Embed Size (px)
DESCRIPTION
Citoesqueleto
Citation preview

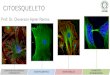
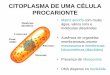
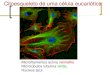
CITOESQUELETO Introdução As células eucariontes possuem uma complexa rede citoplasmática de microtúbulos e microfilamentos situada predominantemente próxima à membrana celular e envolvida em quase todos os aspectos que envolvem a forma e movimento celulares. Essa rede é chamada de CITOESQUELETO. O citoesqueleto desempenha um papel mecânico, de suporte, mantendo a forma celular e a posição de seus componentes. Ele estabelece, modifica e mantém a forma das células. É responsável por movimentos celulares como contração, formação de pseudópodes e deslocamento intracelular de organelas, cromossomos, vesículas e grânulos diversos. O citoesqueleto deve ser considerado como um complexo de túbulos, microfilamentos longos (actina), fi lamentos curtos (miosina), fi lamentos intermediários e macromoléculas protéicas diversas que formam um conjunto dinâmico. Os microtúbulos são estruturas cil índricas de 24 nm, composto por fibrilas longitudinais que contribuem para os movimentos intracelulares; são feixes ordenados de cilindros delgados e longos formados por proteínas (a e b tubulina) que constituem os cílios e flagelos, as fibras do áster e o fuso (na divisão celular), os centríolos e os corpúsculos basais. Os microfilamentos são estruturas de 4 a 6 nm de diâmetro e são formados pela actina, miosina ou outras proteínas contráteis. Participam ativamente nos movimentos celulares. Os filamentos intermediários tem 10 nm de espessura e contém diversas proteínas fibrosas não contráteis que desempenham um papel mecânico. Há vários tipos de filamentos intermediários: filamentos de queratina, neurofi lamentos, filamentos gliais e filamentos heterogêneos. Como o citoesqueleto é muito dinâmico, ele está envolvido em vários movimentos. Estes podem ocorrer em células não-musculares ou em células musculares. Nas células não-musculares os cílios, os flagelos, a ciclose (tipo de movimento de geração de correntes citoplasmáticas por ação de microfilamentos) e o movimento amebóide (que é um tipo de locomoção no qual a célula troca de forma ativamente, enviando pseudópodes citoplasmáticos na direção do movimento) são responsáveis pela locomoção. E nas células musculares a movimentação celular e conseqüente contração muscular ocorre no interior dos sarcômeros. Os sarcômeros são unidades estruturais da miofibrila (estruturas delgadas cilíndricas e contráteis do tecido muscular estriado) onde há a participação de proteínas contráteis como a actina, a miosina e outras. A capacidade que as células eucarióticas possuem de adotar uma variedade de formas e de executar movimentos coordenados e direcionados depende de uma rede complexa de filamentos de proteínas filamentosas que se estendem por todo o citoplasma. Esta rede é chamada de citoesqueleto. O citoesqueleto é composto por vários tipos de fi lamentos de proteínas visíveis ao microscópio eletrônico que cruzam a célula eucariótica formando uma rede tridimensional interligada em todo o citoplasma. É uma estrutura altamente

dinâmica que se reorganiza continuamente sempre que a célula altera a forma, se divide ou responde ao seu ambiente. Ele poderia ser chamado de "citomusculatura", pois é responsável por movimentos como deslocamento de células sobre um substrato e contração muscular; fornecendo também maquinaria necessária para movimentos intracelulares, como o transporte de organelas de um lugar a outro no citoplasma e a segregação dos cromossomos na mitose. O citoesqueleto está ausente nas bactérias e ele pode ter sido um fator crucial na evolução das células eucarióticas. Suas diferentes atividades dependem de três tipos de filamentos protéicos: fi lamentos de actina, microtúbulos e filamentos intermediários que fornecem estrutura, organização e forma celular. Cada um dos tipos do componente do citoesqueleto é composto por subunidades protéicas simples que se polimerizam para formar filamentos de espessura uniforme. Esses filamentos não são estruturas permanentes, pois sofrem constantes desmontagens em suas subunidades monoméricas e remontagens em fi lamentos. Sua localização na célula não é rigidamente fixa e pode alterar-se com a mitose, citocinese ou alterações na forma da célula. Todos os tipos de filamentos se associam com outras proteínas que interligam fi lamentos entre si ou a outros, influenciam na montagem e desmontagem ou movem organelas citoplasmáticas ao longo dos filamentos. Particularmente, os filamentos de actina e microtúbulos são os principais produtores da movimentação das organelas e de toda a célula. A actina e miosina são duas proteínas geradoras de forças. A tropomiosina e troponina C ( que ocorrem no músculo) têm função reguladora. A a actinina participa das interações entre o filamentos de actina e a membrana celular, porém existem outras proteínas fixadoras de actina que podem formar ligações transversais com fi lamentos de actina. Filamentos de actina tendem a formar feixes paralelos com região em que a célula contata com um substrato sólido (isto pode ser visto quando a célula se distende). Esses feixes são chamados de fibras de tensão ou estresse. 2 - Organização Estrutural dos Elementos do Citoesqueleto 2.1 – Microtúbulos Os microtúbulos são estruturas presentes no citoplasma de todas as células eucariontes. As primeiras observações dessas estruturas foram realizadas por De Robertis e Franchi em 1953, no axoplasma de fibras nervosas. Devido ao fato de que a maioria dos microtúbulos são muito lábeis e não resistentes ao efeito de muitos fixadores, só a partir de 1963, com a introdução do glutaldeído com fixação para a microscopia eletrônica, a sua presença pode ser identificada. Os microtúbulos citoplasmáticos apresentam dimensões uniformes por volta de 25 nm de diâmetro externo e vários micrômeros de comprimento e são notavelmente retilíneos. Em corte transversal apresentam uma parede densa de aproximadamente de 6 nm de espessura, e uma região central mais clara. A parede do microtúbulo está constituída por

