Embed Size (px)
Citation preview

1
UNIVERSIDADE FEDERAL DE MINAS GERAIS ESCOLA DE VETERINÁRIA
Colegiado dos Cursos de Pós-Graduação
CÉLULAS TRONCO MESENQUIMAIS DE CÃES ASSOCIADAS OU NÃO À VIDRO BIOATIVO, DIFERENCIAÇÃO OSTEOGÊNICA IN VITRO
E TRATAMENTO DE DEFEITO ÓSSEO CRÍTICO
ENDRIGO GABELLINI LEONEL ALVES
BELO HORIZONTE ESCOLA DE VETERINÁRIA – UFMG
2013

2
ENDRIGO GABELLINI LEONEL ALVES
CÉLULAS TRONCO MESENQUIMAIS DE CÃES
ASSOCIADAS OU NÃO À VIDRO BIOATIVO,
DIFERENCIAÇÃO OSTEOGÊNICA IN VITRO
E TRATAMENTO DE DEFEITO ÓSSEO CRÍTICO
Belo Horizonte
Escola de Veterinária da UFMG
2013
Tese apresentada à Escola de Veterinária da Universidade Federal de Minas Gerais como requisito parcial para obtenção do grau de Doutor em Ciência Animal Área de concentração: Medicina e Cirurgia Veterinárias Orientadora:
Profª Dra. Cleuza Maria de Faria Rezende Co-orientadores: Profa. Dra. Rogéria Serakides
Prof. Dr. Humberto Pereira Oliveira

3
Alves, Endrigo Gabellini Leonel, 1980- A474c Células tronco mesenquimais de cães associadas ou não à vidro bioativo, Diferenciação osteogenica in vitro e tratamento de defeitos ósseos críticos / Endrigo Gabellini Leonel Alves. – 2013. 108 p. : il. Orientadora: Cleuza Maria de Faria Rezende Co-orientadores: Rogéria Serakides, Humberto Pereira Oliveira Tese (doutorado) – Universidade Federal de Minas Gerais, Escola de Veterinária. Inclui bibliografia.
1. Cão – Doenças – Teses. 2. Células-tronco – Teses. 3. Ossos – Regeneração Teses. 4. Ortopedia veterinária – Teses. I. Rezende, Cleuza Maria de Faria. II. Serakides, Rogéria. III. Oliveira, Humberto Pereira. IV. Universidade Federal de Minas Gerais. Escola de Veterinária. V. Título.
CDD- 636.708 973

4
Tese defendida e aprovada em 27 de maio de 2013, pela Comissão Examinadora constituída
por:

5

6
Por todo apoio, carinho e amizade, dedico esse trabalho a minha família
Especialmente
À minha esposa Isabel
Aos meus pais José Walter e Beatriz
Aos meus avos Wilson e Aparecida
Ao meu irmão Enzo
E ao meu afilhado Gabriel

7
Agradecimentos
Agradeço a Deus força maior que rege o equilíbrio de nossas vidas. Obrigado pela paciência e
perseverança.
À minha orientadora profa. Cleuza Maria de Faria Rezende por sua dedicação, amizade e pelo
inestimável conhecimento transmitido.
À minha coorientadora profa. Rogéria Serakides pela oportunidade de trabalhar com células
tronco e por sua orientação e participação ativa nesse trabalho durante todas as fases, da
elaboração do projeto às correções finais da tese.
Ao meu coorientador prof. Humberto Pereira Oliveira pela ajuda nesse estudo, pelos
ensinamentos e pelas correções sempre pertinentes.
À professora Marivalda de Magalhães Pereira e ao Professor Alfredo Miranda de Goes pela boa
vontade e prontidão em ajudar sempre que solicitado. Considero esses dois professores meus
coorientadores extraoficiais, sem a ajuda deles não seria possível desenvolver esse trabalho.
Aos professores Duvaldo Eurides, Betânia Souza Monteiro, Natália de Melo Ocarino por
aceitarem participar da banca de defesa desta tese.
À minha esposa Isabel Rodrigues Rosado pela participação ativa durante todos os experimentos,
pelo carinho, paciência e companheirismo.
À amiga profa. Jankerle Neves Boeloni, pessoa que me ensinou tudo que sei sobre cultura
celular e técnicas de biologia molecular, sempre com boa vontade e disposição para trabalhar.
Aos amigos e colegas de pós graduação prof. Omar Leonardo Aristizabal Paez, Jéssica
Alejandra Castro Varón, Felipe Nemer Machado e prof. Rau Fernando Silva Molano, pela
participação ativa nas cirurgias. Sem vocês não seria possível realizar esse trabalho.
A amiga e colega de pós graduação profa. Fabíola Bono Fukushima pelas anestesias dos cães e
por planejar brilhantemente o estudo anestésico.
À profa. Eliane Gonçalves de Melo pela disposição em ajuda na solução dos problemas
referentes ao hospital veterinário e ao centro de experimentação de pequenos animais. Também
por conseguir todos os exames de Leishimaniose para os cães do experimento.
A profa. Natália de Melo Ocarino por ajudar sempre que solicitado.
Ao prof. Roberto Baracat de Araújo por doar matérias para a realização do experimento.
À aluna de iniciação científica Natália Oliveira Freire pelo apoio no experimento e pelos
cuidados com os cães.
Aos colegas de pós graduação Rodrigo dos Santos Horta e Mariana da Silva Figueiredo, pelos
cuidados analgésicos pós operatórios dos cães.
Ao Professor Mario Jefferson Quirino Louzada pela orientação quanto às análises
densitométrica e por disponibilizar o Laboratório de Biofísica da UNESP Aracatuba, para
realização dessas análises.

8
Aos amigos e colegas de pós-graduação Júneo de Freitas Silva e Pablo Herthel de Carvalho por
ajudar sempre que solicitado.
Às doutoras Alexandra Rodrigues Pereira da Silva e Agda Aline Rocha de Oliveira pela grande
ajuda com a produção do biomaterial utilizado no experimento.
As colegas de pós graduação Ana Cláudia Chagas de Paula, Alessandra Arcoverde Cavalcanti
Zonar e Juliana Lott Carvalho pelo auxílio na caracterização fenotípica das células estudadas
nessa tese.
A todos os demais professores da pós-graduação que com certeza me ajudaram de alguma forma
para minha formação.
Aos meus pais José Walter Leonel Alves e Beatriz Gabellini Alves pelo amor incondicional e
grande incentivo aos estudos. Também por cuidarem dos cães até que eles atingissem a idade
para entrar no experimento.
Aos meus avós Wilson Gabellini e Apparecida Straiotto Gabellini pela amizade, amor e pelos
sábios ensinamentos. Também por cuidarem dos cães até que eles atingissem a idade para entrar
no experimento.
Aos meus sogros Sebastião Carlo da Silva Rosado e Ivonilde Rodrigues Rosado por cuidarem
dos cães até que eles atingissem a idade para entrar no experimento.
Ao grande amigo e colega de profissão Marcus Vinícius Caetano de Souza por conseguir os
cães do experimento e por acompanha-los durante todo o período pré experimental cuidando e
medicando sempre que necessário.
Aos funcionários do hospital veterinário Carlos, Tamires, Terezinha, Ronaldo, Creide, Liu e
Cleiton, que sempre estiveram prontos para ajudar.
Às Enfermeiras do Hospital Veterinário Ana e Sílvia pela colaboração na execução deste
trabalho junto ao bloco cirúrgico e setor de esterilização.
Aos funcionários do setor de patologia Leimar e Adão que sempre ajudaram quando solicitado.
Às funcionárias da secretaria Eliane, Rosangela, Lurdes, que muito me ajudaram com as tarefas
burocráticas.
Aos técnicos em radiologia Elias, Eduardo e Eli pela cooperação na realização dos exames
radiográficos.
À empresa OUROFINO AGRONEGÓCIOS, em especial, à Lídia Marinho Silva Lima
supervisora de registro de produtos pela doação de medicamentos e produtos veterinários para a
utilização nesse estudo.
À empresa PIONEIRA, em especial, à Germano Jeycic pela doação de medicamentos e
produtos medico hospitalares para utilização no experimento.
À empresa CENTRALVET TERAPEÊUTICA INTELIGENTE, em especial, a Ronald
Glanzmann, pela doação de todo o propofol utilizado no presente estudo.
À empresa ROYAL CANNIN, em especial, a Luciana Domingues de Oliveira promotora de
vendas dessa empresa, pela doação de 2420 kg de ração para os animais do presente estudo.

9
À empresa KOWALSKI ALIMENTOS, em especial, a Cristino Motta supervisor de vendas
dessa empresa, pela doação de 600 Kg de ração para os animais do presente estudo.
À empresa LUPUS ALIMENTOS, em especial, a Vivian Aro técnica dessa empresa, pela
doação de 1000 Kg de ração para os animais do presente estudo.
À empresa DB, em especial, a Reinaldo Bispo executivo de vendas dessa empresa, pela doação
de seringas e agulhas para serem utilizadas nesse estudo.
À empresa CRISTÁLIA, pela doação de todo o isofluorano utilizado no presente estudo.
À empresa BIOCONECTVET, em especial Vanderlei A. Pimenta pela doação dos preparadores
de rosca utilizados no presente estudo.
À empresa HERTAPE CALIER, pela doação de 160 doses da vacina MULTI-DOG para a
imunização dos cães desse estudo.
À FAPMIG pelo financiamento do estudo.
Ao CNPq pela concessão da bolsa de estudos.
Aos animais utilizados neste experimento pela doçura mesmo em momentos desconfortáveis.
Ao meu irmão Enzo Gabellini Leonel Alves por ser meu melhor amigo e companheiro de todas
as horas, me apoiando sempre que preciso.

10
Lista de abreviações
BCIP - 5-bromo-4-chloro-3’-indolylphosphate p-toluidine salt
BMP – Proteína ósseas morfogênicas
BSA – Albumina sérica bovina
BSP – Sialoproteina óssea
CaSR - Calcium sensing receptor
CTM- Células tronco mesenquimais
CTM-AD – Células tronco mesenquimais derivadas do tecido adipose
CTM-MO – Células tronco mesenquimais derivadas da medula óssea
DMEM – Dulbecco’s modiffed eagle medium ou meio basal
DMEM PI – Meio basal suplementado como produto iônico
FA – Fosfatase alcalina
FACS - Fluorescence-activated cell sorting
FoxO - Forkhead homeobox tipo O
GAPDH - Gliceraldeído-3-fosfato desidrogenase
HA - Hidroxiapatita
MAPK - Proteína quinase ativada por mitógeno
MOD – Matriz óssea desmineralizada
MTT - Brometo de [3-(4,5-dimetiltiazol-2yl)-2,5-difenil tetrazolium]
NBT - Nitro-blue tetrazolium chloride
NCC – Número de células por campo
NM – Nódulos de mineralização
OC – Osteocalcina
ON- Osteonectina
OST – Meio de diferenciação osteogênica
OST PI – Meio de diferenciação osteogênica suplementado com o produto iônico
OSX- Osterix
PBS - Solução tampão de fosfato padrão
PRP – Plasma rico em plaquetas
RT-PCR – Reação em cadeia da polimerase em tempo real
SDS - Dodecil sulfato de sódio
SFB – Soro fetal bovino
TEOS- Tetraetilortosilicato
TEP - Trietilfosfato

11
Sumário
Lista de abreviações............................................................................................................... 9
Lista de tabelas....................................................................................................................... 11
Lista de figuras....................................................................................................................... 12
Resumo................................................................................................................................... 18
Abstract................................................................................................................................... 19
Introdução............................................................................................................................... 20
Objetivos................................................................................................................................. 20
Capítulo 1: Revisão de literatura........................................................................................ 21
Defeitos ósseos e alternativas para o seu tratamento............................................................. 18
Células tronco e sua aplicação na ortopedia........................................................................... 25
Potencial proliferativo e de diferenciação osteogênicadas CTM......................................... 31
Capítulo 2: Estudo comparativo da diferenciação osteogênica de células tronco
mesenquimais da medula óssea e de tecido adiposo de cães adultos
35
Introdução............................................................................................................................... 35
Material e métodos................................................................................................................. 35
Resultados............................................................................................................................... 42
Discussão................................................................................................................................ 51
Conclusão............................................................................................................................... 53
Capítulo 3: Efeito do produto iônico do biovidro 60S na diferenciação osteogênica de
células tronco mesenquimais do tecido adiposo de cães
54
Introdução............................................................................................................................... 54
Material e métodos................................................................................................................. 54
Resultados............................................................................................................................... 61
Discussão................................................................................................................................ 65
Conclusão............................................................................................................................... 68
Capítulo 4: Tratamento de defeitos ósseos críticos em rádios de cães com matriz
porosa do biovidro 60S associada ou não a células tronco mesenquimais do tecido
adiposo
69
Introdução............................................................................................................................... 69
Material e Métodos................................................................................................................. 70
Resultados............................................................................................................................... 76
Discussão................................................................................................................................ 86
Conclusão............................................................................................................................... 91
Referências Bibliográficas...................................................................................................... 91
Anexos ................................................................................................................................... 108
Protocolo de aprovação pelo comitê de ética ........................................................................ 108

12
Lista de tabelas
Capítulo 2. Estudo comparativo da diferenciação osteogênica de células tronco
mesenquimais da medula óssea e de tecido adiposo de cães adultos
35
Tabela 1. Lista de genes e sequência de nucleotídeos dos iniciadores para RT-PCR
em tempo real
42
Capítulo 3. Efeito do produto iônico do biovidro 60S na diferenciação osteogênica
de células tronco mesenquimais do tecido adiposo de cães
54
Tabela 1. Lista de genes e sequência de nucleotídeos dos iniciadores para RT-PCR
em tempo real
60
Tabela 2. Concentração média dos íons Si e Ca nas amostras dos meios de cultivo
celular basal (DMEM), meio osteogênico (OST), produto iônico do
biovidro 60S em meio basal (DMEM PI) e em osteogênico (OST PI)
61
Capítulo 4. Tratamento de defeitos ósseos críticos em rádios de cães com matriz
porosa do biovidro 60S associada ou não a células tronco
mesenquimais do tecido adiposo
69
Tabela 1. Frequências de ocorrência de reação periosteal, calos periosteal, medular e
intercortical e de continuidade cortical e medular em defeitos ósseos
críticos em rádios de cães tratados com osso autógeno, observadas aos 0,
15, 30, 45, 60, 75 e 90 dias de pós operatório
80

13
Lista de figuras
Capítulo 2. Estudo comparativo da diferenciação osteogênica de células tronco
mesenquimais da medula óssea e de tecido adiposo de cães adultos
36
Figura 1. Fotomicroscopia de contraste de fase das células tronco mesenquimais
extraídas de medula óssea (A) e do tecido adiposo de cães (B). Observar a
morfologia alongada e fusiforme das células. Barra = 70 µm
43
Figura 2. Histogramas da expressão de CD45, CD34, CD90 e CD29 por citometria
de fluxo em células tronco mesenquimais da medula óssea de cães,
mantidas em meio basal (DMEM) após o terceiro repique e confluência de
80 a 90%. Os histogramas mostram a escala de fluorescência no eixo x
considerada positiva quando o pico de células está acima de 101 ou seja,
toda a área do gráfico que se encontra no intervalo da barra M1 é positiva
43
Figura 3. Histogramas da expressão de CD45, CD34, CD90 e CD29 por citometria
de fluxo em células tronco mesenquimais do tecido adiposo de cães,
mantidas em meio basal (DMEM) após o terceiro repique e confluência de
80 a 90%. Os histogramas mostram a escala de fluorescência no eixo x
considerada positiva quando o pico de células está acima de 101 ou seja,
toda a área do gráfico que se encontra no intervalo da barra M1 é positiva
44
Figura 4. Cultura de células tronco mesenquimais extraídas da medula (1) e do
tecido adiposo (2) de cães submetida à diferenciação osteogênica (A),
adipogênica (B) e condrogênica (C) por 21 dias. Observar nas imagens 1A
e 2A os nódulos de mineralização corados em vermelho pela técnica de
Von Kossa; nas imagens 1B e 2B as vesículas lipídicas com aspecto
birrefringente e coloração avermelhada no interior das células coradas
pela técnica Oil Red e nas imagens 1C e 2C os proteoglicanos corados em
violeta vivo pela técnica de PAS. Barras das imagens 1A e 2A = 350 µm e
das imagens 1B, 2B, 1C e 2C = 8,75 µm.
45
Figura 5. Média e desvio padrão da conversão do MTT em cristais de formazan em
culturas de CTM-MO e CTM-AD de cães, mantidas em meio basal
(DMEM) e meio osteogênico (OST) por sete, 14 e 21 dias.
45
Figura 6. Porcentagem média e desvio padrão da área celular por campo em culturas
de CTM-MO e CTM-AD de cães, mantidas em meio basal (DMEM) e
meio osteogênico (OST) por sete, 14 e 21 dias.
46
Figura 7. Média e desvio padrão da atividade da fosfatase alcalina em culturas de
CTM-MO e CTM-AD de cães, mantidas em meio basal (DMEM) e meio
osteogênico (OST) por sete, 14 e 21 dias.
46
Figura 8. Média e desvio padrão do colágeno em culturas de CTM-MO e CTM-AD
de cães, mantidas em meio basal (DMEM) e meio osteogênico (OST) por
sete, 14 e 21 dias.
47

14
Figura 9. Cultura de células tronco mesenquimais extraídas da medula (1) e do
tecido adiposo (2) de cães submetida à diferenciação osteogênica por sete
(A), 14 (B) e 21 (C) dias. Observar aumento crescente da área
mineralizada durante o cultivo. Os nódulos de mineralização foram
corados pela técnica de Von Kossa. O grupo células tronco extraídas do
tecido adiposo (2) apresentou maior área mineralizada quando comparado
com o grupo células tronco extraídas da medula óssea (1) durante todos os
tempos estudados. Barra = 350 µm
48
Figura 10. Média e desvio padrão da área mineralizada por campo em culturas de
CTM-MO e CTM-AD de cães, mantidas em meio basal (DMEM) e meio
osteogênico (OST) por sete, 14 e 21 dias.
48
Figura 11. Média e desvio padrão da quantificação relativa do transcrito gênico para
osterix pela técnica de RT-PCR em tempo real em culturas de CTM-MO e
CTM-AD de cães, mantidas em meio basal (DMEM) e meio osteogênico
(OST) por sete, 14 e 21 dias. Os dados estão expressos em relação ao
osteoblasto canino mantido em meio basal (DMEM). (linha tracejada).
49
Figura 12. Média e desvio padrão da quantificação relativa do transcrito gênico para
sioloproteína óssea (BSP) pela técnica de RT-PCR em tempo real em
culturas de CTM-MO e CTM-AD de cães, mantidas em meio basal
(DMEM) e meio osteogênico (OST) por sete, 14 e 21 dias. Os dados estão
expressos em relação ao osteoblasto canino mantido em meio basal
(DMEM), o valor de expressão do osteoblasto foi considerado igual a um.
49
Figura 13. Média e desvio padrão da quantificação relativa do transcrito gênico para
osteonectina (ON) pela técnica de RT-PCR em tempo real em culturas de
CTM-MO e CTM-AD de cães, mantidas em meio basal (DMEM) e meio
osteogênico (OST) por sete, 14 e 21 dias. Os dados estão expressos em
relação ao osteoblasto canino mantido em meio basal (DMEM). (linha
tracejada).
50
Figura 14. Média e desvio padrão da quantificação relativa do transcrito gênico para
osteocalcina (OC) pela técnica de RT-PCR em tempo real em culturas de
CTM-MO e CTM-AD de cães, mantidas em meio basal (DMEM) e meio
osteogênico (OST) por sete, 14 e 21 dias. Os dados estão expressos em
relação ao osteoblasto canino normal mantido em meio basal (DMEM).
(linha tracejada).
50
Capítulo 3 Efeito do produto iônico do biovidro 60S na diferenciação osteogênica
de células tronco mesenquimais do tecido adiposo de cães
54
Figura 1. Redução do MTT em cristais de formazan (média e desvio padrão) em
culturas de CTM-AD caninas mantidas em meio osteogênico (OST) e em
produto iônico em meio osteogênico (OST PI) por sete, 14 e 21 dias.
60

15
Figura 2. Atividade da fosfatase alcalina (média ± desvio padrão) em culturas de
CTM-AD caninas mantidas em meio osteogênico (OST) e produto iônico
em meio osteogênico (OST PI) por sete, 14 e 21 dias.
62
Figura 3. Média e desvio padrão da quantidade de colágeno em culturas de CTM-
AD caninas cultivadas em meio osteogênico (OST) e produto iônico em
meio osteogênico (OST PI) por sete, 14 e 21 dias.
62
Figura 4. Cultura de CTM-AD caninas mantidas em meio osteogênico (OST) (1) e
produto iônico em meio osteogênico (2) por sete (A), 14 (B) e 21 (C) dias.
Observar os nódulos de mineralização corados pela técnica de Von Kossa.
O grupo com produto iônico do biovidro 60S OST PI (2) mostrou maior
área de mineralização por campo em relação ao controle OST (1). Barra =
350 µm
63
Figura 5. Percentagem média e desvio padrão da área de mineralização por campo
em culturas de CTM-AD caninas cultivadas em meio osteogênico (OST) e
produto iônico em meio osteogênico (OST PI) por sete, 14 e 21 dias.
63
Figura 6. Média e desvio padrão da quantificação relativa do transcrito gênico para
osterix pela técnica de RT-PCR em tempo real em culturas de CTM-AD
caninas mantidas em meio osteogênico (OST) e produto iônico em meio
osteogênico (OST PI) por sete, 14 e 21 dias. Os dados estão expressos em
relação ao osteoblasto canino mantido em meio basal (DMEM)
representado pela linha pontilhada.
64
Figura 7. Média e desvio padrão da quantificação relativa do transcrito gênico para
sialoproteína óssea (BSP) pela técnica de RT-PCR em tempo real em
culturas de CTM-AD caninas mantidas em meio osteogênico (OST) e
produto iônico em meio osteogênico (OST PI) por sete, 14 e 21 dias. Os
dados estão expressos em relação ao osteoblasto canino mantido em meio
basal (DMEM).
64
Figura 8. Média e desvio padrão da quantificação relativa do transcrito gênico para
osteonectina (ON) pela técnica de RT-PCR em tempo real em culturas de
CTM-AD caninas mantidas em meio osteogênico (OST) e produto iônico
em meio osteogênico (OST PI) por sete, 14 e 21 dias. Os dados estão
expressos em relação ao osteoblasto canino mantido em meio basal
(DMEM), representado pela linha pontilhada.
65
Figura 9. Média e desvio padrão da quantificação relativa do transcrito gênico para
osteocalcina (OC) pela técnica de RT-PCR em tempo real em culturas de
CTM-AD caninas mantidas em meio osteogênico (OST) e produto iônico
em meio osteogênico (OST PI) por sete, 14 e 21 dias. Os dados estão
expressos em relação ao osteoblasto canino mantido em meio basal
(DMEM), representado pela linha pontilhada.
65

16
Capítulo 4 Tratamento de defeitos ósseos críticos em rádios de cães com matriz
porosa do biovidro 60S associada ou não a células tronco
mesenquimais do tecido adiposo
69
Figura 1. Sequencia e fotos da cirurgia de implantação do biovidro 60S, associado
ou não das CTM-AD para tratamento de defeitos ósseos críticos em rádios
de cães. Observar nas imagens A - ostectomia; B – posicionamento da
placa cranial ao osso; C e D – matriz do biovidro 60S com forma
semelhante ao segmento osteotomisado; E – deposição das células na
matriz do biovidro; F - defeito ósseo sem preenchimento (controle
negativo); G – defeito ósseo preenchido com enxerto autógeno (controle
positivo); H – defeito ósseo preenchido com a matriz do biovidro 60S e I -
defeito ósseo preenchido com a matriz do biovidro associada com as
CTM-AD.
74
Figura 2. Imagens radiográficas de defeitos ossos críticos em rádios de cães na
projeção médio-lateral, processadas pelo software Digimizer Image
Analysis para mensuração das áreas de preenchimento ósseo (áreas
contornadas em azul na imagem A), área de preenchimento com
biomaterial (área contornada em verde na imagem A), área total do defeito
(áreas contornadas em vermelho na imagem A), áreas de calo periosteal
(áreas contornadas em azul na imagem B) e medular (áreas contornadas
em vermelho na imagem B)
75
Figura 3. Micrografia uma amostra da matriz porosa do biovidro 60S obtida por
microscopia eletrônica de varredura. Observa a presença de poros
interconectados com diâmetro variado
76
Figura 4. Imagens radiográficas de defeitos ossos críticos em rádios de cães não
tratados (controle negativo – A) e tratados com osso autógeno (controle
positivo – B), obtidas aos 90 dias de pós-operatório. Observar na imagem
A o defeito ósseo parcialmente preenchido com osso esponjoso sem a
formação de ponte óssea entre os fragmentos e na imagem B a completa
regeneração óssea, com continuidade das corticais e do canal medular
77
Figura 5. Percentagem média e desvio padrão da área de biomaterial em defeitos
ósseos críticos em rádios de cães, tratados com matriz porosa de biovidro
60S associada (BV+CTM) ou não (BV) às CTM-AD diferenciadas, ao
longo de 90 dias de avaliação pós-operatória.
77
Figura 6. Imagens radiográficas de defeitos ossos críticos em rádios de cães,
tratados com matriz porosa de biovidro 60S (A) e com a associação da
matriz porosa de biovidro 60S à CTM-AD diferenciadas (B), obtidas aos
90 dias de pós-operatório. Observar a ausência de biovidro na região
correspondente ao defeito ósseo em ambos os casos.
78

17
Figura 7. Percentagem média e desvio padrão da área de preenchimento ósseo em
defeitos críticos de rádios de cães, não tratados (C-) e tratados com CTM-
AD diferenciadas (CTM), matriz porosa de biovidro 60S (BV), matriz
porosa de biovidro 60S associada à CTM-AD diferenciadas (BV+CTM) e
osso autógeno (C+), ao longo de 90 dias de avaliação pós-operatória
78
Figura 8. Imagens radiográficas de defeitos ossos críticos em rádios de cães,
tratados com CTM-AD diferenciadas (A), matriz porosa de biovidro 60S
(B) e matriz porosa de biovidro 60S associada à CTM-AD diferenciadas
(C), obtidas aos 90 dias de pós-operatório. Observar em todas as imagens
o preenchimento parcial do defeito com osso esponjoso das extremidades
em direção ao centro e nas imagens B e C, fragmentos de biomaterial no
centro dos defeitos.
79
Figura 9. Percentagem média e desvio padrão da área de preenchimento ósseo em
defeitos críticos em rádios de cães, não tratados (C-) e tratados com CTM-
AD diferenciadas (CTM), matriz porosa de biovidro 60S (BV), matriz
porosa de biovidro 60S associada à CTM-AD diferenciadas (BV+CTM) e
osso autógeno (C+), ao longo de 90 dias de avaliação pós-operatória.
79
Figura 10. Percentagem média e desvio padrão da área dos calos periosteais e
medulares adjacentes às linhas de osteotomia distal e proximal de defeitos
ósseos críticos em rádios de cães tratados com osso autógeno (C+), ao
longo de 90 dias de avaliação pós-operatória
81
Figura 11. Densidade média e desvio padrão de defeitos ósseos críticos em rádios de
cães, não tratados (C-) e tratados com CTM-AD diferenciadas (CTM),
matriz porosa de biovidro 60S (BV), matriz porosa de biovidro 60S
associada à CTM-AD diferenciadas (BV+CTM) e osso autógeno (C+),
aos 45 e 90 dias de pós-operatório. A densidade está expressa em
percentagem relação à densidade normal do rádio de cada animal.
81
Figura 12. Fragmentos ósseos correspondentes a defeitos ósseos críticos em rádios de
cães, não tratados (1A e 1B) e tratados com matriz porosa de biovidro 60S
(2A e 2B), matriz porosa de biovidro 60S associada à CTM-AD
diferenciadas (3A e 3B), osso autógeno (4A e 4B) e CTM-AD
diferenciadas (5), aos 45 (1A, 2A, 3A e 4A) e 90 (1B, 2B, 3B e 4B) dias
de pós-operatório. Notar nas imagens 1A, 1B, 2A, 2B, 3A, 3B e 5, o
preenchimento parcial do defeito com ósseo esponjoso das extremidades
em direção ao centro (regiões com asteriscos vermelhos); nas imagens 2A,
2B e 3A, a matriz porosa do biovidro 60S no centro do defeito ósseo
envolvida por tecido mole (regiões com estrelas verdes); na imagem 2B,
ausência de biomaterial no centro do defeito; na imagem 4A, os calos
periosteais (apontados pelas setas vermelhas), os calos medulares (nas
regiões com asterisco azul), a presença de calo intercortical na área
circulada em vermelho e a ausência na área circulada em preto; e na
imagem 4B, a continuidade das corticais e da medular.
82

18
Figura 13. Micrografia do osso neoformado nas extremidades de defeitos ósseos
críticos em rádios de cães, não tratados (A) e tratados com matriz porosa
de biovidro 60S (B), matriz porosa de biovidro 60S associada à CTM-AD
diferenciadas (C) e CTM-AD diferenciadas (D). Observar que o osso
neoformado é trabecular, com trabéculas espessas. Barra = 93µm
83
Figura 14. Percentagem média e desvio padrão da área de neoformação óssea em
defeitos críticos em rádios de cães, não tratados (C-) e tratados com CTM-
AD diferenciadas (CTM), matriz porosa de biovidro 60S (BV), matriz
porosa de biovidro 60S associada à CTM-AD diferenciadas (BV+CTM) e
osso autógeno (C+), aos 45 e 90 dias de avaliação pós-operatória (medidas
obtidas por histomorfometria).
84
Figura 15. Micrografia do centro de defeitos ósseos críticos em rádios de cães, não
tratados (A) e tratados com CTM-AD diferenciadas (B), matriz porosa do
biovidro 60S (C), matriz porosa do biovidro 60S associada à CTM-AD
diferenciadas (D), 90 dias após a cirurgia. Observar em todas as imagens a
presença de tecido conjuntivo fibroso e a ausência de tecido ósseo. A
barra corresponde a 93µm.
85
Figura 16. Percentagem média e desvio padrão do número de vasos por campo no
centro de defeitos críticos em rádios de cães, não tratados (C-) e tratados
com CTM-AD diferenciadas (CTM), matriz porosa de biovidro 60S (BV),
matriz porosa de biovidro 60S associada à CTM-AD diferenciadas
(BV+CTM) e osso autógeno (C+), aos 45 e 90 dias de avaliação pós-
operatória (medidas obtidas por histomorfometria).
85
Figura 17. Percentagem média e desvio padrão da área de biomaterial no centro de
defeitos críticos em rádios de cães, tratados com matriz porosa de biovidro
60S (BV) e matriz porosa de biovidro 60S associada à CTM-AD
diferenciadas (BV+CTM), aos 45 e 90 dias de avaliação pós-operatória
(obtido por histomorfometria).
86

19
Resumo
O objetivo deste estudo foi comparar o potencial osteogênico das células tronco mesenquimais
extraídas do tecido adiposo (CTM-AD) com as extraídas da medula óssea (CTM-MO) de cães
adultos, avaliar o efeito do produto iônico do biovidro 60S (BV) na diferenciação osteogênica
das CTM-AD e avaliar o efeito da matriz porosa do BV associada ou não a células tronco em
defeitos ósseos críticos de cães adultos. As CTM-AD e CTM-MO foram submetidas à
diferenciação osteogênica e foram avaliados MTT, fosfatase alcalina (FA), colágeno, número de
células por campo (NCC), nódulos de mineralização (NM) e expressão de osterix (OSX),
sialoproteína óssea (BSP), osteonectina (ON) e osteocalcina (OC). O MTT, a FA, o colágeno, o
NCC e a expressão de ON foram maiores nas CTM-AD o que determinou maior osteogênese in
vitro em relação às CTM-MO. As CTM-AD foram então submetidas à diferenciação
osteogênica na presença e na ausência do produto iônico do BV. Avaliou-se o MTT, a FA, o
colágeno, os NM e a expressão de OSX, BSP, ON e OC. As CTM-AD diferenciadas na
presença do produto iônico do BV mostraram maior conversão de MTT e maior expressão de
BSP o que determinou maior osteogênese in vitro. A matriz porosa do BV, associada ou não as
CTM-AD, foi usada no tratamento de defeitos ósseos críticos em rádios de cães adultos.
Distribuiu-se os cães em três grupos de tratamento (BV, CTM-AD e BV+CTM-AD) e dois
controles, negativo e positivo. A regeneração óssea foi avaliada clínica, radiográfica,
densitometrica e histologicamente, ao longo de 90 dias. Os defeitos tratados com BV,
BV+CTM-AD e CTM-AD mostraram regeneração óssea parcial, semelhante entre si, e inferior
aos defeitos tratados com osso autógeno. Conclui-se que as culturas de CTM-AD apresentam
maior potencial osteogênico que as de CTM-MO “in vitro”, que o produto iônico do biovidro
60S favorece a osteogênese “in vitro” de culturas CTM-AD de cães, que a matriz porosa do BV
associada ou não as CTM-AD não é eficiente no tratamento de defeitos ósseos críticos em
rádios de cães adultos e que as CTM-AD favorecem a neovascularização nesses defeitos.
Palavras Chaves: Células tronco, biomateriais, diferenciação osteogênica, regeneração
óssea, ortopedia, cão.

20
Abstract
This thesis aimed to compare the osteogenic potential of mesenchymal stem cells derived from
adipose tissue (AT-MSC) to those extracted from bone marrow (BM-MSC) of adult dogs, to
evaluate the effect of the ionic product of 60S bioglass (BG) in osteogenic differentiation of
AT-MSC, and to study the effect of BG porous matrix whether or not associated to stem cells in
critical bone defects in adult dogs. AT-MSC and BM-MSC underwent osteogenic
differentiation and MTT, alkaline phosphatase (ALP), collagen, number of cells per field
(NCF), nodules of mineralization (NM), and expression of osterix (OSX), sialoprotein bone
(BSP), osteonectin (ON), and osteocalcin (OC) were evaluated. The MTT, ALP, collagen, NCF,
and the expression of ON were higher in AT-MSC, which showed improved in vitro
osteogenesis compared to BM-MSC. AT-MSC was then subjected to osteogenic differentiation
in the presence and absence of the ionic product of BG. MTT, ALP, collagen, NM, as well as
the expression of the OSX, BSP, ON, and OC were evaluated. AT-MSC differentiated in the
presence of the ionic product of BG showed higher MTT conversion and increased expression
of BSP, which showed improved in vitro osteogenesis. The BG porous matrix, whether or not
associated to AT-MSC, was employed in the treatment of critical bone defects in radius of adult
dogs. Dogs were distributed into three treatment groups (BG, AT-MSC, and BG+AT-MSC) and
two controls, positive and negative. Over 90 days, bone regeneration was clinically,
radiographically, densitometrically, and histologically evaluated. The defects treated with BG,
AT-MSC and BG+AT-MSC showed partial bone regeneration, similar to each other, and lower
to defects treated with autogenous bone. In conclusion, cultures of AT-MSC exhibit higher in
vitro osteogenic potential than in vitro BM-MSC; the ionic product of BG improves in vitro
osteogenesis in AT-MSC cultures of dogs; the BV porous matrix, whether or not associated to
AT-MSC, is not effective in the treatment of critical bone defects in radius of adult dogs; and,
finally, AT-MSC improves neovascularization in these defects.
Keywords: Stem cells, biomaterials, osteogenic differentiation, bone regeneration,
orthopedics, dog.

21
Introdução
Um dos desafios da ortopedia veterinária e
humana é o tratamento de defeitos ósseos
críticos, caracterizados por grandes perdas
ósseas incapazes de se regenerarem
naturalmente sem algum tipo de tratamento
adicional à fixação (Drosse et al., 2008;
Reichert et al., 2009, ASTM 2010). Os
defeitos ósseos críticos são tratados pela
associação de métodos de fixação com
enxertos ósseos, preferencialmente
autógeno. A fixação tem como função
impedir a movimentação dos fragmentos
ósseos no foco da fratura, propiciando
assim um microambiente que permita a
regeneração óssea e, o enxerto favorece a
regeneração do tecido ósseo por meio dos
processos de osteocondução e osteogênese.
Alguns inconvenientes como maior tempo
cirúrgico, complicações no local da
colheita, fragilização do local e a
quantidade limitada de osso coletado estão
associados à utilização dos enxertos
autógenos (Santos e Rahal, 2004;
Piermattei et al., 2006; Sanders, 2007). A
produção de matrizes sintéticas a partir de
biomateriais parece ser a alternativa mais
viável e promissora, especialmente quando
combinada com as terapias celulares e
fatores de crescimento (De Long Jr, 2007;
Dutra et al., 2008; Hench, 2009). Diferentes
tipos de biomaterias podem ser utilizados
para produção das matrizes sintéticas, mas
o grupo das biocerâmicas se destaca, visto
que favorecem a regeneração óssea por
osteocondução (Bruder et al., 1998;
Arinzeh et al., 2003; Yuan et al., 2010) e,
em alguns casos, por osteoindução (Xynos
et al., 2000; Xynos et al., 2001; Habibovic
et al., 2006; Hench, 2009; Hopp et al.,
2011). As matrizes tridimensionais
sintéticas podem ser produzidas com uma
arquitetura que mimetiza o osso esponjoso e
podem ser colonizada “in vitro” por células
de interesse, como células tronco e
osteoblastos (Wang, 2003; Brodke et al.,
2006). A vantagem da terapia com células
tronco mesenquimais (CTM) consiste na
utilização de células do próprio indivíduo
sob tratamento, sem risco de rejeição
(Kraus e Kirker-Head, 2006). As CTM são
células multipotentes, encontradas entre as
células diferenciadas de um tecido e com
função de se auto-renovar e de se
diferenciar, originando os tipos celulares do
tecido no qual residem (Presnell et al.,
2002; Grove et al., 2004). Vários tecidos
são fontes de CTM, em maior ou menor
número mas, a medula óssea e o tecido
adiposo são considerados os principais
sítios de CTM devido a maior quantidade e
facilidade de obtenção dessas células
(Payushina et al., 2006; Rebelatto et al.,
2008). No entanto, ainda existem
controvérsias quanto à melhor fonte de
células tronco para terapia de regeneração
óssea em cães. Dessa forma, o presente
estudo avaliou a matriz porosa do biovidro
60S associada à CTM como terapia de
regeneração óssea em defeitos críticos em
cães. Para isso o estudo foi subdividido em
três etapas. Na primeira, o potencial
osteogênico das células tronco
mesenquimais extraídas da medula óssea
(CTM-MO) e do tecido adiposo (CTM-AD)
foi comparado para se identificar a melhor
fonte de CTM para terapias de regeneração
óssea em cães. Na segunda etapa, foi
avaliado o efeito do produto iônico do
biovidro 60S na diferenciação osteogênica
das CTM oriundas da melhor fonte e na
terceira etapa, avaliou-se o efeito da matriz
porosa do biovidro 60S associado às CTM
no tratamento de defeitos ósseos críticos em
cães.
Objetivos
Comparar o potencial osteogênico das
células tronco mesenquimais extraídas da
medula óssea com as extraídas do tecido
adiposo de cães adultos.
Avaliar in vitro, o efeito do produto iônico
do biovidro 60S nas células tronco
mesenquimais extraídas do tecido adiposo
de cães adultos.
Avaliar o efeito da matriz porosa do
biovidro 60S em associação com as CTM
que obtiveram o melhor potencial
osteogênico no tratamento de defeitos
ósseos críticos em cães.

