
unifoliatos laterais aos feridos.
C F LF
A
C F LF

55
5.6– Análise da atividade indutora e caracterização de esxudado de floema
5.6.1 - Teste de indução de PFOs por esxudado de floema
As medições do potencial do esxudado de floema em induzir atividade/síntese
de PFOs foram feitas, tendo-se três parâmetros de comparação. O primeiro é
representado como o controle, onde plantas de feijão-de-corda foram inoculadas
inicialmente com um meio contendo apenas o tampão de esxudação. O segundo
onde plantas foram inoculadas com esxudado de floemas de plantas não feridas, no
intuito de verificar se o floema extraído destas plantas era capaz de induzir alguma
resposta de defesa. Por fim a terceira medição foi feita a partir da inoculação de
plantas com esxudado de floema de plantas lesionadas. Os resultados obtidos em
tais ensaios são apresentados na figura 22, onde é mostrado que ocorreu um
aumento de indução de PFOs em plantas inoculadas com esxudado de plantas
controle (ExuNfer) e de plantas feridas (Exufer), quando comparadas a plantas
somente inoculadas com tampão de extração de floema. Além disso, não houve
diferença significativa entre as respostas para os inóculos de esxudados de plantas
submetidas às duas condições ExuNfer ou Exufer.
5.6.2 – Detecção da presença de AJ em esxudado de floema por
espectrometria de massas
A análise da composição molecular do esxudado de floema extraído de
plantas de feijão-de-corda, por espectrometria de massas (Figura 23), revelou a
existência de componentes moleculares que diferem em ocorrência entre esxudados
de plantas feridas (Exufer) e não feridas (ExuNfer). No entanto foi detectável a
possível ocorrência de ácido jasmônico AJ em esxudado de ambas plantas, feridas
e não feridas ao nível dos unifoliatos.
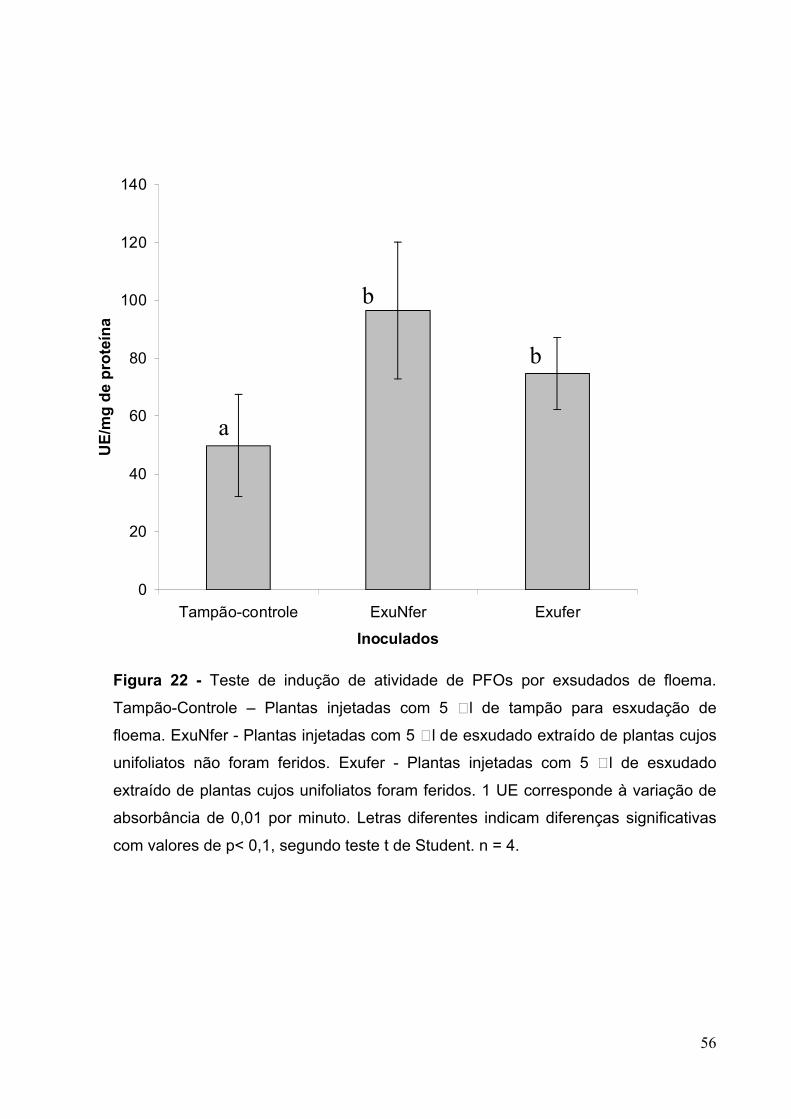
56
Figura 22 - Teste de indução de atividade de PFOs por exsudados de floema.
Tampão-Controle – Plantas injetadas com 5 �l de tampão para esxudação de
floema. ExuNfer - Plantas injetadas com 5 �l de esxudado extraído de plantas cujos
unifoliatos não foram feridos. Exufer - Plantas injetadas com 5 �l de esxudado
extraído de plantas cujos unifoliatos foram feridos. 1 UE corresponde à variação de
absorbância de 0,01 por minuto. Letras diferentes indicam diferenças significativas
com valores de p< 0,1, segundo teste t de Student. n = 4.
0
20
40
60
80
100
120
140
Tampão-controle ExuNfer Exufer
Inoculados
UE
/mg
de
pro
teín
a
a
b
b

57
Figura 23 – Análise comparativa entre dados de espectrometria de massas obtidos
a partir de amostras de esxudados de unifoliatos de feijão-de-corda. Tamp - Tampão
de extração de exsudato, considerado branco de leitura; ExuNfer - esxudado
extraído de plantas cujos unifoliatos não foram feridos; Exufer - esxudado extraído
de plantas cujos unifoliatos foram feridos. A seta indica sinais relativos às massas
moleculares iguais a do ácido jasmônico (AJ).
modo positivo tampao e solvente mais amostra
211 212
212.6356210.7310
210.6531212.3618211.7915211.2063
210.7933
210.7154
210.6765
210.6609
210.6298
210.6142
210.5519
210.7388
212.6277211.7056210.7544
211.0193210.7699
210.9179211.6744
211.4247211.0738
211.7525
212.5261212.4557211.9477
210.6531
210.6375212.3462212.2837211.8930210.7544
C12H18O3
Tamp
ExuNfer
Exufer

58
6- Discussão
6.1 – Análise da indução e caracterização de PFOs
Plantas de V. unguiculata, quando submetidas a injuria mecânica, são
capazes de aumentar a atividade de PFOs tanto a nível local como sistemicamente.
Os dados mostrados na figura 8 revelaram que o perfil de indução por ferimento de
PFOs em unifoliatos de feijão-de-corda apresentou uma estreita similaridade com a
de plantas de Lycopersicum esculentum, quando submetidas ao mesmo tratamento,
sendo a atividade destas aumentada em aproximadamente 18 vezes (Constabel et
al., 1995). Em contraste, o ferimento de batata, em folhas do 6o ao 8o nodo, resultou
em um menor aumento de apenas 1,7 vezes na atividade polifenol oxidásica, 48
horas após o estímulo (Thipyapong et al., 1995). Em ambas as folhas feridas e não
feridas de plantas de tabaco, a atividade de PFO aumentou em 4 e 6 vezes,
respectivamente, após 24 horas, e mais de 10 vezes em ambos unifoliatos, no
tempo de 48 horas após a lesão. Resultados similares foram obtidos em plantas
híbridas de choupo com dois meses de idade, onde se ferindo as folhas inferiores
houve um aumento de duas e quatro vezes na atividade de PFOs nos tempos de 48
e 96 horas, respectivamente, em ambas as folhas feridas e não feridas (Constabel &
Ryan, 1998). Nos resultados desta tese, os níveis de atividade solúvel de PFO foram
significativamente aumentados, enquanto nenhuma diferença foi detectada em
fração insolúvel de proteínas tanto de unifoliatos feridos como laterais aos feridos
(tabela 1). Uma vez que mais de 90 % da atividade total de PFO dos unifoliatos está
concentrada na fração insolúvel (quando consideradas frações sem ativação por
SDS), a qual foi resultante da re-extração do precipitado obtido da primeira extração
com solução de Triton X-100 1%, este trabalho alerta para os métodos empregados
quando se investigando a indução de PFOs em tecidos vegetais submetidos a
estresses. O aumento de atividade de PFO restrito a frações de proteínas solúveis
levanta questionamentos sobre análises de indução de PFOs de trabalhos
previamente publicados, como por exemplo a sondagem desenvolvida por Constabel
& Ryan (1998), na qual foi empregado 0,1% de Triton X-100, para extração protéica.

