Embed Size (px)
Citation preview

1
UNIVERSIDADE FEDERAL DE MATO GROSSO FACULDADE DE AGRONOMIA E MEDICINA VETERINÁRIA
Programa de Pós-graduação em Agricultura Tropical
COMUNIDADE MICROBIANA DO SOLO, BACTÉRIAS DIAZOTRÓFICAS E FUNGOS MICORRÍZICOS
ARBUSCULARES ASSOCIADOS AO ALGODOEIRO, EM DIFERENTES SISTEMAS DE CULTIVO
PATRICIA MARIA COURY DE ANDRADE VILELA
CUIABÁ - MT 2006

2
UNIVERSIDADE FEDERAL DE MATO GROSSO FACULDADE DE AGRONOMIA E MEDICINA VETERINÁRIA
Programa de Pós-graduação em Agricultura Tropical
COMUNIDADE MICROBIANA DO SOLO, BACTÉRIAS DIAZOTRÓFICAS E FUNGOS MICORRÍZICOS
ARBUSCULARES ASSOCIADOS AO ALGODOEIRO, EM DIFERENTES SISTEMAS DE CULTIVO
PATRICIA MARIA COURY DE ANDRADE VILELA
Engenheira Agrônoma
Orientadora: Profa Dra MARIA DE FÁTIMA LOUREIRO
Co-orientador: Pesq. Dr. ARNALDO COLOZZI FILHO
Dissertação apresentada à Faculdade de
Agronomia e Medicina veterinária da
Universidade Federal de Mato Grosso
para obtenção do título de Mestre em
Agricultura Tropical, área de
concentração em Recursos Naturais.
CUIABÁ - MT
2006

3iii
UNIVERSIDADE FEDERAL DE MATO GROSSO FACULDADE DE AGRONOMIA E MEDICINA VETERINÁRIA
Programa de Pós-Graduação em Agricultura Tropical
CERTIFICADO DE APROVAÇÃO
Título: COMUNIDADE MICROBIANA DO SOLO, BACTÉRIAS
DIAZOTRÓFICAS E FUNGOS MICORRÍZICOS ARBUSCULARES
ASSOCIADOS AO ALGODOEIRO, EM DIFERENTES SISTEMAS DE
CULTIVO.
Autora: Patrícia Maria Coury de Andrade Vilela
Orientadora: Dra. Maria de Fátima Loureiro
Co-orientador: Dr. Arnaldo Colozzi Filho
Aprovada em 28 de abril de 2006.
Comissão Examinadora:
___________________________ _______________________
Dra. Maria de Fátima Loureiro Dr. Arnaldo Colozzi Filho
(FAMEV/UFMT) (Orientadora) (IAPAR) (Co-Orientador)
___________________________ _________________________
Dr. Jean Louis R. Belot Dr. Joadil Gonçalves de Abreu
(CIRAD/COODETEC) (FAMEV/UFMT)

4iv
DEDICO
A Deus
Aos meus pais
Edson e Rosina pelo apoio e exemplo de luta
Ao meu esposo
Marcelo, pelo apoio e paciência nos momentos mais difíceis
Aos meus irmãos
Gabrielle e Edson Junior, pela paciência e ajuda imprescindível
A minha sobrinha
Maria Lúcia

5 v
AGRADECIMENTOS
A Universidade Federal de Mato Grosso, Faculdade de Agronomia e
Medicina Veterinária, UFMT/FAMEV, pela oportunidade de realização do
curso.
Ao Instituto Agronômico do Paraná, IAPAR – Londrina, por ter cedido
o laboratório para o desenvolvimento de parte do trabalho.
A Professora Dra. Maria de Fátima Loureiro pela orientação e apoio.
Ao Pesquisador Dr. Arnaldo Colozzi Filho, pela co-orientação do
trabalho e gentil atenção.
Ao Professor Dr. Joadil Gonçalves de Abreu pela colaboração nas
análises estatísticas e sugestões.
A todos os funcionários e estagiários do laboratório de Microbiologia
do solo – IAPAR, em especial ao Osvaldo Machineski, pela colaboração e
auxilio nas análises laboratoriais lá realizadas.
A todos os colegas, funcionários e estagiários do laboratório de
Microbiologia do solo da FAMEV/UFMT, em especial ao Edson Andrade
Junior, Marcianne Quixabeira e Valéria Pires, pela colaboração e execução
dos trabalhos.
Ao Fundo de apoio à cultura do algodão – FACUAL, pelo
financiamento dessa pesquisa.
A diretoria e funcionários da Coodetec pelo apoio e incentivo, em
especial a Tatiane e Arlindo.
Ao pesquisador Dr Jean Louis Belot pelo apoio, incentivo, paciência e
sugestões.
Ao Marcos Pinheiro e Ronier Rezende pelo apoio no momento das
coletas de campo.
Aos professores das disciplinas cursadas pelos conhecimentos
transmitidos, meus colegas de curso pela amizade e as funcionárias da
secretaria pelo atendimento prestado.
A todos que de alguma forma contribuíram para a realização deste
trabalho.

6 vi
SUMÁRIO
LISTA DE FIGURAS...................................................................................... vii
Página
LISTA DE TABELAS.......................................................................................ix
Resumo ..........................................................................................................1
Abstract...........................................................................................................2
1. INTRODUÇÃO............................................................................................3
2. REVISÃO DE LITERATURA.......................................................................5
2.1 A cultura do algodoeiro .............................................................................5
2.2. Sistemas de cultivo ..................................................................................6
2.3. Atividade microbiana nos solos do cerrado .............................................9
2.4. Micorrizas ..............................................................................................10
2.5. Bactérias diazotróficas...........................................................................13
CAPITULO I: Micorrização do algodoeiro em diferentes sistemas de cultivos
em Campo Verde - MT .................................................................................16
Resumo: .......................................................................................................17
Abstract.........................................................................................................18
3.1. INTRODUÇÃO.......................................................................................19
3.2. MATERIAL E MÉTODOS ......................................................................20
3.3. RESULTADOS E DISCUSSÃO .............................................................24
3.4 CONCLUSÃO .........................................................................................31
CAPITULO II: Populações de bactérias e fungos e ocorrência de
Azospirillum em solo sob diferentes sistemas de cultivos do algodoeiro em
Campo Verde - MT ...................................................................................... 32
Resumo ........................................................................................................33
Abstract.........................................................................................................34
4.1. INTRODUÇÃO:......................................................................................35
4.2. MATERIAL E MÉTODOS ......................................................................36
4.3. RESULTADOS E DISCUSSÃO .............................................................38
4.4. CONCLUSÃO ........................................................................................43
5. CONCLUSÕES GERAIS ..........................................................................44
6. REFERÊNCIAS BIBLIOGRÁFICAS..........................................................45
APÊNDICES .................................................................................................57

7vii
LISTA DE FIGURAS
Página
Figura 1. Estádios fenológico do ciclo do algodoeiro e épocas das coletas das amostras, em dias.
20
Figura 2. Fungos micorrízicos arbusculares em raízes do algodoeiro cultivado em sistema plantio direto em rotação bianual com soja seguida de sorgo consorciado com braquiária, em Campo Verde-MT. (Aumento de 40 vezes).
25
Figura 3. Colonização micorrízica (%) do algodoeiro, cultivado em monocultura em sistema convencional (T1), em monocultura em sistema semi-direto sobre palhada de milheto (T2) e em plantio direto em rotação bianual com soja seguida de sorgo consorciado com braquiária (S3), em dois níveis de fertilização, no município de Campo Verde-MT. Média de duas repetições.
25
Figura 4. Esporos de fungos micorrízicos arbusculares isolados da rizosfera do algodoeiro, cultivado em sistema convencional (T1), em Campo Verde-MT. (Aumento de 40 vezes).
28
Figura 5. Densidade de esporos de fungos micorrízicos em solo cultivado com algodoeiro, em monocultura em sistema convencional (T1), em monocultura em sistema semi-direto sobre palhada de milheto (T2) e em plantio direto em rotação bianual com soja seguida de sorgo consorciado com braquiária (S3), nas quatro épocas de coleta, na fertilização reduzida, no município de Campo Verde-MT. Média de seis repetições.
29
Figura 6. Densidade de esporos de fungos micorrízicos em solo cultivado com algodoeiro, em monocultura em sistema convencional (T1), em monocultura em sistema semi-direto sobre palhada de milheto (T2) e em plantio direto em rotação bianual com soja seguida de sorgo consorciado com braquiária (S3), nas quatro épocas de coleta, na fertilização padrão, no município de Campo Verde-MT. Média de seis repetições.
30
Figura 7. População de fungos em solo cultivado com algodoeiro, em monocultura em sistema convencional (T1), em monocultura em sistema semi-direto sobre palhada de milheto (T2) e em plantio direto em rotação bianual com soja seguida de sorgo consorciado com braquiária (S3), em dois níveis de fertilização, no município de Campo Verde-MT. Média de oito repetições.
39

8viii
Figura 8. População de bactérias em solo cultivado com algodoeiro, em monocultura em sistema convencional (T1), em monocultura em sistema semi-direto sobre palhada de milheto (T2) e em plantio direto em rotação bianual com soja seguida de sorgo consorciado com braquiária (S3), em dois níveis de fertilização, no município de Campo Verde-MT. Média de oito repetições
40

9ix
LISTA DE TABELAS
Página
Tabela 1. Fertilização (Kg/ha) aplicada na base e em cobertura, no algodoeiro conduzido em sistemas convencional (T1), semi-direto (T2) e plantio direto (S3), no município de Campo Verde-MT.
21
Tabela 2. Análise química de solo cultivado com algodoeiro nos sistemas de cultivo convencional (T1), semi-direto (T2) e plantio direto, com rotação bianual de culturas (S3), em diferentes níveis de fertilização, em quatro épocas, no município de Campo Verde-MT (média de seis repetições).
22
Tabela 3. Densidade de esporos de fungos micorrízicos (no./50 mL de solo) em monocultura em sistema convencional (T1), em monocultura em sistema semi-direto sobre palhada de milheto (T2) e em plantio direto em rotação bianual com soja seguida de sorgo consorciado com braquiária (S3), nas quatro épocas de coleta, em dois níveis de fertilização, no município de Campo Verde-MT. Média de seis repetições.
27
Tabela 4. População de bactérias diazotróficas (NMPx105/g de solo) em solo cultivado com algodoeiro, em monocultura em sistema convencional (T1), em monocultura em sistema semi-direto sobre palhada de milheto (T2) e em plantio direto em rotação bianual com soja seguida de sorgo consorciado com braquiária (S3), em dois níveis de fertilização, no município de Campo Verde-MT. Média de seis repetições.
42

1
COMUNIDADE MICROBIANA DO SOLO, BACTÉRIAS DIAZOTRÓFICAS E FUNGOS MICORRÍZICOS ARBUSCULARES ASSOCIADOS AO
ALGODOEIRO, EM DIFERENTES SISTEMAS DE CULTIVO
Resumo: Os sistemas de cultivos com seus respectivos manejos causam
grandes impactos na comunidade microbiana do solo, bactérias diazotróficas
e fungos micorrízicos arbusculares. Com isso, o objetivo desse trabalho foi
avaliar os fungos micorrízicos arbusculares, a comunidade microbiana e
bactérias diazotróficas, associados ao algodoeiro, em diferentes sistemas de
cultivo e em dois níveis de fertilização. O delineamento experimental
utilizado foi inteiramente casualizado, com seis tratamentos e seis
repetições, os tratamentos foram dispostos em esquema de parcelas
subdivididas, sendo nas parcelas os dois níveis de fertilização (padrão e
reduzida) e nas sub-parcelas os três sistemas de cultivo [monocultivo de
algodoeiro em preparo convencional (T1), monocultivo de algodoeiro em
plantio semi-direto, sobre palhada de milheto (T2) e algodoeiro em plantio
direto, em rotação bianual, com soja seguido de sorgo consorciado com
Brachiaria ruzizienses (S3)]. Avaliou-se a densidade de esporos do solo, a
colonização radicular do algodoeiro, as populações de bactérias, fungos e
bactérias diazotróficas. O sistema de plantio semi-direto (T2) apresentou
uma tendência a ter uma maior colonização micorrízica e obteve maior
população de bactérias totais, para ambas as fertilizações. O sistema de
plantio direto (S3), apresentou maior densidade de esporos. De modo geral
os sistemas de cultivo e a fertilização do solo atuaram sobre a simbiose
micorrízica-arbuscular. Não houve efeito das fertilizações na população de
fungos totais para os sistemas semi-direto (T2) e plantio direto (S3). A
população de Azospirillum sp., foi influenciada pelas épocas de coleta e
provavelmente pelos tratos culturais, sendo que o sistema semi-direto (T2)
teve melhor efeito sobre essa população.
Palavras chaves: Fungos micorrízicos, colonização radicular, Azospirillum
sp, comunidade microbiana, sistemas de cultivo.

2
SOIL MICROBIAL COMMUNITY, DIAZOTROPHIC BACTERIA, AND ARBUSCULAR MYCORRHIZAL FUNGI ASSOCIATED WITH COTTON IN
DIFFERENT CULTIVATION SYSTEMS
Abstract: Different cultivation systems, with their corresponding
management practices, have a great impact on the soil microbial community,
diazotrophic bacteria, and arbuscular mycorrhizal fungi. The objective of this
work was therefore to evaluate the arbuscular mycorrhizal fungi, microbial
community and diazotrophic bacteria associated with cotton in different
cultivation systems at two fertilization levels. A completely randomized
experimental design was adopted, with six treatments and six replicates. The
treatments were organized in a split-plot arrangement where the two
fertilization levels (standard and reduced) were assigned to plots and three
cultivation systems were assigned to sub-plots [cotton monoculture under
conventional tillage (T1), cotton monoculture under semi-direct planting on
millet trash (T2), and no-till cotton, in a biennial rotation, with soybean
followed by sorghum intercropped with Brachiaria ruzizienses (S3)].
Evaluations included spore density in the soil, cotton root colonization, and
populations of bacteria, fungi, and diazotrophic bacteria. The semi-direct
planting system (T2) showed a tendency to have higher mycorrhizal
colonization and obtained higher total bacterial populations, at both
fertilization levels. The no-till system (S3) showed higher spore density. In
general, the cultivation systems and soil fertilization acted upon the
arbuscular mycorrhizal symbiosis. There was no effect of fertilization on total
fungal population in the semi-direct (T2) and no-till (S3) systems. The
Azospirillum sp. population was influenced by collection season and probably
by management practices. The semi-direct planting system (T2) had the best
effect on this population.
Keywords: Mycorrhizal fungi, root colonization, Azospirillum sp, microbial
community, cultivation systems.

3
1. INTRODUÇÃO O Algodão é uma das fibras vegetais cultivadas mais antigas do
mundo. Sendo uma cultura de grande importância, seu principal produto é a
fibra, que possui diversas aplicações na indústria (Corrêa, 1989). A semente
(caroço) representa aproximadamente 65% do peso da produção e a fibra
35%.
A cotonicultura já atingiu altos níveis de produtividade, mas, para sua
consolidação no cenário nacional, é necessário chegar a sustentabilidade
dos sistemas de produção que depende, entre outros fatores, do potencial
de impacto das tecnologias sobre os agroecossistemas em seus vários
aspectos sociais, ambientais e econômicos.
Nos agroecossistemas só é possível manter o processo em equilíbrio
dinâmico, desde que haja igualdade, estabilidade e produtividade
contempladas dentro das dimensões ecológicas, econômicas e sociais.
Para assegurar a viabilidade do cultivo algodoeiro, é preciso reduzir
os riscos econômicos e ecológicos e aumentar a rentabilidade,
principalmente mediante a redução dos custos de produção, que atingem
níveis muito elevados e, portanto, tornam este cultivo arriscado. A introdução
de variedades resistentes a viroses e bacteriose, e mais tolerantes a
doenças foliares, contribui para o processo de redução de custos e de
consolidação da cotonicultura nos cerrados. Mas existem vias
complementares de redução de custos, em particular tratando da associação
entre microrganismos e variedades no sistema de cultivo.
Dentre os sistemas de cultivo, o plantio direto e o cultivo mínimo são
os mais utilizados atualmente, porém há muitas lavouras que ainda utilizam
o plantio convencional.
A produtividade agrícola é determinada em parte pela disponibilidade
de nitrogênio previamente fixado, seja pelo processo biológico ou pelo
processo químico industrial.
Os principais organismos que podem utilizar o nitrogênio gasoso
atmosférico pertencem ao reino do procariotos e a reação básica parece a
mesma em todos os casos: redução do nitrogênio elementar (N2) à amônia

4
(NH3), catalisada pelo complexo enzimático da nitrogenase. Esses
organismos sejam em simbiose ou em vida livre, como, por exemplo,
Azospirillum, que é uma bactéria fixadora associativa, demonstram grande
potencial em reduzir a dependência de fertilizantes nitrogenados,
reconhecidamente comprometedores da qualidade da água e do ar quando
usado inadequadamente. Assim, todas as possibilidades de incremento da
fixação biológica de nitrogênio na agricultura devem ser exploradas, não
somente como alternativa econômica, mas, também ecológica.
Fungos micorrízicos arbusculares (FMA) são microrganismos que
formam associações simbiotróficas mutualistas (micorrizas arbusculares)
com as raízes de vegetais. Estes fungos ocorrem normalmente na natureza
(Allen, 1991) propiciando às plantas melhor eficiência na absorção de
fósforo. As micorrizas contribuem para o estabelecimento e longevidade dos
agrossistemas (Harley & Smith, 1983; Sieverding, 1990; Abbot & Robson,
1991).
Ainda que sejam organismos de ocorrência generalizada, algumas
perturbações no ecossistema que acarretem mudanças nas propriedades
dos solos poderão contribuir para redução ou eliminação dos propágulos de
FMA em um determinado ambiente. Entre estas perturbações, fatores como
o tipo de preparo de solo e de plantio (preparo convencional), sucessão de
culturas, cultivo de plantas não hospedeiras, pousio prolongado, eliminação
da vegetação de cobertura, fogo, poluição, erosão, compactação, inundação,
uso de fertilizantes em quantidades excessivas, uso de agrotóxicos
sistêmicos (Abbott & Robson, 1991), são responsáveis pela redução do
número de propágulos viáveis de FMA.
Os efeitos das práticas de manejo do solo sobre a percentagem de
colonização de raízes por fungos micorrízicos já foram estudados,
evidenciando efeitos significativos em cultivos de milho e trigo.
Este trabalho teve como objetivo avaliar a comunidade microbiana,
bactérias diazotróficas e fungos micorrízicos arbusculares, associados ao
algodoeiro, em diferentes sistemas de cultivo e em dois níveis de fertilização.