estruturas filamentosas, lineares ou em espiral, que apresentam um diâmetro aproximado de 5 nm. Em corte transversal observam-se geralmente 13 filamentos, separados por uma distância de 4,5 nm de centro a centro. Os microtúbulos são compostos por unidades protéicas bastante semelhantes, mesmo quando diferentes tipos celulares são analisados. O termo tubulina é utilizado para designar a principal proteína dos cílios, flagelos e microtúbulos citoplasmáticos. A tubulina é um dímero de 110.000 a 120.000 daltons. Na maioria dos casos, podem ser identificados dois monômeros de tipos diferentes de tubulina, as a e ß tubulinas embora apresentem pesos semelhantes. O espaço de 8 nm que se observa ao longo do eixo longitudinal do microtúbulos reflete provavelmente o pareamento dos dois tipos de monômeros de tubulina. Cada dímero de tubulina pode-se ligar a uma molécula de cochicina, propriedade esta que permite fazer a determinação da proteína tubulina. A montagem dos microtúbulos a partir dos dímeros de tubulina é um processo especificamente orientado e programado. A quantidade de tubulina a ser polimerizada varia conforme as necessidades da célula. Em condições normais, os microtúbulos se encontram em equilíbrio com a quantidade de tubulina livre. A polimerização dos microtúbulos é favorecida por uma proteinoquinase dependente de AMP cíclico que promove a fosforilação dos monômeros de tubulina. A montagem de monômeros de tubulina é um fenômeno polarizado, isto é, dímeros se unem a um dos extremos dos microtúbulos enquanto se desprendem um do outro. Quando a colchicina se fixa a tubulina, o processo de montagem é inibido, porém a desmontagem ou despolimerização continua ocorrendo. Deste modo, eventualmente, os microtúbulos podem ser completamente despolimerizados. A vimblastina, atua despolimerizando os microtúbulos já formados e, numa segunda etapa, agrega as subunidades de tubulina formando estruturas paracristalinas. Os microtúbulos são polímeros rígidos formados por moléculas de tubulina. Eles se organizam pela adição, às suas extremidades livres, de moléculas de tubulina contendo GTP, sendo que uma das extremidades (extremidades "mais") cresce mais rapidamente do que a outra. A hidrólise do GTP em GDP acontece depois da ligação da tubulina e enfraquece as pontes que mantém os microtúbulos unidos. Os microtúbulos de crescimento lento, são instáveis e sujeitos a desagregação "catastrófica", pela colchicina, mas, podem ser estabilizados por associação com outras estruturas que bloqueiam suas duas extremidades. Os centros organizadores de microtúbulos, tais como os centrossomos, protegem as extremidades "menos" e iniciam continuamente a formação de novos microtúbulos, que crescem em direções aleatórias. Qualquer microtúbulo que, casualmente, encontrar uma estrutura que estabilize sua extremidade livre será seletivamente mantido, enquanto os demais despolimerizarão. Este processo seletivo determinaria a posição dos conjuntos de microtúbulos numa célula. A falta de GDP detém a agregação da tubulina. Em situação in vivo, a polimerização da tubulina está regulada pela cálcio e pela proteína fixadora de cálcio, a calmodulina. Quando ambos formam um agregado, a polimerização é inibida.

As subunidades de tubulina nos microtúbulos (tubulinas alfa e beta) seletivamente estabilizados , são modificadas por acetilação e detirosinação. Estas alterações marcariam os microtúbulos como "maduros" e produziriam sítios para a ligação de proteínas específicas associadas a microtúbulos (MAPs), que promovem uma estabilização adicional contra a desagregação. As proteínas motoras constituem uma classe importante de MAPs que usam a energia da hidrólise do ATP para se mover unidirecionalmente ao longo dos microtúbulos, deslocando um carregamento específico . Em geral, as dineínas movem o seu carregamento em direção da extremidades "mais", enquanto a maioria das quinesinas se movem com o carregamento em direção da extremidade "menos". Tais proteínas motoras são responsáveis, em grande parte, pela organização espacial e pelos movimentos direcionados das organelas no citoplasma. Na célula, existem sítios de organização, tais como o centríolo e os corpúsculos basais dos cílios, a partir dos quais o processo de polimerização é de certa maneira orientado. Freqüentemente os microtúbulos se dispõem radialmente a partir do núcleo da célula e aparecem como fi lamentos retos ou curvos que parecem terminar perto da superfície celular. Esses filamentos desaparecem após o tratamento com Colcemid (um derivado da colchicina) ou por esfriamento, reaparecendo se situação é revertida. Estas observações demonstraram que os microtúbulos se originam das proximidades dos núcleos, a partir de um dos pontos focais correspondentes à região do cromossoma (ou seja, que contém o centríolo) ou centrosfera. Muitos dessas observações sugeriram que a centrosfera é o principal centro organizador dos microtúbulos. Nas células prestes a entrar em mitose, os microtúbulos citoplasmáticos desaparecem e são substituídos por aqueles que integram o fuso e os ásteres. Em células transformadas por um vírus ou por certas substâncias químicas, os microtúbulos estão desorganizados e às vezes desaparecem por completo. A participação dos microtúbulos no citoesqueleto foi bem estudada, tomando-se como modelo os heliozoários. Estes protozoários são envoltos por expansões muito finas do citoplasma, os axonemas, que se dispõem radialmente na célula. Nas micrografias eletrônicas, observavam-se que os axonemas contém numerosos microtúbulos, dispostos ordenadamente em espiral. Interessante é que, submetendo-se esses protozoários à ação da uréia, os axonemas entram em colapso e se retraem. Retirando a uréia do meio, os axonemas regeneram-se em menos de meia hora. O estudo deste fenômeno com o microscópio eletrônico mostrou que, durante o colapso dos axonemas, ocorre uma despolimerização dos microtúbulos, com acúmulo das moléculas globosas de tubulina no citoplasma. Com a retirada da uréia ocorre a repolimerização dos microtúbulos e o reaparecimento dos axonemas. As funções dos microtúbulos são as seguintes: Função Mecânica: Tem-se relacionado a forma de algumas protuberâncias ou prolongamentos celulares com a orientação e distribuição dos microtúbulos. Os microtúbulos são considerados como uma armação que determina a forma e distribuição dos constituintes da célula. A integridade dos microtúbulos é necessária para manter a forma característica de muitas células e a rigidez das estruturas mais amplas. Como exemplo de tais tipos de estruturas temos os axônios e os dendritos dos neurônios. Morfogênese: O papel que os microtúbulos desempenham na aquisição da forma durante