59
Como PFOs são proteínas fracamente aderidas a membranas, estas são facilmente
liberadas por soluções de detergente (Triton X-100) e quantificação de atividade total
(atividade de PFO solúveis somadas a PFO insolúveis) poderia mascarar variações
de isoformas que apresentam diferentes solubilidades.
Isoformas de PFO de muitas espécies de plantas apresentam uma variação
de tamanho de 32 a mais de 200 kDa, sendo que a maioria apresenta Mr entre 35-
70 (Flurkey, 1986; Sherman et al., 1991; Steffens et al., 1994; Fraignier et al., 1995;
Van Gelder et al., 1997; Yang et al., 2000). Estas massas diferem entre as formas
imaturas e maduras e ativas da enzima. Uma PFO de massa equivalente a 67 kDa
foi definida como a forma madura de uma enzima que perdeu seu peptídeo de
importação para o lúmen do cloroplasto (Anderson & Morris, 2003). Neste caso, uma
proteína de 83 kDa foi considerada como uma pró-PFO imatura, não processada,
contendo ainda um fragmento transitório de 14-16 kDa. Em adição, sementes
imaturas de trigo contêm múltiplas isoformas de PFOs mas estas aparentam estar
inibidas (inativas) a menos que sejam tratadas com SDS ou pelo envelhecimento.
Após sofrer clivagem proteolítica e processamento, uma enzima de 40 kDa foi
observada como a forma ativa e madura de uma PFO de uva, originária de uma
proteína precursora completa de 67 kDa, a qual é composta por uma seqüência de
10,6 kDa de um peptídeo transitório de direcionamento para cloroplasto, uma
unidade catalítica de 40,5 kDa contendo duas ligações a cobre e uma extensão C-
terminal de 16,2 kDa (Dry & Robinson, 1994). Outro trabalho tem definido PFOs com
massas moleculares tão baixas quanto 35 a 40 kDa, as quais seriam produtos de
degradação proteolítica, ocorrida durante a extração, ou até mesmo durante o
processamento fisiológico vegetal (Shin et al., 1997). De acordo com Yoruk &
Marshall (2003) e os dados dos trabalhos aqui citados, é relevante mencionar que a
atribuição da multiplicidade de PFOs baseada em estimativas de massa molecular
deve ser interpretada com cautela, uma vez que interconversões de várias formas de
PFOs, sob condições eletroforéticas desnaturantes ou parcialmente desnaturantes,
têm sido amplamente publicadas na literatura (Flurkey, 1990; Cary et al., 1992;
Robinson & Dry, 1992; Fraignier et al., 1995; Marques et al., 1995).
Como pode ser visto na Tabela 2, os procedimentos de isolamento de PFO
resultaram em enriquecimento de atividade de 11,18 vezes, indo de uma atividade
específica de 1,04 do extrato bruto até 11,63 do pico eluído com 0,2 M NaCl via
cromatografia. Similar eficiência de purificação (11,59 vezes) também foi vista em

60
outro trabalho descrito por Gandia-Herrero et al. (2004) para uma PFO solúvel de
beterraba. Estes resultados mostram que apesar do processo de purificação não ter
atingido um rendimento tão grande quanto o de outros mencionados na literatura, o
material estava suficientemente enriquecido em sua atividade polifenol oxidásica por
cromatografia em DEAE-celulose, tendo sido possível a caracterização cinética
desta enzima.
A estabilidade e ativação de F-0,2 por SDS foram muito similares às
observadas para uma PFO purificada de uma variedade de uva (uva Napoleão),
descrito por Núñez-Delicado et al. (2007). Os níveis de ativação de PFOs por SDS é
algo que varia enormemente, dependendo da espécie em estudo: em Vicia faba
aumentam de 65 a 119 vezes (Sanchez-Ferrer et al., 1990; Jiménez & García-
Carmona, 1996); 19 vezes para PFO de uva (Sanchez-Ferrer et al., 1989a); 25
vezes para PFO de pêssego (Laveda et al. 2000); dez vezes para PFO de banana
(Sojo et al., 1998); 4 vezes para PFO de folhas de batata (Sanchez-Ferrer et al.,
1993); 7 vezes para PFO de manga (Robinson et al., 1993). Todas estas atividades
foram medidas sob condições experimentais similares as aqui descritas. Entretanto,
concentrações mais altas deste detergente inibiram significativamente a atividade
desta classe de enzimas (Robb et al., 1964; Sanchez-Ferrer et al., 1989b, 1993). A
PFO de V. unguiculata aqui observada aumentou sua atividade em 10 vezes em
relação a seu estado latente e manteve a atividade em patamares próximos a 50 %,
mesmo em concentrações superiores a 140 mM de SDS (figura 13). Tal
característica sugere que a enzima em estudo apresenta um comportamento
incomum em relação a outras, visto que a maioria das enzimas deste tipo não possui
grande resistência térmica.
Além da elevada resistência a SDS, esta enzima também mostrou ter uma
alta estabilidade térmica. Consta na literatura uma infinidade de trabalhos relatando
sobre a energia de desnaturação de PFO, e um grande leque de valores de
estabilidade foi revisado por Yoruk & Marshall (2003). De forma similar aos dados
obtidos acerca de ativação por SDS e estabilidade da enzima em presença deste
detergente, a estabilidade térmica de F-0,2 foi muito similar a encontrada para PFO
de uvas da variedade Napoleão (Núñez-Delicado et al., 2007), embora esta
apresente uma estabilidade um pouco inferior a encontrada aqui para V. unguiculata
(80 % de atividade residual a 70 oC) (figura 14).

61
Em relação ao efeito do pH sobre a atividade enzimática, o perfil encontrado
para atividade de F-0,2 de V. unguiculata foi muito similar ao visto para PFO de
frutos de nespereiras (Sellés-Marchart et al., 2006), uma PFO latente de raízes de
beterraba (Gandía-Herrero et al., 2004) e uma extraída de flores de brócolis (Gawlik-
Dziki et al., 2007). O pH ótimo de PFOs é algo que varia amplamente de acordo
com cada espécie vegetal, mas em geral a variação fica entre os valores 4,0 e 8,0
(Yoruk & Marshall, 2003), o que está de acordo com os dados apresentados para F-
0,2, com pH ótimo de 6,0, evidenciando que esta enzima tem um perfil de atividade
comum comparada as demais da mesma classe enzimática (figura 15).
Os dados referentes à afinidade por substratos indicam que a enzima é
pertence a família das catecolases, uma característica comum de muitas PFOs de
folhas de plantas. Além deste fato, uma maior especificidade contra os substratos 4-
metil-catecol, catecol e pirogalol (tabela 3) indica uma melhor afinidade para
moléculas pequenas de substratos. Este tipo de preferência por substratos
encontrada para F-0,2 também foi encontrada em PFOs de híbridos de choupo
(Populus trichocarpa e P. deltoides) que é induzida por ferimento e por metil-
jasmonato (Constabel et al., 2000; Wang & Constabel, 2003). Tal maior afinidade
contra os substratos catecol e 4-metil-catecol também foi detectada para PFO de
cloroplasto de folhas de Vigna radiata (Shin et al., 1997), uma espécie do mesmo
gênero Vigna aqui pesquisado. No entanto, os valores de atividade, encontrados
para L-DOPA e ácido clorogênico, diferem em muito dos encontrados nesta tese
para F-0,2. Acrescentando a este fato, diferentemente da enzima aqui descrita, a
PFO de Vigna radiata (Shin et al., 1997) não é uma enzima latente.
Os valores de Km aparentes encontrados aqui para o substrato 4-metil-
catecol diferem significativamente dos medidos para PFO de V. radiata (Shin et al.,
1997). No entanto a atividade relativa contra 4-metil-catecol, neste caso, também é
maior que a encontrada contra catecol, sugerindo uma origem cloroplastídica similar
para ambas, embora a PFO de V. radiata seja diferente por se encontrar em seu
estado ativo. Outra observação é a de que tal enzima foi purificada a partir de
frações de cloroplastos, extraídas com tampão contendo Triton X-100, ou seja,
provavelmente trata-se de enzima contida na fração não solúvel, diferentemente da
F-0,2 extraída de frações solúveis.
Consta na literatura uma infinidade de trabalhos caracterizando a cinética de
inibição de atividade de PFOs, usando-se diversos tipos de inibidores para isto.

