Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE
CENTRO DE BIOCIÊNCIAS
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS BIOLÓGICAS
RIQUEZA DE FUNGOS MICORRÍZICOS ARBUSCULARES EM REMANESCENTE
DE MATA ATLÂNTICA DE TABULEIRO COSTEIRO DO RIO GRANDE DO
NORTE
Aluna: Xochitl Margarito Vista
Orientador: Prof. Dr. Bruno Tomio Goto
Linha de pesquisa: Diversidade Biológica
NATAL - RN
2016

XOCHITL MARGARITO VISTA
RIQUEZA DE FUNGOS MICORRÍZICOS ARBUSCULARES EM REMANESCENTE
DE MATA ATLÂNTICA DE TABULEIRO COSTEIRO DO RIO GRANDE DO
NORTE
NATAL/RN
2016
Dissertação apresentada como requisito parcial à
obtenção do título do mestre, pelo curso de Pós-
graduacão em Ciências Biológicas, do Centro de
Biociências da Universidade Federal do Rio
Grande do Norte.
Área de concentração: Biodiversidade
Orientador: Profr. Dr. Bruno Tomio Goto

Universidade Federal do Rio Grande do Norte - UFRN
Sistema de Bibliotecas - SISBI
Catalogação de Publicação na Fonte. UFRN - Biblioteca Setorial Prof. Leopoldo Nelson - Centro de
Biociências – CB
Vista, Xochitl Margarito.
Riqueza de fungos micorrízicos arbusculares em
remanescente de Mata Atlântica de tabuleiro costeiro do Rio Grande do Norte / Xochitl Margarito Vista. - Natal,
RN, 2016.
84 f.: il.
Dissertação (Mestrado)- Universidade Federal do Rio
Grande do Norte. Centro de Biociências. Programa de
Pós-Graduação em Ciências Biológicas. Orientador: Prof. Dr. Bruno Tomio Goto.
1. Conservação - Dissertação. 2. Diversidade -
Dissertação. 3. Micorríza - Dissertação. 4. Santuário
Ecológico de Pipa - Dissertação. 5. Taxonomia -
Dissertação. I. Goto, Bruno Tomio. II. Título.
RN/UF/BSE-CB CDU
502.1
Elaborado por KATIA REJANE DA SILVA - CRB-15/351

XOCHITL MARGARITO VISTA
RIQUEZA DE FUNGOS MICORRÍZICOS ARBUSCULARES EM REMANESCENTE
DE MATA ATLÂNTICA DE TABULEIRO COSTEIRO DO RIO GRANDE DO
NORTE
Aprovada em 17/08/2016
BANCA EXAMINADORA:
_____________________________________________
Dr. Bruno Tomio Goto
Universidade Federal do Rio Grande do Norte
(Orientador)
_____________________________________________
Dra. Vilma Maria dos Santos
Universidade Federal do Piauí
_____________________________________________
Dr. Iuri Goulart Baseia
Universidade Federal do Rio Grande do Norte
Dissertação apresentada como requisito parcial à
obtenção do título do mestre, pelo curso de Pós-
graduacão em Ciências Biológicas, do Centro de
Biociências da Universidade Federal do Rio
Grande do Norte.
Área de concentração: Biodiversidade

AGRADECIMENTO
Agradeço,
Ao professor Dr. Bruno Goto, orientador, você me deu uma oportunidade quando eu mais
precisei, acreditou em mim e nas minhas capacidades, e por tudo isso lhe agradeço. Por
compartilhar seus conhecimentos comigo e meus colegas do laboratório, por tudo que me
ensinou, pela paciência e humildade que demonstrou no início e por nunca ter desistido de
mim. Pela dedicação, pelo desempenho, pelo profissionalismo e confiança. E tenha certeza de
que tudo o que aprendemos, vamos levar por toda a nossa vida. Você é uma inspiração e um
grande exemplo de vida e para sempre terá minha gratidão.
Ao professor Dr. Iuri Goulart, pela oportunidade de aprender sobre outros grupos de fungos,
também pelas palavras de motivação que sempre me deu e fez incentivar-me, e agradeço a
amizade o carinho e compreensão.
Ao professor Dr. Umberto Laino Fulco, coordenador do programa em ciências Biológicas, por
sempre estar ao pendente de mim no programa, também pela aprendizagem de suas aulas,
pela confiança e o incentivo.
A Louise da Mata, secretária do PPCB, pela paciência e disponibilidade de ajudar a todos os
alunos do programa.
Aos professores, Dra. Vanessa, Dra. Cristina, Dr. Fulvio, Dr. Bruno Bellini, Dr. Renato. Pelas
contribuições realizadas durante a disciplina e pela oportunidade de aprendizados.
Aos professores de português como língua estrangeira, Dr. Daniel Santos, Manoela Salles.
Por ter sempre a gentileza de me ajudar nas disciplinas e pelo apoio e atenção dispensados.
E principalmente a David Hassett, Coordenador do Santuário Ecológico de Pipa (SEP), por
permitirem e apoiar na realização desta pesquisa.
Aos colegas do Laboratório de Biologia de Micorrizas e o Laboratório de Biologia de Fungos:
Marcus, Khadija, Juliana, Jessica, Mariana, Emerson, Sthephania, Ruy, Kássia, Aretha, Alder,
Amanda, Bianca, Donis, Freitas, Rubson, Luana, Julieth, Ana Clarissa, Nathalia, Miguel,
Thiago, Jeferson. Por me permitir formar parte de sua equipe de pesquisa, e apoio durante a
realização das coletas e pela amizade.
Aos amigos que conheci durante a jornada acadêmica, Thiago, Felipe, Michelle, Tanyria,
Marcel, Cristiane e Felipe Marinho pelo coleguismo e oportunidade de saberes em nossas
diferentes áreas e linhas de pesquisa e por sempre ter essa disponibilidade em ajudar-me.

Aos amigos que conheci no Brasil, Martha, Alcidia, Dilaila, Ana, Naudiane, Débora, Esteban,
Cícero, Flodobaldo, Cristina, Ana nemocón, Erika Priscila, Erika nascimento, Gilvan, Paulo,
André, pela hospitalidade de me acolher e ajudar em todo momento, pelo carinho e amizade.
A minha amiga Rosario Orozco e seus filhos, pela confiança, amizade e carinho e sempre tem
a disposição em ajudar.
Aos meus pais, Mariana e Antonio, por acreditarem em mim e me ajudarem, pelo amor
incondicional, apoio constante e paciência por tantas ausências e ainda assim estão sempre a
meu lado em todas as circunstancias.
Aos meus irmãos Yolotl, Gladys, Nereyda, Yael, Giovanni, Mario, pelo amor, carinho e apoio
e pelas palavras de motivação. Também a meus sobrinhos Diego e Yamilet.
Aos membros de diferentes instituições, Consejo Nacional de Ciencia y Tecnologia
(CONACYT), ao Grupo Coímbra, ao TecNM e a Embaixada do México no Brasil. Pelo
suporte financeiro para o desenvolvimento desse trabalho.
Enfim, a todos que de alguma forma contribuíram para a realização deste trabalho e a
finalização de mais essa etapa, muito obrigada.

RESUMO
Os Tabuleiros Costeiros constituem uma unidade geoambiental distribuída por todo o litoral
brasileiro, cuja principal formação vegetal é a Mata Atlântica. Fungos Micorrízicos
Arbusculares (FMA) são simbiontes obrigatórios de plantas, os quais representam um
importante componente da microbiota do solo em ecossistemas naturais e agrícolas. Estudo de
diversidade em ambientes costeiros são relevantes, apresentando peculiaridades bióticas e
abióticas. Além disso, são limitados os inventários de FMA em sistemas costeiros conduzidos
em território brasileiro. São necessárias áreas protegidas, para assegurar a conservação de
habitats onde os FMA ocorrem naturalmente e evoluíram, representando um lugar apropriado
para a conservação in situ. O objetivo deste estudo foi avaliar a diversidade de um complexo
de tabuleiros costeiro do dominio Mata Atlântica do Rio Grande do Norte, a fim de ampliar o
conhecimento sobre a composição e distribuição deste grupo de organismos. Para isso, foram
estudadas nove amostras de solo para identificação taxonômica, três amostras pertecem ao
período chuvoso e seis de estiagem. Foram encontradas 47 espécies de FMA (30 no período
chuvoso e 26 na estiagem), distribuídas em nove famílias: Acaulosporaceae (9 spp.),
Ambisporaceae (3 spp.), Dentiscutataceae (4 spp.), Diversisporaceae (4 spp.),
Entrophosporaceae (4 spp.), Gigasporaceae (3 spp.), Glomeraceae (15 spp.), Racocetraceae (2
sp.), Scutellosporaceae (3 spp.) e 17 gêneros. Das 138 espécies, registrados no bioma Mata
Atlântica, 35%, correspondem aos FMAs em tabuleiros costeiros de remanescentes de Mata
Atlântica estudado. O que representa 16% de espécies conhecidas para o filo. Para o Brasil
representa 30% das espécies. Para Nordeste, representa 38%. Para o RN havia apenas 41
espécies registradas, porém este número foi ampliado para 55 com os dados obtidos neste
presente trabalho. Além disso, a área apresenta 37% das espécies reportadas em áreas
protegidas globalmente. No entanto, a composição de espécies mudou entre os períodos, com
maior número de espécies no período chuvoso. Além disso, a riqueza de espécie foi mais
representativa no período chuvoso do que no de estiagem, mesmo que a força amostral tenha
sido menor no período chuvoso. Os gêneros Acaulospora e Glomus foram as mais
representativos, além disso foi acrecido o registro de Glomus spinuliferum Sieverd. & Oehl,
para o Brasil e para o bioma Mata Atlântica. Os resultados obtidos demonstram o potencial
em diversidade de FMA presentes em áreas de tabuleiros costeiros em remanescentes de Mata
Atlântica.
Palavras-chave: conservação, diversidade, micorríza, santuário ecológico de pipa,
taxonomia

ABSTRACT
The Coastal tablelands constitute a geoenvironmental unit distributed throughout the Brazilian
coast, whose main vegetation is the Atlantic Forest. Arbuscular Mycorrhizal Fungi (AMF) are
obligatory symbionts of plants, which represent an important component of the soil
microbiota in natural and agricultural ecosystems. Study of diversity in coastal environments
are relevant, presenting biotic and abiotic peculiarities. In addition, inventories of AMF in
coastal systems conducted in Brazil are limited. Protected areas are needed to ensure the
conservation of habitats where the AMF occur naturally and have evolved, representing an
appropriate place for in situ conservation. The objective of this study was to evaluate the
diversity of a coastal tablelands complex of the Atlantic Forest domain of Rio Grande do
Norte, in order to increase knowledge about the composition and distribution of this group of
organisms. For this, nine soil samples were studied for taxonomic identification, three
samples of the rainy period and six of dry. We found 47 species of AMF (30 in the rainy
season and 26 in the dry season), distributed in nine families: Acaulosporaceae (9 spp),
Ambisporaceae (3 spp.), Dentiscutataceae (4 spp.), Diversisporaceae (4 spp.),
Entrophosporaceae (4 spp.), Gigasporaceae (3 spp.), Glomeraceae (14 spp.), Racocetraceae (2
sp.), Scutellosporaceae (3 spp.) and 17 genera. Of the 138 species, recorded in the Atlantic
Forest biome, 35% correspond to the AMF in coastal tablelands of Atlantic Forest remnants
studied. This represents 16% of species known for Phyllum. For Brazil it represents 30% of
the species. For the Northeast, it represents 38%. For the RN there were only 41 registered
species, however this number was increased to 55 with the data obtained in this present study.
In addition, the area presents 37% of the species reported in globally protected areas.
However, species composition changed between the periods, with the highest number of
species in the rainy season. In addition, species richness was more representative in the rainy
period than in the dry season, even though the sample strength was lower in the rainy season.
The genera Acaulospora and Glomus were the most representative, in addition, the
registration of Glomus spinuliferum Sieverd & Oehl, was increased for Brazil and for the
Atlantic Forest biome. The results obtained demonstrate the diversity potential of AMF
present in coastal tablelands areas in remnants of Atlantic Forest.
Keywords: mycorrhizae, diversity, taxonomy, conservation

LISTA DE FIGURAS
Figura 1. Árvore filogenética proposta por Oehl et at. (2011a), incluindo taxa
adicionais proposta por Blaszkowski (2012, 2014), Goto et al. (2012), Marinho et al.
(2014), Oehl et al. (2014) .................................................................................................
14
Figura 2. Mapa de biomas do Brasil................................................................................. 15
Figura 3. Representatividade de espécies, família e gênero de Glomeromycota
descritas e reportadas no Brasil.........................................................................................
25
Figura 4. Representatividade de FMA por bioma brasileiro............................................ 26
Figura 5. Mapa dos remanescentes florestais da Mata Atlântica 2014-2015…............... 27
Figura 6. Figura 6. Proporção de famílias de FMA que ocorrem na Mata Atlântica....... 35
Figura 7. Delimitação de tabuleiros costeiros no Brasil................................................... 40
Figura 8. Pontos de coletas no Santuário Ecológico de Pipa........................................... 43
Figura 9. Mostras do solo (50g)....................................................................................... 44
Figura 10. Peneiramento úmido (Gerdemann & Nicolson, 1963).................................... 44
Figura 11. Centrifugação em água e sacarose (50%), (Jenkins, 1964)............................. 44
Figura 12. Isolamento de glomerosporos......................................................................... 44
Figura 13. Montagem de lâminas com PVLG e PVLG + Melzer................................... 45
Figura 14. Avaliação taxonômica..................................................................................... 45
Figura 15. Famílias de FMA que foram registrada em remanecente de Mata Atlântica
de tabuleiro costeiro, Pipa, RN..........................................................................................
50
Figura 16. Representatividade de gêneros registradas em remanecente de Mata
Atlântica de tabuleiro costeiro, Pipa, RN..........................................................................
51
Figura 17. A-D. Espécies de FMA encontradas na estação chuvosa em tabuleiro
costeiro do SEP..................................................................................................................
53
Figura 18. E-H. Espécies de FMA encontradas na estação seca em tabuleiro costeiro
do SEP...............................................................................................................................
54

LISTA DE TABELAS
Tabela 1. Espécies de Glomeromycota que ocorrem no Brasil...................................... 17
Tabela 2. Espécies de FMA registradas em Mata Atlântica........................................... 28
Tabela 3. Espécies de FMA registradas em localidades de Mata Atlântica do Estado
do Rio Grande do Norte...................................................................................................
36
Tabela 4. Ocorrência de espécies de FMA em tabuleiros costeiros de remanescente
de Mata Atlântica do Santuário Ecológico de Pipa no período chuvoso e de estiagem..
46
Tabela 5. Frequência de espécies de FMA por período de coleta em remanecente de
Mata Atlântica de tabuleiro costeiro, Pipa, RN...............................................................
51
Tabela 6. Ocorrência de espécies de FMA em remanecente de Mata Atlântica de
tabuleiro costeiro por amostras no período chuvoso.......................................................
55
Tabela 7. Ocorrências de espécies de FMA em remanecente de Mata Atlântica de
tabuleiro costeiro por amostras no período de estiagem..................................................
56
Tabela 8. Riqueza de espécies de FMA e índices de diversidade, dominância e
equitatividade por período de coleta e amostras em remanescente de Mata Atlântica
de tabuleiro costeiro.........................................................................................................
57
Tabela 9. Propriedades físico-químicas do solo em remanescente de Mata Atlântica
de tabuleiro costeiro do RN.............................................................................................
61

SUMÁRIO
1.INTRODUÇÃO.......................................................................................................... 9
2. REVISÃO DE LITERATURA................................................................................. 11
2.1. Fungos Micorrízicos Arbusculares (FMA).......................................................... 11
2.2. Taxonomia e Sistemática de Fungos Micorrízicos Arbusculares......................... 13
2.3. Diversidade de Fungos Micorrízicos no Brasil.................................................... 15
2.4. Fungos Micorrízicos em Mata Atlântica.............................................................. 26
2.5. Fungos Micorrízicos do Rio Grande do Norte..................................................... 36
2.6. Tabuleiros costeiros.............................................................................................. 39
3. OBJETIVOS.............................................................................................................. 42
3.1. Objetivo Geral...................................................................................................... 42
3.2. Objetivos específicos............................................................................................ 42
4. MATERIAIS E MÉTODOS..................................................................................... 42
4.1. Áreas de estudo..................................................................................................... 42
4.2. Obtenção das amostras......................................................................................... 43
4.3. Avaliação para extração de glomerosporos.......................................................... 43
4.4. Identificação taxonômica...................................................................................... 45
4.5. Análises ecológicas.............................................................................................. 45
5. RESULTADOS E DISCUSSÃO.............................................................................. 46
5.1. Influência de fatores químicos do solo................................................................. 61
6. CONCLUSÕES ......................................................................................................... 63
7. CONSIDERAÇÕES FINAIS.................................................................................... 63
8. REFERÊNCIAS BIBLIOGRÁFICAS.................................................................... 63

12
1. INTRODUÇÃO
Os Fungos Micorrízicos Arbusculares (FMA), atualmente compreendem os representantes
do filo Glomeromycota (Schüßler et al., 2001), e formam um grupo monofilético com três
classes (Archaeosporomycetes, Glomeromycetes e Paraglomeromycetes), cinco ordens
(Archaeosporales, Diversisporales, Gigasporales, Glomerales e Paraglomerales), 15 famílias e
aproximadamente 300 espécies (Oehl et al., 2011; LBM, 2016; Öpik et al., 2013). Esses
fungos são biotróficos obrigatório e se associam com raízes de plantas vasculares terrestres,
epífitas, aquáticas e também com rizoides e talos de briófitas, além de outros vegetais basais,
formando a relação simbiótica mutualística denominada micorriza arbuscular (MA) e
micotalia, para vegetais com e sem raízes, respectivamente (Schüßler et al., 2001).
Os FMA representam um importante componente da microbiota do solo em ecossistemas
naturais e agrícolas, estabelecendo relações com aproximadamente 80% das espécies vegetais
(Siqueira et al., 2002). Esses fungos auxiliam na translocação de nutrientes, disponibilizando-
os para as células do córtex de raízes de plantas, ampliam a capacidade de absorção de água e
aumentam a resistência do sistema radicular ao ataque de patógenos (Jeffries et al., 2003;
Moreira & Siqueira, 2006). Participam do funcionamento dos ecossistemas, devido ao seu
efeito sobre a diversidade e produtividade das comunidades vegetais (van der Heijden et al.,
1998), e são ferramenta importante na recuperação e restabelecimento da vegetação em
ecossistemas frágeis ou degradados (Dandan e Zhiwei, 2007; Souza et al., 2013).
Os FMA atuam como agentes potenciais de controle biológico, amenizando os efeitos ou
danos causados por fitopatógenos, provavelmente por meios indiretos, pois promovem melhor
nutrição das plantas e aumento da resistência do sistema radicular. Devido a sua importância
na ecologia ambiental estão entre os organismos ecologicamente mais significativos no
planeta (Fitter et al., 2011). Considerados como importantes indicadores de qualidade do solo
(Schloter et al., 2003).
A Floresta Atlântica é considerada a segunda maior floresta neotropical depois da Floresta
Amazônica (Campanili e Prochnow, 2006). Aproximadamente 95% de sua extensão encontra-
se em território brasileiro e o restante na Argentina e no Paraguai (Stehmann et al., 2009).
Apresenta aproximadamente 7% de remanescentes de cobertura original, com distribuição
fragmentada nas regiões costeiras, no interior das regiões Sul e Sudeste, nos remanescentes
dos estados de Goiás, Mato Grosso do Sul e no interior de alguns estados do Nordeste
(Zangaro e Moreira, 2010). Segundo bioma mais ameaçado de extinção do mundo, a Floresta
Atlântica, perde apenas para as quase extintas florestas da ilha de Madagascar na costa de

13
África (Campanili e Prochnow, 2006) sendo considerado um dos 25 hotspots mundiais de
biodiversidade (Tabarelli et al.,2005).
O bioma Mata Atlântica é um conjunto de ecossistemas de grande importância, que inclue
as faixas litorâneas do atlântico, com seus manguezais e restingas, florestas de baixada e de
encosta da serra do mar, florestas interioranas, as matas de araucária, os campos de altitude, e
os tabuleiros costeiros onde há uma parcela significativa da diversidade biológica do Brasil e
do mundo (Renato et al., 2009).
Os tabuleiros costeiros constituem uma unidade geoambiental, cuja área no Brasil é
estimada em 20,0 milhões de hectares, que se distribuem desde o Estado do Amapá até o Rio
de Janeiro, sendo considerado a maior ocorrência de formação de sedimentos do período
terciário, dentro de um único país (Jacomine, 1996; Rezende, 2000; Nascimento, 2001). Os
principais tipos de solos encontrados os tabuleiros são os Latossolos Amarelos e Argissolos
Amarelos, caracterizam-se como profundos, ácidos, álicos, com baixa capacidade de troca
catiônica e presença de horizontes coesos (Sobral et al., 2008) e a vegetação nativa da Mata
Atlântica.
O estudo da diversidade em áreas protegidas contribui para o reconhecimento desses
locais como reservas genéticas e também para manutenção e conservação a longo-prazo dos
simbiontes micorrízicos em seus habitats naturais. A proteção dessas áreas é necessária para
assegurar a conservação de habitats onde os FMA ocorrem naturalmente representando
alternativa para a conservação in situ de FMA (Turrini et al., 2010; Turrini e Giovannetti,
2011).
O estudo dos FMA é fundamental para a compreensão da interação com as comunidades
vegetais e novas descobertas podem ser úteis para melhor entender e delimitar os
representantes desse filo (Souza, 2003; Helgason e Fitter, 2005; Goto, 2009). Além disso,
existem poucos inventários da comunidade de FMA em tabuleiros costeiros do dominio Mata
Atlântica, demandando urgentemente a realização de inventários nessas áreas, pois estes
potencialmente abrigam espécies de FMA ainda não descritas pela ciência (Souza et al., 2007;
Souza et al., 2010; Ohsowski et al., 2014).