5
2. REVISÃO DE LITERATURA
2.1 A cultura do algodoeiro O algodoeiro é uma dicotiledônea da família das Malváceas. O gênero
Gossypium é constituído de 52 espécies distribuídas nos continentes: Ásia,
África, Austrália e América, sendo que destas apenas quatro são cultivadas.
O Gossypium arboreum L., cultivado na Índia é importante comercialmente e
o Gossypium herbaceum L. que já teve maior importância no passado,
atualmente é plantado apenas em algumas regiões secas da África e Ásia.
Finalmente, cerca de 90% da produção mundial de algodão é de Gossypium
hirsutum L. e 8% de Gossypium barbadense L. (Lee, 1984).
A origem dos algodoeiros cultivados é de difícil determinação.
Segundo Phillips (1963) e Fryxell (1965) o gênero Gossypium é muito antigo,
tendo os tetraplóides, provavelmente, se originado há aproximadamente 1,5
milhão de anos. Saunders (1961) propõe que o centro de origem do gênero
Gossypium seja a África Central, pois quatro dos sete grupos genômicos
diplóides ocorrem neste continente.
Na Índia, em 1500 a.C., o algodoeiro já era cultivado visando a
fabricação de tecidos. Mil anos depois foram os chineses que iniciaram o
cultivo, que só chegou a Europa no século IV a.C. Na América, o algodão já
era utilizado pelos índios. Portanto, quando o Brasil foi descoberto o algodão
já era cultivado.
Até o final do século XVIII e início do século XIX, a cultura do algodão
no Brasil era basicamente do tipo perene (Gossypium barbadense e
Gossypium hirsutum raça Marie Galante), concentrando-se, principalmente,
na região Nordeste. A partir de 1860 a Inglaterra introduziu a variedade
herbácea (Gossypium hirsutum L. r. Latifolium, tipo upland) no Brasil com o
objetivo de incentivar a produção brasileira e diminuir sua dependência com
relação aos Estados Unidos. Em 1870/71, a cultura do algodão entra em
decadência profunda, somente em 1880/81 ocorre à recuperação.
Avançando rapidamente, em 1940, a cotonicultura brasileira viveu a
fase áurea de expansão de área cultivada. O fim dos anos 80 e início dos 90

6
é marcado por uma forte crise da cotonicultura, sendo que a partir de 1999,
com expansão da cotonicultura para a região Centro-Oeste, principalmente,
Mato Grosso, a situação se modifica e o país volta a ter elevados ganhos de
produtividade e qualidade. Em resumo, a cotonicultura no Brasil manteve
sua área estável nos últimos setes anos (safras 97/98 a 02/03), em 03/04 e
04/05 houve um pequeno aumento e a previsão para a safra 05/06 é de que
tenha uma redução de 32,6% da área plantada a nível nacional (CONAB,
2006).
2.2. Sistemas de cultivo Os solos mais comuns na região dos cerrados são os latossolos,
correspondendo com 46% da área (UFLA, 2006). Esses solos são antigos,
profundos e bem drenados, são ácidos e de baixa fertilidade com altos níveis
de Fe e Al (Klink, 2006). Entretanto, apresentam uma boa estrutura física,
aliada ao relevo predominantemente plano, favorecendo a agricultura
mecanizada (Pivello, 2006).
O algodoeiro tem-se mostrado uma cultura de larga adaptação nos
solos do cerrado, no que se refere às condições edáficas, desde que sejam
efetuadas as devidas correções, passando a apresentar características
suficientes para atender às necessidades básicas ao seu pleno
desenvolvimento (Portalbrasil, 2006).
Os sistemas de cultivo nada mais são do que um conjunto de
operações técnicas utilizadas de maneira idêntica na condução de uma
cultura. Sendo definido pelo tipo de cultura, pela ordem de sucessão das
culturas à nível de parcela assim como o itinerário técnico (Sebillotte, 1990).
Dependendo das operações técnicas utilizadas podem ser classificados
como: plantio direto, plantio convencional, cultivo mínimo, entre outros.
A técnica que inclui em seu preparo a passagem do arado e da grade
niveladora, sem cobertura passou a ser denominada plantio convencional a
fim de distinguir-se do plantio direto, surgido posteriormente, que é a prática
onde é dispensado o uso de cultivos mecânicos (arados, grades, entre
outros) e apresenta uma boa cobertura de palha no solo (Ferraz et al.,

7
2004). O outro sistema, o cultivo mínimo poderia ser um intermediário, onde
se faz o uso de arado, grade e posteriormente planta-se uma cobertura para
o solo.
O plantio direto teve seu inicio na década de 60, com os trabalhos de
Shirley H. Phillips e Harry M. Young Jr., em Kentucky-EUA (Saturnino,
2001). Todavia, o seu desenvolvimento ocorreu a partir da década de 70,
com o surgimento do herbicida paraquat que veio substituir o preparo do
solo no controle de plantas daninhas.
A adaptação do plantio direto às condições do Mato Grosso, evidencia
o fato de que a cultura algodoeira, com altíssimo nível de insumos químicos,
não está sendo sustentável nas suas práticas atuais. Sabe-se que a
produtividade do algodoeiro reduz, influenciada ao mesmo tempo, pelas
práticas excessivas da monocultura e a volta das gradagens, sendo esta
com o objetivo de destruir a soqueira, medida profilática obrigatória por lei
para controle de algumas pragas e doenças (Séguy et al., 2004).
Existe, atualmente, um consenso sobre a exigência de reduzir o risco
econômico e custos de produção para consolidar a cotonicultura no Brasil
Central, principalmente em Mato Grosso, que tem os custos mais altos do
país, cerca de U$ 1.500. Um dos caminhos mais eficientes e seguros para
conseguir esse objetivo passa pela adoção do sistema plantio direto (SPD),
que propicia solos protegidos contra a erosão, com excelentes propriedades
físicas, biologicamente sadios e muito favoráveis à cultura algodoeira,
propiciando menor susceptibilidade a qualquer stress (Séguy et al., 1998 e
2003a). A definição do SPD proposta por Séguy (1998), fica simbolicamente
resumida na seguinte fórmula: SPD = ABC = A (ausência de trabalho do
solo) + B (biodiversidade, opondo-se a monocultura) + C (coberturas
necessárias para cobrir o solo e colonizar o perfil).
Os motivos da adoção do sistema de plantio direto na palha foram,
além da melhoria no controle da erosão, vantagens econômicas como a
redução dos custos de preparo e plantio, e o alívio do calendário operacional
(Landers, 1998). Para gerar fertilidade organo-biológica e reduzir
expressivamente as necessidades em herbicidas e fertilizantes, são

8
necessárias importantes biomassas integradas em SPD diversificados, que
incluem plantas de coberturas e as safrinhas de grãos (Séguy et al., 2003b).
Uma das condições para se implantar o sistema de plantio direto é a
cobertura do solo com uma camada de palha, a fim de proporcionar um
ambiente extremamente favorável às condições químicas, físicas e
biológicas, contribuindo para o controle de plantas daninhas, estabilização
de produção e manutenção da qualidade do solo (Heckler et al., 1998;
Alvarenga et al., 2001). Plantas produtoras de grande biomassa (Eleusine
coracana, sorgo ou milheto consorciado com Brachiaria ruzizienses)
permitem construir uma fertilidade de origem organo-biológica, de grande
importância, pois aumenta a capacidade do solo em produzir com o passar
do anos (Séguy et al., 2001).
O plantio direto com cobertura vegetal permanente funciona como o
ecossistema florestal, em circuito fechado, sem perda marcante de
nutrientes, e neutraliza eficientemente os efeitos nocivos da acidez (Al) nas
culturas sensíveis, constatando mais uma vez a importância desse sistema
(Séguy et al., 2004).
No sistema de plantio direto os microrganismos são beneficiados
pelas menores oscilações térmicas e temperaturas máximas inferiores que
ocorrem nesse sistema em relação ao plantio convencional (Lal e Kharnna,
1993) refletindo positivamente nos microrganismos de importância agrícola
que são termosensíveis.
O teor de matéria orgânica também é maior no plantio direto nos
primeiros centímetros do solo, resultando em maior disponibilidade de
nutrientes para os microrganismos e maior retenção de água no solo,
também importante para a microbiota (Hungria et al., 1995).
Os sistemas de manejo do solo podem favorecer diversos
microrganismos benéficos, entre eles os fungos micorrízicos, entretanto
algumas práticas de manejo mecânico utilizado no sistema convencional
podem reduzir colonização micorrízica nas raízes das plantas, destruindo a
rede de hifas no solo, reduzindo o potencial de inóculo, uma vez que a

9
translocação dos assimilados da planta dentro da hifa é interrompido (Evans
& Miller, 1988).
2.3. Atividade microbiana nos solos do cerrado
A partir de estimativas da diversidade biológica mundial e dos
Cerrados considera-se que a riqueza mínima da região dos Cerrados é da
ordem de 320.000 espécies (Assad, 1996).
A biomassa microbiana do solo é constituída por fungos, bactérias e
actinomicetos que atuam em processos que vão desde a origem do solo,
formação e manutenção da sua estrutura até a decomposição de resíduos
orgânicos, ciclagem de nutrientes, biorremediação de poluentes e metais
pesados (Mendes e Vivaidi, 2001). Também pode ser definida como a parte
da matéria orgânica constituída pelos organismos vivos com volume menor
que 5 a 10 μ m3 (Moreira e Siqueira, 2002).
A atividade microbiana dos solos inclui todas as reações metabólicas
celulares, suas interações e seus processos bioquímicos mediados ou
conduzidos pelos organismos do solo (Matsuoka, 2001).
O aumento da biomassa microbiana e sua atividade no solo têm
implicações práticas importantes na agricultura. Biomassa microbiana maior
implica em maior mobilização temporária de C, N e outros nutrientes e,
conseqüentemente, menor perda de nutrientes do sistema solo-planta
(Colozzi Filho et al., 1999).
A biomassa microbiana pode contribuir significativamente para a
disponibilização do nitrogênio (N) do solo para as plantas, a quantidade de N
inorgânico depende da taxa de decomposição da matéria orgânica do solo
pela biomassa, da imobilização, da desnitrificação e da lixiviação (Lovell &
Hatch, 1998).
Os diferentes sistemas de manejo do solo podem determinar
profundas alterações, provocando mudanças qualitativas e quantitativas nos
microrganismos e suas atividades (Balota, 1997). Isto ocorre porque o
conjunto de práticas agrícolas utilizadas em cada agrossistema condiciona o

10
sistema solo-planta a uma dinâmica própria que pode influenciar a atividade
biológica de maneira diferenciada.
O cultivo convencional reduziu significativamente a diversidade de
bactérias na cultura do trigo, por exemplo, aumentando a predominância de
poucos grupos de microrganismos e reduzindo a fertilidade do solo (Balota,
1997).
Em sistemas onde se proporciona maior cobertura e menor
revolvimento do solo, como plantio direto (Carneiro et al., 1999; Ananyeva et
al., 1999) e sistemas de cultivo mínimo e reduzido (Deng & Tabatabai, 1997;
Kandeler et al., 1999), aumentos na biomassa microbiana e em sua
atividade têm sido observados (Matsuoka, 2001).
O efeito dos sistemas de manejo sobre a microbiota, porém, não se
restringe a aspectos quantitativos, como as alterações na biomassa.
Também a composição da comunidade microbiana pode ser alterada, pois
os grupos microbianos podem ser afetados diferentemente pelas práticas de
manejo (Vargas et al., 2004).
2.4. Micorrizas A primeira ilustração de uma micorriza foi publicada em 1840, quando
Robert Hartig ilustrou raízes finas de pinus. Porém, ele não reconheceu o
componente fúngico separadamente. Em 1847, S. Reissek descreveu as
células do fungo associadas com orquídeas e em 1881, F. Kamienski
mostrou que algumas raízes de árvores tinham uma capa de fungos que as
cercavam e que os nutrientes para serem translocados pela planta
necessitavam penetrar nesta capa (Paul & Clark, 1996).
O termo micorriza (grego “mykes” = fungo, “rhiza” = raiz) foi proposto
em 1885, pelo alemão, Albert Bernard Frank (Sylvia, 1998). Ele caracterizou
o fungo formado externamente em raízes de árvores. Essa associação foi
descrita como endotrófica (Paul & Clark, 1996). O termo trófica foi eliminado
no final da década de 60, passando a serem designadas ectomicorrizas e
endomicorrizas (Siqueira & Klauberg Filho, 2000).

11
Os FMAs estão distribuídos de forma generalizada no ambiente
terrestre, desde regiões árticas até os trópicos (Brundrett, 1991), ocorrendo
em quase todas as famílias de plantas, com exceção das que formam
ectomicorrizas, das que possuem associações específicas e algumas
famílias das monocotiledôneas e dicotiledôneas, como Brassicaceae
(Silveira, 1998).
Os FMAs são simbiontes obrigatórios, ou seja, necessitam da planta
hospedeira para poder completar seu ciclo vital. No córtex da raiz da planta
hospedeira, as hifas dos FMAs percorrem inter e intracelularmente as
células corticais, formando no seu interior, estruturas arboriformes
(arbúsculos) e/ou saculares (vesículas) (Powell & Bagyaraj, 1984).
Silveira (1992) diz que o desenvolvimento do FMA apresenta cinco
estágios: pré-infecção; infecção primária; formação de arbúsculos e/ou
vesículas; desenvolvimento do fungo na raiz e na rizosfera e dispersão do
fungo no solo.
Estabelecida a simbiose, os fungos formadores de micorrizas,
estabelecem uma série de inter-relações biotróficas. A raiz é perfurada pela
hifa e a colonização instala-se apenas nas células do córtex. As hifas
percorrem as células do hospedeiro e nos espaços intercelulares são
formados os arbúsculos e/ou as vesículas. As espécies de FMAs que
formam vesículas intra-radicais possuem células auxiliares no micélio extra-
radical (Silveira, 1992).
As vesículas são estruturas globulares esféricas relacionadas com
armazenamentos de substâncias de excreção ou reserva, como óleos, e
ocorrem inter e intracelularmente às raízes como também no solo (Siqueira
et al., 1986). Os arbúsculos, originados de ramificações dicotômicas de
hifas, ocorrem no interior de células corticais e apresentam circunvoluções
que lhes aumentam a superfície, tendo como função, as trocas nutricionais
entre os organismos simbiontes (Scannerini & Bofante–Fasolo, 1983).
As hifas funcionam como extensão do sistema radicular, aumentando
sua capacidade em explorar maior volume de solo e aumentar a superficie
de contato com o solo e os sítios de absorção de nutrientes, melhorando o

12
estado nutricional da planta através da maior absorção de nutrientes,
principalmente do P, Cu e Zn, que apresentam baixa mobilidade. Dessa
forma, esta simbiose é de grande interesse, principalmente, em condições
tropicais, onde os solos apresentam baixos teores de fósforo disponíveis
devido a sua alta capacidade de fixação (Colozzi Filho & Balota, 1994).
De acordo com Allen (1992), os FMAs contribuíram para a evolução
das plantas, e muitas delas, são extremamente dependentes da associação
micorrízica para a sobrevivência.
A dependência micorrízica pode ser definida como o grau em que a
planta hospedeira é dependente à condição micorrízica para crescimento
máximo em um dado nível de fertilidade do solo (Gerdemann, 1975). As
plantas podem ser dependentes obrigatórias ou facultativas de FMA. As
espécies micotróficas facultativas podem sobreviver e crescer sem micorriza
arbuscular (MA) em um dado nível de fertilidade do solo (Sieverding, 1991).
A alta dependência de MA está relacionada com o requerimento de P e à
baixa capacidade da planta em absorver este nutriente quando não
micorrizadas e com o tipo de sistema radicular da planta, sendo altamente
dependente aquelas espécies com o sistema radicular pouco desenvolvido,
como a mandioca, citros, café, soja e muitos legumes tropicais. Plantas com
alta densidade de raízes e pêlos radiculares longos são menos dependentes
de MA (Sieverding, 1991).
Informações sobre a diversidade e a dinâmica de fungos micorrízicos
em condições de campo são decorrência de estudos quantitativos
determinando a colonização radicular e a densidade de esporos no solo, e
qualitativas, em que a presença de diferentes fungos é determinada para
gênero e/ou espécies, em termos de freqüência e abundância relativa
(Siqueira & Klauberg Filho, 2000).
Vários trabalhos mostram os benefícios da associação com FMAs,
como em mudas de acerola, onde foi observado maior crescimento e
absorção de fósforo, potássio, cobre e zinco (Oliveira & Leal, 2000); para
mudas de maracujazeiro, a inoculação com FMAs proporcionou aumentos
no crescimento e nos teores de nutrientes da parte aérea na fase de

13
produção de mudas (Soares & Martins, 2000). Para a soja, a micorrização
favoreceu de forma significativa o crescimento, nodulação e o acúmulo de
nutrientes (Minhoni et al., 1993) e as plantas de soja micorrizadas
apresentaram maiores teores de fósforo e cálcio nas folhas e menores de
potássio, manganês, zinco e cobre do que aqueles não micorrizadas
(Siqueira et al., 1986).
Siqueira et al. (1986), verificou que o algodoeiro crescendo em solos
deficientes em P exibiu elevada dependência micorrízica e alto grau de
micotrofismo, não apresentando especificidade em nível de colonização.
Entretanto, a efetividade simbiótica foi diferenciada, confirmando a
universalidade da simbiose e sua importância para o crescimento do
algodoeiro em condições de deficiência de P (Davis et al., 1979; Smith et al.,
1986).
O uso de microrganismos com a finalidade de melhorar a
disponibilidade de nutrientes às plantas é uma prática de grande importância
e necessária para a agricultura. Estudos relacionados à ecologia dos FMAs
são essenciais para se planejar o uso efetivo desses fungos na agricultura
comercial ou conhecer sua atividade como componente do ecossistema
como um todo (Fitter, 1991).
2.5. Bactérias diazotróficas Com a exceção da água, o nitrogênio é geralmente considerado o
nutriente mais limitante para o crescimento de plantas no seu ambiente
natural (Franco & Dobereiner, 1994). Apesar da abundância de N2 na
atmosfera, somente os organismos procariontes conseguem converter ou
reduzir enzimaticamente o nitrogênio da atmosfera em amônia. Estes
organismos são denominados diazotróficos e o mecanismo responsável pela
incorporação de N à biomassa é chamado de fixação biológica de nitrogênio
(Marin et al., 1999).
Evans & Burris (1992), caracterizaram as bactérias fixadoras de
nitrogênio ou diazotrofos, em três grupos: diazotrofos de vida livre,
diazotrofos associativos e diazotrofos simbióticos.

14
Os primeiros a serem reconhecidos foram os diazotrofos de vida livre,
que fixam nitrogênio para o seu próprio uso, como é o caso de Beijerinkia
fluminensis e Beijerinkia indica, isoladas da rizosfera de plantas de cana-de-
açúcar crescidas em solos tropicais, sendo demonstrado o seu potencial na
associação com gramíneas (Dobereiner & Ruschel, 1958; Dobereiner e
Alvahydo, 1959). Já os diazotrofos simbióticos, são aqueles que
estabelecem uma interação muito estreita entre o macro e microsimbionte,
em alguns casos formam-se estruturas diferenciadas denominadas nódulos.
Nesse grupo de microrganismos estão os gêneros Azorhizobium,
Bradyrhizobium, Rhizobium, Sinorhizobium, Mesorhizobium, entre outros
(Marin et al., 1999).
Dentro do grupo dos diazotrofos associativos, podemos dividir em
endofíticos facultativos (que podem colonizar tanto a rizosfera como o
interior das raízes) e os endofíticos obrigatórios (que colonizam o interior das
raízes), como proposto por Baldani et al., 1997. Entre os endofíticos
obrigatórios estão: Acetobacter diazotrophicus (Cavalcante & Dobereiner,
1988), Azoarcus spp. (Reinhold-Hurek et al., 1993), Herbaspirillum
seropedicae (Baldani et al., 1986), Herbaspirillum rubrisubalbicans (Baldani
et al., 1996) e Burkholderia spp. (Yabuuchi et al., 1992).
Para endofíticos facultativos, o grupo funcional predominante é do
gênero Azospirillum, redescoberto por Dobereiner & Day (1975), isoladas a
partir de raízes de Digitaria. Estudos recentes relatam Azospirillum sp., como
sendo o grupo funcional mais promissor associado a gramíneas e não
leguminosas, devido a sua capacidade de colonizar todo o interior da planta,
localizando-se dentro de habitats protegidos do oxigênio (Baldani et al.,
1997).
Azospirillum spp. pode ser caracterizado como uma bactéria gram
negativa, tipo bastonete, bastante móvel. São aeróbios típicos, quando
supridos com fonte de nitrogênio combinado, e microaerofílicos, quando
crescem dependentes da fixação de N2. Em meio sólido formam uma
película delgada em forma de véu, abaixo da superfície do meio, onde a

15
concentração de O2 permite a fixação de nitrogênio para iniciar seu
crescimento, movendo-se em direção a superfície (Dobereiner et al., 1995).
A distribuição ecológica de Azospirillum spp. é extremamente ampla
podendo ser considerada uma bactéria universal encontrada colonizando
plantas crescidas em diferentes habitats (Dobereiner et al., 1976 ;Dobereiner
& Pedrosa, 1987). Estirpes tem sido encontradas em associação com
plantas monocotiledôneas, por exemplo: milho, arroz, cana-de-açúcar, entre
outras (Dobereiner et al., 1976; Haahtela et al., 1981) e com as
dicotiledôneas (Rao & Vankateswarlu, 1982).
Cinco espécies de Azospirillum têm sido descritas até o momento: A.
brasilense; A. lipoferum; A. amazonense; A. halopraeferens e A. irakense.
Um dos principais mecanismos de desenvolvimento de plantas relacionados
ao Azospirillum tem sido atribuído à sua capacidade de produzir substâncias
de promoção de crescimento vegetal. Azospirillum estimula a densidade e o
comprimento dos pêlos radiculares, a freqüência de raízes laterais e o
aumento da superfície da raiz. Tais efeitos na morfologia e fisiologia da raiz
resultam em aumento de absorção de água e nutrientes (Okon & Labandera-
Gonzales, 1994). Além de cana-de-açúcar, bactérias endofíticas fixadoras de
nitrogênio têm sido associadas a cereais, palmeiras, gramíneas forrageiras,
frutíferas e outras plantas de importância agronômica. Devido ao potencial
para utilização em grande escala no campo, muito mais estudos
taxonômicos, fisiológicos e genéticos necessitam ser realizados sobre a
interação planta-diazotróficos endofíticos.