22
Capítulo 1: Revisão de literatura
Defeitos ósseos e alternativas para o seu
tratamento
Os defeitos ósseos críticos ainda
representam um grande desafio quanto ao
tratamento (Drosse et al., 2008; Reichert et
al., 2009, ASTM 2010). Para se classificar
um defeito ósseo como crítico deve-se
considerar fatores como espécie, idade,
localização anatômica da lesão, estabilidade
dos fragmentos ósseos e tipo de fixação
(Adan, 1999; Arnold e Adan, 1999; Arnold,
2001; ASTM 2010). Em diáfises de ossos
longos de cães adultos hígidos, consideram-
se críticos aqueles defeitos com extensão
igual ou superior a 1,5 vezes o maior
diâmetro da região óssea acometida (ASTM
2010).
Os enxertos ósseos e seus substitutos são
alternativas para o tratamento das falhas
ósseas críticas, como fraturas cominutivas
ou com grande perda tecidual,
complicações da reparação óssea como
união retardada, não união, má união e
sequestros ósseos. Os enxertos ósseos e
seus substitutos sintéticos favorecem a
regeneração óssea por mecanismos de
osteocondução, osteoindução e osteogênese
(Watson, 2005; Giannoudis et al., 2005). A
osteocondução se refere à capacidade de um
biomaterial ou enxerto fornecer uma matriz
passível de ser colonizada, invadida e/ou
substituída por células osteoprogenitoras,
permitindo assim a formação de um novo
tecido ósseo. O termo osteogênese deve ser
utilizado apenas para aqueles enxertos ou
preparações que possuam células ósseas
viáveis com capacidade de se multiplicar e
produzir novo tecido ósseo, e a
osteoindução é a capacidade de um
biomaterial ou enxerto de estimular a
multiplicação e diferenciação de células
tronco mesenquimais em células
osteoprogenitoras e osteoblastos (Geissler,
2006; De Long Jr et al., 2007).
Os enxertos ósseos são classificados de
acordo com sua origem em autógenos,
homólogos ou heterólogos. Enxerto
autógeno é aquele transplantado de uma
localização anatômica para outra no mesmo
animal. É considerado o mais eficiente na
reparação óssea por ser biocompatível, não
induzir rejeição tecidual e apresentar efeitos
osteocondutor, osteoindutor e osteogênico
(Eppley, 2005; Sanders, 2007). Como
desvantagens, tem-se maior tempo
cirúrgico, complicações no local de colheita
como hematomas, seromas, feridas, dor e
fragilização óssea e limitada quantidade de
material coletado (Santos e Rahal, 2004;
Piermattei et al., 2006; Sanders, 2007).
No aloenxerto ou enxerto homólogo o osso
coletado em um animal é transplantado para
outro da mesma espécie (Watson, 2005).
São indicados para fraturas com grande
perda óssea e possuem osteocondutividade
e osteoindutibilidade, sendo uma alternativa
para substituição do autoenxerto, quando
este não for viável. Apresentam a
desvantagem de risco de transmissão de
doenças e de rejeição, sendo indicada a
realização da triagem dos doadores para que
os riscos sejam minimizados (Geissler,
2006; De Long Jr. et al., 2007).
O enxerto heterólogo ou xenoenxerto é
coletado em uma espécie e transplantado
para outra. Esse tipo de material apresenta
características semelhantes aos aloenxertos,
possui maior potencial imunogênico e
maior probabilidade de rejeição (Santos e
Rahal, 2004; Piermattei et al., 2006).
Geralmente os alo e xenoenxertos são
enxertos corticais constituídos por osso
compacto, denso e de baixa celularidade,
conferindo bom efeito mecânico. Os
enxertos esponjosos são formados por osso
trabecular, poroso e com alta celularidade,
sendo quase sempre autoenxertos. O
enxerto córtico-esponjoso é composto por
osso cortical e esponjoso e possui
característica intermediária (Santos e Rahal,
2004; Giannousdis et al., 2005).
Os enxertos orgânicos são todos aqueles
originados a partir de um tecido animal
(autoenxertos, aloenxertos, xenoenxertos,
concentrados de plaquetas e matriz óssea
desmineralizada), e os inorgânicos ou
aloplásticos são aqueles de origem sintética
como biocerâmicas e biopolímeros (Eppley
et al., 2005).
Devido às limitações da enxertia óssea há
uma incessante busca por inovações nessa
área, principalmente quanto ao
desenvolvimento de novos materiais que
possam substituir a enxertia óssea. A
produção de matrizes sintéticas a partir de
biomateriais parece ser a alternativa mais
viável e promissora, especialmente quando

23
combinada com as terapias celulares e
fatores de crescimento (De Long Jr, 2007;
Dutra et al., 2008).
O termo biomaterial foi definido na
Conferência do Instituto Nacional de
Desenvolvimento do Conselho de Saúde
nos EUA em 1982, como qualquer
substância que não seja droga ou uma
combinação de substâncias, sintéticas ou de
origem animal, que possa ser utilizada por
um período de tempo, como parte de um
sistema que trate, aumente ou substitua
qualquer tecido, órgão ou função do corpo
(Jones e Hench, 2003).
A biocompatibilidade é a habilidade de um
material desempenhar uma função com
resposta apropriada do tecido hospedeiro
em uma aplicação específica, sem agressão
deste. Há algumas décadas, era considerado
biocompatível o material totalmente inerte
implantado no tecido animal, cuja presença
não estimularia nenhuma resposta orgânica.
No entanto, esse conceito alterou-se ao se
reconhecer que qualquer biomaterial
elucida uma resposta do tecido receptor e o
mesmo biomaterial pode mostrar-se
biocompatível para uma aplicação ou para
um tipo de paciente e incompatível para
outras aplicações biomédicas ou para
pacientes de diferentes espécies, raças,
faixas etárias, sexo, estado geral de saúde
(Rezende, 2006).
Dentre os biomateriais com características
favoráveis para produção de substitutos
ósseos encontram-se as biocerâmicas, os
polímeros biodegradáveis e os materiais
compósitos, produzidos pela combinação
dos anteriores (Eppley et al., 2005;
Gianoudis et al., 2005). As matrizes
poliméricas reabsorvíveis e com estrutura
porosa semelhante ao osso trabecular tem
sido produzidas, entretanto, a resposta
biológica aos polímeros quando
implantados no organismo não favorece a
regeneração óssea. Os polímeros não
possuem capacidade de se ligar
quimicamente ao tecido ósseo o que leva a
formação de uma interface fibrosa pouco
coesa. Eles não são capazes de estimular a
atividade osteoblástica mas apresentam
plasticidade e ductibilidade o que favorece
a confecção de matrizes e permite que elas
sofram deformação sem se romper (Jones e
Hench, 2003; Orefice et al., 2006; Alves et
al., 2010a,b). O módulo de elasticidade dos
polímeros é menor que o do tecido ósseo, o
que inviabiliza sua utilização quando há
necessidade de suportar grandes cargas
mecânicas (Alves et al., 2010a,b).
Entretanto, os polímeros podem ser
combinados com outros compostos
terapêuticos como antibióticos, favorecendo
a reparação óssea em casos de infecção,
com fatores de crescimento ósseo e com
cerâmicas bioativas, contribuindo com a
regeneração óssea precoce, estimulando a
proliferação celular e a osteointegração
(Wang, 2003).
Dentre os polímeros biodegradáveis
destacam-se o polihidroxibutirato (PHB),
ácido polilático (PLA), ácido poliglicólico
(PGA) e policaprolactona (PCL) (Wang,
2003). Esses polímeros apresentam
degradação lenta e resistência suficiente
para utilização como matéria prima na
fabricação de substitutos ósseos,
principalmente quando reforçados com
partículas de biocerâmicas (Luklinska e
Schluckwerder, 2003; Alves, et al., 2011).
A utilização da cerâmica como substituto
ósseo data de 1894, quando pela primeira
vez Dreesman et al. utilizaram o gesso
(CaSO4) para essa finalidade. Desde então
as cerâmicas são estudadas e aprimoradas
para serem utilizadas como substitutos
ósseos. Existe atualmente um grupo
específico de cerâmicas bioativas,
consideradas as mais apropriadas para
substituição óssea. Esse grupo é composto
pelos fosfatos de cálcio, biovidros e
vidrocerâmicas. As cerâmicas bioativas são
compatíveis com os tecidos do corpo
humano e animal por possuírem em sua
composição química íons normalmente
encontrados no organismo animal (Wang,
2003; Eppley et al., 2005). Favorecem a
regeneração óssea e a osteointegração dos
implantes por possuírem semelhança
química e física com a matriz óssea mineral
(Hidaka et al., 2006; Vallet-Regí, 2006). As
biocerâmicas têm sido empregadas na
produção de matrizes capazes de absorver
fármacos e fatores de crescimento e
promover sua liberação controlada (Vallet-
Regí, 2006; Pereira et al., 2006; Sanders,
2007). A confecção de matrizes
tridimensionais sintéticas, cuja arquitetura
mimetiza o osso esponjoso, é uma maneira
eficiente de promover a reparação de
grandes falhas ósseas. Por suas

24
propriedades osteocondutivas, as cerâmicas
permitem a migração e adesão de células
tronco mesenquimais (CTM), que podem se
diferenciar em células osteoprogenitoras e
favorecer a regeneração óssea (Watson,
2005: Vallet-Regí, 2006; Mygind et al.,
2007). O tamanho do poro da cerâmica
deve ser semelhante ao do osso esponjoso
(Watson, 2005) para permitir a invasão
vascular, colonização celular e substituição
por tecido ósseo (Geissler, 2006). Chen et
al., (2001) recomendam o diâmetro maior
que 100-150µm e Geissler (2006) cita que o
diâmetro dos poros deve variar entre 150 e
500µm, mas salienta que poros muito
amplos podem enfraquecer o implante. De
Long Jr. et al (2007) relatam que o tamanho
ideal dos poros deve ser de 300 a 500µm,
pois permite maior colonização do
implante. Os autores advertem ainda que, a
presença de interconectividade entre os
poros é tão importante quanto seu tamanho
(Chen et al., 2001; Watson, 2005; Geissler,
2006; Vallet-Regí, 2006; De Long Jr. et al
2007), pois influencia diretamente na tensão
de oxigênio, fator de importância relevante
na diferenciação das células tronco
mesenquimais em células osteoprogenitoras
e dessas em osteoblastos (Chung et al.,
2012). Ao estudar a colonização e
diferenciação de células tronco
mesenquimais em arcabouço de
hidroxiapatita, Mygind et al. (2007) relatam
uma maior colonização nas amostras cujo
diâmetro médio do poro era de 500µm e
uma maior diferenciação celular nas
amostras cujo diâmetro médio era 200µm.
As biocerâmicas possuem diferentes
características químicas que refletem
diretamente na sua reabsorção. A
cristalinidade da cerâmica representa o grau
de organização e de ligação das moléculas,
sendo que quanto mais cristalina a amostra,
maior é a ligação entre as moléculas e
menor a taxa de dissolução,
consequentemente mais lenta é a reabsorção
(Watson, 2005; Pereira et al., 2006; De
Long Jr. et al., 2007). A taxa de reabsorção
difere entre as cerâmicas, sendo que a maior
reabsorção é observada nos vidros
bioativos, seguida pelos fosfatos de cálcio e
hidroxiapatita em ordem decrescente. O
termo fosfato de cálcio é utilizado
genericamente para definir um extenso
grupo de substâncias que possuem essa
composição em suas moléculas. Dentre eles
estão a hidroxiapatita (HA), o fosfato
tricálcico (TCA) e o fosfato de cálcio
bifásico (formado pela mistura de TCA com
HA) (Eppley et al., 2005; Vallet-Regí,
2006). A capacidade desses materiais de se
ligarem quimicamente ao osso através da
superfície do implante ocorre pela formação
de microcristais de apatita semelhante à
apatita óssea, que permitem o crescimento
de tecido ósseo e a osteointegração (Pereira
et al., 2006). Alterações teciduais no
microambiente de implantação da cerâmica
como aumento da acidez, edema e aumento
de temperatura local podem também
acelerar a reabsorção da cerâmica (Eppley
et al., 2005).
A hidroxiapatita (Ca10[PO4]6[OH]2) é uma
das cerâmicas mais difundidas e utilizadas
em cirurgias ortopédicas para
preenchimento de falhas ósseas e na
confecção de compósitos para emprego
ortopédico (Mygind et al., 2007). Ela é o
principal constituinte da matriz óssea
mineral, representando cerca de 70 a 95%
da composição mineral do osso (Eppley et
al., 2005; Pereira et al., 2006). É lentamente
biodegradável e pode ser produzida na
forma porosa, o que provê uma estrutura
para invasão do tecido receptor,
aumentando a ligação osso-implante na sua
interface (Wang et al., 2003). Sua
reabsorção pode ocorrer por dissolução e
por fagocitose (Pereira et al., 2006).
A hidroxiapatita assim como as demais
biocerâmicas é atóxica, pode ser produzida
no tamanho e na quantidade desejada, pode
ser recortada, esterilizada e guardada por
longos períodos de tempo à temperatura
ambiente (Manjubala et al., 2002; Pereira et
al., 2006). A hidroxiapatita porosa permite
a invasão e o crescimento interno de tecido
ósseo, mas apresenta baixa taxa de
dissolução e lenta reabsorção (Geissler,
2006). Dentre as biocerâmicas, a
hidroxiapatita é a que apresenta menor
bioatividade, não apresenta capacidade de
osteoindução e nem se liga aos tecidos
moles, mas é um material osteocondutor
capaz de se ligar ao osso adjacente
promovendo a osteointegração (Geissler,
2006; Pereira et al., 2006).
Os biovidros e as vidro-cerâmicas são os
biomateriais que apresentam a maior
bioatividade quando implantados no tecido

25
ósseo (Hu e Zhong, 2009). São materiais
compostos de sílica amorfa e óxidos de
cálcio, fósforo e sódio, mas também podem
existir, em pequenas quantidades, outros
elementos como zinco, estrôncio, magnésio,
cobre e boro (Hoppe et al., 2011). Os
biovidros são capazes de se ligar
rapidamente ao tecido ósseo e aos tecidos
moles por meio da adesão das fibras de
colágeno à sua superfície (Hench, 1997;
Barrere et al., 2006; Hoppe et al., 2011).
Quando implantado no tecido ósseo o
biovidro reage com o fluido intersticial
desencadeando uma cascata de reações
físico-químicas superficiais e eventos
celulares que culminam com a formação de
uma camada de hidroxiapatita biológica,
deposição e mineralização de osteóide em
sua superfície, o que favorece sua ligação
ao osso e promove uma interface coesa e
resistente (Jones e Hench, 2003; Vallet-
Regí, 2006; Hench, 2009). Os vidros
bioativos podem ser produzidos de duas
formas distintas. No método tradicional as
matérias primas do vidro (óxido de sílica –
SiO2; carbonato de cálcio – CaCO3;
carbonato de sódio – Na2CO3; óxido de
fosforo – P2O5) são misturadas e fundidas a
altas temperaturas, e o vidro é resfriado e
triturado para utilização na forma de
grânulos (Jones e Hench, 2003; Wang et al.,
2003). Os métodos modernos de síntese são
realizados pela rota sol-gel que consiste
basicamente na produção de uma solução
coloidal contendo as partículas desejadas na
composição do material e em seguida essa
solução é gelificada por meio da adição de
um catalizador, o material é seco e tratado
termicamente, tornando-se disponível para
a utilização (Pereira et al., 2005). A rota
sol-gel permite a produção de matrizes
tridimensionais com poros interconectados,
semelhante ao osso trabecular o que torna
esse material uma opção para substituição
da enxertia óssea. Os materiais produzidos
pela rota sol-gel apresentam maior
bioatividade e são reabsorvidos mais
rapidamente (Jones e Hench, 2003; Wang et
al., 2003; Pereira et al., 2006). Os efeitos
orgânicos dos biovidros são mediados
principalmente pelos íons liberados por
dissolução do material após sua
implantação no organismo (Xynos et al.,
2000; Xynos et al., 2001, Hench, 2009). No
entanto, não se sabe ao certo qual a melhor
composição química para a liberação da
concentração ideal de íons para
maximização do processo de regeneração
óssea e nem o exato mecanismo de ação de
cada íon no processo de regeneração óssea
(Hoppe et al., 2011; Rahaman et al., 2011).
Sabe-se que dentre os íons mais
importantes estão o cálcio (Chai et., 2012) e
o silício (Shie et al., 2011). Alguns estudos
já mostraram que os íons Ca e Si favorecem
a regeneração óssea por aumentarem a
proliferação (Maeno et al., 2005; Barradas
et al., 2010; McCullen et al., 2010) e a
diferenciação osteogênica “in vitro”(Jung e
Park, 2010; McCullen et al., 2010).
Os biovidros, assim como as demais
biocerâmicas, podem ser associados a
outros compostos com propriedades
osteoindutivas e osteogênicas tais como
fatores de crescimento osteogênico,
concentrado autógeno de plaquetas (Dutra
et al., 2008), matriz óssea desmineralizada,
aspirado de medula óssea (Kraus e Kirker-
Heard, 2006) e células tronco
mesenquimais (Arinzed et al., 2003). A
associação da matriz cerâmica com
concentrado de plaquetas autógeno (Dutra
et al., 2008) e com células tronco alógenas
mostrou-se mais eficiente na reparação de
falhas ósseas quando comparada com o uso
da cerâmica pura (Arinzed et al., 2003).
O colágeno tipo I é o principal responsável
pela resistência óssea e compõe cerca de
90% da matriz, fato que levou sua
utilização como substituto ósseo. A matriz à
base de colágeno pode contribuir para
deposição de minerais, pode ser
vascularizada e promover meio favorável à
regeneração óssea (Geissler, 2006; Donzelli
et al., 2007). Entretanto, esse biomaterial
pode estimular a reação imune e propicia
fraca estrutura para suporte mecânico. O
colágeno é utilizado para a produção de
esponjas que funcionam como sistema de
liberação controlada para produtos
osteocondutores, osteoindutores e
osteogênicos. Esse biomaterial é utilizado
em associação com biocerâmicas e aspirado
de medula óssea, apresentando a função de
carreador e expansor volumétrico
(Giannousdis et al., 2005).
A matriz óssea desmineralizada (MOD) é
produzida a partir de aloenxertos e é
composta por colágeno, proteínas não
colagênicas e fatores de crescimento ósseo.

26
A MOD facilita o processo de formação
óssea por aumentar a superfície de adesão
celular (osteocondução) e estimular a
formação óssea por fatores de crescimento
(osteoindução). Sua desvantagem é a falta
de padronização quanto à sua composição,
uma vez que são produzidas a partir de
vários doadores. Outro fator é que, durante
o processamento do material, pode haver
danos às proteínas, fazendo com que
algumas percam sua função osteoindutora,
diminuindo o efeito benéfico do produto
final (Watson, 2005; Geissler, 2006).
O concentrado autógeno de plaquetas,
também denominado plasma rico em
plaquetas (PRP) é definido como um
volume de plasma autógeno com
concentração adequada de plaquetas e
outras células sanguíneas que permite altos
níveis de liberação dos fatores de
crescimento em áreas ósseas a serem
recuperadas (Ogino et al. 2005; Silva et al.,
2011). Ao depositar o concentrado
plaquetário em uma lesão óssea há
degranulação das plaquetas e liberação de
fatores de crescimento derivado de
plaquetas (PDGF); transformador beta
(TGF-β1 e β2); semelhante à insulina (IGF-
1 e 2); de endotélio vascular (VEGF);
epidermal (EGF); básico de fibroblastos
(FGF-2); e proteínas morfogênicas (BMPs)
(Hidaka et al., 2006; Nishimura et al.,
2012). Alguns destes fatores promovem
efeito quimiotático e mitogênico para
osteoblastos e pré-osteoblastos,
estimulando um maior aporte e
multiplicação de células ósseas no local de
deposição (Watson, 2005).
A utilização de fatores de crescimento
ósseo para o tratamento de defeitos ósseos
pode ser feita isoladamente, mas os
melhores resultados são obtidos quando se
associa materiais osteocondutores como as
cerâmicas (Notodihardjo et al., 2011).
Reddi & Huggins em 1972, foram os
primeiros pesquisadores a estudar as
proteínas ósseas morfogenéticas (BMPs) e a
associá-las à formação óssea (Geissler,
2006). Atualmente já foram identificadas
mais de 15 BMPs capazes de se ligarem às
células-tronco e desencadear mecanismos
que estimulem a proliferação e
diferenciação celular, resultando em
regeneração óssea (De Long Jr et al., 2007).
As BMPs são proteínas osteoindutoras não
imunogênicas e não espécie-específica que,
ao serem liberadas na corrente sanguínea,
são capazes de estimular as células
perivasculares e mesenquimais
indiferenciadas a se transformarem em
células da linhagem osteoprogenitora.
Algumas destas proteínas, como a BMP-2 e
BMP-7 podem ser produzidas por
tecnologia recombinante e já estão
disponíveis para utilização clínica (Sanders,
2007; Notodihardjo et al., 2012). O fator de
crescimento de endotélio vascular (VEGF)
desempenha uma importante função na
ossificação endocondral, ele atua na
maturação final do condrócito e promove a
invasão vascular da cartilagem o que
permite a migração de células
osteoprogenitoras e osteoblastos
promovendo assim a substituição da
cartilagem por osso (Ballock e O´Keefe,
2003a,b; Geiger et al., 2005; Long, 2012).
Células tronco e sua aplicação na
ortopedia
A presença de células osteoprogenitoras e
células tronco mesenquimais (CTM)
capazes de se transformar em células ósseas
é provavelmente, o fator que mais contribui
para o aumento da atividade osteogênica e
consequentemente, para a reparação de uma
lesão óssea. Quando células tronco são
adicionadas em um sítio de lesão óssea, elas
podem se diferenciar em osteoblastos e
aumentar a síntese de matriz óssea (Arinzeh
et al., 2006; Bielby et al., 2007; Mizuno,
2009). As CTM possuem a capacidade de
se diferenciar in vitro em qualquer célula de
origem mesenquimal como tenócitos,
mioblastos, adipócidos, condrócitos e
osteoblastos (Pittenger et al., 1999;
Shenfield et al., 2002). Para maximizar sua
diferenciação em osteoblastos, pode-se
associá-las a BMPs, matriz óssea
demineralizada, ou a qualquer outro tipo de
substituto osteoindutor (Bruder et al.,
1998a,b; Brodke et al., 2006; Hidaka et al.,
2006; Yuan et al., 2007; Liao et al., 2010).
Existem basicamente três formas de se
utilizar as CTM para o tratamento de lesões
ósseas. A forma mais rápida e simples, já
utilizada clinicamente há anos, trata-se da
injeção de aspirado de medula óssea no
foco da lesão. No entanto, o aspirado de
medula contém pequena quantidade de

27
CTM e vários outros tipos celulares como
hemácias, células tronco hematopoiéticas e
leucócitos, logo o efeito de sua utilização
não pode ser atribuído exclusivamente às
CTM (Kraus e Kirker-Head, 2006; Pountos
et al., 2007; Arthur et al., 2009). O número
de CTM na medula óssea diminui com o
envelhecimento, o que torna essa técnica
menos eficiente em animais geriátricos. A
segunda alternativa é a utilização de CTM
alógenas advindas de bancos de culturas
celulares. É necessário portanto, a
construção de uma banco de CTM que
permaneçam congeladas. Para seu uso, um
pool de células é descongelado e injetado
no foco de lesão. Essa técnica tem a
vantagem de fornecer uma grande
quantidade de CTM em um curto período
de tempo, mas apresenta o inconveniente da
possibilidade de rejeição, uma vez que
utiliza células homólogas e não autógenas
(Arinzeh et al., 2003; Yamada et al., 2003).
A terceira forma de utilização das CTM é
pelo isolamento, cultivo e expansão in vitro
de CTM autógenas (Bruder et al., 1998a;
Brodke et al., 2007; Yuan et al., 2007). Essa
é a técnica que propicia o melhor potencial
de regeneração, pois não há risco de
rejeição e as CTM podem ser diferenciadas
em osteoblastos antes de sua utilização
(Boo et al., 2002). Os inconvenientes estão
relacionados principalmente com o maior
tempo requerido para isolamento e
expansão in vitro das CTM, enquanto as
CTM oriundas do aspirado de medula ou do
banco de células permitem a utilização
imediata (Kraus e Kirker-Head, 2006;
Arthur et al., 2009).
O termo célula tronco (CT) foi criado para
facilitar o entendimento do que é essa
célula, fazendo uma analogia entre uma
árvore e o organismo. A árvore possui um
único tronco de onde se originam vários
galhos, ramos e folhas. Analogamente a
célula tronco é capaz de se dividir e se
diferenciar originando vários tipos de
células especializadas. As células tronco
podem ser classificadas de acordo com seu
potencial de diferenciação em: células
totipotentes, capazes de se diferenciar em
qualquer tipo de célula do organismo
adulto; células pluripotentes, capazes de se
diferenciar em quase todos os tipos de
tecido com exceção da placenta e dos
anexos fetais; células multipotente, capazes
de se diferenciar em um número menor de
linhagens celulares; e células unipotentes
que são capazes de diferenciar em apenas
um tipo de célula especializada (Rao, 2004;
Wagers e Weisman, 2004)
As células tronco são células
indiferenciadas, com alta capacidade de
multiplicação e diferenciação em outros
tipos celulares mediante estímulos
adequados. Elas podem ser divididas em
células tronco embrionárias (CTE), que são
obtidas do blastocisto, ou células tronco
adultas (CTA) que são obtidas após a fase
de blastocisto em feto, filhotes ou do
indivíduo adulto (Grove et al., 2004). As
células tronco mesenquimais (CTM) são
um tipo de celula tronco adulta que podem
ser encontradas em praticamente todos os
tecidos do indivíduo e desempenham a
função de se auto-renovar, manter ou
reparar os tecidos nos quais residem
(Shenfield et al., 2002; Baksh et al., 2004).
As CTE e as CTM tem sido empregadas em
terapias regenerativas, mas cada um dos
tipos celulares tem suas particularidades. As
CTE são mais indiferenciadas que as CTM,
podem facilmente se diferenciar em
qualquer tipo celular (Thomson et al., 1998)
e podem ser expandidas indefinidamente in
vitro, sem a perda de seu potencial de
diferenciação. Mas apresentam também
desvantagens como poder desencadear uma
resposta imunológica quando implantadas
no organismo, visto que são alogênicas e
não autólogas e sua maior indiferenciação e
capacidade mitótica pode levar a
proliferação descontrolada e a desvios na
diferenciação, resultando na formação de
teratomas e de outros tumores (Shenfield et
al., 2002; Rao, 2004). Como as CTM
podem ser colhidas do próprio paciente,
elas podem ser expandidas in vitro e depois
reintroduzidas sem o risco de rejeição
(Baksh et al., 2004; Kraus e Kirker-Head,
2006). Há menor risco de proliferação
desordenada, visto que as CTM são menos
indiferenciadas (Bobis et al., 2006). Mas,
assim como as CTE, elas também
apresentam desvantagens como a
diminuição da capacidade proliferativa e de
diferenciação com a idade, o que limita o
seu transplante autólogo em pacientes
idosos (Heng et al., 2004). As CTM sofrem
senescência quando mantidas em cultivo,
limitando o número de repiques e seu

28
tempo de permanência em cultivo
(Payshina et al., 2006).
As CTM já foram isoladas de vários tecidos
como medula óssea (Boeloni et al, 2009;
Ocarino et al., 2010; Spencer et al., 2012),
tecido adiposo (Zuk et al., 2001; Vieira al.,
2010), derme (Toma et al., 2001), músculo
esquelético, membrana sinovial, pulmão
(Tae et al., 2006), fígado, sangue periférico,
intestino (Presnell et al., 2002), sistema
nervoso periférico, sistema nervoso central,
miocárdio (Beltrami et al., 2003), polpa
dentária (Meirelles et al., 2006), líquido
amniótico, placenta (Bobis et al., 2006;
Filioli Uranio et al., 2011), sangue do
cordão umbilical (Rebelato et al., 2009; Seo
et al., 2009) vasos sanguíneos, córnea e
retina (Meirelles et al., 2006). Devido a
maior facilidade de isolamento, grande
capacidade de expansão in vitro e amplo
potencial de diferenciação, as CTM
oriundas da medula óssea e do tecido
adiposo são as mais estudadas e
empregadas na medicina regenerativa (Zuk
et al., 2001; Payushina et al., 2006; Tae et
al., 2006; Zhu et al., 2008). A diferenciação
das CTM em osteoblastos, condroblastos,
adipócitos e miócitos, ocorre mediante
ativação de fatores de transcrição
específicos ligados aos genes
Runx2/Osterix/Lrp5, SOX5/6/7, PPARγ e
MyoD family, respectivamente (Katagiri e
Takahashi, 2002, Kassem et al., 2007; Liu e
Lee, 2012; Long, 2012; Zhang, 2012).
As células tronco mesenquimais extraídas
da medula óssea (CTM-MO) são também
denominadas células do estroma da medula,
células tronco estromais da medula óssea,
células precursoras do estroma, células
tronco esqueléticas e células progenitoras
adultas (Chen et al., 2008). Elas foram
primariamente descritas em suínos por
Fridenstein e colaboradores em 1970 como
unidades formadoras de colônias de
fibroblastos (Tae et al., 2006; Liu et al.,
2009), mas atualmente já foram isoladas da
medula óssea de diversas espécies como
camundongo, ratos (Boeloni et al, 2009),
gatos (Martins et al., 2002), cães (Chung et
al., 2012), cavalos, coelhos, suínos e seres
humanos (Chamberlain et al., 2007). As
CTM-MO são facilmente isoladas in vitro,
pois possuem a capacidade de aderir à
superfície plástica, característica que não é
compartilhada pelas células tronco
hematopoiéticas. Para o isolamento das
CTM-MO, uma amostra de medula óssea é
cultivada em garrafas plásticas, o que
permite a rápida aderência dessas células.
Transcorrido o tempo de 24 a 48 horas de
cultivo, as garrafas são lavadas com solução
salina isotônica retirando as células
contaminantes não aderidas (hemácias,
leucócitositos e células tronco
hematopoiéticas) e posteriormente são
realizadas trocas sistemáticas do meio de
cultura a cada 72 horas para eliminação
total das células contaminantes (Baksh et
al., 2004; Ocarino et al., 2007; Boeloni et
al, 2009)
As células tronco mesenquimais extraídas
do tecido adiposo (CTM-AD) podem
também ser denominadas células
multipotentes derivadas do tecido adiposo,
pré-adipócitos, células mesenquimais
derivadas do tecido adiposo, células tronco
adultas derivadas do tecido adiposo, células
estromais aderentes derivadas do tecido
adiposo, células tronco mesenquimais
derivadas do tecido adiposo e células
estromais derivadas do tecido adiposo,
sendo essa última nomenclatura definida
como a mais adequada na “Second Annual
International Fat Applied Technology
Society Meeting” (Levi e Longaker, 2011).
As CMT-AD foram primariamente isoladas
e descritas em humanos por Zuk et al., em
2001, mas atualmente já foram isoladas de
diversas espécies como ratos (Zaminy et al.,
2008), camundongos, coelhos, cães
(Neupane et al., 2008; Vieira et al., 2010) e
suínos (Qu et al., 2007; Arrigoni et al.,
2009)
As CTM-AD podem ser extraídas do tecido
adiposo intra-abdominal ou subcutâneo
(Locke et al., 2009; Rada et al., 2009). O
método de isolamento mais empregado é o
enzimático e a colagenase tipo I, a enzima
mais utilizada. Inicialmente é colhida uma
amostra de tecido adiposo, que pode ser
realizada por biópsia ou por lipoaspiração.
A amostra é lavada com solução salina para
remover as células sanguíneas e possíveis
contaminantes como os anestésicos locais
no caso de coleta por lipoaspiração.
Posteriormente a amostra é digerida com
uma solução de colagenase e as frações são
separadas por centrifugação. A fração
estromal que contém as CTM-AD é
cultivada nas mesmas condições das CTM-

29
MO (Zuk et al., 2001; Locke et al., 2009;
Mizuno, 2009). Durante o cultivo, as CTM-
AD também se aderem às placas ou às
garrafas de cultivo (Levi e Longaker, 2011).
Com o objetivo de facilitar a identificação
da CTM e viabilizar sua utilização clínica,
vários estudos tem avaliado a expressão de
proteínas de superfície destas células. As
culturas de CTM podem ser identificadas e
caracterizadas pela expressão ou ausência
de marcadores fenotípicos (Payushina et al.,
2006). The International Society for
Cellular Therapy padronizou critérios
mínimos para classificar uma célula como
célula tronco mesenquimal humana. Ficou
estabelecido que para ser considerada uma
CTM, a célula deve apresentar morfologia
semelhante a fibroblasto, possuir
capacidade de aderir ao plástico e de se
diferenciar em no mínimo três tipos
celulares: osteoblastos, condrócitos e
adipósitos, devem apresentar um perfil de
expressão de proteínas de superfície onde
são expressos os marcadores CD105, CD73
e CD90 e não os marcadores de células
hematopoiéticas CD 45 e CD34 (Dominici,
et al., 2006). Muitos outros marcadores de
superfície já foram estudados em CTM e foi
observado o seguinte perfil de expressão em
CTMs humanas: moléculas de adesão
CD44 (receptor de hialuronato), CD50
(molécula de adesão intercelular 3), CD54
(molécula de adesão intercelular 1), CD56
(molécula de adesão de células neuronais),
CD58 (antígeno 3 associado a função
linfocitária), CD62L (selectina-L),
CD102(molécula de adesão intercelular 2),
CD106(molécula 1 de células de adesão
vascular), CD166(molécula de adesão
ativada por leucócitos) (Strem et al., 2005;
Pountos et al., 2007). Algumas integrinas
CD29(integrina β1), CD49(a,b,c,e,f)
(integrinas α), CD61, CD104 (Lindner et
al., 2010), citocinas e fatores de
crescimento: CD71 (receptor da
transferina), CDw119 (receptor do
interferon γ), CD120 (Receptor do fatores
de necrose tumoral α), CD121 (Receptor de
interleucina 1), CD123 (Receptor de
interleucina 3), CD124 (Receptor de
interleucina 4), CD126 (Receptor de
interleucina 6), CD127 (Receptor de
interleucina 7), CD140a (Receptor do fator
de crescimento derivado das plaquetas),
FGFR (Receptor do fator de crescimento
fibroblástico) e CD271 (receptor do fator de
crescimento neural). Outros marcadores
também são expressos: CD9, CD13
(aminopeptidase N), CD73
(ectonucleotidase 5´), CD90 (Glicoproteina
Thy-1), CD105, CD146, CD157 (antígeno
1 de células estromais da medula óssea),
SH3, D7-FIB, STRO-1, HLA-A,B,C
(moléculas apresentadores de antígeno),
SSEA-3,4 (Pountos et al., 2007; Schaffler e
Buchler, 2007). Marcadores de células
hematopoiéticas como CD1a, CD14
(receptor de lipopolisacarídeo), CD33,
CD34, CD38, CD45 (antígeno comum em
leucócitos) e CD133 (AC133) não são
expressos pelas CTM (Pountos et al., 2007;
Choi et al, 2008).
Alguns marcadores de CTM foram
avaliados em cães, mas grande parte dos
que estão disponíveis no mercado não são
espécie específicos para cães, o que
dificulta a interpretação dos resultados, pois
não se sabe se as proteínas não são
realmente expressas ou se o anticorpo
utilizado não apresenta reação cruzada com
a espécie canina. Dentre os marcadores de
CTM que apresentaram positividade em
CTM de cães estão CD44 (Seo et al., 2009;
Vieira et al., 2009; Lee et al., 2011;
Martinello et al., 2011), CD29 (Seo et al.,
2009; Vieira et al., 2009; Lee et al., 2011),
CD90 (Vieira et al., 2009; Lee et al., 2011;
Martinello et al., 2011), CD33, CD105,
CD184, OCT4 (Seo et al., 2009), CD140a,
CD117 (Martinello et al., 2011), CD56,
CD146, CD271(Rozemuller et al., 2010).
Alguns estudos não obtiveram positividade
para os marcadores CD13 (Vieira et al.,
2009), CD105 (Vieira et al., 2009; Lee et al,
2011) e CD73 (Seo et al., 2009; Vieira et
al., 2009 ), CD90 (Seo et al., 2009)
sabidamente positivos para a espécie
humana, entretanto, a dúvida quanto a sua
expressão permanece devido a falta de
especificidade dos anticorpos utilizados.
Em um estudo recente os marcadores
humanos para CD73, CD90, e CD105 não
apresentaram reação cruzada para a espécie
canina (Rozemuller et al., 2010). Os
marcadores de células hematopoiéticas
CD14 (Seo et al., 2009; Vieira et al., 2009;
Lee et al., 2011), CD34, CD45 (Seo et al.,
2009; Vieira et al., 2009; Lee et al., 2011;
Martinello et al., 2011), CD3 (Lee et al.,
2011), CD117 (Vieira et al., 2009), CD4,

30
CD8a, CD10, CD20, CD24, CD31, CD38,
CD41a, CD49b, CD51, CD62p, CD133,
HLA-DR não são expressos pelas CTM
caninas (Seo et al., 2009). Outro
inconveniente relacionado à caracterização
fenotípica das CTM é que alguns
marcadores como CD44, CD29, CD90
podem também ser expressos por
fibroblastos (Ishii et al., 2005) o que
dificulta a diferenciação das CTM e dos
fibroblastos que podem estar presentes em
uma mesma cultura (Bobis et al., 2006).
Utilizando a técnica de PRC, Lee et al
(2011) constataram que os marcadores de
células tronco CD10, CD29, CD73 e
CD106 não são expressos pelo fibroblasto
canino, o que torna esses marcadores
preferíveis nessa espécie.
Vários meios de cultura como IMDM -
Iscove’s Modified Dulbecco’s Medium
(Martins et al., 2002; Rebelatto et al., 2008;
Lee et al., 2011), RPMI – Roswell Park
Memorial Intitute (Chung et al., 2012),
MEM - Minimal Essential Médium Eagle
(Yamada et al., 2003), α-MEM - Minimal
Essential Médium Eagle Alpha
Modification (Deliloglu-Gurhan et al.,
2006; Bunnel et al., 2008) podem ser
utilizados para isolamento e cultivo de
CTM, entretanto, o meio mais utilizado é o
DMEM - Dulbecco’s modified Eagle
medium enriquecido com antibióticos,
antifúngicos e 10% de soro fetal bovino
(SFB) (Pittenger et al., 1999; Ocarino et al.,
2008; Boeloni et al., 2009; Seo et al., 2009;
Martinello et al., 2011). A suplementação
dos meios de cultivo com SFB é
indispensável para a função celular e
sucesso do cultivo e diferenciação das
células. O SFB fornece substâncias como
fatores de crescimento, citocinas e outras
moléculas orgânicas essenciais para a
sobrevivência e desenvolvimento celular e
essas moléculas não estão presentes mesmo
nos meios de cultura comerciais mais
completos. Já foi comprovado que na
ausência de SFB, as células apresentam
baixo índice mitótico, pouca adesão e
elevada taxa de apoptose (Heng et al.,
2004). Um estudo recente mostrou melhor
resultado no cultivo e diferenciação de
células tronco mesenquimais humanas
quando se substituiu o SFB pelo soro da
mesma espécie (Paula et al., 2012).
Durante o isolamento e cultivo as CTM
aderem à superfície plástica das placas e
garrafas de cultivo e se proliferam
expandindo seu número in vitro por um
longo período (Tae et al., 2006). Com o
passar do tempo e com o aumento do
número de passagens, as CTM se tornam
senescentes e diminui seu potencial
proliferativo (Stenderup et al., 2003). Em
humanos isso ocorre após a trigésima
passagem (Kim et al., 2008). Outros fatores
como idade, presença de doenças sistêmicas
e de alterações metabólicas podem
influenciar o potencial proliferativo e de
diferenciação das CTM (Liu et al., 2009).
Para diferenciação in vitro, o meio basal,
geralmente DMEM acrescido de
antibióticos, antifúngicos e SFB, é
suplementado com substâncias que
estimulam a diferenciação. No caso da
diferenciação osteogênica as substâncias
mais utilizadas são a dexametasona, o ácido
ascórbico e o ß-glicerofosfato (Jackson et
al., 2007; Panetta et al., 2009). A vitamina
D3 também pode ser utilizada nesse
protocolo, associada às demais substâncias
ou em substituição à dexametasona (Zhou
et al., 2006). A dexametasona e a vitamina
D3 atuam no estágio inicial de
diferenciação, desencadeando o processo
(Payushina et al., 2006; Zhou et al., 2006).
Sabe-se que a dexametasona, assim como
outros glicocorticoides, é capaz de
estimular a expressão de runx2 nas CTMs,
iniciando assim o processo de diferenciação
osteogênica (Liu e Lee, 2012) mas, sua
ação prolongada pode induzir apoptose
osteoblástica via inibição de Wnts (Marie,
2008). A vitamina D age nas CTMs
estimulando a expressão de runx2 e
inibindo a expressão de PPARγ2 importante
fator de diferenciação adipogênica, o que
favorece ainda mais o processo de
diferenciação osteogênica (Marie, 2008). O
ácido ascórbico e o β-glicerofosfato são
essenciais nos estágios tardios da
diferenciação osteogênica quando ocorrem
a síntese e mineralização da matriz
extracelular (Payushina et al., 2006). Para
produção da matriz, o osteoblasto sintetiza
grande quantidade de colágeno e o ácido
ascórbico é o cofator indispensável nesse
processo, o que justifica a necessidade de
sua suplementação no meio de
diferenciação osteogênica (Franceschi et a.,

31
2009). O β-glicerofosfato atua
disponibilizando o fosfato necessário no
processo de mineralização da matriz
extracelular (Bruedigam et al., 2011). Após
estímulo para diferenciação, as CTM
sofrem uma divisão celular assimétrica, ou
seja, uma célula mãe origina duas células
filhas, sendo que uma célula filha é idêntica
à célula mãe e a outra inicia o processo de
diferenciação, adquirindo características de
uma célula precursora de determinada
linhagem celular. Essa divisão assimétrica
vai ocorrendo sucessivamente nas células
filhas até que a diferenciação se complete
originado uma célula especializada. O
processo de diferenciação ainda é muito
pouco compreendido mas, estima-se que
cerca de 914 genes estejam envolvidos na
diferenciação osteogênica (Baksh et al
.,2004). O processo de diferenciação
osteogênica é mediado por uma série de
fatores de transcrição e os mais importantes
e conhecidos são o runx2 (core-binding
fator A1- Cbfa), o osterix (Osx) e o ATF4
(cAMP response elements binding protein 2
- CREB2) (Yang e Karsenty, 2002;
Karsenty, 2008; Franceschi et al., 2009). O
runx2 é o fator de transcrição mais
importante e estudado no processo de
diferenciação osteogênica, atuando em
todas as etapas do processo. Ele é
responsável pelo início da diferenciação,
estimulando a transformação de CTMs em
células osteoprogenitoras, mas tem ação
também nos estágios intermediários e finais
da diferenciação, bem como, na síntese e
mineralização da matriz pelo osteoblasto
maduro (Karsenty, 2008; Marie, 2008;
Franceschi et al., 2009; Liu e Lee, 2012). O
runx2 tambem é importante no processo de
diferenciação condrogênica uma vez que
seu estímulo transforma uma CTM em uma
célula progenitora da linhagem
osteocondral, ou seja, após essa etapa a
célula pode diferenciar-se tanto em um
osteoblasto quanto em um condrócito (Liu e
Lee, 2012; Nishimura et al., 2012). O OSX
é o segundo fator de transcrição mais
importante no processo de diferenciação
osteogênica. Ele é imprescindível no
processo e atua na transformação dos pré-
osteoblastos em osteoblastos maduros,
finalizando assim o processo de
diferenciação osteogênica. O OSX é
fundamental na inibição do processo de
diferenciação condrogênica (Nakashima et
al., 2002; Datta et al., 2008; Long, 2012;
Zhang, 2012). Assim como o runx2, o OSX
continua sendo expresso após a
diferenciação osteogênica e atua na
atividade dos osteoblastos controlando a
síntese e a mineralização da matriz
extracelular por meio do controle da
expressão do receptor intranuclear da
vitamina D (Long, 2012; Zhang, 2012).
Muitos outros fatores estão envolvidos no
processo de diferenciação osteogênica
entretanto, a função exata de cada um deles
ainda é pouco conhecida. Alguns fatores
como o ZNF145 (promyelotic leucemia zinc
finger) e o OMD (osteomodulin) participam
de todo o processo de diferenciação, mas
outros fatores como FKBP5 (FK506-
binding protein 5) e CPM
(carboxypeptidase M) atuam apenas no
início do processo e o fator APOD
(apolipoprotein D), nos estágios finais de
diferenciação e maturação osteoblástica. O
fator OMD é um pequeno proteoglicano
rico em leucina importante nos processos de
biomineralização (Liu et al., 2007). O fator
Smad atua na atividade de fosfatase alcalina
e na síntese de osteocalcina pelos
osteoblastos (Katagiri e Takahashi, 2002).
O fator ZNF145 estimula a expressão de
Runx2 que por sua vez, regula a expressão
de vários genes como os referentes ao
colágeno I, fosfatase alcalina, osteopontina,
osteocalcina, sialoproteína óssea (Ducy et
al., 1997; Ducy e Karsenty, 1998; Datta et
al., 2008; Franceschi et al., 2009) Durante e
após a diferenciação osteogênica as células
mudam seu perfil de expressão e passam a
apresentar maior quantidade de marcadores
da diferenciação osteogênica. Dentre os
principais marcadores estão o colágeno tipo
I, a fosfatase alcalina, proteínas não
colagênicas da matriz óssea (osteopontina,
osteonectina, osteocalcina e sialoproteína
óssea), runx2, OSX e BMPs (Marie, 2001;
Halvorsen et al., 2001; Yang e Karsenty,
2002; Nakashima et al., 2002; Payushina et
al., 2006; Kassem et al., 2008). O colágeno
tipo I é o principal constituinte da matriz
óssea orgânica representando cerca de 90%
de sua composição. É considerado um
marcador precoce da diferenciação
osteogênica e pode ser expresso por células
osteoprogenitoras, pré-osteoblastos e
osteoblastos (Nakashima et al., 2002). A

