Embed Size (px)
Citation preview

Di r ecci ó n:Di r ecci ó n: Biblioteca Central Dr. Luis F. Leloir, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires. Intendente Güiraldes 2160 - C1428EGA - Tel. (++54 +11) 4789-9293
Co nta cto :Co nta cto : [email protected]
Tesis Doctoral
Interacción del virus Junín con elInteracción del virus Junín con elcitoesqueleto y la membrana de lacitoesqueleto y la membrana de la
célula huéspedcélula huésped
Cordo, Sandra M.
2004
Tesis presentada para obtener el grado de Doctor en CienciasQuímicas de la Universidad de Buenos Aires
Este documento forma parte de la colección de tesis doctorales y de maestría de la BibliotecaCentral Dr. Luis Federico Leloir, disponible en digital.bl.fcen.uba.ar. Su utilización debe seracompañada por la cita bibliográfica con reconocimiento de la fuente.
This document is part of the Master's and Doctoral Theses Collection of the Central LibraryDr. Luis Federico Leloir, available in digital.bl.fcen.uba.ar. It should be used accompanied bythe corresponding citation acknowledging the source.
Cita tipo APA:
Cordo, Sandra M.. (2004). Interacción del virus Junín con el citoesqueleto y la membrana de lacélula huésped. Facultad de Ciencias Exactas y Naturales. Universidad de Buenos Aires.http://hdl.handle.net/20.500.12110/tesis_n3725_Cordo
Cita tipo Chicago:
Cordo, Sandra M.. "Interacción del virus Junín con el citoesqueleto y la membrana de la célulahuésped". Tesis de Doctor. Facultad de Ciencias Exactas y Naturales. Universidad de BuenosAires. 2004. http://hdl.handle.net/20.500.12110/tesis_n3725_Cordo

FACULTAD DE CIENCIAS EXACTAS Y NATURALES
UNIVERSIDAD DE BUENOS AIRES
“INTERACCIÓNDEL VIRUSjUNÍN CON
EL CITOESQUELETO Y
LA MEMBRANA DE LA CÉLULAHUÉSPE ”
“ Junin Virus interaction with the cytoskeletonand the cellular membrane”
AUTOR:Lic. SANDRA M. CORDO
' DIRECTOR:Dra. NELIDA A. CANDURRA
LABORATORIO DE VIROLOGÍADEPARTAMENTO DE QUÍMICA BIOLÓGICA
FACULTAD DE CIENCIAS EXACTAS Y NATURALESUNIVERSIDAD DE BUENOS AIRES
TESIS PARAOPTAR POR EL TÍTULO DEDOCTOR DE LA UNIVERSIDAD DE BUENOS AIRES
ZMM

Resumen

EI virus Junín (JUNV), agente etiológico de la fiebre hemorrágica argentina, es
un miembro de la familia Arenavirídae. Es un virus envuelto de genoma ARN
segmentado cuyo ciclo de replicación ha sido caracterizado en varios aspectos. La
entrada a las células blanco se realiza vía endocitosis mediada por receptor y posterior
fusión dependiente de PH. Una vez que la nucleocápside se encuentra dentro del
citoplasma se expresan 5 proteínas estructurales, mediante una estrategia de
codificación ambisense. La proteína mayoritaria asociada a la nucleocápside es
denominada NP. A partir de un único precursor glicoproteico (GPC), sintetizado en el
retículo y exportado a la membrana plasmática de la célula, se obtienen dos
glicoproteinas virales denominadas GP1 y GP2. Ambas conforman las estructurasclavifon'nes encontradas en la envoltura viral. Se desconocen hasta el momento los
mecanismos detallados y las estructuras celulares involucradas en el transporte de las
distintas proteínas virales hacia el sitio de brotación. El ensamblado de las partículas
virales se lleva a cabo en Ia membrana plasmática de las células infectadas.
En este trabajo se estudiaron algunos aspectos de la interacción virus-célula
huésped, caracterizando la asociación de NP, GPC y GP1 a distintas estructuras
celulares. En primer lugar se estudio la interacción del virus con la red de filamentos
intermedios (FI) del citoesqueleto. Se utilizó el compuesto acrilamida capaz de
desorganizar especificamente los Fl. La producción de virus infeccioso disminuyó de
manera dosis-dependiente en presencia del mismo. Los resultados, observados en
celulas Vero, cultivos de fibroblastos humanos y astrocitos murinos, fueron
comparables. Se analizó luego el efecto del compuesto sobre diferentes pasos del
ciclo de multiplicación. De esta manera se determinó que la integridad de los FI es
crucial al menos durante los pasos previos a la síntesis de proteínas virales y
posteriores a la intemalización. Mediante extracciones específicas realizadas con
detergentes no iónicos se encontró que NP se asocia a la fracción celular
correspondiente al citoesqueleto, aunque no se determinó a que componente
correspondería esta interacción. Las glicoproteinas no mostraron asociación especifica
a estas fracciones celulares en iguales condiciones.
Por otro lado, modificaciones en el contenido de colesterol de las células
infectadas y tratadas con inhibidores de la enzima HMG-CoAreductasa, disminuyeron
la infectividad y localización de las glicoproteinas en la membrana de las mismas.
Estos resultados llevaron al análisis de la asociación de GPC y GP1 a estructuras de
membrana ricas en colesterol denominadas rafts. Los ensayos de fraccionamiento
celular específicos para estos microdominios de membrana mostraron la localización
de las glicoproteinas en rafts. Esta interacción fue dependiente de la temperatura y de
la presencia de colesterol en los cultivos infectados. Además, la asociación de las

glicoproteínas a las fracciones rafts fue estabilizada a 37°C por Ia adición de
anticuerpos específicos. Estos resultados sugieren la utilización de esta vía como
mecanismo de transporte para GPC y GP1 a la membrana celular de las células
infectadas. Asimismo la localización de las glicoproteínas en los microdominios rafts
podría representar una estrategia para el reclutamiento de los componentes virales
previo al paso de liberación de Ia progenie en las células infectadas.
Palabras claves: Virus Junín, Filamentos Intermedios, Fraccionamiento celular,
Microdominios de membrana.

Abstract

Junin virus (JUNV), the etiological agent of argentine hemorrhagic fever, is a
member of the Arenavin'dae family. The viral cycle multiplication of this RNA virus has
been charactenzed in different aspects. The entrance in the target cells is performed by
receptor-mediated endocytosis following pH dependant fusion step. Once the
nucleocapsids are within the cytoplasm the ambisense strategy allows expression of
five structural proteins. The major nucleocapsid associated protein is named NP. As a
result of the glycoprotein precursor synthesis and after its maturation and exocytic
transport to plasma membrane of infected cells, two glycoproteins are obtained. Both
proteins. named GP1 and GP2, form the claviform structures found in the viral
envelope. At this moment, the detailed mechanisms and cellular structures involved in
the transport of different viralproteins towards the budding site are not known although
the fact that viral assembly is carried out in plasma membrane of infected cells is
currently accepted.
ln this work we have studied some aspects of virus-cell interaction
characten'zing the association of NP, GPC y GP1 to cellular structures. First. we have
studied the interaction between virus and cytoskeletal intermediate filaments (IF). Inthe
presence of acrylamide, compound that selectiver disrupt IF, viral yield was
diminished. The results were similar in Vero cells, human fibroblast cultures and murine
astrocytes. Then, we analyzed the effect of acrylamide on different steps of the
multiplicationcycle. lt was determined that IF integn'tyis essential at least between the
step of internalization and viral protein synthesis. Detergent extractions of infected
cultures demonstrated that NP associates to cellular cytoskeletal fraction although it
was not possible to identify the specific cytoskeletal framework involved in that
association. Glycoproteins did not show specific association to these cytoskeletal
fractions under similarconditions assayed.
On the other hand, modifications in the cholesterol contents of infected cells
induced by HMG-CoA reductase inhibitors. diminished infectivity and glycoproteins
membrane Iocalization. This result leded us to analyse the association of GPC and
GP1 to membrane cholesterol enriched structures named rafts. Specific cellular
fractionation assays showed glycoproteins associated to raft microdomains. This
interaction was temperature and cholesterol dependant. Moreover, glycoprotein
association to membrane rafts was stabilized at 37°C by specific antibody addition.
These results suggest that GPC and GP1 may utilizethe exocytic raft pathway to reach
the plasma membrane of infected cells. Finally, glycoproteins raft association may
represent the strategy to recruit viral proteins before releasing progeny from the cells.

FÜEy 5a?ÉÉÉIMII
FACULTAD DE CIENCIAS EXACTASY NATURALES
UNIVERSIDAD DE BUENOS AIRES
“INTERACCIÓNDEL VIRUSjUNÍN CON
EL CITOESQUELETO Y
LA MEMBRANA DE LA CÉLULAHUÉSPED”
AUTOR:Lic. SANDRA M. CORDO
DIRECTOR:Dra. NELIDA A. CANDURRA
LABORATORIO DE VIROLOGÍADEPARTAMENTO DE QUÍMICA BIOLOGICA
FACULTAD DE CIENCIAS EXACTASY NATURALESUNIVERSIDAD DE BUENOS AIRES
TESIS PARAOPTAR POR EL TÍTULO DEDOCTOR DE LA UNIVERSIDAD DE BUENOS AIRES
2004
:i l.
v, r4 i

“La ciencia tiene la fascinación de la aventura porque, por encima de todo,es una exploración a lo desconocido... el motor de la ciencia es la curiosidaddel hombre”
César Nfilstein (diciembre de 1999)

AGRADECIMIENTOS
Mientras concluye esta etapa tan importante de mi vida personal y profesional en mimemoria desfilan decenas de momentos vividos. Cada uno de ellos se encuentra plagado degente conocida, gente compañera, amiga y consejera. No importa si el momento fue triste,alentador, decepcionante o glorioso nunca estuve sola en este largo camino recorrido y hoyluego de tantos meses de trabajo quiero compartir con cada uno de ustedes la felicidad dehaber llegado a la meta tan largamente añorada.
Quisiera agradecer en primer lugar a mi directora la Dra. Nélida Candurra. Por estos añoscompartidos codo a codo, por sus enseñanzas diarias, su compañia y cordial en estearduo camino científico. Por sus palabras tranquilizadoras frente a mi desesperación, porsus consejos cuando todo parecía duda y por su mirada atenta de todos los días. Pero sobretodo quiero agradecerle 1acandidez de su apoyo y su trato personal.
A la Dra. Elsa Damonte porque sin su apoyo en el momento indicado no habría tenido laoportunidad de realizar este trabajo. Hoy quiero agradecer formalmente su confianza. Porhaberme elegido para formar parte de su grupo de investigación y por apoyarme de muydiversas formas, profesionales y personales, hasta el final.
A1Dr. Luis Scolaro por ser un compañero ideal, por su buen humor y su predisposición aprueba de todo. Por ser un modelo tanto científico como humano. Por todas lasmaravillosas conversaciones compartidas y por el aguante durante los momentos de mayornerviosismo... y por un abrazo tan importante en mi vida.
A la Dra. Viviana Castilla por su dulzura constante, porque en el transcurso de estos añoscreció una amistad inesperada y maravillosa. Por su cariño, su disposición y su temple. Pormostrarme que también es posible conservar los valores y convicciones humanos en estaprofesión.
Quiero agradecer especialmente a mi compañera de cucha Cybele García. Sin saberlocomenzamos juntas este camino virológico y sin planearlo tampoco estamos juntasnuevamente al final de esta historia. Con el correr de los años desarrollamos esta granhermandad y, pensando distinto a veces y coincidiendo otras tantas, construimos estaamistad en la que siempre aprendemos algo. Por sus consejos cientificos y por el apoyoenorme en estos últimos meses de histeria, llanto y diversión. Por el futuro de ambas paraque cada una poco a poco logre alcanzar las nuevas metas.
A Paula Ellenberg quiero agradecer la gran cantidad de momentos compartidos, porque enpoco tiempo me brindó su amistad, me contagió su alegría y su espíritu renovador. Por seruna compañera y amiga solidaria, por ayudarme a ver el lado positivo de las cosas y porhaber soportado estos meses de locuras y mal humor.
Muchas gracias a la Dra. Andrea Barquero con quien compartimos una amistad muyespecial. Muchos momentos quedan en mi recuerdo de las aventuras vividas y todos ellostienen el valor de lo compartido desde el corazón. Gracias por ayudarme a ejercitar loinesperado y por soportar las vetas más desdeñables de mi carácter. Mi deseo es quenuestros caminos sigan unidos más allá de las grandes y maravillosas vueltas de la vida,porque lo realmente importante siempre perdura.

A los amigos “de al lado”, el Dr. Carlos Pujol compañero de docencia durante tantoscuatrimestres, por las charlas entretenidas y la buena onda siempre. A la Dra. josefinaCarlucci por su sincera amistad, hoy muy lejos de aquí pero siempre en los recuerdos masalegres y divertidos. A Erina Petrera por su desinteresada amistad, por los mates y por susmuchas asesorías en el extraño mundo del diseño y la moda. A la Dra. Mónica W’achsmanpor preguntar, por interesarse, por brindar siempre una sonrisa y por ser compinche dealgunas aventuras. A Laura Talarico por su estilo Cándido y amable. A Isabel Paz por estarsiempre atenta y dispuesta ayudarme con las pequeñas cosas.
A la Dra. Celia Coto por sus palabras de aliento, por las anécdotas de y por lascharlas siempre interesantes dispuestas a cubrir mis momentos constantes de curiosidad.
A todo el gran mundo que representa el laboratorio de Virología, porque el constantedesfilar de sus caras y vidas le dieron a esta historia todas las tonalidades necesarias paraconsiderarla una historia real.
A los compañeros del departamento de QB que involuntariamente prestaron su oído atanta locura pre-Tesis. Gracias julia, gracias Nancy por soportar con una sonrisa tantasinterrupciones.
A Jimena Ruiz, por sus ganas, por las charlas circulares en los momentos de terapiacompartida. Sobre todo por demostrarrne que nunca es tarde para iniciar amistades.
Debo unas palabras de agradecimiento a la Ligia por ser un lugar donde compartirsentimientos acallados de tanto ejercitar el pensamiento cientifico. Por las risas yacompartidas _\'por los momentos venideros en tan buena compañia.
Gracias a Alejandra Donato por brindarme su amistad y cariño, por enseñarme tantas cosasy por ser mi modelo docente. A Eduardo de Navarrete por ponerle tanto humor a las horasy anécdotas compartidas. Gracias especiales por las caricaturas de mi vida y las que estan enesta Tesis.
Mis amigos, mis fieles e irremplazables amigos, no tengo palabras para agradecer laspruebas de amistad ofrecidas durante estos años. Mariano, Vane formar parte de sus vidases un regalo maravilloso, gracias por aguantarme y ser siempre ese respaldo al que puedoacudir en mis momentos de flaqueza. Gracias por confiarme los dos frutos de su amor:Mariel _\'Malena. Mil gracias a los que hoy están lejos: a Facu por sus palabras cariñosas ypor su entrega siempre. A Martin y Mariana por todos estos años de amistad compartida,por su apoyo incondicional también en esta última etapa. Gonzalo, Valeria gracias poralentarme constantemente, por darme fuerzas y confianza en mi misma. A Guillo por sumano amiga y honesta siempre dispuesta a ayudar.
Finalmente quiero agradecer a mi familia, a mis padres y hermanos, porque son ellos con suamor incondicional los que me dan fuerzas para alcanzar mis objetivos y plantearrnenuevos. Porque siempre estuvieron y estaran a mi lado diciéndome: vos podes, adelante.
Cuando tenía 18 años y me inscribí en la facultad nunca pensé que iba a terminarrecibiéndome, al recibirme la idea de vivir de mi profesión me parecía un imposible. Hoyque estoy alcanzando el grado académico mas esperado por un profesional me parece tanincierto mi futuro... sin embargo aprendí que la vida es siempre ese paso adelante, tandifícil de dar, pero tan necesario para seguir caminado.

Resultados de esta tesis fueron publicados en el siguiente trabajo:
"Intermediate Filament lntegrity ¡s Required for Junin Virus Replication"
Cordo, S. M. and Candurra, N. A. (2003) Virus Research, 97, 47-55.

ABREVIATURAS:
ADN
AM
ARN
ARNmCsClFHA
FI
GFAP
GP1 y GP2GPCGPI
glmlHA
hs.IFl
JUNV
kbkDa
L/M
LCMV
MCD
MEM
MF
MOI
MT
uCÍ
uMNaCI
NP
nm
p.i.PBSPH
SDSTX-1OO
UFPVSV
WB
X 9
ácido desoxiribonulceicoastrocitos murinosácido ribonulceico
ácido ribonulceico mensajerocloruro de cesio
Fiebre Hemorrágica ArgentinaFilamentos Intermedios
proteina gliofibrilarácida
glicoproteina de envoltura 1 y 2precursor glicoproteicoglicosil fosfatidil inositol
gramos/mililitro
hemaglutininahorasinmunofluorescencia indirectavirus Junínkilobaseskilo Dalton/s
lovastatina/mevalonato
virus de la coriomeningitis IinfocitarimetiI-ciclodextrina
medio esencial mínimomicrofilamentos
multiplicidad de infecciónmicrotúbulosmicro Courie/smicro litro/s
micro Molar
cloruro de sodio
nucleoproteínanano metros
post infecciónbuffer fosfato salinofibroblastos humanossodio dodecil sulfatoTritón X -100
unidades formadoras de placasvirus de la estomatits vesiclularwestern blot
multiplicado por fuerza de gravedad

INDICE PAG
RESUMEN 2
ABSTRACT 5
INTRODUCCIÓN
I. Los Arenavirus 7
II. Fiebre Hemorra'gica Argentina 11
lll. Virus Junin 12
III.1.Vacunas 14
lll.2. Morfología del virión 15
III.3. Estructura Molecular 15
Ill.4. Proteínas virales 16
II|.5. Ciclo de replicación 18
IV. Interacción virus-célula huésped 22
V. Citoesqueleto 25
V.1. Microtúbulos 26
V.2. Microfilamentos 26
V.3. Filamentos Intermedios 27
VI. Microdominios de membrana rafts 31
OBJETIVOS 36
MATERIALES Y MÉTODOS
1. Virus 38
2. Cultivos celulares
2.1. Línea celular Vero 38
2.2. Linea diploide de fibroblastos humanos 39
2.3. Cultivo priman'o de astrocitos murinos 39
2.4. Linea celular MDCK 39
3. Compuestos utilizados 39
4. Obtención de virus purificado y marcado radiactivamente 40
5. Ensayo de viabilidad celular: Método de M'IT 40
6. Curvas de crecimiento 41
7. Síntesis de proteinas 41
8. Titulación de la infectividad viral por el método de UFP
8.1. Virus Junín 42
8.2. Virus Influenza 42
9. Ensayo de inhibición del rendimiento viral 43
10. Ensayo de acción virucida 43

11. Acción del compuesto acrilamida a distintos tiempos
11.1. Adición post-infección 43
11.2. Adición por pre-tratamiento 44
12. Acción de Ia acrlilamida sobre Ia adsorción e internalización del virus 44
13. Estudio de Ia expresión de proteínas
13.1. Inmunofluorescenci indirecta 44
13.2. Marcado metabólico, inmunoprecipitación y SDS-PAGE 46
13.3. Western blot 47
14. Reducción del colesterol y su efecto sobre la multiplicación viral
14.1. Acción de hidroxiesteroles inhibidores dela HMGCoA reductasa. 47
14.2. Lovastatina y mevalonato + metil-B-ciclodextrina 47
15. Cuantificación de colesterol en extractos celulares 48
16. Fraccionamiento en TX-100
16.1. Buffer PIPES/TX-100 48
16.2. Buffer TNE/TX-100 48
17. Estudio de microdominios de membrana rafts y su flotabilidad en gradientesde sacarosa
17.1. Virus Junin 49
17.2. Virus Influenza 50
18. Estudio dela estabilidad de microdominios rafts
18.1. En presencia de metiI-B-ciclodextrina 50
18.2. En presencia de anticuerpos especificos 50
19. Análisis de datos 51
RESULTADOS
Capitqu I
A. Estudio de la interacción de JUNV con los Filamentos Intermedios
1. Caracterización de los sistemas celulares utilizados
1.1. Infección y productividad viral 55
1.2. Determinación de viabilidad celular en presencia de acrilamida 57
1.3. Efecto de Ia acrilamida sobre el citoesqueleto 59
2. Caracterización de la acción de la acrilamida sobre cultivos infectados
2.1. Acción de Ia acrilamida sobre las partículas virales infecciosas 61
2.2. Efecto de Ia acrilamida sobre la producción de particulas infecciosas 62
2.3. Acción de la acrilamida sobre distintas etapas del ciclo de multiplicación 63
2.4. Expresión de los antígenos virales en presencia de la acrilamida 69
2.5. Sintesis de proteína en presencia de acrilamida 71
2.6. Interacción filamentos de vimentina-proteinas virales 72

B. Fraccionamiento celular y proteinas virales
1. Fraccionamiento con dtergentes no-iónicos 75
1.1. Localización de las proteínas virales 75
Capitqu I/
A. Estudio del colesterol y su relación con el ciclo de multiplicación viral
1. Colesterol e hidroxiesteroles
1.1. Acción sobre la viabilidad celular 81
1.2. Acción sobre Ia multiplicación viral 82
1.3. Efecto sobre Ia expresión de proteínas virales 83
2. Reducción en la síntesis de colesterol por acción de la lovastatina
2.1. Acción sobre la viabilidad celular 85
2.2. Acción sobre la producción viral 87
3. Extracción directa de colesterol
3.1. Metilciclodextrina y su acción sobre las particulas infectivas 88
3.2. Metilciclodextrina y expresión de proteinas virales
2.3. Expresión de proteínas virales 88
B. Estudio dela asociación de JUNV a microdominios de membrana rafls
1. Fraccionamiento de membranas en TX-100 90
2. Flotabilidaden gradientes de sacarosa 91
DISCUSIÓN y CONCLUSIONES 98
BIBLIOGRAFÍA 107

Resumen

El virus Junín (JUNV), agente etiológico de la fiebre hemorrágica argentina, es
un miembro de la familia Arenavin'dae. Es un virus envuelto de genoma ARN
segmentado cuyo ciclo de replicación ha sido caracterizado en varios aspectos. La
entrada a las células blanco se realiza vía endocitosis mediada por receptor y posterior
fusión dependiente de pH. Una vez que la nucleocápside se encuentra dentro del
citoplasma se expresan 5 proteinas estructurales, mediante una estrategia de
codificación ambisense. La proteína mayoritaria asociada a la nucleocápside es
denominada NP. A partir de un único precursor glicoproteico (GPC), sintetizado en el
reticulo y exportado a la membrana plasmática de la célula, se obtienen dos
glicoproteinas virales denominadas GP1 y GP2. Ambas conforman las estructurasclaviformes encontradas en la envoltura viral. Se desconocen hasta el momento los
mecanismos detallados y las estructuras celulares involucradas en el transporte de las
distintas proteínas virales hacia el sitio de brotación. El ensamblado de las particulas
virales se lleva a cabo en la membrana plasmática de las células infectadas.
En este trabajo se estudiaron algunos aspectos de la interacción virus-célula
huésped, caracterizando la asociación de NP, GPC y GP1 a distintas estructuras
celulares. En primer lugar se estudio la interacción del virus con la red de filamentos
intermedios (FI) del citoesqueleto. Se utilizó el compuesto acrilamida capaz de
desorganizar especificamente los FI. La producción de virus infeccioso disminuyó de
manera dosis-dependiente en presencia del mismo. Los resultados, observados en
células Vero, cultivos de fibroblastos humanos y astrocitos murinos. fueron
comparables. Se analizó luego el efecto del compuesto sobre diferentes pasos del
ciclo de multiplicación. De esta manera se determinó que la integridad de los FI es
crucial al menos durante los pasos previos a la sintesis de proteínas virales y
posteriores a la internalización. Mediante extracciones especificas realizadas con
detergentes no iónicos se encontró que NP se asocia a la fracción celular
correspondiente al citoesqueleto, aunque no se determinó a que componente
correspondería esta interacción. Las glicoproteinas no mostraron asociación específica
a estas fracciones celulares en iguales condiciones.
Por otro lado, modificaciones en el contenido de colesterol de las células
infectadas y tratadas con inhibidores de la enzima HMG-CoAreductasa, disminuyeron
la infectividad y localización de las glicoproteinas en la membrana de las mismas.
Estos resultados llevaron al análisis de la asociación de GPC y GP1 a estructuras de
membrana ricas en colesterol denominadas rafts. Los ensayos de fraccionamiento
celular especificos para estos microdominios de membrana mostraron la localización
de las glicoproteinas en rafts. Esta interacción fue dependiente de la temperatura y de
la presencia de colesterol en los cultivos infectados. Además, la asociación de las

glicoproteínas a las fracciones rafts fue estabilizada a 37°C por la adición de
anticuerpos específicos. Estos resultados sugieren Ia utilización de esta vía como
mecanismo de transporte para GPC y GP1 a la membrana celular de las células
infectadas. Asimismo la localización de las glicoproteínas en los microdominios rafis
podria representar una estrategia para el reclutamiento de los componentes virales
previo al paso de liberación de la progenie en las células infectadas.

Abstract

Junín virus (JUNV), the etiological agent of argentine hemorrhagic fever, is a
member of the Arenaviridae family. The viral cycle multiplication of this RNA virus has
been characterized in different aspects. The entrance in the target cells is performed by
receptor-mediated endocytosis following pH dependant fusion step. Once the
nucleocapsids are within the cytoplasm the ambisense strategy allows expression of
five structural proteins. The major nucleocapsid associated protein is named NP. As a
result of the glycoprotein precursor synthesis and after its maturation and exocytic
transport to plasma membrane of infected cells. two glycoproteins are obtained. Both
proteins, named GP1 and GP2, form the claviform structures found in the viral
envelope. At this moment, the detailed mechanisms and cellular structures involved in
the transport of different viral proteins towards the budding site are not known although
the fact that viral assembly is carried out in plasma membrane of infected cells is
currently accepted.
In this work we have studied some aspects of virus-cell interaction
characterizing the association of NP, GPC y GP1 to cellular structures. First, we have
studied the interaction between virus and cytoskeletal intermediate filaments (IF). In the
presence of acrylamide, compound that selectiver disrupt IF, viral yield was
diminished. The results were similar in Vero cells, human fibroblast cultures and murine
astrocytes. Then, we analyzed the effect of acrylamide on different steps of the
multiplication cycle. lt was determined that IF integrity is essential at least between the
step of internalization and viral protein synthesis. Detergent extractions of infected
cultures demonstrated that NP associates to cellular cytoskeletal fraction although it
was not possible to identify the specific cytoskeletal framework involved in that
association. Glycoproteins did not show specific association to these cytoskeletal
fractions under similar conditions assayed.
On the other hand, modifications in the cholesterol contents of infected cells
induced by HMG-CoA reductase inhibitors, diminished infectivity and glycoproteins
membrane Iocalization. This result Ieded us to analyse the association of GPC and
GP1 to membrane cholesterol enriched structures named rafts. Specific cellular
fractionation assays showed glycoproteins associated to raft microdomains. This
interaction was temperature and cholesterol dependant. Moreover, glycoprotein
association to membrane rafts was stabilized at 37°C by specific antibody addition.
These results suggest that GPC and GP1 may utilize the exocytic raft pathway to reach
the plasma membrane of infected cells. Finally, glycoproteins raft association may
represent the strategy to recruit viral proteins before releasing progeny from the cells.