62
Nesta tese foram escolhidos os inibidores: Resorcinol, EDTA, tropolona e azida
sódica. Resorcinol é um inibidor que age diretamente sobre o centro ativo desta
enzima, formando uma inibição do tipo competitiva. Porém o uso deste inibidor
reduziu somente 40% da atividade inicial de F-0,2. O agente quelante EDTA e a
azida sódica reduziram a 15 e 100 % da atividade inicial, respectivamente, quando
usados na concentração máxima de 10 mM. Estes dados, especialmente quanto a
inibição por EDTA sugerem uma forte ligação do íons Cu++ no centro ativo da
enzima. Tropolona, um competidor de Cu2+ descrito como um dos mais potentes
inibidores específicos para PFOs (Khan & Andrawis, 1985), reduziu em 50 % a
atividade de F-0,2, quando usado na baixa concentração de 0,01 mM (tabela 5), a
menor concentração efetiva, dentre todos os inibidores testados.
6.2 – Análise da indução, caracterização e efeito de cistatinas em unifoliatos de
V. unguiculata
Consta na literatura a detecção de cistatinas constitutivas de uma enorme
variedade de espécies vegetais, como por exemplo: as cistatinas I e II de arroz (Abe
et al., 1987;1989; Kondo et al., 1991), das cistatinas I e II de milho (Abe e Whitaker,
1988; Abe e Arai, 1991; Abe et al., 1996; Irie et al., 1996), cistatina de sementes de
feijão–de-corda (Fernandes et al., 1993), cistatinas de frutos de abacate (Kimura et
al., 1995), cistatinas de botões florais de Brassica campetris (Lim et al., 1996),
cistatina de sementes de soja (Hines et al., 1991; Misaka et al., 1996), de sementes
de Phaseolus vulgaris (Santino et al., 1998; Brzin et al., 1998), além de muitas
outras. No entanto, ainda se conhece pouco a respeito do papel destas proteínas
nos mecanismos induzidos de defesa em plantas, frente à ação de estresses
bióticos ou abióticos, apesar terem sido detectadas há algum tempo (Barrett &
Kirschke,1981). A detecção, purificação, seqüenciamento e caracterização de
cistatinas expressas de forma constitutiva, em sementes de plantas de feijão-de-
corda, são alvos perseguidos há muito tempo (Flores et al., 2001). Recentemente a
indução de uma multicistatina foi detectada em folhas desta mesma espécie, frente
ao estresse de desidratação (Diop et al. 2004). Neste trabalho uma seqüência de
cDNA codificadora para cistatinas foi isolada de uma biblioteca de cDNA produzida a
partir de plantas de feijão-de-corda sob estímulo de estresse hídrico. Após clonagem
e seqüenciamento foi possível constatar que esta cistatina apresentava um duplo

63
domínio, sendo uma proteína de massa molecular de 22 kDa, e outra suspeita de ter
um triplo domínio com 39 kDa aproximadamente, diferente da cistatina de baixo
peso, de um único domínio, previamente descrita em sementes desta espécie
(Flores et al., 2001). Foi constatado também neste trabalho que só ocorreu a
indução dessas proteínas quando as plantas estavam submetidas a estresse hídrico,
e a expressão de seus RNAm não foi muito aumentada pela ação de ABA. Foi
proposto pelos autores que tais cistatinas induzidas pudessem apresentar um papel
protetor contra as proteases endógenas, evitando a autólise em período de estresse,
relembrando inclusive um papel multifuncional sugerido para cistatinas de variadas
massas moleculares, descrito anteriormente (Flores et al., 2001). Em nossos
estudos foi visto que uma cistatina de massa molecular de 22 kDa, muito similar à
encontrada no trabalho de Diop et al. (2004), é induzida em plantas de feijão-de-
corda tratadas com estresse de ferimento, principalmente em unifoliato ferido (figura
18 B), mas observando-se também a presença de cistatinas constitutivas, que não
mudaram seu perfil de indução, e uma de cistatina de 29 kDa induzida somente em
unifoliatos que sofreram a ação sistêmica do ferimento. A diferente expressão de
cistatinas frente à ação de ferimento também foi vista em soja (Zhao et al. 1996),
onde duas cistatinas tiveram sua expressão aumentada somente em plantas feridas
e mostravam KIs de 500 e 1000 vezes menores que o apresentado por uma cistatina
constitutiva, cuja expressão não foi alterada com o tratamento, mostrando que a
afinidade de cistatinas por suas proteinases-alvo pode variar amplamente de acordo
com seu papel. Tais dados evidenciam a participação destes inibidores em funções
bem diferenciadas no corpo vegetal, não se limitando a um papel de defesa.
Proteinases cisteínicas são as principais enzimas proteolíticas envolvidas com
a senescência de pétalas de flor de íris, podendo inclusive, ser possível prevenir ou
adiar os sintomas de senescência fazendo-se uso de um tratamento a base de
inibidores de proteinases sintéticos (Pak & van Doorn, 2004). Em pétalas de cravo
foi evidenciado que o aumento da expressão de um inibidor de caspases,
proteinases cisteínicas envolvidas com morte celular programada, promovia a
proteção da flor ao processo de senescência e murchamento. Além disso, foi
destacado que a atividade proteolítica estava presente de forma uniforme, durante
todo o período de senescência em diferentes partes da flor, sendo que a proteção
contra os efeitos deletérios das caspases de partes vitais, como, por exemplo,
tecidos ovarianos, foi atribuída à expressão de inibidores destas proteinases, ou

64
seja, cistatinas (Sugawara et al., 2002). Neste sentido, buscamos correlacionar
nossos resultados de detecção de cistatina induzida por ferimento, com um provável
papel de regulação de proteólise endógena nestes tecidos. A detecção do aumento
de expressão da cistatina de 22 kDa majoritariamente em unifoliatos feridos, chama
a atenção ao papel desta em processos fisiológicos não relacionados com defesa,
uma vez que a expressão de proteínas de defesa ocorre tão intensamente a nível
local como de forma sistêmica (Ryan, 2000), sendo em muitos casos, induzida mais
intensamente a nível sistêmico do que em nível local (Thypiapong et al., 1996).
Complementando estes dados também foi visto, neste trabalho, que uma protease
cisteínica minoritária, com Mr de 50 kDa aproximadamente, teve sua atividade
fortemente reduzida somente em unifoliatos de plantas feridas, a nível local e
sistêmico enquanto outra protease majoritária mostrou sua atividade levemente
reduzida somente em unifoliatos feridos. Os dados indicam uma possível
especificidade da cistatina induzida contra proteases endógenas de uniofoliatos de
V. unguiculata, caso esta esteja mesmo exercendo inibição in vivo (figura 20). Um
outro dado complementar mostrado na figura 21, é a perceptível redução do efeito
de senescência, promovida pelo ferimento de unifoliatos, tanto a nível local como
sistêmico. Todos os dados obtidos em relação à indução de cistatinas, redução de
atividade proteolítica cisteínica e inibição de senescência apontam para uma estreita
correlação entre estes, fundamentada por outros anteriores, descritos na literatura.
Baseando-se nos dados aqui mencionados, é proposta a idéia de cistatinas estarem
participando na redução ou supressão de senescência em unifoliatos de plântulas de
feijão-de-corda. No entanto, a idéia não é nova. Há mais de vinte anos, um trabalho
feito com plântulas de aveia revelou que a senescência era inibida por ferimento
(Giridhar & Thimann, 1984). Os autores destacaram a possibilidade de inibidores de
proteases estarem atuando sobre a inibição de proteases envolvidas no processo.
Porém, não seriam os mesmos induzidos como resposta de defesa, uma vez que a
inibição enzimática por eles atribuída, não necessitava de luz, fator que para Ryan e
seu grupo de pesquisa, era imprescindível para indução de inibidores de proteases
relacionados com defesa (Green & Ryan, 1973).
Recentemente foi registrado, na literatura, que plantas de Phaseolus vulgaris
aumentam seus níveis de proteases serínicas, em folhas senescentes, frente à ação
de estresse hídrico (Hieng et al., 2004). Cultivares resistentes desta espécie não
apresentaram aumento da atividade proteásica. Embora as proteases neste caso