14
2. REVISÃO DE LITERATURA
2.1. Fungos Micorrízicos Arbusculares (FMA)
Micorriza (mykes= fungo; rhiza= raiz) é uma associação simbiótica mutualística
obrigatória, que ocorre entre alguns tipos de fungos do solo e raízes da maioria das plantas
vasculares (Sieverding, 1991). A associação é caracterizada pela formação de um grupo
funcional, através do qual ocorre o movimento de fotoassimilados da planta para fungo e de
nutrientes absorvidos do solo pelo fungo para a planta (Sylvia et al., 1998).
Arbert Frank (1877), patologista florestal na Alemanha, empregou pela primeira vez o
termo simbiose em 1885, e foi um dos primeiros pesquisadores a demonstrar que a
colonização das raízes das plantas, pelos fungos, resultava em micélio abundante na rizosfera,
e que este ajudava na absorção de nutrientes do solo. Além disso, também afirmou que o
fungo é incapaz de atacar, injuriar ou causar qualquer disfunção nas raízes, caracterizando a
natureza mutualista da associação, propondo o termo micorrizas.
Atualmente, as micorrizas estão categorizadas em sete tipos distintos: Micorrízas
Arbusculares, Ectomicorrizas, Ectoendomicorrizas, Arbutoide, Monotropoide, Ericoide e
Orquidóide (Allen, 1992). Esta categorização tem como parâmetros as características do
fungo (hifas septadas ou não septadas), a forma de penetração e colonização das células
hospedeiras, presença ou ausência do manto fúngico, alteração da morfologia da raiz,
especificidade do hospedeiro, o tipo do fungo e da planta envolvidos na associação (Smith e
Read 1997 e Brundrett et al., 1995). Aproximadamente 80% das espécies de plantas e 92%
das famílias vegetais são reconhecidas por formar micorríza (Wang e Qiu, 2006).
Considerando a amplitude dessa associação, J.L. Harley afirmou que “plantas não têm
raízes, tem micorrízas”, alertando para o fato de que a condição da raiz não micorrizada é a
exceção na natureza, sendo encontradas nos mais diversos ecossistemas, como florestas
tropicais e temperadas, savanas, desertos, pradarias, dunas, áreas degradadas e também em
sistemas agrícolas (Harley, 1989; Stürmer e Siqueira, 2013). Apenas seis famílias de plantas
são reconhecidamente não micotróficas (Amaranthaceae, Brassicaceae, Caryophillaceae,
Chenopodyaceae, Cyperaceae e Juncaceae), contudo, cada uma dessas famílias possui
representantes que podem estabelecer micorrizas ou cujo status de micotrofismo é fortemente
influenciado pelas condições ambientais (Muthukumar et al., 2004).
De acordo com Kenrick e Crane (1997), as primeiras plantas colonizaram o ambiente
terrestre há 480 milhões de anos. Entretanto, relato de arbúsculos em fósseis de mais de 400

15
milhões de anos (período devoniano) (Doztler et al., 2006, 2009, Redecker et al.,2000a, Remy
et al., 1994), indicaria a presença da simbiose micorrízica nas primeiras plantas vasculares,
demonstrando que os FMA muito provavelmente foram a mais antiga simbiose entre essas
plantas e fungos (Redecker et al., 2000a) e que estes organismos ajudaram os seus
hospedeiros a colonizar o novo ambiente (Redecker, 2002). As micorrizas só foram
reconhecidas e tratadas cientificamente em meados do século XIX, quando foram publicados
os primeiros relatos detalhados da associação entre células radiculares e micélios fúngicos
(Siqueira, 1986).
Em 1845 Tulasne & Tulasne haviam descrito o primeiro gênero (Glomus Tul. & Tul.)
de FMA. Inicialmente com apenas duas espécies (Glomus macrocarpum e G. microcarpum),
Glomus é atualmente considerado o gênero com maior diversidade no filo Glomeromycota.
Sclerocystis Berk. & Broome viria a ser descrito apenas 28 anos depois de Glomus (Berkeley
e Broome, 1873), e a primeira revisão de Endogonaceae, grupo onde se incluía os FMA, foi
publicada apenas em 1922.
Os FMA representam um importante componente da microbiota do solo em
ecossistemas naturais e agrícolas, agindo como uma extensão do sistema radicular das plantas,
contribui para a maior absorção e utilização dos nutrientes do solo (Siqueira et al., 2002),
notadamente o fósforo (P), que é o nutriente mais limitante para a produção agrícola nos
trópicos, dado a sua baixa disponibilidade no solo e a natureza não renovável desse recursos
(Souza et al. 2007). Esses fungos auxiliam na translocação de nutrientes, disponibilizando-os
para as células do córtex de raízes de plantas, favorecendo a nodulação e a fixação de N em
leguminosas, ampliam a capacidade de absorção de água e aumentando a resistência do
sistema radicular da planta ao ataque de patógenos (Jeffries et al., 2003; Moreira e Siqueira,
2006).
Tem sido demonstrado que os FMA têm participação no funcionamento dos
ecossistemas, devido ao seu efeito sobre a diversidade e produtividade das comunidades
vegetais (van der Heijden et al., 1998), promovendo o incremento no crescimento da planta e
tolerância a estresses bióticos e abióticos (Souza et al., 2007) sendo um componente
importante na recuperação e no restabelecimento da vegetação em ecossistemas frágeis ou
degradados (Dandan e Zhiwei, 2007; Souza et al., 2013).

16
2.2. Taxonomia e Sistemática de Fungos Micorrízicos Arbusculares
Atualmente, estudos moleculares indicam que os FMA, do filo Glomeromycota
(Schüßler et al. 2001) formam um grupo monofilético, classificados em três classes
(Archaeosporomycetes, Glomeromycetes e Paraglomeromycetes), cinco ordens
(Archaeosporales, Diversisporales, Gigasporales, Glomerales e Paraglomerales), 15 famílias,
38 gêneros e aproximadamente 300 espécies (Błaszkowski et al., 2012, 2014a; Goto et al.,
2012; Oehl et al., 2011, 2015; Sieverding et al., 2014, Öpik et al. 2013, LBM, 2016).
O filo compreende Geosiphon piriformes (Kütz.) F. Wettst., único representante do
filo que forma associação com algas do gênero Nostoc (Schüßler et al., 2001; Wettstein,
1915). Consistem em organismos que necessitam estar associados a uma raiz fisiologicamente
ativa que lhes fornecem carboidratos e outros fatores para que eles possam crescer, esporular
e, assim, completar seu ciclo de vida, por isso, são considerados biotróficos obrigatórios na
natureza (Stürmer e Siqueira, 2013).
Os esporos de FMA, os maiores da linhagem evolutiva do Reino Fungi, anteriormente
eram chamados clamidósporos ou zigosporos por não serem produzidos em estruturas
sexuadas típicas do filo Zygomycota. Walker e Sanders (1986) sugeriram a utilização do
termo genérico esporo, pois não havia dados suficientes sobre a natureza das estruturas
reprodutivas desses fungos. Com a criação do filo Glomeromycota (Figura 1) era pertinente a
criação de um termo específico para designar os esporos de FMA, assim Goto e Maia (2006)
propuseram o termo glomerosporos, nomenclatura atualmente aceita e utilizada pela
comunidade científica.
Os FMA beneficiam o hospedeiro de forma direta, por melhorar a sua nutrição, e
indiretamente, promovendo tolerância a condições restritivas de origem biótica ou abiótica
(Córdoba et al., 2002). Uma importante característica dos FMA é que eles possuem baixa
especificidade de hospedeiro, podendo colonizar o córtex radicular de diversas espécies de
plantas em um ecossistema, pertencentes a vários grupos (briófitas, pteridófitas,
gimnospermas e angiospermas), em diferentes ecossistemas terrestres que abrangem desde os
trópicos até o ártico (Smith e Read 2008; Stürmer e Siqueira, 2013).
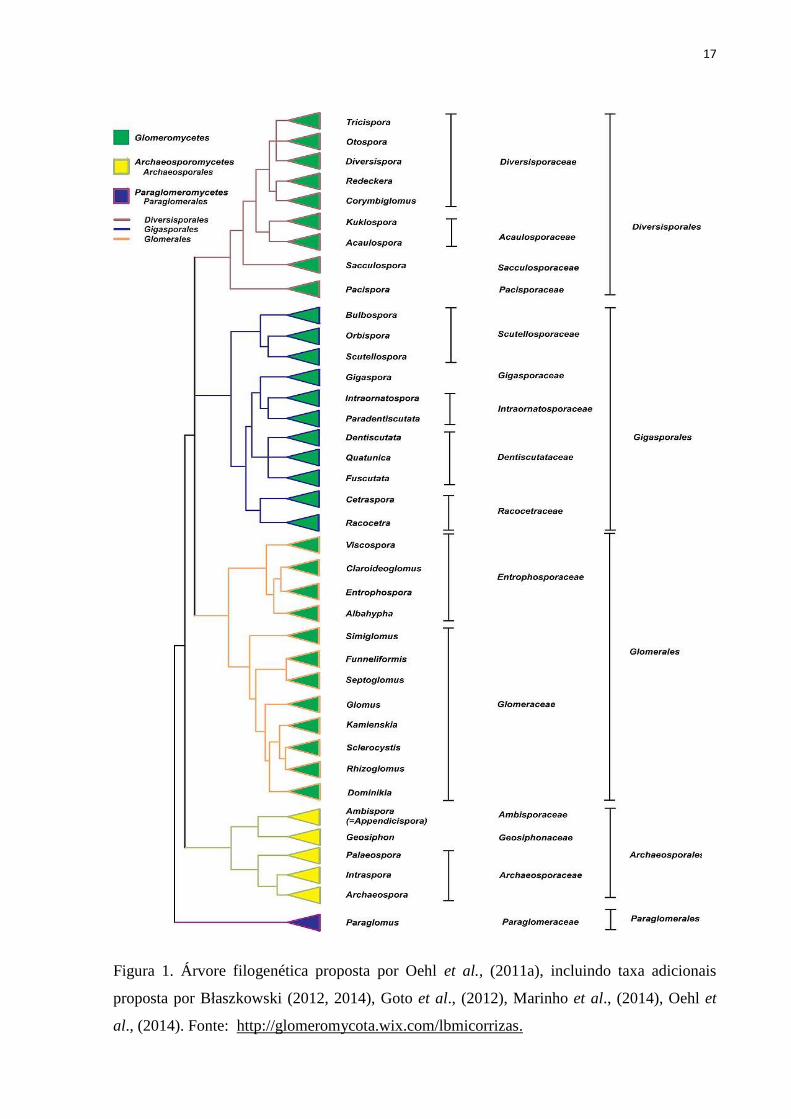
17
Figura 1. Árvore filogenética proposta por Oehl et al., (2011a), incluindo taxa adicionais
proposta por Błaszkowski (2012, 2014), Goto et al., (2012), Marinho et al., (2014), Oehl et
al., (2014). Fonte: http://glomeromycota.wix.com/lbmicorrizas.

18
2.3. Diversidade de Fungos Micorrízicos no Brasil
O Brasil consiste em um dos países mais ricos do mundo, considerado megadiverso e
apresentando um enorme potencial biológico a ser explorado (Stehmann et al., 2009;
Mittermeier et al., 2005; Póvoa et al., 2006; Lino, 1992). Pela sua localização geográfica e
seu tamanho continental abriga seis biomas, segundo o Instituto Brasileiro de Geografia e
Estatística (IBGE): Amazônia, Mata Atlântica, Caatinga, Cerrado, Pantanal, Campos sulinos e
zona costeira (figura 2), (IBGE, 2016).
Os primeiros relatos sobre a ocorrência de fungos micorrízicos no Brasil datam do
início do século XIX (1906–1937) quando J. Rick encontrou, em florestas de Pinus, no Rio
Grande do Sul, fungos do gênero Amanita, caracteristicamente ectomicorrízicos (Siqueira et
al., 2010). Em 1922, Thaxter realizou o primeiro registro de FMA, que consistia em uma
espécie de Redeckera fulva (descrita na época como Endogone fulva). Na década de 1950,
foram relatadas micorrízas em pinheiros do Paraná (Araucária), por Milanez e Monteiro-Neto.
Sacco, publicou observações sobre Sesbania punicea, Benth. Entre 1950 e o final da década
de 1960, Went e Stark, discutiram o papel das micorrizas em sistemas tropicais, fazendo
referência a importância destas no funcionamento da floresta amazônica. A Mata Atlântica,
juntamente com a Amazônia e o Cerrado consistem nos primeiros biomas brasileiros a serem
contemplados em iniciativas de pesquisa (Trufem, 1996).
Figura 2. Mapa de biomas do brasil; fonte:http://www.ibge.gov.br/biomas

19
Contudo, a década de 70 foi o período mais importante na evolução da micorrízologia
brasileira, marcado por estudos restritos a áreas agrícolas, de caráter eminentemente
tecnológico e voltado para aplicações na agricultura, em decorrência do impacto da
consolidação do conhecimento sobre o papel dos FMA na absorção de nutrientes, ocorrida na
década anterior (Zangaro e Moreira, 2010).
Na década de 1980, viveu-se grande empolgação, e interesse renovado à expansão do
treinamento e da pesquisa micorrízica, gerando publicações reveladoras do potencial destes
fungos e sua simbiose com as plantas tropicais (Siqueira et al., 2010). As primeiras revisões
sobre o tema foram publicadas em veículos nacionais, as quais se tornaram referências no
país, como Lopes et al., (1983) e Zambolim e Siqueira, (1985). Nas décadas subsequentes
trabalhos sisando o entendimento do papel dos FMA em ecossistemas naturais foram
desenvolvidos, levando-se em consideração a avaliação da ocorrência e realização de
inventários taxonômicos nos diferentes ecossistemas brasileiros, destacando-se como
importantes fontes de diversidade de FMAs (Stürmer e Siqueira, 2005).
Os Estados que mais se destacaram no estudo de FMAs nessa década foram São Paulo
e Minas Gerais, onde há predominância em avaliações quantitativas e qualitativas das
populações fúngicas, eficiência simbiótica, e distribuição em função de características
climáticas e edáficas em ecossistemas naturais (dunas, cerrado) e agrícolas (café, citrus, soja,
banana, cana, feijão, etc.) (Siqueira e Klauberg Filho, 2000). Na região Sul e em alguns
Estados do Nordeste e nas regiões Norte e Centro Oeste poucos trabalhos (Stürmer e Siqueira,
2005). No entanto, recentemente Silva et al., 2014, registraram a ocorrência de 125 espécies
de FMA distribuídos em 28 gêneros e 14 famílias para Nordeste, indicando os avanços no
conhecimento sobre a diversidade desses fungos em áreas naturais da região.
A estimativa, amplamente aceita, da diversidade de fungos no mundo é de 5,1 milhões
de espécies (Blackwell, 2011). Souza et al., (2008), estima que a diversidade real de FMA
pode estar entre 37.000 e 78.000 espécies.
Atualmente, 156 espécies de FMA foram catalogadas no Brasil, número que
representa 52% do filo Glomeromycota, distribuídas em 14 famílias e 29 gêneros (Tabela 1),
(LBM, 2016). Das 15 famílias e 38 gêneros atualmente descritos para o filo, o Brasil registra
mais de 50% das famílias e gêneros distribuídos entre as cinco ordens do filo Glomeromycota
(Figura 3). Gigasporales e Diversisporales constituem as ordens mais representadas no Brasil,
contudo, tem sido reportadas poucas linhagens basais no país.

20
Tabela 1. Espécies de Glomeromycota que ocorrem no Brasil.
Família Espécie
Acaulosporaceae Acaulospora bireticulata F.M. Rothwell & Trappe
Acaulospora capsicula Błaszk.
Acaulospora cavernata Błaszk.
Acaulospora colossica Schultz, Bever & Morton
Acaulospora delicata C. Walker, C.M. Pfeiffer &
Bloss
Acaulospora denticulata Sieverd. & S. Toro
Acaulospora dilatata J.B. Morton
Acaulospora elegans Trappe & Gerd.
¹Acaulospora endographis B.T. Goto
Acaulospora excavata Ingleby & C. Walker
Acaulospora foveata Trappe & Janos
¹Acaulospora herrerae Furrazola, B.T. Goto, G.A.
Silva, Sieverd. & Oehl
¹Acaulospora ignota Błaszk., Góralska, Chwat &
Goto
Acaulospora koskei Błaszk.
Acaulospora lacunosa J.B. Morton
Acaulospora laevis Gerd. & Trappe
Acaulospora longula Spain & N.C. Schenck
²Acaulospora mellea Spain & N.C. Schenck
Acaulospora morrowiae Spain & N.C. Schenck
Acaulospora nicolsonii C. Walker, L.E. Reed & F.E.
Sanders
Acaulospora paulinae Błaszk.
Acaulospora papillosa C.M.R. Pereira & Oehl
Acaulospora rhemii Sieverd. & S. Toro
Acaulospora reducta Oehl, B.T. Goto & C.M.R.
Pereira
Acaulospora rugosa J.B. Morton

21
Acaulospora scrobiculata Trappe
Acaulospora sieverdingii Oehl, Sýkorová & Błaszk.
Acaulospora spinosa C. Walker & Trappe
Acaulospora tuberculata Janos & Trappe
Ambisporaceae Ambispora appendicula (Spain, Sieverd., N.C.
Schenck) C. Walker
¹Ambispora brasiliensis B.T. Goto, L.C. Maia &
Oehl
Ambispora fecundispora (N.C. Schenck & G.S. Sm.)
C. Walker
Ambispora gerdemannii (S.L. Rose, B.A. Daniels &
Trappe) C. Walker, Vestberg & A.
Ambispora jimgerdemannii (N.C. Schenck & T.H.
Nicolson) C. Walker
Ambispora leptoticha (N.C. Schenck & T.H.
Nicolson) Walker, Vestberg & A. Schüssler
Kuklospora colombiana (Spain & N.C. Schenck)
Oehl & Sieverd.
Kuklospora kentinensis (Wu & Liu) Oehl & Sieverd.
Archaeosporaceae Archaeospora myriocarpa (Spain, Sieverd. & N.C.
Schenck) Oehl, G.A. Silva, B.T. Goto & Sieverd.
Archaeospora trappei (R.N. Ames & Linderman)
J.B. Morton & D. Redecker
Archaeospora undulata (Sieverd.) Sieverd., G.A.
Silva, B.T. Goto & Oehl
Dentiscutataceae Dentiscutata biornata (Spain, Sieverd. & S. Toro)
Sieverd., F.A. de Souza & Oehl
¹Dentiscutata cerradensis (Spain & J. Miranda)
Sieverd., F.A. de Souza & Oehl
¹Dentiscutata colliculosa B.T. Goto & Oehl
Dentiscutata hawaiiensis (Koske & Gemma)
Sieverd., F.A. Souza & Oehl
Dentiscutata heterogama (T.H. Nicolson & Gerd.)

22
Sieverd., F.A. Souza & Oehl
Dentiscutata nigra (J.F. Redhead) Sieverd., F.A.
Souza & Oehl
¹Dentiscutata scutata (C. Walker & Dieder.)
Sieverd., F.A. de Souza & Oehl
Dentistutata reticulata (Koske, D.D. Miller & C.
Walker) Sieverd., F.A. de Souza & Oehl
¹Fuscutata aurea Oehl, C.M. Mello & G.A. Silva
¹Fuscutata heterogama Oehl, F.A. Souza, L.C. Maia
& Sieverd.
¹Fuscutata rubra (Stürmer & J.B. Morton) Oehl, F.A.
de Souza & Sieverd.
Fuscutata savannicola (R.A. Herrera & Ferrer) Oehl,
F.A. Souza & Sieverd.
Quatunica erythropus (Koske & C. Walker) F.A. de
Souza, Sieverd. & Oehl
Diversisporaceae Corymbiglomus tortuosum (N.C. Schenck & G.S.
Sm.) Błaszk. & Chwat
Diversispora insculpta (Błaszk.) Oehl, G.A. Silva &
Sieverd.
Diversispora spurca (C.M. Pfeifer, C. Walker &
Bloss) C. Walker & Schüssler
Diversispora versiformis (P. Karst.) Oehl, G.A. Silva
& Sieverd.
Redeckera fulva (Berk. & Broome) C. Walker & A.
Schüssler
Entrophosporaceae Claroideoglomus claroideum (N.C. Schenck & G.S.
Sm.) C. Walker & A. Schüssler
Claroideoglomus etunicatum (W.N. Becker & Gerd.)
C. Walker & A. Schüssler
Claroideoglomus lamellosum (Dalpé, Koske &
Tews) C. Walker & A. Schüssler
Claroideoglomus luteum (L.J. Kenn., J.C. Stutz &

23
J.B. Morton) C. Walker & A. Schüssler
Entrophospora infrequens (I.R. Hall) R.N. Ames &
R.W. Schneid.
Viscospora viscosa (T.H. Nicolson) Sieverd., Oehl &
F.A. Souza
Gigasporaceae Gigaspora albida N.C. Schenck & G.S. Sm.
Gigaspora decipiens I.R. Hall & L.K. Abbott
Gigaspora gigantea (T.H. Nicholson & Gerd.) Gerd.
& Trappe
Gigaspora margarita W.N. Becker & I.R. Hall
¹Gigaspora ramisporophora Spain, Sieverd. & N.C.
Schenck
Gigaspora rosea T.H. Nicolson & N.C. Schenck
Glomeraceae Funneliformis caledonium (T.H. Nicolson & Gerd.)
C. Walker & A. Schüssler
Funneliformis geosporum (T.H. Nicolson & Gerd.)
C. Walker & A. Schüssler
Funneliformis halonatum (S.L. Rose & Trappe)
Oehl, G.A. Silva & Sieverd.
Funneliformis monosporus (Gerd. & Trappe) Oehl,
G.A. Silva & Sieverd.
Funneliformis mosseae (T.H. Nicolson & Gerd.) C.
Walker & A. Schüssler
Funneliformis verruculosum (Błaszk.) C. Walker &
A. Schüssler
Funneliformis vesiculiferum (Thaxt.) C. Walker & A.
Schüssler
Glomus albidum C. Walker & L.H. Rhodes
Glomus ambisporum G.S. Sm. & N.C. Schenck
Glomus australe (Berk.) S.M. Berch
Glomus arborense McGee
²Glomus brohultii R.A. Herrera, Ferrer & Sieverd.
Glomus clavisporum (Trappe) R.T. Almeida & N.C.

24
Schenck
Glomus diaphanum J.B. Morton & C. Walker
Glomus dimorphicum Boyetchko & J.P. Tewari
Glomus formosanum C.G. Wu & Z.C. Chen
Glomus fuegianum (Speg.) Trappe & Gerd.
Glomus globiferum Koske & C. Walker
Glomus glomerulatum Sieverd.
Glomus heterosporum G.S. Sm. & N.C. Schenck
Glomus lacteum S.L. Rose & Trappe
Glomus macrocarpum Tul. & C. Tul.
Glomus maculosum D.D. Mill. & C. Walker
Glomus magnicaule I.R. Hall
Glomus microcarpum Tul. & C. Tul.
Glomus multicaule Gerd. & B.K. Bakshi
Glomus multisubstensum Mukerji, Bhattacharjee &
J.P. Tewari
Glomus pallidum I.R. Hall
Glomus pansihalos S.M. Berch & Koske
Glomus pellucidum McGee & Pattinson
Glomus nanolumen Koske & Gemma
Glomus reticulatum Bhattacharjee & Mukerji
Glomus rubiforme (Gerd. & Trappe) R.T. Almeida &
N.C. Schenck
Glomus sinuosum (Gerd. & B.K. Bakshi)
R.T.Almeida & N.C. Schenck
Glomus taiwanense (C.G. Wu & Z.C. Chen) R.T.
Almeida & N.C. Schenck ex Y.J. Yao
Glomus tenebrosum (Thaxt.) S.M. Berch
Glomus tenue (Greenall) I.R. Hall
¹Glomus trufemii B.T. Goto, G.A. Silva & Oehl
Glomus vesiculifer (Thaxt.) Gerd. & Trappe
Sclerocystis coremioides Berk. & Broome
Sclerocystis sinuosa Gerd. & B.K. Bakshi

25
Septoglomus constrictum (Trappe) Sieverd., G.A.
Silva & Oehl
Septoglomus deserticola (Trappe, Bloss & J.A.
Menge) G.A. Silva, Oehl & Sieverd.
Septoglomus furcatum Błaszk., Chwat & Kovács,
Ryszka
¹Septoglomus titan B.T. Goto & G.A. Silva
Simiglomus hoi (S.M. Berch & Trappe) G.A. Silva,
Oehl & Sieverd.
Rhizoglomus aggregatum (N.C. Schenck & G.S.
Sm.) Sieverd., G.A. Silva & Oehl
Rhizoglomus clarum (T.H. Nicolson & N.C.
Schenck) Sieverd., G.A. Silva & Oehl
Rhizoglomus fasciculatum (Thaxt.) Sieverd., G.A.
Silva & Oehl
Rhizoglomus intraradices (N.C. Schenck & G.S.
Sm.) Sieverd., G.A. Silva & Oehl
Rhizoglomus invermaium (I.R. Hall) Sieverd., G.A.
Silva & Oehl
Rhizoglomus manihotis (R.H. Howeler, Sieverd. &
N.C. Schenck) Sieverd., G.A. Silva & Oehl
Rhizoglomus microaggregatum (Koske, Gemma &
P.D. Olexia) Sieverd., G.A. Silva & Oehl
¹Rhizoglomus natalense (Błaszk., Chwat & B.T.
Goto) Sieverd., G.A. Silva & Oehl
Intraornatosporaceae ¹Intraornatospora intraornata (B.T. Goto & Oehl)
B.T. Goto, Oehl & G.A. Silva
¹Paradentiscutata bahiana Oehl, Magna, B.T. Goto
& G.A. Silva
¹Paradentiscutata maritima B.T. Goto, D.K. Silva,
Oehl & G.A. Silva
Pacisporaceae Pacispora chimonobambusae (C.G. Wu & Y.S. Liu)
Sieverd. & Oehl ex C. Walker, Vestberg &

26
Schuessler
Pacispora robigina Sieverd. & Oehl
Pacispora scintillans (S.L. Rose & Trappe) Sieverd.
& Oehl ex C. Walker, Vestberg & A. Schüssler
Paraglomeraceae Paraglomus albidum (C. Walker & L.H. Rhodes)
Oehl, F.A. Souza, G.A. Silva & Sieverd.
Paraglomus bolivianum (Sieverd. & Oehl) Oehl &
G.A. Silva
¹Paraglomus brasilianum (Spain & J. Miranda) J.B.
Morton & D. Redecker
Paraglomus occultum (C. Walker) J.B. Morton & D.
Redecker
¹Paraglomus pernambucanum Oehl, C.M. Mello,
Magna & G.A. Silva
Racocetraceae ¹Cetraspora auronigra Oehl, L.L. Lima, Kozovits,
Magna & G.A. Silva
Cetraspora gilmorei (Trappe & Gerd.) Oehl, F.A. de
Souza & Sieverd.
Cetraspora nodosa (Błaszk.) Oehl, G.A. Silva, B.T.
Goto & Sieverd.
Cetraspora pellucida (T.H. Nicolson & N.C.
Schenck) Oehl, F.A. de Souza & Sieverd.
Racocetra castanea (C. Walker) Oehl, F.A. de Souza
& Sieverd.
Racocetra coralloidea (Trappe, Gerd. & I. Ho) Oehl,
F.A. de Souza & Sieverd.
Racocetra fulgida (Koske & C. Walker) Oehl, F.A.
de Souza & Sieverd.
Racocetra gregaria (N.C. Schenck & T.H. Nicolson)
Oehl, F.A. de Souza & Sieverd.
Racocetra minuta (Ferrer & R.A. Herrera) Oehl, F.A.
Souza & Sieverd.
Racocetra persica (Koske & C. Walker) Oehl, F.A.