16
CAPITULO I: Micorrização do algodoeiro em diferentes sistemas de cultivos em Campo Verde - MT

17
MICORRIZAÇÃO DO ALGODOEIRO EM DIFERENTES SISTEMAS DE CULTIVO EM CAMPO VERDE - MT
Resumo: O objetivo deste trabalho foi avaliar os fungos micorrízicos
arbusculares, associados ao algodoeiro, em diferentes sistemas de cultivo e
dois níveis de fertilização. O experimento foi conduzido na Fazenda Mourão,
no município de Campo Verde-MT. O delineamento experimental utilizado foi
inteiramente casualizado, com seis tratamentos e seis repetições, os
tratamentos foram dispostos em esquema de parcelas subdivididas, sendo
nas parcelas os dois níveis de fertilização (padrão e reduzida) e nas sub-
parcelas os três sistemas de cultivo [monocultivo de algodoeiro em preparo
convencional (T1), monocultivo de algodoeiro em plantio semi-direto, sobre
palhada de milheto (T2) e algodoeiro em plantio direto, em rotação bianual,
com soja seguido de sorgo consorciado com Brachiaria ruzizienses (S3)].
Avaliou-se a densidade de esporos do solo e a colonização radicular do
algodoeiro. Foi observado que em todas as amostras de raízes e solo
ocorreram à presença de fungos micorrízicos arbusculares. O sistema de
plantio semi-direto (T2) apresentou uma tendência a ter uma maior
colonização micorrízica. O sistema de plantio direto (S3), apresentou maior
densidade de esporos. De modo geral os sistemas de cultivo e a fertilização
do solo atuaram sobre a simbiose micorrízica-arbuscular.
Palavras chaves: Fungos micorrízicos, colonização radicular, esporos,
densidade de esporos no solo.

18
COTTON MYCORRHIZATION IN DIFFERENT CULTIVATION SYSTEMS IN CAMPO VERDE - MT
Abstract: The objective of this work was to evaluate arbuscular mycorrhizal
fungi associated with cotton in different cultivation systems under two
fertilization levels. The experiment was conducted at Fazenda Mourão, in the
municipality of Campo Verde - MT, Brazil. A completely randomized
experimental design was adopted, with six treatments and six replicates. The
treatments were organized in a split-plot arrangement where the two
fertilization levels (standard and reduced) were assigned to plots and three
cultivation systems were assigned to sub-plots [cotton monoculture under
conventional tillage (T1), cotton monoculture under semi-direct planting on
millet trash (T2), and no-till cotton, in a biennial rotation, with soybean
followed by sorghum intercropped with Brachiaria ruzizienses (S3)]. Spore
density in the soil and cotton root colonization were evaluated. It was
observed that the presence of arbuscular mycorrhizal fungi occurred in all
root and soil samples. The semi-direct planting system (T2) showed a
tendency to have higher mycorrhizal colonization. The no-till system (S3)
showed higher spore density. In general, the cultivation systems and soil
fertilization acted upon the arbuscular mycorrhizal symbiosis.
Keywords: Mycorrhizal fungi, root colonization, spores, spore density in the
soil.

19
3.1. INTRODUÇÃO As micorrizas arbusculares (MAs) são uma associação simbiotrófica
mutualista importante entre fungos Zigomicetos da ordem Glomales e a
maioria das raízes de plantas, e ocorrem nos mais variados ecossistemas.
Muitos são os benefícios desta simbiose, tanto para as plantas quanto para
o fungo, sejam eles nutricionais ou não (Siqueira e Klauberg Filho, 2000).
O algodoeiro é facilmente micorrizado em condições de campo
(Colozzi Filho et al., 1985). Quando se desenvolve em solos deficientes em
fósforo, apresenta alto grau de micotrofismo e elevada dependência
micorrízica (Siqueira et al., 1986).
Apesar dos FMAs mostrarem-se, na maioria das vezes, benéficos às
plantas, os efeitos da micorrização são resultados da interação fungo planta
e que pode ser alterada por fatores que modifiquem um ou mais desses
componentes (Varma & Hock, 1995).
O sistema de cultivo interfere na população do FMAs, pois a
desestruturação do solo pela aração e gradagem pode destruir as hifas
fúngicas (Colozzi Filho et al., 1993). Os mesmos autores relatam ainda que o
revolvimento do solo, provocado pela aração, pode expor os propágulos
(esporos, hifas e fragmentos de raízes colonizadas) na superfície do solo,
inviabilizando-os devido à ação de altas temperaturas e de predadores. Eles
afirmam que o plantio direto e a rotação de culturas podem aumentar o
potencial de inóculo natural desses fungos no solo.
Devido a grande importância dos FMAs, poucas informações
disponíveis sobre a micorrização do algodoeiro e a necessidade de se
estudar alternativas que venham contribuir para a cotonicultura, o presente
trabalho teve por objetivo avaliar os fungos micorrízicos arbusculares
associados ao algodoeiro, em diferentes sistemas de cultivo, em dois níveis
de fertilização.

20
3.2. MATERIAL E MÉTODOS Localização da área:
O estudo foi realizado na área de pesquisa da COODETEC/CIRAD,
Fazenda Mourão, no município de Campo Verde – MT (15o28’73’’S;
54o54’30’’W e 680 m sobre o nível do mar), situada às margens da BR 070,
em uma área de 15 ha cultivada com algodoeiro em diferentes sistemas de
cultivo e dois níveis de fertilização, implantados há quatro anos.
Coletas de amostras no campo:
As amostras de solo rizosférico foram coletadas em quatro épocas e a
das raízes iniciou a partir da segunda época (22/12/04, 05/03/05, 07/05/05 e
18/06/05), que coincidiram com estádios do ciclo fenológico do algodoeiro
(Figura 1).
Dias após a germinação
FIGURA 1. Estádios fenológico do ciclo do algodoeiro e épocas das coletas das amostras, em dias. O delineamento experimental utilizado foi inteiramente casualizado
com seis tratamentos e seis repetições. Os tratamentos foram dispostos em
esquema de parcelas subdivididas, sendo nas parcelas os dois níveis de
fertilização (padrão e reduzida – Tabela 1) e nas sub-parcelas os três
sistemas de cultivo [monocultivo algodoeiro em preparo convencional (T1),
monocultivo algodoeiro em plantio semi-direto, sobre palhada de milheto
(T2); e algodoeiro em plantio direto, em rotação bianual com soja seguida de
sorgo consorciado com Brachiaria ruzizienses (S3)]. O tamanho de cada
sub-parcela foi de 3.600m2 (36x100m).

21
TABELA 1. Fertilização (Kg/ha) aplicada na base e em cobertura, no
algodoeiro conduzido em sistemas convencional (T1), semi-direto (T2) e
plantio direto (S3), no município de Campo Verde-MT.
Tipo N P2O5 K2O Boro
Padrão 130 146 180 2
Reduzida 65 73 90 1
O plantio do algodoeiro foi realizado no dia 03/12/2004, nos três
sistemas descritos acima, sendo que no T2 e S3 foram sobre palhada de
milheto e braquiária, respectivamente. O espaçamento foi de 0,90m, com
densidade de 10 a 12 plantas/metro. Os tratos culturais realizados durante
toda a safra encontram-se no apêndice 1.
Em cada repetição (dentro de cada sub-parcela) foi aberta uma
trincheira onde foi coletada uma amostra de solo na profundidade de 0-
20cm, que permaneceram marcadas durante todas as coletas, perfazendo
um total de 36 amostras (3 sistemas x 2 níveis de fertilização x 6 repetições).
As amostras de raízes foram retiradas de três plantas coletadas nas
sub-parcelas que compuseram uma amostra. Essas plantas foram
selecionadas ao acaso. A retirada de amostras das raízes iniciaram a partir
da coleta 2.
Após a coleta, as amostras de solo e raiz foram embaladas em sacos
plásticos previamente identificados e acondicionadas em caixas de isopor e
transportadas, para o Laboratório de Microbiologia de Solo da
FAMEV/UFMT, no prazo máximo de 24 horas.
No laboratório, uma parte dessas amostras foram separadas e
enviadas para a análise química do solo (Tabela 2) e outra parte foi retirada
para a determinação de umidade (apêndice 2), sendo que no restante do
solo procedeu-se as análises microbiológicas.
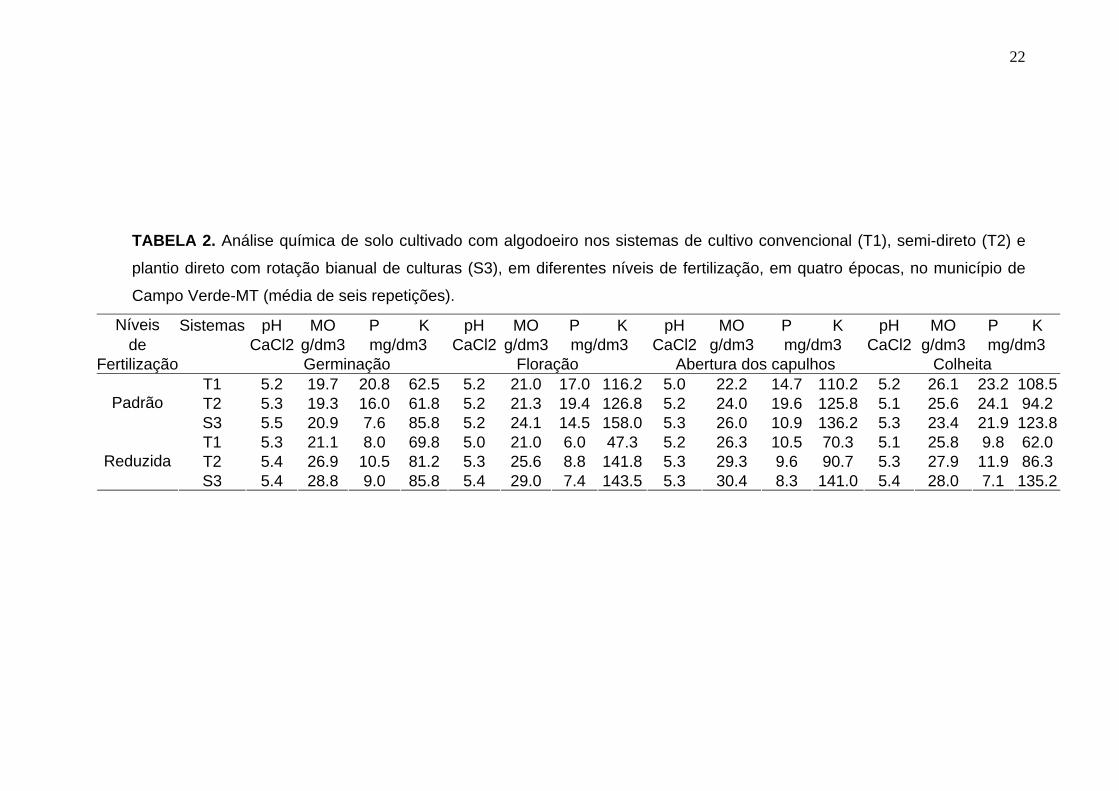
22
TABELA 2. Análise química de solo cultivado com algodoeiro nos sistemas de cultivo convencional (T1), semi-direto (T2) e
plantio direto com rotação bianual de culturas (S3), em diferentes níveis de fertilização, em quatro épocas, no município de
Campo Verde-MT (média de seis repetições).
Níveis Sistemas pH MO P K pH MO P K pH MO P K pH MO P K de CaCl2 g/dm3 mg/dm3 CaCl2 g/dm3 mg/dm3 CaCl2 g/dm3 mg/dm3 CaCl2 g/dm3 mg/dm3
Fertilização Germinação Floração Abertura dos capulhos Colheita T1 5.2 19.7 20.8 62.5 5.2 21.0 17.0 116.2 5.0 22.2 14.7 110.2 5.2 26.1 23.2 108.5 T2 5.3 19.3 16.0 61.8 5.2 21.3 19.4 126.8 5.2 24.0 19.6 125.8 5.1 25.6 24.1 94.2 Padrão
S3 5.5 20.9 7.6 85.8 5.2 24.1 14.5 158.0 5.3 26.0 10.9 136.2 5.3 23.4 21.9 123.8 T1 5.3 21.1 8.0 69.8 5.0 21.0 6.0 47.3 5.2 26.3 10.5 70.3 5.1 25.8 9.8 62.0 T2 5.4 26.9 10.5 81.2 5.3 25.6 8.8 141.8 5.3 29.3 9.6 90.7 5.3 27.9 11.9 86.3 Reduzida
S3 5.4 28.8 9.0 85.8 5.4 29.0 7.4 143.5 5.3 30.4 8.3 141.0 5.4 28.0 7.1 135.2

23
Avaliação da colonização das raízes por FMAs:
Realizou-se, primeiramente, no Laboratório de Microbiologia do Solo
da FAMEV/UFMT, a separação das raízes mais finas do algodoeiro, sendo
acondicionadas em frascos e enviadas ao Laboratório de Microbiologia do
Solo do IAPAR em Londrina/PR, onde foi avaliada a colonização. No IAPAR,
as amostras, passaram por mais uma separação, novamente as raízes mais
finas. Posteriormente, essas raízes foram colocadas em placas de
polietileno, imersas em becker de vidro com solução de KOH 10% para a
clarificação; as raízes mais escuras foram clareadas com solução de H2O2,
sendo em seguida acidificadas em HCl 1% e coloridas com ácido glicerol
contendo azul de tripano (para a coloração das estruturas fúngicas internas),
segundo metodologia de Koske e Gemma (1989). A porcentagem do
comprimento de raízes colonizadas foi avaliada pelo método de intersecção
em quadrantes descrito por Giovanetti e Mosse (1980), onde as raízes
coradas foram espalhadas em uma placa de Petri com fundo quadriculado. A
avaliação foi realizada com o auxílio de microscópio estereoscópico, onde
foram observados a presença e ausência das estruturas fúngicas (hifas,
arbúsculos e/ou vesículas) nos pontos de intersecção das raízes com as
linhas quadriculadas.
Avaliação da densidade de esporos de FMAs:
Os esporos foram extraídos do solo pelo método do peneiramento
úmido de Gerdemann e Nicolson (1963), combinado com a técnica de
centrifugação em sacarose (Sieverding, 1991). Cada amostra de 50 mL de
solo foi homogeneizada em 1000 mL de água, desfazendo-se os torrões
com o auxílio do bastão de vidro. Esperou-se cerca de 30 segundos para a
decantação das partículas de solo e suspensão dos esporos, vertendo o
sobrenadante em duas peneiras com malhas de abertura de 71 e 53 mesh
(processo repetido por três vezes), sendo recolhido o filtrado retido na
peneira de 53 mesh.
O material recolhido foi primeiramente centrifugado em água por 3
minutos a 3000 rpm, desprezando-se o sobrenadante. Foi adicionada uma

24
solução de água e sacarose a 50% e após agitação com o bastão de vidro, a
amostra foi novamente centrifugada por 2 minutos a 2000 rpm. Sendo o
sobrenadante recolhido em peneira de 53 mesh, lavado em água corrente,
seguido de lavagem com água destilada e por fim, colocado em uma placa
de Petri, onde procedeu-se a contagem de esporos utilizando o microscópio
estereoscópico (aumento de 10 a 40 x).
Análise dos dados:
Os dados de colonização de raízes foram submetidos à estatística
descritiva (medidas de posição e dispersão).
Os dados de densidade de esporos de FMAs foram submetidos a
análise de variância (apêndice 3) e teste de médias, conforme metodologia
descrita em Banzatto & Kronka (1992), utilizando o software SAEG (Ribeiro
Junior, 2001).
3.3. RESULTADOS E DISCUSSÃO Em todas as amostras de solo e raízes analisadas foram encontradas
estruturas de FMAs.
A colonização das raízes por FMAs (Figura 2) pode ser considerada
inferior (Tabela 3), quando comparada com as relatadas em outros trabalhos
com algodão - 48,8% (Siqueira et al., 1986), mandioca - 47,9% (Balota et al.,
1999) e com milho - 44% e soja - 38%, após bracquiária (Cordeiro et al.,
2005).