65
tenham sido serínicas, também observa-se, como foi acima mencionado para
proteases cisteínicas, a estreita relação entre a redução da atividade proteolítica, e o
aumento da resistência à senescência foliar causada por estresse hídrico.
Medições diretas da expressão de cistatinas são muitas vezes dificultadas
pelas suas baixas concentrações em tecidos foliares, tal como o que ocorre em
trabalhos semelhantes ao de Diop et al. (2004), no qual foi claramente possível
observar uma grande diferença entre os níveis de mRNA de cistatinas e os seus
respectivos perfis de expressão protéica, justificada por eles como decorrente de
processos pós-transcricionais. O protocolo desenvolvido nesta tese permitiu a
detecção de cistatinas, não somente por eliminar contaminantes indesejáveis ao
ensaio (pigmentos, fenóis), mas também por permitir concentrar as proteínas alvo de
análise nos extratos finais clarificados. É sugerido aqui que este método poderia ser
útil inclusive na detecção da cistatinas em tecido foliar, em outras espécies de
plantas.
6.3 – Análise da composição e efeito eliciador de defesa de esxudado de
floema
Com os dados apresentados na figura 22 é possível observar um aumento dos
níveis de atividade de PFOs em plantas inoculadas com esxudado de floema obtido
de plantas de feijão-de-corda cujos unifoliatos não foram feridos; um efeito indutor
menos pronunciado também é visto quando se inoculou o esxudado de floema de
plantas cujos unifoliatos haviam sido previamente feridos. Estes resultados podem
parecer paradoxais, uma vez que não era esperada a indução de PFO por um
esxudado de floema de plantas não feridas nos unifoliatos. No entanto tal com foi
citado por Buchannan et al. (figura 4), somente o fato da coleta de esxudado ser
feita via corte do tecido vegetal, no caule, seria suficiente para promover uma grande
alteração dos seus componentes. Tais alterações devem ter ocorrido em nossos
experimentos, e um fator de indução de defesa pode ter sido liberado em nossas
amostras. Quanto à menor indução de atividade via inoculação de esxudado de
plantas feridas ao nível de unifoliatos, acredita-se que como foi visto na análise de
espectrometria de massas (figura 23), um sinal correspondente ao ácido jasmônico
aparece em esxudados de plântulas feridas ao nível de unifoliatos, e em plantas
controle, contendo apenas o ferimento no caule, feito para exudação do floema.

66
Estes dados indicam que aparentemente apenas o ferimento do caule é suficiente
para liberação do hormônio AJ, e consequentemente para indução de PFOs, como
foi constatada na figura 22. Tais informações são contrárias aos dados apresentados
em um trabalho anterior similar, feito com plantas de tomate (Ryan,1974), onde foi
visto que em plantas de tomate, feridas ao nível do pecíolo foliar ou caule, a
expressão de inibidores de proteinases serínicas (a proteína de defesa teste em
questão na época) era fracamente estimulada. Entretanto, diante dos dados obtidos
para feijão-de-corda, o ferimento feito no caule promoveu uma considerável
alteração na expressão de PFOs. Pelo menos para a espécie V. unguiculata, nas
definidas condições de estudo, a premissa de que ferimento no caule não induziria
resposta de defesa não é verdadeira. A análise por espectrometria de massas,
portanto, revelou a possível presença de AJ como componente somente presente
em esxudado de plantas feridas ao nível de unifoliatos e caule e plantas controle
feridas somente ao nível de caule, o qual pode ter sido responsável pela indução de
PFO em plantas de feijão-de-corda. A medição do aparecimento de AJ em floema de
plantas feridas é um dado que está de acordo com os mencionados previamente na
literatura e apontam para um uso promissor da técnica mencionada, no intuito de
detectar e caracterizar fitormônios circulantes no floema. Além disso, o esxudado de
unifoliatos de plantas não feridas também foi capaz de induzir a atividade de PFO
quando inoculado (figura 22), o que está de acordo com a análise de espectrometria
de massas (figura 23) onde o AJ também pode estar presente em plantas que não
tiveram os unifoliatos feridos. No entanto, a presença de outras possíveis moléculas
sinalizadoras de defesa, liberadas somente pela lesão feita no caule durante o
processo de sua extração (figura 8), também deve ser considerada.
Além deste sinal, vários outros revelaram a ocorrência de substâncias
presentes em esxudado de plantas feridas, mas não em plantas controle. A análise
de componentes minoritários em esxudado de floema, tais como hormônios
vegetais, ou até mesmo proteínas de defesa, é dificultada não somente pela baixa
concentração destes neste meio vegetal (figura 6), como também pela própria
dificuldade de coleta de esxudado de floema da maioria das espécies, já que ocorre
formação de calose no local do ferimento, pouco tempo após o corte dos elementos
de tubo crivados. Esta é uma defesa natural do organismo vegetal ao
extravasamento deste componente. Para obtenção de quantidade maiores de
exsudato, emprega-se um tampão de extração de pH próximo a 7,0, contendo um

67
agente quelante de cálcio (EDTA) para evitar a formação de calose (Narváez-
Vasquéz et al.,1995). Em poucas espécies, como Ricinus comunis e Curcubita
maxima, é possível a fácil extração e análise do conteúdo floemático. Isto ocorre
porque a concentração de proteínas no floema destas espécies é bem superior às
demais, podendo em certos casos chegar a níveis 10 vezes maiores (Taiz & Zeiger,
2004). Estas características permitiram a execução de uma análise do proteoma de
defesa da espécie C. maxima (Walz et al. 2004). A utilização da espectrometria de
massa na busca de um hormônio vegetal liberado em floema não é inédita.
Hoffmann-Benning et al. (2002) já a utilizaram para identificação de um hormônio
peptídico, responsável por ativar o florescimento de Perilla ocymoides, denominado
florígeno. Esta tese utilizou-se de tal metodologia, pela primeira vez, para estudo
sobre de moléculas sinalizadoras direcionadas à defesa vegetal. Os dados obtidos
até o momento apontam a possível presença destas substâncias em uma fração de
esxudado de floema de plantas de feijão-de-corda; no entanto, esta fração necessita
de etapas posteriores de purificação, para ser possível qualquer outra conclusão a
respeito da natureza molecular deste agente indutor de defesa.

68
7 - CONCLUSÕES
Plantas de feijão-de-corda aumentam seus níveis de atividade de PFOs quando
submetidas a estresse por lesão mecânica.
O aumento de atividade de PFOs em plantas de feijão-de-corda é um processo
sistêmico, ou seja, também foi observado em folhas laterais às feridas.
O aumento de atividade de PFOs só foi observado na fração solúvel desta
enzima; a atividade de frações insolúveis de PFOs não variou como resposta ao
ferimento.
O aumento máximo de atividade de PFOs ocorreu em unifoliatos de plantas, 48
horas após o ferimento mecânico, tanto em unifoliatos feridos quanto em unifoliatos
laterais a estes.
PFOs induzidas em plantas de feijão-de-corda são enzimas latentes, ativadas na
presença de SDS, sendo a concentração de 5mM determinada como ótima para
suas atividades. Apresentaram Mrs de 58 e 73 respectivamente, são termo-
resistentes perdendo a atividade apenas em temperaturas maiores que 70 oC;
possuem pH ótimo em torno de 6,0; melhores substratos catecol e 4-metil-catecol e
são inibidas por tropolona.
Plantas de feijão-de-corda aumentam seus níveis de cistatinas quando
submetidas a estresse por lesão mecânica tanto localmente como de forma
sistêmica.
Diferentes isoformas de cistatinas são expressas frente a ação de ferimento,
sendo uma proteína de massa molecular de 22 kDa, com expressão aumentada
local e sistemicamente, e outra de 29 kDa, induzida somente em unifoliatos laterais
aos feridos.
Uma protease cisteínica de 50 kDa tem sua atividade reduzida em unifoliatos de
plântulas feridas de feijão-de-corda.