INTRODUCCIÓN

n ïTPÓDIIrrïr'n í
I. LOS ARENAVIRUS
La familia Arenaviridae, establecida en el año 1970, comprende un grupo
de virus envueltos con genoma de ARN segmentado. El primer miembro,
descripto en el año 1925, producía una infección humana definida clínicamente
como meningitis aséptica aguda. Esta enfermedad caracterizada por un
aumento de linfocitos en el fluído cerebroespinal libre de bacterias, 10 años
mas tarde fue nombrada como coriomeningitis Iinfocitariay su agente etiológico
denominado LCMV(Amstrong y col., 1934; Traub y co|., 1935). Por más de tres
décadas este virus permaneció huérfano hasta que en 1956 y en 1958 en
Trinidad y en Argentina respectivamente se identificaron 2 nuevos agentes
infecciosos capaces de enfermar ratones. El primero de ellos, el virus Tacaribe
(Downs y col., 1963) fue aislado de murciélagos fructívoros. Aunque
inicialmente parecía no haber motivo para su relación con el virus Junín
descripto en nuestro país por el Dr. Parodi y col. (1958), pronto ambos virus
mostraron características serológicas comunes (Mettler y col., 1963).
Rápidamente otro virus de similares características y patógeno para humanos
fue descripto en Bolivia con el nombre de Machupo (Johnson y col., 1965)
surgiendo así el grupo llamado Complejo Tacaribe (Johnson y col., 1973).
Estudios de microscopía electrónica mostraron por primera vez un
arenavirus en el año 1969 (Bergold y col.) las partículas pleomórficas con
espículas proyectadas hacia el exterior poseían gránulos electro-densos en su
interior (Figura 1). Esta última característica se debe a la incorporación de
ribosomas en los viriones y debido a ella se acuñó el nombre “arena”.
Figura 1. Microfotografía electrónica de una partícula viral perteneciente a la familiaArenaviridae,

n ¡mom VY‘IÓli
Los estudios morfológicos junto con los serológicos entre estos virus y
el inicialmente descripto LCMV condujeron al establecimiento de una nueva
familia: Arenaviridae, propuesta por Rowe y col. en el año 1970. El virus Lassa
causante de fiebre hemorrágica en África (Speir y col, 1970) fue luego incluido
dentro de esta familia.
La familia Arenaviridae agrupa actualmente miembros de diferente
origen geográfico. Su distribución mundial se muestra en la Figura 2 (Clegg y
col., 2000).
4574?: la.
#fiwásafl ¿gm:í‘VwMH ¡s <
l LCM I Ippy l Tacaribe I LatinoI Lassa I Junin m PmaPañ I Parana
I Mopeia E] hlhchupo I Flexal I TamiamiI Mahal: I Guanafito I Pichinde
Figura 2. Distribución geográfica de los principales miembros de la familia Arenaviridae.
Todos sus miembros con excepción del virus Tacaribe son parásitos de
roedores. Dos grandes grupos fueron determinados por fijación de
complemento e inmunofluorescencia y denominados Complejo del Viejo Mundo
y del Nuevo Mundo (Howard y col., 1993). Como se muestra en la Tabla I
dentro del primer grupo se encuentra el virus de la Coriomeningitis Linfocitaria,
especie prototipo de la familia y de distribución universal, junto con otros virus
aislados de África.

n rmnmirrrr'u '.
Dentro del Complejo del Nuevo Mundo o anteriormente llamado
Complejo Tacaribe se encuentran todos los virus americanos descriptos hastael momento.
Especie Reservorio natural Región
Ippy virus Arvicanthis sp. Africa Central
Lassa virus Mástomys sp. Africa Oriental
Viejo Mundo LCMvirus Mus muscu/us Europa y América
Mobala virus Praomys sp. Africa Central
Mopeia virus Mástomys natalensis Mozambique,Zimbabwe
Amapari virus Oryzomys capito BrasilNeacomys guianae
Flexal virus Oryzomys sp. Brasil
Guanarito virus Zygodontomys brevicauda Venezuela
Junín virus Calomys muscu/¡nus Argentina
Latino virus Calomys callosus Bolivia
Machupo virus Calomys callosus Bolivia
Paraná virus Oryzomys buccinatus Paraguay
Nuevo Mundo Pichinde virus Oryzomys albigulan's Colombia
Piritalvirus Sigmodon a/stoni Venezuela
Oliveros virus Bolomis obscurus Argentina
Sabiá virus Desconocido Brasil
Tacaribe virus Artibeus sp. Trinidad
Tamiami virus Sigmodon hispidus Florida, EEUU
Whitewater Arroyo virus
Pampa virus
Neotoma albigula
Bolomis obscurus
Nuevo Mexico,EEUUArgentina
Tabla l. Arenavirus del Viel'oy Nuevo Mundo, su resfiivo huésped v área de incidencia.
Recientemente el análisis exhaustivo del segmento S del ARN genómico
permitió establecer relaciones filogenéticas entre los arenavirus (Bowen y col.,
1996). Estudios comparativos de secuencias parciales de la nucleoproteína de

IT T'I’IU)l‘)I WY‘IÓ‘H
distintos arenavirus confirmó la clasificación en los dos grupos anteriormente
mencionados. Por otro lado permitió establecer tres Iinajes distintos dentro del
Complejo Nuevo Mundo. El árbol filogenético resultante y los Iinajes A, B y C se
muestran en la Figura 3.
VIEJOmuuoo‘m
m TamiamiI Whitewater Arrovo Im
OfivemsxmParanáC
NUEVO MUNDO
Figura 3. Relación filogenética entre los arenavirus. Los cuadros sombreados indican las especies
causantes de fiebres hemorrágicas en humanos.
Una característica importante que presenta la familia Arenaviridae es la
propiedad de varios de sus miembros de causar fiebres hemorrágicas en
humanos. Mientras que en el Viejo Mundo solo el virus Lassa es responsable
de este tipo de enfermedad, en el Nuevo mundo cuatro especies producen
fiebre hemorrágica y todas se encuentran dentro del linaje B. Evolutivamente
este hecho podría indicar que el fenotipo hemorrágico surgió al menos por dos
vías independientes en esta familia. Sin embargo y debido a que aún se
desconocen los mecanismos moleculares por los cuales estos virus producen

n mmm irr‘ir'iïi
una patología hemorrágica en humanos no puede descartarse completamente
el hecho de que algún otro miembro posea esta capacidad aún no elucidada.
El estudio de fiebres hemorrágicas ha sido de gran interés en las últimas
décadas sin embargo muy poco se sabe acerca de la patogenia producida por
arenavirus. EI virus presumiblemente entra al organismo por inhalación y en la
mayoría de los casos se deposita en los bronquiolos. AI alcanzar el sistema
linfático se esparce sistémicamente sin dejar un foco neumónico aparente
(Kenyon y col., 1992). EI virus es capaz de infectar endotelios tanto in vitro
como ¡n vivo (Andrews y coI., 1978; Zaki y col., 1997). Estudios realizados en
cobayos infectados con una variante de Pichinde mostraron la activación de
moléculas mediadoras de shock tales como leucotrienos, plaquetas y
endorfinas (Liuy col., 1986; Peters y co|., 1987, 1989 y 1997). Estas sustancias
aumentaron en respuesta a los altos valores de TNF-a encontrados en dichos
cobayos (Aronson y co|., 1995).
El sangrado profuso, visto a menudo en Ia patología hemorrágica, es
presumiblemente consecuencia del daño vascular ocasionado por el aumento
de citoquinas y virus combinado con una marcada trombocitopenia (Molinas y
col., 1989; Heller y col., 1995). La severidad del cuadro, por otro lado, varía de
paciente en paciente involucrando factores del huésped aún no conocidos en la
recuperación de la enfermedad. El periodo de convalecencia se caracteriza por
la producción pasiva de anticuerpos tanto en pacientes como en modelosanimales.
II. FIEBRE HEMORRÁGICA ARGENTINA (FHA)
La fiebre hemorrágica argentina o “mal de los rastrojos" es una
enfermedad endemo-epidémica del área central de la Pampa húmeda. Esta
enfermedad caracterizada por un conjunto de sintomas iniciales que incluyen
alteraciones hematológicas, cardiovasculares, renales, inmunológicas y
neurológicas dependiendo de sus formas leves o graves, fue detectada por
primera vez en los años 50 en trabajadores rurales (Arribalzaga y col., 1955).
Los estudios patológicos de casos fatales muestran que Ia mayoria de las
lesiones consisten en una vasocongestión generalizada con múltiples
hemorragias en diferentes órganos. La ruptura de capilares es la responsable

de las hemorragias observadas, produciendo el colapso del sistema vascular y
el consiguiente shock hipotenso. Aunque no ha sido demostrada hasta el
momento la replicación viral en el tejido vascular, se ha encontrado que
principalmente linfocitos y monocitos de sangre periférica constituyen el sitio
primario de replicación del virus (Ambrosio y col., 1986). Alternativamente en
algunos pacientes la respuesta inmune resulta muy efectiva en la eliminación
del virus, cursando la enfermedad con los síntomas más leves.
III. VIRUS JUNIN
El virus Junin, agente causal de la FHA, fue aislado e identificado en el
año 1958 por el Dr. Parodi en el Hospital Regional de la ciudad de Junín,
provincia de Buenos Aires. Los primeros aislamientos de este virus se
realizaron a partir de muestras de sangre y órganos provenientes de pacientes.
El principal reservorio natural del virus Junin es el roedor de hábitos
rurales Calomys muscu/¡nus en el cual causa infecciones crónicas que le
permiten la persistencia en la naturaleza. También se ha aislado de Calomys
laucha, Akodon azarae y Oryzomys flavescens (Maiztegui y col., 1975) y en
roedores urbanos como Mus musculus (Sabattini y col., 1977). La enfermedad
presenta una incidencia estacional en la que las cosechas agricolas coinciden
con un incremento de la población de roedores, aumentando asi el contacto
entre el trabajador rural y el reservorio infectado. Las posibles rutas de
infección para los humanos son la cutánea, a través de Iaceraciones en la piel
o mucosas, y respiratoria por medio de la inhalación de aerosoles
contaminados con fluidos (orina, sangre o saliva) de ratones infectados
(Maiztegui y col., 1986).
Si bien la patogénesis de JUNV se ha estudiado en ratones lactantes y
cobayos mediante infecciones experimentales (Buchmeier y col., 1980), es en
primates rhesus macaques donde mejor se reproducen las caracteristicas
patológicas de la enfermedad en humanos (McKee y col., 1985). Debido, sin
embargo, a los inconvenientes obvios en el trabajo con primates se han
realizado y realizan la mayoria de los estudios en ratones utilizando diferentes
vías de inoculación y edad del animal.

ll l'l‘ROIïll(‘('IÓÏl
Área histórica 1955 al 1987Area autolimitada (3.700.000 hab.)Area nueva (1.000.000 hab.)
Fi ura 4. Localización del área endémica ara el virus Junín en la Re ública Ar entina.
La zona endemo-epidémica de la FHA es el centro de la República
Argentina, un área pequeña en comparación a la superficie total del territorio,
pero donde se encuentra una alta densidad de población y un desarrollo rural
fundamental para la economía del país (Figura 4).
Por su significancia económica se consideró de suma importancia el
estudio de esta enfermedad y del agente que la produce (Millsy col., 1991 y
1992; Garcia y col., 1996). EI control del reservorio animal de JUNV ha sido
impracticable debido a las grandes zonas geográficas implicadas. Por otro lado
es evidente el progresivo aumento de áreas afectadas. En este sentido todos
los estudios sobre el virus y su multiplicación en el huésped han sido dirigidos
al desarrollo de una vacuna eficaz. Asimismo, los estudios conducentes a la
caracterización biológica y molecular del ciclo de replicación viral representan
un aporte valioso para el conocimiento de Ia familia Arenaviridae en su totalidad
y la posible elaboración de estrategias preventivas para los virus de esta
familia.

III.1.Vacunas
AI presente Ia única vacuna desarrollada contra un arenavirus es
Candid #1 para la prevención de Ia FHA. A fines de lo década del 70 un
programa de colaboración entre el gobierno argentino, la Organización
Panamericana de la Salud y organismos de los Estados Unidos condujo a la
elaboración de dicha vacuna a virus vivo atenuado y que fue aprobada por la
Federal Drug Administration.
La cepa Candid #1, derivada de la cepa XJ #44, desarrollada con este
propósito mostró una virulencia sobre cobayos marcadamente menor a otras
cepas atenuadas de JUNV (Barrera Oro y col., 1991). En el año 1988
comenzaron las fases de estudios clínicos en humanos con voluntarios tanto en
Argentina como en los Estados Unidos. La vacuna mostró una eficacia del 95%
en la protección contra la FHA. Además en el 90% de los voluntarios se
registraron anticuerpos neutralizantes hasta 10 años luego de su vacunación
(Maiztegui y col., 1998). En el período 1992-1999 la efectividad de la vacuna
aplicada en zonas de alto riesgo fue del 98.1% (Enria y col., 1999).
A pesar de las contraindicaciones para la administración de vacunas a
virus vivo, especialmente las relacionadas con el alto riesgo que representa la
replicación del virus dentro del huésped, Candid #1 mostró ser segura,
altamente inmunogénica y efectiva para la prevención de FHA en zonas
geográficamente limitadas y de alto riesgo para esta enfermedad. Por otro lado,
estudios realizados en cobayos y monos rhesus sugieren su utilizaciónpara la
prevención de la fiebre hemorrágica boliviana producida por el virus Machupo
(Barrera Oro y col., 1982 y 1988; Lupton y col., 1988, Jarhling y col., 1988).
La caracterización molecular de Candid #1 se realizó secuenciando el
segmento S del ARN genómico. Estudios comparativos de la secuencia del
precursor glicoproteico de esta cepa y sus cepas parentales muestran doscambios aminoacídicos localizados en el extremo amino terminal. Estos
estudios sugieren una participación de la glicoproteina en el desarrollo del
fenotipo atenuado de virulencia, aunque aún no esta del todo comprobado.
Otros estudios sugieren que una disminución en la carga de la nucleoproteína
podría relacionarse con la atenuación (Albariño y col., 1997; Ghiringhelli y coI.,
1997). Una vez más se evidencia la importancia del estudio y caracterización

li Í'I'RÜI‘A TH‘IÓTÍ
de este agente infeccioso para el desarrollo o mejora en el futuro de nuevas
vacunas y estrategias de prevención.
III.2.Morfología del virión
Como se mencionó anteriormente los arenavirus son virus envueltos de
morfología variable. Con un diámetro aproximado entre 50 y 300 nm (Compans
y col., 1993) las partículas de menor tamaño suelen presentar forma esférica
mientras que las más grandes son pleomórficas. En el caso del virus Junín se
ha descripto un morfología redonda u oval con diámetros entre 110 y 130 nm
(Lascano y coI., 1971 y 1974). La densidad de Ia partícula viral calculada en
gradientes de CsCI es de 1.20 g/mI (Romanowski y col., 1981).
Microfotografías electrónicas muestran a los viriones recubiertos por una
membrana electrodensa desde la cual emergen proyecciones de estructura
claviforme correspondientes a las glicoproteínas. En su interior se observan
gránulos electrodensos indentificados como ribosomas.
III.3.Estructura molecular
Las partículas virales están compuestas por una envoltura Iipídica,en su
interior se encuentra el genoma el cual consta de dos moléculas de cadena
simple de ARN. La Figura 5 muestra el esquema de una partícula viral y sus
principales componentes.
Envoltura Iipídica
Ribosoma
Figura 5. Esquema de una partícula viraly sus principales componentes.

n mmm irrióï '.
Cada una de las moléculas de ARN difiere en tamaño y secuencia y
posee 2 marcos de lectura abiertos, no superpuestos y de polaridad opuesta.
Esto representa una de las características más sobresalientes de la familia,
determinando además su estrategia tan particular de replicación.
La molécula L de ARN tiene una longitud de 7 kb y en ella se encuentran
los genes para la sintesis de la ARN polimerasa viral (L) y una proteina
denominada Z por su capacidad de asociarse a moléculas de Zinc. La molécula
S de ARN es de aproximadamente 3.5 kb y posee los marcos de lectura para la
expresión de las glicoproteínas (GP1 y GP2) y la nucleoproteína (NP), todas
presentes en la partícula viral madura. Las moléculas de ARN se encuentran
conformando un complejo ribonucleoproteico o nucleocápside en el interior del
virión, constituido principalmente por el ARN correspondiente y la
nucleoproteína NP. En menor medida L y Z se encuentran asociadas a las
nucleocápsides.
III.4.Proteinas virales
Todas las proteinas virales descriptas hasta el momento para JUNV son
estructurales aunque solo algunas son mayoritarias en la partícula viral
madura. La nucleoproteína NP codificada en el extremo 3' del segmento ARN
S representa el constituyente principal de la nucleocápside. Esta proteina fue
descripta por Martinez Segovia y col. (1977) como asociada al ARN genómico,
encontrándose en purificados de JUNV tratados con Tritón X-100 2% y
resueltos en gradientes de CsCI. Estudios serológicos entre distintos arenavirus
demuestran un alto grado de antigenicidad cruzada para NP entre los
miembros del Nuevo Mundo, no asi entre estos y los pertenecientes al Viejo
Mundo (Rowe y col., 1970).
Estudios realizados con LCMV indican que la nucleoproteína de este
virus es requerida en trans para la síntesis de ARN (Lee y col., 2000). Tortorici
y col. (2001) describieron para NP una función antiterminadora de la replicación
¡n vivo.Cuando se procesan cultivos de células Vero infectados con JUNV esta
proteina puede visualizarse por radioinmunoprecipitación o WB con anticuerpos
monoclonales especificos (Sanchez y coI., 1989) como una banda de
aproximadamente 60 kDa. En las mismas preparaciones es frecuente encontrar
dos productos de degradación de 45 y 25 kDa respectivamente. Recientes

n rmnm Irrir'n i
estudios postulan mas funciones regulatorias para NP y sus productos de
degradación en el mantenimiento de cultivos persistentemente infectados
(Ellenberg y col., 2002).
JUNV posee dos glicoproteínas GP1 y GP2 provenientes de un único
precursor GPC que es procesado proteolíticamente durante su caminoexocítico hacia la membrana. Los estudios de la secuencia de bases del ARN S
predicen un peso aproximado de 52 kDa para GPC, sin embargo y debido al
variado grado de glicosilación, este precursor que es detectado muycomunmente en SDS-PAGE muestra una movilidad de 66-72 kDa
aproximadamente. Ocho sitios potenciales de glicosilación hallados en la
secuencia de GPC de JUNV avalan Io antes dicho (Buchmeier y co|., 1987). La
misma secuencia indica el sitio de clivaje para GPC en la posición 306-307 de
la cadena polipeptidica. Tanto GPC como GP1 son fácilmente evidenciables en
muestras inmunoprecipitadas, sin embargo la presencia de GP2 no se
evidencia por esta técnica.
Con un tamaño estimado de 38 kDa GP1 demostró estar expuesta en la
superficie de los viriones de JUNV (Mersich y col.. 1988). Además esta
glicoproteína es la responsable de la producción de anticuerpos neutralizantes
indicando su participación directa en el reconocimiento virus-célula. Estudios
realizados sobre JUNV con inhibidores del procesamiento post-traduccional de
GPC han demostrado que si bien la N-glicosilación es indispensable para la
multiplicación viral no son necesarias las formas complejas de glicosilación
para la infectividad de los viriones (Martinez Peralta y col., 1979; Padula y col..
1984; Silber y col., 1993). Los datos de secuenciación disponibles y estudios
realizados con inhibidores específicos de la miristilación sugieren la acilación
del precursor glicoproteico (Cordo y co|., 1999).
Por otro lado GP2, una glicoproteína integral de membrana, tendría
capacidad fusogénica requerida para el ingreso de las nucleocapsides al
citoplasma. Debido a Ia falta de anticuerpos específicos contra esta proteina
solo es evidenciable su presencia mediante ensayos de fusión a pH ácido
(Castilla y col., 1996).
Dado que las glicoproteínas se encuentran en cantidades equimolares
en el virión, Burns y col. (1991) proponen un modelo para los arenavirus donde
GP1 y GP2 formarían octámeros mediante interacciones iónicas. En conjunto

IïI'nmnurqóï ï
formarian la estructura glicoproteica observada en preparaciones al
microscopio electrónico como imágenes claviformes de 5 -10 nm de longitud.
Por analogía con otros virus de cadena negativa se ha asumido que Les
la ARN polimerasa ARN dependiente viral aunque hay pocos datos al respecto.
Esta proteína codificada en el extremo 3' del segmento de ARN L es un
polipéptido no glicosilado con un peso aproximado de 250 kDa. La
concentración intracelular de L aumenta dramáticamente durante el curso de
una infección aguda con LCMVsin embargo, la actividad polimarasa disminuye
con el tiempo de infección (Fuller-Pace y col.. 1989). Estos hallazgos proponen
una función regulatoria de L en tiempos tardíos de infección que podria afectar
el balance entre transcripción y replicación (Southern y coI.. 1987). Lee y col.
(2000) demuestran en su trabajo que L, junto con NP, es un factor minimo
necesario para Ia replicación del ARN.
La proteína Z se encuentra codificada en el extremo 5’ del segmento L
de ARN. Este polipéptido de un peso aproximado de 11 kDa se ha
caracterizado por su dominio RING finger de unión al metal Zinc (Salvato y col.,
1989). En los últimos años muchos grupos de investigación se han abocado al
estudio de esta pequeña proteína y, aunque su función específica aun no ha
sido determinada, numerosos trabajos muestran su participación en la
replicación viral. Garcin y col. (1993) proponen, en base a experimentos de
transcripción in vitro, que Z es necesaria tanto para la replicación como para la
síntesis de los ARNm virales. Por otro lado para LCMVse observó un efecto
inhibitorio de Z sobre los procesos antes mencionados (Lee y col., 2000).
Estudios con compuestos que alteran específicamente Ia estructura RING
finger demuestran el posible rol de Z en las etapas finales del ciclo de
replicación de JUNV (Garcia y coI., 2003). Las interacciones estructurales
descriptas entre Z y NP son al presente motivo del estudio del posible rol de Z
como proteína de matriz en los arenavirus.
III.5.Ciclo de replicación
La Figura 6 muestra un esquema del ciclo de replicación de un
arenavirus. La entrada del virión a la célula se produce por un mecanismo de
endocitosis mediada por receptor (Borrow y col., 1994). Recientemente la
proteína de membrana a-dystroglycano se ha identificado como receptor celular

nmmm Irrióï l
de LCMV(Cao y col., 1998). Esta proteina, mediadora de interacciones con Ia
matriz extracelular y presente en gran variedad de tejidos, parece también
tener afinidad por otros arenavirus tales como Lassa, Oliveros y Mobala. Sin
embargo, se desconoce aún su relación con JUNV.
El complejo glicoproteico presente en los viriones es el responsable de
las interacciones iniciales con la superficie celular. Anticuerpos dirigidos contra
GP1 pueden neutralizar la adsorción del virus a la célula indicando su papel en
la infección de las células susceptibles.
RibosomasyRNALy 5
EnvolturaLlheracmn viralpor bmtacion
mRNAZ
í _,, GP-1
Gp'z (I-distrogltcano-dlst licanoí/‘r B ’09
Adsorción ypenetración
del citoplasma
Reticulo ' 1endoplasmático
Fusión y liberación:del virión dentro
mRNAZM ORNP
Ribosomas ‘Mo
¿“vai/É
J r' Golgi
GPC+Z
Í Transcripción y Traducción
\ RNP\ intracelular\‘l
É Replicaclonl
NP + L (polimerasa)Z 7
(SP-2
GP-t
Figura 6. Ciclo de multiplicación de un arenavirus.
La entrada de JUNV es inhibida por agentes Iisosomotrópicos y
representó uno de los elementos de prueba del ingreso dependiente de pH de
los viriones al interior de la células (Castilla y col., 1991). Estudios con otros
arenavirus han demostrado además el efecto irreversible del pH sobre la

estructura conformacional de GP1 y su estabildad en la envoltura viral (Di
Simone y co|., 1994).
En el citoplasma ocurre la transcripción y replicación del genoma aunque
aún no se ha esclarecido la necesidad de algún componente celular para laadición de estructuras CAP al extremo 5' de los ARNms. Un inhibidor de la
ARN polimerasa ll como la actinomicina D no impide completamente la
replicación de LCMVpero disminuye notablemente los niveles de producción
viral por razones aun no elucidadas.
Aún cuando se desconocen los detalles para el inicio y los requisitos
exactos de la misma, la estrategia de replicación ambisense (Figura 7) ha sido
profundamente estudiada y descripta por Auperin y col. (1984). Este término
alude al hecho de que cada ARN tiene una región positiva y otra negativa.
Como se mencionó anteriormente cada molécula de ARN codifica para 2
proteínas, uno de los genes se encuentra hacia el extremo 5' y el otro hacia el
3' separados por una región no codificante intergénica de secuencias
autocomplementarias. De acuerdo a esta última caracteristica los ARN pueden
formar estructuras secundarias muy estables conocidas como horquillas.
Aunque el extremo 5' del ARN S contiene la secuencia codificante para
GPC este precursor glicoproteico solo es traducido luego de una ronda de
replicación genómica y a partir del transcripto subgenómico fabricado en la
célula. En el caso de la molécula de ARN L, la proteína Z ubicada en su
extremo 5' es sintetizada por igual mecanismo que GPC. De esta manera los
genes ubicados en la region 5' del ARN genómico tiene en realidad una
polaridad pseudo-positiva.
La replicación viral comienza entonces en los extremos 3' con Ia
transcripción de los genes N y L del ARN S y ARN L respectivamente. Los
mensajeros así fabricados son traducidos por la maquinaria celular
conduciendo a la síntesis de las proteinas NP y L. La síntesis de los
intermediarios antigenómicos completos permite la replicación del genoma para
la nueva progenie así como también para la síntesis de los ya mencionados
ARNm correspondientes a GPC y Z. Varias evidencias sugieren que los
niveles de NP serian los determinantes en el equilibrio entre transcripción y
replicación viral dentro de la célula infectada (Franze-Fernandez y col., 1987).
20

n mmm irrïr'n 1
3a NP mRNA 5!
TRADUCCÍON
l SEGMENTO COMPLEMENTARJO DEL.RNAvS
5/ GP mRNA ar
GP-1 l GP-ZTRADUCCION
Figura 7. Estrategia de replicación ambísense.
No existe mucha información respecto de las etapas finales del ciclo de
replicación. A partir del procesamiento de las glicoproteinas en el reticqu y
aparato de Golgi, el ensamblado y brotación de los viriones se produce en Ia
membrana celular pero se desconocen los eventos que determinan Ia salida del
virus. En general se asume que el ensamblado de las partículas virales es
iniciado por interacciones entre el dominio citoplasmático de una proteina viral
de membrana y los componentes internos de la partícula. En consecuencia el
proceso comienza tan pronto como las proteinas virales de membrana y
citoplasmáticas se encuentran en concentración suficiente dentro de la célula
infectada. Estudios realizados con LCMV(Burns y co|., 1991) han demostrado
la capacidad de GP2 de interactuar con NP a través de su extremo
citoplasmático. Estudios muy recientes estan indicando a la proteina Z como
posible responsable del reclutamiento en la membrana, funcionando como una
proteina matriz en este proceso (Strecker y co|., 2003).
Por último, la salida del virus de la célula infectada esta íntimamente
acoplada al proceso de brotación por el cual el virus adquiere la envoltura
Iipídica. AI alcanzar el medio extracelular la progenie posee la capacidad
necesaria para la infección de una célula blanco y el iniciode un nuevo ciclo de
replicación.
21

IV. INTERACIÓN VIRUS-CÉLULA HUÉSPED
Dado que un virus es incapaz de replicarse por si mismo la entrada a
una célula y el uso de Ia maquinaria y energía celular, en mayor o menor
medida. resulta indispensable para su subsistencia y persistencia en la
naturaleza. De este modo el estudio de Ia multiplicación viral es de muchas
maneras el estudio de la interacción entre las distintas estructuras y proteínas
virales con las de la célula huésped. Los mecanismos por los cuales un virus
puede infectar una célula determinada, multiplicarse eficazmente y producir su
progenie son objeto de estudio de la Virología y biologia celular básica.
conduciendo al conocimiento detallado de cada uno de los pasos del ciclo de
multiplicación viral. Este conocimiento representa el punto de partida para
todos los estudios conducentes al diseño de estrategias antivirales que
permitan combatir la replicación viral. El diseño de vacunas es tambien en este
sentido dependiente del conocimiento de la funcionalidad de las distintas
proteínas virales, su expresión y su interacción con el sistema inmunológico.
Por otro lado, la caracterización de las distintas interacciones entre el virus y la
célula huésped permite la mejor comprensión de los daños citopáticos siendo
una herramienta para la explicación de las patologías producidas por un virus
en los distintos órganos infectados y su manifestación como una enfermedad
con características definidas y propias.
Asimismo es importante señalar la importancia que ha cobrado la
utilización de los sistemas virales como herramienta para el estudio y
caracterización de los procesos básicos desarrollados en una célula eucariota.
Los sistemas de cultivos de células son la principal herramienta para el
estudio de las interacciones virus-célula. Su desarrollo y caracterización
resultan indispensables para el uso de los mismos como un sistema
experimental altamente controlable y reproducible.
El virus Junin es capaz de crecer ¡n vitro en una amplia variedad de
líneas celulares de distintos origenes. En general la replicación es poco
eficiente y no se observa citopatogenicidad, sin embargo en sistemas celulares
como Vero y MRC-5 se observa un efecto citopático característico
(Weissenbacher y coI., 1983). La infección de monocapas con JUNV presenta
dos etapas marcadamente diferentes. La etapa aguda o iniciales aquella en la