27
de Souza & Sieverd.
¹Racocetra tropicana Oehl, B.T. Goto & G.A. Silva
²Racocetra verrucosa (Koske & C. Walker) Oehl,
F.A. de Souza & Sieverd.
Racocetra weresubiae (Koske & C. Walker) Oehl,
F.A. de Souza & Sieverd.
Sacculosporaceae Sacculospora baltica (Błaszk., Madej & Tadych)
Oehl, Palenz., IC. Sánchez Castro, B.T. Goto, G.A
Silva & Sieverd.
Scutellosporaceae ¹Bulbospora minima Oehl, Marinho, B.T. Goto &
G.A. Silva
¹Orbispora pernambucana (Oehl, D.K. Silva, N.
Freitas, L.C. Maia) Oehl, G.A.Silva & D.K. Silva
¹Scutellospora alterata Oehl, J.S. Pontes, Palenz.,
Sánchez-Castro & G.A. Silva
Scutellospora aurigloba (I.R. Hall) C.Walker & F.E.
Sanders
Scutellospora calospora (T.H. Nicolson & Gerd.) C.
Walker & F.E. Sanders
Scutellospora dipapillosa (C. Walker & Koske) C.
Walker & F.E. Sanders
Scutellospora dipurpurescens J.B. Morton & Koske
1. Espécies descritas originalmente a partir de material tipo brasileiro; 2. Espécies em que a
descrição original considera a análise de amostra proveniente do Brasil como material
suplementar para o diagnóstico taxonômico. Fonte:
http://glomeromycota.wix.com/lbmicorrizas.
Das 156 espécies que ocorrem no Brasil, outras 27 foram descritas incluindo material
brasileiro dentre essas 24 foram exclusivamente descritas a partir de material tipo do Brasil. A
Caatinga, a Mata Atlântica e o Cerrado correspondem aos biomas onde foram coletadas essas
espécies novas, os biomas Mata Atlântica, Caatinga e o Cerrado detém maior percentual da
contribuição.
28
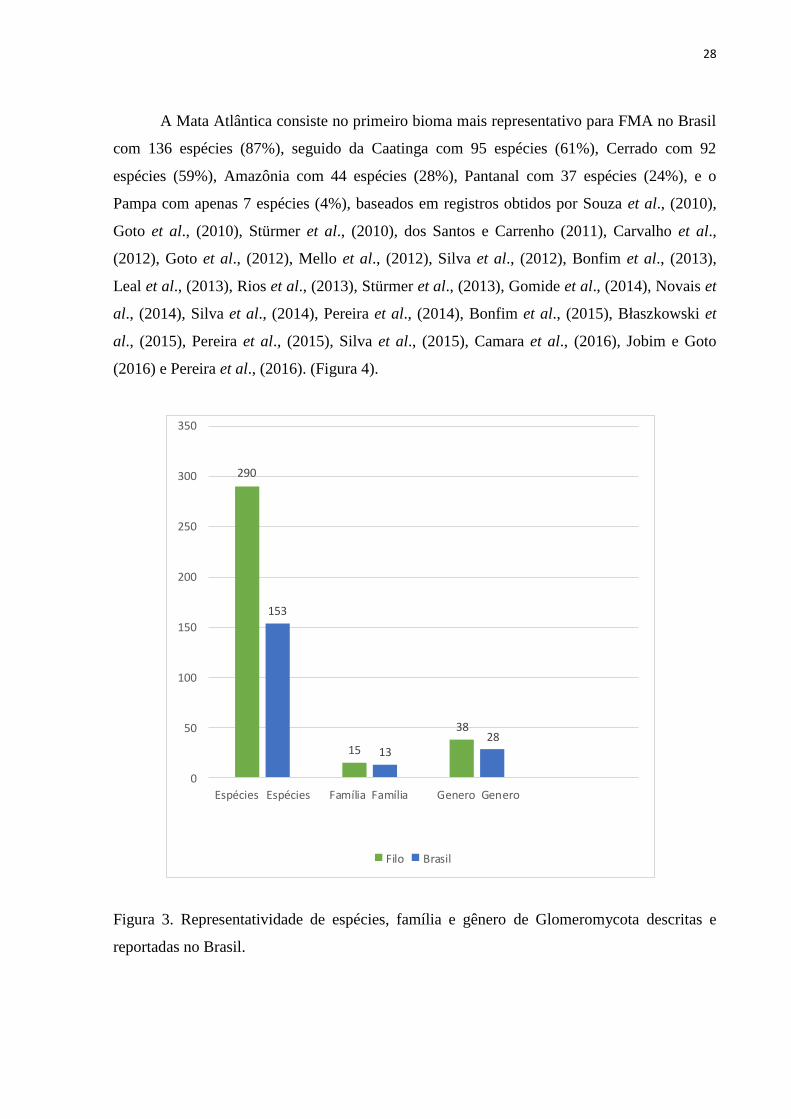
A Mata Atlântica consiste no primeiro bioma mais representativo para FMA no Brasil
com 136 espécies (87%), seguido da Caatinga com 95 espécies (61%), Cerrado com 92
espécies (59%), Amazônia com 44 espécies (28%), Pantanal com 37 espécies (24%), e o
Pampa com apenas 7 espécies (4%), baseados em registros obtidos por Souza et al., (2010),
Goto et al., (2010), Stürmer et al., (2010), dos Santos e Carrenho (2011), Carvalho et al.,
(2012), Goto et al., (2012), Mello et al., (2012), Silva et al., (2012), Bonfim et al., (2013),
Leal et al., (2013), Rios et al., (2013), Stürmer et al., (2013), Gomide et al., (2014), Novais et
al., (2014), Silva et al., (2014), Pereira et al., (2014), Bonfim et al., (2015), Błaszkowski et
al., (2015), Pereira et al., (2015), Silva et al., (2015), Camara et al., (2016), Jobim e Goto
(2016) e Pereira et al., (2016). (Figura 4).
Figura 3. Representatividade de espécies, família e gênero de Glomeromycota descritas e
reportadas no Brasil.
290
15
38
153
1328
0
50
100
150
200
250
300
350
Espécies Espécies Família Família Genero Genero
Filo Brasil
29
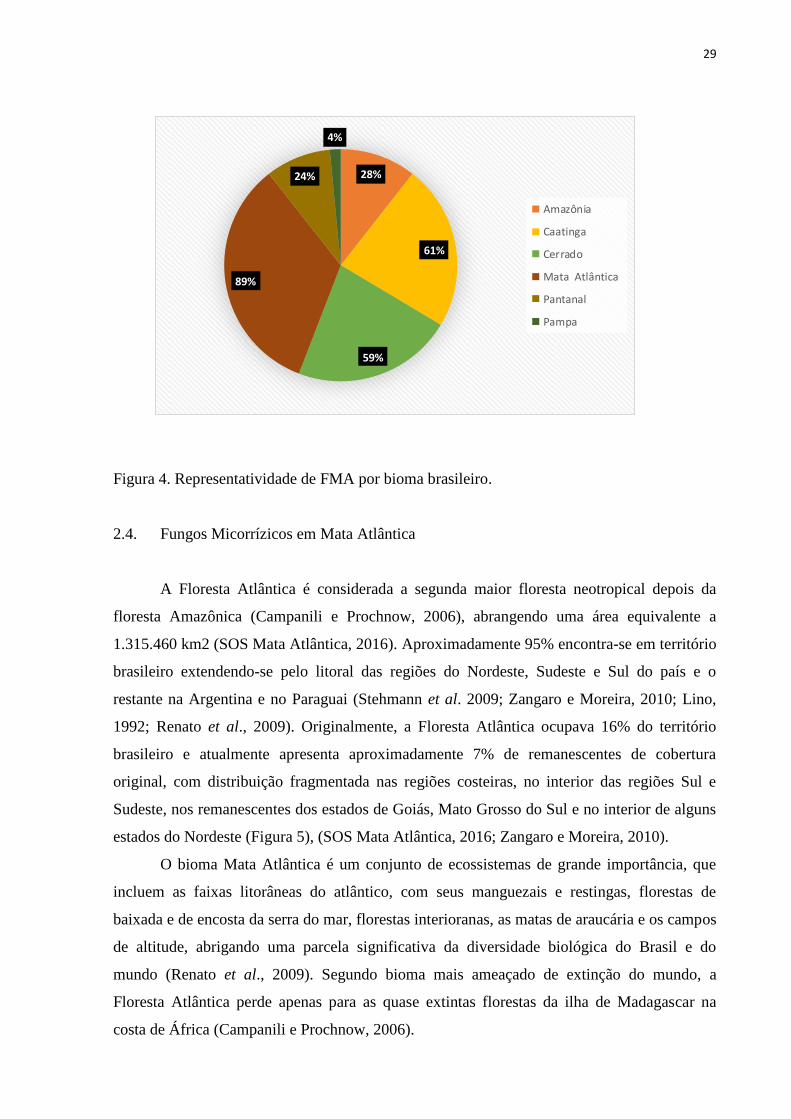
Figura 4. Representatividade de FMA por bioma brasileiro.
2.4. Fungos Micorrízicos em Mata Atlântica
A Floresta Atlântica é considerada a segunda maior floresta neotropical depois da
floresta Amazônica (Campanili e Prochnow, 2006), abrangendo uma área equivalente a
1.315.460 km2 (SOS Mata Atlântica, 2016). Aproximadamente 95% encontra-se em território
brasileiro extendendo-se pelo litoral das regiões do Nordeste, Sudeste e Sul do país e o
restante na Argentina e no Paraguai (Stehmann et al. 2009; Zangaro e Moreira, 2010; Lino,
1992; Renato et al., 2009). Originalmente, a Floresta Atlântica ocupava 16% do território
brasileiro e atualmente apresenta aproximadamente 7% de remanescentes de cobertura
original, com distribuição fragmentada nas regiões costeiras, no interior das regiões Sul e
Sudeste, nos remanescentes dos estados de Goiás, Mato Grosso do Sul e no interior de alguns
estados do Nordeste (Figura 5), (SOS Mata Atlântica, 2016; Zangaro e Moreira, 2010).
O bioma Mata Atlântica é um conjunto de ecossistemas de grande importância, que
incluem as faixas litorâneas do atlântico, com seus manguezais e restingas, florestas de
baixada e de encosta da serra do mar, florestas interioranas, as matas de araucária e os campos
de altitude, abrigando uma parcela significativa da diversidade biológica do Brasil e do
mundo (Renato et al., 2009). Segundo bioma mais ameaçado de extinção do mundo, a
Floresta Atlântica perde apenas para as quase extintas florestas da ilha de Madagascar na
costa de África (Campanili e Prochnow, 2006).
28%
61%
59%
89%
24%
4%
Amazônia
Caatinga
Cerrado
Mata Atlântica
Pantanal
Pampa
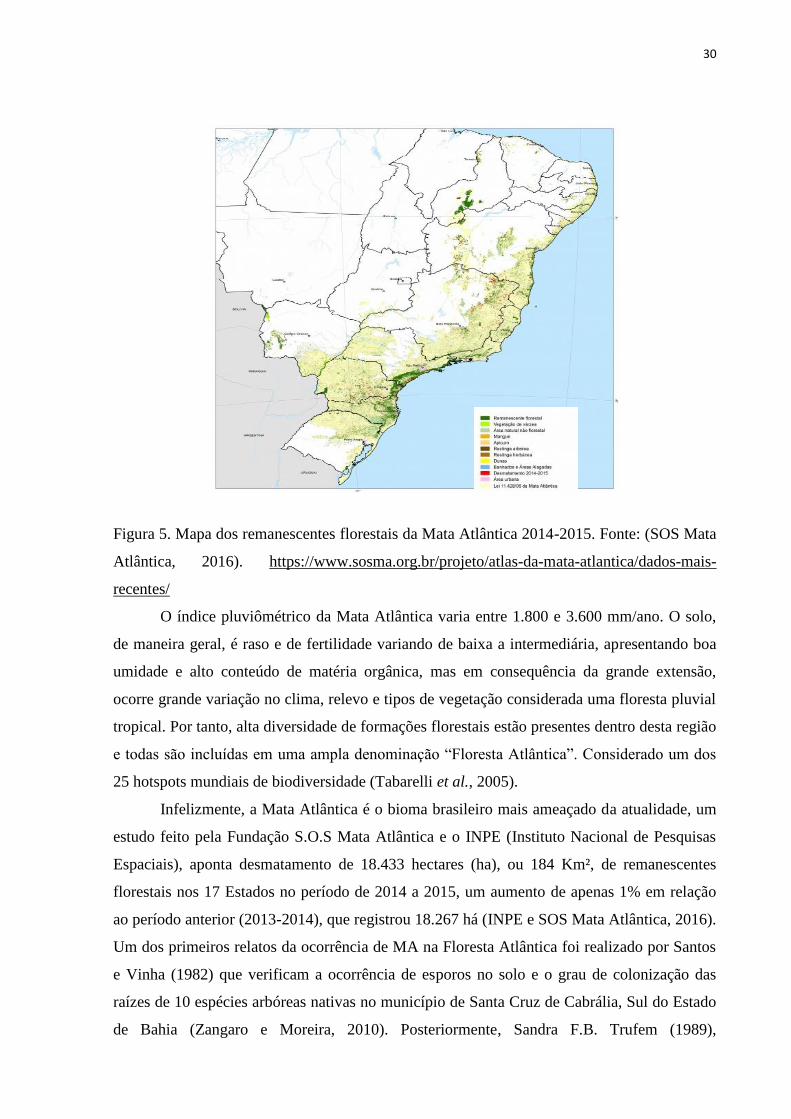
30
Figura 5. Mapa dos remanescentes florestais da Mata Atlântica 2014-2015. Fonte: (SOS Mata
Atlântica, 2016). https://www.sosma.org.br/projeto/atlas-da-mata-atlantica/dados-mais-
recentes/
O índice pluviômétrico da Mata Atlântica varia entre 1.800 e 3.600 mm/ano. O solo,
de maneira geral, é raso e de fertilidade variando de baixa a intermediária, apresentando boa
umidade e alto conteúdo de matéria orgânica, mas em consequência da grande extensão,
ocorre grande variação no clima, relevo e tipos de vegetação considerada uma floresta pluvial
tropical. Por tanto, alta diversidade de formações florestais estão presentes dentro desta região
e todas são incluídas em uma ampla denominação “Floresta Atlântica”. Considerado um dos
25 hotspots mundiais de biodiversidade (Tabarelli et al., 2005).
Infelizmente, a Mata Atlântica é o bioma brasileiro mais ameaçado da atualidade, um
estudo feito pela Fundação S.O.S Mata Atlântica e o INPE (Instituto Nacional de Pesquisas
Espaciais), aponta desmatamento de 18.433 hectares (ha), ou 184 Km², de remanescentes
florestais nos 17 Estados no período de 2014 a 2015, um aumento de apenas 1% em relação
ao período anterior (2013-2014), que registrou 18.267 há (INPE e SOS Mata Atlântica, 2016).
Um dos primeiros relatos da ocorrência de MA na Floresta Atlântica foi realizado por Santos
e Vinha (1982) que verificam a ocorrência de esporos no solo e o grau de colonização das
raízes de 10 espécies arbóreas nativas no município de Santa Cruz de Cabrália, Sul do Estado
de Bahia (Zangaro e Moreira, 2010). Posteriormente, Sandra F.B. Trufem (1989),

31
desenvolveu estudos em áreas de Mata Atlântica no litoral da Ilha do Cardoso, São Paulo, nas
investigações foram registrados um total de 46 espécies, distribuídos entre os gêneros
Acaulospora, Gigaspora, Glomus, Sclerocystis e Scutellospora (Trufem et al., 1989, 1994,
Trufem, 1990, 1995).
Em compilação de registros de espécies de FMA realizados por Zangaro e Moreira
(2010), a Mata Atlântica detinha 78 registros de FMA. Atualmente, o bioma é representado
por 136 espécies, distribuídas em 13 famílias e 27 gêneros, um aumento de 74% obtido em
pouco menos de uma década (Tabela 2). Além disso, a Mata Atlântica detém 87% de
representatividade das espécies que ocorrem no Brasil, um valor em concordância com o
esperado para um bioma que tem sido historicamente mais investigado. As famílias mais
representadas em número de espécies da Mata Atlântica corresponde a Acaulosporaceae e
Glomeraceae, com os gêneros Acaulospora e Glomus (Figura 6). Esse padrão tem sido
documentados em vários estudos de diversidade na Mata Atlântica (Aidar et al., 2004;
Carrenho et al., 2001; Moreira et al., 2009; Stürmer et al., 2006; Trufem et al., 1990).
Tabela 2. Espécies de FMA registradas na Mata Atlântica¹.
Família Espécie
Acaulosporaceae
Acaulospora bireticulata F.M. Rothwell &
Trappe
A. cavernata Błaszk.
A. colossica P.A. Schultz, Bever & J.B. Morton
A. delicata C. Walker, C.M. Pfeiffer & Bloss
A. denticulata C. Walker, C.M. Pfeiffer &
Bloss
A. elegans Trappe & Gerd.
A. excavata Ingleby & C. Walker
A. foveata Trappe & Janos
A. herrerae Furrazola,
B.T. Goto, G.A. Silva, Sieverd. & Oehl
A. ignota Błaszk., Góralska, Chwat & B.T.
Goto
Acaulospora koskei Błaszk.

32
A. lacunosa J.B. Morton
A. laevis Gerd. & Trappe
A. longula Spain & N.C. Schenck
A. mellea Spain & N.C. Schenck
A. minuta Oehl, Tchabi, Hount., Palenz., I.C.
Sánchez & G.A. Silva
A. morrowiae Spain & N.C. Schenck
A. myriocarpa (Spain, Sieverd. & N.C.
Schenck) Oehl, G.A. Silva, B.T. Goto &
Sieverd.
A. papillosa C.M.R. Pereira & Oehl
A. reducta Oehl, B.T. Goto & C.M.R. Pereira.
A. rehmii Sieverd. & S. Toro
A. rugosa J.B. Morton
A. scrobiculata Trappe
A. sieverdingii Oehl, Sýkorová & Błaszk.
A. spinosa C. Walker & Trappe
A. splendida Sieverd., Chaverri & I. Rojas
A. tuberculata Janos & Trappe
Kuklospora colombiana (Spain & N.C.
Schenck) Oehl & Sieverd.
Ambisporaceae Ambispora appendicula (Spain, Sieverd., N.C.
Schenck) C. Walker
A. gerdemannii (S.L. Rose, B.A. Daniels &
Trappe) C. Walker, Vestberg & A. Schüssler
A. fecundispora (N.C. Schenck &
G.S. Sm.) C. Walker, Vestberg & A. Schüssler
A. leptoticha (N.C. Schenck & T.H. Nicolson)
Walker, Vestberg & A. Schüssler
Archaeosporaceae Archaeospora trappei (R.N. Ames &
Linderman) J.B. Morton & D. Redecker
Dentiscutataceae Dentiscutata biornata (Spain, Sierverd. & S.
Toro

33
D. cerradensis (Spain & J. Miranda) Sieverd.,
F.A. de Souza & Oehl
D. colliculosa B.T. Goto & Oehl
D. hawaiiensis (Koske & Gemma) Sieverd.,
F.A. de Souza & Oehl
D. heterogama (T.H. Nicolson & Gerd.)
Sieverd., F.A. de Souza & Oehl
D. nigra (J.F. Readhead) Sieverd., F.A. de
Souza & Oehl
D. reticulata (Koske, D.D. Miller & C. Walker)
Sieverd., F.A. de Souza & Oehl
D. scutata (C. Walker & Dieder.) Sieverd.,
F.A. de Souza & Oehl
Fuscutata aurea (Oehl & Sieverd.) Błaszk.
Chwat, G.A. Silva & Oehl
F. heterogama Oehl, F.A. Souza. L.C. Maia &
Sieverd
F. rubra (Stürmer & J.B. Morton) Oehl, F.A.
de Souza & Sieverd.
F. savannicola (R.A. Herrera & Ferrer) Oehl,
F.A. de Souza & Sieverd.
Quatunica erythropa (Koske & C. Walker)
F.A. de Souza, Sieverd. & Oehl
Diversisporaceae Corymbiglomus globiferum (Koske & C.
Walker) Błaszk. & Chwat.
C. tortuosum (N.C. Schenck & G.S. Sm.)
Błaszk. & Chwat
Diversispora spurca (C.M. Pfeifer, C. Walker
& Bloss) C. Walker & Schüssler
D. trimurales (Koske & Halvorson) C. Walker
& A. Schüssler
D. versiformis (P. Karst.) Oehl, G.A. Silva &
Sieverd.

34
Redeckera fulva (Berk. & Broome) C. Walker
& A. Schüssler
Entrophosporaceae Entrophospora infrequens (I.R. Hall) R.N.
Ames & R.W. Schneid.
Claroideoglomus claroideum (N.C. Schenck &
G.S. Sm.) C. Walker & A. Schüssler
C. etunicatum (W.N. Becker & Gerd.) C.
Walker & A. Schüssler
Viscospora viscosa (T.H. Nicolson) Sieverd.,
Oehl & F.A. Souza
Gigasporaceae Gigaspora albida N.C. Schenck & G.S. Sm.
G. decipiens I.R. Hall & L.K. Abbott
G. gigantea (T.H. Nicholson & Gerd.) Gerd. &
Trappe
G. margarita W.N. Becker & I.R. Hall
G. ramisporophora Spain, Sieverd. & N.C.
Schenck
G. rosea T.H. Nicolson & N.C. Schenck
Glomeraceae Funneliformis geosporus (T.H. Nicolson &
Gerd.) C. Walker & A. Schüssler
F. halonatus (S.L. Rose & Trappe) Oehl, G.A.
Silva & Sieverd.
F. mosseae (T.H. Nicolson & Gerd.) C. Walker
& A. Schüssler
F. monosporus (Gerd. & Trappe) Oehl, G.A.
Silva & Sieverd.
Glomus ambisporum G.S. Sm. & N.C. Schenck
G. arborense McGee
G. australe (Berck.) S.M. Berch
G. botryoides Rothwell & Victor
G. brohultii Sieverd. & Herrera
G. claroideum Schenck & Smith
G. clavisporum (Trappe) R.T. Almeida & N.C.