32
fosfatase alcalina é uma enzima presente na
membrana do osteoblasto e participa da
síntese e mineralização da matriz óssea. No
processo de diferenciação osteogênica sua
expressão pode ser aumentada em até 27
vezes (Kassem et al., 2008) e é considerada
um marcador precoce de diferenciação
osteogênica, sendo expressa por pré-
osteoblastos e osteoblastos (Kassem et al.,
2008). In vitro, a fosfatase alcalina age no
β-glicerofosfato liberando os fosfatos e
aumentando sua concentração local que
culmina com a rápida mineralização da
matriz extracelular rica em colágeno
secretada pelo osteoblasto (Bruedigam et
al., 2011). As proteínas não colagênicas da
matriz são marcadores fidedignos da
diferenciação osteogênica, e estão
relacionadas com o processo de
mineralização da matriz. Algumas são
expressas em maior quantidade no início do
processo de mineralização, outras nas fases
intermediárias e outras nas fases finais de
maturação da matriz óssea. A osteopontina
é uma fosfoproteína que possui domínios
ligantes de cálcio e está relacionada com a
proliferação celular e com a mineralização
da matriz óssea. É considerada um
marcador precoce de diferenciação
osteogênica e é expressa por células
osteoprogenitoras, pré-osteoblastos e
osteoblastos (Donzelli et al., 2007). A
sialoproteina óssea (BSP) é responsável
pelo início do processo de nucleação e
deposição de hidroxiapatita na matriz,
iniciando assim a mineralização da matriz
óssea. Reflete uma fase final de
diferenciação e pode ser expressa por pré-
osteoblastos e osteoblastos (Nefussi et al.,
1997; Zhou et al., 2006). A osteocalcina é
uma glicoproteína específica do tecido
ósseo que atua promovendo a mineralização
da matriz. Ela é expressa apenas por
osteoblastos maduros e reflete o final da
diferenciação e maturação da matriz óssea
(Nefussi et al., 1997; Nakashima et al.,
2002; Zhou et al., 2006; Nakamura et al.,
2009). A osteonectina também está
relacionada com a mineralização da matriz
no estágio final de diferenciação (Strauss et
al., 1990; Nefussi et al., 1997; Nakashima
et al., 2002; Karaoz et al., 2009).
Além das mudanças no perfil de expressão
gênica, as CTM mudam sua morfologia,
durante a diferenciação osteogênica,
adquirindo forma cuboidal ou poligonal
semelhante aos osteoblastos e se agrupam
concentricamente, formando nódulos
mineralizados (Payushina et al., 2006; Tae
et al., 2006). Esse nódulos podem ser
visibilizados pelas técnicas de coloração
Alizarin red ou Von kossa (Chamberlain et
al., 2007), bem como por microscopia
eletrônica e apresentam duas zonas, uma
periférica formada por camadas de células
alongadas separadas por pouca matriz e
uma zona central contendo células com
morfologia variada (alongadas, triangulares
e esféricas), localizadas em lacunas
separadas por grande quantidade de matriz.
Na zona periférica há predomínio de células
e na zona central, de matriz (Eslaminejad e
Taghiyar, 2010). A síntese e mineralização
da matriz extracelular são consideradas
marcadores finais de diferenciação
osteogênica pois refletem toda a atividade
metabólica do osteoblasto (Bruedigam et
al., 2011).
Potencial proliferativo e de diferenciação
osteogênica das CTM
Existem diferentes populações de CTM,
com distintas capacidades proliferativa e de
diferenciação no mesmo indivíduo (Jiang et
al., 2002; Crisan et al., 2008). A
distribuição dessas populações nos
diferentes tecidos (Im et al., 2005; Hayashi
et al., 2007; Yoshimura et al., 2007; Peng et
al., 2008; Zaminy et al., 2008; Filioli-
Uranio et al., 2011; Shafiee et al., 2011;
Dmitrieva et al., 2012; Park et al., 2012;
Monaco et al., 2012; Al-Nbaheen et al.,
2013; Wen et la., in press) e dentro de cada
tecido (Aksu et al., 2008; Neupane et al.,
2008), determina maior ou menor potencial
proliferativo e de diferenciação das CTM
isoladas de locais diferentes no mesmo
indivíduo (Hass et al., 2011; Strioga et al.,
2012). Estudos já mostraram também que
os potenciais proliferativos e de
diferenciação das CTM podem variar de
acordo com a espécie animal (Arrigoni et
al., 2009; Levi et al., 2011), gênero
(Muschler et al., 2001; Aksu et al., 2008;
Jiang et al., 2008) e idade (Zhang et al.,
2009; Hell et al., 2011; Veronesi et al.,
2011; Al-Nbaheen et al., 2012; Kim et al.,
2012; Zaim et al., 2012). Com isso muitos
estudos têm sido realizados com o objetivo

33
de se identificar a melhor fonte de células
tronco para utilização terapêutica na
condição específica de cada paciente
(Mosna et al., 2010; Strioga et al., 2012).
Um dos focos desses estudos está voltado
para a identificação da melhor fonte de
células tronco para a terapia de regeneração
óssea.
Atualmente a medula óssea (Im et al., 2005;
Lee et al., 2011) e o tecido adiposo
(Neupane et al., 2008; Viera et al., 2010)
são considerados os principais sítios para
obtenção dessas células, devido à maior
quantidade de tecido disponível para
colheita, bom rendimento de CTM e
facilidade de obtenção. No entanto, ainda
há dúvidas sobre qual deles é a melhor
fonte de CTM para terapias de regeneração
óssea. Vários estudos a respeito já foram
realizados em humanos (De Ugarte et al.,
2003; Lee et al., 2004; Im et al., 2005;
Skaguchi et al., 2005; Izadpanah et al.,
2006; Kern et al., 2006; Egusa et al., 2007;
Liu et al., 2007; Niermeyer et al., 2007;
Noel et al., 2008; Reberatto et al., 2008;
Zhu et al., 2008; Pachon-Pena et al., 2011;
Shafiee et al., 2011; Adegani et al., 2012;
Al-Nbaheen et al., 2012; Dmitrieva et al.,
2012; Ertas et al., 2012; Park et al., 2012;
Wen et al., in press) e ratos (Yoshimura et
al., 2007; Peng et al., 2008; Hayashi et al.,
2008; Zaminy et al., 2008; Nakanishi et al.,
2011) e alguns em equinos (Ranera et al.,
2012), macacos (Izadpanah et al., 2006),
coelhos (Lin et al., 2009), suínos (Monaco
et al., 2012), mas pouco se sabe sobre o
potencial osteogênico dessas células na
espécie canina (Chung et al., 2012; Spencer
et al., 2012). Alguns desses estudos em
coelhos (Lin et al., 2009), cães (Chung et
al., 2012) e humanos (Izadpanah et al.,
2006) mostraram que as células tronco
mesenquimais oriundas do tecido adiposo
(CTM-AD) apresentam maior osteogênese
in vitro que as oriundas da medula óssea
(CTM-MO). Outros em humanos (Im et al.,
2005; Sakaguchi et al., 2005; Liu et al.,
2007; Noel et al., 2008; Shafiee et al., 2011;
Park et al., 2012), macacos (Izadpanah et
al., 2006), suínos (Monaco et al., 2012) ,
coelhos (Lin et al., 2009) e ratos
(Yoshimura et al., 2007; Hayashi et al.,
2008; Zaminy et al., 2008; Nakanishi et al.,
2011) mostraram que as CTM-MO não só
apresentam uma maior osteogênese in vitro
(Im et al., 2005; Izadpanah et al., 2006; Liu
et al., 2007; Hayashi et al., 2008; Skaguchi
et al., 2005; Zaminy et al., 2008; Shafiee et
al., 2011; Park et al., 2012) mas também,
maior expressão de genes e marcadores de
diferenciação osteogênica (Im et al., 2005;
Liu et al., 2007; Noel et al., 2008;
Yoshimura et al., 2007; Hayashi et al.,
2008; Zaminy et al., 2008; Lin et al., 2009;
Nakanishi et al., 2011; Shafiee et al., 2011;
Park et al., 2012; Monaco et al., 2012)
quando submetidas aos protocolo de
diferenciação osteogênica in vitro.
Entretanto, existem também estudos em
humanos (De Ugarte et al., 2003; Noel et
al., 2008; De Ugarte et al., 2003; Noel et
al., 2008; Reberatto et al., 2008; Pachon-
Pena et al., 2011), equinos (Ranera et al.,
2012), cães (Chung et al., 2012; Spencer et
al., 2012) e ratos (Peng et al., 2008) que não
mostram diferenças significativas na
osteogênese “in vitro’ (De Ugarte et al.,
2003; Lee et al., 2004; Noel et al., 2008;
Reberatto et al., 2008; Pachon-Pena et al.,
2011; Wen et al., in press) e nem na
expressão de marcadores de diferenciação
osteogênica (De Ugarte et al., 2003; Lee et
al., 2004; Peng et al., 2008; Reberatto et al.,
2008; Pachon-Pena et al., 2011; Chung et
al., 2012; Ranera et al., 2012; Spencer et al.,
2012; Wen et al., in press) entre as CTM-
MO e CTM-AD. Essa divergência entre os
resultados pode ser justificada pelas
variações na metodologia de cada estudo,
pois vários fatores relacionados ao cultivo e
diferenciação “in vitro” da CTM como
protocolo de isolamento das CTM
(principalmente ligados ao método de
dissociação celular) (Sethe et al., 2006),
meio de cultura utilizado (Schwarz et al.,
2012), suplementos utilizados no meio de
cultura para diferenciação celular (Bellows
et al., 2003; Fiorentini et al., 2011),
protocolos de transfecção (Lin et al., 2009)
e outros fatores relacionados com o
microambiente de cultivo como a presença
de biomateriais (Makhluf et al., 2000),
tensão de oxigênico (Chung et al., 2012),
pressão (Park et al., 2012) podem alterar os
potenciais proliferativos e osteogênico das
CTM.
O potencial proliferativo das CTM é um
importante fator para a osteogênese “in
vitro” e “in vivo”, pois uma célula mais
proliferativa pode originar um maior

34
número de células diferenciadas e assim
produzir mais matriz extracelular e
favorecer a osteogênese e a regeneração
óssea quando utilizada clinicamente para
esse fim. Estudos comparativos entre o
potencial proliferativo das CTM-MO e das
CTM-AD em humanos (De Ugarte et al.,
2003; Skaguchi et al., 2005; Lee et al.,
2004; Kern et al., 2006; Izadpanah et al.,
2006; Shafiee et al., 2011; Dmitrieva et al.,
2012; Wen et al., in press), equinos (Ranera
et al., 2012), macacos (Izadpanah et al.,
2006), cães (Chung et al., 2012), coelhos
(Lin et al., 2009) e ratos (Yoshimura et al.,
2007; Peng et al., 2008; Nakanishi et al.,
2011) são encontrados na literatura recente.
A maioria deles mostrou que as CTM-AD
apresentam um maior potencial
proliferativo que as CTM-MO (Lee et al.,
2004; Izadpanah et al., 2006; Kern et al.,
2006; Yoshimura et al., 2007; Peng et al.,
2008; Lin et al., 2009; Chung et al., 2012;
Dmitrieva et al., 2012; Wen et al., in press),
no entanto, alguns observaram maior
potencial proliferativo nas CTM-MO
(Skaguchi et al., 2005; Izadpanah et al.,
2006; Nakanishi et al., 2011) e outros não
encontraram diferença significativa (De
Ugarte et al., 2003; Shafiee et al., 2011;
Ranera et al., 2012).
Fatores como a idade (Zhang et al., 2009;
Veronesi et al., 2011; Kim et al., 2012;
Zaim et al., 2012), o gênero (Muschler et
al., 2001; Aksu et al., 2008; Jiang et al.,
2008) e os hábitos alimentares do doador
(Zhou et al., 2010; Ko et al., 2011), o uso
de alguns medicamentos como os anti-
inflamatórios (Molinuevo et al., 2010;
Choi et al., 2011; Muller et al., 2011) e
doenças (Jiang et al., 2008; Veronesi et al.,
2011) podem reduzir o potencial
osteogênico das CTM mas, a senescência
(idade in vitro) é considerada a principal
causa de redução e perda do potencial
osteogênico das CTM “in vitro” (Bonab et
al., 2006).
A senescência é a perda da capacidade
proliferativa que ocorre fisiologicamente
com o aumento do número de passagens
durante o cultivo (Mosna et al., 2010). Com
o aumento do tempo de cultura, as CTM
passam a acumulam vacúolos lipídicos no
seu interior, diminuem a taxa de
crescimento até perderem totalmente sua
capacidade mitótica e entrarem em
apoptose (Mosna et al., 2010). O principal
mecanismo envolvido na senescência
celular parece ser o encurtamento do
telômero. A cada replicação a célula perde
um segmento do telômero até chegar a um
comprimento mínimo e quando isso ocorre,
a célula perde sua capacidade mitótica
(Sethe et al., 2006). A atividade de uma
enzima denominada telomerase está
diretamente relacionada ao processo de
manutenção ou encurtamento do telômero
pois essa enzima é capaz de sintetizar novos
segmentos de telômero reduzindo assim seu
encurtamento durante a replicação celular
(Sethe et al., 2006). Essa enzima pode ser
usada como parâmetro para avaliação da
senescência das CTM. A perda gradual do
telômero com o aumento do número de
passagens já foi comprovada em CTM
humanas mantidas “in vitro” (Stenderup et
al., 2003; Bonab et al., 2006). Alguns
estudos em humanos (De Ugarte et al.,
2003; Izadpanah et al., 2006; Kern et al.,
2006; Dmitrieva et al., 2012), macacos
(Izadpanah et al., 2006), cães (Chung et al.,
2012) e ratos (Peng et al., 2008) avaliaram
parâmetros relativos à senescência das
CTM-AD e CTM-MO. A maioria deles
mostrou que as que as CTM-AD são mais
resistentes à senescência que as CTM-MO
(Izadpanah et al., 2006; Kern et al., 2006;
Peng et al., 2008; Chung et al., 2012;
Dmitrieva et al., 2012), no entanto, em um
estudo em macacos observou-se maior
resistência à senescência nas CTM-MO
(Izadpanah et al., 2006) e outro em
humanos não verificou-se diferença
significativa entre as duas cultura (De
Ugarte et al., 2003). As CTM-AD e CTM-
MO também apresentam diferenças quanto
a resistência à apoptose quando cultivadas
“in vitro” (Ranera et al., 2012) e quando
submetidas a situações adversas como
estresse oxidativo (Ertas et al., 2012) e
privação de nutrientes como o soro fetal
bovino (Peng et al., 2008; Ertas et al.,
2012). A maioria dos estudos mostra que as
a CTM-AD são mais resistentes à apoptose
quando mantidas em cultura “in vitro”
(Ranera et al., 2012) e quando submetidas
ao estresse oxidativo (Ertas et al., 2012) e à
privação sérica (Peng et al., 2008; Ertas et
al., 2012). Entretanto, outro estudo com
CTM de ratos observou maior resistência
das CTM-MO quando submetidas ao

35
estresse oxidativo (Peng et al., 2008). As
CTM-AD apresentam também maior
resistência a hipóxia que as CTM-MO
(Chung et al., 2012) e são genética
(Adegani et al., 2012) e morfologicamente
mais estáveis ao longo da cultura, o que
mantem o potencial de diferenciação dessas
células por um período mais prolongado
(Izadpanah et al., 2006). Um estudo sobre a
capacidade imunomoduladora das CTM-
AD e CTM-MO não mostrou diferença
significativa entre os dois tipos celulares
(Niermeyer et al., 2007). O rendimento de
CTM por grama de tecido é outro fator a ser
considerado quando se pesquisa a melhor
fonte de CTM para utilização clínica. A
maioria dos estudos mostra maior
rendimento de CTM nos tecido adiposo em
comparação com a medula óssea
(Yoshimura et al., 2007; Hayashi et al.,
2008; Chung et al., 2012; Hass et al., 2011).

36
Capítulo 2: Estudo comparativo da
diferenciação osteogênica das células
tronco mesenquimais da medula óssea e
do tecido adiposo de cães adultos.
Introdução
O cultivo celular bem sucedido e o
subsequente uso das células para tratamento
de diferentes alterações é o propósito das
pesquisas na área de terapia celular e, por
ser um tema relativamente recente, muitos
questionamentos ainda carecem de
respostas. Sabe-se que existem diferentes
populações de células tronco mesenquimais
(CTM), com distintas capacidades
proliferativa e de diferenciação no mesmo
indivíduo (Jiang et al., 2002; Crisan et al.,
2009). Essas populações são também
desigualmente distribuídas nos tecidos, o
que determina maior ou melhor potencial
proliferativo e de diferenciação das CTM
isoladas de locais diferentes no mesmo
indivíduo (Hass et al., 2011; Strioga et al.,
2012). Estudos já mostraram que o
potencial osteogênico das CTM pode variar
de acordo com o tecido de origem (Im et
al., 2005; Hayashi et al., 2007; Yoshimura
et al., 2007; Peng et al., 2008; Zaminy et
al., 2008; Filioli-Uranio et al., 2011;
Shafiee et al., 2011; Al-Nbaheen et al.,
2012; Dmitrieva et al., 2012; Park et al.,
2012; Monaco et al., 2012; Wen et la., in
press), com o local de colheita (Aksu et al.,
2008; Neupane et al., 2008), com a espécie
animal (Arrigoni et al., 2009; Levi et al.,
2011), com o gênero (Muschler et al., 2001;
Aksu et al., 2008; Jiang et al., 2008) e com
a idade (Zhang et al., 2009; Veronesi et al.,
2011; Al-Nbaheen et al., 2012; Kim et al.,
2012; Zaim et al., 2012). Portanto, para
terapias de regeneração óssea é importante
se conhecer a melhor fonte de CTM.
A medula óssea (Im et al., 2005; Lee et al.,
2011) e o tecido adiposo (Neupane et al.,
2008; Viera et al., 2010) são considerados
os principais sítios para obtenção dessas
células, devido à maior quantidade e
facilidade de colheita e isolamento. No
entanto, ainda não se sabe ao certo qual
delas é a melhor fonte de CTM para
terapias de regeneração óssea. Muitos
estudos a respeito já foram realizados em
humanos (De Ugarte et al., 2003; Lee et al.,
2004; Im et al., 2005; Skaguchi et al., 2005;
Izadpanah et al., 2006; Liu et al., 2007;
Noel et al., 2008; Reberatto et al., 2008;
Pachon-Pena et al., 2011; Shafiee et al.,
2011; Park et al., 2012; Wen et al., in press)
e ratos (Yoshimura et al., 2007; Hayashi et
al., 2008; Peng et al., 2008; Zaminy et al.,
2008; Nakanishi et al., 2011) mas, pouco se
sabe sobre o potencial osteogênico dessas
células em outras espécies como o cão
(Chung et al., 2012; Spencer et al., 2012).
Alguns desses estudos sugerem que as
CTM oriundas da medula óssea (CTM-MO)
possuem maior potencial osteogênico que
as CTM do tecido adiposo (CTM-AD) (Im
et al., 2005; Skaguchi et al., 2005; Liu et
al., 2007; Yoshimura et al., 2007; Hayashi
et al., 2008; Zaminy et al., 2008; Nakanishi
et al., 2011; Shafiee et al., 2011; Park et al.,
2012; Monaco et al., 2012), outros indicam
maior potencial osteogênico nas CTM-AD
(Lin et al., 2009 ; Chung et al., 2012) e
existem aqueles que mostram potencial
osteogênico semelhante entre os dois tipos
celulares (De Ugarte et al., 2003; Lee et al.,
2004; Noel et al., 2008; Peng et al., 2008;
Reberatto et al., 2008; Pachon-Pena et al.,
2011; Spencer et al., 2012; Ranera et al.,
2012; Wen et al., in press). Assim, o
objetivo do presente estudo foi avaliar
quantitativamente o potencial osteogênico
das CTM-MO e das CTM-AD de cães
adultos jovens, visando identificar a melhor
fonte de CTM para terapias de regeneração
óssea em cães.
Material e métodos
Foram utilizadas as bases físicas e a infra-
estrutura do Centro de Experimentação de
Pequenos Animais e do bloco cirúrgico do
Hospital Veterinário, do Núcleo de Células
Tronco e Terapia Celular e dos
Laboratórios de Biologia Molecular e de
Histopatologia e Imunoistoquímica do
Departamento Clínica e Cirurgia
Veterinárias da Escola de Veterinária da
UFMG, do Laboratório de Biologia
Molecular do Departamento de Imunologia
e Bioquímica do Instituto de Ciências
Biológicas da UFMG. O projeto foi
aprovado pelo Comitê de Ética em
Experimentação Animal (CETEA) da
UFMG1.
1 Número de protocolo no CETEA 157/2009

37
Animais
Foram utilizados três cães, machos, não
castrados, sem raça definida com dois anos
de idade e massa corporal média de 18 Kg.
Os cães foram recolhidos em fazendas da
zona rural do município de Lavras quando
filhotes e criados até os dois anos de idade
para realização do presente estudo. No
período experimental os cães foram
mantidos em canis individuais tipo solário,
com área de 4,5m2, pertencentes ao Centro
de Experimentação de Pequenos Animais
do Hospital Veterinário da Escola de
Veterinária da UFMG, onde receberam
ração comercial2 e água a vontade. Todos
os animais passaram por um período
mínimo de 30 dias de adaptação às novas
instalações. Nessa fase eles foram avaliados
clinicamente e por meio de exames
radiográficos e laboratoriais (hemograma,
pesquisa de hemoparasitas e análise
sorológica para leishmaniose), assegurando-
se que todos estavam saudáveis e aptos para
serem utilizados no presente estudo. Todos
os animais foram desverminados, vacinados
e receberam medicação ectoparasiticida.
Colheita das amostras de medula óssea e
do tecido adiposo
As colheitas das amostras de medula óssea
e de tecido adiposo foram realizadas no
centro cirúrgico do Hospital Veterinário da
Escola de Veterinária da Universidade
Federal de Minas Gerais (UFMG).
Para a colheita das amostras, os cães foram
submetidos apenas ao jejum sólido de oito
horas. Como medicação pré-anestésica,
foram empregados 1mg/kg IM de xilazina3
e 15mg/kg IM de quetamina4, na mesma
seringa. O membro pélvico direito foi
submetido à tricotomia desde a coluna
lombo-sacra até o tarso e preparado para
cirurgia asséptica. A veia cefálica foi
canulada e os animais receberam propofol5
(3mg/kg IV) para intubação e manutenção
anestésica. Como analgésico e anti-
inflamatório foi administrado meloxicam6
2 Canitos, Kowalski, Apucarana, PR, Brasil 3 Calmiun, Agener União, Embu-Guaçu, SP, Brasil 4 Vetanarcol, Konig, Santana do Parnaíba, SP, Brasil 5 Fresofol, Fresenius Kabi, Campinas, SP, Brasil 6 Maxicam, Ouro Fino, Cravinhos, SP, Brasil
(0,2mg/kg IM) imediatamente após a
indução anestésica.
A coleta da medula óssea foi realizada por
punção aspirativa no platô tibial, pelo
acesso medial, entre o ligamento patelar e o
ligamento colateral medial, com agulha
16G com mandril, acoplada a uma seringa
de 10 mL contendo 0,5 mL de heparina
sódica7 na concentração de 5000 UI/mL.
Foi coletado 1 mL de medula óssea de cada
animal, totalizando 3mL de amostra final.
As amostras de tecido adiposo foram
coletadas cirurgicamente, removendo tecido
adiposo subcutâneo da região glútea
imediatamente acima do trocânter maior.
Foi coletado aproximadamente 1cm3 de
tecido adiposo de cada animal totalizando
cerca de 3cm3 de amostra final.
Imediatamente após a coleta as amostras de
medula e de tecido adiposo foram colocadas
separadamente em tubos falcon de 50 mL
contendo 20 mL de meio de cultura
Dulbecco’s Modiffed Eagle Medium baixa
glicose8 (DMEM), à temperatura ambiente
e encaminhadas para sala de cultura para
realização dos protocolos de extração para
cada tipo de tecido. O intervalo entre a
colheita e o processamento foi de cerca de
60 minutos.
As amostras de tecido adiposo e de medula
óssea foram processadas separadamente
para a obtenção de células tronco
mesenquimais, constituindo-se dois grupos,
células tronco mesenquimais extraídas da
medula óssea de cães (CTM-MO) e células
tronco mesenquimais extraídas do tecido
adiposo de cães (CTM-AD).
Posteriormente, foi determinado o potencial
osteogênico das CTM-AD e CTM-MO.
Cada tipo de célula foi cultivado em
DMEM e em meio de diferenciação
osteogênica por sete, 14 e 21 dias. Após
esses períodos, foram realizados os testes
de conversão do brometo de [3-(4,5-
dimetiltiazol-2yl)-2,5-difenil tetrazolium]
(MTT) em cristais de formazan, atividade
da fosfatase alcalina (FA), síntese de
colágeno, quantificação da área
mineralizada por campo e avaliação da
expressão relativa de osterix (OSX),
sialoproteína óssea (BSP), osteonectina
(ON) e osteocalcina (OC) por RT-PCR em
7 Parinex, Hipolabor, Belo Horizonte, MG, Brasil 8 DMEM, Gibco, Grand Island, NY, USA

38
tempo real. Os protocolos estão descritos
detalhadamente a seguir.
Isolamento, cultivo e expansão de células
tronco mesenquimais
Extração e cultivo de células tronco
mesenquimais da medula óssea e do
tecido adiposo
No Núcleo de Células Tronco da Escola de
Veterinária da UFMG a amostra de medula
óssea foi centrifugada por 10 minutos a
1400rpm e levada para capela de fluxo
laminar para manipulação asséptica. O
sobrenadante foi desprezado e o pellet de
células foi ressuspenso em 20 mL de meio
de cultura para células tronco
mesenquimais (meio basal), DMEM com
baixa glicose enriquecido com gentamicina
(60 μg/L), penicilina (100 U/mL),
estreptomicina (100μg/mL), anfotericina
(25μg/mL)9 e 10% de soro fetal bovino
10.
Uma alíquota de 2 mL do meio com as
células ressuspensas foi transferida para
cada garrafa de cultivo celular T7511
e 8 mL
de meio foram adicionados em cada
garrafa. Foram feitas dez garrafas.
Realizou-se homogeneização e as garrafas
foram incubadas em estufa a 37oC e 5% de
CO2. Após 48h as células foram lavadas
duas vezes com PBS 0,15 molar para
remoção das hemácias e demais células não
aderidas. O meio de cultivo foi trocado duas
vezes por semana.
A extração das células tronco
mesenquimais de tecido adiposo foi
realizada segundo o protocolo descrito a
seguir que foi adaptado de protocolos
estabelecidos por Neupane et al. (2008) e
Vieira et al. (2010).
As amostras de tecido adiposo foram
também manipuladas na capela de fluxo
laminar. As amostras foram lavadas duas
vezes com PBS 0,15 molar para retirada de
restos de sangue e debris celulares. Para
digestão do tecido conjuntivo e liberação
das células, as amostras foram colocadas
em outro tubo falcon de 50 mL contendo 20
mL de uma solução estéril de colágenase
9 PSA, Sigma-Aldrich, St. Louis, MO, USA 10 Soro fetal bovino, Sorali, Campo Grande, MS, Brasil 11 Garrafa T75, TPP (Techno Plastic Products in
Trasadingen), Switzerland, Germany
B12
0,1% P/V. As amostras foram
fragmentadas a partículas menores com
tesoura cirúrgica estéril. A tesoura foi
introduzida no tubo falcon de 50mL e com
movimentos de abrir e fechar dos ramos o
tecido foi fragmentado ao máximo possível.
Em seguida a amostra foi encubada em
estufa a 37oC e 5% de CO2 durante 45
minutos e a cada 15 minutos a amostra foi
vigorosamente agitada para facilitar o
processo de digestão. Transcorrido o tempo
de digestão, a amostra foi centrifugada por
10 minutos a 1400rpm, o sobrenadante
contendo a fração adiposa foi descartado e
o pellet foi ressuspenso em 10 mL PBS
0,15 molar para lavagem e retirada dos
resíduos de colagenase. A amostra foi
colocada em outro tubo falcon para reduzir
a contaminação por gordura retida na
parede do tubo e novamente centrifugada
por 10 minutos a 1400rpm. Após a
centrifugação o sobrenadante foi novamente
descartado e o pellet, fração estromal, foi
ressuspenso em 20 mL de meio de cultura
para células tronco, da mesma forma como
descrito para as células extraídas da
medula. Uma alíquota de 2 mL do meio
com as células ressupensas foi transferida
para cada garrafa de cultivo celular T75 e 8
mL de meio foram adicionados em cada
garrafa. Foram feitas dez garrafas no total.
Realizou-se homogeneização e as garrafas
foram incubadas em estufa a 37oC e 5% de
CO2. Após 48h foi realizada troca parcial
do meio de cultura, renovando apenas 5
mL. Após mais 48h as células foram
lavadas duas vezes com PBS 0,15 molar
para remoção de debris e células não
aderidas. O meio de cultivo foi trocado duas
vezes por semana.
Assim que foi atingido 80 a 90% de
confluência, as células de cada grupo foram
separadamente repicadas. Para
desprendimento das células, as garrafas
foram lavadas com 10 mL de PBS 0,15
molar para retirada dos restos de meio de
cultura. Após a lavagem, foram adicionados
1,5 mL de tripsina e a garrafa foi incubada
por 10 minutos em estufa a 37oC e 5% de
CO2. Transcorrido o tempo de ação da
tripsina a mesma foi inativada com 5 mL de
DMEM enriquecido com 10% de soro fetal
12 Colagenase B, Roche Applied Science, Penzberg,
Germany

39
bovino. As garrafas foram então lavadas
com o próprio meio para retirada das
células e uma alíquota de 1 mL foi repicada
para outra garrafa T75 com mais 9 mL de
meio de cultura. Após o terceiro repique
ambos os tipos celulares foram
caracterizados fenotipicamente quanto à
expressão de CD90, CD29, CD45 e CD34 e
submetidas à diferenciação osteogênica por
sete, 14 e 21 dias.
Caracterização Fenotípica
As células de cada grupo foram submetidas
separadamente à ação da tripsina e
ressuspensas em 1 mL de solução PBS
0,15M + 10% SFB + 1% azida sódica à
temperatura de 4oC para fixação, evitando a
internalização dos epítopos de interesse.
Elas foram contadas em câmara de
Neubauer e plaqueadas na concentração de
1x105
células por poço em placas de 96
poços de fundo redondo. Foram feitos dois
poços para controle negativo, dois poços
para controle de marcação inespecífica só
com anticorpo secundário conjugado com
FITC13
e dois poços para cada marcador
CD9014
, CD29-PE15
, CD4516
e CD34-PE17
.
Após o plaqueamento, as células foram
centrifugadas a 1400 rpm durante 10
minutos, o sobrenadante foi descartado e
adicionou-se 100µL de anticorpo primário
para cada marcador. A placa foi incubada
protegida da luz, a 4oC por 30 minutos. Nos
controles negativos e para marcação
inespecífica foram adicionados 100µL de
PBS 0,15M + 3% BSA. Utilizou-se as
diluições de 1/5 para o CD29-PE, 1/10 para
o CD34-PE e 1/500 para o CD90 e CD45.
Esta diluição foi feita com solução de PBS
0,15M + 3% BSA. Transcorrido o período
de incubação, as células foram
centrifugadas por 10 min a 1400 rpm e o
sobrenadante foi descartado. As células
foram então submetidas a três lavagens com
200µL de PBS 0,15M + 3% BSA a 4oC
13 Anticorpo secundário policlonal de coelho anti IgG de
rato conjugado com FITC (ab6730), abcam, Cambridge,
Estados Unidos. 14 Anticorpo monoclonal de rato antiCD90 (ab22541),
abcam, Cambridge, Estados Unidos. 15 Anticorpo monoclonal de camundongo antiCD29
(ab64629), abcam, Cambridge, Estados Unidos. 16 Anticorpo monoclonal de rato antiCD45 (ab22514), abcam, Cambridge, Estados Unidos. 17 Anticorpo monoclonal de rato antiCD34 (ab42902),
abcam, Cambridge, Estados Unidos.
para retirada dos anticorpos não ligados.
Em seguida adicionou-se 100µL do
anticorpo secundário conjugado com FITC
nos poços referentes ao controle de ligação
inespecífica, e anticorpos primários não
conjugados com fluoróforo CD90 e CD45.
Os demais poços, controle negativo, CD29-
PE e CD34-PE receberam 100µL de PBS
0,15M + 3% BSA a 4oC. As células foram
incubadas por mais 30 minutos a 4oC,
protegidas da luz. Após o período de
incubação as células foram centrifugadas
por 10 min a 1400 rpm, o sobrenadante foi
descartado e as células foram submetidas a
mais três lavagens com 200µL de PBS
0,15M + 3% BSA a 4oC para retirada dos
anticorpos não ligados. Em seguida as
células foram ressuspensas em 200µL de
PBS 0,15M + 10% SFB + 1% azida sódica
a 4oC, por poço, e imediatamente
encaminhadas para leitura no citômetro de
fluxo FACScan (Fluorescence Activated
Cell Analyser), empregando-se o software
Cell Quest, com aquisição de 20000
eventos, tendo como parâmetros FSC e SSC
em escala linear e FL1 (para FIFC) e FL2
(para PE) em escala logarítmica. Os dados
foram analisados pelo programa WinMDI
por gráficos dot plot.
Osteoblastos caninos
Como controle positivo da diferenciação
osteogênica foi utilizado uma cultura
primária de osteoblastos caninos18
. A
cultura foi descongelada conforme
protocolo sugerido pelo fabricante, os
osteoblastos foram cultivados em garrafas
T75, com os mesmos meios e sob as
mesmas condições das CTM-MO e CTM-
AD. Após o quarto repique e obtenção de
80 a 90 % de confluência as osteoblastos,
foram utilizados para extração do RNA
total e posterior análise da expressão de
osterix, sialoproteína óssea, osteonectina e
osteocalcina pela técnica de RT-PCR em
tempo real.
18 CnOb - canine osteoblasts, Cell Application , San Diego,
CA , USA

40
Teste de viabilidade celular pelo azul de
Tripan
Após a caracterização fenotípica e antes do
cultivo em meio de diferenciação, as CTM
de cada grupo experimental foram avaliadas
quanto à viabilidade celular pelo azul de
Tripan. As CTM foram cultivadas em
garrafas T75 (1x104 células/cm
2) com
DMEM e no momento do teste foram
lavadas com PBS (0,15M) e submetidas à
ação da tripsina. O sobrenadante contendo
as células foi colhido e centrifugado a
1400g por 10 minutos, as células foram
ressuspensas em meio basal e coradas pelo
azul de Tripan. As células viáveis
(transparentes) e inviáveis (em azul) de
cada grupo foram quantificadas em câmara
de Neubauer.
Cultivo de células tronco mesenquimais
da medula óssea e do tecido adiposo em
meio de diferenciação adipogênica
Após o terceiro repique as células de cada
grupo foram submetidas separadamente à
ação da tripsina e plaqueadas na densidade
de 1x104 células/cm
2, em seis repetições em
placas de seis poços. Após a obtenção de 60
a 70% de confluência celular o meio basal
foi substituído por meio adipogênico que é
constituído do DMEM enriquecido com
soro fetal bovino (10%), dexametasona
(1M), insulina (10L/mL), indometacina
(100mM) e isobutilmetilxantina (500mM).
Em seguida as células foram cultivadas a
37oC e 5% de CO2 por 21 dias e o meio de
cultura trocado a cada quatro dias.
Transcorrido esse período as células foram
fixadas em formalina 10% por 60 minutos,
coradas pela técnica de Oil Red e avaliados
por microscopia óptica, para confirmação
da diferenciação adipogênica.
Cultivo de células tronco mesenquimais
da medula óssea e do tecido adiposo em
meio de diferenciação condrogênica
Após o terceiro repique as células de cada
grupo foram submetidas separadamente à
ação da tripsina e 1x105 células de cada
grupo foram colocadas em tubos falcon de
15 ml e cultivadas em meio basal por 24
horas. Após esse período o meio basal foi
substituído por meio condrogênico que é
constituído do DMEM enriquecido com
soro fetal bovino (1%), albumina sérica
bovina (0,0125g/mL), piruvato (100M),
insulina (6,25g/mL), transferrina
(6,25g/mL), ácido ascórbico (50gM),
dexametasona (100nM) e TGF-β1
(10ng/mL). Em seguida as células foram
cultivados a 37oC e 5% de CO2 por 21 dias
e o meio de cultura trocado a cada quatro
dias. Transcorrido esse período os pellets
celulares foram fixados em formalina 10%
por 60 minutos e processados pela técnica
rotineira de inclusão em parafina. Cortes
histológicos com 5m de espessura foram
corados pela técnica de PAS e avaliados por
microscopia óptica, para confirmação da
diferenciação condrogênica.
Cultivo de células tronco mesenquimais
da medula óssea e do tecido adiposo em
meio de diferenciação osteogênica
Após o terceiro repique as células de cada
grupo foram submetidas separadamente à
ação da tripsina e plaqueadas na densidade
de 1x104 células/cm
2, em quatro repetições,
em garrafas T25 e em seis repetições em
placas de seis e 24 poços19
de acordo com a
especificação de cada teste. Após a
obtenção de 60 a 70% de confluência
celular o meio basal foi substituído por
meio osteogênico que é enriquecido com
ácido ascórbico (50 g/mL), ß-
glicerofosfato (10 mM)20
e dexametasona
(0,1 M)21
, acrescido de 10% de soro fetal
bovino. Em seguida as células foram
cultivadas a 37oC e 5% de CO2 por sete, 14
e 21 dias. Transcorridos esses períodos
foram avaliados a conversão do MTT em
cristais de formazan, a atividade da
fosfatase alcalina, a síntese de colágeno, a
área celular e mineralizada por campo e a
expressão relativa dos transcritos gênicos
para osterix (OSX), sialoproteina óssea
(BSP), osteonectina (ON) e osteocalcina
(OC) por RT-PCR em tempo real.
19 TPP (Techno Plastic Products in Trasadingen), Switzerland, Germany 20 Sigma-Aldrich, St. Louis, MO, USA 21 Aché, Guarulhos, SP, Brasil

41
Teste de conversão do MTT em cristais
de formazan
Para o teste de conversão do MTT as CTM
de cada grupo foram cultivadas
separadamente em placas de 24 poços com
DMEM e meio osteogênico e ao término de
cada período, submetidas ao teste de
conversão do MTT {brometo de [3-(4,5-
dimetiltiazol-2yl)-2,5-difenil tetrazolium]}
em cristais de formazan. O meio foi
substituído por 210L de meio osteogênico
com soro fetal bovino em cada poço e
170L de MTT (5mg/mL)22
. A placa foi
incubada por duas horas em estufa a 37oC e
5% de CO2. Os cristais de formazan foram
observados ao microscópio antes do
acréscimo de 210L de SDS (sódio dodecil
sulfato)-10% HCl que permaneceu
overnight em estufa a 37oC e 5% de CO2.
Posteriormente, 100L de cada poço foram
transferidos para placas de 96 poços para
análise na leitora de placas com
comprimento de onda de 595nm de acordo
com Valério et al. (2004).
Avaliação da atividade da fosfatase
alcalina
Para avaliação da atividade da fosfatase
alcalina as CTM de cada grupo foram
cultivadas separadamente em placas de 24
poços com DMEM e meio osteogênico. Ao
término de cada período, as culturas foram
lavadas com PBS (0,15 molar). Em cada
poço, foram acrescentados 200L de
solução de BCIT/NBT23
(1mL de tampão
da fosfatase alcalina, 4,4L de NBT {nitro-
blue tetrazolium chloride} e 3,3L de BCIP
{5-bromo-4-chloro-3’-indolylphosphate p-
toluidine salt}). As amostras ficaram duas
horas na estufa a 37oC e 5% de CO2 e foram
observadas ao microscópio óptico antes do
acréscimo de 200L de SDS (sódio dodecil
sulfato)-10% HCl. Estas amostras
permaneceram overnight em estufa a 37oC e
5% de CO2. Posteriormente, 100L de cada
poço foram transferidos para placas de 96
poços para leitura em espectrofotômetro
com comprimento de onda de 595nm de
acordo com Ocarino et al. (2008) e Boeloni
et al. (2009).
22 Invitrogen, Carlsbad, CA, USA 23 Zymed Laboratories, CA, USA
Avaliação da síntese de colágeno
Para dosagem do colágeno as CTM de cada
grupo foram cultivadas separadamente em
placas de 24 poços com DMEM e meio
osteogênico. Ao término de cada período,
as culturas foram lavadas com PBS (0,15
molar). Adicionou-se em seguida, 1ml de
Bouin em cada poço, para fixação e a placa
foi incubada por duas horas em estufa a
37oC e 5% de CO2. As placas foram então
retiradas da estufa e colocada overnight a
6oC em geladeira. Transcorrido esse
período as placas foram lavadas quatro
vezes com água osmose reversa e secas
para posterior coloração com Sirius Red
durante 30 minutos à temperatura ambiente.
O excesso do corante foi removido e as
células foram lavadas três vezes com
solução de HCl 0,01N e secas.
Posteriomente foram adicionados 300L de
NaOH 0,5M e a placa foi incubada por mais
30 minutos. Em seguida, 100L de cada
poço foram transferidos para placas de 96
poços para leitura em espectrofotômetro
com comprimento de onda de 540nm de
acordo com Ocarino et al. (2008) e Boeloni
et al. (2009).
Determinação da porcentagem de células
por campo
Para a determinação da porcentagem de
células por campo as CTM de cada grupo
foram cultivadas separadamente em placas
de seis poços com lamínulas (22x22mm)
estéreis, com DMEM e meio osteogênico.
Ao término desse período, as culturas foram
fixadas em álcool 70% e submetidas à
coloração por hematoxilina-eosina (Prophet
et al., 1992). Foi determinada a
porcentagem de células por campo com o
auxílio de uma ocular micrométrica,
contendo uma gratícula com 121 pontos,
em 25 campos, com objetiva de 4.
Avaliação dos nódulos de mineralização e
determinação da área mineralizada por
campo
Para a determinação da porcentagem de
área mineralizada por campo, as CTM de
cada grupo foram cultivadas separadamente
em placas de seis poços com lamínulas

42
(22x22mm) estéreis, com DMEM e meio
osteogênico. Após esse período, as CTM
foram lavadas com PBS 0,15M, fixadas
com álcool 70% por 24 horas e coradas pelo
método de Von Kossa (adaptado de Prophet
et al., 1992) para avaliação da porcentagem
de área mineralizada por campo com o
auxílio de uma ocular micrométrica,
contendo uma gratícula com 121 pontos,
em 25 campos, com objetiva de 4.
Quantificação relativa dos transcritos
gênicos para osterix, sialoproteína óssea,
osteonectina e osteocalcina por RT-PCR
em tempo real
A extração do RNA total das células foi
feita em quatro garrafas T25 por grupo com
o emprego do Trizol24
. O método de
extração consistiu de uma etapa inicial de
lise e homogeneização da monocamada de
células por cinco minutos à temperatura
ambiente, para completa dissociação dos
complexos nucleoprotéicos. O lisado foi
transferido para um microtubo de 1,5 mL e
foram adicionados 0,2 mL de clorofórmio,
seguido de 15 segundos de
homogeneização, três minutos de incubação
à temperatura ambiente e centrifugação a
12.000 g por 15 minutos à 4ºC, para
separação em três fases, onde a fase incolor
superficial (aquosa) continha o RNA. Na
terceira etapa, a fase aquosa foi transferida
para um novo tubo, acrescido de 0,5mL de
álcool isopropílico e incubada por 30
minutos à temperatura -80ºC. Em seguida, a
solução foi descongelada e centrifugada a
12.000g por 10 minutos a 4ºC para
precipitação do RNA. O pellet foi então
lavado com 1 mL de etanol a 75%,
homogeneizado e centrifugado a 7.500g por
cinco minutos a 4ºC. O RNA foi
solubilizado em água DEPC25
livre de
RNAse e imediatamente armazenado a -
80ºC. A concentração de RNA foi
determinada pela leitura da absorbância a
260/280 nm, por espectrofotometria. Foram
realizadas as reações de transcrição reversa
utilizando-se Kit comercial26
. Utilizou-se 1
µg de RNA total para a síntese de cDNA
24 Invitrogen, USA 25 Água tratada com dimetil pirocarbonato (DEPC), Invitrogen, USA 26 SuperScriptTM III Platinum® Two-Step qRT-PCR Kit
with SYBR Green, Invitrogen, CA, USA
com um volume final de 20 µL. As reações
de PCR foram feitas em tempo real,
utilizando-se 2µg de cDNA, 5pM de cada
iniciador e 12,5µL do reagente syber
Green27
em um volume final de 25 µL de
reação em um tubo28
, no aparelho
SmartCycler System29
. Os parâmetros
utilizados para amplificação foram: 50°C
por 120 segundos, 95°C por 150 segundos e
45 ciclos, 95°C por 15 segundos e 60°C por
30 segundos. Os iniciadores foram
pesquisados na literatura ou delineados com
base na sequência do mRNA Canis
familiares (Tab. 1). A expressão gênica foi
calculada usando o método 2-∆∆CT
, onde os
resultados obtidos para cada grupo foram
comparados quantitativamente após a
normalização baseada na expressão de
gliceraldeído-3-fosfato desidrogenase
(GAPDH) Canis familiares. Os níveis de
expressão obtidos nas culturas de
osteoblastos foram utilizados como controle
positivo da diferenciação osteogênica e
como padrão de expressão no cálculo da
expressão relativa de cada transcrito.
Análise estatística
Realizou-se análise de variância (ANOVA)
e para cada variável foram determinados a
média e o desvio padrão. As médias foram
comparadas pelo teste SNK utilizando o
programa Graphpad Instat 330
. Diferenças
foram consideradas significativas para
p0,05 (Sampaio, 1998).
27 SuperScriptTM III Platinum® Two-Step qRT-PCR Kit
with SYBR Green, Invitrogen, CA, USA 28 SmartCycler® Tube-25µL, Cepheid, Sunnyvale, CA,
USA 29 SmartCycler® System, Cepheid, Sunnyvale, CA, USA 30 GraphPad Software Inc., San Diego, USA

43
Tabela 1: Lista de genes e sequência de nucleotídeos dos iniciadores para RT-PCR em tempo real.
Genes
Referência ou
No. de acesso
Oligonucleotídios iniciadores
(sequências de nucleotídeos 5’ a 3’)
Temperatura de
anelamento (◦C)
Tamanho do
produto
(pares de base)
Osterix (OSX)
(Neupane et al.,
2008)
F- ACGACACTGGGCAAAGCAG
R- CATGTCCAGGGAGGTGTAGAC
60 285
Sialoproteina
óssea (BSP)
(Vieira et al.,
2010)
F- TTGCTCAGCATTTTGGGAAT
R- AACGTGGCCGATACTTAAAGAC
60 295
Osteonectina
(ON)
(XM_849889.1)
F- GCCTTGGCAGCCCCTCAACA
R- CACCTGCACGGGGTTGGCTC
60 108
Osteocalcina
(OC)
(Neupane et al.,
2008)
F- GAGGGCAGCGAGGTGGTGAG
R- TCAGCCAGCTCGTCACAGTTGG
62 134
GAPDH
(Vieira et al.,
2010)
F- CCATCTTCCAGGAGCGAGGAT
R- TTCTCCATGGTGGTGAAGAC
60 97
Resultados
Isolamento, cultivo e expansão das
células tronco mesenquimais extraídas
da medula óssea e do tecido adiposo
Os protocolos utilizados para isolamento,
cultivo e expansão das CTM-MO e CTM-
AD foram eficientes e permitiram a
manutenção das culturas até a sexta
passagem quando as células foram
descartadas. As CTM-MO e as CTM-AD
apresentaram morfologia alongada,
fusiforme, semelhante à morfologia de
fibroblastos (Fig.1).
Caracterização Fenotípica
A caracterização fenotípica por citometria
de fluxo das CTM-MO mostrou baixa
expressão de marcadores de células
hematopoiéticas CD45 (1,45%) e CD34
(1,53%) e alta expressão de marcadores de
células tronco CD90 (80,04%) e CD29
(96%) (Fig.2).
Resultado semelhante foi obtido após a
caracterização fenotípica por citometria de
fluxo das CTM-AD, isso é, baixa expressão
de marcadores de células hematopoiéticas
CD45 (1,54%) e CD34 (0,88%) e alta
expressão de marcadores de células tronco
CD90 (60,94%) e CD29 (77,08%) (Fig.3).
Cultivo de células tronco mesenquimais
da medula óssea e do tecido adiposo em
meio de diferenciação osteogênica,
adipogênica e condrogênica.
As CTM-AD e CTM-MO apresentaram a
formação de nódulos de mineralização após
21 dias de cultivo em meio osteogênico,
caracterizando sucesso na diferenciação
osteogênica (Fig.4). Verificou-se a
formação de vesículas lipídicas com
aspecto birrefringente e coloração
avermelhada no interior do citoplasma das
CTM-MO e CTM-AD após 21 dias de
cultivo em meio adipogênico o que
caracteriza a diferenciação adipogênica
dessas células (Fig.4). Após 21 dias de
cultivo em meio condrogênico foi possível
observar a diferenciação das CTM-MO e
CTM-AD em condrócitos com o acúmulo
de proteoglicanos corados em violeta vivo
no interior do citoplasma e na matriz
extracelular (Fig.4).