ITï'l'l’f‘ilïllf‘f‘lf'fiï
cual los niveles de producción viral son máximos y la destrucción celular
marcada. Entre el tercer y quinto dia se observa el efecto citopático
acentuándose los focos de células redondeadas que con su posterior
desprendimiento originarán las denominadas placas bajo medio semisólido
(Tkaczenski y coI., 1970). En la segunda etapa, a partir de los 8-10 días p.i., las
células sobrevivientes comienzan a repoblar el cultivo estableciéndose una
acción moderada del virus sobre éstas. Esta etapa es también llamada de
persistencia y su establecimiento es una de las característica de los arenavirus
que los ha hecho objeto de estudio en este aspecto.
El reconocimiento de la célula huésped es el primer paso en la
interacción virus-célula. Esta etapa, que requiere la especificidad de interacción
entre la membrana del virus envuelto y la membrana celular, conducirá al inicio
de la infección determinando además el tropismo del virus. Un virus puede
utilizar una amplia variedad de moléculas de superficie para su ingreso a la
célula. Proteinas receptoras asi como carbohidratos o glicolipidos son en
general los que permiten la adsorción inicial a la membrana celular.
Usualmente otras moléculas co-receptoras asi como proteinas virales
específicas pueden desencadenar y facilitar el evento de fusión. Una vez
dentro de la célula los eventos de replicación y síntesis de proteinas son
llevados a cabo en su mayoría por la maquinaria celular.
La maduración y transporte de las proteínas virales en las etapas tardías
del ciclo viral son determinantes en la producción de las nuevas particulas
infectivas. A este respecto las glicoproteínas virales, especialmente la
hemaglutinina (HA) del virus Influenza asi como la glicoproteína G del virus de
la estomatitis vesicular (VSV) han sido utilizadas para el estudio de la
biogénesis, transporte y localizaciónde proteinas de membrana. De esta forma
ambas glicoproteínas estan ampliamente caracterizadas y estudiadas en este
aspecto siendo referentes para el estudio de muchas otras proteinas virales.
Sin embargo, cabe destacar que la elección de Influenza y VSV como modelo
para el estudio de estos procesos ha sido favorecida en parte por su alta
capacidad replicativa que facilitael abordaje experimental.
Los virus con la propiedad de producir infecciones persistentes en sus
hospedadores son de particular interés debido a Ia capacidad de replicarse en
una célula sin producir su muerte o un efecto muy dañino sobre sus
23

Iïmmm irr‘róï '.
estructuras. Esta es la base para la explicación del establecimiento de
infecciones crónicas en la naturaleza por los arenavirus y otros virus con igual
propiedad (Howard y col., 1986; Lehmann-Grube y col., 1972 y 1984; Rawls y
coL,1981)
El hecho de que un virus pueda establecer infecciones muy poco
dañinas para la célula es una muestra tambien de un alto grado de co
evolución donde la relación entre el virus y la célula es aún mas estrecha.
demostrando la íntima dependencia del primero por el segundo. Este aspecto
representa, sin embargo, un desafío experimental debido a las obvias
implicancias de estudiar procesos virales que producen una pobremanifestación celular. En estos casos es frecuente recurrir a la alteración de
algún mecanismo o estructura celular para observar sus efectos sobre la
normal multiplicación del virus. Los resultados observados permiten el estudio
de dichas interacciones virus-célula aunque frecuentemente es necesario un
abordaje molecular para determinar asociaciones más específicas. Por otro
lado la utilización de técnicas moleculares, como la expresión individual de
proteinas virales en sistemas transfectados, presentan también dificultades a lahora de las conclusiones debido a Ia ausencia total de las interacciones con el
resto de las proteínas virales que se dan naturalmente en una infección con
virus entero.
En este trabajo de tesis se han caracterizado parte de los pasos que
llevan al transporte y localización de las proteínas del virus Junin en la
membrana celular y su importancia en la infectividad de Ia progenie. En primer
lugar se estudió la participación de los filamentos intermedios en el ciclo de
multiplicaciónviral. Los resultados obtenidos junto con datos anteriores a este
trabajo nos condujeron al segundo aspecto estudiado: Ia localización y
transporte de las proteinas virales en membranas enriquecidas en colesterol
denominadas rafts. A continuación se detalla una breve introducción y reseña
de estas estructuras celulares para la mejor comprensión de estos procesos y
sus posibles implicancias en el conocimiento del ciclo de multiplicación de
JUNV.
24

ni'i‘imm ir‘rir'm
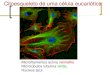
V. CITOESQUELETO
Las células eucariotas poseen tres tipos de redes proteicas filamentosas
perfectamente diferenciables y especialmente distribuidas en el interior del
citoplasma. Conformado por microtúbulos, microfilamentos y filamentos
intermedios el citoesqueleto constituye una intrincada red funcional y
estructural (Figura 8).
En su conjunto estos polímeros de variada longitud y calibre conforman
el esqueleto celular cuya función es la de mantener la estructura y morfología
general de la célula, así como transportar y mantener la localización de las
organelas en el citoplasma. Cada red muestra un arreglo y función particular,
sin embargo usualmente sus funciones dependen de proteínas accesorias
motoras, proteínas que permiten la asociación entre los distintos elementos del
citoesqueleto y aquellas que conectan el citoequeleto con cualquier otraestructura celular.
TubuünaMICROTUBULO
25 nm i'L y
A.
7 nm
FILAMENTO
8-10 nm C INTERMEDIO
Figura 8. Componentes del citoesqueleto.
25

V.1. Microtúbulos (MT)
Los microtúbulos, estructuras huecas de 25 nm de diámetro, son el
resultado de la polimerización de proteínas denominadas tubulina. Se
encuentran distribuidos en el citoplasma irradiando de una zona cercana al
núcleo (centrosoma) hacia la periferia de la célula. Son estructuras polares
dado que su polimerizaciónes más rápida y estable hacia uno de los extremos.
Además, extremadamente dinámicos, pueden renovarse según los
requerimientos celulares y mediante la regulación de la polimerización
despolimerización de los mismos.
Entre las funciones que cumplen los MT es de gran importancia la
localización intracelular especifica de las organelas. Tanto el reticulo
endoplasmático como el aparato de Golgi cambian su patrón de localización al
tratar las células con agentes depolimerizantes de MT. Asimismo el
desplazamiento de organelas dentro del citoplasma depende de la red de MTy
sus proteínas motoras asociadas. Mitocondrias, granulos de secreción y
vesículas trasportadoras han sido decriptas como asociadas a MT en su
desplazamiento dentro del citoplasma. Los MT son responsables del sostén
interno y funcionamiento de cilios y flagelos. En estas estructuras los
microtúbulos se organizan de manera diferente formando asociaciones
macromoleculares de alta complejidad que permiten el movimiento coordinado
de la célula. Por ultimo otra función de gran importancia y una de las primeras
en ser descriptas es la participación de los MTen la división celular. Formando
parte del huso mitótico los MT participan activamente en la segregación de los
cromosomas duplicados.
V.2.Microfilamentos (MF)
Los microfilamentos están formados por una subunidad proteica globular
de aproximadamente 45 kDa llamada actina. La polimerización de estas
subunidades requiere energia y produce filamentos de un diámetro aproximado
de 6 a 10 nm. Aunque los filamentos de actina pueden encontrarse distribuidos
en todo el citoplasma su localización típica es bajo la membrana plasmática
formando el cortex celular o actina cortical. Esta red, formada por una gran
cantidad de filamentos de actina paralelos entre si, se encuentra además
asociada a polímeros de miosina los cuales permiten la contractibilidad del
26

sistema. La localización y nucleación de los MF bajo la membrana plasmática
es dinámica y esta controlada por señales extracelulares que pueden inducir la
re-estructuración local de la actina cortical. De manera inversa una re
organización del con‘ex puede influenciar el comportamiento de la membrana
asociada a este. Estructuras especiales como las microvellosidades de las
células epiteliales se encuentran sostenidas internamente por MFs. La emisión
de filopodiosen células como los macrófagos es también dependiente de la red
de MFs. Por último, durante la división celular la estrangulación del citoplasma
depende de la re-estructuración y acción coordinada de MFs y filamentos demiosina.
V.3.Filamentos intermedios (FI)
Los filamentos intermedios representan el tercer tipo de polímero
perteneciente al citoesqueleto. Con un diámetro de 8 a 10 nm estos polímeros
presentes en organismos pluricelulares son particularmente abundantes en el
citoplasma de células sujetas a estrés mecánico dado que su función principal
es aportar resistencia. Células epiteliales y nerviosas así como musculares
presentan este tipo característico de filamentos. A diferencia de los MT y MF
altamente conservados en la evolución de los ecucariontes, los FI están
constituidos por una gran variedad de proteinas relacionadas tales como
vimentina, laminina o queratina.
Distintos tipos celulares difieren en el tipo particular de Fl expresado. Los
Fl están compuestos de subunidades proteicas de estructura fibrosa y no
globular como Io son los monómeros de actina y tubulina. Su estructura básica
consta de una cadena polipeptidica con un dominio central helicoidal.
presentando en los extremos amino y carboino terminal dominios globulares. AI
parecer sin gasto de energía estos monómeros se asocian en dímeros y luego
en tetrámeros que conforman la subunidad estructural de la que se componen
los filamentos. Los tetrámeros libres en el citoplasma son difíciles de hallar
encontrándose, en la mayoría de los casos, polimerizados en el estado
filamentoso. Sin embargo bajo ciertas circunstancias la célula puede regular el
ensamblado de estas unidades como es el caso de la laminina nuclear cuya
despolimerizaciónes desencadenada por la fosforilaciónde sus subunidades.

Los filamentos intermedios presentes en células de vertebrados pueden
agruparse según se muestra en la Tabla ll.
La lámina nuclear presente en la cara interna del núcleo de las células
esta compuesta por Iamininas. Estas proteínas que son dirigidas desde el
citoplasma a su destino gracias a una señal de localización nuclear propia
forman una malla en asociación con otras proteínas. En condiciones normales,
esta malla de FI interrumpida solo por los poros nucleares, se asocia tanto a la
cromatina como a la membrana nuclear.
Los FI presentes en células de invertebrados son similares a la laminina
sugiriendo que esta pudo ser la primera en su tipo a partir de la cual surgieronotras formas de FI.
Clase Composición del polímero Localización celular
Lámina . . '
nuclear LamInIna A, B y C Nucleo
Queratinas Queratina I (ácida) y II(básica/neutra) Células epiteliales y sus derivados
(pelo, uñas)
Vimentinas Vimentina Células de origen mesenquimáticoDesmina Células musculares
Proteína ácida gliofibrilar(GFAP) Células de la glía (astrocitos y
células de Schwann)
Filamentos Neurofilamentos Neuronas
Intermedios
neuronales
Tabla ll. Clasificación de filamentos intermedios en células de vertebrados.
La familia de queratinas es la más grande y diversa dentro de los
filamentos intermedios. Su localización por excelencia son las células
epiteliales, encontrándose alrededor de 20 tipos distintos en el epitelio humano.
En una misma célula una gran variedad de queratinas pueden polimerizar para
formar el sistema de FI. La combinación particular de queratinas dependerá del
tipo celular y su localización. En el órgano de la piel, por ejemplo, las distintas
capas de la epidermis expresan diferentes grupos de queratinas. Esta

n i'I'RODlHY‘IÍH '.
heterogenicidad es sumamente útil para el diagnóstico e identificación decarcinomas.
Los filamentos de vimentina y los relacionados a ésta forman en general
polímeros de un solo tipo de proteína. En el citoplasma de células de origen
mesodérmico incluyendo fibroblastos, células endoteliales, glóbulos blancos y
en líneas celulares de otros orígenes la vimentina representa el tipo principal de
FI. La desmina se encuentra principalmente en células pertenecientes al
músculo liso, esquelético o estriado observándose, en estos casos, una
asociación importante de los filamentos de actina y miosina con los de
desmina. Células de la glía como astrocitos y células de Schwann presentan un
tercer tipo proteico llamado proteína ácida gliofibrilar (GFAP). Los filamentos
gliales pueden estar formados por distintos tipos de proteínas ácidas e incluso
pueden formar co-polímeros con vimentina. Sin embargo células que expresan
tanto vimentina como queratina no muestran polímeros mixtos sino que cada
proteína forma su propio filamento.
Por último las células nerviosas presentan neurofilamentos localizados a
lo largo del axón. Estos polímeros usualmente conformados por tres tipos
diferentes de proteínas se asocian entre si en distintos puntos y a lo largo de su
eje mayor. Esta disposición representaría el soporte mecánico básico de
prolongaciones de membranas tan largas y delgadas como los axonesneuronales.
Experimentalmente el citoesqueleto ha sido clásicamente definido como
una matriz proteica insoluble en detergentes no iónicos (Osbom y col., 1977).Cuando cultivos celulares son tratados con NP-4O o TX-100 la fracción
resistente a la extracción se encuentra enriquecida en MTy MF como en FI.
Estudios realizados en células Ptk1 demostraron la presencia de
ribonuceloproteínas llamadas prosomas asociadas a los filamentos intermedios
(OIink-Couxy col., 1992, Arcangelettí y col., 1992 y 1997b). Mas tarde estudios
exhaustivos acerca de Ia naturaleza estructural de los prosomas y su
asociación con ARN mensajeros demostró la relación funcional entre estos y
las redes de vimentina y queratina. Los prosomas son complejos constituidos
por ARNm y proteínas de bajo peso molecular (19-36 kDa). La composición
proteica de una partícula individual está en relación con el tipo celular y la
población de ARNm. En algunos casos se encuentran también pequeños ARN

lï ïTp0m irmúï 1
de 50 a 150 nucleótidos de largo cuya función parece ser la estabilización de
los ARNm. Los complejos, de una ultraestructura cilindrica, llegan a un peso
molecular de 720 kDa y presentan en su interior una función proteasa.
Dada la cantidad de estudios que muestran la asociación de prosomas a
ARNm no traducidos y su ausencia total en complejos de poliribosomas (Spohr
y col., 1970; Grossi de Sa y col., 1988a y b; Olink-Coux y col., 1992) fueron
propuestos como marcadores citológicos de ARNm en estado inactivo o no
traducido (Scherrer y col., 1994). Además esto sugirió su posible función en la
represión de los mensajeros. Los prosomas podrian actuar como un factor en
trans capaz de regular el pasaje de un ARNm de su forma inactiva a su forma
activa asociándose o no a los mismos. En este sentido la traducción in vitrode
ARNmvirales (adenovirus. virus de mosaico del tabaco) mostró ser inhibida por
prosomas en condiciones en que la traducción del ARNm de globina no fueafectado.
Los prosomas no solo resisten la extracción con detergentes no iónicos
sino también bajas concentraciones de SDS, deoxicolato. RNasa y alta fuerza
iónica (Schmid y col., 1984). Estudios con anticuerpos monoclonales mostraron
la co-Iocalización de prosomas y filamentos intermedios. Tanto en tipos
celulares expresando vimentina y desmina así como queratina se demostró un
alto porcentaje de superposición de imágenes. Aunque aún no se realizaron
estudios similares en células nerviosas ya se ha demostrado la presencia de
prosomas específicos en las mismas. De esta manera Scherrer y col. (1994)
proponen que los FI transportan los prosomas y por Io tanto los ARNm
inactivos, siendo asi la red de filamentos intermedios la responsable de la
citodistribución selectiva de mensajeros previa a la traducción. Los estudios
dinámicos realizados con drogas como Ia acrilamida, una neurotoxina que
induce selectivamente el colapso de los FI sin alterar las otras redes del
citoesqueleto, indicaron Ia asociación directa entre prosomas y diferentes tipos
de FI. Durante el colapso ambos tipos de antígenos permanecen asociados
mostrando una íntima interacción (Olink-Coux, 1992).
30

VI.MICRODOMINIOS DE MEMBRANA RAFTS
Las membranas celulares contienen diferentes tipos de lípidos tales
como glicerofosfolipidos, esfingolipidos y colesterol. Asociados en
microdominios cada uno de estos componentes posee diferentes
caracteristicas fisico-químicas y biológicas coexistiendo en la membrana
plasmática de células eucariotas. En los últimos años se han estudiado
microdominios de membrana de composición específica denominados rafts.
Los rafts son estructuras complejas que segregan del resto de la membrana
plasmática caracterizadas por una alta concentración de colesterol y
esfingolípidos en la capa extracelular de la membrana. Debido a su
composición son caracterizados por su resistencia a la extracción con TX-100 a
bajas temperaturas y aislados como membranas de baja densidad en
gradientes de flotación en sacarosa (Brown y coI., 2000). Un modelo para la
estructura de estos microdominios fue propuesto por Simons y col. (1997) y
ampliamente aceptado. En él se propone una estrecha asociación lateral entre
los glicolípidosdebido a sus largas cadenas de ácidos grasos saturados. Las
moléculas de colesterol intercaladas entre los lípidos serían los determinantes
en la estabilización de estas estructuras o microdominios. Se ha definido este
estado particular como fase líquido-ordenada debido a su comportamiento
físico-químico dentro del resto de la membrana plasmática considerada, en el
mismo contexto teórico. como fase líquido-desordenada. De esta manera se
explica la tendencia de los rañ‘s a permanecer como una unidad dentro del
resto de la membrana plasmática y su relativa estabilidad en detergentes no
iónicos. El lado citoplasmático de estos microdominios esta pobremente
caracterizado, su composición incluiría fosfolípidos de cadena saturada y
colesterol (Fridriksson y col., 1999). Aunque no se conoce como estarían
acoplados el lado citoplasmático con el extracelular se ha sugerido que las
largas cadenas carbonadas pertenecientes a los esfingolipidos en el lado
exoplásmico podrían acoplar el citoplasmático por interdigitación (Simons y
col., 2000).
Otra propiedad de estos microdominios que ha despertado un interés
aun mayor es la capacidad de incluir proteínas específicas dentro de su
3]

Ii i'l'lU )I‘)lVTIÓH
estructura. Estas proteínas podrian además colaborar en al acoplamiento de
ambas caras de Ia membrana.
Distintas clase de proteínas se han definido como asociadas o afines a
rafts (Figura 9). Proteínas integrales, como caveolina 1, contienen varias
señales de palmitilaciónque favorecen Ia asociación a la membrana. Proteínas
periféricas, como Ia subunidad ocde Ia proteína G o la familia de quinasas Src,
localizan en estos microdominios (Mukherjee y col., 2003). Estas proteínas
asociadas al lado citoplasmático de Ia membrana se encuentran doblemente
aciladas, por palmitilación o miristilación (Resh y coI., 1999). Un tercer tipo de
proteína se ha descripto asociada a rafts y son las ubicadas del lado externo de
la membrana. Aunque estas son inicialmente proteínas transmembrana,
posteriormente se clivan y modifican con un incoIípido. De esta forma Ia
asociación de Ia proteína a la membrana es vía una cadena acílica saturada,
usualmente perteneciente a la molécula de glicofosfatidilinositolo GPI (Hooper
y co|., 1999). Esta topología determina que este tipo de proteínas asociadas a
rafts no puedan unirse al citoesqueleto aún cuando sean insolubles en TX-100.
Los dominios extracelulares sin embargo podrían interactuar con proteínas
transmembranas ligadas directamente con algún elemento del citoesqueleto.
MEDIOEXTRACELULAR
ClTOPLASMA
Figura 9. Proteínas de membrana asociadas a microdominiosrafts.
Las proteínas asociadas a rafts vía GPI pueden, oportunamente, ser
liberadas por fosfolipasas. De manera similar las proteínas integrales o
32

17ï'nmm irrjr'n 1
asociadas a la cara citoplasmática de la membrana pueden ser controladas por
la palmitilación reversible de sus cadenas polipeptídicas. Estos mecanismos
sugieren Ia flexibilidadde la asociación entre proteinas y rafts y su potencial rol
biológico.
La distribución de los rafts en la membrana depende del tipo celular. En
células epiteliales o células nerviosas donde existe una polarización de la
membrana estos dominios se localizan preferencialmente en el lado apical o en
la membrana axonal respectivamente. En células no polarizadas como
linfocitos o fibroblastos los rafts se distribuyen a lo largo de toda la superficie
celular sin obvias preferencias.Dado el constante intercambio de membranas en una célula no es de
extrañar el hecho de que los rafts se encuentren en membranas del camino
biosintético y exocítico celular. Mientras el colesterol es sintetizado en el
reticulo endoplasmático, los esfingolípidos y sus modificaciones se completan
en el aparato de Golgi (van Meer, 1989). De esta manera se ha sugerido que el
compartimiento de ensamblado del raft sería el aparato de Golgi (Brown y col.,
1998). Las vesículas internas así constituidas viajarian principalmente a Ia
membrana plasmática enriqueciéndose en sus componentes vía un mecanismo
retrógrado de exclusión de otros componentes hacia el reticulo. La inclusión de
proteínas en el camino exocítico de estas vesículas representa el principal
mecanismo para la distribución polarizada de proteinas en células epiteliales
(Keller y col., 1997). Por otro lado una vez en la membrana plasmática los rafts
son continuamente endocitados (Mukherjee y col., 2000). Su reciclado puede,
finalmente, llevarlos al Golgi vía endosomas o devolverlos a Ia membrana
celular (Puri y col., 1999).
Una de las razones por las cuales ha sido dificultoso probar la existencia
de estos dominios se relaciona con su pequeño tamaño y la consecuente
dificultad para ser resueltos por la microscopía clásica. Estudios de
entrecruzamiento con anticuerpos y Iectinas en células vivas permitieron
sortear el problema y una vez mas confirmar su existencia (Janes y col., 1999;
Harder y col., 1998). EI agrupamiento de un número variable de rafts inducido
por anticuerpos especificos produce dominios visibles al microscopio óptico.
Las imágenes en forma de parches sobre la membrana celular corresponden aestructuras del orden del micrón. En fibroblastos se han caracterizado

microdominios de aproximadamente 50 nm de diámetro mediante la utilización
de anticuerpos (Pralle y col., 2000).
Diferentes virus envueltos se ensamblan y brotan de membranas
intracelulares especificas tales como el retículo endoplásmico o diferentes
compartimentos del Golgi. Asimismo los virus que brotan de la membrana
plasmática celular no Io hacen al azar. Como Io demuestran muchos estudios
diferentes virus se ensamblan y brotan asimétricamente de la superficie de
células epiteliales polarizadas. Algunos Io hacen desde la membrana apical
mientras otros brotan de la membrana basolateral de las mismas (Compans y
col., 1995; Barman y co|., 2000).
El rol de los rafts en el proceso de ensamblado y brotación viral es objeto
de estudio desde hace varios años. Muchas proteinas virales se han descripto
como residentes de rafts y su función asociada con el reclutamiento y
ensamblado de los componentes del virión. Por otro lado, el mecanismo por el
cual la envoltura viral excluye Ia mayoría de las proteinas celulares o en
algunos casos incluye selectivamente algunas proteinas específicas podría ser
explicado en el contexto de la teoria de rafts.

OBJETIVOS

Estudios realizados por nuestro grupo de trabajo han demostrado la
participación del citoesqueleto en eI transporte de las glicoproteínas de JUNV a
la superficie celular. Tanto la integridad de microtúbulos como microfilamentos
es necesaria para Ia replicación viral. La expresión de las glicoproteínas virales
en la membrana celular fue disminuida cuando se alteró la red de MT o MF.
Asimismo el análisis por inmunofluorescencia de cultivos infectados y extraídos
con TX-100 mostró que las glicoproteínas permanecían mayoritariamente
unidas a la fracción resistente al detergente (Candurra y col., 1999).
Quedaba por explorar Ia participación de los filamentos intermedios en el
ciclo de multiplicación de JUNV y su relación con la localización de la
glicoproteina en las distintas fracciones celulares. Por otro lado la posible
miristilación del precursor glicoproteico de JUNV (Cordo y col., 1999) sugería
un mecanismo para su asociación especifica a microdominios de membrana
enriquecidos en colesterol.
El trabajo de tesis que a continuación se presenta se llevó acabo con el
objetivo de determinar 1) Ia participación del citoesqueleto, particularmente la
red de filamentos intermedios, en el ciclo de replicación de JUNV y 2) la
asociación de las proteínas virales a membrana de la célula huésped, con la
finalidad de esclarecer el comportamiento frente a detergentes no iónicos.

MATERIALES Y MÉTODOS

1. Virus
Se utilizó Ia cepa IV4454 del virus Junín (JUNV) atenuada para cobayo.
La misma fue purificada por dos clonados en células Vero. Se prepararon los
stocks de trabajo infectando las células con una multiplicidad de infección
(MOI)de 0.1 UFP/célula. Se recogieron los sobrenadantes a los 4-5 días post
infección (p.i.), se titularon por el método de unidades formadoras de placas
(UFP) en células Vero y se almacenaron en alícuotas a -80°C. Los títulos de los
stocks fueron en todos los casos del orden de 106UFP/ml.
Se utilizó virus Influenza A cepa A/Texas/36/91 cedida por eI Área de
Virus Respiratorios del Instituto Malbrán. La semilla se amplificó en células
MDCK. Las células se infectaron (MOI = 1) con un inóculo en medio de cultivo
sin suero y suplementado con 2 pg/ml tripsina tratada con tiolsulfoniI-fenilalanil
clorometiI-ketona (TPKC). Luego de 1 hora de adsorción a temperatura
ambiente se retiró el inóculo y se incubó con el mismo medio. Los
sobrenadantes se cosecharon a las 48 horas p.i. y se titularon por el método de
UFP en la línea celular MDCK con las modificaciones que se indican en la
sección 8.2.
2. Cultivos celu/ares
2.1. Linea celu/ar Vero (ATCC CCL 81)
Proveniente de riñón de mono verde africano (Cercopithecus aethiops).
Los cultivos se crecieron en medio esencial mínimo de Eagle (MEM)
suplementado con 5 % de suero de ternera (ST) inactivado y 50 pg/ml de
gentamicina. Las monocapas se subcultivaron periódicamente y luego de
alcanzada la confluencia se mantuvieron en MEMsuplementado con 1,5 % de
ST. Cuando estos cultivos se utilizaron para plaquear virus Junin se
mantuvieron en medio semisólido de plaqueo (MEM2X: 4% de ST inactivado,
100 pg/ml de gentamicina) en partes iguales con metilcelulosa 1.4%. En los
casos en los que los cultivos se incubaron en estufa con 4% de 002 los medios
se suplementaron con 20 pM de buffer HEPES.
38

2.2. Linea diploide de flbrob/astos humanos (PH)
Cedida gentilmente por la Dra. G. Carballal (CEMIC, Bs. As., Argentina). Los
cultivos se crecieron y propagaron en MEM suplementado con 10% de suero
fetal bovino inactivado y 50 pg/ml de gentamicina. Las monocapas se
subcultivaron semanalmente y se utilizarondurante 10 pasajes.
2.3. Cultivo priman'o de astrocitos murinos (AM)
Los cultivos se obtuvieron a partir de cerebros de ratones OF1 (lffa
Credo, Lyon, Francia) de 1-3 dias de vida. El material se extrajo en esterilidad,
se resuspendió en bufferfosfato salino (PBS), se disgregó en forma mecánica y
luego enzimática con tripsina 0.02% durante 15 minutos a 37°C. Se centrifugó
a 1000 xg y las células fueron resuspendidas en MEM 10% suero fetal bovino
a una densidad de 1x106 células/ml. Luego de formada la monocapa los
cultivos se subcultivaron y se incubaron a 37°C hasta el momento de Ia
infección.
2.4. Linea celular MDCK (ATCC CCL 34)
Provenientes de riñón de perro (Canis famílian's). Los cultivos se
crecieron en MEM suplementado con 10% de SF inactivado y 50 pg/ml de
gentamicina. Las monocapas se subcultivaron periódicamente y luego de
alcanzada la confluencia se mantuvieron en MEM suplementado con 3% de
SF. Cuando estos cultivos se utilizaron para plaquear virus Influenza se
crecieron en medio semisólido de plaqueo consistente en MEMsuplementado
con tripsina tratada con TPCK 2 pg/ml, DEAE-dextrán 300 ug/ml y agarosa 1%.
3. Compuestos utilizados
Los compuestos fueron adquiridos en Sigma-Aldrich USA. Las
soluciones madres se fraccionaron apropiadamente y se mantuvieron en -80°C.Acrilamida: solución madre de 100mM en MEM.
Hidroxiesteroles: soluciones madre 100 mMen etanol.
Mevalonato: solución madre 100 mM en etanol.
MetiI-B-ciclodextrina:solución madre 500 mM en agua.
Lovastatina: solución madre 10 mM en DMSO.
39