35
Schenck
G. constrictum Trappe
G. diaphanum J.B. Morton & C. Walker
G. deserticola Trappe, Bloss & Menge
G. diaphanum Morton & Walker
G. etunicatum Becker & Gerdemam
G. formosanum C.G. Wu & Z.C. Chen
G. globiferum (Koske & C. Walker) Błaszk. &
Chwat
G. geosporum (Nicol. & Gerd.) C. Walker
G. glomerulatum Sieverd.
G. heterosporum G.S. Sm. & N.C. Schenck
G. invermaium Hall
G. macrocarpum Tul. & C. Tul.
G. maculosum D.D. Mill & C. Walker
G. microaggregatum Koske, Gemma & Olexia
G. microcarpum Tul. & C. Tul.
G. mosseae (Nicol. & Gerd.) Gerd. & Trappe
G. multicaule Gerd. & B.K. Bakshi
G. pallidum I.R. Hall
G. pansihalos S.M. Berch & Koske
G. pachycaule (C.G. Wu & Z.C. Chen)
Sieverd. & Oehl
G. reticulatum Bhattacharjee & Mukerji
G. rubiforme (Gerd. & Trappe) R.T. Almeida
& N.C. Schenck
G. sinuosum (Gerd. & B.K. Bakshi) R.T.
Almeida & N.C. Schenck
G. taiwanense (C.G. Wu & Z.C. Chen) R.T.
Almeida & N.C. Schenck ex Y.J. Yao
G. tenebrosum (Thaxt.) S.M. Berch
G. trufemii B.T. Goto, G.A. Silva & Oehl
G. vesiculiferum (Thaxt.) Gerd. & Trappe

36
Rhizoglomus aggregatum (N.C. Schenck &
G.S. Sm.) Sieverd., G.A. Silva & Oehl
R. clarum (T.H. Nicolson & N.C. Schenck)
Sieverd., G.A. Silva & Oehl
R. fasciculatum (Thaxt.) Sieverd., G.A. Silva &
Oehl
R. intraradices (N.C. Schenck & G.S. Sm.)
Sieverd., G.A. Silva & Oehl
R. invermaium (I.R. Hall) Sieverd., G.A. Silva
& Oehl
R. manihotis (R.H. Howeler, Sieverd. & N.C.
Schenck) Sieverd., G.A. Silva & Oehl
R. microaggregatum (Koske, Gemma & P.D.
Olexia) Sieverd., G.A. Silva & Oehl
R. natalense (Błaszk. Chwat & B.T. Goto)
Sieverd., G.A. Silva & Oehl
Sclerocystis coremioides Berk. & Broome
S. sinuosa Gerd. & B.K. Bakshi
Septoglomus constrictum (Trappe) Sieverd.,
G.A. Silva & Oehl
S. deserticola (Trappe, Bloss & J.A. Menge)
G.A. Silva, Oehl & Sieverd.
Simiglomus hoi (S.M. Berch & Trappe) G.A.
Silva, Oehl & Sieverd.
Intraornatosporaceae Intraornatopsora intraornata (B.T. Goto &
Oehl) B.T. Goto, Oehl & G.A. Silva
Paradentiscutata bahiana Oehl, Magna, B.T.
Goto & G.A. Silva
P. maritima B.T. Goto, D.K. Silva, Oehl &
G.A. Silva
Paraglomeraceae Paraglomus albidum (C. Walker & L.H.
Rhodes) Oehl, F.A. Souza, G.A. Silva &
Sieverd.

37
P. bolivianum (Sieverd. & Oehl) Oehl & G.A.
Silva
P. occultum (C. Walker) J.B. Morton & D.
Redecker
P. pernambucanum Oehl, C.M. Mello, Magna
& G.A. Silva
Racocetraceae Cetraspora gilmorei (Trappe & Gerd.) Oehl,
F.A. de Souza & Sieverd.
C. pellucida (T.H. Nicolson & N.C. Schenck)
Oehl, F.A. de Souza & Sieverd.
Racocetra castanea (C. Walker) Oehl, F.A. de
Souza & Sieverd.
R. coralloidea (Trappe, Gerd. & I. Ho) Oehl,
F.A. de Souza & Sieverd.
R. fulgida (Koske & C. Walker) Oehl, F.A. de
Souza & Sieverd.
R. gregaria (N.C. Schenck & T.H. Nicolson)
Oehl, F.A. de Souza & Sieverd.
R. persica (Koske & C. Walker) Oehl, F.A. de
Souza & Sieverd.
R. tropicana Oehl, B.T. Goto & G.A. Silva
R. verrucosa (Koske & C. Walker) Oehl, F.A.
de Souza & Sieverd.
R. weresubiae (Koske & C. Walker) Oehl, F.A.
de Souza & Sieverd.
Sacculosporaceae Sacculospora baltica (Błaszk., Madej &
Tadych) Oehl, Palenz., IC. Sánchez-Castro,
B.T. Goto, G.A. Silva & Sieverd.
Scutellosporaceae Orbispora pernambucana (Oehl, D.K. Silva, N.
Freitas, L.C. Maia) Oehl, G.A. Silva & D.K.
Silva
Scutellospora aurigloba (I.R. Hall) C.Walker
& F.E. Sanders
38
S. calospora (T.H. Nicolson & Gerd.) C.
Walker & F.E. Sanders
S. dipapillosa (C. Walker & Koske) C. Walker
& F.E. Sanders
S. dipurpurescens J.B. Morton & Koske
1. Lista baseada em registros obtidos por Souza et al., (2010), Dos Santos e Carrenho (2011),
Goto et al., (2012), Mello et al., (2012), Silva et al., (2012), Bonfim et al., (2013), Stürmer et
al., (2013), Novais et al., (2014), Silva et al., (2014), Pereira et al., (2014), Bonfim et al.,
(2015), Blaszkowski et al., (2015), Pereira et al., (2015), Silva et al., (2015), Camara et al.,
(2016), Jobim e Goto, (2016) e Pereira et al., (2016).
A Mata Atlântica é o bioma com o maior número de espécies novas descritas no
Brasil: Acaulospora reducta, A. endographis, A. herrerae, A. ignota, A. papillosa, Dentiscuta
colliculosa, Fuscutata aurea, Glomus trufemii, Intraspora intraornata, Orbispora
pernambucana, Paradentiscutata bahiana, P. maritima, Racocetra tropicana e Rhizoglomus
natalense são espécies descritas originalmente para o bioma (Błaszkowski, 2014;
Błaszkowski et al., 2015; Furrazola et al., 2013; Goto et al., 2010, 2011, 2012a, b, 2013;
Mello et al., 2012; Oehl et al., 2011d; Pereira et al., 2015; Pereira et al., 2016).
20%
3%0,70%
9%
4%
3%
4%
38%
2% 3%
7%
0,70%
5%Acaulosporaceae
Ambisporaceae
Archaeosporaceae
Dentiscutataceae
Diversisporaceae
Entrophosporaceae
Gigasporaceae
Glomeraceae
Intraornatosporaceae
Paraglomeraceae
Racocetraceae
Sacculosporaceae
Scutellosporaceae

39
Figura 6. Proporção de famílias de FMA que ocorrem na Mata Atlântica.
2.5. Fungos Micorrízicos no Rio Grande do Norte
O Rio Grande do Norte possui 41 espécies, 19 gêneros e 11 famílias registradas. (Lista
baseada em registros obtidos por Jobim e Goto, 2016, Błaszkowski et al. 2015, Silva et al.,
2014, Błaszkowski et al. 2014, Goto et al. 2012, Furrazola et al. 2011) (Ver tabela 3). Além
disso, apresenta poucos inventários taxonômicos de FMA, contudo, os estudos conduzido
nesse estado são registradas cinco espécies novas para a ciência (A. herrerae Furrazola, B.T.
Goto, G.A. Silva, Sieverd. & Oehl, A. ignota Błaszk., Góralska, Chwat & B.T. Goto, G.
trufemii B.T. Goto, G.A. Silva & Oehl, P. maritima B.T. Goto, D.K. Silva, Oehl & G.A. Silva
e R. natalense Błaszk., Chwat & B.T. Goto), além de uma nova família (Intraornatosporaceae)
e um novo gênero (Paradentiscutata), todas os referidos táxons descritos originalmente para
ecossistemas dunares sob influência marítima, revelando um cenário taxonomicamente
promissor.
Tabela 3. Espécies de FMA registradas em localidades de Mata Atlântica do Estado do
Rio Grande do Norte.
Família Espécie Área
Acaulosporaceae
Acaulospora cavernata Błaszk. PEDG¹
Acaulospora foveata Trappe & Janos BI², PEDN³,
PEDG
Acaulospora herrerae Furrazola, B.T. Goto,
G.A. Silva, Sieverd. & Oehl
PEDN
Acaulospora ignota Błaszk., Góralska, Chwat
& B.T. Goto
PEDN
Acaulospora lacunosa J.B. Morton PEDG
Acaulospora morrowiae Spain & N.C. Schenck BI, PEDG
Acaulospora scrobiculata Trappe. PEDG
Acaulospora spinosa C. Walker & Trappe BI
Ambisporaceae

40
Ambispora appendicula (Spain, Sieverd., &
N.C. Schenck) C. Walker
PEDN, PEDG
Ambispora gerdemanii (S.L. Rose, B.A.
Daniels & Trappe) C. Walker, Vestberg & A.
Schüssler
BI, PEDG
Racocetraceae
Cetraspora gilmorei (Trappe & Gerd.) Oehl,
F.A. de Souza & Sieverd.
BI, PEDG
Racocetra gregaria (N.C. Schenck & T.H.
Nicolson) Oehl, F.A. de Souza & Sieverd.
BI, PEDG
Racocetra verrucosa (Koske & C. Walker)
Oehl, F.A. de Souza & Sieverd.
BI
Racocetra weresubiae (Koske & C. Walker)
Oehl, F.A. de Souza & Sieverd.
PEDG
Entrophosporaceae
Claroideoglomus etunicatum (W.N. Becker &
Gerd.) C. Walker & A. Schüssler
BI
Diversisporaceae
Corymbiglomus tortuosum (N.C. Schenck &
G.S. Sm.) Błaszk. & Chwat
BI, PEDG
Dentiscutataceae
Dentiscutata biornata (Spain, Sieverd. & S.
Toro) Sieverd., F.A. de Souza & Oehl
PEDG
Dentiscutata colliculosa B.T. Goto & Oehl PEDG
Dentiscutata scutata Sieverd., F.A. Souza &
Oehl
PEDN
Fuscutata heterogama (T.H. Nicolson & Gerd.)
Sieverd., F.A. de Souza & Oehl
PEDG
Fuscutata rubra (Stürmer & J.B. Morton)
Oehl, F.A. de Souza & Sieverd.
BI, PEDG
Glomeraceae
Funneliformis halonatus (S.L. Rose & Trappe)
Oehl, G.A. Silva & Sieverd.
PEDN
41
Septoglomus furcatum Błaszk., Chwat &
Kovács & Ryszka
BI
Glomus australe (Berck.) S.M. Berch BI
Glomus coremioides (Berk. & Broome) D.
Redecker & J.B. Morton
PEDG
Glomus fulvum (Berk. & Broome) C. Walker &
A. Schüssler
PEDG
Glomus glomerulatum Sieverd. BI, PEDN,
PEDG
Glomus pachycaule (C.G. Wu & Z.C. Chen)
Sieverd. & Oehl
PEDG
Glomus sinuosum (Gerd. & B.K. Bakshi) R.T.
Almeida & N.C. Schenck
PEDG
Glomus taiwanensem (C.G. Wu & Z.C. Chen)
R.T. Almeida & N.C. Schenck ex Y.J. Yao
PEDG
Glomus trufemii B.T. Goto, G.A. Silva & Oehl PEDN, BI
Rhizoglomus natalense Błaszk., Chwat & B.T.
Goto
PEDN
Gigasporaceae
Gigaspora albida N.C. Schenck & G.S. Sm. BI, PEDG
Gigaspora decipiens I.R. Hall & L.K. Abbott BI, PEDG
Redeckera fulva (Berk. & Broome) C. Walker
& A. Schüssler
PEDG
Intraornatosporaceae
Intraornatospora intraornata (B.T. Goto &
Oehl) B.T. Goto, Oehl & G.A. Silva
PEDG
Sclerocystis coremioides Berk. & Broome PEDG
Sclerocystis sinuosa Gerd. & B.K. Bakshi PEDG
Paradentiscutata marítima B.T. Goto, D.K.
Silva, Oehl & G.A. Silva
BI, PEDG
Scutellosporaceae

42
Orbispora pernambucana (Oehl, D.K. Silva, N.
Freitas, L.C. Maia) Oehl, G.A.Silva & D.K.
Silva
BI, PEDG
Sacculosporaceae
Sacculospora baltica (Błaszk., Madej &
Tadych) Oehl, Palenzuela, I.C. Sánchez, B.T.
Goto, G.A. Silva & Sieverd.
PEDG
1. PEDG = Parque Ecológico Dunas de Genipabu; 2. BI = Barreira do Inferno; 3. PEDN =
Parque Estadual das Dunas de Natal.
2.6. Tabuleiros costeiros
Os tabuleiros costeiros apresentam feições geomorfológicas de superfície do tipo tabular,
dissecada por vales poucos profundos de encostas com forte declividade em forma de ¨U¨,
próximas ao oceano Atlântico. Estão relacionados com os sedimentos da Formação Barreiras
a deposição desses sedimentos ocorreu no período Terciário, sob um clima árido ou semiárido
e terminou durante o pleistoceno, com a mudança para um clima quente e úmido (SUGUIU et
al., 1985). Cuja área no Brasil é estimada em 20,0 milhões de hectares, que se distribuem
desde o Estado do Amapá até o Rio de Janeiro, constituindo uma unidade geoambiental
(Figura 7). O revelo em sua maioria é suave ondulados, ondulados ou até fortemente ondulada
com elevações de topos planos (chãs), com altitudes que variam de 20,0 a 220,0 metros, e
grande variacão climática e vegetativa. É considerada a maior ocorrência de formação de
sedimentos do período terciário (últimos 65 milhões de anos), dentro de um único país
(Jacomine, 1996; Rezende, 2000; Nascimento, 2001).
No Nordeste abrange uma área aproximada de 8.420.00 hectares. É formado de platôs
sedimentares, variando sua altitude entre 30 a 150 metros. Com relação ao relevo nota-se uma
variação entre vales estreitos e encostas íngremes ou vales abertos constituídos de encostas
suaves ou podendo apresentar-se fundo com amplas várzeas. A vegetação nativa é a Mata
Atlântica. De forma geral, os solos são de baixa fertilidade natural, possuindo pouca aptidão
para acúmulo de água, são ácidos, com baixo estoque de matéria orgânica, baixa capacidade
de troca catiônica, devido á presença de caulinita na fração argila estes solos contém alumínio
trocável e prevalece solos que apresenta saturação por alumínio igual ou maior que 50% e em
menor porcentagem apresentam-se também solos distróficos, ou seja solos que apresenta

43
saturação por bases e por alumínio menor que 50%. Verifica-se também que estes solos
apresentam baixos teores de fósforo e potássio e a matéria orgânica é quem fornece a maior
quantidade de nitrogênio do solo (Sobral et al., 2008).
Os principais solos do ecossistema dos tabuleiros (Latossolos Amarelos e Argissolos
Amarelos) caracterizam-se como profundos, ácidos, álicos, com baixa capacidade de troca
catiônica e presença de horizontes coesos (Jacomine, 1996; Ribeiro, 1998; Rezende, 2000;
Nascimento, 2001; e Lima et al., 2004). O termo “coeso” tem sido utilizado com significado
de tenaz (Rezende, 2000) para caracterizar horizontes minerais subsuperficiais do solo que
apresentam aumento de coesão entre as suas partículas, tornando-se duro, muito duro ou
extremamente duro quando seco e friável quando úmido (Jacomine, 1996). A densidade do
solo desse horizonte é maior que os horizontes subjacentes e não apresentam uma organização
estrutural visível, são maciços (Embrapa, 1999). Os solos com essas características são
denominados no Brasil como “solos coesos” ou “solos com horizontes coesos” (Lima et al,
2004).
Figura 7. Delimitação de tabuleiros costeiros no Brasil. Fonte: Embrapa (2014) modificado.
No atual Sistema Brasileiro de Classificação de Solos (SiBCS), o caráter coeso já foi
incluído como característica diagnóstica atribuída somente à classe dos Latossolos Amarelos
do 3º nível categórico (Embrapa, 1999). Embora o SiBCS considere o termo “coeso” somente
Tabuleiros Costeiros

44
para a classe dos Latossolos Amarelos, há ocorrências da presença desse comportamento em
horizontes subsuperficiais em Argissolos Acinzentados e Argissolos Amarelos (Jacomine,
2001). Encontram-se ainda, em menor expressão, Neossolos Quartzarênicos, Plintossolos e
Espodossolos (Jacomine, 1996; Nascimento, 2001; Jacomine, 2001; Ribeiro, 2001). Esses
solos apresentam disponibilidade de nutrientes limitada, com baixos valores de soma de bases
e de capacidade de troca catiônica, valores elevados de alumínio trocável e de saturação de
alumínio. Valores de capacidade de troca catiônica potencial CTC inferiores a 7,9 cmolc kg-1,
refletem a mineralogia caulinítica do material de origem dos solos (Ribeiro,1986; Jacomine,
1996; Silva; Lima Neto et al., 2009).
Segundo Cintra et al. (2004) os principais solos presentes nos tabuleiros costeiros são
os Latossolos Amarelos e Argissolos Amarelos, além das características anteriormente citadas
estes solos apresentam horizontes coesos. Esta denominação dar-se devido à necessidade de
se denominar o adensamento de horizontes superficiais associada a diferentes graus de
coesão, ou seja, aumento da coesão entre as partículas do solo tornando assim o solo mais
denso. Devido ao processo de erosão dos solos submetidos, estes horizontes podem aparecer
próximos a superfície, geralmente nos primeiros 10 a 20 cm, já em solos sob floresta nativa,
estes horizontes estão localizados em profundidades diferenciadas, normalmente coincidindo-
se com os horizontes AB e/ou BA. Nestes solos a umidade é um fator relevante visto que
quando secos eles possuem uma consistência dura e quando úmido apresenta uma
consistência friável.
A origem dos horizontes coesos ainda é questionável, segundo Pacheco (2010) esta
origem pode estar relacionada a diversos fatores, como a presença de compostos orgânicos
pouco polimerizados, perda de argilas para camadas superiores, presença de ferro e argila nos
micrósporos, utilização intensa de cultivo, mudanças sucessivas de ciclos de umedecimento e
secagem. Outros fatores contribuem para a formação destes horizontes a exemplo dos
processos geomorfológicos. Também pode influenciar neste processo a obstrução dos
micrósporos do solo pelo deslocamento de máteria leve para horizonte inferiores. Esta coesão
produz efeito negativo no fluxo de ar, água, nutrientes, temperatura, microorganismos e no
desenvolvimento radicular devido á diminuição do diâmetro dos poros do solo, diminuindo-se
assim a produção vegetal.
Lima Neto et al., (2010) pesquisando os atributos químicos, minerológicos e
micromorfólogicos de horizontes coesos em tabuleiros verificaram que a gênese dos
horizontes coesos não estão relacionados aos agentes cimentantes, segundo estes autores a sua
gênese apresenta duas fases distintas, a iluviação de argila fina, provocando o estupimento dos

45
poros do solo e em seguida perda de ferro na parte superior fazendo com que a estructura
entre em colapso.
3.OBJETIVOS
3.1. Objetivo Geral
Avaliar a diversidade de um complexo de tabuleiros costeiro do dominio Mata Atlântica do
Rio Grande do Norte, a fim de ampliar o conhecimento sobre a distribuição deste grupo de
organismos.
3.2. Objetivos específicos
Coletar especimes no período chuvoso e estiagem;
Identificar e inventariar espécies de FMA na área escolhida;
Avaliar abundancia de glomerosporos de FMA no solo
Comparar a composição de espécies de FMA entre o período chuvoso e estiagem;
Avaliar a Riqueza de espécies de FMA entre as duas estações (chuva e estiagem);
4. MATERIAIS E MÉTODOS
4.1. Área de estudo
O presente estudo foi realizado na Unidade de Conservação (UC) do Estado do Rio
Grande do Norte: o Santuário Ecológico de Pipa (SEP).
O SEP foi criado em 1986, e trata-se de, uma área particular, localizada na Praia de Pipa,
entre as coordenadas (06°11’00’’- 06°17’30’’S e 35°17’30’’- 35°12’30’’W), com altitude
aproximada de 30 metros (BRASIL, 1985). O SEP está localizado no município de Tibau do
Sul, Litoral Sul do Estado, ocupa aproximadamente 60 ha., sendo constituído de elementos
representativos da Mata Atlântica. A área apresenta clima tropical úmido (As’), seguindo o
sistema de classificação de Köppen (1948), caracterizando-se por chuvas de inverno e verão
seco. A precipitação média anual é de 1500 mm (Silva, 1997), a estação seca ocorre entre os
meses de agosto a janeiro e a estação chuvosa ocorre entre os meses fevereiro a julho
(Jacomine et al., 1971). A cobertura vegetal é composta por floresta de tabuleiro (em seu
estado original), no entanto várias formações vegetais podem ser encontradas na área como as
restingas e dunas, ecossistemas costeiros estão incluidas no Bioma Mata Atlântica, ocupando

46
cerca de 80% da costa brasileira (Lacerda et al., 1993). O Santuário é um posto Avançado da
Reserva da Biosfera da Mata Atlântica, título concedido pela UNESCO em 1994 (Moraes,
2016). O SEP estão descritas 168 espécies de plantas, distribuídas em 62 famílias. Dentre as famílias
mais representativas, quanto ao número de espécies, destacaram-se Cyperaceae (12 spp.), seguida de
Myrtaceae e Poaceae (11 spp.), Asteraceae (10 spp.), Fabaceae e Rubiaceae (8 spp.), Caesalpiniaceae e
Malpighiaceae (6 spp.), Boraginaceae e Euphorbiaceae (5 spp.), Polygonaceae, Sapindaceae e
Verbenaceae (4 spp.), Apocynaceae, Chrysobalanaceae, Mimosaceae e Passifloraceae (3 spp.),
contribuindo com 63,09% do total das espécies (Almeida et al., 2006).
4.2. Obtenção das amostras
As amostras do solo foram coletadas durante duas épocas do ano: período chuvoso e
de estiagem (Junho e Outubro de 2015). As amostras foram coletadas selecionando libremente
cinco pontos: (placa 2, mirante prainha, mirante das tartarugas, mirante dos golfinhos e trilha
da peroba) (Figura 8). 10 amostras coletadas no ponto Placa 2, mirante prainha, ponto mirante
das tartarugas e mirante dos golfinhos e outras 10 amostras foram coletadas na trilha da
peroba. Totalizando 20 amostras para cada período de coleta retiradas na profundidade de 0-
20 cm. Das 20 amostras coletadas apenas foram tratadas 9 amostras de solo neste estudo (3
são do período chuvoso e 6 do período de estiagem). As amostras obtidas foram
acondicionadas em sacos plásticos e transportadas para o Laboratório de Biologia de
Micorrízas da Universidade Federal do Rio Grande do Norte (UFRN), onde foram mantidos a
temperatura ambiente de (26°C), para avaliação da comunidade de FMA. Parte do material
coletado foi encaminhada para a EMPARN para realização de análises físico-químicas (macro
e micronutrientes).