a diferenciação celular está relacionado com a sua função mecânica. Um exemplo interessante é proporcionado pelas mudanças morfogenéticas que ocorrem durante a espermiogênese. O aumento do núcleo da espermátide ocorre simultaneamente com a produção de um conjunto ordenado de microtúbulos que se dispõe em torno do mesmo. Polarização e motilidade celulares: A determinação da polaridade intrínseca de certas células também está relacionada com a função mecânica dos microtúbulos. O tratamento com Colcemid determina mudanças na motil idade. Quando algum tipo de movimento persiste, o deslocamento direcionado da célula é substituído, por exemplo, por um movimento ao acaso. Circulação e transporte: Os microtúbulos podem intervir também no transporte de macromoléculas no interior da célula. É provável que para tal finalidade formem "canais" no citoplasma. Um exemplo da correlação entre os microtúbulos e o transporte de partículas é fornecido pelo melanócito, no qual se observa o movimento dos grânulos de melanina entre os canais criados, na matriz citoplasmática, pelos microtúbulos. Transudação sensorial: Nos receptores sensoriais encontram-se com freqüência conjuntos de microtúbulos organizados de maneira bastante regular. Postula-se sua possível função na transudação de energia. 2.2 – Microfilamentos Os microfilamentos são estruturas delgadas formados por duas cadeias em espiral de monômeros globosos da proteína actina G (lembrando dois colares de pérolas enroladas) que se associam formando uma estrutura quaternária fibrosa (actina F). Observados ao Microscópio Eletrônico, mostram-se como estruturas finas de 6 a 8 nm de diâmetro. Subunidades individuais de actina se polimerizam para formar fi lamentos de actina. A proteína fi lamina segura dois microfilamentos quando eles se cruzam em ângulo reto. Os filamentos são unidos através de ligações cruzadas por uma outra proteína, a fodrina, formando agregados ou feixes lado-a-lado. Os microfilamentos de actina são muito abundantes em células musculares e também existem em plantas (apesar da ausência de músculos). A actina constitui 5 a 30% de todas as proteínas do citoplasma. Diferentes estudos têm sugerido que esses filamentos representem sistemas contráteis. Este conceito tem sido confirmado pela demonstração de que as proteínas contráteis do músculo (actina e miosina) se encontram amplamente distribuídas na maioria das células. Abaixo da membrana plasmática há feixes de filamentos constituídos por actina, os quais estão em continuidade com um retículo de filamentos semelhantes que atravessam o citosol. Os canais ou espaços criados nesse retículo ou trama trabecular são de 50 a 100 nm e, na célula viva, permitem a difusão de líquidos e metabóli tos por todo o citosol. Os filamentos dessa trama se encontram em contato com as vesículas do retículo endoplasmático, com microtúbulos e com polissomas, os quais parecem estar todos contidos e sustentados por essa trama. O fato de que essa trama pode ser armada e desarmada facil itaria também pequenos movimentos das organelas contidas no citoesqueleto.

A exemplo dos microtúbulos, que são despolimerizados pela colchicina, os microfilamentos são despolimerizados pela citocolasina B. Tem sido demostrado que essa droga inibe diversas atividades celulares onde participam alguns tipos de microfilamentos, por exemplo, a contração do músculo liso, a migração celular, a endocitose, a exocitose e outros processos, destruindo a organização regular dos microfilamentos associados com algumas dessas funções. Essa inibição pode ser tanto pela destruição quanto pela estabilização de microfilamentos, mostrando que o dinamismo entre os monômeros de actina dos microfilamentos e os do pool citoplasmático é essencial para seu funcionamento. Sem dúvida, nem todos os microfilamentos são sensíveis a citocolasina B. Conclui-se que os filamentos sensíveis a citocolasina B constituem a maquinaria contrátil das células não musculares. No músculo estriado os microfilamentos de actina reagem com os filamentos de miosina promovendo a contração muscular. Nesse tipo de músculo, os fi lamentos de actina e os de miosina apresentam um grau muito elevado de organização espacial. Nas células musculares lisas encontradas, por exemplo, no tubo digestivo , nos vasos sangüíneos, no útero, etc, a contração também se deve à interação actina-miosina. Os microfilamentos de actina são muito abundantes nos músculos e são encontrados, embora em menor número, no citoplasma de todas as células mas filamentos longos de miosina só foram encontrados nas células musculares estriadas. Nas células musculares lisas e nos demais tipos celulares, a miosina está presente como filamentos curtos ou bastonetes finos que, em cooperação com o microfilamento de actina são responsáveis pela contração. Portanto, a contração celular tem como base a interação actina- miosina. Nas células musculares estriadas, que são especializadas para contração rápida e forte, a organização das duas proteínas (actina-miosina) atinge um grau muito elevado, o que não acontece e nem é necessário para outras células. Muitos movimentos celulares como a citocinese (divisão citoplasmática), a endocitose, fenômenos como o fenômeno do capuz, a fusão celular e a mobilização dos receptores requerem a interação dos microfilamentos com a membrana celular. Essa interação se estabelece através de ligações estáveis como o ATP e por 5 proteínas especiais, como a vinculina. Alguns distúrbios nas células podem provocar a fosforilação da vinculina contribuindo para a desorganização do citoesqueleto unido à membrana celular. As desorganizações do microfilamento alteram a forma da célula. 2.3 - Filamentos Intermediários Os filamentos intermediários são polímeros fortes parecidos a cabos e são constituídos de polipeptídios fibrosos resistentes ao estiramento, desempenhando um papel estrutural na célula. Sua principal função é a de resistir à tensão mecânica, ou seja, proporcionar estabilidade mecânica às células. São chamados de "intermediários" porque seu diâmetro aparente (10 nm de espessura) está entre o dos filamentos de actina e o dos filamentos de miosina; sendo seu diâmetro