69
Plântulas de feijão-de-corda feridas são mais resistentes à senescência que
plântulas controle, não lesionadas.
Indução de cistatinas, redução de atividade proteinásica do tipo cisteínica
específica e a resistência à senescência são eventos correlacionados à resposta a
ferimento em plântulas de feijão-de-corda.
Esxudatos de floema de plantas feridas ao nível de unifoliatos e ao nível de caule
foram capazes de aumentar a expressão de PFOs quando inoculados em plantas
teste, evidenciando a presença de moléculas indutoras de defesa nestas amostras.
Através de espectrometria de massas foi observada a possível presença de ácido
jasmônico em esxudado de plântulas feridas de feijão-de-corda.

70
8- REFERÊNCIAS BIBLIOGRÁFICAS
Abe, M. & Whitaker, J. R. (1988). Purification and characterization of a
cysteine proteinase inhibitor from the endosperm of corn. Agric. Biol. Chem., 52:
1583-1584.
Abe, K., Emori, Y., Kondo, H., Susuki, K. & Arai, S. (1987). Molecular
cloning of a cysteine proteinase inhibitor of rice (Oryzacystatin). Homology with
animal cystatins and transient expression in the ripening process of rice seeds. J.
Biol. Chem. 262:16793 – 16797.
Abe, K., Kondo, K., Watanabe, H., Emori, Y. & Arai, S. (1991). Oryzacystatins
as the first well-defined cystatins of plant origin and their target proteinases in rice
seeds. Biomed. Biochim. Acta. 50: 637-641.
Abe, M., Domoto, C., Watanabe, H., Abe, K. & Arai, S. (1996). Structural
organization of the gene encoding corn cystatin. Biosci. Biotech. Biochem., 60: 1173-
1175.
Anderson, J. V. & Morris, C. F. (2003) Purification and analysis of wheat grain
polyphenol oxidase (PFO) protein. Cereal Chem. 80:135-143.
Aldridge, D. C., Galt, S., Giles, D. & Turner, W. B. (1971) Metabolites of
Lasiodiplodia theobromae. J. Chem. Soc. 1623–1627.
∙ Aquino-Bolaños, E. N. & Mercado-Silva, E. (2004) Effects of polyphenol
oxidase and peroxidase activity, phenolics and lignin content on the browning of cut
jicama. Postharvest Biol. Technol. 33: 275-283.
Aydemir, T. (2004) Partial purification and characterization of polyphenol
oxidase from artichoke (Cynara scolymus L.) heads. Food Chem. 87:59–67.
Aydin S., Geckil, H., Zengin, F., Ozercan, H. I., Karatas, F., Aydin, S, Dilek, T-
B., Yusuf O., Ferda D. & Venhar C. (2006) Ghrelin in plants: What is the function of
an appetite hormone in plants? Peptides. 1597-1602.

71
Barrett, A.J. & Kirschke, H. (1981) Cathepsin B, cathepsin H, and cathepsin L.
Methods Enzymol. 80:535–561.
Barrett, A. J. (1987) The cystatins: A new class of peptidase inhibitors. Trends
Biochem. Sci., 12:193 – 196.
Belenghi B, Acconcia F, Trovato M, Perazzolli M, Bocedi A, Polticelli F,
Ascenzi P. & Delledonne M. (2003) AtCYS1, a cystatin from Arabidopsis thaliana,
suppresses hypersensitive cell death. European Journal of Biochemistry. 270:2593–
2604.
Bertrand, G., (1896) Sur une nouvelle oxydase, ou ferment soluble oxidant,
d’origine végétale. C. R. Acad. Sci. (Paris) 122:1215–1217.
Boonsiri, K., Ketsa, S. & van Doorn, W. G. (2007) Seed browning of hot
peppers during low temperature storage. Postharvest Biol. Technol. 45:358-365.
Bowles, D. (1998) Signal transduction in the wound response of tomato plants.
Phil. Trans. R. Soc. Lond. B. 353:1495-1510.
Bruxelles, G. L. & Roberts, M. R. (2001) Signals Regulating Multiple
Responses to Wounding and Herbivores Critical Reviews in Plant Sciences. 20:487-
521.
Bradford, J. D. (1976) A rapid and sensitive method for quantitation of
microgram quantities of protein utilizing the principle of dye binding. Analitichal
Biochemistry. 72:248-254.
Brzin, J., Popovic, T., Ritonja, A., Puizdar, V. & Kidric, M. (1998) Related
cystatin inhibitors from leaf and seed of Phaseolus vulgaris L. Plant Science. 138: 17-
26.

72
Buchana, B. B., Gruissem, W. & Jones, R. L. (2000) Biochemistry & Molecular
Biology of Plants, first edition, Chapter 15 : 730 – 784.
Carasco, J. F., Xavier-Filho, J. (1981) Sequential expression of trypsin
inhibitors in developing fruit of cowpea (Vigna unguiculata (L) Walp.), Annals of
Botany Company. 47: 259 - 266.
Carvalho, A. O., Machado, O. L. T., Da Cunha, M., Santos, I. S. & Gomes, V.
M. (2001) Antimicrobial peptides and immunolocalization of a LTP in Vigna
unguiculata seeds. Plant Physiol. Biochem. 39, 137-146.
Cary J.W., Lax A.R. & Flurkey, W.H. (1992) Cloning and characterization of
cDNAs coding for Vicia faba polyphenol oxidase. Plant Mol.Biol. 20:245-253.
Chazarra, S., Cabanes, J., Escribano J. & García-Carmona, F. (1997) Kinetic
study of the suicide inactivation of latent polyphenoloxidase from iceberg lettuce
(Lactuca sativa) induced by 4-tert-butylcatechol in the presence of SDS. Biochim.
Biophys. Acta 1339, 297-303.
Cloutier, C., Jean, C., Fournier, M. Yelle, S. & Michaud, D. (2000) Adult
Colorado potato beetles, Leptinotarsa decemlineata compensate for nutritional stress
on oryzacystatin I-transgenic potato plants by hypertrophic behavior and over-
production of insensitive proteases, Arch. Insect Biochem. Physiol. 44:69–81
Concellón, A., Añón, M. C. & Chaves, A. R. (2004) Characterization and
changes in polyphenol oxidase from eggplant fruit (Solanum melongena L.) during
storage at low temperature. Food Chem. 88:17-24.
Constabel, C.P. & Ryan, C.A., (1998) A survey of wound- and methyl
jasmonate-induced leaf polyphenol oxidase in crop plants. Phytochemistry. 47:507-
511.

73
Constabel, C.P., Bergey, D.R. & Ryan, C. A. (1995) Systemin activates
synthesis of wound-inducible tomato leaf polyphenol oxidase via the octadecanoid
defense signaling pathway. Proc. Natl. Acad. Sci. U.S.A. 92:407–411.
Constabel, C.P., Yip, L., Patton, J.J. & Christopher, M.E. (2000) Polyphenol
oxidase from hybrid poplar. Cloning and expression in response to wounding and
herbivory. Plant Physiol. 124, 285-295.
Demeke, T. & Morris, C.F. (2002) Molecular characterization of wheat
polyphenol oxidase (PFO). Theor. Appl. Genet. 104: 813–818.
Diop, N. N., Kidric, M., Repellin, A., Gareil M., d'Arcy-Lameta, A., Thi, A. T. P.
& Zuily-Fodil, Y. A. (2004) A multicystatin is induced by drought-stress in cowpea
(Vigna unguiculata (L.) Walp.) leaves. FEBS Lett. 577: 545-550.
Doan, A., Ervin, G. & Felton, G. (2004) Temporal effects on jasmonate
induction of anti-herbivore defense in Physalis angulata: seasonal and ontogenetic
gradients. Biochem. Syst. Ecol. 32:117-126.
Dry, I.B. & Robinson, S.P. (1994) Molecular cloning and characterization of
grape berry polyphenol oxidase. Plant Mol. Biol. 26: 495-502.
Duffey, S. S. & Felton, G.W., (1991) Enzymatic antinutritive defenses of the
tomato against insects. In: P.A. Hedin, (Ed.), Naturally Occurring Pest Bioregulators,
American Chemical Society, Washington. 166-197.
Farmer, E. E.; Ryan, C. A. (1990) Interplant communication: airborn methyl
jasmonate induces synthesis of proteinase inhibitors in plant leaves. Proceedings
National Academy of Sciences, Washington. 87: 7713-7716.
Farmer, E.E. and Ryan C.A., (1992) Octadecanoid precursors of jasmonic acid
activate the synthesis of wound-inducible proteinase inhibitors, Plant Cell 4. 129–
134.