25
FIGURA 2. Fungos micorrízicos arbusculares em raízes do algodoeiro
cultivado em sistema plantio direto em rotação bianual com soja seguida de
sorgo consorciado com braquiária, em Campo Verde-MT. (Aumento de 40
vezes).
Não houve grande variabilidade na colonização micorrízica entre os
sistemas de cultivo (figura 3). Porém, há uma tendência de que a fertilização
reduzida tenha influenciado na colonização. Constatando que a colonização
micorrízica pode ser afetada por inúmeros fatores como manejo do solo,
densidade de raízes, idade da planta, a espécie vegetal, entre outros,
corroboram com isso Afek et al. (1990).
0
5
10
15
20
25
30
35
T1 T2 S3 T1 T2 S3
Fertilização padrão Fertilização reduzida
FIGURA 3. Colonização micorrízica (%) do algodoeiro cultivado em
monocultura em sistema convencional (T1), em monocultura em sistema
semi-direto sobre palhada de milheto (T2) e em plantio direto em rotação
bianual com soja seguida de sorgo consorciado com braquiária (S3), em dois

26
níveis de fertilização, no município de Campo Verde-MT. Média de duas
repetições.
Primeiramente, há que considerar certas particularidades
metodológicas, que nessa situação usou-se o perfil de 0-20cm para as
coletas das amostras, independente dos sistemas de cultivo, o que pode ter
influenciado em alguns dados observados. Pois se sabe que há diferenças
entre o sistema radicular do algodoeiro em função do sistema de cultivo e
também quanto à ocorrência do microrganismo do solo ao longo do perfil.
Corroboram resultados obtidos por Carneiro et al. (2004), onde constataram
que na profundidade de 0-5cm a diferença significativa entre o sistema de
plantio direto e plantio convencional, para a população microbiana do solo.
No plantio, os esporos estavam presentes em todas as amostras
analisadas. Na fertilização reduzida o sistema S3 apresentou maior
densidade de esporos 66,57/ 50mL de solo. Já na fertilização padrão, não
houve diferença entre a densidade de esporos de FMAs nos três sistemas
avaliados (tabela 3).
Na floração, o sistema S3 na fertilização reduzida apresentou maior
número de esporos quando comparado aos demais sistemas (T1 e T2)
(tabela 3).
Na abertura dos capulhos, em todos os sistemas (T1, T2 e S3) na
fertilização padrão houve maior densidade de esporos, quando comparado
com a fertilização reduzida (tabela 3).
Na colheita, a maior densidade de esporos foi observada na
fertilização padrão quando comparada com a fertilização reduzida, para os
sistemas T1 e S3. Já para o T2 não houve diferenças significativas entre as
fertilizações padrão e reduzida (tabela 3).
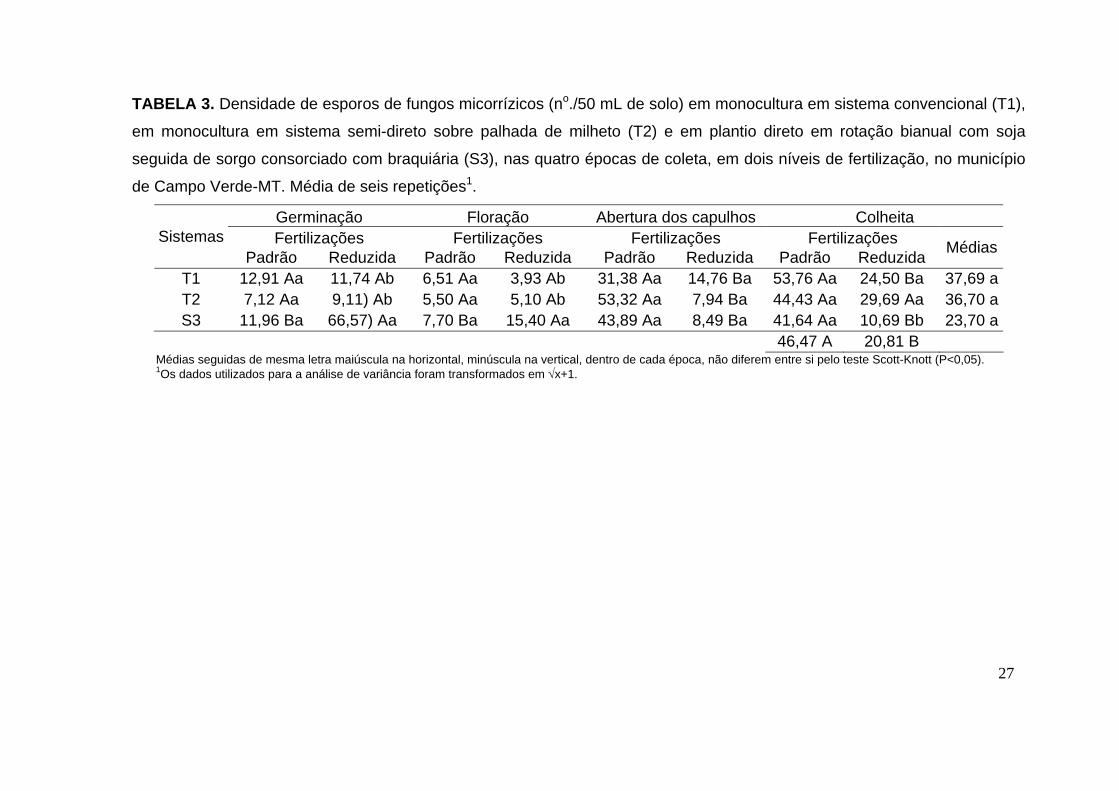
TABELA 3. Densidade de esporos de fungos micorrízicos (no./50 mL de solo) em monocultura em sistema convencional (T1),
em monocultura em sistema semi-direto sobre palhada de milheto (T2) e em plantio direto em rotação bianual com soja
seguida de sorgo consorciado com braquiária (S3), nas quatro épocas de coleta, em dois níveis de fertilização, no município
de Campo Verde-MT. Média de seis repetições1.
27
Germinação Floração Abertura dos capulhos Colheita Fertilizações Fertilizações Fertilizações Fertilizações Sistemas
Padrão Reduzida Padrão Reduzida Padrão Reduzida Padrão Reduzida Médias
T1 12,91 Aa 11,74 Ab 6,51 Aa 3,93 Ab 31,38 Aa 14,76 Ba 53,76 Aa 24,50 Ba 37,69 a T2 7,12 Aa 9,11) Ab 5,50 Aa 5,10 Ab 53,32 Aa 7,94 Ba 44,43 Aa 29,69 Aa 36,70 a S3 11,96 Ba 66,57) Aa 7,70 Ba 15,40 Aa 43,89 Aa 8,49 Ba 41,64 Aa 10,69 Bb 23,70 a
46,47 A 20,81 B Médias seguidas de mesma letra maiúscula na horizontal, minúscula na vertical, dentro de cada época, não diferem entre si pelo teste Scott-Knott (P<0,05). 1Os dados utilizados para a análise de variância foram transformados em √x+1.

28
Pode-se verificar a diversidade de esporos encontrados nas amostras
de solo, onde cada forma, tamanho e coloração indicam uma espécie
diferente (figura 4). Porém, não foi realizada a identificação das espécies,
sendo difícil concluir que uma maior densidade de esporos significa uma
melhor eficiência. Estudos realizados por Howeler et al. (1987) salientam
que nem todas as espécies associadas naturalmente a cultura são
eficientes, pois há espécies de fungos micorrízicos que apresentam efeito
diferenciado nas plantas hospedeiras.
FIGURA 4. Esporos de fungos micorrízicos arbusculares isolados da
rizosfera do algodoeiro, cultivado em sistema plantio convencional (T1), em
Campo Verde-MT. (Aumento de 40 vezes).
Apesar de maior densidade de esporos no solo indicar condição
desfavorável aos FMAs, sendo o revolvimento do solo um dos fatores
principais e a tendência no plantio convencional haver maior esporulação do
que em sistemas de plantio direto, vem sendo observado em algumas
situações, o inverso. Corrobora com essa afirmação os resultados de Silva
(2004), onde na maioria das situações, a densidade de esporos foi mais
elevada em sistema de plantio direto quando comparado ao plantio
convencional, com cultivo de soja.
Com os resultados da presente pesquisa, onde nas épocas de plantio
e floração o sistema S3 apresentou as maiores densidades de esporos, na
fertilização reduzida (figura 5), deve-se levar em consideração que no
sistema S3 antes do plantio algodoeiro, era cultivado braquiária que é um
hospedeiro multiplicador de FMAs, apresentando alta dependência
micorrízica, o que conseqüentemente, acarretou o aumento do inóculo no
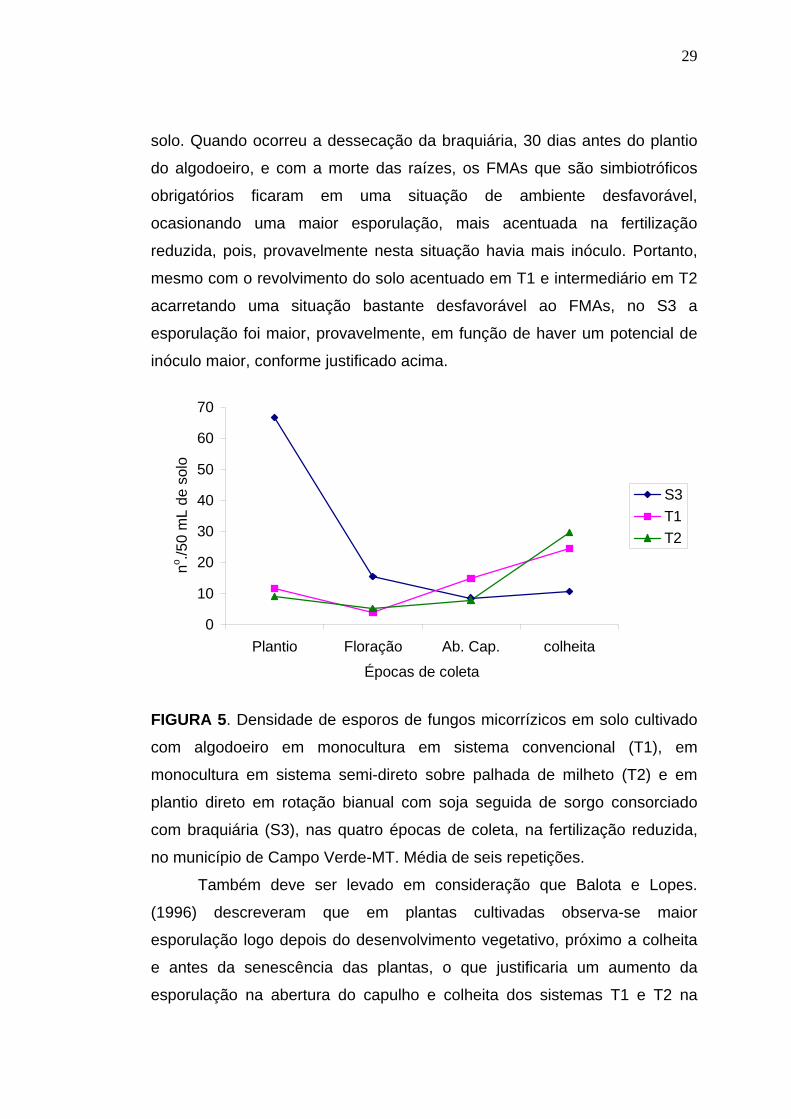
29
solo. Quando ocorreu a dessecação da braquiária, 30 dias antes do plantio
do algodoeiro, e com a morte das raízes, os FMAs que são simbiotróficos
obrigatórios ficaram em uma situação de ambiente desfavorável,
ocasionando uma maior esporulação, mais acentuada na fertilização
reduzida, pois, provavelmente nesta situação havia mais inóculo. Portanto,
mesmo com o revolvimento do solo acentuado em T1 e intermediário em T2
acarretando uma situação bastante desfavorável ao FMAs, no S3 a
esporulação foi maior, provavelmente, em função de haver um potencial de
inóculo maior, conforme justificado acima.
0
10
20
30
40
50
60
70
Plantio Floração Ab. Cap. colheita
Épocas de coleta
no ./50
mL
de s
olo
S3T1T2
FIGURA 5. Densidade de esporos de fungos micorrízicos em solo cultivado
com algodoeiro em monocultura em sistema convencional (T1), em
monocultura em sistema semi-direto sobre palhada de milheto (T2) e em
plantio direto em rotação bianual com soja seguida de sorgo consorciado
com braquiária (S3), nas quatro épocas de coleta, na fertilização reduzida,
no município de Campo Verde-MT. Média de seis repetições.
Também deve ser levado em consideração que Balota e Lopes.
(1996) descreveram que em plantas cultivadas observa-se maior
esporulação logo depois do desenvolvimento vegetativo, próximo a colheita
e antes da senescência das plantas, o que justificaria um aumento da
esporulação na abertura do capulho e colheita dos sistemas T1 e T2 na
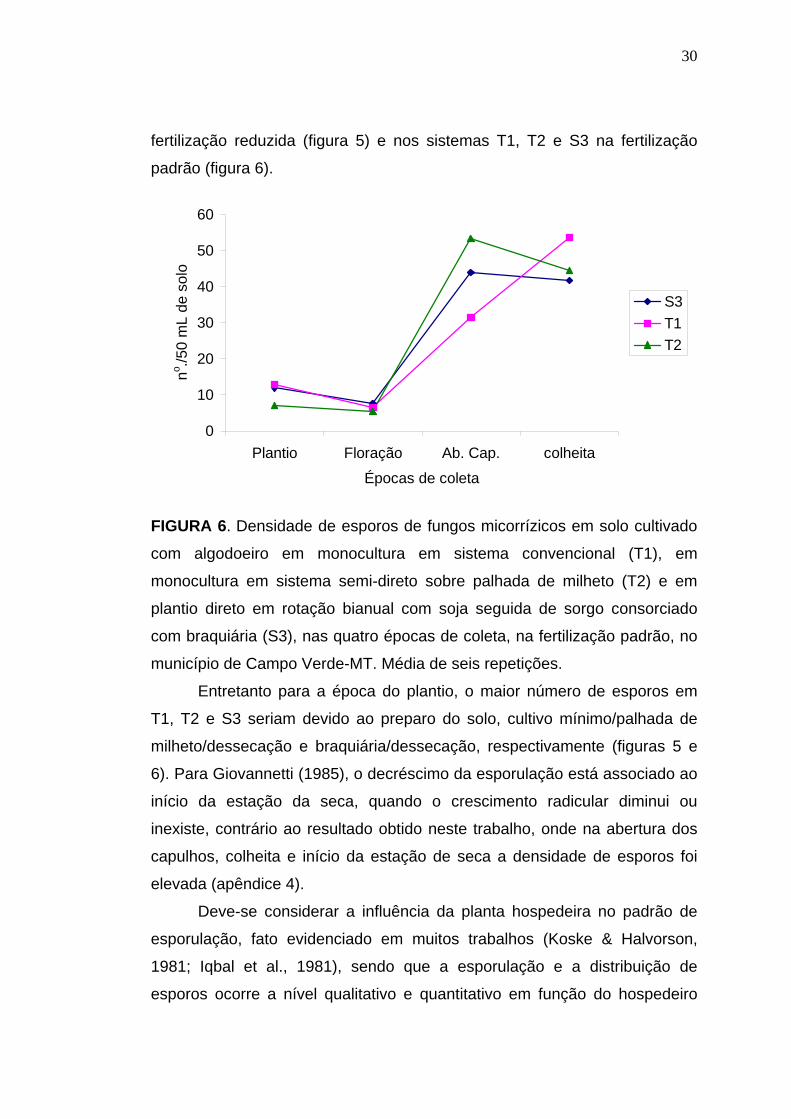
30
fertilização reduzida (figura 5) e nos sistemas T1, T2 e S3 na fertilização
padrão (figura 6).
0
10
20
30
40
50
60
Plantio Floração Ab. Cap. colheita
Épocas de coleta
no ./50
mL
de s
olo
S3T1T2
FIGURA 6. Densidade de esporos de fungos micorrízicos em solo cultivado
com algodoeiro em monocultura em sistema convencional (T1), em
monocultura em sistema semi-direto sobre palhada de milheto (T2) e em
plantio direto em rotação bianual com soja seguida de sorgo consorciado
com braquiária (S3), nas quatro épocas de coleta, na fertilização padrão, no
município de Campo Verde-MT. Média de seis repetições.
Entretanto para a época do plantio, o maior número de esporos em
T1, T2 e S3 seriam devido ao preparo do solo, cultivo mínimo/palhada de
milheto/dessecação e braquiária/dessecação, respectivamente (figuras 5 e
6). Para Giovannetti (1985), o decréscimo da esporulação está associado ao
início da estação da seca, quando o crescimento radicular diminui ou
inexiste, contrário ao resultado obtido neste trabalho, onde na abertura dos
capulhos, colheita e início da estação de seca a densidade de esporos foi
elevada (apêndice 4).
Deve-se considerar a influência da planta hospedeira no padrão de
esporulação, fato evidenciado em muitos trabalhos (Koske & Halvorson,
1981; Iqbal et al., 1981), sendo que a esporulação e a distribuição de
esporos ocorre a nível qualitativo e quantitativo em função do hospedeiro

31
(Trufem & Banoni, 1985; Sieverding, 1991). E também a influência dos
sistemas de cultivo, que determinam profundas alterações, provocando
mudanças qualitativas e quantitativas nos microrganismos e suas atividades
(Balota, 1997). Pois o conjunto de práticas agrícolas utilizadas condiciona
um sistema solo-planta próprio, com atividade biológica diferenciada.
3.4 CONCLUSÃO Os sistemas de cultivo e a fertilização do solo atuam sobre a simbiose
micorrízica-arbuscular.
Houve uma tendência de que plantas de algodoeiro cultivadas em
sistema semi-direto (T2) apresentassem uma maior colonização.
A diversidade de esporos de fungos micorrízicos arbusculares no solo
foi maior no tratamento com fertilização padrão.
No sistema de cultivo do algodoeiro em plantio direto (S3), apresentou
uma densidade de esporos de FMAs maior.

32
CAPITULO II: Populações de bactérias e fungos e ocorrência de Azospirillum em solo sob diferentes sistemas de cultivos
do algodoeiro em Campo Verde - MT

33
POPULAÇÕES DE BACTÉRIAS E FUNGOS E OCORRÊNCIA DE AZOSPIRILLUM EM SOLO SOB DIFERENTES SISTEMAS DE CULTIVO
DO ALGODOEIRO EM CAMPO VERDE - MT Resumo: O objetivo deste trabalho foi avaliar a comunidade microbiana e
bactérias diazotróficas, associadas ao algodoeiro, em diferentes sistemas de
cultivo, em dois níveis de fertilização. O experimento foi conduzido na
Fazenda Mourão, no município de Campo Verde-MT. O delineamento
experimental utilizado foi inteiramente casualizado, com seis tratamentos e
seis repetições, os tratamentos foram dispostos em esquema de parcelas
subdivididas, sendo nas parcelas os dois níveis de fertilização (padrão e
reduzida) e nas sub-parcelas os três sistemas de cultivo [monocultivo
algodoeiro em preparo convencional (T1), monocultivo algodoeiro, em
plantio semi-direto, sobre palhada de milheto (T2) e algodoeiro em plantio
direto, em rotação bianual, com soja seguido de sorgo consorciado com
Brachiaria ruzizienses (S3)]. Avaliou-se a população de bactérias e fungos
de solo, pelo método do isolamento em meio seletivo e contagem das
unidades formadoras de colônias em placas, e a população de bactérias
diazotróficas pelo método do número mais provável, baseado na presença
ou ausência de película formada em meio semi-sólido. Não houve efeito das
fertilizações no número de fungos totais para os sistemas T2 e S3. O
sistema T2 tem maior número de bactérias totais, igual ao observado para o
nível de fertilização reduzida. A população de Azospirillum sp foi influenciada
pelas épocas de coleta e pelos tratos culturais, sendo que o sistema T2 foi o
que teve o melhor efeito sob a população de Azospirillum sp, considerando
as 4 épocas de coleta.
Palavras chaves: Azospirillum sp, sistemas de cultivo, fungos totais,
bactérias totais.

34
BACTERIAL AND FUNGAL POPULATIONS AND OCCURRENCE OF AZOSPIRILLUM IN A SOIL UNDER DIFFERENT COTTON CULTIVATION
SYSTEMS IN CAMPO VERDE - MT
Abstract: The objective of this work was to evaluate the microbial community
and diazotrophic bacteria associated with cotton in different cultivation
systems and two fertilization levels. The experiment was conducted at
Fazenda Mourão, in the municipality of Campo Verde - MT, Brazil. A
completely randomized experimental design was adopted, with six
treatments and six replicates. The treatments were organized in a split-plot
arrangement where the two fertilization levels (standard and reduced) were
assigned to plots and three cultivation systems were assigned to sub-plots
[cotton monoculture under conventional tillage (T1), cotton monoculture
under semi-direct planting on millet trash (T2), and no-till cotton, in a biennial
rotation, with soybean followed by sorghum intercropped with Brachiaria
ruzizienses (S3)]. The soil bacterial and fungal populations were evaluated
by the isolation method in selective medium and the counting of colony-
forming units in plates; the diazotrophic bacterium population was evaluated
by the most-probable-number method, based on the presence or absence of
a skin membrane formed on a semi-solid medium. There was no effect of
fertilization on the total number of fungi in systems T2 and S3. System T2
showed a higher number of total bacteria, equal to the number observed at
the reduced fertilization level. The Azospirillum sp population was influenced
by collection season and by management practices. System T2 had the best
effect under the Azospirillum sp population, considering the 4 collection
seasons.
Keywords: Azospirillum sp, cultivation systems, total fungi, total bacteria.