47
Figura 8. Pontos de coletas no Santuário Ecológico de Pipa, litoral sul do RN.
4.3. Avaliação para extração de glomerosporos
Os glomerosporos foram extraídos a partir de 50g de solo rizosférico de campo pela
técnica de peneiramento úmido (Gerdemann and Nicolson, 1963). E centrifugação em água e
sacarose (50%) (Jenkins, 1964) utilizando peneiras com abertura de 710,150 e 63 μm (Figuras
9-12).
Fonte: http://glomeromycota.wixsite.com
4.4. Identificação taxonômica
Após a extração, os glomerosporos foram separados em morfotipos (tamanho e cor)
com auxílio de estereomicroscópio e montadas em lâminas com PVLG (ácido polivinílico
lacto-glicerol) e PVLG + reagente de Melzer, para posteriormente visualização no
Figura 10. Peneiramento úmido
(Gerdemann & Nicolson, 1963).
Figura 9. Mostras de solo (50g).
Figura 11. Centrifugação em água e
sacarose (50%), (Jenkins, 1964).
Figura 12. Isolamento dos glomerosporos

48
microscópio óptico e avaliação taxonômica. Para a identificação das espécies de FMA foram
consultados manuais de identificação e chaves dicotômicas proposta por Schenck e Pérez
(1990), Goto (2009), Błaszkowski (2012) e confronto com descrições disponíveis em
coleções internacionais (http://www.zor.zut.edu.pl/Glomeromycota/, http:invam.wvu.edu/ e
http://glomeromycota.wix.com/lbmicorrizas) e demais literaturas pertinentes (Figura 13 e 14),
considerando a classificação proposta por Oehl et al., (2011a) e táxons adicionais propostos
por Błaszkowski (2012, 2014), Goto et al., (2012), Marinho et al., (2014) e Oehl et al.,
(2014).
4.5. Análises Ecológicas
Após a identificação de todas as espécies da comunidades de FMA forão avaliadas em
termos quantitativos e qualitativos a partir de dados populacionais (abundância e frequência
de ocorrência) e sua estruturação analisada por meio de índices ecológicos (riqueza,
dominância, diversidade e equitatividade). Forão determinados: 1) a riqueza de espécies (R),
avaliada pela razão entre o número de espécies presentes em 50g de solo e o número de
amostras, 2) o índice da dominância de Simpson (Is) [Is = 1-L], onde L = ni (n-1) / N (N-1),
ni = número de esporos de cada espécie de FMA “i”, N = número total de espécies de fungos
micorrízicos, e 3) o índice de diversidade se Shannon (H) [H = - (pi log pi), onde pi = ni/N;
ni = número de cada espécie de FMA, N = número total de espécies e 4) equitabilidade de
Polieu (Jʼ= Hʼ/ log2S) foram obtidos com auxílio do software PAST versão 2.17 (Hammer et
al., 2001).
Figura 13. Montagem de lâminas com
PVLG e PVLG + Melzer
Figura 14. Avaliação taxonômica

49
5. RESULTADOS E DISCUSSÃO
Foram identificadas 47 espécies de FMA (30 no período chuvoso e 26 no de estiagem
e apenas 9 espécies ocorrendo em ambas estações), distribuídas em 9 famílias e 17 gêneros
(Tabela 4). Das aproximadamente 300 espécies, atualmente reconhecidos para o filo, os
tabuleiros costeiros do presente trabalho apresenta 16%, das espécies, reconhecidos
globalmente, assim como 30%, daqueles citados para o Brasil. No entanto, das 136 espécies,
registradas no bioma Mata Atlântica, 35% das espécies, são representados em tabuleiros
costeiros do presente trabalho, respectivamente as 125 espécies, reportados para o Nordeste
representa 38%, das espécies, distribuídos entre as cinco ordens do filo Glomeromycota.
Gigasporales, Glomerales e Diversisporales constituem as ordens mais representadas em
tabuleiros costeiros de remanescentes de Mata Atlântica, no entanto Archaeosporales e
Paraglomerales têm sido pouco documentadas. Para o Brasil, foram acrescidos o registro de
Glomus spinuliferum Sieverd. & Oehl, aumentando para 157 o número de espécie registradas
no país e 137 para o bioma Mata Atlântica. Além disso, 10 espécies encontradas são
potencialmente novas para a ciência.
Tabela 4. Ocorrência de espécies de FMA em remanescente de Mata Atlântica de tabuleiros
costeiros do Santuário Ecológico de Pipa no período chuvoso e de estiagem.
Espécie
Período
Chuvoso Estiagem
Ambisporaceae
Ambispora
Ambispora callosa (Sieverd.) C. Walker, Vestberg
& A. Schüssler
X
³Ambispora reticulata Oehl & Sieverd. X X
Ambispora sp.nov.1 X
Acaulosporaceae
Acaulospora

50
Acaulospora cavernata Błaszk. X
Acaulospora aff. herrerae Furrazola, B.T.Goto,
G.A.Silva, Sieverd. & Oehl
X
Acaulospora reducta Oehl, B.T. Goto & C.M.R.
Pereira
X
Acaulospora aff. tortuosa Palenz., Oehl, Azcon-
Aguilar & G.A.Silva
X
Acaulospora foveata Trappe & Janos X
³Acaulospora herrerae Furrazola, B.T. Goto, G.A.
Silva, Sieverd. & Oehl
X X
Acaulospora ignota Błaszk., Góralska, Chwat &
Goto
X
Acaulospora scrobiculata Trappe X
Acaulospora sp. nov.2 X
Dentiscutataceae
Dentiscutata
³Dentiscutata sp. nov. 3 X X
Fuscutata
Fuscutata rubra (Stürmer & J.B. Morton) Oehl,
F.A. de Souza & Sieverd.
X
Fuscutata aurea Oehl, C.M. Mello & G.A. Silva X
Fuscutata heterogama Oehl, F.A. Souza, L.C.
Maia & Sieverd.
X
Diversisporaceae
Diversispora
³Diversispora aff. peridiata Błaszk., Chwat,
Kovács & Góralska
X X
Diversispora aff. varaderana Błaszk., Chwat,
Kovács & Góralska
X
Diversispora sp. nov. 4 X
Corymbiglomus
Corymbiglomus sp. nov. 5 X
Racocetraceae

51
Cetraspora
Cetraspora gilmorei (Trappe & Gerd.) Oehl, F.A.
de Souza & Sieverd.
X
Redeckera
Redeckera fulva (Berk. & Broome) C. Walker &
A. Schüssler
X
Entrophosporaceae
Claroideoglomus
Claroideoglomus aff. etunicatum (W.N. Becker &
Gerd.) C. Walker & A. Schüssler
X
Claroideoglomus hanlinii Błaszk., Chwat &
Góralska
X
Claroideoglomus lamellosum (Dalpé, Koske &
Tews) C. Walker & A. Schüssler
X
Claroideoglomus etunicatum (W.N. Becker &
Gerd.) C. Walker & A. Schüssler
X
Gigasporaceae
Gigaspora
Gigaspora gigantea (T.H. Nicholson & Gerd.)
Gerd. & Trappe
X
Gigaspora decipiens I.R. Hall & L.K. Abbott X
³Gigaspora sp. nov. 6 X X
Glomeraceae
Funneliformis
Funneliformis halonatum (S.L. Rose & Trappe)
Oehl, G.A. Silva & Sieverd.
X
Glomus
Glomus brohultii R.A. Herrera, Ferrer & Sieverd. X
³Glomus glomerulatum Sieverd. X X
Glomus sinuosum (Gerd. & B.K. Bakshi) R.T.
Almeida & N.C. Schenck
X
²ʼ³Glomus spinuliferum Sieverd. & Oehl X X
Glomus trufemii B.T. Goto, G.A. Silva & Oehl X

52
Glomus sp. nov. 7 X
³Glomus aff. brohultii Sieverd. & Herrera X X
³Glomus aff. trufemii B.T. Goto, G. A. Silva &
Oehl
X X
Rhizoglomus
Rhizoglomus aggregatum (N.C. Schenck & G.S.
Sm.) Sieverd., G.A. Silva & Oehl
X
Rhizoglomus aff. aggregatum (N.C. Schenck &
G.S. Sm.) Sieverd., G.A. Silva & Oehl
X
Rhizoglomus clarum (T.H. Nicolson & N.C.
Schenck) Sieverd., G.A. Silva & Oehl
X
Septoglomus
Septoglomus titan B.T. Goto & G.A. Silva X
Septoglomus sp. nov. 8 X
Simiglomus
Simiglomus sp. nov. 9 X
Scutellosporaceae
Orbispora
Orbispora pernambucana (Oehl, D.K. Silva, N.
Freitas & L.C. Maia) Oehl, G.A. Silva & D.K.
Silva
X
Scutellospora
Scutellospora sp. nov. 10 X
Scutellospora calospora (T.H. Nicolson & Gerd.)
C. Walker & F.E. Sanders
X
1-10. São espécies novas para a ciência; ²novo registro para o Brasil e para o bioma Mata
Atlântica; e ³espécies que se repetem nos dois períodos.
Enquanto que o RN com apenas 41 espécies registradas até o momento, foi
acrescentado 14 espécies, permitindo o aumento desse número para 55 espécies com os
registros das espécies, Ambispora callosa, A. reticula, Acaulospora reducta, F. aurea, R.
fulva, C. hanlinii, C. lamellosum, G. gigantea, G. brohultii, G. spinuliferum, R. aggregatum,
R. clarum, R. titan e S. calospora, correspodem a novos registros para o estado. Esse fato
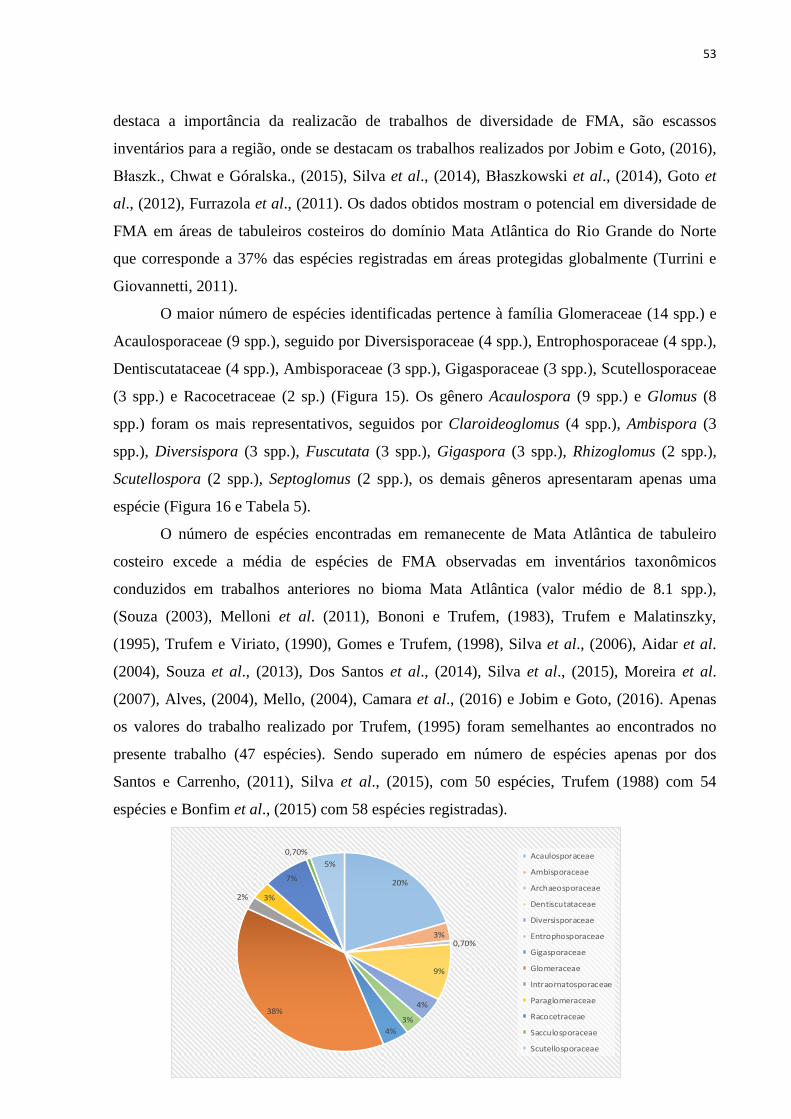
53
destaca a importância da realizacão de trabalhos de diversidade de FMA, são escassos
inventários para a região, onde se destacam os trabalhos realizados por Jobim e Goto, (2016),
Błaszk., Chwat e Góralska., (2015), Silva et al., (2014), Błaszkowski et al., (2014), Goto et
al., (2012), Furrazola et al., (2011). Os dados obtidos mostram o potencial em diversidade de
FMA em áreas de tabuleiros costeiros do domínio Mata Atlântica do Rio Grande do Norte
que corresponde a 37% das espécies registradas em áreas protegidas globalmente (Turrini e
Giovannetti, 2011).
O maior número de espécies identificadas pertence à família Glomeraceae (14 spp.) e
Acaulosporaceae (9 spp.), seguido por Diversisporaceae (4 spp.), Entrophosporaceae (4 spp.),
Dentiscutataceae (4 spp.), Ambisporaceae (3 spp.), Gigasporaceae (3 spp.), Scutellosporaceae
(3 spp.) e Racocetraceae (2 sp.) (Figura 15). Os gênero Acaulospora (9 spp.) e Glomus (8
spp.) foram os mais representativos, seguidos por Claroideoglomus (4 spp.), Ambispora (3
spp.), Diversispora (3 spp.), Fuscutata (3 spp.), Gigaspora (3 spp.), Rhizoglomus (2 spp.),
Scutellospora (2 spp.), Septoglomus (2 spp.), os demais gêneros apresentaram apenas uma
espécie (Figura 16 e Tabela 5).
O número de espécies encontradas em remanecente de Mata Atlântica de tabuleiro
costeiro excede a média de espécies de FMA observadas em inventários taxonômicos
conduzidos em trabalhos anteriores no bioma Mata Atlântica (valor médio de 8.1 spp.),
(Souza (2003), Melloni et al. (2011), Bononi e Trufem, (1983), Trufem e Malatinszky,
(1995), Trufem e Viriato, (1990), Gomes e Trufem, (1998), Silva et al., (2006), Aidar et al.
(2004), Souza et al., (2013), Dos Santos et al., (2014), Silva et al., (2015), Moreira et al.
(2007), Alves, (2004), Mello, (2004), Camara et al., (2016) e Jobim e Goto, (2016). Apenas
os valores do trabalho realizado por Trufem, (1995) foram semelhantes ao encontrados no
presente trabalho (47 espécies). Sendo superado em número de espécies apenas por dos
Santos e Carrenho, (2011), Silva et al., (2015), com 50 espécies, Trufem (1988) com 54
espécies e Bonfim et al., (2015) com 58 espécies registradas).
20%
3%0,70%
9%
4%
3%
4%
38%
2% 3%
7%
0,70%
5%Acaulosporaceae
Ambisporaceae
Archaeosporaceae
Dentiscutataceae
Diversisporaceae
Entrophosporaceae
Gigasporaceae
Glomeraceae
Intraornatosporaceae
Paraglomeraceae
Racocetraceae
Sacculosporaceae
Scutellosporaceae
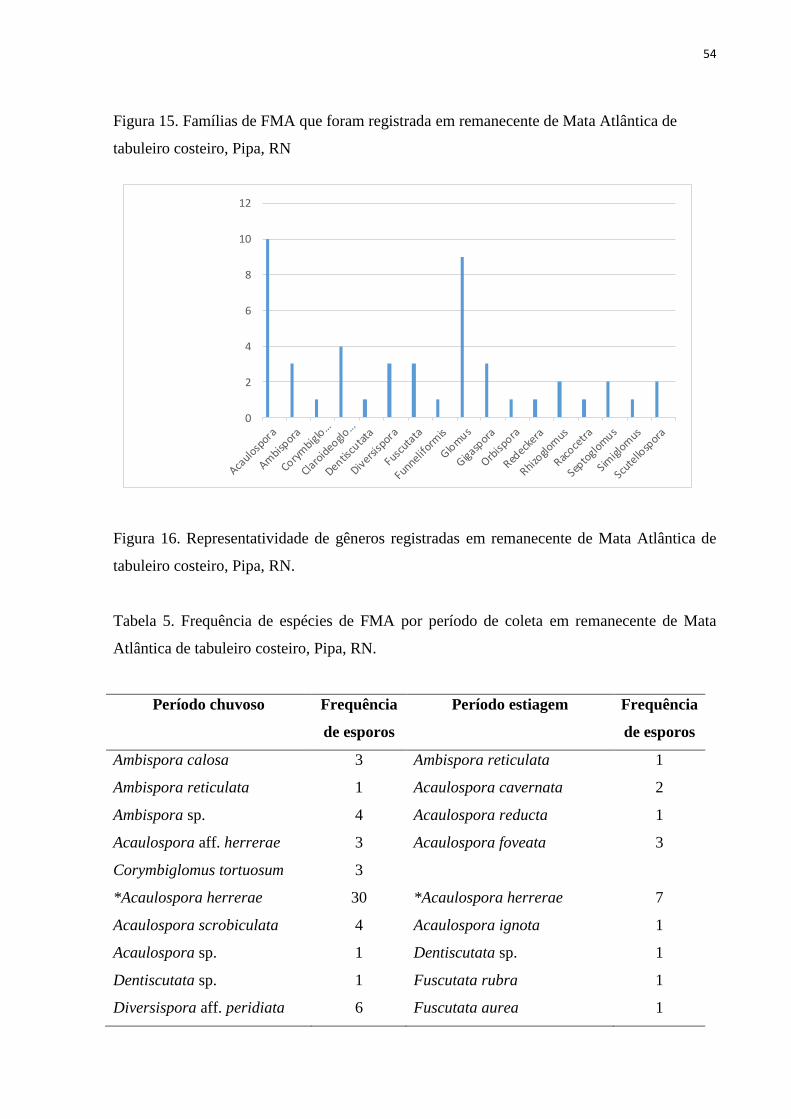
54
Figura 15. Famílias de FMA que foram registrada em remanecente de Mata Atlântica de
tabuleiro costeiro, Pipa, RN
Figura 16. Representatividade de gêneros registradas em remanecente de Mata Atlântica de
tabuleiro costeiro, Pipa, RN.
Tabela 5. Frequência de espécies de FMA por período de coleta em remanecente de Mata
Atlântica de tabuleiro costeiro, Pipa, RN.
Período chuvoso Frequência
de esporos
Período estiagem Frequência
de esporos
Ambispora calosa 3 Ambispora reticulata 1
Ambispora reticulata 1 Acaulospora cavernata 2
Ambispora sp. 4 Acaulospora reducta 1
Acaulospora aff. herrerae 3 Acaulospora foveata 3
Corymbiglomus tortuosum 3
*Acaulospora herrerae 30 *Acaulospora herrerae 7
Acaulospora scrobiculata 4 Acaulospora ignota 1
Acaulospora sp. 1 Dentiscutata sp. 1
Dentiscutata sp. 1 Fuscutata rubra 1
Diversispora aff. peridiata 6 Fuscutata aurea 1
0
2
4
6
8
10
12

55
*Diversispora aff.
varaderana
10 Fuscutata heterogama 1
Diversispora sp. 6 *Diversispora aff. peridiata 17
Corymbiglomus sp. 1 Cetraspora gilmorei 1
Claroideoglomus aff.
etunicatum
1 *Claroideoglomus aff.
hanlinii
16
Gigaspora sp. 2 Claroideoglomus aff.
lamellosum
4
Funneliformis halonatum 2 Claroideoglomus etunicatum 1
Glomus brohultii 1 Gigaspora gigantea 1
Glomus glomerulatum 4 Gigaspora decipiens 1
Glomus sinuosum 1 Gigaspora sp. 4
*Glomus spinuliferum 16 *Glomus glomerulatum 86
Glomus trufemii 4 Glomus spinuliferum 1
Glomus sp. 1 *Glomus aff. brohultii 36
Glomus brohultii 2 *Glomus aff. trufemii 11
*Glomus aff. trufemii 56 Rizoglomus aff. aggregatum 19
*Rhizoglomus aggregatum 25 Orbispora pernambucana 1
Rhizoglomus clarum 1 Septoglomus sp. 1
Redeckera fulva 1 *Simiglomus sp. 102
Septoglomus titan 1
Scutellospora sp. 1
Scutellospora calospora 1
*Espécies mais frequentes.
De acordo com os dados obtidos as espécies mais frequentes no período chuvoso
foram Glomus aff. trufemii com (56 ocorrências), seguido por Acaulospora herrerae (30),
Rhizoglomus aggregatum (25), Glomus spinuliferum (16), Diversispora aff. varaderana (10),
no entanto Diversispora aff. peridiata e Diversispora sp. nov. apresentam (6), Ambispora aff.
callosa, Acaulospora aff. herrerae e Acaulospora aff. tortuosa apresentam (3), Gigaspora sp.
nov., Funneliformis halonatum, Glomus aff. brohultii apresentam (2), as seguintes espécies
ocorreram apenas uma vez: Ambispora reticulata, Acaulospora sp. nov., Dentiscutata sp.
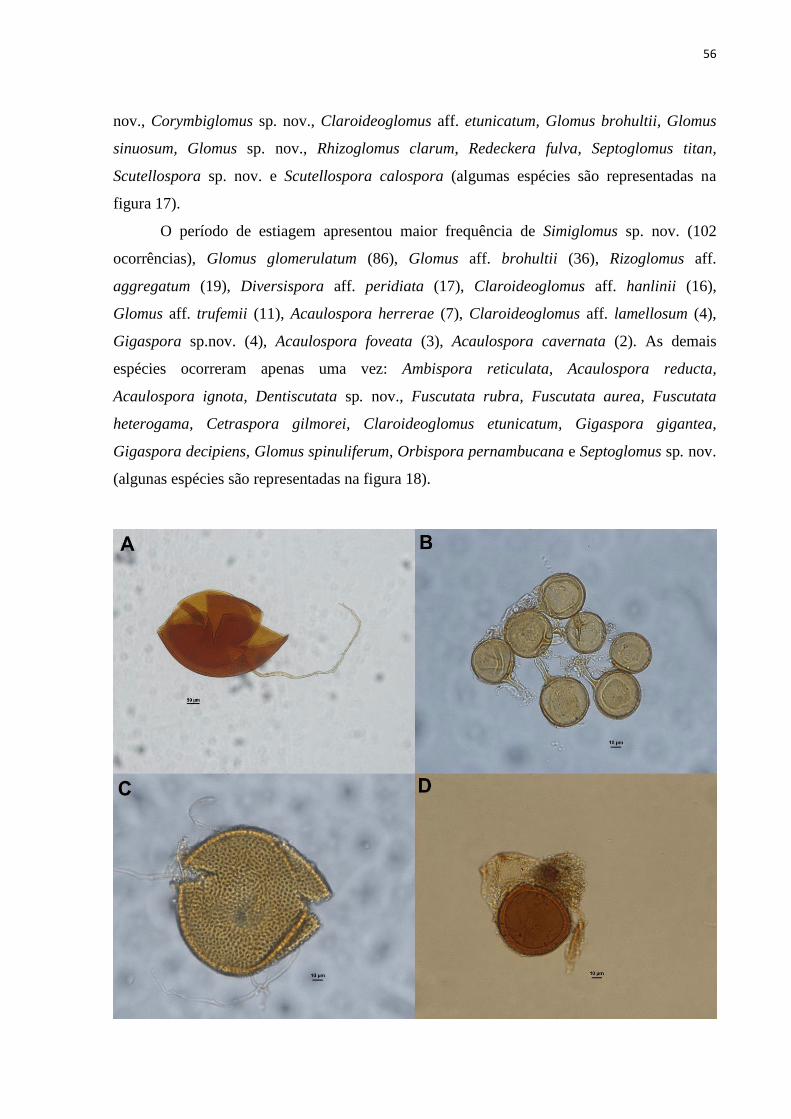
56
nov., Corymbiglomus sp. nov., Claroideoglomus aff. etunicatum, Glomus brohultii, Glomus
sinuosum, Glomus sp. nov., Rhizoglomus clarum, Redeckera fulva, Septoglomus titan,
Scutellospora sp. nov. e Scutellospora calospora (algumas espécies são representadas na
figura 17).
O período de estiagem apresentou maior frequência de Simiglomus sp. nov. (102
ocorrências), Glomus glomerulatum (86), Glomus aff. brohultii (36), Rizoglomus aff.
aggregatum (19), Diversispora aff. peridiata (17), Claroideoglomus aff. hanlinii (16),
Glomus aff. trufemii (11), Acaulospora herrerae (7), Claroideoglomus aff. lamellosum (4),
Gigaspora sp.nov. (4), Acaulospora foveata (3), Acaulospora cavernata (2). As demais
espécies ocorreram apenas uma vez: Ambispora reticulata, Acaulospora reducta,
Acaulospora ignota, Dentiscutata sp. nov., Fuscutata rubra, Fuscutata aurea, Fuscutata
heterogama, Cetraspora gilmorei, Claroideoglomus etunicatum, Gigaspora gigantea,
Gigaspora decipiens, Glomus spinuliferum, Orbispora pernambucana e Septoglomus sp. nov.
(algunas espécies são representadas na figura 18).