74
Felton G.W., Donato, K.K., Broadway, R.M., Duffey, S.S. (1992) Impact of
oxidized plant phenolics on the nutritional quality of dietary protein to a noctuid
herbivore Spodoptera exigua. J. Insect Physiol. 38: 277-285.
Felton, G.W., Donato, K.K., Del Vecchio, R.J., Duffey, S.S. (1989) Activation of
plant foliar oxidases by insect feeding reduces nutritive quality of foliage for noctuid
herbivores. J. Chem. Ecol. 15:2667-2694.
Fernandes, K. V. S., Sabelli, P. A., Barratt, D. H., Richardson, M., Xavier-
Filho, J., Shewry, P. R. (1993) The resistance of cowpea seeds to bruchid beetles is
not related to levels of cysteine proteinase inhibitors. Plant Mol. Biol. 23:215-219.
Firmino, F., Fernandes, K. V. S., Sales, M. P., Gomes, V.M., Miranda, M. R.
A., Domingues, S. J. S., Xavier-Filho, J. (1996) Cowpea (Vigna unguiculata), vicilins
associate with putative chitinous structures in midgut and feces of the bruchid beatles
Callosobruchus maculatus and Zabrotes subfasciatus. Braz. J. Med. Biol. Res. 29:
749-756.
Flurkey, W.H. (1986) Polyphenoloxidase in higher plants. Plant Physiol. 81:
614-618.
Flurkey, W.H. (1990) Electrophoretic and molecular weight anomalies
associated with broad bean polyphenoloxidase in SDS-PAGE electrophoresis.
Phytochemistry. 29:387-391.
Fraignier, M., Marques, L., Fleuriet, A., Mmacheix, J. (1995) Biochemical and
immunochemical characteristics of polyphenol oxidase from different fruits of Prunus.
J. Agric. Food Chem. 43:2375-2380.
Gandía-Herrero, F., García-Carmona, F., Escribano, J. (2004) Purification and
characterization of a latent polyphenol oxidase from beet root (Beta vulgaris L.). J.
Agric. Food Chem. 52:609–615.

75
Gawlik-Dziki, U., Szymanowska, U., Baraniak, B., 2007. Characterization of
polyphenol oxidase from broccoli (Brassica oleracea var. botrytis italica) florets. Food
Chem. 105: 1047-1053.
Giridhar, G. and Thimann, K. V. (1985) Interaction between Senescence and
Wounding in Oat Leaves. Plant Physiol. 78: 29-33.
Green, T. R. and Ryan, C. A. (1972) Wound-induced proteinase inhibitor in plant
leaves: A possible defense mechanism against insect. Science. 175:776-777.
Green T.R and Ryan CA. (1973) Wound-induced Proteinase Inhibitor in Tomato
Leaves: Some Effects of Light and Temperature on the Wound Response 1. Plant
Physiol. 51(1):19–21.
Geelman, R. A., Tierney, M. L., and Mullet, J. E. (1992) Jasmonic and methyl
jasmonate accumulate in wounded soybean and modulate wound gene
expression. Proc. Natl. Acad. Sci. USA. 89:4938–4941.
Gomes, V. M. and Xavier-Filho, J., (1994) Biochemical defenses of plants.
Arq. Biol. Tecnol. 37:371-383.
∙ Gomes, V.M., Oliveira, A.E.A., Xavier-Filho, J. (1996) A chitinase and a β-1,3-
glucanase isolated from the seeds of cowpea (Vigna unguiculata L.Walp) inhibit the
growth of fungi and insect pests of the seeds. J. Sci. Food Agric. 72:86–90.
Groβ. N., Wasternack, C., Köck, M. (2004) Wound-induced RNaseLE
expression is jasmonate and systeminin independent and occurs only locally in
tomato (Lycopersicon esculentum cv. Lukullus). Phytochemistry. 65:1343–1350.
Gutierrez-Campos, R., Torres-Acosta, J.A., Saucedo-Arias, L.J. and Gomez-
Lim, M.A. (1999) The use of cysteine proteinase inhibitors to engineer resistance
against potyviruses in transgenic tobacco plants. Nature Biotechnology. 17:1223 –
1226.

76
Hause, B., Hause, G., Kutter, C., Miersch, O. and Wasternack, C. (2003)
Enzymes of jasmonate biosynthesis occur in tomato sieve elements, Plant Cell
Physil. 44:643–648.
Haq, S. K., Atif, S. M. and Khan, R. H. (2004) Protein proteinase inhibitor
genes in combat against insects, pests, and pathogens: natural and engineered
phytoprotection Archives of Biochemistry and Biophysics. 431:145-159.
Hieng, B. Ugrinovic, K. Sustar-Vozlic J. and Kidric, M. (2004) Different classes
of proteases are involved in the response to drought of Phaseolus vulgaris L,
cultivars differing in sensitivity. J. Plant Physiol.161:519–530.
Hines, M.E., Osuala, C.I., & Nielsen, S.S. (1991) Isolation and partial
characterization of a soybean cystatin cysteine proteinase inhibitor of coleopteran
digestive proteolytic activity. J. Agric. Food Chem. 39:1515-1520.
Hoffmann-Benning, S., Gage, D.A., McIntosh, L., Kende, H. and Zeevaart,
J.A.D. (2002) Comparison of peptides in the phloem sap of flowering and non-
flowering Perilla and lupine plants using microbore HPLC followed by matrix-assisted
laser desorption/ionization time-of-flight mass spectrometry. Planta 216:140–147.
Irie, K., Hosoyama, H., Tabeuchi, T., Iwabuchi, K., Watanabe, H., Abe, M.,
Abe, K. & Arai, S. (1996) Transgenic rice established to express corn cystatin
exhibits strong inhibitory activity against insect gut proteinases. Plant Mol. Biol. 30:
149-157.
Jiménez, M. and García-Carmona, F. (1996) The effect of sodium dodecyl
sulphate on polyphenol oxidase. Phytochemistry. 42:150-1509.
Laemmli, U.K. (1970) Cleavage of structural proteins during assembly of the
head of bacteriophage, Nature. 227:680-685.

77
Jongsma M.A., Bakker P.L., Peters J, Bosch D. and Stiekema W. J. (1995)
Adaptation of Spodoptera exigua larvae to plant proteinase inhibitors by induction of
gut proteinase activity insensitive to inhibition. Proc Natl Acad Sci USA. 92: 8041–
8045.
Khan, V. and Andrawis, A. (1985) Inhibition of mushroom tyrosinase by
tropolone. Phytochemistry. 24. 905–908.
Kimura, M., Ikeda, T., Fukumoto, D., Yamasaki, N. & Yokemura, M. (1995).
Primary structure of cysteine proteinase inhibitor from the fruit of avocado (Persea
americana Mill). Biosci. Biotech. Biochem., 59:2328-2329.
Kondo H., Abe K., Emori Y. & Arai S. (1991). Gene organization of
oryzacystatin-II, a new cystatin superfamily member of plant origin, is closely related
to that of oryzacystatin-I but different from those of animal cystatins. Febs Lett., 278:
87 – 90.
Kouzuma, Y., Kawano, K., Kimura, M., Yamasaki, N., Kadowaki T.,
Yamamoto, K. (1996) Purification, characterization and sequencing of two cysteine
proteinase inhibitors, Sea and Scb, from sunflower (Heliantus annuus) seeds. J.
Biochem. 119:1106-1113.
Krumm, T., Bandemer, K. and Boland, W. (1995) Induction of volatile
biosynthesis in the Lima bean (Phaseolus lunatus) by leucine- and isoleucine
conjugates of 1-oxo-and 1-hydroxyindan-4-carboxylic acid: evidence for amino acid
conjugates of jasmonic acid as intermediates in the octadecanoid signalling pathway.
FEBS Lett. 377:523–529.
Laemmli, U.K. (1970) Cleavage of structural proteins during assembly of the
head of bacteriophage. Nature. 227: 680-685.
Laudert, D. and Weiler, E. W. (1998) Allene oxide synthase: a major control
point in Arabidopsis thaliana octadecanoid signalling. Plant J. 15:675-684.

