Embed Size (px)
Citation preview

Universidade Federal do Rio de Janeiro Centro de Ciências da Saúde Instituto de Ciências Biomédicas
CARACTERIZAÇÃO MORFO-FUNCIONAL E MOLECULAR DAS CÉLULAS MESENQUIMAIS DO ESTROMA DO TECIDO ADIPOSO E SUA POSSÍVEL IDENTIDADE COM O ESTROMA DA MEDULA ÓSSEA
LEANDRA SANTOS BAPTISTATESE SUBMETIDA AO PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS
MORFOLÓGICAS DA UNIVERSIDADE FEDERAL DO RIO DE JANEIRO,
VISANDO A OBTENÇÃO DO GRAU DE DOUTOR.
ORIENTADORES: PROFA. MARIA ISABEL DORIA ROSSI - PROF. RADOVAN BOROJEVIC
Março de 2008

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

i
Examinadores:
Profa. Nance Nardi (Dept. de Genética, UFRGS)
Prof. José Mauro Granjeiro (Centro de Estudos Gerais, Instituto de Biologia, UFF)
Profa. Tatiana Lobo Coelho de Sampaio (Dept. Histologia e Embriologia, UFRJ)
Prof. César Cláudio-da-Silva (Dept. de Clínica Médica, Faculdade de Medicina, UFRJ)
como suplente externo.
Prof. Leonardo Rodrigues Andrade (Dept. Histologia e Embriologia, UFRJ), como
revisor e suplente interno.
Orientadores:
Profa. Maria Isabel Doria Rossi (Dept. Histologia e Embriologia, UFRJ).
Prof. Radovan Borojevic (Dept. Histologia e Embriologia, UFRJ).
Prof. Vivaldo Moura Neto (Coordenador do Curso de Pós-graduação em Ciências
Morfológicas (PCM).

ii
Baptista, Leandra Santos. Caracterização Morfo-Funcional e Molecular das Células Mesenquimais do Estroma do Tecido Adiposo e sua possível Identidade com o Estroma da Medula Óssea/ Leandra Santos Baptista. Rio de Janeiro: UFRJ/Pós-graduação em ciências morfológicas, 2008. xii 113p. Tese – Universidade Federal do Rio de Janeiro, Pós-graduação em ciências morfológicas, 2008. 1. Células-tronco 2. Tecido adiposo 3. Multipotencialidade – Tese.

iii
"O prazer no trabalho aperfeiçoa a obra." Aristóteles
"Procure ser um homem de valor em vez de ser um homem de sucesso."
Albert Einstein
“A vida não passa de um instante, mas basta este instante para empreendermos coisas eternas.”
E. Bersot.

iv
Aos meus pais, Divino e Sônia Baptista, pela eterna dedicação aos filhos.

v
AGRADECIMENTOS Aos meus pais por todo apoio e fé, a vida sem vocês com certeza seria uma
jornada mais difícil. AMO MUITO VOCÊS!
Ao meu irmão e melhor amigo, Raphael, por todo carinho. TAMBÉM TE AMO
MUITO!
Ao meu marido Daniel, primeiro por ouvir todos os dias as minhas derrotas e
vitórias; alegrias e tristezas; e segundo por me ajudar nos layouts dos meus trabalhos
com o seu talento inconfundível. NEM PRECISO DIZER QUE TE AMO!
A minha orientadora e amiga, Isabel, que me acolheu desde as minhas
primeiras “pipetadas” e me acompanhou com muita dedicação e carinho até aqui, me
ensinando muito mais do que ciência. Nunca vou me esquecer das nossas conversas
bem humoradas, científicas ou não. Tenho muito orgulho de ter aprendido com você.
Ao meu orientador Prof. Radovan, por ser um grande exemplo de professor,
pesquisador e pessoa.
AOS DOIS MUITO OBRIGADA PELA CONFIANÇA!
As minhas primeiras e únicas alunas, Carolina e Karina. Acho que não tenho
palavras para agradecer tamanha dedicação. Desejo a vocês toda felicidade e
sucesso do mundo; e saibam que ganharam uma grande amiga para todas as horas!
JÁ ESTOU COM SAUDADES ...
Ao meus grandes amigos e colaboradores, Cesar e Marcelo, sem vocês
simplesmente este trabalho não teria acontecido!
Ao meu revisor, Prof. Leonardo Andrade, pela tese, trabalhos desenvolvidos
em colaboração e companheirismo.
As minhas amigas do banco de células, Daiana, Danielle, Carolina, Karina,
Natália (a equipe exclusivamente feminina da Prof. Isabel) pela grande ajuda e
principalmente pelos grandes congressos e nights! Ah, não posso esquecer da
Karlinha, que apesar de não ser do banco, também é do nosso grupo de tecido
adiposo. Ainda teremos muitas estórias!
A equipe do banco de células e do transplante de medula óssea pelos vários
quebra-galhos, em especial ao Prof. Hélio Dutra.
Ao pessoal do laboratório de proliferação e diferenciação celular e de patologia
celular, em especial a Virgínia e a Prof. Valéria pela grande ajuda.
Ao pessoal da excellion, em especial ao Hamilton e a Kelly, pelos anticorpos e
análises de citometria.
A Edna e Gorete grandes desatadoras de nós! Muito obrigada.
A todos aqueles que de alguma maneira colaboraram para este trabalho.

vi
RESUMO As células-tronco mesenquimais (MSCs) podem ser isoladas de praticamente
todos os tecidos adultos e fetais com posterior expansão in vitro. O tecido adiposo
subcutâneo está principalmente envolvido na estocagem de energia além de ser
considerado como um órgão endócrino em adultos. Sua fração estromal-vascular
contém células-tronco multipotentes que dividem com a MSC da medula óssea a
capacidade de diferenciação em diferentes linhagens mesenquimais além do efeito
imunomodulador. Em vista disso, o tecido adiposo pode representar uma fonte
abundante de MSC e o potencial de aplicação destas células como uma fonte
alternativa as MSCs da medula óssea é de grande interesse. O objetivo deste trabalho
foi isolar e avaliar a multipotencialidade das MSC do tecido adiposo assim como as
suas similaridades com as MSCs da medula óssea. As MSCs do tecido adiposo foram
isoladas de lipoaspirados ou do tecido adiposo subcutâneo por digestão enzimática e
caracterizadas por citometria de fluxo. Também foram investigados o potencial de
diferenciação para as linhagens mesodérmicas e a capacidade de sustentação da
hematopoiese in vitro. A população de células derivada do tecido adiposo era
heterogênea, constituída por pré-adipócitos, MSCs, células endoteliais maduras e
células hematopoiéticas. O potencial de diferenciação em adipócitos, osteoblastos,
condroblastos e células endoteliais foi confirmado in vitro, embora um aparente
comprometimento para a linhagem adipogênica tenha sido observado através de
análises morfológicas e pela expressão constitutiva do gene PPARγ2. Além disso,
assim como as MSCs da medula óssea, as de tecido adiposo foram capazes de
sustentar a hematopoiese in vitro, embora uma tendência de geração de macrófagos
a partir de progenitores hematopoiéticos tenha sido observada, o que se assemelha ao
desenvolvimento do quadro de obesidade. Contudo, quando as MSC do tecido
adiposo foram induzidas para o fenótipo osteoblástico elas foram capazes de sustentar
a célula-tronco hematopoiética (HSC) in vitro. Estes resultados sugerem que o
comprometimento adipogênico, provavelmente ditado pelo microambiente do tecido
adiposo, pode ser superado pela indução osteogênica.

vii
ABSTRACT Mesenchymal stem cells (MSC) can be isolated from virtually all adult and fetal
tissues and expanded in vitro. Adipose subcutaneous tissue is mainly involved in
energy storage and also considered as an endocrine organ in adults. The stromal
vascular fraction contains multipotent stem cells that share with bone marrow
mesenchymal stem cell (BM-MSC) not only the capacity to differentiate into different
mesenchymal lineages, but also an immunomodulatory effect. In view of this, adipose
tissue could represent an abundant source of MSCs and the potential application of
these cells as an alternative source to bone marrow MSCs has raised great interest.
The objective of the present work was to isolated and evaluate the multipotentiality of
MSCs from adipose tissue as well as their similarities with MSCs from bone marrow.
Adipose MSCs were isolated from lipoaspirates or subcutaneous fat tissue by
enzymatic digestion and characterized by flow cytometry. The potential to differentiate
in vitro towards mesodermal lineages and the capacity to support hematopoietic
progenitors were investigated. Adipose tissue-derived cell population was
heterogeneous and contained preadipocytes, MSCs, mature endothelial cells and
hematopoietic cells. The potential to differentiate into adipocytes, osteoblasts,
condroblasts and endothelial cells was confirmed in vitro, although an apparent
commitment to adipogenic lineage was observed in morphological analyses and by the
constitutive expression of PPARγ2 gene. Besides, as bone marrow MSCs, adipose
tissue-derived MSCs were able to support hematopoiesis in vitro, although a trend to
induce proliferation and differentiation of hematopoietic progenitor cells toward
macrophages were observed, resembling the obesity scenario. However, when
adipose tissue MSCs were induced to the osteoblast phenotype they were able to
sustain the hematopoietic stem cells (HSC) in vitro. These results suggest that the
adipogenic commitment, probably dictated by the adipose tissue microenvironment,
could be bypassed by osteogenic induction.

viii
LISTA DE ABREVIATURAS AGM = (aorta-gonad-mesonephros)
α-SMA = (alpha smooth muscle actin)
Ang = (Angiopoietina)
BMP (bone morphogenetic protein)
CD = (cluster of differentiation)
CFU-F = (colony-forming units–fibroblastic)
CFU-GM = (colony unit formation of granulocytes and macrophages)
CFU-S = (colony unit formation of spleen)
CLP = (commun lymphoid progenitors)
CMP = (commun myeloid progenitors)
Flk-1 = (Vascular endothelial growth factor receptor)
GM-CSF = (Granulocyte-macrophage colony-stimulating factor)
G-CSF = (Granulocyte colony-stimulating factor)
GvHD = (Graft-versus-host disease)
HSC = (Hematopoietic stem cell)
HUVEC = (human umbilical cord endothelial cells)
IL = (Interleukin)
KDR = (Vascular endothelial growth factor receptor 2)
LIF = (leukemia inhibitory factor)
LTBMC = (long-term bone marrow culture)
LTC-IC = (long term clonogenic- initiating cell)
MAPC = (multipotent adult progenitor cells)
MCP-1 = (monocyte chemotactic protein-1)
M-CSF (Macrophage colony-stimulating factor)
MGG = (May-Grünwald – Giemsa)
MO = (medula óssea)
MSC = (Mesenchymal stem cell)
OPG = (osteoprotegerin)
PDGF = (platelet-derived growth factor)
PECAM-1 = (platelet endothelial cell adhesion molecule-1)
PPARγ2 = (peroxisome proliferator-activated receptor-gamma-2)
RANKL = (Receptor activator of nuclear factor kappa B ligand)
SCF = (stem cell factor)
SDF-1α = (Stromal derived factor- 1alpha)
TA = (tecido adiposo)

ix
TAB = (tecido adiposo branco)
TAM = (tecido adiposo marrom)
Tal-1 = (T-cell acute lymphocytic leukemia 1)
TGFβ = (Transforming growth factor-beta)
TNF-α = (tumor necrosis factor-alpha)
Tie-2 = (Angiopoietin-1 receptor)
VCAM = (vascular cell adhesion molecule)
VE (vascular–endothelial)-caderina VEGF = (Vascular endothelial growth factor)
vWf = (von-Willebrand factor)

x
1. INTRODUÇÃO 1.1. CÉLULAS – TRONCO .............................................................................................1 1.1.a. CÉLULAS – TRONCO PLURIPOTENTES ...........................................................1 1.1.b. POTENCIAL DE DIFERENCIAÇÃO DAS CÉLULAS – TRONCO NO ORGANISMO ADULTO ..................................................................................................2 1.2. MEDULA ÓSSEA (MO) ...........................................................................................4 1.2.a. MICROAMBIENTE MEDULAR .............................................................................4 1.2.b.HEMOPOIETINAS, QUIMIOCINAS E MATRIZ EXTRACELULAR........................7 1.2.c. CÉLULAS-TRONCO HEMATOPOIÉTICAS (HSC) ..............................................9 1.2.d. CÉLULAS-TRONCO MESENQUIMAIS (MSC) ..................................................10 1.3. TECIDO ADIPOSO (TA) ........................................................................................12 1.3.a. TECIDO ADIPOSO (TA) COMO UM ÓRGÃO ENDÓCRINO ............................12 1.3.b. MSC DE TECIDO ADIPOSO (TA) .....................................................................14 1.4. ONTOGENIA DAS MSC ........................................................................................16 1.4.a. PERICITOS ........................................................................................................16 1.4.b. HEMANGIOBLASTOS E CÉLULAS DO SOMITO .............................................16 1.4.c. MESANGIOBLASTO ..........................................................................................19 1.4.d.CÉLULAS DA CRISTA NEURAL ........................................................................20 1.5. MSC NO ORGANISMO ADULTO .........................................................................21 1.5.a. MARCADORES DE SUPERFÍCIE .....................................................................22 1.5.b. DIFERENCIAÇÃO ORTODOXA ........................................................................23 1.5.c. REGULAÇÃO DAS VIAS OSTEOGÊNICA E ADIPOGÊNICA ...........................24 1.5.d. O POTENCIAL DE DIFERENCIAÇÃO DAS DIFERENTES FONTES DE MSC ..............................................................................................................................25 1.5.e. DIFERENCIAÇÃO NÃO-ORTODOXA ...............................................................26 1.5.f. POTENCIAL IMUNOMODULADOR ....................................................................27 1.6. ENSAIOS CLÍNICOS COM MSCs ........................................................................28 2. OBJETIVOS 2.1. OBJETIVO GERAL ................................................................................................30 2.2. OBJETIVOS ESPECÍFICOS .................................................................................30 3. METODOLOGIA 3.1. AMOSTRAS E TÉCNICAS DE ISOLAMENTO .....................................................31 3.1.a. AMOSTRAS E CÉLULAS ..................................................................................31 3.1.b. ISOLAMENTO DE MSCs ...................................................................................31 3.1.c. OBTENÇÃO DE CÉLULAS DO CORDÃO E DO SANGUE DE CORDÃO UMBILICAL ...................................................................................................................32 3.2. ENSAIOS FUNCIONAIS .......................................................................................33 3.2.a. INDUÇÃO PARA A VIA OSTEOGÊNICA ..........................................................33 3.2.b. INDUÇÃO DE MSCS PARA A VIA ADIPOGÊNICA ..........................................34 3.2.c. INDUÇÃO DE MSCS DE TA PARA A VIA CONDROGÊNICA ..........................34 3.2.d. INDUÇÃO DE MSCS DE TA PARA A VIA ENDOTELIAL .................................35 3.2.e. SUSTENTAÇÃO DA HEMATOPOESE EM CULTURAS DE LONGA DURAÇÃO ...................................................................................................................36

xi
3.3. TÉCNICAS EXPERIMENTAIS ..............................................................................37 3.3.a. CITOMETRIA DE FLUXO ..................................................................................37 3.3.b. SELEÇÃO DAS CÉLULAS DE TA .....................................................................38 3.3.c. HISTOLOGIA, IMUNOFLUORESCÊNCIA E IMUNOHISTOQUÍMICA ..............38 3.3.d. MICROSCOPIA ELETRÔNICA DE VARREDURA (MEV)..................................39 3.3.e. EXTRAÇÃO DE RNA TOTAL E SÍNTESE DE cDNA ........................................40 3.3.f. REAÇÃO DE POLIMERASE EM CADEIA COM TRANSCRIÇÃO REVERSA (RT– PCR) ....................................................................................................................40 3.3.g. ANÁLISE ESTATÍSTICA ....................................................................................40 4. RESULTADOS 4.1. MORFOLOGIA E FENÓTIPO DAS MSC DE TA. .................................................42 4.1.a. ANÁLISE DA POPULAÇÃO TOTAL (FIG. 12 – 15) ...........................................42 4.1.b. ANÁLISE DE POPULAÇÕES CELULARES SELECIONADAS (FIG. 16 – 20) ...............................................................................................................43 4.2. MULTIPOTENCIALIDADE ....................................................................................50 4.2.a. DIFERENCIAÇÃO PARA AS TRÊS LINHAGENS MESODÉRMICAS: ADIPOGÊNICA, OSTEOGÊNICA E CONDROGÊNICA (FIG. 21 – 31) ..............................................................................................................50 4.2.b. DIFERENCIAÇÃO PARA A LINHAGEM ENDOTELIAL (FIG. 32 – 35) .............58 4.3. POTENCIAL DE SUSTENTAÇÃO HEMATOPOIÉTICA........................................62 4.3.a. CO-CULTURA COM CÉLULAS MONONUCLEARES DE SANGUE DE CORDÃO UMBILICAL (FIG. 36 – 41) ..........................................................................62 4.3.b. ANÁLISES DO CO-CULTIVO COM CÉLULAS CD34+ DE SANGUE DE CORDÃO UMBILICAL (FIG. 42 – 45) ..........................................................................70 5. DISCUSSÃO 5.1. FENÓTIPO DAS MSC DE TA................................................................................73 5.1.a. A FRAÇÃO ADERENTE DO TA É CONSTITUÍDA POR CÉLULAS DE ORIGEM ESTROMAL E ENDOTELIAL .......................................................................................73 5.1.b. AS CÉLULAS CD34+ DA FRAÇÃO ADERENTE DO TA ADQUIREM O MARCADOR CD105 IN VITRO ....................................................................................76 5.2. MULTIPOTENCIALIDADE ....................................................................................77 5.2.a. AS MSC DE TA SOB INDUÇÃO ADQUIREM FENÓTIPOS OSTEOGÊNICO, ADIPOGÊNICO E CONDROGÊNICO IN VITRO .........................................................77 5.2.b. AS MSC DE TA SÃO MORFOLOGICA E GENETICAMENTE DIFERENTES DAS MSC DE MO ........................................................................................................78 5.2.c. AS MSC DE TA APRESENTAM UM POTENCIAL ADIPOGÊNICO DIFERENTE DAS MSC DE MO ........................................................................................................79 5.2.d. AS MSC DE TA PARECEM SER COMPROMETIDAS COM A VIA ENDOTELIAL...............................................................................................................81 5.3. POTENCIAL DE SUSTENTAÇÃO HEMATOPOIÉTICA........................................84 5.3.a. AS MSC DE TA APRESENTAM SIMILARIDADES COM AS DE MO ................84 5.3.b. AS MSCS PRÉ-INDUZIDAS PARA AS VIAS ADIPOGÊNICA E OSTEOGÊNICA APRESENTAM PERFIS DE GERAÇÃO DE CÉLULAS HEMATOPOIÉTICAS DISTINTOS ..................................................................................................................85 5.3.c. AS MSCS DE TA QUANDO INDUZIDAS PARA A LINHAGEM OSTEOBLÁSTICA SÃO CAPAZES DE MANTER AS HSCs .......................................87

xii
5.4.CONSIDERAÇÕES FINAIS ...................................................................................89 6. CONCLUSÕES 6.1.FENÓTIPO..............................................................................................................91 6.2.MULTIPOTENCIALIDADE......................................................................................92 6.3.POTENCIAL DE SUSTENTAÇÃO HEMATOPOIÉTICA.........................................92 6.4.CONCLUSÃO FINAL...............................................................................................92 7. REFERÊNCIAS BIBLIOGRÁFICAS ........................................................................93

1
1. INTRODUÇÃO 1.1. CÉLULAS – TRONCO 1.1.a. Células – tronco pluripotentes
As células-tronco são consideradas as unidades responsáveis pelos processos
de geração e de regeneração do organismo adulto (WEISSMAN, 2000). As células-
tronco adultas são definidas pela capacidade de auto-renovação e pela geração de
células maduras de um tecido particular ao longo da diferenciação (REYA et al., 2001).
Mais recentemente, a hipótese de que estas células localizam-se em regiões
específicas, denominadas nichos, nos diferentes tecidos foi amplamente demonstrada
(KAPLAN et al., 2007). Acredita-se que o nicho em que essas células residem é
determinante na regulação das suas propriedades de células-tronco (SPRADLING et
al., 2001).
As células-tronco pluripotentes, teoricamente, podem gerar todos os tipos
celulares e possuem uma alta capacidade de proliferação em um estado
indiferenciado por um período prolongado em cultura. Estas células são derivadas
preferencialmente de tecidos embrionários, principalmente da massa interna do
blastocisto (DONOVAN & GEARHART, 2001). Recentemente, células pluripotentes
têm sido identificadas na medula óssea (MO) adulta (REYES et al., 2001; KUCIA et al.,
2007) e também no sangue de cordão (KUCIA et al., 2007). Células-tronco com estas
características, denominadas de células adultas progenitoras multipotentes (MAPC,
Multipotent Adult Progenitor Cells), expressam marcadores de células embrionárias
(Oct-4, Rex-1, SSEA-1) e quando implantadas no blastocisto foram capazes de gerar
todos os tipos celulares (JIANG et al., 2002). Foi demonstrado, ainda, que as MAPC
também possuem capacidade de reconstituição de longa duração do sistema
hematopoiético (SERAFINI et al., 2007), característica anteriormente atribuída
somente às células-tronco hematopoiéticas (HSC, Hematopoietic Stem Cell)
(WEISSMAN, 2000). Porém, esses trabalhos vêm sendo de difícil reprodução pela
comunidade científica (CHECK, 2007).
As células-tronco embrionárias representam uma importante ferramenta de
estudo do desenvolvimento em diversos tecidos (HIRASHIMA et al., 1999; KEHAT et
al., 2001). Muitos sistemas de diferenciação in vitro têm sido desenvolvidos para as
células-tronco embrionárias murinas, incluindo métodos reprodutíveis de diferenciação
em precursores hematopoiéticos e neurais, cardiomiócitos, células secretoras de
insulina, células endoteliais e vários outros tipos celulares (AMIT & ITSKOVITZ-
ELDOR, 2002).

2
A habilidade das células-tronco pluripotentes de dar origem a um enorme
repertório de células diferenciadas é a razão pela qual elas podem vir a ser úteis em
protocolos de terapias celulares (DONOVAN & GEARHART, 2001). Muitos esforços
têm sido feitos na derivação de novas linhagens de células-tronco embrionárias
humanas (LEVENSTEIN et al., 2006), mas devido a problemas éticos, ao potencial
oncogênico e a rejeição pelo hospedeiro, outras fontes para o uso em terapias
celulares têm sido pesquisadas (WANG et al., 2000).
1.1.b. Potencial de diferenciação das células – tronco no organismo adulto
As células-tronco adultas estão presentes em praticamente todos os tecidos de
animais adultos e contribuem para os processos de reparo e da homeostasia tecidual.
Tais células podem ser multipotentes, gerando várias linhagens de células
diferenciadas, ou oligopotentes, gerando somente uma ou poucas linhagens. Em
tecidos adultos, a população de células-tronco é rara (GOODELL, 2001; KRAUSE,
2002). Estudos recentes (ver abaixo) surpreenderam a comunidade científica, ao
demonstrarem que células-tronco adultas podem adotar perfis de expressão e fenótipo
funcional de células de outros tecidos. Esta propriedade foi denominada plasticidade
(HERZOG et al., 2003).
A primeira evidência de plasticidade das células-tronco adultas veio de estudos
de transplantes de MO em modelos humanos e animais, em que foi possível distinguir
através de diferentes métodos, entre as células do doador e as do recipiente
(GOODELL, 2001). A análise de diferentes órgãos revelou a presença de células
derivadas do doador em tecidos não-hematopoiéticos, incluindo tecidos epiteliais
(KRAUSE et al., 2001), coração (QUAINI et al., 2002), trato gastrointestinal
(OKAMOTO et al., 2002) e músculo esquelético, onde permaneceram enxertadas por
uma década na medula e músculo esquelético do hospedeiro (GUSSONI et al., 2002).
Diversos mecanismos podem ser especulados para a plasticidade de células-
tronco derivadas da MO (Fig. 1): (1) Células-tronco não comprometidas com a
linhagem hematopoiética poderiam se diferenciar nos diversos tipos celulares; (2)
células-tronco já comprometidas poderiam assumir novos fenótipos pelo mecanismo
de transdiferenciação que pode ser direto; (3) ou indireto, onde primeiro a célula-
tronco já comprometida deve passar pelo processo de desdiferenciação; (4) e por
último, as células-tronco poderiam fusionar com as células do tecido residente,
assumindo assim, o novo fenótipo (HERZOG et al., 2003).
Em todos estes casos, o nicho tem papel decisivo no novo comprometimento
dessas células. As células-tronco são capazes de responder a estímulos externos,

3
como por exemplo, ligação a proteínas da matriz específicas para cada tipo de tecido,
que levam ao desencadeamento de diversas vias, que por sua vez poderiam levar a
diferenciação, proliferação ou quiescência (SPRADLING et al., 2001). Além da
composição protéica, a elasticidade da matriz também pode regular as vias de
diferenciação das células-tronco. Foi demonstrado que células-tronco mesenquimais
do estroma da medula quando cultivadas sobre matrizes mais resistentes e pouco
elásticas diferenciam-se para a linhagem osteogênica, enquanto o contato com
matrizes mais flexíveis e maleáveis favorece a diferenciação para a linhagem
neurogênica (ENGLER et al., 2006). Este fenômeno pode estar associado a
modificações da forma celular e do citoesqueleto, como veremos adiante.
Figura 1. Possíveis mecanismos de plasticidade das células-tronco em resposta ao nicho. Fonte: HERZOG et al., 2003.
Vários ensaios in vivo de regeneração tecidual em modelos animais após
trauma ou lesão induzida, obtiveram resultados ambíguos em relação à plasticidade
das células-tronco de MO. Esta discordância de resultados pode ser devida a
problemas técnicos, uma vez que um conjunto de análises e controles experimentais
apropriados são necessários para melhor avaliar os resultados (GROVE et al., 2004).
Ensaios clínicos em protocolos de medicina regenerativa, principalmente na
área de cardiologia, relataram o efeito benéfico das células-tronco de MO na fisiologia
tecidual. Esse efeito pode ter sido alcançado, a exemplo do descrito anteriormente,
através da secreção de fatores, como por exemplo, fatores pró-angiogênicos,
estimulando assim a regeneração tecidual de forma indireta, ou por mecanismos de
diferenciação, transdiferenciação ou fusão celular. Nenhum desses mecanismos foi
comprovado de maneira irrefutável e a proposta mais provável seria que a
regeneração tecidual se daria por uma combinação destes (DIMMELER et al., 2005).

4
1.2. MEDULA ÓSSEA 1.2.a. Microambiente medular
Na MO residem ao menos dois tipos de células-tronco: as HSCs e as células-
tronco mesenquimais (MSC, Mesenchymal Stem Cell); estes dois tipos celulares co-
existem funcional e espacialmente. In vivo, as células hematopoiéticas são
continuamente formadas enquanto as células estromais são mais quiescentes e estão
ligadas à matriz extracelular, formando estruturas complexas, multicelulares (Fig.2)
(BIANCO & ROBEY, 2000).
Figura 2. Estrutura do nicho. (A) Células-tronco quiescentes são estocadas em contato com células do estroma (em azul) responsáveis por sua manutenção. (B) Quando estas células-tronco entram em contato com outros tipos de células estromais (em verde) os mecanismos de diferenciação e/ou auto-renovação são acionados. Adaptado de Wilson & Trumpp 2006.
As células estromais são tanto de origem mesenquimal como hematopoiética e
incluem osteoblastos, células reticulares ou adventiciais, células acumuladoras de
gordura, miofibroblastos, células endoteliais, células musculares lisas da parede
vascular, células dendríticas e macrófagos (BIANCO et al., 2001; VERFAILLIE, 2000).
A primera evidência de um papel importante das células estromais na sustentação da
hematopoese veio da observação de FRIEDENSTEIN et al. (1966), que através do
transplante de células estromais na cápsula renal, mostraram a formação de ossículos
com células hematopoiéticas derivadas do hospedeiro.

5
Dentro da população de células estromais estão as MSCs, que são capazes
de auto-renovação, bem como de diferenciação em muitos tecidos derivados do
mesênquima. As características comuns entre as células referidas por diferentes
pesquisadores como MSC é que elas crescem em cultura como células aderentes e
possuem capacidade de diferenciação assim como as HSCs (KRAUSE, 2002). Um
dos principais mecanismos pelo qual as MSCs regulam a hematopoiese é pela
produção de citocinas, mas não está claro se MSCs de diferentes fontes possuem
perfis de citocinas específicos na regulação de proliferação e diferenciação das células
hematopoiéticas (DAZZI et al., 2006). Acreditava-se que a hierarquia do componente estromal fosse como a do
sistema hematopoiético, rígida, ou seja, o comprometimento das células precursoras
parecia ser progressivo e irreversível. Porém, modelos mais recentes de plasticidade,
sugerem que as MSCs, uma vez que tenham se transformado em células
acumuladoras de gordura, podem reverter o seu fenótipo para um estroma de
sustentação de hematopoiese dependendo das condições fisiológicas do organismo
(BIANCO & ROBEY, 2000).
Durante o desenvolvimento embrionário, o estabelecimento da hematopoese
definitiva está ligado às células endoteliais na aorta dorsal (PIETILÄ & VAINIO, 2005).
A chegada das HSCs ocorre somente depois do desenvolvimento de estruturas
específicas organizadas pelas células vasculares (CHARBORD et al., 1996). Após a
sua formação, as células estromais ainda são responsáveis por prover a maioria dos
fatores requeridos para o ordenado desenvolvimento das células progenitoras
hematopoiéticas (VERFAILLIE, 2000).
A caracterização do papel do microambiente da MO no processo
hematopoiético tornou-se possível com o estabelecimento de culturas dependentes de
estroma, inicialmente desenvolvidas por DEXTER et al. (1985), também nomeadas de
culturas de longa duração de MO (LTBMC, long-term bone marrow culture). Quando a
MO é examinada a partir de aspirados ou biópsia, células hematopoiéticas em
diferentes estágios de diferenciação podem ser encontradas em áreas distintas do
espaço medular (QUESENBERRY et al., 1994). O microambiente hematopoiético
produz e concentra citocinas hematopoiéticas que podem induzir ou inibir a
proliferação e diferenciação de células progenitoras (VERFAILLIE, 2000). As células
hematopoiéticas ao longo do seu comprometimento em células diferenciadas se
encaminham para a parte mais central da MO, caindo nos sinusóides e alcançando a
corrente sanguínea (CHARBORD et al., 1996).
A localização diferenciada dos progenitores hematopoiéticos na MO tem levado
ao conceito de um nicho específico para as HSCs. Por exemplo, as células mais

6
precursoras podem ser encontradas na região subendosteal em proximidade com os
osteoblastos (Fig. 3) (QUESENBERRY et al., 1994). O contato de células primitivas
com um microambiente de sustentação de células-tronco aumenta as divisões
assimétricas de progenitores, tanto primitivos como comprometidos (PUNZEL et al.,
2003).
Dois estudos recentes comprovaram que as células osteoblásticas
representam um componente regulatório do microambiente da MO (Fig. 2). As células
osteoblásticas funcionam como um componente chave na organização dos nichos de
HSC, regulando o tamanho da população de HSC (CALVI et al., 2003; ZHANG et al.,
2003). Na presença de osteoblastos, a maioria dos progenitores hematopoiéticos se
mantém em um estágio imaturo (BALDUÍNO et al., 2005; TAICHMAN et al., 2005).
Além disso, na MO a expressão de osteopontina, uma molécula associada à
manutenção do fenótipo imaturo das células hematopoiéticas, é restrita à superfície
endosteal do osso e as células hematopoiéticas primitivas demonstram adesão
específica a esta proteína (NILSSON et al., 2005). Porém, o nicho subendosteal não é
o único para as HSCs. Estas células também interagem com as células endoteliais dos
sinusóides da MO e de sítios extramedulares. As células endoteliais expressam
fatores que regulam a manutenção e função das HSCs (KIEL et al., 2005), embora
existam evidências de diferenças importantes entre os dois possíveis nichos de HSC
na capacidade de manutenção desta população (HAYLOCK et al., 2007).
Figura 3. Microambiente da MO. Fonte: Dazzi et al., 2006.

7
1.2.b. Hemopoietinas, quimiocinas e matriz extracelular
Desenvolvimento, homeostasia, circulação e capacidade de resposta do
sistema hematopoiético são altamente regulados por uma rede complexa de
comunicações que é mediada por sinais intracelulares. Estes sinais são
desencadeados por contato direto célula-célula ou célula-matriz ou ainda pela
liberação de citocinas solúveis (KURZROCK, 2000). Dentre as vias de sinalização
responsáveis pela manutenção da capacidade tronco do sistema hematopoiético, as
proteínas da via de Wnt (VAN DEN BERG et al., 1998), assim como a via de notch
com o seu ligante delta-1 são potencialmente importantes, tanto na expansão dos
progenitores mielóides como dos linfóides (OHISHI et al., 2002). O papel de Wnt na
hematopoiese foi mais bem descrito pela purificação da proteína Wnt3a, a qual induz a
auto-renovação das HSCs (WILLERT et al., 2003).
Muitos destes fatores de crescimento hematopoiético são produzidos pelas
células estromais da MO, as quais residem em proximidade com os precursores
hematopoiéticos (KURZROCK, 2000). Em geral, dois ou mais tipos de citocinas são
necessárias para cada linhagem hematopoética completar seu potencial de
desenvolvimento (WEISSMAN, 2000). As células estromais produzem fatores de
crescimento a partir da adesão de progenitores. Esta adesão pode ocorrer através da
expressão de ligantes na superfície da célula, tais como a molécula de adesão celular
vascular (VCAM-1, vascular cell adhesion molecule-1), com a expressão na superfície
celular de fatores de crescimento, como, por exemplo, fator de crescimento de células
tronco (SCF, stem cell factor), ou por componentes da matriz extracelular, secretados
pelas células de estroma (KOVACH et al., 1995). Em culturas de MO de longa
duração, colágeno, fibronectina e laminina são secretados inicialmente, e a deposição
extracelular destas proteínas coincide com uma hematopoiese ativa (ABBOUD &
LICHTMAN, 2001).
Os osteoblastos humanos expressam várias moléculas conhecidamente
capazes de modular a hematopoiese, como o fator de estimulação de granulócitos (G-
CSF-granulocyte colony-stimulating factor), o fator de estimulação de granulócitos e de
macrófagos (GM-CSF-granulocyte-macrophage colony-stimulating factor), o fator de
estimulação de macrófagos (M-CSF-macrophage colony-stimulating factor), a
interleucina 1beta (IL-1β-interleukin-1beta), a interleucina 6 (IL-6-interleukin-6), a
interleucina 7 (IL-7-Interleukin-7), o fator inibitório de leucemia (LIF-leukemia inhibitory
factor), a osteoprotegerina (OPG-osteoprotegerin), o receptor ativador do ligante do
fator nuclear kappa B (RANKL-receptor activator of nuclear factor kappa B ligand), o
fator derivado de estroma (SDF-1α-stromal derived factor- 1alpha), o fator de necrose

8
tumoral alpha (TNF-α-Tumor necrosis factor-alpha) e o fator de crescimento vascular
endotelial (VEGF-vascular endothelial growth factor) (GREENFIELD et al., 1996;
TAICHMAN & EMERSON, 1998; HOLFBAUER et al., 2000; PONOMARYOV et al.,
2000). A quimiocina SDF-1α é particularmente importante na interação HSC-
osteoblasto, além de ser responsável pela retenção dos progenitores hematopoéticos
na medula (HAMADA et al., 1998; WANG et al., 1998; PELED et al., 1999).
A célula acumuladora de gordura é a célula estromal mais abundante na MO
adulta, onde o seu verdadeiro papel ainda gera controvérsias. Existem algumas
hipóteses para sua função: (1) As células acumuladoras de gordura possuem um
papel passivo, ocupando o excesso de espaço na cavidade medular (GIMBLE et al.,
1996); (2) podem possuir um papel ativo no metabolismo lipídico sistêmico; (3) são
responsáveis por prover um reservatório energético localizado na MO (YOKOTA et al.,
2002); e (4) podem contribuir diretamente na promoção da hematopoiese e influenciar
a osteogênese (GIMBLE et al., 1996).
Uma das principais proteínas secretada pelas células acumuladoras de gordura
é a leptina, que promove a formação de osteoblastos e hematopoiese, ao mesmo
tempo em que inibe a adipogênese (UMEMOTO et al., 1997; LAHARRAGUE et al.,
1998; THOMAS et al., 1999). Outras proteínas produzidas por essas células, com
papel ativo na hematopoiese, são as adiponectinas (DIMASCIO et al., 2007), responsáveis por bloquear a mielopoiese em ensaios clonais de precursores de
células hematopoiéticas (LAHARRAGUE et al., 2000; YOKOTA et al., 2000).
Adicionalmente, foi descrito que as HSC expressam duas classes de receptores para a
proteína adiponectina e a ligação destes promove a proliferação das HSC, avaliado
por ensaios in vitro e in vivo (DIMASCIO et al., 2007).
A diferenciação para a via adipogênica se acompanha de redução da
expressão constitutiva de M-CSF, SCF, IL-6, LIF, osteopontina e osteocalcina
(UMEZAWA et al., 1991; GIMBLE et al., 1992; DORHEIN et al., 1993), e o seu
potencial de sustentação hematopoiética pode estar relacionado com a diferenciação
para as linhagens mieloíde e linfóide (células B) de progenitores hematopoiéticos
CD34+ (CORRE et al., 2004).
Curiosamente, células fibroblastóides derivadas de órgãos não hematopoéticos
liberam frequentemente os mesmos tipos de citocinas, mas não são capazes de
manter a hematopoiese, sugerindo que as células fibroblastóides derivadas de órgãos
hematopoéticos podem expressar moléculas importantes para a hematopoese que
ainda não foram descritas (VERFAILLIE, 2000). Por exemplo, fibroblastos da derme
não expressam VCAM-1 (CD106), e isto é responsável pela incapacidade destes
fibroblastos de manter a hematopoese de células B (Burger et al., 2001).

9
1.2.c. Células-tronco hematopoiéticas
Todos os tipos de células sanguíneas derivam de um precursor comum, a
HSC. Estas células são definidas como as únicas células capazes de reconstituição de
longo prazo de todo o sistema hematopoiético, após uma irradiação letal (BONIFER et
al., 1998). A reconstituição de longo prazo de hospedeiros irradiados e os posteriores
transplantes para hospedeiros secundários representam os principais critérios para a
demonstração das propriedades das HSC; que são a auto-renovação e a
diferenciação para as diferentes linhagens do sangue (KRAUSE et al., 2001). No
sangue circulante existe uma pequena proporção destas células (tronco e
progenitoras), que podem ser isoladas por técnicas especiais (QUESENBERRY &
COLVIN, 2001).
TILL & MCCULLOUGH (1961) comprovaram a existência das HSCs através de
um ensaio in vivo de colônias capaz de identificar células-tronco multipotentes, a
unidade formadora de colônias do baço (CFU-S-colony unit formation of spleen).
Posteriormente, foram descritos ensaios de colônia in vitro para células progenitoras
hematopoiéticas (BRADLEY & METCALF, 1966), como a unidade formadora de
colônias de granulócitos e de macrófagos (CFU-GM-colony unit formation of
granulocytes and macrophages). Os ensaios clonogênicos levaram à descrição de
moléculas que inibiam ou estimulavam o crescimento de células hematopoiéticas
(PLUZNIK & SACHS, 1965). Outro ensaio in vitro, denominado de célula iniciadora de
culturas de longo prazo (LTC-IC-Long Term Culture-Initianting Cell), baseado nas
culturas de longa duração desenvolvidas por Dexter et al., (1965) e que se baseia na
capacidade de manter a produção de células hematopoiéticas por longos períodos,
permitiu caracterizar a população de HSC humana (CROISILLE et al., 1994). O
desenvolvimento destas técnicas levou ao desenho de um modelo hierárquico para o
sistema hematopoiético, onde as HSC de reconstituição de longo prazo dão origem a
HSC de reconstituição de curto prazo, que por sua vez dão origem a progenitores
multipotentes, os quais irão gerar progenitores comprometidos com linhagens únicas
(Fig. 4) (REYA et al., 2001). A diferenciação dos progenitores hematopoiéticos é regulada por fatores de
transcrição; não apenas pela ausência ou expressão do fator, mas em alguns casos
pela baixa ou alta expressão do mesmo, como o que ocorre com o fator de trancrição
hematopoiético - PU.1, que, em baixas concentrações induz a diferenciação de
progenitores jovens para linfócitos B e em altas concentrações para macrófagos
(DEKOTER & SINGH, 2000).

10
1.2.d. Células-tronco mesenquimais No estroma da MO existe uma subpopulação de células não-hematopoiéticas
que foram denominadas - MSCs ou células-tronco estromais. Estas células podem ser
expandidas ex vivo e induzidas tanto in vitro como in vivo, para fenótipos
terminalmente diferenciados que incluem osteoblastos, condrócitos, adipócitos,
tenócitos (células do tendão), células musculares e para um estroma sustentador de
hematopoiese (MINGUELL et al., 2001). Ainda não há um consenso entre os
pesquisadores sobre a definição da MSC ou sobre como elas devem ser isoladas e
expandidas in vitro. Além disso, diferentemente das HSCs, nenhum perfil específico de
expressão de marcadores de superfície capazes de identificar em definitivo tal célula
foi descrito (HERZOG et al., 2003).
Figura 4. As HSC podem ser subdivididas em células de auto-renovação de longo prazo, de curto-prazo e em progenitores multipotentes (as setas vermelhas indicam a auto-renovação). Os progenitores multipotentes dão origem a progenitores linfóides e a progenitores mielóides. Fonte: Reya et al., 2001. As células-tronco estromais da MO foram primeiramente isoladas e
caracterizadas através dos trabalhos de FRIEDENSTEIN et al. (1966 e 1968). Eles
caracterizaram essas células como aderentes, clonogênicas, não-fagocíticas, com
morfologia fibroblastóide, definida como a unidade formadora de colônias
fibroblastóides (CFU-F-colony forming units fibroblastic). Essas colônias
fibroblastóides, sob condições experimentais apropriadas dão origem a um grande
espectro de células diferenciadas de tecidos conjuntivos (FRIEDENSTEIN et al.,
1974).