57
Figura 17. A-D. Espécies de FMA encontradas no período chuvoso em tabuleiro costeiro do
SEP. A) Gigaspora sp., B) Rhizoglomus aggregatum, C) Acaulospora scrobiculata e D)
Glomus trufemii.
Figura 18. A-D. Espécies de FMA encontradas no período de estiagem em tabuleiro costeiro
do SEP. A) Orbispora pernambucana, B) Glomus glomerulatum, C) Dentiscutata sp. e D)
Acaulospora cavernata.
De forma geral, espécies dos gêneros Glomus e Acaulospora são mais comuns tanto
em ambientes naturais como manejados (Aidar et al., 2004; Jefwa et al., 2012; Oehl et al.,
2003). Os gêneros Glomus e Acaulospora têm sido detectados como frequentes na área
estudadas de Mata Atlântica, ocorrendo com maior densidade de esporos e números de
espécies em relação aos demais gêneros (Tabela 6 e 7). Já que isso é decorrente do maior

58
número de espécies descritas para esses gêneros e da disseminação e adaptabilidade dessas
espécies (Daniell et al., 2001). Nos solos estudados espécies de Acaulospora e Glomus
ocorreram com maior freqüência, fato também relatado para outros ambientes de floresta
tropical (Guadarrama & Alvaréz-Sanchéz, 1999; Zhaon et al., 2001; Muthukumar et al.,
2003).
As espécies de Acaulospora apresentam melhor estabelecimento em solos com faixas
de pH ácido (Sieverding, 1991), como o das regiões tropicais. Algumas espécies de Glomus
parecem ser afetadas pelo tipo de uso do solo, como as esporocárpicas (Tchabi et al., 2008).
Espécies do gênero Glomus parecem pouco competitivas por não apresentarem agressividade
na colonização de determinadas áreas, em decorrência da dormência prolongada dos esporos
(Santos e Carrenho, 2011).
Segundo Carrenho (1998), estes gêneros Acaulospora e Glomus apresentam maior
capacidade de adaptação a solos submetidos a diferentes variações nos teores de matéria
orgânica, calagem, textura, entre outros fatores, demostrando que as espécies são tolerantes a
perturbações ambientais. A ordem Gigasporales podem dominar a comunidade nos ambientes
que sofrem grandes impactos e são naturalmente estressados, como na vegetação das dunas e
restingas.
Tabela 6. Ocorrência de espécies de FMA em remanecente de Mata Atlântica de tabuleiro
costeiro por amostras no período chuvoso.
Chuva 1P01 1P02 1P03
Ambispora callosa 2 1 0
Acaulospora aff. herrerae 1 1 1
Acaulospora sp. 1 0 0
Acaulospora herrerae 2 1 27
Corymbiglomus sp. 1 0 0
Diversispora aff. varaderana 10 1 0
Diversispora sp. 6 0 0
Diversispora aff. peridiata 2 4 0
Funneliformis halonatum 1 0 2
Gigaspora sp. 1 0 1
Glomus sp. 1 0 0
Glomus brohultii 1 0 0
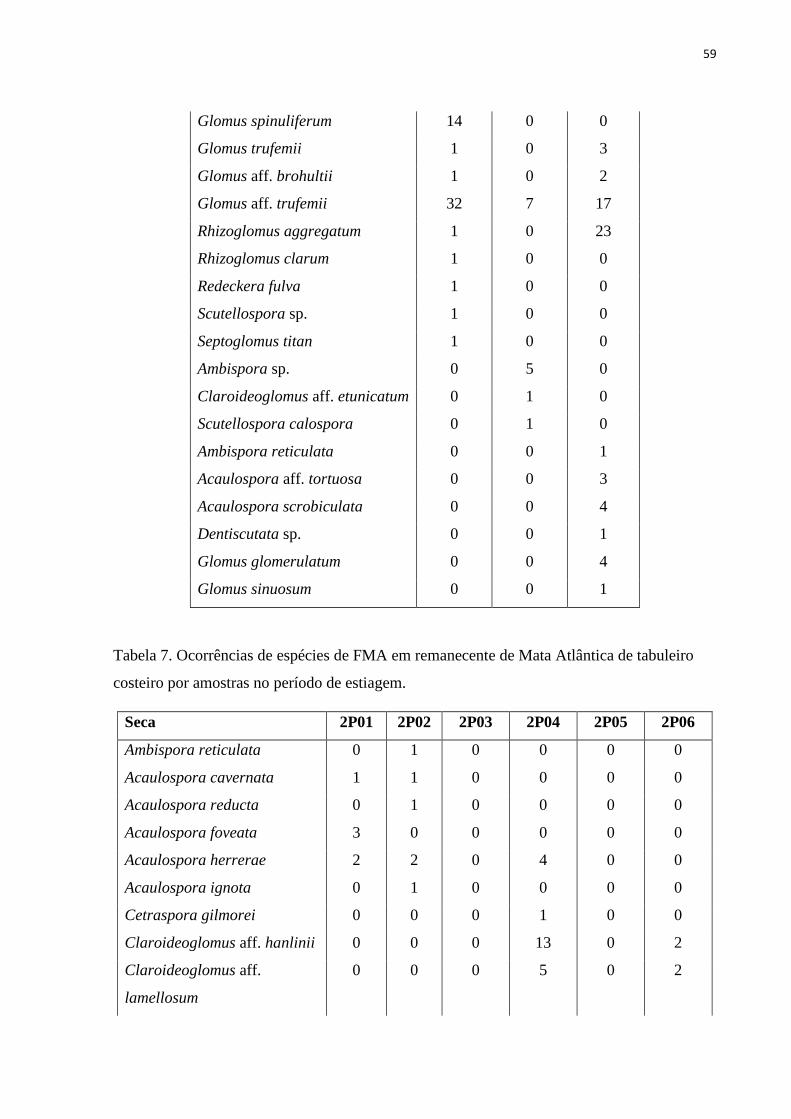
59
Glomus spinuliferum 14 0 0
Glomus trufemii 1 0 3
Glomus aff. brohultii 1 0 2
Glomus aff. trufemii 32 7 17
Rhizoglomus aggregatum 1 0 23
Rhizoglomus clarum 1 0 0
Redeckera fulva 1 0 0
Scutellospora sp. 1 0 0
Septoglomus titan 1 0 0
Ambispora sp. 0 5 0
Claroideoglomus aff. etunicatum 0 1 0
Scutellospora calospora 0 1 0
Ambispora reticulata 0 0 1
Acaulospora aff. tortuosa 0 0 3
Acaulospora scrobiculata 0 0 4
Dentiscutata sp. 0 0 1
Glomus glomerulatum 0 0 4
Glomus sinuosum 0 0 1
Tabela 7. Ocorrências de espécies de FMA em remanecente de Mata Atlântica de tabuleiro
costeiro por amostras no período de estiagem.
Seca 2P01 2P02 2P03 2P04 2P05 2P06
Ambispora reticulata 0 1 0 0 0 0
Acaulospora cavernata 1 1 0 0 0 0
Acaulospora reducta 0 1 0 0 0 0
Acaulospora foveata 3 0 0 0 0 0
Acaulospora herrerae 2 2 0 4 0 0
Acaulospora ignota 0 1 0 0 0 0
Cetraspora gilmorei 0 0 0 1 0 0
Claroideoglomus aff. hanlinii 0 0 0 13 0 2
Claroideoglomus aff.
lamellosum
0 0 0 5 0 2
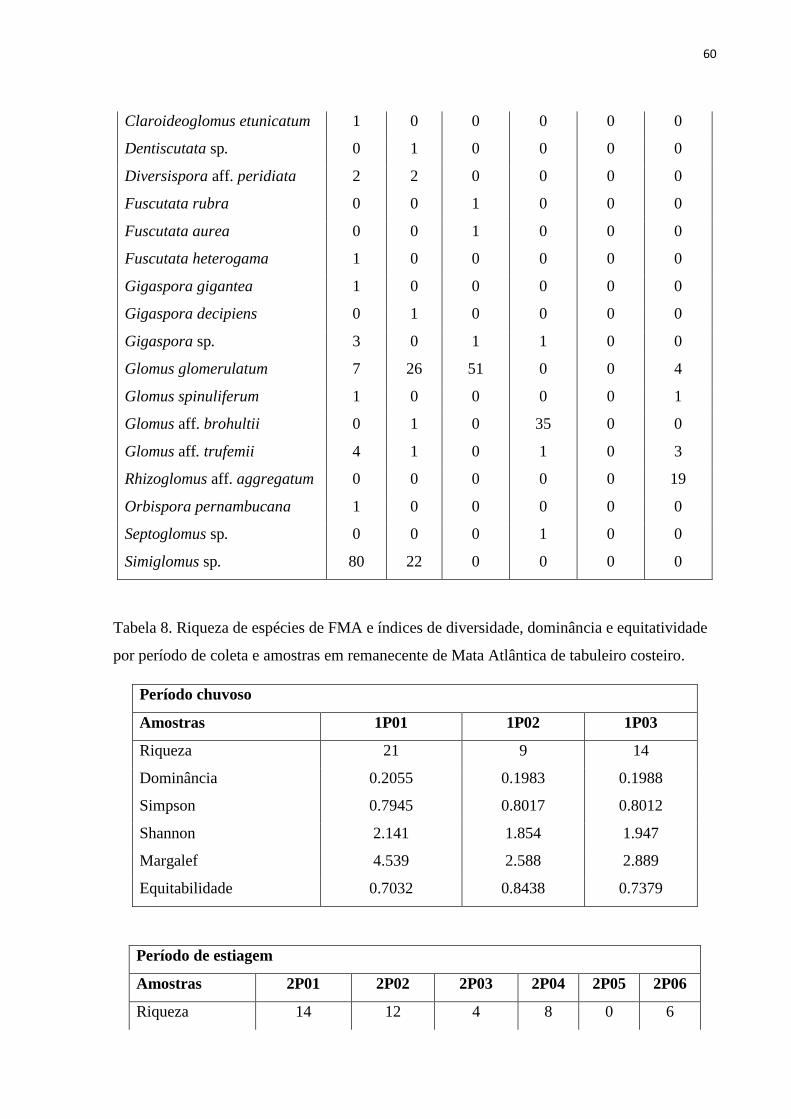
60
Claroideoglomus etunicatum 1 0 0 0 0 0
Dentiscutata sp. 0 1 0 0 0 0
Diversispora aff. peridiata 2 2 0 0 0 0
Fuscutata rubra 0 0 1 0 0 0
Fuscutata aurea 0 0 1 0 0 0
Fuscutata heterogama 1 0 0 0 0 0
Gigaspora gigantea 1 0 0 0 0 0
Gigaspora decipiens 0 1 0 0 0 0
Gigaspora sp. 3 0 1 1 0 0
Glomus glomerulatum 7 26 51 0 0 4
Glomus spinuliferum 1 0 0 0 0 1
Glomus aff. brohultii 0 1 0 35 0 0
Glomus aff. trufemii 4 1 0 1 0 3
Rhizoglomus aff. aggregatum 0 0 0 0 0 19
Orbispora pernambucana 1 0 0 0 0 0
Septoglomus sp. 0 0 0 1 0 0
Simiglomus sp. 80 22 0 0 0 0
Tabela 8. Riqueza de espécies de FMA e índices de diversidade, dominância e equitatividade
por período de coleta e amostras em remanecente de Mata Atlântica de tabuleiro costeiro.
Período chuvoso
Amostras 1P01 1P02 1P03
Riqueza 21 9 14
Dominância 0.2055 0.1983 0.1988
Simpson 0.7945 0.8017 0.8012
Shannon 2.141 1.854 1.947
Margalef 4.539 2.588 2.889
Equitabilidade 0.7032 0.8438 0.7379
Período de estiagem
Amostras 2P01 2P02 2P03 2P04 2P05 2P06
Riqueza 14 12 4 8 0 6

61
Dominância 0.5571 0.3267 0.893 0.3867 0 0.411
Simpson 0.4429 0.6733 0.107 0.6133 0 0.589
Shannon 1.172 1.503 0.2759 1.301 0 1.255
Margalef 2.777 2.687 0.7521 1.703 0 1.456
Equitabilidade 0.4441 0.6048 0.1988 0.6259 0 0.7003
O conhecimento sobre o papel que desempenham e a importância da diversidade dos
FMA para o funcionamento dos ecossistemas tem levado a grande esforço para identificar as
espécies que colonizam as plantas nos sistemas naturais (Rodriguez-Echeverria e Freitas,
2006). Além disso, dados sobre abundancia, diversidade, distribuição espacial e temporal de
espécies de FMA são relevantes para o entendimento da interação entre esses fungos e a
vegetação associada (Sturmer e Bellei, 1994).
Os índices de diversidade por ponto e períodos de amostragem encontram-se
expressos na tabela 8. No período chuvoso, a amostra 1P01 apresentou maiores valores para a
riqueza de espécies (21), seguida pela amostra 1P03 (14) e amostra 1P02 (9). Isto também
deve-se a que o numero amostral foi menor (apenas 3 amostras). Já no período seco, o número
amostral foi maior (6 amostras) a amostra 2P01 apresentou riqueza equivalente a 26 espécies,
seguida pela amostra 2P02 (12), 2P04 (8), 2P06 (6), 2P03 (4) e a amostra 2P05 não
apresentou nenhuma espécie, portanto, não houve representatividade em riqueza.
Comparando-se com outros trabalhos realizados por Souza et al., (2013) onde se
obteve uma maior riqueza (28) foi encontrada nas áreas revegetadas, em comparação com a
área de floresta que apresentou 10 espécies. Moreira et al., (2007) deteve maior riqueza no
Parque Estadual Campos de Jordao (18) do que no Parque Estadual Turístico de Alto de
Ribeiro (7), sendo apenas cinco espécies comuns a ambos parques.
Vários fatores podem influenciar a ocorrência de FMA, entre eles o hospedeiro, a
sazonalidade e os fatores edáficos. A sazonalidade parece ter influenciado o número de
esporos do solo e a diversidade de espécies de FMA nos diferentes ambientes da Mata
Atlântica. Bononi e Trufem (1983), Trufem e Viriato (1990), Santos (2001), Aidar et al.,
(2004) e Silva et al., (2006) verificaram maior densidade de esporos e riquezas de espécies no
período chuvoso do que na estiagem em floresta secundária.
O número de esporos diferiu entre os períodos avaliados na riqueza de espécies por
amostras, no entanto a composição de espécies de FMA mudou com os períodos, porém foi
detectada uma tendência de aumento de número de espécies no período chuvoso, que no
periodo de estiagem. Das 47 espécies de FMA identificadas (ver tabela 46), 30 são exclusivas

62
no período chuvoso e 26 no de estiagem, onde apenas 9 destas espécie ocorreram em ambas
estações.
O clima controla a formação da associação e estabelecimento das comunidades de
FMA tanto diretamente, pela maior disponibilidade hídrica e altas temperaturas, quanto
indiretamente, pela maior necessidade nutricional e hídrica das plantas em certas épocas do
ano.
Segundo Trufem e Bononi (1985) existem espécies que ocorrem na rizosfera da
maioria das plantas hospedeiras durante todo o ano; algumas espécies ocorrem com grande
abundancia, porém são restritas a certas espécies de plantas e épocas do ano; outras
apresentam baixa abundância e ausência de restrições em relação ao hospedeiro ou épocas do
ano; algumas espécies de FMA mostram baixas abundancias e restrições para hospedeiro e
época do ano.
Os valores registrados por Trufem (1995), semelhantes aos registados em fragmentos
de Mata Atlântica de tabuleiro costeiro, mostram a ocorrência de 47 espécies de FMA em solo
rizosferico de restinga de 51 fitosimbiontes no Parque Estadual da Ilha do Cardoso em São
Paulo. E aproximados aos valores encontrados por Jobim e Goto, 2016, considerando o único
inventário de diversidade do RN, registraram 46 espécies de FMA em dunas marítimas no
Parque Ecológico Dunas de Genipabu, a maior riqueza e diversidade foram associados ao
período de estiagem.
Diversos autores detetaron valores elevados de diversidade de FMA em diferentes
fitofisionomia da Mata Atlântica dunas, restinga, florestas ombrofila densa, floresta ombrófila
mixta ou floresta de auracaria tais como (Dos Santos e Carenho, 2011; Silva et al., 2015;
Bonfim et al., 2015; Camara et al., 2016). Por outro lado, outros autores registraron baixa
diversidade de FMA em diversas fitofisionomias da Mata Atlântica (Bononi e Trufem, 1983;
Trufem e Viriato, 1990; Gomes e Trufem, 1998; Trufem e Malatinszky, 1995; Souza, 2003;
Aidar et al., 2004; Alves, 2004; Melo, 2004; Silva et al., 2006, Moreira et al., 2007; Melloni et
al., 2011; Souza et al., 2013; Santos et al., 2014; Silva et al., 2015).
Enquanto a valores encontrados na avaliação de glomerosporos na rizosfera, Araújo et
al., (2003) verificaram entre o número 52 e 132 glomesporos de FMA em Melastomataceae
de fragmento de Mata Atlântica, no Parque Metropolitano de Pituaçu, Salvador, Bahia,
apresentam abundância de gêneros de 10% para Acaulospora, de 11% para Kuklospora.
Em otros países como na floresta tropical úmido do México, Guadarrama e Álvarez-
Sánchez (1999) observaram o contrario deste trabalho a riqueza de espécies e o número de
glomesporos aumentam no período de estiagem e decresceram no chuvoso. Observaram

63
também que durante a estiagem a taxa de crescimento das raízes diminuía e o número de
esporos no solo aumentava, enquanto no período chuvoso a taxa de crescimento das raízes
aumentava, influenciando o aumento da germinação dos glomesporos e na colonização das
raízes, com consequência na diminuição da esporulação. E outros trabalhos mostram todo o
contrario como o trabalho realizado por Brundrett e Ashwath, (2013) sobre associações
micorrízicos arbusculares (FMA) em habitats não perturbadas e perturbados nas proximidades
do Parque Nacional de Kakadu, na Austrália tropical, mostra um número menor de esporos
em floresta tropical em comparação em outros dois sítios de amostragem.
De acordo com o estudo realizado por Turrini e Giovannetti (2011), em 96 áreas
protegidas em todo o mundo, registraram um total de 127 espécies de FMA. Dados que
confrontan a diversidade de FMA em áreas protegidas e a importância destas como áreas
apropriadas para a conservação in situ da FMA, confrontando com os dados encontradas em
tabulerios costeiros do SEP respresenta um total de 37% de espécies de FMA.
Segundo Bever et al., (2001) existem duas hipóteses que podem explicar a
manutenção de alta diversidade de espécies de FMA em uma comunidade. A primeira sugere
que as espécies podem ser equivalentemente competitivas dentro de um simples nicho, como
o córtex das raízes finas das plantas hospedeiras. Neste caso, a diversidade é sustentada por
um processo direcionado de maneira aleatória. Esta possibilidade se apoia no fato de que uma
certa espécie de planta pode ser colonizada por muitas espécies de FMA. A segunda hipótese
propõe que as espécies de fungos são ecologicamente distintas e ocupam diferentes nichos.
Um fungo individual será competitivamente superior em seu nicho específico e a presença de
múltiplos nichos em um hábitat resultará em ativa manutenção de uma elevada diversidade na
comunidade de FMA. Sugere também que fatores bióticos e abióticos podem estar envolvidos
na manutenção da elevada diversidade em uma comunidade de FMA.
De acordo com a hipótese proposta por Connell (1978), a alta diversidade, comumente
observada em espécies arbóreas em florestas tropicais úmidas, é reflexo de um estado natural
de desequilíbrio ambiental, a diversidade assume níveis baixos tantos para distúrbios
ambientais mínimos como máximos, se alcança um valor máximo de diversidade em
dosagens intermediárias de perturbação, caso não haja distúrbios, o sistema irá encaminhar
para um estado em equilíbrio com baixos índices de diversidade.
Segundo Abbott e Gazey (1994), ainda não se sabe o quanto a teoria de Connel se
aplica aos FMA. Entretanto, ao discutirem os efeitos dos distúrbios ambientais sobres estes
organismos, os autores afirmam que um aumento inicial da diversidade de FMA pode
acontecer como repostas a uma entrada de propágulos oriundo de solos vizinhos a região

64
impactada. A depender da composição das espécies presentes inicialmente no ambiente e das
espécies imigrantes, interações competitivas ou positivas podem atuar, aumentando o
diminuindo a abundância de determinadas espécies ou a diversidade do ambiente.
5.1. Influência de fatores químicos do solo
Quanto a caracterização física, no período de estiagem foram encontradas maiores
valores de pH, P, Zn e Mn relação ao período chuvoso, que apresentou maior valor de H+Al,
e Fe caracterizando um solo mais ácido (Tabela 9).
Tabela 9. Propriedades físico-químicas do solo em remanescente de Mata Atlântica de
tabuleiro costeiro do RN.
VARIÁVEIS CHUVA ESTIAGEM
MACRONUTRIENTES
pH em água (1:2,5) 4,93 5,40
Alumínio (cmolc.dm-3) 0,14 0,14
Fósforo (mg.dm-3) 6 7
Potássio (mg.dm-3) 78 88
Cálcio (cmolc.dm-3) 1,36 2,76
Magnésio (cmolc.dm-3) 0,90 1,05
Hidrogenio + alumínio (cmolc.dm-3) 3,33 3,01
Sódio (mg.dm-3) 53 57
MICRONUTRIENTES
Ferro (mg.dm-3) 14,42 10,70
Zinco (mg.dm-3) 1,09 1,20
Manganês (mg.dm-3) 7,76 16,65
Cobre (mg.dm-3) 0,07 0,06
A estrutura das comunidades de FMA pode ser afetada por fatores físicos, químicos e
biológicos, entre os quais: composição vegetal, temperatura, umidade e pH do solo,
disponibilidade de nutrientes e estresse antropogênico, incluindo compactação do solo e
presença de metais pesados (Entry et al., 2012).
Temperatura e umidade também são fatores limitantes para o desenvolvimento das
espécies de FMA, pois interferem diretamente em seu comportamento fisiológico. Cada

65
espécie de FMA possui uma temperatura ótima de germinação, que pode variar de 18-25ºC.
Em geral, a umidade é próxima à capacidade de campo (Siqueira et al., 1985). Fora desses
limites, apenas as espécies mais resistentes passam a colonizar os ambientes. Há também
restrição ao estabelecimento de espécies mais sensíveis de FMA em ambientes com
compactação ou erosão do solo, visto que a camada superficial do solo é um dos principais
habitats desses fungos (Brundrett et al., 1996).
O pH do solo pode afetar a diversidade micorrízica de forma direta, atingindo os
fungos, ou de forma indireta, comprometendo a disponibilidade de nutrientes para o fungo ou
para a planta. A germinação dos glomerosporos, por exemplo, está altamente relacionada ao
pH do solo e, aparentemente, espécies de Acaulospora, Gigaspora e Scutellospora “lato
sensu” são favorecidas em ambientes com pH de 4 a 6, enquanto espécies de Glomus “lato
sensu” são favorecidas na faixa de 6 a 8 (Zhu et al., 2007). Entretanto, Trufem et al. 1994,
observaram que rizosferas de plantas do litoral arenoso do parque estadual da Ilha do Cardoso
em São paulo, são pobres em P (lug/cm3) e em matéria orgânica (0,15%), com pH levemente
ácido (pH =6,3), portanto, com condições ecológicas que justificam e até mesmo facilitam a
ocorrência de FMA. Outro estudo também realizado por Trufem, (1995), demonstrou que à
medida que se caminha do litoral para o interior da Ilha do Cardoso, as condições de solo se
modificam, aumentando os teores de fósforo, matéria orgânica, potássio, cálcio, magnésio e
enxofre. O pH deixa de ser tão ácido e os teores de alumínio passam por oscilações. Já Souza
et al., (2003) e Trufem (1990) definem que a distribuição de espécies de Acaulosporaceae e
Glomeraceae são favorecidos em solos de baixo pH (3,5–5,8) e baixos teores de fósforo (4-5
mg.dm-3), o que corroboram com os resultados obtidos em remanescente de Mata Atlântica
sob tabuleiro costeiro, possivelmente sendo a disponibilidade de elevada de fósforo no
período de estiagem o fator que justificaria a menor diversidade destes gêneros ocorrendo até
o momento nesta região (ver tabela 9). Em solos com pH ácido entre 4,5 e 4,8, a ocorrência de
algumas espécies de FMA pode ser favorecida, como é o caso de espécies do gênero Glomus
(Borba & Amorim, 2007). Contudo, este resultado divergiu do encontrado em dunas fixas na
Praia da Joaquina en Santa Catarina, onde predominaram espécies de Glomus, apesar do pH
5,9 e, portanto, consideravelmente menos ácido (Cordoba et al., 2001). Os FMA também são
afetados pelo excesso ou escassez de nutrientes presentes no solo, principalmente o fósforo.
Em geral, alta concentração de P disponível no solo restringe a colonização
micorrízica, com queda na contribuição do fungo para absorção desse nutriente e consequente
redução da densidade de FMA, isso ocorre porque quando as plantas estão supridas de P,
torna-se desnecessário desperdiçar energia investindo na associação micorrízica (Diniz,

66
2006). Siqueira (1994) menciona que espécies de Gigasporales apresentam preferência por
solos com grandes concentrações de fósforo (P > 40 mg.dm-3) e ácidos, com pH variando
entre 5,5 e 4,8. Entretanto, solos com valores de pH menores que os testados por Siqueira
(1994), como no período chuvoso (pH = 4,93) apresentou diversidade maior em relação aos
solos do período de estiagem (pH = 5,40), mesmo o período chuvoso apresentado menor
disponibilidade de fósforo no solo de 6 mg.dm-3 em relação aos 7 mg.dm-3 do período de
estiagem, possivelmente os solos do período chuvoso já acidos demais para Gigasporales,
desfavorecendo este grupo. Por outro lado em condições de maiores teores de P no solo, tende
a ser menor a formação de micorriza e, portanto, é menor a presença de FMA no solo
(Moreira & Siqueira, 2006).
6. CONCLUSÕES
Os remanescente do domínio Mata Atlântica em tabuleiros costeiros apresentam alta
diversidade de FMA, mostrando que a riqueza e composição de espécies de FMA se
presentaram nas duas estações, mesmo tendo um numero amostral menor (9 amostras de
solo). Os gêneros Acaulospora e Glomus foram as mais representativos e a espécie Glomus
spinuliferum como novo registro para a Mata Atlântica.
7. CONSIDERAÇÕES FINAIS
Os remanescentes de Mata Atlântica em tabuleiros costeiros apresentam um alto
número de espécies, despontando como uma área biologicamente rica e taxonomicamente
promissora para novas espécies. Vale confrontar que a diversidade encontrada no estudo é
fruto de dados de coleta limitado, se comparando aos trabalhos de Trufem (1995), Souza
(2003), dos Santos e Carrenho (2011). De qualquer modo, este ainda pode ser considerado
uma importante contribuição para o conhecimento da diversidade de FMA no estado do Rio
Grande do Norte e como primero inventário para o litoral sul do RN. Apesar dos estudos
sobre estes fungos no estado, muitos são de descripção de novos taxa, onde se destacam os
trabalhos de Goto et al., (2010, 2011, 2012) e Błaszkowski et al., (2014; 2015). No entanto,
em quanto a inventários, no RN, apenas existe um único inventario de diversidade, realizado
por Jobim e Goto (2016), mostrando que realmente são restritos.
8. REFERÊNCIAS BIBLIOGRÁFICAS

67
ABBOTT, L.K.; GAZEY, C. 1994. An ecological view of the formation of VA
mycorrhizas. Plant and soil 159 (1): 69-78.
AIDAR, M.P.M.; CARRENHO, R.; JOLY, C.A. 2004. Aspects of arbuscular mycorrhizal
fungi in na Atlanctic Forest chronosequene in Parque Estadual Turístico do Alto Ribeira
(PETAR), SP. Biota Neotropica 4: 1- 5.
ALLEN, M.F. 1992. Mycorrhizal functioning. London: Chapman Hall 515 p.
ALMEIDA JR, E. B.; ZICKEL, C. S. MENDONÇA P. R. M. 2006. Caracterização e espectro
biológico da vegetação do litoral arenoso do Rio Grande do Norte. Revista de Geografia
(Recife), 23(3), 66-85p.
ALVES, L.J. 2004. Efeito da fragmentação florestal sobre as comunidades de fungos
micorrízicos arbusculares da floresta Atlântica do extremo sul da Bahia. Dissertação de
Mestrado em Ecologia e Biomonitoramento. Universidade Federal da Bahia. Salvador,
Bahia. 84 p.
ARAÚJO, C.V.M.; SANTOS, O.M.; ALVES, L.J.; MUNIZ, C.R.R. 2003. Fungos
micorrízicos arbusculares em espécies de Melastomataceae no Parque Metropolitano de
Pituaçu, Salvador-Bahia-Brasil. Sitientibus série Ciências Biológicas 3: 115-119.
BLACKWELL, M. 2011. The Fungi: 1, 2, 3… 5.1 million species?. American journal of
botany 98(3): 426-438.
BERKELEY, M.J.; BROOME, C.E. 1873. Enumeration of the Fungi of Ceylon. Part II.
Containing the remainder of the Hymenomycetes, with the remaining established tribes of
Fungi. Journal of the Linnean Society of London, Botany 14 (73): 29-64.
BEVER, J.D.; SCHULTZ, P.A.; PRINGLE, A.; MORTON, J.B. 2001. Arbuscular
mycorrhizal fungi: more diverse than meets the eye, and the ecological tale of why. Ecology.
Bioscience 51(11): 923-931.