4. Obtención de virus purificada v marcado radioactivamente
Monocapas de células Vero crecidas a confluencia en botellas T75 se
infectaron con JUNV (MOI = 0.1) y se incubaron a 37°C durante 1 hora para
permitir la adsorción del virus. Luego se agregó MEMsuplementado con 1.5 %
de ST y se incubó durante 72 hs. p.i. AI concluir este período se colocó medio
de ayuno (MEMdepletado de los aminoácidos metionina y cisteína) durante 1.5
horas. Luego se incorporó directamente en el medio de ayuno una solución
radioactiva conteniendo Sss-metionina y S35-cisteína (Expre S35835, 100uCi/ml.
NEN, USA) y se incubó por un período de 24 hs. a 37°C. Por último se cosechó
el sobrenadante y se clarificóa 10000 x g durante 15 minutos.
Para la concentración se ultracentrifugó durantes 1.5 hs. a 120000 x g y
se resuspendió el pellet por sonicación. La muestra recuperada se purificó en
un gradiente de sacarosa discontinuo (15%-60°/o),recolectando la interfase con
pipeta pasteur y ultracentrifugando nuevamente. Las muestras fueron diluídas
en buffer TES (Tris 0.01M, NaCl 0.1 M, EDTA 0.001M, PH 7.4).
Se determinó la radioactividad asociada en un contador de centelleo y el
titqu infectivo para el virus purificado siendo 5.7 x107 dpm/ml y 4.3 x 107
UFP/ml respectivamente.
5. Ensayo de viabilidad celular: Método del MTT
Se utilizó el método colorimétrico descripto por Mosmann y col. (1983).
Monocapas de células Vero crecidas en microplacas de 96 cavidades se
trataron con distintas concentraciones del compuesto. Luego de 24 horas de
tratamiento se agregan 10 pl de una solución 5 mg/ml de M1T (3-4,5
dimetiltiazol-2-yl)-2,5-difenil bromuro de tetrazolio, Sigma-Aldrich, USA) en
cada cavidad.
EI compuesto MTI' es clivado en presencia de la enzima mitocondrial
succinato deshidrogenasa originando un subproducto coloreado (cristales
azules de formazán).
Transcurridas 2 horas a 37°C se retiran los sobrenadante, se colocan
200 pl de etanol 95%, se resuspende el precipitado y se mide la absorbancia
4o

M "xTFRI r‘il F‘: T ixfl:"i'(‘¡I‘¡ (3‘:
en un lector de ELISA a una longitud de onda de prueba de 492 nm y una
longitud de referencia de 690 nm.
Teniendo en cuenta que la adsorbancia de la solución en las muestras
es proporcional al número de células viables presentes al momento del ensayo
se determina la viabilidadde las monocapas. Cada concentracion se realizó por
triplicado. Se calculó la CC50,concentración del compuesto que reduce el 50%
de la viabilidad celular, con el promedio de las de absorbancia tomando como
control monocapas tratadas solo con el disolvente del compuesto.
6. Curvas de crecimiento
Cultivos de células Vero, PH o AM crecidos en monocapa se infectaron
con JUNV (MOI = 0.1) al alcanzar confluencia. A distintos días p.i. se
cosecharon los sobrenadantes y se titularon por el método de UFP en célulasVero.
7. Síntesis de groteínas
Para el estudio de la sintesis de proteínas celulares los cultivos de
células Vero se incubaron con distintas concentraciones de acrilamida en un
rango de 1 a 3 mM. Luego de 24 hs. de tratamiento los cultivos se Iavaron con
PBS y se realizó un pulso de marcación de 1 hora con aminoácidos
radioactivos (EXPRE835835, 1pCi/ml, NEN. USA) en presencia del compuesto.
Finalmente los cultivos se Iavaron con PBS frío y se precipitaron las proteínas
con ácido perclórico (PCA) 1.5%. Los cultivos se lisaron en SDS 1% - NaOH
0.1 M y se agregó a las muestras líquido de centelleo. La radiactividad
asociada se midió en contador WALLAC1400 DSA.
Para la determinación de la síntesis de proteínas virales los cultivos
infectados y tratados con acrilamida se marcaron metabolicamente, se
inmunopreciptaron y resolvieron en geles SDS-PAGE como se indica en la
sección 12.2.
4l

8. Titulacióndela infectividad vira/ por e/ método de UFP
8.1. Virus Junín
Se prepararon diluciones seriadas al décimo en MEM (1.5% ST) del
stock viral o de la muestra a titular. Monocapas de células Vero, crecidas en
microplacas de 24 cavidades, fueron infectadas con un volumen de inóculo de
100 pl por duplicado de cada dilución. Se deja adsorber el virus durante 1 hora
a 37° C, se retira el inóculo y se agrega medio semisólido de plaqueo. Las
microplacas se incuban por 7 días en estufa de C02. Transcurrido este periodo
se fijan las células en formol 10% por 20 minutos, se lavan con agua destilada
y se revelan las placas agregando cristal violeta al 1%.
Se determinó el título según la fórmula: UFP/mi = x / (dil. vol)
8.2. Virus Influenza
Se prepararon diluciones seriadas al décimo en MEM sin suero y
suplementado con 2 pg/ml tripsina tratada con TPKC. Luego de 1 hora de
adsorción a temperatura ambiente se retiró el inóculo y se incubó con medio de
plaqueo como se indica en Ia sección 2.4. La microplacas se incuban invertidas
a 33°C en estufa de C02 durante 72 horas. Transcurrido este período se fijan
las células en formol 10% por 20 minutos, se lavan con agua destilada y se
revelan las placas agregando cristal violeta al 1%. Los títulos se determinaron
como se indica en la sección 7.1.
9. Ensayo de inhibición del rendimiento viral
Monocapas de células crecidas en microplacas de 24 cavidades fueron
infectadas con virus Junín (MOI = 0.1) y luego de una hora de adsorción se
retiró el inóculo. Al tiempo adecuado las células fueron tratadas, por duplicado.
con distintas concentraciones del compuesto a evaluar y se mantuvieron a
37°C en estufa de COz hasta el momento de realizar la cosecha. Los
sobrenadantes se titulan por el método de UFP para el cálcqu de la infectividad
remanente. EI porcentaje de inhibición se calcula con respecto a los controles
infectados y sin tratar. Se calcula, además, la Clso como la concentración del
compuesto que reduce el 50% de la infectividad viral.

Í\L"\'I'Í":RI -\l 1-": T Mil-"l'fil‘fl‘fï
Para la titulación de infectividad asociada a célula se procesan las
monocapas, una vez cosechado el sobrenadante, lavando tres veces con PBS
y agregando MEM (1.5% ST). Luego se realizan dos ciclos de congelado y
descongelado y se centrífuga a 10000 x g por 10 minutos para separar los
restos celulares. Con el sobrenadante obtenido se procede a titular por UFP de
igual forma que con el sobrenadante extracelular.
10. Ensayo de acción virucida
Volúmenes iguales de una suspensión de JUNV conteniendo 1x106UFP
y una concentración adecuada del compuesto a evaluar se incubaron a 37°C
durante 90 minutos. Como control se incubó, en las mismas condiciones, una
suspensión viral con igual volumen de MEM.Luego se realizaron diluciones de
las distintas mezclas y se procedió a su titulación por UFP en células Vero,
tomando en cuenta que en las diluciones ensayadas sobre las células los
compuestos no ejercieran efecto antiviral. Se calculó para cada compuesto el
porcentaje de infectividadremanente respecto del control.
11. Acción del compuesto acrilamida a distintos tiemgos
11.1. Adición post-infección
Cultivos de células fueron crecidas en monocapas hasta confluencia y
luego infectados con JUNV (MOI = 0.1). A distintas horas p.i. se colocó MEM
conteniedo 2 mM de acrilamida. Se cosecharon los distintos sobrenadantes a
las 24 hs. p.i. se titularon por el método de UFP. Sobrenadantes de células
infectadas y sin tratar fueron cosechadas como control.
Otra serie de cultivos fueron infectados y luego del período de adsorción
se colocó MEMconteniendo acrilamida 2 mM. A distintos tiempos p.i. se retiró
el medio. se lavó con PBS y se colocó medio fresco. Todos los sobrenadantes
se cosecharon a las 24 hs. p.i. para su titulación por el método de UFP.
11.2. Adición por pre-tratamiento (PT)
Cultivos de células crecidas en monocapas hasta confluencia fueron
incubados con MEM(1.5% ST) conteniendo 2 mM de acrilamida durante 24, 8,
43

M ¿TFRI -‘\I I-“ï T ¡till-"I'OI‘K7‘:
6, 4 o 2 horas antes de ser infectados. Luego del período de pre-tratamiento seinfectaron los cultivos adsorbiendo el virus durante una hora a 37°C
(MOI = 0.1). Finalmente se retiró el inóculo se cubrieron las monocapas con
medio fresco y se cosecharon los sobrenadantes correspondientes a las 24 hs.
p.i. para ser titulados por el método de UFP.
12. Acción dela acrilamida sobre la adsorción e intema/¡zación del virus
Cultivos de células Vero crecidas en monocapa fueron infectadas con un
stock de JUNV marcado radiactivamente como se indica en la sección 4. Para
ello 100 ul de la suspensión (200000 dpm) se adsorbieron al cultivo, en
presencia o ausencia del compuesto, incubando a 4°C durante 1 hora. Luego
se retiró el inóculo, se lavó tres veces con PBS-BSA 0.2% y se lisaron los
cultivos con una solución de NaOH-SDS (0.1 N, 1% respectivamente) durante
30 minutos a temperatura ambiente. Se recogieron las muestras y se
resuspendieron en 2 ml de liquido de centelleo para cuantificar la radioactividadasociada.
Cuando se evaluó la internalización se procedió a adsorber eI inóculo y
lavar las monocapas como se mencionó anteriormente. Luego de los lavados
se incubaron los cultivos durante 1 hora a 37°C en PBS-BSA 0.2% para
permitir la internalización de los viriones marcados radiactivamente. Finalizado
el tiempo se retiró el medio y se adicionó una solución de proteinasa K en PBS
(1mg/ml) para eliminar el virus adsorbido pero no internalizado. Se incubó
durante 45 minutos a 4°C y se detuvo la reacción con 0.5 mI de una solución
PBS-BSA 3% adicionada con 2 mM de feniI-metiI-sulfonil-fluoruro (PMSF). Se
centrifugaron las muestras, se resuspendieron en 0.2 ml de NaOH-SDS y se
agregó liquido de centelleo para cuantificar la radioactividad como se mencionóanteriormente.
13. Estudio dela expresión de proteínas
13.1. Inmunofluorescencia indirecta (IFI)
Monocapas de células crecidas sobre cubreobjetos en microplacas de
24 cavidades fueron infectados con JUNV (MOI = 0.1). Luego de una hora de
44

adsorción a 37°C se retiró el inóculo y se trató con distintas concentraciones o
a distintos tiempos con una concentración fija de los compuestos. A las 24
horas p.i. se cosecharon los sobrenadantes para su titulaciónpor UFP y se Iavó
tres veces con PBS. Para realizar IFItotal o citoplasmática se fijaron las células
con metanol durante 10 minutos a una temperatura de -20°C. Para IFI de
superficie o membrana se fijó 10 minutos con formaldehído a temperaturaambiente.
Luego del período de fijación se realizan tres lavados con PBS, en
ambos casos. y se agrega 50 pg/ml de Ig de cabra por 15 minutos a 37°C como
bloqueante. Para revelar el antígeno viral se utilizaron anticuerpos
monoclonales reactivos contra GP1 y su precursor GPC o reactivos contra NP
(GBOB-BEOBo IC06-BE10 respectivamente, Sanchez et al. 1989). Para revelar
filamentos intermedios se utilizaron anticuerpos IgG anti-GFAP (Sigma-Aldrich,
USA) desarrollados en conejo o anticuerpos monoclonales anti-vimentina (clon
V9, Sigma-Aldrich, USA). Para Ia IFl de microtúbulos se utilizaron anticuerpos
monoclonales anti-a-tubulina (clon B512, Sigma-Aldrích, USA). Por último se
realizó una incubación con suero anti-lgG de ratón o conejo desarrollados en
cabra, conjugados con fluoresceina (Sigma-Aldrich, USA), por 30 minutos a
37°C.
Solo en los casos en los que se marcaron los filamentos de actina se
utilizó faloídína-FICT (Sigma-Aldrich, USA) y la técnica de inmunofluorescencia
directa. En todos los casos luego de un lavado final con agua destilada las
células fueron montadas en una solución tamponada de glicerol conteniendo
2.5 % de DABCO (1,4-diazabiciclo octano) como amplificador de la señal
fluorescente.
Los preparados se examinaron en un microscopio óptico Olympus
BX51. El porcentaje de células flourescentes en cada cubreobjeto fue calculado
de 20 campos ópticos seleccionados al azar, tomando como control las
monocapas infectadas y sin tratar.
En los ensayo de co-Iocalización los cultivos se incubaron
simultáneamente con el anticuerpo monoclonal para revelar el antígeno
vimentina y suero de cobayo policlonal monoespecifico reactivo contra Ia
glicoproteina viral. En Ia segunda incubación simultáneamente se agregaron
45

M -'\'I'FRI MJ“: Y MÚ'I'OI‘K‘)‘:
los anticuerpos específicos anti-ratón y anti-cobayo correspondientes. Los
preparados se observaron en un microscopio confocal Zeiss LSM510.
13.2. Marcado metabólico, inmunoprecipitación y SDS-PAGE
Monocapas de células Vero crecidas en microplacas de 24 cavidades
fueron infectadas con JUNV (MOI = 1). Finalizada la hora de adsorción se
incubó en MEM adicionado con 1.5% de ST. A las 48 hs. p.i. se colocó medio
de ayuno (MEMdepletado de los aminoácidos metionina y cisteína) durante
1.5 horas. Luego se incorporó una solución radioactiva conteniendo
Sas-metionina y Sas'cisteína (EXPRES35835, 100pCi/ml, NEN, USA) y se incubó
por un periodo de 3 hs. a 37°C. Finalmente las células se Iavaron con PBS, se
Iisaron y resuspendieron con 75 pl de buffer RIPA (NaCI 0.15M, SDS 0.1%,
Triton X-100 1%, PMSF 0.4 mM, aprotinina 10 pg/ml, deoxicolato de sodio 1%,
Tris-HCI 0.01M pH 7.4). Las muestras fueron almacenadas a -80°C hasta el
momento de ser utilizadas.
Para realizar la inmunoprecipitación se sonicaron los Iisados durante 5
minutos y luego se adicionaron los anticuerpos monoclonales correspondientes
a una dilución 1/100 incubando 30 minutos a 37°C y 90 minutos a 4 °C. Se
utilizó como medio de arrastre para los complejos inmunes una solución de
proteína A-sefarosa (100 mg/ml RIPA) y se incubó nuevamente 30 minutos a
37°C y 90 minutos a 4°C con agitación. Finalmente se realizaron tres ciclos de
lavado con buffer RIPA, se resuspendieron los inmunoprecipitados en buffer
muestra 4X (Tris-HCI 0.06M pH 6.8, SDS 5%, glicerol 10%, 2-[3-mercaptoetanol
2 %, azul de bromofenol 0.005%) y se incubó 1 minuto a 100°C conservando el
sobrenadante.
Para la resolución de las proteinas en minigeles de poliacrilamida
desnaturalizantes (SDS-PAGE)se sembraron las muestras en un concentrador
(4% de acrilamida en buffer Tris-HCl 0.5 M pH 6.8, SDS 10%) junto con
soluciones patrones de peso molecular. Las corridas en el gel separador (10 %
de acrilamida en buffer Tris-HCI 1.5 M pH 8.8, SDS 10%), se llevaron a cabo
durante 1.5 horas a 150 V y 90 mA. Una vez finalizada la electroforesis, los
geles se tiñeron con una solución de Coomassie Blue R250 al 0.25 °/oen
metanol: agua: ácido acético glacial (50:50:10) Luego de decolorar los geles se
procesan, para su posterior fluorografia, realizando lavados de 1 hora en
46

concentraciones crecientes de DMSO. Finalmente los mismos se incubaron
durante 3 horas en una solución de PPO (2,5-difeniloxazol, Sigma-USA)
amplificadora de señal, se secaron y se expusieron sobre una película AgfaCurix RPI a -80°C hasta su revelado.
13.3. Western b/ot (WB)
Las muestras a procesar por esta técnica fueron sonicadas durante 5
minutos y resuspendidas en buffer muestra SDS-PAGE 4X previamente a ser
resueltas en geles 10% como se describió en la sección 11.2. Luego setransfirió durante 1.5 horas a 30 mA sobre una membrana de PVDF
(Polyscreen, PerkinElmer Life Sciences Inc.) y se bloqueó durante una noche
en una solución de leche 5% en PBS a 4°C. Para revelar la presencia de la NP
del virus Junín se incubó con anticuerpos monoclonales específicos (|006
BE10) durante 1 hora a temperatura ambiente. Para revelar la presencia de la
HA de Influenza se utilizó un anticuerpo monoclonal H1 cedido gentilmente por
el Dr. Jorge Cámara (Instituto de Virología J. M. Vanella, Córdoba).
Finalmente se realiza la incubación con una dilución 1/1000 del segundo
suero anti-ratón peroxidasa (ICN) en iguales condiciones y se reveló con el
sistema de quimioluminiscencia ECL (Amershan-Pharmacia).
14. Reducción de/ colesterol y su efecto sobre la multiQ/icaciónviral
14.1. Acción de hidroxiesteroles inhibidores dela HMG CoA reductasa
Cultivos de células Vero se infectaron con JUNV (MOI = 0.1). Luego de
la hora de adsorción se agregó MEMconteniendo distintas concentraciones de
compuesto a evaluar y se incubó durante 24 hs. Se cosecharon los
sobrenadantes y el virus asociado a célula para su titulación por el método de
UFP.
14.2. Acción de Iovastatina y meva/onato + metiI-B-ciclodextn'na
Cultivos de células Vero crecidos hasta confluencia se incubaron con
una combinación de Iovastatina/mevalonato (L/M). Se incubaron durante 24
horas y luego se infectaron con JUNV (MOI = 0.1). AI finalizar el periodo de
47

adsorción se agregaron nuevamente los compuestos durante 24 hs p.i. y se
cosecharon los sobrenadantes para titular por el método de UFP.
En los casos indicados, luego de la cosecha, se realizó un pulso de
extracción con el compuesto metiI-B-ciclodextrina (MCD) 10 mM durante 30
minutos a 37°C. Finalmente los cultivos fueron fijados para IFI citoplasmática y
de membrana según se describe en la sección 13.1.
15. Cuantiflcacíón de colesterol en extractos celulares
Monocapas de células Vero crecidas en microplacas de 6 cavidades
fueron tratadas con los compuestos indicados o incubadas en ausencia de los
mismos por igual período de tiempo como control. Luego del tratamiento las
monocapas se lavaron con PBS y se levantaron las células con 250 ul de agua
estéril en cada cavidad y se colectaron por centrifugación.
Los lípidos totales fueron extraídos en todos los casos por el método de
Bligh y Dyer (1959). La muestras se homogeneizaron con una mezcla de
cloroformo: metano (1:2) hasta formar un sistema miscible con el agua de las
células. El homogenato fue diluido con cloroformo y agua (1: 0.8) formándose
dos fases. Después de la centrifugación la fase metanólica fue descartada y la
fase clorofórmica conteniendo los lípidos fue evaporada a sequedad con
nitrógeno. El colesterol se cuantificó en cromatografía de capa delgada. Este
estudio se realizo en el laboratorio Dr. R. lgaI de la Facultad de Ciencias
Médicas de la Universidad de La Plata.
16. Fraccionamiento en TX-100
16.1. Buffer PlPES/TX-100
Cultivosde células Vero crecidas en microplacas de 24 cavidades fueron
infectadas con JUNV (MOI = 1). Luego de la hora de adsorción se retiró el
inóculo y se incubo en MEM (1.5 °/oST) a 37°C. A las 48 horas p.i. se retiró el
medio, se Iavó con PBS, se realizó el ayuno y el marcado metabólico como se
indica en la sección 11.2. Las monocapas se extrajeron sobre hielo durante 10
minutos en buffer de extracción (PIPES 10 mM pH 6.8, KCI 100 mM, MgClz 2.5
48

M "x'I'FJZI -\I F‘: Y RÜZ'TOI‘K‘)‘:
mM, C8CI2 1 mM, EGTA 2 mM, sacarosa 0.3 M. PMSF 1 mM) adicionado con
TX-1OO 1%.
El material remanente sobre la microplaca se levantó con buffer RIPA. El
sobrenadante de la extracción se colectó y se le adicionaron los componentes
necesarios para igualarlo a buffer RIPA. Ambos materiales se
inmunoprecipitaron y resolvieron en geles SDS-PAGE como se indica en la
sección 13.2. para el revelado de las proteínas virales.
16.2. Buffer TNE/TX-100
Cultivos de células Vero crecidas en microplacas de 24 cavidades fueron
infectadas con JUNV (MOI = 1). Luego de la hora de adsorción se retiró el
inóculo y se incubo en MEM (1.5 % ST) a 37°C. A las 48 horas p.i. se retiró el
medio, se Iavó con PBS, se realizó el ayuno y el marcado metabólico como se
indica en la sección 13.2.
Las monocapas se extrajeron sobre hielo durante 30 minutos en buffer
de extracción TNE/TX-100 (Tris-HCI 25 mM pH 7.5, NaCI 150 mM, EDTA 5
mM, TX-100 1%, PMSF 0.4 mM, aprotinina 10 ug/ml, previamente enfriado)
durante 30 minutos a 4°C. Se recogió el lisado y se centrifugó durante 15
minutos a 12000 xg. El sobrenadante asi como el pellet se inmunoprecipitaron
y resolvieron en geles SDS-PAGE como se indica en la sección 13.2.
17. Estudio de microdominios de membrana rafts y su flotabilidad en
gradientes de sacarosa
17.1. Virus Junín
Se infectaron cultivos de células Vero en botellas T75 con JUNV
(MOI = 1), luego de la hora de adsorción se retiró el inóculo y se incubo en
MEM (1.5 °/oST) a 37°C. A las 48 horas p.i. se retiró el medio, se lavó con PBS,
se realizó el ayuno y el marcado metabólico como se indica en la sección 11.2.
Luego se retiró el medio, se lavó con PBS frio. se retiró el excedente
cuidadosamente y se extrajo con 600 ul de buffer de extracción TNEfTX-100
durante 30 minutos a 4°C o a 37°C (Manié y col., 2000) según el caso. Se
recogió el lisado y se llevóa 40% de sacarosa. Seguidamente se armó el resto
del gradiente con soluciones de 30% y 4% de sacarosa en TNE. Se
49

M "x'l'l-ZRIRU“: Y l’xflÏ'TODÜ‘:
ultracentrifugó a 200000 x g durante 21 horas a 4°C y se desarmó el gradiente
en 8 fracciones de 500u| desde la porción superior al fondo del tubo
(numeradas 1 a 8 respectivamente). Alícuotas iguales de cada muestra se
inmunoprecipitaron y sembraron en geles SDS-PAGE como se indica en la
seción 13.2. para revelar la presencia de las proteinas virales.
17.2. Virus Influenza
Cultivos de células MDCK crecidos en botellas T75 se infectaron
(MOI = 1) con la cepa AfTexas/36/91 de virus Influenza. Luego de la hora de
adsorción en MEM(suplementado con tripsina tratada con TPCK ) se retiró el
inóculo y se incubo a 37°C. A las 24 horas p.i. se retiró el medio, se Iavó con
PBS frio, se retiró el excedente cuidadosamente y se extrajo con 600 ul de
buffer TNE/TX-100a 4°C o a 37°C. Se sembró en gradiente y se ultracentrifugó
como se indica en Ia sección 17.1. Luego de desarmar el gradiente las
fracciones se analizaron por WB como se indica en la sección 13.3.
18. Estudio de la estabilidad de microdominios rafts
18.1. En presencia de metil-B-ciclodextrina
Se infectaron cultivos de células Vero en botellas T75 con JUNV
(MOI = 1), luego de la hora de adsorción se retiró el inóculo y se incubó en
MEM(1.5 °/oST) a 37°C. A las 48 horas p.i. se retiró el medio, se lavó con PBS,
se realizó el ayuno y el marcado metabólico como se indica en la sección 13.2.
Luego agregó al medio MCD 10 mM durante 30 minutos a 37°C.
Los cultivos se extrajeron con buffer TNE/TX-100 a 4°C y se sembraron
en gradiente de sacarosa como se indica en la sección 17.1. Las fracciones se
analizaron por inmunoprecipitación y resolución en minigeles SDS-PAGE como
en la sección 13.2.
18.2. En presencia de anticuerpos específicosSe infectaron cultivos de células Vero crecidos en botellas T75 con
JUNV (MOI = 1), luego de la hora de adsorción se retiró el inóculo y se incubó
en MEM (1.5 °/oST) a 37°C. A las 48 horas p.i. se retiró el medio, se lavó con
PBS, se realizó el ayuno y el marcado metabólico como se indica en la sección

KL-‘x'l'l-‘RI-‘\I F‘: Y MIÏ'IÏ ‘¡Df 3‘:
13.2. Luego agregaron anticuerpos monoclonales especificos para GP1 y GPC
(1/100), se incubó 30 minutos a 37°C y 15 minutos a 4°C. Finalmente se retiró
el medio, se Iavó con PBS frío, se extrajo con 600 pl de buffer TNE/TX-100 a
4°C o a 37°C y se sembró en gradiente de sacarosa según se detalla en Ia
sección 17.1. Las fracciones se analizaron por inmunoprecipitación y resolución
en geles SDS-PAGE como se indica en la sección 13.2.
19. Análisis de datos
Todos los experimentos fueron realizados por duplicado y repetidos almenos 3 veces. Los datos obtenidos fueron analizados estadísticamente
calculando las medias aritméticas y los errores estándar.