44
Figura 1: Fotomicroscopia de contraste de fase das células tronco mesenquimais extraídas de medula
óssea (A) e do tecido adiposo de cães (B). Observar a morfologia alongada e fusiforme das células. Barra
= 70 µm
Figura 2: Histogramas da expressão de CD45, CD34, CD90 e CD29 por citometria de fluxo em células
tronco mesenquimais da medula óssea de cães, mantidas em meio basal (DMEM) após o terceiro repique
e confluência de 80 a 90%. Os histogramas mostram a escala de fluorescência no eixo x considerada
positiva quando o pico de células está acima de 101 ou seja, toda a área do gráfico que se encontra no
intervalo da barra M1 é positiva.

45
Figura 3: Histogramas da expressão de CD45, CD34, CD90 e CD29 por citometria de fluxo em células
tronco mesenquimais do tecido adiposo de cães, mantidas em meio basal (DMEM) após o terceiro
repique e confluência de 80 a 90%. Os histogramas mostram a escala de fluorescência no eixo x
considerada positiva quando o pico de células está acima de 101 ou seja, toda a área do gráfico que se
encontra no intervalo da barra M1 é positiva.

46
Figura 4: Cultura de células tronco mesenquimais extraídas da medula (1) e do tecido adiposo (2) de cães
submetida à diferenciação osteogênica (A), adipogênica (B) e condrogênica (C) por 21 dias. Observar nas
imagens 1A e 2A os nódulos de mineralização corados em vermelho pela técnica de Von Kossa; nas
imagens 1B e 2B as vesículas lipídicas com aspecto birrefringente e coloração avermelhada no interior
das células coradas pela técnica Oil Red e nas imagens 1C e 2C os proteoglicanos corados em violeta
vivo pela técnica de PAS. Barras das imagens 1A e 2A = 350 µm e das imagens 1B, 2B, 1C e 2C = 8,75
µm.
Avaliação da diferenciação osteogênica
Avaliação da conversão do MTT em
cristais de formazan
Em todos os grupos houve um aumento da
capacidade de conversão de MTT em
cristais de formazan ao longo do tempo de
cultivo, independente do meio utilizado
(Fig.5). As CTM mantidas em meio basal
mostraram maior capacidade de conversão
de MTT que as mantidas em meio
osteogênico, aos 14 e 21 dias de cultivo, já
aos sete dias, as CTM mantidas em meio
osteogênico mostraram maior conversão de
MTT que as mantidas em meio basal
(Fig.5). Quando mantidas em meio basal, as
CTM-AD mostraram maior conversão de
MTT que as CTM-MO, em todos os tempos
estudados (Fig.5). Em meio osteogênico, as
CTM-AD mostraram maior conversão de
MTT que as CTM-MO aos sete e 21 dias de
cultivo e nenhuma diferença foi observada
aos 14 (Fig.5).
Figura 5. Média e desvio padrão da conversão do MTT em cristais de formazan em culturas de CTM-MO
e CTM-AD de cães, mantidas em meio basal (DMEM) e meio osteogênico (OST) por sete, 14 e 21 dias. *
Representa diferença estatística (P<0,05).

47
Avaliação da área celular por campo
Em todos os grupos houve um aumento da
percentagem de células por campo, ao
longo do cultivo, independente do meio
utilizado. As CTM-MO mantidas em meio
basal mostraram maior celularidade que as
mantidas em meio osteogênico, aos 21 dias
de cultivo, já aos sete e 14, nenhuma
diferença foi observada (Fig.6). As CTM-
AD mantidas em meio basal mostraram
maior celularidade que as mantidas em
meio osteogênico, em todos os tempos
estudados (Fig.6). As CTM-ADs mostraram
maior celularidade que as CTM-MOs em
todos os tempos estudados independente do
meio de cultura utilizado (Fig.6).
Figura 6: Porcentagem média e desvio padrão da área celular por campo em culturas de CTM-MO e
CTM-AD de cães, mantidas em meio basal (DMEM) e meio osteogênico (OST) por sete, 14 e 21 dias. *
Representa diferença estatística (P<0,05).
Avaliação da atividade da fosfatase
alcalina
As CTM mantidas em meio osteogênico
mostraram maior atividade da fosfatase
alcalina que as mantidas em meio basal, em
todos os tempos estudados (Fig.7).
Independente do meio de cultura utilizado,
as CTM-AD mostraram maior atividade da
fosfatase alcalina que as CTM-MO, em
todos os tempos estudados (Fig.7).
Figura 7: Média e desvio padrão da atividade da fosfatase alcalina em culturas de CTM-MO e CTM-AD
de cães, mantidas em meio basal (DMEM) e meio osteogênico (OST) por sete, 14 e 21 dias. * Representa
diferença estatística (P<0,05).
Avaliação da síntese de colágeno
Em todos os grupos houve um aumento na
síntese de colágeno ao longo do tempo de
cultivo, independente do tipo de meio
utilizado. As CTM-MO mantidas em meio
basal mostraram maior dosagem de
colágeno que as mantidas em meio
osteogênico, aos sete e 21 dias de cultivo, já
aos 14 dias, maior síntese de colágeno foi
observada nas CTM-MO mantidas em meio
osteogênico (Fig. 8). As CTM-AD mantidas
em meio basal mostraram maior síntese de
colágeno que as mantidas em meio
osteogênico, aos 14 e 21 dias de cultivo, já
aos sete dias, maior síntese de colágeno foi

48
observada nas CTM-AD mantidas em meio
osteogênico (Fig.8). As CTM-AD
mostraram maior síntese de colágeno que as
CTM-MO em todos os tempos estudados,
independente do meio de cultivo utilizado
(Fig.8).
Figura 8: Média e desvio padrão do colágeno em culturas de CTM-MO e CTM-AD de cães, mantidas em
meio basal (DMEM) e meio osteogênico (OST) por sete, 14 e 21 dias. * Representa diferença estatística
(P<0,05).
Avaliação da formação de nódulos e da
área mineralizada por campo
As CTM-MO submetidas à diferenciação
osteogênica formaram apenas um nódulo de
mineralização aos 21 dias de cultivo em
todos os campos avaliados (Fig.4, 9 e 10).
As CTM-AD submetidas à diferenciação
osteogênica formaram nódulos de
mineralização em todos os tempos
estudados e mostraram um aumento na
porcentagem de área mineralizada por
campo, ao longo do tempo de cultivo (Fig.9
e 10). Não houve, em nenhum dos tempos
estudados, formação de nódulos de
mineralização nas CTM-AD e CTM-MO
mantidas em meio basal (Fig.9 e 10). As
CTM-AD em meio osteogênico mostraram
maior área mineralizada por campo que as
CTM-MO sob as mesmas condições, em
todos os tempos estudados (Fig.9 e 10).
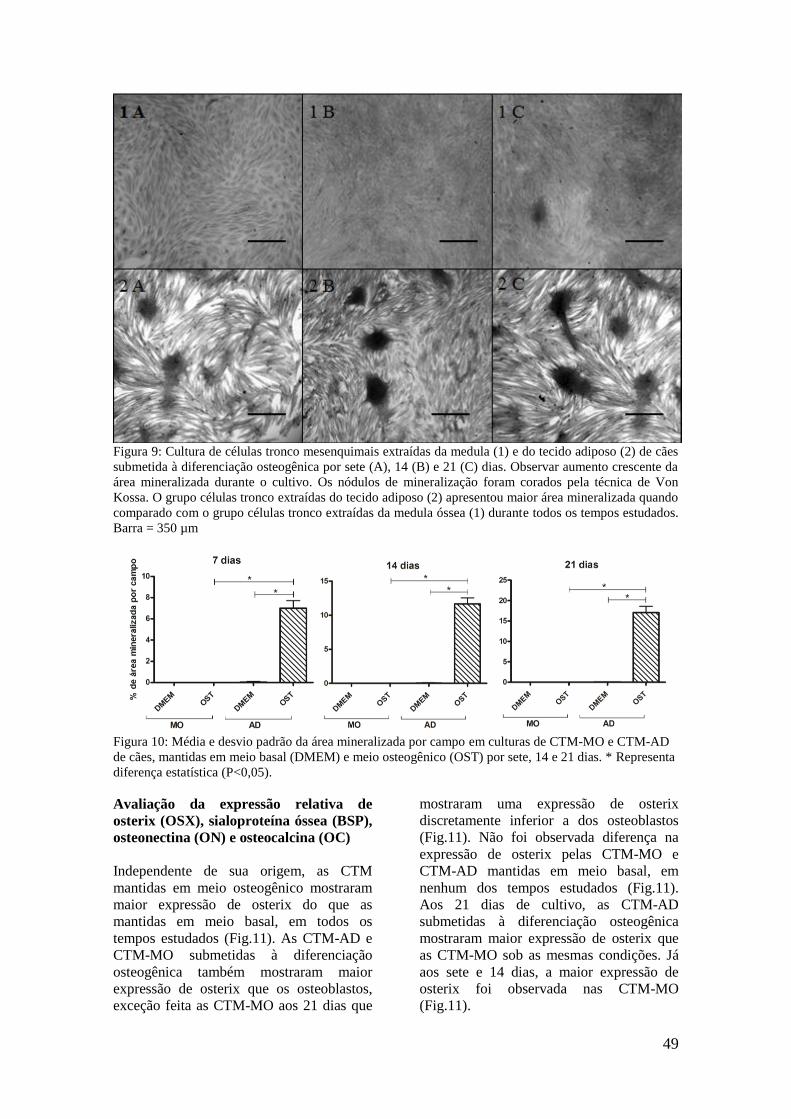
49
Figura 9: Cultura de células tronco mesenquimais extraídas da medula (1) e do tecido adiposo (2) de cães
submetida à diferenciação osteogênica por sete (A), 14 (B) e 21 (C) dias. Observar aumento crescente da
área mineralizada durante o cultivo. Os nódulos de mineralização foram corados pela técnica de Von
Kossa. O grupo células tronco extraídas do tecido adiposo (2) apresentou maior área mineralizada quando
comparado com o grupo células tronco extraídas da medula óssea (1) durante todos os tempos estudados.
Barra = 350 µm
Figura 10: Média e desvio padrão da área mineralizada por campo em culturas de CTM-MO e CTM-AD
de cães, mantidas em meio basal (DMEM) e meio osteogênico (OST) por sete, 14 e 21 dias. * Representa
diferença estatística (P<0,05).
Avaliação da expressão relativa de
osterix (OSX), sialoproteína óssea (BSP),
osteonectina (ON) e osteocalcina (OC)
Independente de sua origem, as CTM
mantidas em meio osteogênico mostraram
maior expressão de osterix do que as
mantidas em meio basal, em todos os
tempos estudados (Fig.11). As CTM-AD e
CTM-MO submetidas à diferenciação
osteogênica também mostraram maior
expressão de osterix que os osteoblastos,
exceção feita as CTM-MO aos 21 dias que
mostraram uma expressão de osterix
discretamente inferior a dos osteoblastos
(Fig.11). Não foi observada diferença na
expressão de osterix pelas CTM-MO e
CTM-AD mantidas em meio basal, em
nenhum dos tempos estudados (Fig.11).
Aos 21 dias de cultivo, as CTM-AD
submetidas à diferenciação osteogênica
mostraram maior expressão de osterix que
as CTM-MO sob as mesmas condições. Já
aos sete e 14 dias, a maior expressão de
osterix foi observada nas CTM-MO
(Fig.11).

50
Figura 11: Média e desvio padrão da quantificação relativa do transcrito gênico para osterix pela técnica
de RT-PCR em tempo real em culturas de CTM-MO e CTM-AD de cães, mantidas em meio basal
(DMEM) e meio osteogênico (OST) por sete, 14 e 21 dias. Os dados estão expressos em relação ao
osteoblasto canino mantido em meio basal (DMEM) (linha tracejada). *Representa diferença estatística
(P<0,05).
As CTM mantidas em meio osteogênico
mostraram maior expressão de sialoproteína
óssea do que as mantidas em meio basal,
exceção feita as CTM-MO aos 14 dias que
não mostraram diferença na expressão de
sialoproteína quando cultivadas em meio
basal e em meio osteogênico (Fig.12). Os
níveis de expressão de sialoproteína óssea
apresentados pelas CTM-MO e CTM-AD
submetidas à diferenciação osteogênica não
alcançaram os níveis osteoblásticos de
expressão de sialoproteína óssea, em
nenhum dos tempos estudados (Fig.12). As
CTM-MO mantidas em meio basal
mostraram maior expressão de sialoproteína
óssea que as CTM-AD sob mesmas
condições, aos sete dias de cultivo, mas
nenhuma diferença foi observada aos 14 e
21 (Fig.12). Aos 14 dias de cultivo, as
CTM-AD submetidas à diferenciação
osteogênica mostraram maior expressão de
sialoproteía óssea que as CTM-MO sob
mesmas condições, já aos sete e 21 dias, a
maior expressão de sialoproteía óssea foi
observada nas CTM-MO (Fig.12).
Figura 12: Média e desvio padrão da quantificação relativa do transcrito gênico para sioloproteína óssea
(BSP) pela técnica de RT-PCR em tempo real em culturas de CTM-MO e CTM-AD de cães, mantidas em
meio basal (DMEM) e meio osteogênico (OST) por sete, 14 e 21 dias. Os dados estão expressos em
relação ao osteoblasto canino mantido em meio basal (DMEM), o valor de expressão do osteoblasto foi
considerado igual a um. *Representa diferença estatística (P<0,05).

51
Independente de sua origem, as CTM
mantidas em meio osteogênico mostraram
maior expressão de osteonectina que as
mantidas em meio basal, em todos os
tempos estudados. As CTM-AD e CTM-
MO submetidas à diferenciação osteogênica
também mostraram maior expressão de
osteonectina que os osteoblastos, exceção
feita as CTM-MO aos 21 dias que
mostraram uma expressão de osteonectina
inferior a dos osteoblastos (Fig.13). Não foi
observada diferença na expressão de
osteonectina pelas CTM-MO e CTM-AD
mantidas em meio basal, em nenhum dos
tempos estudados (Fig.13). As CTM-AD
submetidas à diferenciação osteogênica
mostraram maior expressão de osteonectina
que as CTM-MO sob as mesmas condições,
em todos os tempos estudados (Fig.13).
Figura 13: Média e desvio padrão da quantificação relativa do transcrito gênico para osteonectina (ON)
pela técnica de RT-PCR em tempo real em culturas de CTM-MO e CTM-AD de cães, mantidas em meio
basal (DMEM) e meio osteogênico (OST) por sete, 14 e 21 dias. Os dados estão expressos em relação ao
osteoblasto canino mantido em meio basal (DMEM). (linha tracejada). *Representa diferença estatística
(P<0,05).
Independente de sua origem, as CTM
mantidas em meio osteogênico mostraram
maior expressão de osteocalcina que as
mantidas em meio basal e que os
osteoblastos, em todos os tempos estudados
(Fig.14). Não foi observada diferença na
expressão de osteocalcina pelas CTM-MO e
CTM-AD mantidas em meio basal, em
nenhum dos tempos estudados (Fig.14). As
CTM-MO submetidas a diferenciação
osteogênica mostraram maior expressão de
osteocalcina que as CTM-AD sob as
mesmas condições de cultivo, em todos os
tempos estudados (Fig.14).
Figura 14: Média e desvio padrão da quantificação relativa do transcrito gênico para osteocalcina (OC)
pela técnica de RT-PCR em tempo real em culturas de CTM-MO e CTM-AD de cães, mantidas em meio
basal (DMEM) e meio osteogênico (OST) por sete, 14 e 21 dias. Os dados estão expressos em relação ao
osteoblasto canino normal mantido em meio basal (DMEM). (linha tracejada). *Representa diferença
estatística (P<0,05).

52
Discussão
A forma alongada semelhante a fibroblasto
apresentada pelas CTM-MO e CTM-AD
(Fig.1) e sua capacidade de diferenciação
em osteoblastos, adipócitos e condrócitos
(Fig.4), mostra sucesso no isolamento das
células tronco mesenquimais (CTM), pois
esses são os critérios definidos como padrão
pela The International Society for Cellular
Therapy (Dominici, et al., 2006) e relatada
por outros pesquisadores (Martinello et al.,
2011; Chung et al., 2012; Spencer et al.,
2012; Takemitsu et al., 2012). A alta
expressão de marcadores de células tronco e
baixa de marcadores de células
hematopoiéticas observadas no presente
estudo (Fig.2 e 3), confirmam o sucesso no
isolamento das CTM a partir de amostras de
medula óssea e de tecido adiposo. Esse
padrão de expressão é mundialmente aceito
como característico de células tronco
mesenquimais (Dominici, et al., 2006; Seo
et al., 2009; Rozemuller et al., 2010; Vieira
et al., 2010; Lee et al., 2011; Martinello et
al., 2011; Spencer et al., 2012). A sociedade
internacional de terapia celular (The
International Society for Cellular Therapy)
definiu que as CTM devem expressar
marcadores como CD105, CD73 e CD90 e
não expressar os marcadores de células
hematopoiéticas (Dominici, et al., 2006).
No presente estudo, tanto as CTM-MO
quanto as CTM-AD mostraram uma alta
expressão de CD90, marcador característico
de células tronco mesenquimais (Vieira et
al.,2010; Lee et al., 2011; Martinello et al.,
2011; Takemitsu et al., 2012) e baixa
expressão de CD45 e CD34, marcadores de
células hematopoiéticas (Dominici, et al.,
2006; Takemitsu et al., 2012), excluindo a
possibilidade de isolamento de células
tronco hematopoiética especialmente na
cultura extraída da medula óssea. As
culturas de CTM isoladas no presente
experimento apresentaram uma alta
expressão de CD29, outro marcador de
células tronco (Seo et al., 2009; Vieira et
al.,2010) que não é expresso por
fibroblastos caninos (Lee et al., 2011),
excluindo assim a possibilidade do
isolamento de fibroblastos principalmente,
na cultura extraída do tecido adiposo.
Outros marcadores de CTM podem também
ser utilizados em cães como CD44 (Seo et
al., 2009; Vieira et al., 2010; Lee et al.,
2011; Martinello et al., 2011; Takemitsu et
al., 2012), CD33, CD105, CD184, OCT4
(Seo et al., 2009), CD140a, CD117
(Martinello et al., 2011), CD56, CD146,
CD271(Rozemuller et al., 2010), mas não
foram utilizados nesse estudo, pela falta de
especificidade dos anticorpos disponíveis
no mercado para células de cão, na ocasião
em que o estudo foi conduzido.
A menor conversão de MTT em cristais de
formazan observada nas CTM submetidas à
diferenciação osteogênica no presente
estudo (Fig.5), sugere menor atividade
mitocondrial nos grupos submetidos à
diferenciação o que também foi observado
em outros estudos semelhantes em ratos e
humanos (Baksh et al .,2004; Li e Sun,
2011). Isso pode ter ocorrido devido ao
menor metabolismo dos osteoblastos
quando comparado com as CTM
indiferenciadas (Payushina et al., 2006;
Kassem et al., 2008; Datta et al., 2008) mas
também, pela menor quantidade de células
presente nos grupos submetidos à
diferenciação osteogênica como observado
no presente estudo. Com a progressão dos
estágios de diferenciação há uma redução
da capacidade proliferativa da célula e um
deslocamento da atividade celular para
síntese de proteínas específicas da célula
diferenciada e dos componentes da matriz
óssea extracelular (Baksh et al ., 2004;
Payushina et al., 2006; Datta et al., 2008).
A maior conversão de MTT em cristais de
formazan (Fig.5) e maior celularidade
(Fig.6) observadas nas CTM-AD quando
submetidas à diferenciação, sugere uma
maior atividade dessas células em relação
às CMT-MO sob as mesmas condições.
Esta pode ser uma das vantagens do uso das
CTM-AD visto que uma célula mais ativa
pode produzir mais matriz extracelular,
favorecendo assim a regeneração óssea
quando utilizada clinicamente para esse
fim.
O aumento gradual e progressivo da
atividade da fosfatase alcalina (FA) durante
o processo de diferenciação osteogênica em
ambos os tipos celulares CTM-MO e CTM-
AD, observado no presente estudo (Fig.7),
sugere sucesso na diferenciação osteogênica
dessas células. A maior atividade da FA
observada nas CTM-AD em relação às
CTM-MO sugere um maior potencial

53
osteogênico das CTM-AD, visto que a FA é
um dos marcadores mais utilizados na
diferenciação osteogênica in vitro
(Eslaminejad e Taghiyar , 2010; Li e Sun,
2011). A FA é uma enzima presente na
membrana do osteoblasto que participa da
síntese e mineralização da matriz óssea
(Datta et al., 2008; Franceshi et al., 2009).
No processo de diferenciação osteogênica
sua expressão pode ser aumentada em até
27 vezes (Kassem et al., 2008). Nefussi et
al (1997) relataram que os locais de maior
expressão da FA no início do processo de
diferenciação são aqueles onde futuramente
se formarão os nódulos de mineralização.
Ela é considerada um marcador precoce de
diferenciação osteogênica, sendo expressa
por pré-osteoblastos e osteoblastos (Kassem
et al., 2008).
A maior síntese de colágeno (Fig.8) e maior
área mineralizada por campo (Fig.9 e 10)
observadas nas CTM-AD em relação às
CTM-MO mantidas em meio osteogênico,
mostra maior capacidade de produção de
matriz extracelular das CTM-AD em
comparação com as CTM-MO sob as
mesmas condições de cultivo. O colágeno
tipo I é o principal constituinte da matriz
óssea orgânica representando cerca de 90%
de sua composição e é considerado um
marcador precoce da diferenciação
osteogênica, podendo ser expresso por
células osteoprogenitoras, pré-osteoblastos
e osteoblastos (Nakashima et al., 2002). A
formação de nódulos de mineralização a
partir do sétimo dia de cultivo nas CTM-
AD mantidas em meio osteogênico (Fig.9),
confirma a diferenciação dessas células.
Após a diferenciação osteogênica as CTM
adquirem forma cuboidal ou poligonal
semelhante aos osteoblastos e se agrupam
concentricamente formando nódulos
mineralizados (Payushina et al., 2006; Tae
et al., 2006; Eslaminejad & Taghiyar,
2010). Esses nódulos são considerados
indicadores de diferenciação osteogênica
“in vitro” (Eslaminejad & Taghiyar, 2010;
Li e Sun, 2011; Martinello et al., 2011;
Chung et al., 2012; Spencer et al., 2012) e
sua quantificação é a forma mais fidedigna
de avaliar a osteogênese “in vitro”, visto
que a produção de matriz mineralizada, não
indica apenas fase final de diferenciação
osteogênica, mas também a atividade
metabólica das células diferenciadas
(Bruedigam et al., 2011)
O aumento na expressão de osterix
encontrado nas CTM-MO e CTM-AD
submetidas ao protocolo de diferenciação
(Fig.11) é outro importante indicativo de
diferenciação osteogênica. O osterix é um
fator de transcrição imprescindível no
processo de diferenciação osteogênica e
atua na transformação dos pré-osteoblastos
em osteoblastos maduros, finalizando assim
o processo de diferenciação (Nakashima et
al., 2002; Datta et al., 2008; Franceschi et
al., 2009). O aumento da expressão de
osterix pelas CTM submetidas à
diferenciação osteogênica já foi relatado em
outros estudos semelhantes em ratos e
humanos e é considerado um marcador
fidedigno da diferenciação osteogênica em
cães (Neupane et al., 2008; Chung et al.,
2012)
O aumento da expressão de sialoproteina
óssea (BSP) (Fig.12), osteonectina (ON)
(Fig.13) e osteocalcina (OC) (Fig.14)
durante o processo de diferenciação
observado no presente estudo, comprova o
estágio final de diferenciação osteogênica
das CTM-MO e CTM-AD. As proteínas
não colagênicas da matriz são consideradas
marcadores fidedignos de diferenciação
osteogênica e estão intimamente
relacionados com a produção e
mineralização da matriz extracelular. A
BSP é uma das proteínas responsáveis pelo
início do processo de nucleação e deposição
de hidroxiapatita na matriz óssea. O
aumento na sua expressão reflete uma fase
final de diferenciação e início de
mineralização da matriz. A sialoproteina
óssea pode ser expressa por pré-
osteoblastos e osteoblastos (Zhou et al.,
2006). O aumento de sua expressão também
já foi relatado durante o processo de
diferenciação osteogênica de CTM-AD em
cães (Neupane et al., 2008; Vieira et al.,
2010). A osteocalcina é uma glicoproteína
específica do tecido ósseo que atua
promovendo a mineralização da matriz. Ela
é expressa por osteoblastos e reflete
também o final da diferenciação e
maturação da matriz óssea e é considerada
o marcador tardio mais específico da
diferenciação osteogênica (Zhou et al.,
2006; Nakashima et al., 2002). O aumento
crescente de sua expressão é característico

54
do processo de diferenciação osteogênica
de CTM-MO e CTM-AD (Chung et al.,
2012). A osteonectina também está
relacionada com a mineralização da matriz
no estágio final de diferenciação (Strauss et
al., 1990; Nefussi et al., 1997; Nakashima
et al., 2002; Karaoz et al., 2009).
Embora as CTM-MO tenham mostrado um
predomínio de maior expressão de osterix
(Fig.11), sialoproteina óssea (Fig.12) e
osteocalcina (Fig.14) em relação às CTM-
AD, isso não promoveu maior osteogênese
“in vitro” (Fig.9 e 10). A quantificação dos
transcritos gênicos para determinadas
proteínas como a sialoproteína óssea e a
osteocalcina é um importante indicativo de
sua síntese, no entanto, não garante que
essas proteínas estejam de fato sendo
sintetizadas. Fatores relacionados com a
exportação do RNA mensageiro (mRNA)
do núcleo para o citoplasma, com a
estabilização mRNA para que ele chegue
íntegro no local de tradução e fatores
relacionados diretamente com a tradução
podem fazer com que um mRNA não seja
traduzido em proteína (Fabian et al., 2010).
Estudos recentes descobriram microRNAs
capazes de inibir a expressão de genes após
sua transcrição (Kim et al., 2012). Existem
evidências de que esses microRNAs estão
envolvidos em uma série de processos
biológicos e dentre eles, a proliferação
(Mizuno et al., 2008; Wagner et al., 2008) e
diferenciação osteogênica (Kim et al.,
2009). Alguns microRNAs como miR-29c,
miR-369-5p, miR-371 e miR-499 tem sua
expressão aumentada com o aumento do
número de passagens e senescência celular
“in vitro” o que sugere que eles regulam a
proliferação celular (Wagner et al., 2008).
Outros como miR-204 e miR-211 são
comprovadamente reguladores do runx-2,
gene mais importante da diferenciação
osteogênica, sendo que a superexpressão
desses microRNAs reduzem a resposta
desencadeada pelo runx-2 (Kim et al.,
2012). Também já foram estudados os
microRNAs miR-125b e miR-196a que
regulam a diferenciação osteogênica
levando a redução da proliferação celular e
inibição da diferenciação osteogênica,
respectivamente (Mizuno et al., 2008; Kim
et al., 2009).
Conclusão
Nas condições em que foi realizado este
experimento pode-se dizer que:
As culturas de CTM-AD apresentam maior
potencial osteogênico que as de CTM-MO
“in vitro” com maior área mineralizada por
campo, maior atividade da fofatase alcalina,
maior produção de colágeno e maior
expressão de osteonectina, importantes
indicadores de osteogênese.

55
Capítulo 3: Efeito do produto iônico do
biovidro 60S na diferenciação
osteogênica de células tronco
mesenquimais do tecido adiposo de cães
Introdução
Dentre os biomateriais disponíveis para
produção de matrizes sintéticas e sua
utilização como substituto ósseo
encontram-se os biovidros que são
considerados os mais bioativos e favoráveis
à regeneração óssea (Hench, 2009). Seu
efeito biológico pode variar de acordo com
a composição química, grau de organização
das moléculas (cristalinidade), topografia e
microestrutura da matriz bem como, com as
condições do microambiente que podem
determinar maior ou menor dissolução do
biomaterial e alterar consequentemente, seu
efeito biológico (Hoppe et al., 2011;
Rahaman et al., 2011). Acredita-se que o
efeito biológico dos biovidros seja mediado
principalmente pelos íons cálcio (Chai et.,
2012) e silício (Shie et al., 2011) liberados
na sua dissolução. Já foi comprovado que
esses íons podem favorecem a proliferação
(Maeno et al., 2005; Barradas et al., 2010;
McCullen et al., 2010) e diferenciação
osteogênica “in vitro”(Jung e Park, 2010;
McCullen et al., 2010). Mas não se sabe ao
certo qual a composição química ideal do
biovidro para que quando implantado no
organismo libere a combinação e a
concentração ideal de íons para
maximização do processo de regeneração
óssea. A resposta biológica a um
biomaterial pode variar de acordo com o
tecido no qual será implantado (Rahaman et
al., 2011) e com a espécie (Bielby et al.,
2004; Reilly et al., 2007). Em um estudo
“in vitro” com osteoblastos de ratos, o
biovidro 60S de composição molar 4%
P2O5; 36% CaO; 60% SiO2 mostrou taxas
satisfatórias de liberação de íons cálcio e
silício, que favoreceram a proliferação
celular (Valerio et al., 2004), mas nada se
sabe dos efeitos desse biomaterial sobre a
diferenciação osteogênica de células tronco
mesenquimais de cães. Portanto, o objetivo
do presente estudo é avaliar o efeito do
produto iônico do biovidro 60S na
diferenciação osteogênica de células tronco
mesenquimais extraídas do tecido adiposo
de cães, com vistas a futura utilização desse
biomaterial associado às CTM-AD para
tratamento de defeitos ósseos em cães.
Material e métodos
Foram utilizadas as bases físicas e a infra-
estrutura dos laboratórios de Cultivo de
Células Tronco e Terapia Celular, de
Biologia Molecular e de Histopatologia e
Imunoistoquímica do Departamento Clínica
e Cirurgia Veterinárias da Escola de
Veterinária da UFMG, do Laboratório de
Biologia Molecular do Departamento de
Imunologia e Bioquímica do Instituto de
Ciências Biológicas da UFMG e do
Laboratório de Biomateriais do
Departamento de Engenharia Metalúrgica e
de Materiais da UFMG. O projeto foi
aprovado pelo Comitê de Ética em
Experimentação Animal (CETEA) da
UFMG31
.
Produção do biovidro 60S
A produção do biovidro 60s de composição
molar 4% P2O5; 36% CaO; 60% SiO2 foi
realizada no Laboratório de Biomateriais do
Departamento de Engenharia Metalúrgica e
de Materiais da UFMG conforme descrito a
seguir: foram adicionados em um bequer,
133,8 ml de tetraetilortosilicato (TEOS) e
13,6 ml de trietilfosfato (TEP) e em outro,
97,9 ml de água deionizada e 16,3 ml de
solução de ácido nítrico 2 N, as soluções
foram homogeneizadas e reservadas. Em
seguida as duas soluções foram misturadas
em um terceiro bequer maior e mantidas
sob agitação magnética por 60 minutos com
o objetivo de hidrolisar o TEOS.
Transcorrido este período, 85,01g de nitrato
de cálcio [Ca(NO3)2 4H2O] foi
gradativamente adicionado à solução que
permaneceu sob agitação magnética por
mais 30 minutos. Em seguida a solução foi
vertida em um recipiente de polipropileno
que foi hermeticamente fechado e incubado
a 60◦C por 72 horas para gelação e
maturação. Transcorrido esse período a
tampa do recipiente foi trocada por outra
com múltiplos orifícios e este foi então
submetido a um protocolo de secagem,
permanecendo a 60◦C por 72 horas,
seguindo-se um aumento gradual da
31 Número de protocolo no CETEA (157/2009)

56
temperatura em 10◦C a cada 24 horas, até
atingir a temperatura de 120◦C (Coelho et
al., 2003; Coelho et al., 2005). Após a
secagem a amostra foi moída manualmente
em um gral de quartzo e foi separada a
fração com granulometria menor que
137µm por meio de uma peneira 100 mesh.
A eliminação de possíveis resíduos foi
realizada por queima na mufla a 700◦C por
seis horas quando foi utilizada uma taxa de
aquecimento e de resfriamento de 1◦C por
minuto.
Preparação do meio produto iônico do
biovidro 60S
Para obtenção do produto iônico uma
amostra de 6 g do biovidro 60S foi
adicionada a 1 L de meio basal [DMEM
com baixa glicose enriquecido com
gentamicina (60 μg/L), penicilina (100
UI/ml), estreptomicina (100μg/mL),
anfotericina (25μg/mL)32
] e a mistura foi
vigorosamente homogeneizada, seguindo-se
um período de incubação de 12 horas a 6◦C.
Transcorrido o tempo de incubação, a
suspensão foi filtrada em membrana de
22µm e o pH normalizado para 7.2,
concluindo-se o processo de produção do
produto iônico do biobidro 60S.
Para avaliação do efeito do produto iônico
nas células tronco mesenquimais de origem
adiposa (CTM-AD) foram constituídos dois
meios de cultura expressos a seguir:
1- Produto iônico em meio de diferenciação
osteogênica, denominado OST PI –
Constituiu-se do produto iônico acrescido
de 50 µg/ml de vitamina C, 10mM de β
glicerolfosfato, 0,1µM de dexametasona e
10% de soro fetal bovino;
2- Meio de diferenciação osteogênica
(controle), denominado OST – Constituiu-
se do DMEM com baixa glicose
enriquecido com gentamicina (60 μg/L),
penicilina (100 U/ml), estreptomicina
(100μg/mL), anfotericina (25μg/mL)33
,
10% de soro fetal bovino, 50 µg/ml de
vitamina C, 10mM de β glicerolfosfato e
0,1µM de dexametasona.
32 PSA, Sigma-Aldrich, St. Louis, MO, USA 33 PSA, Sigma-Aldrich, St. Louis, MO, USA
Avaliação das concentrações dos íons Si e
Ca nos meios de cultivo celular.
Previamente ao uso dos meios de cultura
nas CMT-AD caninas determinou-se a
concentração dos íons Si e Ca nos mesmos.
Utilizou-se a técnica de espectrometria de
absorção atómica (GBC-Avanta) pelo
método de adição padrão.
Animais
Foram utilizados três cães, machos, não
castrados, sem raça definida com dois anos
de idade e massa corporal média de 18 Kg.
Os cães foram recolhidos em fazendas da
zona rural do município de Lavras quando
filhotes e criados até os dois anos de idade
para realização do presente estudo. No
período experimental os cães foram
mantidos em canis individuais tipo solário,
com área de 4,5m2, pertencentes ao Centro
de Experimentação de Pequenos Animais
do Hospital Veterinário da Escola de
Veterinária da UFMG, onde receberam
ração comercial34
e água a vontade. Todos
os animais passaram por um período
mínimo de 30 dias de adaptação às novas
instalações. Nessa fase eles foram avaliados
clinicamente e por meio de exames
radiográficos e laboratoriais (hemograma,
pesquisa de hemoparasitas e análise
sorológica para leishmaniose), assegurando-
se que todos estavam saudáveis e aptos para
serem utilizados no presente estudo. Todos
os animais foram desverminados, vacinados
e receberam medicação ectoparasiticida.
Obtenção, cultivo e expansão das células
tronco mesenquimais do tecido adiposo
de cães
A colheita das amostras de tecido adiposo
foi realizada no centro cirúrgico do Hospital
Veterinário da Escola de Veterinária da
Universidade Federal de Minas Gerais
(UFMG). Para colheita das amostras, os
cães foram submetidos ao jejum sólido de
oito horas. Como medicação pré-anestésica,
foram empregados 1mg/kg IM de xilazina35
e 15mg/kg IM de quetamina36
, na mesma
seringa. A região glútea direita foi
34 Canitos, Kowalski, Apucarana, PR, Brasil 35 Calmiun, Agener União, Embu-Guaçu, SP, Brasil 36 Vetanarcol, Konig, Santana do Parnaíba, SP, Brasil