11
Figura 5. Durante o desenvolvimento na medula óssea, as MSCs se tornam comprometidas com a linhagem osteogênica sob influência do fator de transcrição CBFA-1, de modo que as células adquirem um fenótipo de osteoblastos. Quando há formação óssea, estas células se diferenciam em um estroma medular primitivo de sustentação da hematopoese. Na vida pós-natal as MSCs se diferenciam em células acumuladoras de gordura, podendo reverter seu fenótipo para ambos os tipos celulares de acordo com a necessidade do organismo. Fonte: Bianco & Robey, 2000.
Uma única colônia de CFU-F gera ossículos idênticos aos gerados por
linhagens estromais não-clonais, as quais provavelmente representam células de
fenótipos múltiplos e diferenciados. Porém CFU-Fs são heterogêneos em relação à
capacidade osteogênica. Nesse estudo, em torno de 50% das colônias demonstraram
potencial osteogênico, onde algumas desenvolveram osso e tecido hematopoiético,
outras desenvolveram somente osso sem hematopoiese, e até nenhum osso, avaliado
por ensaios in vivo (KUZNETSOV et al., 1997). Baseado na diversidade de clones de
CFU-F, foi desenvolvido o conceito de que cartilagem, osso, e outros tecidos
conjuntivos derivam de um ancestral comum, a célula-tronco estromal ou MSC. Essas
células-tronco permanecem no organismo pós-natal e residem no organismo adulto
(Fig. 5) (BIANCO & ROBEY, 2000). No entanto, só recentemente, a propriedade de
auto-renovação, uma característica de células-tronco, das MSC de MO foi
demonstrada. SACHETTI et al. (2007) demonstraram que células da MO humana,
isoladas de CFU-F e que expressam CD146, um marcador característico de células
perivasculares, têm propriedade de auto-renovação e multipotencialidade.

12
1.3. TECIDO ADIPOSO (TA)
Na maioria dos mamíferos, estão presentes dois tipos de tecido adiposo:
marrom (TAM) e branco (TAB). Os dois tecidos possuem papel ativo na regulação da
homeostase energética através do metabolismo (HAHN & NOVAK, 1975) e da
liberação de várias moléculas bioativas, chamadas de adipocinas (TRAYHURN, 2005).
Particularmente em humanos, o único tipo de TA que persiste ao longo da vida pós-
embrionária é o TAB. Este ainda pode ser dividido em dois tipos: subcutâneo e
visceral (WAJCHENBERG et al., 2000). As diferentes localizações do TAB também
correspondem a diferentes propriedades metabólicas e funções, o que representa um
importante foco no mundo científico atual (PRUNET-MARCASSUS et al., 2006). O
TAB visceral participa da patogênese de desordens metabólicas (WAJCHENBERG et
al., 2000) enquanto que no subcutâneo foi demonstrada a presença de células com
múltiplo potencial de diferenciação (ZUK et al., 2001; BAPTISTA et al., 2007).
O desenvolvimento do TAB em humanos tem início no segundo terço do
período gestacional com a agregação de uma densa massa de células mesenquimais
indiferenciadas, orquestrada com o início da organização vascular. As células desses
agregados diferenciam para células adiposas primitivas, adjacentes a capilares
(HAUSMAN & RICHARDSON, 2004). Estágios mais avançados do desenvolvimento
do TAB estão relacionados ao crescimento dos agregados e a maturação das células
adiposas. Assim como a medula óssea, o TAB é um derivado do mesoderma
embrionário e gera, quando colocado em cultura, uma fração celular aderente, um
estroma (AILHAUD et al., 1992).
A fração estromal do TAB é constituída de pré-adipócitos, células endoteliais,
macrófagos e fibroblastos (CASTEILLA et al., 2004). Existem diversas evidências de
relações parácrinas e/ou autócrinas entre as células endoteliais e os pré-adipócitos
(HAUSMAN & RICHARDSON, 2004). Os pré-adipócitos progressivamente acumulam
lipídios e adquirirem características de adipócitos maduros (AILHAUD et al., 1992).
Esse tipo celular, pré-adipócito, é pouco caracterizado, não existindo nenhum
marcador antigênico específico, por isso sua localização no TAB ainda é desconhecida
(CASTEILLA et al., 2004). 1.3.a. Tecido Adiposo como um órgão endócrino
O conceito do TAB como um órgão endócrino é recente e talvez a simplicidade
aparente deste tecido (constituído de adipócitos em sua maioria) tenha sido o principal
motivo para que muitos pesquisadores o tenham tido somente como um local de

13
reserva, síntese e quebra de lipídios (HAHN & NOVAK, 1975; TRAYHURN, 2005). O
TAB é responsável pela secreção de inúmeros fatores envolvidos em processos
fisiológicos e metabólitos – as adipocinas. Estes fatores possuem um papel relevante
nas patologias associadas à obesidade (FRÜHBECK et al., 2001; RAJALA &
SCHERER, 2003). As adipocinas incluem citocinas clássicas (TNF-α e IL-6), fatores
de crescimento tais como o fator de crescimento transformador beta (TGF-β -
transforming growth factor-beta), o hormônio leptina, proteínas envolvidas na
homeostase vascular (ativador do inibidor do plasminogênio-1), na regulação da
pressão sanguínea (angiotensinogênio), metabolismo lipídico (proteína ligada a retinol)
e na homeostase de glicose (adiponectina), além de fatores angiogênicos (VEGF).
Desta forma, se torna evidente que o TAB é um órgão secretório complexo integrado
com outros tecidos tais como cérebro, músculo esquelético (TRAYHURN, 2005), osso
e o sistema vascular (KOSHIYAMA et al., 2006).
Estas características fazem do TAB um excelente cenário para eventos
inflamatórios. Durante o desenvolvimento da obesidade, além de ocorrer hiperplasia e
hipertrofia de células adipóciticas (LEHRKE & LAZAR, 2004), mudanças inflamatórias
são geradas pelos adipócitos, células endoteliais e os pré-adipócitos (WELLEN &
HOTAMISLIGIL, 2003). A secreção de adipocinas irá levar a um quadro de inflamação
crônica (LEHRKE & LAZAR, 2004), que é agravado pela infiltração macrofágica (Fig.
6) (WEISBERG et al., 2003; XU et al., 2003).
Figura 6. O tecido adiposo obeso é caracterizado pela inflamação e progressiva infiltração por macrófagos concomitante ao desenvolvimento da obesidade. Mudanças no tamanho do adipócito e gordura leva a mudanças físicas na área adjacente e modificações da função parácrina do adipócito, células endoteliais e pré-adipócitos. Fonte: Wellen & Hotamisligil, 2003.

14
As MSCs que geram os pré-adipócitos no TAB participam de todas as etapas
do processo de obesidade, seja de maneira direta, pelo acúmulo lípidico
intracitoplasmático (LEHRKE & LAZAR, 2004) e pela transdiferenciação em
macrófagos (CHARRIÈRE et al., 2003), ou de maneira indireta, através da secreção
de adipocinas, que além de estimular a angiogênese e a adipogênese no TAB (ZHOU
et al., 2007) estimulam a infiltração monocítica (CURAT et al., 2004) e macrofágica
(ZHOU et al., 2007).
1.3.b. MSC de Tecido Adiposo
Estudos recentes demonstraram que no TA subcutâneo assim como na MO,
residem células progenitoras mesodermais multipotentes (ZUK et al., 2001), sendo
assim, excelentes candidatas para protocolos de medicina regenerativa de osso,
cartilagem, distrofias musculares e reconstrução de tecidos moles (SCHÄFFLER &
BÜCHLER, 2007). Esses progenitores multipotentes estão presentes em praticamente
todos os órgãos do organismo adulto (DA SILVA MEIRELLES et al., 2006) e, por isso,
podem ser a razão da integração do TAB com os outros órgãos (KOSHIYAMA et al.,
2006). Fatores transcricionais expressos por estas células, como o receptor
peroxissomo de ativação de proliferação (PPARγ-peroxisome proliferator-activated
receptor-gamma) e CBFA-1, regulam a sua diferenciação para as vias adipogênica e
osteogênica, respectivamente, podendo desta forma estar envolvidos em patologias
(osteoporose e obesidade visceral) em que esse balanço é rompido (NUTTALL &
GIMBLE, 2004; KOSHIYAMA et al., 2006). O procedimento cirúrgico simples, o fácil e repetitivo acesso ao TA subcutâneo
assim como a baixa complexidade dos procedimentos enzimáticos de isolamento
fazem desta fonte de MSC a mais atrativa para pesquisadores e clínicos (CASTEILLA
et al., 2005). O ponto crítico é a ausência de uma caracterização tanto molecular como
celular das células-tronco multipotentes dentro do estroma do TA e apesar de terem
surgido muitos trabalhos visando alcançar esse objetivo, ainda não há um consenso
sobre a verdadeira identidade das MSC do TA (SCHÄFFLER & BÜCHLER, 2007).
Muitos pesquisadores acreditam que essas células são bastante semelhantes às MSC
de MO (ZUK et al., 2001; WAGNER et al., 2005; IZADPANAH et al., 2006; KERN et
al., 2006), e alguns acreditam em suas propriedades únicas, principalmente em
relação ao potencial adipogênico (CASTEILLA et al., 2004; LIU et al., 2007; BAPTISTA
et al., 2007).
CASTEILLA et al. (2004) defendem a hipótese de que as MSC do TA são na
verdade pré-adipócitos plásticos. Esses pré-adipócitos poderiam assumir fenótipos de

15
osso, cartilagem, músculo esquelético (ZUK et al., 2001) e até de células estromais
capazes de sustentar a hematopoese in vitro (CORRE et al., 2006; BAPTISTA et al.,
2007), características marcantes de progenitores estromais da MO (BIANCO &
ROBEY, 2000). O verdadeiro papel dos pré-adipócitos na homeostase do TA seria o
comprometimento e posterior diferenciação tanto em células endoteliais como em
adipócitos (Fig. 7) (PLANAT-BENARD et al., 2004a), ambos os tipos celulares
necessários no processo de desenvolvimento da obesidade (WELLEN &
HOTAMISLIGIL, 2003). Alguns trabalhos também demonstram a diferenciação desses
pré-adipócitos em macrófagos (COUSIN et al., 1999; CHARRIÈRE et al., 2003;
SCHÄFFLER et al., 2007), o que não é surpreendente já que o próprio pré-adipócito
possui características macrofágicas (CASTEILLA et al., 2004) e expressam
constitutivamente receptores toll-like, que desempenham um papel crucial na
imunidade (SCHÄFFLER et al., 2007). Essa possível transdiferenciação aumentaria o
quadro inflamatório estabelecido na obesidade (COUSIN et al., 2001).
Figura 7. Plasticidade das linhagens de células adipocíticas em células endoteliais. Progenitores adipocíticos possuem a habilidade de se diferenciar em adipócitos e também de adquirir um fenótipo endotelial funcional. Adipócitos maduros podem se de-diferenciar e reverter para um fenótipo bi-potente. A identificação de vias que regulam esses processos poderia levar ao desenvolvimento de novas estratégias terapêuticas para o controle do desenvolvimento do tecido adiposo e para angiogênese terapêutica. Fonte: Planat-Benard et al., 2004.

16
1.4. ONTOGENIA DAS MSC 1.4.a. Pericitos
O estroma medular primitivo é estabelecido no desenvolvimento através de
uma série de eventos complexos que se iniciam com a diferenciação de células
osteogênicas primitivas que irão formar rudimentos ósseos, estes rudimentos são
posteriormente invadidos por vasos sanguíneos (CHARBORD et al., 1996). No
organismo pós-natal, a rede vascular medular, assim como no sistema circulatório de
outros órgãos, é delimitada por uma camada contínua de células endoteliais e pericitos
subendoteliais. As células estromais da MO, com características de MSC, residem na
orientação abluminal dos sinusóides e podem ser classificadas como pericitos
especializados (BIANCO et al., 2001; SACCHETTI et al., 2007). O nicho perivascular
também foi descrito especificamente em MSC de polpa dentária e de tecido adiposo
(SHI & GRONTHOS, 2003; ZANNETTINO et al., 2008) e, em praticamente todos os
órgãos no organismo adulto (SILVA MEIRELLES et al., 2006; ZENGIN et al., 2006),
podendo ser prontamente mobilizados em casos de lesão tecidual (SILVA
MEIRELLES et al., 2006; LAMAGNA et al., 2006). Porém sua função melhor descrita
na literatura é a estabilização mecânica de microvasos. Através da emissão de
prolongamentos citoplasmáticos, os pericitos estabelecem contatos focais com as
células endoteliais (VON TELL et al., 2006).
Os pericitos isolados da microvasculatura da retina bovina retêm um fenótipo
de célula-tronco multipotente para as linhagens osteogênica (SCHOR et al., 1990),
condrogênica e adipogênica (FARRINGTON-ROCK et al., 2004) e também para um
estroma sustentador de hematopoese (TINTUT et al., 2003). Outra característica
comum entre as células-tronco estromais da MO e os pericitos é a positividade para
fosfatase-alcalina e para alpha actina de músculo liso (α-SMA-alpha smooth muscle
actin) (BIANCO et al., 2001); apesar da expressão deste último marcador ser
heterogênea nos diversos tecidos (NEHLS et al., 1991).
1.4.b. Hemangioblastos e células do somito
Um dos primeiros passos durante o desenvolvimento vascular é a
diferenciação de células-tronco pluripotentes em células endoteliais. Os fatores de
transcrição específicos que medeiam esses eventos ainda não foram identificados,
mas a expressão de marcadores específicos para as células endoteliais tais como
CD31 ou molécula de adesão celular endotélio plaquetas-1 (PECAM-1 - platelet
endothelial cell adhesion molecule-1) é determinante para o início do processo

17
(OETTGEN, 2001). As células endoteliais se diferenciam dos angioblastos no embrião,
mas também dividem uma origem comum com as células hematopoiéticas (Fig. 8)
(CARMELIET, 2003).
Experimentos de transplante de células de codorna para pintos demonstraram
que as células endoteliais embrionárias se originam de duas linhagens mesodermais.
Uma do mesoderma da esplancnopleura, que também exibe um potencial
hematopoiético e dá origem à rede endotelial dos órgãos viscerais; e a outra dos
somitos, que é puramente endotelial e dá origem a vasculatura das paredes do corpo
(PARDANAUD et al., 1996).
A possível existência de um progenitor comum para as linhagens endoteliais e
hematopoiéticas, o hemangioblasto, foi originalmente sugerida por causa da íntima
associação das células hematopoiéticas com as células endoteliais nas chamadas
“blood islands” (ilhotas sanguíneas) do embrião em desenvolvimento (PIETILÄ &
VAINIO, 2005). Essas ilhotas sanguíneas derivam de agregados de células
mesodermais que colonizam o saco vitelino aproximadamente sete dias pós-
fecundação. As células do centro desses agregados dão origem a células
hematopoiéticas embrionárias enquanto a população periférica se diferencia em
células endoteliais, formando a primeira estrutura vascular (WAGNER, 1980).
Figura 8. Formação da rede vascular. Os progenitores endoteliais se diferenciam em células endoteliais arteriais e venosas, formando um capilar primitivo. Os vasos emitem brotamentos e são estabilizados pelos pericitos, que se diferenciam dos seus próprios progenitores. HSCs contribuem para a angiogênese indiretamente pela diferenciação para leucócitos ou plaquetas. Flk-1, Vascular endothelial growth factor receptor 2; Tie-2, Angiopoietin-1 receptor; Ang-1 ou 2, Angiopoietinas; Shh, Sonic hedgehog; Grdl, Gridlock; Mφ, macrófago. Fonte: Carmeliet, 2003.

18
A existência do hemangioblasto ganhou maior suporte a partir de observações
de que as linhagens hematopoiéticas e endoteliais compartilham a expressão de
alguns genes (ASAHARA et al., 1997). Posteriormente essa célula foi identificada em
estudos in vitro a partir de células-tronco embrionárias (CHOI et al., 1998) e também
no organismo pós-natal, pela expressão dos marcadores de superfície CD34 e do
receptor de fator de crescimento endotelial vascular 2 (KDR-Vascular endothelial
growth factor receptor 2) (PELOSI et al., 2002). A hematopoiese gerada pelo saco vitelino, um sítio extra-embrionário, não é
definitiva e sim transitória. HSCs derivadas do saco vitelino não são capazes de
repopular a MO adulta (PIETILÄ & VAINIO, 2005). A verdadeira fonte da
hematopoese definitiva no adulto é a região da aorta-gônada-mesonefros (AGM -
aorta-gonad-mesonephros), a qual em estágios mais anteriores do desenvolvimento é
representada pela esplancnopleura (MEDVINSKY & DZIERZAK, 1996; GODIN &
CUMANO, 2002). As HSCs depois de colonizar a AGM se localizam especificamente
adjacentes a camada endotelial da parede da aorta dorsal (DE BRUIJN et al., 2002) e
adquirem o potencial para repopular o fígado e a MO adulta (PIETILÄ & VAINIO,
2005). A origem das células endoteliais a partir dos somitos está relacionada com a
aorta da região da AGM, que é um dos primeiros vasos a se formar no embrião. Antes
da sua fusão, as células endoteliais que a compõem se originam da esplancnopleura.
Logo após a sua fusão, o teto da aorta começa a ser colonizado por células endoteliais
que se originam nos somitos enquanto as das paredes e do chão permanecem como
células endoteliais da esplancnopleura (POUGET et al., 2006). As da esplancnopleura,
como já mencionado, vão então gerar um sítio hematopoiético embrionário, onde as
células endoteliais perdem os seus marcadores e adquirem marcadores
hematopoiéticos (JAFFREDO et al., 1998). Quando a hematopoiese é completada, as
células endoteliais originárias dos somitos substituem as da esplancnopleura
(POUGET et al. , 2006).
A subseqüente maturação do vaso requer o recrutamento de células
mesenquimais ou pericitos e sua posterior diferenciação em células musculares lisas.
Isto ocorre através da liberação de fatores de crescimento específicos, sendo o
principal o fator de crescimento derivado de plaqueta (PDGF-platelet-derived growth
factor) (OETTGEN, 2001). Ainda não está comprovada a origem dessas células
perivasculares. Uma das hipóteses é que elas possam ter origem nos somitos, pois as
células recrutadas durante o desenvolvimento da aorta são α-SMA positivas (POUGET
et al., 2006).

19
1.4.c. Mesangioblasto
A noção de que as HSCs se originam de um local específico, constituída por
progenitores endoteliais da esplancnopleura (JAFFREDO et al., 1998), levam a
algumas hipóteses especulativas de que outros progenitores/células-tronco possam
ser gerados na mesma região (COSSU & BIANCO, 2003). Análises clonais de
rudimentos de órgãos explantados levam a uma identificação positiva, na aorta dorsal
embrionária, de uma população celular que expressa simultaneamente marcadores de
origem endotelial e miogênica e ainda são capazes de diferenciação miogênica in vitro
(DE ANGELIS et al., 1999).
Transplantes de células não cultivadas da aorta dorsal de codorna, no broto da
asa de embriões de galinha, demonstram a geração de músculo esquelético com as
células do doador. Além disso, células diferenciadas de diversos tecidos conjuntivos
também foram observadas nesses experimentos, levando a possibilidade de que
esses progenitores associados com a aorta dorsal não são somente miogênicos.
Quando esses progenitores foram co-cultivados in vitro com células hematopoiéticas
totais da MO, há uma expansão de células CD45+ (MINASI et al., 2002), o que revela
mais uma característica de progenitor mesenquimal (BIANCO & ROBEY, 2000). Esses
progenitores também foram encontrados em organismos pós-natal (COSSU &
BIANCO, 2003). As células progenitoras identificadas na região da aorta dorsal foram nomeadas
de “mesangiblastos”, nome escolhido para indicar um progenitor comum para os
derivados vasculares e extra-vasculares mesodermais (Fig. 9) (COSSU & BIANCO,
2003). Os clones de mesangioblastos recém isolados em cultura expressam diversos
marcadores endoteliais tais como Flk-1, CD34, VE (vascular–endothelial)-caderina
(MINASI et al., 2002). Mas, diferentemente do hemangioblasto, esses progenitores
não geram células hematopoiéticas (COSSU & BIANCO, 2003).
Curiosamente, os mesangioblastos expressam receptores da família do TGFβ
/BMP (bone morphogenetic protein) e se diferenciam de maneira eficiente em células
do músculo liso, em resposta a TGFβ, e em osteoblastos em resposta a BMP
(TAGLIAFICO et al., 2004). Esses dados se correlacionam com a grande proporção de
mesangioblastos recém-isolados que expressam α-SMA em níveis altos, característica
de células precursoras de músculo liso e pericitos (COSSU & BIANCO, 2003). Recentemente, também foram identificadas MSCs na região da AGM que
surgem de forma simultânea às HSCs (MENDES et al., 2005; DURAND et al., 2006).
Contudo, neste estágio de desenvolvimento, a presença das MSC é independente da
atividade das HSC. Elas aumentam numericamente durante o desenvolvimento até

20
encontrar um platô na MO adulta (MENDES et al., 2005). As MSCs da região da AGM
possuem diversos potenciais de diferenciação, tornando o microambiente altamente
complexo (DURAND et al., 2006). Por exemplo, progenitores osteogênicos e
condrogênicos estão ambos localizados na aorta dorsal, mesênquima e na região
urogenital, enquanto os progenitores adipogênicos estão presentes somente na região
urogenital (MENDES et al., 2005). Estes dados corroboram com alguns estudos que
demonstraram uma independência precoce na ontogenia da linhagem mesenquimal
adipogênica, quando comparada com as osteogênica e condrogênica (BANFI et al.,
2000).
Figura 9. Esquema que sugere uma possível origem do mesangioblasto. Influências ventralizantes do ambiente direcionam a um potencial hematopoiético nos angioblastos. Influências desconhecidas preservam um potencial para a diferenciação em linhagens mesodermais em outra população de angioblastos. Divisões assimétricas segregam a progênie hematopoiética do hemangioblasto na espaço luminal e a progênie de mesoangioblastos no subendotelial, espaço abluminal. Flk-1 – progenitor mesodermal; Tal-1(T-cell acute lymphocytic leukemia 1) – progenitores hematopoiéticos. Fonte: COSSU & BIANCO, 2003. 1.4.d. Células da Crista Neural
A crista neural produz células pluripotentes dotadas de propriedades
migratórias (LE DOUARIN et al., 2004). Essas células migram para a região da AGM
através de influências dorsalizantes e podem secretar sinais que influenciam o
desenvolvimento das HSCs (PIETILÄ & VAINIO, 2005). As células da crista neural

21
dão origem a neurônios, glia, melanócitos, células endócrinas e a diversos derivados
mesenquimais (LE DOUARIN et al., 2004).
A crista neural cefálica provê pericitos e células do músculo liso para todos os
vasos sanguíneos da face e do cérebro anterior (ETCHEVERS et al., 2001). O
mesênquima do mesoderma cefálico é o sítio de ondas sucessivas de determinação e
diferenciação celular. Células expressando Flk-1 se tornam as células endoteliais dos
vasos em desenvolvimento. Deste estágio em diante, elas se associam a células da
crista neural que se diferenciam em pericitos e em tecido muscular e conjuntivo da
face externa das paredes dos vasos (COULY et al., 1996).
Curiosamente, MSCs da polpa dentária exibem características fenótipicas
diferentes das de MO, remetendo ou a duas origens diferentes na ontogenia, ou a
especificações ditadas pelos diferentes microambientes (SHI & GRONTHOS, 2003).
Recentemente, também foi demonstrado que MSCs da MO e do maxilar/mandíbula,
do mesmo indivíduo, são diferentes quanto ao tipo de osso que elas formam após
transplantes in vivo (AKINTOYE et al., 2006). As MSCs da polpa dentária assim como
as do maxilar e mandíbula poderiam ter origem a partir de células da crista neural, e
as da MO a partir de progenitores mesodermais da região AGM (ROBEY & BIANCO,
2006).
1.5. MSC NO ORGANISMO ADULTO MSCs têm sido descritas por estarem presentes principalmente na MO
(PROCKOP, 1997), mas também têm sido isoladas de diferentes tecidos tais como o
TA (ZUK et al., 2001), na articulação sinovial (DE BARI et al., 2001), no músculo
esquelético (DELLAVALLE et al., 2007), na polpa dentária e no ligamento periodontal
(SHI & GRONTHOS, 2003), no maxilar e na mandíbula (AKINTOYE et al., 2006), na
placenta (KÖGLER et al., 2004), no couro cabeludo humano (SHIH et al., 2005) e em
baixo número no sangue de cordão umbilical (WANG et al., 2004) e no sangue
periférico (KUZNETSOV et al., 2001). Se as MSCs derivadas dessas diferentes fontes
representam fundamentalmente tipos celulares similares, ainda não está claro.
Também foi demonstrado que as MSCs podem residir em todos os tecidos do
organismo adulto (SILVA MEIRELLES et al., 2006). Todas as fontes de MSC possuem
similaridades e diferenças em relação à expressão de marcadores de superfície.
Porém, diferentemente das HSCs humanas que são definidas principalmente pela
expressão de CD34 (KRAUSE et al., 2001), ainda não há um marcador único que
defina a população de MSC. A característica fenótipica em comum das diferentes
fontes de MSC é a adesão ao plástico de cultura (JAVAZON et al., 2004).

22
1.5.a. Marcadores de superfície
Um dos primeiros marcadores descritos para a identificação de células
precursoras estromais foi o STRO-1 (SIMMONS & TOROK-STORB, 1991a), que por
outro lado é expresso por 5% da população de CFU-F positiva para CD34 (SIMMONS
& TOROK-STORB, 1991b). As diferentes populações STRO-1+ ou STRO-1- também
exibem características intrínsecas na sustentação da hematopoese (BENSIDHOUM et
al., 2004). Atualmente sua expressão ainda é reportada (GONÇALVES et al., 2006;
ZANNETTINO et al., 2008) ou não (COLTER et al., 2001; DE UGARTE et al., 2003) em MSC, sendo variável também para diferentes fontes (GRONTHOS et al., 2001).
As características que definem a MSC são inconsistentes entre os
pesquisadores. Muitos laboratórios têm desenvolvido métodos para o isolamento e
expansão das MSCs, os quais invariavelmente possuem diferenças sutis e,
ocasionalmente, diferenças significantes (JAVAZON et al., 2004). Para resolver essas
diferenças a Sociedade Internacional para Terapia Celular (ISCT) propôs uma série de
padrões que definem uma MSC. Além da aderência ao plástico e o potencial de
diferenciação multipotente para as linhagens mesodérmicas, as MSC devem
expressar os marcadores de superfície: CD105, CD73 e CD90; e não devem
expressar CD45, CD34, CD14 ou CD11b, CD79α ou CD19 e HLA de classe II
(DOMINICI et al., 2006). A última coleção de marcadores representa
preferencialmente células de origem hematopoiética (LODIE et al., 2002; LEE et al.,
2004). Apesar da expressão de CD105 ser um consenso para a maioria dos
pesquisadores (GRONTHOS et al., 2001; LODIE et al., 2002; BANAS et al., 2007;
FALANGA et al., 2007; ZANNETTINO et al., 2008), o mesmo não ocorre em relação à
expressão de CD34. A maioria dos autores demostraram que ele não é expresso,
principalmente nas MSC de MO e sangue de cordão umbilical (LODIE et al., 2002 e
LEE et al., 2004; TONDREAU et al., 2005; FALANGA et al., 2007). Outros trabalhos
demonstram a expressão deste marcador somente em MSC de TA (GRONTHOS et
al., 2001; DE UGARTE et al., 2003; PLANAT-BENARD et al., 2004a; BAPTISTA et al.,
2007), onde esse marcador possivelmente representa um fenótipo de células
progenitoras endoteliais, em contraposição a um fenótipo de endotélio maduro,
representado pela falta de expressão de CD31 (PLANAT-BENARD et al., 2004a). Estudos recentes têm demonstrado que MSCs positivas para os marcadores
de superfície citados acima e que são multipotentes para as linhagens mesodérmicas,
podem ser enriquecidas do sangue de cordão umbilical e periférico pela seleção de
CD133 (TONDREAU et al., 2005) e da MO pela seleção para SSEA-1 (ANJOS-

23
AFONSO & BONNET, 2007), SSEA-4 (GANG et al., 2007), ou para o receptor do fator
de crescimento de nervos (CD271) (QUIRICI et al., 2002).
Outros estudos encontraram marcadores específicos de pericitos tais como
CD146 e 3G-5 em MSC derivadas de MO (SHI & GRONTHOS, 2003), o que é consistente com a hipótese de que pericitos vasculares especializados na MO
representam a população de MSC in vivo (BIANCO et al., 2001; BRACHYOGEL et al.,
2005). Recentemente, foi demonstrado que MSC de TA também expressam esses
marcadores de células perivasculares (ZANNETTINO et al., 2007). A molécula de adesão ALCAM – CD166 também foi sugerida como marcador
de MSC derivadas do pericôndrio da MO de membros fetais em camundongos,
participando ativamente na formação desse órgão hematopoiético (ARAI et al., 2002).
Esta molécula também parece participar na vasculo-angiogênese e na sustentação da
hematopoiese embrionárias (OHNEDA et al., 2001), sendo também expressa em MSC
de TA (GRONTHOS et al., 2001) e de MO (MARESCHI et al., 2006).
As MSCs de MO assim como as HSCs também expressam o receptor para a
quimiocina SDF-1α, CXCR4 (PONTE et al., 2007). Estas células pós-transplante
migram especificamente para a MO através da ativação desses receptores (WYNN et
al., 2004).
1.5.b. Diferenciação Ortodoxa As MSCs podem assumir diferentes fenótipos tais como condrócitos,
osteoblastos, adipócitos ou células endoteliais. Este comportamento, ambos in vitro e
in vivo, foi o primeiro vislumbre da propriedade que agora é referida como plasticidade
(GOODELL, 2001). Um tecido derivado do mesoderma pode conter alguns desses
tipos celulares em diferentes estágios de comprometimento, ou até todos, como é o
caso da MO. A noção de que cada um desses fenótipos celulares pode mudar para
outro sob circunstancias específicas é consistente com o desenvolvimento e a
manutenção do órgão do qual estas células derivam. Desta forma este tipo de
plasticidade é ortodoxa, significando que ela permanece dentro do contexto da
homeostasia tecidual (BIANCO et al., 2001).
Já foi demonstrado, por exemplo, que linhagens clonais de células
acumuladoras de gordura da MO podem ser direcionadas para uma diferenciação
osteogênica e formam ossos genuínos em ensaios in vivo (BENNETT et al., 1991;
BERESFORD et al., 1992). Anteriormente, também já havia sido notada a habilidade
de células estromais, responsáveis pela sustentação da hematopoese, de se converter
em células acumuladoras de gordura (BIANCO et al., 1988). Os condrócitos também
podem se desdiferenciar em cultura para um fenótipo osteogênico (GENTILI et al.,

24
1993). Todos esses resultados realçam a natureza não irreversível da diferenciação
desses tipos celulares, levando à noção de que o comprometimento e a diferenciação
possivelmente não são processos unidirecionais e terminais, como é mostrado em
sistemas de multi-linhagens dependentes de uma única célula-tronco (BIANCO et al.,
2001).
Cada via de diferenciação ativa a expressão de genes e proteínas particulares.
O processo de diferenciação osteogênica pode ser dividido em três estágios distintos:
(1) Proliferação celular; (2) Maturação da matriz e (3) Mineralização. Um número de
genes, incluindo fosfatase alcalina, colágeno do tipo I, sialoproteína, osteopontina e
osteocalcina, são expressos em altos níveis por períodos determinados durante a
diferenciação (BECK, 2003). Os eventos que definem a maturação de condrócitos
dividem características comuns com a osteogênese, tais como a expressão de
fosfatase alcalina, a formação de matriz, a deposição do mineral hidroxiapatita e o
requerimento do fosfato inorgânico (WUTHIER, 1993). As moléculas específicas
expressas nessa via de diferenciação compreendem as proteínas de matriz
extracelular tais como colágeno do tipo II, agrecan e decorina (WINTER et al., 2003).
A primeira marca do processo adipogênico é a alteração dramática no formato
celular, que é convertido de um aspecto fibroblastóide para um formato esférico
(MCBEATH et al., 2004). Os eventos requeridos para essa alteração morfológica
incluem a degradação proteolítica da matriz extracelular pelos pré-adipócitos, a
expressão de gens adipocíticos, o acúmulo de lipídeos (GREGOIRE, 2001) e
alterações no citoesqueleto (MCBEATH et al., 2004). Os fatores de transcrição críticos
na adipogênese incluem C/EBPα e/ ou PPARγ (GREGOIRE, 2001).
1.5.c. Regulação das vias osteogênica e adipogênica
As vias osteogênica e adipogênica são balanceadas fisiologicamente através
da expressão de fatores de transcrição (NUTTALL & GIMBLE, 2004) e por proteínas
do citoesqueleto (MCBEATH et al., 2004). Desordens nesse balanço podem levar a
quadros patológicos, como a osteoporose. Os mecanismos transcricionais na
regulação da adipogênese e osteogênese dependem em parte, de receptores
nucleares hormonais e seus ligantes. Um destes receptores é o PPARγ2, cujos
ligantes incluem ácidos graxos de cadeia longa, derivados oxidados dos ácidos
graxos, e compostos de thiazolidinediona (NUTTALL & GIMBLE, 2004).
Os ligantes para PPARγ2 promovem adipogênese nas células estromais da MO
e inibem a osteogênese (Fig. 10). Estes agentes não ativam somente o receptor
nuclear PPARγ2, mas também suprimem as ações da proteína regulatória

25
transcricional – CBFA-1 (LECKA-CZERNIK et al., 2002). Em modelos animais o mais
potente ligante para PPARγ2 - rosiglitazona, aumenta o volume do tecido adiposo na
MO enquanto reduz a densidade mineral óssea e a área trabecular do osso (RZONCA
et al., 2004).
Os osteoblastos também são responsáveis pela secreção de adiponectinas
(BERNER et al., 2004), proteínas antes atribuídas somente aos adipócitos do TA
subcutâneo (MAEDA et al., 1996). As adiponectinas estão presentes na MO e regulam
negativamente a diferenciação adipogênica (YOKOTA et al., 2002) assim como a
diferenciação de progenitores mielóides (YOKOTA et al., 2000). Este último dado é
conflitante com a observação de que essas proteínas promovem a proliferação de
células osteoblásticas (BERNER et al., 2004), já que o principal nicho das HSCs é o
subendosteal (TAICHMAN, 2005). Novas avaliações do papel das adiponectinas na
MO e no balanço das vias adipogênicas e osteogênicas são necessárias, já que
recentemente também foi descoberto que as HSCs expressam receptores para essas
proteínas (DIMASCIO et al., 2007).
Figura 10. Modelo proposto para as mudanças na MSC no comprometimento para as vias adipogênica e osteogênica. Quando fatores de transcrição da família dos PPARγ são ativados, a diferenciação osteogênica cessa, sendo substituída pela via adipogênica. Adaptado de McBeath et al., 2004.
1.5.d. O potencial de diferenciação das diferentes fontes de MSC Mesmo na MO existem populações de MSC funcionalmente heterogêneas
(MURAGLIA et al., 2000; COLTER et al., 2001) e que contêm progenitores não
diferenciados com potenciais de diferenciação variados (KUZNETSOV et al., 1997).
Também no TA subcutâneo foi demonstrado MSC em diferentes estágios de
diferenciação (ZUK et al., 2002). As diferenças entre sub-populações encontradas no

26
mesmo órgão também podem refletir em diferenças encontradas entre diversos
órgãos. As diferentes fontes de MSC anteriormente mencionadas possuem
populações de MSC que não são funcionalmente equivalentes em relação ao potencial
de diferenciação (PHINNEY & PROCKOP, 2007).
Além da MO, as principais fontes de MSC são o TA e o sangue de cordão
umbilical. Em 2005, através de ensaios de avaliação da expressão global de genes
(RNA-microarray), foram detectadas diferenças na expressão de genes mesodérmicos
entre as MSC das fontes citadas, o que pode estar correlacionado com diferentes
ontogenias. Curiosamente, as MSC de sangue de cordão, quando comparadas com as
de MO e TA, apresentam um baixo (WAGNER et al., 2005) ou nenhum potencial
adipogênico, além de um baixo rendimento no momento do isolamento (KERN et al.,
2006).
Ainda em relação ao potencial adipogênico, as MSC de TA são mais
competentes, enquanto as MSC de MO são mais comprometidas para as vias
osteogênica e condrogênica (LIU et al., 2007), característica que pode estar
relacionada com o microambiente tecidual em que essas células residem (BAPTISTA
et al., 2007). As MSC de TA possuem expressão constitutiva para subtipos do gene
PPARγ (ZUK et al., 2002; BAPTISTA et al., 2007) e somente as MSC de MO, quando
cultivadas em um sistema tridimensional induzidas para a via condrogênica, modulam
os genes relacionados de forma eficiente (WINTER et al., 2003). Apesar do seu
comprometimento com a via adipogênica, as MSC de TA, quando cultivadas em
biomateriais de hidroxiapatita para implantação in vivo, são capazes de formar uma
matriz osteóide (HICOK et al., 2004).
1.5.e. Diferenciação Não-Ortodoxa Vários estudos têm indicado ou implicado que progenitores com um aparente
potencial de diferenciação não-ortodoxo podem ser encontrados em tecidos no
organismo adulto (BIANCO et al., 2001). Primeiro foi mostrado que a MO contém
progenitores miogênicos sistematicamente transplantáveis (FERRARI et al., 1998), em
seguida que células da MO poderiam gerar hepatócitos (LAGASSE et al., 2000) e por
último, que as MSC da MO poderiam ter um potencial neurogênico in vivo (KOPEN et
al., 1999).
Em relação ao potencial neurogênico, diversos grupos obtiveram resultados
positivamente semelhantes tanto para as MSC de MO (GROVE et al., 2004), quanto
para as de TA (SCHÄFFLER & BÜCHLER, 2007). Porém, os métodos utilizados para
promover a diferenciação neural in vitro e acessar a biologia das células diferenciadas

27
são fragmentados e inconsistentes (PHINNEY & PROCKOP, 2007). Os agentes
indutores frequentemente promovem o rompimento da rede de filamentos de actina
levando a uma retração citoplasmática, que se assemelha ao brotamento de neuritos
(NEUHUBER et al., 2004). Porém, na presença de matrizes frouxas, ou seja com
menor elasticidade, que mimetizam o microambiente do tecido nervoso, as MSC de
MO emitem prolongamentos e a dinâmica do brotamento é claramente o oposto da
retração do corpo celular induzido por DMSO, um agente frequentemente utilizado
como indutor neurogênico (ENGLER et al., 2006).
Recentemente, foram encontrados em uma baixa freqüência clones na
população de MSC de TA cultivadas em matriz semi-sólida de metilcelulose, com
potencial de diferenciação espontânea para cardiomiócitos funcionalmente
competentes (PLANAT-BÉNARD et al., 2004b). Agentes demetilantes também são
capazes de induzir mudanças nas propriedades eletrofisiológicas das MSC de MO,
mas não há a geração de cardiomiócitos funcionais (BALANA et al., 2006).
As MSC de MO após uma administração sistêmica, são capazes de se
diferenciar em vários tipos celulares do epitélio (KRAUSE et al., 2001). Essas células
quando co-cultivadas com células epiteliais das vias aéreas adquirem a expressão de
diversos marcadores epiteliais (SPEES et al., 2003).
Em termos de ontogenia não é surpreendente que esses progenitores
mesodérmicos possuam potencial epitelial, pois todo o mesênquima é derivado do
epitélio (PÉREZ-POMARES & MUÑOZ-CHÁPULI, 2002). Já em relação ao potencial
miogênico, foi demonstrada a colonização da aorta da região da AGM pelas células do
somito (POUGET et al., 2006), local onde também foram descritos os mesangioblastos
(MINASI et al., 2002), o que torna bem provável a diferenciação em células do
músculo esquelético.
Como mencionado anteriormente, as células da crista neural dão origem tanto
a componentes celulares do sistema nervoso como a componentes do sistema
cardiovascular (LE DOUARIN et al., 2004). Como a origem das células perivasculares
não é unilateral (ROBEY & BIANCO, 2006), é possível que os potenciais
cardiogênicos e neurogênicos variem de acordo com a fonte de MSCs.
1.5.f. Potencial Imunomodulador As MSCs além da multipotencialidade para linhagens mesodérmicas, têm
propriedade imunomoduladora, o que pode auxiliar no tratamento de doenças auto-
imunes e em transplantes de MO (ZHAO et al., 2004). A associação de MSCs com a
MO total a ser transplantada tem sido utilizada na clínica e além de reduzir o tempo de