78
Laveda, F., Núñez-Delicado, E., García-Carmona F., Sánchez-Ferrer, A.,
(2000) Reversible sodium dodecyl sulfate activation of latent peach polyphenol
oxidase by cyclodextrins. Arch. Biochem. Biophys. 379:1-6.
Laveda, F., Núñez-Delicado, E., García-Carmona, F. and Sánchez-Ferrer, A.
(2001) Proteolytic activation of latent paraguaya peach PFO. Characterization of
monophenolase activity. J. Agric. Food Chem. 49 :1003-1008.
Lawrence, P. K. and Koundal, K. R. (2002) Plant protease inhibitors in control
of phytophagous insects. Electronic Journal of Biotechnology. 5:93–106.
Lax, A.R., Vaughn K. C. and Templeton G. E. (1984) cDNA cloning and
expression of potato polyphenol oxidase. J. Hered. 21:285-287.
Lee, G.I. and Howe, G.A. (2003) The tomato mutant spr1 is defective in
systemin perception and the production of a systemic wound signal for defense gene
expression. Plant J. 33:567–576.
Lim, C.O., Lee, S.I., Chung, W.S., Park, S.H., Hwang, I. and Cho, M.J. (1996).
Characterization of a cDNA encoding cysteine proteinase inhibitor from Chinese
cabbage (Brassica campestris L. ssp. Pekinensis) flower buds. Plant Mol. Biol., 30:
373-379.
Lindsey, K., Casson, S. and Chilley, P. (2001) Peptides: new signalling
molecules in plants. Trends in Plant Science. 7:78-83.
Macedo, M.L.R., Andrade L. B. S., Moraes R. A. and Xavier-Filho J. (1993)
Vicilin variants and the resistance of cowpea (Vigna unguiculata) seeds to the
cowpea weevil (Callosobruchus maculatus). Comp. Biochem. Physiol.105:89-94.
Margis, R., Reis, E.M. and Villeret, V. (1998) Strutural and phylogenetic
relationships among plant and animal cystatins. Arch. Biochem. Biophys. 359:24-30.

79
Manivannan, P., Jaleel, C. A., Kishorekumar, A., Sankar, B., Somasundaram,
R., Sridharan, R. and Panneerselvam, R. (2007) Changes in antioxidant metabolism
of Vigna unguiculata (L.) Walp. by propiconazole under water deficit stress. Colloids
Surf. B. 57:69-74.
Marques, L., Fleuriet, A., Macheix, J., (1995) Characterization of multiple
forms of polyphenoloxidase from apple fruit. Plant Physiol. Biochem. 33:193-200.
Mayer, A.M. (2006) Polyphenol oxidases in plants and fungi: going places? A
review. Phytochemistry. 67: 2318-2331.
Machleidt, W., Thiele, U., Laber, B., Assfalg-Machleidt, I., Ester1, A., Wiegand,
G., Kos, J., Turk, V. and Bode, W. (1989) Mechanism of inhibition of papain by
chicken egg white cystatin. FEBS Lett. 243:234-238.
Mahanil, S., Attajarusit, J., Stout, M. J. and Thipyapong., P. (2008)
Overexpression of tomato polyphenol oxidase increases resistance to common
cutworm. Plant Science. 174:456-466.
Mazzafera, P., Robinson, S. P. (2000) Characterization of polyphenol oxidase
in coffee. Phytochemistry. 55:285-296.
McFarland, D. and Ryan C. A. (1974) Protein inhibitor-inducing factor in plant
leaves: A phylogenetic survey. Plant Physiology. 54:706-708.
Michaud, D.; Nguyen-Quoc, B. and Yelle, S. (1993) Selective inhibition of
Colorado potato beetle cathepsin H by oryzacystatins I and II. FEBS Letters. 331:
173 -176.
Michaud, D. Cantin, L. and Vrain, T. C. (1995) Carboxy-terminal truncation of
oryzacysatin II by oryzacytatin-insensitive insect digestive proteinases. Archives of
Biochemistry and Biophysics. 322:469-474.

80
Misaka, T., Kuroda, M., Iwabuchi, K. and Arai, S. (1996) Soyacystatin, a novel
cysteine proteinase inhibitor in soybean, is distinct in protein structure and gene
organization from other cystatins of animal and plant origin. Eur. J. Biochem. 240:
609-614.
Mohammadi, M. and Kazemi, H. (2002) Changes in peroxidase and
polyphenol oxidase activities in susceptible and resistant wheat heads inoculated
with Fusarium graminearum and induced resistance. Plant Science 162: 491–498.
Moore, B. M. and Flurkey, W. H. (1988) Mini-chromatofocusing of plant and
fungal polyphenoloxidases. Anal. Biochem. 172: 504-508.
Narváez-Vásquez, J.; Pearce G., Orozco-Cardenas, M. L.; Franceschi. V. R.
and Ryan C. A.; (1995) Autoradiographic and biochemical evidence for the systemic
translocation of systemin in tomato plants. Planta, 195,593-600.
Núñez-Delicado, E., Serrano-Megias, M., Perez-Lopez, A. J., Lopez-Nicolas,
J. M. (2007) Characterization of polyphenol oxidase from Napoleon grape. Food
Chem.100 :108-114.
Orozco-Cárdenas, M.L., Narváz-Vásquez, J. and Ryan, C.A. (2001) Hydrogen
peroxide acts as a second messenger for the induction of defense genes in tomato
plants in response to wounding, systemin, and methyl jasmonate. Plant Cell. 13:179–
191.
Pak, C. and van Doorn, W. G. (2005) Delay of Iris lower senescence by
protease inhibitors. New Phytologist. 165: 473–480.
Pearce, G., Strydom, D., Johnson, S. and Ryan, C.A., (1991) A polypeptide
from tomato leaves induces wound-inducible proteinase inhibitor proteins. Science.
253: 895–898.
Pearce, G., Narvaez-Vasquez, J. and C. Ryan A. 2005 Systemins Handbook
of Biologically Active Peptides, 49-53 .

81
Reymond, P., Weber, H., Damond, M., and Farmer, E. E. (2000) Differential
gene expression in response to mechanical wounding and insect feeding in
Arabidopsis. Plant Cell. 12:707-719.
Robb, D.A., Mapson, L.W., Swain, T., 1964. Activation of the latent tyrosinase
of broad bean. Nature 201, 503-504.
Roberts, K. (1992) Potential awareness of plants. Nature. 360 : 14 –15.
Robinson, S.P. and Dry, I.B. (1992)Broad bean leaf polyphenol oxidase is a 60
Kilodalton protein susceptible to proteolytic cleavage. Plant Physiol. 99:317-323.
Robinson, S.P., Loveys, B.R., Chacko, E.K., 1993. Polyphenol oxidase
enzymes in the sap and skin of mango fruit. Aust. J. Plant Physiol. 20, 99-107.
Rogelj, B., Popovic T., Ritonja, A., Strukelj, B. & Brzin, J. (1998)
Chelidocystatin, a novel phytocystatin from Chelidonium majus. Phytochemistry. 49:
1645-1649.
Rojo, E., Léon, J. & Sánchez-Serrano, J. J. (1999) Cross-talk between wound
signalling pathways determines local versus systemic gene expression in Arabidopsis
thaliana. Plant J. 20:135-142.
Ryan, C. A. (1974) Assay and Biochemical properties of the proteinase
inhibitor inducing factor, a wound hormone. Plant Physiology, 54: 328-332.
Ryan C. A. (1990) Proteinase inhibitors in plants: Genes for improving
defenses against insects and pathogens. Annual Review of Phytopathology. 28:425-
449.
Ryan C.A., Pearce, G.L. & McGurl, B. (1995) Systemin, an inducer of plant
defense proteins, and methods of use. Biotechnology Advances, 13, 728.