57
submetida à tricotomia e preparada para
cirurgia asséptica. A veia cefálica foi
canulada e os animais receberam propofol37
(3mg/kg IV) para intubação e manutenção
anestésica. Como analgésico e anti-
inflamatório foi administrado meloxicam38
(0,2mg/kg IM) imediatamente após a
indução anestésica.
As amostras de tecido adiposo foram
colhidas cirurgicamente do tecido adiposo
subcutâneo da região glútea imediatamente
acima do trocânter maior. Foi colhido
aproximadamente 1cm3 de tecido adiposo
de cada animal totalizando cerca de 3cm3
de amostra final. Imediatamente após a
colheita, as amostras de tecido adiposo
foram colocadas em tubos falcon de 50 mL
contendo 20 mL de meio de cultura
Dulbecco’s Modiffed Eagle Medium baixa
glicose39
(DMEM), à temperatura ambiente.
Em seguida as amostras foram
encaminhadas ao Núcleo de Células Tronco
da Escola de Veterinária da UFMG, onde as
CTM foram extraídas e cultivadas segundo
o protocolo descrito a seguir que foi
adaptado de protocolos estabelecidos por
Neupane et al. (2008) e Vieira et al. (2010).
As amostras de tecido adiposo foram
manipuladas de forma asséptica na capela
de fluxo laminar. Elas foram lavadas duas
vezes com PBS 0,15 molar para retirada de
restos de sangue e debris celulares. Para
digestão do tecido conjuntivo e liberação
das células, as amostras foram colocadas
em outro tubo falcon de 50 mL contendo 20
mL de uma solução estéril de colagenase
B40
0,1% P/V. As amostras foram então
fragmentadas a partículas menores com
auxílio de uma tesoura cirúrgica estéril. A
tesoura foi introduzida no tubo falcon de
50mL e com movimentos de abrir e fechar
dos ramos o tecido foi fragmentado ao
máximo possível. Em seguida a amostra foi
encubada em estufa a 37oC e 5% de CO2
durante 45 minutos e a cada 15 minutos a
amostra foi vigorosamente agitada para
facilitar o processo de digestão.
Transcorrido o tempo de digestão a amostra
foi centrifugada por 10 minutos a 1400rpm,
o sobrenadante contendo a fração adiposa
37 Fresofol, Fresenius Kabi, Campinas, SP, Brasil 38 Maxicam, Ouro Fino, Cravinhos, SP, Brasil 39 DMEM, Gibco, Grand Island, NY, USA 40 Colagenase B, Roche Applied Science, Penzberg,
Germany
foi descartado e o pellet foi ressuspenso em
10 mL PBS 0,15 molar para lavagem e
retirada dos resíduos de colagenase. A
amostra foi colocada em outro tubo falcon
para reduzir a contaminação por gordura
retida na parede do tubo e novamente
centrifugada por 10 minutos a 1400rpm.
Após a centrifugação o sobrenadante foi
novamente descartado e o pellet, fração
estromal, foi ressuspenso em 20 mL de
meio de cultura para células tronco (meio
basal), DMEM com baixa glicose,
enriquecido com gentamicina (60 μg/L),
penicilina (100 UI/mL), estreptomicina
(100μg/mL), anfotericina (25μg/mL)41
e
10% de soro fetal bovino42
. Uma alíquota
de 2 mL do meio com as células
ressupensas foi transferida para cada
garrafa de cultivo celular T75 e 8 mL de
meio foram adicionados em cada garrafa.
Foram feitas dez garrafas no total.
Realizou-se uma cuidadosa
homogeneização e as garrafas foram
incubadas em estufa a 37oC e 5% de CO2.
Após 48h foi realizada troca parcial do
meio de cultura, renovando apenas 5 mL.
Após mais 48h as células foram lavadas
duas vezes com PBS 0,15 molar para
remoção de debris e células não aderidas. O
meio de cultivo foi trocado duas vezes por
semana a fim de se eliminar células não
aderidas garantindo a obtenção de células
tronco mesenquimais.
Assim que foi atingido 80 a 90% de
confluência, as células foram repicadas.
Para desprendimento das células, as
garrafas foram lavadas com 10 mL de PBS
0,15 molar para retirada dos restos de meio
de cultura. Após a lavagem, foi adicionado
1,5 mL de tripsina e a garrafa foi incubada
por 10 minutos em estufa a 37oC e 5% de
CO2. Transcorrido o tempo de ação da
tripsina a mesma foi inativada com 5 mL de
DMEM enriquecido com 10% de soro fetal
bovino. As garrafas foram então lavadas
com o próprio meio para retirada das
células e uma alíquota de 1 mL foi repicada
para outra garrafa T75 com mais 9 mL de
meio de cultura. Após o terceiro repique as
células foram então avaliadas quanto a
viabilidade celular e utilizadas para
41 PSA, Sigma-Aldrich, St. Louis, MO, USA 42 Soro fetal bovino, Sorali, Campo Grande, MS, Brasil

58
avaliação do efeito do produto iônico do
biovidro 60S.
Osteoblastos caninos
Como controle positivo da diferenciação
osteogênica foi utilizado uma cultura
primária de osteoblastos caninos43
. A
cultura foi descongelada conforme
protocolo sugerido pelo fabricante, os
osteoblastos foram cultivados em garrafas
T75, com DMEM com baixa glicose,
enriquecido com gentamicina (60 μg/L),
penicilina (100 UI/mL), estreptomicina
(100μg/mL), anfotericina (25μg/mL)44
e
10% de soro fetal bovino45
, sob as mesmas
condições das CTM-AD. Após o quarto
repique e obtenção de 80 a 90 % de
confluência os osteoblastos, foram
utilizados para extração do RNA total e
posterior análise da expressão de osterix,
sialoproteína óssea, osteonectina e
osteocalcina pela técnica de RT-PCR em
tempo real.
Avaliação da viabilidade das CTM-AD
pelo azul de Tripan
Para avaliação da viabilidade celular a
cultura de CTM-AD foi lavada com PBS
(0,15M) e submetida à ação da tripsina. O
sobrenadante contendo as células foi
colhido e centrifugado a 1400rpm por 10
minutos, as CTM-AD foram ressuspensas
em meio basal e coradas pelo azul de
Tripan. As CTM-AD viáveis
(transparentes) e inviáveis (em azul) foram
quantificadas em câmara de Neubauer.
Avaliação do efeito do produto iônico do
biovidro 60S nas CTM-AD caninas
Após o terceiro repique e avaliação da
viabilidade celular as CTM-AD foram
submetidas à ação da tripsina e plaqueadas
na densidade de 1x104 células/cm
2, em
quatro repetições, nas garrafas T25 e em
seis repetições nas placas de seis e 24
poços46
de acordo com a especificação de
43 CnOb - canine osteoblasts, Cell Application , San Diego, CA , USA 44 PSA, Sigma-Aldrich, St. Louis, MO, USA 45 Soro fetal bovino, Sorali, Campo Grande, MS, Brasil 46 TPP (Techno Plastic Products in Trasadingen), Switzerland, Germany
cada teste detalhadas a seguir. Após a
obtenção de 60 a 70% de confluência
celular o meio basal foi substituído pelos
meios a serem avaliados OST PI e OST.
Em seguida as CTM-AD foram cultivadas a
37oC e 5% de CO2 por sete, 14 e 21 dias.
Transcorridos esses períodos, avaliou-se em
cada grupo a conversão do MTT em cristais
de formazan, a atividade da fosfatase
alcalina (FA), a síntese de colágeno, a área
celular e mineralizada por campo e as
expressões relativas de osterix (OSX),
sialoproteina óssea (BSP), osteonectina
(ON) e osteocalcina (OC) por RT-PCR em
tempo real.
Avaliação da conversão do MTT em
cristais de formazan
Para avaliação da conversão de MTT em
cristais de formazan as CTM-AD foram
cultivadas em placas de 24 poços. Ao
término de cada período de cultivo os meios
(OST PI e OST) foram substituídos por
210L de meio basal em cada poço e
170L de MTT (5mg/mL)47
. A placa foi
incubada por duas horas em estufa a 37oC e
5% de CO2. Após esse período os cristais
de formazan foram observados ao
microscópio óptico e em seguida,
adicionou-se 210L de SDS (sódio dodecil
sulfato)-10% HCl em cada poço. A placa
foi então incubada overnight em estufa a
37oC e 5% de CO2 e posteriormente, 100L
de cada poço foram transferidos para placas
de 96 poços para análise na leitora de placas
com comprimento de onda de 595nm de
acordo com Valério et al. (2004).
Avaliação da atividade da fosfatase
alcalina
Para avaliação da atividade da fosfatase
alcalina as CTM-AD foram cultivadas em
placas de 24 poços. Ao término de cada
período, as culturas foram lavadas com PBS
(0,15 molar) e foi adicionado 200L de
solução de BCIT/NBT48
(1mL de tampão
da fosfatase alcalina, 4,4L de NBT {nitro-
blue tetrazolium chloride} e 3,3L de BCIP
{5-bromo-4-chloro-3’-indolylphosphate p-
toluidine salt}), em cada poço. As amostras
47 Invitrogen, Carlsbad, CA, USA 48 Zymed Laboratories, CA, USA

59
permaneceram duas horas na estufa a 37oC
e 5% de CO2 e foram observadas ao
microscópio óptico. Em seguida, adicionou-
se 200L de SDS (sódio dodecil sulfato)-
10% HCl em cada poço e a amostra foi
incubada overnight em estufa a 37oC e 5%
de CO2. Posteriormente, 100L de cada
poço foram transferidos para placas de 96
poços para leitura em espectrofotômetro
com comprimento de onda de 595nm de
acordo com Ocarino et al. (2008) e Boeloni
et al. (2009).
Avaliação da síntese de colágeno
Para avaliação da síntese de colágeno
CTM-AD foram cultivadas em placas de 24
poços. Ao término de cada período, as
culturas foram lavadas com PBS (0,15
molar). Em seguida adicionou-se 1ml de
Bouin em cada poço, para fixação e a placa
foi incubada por duas horas em estufa a
37oC e 5% de CO2. As placas foram então
retiradas da estufa e colocadas overnight a
6oC em geladeira. Transcorrido esse
período as placas foram lavadas quatro
vezes com água osmose reversa e secas
para posterior coloração com Sirius Red
durante 30 minutos à temperatura ambiente.
O excesso do corante foi removido e as
células foram lavadas três vezes com
solução de HCl 0,01N e mantidas a
temperatura ambiente para secagem.
Posteriormente foram adicionados 300L
de NaOH 0,5M e a placa foi incubada em
repouso por mais 30 minutos.
Posteriormente, 100L de cada poço foram
transferidos para placas de 96 poços para
leitura em espectrofotômetro com
comprimento de onda de 540nm de acordo
com Ocarino et al. (2008) e Boeloni et al.
(2009).
Avaliação dos nódulos de mineralização e
determinação da área mineralizada por
campo
Para avaliação da formação de nódulos de
mineralização e determinação da
porcentagem de área mineralizada por
campo, as CTM-AD foram cultivadas em
placas de seis poços com lamínulas49
estéreis. Ao término de cada período de
49 Lamínula de plástico, Newton, NC, USA.
avalição, as culturas foram lavadas com
PBS 0,15M, fixadas com álcool 70% por 24
horas e coradas pelo método de Von Kossa
(adaptada de Prophet et al., 1992). Foi então
determinada a porcentagem de área
mineralizada por campo com o auxílio de
uma ocular micrométrica, contendo uma
gratícula com 121 pontos, em 25 campos,
com objetiva de 4.
Quantificação relativa dos transcritos
gênicos para osterix, sialoproteína óssea,
osteonectina e osteocalcina por RT-PCR
em tempo real
Realizou-se, em todos os grupos, a
quantificação relativa da expressão de
osterix (OSX), sialoproteína óssea (BSP),
osteonectina (ON) e osteocalcina (OC) nos
três períodos estudados, pela técnica de RT-
PCR em tempo real. A extração do RNA
total das células foi feita em quatro garrafas
T25 por grupo, com o emprego do Trizol50
.
O método de extração consistiu de uma
etapa inicial de lise e homegenização da
monocamada de células por cinco minutos à
temperatura ambiente para completa
dissociação dos complexos nucleoprotéicos.
O lisado foi transferido para um microtubo
de 1,5mL e foram adicionados 0,2mL de
clorofórmio, seguido de 15 segundos de
homogeneização, três minutos de incubação
à temperatura ambiente e centrifugação a
12.000g por 15 minutos à 4ºC, para
separação em três fases, onde a fase incolor
superficial continha o RNA. Na terceira
etapa, a fase aquosa foi transferida para um
novo tubo, com a adição de 0,5mL de
álcool isopropílico e incubação por 30
minutos à temperatura -80ºC. Em seguida a
solução foi descongelada e centrifugada a
12.000g por 10 minutos a 4ºC para
precipitação do RNA. O pellet foi então
lavado com 1 mL de etanol a 75%,
homogeneizado e centrifugado a 7.500g por
cinco minutos a 4ºC. O RNA foi
solubilizado em água DEPC51
livre de
RNAse e imediatamente armazenado a -
80ºC. A concentração de RNA foi
determinada pela leitura da absorbância a
260/280 nm por espectrofotometria. Foram
realizadas as reações de transcrição reversa
50 Invitrogen, USA 51 Água tratada com dimetil pirocarbonato (DEPC), Invitrogen, USA

60
com um Kit comercial52
, utilizando-se 1µg
de RNA total para a síntese de cDNA com
um volume final de 20µL. Para as reações
de PCR em tempo real foi utilizado 2µg de
cDNA, 5pM de cada iniciador e 12,5µL do
reagente syber Green53
em um volume final
de 25 µL de reação em um tubo54
, no
aparelho SmartCycler System55
. Os
parâmetros utilizados para amplificação
foram de 50°C por 120 segundos, 95°C por
150 segundos e 45 ciclos de 95°C por 15
segundos e 60°C por 30 segundos. Os
iniciadores foram pesquisados na literatura
ou delineados com base na sequência do
mRNA Canis familiares (Tab. 1). A
expressão gênica foi calculada usando o
método 2-∆∆CT
, onde os resultados obtidos
para cada grupo foram comparados
quantitativamente após a normalização
baseada na expressão de gliceraldeído-3-
fosfato desidrogenase (GAPDH) Canis
familiares. Os níveis de expressão obtidos
nas culturas de osteoblastos foram
utilizados como controle positivo da
diferenciação osteogênica e como padrão de
expressão no cálculo da expressão relativa
de cada transcrito.
52 SuperScriptTM III Platinum® Two-Step qRT-PCR Kit with SYBR Green, Invitrogen, CA, USA 53 SuperScriptTM III Platinum® Two-Step qRT-PCR Kit with SYBR Green, Invitrogen, CA, USA 54 SmartCycler® Tube-25µL, Cepheid, Sunnyvale, CA, USA 55 SmartCycler® System, Cepheid, Sunnyvale, CA, USA

61
Tabela 1: Lista de genes e sequência de nucleotídeos dos iniciadores para RT-PCR em tempo real.
Genes
Referência ou
No. de acesso
Oligonucleotídios iniciadores
(sequências de nucleotídeos 5’ a 3’)
Temperatura de
anelamento (◦C)
Tamanho do
produto
(pares de bases)
Osterix (OTX)
(Neupane et al.,
2008)
F- ACGACACTGGGCAAAGCAG
R- CATGTCCAGGGAGGTGTAGAC
60 285
Sialoproteina
óssea (BSP)
(Vieira et al.,
2010)
F- TTGCTCAGCATTTTGGGAAT
R-AACGTGGCCGATACTTAAAGAC
60 295
Osteonectina
(ON)
(XM_849889.1)
F- GCCTTGGCAGCCCCTCAACA
R- CACCTGCACGGGGTTGGCTC
60 108
Osteocalcina
(OC)
(Neupane et al.,
2008)
F- GAGGGCAGCGAGGTGGTGAG
R- TCAGCCAGCTCGTCACAGTTGG
62 134
GAPDH
(Vieira et al.,
2010)
F- CCATCTTCCAGGAGCGAGGAT
R- TTCTCCATGGTGGTGAAGAC
60 97
Análise estatística
Realizou-se análise de variância (ANOVA)
e para cada variável foram determinados a
média e o desvio padrão. As médias foram
comparadas pelo teste T de Student
utilizando o programa Graphpad Instat 356
.
Diferenças foram consideradas
significativas se p0,05 (Sampaio, 1998).
Resultados
As partículas do biovidro 60S apresentaram
dissolução parcial após serem adicionadas
aos meios de cultivo celular, causando um
aumento significativo na concentração dos
íons Si e Ca (Tab.3). A concentração do íon
Si foi próxima de zero no meio que não
entrou em contato com o biovidro (OST) e
no meio que recebeu o produto iônico (OST
PI), sua concentração foi de 41,28 mg/L. A
concentração do íon Ca foi de 87,5 mg/L no
meio que não entrou em contato com o
biovidro (OST) e no meio que recebeu o
produto iônico (OST PI), sua concentração
foi de 156,5 mg/L, representando uma
aumento na ordem de 78,85% (Tab.2).
56 GraphPad Software Inc., San Diego, USA

62
Tabela 2: Concentração média dos íons Si e Ca nas amostras dos meios de cultivo celular basal (DMEM),
meio osteogênico (OST), produto iônico do biovidro 60S em meio basal (DMEM PI) e em osteogênico
(OST PI).
Meio de cultura Concentração de Si Concentração de Ca
OST < 1 mg/L 87,5 mg/L
OST PI 41,28 mg/L 156,5 mg/L
Avaliação da conversão do MTT em
cristais de formazan
A suplementação do meio osteogênico com
produto iônico (OST PI) aumentou a
conversão de MTT em cristais de formazan
aos 21 dias de cultivo, reduziu aos sete e
nenhum efeito foi observado aos 14 (Fig.1).
Embora não tem sido quantificada,
verificou-se uma maior densidade celular
nas culturas que receberam o meio
osteogênico suplementado com produto
iônico (OST PI) em relação as que
receberam o meio osteogênico, aos 14 e 21
dias de cultivo.
Figura 1: Redução do MTT em cristais de formazan (média e desvio padrão) em culturas de CTM-AD
caninas mantidas em meio osteogênico (OST) e em produto iônico em meio osteogênico (OST PI) por
sete, 14 e 21 dias. *Representa diferença estatística (P<0,05).
Avaliação da atividade da fosfatase
alcalina
As CTM-AD mantidas em meio
osteogênico suplementado com produto
iônico (OST PI) mostraram menor atividade
da fosfatase alcalina que as mantidas em
meio osteogênico (OST) aos 14 e 21 dias de
cultivo. Aos sete, nenhuma diferença
estatística foi observada (Fig. 2).
Figura 2: Atividade da fosfatase alcalina (média ± desvio padrão) em culturas de CTM-AD caninas
mantidas em meio osteogênico (OST) e produto iônico em meio osteogênico (OST PI) por sete, 14 e 21
dias. *Representa diferença estatística (P<0,05).

63
Avaliação da síntese de colágeno
A suplementação do meio osteogênico com
produto iônico (OST PI) reduziu a síntese
de colágeno aos sete e 21 dias de cultivo e
nenhum efeito foi observado aos 14 (Fig.
3).
Figura 3: Média e desvio padrão da quantidade de colágeno em culturas de CTM-AD caninas cultivadas
em meio osteogênico (OST) e produto iônico em meio osteogênico (OST PI) por sete, 14 e 21 dias.
*Representa diferença estatística (P<0,05).
Avaliação dos nódulos de mineralização e
determinação da área mineralizada por
campo
As CTM-AD mantidas em meio
osteogênico (OST) e meio osteogênico
suplementado com produto iônico (OST PI)
formaram nódulos de mineralização em
todos os tempos estudados (Fig.4 e 5). A
área mineralizada por campo aumentou
com o tempo de cultivo nas culturas
mantidas em ambos os meios. Verificou-se
maior área mineralizada por campo nas
culturas de CTM-AD mantidas em meio
osteogênico suplementado com produto
iônico (OST PI), em relação as mantidas em
meio osteogênico (OST), em todos os
tempos estudados (Fig.4 e 5).
Figura 4: Cultura de CTM-AD caninas mantidas em meio osteogênico (OST) (1) e produto iônico em
meio osteogênico (2) por sete (A), 14 (B) e 21 (C) dias. Observar os nódulos de mineralização corados
pela técnica de Von Kossa. O grupo com produto iônico do biovidro 60S OST PI (2) mostrou maior área
de mineralização por campo em relação ao controle OST (1). Barra = 350 µm

64
Figura 5: Percentagem média e desvio padrão da área de mineralização por campo em culturas de CTM-
AD caninas cultivadas em meio osteogênico (OST) e produto iônico em meio osteogênico (OST PI) por
sete, 14 e 21 dias. *Representa diferença estatística (P<0,05).
Avaliação da expressão relativa dos
transcritos gênicos para osterix,
sialoproteína óssea, osteonectina e
osteocalcina
As CTM-AD mantidas em meio
osteogênico (OST) e em meio osteogênico
suplementado com produto iônico (OST PI)
mostraram expressão de osterix semelhante
e superiores à expressão osteoblástica, em
todos os tempos estudados (Fig.6).
Figura 6: Média e desvio padrão da quantificação relativa do transcrito gênico para osterix pela técnica de
RT-PCR em tempo real em culturas de CTM-AD caninas mantidas em meio osteogênico (OST) e produto
iônico em meio osteogênico (OST PI) por sete, 14 e 21 dias. Os dados estão expressos em relação ao
osteoblasto canino mantido em meio basal (DMEM) representado pela linha pontilhada. *Representa
diferença estatística (P<0,05).
As culturas de CTM-AD mantidas em meio
osteogênico suplementado com o produto
iônico (OST PI) mostraram maior
expressão de sialoproteína óssea que as
mantidas em meio osteogênico (OST), aos
14 e 21 dias. Nenhuma diferença foi
observada aos sete. Independente do meio
de cultura utilizado as CTM-AD mostraram
níveis de expressão sialoproteína óssea
inferiores aos níveis osteoblásticos, em
todos os tempos estudados (Fig.7).

65
Figura 7: Média e desvio padrão da quantificação relativa do transcrito gênico para sialoproteína óssea
(BSP) pela técnica de RT-PCR em tempo real em culturas de CTM-AD caninas mantidas em meio
osteogênico (OST) e produto iônico em meio osteogênico (OST PI) por sete, 14 e 21 dias. Os dados estão
expressos em relação ao osteoblasto canino mantido em meio basal (DMEM). *Representa diferença
estatística (P<0,05).
Verificou-se menor expressão de
osteonectina nas culturas de CTM-AD
mantidas meio osteogênico suplementado
com produto iônico (OST PI) em relação às
mantidas em meio osteogênico (OST), aos
14 dias de cultivo, mas nenhuma diferença
significativa foi observada nos demais
tempos avaliados (Fig.8).
Figura 8: Média e desvio padrão da quantificação relativa do transcrito gênico para osteonectina (ON)
pela técnica de RT-PCR em tempo real em culturas de CTM-AD caninas mantidas em meio osteogênico
(OST) e produto iônico em meio osteogênico (OST PI) por sete, 14 e 21 dias. Os dados estão expressos
em relação ao osteoblasto canino mantido em meio basal (DMEM), representado pela linha pontilhada.
*Representa diferença estatística (P<0,05).
As CTM-AD mantidas em meio
osteogênico (OST) e em meio osteogênico
suplementado com o produto iônico (OST
PI) não mostraram diferenças significativas
na expressão de osteocalcina em nenhum
dos tempos estudados (Fig.9).
Figura 9: Média e desvio padrão da quantificação relativa do transcrito gênico para osteocalcina (OC)
pela técnica de RT-PCR em tempo real em culturas de CTM-AD caninas mantidas em meio osteogênico
(OST) e produto iônico em meio osteogênico (OST PI) por sete, 14 e 21 dias. Os dados estão expressos
em relação ao osteoblasto canino mantido em meio basal (DMEM), representado pela linha pontilhada.
*Representa diferença estatística (P<0,05).

66
Discussão
O aumento na concentração do íon Si até
41,28 ppm decorrentes da dissolução do
biovidro 60S (Tab.2) era esperado e foi
significativamente superior ao observado
em alguns biovidros produzidos por fusão
(derretimento) (4,36 a 19,4 ppm) (Xynos et
al., 2000; Xynos et al., 2001; Qiu et al.,
2009; Tsigkou et al., 2009), mas inferior ao
observado em outros produzidos pela rota
sol-gel (50,2 a 203,11 ppm) (Bielby et al.,
2004; Christodoulou et al., 2005,
Christodoulou et al., 2006). Esta variação
pode ser explicada pela diferença de
composição química entre esses
biomateriais, mas também pela diferença na
organização das moléculas que pode
determinar maior ou menor dissolução do
material quando exposto a um meio aquoso
(Pereira et al., 2005). Da mesma forma, o
aumento na concentração do íon Ca até a
156,5 ppm decorrentes da dissolução do
biovidro 60S (Tab.2) era também esperado,
mas foi significativamente superior ao
observado em outros biovidros produzidos
por fusão (derretimento) (62,7 a 88,35 ppm)
(Xynos et al., 2000; Xynos et al., 2001; Qiu
et al., 2009; Tsigkou et al., 2009) e pela rota
sol-gel (47,06 a 95,8 ppm) (Bielby et al.,
2004; Christodoulou et al., 2005,
Christodoulou et al., 2006). Isto implica que
o mesmo tipo de biomaterial pode fornecer
diferentes resultados segundo sua
composição específica e grau de
organização das moléculas, o que justifica
as diferentes respostas encontradas na
literatura.
O aumento da conversão de MTT em
cristais de formazan nas CTM-AD mantidas
em meio osteogênico suplementado com
produto iônico (OST PI) em relação às
mantidas em meio osteogênico (OST), aos
21 dias de cultivo (Fig.1), mostra que os
níveis de silício (41,28 ppm) e cálcio (41,28
ppm) observados no presente estudo são
seguros para as CTM-AD de cães. Um
estudo recente com cultura de osteoblastos
mostrou que a concentração de silício é
tóxica para essas células a partir da
concentração de 169,35 ppm, quando
ocorre uma redução da viabilidade celular e
aumento da apoptose (Shie et al., 2011). No
presente estudo, a concentração de silício
(41,28 ppm) permaneceu bem abaixo dos
níveis tóxicos relatados por Shie et al.
(2011). Quanto ao cálcio, vários estudos já
avaliaram seu efeito na viabilidade celular e
mostraram que os níveis de cálcio
extracelular não devem ultrapassar a
concentração de 240 ppm em culturas de
osteoblastos imortalizados (Nakamura et
al., 2010), 300 ppm em culturas primárias
de osteoblastos de ratos (Valerio et al.,
2004), 312,60 ppm em cultura de CTM-MO
humanas (Barradas et al., 2012) e de ratos
(Wen et al., 2012a), 400,78 ppm em
culturas de osteoblastos de camundongos
(Maeno et al., 2005), 641,24 ppm em
culturas de CTM-AD humanas (McCullen
et al., 2010) e 801,56 ppm em culturas de
cementoblastos imortalizados de
camundongos (Kanaya et al., 2010).
Embora os níveis de cálcio liberados pelo
biovidro 60S (156,5 ppm) sejam superiores
aos liberados por outros biomateriais
semelhantes, eles estão muito abaixo dos
níveis considerados tóxicos.
O maior MTT observado aos 21 dias nas
CTM-AD suplementadas com o produto
iônico (Fig.1) mostra que os íons liberados
pelo biovidro 60S favorecem a atividade
dessas culturas por aumentarem a atividade
mitocondrial e/ou o número de células na
cultura, o que de fato foi observado no
presente estudo embora não tenha sido
quantificado. Tanto o aumento do número
de células quanto o aumento da atividade de
cada uma delas favorecem a produção de
matriz extracelular o que é uma vantagem
para a regeneração óssea. O teste de MTT
baseia-se na capacidade da enzima
succinato desidrogenase converter o sal
tetrazolium (MTT), que é hidrossolúvel e
de cor amarela, em cristais de formazan,
que são de cor azul escura. A enzima
succinato desidrogenase participa do ciclo
de Krebs e está presente principalmente, em
mitocôndrias viáveis (Mossmann, 1983,
Hansen et al., 1989).
O significativo aumento da área
mineralizada (Fig.6) observados nas
culturas que receberam o produto iônico
mostra que os íons liberados na dissolução
do biovidro 60S favorecem a osteogênese
“in vitro” e leva a crer que esse biomaterial
possa apresentar resultado semelhante
também “in vivo”. Sabe-se que o produto
de dissolução de algumas biocerâmicas
pode favorecer a proliferação (Xynos et al.,

67
2000; Xynos et al., 2001; Qiu et al., 2009;
Sun et al., 2009 b; Tsigkou et al., 2009) e a
diferenciação osteogênica (Qiu et al., 2009;
Sun et al., 2009 a; Sun et al., 2009 b;
Tsigkou et al., 2009; Jung e Park, 2010; Li
e Sun, 2011), bem como a osteogênese “in
vitro” (Sun et al., 2009 a; Tsigkou et al.,
2009; Li e Sun, 2011), mas o presente
estudo é o primeiro a mostrar os efeitos
benéficos do produto iônico do biovidro
60S nas diferenciação osteogênica de CTM-
AD de cães. Esses efeitos benéficos são
atribuídos principalmente aos íons silício e
cálcio, liberados durante o processo de
dissolução dos biovidros (Shie et al., 2011;
Varanasi et al., 2012). Estudos com cultivo
de osteoblastos de rato já mostraram que o
silício é capaz de aumentar a atividade e
proliferação celular (Shie et al., 2011), a
expressão de genes ligados a diferenciação
osteogênica (Varanasi et al., 2012) e de
favorecer a mineralização da matriz
extracelular (Shie et al., 2011). Sabe-se
também que o cálcio extracelular é capaz de
aumentar a atividade e proliferação celular
em culturas de osteoblastos de rato (Maeno
et al., 2005) e em culturas de CTM-MO
(Barradas et al., 2010) e CTM-AD de
humanos (McCullen et al., 2010). Já foi
também mostrado que o cálcio extracelular
é capaz de aumentar a expressão de genes
ligados a diferenciação osteogênica em
osteoblastos de roedores (Maeno et al.,
2005; Jung e Park, 2010) e em CTM-MO
humanas (Barradas et al., 2010), bem como
de favorecer a síntese e mineralização da
matriz extracelular por osteoblastos
(Nakamura et al., 2010) e por CTM-AD
humanas diferenciadas (McCullen et al.,
2010). Os estudos previamente citados
mostram que o cálcio é capaz de favorecer a
diferenciação osteogênica e a osteogênese
“in vitro”, mas pouco se conhece a respeito
dos mecanismos envolvidos. Sabe-se que o
cálcio extracelular é capaz de controlar a
aposição e reabsorção de matriz óssea, por
meio do controle da secreção de
paratormônio (PTH) “in vivo” (Marie et al.,
2010). Os receptores CaSR atuam no
controle da proliferação, diferenciação e
apoptose, por meio de diferentes vias de
sinalização. O aumento do cálcio
extracelular pode agir nos receptores CaSR,
ativando a via Extracellular Signal
Regulated Kinase – Mitogen-Activated
Protein Kinase (ERK-MAPK) e aumentando
a proliferação osteoblástica (Huang et al.,
2001), mas podem também ser ativadas
outras vias como a Akt (também conhecida
como Protein Kinase B- PKB), que induz a
apoptose celular (Fromigué et al., 2009).
Sabe-se também que os CaSR são
moduladores importantes da diferenciação
osteogênica in vivo (Chang et al., 2008) e
que a via (ERK-MAPK) controla a
diferenciação osteogênica pelo controle da
expressão Runx2 (Ge et al., 2007). A via
ERK-MAPK é considerada o principal meio
de comunicação da superfície celular com o
núcleo que controla a diferenciação celular
(Ge et al., 2007; Marie et al., 2010). Já foi
comprovado que o aumento de cálcio
extracelular é capaz de aumentar a atividade
da fosfatase alcalina e as expressões de
osteopontina (OPN) e osteonectina (BSP)
em osteoblastos de rato e que esse efeito é
mediado pelos canais de cálcio tipo-L (L-
type Calcium Channel) e pela via de
sinalização calcium/calmodulin-dependente
da protein kinase 2 (CaM-CaMK2) (Jung e
Park, 2010). Um estudo com CTM-MO
humanas mostrou que o cálcio extracelular
é capaz de aumentar a proliferação celular e
estimular a diferenciação osteogênica pelo
aumento da expressão de BMP-2 e que esse
aumento da expressão de BMP-2 está
diretamente relacionado com os canais de
cálcio tipo L voltagem dependente (Type L
Voltage-Gated Ca2+
Channels-L-VGCCs) e
com a via de sinalização Extracellular
Signal Reguleted Kinase 1 and 2 (MEK1/2).
Esse mesmo estudo mostrou que o aumento
da expressão de BMP-2 não é mediado
pelos receptores CaSR e nem pelos
Metabotropic Glutamate Receptor Type 1
(mGlrR1) e que não há um efeito direto do
cálcio na transcrição gênica de BMP-2
(Barradas et al., 2010). Recentemente outro
estudo reafirmou a importância dos canais
de cálcio tipo L voltagem dependente (Type
L Voltage-Gated Ca2+
Channels-L-VGCCs)
na proliferação e diferenciação osteogênica
de CTM-MO de ratos mediada pelo cálcio.
Entretanto, esse estudo mostrou também
que o bloqueio desses canais não anula
totalmente os efeitos do cálcio na
proliferação e diferenciação osteogênica, o
que sugere o envolvimento de outros
mecanismos nesse processo (Wen et al.,
2012).

68
Muito embora o produto iônico do biovidro
60S tenha reduzido a síntese de colágeno,
aos sete e 21 dias (Fig.4) e a atividade da
fosfatase alcalina aos 14 e 21 dias de
cultivo (Fig.3), isso não interferiu na síntese
e mineralização da matriz extracelular pelas
CTM-AD de cães (Fig.5 e 6). Estudos com
culturas de osteoblastos de ratos (Valerio et
al., 2004) e de humanos (Tsigkou et al.,
2009) mostraram efeito estimulatório do
biovidro na síntese de colágeno (Valerio et
al., 2004; Tsigkou et al., 2009) e na
atividade da fosfatase alcalina (Tsigkou et
al., 2009). No entanto, esses estudos
trabalharam com espécie e tipo celular
diferentes dos utilizados no presente estudo,
o que pode justificar a diferença de resposta
observada.
A maior expressão de sialoproteina óssea e
maior área mineralizada por campo
observadas nas CTM-AD mantidas meio
osteogênico suplementado com produto
iônico (OST PI) em relação às mantidas em
meio osteogênico (OST) (Fig.8) mostram
que o produto iônico do biobidro 60S
favorece a osteogênese in vitro. Sugere
também que esse aumento na osteogênese
seja mediado pelo aumento da expressão da
sialoproteína óssea. A BSP é uma das
proteínas responsáveis pelo início do
processo de nucleação e deposição de
hidroxiapatita na matriz óssea. O aumento
na sua expressão reflete a fase final de
diferenciação e início de mineralização da
matriz. A sialoproteina óssea pode ser
expressa por pré-osteoblastos e osteoblastos
(Zhou et al., 2006).
A insignificante variação na expressão de
osterix (Fig.7), osteonectina (Fig.9) e
osteocalcina (Fig.10) observada nas CTM-
AD que receberam o produto iônico, sugere
que o efeito do produto de dissolução do
biovidro 60S não é mediado pelas
expressões dessas proteínas. O osterix é um
fator de transcrição imprescindível no
processo de diferenciação osteogênica e
atua na transformação dos pré-osteoblastos
em osteoblastos maduros, finalizando assim
o processo de diferenciação osteogênica
(Nakashima et al., 2002; Datta et al., 2008;
Long, 2012; Zhang, 2012). Sabe-se também
que o osterix pode influenciar a síntese e
mineralização da matriz extracelular por
meio do controle da expressão do receptor
intranuclear da vitamina D (Long, 2012;
Zhang, 2012). A osteonectina e a
osteocalcina são proteínas não colagênicas
da matriz e estão intimamente relacionadas
com o processo de síntese e mineralização
da matriz óssea e com o estágio final de
diferenciação osteogênica (Strauss et al.,
1990; Nefussi et al., 1997; Nakashima et
al., 2002; Karaoz et al., 2009).
Conclusão
Nas condições em que foi realizado este
experimento pode-se dizer que:
O produto iônico do biovidro 60S favorece
a osteogênese “in vitro” de culturas CTM-
AD de cães com aumento da conversão de
MTT, da área mineralizada por campo e da
expressão de sialoproteína óssea.

69
Capítulo 4: Tratamento de defeitos
ósseos críticos em rádios de cães com
matriz porosa do biovidro 60S associada
ou não a células tronco mesenquimais do
tecido adiposo.
Introdução
Um dos grandes desafios da ortopedia
veterinária e humana é o tratamento de
defeitos ósseos críticos, caracterizados por
grandes perdas incapazes de se regenerarem
naturalmente sem algum tipo de tratamento
adicional à fixação (Drosse et al., 2008;
Reichert et al., 2009, ASTM 2010).
Geralmente, os defeitos ósseos críticos são
tratados pela associação de um método de
fixação com enxertos ósseos,
preferencialmente autógeno. A fixação tem
como função impedir a movimentação dos
fragmentos ósseos no foco da fratura,
propiciando assim um microambiente que
permita a regeneração óssea e o enxerto
favorece a regeneração do tecido ósseo por
meio dos processos de osteocondução,
osteoindução e osteogênese (Tseng et al.,
2008). Entretanto, alguns inconvenientes
como maior tempo cirúrgico, complicações
no local da colheita, fragilização do local e
a quantidade limitada de osso coletado
estão associados à utilização dos enxertos
autógenos, considerado o melhor até o
presente momento (Santos e Rahal, 2004;
Piermattei et al., 2006; Sanders, 2007;
Strioga et al., 2012). Devido a isso, muito
se tem estudado em busca de alternativas
eficientes para substituição dessa forma de
enxertia e a produção de matrizes sintéticas
a partir de biomateriais parece ser a
alternativa mais viável e promissora,
especialmente quando combinada com as
terapias celulares e fatores de crescimento
(De Long Jr, 2007; Dutra et al., 2008;
Hench, 2009). Diferentes tipos de
biomaterias podem ser utilizados para
produção dessas matrizes sintéticas, mas o
grupo das biocerâmicas se destaca, uma vez
que favorece a regeneração óssea por
osteocondução (Bruder et al., 1998;
Arinzeh et al., 2003; Yuan et al., 2010) e,
em alguns casos, por osteoindução (Xynos
et al., 2000; Xynos et al., 2001; Habibovic
et al., 2006; Hench, 2009; Hopp et al.,
2011). Dentre as biocerâmicas, os vidros
são considerados os mais ativos e
favoráveis à regeneração óssea (Hench,
2009). Quando implantados no osso reagem
com o fluido intersticial desencadeando
uma cascata de reações físico-químicas e
eventos celulares que culminam com a
formação de uma camada de hidroxiapatita
biológica, deposição e mineralização de
osteóide em sua superfície, o que favorece
sua ligação ao osso e promove uma
interface coesa e resistente (Jones e Hench,
2003; Vallet-Regí, 2006; Hench, 2009). O
efeito do biovidro na regeneração óssea
pode variar de acordo com a composição
química, grau de organização das
moléculas, topografia e microestrutura da
matriz bem como, com as condições do
microambiente de implantação que podem
determinar maior ou menor dissolução do
biomaterial e consequentemente alterar seu
efeito biológico (Hoppe et al., 2011;
Rahaman et al., 2011). O biovidro 60S de
composição molar 4% P2O5; 36% CaO;
60% SiO2 é um desses biomateriais e já
mostrou efeitos benéficos na regeneração
óssea in vitro favorecendo a proliferação de
osteoblastos de ratos (Valerio et al., 2004) e
aumentando a osteogênese durante o
processo de diferenciação osteogênica de
CTM-AD de cães (capítulo 3 do presente
estudo) e in vivo, propiciando a osteogênese
em defeitos ósseos não críticos em
mandíbulas de cães (Dutra et al., 2008).
A matriz do biovidro 60S pode ser
produzida de forma porosa com uma
arquitetura que mimetiza o osso esponjoso e
pode ser associada a células tronco e
osteoblastos (Wang, 2003; Brodke et al.,
2006). As células tronco mesenquimais
(CTM) são células multipotentes,
encontradas entre as células diferenciadas
de um tecido e com função de se
autorrenovar e se diferenciar, originando os
tipos celulares do tecido no qual residem
(Presnell et al., 2002; Grove et al., 2004).
Vários tecidos são fontes de CTM, em
maior ou menor número (Payushina et al.,
2006) mas, o tecido adiposo é considerado a
melhor fonte dessas células para utilização
clínica pela grande quantidade de CTM
presente nesse tecido, facilidade de
obtenção do tecido adiposo e de isolamento
dessas células (Zuk et al., 2001; Neupane et
al., 2008; Vieira al., 2010). As CTM de
origem adiposa (CTM-AD) apresentam
maior potencial osteogênico que as CTM

70
originadas da medula óssea (CTM-MO)
(Lin et al., 2009), outra importante fonte de
CTM para utilização terapêutica (Bruder et
al., 1998; Arinzeh et al., 2003). Esse maior
potencial osteogênico das CTM-AD em
relação às CTM-MO já foi comprovado na
espécie canina (capítulo 2 do presente
estudo). Para garantir a diferenciação
osteogênica das CTM-AD essa etapa pode
ser realizada in vitro antes da sua utilização
clínica, embora um estudo recente não
tenha mostrado vantagens para a
regeneração óssea quanto se utilizou as
CTM diferenciadas em relação às
indiferenciadas (Beloti et al., 2012). Dessa
forma, o objetivo do presente estudo foi
avaliar a matriz porosa do biovidro 60S
associada ou não as CTM-AD diferenciadas
no tratamento de defeitos ósseos críticos em
rádios de cães.
Material e métodos
Foram utilizadas as bases físicas e a infra-
estrutura do Centro de Experimentação de
Pequenos Animais, do bloco cirúrgico e do
setor de diagnóstico por imagem do
Hospital Veterinário, do Núcleo de Células
Tronco e Terapia Celular e dos
Laboratórios de Histopatologia e
Imunoistoquímica do Departamento de
Clínica e Cirurgia Veterinárias da Escola de
Veterinária da UFMG, do Laboratório de
Biomateriais do Departamento de
Engenharia Metalúrgica e de Materiais da
UFMG e do Laboratório de Densitometria
da Faculdade de Medicina Veterinária da
Universidade Estadual Paulista “Júlio de
Mesquita Filho” campus de Araçatuba. O
projeto foi aprovado pelo Comitê de Ética
em Experimentação Animal da UFMG57
.
Produção da matriz porosa do biovidro
60S
A produção do biovidro 60S de composição
molar 4% P2O5; 36% CaO; 60% SiO2 foi
realizada no Laboratório de Biomateriais do
Departamento de Engenharia Metalúrgica e
de Materiais da UFMG conforme descrito a
seguir: foram adicionados em um bequer,
133,8 mL de tetraetilortosilicato (TEOS) e
57 Número de protocolo no CETEA (157/2009)
13,6 mL de trietilfosfato (TEP) e em outro,
97,9 mL de água deionizada e 16,3 mL de
solução de ácido nítrico 2N, as soluções
foram homogeneizadas e reservadas. Em
seguida as duas soluções foram misturadas
em um terceiro bequer maior e mantidas
sob agitação magnética por 60 minutos com
o objetivo de hidrolisar o TEOS.
Transcorrido este período, 85,01g de nitrato
de cálcio [Ca(NO3)2 4H2O] foi
gradativamente adicionado à solução que
permaneceu sob agitação magnética por
mais 30 minutos. Para permitir aeração e
formação da espuma do biovidro 60S, foi
adicionado 1mL de surfactante para cada
80mL solução. Em seguida 4mL de solução
aquosa de ácido fluorídrico (HF) 10% foi
acrescida a 100mL da solução final para
catalisar a sua gelificação. A solução foi
então aerada por dois minutos com o
auxílio de uma batedeira elétrica e após
esse período a espuma de biovidro foi
vertida em um recipiente de polipropileno
cujo interior possuía forma cilíndrica com
2cm de diâmetro e 3cm de altura. O
recipiente foi então hermeticamente
fechado e incubado a 60◦C em estufa por 72
horas para gelação e maturação.
Transcorrido esse período a tampa do
recipiente foi trocada por outra com
múltiplos orifícios e o biovidro foi
submetido a um protocolo de secagem em
estufa, permanecendo a 60◦C por 72 horas,
seguindo-se um aumento gradual da
temperatura em 10◦C a cada 24 horas, até
atingir a temperatura de 120◦C. Após a
secagem as amostras do biovidro 60S foram
tratadas termicamente na mufla a 700◦C por
6 horas quando foi utilizada uma taxa de
aquecimento e de resfriamento de 1◦C por
minuto (Coelho et al., 2003; Coelho et al.,
2005). Após o tratamento térmico as
amostras foram embaladas separadamente
em recipientes plásticos hermeticamente
fechados e esterilizadas por radiação gama.
Avaliação da porosidade da matriz do
biovidro 60S
A porosidade total da matriz do biovidro
60S foi calculada pela seguinte fórmula:
Porosidade total (%) =
(
)x 100
Onde a densidade verdadeira foi medida por
picnometria de Hélio (2,7 g/cm3) e a