28
enxerto, diminui a incidência da doença enxerto versus hospedeiro – GvHD (Graft-
versus-host disease) (LE BLANC & RINGDÉN, 2006).
As principais fontes de MSC já testadas tanto em modelos animais de GvHD
(FANG et al., 2006; YAÑEZ et al., 2006), quanto no tratamento de pacientes
(RINGDÉN et al., 2006) demonstram resultados promissores. Exceto as MSCs de
sangue de cordão, que parecem não ter efeito imunomodulador (TISATO et al., 2007).
Os mecanismos de imunomodulação exercidos pelas MSCs ainda não foram
totalmente elucidados, mas já foi descrito que essas células alteram o perfil de
secreção de citocinas de células dendríticas, células T efetoras e células Natural Killer
para um fenótipo mais anti-inflamatório ou tolerante (AGGARWAL & PITTENGER,
2005).
1.6. ENSAIOS CLÍNICOS COM MSCs
Devido ao grande potencial terapêutico das MSC na medicina regenerativa, o
Comitê de células mesenquimais e células tronco teciduais da Sociedade Internacional
para Terapia Celular, propôs recentemente critérios mínimos para a definição das
MSC humanas. Três características principais definem a MSC: (1) Aderência ao
plástico in vitro; (2) Expressão de antígenos de superfície específicos; (3) Potencial de
diferenciação para as três principais linhagens mesodermais: osteoblastos, adipócitos
e condrócitos (DOMINICI et al., 2006).
As MSCs de MO foram primeiramente utilizadas em ensaios clínicos no
tratamento da osteogenesis imperfecta (HORWITZ et al., 2001) em
mucopolisacaridoses e leucodistrofias metacromáticas, as quais levam a defeitos
ósseos e neurológicos (KOÇ et al., 2002), em GvHD (RINGDÉN et al., 2006), em
esclerose múltipla (MOHYEDDIN BONAB et al., 2007), no infarto do miocárdio
(SCHÄCHINGER et al., 2006) e em outras doenças. Alguns trabalhos sugerem o
potencial neurogênico das MSC (PHINNEY & PROCKOP, 2007) e o seu uso clínico
em pacientes com autismo têm sido defendido, pois esta patologia está
frequentemente associada a anormalidades do sistema imunológico e a hipoperfusão
neural (ICHIM et al., 2007).
Em humanos, as limitações biológicas para a regeneração cardíaca criaram
oportunidades para novos ensaios clínicos. A área cardíaca foi uma das pioneiras em
protocolos de terapias celulares com células de MO (DIMMELER et al., 2005), sendo
primeiramente utilizadas células mononucleares (STRAUER et al., 2002; PERIN et al.,
2003; WOLLERT et al., 2004). A partir desses ensaios foram propostas as primeiras
hipóteses para os mecanismos de ação das células de MO, mas provavelmente ocorre

29
uma sinergia entre os diversos mecanismos (Fig. 11) (DIMMELER et al., 2005).
Posteriormente, com a descoberta da multipotencialidade das MSC e seu potencial de
diferenciação também para cardiomiócitos e vasos sanguíneos (PITTENGER &
MARTIN, 2004), essa fonte de células da MO passou a ser utilizada em ensaios
clínicos (SCHÄCHINGER et al., 2006).
As MSCs de MO, devido ao seu grande potencial osteogênico, é a fonte mais
indicada para tratamento de defeitos críticos ósseos (ROBEY & BIANCO. 2006), mas
as MSCs de TA também podem ser utilizadas neste e em outros protocolos em
medicina regenerativa (SCHÄFFLER & BÜCHLER, 2007). O primeiro ensaio clínico a
utilizar essa nova fonte de MSC, foi o tratamento de fístulas geradas pela doença de
Chron, uma patologia essencialmente de caráter auto-imune (GARCÍA-OLMO et al.,
2005). As propriedades imunossupressoras das MSCs de TA permitem a sua
utilização em diversos ensaios clínicos, tais como no tratamento de GvHD, que
demonstra excelentes resultados tanto em modelos animais (YAÑEZ et al., 2006),
como em um estudo piloto em um paciente em estágios iniciais da doença (FANG et
al., 2006).
Figura 11: Mecanismos de ação. Células progenitoras podem facilitar a recuperação funcional do miocárdio infartado por vários mecanismos, incluindo o aumento da neovascularização de maneira direta ou indireta. Fatores parácrinos liberados pelas células progenitoras podem inibir a apoptose dos cardiomiócitos, afetar o remodelamento, ou aumentar o reparo endógeno (por exemplo, através de progenitores residentes). A diferenciação em cardiomiócitos pode contribuir para a regeneração cardíaca. A extensão pelas quais esses diferentes mecanismos estão ativos pode depender criticamente do tipo de injúria tecidual, tais como aguda ou crônica. Fonte: Dimmeler et al., 2005.
O uso das MSCs também tem sido proposto em protocolos de bioengenharia
de pele (BURD et al., 2007), onde modelos animais revelam que somente as MSCs de
MO e não as HSCs, são capazes de adquirir marcadores de superfície específicos de

30
queratinócitos e de formar estruturas glandulares, sugerindo uma contribuição direta
para a regeneração cutânea (WU et al., 2007). Resultados semelhantes foram
observados na regeneração de feridas de camundongos diabéticos (JAVAZON et al.,
2007) e o uso das MSC de MO associadas à biomateriais de colágeno também já foi
testado em pacientes diabéticos (VOJTASSÁK et al., 2006).
As MSC de TA já foram utilizadas no tratamento de rítides (STASHOWER et
al., 1999) e o nosso grupo tem tido excelentes resultados para preenchimentos
cutâneos com a associação dessas células ao ácido hialurônico (dados não
publicados). O uso dessa nova fonte de MSC em protocolos de terapias celulares
ainda é restrito, mas sua multipotencialidade e similaridades com as MSC de MO faz
dessa fonte de fácil acesso, uma excelente candidata para protocolos em medicina
regenerativa (FRASER et al., 2006). Desta forma, é importante aprofundar o
conhecimento sobre a biologia das MSCs de TA, assim como testar sua semelhança
com as MSCs de MO, para que cada tipo celular seja indicado da melhor maneira para
os diversos protocolos de terapias celulares.
2.OBJETIVOS
2.1. Objetivo Geral Analisar os componentes celulares do tecido adiposo subcutâneo humano,
avaliando a multipotencialidade da sua fração mesenquimal in vitro.
2.2. Objetivos Específicos 1 – estabelecimento da técnica e cultivo das MSCs de TA subcutâneo humano.
2 – caracterização morfológica e fenotípica das MSCs de TA, através de
técnicas de microscopia e citometria de fluxo.
3 – avaliação in vitro do potencial de diferenciação paras as vias adipogênica,
osteogênica, condrogênica e endotelial das MSCs de TA, através de ensaios
funcionais e expressão de marcadores específicos.
4 – avaliação do potencial de sustentação hematopoiética das MSCs de TA
induzidas ou não para as vias adipogênica e osteogênica, em comparação com as
MSCs de MO, através de ensaios de co-cultivo, expressão gênica e de quantificação
de progenitores hematopoiéticos.

31
3. METODOLOGIA
3.1. AMOSTRAS E TÉCNICAS DE ISOLAMENTO.
3.1.a. Amostras e células.
As amostras de MO foram obtidas de aspirados da crista ilíaca de doadores
voluntários da Unidade de Transplante de Medula Óssea do Serviço de Hematologia
do Hospital Universitário Clementino Fraga Filho (HUCFF), da Universidade Federal
do Rio de Janeiro (UFRJ). Amostras de TA subcutâneo ou de lipoaspirado, ambos da
parede abdominal, foram obtidas de pacientes submetidos à cirurgia estética no
HUCFF. A mediana da idade dos pacientes foi de 41,8 ± 2,1, totalizando vinte, todos
do sexo feminino. Os procedimentos foram aprovados pelo Comitê de Ética em
Pesquisa (CEP) deste hospital – portaria: 088/04. Todos os pacientes leram,
concordaram e asinaram o termo de consentimento livre e esclarecido.
As amostras de cordão e de sangue de cordão umbilical humanos foram
coletadas em solução salina tamponada com heparina (20 UI/mL; Liquemine, Roche),
na Maternidade-escola Pró-Matre no Rio de Janeiro. A obtenção destas amostras foi
aprovada pelo comitê de ética desta instituição.
Fibroblastos de pele humana (FPH) e a linhagem estromal de MO murina –
MS5 foram fornecidos pelo Banco de Células do Rio de Janeiro (BCRJ), HUCFF,
UFRJ.
3.1.b. Isolamento de MSCs.
Obtenção de MSCs de MO.
Os aspirados de MO foram submetidos à hemossedimentação com Hespan®
(hidroxietil de amido McGaiy Inc., Irvine, CA) na diluição de 1:6. Em seguida, as
células foram lavadas com tampão fosfato salino (PBS), centrifugadas a 500 g
(gravidades) por 15 minutos, ressuspensas em Iscove´s (Gibco BRL, Rockville, MD)
ou Dulbecco’s modified Eagle Medium-Low Glicose (DMEM, LGC, Cotia, SP)
suplementado com 10% de soro fetal bovino (SFB, Cultilab, Campinas, SP), 100 U/ml
de penicilina sódica e 100 μg/mL de estreptomicina) e 10 mL de suspensão com 1-2 x
106 células/mL foram distribuídos em garrafas de cultura de 25 cm2. As células foram
mantidas em estufa a 37ºC com 5% de CO2 por três a quatro dias. Após este período
as células não aderentes foram retiradas e a monocamada foi lavada com PBS e
mantida nas mesmas condições para expansão. Quando confluentes, as células foram
dissociadas com solução de tripsina 0,125% (Gibco) e 0,78 mM de EDTA (Invitrogen,

32
Gibco) e distribuídas numa densidade de 0,8 – 1,3 x 104 células/cm2 para expansão. A
dissociação enzimática da monocamada confluente seguida de redistribuição para
expansão foi denominada, neste trabalho, de passagem.
Obtenção de MSCs de TA.
Pequenos fragmentos de TA foram obtidos após a retirada de vasos
sanguíneos e do excesso de tecido conjuntivo. Estes fragmentos ou o material obtido
da lipoaspiração foram submetidos à digestão enzimática com 1 mg/mL de colagenase
IA (Sigma Chemical Co, St Louis, MO.) por 20 minutos a 37ºC. A suspensão obtida foi
filtrada em malha de nylon com poro de 150 μm, centrifugadas a 300 g por 10 minutos
e lavada duas vezes com PBS. As células foram ressuspensas em DMEM com 10%
de SFB e antibiótico e 5 mL de suspensão com 105 células/cm2 foram distribuídos em
garrafas de cultura de 25 cm2, que foram mantidas em estufa a 37ºC com 5% de CO2.
Após aproximadamente 16 horas, as células não aderentes foram retiradas e a fração
aderente foi lavada com PBS, sendo mantida a 37ºC com 5% de CO2 para expansão.
Quando confluentes, as células foram dissociadas como descrito, sendo redistribuídas
em uma densidade de 0,8 – 1,3 x 104 células/cm2 para expansão, caracterizando uma
passagem.
3.1.c. Obtenção de células do cordão e do sangue de cordão umbilical.
Obtenção de células endoteliais maduras do cordão umbilical – HUVEC (human
umbilical cord endothelial cells). A veia do cordão umbilical foi canulada em ambas as extremidades. Por
intermédio das cânulas o cordão foi lavado com PBS pelo menos duas vezes e
incubado com a solução de colagenase IA (Sigma Chemical Co.) 1 mg/mL por 10
minutos a 37ºC. A suspensão foi então recolhida e centrifugada a 300 g por 10
minutos. As células de cada cordão foram ressuspensas em 5 mL de Endothelial Cell
Basal Medium 2 (EBM-2 - Cambrex Biosciesnce, Walkersville, MD) suplementado com
Endothelial Growth Medium 2 (EGM-2 – Cambrex Bioscience), distribuídas em uma
garrafa de cultura de 25 cm2 e mantidas em estufa a 37ºC com 5% de CO2. Após
aproximadamente 16 horas, as células não aderentes foram retiradas e a fração
aderente foi lavada com PBS, sendo mantida a 37ºC com 5% de CO2 para expansão.
Quando confluentes, as células foram dissociadas como descrito, e utilizadas para os
experimentos.

33
Obtenção de células mononucleares e CD34+ do sangue de cordão umbilical.
As células mononucleadas foram obtidas por centrifugação em gradiente de
Ficoll-Paque TM Plus (Amersham Biosciences, Auckland, CA). Para tanto, as amostras
foram diluídas em PBS na proporção de 1:4, aplicadas lentamente sobre a solução de
Ficoll-Paque e, em seguida, centrifugadas a 500 g por 15 minutos. O anel de células
mononucleares foi retirado e lavado com PBS. As células foram ressuspensas em
DMEM com 10% de SFB e antibiótico para quantificação em hemocitômetro,
utilizando-se líquido de Turk. Após a quantificação, as células foram novamente
centrifugadas e ou foram congeladas em meio de criopreservação, ou seja, DMEM
contendo 10% de Dimetilsulfóxido (DMSO, Sigma Chemical Co) e 50% de SFB ou
procedeu-se o isolamento de células CD34+ por microesferas magnéticas (CD34
Progenitor Cell Selection System – Dynal Biotech, Bromborough, Wirral). Para tal, foi
seguido o protocolo recomendado pelo fabricante. O grau de pureza dos isolamentos
variou na faixa de 70 – 90%, avaliado por citometria de fluxo. As células CD34+ foram
então congeladas ou utilizadas para os experimentos.
3.2. ENSAIOS FUNCIONAIS.
3.2.a. Indução para a via osteogênica.
A diferenciação osteogênica das células expandidas até a terceira passagem
foi induzida através de cultura em meio osteoindutor, ou seja, DMEM suplementado
com 10% SFB, 5 X 10-6M de ácido ascórbico (Sigma Chemical Co), 10-8M de
dexametasona (Sigma Chemical Co) e 10-2M de β-glicerofosfato (Sigma Chemical Co)
por até quatro semanas. Controles pareados foram mantidos em meio não indutor
(DMEM suplementado com 10% SFB e antibióticos). Esta diferenciação foi
acompanhada pela revelação de depósitos de cálcio, através da coloração de Von
Kossa modificado (ZUK et al., 2001). As análises foram feitas em microscópio invertido
(Leika DC300). Durante o período de indução, amostras (106 células) foram retiradas e
as células foram lisadas em 1 mL de TRIZOLTM Reagent (Invitrogen) para posterior
isolamento do RNA total. As MSC induzidas por apenas uma semana foram
denominadas neste trabalho de pré-osteoblásticas.
As MSCs também foram distribuídas, na concentração de 2 X 105 células,
sobre pastilhas ou amostras porosas de hidroxiapatita, com área aproximada de 1,88
cm2, que foram fornecidas pelos Dr. Marcelo Prado do Instituto Militar de Engenharia
(IME) e Dr. Alexandre Rossi do Centro Brasileiro de Pesquisas Físicas (CBPF). As
células foram mantidas por até uma semana ou em DMEM com 10% de SFB e

34
antibióticos ou em meio osteoindutor, e posteriormente fixadas para análise por
microscopia eletrônica de varredura (MEV). A expressão de osteopontina foi avaliada
por imunofluorescência em microscópio confocal a laser (LSM META 510 - Zeiss).
3.2.b. Indução de MSCs para a via adipogênica.
A diferenciação adipogênica foi induzida através da incubação das células
expandidas até a terceira passagem em DMEM com 10% de SFB, suplementado com
10 μM de Insulina (Biohulin®, Montes Claros, MG), 0,5mM de Isobutilmetilxantina
(IBMX, Sigma Chemical Co), 1 μM de dexametasona (Sigma Chemical Co) e 200 μM
de Indometacina (Sigma Chemical Co) por até seis semanas. Controles pareados
foram mantidos em meio não indutor (DMEM suplementado com 10% SFB e
antibióticos). A presença de acúmulos lipídicos citoplasmáticos foi evidenciada através
da coloração de Oil Red O (PROPHET et al., 1994). Em seguida, as lamínulas foram
contracoradas com Hematoxilina de Harris (Merck) e seladas em glicerol (Sigma
Chemical Co) para posterior análise morfológica ao microscópio óptico (Nikon Eclipse
E400). Durante o período de indução, amostras (106 células) foram retiradas e as
células foram lisadas em 1 mL de TRIZOLTM Reagent (Invitrogen) para posterior
isolamento do RNA total. As MSC induzidas por apenas uma semana foram
denominadas neste trabalho de pré-adipocíticas.
3.2.c. Indução de MSCs de TA para a via condrogênica.
As MSCs de TA expandidas até a passagem três foram cultivadas em
condições padrão para a formação de esferóides (SCHIPANI et al., 2001; ROSSI et
al., 2005, INDOVINA et al., 2007). Em seguida, os esferóides foram mantidos em
DMEM com 10% de SFB suplementado com 2,5 x10-4M de ácido ascórbico (Sigma
Chemical Co), 10-7M de dexametasona (Decadron, Roche, Rio de Janeiro, RJ) e 10
ng/ml de TGF-β1 (Transforming Growth Factor-β; R & D Systems, Minneapolis, MN). O
meio foi renovado semanalmente, com reposição de ácido ascórbico a cada três ou
quatro dias, durante 28 dias. Controles pareados foram realizados mantendo os
esferóides em meio não indutor (DMEM suplementado com 10% SFB). Após este
período os esferóides foram fixados e incluídos em parafina. Os cortes de esferóides
foram corados pelo método de Alcian Blue (Merck, Whitehouse Station, NJ) em pH 1,0
(BANCROFT & STEVENS, 1996).

35
3.2.d. Indução de MSCs de TA para a via endotelial.
Ensaios em sistema de esferóides.
As MSC de TA foram cultivadas em sistema de esferóides em EBM-2
suplementado com EGM-2 ou em DMEM com 10% de SFB e antibióticos, por até dez
dias sem troca de meio. Após este período, os esferóides foram retirados para cultura
em gel de colágeno ou para análise morfológica. Neste caso, foram fixados e
processados para inclusão em parafina ou congelados. A diferenciação para a
linhagem endotelial foi acompanhada pela expressão de CD31, VEGF e vWF (fator de
von Willebrand), avaliada por imunofluorescência de cortes congelados ou por
imunohistoquimica. Para verificar a expressão de genes envolvidos na diferenciação
endotelial, cerca de dez esferóides foram lisados em 0,5 mL de TRIZOLTM Reagent
(Invitrogen) para posterior isolamento do RNA total.
Co-cultivo com HUVECs.
As MSC de TA e HUVECs foram cultivadas simultaneamente em igual
concentração de células – 104 células/poço em placas de 24 poços, e acompanhadas
por até dez dias. Após este período, as células foram fixadas em paraformaldeído a
4% tamponado por uma hora a temperatura ambiente e a expressão de CD31 foi
avaliada por imunofluorescência. A análise foi realizada em microscópio de
imunofluorescência (Nikon Eclipse E400).
Para avaliar a interação das MSCs de TA com as HUVECs em modelo
tridimensional, que favorece a formação de estruturas tubulares (Kelm et al., 2005), as
células foram cultivadas em modelo de esferóide multicelular. Após a formação de
esferóides de TA, as HUVECs foram distribuídas sobre estes, mantendo-se uma
relação destas células de 1:2 (esferóides mistos). Após dez dias, foi seguido o
protocolo de fixação e inclusão em parafina. Os cortes foram analisados em
microscópio óptico (Nikon Eclipse E400) através da coloração de Hematoxilina e
Eosina (H-E).
Brotamento em Gel de Colágeno.
Após a formação dos esferóides de MSC de TA, HUVEC e mistos, estes foram
inseridos em gel de colágeno do tipo I extraído da cauda de rato em pH 3,85 na
concentração de 14,3 mg/mL e posteriormente diluído em ácido acético (Merck) 3,5 X
10-13 N para uma concentração final de 0,6 mg/mL. Para verificar a formação de
brotamentos a partir dos esferóides, como descrito (KORFF & AUGUSTIN, 1999),
estes foram mantidos na presença de VEGF (Vascular Endothelial Growth Factor)
recombinante humano (Sigma Chemical Co) na concentração final de 2,5 ng/mL ou

36
uma monocamada de MSC de TA na concentração de 5 X 103 células/poço. O padrão
de brotamento dos esferóides foi analisado por até três dias em microscópio invertido
(Leica DC300).
3.2.e. Sustentação da Hematopoese em Culturas de Longa Duração.
Co-cultivo com células mononucleares de sangue de cordão.
Culturas de longa duração foram estabelecidas pela modificação do método de
Dexter (DEXTER & TESTA, 1976). As MSC de MO e de TA induzidas ou não por uma
semana para as vias adipogênica e osteogênica, e os FPH foram cultivadas em
garrafas de 25 cm2, em triplicata. Após o estabelecimento da monocamada, as células
mononucleares de sangue de cordão foram distribuídas sobre os estromas na
concentração de 5 X 105 células/mL. As co-culturas foram mantidas em estufa a 37ºC
com 5% de CO2 por até 32 dias em meio Iscove’s suplementado com 12,5% de SFB e
12,5% de soro de cavalo (Sigma Chemical Co), 10-6 M de hidrocortisona (Sigma
Chemical Co) e antibióticos. A cada três dias, alíquotas do sobrenadante foram
retiradas, quantificadas em hemocitômetro com azul de Trypan, sendo que parte das
células foi usada para a confecção de citocentrifugados, corados pelo método de May
Grunwald-Giemsa (MGG).
Semanalmente, metade do meio era trocado, retirando-se as células do
sobrenadante, que foram utilizadas em ensaios de formação de colônia de granulócito
e macrófago em meio semi-sólido (CFU-GM, granulocyte-macrophage colony forming
unit) (DUTRA et al., 1997). Resumidamente, o meio semi-sólido consistia de duas
camadas com os seguintes reagentes: 20% de Iscove`s Modified Dulbecco`s Medium
1X (IMDM - GIBCO®) + 30% de IMDM 2X + 20% de SFB + 30% de solução de ágar
1,8% em água destilada (Sigma Chemical Co). Após o plaqueamento e polimerização
da primeira camada, foi adicionada a segunda acrescida das células hematopoiéticas
(5 X 104 células) e do sobrenadante da linhagem de carcinoma de bexiga (5637) –
enriquecido em hemopoietinas, principalmente GM-CSF. Ao final de 12 dias de
incubação em estufa a 37ºC com 5% de CO2, as placas foram fixadas em vapor de
formol 10% (Merck) e o número de colônias (grupo de células com mais de 50 células)
formadas foi quantificada com o auxílio de microscópio invertido (Nikon - TMS).
Co-cultivo com células CD34+ de sangue de cordão. As MSC de MO não induzidas e as de TA induzidas ou não por duas semanas
para a via osteogênica, foram distribuídas em placas de 24 poços em triplicatas e as

37
células CD34+, isoladas magneticamente, como descrito anteriormente, do sangue de
cordão, foram distribuídas em uma concentração de 2 X 104 células/poço. As co-
culturas foram mantidas em estufa a 37ºC com 5% de CO2 por até 21 dias em Alpha-
Medium (α-MEM - GIBCO®) suplementado com 12,5% de SFB e 12,5% de soro de
cavalo (Sigma Chemical Co), 10-4 M de β-mercaptoetanol (Merck) e antibióticos
(CROISILLE et al., 1994). A cada três dias, as células hematopoiéticas eram retiradas
dos poços para a quantificação em hemocitômetro com azul de Trypan, sendo que
parte das células foi usada para a confecção de citocentrifugados e a outra parte para
análise em citometria de fluxo. Após uma semana, parte das células foi utilizada em
ensaios de LTC-IC (long term clonogenic- initiating cell) e de CFU-GM. Os ensaios de
LTC-IC foram realizados como previamente descrito (ISSAAD et al., 1993).
Resumidamente, as células hematopoiéticas foram retiradas e distribuídas, em
diluição limitante (50 a 400 células/poço), sobre monocamada de linhagem de estroma
medula murino, a MS5, preparada 24h antes em placa de 96 poços, tendo sido
utilizados 30 poços/diluição. As placas foram mantidas em estufa a 33ºC com 5% de
CO2 por até quatro semanas, com troca semanal de meio de cultura. Todos os poços
foram examinados pelo menos uma vez por semana e a presença de colônias de
células hematopoiéticas foi assinalada.
3.3. TÉCNICAS EXPERIMENTAIS.
3.3.a. Citometria de fluxo.
As células em suspensão foram lavadas em tampão de FACS (PBS contendo
3% de SFB e 0,1% de Azida Sódica) e incubadas por 30 minutos a 4ºC com
anticorpos monoclonais conjugados aos fluorocromos FITC, PE, PerCP, PE-Cy5 e
APC. Foram analisados os seguintes marcadores de superfície: CD105 (clone 166707;
R & D Systems), CD34 (clone 8G12; Becton & Dickinson, Franklin Lakes, NJ), CD45
(clone H130; Becton & Dickinson), CD13 (clone TÜK 1; Caltag Lab), CD14 (clone TÜK
4; Caltag Lab), CD146 (clone P1H12; Becton & Dickinson), CD31 (clone L133.1;
Becton & Dickinson), CD90 (clone 5E10; Becton & Dickinson), CD44 (clone G44-26;
Becton & Dickinson) e CD49d (clone 9F10; Becton & Dickinson). Após a incubação
com os anticorpos, quando necessário, as hemácias foram lisadas pela incubação, por
15 minutos a temperatura ambiente com a solução de lise (FACSTM lysing solution -
Becton & Dickinson) conforme as recomendações do fabricante, e a suspensão celular
foi lavada com tampão de FACS e adquirida em citômetro FACScalibur, equipado com
o programa Cell Quest 3.1 (Becton & Dikinson) ou em citômetro FACScanto, equipado
com o programa FACS Diva Software 4.0.

38
3.3.b. Seleção das células de TA.
As células de TA recém-isoladas foram incubadas, como descrito acima, com
os anticorpos monoclonais CD105, CD34 e CD45, conjugados diretamente com os
fluorocromos FITC, PE e PE-Cy5, respectivamente. A suspensão foi lavada e as
células sorteadas em Mo-Flo Fluorescence Activated Cell Sorter (FACS) - Dako
Cytomation equipado com o programa Summit 4.3 para aquisições e análises. Após a
seleção, foi feita avaliação da pureza das populações e da viabilidade, pelo Azul de
Trypan. As populações celulares selecionadas foram distribuídas separadamente em
placas de cultura com DMEM com 10% SFB e antibióticos e mantidas em estufa a
37ºC com 5% de CO2.
3.3.c. Histologia, Imunofluorescência e Imunohistoquímica.
Para análise histológica dos esferóides, estes foram fixados em
paraformaldeído 4% em PBS por pelo menos seis horas. Em seguida, parte dos
esferóides foi incubada em soluções seriadas de sacarose 15% e 30% em água
destilada por 6h cada, incluída em OCT (Optimal Cutting Temperature – Tissue-Tek Ò
– Gentaur, Bruxelas) e congelada. Os blocos foram armazenados em freezer -70°C e
posteriormente cortados com espessura de 3 – 5 μm em criostato (Leica CM 1850).
Outra parte dos esferóides foi processada rotineiramente para inclusão em
parafina. Foram realizados cortes com 5 μm de espessura em micrótomo (LUPE, 820
Spencer) que foram corados por Hematoxilina e Eosina (H-E) e selados em Entellan
(Merck) para posterior análise ao microscópio óptico (Nikon Eclipse E400).
As análises por imunofluorescência foram realizadas em cortes congelados ou
em citocentrifugados ou, ainda, em células cultivadas sobre hidroxiapatita. A
diferenciação para a linhagem endotelial em sistema de esferóides foi acompanhada
por com o anticorpo primário de camundongo anti-CD31 humano (Dakocytomation,
Fort Collins, Colorado) na diluição de 1:40 seguido da incubação com o secundário de
cabra anti-IgG conjugado a FITC (CALTAG, Invitrogen) na diluição de 1:100. Para a
marcação nuclear foi utilizada solução de DAPI 1:5000 em PBS. As observações
foram realizadas em microscópio confocal a laser (LSM META 510 - Zeiss). Já nos
citocentrifugados a marcação para CD31 foi acompanhada com o anticorpo primário
de cabra humano (Sigma Chemical Co) na diluição de 1:100 seguido da incubação
com o secundário de camundongo anyi-IgG conjugado a Rodamina (Sigma Chemical
Co). As observações foram realizadas em microscópio óptico de fluorescência (Nikon
Eclipse E400). Ensaios de imunofluorescência também foram realizados nas MSC
cultivadas em arcabouços porosos de hidroxiapatita. Para o acompanhamento da

39
diferenciação osteogênica, foi utilizado o anticorpo primário policlonal anti-
osteopontina (Santa Cruz, Santa Cruz, CA) na diluição 1:500 incubado a 4°C por 12
horas após o bloqueio com PBS-3%BSA. Em seguida as amostras foram incubadas
com o anticorpo secundário conjugado a TRITC na diluição 1:1000 por uma hora, para
a revelação da marcação. Os fragmentos do arcabouço de hidroxiapatita foram
analisados em microscópio confocal a laser (LSM META 510 - Zeiss).
A reação imunohistoquímica foi realizada após desparafinização dos cortes,
seguida da recuperação antigênica com solução de ácido cítrico pH 6,0 em panela de
pressão por cinco minutos. Após dois bloqueios seguidos, o primeiro com cloreto de
amônio 50 mM pH 8,0 por 30 minutos e o segundo para sítios inespecíficos, com uma
solução de PBS/BSA 5% por 1 hora, os anticorpos primários de coelho anti-vWF
(DakoCytomation) na diluição de 1:50 e de camundongo anti-VEGF (Santa Cruz) na
diluição de 1:100 foram adicionados sobre os cortes que foram incubados a 4°C por 12
horas. Depois de sucessivas lavagens com PBS/Tween 0,25% por 5 minutos, foi
realizada a inativação da peroxidase endógena com peróxido de hidrogênio 70% em
metanol por 15 minutos (protegido da luz). Os cortes foram então novamente lavados,
revelados com o kit de estreptavidina (LSAB, DakoCytomation) por uma hora e meia,
contracorados com hematoxilina, desidratados rapidamente e selados em Entellan
(Merck) para posterior análise ao microscópio óptico (Nikon Eclipse E400).
3.3.d. Microscopia eletrônica de varredura (MEV).
As células cultivadas em arcabouços porosos de hidroxiapatita foram, após a
retirada do meio de cultura e de sucessivas lavagens com PBS, fixadas em
glutaraldeído a 2,5% (Ted Pella Inc, Redding, CA) em tampão cacodilato de sódio
(Sigma Chemical Co) 0,1M por duas horas. Após três lavagens com o mesmo tampão,
o material foi pós-fixado em Tetróxido de Ósmio 1% (E.M.S) por 40 minutos, na
ausência de luz, e novamente lavado para a desidratação com banhos sucessivos de
10 minutos, cada, com álcool 35%, 50%, 70%, 95% e 100%. Em seguida, o material
foi seco pelo método de ponto critico e posteriormente recoberto (BAL-TEC/ SCD
O5O) com fina camadas de ouro para a metalização. Finalmente, as amostras foram
transferidas para suportes de alumínio (“STUBS”) sobre fita de carbono para a
visualização em microscópio eletrônico de varredura (Jeol 5310), operado em 20 kv.
Foram obtidas imagens digitais com programa SEMafore com resolução de 1024 X
960 Pixels.

40
3.3.e. Extração de RNA Total e Síntese de cDNA.
As amostras foram lisadas com TRIZOLTM Reagent e o RNA total foi extraído
seguindo-se as recomendações do fabricante. O RNA total foi diluído em água com
0,1% de DEPC (Dietil-pirocarbonato) e a relação entre ácido nucléico e proteína foi
determinada em espectrofotômetro, em comprimento de onda de 260 nm (nucleotídeo)
e 280 nm (proteínas). As amostras que apresentaram uma relação entre RNA e
proteínas acima de 1,6 foram utilizadas na síntese de cDNA. Para a síntese de 20 µL
de cDNA, 2 μg de RNA foram utilizados, sendo o volume ajustado com água DEPC,
para 11 µL. A este volume foi adicionado 1 μL de Oligo DT primer 500 μg/mL (Gibco) e
a mistura foi incubada a 68°C por 4 minutos. Após este período, adicionou-se 4 μL do
tampão da enzima transcriptase reversa (Invitrogen), 2 μL de 0,1 M de DTT
(Invitrogen), 1 μL de 10 mM de mistura de nucleotídeos (Invitrogen) e 1μL da enzima
de transcrição reversa, (M-MLV RT 200 U/mL; Invitrogen). A mistura foi incubada a
39°C por 2 h. Em seguida, foi colocada a 90 °C por 10 minutos e armazenada a –
20°C.
3.3.f. Reação de polimerase em cadeia com transcrição reversa (RT– PCR), Reverse Transcriptase-Polymerase Chain Reaction).
Para as reações de RT-PCR, 2 μL de cDNA foram adicionados a 20 μL de uma
solução contendo os primers (mix de PCR, anexo) e 0,3 μL de Taq Polimerase 5U/µL
(Cenbiot Enzimas, Campus do Vale da UFRGS). Após 30 segundos a 98ºC, foram
realizados 34 ciclos de 96°C por 45 segundos seguidos de 1 minuto a temperatura
média de anelamento para cada primer (Tab.1) e 72°C por 1 minuto para
alongamento. Ao final, um ciclo a 75ºC por 5 minutos. Os primers utilizados estão
listados na tabela 1. O produto amplificado foi submetido a eletroforese em gel de
agarose 1,2% e posterior coloração com brometo de etídeo 0,17 µg/mL de TAE (Tris-
Acetato-EDTA). A visualização foi realizada com o auxílio de um transiluminador.
3.3.g. Análise Estatística.
Foi utilizado teste não paramétrico, não pareado, de Mann-Whitney para a
análise de significância estatística nos experimento de CFU-GM. Foi considerado
significativo p< 0,05.
A freqüência de LTC-IC foi calculada usando o método estatístico de Poisson
(SUTHERLAND et al., 1990).

41
Tabela 1. Primers utilizados, temperatura média de anelamento e tamanho do produto amplificado (pb, pares de bases).
Sequência TM Produto
ANG-1 5’GTGGAATCTGTCATATACTGTGAA3’ 3’ACTGTGCAGATGTATATGAAGC5’ 54°C 326pb
Delta-1 5’CCACGCAGATCACGAACACC3’ 3’TTGCTATGACGCACCATCC5’ 55°C 335pb
FLT3L 5’AACAACCTATCTCCTCCTGCT3’ 3’GGCACATTTGGTGACAAAGTG5’ 55°C 306pb
G-CSF 5’GCCAGCTCCCTGCCCCAGAGCTTC3’ 3’GTGTGTCCAAGGTGGGACCCAACT5’ 64°C 300pb
GM-CSF 5’CAGGAGGCCCGGCGTCTCCTGAAC3’ 3’ACAAGCAGAAAGTCCTTCAGGTTC5’ 64°C 289pb
HGF 5’GAGGGACATAAGAAAAGAAGA3’ 3’GTGTGGTATCATGGAACTCCA5’ 55°C 386pb
IL-6 5’AACTCCTTCTCCACAAGCGCCTT3’ 3’CCGAAGAGCCCTCAGGCTGG5’ 64°C 626pb
IL-7 5’ATGAGAGTGTTCTAATGGTC3’ 3’GGAGCATTCAGTTTCCATTG5’ 51°C 619pb
KL/SCF 5’GACAGC TTGACTGATCTTCTGGAC3’ 3’ACTGCTGTCATTCCTAAGGGAGCT5’ 58°C 359pb
LIF 5’TGAACCAGATCAGGAGCCAA3’ 3’TCGGTTCACAGCACACTTCA5’ 55°C 505pb
M-CSF 5’TGTTGTTGGTCTGTCTCCTGG3’ 3’TTCTCCAGCAACTGGAGAGGT5’ 58°C 384pb
SDF-1 5’CCGCGCTCTGCC TCAGCGACGGGAAG3’ 3’CTTGTTTAAAGCTTTCTCCAGGTACT5’ 64°C 226pb
TGF- β1 5’TGAACCGGCCTTTCCTGCTTCTCATG3’ 3’GCGGAAGTCAATGTACAGCTGCCGC5’ 64°C 151pb
TNF- α 5’ACAAGCCTGTAGCCCATGTT3’ 3’AAAGTAGACCTGCCCAGACT5’ 55°C 427pb
TPO 5’TGCTCCGAGGAAAGGTGCGTT3’ 3’GGGAAGAGCGTATACTGTCCA5’ 59°C 459pb
VEGF 5’AGACCCTGGTGGACATCT TC3’ 3’TGCATTCACATTTGTTGTGC5’ 51°C 224pb
CBFA-1 5’ CTCACTACCACACCTACCTG3’ 3’CAATATGGTCGCCAAACAGATTC5’
57ºC 319 pb
Osteocalcina (OC) 5’ATGAGAGCCCTCACACTCCTC3’ 3’GCCGTAGAAGCGCCGATAGGC5’
64ºC 294 pb
Osteopontina (OP) 5’CCCTTCCAAGTAAGTCCAACGAAAGC3’ 3’CTGGATGTCAGGTCTGCGAAACTTC5’
62ºC 323 pb
PPARγ2 5’GCTGTTATGGGTGAAACTCTG3’ 3’ATAAGGTGGAGATGCAGG5’
53ºC 330 pb
Leptina 5’GGCTTTGGCCCTATCTTTTC3’ 3’GCTCTTAGAGAAGGCCAGCA5’
56ºC 316 pb
GAPDH 5’ATCACCATCTTCCAGGAGCG3’ 3’CCTGCTTCACCACCTTCTTG5’
57,5ºC 587 pb

42
4. RESULTADOS 4.1. MORFOLOGIA E FENÓTIPO DAS MSC DE TA. 4.1.a. Análise da população total.
Após a digestão enzimática do TA, a população celular obtida foi heterogênea
(Fig. 12), com presença de células hematopoiéticas, evidenciadas pela expressão de
CD45. As células CD45 representam a população hematopoiética residente e/ou
circulante do TA. Ensaios clonogênicos de CFU-GM foram realizados com a fração
celular imediatamente pós-digestão, porém não foram encontradas colônias positivas,
o que pode indicar células hematopoiéticas no TA em estágios mais avançados de
diferenciação (dados não mostrados). A população de células hematopoiéticas foi
excluída e os outros marcadores foram analisados na população da região 3 (Fig.
12D). Verificou-se que a população CD45- é heterogênea quanto a expressão de
CD34, CD105, CD146, CD31 e CD90 (Fig. 12 E-K). A maioria das células CD45-
expressa CD34 com intensidade variável e uma pequena população (de 7 a 10%)
destas expressa também CD105 (endoglina) (Fig. 12E). A análise da expressão de
CD146 e CD105 (Fig. 12F) demonstrou a presença de células CD146+CD105-,
CD146+CD105+ e CD146-CD105+. Ainda, a análise da expressão de CD146 e CD34
na população CD45- (Fig. 12G) permitiu subdividir esta população em células
CD146+CD34-, CD146+CD34+ e CD146-CD34+, sendo que esta incluía a população
CD34Hi. Deve ser notada que intensidade de fluorescência para CD146 variou, ou
seja, pode-se observar uma população com alta expressão (CD146Hi) e outra com
baixa expressão (CD146Lo) deste marcador. Embora não tenha sido possível verificar
a co-expressão de CD146, CD105 e CD34 em função dos fluorocromos conjugados a
estes anticorpos, a intensidade de fluorescência permite supor que as células CD34Hi
constituem uma população negativa tanto para CD105 quanto para CD146. Além
disto, com o auxílio do marcador CD31 (PECAM-1), foi observado que as células
endoteliais maduras estão presentes na população CD34+ e CD105+, e representam
cerca de 10% da população CD34+ (Fig. 12H) e 7% da população CD105+ (Fig. 12I).
No entanto, o percentual de células CD31+ pode variar de acordo com a amostra
analisada (Fig. 12J). Além disto, a população CD34+ é homogeneamente positiva para
CD90 (Fig. 12K), sendo que duas subpopulações de células, que variavam quanto a
intensidade de fluorescência para CD44 (Fig. 12L) foram encontradas nesta fração.
Após adesão em plástico por 18 horas, as células hematopoiéticas são
perdidas, restando células aderentes pequenas, que ainda não espraiaram (Fig. 13A).
Esta morfologia é mantida por até três dias e está relacionada com a manutenção da
expressão de CD34 (Fig. 14D). Após expansão, na fase de pré-confluência, as células