68
BŁASZKOWSKI, J. 2012. Glomeromycota. W. Szafer Institute of Botany Polish Academy of
Sciences, Kraków. ISBN: 978-83-89648-82-2. 304 p.
BŁASZKOWSKI, J.; CHWAT, G.; GÓRALSKA, A.; PRZEMYSLAW, R.; KOVÁCS, G.M.
2014. Two new genera, Dominikia and Kamienskia, and D. disticha sp. nov. in
Glomeromycota. Nova Hedwigia 100: 225- 238.
BŁASZKOWSKI, J.; CHWAT, G.; GÓRALSKA, A. 2015. Acaulospora ignota and
Claroideoglomus hanlinii, two new species of arbuscular mycorrhizal fungi (Glomeromycota)
from Brazil and Cuba. Mycological Progress 14(4), 1-11.
BONFIM, J.A.; VASCONCELLOS, R.L.F.; STÜRMER, S.L.; CARDOSO, E.J.B.N. 2013.
Arbuscular mycorrhizal fungi in the Brazilian Atlantic Forest: a gradient of environmental
restoration. Applied Soil Ecology 71: 7-14.
BONFIM, J.A.; VASCONCELLOS, R.L.F.; GUMIERE, T.; MESCOLOTTI, D.D.L.C.;
OEHL, F. CARDOSO, E.J.B.N. 2015. Diversity of Arbuscular Mycorrhizal Fungi in a
Brazilian Atlantic Forest Toposequence. Microbial ecology 71(1): 164-177.
BONONI, V.L.R.; TRUFEM, S.F.B. 1983. Endomicorrizas vesículo-arbusculares do cerrado
da Reserva Biológica de Moji-Guaçú, SP, Brasil. Rickia 10: 55-84.
BORBA, M.F.; AMORIM, S.M.C. 2007. Fungos micorrízicos arbusculares em sempre-vivas:
subsídio para cultivo e replantio em áreas degradadas. Revista de Biologia e Ciências da
Terra.7 (2): 20-27p.
BRASIL. 1985. Anuário Estatístico do Rio Grande do Norte, Natal, IDEC, v. 12: 266.
BRUNDETT, M.C.; ASHWATH, N.; JASPER, D.A. 1995. Mycorrhizas in the kakadu region
of tropical Australia. Plant and Soil, Dordrecht, v. 184, n. 1: 173-184.
BRUNDRETT, M.; BOUGHER, N.; DELL, B., GROVE, T., MALAJCZUK, N. 1996.
Working with Mycorrhizas in Forestry and agriculture. ACIAR Monograph 3: 141-145p.

69
BRUNDRETT, M.C.; ASHWATH, N. 2013. Glomeromycotan mycorrhizal fungi from
tropical Australia III. Measuring diversity in natural and disturbed habitats. Plant and
soil 370 (1-2): 419-433.
CAMARA, R.; PEREIRA, M.G.; SILVA, C.F.D.; PAULA, R.R.; SILVA, E.M.R.D. 2016.
Arbuscular Mycorrhizal Fungi in Two Forest Fragments of Periodically Floodable Restinga at
Marambaia, RJ. Floresta e Ambiente, (AHEAD) 23(1): 33-42
CAMPANILI, M.; PROCHNOW, M. 2006. Atlantic Forest: a network for the forest, Brasília:
RMA, 332 p.
CARRENHO, R.; TRUFEM, S.F.B.; BONONI, V.L.R. 1998. Arbuscular mycorrhizal fungi
in Citrus sinensis/ C. limon trated with Fosety-Al and Metalaxyl. Mycological Research 102:
677-682.
CARRENHO, R.; TRUFEM, S.F.B.; BONONI, V.L.R. 2001. Fungos micorrízicos
arbusculares em rizosferas de três espécies de fitobiontes instaladas em área de mata ciliar
revegetada. Acta Botanica Brasilica 15: 115-124.
CARRENHO, R. C.; BALOTA, S.M.G.; COLOZZI-FILHO, E.L.A. 2010. Fungos
micorrízicos arbusculares em agroecossistemas brasileiros In: Siqueira, J.O., Souza, F.A.,
Cardoso, E.J.B.N., Tsai, S.M., (ed.) Micorrizas: 30 anos de pesquisa no Brasil. Lavras,
Editora UFLA. 215-250p.
CARVALHO, F.; SOUZA, F.A.; CARRENHO, R.; SOUZA, F.M.M.; JESUS, E.C.;
FERNANDES, G.W. 2012. The mosaic of habitats in the high-altitude Brazilian rupestrian
fields is a hotspot for arbuscular mycorrhizal fungi. Appl. Soil Ecology 52: 9–19.
CINTRA, F.L.D.; PORTELA, J.C.; NOGUEIRA, L.C. 2004. Caracterização física e hídrica
em solos dos tabuleiros costeiros no distrito de irrigação Platô de Neópolis. Revista
Brasileira de Engenharia Agrícola e Ambiental, v. 08, n. 01. 45-50p.

70
CORDOBA, A.S.; MENDONCA, M.M.; STURMER, S.L.; RYGLEWICZ, P.T. 2001.
Diversity of arbuscular mycorrhizal fungi along a sand dune stabilization gradient: a case
study at Praia de Joaquina, Ilha de Santa Catarina, South Brazil. Mycoscience 42:379-387p.
CORDOBA, A.S.; MENDONCA, M.M.; ARAUJO, E.F. 2002. Avaliacao da diversidade
genética de fungos micorrizicos arbusculares em tres estadios de estabilizacao de dunas.
Revista Brasileira de Ciência do Solo 26: 931-937.
DANDAN, Z.; ZHIWEI, Z. 2007. Biodiversity of arbuscular mycorrhizal fungi in the hot-dry
valley of the Jinsha River, southwest China. Applied Soil Ecology 37:118-128.
DA SILVA K.J.D. 2013. Fungos micorrízicos arbusculares (Glomeromycota) em áreas de
tabuleiro costeiro, do centro de lançamento barreira do inferno, Parnamirim, RN. Monografia
TCC em Ciências Biológicas. Universidade Federal do Rio Grande do Norte, Natal, RN. 45
p.
DINIZ, V. M. 2006. Absorção de fósforo e nitrogênio por espécies arbóreas da Caatinga
nordestina inoculadas com fungos micorrízicos. Dissertação. Universidade Federal de
Campina Grande. Patos, Paraíba.
DOS SANTOS, F.E.F.D.; CARRENHO, R. 2011. Diversity of arbuscular mycorrizal fungi in
an impacted forest remnant-Parque Cinquentenário, Maringá, Paraná, Brazil. Acta Botanica
Brasilica 25(2): 508-516.
DOS SANTOS, R.S.; BARRETO, P.A.B.; SCORIZA, R.N. 2014. Efeito da sazonalidade na
comunidade de fungos micorrízicos arbusculares em um fragmento de mata de cipóem Vitória
da Conquista, Bahia. Revista Brasileira de Biociências 12(1): 46.
DOTZLER, N.; KRINGS, M.; TAYLOR, T.N.; AGERER, R. 2006. Germination Shields in
Scutellospora (Glomeromycota: Diversisporales, Gigasporaceae) from the 400 million-year-
old Rhynie chert. Mycological Progress 5: 178-184.

71
EMBRAPA -.Empresa Brasileira de Pesquisa Agropecuária Centro Nacional de Pesquisa de
Solos. 1999. EMBRAPA - CNPS. Sistema Brasileiro de Classificação de Solos. Brasília:
412p.
ENTRY, J.A.; RYGIEWICZ, P.T.; WATRUD, L.S.; DONNELLY, P.K. 2012. Influence of
adverse soil conditions on the formation and function of Arbuscular mycorrhizas. Advances
in Environmental Research 7: 123-138p.
FRANK, A.B. 1885. Ueber die auf Wurzelsymbiose beruhende Ernährung gewisser Bäume
durch unterirdische Pilze. Berichte der Deutschen Botanischen Gesellschaft 3: 128–145.
FRANK, A.B. 1877. Über die biologischen Verhältnisse des Thallus einiger Krustflechten.
Cohn Beitr Biol Pflanzen 2:123–200.
FITTER, A.H.; HELGASON, T.; HODGE, A. 2011. Nutritional exchanges in the arbuscular
mycorrhizal symbiosis: implications for sustainable agriculture. Fungal Biology
Reviews 25(1): 68-72.
FURRAZOLA, E.; GOTO, B.T.; SILVA, G.A.; TORRES-ARIAS, Y.; MORAIS, T.; LIMA,
C.E.P.; FERREIRA, A.C.A.; MAIA, L.C.; SIEVERDING, E.; OEHL, F. 2013. Acaulospora
herrerae, a new pitted species in the Glomeromycetes from Cuba and Brazil. Nova Hedwigia
97: 401-413.
FUNDAÇÃO SOS MATA ATLÂNTICA & INPE - INSTITUTO NACIONAL DE
PESQUISAS ESPACIAIS. 2016. Atlas dos remanescentes florestais da Mata Atlântica
período 2014 – 2015. Relatório Parcial. Fundação SOS Mata Atlântica / Instituto
Nacional de Pesquisas Espaciais, São Paulo, 156p. https://www.sosma.org.br/nossa-
causa/floresta/a-mata-atlantica> Acesso em: 21 de junho, 2016.
GERDEMANN, J.W.; NICOLSON, T.H. 1963. Spores of mycorrhizal Endogone species
extracted from soil by wet-sieving and decanting. Transactions of the British Mycological
Society 46: 235-244.

72
GIOVANNETTI, M.; GIANINAZZI-PEARSON, V. 1994. Biodiversity in arbuscular
mycorrhizal fungi. Mycological Reserch 98:705-715.
GIOVANNETTI, M.; SBRANA, C.; AVIO, L.; STRANI, P. 2004. Patterns of belowground
plant interconnections established by means of arbuscular mycorrhizal networks. New Phytol
164:175–181.
GOMES, S.D.P.; TRUFEM, S.F.B. 1998. Fungos micorrízicos arbusculares (Glomales,
Zygomycota) na Ilha dos Eucaliptos, Represa do Guarapiranga, São Paulo, SP. Acta
Botanica Brasilica.
GOMIDE, P.H.O.; SILVA, M.L.N.; SOARES, C.R.F.S.; CARDOSO, E.L.; CARVALHO, F.;
LEAL, P.L.; MARQUES, R.M.; STÜRMER, S.L. 2014. Fungos micorrízicos arbusculares em
fitofisionomias do pantanal da Nhecolândia, Mato Grosso do Sul. Revista Brasileira de
Ciência do Solo 38: 114-1127.
GOTO, B.T. 2009. Taxonomia de Glomeromycota: revisão morfológica, chaves dicotômicas
e descrição de novos táxons. Tese de doutorado em Biologia de Fungos. Recife.
Universidade Federal de Pernambuco, 358 p.
GOTO, B.T.; MAIA, L.C. 2006. Glomerospores: a new denomination for the spores of
Glomeromycota, a group molecularly distinct from the Zygomycota. Mycotaxon 96: 129-
132.
GOTO, B.T.; SILVA, G.A.; YANO MELO, A.M.; MAIA, L.C. 2010. Checklist of the
arbuscular mycorrhizal fungi (Glomeromycota) in the Brazilian semiarid. Mycotaxon 113:
251-254.
GOTO, B.T.; SILVA, G.A..; MAIA, L.C.; OEHL, F. 2010. Dentiscutata colliculosa, a new
species in the Glomeromycetes from Northeastern Brazil with colliculate spore
ornamentation. Nova Hedwigia 90(3-4): 383-393.

73
GOTO, B.T.; SILVA, G.A.; MAIA, L.C.; SOUZA, R.G.; COYNE, D.; TCHABI, A.;
LAWOUIN, L.; HOUNTONDJI, F.; OEHL, F. 2011. Racocetra tropicana, a new species in
the Glomeromycetes from tropical areas. Nova Hedwigia 92: 69–82.
GOTO, B.T.; SILVA, G.A.; ASSIS, D.M.A.; SILVA, D.K.A.; SOUZA, R.G.; FERREIRA,
A.C.A.; JOBIM, K.; MELLO, C.M.A.; VIEIRA, H.E.E.; MAIA, L.C.; OEHL, F. 2012.
Intraornatosporaceae: (Gigasporales), a new family with two new genera and two new
species. Mycotaxon 119: 117–132.
GOTO, B.T.; JARDIM, J.G.; SILVA, G.A.; FURRAZOLA, E.; TORRES-ARIAS, Y.;
OEHL, F. 2012b. Glomus trufemii (Glomeromycetes), a new sporocarpic species from
Brazilian sand dunes. Mycotaxon 120: 1-9.
GOTO, B.T.; SILVA, G.A.; ASSIS, D.M.A.; SILVA, D.K.A.; SOUZA, R.G.; FERREIRA,
A.C.A.; JOBIM, K.; MELLO, C.M.A.; VIEIRA, H.E.E.; MAIA, L.C.; OEHL, F. 2012.
Intraornatosporaceae: (Gigasporales), a new family with two new genera and two new
species. Mycotaxon 119: 117–132.
GOTO, B.T.; PEREIRA, C.M.; NOBRE, C.P.; ZATORRE, N.P.; COVACEVICH, F.;
BERBARA, R.L.L.; MAIA, L.C. 2013. Acaulospora endographis (Glomeromycetes), a new
fungus with a complex spore wall. Mycotaxon 123: 403-408.
GOEDERT, W.J. 1980. Uso e manejo dos recursos naturais do cerrado: solo e clima. In anais
do V simpósio sobre o cerrado.Cerrado: uso e manejo (D. Marchetti e A.D. Machado, eds.).
Editerra, Brasília. 475-498p.
GUADARRAMA, P.; ÁLVAREZ-SÁNCHEZ, F.J. 1999. Abundance of arbuscular
mycorrhizal fungi spores in different environments in a tropical rain forest, Veracruz,
Mexico. Mycorrhiza 8(5): 267-270.
HAMMER, Ø.; HARPER, D.A.T.; RYAN, P.D. 2001. PAST: Paleontological statistics
software package for education and data analysis. Palaeontologia Electronica 4(1): 9p.
Version 2.17, Manual. http://palaeo-electronica.org/2001_1/past/issue1_01.htm
HARLEY, J.L. 1989. The significance of mycorrhiza. Mycological Research 92: 129-139.

74
HAWKSWORTH, D.L. 1991. The fungal dimension of biodiversity: magnitude, significance,
and conservation. Mycological Research 95 (6): 641-655.
HELGASON, T.; DANIEL, T.; HUSBAND, R.; FITTER, A.; YOUNG, J. 1998. Ploughing
up the Wood-wide web?. Nature 394: 431.
HELGASON, T.; FITTER, A. 2005. The ecology and evolution of the arbuscular mycorrhizal
fungi. Mycologist 19: 96-101.
IBGE - Instituto Brasileiro de Geografia e Estatística. 2016. Mapa de Biomas e Vegetacão do
Brasil. Rio de Janeiro: IBGE. Disponível em:< http://www.ibge.gov.br/biomas >. Acesso em:
10 de junho de 2016.
JACOMINE, P.K.T. 2001. Evolução do conhecimento sobre solos coesos no brasil. in:
workshop coesão em solos dos tabuleiros costeiros, aracaju. anais... aracaju: embrapa
tabuleiros costeiros, 19-46p.
JACOMINE, P.K.T. 1996. Distribuição geográfica, características e classificação dos solos
coesos dos Tabuleiros Costeiros. In: Reunião Técnica sobre Solos dos Tabuleiros Costeiros.
Anais... EMBRAPA/UFBA. 13-26p
JACOMINE, P.K.T.; SILVA, F.B.R.; FORMIGA, R.A.; ALMEIDA, J.C.; BELTRÃO, V.
D.A.; PESSOA, S.C.P.; FERREIRA, R.C. 1971. Levantamento exploratório-Reconhecimento
de solos do Estado do Rio Grande do Norte. Recife, SUDENE/ Divisão de pesquisa
pedológica. Recife, v.1. Boletim Técnico, n.21, (serie pedológica, n.9). DPP/DA. Convênio
MA/DNPEA/SUDENE/DRN, MA/CONTAP/USAID/BRASIL, p.531.
JEFFRIES, P.; GIANINAZZI, S.; PEROTTO, S.; TURNAU, K.; BAREA, J. M. 2003. The
contribution of arbuscular mycorrhizal fungi in sustainable maintenance of plant health and
soil fertility. Biology and fertility of soils 37(1): 1-16.
JENKINS, W.R. 1964. A rapid centrifugal – flotation technique for separating nematodes
from soil. Plant Diasease Report 48: 692.

75
JOBIM, K.; GOTO B.T. 2016. Diversity of arbuscular mycorrhizal fungi (Glomeromycota) in
maritime sand dunes of Brazilian northeast. Studies in Fungi 1(3): 43–55.
KENRICK, P.; CRANE, P.R. 1997. The origin and early evolution of plants on
land. Nature 389(6646): 33-39.
KÖPPEN, W. 1948. Climatologia: com un estudio de los climas de la tierra. Fondo de
Cultura Economica. Mexico.
LACERDA, L.D.; ARAÚJO, D.S.D.; MACIEL, N.C. 1993. Dry coastal ecosystems of the
tropical Brazilian coast. In: van der Maarel E. (ed). Dry coastal ecosystems: Africa, America,
Asia, Oceania. Elsevier, Amsterdam 477-493 p.
LABORATÓRIO DE BIOLOGIA DE MICORRIZAS. 2016. Lista global das espécies do
filo Glomeromycota. Disponível em: http://glomeromycota.wix.com/lbmicorrizas>. Acesso
em: 9 de junho de 2016.
LEAL, P.L.; SIQUEIRA, J.O.; STÜRMER, S.L. 2013. Switch of tropical Amazon forest to
pasture affects taxonomiccomposition but not species abundance and diversity of
arbuscularmycorrhizal fungal community. Applied Soil Ecology 71: 72– 80.
LIMA, H.V.; SILVA, A.P.; JACOMINE, P.T.K.; ROMERO, R.E.; LIBARDI, P.L. 2004.
Identificação e caracterização de solos coesos no estado do Ceará. R. Bras. Ci. Solo, v. 28.
467-476p.
LIMA, R.A.A.D. 2013. Dinâmica sazonal de fungos micorrízicos arbusculares
(Glomeromycota) em cerradão e brejo de altitude na Chapada do Araripe, CE. Dissertação de
Mestrado em Sistematica e Evolução. Universidade Federal do Rio Grande do Norte. Natal,
RN. 79 p.
LIMA NETO, J. de A.; RIBEIRO, M.R.; CORRÊA, M.M.; SOUZA JÚNIOR, V.S. de;
ARAÚJO FILHO, J.C. de.; LIMA, J.F.W.F. 2010. Atributos químicos, mineralógicos e

76
micromorfológicos de horizontes coesos de Latossolos e Argissolos dos Tabuleiros Costeiros
do Estado de Alagoas. Revista Brasileira de Ciência do Solo, Viçosa, v. 34, n. 2, 473-486p.
LIMA NETO, J. de A.; RIBEIRO, M.R.; CORRÊA, M.M.; SOUZA JÚNIOR, V.S. de;
LIMA, J.F.W.F.; FERREIRA, R.F.A.L. 2009. Caracterização e gênese do caráter coeso em
latossolos amarelos e argissolos dos tabuleiros costeiros do estado de Alagoas. Revista
Brasileira de Ciência do Solo, Viçosa, v. 33, n. 4, 1001-1011p.
LINO, F.C. 1992. Reserva da Biosfera da Mata Atlântica. Consórcio Mata Atlântica-
Universidade Estadual de Campinas (1): 1 - 101.
LOPES, E.S.; OLIVEIRA, E.D.; DIAS, R., SCHENCK, N.C. 1983. Occurrence and
distribution of vesicular-arbuscular mycorrhizal fungi in coffee (Coffea arabica L.)
plantations in Central Sao Paulo State, Brazil. Turrialba, 33(4):417-422, 1983.
LOPES, A.S. 1984. Solos sob cerrado. Características, propiedades e manejo. Asociação
Brasileira de Pesquisa da Postassa e do Fosfato, Piracicaba.
MALAVOLTA, E.1980. Elementos de nutrição mineral de plantas. São Paulo, Ceres.
251p.
MARINHO, F.; SILVA, G.A.; FERREIRA, A.C.A.; VERAS, J.S.N.; SOUSA, N.M.F.;
GOTO, B.T.; MAIA, L.C.; OEHL, F. 2014. Bulbospora minima, a new genus and a new
species from semi-arid Northeast Brazil. Sydowia 66: 313-323
MARTÍNEZ-GARCÍA, L.B.; DE DIOS MIRANDA, J.; PUGNAIRE, F.I. 2012. Impacts of
changing rainfall patterns on mycorrhizal status of a shrub from arid environments. European
Journal of Soil Biology, 50: 64-67.
MELO, A.M.M. 2004. Fungos micorrízicos arbusculares (Glomeromycota) em fragmentos de
Mata Atlântica no centro de endemismo Pernambuco. Dissertação de Mestrado em Biologia
de Fungos. Universidade Federal de Pernambuco, Recife, Pernambuco. 49 p.