71
densidade volumétrica foi calculada
dividindo-se a massa da matriz (g) pelo seu
volume (cm3)
Caracterização morfológica da matriz
porosa do biobidro 60S
Para análise morfológica da estrutura
porosa, as matrizes foram recobertas com
carbono (SPI/Supplies-EUA) e analisadas
por microscopia eletrônica de varredura
(MEV-FEI-Inspect-S50/Republica Tcheca).
Obtenção, cultivo e expansão das células
tronco mesenquimais do tecido adiposo
de cães
A colheita das amostras de tecido adiposo
foi realizada no centro cirúrgico do Hospital
Veterinário da Escola de Veterinária da
Universidade Federal de Minas Gerais
(UFMG). Foram utilizados três cães,
machos, não castrado, sem raça definida
com dois anos de idade e massa corporal
média de 18 Kg. Para colheita das amostras,
os cães foram submetidos ao jejum sólido
de oito horas. Como medicação pré-
anestésica, foram empregados 1mg/kg, IM,
de xilazina58
e 15mg/kg, IM, de
quetamina59
, na mesma seringa. A região
glútea direita foi submetida à tricotomia e
preparada para cirurgia asséptica. A veia
cefálica foi canulada e os animais
receberam propofol60
(3mg/kg IV) para
intubação e manutenção anestésica. Como
analgésico e anti-inflamatório foi
administrado meloxicam61
(0,2mg/kg IM)
imediatamente após a indução anestésica.
As amostras de tecido adiposo foram
colhidas cirurgicamente do tecido adiposo
subcutâneo da região glútea imediatamente
acima do trocânter maior. Foi colhido
aproximadamente 1cm3 de tecido adiposo
de cada animal totalizando 3cm3 de amostra
final. Imediatamente após a colheita, as
amostras de tecido adiposo foram colocadas
em tubos falcon de 50 mL contendo 20 mL
de meio de cultura Dulbecco’s Modiffed
Eagle Medium baixa glicose62
(DMEM), à
temperatura ambiente. Em seguida as
58 Calmiun, Agener União, Embu-Guaçu, SP, Brasil 59 Vetanarcol, Konig, Santana do Parnaíba, SP, Brasil 60 Fresofol, Fresenius Kabi, Campinas, SP, Brasil 61 Maxicam, Ouro Fino, Cravinhos, SP, Brasil 62 DMEM, Gibco, Grand Island, NY, USA
amostras foram encaminhadas ao Núcleo de
Células Tronco da Escola de Veterinária da
UFMG, onde as CTM foram extraídas e
cultivadas segundo o protocolo descrito a
seguir que foi adaptado de protocolos
estabelecidos por Neupane et al. (2008) e
Vieira et al. (2010).
As amostras de tecido adiposo foram
manipuladas de forma asséptica na capela
de fluxo laminar. Elas foram lavadas duas
vezes com PBS 0,15 molar para retirada de
restos de sangue e debris celulares. Para
digestão do tecido conjuntivo e liberação
das células, as amostras foram colocadas
em outro tubo falcon de 50 mL contendo 20
mL de uma solução estéril de colágenase
B63
0,1% P/V. As amostras foram então
fragmentadas a partículas menores com
auxílio de uma tesoura cirúrgica estéril. A
tesoura foi introduzida no tubo falcon de
50mL e com movimentos de abrir e fechar
dos ramos o tecido foi fragmentado ao
máximo possível. Em seguida a amostra foi
encubada em estufa a 37oC e 5% de CO2
durante 45 minutos e a cada 15 minutos a
amostra foi vigorosamente agitada para
facilitar o processo de digestão.
Transcorrido o tempo de digestão a amostra
foi centrifugada por 10 minutos a 1400rpm,
o sobrenadante contendo a fração adiposa
foi descartado e o pellet foi ressuspenso em
10 mL PBS 0,15 molar para lavagem e
retirada dos resíduos de colagenase. A
amostra foi colocada em outro tubo falcon
para reduzir a contaminação por gordura
retida na parede do tubo e novamente
centrifugada por 10 minutos a 1400rpm.
Após a centrifugação o sobrenadante foi
novamente descartado e o pellet, fração
estromal, foi ressuspenso em 20 mL de
meio de cultura para células tronco (meio
basal), DMEM com baixa glicose,
enriquecido com gentamicina (60 μg/L),
penicilina (100 UI/mL), estreptomicina
(100μg/mL), anfotericina (25μg/mL)64
e
10% de soro fetal bovino65
. Uma alíquota
de 2 mL do meio com as células
ressupensas foi transferida para cada
garrafa de cultivo celular T75 e 8 mL de
meio foram adicionados em cada garrafa.
Foram feitas dez garrafas no total.
63 Colagenase B, Roche Applied Science, Penzberg, Germany 64 PSA, Sigma-Aldrich, St. Louis, MO, USA 65 Soro fetal bovino, Sorali, Campo Grande, MS, Brasil

72
Realizou-se uma cuidadosa
homogeneização e as garrafas foram
incubadas em estufa a 37oC e 5% de CO2.
Após 48h foi realizada troca parcial do
meio de cultura, renovando apenas 5 mL.
Após mais 48h as células foram lavadas
duas vezes com PBS 0,15 molar para
remoção de debris e células não aderidas. O
meio de cultivo foi trocado duas vezes por
semana a fim de se eliminar células não
aderidas garantindo a obtenção de células
tronco mesenquimais.
Assim que foi atingido 80 a 90% de
confluência, as células foram repicadas.
Para desprendimento das células, as
garrafas foram lavadas com 10 mL de PBS
0,15 molar para retirada dos restos de meio
de cultura. Após a lavagem, foi adicionado
1,5 mL de tripsina e a garrafa foi incubada
por 10 minutos em estufa a 37oC e 5% de
CO2. Transcorrido o tempo de ação da
tripsina a mesma foi inativada com 5 mL de
DMEM enriquecido com 10% de soro fetal
bovino. As garrafas foram então lavadas
com o próprio meio para retirada das
células e uma alíquota de 1 mL foi repicada
para outra garrafa T75 com mais 9 mL de
meio de cultura.
Após o terceiro repique as células foram
então avaliadas quanto à viabilidade celular
pelo azul de Tripan e plaqueadas na
densidade de 1x104 células/cm
2 em garrafas
T75. Quando as garrafas atingiam 60 a 70%
de confluência celular o meio basal foi
substituído por meio osteogênico que é
constituído do meio basal enriquecido com
ácido ascórbico (50 g/mL), ß-
glicerofosfato (10 mM)66
e dexametasona
(0,1 M)67
. As células foram então
cultivadas a 37oC e 5% de CO2 por 21 dias
para diferenciação osteogênica.
Transcorrido esse período as células foram
desprendidas da superfície das garrafas de
cultivo por meio da ação da tripsina e
lavadas duas vezes com PBS 0,15 molar
para retirada dos resíduos de soro fetal
bovino. As células foram então
ressuspensas em DMEM puro, contadas em
câmara de Neubauer e utilizadas para o
tratamento de defeitos ósseos associadas ou
não a matriz porosa do biovidro 60S na
densidade 3x106 células por cm
3 de defeito
66 Sigma-Aldrich, St. Louis, MO, USA 67 Aché, Guarulhos, SP, Brasil
assim como o utilizado por Bruder et al.
(1998) e Arinzeh et al. (2003).
Implantação da matriz porosa do
biovidro 60S associado ou não as CTM-
AD diferenciadas para tratamento de
defeitos ósseos críticos
As intervenções cirúrgicas para realização
dos defeitos ósseos críticos em rádios de
cães e seu preenchimento de acordo com
cada grupo experimental, foram realizadas
no centro cirúrgico do Hospital Veterinário
da Escola de Veterinária da Universidade
Federal de Minas Gerais (UFMG). Foram
utilizados 35 cães, machos, intactos, sem
raça definida com dois anos de idade e
massa corporal média de 25 Kg. Os cães
foram recolhidos em fazendas da zona rural
do município de Uberaba quando filhotes e
criados até os dois anos de idade para
realização do presente estudo. No período
experimental os cães foram mantidos em
canis individuais tipo solário, com área de
4,5m2, pertencentes ao Centro de
Experimentação de Pequenos Animais do
Hospital Veterinário da Escola de
Veterinária da UFMG, onde receberam
ração comercial68
e água a vontade. Todos
os animais passaram por um período
mínimo de 30 dias de adaptação às novas
instalações. Nessa fase eles foram avaliados
clinicamente e por meio de exames
radiográficos e laboratoriais (hemograma,
pesquisa de hemoparasitas e análise
sorológica para leishmaniose), assegurando-
se que todos estavam saudáveis e aptos para
serem utilizados no presente estudo. Todos
os animais foram desverminados, vacinados
e receberam medicação ectoparasiticida. Os
cães foram então aleatoriamente
distribuídos em quatro grupos
experimentais como descrito a seguir:
grupo controle negativo (C-) – animais
com defeitos ósseos críticos sem
preenchimento (n=10); grupo tratado com
biovidro (BV) - animais com defeitos
ósseos críticos preenchidos com a matriz
porosa do biovidro 60S (n=10); grupo
tratado com a associação do biovidro
com as CTM-AD diferenciadas
(BV+CTM) - animais com defeitos ósseos
críticos preenchidos com a matriz porosa do
68 Canitos, Kowalski, Apucarana, PR, Brasil

73
biovidro 60S associada às CTM-AD
diferenciadas (n=10) e grupo tratado com
as CTM-AD diferenciadas (CTM) -
animais com defeitos ósseos críticos
tratados pela injeção local das CTM-AD
diferenciadas (n=5). O controle positivo
(C+) foi realizado em todos os animais
utilizando-se o rádio contralateral, com
defeito crítico preenchido com o próprio
fragmento ósseo retirado da ostectomia.
Para realização das cirurgias, os cães foram
submetidos ao jejum sólido de oito horas e
receberam como medicação pré-anestésica
atropina69
(0,02 mg/kg SC) e 10 minutos
após, acepromazina70
(0,03 mg/kg IM) e
morfina71
(0,8 mg/kg IM) na mesma
seringa. A veia safena lateral foi canulada e
os membros torácicos foram submetidos à
tricotomia, que se estendeu da região dos
metacarpos até a coluna. A indução
anestésica foi realizada com propofol72
(3
mg/kg IV) e diazepam73
(0,3 mg/kg IV),
seguindo-se a intubação endotraqueal e
manutenção anestésica por meio da
inalação espontânea de isofluorano74
diluído em oxigênio puro, em circuito
semifechado de baixo fluxo. O animal foi
então posicionado em decúbito esternal e os
membros torácicos foram preparados para
cirurgia asséptica com solução degermante
a base de iodo e álcool iodado. Para
controle da dor e inflamação foram
administrados tramadol75
(3mg/kg SC TID)
e meloxicam76
(0,2 mg/kg IM SID) durante
cinco dias consecutivos iniciando-se a
primeira aplicação imediatamente após o
término da cirurgia, bem como
antibioticoterapia profilática com cefalexina
(30 mg/kg VO, BID) durante sete dias
consecutivos, iniciando se a primeira dose
no pré-operatório imediato via intravenosa.
Para a exposição óssea, foi realizada uma
incisão de pele crânio-medial ao rádio,
divulsão romba do tecido subcutâneo e
afastamento lateral da musculatura
extensora (Fig.1), segundo Piermattei e
Johnson (2004). O periósteo foi afastado e
69 Pasmodex, Isofarma, Eusébio, CE, Brasil 70 Acepran 0,2%, Vetnil, São Paulo, SP, Brasil 71 Dimorf, Cristália, Itapira, SP, Brasil 72 Fresofol, Fresenius Kabi, Campinas, SP, Brasil 73 Diazepam, União Química, Embu-Guaçu, SP, Brasil 74 Isoforine, Cristália, Itapira, SP, Brasil 75 Cloridrato de Tramadol, Teuto, Anápolis, GO, Brasil 76 Maxicam injetável 0,2%, Ouro Fino, Cravinhos, SP,
Brasil
o diâmetro do terço médio do rádio no
sentido médio-lateral foi mensurado com
auxílio de um paquímetro estéril. Foi então
realizada uma ostectomia transversal do
terço médio do rádio com extensão
correspondente a uma vez e meio o
diâmetro do osso (Fig.1), assegurando a
criação de um defeito ósseo crítico (Millis e
Jackson, 2003). Para realização desse
procedimento foi utilizada uma serra óssea
oscilatória pneumática, sob irrigação
constante com solução fisiológica. A
estabilização óssea foi realizada com uma
placa de aço F318 em ponte, posicionada na
superfície cranial do rádio e fixada com seis
parafusos bicorticais, sendo três no
segmento proximal e três no segmento
distal (Fig.1). As perfurações ósseas foram
realizadas com broca de diâmetro igual ao
do centro do parafuso, utilizando-se
furadeira pneumática com rotação
controlável e irrigação constante. A
profundidade de cada orifício foi
mensurada, a rosca foi preparada e os
parafusos apertados manualmente.
Inicialmente foram colocados os parafusos
adjacentes à falha óssea e depois os das
extremidades da placa, seguindo-se os
demais alternadamente. Após a fixação
óssea o defeito foi tratado de acordo com
cada grupo experimental (Fig.1) e a síntese
tecidual foi realizada de forma rotineira
sendo o tecido subcutâneo aproximado em
padrão simples contínuo com caprofil 3-0 e
a pele com simples separado com náilon
nomofilamentar 3-0. Após a cirurgia foram
utilizadas bandagens acolchoadas em
ambos os membros torácicos para
prevenção do edema pós-operatório e
proteção da ferida cirúrgica, mantidas
durante sete dias.

74
Figura 1: Sequencia e fotos da cirurgia de implantação do biovidro 60S, associado ou não das CTM-AD
para tratamento de defeitos ósseos críticos em rádios de cães. Observar nas imagens A - ostectomia; B –
posicionamento da placa cranial ao osso; C e D – matriz do biovidro 60S com forma semelhante ao
segmento osteotomisado; E – deposição das células na matriz do biovidro; F - defeito ósseo sem
preenchimento (controle negativo); G – defeito ósseo preenchido com enxerto autógeno (controle
positivo); H – defeito ósseo preenchido com a matriz do biovidro 60S e I - defeito ósseo preenchido com
a matriz do biovidro associada com as CTM-AD.
Avaliações clínica e radiográfica
As avaliações clínicas de todos os animais
foram realizadas às 1, 3, 5, 8 e 24 horas
após a cirurgia e diariamente até o 10◦ dia
de pós-operatório com o objetivo de
monitorar a analgesia e eventuais
intercorrências.
As avalições radiográficas foram realizadas
no setor de diagnóstico por imagem do
Hospital Veterinário da Escola de
Veterinária da UFMG, utilizando-se um
aparelho de raio-x digital Regius modelo
11077
. Foram realizadas as projeções
radiográficas médio-lateral e crânio-caudal
de ambos os rádios no pré e pós-operatórios
imediatos e aos 15, 30, 45, 60, 75 e 90 dias
após a intervenção cirúrgica em cinco
animais de cada grupo. Os outros cinco dos
grupos, C-, BV e BV+CTM foram
avaliados radiograficamente até o 45o dia de
pós-operatório quando foram submetidos à
eutanásia para avaliação histológica. A
projeção crânio-caudal foi utilizada para
monitoração do alinhamento ósseo e a
77 Konica Minolta, Tóquio, Japão

75
projeção médio-lateral, para mensuração
das áreas de crescimento ósseo nas linhas
de osteotomia, áreas compostas pelo
biomaterial, áreas de calo periosteal e
medular. As mensurações foram realizadas
com auxílio do software Digimizer Image
Analysis. As áreas correspondentes ao
crescimento ósseo e ao biomaterial foram
expressas em percentagem em relação à
área total da falha óssea de cada animal
considerada 100% (Fig.2). As áreas dos
calos periosteais e medulares também
foram expressas em percentagem e a maior
área de cada calo para cada animal foi
considerada 100% (Fig.2). As frequências
de reação periosteal, de calo periosteal, de
calo medular, de calo intercortical, de
continuidade cortical e de continuidade
medular foram também avaliadas em cada
tempo.
Figura 2. Imagens radiográficas de defeitos ossos críticos em rádios de cães na projeção médio-lateral,
processadas pelo software Digimizer Image Analysis para mensuração das áreas de preenchimento ósseo
(áreas contornadas em azul na imagem A), área de preenchimento com biomaterial (área contornada em
verde na imagem A), área total do defeito (áreas contornadas em vermelho na imagem A), áreas de calo
periosteal (áreas contornadas em azul na imagem B) e medular (áreas contornadas em vermelho na
imagem B).
Avaliações densitométrica e histológica
As avaliações densitométricas e
histológicas foram realizadas aos 45 dias de
pós-operatório em cinco animais dos grupos
C-, BV e BV+CTM e aos 90 dias de pós-
operatório nos demais cinco animais dos
seguintes grupos C-, BV, BV+CTM e
CTM. Essas avaliações foram feitas
utilizando-se apenas os ossos rádios após
colheita e fixação em formalina tamponada
10%. Para isso os animais foram
submetidos à eutanásia por meio da
aplicação de xilazina (1mg/kg) e quetamina
(15mg/kg) na mesma seringa via
intramuscular, seguida da canulação da veia
cefálica e injeção de propofol até parada
cardiorrespiratória, seguida da injeção de 40
mL de cloreto de potássio via intravenosa.
Os rádios e ulnas direitos e esquerdos foram
dissecados, colhidos e fixados em formalina
tamponada 10% por 30 dias.
A avaliação densitométrica foi realizada no
Laboratório de Densitometria da Faculdade
de Medicina Veterinária da Universidade
Estadual Paulista “Júlio de Mesquita
Filho”, campus de Araçatuba, utilizando-se
um densitômetro de dupla emissão de raios-
x78
. A densidade mineral óssea (g/cm2) foi
mensurada na região correspondente às
falhas ósseas com incidência na projeção
médio-lateral. Os valores de densidade
obtidos de cada falha, de cada animal foram
convertidos em percentagem da densidade
óssea normal do rádio de cada animal,
considerada 100%. A densidade óssea
considerada normal foi obtida na região
78 Densitômetro DXA, modelo DPX-Alpha Lunar

76
correspondente aos defeitos tratados com o
autoenxerto (controle +).
A avaliação histológica e histomorfométrica
foi realizada no laboratório de
Histopatologia e Imunoistoquímica do
Departamento de Clínica e Cirurgia
Veterinárias da Escola de Veterinária da
UFMG. Os rádios e ulnas foram
transversalmente seccionados na altura dos
parafusos adjacentes à falha. O segmento
ósseo central, contendo a falha, foi
submetido ao protocolo de
desmineralização com solução aquosa de
ácido fórmico 50% v/v e citrato de sódio
9% p/v por 90 dias, sendo trocada a cada
quatro dias. Após esse período foi realizada
uma secção longitudinal do segmento ósseo
correspondente à falha e as metades foram
avaliadas macroscopicamente e processadas
pela técnica rotineira de inclusão em
parafina. Cortes histológicos de 5 μm foram
obtidos de cada bloco e corados pela
técnica da hematoxilina-eosina. As lâminas
foram avaliadas descritivamente por
microscopia óptica. A área de neoformação
óssea foi quantificada com o auxílio de uma
ocular micrométrica, contendo uma
gratícula com 121 pontos e com objetiva de
4 percorrendo-se toda a extensão do
defeito. A área de neoformação óssea foi
expressa em relação à área total do defeito
de cada animal. Também foi quantificado o
número de vasos por campo em 12 campos
tomados na área do defeito com objetiva de
20.
Análise estatística
Foi utilizado delineamento inteiramente
casualizado com controle positivo pareado.
Os dados não mensuráveis foram analisados
descritivamente. Os dados paramétricos
foram submetidos à análise de variância
(ANOVA) e as médias comparadas pelo
teste Student Newman Keuls (SNK)
utilizando o programa Graphpad Instat 379
.
Diferenças foram consideradas
significativas se p0,05 (Sampaio, 1998).
79 GraphPad Software Inc., San Diego, USA
Resultados
Avaliação da matriz porosa do biovidro
60S
A matriz do biovidro 60S apresentou
porosidade média de 97,46%, variando de
97,01 a 97,8%. A microscopia eletrônica
mostrou que a matriz apresentava poros
interconectados com diâmetro médio de
507µm variando de 300 a 700µm (Fig.3).
Figura 3. Micrografia de uma amostra da matriz
porosa do biovidro 60S obtida por microscopia
eletrônica de varredura. Observa a presença de
poros interconectados com diâmetro variado.
Avaliações clínica e radiográfica
Os animais adaptaram-se bem aos canis e
não apresentaram intercorrências clínicas
que pudessem comprometer os resultados
da pesquisa. Todos os cães foram capazes
de deambular 24 horas após a cirurgia e o
protocolo analgésico utilizado foi eficiente,
mantendo os animais sem sinais de dor no
período pós-operatório.
A avaliação radiográfica pré-operatória
possibilitou a escolha prévia das placas e
parafusos de dimensões adequadas para
cada animal, permitiu estimar previamente
a extensão do defeito ósseo e as dimensões
da matriz de biovidro para preenchimento
total do defeito. O cálculo prévio da área do
defeito possibilitou também definir a
quantidade de células tronco necessária
para cada animal.
A extensão do defeito de uma vez e meia o
diâmetro ósseo atingiu o objetivo proposto
de criação de defeitos ósseos críticos em
rádios de cães. Os controles negativos (C-)

77
mostraram preenchimento ósseo parcial da
falha de no máximo 56,68% sem a
formação de ponte óssea entre as
extremidades e os controles positivos (C+)
mostraram regeneração óssea completa em
todos os animais aos 90 dias (Fig.4).
Figura 4: Imagens radiográficas de defeitos ossos críticos em rádios de cães não tratados (controle
negativo – A) e tratados com osso autógeno (controle positivo – B), obtidas aos 90 dias de pós-operatório.
Observar na imagem A o defeito ósseo parcialmente preenchido com osso esponjoso sem a formação de
ponte óssea entre os fragmentos e na imagem B a completa regeneração óssea, com continuidade das
corticais e do canal medular.
As avaliações radiográficas pós-operatórias
permitiram o acompanhamento da
regeneração óssea e da evolução de
reabsorção da matriz porosa do biovidro
60S durante os 90 dias subsequentes a sua
implantação nos defeitos ósseos. A área
correspondente ao biomaterial mostrou
rápida reabsorção após sua implantação
com redução média de 12,62% e 13,17% a
cada 15 dias nos grupos BV e BV+CTM
respectivamente (Fig.5). O grupo BV+CTM
mostrou reabsorção do biovidro
estatisticamente inferior, aos 15 dias de
implantação, mas essa diferença não foi
observada nos demais tempos estudados
(Fig.5). A matriz porosa do biovidro 60S
foi completamente reabsorvida em 40% e
60% dos animais aos 90 dias nos grupos
BV e BV+CTM, respectivamente (Fig.6).
Figura 5: Percentagem média e desvio padrão da área de biomaterial em defeitos ósseos críticos em rádios
de cães, tratados com matriz porosa de biovidro 60S associada (BV+CTM) ou não (BV) às CTM-AD
diferenciadas, ao longo de 90 dias de avaliação pós-operatória (medidas obtidas radiograficamente).
*Representa diferença estatística (P<0,05).
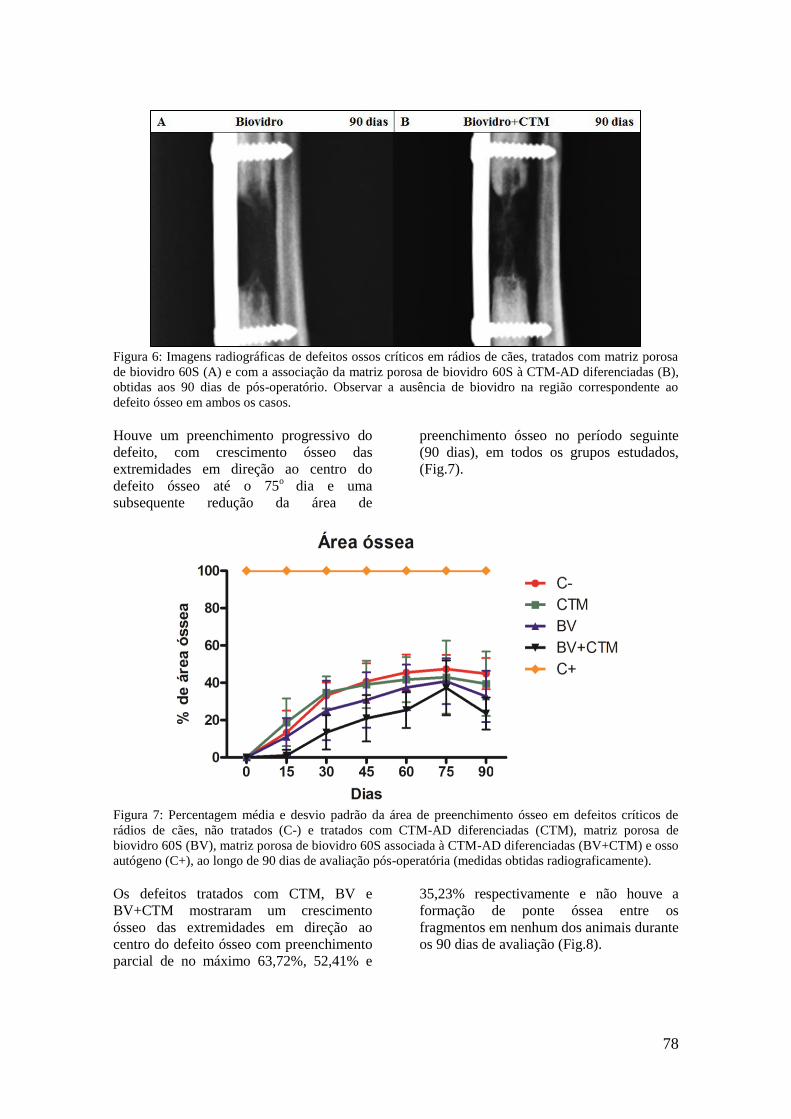
78
Figura 6: Imagens radiográficas de defeitos ossos críticos em rádios de cães, tratados com matriz porosa
de biovidro 60S (A) e com a associação da matriz porosa de biovidro 60S à CTM-AD diferenciadas (B),
obtidas aos 90 dias de pós-operatório. Observar a ausência de biovidro na região correspondente ao
defeito ósseo em ambos os casos.
Houve um preenchimento progressivo do
defeito, com crescimento ósseo das
extremidades em direção ao centro do
defeito ósseo até o 75o
dia e uma
subsequente redução da área de
preenchimento ósseo no período seguinte
(90 dias), em todos os grupos estudados,
(Fig.7).
Figura 7: Percentagem média e desvio padrão da área de preenchimento ósseo em defeitos críticos de
rádios de cães, não tratados (C-) e tratados com CTM-AD diferenciadas (CTM), matriz porosa de
biovidro 60S (BV), matriz porosa de biovidro 60S associada à CTM-AD diferenciadas (BV+CTM) e osso
autógeno (C+), ao longo de 90 dias de avaliação pós-operatória (medidas obtidas radiograficamente).
Os defeitos tratados com CTM, BV e
BV+CTM mostraram um crescimento
ósseo das extremidades em direção ao
centro do defeito ósseo com preenchimento
parcial de no máximo 63,72%, 52,41% e
35,23% respectivamente e não houve a
formação de ponte óssea entre os
fragmentos em nenhum dos animais durante
os 90 dias de avaliação (Fig.8).

79
Figura 8: Imagens radiográficas de defeitos ossos críticos em rádios de cães, tratados com CTM-AD
diferenciadas (A), matriz porosa de biovidro 60S (B) e matriz porosa de biovidro 60S associada à CTM-
AD diferenciadas (C), obtidas aos 90 dias de pós-operatório. Observar em todas as imagens o
preenchimento parcial do defeito com osso esponjoso das extremidades em direção ao centro e nas
imagens B e C, fragmentos de biomaterial no centro dos defeitos.
As áreas de preenchimento ósseo nos
grupos CTM, BV e BV+CTM foram
significativamente inferiores ao controle
positivo (C+) em todos os tempos
estudados (Fig.9). Os grupos CTM e BV
mostraram área de preenchimento ósseo
semelhante ao controle negativo (C-) em
todos os tempos estudados (Fig.9). O grupo
BV+CTM mostrou valores de
preenchimento ósseo inferiores aos demais
grupos aos 15 e 30 dias, sua área de
preenchimento também foi inferior aos
grupos C- e CTM aos 30 e 45 dias e ao
grupo C- aos 60 e 90 dias de pós-operatório
(Fig.9).
Figura 9: Percentagem média e desvio padrão da área de preenchimento ósseo em defeitos críticos em
rádios de cães, não tratados (C-) e tratados com CTM-AD diferenciadas (CTM), matriz porosa de
biovidro 60S (BV), matriz porosa de biovidro 60S associada à CTM-AD diferenciadas (BV+CTM) e osso
autógeno (C+), ao longo de 90 dias de avaliação pós-operatória (medidas obtidas radiograficamente).
*Representa diferença estatística (P<0,05).

80
Os controles positivos se regeneram de
forma indireta, com a formação de
pequenos calos periosteais, medulares e
intercorticais, com completa consolidação
óssea aos 90 dias em100% dos animais. As
frequências de observação de reação
periosteal, medular e intercortical e da
continuidade cortical e medular, em cada
tempo, estão apresentadas na tabela 1 e a
dinâmica de crescimento e regressão dos
calos na figura 10.
Tabela 1: Frequências de ocorrência de reação periosteal, calos periosteal, medular e intercortical e de
continuidade cortical e medular em defeitos ósseos críticos em rádios de cães tratados com osso autógeno,
observadas aos 0, 15, 30, 45, 60, 75 e 90 dias de pós operatório.
Dias
0 15 30 45 60 75 90
Reação periosteal proximal
0% 64,29% 25% 0% 0% 0% 0%
Reação periosteal distal
0% 50% 28,57% 0% 0% 0% 0%
Calo periosteal proximal
0% 3,57% 67,86% 71,43% 31,25% 25% 6,25%
Calo periosteal distal
0% 3,57% 57,14% 60,71% 50% 18,75% 0%
Calo medular proximal
0% 14,29% 85,71% 82,14% 43,75% 31,25% 6,25%
Calo medular distal
0% 10,71% 92,86% 82,14% 62,5% 43,75% 6,25%
Calo intercortical proximal
adjacente à ulna
0% 0% 50% 85,71% 100% 100% 100%
Calo intercortical proximal
adjacente à placa
0% 0% 35,71% 82,14% 100% 100% 100%
Calo intercortical distal
adjacente à ulna
0% 0% 53,57% 89,29% 100% 100% 100%
Calo intercortical distal
adjacente à placa
0% 0% 35,71% 85,71% 100% 100% 100%
Continuidade cortical
proximal adjacente à ulna
0% 0% 7,14% 10,71% 43,75% 75% 100%
Continuidade cortical
proximal adjacente à placa
0% 0% 0% 3,57% 31,25% 56,25% 100%
Continuidade cortical distal
adjacente à ulna
0% 0% 0% 7,14% 50% 68,75% 100%
Continuidade cortical distal
adjacente à placa
0% 0% 0% 0% 37,5% 62,5% 100%
Continuidade medular
proximal
0% 0% 0% 10,71% 56,25% 68,75% 93,75
%
Continuidade medular distal 0% 0% 0% 25% 43,75% 62,5% 100%

81
Figura 10: Percentagem média e desvio padrão da área dos calos periosteais e medulares adjacentes às
linhas de osteotomia distal e proximal de defeitos ósseos críticos em rádios de cães tratados com osso
autógeno (C+), ao longo de 90 dias de avaliação pós-operatória.
Avaliação densitométrica
Todos os grupos apresentaram menor
densidade na região do defeito ósseo em
relação aos controles positivo (C+), aos 45
e 90 dias (Fig.11). Aos 45 dias, os grupos
BV e BV+CTM mostraram densidade
semelhante entre si e superiores ao grupo
controle negativo (C-) e aos 90 dias,
nenhuma diferença foi observada entre os
grupos C-, CTM, BV e BV+CTM (Fig.11).
Figura 11: Densidade média e desvio padrão de defeitos ósseos críticos em rádios de cães, não tratados
(C-) e tratados com CTM-AD diferenciadas (CTM), matriz porosa de biovidro 60S (BV), matriz porosa
de biovidro 60S associada à CTM-AD diferenciadas (BV+CTM) e osso autógeno (C+), aos 45 e 90 dias
de pós-operatório. A densidade está expressa em percentagem relação à densidade normal do rádio de
cada animal. *Representa diferença estatística (P<0,05).
Avaliação macroscópica
A avaliação macroscópica dos fragmentos
ósseos desmineralizados, após secção
longitudinal revelou o preenchimento
parcial dos defeitos com tecido ósseo
esponjoso, com crescimento das
extremidades ósseas em direção ao centro,
nos grupos C-, CTM, BV e BV+CTM aos
45 e 90 dias (Fig.12). Nos grupos BV e
BV+CTM foi possível observar parte da
matriz porosa do biovidro 60S no centro do
defeito ósseo, envolvida por tecido mole,
aparentemente conjuntivo (Fig.12). Não foi

82
possível observar a presença do biomaterial
em 40% e 60% das amostras dos grupos
BV e BV+CTM respectivamente (Fig.12).
Verificou-se nas amostras do controle
positivo, aos 45 dias, a formação de calos
periosteais, medulares e intercorticais
(Fig.12). Aos 90 dias, observou-se
continuidade cortical e medular no
segmento correspondente ao defeito ósseo
(Fig.12).
Figura 12: Fragmentos ósseos correspondentes a defeitos ósseos críticos em rádios de cães, não tratados (A1 e A2) e tratados com
matriz porosa de biovidro 60S (B1 e B2), matriz porosa de biovidro associada à CTM-AD diferenciadas (C1 e C2), osso autógeno (D1 e D2) e CTM-AD diferenciadas (E), aos 45 (A1, B1, C1 e D1) e 90 (A2, B2, C2, D2 e E) dias de pós-operatório. Notar nas
imagens A1, A2, B1, B2, C1, C2 e E, o preenchimento parcial do defeito com ósseo esponjoso das extremidades em direção ao
centro (regiões com asteriscos vermelhos); nas imagens B1, C1 e C2, a matriz porosa do biovidro no centro do defeito ósseo envolvida por tecido mole (regiões com estrelas verdes); na imagem B2, ausência de biomaterial no centro do defeito; na imagem
D1, os calos periosteais (apontados pelas setas vermelhas), os calos medulares (nas regiões com asterisco azul), a presença de calo intercortical na área circulada em vermelho e a ausência na área circulada em preto; e na imagem D2, a continuidade das corticais e
da medular.

83
Avaliação histológica e
histomorfométrica
A avaliação histológica dos grupos controle
negativo (C-), biovidro (BV), biovidro mais
células (BV+CTM) e célula (CTM),
mostrou neoformação óssea das
extremidades do defeito em direção ao
centro, sem união entre elas. O osso
neoformado em todos os defeitos dos
grupos C-, BV, BV+CTM e CTM foi
esponjoso, com trabéculas espessas
predominantemente recobertas por
osteoblastos volumosos. Verificou-se
também uma predominância de osteócitos
ativos em lacunas alargadas (Fig.13). Aos
45 dias, observou-se resquícios de tecido
cartilaginoso em meio ao osso neoformando
em todos os defeitos do grupo controle
negativo.
Verificou-se, pela avaliação
histomorfométrica, aos 45 e 90 dias, que as
áreas de neoformação óssea nos grupos
CTM, BV e BV+CTM foram
significativamente inferiores ao controle
positivo (Fig.14). Os grupos C-, BV e CTM
mostraram área de neoformação óssea
semelhante, aos 45 e 90 dias. O grupo
BV+CTM mostrou menor área de
neoformação óssea que o controle negativo
(C-) em todos os tempos estudados
(Fig.14).
Figura 13: Micrografia do osso neoformado nas extremidades de defeitos ósseos críticos em rádios de
cães, não tratados (A) e tratados com matriz porosa de biovidro 60S (B), matriz porosa de biovidro 60S
associada à CTM-AD diferenciadas (C) e CTM-AD diferenciadas (D). Observar que o osso neoformado é
trabecular, com trabéculas espessas predominantemente recobertas por osteoblastos ativos. Hematoxilina
eosina. Barra = 93µm.

84
Figura 14: Percentagem média e desvio padrão da área de neoformação óssea em defeitos críticos em
rádios de cães, não tratados (C-) e tratados com CTM-AD diferenciadas (CTM), matriz porosa de
biovidro 60S (BV), matriz porosa de biovidro 60S associada à CTM-AD diferenciadas (BV+CTM) e osso
autógeno (C+), aos 45 e 90 dias de avaliação pós-operatória (medidas obtidas por histomorfometria).
*Representa diferença estatística (P<0,05).
Verificou-se, que o centro dos defeitos dos
grupos C- e CTM era formado por tecido
conjuntivo fibroso desorganizado, mas nos
defeitos do grupo CTM o tecido conjuntivo
era menos denso, haviam agrupamentos
celulares com características de endotélio
vascular e a neovascularização era muito
mais intensa que no grupo C- (Fig.15).
O centro dos defeitos dos grupos (BV e
BV+CTM) era formado pela matriz do
biovidro 60S com tecido conjuntivo no
interior dos poros e em torno da matriz
(Fig.15). Foi observada a presença de um
tecido semelhante ao mixomatoso em
algumas áreas dos defeitos do grupo
BV+CTM, em outras, verificou-se a
presença de aglomerados de células
semelhantes às endoteliais e a
neovascularização. Verificou-se, aos 90
dias, maior neovascularização nos grupos
CTM e BV+CTM em relação aos grupos
controle negativo e BV, respectivamente
(Fig. 16).

85
Figura 15: Micrografia do centro de defeitos ósseos críticos em rádios de cães, não tratados (A) e tratados
com CTM-AD diferenciadas (B), matriz porosa do biovidro 60S (C), matriz porosa do biovidro 60S
associada à CTM-AD diferenciadas (D), 90 dias após a cirurgia. Observar em todas as imagens a
presença de tecido conjuntivo fibroso e a ausência de tecido ósseo. Hematoxilina eosina. Barra = 93µm.
Figura 16: Percentagem média e desvio padrão do número de vasos por campo no centro de defeitos
críticos em rádios de cães, não tratados (C-) e tratados com CTM-AD diferenciadas (CTM), matriz porosa
de biovidro 60S (BV), matriz porosa de biovidro 60S associada à CTM-AD diferenciadas (BV+CTM) e
osso autógeno (C+), aos 45 e 90 dias de avaliação pós-operatória (medidas obtidas por histomorfometria).
*Representa diferença estatística (P<0,05).