43
adquirem morfologia fibroblastoíde (Fig. 13B) e há uma diminuição na expressão de
CD34 que é concomitante ao aumento da expressão de CD105 (Figs. 14D e E). A
fração aderente torna-se homogeneamente positiva para CD49d, CD13 (Figs. 14G e
H), CD105, CD90 (Figs. 15C e D) e negativa para CD34 somente após a terceira
passagem (Fig. 14F), além de aumentar de tamanho (Figs. 14A e B). Esses dados
permitem inferir que a morfologia fibroblastóide está relacionada a uma mudança de
expressão entre os marcadores CD34 e CD105. Não se pode afirmar, no entanto, que
a população CD34+, adquiriu a expressão de CD105, pois pode ter ocorrido apenas
uma seleção populacional in vitro. Parte da população CD105 também é positiva para
o marcador CD146 (Fig. 15B), mas somente a população com alta expressão para
este marcador é CD90 negativa (Fig. 15C). De acordo com o padrão de expressão
para CD146, a população CD90-, possui o mesmo perfil da população CD34+CD146+
pós-digestão (Fig. 15C).
4.1.b. Análise de Populações Celulares Selecionadas
Com o objetivo de averiguar se a perda do marcador CD34 e o ganho de
CD105 nas células aderentes ocorria em função de seleção in vitro, as células obtidas
após digestão enzimática de TA foram sorteadas para esses marcadores,
selecionando-se duas populações: CD34+CD105+CD45- (Fig. 16B) e CD34+CD105-
CD45- (Fig. 16C). A análise após o procedimento mostrou bom grau de pureza, 90% e
95%, respectivamente (Figs. 16B e C), com boa viabilidade celular (<90%) (não
mostrado).
As duas populações foram distribuídas em placas de cultura, mostrando
morfologia arredondada (Fig. 17). A população CD34+CD105+CD45- não proliferou in
vitro, mas a população CD34+CD105-CD45- apresentou um padrão morfológico com
três dias de cultura semelhante à fração aderente do TA (Fig. 17B). Após sete dias, a
população CD34+CD105+CD45- morreu e a CD34+CD105-CD45- atingiu a confluência.
Análises por citometria de fluxo revelaram que 73,05% da população CD34+CD105-
CD45- adquiriu o marcador CD105 (Fig. 18B) durante a expansão. As células CD34+CD105+ são maiores (Figs. 18C e D), quando comparadas com as células
CD34+ (Figs. 18E e F). Esses dados comprovam que a mudança morfológica se deve
a uma modulação da expressão de CD34 e CD105 pelas células durante sua
expansão e não por seleção de uma subpopulação.
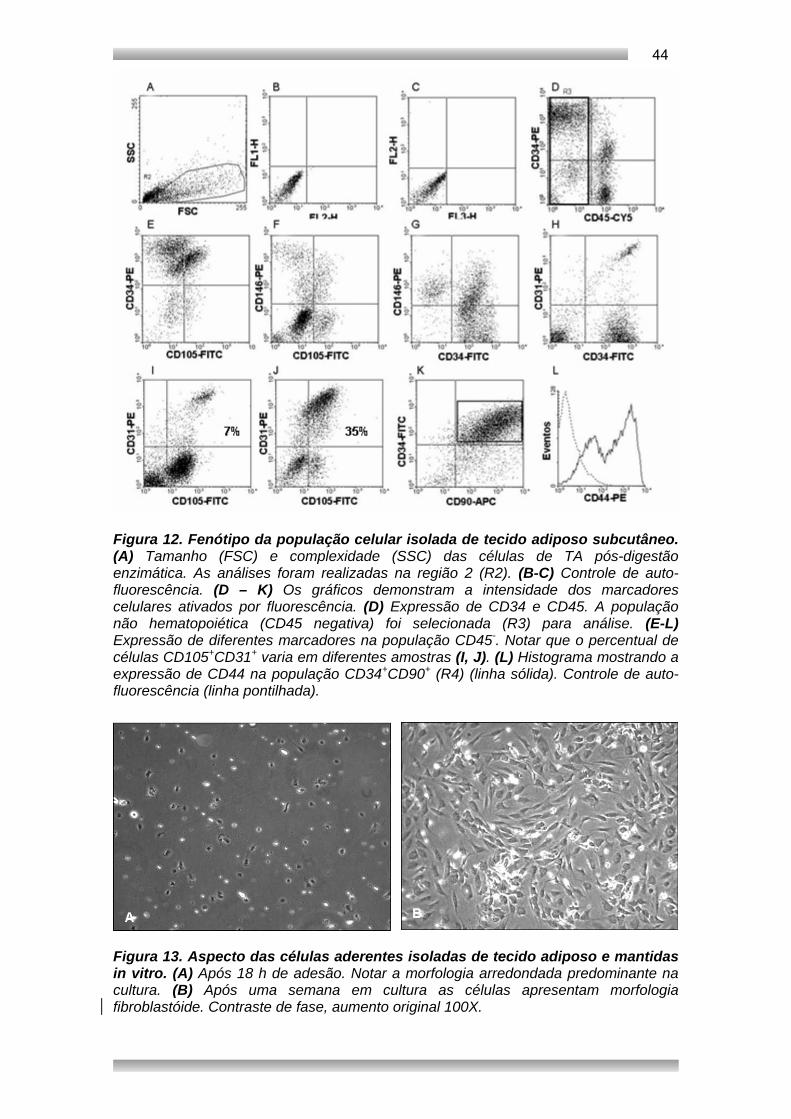
44
Figura 12. Fenótipo da população celular isolada de tecido adiposo subcutâneo. (A) Tamanho (FSC) e complexidade (SSC) das células de TA pós-digestão enzimática. As análises foram realizadas na região 2 (R2). (B-C) Controle de auto-fluorescência. (D – K) Os gráficos demonstram a intensidade dos marcadores celulares ativados por fluorescência. (D) Expressão de CD34 e CD45. A população não hematopoiética (CD45 negativa) foi selecionada (R3) para análise. (E-L) Expressão de diferentes marcadores na população CD45-. Notar que o percentual de células CD105+CD31+ varia em diferentes amostras (I, J). (L) Histograma mostrando a expressão de CD44 na população CD34+CD90+ (R4) (linha sólida). Controle de auto-fluorescência (linha pontilhada).
Figura 13. Aspecto das células aderentes isoladas de tecido adiposo e mantidas in vitro. (A) Após 18 h de adesão. Notar a morfologia arredondada predominante na cultura. (B) Após uma semana em cultura as células apresentam morfologia fibroblastóide. Contraste de fase, aumento original 100X.
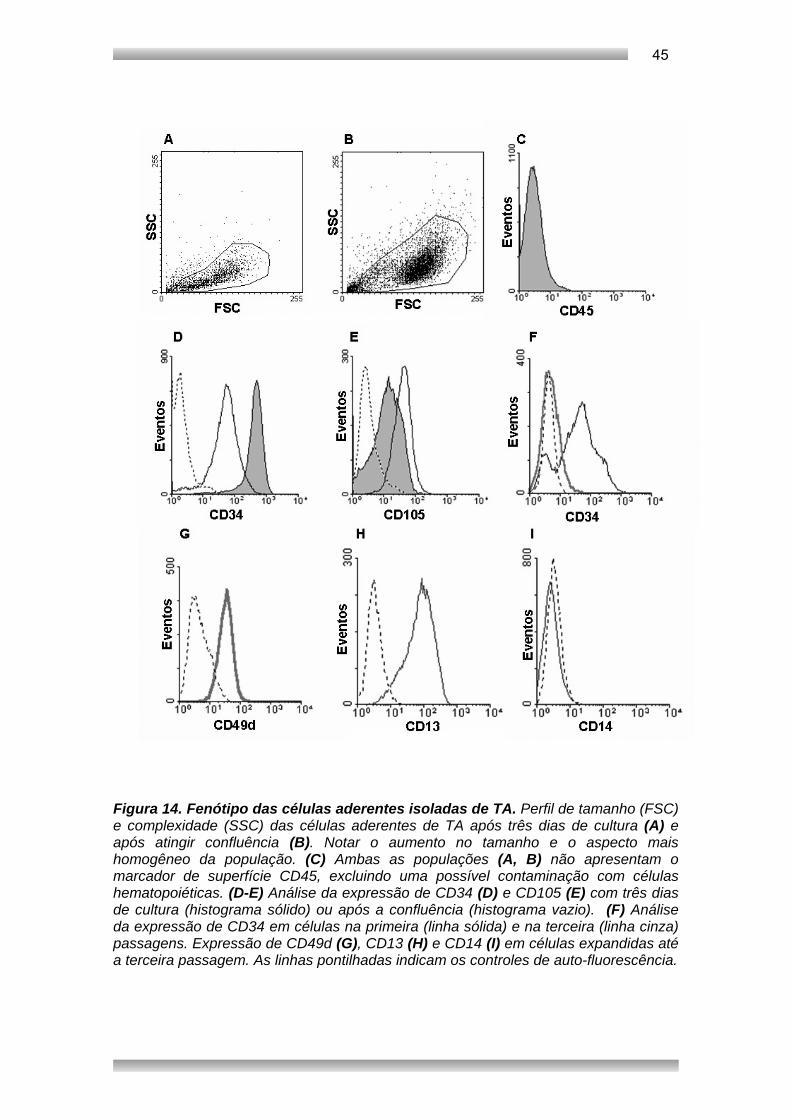
45
Figura 14. Fenótipo das células aderentes isoladas de TA. Perfil de tamanho (FSC) e complexidade (SSC) das células aderentes de TA após três dias de cultura (A) e após atingir confluência (B). Notar o aumento no tamanho e o aspecto mais homogêneo da população. (C) Ambas as populações (A, B) não apresentam o marcador de superfície CD45, excluindo uma possível contaminação com células hematopoiéticas. (D-E) Análise da expressão de CD34 (D) e CD105 (E) com três dias de cultura (histograma sólido) ou após a confluência (histograma vazio). (F) Análise da expressão de CD34 em células na primeira (linha sólida) e na terceira (linha cinza) passagens. Expressão de CD49d (G), CD13 (H) e CD14 (I) em células expandidas até a terceira passagem. As linhas pontilhadas indicam os controles de auto-fluorescência.
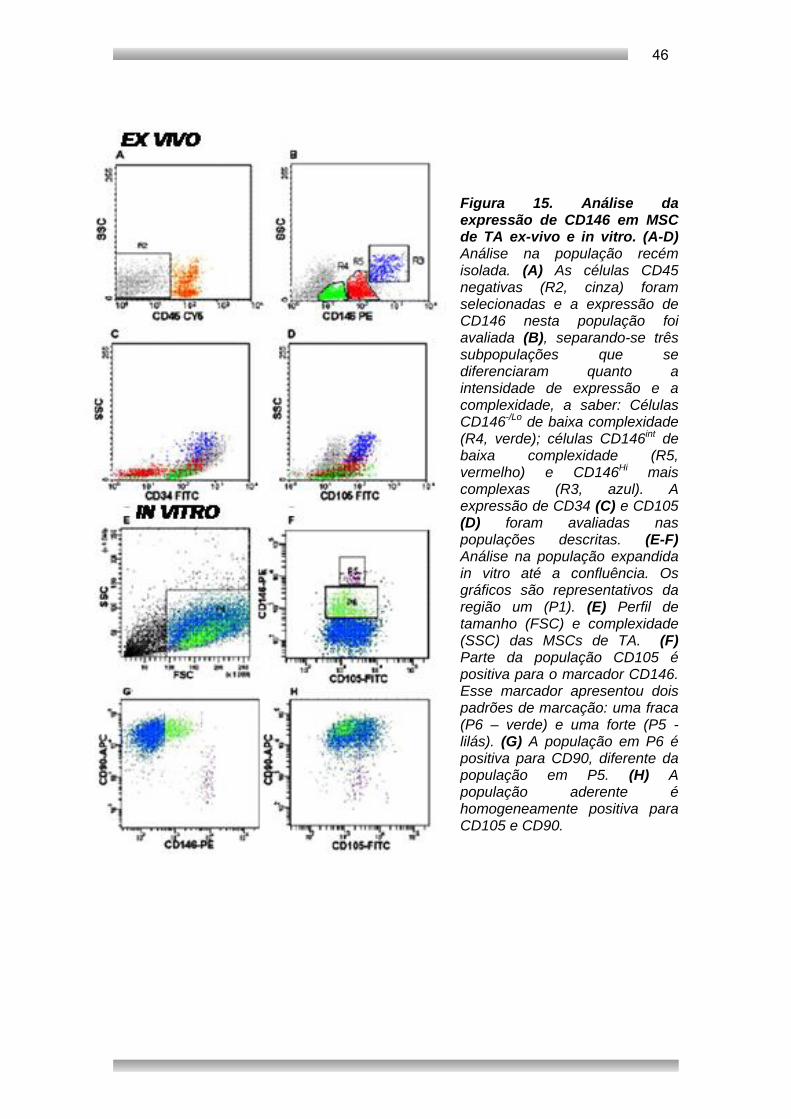
46
Figura 15. Análise da expressão de CD146 em MSC de TA ex-vivo e in vitro. (A-D) Análise na população recém isolada. (A) As células CD45
negativas (R2, cinza) foram selecionadas e a expressão de CD146 nesta população foi avaliada (B), separando-se três subpopulações que se diferenciaram quanto a intensidade de expressão e a complexidade, a saber: Células CD146-/Lo de baixa complexidade (R4, verde); células CD146int de baixa complexidade (R5, vermelho) e CD146Hi mais complexas (R3, azul). A expressão de CD34 (C) e CD105 (D) foram avaliadas nas populações descritas. (E-F) Análise na população expandida in vitro até a confluência. Os gráficos são representativos da região um (P1). (E) Perfil de tamanho (FSC) e complexidade (SSC) das MSCs de TA. (F) Parte da população CD105 é positiva para o marcador CD146. Esse marcador apresentou dois padrões de marcação: uma fraca (P6 – verde) e uma forte (P5 - lilás). (G) A população em P6 é positiva para CD90, diferente da população em P5. (H) A população aderente é homogeneamente positiva para CD105 e CD90.

47
Figura 16. Sorteamento de células de TA obtidas pós-digestão enzimática. (A) Aspecto da população total: (A.1) FSC e SSC; (A.2) CD34-PE e CD45-PECy5; (A.3) CD34-PE e CD105-FITC nas células CD45- (R3). (B) Análise após seleção da população CD34+CD105+ (R5 em A.3): (B.1) FSC e SSC; (B.2) CD34-PE e CD45-PECy5; (B.3) CD34-PE e CD105-FITC. (C) Análise após seleção da população CD34+CD105- (R4 em A.3): (C.1) FSC e SSC; (C.2) CD34-PE e CD45-PECy5; (C.3) CD34-PE e CD105-FITC.
Figura 17. Morfologia das células sorteadas em cultura. Após três dias em cultura as populações CD34+CD105+CD45- (A) e CD34+CD105-CD45- (B). Somente em (B) podemos observar células aderidas ao plástico. Notar a presença de abundantes restos celulares nas culturas da população CD105+ (A-setas). Contraste de fase, aumento original 200X.
PRÉ-SELEÇÃO
PÓS-SELEÇÃO

48
Na segunda passagem as células CD34+CD105- já são homogeneamente
positivas para CD105 (Figs. 19 B e C), perfil também encontrado em células não
sorteadas do mesmo paciente, na mesma passagem (Figs. 19D e E). A morfologia das
duas culturas também é bastante semelhante neste momento (Figs. 20A e B).
Figura 18. As células CD34+CD105- adquirem CD105 após sete dias em cultura. (A) Perfil de tamanho (FSC) e complexidade (SSC) da população CD34+CD105-CD45- após sete dias de cultura. As células CD34+ adquiriram o marcador CD105 in vitro (D). As células menores (região marcada em B) são predominantemente CD34+CD105- (E), enquanto as células maiores e mais complexas (região marcada em C) são CD34+CD105+ (F).
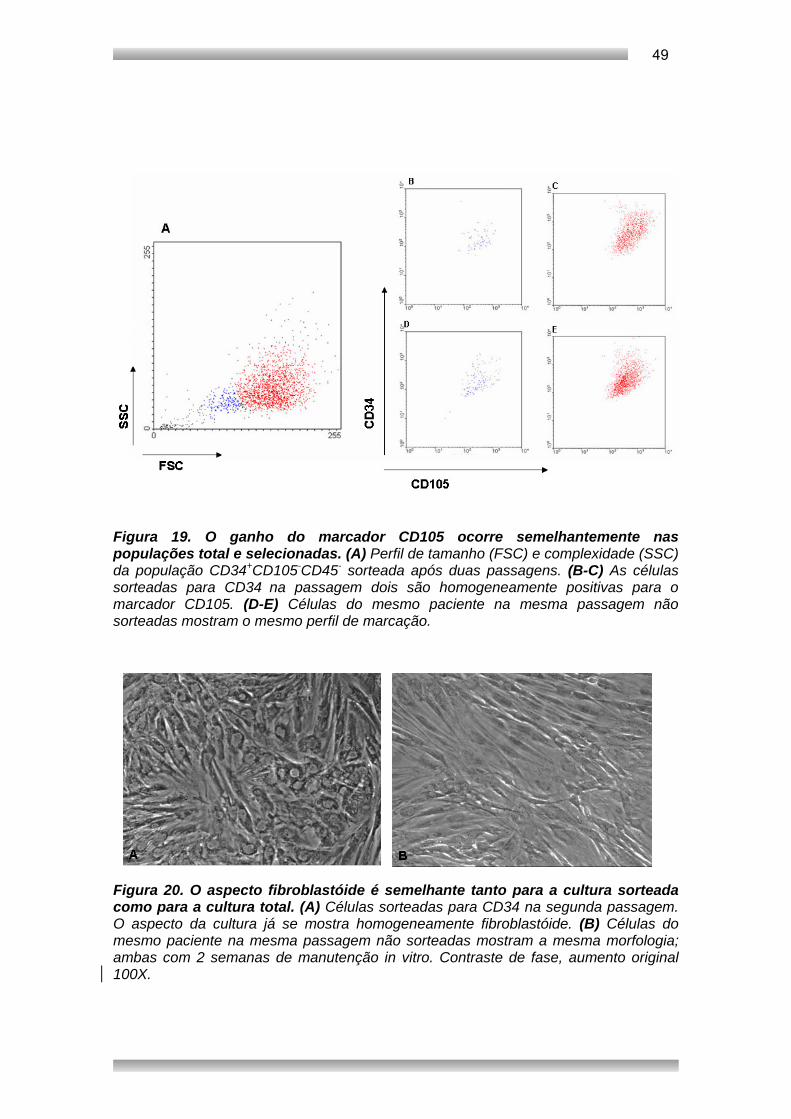
49
Figura 19. O ganho do marcador CD105 ocorre semelhantemente nas populações total e selecionadas. (A) Perfil de tamanho (FSC) e complexidade (SSC) da população CD34+CD105-CD45- sorteada após duas passagens. (B-C) As células sorteadas para CD34 na passagem dois são homogeneamente positivas para o marcador CD105. (D-E) Células do mesmo paciente na mesma passagem não sorteadas mostram o mesmo perfil de marcação.
Figura 20. O aspecto fibroblastóide é semelhante tanto para a cultura sorteada como para a cultura total. (A) Células sorteadas para CD34 na segunda passagem. O aspecto da cultura já se mostra homogeneamente fibroblastóide. (B) Células do mesmo paciente na mesma passagem não sorteadas mostram a mesma morfologia; ambas com 2 semanas de manutenção in vitro. Contraste de fase, aumento original 100X.

50
4.2. MULTIPOTENCIALIDADE 4.2.a. Diferenciação para as três linhagens mesodérmicas: adipogênica, osteogênica e condrogênica
Após o estabelecimento da monocamada de células do estroma do tecido
adiposo (MSCs de TA), as células foram expandidas até a terceira passagem e foram
realizados ensaios de diferenciação in vitro. As células foram cultivadas com os meios
indutores específicos (vide metodologia) para a diferenciação nas três linhagens
mesodérmicas – osso, adipócitos e cartilagem. Os ensaios para a via osteogênica e
adipogênica foram realizados em cultura convencional, bidimensional, ou seja, na
monocamada aderida ao plástico, e, após uma semana, as células já apresentavam
mudanças morfológicas (Figs. 21 A, B e C). As células induzidas para a via
adipogênica mudaram sua morfologia para um aspecto menos fusiforme (Fig. 21B),
em comparação às células induzidas para a via osteogênica (Fig. 21C), que por sua
vez se mostraram semelhantes ao controle (Fig. 21A). Na indução adipogênica foi
possível observar uma menor densidade celular, provavelmente acarretada pela
inibição da proliferação. Após quatro semanas de indução, verificou-se a presença de
células contendo vacúolo único com afinidade pelo corante Oil Red O (células
univacuoladas) na diferenciação adipogênica (Fig. 22A) e depósitos de cálcio,
evidenciados pela coloração de Von Kossa, na diferenciação osteogênica (Fig. 22B). A
indução para cartilagem foi realizada em um sistema de cultura tridimensional, do tipo
esferóide (vide metodologia). Após a formação dos esferóides, estes foram incubados
em meio indutor, contendo TGF-β, por até quatro semanas. A análise histológica dos
esferóides mostrou deposição de matriz extracelular, revelada pelo corante alcian blue
(pH ácido), que possui afinidade por glicosaminoglicanos e proteoglicanos ácidos (Fig.
22C).
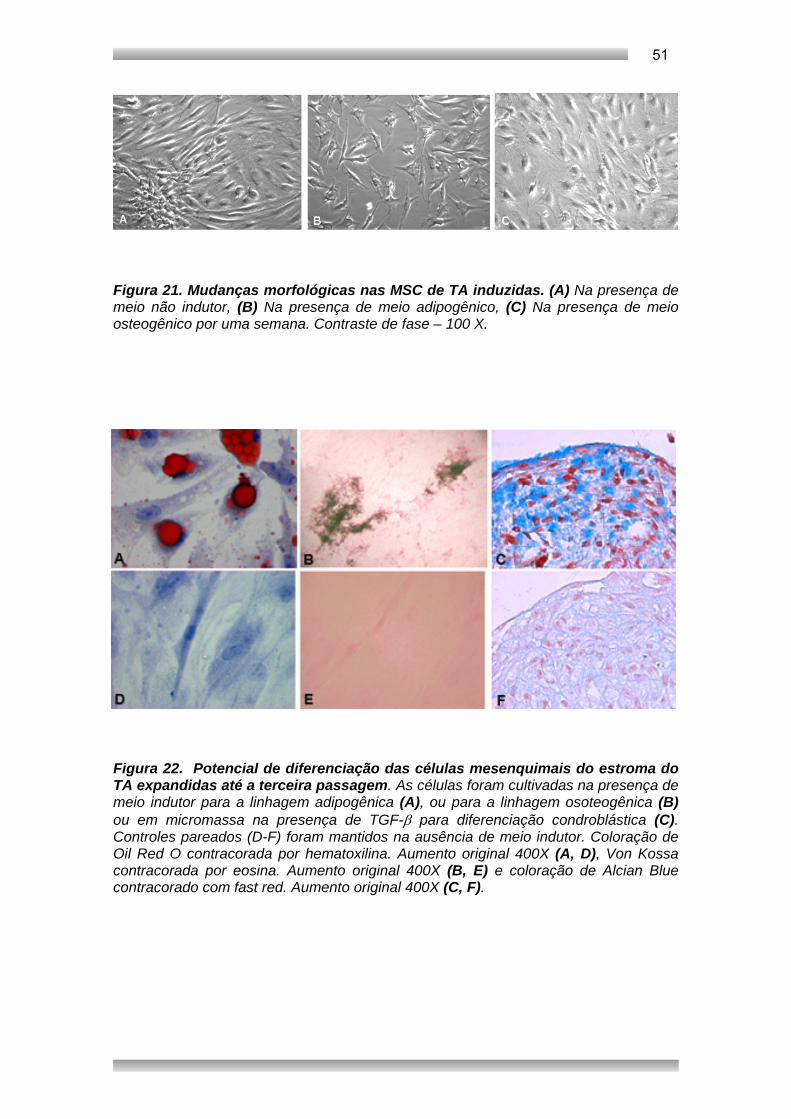
51
Figura 21. Mudanças morfológicas nas MSC de TA induzidas. (A) Na presença de meio não indutor, (B) Na presença de meio adipogênico, (C) Na presença de meio osteogênico por uma semana. Contraste de fase – 100 X.
Figura 22. Potencial de diferenciação das células mesenquimais do estroma do TA expandidas até a terceira passagem. As células foram cultivadas na presença de meio indutor para a linhagem adipogênica (A), ou para a linhagem osoteogênica (B) ou em micromassa na presença de TGF-β para diferenciação condroblástica (C). Controles pareados (D-F) foram mantidos na ausência de meio indutor. Coloração de Oil Red O contracorada por hematoxilina. Aumento original 400X (A, D), Von Kossa contracorada por eosina. Aumento original 400X (B, E) e coloração de Alcian Blue contracorado com fast red. Aumento original 400X (C, F).

52
Durante a expansão in vitro das MSC de TA, verificou-se que sua morfologia
era bem semelhante à de MO em baixa confluência (Figs. 23 A e B). Porém, após a
confluência notou-se que algumas células da monocamada de MSCs de TA
acumulavam lipídeos espontaneamente (Fig. 23C). Este acúmulo lipídico foi
acompanhado de mudanças morfológicas na monocamada, observadas pela
microscopia eletrônica de varredura. Nas áreas de maior confluência, algumas células
de TA assumiram uma morfologia mais arredondada, característica de adipócitos (Fig.
23E), enquanto as MSC de MO permaneceram fibroblastóides (Fig. 23F). As análises por RT-PCR revelaram diferenças na expressão de genes
envolvidos na diferenciação adipogênica entre as MSCs de TA e de MO. Mesmo sem
indução para a via adipogênica, as MSCs de TA expressaram o gene PPARγ2
(peroxisome proliferator-activated receptor-gamma-2), além dos genes envolvidos com
a diferenciação osteogênica, que são constitutivamente expressos também pelas
células de MO (Fig. 24). Portanto, ou as MSCs de TA apresentam um maior
comprometimento com a via adipogênica, ou a monocamada é heterogênea, com
presença de células comprometidas com a linhagem adipogênica, possivelmente pré-
adipócitos, e células mais indiferenciadas multipotentes.
Para testar se o potencial de diferenciação para a linhagem adipogênica e
osteogênica diferia entre as MSCs de TA e de MO, foi feita uma análise comparativa.
Assim, durante a indução adipogênica, a intervalos semanais, a monocamada foi
fixada, corada pelo Oil Red O e o percentual de células, multivacuolares e
univacuolares, acumuladoras de lipídeos foi determinado (Figs. 25 e 26). O potencial
osteogênico foi também determinado em monocamada (Fig. 28) ou sobre
hidroxiapatita (Fig. 30). Ao mesmo tempo, foram retiradas amostras para análise, por
RT-PCR, da expressão dos genes envolvidos na diferenciação para estas linhagens
(Figs 27 e 29).
Em relação ao potencial adipogênico, foi possível notar uma rápida
diferenciação das MSCs de TA em relação as da MO (Figs. 25 e 26), pois após quatro
semanas já se notava a presença de mais de 50% de células univacuoladas (Fig.
26B). Já nas MSC de MO, o estágio final da diferenciação adipocítica só foi alcançado
em seis semanas. Esta diferença na resposta ao meio indutor pode ser explicado pela
expressão de PPARγ2, que, nas MSCs de MO, ocorreu somente após uma semana de
indução (Fig. 27A). Curiosamente, observou-se uma expressão constitutiva de leptina
nas células de MO, enquanto que nas MSCs de TA a expressão deste gene só
ocorreu após quatro semanas de indução (Fig. 27B).

53
Figura 23. MSC de TA e de MO em cultura sem agentes indutores. (A, C, E) MSC de TA em subconfluência, e em hiperconfluência coradas por Oil Red O e hematoxilina e analisadas por microscopia eletrônica de varredura, respectivamente. Setas indicam acúmulos lipídicos em C. (B, D, F) MSC de MO nas mesmas condições. (A - D) Contraste de fase – aumento original 100 X. Figura 24. Expressão de CBFA-1/RUNX-2, osteocalcina, osteopontina, e PPARγ2 nas MSC de MO e de TA. O RNA total das células expandidas foi isolado, e o cDNA amplificado com primers específicos. Os produtos do RT-PCR foram analisados por gel de agarose corado por brometo de etídeo. O tamanho do produto amplificado se encontra abaixo dos genes respectivamente.
TA MOTA MO
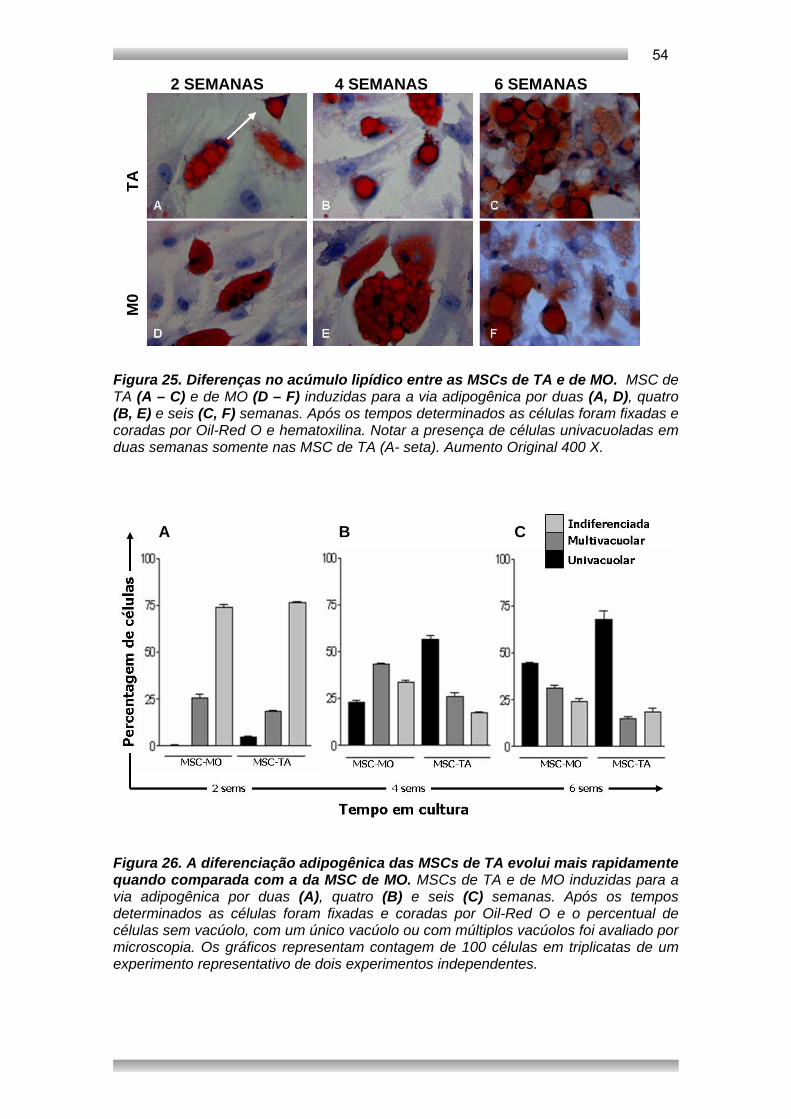
54
Figura 25. Diferenças no acúmulo lipídico entre as MSCs de TA e de MO. MSC de TA (A – C) e de MO (D – F) induzidas para a via adipogênica por duas (A, D), quatro (B, E) e seis (C, F) semanas. Após os tempos determinados as células foram fixadas e coradas por Oil-Red O e hematoxilina. Notar a presença de células univacuoladas em duas semanas somente nas MSC de TA (A- seta). Aumento Original 400 X.
Figura 26. A diferenciação adipogênica das MSCs de TA evolui mais rapidamente quando comparada com a da MSC de MO. MSCs de TA e de MO induzidas para a via adipogênica por duas (A), quatro (B) e seis (C) semanas. Após os tempos determinados as células foram fixadas e coradas por Oil-Red O e o percentual de células sem vacúolo, com um único vacúolo ou com múltiplos vacúolos foi avaliado por microscopia. Os gráficos representam contagem de 100 células em triplicatas de um experimento representativo de dois experimentos independentes.
A B C
M0
T
A
2 SEMANAS 4 SEMANAS 6 SEMANAS

55
Figura 27. Análise da expressão de mRNA de genes ligados à diferenciação adipogênica. (A) PPARγ2, 330pb; (B) Leptina, 326pb; (C) GAPDH, 587pb. (A e B): Linha 1: Padrão de peso polecular 100pb; Linha 2: controle da reação de PCR(cPCR); Linha 3: MSC de MO não induzida; Linha 4 e 5: células de MO induzidas por 1 e 4 semanas, respectivamente; Linha 6: MSC de TA não induzida; Linhas 7 e 8: MSCs de TA induzidas por uma e quatro semanas, respectivamente. (C): Linha 1: Padrão de Peso Molecular 100pb; Linha 2: MSC de MO não induzida; Linha 3 e 4: Células de MO induzidas por uma e quatro semanas, respectivamente; Linha 5: MSC de TA não induzida; Linhas 6 e 7: Células de TA induzidas por uma e quatro semanas, respectivamente.
Quanto ao potencial osteogênico, verificou-se que as MSCs de TA e de MO,
expandidas até a terceira passagem e mantidas por até três semanas na presença de
meio osteoindutor, mostram depósitos de cálcio na monocamada, de forma muito
semelhante (Fig. 28). A análise da expressão gênica também revelou padrões
semelhantes, a não ser pela perda de osteopontina pelas MSC de TA em três
semanas (Fig. 29). As MSCs de TA também foram cultivadas por até duas semanas
em esponjas de hidroxiapatita, um biomaterial osteocondutor utilizado em diversos
protocolos de bioengenharia óssea (ROBEY & BIANCO, 2007). Após uma semana, foi
possível notar a intensa proliferação das células de MO (Fig. 30B), quando comparado
com as de TA (Fig. 30A), pois o número de células iniciais foi o mesmo. A morfologia
também se apresentou ligeiramente diferente, notando-se, em especial, uma
morfologia mais miofibroblastóide nas células de MO (Fig. 30 C e D). O aspecto
morfológico encontrado nas esponjas de hidroxiapatita foi bem similar ao encontrado
na monocamada confluente (Fig. 23E-F), ambos analisados por microscopia eletrônica
de varredura. Análises por microscopia confocal revelaram marcação positiva para
osteopontina, idêntica para ambas as células (Fig. 30E-F).
Para verificar se a população celular aderente recém-isolada do TA era
heterogênea quanto ao potencial de diferenciação para a linhagem adipogênica e
osteogênica, estas foram cultivadas em meio indutor misto (vol/vol de meio
osteogênico e adipogênico) por até dez dias e a monocamada foi corada com Oil Red
O e Von Kossa. Verificou-se a presença tanto de depósitos de cálcio, quanto de
células acumuladoras de lipídeos (Fig. 31A). No entanto, foi observado um maior
número de células induzidas para a via adipogênica (Fig. 31B).

56
Figura 28. As MSCs de TA e de MO apresentam depósitos de cálcio de forma semelhante. MSCs de TA (A) e de MO (B) induzidas para a via osteogênica por três semanas. As células foram fixadas e coradas pelo método de Von Kossa. Aumento Original 400 X.
Figura 29. Análise da expressão de mRNA de genes ligados à diferenciação osteogênica. (A) CBFA-1, 319pb; (B) Osteocalcalcina, 294pb; (C) Osteopontina, 323pb; (D) GAPDH, 587pb. (A-C): Linha 1: Padrão de peso molecular 100pb; Linha 2: cPCR; Linha 3: MSC de MO não induzida; Linha 4 e 5: células de MO induzidos por 1 e 4 semanas, respectivamente; Linha 6: MSC de TA não induzida; Linhas 7 e 8: Células de TA induzidas por uma e quatro semanas, respectivamente. (D): Linha 1: Padrão de Peso Molecular 100pb; Linha 2: MSC de MO não induzida; Linha 3 e 4: Células de MO induzidas por uma e quatro semanas, respectivamente; Linha 5: MSC de TA não induzida; Linhas 6 e 7: Células de TA induzidas por uma e quatro semanas, respectivamente. Figura 30. Cultura de MSCs de TA e de MO em arcabouços de hidroxiapatita. MSC de TA (A, C e E) e de MO (B, D e F) foram distribuídas sobre arcabouços porosos de hidroxiapatita e mantidas em cultura por uma semana. Notar o maior recobrimento da superfície do biomaterial pelas células de MO (B) quando comparadas com as de TA (A). Detalhe do aspecto miofibroblastóide das células de MO (D) na hidroxiapatita. Expressão de osteopontina nas células de TA (E) e de MO (F) cultivadas em hidroxiapatita. (A-D) Microscopia eletrônica de varredura. Barra = 500μm (A E B) e 10μm (C e D). (E-F) Microscopia confocal. Dimensões = 230,34 μm x 230,34 μm. Figura 31. MSC de TA na presença de meio indutor misto diferenciam para as linhagens osteogênica e adipogênica. No momento do estabelecimento da cultura, as células foram incubadas em meio indutor misto por até 10 dias, fixadas e coradas pelos métodos de Oil Red e Von Kossa e contracoradas por hematoxilina. (A) Notar a presença de depósitos de cálcio expressivos – seta, circundados por células com acúmulos lipídicos multivacuolados. (B) Representação de vários campos, onde o número de células comprometidas para a linhagem adipogênica é expressivamente maior do que para a linhagem osteogênica. Aumento original 200X.
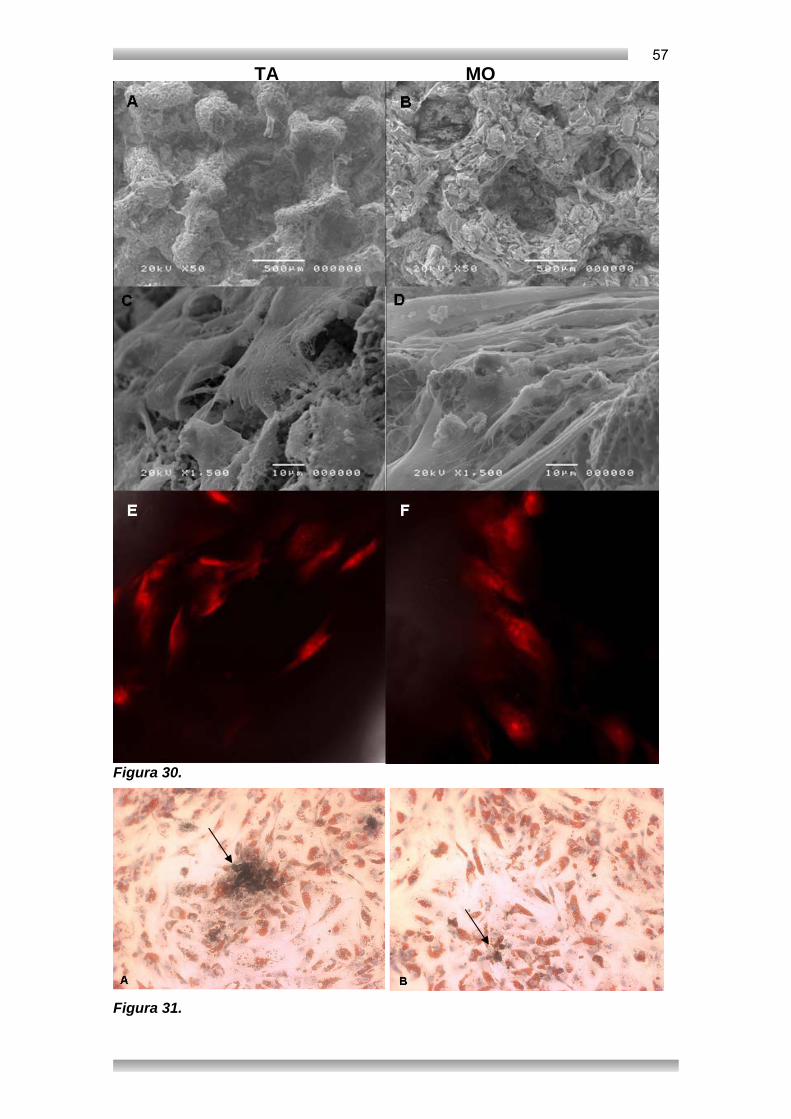
57
Figura 30. Figura 31.
TA MO

58
4.2.b. Diferenciação para a linhagem endotelial O papel das MSCs de TA na angiogênese tem sido amplamente relatada
(FUKUMURA et al., 2003; KUBIS et al., 2007; MIRANVILLE et al., 2004; MOON et al.,
2006, PLANAT-BENARD et al., 2004a; SUMI et al., 2007; WOSNITZA et al., 2007),
mas, embora não haja dúvida sobre o efeito estimulador destas células na
angiogênese, a observação de uma diferenciação direta para a linhagem endotelial é
controversa. Para testar a capacidade das MSCs isoladas de TA humano de se
diferenciarem em células endoteliais ou de interagirem com endotélio maduro, à
semelhança de pericitos, modelos de cultura tridimensional do tipo esferóide foram
desenvolvidos. Os esferóides são agregados de células responsáveis tanto pela
geração de um centro de hipóxia, como pela síntese de proteínas da matriz
extracelular. Desta forma este sistema é descrito como modulador positivo da
agiogênese em células endotelais maduras ou ainda em células comprometidas com
esa via (KELM et al., 2005).
O sistema de cultivo em esferóide utilizado permitiu a diferenciação das MSCs
de TA para a linhagem endotelial, mesmo na ausência do meio próprio para células
endoteliais. No interior dos esferóides foram observadas estruturas que se
assemelhavam a vasos (Figs. 32A e B) e que serão investigadas posteriormente
através de microscopia eletrônica de transmissão. Os esferóides também
apresentaram células positivas para CD31 (Figs. 33A e B) e vWf (Fig. 32D) e secretam
a proteína VEGF (Fig. 32C). O mesmo não ocorreu para as MSC de TA cultivadas em
monocamada (Figs. 33C, D e E). Curiosamente, este sistema de cultivo tridimensional
induziu a expressão gênica de angiopoietina-1 (Ang-1) pelas MSCs de TA. Este gene
possui um papel importante na formação de vasos (SURI et al., 1996) e não é
expresso pelas MSC de TA cultivadas em monocamada (Fig. 33F). A expressão de
VEGF (Vascular Endothelial Growth Factor) e SDF-1α (Stromal Derived Factor-1), que
também têm papel importante na angiogênese (FIEDLER & AUGUSTIN, 2006), parece
ser constitutiva, tendo sido observada tanto nas células cultivadas em modelo de
esferóide, como nas mantidas em monocamada (Figs. 33G e H).
Devido à diferenciação obtida para a via endotelial em sistema de esferóides,
foi investigado se as MSCs de TA seriam capazes de modular células endoteliais
maduras humanas (HUVEC). As MSCs de TA foram co-cultivadas com as HUVEC em
monocamada sem meio de cultura endotelial por até quatro dias. Após 24 horas de
cultura, já foi possível observar a emissão de prolongamentos das MSCs de TA em
torno das HUVEC, que, caracteristicamente, formam grupamentos de células
poligonais (Fig. 34A). Esta interação parece ter favorecido a manutenção das