também intermediário ao dos filamentos de actina e dos microtúbulos . Esses filamentos intermediários são muito heterogêneos quando se consideram suas propriedades bioquímicas, porém, quanto à morfologia e localização, é possível agrupá-los em apenas 4 tipos principais. Os filamentos intermediários formam retículos que interconectam o núcleo à superfície celular. Inserem-se no envoltório nuclear formando uma espécie de rede ao redor do núcleo, irradiando-se pelo citoplasma. Todos os filamentos intermediários são constituídos pela agregação de moléculas alongadas, cada uma formada por 3 cadeias polipeptídicas enroladas em hélice. A presença dos filamentos intermediários se dá em grande número no epitélio ligando as células entre si por junções especializadas, nas células nervosas ao longo do comprimento dos axônios e em todos os tipos de células musculares. Os tipos de filamentos intermediários são: Filamentos de queratina: São encontrados exclusivamente em células epiteliais e estruturas derivadas como pêlos, unhas e chifres. Denominados também de tonofilamentos, filamentos de pré- queratina ou de citoqueratina constituem a classe mais complexa de filamentos intermediários. Esses filamentos são ancorados à superfície da célula onde tendem a convergir sobre os desmossomas. As citoqueratinas de mamíferos são proteínas alfa fibrosas que são sintetizadas nas camadas vivas da epiderme e formam a maior parte da camada morta de extrato córneo. Neurofilamentos: Junto com os microtúbulos são os principais elementos estruturais dos axônios, dos dendritos e do pericário dos neurônios. Contém 3 polipeptídeos e são muito sensíveis à presença do cálcio. Formam um emaranhado tridimensional que faz do axoplasma um gel altamente estruturado. Filamentos Gliais: Encontram-se dispersos no citoplasma dos astócitos por uma proteína que apresenta propriedades muito ácidas. A glia (ou neuróglia) é encontrada no tecido nervoso do sistema nervos central e é constituída por alguns tipos celulares, todos com ramificações. Filamentos Heterogêneos: Incluem-se os filamentos intermediários de morfologia e localizações similares, porém que contém diferentes proteínas. Como a Desmina, a Vimentina e a Sinemina (peso molecular 55000) são as mais freqüentes, encontrados nos fibroblastos e em todas as células de origem mesenquimal. Por meio de imunofluorescência é possível observar que apresentam uma disposição ondulada (vimentus-ondeado). Os filamentos de DESMINA (peso molecular 55000) são encontrados nas células musculares lisas e nas linhas Z e M das células estriadas que constituem os músculos esqueléticos e a musculatura cardíaca. As micrografias têm mostrado que os filamentos intermediários podem ligar-se aos microtúbulos através de pontes delgadas de um material ainda desconhecido. Isto é particularmente evidente em certos prolongamentos (axônio) das células nervosas. Esta associação também foi demonstrada por imunocitoquímica. A dissolução dos microtúbulos de fibroblastos por meio de colchicina modifica a posição dos filamentos de vimentina. Enquanto os microtúbulos estão

intactos, os filamentos de vimentina percorrem todo o citoplasma, mas após a desmontagem dos microtúbulos pela colchicina, os filamentos de vimentina, aglutinam-se em volta do núcleo de fibroblasto. Os monômeros de vários tipos de filamentos intermediários diferem na seqüência de aminoácidos e possuem pesos moleculares muito diferentes. Porém, todos possuem um domínio central homólogo, na forma de um bastão, que apresenta uma estrutura estendida e enrolada quando a proteína forma dímero. Dois dímeros se associam entre si e formam um tetrâmero simétrico, organizando-se em grandes conjuntos sobrepostos formando filamentos intermediários não polimerizado. Os Domínios em forma de bastão das subunidades constituem a estrutura central do filamento intermediário, enquanto os domínios de ambas as extremidades projetam-se para fora. Uma função dos domínios terminais variáveis pode se permitir que cada tipo de filamento se associe com outros componentes específicos, a fim de posicioná-lo de forma adequada para cada tipo de célula. 3 - Estruturas Compostas por Microtúbulos Várias estruturas celulares derivam de grupos especiais de microtúbulos. Alguns deles, com ásteres do fuso, constituem organelas transitórias que surgem e desaparecem de acordo com os ciclos mitóticos e meióticos. Outros, tais como os cílios e flagelos, corpos (ou corpúsculos) basais e centríolo, são do tipo mais permanente. 3.1 - O aparelho mitótico O áster aparece como um grupo de microtúbulos radiais que convergem para o centríolo, em volta do qual se observa freqüentemente uma zona clara denominada de microcentro ou centrossoma. Pouco se sabe da função dos ásteres, além de que nas células animais geralmente o fuso se forma entre eles, durante a mitose. O fuso também é composto por microtúbulos, podendo ser convenientemente estudado através da microscopia eletrônica. (O fato de que tanto os microtúbulos das áster como os do fuso apresentam uma propriedade física denominada birrefringência positiva permite observá-los ao microscópio de polarização e de interferência, em células vivas). Tem-se reconhecido dois tipos de aparelho mitótico; um que depende da presença de ásteres (fuso astral) e outro que independem desses (fuso anastral). Nas células vegetais não se encontram nem centríolos nem ásteres. Nestas células, a formação dos microtúbulos não está relacionada com os pólos, mas sim com os cinetócoros, que são regiões especiais dos centrômeros dos cromossomas. Estudos imunocitoquímicos têm mostrado que os centríolos, o fuso e o cinetocoros contém tubulina, sugerindo que estas estruturas podem ser intermediárias na montagem da tubulina em microtúbulos. Inicialmente, as fibras do fuso foram dividas em 3 tipos: cromossômicas, que unem os cromossomas aos pólos; contínuas, que se estendem de pólo a pólo; interzonais, que são observadas na anáfase e na telófase entre os cromossomas filhos. Entretanto, a microscopia eletrônica demonstrou a existência de outros tipos: microtúbulos cinetocóricos, que como o nome indica, se originam nessa região dos cromossomas;