35
4.1. INTRODUÇÃO: A comunidade microbiana do solo é formada por fungos, bactérias e
actinomicetos, sendo influenciada pela temperatura, umidade e aeração do
solo, disponibilidade de nutrientes e substratos orgânicos. Estes fatores
podem ser modificados pelos sistemas de manejo, em razão da forma como
os resíduos das culturas anteriores são depositados e do grau de
revolvimento do solo (Vargas & Scholles, 2000).
A atividade microbiana pode ser dividida em dois tipos: geral e
especifica. A geral é aquela proveniente de todos ou quase todos os
microrganismos do solo, como a respiração e produção de calor, por
exemplo. Já a específica é realizada por grupos específicos e/ou funcionais
como os fixadores de nitrogênio e os nitrificadores, entre outros.
O grupo funcional dos organismos fixadores de nitrogênio
(diazotróficos) é mediado por uma parcela dos procariotos que, apesar de
relativamente pequena, apresenta alta diversidade morfológica, fisiológica,
genética e filogenética.
Os microrganismos diazotróficos podem desempenhar importante
papel na reabilitação e sustentabilidade dos ecossistemas, uma vez que
incorporam N por meio da fixação biológica e produzem e liberam
substâncias reguladoras do crescimento vegetal, as quais contribuem para
melhorar a nutrição mineral e utilização de água pelas plantas (Bazzicalupo
& Okon, 2000).
Durante os últimos anos novas bactérias fixadoras de nitrogênio foram
identificadas, incluindo espécies dos gêneros Azospirillum, Herbaspirillum,
Acetobacter, Azoarcus e Burkholderia (Tarrand et al., 1978; Baldani et al.,
1986; Gillis et al., 1991; Cavalcante & Döbereiner, 1988; Reinhold-Hurek et
al., 1993; Hartmann et al., 1995), despertando um crescente interesse no
isolamento de bactérias redutoras de nitrogênio no interior de plantas
(Boddey, 1995; Triplett, 1996; Boddey et al., 1997; Hallmann et al., 1997). O
estabelecimento associativo na rizosfera, ou endofítico em raízes ou partes
aéreas, de bactérias diazotróficas com plantas da família Graminae, apesar
da não existência de evidências diretas, tem sido considerado uma

36
explicação plausível para a fixação biológica de nitrogênio observada em tais
plantas.
Nos diferentes agroecossistemas, a fixação biológica de N2 (FBN)
associada a diversas espécies de plantas é uma etapa importante no ciclo
de N, visto que bactérias diazotróficas podem contribuir no suprimento de
parte do N necessário às plantas (Chalk, 1991).
Devido a grande importância da comunidade microbiana e das
bactérias diazotróficas e a necessidade de buscar alternativas que venham
contribuir para a cotonicultura, o presente trabalho teve por objetivo verificar
o efeito de três tipos de sistemas de cultivo do algodoeiro na população de
bactérias, fungos e bactérias diazotróficas do solo.
4.2. MATERIAL E MÉTODOS Localização da área:
O estudo foi realizado na área de pesquisa da COODETEC/CIRAD,
Fazenda Mourão, no município de Campo Verde – MT (15o28’73’’S;
54o54’30’’W e 680 m sobre o nível do mar), situada às margens da BR 070,
em uma área de 15 ha cultivada com algodoeiro em diferentes sistemas de
cultivo e dois níveis de fertilização, implantados há quatro anos.
Coletas de amostras no campo:
As amostras de solo rizosférico foram coletadas em quatro épocas e a
das raízes iniciou a partir da segunda época (22/12/04, 05/03/05, 07/05/05 e
18/06/05), que coincidiram com estádios do ciclo fenológico do algodoeiro
(Figura 1).
O delineamento experimental utilizado foi inteiramente casualizado
com seis tratamentos e seis repetições. Os tratamentos foram dispostos em
esquema de parcelas subdivididas, sendo nas parcelas dois níveis de
fertilização (padrão e reduzida – Tabela 1) e nas sub-parcelas três sistemas
de cultivo [monocultivo algodoeiro em preparo convencional (T1),
monocultivo algodoeiro em plantio semi-direto, sobre palhada de milheto
(T2); e algodoeiro em plantio direto, em rotação bianual com soja seguida de

37
sorgo consorciado com Brachiaria ruzizienses (S3)]. O tamanho de cada
sub-parcela foi de 3.600m2 (36x100m).
O plantio do algodoeiro foi realizado no dia 03/12/2004, nos três
sistemas descritos acima, sendo que no T2 e S3 foram sobre palhada de
milheto e braquiária, respectivamente. O espaçamento foi de 0,90m, com
densidade de 10 a 12 plantas/metro. Os tratos culturais realizados durante
toda a safra encontram-se no apêndice 1.
Em cada repetição (dentro de cada sub-parcela) foi aberta uma
trincheira onde foi coletada uma amostra de solo na profundidade de 0-
20cm, que permaneceram marcadas durante todas as coletas, perfazendo
um total de 36 amostras (3 sistemas x 2 níveis de fertilização x 6 repetições).
Após a coleta, as amostras de solo foram embaladas em sacos
plásticos previamente identificados e acondicionadas em caixas de isopor e
transportadas, para o Laboratório de Microbiologia de Solo da
FAMEV/UFMT, no prazo máximo de 24 horas.
No laboratório, uma parte dessas amostras foram separadas e
enviadas para a análise química do solo (Tabela 2) e outra parte foi retirada
para a determinação de umidade (apêndice 2), sendo que no restante do
solo procedeu-se as análises microbiológicas.
Avaliação da população de bactérias e fungos do solo
Para a determinação da população de fungos e bactérias do solo foi
utilizado o método do isolamento em meio seletivo e contagem das unidades
formadoras de colônias (UFC) em placa.
Após peneiramento do solo (peneira < 2 mm), foram pesados 10g de
solo/amostra, e acondicionados em frascos contendo 90 mL de água
destilada e esterilizada. Os frascos foram agitados durante 20 minutos a 120
rpm. Em seguida, foi realizada a diluição decimal (10-2 para fungos e 10-4
para bactérias) e posteriormente a inoculação na superfície dos meios
sólidos específicos (100 μL.placa-1). Foram utilizados meios sólidos
específicos, Martin para fungos e Agar nutrientes para bactérias. As placas

38
foram, então, incubadas a 30oC por 72 e 24 horas para fungos e bactérias,
respectivamente.
Após esse período foram feitas as contagens de unidades formadoras
de colônias (UFC).
Isolamento e contagem de bactérias diazotróficas
Para o isolamento das bactérias diazotróficas do solo rizosférico,
utilizou-se o meio de cultura NFb (Azospirillum spp.), segundo Döbereiner et
al. (1995).
Foram usados 10 g de solo de cada amostra, homogeneizados em 90
mL de solução salina, durante 20 minutos. Em seguida, foi realizada a
diluição seriada das amostras (10-2 a 10-7). A inoculação de 0,1 mL de cada
diluição foi feita no centro do meio específico, sendo inoculados três frascos
para cada diluição. Os frascos de vidro já inoculados foram levados para a
sala de crescimento, onde permaneceram por 72 horas. Posteriormente foi
efetuada a quantificação dos diazotróficos, através do método do número
mais provável (NMP), baseado na presença ou ausência da película formada
no meio semi-sólido, conforme a tabela McCrady.
Análise dos dados:
Os dados de população de fungos e bactérias do solo (UFC.g de solo-
1) foram submetidos a estatística descritiva (medidas de posição e
dispersão).
Os dados de contagem de bactérias diazotróficas foram submetidos a
análise de variância (apêndice 5) e teste de médias conforme metodologia
descrita em Banzatto e Kronka (1992), utilizando o software SAEG (Ribeiro
Junior, 2001).
4.3. RESULTADOS E DISCUSSÃO
A população de fungos totais nos sistemas semi-direto (T2) e direto
(S3) não foram afetados com a fertilização, demonstrando um certo
equilíbrio desses sistemas, diferente do que ocorreu no sistema
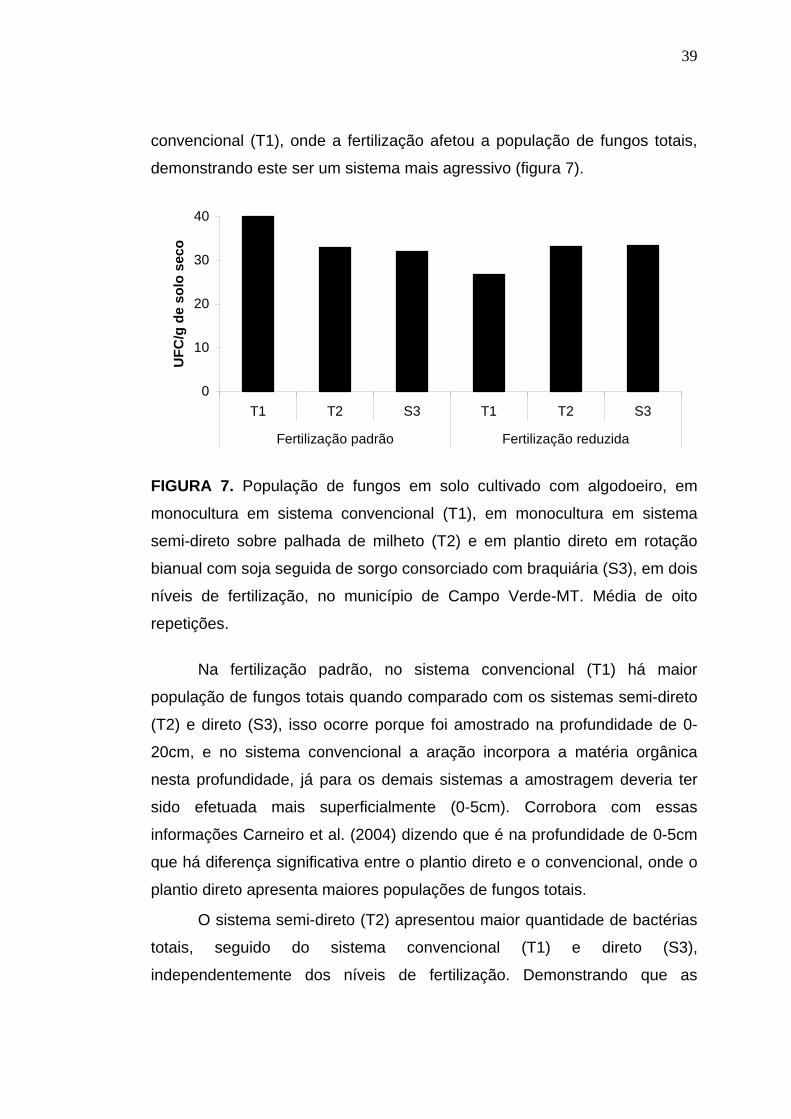
39
convencional (T1), onde a fertilização afetou a população de fungos totais,
demonstrando este ser um sistema mais agressivo (figura 7).
0
10
20
30
40
T1 T2 S3 T1 T2 S3
Fertilização padrão Fertilização reduzida
UFC
/g d
e so
lo s
eco
FIGURA 7. População de fungos em solo cultivado com algodoeiro, em
monocultura em sistema convencional (T1), em monocultura em sistema
semi-direto sobre palhada de milheto (T2) e em plantio direto em rotação
bianual com soja seguida de sorgo consorciado com braquiária (S3), em dois
níveis de fertilização, no município de Campo Verde-MT. Média de oito
repetições.
Na fertilização padrão, no sistema convencional (T1) há maior
população de fungos totais quando comparado com os sistemas semi-direto
(T2) e direto (S3), isso ocorre porque foi amostrado na profundidade de 0-
20cm, e no sistema convencional a aração incorpora a matéria orgânica
nesta profundidade, já para os demais sistemas a amostragem deveria ter
sido efetuada mais superficialmente (0-5cm). Corrobora com essas
informações Carneiro et al. (2004) dizendo que é na profundidade de 0-5cm
que há diferença significativa entre o plantio direto e o convencional, onde o
plantio direto apresenta maiores populações de fungos totais.
O sistema semi-direto (T2) apresentou maior quantidade de bactérias
totais, seguido do sistema convencional (T1) e direto (S3),
independentemente dos níveis de fertilização. Demonstrando que as
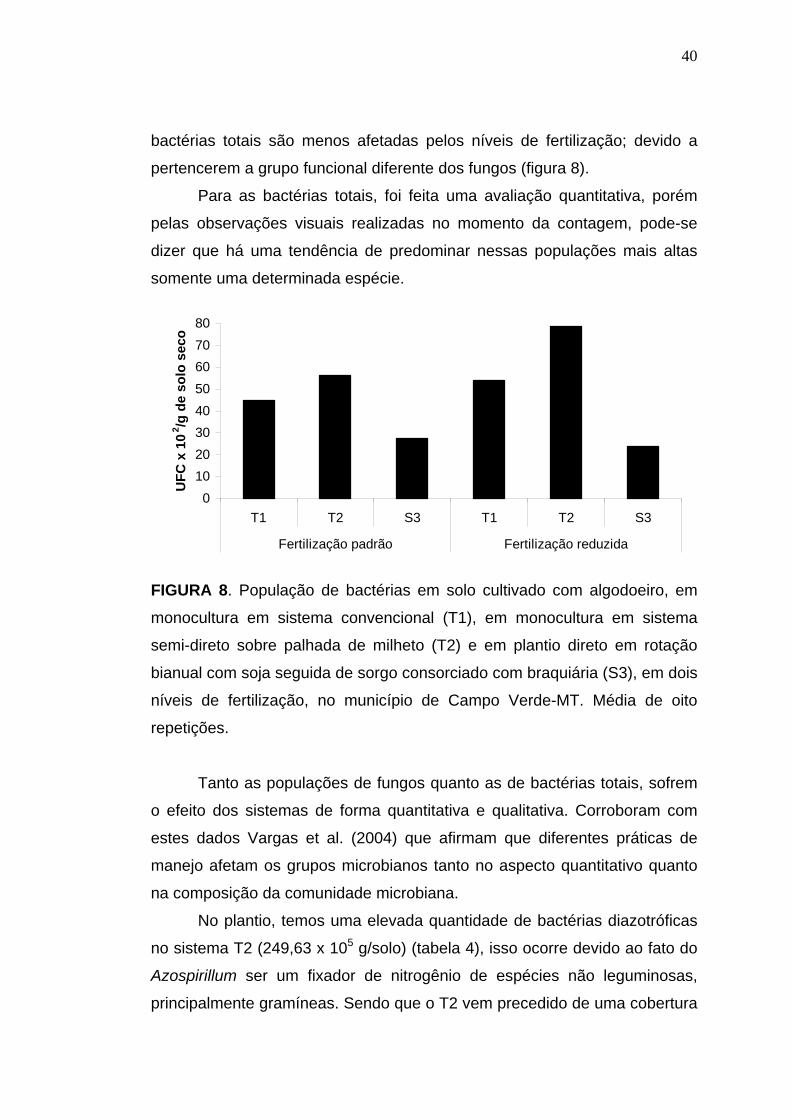
40
bactérias totais são menos afetadas pelos níveis de fertilização; devido a
pertencerem a grupo funcional diferente dos fungos (figura 8).
Para as bactérias totais, foi feita uma avaliação quantitativa, porém
pelas observações visuais realizadas no momento da contagem, pode-se
dizer que há uma tendência de predominar nessas populações mais altas
somente uma determinada espécie.
01020304050607080
T1 T2 S3 T1 T2 S3
Fertilização padrão Fertilização reduzida
UFC
x 1
0 2/g
de
solo
sec
o
FIGURA 8. População de bactérias em solo cultivado com algodoeiro, em
monocultura em sistema convencional (T1), em monocultura em sistema
semi-direto sobre palhada de milheto (T2) e em plantio direto em rotação
bianual com soja seguida de sorgo consorciado com braquiária (S3), em dois
níveis de fertilização, no município de Campo Verde-MT. Média de oito
repetições.
Tanto as populações de fungos quanto as de bactérias totais, sofrem
o efeito dos sistemas de forma quantitativa e qualitativa. Corroboram com
estes dados Vargas et al. (2004) que afirmam que diferentes práticas de
manejo afetam os grupos microbianos tanto no aspecto quantitativo quanto
na composição da comunidade microbiana.
No plantio, temos uma elevada quantidade de bactérias diazotróficas
no sistema T2 (249,63 x 105 g/solo) (tabela 4), isso ocorre devido ao fato do
Azospirillum ser um fixador de nitrogênio de espécies não leguminosas,
principalmente gramíneas. Sendo que o T2 vem precedido de uma cobertura

41
de milheto, que contribuiu para aumentar a população, também ocorreu a
mesma situação no S3, porém em menor proporção já que antes da
braquiária era soja, uma leguminosa que possui um fixador eficiente
(Rhizobium), que inibiu um pouco a população de Azospirillum.
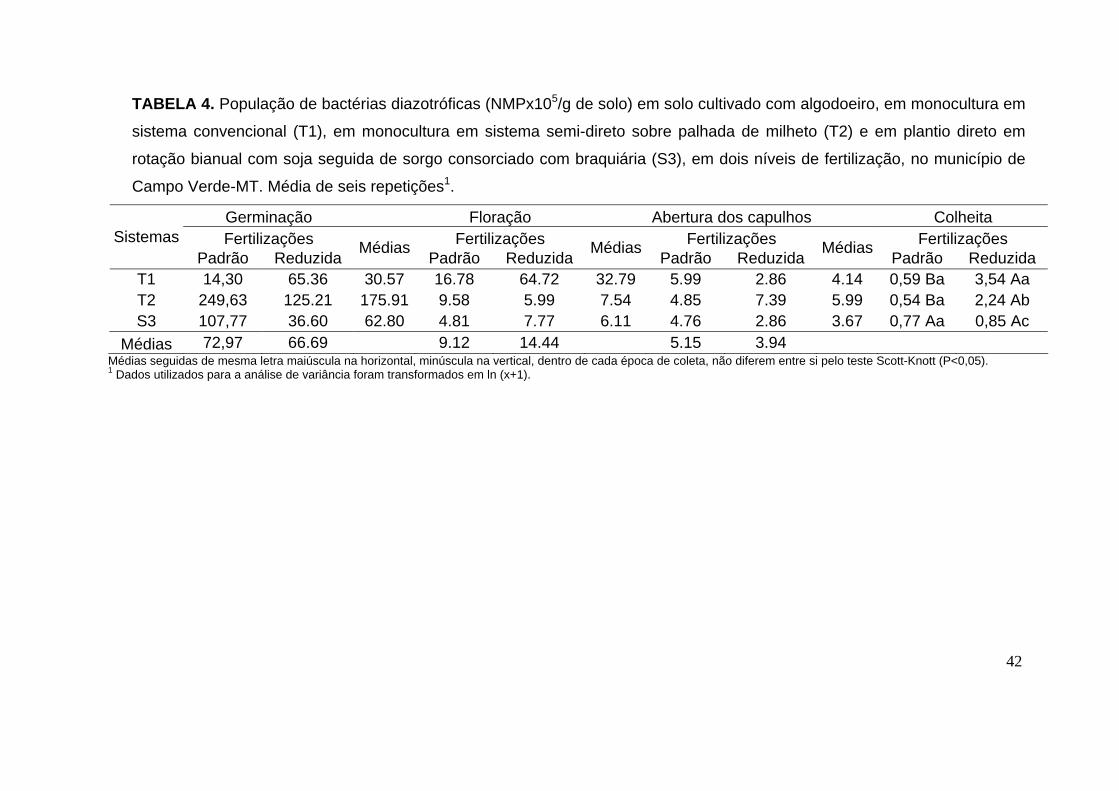
Germinação Floração Abertura dos capulhos Colheita Fertilizações Fertilizações Fertilizações Fertilizações Sistemas
Padrão Reduzida Médias Padrão Reduzida Médias Padrão Reduzida Médias Padrão Reduzida T1 14,30 65.36 30.57 16.78 64.72 32.79 5.99 2.86 4.14 0,59 Ba 3,54 Aa T2 249,63 125.21 175.91 9.58 5.99 7.54 4.85 7.39 5.99 0,54 Ba 2,24 Ab S3 107,77 36.60 62.80 4.81 7.77 6.11 4.76 2.86 3.67 0,77 Aa 0,85 Ac
Médias 72,97 66.69 9.12 14.44 5.15 3.94
42
TABELA 4. População de bactérias diazotróficas (NMPx105/g de solo) em solo cultivado com algodoeiro, em monocultura em
sistema convencional (T1), em monocultura em sistema semi-direto sobre palhada de milheto (T2) e em plantio direto em
rotação bianual com soja seguida de sorgo consorciado com braquiária (S3), em dois níveis de fertilização, no município de
Campo Verde-MT. Média de seis repetições1.
Médias seguidas de mesma letra maiúscula na horizontal, minúscula na vertical, dentro de cada época de coleta, não diferem entre si pelo teste Scott-Knott (P<0,05). 1 Dados utilizados para a análise de variância foram transformados em ln (x+1).