RESULTADOS

CAPÍTULO I

Participación del Citoesqueleto
en e] Ciclo de Multiplicación
de] Virusjum'n

A. Estudio dela interacción de JUNVcon los FiIamentos Intermedios
1. Caracterización delos sistemas celulares utilizados
1.1. Infección y productividad viral
Para cumplir con el primer objetivo de trabajo y estudiar la interacción
JUNV- FI se emplearon tres sistemas celulares. La línea celular Vero, utilizada
de rutina para la producción del virus Junín y la mayoria de los arenavirus, fue
el principalsistema utilizado en este trabajo. Esta linea donde se han realizado
la mayoría de las caracterizaciones del ciclo de multiplicación ¡n vitro es
fácilmente cultivable y en ella se alcanzan valores de producción viral
altamente aceptables. En estos cultivos se expresa el antígeno vimentina como
red principal de Fl. Se utilizó además un cultivo dipliode de fibroblastos
humanos (PH) con el propósito de ampliar los resultados obtenidos en otro
sistema celular que expresara el mismo tipo de filamento intermedio. Asimismo
cultivos primarios de astrocitos murinos (AM) caracterizados por la expresión
de GFAP fueron desarrollados con el propósito de comparar la funcionalidad de
dos tipos diferentes de Fl en su relación con el mismo virus.
Los dos sistemas antes mencionados no habían sido caracterizados
hasta el presente como sistema de replicación para JUNV por lo que
inicialmente se determinó la permisividad y productividad de los mismos
mediante curvas de crecimiento. Se observó además la morfología de Ia
monocapa durante el transcurso de los experimentos para Ia determinación del
grado de acción citopática (ACP). Todas las observaciones se realizaron
tomando como referencia cultivos controles sin infectar y con igual tiempo de
crecimiento en monocapa.
En todos los casos los cultivos se infectaron a una MOI= 0.1 para evitar
la presencia de particulas interferentes características y abundantes en las
infecciones con alta relación virus-célula. Luego de la adsorción los cultivos se
mantuvieron a 37°C en estufa de C02. A distintos tiempos p.i. se cosecharon
los sobrenadates y se evaluó la productividad por el método de UFP.
Como se observa en la Figura 10 todos los cultivos fueron sensibles a la
infección y productores de partículas infecciosas. Los valores de infectividad

lll-"lll! TUN)“ - ("KI’ÍTUI (‘i I
alcanzados en células Vero fueron concordantes con los descriptos en
bibliografía, estableciéndose el iniciode ACP al tercer día p.i.
La linea diploide PH mostró una curva similar al cultivo patrón, si bien
con titulos similares en los primeros días, los valores máximos de producción
viral alcanzados fueron menores. El aspecto de Ia monocapa fue siempre muy
denso. las células mostraron una morfología alargada orientándose en grupos
numerosos y paralelas a su eje mayor. No se observó un ACP característico o
definido en este tipo de cultivo.
10’ _10°; /./I\10‘1: '10H: \!:1o“! 9/910’];
Infectividad(UFPImI) 10"!10°' , . . . . fi r
t 1 2 3 4 5 6 7
Tiempo (dias p.i.)
Figura 10. Multiplicacióndel virus Junín en distintos sistemas celulares. Cultivos de células Vero
(A), astrocitos murinos (o) y fibroblastos humanos (o) fueron infectados con JUNV (MOI = 0.1). A
distintos dias p.i. se cosecharon los sobrenadantes y se titularon por el método de UFP.
Los cultivos de AM alcanzaron una buena confluencia al tercer dia de
sembrados, las células de aspecto irregular mostraron algunas prolongaciones
poco definidas. La curva de crecimiento viral en el cultivo primario de AM
mostró diferencias significativas con los dos sistemas antes descriptos. El
estado productivo alcanzado a las 24 hs. p.i. fue un orden menor al observado
en células Vero, asimismo fue mas lenta la evolución del crecimiento viral
manteniéndose constante el título del virus en el medio extracelular durante los
primeros días p.i. Al quinto dia se alcanzó el máximo de producción en el
sobrenadante y, aunque el título (1x105 UFP/ml) fue significativamente mayor
que los observados días anteriores, también fue dos órdenes de diferencia
menor que el alcanzado en el cultivo de células patrón Vero. Algunas
alteraciones morfológicas como redondeamiento celular fueron observadas en
56

RFCIII 'l'-\I‘¡(‘¡‘ï - ("KFÍTUI (31
el cultivo primario al tercer dia luego de la infección, variando este proceso muy
lentamente en los subsiguientes días. AI séptimo día se realizó Ia última
cosecha debido a que tanto los cultivos sin infectar como los infectados
mostraban un porcentaje de desprendimiento celular cercano al 30% de la
monocapa. Esta alteración inespecifica debido probablemente al
envejecimiento del cultivo aumentó con el correr de los días hasta el
desprendimiento total de la monocapa entre los días 9 a 11 p.i.
1.2. Determinación de viabilidad celular en presencia de acrilamida
Para el inicio del estudio de la participación de los FI en el ciclo de
multiplicación de JUNV se indujo la desorganización de esta red del
citoesqueleto utilizando el compuesto acrilamida. Como paso previo, sobre
cada sistema virus-célula descripto en la sección anterior, se determinó la
viabilidad celular a distintas concentraciones de acrilamida. La actividad
metabólica de los cultivos se tomó como signo de viabilidad evaluándose por el
método del M1T. Adicionalmente se observaron los cultivos al microscopio
óptico durante todos los períodos de tratamiento y en cada uno de los
experimentos. Los cultivos sembrados y crecidos hasta alcanzar confluenciafueron tratados durante 24 hs. con concentraciones crecientes de acrilamida en
un rango de 0 - 20 mM y teniendo en cuenta datos bibliográficos anteriores
(Arcangeletti y col., 1997). La Tabla lll muestra los valores de viabilidad para la
línea celular Vero. correspondientes a cada una de las concentraciones de
acrilamidaensayadas.
Acrilamida (mM)
-- 2 4 6 8 12 16 20
Absorbancia 0.833 0.774 0.615 0.325 0.255 0.236 0.212 0.206
Viabilidad (%) 100 93 70 37 29 24 27 23
'l’abla Ill. Viabilidad celular. Cultivos de células Vero crecidos en monocapas fueron tratados durante 24
horas con distintas concentraciones del compuesto acrilamida. AIfinalizar el tratamiento se determinó Ia
viabilidad por el método de MTT en un lector de ELISA. Los valores de absorbancia corresponden al
promedio de triplicados. El porcentaje se calculó con respecto a controles sin tratar.
57

[11"‘ïlll'l'-'\I‘¡(‘¡‘ï- (‘ KFÍ'I'III (‘1I
Como se puede observar la viabilidaddisminuye de manera dependiente
de la dosis de acrilamida. Los cultivos mostraron sensibilidad al compuesto
produciéndose redondeamiento celular a partir de la concentración 6 mM. Con
estos datos se obtuvo una CCso= 5.2 mM para este cultivo.
Teniendo en cuenta el efecto del compuesto sobre los cultivos de células
Vero se ensayó sobre el cultivo primario de AM y la línea PH un rango mas
acotado de concentraciones. Mediante el mismo ensayo de viabilidad
anteriormente utilizado (MTF) se determinó el efecto del compuesto sobre
estos cultivos. Los resultados se observan en la Tabla IV. Se muestra en la
misma que la viabilidad disminuye con Ia concentración del compuesto para
ambos cultivos, sin embargo, mientras la viabilidadde los astrocitos permanece
por encima del 70 °/o a las dosis ensayadas los cultivos de células PH
resultaron significativamente mas sensibles. A la concentración máxima
ensayada la viabilidad cayó al 50% con respecto a los cultivos controles. En
estas condiciones las cultivos presentaron una morfologíaalterada típicamente
redondeada, además alrededor de un 10 % de la monocapa se encontró
desprendida. En Ia máxima concentración ensayada ambos efectos se
pronunciaron significativamente como se corrobora por el valor de viabilidad
(37 °/o).Finalmente se muestran en Ia tabla los valores calculados para la CC50
en cada caso.
Acrilamida (mM)CCso
-- 0.25 0.5 1 2 3 4 6
Absorbancia 0.697 0.595 0.576 0.602 0.586 0.567 0.535 0.496E >6 mM< Viabilidad(%) 100 85 83 86 a4 81 77 71
Absorbancia 0.852 0.849 0.853 0.334 0.800 0.792 0.426 0.315
E 4mMViabilidad(%) 100 100 100 98 94 93 50 37
Tabla IV. Viabilidad celular. Cultivos primarios de AM y células PH crecidos en monocapas fueron
tratados durante 24 horas con distintas concentraciones del compuesto acrilamida. Al finalizar el
tratamiento se determinó la viabilidad por el método de MTT en un lector de ELISA. Los valores de
absorbancia corresponden al promedio de triplicados. El porcentaje se calculó con respecto a controlessin tratar.

FF‘ÏUÏ TADO‘: —F ‘ T'ÍTUÏ O I —
1.3. Efecto de la acrilamida sobre el citoesqueleto
La acción de la acrilamida sobre los elementos del citoesqueleto se ha
descripto en trabajos anteriores, sin embargo para corroborar y caracterizar su
efecto en los sistemas utilizados se determinó la expresión de los distintos
anfígenos mediante la tecnica de IFl.
Figura 11. Acción de Ia acrilamida sobre los filamentos intermedios.
Cultivos de células Vero (A y B), PH (C y D) o astrocitos murinos (E y F) fueron incubados durante 24 hs.
en ausencia (A, C y E) o presencia de acrilamida 2 mM (B, D y F). Los cultivos se lavaron con PBS, y se
fijaron con metanol durante 10 minutos a -20°C. Los filamentos intermedios se revelaron con anticuerpos
monoclonales específicos según se detalla en materiales y métodos. Aumentos x100.
59

Los cultivos controles y tratados con una concentración de acrilamida 2
mM fueron crecidos sobre cubreobjetos y fijados luego del tratamiento. Se
reveló luego con anticuerpos monoclonales específicos para FI.
En la Figura 11 se muestra las fotografias correspondientes. Como
puede observarse los delgados filamentos de vimentina forman una malla
perinuclear que se extiende hacia la periferia de la célula. Tanto en células
Vero como en PH (panel A y C) el patrón del antígeno es similar y comparable
a los descriptos para otros sistemas celulares.
Similares caracteristicas presentaron los filamentos de GFAP en los
cultivos de astrocitos. Aunque la red se muestra mucho mas definida en el
citoplasma pueden apreciarse también delgadas fibras en los procesos de
membrana o extenciones (panel E). Cuando los cultivos se trataron con el
compuesto el patrón de lFI de los antígenos se alteró completamente. La
Figura 11 (panel B, D y F) muesta un evidente acortamiento y desorganización
de los filamentos intermedios. Luego de 24 hs. en presencia de acrilamida
estos se asocian lateralmente formando acúmulos en el citoplasma.
Distintos trabajos mostraron la especificidad de la acción de este
compuesto sobre los FI, sin embargo para corroborarlo se analizó por
inmunofluorescencia el patrón de microtúbulos y filamentos de actina en
cultivos de células Vero.
Los cultivos fueron crecidos a confluencia y, luego de 24 hs. en
presencia de la concentración del compuesto que alteró Ia estructura del
antígeno vimentina, se procesaron y revelaron con los reactivos específicos
que se indican en materiales y métodos. Como se observa en la Figura 12 no
hay diferencias entre el patrón de MF en las condiciones control y los tratados
con acrilamida 2 mM (paneles A y B). Cuando se analizó el efecto del
compuesto sobre los MT (paneles C y D) tampoco se encontraron diferencias
en presencia o ausencia del compuesto. Para todos los campos examinados
tanto los microfilamentos como los microtúbulos se observaron definidos y
estructuralmente íntegros.

mmm TADO‘Z- rAPiTm o l —
Figura 12. Acción de la acrilamida sobre microtúbulos y microfilamentos.
Cultivos de células Vero controles (A y C) y tratados con acrilamida 2 mM (B y D). Luego de 24 hs. se
lavaron con PBS y se fijaron para su análisis por IFI. Se utilizó faloidina-FICT para el marcado de
microfilamentos (A Y B) y anticuerpos monoclonales para el marcado de microtúbulos (C y D) como se
indica en materiales y métodos. Aumentos x100.
2. Caracterizacióndela acción dela acrilamida sobre cultivos infectados
2.1. Acción dela acrilamida sobre las Qan‘ículasvirales infecciosas
Como primer paso a la utilización de un compuesto en un sistema viral
es necesario determinar su actividad virucida. La acción directa de un
compuesto sobre las partículas virales puede producir su inactivación debido a
cambios en la estructura del virión que luego modifiquen la infectividad del
mismo.
Dado que en los ensayos de 24 hs. las particulas se acumulan en el
sobrenadante celular y están en contacto directo con el compuesto durante ese
período se analizó su efecto sobre las mismas. Se realizó un ensayo virucida
exponiendo un alícuota del stock viral a una concentración 100 mM de
61

acrilamida durante 1.5 hs a 37°C. Luego de este período se tituló la infectividad
remanente por el método de UFP.
Los resultados indicaron que en las condiciones ensayadas no hubo un
efecto significativo del compuesto sobre los viriones. Los titulos del stock
control (5.1x105 UFP/ml) fueron comparables a los correspondientes al stock
tratado con acrilamida (4.8x 1o5UFP/ml).
2.2. Efecto dela acrilamida sobre la producción de partículas infecciosas
Una vez determinado el efecto del compuesto sobre los distintos
elementos del citoesqueleto teniendo en cuenta su efecto sobre la viabilidad
celular y su acción no virucida se ensayó la acrilamida sobre el sistema viral en
el rango de concentración de 0 a 4 mM. Para determinar el efecto de disrupción
de los FI sobre la producción viral se infectaron monocapas y luego se
incubaron durante 24 hs. en presencia del compuesto. Tanto en el caso de los
cultivos de células Vero como en los de PH el compuesto fue agregado
inmediatamente luego de la adsorción. Debido a los bajos valores de
infectividad obtenidos durante los primeros días p.i. en los cultivos primarios de
AM se colocó el compuesto sobre los mismos al tercer día p.i. La producción
viral alcanzada en los AM en este día es comparable a la producida en las
primeras 24 hs. de infección en el cultivo patrón de células Vero (Figura 10).
Luego del período de tratamiento los sobrenadantes celulares asi como
el virus asociado a célula fueron cosechados y titulados por el método de UFP.
La Figura 13 muestra el porcentaje de producción viral luego de 24 hs.
de tratamiento con respecto a las monocapas infectadas y sin tratar. Para todos
los sistemas ensayados la producción de particulas virales infecciosas
disminuye de manera dependiente de la dosis, alcanzándose una inhibicióndel90 % a la concentración de acrilamida 2 mM.
En estas condiciones la viabilidad de los cultivos fue superior al 90 %.
Los valores de inhibición de la multiplicación viral para el virus asociado a
célula fueron comparables a los del virus extracelular.

,_chf_wvr "a Wa__,n,fia, ,,mt V___v .7 l¡l“ll I\Ifll“ ("Pillilvi
—100
—60
Infectividad(%control)
(|011U°°°/o)pepiuqem
Acrilamida (mM)
Figura 13. Acción de la acrilamida sobre Ia producción de JUNV.
Cultivos de células Vero (A), astrocitos murinos (B) y fibroblastos humanos (C) fueron infectados con
JUNV. Luego de Ia adsorción se trató con distintas concentraciones de acrilamida. A las 24 hs. p.i. se
cosecharon los sobrenadantes y se tituló por el método de UFP. Paralelamente se determinó la viabilidad
celular en presencia de acrilamida por el método de MTT(o)
2.3. Acción de la acrilamida sobre distintas etapas del ciclo demultiplicación
En acuerdo con los resultados anteriores y para iniciar los estudios
tendientes a determinar la etapa del ciclo de replicación dependiente de la
integridad de los FI se seleccionó la concentración 2 mM de acrilamida dado
63

Rl5‘ïl l] 'l' “30‘: ‘(‘ \17Í'l‘lll () I —
que producía un efecto inhibitorio igual o mayor al 90% sobre la producción
viral (dependiendo del sistema celular analizado). Esta inhibición resultó ser
especifica dado que, en estas condiciones, los valores de viabilidad siempre se
mantuvieron por encima del 95 %. Asimismo, bajo estas condiciones, las
monocapas no mostraron signos morfológicosanómalos o no esperados.
Para analizar el efecto del compuesto sobre las diferentes etapas delciclo viral los cultivos de células Vero se trataron con acrilamida a distintos
tiempos p.i. Los sobrenadantes fueron cosechados a las 24 hs. y se titularon
por el método de UFP. La Figura 14 muestra la inhibición de la multiplicación
viral cuando Ia droga fue agregada a las 0, 2, 4, 6, 8, 12, 14, 17 y 21hs. p.i. y
los sobrenadantes de los cultivos se cosecharon a las 24 hs p.i. Los valores se
expresan como el porcentaje del control infectado y sin droga cosechado al
mismo tiempo.
100
Inhibición(%delcontrol)
0 2 4 6 8 12 14 17 21
Tiempo de adición (horas p.i.)
Figura 14. Efecto del tiempo de adición de acrilamida sobre la multiplicación de JUNV.
Monocapas de células Vero fueron infectadas con JUNV (MOI = 0.1). Luego de la adsorción y a
distintos tiempos p.i. se adicionó al medio acrilamida 2 mM. Se incubaron los cultivos hasta las 24
hs. p.i. para titular Ia infectividad por el método de UFP. Los datos se expresan como % del control
infectado, sin tratar y titulado a las 24 horas p.i. cuyo valor de inhibición corresponde al 0%.
Los resultados indican que la inhibición de la producción viral fue
máxima (96 0/o)cuando el compuesto fue agregado durante 24 hs. siendo estos
valores similares a los obtenidos en las curvas de inhibición realizadas
previamente. Sin embargo el efecto inhibitoriofue menor, alrededor del 50 °/o,al
ser agregado a partir de las 8 hs. p.i. Estos resultados podían estar indicando
64

RIWIH'l‘\13()‘3-(‘\|’Í'l'lll(Hi
que la integridad de los Fl era necesaria para la replicación normal del virus en
los primeros estadios del ciclo viral.
En otro experimento se realizó el ensayo inverso. Los cultivos fueron
infectados e inmediatamente luego de la adsorción se agregó acrilamida 2 mM
según se detalla en materiales y métodos. El compuesto fue retirado a distintas
horas, los cultivos lavados con PBS y cosechados los sobrenadantes a las 24
hs. p.i. Adicionalmente se realizaron tratamientos por distintos lapsos de tiempo
previo a Ia infección de las células (PT), cosechando luego el virus en el medio
extracelular a las 24 hs. p.i. La Figura 15 resume los resultados obtenidos.
La acción de la acrilamida mostró ser independiente del momento de
adición sobre los cultivos.
4nn¡vu
60
40
lnhibición(%delcontrol)
O O O N v (D oo\ \ \ \ \ x0'] C) O O O5; Y-8/0 0/24
-24/0
Tratamiento (horas)
Figura 15. Efecto de la acrilamida sobre distintas etapas del ciclo de multiplicación de JUNV.
Cultivos de células Vero fueron tratados por cortos períodos de tiempo con acrilamida 2 mM. El
compuesto se adicionó previo a la infección (PT), inmediatamente luego de la adsorción o a tiempos
tardíos. Luego del tratamiento se retiró el medio, se Iavó con PBS y se incubó hasta las 24 hs. p.i. Los
sobrenadantes se cosecharon luego de ese período para su titulación por el método de UFP.
Cuando el compuesto fue agregado durante cortos períodos de tiempo
durante las etapas iniciales de Ia infección o previo a ésta el efecto fue
maracadamente reducido. A modo de ejemplo los valores de inhibicióncuando
la acrilamida fue agregada 4 hs. antes de la infección o durante las 4 horas
subsiguientes a la misma son cercanos al 20 %. Estos datos sugirieron que la
presencia del compuesto durante un tiempo prolongado era necesaria para
65

interferir marcadamente con la producción de la progenie, indicando un tiempo
mínimo (22 a 24 horas de tratamiento) para producir valores de inhibición del
orden del 90 %. Se analizó el patrón de vimentina en cultivos de células Vero
tratados durante distintos tiempos con acrilamida 2 mM. Los ensayos de IFI
mostraron la agregación y alteración de la vimentina a partir de las 20 horas de
tratamiento. Estos datos muestran que se requiere un prolongado período de
tiempo para colapsar la red de Fl y consecuentemente reducir la multiplicación
de JUNV.
Solo alrededor de un 20 a 30 % de inhibiciónfue alcanzado en tiempos
breves antes y después de la infección de los cultivos (Figura 15), sin embargo
no fue posible descartar algún efecto directo sobre las primeras etapas del ciclo
de multiplicación del virus.
Para caracterizar el efecto de la acción prolongada y reversibilidad de la
arcrilamida sobre los cultivos infectados se realizó un ensayo de reversibilidad.
Se trataron los cultivos de células Vero durante 24 hs. con el compuesto,
lavando con PBS luego de este período y reemplazando por medio fresco para
continuar la incubación por otras 24 horas. Se realizó el tratamiento inverso y
además se incubó en presencia de acrilamida durante 48 hs (Tabla V).
Tratamiento (tiempo p.i.) Infectividad (UFP/ml) Inhibición (%control)
— 1.1 x106 —
Acrilamida 0 - 48 hs. 63 x102 99'9 0/0
Acrilamida24 - 48 hs 5.0 x104 955 %
Acrilamida0 - 24 hs. x105 33.7 %
Tabla V. Reversibilidad dela acción de Ia acrilamida.
Cultivos de células Vero fueron infectados con JUNV (MOI= 0.1). Luego de la adsorción se agregó MEM
o MEMconteniendo acrilamida 2 mM de acuerdo a los tiempos y tratamientos indicados. En todos los
casos se determinó la infectividad por el método de UFP a las 48 hs. p.i. Los valores de inhibición se
expresan respecto al control sin tratar.
En cada uno de estos puntos se procedió a la titulación de Ia infectividad
por el método de UFP. Cuando el compuesto se agrega desde las 24 a las 48
66

lll-"WII TKDÜ‘: - ("KPÍ'I'IJI f) I
hs. p.i. la inhibición resulta comparable a los tratamientos durante las primeras
24 hs. de infección. Los valores de inhibición luego de 24 hs. de retirado el
compuesto fueron cercanos al 34% indicando una reversibilidad del 66%.
El mismo ensayo se realizó sobre los cultivos de AM y células PH. En
ambos casos el efecto fue reversible con valores ligeramente menores a los
observados para los cultivos de células Vero (Tabla VI). La reversibilidad luego
de 24 hs. en ausencia del compuesto fue del 45% y 35% para PH y AM
respectivamente.
Tratamiento (tiempo lnfectividad (% inhibición)
P-i.) PH AM
1.1 x105 UFP/ml 2.5 x104 UFP/
Acrilamida o - 4a hs. 1-0 x103 UFP/ml (99.1%) 4.o x1o‘ UFP/ml (99.8%)
Acñlamma 24 _48 ha 7.9 x103 UFP/ml (92.8%) 4.9 x102 UFP/ml (98.0%)
Acruamma o _24 ha 5.ox1o‘ UFP/ml (54.5%) 8.7 x103 UFP/ml (65.2%)
Tabla VI. Reversibilidad de Ia acción de la acrilamida.
Cultivos de fibroblastos humanos (PH) o astrocitos mun'nos (AM) fueron infectados con JUNV (MO|=
0.1). Luego de la adsorción se agregó MEM o MEM conteniendo acrilamida 2 mM de acuerdo a los
tiempos y tratamientos indicados. En todos los casos se determinó la infectividad por el método de UFP
a las 48 hs. p.i. Los valores de inhibición se expresan respecto al control sin tratar.
Como siguiente paso se evaluó el requerimiento de la integridad de los
FI durante las etapas de adsorción e internalización del virus mediante ensayos
directos descriptos detalladamente en materiales y métodos. Se infectaron
cultivos de células Vero con virus marcado radioactivamante y luego se
determinó la marca asociada a células. Los cultivos se pre-trataron durante 24
y 4 hs. con acrilamida 2 mM.
En la Figura 16 se resumen los valores registrados en las distintas
condiciones. Como se puede observar los valores de adsorción en presencia
del compuesto son comparables a los valores control y a los obtenidos en los
cultivos pre-tratados. Asimismo. independientemente del tiempo de pre
67

Rlï‘ïlll 'l'\l‘)()‘3s(‘ \I7Í'l‘lll014
tratamiento el efecto sobre la integridad de los FI no demostró afectar el
reconocimiento virus-célula, paso inicial de la infección. Dado que la adsorción
no mostró requerir de una normal red de Fl la siguiente etapa analizada en el
ciclo de multiplicación fue la internalización de los viriones adsorbidos.
2oooo_
15000_
1oooo_
sooo_
3SS-JUNVasociado
¡O
control PT 24 hs PT 4 hs
Figura 16. Efecto del compuesto acrilamida sobre las etapas iniciales de la infección.
Cultivos de células Vero fueron pre-tratados (PT) con acrilamida durante 24 o 4 hs. previo a la
infección. Luego de este período se Iavaron los cultivos y se infectaron con 35S-JUNV (MOl = 0.1)
como se indica en materiales y métodos. Para determinar la adsorción (I) e internalización (I) se
lisaron los cultivos y se midió la radioactividad asociada. Cultivos sin tratar se infectaron y
procesaron de igual modo como control. Los valores se expresan como dpm promedio i SD.
Utilizando las mismas condiciones antes analizadas se determinó la
radioactividad asociada a los cultivos luego de una incubación a 37°C durante
1.5 hs. post adsorción. Como se observa en la Figura 16 la internalización de
los viriones en las células no fue afectada en presencia del compuesto. Los
valores se expresan en porcentaje del total de dpm adsorbidas en las
condiciones control. Alrededor de un 30% del virus que es adsorbido ingresa al
interior de la célula durante las primeras horas, en condiciones normales y a la
temperatura permisiva de 37°C. AI tratar los cultivos con el compuesto y
permitir la internalización del virus los valores de radioactividad incorporada no
variaron respecto de los controles. En presencia de Ia acrilamida el virus que
ingresó a la célula también representa un 30% del virus adsorbido.
68

lil“! ll 'l' \I)< T: —(‘ \I‘Í'l‘l ll (Hi
2.4. Expresión de los antígenos virales en gresencia de la acrilamida.
Figura 17. Acción de la acrilamida sobre las proteínas virales.
Cultivos de células Vero (C - H), PH (A y B) fueron incubados durante 24 hs. en ausencia (A, C, E y
G) o presencia de acrilamida 2 mM (B, D, F y H). Los cultivos se Iavaron con PBS, y se fijaron para
IFI. Las proteínas virales se revelaron con anticuerpos monoclonales específicos segun se detalla en
materiales y métodos. Nucleoproteína: A -D. Glicoproteinas: E —H. Aumentos x 40.
69

RI-“ïlll T-\D(‘)‘: —("KPÍ'I'HI (31
Dado que la acrilamida mostró no afectar el ingreso del virus a las
células se realizaron estudios para analizar las consecuencias de la
desorganización estructural de los FI sobre la expresión de antígenos virales
en cultivos infectados. Para ello se realizó IFI citoplasmática en todos los
cultivos tratados durante 24 hs. con acrilamida y luego fijadas para ser
revelados con anticuerpos monoclonales específicos.
En las condiciones control las células muestran el patrón de proteínas
virales característico. La NP presenta una típica fluorescencia punteada y
distribuida uniformemente en todo el citoplasma (Figura 17, panel A y C). Por
el contrario, la glicoproteína muestra un patrón difuso citoplasmático (panel E).
Cuando los cultivos fueron tratados se observó una disminución en el
número de células fluorescentes para el antígeno viral NP, esta disminución en
la expresión se observó tanto en los cultivos de células Vero (panel D) como en
los de PH (panel B). La reducción en la expresión de las glicoproteínas fue
similar en los cultivos procesados para inmunofluorescencia citoplasmática
(panel F) como de superficie (panel H). Se observa, entonces, que la expresión
de ambos antígenos virales está notablemente disminuida en presencia de la
dosis de acrilamida que modifica la estructura de los filamentos de vimentina.
Todos estos resultados fueron similares en los cultivos primarios de AM.
En la Tabla Vll se resume la cuantificación de células fluorescentes para
cada antígeno viral en cultivos de células Vero.
N° de células positivas“
Tratamiento IFIcitoplasmática IFIde superficie
Nucleoproteína Glicoproteína Glicoproteína
--- 72.4 :t 9.0 60.8 i11.5 66.3 :l:6.1
acrilamida 41.8 i 4.1 35.9 :t:5.2 38.0 i 2.8
(42 %)b (41 %)b (42 %)"
Tabla Vll. Cuantiticación del efecto de la acrilamida sobre Ia expresión de las proteinas virales de JUNV.
Células Vero infectadas y tratadas durante 24 hs. con el compuesto se procesaron para IFIy se revelaron
las proteínas virales indicadas con anticuerpos monoclonales específicos.
° Número (promedio :t SD) de células positivas contabilizadas de 20 campos para cada muestra
bPorciento de inhibición respecto de controles sin tratamiento.
70

RF‘ïllI 'I'-\I‘)(‘¡‘: —(‘RPÍTHI (‘i l
La disminución en el número de células positivas para NP y GP1, tanto
en el citoplasma como en membrana, fue cercano al 40 % indicando que la
perturbación de la integridad de los FI afecta la expresión normal de las
principales proteínas de JUNV. Sin embargo Ia alteración de esta red del
citoesqueleto no parece afectar el transporte a membrana de las glicoproteinas
como sí fue descripto para la alteración de MTy MF (Candurra y co|., 1999).
2.5. Síntesis de proteínas en presencia de acrilamida.
Debido a la disminución en la expresión de las principales proteinas del
virus, observada por el ensayo de IFI, se quiso determinar si el tratamiento con
el compuesto podia tener un efecto sobre la síntesis de proteinas celulares y/o
virales. Para evaluar Ia sintesis de proteínas celulares se marcaronmetabolicamente cultivos de células Vero tratados con concentraciones
crecientes de acrilamida. Luego los Iisados se precipitaron con PCA y se
determinó Ia radioactividad asociada a los mismos. Como se indica en la
Tabla VIIIlos valores de dpm asociados a los cultivos crecidos en presencia y
o ausencia de acrilamida no difieren significativamente.
Acrilamida (mM)
-- 1 2 3
dpm ( X i SD) 43299 i 4765 44661 i 5505 42230 i 4765 46027 i 1812
Tabla Vlll. Síntesis de proteinas celulares. Cultivos de células Vero fueron marcados metabolicamente en
presencia de distintas concentraciones de acrilamida o en ausencia de Ia misma durante 24 horas. Luego
de este periodo se lavaron los cultivos y se los proceso para determinar la sintesis de proteínas por el
método de precipitación con PCA.
El virus Junín no produce shutt-off celular y, si bien posee su propia
polimerasa para el inicio del ciclo replicativo, hasta el momento no se ha
descripto que posea la capacidad de regular la síntesis de sus propias
proteínas de manera independiente de la síntesis celular. Sin embargo, para
descartar el hecho de que la desorganización de los FI pudiera tener algún
efecto especifico y desconocido sobre la síntesis de proteinas virales, se
realizó un ensayo de marcado metabólico en presencia del compuesto. Para
7l

Rlï‘ïlll 'I'\I‘)(‘)‘1 AF \l’Í'l'lll l) l —
ello se trataron cultivos infectados con acrilamida 2 Mm. Luego de 48 hs. p.i se
marcaron radioactivamente y se procesaron los Iisados celulares para su
posterior inmunoprecipitación con anticuerpos monoclonales. Las muestras se
separaron en SDS-PAGE 10%, en Ia Figura 18 se muestran las fluorografías
correspondientes.
1 2 3 4I66> 1 GPCa, É NP45> v” r, aq #43
. <2: GP1
25> - - “5
Figura 18. Sintesis de proteinas virales.
Cultivos de células Vero se infectaron con JUNV (MOI = 1) y se incubaron durante 24 hs. En
ausencia (calles 1 y 2) o presencia de acrilamida 2 mM (calles 3 y 4). A las 48 hs. p.i. se realizó un
maracado metabólico. Los Iisados correspondientes se inmunoprecipitaron con anticuerpos
monoclonales especificos para el revelado de las glicoproteinas (calle 1 y 3) o la nucleoproteina
(calle 2 y 4).
Como se puede observar el patrón de bandas correspondientes a la
nucleoproteina y sus productos de degradación (43 y 25 kDa) fue comparable
entre las muestras tratadas (calle 4) y las muestras control (calle 2). Por esta
metodología tampoco se observan efectos significativos sobre la síntesis de
GP1 y su precursor GPC (calles 1y 3) en presencia del compuesto.
2.6. Interacción filamentos de vimentina-Qroteínas virales.
Dado que el compuesto acrilamida no mostró efectos sobre la síntesis
de proteínas virales o celulares y sin embargo se observó que la
72

RI-‘QUI'I‘ADÜ‘:-(‘-\I7ÍTUI (‘11
desorganización de los FI produjo una disminución de Ia expresión de la
glicoproteína viral en la superficie de las células infectadas se quiso explorar la
posibilidad de que algún paso en el transporte de esta proteína hacia la
membrana dependiese de la red de FI.
Varios trabajos describen la asociación de FI con complejos proteicos
relacionados con el transporte, transcripción y/o traducción de ARN
mensajeros. Se ha descripto además Ia interacción, debido a una asociación
directa, entre proteínas virales y elementos del citoesqueleto. Principalmente
varias glicoproteínas han sido descriptas en su asociación con el citoesqueleto
durante el transporte a membrana. Para analizar la posibilidad de que GPC o
aún GP1 se asociara en estadios tempranos o tardíos del ciclo de replicación
viral a los filamentos de vimentina se realizaron ensayos de co
inmunoprecipitación. Los cultivos se infectaron, a las 48 hs. p.i. se marcaron
metabolicamente y luego se inmunoprecipitaron con los anticuerpos anti
viementina, anti-GPC y GP1 o anti-NP utilizados en los experimentos de IFI.
Las muestras fueron sembradas en geles SDS-PAGE para su resolución, sin
embargo, en las fluorografías correspondientes no pudo observarse ninguna
banda adjudicable a la vimentina en presencia del suero especifico. Aún
cuando se variaron las concentraciones de anticuerpo y algunas condiciones
de inmunoprecipitación, no pudo evidenciarse la banda control correspondiente
a ésta proteína. Las muestras inmunoprecipitadas con anticuerpos reactivos
contra las proteinas virales tampoco co-precipitaron bandas en el rango de lavimentina.
Se realizaron además extracciones en buffers específicos para el
enriquecimiento del citoesqueleto en Fl. Los cultivos extraídos se analizaron
por la técnica de IFI sin poder evidenciar proteínas virales en los mismos.
lguales resultados se observaron cuando las mismas muestras fueron
analizadas por inmunoprecipitación con anticuerpos específicos para las
proteínas virales.
Finalmente se realizaron ensayos de co-inmunomarcación como otra
alternativa para determinar alguna posible asociación entre las glicoproteínas
virales y los FI.

