Embed Size (px)
Citation preview

0
U
F
R
N
U
F
R
N
MARCELO ANDERSON BARBOSA NASCIMENTO
ESTUDO DA EXPRESSÃO IMUNO-
HISTOQUÍMICA DE SOX2, FGF-10 E WNT-1 EM
LESÕES ODONTOGÊNICAS EPITELIAIS
BENIGNAS
NATAL-RN
2018

1
MARCELO ANDERSON BARBOSA NASCIMENTO
ESTUDO DA EXPRESSÃO IMUNO-HISTOQUÍMICA DE SOX2, FGF-10
E WNT-1 EM LESÕES ODONTOGÊNICAS EPITELIAIS BENIGNAS
NATAL/RN
2018

2

3
UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE
CENTRO DE CIÊNCIAS DA SAÚDE
PROGRAMA DE PÓS-GRADUAÇÃO EM PATOLOGIA ORAL
ESTUDO DA EXPRESSÃO IMUNO-HISTOQUÍMICA DE SOX2, FGF-10 E WNT-1
EM LESÕES ODONTOGÊNICAS EPITELIAIS BENIGNAS
NATAL/RN
2018
Tese apresentada ao Programa de Pós-Graduação em
Patologia Oral da Universidade Federal do Rio Grande
do Norte, como parte dos requisitos para a obtenção do
Título de Doutor em Patologia Oral.
Doutorando: Marcelo Anderson Barbosa Nascimento
Orientadora: Profª. Drª. Lélia Batista de Souza

4

5

6
Dedicatória

7
Aos meus pais, Joaquim Marcelino (in memoriam) e Luiza Maria, pela vida e pela educação que me fez alcançar além do que eu sonhava.
Às minhas irmãs, Aninha e Iara, que sonham comigo apesar da distância.
Ao meu irmão Marcieudo, pela amizade e apoio.
Ao meu sobrinho, Enzo, por encher meus dias de alegria e esperança.
A Kleyber, parceiro paciente nestes anos.
A todos os meus professores, do “pré-escolar” até agora, vocês são minha inspiração.
“A verdadeira alegria vem da harmonia profunda entre as pessoas, que todos experimentam no seu coração e que nos faz sentir a beleza de estar juntos, de apoiar-se mutuamente no
caminho da vida.”
Jorge Mario Bergoglio

8
Agradecimentos

9
Àquele que me feriu de Amor, que me constrói quando me desfaço, que me
faz ir avante quando temo sair do lugar, minha constante e eterna gratidão.
À minha orientadora, Profª Drª Lélia Batista de Souza, pela desmedida
paciência comigo durante esses anos de Pós-Graduação, aproveito para pedir
perdão pela minha falta de dedicação. E agradeço pela sua dedicação de
vida em favor da Odontologia e pesquisa científica.
Aos professores do Programa de Pós-Graduação em Patologia Oral Prof.
Dr. Leão Pereira Pinto, Profª Drª Roseana de Almeida Freitas, Profª Drª
Hébel Cavalcante Galvão, Profª Drª Lélia Maria Guedes Queiroz, Prof Dr
Antônio de Lisboa Lopes Costa, Profª Drª Ana Myriam Costa de Medeiros,
Profª Drª Márcia Cristina da Costa Miguel, Profª Drª Érika Janine Dantas
da Silveira, Profª Drª Patrícia Teixeira de Oliveira, Prof Dr Carlos Augusto
Galvão Barbosa que por caminhos interessantes nos encontram… e neste
encontro, desenterram-nos, nos dá polimento, põe-nos no presente. Aos
poucos lapidam-nos, aos toques, com jeito, a um olhar atento para nos
tornar de uma pedra rústica à uma pedra polida, preciosa.
Profª Drª Karuza, Prof Fábio Wildson, Profª Ivana, Profª Patrícia
Teixeira, Prof Gustavo, Prof Manoel, Prof Cassiano, Prof Pedro Paulo,
Prof Dácio, Prof Alberto, Prof Cícero, Profª Rejane, Profª Jovanka, Profª
Juacema, Profª Raquel, Profª Aparecida, Profª Maria Alice, obrigado pela
inspiração, pelo apoio e incentivo, tenham a certeza que se estou aqui foi
para tentar ser o professor que vocês são para mim.
À minha turma de mestrado e doutorado, Manoela, Thais, Malu, Luciana,
Vivi, Andréia, Tiago, Luiz, Nice e Laura, com a amizade que foi sendo
construída com vocês o caminho foi mais leve e prazeroso.
Aos meus irmãos de orientação, Keila, Adriana, Jamile, Vilson, Leorik,
Dáurea, obrigado pelo apoio intelectual e pela cumplicidade.

10
À Denise (amor meu), e demais xuxus, Ana Luiza, Rose, Melka (mestra),
Clarissa, Bárbara, Natália, pela imensa contribuição no início e ao longo de
nossa jornada patológica.
Aos demais colegas, Cyntia, Maiara, Stefânia, Joabe, Emeline, Felipe,
Fernanda, Jadson, Eduardo, Emília, Salomão, Rafaela, Luiz Arthur,
Amanda, Mara, Hugo, Jefferson, Marianna, Patrícia, Rodrigo, Thalita e
Angélica, obrigado pelas conversas, conselhos, risadas, ensino e companhia
carinhosa.
Aos meus coordenadores da Escola Técnica Potiguar, Disraeli e Lidiane, e
demais colegas e professores, obrigado pela confiança e compreensão durante
este período.
À Gracinha, Lourdinha, Hévio, Sandrinha e Ricardo, pelo auxílio e
dedicação desmedida para conosco.
À minha família pelo apoio e torcida.
À Fernanda Duarte, minha amiga-mana pela sua e amizade, confiança,
irmandade e tremenda paciência em meu auxílio. À Karlla Luah pela
amizade e por aguentar minhas loucuras.
Aos meus amigos (me perdoem se alguém não for citado) Filipe, Tiago, Jean
(juntos formamos aquele quarteto), Hudson, Dani (amada), Luis Carlos,
Carla Paiva, Henrique, Alberto, Carla Costa, Vanessa do Carmo, Suelanne,
Joelia, Rafael, Moniquinha, Derson, Tati (pqna), Gustavo e Ana Sampaio,
Genianny, Rafaela Nobrega, Giselle, Janaides, Belinha, Mienda e Andrew
(pela ajuda na pesquisa) que deposito meu amor e que me retribuem da
mesma forma, como gostaria de abraçá-los e agradecer por acreditarem em
mim.
Aos meus pacientes e alunos, pela confiança e compreensão.

11
Ao Programa de Pós-Graduação em Patologia Oral – PPGPO e a
Universidade Federal do Rio Grande do Norte, pela disponibilidade de
infraestruturas de material e de recursos humanos.
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq)
pelo auxílio financeiro na minha formação e desenvolvimento deste
trabalho.

12
Para vir ao que não sabes – hás de ir por onde não sabes.
Para vir a saber tudo – não queiras saber algo em nada.
João da Cruz.

13
Resumo

14
RESUMO
Os dentes desenvolvem-se a partir de interações sequenciais entre o epitélio e o mesênquima
derivado da crista neural em diferentes estágios de histodiferenciação e morfodiferenciação.
Ao final da odontogênese, espera-se que as estruturas que participaram da formação destes
tecidos desapareçam ou permaneçam quiescentes. Não é incomum que os remanescentes
epiteliais da odontogênese originem lesões, como cistos e tumores odontogênicos. No
desenvolvimento dentário precoce, a manutenção das células-tronco é regulada por uma série
de fatores de transcrição específicos, que inclui OCT-4, SOX-2, Nanog, Stat-3 e c-Myc e
diversos outros genes Homeobox e vias de transcrição (SHH, Wnt/β-catenina, FGF, BMP)
contribuem para o destino e diferenciação celular. No entanto, há a participação destes genes
e vias na patogênese de vários tipos de tumores. O objetivo do presente estudo foi avaliar a
imunoexpressão de SOX2, FGF-10 e Wnt-1 em uma série de casos de lesões odontogênicas e
alguns espécimes de germes dentários. A amostra consistiu de 20 Ceratocistos Odontogênicos
(CO), 20 Ameloblastomas sólidos (AM), 20 Tumores odontogênicos adenomatoides (TOA),
10 Tumores odontogênico epitelial calcificante (TOEC) e 05 casos de germes dentários
usados comparativamente. A imunoexpressão foi avaliada de acordo com o percentual de
células epiteliais imunomarcadas e intensidade de células positivas resultando na pontuação
de imunomarcação total (PIT) que variou de 0 a 7. A análise da imunoexpressão da SOX2
revelou positividade na maioria dos casos das lesões estudadas. A pontuação de
imunomarcação para SOX2 revelou haver diferença estatisticamente significativa entre os
grupos de lesões estudadas, com maior frequência em CO e TOEC (p <0,001). Após o
pareamento, observou-se diferença significativa entre AM e CO, AM e TOEC, CO e TOA,
CO e TOEC e, TOA e TOEC (p <0,05). A análise da imunoexpressão da FGF-10 e Wnt-1
revelou positividade em todos os casos das lesões estudadas, mas sem diferença
estatisticamente significativa entre os grupos de lesões estudadas (p = 0,628). Houve
diferença significativa em relação aos escores de positividade para Wnt-1 (p <0,001) com
maior frequência em CO e TOA. Após o pareamento, observou-se existir diferença
estatisticamente significativa entre AM e CO, AM e TOEC, CO e TOEC e, TOA e TOEC (p
<0,05). O padrão de expressão de SOX2, FGF-10 e Wnt-1, em germes dentários e nas lesões
odontogênicas aqui avaliadas, confirma a participação destas vias na odontogênese e também
no desenvolvimento das lesões odontogênicas.
Palavras-chave: Células-tronco, Lesões Odontogênicas Epiteliais Benignas; Imuno-
histoquímica; SOX2; FGF10; Wnt-1.

15
Abstract

16
ABSTRACT
Dental development occurs from sequential interactions between the epithelium and the
mesenchyme derived from the neural crest at different stages of histodifferentiation and
morphodifferentiation. At the end of tooth development, the structures that participated in the
formation of these tissues are expected to disappear or remain quiescent. It is not uncommon
that the epithelial remnants of the tooth development originate lesions such as odontogenic
cysts and tumors. In early tooth development, stem cell maintenance is regulated by specific
transcription factors, which includes OCT-4, SOX-2, Nanog, Stat-3 and c-Myc and several
other Homeobox genes and transcription pathways (SHH, Wnt/β-catenin, FGF, BMP)
contribute to cell fate and differentiation. However, there is involvement of these genes and
pathways in the pathogenesis of several types of tumors. The aim of the present study was to
evaluate the immunoexpression of SOX2, FGF-10 and Wnt-1 in a case series of odontogenic
lesions and some specimens of dental germs. The sample consisted of 20 Odontogenic
Keratocysts (OK), 20 solid ameloblastomas (AM), 20 adenomatoid odontogenic tumors
(AOT), 10 calcifying epithelial odontogenic tumors (CEOT) and 5 dental gerns for
comparison. Immunoexpression was evaluated according to the percentage of immunostained
epithelial cells and intensity of the positive cells resulting in total immunostaining score (PIT)
ranging from 0 to 7. The analysis of SOX2 immunoexpression revealed positivity in most
cases of the lesions studied. The immunostaining score for SOX2 revealed a statistically
significant difference between the groups of lesions studied, with a higher frequency in OK
and CEOT (p < 0.001). After pairing, we observed a significant difference between AM and
OK, AM and CEOT, OK and AOT, OK and CEOT, and AOT and CEOT (p <0.05). Analysis
of the FGF-10 and Wnt-1 immunoexpression revealed positivity in all cases of the lesions
studied, with no statistically significant difference between the groups of lesions studied (p =
0.628). There was a significant difference in relation to the positivity scores for Wnt-1 (p
<0.001) with higher frequency in OK and AOT. After pairing, there was a statistically
significant difference between AM and OK, AM and CEOT, OK and CEOT and, AOT and
CEOT (p <0.05). The expression pattern of SOX2, FGF-10 and Wnt-1 in dental germs and
odontogenic lesions evaluated here confirms the participation of these pathways in the tooth
development as well as in the development of odontogenic lesions.
Keywords: Stem cells, Benign Odontogenic Epithelial Lesions; Immunohistochemistry;
SOX2; FGF10; Wnt-1.

17
Lista de Ilustrações

18
LISTA DE ILUSTRAÇÕES
Figura 1. Fotomicrografia evidenciando a fraca marcação nuclear de SOX2 em
Ameloblastoma com evidente marcação em vasos sanguíneos no estroma
(Panoramic Viewer, Advance).........................................................................70
Figura 2. Fotomicrografia evidenciando a fraca marcação nuclear e citoplasmática de
SOX2 em Tumor Odontogênico Epitelial Calcificante (Panoramic Viewer,
Advance)..........................................................................................................70
Figura 3. Fotomicrografia evidenciando a fraca marcação citoplasmática de SOX2 em
Tumor Odontogênico Epitelial Calcificante com escassa marcação no material
amiloide e anéis de Liesegang (Panoramic Viewer, Advance)........................71
Figura 4. Fotomicrografia evidenciando a fraca marcação citoplasmática de SOX2 em
Tumor Odontogênico Adenomatoide (Panoramic Viewer, Advance).............71
Figura 5. Fotomicrografia evidenciando marcação nuclear de SOX2 em Ceratocisto
Odontogênico (Panoramic Viewer, Advance).................................................72
Figura 6. Fotomicrografia evidenciando a marcação nuclear e citoplasmática de SOX2
em germe dentário na fase de campânula (Panoramic Viewer, Advance)......72
Figura 7. Fotomicrografia evidenciando a marcação citoplasmática de FGF-10 em
Ameloblastoma (Panoramic Viewer, Advance)..............................................73
Figura 8. Fotomicrografia evidenciando marcação nuclear e citoplasmática de FGF-10
em Tumor odontogênico epitelial calcificante (Panoramic Viewer,
Advance)..........................................................................................................73
Figura 9. Fotomicrografia evidenciando marcação citoplasmática de FGF-10 em Tumor
odontogênico adenomatoide (Panoramic Viewer, Advance)...........................74
Figura 10. Fotomicrografia evidenciando a marcação de FGF-10 em Ceratocisto
Odontogênico, com evidente marcação em vasos sanguíneos no estroma
(Panoramic Viewer, Advance).........................................................................74

19
Figura 11. Fotomicrografia evidenciando a marcação nuclear e citoplasmática de FGF-10
em germe dentário na fase de campânula (Panoramic Viewer, Advance).......75
Figura 12. Fotomicrografia evidenciando a moderada marcação citoplasmática de Wnt-1
em Ameloblastoma com evidente marcação em vasos sanguíneos no estroma
(Panoramic Viewer, Advance).........................................................................75
Figura 13. Fotomicrografia evidenciando a fraca marcação citoplasmática de Wnt-1 em
Tumor Odontogênico Epitelial Calcificante com escassa marcação no material
amilóide (Panoramic Viewer, Advance)..........................................................76
Figura 14. Fotomicrografia evidenciando a marcação citoplasmática de Wnt-1 em Tumor
odontogênico adenomatoide (Panoramic Viewer, Advance)...........................76
Figura 15. Fotomicrografia evidenciando a marcação citoplasmática de Wnt-1 em
Ceratocisto Odontogênico com evidência em suprabasal e superficial
(Panoramic Viewer, Advance).........................................................................77
Figura 16. Fotomicrografia evidenciando a marcação nuclear e citoplasmática de Wnt-1
em germe dentário na fase de campânula, com fraca marcação nas regiões das
alças cervicais (Panoramic Viewer, Advance).................................................77
Figura 17. Fotomicrografia evidenciando a marcação nuclear de SOX2
predominantemente na camada basal e suprabasal em mucosa oral subjacente
a área do parênquima de um caso de Ameloblastoma (Panoramic Viewer,
Advance)..........................................................................................................78

20
Lista de Quadro e Tabelas

21
LISTA DE QUADROS E TABELAS
QUADRO
Quadro 1 Especificidade, clone, fabricante, diluição e incubação..................................56
TABELAS
Tabela 1. Distribuição absoluta e relativa dos casos de acordo com a pontuação de
imunomarcação total (PIT) para SOX-2 no parênquima das lesões
odontogênicas estudadas. Natal – RN, 2018……………………………...62
Tabela 2. Distribuição absoluta e relativa dos casos acordo com os escores de
positividade para SOX-2 no parênquima das lesões odontogênicas estudadas.
Natal – RN, 2018………………………………..………………………….62
Tabela 3. Tamanho da amostra, mediana, quartis 25 e 75, média dos postos, estatística
KW e significância estatística (p) para os escores de positividade para SOX2
em relação aos grupos de lesões odontogênicas estudadas. Natal – RN,
2018............………………………….………………………….………….63
Tabela 4. Avaliação da imunoexpressão de SOX2 de acordo com as lesões. Natal –
RN, 2018..………………………………………….……………………….63
Tabela 5. Distribuição dos casos de acordo com a pontuação de imunomarcação total
(PIT) para FGF-10 no parênquima das lesões odontogênicas estudadas. Natal
– RN, 2018.....................................................................................................64
Tabela 6. Distribuição absoluta e relativa dos casos acordo com os escores de
positividade para FGF-10 no parênquima das lesões odontogênicas
estudadas. Natal – RN, 2018. ……………………………..………………..65
Tabela 7. Tamanho da amostra, mediana, quartis 25 e 75, média dos postos, estatística
KW e significância estatística (p) para os escores de positividade para FGF-
10 em relação aos grupos de lesões odontogênicas estudadas. Natal – RN,
2018.....................................………………………………………………..65

22
Tabela 8. Distribuição dos casos de acordo com a pontuação de imunomarcação total
(PIT) paraWnt-1 no parênquima das lesões odontogênicas estudadas. Natal –
RN, 2018........................................................................................................66
Tabela 9. Distribuição absoluta e relativa dos casos acordo com os escores de
positividade para Wnt-1 no parênquima das lesões odontogênicas estudadas.
Natal – RN, 2018.....................................…………………………………..67
Tabela 10. Tamanho da amostra, mediana, quartis 25 e 75, média dos postos, estatística
KW e significância estatística (p) para os escores de positividade para Wnt-1
em relação aos grupos de lesões odontogênicas estudadas. Natal – RN,
2018.............................................…………………………………………..67
Tabela 11. Avaliação da imunoexpressão de Wnt-1 de acordo com as lesões. Natal –
RN, 2018.....................................…………………………………………..68
Tabela 12. Tamanho da amostra, comparação, coeficiente de correlação de Spearman (r)
e significância estatística (p) para os escores de positividade para SOX2,
FGF-10 e Wnt-1 em relação aos grupos de lesões odontogênicas estudadas.
Natal – RN, 2018...........................................................................................69

23
Abreviaturas e Siglas

24
LISTA DE SIGLAS E ABREVIATURAS
AM Ameloblastoma
APC Do inglês Adenomatous Polyposis Coli
BIO Do inglês Bromoindirubin oxime
BMI-1 Do inglês B-cell-specific moloney murine leukemia virus integration
site 1
BMP Do inglês Bone morphogenetic protein
BRAF Do inglês v-raf murine sarcoma viral oncogene homolog B1
BSA Do inglês Bovine Serum Albumin
CES Células-tronco embrionárias
c-MYC Do inglês myelocytomatosis oncogene
CNS Conselho Nacional de Saúde
CO Ceratocisto Odontogênico
DAB Diaminobenzidina
DKK1 Do inglês Dickkopf-related protein 1 precursor
DLX Do inglês Distal-less homeobox
DNA Do inglês desoxyribonucleic acid
Dvl Do inglês dishevelled segment polarity protein 1
EGF Do inglês epidermal growth factor
Erk Cinases reguladas por sinal extracelular
FGF Do inglês Fibroblast growth factor
FGFR Receptor de Fibroblast growth factor
FoxP1 Do inglês Forkhead box P1
GSK3β Glicogênio Sintase Quinase 3β
hESC Células-tronco embrionárias humanas
HMG Do inglês High mobility group
HoxB4 Do inglês Homeobox B4
IRX Do inglês Iroquois homeobox
ISL Do inglês insulin I gene