43
Quando comparamos as coletas, observa-se redução acentuada na
população de Azospirillum do plantio para a colheita, isso pode ser devido ao
fato de que a partir da floração inicia a adubação de cobertura, que contém
nitrogênio, o que inibe a população de Azospirillum. Também deve ser
considerada a grande quantidade de aplicações de defensivos.
O comportamento dos sistemas de cultivo é diferenciado, porém há
efeito dos mesmos sob os microrganismos do solo, indo de encontro com a
afirmação de Balota (1997), onde os diferentes sistemas determinam
profundas alterações nos microrganismos e nas suas atividades,
influenciando-as de maneira diferenciada.
4.4. CONCLUSÃO Não houve interferência das fertilizações no número de fungos totais
para os sistemas semi-direto (T2) e plantio direto (S3).
O sistema semi-direto (T2) tem maior população de bactérias totais,
mesma situação quando referimos ao nível de fertilização reduzida.
A população de Azospirillum sp foi influenciada pelas épocas de
coleta e pelos tratos culturais.
O sistema semi-direto (T2) foi o que teve a maior a população de
Azospirillum sp, considerando as 4 épocas de coleta.

4444
5. CONCLUSÕES GERAIS
Avaliou-se a comunidade microbiana, bactérias diazotróficas e fungos
micorrízicos arbusculares, associados ao algodoeiro, em diferentes sistemas
[monocultivo algodoeiro em preparo convencional (T1), monocultivo
algodoeiro em plantio semi-direto, sobre palhada de milheto (T2); e
algodoeiro em plantio direto, em rotação bianual com soja seguida de sorgo
consorciado com Brachiaria ruzizienses (S3)] e em dois níveis de fertilização
(padrão e reduzida).
Foi constatada a presença dos microrganismos de solo e raízes em
todas as amostras.
Houve efeito dos sistemas de cultivo e da fertilização sobre a
comunidade microbiana, bactérias diazotróficas e fungos micorrízicos
arbusculares, porém de forma diferenciada.
O sistemas semi-direto (T2) foi o que obteve melhor resposta dos
microrganismos, ou seja, as maiores populações, quando analisado todas as
variáveis avaliadas. Porém o sistema plantio direto (S3) mostrou-se bem
equilibrado e isso refletiu no campo por ter apresentado a maior
produtividade (média de 284@/ha de algodão em caroço) quando
comparado aos demais sistemas.

45
6. REFERÊNCIAS BIBLIOGRÁFICAS
ABBOTT, L. K.; ROBSON, A. D. Factores influencing the occurrence of vesicular arbuscular mycorrhizas. Agriculture, Ecossystems and Environment, v. 35, p. 125-150, 1991.
AFEK, U.; RINALDELLI, J. A.; MENGE, E. L.; et al. Mycorrhizal inoculum influence colonization of cotton, onion and pepper seedlings. Soc. Hort. Sci., v.115, n.1, p. 938-942, 1990.
ALLEN, M. F. The Ecology of Mycorrhizae. Cambridge Studies in Ecology. Cambridge: Cambridge University Press, 1991, 184 p.
ALLEN, M. F. ed. Mycorrhizal functioning. London: Chapman Hall, 1992, 515 p.
ALVARENGA, R. C.; CABEZAS, W. A. L.; CRUZ, J. C.; et al. Plantas de cobertura de solo para sistema plantio direto. In: Informe Agropecuário, v.22, n. 208, p. 25-42, 2001.
ANANYEVA, N. D.; DEMKINA, T. S.; JONES, W. J.; et al. Microbial biomass in solis of Rússia under long-term management practices. Biology Fertility Soils, v.29, p.291-299, 1999.
ASSAD, Maria Leonor R. C. L. Recursos Biológicos: Ocorrência e variabilidade. VIII Simpósio do Cerrado, 1996.
BALDANI, J. I.; BALDANI, V. L. D.; SELDIN, L.; et al. Characterization of Herbaspirillum sepopedicae gen. nov sp. Nov., a root-associated nitrogen-fixing bacterium. International Journal of Systematic Bacteriology, v. 36, n.1, p.86-93, 1986.
BALDANI, J. I.; POT, B.; KIRCHHOF, G.; et al. Emended description of Herbaspirillum; inclusion of [Pseudomonas] rubrisubalbicans, a mild plnat pathogen, as Hesbaspirillum rubrisubalbicans comb. Nov.; and classification of a group of clinical isolates (EF group 1) as Herbaspirillum species 3. International Journal of Systematic Bacteriology, Baltimore, v.46, n.3, p.802-810, 1996.

4646
BALDANI, J. I.; CARUSO, L.; BALDANI, V. L. D.; et al. Recent advances in BNF with non-legume plants. Soil Biology and Biochemistry, Oxford, v.29, n.5/6, p.911-928, 1997.
BALOTA, E. L.; LOPES, E. S. Introdução de fungo micorrízico arbusculares no cafeeiro em condições de campo: II. Flutuação sazonal de raízes, de colonização e de fungos micorrízicos arbusculares associados. Revista Brasileira de Ciência do Solo, v.20, p.225-232, 1996.
BALOTA, E. L. Alterações microbiológicas em solo cultivado sob o plantio direto. In: PEIXOTO, R.T.G. dos; ARKENS, D.C.; SAMAHA, M.J. (ed) Plantio Direto: o caminho para uma agricultura sustentável. Palestras do 10 Congresso Brasileiro de Plantio Direto para uma agricultura sustentável. Ponta Grossa, 1997, 257p.
BALOTA, E. L.; LOPES, E. S.; HUNGRIA, M.; et al. Ocorrência de bactérias diazotróficas e fungos micorrízicos arbusculares na cultura da mandioca. Revista Brasileira de Ciência do Solo, v.34, n.7, p.1265-1276, 1999.
BANZATTO, D. A.; KRONKA, S. N. Experimentação Agrícola, Jaboticabal: FUNEP, 1992, 247p.
BAZZICALUPO, M.; OKON, Y. Associative and endophytic symbiosis. In: PEDROSA, F.; HUNGRIA, M.; YATES, M.G.; NEWTON, W.E., eds. Nitrogen fixation: from molecules to crop productivity. Dordrecht, Kluwer Academic Publishers, p.409-410, 2000.
BODDEY, R. M. Biological nitrogen fixation in sugar cane – a key to energetically viable biofuel production. CRC Critical Reviews in Plant Sciences, v.14, p.263-279,1995.
BODDEY, R. M., De OLIVEIRA, O C.; URQUIACA, S.; et al. Biological nitrogen fixation associated with sugar-cane and rice: contribuitions and prospects for improvement. Plant and Soil, v.174, p.195-209, 1997.
BRUNDRETT, M. C. Mycorrhizas in natural ecossystems. In: MACFAYDEN, A.; BEGON, M.; FITTER A. H. Advances in Ecological Research. Academic Press, v. 21, p.171-313, 1991.

47
CARNEIRO, R. G.; MENDES, I. C.; CARVALHO, A. M.; et al. Dinâmica de variáveis biológicas associadas ao ciclo do fósforo em solo de cerrado sob diferentes sistemas de manejo. Embrapa Cerrados, n.36, p.1-5, 1999 (Pesquisa em andamento).
CARNEIRO, R. G.; MENDES, I. C.; LOVATO, P. E.; et al. Indicadores biológicos associados ao ciclo do fósforo em solos de Cerrado sob plantio direto e plantio convencional. Pesquisa Agropecuária Brasileira, Brasília, v.39, n.7, p.661-669, 2004.
CAVALCANTE, V. A.; DÖBEREINER, J. A new acid-tolerant nitrogen-fixing bacterium associated with sugar-cane. Plant and Soil, Dordrecht, v.108, n.1, p.23-31, 1988.
CHALK, P. M. The contribution of associative and symbiotic nitrogen fixation to the nitrogen nutrition of non-legumes. Plant Soil, n.132, p.29-39, 1991.
COLOZZI FILHO, A.; SIQUEIRA, J. O.; OLIVEIRA, E. Ocorrência de micorriza vesicular-arbusculares em alguns ecossistemas do Estado de Minas Gerais. In: Reunião Brasileira sobre Micorrizas, Lavras. Programa e Resumos, Edição FAEPE, 1985, 68p.
COLOZZI FILHO, A.; ANDRADE, D. S.; BALOTA, E. L. Esporulação de fungos micorrízicos vesículo-arbusculares sem solo sob plantio direto ou convencional e rotação de culturas. In: Encontro Latino Americano sobre Plantio Direto na Pequena Propriedade, Ponta Grossa, 1993.
COLOZZI FILHO, A.; BALOTA, E. L. Micorrizas arbusculares. In: HUNGRIA, M.; ARAUJO, R.S. Manual de métodos empregados em estudos de microbiologia agrícola. EMBRAPA-CNPSo-CNPAF, 1994, 542p.
COLOZZI FILHO, A.; BALOTA, E. L.; ANDRADE, D. S. Microrganismos e processos biológicos no sistema de plantio direto. In: SIQUEIRA J.O.; MOREIRA F.M.S.; A.S. LOPES; GUILHERME L.R.G.; FAQUIM V.; FURTINI NETO A.E.,; . CARVALHO J.G. Soil Fertility, Soil Biology and Plant Nutrition Interrelationships, v. 1, p. 487-508, 1999.
CONAB, 2006.<http:/www.conab.org.br> Acesso em 07/03/2006.

48
CORDEIRO, M. A. S.; CARNEIRO, M. A. C.; PAULINO, H. B.; et al. Colonização e densidade de esporos de fungos micorrízicos em dois solos do cerrado sob diferentes sistemas de manejo. Pesquisa Agropecuária Tropical, v.35, n.3, p.147-153, 2005.
CORRÊA, J. R. V. Algodoeiro: informações básicas para o seu cultivo. EMBRAPA-UEPAE Belém, 1989, 29p.
DAVIS, R. M.; MENGE, J. A.; ERWIN, D. C. Influence of Glomus fasciculatus and soil phosphorus on verticillium wilt of cotton. Phytopathology, v.69, p.453-456, 1979.
DENG, S. P.; TABATABAI, M. A. Effect of tillage and residue management on enzyme activities in soils. III. Phosphatases and Arylsulfatase. Biology Fertility Soils, v.24, p.141-146, 1997.
DÖBEREINER, J.; RUSCHEL, A. P. Uma nova espécie de Beijerinkia. Revista de Biologia. v.1, p.261-272, 1958.
DÖBEREINER, J.; ALVAHYDO, R. Sobre a influência da cana-de-açúcar na ocorrência de Beijerinckia no solo. II-Influência das diversas partes do vegetal. Revista Brasileira de Biologia, Rio de Janeiro, v.19, n.4, p.401-412, 1959.
DÖBEREINER, J.; DAY, J. M. Associative symbioses in tropical grasses: characterization of microorganisms and nitrogen-fixing sites. In: NEWTON, W.E.; NYMAN, C.J., ed. Nitrogen fixation; proceedings of the 1st International Symposium on Nitrogen Fixation. Pullman: Washington State University, v.2, p.518-538, 1975.
DÖBEREINER, J.; MARRIEL, I. E.; NERY, M. Ecological distribution of Spirillum lipoferum Beijerinck. Canadian Journal of Microbiology, Ottawa, v.22, n.10, p.1464-1473, 1976.
DÖBEREINER, J.; PEDROSA, F. O. Nitrogen-fixing bactéria in nonleguminous crop plants. Madison: Springer-Verlag, 1987, 155p.

49
DÖBEREINER, J., BALDANI, J. I., BALDANI, V. L. D. Como isolar e identificar bactérias diazotróficas de plantas não leguminosas. Brasília: EMBRAPA, SPI.; Itaguaí: EMBRAPA, CNPAB, 1995, 60p.
EVANS, D. G., MILLER M. H. Vesicular-arbuscular – mycorrhizal, and the soil disturbance-induced reduction of nutrient absorption in maize. Casual relations. New Phytol. v. 110, p.67-74, 1988.
EVANS, H. J. BURRIS, R. H. Highlights in biological nitrogen fixation during the last 50 years. In: STACEY, G.; BURRIS, R. H.; EVANS, H. J. eds. Biological Nitrogen Fixation. New York: Chapman and Hall, p.1-42, 1992.
FERRAZ, F. M.; MÔNACO, G. M.; PUSCH, J.A. Plantio direto avança no Sul e nos cerrados. Agrianual, p.23-26, 2004.
FITTER, A. H. Costs and benefits of mycorrhizas: Implications for functioning under natural conditions. Experientia, v.47, p.350-355, 1991.
FRANCO, A. A.; DÖBEREINER, J. A biologia do solo e a sustentabilidade dos solos tropicais. Summa Phytopathológica, São Paulo, v.20, n.1, p.68-74, 1994.
FRYXELL, P. A. Stages in the evolution of Gossypium. Advanced Frontiers Plant Science, v.10, p.31-56, 1965.
GERDEMANN, J. W. Vesicular-arbuscular mycorrhizae. In: TORREY, J. G.; CLARKSON, D. T.(Ed), The development and function of roots. Academic Press, p.575-91, 1975.
GERDEMANN, J. W.; NICOLSON, T. H. Spores of mycorrhizal endogone species extracted from soil by wet sieving and decating. Transactions of the British Mycological Society, v.46, p. 235-44, 1963.
GILLIS, M.; DÖBEREINER, J.; POT, B.; et al. Taxonomic relationships between (Pseudomonas) rubrisubalbicans, some-clinical isolates (EF Group 1), Herbaspirillum seropedicae and (Azospirillum ) autotrophicum. In: POLSINELLI, M.; MATERASSI, R.; VINCENZI, M. (Ed.) Nitrogen Fixation. Dordrecht: Kluwer Academic Publishers, 1991. 292p.

50
GIOVANETTI, M. Seasonal variations of vesiular-arbuscular mycorrhizas and Endogonaceous spores in a maritime sand dune. Trans. Br. Mycol. Soc., v.84, p.679-684, 1985.
GIOVANNETTI, M.; MOSSE, B. An evaluation of techniques for measuring vesicular-arbuscular micorrhizal infection in roots. New Phytologist, v. 84, p. 484-500, 1980.
HAAHTELA, K.; WAARTIOVAARA, T.; SUNDMAN, V.; et al. Root-associated N2 fixation (acetylene reduction) by Enterobacteriaceae and Azospirillum strains in cold climate spodosols. Applied and Environmental Microbiology, Washinngton, v.41, n.1, p.203-206, 1981.
HALLMANN, J.; QUADT-HALMANN, A.; MAHAFFEE, W. F; KLOEPPER, J.W. Bacterial endophytes in agricultural crops. Canadian Journal of Microbiology, v.43, p.895-914, 1997.
HARLEY, J. L.;SMITH, S. T. Mycorrhizal Simbiosis. NeW YorK : Academic Press, 1983, 483p.
HARTMANN, A.; BALDANI, J. I.; KIRCHROF, G.; et al. Taxonomic and ecologic studies of diazotrophic rhizosphere bacteria using phylogenetic probes. In: FENDRIK, I.; DEL GALLO, M.; VANDERLEYDEN, J.; ZANARICZY, M. (Ed.) Azospirillum VI and related microorganisms. Berlin: Springer-Verlag. 1995. 415p.
HECKLER. J. C.; HERMANI, L. C.; PITOL, C. Palha. In: SALTON, J.C.; HERMANI, L.C.; FONTES, C. Z. (Org) Sistema Plantio direto: o produtor pergunta a EMBRAPA responde. EMBRAPA – CPAO, p. 37 – 49, 1998.
HOWELER, R. H.; SIEVERDING, E.; SAIF, R. S. Pratical aspects of mycorrhizal technology tropical crops and pasture. Plant and Soil, Dordrecht, v.100, p.249-283, 1987.
HUNGRIA, M.; ANDRADE, D. S.; COLOZZI FILHO, A.; et al. Ecologia microbiana em solos sob cultivo na Região Sul do Brasil. In: HUNGRIA, M.; BALOTA, E. L.; COLOZZI FILHO, A.; ANDRADE, D. S., EDS. Microbiologia do solo: desafios para o século XXI. Londrina: IAPAR/EMBRAPA-CNPSo, p.234-270, 1995.

51
IQBAL, S. H. et al. A field survy of micorrhizal associations in terms of Pakistan. New Phytalogist, v.87, p.69-79, 1981.
KANDELER, E.; TSCHERKO, D.; SPIEGEL, H. Long-term monitoring of microbial biomass, N mineralisation and enzyme activities of a Chernozem under different tillage management. Biology Fertility Soils, v.28, p.343-351, 1999.
KLINK, C. A.; MIRANDA, H. S.; GONZALES; M. I. et al. O bioma cerrado site 3. Disponível em: http://www.icb.ufmg.br/rpeld/port_site03.pdf. Acesso em: 10 de maio de 2006.
KOSKE, R. E.; HALVORSON, W. L. Ecological studies of vesicular-arbuscular mycorrhizae in a barrier sand dune. Canadian Journal of Botany, v.59, p.1413-1422, 1981.
KOSKE, R. E; GEMMA, J. N. A modified procedure for staining roots to detect VA micorrhizas. Mycological Research, v.4, n.92, p. 486-488, 1989.
LAL, I. L.; KHANNA, S. Renadulotin and nitrogen fixing potential of Acacia nilotica inoculated with Rhizobium isolates. Canadian Journal of microbiology, v.39, p.87-91, 1993.
LANDERS, J. N. Technology transfer mechanisms for the new direct sowing techniques in the savannahs of central Brazil. In: Rasolo, F. et Raunet, M., (eds). Gestion agrobiologique des sols et des systèmes de culture. Actes de l´atelier international, Antsirabe, Madagascar, 1998.
LEE, J. A. Cotton as a world crop. In: RHOEL, R. J.; LEWIS, C. F. (eds). Cotton. American Society of Agronomy, p.1-16, 1984.
LOVELL, R. D.; HACTCH, D. J. Stimulation of microbial activity following spring applications of nitrogen. Biology and Fertility of Solis, v.26, p.28-30, 1998. MARIN, V. A.; BALDANI, V. L. D.; TEIXEIRA, K. R. S.; et al. Fixação biológica de nitrogênio: Bactérias fixadoras de nitrogênio de importância para a Agricultura Tropical. Série documentos, EMBRAPA-CNPAB, abril, p.4-34, 1999.