RI<"3l‘l'I‘ \l)()‘3 —(‘\I‘Í'l'l'l (>17
Los cultivos se infectaron y a las 24 hs. p.i. se fijaron para realizar
inmunoflourescencia de doble marcación. Se observa en Ia Figura 19 el patrón
tipico de filamentos de vimentina (panel B) en células Vero. En ei panel A se
muestra el patrón de IFI correspondiente a la glicoproteína viral. En Ia imagen
superpuesta de los dos fluorocromos no se observa co-Iocalización de los
antígenos por Io que no habría interacción entre ambos tipos de proteínas, al
menos en los tiempos y condiciones ensayados en este trabajo.
Figura 19. Inmunocoiocalización de proteínas virales y vimentina.
Cultivos de células Vero se infectaron con JUNV (MOI= 0.1) y luego de 24 hs. se procesaron
para IFI con anticuerpos específicos para el revelado de las glicoproteínas (panel A) o de
vimentina (panel B). Panel C: superposición de imágenes.
74

B. Fraccionamiento celular y proteinas virales
1. Fraccionamiento con detergentes no iónicos
1.1. Localización de las proteínas virales
En los últimos años se ha descripto Ia asociación de distintas proteinas
virales a fracciones resultantes de extraer los cultivos celulares con diferentes
detergentes, condiciones de salinidad y/o temperatura.
Las fracciones celulares que presentan insolubilidad a detergentes no
iónicos se encuentran enriquecidas en material perteneciente al citoesqueleto.
Con el objetivo de determinar la distribución de las proteinas virales en este
tipo de fracciones se realizaron extracciones a 4°C en buffer PIPES/TX-100 1%
adicionado con agentes estabilizantes del citoesqueleto.
Los cultivos fueron infectados con JUNV y marcados metabolicamente a
las 48 hs. p.i. Luego se extrajeron durante 10 minutos recogiendo el
sobrenadante (soluble). EI material remanente sobre Ia microplaca se levanto
en buffer RIPA (insoluble) y se inmunoprecipitaron para la determinación de las
proteinas virales en ambos tipos de muestras como se indica en materiales ymétodos.
La Figura 20 muestra los resultados obtenidos luego de la resolución de
los inmunoprecipitados en SDS-PAGE y su posterior fluorografia. Las calles 1 y
2 corresponden a muestras provenientes de cultivos sin infectar y las calles 3 y4 a las muestras infectadas con JUNV. Se observan tanto en el S como en l
algunas bandas celulares arrastradas por los anticuerpos monoclonalesutilizados.
Las condiciones del buffer de extracción mostraron la estabilización del
citoesqueleto como se puede observar por la presencia de la banda
correspondiente a la actina (45 kDa) en las muestras resistentes a la extracción
(calles 2 y 4). En las condiciones del ensayo se puede observar que la
nucleoproteína viral se encuentra asociada completamente a Ia fracción
insoluble en TX-100. Sus productos de degradación usualmente detectados
con esta técnica también se encuentran preferencialmente en la fracción
75

Rlï‘ïl Il 'l' \I)( )‘3—(' \PÍ'l'lll () l —
resistente a la extracción (panel A, calle 4). Por el contrario GP1 y su precursor
GPC se solubilizaron en estas condiciones (panel B, calle 3).
Figura 20. Fraccionamiento en buffer PIPES/ TX-100.
Cultivos de células Vero sin infectar (calles 1 y 2) e infectados (calles 3 y 4) con JUNV (MOI = 1) se
marcaron metabolicamente a las 48 hs. p.i. Luego de lavar las monocapas con PBS se extrajeron
durante 10 minutos sobre hielo en buffer adicionado con TX-1OO1%. El material en el sobrenadante
(S) asi como el material remanente luego de Ia extracción (I) se inmunoprecipitó con anticuerpos
específicos para el revelado de las proteínas virales. Panel A: anticuerpos anti-nucleoproteína. Panel
B: anticuerpos anti-glicoproteínas.
La localización de una proteina en la fracción resistente a la extracción
es un fuerte indicativode que la misma se encuentra asociada al citoesqueleto.
Sin embargo, para corroborar que condiciones podian alterar esta asociación
se realizaron extracciones a distinta temperatura o con la adición de alta fuerza
iónica. En ambas condiciones los Iisados se fraccionaron luego de Ia extracción
por centrifugado a 10000 xg. Las muestras se procesaron para WB utilizando el
mismo anticuerpo que en los experimentos de inmunoprecipitación para el
76

¡“ml -¡'i' \l)r rr . (‘ wí'i'i 'u) l
revelado de NP. Como se observa en Ia Figura 21 tanto las condiciones de alta
salinidad como temperatura solubilizaron parcialmente a NP sugiriendo
nuevamente su asociación al citoesqueleto.
NaCI 37°C 4°C
Figura 21. Fraccionamiento de NP en distintas condiciones de extracción.
Cultivos de células Vero infectados con JUNV (MOI = 1) durante 48 hs. se extrajeron con buffer
PIPESfl'X-100 1% durante 10 minutos en condiciones de alta salinidad (NaCl 1M), y a distintas
temperaturas (37° o 4°C). En todos los casos los Iisados obtenidos se centrifugaron a 10000 xg
separándose el sobrenadante (S) del pellet (P). Todas las muestras se revelaron por la técnica de WB
con anticuerpos especificos para Ia nucleoproteina viral.
La localización de una proteína en Ia fracción soluble a TX-100 puede
indicar su presencia en la fracción celular Iipídica o simplemente en el soluble
celular. Sin embargo Ia localización en Ia fracción soluble de una proteína no es
tampoco prueba de su no asociación al citoesqueleto. Su interacción puede no
estar estabilizada o favorecida en las condiciones de extracción. Candurra y
col. (1999) hallaron a GP1 preferencialmente en Ia fracción insoluble de
extractos de células Vero. Dado que las soluciones de extracción utilizadas en
este trabajo no fueron las mismas y tanto GP1 como GPC mostraron estar
presentes en Ia fracción soluble se realizó un experimento comparable al
realizado para NP con el objetivo de explorar el comportamiento de las
glicoproteínas en distintas condiciones de extracción y su relación con la
localización celular.
Cultivos de células Vero fueron infectados con JUNV y marcados
metabolicamente a las 48 hs. p.i. Luego de este período se extrajeron en
77

RI‘QI lI 'l' \l)( )Íï —(‘M‘Í'l‘l ll () I —
condiciones de alta salinidad (NaCl 1 M) y diferentes temperaturas (4 o 37°C).
Las muestras se inmunoprecupitaron con anticuerpos reactivos contra GPC y
GPl y luego se resolvió en SDS-PAGE. La Figura 13 muestra las fluorografías
correspondientes. En todas las condiciones ensayadas las glicoproteínas de
JUNV mostraron una distribución similar. Ambas proteínas se encuentran
preferencialmente en el sobrenadante de extracción.
25)
NaCI 37°C 4°C
Figura 22. Fraccionamiento de GPC y GP1 en distintas condiciones de extracción.
Cultivos de células Vero infectados con JUNV (MOI = 1) durante 48 hs. se extrajeron con buffer
PIPES/TX-1OO 1% durante 10 minutos en condiciones de alta salinidad (NaCI 1M), y a distintas
temperaturas (37° o 4°C). En todos los casos los Iisados obtenidos se centrifugaron a 10000 xg
separándose el sobrenadante (S) del peIIet (P). Todas las muestras se revelaron por Ia técnica de
inmunoprecipitación con anticuerpos específicos para las glicoproteínas GPC y GP1.
78

CAPÍTULO II

Interacción de] Virusjunín
con Ia Membrana de 12Célula Huésped

A. Estudio del colesterol y su relación con el ciclo de multiplicación viral
1. Colesterol e hidroxiesteroles
1.1. Acción sobre la viabilidad celular
Para la utilización de los hidroxiesteroles sobre los cultivos de células se
determinó primero su efecto sobre la viabilidad celular. Se trataron monocapas
de células Vero con concentraciones crecientes de los compuestos y luego de
24 hs. se determinó la viabilidad por el método de MTI'.
Como se observa en la Tabla IX los hidroxiesteroles no mostraron
efectos significativos sobre los cultivos en los rangos de concentración
ensayados así como tampoco concentraciones semejantes de colesterol.
Concentración (pM)
-- 25 50 100 150 200 Compuesto (CCso)
Absorbancia 0.560 0.557 0.552 0.562 0.548 0.541 -,_OH
Viabilidad (%) 100 99 98 100 97 96 ( > 200 llM)
Absorbancia 0.560 0.548 0.530 0.515 0.487 0.314 200"
Viabilidad (%) 100 94 94 92 87 56 ( > 200 PM)
Absorbancia 0.560 0.564 0.685 0.532 0.612 0.562 250...
Viabilidad (%) 100 100 100 95 100 100 ( > 200 ilM)
Absorbancia 0.560 0.499 0.543 0.507 0.459 0.514 cueste,“
Viabilidad (%) 100 89 97 90 82 91 ( > 200 llM)
Tabla IX.Acción de hidroxiesteroles sobre la viabilidad celular.
Cultivos de células Vero fueron tratados durante 24 hs. con distintas concentraciones de un hidroxiesterol.
Luego de este periodo se determinó la viabilidad celular por el método de M'I'i'. Los valores de viabilidad se
expresan como porcentaje del control correspondiente a cultivos expuestos a la concentración de solvente
(etanol) presente en la máxima concentración de compuesto analizada. Los valores de absorbancia
corresponden al promedio de triplicados.
81

mmm 'r,\r,(‘,'= . (‘-‘\PÍ'I'lil (‘i ll
Asimismo cuando se observaron los cultivos al microscopio óptico no se
detectaron cambios morfológicos significativos con respecto a los cultivos
controles, durante todos los períodos de tratamiento y para cada uno de los
compuestos utilizados. En todos los casos no pudo determinarse la C050 hasta
las concentraciones ensayadas
1.2. Acción sobre la multiplicación viral.
Una vez determinada Ia viabilidad de los compuestos sobre las células
se evaluó el efecto de los derivados hidroxilados del colesterol 20 a, 7B y 25
OH colesterol sobre la multiplicación de JUNV.
Los hidroxiesteroles fueron agregados al medio de cultivo luego de la
adsorción viral en concentraciones entre 5 - lOOuM. A las 24 hs. p.i. se
cosecharon los sobrenadantes y se determinó la infectividad por el método de
UFP (Figura 23). Para el 20 a-OH como para el 25-OH colesterol se obtuvo
una reducción dosis-dependiente alcanzándose inhibiciones del 85% en la
infectividadviral para las mayores concentraciones ensayadas, con Clsoentre
20 —40 uM.
El virus Junin no posee formas intracelulares infectivas sin embargo al
titular el virus asociado a célula en condiciones control usualmente se observan
valores de infectividad 1 logaritmo inferior a los encontrados en el
sobrenadante. EI virus asociado a célula representaría Ia proporción de virus
que se encuentra brotando de la membrana celular pero aún no se ha
separado físicamente de ella. Cuando se tituló el virus asociado a célula en
presencia de los hidroxiesteroles no se observó una disminución en la
infectividad del virus asociado a célula en el rango de concentraciones
ensayado sugiriendo que en presencia de los compuestos se altera el paso debrotación.
El compuesto 7B-OH colesterol no mostró una actividad antiviral
significativa en las condiciones del ensayo. Por último tampoco el colesterol por
si mismo no mostró tener un efecto negativo sobre la infectividad de JUNV.
82

Infectividad(logUFP/ml)
Infectividad(logUFP/ml)
Rlï‘ïl ll 'l"\I‘)(‘)‘z —(‘ \PÍ'l‘l Jl Ü ll —
-100
-100
I I I l I l l I I I l
._._. —100 o—.\.I\ <CI\«ox.I- —
m É '\.5,5- -80 n- 5‘5- \. -80\ a “5 \m- .\ _eoE: g! I \ -60\I— O _ \'_'- \ v 5'0_30 l\\\‘ ' l ÉL 1: \\\\I ¡————-———————————=N
- 4o g E —4oo >45- g É: 45
» 20 q. 8 —20A g “E BV _
A n n A fl ni | l | | l I i I I l l l I
o 20 4o so so 100 o 20 4o so so 100
20-OH colesterol (pM) 25-OH colesterol (pM)
E n 1: nv I I I I I ' l v l I ' l ' l l r._._.\'/ -100 2 ._._.\/,/'\.,——I/.\' < E I /II -' \s a / -\-\.
5,5 — - 80 :: u. 5‘5_ I - soa: DN m
-5° 2 3, -eo
&0— ,zr""'_.\\\\\\\\\ se 15 ío‘I.——___________——I CL N \' _4o(_D_ I'\./'\I/. _40
o .245v g 13 45
—2o :- 2 —20C °_ E DV
4 n ' ' ' ' ' ' n n l I I I I I n° 2° 4° 50 80 100 o 2o 4o 60 so 100
7B-OHcolesterol colesterol (pM)
Figura 23. Efecto de hidroxiesteroles sobre la infectividad del virus Junín.
Cultivos de células Vero fueron infectados con JUNV (MOl = 0.1) y tratados con distintas concentraciones
de 20-OH colesterol (A), 25-OH colesterol (B), 76-OH colesterol (C) o colesterol (D). Luego de 24 hs. se
cosecharon los sobrenadantes y se titularon por el método de UFP (I). Para el virus asociado a célula se
determino la infectividad por igual método (I). Otro grupo de cultivos en iguales condiciones fueron
utilizados para determinar la viabilidad celular (I).
1.3. Efecto sobre la expresión de proteínas virales
Cultivos de células Vero se infectaron con JUNV y luego del tratamiento
con los hidroxiesteroles se procesaron para realizar un ensayo de
inmunofluorescencia indirecta. Las fotografías correspondientes al compuesto
20-OH colesterol se pueden observar en la Figura 24. Para el marcado de la
IFI
monoclonal específico (paneles A y B). En este caso no se observan
nucleoproteína se realizó citoplasmática utilizando un anticuerpo
diferencias entre los cultivos controles y los tratados.
83
(Ionuoolap%)pep"!qu
(Ionuoolap%)pepuiqem

RF‘ïlJIT mas - (“APÍTIIÏo n
Los cultivos revelados con anticuerpos monoclonales reactivos contra la
glicoproteína viral no mostraron diferencias significativas en la flourescencia
citoplasmática. Sin embargo al analizar los preparados fijados para IFl de
membrana se observa una reducción en el número de células fluorescentes
con respecto a los controles sin tratar (paneles C y D). La disminución se
observa tanto en los tratamientos con 25-OH como con 20-OH colesterol.
Figura 24. Expresión de la glicoproteína y la nucleoproteina viral en presencia de hidroxiesteroles.
Cultivos de células Vero se infectaron con JUNV (MOI= 0.1) y luego de la adsorción cultivos controles (A
y C) y tratados con 20-OH colesterol (B y D) se incubaron durante 24 hs. Los cultivos se fijaron para IFI
de membrana (C y D) o citoplasmática (A y B) como se indica en materiales y métodos. Para el revelado
de las proteínas virales se utilizaron anticuerpos monoclonales específicos. Aumentos x100 (A y B), x40
(C y D).
Para Ia cuantificación del efecto de los hidroxiesteroles sobre las
proteínas virales se contabilizó en cada caso el número de células
fluorescentes. En la Tabla X se resumen los resultados obtenidos por campo y
los porcentajes respecto al control en ausencia de los compuestos.
84

PF‘ïl Il T “30‘: —(‘ M’ÍTIII 0 Il
Como se puede observar la expresión de la nucleoproteina viral, en
presencia de los compuestos, no fue significativamente alterada mientras que
Ia glicoproteína disminuye su expresión en Ia membrana plasmática.
Glicoproteina Nucleoproteína
n° de células n° de célulasCompuesto % control % control
fluorescentes fluorescentes
— 64.2 :t 5.5 100 59.2 i 8.3 100
20-0H colesterol 28.3 i 6.3 44 52.5 :t 7.3 100
25-0H colesterol 27.0 i 3.5 42 53.8 :t 3.4 100
Tabla x. Expresión de proteínas virales determinado por inmunofluorescencia indirecta.
Cultivos de células Vero se infectaron con JUNV (MOI = 0.1) luego se trataron durante 24 hs. con el
hidroxiesterol correspondiente. Las monocapas se fijaron para IFI total o de superficie y se determinó la
presencia de las proteinas virales con anticuerpos monoclonales especificos. Las muestras se observaron
en microscopio óptico y se contabilizó, para cada caso, el número de células fluorescentes de 20 campos
seleccionados al azar. Los resultados se expresan como el promedio :l: SD provenientes de al menos 3
experimentos independientes.
2. Reducción en Ia síntesis de colesterol por acción de la lovastatina
2.1. Acción sobre la viabilidad celular
Con el objeto de reducir de manera significativa la cantidad de colesterol
en Ia membrana plasmática se utilizó lovastatina, droga descripta como
inhibidora específica de la enzima 3-HMG-COAreductasa. Previamente a la
infección de los cultivos se realizó un ensayo de viabilidad celular.
Debido a que las células animales no pueden prescindir completamente
del colesterol se combinó el tratamiento de lovastatina con el compuesto
mevalonato, precursor de la síntesis del mismo. De esta manera aunque las
concentraciones de colesterol pueden reducirse significativamente en el
sistema siempre existe una cantidad compatible con la viabilidad celular.
Para determinar las concentraciones adecuadas de cada componente ennuestro sistema se sometieron distintos cultivos a una serie de combinaciones
85

de tratamientos con lovastatina en un rango de 0-80 pM y mevalonato de 0
0.25 mM. Se siguió la evolución de las monocapas por microscopio óptico
durante 48 hs a fin de determinar Ia presencia de cambios morfológicos
anormales en las mismas y se determinó la viabilidad de los cultivos por el
método de MTT. Como se muestra en Ia Tabla XI la viabilidad celular en el
rango de concentraciones de lovastatina disminuye hasta un 60 % solo cuando
no hay presente mevalonato en el medio. En el resto de los casos se mantiene
siempre por encima del 90 %.
Lovastatina (pM)Mevalonato
5 10 20 40 80(mM)
-- 0.656 i 0.05 0.507 i 0.02 0.499 d:0.05 0.467 i 0.02 0.431i 0.08 0.407 i: 0.02
(100 %) (77 %) (76 %) (71 %) (65 %) (62 %)
0_125 0.659 :L-0.02 0.618 :l:0.02 0.630 :t 0.02 0.615 i 0.05 0.612 :l:0.07 0.594 á:0.08
(100 %) (94 %) (96 %) (94 %) (93 %) (90 %)
0250 0.650 :i:0.08 0.651 :t:0.05 0.648 t 0.03 0.646 i 0.02 0.641 :t 0.08 0.633 :t 0.05
(100 %) (99 %) (99 %) (98 %) (98 %) (96 %)
Tabla XI.Viabilidadcelular en presencia de compuestos inhibidores de la sintesis del colesterol.
Cultivode células Vero se trataron con concentraciones crecientes de lovastatina y mevalonato durantes 48
hs. luego de este período se determinó la viabilidad celular por el método de MTT.Los resultados se expresan
como la absorbancia promedio de triplicados :t SD. El porcentaje viabilidad celular calculado respecto a los
controles sin tratar se muestra entre paréntesis.
En cuanto a los cambios morfológicos solo se observaron a partir de las
24 hs. de incubación células redondeadas en las concentraciones mayores delovastatina en ausencia de mevalonato. Estas alteraciones aumentaron
progresivamente hasta las 48 hs. comprometiendo hasta un 30% de la
monocapa. Teniendo en cuenta datos anteriores y los propios resultados de
viabilidad celular se decidió utilizar para el resto de los experimentos la
concentración de lovastatina 10 uM combinada con la menor concentración de
mevalonato ensayada (0.125 mM).

FF‘ïIlI TADÓ‘Ï —(‘KT’ÍTUI O II
2.2. Acción sobre la producción viral
Se ensayó la combinación de Iovastatina/mevalonato (L/M) elegida
anteriormente sobre cultivos infectados y se evaluó su efecto sobre la
producción de partículas infecciosas. Los cultivos se trataron con L/Mdurante
24 hs., luego se los infectó y se realizó el tratamiento durante las siguientes 24
hs. p.i. al término de este tiempo se cosecharon los sobrenadantes y se tituló la
infectividad en los mismos por el método de UFP. Los títulos calculados
mostraron una inhibición del (68 %) en la producción de partículas infecciosas
en presencia de L/M (1.8 x 105 UFP/ml) en relación con los cultivos control
(5.7 x 1o5 UFP/ml).
2.3. Expresión de proteínas virales
Para analizar la expresión de las proteínas virales en presencia delosvastatina se realizó IFI de cultivos sometidos al tratamiento L/M antes
descripto. La Figura 25 muestra las fotografías correspondientes. La expresión
de la glicoproteína viral en las células infectadas disminuye en presencia de los
compuestos. Los sobrenadantes correspondientes a los cultivosobservados en
esta figura fueron titulados obteniéndose los valores de reducción antes
determinados. Bajo estas condiciones la expresión de la nucleoproteina viral no
mostró diferencias con respecto a los controles.
Figura 25. Expresión de la glicoproteína viral.
Cultivos de células Vero se infectaron con JUNV (MOI = 0.1) y se trataron con Iovastatina y
mevalonato como se describe en materiales y métodos (B). Luego de 48 hs. de tratamiento los
cultivos se Iavaron y se procesaron para IFIjunto con cultivos controles (A).

FF‘ÍIIÏTmm -mmmn o II
3. Extracción directa de colesterol
3.1. MetiI-fi-cic/odextrinay su acción sobre las particulas ¡nfectivas
A fin de reducir de manera directa la cantidad de colesterol en la
membrana celular se utilizó el compuesto metil-B-ciclodextrina (MCD) cuya
propiedad es secuestrar especificamente esta molécula. Se utilizó la
concentración 10 mM para realizar extracciones de cultivos infectados y
previamente tratados como en la sección anterior con L/M.
Para determinar inicialmente el efecto del compuesto sobre el virus se
expusieron partículas infecciosas a la acción directa de este compuesto en un
ensayo de acción virucida como se indica en materiales y métodos. Luego de
1.5 hs. a 37°C se tituló la infectividad remanente de la muestra tratada y la
control por el método de UFP. Los datos obtenidos mostraron una acción
directa del compuesto sobre las partículas disminuyendo su infectividad 2
logaritmos (5.2 x103 UFP/ml) respecto de los controles (6.3 x 105UFP/ml).
3.2. Metil-[i-cic/odextrinay expresión de proteínas virales
Figura 26. Expresión de la glicoproteína en cultivos extraídos con MCD.
Cultivos de células Vero se infectaron con JUNV (MOI = 0.1) y se trataron con L/Mcomo se describe en
materiales y métodos. Luego de 48 hs. de tratamiento se realizó una extracción con MCD durante 40
minutos (B). Los cultivos se lavaron y procesaron para IFIjunto a cultivos controles (A).
Se trataron cultivos infectados con L/M según se detalla en la sección
2.2 y luego se realizó un pulso de extracción de 40 minutos con el compuesto
MCD 10 mM. La expresión de la glicoproteína viral en la membrana celular se
88

mmm T.\[)(‘,C - (‘APÍ’I‘UID II
determinó mediante IFI. Como se puede observar la extracción del colesterol
produce una reducción visiblemente significativa de la glicoproteína en la
membrana celular (Figura 26). No se observaron diferencias en la expresión de
la nucleoproteína en los cultivos tratados.
Para determinar la disminución de la glicoproteína observada por IFI se
cuantificó el número de células fluorescentes en los experimentos
correspondientes a las Figuras 16 y 17.
Adicionalmente y en cultivos paralelos se determinó el contenido lipidico
de las membranas celulares. Se extrajeron los lípidos por el método de Bligh 8.
Dyer y se determinó la cantidad de colesterol remanente por cromatografía en
capa delgada. En la Tabla XIIse resumen los resultados de la cuantificación de
colesterol y los porcentajes de células fluorescentes luego de cada tratamiento.
Colesterol Glicoproteína (IFI)
n° de célulasTratamiento pg (x :t s) % control % control
fluorescentes
- l -- 2.07 :t 0.29 100 31.1 :t 3.8 100
lovastatina l mevalonato 1.35 :t 0.32 65 13.1 :t 3.4 42
lovastatina l mevalonato0.59 i 0.08 29 1.8 :l:0.8 5
+ metil-B-ciclodextrina
Tabla XII. Cuantificación de la expresión de proteinas virales y contenido lipidico de las membranascelulares.
Se cuantificó el n° de células fluorescentes correspondientes a los ensayos de las figuras 16 (L/M) y 17
(L/M + MCD) tomando 20 campos ópticos elegidos al azar en cada uno de los casos. Paralelamente se
determinó el contenido de colesterol en las mismas membranas mediante cromatografía en capa delgada.
Se expresan los porcentajes respecto de los controles sin tratamiento.
Como se puede observar el contenido lipidico de las membranas
tratadas disminuye respecto de los controles. Cuando luego del tratamiento se
extrajeron los cultivos con MCD el contenido de colesterol en los mismos se
redujo hasta un 70% confirmando la acción del compuesto. En estas
condiciones la expresión de Ia glicoproteína en la membrana plasmática fue el
5% del valor en los cultivos normales.

Rlï‘ll ll 'l' \I‘)(‘)‘3 - (‘ \I’Í'l‘lll O II —
B. Estudio de la asociación del virus Junín a microdominios de membrana
1. Fraccionamiento de membranas en TX-100
La disminución de colesterol en los cultivos infectados afectó la
producción viral y presencia de glicoproteinas virales en la membrana
plasmática de las células infectadas. Por otro lado dado que las glicoproteinas
virales no mostraron asociación al citoesqueleto, su localización en los
sobrenadantes de extracciones con TX-100 (Figura 20) podría estar
relacionada con la asociación específica a microdomios rafts. Para abordar
este aspecto se realizaron extracciones a 4°C durante 30 minutos en buffer
Tris-EDTA salino adicionado con TX-100 1% y en ausencia de agentes
estabilizantes del citoesqueleto. Luego se procesó el sobrenadante (S) y el
material pelleteable (P). Cuando las muestras fueron inmunoprecipitadas se
observó una distribución diferencial de las proteínas virales.
S P S P
""1! l o La
GPC:>NP
GP1 :1)
Figura 27. Fraccionamiento en buffer TNE/ TX-100.
Cultivos de células Vero se infectaron con JUNV (MOI = 1) y se marcaron metabolicamente a las 48 hs.
p.i. Luego de lavar las monocapas con PBS se extrajeron durante 10 minutos sobre hielo en buffer
adicionado con TX-100 1%. Los Iisados se centrifugaron a 10000 xg para separar el sobrenadante (S) del
pellet (P). Ambos materiales se inmunoprecipitaron con anticuerpos especificos para el revelado de las
proteínas virales. Calles 1 y 2: anticuerpos anti-glicoproteinas. Calles 3 y 4: anticuerpos anti-NP.
90

¡“mln TADQS . (‘-\I’ÍI'UI (311
La Figura 27 muestra las fluorografías correspondientes a las muestras
inmunoprecipitadas con anticuerpos monoclonales para GP1 y GPC (líneas 1 y
2) y para NP (líneas 3 y 4). Como se puede observar NP se encuentra
mayoritariamente en la fracción remanente luego de la extracción mientras que
la glicoproteína GP1 esta asociada a la fracción del sobrenadante. EI
fraccionamiento de las proteinas virales en este buffer de extracción fue similar
al observado en la Figura 20.
2. Flotabilidaden gradientes de sacarosa
Para determinar la naturaleza del material en el sobrenadante de las
extracciones se sembraron los Iisados en gradientes isopícnicos.
Cultivos de células Vero infectados y marcados radioactivamente fueronextraídos como se mencionó anteriormente. Los Iisados se sembraron en
gradientes de sacarosa 5 - 40%, se ultracentrifugaron durante 21 horas y se
desarmaron obteniéndose 8 fracciones. Volúmenes iguales de cada fracción
fueron inmunoprecipitadas con anticuerpos monoclonales contra la
glicoproteína GP1 y su precursor y luego resueltos en geles de poliacrilamida.
La nucleoproteína del virus fue revelada en cada una de las fracciones con
anticuerpos específicos por la técnica de WB.
Las proteínas con localización en rafts no son solubilizadas por el
detergente a baja temperatura y, por lo tanto, flotan en los gradientes
ubicándose en las primeras fracciones o menos densas. Una de las proteinas
virales mejor descriptas como asociadas este tipo de fracciones es la
glicoproteína hemaglutinina (HA) del virus Influenza. Se utilizó este sistema,
entonces, como control positivo. En similares condiciones a las utilizadas para
JUNV se procesaron cultivos infectados con virus Influenza para la posterior
detección por WB de la HA. Los resultados de las extracciones a 4°C se
muestran en la Figura 28.
Como se puede observar y de acuerdo a Io esperado Ia HA del virus
Influenza se encuentra principalmente en las fracciones menos densas
correspondientes a los microdominios enriquecidos en colesterol (panel A). La
presencia de pequeñas cantidades de HAen las fracciones 6-8 probablemente
representan complejos completamente solubilizados por el detergente. Bajo las
91

Rlïcl ll T \l‘)()‘3 - (‘ \PÍTllI O II —
mismas condiciones Ia glicoproteína del virus Junín se encontró asociada a
fracciones de baja densidad (panel B). Se observa tanto el precursor como Ia
glicoproteína GP1 mayoritariamente distribuidos en las fracciones superiores.
Se analizó la localización de Ia nucleoproteína en las fracciones
mediante un ensayo de WB como se describe en materiales y métodos. Se
puede observar en el panel C que NP se distribuye a Io largo de todo el
gradiente sin mostrar una localización preferencial por alguna de las fracciones.
HA w¡C
GPCE> omita-h. ri“. 4664 45
GP1l:> ¿a b
NPI > ¡jqm fl r‘ lC
Figura 28. Distribuciónde proteínas virales en gradientes de sacarosa.
Cultivos de células Vero y MDCK se infectaron con JUNV o Influenza respectivamente. Se
extrajeron en TX-1OO1% a 4°C y durante 30 minutos. Sobre los Iisados obtenidos se armó un
gradiente de sacarosa según se indica en materiales y métodos. Se ultracentrifugó y se
inmunoprecipitaron las fracciones obtenidas (panel B) o se revelaron por WB (paneles A y C) con
anticuerpos específicos.
Los microdominios enriquecidos en colesterol presentan una gran
estabilidad 4°C sin embargo a temperaturas mayores la insolubilidad en TX-100
comienza a disminuir significativamente. Para examinar el comportamiento de
las proteínas de JUNV en estas condiciones se realizó el mismo ensayo de
extracción pero a una temperatura de 37°C. Nuevamente se incluyó la
92

v
.v'vv“r
v‘
lllmlll T “70‘: - (‘ \PÍ'l'lll 0 Il —
extracción de HA como control. En la Figura 29 se muestran como la
glicoproteína de JUNV y su precursor se distribuye a Io largo de todo el
gradiente (panel B). Asimismo en estas condiciones la HA de Influenza
incrementó su solubilidad mostrando una distribución equitativa en las fraciones
(panel C).
HA :9 gave“;GPCI:.’> 466
4 45
GP1(:>
NPI:í>C
Figura 29. Distribuciónde proteínas virales en gradientes de sacarosa.
Cultivos de células Vero y MDCKse infectaron con JUNV o Influenza respectivamente. Se extrajeron en
TX-100 1% a 37°C y durante 30 minutos. Sobre los Iisados obtenidos se armó un gradiente de sacarosa
según se indica en materiales y métodos. Se ultracentrifugó y se inmunoprecipitaron las fracciones
obtenidas (panel B) o se revelaron por WB (paneles A y C) con anticuerpos específicos.
La localización de NP no vario significativamente a 37°C como puede
observarse en el panel C.
Una de las moléculas de mayor relevancia en la composición de los
microdominios rafts es el colesterol. Intercalándose entre los fosfolípidos es
determinante en la estabilidad de dichas estructuras. Para comprobar la
asociación especifica de la glicoproteína viral a las fracciones enriquecidas en
colesterol los cultivos infectados con JUNV se trataron con metiI-B-ciclodextrina
y luego se procesaron con TX-100 1% a 4°C como se describió anteriormente.
93

Rlï‘ïl ll 'l' “30‘: —r \I’Í'l'lll () II —
Nuevamente en las condiciones control GP1 y GPC se encuentran
preferentemente en las fracciones superiores 4 y 5 (Figura 30, panel A). Al
secuestrar de las membranas las moléculas de colestrol la glicoproteina del
virus se encontró distribuida en todas las fracciones del gradiente (panel B).
Estos resultados demostraron que la presencia de GP1 y su precursor
en las fracciones de menor densidad se debe a su asociación específica a los
microdominios de membrana enriquecidos en colesterol.
GPC I:I> I 4 66
4 45
GP1:>
GPC |:> 4 66
445
GP1:>
Figura 30. Distribuciónde proteinas virales en gradientes de sacarosa.
Cultivos de células Vero infectaron con JUNV. A las 48 hs. p.i. cultivos controles (panel A) y tratados con
MCD durante 30 minutos (panel B) se extrajeron en TX-1OO1% a 4°C y durante 30 minutos. Sobre los
Iisados obtenidos se armó un gradiente de sacarosa según se indica en materiales y métodos. Se
ultracentrifugó y se inmunoprecipitaron las fracciones obtenidas para el revelado de GP1 y GPC.
El entrecruzamiento con anticuerpos específicos mostró favorecer
también la estabilidad de los microdominios rafts aumentando su tamaño y
asociación por dimerización de proteínas contenidas en su estructura.
Se realizó a continuación un ensayo de entrecruzamiento utilizando
anticuerpos específicos para GP1 y su precursor. Los cultivos se incubaron con
94

RFQIIÏ TADÜ‘z - (‘APÍTUI 0 II
los anticuerpos durante 30 minutos y luego se realizó la extracción a 4°C o a
37°C. Los lisados obtenidos se llevaron un gradiente como en los experimentos
anteriores. En la Figura 31 se muestran las fluorografías obtenidas.
GPC «:9 4 664 45
GP1 i:>
GPC iz) 4 66
4 45
GP1 :1)
Figura 31. Estabilización de microdominiospor entrecruzamiento con anticuerpos.
Cultivos de células Vero infectaron con JUNV (MOI = 1). A las 48 hs. p.i. los cultivos se incubaron oon
anticuerpos específicos para GPC y GP1 durante 30 minutos a 37°C. Luego de este período se
extrajeron todos los cultivos en TX-100 1% a 4°C (panel A) o a 37°C (panel B) durante 30 minutos. Sobre
los lisados obtenidos se armó un gradiente de sacarosa según se indica en materiales y métodos. Se
ultraoentn'fugóy se inmunoprecipitaron las fracciones obtenidas para el revelado de GP1 y GPC.
En el panel A al igual que en las Figuras 19 y 21 correspondientes a la
extracción a 4°C se observan las glicoproteínas en las fracciones superiores
debido a su baja densidad. Como se mostró anteriormente al realizar la
extracción a 37°C (Figura 20) los microdominios aumentan su solubilidad en
TX-100 perdiendo su característica flotabilidad en las fracciones superiores del
95

gradiente. En la Figura 22 se observa que al extraer a 37°C los cultivos
previamente incubados con anticuerpos específicos. las glicoproteinas GPC y
GP1 se localizan en las fracciones 5 y 6 del gradiente. Estos resultados
muestran la estabilización de los microdominios rafts a 37°C por la adición
anticuerpos especificos para las glicoproteinas virales.

DISCUSIÓN Y CONCLUSIONES

I‘JL‘:(‘II(:I(‘)Ï ï \-(‘(‘¡i N] U‘ïlf‘ri ïF‘:
El virus Junín es un virus envuelto perteneciente a Ia familia Arenavin'dae
y representa uno de los 5 miembros capaces de producirfiebres hemorrágicas
en humanos. Distintos grupos de investigación han abordado el estudio de esta
patogenia y sus respectivos agentes etiológicos con el objetivo de desentrañar
las bases celulares y moleculares subyacentes. Hasta el momento es poco lo
que se sabe sobre el ciclo de replicación de JUNV, su interacción con la célula
huésped y con el sistema inmunológico del organismo infectado. Sin embargo,
la única vacuna desarrollada en el mundo para un arenavirus es la vacuna
candid #1 para la prevención de la FHA. La misma mostró una alta eficiencia
aunque debido a factores económicos y al hecho de que es una vacuna
desarrollada a virus atenuado solo es administrada en zonas de alto riesgo.
En conjunto el conocimiento íntimo de un ciclo de replicación viral es el
punto de partida para el desarrollo racional de estrategias tendientes al control,
cura y/o prevención de las enfermedades virales. La sintesis, modificación y
transporte de proteínas virales a la membrana de una célula infectada
representan una serie de eventos determinantes en el ciclo de replicación viral.
La infectividad de la progenie, y su consecuente potencial patogénico,
dependen de la adecuada interacción virus-célula huésped. La oportuna
utilización de los recursos celulares es determinante para la supervivencia y
prevalencia del virus en la naturaleza. Cada paso del ciclode replicación posee
requerimientos específicos y cada estructura celular utilizada representa un
punto de estudio básico para la virología.
En este trabajo se estudió la interacción virus-célula huésped con
particular interés en dos estructuras celulares: el citoesqueleto y las
membranas lipídicas. Se determinó la localización preferencial de NP a Ia
fracción celular correpondiente al citoesqueleto y Ia asociación de las
glicoproteínas virales a los microdominios rafts de membrana.
La identificación de moléculas de actina asociada a viriones purificados
de JUNV condujo a la hipótesis de que la misma podia participar activamente
en su ciclo de replicación (Pasian y col.. 1983). Estudios posteriores en este
sentido demostraron que drogas desestabilizantes de la red de microfilamentos
ejercían un efecto inhibitoriosobre la multiplicación de JUNV y otros arenavirus
(Castilla y col., 1994; Candurra y col., 1996). Recientemente se demostró que
la perturbación específica tanto de la red de microtúbulos como la de
98

microfilamentos producía una disminución general de la producción viral
(Candurra y coI., 1999). En el mismo trabajo se demostró que dicha
perturbación tenía efectos sobre las etapas tardías del ciclo de multiplicación
alterando el trasporte de la glicoproteínas y la consecuente producción de
partículas infecciosas. En este trabajo se estudió ia participación en el ciclo de
multiplicación de JUNV de la tercera red componente del citoesqueleto: los
filamentos intermedios. Una de las particularidades de los FI es su expresión
variada en células de diferentes tejidos u origen embriológico. Por este motivo
se utilizaron en este estudio 2 cultivos con expresión de vimentina (una línea
celular y un cultivo diploide) y un cultivo primario de astrocitos con expresión
preferencial de GFAP.
El requerimiento de los FI en las distintas etapas del ciclo de replicación
se exploró utilizando el compuesto acrilamida capaz de producir Ia alteración
específica de redes de vimentina, citoqueratina y neurofilamentos (OIink-Couxy
col., 1992; Sager y coI., 1989).
Estudios realizados con influenza A mostraron la reorganización de
filamentos de vimentina y citoqueratina producida durante el curso de la
infección en células LLC-MK2.La alteración en el patrón de expresión de estos
antígenos producida por el virus fue comparable al efecto de el compuesto
acrilamida sobre células sin infectar (Arcangeletti y coI._1997a). Los cultivos de
células Vero tradicionalmente utilizados para Ia multiplicación del virus Junín no
muestran alteraciones específicas de ninguna de las redes de citoesqueleto
durante el transcurso de Ia infección. Asimismo los cultivos diploides como los
cultivos primarios utilizados en este trabajo no mostraron modificaciones en la
estructura del citoesqueleto al ser infectados con JUNV. El análisis del
requerimiento de esta estructura celular durante el ciclo de multiplicación viral
se realizó, por Iotanto, utilizando el compuesto acrilamida.
Se determinó que la perturbación de los FI produce una disminución en
la producción general de virus infeccioso en los tres tipos de cultivos utilizados
y tratados con acrilamida durante 24 hs. Cuando se evaluó el efecto del
compuesto durante distintos periodos de tratamiento no se observó acción
directa sobre los pasos iniciales de adsorción e internaiización del virus a la
célula (Figura 16). Se analizaron etapas tardías de la infección y la expresión
99

DI‘W‘IÏQVIHï \-(‘(‘¡ï irjjl‘ïlñï il":
de las proteínas virales analizada por IFI mostró una disminución en ambas
GP1 y NP luego de 24 hs. de tratamiento (Figura 17).
Arcangeletti y col. (1997a) observaron disminución en Ia síntesis de
proteínas celulares y virales en cultivos LLC-MK2infectados con virus Influenza
A y luego de un tratamiento con acrilamida 5 mM. Para descartar un efecto
directo del compuesto sobre la síntesis de proteínas en el sistema utilizado se
realizó un marcado metabólico en presencia de acrilamida. El análisis de los
cultivos no mostró efectos significativos sobre la incorporación de aminoácidos
radiactivos en las 4 horas del ensayo. La síntesis de proteínas celulares así
como las virales no fue afectada cuando se desorganizó la red de FI con unaconcentración de acrilamida de 2 mM.
Los resultados obtenidos en este trabajo muestran que la integridad de
los Fl no es requerida durante los pasos de adsorción e internalización del
virus. La síntesis de proteínas tampoco mostró ser directamente dependiente
de esta red sin embargo. y debido a que la expresión de las proteínas virales a
las 24 hs. p.i se encuentra disminuida, un paso posterior al ingreso del virus y
anterior a la síntesis podría ser el afectado al desorganizar los FI.
Existe una amplia evidencia que demuestra la asociación del
citoesqueleto con complejos proteicos denominados prosomas. Estas
estructuras. que se acomplejan en trans a ARNm en estado inactivo, se
encuentran específicamente asociadas a las redes de filamentos intermedios
en distintos tipos celulares como fibroblastos, células musculares y epiteliales
(Grossi de Sa y col., 1988a y b; Olinx-Coux y col., 1992; Arcangeletti y col.,
2000). En vista de esta asociación se asume que los FI podrian servir como
guía para el transporte y localización de los ARNm dentro del citoplasma
celular. La perturbación de los FI con acrilamida no produce la disociación entre
Ia red de citoqueratina y el antígeno prosomal p27 (Arcangeletti y col., 1997a).
Katze y col. (1989) proponen en base a sus estudios que los prosomas
serian necesarios, aunque no suficientes, para que ARNm virales y celulares
sean traducidos en proteínas. Aunque en el presente trabajo no se realizaron
estudios de localización de ARNm podría sugerirse que la disminución en la
expresión de las proteínas virales en presencia de la acrilamida afecta el
destino de los complejos ARNm-prosomas impidiendo su activación y posterior
traducción. Si los prosomas permanecen unidos a la red de vimentina en las
lOO

DI‘ICUIÍJÓÏ i \-(‘(‘)ï ir] lI‘ÏIGT iF‘ï
células tratadas esto mantendría a los mensajeros en su estado inactivo
produciendo la disminución observada en la expresión de proteinas virales. En
este sentido los ensayos realizados para determinar la síntesis de proteínas en
células tratadas con acrilamida reflejan el correcto funcionamiento de este
proceso durante el lapso de incorporación de aminoácidos marcados y no la
acumulación de proteínas sintetizadas durante todo el tratamiento. En el futuro
deberán realizarse estudios relacionados con el transporte y localización de los
ARNm virales de JUNV para determinar con mayor certeza su relación con los
Fl de la célula infectada.
Se estudiaron los patrones de IFI de los filamentos de vimentina y las
proteinas virales mediante ensayos de co-Iocalización. Por esta técnica no
pudo observarse una asociación directa entre los FI y la glicoproteína viral.
Estudios realizados con el virus respiratorio sincicial en células Hep-2
mostraron su capacidad para producir una desorganización especifica de la red
de FI. En este trabajo cuando se realizaron extracciones específicas para
citoesqueleto solo la nucleoproteína viral mostró una asociación univoca a la
correspondiente fracción. Esta asociación fue parcialmente desestabilizada en
condiciones de alta salinidad (García Barreno y col., 1988).
Debido a que en las extracciones anteriores la glicoproteína de JUNV
fue localizada en la fracción del sobrenadante y estudios anteriores de nuestro
grupo de investigación reportaban su presencia preferencial en el material
correspondiente al citoesqueleto, se realizaron estudios tendientes a
determinar la naturaleza de este comportamiento.
La presencia de una proteina en la fracción resistente a la extracción con
detergentes no iónicos es un fuerte indicativo de su asociación con el
citoesqueleto. Sin embargo la localizaciónen el sobrenadante puede deberse a
diferentes circunstancias. Estas podrían ser que la asociación con el
citoesqueleto no se encuentra estabilizada en las condiciones del ensayo dado
que algunas interacciones requieren la presencia de agentes quelantes de Ca++
para la estabilización del citoesqueleto. Por otro lado las proteínas podrian
encontrarse en el sobrenadante debido a su asociación a fracciones Iipídicasde diferente naturaleza entre las cuales se encuentran los rafts.
En los últimos años se han descripto los microdominios de membrana
ricos en colesterol denominados rafts. Si bien estos son insolubles en TX-100
10]

leV‘II‘ïlÚï ï x-(T‘fi'I(‘Ill‘ïlf‘lï il“:
no necesariamente permanecen en la fracción remanente a la extracción.
Inicialmente las fracciones insolubles en detergentes no iónicos se han definido
en base al material remanente que permanece retenido sobre el sustrato luego
de la extracción, o a la fracción pelleteable a 10000 xg. Si bien esto es válido
para el material asociado a citoesqueleto, que es en general de gran peso
molecular y exclusivamente de origen proteico, no necesariamente lo es para
los rafts de tamaño tan pequeño y que, aún siendo insolubles, podrían también
particionar en el sobrenadante de las extracciones.Cuando se comenzaron a estudiar los microdominios rafts se
establecieron técnicas más especificas para su determinación en los Iisados
celulares provenientes de extracciones con detergentes. Su caracterización se
basa en su flotabilidad en gradientes de sacarosa (Brown y coI., 1997).
Estudios realizados con HIV demostraron la asociación específica de Ia
glicoproteína Gag a microdominios rafts, en las fracciones menos densas de
gradientes de sacarosa, provenientes de sobrenadante postnuclear (Ono y coI.,
2001).
Análisis sobre la composición lipídica de la envoltura del virus Junin
mostraron una proporción del 60% de colesterol respecto de las membranas
celulares de las que brota (Bartollota, comunicación personal). Teniendo en
cuenta este antecedente y el hecho de que no estuviera aún definido
completamente el comportamiento de Ia glicoproteína viral en TX-100 se
iniciaron estudios tendientes a analizar la relación del virus con las membranas
enriquecidas en colesterol.
Para estudiar la importancia del colesterol en el ciclo de multiplicación
del JUNV se utilizaron análogos hidroxilados del mismo. Estos compuestos han
sido previamente caracterizados como moduladores negativos de la enzima
HMG CoA reductasa produciendo, en cultivos expuestos a ellos, una
disminución en Ia cantidad total de colesterol en las células. El colesterol es
una molécula de caracteristicas hidrófobicas y de gran importancia en las
membranas de células animales, reducir sus valores intracelulares puede por lo
tanto provocar cambios en las estructuras asociadas a éste, influyendo en
procesos como el transporte y localización de proteínas de membrana.
El primer abordaje experimental se llevó a cabo determinando el efecto
de los hidroxesteroles sobre la producción viral. Dos análogos mostraron
l02

13191] J‘ZIÓÏ ï \' (Y‘JI ¡("l ll‘ÏIÓÏ ïF‘:
actividad antiviral sobre JUNV. También se determinó que en presencia de los
mismos solo la expresión de las glicoproteínas estaba reducida en membrana
sugiriendo un efecto de los compuestos sobre el transporte hacia la misma.
Utilizando el compuesto Iovastatina, inhibidor de Ia sintesis del
colesterol, se demostró nuevamente la necesidad de concentraciones normales
del mismo para la correcta localización de GP1 en la membrana celular.
Finalmente se redujo de manera directa la cantidad de colesterol en las
membranas utilizando el compuesto MCD. En estas condiciones fue máxima la
reducción de la glicoproteína en la membrana de las células infectadas, sin
afectar la presencia y localización de la nucleoproteína viral.
Habiendo determinado la importancia de la presencia de colesterol en la
localización y expresión normal de la glicoproteína viral se exploró la posibilidad
de que Ia misma fuese incorporada en microdominiosde membrana rafts.
Se realizaron extracciones con TX-100 y se sometieron los Iisados a
ultracentrifugación en gradiente de sacarosa. El análisis de las fracciones
obtenidas demostró la presencia de las glicoproteínas en la zona superior o de
menor densidad dentro del gradiente. La asociación especifica de GPC asi
como de GP1 a dichas fracciones mostró ser dependiente de la temperatura y
de la presencia de colesterol en las membranas. La adición de anticuerpos
especificos estabilizó Ia presencia de las proteinas en las de menor densidad a
37°C. Todas estas propiedades en conjunto demuestran la incorporación de las
glicoproteínas virales GPC y GP1 en microdominios de membrana rafts.
Distintos trabajos permitieron Ia caracterización de proteinas virales
como residentes de microdominios rafts. Estudios realizados con el virus
Influenza han demostrado la asociación de la glicoproteína HA en estos
microdominios. La presencia de colesterol fue crítica para la insolubilidad de
esta proteina en TX-100. La remoción de colesterol producida por el
tratamiento con Iovastatina o MCD produjo un incremento en la solubilidad de
HAacompañada por una disminución de su presencia en la membrana y en las
fracciones rafts (Keller y coI.. 1998). Este virus, además, mostró una salida
preferencial por Ia cara apical de cultivos MDCK polarizados aportando
evidencias biológicas a su localización en rafts. A este respecto estudios
realizados con JUNV indican la salida preferencial del mismo por la cara apical
de cultivos polarizados Vero C1008 (Cesio y Acuña. comunicación personal).
103

La utilización de proteínas quimera demostraron que el dominio
transmembrana de HA resulta determinante para esta localización. El dominio
transmembrana de la proteina G de VSV o el ectodominio de HAno permitieron
la localización de HA en las fracciones insolubles a TX-100 (Scheiffele y col.,
1997).
Si bien no se analizaron los determinantes moleculares relacionados con
la asociación de GP1 a los microdominios rafts dos posibles y no excluyentes
circunstancias podrian colaborar en ello. Aunque GP1 es de naturaleza
periférica su estrecha asociación con GP2, una glicoproteina transmembrana,
podría estar dirigiendo su camino a las membranas enriquecidas en colesterol.
Desafortunadamente la falta de antigenicidad de GP2 y la consecuente falta de
anticuerpos contra la misma dificulta su estudio.
Otro factor que podria estar colaborando y favoreciendo la unión de GP1
a los rafts podría ser su miristilación. Nguyen y col., (2000) mostraron para HIV
la asociación preferencial de la forma miristilada de Gag a los microdominios
rafts. Estudios realizados con JUNV basados en la presencia de sitio de
miristilación en la secuencia de GP1 y la actividad antiviral demostrada para
análogos competitivos del ácido mirísitico han sugerido Ia modificación de la
glicoproteina con este ácido graso (Cordo y col.,1999).
La incorporación de las proteinas virales en los microdominios rafts no
solo dirigiría su transporte hacia la membrana sino que además facilitaria el
proceso de brotación desde las mismas. La naturaleza viscosa y poco fluida de
la fase liquido-ordenada respecto del resto de la membrana podría inducir la
curvatura o protrución favoreciendo el egreso de la partícula viral. Por otro lado
estudios recientes demostraron que HA no es suficiente para la brotación de
viriones. La presencia de Ia proteina de matriz M1 es necesaria para este
proceso sugiriendo la participación de otras estructuras celulares tales como el
citoesqueleto para el evento de brotación (Gomez-Puertas y col., 2000).
Cultivos infectados con el virus de sarampión mostraron la localización
de sus proteínas F y H en las fracciones de gradientes correspondientes a rafts
(Blau y coI., 1995). Sin embargo cuando estas proteinas fueron expresadas
separadamente solo F se asoció con los microdominios rafts. Estos resultados
muestran que algunas proteínas no poseen por si mismas esta capacidad sino
104

que dependen de la interacción con otras para facilitar su asociación a los
microdominios (Vincent y col., 2000).
Estudios realizados sobre la proteina Z de varios arenavirus postulan
distintas funciones regulatorias (Garcin y col., 1993; Borden y co|., 1998a y b;
Campbell Dwyer y col.. 2000). Asimismo, debido a su caracterización como
proteína estructural de LCMV,se ha sugerido su rol como proteína de matriz
(Salvato y col., 1992). En un estudio reciente Strecker y col. (2003)
demostraron tambien Ia presencia de Z en las partículas de virus Lassa y su
probable rol como proteína de matriz. Esta proteína se encontró fuertementeasociada a membranas insolubles en TX-100. Además mostró ser suficiente
para la liberación de partículas en células transfectadas solo con el gen
correspondiente a Z. Aunque aún no se ha determinado la presencia de Z en
microdominios rafts, la misma podria colaborar en la unión de las glicoproteínas
a estos dominios favoreciendo el encuentro del resto de los componentes
virales para el ensamblado y posterior liberación de la progenie.
Durante el curso de una infección cada componente de Ia partícula viral
es sintetizado en distintos compartimentos celulares, luego, cada proteina es
transportada hacia un lugar común donde interacciones de diverso origen
promoverán la morfogénesis y liberación de la progenie viral. Este trabajo
permitió determinar que la integridad de los Fl es crucial al menos durante los
pasos previos a la sintesis de proteínas virales, luego de la internalización del
virus. La nucleoproteína de JUNV se asocia a Ia fracción celular
correspondiente al citoesqueleto y, aunque no se determinó a que componente
correspondería, ésta interacción podría facilitar el trasporte de NP a la
membrana plasmática. La glicoproteína viral no parece interaccionardirectamente con los filamentos de vimentina. Por otro lado se determinó la
asociación específica de GP1 y GPC a los microdominios rafts sugiriendo la
utilizaciónde esta via como mecanismo de transporte a la membrana celular delas células infectadas.
ll
/ I‘ll/M41CWW“gwbwi combo
105

BIBLIOGRAFÍA

mm [OCR RFÍ x
Albariño, C. G., P. D. Ghiringhelli, et al. (1997). "Molecular characterization ofattenuated Junin virus strains." J Gen Virol78 (Pt 7): 1605-10.
Ambrosio, A. M., D. A. Enria, et al. (1986). "Junin virus isolation from lymphomononuclear cells of patients with Argentine hemorrhagic fever." Intervirology25(2): 97-102.
Andrews, B. S., A. N. Theofilopoulos, et al. (1978). "Replication of dengue andjunin viruses in cultured rabbit and human endothelial cells." Infect lmmun20(3): 776-81.
Arcangeletti, C., Olink-Coux, M., Minisini, R., Huesca, M., Chezzi, C., Scherrer,K. (1992) "Patterns of cytodistribution of prosomal antigens on the vimentin andcytokeratin networks of monkey kidney cells". Eur J Cell Biol. 59, 464-76.
Arcangeletti, M. C., Pinardi, F., Missorini, S., De Conto, F., Conti, G.,Portincasa, P., Scherrer, K. Chezzi, C. (1997a) "Modification of cytoskeletonand prosome networks in relation to protein synthesis in influenza A virusinfected LLC-MK2cells" Virus Res. 51, 19-34.
Arcangeletti, C., Sutterlin, R., Aebi, U., De Conto, F., Missorini, S., Chezzi, C. &Scherrer, K. (1997b) "Visualization of prosomes (MCP-proteasomes),intermediate filament networks by "instantaneous fixation" preserving thecytoskeleton" J Struct Biol. 119, 35-58.
Arcangeletti, C., De Conto, F., Sutterlin, R., Pinardi, F., Missorini, S., Geraud,G., Aebi, U., Chezzi, C. & Scherrer, K. (2000) "Specific types of prosomesdistribute differentially between intermediate and actin filaments in epithelial,fibroblastic and muscle cells" Eur J Cell Biol. 79, 423-37.
Armstrong, C. L. R. (1934). "Experimental lymphocytic choriomeningitis ofmonkeys and mice produced by a virus encountered in studies of the 1933 St.Louis encephalitis epidemic." Public Health Republic 49: 1019-1027.
Aronson, J. F., N. K. Herzog, et al. (1995). "Tumor necrosis factor and thepathogenesis of Pichinde virus infection in guinea pigs." Am J Trop Med Hyg52(3): 262-9.
Arribalzaga, R. A. (1955). "New epidemic disease due to unidentified germ:nephrotoxic, leukopenic and enanthematous hyperthermia." Dia Med 27(40):1204-10.
Auperin, D. D., V. Romanowski, et al. (1984). "Sequencing studies of pichindearenavirus S RNA indicate a novel coding strategy, an ambisense viral S RNA."J Virol 52(3): 897-904.
Barman, S. Nayak, D. P. (2000) "Analysis of the transmembrane domain ofinfluenza virus neuraminidase, a type IItransmembrane glycoprotein, for apicalsorting and raft association" J Virol.74, 6538-45.
107

nm] man «r-í-\
Barrera Oro, J. G. Eddy, G. A. (1982) "Characteristics of Candidate liveattenuated Junin virus vaccine" IV International Conference on ComparativeVirology. Banff, Alberta, Canada. Abstract 84-10.
Barrera Oro, J. G., Lupton, H. W., Jahrling, P. B. et al. (1988) "Cross protectionagainst Machupo virus with Candid #1 live-attenuated Junin virus vaccine. l.The post-vaccination prechallenge immune response" Second InternationalConference on Comparative Virology, Mar del Plata, Argentina. Abstract P.E.1.
Barrera Oro, J. G. and K. T. McKee, Jr. (1991). "Toward a vaccine againstArgentine hemorrhagic fever." Bull Pan Am Health Organ 25(2): 118-26.
Bergold, G. H., L. A. Gonzalez Marquez, et al. (1969). "Structure of ninearboviruses." J Gen Virol 5(1): 135-40.
Blau, D. M., Compans, R. W. (1995) "Entry and release of measles virus arepolarized in epithelial cells" Virology. 210, 91-9.
Bligh, E. G. & Dyer, W. J. (1959) A rapid method of total Iipidextraction andpurification, Can J Med Sci. 37, 911-7.
Borden, K. L., Campbell Dwyer, E. J., Salvato, M. S. (1998a) "An arenavirusRING (zinc-binding) protein binds the oncoprotein promyelocyte Ieukemiaprotein (PML) and relocates PML nuclear bodies to the cytoplasm" J Virol. 72,758-66.
Borden, K. L., Campbell Dwyer, E. J., Carlile, G. W., Djavani, M., Salvato, M. S.(1998b) "Two RING finger proteins, the oncoprotein PML and the arenavirus Zprotein, colocalize with the nuclear fraction of the ribosomal P proteins" J Virol.72, 3819-26.
Borrow, P. and M. B. Oldstone (1994). "Mechanism of Iymphocyticchoriomeningitis virus entry into cells." Wrology 198(1): 1-9.
Bowen, M. D., C. J. Peters, et al. (1996). "The phylogeny of New World(Tacaribe complex) arenaviruses." Virology219(1): 285-90.
Brown, D. A., London, E. (1997) "Structure of detergent-resistant membranedomains: does phase separation occur in biological membranes?" BiochemBiophys Res Commun. 240, 1-7.
Brown, D. A. and E. London (1998). "Functions of Iipid rafts in biologicalmembranes." Annu Rev Cell Dev Biol 14: 111-36.Brown, D. A. and E. London (2000). "Structure and function of sphingolipid- andcholesterol-rich membrane rafts." J BiolChem 275(23): 17221-4.
Buchmeier, M. J., Welsh, R. M., Dutko, F. J. & Oldstone, M. B. (1980) "TheVirologyand immunobiology of Iymphocytic choriomeningitis virus infection" AdvImmunol. 30, 275-331.
l08

mm man «Fi-\
Buchmeier, M. J., P. J. Southern, et al. (1987). "Site-specific antibodies define acleavage site conserved among arenavirus GP-C glycoproteins." J Virol61(4):982-5.
Burns, J. W. and M. J. Buchmeier (1991). "Protein-protein interactions inlymphocytic choriomeningitis virus." Virology 183(2): 620-9.
Campbell Dwyer, E. J., Lai, H., MacDonald, R. C., Salvato, M. S. & Borden, K.L. (2000) The lymphocytic choriomeningitis virus RING protein Z associateswith eukaryotic initiation factor 4E and selectiver represses translation in aRING-dependent manner, J Virol.74, 3293-300.
Cao, W., M. D. Henry, et al. (1998). "Identification of alpha-dystroglycan as areceptor for lymphocytic choriomeningitis virus and Lassa fever virus." Science282(5396): 2079-81.
Candurra, N. A., Maskin, L. & Damonte, E. B. (1996) "Inhibition of arenavirusmultiplication in vitro by phenotiazines" Antiviral Res. 31, 149-58.
Candurra, N. A., Lago, M. J., Maskin, L. & Damonte, E. B. (1999) "Involvementof the cytoskeleton in Junin virus multiplication" J Gen Virol. 80 ( Pt 1), 147-56
Castilla, V., S. E. Mersich, et al. (1991). "Lysosomotropic compounds inhibitingthe multiplication of Junin virus" Rev Argent Microbiol23(2): 86-9.
Castilla, V., Mersich, S. E., Candurra, N. A. & Damonte, E. B. (1994) The entryof Junin virus into Vero cells, Arch Virol. 136, 363-74.
Castilla, V. & Mersich, S. E. (1996) Low-pH-induced fusion of Vero cellsinfected with Junin virus, Arch Virol. 141, 1307-17.
Clegg JCS, B. M., Buchmeier MJ, Gonzalez J-P, Peters CJ, Rico-Hesse R,RomanowskiV (2000). "Family Arenaviridae. ln: Regenmortel MHV van,Fauquet CM, Bishop DHL, Carstens EB, Estes MK, Lemon SM, Maniloff J,Mayo MA, McGeoch DJ, Pringle CR, Vchner RB (eds)." Virus Taxonomy.(Seventh report of International Committee on Taxonomy of Viruses. Academic,Orlando).
Compans, R. (1993) Arenavirus ultrastructure and morphogenesis. In TheArenaviridae. Salvato (eds). Plenum Press, New York, pp 51-83.
Compans, R. W. (1995). "Virus entry and release in polarized epithelial cells."Curr Top Microbiol lmmunol 202: 209-19.
Cordo, S. M., Candurra, N. A. & Damonte, E. B. (1999)" Myristic acid analogsare inhibitors of Junin virus replication" Microbes Infect. 1, 609-14.
Di Simone, C., M. A. Zandonatti, et al. (1994). "Acidic pH triggers LCMVmembrane fusion activity and conformational change in the glycoprotein spike."Virology 198(2): 455-65.
109

mm man wi a
Downs, W. G., Anderson, C. R., Spence, L., Aitken, T. H. G. & Greenhal, A. G.(1963) "Tacaribe virus, a new agent isolated from Artibeus bats and mosquitoesin Trinidad, West Indies" Am J Trop Med Hyg. 12, 640.
Ellenberg, P., M. Edreira, et al. (2002). "Synthesis and expression of viralantigens in Vero cells persistently infected with Junin virus." Arch Virol 147(8):1543-57.
Enria, D., Feuillade, M.. Levis, S. & al., e. (1999) "Impact of vaccination of ahigh risk population for Argentine hemorrhagic fever with a live attenuated Juninvaccine" ln: Saluzzo JF, Dodet B (eds) Factors in the Emergence and Controlof Rodent-borne Viral Diseases. Elsevier SAS, Paris, France, pp 273-280.
Franze-Fernandez, M. T., Zetina, C., lapalucci, S., Lucero, M. A., Bouissou, C.,Lopez, R., Rey, 0., Daheli, M., Cohen, G. N. & Zakin, M. M. (1987) "Molecularstructure and early events in the replication of Tacaribe arenavirus S RNA"Virus Res. 7, 309-24.
Fridriksson, E. K., Shipkova, P. A., Sheets, E. D., Holowka, D., Baird, B. &McLafferty, F. W. (1999) "Quantitative analysis of phospholipids in functionallyimportant membrane domains from RBL-2H3 mast cells using tandem highresolution mass spectrometry" Biochemistry. 38, 8056-63.
Fuller-Pace, F. V. and P. J. Southern (1989). "Detection of virus-specific RNAdependent RNA polymerase activity in extracts from cells infected withlymphocytic choriomeningitis virus: in vitro synthesis of full-length viral RNAspecies." J Virol63(5): 1938-44.
Garcia, J., Calderon, G., Sabattini, M. & al., e. (1996) "Infección por virus Junin(VJ) de Calomys musculinus (Cm) en áreas con diferente situaciónepidemiológica para la Fiebre Hemorrágica Argentina (FHA)" Medicina (BAires). 56, 624.
Garcia, C. C., B. N. Brousse, et al. (2003). "Inhibitory effect ofthiosemicarbazone derivatives on Junin virus replication in vitro." Antivir ChemChemother 14(2): 99-105.
Garcia-Barreno, B.. Jorcano, J. L., Aukenbauer, T., Lopez-Galindez, C. &Melero, J. A. (1988) "Participation of cytoskeletal intermediate filaments in theinfectious cycle of human respiratory syncytial virus (RSV)" Virus Res. 9, 30721.
Garcin, D., Rochat, S. & Kolakofsky, D. (1993) "The Tacaribe arenavirus smallzinc finger protein is required for both mRNA synthesis and genome replication"J Virol. 67, 807-12.
Ghiringhelli, P. D., C. G. Albarino, et al. (1997). "The glycoprotein precursorgene of the attenuated Junin virus vaccine strain (Candid #1)." Am J Trop MedHyg 56(2): 216-25.
llO

Gomez-Puertas, P., Albo, C., Perez-Pastrana, E., Vivo, A. & Portela, A. (2000)"Influenza virus matrix protein is the major driving force in virus budding" J Virol.74, 11538-47.
Grossi de Sa, M. F., Martins de Sa, C., Harper, F., OIink-Coux, M., Huesca, M.& Scherrer, K. (1988a) "The association of prosomes with some of theintermediate filament networks of the animal cell "J Cell Biol. 107, 1517-30.
Grossi de Sa, M. F., Martins de Sa, C., Harper, F., Coux, 0., Akhayat, 0., Pal,J. K., Florentin, Y. & Scherrer, K. (1988b) "Cytolocalization of prosomes as afunction of differentiation" J Cell Sci. 89 (Pt 2), 151-65.
Harder, T., P. Scheiffele, et al. (1998). "Lipid domain structure of the plasmamembrane revealed by patching of membrane components." J Cell Biol 141(4):929-42.
Howard, C. R. (1986) "Arenaviruses" In: Perspectives in medical virology. Vol 2.Amsterdam: Elsevier, 1986.
Howard, C. R. (1993). "Antigenic diversity amog the arenaviruses." In: SalvatoMS (ed) The Arenaviridae. Plenum, New York.
Heller, M. V., Marta, R. F., Sturk, A., Maiztegui, J. l., Hack, C. E., Cate, J. W. &Molinas, F. C. (1995) "Early markers of blood coagulation and fibrinolysisactivation in Argentine hemorrhagic fever" Thromb Haemost. 73, 368-73.
Hooper, N. M. (1999) "Detergent-insoluble glycosphingolipid/cholesterol-richmembrane domains, Iipid rafts and caveolae" (review), Mol Membr Biol. 16,145-56.
Jahrling, P. B., Trotter, R. W., Barrera Oro, J. G. 8. aI., e. (1988) "Crossprotection against Machupo virus with Candid #1 Junin vaccine: Ill, postchallenge clinical findings". Second International Conference on the Impact ofViral Diseases on the Development of Latin American Countries and theCaribbean Region, Mar del Plata, Argentina. Abstract PE3.
Janes, P. W., S. C. Ley, et al. (1999). "Aggregation of Iipid rafts accompaniessignaling via the T cell antigen receptor." J Cell Biol 147(2): 447-61.
Johnson, K. M., V\ñebenga, N. H., Mackenzie, R. B., Kuns, M. L., Tauraso, N.M., Shelokov, A., Webb, P. A., Justines, G. & Beye, H. K. (1965) "Wrusaisolations from human cases of hemorrhagic fever in Bolivia"Proc Soc ExpBiol Med. 118, 113.
Johnson, K. M., Webb, P. A. & Justines, G. (1973) "Biology of Tacaribecomplex viruses" In Lymphocytic Choriomeningitis Virus and OtherArenaviruses (F. lehmann-Grube, ed). Springer-Verlag, Viena, pp 241-258.
lll

mm man wi a
Katze, M. G., Lara, J. & Wambach, M. (1989) "Nontranslated cellular mRNAsare associated with the cytoskeletal framework in influenza virus or adenovirusinfected cells" Virology. 169, 312-22.
Keller, P. & Simons, K. (1997) "Post-Golgi biosynthetic trafficking" J Cell Sci.110 (Pt 24), 3001-9.
Keller, P. & Simons, K. (1998) "Cholesterol is required for surface transport ofinfluenza virus hemagglutinin" J Cell Biol. 140, 1357-67.
Kenyon, R. H., K. T. McKee, Jr., et al. (1992). "Aerosol infection of rhesusmacaques with Junin virus." Intervirology 33(1): 23-31.
Lascano, E. F. & Berria, M. I. (1971) "Electron microscopy of the Junin vius inprimary rabbit and hamster kidney cultures" Medicina (B Aires). 31, 1-5.
Lascano, E. F. & Berria, M. I. (1974) "Ultrastructure of Junin virus in mousewhole brain and mouse brain tissue cultures" J Virol. 14, 965-74.
Lee, K. J., I. S. Novella, et al. (2000). "NP and L proteins of Iymphocyticchoriomeningitis virus (LCMV) are sufficient for efficient transcription andreplication of LCMVgenomic RNA analogs." J Virol 74(8): 3470-7.
Lehmann-Grube, F. (1972). "Persistent infection of the mouse with the virus ofIymphocytic choriomeningitis." J Clin Pathol Suppl (R Coll Pathol) 6: 8-21.
Lehmann-Grube, F. (1984). "Portraits of viruses: arenaviruses." Intervirology22(3): 121-45.
Liu, C. T., P. B. Jahrling, et al. (1986). "Evidence for the involvement ofsulfidopeptide leukotrienes in the pathogenesis of Pichinde virus infection instrain 13 guinea pigs." Prostaglandins Leukot Med 24(2-3): 129-38.
Lupton, H. W., Jahrling, P. B., Barrera Oro, J. G. & a|., e. (1988) "Crossprotection against Machupo virus with Candid #1 Junin vaccine: II, postchallenge virological and immunological findings" Second InternationalConference on the Impact of Viral Diseases on the Development of LatinAmerican Countries and the Caribbean Region, Mar del Plata, Argentina.Abstract PE2.
Maiztegui, J., M. Feuillade, et al. (1986). "Progressive extension of the endemicarea and changing incidence of Argentine Hemorrhagic Fever." Med Microbiollmmunol (Berl) 175(2-3): 149-52.
Maiztegui, J. I. (1975). "Clinical and epidemiological patterns of Argentinehaemorrhagic fever." BullWorld Health Organ 52(4-6): 567-75.
Maiztegui, J. I., K. T. McKee, Jr., et al. (1998). "Protective efficacy of a liveattenuated vaccine against Argentine hemorrhagic fever. AHF Study Group." JInfect Dis 177(2): 277-83.

Manié, S.N., Debreyne, S., Vincent, S., Gerlier, D. (2000) "Measles virusstructural components are enriched into Iipid rafts microdomains: a potentialcellular location for virus assembly" J Virol, 74 (1) 305-311.
Martinez Peralta, L. A., Leon, M. E., Coto, C. E. & Laguens, R. P. (1979) "Effectof glucosamine on the replication of the arenavirus Junin in Vero cells"Intervirology. 11, 188-90.
Martinez Segovia, Z. M. and M. I. De Mitri (1977). "Junin virus structuralproteins." J Virol21(2): 579-83.
McKee, K. T., Jr., B. G. Mahlandt, et al. (1985). "Experimental Argentinehemorrhagic fever in rhesus macaques: viral strain-dependent clinicalresponse." J Infect Dis 152(1): 218-21.
Mersich, S. E., V. Castilla, et al. (1988). "Lectin affinity of Junin virusglycoproteins." Ann Inst Pasteur Virol 139(3): 277-83.
Mettler, N., Casals, J. & Shope, R. (1963) "Study of antigenic relationshipbetween Junín virus, the etiological agent of Argentine hemorahagic fever, andother arthropode borne viruses" Am J of Trop Med Hyg. 12, 647-652.
Mills, J. N., B. A. Ellis, et al. (1991). "Junin virus activity in rodents from endemicand nonendemic loci in central Argentina." Am J Trop Med Hyg 44(6): 589-97.
Mills, J. N., Ellis, B. A., McKee, K. T., Jr., Calderon, G. E., Maiztegui, J. I.,Nelson, G. 0., Ksiazek, T. G., Peters, C. J. & Childs, J. E. (1992) A longitudinalstudy of Junin virus activity in the rodent reservoir of Argentine hemorrhagicfever, Am J Trop Med Hyg. 47, 749-63.
Molinas FC, B. M. d., Maiztegui JI (1989). "Hemostasis and the complementsystem in Argentine hemorrhagic fever." Rev Infect Dis 11((Suppl 4)): 762-767.
Mosmann, T. (1983). "Rapid colorimetric assay for cellular growth and survival:application to proliferation and cytotoxicity assays." J lmmunol Methods 65(1-2):55-63.
Mukherjee, S. & Maxfield, F. R. (2000) Role of membrane organization andmembrane domains in endocytic Iipidtrafficking, Traffic. 1, 203-11.
Mukherjee, A., L. Arnaud, et al. (2003). "Lipid-dependent recruitment ofneuronal Src to Iipidrafts in the brain." J Biol Chem 278(42): 40806-14.
Nguyen, D. H. & Hildreth, J. E. (2000) "Evidence for budding of humanimmunodeficiency virus type 1 selectiver from glycolipid-enriched membraneIipid rafts" J Virol. 74, 3264-72.
ll3

OIink-Coux, M., Huesca, M. & Scherrer, K. (1992) "Specific types of prosomesare associated to subnetworks of the intermediate filaments in PtK1cells" Eur J Cell Biol. 59, 148-59.
Ono, A. & Freed, E. O. (2001) "Plasma membrane rafts play a critical role inHIV-1assembly and release" Proc Natl Acad Sci U S A. 98, 13925-30.
Osborn, M. & Weber, K. (1977) "The detergent-resistant cy'toskeleton of tissueculture cells includes the nucleus and the microfilament bundles" Exp Cell Res.106, 339-349.
Padula, P. J. & de Martinez Segovia, Z. M. (1984) "Replication of Junin virus inthe presence of tunicamycin" lntervirology. 22, 227-31.
Parodi, A. S., D. J. Greenway, et al. (1958). "Concerning the epidemic outbreakin Junin." Dia Med 30(62): 2300-1.
Pasian, E. L., Fernandez Cobo, M., Padula, P., De Mitri, M. I. & MartinezSegovia, Z. M. (1983) "Presence of actin in Junin virus" ComunicacionesBiológicas. 2, 25-30.
Peters, C. J., Jahrling PB (1987). "Experimental studies of arenaviralhemorrhagic fevers." (In: Oldstone MB (ed) Current Topics in Microbiology andInmmunology. Vol 134. Arenaviruses: epidemiology and inmmunoterapy.Springer Verlag, Berlin Heidelberg New York): pp 5-68.
Peters, C. J., C. T. Liu, et al. (1989). "Pathogenesis of viral hemorrhagic fevers:Rift Valley fever and Lassa fever contrasted." Rev Infect Dis 11 Suppl 4:S 743-9.
Peters, C. J. (1997). "Pathogenesis of viral hemorrhagic fevers." (In: NathansonN, Ahmed R, Gonzalez-Scarano F, Griffin D, Holmes KV, Murphy FA, RobinsonHL(eds) Viral pathogenesis. Lippincott-raven, Philadelphia): pp 779-799.
Pralle, A., P. Keller, et al. (2000). "Sphingolipid-cholesterol rafts diffuse as smallentities in the plasma membrane of mammalian cells." J Cell Biol 148(5): 9971008.
Puri, V., Watanabe, R., Dominguez, M., Sun, X., Wneatley, C. L., Marks, D. L. &Pagano, R. E. (1999) "Cholesterol modulates membrane traffic along theendocy‘tic pathway in sphingolipid-storage diseases" Nat Cell Biol. 1, 386-8.
Rawls, W. E., M. A. Chan, et al. (1981). "Mechanisms of persistence inarenavirus infections: a brief review." Can J Microbiol27(6): 568-74.
Resh, M. D. (1999) "Fatty acylation of proteins: new insights into membranetargeting of myristoylated and palmitoylated proteins" Biochim Biophys Acta.1451, 1-16.
H4

mnunan«m
Romanowski, V. (1981) "Estructura bioquímica del virus Junín" Tesis doctoral.Facultad de Ciencias Exactas, Universidad de la Plata. Argentina.Rowe, W. P., F. A. Murphy, et al. (1970). "Arenoviruses: proposed name for anewly defined virus group." J Virol 5(5): 651-2.
Sabattini, M., Gonzalez de Rios L., Diaz G., Vega V. (1977). "Infección natural yexperimental de roedores con virus Junín." Medicina (Buenos Aires) 37: 149161
Sager. P. R. (1989) "Cytoskeletal effects of acrylamide and 2,5-hexanedione:selective aggregation of vimentin filaments" Toxicol Appl Pharmacol. 97, 14155.
Salvato. M. S., Schweighofer, K. J., Burns, J. 8. Shimomaye, E. M. (1992)"Biochemical and immunological evidence that the 11 kDa zinc-binding proteinof Iymphocytic choriomeningitis virus is a structural component of the virus"Virus Res. 22, 185-98.
Salvato, M. S. and E. M. Shimomaye (1989). 'The completed sequence ofIymphocytic choriomeningitis virus reveals a unique RNA structure and a genefor a zinc finger protein." Virology 173(1): 1-10.
Sanchez, A., D. Y. Pifat, et al. (1989). "Junin virus monoclonal antibodies:characterization and cross-reactivity with other arenaviruses." J Gen Virol 70 (Pt 5): 1125-32.
Scheiffele, P., Roth, M. G. & Simons, K. (1997) "Interaction of influenza virushaemagglutinin with sphingolipid-cholesterol membrane domains via itstransmembrane domain" Embo J. 16, 5501-8.
Scherrer, K. and F. Bey (1994). "The prosomes (multicatalytic proteinases;proteasomes) and their relationship to the untranslated messengerribonucleoproteins, the cytoskeleton, and cell differentiation." Prog Nucleic AcidRes Mol Biol 49: 1-64.
Schmid, H. P., Akhayat, 0., Martins De Sa, C., Puvion, F., Koehler, K. &Scherrer, K. (1984) "The prosome: an ubiquitous morphologically distinct RNPparticle associated with repressed mRNPs and containing specific ScRNA anda characteristic set of proteins" Embo J. 3, 29-34.
Silber, A. M., Candurra, N. A. & Damonte, E. B. (1993) "The effects ofoligosaccharide trimming inhibitors on glycoprotein expression and infectivityofJunin virus" FEMS Microbiol Lett. 109, 39-43.
Simons. K. and E. Ikonen (1997). "Functional rafts in cell membranes." Nature387(6633): 569-72.
Simons, K. and D. Toomre (2000). "Lipidrafts and signal transduction." Nat RevMol Cell Biol 1(1): 31-9.
115

Southern, P. J., M. K. Singh, et al. (1987). "Molecular characterization of thegenomic S RNA segment from Iymphocytic choriomeningitis virus." Virology157(1): 145-55.
Speir, R. W., Wood, 0., Liebhaber, H. & Buckley, S. M. (1970) "Lassa fever, anew virus disease of man from West Africa. IV.Electron microscopy of Vero cellcultures infected with Lassa virus" Am J Trop Med Hyg. 19, 692-4.
Spohr, G., N. Granboulan, et al. (1970). "Messenger RNA in HeLa cells: aninvestigation of free and polyribosome-bound cytoplasmic messengerribonucleoprotein particles by kinetic labelling and electron microscopy." Eur JBiochem 17(2): 296-318.
Strecker, T., Eichler, R., Meulen, J., Weissenhorn, W., Dieter Klenk, H., Garten,W. & Lenz, O. (2003) "Lassa virus Z protein is a matrix protein and sufficient forthe release of virus-like particles" J Virol. 77, 10700-5.
Tkaczenski, L. Z. & Boxaca, M. C. (1970) "Cambios citoquímicos en célulasVero infectadas con alguno virus del grupo Tacaribe" Medicina (B Aires). 30,28-37.
Tortorici, M. A., Ghiringhelli, P. D., Lozano, M. E., Albarino, C. G. &Romanowski, V. (2001) "Zinc-binding properties of Junin virus nucleocapsidprotein" J Gen Virol. 82, 121-8.
Traub, E. (1935). "A filterable virus recoveres from withe mice." Science 81:198-299.
van Meer, G. (1989) "Lipidtraffic in animal cells" Annu Rev Cell Biol. 5, 247-75.
Vincent, S., Gerlier D., Manié, S. (2000) "Measles Virus Assembly withinMembrane Rafts" J ViroI74 (21) 9911-9915
Weissenbacher, M. C., M. A. CaIeIIo, et al. (1983). "Junin virus infection ingenetically athymic mice." Intervirology 19(1): 1-5.
Zaki SR, P. C. (1997). "Viral hemorrhagic fevers." (ln: Connor DH, ChandlerFW, Schwartz DA. Manz HJ, Lack EE (eds) The pathology of infectiousdisease. Appleton and Lange, NonNalk,Conn.,): pp 347-364.
116