25
Klf4 Factor de Krüppel
LEF Do inglês Lymphoid enhancer-binding factor
LRP5/6 Do inglês Low-density lipoprotein receptor-related protein 5 e 6
MAPK Do inglês mitogen-activated protein kinase
Msx Do inglês Msh homeobox
Nanog Do inglês Nanog homeobox
OCT4 Fator de transcrição de ligação Octamer 4
OMS OrganizaçãoMudial da Saúde
OSX Do inglês Osterix
p63 Do inglês tumor protein p63
Pax Do inglês Paired box
PBS Do inglês phosphate buffered saline
PCP Do inglês Planar Cell Polarity
PIT Pontuação de imunomarcação total
Pitx2 Do inglês pituitary homeobox 2
PTCH Do inglês Protein patched homolog
RNA Do inglês ribonucleic acid
RTK Do inglês Receptor tyrosine kinases
RUNX Do inglês Runt-related transcription factor
Sca-1 Do inglês Stem cells antigen-1
SHH Do inglês Sonic Hedgehog
shRNA Do inglês short hairpin RNA
SISNEP Sistema Nacional de Ética em pesquisa
Smad Do inglês Sma and Mad related proteins
SMO Do inglês Smoothened
SNC Sistema nervoso central
SOX2 Região de determinação do sexo Y-box2
SPSS Do inglês Statistical Package for the Social Sciences
Sry Do inglês sex-determining region Y

26
Stat-3 Transdutor de sinal e ativador da transcrição 3
TBX Do inglês T-box
TCF Fator de células T
TGF Fator de crescimento tumoral
TOs Tumores odontogênicos
TOA Tumor odontogênico adenomatoide
TOEC Tumor odontogênico epitelial calcificante
UFRN Universidade Federal do Rio Grande do Norte
Wif1 Do inglês Wnt inhibitory factor 1
Wnt Do inglês Wingless-related integration site

27
Sumário

28
SUMÁRIO
1 INTRODUÇÃO ................................................................................................................... 31
2 REVISÃO DE LITERATURA ........................................................................................... 34
2.1 ODONTOGÊNESE ........................................................................................................ 34
2.2 CÉLULAS-TRONCO E ODONTOGÊNESE ................................................................ 36
2.3 FATOR DE TRANSCRIÇÃO SOX2 ............................................................................. 38
2.4 FGF-10 ............................................................................................................................ 41
2.5 WNT-1 ............................................................................................................................ 43
2.6 CÉLULAS-TRONCO E LESÕES ODONTOGÊNICAS .............................................. 45
2.7 AMELOBLASTOMA .................................................................................................... 46
2.8 TUMOR ODONTOGÊNICO EPITELIAL CALCIFICANTE ...................................... 48
2.9 TUMOR ODONTOGÊNICO ADENOMATOIDE ........................................................ 49
2.10 CERATOCISTO ODONTOGÊNICO .......................................................................... 50
3 PROPOSIÇÃO .................................................................................................................... 53
4 MATERIAIS E MÉTODOS ............................................................................................... 55
4.1 CONSIDERAÇÕES ÉTICAS ........................................................................................ 55
4.2 CARACTERIZAÇÃO DO ESTUDO ............................................................................ 55
4.3 POPULAÇÃO ................................................................................................................ 55
4.4 AMOSTRA ..................................................................................................................... 55
4.5 CRITÉRIOS DE SELEÇÃO DA AMOSTRA ............................................................... 56
4.5.1 Critérios de inclusão............................................................................................... 56
4.5.2 Critérios de exclusão .............................................................................................. 56
4.6 ESTUDO MORFOLÓGICO .......................................................................................... 56
4.7 ESTUDO IMUNO-HISTOQUÍMICO ........................................................................... 57
4.8 ANÁLISE DO PERFIL IMUNO-HISTOQUÍMICO ..................................................... 58
4.9 ANÁLISE ESTATÍSTICA ............................................................................................. 59
5 RESULTADOS .................................................................................................................... 61

29
5.1 ANÁLISE MORFOLÓGICA ......................................................................................... 61
5.2 RESULTADOS IMUNO-HISTOQUÍMICOS ............................................................... 62
5.2.1 Análise da imunoexpressão de SOX-2 .................................................................. 62
5.2.2 Análise da imunoexpressão de FGF-10 ................................................................ 66
5.2.3 Análise da imunoexpressão de Wnt-1................................................................... 68
5.2.4 Correlação das imunoexpressões de SOX2, FGF-10 e Wnt-1 ............................ 70
6 DISCUSSÃO ........................................................................................................................ 82
7 CONCLUSÕES .................................................................................................................... 90
8 REFERÊNCIAS .................................................................................................................. 92
ANEXO .................................................................................................................................. 101

30
Introdução

31
1 INTRODUÇÃO
Os dentes desenvolvem-se a partir de interações sequenciais entre o epitélio e o
mesênquima derivado da crista neural em um processo denominado de odontogênese, que
possui diferentes estágios de histodiferenciação e morfodiferenciação. O tamanho, forma,
estrutura dos dentes e também a posição dos dentes são determinados por genes associados a
interações e processos principais, presentes na região durante o desenvolvimento e maturação
dentária (NANCI, 2013; LI; PARADA; CHAI, 2017). Estes genes também são reguladores
centrais do desenvolvimento e estão associados às interações entre as células. Ao final da
odontogênese, ocorre uma desagregação das estruturas epiteliais que participaram na
formação dos tecidos dentários e espera-se que, com o desenvolvimento, essas células
desapareçam ou permaneçam quiescentes (ISHIKAWA et al., 2017). Durante esse processo
não é incomum que os remanescentes epiteliais da odontogênese originem lesões, como cistos
e tumores odontogênicos (AGARWAL et al., 2016; JIANG et al., 2016).
As lesões odontogênicas constituem um grupo heterogêneo de patologias que
acometem os maxilares, com origem a partir dos remanescentes epiteliais ou mesenquimais da
embriogênese dentária. Essas lesões podem variar de cistos a neoplasias verdadeiras, que por
vezes exibem manifestações clínicas variadas. Os cistos odontogênicos são patologias
relativamente frequentes na prática odontológica, sendo que os mais prevalentes são os cistos
residuais, dentígero e o Ceratocisto odontogênico (CO) (DE SOUZA et al., 2010). Essas
lesões geralmente apresentam uma evolução lenta e benigna. Entretanto, alguns cistos podem
atingir um tamanho significativo se não forem diagnosticados e tratados de forma apropriada,
resultando em importantes transtornos para os pacientes (SELVAMANI; DONOGHUE;
BASANDI, 2012).
As neoplasias derivadas dos tecidos epiteliais, ectomesenquimais ou mesenquimais
que dão origem aos dentes são denominadas de tumores odontogênicos. Algumas dessas
lesões podem apresentar um comportamento maligno, enquanto que outras possuem uma
evolução tão benigna, que parecem representar apenas malformações semelhantes a um tumor
(hamartomas) (RAMOS et al., 2013). O Ameloblastoma é uma neoplasia benigna que se
desenvolve a partir de elementos celulares epiteliais odontogênicos; caracterizado por
crescimento lento, localmente invasivo e se infiltrar entre as trabéculas ósseas, apresentando
altas taxas de recorrência (CHAUDHARY et al., 2011). O Tumor odontogênico
adenomatoide (TOA) é uma lesão de origem epitelial odontogênica evidenciando padrões
arquiteturais variados em meio a um escasso estroma de tecido conjuntivo maduro. É um

32
tumor benigno, indolente, de crescimento lento, limitado e tratado por excisão local e raras
recorrências (BASKARAN et al., 2011). O Tumor odontogênico epitelial calcificante (TOEC)
é um tumor benigno raro, de comportamento localmente invasivo, que representa menos de
1% de todos os tumores odontogênicos (EL-NAGGAR et al., 2017).
A regulação de células-tronco é fundamental para a homeostase e função dos tecidos.
Distúrbios na sua regulação pode levar à proliferação excessiva de células e muitos tumores
são originados e/ou mantidos por células-tronco tumorais. Marcadores de células-tronco são
expressos em muitas células tumorais e estão envolvidos em várias etapas de formação de
tumores. Nos tumores odontogênicos, a contribuição de células-tronco dentárias para a
geração de tumores odontogênicos ainda não é bem conhecida (SARKAR;
HOCHEDLINGER, 2013; MIRAN; MITSIADIS; PAGELLA, 2016).
No desenvolvimento dentário precoce, a manutenção das células-tronco é regulada por
uma série de fatores de transcrição específicos, que inclui OCT-4, SOX-2, Nanog, Stat-3 e c-
Myc (RAMANATHAN et al., 2018). E diversos outros genes Homeobox e vias de transcrição
(SHH, Wnt/β-catenina, FGF, BMP) contribuem para diferenciação celular. Porém, em
diversos tipos de tumores há a participação destes genes e vias em sua patogênese (HUANG;
THESLEFF, 2013; SURYADEVA; KHAN, 2015; PUTHIYAVEETIL et al., 2016). E a analise
destes genes se faz importante no estudo da patogênese de cistos e tumores odontogênicos,
considerando que estes se originam de alterações que podem ocorrer nestas vias durante a
odontogênese.
Tendo em vista os escassos estudos acerca da participação dos fatores de
pluripotência, e das vias de transcrição relacionadas a estes fatores na patogênese das lesões
odontogênicas, o objetivo de nosso estudo foi avaliar a imunoexpressão de SOX2, FGF-10 e
Wnt-1 em uma série de casos de Ceratocisto Odontogênico, Ameloblastoma, Tumor
odontogênico adenomatoide e Tumor odontogênico epitelial calcificante além de alguns
germes dentários para efeito comparativo, visando fornecer subsídios para a melhor
compreensão da participação destas proteínas no desenvolvimento e comportamento biológico
das lesões odontogênicas estudadas.

33
Revisão de Literatura

34
2 REVISÃO DE LITERATURA
2.1 ODONTOGÊNESE
A odontogênese tem início no embrião, entre a sexta e a sétima semana de vida intra-
uterina, quando células epiteliais invaginam para o ectomesênquima e começam a sofrer uma
condensação dividindo-se em dois processos: um direcionado para a região anterior,
recebendo a denominação de lâmina vestibular e outro direcionado mais profundamente (para
lingual ou palatina) conhecido como lâmina dentária. É nesta última que ocorre uma contínua
e progressiva histodiferenciação das células epiteliais e do ectomesênquima adjacente dando
origem aos germes dentários e futuros dentes (NANCI, 2013).
Os dentes desenvolvem-se a partir de interações sequenciais entre o epitélio e o
mesênquima derivado da crista neural. O primeiro sinal morfológico do desenvolvimento
dentário é um espessamento epitelial no primeiro arco branquial evidente na 6ª semana de
vida embrionária. Em seguida, assume progressivamente as fases de botão, capuz e
campânula. Os nós de esmalte primários aparecem no epitélio interno do órgão do esmalte no
estágio inicial da fase de capuz, mas desaparecem no estágio final. Posteriormente, as células
epiteliais se diferenciam em ameloblastos e os odontoblastos diferenciam-se a partir das
células mesenquimais da papila dental. Concomitantemente, fatores de crescimento que são
vitais ao desenvolvimento durante a embriogênese, como o Fator de Crescimento de
Fribroblasto (FGF), o Fator de Crescimento Epidérmico (EGF), Fator de Crescimento
Transformante (TGF) e Proteínas Ósseas Morfogenéticas (BMP), também estão presentes na
odontogênese e desempenham papéis críticos na regulação do desenvolvimento dentário e
direcionam a coordenação precisa da diferenciação celular, crescimento, migração, indução,
fusão e desintegração (KAWASAKI et al., 2015).
O tamanho, forma, estrutura dos dentes e também a posição dos dentes são
determinados por genes associados a interações e processos principais presentes na região
durante o desenvolvimento e maturação dentária. Estes genes também são reguladores
centrais do desenvolvimento e estão associados às interações entre as células. As vias incluem
genes que codificam os sinais reais, seus receptores, mediadores das vias de sinalização e
fatores de transcrição. Dentre estes, os genes Homeobox são genes reguladores que codificam
proteínas nucleares que atuam como fatores de transcrição e regulam aspectos da

35
morfogênese e diferenciação celular durante o desenvolvimento embrionário normal
(RAMANATHAN et al., 2018).
O gene homeobox de segmento muscular (Msx) no mesênquima dentário,
especificamente o Msx-1 e Msx-2, são induzidos por BMP e participam da regulação do ciclo
celular das células mesenquimais dentárias e posteriormente pela inibição da expressão de
BMP-2 e BMP-4 durante a fase de capuz. A família gênica que apresenta uma estrutura de
ligação ao DNA chamada de paired box (Pax), também funciona como fator de transcrição
presente no mesênquima. O fator de transcrição Pax-9 regula a expressão de moléculas
mesenquimais odontogênicas e a mutação neste gene pode causar agenesia dentária. O gene
Sonic Hedgehog (Shh), expresso na primeira fase da morfogênese, regula as células da papila
dentária e o tamanho do dente, e a regulação deste gene pode ser controlada pelo FGF. A via
de sinalização de Wnt/β-catenina se faz presente ao longo do epitélio e no mesênquima
odontogênico durante o desenvolvimento dentário e participa no processo de diferenciação
dos odontoblastos (THESLEFF, 2006; HUANG; THESLEFF, 2013; SURYADEVA; KHAN,
2015; PUTHIYAVEETIL et al., 2016).
A expressão de FGF, durante a morfogênese dentária, está associada a eventos de
iniciação que induzem a expressão de outros genes estimulando a proliferação celular no
epitélio. O aumento da expressão de FGF-10, que pode ser induzida pela via Wnt/β-Catenina,
mantém um compartimento de células-tronco que forneceria células epiteliais ao epitélio
dentário interno, que podem sofrer diferenciação em células do estrato intermediário ou em
ameloblastos. A diminuição ou ausência da expressão de FGF-10 interrompe a formação da
coroa e leva ao desenvolvimento da raiz (ENNES et al., 2012; AURREKOETXEA et al.,
2016).
O desenvolvimento das raízes dentárias é direcionado pela bainha epitelial de Hertwig
e durante o desenvolvimento dentário inicial, as interações recíprocas e sequenciais entre o
epitélio e o mesênquima levam à formação de dentina radicular, cemento e tecidos
periodontais. As principais vias de sinalização envolvidas nesses processos são as vias Tgfβ /
Bmp, Wnt, Fgf e Shh, que funcionam conjuntamente com múltiplos fatores de transcrição
para mediar as interações que orientam o desenvolvimento da raiz (LI; PARADA; CHAI,
2017).

36
Ao final da odontogênese, ocorre uma desagregação das estruturas epiteliais que
participaram na formação dos tecidos dentários e espera-se que, com o desenvolvimento,
essas células desapareçam ou permaneçam quiescentes (ISHIKAWA et al., 2017). Durante
esse processo não é incomum que os remanescentes epiteliais da odontogênese originem
lesões, como cistos e tumores odontogênicos. O espectro do comportamento biológico destas
lesões é amplo, onde podemos encontrar proliferações hamartomatosas, tumores benignos não
agressivos, tumores benignos agressivos e tumores malignos, por vezes com capacidade
metastática (AGARWAL et al., 2016; JIANG et al., 2016).
2.2 CÉLULAS-TRONCO E ODONTOGÊNESE
Mesmo após a odontogênese, ainda há células-tronco nos tecidos dentários. Células-
tronco mesenquimais humanas podem ser isoladas de dentes decíduos e permanentes e são
capazes de regenerar tecidos dentários específicos e se diferenciar em outros tecidos
(RODRÍGUEZ-LOZANO et al, 2011). As células-tronco têm o potencial de se desenvolver
em vários tipos de células diferentes no corpo durante a embriogênese e desenvolvimento
humano. Além disso, em muitos tecidos servem como um sistema de reposição celular,
dividindo-se para reabastecer outras células. Quando uma célula-tronco se divide, cada célula
tem potencial para permanecer uma célula-tronco ou se diferenciar em outro tipo de célula
com uma função mais especializada. Embora várias moléculas tenham sido implicadas na
biologia das células-tronco ao longo dos últimos anos, os modos de ação e comportamento
biológico destas moléculas permanecem incompletamente compreendidos (SARKAR;
HOCHEDLINGER, 2013).
Os dentes exibem diversas formas de reparo em resposta a danos e as células-tronco da
polpa dentária fornecem uma fonte de células para substituir as danificadas e facilitar o reparo
(MIRAN; MITSIADIS; PAGELLA, 2016). As células-tronco estão presentes em várias partes
do dente decíduo ou permanente, como o ligamento periodontal, polpa, folículo, as raízes em
crescimento, assim como em mucosa oral, e desempenham papéis dinâmicos no
desenvolvimento e suporte dos dentes. As células-tronco mesenquimais da polpa de dentes
decíduos esfoliados podem ser obtidas com facilidade, tornando-as uma fonte de células-
tronco autólogas para uso potencial na restauração de tecido pulpar vital removido devido à
infecção, na regeneração do ligamento perdido na doença periodontal e na geração de

37
estruturas dentárias parciais ou completas para formar implantes biológicos (JUURI et al.,
2013b; ABDUWELI et al., 2014; ORSINI et al., 2015; SHARPE, 2016).
Um nicho de células-tronco é definido como o microambiente em que estas células
estão situadas nos tecidos e órgãos. A interação com diversos tipos de células pode regular a
diferenciação celular nestes nichos. Assim, no desenvolvimento embrionário precoce, a
manutenção das células-tronco e o destino das células é regulada por uma série de fatores de
transcrição específicos, que inclui o fator de transcrição de ligação Octamer 4 (OCT-4), a
região de determinação do sexo Y-box2 (SOX-2), a proteína homeobox Nanog, o transdutor
de sinal e ativador da transcrição 3 (Stat-3) e alguns outros fatores, por exemplo, c-Myc e
factor de Krüppel (Klf4) (RAMANATHAN et al., 2018).
Alguns estudos avaliaram o padrão de expressão dos fatores de transcrição de células-
tronco pluripotentes, tais como o Oct-4, SOX2, Nanog e Stat-3, bem como o seu potencial
envolvimento na regulação dos nichos de células-tronco em vários estágios da morfogênese
inicial in situ de dentes humanos. De um modo geral, observou-se que todos os fatores de
transcrição analisados estavam ativos durante a morfogênese fetal dos dentes humanos.
Portanto, a inter-relação entre esses fatores de transcrição é complexa e está relacionada à
formação do nicho de células-tronco e que regulam a diferenciação em ameloblastos e
odontoblastos (ZHANG et al., 2012; CUNHA et al., 2013).
Juuri et al. (2013b) avaliaram germes dentários em distintas fases do
desenvolvimento, obtidos a partir de mamíferos e répteis. Foi observado que a expressão de
SOX-2 apresentava um padrão similar em células dentárias epiteliais, assim como no epitélio
oral em todas as espécies, e em cortes histológicos sucessivos da lâmina dentária em
mamíferos e répteis. As células do epitélio dental positivas para SOX-2 apresentavam
competência para formação de dentes sucessores e que esse fator de transcrição pode ser um
marcador de células epiteliais odontogênicas progenitoras que não finalizaram seu processo
de diferenciação. Após a supressão condicional de SOX2 em ratos, observou-se o crescimento
anormal do epitélio responsável pela formação dos molares posteriores, o epitélio oral foi
marcadamente hiperplásico e a lâmina dental apresentou-se alongada.

38
2.3 FATOR DE TRANSCRIÇÃO SOX2
Durante o desenvolvimento, regulação, destino, decisão e diferenciação de células-
tronco progenitoras o gene SRY (região Y determinante do sexo) - box 2 (SOX2) e seus
fatores de transcrição desempenham inúmeras funções. Proteínas Sox se ligam a regiões no
DNA e podem formar complexos com outros fatores de transcrição, que são regulados em
vários níveis e agem de uma forma dose-dependente. Vinte genes Sox que compartilham um
domínio DNA altamente conservado HMG (do inglês High mobility group) foram
identificados. O domínio de HMG medeia a ligação no DNA e foi originalmente identificado
no gene do fator determinante testicular Sry (região determinante de sexo Y) em mamíferos.
Com base nas semelhanças no domínio HMG, genes Sox são divididos em subgrupos de A a
H. Os membros do mesmo subgrupo compartilham propriedades bioquímicas semelhantes e
têm funções sobrepostas e redundantes (JUURI et al., 2013b).
A proteína SOX2 foi conhecida inicialmente por regular a autorrenovação de células-
tronco embrionárias em ratos e humanos (CES). É também importante para a manutenção de
células-tronco em vários tecidos adultos, incluindo o cérebro e traqueia e é um dos principais
fatores de transcrição para o estabelecimento de células-tronco pluripotentes induzidas.
Recentemente, a amplificação do gene de SOX2 tem sido associada com o desenvolvimento
de carcinoma de células escamosas em vários sítios tais como o pulmão e o esôfago. Esses
diferentes papéis para SOX2 envolvem complicadas redes de regulação que consistem em
microRNAs, quinases e moléculas de sinalização. Embora os níveis de SOX2 sejam
modulados transcricionalmente e translacionalmente, a modificação pós-translacional também
é importante para as várias funções do SOX2. Em diversos casos de neoplasias malignas,
altos níveis de SOX2 estão correlacionados com pior prognóstico e aumento da proliferação
de células-tronco tumorais. Portanto, SOX2 pode ser potencialmente explorada como uma
nova via terapêutica para o tratamento de câncer (LIU et al. 2013). SOX2, juntamente com
Oct4 e Nanog, mantem a célula em estado de célula-tronco; estes genes são induzidos por
diversos fatores como FGF, β-catenina, Erk ½ e Smad 2 e 3. Porém, outros fatores (BMP,
Wnt) induzem estas células a diferenciação. São reconhecidos 20 tipos de genes Sox que
foram descobertos em ratos e humanos (SCHEPERS; TEASDALE; KOOPMAN, 2002;
SARKAR; HOCHEDLINGER, 2013).
Em sua maioria, cada gene possui funções específicas, porém, diferentes fatores Sox
podem ter diferentes funções biológicas em um mesmo domínio de ligação no DNA.