52
MATSUOKA, M. Características biológicas de solos sob vegetação nativa e sistemas agrícolas anuais e perenes na região de Primavera do Leste. 2001. 123f. Dissertação (Mestrado em Agricultura Tropical) – Faculdade de Agronomia e Medicina Veterinária, Universidade Federal de Mato Grosso, Cuiabá-MT, 2001.
MENDES, I. C.; VIVAIDI, L. A.; Dinâmica da Biomassa e atividade microbiana em uma área sob mata de galeria na região do Distrito Federal. In:RIBEIRO, J. F.; FONSECA, C. E.; SOUSA – SILVA, J. C. Cerrado: caracterização e recuperação de matas de galeria. Embrapa Cerrados, 2001.
MINHONI M. T. A.; CARDOSO E. B. J. N.; EIRA A. F. Efeitos da interação de fosfato de rocha, matéria orgânica e fungo micorrízico no crescimento e na absorção de nutrientes pela soja. Revista Brasileira de Ciência do Solo v.17 p. 165-171, 1993.
MOREIRA, F. M. S.; SIQUEIRA, J. O. Microbiologia e bioquímica do solo, 2002, 626 p.
OKON, Y.; LABANDERA-GONZALEZ, C. A. Agronomic applications of Azospirillum: an evaluation of 20 years worldwide field inoculation. Soil Biology and Biochemistry, v.26, p.1591-1601, 1994.
OLIVEIRA, A. A. R.; LEAL, L. C. de. Influência de fungos micorrízicos no desenvolvimento de mudas de acerola. In: Reunião brasileira de fertilidade do solo e nutrição de plantas, 24. Reunião brasileira sobre micorrizas, 8. Simpósio brasileiro de microbiologia do solo, 6. Reunião brasileira de biologia do solo, 2000. CD-rom.
PAUL, E. A.; CLARK, F. E. Mycorrhizal relationships. In: PAUL, E. A.; CLARK, F. E. Soil Microbiology and Biochemistry. Academic Press, p.245-263, 1996.
PHILLIPS, L. L. The cytogenetics of gossypium and the origin of new world cottons. Evolution, v.17, p.460-469, 1963.

53
PIVELLO, V. R. Invasões biológicas no cerrado brasileiro: Efeitos da introdução de espécies exóticas sobre a biodiversidade. Ecologia Informativo, n.33, 2006.
PORTALBRASIL, 2006 <www.portalbrasil.net/cerrado_climarelevo.htm > Acesso em: 10 de maio de 2006.
POWELL, C. L.; BAGYARAJ, D. J. VA Mycorrhiza. Boca Raton, 1984, 234p.
RAO, A. V.; VANKATESWARLU, B. Associative symbiosis of Azospirillum lipoferum with dicotyledonous succulent plants of the Indian desert. Canadian Journal of Microbiology, Ottawa, v.28, n.7, p.778-782, 1982.
REINHOLD-HUREK, B.; HUREK, T.; GILLS, M.; et al. Azoarcus gen. nov. nitrogen-fixing protobacteria associated with roots of Kallar grass (Leptochloa fusca (L.) Kunth), and description of two species, Azoarcus indigens sp. Nov. and Azoarcus communis nov. International Journal of Systematic Bacteriology, v.43, p.574-584, 1993.
RIBEIRO JUNIOR, J. I. Análises estatísticas no SAEG. Viçosa: UFV, 2001. 301p.
SATURNINO, H. M. Evolução do plantio direto e as perspectivas nos cerrados. In: Informe Agropecuário. Belo Horizonte: EPAMIG, v.22, n.208, p.5-12, 2001.
SAUNDERS, J. H. The wild species of Gossypium and their evolutionary history, 1961.
SCANNERINI, S.; BOFANTE-FASOLO, P. Comparative ultra estrutural analysis of mycorrhizal associations. Canadian Journal of Botany, v.61, p.917-943, 1983.
SEBILLOTTE, M. Système de culture, un concept opératoire pour les agronomes. IN: Combe, L. & Piccard, D. (Ed.). Les systèmes de culture. Paris, INRA, 1990, 196p.

54
SEGUY, L.; BOUZINAC, S.; MAEDA, E.; MAEDA, N. Large scale mechanized direct drilling of cotton in Brazil. p.11-17. The Icac Recorder, 1998.
SEGUY, L.; BOUZINAC, S.; MARONEZZI, A. C. Un dossier du semis direct: Systèmes de cultura et dynamique de la matière organique. Montpellier: CIRAD-CA/GEC, 2001. 203p.
SEGUY, L.; BOUZINAC, S.; MARONEZZI, A. C.; SCOPEL, E.; BELOT, J. L.; MARTIN, J. The success of no-tillage with cover-crops for Savannah regions. In : II Congresso mundial sobre agricultura conservacionista. Produzindo em harmonia com a natureza. Anais. Volume I. Palestras. FBPDP, Ponta Grossa p.153-155, 2003 (a).
SEGUY, L.; BOUZINAC, S.; SCOPEL, E.; RIBEIRO, F. New concepts for sustainable management of cultivated soils through direct seeding mulch based systems : The Cirad experience, partnership and networks. In : II Congresso mundial sobre agricultura conservacionista. Produzindo em harmonia com a natureza. Anais. Volume I. Palestras. FBPDP, Ponta Grossa. p.142-144, 2003 (b).
SEGUY, L.; BOUZINAC, S.; BELOT, J. L.; MARTIN, J. Sistema de produção sustentavel de algodão para os cerrados úmidos do Brasil Central. In: ZAMBOLIM, L.; SILVA, A. A.; AGNES E. L. Integração agricultura-pecuária. Viçosa, p.385-397, 2004.
SIEVERDING, E. Ecology of VAM fungi. In tropical agrosystems. Agriculture, Ecosystems and Enviroment, v. 29, p. 369-390, 1990.
SIEVERDING, E. Vesicular-Arbuscular mycorrhizae management in tropical agrosystems. Eschborn, 1991, 371p.
SILVA, A. P. Micorrizas arbusculares no agrossistema soja no estado de Mato Grosso. 2004. 118f. Dissertação (Mestrado em Agricultura Tropical) – Faculdade de Agronomia e Medicina Veterinária, Universidade Federal de Mato Grosso, Cuiabá-MT, 2004. SILVEIRA, A. P. D. Microbiologia do Solo. Campinas. Sociedade Brasileira de Ciências do Solo, 1992, 282p.

55
SILVEIRA, A. P. D. da. Ecologia de fungos micorrízicos arbusculares. In: MELO, I. S.; AZEVEDO, J. L. Ecologia Microbiana. EMBRAPA-CNPMA, 1998, 488p.
SIQUEIRA, J. O.; COLOZZI FILHO, A.; FARIA, F. H. S; OLIVEIRA, E. Efetividade simbiótica de fungos micorrízicos vesículo-arbusculares para o algodoeiro. Revista Brasileira de Ciência do Solo, v.10, p.213-218, 1986.
SIQUEIRA, J. O.; KLAUBERG FILHO, O. Micorrizas arbusculares: A pesquisa brasileira em perspectiva. In: NOVAIS, F. R. de; ALVAREZ, V. H. V; SCHAEFER, C. E. G. R. (eds). Tópicos em Ciência do Solo, v.1, 2000, 352p.
SMITH, G. S.; RONCADORI, R. W.; HUSSEY, R. S. Interaction of endomycorrhizal fungi, superphosphate, and Meloidogyne incognita on cotton in microplot and field studies. Journal Nematology, v.18, p.208-216, 1986.
SOARES, A. C. F.; MARTINS, M. A. Influência de fungos micorrízicos arbusculares associados à adição de compostos fenólicos, no crescimento de mudas de maracujazeiro amarelo (Passiflora edulis f. flavicarpus). Revista Brasileira de Ciência do Solo, v.24, p.731-740, 2000.
SYLVIA, D. M. Mycorrhizal Symbioses. In: SYLVIA, D. M.; FUHRMANN, J. J.; HARTEL, P. G.; ZUBERER, D. A. Principles and apllications of soil microbiology. Prentice Hall, p.409-426, 1998.
TARRAND, J. J.; KRIEG, N. R.; DÖBEREINER, J. A taxonomic study of the Spirillum lipoferum grup., with description of a new genus, Azospirillum gen. nov and two species. Azospirillum lipoferum (Beijerinck) com. nov and Azospirillum brasilense sp. nov. Canadian Journal of Microbiology, v.8 p.967-980, 1978.
TRIPLETT, E. W. Diazotrophic endophytes progress and prospects for nitrogen fixation in monocots. Plant and Soil, v.186, p.2938, 1996.
TRUFEM, G.B.; BANONI, V.L.R. Micorrizas vesiculo arbusculares de culturas introduzidas em áreas de cerrado. Rickia, v.12, p.165-187, 1985.

56
UFLA, 2006 <hppt://www.dcs.ufla.br/cerrados/portugues/CIntrol.htm> Acesso em: 10 de maio de 2006.
VARGAS, E. D.; SCHOLLES, D. Biomassa microbiana e produção de C-CO2 e N mineral de um Podzólico Vermelho-Escuro submetido a diferentes sistemas de manejo. Revista Brasileira de Ciência do Solo. Viçosa, v.24, p.35-42, 2000.
VARGAS, L. K.; SELBACH, P. A.; SÁ, E. L. S. Alterações microbianas no solo durante o ciclo do milho nos sistemas plantio direto e convencional. Pesquisa Agropecuária Brasileira, v.39, n.8, p.749-755, 2004.
VARMA, A.; HOCK, B. Mycorrhiza: structure, function, molecular biology and biotechnology. SPRING-VERLAG, 1995, 747p.
YABUUCHI, E.; KOSAKO, Y.; OYAIZU, H.; et al. Proposal of Burkholderia gen. Nov. and transfer of seven species of the genus Pseudomonas homology group II to the new genus, with the type species Burkholderia cepacia (Palleroni and Holmes, 1981) comb. Nov. Microbiology and Immunology, Tokyo, v.36, p.1251-1275, 1992.

57
APÊNDICES
57
Apêndice 1. Tratos culturais realizados no sistema convencional (T1), sistema semi-direto sobre palhada de milheto (T2) e em
plantio direto em rotação bianual com soja seguida de sorgo consorciado com braquiária (S3), em dois níveis de fertilização
(padrão - FP e reduzida – FR), no município de Campo Verde-MT.
Fertilização Sistema Produtos Dose/ha Data FP T1 Metamidofos, Saurus, Agrex 0.7, 0.125 e 0.250 29/12/04 FP T2 Metamidofos, Saurus, Agrex 0.7, 0.125 e 0.250 29/12/04 FP S3 Metamidofos, Saurus, Agrex 0.7, 0.125 e 0.250 29/12/04 FR T1 Metamidofos, Saurus, Agrex 0.7, 0.125 e 0.250 29/12/04 FR T2 Metamidofos, Saurus, Agrex 0.7, 0.125 e 0.250 29/12/04 FR S3 Metamidofos, Saurus, Agrex 0.7, 0.125 e 0.250 29/12/04 FP T1 Dissulfan e Saurus 2.0 e 0.125 14/01/05 FP T2 Dissulfan e Saurus 2.0 e 0.125 13/01/05 FP S3 Dissulfan, Saurus e Gallant 2.0, 0.15 e 0.40 14/01/05 FR T1 Dissulfan, Saurus e Gallant 2.0, 0.15 e 0.40 14/01/05 FR S3 Dissulfan, Saurus e Gallant 2.0, 0.15 e 0.40 14/01/05 FP T1 Mentox, Marshal 400, Priori e Nimbus 1.0, 0.5, 0.2 e 0.2 19/01/05 FP T2 Mentox, Marshal 400, Priori e Nimbus 1.0, 0.5, 0.2 e 0.2 19/01/05 FP S3 Mentox, Marshal 400, Priori e Nimbus 1.0, 0.5, 0.2 e 0.2 19/01/05 FR T1 Mentox, Marshal 400, Priori e Nimbus 1.0, 0.5, 0.2 e 0.2 19/01/05 FR T2 Mentox, Marshal 400, Priori e Nimbus 1.0, 0.5, 0.2 e 0.2 19/01/05 FR S3 Mentox, Marshal 400, Priori e Nimbus 1.0, 0.5, 0.2 e 0.2 19/01/05 FP T1 Lanate, Rimon, Gallant, Miner oil e Saurus 1.0, 0.15, 0.4, 0.4 e 0.15 02/02/05 FP T2 Lanate, Rimon, Gallant, Miner oil e Saurus 1.0, 0.15, 0.4, 0.4 e 0.15 02/02/05 FP S3 Lanate, Rimon, Gallant, Miner oil e Saurus 1.0, 0.15, 0.4, 0.4 e 0.15 02/02/05 FR T1 Lanate, Rimon, Gallant, Miner oil e Saurus 1.0, 0.15, 0.4, 0.4 e 0.15 02/02/05 FR T2 Lanate, Rimon, Gallant, Miner oil e Saurus 1.0, 0.15, 0.4, 0.4 e 0.15 02/02/05 FR S3 Lanate, Rimon, Gallant, Miner oil e Saurus 1.0, 0.15, 0.4, 0.4 e 0.15 02/02/05 FP T1 Tuval, Saurus e Match 0.1, 0.2 e 0.2 04/02/05
58