microtúbulos polares, que se originam nos pólos; e microtúbulos livres. Estudos quantitativos excluíram a existência das chamadas fibras contínuas, uma vez que geralmente os microtúbulos não se estendem de um pólo ao outro. Um dos aspectos que mais chamam a atenção no fuso é a facili dade com que se arma e desarma, tanto "in vivo" como em respostas a tratamentos diversos, pois normalmente a célula contém um gene "pool") de monômeros de tubulina que se encontram em uma espécie de equil íbrio dinâmico com microtúbulos polimerizados. Durante a mitose , também é produzido um ciclo de polimerização e despolimerização tanto nos microtúbulos citoplasmáticos quanto naqueles incorporados ao fuso. Acredita-se que a concentração intracelular de cálcio tenha um importante papel nesse ciclo. Recentemente estabeleceu-se uma relação entre a função desse íon e uma proteína de baixo peso molecular, fixadora de cálcio. 3.2 - Cílios e Flagelos Os cílios são estruturas com aspecto de pequenos pêlos com 0,25 micrômeros de diâmetros constituídos por um feixe de microtúbulos dispostos paralelamente e envoltos por uma membrana. Os cílios são curtos, múltiplos. Estendem-se a partir da superfície de muitos tipos de células e são encontrados na maioria das espécies animais, em muitos protozoários e em algumas plantas inferiores. Têm como função primária a de movimentar fluido sobre a superfície celular ou deslocar células isoladas através de um fluido. Os flagelos dos espermatozóides de muitos protozoários possuem grande semelhança com cílios, porém muito mais longos. Os flagelos são geralmente únicos e bem longos. No corpo humano estão situados apenas nos espematozóides. Na cauda do espermatozóides já foram verificadas mais de 200 tipos de proteínas. Os corpúsculos basais se reproduzem por mecanismo pouco conhecido. Estes processos em geral iniciam-se por aglomeração de substâncias eletrondensas, o material pericentriolar, que podem ocorrer perto de centríolos preexistentes ou, então, livre no citoplasma, independentemente de centríolos. O movimento de um cílio ou de um flagelo é produzido pela curvatura de seu núcleo chamado axonema. O axonema é composto por microtúbulos e suas proteínas associadas. Tanto cílios como flagelos são feixes de microtúbulos. Os microtúbulos estão dispostos nesta forma: nove microtúbulos duplos especiais formam um anel ao redor de um par de microtúbulos simples. Este arranjo é característico de quase todas as formas de cílios e flagelos eucarióticos desde protozoários até humanos. Enquanto o par central é formado por microtúbulos completos e separados; os pares externos ou periféricos são compostos por um microtúbulo completo e outro parcial, mantidos unidos . Descrevem-se ainda , nos cílios e flagelos , pequenos braços que partem de um dos microtúbulos dos pares periféricos ligando-se aos pares vizinhos: são formados por uma proteína , a nexina. Além desta estrutura , descrevem-se pequenas pontes que unem-se aos pares de microtúbulos periféricos. Sabe-se que os pequenos braços são constituídos por uma proteína de pequena atividade ATPásica chamada dineína. Uma série de

experiências realizadas com flagelos isolados de cauda de espermatozóide demonstraram que os pares de microtúbulos deslizam entre si durante a contração e que a força motriz para este movimento deriva da interação dos braços de dineína com microtúbulos vizinhos. A dineína é uma molécula grande (400.000 dáltons). Ela estabelece contato com a tubulina dos microtúbulos vizinhos e gera força da mesma intensidade que a miosina com a actina. Essas forças promovem movimentos de deslizamento entre pares de microtúbulos vizinhos, provocando o deslizamento de um par em relação ao outro. Este deslizamento é limitado por proteínas que prendem os pares de microtúbulos aos outros. O resultado da ação destas forças contidas leva a um dobramento dos cílios. Portanto é a dineína que dirige os movimentos dos cílios e flagelos em células não-musculares. Várias observações sugerem que o ATP fornece energia para o movimento cili ar e flagelar. Por exemplo, a queda do teor de ATP nos espermatozóides diminui sua motil idade: a adição de ATP a células ciliadas e flagelos isolados e previamente tratados por detergentes ( para remover a membrana e facil itar a entrada de ATP) promove rigorosos movimentos dos cílios e flagelos. Nas células do epitélio cil iado, as mitocôndrias dispõem-se principalmente no pólo apical, em situação ideal para fornecer aos cílios ATP por elas induzido. Fenômeno análogo ocorre nos espermatozóides dos mamíferos, cujos flagelo é envoltos por uma espiral de mitocôndrias na sua porção inicial. Os cílios e flagelos são estruturas complexas constituídas por mais de 30 proteínas diferentes, várias das quais são imprescindíveis para a sua movimentação. Recentemente , estudando os flagelos de pessoas estéreis, verificou-se que a doença de Manes-Kartagener descrita há várias décadas e caracterizada por esteril idade e infecção respiratória crônica é causada pela ausência de braços de dineína nos cílios e flagelos, o que impede a sua movimentação impossibil itando o movimento de espermatozóides e o batimento celular responsáveis pela eliminação contínua de poeiras que penetram na árvore respiratória. Recentemente, foram descritas outras doenças de sintomatologia semelhante, causadas pela ausência de outras proteínas dos cílios e flagelos que também participam de sua movimentação, criando assim um complexo de enfermidades: síndrome dos cílios imóveis. Os flagelos e cílios crescem a partir de corpúsculos basais ou cinetossomas semelhantes aos centríolos que possuem nove grupos de três microtúbulos fundidos em tripletes e formam a parede do mesmo. Durante a formação ou regeneração de um cílio cada par de microtúbulos do axonema se forma a partir de dois dos microtúbulos do triplete do corpúsculo basal. Freqüentemente os corpúsculos basais apresentam prolongamentos de estriações transversais que se dirigem para o centro do citoplasma, formando as chamadas raízes dos cílios, que teriam a função de sustentar e ancorar os cílios na célula. 3.3 - Centríolos e Corpúsculos Basais Os centríolos são estruturas cil índricas que medem 0,2 mm x 0,5 mm, podendo alcançar até 2 mm no eixo longitudinal. Apresentam as extremidades abertas, a menos que o

centríolo carregue um cílio. Nesse caso, as duas estruturas se encontram separadas por uma placa ciliar. A parede do centríolo possui 9 grupos de microtúbulos dispostos em círculo. Cada grupo é formado por um conjunto de 3 microtúbulos (em vez de 2 observados nos cílios), os quais estão inclinados em direção ao centro. Os túbulos são designados como subfibrilas A, B e C, a partir do centro para a periferia. Tanto a subfibrila A como a B atravessam a placa cil iar e se continuam como as subfibrilas correspondentes do axonema; a subfibrila C termina próximo da placa cili ar. Nos centríolos não existem microtúbulos centrais nem braços especiais. Os centríolos se apresentam nas células em par. Os centríolos-irmãos deste par dispõem-se perpendicularmente entre si e um par já duplicado de cada centríolo (consistindo em um centríolo recém-formado e o seu genitor, se acha em cada pólo do fuso da metáfase. Os dois membros de cada par vão se separar no período da intérfase subseqüente. O centríolo cresce a partir do extremo distal e, no caso dos cinetossomas (corpúsculos basais), os cílios se formam a partir desta extremidade. Os procentríolos, que se formam em ângulo reto em relação ao centríolo, estão localizados perto da extremidade proximal. Existem atualmente vários indícios de que os centríolos poderiam se originar por diferentes mecanismos. Os centríolos destinados a formar fusos mitóticos se originam diretamente da parede do centríolo preexistente. Os centríolos filhos aparecem primeiro sob a forma de estruturas anulares (procentríolos) que crescem até se converter em cilindros. Os grupos de 3 túbulos originam-se em grupos simples ou duplos que aparecem primeiro na base do procentríolo. Quando atingem metade do seu crescimento, os centríolos filhos são liberados no citoplasma, onde se completa a sua maturação. O outro mecanismo de formação do centríolo dá origem aos centríolos que são destinados a se transformar em corpúsculos basais, como no caso do epitélio cili ado. Essas estruturas se organizam progressivamente a partir de um material fibrogranular precursor localizado no citoplasma apical. Os cinetossomas recém formados se aliam em fileiras abaixo da membrana celular, e cada um deles pode então produzir satélites ao seu lado, uma raiz a partir da base e um cílio a partir do ápice. O desenvolvimento do cílio começa com o surgimento de uma vesícula que adere ao extremo distal do cinetossoma. Ao crescer, o corpo do cílio empurra para fora a parede da vesícula, a qual se constitui em uma bainha ciliar temporária até que se forme a permanente. 4 - Papéis Funcionais em Células Não-Musculares A base molecular da contração nos sistemas não-musculares se encontra ainda no terreno das hipóteses, porém o modelo que está se desenvolvendo é, de certa maneira, semelhante ao do músculo, uma vez que se baseia na interação dos fi lamentos de actina e miosina, como conseqüente produção de uma força de deslocamento. O sistema de contração em células não-musculares é muito lento quando comparado as células musculares. Provavelmente isto ocorre porque as proteínas que constituem o sistema não-muscular apresentam uma distribuição mais ao acaso e também por possuírem uma concentração muito menor de miosina.

Para que possa ocorrer vários dos movimentos das células não-musculares (extensão e troca na forma das células, locomoção, formação de pseudópodes) a superfície celular e o sistema contrátil devem estar intimamente acoplados. Esse acoplamento poderia ser estabelecido pelas moléculas de alfa actina que ligam a actina à membrana celular . Diferente dos músculos que possuem os filamentos de actina estáveis, a maioria das células não-musculares contém filamentos ou estruturas transitórias que podem aparecer e desaparecer em menos de um minuto. Portanto deve haver um mecanismo que controla a montagem e desmontagem dos monômeros de actina e a formação de fibras de tensão. Alguns exemplos dessas estruturas transitórias podem ser: o anel contrátil , um feixe de filamentos de actina e de filamentos de miosina II em forma de cinturão. Este anel aparece sob a membrana plasmática durante a fase M do ciclo celular; forças geradas por ele buscam a membrana para dentro e assim contraem a parte mediana da célula, levando a separação de duas células filhas, num processo conhecido como citocinese. Outro exemplo de estruturas contráteis temporárias dos filamentos de actina e miosina II são as fibras tensionais, componentes importantes do citoesqueleto de fibroblastos. Mas nem todas as estruturas contráteis de fi lamentos de actina e miosina encontradas nas células não-musculares são transitórias. Aquelas associadas com junções intercelulares chamadas cinturões de adesão, por exemplo, são freqüentemente mais duradouras. Os cinturões de adesão são encontrados próximos à superfície apical de células epiteliais. Entre suas funções parecem ter um papel importante no dobramento das camadas epiteliais durante a embriogênese. Proteínas fixadoras de actina: São proteínas que possuem interação com as proteínas de actina. Baseando-se em sua possível função, pode-se classificar essas proteínas em: (A) aquelas que tendem a produzir uniões transversais, favorecendo a formação de géis (geli ficação); (B) aquelas que produzem o efeito oposto (solação), ou seja a diminuição da viscosidade, fragmentando os fi lamentos de actina e formando um capuz "cap", e (C) aquelas que tendem a estabilizar a actina em sua formação monomérica (actina G) e que também levam à formação do estado sol. A calmodulina é uma proteína com afinidade para o íon Ca?² e quando forma um complexo com o mesmo (complexo calmodulina) ativa a cinase da cadeia leve da miosina, enzima que catalisa a fosforilação da miosina. Em conseqüência muda a conformação das cabeças de miosina, resultando no deslizamento dos microfilamentos de actina adjacentes, causando a contração nas células não-musculares. Existem basicamente 2 tipos de motilidade celular, nos quais ocorrem com a participação dos microfilamentos e as interações entre a actina e miosina. A ciclose é um tipo de movimento das células não-musculares. Esse tipo de movimento é caracterizado pela geração de correntes citoplasmáticas provocadas por ação de microfilamentos e que através dessas correntes contínuas deslocam grânulos citoplasmáticos. A corrente citoplasmática ou ciclose pode ser facilmente observada em células vegetais. Nestas células, em geral, o citoplasma está circunscrito a uma capa próxima à parede celulósica e as finas trabéculas que atravessam o grande vacúolo central e é possível observar-se correntes contínuas que deslocam os cloroplastos e outros grânulos citoplasmáticos. O experimento clássico sobre ciclose foi realizado em grandes células cilíndricas de

Niella, as quais possuem uma capa protoplasmática delgada rodeando o vacúolo central. Esta capa protoplasmática está subdividida em uma região cortical de citoplasma estruturado, no qual estão incluídos cloroplastos imóveis, e uma capa isolada de citoplasma (endoplasma) onde a ciclose ocorre. O conjunto está rodeado pela membrana celular, abaixo da qual se observa uma faixa única de microtúbulos. Na região cortical, abaixo dos cloroplastos, existem feixes de filamentos de actina cuja direção é paralela à direção da ciclose. Acredita-se que a força impulsora inserida dada pelo mecanismo de interação de actina - miosina que ocorreria na região de limite entre o citoplasma estático e o endoplasma móvel. O movimento amebóide é um outro tipo de sistema de contração das células não-musculares. Este tipo de movimento está presente nos protozoários (amebas) e em células animais (leucócitos, fibroblastos, macrófagos). Ele se realiza através da extensão, pela célula, de prolongamento da camada cortical do citoplasma muito rica em actina. Esse prolongamento é denominado pseudópodos, e, ao se fixar num substrato, parece puxar o resto da célula em sua direção. Embora esta forma de locomoção possa ser melhor observada na amebas, também existem muitos outros tipos celulares, como, por exemplo, nos leucócitos. Em um cultivo formado por uma monocamada de células observa-se, às vezes, um tipo de movimento amebóide ruffling (folho, pregueado). Este movimento é feito através da emissão de prolongamentos da superfície, que estabelece contato intermitente com a superfície no vidro. Por meio da aderência e separação alternadas deste prolongamento pregueado, a célula consegue deslocar-se sobre uma superfície de suporte e movimentar-se ativamente até a zona de migração do cultivo. Acredita-se geralmente que os microfilamentos participem no movimento amebóide. Têm sido observados, em amebas, filamentos de actina e de miosina mais espessos, e parece não haver dúvidas de que a interação actina-miosina proporciona a força motora. Entretanto, as opiniões estão divididas no que se refere ao local mais provável da contração. Enquanto que alguns pesquisadores consideram a região posterior da ameba que avança como a mais ativa, outros dão maior importância maior à extremidade anterior. As microvilosidades são projeções digitiformes encontradas na superfície de muitas células animais. São particularmente abundantes em algumas células epiteliais que necessitam de uma grande área de superfície para desempenhar eficientemente suas funções. O estudo destas microvilosidades por microscopia eletrônica revelou a presença de actina, miosina e alfa-actina, proteínas relacionadas com a contração, mas que nas microvilosidades possuem a função de sustentação da forma alongada das microvilosidades. Estão presentes 40 microfilamentos de actina no interior das microvilosidades. Estes microfilamentos se prendem em espessamentos que ocorrem nas extremidades dos microvilos e que se ligam entre si e com a membrana plasmática por meio de várias proteínas. As células ciliares presentes no organismo humano (na árvore respiratória e trompa uterina) encontram-se associadas a células que secretam muco e têm como conseqüência o transporte unidirecional de uma camada delgada de muco que reveste a superfície

interna destas estruturas. Desta maneira a poeira que atinge a árvore respiratória é captada pelo muco e transportada para a cavidade bucal, enquanto que na trompa ocorre um fluxo de muco para o útero, o que facil ita o transporte dos óvulos. Já nos espermatozóides seu movimento flagelar ocorre por um abalo tipo vaivém devido a uma contração na sua base que se transmite ao longo de sua extensão em uma região rica em mitocôndrias, perto do núcleo do espermatozóide. A atividade do flagelo movimenta o espermatozóide para frente. Como já foi estudado na parte 3.2 Cílios e Flagelos, estes tem grande participação nos processos de movimentação de células não-musculares. Existe também microtúbulos não só nos cílios e flagelos mas também no citoplasma. Os microtúbulos citoplasmáticos se encontram em constante reorganização, crescendo em ema parte graças a polimerização local dos dímeros de tubulina (parte "mais") e diminuindo em outra graças a despolimerização da tubulina (parte "menos"). As principais funções desses microtúbulos são: processos de mudança de forma celular, transporte intra-citoplasmático de partículas e para os movimentos cromossômicos na mitose. Os microtúbulos também participam do transporte intracelular de partículas em células não-musculares, como no caso do fluxo que ocorre nos axônios e no transporte e exocitose dos grânulos de secreção nas células glandulares. Nestas situações, os microtúbulos não se associam como ocorre nos cílios e flagelos, não apresentam os braços de dineína e esta proteína é substituída por outra chamada cinesina (Fig 3.2.G), que também apresenta atividade ATPásica. Na realidade, o funcionamento dos microtúbulos é extremamente complexo, já que apresentam inúmeras proteínas que se associam a eles. 5 - Papéis Funcionais em Células Musculares O tecido muscular é responsável pelos movimentos corporais. É constituído por células alongadas caracterizado pela presença de grande quantidade de filamentos citoplasmáticos responsáveis pela contração. De acordo com sua classificação morfológica é dividido em músculo liso, músculo estriado esquelético e estriado cardíaco. As células musculares são tão diferenciadas que seus componentes receberam nomes especiais. A membrana é chamada de sarcolema, o citoplasma (preenchido principalmente de fibrilas paralelas - as miofibrilas) recebe o nome de sarcoplasma, o retículo endoplasmático é chamado de retículo sarcoplasmático, e as mitocôndrias recebem o nome de sarcossomas. A célula muscular estriada apresenta um alto grau de especialização para realizar movimentos. Na vida embrionária, as células musculares precursoras se fundem formando as fibras musculares estriadas esqueléticas, que agrupam-se em feixes cujas extremidades são presas a tendões inseridas nos ossos. Estas fibras contêm um feixe de delgadas estruturas cilíndricas denominadas miofibri las. As miofibrilas são cil índricas, apresentam um diâmetro de 1 a 2 mm e correm longitudinalmente à fibra muscular,

preenchendo quase que completamente seu interior. Cada miofibrila apresenta alternadamente bandas I e bandas A. As miofibrilas são formadas por unidades repetidas, denominadas sarcômeros. Cada unidade desta (sarcômero) é limitada por duas estrias finas e eletrondensas denominadas estrias Z. O sarcômero é constituído por filamentos de dois tipos. Em um dos tipos os filamentos são finos, dirigem-se medialmente, porém não atingem o centro do sarcômero e são constituídas por actina. No outro tipo os filamentos são grossos, encontram-se no centro do sarcômero e são constituídos por proteínas fibrilares chamadas miosina . As miofibrilas do tecido muscular estriado contém basicamente 4 tipos básicos de proteínas: miosina, actina, troponina e tropomiosina. MIOSINA: É uma molécula grande tendo forma de bastão sendo formada por 2 peptídeos enrolados em hélice. Em uma de suas extremidades a miosina apresenta uma saliência globular ou cabeça, que possui locais específicos para combinação com o ATP e é dotada de atividade ATPásica. Quando submetida a ligeira proteólise, a molécula de miosina pode ser dividida em 2 fragmentos: meromiosina leve e meromiosina pesada. O fragmento leve corresponde à maior parte da porção do bastão da molécula, enquanto que a pesada contém a saliência globular (cabeça). ACTINA: Apresenta-se sob forma de polímeros longos (actina F) formados por duas cadeias de monômeros globulares (actina G) torcidas uma sobre a outra. Uma característica importante das moléculas de actina G é a sua assimetria estrutural. Quando elas se polimerizam para formar a actina F, frente a um monômero combina-se com a parte posterior do outro produzindo um fi lamento polarizado. TROPONINA: É um complexo por 3 subunidades: TnT, que se liga fortemente a tropomiosina; TnC, que têm grande afinidade para os íons cálcio; e TnI, que cobre o sítio ativo da actina onde ocorre a interação entre a actina e a miosina. Cada molécula de tropomiosina tem um local específico onde se prende um complexo (três subunidades) de tropomiosina. TROPOMIOSINA: É uma molécula longa e fina , com cerca de 40 nm de comprimento, contendo duas cadeias polipeptídicas, uma enrolada na outra. As moléculas de tropomiosina unem-se umas às outras pelas extremidades, para formar filamentos longos que se localizam ao longo do sulco existente entre os dois filamentos de actina F. O sarcômero em repouso consiste em fi lamentos finos(actina) e grossos(miosina) que se sobrepõem parcialmente. Durante o ciclo de contração os dois tipos de filamentos conservam seus comprimentos originais. Se a contração não é devido ao encurtamento dos fi lamentos, deve ser conseqüência de um aumento na zona de sobreposição entre os filamentos. A principal função desempenhada pelas estruturas do citoesqueleto nas células musculares é a movimentação celular, que resulta na contração muscular. A contração muscular ocorre pelo deslizamento dos fi lamentos de actina sobre os filamentos de miosina para o interior do sarcômero, isso provoca a aproximação das estrias Z. Comparando-se o músculo contraído com o distendido, nota-se que no contraído os filamentos finos de actina ficam menos visíveis graças ao seu deslizamento por entre os filamentos grossos de miosina.

A contração se inicia na faixa A, onde os filamentos finos e grossos se sobrepõem. Durante o ciclo de contração a actina e miosina interagem da seguinte maneira: Durante o repouso, ATP liga-se à ATPase das cabeças da miosina. Para atacar a molécula de ATP e liberar energia, a miosina necessita da actina que atua como um cofator. No músculo em repouso a miosina não pode associar-se a actina devido à repressão do local de ligação do complexo troponina-tropomiosina fixado sobre o filamento de actina F. Todavia, quando há disponibilidade dos íons cálcio estes combinam-se com a unidade TnC da troponina. Isto muda a configuração espacial das três subunidades de troponina e empurra a molécula de tropomiosina mais para dentro do sulco da hélice de actina. Em conseqüência , ficam expostos os locais de ligação dos componentes globulares da actina e esta fica livre para interagir com a cabeça da molécula de miosina. A combinação dos íons cálcio com a TnC corresponde à fase em que o complexo miosina-ATP é ativado. Como resultado da ponte entre a cabeça da miosina e a subunidade de actina G do filamento fina, o ATP é convertido em ADP+P e energia. Esta atividade leva a uma deformação da cabeça e de parte do segmento em bastão da miosina, aumentando a curvatura da cabeça. Como a actina está combinada com a miosina , o movimento da cabeça da miosina empurra o filamento da actina, promovendo seu deslizamento sobre o fino fi lamento de miosina. Embora os filamentos grossos possuam um elevado número de cabeças de miosina, em cada momento da contração apenas um pequeno número de cabeça alinha-se com os locais de combinação da actina. À medida em que as cabeças de miosina movimentam a actina, novos locais para formação das pontes actina- miosina aparecem. Não existindo o ATP, o complexo actina-miosina torna-se estável; isto explica a rigidez muscular muito intensa que ocorre após a morte (rigor motriz).