59
HUVECs, pois após quatro dias os ninhos de células endoteliais permaneceram
estáveis (Fig. 34C), o mesmo não ocorrendo quando as HUVEC foram cultivadas na
ausência de meio de endotélio ou de MSCs de TA (Fig. 34B, D). A interação das MSC
de TA com as HUVEC aparentemente não foi capaz de modular positivamente a
expressão de CD31 nas MSCs de TA, pois a marcação ficou restrita a ílhotas
pequenas de células poligonais (Fig. 34E), diferentemente do que foi visto em sistema
de esferóides. Nos esferóides mistos notou-se uma segregação das duas populações
celulares. As HUVECS formaram um esferóide que ficou excentricamente disposto,
mas circundado pelo esferóide de MSCs de TA (Fig. 34F).
Além disto, as MSCs de TA também foram capazes de estimular o brotamento
de esferóides de HUVEC em gel de colágeno (Fig. 35A) com padrão semelhante ao
estímulo de VEGF (Fig. 35B). Os esferóides de MSC de TA também responderam ao
VEGF, mas de maneira contrária aos de HUVEC. Ou seja, seu brotamento
espontâneo (Fig. 35C) cessou na presença do fator (Fig. 35D), o mesmo sendo
observado quando os esferóides mistos foram colocados em gel de colágeno (Fig.
35E).
Figura 32. Os esferóides de MSC de TA apresentam estruturas semelhantes a vasos. Cortes de parafina de esferóides de MSC de TA expandidas até a terceira passagem sem indução. (A, B) Aspecto histológico dos esferóides de MSCs de TA, mostrando detalhe (B) de estrutura com luz central (seta). H-E, aumentos originais de 400X e 1000X, respectivamente. (C) Imunohistoquímica para VEGF. Notar a presença do fator de crescimento VEGF preferencialmente no interior dos esferóides. Aumento original 200 X. (D) Imunohistoquímica para vWf. Aumento original 400 X.
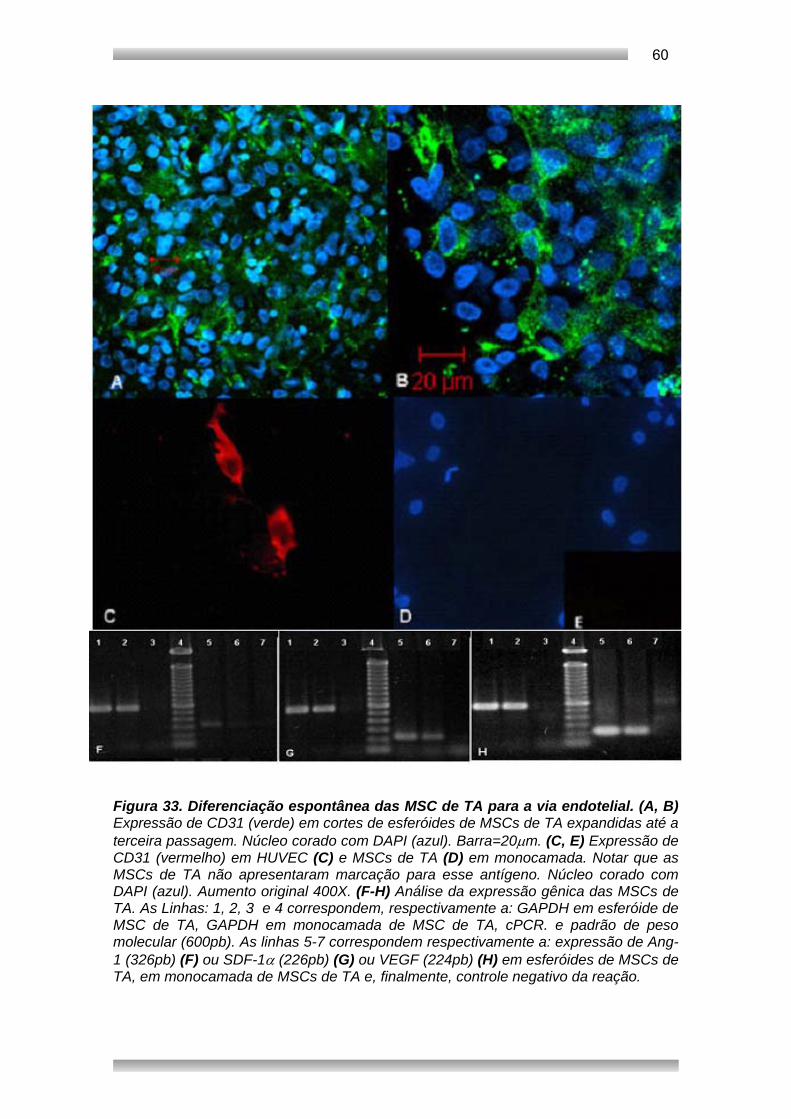
60
Figura 33. Diferenciação espontânea das MSC de TA para a via endotelial. (A, B) Expressão de CD31 (verde) em cortes de esferóides de MSCs de TA expandidas até a terceira passagem. Núcleo corado com DAPI (azul). Barra=20μm. (C, E) Expressão de CD31 (vermelho) em HUVEC (C) e MSCs de TA (D) em monocamada. Notar que as MSCs de TA não apresentaram marcação para esse antígeno. Núcleo corado com DAPI (azul). Aumento original 400X. (F-H) Análise da expressão gênica das MSCs de TA. As Linhas: 1, 2, 3 e 4 correspondem, respectivamente a: GAPDH em esferóide de MSC de TA, GAPDH em monocamada de MSC de TA, cPCR. e padrão de peso molecular (600pb). As linhas 5-7 correspondem respectivamente a: expressão de Ang-1 (326pb) (F) ou SDF-1α (226pb) (G) ou VEGF (224pb) (H) em esferóides de MSCs de TA, em monocamada de MSCs de TA e, finalmente, controle negativo da reação.
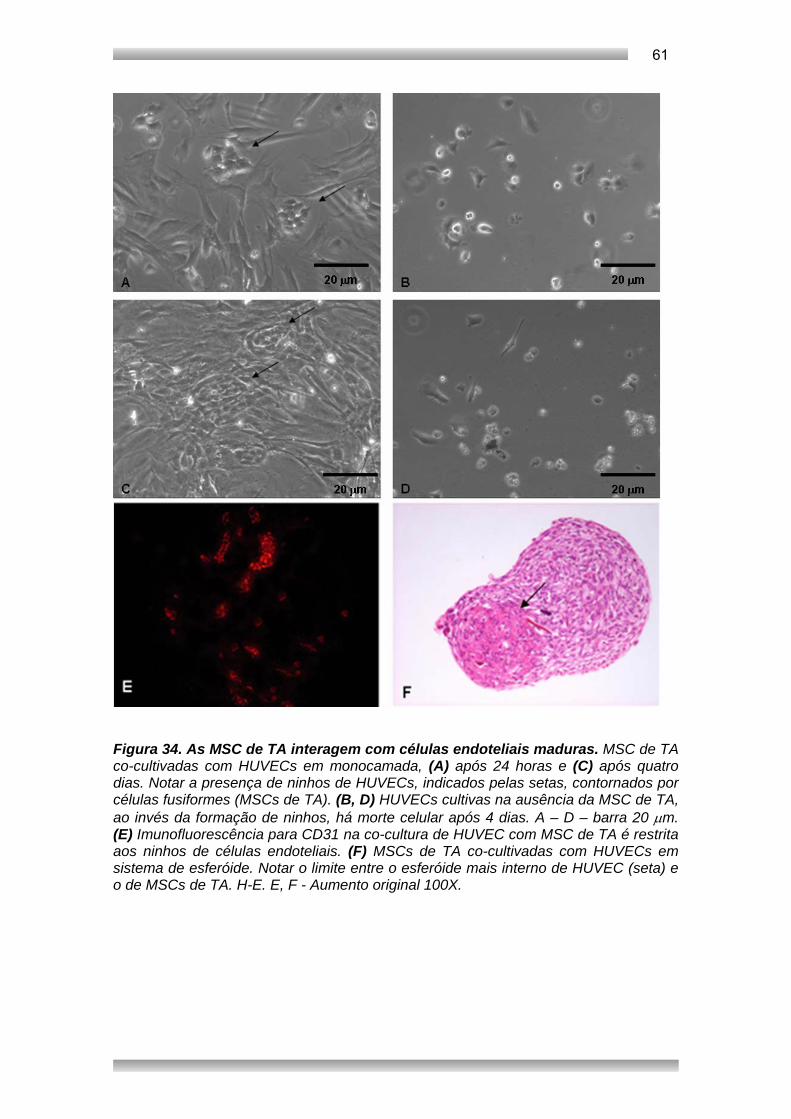
61
Figura 34. As MSC de TA interagem com células endoteliais maduras. MSC de TA co-cultivadas com HUVECs em monocamada, (A) após 24 horas e (C) após quatro dias. Notar a presença de ninhos de HUVECs, indicados pelas setas, contornados por células fusiformes (MSCs de TA). (B, D) HUVECs cultivas na ausência da MSC de TA, ao invés da formação de ninhos, há morte celular após 4 dias. A – D – barra 20 μm. (E) Imunofluorescência para CD31 na co-cultura de HUVEC com MSC de TA é restrita aos ninhos de células endoteliais. (F) MSCs de TA co-cultivadas com HUVECs em sistema de esferóide. Notar o limite entre o esferóide mais interno de HUVEC (seta) e o de MSCs de TA. H-E. E, F - Aumento original 100X.
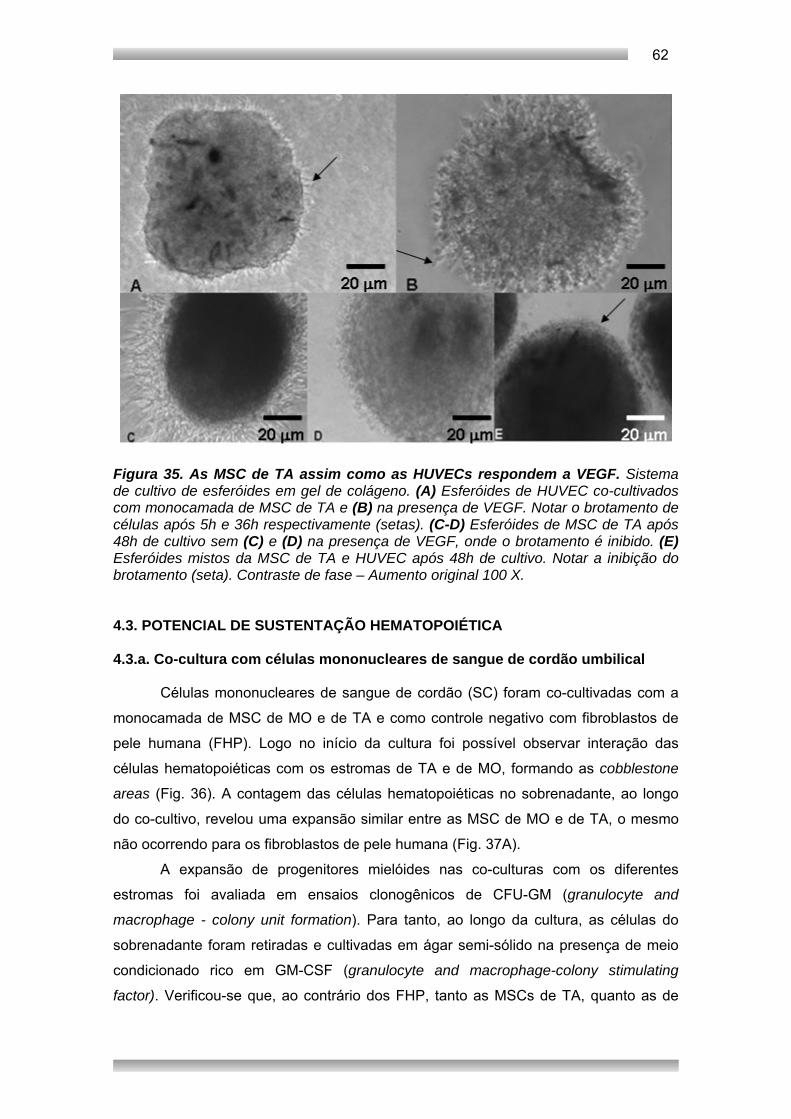
62
Figura 35. As MSC de TA assim como as HUVECs respondem a VEGF. Sistema de cultivo de esferóides em gel de colágeno. (A) Esferóides de HUVEC co-cultivados com monocamada de MSC de TA e (B) na presença de VEGF. Notar o brotamento de células após 5h e 36h respectivamente (setas). (C-D) Esferóides de MSC de TA após 48h de cultivo sem (C) e (D) na presença de VEGF, onde o brotamento é inibido. (E) Esferóides mistos da MSC de TA e HUVEC após 48h de cultivo. Notar a inibição do brotamento (seta). Contraste de fase – Aumento original 100 X. 4.3. POTENCIAL DE SUSTENTAÇÃO HEMATOPOIÉTICA 4.3.a. Co-cultura com células mononucleares de sangue de cordão umbilical Células mononucleares de sangue de cordão (SC) foram co-cultivadas com a
monocamada de MSC de MO e de TA e como controle negativo com fibroblastos de
pele humana (FHP). Logo no início da cultura foi possível observar interação das
células hematopoiéticas com os estromas de TA e de MO, formando as cobblestone
areas (Fig. 36). A contagem das células hematopoiéticas no sobrenadante, ao longo
do co-cultivo, revelou uma expansão similar entre as MSC de MO e de TA, o mesmo
não ocorrendo para os fibroblastos de pele humana (Fig. 37A).
A expansão de progenitores mielóides nas co-culturas com os diferentes
estromas foi avaliada em ensaios clonogênicos de CFU-GM (granulocyte and
macrophage - colony unit formation). Para tanto, ao longo da cultura, as células do
sobrenadante foram retiradas e cultivadas em ágar semi-sólido na presença de meio
condicionado rico em GM-CSF (granulocyte and macrophage-colony stimulating
factor). Verificou-se que, ao contrário dos FHP, tanto as MSCs de TA, quanto as de

63
Mo foram capazes de expandir significativamente (p<0,01) os progenitores mielóides
presentes no sangue de cordão (Fig. 37B). A diferença no número de colônias gerado
pelas monocamadas de MSCs de MO e de TA nos dois pontos analisados não foi
significativa (p>0,05).
A análise diferencial dos citocentrifugados revelou uma geração de células
hematopoiéticas pelas MSCs de TA com a mesma dinâmica das células de MO (Figs.
37C e D). No entanto, observou-se uma diferença no perfil hematopoiético gerado por
essas duas monocamadas no final do co-cultivo (28 dias), caracterizada pelo pico
expressivo de células mono-macrofágicas nas MSCs de TA (Fig. 37C). Estas células
são provavelmente derivadas de precursores monocíticos que podem ser detectados
após 10 dias de co-cultura com as MSCs de TA (Fig. 38D). No mesmo tempo de co-
cultivo, as MSCs de MO geraram um pico de polimorfonucleares (Fig. 38C).
No microambiente da medula óssea, as diferentes populações estromais
derivadas das MSCs parecem ter papel específico. Com o objetivo de investigar esses
dados, as monocamadas de ambas as células (MSCs de TA e de MO) foram induzidas
por uma semana para as linhagens adipocítica ou osteogênica, denominadas neste
trabalho de pré-adipócitos e pré-osteoblastos. Através da contagem das células
hematopoiéticas geradas nessas co-culturas foi possível observar uma expansão
destas em contato com as monocamadas de pré-adipócitos de TA e de MO,
contrastando com a manutenção das células hematopoiéticas nas co-culturas com
pré-osteoblastos derivados de MSCs de TA (Fig. 28A) e de MO (Fig. 39B). Esses
dados são sustentados pelas contagens de colônias CFU-GM, que revelaram uma
expansão de progenitores mielóides nos estromas de pré-adipócitos, novamente de
maneira semelhante para as MSCs de TA e de MO (Fig. 39C), sendo que o maior
número de colônias geradas foi nos estromas induzidos para a linhagem adipogênica.
A análise diferencial dos citocentrifugados demonstrou uma expansão
preferencial de progenitores comprometidos com a série mielóide após oito dias nos
estromas de pré-adipócitos. Essa expansão foi acompanhada por um aumento no
percentual das células macrofágicas e diminuição de blastos (Figs. 40 A e B). Essas
observações são complementares a contagem de CFU-GM, que foi relativamente
expressiva em oito dias (Fig. 39C). Outro dado importante foi o aumento, em torno de
10%, no percentual de macrófagos após 14 dias de co-cultura com estroma de pré-
adipócitos derivados de MSCs de MO (Fig. 40B).
Nos estromas pré-osteoblásticos, o perfil de geração de células
hematopoiéticas é completamente diferente dos estromas pré-adipocíticos. Em oito
dias há uma discreta manutenção de células blásticas na MSC de TA (Fig. 40C) e
expansão das mesmas nas células de MO (Fig. 40D). Em nenhum momento de co-

64
cultivo há a expansão de progenitores mieloídes, correlacionando com a escassez de
colônias de CFU-GM nesse co-cultivo (Figs. 39 C e D). Apesar da falta de expansão
de progenitores, no final do co-cultivo há a geração de células hematopoiética
terminalmente diferenciadas – os polimorfonucleares (Figs. 40 C e D).
Análises por RT-PCR de genes envolvidos em hematopoiese revelam perfis
muito semelhantes para as MSs de TA e de MO, ambas sem indução. A indução para
as duas vias, adipogênica e osteogênica, são capazes de modular as hemopoietinas
de maneira particular (Fig. 41 e Tab. 2). Dentre elas, GM-CSF (Granulocyte-
macrophage colony-stimulating factor) é regulada positivamente na indução
osteogênica e LIF (leukemia inhibitory factor) negativamente na indução adipogênica
(Fig. 41).
Figura 36. Interação das células hematopoiéticas com estromas de TA e de MO. Após quatro dias de co-cultivo as células mononucleares de sangue de cordão interagiram com as MSCs de TA (A) e de MO (B), formando cobblestone areas. Contraste de fase. Aumento Original 200 X.
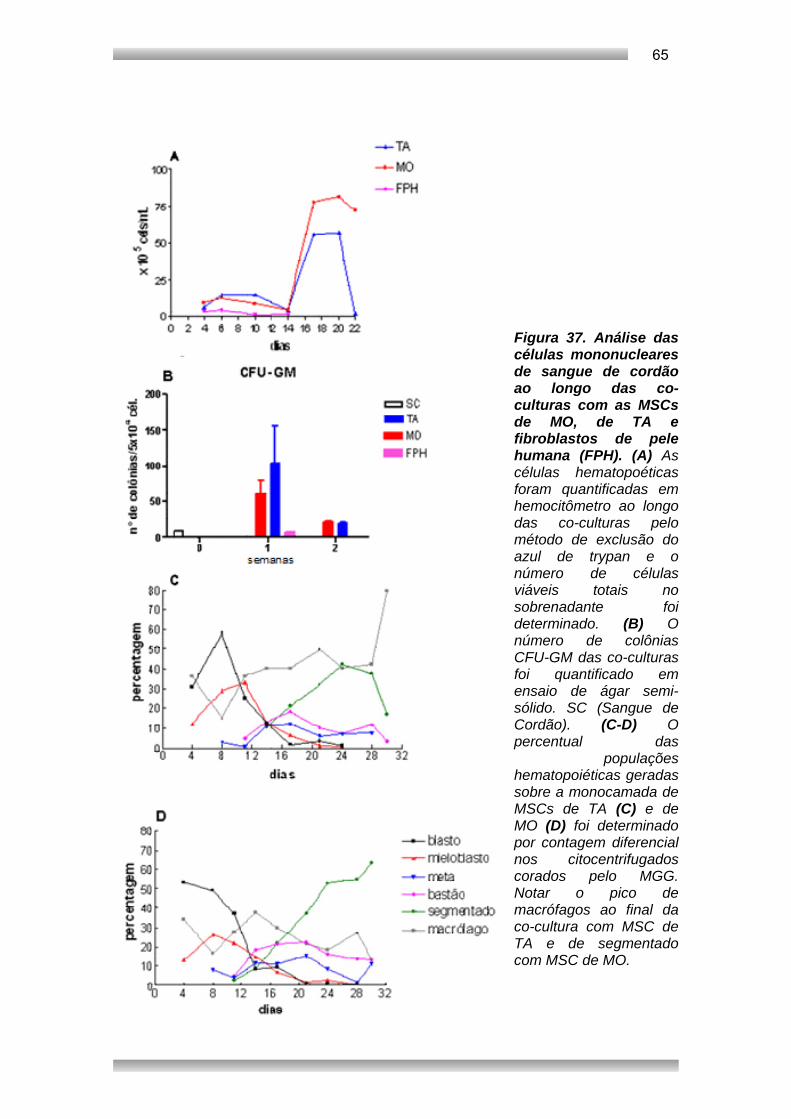
65
Figura 37. Análise das células mononucleares de sangue de cordão ao longo das co-culturas com as MSCs de MO, de TA e fibroblastos de pele humana (FPH). (A) As células hematopoéticas foram quantificadas em hemocitômetro ao longo das co-culturas pelo método de exclusão do azul de trypan e o número de células viáveis totais no sobrenadante foi determinado. (B) O número de colônias CFU-GM das co-culturas foi quantificado em ensaio de ágar semi-sólido. SC (Sangue de Cordão). (C-D) O percentual das
populações hematopoiéticas geradas sobre a monocamada de MSCs de TA (C) e de MO (D) foi determinado por contagem diferencial nos citocentrifugados corados pelo MGG. Notar o pico de macrófagos ao final da co-cultura com MSC de TA e de segmentado com MSC de MO.
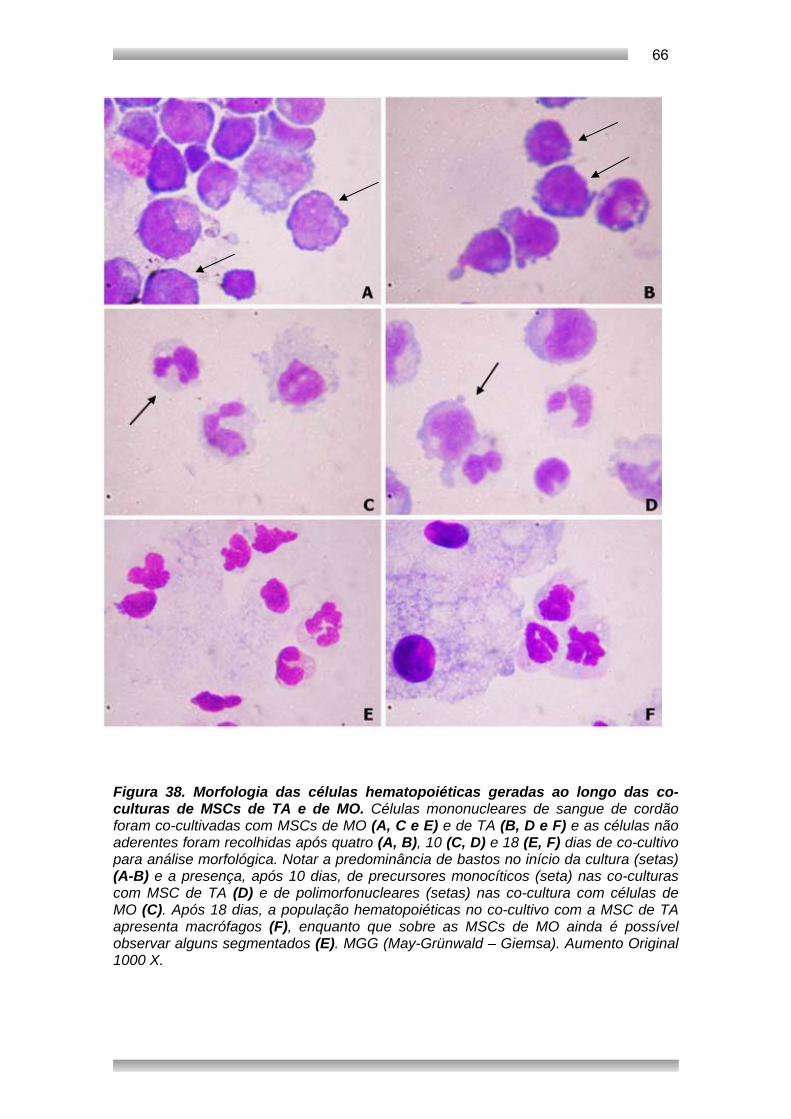
66
Figura 38. Morfologia das células hematopoiéticas geradas ao longo das co-culturas de MSCs de TA e de MO. Células mononucleares de sangue de cordão foram co-cultivadas com MSCs de MO (A, C e E) e de TA (B, D e F) e as células não aderentes foram recolhidas após quatro (A, B), 10 (C, D) e 18 (E, F) dias de co-cultivo para análise morfológica. Notar a predominância de bastos no início da cultura (setas) (A-B) e a presença, após 10 dias, de precursores monocíticos (seta) nas co-culturas com MSC de TA (D) e de polimorfonucleares (setas) nas co-cultura com células de MO (C). Após 18 dias, a população hematopoiéticas no co-cultivo com a MSC de TA apresenta macrófagos (F), enquanto que sobre as MSCs de MO ainda é possível observar alguns segmentados (E). MGG (May-Grünwald – Giemsa). Aumento Original 1000 X.
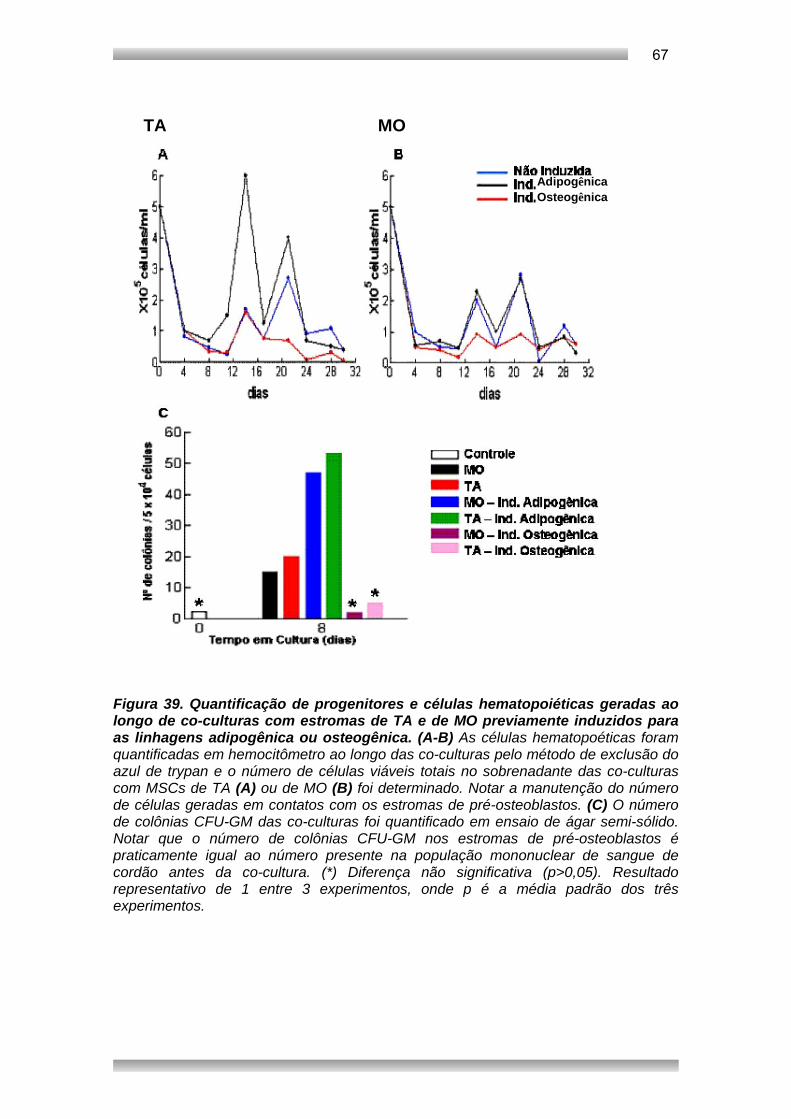
67
Figura 39. Quantificação de progenitores e células hematopoiéticas geradas ao longo de co-culturas com estromas de TA e de MO previamente induzidos para as linhagens adipogênica ou osteogênica. (A-B) As células hematopoéticas foram quantificadas em hemocitômetro ao longo das co-culturas pelo método de exclusão do azul de trypan e o número de células viáveis totais no sobrenadante das co-culturas com MSCs de TA (A) ou de MO (B) foi determinado. Notar a manutenção do número de células geradas em contatos com os estromas de pré-osteoblastos. (C) O número de colônias CFU-GM das co-culturas foi quantificado em ensaio de ágar semi-sólido. Notar que o número de colônias CFU-GM nos estromas de pré-osteoblastos é praticamente igual ao número presente na população mononuclear de sangue de cordão antes da co-cultura. (*) Diferença não significativa (p>0,05). Resultado representativo de 1 entre 3 experimentos, onde p é a média padrão dos três experimentos.
TA MO
Osteogênica Adipogênica

68
Figura 40. Os estromas pré-adipocíticos e pré-osteoblásticos induziram perfis hematopoiéticos distintos. Contagem diferencial dos citocentrifugados corados pelo método de MGG (May-Grünwald – Giemsa). (A, B) O perfil das células hematopoiéticas geradas ao longo do co-cultivo pré-adipocítico é preferencialmente macrofágica. (C, D) Os estromas pré-osteoblásticos não expandem células comprometidas com a linhagem mielóide. Figura 41. Análise da expressão de mRNA de genes ligados à hematopoiese. O RNA total das células expandidas foi isolado, e o cDNA amplificado com primers específicos. Os produtos do RT-PCR foram analisados por gel de agarose corado por brometo de etídeo. O tamanho do produto amplificado está respectivamente ao lado dos genes. Linhas 1 e 2: MSC de MO e de TA sem indução, respectivamente. Linhas 3 e 4: MSC de MO e de TA pré-osteoblásticas, respectivamente. Linhas 5 e 6: MSC de MO e de TA pré-adipocíticas, respectivamente.
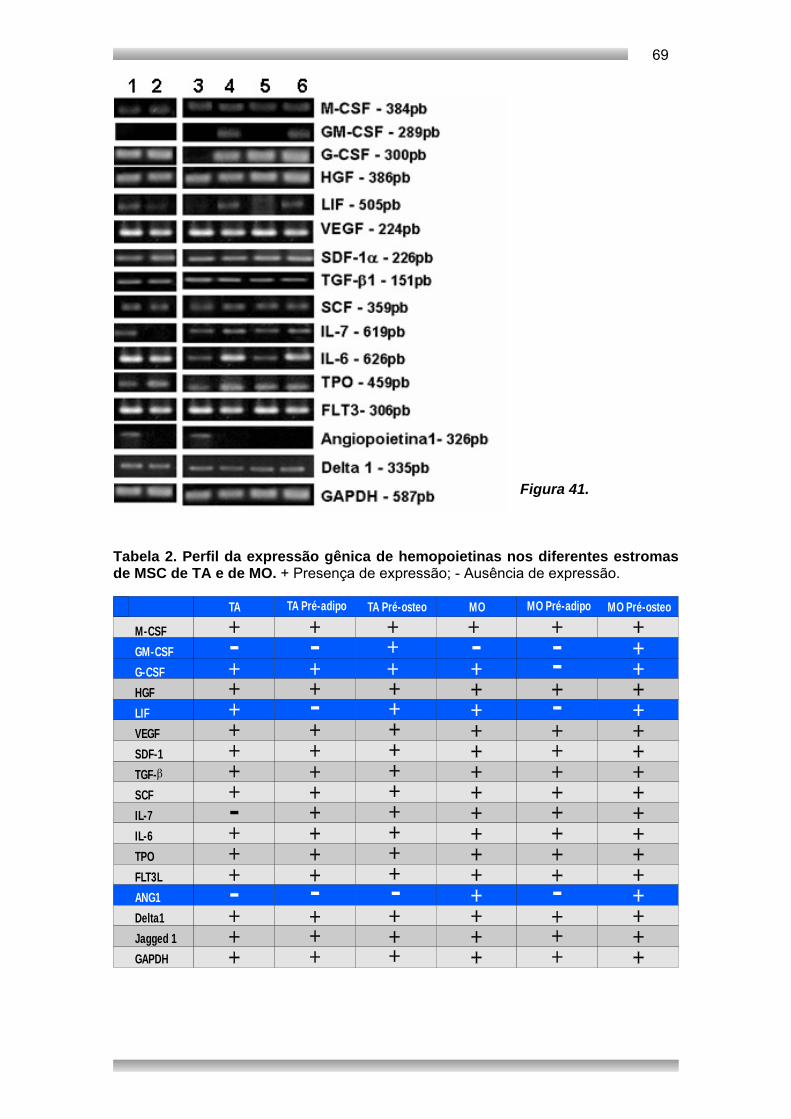
69
Figura 41.
Tabela 2. Perfil da expressão gênica de hemopoietinas nos diferentes estromas de MSC de TA e de MO. + Presença de expressão; - Ausência de expressão.
M-CSF
HGF
VEGFSDF-1TGF-SCFIL-7IL-6TPOFLT3L
Delta1Jagged 1GAPDH
GM-CSFG-CSF
LIF
ANG1
TA TA Pré-adipo TA Pré-osteo MO MO Pré-adipo MO Pré-osteo

70
4.3.b. Análises do co-cultivo com células CD34+ de sangue de cordão umbilical Com o objetivo de investigar se o fenótipo osteoblástico induzido nas MSCs de
TA era capaz de levar à manutenção das HSCs, mimetizando o nicho da MO in vivo
(TAICHMAN, 2005), a monocamada foi induzida por até duas semanas para posterior
co-cultura com células CD34+. Periodicamente, as células hematopoiéticas geradas
nas co-culturas foram quantificadas e analisadas por citometria de fluxo. Quando
comparadas com a MO, as MSCs de TA apresentaram um número absoluto
semelhante de células CD34+, assim como de células totais, geradas ao longo da co-
cultura (Fig. 42). Em sete dias, o percentual de células comprometidas com a linhagem
mielóide, identificadas pelo marcador de superfície CD13, foi maior nas MSC de TA
(Tab. 3). Nas co-culturas com MSC de TA, somente 17,5% das células mielóides
expressavam CD34, enquanto que nas co-culturas com MSC de MO, este percentual
foi de 40,9%, sugerindo uma diferenciação precoce dos progenitores mielóides em
contato com as MSC de TA. Já o percentual de células comprometidas com a
linhagem monocítica/macrofágica (CD14) foi semelhante (dado não mostrado). No
entanto, uma diferença na intensidade de fluorescência deste marcados foi observada
(Tab. 3). As células geradas nas co-culturas com MSCs de MO apresentavam uma
intensidade de fluorescência para CD14 maior, possivelmente indicando um outro tipo
de população de células CD14+.
A análise das co-culturas de células CD34+ sobre MSCs de TA induzidas por
duas semanas para a linhagem osteogênica mostrou um perfil semelhante em relação
ao percentual de células CD34+ (Fig. 43A). Porém, verificou-se um aumento no
número absoluto de células CD34+ em quatro dias nas MSCs de TA induzidas, que,
em sete dias, se equiparou ao observado nas MSCs não induzidas (Fig. 43B). A
análise diferencial, nos citocentrifugados de células não-aderentes produzidas nestas
culturas, evidenciou uma diferença marcante na população de blastos e mieloblastos
após sete dias, ressaltando-se a predominância de mieloblastos nas co-culturas com
MSCs de TA não induzidas (Fig. 44).
Para verificar a freqüência de células com propriedade de HSC, foi realizado o
ensaio de LTC-IC (Long Term Culture- Initiating Cell), que permite quantificar in vitro
células capazes de reconstituição hematopoética de longo prazo (CROISILLE et al.,
1994), ou seja com características de HSC. Assim, as células hematopoiéticas
geradas nos co-cultivos após sete dias foram recolhidas, quantificadas e distribuídas,
em diluição limitante, sobre monocamada de uma linhagem de estroma murino, a MS-
5. Utilizando o método estatístico de Poisson (SUTHERLAND et al., 1990), os
resultados preliminares permitiram verificar que, após quatro semanas, a freqüência
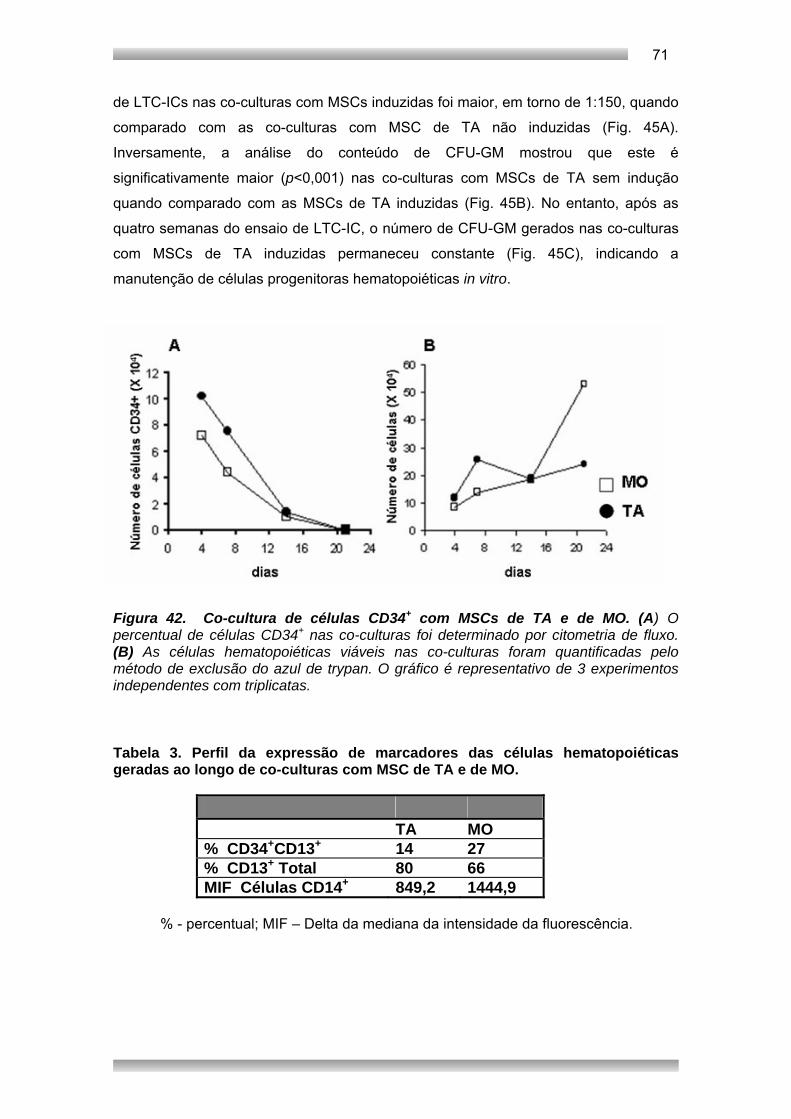
71
de LTC-ICs nas co-culturas com MSCs induzidas foi maior, em torno de 1:150, quando
comparado com as co-culturas com MSC de TA não induzidas (Fig. 45A).
Inversamente, a análise do conteúdo de CFU-GM mostrou que este é
significativamente maior (p<0,001) nas co-culturas com MSCs de TA sem indução
quando comparado com as MSCs de TA induzidas (Fig. 45B). No entanto, após as
quatro semanas do ensaio de LTC-IC, o número de CFU-GM gerados nas co-culturas
com MSCs de TA induzidas permaneceu constante (Fig. 45C), indicando a
manutenção de células progenitoras hematopoiéticas in vitro.
Figura 42. Co-cultura de células CD34+ com MSCs de TA e de MO. (A) O percentual de células CD34+ nas co-culturas foi determinado por citometria de fluxo. (B) As células hematopoiéticas viáveis nas co-culturas foram quantificadas pelo método de exclusão do azul de trypan. O gráfico é representativo de 3 experimentos independentes com triplicatas. Tabela 3. Perfil da expressão de marcadores das células hematopoiéticas geradas ao longo de co-culturas com MSC de TA e de MO.
TA MO % CD34+CD13+ 14 27 % CD13+ Total 80 66 MIF Células CD14+ 849,2 1444,9
% - percentual; MIF – Delta da mediana da intensidade da fluorescência.
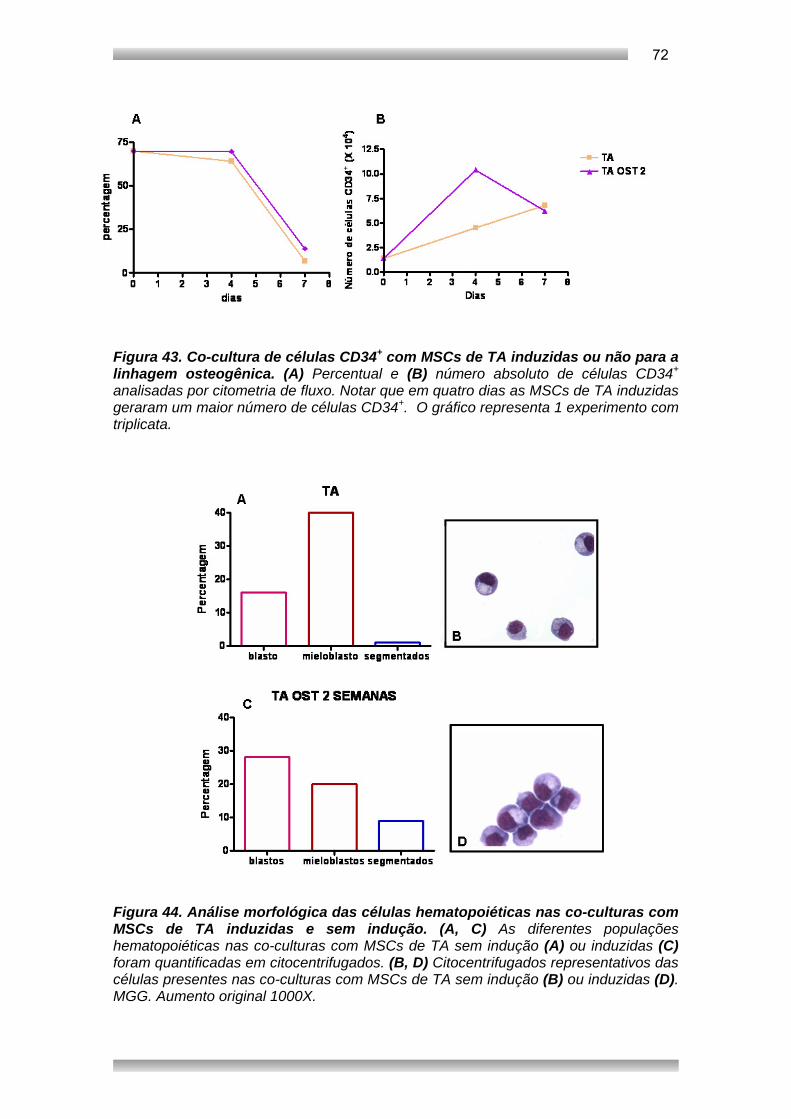
72
Figura 43. Co-cultura de células CD34+ com MSCs de TA induzidas ou não para a linhagem osteogênica. (A) Percentual e (B) número absoluto de células CD34+ analisadas por citometria de fluxo. Notar que em quatro dias as MSCs de TA induzidas geraram um maior número de células CD34+. O gráfico representa 1 experimento com triplicata.
Figura 44. Análise morfológica das células hematopoiéticas nas co-culturas com MSCs de TA induzidas e sem indução. (A, C) As diferentes populações hematopoiéticas nas co-culturas com MSCs de TA sem indução (A) ou induzidas (C) foram quantificadas em citocentrifugados. (B, D) Citocentrifugados representativos das células presentes nas co-culturas com MSCs de TA sem indução (B) ou induzidas (D). MGG. Aumento original 1000X.
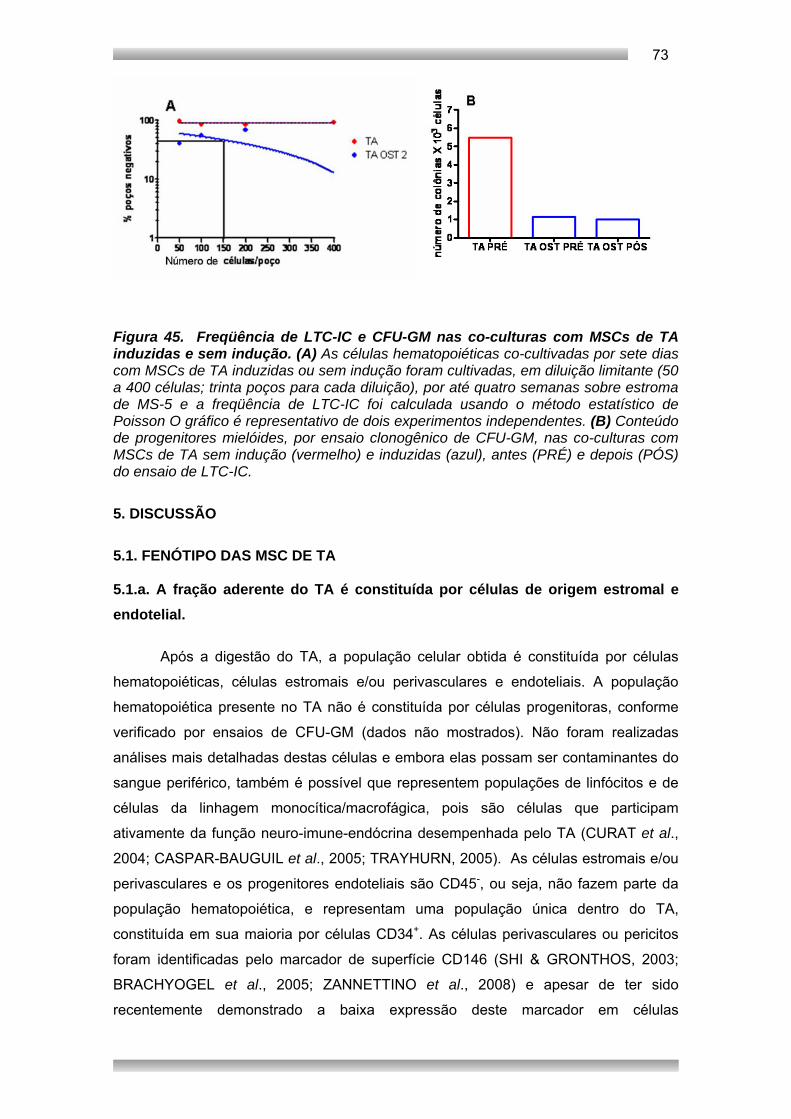
73
Figura 45. Freqüência de LTC-IC e CFU-GM nas co-culturas com MSCs de TA induzidas e sem indução. (A) As células hematopoiéticas co-cultivadas por sete dias com MSCs de TA induzidas ou sem indução foram cultivadas, em diluição limitante (50 a 400 células; trinta poços para cada diluição), por até quatro semanas sobre estroma de MS-5 e a freqüência de LTC-IC foi calculada usando o método estatístico de Poisson O gráfico é representativo de dois experimentos independentes. (B) Conteúdo de progenitores mielóides, por ensaio clonogênico de CFU-GM, nas co-culturas com MSCs de TA sem indução (vermelho) e induzidas (azul), antes (PRÉ) e depois (PÓS) do ensaio de LTC-IC.
5. DISCUSSÃO
5.1. FENÓTIPO DAS MSC DE TA 5.1.a. A fração aderente do TA é constituída por células de origem estromal e endotelial. Após a digestão do TA, a população celular obtida é constituída por células
hematopoiéticas, células estromais e/ou perivasculares e endoteliais. A população
hematopoiética presente no TA não é constituída por células progenitoras, conforme
verificado por ensaios de CFU-GM (dados não mostrados). Não foram realizadas
análises mais detalhadas destas células e embora elas possam ser contaminantes do
sangue periférico, também é possível que representem populações de linfócitos e de
células da linhagem monocítica/macrofágica, pois são células que participam
ativamente da função neuro-imune-endócrina desempenhada pelo TA (CURAT et al.,
2004; CASPAR-BAUGUIL et al., 2005; TRAYHURN, 2005). As células estromais e/ou
perivasculares e os progenitores endoteliais são CD45-, ou seja, não fazem parte da
população hematopoiética, e representam uma população única dentro do TA,
constituída em sua maioria por células CD34+. As células perivasculares ou pericitos
foram identificadas pelo marcador de superfície CD146 (SHI & GRONTHOS, 2003;
BRACHYOGEL et al., 2005; ZANNETTINO et al., 2008) e apesar de ter sido
recentemente demonstrado a baixa expressão deste marcador em células