58
FP T2 Tuval, Saurus e Match 0.1, 0.2 e 0.2 04/02/05 FP S3 Tuval, Saurus e Match 0.1, 0.2 e 0.2 04/02/05 FR T1 Saurus e Match 0.2 e 0.2 07/02/05 FR S3 Tuval, Saurus e Match 0.15, 0.2 e 0.2 07/02/05 FP T1 Eminent, Rimon, Polo e Pirefos 0.4, 0.1, 0.5 e 0.6 11/02/05 FP T2 Eminent, Rimon, Polo e Pirefos 0.4, 0.1, 0.5 e 0.6 11/02/05 FP S3 Eminent, Rimon, Polo e Pirefos 0.4, 0.1, 0.5 e 0.6 11/02/05 FR T1 Eminent, Rimon, Polo e Pirefos 0.4, 0.1, 0.5 e 0.6 11/02/05 FR T2 Eminent, Rimon, Polo e Pirefos 0.4, 0.1, 0.5 e 0.6 11/02/05 FR S3 Eminent, Rimon, Polo e Pirefos 0.4, 0.1, 0.5 e 0.6 11/02/05 FP T1 Pirephos, Fury400, Miner oil, Eminent e Abamectin 0.6, 0.15, 0.2, 0.5 e 0.5 16/02/05 FP T2 Pirephos, Fury400, Miner oil, Eminent e Abamectin 0.6, 0.15, 0.2, 0.5 e 0.5 16/02/05 FP S3 Pirephos, Fury400, Miner oil, Eminent e Abamectin 0.6, 0.15, 0.2, 0.5 e 0.5 16/02/05 FR T1 Pirephos, Fury400, Miner oil, Eminent e Abamectin 0.6, 0.15, 0.2, 0.5 e 0.5 16/02/05 FR T2 Pirephos, Fury400, Miner oil, Eminent e Abamectin 0.6, 0.15, 0.2, 0.5 e 0.5 16/02/05 FR S3 Pirephos, Fury400, Miner oil, Eminent e Abamectin 0.6, 0.15, 0.2, 0.5 e 0.5 16/02/05 FP T1 Sulf.Manganes, Uréia, Ac. Borico, Dissulfan e Saurus 0.6, 10, 0.4, 2 e 0.15 19/02/05 FP T2 Sulf.Manganes, Uréia, Ac. Borico, Dissulfan e Saurus 0.6, 10, 0.4, 2 e 0.15 19/02/05 FP S3 Sulf.Manganes, Uréia, Ac. Borico, Dissulfan e Saurus 0.6, 10, 0.4, 2 e 0.15 19/02/05 FR T1 Sulf.Manganes, Uréia, Ac. Borico, Dissulfan e Saurus 0.6, 10, 0.4, 2 e 0.15 19/02/05 FR T2 Sulf.Manganes, Uréia, Ac. Borico, Dissulfan e Saurus 0.6, 10, 0.4, 2 e 0.15 19/02/05 FR S3 Sulf.Manganes, Uréia, Ac. Borico, Dissulfan e Saurus 0.6, 10, 0.4, 2 e 0.15 19/02/05 FP T1 Dissulfan, Saurus, Ureia, Mancozin Sulf. Manganes e Ac. Borico 2, 0.2, 10, 1.5, 0.6 e 0.4 23/02/05 FP T2 Dissulfan, Saurus, Ureia, Mancozin Sulf. Manganes e Ac. Borico 2, 0.2, 10, 1.5, 0.6 e 0.4 23/02/05 FP S3 Dissulfan, Saurus, Ureia, Mancozin Sulf. Manganes e Ac. Borico 2, 0.2, 10, 1.5, 0.6 e 0.4 23/02/05 FR T1 Dissulfan, Saurus, Ureia, Mancozin Sulf. Manganes e Ac. Borico 2, 0.2, 10, 1.5, 0.6 e 0.4 23/02/05 FR T2 Dissulfan, Saurus, Ureia, Mancozin Sulf. Manganes e Ac. Borico 2, 0.2, 10, 1.5, 0.6 e 0.4 23/02/05 FR S3 Dissulfan, Saurus, Ureia, Mancozin Sulf. Manganes e Ac. Borico 2, 0.2, 10, 1.5, 0.6 e 0.4 23/02/05 FP T1 Mentox, Score e Abamectin 1, 0.5 e 0.5 3/3/2005 FP T2 Mentox, Score, Abamectin, Miner oil e Match 1, 0.5 , 0.5, 0.5 e 0.3 3/3/2005
59
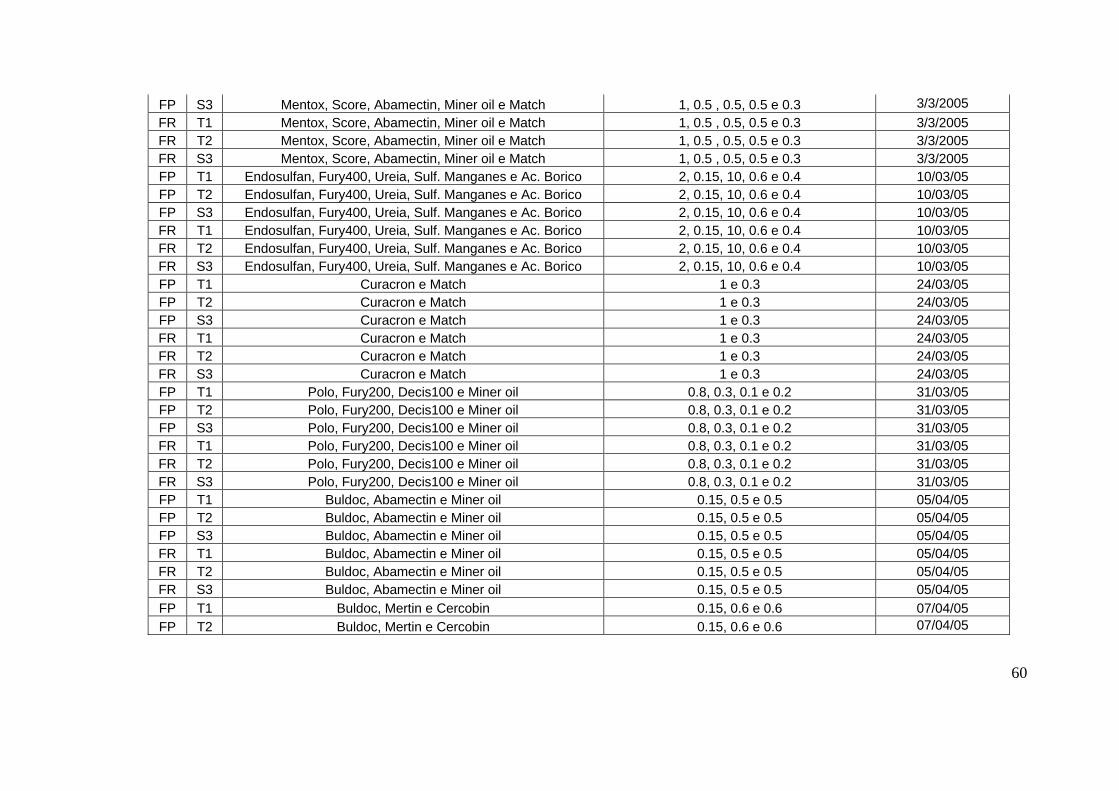
59
FP S3 Mentox, Score, Abamectin, Miner oil e Match 1, 0.5 , 0.5, 0.5 e 0.3 3/3/2005 FR T1 Mentox, Score, Abamectin, Miner oil e Match 1, 0.5 , 0.5, 0.5 e 0.3 3/3/2005 FR T2 Mentox, Score, Abamectin, Miner oil e Match 1, 0.5 , 0.5, 0.5 e 0.3 3/3/2005 FR S3 Mentox, Score, Abamectin, Miner oil e Match 1, 0.5 , 0.5, 0.5 e 0.3 3/3/2005 FP T1 Endosulfan, Fury400, Ureia, Sulf. Manganes e Ac. Borico 2, 0.15, 10, 0.6 e 0.4 10/03/05 FP T2 Endosulfan, Fury400, Ureia, Sulf. Manganes e Ac. Borico 2, 0.15, 10, 0.6 e 0.4 10/03/05 FP S3 Endosulfan, Fury400, Ureia, Sulf. Manganes e Ac. Borico 2, 0.15, 10, 0.6 e 0.4 10/03/05 FR T1 Endosulfan, Fury400, Ureia, Sulf. Manganes e Ac. Borico 2, 0.15, 10, 0.6 e 0.4 10/03/05 FR T2 Endosulfan, Fury400, Ureia, Sulf. Manganes e Ac. Borico 2, 0.15, 10, 0.6 e 0.4 10/03/05 FR S3 Endosulfan, Fury400, Ureia, Sulf. Manganes e Ac. Borico 2, 0.15, 10, 0.6 e 0.4 10/03/05 FP T1 Curacron e Match 1 e 0.3 24/03/05 FP T2 Curacron e Match 1 e 0.3 24/03/05 FP S3 Curacron e Match 1 e 0.3 24/03/05 FR T1 Curacron e Match 1 e 0.3 24/03/05 FR T2 Curacron e Match 1 e 0.3 24/03/05 FR S3 Curacron e Match 1 e 0.3 24/03/05 FP T1 Polo, Fury200, Decis100 e Miner oil 0.8, 0.3, 0.1 e 0.2 31/03/05 FP T2 Polo, Fury200, Decis100 e Miner oil 0.8, 0.3, 0.1 e 0.2 31/03/05 FP S3 Polo, Fury200, Decis100 e Miner oil 0.8, 0.3, 0.1 e 0.2 31/03/05 FR T1 Polo, Fury200, Decis100 e Miner oil 0.8, 0.3, 0.1 e 0.2 31/03/05 FR T2 Polo, Fury200, Decis100 e Miner oil 0.8, 0.3, 0.1 e 0.2 31/03/05 FR S3 Polo, Fury200, Decis100 e Miner oil 0.8, 0.3, 0.1 e 0.2 31/03/05 FP T1 Buldoc, Abamectin e Miner oil 0.15, 0.5 e 0.5 05/04/05 FP T2 Buldoc, Abamectin e Miner oil 0.15, 0.5 e 0.5 05/04/05 FP S3 Buldoc, Abamectin e Miner oil 0.15, 0.5 e 0.5 05/04/05 FR T1 Buldoc, Abamectin e Miner oil 0.15, 0.5 e 0.5 05/04/05 FR T2 Buldoc, Abamectin e Miner oil 0.15, 0.5 e 0.5 05/04/05 FR S3 Buldoc, Abamectin e Miner oil 0.15, 0.5 e 0.5 05/04/05 FP T1 Buldoc, Mertin e Cercobin 0.15, 0.6 e 0.6 07/04/05 FP T2 Buldoc, Mertin e Cercobin 0.15, 0.6 e 0.6 07/04/05
60
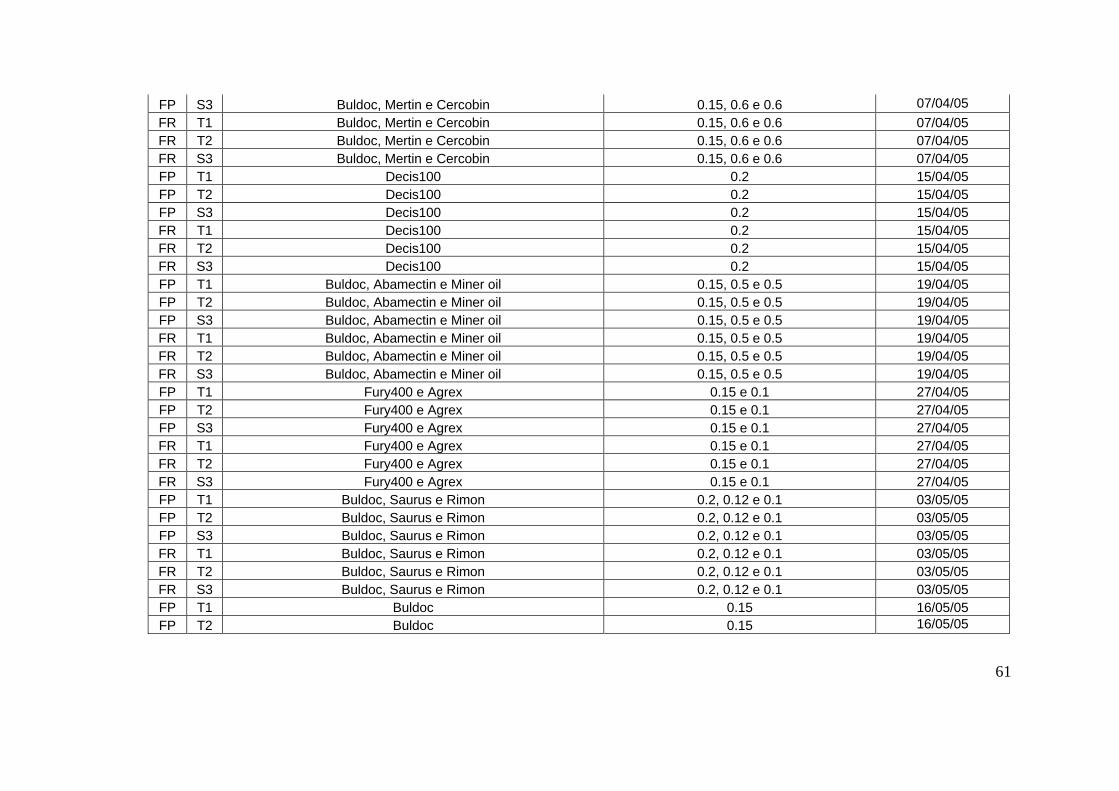
60
FP S3 Buldoc, Mertin e Cercobin 0.15, 0.6 e 0.6 07/04/05 FR T1 Buldoc, Mertin e Cercobin 0.15, 0.6 e 0.6 07/04/05 FR T2 Buldoc, Mertin e Cercobin 0.15, 0.6 e 0.6 07/04/05 FR S3 Buldoc, Mertin e Cercobin 0.15, 0.6 e 0.6 07/04/05 FP T1 Decis100 0.2 15/04/05 FP T2 Decis100 0.2 15/04/05 FP S3 Decis100 0.2 15/04/05 FR T1 Decis100 0.2 15/04/05 FR T2 Decis100 0.2 15/04/05 FR S3 Decis100 0.2 15/04/05 FP T1 Buldoc, Abamectin e Miner oil 0.15, 0.5 e 0.5 19/04/05 FP T2 Buldoc, Abamectin e Miner oil 0.15, 0.5 e 0.5 19/04/05 FP S3 Buldoc, Abamectin e Miner oil 0.15, 0.5 e 0.5 19/04/05 FR T1 Buldoc, Abamectin e Miner oil 0.15, 0.5 e 0.5 19/04/05 FR T2 Buldoc, Abamectin e Miner oil 0.15, 0.5 e 0.5 19/04/05 FR S3 Buldoc, Abamectin e Miner oil 0.15, 0.5 e 0.5 19/04/05 FP T1 Fury400 e Agrex 0.15 e 0.1 27/04/05 FP T2 Fury400 e Agrex 0.15 e 0.1 27/04/05 FP S3 Fury400 e Agrex 0.15 e 0.1 27/04/05 FR T1 Fury400 e Agrex 0.15 e 0.1 27/04/05 FR T2 Fury400 e Agrex 0.15 e 0.1 27/04/05 FR S3 Fury400 e Agrex 0.15 e 0.1 27/04/05 FP T1 Buldoc, Saurus e Rimon 0.2, 0.12 e 0.1 03/05/05 FP T2 Buldoc, Saurus e Rimon 0.2, 0.12 e 0.1 03/05/05 FP S3 Buldoc, Saurus e Rimon 0.2, 0.12 e 0.1 03/05/05 FR T1 Buldoc, Saurus e Rimon 0.2, 0.12 e 0.1 03/05/05 FR T2 Buldoc, Saurus e Rimon 0.2, 0.12 e 0.1 03/05/05 FR S3 Buldoc, Saurus e Rimon 0.2, 0.12 e 0.1 03/05/05 FP T1 Buldoc 0.15 16/05/05 FP T2 Buldoc 0.15 16/05/05
61
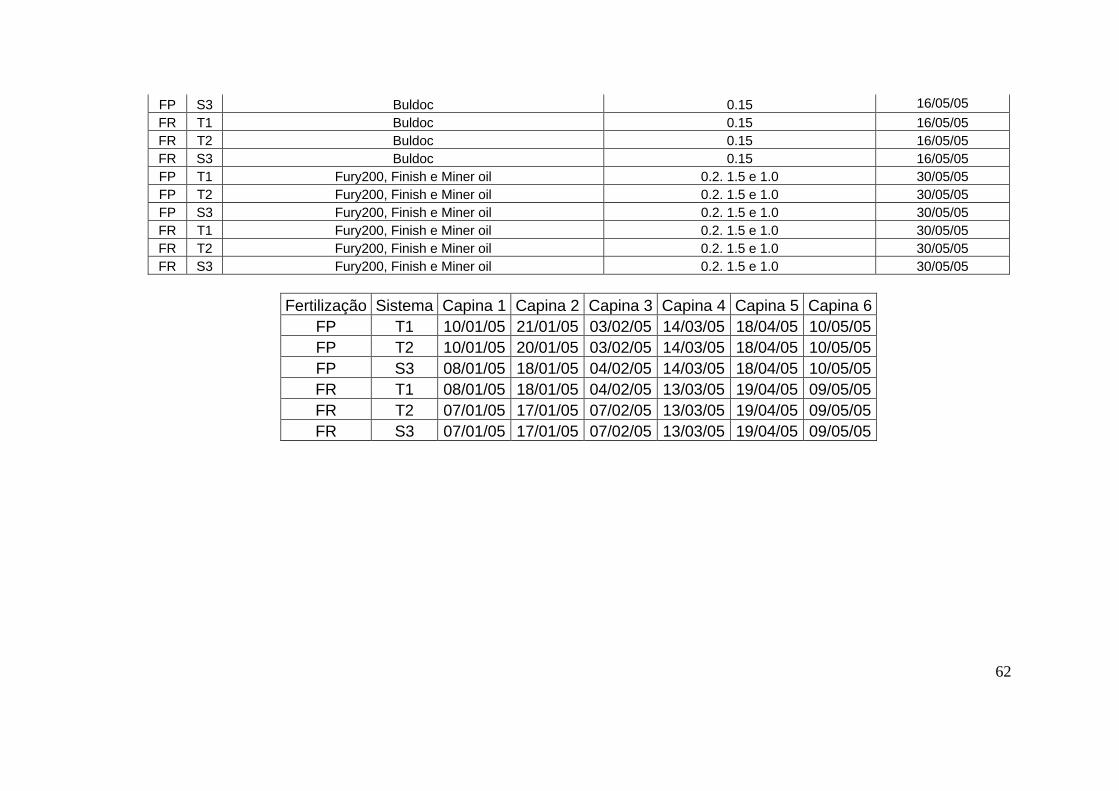
62
61
FP S3 Buldoc 0.15 16/05/05 FR T1 Buldoc 0.15 16/05/05 FR T2 Buldoc 0.15 16/05/05 FR S3 Buldoc 0.15 16/05/05 FP T1 Fury200, Finish e Miner oil 0.2. 1.5 e 1.0 30/05/05 FP T2 Fury200, Finish e Miner oil 0.2. 1.5 e 1.0 30/05/05 FP S3 Fury200, Finish e Miner oil 0.2. 1.5 e 1.0 30/05/05 FR T1 Fury200, Finish e Miner oil 0.2. 1.5 e 1.0 30/05/05 FR T2 Fury200, Finish e Miner oil 0.2. 1.5 e 1.0 30/05/05 FR S3 Fury200, Finish e Miner oil 0.2. 1.5 e 1.0 30/05/05
Fertilização Sistema Capina 1 Capina 2 Capina 3 Capina 4 Capina 5 Capina 6FP T1 10/01/05 21/01/05 03/02/05 14/03/05 18/04/05 10/05/05FP T2 10/01/05 20/01/05 03/02/05 14/03/05 18/04/05 10/05/05FP S3 08/01/05 18/01/05 04/02/05 14/03/05 18/04/05 10/05/05FR T1 08/01/05 18/01/05 04/02/05 13/03/05 19/04/05 09/05/05FR T2 07/01/05 17/01/05 07/02/05 13/03/05 19/04/05 09/05/05FR S3 07/01/05 17/01/05 07/02/05 13/03/05 19/04/05 09/05/05
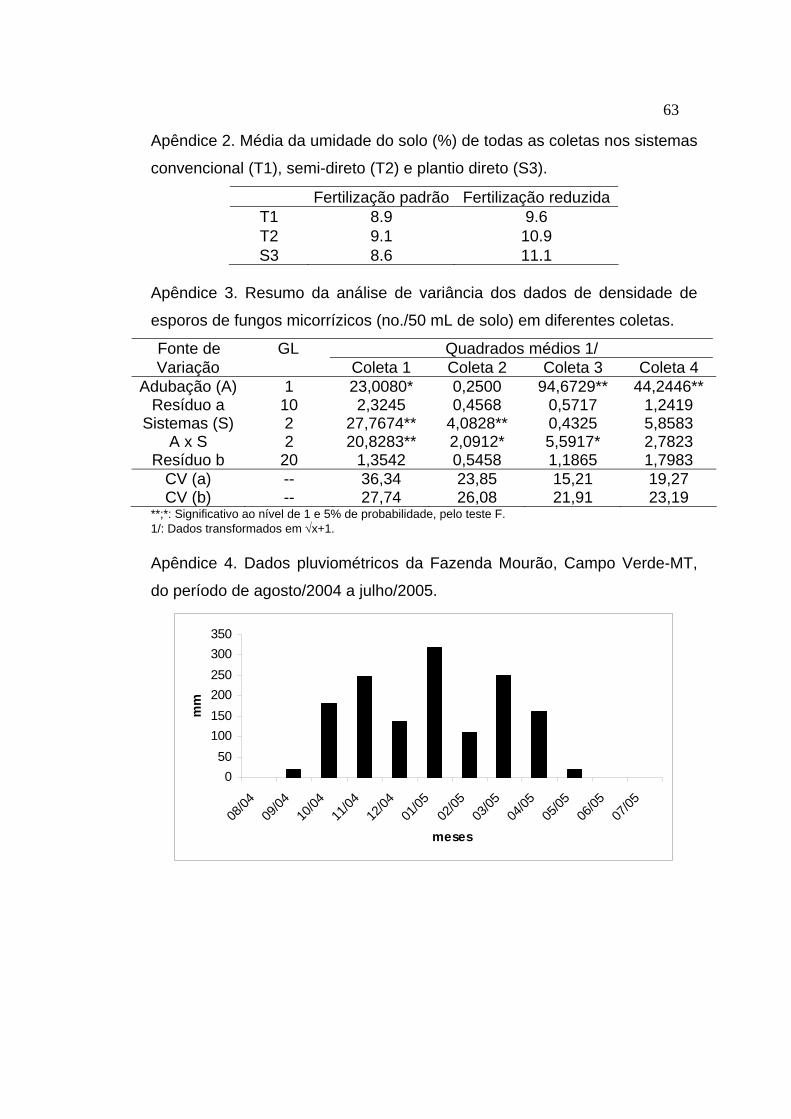
Apêndice 2. Média da umidade do solo (%) de todas as coletas nos sistemas
convencional (T1), semi-direto (T2) e plantio direto (S3).
63
Fertilização padrão Fertilização reduzida T1 8.9 9.6 T2 9.1 10.9 S3 8.6 11.1
Apêndice 3. Resumo da análise de variância dos dados de densidade de
esporos de fungos micorrízicos (no./50 mL de solo) em diferentes coletas.
Fonte de GL Quadrados médios 1/ Variação Coleta 1 Coleta 2 Coleta 3 Coleta 4
Adubação (A) 1 23,0080* 0,2500 94,6729** 44,2446** Resíduo a 10 2,3245 0,4568 0,5717 1,2419
Sistemas (S) 2 27,7674** 4,0828** 0,4325 5,8583 A x S 2 20,8283** 2,0912* 5,5917* 2,7823
Resíduo b 20 1,3542 0,5458 1,1865 1,7983 CV (a) -- 36,34 23,85 15,21 19,27 CV (b) -- 27,74 26,08 21,91 23,19
**;*: Significativo ao nível de 1 e 5% de probabilidade, pelo teste F. 1/: Dados transformados em √x+1. Apêndice 4. Dados pluviométricos da Fazenda Mourão, Campo Verde-MT,
do período de agosto/2004 a julho/2005.
050
100150200250300350
08/04
09/04
10/04
11/04
12/04
01/05
02/05
03/05
04/05
05/05
06/05
07/05
meses
mm

64
Apêndice 5. Resumo da análise de variância dos dados de número mais
provável de bactérias diazotróficas (no.x105/50 mL de solo) em diferentes
coletas.
Fonte de GL Quadrados médios Variação Coleta 1 1/ Coleta 2 1 / Coleta 3 1/ Coleta 4 2/
Adubação (A) 1 0,0608 1,8632 0,6778 2,1218** Resíduo a 10 6,9044 1,9832 0,8433 0,1385
Sistemas (S) 2 9,3568 10,1012 0,7732 0,3606** A x S 2 5,8771 2,4720 1,1387 0,5501**
Resíduo b 20 3,8400 3,4395 1,0057 0,0581 CV (a) -- 61,83 57,61 60,65 19,27 CV (b) -- 46,11 75,86 66,53 23,19
**: Significativo ao nível de 1% de probabilidade, pelo teste F. 1/: Dados transformados em ln (x+1). 2/: Dados transformados em √x+1.





