77
MELLO, C.M.A.; SILVA, G.A.; VIEIRA, H.E.E.; SILVA, I.R.; MAIA, L.C.; OEHL. F.
2012. Fuscutata aurea, a new species in the Glomeromycetes from cassava and maize fields
in the Atlantic rainforest zone of Northeastern Brazil. Nova Hedwigia 1: 267 – 275.
MELLONI, R.; DA COSTA GUIDA, E.; DE ANDRADE, M.R.; MELLONI, E.G.P. 2011.
Fungos micorrízicos arbusculares em solos da Reserva Biológica Municipal Serra dos
Toledos, Itajubá/MG. Ciência Florestal 21(4): 799-809.
MITTERMEIER, R.A.; FONSECA, G.A.B.; RYLANDS, A.B.; BRANDON, K.A. 2005.
Brief History of Biodiversity Conservation in Brazil. Conservation Biology 19: 601 – 607.
MORAES, J.E.P. 2016. Estudo dos ecossistemas e unidades de conservação da bacia potiguar
e análise dos impactos ambientais das atividades petrolíferas na região costeira do Rio Grande
do Norte. 56 edicao. Revista Educação Ambiental em Ação. Disponível em:
http://www.revistaea.org. Acesso em: 10 de junho de 2016.
MOREIRA, F.S.; SIQUEIRA, J.O. 2006. Microbiologia e Bioquímica do Solo. Editora
UFLA, Lavras, MG, Brasil.
MOREIRA, M.; NOGUEIRA, M.A.; TSAI, S.M.; GOMES-DA-COSTA, S.M.; CARDOSO,
E.J. 2007. Sporulation and diversity of arbuscular mycorrhizal fungi in Brazil Pine in the field
and in the greenhouse. Mycorrhiza 17(6): 519-526.
MOREIRA, M.; BARETTA, D.; TSAI, S.M.; CARDOSO, E.J.B.N. 2009. Arbuscular
mycorrhizal fungal communities in native and in replanted Araucaria forest. Scientia
Agricola 66: 677-684.
MOSSE, B.; STRIBLEY, D.P.; LETACON, F. 1981. Ecology of mycorrhizal fungi. Adv.
Microb. Ecol. 5: 137-210p.
MUTHUKUMAR, T.; UDAIYAN, K.; SHANMUGHAVEL, P. 2004. Mycorrhiza in sedges -
an overview. Mycorrhiza 14: 65-77.

78
NASCIMENTO, G.B. 2001. Caracterização dos solos e avaliação de propriedades edáficas
em ambientes de Tabuleiros Costeiros da região Norte Fluminense (RJ). Rio de Janeiro, RJ:
UFRRJ. Dissertação (Mestrado) – Universidade Federal Rural do Rio de Janeiro.
NOVAIS, C.B.; BORGES, W.L.; JESUS, E.C.; SAGGIN-JÚNIOR, O.J.; SIQUEIRA, J.O.
2014. Inter- and intraspecific functional variability of tropical arbuscular mycorrhizal fungi
isolates colonizing corn plants. Applied Soil Ecology 76: 78– 86.
OEHL, F.; SILVA, G.A.; GOTO, B.T.; MAIA, L.C.; SIEVERDING, E. 2011.
Glomeromycota: two new classes a new ordem. Mycotaxon 116: 365-379.
OEHL, F.; SIEVERDING, E.; PALENZUELA, J.; INEICHEN, K.; SILVA, G.A. 2011a.
Advances in Glomeromycota taxonomy and classification. IMA Fungus 2: 191–199.
OEHL, F.; SILVA, D.K.A.; MAIA, L.C.; SOUSA, N.M.F.; VIEIRA, H.E.E.; SILVA, G.A.
2011d. Orbispora gen. nov., ancestral in the Scutellosporaceae (Glomeromycetes).
Mycotaxon 116: 161–169.
OEHL, F.; SÁNCHEZ-CASTRO, I.; PALENZUELA, J.; SILVA, G.A. 2014. Palaeospora
spainii, a new arbuscular mycorrhizal fungus from Swiss agricultural soils. Nova Hedwigia
101: 89-102.
OHSOWSKI, B.M.; ZAITSOFF, P.D.; ÖPIK, M.; HART, M.M. 2014. Where the wild things
are: looking for uncultured Glomeromycota. New Phytologist 204(1): 171-179.
ÖPIK, M.; DAVISON, J.; MOORA, M.; ZOBEL, M. 2013. DNA-based detection and
identification of Glomeromycota: the virtual taxonomy of environmental sequences
1. Botany 92(2): 135-147.
PACHECO, E. P.2010. Estudo da compressibilidade e qualidade de um Argissolo Amarelo
cultivado com cana-de-açúcar nos Tabuleiros Costeiros de Alagoas. 2010. Tese (Doutorado
em Ciência do Solo) - Universidade Federal Rural de Penambuco, Recife – PE. 118p.

79
PEREIRA, C.M.R.; SILVA, D.K.A.; FERREIRA, A.C.A.; GOTO, B.T.; MAIA, L.C. 2014.
Diversity of arbuscular mycorrhizal fungi in Atlantic forest areas under different land
uses.Agriculture, Ecosystems and Environment 185: 245–252.
PEREIRA, C.M.R.; GOTO, B.T.; SILVA, D.K.A.; FERREIRA, A.C.A.; SOUZA, F. A.;
SILVA, G.A.; MAIA, L.C.; OEHL, F. 2015. Acaulospora reducta sp. nov. and Acaulospora
excavata, two glomeromycotan fungi with pitted spores from Brazil. Mycotaxon 130(4): 983-
995.
PEREIRA, C.M.; MAIA, L.C.; SÁNCHEZ-CASTRO, I.V.Á.N.; PALENZUELA, J.; SILVA,
D.K.; SUDOVÁ, R.; SILVA, G.A. 2016. Acaulospora papillosa, a new mycorrhizal fungus
from NE Brazil, and Acaulospora rugosa from Norway. Phytotaxa 260(1): 14-24.
PÓVOA, I.C.F.; SILVA, N.S.; AQUINO-SILVA, M.R. 2006. Biodiversidade e
Desenvolvimento Sustentável. In: X Encontro Latino Americano De Iniciação Científica E Vi
Encontro Latino Americano De Pós-Graduação–Universidade Do Vale Do Paraíba. São
Paulo: Revista Univap: 96.
R DEVELOPMENT CORE TEAM. R. 2011. A language and environment for statistical
computing. Vienna: R Foundation for Statistical Computing. Disponível em:
http://www.R-project.org/.
REDECKER, D.; KODNER, R.; GRAHAM, L.E., 2000a. Glomalean fungi from the
Ordovician. Science 289: 1920-1921.
REDECKER, D.; KODNER, R.; GRAHAM, L.E. 2002. Palaeoglomus grayi from the
Ordovincian. Mycotaxon 84: 33 – 37.
REMY, W.; TAYLOR, T.N.; HASS, H.; KERP, H. 1994. Four Hundredmillion-year-old
vesicular arbuscular mycorrhizae. Proceedings of the National Academy of Sciencies, USA
91: 11841-11843.
REZENDE, J. de O. 2000. Solos coesos dos Tabuleiros Costeiros: Limitações agrícolas e
manejo. Salvador: SEAGRI, SPA. Série Estudos Agrícolas, 1. 117p.

80
RIBEIRO, L.P. 2001. Evolução da cobertura pedológica dos tabuleiros costeiros e a gênese
dos horizontes coesos. in: workshop coesão em solos dos tabuleiros costeiros. Aracaju.
Anais... Aracaju: EMBRAPA Tabuleiros Costeiros, 93-121p.
RIBEIRO, L.P. 1998. Os Latossolos Amarelos do recôncavo Baiano: gênese, evolução e
degradação. Salvador, SEPLANTEC/CADCT. 99p.
RIBEIRO, L.P. 1986. Sílica em horizontes coesos de solos da Bahia. Universitas, n.38. 59-
80p.
RIOS, T.; SOUZA, R.G.D.; MAIA, L.C.; OEHL, F.; LIMA, C.E.P. 2013. Arbuscular
mycorrhizal fungi in a semi-arid, limestone mining-impacted area of Brazil. Acta Botanica
Brasilica, 27(4): 688-693.
RODRÍGUEZ-ECHEVERRÍA, S.; FREITAS, H. 2006. Diversity of AMF associated with
Ammophila arenaria ssp. Arundinacea in Portuguese sand dunes. Mycorrhiza 16(8): 543-
552.
SANTOS, I.S. 2001. Fungos micorrízicos arbusculares em ambiente de Mata Atlântica e de
Eucaliptos na região de Entre Rios, Bahia. Salvador, Dissertação de Mestrado. Universidade
Federal da Bahia. 69p.
SANTOS, O.M.; VINHA, S.G. 1982. Ocorrência de micorrizas em árvores nativas do Sul da
Bahia. 1. Estação Ecológica do Pau Brasil. Theobroma 12: 261 – 265.
SCHENCK N.C.; PÉREZ Y. 1900. Manual for the identification of VA mycorrhizal fungi.
Gainesville: Synergistic Publications. 286 p.
SCHLOTER, M.; DILLY, O.; MUNCH, J.C. 2003. Indicators for evaluating soil quality.
Agriculture, Ecosystems and Environment 98: 255-262.
SCHÜßLER, A.; SCHWARZOTT, D.; WALKER, C. 2001. A new phylum, the
Glomeromycota: phylogeny and evolution. Mycological Research 105: 1413 – 1421.

81
SIEVERDING, E. 1991. Vesicular-arbuscular mycorrhiza management in tropical
agrosystems. Eschborn: Technical Cooperation, Germany. 371 p.
SIEVERDING, E.; SILVA, G.A.; BERNDT, R.; OEHL, F. 2014. Rhizoglomus, a new genus
of the Glomeraceae. Mycotaxon 129: 373-386.
SILVA, M.M.F. 1997. Impactos Ambientais no Município de Tibau do Sul-RN. Monografia
em Ciências Biológicas. Universidade Federal Rural de Pernambuco, Recife.
SILVA, C.F.; PEREIRA, M.G.; DA SILVA, E.M.R.; CORREIA, M.E.F.; SAGGIN-JÚNIOR,
O.J. 2006. Fungos micorrízicos arbusculares em áreas no entorno do Parque Estadual da Serra
do Mar em Ubatuba (SP). Revista Caatinga 19(1).
SILVA, D.K.A.; PEREIRA, C.M.R.; SOUZA, R.G.; SILVA, G.A.; OEHL, F.; MAIA, L.C.
2012. Diversity of arbuscular mycorrhizal fungi in resting and dunes areas in Brazilian
Northeast. Biodiversity and Conservation 21: 2361–2373.
SILVA, I.R.; DE MELLO, C.M.A.; NETO, R.A.F.; DA SILVA, D.K.A.; DE MELO, A.L.;
OEHL, F.; MAIA, L.C. 2014. Diversity of arbuscular mycorrhizal fungi along an
environmental gradient in the Brazilian semiarid. Applied Soil Ecology, 84: 166-175.
SILVA, D.K.A.; GOTO, B.T; OEHL, F.; DA SILVA, G.A.; NOBRE, C.P.; PEREIRA
C.M.R.; MELLO, C.M.A.; ASSIS, D.M.A.; MARINHO, F.; SILVA, I.R.; PONTES, J.S.P.;
JOBIM, K.; VIEIRA, L.C.; SOUZA, N.M.F.; LIMA, R. A.A.; MAIA, L.C. 2014. Arbuscular
mycorrhizal fungi: new records in Northeast of Brazil. Boletim do Museo de Biologia Mello
Leitão (N. Sér.) 36:35-50.
SILVA, D.K.A.; COUTINHO, F.P.; ESCOBAR, I.E.C.; DE SOUZA, R.G.; OEHL, F.,
SILVA, G.A.; MAIA, L.C. 2015. The community of arbuscular mycorrhizal fungi in natural
and revegetated coastal areas (Atlantic Forest) in northeastern Brazil. Biodiversity and
Conservation, 24(9): 2213-2226.

82
SILVA, D.K.A.; DE SOUZA, R.G.; DE ALENCAR VELEZ, B.A.; DA SILVA, G.A.;
OEHL, F.; MAIA, L.C. 2015. Communities of arbuscular mycorrhizal fungi on a vegetation
gradient in tropical coastal dunes. Applied Soil Ecology, 96: 7-17.
SIQUEIRA, J. O.; HUBBELL. D. H.; SCHENCK. N. C. 1982. Spore germination and germ
tube growth of a vesicular-arbuscular mycorrhizal fungus in vitro. In: D. K. ARORA, B.
RRAI, K. G. MUKERJI & G. R. KNUDSEN. Handbook of applied mycology: soil and
plants. Varanasi, Banaras Hindu University, 1992, 3-34p.
SIQUEIRA, J.O.; SYLVIA, D.; GIBSON, J.; HUBBEL, D. 1985. Spore, germination, and
germ tubes of vesicular arbuscular mycorrhizal fungi. Canadian Journal of Microbiology
31: 965-997p.
SIQUEIRA, J.O. Micorrizas: forma e função. 1986. In: REUNÃO BRASILEIRA SOBRE
MICORRIZAS, 1. Lavras. Anais.Lavras: FAEPE. 5- 32p.
SIQUEIRA, J.O. 1994. Micorrizas arbusculares. In Microrganismos de importancia
agrícola. (R.S. Araújo & M. Hungria, eds.). Embrapa-SPI, Brasília. 235-249p.
SIQUEIRA, J.O.; KLAUBERG FILHO, O. 2000. Micorrizas arbusculares: a pesquisa
brasileira em perspectiva. In: NOVAIS, R.F. de; ALVAREZ, V.H.; SCHAEFER, C.E.G.R.
(Ed.). Tópicos em ciência do solo. Viçosa: UFV, v. 1, 235-264 p.
SIQUEIRA, J.S.; LAMBAIS, M.R.; STÜRMER, S.L. 2002. Fungos Micorrízicos
Arbusculares: Características, associação simbiótica e aplicação na agricultura.
Biotecnologia Ciência e Desenvolvimento 25: 12-21.
SOBRAL, L. F.; SILVEIRA, K. R. da; VIANA, R. D. 2008. Correlações entre os métodos
Mehlich 1, Mehlich 3 e resina para análise de P e K em solos dos Tabuleiros Costeiros.
Boletim de Pesquisa e Desenvolvimento 33. Aracaju: Embrapa Tabuleiros Costeiros.
SOUZA, R.G.; MAIA, L.C.; SALES, M.; TRUFEM, S.F.B. 2003. Diversidade e potencial de
infectividade de fungos micorrízicos arbusculares em área de caatinga, na Região de Xingó,
Estado de Alagoas, Brasil. Revista Brasileira de Botânica 26 (1): 49-60.

83
SOUZA, F.A.; SILVA, I.C.L.; BERBARA, R.L.L. 2007. Fungos Micorrízicos Arbusculares:
muito mais diversos do que se Imaginava. Biodiversidade 5: 551-556.
SOUZA, F.A.; SILVA, I.C.L.; BERBARA, R.L.L. 2008. Fungos micorrízicos arbusculares:
muito mais diversos do que se imaginava. In: Moreira, F. M. S.; Siqueira, J. O.; Brussaard, L.
(Ed.) Biodiversidade do solo em ecossistemas brasileiros. Lavras: Editora UFLA. 483-536
p. Parceria: UFRJ; UFRRJ.
SOUZA, F.A.; STÜRMER, S.L.; CARRENHO, R.; TRUFEM, S.F.B. 2010. Classificação e
taxonomia de fungos micorrízicos arbusculares e sua diversidade e ocorrência no Brasil. In:
SIQUEIRA, J.O.; SOUZA, F.A.; CARDOSO, E.J.B.N.; TSAI, S.M. (Org.). Micorrizas: 30
anos de pesquisas no Brasil. Lavras: Editora UFLA, 15–73 p.
SOUZA, R.G.; SILVA, D.K.A.; MELLO, C.M.A.; GOTO, B.T.; SILVA, F.S.B.; SAMPAIO,
E.V.S.B.; MAIA, L.C. 2013. Arbuscular mycorrhizal fungi in revegetated mined dunes. Land
Degradation & Development 24(2): 147-155.
SUGUIO, K.; MARTIN, L.; BITTENCOURT, A. C. S. P.; DOMINGUES, J.M.L.; FLEXOR,
J.; AZEVEDO, A.E.G. 1985. Flutuações relativas do mar durante o Quaternário Superior ao
longo do litoral brasileiro e suas implicações na sedimentação costeira. Revista Brasileira de
Geociências, São Paulo, v. 15, n. 4, 273-286p.
SMITH, S.E.; READ, D.J. The symbiontes forming VA mycorrhizas. 1997. In: SMITH, S. E.;
READ, D.J. (Ed.). Mycorrhizas symbiosis. San Diego: Academic Press: 33-80.
SMITH, S.E.; READ, D. 2008. Mycorrhizal Symbiosis. 3ª ed. San Diego: Academic Press.
STEHMANN, J.R.; FORZZA, R.C; SALINO, A.; SOBRAL, M.; DA COSTA D.P.;
KAMINO L.H.Y. 2009. Diversidade taxonômica na Floresta Atlântica. Plantas da floresta
atlântica (Vol. 1). Rio de Janeiro: Jardim Botânico do Rio de Janeiro, 3-12.
STÜRMER, S.L.; BELLEI, M.M. 1994. Composition and seasonal variation of spore
populations of arbuscular mycorrhizal fungi in dune soils on the island of Santa Catarina,
Brazil. Canadian Journal of Botany 72(3): 359-363.

84
STÜMER, S.L.; SIQUEIRA, J.O. 2005. Diversity of Arbuscular Mycorrhizal Fungi in
Selected Ecosystems. In: MOREIRA, F.M.S.; SIQUEIRA, J.O.; 63 BRUSSARD, L. (Ed.).
Soil biodiversity in Amazonian and other brazilian ecosystems. Wallingford: CABI
Publishing.
STÜRMER, S.L.; KLAUBERG, F.; QUEIROZ, M.H.D.; MENDONÇA, M.M.D. 2006.
Ocurrence of arbuscular mycorrhizal fungi in soils of early stages f a secondary sucession of
Atlantic Forest in South Brazil. Acta Botanica Brasílica 20: 513 – 531.
STÜRMER, S.L.; SIQUEIRA, J.O. 2010. Species richness and spore abundance of arbuscular
mycorrhizal fungi across distinct land uses in Western Brazilian Amazon. Mycorrhiza 21(4):
255-267.
STÜRMER, S.L.; SIQUEIRA, J.O. 2013. Fungos Micorrízicos. In: MOREIRA, F.M.S.;
CARES, J.E.; ZANETTI, R.; STÜRMER, S.L. (Eds.). O Ecossistema solo: componentes,
relações ecológicas e efeitos na produção vegetal. Lavras: Editora UFLA, p. 289-311.
STÜERMER, S.L.; STÜERMER, R.; PASQUALINI, D. 2013. Taxonomic diversity and
community structure of arbuscular mycorrhizal fungi (Phylum Glomeromycota) in three
maritime sand dunes in Santa Catarina state, south Brazil. Fungal Ecology 6(1): 27-36.
SYLVIA, D.M.; FUHRMANN, J.J.; HARTEL, P.G.; ZUREBER, D.A. 1998. Principles and
applications of the soil microbiology. New Jersey: Prentice Hall, 551p.
TABARELLI, M.; PINTO, L.P.; SILVA, J.M.C.; HIROTA, M.M.; BEDÊ, L.C. 2005.
Desafios e oportunidades para a conservação da biodiversidade na Mata Atlântica brasileira.
Megadiversidade 1: 132 – 138.
THAXTER, R. 1922. A revision of Endogonaceae. Proceedings of the American Academy
of Art and Sciences 57: 291–351.

85
TRUFEM, S.F.B. 1996. Methods for the assessment of diversity in Mycorrhizae. In: Bicudo
CEM, Menezes NA (Eds.). Biodiversity in Brazil: first approach. São Paulo: CNPq, p. 49–
63.
TRUFEM, S.F.B.; BONONI, V.L.R. 1985. Micorrizas vesículo-arbusculares de culturas
introduzidas em áreas de cerrado. Rickia 12; 165-187.
TRUFEM, S.F.B.; OTOMO, H.S.; MALATINSZKY, S.M.M. 1989. Fungos micorrízicos
vesículos arbusculares em rizosferas de plantas em dunas do Parque Estadual da Ilha do
Cardoso, São Paulo, Brasil. Acta Botanica Brasilica 3: 141-152.
TRUFEM, S.F.B. 1990. Aspectos ecológicos de fungos micorrízicos vesículoarbusculares da
mata tropica tropical úmida da Ilha do Cardoso, SP, Brasil. Acta Botanica Brasilica 4: 31-45.
TRUFEM, S.F.B.; VIRIATO, A. 1990. Fungos micorrízicos vesículo-arbusculares da Reserva
Biológica do Alto da Serra de Paranapiacaba, São Paulo, Brasil. Revista brasileira de
Botânica 13; 49-54.
TRUFEM, S.F.B.; MALATINSZKY, S.M.M.; OTOMO, H.S. 1994. Fungos micorrízicos
arbusculares em rizosferas de plantas do litoral arenoso do Parque Estadual da Ilha do
Cardoso, SP, Brasil. Acta Botanica Brasilica 8: 219-229.
TRUFEM, S.F.B.; MALATINSZKY, S.M.M. 1995. Fungos micorrízicos arbusculares de
Melastomataceae e outras plantas nativas resistentes e sensíveis à poluição na Reserva
Biológica do Alto da Serra de Paranapiacaba, SP, Brasil. Hoehnea 22(1/2); 77-89.
TRUFEM S.T.B. 1995. Aspectos ecológicos de fungos micorrizicos arbusculares na rizosfera
de plantas da restinga da Ilha do Cardoso, SP, Brasil. Revista Brasileira de Botânica, vol.
18, n.1. 51-60p.
TULASNE, L.R.; TULASNE, C. 1845. Fungi nonnulli hypogaei, novi v. minus cogniti act.
Nuovo Giornale Botanico Italiano 2: 35–63.

86
TURRINI, A.; SBRANA, C.; STRANI, P.; PEZZAROSSA, B.; RISALITI, R.;
GIOVANNETTI, M. 2010. Arbuscular mycorrhizal fungi of a Mediterranean island
(Pianosa), whitin a UNESCO Biosphere Reserve. Biology and Fertility of Soils 46: 511-520.
TURRINI, A.; GIOVANNETTI, M. 2011. Arbuscular mycorrhizal fungi in national parks,
nature reserves and protected areas worldwide: a strategic perspective for their in situ
conservation. Mycorrhiza 22(2); 81-97.
VAN DER HEIJDEN, M.G.A.; KLIRONOMOS, J.N.; URSIC, M.; MOUTOGLIS, P.;
STREITWOLF-ENGEL, R.; BOLLER, T.; WIEMKEN, A.; SANDERS, I.R. 1998.
Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and
productivity. Nature 396: 69-72.
VAN DER HEIJDEN, M.G.A.; BOLLER, T.; WIEMKEN, A.; SANDERS, I.R. 1998.
Different arbuscular mycorrhizal fungal species are potential determinants of plant
community structure. Ecology 79: 2082-2091.
WALKER, C.; SANDERS, F.E. 1986. Taxonomic concepts in the Endogonaceae. III. The
separation of Scutellospora gen. nov. From Gigaspora Gerd. & Trappe. Mycotaxon.
WANG, B.; QIU, Y.L. 2006. Phylogenetic distribution and evolution of mycorrhizas in land
plants. Mycorrhiza 16; 299-363.
WETTSTEIN, F. 1915. Geosiphon Fr Wettst, eine neue, interessante Siphonee.
Österreichische botanische Zeitschrift 65: 145-56.
ZAMBOLIM, L.; SIQUEIRA, J.O. 1985. Importância e potencial das Associações
micorrizicas para a agricultura. Empresa de Pesquiza Agropecuária de Minas Gerais
(EPAMIG). Serie/Documentos 26. Brasil.
ZANGARO, W.; MOREIRA, M. 2010. Micorrizas Arbusculares nos Biomas Floresta
Atlântica e Floresta de Araucária. In: SIQUEIRA, J.O.; SOUZA, F.A.; CARDOSO, E.J.B.N.;
TSAI, S.M. (Org.). Micorrizas: 30 anos de pesquisas no Brasil. Lavras: Editora UFLA, p
279 - 310.

87
ZHU, H.H.; YAO, Q.; SUN, X.T.; HU, Y.L. 2007. Colonization, ALP activity and plant
growth promotion of native and exotic arbuscular mycorrhizal fungi at low pH. Soil Biology
& Biochemistry 39: 942–950p.





