74
perivasculares do músculo esquelético (DELLAVALLE et al., 2007), na MO são
responsáveis pela organização do microambiente hematopoiético (SACCHETTI et al.,
2007). No TA, células positivas para este marcador estão presentes em ambas as
frações celulares: CD34+CD45- e CD34-CD45- e são homogeneamente positivas para
CD105. As células endoteliais maduras identificadas pelo marcador CD31, assim
como as células CD146+, estão presentes na população CD34+CD105+, mas o seu
percentual de expressão é bastante variável de acordo com a amostra analisada.
A população CD34+ também é homogeneamente positiva para CD90, marcador
considerado característico de MSC na MO (DOMINICI et al., 2006). Desta forma, no
TA subcutâneo reside uma população de MSC identificada pelos marcadores CD105 e
CD90, mas diferente da MO, esta população também é positiva para CD34.
Recentemente foi identificada uma pequena população de MSC na MO oriundas da
fração CD34+CD45+, embora a maioria tenha sido obtida da fração duplamente
negativa (KAISER et al., 2007). Em nosso trabalho, na população de células CD34+,
foram identificadas duas subpopulações CD44+ (Fig. 12L). Esta molécula de adesão já
foi identificada em diferentes fontes de MSC (HUANG et al., 2007; JO et al., 2007;
JONES et al., 2007; KAISER et al., 2007; ZANNETTINO et al., 2008), e também
parece estar relacionada à população de células estromais e/ou perivasculares, assim
como CD105 e CD90 (ZANNETTINO et al., 2007). Como em nosso trabalho foi
encontrada uma expressão heterogênea para este marcador, e células progenitoras
mesodermais do TA expressam duplamente CD44 e CD34 (GESTA et al., 2007),
talvez essas subpopulações possam indicar células com multipotencialidade distintas.
Outro marcador analisado que revelou heterogeneidade populacional, foi a molécula
de CD146. Todas as três subpopulações identificadas são CD105+ (Fig. 15D), mas
somente a população com alta expressão para CD146 é homogeneamente positiva
para CD34 (Fig. 15C). Esta observação talvez seja mais um indício de que no TA
possa existir progenitores tantos de origem endotelial (CD146+CD34+, CD146-CD34+),
como estromal e/ou perivascular (CD146+CD105+). Análises de multipotencialidade e
auto-renovação destas diferentes populações são necessárias para tentar definir a
possível identidade das MSC no TA.
A partir das análises fenotípicas descritas acima, podemos definir que a fração
aderente do TA é constituída essencialmente por duas subpopulações de células
CD45-CD90+CD44+, que se distinguem pela expressão de CD34 e CD105: as células
CD34+CD105-CD146- e as CD34+CD105+CD146+. Antes de atingir a confluência, a
morfologia é predominantemente arredondada, diferente das células estromais de MO
que possuem morfologia fibroblastoíde desde o estabelecimento do cultivo
(FRIEDENSTEIN et al., 1974; BIANCO & ROBEY, 2000). A morfologia fibroblastoíde é

75
alcançada pela fração aderente do TA somente após a confluência e se mantêm ao
longo das passagens. A mudança para uma morfologia mais fibroblastóide foi
acompanhada pela perda gradual da expressão de CD34 e o concomitante aumento
na expressão de CD105 (Figs. 14D e E), até que após a terceira a passagem, a fração
aderente se torna homogeneamente CD105+CD49+CD13+. O aumento de marcadores
estromais juntamente com a perda de CD34 também foi observado em outro modelo
de MSC de TA (MITCHELL et al., 2006) e demonstrado em progenitores vasculares da
parede da aorta humana (ZENGIN et al., 2006). Os progenitores da adventícia da
aorta média e de coração de murinos também são morfologicamente heterogêneos,
com células arredondadas e fibroblastóides (HU et al., 2004). Essas duas
características, morfológicas e fenotípicas, inerentes às células de TA e similares a
progenitores vasculares residentes, nos levaram a investigar um possível
comprometimento angiogênico dessas células e também a idéia de uma localização
perivascular desses progenitores no TA, posteriormente confirmada por outros
pesquisadores (ZANNETTINO et al., 2007). A perda do marcador de superfície CD34 também é acompanhada por um
discreto aumento na população CD146 e CD90 positivas. Esses dois marcadores
estão presentes na população CD105+, mas uma subpopulação com uma maior
expressão para CD146 é CD90 negativa (Fig. 15G). Como já anteriormente
mencionado, o marcador de superfície CD146 foi recentemente utilizado para a
identificação de uma população de MSC na MO, com capacidade de auto-renovação
(SACCHETTI et al., 2007). Já a molécula de CD90 além de ter sido proposta como um
dos marcadores característicos de MSCs (DOMINICI et al., 2006), representa a
molécula de Thy-1, uma glicoproteína de membrana, inicialmente descrita em
populações de HSCs capazes de reconstituição hematopoiética de longa duração
(MULLER-SIEBURG et al., 1986). Uma população de MSCs duplamente positiva para
os marcadores CD146 e CD90 também foi encontrada na MO humana (MÜLLER et
al., 2006), mas não há relatos de uma subpopulação CD146+CD90-. Quando
comparada com outras fontes de MSC, somente a MO possui uma população
homogênea com alta expressão para CD146 sem a presença de populações com
menor expressão (SACCHETTI et al., 2007), diferente do que foi encontrado em nosso
trabalho.
A partir das análises fenotípicas das células obtidas pós-digestão enzimática e
da fração aderente in vitro do TA e também da comparação com os trabalhos que
utilizam MSC de MO, podemos concluir que, apesar das similaridades entre essas
duas fontes de MSC, no TA existem subpopulações que diferem do perfil fenótipo
encontrado na MO. Somente no TA foi observada a presença de uma população

76
positiva para marcadores tanto de células estromais e/ou perivasculares (CD105,
CD146, CD90 e CD44) como de origem endotelial (CD34 e CD31), o que nos remete a
progenitores essencialmente endoteliais neste tecido (ASAHARA et al., 1997; ZENGIN
et al., 2006).
5.1.b. As células CD34+ da fração aderente do TA adquirem o marcador CD105 in
vitro.
Os marcadores que melhor definiram duas sub-populações distintas na fração
aderente do TA, CD34+CD105-CD45- e CD34+CD105+CD45-, foram utilizados para a
sua seleção imediatamente após a digestão do tecido. As células foram cultivadas por
três dias, após os quais a população CD34+CD105-CD45- se apresentou
morfologicamente semelhante as culturas de TA não selecionadas (Fig. 17B), diferente
da população CD34+CD105+CD45-, que mostrou uma morfologia não usual para
células aderentes (Fig. 17A). Curiosamente, Ogawa e colaboradores em 2006,
relataram que células fibroblastóides, capazes de geração de CFU-F, poderiam ser
derivadas de HSCs da MO (EBIHARA et al., 2006; LARUE et al., 2006) e uma
subpopulação de progenitores endoteliais KDR+, mobilizados do sangue periférico de
pacientes tratados com G-CSF, expressam simultaneamente marcadores monocíticos
(CD45 e CD14) e endoteliais (CD31 e CD105) (BRUNO et al., 2007). Também
recentemente alguns trabalhos mostraram que as células monocíticas dividem alguns
marcadores com progenitores endoteliais. Esses marcadores podem ser modulados in
vitro em condições angiogênicas, com perda simultânea dos marcadores monocíticos,
mas essas células não são capazes nem de formar redes vasculares (ROHDE et al.,
2006) e nem de gerar colônias de progenitores endoteliais (LÓPEZ-HOLGADO et al.,
2007). Em nosso trabalho, as células com marcadores endoteliais e de origem não-
hematopoiética (CD34+CD105+CD45-) apresentaram uma morfologia de célulasainda
não descrita, mas a cultura morreu rapidamente não permitindo a caracterização desta
população. Não é possível, neste momento, afirmar se a morte da cultura se deveu ao
fato desta população celular ser terminalmente diferenciada, ou seja, com baixa
capacidade proliferativa ou ao processo de seleção utilizado (sorteamento em alta
pressão).
A população CD34+CD105-CD45- foi expandida em cultura e também
apresentou uma cinética de ganho do marcador CD105. As células CD105+
representam a população de maior tamanho, enquanto as CD34+ as menores (Fig.
18). Essa observação é consistente com a hipótese da relação entre o ganho do
marcador CD105 e a mudança para uma morfologia mais fibroblastoíde, pois as

77
células com aspecto fibroblastoíde são maiores do que as com aspecto arredondado
do início da cultura. Assim como na cultura com a população total do TA, após duas
passagens, a população sorteada para CD34 se torna homogeneamente positiva para
CD105 e adquire uma morfologia fibroblastoíde. Essa regulação positiva do marcador
CD105 in vitro também foi descrita para progenitores vasculares da parede da aorta
humana, mas não foram investigadas correlações funcionais atribuídas ao ganho
deste marcador (ZENGIN et al., 2006). O ganho do marcador CD105 na população
CD34+ ressalta o que nós e outros pesquisadores (MITCHELL et al., 2006) já
havíamos observado em relação à regulação positiva de marcadores estromais in vitro
nas MSC de TA. São necessários testes funcionais de diferenciação in vitro e in vivo,
para avaliar se a verdadeira população de MSC in vivo é CD105-; sendo assim este
marcador seria somente um artefato adquirido in vitro.
5.2. MULTIPOTENCIALIDADE
5.2.a. As MSC de TA sob indução adquirem fenótipos osteogênico, adipogênico e condrogênico in vitro.
A partir do estabelecimento da monocamada derivada de células do TA, foram
realizados testes da sua multipotencialidade ortodoxa. As MSC de TA com apenas
uma semana de indução já apresentaram mudanças na morfologia. A indução
adipogênica acarretou em uma morfologia mais arrendondada, ao contrário da
osteogênica em que as células se apresentaram com um aspecto morfológico
semelhante as células não induzidas. Esta mudança de morfologia está de acordo
com os estudos de diferenciação de MSCs que demonstraram a necessidade da
ativação da via de RhoA para o comprometimento para a via osteogênica, e a sua
desativação para a via adipogênica. O formato celular mais arredondado é o principal
responsável pela inibição da via de RhoA (MCBEATH et al., 2004). Após quatro
semanas de indução, parte das células MSC de TA se tornam terminalmente
diferenciadas para ambas as vias. A ausência de diferenciação em grande parte da
monocamada é explicada pela heterogeneidade dos estromas tanto de MO como de
TA. O estroma de ambas as fontes é composto por células multipotentes,
oligopotentes e unipotentes para as diversas vias de diferenciação (KUZNETSOV et
al., 1997; ZUK et al., 2002).
A indução condrogênica foi realizada em sistema de esferóides, que, em
função da geração de um centro de hipóxia e do seu arcabouço em três dimensões,
facilita a diferenciação das MSCs em condroblastos através da expressão de

78
moléculas específicas e também de mecanoreceptores (SCHIPANI et al., 2001). Após
quatro semanas, foi possível notar um aumento no acúmulo de matriz extracelular e
proteoglicanos, porém sem mudanças morfológicas. As MSC de MO quando induzidas
para essa via em sistema tridimensional, apresentam mudanças morfológicas para
células condroblásticas e também marcação para colágeno do tipo II, específico para o
tecido cartilaginoso (MURAGLIA et al., 2003). Os esferóides de TA em nosso trabalho
também apresentaram marcação para colágeno do tipo II, porém pouco expressiva
(dados não mostrados). Outros trabalhos comparando a diferenciação de MSC de MO
e de TA relatam uma ativação da expressão de genes que caracterizam uma
diferenciação completa para a via cartilaginosa somente nas células de MO (WINTER
et al., 2003; HENNIG et al., 2007). A capacidade condrogênica reduzida das MSC de
TA parece ser compensada pela aplicação exógena de fatores, tais como BMP-6
(HENNIG et al., 2007).
5.2.b. As MSC de TA são morfologica e geneticamente diferentes das MSC de MO.
Em baixa densidade, a monocamada de células já expandida dessas duas
diferentes fontes, MO e TA, é morfologicamente semelhante. Porém, quando o estágio
de confluência é atingido, as MSC de TA começam a acumular gotículas de lipídeos
espontaneamente (Fig. 23C). Análises de microscopia de varredura demonstraram um
formato mais arrendondado nas áreas de maior confluência da monocamada de TA,
ao contrário do aspecto miofibroblastoíde característico das células estromais de MO
(Fig. 23E, F). Um dos primeiros passos para a diferenciação adipogênica é a mudança
para uma morfologia mais arredondada (GREGOIRE, 2001), que in vitro é facilitada
pela confluência (MCBEATH et al., 2004). Alguns estudos discutem a hipótese de que
a alta densidade da monocamada de MSC acarreta em uma perda da sua
multipotencialidade (COLTER et al., 2001). Desta forma, as células poderiam se
diferenciar espontaneamente na linhagem para a qual se encontram mais
comprometidas. As análises moleculares revelaram um padrão de expressão gênica
semelhante para genes envolvidos na osteogênese, apesar das MSCs de MO
aparentarem uma maior expressão. A diferença marcante foi em relação à
adipogênese, já que somente as MSCs de TA expressam o gene PPARγ2 (Fig. 24),
principal responsável pelo comprometimento de células para essa via. Isto explica em
parte, o acúmulo espontâneo de microvesículas de lipídeos. Alguns trabalhos, através
de análises de microarray, revelaram um comprometimento osteogênico e
condrogênico das MSC de MO, e adipogênico das de TA (LIU et al., 2007). O grau de

79
comprometimento dessas diferentes fontes de MSC pode estar diretamente
relacionado com o microambiente em que elas estão inseridas in vivo. O
microambiente hematopoiético é complexo, onde o osso é constantemente
remodelado e possui um papel determinante na manutenção das HSCs (TAICHMAN,
2005), assim como as células do estroma subendotelial as quais são as maiores
secretoras de Angiopoietina-1 (SACCHETTI et al., 2007). Já o microambiente do TA é
constituído basicamente por pré-adipócitos, adipócitos e células endoteliais, sendo a
angiogênese um processo determinante na expansão e/ou retração desse tecido
(CASTEILLA et al., 2004). Se realmente essas duas diferentes fontes de MSC
possuem uma origem embriológica comum (COSSU & BIANCO, 2003), é razoável
atribuir essa diferença de comprometimento observada em nosso trabalho e por outros
pesquisadores, aos diferentes microambientes. Além disso, seguindo uma hipótese de
heterogeneidade da população de MSC no TA, junto com o fato destas células
apresentarem potencial osteogênico, talvez a expressão gênica de PPARγ2 esteja
somente em parte desta população. Ensaios clonais com análises posteriores de
diferenciação e expressão gênica estão sendo realizados para a comprovação desta
hipótese.
5.2.c. As MSC de TA apresentam um potencial adipogênico diferente das MSC de MO.
A adipogênese nas MSCs de TA apresentou uma evolução mais rápida,
quando comparada com as MSCs de MO. Em duas semanas, as células de TA que
entraram nessa via de diferenciação, já haviam fundido suas gotículas lipídicas,
alcançando assim o fenótipo de adipócito maduro (Figs. 25A e 26A). Já as MSCs de
MO não apresentaram células univacuoladas em duas semanas (Figs. 25D e 26A), e
em quatro semanas a maioria das células ainda era multivacuolada, com o citoplasma
bastante hipertrofiado (Figs. 25E e 26B). A expressão de PPARγ2, diferente da
expressão constitutiva do TA, nas MSC de MO é regulada positivamente somente
após uma semana de indução. Isto pode explicar o atraso na diferenciação em células
univacuoladas. Já a expressão de leptina é constitutiva nas células de MO e parece
aumentar após quatro semanas de indução. A leptina aumenta a proliferação das
HSCs e consequentemente a hematopoiese (LAM & LU, 2007), desta forma é
coerente sua expressão constitutiva nas MSC de MO. Nas MSC de TA a expressão de
leptina é regulada positivamente somente após quatro semanas de indução
adipogênica, provavelmente por causa do grande aumento em células univacuoladas.
No TA, os adipócitos são os grandes responsáveis pela secreção de leptina (CAO,

80
2007), que, além de estimular a proliferação de monócitos e macrófagos, é um dos
fatores responsáveis pelos eventos pró-inflamatórios desencadeados no
desenvolvimento da obesidade (LAM & LU, 2007). Em seis semanas a maioria das
MSC de TA era univacuolada, mimetizando o microambiente deste tecido in vitro. Já
as células estromais de MO ainda apresentavam mais da metade das células ou
multivacuoladas ou não diferenciadas. As diferenças morfológicas observadas na
adipogênese desses dois tipos de células, ainda não foram descritas na literatura. Os
trabalhos de comparação dessa via de diferenciação não reportam diferenças entre
essas duas fontes de MSC. Isto pode ser explicado em alguns casos pela falta de
acompanhamento da adipogênese in vitro (GRONTHOS et al., 2001; LEE et al., 2004),
ou ainda porque análises funcionais bioquímicas não demonstram diferenças entre
esses dois tipos de células acumuladoras de gordura (DICKER et al., 2005). Talvez a
principal e única diferença seja mesmo na evolução da adipogênese, que é mais
rápida nas MSC de TA constitutivamente PPARγ2 positivas.
Ambas as MSC apresentaram um padrão semelhante de calcificação in vitro
(Fig. 28), assim como o perfil de expressão gênica para a via osteogênica (Fig. 29). As
MSCs de TA parecem apresentar níveis menores de expressão, quando comparadas
com as de MO, porém ensaios quantitativos são necessários para a confirmação
desses resultados. Curiosamente, a expressão de osteopontina foi regulada
negativamente após quatro semanas de indução somente nas MSC de TA, e, além
disso, estas células mesmo sem indução osteogênica expressam CBFA-1, o gene
indispensável para a via osteogênica, junto com a expressão de PPARγ2. Esta
expressão simultânea é curiosa, já que agentes indutores de adipogênese,
responsáveis pela ativação da transcrição de PPARγ, inibem a expressão e as ações
de CBFA-1 (NUTTALL & GIMBLE, 2004). A partir de modelos murinos, foi descrito
uma cinética temporal e espacial na secreção de proteínas envolvidas na
osteogênese. A presença de ácido ascórbico, um dos agentes indutores desta via,
aumenta a síntese de colágeno e durante este estágio, outras proteínas acumulam na
matriz recém-sintetizada tais como a sialoproteína do osso, osteopontina e
osteocalcina. O aumento nos níveis de osteocalcina regula negativamente os de
osteopontina e vice-versa (BECK, 2003). Muito provavelmente não foi isto que ocorreu
na regulação negativa de osteopontina nas MSC de TA, já que os níveis de
osteocalcina permaneceram constantes e essa regulação não foi observada nas
MSCs de MO. Zuk et al. (2002) também não observaram essa expressão temporal de
proteínas em ambas as fontes de MSC humanas. Além disso, a osteopontina não é
uma proteína específica de osteogênese; seus níveis também aumentam durante
processos de cicatrização, inflamação (GIACHELLI & STEITZ, 2000), obesidade

81
(NOMIYAMA et al., 2007) e esta proteína ainda é responsável por compor o nicho
subendosteal da MO (NILSSON et al., 2005).
Como não foram vistas diferenças marcantes na osteogênese, tanto a nível
celular como molecular entre as duas fontes de MSC, ambas as células tiveram seu
potencial testado na presença de um biomaterial ósseo composto de hidroxiapatita.
Após sete dias de cultura, foi possível notar intensa proliferação das MSCs de MO na
superfície do arcabouço de hidroxiapatita, ao contrário das MSCs de TA (Figs. 30A e
B). No interior do material, as MSCs de TA, na maioria das áreas analisadas,
encontrava-se com um aspecto menos espalhado enquanto as de MO tinham aspecto
miofibroblastoíde (Fig. 30C, D). Apesar das diferenças morfológicas, ambas as células
marcaram positivamente para osteopontina, o que novamente pode indicar a falta de
especificidade desta proteína para a via osteogênica. É importante ressaltar que
essas diferenças morfológicas e proliferativas encontradas entre as células na
presença de um material com propriedades osteocondutoras e osteoindutoras
(SPIVAK & HASHARONI, 2001; HASEGAWA et al., 2007; INANÇ et al., 2007), reforça
a hipótese do maior comprometimento das MSCs de TA para a via adipogênica e das
de MO para a via osteogênica. Para que as MSCs de TA demonstrem mais afinidade
com o biomaterial ósseo, talvez seja necessária uma indução osteogênica prévia, ou
ainda aumentar o tempo de contato com biomateriais de hidroxiapatita.
Com o objetivo de ratificar o maior comprometimento adipogênico das MSCs
de TA, a monocamada recém-isolada foi incubada por até dez dias em meio indutor
misto para as linhagens adipogênica e osteogênica. Foi possivel observar pequenas
áreas de deposição de cálcio, circundadas por células com acúmulos lipídicos
citoplasmáticos (Fig. 31). Apesar de o número de células diferenciadas para a via
adipogênica não ter sido quantificada, as áreas com marcação para Oil Red O foram
maiores do que as com depósito de cálcio, o que indiretamente revela uma população
heterogênea com um maior número de células comprometidas para adipócitos.
Provavelmente, essa população de células é responsável pela expressão constitutiva
de PPARγ2 nos cultivos de MSC de TA.
5.2.d. As MSC de TA parecem ser comprometidas com a via endotelial Além da expressão de marcadores de superfície de progenitores de células
endoteliais, as MSC de TA quando cultivadas em sistema de esferóides regulam
positivamente e de maneira rápida marcadores mais tardios para essa via. Após dez
dias de incubação em meio não indutor, os esferóides de MSC de TA apresentaram
marcação para CD31; o mesmo não ocorrendo quando essas células são mantidas

82
em monocamada. Apesar de uma fração das células de TA apresentarem este
marcador endotelial no momento do isolamento, ou ele é regulado negativamente,
possivelmente juntamente com o CD34, após a expansão in vitro, ou esta população
representa células endoteliais maduras que não se expandem no sistema de cultura
convencional. O sistema tridimensional também é responsável pela regulação positiva
do gene angiopoietina-1, diretamente envolvido em processos angiogênicos (FIEDLER
& AUGUSTIN, 2006) e regulador do nicho hematopoiético na MO (SACCHETTI et al.,
2007); e também pela expressão gênica e síntese de VEGF; e de vWf. Alguns
trabalhos também já haviam relatado essa diferenciação aparentemente espontânea,
porém em sistema de matrigel, o qual é composto por diversos fatores desconhecidos
incluindo angiogênicos (PLANAT-BENARD et al., 2004a; MOON et al., 2006). Também
em sistema de matrigel, células da MO CD34-CD146+ adotam preferencialmente um
fenótipo de células perivasculares ao invés de endoteliais (SACCHETTI et al., 2007). A
regulação positiva de angiopoietina-1 é consistente com o fenótipo de células
perivasculares, pois sua ligação com o receptor Tie-2, comumente expresso em
células endoteliais, ativa a formação de vasos sanguíneos (SURI et al., 1996).
Os esferóides de TA também apresentaram estruturas semelhantes a vasos
(Figs. 33A e B), já observadas em outro trabalho que utilizou células tumorais de
mama (TIMMINS et al., 2004). O mais intrigante é que de acordo com a literatura, as
células perivasculares não dão origem a células endoteliais, pois possuem origem
embriológica distinta (OETTGEN, 2001). Isto nos leva a duas hipóteses: (1) As MSCs
de TA somente apresentam marcadores de células perivasculares, mas não são
pericitos funcionais; (2) ou podem ser pericitos em um estágio de ativação distinto dos
já descritos, particular para o TA subcutâneo. Esta última hipótese é sustentada por
um trabalho recente que demonstrou que as células perivasculares podem ser
derivadas de progenitores de células endoteliais in vitro, e que não somente
estabilizam os capilares, mas também participam do remodelamento vascular in vivo
(HAGEDORN et al., 2004).
9 Com o objetivo de investigar sua possível identidade com as células
perivasculares, o efeito das MSCs de TA em células endoteliais maduras (HUVEC)
também foi analisado. As MSC de TA quando co-cultivadas em monocamada com as
HUVEC, interagem intimamente com estas células formando estruturas similares as de
células perivasculares (Figs. 34A e C). Além disso, são capazes de promover a
sobrevivência das HUVEC, já que quando estas células são cultivadas sem a
presença das MSC de TA e nas mesmas condições de cultivo, predomina intensa
morte celular. A secreção de fatores angiogênicos e anti-apoptóticos já havia sido
descrita para as MSCs de TA (REHMAN et al., 2004), porém acreditamos que além

83
dessa regulação parácrina, haja moléculas de adesão específicas envolvidas dada a
interação particular dessas células com as HUVEC. Este padrão de interação se
repetiu no sistema de esferóides, onde as MSC de TA, através da emissão de
prolongamentos, circundou o ninho de HUVEC formado (Fig. 34F). É importante
ressaltar que este padrão de interação não é comumente descrito na literatura.
Normalmente, em sistema de co-cultivo com HUVEC em esferóides, essas células
tendem a se localizar somente na superfície externa (KELM et al., 2005). Somente um
trabalho descreveu um padrão de interação semelhante ao nosso, e este foi atribuído
indiretamente a maior produção de proteínas envolvidas com o processo de hipóxia, e
diretamente a ligação diferenciada dessas células a proteínas da matriz extracelular,
as quais permanecem desconhecidas (TIMMINS et al., 2004). Células de MO CD34-
CD146+ em matrigel também apresentam um padrão de interação com HUVEC similar
as de células perivasculares (SACCHETTI et al., 2007). Porém, essas duas fontes de
MSC, MO e TA, diferem na expressão de CD34, e talvez esta molécula possa estar
relacionada à diferenciação endotelial alcançada somente pelas células de TA. A
expressão da molécula de CD31 novamente não foi regulada positivamente em
monocamada, ainda que na presença da HUVEC, o que reafirma o papel de fatores
liberados durante a condição de hipóxia nos esferóides de TA para a diferenciação
endotelial.
O papel da liberação de fatores angiogênicos também foi avaliado em gel de
colágeno, frequentemente utilizado para a quantificação de brotamento de capilares
em esferóides (JAKOBSSON et al., 2007). Na presença de VEGF os esferoídes de
HUVEC emitem brotamentos após três dias (Fig. 35B), enquanto na presença da
monocamada das MSC de TA foi possível observar brotamentos semelhantes em
apenas cinco horas (Fig. 35A). Estes dados confirmam a hipótese de secreção de
fatores angiogênicos e anti-apoptóticos pelas MSC de TA, e que estes fatores são
capazes de estimular células endoteliais maduras. Diversos trabalhos demonstram a
importância de fatores secretados pelos pericitos na promoção da migração de células
endoteliais e também na morfogênese durante os estágios iniciais da
neovascularização (WATANABE et al., 1997; FUKUSHI et al., 2004). Estes dados
estão de acordo com os nossos resultados, e somente um trabalho observou um efeito
inibitório promovido por estas células perivasculares (MCILROY et al., 2006). Os
esferóides das MSC de TA em condições padrões de cultura, são capazes de emitir
prolongamentos celulares em gel de colágeno, porém tanto na presença de VEGF
quanto na de HUVEC, esses prolongamentos cessam. Essa capacidade migratória
das MSC de TA pode ser atribuída à expressão de CD34 por dois motivos: (1) em
células-tronco embrionárias o brotamento em gel de colágeno pode ser atribuído

84
diretamente a este marcador (MCCLOSKEY et al., 2006); (2) as MSC de MO, que não
expressam este marcador, quando plaqueadas nas mesmas condições, não são
capazes de emitir prolongamentos (dados não mostrados). A inibição da emissão de
prolongamentos só permite inferir que tanto fatores angiogênicos, quanto contatos
celulares com células endoteliais inibem a característica migratória das MSC de TA.
Apesar de já ter sido identificado um inibidor de migração de células murais
especificamente produzido por células endoteliais, a VE-estatina (SONCIN et al.,
2003), normalmente o VEGF é responsável pela migração de pericitos (LIU et al.,
2005). De acordo com esses dados preliminares, só podemos concluir que, se
realmente as MSCs de TA representam uma população de pericitos, estes pericitos
são distintos dos já descritos. Ensaios in vivo são necessários para a descoberta da
verdadeira identidade das células progenitoras do TA.
5.3. POTENCIAL DE SUSTENTAÇÃO DA HEMATOPOIESE. 5.3.a. As MSC de TA apresentam similaridades com as de MO.
Após quatro dias de co-cultivo com células mononucleares de sangue de
cordão, já foi possível observar “cobblestone areas”, interação característica de
progenitores hematopoiéticos com o estroma. Seguido desta interação, houve a
expansão de células hematopoiéticas, somente nos estromas de MSC de MO e de TA.
Nas co-culturas com FPH, essas células se esgotam em apenas quatorze dias. O
esgotamento das células foi confirmado por ensaios clonogênicos de CFU-GM, onde,
comparada com as células mononucleares antes do cultivo, o FPH não foi capaz de
expandir clones comprometidos para as linhagens granulocítica e macrofágica, ao
contrário das MSC de MO e de TA (Fig. 37B). A diferença no número de colônias
geradas em uma semana de cultivo entre as MSCs não foi estaticamente significativa.
Após três semanas, não houve geração de colônias CFU-GM nesses cultivos (dados
não mostrados), o que correlaciona diretamente com o perfil de geração de células
hematopoiéticas. Nestes cultivos a diminuição significativa de células progenitoras
comprometidas com a linhagem mielóide (mieloblastos) acontece de forma simultânea
ao aumento de células diferenciadas (meta, bastão e segmentados) entre quinze e
vinte dias (Fig. 37C, D).
Curiosamente, após aproximadamente vinte dias de cultivo, particularmente
nas MSC de TA, houve uma geração preferencial de células macrofágicas em
detrimento de segmentados, provavelmente derivadas de precursores monocíticos
detectados em dez dias de co-cultivo (Fig. 38D). O macrófago é um tipo celular
funcionalmente integrado com o osso, sistema vascular e o TA. Esta célula pode se

85
diferenciar em osteoclastos no tecido ósseo e também é capaz de infiltrar o sistema
vascular e o TAB, levando a quadros de inflamação crônica, os quais podem causar
ateroesclerose ou resistência a insulina secundária à obesidade (KOSHIYAMA et al.,
2006). No caso da obesidade, o papel do macrófago foi recentemente descoberto
(WEISBERG et al., 2003; XU et al., 2003) e, atualmente, foi evidenciado o
envolvimento do fator transcricional PPARγ na ativação destas células e na indução de
resistência a insulina (ODEGAARD et al., 2007). As MSC de TA, diferentemente das
de MO, expressam constitutivamente o gene PPARγ2 e, provavelmente, esta seja a
causa da expansão preferencial de células macrofágicas. Durante o desenvolvimento
da obesidade, a infiltração do TA por macrófagos pode se dever a mobilização de
monócitos com posterior diferenciação em macrófagos in situ (CURAT et al., 2004) ou
a transdiferenciação dos pré-adipócitos diretamente em células macrofágicas
(CHARRIÈRE et al., 2003). A migração de monócitos para o TA durante a obesidade
parece ser mediado pela secreção de MCP-1, sintetizado pelos adipócitos
hipertrofiados (WELLEN & HOTAMISLIGIL, 2003). No entanto, nossos dados estão de
acordo com trabalho recente que mostrou que as células progenitoras do TA,
CD34+CD45-, é que secretam altos níveis da quimiocina MCP-1 (monocyte
chemotactic protein-1), diretamente envolvida no recrutamento de macrófagos (ZHOU
et al., 2007). Devido à propriedade destas células de gerar macrófagos a partir de
células hematopoiéticas in vitro, talvez esta diferenciação possa ocorrer no TA in situ
em situações fisiológicas e ser bastante agravada em quadros de obesidade. Com
isso as MSC teriam participação ativa no desenvolvimento do quadro inflamatório da
obesidade.
5.3.b. As MSCs pré-induzidas para as vias adipogênica e osteogênica apresentam perfis de geração de células hematopoiéticas distintos.
Com o objetivo de melhor investigar as propriedades de sustentar a
hematopoiese das MSC de TA, comparando-as às de MO, as duas populações
celulares foram induzidas para os fenótipos adipocítico e osteoblástico, ambos
constituintes do estroma medular. Os estromas pré-adipocíticos foram responsáveis
por uma expansão significativa de células hematopoiéticas, ao contrário dos pré-
osteoblásticos. As análises de CFU-GM também revelaram um maior número de
colônias geradas pelos estromas pré-adipocíticos novamente para ambas as fontes de
células (Fig. 39). Os estromas pré-osteoblásticos não foram capazes de expansão de
colônias de granulócitos e macrofágos, propriedade que foi correlacionada com a
inibição de geração de mieloblastos e a discreta manutenção blástica por mais de

86
vinte dias. Os dados obtidos em nosso trabalho estão de acordo com o papel de
células osteoblásticas na regulação das HSC (CALVI et al., 2003; ZHANG et al., 2003;
BALDUINO et al., 2005) e de células acumuladoras de lipídeos na cavidade medular,
cujo papel ainda é obscuro, mas que parecem estimular a hematopoiese, contribuindo,
principalmente, na geração de células diferenciadas, pois não seriam capazes de
manter as HSC (CORRE et al., 2004). Em nosso trabalho, de fato, o número de
células blásticas caiu precocemente em ambos os estromas pré-adipocíticos, quando
comparados com os pré-osteoblásticos (Fig. 40).
As MSC de TA induzidas para osteoblastos apresentaram um perfil de geração
de células hematopoiéticas semelhante às células de MO, apesar da expressão
constitutiva de PPARγ2 e do microambiente deste tecido não ser constituído por
células osteoblásticas. Porém somente nos cultivos com MSC de MO foi possível
observar uma expansão e manutenção blástica em longo prazo (Fig. 40D). A
plasticidade evidenciada pelas MSC de TA e o fenótipo osteoblástico mais eficiente
das células de MO reforçam a teoria de que estas duas diferentes fontes possuem
uma origem embriológica comum e que suas características intrínsecas seriam
geradas pelos seus respectivos microambientes.
Os estromas diferenciados para a via adipogênica induziram uma diferenciaçãp
preferencial das células hematopoiéticas para a linhagem macrofágica, que foi
novamente mais evidente nas culturas com MSC de TA. Esta maior produção de
macrófagos pelas MSC de MO reforça a idéia de que o fenótipo adipocítico induz
diferenciaçcão preferencial para esta linhagem e é possivelmente também causado
pela expressão de PPARγ2, regulado positivamente após uma semana de indução
adipogênica.
O perfil da expressão gênica de hemopoietinas também é semelhante para
ambas as fontes de MSC, com algumas particularidades (Fig. 41 e Tab. 2). Somente
as MSC de MO expressam o mRNA para angiopoietina-1, que além do seu potencial
angiogênico, é um importante componente dos nichos de HSC (ARAI et al., 2004;
KIEL et al., 2005). Após a indução pré-adipocítica, as MSC de MO regulam
negativamente este fator, o que talvez possa explicar a sua semelhança na geração
de células hematopoiéticas com as MSC de TA sem indução. Outro gene regulado
negativamente durante a pré-indução adipocítica em ambas as fontes de MSC, foi o
LIF. Este fator possui um papel inibitório na proliferação de células hematopoiéticas
(LIEBERMANN & HOFFMAN, 1994), o que, em nosso trabalho, se correlaciona com o
menor número de colônias CFU-GM encontrado nos estromas sem indução e com
indução para a via osteogênica (Fig 39C). Uma possível explicação para a regulação
negativa pós indução adipogênica, seria que a dexametasona em concentração igual à