86
Observou-se uma significativa redução na
quantidade da matriz do biovidro dos 45
para os 90 dias de avaliação nos grupos que
receberam o biomaterial (Fig.17). Aos 90
dias de avaliação foram observados apenas
resquícios na matriz do biovidro, em dois
(40%) defeitos do grupo BV e três do
BV+CTM. Raras células gigantes
multinucleadas foram observadas próximas
a matriz do biovidro nos grupos BV e
BV+CTM.
Figura 17: Percentagem média e desvio padrão da área de biomaterial no centro de defeitos críticos em
rádios de cães, tratados com matriz porosa de biovidro 60S (BV) e matriz porosa de biovidro 60S
associada à CTM-AD diferenciadas (BV+CTM), aos 45 e 90 dias de avaliação pós-operatória (medidas
obtidas por histomorfometria). *Representa diferença estatística (P<0,05).
Não foi observada neoformação óssea no
centro dos defeitos em nenhum dos grupos
C-, BV, BV+CTM e CTM. Também não
foi observada a presença de infiltrado
inflamatório em nenhum dos defeitos de
todos os grupos.
Verificou-se predomínio da regeneração
óssea endocondral nos defeitos tratados
com o osso autógeno (C+), incompleta aos
45 dias e completa aos 90. A regeneração
óssea da cortical adjacente à placa ocorreu
predominantemente via endosteal (região
medular) e da cortical oposta
predominantemente periosteal (calo
periosteal) e com maior volume de osso
neoformado. Observou-se algumas áreas
com lacunas vazias na matriz óssea do
segmento utilizado para o preenchimento da
falha, principalmente na cortical adjacente a
placa.
Discussão
Para o emprego clínico de uma nova terapia
para regeneração óssea é necessário o
estudo experimental prévio em indivíduos
preferencialmente, da mesma espécie em
que será utilizada, uma vez que a resposta
biológica pode variar de acordo com a
espécie não permitindo extrapolação
fidedigna de uma espécie para outra, razão
para a seleção do cão como modelo
experimental. O cão é uma espécie muito
utilizada em estudos de regeneração de
defeitos ósseos críticos (Bruder et al.,
1998; Arinzeh et al., 2003; Hu et al., 2003;
Brodke et al., 2006; Wu et al., 2006; Cui et
al., 2007; He et al., 2007; Yuan et al., 2007;
Zhao et al., 2009; Yuan et al., 2010; Liu et
al., 2013) e é considerado um bom modelo
para estudos pré clínicos quando se propõe
aplicar as técnicas em pacientes humanos

87
(Gugala et al., 2007; Reichert et al., 2009).
Entretanto, outras espécies como o coelho
(Wan et al., 2006; Nather et al., 2010), a
cabra (Liu et al., 2008; Zhu et al., 2010), a
ovelha (Kon et al., 2000; Viateau et al.,
2007) e o rato são também utilizadas em
estudos de regeneração óssea (Levi et al.,
2010).
A metodologia de estudo deve reproduzir o
mais fielmente possível o problema clínico
a ser tratado, razão da seleção do estudo
com defeitos ósseos críticos em rádio de
cães. A maioria dos estudos trabalharam
com modelos de defeitos ósseos críticos em
fêmur (Bruder et al., 1998; Arinzeh et al.,
2003; Brodke et al., 2006), mandíbula (Wu
et al., 2006; He et al., 2007; Yuan et al.,
2007; Zhao et al., 2009; Yuan et al., 2010) e
crânio (Cui et al., 2007; Liu et al., 2013) de
cães, mas o uso do osso rádio como no
presente estudo, reproduz mais
fidedignamente o maior problema da
prática clínica, que é a não união óssea em
rádio de cães, correspondendo, segundo
Millis e Jackson (2003) a 40,6% dos casos
de não união em cães. Essa maior
propensão a não união de fratura de rádio e
ulna está associada principalmente ao
menor suprimento vascular na metáfise
distal de rádios de cães de pequenos porte
(Welch et al., 1997; Jackson e Pacchiana,
2004) e à pequena cobertura muscular do
osso rádio, que prejudicam o aporte
vascular para a região óssea fraturada e
aumenta o risco de problemas na
consolidação óssea (Millis e Jackson, 2003;
Jackson e Pacchiana, 2004; Johnson et al.,
2005).
O preenchimento parcial dos defeitos
ósseos observado aos 90 dias de pós
operatório no grupo controle negativo (C-)
(Fig.4, 7, 9, 12 e 14) comprova que a
extensão do defeito ósseo foi suficiente para
causar um defeito crítico, que necessita de
suporte para regeneração (Drosse et al.,
2008; Reichert et al., 2009, ASTM 2010) e
valida portanto, a metodologia utilizada.
Segundo Millis e Jackson (2003) e a ASTM
(2010) um segmento de defeito ósseo uma
vez e meia o diâmetro diafisário excede a
capacidade regenerativa do osso em cães
com maturidade esquelética e resulta em
não união. Outros pesquisadores, no
entanto, consideram defeitos ósseos críticos
aqueles com extensões iguais ou superiores
a duas ou duas vezes e meia o maior
diâmetro do osso (Gugala, et al., 2007;
Reichert et al., 2009)
A completa regeneração óssea observada
aos 90 dias de pós operatório em todos os
defeitos tratados com o autoenxerto (grupo
C+) (Fig.4, 7, 9 e 12) mostra que todos os
animais apresentaram capacidade de
regeneração óssea normal e que a técnica de
estabilização óssea foi adequada. A fixação
óssea inadequada é uma das principais
causas de problemas na consolidação óssea
(Piermattei et al., 2006) e a causa mais
frequente de não união de fraturas em
pequenos animais (Millis e Jackson, 2003;
Jackson e Pacchiana, 2004; Johnson et al.,
2005). É necessário portanto, garantir uma
estabilização óssea adequada quando se
estuda novas terapias e biomateriais para
substituição óssea. O excesso de
instabilidade no foco de fratura pode levar a
não regeneração óssea mesmo que o
biomaterial ou a terapia apresentem as
melhores características para promoção da
cura.
A maior densidade observada aos 45 dias
de pós operatório nos grupos tratados com a
matriz porosa do biovidro 60S, associada
ou não às CTM (grupo BV e BV+CTM) em
relação ao grupo controle negativo (C-)
(Fig.11) se deve à maior densidade do
biomaterial em relação aos tecidos moles,
uma vez que radiográfica e
histologicamente não foi observada
superioridade desses grupos (BV e
BV+CTM) em relação ao controle
negativo, quanto à regeneração ósseo no
defeito (Fig.7, 9, 12 e 14). Além disso, a
semelhança nos resultados da avaliação
densitométrica dos grupos C-, BV,
BV+CTM e CTM, aos 90 dias de pós
operatório (Fig.11), sugere semelhança no
processo de regeneração óssea nesses
grupos. É valido ressaltar que nesse período
(90 dias) a maior parte da matriz do
biovidro 60S já havia sido reabsorvida
(Fig.5, 12 e 17) pelo organismo, reduzindo
assim a densidade da área do defeito
(Fig.11). A densitometria óssea é uma
técnica que permite avaliação da massa
mineral óssea de forma precisa e não
invasiva, sendo muito utilizada para
diagnóstico e acompanhamento e doenças
osteopênicas e na quantificação da massa
mineral óssea em estudos de novas terapias

88
para regeneração óssea (Louzada et al.,
1990; Almeida Paz e Bruno, 2006). No
entanto, quando se estuda biomateriais
como cerâmicas para o tratamento de
defeitos ósseos, a avaliação densitométrica
não é capaz de diferenciar o aumento de
densidade devido à regeneração óssea
daquele decorrente do preenchimento da
falha com o biomaterial, logo, a
interpretação dos resultados deve ser feita
com cautela, considerando esse fato.
A regeneração óssea incompleta com
preenchimento parcial dos defeitos
observada nos grupos BV e BV+CTM
(Fig.7, 8, 9, 12 e 14) mostra que nenhum
desses tratamentos foi eficiente na
regeneração de defeitos ósseos críticos de
rádios de cães. Acredita-se que a rápida
reabsorção da matriz porosa do biovidro
60S seja a principal causa de insucesso dos
tratamentos BV e BV+CTM pois, se a taxa
de reabsorção do biomaterial for superior a
taxa de neoformação óssea ocorrerá a
formação de uma lacuna entre o osso e o
biomaterial impedindo assim a
osteocondução e a completa regeneração do
defeito. Essa lacuna entre o biomaterial e
osso é na maioria das vezes preenchida por
tecido conjuntivo como o relatado em
estudos semelhantes com outros
biomateriais (Hu et al., 2003; Wu et al.,
2006; Zhu et al., 2006; Yuan et al., 2007;
Liu et al., 2008) e verificado
histologicamente neste trabalho. Em um
estudo prévio com esse mesmo biomaterial
no tratamento de pequenos defeitos em osso
alveolar de mandíbulas de cães, o biovidro
60S favoreceu a regeneração óssea sem a
formação de tecido conjuntivo na interfase
osso implante (Dutra et al., 2008). Acredita-
se que essa divergência de resultados esteja
diretamente relacionada à extensão do
defeito que foi muito superior no presente
estudo e com o tipo e localização do
defeito, que no presente estudo foi crítico
em um segmento completo de osso longo
composto principalmente por osso
osteônico que apresenta capacidade
regenerativa inferior ao osso trabecular de
defeitos alveolares em mandíbulas.
A alta porosidade das matrizes e o baixo
grau de organização das moléculas
(material amorfo) do biovidro 60S (Fig.3)
são características que lhe conferem alta
taxa de reabsorção após sua implantação no
organismo, o que pode justificar o
insucesso nos tratamento BV e BV+CTM.
Quando se considera a composição
química, os biovidros são os biomaterias
que apresentam a maior taxa de reabsorção
dentre todas as biocerâmicas (Pereira et al.,
2006). O grau de organização das
moléculas, também denominado
cristalinidade, interfere na taxa de
dissolução e reabsorção das biocerâmicas,
sendo que quanto maior a cristalinidade das
amostras menor será sua taxa de reabsorção
após implantação no organismo (Hoppe et
al., 2011). As características
microestruturais da matriz como porosidade
e interconectividade dos poros também
influenciam a taxa de reabsorção. Quanto
mais porosa e interconectada a matriz,
menor a densidade e maior a área
superficial sujeita a ação dos líquidos
intersticiais e células, o que ocasiona maior
taxa de reabsorção (Rahaman et al., 2011).
Além disso, as características do
microambiente de implantação como aporte
vascular, aumento de pH, presença de
edema e aumento de temperatura local
podem acelerar a reabsorção das cerâmicas
(Chai et al., 2012).
Por outro lado, a alta porosidade (97%) e o
amplo diâmetro dos poros (350 a 750µm)
interconectados observados nas matrizes
dos biovidro 60S facilitam sua colonização
e neovascularização como observado
histologicamente. O tamanho dos poros da
cerâmica deve ser semelhantes ao do osso
esponjoso (Watson, 2005) para permitir a
invasão vascular, colonização celular e
substituição por tecido ósseo (Geissler,
2006). Chen et al., (2001) recomendam um
diâmetro maior que 100-150µm e Geissler
(2006) cita que o diâmetro dos poros deve
variar entre 150 e 500µm, mas salienta que
poros muito amplos podem enfraquecer a
matriz. De Long Jr. et al (2007) relatam que
o tamanho ideal dos poros deve ser de 300 a
500µm, pois permite maior colonização do
implante. Tão importante quanto o diâmetro
dos poros, é a presença de
interconectividade entre eles (Chen et al.,
2001; Watson, 2005; Geissler, 2006; Vallet-
Regí, 2006; De Long Jr. et al 2007), pois
isso influencia diretamente na tensão de
oxigênio, fator de importância relevante na
diferenciação das células tronco
mesenquimais em células osteoprogenitoras

89
e dessas em osteoblastos (Chung et al.,
2012). Ao estudar a colonização e
diferenciação de células tronco
mesenquimais em matrizes de
hidroxiapatita, Mygind et al. (2007)
observaram maior colonização nas amostras
com poro de diâmetro médio de 500µm e
maior diferenciação celular nas amostras
com diâmetro de 200µm.
Embora a associação das CTM-AD
diferenciadas com a matriz porosa do
biovidro 60S não tenha aumentado a
osteogênese no presente trabalho (Fig.7, 9,
12 e 14), essa é uma prática que comumente
favorece a regeneração óssea, fato já
mostrado em outros estudos semelhantes
com matrizes produzidas a partir de outros
biomaterias de reabsorção mais lenta que o
biovidro 60S como os corais (Zhu et al.,
2006; Cui et al., 2007; Viateau et al., 2007;
Yuan et al., 2010; Zhu et al., 2010; Liu et
al., 2013), os fosfatos tricálcicos (TCP)
(Wu et al., 2006; Yuan et al., 2007; He et
al., 2007; Liu et al., 2008), os fosfatos
bifásicos (TCP/HA) (Bruder et al., 1998;
Arinzeh et al., 2003; Kok et al., 2003), as
hidroxiapatitas (HA) (Kon et al., 2000;
Zhao et al., 2009) e os compósito de
cerâmicas com polímeros (Mylonas et al.,
2007; Levi et al., 2010; Belocio et al.,
2012). A presença de células
osteoprogenitoras e CTM capazes de se
transformar em células ósseas é
provavelmente, o fator que mais contribui
para o aumento da atividade osteogênica e
consequentemente, para a reparação de uma
lesão óssea. Quando CTM são adicionadas
em um sítio de lesão óssea, elas podem se
diferenciar em osteoblastos e aumentar a
síntese de matriz óssea (Arinzeh et al.,
2005; Bielby et al., 2007; Mizuno, 2009).
Por outro lado, as CTM possuem a
capacidade de se diferenciar em qualquer
célula de origem mesenquimal como
tenócitos, mioblastos, adipócidos e
condrócitos (Pittenger et al., 1999;
Shenfield et al., 2002). Para maximizar sua
diferenciação osteogênica recomenda-se
diferencia-las “in vitro” antes da sua
utilização clínica (Yamada et al., 2003;
Boo et al., 2004; Wu et al., 2006; Zhu et
al., 2006; Cui et al., 2007; He et al., 2007;
Yoon et al., 2007; Yuan et al., 2007;
Viateau et al., 2007; Liu et al., 2008; Liao
et al., 2009; Zhao et al., 2009; Yuan et al.,
2010; Belocio et al., 2012; Liu et al., 2013),
assim como o realizado no presente estudo.
Outras técnicas podem também ser
utilizadas como associação a BMPs (Jeon et
al., 2008; Levi et al., 2010; Zhu et al.,
2010), a matriz óssea desmineralizada
(Mauney et al., 2004; Brodke et al., 2006)
e a qualquer outro tipo de biomaterial
osteoindutor (Habibovic et al., 2006;
Tsigkou et al., 2009; Hopp et al., 2011; Lin
e Sun, 2011).
Outra questão ainda controversa refere-se à
melhor forma de colonização das matrizes.
Alguns estudos recomendam a colonização
“in vivo” (Bruder et al., 1998; Kon et al.,
2000; Arinzeh et al., 2003; Kok et al., 2003;
Mauney et al., 2004; Arinzeh et al., 2005;
Wan et al., 2006; Wu et al., 2006; He et al.,
2007; Mylonas et al., 2007; Yuan et al.,
2007; Liao et al., 2009; Zhao et al., 2009;
Levi et al., 2010; Yuan et al., 2010), assim
como o proposto no presente estudo. Nesse
tipo de colonização as CTM são semeadas
na matriz de biomaterial e ambas
implantadas no organismo, ou seja, a
colonização da matriz ocorre após sua
implantação no organismo. A colonização
“in vivo” tem a vantagem de ser uma
técnica de utilização imediata, pois não é
necessário o cultivo da matriz “in vitro”
antes da implantação, possibilitando assim
sua utilização em pacientes que necessitam
de cirurgias emergenciais. Outros estudos
sugerem a colonização “in vitro” (Yamada
et al., 2003; Boo et al., 2004; Zhu et al.,
2006; Cui et al., 2007; Yoon et al., 2007;
Viateau et al., 2007; Jeon et al., 2008; Liu
et al., 2008; Zhu et al., 2010; Belocio et al.,
2012; Liu et al., 2013). As CTM são
semeadas na matriz de biomaterial e
cultivadas em estufa antes da sua
implantação, garantindo a completa
colonização da matriz antes de seu
emprego. No entanto, isso não garante a
sobrevivência e atividade dessas células
após sua implantação no organismo
principalmente quando se trabalha com
matrizes grandes onde as células do centro
da matriz são pouco nutridas devido a
distância física dos vasos sanguíneos,
principalmente nos primeiros dias após a
implantação quando ainda não existe
vascularização do interior da matriz. Além
disso, há dificuldade na colonização interna
de grandes matrizes “in vitro” devido a

90
problemas relacionados com a semeadura e
nutrição das células no centro da matriz
(Korossis et al., 2005; Silva et al., 2013).
Algumas técnicas utilizando centrifugação
(Belocio et al., 2012) e vácuo (Bruder et
al., 1998; Arinzeh et al., 2003; Arinzeh et
al., 2005) tem sido propostas para sanar a
dificuldade da semeadura interna das CTM
e a utilização de biorreatores para melhorar
a perfusão interna das matrizes durante seu
cultivo “in vitro” (Korrossis et al., 2005;
Silva et al., 2013).
Embora não se tenha observado nenhum
indício histológico de rejeição das CTM-
AD, a ineficiência na regeneração óssea
observada nos grupos tratados com essas
células (CTM e BV+CTM) pode estar
relacionada com a utilização de células
alógenas, pois células estranhas ao
organismo podem incitar uma resposta
imunológica prejudicando a regeneração
óssea (Kraus e Kirker-Head, 2006; Mosna
et al., 2010; Strioga et al., 2012). No
entanto, estudos já mostraram que as CTM
alogênicas diferenciadas ou não, possuem a
capacidade de imunomodulação (Niermeyer
et al., 2007) e não são rejeitadas quando
implantadas em indivíduos
imunologicamente compatíveis (Arinzeh et
al., 2003; Kok et al., 2003; Cui et al., 2007;
Mylonas et al., 2007). Estudos mostraram
também sucesso com a utilização de CTM
alogênicas para regeneração óssea (Arinzeh
et al., 2003; Kok et al., 2003; Wan et al.,
2006; Cui et al., 2007; Mylonas et al., 2007;
Belocio et al., 2012). A utilização
terapêutica de CTM alogênicas, assim como
o realizado no presente estudo, apresenta
vantagens em relação à utilização de CTM
autógenas, pois quando se trabalha com
CTM alogênicas é possível a criação de
bancos de células criopreservadas (Kok et
al., 2003; Martinello et al., 2011; Spencer et
al., 2012) o que permite sua pronta
utilização clínica em casos emergenciais
(Kraus e Kirker-Head, 2006). Além disso,
pode-se selecionar doadores jovens em
perfeitas condições de saúde o que propicia
o isolamento de CTM com alto potencial
regenerativo, permitindo assim sua
utilização terapêutica em pacientes
geriátricos e com outras condições que
podem afetar o potencial regenerativo das
CTM autógenas como a presença de
doenças genéticas.
A quantidade das células utilizadas é outro
fator que pode ter contribuído para o
insucesso dos tratamentos que utilizaram as
CTM-AD (CTM e BV+CTM).
Teoricamente quanto maior o número de
células utilizado, maior será a produção de
matriz e melhor será a regeneração óssea,
entretanto, quanto maior o número de
células maior o tempo necessário para
expandi-las “in vitro” antes da sua
utilização terapêutica e com isso maior o
tempo para se realizar o tratamento. Por
isso a necessidade de se descobrir o número
mínimo de células por volume de defeito
ósseo capaz de promover um resultado
efetivo quanto a regeneração óssea. A
maioria dos estudos mostram sucesso com a
semeadura de quantidades da ordem de 106
CTM/cm3 de defeito (Bruder et al., 1998;
Kon et al., 2000; Arinzeh et al., 2003; Kok
et al., 2003; Boo et al., 2004; Mauney et
al., 2004; Arinzeh et al., 2005; Wan et al.,
2006; Zhu et al., 2006; He et al., 2007;
Mylonas et al., 2007; Liu et al., 2008;
Nather et al., 2010; Zhu et al., 2010), assim
como o utilizado no presente estudo. Outros
sugerem quantidades menores da ordem de
105
CTM/cm3 de defeito (Viateau et al.,
2007; Yoon et al., 2007; Levi et al., 2010;
Belocio et al., 2012), mas existem também
aqueles que recomendam quantidades
maiores da ordem de 107
CTM/cm3
(Yamada et al., 2003; Cui et al., 2007;
Yuan et al., 2007; Liao et al., 2009; Zhao et
al., 2009; Yuan et al., 2010; Liu et al.,
2013) e 108
CTM/cm3 para uma efetiva
regeneração óssea (Wu et al., 2006).
A diferenciação osteogênica das CTM-AD
por 21 dias antes da sua utilização no
tratamento dos defeitos ósseos foi
previamente definida com base nos
resultados obtidos no capítulo 3 da presente
tese que mostram maior osteogênese “in
vitro” nesse período. No entanto, vários
outros tempos de diferenciação, como cinco
(He et al., 2007), sete (Zhu et al., 2006;
Yuan et al., 2007; Liu et al., 2008; Yuan et
al., 2010; Liu et al., 2013), 10 (Cui et al.,
2007; Viateau et al., 2007; Liao et al.,
2009), 14 (Yoon et al., 2007; Zhao et al.,
2009) e 20 dias, tem sido empregados com
sucesso na regeneração óssea (Yamada et
al., 2003; Boo et al., 2004). Mas
recentemente um estudo mostrou que as
CTM indiferenciadas e em estágio inicial de

91
diferenciação (sete a 14 dias) são mais
eficientes na regeneração óssea do que
células em estágio final de diferenciação
(21 dias) (Belocio et al., 2012), o que pode
justificar a ineficiência das CTM-AD na
regeneração óssea observada no presente
estudo.
Conclusão
Nas condições em que foi realizado esse
estudo pode-se afirmar que:
A matriz porosa do biovidro 60S associada
ao não as CTM-AD diferenciadas não é
eficiente no tratamento de defeitos ósseos
críticos no rádio de cães adultos.
As células tronco mesenquimais de origem
adiposa favorecerem a neovascularização
de defeitos ósseos críticos no rádio de cães
adultos.
Bibliografia
ABDALLAH, B.M.; HAACK-
SORENSEN, M.; FINK, T.; et al. Inhibition
of osteoblast differentiation but not
adipocyte differentiation of mesenchymal
stem cells by sera obtained from aged
females. Bone.,v.39, p.181-188, 2006.
ADAM, J.A. A simplified model of wound
healing with particular reference to the
critical size defect. Mathematical and
Computer Modelling. v.30, p.23-32, 1999.
ADEGANI, F.J.; LANGROUDI, L.;
AREFIAN, E.; et al. A comparison of
pluripotency and differentiation status of
four mesenchymal adult stem cells. Mol
Biol Rep., 2012.
AKSU, A.E.; RUBIN, J.P.; DUDAS, J.R.;
et al. Role of gender and anatomical region
on induction of osteogenic differentiation of
human adipose –derived stem cells. Ann
Plast Surg., v.60, p.306-322, 2008.
ALMEIDA PAZ, I.C.L.; BRUNO, L.D.G.
Bone mineral density: Review. Rev Bras
Cienc Avic., v.8, p.69-73, 2006.
ALMEIDA, M.; HAN, L.; MARTIN-
MILLAN, M.; et al. Oxidative stress
antagonizes Wnt signaling in osteoblast
precursors by diverting β-catenin from T
cell factor-to forkhead box o-mediated
transcription. J Biol Chem., v.282, p.27298-
27305, 2007.
AL-NBAHEEN, M.; VISHNUBALAJI, R.;
ALI, D.; et al. Human stromal
(mesenchymal) stem cells from bone
marrow, adipose tissue and skin exhibit
differences in molecular phenotype and
differentiation potential. Stem Cells Rev.,
v.9, p.32-43, 2013.
ALNO, N.; JEGOUX, F.; PELLEN-
MUSSI, P.; et al. Development of a thre-
dimensional model for rapid evaluation of
bone substitutes in vitro: Effect of the 45S5
bioglass, J. Biomed. Mater. Res. A, v.95,
p.137-145, 2010.
ALVES, E.G.L.; REZENDE, C.M.F.;
OLIVEIRA, H.P.; et al. Avaliação
mecânica da placa de compósito de
polihidroxibutirado e hidroxiapatita em
modelos ósseos de gato. Arq. Bras. Med.
Vet. Zoote., v.62, n.6, p.1367-1374, 2010.
ALVES, E.G.L.; REZENDE, C.M.F.;
OLIVEIRA, H.P.; et al. Emprego
experimental da placa de compósito poli-
hidroxibutirato/hidroxiapatita na fixação
femoral em gatos. Arq. Bras. Med. Vet.
Zoote., v.62, n.5, p.1128-1134, 2010.
ALVES, E.G.L.; REZENDE, C.M.F.;
SERAKIDES, R.; et al. Orthopedic
implanto f polyhydroxybutirate (PHB) and
hydroxyapatite composite em cats. J
FelineMed Surg., v.13, p.546-552, 2011.
ARINZEH, T.L.; PETER, S.J.;
ARCHAMBAULT, M.P.; et al. Allogeneic
mesenchymal stem cells regenerate bone in
a critical-size canine segmental defect. J.
Bone Joint Surg. Am. v.85, p.1927-1935,
2003.

92
ARINZEH, T.L.; TRAN, T.; MCALARY,
J.; et al. A comparative study of biphasic
calcium phosphate ceramics for human
mesenchymal stem-cell-induced bone
formation. Biomaterials., v.26, p.3631-
3638, 2005.
ARNOLD, J.S. A simplified model of
wound healing III the critical size defect in
three dimensions. Mathematical and
Computer Modelling. v.34, p.385-392,
2001.
ARNOLD, J.S.; ADAM, J.A. A simplified
model of wound healing II the critical size
defect in two dimensions. Mathematical
and Computer Modelling. v.30, p.47-60,
1999.
ARRIGONI, E.; LOPA, S.O.;
GIROLAMO, L. et al. Isolation,
characterization and osteogenic
differentiation of adipose tissue-derived
stem cells: from small to large animal
models. Cell. Tissue Res., v.338, p.401-411,
2009.
ARTHUR, A.; ZANNETTINO, A.;
GRONTHOS, S. The therapeutic
applications of multipotential
mesnchymal/stromal stem cells in skeletal
tissue repair. J. Cel. Physiol., v.218, p.237-
234, 2009.
ASTM F2721-09 Standard guide for pre-
clinical in vivo evaluation in critical size
segmental bone defects, Developed by
subcommittee F04.44. Book of Standards,
v.13.02.
BAKSH, D.; SONG, L. TUAN, R.S. Adult
mesenchymal stem cells: characterization,
differentiation and application in cell and
gene therapy. J. Cell. Mol. Med., v.8, p.
301-316, 2004.
BALLOCK, R.T.; O´KEEFE, R.J.
Physiology and pathophysiology of the
growth plate. Birth Defects Res. C. Embryo
Today, v.69, p.123-143, 2003.
BALLOCK, R.T.; O´KEEFE, R.J. The
biology of the growth plate. J. Bone Joint
Sur., v.85, p.715-126, 2003.
BARRADAS, A.M.C.; FERNANDES,
H.A.M.; GROEN, N.; et al. A calcium-
induced signaling cascade leading to
osteogenic differentiation of human bone
marrow-derived mesenchymal stromal
cells. Biomaterials, v.33, p.3205-3215,
2012.
BARRERE, F.; Van BLITTERSWIJK.
C.A.; GROOT, K. Bone regeneration:
molecular and cellular interactions with
calcium phosphate ceramics. International
Journal of Nanomedicine. v.1, p.317-332,
2006.
BAXTER, M.A.; WYNN, R.F.; JOWITT,
S.N.; et al. Study of telomere length reveals
rapid aging of human marrow stromal cells
following in vitro expansion. Stem Cells.,
v.22, p.675-682, 2004.
BELLOWS, C.G.; PEI, W.; JIA, Y.; et al.
Proliferation, differentiation and self-
renewal of osteoprogenitors in vertebral cell
populations from aged and young female
rats. Mech Ageing dev., v.18, p.747-757,
2003.
BELLOWS, C.G.; REIMERS, S.M.;
HEERSCHE, J.N.M. Expression of
mRNAs for type-I collagen, bone
sialoprotein, osteocalcin, and osteopontin at
different stages of osteoblastic
differentiation and their regulation by 1,25
dihydroxyvitamin D3. Cell Tissue Res.,
v.297, p. 249-259, 1999.
BELOTI, M.M.; SICCHIERI, L.G.;
OLIVEIRA, P.T.; et al. The influence of
osteoblast differentiation stage on bone
formation in autogenously implanted cell-
based poly(lactide-co-glycolide) and
calcium phosphate constructs. Tissue Eng
PartA., v.18, p.999-1005, 2012.

93
BELTRAMI, A.P.; BARLUCCHI, L.;
TORELLA, D. et al. Adult cardiac stem
cells are multipotent and support
myocardial regeneration. Cell, v.114,
p.763-776, 2003.
BIELBY, R.; JONES, E.; McGONAGLE,
D. The role of mesnchymal stem cells in
maintenance and repair of bone, Injury,
v.38s1, p.s26-s32, 2007.
BOBIS, S.; JAROCHA, D.; MAJKA, M.
Mesenchymal stem cells: characteristics
and clinical applications. Folia Histochem.
Cytobiol., v.44, p.215-30, 2006.
BOELONI, J.N.; OCARINO, N.M.;
BOZZI, A. et al. Dose-dependent effects of
triiodothyronine on osteogenic
differentiation of rat bone marrow
mesenchymal stem cells. Horm Res., v.71,
p.88-97, 2009.
BONAB, M.M.; ALIMOGHADDAM, K.;
TALEBIAN, F.; et al. Aging of
mesenchymal stem cell in vitro. BMC Cell
Biol., v.7, p.1-7, 2006.
BONEWALD, L. Osteocytes as
multifunctional cells. J. Musculoskelet.
Neuronal Interact., v.6, p.331-333, 2006.
BOO, J.S.; YAMADA, Y.; OKAZAKI, Y.;
et al. Tissue-engineered bone using
mesenchymal stem cells and a
biodegradable scaffold. J Craniofac Surg.
v.13, p.34-37, 2004.
BOYLE, W.J.; SIMONET, W.S.; LACEY,
D.L. Osteoclast differentiation and
activation. Nature, v.423, p.337-342, 2003.
BRAKSPEAR, K.S.; MASON, D.J.
Glutamate signaling in bone. Front
Endocrinol.v.3, p.1-8, 2012.
BRODKE, D.; PEDROSO, H.A.; KAPUR,
T.A.; et al. Bone grafts prepared with
selective cell retention technology heal
canine segmental defects as effectively as
autograft. Journal of Orthopaedic
Research. v.24, p.857-866, 2006.
BRUDER, S.P.; JAISWALL, N.;
RICALTON, N.S.; et al. Mesenchymal
stem cells in osteobiology and applied bone
regeneration. Clin. Orthop. Relat. Res,.
v.355, p.s247-s256, 1998b.
BRUDER, S.P.; KRAUS, K.R.;
GOLDBERG, V.M.; et al. The effect of
implants loaded with autologous
mesenchymal stem cells on the healing of
canine segmental bone defects. J Bone Joint
Surg Am. v.80, p.985-996, 1998a.
BRUEDIGAN, C.; DRIEL, M.V.;
KOEDAM, M. et al. Basic techniques in
human mesenchymal stem cell cultures:
differentiation into osteogenic and
adipogenic lineages, genetic perturbations,
and phenotypic analyses. Curr Protoc Stem
Cell Biol, v.17, p.1-19, 2011.
BUNNELL, B.A.; FLAAT, M.;
GAGLIARDI, C.; et al. Adipose-derived
stem cells: Isolation, expansion and
differentiation. Methods. v.45, p.115-120,
2008.
CAO, J.; VENTON, L.; SAKATA, T.; et al.
Expression of RANKL and OPG correlates
with age-related bone loss in male C57BL/6
mice. J Bone Miner Res., v.18, p.270-277,
2003.
CHAI, Y.C.; CARLIER, A.; BOLANDER,
J.; Current views on calcium phosphate
osteogenicity and the translation into
effective bone regeneration strategies. Acta
Biomater., v.8, p.3876-3887, 2012.
CHAMBERLAIN, G.; FOX, J.; ASHTON,
B. et al. Concise review: mesenchymal stem
cells: their phenotype, differentiation
capacity, immunological features, and
potential for homing. Stem Cells, v.25,
p.2739-2749, 2007.
CHANG, W.; TU, C.; CHEN, T.H.; et al.
The extracellular calcium-sensing receptor
(CaSR) is a critical modulator of skeletal
development. Sci Signal., v.1, p.1-35, 2008.

94
CHEN, G.; USHIDA, T.; TATEISHI, T.
Development of Biodegradable Porous
Scaffolds for Tissue Engineering. Mater Sci
Eng C. v.17, n.1-2., p.63-69, 2001.
CHEN, Y.; SHAO, J-Z.; XIANG, L-X. et
al. Mesenchymal stem cells: a promising
candidate in regenerative medicine. Int. J.
Biochem. Cell Biol., v.40, p.815-820, 2008.
CHOI, Y.J.; LEE, J.Y.; LEE, S.J.; et al.
Alpha-adrenergic blocker mediated
osteoblastic stem cell differentiation.
Biochem Biophys Res Comun., v.416,
p.232-238, 2011.
CHOI, Y.S.; NOH, S.E.; LIM, S.M.; et al.
Multipotency and growth characteristic of
periosteum-derived progenitor cells for
chondrogenic, osteogenic and adipogenic
differentiation. Biotechnol. Lett, v.30,
p.593-601, 2008.
CHRISTODOULOU, I.; BUTTERY,
L.D.K.; SARAVANAPAVAN, P.; et al.
dose- and time-dependent effect of
bioactive gel-glass ionic- dissolution
products on human fetal osteoblast-specific
gene expression. J Biomed Mater Res Part
B: Appl Biomater., v.74, p.529-537, 2005.
CHRISTODOULOU, I.; BUTTERY,
L.D.; TAI, G.; et al. Characterization of
human fetal osteoblast by microarray
analysis following stimulation with 58S
bioactive Gel-Glass ionic dissolution
products. J. Biomed. Mater. Res. B. Appl.
Biomater., v.77, p.431-446, 2006.
CHUNG, D.; HAYASHI, K.;
TOUPADAKIS, C.A.; et al. Osteogenic
proliferation and differentiation of canine
bone marrow and adipose tissue derived
mesenchymal stromal cells and the
influence of hypoxia. Res. Vet. Sci. v.91,
p.66-75, 2012.
CIVITELLI, R. Cell-cell communication in
the osteoblast/osteocyte lineage. Arch.
Biochem. Biophys., v.473, p.188-192, 2008.
COELHO, M.B.; SOARES, I.R.;
MANSUR, H.S.; et al. Effect of the Type of
Surfactant on Bioactive Glasses Foam
Formation. Key Eng Mater., v. 240-242, p.
257-260, 2003.
COELHO, M. B.; PEREIRA, M. M. Sol-
gel synthesis of bioactive glass scaffolds for
tissue engineering: effect of surfactant type
and concentration. J Biomed Mater Res B
Appl Biomater., v. 75, p. 451-456, 2005.
CRISAN, M.; CHEN, C.W.; CORSELLI,
M.; et al. Perivascular multipotent
progenitor cell in human organs. Ann NY
Acad Sci., v.1176, p.118-123, 2009.
CRISAN, M.; YAP, S. ; CASTEILLA, L. ;
et al., A perivascular origin for
mesenchymal stem cells in multiple human
organs. Cell Strem Cell., v.3, p.301-313,
2008.
CSAKI, C.; MATIS, U.; MOBASHERI, A.;
et al. Co-culture of canine mesenchymal
stem cells with primary bone-derived
osteoblasts promotes osteogenic
differentiation. Histochem Cell Biol., p.251-
266, v. 131, 2009.
CUI, L.; LIU, B.; LIU, G.; et al. Repair of
cranil bone defects with adipose derived
stem cells and coral scaffold in canine
model. Biomaterials., v.28, p.5477-5486,
2007.
DATTA, H.K.; NG, W.F.; WALKER, J.A.
et al. The cell biology of bone metabolism.
J. Clin. Pathol., v.61, p.577-587, 2008.
DE LONG JR, W.G.; EINHORN, T.A.;
KOVAL, K.; et al. Bone Grafts and Bone
Substitutes in Orthopaedic Trauma Surgery.
A Critical Analysis. J Bone Joint Surg.
v.89. n.3, p.649-658, 2007.
DE UGARTE, D,A.; MORIZONO,
K.; ELBARBARY, A.; et al. Comparison
of Multi-lineage cells from human adipose
tissue and bone marrow. Cells Tissues
Organs., v.174, p.101-109, 2003.

95
DELILOGLU-GURHAN, S.I.;
VATANSEVER, H.S.; OZDAL-KURT, F.;
et al. Characterization of osteoblasts
derived from bone marrow stromal cells in
a modified cell culture system. Acta
Histochemica, v.108, p.49-57, 2006.
DMITRIEVA, R.; MINULLINA, I.R.;
BILIBINA, A.A.; et al. Bone marrow-and
subcutaneous adipose tissue-derived
mesenchymal stem cells. Cell Cycle., v.11,
p.377-383, 2012.
DOMINICI, M.L.; Le BLANC, K.;
MUELLER, I.; et al. Minimal criteria for
defining multipotencial mesenquimal
stromal cells. The international society for
cellular therapy position statement.
Cytotherapy. v.8, p.315-317, 2006.
DONZELLI, E.; SALVADÈ, A.; MIMO, P.
et al. Mesenchymal stem cells cultured on a
collagen scaffold: in vitro osteogenic
differentiation. Arch. Oral Biol., v.52, p.63-
73, 2007.
DROSSE, I.; VOLKMER, E.; SEITZ, S.; et
al. Validation of femoral critical size defect
model for orthotopic evaluation of bone
healing: A biomechanical, veterinary and
trauma surgical perspective. Tissue
Engineering., v.14, p.79-88, 2008.
DUCY, P.; KARSENTY, G. Genetic
control of cell differentiation in the
skeleton. Curr. Opin. Cell Biol., v.10,
p.614-619, 1998.
DUCY, P.; ZHANG, R.; GEOFFROY, V.;
et al. Osf2/Cbfa1: a transcriptional activator
of osteoblast differentiation. Cell, v.89,
p.747-754, 1997.
DUTRA, C.E.; PEREIRA, M.M.;
SERAKIDES, R.; et al. In vivo evaluation
of bioactive glass foams associated with
platelet-rich plasma in bone defects. J.
Tissue Eng. Regen. Med. v.4, p.221-227,
2008.
EGUSA, H.; IIDA, K.; KOBAYASHI, M.;
et al. Downregulation of extracellular
matrix-related gene clusters during
osteogenic differentiation of human bone
marrow- and adipose tissue-derived stromal
cells. Tissue Eng., v.13, p.2589-2600, 2007.
EPPLEY, B.L.; PIETRZAK, W.S.;
BLANTON, M.W. Allograft and alloplastic
bone substitutes: A review of science and
technology for the craniomaxillofacial
surgeon. J Craniofac Surg. v.16, n.6, p.981-
989, 2005.
ERTAS, G.; URAL, E.; URAL, D.; et al.
Comparative analyses of apoptotic
resistance of mesenchymal stem cells
isolated from human bone marrow and
adipose tissue. ScientificWorldJournal.,
2012.
ESLAMINEJAD, W.B.; TAGHIYAR, L.
Study of the structure os canine
mesenchymal stem cell osteogenic culture.
Anat. Histol. Embryol. v.39, p.446-455,
2010.
FABIAN, M.R.; SONENBERG, N.;
FILIPOWICZ, W. Regulation of mRNA
translation and stability by microRNAs.
Annu Rev Biochem., v.79, p.351-379, 2010.
FILIOLI-URANIO, M.; VALENTINI,
L.; LANGE-CONSIGLIO, A.; et al.
Isolation, proliferation, cytogenetic and
molecular characterization and in vitro
differentiation potency of canine stem cells
from foetal adnexa: a comparative study of
amniotic fluid, amnion and umbilical cord
matrix. Mol. Reprod. Dev., v.78, p.361-373,
2011.
FIORENTINI, E.; GRANCHI, D.;
LEONARDI, E.; et al. Effects of osteogenic
differentiation inducers on in vitro
expanded adult mesenchymal stromal cells.
Int J Artif Organs., v.34, p.998-1011, 2011.

96
FORMIGUE, O.; HAY, E.; BARBARA,
A.; et al. Calcium sensing recptor-
dependent and receptor-independent
activation of osteoblast replication and
survival by strontium ranelate. J Cell Mol
Med., v.13, p.2189-2199, 2009.
FRANCESCHI, R.T.; GE, C.; XIAO, G.; et
al. Transcriptional regulation of osteoblasts.
Cells Tissues Organs, v.189, p.144-152,
2009.
GE, C.; XIAO, G.; JIANG, D.; et al.
Critical role of the extracellular signal-
regulated kinase-MAPK pathway in
osteoblast differentiation and skeletal
development. J Cell Biol., v.176, p.709-
718, 2007.
GEISSLER, W.B. Bone Graft Substitutes in
the Upper Extremity. Hand Clin. v.22, n.3,
p.329-339, 2006.
GIANNOUDIS, P.V.; DINOPOULOS, H.;
TSIRIDIS, E. Bone substitutes: an update.
Injury. v.36, n.3, p.20-27, 2005.
GONG, Z.; WEZEMAN, F. Inhibitory
effect of alcohol on osteogenic
differentiation in human bone marrow-
derived mesenchymal stem cells. Alcohol
Clin Exp Res., v.28, p.468-479, 2004.
GROVE, J.E.; BRUSCIA, E.; KRAUSE,
D.S. Plasticity of bone marrow-derived
stem cells. Stem Cells, v.22, p.487-500,
2004.
GUGALA, Z.; LINDSEY, R.W.;
GOGOLEWSKI, S. New approaches in the
treatment of critical-size segmental defects
in long bones. Macromol Symp., v.253,
p.147-161, 2007.
HABIBOVIC, P.; YUAN, H.; DOEL,
M.V.D.; et al. Relevance of osteoinductive
biomaterials in critical-sized orthotopic
defect. J Orthop Res.,v.24, p.867-876,
2006.
HADJIDAKIS, D.J.; ANDROULAKIS, I.I.
Bone remodeling. Ann. N.Y. Acad. Sci.,
v.1092, p.385-396, 2006.
HALVORSEN, Y.D.; FRANKLIN, D.;
BOND, A.L.; et al. Extracellular matrix
mineralization and osteoblast gene
expression by human adipose tissue-derived
stromal cells. Tissue Eng., v.7, p.729-741,
2001.
HANSEN, M.B.; NIELSEN, S.E.; BERG,
K. Re-examination and further development
of a precise and rapid dye method for
measuring cell growth/cell kill. J. Immunol.
Methods, v.119, p.203-210, 1989.
HASS, R.; KASPER, C.; BOHM, S.; et al.
Different populations and sources of human
mesenchymal stem cells (MSC): A
comparison of adult and neonatal tissue-
derived MSC. Cell Commun Signal., v.9,
p.1-14, 2011.
HAYASHI, O.; KATSUBE, Y.; HIROSE,
M.; et al. Comparison of osteogenic ability
of rat mesenchymal stem cells from bone
marrow, periosteum, and adipose tissue.
Calcif Tissue Int., v.82, p.238-247, 2008.
HE, Y.; ZHANG, Z.Y.; ZHU, H.G.; et al.
Experimental study on reconstruction of
segmental mandible defects using tissue
engineered bone combined bone marrow
stromal cells with three-dimensional
tricalcium phosphate. J Craniofac Surg.,
v.18, p.800-805, 2007.
HELL, R.C.R.; OCARINO, N.M.;
BOELONI, J.N.; et al. Physical activity
improve age-related decline in the
osteogenic potential of rats` bone marrow-
derived mesenchymal stem cells. Acta
Physiol., v.205, p.292-301, 2012.
HENCH, L.L. Genetic design of bioactive
glass. Journal of European Ceramic
Society. v.29, p.1257-p.1265, 2009.
HENCH, L.L. Sol-gel materials for
bioceramic applications. Current Opinion
in Solid State e Materials Science. v.2,
p.604-610, 1997.
HENG, B.C.; CAO, T.; LEE, E.H.
Directing stem cell differentiation into the
chondrogenic lineage in vitro. Stem Cells,
v.22, p.1152-1167, 2004.

97
HIDAKA, C.; CUNNINGLAM, M.E.;
RODEO, S.A.; et al. Modern biologics use
in orthopaedic surgery. Curr. Opin.
Rheumat., v.18, n.1, p.74-79, 2006.
HOPPE, A.; GÜLDAL, N.S.;
BOCCACCINI, A.R. A review of the
biological response to ionic products from
bioactive glasses and glass-ceramics.
Biomaterials., v.32, p.2757-2774, 2011.
HU, Y.; ZHANG, C.; SHANG, S.; et al.
development of a porous poly(L-lactic
acid)/hydroxyapatite/collagen scaffold as
BMP delivery system and its use in healing
canine segmental bone defect. J Biomed
MaterA., v.67, p.591-598, 2003.
HU, Y.C.; ZHONG, J.P. Osteostimulation
of bioglass. Chin. Med. J. (Engl). v.122,
p.2386-2389, 2009.
HUANG, Z.; CHENG, S.L.;
SLATOPOLSKY, E. Sustained activation
of extracellular signal-regulated kinase
pathway is required for extracellular
calcium stimulation of human osteoblast
proliferation. J Biol Chem., v.276, p.21351-
21358, 2001.
HULSE, D.; HYMAN, D. Fracture biology
and biomechanics. In: SLATTER, D. (Ed).
Textbook of small Animal Surgery. v.1. 3ed.
Philadelphia:Saunders. 2003. p.1785-1792.
IM, G.I.; SHIN, Y.W.; LEE, K.B. Do
adipose tissue-derived mesenchymal stem
cells have tha same osteogenic and
chondrogenic potential as bone marrow-
derived cells?. Osteoarthritis
Cartilage., p.845-853, v.13, 2005.
ISHII, M.; KOIKE, C.; IGARASHI, A. et
al. Molecular markers distinguish bone
marrow mesenchymal stem cells from
fibroblasts. Biochem. Biophys. Res.
Commun., v.332, p.297-303, 2005a.
IZADPANAH, R.; TRYGG, C.; PATEL,
B.; et al. Biologic properties of
mesenchymal stem cells derived from bone
marrow and adipose tissue. J Cell
Biochem., v.99, p.1285-1297, 2006.
JACKSON, L.; JONES, D.R.; SCOTTING,
P. et al. Adult mesenchymal stem cells:
differentiation potential and therapeutic
applications. J. Postgrad. Med., v.53,
p.121-127, 2007.
JACKSON, L.C.; PACCHIANA, P.D.
Common complications of fracture repair.
Clin Tech Small Anim Pract., v.19, p.168-
179, 2004.
JEON, Q.; RHIE, J.W.; KWON, K.; et al.
In vivo bone formation following
transplantation of human adipose-derived
stromal cells that are not differentiated
osteogenically. Tissue Eng PartA., v.14,
p.1285-1294, 2008.
JIANG, Y.; JAHAGIRDAR, B.N.;
REINHARDT, R.L. et al., Pluripotency of
mesenchymal stem cells derived from adult
marrow. Nature, v.418, p.41-49, 2002.
JIANG, Y.; MISHIMA, H.; SAKAI, S.; et
al. Gene expression analysis of major
lineage-defining factors in human bone
marrow cells: Effect of aging, gender, and
age-related disorders. J Orthop Res., v.26,
p.910-917, 2008.
JINGUSHI, S.; JOYCE, M.E.;
BOLANDER, M.E. Genetic expression of
extracelular matrix proteins correlates with
histologic changes durind fracture. J. Bone
Miner. Res. v.7, p.1045-1055, 1992.
JOHNSON, A.L. Fundamental of
orthopedic surgery and fracture
management. In: FOSSUM, T.W. (Ed)
Small animal surgery. 3ed, St.
Louis:Mosby, 3007, p.930-1014.
JOHNSON, A.L.; HOULTON, J.E.F.;
VANNINI, R. AO principles of fracture
management in the dog and cat. New York:
Thieme, 2005, 529p.
JONES, J.R.; HENCHCH, L.L.
Regeneration of Trabecular Bone Using
Porous Ceramics. Curr Opin Solid State
Mater. v.7, n.4-5, p.301-307, 2003.