82
Ryan, C.A., 2000. The systemin signaling pathway: differential activation of
plant defensive genes. Biochim. Biophys. Acta 1477, 112-121.
Sales, M. P., Gomes, V. M., Fernandes, K. V. & Xavier-Filho, J. (1996) Chitin-
binding proteins from cowpea (Vigna unguiculata) seeds, Brazilian Journal of Medical
and Biological Research = Revista Brasileira de Pesquisas Medicas e Biologicas /
Sociedade Brasileira de Biofisica ... [et al.]. 29:319-326.
Sanchez-Ferrer, A., Bru, R. & Garcia-Carmona, F. (1989) Novel procedure for
extraction of a latent grape polyphenoloxidase using temperature-induced phase
separation in Triton X-114. Plant Physiol. 91:1481-1487.
Sanchez-Ferrer, A., Bru, R., Garcia-Carmona, F., 1990. Partial purification of a
thylakoid-bound enzyme using temperature-induced phase partitioning. Anal.
Biochem. 184 : 279-282.
Sanchez-Ferrer, A., Levada, F. & Garcia-Carmona, F. (1993) Substrate
dependent activation of latent potato leaf polyphenol oxidase by anionic surfactants.
J. Agric. Food Chem. 41: 1583-1586.
Sanchez-Ferrer, A., Villalba, J., Garcia-Carmona, F., 1989b. Triton X-114 as a
tool for purifying spinach polyphenol oxidase. Phytochemistry 28 : 1321-1325.
Santino, A., Gallo, A., Leone, A., Poltronieri, P., Zacheo, G. (1998).
Purification and caracterisation of a novel papain inhibitor from common bean
(Phaseolus vulgaris) seed. Physiol. Plant. 104, 293–298.
Schmelz, E. A., Carroll M. J., LeClere, S., Phipps S. M., Meredith, J., Chourey,
P. S., Alborn H. T. & Teal, P. E. A. (2006) Fragments of ATP synthase mediate plant
perception of insect attack. PNAS. 103:8894-8899.
Sellés-Marchart, S., Casado-Vela, J. & Bru-Martínez, R. (2006) Isolation of a
latent polyphenol oxidase from loquat fruit (Eriobotrya japonicaLindl.): kinetic

83
characterization and comparison with the active form. Arch.Biochem. Biophys. 446:
175-185.
Sellés-Marchart, S., Casado-Vela, J. and Bru-Martínez, R. (2007) Effect of
detergents, trypsin and unsaturated fatty acids on latent loquat fruit polyphenol
oxidase: Basis for the enzyme’s activity regulation. Arch.Biochem. Biophys. 464:295 -
305.
Sherman, T.D., Vaughn, K.C. & Duke, S.O. (1991) A limited survey of the
phylogenetic distribution of polyphenol oxidase. Phytochemistry. 30:2499-2506.
Shi, C., Daí, Y., Xu, X., Xie, Y. & Liu, Q. (2002) The purification of polyphenol
oxidase from tobacco. Protein Exp. Purif. 24: 51–55.
Shin, R., Froderman, T. & Flurkey, W. H. (1997) Isolation and characterization
of a mung bean leaf polyphenol oxidase. Phytochemistry. 45:15-21.
Simões-Araújo, J. L., Rodrigues, R. L., De, A. G., Liliane, B., Mondego, J. M. C.,
Alves-Ferreira, M., Rumjanek, N. G. & Margis-Pinheiro, M. (2002) Identification of
differentially expressed genes by cDNA-AFLP technique during heat stress in
cowpea nodules. FEBS Letters. 515: 44-50.
Sojo, M.M., Núñez-Delicado, E., Garcia-Carmona & F., Sanchez-Ferrer, A.
(1998) Partial purification of a banana polyphenol oxidase using Triton X-114 and
PEG 8000 for removal of polyphenols. J. Agric. Food Chem. 46:4924-4930.
Steffens, J. C., Harel E., Hunt, M. D. (1994) Polyphenol oxidase. In: Ellis, B.E.;
Kuroki, G.W.; Stafford, H.A. (Eds.), Recent Advances in Phytochemistry. New York:
Plenum Press. 28:275-312.
Stratmann, J. W. (2003) Long distance run in the wound response – jasmonic
acid is pulling ahead, Trends in Plant Science. 8:247-250.

84
Sugawara, H., Shybuya, K. Yoshioka.T., Hashiba and Satoh, T. (2002) Is a
cysteine proteinase inhibitor involved in the regulation of petal wilting in senescing
carnation (Dianthus cariophyllus L.) Flowers ? Journal of Exper. Botany. 53:407 -
413.
Thipyapong, P., Jeffrey, M., Wolfe D.W., Steffens, J. C., 2004.Suppression of
polyphenol oxidases increases stress tolerance intomato. Plant Sci. 167, 693-703.
Thipyapong, P., Hunt, M.D., Steffens, J.C., 1995. Systemic wound induction
of potato (Solanum tuberosum) polyphenol oxidase.Phytochemistry 40, 673–676.
Towbin, H.,Stachelin, N.T., Gordon, J. (1979) Eletroforetic transfer of protein
from polyacrylamide gels to nitrocelulose sheets, procedures and some aplications.
Proceedings of the National Academy of Sciences of the United States of America,
76: 4350 – 4354.
Turlings, T. C. J., Loughrin, J. H., McCall, P. J., Röse, U. S. R., Lewis, W. J.,
and Tumlinson, J. H. (1995) How caterpillar-damaged plants protect themselves by
attracting parasitic wasps. Proc. Natl. Acad. Sci. U. S. A. 92:4169-4174.
Valentines, M.C., Vilaplana, R., Torres, R., Usall J. & Larrigaudière, C. (2005)
Specific roles of enzymatic browning and lignification in apple disease resistance.
Postharvest Biol. Technol. 36:227-234.
Van Gelder, C.W.G., Flurkey, W.H., Wichers, H.J., (1997) Sequence and
structural features of plant and fungal tyrosinases. Phytochemistry 45, 1309-1323.
Volkov, A. G. and Haack, R. A. (1995) Insect-induced bioelectrochemical
signals in potato plants. Bioelectrochemistry and Bioenergetics. 37: 55-60.
Wang, J. & Constabel, C. P. (2003) Biochemical characterization of two
differentially expressed polyphenol oxidases from hybrid poplar..Phytochemistry. 64:
115-121.

85
Wasternack, Stenze, C., I. l, Hause, B., Hause, G., Kutter C., Maucher, H.,
Neumerkel, J., Feussner I. and Miersch, O. (2006) The wound response in tomato –
Role of jasmonic acid. J. of Plant Physiol. 163: 297-306.
Whitaker, J.R. & Lee, C.Y. (1995) Recent advances in chemistry of enzymatic
browning: an overview. In: Lee, C.Y.,Whitaker, J.R. (Eds.),EnzymaticBrowning and
its Prevention.American Chemical Society, Washington, DC. 2-7.
Wildon, D.C., Thain, J. F. , Minchin, P. E. R., Gubb, I. R., Reilly, A. J., Skipper,
Y.,D., Doherty, H.M., O’Donnel, P. J. & Bowles, D. J. (1992) Electrical signalling and
systemic proteinase inhibitor induction in the wounding plant. Nature. 360: 62 –65.
Xavier-Filho, J., Campos, F. A. P., Ary, M. B., Silva, C. P., Carvalho, M. M. M.,
Macedo, M. L. R., Lemos, F. J. A. & Grant, G. (1989) Poor correlation between the
levels of proteinase inhibitors found in seeds of different cultivars of cowpea (Vigna
unguiculata) and the resistance/susceptibility to predation by Callosobruchus
maculatus. J. Agric. Food Chem. 37:1139-1143.
Yang, C., Fujita, S., Ashrafuzzaman, M., Nakamura, N. & Hayashi, N. (2000)
Purification and characterization of polyphenol oxidase from banana (Musa
sapientum L.) pulp. J. Agric. Food Chem. 48:2732-2735.
Ye, X. Y., Wang, H. X. & Ng, T. B. (2000) Structurally dissimilar proteins with
antiviral and antifungal potency from cowpea (Vigna unguiculata) seeds, Life
Sciences. 67:3199-3207.
Yoruk, R. & Marshall, M. R. (2003) Physicochemical properties and function of
plant polyphenol oxidase: a review. J. Food Biochem. 27:361–422.
Zhao, Y., Botella, M. A., Subramanian, L., Niu, X., Nielsen, S. S., Bressan, R.
A., & Hasegawa, P. M. (1996) Two Wound-lnducible Soybean Cysteine Proteinase
lnhibitors Have Greater lnsect Digestive Proteinase Inhibitory Activities than a
Constitutive Homolog. Plant Physiol. 111:1299-1306.

86
Zhu-Salzman, K., Koiwa, H., Salzman, R.A., Shade, R.E. & Ahn, J.E. (2003)
Cowpea bruchid Callosobruchus maculatus uses a three-component strategy to
overcome a plant defensive cysteine protease inhibitor. Insect Molecular Biology
12:135–145.

1
ANEXO

2