39
Seletividade de genes-alvo por diferentes fatores Sox pode ser alcançado através de afinidade
diferencial para determinadas sequências flanqueadoras próximas aos locais de consenso Sox,
ou através de homo ou heterodimerização entre proteínas Sox, modificações pós-
translacionais de fatores Sox, ou interação com outros co-fatores. Esta versatilidade molecular
pode assim explicar porque os mesmos fatores Sox podem desempenhar papéis moleculares e
funcionais muito diferentes em contextos biológicos distintos (WEGNER, 2010).
A expressão de SOX2 é regulada e modulada extra e intracelularmente. Sinais
extracelulares pertencentes às principais vias de sinalização pode positiva ou negativamente
controlar os níveis de expressão SOX2. Por exemplo, a expressão de SOX2 é induzida pela
sinalização Wnt nas células da papila gustativa em desenvolvimento, enquanto que a
superexpressão de SOX2 em osteoblastos leva à inibição da via de sinalização Wnt canônica.
SOX2 também suprime a diferenciação dos osteoblastos, pela inibição de genes Adenomatous
Polyposis Coli (APC) e Glicogênio Sintase Quinase (GSK3ß), que são reguladores negativos da
via Wnt. Além disso, a sinalização FGF está associada à regulação SOX2: sinalização FGF
regula negativamente SOX2 durante o desenvolvimento do intestino anterior, enquanto que
em progenitores osteoblastos, FGF regula positivamente SOX2 (JUURI et al., 2013b).
O estudo de Seo et al. (2011) mostrou que essas múltiplas funções da SOX2 são
codificadas em domínios distintos. A função de autorrenovação de SOX2 é dependente da sua
atividade de transcrição e requer tanto a sua ligação com o DNA, assim como a ativação de
regiões C-terminais, enquanto apenas a terceira região de transativação C-terminal é
necessária para a ligação com β-catenina, interferindo na transcrição induzida por Wnt. Os
resultados deste estudo com relação a análise de expressão gênica em osteoblastos que não
expressam SOX2 apoiam fortemente a noção de que SOX2 contribui para a manutenção das
células-tronco, o gene SOX2 inibe a diferenciação através da diminuição da sinalização Wnt
por mecanismos trancricionais e pós-transcricionais, e que os fatores APC e GSK3β, que são
reguladores negativos da via Wnt, são alvos diretos de SOX2 em osteoblastos. Vários genes,
tais como o FoxP1 e BMI-1, estão associados com a manutenção das células em um estado de
não diferenciação e encontram-se regulados negativamente com a inativação de SOX2.
As mutações nos componentes da via Wnt, como APC e β-catenina que participam da
sinalização tecidual constitutiva, estão causalmente ligadas ao desenvolvimento de muitos
tipos de câncer. A expressão do gene Sox também é desregulada em uma ampla variedade de
cânceres humanos e há evidências de que os fatores SOX afetem a tumorigênese modulando a

40
atividade da β-catenina e a expressão de genes-alvo de Wnt oncogênicos, como Ciclina-D1 e
c-Myc (BASU-ROY et al.,2012; KORMISH; SINNER; ZORN, 2010).
Banerjee et al. (2016) estudaram a expressão de SOX2 e Oct-4 em folículos dentários,
cisto radicular, cisto dentigero, ceratocisto odontogênico, ameloblastoma, tumor odontogênico
adenomatoide e carcinoma ameloblástico. Os resultados mostraram a presença de células com
perfil de células-tronco nos tecidos normais e de lesões odontogênicas. SOX2 foi o fator mais
consistente e confiável na detecção de células-tronco. As expressões destes fatores nas
células-tronco são mantidas após a transformação tumoral e, provavelmente, sugerem que não
há alteração fenotípica das células-tronco na progressão do estado embrionário normal para o
seu componente tumoral. A quantificação e a localização indicam o provável papel das
células na patogênese das lesões.
Para que ocorra a manutenção de uma neoplasia se faz necessário um reservatório de
células com capacidade de autorrenovação, denominadas de células-tronco tumorais. Basu-
Roy et al. (2012) avaliaram a expressão de SOX2 em linhagens de células de osteossarcoma
em ratos e humanos e observaram uma alta expressão deste gene, assim como em amostras
tumorais. Células de osteossarcoma têm a capacidade aumentada de crescimento em
suspensão e organizam-se em aglomerados denominados de osteoesferas, que exibem alta
expressão de SOX2 e o marcador de células-tronco, Sca-1. A depleção de SOX2 por shRNAs
nas células derivadas de osteossarcoma de roedores reduz drasticamente suas propriedades
transformadoras in vitro e sua capacidade para formar tumores. Células com depleção de
SOX2 perdem a capacidade de formar osteoesferas e diferenciaram-se em osteoblastos
maduros, que exibem uma diminuição da expressão do gene Sca-1 e aumento da via de
sinalização Wnt. Portanto, SOX2 é necessária para a proliferação e autorrenovação de células
em osteossarcoma e antagoniza a ação pró-diferenciação de Wnt, suportando a ideia da ação
de supressão tumoral da via Wnt.
A sinalização Wnt/β-catenina regula o destino celular durante o desenvolvimento de
órgãos e a manutenção do tecido pós-natal, mas sua contribuição para a especificação de
diferentes linhagens epiteliais de pulmão ainda é incerta. Para esclarecer esta questão,
Hashimoto et al. (2012) utilizaram uma abordagem com recombinase Cre (Cre)-LoxP para
ativar a sinalização canônica Wnt ectópica no desenvolvimento endodérmico de pulmão.
Observou-se que a ativação Wnt dentro da endoderme distal de pulmão permitiu o
desenvolvimento normal do epitélio alveolar, mas levou à perda de epitélio bronquiolar e

41
ectasia de vias aéreas distais, levando a expressão ectópica de um fator estimulador linfóide e
um fator de células T (LEF e TCF, respectivamente) e ausência do gene SRY (região Y
determinante do sexo) - box 2 (SOX2) e expressão da proteína p63 (p63) em derivados
proximais. A perda condicional de SOX2 nas vias aéreas resultou em defeitos na
diferenciação epitelial juntamente com a ativação ectópica da via canônica Wnt. Estes dados
sugerem que Wnt regula negativamente um programa de sinalização dependente de SOX2,
necessária para a progressão do desenvolvimento da linhagem bronquiolar.
2.4 FGF-10
A família de proteínas de sinalização de fator de crescimento de fibroblastos (FGF)
inclui 22 membros que foram identificados com base na homologia de sequência de DNA.
Dezoito destes FGFs funcionam como ligantes que interagem com quatro receptores tirosina
quinases (RTKs) em camundongos e humanos. Os quatro FGFs restantes (FGF 11-14) são
proteínas intracelulares que não interagem com os receptores de FGF (FGFRs). Uma quinta
proteína semelhante a FGFR (FGFRL1) também foi identificada que carece de um domínio de
tirosina quinase intracelular e provavelmente regula negativamente os FGFRs ao interagir
com os ligantes. A sinalização FGF, comumente estudada em organismos que vão desde
cnidários até humanos, age pleiotropicamente durante a embriogênese, por possuir
propriedade de determinar mais de uma característica fenotípica, e também regula múltiplas
funções homeostáticas e reparadoras em adultos. Além disso, a ativação patológica das
FGFRs está presente em muitas doenças congênitas e câncer. Várias estratégias terapêuticas
estão atualmente sendo desenvolvidas para modular a sinalização de FGFR em várias
patologias. Compreender os mecanismos que governam a sinalização de FGF é, portanto,
importante para apreciar muitos aspectos da biologia do FGF e seu papel nas patologias
(BREWER; MAZOT; SORIANO, 2016).
A análise filogenética sugere que 22 genes FGF podem ser organizados em sete
subfamílias que contenham dois a quatro membros. Os genes da subfamília Fgf1, Fgf4, Fgf7,
Fgf8 e Fgf9 codificam FGFs canônicos que são secretados, que se ligam e ativam FGFRs
tendo a heparina como cofator. Os membros da subfamília Fgf15/19 codificam FGFs
endócrinos, que se ligam e ativam FGFRs com a proteína da família Klotho como cofator. Os
genes da subfamília Fgf11 codificam FGFs intracelulares, que são proteínas não-sinalizadoras

42
que servem como cofactores para canais de sódio e outras moléculas. Ambos, ligantes e
receptores de FGF, podem se localizar no núcleo celular onde realizam funções de sinalização
que podem ser independentes da atividade de receptor tirosina quinase. FGF1 no núcleo
estimula a síntese de DNA independente de FGFR e FGF2 está associada com a proliferação
de células de glioma. Ainda não é claro se FGFs têm funções de transcrição direta ou exercem
sua atividade no núcleo através de interações com outras moléculas (ORNITZ; ITOH, 2015).
Muitas proteínas de FGF que são secretadas em condições fisiológicas, existem
principalmente como proteína instáveis e sua estabilidade relacionada ao ligante esteja
associada à sua atividade biológica, restringindo-a ao tecido vizinho em que é secretada,
regulando o potencial de sinalização ao restringir o tempo de produção da sinalização. O
sulfato de heparina também influencia o turnover e na estabilidade da estrutura proteica da
maioria de FGFs parácrinos em condições fisiológicas (FGF1-4 / 6-9 / 16-18 / 20/22), com
exceção do FGF10, cuja rápida degradação é independente da presença de heparina
(MIKOLAJCZAK, 2016).
A sinalização do FGF é um componente crítico de genes que regulam o
desenvolvimento durante a embriogênese, determinando a manutenção de populações de
células progenitoras, diferenciação e morfogênese. Vários ligantes de FGF são expressos no
desenvolvimento pulmonar, incluindo Fgf7, Fgf8, Fgf9, Fgf10 e Fgf18. Destes, apenas o
FGF10 é absolutamente necessário para a formação inicial do pulmão. O Fgf10 é
dinamicamente expresso no mesênquima submesotelial em locais onde serão formados brotos
pulmonares futuros. Ele atua sobre o epitélio que expressa Fgfr2b para induzir crescimento de
brotos, provocando uma resposta quimiotáctica e de migração, mantendo as células epiteliais
distais em um estado indiferenciado (VOLCKAERT; DE LANGHE, 2015).
Várias evidências apontam para a importância do papel do FGF durante o
desenvolvimento dentário. A expressão de FGF se restringe inicialmente à banda epitelial
primária e persiste até o início do estágio de broto. Um papel importante deste fator é sua
participação na diferenciação de ameloblastos, como o FGF4 e FGF9, presentes no epitélio
interno órgão do esmalte (PUTHIYAVEETIL et al., 2016).
A sinalização de FGF10 no mesênquima dentário desempenha um papel fundamental
no controle da transição do estágio de formação da coroa para a raiz através da regulação da
formação de HERS durante o desenvolvimento. A ausência da expressão de Fgf10 no

43
mesênquima dentário próximo a alça cervical é necessária para o início da formação de raízes
em molares e a persistência da expressão de Fgf10 é necessária para o crescimento contínuo
de coroas em molares. O aumento da expressão de Fgf10 no estágio de transição de coroa
para raiz pode levar à inibição da formação de HERS. Portanto, sugere-se que Fgf10 controla
a troca entre a formação da coroa e da raiz durante o desenvolvimento dentário ao regular as
interações epitélio e mesênquima (LI; PARADA; CHAI, 2017).
Aurrekoetxea et al. (2016) estudaram a expressão de genes Fgf em amostras de
molares após hiperativação da via Wnt/β-catenina e observaram que alguns membros da
família FGFs, como Fgf4 ou Fgf10, são essenciais para a proliferação celular e para a
formação correta de cúspides dentárias. A expressão de Fgf9 aumentou ligeiramente em
molares tratados por 48h com reagente de Bromoindirubin oxime (BIO), que é um inibidor de
Glicogênio Sintase Quinase (GSK-3) que está relacionado com diversos processos
fisiológicos como a síntese de glicogênio, diferenciação e determinação celular fetal,
proliferação e motilidade celular. No entanto, a expressão de Fgf4 e Fgf10 aumentou
acentuadamente. A mudança de expressão mais importante foi observada para Fgf4, que
normalmente era restrita aos nós de esmalte em amostras controle e cuja presença estendeu-se
ao longo do epitélio dentário interno em amostras tratadas com BIO. A expressão de Fgf10
também foi regulada positivamente, mas esse aumento foi restringido à região mesenquimal.
2.5 WNT-1
A sinalização Wnt representa uma de várias vias, incluindo Notch-Delta, Hedgehog,
TGF-β e BMP, que estão implicadas nos processos de desenvolvimento. Cada uma dessas
vias de sinalização é conservada em sua evolução e genérica em sua atividade.
Fundamentalmente, Wnts são fatores estimuladores do crescimento e induzem à proliferação
celular por controlar o ciclo celular em vários pontos. Diferentemente de outros fatores de
crescimento, a sinalização Wnt possui a capacidade de dar forma aos tecidos em crescimento,
regular a pluripotência das células-tronco e atuar no processo como fator de crescimento
direcional durante a diferenciação celular na embriogênese e reparo tecidual (NUSSE;
CLEVERS, 2017).
Existem 19 genes Wnt identificados no genoma humano e as proteínas secretadas são
glicosiladas que, geralmente, atuam localmente de forma parácrina ou autócrica, ligando-se à

44
célula alvo que expressa um receptor Frizzled, formando um complexo com o co-receptor
LRP5/6 (proteína relacionada ao receptor de lipoproteína de baixa densidade 5/6). Isso
desencadeia a ativação de Disheveled (Dvl) e desfaz um complexo composto por GSK3β,
APC e Axin. Em um modelo em que a via é simplificada nos estados "ativado e não ativado",
o fator de transcrição β-catenina é continuamente degradado pelo complexo GSK3β / APC /
Axin, quando os ligantes Wnt estão ausentes, e a via é inativa. Na presença de ligantes Wnt
ligados aos seus receptores, não ocorre a degradação de β-catenina e esta torna-se livre para
translocar para o núcleo e ocorrer a combinação com fatores de transcrição TCF / LEF para
iniciar a transcrição de genes alvo Wnt (JANSSON; KIM; CHENG, 2015).
Comumente, a via Wnt é dividida em sinalização dependente de β-catenina (canônica),
como citado anteriormente, e independente (não canônica). As vias não canônicas são a
Planar Cell Polarity (PCP), responsável pela reorganização do citoesqueleto e a Wnt
dependente de Cálcio (Wnt/Ca+), que ativam genes envolvidos na migração e destino celular.
A via canônica é ativada pelos ligantes Wnt1, 2, 3, 3A, 8A, 8B, 10A e 10B; a via não
canônica Wnt/Ca+ é ativa por Wnt4, 5B, 6, 7, 7B e 11 enquanto a via PCP é ativada por
Wnt5A e 11 (KATOH; KATOK, 2017).
O gene Wnt1 é altamente expresso em ratos no tubo neural dorsal antes da emigração
da crista neural e tem múltiplos papéis no início do desenvolvimento do mesencéfalo. A
inativação do gene Wnt1 em camundongos resulta na perda de toda a região do mesencéfalo.
A expressão ectópica de Wnt1 ao longo do Sistema Nervoso Central (SNC) sob o controle do
amplificador HoxB4 aumenta dramaticamente a mitogênese, a proliferação e o crescimento
excessivo do mesencéfalo sem alterar o padrão de formação do SNC. Além disso, quando
Wnt1 foi expresso ectopicamente sob o controle do promotor endógeno engrailed 1 em ratos
En1Wnt1
, a formação do padrão no mesencéfalo foi mantida, mas o aumento da região dorso-
caudal foi observado (LEWIS et al., 2013).
O estado de pluripotência das células-tronco embrionárias humanas (hESC) é
caracterizado e mantido pela expressão dos fatores de transcrição Oct4, Nanog e Sox2, que
podem ser inativados pelo padrão de outros fatores e vias como FGF e Wnt (SARKAR;
HOCHEDLINGER, 2013). O perfil de expressão de RNA em uma linhagem de hESC
mostrou que essas células expressam RNA para cada um dos 19 genes Wnt e, além disso,
expressam todos os seus respectivos receptores. Cada gene possui funções especificas na
manutenção de células-tronco ou em sua diferenciação. Por exemplo, Wnt1 está relacionado

45
com a manutenção de células-tronco neurais, mas também está envolvido na diferenciação de
células-tronco mesenquimais, pois a indução da expressão de Wnt-1 em pré-adipócitos leva a
inibição da adipogênese (ROOS et al., 2000; CAMP et al., 2014; LIEN; FUCHS, 2014).
De acordo com Gaete e Tucker (2013) a ativação da via Wnt / β-catenina em órgãos
dentários de cobra, promove mudanças na proliferação e no padrão molecular da lâmina
dentária, resultando em perda do surgimento organizado de germes dentários. Esses resultados
sugerem que os componentes epiteliais são críticos para o arranjo de órgãos que se
desenvolvem em sequência e destacam o papel da sinalização Wnt / β-catenina em tais
processos.
Além de participar da embriogênese e da determinação tecidual, a via sinalização Wnt
também tem sido fortemente associada ao câncer, onde em casos de câncer gastrointestinal,
leucemias e melanomas esta via, seja no aumento da expressão de Wnt ou pela ausência de
seus reguladores, contribui para o desenvolvimento tumoral (ZHAN; RINDTORFF;
BOUTROS, 2017).
2.6 CÉLULAS-TRONCO E LESÕES ODONTOGÊNICAS
Sabe-se que tumores odontogênicos são derivados de tecidos relacionados com o
germe dentário em desenvolvimento, mas o papel das células-tronco na patogênese destes
tumores não é bem estabelecido e são escassos os estudos com marcadores moleculares para
identificar células-tronco (JUURI et al.,2013a). Compreender os mecanismos celulares e
moleculares subjacentes à autorrenovação e diferenciação, das células-tronco epiteliais e
progenitoras dentais, é essencial para o desenvolvimento de novas terapias regenerativas e
desvendar a patogênese dos tumores odontogênicos (KETTUNEN et al., 2000; CUNHA et
al.,2013; GARG et al., 2015).
Estudos imuno-histoquímicos confirmam a presença de proteínas FGF, BMP, β-
catenina e Wnt em diversos tumores odontogênicos (SIRIWARDENA et al., 2009; ALVES
PEREIRA et al., 2010; HAKIM et al., 2011; NASCIMENTO et al., 2017). Estes fatores
podem agir de forma sinérgica para manutenção de um perfil de célula-tronco somática ou
induzir a diferenciação celular nestes tumores (SARKAR; HOCHEDLINGER, 2013).

46
Juuri et al. (2013a) procuraram explorar se a expressão de SOX2 também está
associada com a formação de ameloblastoma. As estruturas epiteliais do ameloblastoma se
assemelham a alça cervical vestibular do incisivo de rato, onde foram detectadas células
tronco SOX2+. Além disso, em ameloblastoma, a produção de células semelhantes a pré-
ameloblastos é semelhante e contínuo como no incisivo de rato. Neste estudo, verificaram que
SOX2 foi expressa no epitélio do tumor em dois padrões diferentes de ameloblastoma, o
folicular e plexiforme. A expressão SOX2 localizava-se nas camadas epiteliais organizadas
em paliçada de ilhas epiteliais, bem como nas células semelhantes ao retículo estrelado do
órgão do esmalte. O mesênquima circundante foi principalmente negativo para SOX2. Uma
vez que ameloblastoma ocorre frequentemente na região do 3º molar inferior, foi realizada em
ratos a fragmentação da lâmina dentária que liga o 3º molar ao segundo molar por cortes
histológicos. Curiosamente, a expressão intensa de SOX2 foi detectada nos fragmentos da
lâmina dentária e na região posterior do 3º molar, que é o local para o desenvolvimento de um
quarto molar. A lâmina dentária de um primeiro molar humano também expressou SOX2. As
células epiteliais que cobrem parte da raiz do dente, denominada bainha epitelial de Hertwig e
os restos epiteliais de Malassez, também têm sido sugeridos como possíveis origens de
ameloblastoma, mas não foi observada expressão de SOX2 nestas células epiteliais.
Entretanto, alguns estudos genéticos e imuno-histoquímicos apontam uma menor
expressão do fator SOX2 em ameloblastoma e maior imunorreatividade em Ceratocisto
odontogênico e Carcinoma Ameloblástico (LEI et al., 2014; HEIKINHEIMO et al., 2015;
BANERJEE et al., 2016).
2.7 AMELOBLASTOMA
O Ameloblastoma é uma neoplasia benigna que se desenvolve a partir de elementos
celulares epiteliais odontogênicos; é considerado um dos tumores mais frequentemente
encontrado nos ossos maxilares, sendo caracterizado por exibir crescimento lento, localmente
invasivo e que pode se infiltrar entre as trabéculas ósseas, apresentando altas taxas de
recorrência (KUMAMOTO; OOYA, 2008; CHAUDHARY et al., 2011). Esses tumores
podem surgir a partir de restos epiteliais da lâmina dentária e de um órgão do esmalte em
desenvolvimento. Acomete principalmente adultos na terceira e quinta década de vida, sem
predileção por sexo e corresponde de 10% a 30% de todos os tumores odontogênico afetando

47
principalmente a região posterior da mandíbula (CHAUDHARY et al., 2011; LEE; KIM,
2013).
De acordo com a Organização Mundial da Saúde (OMS), o ameloblastoma contém
epitélio odontogênico e estroma fibroso sem envolvimento do ectomesênquima, apresentando
quatro variantes clínico-patológicas: o ameloblastoma sólido/multicístico, unicístico e
periférico/extraósseo e metastático (EL-NAGGAR et al., 2017; FREGNANI et al., 2010).
O aumento de volume representa o sintoma clínico mais comum e radiograficamente
esses tumores são caracterizados como lesões expansivas, radiolúcidas, uni ou multiloculares,
podendo apresentar erosão, rompimento das corticais ósseas, além de reabsorção das raízes
dos elementos dentários adjacentes (WAKOH et al., 2011; RICCI et al., 2012).
O Ameloblastoma sólido é o tipo clínico-patológico mais comum, com
comportamento localmente agressivo. Histopatologicamente, pode apresentar diversos
padrões como: folicular, plexiforme, de células granulares, células basais e acantomatoso.
Estes padrões, no entanto, não interferem quanto ao tratamento estabelecido (HUNASGI et
al., 2013).
O tipo folicular apresenta ninhos com células periféricas, semelhantes a ameloblastos,
que podem estar organizadas em paliçada, com polaridade invertida centralmente as células
são organizadas frouxamente semelhantes ao retículo estrelado do órgão do esmalte (LEE;
KIM, 2013).
No tipo plexiforme, o epitélio odontogênico apresenta-se em forma de cordões
anastomosados exibindo células cúbicas ou colunares, semelhantes a ameloblastos e células
centrais frouxamente arranjadas (CASTRO-SILVA et al., 2012; LEE; KIM, 2013).
O ameloblastoma acantomatoso é raro e exibe ninhos sólidos de células epiteliais com
paliçada periférica e metaplasia de células escamosas na porção central; o ameloblastoma de
células granulares também é uma variante rara, em que as células tumorais localizadas na
porção central das ilhas tumorais apresentam citoplasma granular eosinofílico e células
periféricas que se assemelham a ameloblastos (LEE; KIM, 2013). O padrão de células basais,
é o tipo menos comum sendo composto por ninhos de células basalóides uniformes, não
apresentando retículo estrelado na porção central dos ninhos e as células periféricas tendem a
ser cúbicas em vez de colunares (HIROTA et al.,2005).

48
O ameloblastoma unicístico exibe um comportamento menos agressivo que o tipo
sólido e apresenta características clínicas, radiográficas semelhantes a um cisto. O exame
histopatológico mostra epitélio ameloblastomatoso, revestindo uma cavidade cística, em que
as células epiteliais proliferam, com ou sem crescimento luminal por vezes
mural (CHAUDHARY et al, 2011; RICCI et al, 2012; LEE; KIM, 2013).
O padrão desmoplásico foi considerado como subtipo histopatológico do
Ameloblatoma sólido pela OMS, em sua última classificação, sendo caracterizado por
pequenos ninhos e cordões de epitélio odontogênico comprimidos em um estroma
densamente colagenizado (PHILIPSEN, REICHART, 2004; FULCO et al, 2010; RICCI et al,
2012; EL-NAGGAR et al., 2017). Em virtude da capacidade do ameloblastoma se infiltrar
pelas trabéculas ósseas, o tratamento preferencial do tumor é cirúrgico, visando menores
riscos de recidivas e melhores prognósticos (GUNAWARDHANA et al., 2011; HERTOG et
al, 2011).
2.8 TUMOR ODONTOGÊNICO EPITELIAL CALCIFICANTE
O tumor odontogênico epitelial calcificante (TOEC) é um tumor benigno raro, de
comportamento localmente invasivo, que representa menos de 1% de todos os tumores
odontogênicos (EL-NAGGAR et al., 2017). É também conhecido como Tumor de Pindborg e
localiza-se nos ossos maxilares com predileção pela região posterior de mandíbula, podendo
ter localização extra-óssea ou intra-óssea, sendo esta última mais frequente. Quando
localizados no tecido gengival, são menos agressivos do que os intraósseos (CHRCANOVIC;
GOMEZ, 2017).
O TOEC cresce causando expansão da cortical óssea (SLOOTWEG, 2006) e ao exame
radiográfico, mostra área radioluscente unilocular ou multilocular, contendo massas
radiopacas que aumentam de tamanho e radiopacidade com o tempo (DEBONI et al., 2006).
As características histopatológicas do TOEC incluem proliferação de células epiteliais
poliédricas organizadas em ilhotas e lençóis dispersos em estroma abundante e eosinofílico.
As células neoplásicas apresentam bordas citoplasmáticas bem definidas, pontes intercelulares
evidentes e núcleos frequentemente pleomórficos, apesar de exibirem raras mitoses. As
células tumorais produzem material amiloide, homogêneo e eosinofílico, que pode tornar-se

49
calcificado na forma de anéis concêntricos denominados de anéis de Liesegang. A verdadeira
natureza do material amiloide é ainda incerta (TAKATA; SLOOTWEG, 2005).
Devido às características apresentadas pelos casos de TOECs intraósseos ou de
localização periférica, torna-se evidente que outras fontes, além do epitélio reduzido do órgão
do esmalte, devem ser consideradas quando se discute a histogênese da lesão. A localização
periférica sugere a possibilidade de que o tumor se origine de restos da lâmina dentária ou de
células basais do epitélio oral (SLOOTWEG, 2006).
O prognóstico para a maioria dos casos de TOEC é considerado bom, com pequenas
taxas de recidiva e baixíssimas chances de transformação maligna. Há relatos de TOEC com
comportamento mais agressivo, em que foi verificada tumefação dolorosa, com rápido
crescimento da massa tumoral, ruptura da cortical, compressão do nervo alveolar inferior e
infecção secundária (DEBONI et al., 2006).
2.9 TUMOR ODONTOGÊNICO ADENOMATOIDE
O tumor odontogênico adenomatoide (TOA) é uma lesão de origem epitelial
odontogênica composta de epitélio odontogênico evidenciando padrões arquiteturais variados
em meio a um escasso estroma de tecido conjuntivo maduro (BASKARAN et al., 2011).
Muitas vezes é inicialmente diagnosticado, radiograficamente, como cisto odontogênico e
representa cerca de 1% a 9% de todos os tumores odontogênicos (LEE; KIM, 2013;
PHILIPSEN et al., 2007).
O TOA pode se apresentar como uma lesão única com predileção em pacientes do
sexo feminino, na segunda década de vida e no sextante anterior da maxila, podendo ser
sobrepostos a cistos dentígeros ou a dentes não erupcionados, geralmente associados aos
caninos permanentes. Além disso, os casos mostram opacidades dispersas (flocos) dentro de
uma radiolucência unilocular (RICK, 2004; ROBINSON; VINCENT, 2012).
Existem três variantes do TOA: a variante folicular (73%) está associada a um dente
impactado e deslocado, sendo confundido radiograficamente como um cisto dentigero; a
extrafolicular (24%) imita um cisto radicular ao redor do ápice de um dente; e a variante
periférica (3%) apresenta-se como um defeito do osso periodontal ou crescimento ectópico.
Na reavaliação da origem e patogênese do TOA, parece que este tumor, ou hamartoma, é

50
derivado do epitélio odontogênico do complexo da lâmina dentária ou de seus remanescentes
(CHINDASOMBATJAROEN et al., 2012).
Histopatologicamente, a maioria dos casos apresenta um padrão de crescimento
predominantemente sólido ou uma proporção semelhante de padrões sólido e cribriforme.
Material amorfo eosinofílico e espaços semelhantes a ductos são observados em 90% a 100%
dos casos. Estruturas enoveladas e áreas semelhantes ao tumor odontogênico epitelial
calcificante também podem ser encontradas. Quantidades variáveis de material calcificado são
observadas na maioria dos TOAs, enquanto que osteodentina e hialinização perivascular
raramente estão presentes (DE MATOS et al., 2012). O TOA é um tumor benigno, indolente,
de crescimento lento, limitado e por isso, é tratado por excisão local e as recorrências são
extremamente raras (EL-NAGGAR et al., 2017).
2.10 CERATOCISTO ODONTOGÊNICO
O Ceratocisto Odontogênico é uma forma distinta de cisto odontogênico de
desenvolvimento que merece atenção especial devido às suas características histopatológicas,
ao seu comportamento clínico agressivo e a sua alta taxa de recidiva (MYOUNG et al., 2001;
SHEAR, 2003; NEVILLE et al., 2016). Essa lesão pode ser relacionada com a síndrome do
carcinoma nevóide basocelular, também denominada de síndrome de Gorlin (FIGUEROA et
al., 2010; HAKIM et al., 2011).
Acredita-se que o Ceratocisto odontogênico origina-se a partir de remanescentes da
lâmina dentária (TSUKAMOTO et al., 2001), embora alguns autores apontem como origem
as células da camada basal do epitélio oral adjacente à lesão ou ainda, a partir da proliferação
de pequenos harmatomas epiteliais do epitélio gengival (AMORIM et al., 2003;
STOELINGA, 2003; MENON, 2015; JIANG et al, 2016).
Geralmente essa lesão é assintomática, porém quando atinge grandes extensões,
apresenta sintomatologia dolorosa (NEVILLE et al., 2016). Estudos epidemiológicos revelam
que o ceratocisto odontogênico constitui o terceiro tipo mais comum de cisto odontogênico,
com frequências entre 1,3% e 21,5% (MOSQUEDA-TAYLOR et al., 2002; JONES; CRAIG;
FRANKLIN, 2006; GROSSMANN et al., 2007; DE SOUZA et al., 2010). Apresenta
predileção pelo sexo masculino, acometendo indivíduos entre a segunda e terceira década de

51
vida (SHEAR, 2003; NEVILLE et al., 2016). Em relação ao sítio anatômico de 61,5% a
76,5% dos casos ocorrem em mandíbula, sendo a região posterior e o ramo ascendente as
áreas mais comumente afetadas (GONZÁLEZ-ALVA et al., 2008; TORTORICI et al., 2008;
DE SOUZA et al., 2010).
Histopatologicamente, o ceratocisto odontogênico exibe um revestimento epitelial
bastante típico, com 5 a 8 camadas de células, cuja camada basal apresenta células colunares
ou cúbicas dispostas em paliçada, muitas vezes com hipercromatismo nuclear e camada
superficial paraceratinizada e corrugada. A interface epitélio-conjuntivo frequentemente é
plana e frequentemente há destacamento do revestimento epitelial. A cápsula de tecido
conjuntivo é relativamente delgada, friável e bem vascularizada, geralmente sem infiltrado
inflamatório (NEVILLE et al., 2016). Em alguns casos, podem ser observadas a presença de
ilhas de epitélio odontogênico e pequenos cistos satélites (EL-NAGGAR et al., 2017;
NEVILLE et al., 2016).
O ceratocisto odontogênico apresenta um mecanismo de crescimento diferenciado que
pode estar relacionado a fatores desconhecidos, inerentes ao próprio epitélio ou à atividade
enzimática na cápsula fibrosa (LI et al., 1996; NEVILLE et al., 2016). Alguns estudos,
avaliando marcadores de proliferação celular como o PCNA (Antígeno Nuclear de
Proliferação Celular) e Ki-67, têm demonstrado uma imunoexpressão elevada para ambos os
marcadores, sugerindo que o comportamento mais agressivo do ceratocisto pode estar
relacionado à sua elevada atividade proliferativa (KIM et al., 2003; GURGEL et al., 2008).
Portanto, reconhecendo a importância da presença de WNT, FGF, e SOX durante o
processo de manutenção da pluripotência, renovação celular e diferenciação celular durante a
odontogênese, se faz necessário pesquisas que evidenciem o papel destes fatores nas diversas
lesões que possuem origem do aparato odontogênico a fim de elucidar a natureza e os
divergentes comportamentos biológicos de lesões odontogênicas.

52
Proposição

53
3 PROPOSIÇÃO
As células-tronco são fundamentais para homeostase dos tecidos e pesquisas têm
demonstrado a sua participação no processo de formação de dentes. Distúrbios na sua
regulação podem levar à proliferação excessiva de células, culminando na formação de cistos
e tumores. Sabendo-se que as lesões odontogênicas se originam a partir de remanescentes da
embriogênese dentária, nos propomos a avaliar a participação de células com fenótipo de
pluripotência em lesões epiteliais odontogênicas benignas, através da expressão imuno-
histoquímica do fator de transcrição SOX2, como marcador celular de pluripotência e dos
fatores Wnt e FGF-10 que atuam, respectivamente, como indutor e inibidor da SOX2 em
diversos processos biológicos e patológicos. Pretende-se também, correlacionar o padrão de
imunoexpressão dos fatores estudados com os comportamentos biológicos variados das lesões
constantes do estudo.

54
Material e Métodos

55
4 MATERIAIS E MÉTODOS
4.1 CONSIDERAÇÕES ÉTICAS
A presente pesquisa foi registrada no sistema da Base Nacional de Registros de
Pesquisas envolvendo Seres Humanos (Plataforma Brasil) e de acordo com as Diretrizes e
Normas Regulamentadoras de Pesquisas Envolvendo Seres Humanos (Resolução CNS
446/2012), submetida à análise de seu conteúdo pelo Comitê de Ética em Pesquisa da
Universidade Federal do Rio Grande do Norte e aprovado sob o número de parecer
031235/2016.
O presente trabalho foi executado a partir de espécimes teciduais, emblocados em
parafina, pertencentes ao arquivo do Serviço de Anatomia Patológica da Disciplina de
Patologia Oral do Departamento de Odontologia da UFRN.
4.2 CARACTERIZAÇÃO DO ESTUDO
Esta pesquisa consistiu em uma análise descritiva e comparativa da expressão imuno-
histoquímica de SOX2, Wnt-1 e FGF-10 em espécimes de lesões odontogênicas benignas de
origem epitelial e germes dentários.
4.3 POPULAÇÃO
Do total de casos registrados e diagnosticados nos arquivos do Serviço de Anatomia
Patológica da Disciplina de Patologia Oral do Departamento de Odontologia da Universidade
Federal do Rio Grande do Norte/UFRN, foram selecionados os casos de lesões odontogênicas
que constituíram a amostra da presente pesquisa.
4.4 AMOSTRA
A amostra foi constituída por 20 casos de ameloblastoma do tipo sólido, 20 casos de
tumor odontogênico adenomatoide, 10 casos de tumor odontogênico epitelial calcificante e 20
casos de ceratocisto odontogênico isolado, todos emblocados em parafina, diagnosticados no
serviço anteriormente citado. Também, selecionou-se 05 casos de germes dentários em
diversas fases de desenvolvimento, principalmente na fase de campânula, para efeito
comparativo do estudo.

56
4.5 CRITÉRIOS DE SELEÇÃO DA AMOSTRA
4.5.1 Critérios de inclusão
Foram selecionados 20 casos de ameloblastomas, 20 casos de tumor odontogênico
adenomatoide, 10 casos de tumor odontogênico epitelial calcificante e 20 casos de
ceratocistos odontogênicos que, conforme informações contidas nas fichas de
requisição de biópsias, não foram submetidos ao processo de descalcificação;
Foram selecionados e incluídos no estudo os casos das referidas lesões cujos blocos do
material incluído em parafina se apresentaram em boas condições de armazenamento e
continham material suficiente para o estudo pretendido;
Foram incluídos no estudo, 5 germes dentários para efeito comparativo, provenientes
de remoção por indicação ortodôntica.
4.5.2 Critérios de exclusão
Foram excluídos do estudo, todos os casos das lesões selecionadas que tenham sido
submetidas ao processo de descalcificação, o que poderá acarretar perda de
antigenicidade do material;
Foram excluídos do estudo, todos os espécimes de ameloblastomas cujos dados clínicos,
radiográficos e macroscópicos constantes nos arquivos não satisfaçam os requisitos para
que fosse seguramente estabelecido o tipo clínico-patológico;
Casos de CO secundariamente inflamados;
Foram excluídos do estudo os casos cujos blocos não se apresentem com material em
boas condições de armazenamento e contenham material insuficiente para os estudos
pretendidos.
4.6 ESTUDO MORFOLÓGICO
Para o estudo morfológico, foram utilizadas lâminas com cortes de 5μm de espessura
do material incluído em blocos de parafina, corados pela técnica da Hematoxilina/ Eosina,
arquivadas no serviço de Anatomia Patológica da Disciplina de Patologia Oral da UFRN e
examinadas à microscopia de luz, sendo realizada uma análise descritiva dos aspectos
histopatológicos observados nas lesões objeto da presente pesquisa.