98
JONES. J.R.; TSIGKOU, O.; COATES,
E.E.; et al. Extracellular matrix formation
and mineralization on phosphate-free
porous bioactive glass scaffold using
primary human osteoblast.
Biomaterials., v.28, p.1653-1663, 2006.
JUNG, G.Y.; PARK, Y.J. Effects of HA
released calcium ion on osteoblast
differentiation. J Mater Sci: Mater Med,
v.21, p.1649-1654, 2010.
JUSTESEN, J.; STENDERUP, K.;
ERIKSEN, E.F.; et al. Maintenance of
osteoblastic and adipocytic differentiation
potential with age and osteoporosis in
human marrow stromal cells cultures.
Calcif Tissue Int., v.71, p.36-44, 2002.
KANAYA, S.; NEMOTO, E.; EBE, Y.; et
al. Elevated extracelular calcium increases
fibroblast growth fator-2-gene and protein
expression levels via a cAMP/PKA
dependent pathway in cementoblasts.
Bone., v.47, p.564-572, 2010.
KARAOZ, E.; AKSOY, A.; AYHAN, S. et
al. Characterization of mesenchymal stem
cells from rat bone marrow: ultrastructural
properties, differentiation potential and
immunophenotypic markers. Histochem.
Cell Biol., v.132, p.533-546, 2009.
KARSENTY, G. Transcriptional control o
skeletogenesis. Annu Rev Genomics Hum
Genet. v.9, p.183-196, 2008.
KASSEN, M.; ABDALLAH, B.M.;
SAEED, H. Osteoblastic cells:
differentiation and Trans-differentiation.
Arch. Biochem. Biophys., v.473, p.183-187,
2008.
KATAGIRI, T.; TAKAHASHI, N.
Regulatory mechanisms of osteoblast and
osteoclast differentiation. Oral Dis., v.8,
p.147-159, 2002.
KERN, S.; EICHLER, H.; STOEVE, J.; et
al. Comparative analysis of mesenchymal
stem cells from bone marrow, umbilical
cord blood, or adipose tissue. Stem Cells.,
v.24, p.1294-1301, 2006.
KHAN, W.S.; ADESIDA, A.B.; TEW,
S.R.; et al. The epitope charactetisation and
the osteogenic differentiation potential of
human fat pad-derived stem cells is
maintained with ageing in later life. Injury,
v.40, p.150-157, 2009.
KIM, J.; KANG, J.W.; PARK, J.H. et al.
Biological characterization of long-term
cultured human mesenchymal stem cells.
Arch. Pharm. Res., v.32, p.117-126, 2008.
KIM, M.; KIM, C.W.; CHOI, Y.S.; et al.
Age-related alterations in mesenchymal
stem cells related to shift in differentiation
from osteogenic to adipogenic potential:
Implication to age-associated bone diseases
and defects. Mech Ageing Dev., v.133,
p.215-225, 2012.
KIM, Y.J.; BAE, S.W.; YU, S.S.; et al.
miR-196a regulates proliferation and
osteogenic differentiation in mesenchymal
stem cells derived from human adipose
tissue. J Bone Miner Res., v.24, p.816-825,
2009
KO, C.H.; SIU, W.S.; WONG, H.L.; et al.
Pro-bone and antifat effects of grren tea and
its polyphenol, epigallocatechin, in rat
mesenchymal stem cells in vitro. Agric
Food Chem., v.59, p.9870-9876, 2011.
KOK, I.J.D.; PETER, S.J.;
ARCHAMBAULT, M.; et al. Investigation
of allogeneic mesenchymal stem cell-based
alveolar bone formation: oreliminary
findings. Clin Oral Implants Res., v.14,
p.481-489, 2003.
KON, E.; MURAGLIA, A.; CORSI, A.; et
al. Autologous bone marrow stromal cells
loaded onto porous hydroxyapatite ceramic
accelerate bone repair in critical-size
defects of sheep long bones. J Biomed
Mater Res., v.49, p. 328-337, 2000.
KOROSSIS, S.A.; BOLLAND, F.;
KEARNEY, J.N.; et al. Bioreactors in
tissue engineering. In: ASHAMMAKHI,
N.; REIS, R.L. (Ed.) Topics in tissue
engineering. 1.ed. cidade:editora, 2006, 2o
V, Cap.8, p.1-23.

99
KOUSTENI, S. FoxO1, the transcriptional
chief of staff of energy metabolism. Bone.,
v.50, p.437-443, 2012.
KRAUS, K.H.; KIRKER-HEAD, C.
Mesenchymal stem cells and bone
regeneration. Vet. Surg., v.35, n.3, p.232-
242, 2006.
KRETLOW, J.D.; JIN, Y.Q.; LIU, W.; et
al. Donor age and cell passage affects
differentiation potential of murine bone
marrow-derived stem cells. BMC Cell Biol.,
v.9, p.1-13, 2008.
LEE, R.H.; KIM, B.C,; CHOI, I.S.; et al.
Characterization and expression analysis of
mesenchymal stem cells from human bone
marrow and adipose tissue. Cell Physiol
Biochem., v.14, p.311-324, 2004.
LEE, W.S.; SUZUKI, Y.; GRAVES, S.S.;
et al. Canine bone marrow-derived
mesenchymal stromal cells suppress
alloreactive lymphocyte proliferation in
vitro but fail to enhance engraftment in
canine bone marrow transplantation. Biol.
Blood Marrow Transplant., v.4, p.465-475,
2011.
LESKELA, H.V.; RISTELI, J.;
NISKANEN, S.; et al. Osteoblast
recruitment from stem cells does not
decrease by age at late adulthood. Biochen
Biophys Res Commun., v.311, p.1008-1013,
2003.
LEVI, B.; JAMES, A.W.; NELSON, E.R.;
et al. Human adipose derived stromal cells
heal critical size mouse calvarial defects.
Plos One., v.5, p.1-11, 2010.
LEVI, B.; LONGAKER, M.T. Adipose
derived stromal cells for skeletal
regenerative medicine. Stem Cells, v.29,
p.576-582, 2011.
LEVI, B.; NELSON, E.R.; BROWN, K.; et
al. Differences in osteogenic differentiation
of adipose –derived stromal cells from
murine, canine, and human sources in vitro
and in vivo. Plas Reconstr Surg., v.128,
p.373-385, 2011.
LEVI, B.; NELSON, E.R.; BROWN, K.; et
al. differences in osteogenic differentiation
of adipose-derived stromal cells from
murine, canine, and human sources in vitro
and in vivo. Plast. Reconstr. Surg., v.128,
p.373-386, 2011.
LI, H.W.; SUN, J.Y. Effects of dicalcium
silicate coating ionic dissolution products
on human mesenchymal stem-cell
proliferation and osteogenic differentiation.
J. Int. Med. Res., v.39, p.112-128, 2011.
LIAO, H.T.; CHEN, C.T.; CHEN, C.H.; et
al. Combination of guided osteogenesis
with autologous platelet-rich fibrin glue and
mesenchymal stem cells for mandibular
reconstruction. J. Trauma, v.70, p. 228-237,
2011.
LIN, L.; SHEN, Q.; WEI, X.; et al.
Comparison of osteogenic potentials of
BMP2 transduced stem cells from
autologous bone marrow and fat tissue in
rabbit model of calvarial defects. Calcif
Tissue Int., v.85, p.55-65, 2009.
LINDNER, U.; KRAMER, J.;
ROHWEDEL, J.; et al. Mesenchymal stem
or stromal cells: Toward a better
understanding of their biology. Transfus.
Med. Hemother., v.37, p.75-83, 2010.
LIU, G.; ZHANG, Y.; LIU, B.; et al. Bone
regeneration in a canine model using
allogeneic adipose derived stem cells and
coral scaffold. Biomaterials., v.34, p.2655-
2664, 2013.
LIU, G.; ZHAO, L.; ZHANG, W.; et al.
Repair of goat tibial defects with bone
marrow stromal cells and β-tricalcium
phosphate. J Mater Sci: Mater Med., v.19,
p.2367-2376, 2008.
LIU, T.M., LEE, E.H. Transcriptional
regulatory cascades in runx2-dependent
bone development. Tissue Eng PartB Rev,
v., p., 2012. In press

100
LIU, T.M.; MARTINA, M.;
HUTMACHER, D.W.; et al. Identification
of common pathways mediating
differentiation of bone marrow-and adipose
tissue-derived human mesenchymal stem
cells into three mesenchymal lineages. Stem
Cells.v.25, p.750-760, 2007.
LIU, Z.J.; ZHUGE, Y.; VELAZQUEZ, C.
Trafficking and differentiation of
mesenchymal stem cells. J. Cell. Biochem.,
v.106, p.984-991, 2009.
LOCKE, M.; WINDSOR, J.; DUNBAR, R.
Human adipose-derived stem cells:
isolation, characterization and applications
in surgery. ANZ. J. Surg., v.79, p.235-244,
2009.
LONG, F. Building strong bones :
molecular regulation of the osteoblast
lineage. Nat Rev Mol Cell Biol, v.13, p.27-
38, 2012.
LOUZADA, M.J.Q. ; XAVIER, C.A.M. ;
MARQUES NETO, J.F.M. Ação da
calcitonina na resolução de perfuraçoes
ósseas em coelhos: controle
fotodensitométrico e histológico. Rev Bras
Reumatol., v.30, p.111-116, 1990.
LUKLINSKA, Z.B.; SCHLUCKWERDER,
H. In vivo response to HA-
polyhydroxybutyrate/polyhydroxyvalerate
composite. J Mycrosc. v.211, n.2, p.121-
129, 2003.
MACKIE, E.J. Osteoblasts: novel roles in
orchestration of skeletal architecture. Int. J.
Biochem. Cell Biol., v.35, p.1301-1305,
2003.
MAENO, S.; NIKI, Y., MATSUMOTO,
H.; et al. The effect of calcium ion
concentration on osteoblast viability,
proliferation and differentiation in
monolayer and 3D culture. Biomaterials,
v.26, p.4847-4855, 2005.
MAKHLUF, H.A.; MUELLER, S.M.;
MIZUNO, S.; et al. Age-related decline in
osteoprotegerin expression by human bone
marrow cells cultured in three-dimensional
collagen sponges. Biochem Biophys res
Commun., v.268, p.669-672, 2000.
MANJUBALA, I.; SIVAKUMAR, M.;
SURESHKUMAR, R.V.; et al. Bioactivity
and osseointegration study of calcium
phosphate ceramic of different chemical
composition. J Biomed Mater Res. v.63,
n.2, p.200-208, 2002.
MANTOVANI, C.; RAIMONDO, S.;
HANEEF, M.S.; et al. Morphological,
molecular and functional differences of
adult bone marrow- and adipose-derived
stem cells isolated from rats od different
ages. Exp Cell Res., v.318, p.2034-2048,
2012.
MARESCHI, K.; FERRERO, I.;
RUSTICHELLI, D.; et al. Expansion of
mesenchymal stem cells isolated from
pediatric and adult donor bone marrow. J
Cell Biochem., v.97, p.744-754, 2006.
MARIE, P.J. The calcium-sensing receptor
in bone: Apotential therapeutic target in
osteoporosis. Bone., v.46, p.571-576, 2010.
MARIE, P.J. The molecular genetics of
bone formation. Am. J. Pharmacogenomics,
v.1, p.175-187, 2001.
MARIE, P.J. Transcription factors
controlling osteoblastogenesis. Arch
Biochem Biophys, v.473, p.98-105, 2008.
MARTIN, D.R.; COX, N.R.; HATHCOCK,
T.L.; et al. Isolation and characterization of
multipotencial mesenchymal stem cells
from feline bone marrow. Experimental
Hematology. v.30, p.879-886, 2002.
MARTINELLO, T.; BRONZINI, I.;
MACCATROZZO, L.; et al. Canine
adipose-derived-mesenchymal stem cells do
not lose stem features after a long-term
cryopreservation. Res. Vet. Sci. v.91, p.18-
24, 2011.
MAUNEY, J.R.; JAQUIERY, C.;
VOLLOCH, V.; et al. In vitro and in vivo
evaluation of differentially demineralized
cancellous bone scaffolds combined with
human bone marrow stromal cells for tissue
engineering. Biomaterials., v.26, p.3173-
3185, 2005.

101
MCCULLEN, S.D.; ZHAN, J.;
ONORATO, M.L.; et al. Effect of varied
ionic calcium on human adipose-derived
stem cell mineralization. Tissue Eng PartA,
v.16, p.1971-1981, 2010.
MEIRELLES, L.S.; CHAGASTELLES,
P.C.; NARDI, N.B. Mesenchymal stem
cells reside in virtually all post-natal organs
and tissues. J. Cell Sci., v.119, p.2204-
2213, 2006.
MELIGA, E.; STREM, B.M.; DUCKERS,
H.J.; et al. Adipose-derived cells. Cell
Transplant.v.16, p.93-970, 2007.
MILLIS, D.L.; JACKSON, A.M.
Nonunion, delayded union, and malunion.
In: SLATTER, S. Textbook of small animal
surgery. 3.ed. Philadelphia: Saunders, 2003.
p.1846-1861.
MIZUNO, H. Adipose-derived stem cells
for tissue repair and regeneration: ten years
of research and a literature review. J.
Nippon. Med. Sch., v.76, p.56-66, 2009.
MIZUNO, Y.; YAGI, K.; TOKUZAWA,
Y.; et al. miR-125b inhibits osteoblastic
differentiation by down-regulation of cell
proliferation. Biochen Biophys Res
Commun., v.368, p.267-272, 2008.
MOERMAN, E.J.; TENG, K.; LIPSCHITZ,
D.A.; et al. Aging activates adipogenic and
suppresses osteogenic programs in
mesenchymal marrow stromal/stem cells:
the role of PPAR-γ2 transcription factor and
TGF-β/BMP signaling pathways. Aging
Cell., v.3, p.379-389, 2004.
MOLINUEVO, M.S.; SCHUMAN, L.;
MCCARTHY, A.D.; et al. Effect of
metformin on bone marrow progenitor cell
differentiation: in vivo and in vitro studies.
J Bone Miner Res., v.25, p.211-221, 2010.
MONACO, E.; BIONAZ, M.; ZAS, S.R.; et
al. Transcriptomics comparison between
porcine adipose and bone marrow
mesenchymal stem cells during in vitro
osteogenic and adipogenic differentiation.
Plos One., v.7, p.1-19, 2012.
MOSNA, F.; SENSEBE, L.; KRAMPERA,
M. Human bone marrow and adipose tissue
mesenchymal stem cells: A user`s guide.
Stem Cell Dev. v.19, p.1449-1470, 2010.
MOSSMANT, T. Rapid colorimetric assay
for celular growth and survival: applicaion
to proliferation and cytotoxicidade assays.
J. Immunol. Methods,. v.65, p.55-65, 1983.
MULLER, M.; RAABE, O.; ADDICKS,
K.; et al. Effects of non-steroidal anti-
inflammatory drugs on proliferation,
differentiation and migration in equine
mesenchymal stem cells. Cell Biol Int.,
v.35, p.235-248, 2011.
MUSCHLER, G.F.; NITTO, H.; BOEHM,
C.A.; et al. Age-and gender related changes
in the cellularity of human bone marrow
and the prevalence of osteoblastic
progenitors. J Orthop Res., v.19, p.117-125,
2001.
MYGIND, T.; STIEHLER, M.;
BAATRUP, A.; et al. Mesenchymal stem
cell ingrowth and differentiation on
coralline hydroxyapatite scaffolds.
Biomaterials. v.28, n.6, p.1036-1047, 2007.
MYLONAS, D.; VIDAL, M.D.; KOK,
I.J.D.; et al. Investigation of a thermoplastic
polymeric carrier for bone tissue
engineering using allogeneic mesenchymal
stem cells in granular scaffolds. J
Prosthodont., v.16, p.421-430, 2007.
NAKAMURA, A.; DOHI, Y.; AKAHANE,
M. et al. Osteocalcin secretion as an early
marker of in vitro osteogenic differentiation
of rat mesenchymal stem cells. Tissue Eng.
Part C Methods, v.15, p.169-180, 2009.
NAKAMURA, S.; MATSUMOTO, T.;
SASAKI, J.I.; et al. Effect of calcium ion
concentrations on osteogenic differentiation
and hematopoietic stem cell niche-related
protein expression in osteoblasts. Tissue
Eng Part A., v.16, p.2467-2473, 2010.

102
NAKANISHI, C.; NAGAYA, N.;
OHNISHI, S.; et al. Gene and protein
expression analysis of mesenchymal stem
cells derived from rat adipose tissue and
bone marrow. Circ J., v.75, p.2260-2268,
2011.
NAKASHIMA, K.; ZHOU, X.; KUNKEL,
G.; et al. The novel zinc finger-containing
transcription factor osterix is required for
osteoblast differentiations and bone
formation. Cell, v.108, p.17-29, 2002.
NATHER, A.; DAVID, V.; TENG, J.W.H.;
et al. Effect of autologous mesenchymal
stem cells on biological healing of
allografts in critical-sized tibial defects
simulated in adult rabbits. Ann Acad Med
Singapore., v.39, p.599-606, 2010.
NEFUSSI, J.R.; BRAMI, G.;
MODROWSKI, D.; et al. Sequential
expression of bone matrix proteins during
rat calvaria osteoblast differentiation and
bone nodule formation in vitro. J.
Histochem. Cytochem., v.45, p.493-503,
1997.
NEUPANE, M.; CHANG, C.; KIUPEL,
M.; et al. Isolation and characterization of
canine adipose-derived mesenquimal stem
cells. Tissue Engineering: Part A, v.14,
p.1007-1015, 2008.
NG, T.K.; CARBALLOSA, C.M.;
PELAEZ, D.; et al. Nicotine alters
microRNA expression and hinder human
adult stem cell regenerative potential. Stem
Cells Dev., v.22, p.1-10, 2013.
NIERMEYER, P.; KORNACKER, M.;
MEHLHORN, A.; et al. Comparison of
immunological properties of bone marrow
stromal cells and adipose tissue-derived
stem cells before and after osteogenic
differentiation in vitro. Tissue Eng., v.13,
p.111-121, 2007.
NISHIMURA, R.; HATA, K.;
MATSUBARA, T.; et al. Regulation of
bone and cartilage development by network
between BMP signalling and transcription
factors. Biochem,v.151, p.247-254, 2012.
NOEL, D.; CATON, D.; ROCHE, S.; et al.
Cell specific differences between human
adipose-derived and mesenchymal-stromal
cells despite similar differentiation
potentials. Exp Cell Res., v.314, p.1575-
1584, 2008.
NOTODIHARDJO, F.Z.; KAKUDO, N.;
KUSHIDA, S.; et al. Bone regeneration
with BMP-2 and hydroxyapatite in a
critical-size calvarial defect in rats. J
Craniomaxillofac surg. v.40, p.287-291,
2012.
OCARINO, N.M.; BOELONI, J.N.; GOES,
A.M. et al. Osteogenic differentiation of
mesenchymal stem cells from osteopenic
rats subjected to physical activity with and
without nitric oxide synthase inhibition.
Nitric Oxide, v.19, p.320-325, 2008.
OCARINO, N.M.; BOELONI, J.N.;
JORGETTI, V.; et al. Intra-bone marrow
injection of mesenchymal stem cells
improves the femur bone mass of
osteoporotic female rats. Connect. Tissue
Res., n.51, p.426-436, 2010.
OCARINO, N.M.; MARUBAYASHI, U.;
CARDOSO, T.G.S. et al. Physical activity
in osteopenia treatment improved the mass
of bones directly and indirectly submitted
to mechanical impact. J. Musculoskelet.
Neuronal Interact., v.7, p.84-93, 2007.
OGINO, Y.; AYUKAWA, Y.;
TSUKIYAMA, Y.; et al. The effect of
platelet-rich plasma on cellular response of
rat bone marrow cells in vitro. Oral Surg.
Oral Med. Oral Pathol. Oral Radiol.
Endod., v.100, p.302-307, 2005.
ORÉFICE, R.L. Materiais poliméricos –
Ciência e aplicações como biomateriais. In:
ORÉFICE, R.L.; PEREIRA, M.M.;
MANSUR, H.S. (Ed) Biomateriais
Fundamentos e Aplicações. 1ed. Rio de
Janeiro: Cultura Médica. 2006. p.83- 156.
PACHON-PENA, G.; YU, G.; TUCKER,
A.; et al. Stromal stem cells from adipose
tissue and bone marrow of age-matched
female donor display distinct
immunophenotypic profiles. J Cell Physiol.,
v.226, p.843-851, 2011.

103
PANETTA, N.J.; GUPTA, D.M.;
QUARTO, N.; et al. Mesenchymal stem
cells for skeletal tissue engineering.
Panminerva Med., v.51, p.25-41, 2009.
PARK, S.H.; SIM, W.Y.; MIN, B.H.; et al.
Chip-based comparison of the osteogenesis
of human bone marrow-and adipose tissue-
derived mesenchymal stem cells under
mechanical stimulation. Plos One., v.7, p.1-
11, 2012.
PAULA, A.C.C.; ZONARI, A.A.C.;
MARTINS, T.M.M. et al. Human serum is
a suitable supplement for the osteogenic
differentiation of human adipose-derived
stem cells seeded on poly-3-
hydroxibutyrate-co-3-hydroxyvalerate
scaffolds. Tissue Eng Part A. v.19, p.1-13,
2012.
PAYUSHINA, O.V.; DOMARATSKAYA,
E.I.; STAROSTIN, V.I. Mesenchymal stem
cells: sources, phenotype, and
differentiation potential. Cell Biol., v.33,
p.2-18, 2006.
PENG, L.; JIA, Z.; YIN, X.; et al.
Comparative analysis of mesenchymal stem
cells from bone marrow, cartilage, and
adipose tissue. Stem Cells Dev., v.17,
p.761-774, 2008.
PEREIRA, M.M.; JONES, J.R.; OREFICE,
R.L.; et al. Preparation of bioactive glass-
polyvinil alcohol hybrid foams by the sol-
gel method. J Mater Sci Mater Med., v.16,
p.1045-1050, 2005.
PEREIRA, M.M.; VASCONCELOS, W.L.;
ZAVAGLIA, C.A.C. Materiais cerâmicos –
Ciência e aplicações como biomateriais. In:
ORÉFICE, R.L.; PEREIRA, M.M.;
MANSUR, H.S. (Ed) Biomateriais
Fundamentos e Aplicações. 1ed. Rio de
Janeiro: Cultura Médica. 2006. p.39- 58.
PIERMATTEI, D.L.; FLO, G.L.;
DECAMP, C.E. Handbook of small animal
orthopedics and fracture repair. 4ed. St.
Louis:Saunders, 2006. 832p.
PITTENGER, M.F.; MACKAY, A.M.;
BECK, S.C. et al. Multilineage potential of
adult human mesenchymal stem cells.
Science, v.284, p.143-147, 1999.
POUNTOS, I.; CORSCADDEN, D.;
EMERY, P.; et al. Mesenchymal stem cell
tissue engineering: techniques for isolation,
expansion and application. Injury. v.38,
p.s23-s33, 2007.
PRESNELL S.C.; PETERSEN, B.;
HEIDARAN, M. Stem cells in adult tissues.
Cell Dev. Biol., v.13, p.369-76, 2002.
PROPHET, E.B.; MILLS, B.;
ARRINGTON, J.B. et al. Laboratory
Methods in Histotechnology. Washington
D. C.: Armed Forces Institute of Pathology,
1992. 278p.
QIU, Z.; YANG, H.; WU, J.; et al. Ionic
dissolution products of NovaBone®
promote osteoblastic proliferation via
influences on the cell cycle. J. Int. Med.
Res., v.37, p.737-745, 2009.
QU, C.Q.; ZHANG, G.H.; ZHANG, L.J. et
al. Osteogenic and adipogenic potential of
porcine adipose mesenchymal stem cells. In
Vitro Cell Dev. Biol. Anim., v.43, p.95-100,
2007.
RACHED, M.T.; KODE, A.; XU, L.; et al.
FoxO is a positive regulator of bone
formation by favoring protein synthesis and
resistance to oxidative stress in osteoblasts.
Cell Metab., v.11, p.1-25, 2010.
RADA, T.; REIS, R.L.; GOMES, M.E.
Adipose tissue-derived stem cells and their
application in bone and cartilage tissue
engineering. Tissue Eng., v.15, p.113-125,
2009.
RAHAMAN, M.N.; DAY, D.E.; BAL, S.;
et al. Bioactive glass in tissue engineering.
Acta Biomater.,v.7, p.2355-2373, 2011.
RAHN, B.A. Bone healing: Histologic and
physiologic concepts. In: Sumner-Smith, G:
Bone in clinical orrtopedics, 2.ed,
Stuttugard:Thieme, 2002, p.287-325.

104
RAISZ, L.G. Physiology and
pathophysiology of bone remodeling. Clin.
Chem., v.45, p.1353-1358, 1999.
RANERA, B.; ORDOVAS, L.; LYAHYAI,
J.; et al. Comparative study of equine boné
marrow and adipose tissue-derived
mesenchymal stromal cells. Equine Vet J.,
v.44, p.33-42, 2012.
RAO, M.S. Stem sense: a proposal for the
classification of stem cells. Stem Cells Dev.,
v.13, p.452-455, 2004.
REBELATTO, C.K.; AGUIAR, A.M.;
MORETAO, M.P.; et al. Dissimilar
differentiation of mesenchymal stem cells
from bone marrow, umbilical cord blood
and adipose tissue. Exp. Biol. Med., v.233,
p.901-913, 2008.
REICHERT, J.C.; SAIFZADEH, S.;
WULLSCHLEGER, M.E.; et al. The
challenger of establishing preclinical
models for segmental bone defect research.
Biomaterials, v.30, p. 2149-2163, 2009.
REILLY, G.C.; RADIN, S.; CHEN, A.T.;
et al. Differential alkaline phosphatase
responses of rat and human bone marrow
derived mesenchymal stem cells to 45S5
bioactive glass. Biomaterials., v.28, p.4091-
4097, 2007.
REZENDE, C.M.F. Teste in vivo de
Biomateriais e Histotécnicas. In:
ORÉFICE, R.L.; PEREIRA, M.M.;
MANSUR, H.S. (Ed) Biomateriais
Fundamentos e Aplicações. 1ed. Rio de
Janeiro: Cultura Médica. 2006. p.299- 314.
ROBLING, A.G.; CASTILLO, A.B.;
TURNER, C.H. Biomechanical and
molecular regulation of bone remodeling.
Annu. Rev. Biomed. Eng., v.8, p.455-498,
2006.
ROE S. Internal fracture fixation. In:
SLATTER, S. Textbook of small animal
surgery. 3.ed. Philadelphia: Saunders, 2003.
p.1798-1818.
ROZEMULLER, H.; PRINS, H.;
NAAIJKENS, B.; et al. Prospective
isolation of mesenquimal stem cells from
multiples mammalian species using cross-
reaction anti human monoclonal antibodies.
Stem. Cells. Dev. V.19, p.1911-1921, 2010.
SAMPAIO, I.B.M. Estatística aplicada à
experimentação animal. Belo Horizonte:
FEP/MVZ, 1998, 211p.
SANDERS, R. Bone graft substitutes:
separating fact from fiction. The Journal of
Bone and Joint Surgery, v.89, n.3, p.469,
2007.
SANTOS, F.C.; RAHAL, S.C. Enxerto
ósseo esponjoso autólogo em pequenos
animais. Cienc. Rural, v.34, p.1969-1975,
2004.
SCHÄFFLER, A.; BÜCHLER, C. Concise
Review: adipose tissue-derived stromal
cells - basic and clinical implications for
novel cell-based therapies. Stem Cells, v.25,
p.818-827, 2007.
SCHWARZ, C.; LEICHT, U.; ROTHE, C.;
et al. Effects of different media on
proliferation and differentiation capacity of
canine, equine and porcine adipose derived
stem cells. Res. Vet. Sci. v.93, p.457-567,
2012.
SEO, M.; JEONG, Y.; PARK, J.; et al.
Isolation and characterization of canine
umbilical cord blood-derived mesenchymal
stem cells. J. Vet. Sci., v.10, p.181-187,
2009.
SETHE, S.; SCUTT, A.; STOLZING, A.
Aging of mesenchymal stem cells. Agering
Res Rev., v.5, p.91-116, 2006.
SHENFIELD, F.; PENNINGS, G.;
SUREAU, C. et al. Stem cells. Hum.
Reprod., v.17, p.1409-1410, 2002.
SHI, Y.Y.; NACAMULI, R.P.; SALIM, A.;
et al. The osteogenic potential of adipose-
derived mesenchymal cells in maintained
with aging. Plast Reconstr Surg., v.116,
p.1686-1696, 2005.

105
SHIE, M.Y.; DING, S.J.; CHANG, H.C.
The role of silicon in osteoblast-like cell
proliferation and apoptosis. Acta.
Biomater., v.7, p.2604-2614, 2011.
SILVA, A.R.P.; PAULA, A.C.C.;
ZONARI, A.; et al. Development of a
protocol for human adipose stem cell
culture in CO2 independent medium and
perfusion bioreactor. J Tissue Sci Eng.,
S11:006, 2013.
SILVA, R.F.; REZENDE, C.M.F.; PAES-
LEMES, F.O.; et al. Evaluación del método
del tubo para concentrar plaquetas canina:
estúdio celular. Arch. Med. Vet., v.43, p.95-
98, 2011.
SKAGUCHI, Y.; SEKIYA, I.;
YAGISHITA, K.; et al. Comparison of
human stem cells derived from various
mesenchymal tissues: Superiority of
synovium as a cell source. Arthritis
Rheum.,v.52, p.2521-2529, 2005.
SPENCER, N.D.; CHUN, R.; VIDAL,
M.A.; et al. In vitro expansion and
differentiation of fresh and revitalized adult
canine bone marrow-derived and adipose
tissue-derived stromal cells. Vet j., v.191,
p.231-239, 2012.
STENDERUP, K.; JUSTESEN, J.;
CLAUSEN, C. et al. Aging is associated
with decreased maximal life span and
accelerated senescence of bone marrow
stromal cells. Bone, v.33, p.919-926, 2003.
STENDERUP, K.; JUSTESEN, J.;
ERIKSEN, E.F.; et al. Number and
proliferative capacity of osteogenic stem
cells are maintained during aging and in
patients with osteoporosis. J Bone Miner
Res., v.16, p.1120-1129, 2001.
STENDERUP, K.; ROSADA, C.;
JUSTESEN, J.; et al. Age human bone
marrow stromal cells maintaining bone
forming capacity in vivo evaluated using an
improved method of visualization.
Biogerontology., v.5, p.107-118, 2004.
STIFFLER, K.S. Internal fracture fixation.
Clin Tech Small Anim Pract. v.19, n.3,
p.105-113, 2004.
STOLZING, A.; JONES, E.;
MCGONAGLE, D.; et al. Age-related
changes inhuman bone marrow-derived
mesenchymal stem cells: Consequences of
cell therapies. Mech Ageing Dev., v.129,
p.163-173, 2008.
STORZ, P. Forkhead homeobox type O
transcription factors in the responses to
oxidative stress. Antioxid Redox Signal.,
v.14, p. 593-605, 2011.
STRAUSS, P.G.; CLOSS, E.I.; SCHMIDT,
J.; et al. Gene expression during osteogenic
differentiation in mandibular condyles in
vitro. J Cell Biol., v.110, p.1369-1378,
1990.
STREM, B.M.; HICOK, K.C.; ZHU, M.; et
al. Multipotencial differentiation of adipose
tissue-derived stem cells. Keio J. Med.,v.54,
p.132-141, 2005.
STRIOGA, M.; VISWANATHAN, S.;
DARINSKAS, A.; et al. Same or not the
same? Comparison of adipose tissue-
derived versus bone marrow-derived
mesenchymal stem and stromal cells. Stem
Cells Dev., v.21, p.2724-2752, 2012.
SUN, J.; WEI, L.; LIU, X.; et al. Influences
of ionic dissolution products of dicalcium
silicate coating on osteoblastic
proliferation, differentiation and gene
expression. Acta Biomater., v.5, p.1284-
1293, 2009b.
SUN, J.; LI, J.; LIU, X.; et al. Proliferation
and gene expression of osteoblasts cultured
in DMEM containing the ionic products of
dicalcium silicate coating. Biomed.
Pharmacother., v.63, p.650-657, 2009a.
SUN, J.; WEI, L.; LIU, X.; et al. Influences
of ionic dissolution products of dicalcium
silicate coating on osteoblastic
proliferation, differentiation and gene
expression. Acta. Biomater., v.5, p.1284-
1293, 2008.
TAE, S.; LEE, S.; PARK, J. et al.
Mesenchymal stem cells for tissue
engineering and regenerative medicine.
Biomed. Mater., v.1, p.63-71, 2006.

106
TAKEMITSU, H.; ZHAO, D.;
YAMAMOTO, I.; et al. comparison of
bone and adipose tissue-derived canine
mesenchymal. BMC Vet Res.,v.8, p.1-9,
2012.
TATE, M.L.K.; ADAMSON, J.R.; TAMI,
A.E. et al. The osteocyte. Int. J. Biochem.
Cell Biol., v.36, p.1-8, 2004.
THOMPSON, K. Bone and joints. In:
MAXIE, M.G. (Ed) Pathology of domestic
animals. 5ed. London:Saunders. 2007. p.2-
23.
THOMSON, J.A.; ITSKOVITZ-ELDOR,
J.; SHAPIRO, S.S. et al. Embryonic stem
cell lines derived from human blastocysts.
Science, v.282, p.1145-1147, 1998.
TOMA, J.G.; AKHAVAN, M.;
FERNANDES, K.J. et al. Isolation of
multipotent adult stem cells from the
dermis of mammalian skin. Nat. Cell Biol.,
v.3, p.778-784, 2001.
TSENG, S.S.; LEE, M.A.; REDDI, A.H.
Nonunions and the potential of stem cells in
fracture-healing. J Bone Joint Surg Am.,
v.90, p.92-98, 2008.
TSIGKOU, O.; JONES, J.R.; POLAK,
J.M.; et al. Differentiation of fetal
osteoblasts and formation of mineralized
bone nodules by 45S5 Bioglass conditioned
medium in the absence of osteogenic
supplements. Biomaterials, v.30, p.3542-
3550, 2009.
VALÉRIO, P.; PEREIRA, M.M.; GOES,
A.M.; et al. The effect of ionic products
from bioactive glass dissolution on
osteoblast proliferation and collagen
production. Biomaterials., v.25, p.2941-
2948, 2004.
VALLET-REGI, M. Revisiting ceramics
for medical applications. Dalton Trans.
v.28, n.44, p.5211-5220, 2006.
VARANASI, V.G.; LEONG, K.K.;
DOMINIA, L.M.; et al. Si e Ca individually
and combinatorially target enhanced
MC3T3-E1 subclone 4 early osteogenic
marker expression. J Oral Implantol., v.38,
p.325-336, 2012.
VERONESI, F.; TORRICELLI, P.;
BORSARI, V.; et al. Mesenchymal stem
cells in the aging and osteoporotic
population. Crit Rev Eukaryot Gene Expr.,
v.21, p.363-377, 2011.
VIATEAU, V.; GUILLEMIN, G.;
BOUSSON, V.; et al. Long-bone critical-
size defects treated with tissue-engineered
grafts: A study on sheep. J Orthop Res.,
v.25, p.741-749, 2007.
VIEIRA, N.M.; BRANDALISE, V.;
ZUCCONI, E.; et al. Isolation,
characterization, and differentiation
potential of canine adipose-derived stem
cells. Cell Transplant. v.19, p.279-289,
2010.
WAGERS, A.J.; WEISSMAN, I.L.
Plasticity of adult stem cells. Cell, v.116,
p.639-648, 2004.
WAGNER, W.; BORK, S.; HORN, P.; et
al. Aging and replicative senescence have
related effects on human stem and
progenitor cells. Plos One., v.4, p.1-13,
2009.
WAGNER, W.; HORN, P.; CASTOLDI,
M.; et al. Replicative senescence of
mesenchymal stem cells: A continuous and
organized process. Plos One., v.3, p.1-12,
2008.
WAN, C.; HE, Q.; LI, G. Allogenic
peripheral blood derived mesenchymal stem
cells(MSCs) enhance bone regeneration in
rabbit ulna critical-size bone defect model.
J Orthop Res., v.24, p.610-618, 2006.

107
WANG, M. Developing bioactive
composite materials for tissue replacement.
Biomaterials. v.24, n.13, p.2133–2151,
2003.
WATSON, J.T. Overview of Biologics, J
Orthop Trauma. v.19, n.10, p.14-16, 2005.
WELCH, J.A.; BOUDRIEAU, R.J.;
DEJARDIN, L.; et al. The intraosseous
blood supply of the canine radius:
Implications for healing of distal fractures
in small dogs. Vet Sug., v.26, p.57-61,
1997.
WEN, L.; WANG, Y.; WANG, H.; et al. L-
type calcium channels play a crucial role in
the proliferation and osteogenic
differentiation of bone marrow
mesenchymal stem cells. Biochem Biophys
Res Commun., v.424, p.439-445, 2012a.
WEN, Y.; JIANG, B.; CUI, J.; et al.
Superior osteogenic capacity of different
mesenchymal stem cells for bone tissue
engineering. Oral Surg Oral Med Oral
Pathol Oral Radiol., v., p. 2012- IN
PRESS.
WU, W.; CHEN, X.; MAO, T.; et al. Bone
marrow-derived osteoblast seeded into
porous beta-tricalcium phosphate to repair
segmental defect in canine´s mandibular.
Ulus Travma Acil Cerrahi Derg.,v.12,
p.268-276, 2006.
XYNOS, I.D.; EDGAR, A.J.; BUTTERY,
L.D.K.; et al. Gene-expression profiling of
human osteoblasts following treatment with
the ionic products of Bioglass 45S5
dissolution. J. Biomed. Mater. Res.,v.55,
p.151-157, 2001.
XYNOS, I.D.; EDGAR, A.J.; BUTTERY,
L.D.K.; et al. Ionic products of bioactive
glass dissolution increase proliferation of
human osteoblasts and induce insulin-like
growth factor II mRNA expression and
protein synthesis. Biochem Bioph Res
Co.,v.276, p.461-465, 2000.
YAMADA, Y,; BOO, J.S.; OZAWA, R.; et
al. Bone regeneration following injection of
mesenchymal stem cells and fibrin glue
with a biodegradable scaffold. J
CranioMaxillofac Surg. v.31, p.27-33,
2003.
YANG, X.; KARSENTY, G. Transcription
factors in bone: developmental and
pathological aspects. Trends Mol. Med.,
v.8, p.340-345, 2002.
YOON, E.; DHAR, S.; CHUN, D.E.; et al.
In vivo osteogenic potential of human
adipose-derived stem cells/poly lactide-co-
glycolic acid constructs for bone
regeneration in a rat critical-sized calvarial
defect model. Tissue Eng., v.13, p.619-627,
2007.
YOSHIMURA, H.; MUNETA,
T.; NIMURA, A.; et al. Comparison of rat
mesenchymal stem cells derived from bone
marrow, synovium, periosteum, adipose
tissue and muscle. Cell. Tissue. Res., v.327,
p.449-462, 2007.
YUAN, J.; CUI, L.; ZHANG, W.J.; et al.
Repair of canine mandibular bone defects
with bone marrow stromal cells and porous
β-tricalcium phosphate. Biomaterials, v.28,
p.1005-1013, 2007.
YUAN, J.; ZHANG, W.J.; LIU, G.; et al.
Repair of canine mandibular bone defects
with bone marrow stromal cells and coral.
Tissue Eng PartA.,v.16, p.1385-1394, 2010.
ZAIM, M.; KARAMAN, S.; CETIN, G.; et
al. Donor age and long-term culture affect
differentiation and proliferation of human
bone marrow mesenchymal stem cells. Ann
Hematol., v.91, p.1175-1186, 2012.
ZAMINY, A.; KASHANI, I.R.;
BARBARESTANI, M. et al. Osteogenic
differentiation of rat mesenchymal stem
cells from adipose tissue in comparison
with bone marrow mesenchymal stem cells:
melatonin as a differentiation factor. Iran.
Biomed. J., v.12, p.133-141, 2008.

108
ZHANG, C. Molecular mechanisms of
osteoblast-specific transcription factor
osterix effect on bone formation. Beijing
Da Xue Xue Bao. v.44, p.659-665, 2012
ZHANG, Z.H.; TEOH, S.H.; CHONG,
M.S.K.; et al. Superior osteogenic capacity
for bone tissue engineering of fetal
compared with perinatal and adult
mesenchymal stem cells. Stem Cells., v.27,
p.126-137, 2009.
ZHAO, J.; ZHANG, Z.; WANG, S.; et al.
Apatite-coated silk fibroin scaffolds to
healing mandibular border defects in
canines. Bone., v.45, p.517-527, 2009.
ZHOU, S.; GREENBERGER, J.S.;
EPPERLY, M.W.; et al. Age-related
intrinsic changes in human bone marrow-
derived mesenchymal stem cells and their
differentiation to osteoblasts. Aging Cell.,
v.7, p.335-343, 2008.
ZHOU, Y.; GUAN, X.X.; ZHU, Z.L.; et al.
Caffeine inhibits the viability and
osteogenic differentiation of rat bone
marrow-derived mesenchymal stromal
cells. Br J Pharmacol., v.161, p.1542-1552,
2010.
ZHOU, Y.S.; LIU, Y.S.; JIAN-GUO, T. Is
1,25-dihydroxyvitamin D3 an ideal
substitute for dexamethasone for inducing
osteogenic differentiation of human adipose
tissue-derived stromal cells in vitro? Chin.
Med. J. v.119, p.1278-1286, 2006.
ZHU, L.; CHUANCHANG, D.; WEI, L.; et
al. Enhanced Healing of goat femur-defect
using BMP7 gene-modified BMSCs and
load-bearing tissue-engineered bone. J
Orthop Res., v.28, p.412-418, 2010.
ZHU, L.; LIU, W.; CUI, L.; et al. Tissue-
engineered bone repair of goat fêmur
defects with osteogenically induced bone
marrow stromal cells. Tissue Eng., v.12,
p.423-433, 2006.
ZHU, M.; KOHAN, E.; BRADLEY, J.; et
al. The effect of age on osteogenic,
adipogenic and proliferative potential of
female adipose-derived stem cells. J Tissue
Eng Regen Med., v.3, p.290-301, 2009.
ZHU, X.; LIU, T.; SONG, K. et al. Adipose
tissue-derived stem cell: a better stem cell
than BMSC. Cell Biochem. Funct., v. 26,
p.664-675, 2008.
ZUK, P.A.; ZHU, M.; MIZUNO, H. et al.
Multilineage cells from human adipose
tissue: implications for cell-based therapies.
Tissue Eng., v.7, p.211-228, 2001.





