87
utilizada no meio indutor adipocítico, é responsável por um rápido aumento na taxa de
degradação do mRNA do LIF (GROSSET et al., 1999). O mRNA para G-CSF também
é regulado negativamente na indução pré-adipocítica, mas somente nas MSC de MO.
Como esta hemopoietina exerce o seu papel tanto em células hematopoiéticas
progenitoras (WRIGHT et al., 2002), como em células terminalmente diferenciadas
(LIANG, 2003), a sua ausência nas MSCs de MO pré-adipocíticas pode explicar as
diminuições nos percentuais de células blásticas e segmentadas, em comparação
com as MSC pré-osteoblásticas e não induzidas. Já o mRNA de GM-CSF é regulado
positivamente em ambas as MSCs induzidas para o fenótipo osteoblástico. Esta
citocina possui efeitos similares ao G-CSF (LIANG, 2003), logo sua regulação deve
estar diretamente relacionada à manutenção blástica observada nesses estromas.
5.3.c. As MSCs de TA quando induzidas para a linhagem osteoblástica são capazes de manter as HSCs.
Assim como no co-cultivo com células mononucleares, as MSC de TA são
capazes de expandir células hematopoiéticas CD34 purificadas (Fig. 42 e Tab. 3). As
MSC de TA são capazes de manter as células CD34+ por até 15 dias, mas somente as
de MO expandem o número total de células após esse período. A não expansão de
células progenitoras nas MSC de TA se correlaciona com as análises fenotípicas das
células geradas ao longo do co-cultivo. Em sete dias o percentual de células CD34
comprometidas com a linhagem mielóide (CD13+) é quase o dobro nas células de MO
em comparação com as de TA; o contrário ocorre com células mielóides que já
perderam o marcador de HSCs. Essa diferenciação precoce observada no co-cultivo
com MSC de TA se correlaciona com os resultados anteriores que revelaram uma
maior geração de células mielóides nos estromas adipocíticos e a manutenção blástica
nos osteoblásticos. Como neste ensaio as MSC não foram induzidas para estas vias,
novamente nos parece que as MSC de MO são mais comprometidas com a linhagem
osteoblástica e as de TA com a adipocítica.
Curiosamente, ao final do co-cultivo, o perfil, e não o percentual, de células
monocíticas/macrofágicas (CD14+) é ligeiramente diferente entre as MSC. A média da
intensidade de fluorescência desta população de células geradas em contato com as
MSC de MO é significativamente maior do que com as MSC de TA. Alguns trabalhos
identificaram sub-populações de monócitos baseados na expressão de CD14.
Somente as células monocíticas com alta expressão para este marcador possuem
propriedades inflamatórias e estas gerariam células CD14 com baixa expressão que
adquirem CD16, o receptor da fração constante (Fc) da IgG (FcγRIII) (RANDOLPH et

88
al., 2002; TACKE & RANDOLPH, 2006). Provavelmente, as MSC de TA seriam
capazes de promover a diferenciação in situ de monócitos CD14Hi para macrófagos
CD14LoCD16+. Esta hipótese necessita de ensaios funcionais para a sua
comprovação, mas está de acordo com a descoberta de macrófagos com
propriedades anti-inflamatórias, especificamente residentes no TA em condições
fisiológicas normais (ZEYDA & STULNIG, 2007) e está diretamente correlacionada
com a predominante produção de monócitos somente no co-cultivo com as MSCs de
TA (Fig. 38D).
Seguindo a nossa hipótese de que as propriedades adipocíticas das MSC de
TA foram geradas pelo microambiente, e que por esta razão podem ser reversíveis, as
MSC de TA foram induzidas por até duas semanas para a via osteoblástica, e a
sustentação de progenitores hematopoiéticos foi avaliada. O percentual de células
CD34+ mantidas ao longo do co-cultivo é semelhante para ambas as MSC de TA,
induzidas e não induzidas, porém o número absoluto destas células hematopoiéticas
em quatro dias é maior quando as células de TA são induzidas para o fenótipo
osteoblástico (Fig. 43). As análises morfológicas das células hematopoiéticas também
revelaram um maior percentual de progenitores mielóides nas MSC de TA e de blastos
nas MSC de TA osteoblásticas em sete dias de co-cultivo. Apesar do maior percentual
blástico nas MSCs induzidas para a linhagem osteoblástica, com estes dados não
podemos afirmar que este estroma é capaz de manter e/ou gerar HSCs. Uma das
principais propriedades que definem uma HSC é a sua capacidade de auto-renovação.
Logo, as células hematopoiéticas geradas em sete dias em ambos os co-cultivos
foram cultivadas em um ensaio que permite determinar a freqüência de HSC de longa
duração em uma determinada população – o LTC-IC. Após mais quatro semanas de
cultivo, somente as células geradas no co-cultivo com as MSC de TA induzidas por
duas semanas para linhagem osteoblástica foram capazes de manter culturas
positivas, ou seja, apresentando produção de células hematopoiéticas. A freqüência
de LTC-IC foi calculada segundo o método estatístico de Poisson (SUTHERLAND et
al., 1990), que revelou que em cada 150 células, uma pode ser considerada HSC de
longa-duração. Também foram realizados ensaios de CFU-GM antes e depois do
ensaio de LTC-IC. O número de colônias geradas pelas MSC induzidas para a
linhagem osteoblástica foi praticamente o mesmo, nestes dois momentos (Fig. 45).
Podemos então inferir de maneira indireta que o número de progenitores
hematopoiéticos foi mantido em longo prazo. Novamente, as MSC de TA sem indução
apresentaram um maior número de colônias CFU-GM antes do ensaio de LTC-IC,
revelando um maior comprometimento mielóide. Provavelmente em razão deste

89
comprometimento não foi possível detectar colônias LTC-IC positivas após quatro
semanas.
Outros trabalhos também avaliaram o papel da MSC de TA na sustentação da
hematopoese (CORRE et al., 2006; WAGNER et al., 2007). CORRE e colaboradores
(2006), não observaram a manutenção de blastos em seus co-cultivos com MSC de
TA ainda que induzidas para a via osteogênica. Neste estudo, além de não terem sido
utilizados ensaios de LTC-IC, as MSC de TA foram induzidas por quatro semanas, até
o estágio terminal de diferenciação. Somente os osteoblastos imaturos respondem a
fatores pró-osteogênicos e sustentam os eventos de osteoclastogênese in vitro
(ATKINS et al., 2003) e in vivo (CORRAL et al., 1998). As MSCs de MO cultivadas em
meio osteogênico por mais de duas semanas mostram vários focos de mineralização,
mas perdem a capacidade de sustentação de hematopoese (MIURA et al., 2006). O
outro estudo (WAGNER et al., 2007), utilizou a técnica de LTC-IC para a quantificação
de HSCs, porém em MSC de TA sem indução obtendo resultados semelhantes aos
nossos.
As propriedades hematopoiéticas das MSC de TA mesmo em diferentes
condições, nos levaram a especular se uma vez estas células inseridas no
microambiente medular, elas seriam capazes de se integrar funcionalmente. A
comprovação desta hipótese abre novas alternativas para diversos tratamentos, desde
a redução do tempo de reconstituição hematopoiética pós-transplante de MO até para
casos de aplasia medular.
5.4.CONSIDERAÇÕES FINAIS
As MSC de TA são semelhantes às de MO, mas talvez por possuírem
populações fenotipicamente distintas, também são únicas no seu potencial de
diferenciação mesodérmica e nas propriedades hematopoiéticas. As células de TA são
enriquecidas em progenitores endoteliais (células CD34+), e o marcador CD105,
amplamente descrito para as MSC de MO, foi regulado positivamente dentro da
população de células CD34+, após expansão in vitro. Este dado nos leva a hipótese
de que a molécula CD105 não seja um marcador de MSC no TA. Essas células
também possuem propriedades pró-angiogênicas, adquirem marcadores endoteliais
de maneira espontânea e ainda modulam células endoteliais maduras in vitro. A
capacidade de diferenciação endotelial aparentemente exclui a possibilidade de as
MSC de TA serem células perivasculares in vivo. Porém, essas células PPARγ2+
também são mais comprometidas com a via adipogênica e possuem um baixo
potencial condrogênico, e por isso, podem ser células perivasculares em um diferente

90
estágio de ativação das células de MO. O seu potencial de sustentação da
hematopoiese é fortemente representado pela geração de células macrofágicas após
a grande expansão de células mielóides (Fig. 46).
Todas estas características das populações de células de TA encontradas em
nosso trabalho, remetem a uma memória de microambiente (Fig 47). No TA, durante o
desenvolvimento da obesidade, são formados centros de hipóxia durante a hipertrofia
e/ou hiperplasia das células adipocíticas. Esta hipóxia leva a estímulos de
angiogênese e estes vasos recém-formados passam a ser responsáveis pela nutrição
dos novos adipócitos. Sem angiogênese, não há o aumento da massa do TA, e
consequentemente não há o aumento de peso. As MSC de TA possuem papel tanto
na formação de novos vasos, através de transdiferenciação ou pelo estímulo de
células endoteliais maduras, como na adipogênese. Outro componente celular
importante no processo de obesidade é o macrófago. Apesar de já ter sido
demonstrado que grande parte destas células no TA são oriundas da MO, também é
possível que MSC do próprio tecido estejam promovendo a diferenciação de
progenitores hematopoiéticos circulantes em células macrofágicas. Em nosso trabalho
mostramos diferenciação espontânea para endotélio e adipócitos, além da formação
de macrófagos a partir de progenitores hematopoiéticos. Porém, após a indução para
a via osteogênica, as MSCs de TA foram capazes de manter as HSC in vitro, tornando
possível sua aplicação em casos de reconstituição hematopoiética.
No âmbito da terapia celular, podemos utilizar diferentes fontes de células-
tronco para diferentes terapias, mas a partir do conhecimento da biologia destas
células, seremos capazes de modular o seu fenótipo in vitro para posterior aplicação in
vivo.
Figura 46. A MSC do TA e o seu possível papel pleiotrópico na
adipogênese, angiogênese e geração de células macrofágicas. Além das MSC, as células endoteliais, pré-adipócitos e adipócitos também participam da homeostase tecidual através da secreção de fatores de crescimento, citocinas e hormônios.

91
Figura 47. O TA e sua possível diversidade celular. Esquema representando as populações celulares do TAB subcutâneo já descritas, e a possível infiltração de progenitores hematopoiéticos, atribuindo uma nova função a este órgão endócrino. As MSC com localização perivascular, além da multipotencialidade exercem funções proliferativas e de estímulos de diferenciação nas diferentes populações celulares representadas acima. 6. CONCLUSÕES De acordo com a avaliação dos dados descritos anteriormente, podemos
concluir que no TA subcutâneo residem células-tronco multipotentes com
características únicas, provavelmente ditadas pelo microambiente. As nossas
conclusões e hipóteses geradas a partir dos nossos resultados podem ser separadas
em três tópicos principais.
6.1. Fenótipo A fração aderente das células de TA, que de acordo com a literatura representa
a população de MSC, é heterogênea em relação à expressão de marcadores de
superfície. As MSC do TA podem ser divididas em duas populações principais:
(1)CD34+CD105-CD146-CD90+CD44+CD45-. (2)CD34+CD105+CD146+CD90+CD44+CD45-.
Através da seleção destas populações foi possível comprovar a perda do CD34
com o ganho simultâneo do marcador CD105. Este dado nos levou a hipótese de que

92
a molécula de CD105 não represente a verdadeira população de progenitores
mesodermais no TA, sendo desta forma um artefato adquirido in vitro.
6.2. Multipotencialidade Assim como as MSC de MO, as de TA, embora heterogêneas, são capazes de
diferenciação para as três linhagens mesodérmicas, apesar do baixo potencial
condrogênico e do comprometimento com a linhagem adipocítica confirmado pela
expressão constitutiva do gene PPARγ2.
Os nossos resultados para a via endotelial nos levaram à hipótese de um papel
ativo das MSC de TA na angiogênese através de dois mecanismos: (1) Diferenciação em células endoteliais; (2) Ativação direta e indireta das células endoteliais. 6.3. Potencial de sustentação hematopoiética As MSCs de TA, assim como as de MO, são capazes de geração mielóide in
vitro, mas gerando, preferencialmente, monócitos e/ou macrófagos. A indução para o fenótipo adipocítico induz expansão e diferenciação de progenitores mielóides e favorece a produção de monócitos/macrófagos, e a osteogênica, a manutenção de HSCs. 6.4. Conclusão final Como os nossos resultados foram obtidos a partir da população total de MSC
de TA, talvez a população de pré-adipócitos também seja plástica. Na maioria dos
tecidos existem células acumuladoras de gordura em íntimo contato com as células
estromais de cada órgão. É possível então que estas células possuam papéis na
organogênese e angiogênese de vários órgãos, tornando indispensável a busca pelas
verdadeiras identidade e potencialidade das células estromais que residem no tecido
adiposo.

93
7. REFERÊNCIAS BIBLIOGRÁFICAS Abboud CN, Lichtman MA. Structure of the marrow and the hematopoietic microenvironment. In: Beutler E, Litchman MA, Coller BS, Kipps TJ, Seligsohn U, Williams Hematology – Sexta edição, McGraw-Hill – New York. 2001. p35. Aggarwal S, Pittenger MF. Human mesenchymal stem cells modulate allogeneic immune cell responses. Blood. 2005;105:1815-22. Ailhaud G, Grimaldi P, Négrel R. Cellular and molecular aspects of adipose tissue development. Annu Rev Nutr. 1992;12:207-33. Akintoye SO, Lam T, Shi S, Brahim J, Collins MT, Robey PG. Skeletal site-specific characterization of orofacial and iliac crest human bone marrow stromal cells in same individuals. Bone. 2006;38:758-68. Amit M, Itskovitz-Eldor J. Derivation and spontaneous differentiation of human embryonic stem cells. J Anat. 2002;200:225-32. Anjos-Afonso F, Bonnet D. Nonhematopoietic/endothelial SSEA-1+ cells define the most primitive progenitors in the adult murine bone marrow mesenchymal compartment. Blood. 2007;109:1298-306. Arai F, Ohneda O, Miyamoto T, Zhang XQ, Suda T. Mesenchymal stem cells in perichondrium express activated leukocyte cell adhesion molecule and participate in bone marrow formation. J Exp Med. 2002;195:1549-63. Arai F, Hirao A, Ohmura M, Sato H, Matsuoka S, Takubo K, Ito K, Koh GY, Suda T. Tie2/angiopoietin-1 signaling regulates hematopoietic stem cell quiescence in the bone marrow niche. Cell. 2004;118: 149-61. Asahara T, Murohara T, Sullivan A, Silver M, van der Zee R, Li T, Witzenbichler B, Schatteman G, Isner JM. Isolation of putative progenitor endothelial cells for angiogenesis. Science. 1997;275:964-7. Atkins, G. J., Kostakis, P., Pan, B., Farrugia, A., Gronthos, S., Evdokiou, A., Harrison, K., Findlay, D. M., and Zannettino, A. C.. RANKL expression is related to the differentiation state of human osteoblasts. J. Bone Miner Res. 2003;18: 1088-98. Balana B, Nicoletti C, Zahanich I, Graf EM, Christ T, Boxberger S, Ravens U. 5-Azacytidine induces changes in electrophysiological properties of human mesenchymal stem cells. Cell Res. 2006;16:949-60. Balduino A, Hurtado SP, Frazão P, Takiya CM, Alves LM, Nasciutti LE, El-Cheikh MC, Borojevic R. Bone marrow subendosteal microenvironment harbours functionally distinct haemosupportive stromal cell populations. Cell Tissue Res. 2005;319:255-66. Banas A, Teratani T, Yamamoto Y, Tokuhara M, Takeshita F, Quinn G, Okochi H, Ochiya T. Adipose tissue-derived mesenchymal stem cells as a source of human hepatocytes. Hepatology. 2007;46:219-28. Bancroft JD, Stevens A. Theory and Practice of Histological Techniques. Churchill livingstone, Quarta Edição, 1996, chapter 10, p190.

94
Banfi A, Muraglia A, Dozin B, Mastrogiacomo M, Cancedda R, Quarto R. Proliferation kinetics and differentiation potential of ex vivo expanded human bone marrow stromal cells: Implications for their use in cell therapy. Exp Hematol. 2000;28:707-15. Baptista LS, Pedrosa CGS, Silva KR, Otazú IB, Takyia CM, Dutra HS, Cláudio-da-Silva C, Borojevic R, Rossi MID. Bone Marrow and Adipose Tissue-Derived Mesenchymal Stem Cells: How Close Are They? J Stem Cells. 2007;2: issue 2. Beck GR Jr. Inorganic phosphate as a signaling molecule in osteoblast differentiation. J Cell Biochem. 2003;90:234-43. Bennett JH, Joyner CJ, Triffitt JT, Owen ME. Adipocytic cells cultured from marrow have osteogenic potential. J Cell Sci. 1991;99:131-9 Bensidhoum M, Chapel A, Francois S, Demarquay C, Mazurier C, Fouillard L, Bouchet S, Bertho JM, Gourmelon P, Aigueperse J, Charbord P, Gorin NC, Thierry D, Lopez M. Homing of in vitro expanded Stro-1- or Stro-1+ human mesenchymal stem cells into the NOD/SCID mouse and their role in supporting human CD34 cell engraftment. Blood. 2004;103:3313-9. Beresford JN, Bennett JH, Devlin C, Leboy PS, Owen ME. Evidence for an inverse relationship between the differentiation of adipocytic and osteogenic cells in rat marrow stromal cell cultures. J Cell Sci. 1992;102:341-51. Berner HS, Lyngstadaas SP, Spahr A, Monjo M, Thommesen L, Drevon CA, Syversen U, Reseland JE. Adiponectin and its receptors are expressed in bone-forming cells. Bone. 2004;35:842-9. Bianco P, Costantini M, Dearden LC, Bonucci E. Alkaline phosphatase positive precursors of adipocytes in the human bone marrow. Br J Haematol. 1988;68:401-3. Bianco P, Gehron Robey P. Marrow stromal stem cells. J Clin Invest. 2000;105:1663-8. Bianco P, Riminucci M, Gronthos S, Robey PG. Bone marrow stromal stem cells: nature, biology, and potential applications. Stem Cells. 2001;19:180-92. Bonifer C, Faust N, Geiger H, Müller AM. Developmental changes in the differentiation capacity of haematopoietic stem cells. Immunol Today. 1998;19:236-41. Brachyogel B, Moch H, Pausch F, Schlötzer-Schrehardt U, Hofmann C, Hallmann R, von der Mark K, Winkler T, Pöschl E. Perivascular cells expressing annexin A5 define a novel mesenchymal stem cell-like population with the capacity to differentiate into multiple mesenchymal lineages. Development 2005;132:2657-68. Bradley TR, Metcalf D. The growth of mouse bone marrow cells in vitro. Aust J Exp Biol Med Sci. 1966;44:287-99. Bruno S, Bussolati B, Scacciatella P, Marra S, Sanavio F, Tarella C, Camussi G. Combined administration of G-CSF and GM-CSF stimulates monocyte-derived pro-angiogenic cells in patients with acute myocardial infarction. Cytokine. 2006;34:56-65. Burd A, Ahmed K, Lam S, Ayyappan T, Huang L. Stem cell strategies in burns care. Burns. 2007;33:282-91.

95
Burger J.A, Zvaifler N.J, Tsukada N, Firestein G.S, Kipps T.J. Fibroblast- like synoviocytes support B-cell pseudoemperipolesis via a stromal cell- derived factor-1 and CD106 (VCAM- 1)-dependent mechanism. J Clin Invest. 2001;107: 305-15. Calvi LM, Adams GB, Weibrecht KW, Weber JM, Olson DP, Knight MC, Martin RP, Schipani E, Divieti P, Bringhurst FR, Milner LA, Kronenberg HM, Scadden DT. Osteoblastic cells regulate the haematopoietic stem cell niche. Nature. 2003; 425:841- 6. Cao Y. Angiogenesis modulates adipogenesis and obesity. J Clin Invest. 2007 Sep;117:2362-8. Carmeliet P. Angiogenesis in health and disease. Nat Med. 2003;9:653-60. Caspar-Bauguil S, Cousin B, Galinier A, Segafredo C, Nibbelink M, André M, Casteilla L, Pénicaud L. Adipose tissues as an ancestral immune organ: site-specific change in obesity. FEBS Lett. 2005;579:3487-92. Caspar-Bauguil S, Cousin B, André M, Nibbelink M, Galinier A, Periquet B, Casteilla L, Pénicaud L. Weight-dependent changes of immune system in adipose tissue: importance of leptin. Exp Cell Res. 2006;312: 2195-202. Casteilla L, Charrière G, Laharrague P, Cousin B, Planat-Benard V, Péricaud L, Chavoin JP. Tissus adipeux, chirurgie plastique et reconstructrice: le retour aux sources. Ann Chir Plast Esthet. 2004;49:409-18. Casteilla L, Planat-Bénard V, Cousin B, Silvestre JS, Laharrague P, Charrière G, Carrière A, Pénicaud L. Plasticity of adipose tissue: a promising therapeutic avenue in the treatment of cardiovascular and blood diseases? Arch Mal Coeur Vaiss. 2005;98:922-6. Charrière G, Cousin B, Arnaud E, André M, Bacou F, Penicaud L, Casteilla L. Preadipocyte conversion to macrophage. Evidence of plasticity. J Biol Chem. 2003;278:9850-5. Charbord P, Tavian M, Humeau L, Péault B. Early ontogeny of the human marrow from long bones: an immunohistochemical study of hematopoiesis and its microenvironment. Blood. 1996;87:4109-19. Check E. Stem cells: the hard copy. Nature. 2007;446:485-6. Choi K, Kennedy M, Kazarov A, Papadimitriou JC, Keller G. A common precursor for hematopoietic and endothelial cells. Development. 1998;125:725-32. Colter DC, Sekiya I, Prockop DJ. Identification of a subpopulation of rapidly self-renewing and multipotential adult stem cells in colonies of human marrow stromal cells. Proc Natl Acad Sci U S A. 2001;98:7841-5. Corral, D. A., Amling, M., Priemel, M., Loyer, E., Fuchs, S., Ducy, P., Baron, R., and Karsenty, G. Dissociation between bone resorption and bone formation in osteopenic transgenic mice. Proc Natl. Acad. Sci. US. 1998; 95: 13835-40. Corre J, Planat-Benard V, Corberand JX, Pénicaud L, Casteilla L, Laharrague P. Human bone marrow adipocytes support complete myeloid and lymphoid differentiation from human CD34 cells. Br J Haematol. 2004;127:344-7.

96
Corre J, Barreau C, Cousin B, Chavoin JP, Caton D, Fournial G, Penicaud L, Casteilla L, Laharrague P. Human subcutaneous adipose cells support complete differentiation but not self-renewal of hematopoietic progenitors. J Cell Physiol. 2006;208:282-8. Cossu G, Bianco P. Mesoangioblasts--vascular progenitors for extravascular mesodermal tissues. Curr Opin Genet Dev. 2003;13:537-42. Couly G, Grapin-Botton A, Coltey P, Le Douarin NM. The regeneration of the cephalic neural crest, a problem revisited: the regenerating cells originate from the contralateral or from the anterior and posterior neural fold. Development. 1996;122:3393-407. Cousin B, Munoz O, Andre M, Fontanilles AM, Dani C, Cousin JL, Laharrague P, Casteilla L, Pénicaud L. A role for preadipocytes as macrophage-like cells. FASEB J. 1999;13:305-12. Cousin B, André M, Casteilla L, Pénicaud L. Altered macrophage-like functions of preadipocytes in inflammation and genetic obesity. J Cell Physiol. 2001;186:380-6. Croisille L, Auffray I, Katz A, Izac B, Vainchenker W, Coulombel L. Hydrocortisone differentially affects the ability of murine stromal cells and human marrow-derived adherent cells to promote the differentiation of CD34++/CD38- long-term culture-initiating cells. Blood. 1994;84:4116-24. Curat CA, Miranville A, Sengenès C, Diehl M, Tonus C, Busse R, Bouloumié A. From blood monocytes to adipose tissue-resident macrophages: induction of diapedesis by human mature adipocytes. Diabetes. 2004; 53: 1285-92. da Silva Meirelles L, Chagastelles PC, Nardi NB. Mesenchymal stem cells reside in virtually all post-natal organs and tissues. J Cell Sci. 2006;119:2204-13. Dazzi F, Ramasamy R, Glennie S, Jones SP, Roberts I. The role of mesenchymal stem cells in haemopoiesis. Blood Rev. 2006; 20: 161-71. De Angelis L, Berghella L, Coletta M, Lattanzi L, Zanchi M, Cusella-De Angelis MG, Ponzetto C, Cossu G. Skeletal myogenic progenitors originating from embryonic dorsal aorta coexpress endothelial and myogenic markers and contribute to postnatal muscle growth and regeneration. J Cell Biol. 1999;147:869-78.
De Bari C, Dell’Accio F, Tvlzanowski P, and Luyten FP. Multipotent mesenchymal stem cells from adult human synovial membrane. Arthritis Rheum. 2001; 44: 1928-42. de Bruijn MF, Ma X, Robin C, Ottersbach K, Sanchez MJ, Dzierzak E. Hematopoietic stem cells localize to the endothelial cell layer in the midgestation mouse aorta. Immunity. 2002;16:673-83. DeKoter RP, Singh H. Regulation of B lymphocyte and macrophage development by graded expression of PU.1. Science. 2000;288:1439-41. Dellavalle A, Sampaolesi M, Tonlorenzi R, Tagliafico E, Sacchetti B, Perani L, Innocenzi A, Galvez BG, Messina G, Morosetti R, Li S, Belicchi M, Peretti G, Chamberlain JS, Wright WE, Torrente Y, Ferrari S, Bianco P, Cossu G. Pericytes of human skeletal muscle are myogenic precursors distinct from satellite cells. Nat Cell Biol. 2007;9:255-67.

97
De Ugarte DA, Alfonso Z, Zuk PA, Elbarbary A, Zhu M, Ashjian P, Benhaim P, Hedrick MH, Fraser JK. Differential expression of stem cell mobilization-associated molecules on multi-lineage cells from adipose tissue and bone marrow. Immunol Lett. 2003;89:267-70. Dexter TM, Testa NG. Differentiation and proliferation of hemopoietic cells in culture. Methods Cell Biol. 1976;14:387-405. Dexter TM, Whetton AD, Spooncer E, Heyworth C, Simmons P. The role of stromal cells and growth factors in haemopoiesis and modulation of their effects by the src oncogene. J Cell Sci Suppl. 1985;3:83-95. Dicker A, Le Blanc K, Aström G, van Harmelen V, Götherström C, Blomqvist L, Arner P, Rydén M. Functional studies of mesenchymal stem cells derived from adult human adipose tissue. Exp Cell Res. 2005;308:283-90. DiMascio L, Voermans C, Uqoezwa M, Duncan A, Lu D, Wu J, Sankar U, Reya T. Identification of adiponectin as a novel hemopoietic stem cell growth factor. J Immunol. 2007;178:3511-20. Dimmeler S, Zeiher AM, Schneider MD. Unchain my heart: the scientific foundations of cardiac repair. J Clin Invest. 2005;115:572-83. Dominici M, Le Blanc K, Mueller I, Slaper-Cortenbach I, Marini F, Krause D, Deans R, Keating A, Prockop Dj, Horwitz E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy. 2006;8:315-7. Donovan PJ, Gearhart J. The end of the beginning for pluripotent stem cells. Nature. 2001;414:92-7. Dorheim MA, Sullivan M, Dandapani V, Wu X, Hudson J, Segarini PR, Rosen DM, Aulthouse AL, Gimble JM. Osteoblastic gene expression during adipogenesis in hematopoietic supporting murine bone marrow stromal cells. J Cell Physiol. 1993;154:317-28. Durand C, Robin C, Dzierzak E. Mesenchymal lineage potentials of aorta-gonad-mesonephros stromal clones. Haematologica. 2006;91:1172-9. Dutra HS, Rossi MI, Azevedo SP, el-Cheikh MC, Borojevic R. Haematopoietic capacity of colony-forming cells mobilized in hepatic inflammatory reactions as compared to that of normal bone marrow cells. Res Immunol. 1997;148:437-44. Ebihara Y, Masuya M, Larue AC, Fleming PA, Visconti RP, Minamiguchi H, Drake CJ, Ogawa M. Hematopoietic origins of fibroblasts: II. In vitro studies of fibroblasts, CFU-F, and fibrocytes. Exp Hematol. 2006;34:219-29. Engler AJ, Sen S, Sweeney HL, Discher DE. Matrix elasticity directs stem cell lineage specification. Cell. 2006;126:677-89. Etchevers HC, Vincent C, Le Douarin NM, Couly GF. The cephalic neural crest provides pericytes and smooth muscle cells to all blood vessels of the face and forebrain. Development. 2001;128:1059-68.

98
Falanga V, Iwamoto S, Chartier M, Yufit T, Butmarc J, Kouttab N, Shrayer D, Carson P. Autologous bone marrow-derived cultured mesenchymal stem cells delivered in a fibrin spray accelerate healing in murine and human cutaneous wounds. Tissue Eng. 2007;13:1299-312. Fang B, Song YP, Liao LM, Han Q, Zhao RC. Treatment of severe therapy-resistant acute graft-versus-host disease with human adipose tissue-derived mesenchymal stem cells. Bone Marrow Transplant. 2006;38:389-90. Farrington-Rock C, Crofts NJ, Doherty MJ, Ashton BA, Griffin-Jones C, Canfield AE. Chondrogenic and adipogenic potential of microvascular pericytes. Circulation. 2004;110:2226-32. Ferrari G, Cusella-De Angelis G, Coletta M, Paolucci E, Stornaiuolo A, Cossu G, Mavilio F. Muscle regeneration by bone marrow-derived myogenic progenitors. Science. 1998;279:1528-30. Fiedler U, Augustin HG. Angiopoietins: a link between angiogenesis and inflammation. Trends Immunol. 2006;27:552-8. Fraser JK, Wulur I, Alfonso Z, Hedrick MH. Fat tissue: an underappreciated source of stem cells for biotechnology. Trends Biotechnol. 2006;24:150-4. Friedenstein AJ, Piatetzky-Shapiro II, Petrakova KV. Osteogenesis in transplants of bone marrow cells. J Embryol Exp Morphol. 1966; 16: 381-90. Friedenstein AJ, Petrakova KV, Kurolesova AI, Frolova GP. Heterotopic of bone marrow. Analysis of precursor cells for osteogenic and hematopoietic tissues. Transplantation. 1968;6:230-47. Friedenstein AJ, Deriglasova UF, Kulagina NN, Panasuk AF, Rudakowa SF, Luriá EA, Ruadkow IA. Precursors for fibroblasts in different populations of hematopoietic cells as detected by the in vitro colony assay method. Exp Hematol. 1974;2:83-92. Frühbeck G, Gómez-Ambrosi J, Muruzábal FJ, Burrell MA. The adipocyte: a model for integration of endocrine and metabolic signaling in energy metabolism regulation. Am J Physiol Endocrinol Metab. 2001;280: E827-47. Fukumura D, Ushiyama A, Duda DG, Xu L, Tam J, Krishna V, Chatterjee K, Garkavtsev I, Jain RK. Paracrine regulation of angiogenesis and adipocyte differentiation during in vivo adipogenesis. Circ Res. 2003; 93: 88-97. Fukushi J, Makagiansar IT, Stallcup WB. NG2 proteoglycan promotes endothelial cell motility and angiogenesis via engagement of galectin-3 and alpha3beta1 integrin. Mol Biol Cell. 2004;15:3580-90. Gang EJ, Bosnakovski D, Figueiredo CA, Visser JW, Perlingeiro RC. SSEA-4 identifies mesenchymal stem cells from bone marrow. Blood. 2007 Feb 15;109(4):1743-51. García-Olmo D, García-Arranz M, Herreros D, Pascual I, Peiro C, Rodríguez-Montes JA. A phase I clinical trial of the treatment of Crohn's fistula by adipose mesenchymal stem cell transplantation. Dis Colon Rectum. 2005;48:1416-23.

99
Gentili C, Bianco P, Neri M, Malpeli M, Campanile G, Castagnola P, Cancedda R, Cancedda FD. Cell proliferation, extracellular matrix mineralization, and ovotransferrin transient expression during in vitro differentiation of chick hypertrophic chondrocytes into osteoblast-like cells. J Cell Biol. 1993;122:703-12. Gesta S, Tseng YH, Kahn CR. Developmental origin of fat: tracking obesity to its source. Cell. 2007;131:242-56. Giachelli CM, Steitz S. Osteopontin: a versatile regulator of inflammation and biomineralization. Matrix Biol. 2000;19:615-22. Gimble JM, Youkhana K, Hua X, Bass H, Medina K, Sullivan M, Greenberger J, Wang CS. Adipogenesis in a myeloid supporting bone marrow stromal cell line. J Cell Biochem. 1992;50:73-82. Gimble JM, Robinson CE, Wu X, Kelly KA. The function of adipocytes in the bone marrow stroma: an update. Bone. 1996;19:421-8. Gonçalves R, Lobato da Silva C, Cabral JM, Zanjani ED, Almeida-Porada G. A Stro-1(+) human universal stromal feeder layer to expand/maintain human bone marrow hematopoietic stem/progenitor cells in a serum-free culture system. Exp Hematol. 2006; 34:1353-9. Goodell MA. Stem cells: is there a future in plastics? Curr Opin Cell Biol. 2001;13:662-5. Greenfield EM, Horowitz MC, Lavish SA. Stimulation by parathyroid hormone of interleukin-6 and leukemia inhibitory factor expression in osteoblasts is an immediate-early gene response induced by cAMP signal transduction. J Biol Chem. 1996;271:10984-9. Gregoire FM. Adipocyte differentiation: from fibroblast to endocrine cell. Exp Biol Med. 2001;226:997-1002. Gronthos S, Franklin DM, Leddy HA, Robey PG, Storms RW, Gimble JM. Surface protein characterization of human adipose tissue-derived stromal cells. J Cell Physiol. 2001;189:54-63. Grosset C, Taupin JL, Lemercier C, Moreau JF, Reiffers J, Ripoche J. Leukaemia inhibitory factor expression is inhibited by glucocorticoids through post-transcriptional mechanisms. Cytokine. 1999;11:29-36. Grove JE, Bruscia E, Krause DS. Plasticity of bone marrow-derived stem cells. Stem Cells. 2004;22:487-500. Gussoni E, Bennett RR, Muskiewicz KR, Meyerrose T, Nolta JA, Gilgoff I, Stein J, Chan YM, Lidov HG, Bönnemann CG, Von Moers A, Morris GE, Den Dunnen JT, Chamberlain JS, Kunkel LM, Weinberg K. Long-term persistence of donor nuclei in a Duchenne muscular dystrophy patient receiving bone marrow transplantation. J Clin Invest. 2002;110:807-14. Hagedorn M, Balke M, Schmidt A, Bloch W, Kurz H, Javerzat S, Rousseau B, Wilting J, Bikfalvi A. VEGF coordinates interaction of pericytes and endothelial cells during vasculogenesis and experimental angiogenesis. Dev Dyn. 2004;230:23-33.

100
Hahn P, Novak M. Development of brown and white adipose tissue. J Lipid Res. 1975;16:79-91. Hamada T, Möhle R, Hesselgesser J, Hoxie J, Nachman RL, Moore MA, Rafii S. Transendothelial migration of megakaryocytes in response to stromal cell-derived factor 1 (SDF-1) enhances platelet formation. J Exp Med. 1998;188:539-48. Hasegawa S, Neo M, Tamura J, Fujibayashi S, Takemoto M, Shikinami Y, Okazaki K, Nakamura T. In vivo evaluation of a porous hydroxyapatite/poly-DL-lactide composite for bone tissue engineering. J Biomed Mater Res A. 2007;81:930-8. Hausman GJ, Richardson RL. Adipose tissue angiogenesis. J Anim Sci. 2004;82:925-34. Hennig T, Lorenz H, Thiel A, Goetzke K, Dickhut A, Geiger F, Richter W. Reduced chondrogenic potential of adipose tissue derived stromal cells correlates with an altered TGFbeta receptor and BMP profile and is overcome by BMP-6. J Cell Physiol. 2007;211:682-91. Herzog EL, Chai L, Krause DS. Plasticity of marrow-derived stem cells. Blood. 2003;102:3483-93. Hicok KC, Du Laney TV, Zhou YS, Halvorsen YD, Hitt DC, Cooper LF, Gimble JM. Human adipose-derived adult stem cells produce osteoid in vivo. Tissue Eng. 2004;10:371-80. Hirashima M, Kataoka H, Nishikawa S, Matsuyoshi N, Nishikawa S. Maturation of embryonic stem cells into endothelial cells in an in vitro model of vasculogenesis. Blood. 1999;93:1253-63. Hofbauer LC, Khosla S, Dunstan CR, Lacey DL, Boyle WJ, Riggs BL. The roles of osteoprotegerin and osteoprotegerin ligand in the paracrine regulation of bone resorption. J Bone Miner Res. 2000;15:2-12. Horwitz EM, Prockop DJ, Gordon PL, Koo WW, Fitzpatrick LA, Neel MD, McCarville ME, Orchard PJ, Pyeritz RE, Brenner MK. Clinical responses to bone marrow transplantation in children with severe osteogenesis imperfecta. Blood. 2001;97:1227-31. Hu Y, Zhang Z, Torsney E, Afzal AR, Davison F, Metzler B, Xu Q. Abundant progenitor cells in the adventitia contribute to atherosclerosis of vein grafts in ApoE-deficient mice. J Clin Invest. 2004;113:1258-65. Huang T, He D, Kleiner G, Kuluz J. Neuron-like differentiation of adipose-derived stem cells from infant piglets in vitro. J Spinal Cord Med. 2007;30 Suppl 1:S35-40. Ichim TE, Solano F, Glenn E, Morales F, Smith L, Zabrecky G, Riordan NH. Stem cell therapy for autism. J Transl Med. 2007;27:5:30. Inanç B, Eser Elçin A, Koç A, Baloş K, Parlar A, Murat Elçin Y. Encapsulation and osteoinduction of human periodontal ligament fibroblasts in chitosan-hydroxyapatite microspheres. J Biomed Mater Res A. 2007;82:917-26.

101
Indovina P, Collini M, Chirico G, Santini MT. Three-dimensional cell organization leads to almost immediate HRE activity as demonstrated by molecular imaging of MG-63 spheroids using two-photon excitation microscopy. FEBS Lett. 2007;581:719-26. Issaad C, Croisille L, Katz A, Vainchenker W, Coulombel L. A murine stromal cell line allows the proliferation of very primitive human CD34++/CD38- progenitor cells in long-term cultures and semisolid assays. Blood. 1993;81:2916-24. Izadpanah R, Trygg C, Patel B, Kriedt C, Dufour J, Gimble JM, Bunnell BA. Biologic properties of mesenchymal stem cells derived from bone marrow and adipose tissue. J Cell Biochem. 2006;99:1285-97. Jaffredo T, Gautier R, Eichmann A, Dieterlen-Lièvre F. Intraaortic hemopoietic cells are derived from endothelial cells during ontogeny. Development. 1998;125:4575-83. Jakobsson L, Kreuger J, Claesson-Welsh L. Building blood vessels--stem cell models in vascular biology. J Cell Biol. 2007;177:751-5. Javazon EH, Beggs KJ, Flake AW. Mesenchymal stem cells: paradoxes of passaging. Exp Hematol. 2004;32:414-25. Javazon EH, Keswani SG, Badillo AT, Crombleholme TM, Zoltick PW, Radu AP, Kozin ED, Beggs K, Malik AA, Flake AW. Enhanced epithelial gap closure and increased angiogenesis in wounds of diabetic mice treated with adult murine bone marrow stromal progenitor cells. Wound Repair Regen. 2007;15:350-9. Jiang Y, Jahagirdar BN, Reinhardt RL, Schwartz RE, Keene CD, Ortiz-Gonzalez XR, Reyes M, Lenvik T, Lund T, Blackstad M, Du J, Aldrich S, Lisberg A, Low WC, Largaespada DA, Verfaillie CM. Pluripotency of mesenchymal stem cells derived from adult marrow. Nature. 2002;418:41-9. Jo YY, Lee HJ, Kook SY, Choung HW, Park JY, Chung JH, Choung YH, Kim ES, Yang HC, Choung PH. Isolation and characterization of postnatal stem cells from human dental tissues. Tissue Eng. 2007;13:767-73. Jones BJ, Brooke G, Atkinson K, McTaggart SJ. Immunosuppression by placental indoleamine 2,3-dioxygenase: a role for mesenchymal stem cells. Placenta. 2007; 28:1174-81. Kaiser S, Hackanson B, Follo M, Mehlhorn A, Geiger K, Ihorst G, Kapp U. BM cells giving rise to MSC in culture have a heterogeneous CD34 and CD45 phenotype. Cytotherapy. 2007;9:439-50. Kaplan RN, Psaila B, Lyden D. Niche-to-niche migration of bone-marrow-derived cells. Trends Mol Med. 2007 Feb;13:72-81. Kehat I, Kenyagin-Karsenti D, Snir M, Segev H, Amit M, Gepstein A, Livne E, Binah O, Itskovitz-Eldor J, Gepstein L. Human embryonic stem cells can differentiate into myocytes with structural and functional properties of cardiomyocytes. J Clin Invest. 2001;108:407-14. Kelm JM, Diaz Sanchez-Bustamante C, Ehler E, Hoerstrup SP, Djonov V, Ittner L, Fussenegger M. VEGF profiling and angiogenesis in human microtissues. J Biotechnol. 2005;118:213-29.