57
4.7 ESTUDO IMUNO-HISTOQUÍMICO
Do material emblocado em parafina foram obtidos cortes de 3μm de espessura e
estendidos em lâminas de vidro previamente limpas e preparadas com adesivo à base de
organosilano (3-aminopropyltriethoxi-silano, Sigma Chemical Co, St. Louis, MO, USA). O
material foi posteriormente submetido aos anticorpos anti-SOX2, anti-FGF-10 e anti-Wnt-1
(Quadro 1).
A coloração imuno-histoquímica para SOX2 foi realizada em cortes histológicos de 3
μm de espessura das lesões estudadas e dos germes dentários, utilizando o sistema de
detecção ADVANCETM
HRP (Dako, Nort America Inc., Carpinteria,CA, USA). Os cortes
foram submetidos a recuperação antigênica em solução de TrylogyTM
(Cell Marque). A
atividade da peroxidase endógena foi bloqueada através da incubação com 3% de peróxido de
hidrogênio, durante 15 minutos. As lâminas foram incubadas com anticorpo policlonal anti-
humano/rato SOX2 (Y-17, Santa Cruz Biotechnology Inc, Santa Cruz, CA, EUA) a 4 °C
overnight com concentração de 1:40, seguido por três lavagens suaves com tampão e
posteriormente coradas durante 5 min com 3'3 diaminobenzidina (DAB). Os cortes
histológicos foram contracorados com Hematoxilina de Mayer. Células-tronco embrionárias
foram utilizadas como controle positivo para SOX2 (WANG et al., 2009).
Com relação a expressão de FGF-10, os cortes histológicos de 3 µm de espessura
foram desparafinados e imersos em 3% de peróxido de hidrogênio durante 30 minutos e soro
de cabra a 10% preparado em solução salina tamponada com fosfato (PBS) durante 20
minutos. O anticorpo primário policlonal FGF-10 (H-121, Santa Cruz Biotechnology, CA) em
diluição de 1:50, foi aplicado overnight numa câmara húmida a 4 °C. A reatividade do
anticorpo foi posteriormente visualizada com o kit ADVANCETM
HRP (Dako Nort America
Inc, Carpinteria, CA, USA), seguindo as recomendações do fabricante e contracorados com
Hematoxilina de Mayer. Os cortes histológicos foram contracorados com Hematoxilina de
Mayer. Tecido de carcinoma de próstata humana foi utilizado como controle positivo para
FGF-10 (NAKAO et al., 2013).
Para o estudo da expressão de Wnt-1 foram utilizados cortes de 3 µm de espessura
desparafinados e utilizado o anticorpo primário policlonal anti-Wnt-1 (G-19, Santa Cruz
Biotechnology Inc. Santa Cruz CA), com diluição de 1:200. A reatividade do anticorpo foi
posteriormente visualizada com o kit ADVANCETM
HRP (Dako Nort America Inc,
Carpinteria, CA, USA), seguindo as recomendações do fabricante e contracorados com

58
Hematoxilina de Mayer. Extrato de tecido da medula espinhal humana foi utilizado como
controle positivo para Wnt-1 (SONG et al., 2009).
Os controles negativos consistiram na substituição dos anticorpos primários por
albumina de soro bovino a 1% (BSA – Bovine Serum Albumin) em solução tampão.
Especificidade Clone/
Referência Fabricante Diluição
Recuperação
Antigênica* Incubação
SOX2 Y-17 Santa Cruz
Biotechnology 1:40 Trilogy/Pascal
Overnight a
4 °C
Wnt-1 G-19 Santa Cruz
Biotechnology 1:200 Trilogy/Pascal
Overnight a
4 °C
FGF-10 H-121 R&D Systems 1:50 Trilogy/Pascal Overnight a
4 °C
Quadro 1: Especificidade, clone, fabricante, diluição e incubação.
*A recuperação antigênica de cada anticorpo utilizado foi feita de acordo com as
recomendações dos fabricantes e otimizadas no Laboratório de Imuno-histoquímica do
Programa de Pós-graduação em Patologia Oral/UFRN.
4.8 ANÁLISE DO PERFIL IMUNO-HISTOQUÍMICO
Após o processamento dos cortes histológicos, cada espécime foi analisado em sua
totalidade sob microscopia de luz (Olympus CH30, Olympus Japan Co., Tokyo, JPN). As
análises das reações imuno-histoquímicas para SOX2, Wnt-1 e FGF-10 foram feitas por
adaptação dos estudos de Wang et al. (2009) e Halifu et al. (2016). A marcação celular no
núcleo e citoplasma foi considerada como positiva quando apresentasse coloração
acastanhada , sendo avaliada em todo o componente epitelial das lesões constantes do estudo.
A análise da expressão foi feita de forma semi-quantitativa, sendo a expressão dos fatores
estudados determinada de acordo com a intensidade e a porcentagem de células tumorais
imunorreativas; a porcentagem de positividade celular foi categorizada como 0 (≤ 5%), 1 (6-
25%), 2 (26-50%), 3 (5l-75%) e 4 (76-100 %). A intensidade foi classificada como 0 quando
não foram identificadas células positivas; 1, fraco; 2, moderada; e 3, forte.
De acordo com a somatória de intensidade e porcentagem de imunomarcação, obteve-se
a Pontuação de imunomarcação total (PIT), variando de 0 a 7. Os escores finais foram
numerados para cada marcador:

59
SOX2: 1 (soma entre 0-2); 2 (soma entre 3-4); 3 (soma ≥5).
Wnt-1: 1 (soma entre 0-4); 2 (soma de 5); 3 (soma ≥6).
FGF-10: 1 (soma entre 0-3); 2 (soma entre 4-5); 3 (soma ≥6).
4.9 ANÁLISE ESTATÍSTICA
Os dados foram digitados originalmente em planilha eletrônica Excel (Microsoft
Windows Vista Home edition®) e posteriormente exportados para o programa Statistical
Package for the Social Sciences (IBM Corp., Armonk, NY, USA), onde foram feitas as
análises.
Considerando a não normalidade e a independência da amostra foram utilizados os
testes de Kruskal-Wallis e de Mann Whitney para avaliar diferenças significativas. Para
verificar a correlação entre as imunoexpressões dos marcadores foi utilizado o teste de
Spearman. Os valores foram considerados estatisticamente significativos quando p<0,05.

60
Resultados

61
5 RESULTADOS
5.1 ANÁLISE MORFOLÓGICA
A análise microscópica dos casos de Ameloblastomas sólidos revelou proliferação das
células epiteliais em ilhas e cordões que se anastomosavam, e a presença de células periféricas
organizadas em paliçada, hipercromáticas e núcleo com polaridade invertida. Evidenciou-se
que a maioria dos casos constantes de nossa amostra apresentava-se com o padrão
histopatológico plexiforme (60%).
Em oito casos (40%) predominou o padrão folicular, onde as células organizavam-se
em ninhos com células periféricas em paliçada e células centrais organizadas frouxamente
semelhante ao retículo estrelado do órgão do esmalte. Em alguns casos, houve associação do
padrão plexiforme com a variante folicular. O estroma era predominantemente fibroso denso,
por vezes com áreas de densidade variada, com vasos sanguíneos de pequeno calibre
apresentando-se, ocasionalmente congestos.
As características histopatológicas dos casos de TOEC demonstravam proliferação de
células epiteliais poliédricas organizadas em ilhotas e lençóis dispersas em estroma abundante
e eosinofílico. As células neoplásicas apresentaram contornos bem definidos, pontes
intercelulares proeminentes e núcleos frequentemente pleomórficos e hipercromáticos. Foi
evidenciada, também, a presença de material amilóide homogêneo e eosinofílico que, por
vezes, exibia anéis de Liesegang.
Avaliando microscopicamente os casos de COs, observou-se o epitélio cístico de
espessura uniforme exibindo de 6 a 8 camadas de células, com camada superficial de
paraceratina exibindo corrugações. A células da camada basal exibiam hipercromatismo,
núcleos com polaridade invertida e organizavam-se em paliçada. A junção epitélio-conjuntivo
se apresentava plana com destacamento do epitélio em relação ao tecido conjuntivo. A
cápsula de tecido conjuntivo era delgada e por vezes exibia leve a moderado infiltrado
inflamatório mononuclear.
Histopatologicamente, os casos de TOA apresentavam áreas sólidas constituídas por
células epiteliais do parênquima tumoral e escasso estroma. As células neoplásicas eram
predominantemente cúbicas e ovais, exibindo estruturas espiraladas semelhantes a rosetas,
assim como estruturas semelhantes a ductos, circundadas, em sua maioria, por células

62
colunares com núcleo polarizado. Observou-se ainda a presença de estruturas calcificadas. Em
alguns casos, foi evidenciada a presença de cápsula de tecido conjuntivo fibroso denso.
5.2 RESULTADOS IMUNO-HISTOQUÍMICOS
Na avaliação da imunoexpressão dos marcadores utilizados em nossa pesquisa
evidenciamos que no parênquima dos casos de Ameloblastoma, as células da camada
periférica ou camada basal não exibiam marcação ou eram predominantemente fracas ou
moderadas, enquanto as células centrais exibiam fraca ou moderada marcação para SOX2. No
estroma, também se evidenciou positividade para todas as moléculas utilizadas na pesquisa,
principalmente em fibroblastos, células endoteliais e células inflamatórias (Fig. 1).
Nos casos de TOEC, a marcação para SOX-2 foi predominantemente nuclear e
citoplasmática e com intensidade de moderada a forte (Fig. 2 e 3 ) enquanto nos casos de
TOA, observou-se de fraca a moderada expressão citoplasmática e, raramente, nuclear
(Fig.4).Avaliando a imunoexpressão para SOX-2 , com intensidade de moderada a forte nas
camadas de células epiteliais nos casos de CO observou-se marcação na camada basal e
parabasal enquanto as células superficiais não exibiam expressão (Fig.,5)
Com relação a expressão para FGF-10, todos os casos de AM foram positivos, com
variação de intensidade entre os casos avaliados, com marcação predominantemente
citoplasmática e, raramente nuclear(Fig.7). Nos casos de TOEC não foram observadas
diferenças na intensidade de expressão no parênquima tumoral (Fig.8). Quando avaliada a
intensidade de imunomarcação para FGF-10 nas células neoplásicas nos casos de TOA
observou-se moderada marcação citoplasmática e, raramente nuclear (Fig.9). Nos casos de
CO, foi evidenciada marcação citoplasmática moderada em todas as camadas epiteliais
(Fig.10).
Todos os casos de AM foram positivos para Wnt-1 com variação de intensidade entre
os casos avaliados com marcação predominantemente citoplasmática (Fig.12). Para os casos
de TOEC evidenciou-se fraca marcação citoplasmática para Wnt-1 no parênquima tumoral
(Fig.13). Quando avaliada a intensidade de imunomarcação para Wnt-1 nas células
neoplásicas nos casos de TOA observou-se moderada marcação citoplasmática e, raramente
nuclear (Fig.14). A marcação para Wnt-1 foi maior e mais intensa na camada superficial e
fraca ou ausente na camada basal e parabasal nos casos de CO (Fig. 15).

63
Nos 05 casos de germes dentários avaliados, utilizados para fins comparativos, a
marcação foi nuclear e citoplasmática nas diferentes células epiteliais e da papila dentária para
todas as proteínas estudadas. A imunoexpressão para SOX-2 exibiu uma marcação nuclear e
citoplasmática na maioria dos casos dos germes dentários (Fig. 6), enquanto a
imunoexpressão de FGF-10 (Fig. 11) e Wnt-1 exibiu marcação tipicamente citoplasmática,
apresentando variações de intensidade de expressão tanto nas estruturas epiteliais quanto
mesenquimais Nos casos em que havia a presença da bainha epitelial de Hertwig, evidenciou-
se menor imunoexpressão para Wnt-1 (Fig. 16).
Também, foi evidenciada marcação nuclear e citoplasmática para SOX2 nas células da
camada basal e parabasal de epitélio de mucosa oral normal, adjacente a fragmentos das
lesões estudadas (Fig. 17).
5.2.1 Análise da imunoexpressão de SOX-2
A análise da imunoexpressão da SOX2 revelou positividade na maioria dos casos das
lesões constantes do estudo. Especificamente no parênquima dos casos de AM, a
imunoexpressão apresentou maior frequência de PIT 2 e 3 (Tabela1) e maior frequência do
escore 1 e 2 (Tabela 2). Os casos de CO exibiram maiores valores com relação às frequências
de PIT 6 e 7, representando 70% dos casos e, com maior frequência de escore 3 (85%). O
teste não paramétrico de Kruskal-Wallis revelou haver diferença estatisticamente significativa
entre os grupos de lesões em relação às medianas dos escores de positividade para SOX2 (p =
<0,001) (Tabela 3). A fim de identificar esta diferença entre as lesões para a imunoexpressão
de SOX2 foi realizado o teste de Mann-Whitney (Tabela 4), que mostrou maior valor de
mediana em CO e TOEC, e revelou existir diferença estatisticamente significativa, entre AM
e CO (p=<0,001), AM e TOEC (p=0,007), CO e TOA (p=<0,001), CO e TOEC (p=<0,001) e TOA e
TOEC (p=0,024).

64
Tabela 01. Distribuição absoluta e relativa dos casos de acordo com a pontuação de
imunomarcação total (PIT) para SOX-2 nas células epiteliais das lesões estudadas. Natal
– RN, 2018.
Grupo
Pontuação de Imunomarcação Total
0
n (%)
1
n (%)
2
n (%)
3
n (%)
4
n (%)
5
n (%)
6
n (%)
7
n (%)
AM 1 (5,0) 2 (10,0) 4 (20,0) 9 (45,0) 1 (5,0) 3 (15,0) 0 (0,0) 0 (0,0)
CO 0 (0,0) 0 (0,0) 2 (10,0) 0 (0,0) 1 (5,0) 3 (15,0) 8 (40,0) 6 (30,0)
TOA 0 (0,0) 1 (5,0) 7 (35,0) 3 (15,0) 8 (40,0) 0 (0,0) 1 (5,0) 0 (0,0)
TOEC 0 (0,0) 0 (0,0) 1 (10,0) 2 (20,0) 2 (20,0) 1 (10,0) 4 (40,0) 0 (0,0)
Fonte: Programa de Pós-Graduação em Patologia Oral/ UFRN.
Legenda: AM – Ameloblastoma sólido; CO – Ceratocisto odontogênico; TOA – Tumor odontogênico
adenomatoide; TOEC – Tumor odontogênico epitelial calcificante.
Tabela 02. Distribuição absoluta e relativa dos casos acordo com os escores de positividade
para SOX-2 nas células epiteliais das lesões odontogênicas estudadas. Natal –
RN, 2018.
Grupo
Escores de positividade
1- Soma entre 0 e 2
n (%)
2- Soma entre 3 e 4
n (%)
3- Soma ≥5
n (%)
Total
n (%)
AM 7 (35,0) 10 (50,0) 3 (15,0) 20 (100,0)
CO 2 (10,0) 1 (5,0) 17 (85,0) 20 (100,0)
TOA 8 (40,0) 11 (55,0) 1 (5,0) 20 (100,0)
TOEC 1 (10,0) 4 (40,0) 5 (50,0) 10 (100,0)
Fonte: Programa de Pós-Graduação em Patologia Oral/ UFRN.
Legenda: AM – Ameloblastoma sólido; CO – Ceratocisto odontogênico; TOA – Tumor odontogênico
adenomatoide; TOEC – Tumor odontogênico epitelial calcificante.

65
Tabela 03. Tamanho da amostra, mediana, quartis 25 e 75, média dos postos e significância
estatística (p) para os escores de positividade para SOX2 em relação aos grupos
de lesões odontogênicas estudadas. Natal – RN, 2018.
Grupo n Mediana Q25-Q75 Média dos postos p
AM 20 3,00 2,00 – 3,00 23,25 <0,001*
CO 20 6,00 5,00 – 7,00 53,73
TOA 20 3,00 2,00 – 4,00 26,30
TOEC 10 4,50 3,00 – 6,00 41,95
Fonte: Programa de Pós-Graduação em Patologia Oral/ UFRN.
Legenda: AM – Ameloblastoma sólido; CO – Ceratocisto odontogênico; TOA – Tumor odontogênico
adenomatoide; TOEC – Tumor odontogênico epitelial Calcificante.
*: Teste de Kruskal-Wallis.
Tabela 04. Tamanho da amostra, mediana, quartis 25 e 75, média dos postos e significância
estatística (p) para os escores de positividade para SOX2 em relação aos grupos
de lesões odontogênicas estudadas. Natal – RN, 2018.
Grupo n Mediana Q25-Q75 Média dos postos p
AM 20 3,00 2,00 – 3,00 12,40 <0,001*
CO 20 6,00 5,00 – 7,00 28,60
AM 20 3,00 2,00 – 3,00 19,33 0,529*
TOA 20 3,00 2,00 – 4,00 21,68
AM
TOEC
20
10
3,00
4,50
2,00 – 3,00
3,00 – 6,00
12,53
21,45
0,007*
CO
TOA
20
20
6,00
3,00
5,00 – 7,00
2,00 – 4,00
28,35
12,65
<0,001*
CO
TOEC
20
10
6,00
4,50
5,00 – 7,00
3,00 – 6,00
17,78
10,95
0,044*
TOA
TOEC
20
10
3,00
4,50
2,00 – 4,00
3,00 – 6,00
12,98
20,55
0,024*
Fonte: Programa de Pós-Graduação em Patologia Oral/ UFRN.
Legenda: AM – Ameloblastoma sólido; CO – Ceratocisto odontogênico; TOA – Tumor odontogênico
adenomatoide; TOEC – Tumor odontogênico epitelial calcificante.
*: Teste de Mann-Whitney.

66
5.2.2 Análise da imunoexpressão de FGF-10
A análise da imunoexpressão de FGF-10 revelou positividade em todos os casos das
lesões estudadas. Especificamente no componente epitelial dos casos, a imunoexpressão
apresentou maior frequência de PIT 5 a 7 (Tabela 5) e maior frequência do escore 3 (Tabela
6). O teste não paramétrico de Kruskal-Wallis revelou não haver diferença estatisticamente
significativa entre os grupos de lesões em relação às medianas dos escores de positividade
para FGF-10 (p = 0,628) (Tabela 7). A fim de identificar diferenças entre as lesões para a
imunoexpressão de FGF-10 foi realizado o teste de Mann-Whitney, que revelou não existir
diferença estatisticamente significativa após o pareamento entre as lesões (p>0,05).
Tabela 05. Distribuição dos casos de acordo com a pontuação de imunomarcação total (PIT) para
FGF-10 no componente epitelial das lesões odontogênicas estudadas. Natal – RN, 2018.
Grupo
Pontuação de Imunomarcação Total
0
n (%)
1
n (%)
2
n (%)
3
n (%)
4
n (%)
5
n (%)
6
n (%)
7
n (%)
AM 0 (0,0) 0 (0,0) 1 (5,0) 1 (5,0) 4 (20,0) 4 (20,0) 5 (25,0) 5 (25,0)
CO 0 (0,0) 0 (0,0) 0 (0,0) 1 (5,0) 3 (15,0) 6 (30,0) 7 (35,0) 3 (15,0)
TOA 0 (0,0) 0 (0,0) 0 (0,0) 0 (0,0) 1 (5,0) 6 (30,0) 10 (50,0) 3 (15,0)
TOEC 0 (0,0) 0 (0,0) 0 (0,0) 0 (0,0) 1 (10,0) 1 (10,0) 7 (70,0) 1 (10,0)
Fonte: Programa de Pós-Graduação em Patologia Oral/ UFRN.
Legenda: AM – Ameloblastoma sólido; CO – Ceratocisto odontogênico; TOA – Tumor odontogênico
adenomatoide; TOEC – Tumor odontogênico epitelial calcificante.
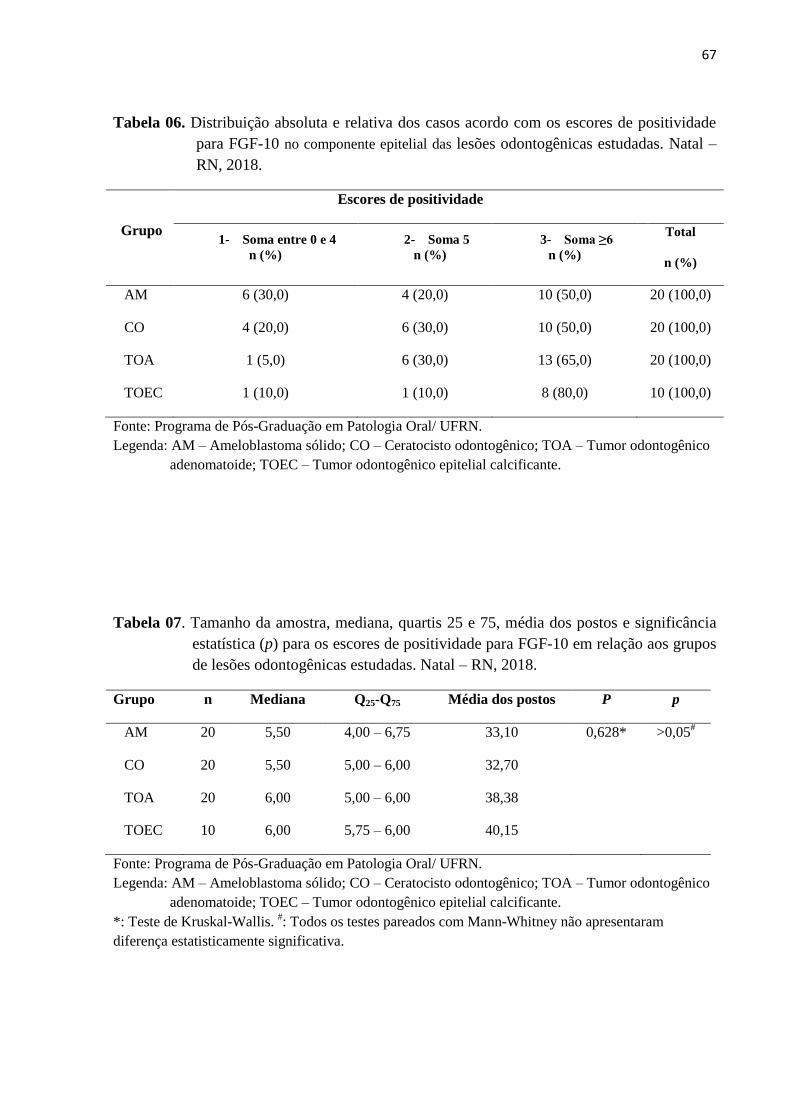
67
Tabela 06. Distribuição absoluta e relativa dos casos acordo com os escores de positividade
para FGF-10 no componente epitelial das lesões odontogênicas estudadas. Natal –
RN, 2018.
Grupo
Escores de positividade
1- Soma entre 0 e 4
n (%)
2- Soma 5
n (%)
3- Soma ≥6
n (%)
Total
n (%)
AM 6 (30,0) 4 (20,0) 10 (50,0) 20 (100,0)
CO 4 (20,0) 6 (30,0) 10 (50,0) 20 (100,0)
TOA 1 (5,0) 6 (30,0) 13 (65,0) 20 (100,0)
TOEC 1 (10,0) 1 (10,0) 8 (80,0) 10 (100,0)
Fonte: Programa de Pós-Graduação em Patologia Oral/ UFRN.
Legenda: AM – Ameloblastoma sólido; CO – Ceratocisto odontogênico; TOA – Tumor odontogênico
adenomatoide; TOEC – Tumor odontogênico epitelial calcificante.
Tabela 07. Tamanho da amostra, mediana, quartis 25 e 75, média dos postos e significância
estatística (p) para os escores de positividade para FGF-10 em relação aos grupos
de lesões odontogênicas estudadas. Natal – RN, 2018.
Grupo n Mediana Q25-Q75 Média dos postos P p
AM 20 5,50 4,00 – 6,75 33,10 0,628* >0,05#
CO 20 5,50 5,00 – 6,00 32,70
TOA 20 6,00 5,00 – 6,00 38,38
TOEC 10 6,00 5,75 – 6,00 40,15
Fonte: Programa de Pós-Graduação em Patologia Oral/ UFRN.
Legenda: AM – Ameloblastoma sólido; CO – Ceratocisto odontogênico; TOA – Tumor odontogênico
adenomatoide; TOEC – Tumor odontogênico epitelial calcificante.
*: Teste de Kruskal-Wallis. #: Todos os testes pareados com Mann-Whitney não apresentaram
diferença estatisticamente significativa.
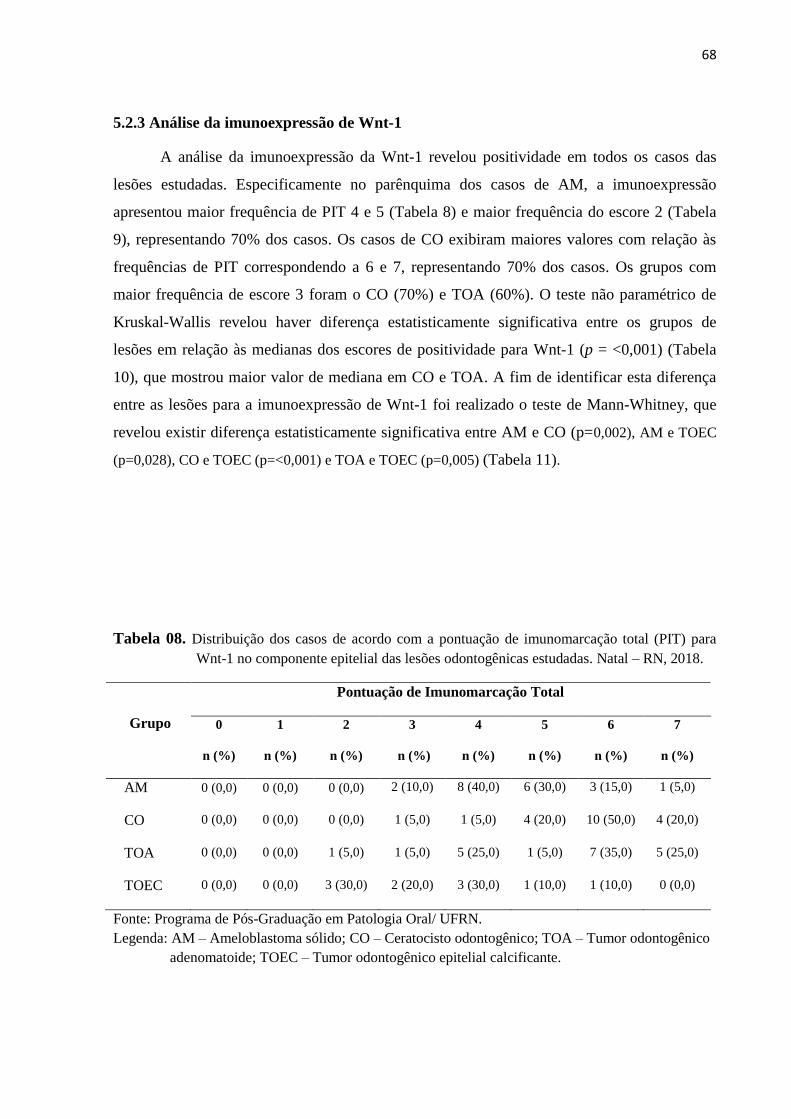
68
5.2.3 Análise da imunoexpressão de Wnt-1
A análise da imunoexpressão da Wnt-1 revelou positividade em todos os casos das
lesões estudadas. Especificamente no parênquima dos casos de AM, a imunoexpressão
apresentou maior frequência de PIT 4 e 5 (Tabela 8) e maior frequência do escore 2 (Tabela
9), representando 70% dos casos. Os casos de CO exibiram maiores valores com relação às
frequências de PIT correspondendo a 6 e 7, representando 70% dos casos. Os grupos com
maior frequência de escore 3 foram o CO (70%) e TOA (60%). O teste não paramétrico de
Kruskal-Wallis revelou haver diferença estatisticamente significativa entre os grupos de
lesões em relação às medianas dos escores de positividade para Wnt-1 (p = <0,001) (Tabela
10), que mostrou maior valor de mediana em CO e TOA. A fim de identificar esta diferença
entre as lesões para a imunoexpressão de Wnt-1 foi realizado o teste de Mann-Whitney, que
revelou existir diferença estatisticamente significativa entre AM e CO (p=0,002), AM e TOEC
(p=0,028), CO e TOEC (p=<0,001) e TOA e TOEC (p=0,005) (Tabela 11).
Tabela 08. Distribuição dos casos de acordo com a pontuação de imunomarcação total (PIT) para
Wnt-1 no componente epitelial das lesões odontogênicas estudadas. Natal – RN, 2018.
Grupo
Pontuação de Imunomarcação Total
0
n (%)
1
n (%)
2
n (%)
3
n (%)
4
n (%)
5
n (%)
6
n (%)
7
n (%)
AM 0 (0,0) 0 (0,0) 0 (0,0) 2 (10,0) 8 (40,0) 6 (30,0) 3 (15,0) 1 (5,0)
CO 0 (0,0) 0 (0,0) 0 (0,0) 1 (5,0) 1 (5,0) 4 (20,0) 10 (50,0) 4 (20,0)
TOA 0 (0,0) 0 (0,0) 1 (5,0) 1 (5,0) 5 (25,0) 1 (5,0) 7 (35,0) 5 (25,0)
TOEC 0 (0,0) 0 (0,0) 3 (30,0) 2 (20,0) 3 (30,0) 1 (10,0) 1 (10,0) 0 (0,0)
Fonte: Programa de Pós-Graduação em Patologia Oral/ UFRN.
Legenda: AM – Ameloblastoma sólido; CO – Ceratocisto odontogênico; TOA – Tumor odontogênico
adenomatoide; TOEC – Tumor odontogênico epitelial calcificante.

69
Tabela 09. Distribuição absoluta e relativa dos casos acordo com os escores de positividade
para Wnt-1 no componente epitelial das lesões odontogênicas estudadas. Natal –
RN, 2018.
Grupo
Escores de positividade
1- Soma entre 0 e 3
n (%)
2- Soma entre 4 e 5
n (%)
3- Soma ≥6
n (%)
Total
n (%)
AM 2 (10,0) 14 (70,0) 4 (20,0) 20 (100,0)
CO 1 (5,0) 5 (25,0) 14 (70,0) 20 (100,0)
TOA 2 (10,0) 6 (30,0) 12 (60,0) 20 (100,0)
TOEC 5 (50,0) 4 (40,0) 1 (10,0) 10 (100,0)
Fonte: Programa de Pós-Graduação em Patologia Oral/ UFRN.
Legenda: AM – Ameloblastoma sólido; CO – Ceratocisto odontogênico; TOA – Tumor odontogênico
adenomatoide; TOEC – Tumor odontogênico epitelial calcificante.
Tabela 10. Tamanho da amostra, mediana, quartis 25 e 75, média dos postos e significância
estatística (p) para os escores de positividade para Wnt-1 em relação aos grupos
de lesões odontogênicas estudadas. Natal – RN, 2018.
Grupo n Mediana Q25-Q75 Média dos postos p
AM 20 4,50 4,00 – 5,00 29,18 <0,001*
CO 20 6,00 5,00 – 6,00 46,13
TOA 20 6,00 4,00 – 6,75 40,80
TOEC 10 3,50 2,00 – 4,25 16,30
Fonte: Programa de Pós-Graduação em Patologia Oral/ UFRN.
Legenda: AM – Ameloblastoma sólido; CO – Ceratocisto odontogênico; TOA – Tumor odontogênico
adenomatoide; TOEC – Tumor odontogênico epitelial calcificante.
*: Teste de Kruskal-Wallis.

70
Tabela 11. Tamanho da amostra, mediana, quartis 25 e 75, média dos postos e significância
estatística (p) para os escores de positividade para Wnt-1 em relação aos grupos
de lesões odontogênicas estudadas. Natal – RN, 2018.
Grupo n Mediana Q25-Q75 Média dos postos p
AM 20 4,50 4,00 – 5,00 14,90 0,002*
CO 20 6,00 5,00 – 6,00 26,10
AM 20 4,50 4,00 – 5,00 17,30 0,086*
TOA 20 6,00 4,00 – 6,75 23,70
AM
TOEC
20
10
4,50
3,50
4,00 – 5,00
2,00 – 4,25
17,98
10,55
0,028*
CO
TOA
20
20
6,00
6,00
5,00 – 6,00
4,00 – 6,75
21,55
19,45
0,583*
CO
TOEC
20
10
6,00
3,50
5,00 – 6,00
2,00 – 4,25
19,48
7,55
<0,001*
TOA
TOEC
20
10
6,00
3,50
4,00 – 6,75
2,00 – 4,25
18,65
9,20
0,005*
Fonte: Programa de Pós-Graduação em Patologia Oral/ UFRN.
Legenda: AM – Ameloblastoma sólido; CO – Ceratocisto odontogênico; TOA – Tumor odontogênico
adenomatoide; TOEC – Tumor odontogênico epitelial calcificante.
*: Teste de Mann-Whitney.
5.2.4 Correlação das imunoexpressões de SOX2, FGF-10 e Wnt-1
Foram analisadas possíveis correlações entre os escores de imunopostividade para
SOX2, FGF-10 e Wnt-1 empregando-se o coeficiente de correlação de Spearman (r) para a
amostra total e não sendo encontradas correlações com significância estatística (p=>0,05).
Observou-se que entre SOX2 e Wnt-1 a correlação foi fraca e positiva (r=0,216; p=0,073),
entre SOX2 e FGF-10 a correlação foi fraca e positiva (r=0,053; p=0,666) e entre Wnt-1 e
FGF-10 a correlação foi fraca e negativa (r= -0,012; p=0,924).
Analisando-se separadamente, as correlações dos marcadores nos grupos de lesões
estudadas, foi observada uma correlação fraca e negativa sem significância estatística entre
FGF-10 e Wnt-1 em CO (r= -0,361; p=0,118) e TOEC (r= -0,258; p=0,471). Uma correlação
fraca e positiva sem significância estatística entre FGF-10 e Wnt-1 em AM (r= 0,051;

71
p=0,831) e TOA (r= 0,285; p=0,223). Ao avaliar SOX2 e Wnt-1 em TOEC foi observada uma
correlação positiva moderada (r=0,626; p=0,053), mas sem significância estatística (Tabela
12).
Tabela 12. Tamanho da amostra, comparação, coeficiente de correlação de Spearman (r) e
significância estatística (p) para os escores de positividade para SOX2, FGF-10 e Wnt-1
em relação aos grupos de lesões odontogênicas estudadas. Natal – RN, 2018.
Grupo n Comparação r p
20 SOX2 x FGF-10 0,157 0,508
AM 20 SOX2 x Wnt-1 -0,199 0,401
20 FGF-10 x Wnt-1 0,051 0,831
20 SOX2 x FGF-10 0,273 0,244
CO 20 SOX2 x Wnt-1 0,029 0,902
20 FGF-10 x Wnt-1 -0,361 0,118
20 SOX2 x FGF-10 0,121 0,612
TOA 20 SOX2 x Wnt-1 0,271 0,247
20 FGF-10 x Wnt-1 0,285 0,223
10 SOX2 x FGF-10 -0,091 0,803
TOEC 10 SOX2 x Wnt-1 0,626 0,053
10 FGF-10 x Wnt-1 -0,258 0,471
Fonte: Programa de Pós-Graduação em Patologia Oral/ UFRN.
Legenda: AM – Ameloblastoma sólido; CO – Ceratocisto odontogênico; TOA – Tumor odontogênico
adenomatoide; TOEC – Tumor odontogênico epitelial calcificante.

72
Figura 2. Fotomicrografia evidenciando a fraca marcação nuclear e citoplasmática de SOX2
em Tumor Odontogênico Epitelial Calcificante (Panoramic Viewer, Advance).
Figura 1. Fotomicrografia evidenciando a fraca marcação nuclear de SOX2 em
Ameloblastoma com evidente marcação em vasos sanguíneos no estroma (Panoramic Viewer,
Advance).

73
Figura 3. Fotomicrografia evidenciando a fraca marcação citoplasmática de SOX2 em Tumor
Odontogênico Epitelial Calcificante com escassa marcação no material amilóide e anéis de
Liesegang (Panoramic Viewer, Advance).
Figura 4. Fotomicrografia evidenciando a fraca marcação citoplasmática de SOX2 em Tumor
Odontogênico Adenomatoide (Panoramic Viewer, Advance).
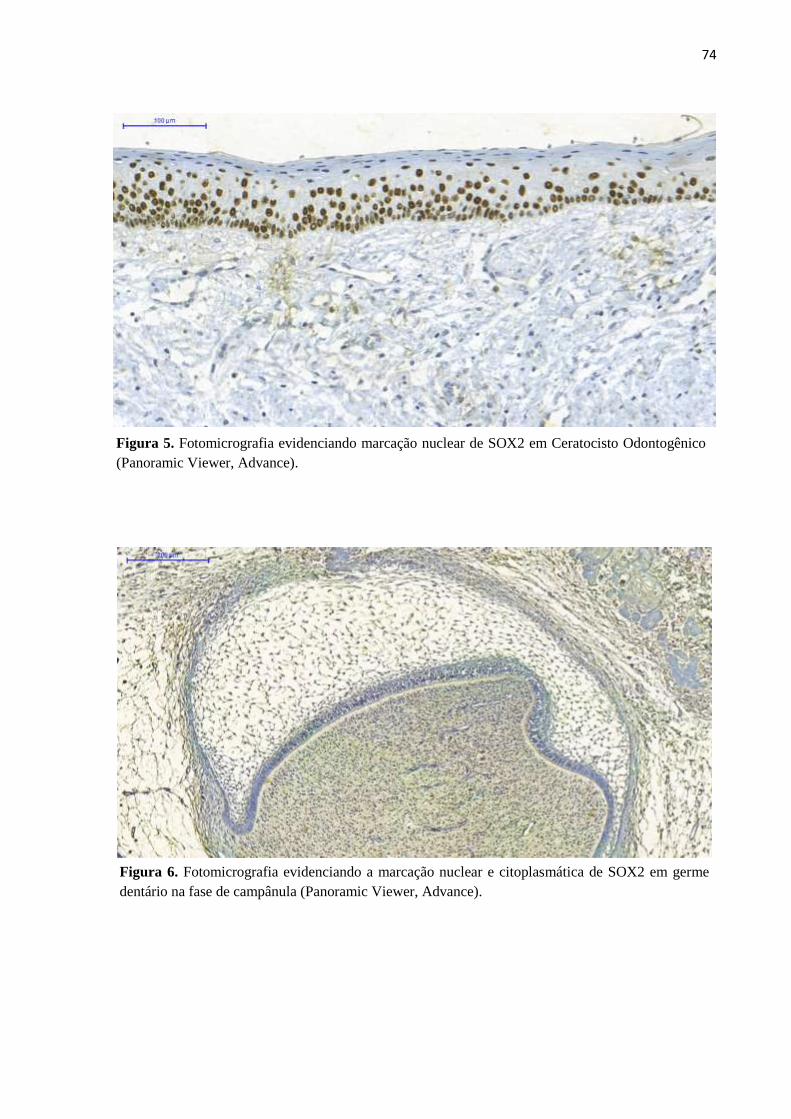
74
Figura 5. Fotomicrografia evidenciando marcação nuclear de SOX2 em Ceratocisto Odontogênico
(Panoramic Viewer, Advance).
Figura 6. Fotomicrografia evidenciando a marcação nuclear e citoplasmática de SOX2 em germe
dentário na fase de campânula (Panoramic Viewer, Advance).

75
Figura 7. Fotomicrografia evidenciando a marcação citoplasmática de FGF-10 em Ameloblastoma
(Panoramic Viewer, Advance).
Figura 8. Fotomicrografia evidenciando marcação nuclear e citoplasmática de FGF-10 em Tumor
odontogênico epitelial calcificante (Panoramic Viewer, Advance).

76
Figura 9. Fotomicrografia evidenciando marcação citoplasmática de FGF-10 em Tumor
odontogênico adenomatoide (Panoramic Viewer, Advance).
Figura 10. Fotomicrografia evidenciando a marcação de FGF-10 em Ceratocisto Odontogênico, com
evidente marcação em vasos sanguíneos no estroma (Panoramic Viewer, Advance).

77
Figura 11. Fotomicrografia evidenciando a marcação nuclear e citoplasmática de FGF-10 em
germe dentário na fase de campânula (Panoramic Viewer, Advance).
Figura 12. Fotomicrografia evidenciando a moderada marcação citoplasmática de Wnt-1 em
Ameloblastoma com evidente marcação em vasos sanguíneos no estroma (Panoramic Viewer,
Advance).

78
Figura 13. Fotomicrografia evidenciando a fraca marcação citoplasmática de Wnt-1 em Tumor
Odontogênico Epitelial Calcificante com escassa marcação no material amilóide (Panoramic
Viewer, Advance).
Figura 14. Fotomicrografia evidenciando a marcação citoplasmática de Wnt-1 em Tumor
odontogênico adenomatoide (Panoramic Viewer, Advance).

79
Figura 15. Fotomicrografia evidenciando a marcação citoplasmática de Wnt-1 em Ceratocisto
Odontogênico com evidência em suprabasal e superficial (Panoramic Viewer, Advance).
Figura 16. Fotomicrografia evidenciando a marcação nuclear e citoplasmática de Wnt-1 em
germe dentário na fase de campânula, com fraca marcação nas regiões das alças cervicais
(Panoramic Viewer, Advance).

80
Figura 17. Fotomicrografia evidenciando a marcação nuclear de SOX2 predominantemente
na camada basal e suprabasal em mucosa oral subjacente a um caso de Ameloblastoma
(Panoramic Viewer, Advance).

81
Discussão

82
6 DISCUSSÃO
As interações epiteliais-mesenquimais, durante a odontogênese, são uma série de
comunicações programadas, sequenciais e recíprocas que dão origem a uma grande variedade
de células e tecidos altamente especializados. As principais vias de sinalização incluem:
BMP, FGF, SHH, WNT e seus respectivos receptores (RAMANATHAN et al., 2018). O gene
Pax9 (Paired box gene 9), atua como um marcador mesenquimal precoce para determinar a
localização exata de formação de germes dentários, que é induzida por Fgf8, mas é regulada
negativamente pelas proteínas BMP2 e BMP4. O fator de transcrição Pitx2 (Paired-like
homeodomain transcription factor 2) é importante na determinação do destino celular na
odontogênese (HUANG; THESLEFF, 2013; SURYADEVA; KHAN, 2015; PUTHIYAVEETIL et
al., 2016).
Durante a odontogênese, as células-tronco (epiteliais e mesenquimais) são essenciais
para a morfogênese dentária. Diversas pesquisas têm utilizado as células-tronco
mesenquimais dentárias por serem facilmente obtidas, enquanto os estudos são escassos com
as células epiteliais odontogênicas por serem perdidas após a conclusão da formação do
esmalte. Ainda na vida adulta, as células-tronco se fazem presentes na polpa dentária e são
importantes para a manutenção e regeneração tecidual (CUNHA et al., 2013; PRIYA te al.,
2015; MIRAN; MITSIADIS; PAGELLA, 2016; SHARPE, 2016;).
Após a formação dos dentes, ocorre uma desagregação das estruturas que participaram
da formação dos tecidos dentários e espera-se que, com o desenvolvimento, essas células
desapareçam ou permaneçam quiescentes (ISHIKAWA et al., 2017). Porém, diversas lesões
podem surgir decorrentes de alterações em remanescentes epiteliais da odontogênese
originando lesões com um comportamento biológico diverso, desde hamartomas a tumores
malignos odontogênicos (AGARWAL et al., 2016; JIANG et al., 2016). Da mesma forma que
a morfogênese dentária não é determinada por um único gene, mas por numerosos fatores
genéticos e epigenéticos, a maioria dos defeitos de desenvolvimento nos dentes geralmente
ocorrem como resultado de mutações em genes que codificam moléculas de sinalização e
fatores de transcrição (SURYADEVA; KHAN, 2015).
A etiologia exata e a patogênese das lesões de origem odontogênica ainda
permanecem incertas e alterações em vários genes e moléculas afetam o desenvolvimento e a
progressão destas lesões. A análise comparativa, entre os mecanismos que ocorrem durante a
odontogênese e nas lesões de origem odontogênicas, é válida pois nos fornece o melhor

83
entendimento da patogênese destas alterações. A capacidade das células destas lesões de
apresentar genes expressos durante a odontogênese é bem conhecida, porém as suas possíveis
alterações genéticas e o padrão de expressão ainda não estão bem elucidados (CUNHA et al.,
2013; HUANG; THESLEFF, 2013; GARG et al., 2015).
O CO possui uma natureza mais agressiva e com altos índices de recorrência
comparado a outros cistos odontogênicos. O AM, apesar de ser uma neoplasia benigna, possui
um comportamento localmente agressivo e diversas alterações genéticas têm sido
evidenciadas neste tumor. O TOA e o TOEC são tumores que, possivelmente possuem a
mesma origem do CO e Ameloblastoma, porém, com comportamentos biológicos indolentes e
formação de tecido mineralizado (REGEZI, 2002; JIANG et al., 2016; CHRCANOVIC;
GOMEZ, 2017). Diversos genes envolvidos na odontogênese estão desregulados e envolvidos
na patogênese destes tumores. Por estes motivos, estas lesões são bastante estudadas e estas
características foram cruciais para nosso estudo comparativo a fim elucidar possíveis
mecanismos que diferem nestas lesões (GARG et al., 2015).
Recentemente, mutações no gene BRAF foram identificadas em uma série de casos de
AM e alterações neste gene contribui para a ativação da sinalização MAPK. Ameloblastoma
benigno e metastatizante apresentam reatividade para SHH e para a proteína SMO
(Smoothened) que é um receptor de ligação à proteína G não clássico que medeia a
sinalização de Sonic Hedgehog (SHH) e é normalmente reprimido por PTCH1 na ausência de
ligantes da via SHH. As mutações SMO foram relatadas entre 16-39% em casos de AMs. A
presença de polimorfismos e as mutações germinativas deletérias em PTCH1 demonstraram
que afetam o risco para o desenvolvimento de AM (SWEENEY et al., 2014; GARG et al.,
2015; JIANG et al., 2016).
Durante os estágios iniciais da embriogênese, SOX2 transloca do citoplasma para o
núcleo para impulsionar a transcrição de seus genes-alvo. Porém, a acetilação de SOX2, por
meio do resíduo de lisina k75 conservado, induz a sua exportação nuclear em células-tronco
embrionárias. Além disso, o aumento da acetilação celular leva à redução nos níveis de SOX2
por ubiquitinação e degradação proteossômica, revogando sua capacidade de impulsionar a
transcrição de seus genes -alvo. Por outro lado, o bloqueio da acetilação mantém SOX2 no
núcleo o que leva a manutenção da célula em estado de pluripotência. A exportação nuclear
mediada por acetilação pode ser um mecanismo regulatório comumente usada para muitos
membros da família Sox (BALTUS et al., 2009; YANG et al., 2014). Diversos estudos

84
apontam que SOX2 é regulada positivamente em diversos tipos de tumores (MÜLLER et al.,
2016; REN; ZHANG; JI, 2016) e em lesões odontogênicas (JUURI et al., 2013a;
HEIKINHEIMO et al., 2015; BANERJEE et al., 2016). Os resultados da presente pesquisa
demonstraram que a via de sinalização SOX2 entre o citoplasma e o núcleo pode participar
no desenvolvimento das lesões avaliadas, porém apresentou maior expressão a nível nuclear e
citoplasmático na camada germinativa em CO, seguido do TOEC, e menor imunoexpressão
de SOX2 em TOA e AM com diferença estatisticamente significativa (p<0,001).
Juuri et al. (2013a) compararam a imunoexpressão de SOX2 de cinco casos de AM
com cortes sequenciais da região de formação de terceiros molares de ratos e observou um
padrão de imunomarcação semelhante entre os espécimes tumorais e fragmentos da lâmina
dentária em ratos, propondo que SOX2 contribui para a manutenção de um padrão de células
com características de pluripotência em AM. Os resultados do presente estudo mostraram
resultados diferentes de imunoexpressão de SOX2 em AM, assim como em TOA, com fraca
marcação ou mesmo ausência de expressão, diferentemente do padrão de expressão em CO
que exibiu escore 3 de marcação na maioria dos casos. Estes resultados corroboram o estudo
de Heikinheimo et al. (2015) que avaliou geneticamente 15 casos de AM e 12 de CO e
constataram que SOX2 estava regulada positivamente com significância em CO ao contrário
do evidenciado em AM. Esta diferença está relacionada com o processo de diferenciação das
células epiteliais do ameloblastoma as quais se diferenciam semelhantes aos ameloblastos,
induzida por fatores de transcrição específicos dos dentes. Ao contrário, o CO possui uma
diferenciação semelhante a queratinócitos, pela indução de genes da via SHH, por marcadores
de células-tronco e marcadores de diferenciação em epitélio escamoso terminal.
Bandyopadhyay et al. (2017) realizaram um estudo imuno-histoquímico com SOX2 sendo
evidenciad expressão em todos os casos de CO e negatividade para os AM. Portanto, pode-se
sugerir que SOX2 está relacionada com o desenvolvimento destas lesões, porém em CO e
TOEC esta proteína está presente ativamente e pode estar envolvida em processos de
manutenção da pluripotência, proliferação e diferenciação das células epiteliais.
Diversos fatores, como Wnt-1, podem induzir a diferenciação após a inativação dos
fatores de pluripotência. No presente estudo, foi observado que, em fragmentos de mucosa
oral adjacente, em alguns espécimes da amostra, o SOX2 estava expresso nas camadas basal e
suprabasal e de forma contrária, a expressão de Wnt-1 nestes mesmos espécimes foi
evidenciada nas camadas mais superficiais. Esta observação pode ser confirmada pelos

85
estudos de Slavik et al. (2007) que afirmam que a presença de Wnt-1 em queratinócitos induz
a diferenciação, estando ausente na camada basal. De acordo com Okubbo, Pevny e Hogan
(2006), a expressão de SOX2 na camada basal é esperada visto a sua participação na
manutenção da pluripotência celular necessária para a homeostase tecidual e que a diminuição
de sua expressão é dose-dependente dos fatores que induzem a diferenciação nas camadas
superficiais do epitélio.
Além disso, a sinalização Wnt possui a capacidade de dar forma aos tecidos em
crescimento, regular a pluripotência das células-tronco e também atua como fator de
crescimento direcional durante a diferenciação celular na embriogênese e reparo tecidual
(NUSSE; CLEVERS, 2017). Wnt pode regular negativamente vias de sinalização dependente
de SOX2, necessárias para a progressão do desenvolvimento de uma linhagem bronquiolar
(HASHIMOTO et al., 2012). A inativação do gene Wnt1 em camundongos resulta na perda
de toda a região do mesencéfalo (LEWIS et al., 2013). Na presente pesquisa, todos os casos
de germes dentários e das lesões odontogênicas foram positivos para Wnt-1, principalmente
em CO e TOA, com expressão tipicamente citoplasmática com diferença estatisticamente
significativa (p=<0,001), levando-nos a crer que este fator de transcrição além de participar da
odontogênese, também participa da patogênese destas lesões.
No mesênquima dentário, a sinalização de FGF-10 desempenha um papel fundamental
no controle da transição do estágio de formação da coroa para a raiz, quando ocorre a
diminuição da sua expressão no mesênquima próximo a região onde ocorre a formação inicial
da bainha epitelial de Hertwig, embora sua contínua expressão se faça necessária, no epitélio
odontogênico, para a formação das coroas dentárias (ENNES et al., 2012; LI; PARADA;
CHAI, 2017). Na presente pesquisa, observamos a expressão de FGF-10 em todas as células
epiteliais nos germes dentários assim como no epitélio de todas as lesões estudadas, embora
sem diferença estatisticamente significativa (p=0,628).
Estudos realizados por Nakao et al. (2013) e Matsuda et al. (2017) em culturas de
células derivadas de AM demonstraram que a indução com a proteína recombinante FGF-10
aumenta a proliferação de células tumorais através da via MAPK que, por sua vez, ativa
fatores de transcrição nucleares. Na presente pesquisa, todos os casos de AM foram positivos
para FGF-10, que em sua maioria exibiu escore de imunoexpressão entre 2 e 3, representando
70% dos casos analisados. De forma semelhante, a maioria dos casos de TOA e TOEC exibiu
escore 3 de imunoexpressão de FGF-10, resultados que corroboram os estudos citados

86
anteriormente de que esta proteína participa do processo de desenvolvimento em casos de
AM, assim como em TOA e TOEC.
No presente estudo, evidenciou-se marcação citoplasmática de FGF-10 nos casos de
CO com escore 3 de imunoexpressão, na maioria dos casos, principalmente nas camadas basal
e parabasal, semelhante ao que ocorreu na avaliação da expressão para SOX2, o que pode
indicar a participação destas proteínas na proliferação e diferenciação celular nestas lesões, de
forma similar ao que ocorre na morfogênese dentária onde FGF-10, regulado positivamente
por meio da via Wnt/β-catenina, induz a manutenção e proliferação de células-tronco
(KETTUNEN et al., 2000; HARADA et al., 2002; AURREKOETXEA et al., 2016).
O papel da via Wnt (Wnt3, 6, 10a, 10b) na morfogênese dentária já é bem estabelecido
(HUANG; THESLEFF, 2013), porém são escassos os estudos avaliando o papel de Wnt-1 em
lesões odontogênicas, embora algumas pesquisas discutam a importância de Wnt-1 na
manutenção da pluripotência de células-tronco (ROSS et al., 2000; CAMP et al., 2014). Choi
et al., (2012) observaram que Wnt1 contribui para a tumorigênese ao conferir propriedades de
células-tronco em células tumorais de câncer de pulmão em um modelo animal e que o
bloqueio da sinalização, especificamente de Wnt-1, pode contribuir para o desenvolvimento
de terapias antitumorais mais eficazes, por diminuir a formação de microesferas e do volume
tumoral.
A via Wnt/ β-catenina também participa do processo de proliferação da bainha
epitelial de Hertwig induzindo a diferenciação em cementoblastos e odontoblastos embora
posteriormente, ocorra a expressão de DKK1 (um antagonista da via Wnt canônica) nas
células epiteliais com o objetivo de regular negativamente a expressão de Wnt (HUANG;
THESLEFF, 2013). Nesta pesquisa, alguns casos de germes dentários revelaram uma menor
expressão de Wnt-1 nas células na região da bainha epitelial de Hertwig,. De acordo com Cao
et al. (2015), essa menor expressão de Wnt ocorre pela regulação de DKK1 sendo o aumento
de DKK1 induzido por OSX (Osterix), que é um fator de transcrição necessário durante a
cementogênese uma vez que este induz a diferenciação dos cementoblastos.
Observou-se, também, nos casos de germes dentários constantes da presente pesquisa,
em que se apresentavam em fases tardias do estágio de campânula, persistência da expressão
de Wnt-1 no epitélio odontogênico e fraca imunoexpressão na região de ectomesênquima. Lee
et al. (2015) constataram que a expressão de Wnt/β-catenina na odontogênese é bastante

87
variável entre o epitélio e o ectomesênquima. No início da fase de campânula, a inibição de
Wnt no epitélio, por meio do fator 1 de inibição de Wnt (Wif1), é crucial para a diminuição da
proliferação na região de determinação do nó de esmalte para a formação das cúspides.
Posteriormente, ocorre o aumento da expressão de Wif1 na região do ectomesênquima e a
consequente diminuição da expressão de Wnt. Ao considerar que todos os casos das lesões
constantes da presente pesquisa exibiam imunoexpressão para Wnt-1 no componente epitelial,
este fato sugere que este fator de transcrição participa da patogênese destas lesões e que
possíveis alterações no mecanismo de regulação desta via podem levar ao aumento de sua
sinalização.
Os resultados da presente pesquisa sugerem que a imunoexpressão de Wnt-1 tanto no
citoplasma quanto no núcleo das células epiteliais das lesões avaliadas pode estar participando
do desenvolvimento e progressão destas lesões, embora tenha se apresentado com maior
expressão a nível citoplasmático nas camadas parabasal e superficial em CO, seguido do
TOA, e menor expressão em AM e TOEC com diferença estatisticamente significativa
(p<0,001). Esta consistente imunoexpressão de Wnt-1 em CO, sugere sua importância na
diferenciação celular nestes casos, diferentemente de SOX2 nas camadas basal e parabasal
que, além de estar envolvido nos processos de diferenciação de ceratinócitos, induz a
proliferação e manutenção da pluripotência (OKUBBO; PEVNY; HOGAN, 2006; SLAVIK
et al., 2007; HEIKINHEIMO et al., 2015).
Ao analisar as correlações dos marcadores em cada grupo de lesões constantes da
presente pesquisa, não foi observada correlação entre FGF-10 e Wnt-1 em AM (r= 0,051;
p=0,831) e TOA (r= 0,285; p=0,223). Além disso, essas duas neoplasias que possuem
diferenciação ameloblástica, exibiram menor frequência de imunoexpressão de SOX2,
possivelmente, pela ação de fatores envolvidos na diferenciação de epitélio odontogênico
(PITX2, MSX2, DLX1,2,3,4, ISL1, TBX1, IRX1, RUNX1 e SOX9) que estão mais presentes
nestes tumores (HEIKINHEIMO et al., 2015; BANERJEE et al., 2016)
Também, foi observada uma correlação fraca e negativa sem significância estatística
entre FGF-10 e Wnt-1 em CO (r= -0,361; p=0,118) e TOEC (r= -0,258; p=0,471). De acordo
com alguns autores (HARADA et al., 2002; SLAVIK et al., 2007) maiores níveis de expressão
de FGF-10 são necessários para que ocorra a manutenção da pluripotência em células
epiteliais, pela indução de SOX2, que consequentemente, diminui a expressão de Wnt-1, que
é responsável pela diferenciação celular.

88
Ao considerarmos o comportamento biológico e os índices de recorrência das lesões
estudadas, sabendo-se que o CO apresenta altas taxas de recorrência, isso poderia estar
relacionado a expressão de SOX2, já que na presente pesquisa esta lesão apresentou altas
frequências de imunoexpressão de SOX2. Porém, não podemos afirmar que isto seja um fato
determinante no comportamento biológico do CO pois, as altas taxas de recorrências estão
muitas vezes relacionadas com o tratamento utilizado para esta lesão. Além disso, o AM que é
uma neoplasia e também possui altas taxas de recorrência, apresentou baixa frequência de
imunoexpressão para SOX2 em comparação com o TOEC, que é localmente invasivo e o
TOA que é conhecidamente uma lesão indolente (REGEZI, 2012; LEE; KIM, 2013). Apesar
destas lesões possuírem a mesma origem a partir de epitélio odontogênico, apresentam
comportamentos biológicos diferentes, porém as diferentes imunoexpressões de FGF-10,
Wnt-1 e SOX2, na presente pesquisa, sugerem que as mesmas não têm relação direta com o
comportamento biológico destas lesões.
Apesar dos resultados desta pesquisa não sugerirem relação entre a expressão de
SOX2 e o comportamento biológico das lesões estudadas, no entanto, o estudo de Lee et al.
(2014) em carcinoma de células escamosas demonstrou que este fator pode ser usado como
valor prognóstico, visto que ele está envolvido com o aumento da proliferação celular,
manutenção da pluripotência e quimioresistência.
Estudos imuno-histoquímicos têm utilizado e nomeado o SOX2 como marcador de
células-tronco (BANERJEE et al., 2016; BANDYOPADHYAY et al., 2017), porém após nossos
resultados em lesões odontogênicas epiteliais benignas, entendemos que este marcador
isoladamente não é suficiente para identificar células-tronco tumorais e assim como Müller et
al. (2016) e Lee et al. (2014), podemos denominar SOX2 como um fator de pluripotência e de
autorrenovação celular nestas lesões.
Desta forma, o estudo das vias de transcrição envolvidas no processo de pluripotência
de células-tronco e de seus reguladores em lesões odontogênicas se faz importante para o
melhor entendimento da sua participação no desenvolvimento e comportamento biológico
destas lesões. Consequentemente, o desenvolvimento de novas terapias mais conservadoras
pode surgir como uma alternativa, já que alguns estudos com linhagens celulares de outros
tipos de tumores demostram que o bloqueio da sinalização de Wnt-1 e SOX2 resulta na
diminuição da proliferação e migração celular neoplásica (POLAKOVA; DUSKOVA;
SMAHEL, 2014; CHANG et al., 2015).

89
Conclusões

90
7. CONCLUSÕES
Diante dos resultados obtidos na presente pesquisa, podemos concluir que:
1. As células epiteliais dos casos de AM, TOA, CO e TOEC foram positivas para SOX2,
FGF-10 e Wnt-1 o que sugere a participação destes fatores no desenvolvimento destas
lesões, porém sem evidente relação com os diferentes comportamentos biológicos.
2. Dentre as células epiteliais das lesões avaliadas, foi evidenciado que CO e TOEC
exibiram maior imunoexpressão nuclear e citoplasmática para SOX-2, com diferença
estatística significativa em relação a marcação em AM e TOA, sugerindo que SOX2
está relacionado com as lesões com diferenciação epitelial em comparação com
tumores com diferenciação ameloblástica.
3. As imunoexpressões nuclear para SOX2 e citoplasmática para FGF-10 nas camadas
basal e parabasal dos COs, indicam o papel destes fatores de transcrição na
manutenção da pluripotência, na diferenciação e proliferação desta lesão.
4. Os casos de CO e TOA exibiram maior imunoexpressão de Wnt-1, com diferença
estatística significativa em relação à marcação evidenciada em AM e TOEC. TBM
5. Não houve correlação estatisticamente significativa entre a imunoexpressão de SOX2,
FGF-10 e Wnt-1 nas lesões estudadas.
6. Os casos de germes dentários analisados no presente estudo foram positivos para
SOX2, FGF-10 e Wnt-1, o que confirma a participação destes fatores de transcrição
durante a odontogênese.

91
Referências

92
8. REFERÊNCIAS
ABDUWELI et al. Tooth replacement and putative odontogenic stem cell niches in
pharyngeal dentition of medaka (Oryzias latipes). Microscopy, v. 63, n. 2, p. 141-53, 2014.
AGARWAL, S. et al. Survival and prognosis for malignant tumors of odontogenic origin.
Otolaryngology-Head and Neck Surgery, v. 155, n. 1, p.113-116, 2016.
ALVES PEREIRA, K. M. et al. Immunohistochemical expression of E-cadherin and β-
catenin in ameloblastomas and tooth germs. Oral Surg. Oral Med. Oral Pathol. Oral
Radiol. Endod., v.109, p.425-431, 2010.
AMORIM, R. F. B. et al. Ceratocisto odontogênico: estudo epidemiológico de 26 casos. Rev
Odonto Ciência, v.18, p. 23-30, 2003.
AURREKOETXEA, M. et al. Wnt/β-catenin regulates the activity of epiprofin/Sp6, SHH,
FGF, and BMP to coordinate the stages of odontogenesis. Front. Cell Dev. Biol., v. 4, n. 25,
p. 1-14, 2016.
BALTUS, G. A. et al. Acetylation of Sox2 induces its nuclear export in embryonic stem cells.
Stem Cells, v. 27, n. 9, p. 2175-2184, 2009.
BANDYOPADHYAY, A. et al. Cancer stem cell markers, SOX 2 and OCT 4 in
ameloblastoma and keratocystic odontogenic tumor: an immunohistochemical study. J Int
Oral Health, v. 9, n. 1, p. 28-32, 2017.
BANERJEE, A. et al. OCT4 and SOX2 are reliable markers in detecting stem cells in
odontogenic lesions. J. Orofac. Sci., v. 8, p. 16-21, 2016.
BASKARAN, P. et al. Adenomatoid odontogenic tumor − A report of two cases with
histopathology correlation. J. Clin. Imaging Sci., v. 1, n. 64, 2011.
BASU-ROY, U. et al. SOX2 maintains self-renewal of tumor initiating cells in
Osteosarcomas. Oncogene, v. 31, n. 18, p. 2270–2282, 2012.
BREWER, J. R.; MAZOT, P.; SORIANO, S. Genetic insights into the mechanisms of Fgf
signaling. Genes & Dev., v. 30, n. 7, p. 751-771, 2016.
CAMP, J. K. V. et al. Wnt signaling and the control of human stem cell fate. Stem Cell Rev.
and Rep., v. 10, n. 2, p. 207-229, 2013.
CAO, Z. et al. Osterix controls cementoblast differentiation through downregulation of Wnt-
signaling via enhancing DKK1 expression. Int. J. Biol. Sci., v. 11, n. 3, p. 335-344, 2015.
CASTRO-SILVA, I. L. et al. Difficulties in the diagnosis of plexiform ameloblastoma. Oral
and Maxillofac. Surg., v.16, p.115-118, 2012.

93
CHANG, L. et al. Identification of a new class of WNT1 inhibitor: cancer cells migration, G-
quadruplex stabilization and target validation. Oncotarget, v. 7, n. 42, p. 67986- 68001, 2015.
CHAUDHARY, Z., et al. Unicystic ameloblastoma: a diagnostic dilemma. Natl J Maxillofac
Surg.v. 2, p. 89-92, 2011.
CHINDASOMBATJAROEN, et al. Calcifying cystic odontogenic tumor and adenomatoid
odontogenic tumor: radiographic evaluation. Oral Surg Oral Med Oral Pathol Oral
Radiol., v. 114, n. 6, p. 796-803, 2012.
CHOI, A. et al. Inhibition of Wnt1 expression reduces the enrichment of cancer stem cells in a
mouse model of breast cancer. Biochemical and Biophysical Research Communications, v.
425, n. 2, p. 436-442, 2012.
CHRCANOVIC, B. R.; GOMEZ, R. S. Calcifying epithelial odontogenic tumor: an updated
analysis of 339 cases reported in the literature. Journal of Cranio-Maxillofacial Surgery, v. 45,
n. 8, p. 1117-1123, 2017.
CUNHA, J. M. et al. Pluripotent stem cell transcription factors during human odontogenesis.
Cell Tissue Res, v. 353, p. 435–441, 2013.
DEBONI, M. C. Z. Clinical, radiological e histological features of calcifying epithelial
odontogenic tumor case report. Braz Dent J, v.17, n.2, p.171-174, 2006.
DE MATOS, F. R. et al. Adenomatoid odontogenic tumor: retrospective study of 15 cases
with emphasis on histopathologic features. Head and Neck Pathology, v. 6, p. 430–437,
2012.
DE SOUZA, L. B. et al. Odontogenic cysts: demographic profile in a Brazilian population
over a 38-year period. Med Oral Patol Oral Cir Bucal, v. 15, n. 4, p. 583-90, 2010.
EL-NAGGAR, A. K. et al. WHO Classification of of head and neck tumours. 4ª edição.
Lyon: IARC Press, 2017.
ENNES, J. P. et al. Immunolocalization of FGF-10 in murine incisors and molar germs. Rev
Gaucha Odontol., v. 60, n. 4, p. 497-502, 2012.
FIGUEROA, A. et al. Keratocystic odontogenic tumor associated with nevoid basal cell
carcinoma syndrome: similar behavior to sporadic type? Otolaryngol Head Neck Surg, v.
142, n. 2, p. 179-83, 2010.
FREGNANI, E. R. et al. Presence of myofibroblasts and expression of matrix
metalloproteinase-2 (MMP-2) in ameloblastomas correlate with rupture of osseous cortical.
Pathol Oncol Res. v.15, p.231-240, 2009.
FULCO, G. M. et al. Solid ameloblastomas-retrospective clinical and histopathologic study of
54 cases. Braz J Otorhinolaryngol. v. 76, n.2, p.172-177, 2010.

94
GAETE, M; TUCKER, A. S. Organized emergence of multiple-generations of teeth in snakes
is dysregulated by activation of Wnt/Beta-catenin signalling. PLoS ONE, v. 8, n. 9, p.
e74484. 2013.
GARG, K. et al. Molecular and genetic aspects of odontogenic tumors: a review. Iran J Basic
Med Sci, v. 18, p. 529‐536, 2015.
GONZÁLEZ-ALVA, P. et al. Keratocystic odontogenic tumor: a retrospective study of 183
cases. J Oral Sci, v. 50, p. 205-12, 2008.
GROSSMANN, S. M. et al. Demographic profile of odontogenic and selected
nonodontogenic cysts in a Brazilian population. Oral Surg Oral Med Oral Pathol Oral
Radiol Endod, v. 104, n. 6, p. 35-41, 2007.
GUNAWARDHANA, K. S. N. D. et al. A clinico-pathological comparison between
mandibular and maxillary ameloblastomas in Sri Lanka. J Oral Pathol Med., v.39, p.236-
241, 2010.
GURGEL, C. A. S. et al. Expression of Ki-67, p53 and p63 proteins in keratocyst
odontogenic tumors: an immunohistochemical study. J Mol Hist, v. 39, p. 311-6, 2008.
HAKIM, S. G. et al. Altered expression of cell-cell adhesion molecules β-catenin/E-cadherin
and related Wnt-signaling pathway in sporadic and syndromal keratocystic odontogenic
tumors. Clin Oral Investig, v. 15, n. 3, p. 321-8, 2011.
HARADA, H. et al. FGF10 maintains stem cell compartment in developing mouse incisors.
Development, v. 129, p. 1533-1541, 2002.
HALIFU, Y. et al. Wnt1 and SRFP1 as potential factors and therapeutic targets in cutaneous
squamous cell carcinoma. Genet Mol Res. v. 15, n. 2, 2016.
HASHIMOTO, Shuichi et al. β-Catenin–SOX2 signaling regulates the fate of developing
airway epithelium. Journal of Cell Science, v. 125, p. 1-11, 2012.
HEIKINHEIMO, K. et al. Early dental epithelial transcription factors distinguish
ameloblastoma from keratocystic odontogenic tumor. Journal of Dental Research, v. 94, n.
1, p. 101–111, 2015.
HERTOG, D. et al. Management of recurrent ameloblastoma of the jaws; a 40-year single
instituition experience. Oral Oncol., v. 47, p. 145-146, 2011.
HIROTA, M. et al. Desmoplastic ameloblastoma featuring basal cell ameloblastoma: a case
report. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod., v. 99, p. 160-164, 2005.
HUANG, G. T.; THESLEFF, I. Stem cells in craniofacial development and regeneration.
1ª edição. Canadá: Wiley-Blackwell, 2013.
HUNASGI, S. Rare giant granular cell ameloblastoma: a case report and an
immunohistochemical study. Case Rep Dent. v. 7, 2013.

95
ISHIKAWA, Y. et al. Quiescent adult stem cells in murine teeth are regulated by Shh
signaling. Cell and Tissue Research, v. 369, n. 3, p. 459-512, 2017.
JANSSON, L.; KIM, G. S.; CHENG, A. G. Making sense of Wnt signaling—linking hair cell
regeneration to development. Frontiers in Cellular Neuroscience, v. 9, n. 66, p. 1-12, 2015.
JONES, A. V.; CRAIG, G. T.; FRANKLIN, C. D. Range and demographics
of odontogenic cysts diagnosed in a UK population over a 30-year period. J Oral Pathol
Med, v. 35, n. 8, p. 500-7, 2006.
JIANG, C. et al. Benign odontogenic tumors: origins, immunophenotypic features, and
genetic alterations. Current Oral Health Reports, v. 3, n. 2, p. 93-101, 2016.
JUURI, E. et al. Expression of the stem cell marker, SOX2, in ameloblastoma and dental
epithelium. Eur J Oral Sci, v. 121, p. 509–516, 2013a.
JUURI, E. et al. SOX2 marks epithelial competence to generate teeth in mammals and
reptiles. Development, v. 140, p. 1424-1432, 2013b.
KATOH, M.; KATOH, M. Molecular genetics and targeted therapy of WNT-related human
diseases (Review). International Journal of Molecular Medicine, v. 40, p. 587-606, 2017.
KAWASAKI, K. et al. Expression of Sox genes in tooth development. Int J Dev Biol., n. 59,
v. 10-12, p. 471-478, 2015.
KETTUNEN, P. et al. Associations of FGF-3 and FGF-10 with signaling networks regulating
tooth morphogenesis. Developmental dynamics, v. 219, p. 322–332, 2000.
KIM, D. K. et al. Comparative Ki-67 expression and apoptosis in the odontogenic keratocyst
associated with or without an impacted tooth in addition to unilocular and multilocular
varieties. Yonsei Med J, v. 44, p. 841-6, 2003.
KORMISH, J. D.; SINNER, D.; ZORN, A. M. Interactions between SOX factors and Wnt/β-
catenin signaling in development and disease. Dev Dyn. v. 239, n. 1, p. 56–68, 2010.
KUMAMOTO, H.; OOYA, K. Immunohistochemical detection of BH3-only protein in
ameloblastic tumors. Oral Dis. v. 14, p. 550-555, 2008.
LEE, M. et al. Roles of Wnt inhibitory factor 1 during tooth morphogenesis. Cell and Tissue
Research, v. 362, n. 1, p. 61-68, 2015.
LEE, S. H. et al. SOX2 regulates self-renewal and tumorigenicity of stem-like cells of head
and neck squamous cell carcinoma. British Journal of Cancer, v. 111, n. 11, p. 2122–2130,
2014.
LEE, S. K.; KIM Y. S. Current concepts and occurrence of epithelial odontogenic tumors: I.
Ameloblastoma and adenomatoid odontogenic tumor. Korean J Pathol. v. 47, p. 191-202,
2013.

96
LEI, Y. et al. Evaluation of SOX2 as a potential marker for ameloblastic carcinoma. Oral
Surg Oral Med Oral Pathol Oral Radiol. v. 117, n. 5, p. 608-616.e1. 2014.
LEWIS, A. E. et al. The widely used Wnt1-Cre transgene causes developmental phenotypes
by ectopic activation of Wnt signaling. Developmental Biology, v. 379, n. 2, p. 229-234,
2013.
LI, J.; PARADA, C.; CHAI, Y. Cellular and molecular mechanisms of tooth root
development. Development, v. 144, n. 3, p. 374-384, 2017.
LI, T. J. et al. p53 expression in odontogenic keratocyst epithelium. J Oral Pathol Med, v.
25, p. 249-55, 1996.
LIEN, W.; FUCHS, E. Wnt some lose some: transcriptional governance of stem cells by
Wnt/ -catenin signaling. Genes & Development, v. 28, n. 14, p. 1517-1532, 2014.
LIU, K. et al. The multiple roles for SOX2 in stem cell maintenance and Tumorigenesis. Cell
Signal, v. 25, n. 5, 2013.
MATSUDA, Y. et al. Distinct role of transforming growth factor-beta 1 and fibroblast growth
factors in human amelobastoma epithelial cell proliferation. Biochemistry and Molecular
Biology, v. 2, n. 1, p. 1-5, 2017.
MENON, S. Keratocystic odontogenic tumours: etiology, pathogenesis and treatment
revisited. J. Maxillofac. Oral Surg., v. 14, n. 3, p. 541–547, 2015.
MIKOLAJCZAK, M. Identification and analysis of a novel nuclear role of Fibroblast
Growth Factor 10 (FGF10). Tese (Doutorado em Fisiologia) - Escola de Ciências
Biológicas, Universidade de East Anglia. Norwich, p. 285. 2016.
MIRAN. S.; MITSIADIS, T. A.; PAGELLA, P. Innovative dental stem cell-based research
approaches: the future of dentistry. Stem Cells International, p. 1-7, 2016.
MOSQUEDA-TAYLOR, A. et al. Odontogenic cysts. Analysis of 856 cases. Med Oral, v. 7,
n. 2, p. 89-96, 2002.
MÜLLER, M. et al. The role of pluripotency factors to drive stemness in gastrointestinal
cancer. Stem Cell Research, v.16, n. 2, p. 349-357, 2016.
MYOUNG, H. et al. Odontogenic keratocyst: review of 256 cases for recurrence and
clinicopathologic parameters. Oral Sur Oral Med Oral Pathol Oral Radiol Endod, v. 91, p.
328-33, 2001.
NAKAO, Y. et al. Fibroblast growth factors 7 and 10 are involved in ameloblastoma
proliferation via the mitogen-activated protein kinase pathway. International journal of
oncology, v. 43, p. 1377-1384, 2013.
NANCI, A. Ten Cate’s oral histology: development, structure, and function. Mosby
Elsevier, St Louis. 2013.

97
NASCIMENTO, M. A. B. et al. Immunoexpression of BMP-2 and BMP-4 and their
receptors, BMPR-IA and BMPR-II, in ameloblastomas and adenomatoid odontogenic tumors.
Archives of Oral Biology, v. 73, p. 223-229, 2017.
NEVILLE, B. W. et al. Patologia Oral e Maxilofacial. 4 ed. Rio de Janeiro: Elsevier; 2016.
NUSSE, R.; CLEVERS, H. Wnt/β-catenin signaling, disease, and emerging therapeutic
modalities. Cell, v. 169, n. 6, p. 985-999, 2017.
OKUBBO, T.; PEVNY, L. H.; HOGAN, B. L. M. Sox2 is required for development of taste
bud sensory cells. Genes & Development, v. 20, n. 19, p. 2654-2659, 2006.
ORNITZ, D. M.; ITOH, N. The fibroblast growth factor signaling pathway. WIREs Dev
Biol., v. 4, n. 3, p. 215-266, 2015.
ORSINI, G. et al. In vivo administration of dental epithelial stem cells at the apical end of the
mouse incisor. Frontiers in Physiology, v. 6, n. 112, p. 1-5, 2015.
PHILIPSEN, H. P., REICHART, P. A. Unicystic ameloblastoma odontogenic tumors and
allied lesions. Quintessence. p. 77-86, 2004.
PHILIPSEN, H. P. et al. An updated clinical and epidemiological profile of the adenomatoid
odontogenic tumour: a collaborative retrospective study. J Oral Pathol Med. v.36, n.7,
p.383-393, 2007.
POLAKOVA, I.; DUSKOVA, M.; SMAHEL, S. Antitumor DNA vaccination against the
Sox2 transcription factor. International Journal of Oncology, v. 45, p. 139-146, 2014.
PRIYA, S. P. et al. Odontogenic epithelial stem cells: hidden sources. Laboratory
Investigation, v. 95, n. 12, p. 1344-1352, 2015.
PUTHIYAVEETIL, J. S. V. et al. Epithelial-mesenchymal interactions in tooth development
and the significant role of growth factors and genes with emphasis on mesenchyme – A
Review. Journal of Clinical and Diagnostic Research, v. 10, n. 9, p. ZE05-ZE09, 2016.
RAMANATHAN, A. et al. Homeobox genes and tooth development: Understanding the
biological pathways and applications in regenerative dental science. Archives of Oral
Biology, v. 85, p. 23-39, 2018.
RAMOS, G. O. et al. Odontogenic tumors: a 14-year retrospective study in Santa Catarina,
Brazil. Braz. oral res., São Paulo, v. 28, n. 1, 2013.
REGEZI, J. A. Odontogenic cysts, odontogenic tumors, fibroosseous, and giant cell lesions of
the jaws. Mod Pathol, v. 15, n. 3, p. 331–341, 2002.
REN, Z.; ZHANG, C.; JI, T. Expression of SOX2 in oral squamous cell carcinoma and the
association with lymph node metastasis. Oncology Letters, v. 11, n. 3, p. 1973-1979, 2016.
RICCI, M. et al. An unusual case of unicystic intramural ameloblastoma and review of the
literature. Contemp Clin Dent. v. 3, p. 233-9, 2012.

98
RICK, G. M. Adenomatoid odontogenic tumor. Oral Maxillofac Surg Clin North Am, v.
16, p. 333–54, 2004.
ROBINSON, R. A.; VINCENT, S. D. Tumors and Cysts of the Jaws. AFIP Atlas of Tumor
Pathology. 4th ser. Silver Spring, MD: American Registry of Pathology; p. 64-68, 2012.
RODRÍGUEZ-LOZANO, F. J. et al. Mesenchymal stem cells derived from dental tissues.
International Endodontic Journal, v. 44, n. 9, p. 800-806, 2011.
ROSS, S. E. et al. Inhibition of adipogenesis by Wnt signaling. Science, v. 289, n. 5481, p.
950-953, 2000.
SARKAR, A.; HOCHEDLINGER, K. The Sox family of transcription factors: versatile
regulators of stem and progenitor cell fate. Cell Stem Cell, v. 12, n. 1, p. 15-30, 2013.
SCHEPERS, G. E., TEASDALE, R. D.; KOOPMAN, P. Twenty pairs of sox: extent,
homology, and nomenclature of the mouse and human sox transcription factor gene families.
Dev. Cell, v. 3, p. 167–170, 2002.
SELVAMANI, M.; DONOGHUE, M.; BASANDI, P. S. Analysis of 153 cases of
odontogenic cysts in a South Indian sample population: a retrospective study over a decade.
Braz. Oral Res., São Paulo, v. 26, n. 4, 2012.
SEO, E. et al. Distinct functions of SOX2 control self-renewal and differentiation in the
osteoblast lineage. Molecular and cellular biology, v. 31, n. 22, p. 4593–4608, 2011.
SHEAR, M. Odontogenic keratocysts: clinical features. Oral Maxillofac Surg Clin North
Am, v. 15, n. 3, p. 335-45, 2003.
SHARPE, P. T. Dental mesenchymal stem cells. Development, v. 143, n. 13, p. 2273-2280,
2016.
SIRIWARDENA, B. S. M. S. et al. Aberrant b-catenin expression and adenomatous
polyposis coli gene mutation in ameloblastoma and odontogenic carcinoma. Oral Oncology,
v. 45, p. 103–108, 2009.
SLAVIK, M. A. et al. Wnt signaling induces differentiation of progenitor cells in organotypic
keratinocyte cultures. BMC Developmental Biology, v. 7, n. 1, p. 1-8, 2007.
SLOOTWEG, P. J. Odontogenic tumours - An update. Curr Diagn Pathol, v. 12, p. 54-65,
2006.
SONG, J. et al. Differentiation and regenerative capacities of human odontoma-derived
mesenchymal cells. Differentiation, v. 77, n. 1, p. 29–37, 2009.
STOELINGA, P. J. W. Etiology and pathogenesis of keratocysts. Oral and Maxillofacial
Surgery Clinics of North America, v. 15, n. 3, p. 317-324, 2003.
SURYADEVA, S; KHAN, M. B. Role of Homeobox genes in tooth morphogenesis: A
Review. Journal of Clinical and Diagnostic Research, v. 9, n. 2, p. ZE09-ZE12, 2015.

99
SWEENEY, R. T. et al. Identification of recurrent SMO and BRAF mutations in
ameloblastomas. Nature Genetics, v. 46, n. 7, p. 722-725, 2014.
TAKATA, T.; SLOOTWEG, P. J. Calcifying epitelial odontogenic tumor. In: BARNES et
al.(Ed.) Pathology & genetics of head and neck tumours. Lyon:IARC Press, 2005. p. 302-
303.
THESLEFF, I. The genetic basis of tooth development and dental defects. Am J Med Genet
Part A, v. 140A, n. 23, p. 2530–2535, 2006.
TORTORICI, S. et al. Prevalence and distribution of odontogenic cysts in Sicily: 1986-2005.
J Oral Sci, v. 50, n. 1, p. 15-8, 2008.
TSUKAMOTO, G. et al. A radiologic analysis of dentigerous cysts and odontogenic
keratocysts associated with a mandibular third molar. Oral Surg Oral Med Oral Pathol
Oral Radiol Endod, v. 91, p. 743-7, 2001.
VOLCKAERT, T.; DE LANGHE, S. P. Wnt and FGF mediated epithelial-mesenchymal
crosstalk during lung development. Developmental Dynamics, v. 244, n. 3, p. 342-366,
2014.
WANG, Q. et al. Oct3/4 and SOX2 are significantly associated with an unfavorable clinical
outcome in human esophageal squamous cell carcinoma. Anticancer res., v. 29, p. 1233-
1242, 2009.
WAKOH, M. et al. Reliance on diagnostic elements in panoramic imaging with focus on
ameloblastoma and keratocystic odontogênica tumor: Psychometric study. Bull Tokyo Dent
Coll.v.52, p. 1-12, 2011.
WEGNER, M. Allpurpose Sox: The many roles of Sox proteins in gene expression. Int. J.
Biochem. Cell Biol. v. 42, p. 381–390, 2010.
YANG, J. et al. Gating pluripotency via nuclear pores. Trends in Molecular Medicine, v.
20, n. 1, p. 1-7, 2014.
ZHANG, L. et al. Expression pattern of SOX2 during mouse tooth development. Gene
Expression Patterns, v. 12, p. 273–281, 2012.
ZHAN, T.; RINDTORFF, N.; BOUTROS, M. Wnt signaling in cancer. Oncogene, v. 36, n.
11, p. 1461-1473, 2016.

100
Anexo

101
ANEXO

102
103

104





