102
Kern S, Eichler H, Stoeve J, Klüter H, Bieback K. Comparative analysis of mesenchymal stem cells from bone marrow, umbilical cord blood, or adipose tissue. Stem Cells. 2006;24:1294-301. Kiel MJ, Yilmaz OH, Iwashita T, Yilmaz OH, Terhorst C, Morrison SJ. SLAM family receptors distinguish hematopoietic stem and progenitor cells and reveal endothelial niches for stem cells. Cell. 2005;121:1109-21. Koç ON, Day J, Nieder M, Gerson SL, Lazarus HM, Krivit W. Allogeneic mesenchymal stem cell infusion for treatment of metachromatic leukodystrophy (MLD) and Hurler syndrome (MPS-IH). Bone Marrow Transplant. 2002;30:215-22.
Kögler G, Sensken S, Airey JA, Trapp T, Müschen M, Feldhahn N, Liedtke S, Sorg R V, Fischer J, Rosenbaum C, Greschat S, Knipper A, Bender J, Degistirici Ö, Gao J, Caplan AI, Colletti EJ, Almeda-Porada G, Müller HW, Zanjani E, Wernet P. A new human somatic stem cell from placental cord blood with intrinsic pluripotent differentiation potential. J Exp Med. 2004; 200:123-35.
Kopen GC, Prockop DJ, Phinney DG. Marrow stromal cells migrate throughout forebrain and cerebellum, and they differentiate into astrocytes after injection into neonatal mouse brains. Proc Natl Acad Sci U S A. 1999;96:10711-6. Korff T & Augustin HG. Tensional forces in fibrillar extracellular matrices control directional capillary sprouting. J Cell Sci. 1999;112: 3249-58. Koshiyama H, Ogawa Y, Tanaka K, Tanaka I. The unified hypothesis of interactions among the bone, adipose and vascular systems: 'osteo-lipo-vascular interactions'. Med Hypotheses. 2006;66:960-3.
Kovach NL, Lin N, Yednock T, Harlan JM, Broudy VC. Stem cell factor modulates avidity of alpha 4 beta 1 and alpha 5 beta 1 integrins expressed on hematopoietic cell lines. Blood. 1995 Jan 1;85(1):159-67. Krause DS, Theise ND, Collector MI, Henegariu O, Hwang S, Gardner R, Neutzel S, Sharkis SJ. Multi-organ, multi-lineage engraftment by a single bone marrow-derived stem cell. Cell. 2001;105:369-77. Krause DS. Plasticity of marrow-derived stem cells. Gene Ther. 2002;9:754-8. Kubis N, Tomita Y, Tran-Dinh A, Planat-Benard V, André M, Karaszewski B, Waeckel L, Pénicaud L, Silvestre JS, Casteilla L, Seylaz J, Pinard E. Vascular fate of adipose tissue-derived adult stromal cells in the ischemic murine brain: A combined imaging-histological study. Neuroimage. 2007; 34(1):1-11. Kucia M, Wysoczynski M, Ratajczak J, Ratajczak MZ. Identification of very small embryonic like (VSEL) stem cells in bone marrow. Cell Tissue Res. 2007 Sep 9; [Epub ahead of print]. Kurzrock, R. Hematopoietic Growth Factors. In: Bast, R.C.; Kufe, D.W.; Pollock, R.E.; Weichselbaum, R.R.; Holland, J.F.; Frei, E. Cancer Medicine. Quinta Edição, BC Decker Inc - Hamilton (Canada): 2000.

103
Kuznetsov SA, Krebsbach PH, Satomura K, Kerr J, Riminucci M, Benayahu D, Robey PG. Single-colony derived strains of human marrow stromal fibroblasts form bone after transplantation in vivo. J Bone Miner Res. 1997;12:1335-47. Kuznetsov SA, Mankani MH, Gronthos S, Satomura K, Bianco P, Robey PG. Circulating skeletal stem cells. J Cell Biol. 2001;153:1133-40. Lagasse E, Connors H, Al-Dhalimy M, Reitsma M, Dohse M, Osborne L, Wang X, Finegold M, Weissman IL, Grompe M. Purified hematopoietic stem cells can differentiate into hepatocytes in vivo. Nat Med. 2000;6:1229-34. Laharrague P, Larrouy D, Fontanilles AM, Truel N, Campfield A, Tenenbaum R, Galitzky J, Corberand JX, Pénicaud L, Casteilla L. High expression of leptin by human bone marrow adipocytes in primary culture. FASEB J. 1998;12:747-52. Laharrague P, Oppert JM, Brousset P, Charlet JP, Campfield A, Fontanilles AM, Guy-Grand B, Corberand JX, Pénicaud L, Casteilla L. High concentration of leptin stimulates myeloid differentiation from human bone marrow CD34+ progenitors: potential involvement in leukocytosis of obese subjects. Int J Obes Relat Metab Disord. 2000;24:1212-6. Lam QL, Lu L. Role of leptin in immunity. Cell Mol Immunol. 2007;4:1-13. Lamagna C, Bergers G. The bone marrow constitutes a reservoir of pericyte progenitors. J Leukoc Biol. 2006;80:677-81. LaRue AC, Masuya M, Ebihara Y, Fleming PA, Visconti RP, Minamiguchi H, Ogawa M, Drake CJ. Hematopoietic origins of fibroblasts: I. In vivo studies of fibroblasts associated with solid tumors. Exp Hematol. 2006;34:208-18. Le Blanc K, Ringdén O. Mesenchymal stem cells: properties and role in clinical bone marrow transplantation. Curr Opin Immunol. 2006;18:586-91. Lecka-Czernik B, Moerman EJ, Grant DF, Lehmann JM, Manolagas SC, Jilka RL. Divergent effects of selective peroxisome proliferator-activated receptor-gamma 2 ligands on adipocyte versus osteoblast differentiation. Endocrinology. 2002;143:2376-84. Le Douarin NM, Creuzet S, Couly G, Dupin E. Neural crest cell plasticity and its limits. Development. 2004;131:4637-50. Lee RH, Kim B, Choi I, Kim H, Choi HS, Suh K, Bae YC, Jung JS. Characterization and expression analysis of mesenchymal stem cells from human bone marrow and adipose tissue. Cell Physiol Biochem. 2004;14:311-24. Lehrke M, Lazar MA. Inflamed about obesity. Nat Med. 2004;10:126-7. Levenstein ME, Ludwig TE, Xu RH, Llanas RA, VanDenHeuvel-Kramer K, Manning D, Thomson JA. Basic fibroblast growth factor support of human embryonic stem cell self-renewal. Stem Cells. 2006;24:568-74. Liang DC. The role of colony-stimulating factors and granulocyte transfusion in treatment options for neutropenia in children with cancer. Paediatr Drugs. 2003;5(10):673-84.

104
Liebermann DA, Hoffman B. Differentiation primary response genes and proto-oncogenes as positive and negative regulators of terminal hematopoietic cell differentiation. Stem Cells. 1994;12:352-69. Liu W, Parikh AA, Stoeltzing O, Fan F, McCarty MF, Wey J, Hicklin DJ, Ellis LM. Upregulation of neuropilin-1 by basic fibroblast growth factor enhances vascular smooth muscle cell migration in response to VEGF. Cytokine. 2005;32:206-12. Liu TM, Martina M, Hutmacher DW, Hui JH, Lee EH, Lim B. Identification of common pathways mediating differentiation of bone marrow- and adipose tissue-derived human mesenchymal stem cells into three mesenchymal lineages. Stem Cells. 2007;25:750-60. Lodie TA, Blickarz CE, Devarakonda TJ, He C, Dash AB, Clarke J, Gleneck K, Shihabuddin L, Tubo R. Systematic analysis of reportedly distinct populations of multipotent bone marrow-derived stem cells reveals a lack of distinction. Tissue Eng. 2002;8:739-51. López-Holgado N, Alberca M, Sánchez-Guijo F, Villarón E, Almeida J, Martín A, Armellini A, García C, Blanco B, Sánchez-Abarca I, Martín S, Pérez-Simón JA, García-Sanz R, San Miguel JF, del Cañizo MC. Short-term endothelial progenitor cell colonies are composed of monocytes and do not acquire endothelial markers. Cytotherapy. 2007;9(1):14-22. Maeda K, Okubo K, Shimomura I, Funahashi T, Matsuzawa Y, Matsubara K. cDNA cloning and expression of a novel adipose specific collagen-like factor, apM1 (AdiPose Most abundant Gene transcript 1). Biochem Biophys Res Commun. 1996; 221:286-9. Mareschi K, Ferrero I, Rustichelli D, Aschero S, Gammaitoni L, Aglietta M, Madon E, Fagioli F. Expansion of mesenchymal stem cells isolated from pediatric and adult donor bone marrow. J Cell Biochem. 2006;97:744-54. McBeath R, Pirone DM, Nelson CM, Bhadriraju K, Chen CS. Cell shape, cytoskeletal tension, and RhoA regulate stem cell lineage commitment. Dev Cell. 2004;6:483-95. McCloskey KE, Smith DA, Jo H, Nerem RM. Embryonic stem cell-derived endothelial cells may lack complete functional maturation in vitro. J Vasc Res. 2006;43:411-21.
Mcilroy M, O'Rourke M, McKeown SR, Hirst DG, Robson T. Pericytes influence endothelial cell growth characteristics: role of plasminogen activator inhibitor type 1 (PAI-1). Cardiovasc Res. 2006;69:207-17.
Medvinsky A, Dzierzak E. Definitive hematopoiesis is autonomously initiated by the AGM region. Cell. 1996;86:897-906. Mendes SC, Robin C, Dzierzak E. Mesenchymal progenitor cells localize within hematopoietic sites throughout ontogeny. Development. 2005;132:1127-36. Minasi MG, Riminucci M, De Angelis L, Borello U, Berarducci B, Innocenzi A, Caprioli A, Sirabella D, Baiocchi M, De Maria R, Boratto R, Jaffredo T, Broccoli V, Bianco P, Cossu G. The meso-angioblast: a multipotent, self-renewing cell that originates from the dorsal aorta and differentiates into most mesodermal tissues. Development. 2002;129:2773-83.

105
Minguell JJ, Erices A, Conget P. Mesenchymal stem cells. Exp Biol Med. 2001;226:507-20. Miranville A, Heeschen C, Sengenès C, Curat CA, Busse R, Bouloumié A. Improvement of postnatal neovascularization by human adipose tissue-derived stem cells. Circulation. 2004 Jul 20;110:349-55. Mitchell JB, McIntosh K, Zvonic S, Garrett S, Floyd ZE, Kloster A, Di Halvorsen Y, Storms RW, Goh B, Kilroy G, Wu X, Gimble JM. Immunophenotype of human adipose-derived cells: temporal changes in stromal-associated and stem cell-associated markers. Stem Cells. 2006;24:376-85. Miura, Y., Gao, Z., Miura, M., Seo, B-M., Sonoyama, W., Chen, W., Gronthos, S., Zhang, L., and Shi, S.. Mesenchymal stem cell-organized bone marrow elements: An alternative hematopoietic progenitor resource. Stem Cells. 2006; 24: 2428-36. Mohyeddin Bonab M, Yazdanbakhsh S, Lotfi J, Alimoghaddom K, Talebian F, Hooshmand F, Ghavamzadeh A, Nikbin B. Does mesenchymal stem cell therapy help multiple sclerosis patients? Report of a pilot study. Iran J Immunol. 2007;4:50-7. Moon MH, Kim SY, Kim YJ, Kim SJ, Lee JB, Bae YC, Sung SM, Jung JS. Human adipose tissue-derived mesenchymal stem cells improve postnatal neovascularization in a mouse model of hindlimb ischemia. Cell Physiol Biochem. 2006;17:279-90. Müller I, Kordowich S, Holzwarth C, Spano C, Isensee G, Staiber A, Viebahn S, Gieseke F, Langer H, Gawaz MP, Horwitz EM, Conte P, Handgretinger R, Dominici M. Animal serum-free culture conditions for isolation and expansion of multipotent mesenchymal stromal cells from human BM. Cytotherapy. 2006;8:437-44. Muller-Sieburg CE, Whitlock CA, Weissman IL. Isolation of two early B lymphocyte progenitors from mouse marrow: a committed pre-pre-B cell and a clonogenic Thy-1-lo hematopoietic stem cell. Cell. 1986;44:653-62. Muraglia A, Cancedda R, Quarto R. Clonal mesenchymal progenitors from human bone marrow differentiate in vitro according to a hierarchical model. J Cell Sci. 2000; 113:1161-6. Muraglia A, Corsi A, Riminucci M, Mastrogiacomo M, Cancedda R, Bianco P, Quarto R. Formation of a chondro-osseous rudiment in micromass cultures of human bone-marrow stromal cells. J Cell Sci. 2003;116:2949-55. Nehls V, Drenckhahn D. Heterogeneity of microvascular pericytes for smooth muscle type alpha-actin. J Cell Biol. 1991;113:147-54. Neuhuber B, Gallo G, Howard L, Kostura L, Mackay A, Fischer I. Reevaluation of in vitro differentiation protocols for bone marrow stromal cells: disruption of actin cytoskeleton induces rapid morphological changes and mimics neuronal phenotype. J Neurosci Res. 2004;77:192-204. Nilsson SK, Johnston HM, Whitty GA, Williams B, Webb RJ, Denhardt DT, Bertoncello I, Bendall LJ, Simmons PJ, Haylock DN. Osteopontin, a key component of the hematopoietic stem cell niche and regulator of primitive hematopoietic progenitor cells. Blood. 2005;106:1232-9.

106
Nomiyama T, Perez-Tilve D, Ogawa D, Gizard F, Zhao Y, Heywood EB, Jones KL, Kawamori R, Cassis LA, Tschöp MH, Bruemmer D. Osteopontin mediates obesity-induced adipose tissue macrophage infiltration and insulin resistance in mice. J Clin Invest. 2007;117:2877-88. Nuttall ME, Gimble JM. Controlling the balance between osteoblastogenesis and adipogenesis and the consequent therapeutic implications. Curr Opin Pharmacol. 2004;4:290-4. Odegaard JI, Ricardo-Gonzalez RR, Goforth MH, Morel CR, Subramanian V, Mukundan L, Eagle AR, Vats D, Brombacher F, Ferrante AW, Chawla A. Macrophage-specific PPARgamma controls alternative activation and improves insulin resistance. Nature. 2007;447:1116-20. Oettgen P. Transcriptional regulation of vascular development. Circ Res. 2001;89:380-8. Ohishi K, Varnum-Finney B, Bernstein ID. Delta-1 enhances marrow and thymus repopulating ability of human CD34(+)CD38(-) cord blood cells. J Clin Invest. 2002;110:1165-74. Ohneda O, Ohneda K, Arai F, Lee J, Miyamoto T, Fukushima Y, Dowbenko D, Lasky LA, Suda T. ALCAM (CD166): its role in hematopoietic and endothelial development. Blood. 2001;98:2134-42. Okamoto R, Yajima T, Yamazaki M, Kanai T, Mukai M, Okamoto S, Ikeda Y, Hibi T, Inazawa J, Watanabe M. Damaged epithelia regenerated by bone marrow-derived cells in the human gastrointestinal tract. Nat Med. 2002;8:1011-7. Pardanaud L, Luton D, Prigent M, Bourcheix LM, Catala M, Dieterlen-Lievre F. Two distinct endothelial lineages in ontogeny, one of them related to hemopoiesis. Development. 1996;122:1363-71. Peled A, Petit I, Kollet O, Magid M, Ponomaryov T, Byk T, Nagler A, Ben-Hur H, Many A, Shultz L, Lider O, Alon R, Zipori D, Lapidot T. Dependence of human stem cell engraftment and repopulation of NOD/SCID mice on CXCR4. Science. 1999;283:845-8. Pelosi E, Valtieri M, Coppola S, Botta R, Gabbianelli M, Lulli V, Marziali G, Masella B, Müller R, Sgadari C, Testa U, Bonanno G, Peschle C. Identification of the hemangioblast in postnatal life. Blood. 2002;100:3203-8. Pérez-Pomares JM, Muñoz-Chápuli R. Epithelial-mesenchymal transitions: a mesodermal cell strategy for evolutive innovation in Metazoans. Anat Rec. 2002;268:343-51. Perin EC, Dohmann HF, Borojevic R, Silva SA, Sousa AL, Mesquita CT, Rossi MI, Carvalho AC, Dutra HS, Dohmann HJ, Silva GV, Belém L, Vivacqua R, Rangel FO, Esporcatte R, Geng YJ, Vaughn WK, Assad JA, Mesquita ET, Willerson JT. Transendocardial, autologous bone marrow cell transplantation for severe, chronic ischemic heart failure. Circulation. 2003;107:2294-302. Phinney DG, Prockop DJ. Concise Review: Mesenchymal Stem/Multi-Potent Stromal Cells (MSCs): The State of Transdifferentiation and Modes of Tissue Repair - Current Views. Stem Cells. 2007; 25: 2896-902.

107
Pietilä I, Vainio S. The embryonic aorta-gonad-mesonephros region as a generator of haematopoietic stem cells. APMIS. 2005;113:804-12. Pittenger MF, Mackay AM, Beck SC, Jaiswal RK, Douglas R, Mosca JD, Moorman MA, Simonetti DW, Craig S, Marshak DR. Multilineage potential of adult human mesenchymal stem cells. Science. 1999;284:143-7. Pittenger MF, Martin BJ. Mesenchymal stem cells and their potential as cardiac therapeutics. Circ Res. 2004;95:9-20. Planat-Benard V, Silvestre JS, Cousin B, André M, Nibbelink M, Tamarat R, Clergue M, Manneville C, Saillan-Barreau C, Duriez M, Tedgui A, Levy B, Pénicaud L, Casteilla L. Plasticity of human adipose lineage cells toward endothelial cells: physiological and therapeutic perspectives. Circulation. 2004a; 109:656-63. Planat-Bénard V, Menard C, André M, Puceat M, Perez A, Garcia-Verdugo JM, Pénicaud L, Casteilla L. Spontaneous cardiomyocyte differentiation from adipose tissue stroma cells. Circ Res. 2004b; 94:223-9. Pluznik DH, Sachs L. The cloning of normal "mast" cells in tissue culture. J Cell Physiol. 1965;66:319-24. Ponomaryov T, Peled A, Petit I, Taichman RS, Habler L, Sandbank J, Arenzana-Seisdedos F, Magerus A, Caruz A, Fujii N, Nagler A, Lahav M, Szyper-Kravitz M, Zipori D, Lapidot T. Induction of the chemokine stromal-derived factor-1 following DNA damage improves human stem cell function. J Clin Invest. 2000;106:1331-9. Ponte AL, Marais E, Gallay N, Langonné A, Delorme B, Hérault O, Charbord P, Domenech J. The in vitro migration capacity of human bone marrow mesenchymal stem cells: comparison of chemokine and growth factor chemotactic activities. Stem Cells. 2007;25:1737-45. Pouget C, Gautier R, Teillet MA, Jaffredo T. Somite-derived cells replace ventral aortic hemangioblasts and provide aortic smooth muscle cells of the trunk. Development. 2006;133:1013-22. Prockop DJ. Marrow stromal cells as stem cells for nonhematopoeitic tissues. Science. 1997; 276:71-4. Prophet EB, Mills B, Arrington JB, Sobin LH. Laboratory Methods in Histotechnology., American Registry of Pathology, Segunda edição, Washington – 1994, chapter 19, p178. Prunet-Marcassus B, Cousin B, Caton D, André M, Pénicaud L, Casteilla L. From heterogeneity to plasticity in adipose tissues: site-specific differences. Exp Cell Res. 2006;312:727-36. Punzel M, Liu D, Zhang T, Eckstein V, Miesala K, Ho AD. The symmetry of initial divisions of human hematopoietic progenitors is altered only by the cellular microenvironment. Exp Hematol. 2003;31:339-47. Quaini F, Urbanek K, Beltrami AP, Finato N, Beltrami CA, Nadal-Ginard B, Kajstura J, Leri A, Anversa P. Chimerism of the transplanted heart. N Engl J Med. 2002;346:5-15.

108
Quesenberry PJ, Crittenden RB, Lowry P, Kittler EW, Rao S, Peters S, Ramshaw H, Stewart FM. In vitro and in vivo studies of stromal niches. Blood Cells. 1994;20:97-104. Quesenberry, P.J, Colvin, G.A. Hematopoietic stem cells, progenitor cells, and cytokines. In: Beutler E, Litchman MA, Coller BS, Kipps TJ, Seligsohn U, Williams Hematology – Sexta edição, McGraw-Hill – New York. 2001. p153. Quirici N, Soligo D, Bossolasco P, Servida F, Lumini C, Deliliers GL. Isolation of bone marrow mesenchymal stem cells by anti-nerve growth factor receptor antibodies. Exp Hematol. 2002;30:783-91. Rajala MW, Scherer PE. Minireview: The adipocyte--at the crossroads of energy homeostasis, inflammation, and atherosclerosis. Endocrinology. 2003;144:3765-73. Randolph GJ, Sanchez-Schmitz G, Liebman RM, Schäkel K. The CD16(+) (FcgammaRIII(+)) subset of human monocytes preferentially becomes migratory dendritic cells in a model tissue setting. J Exp Med. 2002; 196: 517-27. Rehman J, Traktuev D, Li J, Merfeld-Clauss S, Temm-Grove CJ, Bovenkerk JE, Pell CL, Johnstone BH, Considine RV, March KL. Secretion of angiogenic and antiapoptotic factors by human adipose stromal cells. Circulation. 2004;109:1292-8. Reya T, Morrison SJ, Clarke MF, Weissman IL. Stem cells, cancer, and cancer stem cells. Nature. 2001;414:105-11. Reyes M, Lund T, Lenvik T, Aguiar D, Koodie L, Verfaillie CM. Purification and ex vivo expansion of postnatal human marrow mesodermal progenitor cells. Blood. 2001;98:2615-25. Ringdén O, Uzunel M, Rasmusson I, Remberger M, Sundberg B, Lönnies H, Marschall HU, Dlugosz A, Szakos A, Hassan Z, Omazic B, Aschan J, Barkholt L, Le Blanc K. Mesenchymal stem cells for treatment of therapy-resistant graft-versus-host disease. Transplantation. 2006;81:1390-7. Robey PG, Bianco P. The use of adult stem cells in rebuilding the human face. J Am Dent Assoc. 2006;137:961-72. Rohde E, Malischnik C, Thaler D, Maierhofer T, Linkesch W, Lanzer G, Guelly C, Strunk D. Blood monocytes mimic endothelial progenitor cells. Stem Cells. 2006;24:357-67. Rossi MI, Barros AP, Baptista LS, Garzoni LR, Meirelles MN, Takiya CM, Pascarelli BM, Dutra HS, Borojevic R. Multicellular spheroids of bone marrow stromal cells: a three-dimensional in vitro culture system for the study of hematopoietic cell migration. Braz J Med Biol Res. 2005;38:1455-62. Rzonca SO, Suva LJ, Gaddy D, Montague DC, Lecka-Czernik B. Bone is a target for the antidiabetic compound rosiglitazone. Endocrinology. 2004;145:401-6. Sacchetti B, Funari A, Michienzi S, Di Cesare S, Piersanti S, Saggio I, Tagliafico E, Ferrari S, Robey PG, Riminucci M, Bianco P. Self-renewing osteoprogenitors in bone marrow sinusoids can organize a hematopoietic microenvironment. Cell. 2007;131:324-36.

109
Schächinger V, Erbs S, Elsässer A, Haberbosch W, Hambrecht R, Hölschermann H, Yu J, Corti R, Mathey DG, Hamm CW, Süselbeck T, Assmus B, Tonn T, Dimmeler S, Zeiher AM; REPAIR-AMI Investigators. Intracoronary bone marrow-derived progenitor cells in acute myocardial infarction. N Engl J Med. 2006;355:1210-21. Schäffler A, Büchler C. Concise review: adipose tissue-derived stromal cells--basic and clinical implications for novel cell-based therapies. Stem Cells. 2007;25:818-27. Schäffler A, Schölmerich J, Salzberger B. Adipose tissue as an immunological organ: Toll-like receptors, C1q/TNFs and CTRPs. Trends Immunol. 2007;28:393-9. Schipani E, Ryan HE, Didrickson S, Kobayashi T, Knight M, Johnson RS. Hypoxia in cartilage: HIF-1alpha is essential for chondrocyte growth arrest and survival. Genes Dev. 2001;15:2865-76. Schor AM, Allen TD, Canfield AE, Sloan P, Schor SL. Pericytes derived from the retinal microvasculature undergo calcification in vitro. J Cell Sci. 1990;97:449-61. Serafini M, Dylla SJ, Oki M, Heremans Y, Tolar J, Jiang Y, Buckley SM, Pelacho B, Burns TC, Frommer S, Rossi DJ, Bryder D, Panoskaltsis-Mortari A, O'Shaughnessy MJ, Nelson-Holte M, Fine GC, Weissman IL, Blazar BR, Verfaillie CM. Hematopoietic reconstitution by multipotent adult progenitor cells: precursors to long-term hematopoietic stem cells. J Exp Med. 2007;204:129-39. Shi S, Gronthos S. Perivascular niche of postnatal mesenchymal stem cells in human bone marrow and dental pulp. J Bone Miner Res. 2003;18:696-704. Shih DT, Lee DC, Chen SC, Tsai RY, Huang CT, Tsai CC, Shen EY, Chiu WT. Isolation and characterization of neurogenic mesenchymal stem cells in human scalp tissue. Stem Cells. 2005; 23:1012-20. Simmons PJ, Torok-Storb B. Identification of stromal cell precursors in human bone marrow by a novel monoclonal antibody, STRO-1. Blood. 1991a;78:55-62. Simmons PJ, Torok-Storb B. CD34 expression by stromal precursors in normal human adult bone marrow. Blood. 1991b;78:2848-53. Soncin F, Mattot V, Lionneton F, Spruyt N, Lepretre F, Begue A, Stehelin D. VE-statin, an endothelial repressor of smooth muscle cell migration. EMBO J. 2003;22:5700-11. Spees JL, Olson SD, Ylostalo J, Lynch PJ, Smith J, Perry A, Peister A, Wang MY, Prockop DJ. Differentiation, cell fusion, and nuclear fusion during ex vivo repair of epithelium by human adult stem cells from bone marrow stroma. Proc Natl Acad Sci U S A. 2003;100:2397-402. Spivak JM, Hasharoni A. Use of hydroxyapatite in spine surgery. Eur Spine J. 2001;10 Suppl 2:S197-204 Spradling A, Drummond-Barbosa D, Kai T. Stem cells find their niche. Nature. 2001;414:98-104. Stashower M, Smith K, Williams J, Skelton H. Stromal progenitor cells present within liposuction and reduction abdominoplasty fat for autologous transfer to aged skin. Dermatol Surg. 1999;25:945-9.

110
Strauer BE, Brehm M, Zeus T, Köstering M, Hernandez A, Sorg RV, Kögler G, Wernet P. Repair of infarcted myocardium by autologous intracoronary mononuclear bone marrow cell transplantation in humans. Circulation. 2002;106:1913-8. Sumi M, Sata M, Toya N, Yanaga K, Ohki T, Nagai R. Transplantation of adipose stromal cells, but not mature adipocytes, augments ischemia-induced angiogenesis. Life Sci. 2007 Jan 16;80: 559-65. Suri C, Jones PF, Patan S, Bartunkova S, Maisonpierre PC, Davis S, Sato TN, Yancopoulos GD. Requisite role of angiopoietin-1, a ligand for the TIE2 receptor, during embryonic angiogenesis. Cell. 1996; 87:1171-80. Sutherland HJ, Lansdorp PM, Henkelman DH, Eaves AC, Eaves CJ. Functional characterization of individual human hematopoietic stem cells cultured at limiting dilution on supportive marrow stromal layers. Proc Natl Acad Sci U S A. 1990;87:3584-8. Tacke F, Randolph GJ. Migratory fate and differentiation of blood monocyte subsets. Immunobiology. 2006;211:609-18. Tagliafico E, Brunelli S, Bergamaschi A, De Angelis L, Scardigli R, Galli D, Battini R, Bianco P, Ferrari S, Cossu G, Ferrari S. TGFbeta/BMP activate the smooth muscle/bone differentiation programs in mesoangioblasts. J Cell Sci. 2004;117:4377-88. Taichman RS, Emerson SG. The role of osteoblasts in the hematopoietic microenvironment. Stem Cells. 1998;16:7-15. Taichman RS. Blood and bone: two tissues whose fates are intertwined to create the hematopoietic stem-cell niche. Blood. 2005;105:2631-9. Thomas T, Gori F, Khosla S, Jensen MD, Burguera B, Riggs BL. Leptin acts on human marrow stromal cells to enhance differentiation to osteoblasts and to inhibit differentiation to adipocytes. Endocrinology. 1999;140:1630-8. Till u.E, McCulloch, EA. A direct measurement of the radiation sensitivity of normal mouse bone marrow cells. Radiat Res. 1961;14: 213-22. Timmins NE, Dietmair S, Nielsen LK. Hanging-drop multicellular spheroids as a model of tumour angiogenesis. Angiogenesis. 2004;7:97-103. Tintut Y, Alfonso Z, Saini T, Radcliff K, Watson K, Boström K, Demer LL. Multilineage potential of cells from the artery wall. Circulation. 2003;108:2505-10. Tisato V, Naresh K, Girdlestone J, Navarrete C, Dazzi F. Mesenchymal stem cells of cord blood origin are effective at preventing but not treating graft-versus-host disease. Leukemia. 2007;21:1992-9. Tondreau T, Meuleman N, Delforge A, Dejeneffe M, Leroy R, Massy M, Mortier C, Bron D, Lagneaux L. Mesenchymal stem cells derived from CD133-positive cells in mobilized peripheral blood and cord blood: proliferation, Oct4 expression, and plasticity. Stem Cells. 2005;23:1105-12. Trayhurn P. The biology of obesity. Proc Nutr Soc. 2005;64:31-8.

111
Umemoto Y, Tsuji K, Yang FC, Ebihara Y, Kaneko A, Furukawa S, Nakahata T. Leptin stimulates the proliferation of murine myelocytic and primitive hematopoietic progenitor cells. Blood. 1997;90:3438-43. Umezawa A, Tachibana K, Harigaya K, Kusakari S, Kato S, Watanabe Y, Takano T. Colony-stimulating factor 1 expression is down-regulated during the adipocyte differentiation of H-1/A marrow stromal cells and induced by cachectin/tumor necrosis factor. Mol Cell Biol. 1991;11:920-7. Van Den Berg DJ, Sharma AK, Bruno E, Hoffman R. Role of members of the Wnt gene family in human hematopoiesis. Blood. 1998;92:3189-202. Verfaillie, C.M. Anatomy and Physiology of hematopoiesis. In: Hoffman R, Benz EJ, Shattil SJ, Furie B, Cohen HJ, Silberstein LE, McGlave P, Hematology – Basic Principles and Pratice –Terceira Edição, Churchill Livingstone – Philadelphia. 2000. p139. Vojtassák J, Danisovic L, Kubes M, Bakos D, Jarábek L, Ulicná M, Blasko M. Autologous biograft and mesenchymal stem cells in treatment of the diabetic foot. Neuro Endocrinol Lett. 2006;27:134-7. von Tell D, Armulik A, Betsholtz C. Pericytes and vascular stability. Exp Cell Res. 2006;312:623-9. Wagner RC. Endothelial cell embryology and growth. Adv.Microcirc. 1980; 9: 45-75. Wagner W, Wein F, Seckinger A, Frankhauser M, Wirkner U, Krause U, Blake J, Schwager C, Eckstein V, Ansorge W, Ho AD. Comparative characteristics of mesenchymal stem cells from human bone marrow, adipose tissue, and umbilical cord blood. Exp Hematol. 2005;33:1402-16. Wagner W, Roderburg C, Wein F, Diehlmann A, Frankhauser M, Schubert R, Eckstein V, Ho AD. Molecular and secretory profiles of human mesenchymal stromal cells and their abilities to maintain primitive hematopoietic progenitors. Stem Cells. 2007; 25: 2638-47. Wajchenberg BL. Subcutaneous and visceral adipose tissue: their relation to the metabolic syndrome. Endocr Rev. 2000;21:697-738 Wang JF, Liu ZY, Groopman JE. The alpha-chemokine receptor CXCR4 is expressed on the megakaryocytic lineage from progenitor to platelets and modulates migration and adhesion. Blood. 1998;92:756-64. Wang JF, Wang LJ, Wu YF, Xiang Y, Xie CG, Jia BB, Harrington J, McNiece IK. Mesenchymal stem/progenitor cells in human umbilical cord blood as support for ex vivo expansion of CD34(+) hematopoietic stem cells and for chondrogenic differentiation. Haematologica. 2004;89:837-44. Wang JS, Shum-Tim D, Galipeau J, Chedrawy E, Eliopoulos N, Chiu RC. Marrow stromal cells for cellular cardiomyoplasty: feasibility and potential clinical advantages. J Thorac Cardiovasc Surg. 2000;120:999-1005.

112
Watanabe S, Morisaki N, Tezuka M, Fukuda K, Ueda S, Koyama N, Yokote K, Kanzaki T, Yoshida S, Saito Y. Cultured retinal pericytes stimulate in vitro angiogenesis of endothelial cells through secretion of a fibroblast growth factor-like molecule. Atherosclerosis. 1997;130:101-7. Weisberg SP, McCann D, Desai M, Rosenbaum M, Leibel RL, Ferrante AW Jr. Obesity is associated with macrophage accumulation in adipose tissue. J Clin Invest. 2003;112:1796-808. Weissman IL. Translating stem and progenitor cell biology to the clinic: barriers and opportunities. Science. 2000;287:1442-6. Wellen KE, Hotamisligil GS. Obesity-induced inflammatory changes in adipose tissue. J Clin Invest. 2003;112:1785-8. Willert K, Brown JD, Danenberg E, Duncan AW, Weissman IL, Reya T, Yates JR 3rd, Nusse R. Wnt proteins are lipid-modified and can act as stem cell growth factors. Nature. 2003;423:448-52. Wilson A, Trumpp A. Bone-marrow haematopoietic-stem-cell niches. Nat Rev Immunol. 2006; 6:93-106. Winter A, Breit S, Parsch D, Benz K, Steck E, Hauner H, Weber RM, Ewerbeck V, Richter W. Cartilage-like gene expression in differentiated human stem cell spheroids: a comparison of bone marrow-derived and adipose tissue-derived stromal cells. Arthritis Rheum. 2003;48:418-29. Wollert KC, Meyer GP, Lotz J, Ringes-Lichtenberg S, Lippolt P, Breidenbach C, Fichtner S, Korte T, Hornig B, Messinger D, Arseniev L, Hertenstein B, Ganser A, Drexler H. Intracoronary autologous bone-marrow cell transfer after myocardial infarction: the BOOST randomised controlled clinical trial. Lancet. 2004 Jul 10-16;364(9429):141-8. Wosnitza M, Hemmrich K, Groger A, Gräber S, Pallua N. Plasticity of human adipose stem cells to perform adipogenic and endothelial differentiation. Differentiation. 2007; 75:12-23. Wright DE, Bowman EP, Wagers AJ, Butcher EC, Weissman IL. Hematopoietic stem cells are uniquely selective in their migratory response to chemokines. J Exp Med. 2002;195:1145-54. Wu Y, Chen L, Scott PG, Tredget EE. Mesenchymal stem cells enhance wound healing through differentiation and angiogenesis. Stem Cells. 2007;25:2648-59. Wuthier RE. Involvement of cellular metabolism of calcium and phosphate in calcification of avian growth plate cartilage. J Nutr. 1993;123:301-9. Wynn RF, Hart CA, Corradi-Perini C, O'Neill L, Evans CA, Wraith JE, Fairbairn LJ, Bellantuono I. A small proportion of mesenchymal stem cells strongly expresses functionally active CXCR4 receptor capable of promoting migration to bone marrow. Blood. 2004;104:2643-5. Xu H, Barnes GT, Yang Q, Tan G, Yang D, Chou CJ, Sole J, Nichols A, Ross JS, Tartaglia LA, Chen H. Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J Clin Invest. 2003;112:1821-30.

113
Yañez R, Lamana ML, García-Castro J, Colmenero I, Ramírez M, Bueren JA. Adipose tissue-derived mesenchymal stem cells have in vivo immunosuppressive properties applicable for the control of the graft-versus-host disease. Stem Cells. 2006;24:2582-91. Yokota T, Oritani K, Takahashi I, Ishikawa J, Matsuyama A, Ouchi N, Kihara S, Funahashi T, Tenner AJ, Tomiyama Y, Matsuzawa Y. Adiponectin, a new member of the family of soluble defense collagens, negatively regulates the growth of myelomonocytic progenitors and the functions of macrophages. Blood. 2000;96:1723-32. Yokota T, Meka CS, Medina KL, Igarashi H, Comp PC, Takahashi M, Nishida M, Oritani K, Miyagawa J, Funahashi T, Tomiyama Y, Matsuzawa Y, Kincade PW. Paracrine regulation of fat cell formation in bone marrow cultures via adiponectin and prostaglandins. J Clin Invest. 2002;109:1303-10. Zannettino AC, Paton S, Arthur A, Khor F, Itescu S, Gimble JM, Gronthos S. Multipotential human adipose-derived stromal stem cells exhibit a perivascular phenotype in vitro and in vivo. J Cell Physiol. 2008; 214: 413-21. Zeyda M, Stulnig TM. Adipose tissue macrophages. Immunol Lett. 2007; 112: 61-7. Zengin E, Chalajour F, Gehling UM, Ito WD, Treede H, Lauke H, Weil J, Reichenspurner H, Kilic N, Ergün S. Vascular wall resident progenitor cells: a source for postnatal vasculogenesis. Development. 2006;133:1543-51. Zhang J, Niu C, Ye L, Huang H, He X, Tong WG, Ross J, Haug J, Johnson T, Feng JQ, Harris S, Wiedemann LM, Mishina Y, Li L. Identification of the haematopoietic stem cell niche and control of the niche size. Nature. 2003;425:836-41. Zhao RC, Liao L, Han Q. Mechanisms of and perspectives on the mesenchymal stem cell in immunotherapy. J Lab Clin Med. 2004;143:284-91. Zhou HR, Kim EK, Kim H, Claycombe KJ. Obesity-associated mouse adipose stem cell secretion of monocyte chemotactic protein-1 (MCP-1). Am J Physiol Endocrinol Metab. 2007; 293: E1153-8.
Zuk PA, Zhu M, Mizuno H, Huang J, Futrell JW, Katz AJ, Benhaim P, Lorenz HP, Hedrick MH. Multilineage cells from human adipose tissue: Implications for cell-based therapies. Tissue Eng. 2001;7:211-28. Zuk PA, Zhu M, Ashjian P, De Ugarte DA, Huang JI, Mizuno H, Alfonso ZC, Fraser JK, Benhaim P, Hedrick MH. Human adipose tissue is a source of multipotent stem cells. Mol Biol Cell. 2002;13:4279-95.

ANEXO




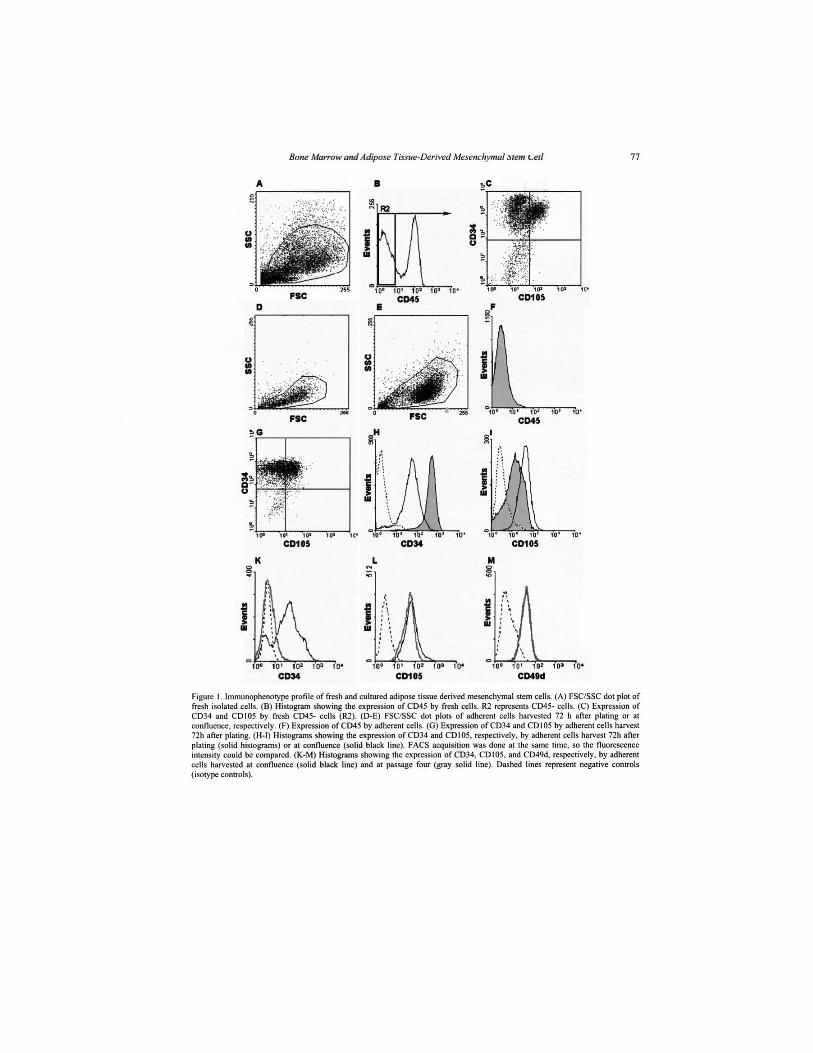












Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo