Embed Size (px)
Citation preview

UNIVERSIDADE DO ALGARVE Faculdade de Ciências e Tecnologia LICENCIATURA EM BIOQUÍMICA
Relatório de Estágio
Estudo da Incorporação de Subunidades Durante a Montagem do Proteassoma 20S
Autor: António José L. Marques Orientador: Prof. Doutora Paula C. Ramos
Faro, Dezembro 2001

"There must be no barriers for freedom of inquiry.
There is no place for dogma in science. The
scientist is free, and must be free to ask any
question, to doubt any assertion, to seek for any
evidence, to correct any errors."
Robert Oppenheimer

Agradecimentos 3
Agradecimentos
No final destes cinco anos de licenciatura quero expressar o meu mais profundo agradecimento, e sabendo que a retribuição de forma alguma será alcançada, ...
... a todo o corpo docente do curso de Bioquímica pelos conhecimentos e experiências transmitidos, não só a nível profissional como pessoal e em especial à Professora Doutora Paula Ramos, minha orientadora de estágio, pela oportunidade que me apresentou ao aceitar-me como orientando e iniciar assim a minha passagem pelo mundo da ciência e investigação.
... a toda a minha família, em especial aos meus Pais por todo o seu
amor, dedicação e esforço, não só no tempo da minha passagem pela Universidade, mas durante todo o tempo anterior. Também por toda a educação que, na sua forma simples de a transmitirem, me levaram a ser o eu de hoje. E não me poderia esquecer do gosto pela incessante busca por conhecimento e novas experiências que desde cedo me incutiram.
... à Bárbara, por toda a sua amizade e amor e por ser ela própria em
todas as alturas. Por tudo o que me deu e transmitiu ao longo destes dois anos, por todas as experiências e momentos partilhados.
... a todos os meus amigos que, perto ou longe, me apoiaram ao longo
destes cinco anos de passagem por esta academia, sempre com a sua incondicional amizade, nos momentos melhores e piores, com a força que só um verdadeiro amigo pode dar.
... à Professora Doutora Deborah Power, pela disponibilidade mostrada
quanto à utilização do seu equipamento. ... ao Professor Doutor Jurgen Dohmen, pelas ideias e conhecimentos
transmitidos, para além das estirpes de levedura por ele construídas, sem as quais este trabalho estaria muito mais dificultado.
... ao Markus London, pela amável cedência da estirpe JD305. ... à Fundação para a Ciência e Tecnologia pela bolsa cedida. ... a todos os outros não mencionados que, directa ou indirectamente,
contribuiram para a conclusão da minha licenciatura. O meu obrigado a todos vocês... Independentemente da proximidade ou
distância, as recordações ficarão para sempre...

Abreviaturas 4
Abreviaturas Ab - Anticorpo, do inglês Antibody
Ag - Antigénio, do inglês Antigene
Ala - Alanina, do inglês Alanine
Asp - Aspartato, do inglês Aspartate
ATP - Adenosina-5’-Trifosfato, do inglês Adenosine-5’-TriPhosphate
bicina - NN-Bis-(2-hidroxietil)-glicina
bisacrilamida - N’,N’ – metilenobisacrilamida
bp - Par de bases, do inglês base pair
BSA - Albumina de Soro Bovino, do inglês Bovine Serum Albumin
CBB - Azul de Coomassie, do inglês Coomassie Brilliant Blue
CE - Extracto Cru, do inglês Crude Extract
Cys - Cisteína, do inglês Cysteine
Da - Unidade de massa molecular, correspondente à massa do átomo
de hidrogénio
DCE - Extracto Cru Dessalinizado, do inglês Desalted Crude Extract
DNA - Ácido Desoxirribonuleico, do inglês Desoxirribonucleic Acid
DMSO - Dimetilsulfóxido
DTT - Ditiotreitol
EDTA - Ácido EtilenoDiaminoTetraAcético
FLAG - Epítopo composto pela sequência de aminoácidos DYKDDDDK
FT - Fracção não ligada, do inglês Flow Through
Glu - Glutamato, do inglês Glutamate
Gly - Glicina, do inglês Glycine
ha - Antigénio composto pela sequência de aminoácidos
YPYDVPDYA
HEPES - N-(2-Hidroxietil)piperazina-N’-(2-ácido etanosulfónico)
Hidrolase Ntn - Hidrolases Nucleófilas de Terminal N, do inglês N terminal
nucleophile hidrolase
His - Histidina, do inglês Histidine
IgG - Imunoglobulina G
LB - Meio de cultura Luria-Brot

Abreviaturas 5
LB-Amp - Meio de cultura LB com Ampicilina
Leu - Leucina, do inglês Leucine
LiAc - Acetato de Lítio, do inglês Lithium Acetate
log - Logarítmo de base 10
Lys - Lisina, do inglês Lysine
MOPS - 3-[N-Morpholino]Propane-Sulfonic Acid
MW - Massa molecular, do inglês Molecular Weight
NaCl - Cloreto de Sódio, do alemão NatriumChlorid
OD600nm - Densidade óptica a 600 nm, do inglês Optical Density
PEG - Polietilenoglicol
PCR - do inglês, Polymerase Chain Reaction
PSA - Persulfato de Amónio
PVDF - Fluoreto de polivinilideno, do inglês Polyvinylidene Fluoride
RNase - Hidrolase de Ácido Ribonucleico, do inglês Ribonucleic Acid
Hidrolase
rpm - Unidade de rotação, correspondente a uma rotação por minuto
SB - Tampão amostra desnaturante, do inglês Sample Buffer
SD - Meio de cultura mínimo para levedura
SDS - Dodecil Sulfato de Sódio, do inglês Sodium Dodecyl Sulfate
Ser - Serina, do inglês Serine
SN - Tampão amostra nativo, do inglês Sample Buffer for Natives
Thr - Treonina, do inglês Threonine
TEMED - N,N,N’,N’-tetrametiletilenodiamina Tris - Tris-(hidroximetil)-aminometano
Ub - Ubiquitina
Ura - Uracilo, do inglês Uracil
UV - Ultravioleta
V0 - Volume nulo
Ve - Volume de eluição
Vt - Volume total
YPD - Meio de cultura completo para levedura, do inglês Yeast extract,
Peptone, Dextrose

Resumo 6
Resumo
A via de degradação proteica dependente de ubiquitina/proteassoma é
responsável pela maior parte da proteólise não lisossomal em células
eucariotas, que inclui a degradação de proteínas com semi-vida curta, proteínas
mal formadas, factores de transcrição, reguladores celulares, geração de
péptidos antigénicos e proteínas afectadas pelo efeito de vários tipos de stress.
Em eucariotas, o proteassoma 20S é caracterizado por possuir 14
subunidades diferentes na sua constituição, sete α e sete β, organizadas em
quatro anéis heptaméricos na forma α7β7β7α7.
Apesar da estrutura cristalina do proteassoma 20S estar resolvida e de os
passos finais na montagem deste complexo estarem caracterizados, pouco se
sabe sobre os passos iniciais deste processo. Com o objectivo de alargar o
conhecimento científico existente sobre a biogénese do proteassoma, foram
feitos estudos utilizando estirpes da levedura Saccharomyces cerevisiae com as
subunidades α7/Prs1p e β4/Pre1p e o factor de maturação Ump1p marcados
com epítopos detectáveis imunoquimicamente.
Observou-se que cádmio e choque térmico interferem no crescimento das
culturas mas não provocam aumento na expressão proteica do proteassoma
nem interferem na sua actividade catalítica. A delecção da subunidade α3/Y13
afecta a actividade quimotríptica do proteassoma 20S, e também interfere com a
distribuição normal da subunidade Prs1p ao longo do processo de montagem.
A comparação da distribuição das subunidades β4/Pre1p e α7/Prs1p em
complexos proteassomais indicou-nos que a β4/Pre1p parece ser incorporada
nos complexos pre-proteassomais numa fase mais tardia em relação à
α7/Prs1p, sugerindo que a α7/Prs1p é uma subunidade envolvida nos passos
iniciais da montagem do proteassoma 20S in vivo.

Abstract 7
ABSTRACT
The ubiquitin/proteasome pathway is responsible for most of the non
lysossomal proteolysis in eukaryotic cells, including degradation of short-lived
and abnormal proteins, transcription factors, cellular regulators, peptidic antigen
generation and proteins affected by several kinds of stress.
The eukaryotic 20S proteasome is characterized by the presence of 14
different subunits, seven α-type and seven β-type, organized in four heptameric
rings with the arrangement α7β7β7α7.
Although the crystal structure of the 20S proteasome is known and the
final steps in its assembly are characterized, little is known about the initial steps
of this process. In order to understand better the proteasome biogenesis, several
studies, using yeast strains with the subunits α7/Prs1p and β4/Pre1p and the
maturation factor Ump1p tagged for immunological detection were performed.
It was observed that cadmium and heat shock interfere in cell culture grow
but do not increase protein expression of the proteasome nor interfere with its
catalytic activity. In spite of that, the chymotryptic-like activity is altered by the
deletion of the α3/Y13 subunit, such deletion also influences the wild type
distribution of subunit Prs1p during the assembly process.
Comparison of the distribution of the subunits β4/Pre1p and α7/Prs1p in
different assembly complexes showed that the β4/Pre1p seems to be included in
the pre-proteasomal complexes in a later stage than α7/Prs1p, suggesting that
α7/Prs1 can be a subunit involved in earlier assembly steps of the proteasome in
vivo.

Índice 8
Índice
Agradecimentos 3 Sinónimos e Acrónimos 4 Resumo 6 Abstract 7 1. Introdução 12 1.1 – Prespectiva Histórica 12 1.2 – Ubiquitina em Levedura 13 1.2.1 – Enzima de Activação de Ubiquitina (E1) 16 1.2.2 – Enzimas de Conjugação de Ubiquitina (E2) 16 1.2.3 – Enzimas de Ligação Ubiquitina-Proteína (E3) 17 1.3 – Estrutura do Proteassoma 17 1.3.1 - Estrutura Cristalina do Proteassoma 20S da Arqueobactéria
Thermoplasma acidophilum
18
1.3.2 – Estrutura Cristalina do Proteassoma 20S de S. cerevisiae 19 1.4 – Identificação dos Centros Catalíticos 20 1.5 – Montagem do Proteassoma 20S 22 1.5.1 – Montagem do Proteassoma 20S em Thermoplasma 22 1.5.2 – Montagem do Proteassoma 20S em Rhodococcus 23 1.5.3 – Montagem do Proteassoma 20S em Eucariotas 23 1.5.4 – Propeptídios no Proteassoma 20S 26 1.6 – Objectivo 27 2. Materiais e Métodos 28 2.1 – Material Biológico 28 2. 2 – Métodos 28 2.2.1 – Cultura de células de E. coli 28 2.2.2 – Cultura de Células de S. cerevisiae 29
2.2.3 – Marcação da Subunidade α7/Prs1p do Proteassoma com o Epítopo ha 30 2.2.3.1 – Produção de Células Competentes de E. coli (JM105) 30 2.2.3.2 – Transformação de Células Competentes de E. coli (JM105) 31 2.2.3.3 – Purificação de DNA Plasmídico 32

Índice 9
2.2.3.4 – Linearização do Plasmídio pCR8 para Integração em Levedura 33 2.2.3.5 – Produção e Transformação de Células Competentes de Levedura 33 2.2.3.6 – Polymerase Chain Reaction (PCR) 35 2.2.4 – Extractos Proteicos 36 2.2.4.1 – Extracção por Maceração das Células 36 2.2.4.2 – Extracção com Esferas de Vidro 37 2.2.5 – Quantificação Proteica 37 2.2.6 – Cromatografia de Filtração em Gel 39 2.2.6.1 – Cromatografia de Filtração em Gel em Superose 6 40 2.2.6.2 – Cromatografia de Filtração em Gel para Dessalinização 41 2.2.6.2.1 – Cromatografia de Filtração em Gel em PD-10 42 2.2.7 – Electroforese de Biomoléculas 42 2.2.7.1 – Electroforese de Ácidos Nucleicos em Géis de Agarose 43 2.2.7.2 – Electroforese de Proteínas em Géis de Poliacrilamida
2.2.7.2.1 – Electroforese de Proteínas em Condições Desnaturantes
44 45
2.2.7.2.2 – Electroforese de Proteínas em Condições Nativas 47 2.2.8 – Detecção de Polipéptidos em Géis de Poliacrilamida 48 2.2.8.1 – Coloracão de Proteínas com Azul de Coomassie 48 2.2.8.2 – Coloração de Proteínas com Prata 49 2.2.9 – “Immunoblot” 50 2.2.9.1 – Electrotransferência de Proteínas 51 2.2.9.2 – Fixação das Proteínas à Membrana de PVDF 52 2.2.9.3 – Bloqueamento da Membrana de PVDF 53 2.2.9.4 – Incubação com Anticorpos 55 2.2.9.5 – Revelação do “Immunoblot” 56 2.2.10 – Cromatografia de Metalo-Afinidade 57 2.2.11 – Medição da Actividade Quimotríptica do Proteassoma 59 3. Resultados 60 3.1 – Efeito do Cádmio e do Choque Térmico no Crescimento de S.
cerevisiae e na Expressão da Subunidade Proteassomal α7/Prs1p
60

Índice 10
3.2 – Marcação da Subunidade α7/Prs1p com o Epítopo ha 63
3.2.1 – Estratégia Usada para a Marcação da Subunidade α7/Prs1p com ha 63
3.2.2 – Análise da Linearização do Plasmídio pCR8 65 3.2.3 – Análise da Integração por PCR e Immunoblot 65
3.2.4 – Observação do Fenótipo da Estirpe Mutante (Y13∆) sob Choque
Térmico
66
3.3 – Presença de Várias Subunidades Proteassomais ao Longo das Fracções
Obtidas por Filtração em Gel
66 3.3.1 – Análise da Actividade Quimotríptica em JD183 e AM1 66 3.3.2 – Presença de Várias Subunidades Marcadas ao Longo do Padrão de
Separação por Filtração em Gel
68
3.4. – Análise de Complexos Contendo a Subunidade β4/Pre1p e o factor de
maturação Ump1p
69
3.5 – Análise de complexos proteassomais contendo β4/Pre1p, α7/Prs1p e
Ump1p
71
3.5 – Purificação de Complexos contendo α7/Prs1pFLAG-6His 73
4 – Discussão 75 4.1 - Efeito do Cádmio e do Choque Térmico no Crescimento de S.
cerevisiae e na Expressão da Subunidade Proteassomal α7/Prs1p
75
4.1.1 – O Cádmio Influencia o Crescimento Celular em Levedura 75 4.1.2 – O Cádmio e Choque Térmico não Afectam os Níveis de Expressão
nem a Actividade Quimotríptica do Proteassoma
76
4.2 – Marcação da Subunidade α7/Prs1p com o Epítopo ha 76
4.2.1 – Verificação da Linearização do pCR8 por Cla I 76 4.2.2 – Verificação da Integração por PCR e Immunoblot 77
4.2.3 – A Estirpe Mutante JD305 (Y13∆) é Sensível a Stress Provocado por
Choque Térmico
77
4.3 – Existem Diferenças na Distribuição das Várias Subunidades ao Longo de
Complexos Separados por Exclusão Molecular
78
4.3.1 – A Estirpe Mutante AM1 Apresenta uma Actividade Catalítica
Semelhante à Estirpe Selvagem no Proteassoma 26S, mas mais

Índice 11
Reduzida no Proteassoma 20S 79
4.3.2 – A Subunidade α7/Prs1p Está Presente ao Longo de Todas as
Fracções Obtidas por Filtração em Gel
79
4.3.3 – A Delecção da Subunidade α3/Y13 Provoca Alterações na Distribuição
da Subunidade α7/Prs1p
82
4.4 – Purificação de Complexo contendo α7Prs1pFLAG-6His 83
5 – Conclusões 85 6 – Perspectiva Futura 86 7 – Bibliografia 87 8 – Anexos 96

Introdução 12
1 – Introdução 1.1 – Perspectiva Histórica O lisossoma tem sido considerado como o local de “despejo” da célula
pela sua capacidade de degradar proteínas, devido ao grande número de
proteases não específicas presentes, e de digerir outros compostos não
necessários. No entanto, como este processo de degradação não é específico
(Bohley et al., 1992) não era previsível que fosse responsável pela degradação
selectiva de proteínas, requisito necessário ao mecanismo de regulação celular.
Em 1980 foi isolada e caracterizada uma endopeptidase que, devido à
existência de várias subunidades na sua composição, foi designada por
complexo proteolítico multicatalítico (“multicatalytic protease complex”). Este
complexo, de elevada massa molecular, com cerca de 700 kDa, possui várias
actividades proteolíticas, nomeadamente actividade tríptica, quimotríptica e pós-
acídica (Wilk e Orlowski, 1980, 1983; Orlowski e Wilk, 1981). Aproximadamente
na mesma altura, foi também descoberto um complexo proteolítico alcalino
existente no músculo da carpa, com cerca de 600 kDa, com 15 nm de
comprimento e 10 nm de diâmetro, composto por quatro anéis (Hase et al.,
1980). Complexos proteolíticos de massa molecular elevada foram, também,
descritos noutros tipos de células como eritrócitos (Edmunds e Pennington,
1982) e células do músculo esquelético de rato (Dahlmann et al., 1983). Em
1984 foi descoberto um mesmo complexo semelhante em levedura (Achstetter
et al., 1984). Após o reconhecimento da sua função nas diversas espécies, esta
protease multicatalítica passou a ser denominada por proteassoma (Arrigo et al.,
1988).
No final dos anos oitenta, a identificação de um complexo maior contendo
o núcleo proteolítico previamente identificado, levou à distinção dos dois
complexos com base na sua constante de sedimentação: o proteassoma 26S e
o proteassoma 20S (Hough et al., 1987; Eytan et al., 1989; Discoll e Goldberg,
1990).

Introdução 13
A associação do proteassoma com a via proteolítica dependente da
ubiquitina teve início em 1986 com a descoberta de uma protease dependente
de ATP que degradava lisozima ubiquitilada (Hough et al., 1986). No ano
seguinte foi identificada esta protease como sendo o proteassoma 26S (Hough
et al., 1987). Heinemeyer et al., 1991 demonstrou que mutações no
proteassoma que originavam uma actividade quimotríptica baixa levava à
acumulação de proteínas ubiquitiladas na célula, demonstrando claramente a
participação do proteassoma no sistema proteolítico dependente de ubiquitina.
Desde a degradação de proteínas com semi-vida curta, proteínas com
erros de síntese ou desnaturadas, degradação de factores de transcrição,
geração de péptidos antigénicos para serem apresentados pela classe I do
complexo MHC (“major histocompatibility complex”), regulação do ciclo celular,
envolvimento no controlo de qualidade das proteínas no retículo endoplasmático,
muitas foram as funções já associadas a este complexo multicatalítico que é o
proteassoma.
1.2 – Ubiquitina em Levedura
A ubiquitina (Ub) é uma proteína com 76 resíduos de aminoácidos,
altamente conservada (entre levedura e mamíferos existe apenas uma diferença
de três resíduos de aminoácidos) e presente em todas as células eucariotas
(Özkaynak et al., 1987; Hershko et al., 1998) (figura 1).
Figura 1 – Representação esquemática da estrutura terciária da ubiquitina de levedura, determinada por cristalografia de raio-X a 1,8 Å de resolução. Destaque para o resíduo Lys48 (amarelo) e Gly76 (azul). Baseado na estrutura da Ub determinada por Vijay-Kumar et al., 1987

Introdução 14
A Ub está associada ao principal sistema proteolítico extra-lisossomal
presente nas células eucariotas (a via proteolítica dependente de
Ub/proteassoma). Esta via está envolvida na regulação da divisão e ciclo celular,
diferenciação e desenvolvimento, mecanismos de resposta celular a stress e
modeladores extracelulares, modulação de receptores da superfície membranar,
canais iónicos e mecanismos de secreção, reparação genómica a nível dos
ácidos desoxirribonucleicos (DNA) e biogénese de organelos (Hilt e Wolf, 2000).
A Ub, no seu estado livre, não possui qualquer actividade enzimática e,
quando ligada de forma covalente a outras proteínas, funciona como marcador,
seleccionando as proteínas para destruição pelo proteassoma 26S
(Hochstrasser, 1996; Smith, 1996; Pickart, 1997).
As proteínas-substrato que sofrem ubiquitilação são proteínas com erros
de síntese ou estruturalmente deformadas. No entanto, são também marcados
para degradação reguladores celulares, muitos dos quais com tempos de semi-
vida curtos, em resposta a sinais específicos ou em determinados passos do
ciclo celular.
A degradação de uma proteína pelo mecanismo proteolítico dependente
de Ub/proteassoma pode ser resumida em dois passos. O primeiro envolve a
ligação de várias moléculas de Ub ao substrato. O segundo leva ao
reconhecimento e degradação da proteína marcada pelo proteassoma 26S (Hilt
e Wolf, 2000).
A conjugação da Ub à proteína com vista a ser degradada é feita em três
passos: inicialmente a Ub é activada no seu terminal carboxílico (resíduo de
glicina (-Gly)) pelo enzima de activação de Ub, E1. Seguidamente, um dos
enzimas de conjugação de Ub (E2), transfere a Ub activada para um dos
enzimas do grupo E3, enzimas de ligação de Ub que, por sua vez, selecciona a
proteína a degradar através da interacção com um sinal de degradação
existente na proteína alvo. A conjugação da Ub à proteína substrato é efectuada
por este grupo de enzimas, que catalisam o último passo da conjugação, a
ligação Ub-proteína alvo. Forma-se então uma ligação isopeptídica entre o grupo

Introdução 15
carboxílico activado do terminal C da Ub e o grupo ε-NH2 de um resíduo de lisina
(Lys) existente na proteína alvo. Sucedem-se vários passos de enlongamento da
cadeia de Ub, com a ligação de um certo número destas proteínas à Ub já ligada
à proteína alvo. O crescimento desta cadeia poliubiquitilada é efectuado pela
ligação de outras Ub activadas ao resíduo Lys48 da Ub que lhes precede (figura
2).
Estas cadeias de poli-Ub ligadas à proteína alvo formam um conjunto
que é reconhecido por um ou mais constituintes da partícula reguladora 19S do
proteassoma (Pickart et al., 1997) e iniciar assim o processo de degradação por
parte deste.
Figura 2 – Mecanismo de conjugação da Ub à proteína a degradar. Activação da Ub pela E1, transporte até à proteína alvo ligada à E3 pela E2, conjugação da Ub à proteína alvo pela E3, formação da cadeia de poli-Ub e reconhecimento e degradação da proteína alvo pelo proteassoma. As Ub, através de isopeptidases, são libertadas e recicladas, podendo ser utilizadas novamente após activação (estrutura do proteassoma 26S adaptada de Voges et al., 1999).

Introdução 16
Na levedura, a Ub é codificada por quatro genes, UBI1, UBI2, UBI3 e
UBI4. Os três primeiros codificam formas precursoras de Ub que, após a
tradução, são rapidamente processadas para formar Ub livre em condições
normais de crescimento (Hochstrasser, 1996). O quarto gene, UBI4, é apenas
expresso em condições de stress e é constituído por cinco repetições da
sequência que codifica para uma molécula de Ub.
1.2.1 – Enzima de Activação de Ubiquitina (E1) O enzima de activação de ubiquitina é essencial para o correcto
funcionamento da via proteolítica dependente de Ub/proteassoma e mesmo para
a viabilidade das células, uma vez que a delecção do gene de S. cerevisiae,
UBA1, que codifica para este enzima de 114 kDa conduz à morte das células
(McGrath et al., 1991).
O E1 é fosforilado e aparenta ter como única função a catálise das
modificações necessárias à activação das moléculas de Ub. O enzima hidrolisa
ATP durante a formação de um éster de tiol entre o grupo carboxílico da glicina
(Gly76) terminal da Ub e um resíduo de cisteína do enzima, dando origem a uma
ligação tioéster E1-Ub de alta energia (McGrath et al., 1991).
1.2.2 – Enzimas de Conjugação de Ubiquitina (E2) No genoma de S. cerevisiae foram detectados treze genes que codificam
enzimas de conjugação de Ub. Estes enzimas caracterizam-se pela presença de
um domínio conservado de aproximadamente 150 resíduos de aminoácidos, o
domínio de conjugação de Ub (domínio UBC). Na região central deste domínio
encontra-se o resíduo de Cys necessário para a ligação tioéster com a Ub
(Jentsch et al., 1990).
Alguns destes enzimas apresentam domínios diferentes entre si, que
poderão estar relacionados com a especificidade para o substrato e para o
enzima E3 que lhe corresponde (Jentsch, 1992).

Introdução 17
Uma das principais funções até agora atribuídas a este conjunto de
enzimas é o transporte e transferência da Ub activada para o complexo E3-
substrato.
1.2.3 – Enzimas de Ligação Ubiquitina-Proteína (E3) Apesar de se aceitar que os enzimas do conjunto E3 têm uma forte
participação no reconhecimento de substratos (as proteínas para degradar) o
nível de conhecimento sobre estes enzimas é ainda muito baixo (Peters et al.,
1998). Uma das maiores dificuldades na identificação e caracterização de novos
enzimas desta família deve-se ao facto da existência de baixa homologia entre
diferentes tipos de E3, bem como os diferentes tipos de funcionamento destes
enzimas (Hershko e Ciechanover, 1998).
Em alguns casos de E3, o substrato tem uma interacção directa com o
enzima, enquanto que noutros casos, essa interacção é efectuada via uma
molécula adaptadora. O método de transferência da Ub activada para a proteína
alvo também varia consoante a E3 em causa, podendo haver a transferência da
ligação tioéster da E2 directamente para a E3 e daí para o substrato ou a ligase
participar apenas como ajudante na transferência da Ub da E2 directamente
para o substrato, ocorrendo neste caso a formação de um complexo E3-E2-
substrato (Hershko e Ciechanover, 1998).
A definição de enzima E3 pode então ser apresentada como a de um
enzima que liga, directa ou indirectamente, um substrato específico e promove a
transferência da Ub, directa ou indirectamente, de um intermediário tioéster para
uma ligação amida com proteínas ou cadeias poli-Ub.
1.3 – Estrutura do Proteassoma O proteassoma 26S é composto por um núcleo proteolítico, o
proteassoma 20S e por dois complexos regulatórios, os complexos 19S, também
designados por PA700 (Peters et al., 1993). Pensa-se que estes complexos

Introdução 18
regulatórios, dependentes de ATP, estão envolvidos no reconhecimento das
cadeias de poli-Ub ligadas a proteínas para degradação e no desenrolamento e
translocação dos substratos para o interior da câmara catalítica localizada no
proteassoma 20S (Tanaka et al., 1997; Seeger et al., 1997). Os complexos 19S
podem ser substituídos por outros complexos activadores da proteólise,
independentes de ATP, os complexos PA28 (Chu-Ping et al., 1992).
1.3.1 - Estrutura Cristalina do Proteassoma 20S da Arqueobactéria Thermoplasma acidophilum
Imagens obtidas por microscopia electrónica do proteassoma 20S de
arqueobactérias (Hegerl et al., 1991) e de eucariotas (Baumeister et al., 1988)
são praticamente indistinguíveis. Ambas mostram uma partícula cilíndrica com
aproximadamente 150 Å de comprimento e 110 Å de diâmetro.
Ao contrário dos eucariotas, as arqueobactérias possuem um
proteassoma composto por apenas duas subunidades diferentes, denominadas
por α e β (Dahlmann et al., 1989), com elevada homologia da sua estrutura
primária e terciária, organizadas num complexo com quatro anéis, cada qual
com sete subunidades e em que os dois anéis formados por subunidades β se
encontram entre os dois anéis formados por subunidades α (Grziwa et al., 1991)
(figura 3).
A estrutura cristalina do proteassoma de T. acidophilum veio, ainda,
demonstrar que os anéis formam um complexo cujo interior é dividido em três
compartimentos: duas antecâmaras e uma câmara central (Jap et al., 1993;
Löwe et al., 1995).
Figura 3 – Estrutura cristalina do proteassoma de T. acidophilum. A vermelho estão representadas as subunidades β entre as subunidades α, a azul (Löwe et al, 1995).

Introdução 19
1.3.2 – Estrutura Cristalina do Proteassoma 20S de S. cerevisiae
Na levedura S. cerevisiae, as subunidades α e β presentes divergiram
das que se encontram em arqueobactérias dando origem a sete formas
diferentes para cada um dos tipos. A estrutura α7β7β7α7 mantém-se, ocupando
agora, cada uma das catorze subunidades diferentes, uma posição específica e
dando origem a uma simetria do tipo C2, como observado na sua estrutura
cristalina (Groll et al., 1997) (figura 4).
A estrutura tridimensional de proteassomas de mamíferos não é ainda
conhecida embora a topologia exacta das subunidades vizinhas em cada anel
do proteassoma humano tenha sido elucidada utilizando métodos de
“crosslinking”, e à excepção da subunidade β7 que se encontra na posição de β2
o rearranjo das subunidades é idêntico ao encontrado para o proteassoma de
levedura (Kopp et al., 1997). No entanto, três das subunidades β podem ser
substituídas por outras subunidades induzidas por citoquinas, introduzindo assim
o funcionamento do proteassoma no sistema imunitário.
A entrada para o canal central do proteassoma 20S de levedura está
obstruída por uma rede formada pelos resíduos pertencentes ao terminal N de
cada subunidade α (figura 5).
Figura 4 – Estrutura cristalina do proteassoma 20S de S. cerevisiae. Em tons de azul são apresentadas as subunidades α e em tons de verde as subunidades β (Groll et al., 1997).

Introdução 20
No caso das arqueobactérias, esta rede não é tão complexa como nos
eucariotas, podendo ser esta uma justificação para a existência de activadores
nos eucariotas e não nos procariotas.
Um aspecto interessante da estrutura das subunidades α do proteassoma
20S de S. cerevisiae está no tamanho dos terminais C, que se encontram na
periferia da molécula. Algumas subunidades β também possuem longos teminais
carboxílicos que por sua vez interagem com outras subunidades vizinhas e são
o factor determinante para a posição específica das subunidades na estrutura
(Groll et al., 1997).
1.4 – Identificação dos Centros Catalíticos Estudos efectuados usando mutagénese dirigida (Seemüller et al., 1995;
Seemüller et al., 1995) e co-cristalografia das subunidades β de Thermoplasma
com inibidores mostraram que o resíduo de treonina (Thr1) é o nucleófilo
necessário para iniciar a hidrólise de ligações peptídicas. A substituição deste
resíduo por alanina (Ala) origina proteassomas inactivos (Seemüller et al., 1995).
Esta função do Thr1 como centro catalítico formado por um único resíduo
colocou o proteassoma numa nova família de proteínas designada por
Figura 5 – Entrada do canal central do proteassoma 20S de T. acidophilum (esquerda, (Löwe et al.,
1995)) e do proteassoma 20S de S. cerevisiae (direita, (Groll et al., 1997)).

Introdução 21
hidrolases nucleófilas do terminal N (hidrolases Ntn) (Brannigan et al., 1995).
Outros resíduos que se revelaram essenciais para a actividade catalítica do
proteassoma no Thermoplasma foram o resíduo de glutamato 17 (Glu17), o
resíduo de aspartato 166 (Asp166) e o resíduo de lisina 33 (Lys33) (Seemüller et
al., 1995).
Em levedura, nos três pares de subunidades β com actividade catalítica,
estão presentes os mesmos resíduos considerados essenciais no
Thermoplasma, com excepção do Glu17 que se encontra substituído por um
resíduo de aspartato. Na zona envolvente ao centro catalítico situam-se os
resíduos de Lys33, Asp17, Ser129 e Asp166 que formam um sistema de ligações de
hidrogénio e pontes salinas que levam à polarização do carbono do grupo
carbonilo da ligação peptídica a ser clivada, facilitando assim o ataque
nucleofílico do Thr1 (Ditzel et al., 1998). A actividade peptidásica do
proteassoma 20S foi testada utilizando como substrato vários péptidos sintéticos
cuja degradação originava um produto fluorogénico, permitindo a caracterização
da especificidade das diferentes subunidades catalíticas. Assim, à subunidade
β1/Pre3p corresponde a actividade pós-acídica, a subunidade β2/Pup1p
apresenta actividade tríptica e a actividade quimotríptica é catalisada pela
subunidade β5/Pre2p (Chen et al., 1996; Arendt et al., 1997; Heinemeyer et al.,
1997). A integridade dos sítios catalíticos é sensível a alterações
conformacionais nas suas regiões mais próximas já que mutações noutras
subunidades β que não as três catalíticas e mesmo em subunidades α, apesar
da distância, levam a uma quebra na actividade de proteólise (Heinemeyer et al.,
1991; Hilt et al., 1993; Chen et al., 1996), ficando em aberto a participação das
outras subunidades proteassomais na degradação de substratos. Foi ainda
sugerida a hipótese de cooperatividade entre as subunidades catalíticas perante
um mesmo substrato (Djaballah et al., 1992; Stein et al., 1996).
A degradação de substratos proteicos pelo proteassoma 20S resulta
numa mistura de fragmentos com cerca de 3 a 30 resíduos de aminoácidos
(Wenzel et al., 1994; Niedermann et al., 1997).

Introdução 22
1.5 – Montagem do Proteassoma 20S A montagem e maturação do proteassoma 20S em eucariotas, requer
uma coordenação precisa de eventos de forma que as 14 subunidades
diferentes encontrem a sua localização correcta no complexo activo constituído
por 28 subunidades (Schmidt et al., 1997; Gerards et al., 1998).
As subunidades β do complexo proteassomal em arqueobactérias
(Thermoplasma) (Zwickl et al., 1992), eubactérias (Rhodococcus) (Zühl et al.,
1997) e eucariotas são sintetisadas na forma de precursores cataliticamente
inactivos, com uma sequência de aminoácidos no terminal N (propeptídio) cuja
clivagem torna as subunidades activas.
1.5.1 – Montagem do Proteassoma 20S em Thermoplasma
A expressão, em separado, das subunidades α e β do proteassoma de
Thermoplasma, em E. coli, mostrou que apenas a primeira tem a capacidade
de formar uma estrutura polimérica na forma de um anel duplo com sete
monómeros cada (Zwickl et al., 1992, 1994). Quanto às subunidades β, não
houve qualquer tipo de oligomerização, nem maturação das suas proformas. A
expressão de subunidades β sem o propeptídio respectivo, na ausência de
subunidades α, revelou-as inactivas cataliticamente. No entanto, co-expressão
dos monómeros α e β deu origem a proteassomas funcionais,
independentemente da presença ou ausência dos propeptídios. Estes resultados
sugeriram que um anel α poderá servir de local de ligação das subunidades β e
que, alterações conformacionais nas subunidades β, originadas pela ligação
destas ao anel α, permitam a junção de duas metades do proteassoma,
montadas separadamente, e levem à degradação dos propeptídios tornando as
subunidades β cataliticamente activas.

Introdução 23
1.5.2 – Montagem do Proteassoma 20S em Rhodococcus O proteassoma 20S de eubactéria Rhodococcus erythropolis possui na
sua constituição duas subunidades α e duas β diferentes, com grande homologia
entre elas, organizadas na estrutura α7β7β7α7, característica do proteassoma
20S (Tanamura et al., 1995).
A expressão de qualquer das combinações possíveis entre as
subunidades α e β do proteassoma 20S de Rhodococcus em E. coli originou
proteassomas funcionais, sem a necessidade de qualquer cofactor adicional
(Zühl et al., 1997). A expressão das subunidades α sem as β, contrariamente ao
que se observou para as subunidades de Thermoplasma, não levou à
oligomerização, permanecendo as subunidades na sua forma monomérica,
sugerindo que os primeiros complexos deste proteassoma 20S são formados
por heterodímeros α/β (Zühl et al., 1997).
Apesar da formação de proteassomas funcionais não necessitar do
propeptídio das subunidades β (mais longos que o das subunidades β de
Thermoplasma), a sua ausência torna a biogénese um processo mais moroso. A
adição do propeptídio em trans leva a uma aceleração do processo, o que
sugere que a autólise do propeptídio é um dos passos limitantes na formação do
proteassoma 20S (Hilt e Wolf, 2000). A utilização de mutantes com deficiências
no processamento dos propeptídios permitiu a caracterização do proteassoma
nesta fase da montagem, revelando uma estrutura com aproximadamente 100
kDa adicionais, relativamente ao 20S processado, correspondentes aos 14
propeptídios. A estrutura foi observada ao microscópio electrónico revelando que
os propeptídios ocupam a cavidade central e podendo estender-se até às duas
antecâmaras (Mayr et al., 1998).
1.5.3 – Montagem do Proteassoma 20S em Eucariotas

Introdução 24
A montagem do proteassoma 20S em eucariotas é um processo ainda
pouco conhecido.
Três subunidades α de origem humana foram expressas individualmente
em E. coli, e apenas a subunidade α7/HsC8 revelou a formação de um anel
duplo semelhante ao obtido em Thermoplasma (Gerards et al., 1997). As
subunidades α6/HsC2 e α1/Pros27 não apresentaram qualquer tipo de
oligomerização quando expressas individualmente, mas foram incorporadas nos
anéis formados quando coexpressas com α7/HsC8. A incorporação não se
mostrou dependente de qualquer tipo de interacção α7/α6 ou α7/α1 uma vez
que a composição das estruturas obtidas foi bastante heterogénia (Gerards et
al., 1998).
Em mamíferos foram descritos intermediários 13-16S (Frentzel et al.,
1994) e 15S (Yang et al., 1995), proteoliticamente inactivos e que exibiam
padrões de subunidades diferentes do encontrado no proteassoma 20S. Estes
complexos apresentavam todas as subunidades α mas estavam em falta
algumas subunidades β e as presentes apresentavam-se ainda na forma
precursora (Frentzel et al., 1994). O complexo 15S foi descrito como precursor
da maturação do proteassoma 20S (Yang et al., 1995). A forma 13S, de 300
kDa, foi identificada como meio proteassoma contendo subunidades β não
processadas. A forma 16S possuía uma massa total de 600 kDa (Schmidtke et
al., 1997).
Apesar das semelhanças de constituição dos dois complexos 16S e 20S,
a diferença no factor de sedimentação pode dever-se a diferenças
conformacionais entre os dois complexos que, por arranjos estruturais
posteriores e assistidos por chaperones, leva à maturação do complexo 16S em
proteassoma 20S (Schmidtke et al., 1997).
Em rato, foram também foram encontrados estes complexos contendo
todas as subunidades α e um conjunto de subunidades β (β2, β3 e β4), o que
compromete a hipótese de os complexos percursores serem formados por
dímeros α/β como na eubactéria Rhodococcus (Zühl et al., 1997). De notar que

Introdução 25
estas subunidades β não estabelecem contacto directo com as subunidades β
correspondentes do anel β simétrico. A entrada das restantes subunidades β
aparenta ser um processo rápido, uma vez que só foram encontradas em
complexos com um curto tempo de existência (Nandi et al., 1997). As
subunidades β5, β6 e β7 possuem extensões no terminal C direccionadas para
o anel β oposto (Groll et al., 1997) que poderão servir para aumentar a
especificidade do posicionamento na junção de dois meios proteassomas.
A montagem correcta de dois complexos correspondentes a meio
proteassoma depende de um factor adicional, o Ump1p (Ramos et al., 1998).
Este factor encontra-se presente em complexos precursores que se julgam
corresponder a metades proteassomais e é enclausurado quando duas dessas
estruturas se juntam para formar a partícula 20S. Após a maturação das
subunidades β o Ump1p é degradado tornando-se o primeiro substrato do
complexo (figura 6). Este factor não é essencial para a maturação proteassomal
como verificado pela viabilidade celular de células cujo gene UMP1 foi
delectado, mas em sua ausência, o processo de maturação e montagem do
proteassoma é ineficiente resultando na acumulação de precursores
proteassomais e na reduzida actividade catalítica dos proteassomas formados
(Ramos et al., 1998).
~15S – Complexo Ump1p Estruturas intermediárias proteassoma 20S
Figura 6 – Modelo da função do Ump1p na maturação do proteassoma 20S. Dois complexos 15S (A) contendo o factor Ump1p unem-se (1) para dar origem a uma estrutura intermediária (B) onde ocorrem arranjos conformacionais que conduzem à autocatálise dos propeptídios (2) tornando assim as subunidades proteolíticas activas (C). Esta activação permite a degradação de substratos, incluindo o próprio Ump1p (3) , com o proteassoma 20S a atingir a sua forma funcional (D); adaptado de Ramos et al., 1998

Introdução 26
1.5.4 – Propeptídios no Proteassoma 20S
À semelhança de outras proteinases, os propeptídios das subunidades β
catalíticas do proteassoma 20S aparentam ter uma função inibitória das mesmas
de modo a impedir a função proteolítica prematura das subunidades. Além disso,
é possível que funcionem como ajudantes na obtenção do enrolamento e
posicionamento correctos das subunidades na estrutura.
A delecção do propeptídio da subunidade β5/Pre2p em levedura verificou-
se ser letal (Chen et al., 1996). A coexpressão da subunidade β5/Pre2p com a
subunidade β5/Pre2p mutante à qual faltava o propeptídio mostrou não haver
incorporação da última na estrutura proteassomal, provando assim a
necessidade do propeptídio de 75 resíduos para a correcta incorporação desta
subunidade. A adição do propeptídio na forma trans compensou plenamente a
falta do mesmo na subunidade β5 mutada (Chen et al., 1996).
A função do propeptídio da subunidade β5/Pre2p foi salientada pela
descoberta do factor adicional Ump1p na montagem do proteassoma (Ramos et
al., 1998). No entanto, apesar da presença deste propeptídio ser essencial
(fornecido em trans ou na subunidade não processada) em presença de Ump1p,
quando o gene codificante para este factor foi delectado, o propeptídio tornou-se
dispensável, dando origem a células viáveis (Ramos et al., 1998). Estes
resultados excluem a hipótese da necessidade do propeptídio da subunidade
β5/Pre2p para a introdução desta subunidade na estrutura e, no entanto,
mostram-no essencial na presença do factor Ump1p, sugerindo uma interacção
entre este e o propeptídio, que promove alterações conformacionais necessárias
para a correcta maturação do proteassoma 20S em levedura (Ramos et al.,
1998).
Os propeptídios das subunidades β1/Pre3p, β2/Pup1p e β7/Pre4p não
são essenciais para a viabilidade celular (Jäger et al., 1999). No entanto, foram
encontradas anormalidades no processamento da subunidade β5/Pre2p na
ausência destes propeptídios, demonstrando assim que o processo de
maturação é afectado (Arendt et al., 1999). A expressão do propeptídio da

Introdução 27
subunidade β2/Pup1p na forma trans quando esta é expressa sem ele diminuiu
os defeitos de processamento (Jäger et al., 1999).
A análise dos propeptídios da subunidade β5/Pre2p das várias espécies
estudadas permite observar que existe grande uniformidade no seu tamanho,
em contraste com a diversividade da sua sequência primária, o que leva a
suspeitar de uma co-evolução entre estes e os vários homólogos de Ump1p
encontrados em eucariotas (Hilt e Wolf, 2000).
As diferentes contribuições dos propeptídios das várias subunidades β
para a montagem e maturação proteassomais reflecte a conjugação e
interdependência do processamento dos propeptídios.
1.6 – Objectivo Este trabalho de conclusão de licenciatura em Bioquímica teve como
objectivo científico aumentar o conhecimento actual sobre a montagem do
proteassoma 20S, um dos elementos chave do principal processo de
degradação selectiva na célula, a via de degradação dependente de
ubiquitina/proteassoma, em S. cerevisiae, nomeadamente ao nível dos
complexos precursores da montagem, através da identificação e caracterização
dos mesmos.

Materiais e Métodos 28
2 – Materiais e Métodos 2.1 – Material Biológico A realização deste trabalho envolveu a utilização de várias estirpes de
Saccharomyces cerevisiae (levedura de padeiro) e Escherichia coli, com
características específicas, como descrito na Tabela I.
Tabela I – Material biológico utilizado, com destaque para as suas características genómicas
relevantes;
Estirpes de S. cerevisiae Estirpe Genótipo relevante Fonte Comentário
JD47-13C MATa his3-∆200 leu2-3, 112 lys2-801 trp1-∆63 ura3-52 Dohmen J Dohmen et al., 1995
JD125 MATa PRE1-ha::YIplac 211 Dohmen J derivada de JD47-13C
JD127 MATa UMP1-ha::Ylplac128 PRE1-ha::Ylplac211 Dohmen J derivada de JD47-13C
JD129 MATa UMP1-ha::YIplac 128 Dohmen J derivada de JD47-13C
JD183 MATa PRS1-ha::YIplac211 Ramos P derivada de JD47-13C JD184 MATa PRS1-FLAG-6his::YIplac211 Ramos P derivada de JD47-13C JD228 MATa PRS1-FLAG-6His Ramos P derivada de JD47-13C JD305
AM1
MATa Y13∆::HIS3
MATa Y13∆::HIS3 PRS1-ha::YIplac211
London M
Este trabalho
derivada de JD47-13C
derivada de JD305
Estirpes de E. coli Estirpe Genótipo relevante Fonte JM105 end A1, thi, rps L, sbc B15, hsd R4, ∆ (lac-proAB),[F’,
traD 36, proAB, lac IqZ ∆M15]
Yanisch et al., 1985
2. 2 – Métodos 2.2.1 – Cultura de células de E. coli
Um óptimo desenvolvimento das células ocorre quando estas se
encontram num meio que contenha todos os nutrientes necessários. No caso de
meios selectivos, devem conter ou excluir a substância de selecção.

Materiais e Métodos 29
As culturas de E. coli foram efectuadas em meio Luria-Brot (LB) contendo
0,5% (m/v) extracto de levedura (GIBCOBRL), 1% (m/v) Bacto-Triptona (DIFCO)
e 0,5% (m/v) NaCl. O meio selectivo com ampicilina (LB-Amp) foi obtido
adicionando 0,12% (m/v) deste antibiótico. Os meios sólidos foram conseguidos
pelo acréscimo de 2% (m/v) de agar. Os meios foram esterilizados no autoclave
a 120 ºC, 0,2 MPa, durante 20 minutos. A solução stock de ampicilina (150mM)
foi esterilizada através de um filtro de 0,2 µm.
As células foram plaqueadas a partir de um stock em 15% (v/v) de glicerol
armazenado a –80 ºC, retirando-se, com um palito estéril, uma pequena
quantidade e espalhando-se ao longo da placa de forma a obter colónias
isoladas. As culturas líquidas foram obtidas pela inoculação de uma determinada
quantidade de meio líquido, normalmente 5 mL, com uma colónia proveniente de
uma placa contendo meio sólido. Todo o procedimento foi efectuado em redor
de uma chama de bico de Bunsen de forma a manter as condições estéreis.
As células foram deixadas a crescer numa incubadora (Orbital Incubator
SI50 – STUART SCIENTIFIC) a 37 ºC, durante a noite. No caso de culturas em
meio líquido foi mantida agitação orbital de 200 rpm.
2.2.2 – Cultura de Células de S. cerevisiae
As estirpes de levedura utilizadas neste trabalho derivam da estirpe JD47-
13C cujo genótipo se encontra descrito na Tabela I. Estas células são
auxotróficas para os aminoácidos histidina, leucina, lisina, triptofano e para o
uracilo, isto é, só crescem em meios contendo estes aminoácidos uma vez que
genes que codificam para enzimas das vias de síntese destes compostos se
encontram mutados.
As estirpes JD47-13C, JD305 e JD228 foram cultivadas em meio
completo YPD (1% (m/v) de extracto de levedura, 2% (m/v) de peptona, 2%
(m/v) de glucose (MERCK)). O meio YPD sólido foi obtido pela adição de 2%
(m/v) de agar antes da esterilização da solução, em autoclave, nas condições
acima mencionadas.

Materiais e Métodos 30
O meio mínimo é composto por 2% (m/v) de glucose, 0,25% (m/v) de SD
(DIFCO), 0,002% (m/v) de adenina, 0,004% (m/v) de uracilo, 0,002% (m/v) de
arginina, 0,001% (m/v) de histidina, 0,006% (m/v) de isoleucina, 0,006% (m/v) de
leucina, 0,004% (m/v) de lisina, 0,001% (m/v) de metionina, 0,006% (m/v) de
fenilalanina, 0,005% (m/v) de treonina e 0,004% (m/v) de triptofano. No caso de
meios selectivos foi excluído o composto de selecção. No caso de meios sólidos,
preparou-se uma solução de 2% (m/v) de agar e, no seu estado líquido,
adicionaram-se os restantes componentes do meio, devidamente estéreis, nas
concentrações respectivas.
As estirpes JD129, JD183, JD184 e AM1 foram cultivadas em meio SD-
Ura (meio mínimo sem uracilo). A estirpe JD125 foi cultivada em meio SD-Leu
(meio mínimo sem leucina). A estirpe JD127 foi cultivada em meio SD-Ura-Leu
(meio mínimo com exclusão de leucina e uracilo).
A inoculação dos meios sólido e líquido realizou-se de modo semelhante
ao descrito para E. coli.
As células foram colhidas quando apresentavam densidade óptica a
600 nm (OD600nm) de 1,0, 3,0 ou 6,0 ± 0,2 através de centrifugação a 3000 g,
4 ºC, durante 5 minutos. Os sedimentos foram lavados uma vez com água, para
eliminar resíduos do meio de cultura.
As células cujos extractos proteicos se destinaram à análise e purificação
de complexos proteasomais foram ressuspensas em 2 mL de tampão de
extracção por grama de massa fresca e congeladas em azoto líquido sendo
depois armazenadas a –80 ºC até serem utilizadas.
2.2.3 – Marcação da Subunidade α7/Prs1p do Proteassoma com o Epítopo
ha em JD305 2.2.3.1 – Produção de Células Competentes de E. coli (JM105) A obtenção de células competentes de E. coli, ou seja, células com a
capacidade de receber material genético, nomeadamente plasmídios, e de o

Materiais e Métodos 31
replicar durante a sua multiplicação, realizou-se de acordo com uma variante do
método desenvolvido por Cohen et al. (1972).
Todo o procedimento foi executado utilizando material e soluções
estéreis, previamente colocados a 4 ºC.
Inocularam-se 5 mL de LB com as células de E. coli (JM105) e deixou-se
a crescer a 37 ºC durante a noite. Mediu-se a OD600nm desta cultura e procedeu-
se à diluição necessária para começar uma nova cultura de 100 mL com
OD600nm=0,2. Quando a cultura atingiu a OD600nm=0,6 colocou-se o frasco em
gelo e agitou-se suavemente para garantir um arrefecimento rápido. Colheram-
se as células a 3840 g (JA-14 – Beckman), durante 10 minutos, a 4 ºC.
Ressuspendeu-se o sedimento por agitação, em gelo, em 20 mL de TfB I (30
mM de acetato de potássio, 50 mM de cloreto de manganésio (MERCK), 100mM
de cloreto de potássio, 10mM de cloreto de cálcio e 15% (v/v) de glicerol).
Repetiu-se a centrifugação e, da mesma forma, ressuspendeu-se o sedimento
em 4 mL de TfB II (10mM de Na-MOPS a pH 7, 75mM de cloreto de cálcio,
10mM de cloreto de potássio e 15% (v/v) de glicerol). A suspensão foi dividida
em alíquotas de 100 µL, congeladas em azoto líquido e armazenadas a –80 ºC.
2.2.3.2 – Transformação de Células Competentes de E. coli (JM105) Com este processo pretendeu-se introduzir o plasmídio pCR8 (figura 7)
nas células JM105 de modo que, durante a sua multiplicação possa ocorrer
amplificação do plasmídio que posteriormente pode ser extraído e purificado.
Assim, utilizando uma pequena quantidade de um plasmídio purificado é
possível produzir grandes quantidades desse mesmo material.
Removeu-se uma alíquota de 100 µL de células competentes de E. coli
(JM105) armazenadas a –80 ºC e colocou-se em gelo até descongelar.
Adicionou-se 10 µL do plasmídio pCR8 e manteve-se o tubo em gelo durante 30
minutos. Para facilitar a entrada do plasmídio nas células, induziu-se um choque
térmico colocando as células num banho a 42 ºC, durante 5 minutos. As células
foram então diluídas em 1 mL de meio LB e deixadas a crescer durante 1 hora, a

Materiais e Métodos 32
37 ºC, tendo sido, de seguida, centrifugadas durante 30 segundos a 16100 g
(Ependorff Centifuge 5145D). Removeu-se cerca de 0,75 mL do sobrenadante,
ressuspendeu-se o sedimento no sobrenadante restante e plaqueou-se em meio
sólido selectivo, LB-Amp. Incubou-se a placa a 37 ºC, durante a noite.
O plasmídio usado, pCR8 (figura 7), contém um gene que promove
resistência ao antibiótico ampicilina (AmpR). Deste modo, apenas as células que
foram transformadas conseguem crescer no meio de crescimento selectivo.
2.2.3.3 – Purificação de DNA Plasmídico A extracção e purificação do plasmídio pCR8 realizou-se com o auxílio do
QIAPREP SPIN MINIPREP KIT (QIAGEN), seguindo-se todo o protocolo
recomendado pelo fabricante (QIAGEN, 1998).
Inocularam-se 5 mL de meio LB-Amp com uma colónia de JM105
transformada com pCR8 e deixou-se a crescer, durante a noite a 37 ºC.
Colheram-se as células numa centrífuga Eppendorf a 4500 g e ressuspendeu-se
o sedimento em tampão P1 com RNase. Adicionaram-se 250 µL de tampão P2 e
agitou-se o eppendorf por inversão até a solução se aparentar viscosa e
ligeiramente translúcida. Adicionaram-se 350 µL de tampão N3 e agitou-se
Figura 7 – Mapa do plasmídio pCR8 amplificado em E. coli, e utilizado para transformar células competentes de levedura; HA – epítopo (YPYDVPDYA) CYC1 – terminador do citocromo c URA3 – via de síntese do uracilo AmpR – resistência à ampicilina Ori – Origem da replicação
4612 bp
AmpR

Materiais e Métodos 33
rapidamente, por inversão, 6 vezes. Centrifugou-se a 10400 g, durante 10
minutos, e transferiu-se o sobrenadante para a coluna QIAprep, montada num
tubo de colheita de 2 mL. Centrifugou-se o conjunto durante 1 minuto e
desprezou-se o volume eluído. Lavou-se a coluna com 0,5 mL de tampão PB e
repetiu-se o passo de centrifugação, novamente desprezando o volume eluído.
Adicionou-se 0,75 mL de tampão PE à coluna e repetiu-se a centrifugação, mais
uma vez desprezando o volume eluído. Centrifugou-se novamente para eliminar
quaisquer resíduos de tampão PE ainda existentes na coluna. Colocou-se a
coluna num eppendorf limpo e eluiu-se o pasmídio da coluna pela adição de 50
µL de tampão EB (10mM Tris, pH 8,5), esperando cerca de 1 minuto antes de
voltar a centrifugar.
2.2.3.4 – Linearização do Plasmídio pCR8 para Integração em Levedura
O único sítio de restrição para o enzima Cla I (AT/CGAT) existente no
plasmídio pCR8 encontra-se dentro da região do gene PRS1 inserida neste
plasmídio, sendo portanto possível linearizar o DNA recorrendo a este enzima.
A 5 µL da solução de DNA plasmídio purificado anteriormente adicionou-
se 2 µL de tampão (tampão H, ROCHE), 1 µL de Cla I (ROCHE) e 12 µL de
água. Agitou-se ligeiramente a solução para a homogeneizar e colocou-se num
banho a 37 ºC, durante 2 horas. O sucesso da digestão foi verificado por
electroforese em gel de agarose.
2.2.3.5 – Produção e Transformação de Células Competentes de Levedura A produção de células competentes de levedura tem como objectivo obter
células com a capacidade de interiorizarem material genético que não o seu e,
dependendo do material genético inserido, promoverem a sua integração ou
simplesmente iniciarem a transcrição deste.
Foram usados dois métodos diferentes para promover a competência e
transformação das células. O primeiro tem a vantagem de separar os passos de

Materiais e Métodos 34
indução de competência e transformação, permitindo assim o armazenamento
de células competentes para transformações futuras (Dohmen et al., 1991). No
entanto, é um processo mais demorado que o segundo (Agatep et al., 1998,
Gietz et al., 1994), em que a indução de competência e transformação são
efectuadas de um modo consecutivo.
Na realização do primeiro processo foi preparada uma cultura de 10 mL
de células (JD305) em meio completo (YPD) até atingir uma OD600nm
aproximadamente de 0,6. Centrifugou-se a 3000 g, durante 5 minutos, a 4 ºC, e
lavou-se o sedimento em 5 mL de solução A (1M de sorbitol (KOSH-LIGHT),
10mM de bicina (BDH CHEMICALS), 3% (v/v) de etilenoglicol (MERCK), 5%
(v/v) de DMSO (RIEDEL), pH 8,35). Voltou-se a centrifugar e ressuspendeu-se o
sedimento em 0,2 mL de solução A. A suspensão foi então congelada a –80 ºC.
Para a transformação adicionaram-se 5 µL do plasmídio pCR8 anteriormente
purificado e digerido com Cla I e agitou-se num vortex à velocidade máxima, a
37 ºC, durante 5 minutos. Adicionou-se 1 mL de solução B (40% ((m/v) de
polietilenoglicol 1000 (SIGMA), 200 mM de bicina, pH 8,35) e resuspendeu-se
por inversão. As células foram, então, incubadas durante 1 hora, a 30 ºC.
Centrifugou-se a suspensão a 800 g, durante 5 minutos à temperatura ambiente
e lavaram-se as células com 1 mL de solução C (150 mM de NaCl, 10 mM de
bicina, pH 8,35). Centrifugou-se de novo a suspensão e ressuspendeu-se o
sedimento em 200 µL de solução C. As células em suspensão foram plaqueadas
em meio sólido SD-Ura a fim de seleccionar as células transformadas. A placa
foi colocada a 30 ºC, durante 3 dias. Das colónias obtidas seleccionaram-se oito
que foram cultivadas numa nova placa SD-Ura para obenção de colónias
isoladas.
O segundo processo de indução de competência e transformação foi
realizado utilizando 10 mL de uma cultura de células JD305 crescidas em YPD,
durante a noite. Centrifugou-se a suspensão e lavaram-se as células com 1 mL
de água. Centrifugou-se de novo e ressuspendeu-se o sedimento em 1 mL de
100 mM de acetato de lítio (LiAc), incubou-se a 30 ºC, durante 5 minutos. A
cultura foi centrifugada a 16000 g, durante 1 minuto e adicionou-se ao

Materiais e Métodos 35
sedimento, por esta ordem, 240 µL de 50% ((m/v) de PEG 3350, 36 µL de 1 M
de LiAc, 5 µL de “calf thymus” DNA, 5 µL de pCR8 purificado e linearizado como
descrito anteriormente e 60 µL de água. Agitou-se a mistura com a ajuda de um
vortex na velocidade máxima, durante 1 minuto e provocou-se de imediato um
choque térmico num banho a 42 ºC, durante 20 minutos. Centrifugou-se durante
15 segundos, a 16000 g, e removeu-se o sobrenadante com a ajuda de uma
micropipeta. Ressuspendeu-se em 200 µL de água e plaqueou-se a suspensão
em meio sólido selectivo SD-Ura, tendo-se deixado a crescer a 30 ºC durante 3
dias. As colónias visíveis foram marcadas e de novo plaqueadas em SD-Ura. Só
as células que sofreram recombinação homóloga e, portanto, possuem o
plasmídio pCR8 integrado no seu genoma têm capacidade de se desenvolverem
em meio sem uracilo, uma vez que possuem o gene URA3 que codifica para um
enzima da via biossintética do uracilo.
2.2.3.6 – Polymerase Chain Reaction (PCR)
A técnica de PCR permite a amplificação selectiva de material genético.
Uma das vantagens apresentadas por esta técnica reside no facto de necessitar
de uma baixa concentração de moléculas “molde” para efectuar a amplificação.
Neste estudo, esta técnica foi aplicada para amplificar uma parte do
plasmídio inserido em levedura e assim verificar a sua integração. Foi
desenhado um oligonucleótido contendo a sequência do início da ORF (“open
reading frame”) de PRS1, e portanto, fora do fragmento do gene PRS1
(5’∆ PRS1) clonado no plasmídeo pCR8 (5’-
GCGCATATGACATCAATTGGTACTGGT-3’) e um oligonucleótido contendo
uma sequência presente no terminador do citicromo c (CYC1) existente no
plasmídio introduzido (5’-GCCTTCGAGCGTCCAAAAACCTTC-3’). Deste modo,
só no caso de ter havido uma integração correcta se iria obter produto de PCR.
Foram removidas células das colónias a testar a partir de uma cultura em
meio sólido, para além de células de JD47-13C (estirpe não marcada) utilizada
como controlo negativo e de células de JD183, controlo positivo, e colocadas no

Materiais e Métodos 36
fundo de tubos de PCR de 0,2 mL. Seguidamente, provocou-se a ruptura da
membrana celular por exposição a microondas, durante 2 minutos, a 700 Watts
de potência. Fez-se uma solução contendo 20 µL do tampão de polimerase
(ROCHE), 8 µL de cloreto de magnésio a 25mM, 8 µL de uma solução a 10 mM
de nucleótidos, 4 µL de cada oligonucleótido, 156 µL de água estéril e 1 µL de
polimerase (Expand High Fidelity PCR System, ROCHE). A cada tubo contendo
as células foram adicionados 25 µL desta solução. A amplificação foi feita num
termociclador (Personal Cycler, BIOMETRA) com o seguinte programa:
Passo Descrição Tempo (segundos)
Temperatura (ºC)
1 150 94
2 desnaturação 30 94
3 annealing 60 54
4 enlongamento 90 72
5 300 72
2.2.4 – Extractos Proteicos
2.2.4.1 – Extracção por Maceração das Células As pastilhas de células no tampão de extracção desejado (tampão 26S
(50mM Tris, 5mM cloreto de magnésio, 2mM ATP (BOEHRINGER MANNHEIM),
1mM DTT (ACROS), 15% (v/v) glicerol, pH 7,5) ou tampão MA (50mM Tris-HCl,
15% (v/v) glicerol, 5mM cloreto de magnésio, pH7,5)), previamente congeladas e
armazenadas a –80 ºC foram transferidas para um almofariz contendo azoto
líquido e com o auxílio do pilão procedeu-se à sua maceração, renovando o
azoto líquido de modo a nunca deixar o homogenato descongelar. Depois da
completa maceração e redução das células a um pó muito fino, deixou-se o
homogenato descongelar e adicionaram-se inibidores de proteases (Complete,
MINI, EDTA Free – BOEHRINGER MANNHEIM) a partir de uma solução sete
33 ciclos

Materiais e Métodos 37
vezes concentrada, em tampão de extracção, para uma concentração final de
uma vez. Centrifugou-se o extracto a 31200 g (JA 18.1), durante 10 minutos, a 4
ºC. Desprezou-se o sedimento e centrifugou-se o sobrenadante a 60000 g, na
ultracentrífuga de bancada TLA-100.4 (BECKMAN TLX), durante 30 minutos, a 2
ºC. Desprezou-se o sedimento e congelou-se o sobrenadante, correspondente
ao extracto cru (CE), em azoto líquido.
Este método foi o utilizado para a maioria das extracções efectuadas uma
vez que se obtém extractos com uma elevada concentração proteica e, quando
foi necessário extrair proteínas a partir de uma grande quantidade de células.
2.2.4.2 – Extracção com Esferas de Vidro Este método de extracção foi aplicado quando foram necessárias
pequenas quantidades de proteína, não sendo por isso a análise afectada pelo
baixo rendimento de extracção conseguido com este método.
A um sedimento de células colhidas, com um volume aproximado de 200
µL, foi adicionado igual volume de esferas de vidro e tampão de lise (50mM de
HEPES, 1mM de EDTA, 1% (v/v) de Triton X-100). A mistura foi deixada em gelo
durante pelo menos 1 minuto. Depois de arrefecida, a mistura foi agitada no
vortex à velocidade máxima durante 30 segundos e de novo colocada em gelo
durante 30 segundos. O extracto foi centrifugado a 40600 g (JA 18.1). O
sobrenadante obtido corresponde ao extracto proteico (CE) a analisar.
2.2.5 – Quantificação Proteica

Materiais e Métodos 38
As concentrações de proteínas foram determinadas através o método de
Bradford. Este método colorimétrico utiliza como corante o Coomassie Brilliant
Blue G-250 (CBB G-250) (figura 8) que, devido à sua carga negativa se liga aos
resíduos básicos na proteína e, em solução acídica, sofre uma alteração do seu
máximo de absorção de 465 nm (apresenta cor vermelha) para 595 nm
(apresenta cor azul) quando a ligação ocorre (Robyt et al., 1990), apresentando
assim diferenças de cor em resposta a diferentes concentrações proteicas
(Bradford, 1976).
Uma das grandes vantagens deste método, para além da sua
sensibilidade (0 – 20 µg de proteína), é a sua baixa susceptibilidade a
interferência por outras substâncias. Além disso, devido à similaridade nas
curvas de resposta, uma grande gama de proteínas padrão pode ser utilizada
para a calibração (Robyt et al., 1990). No presente trabalho a proteína padrão
utilizada foi a albumina de soro bovino (BSA). Partindo-se de uma solução stock
de BSA a 0,5 mg/mL foram feitas diluições em água para obter soluções com 2,5
µg, 5,0 µg, 10 µg, 15 µg e 20 µg de BSA, num volume total de 800 µL. A estas
soluções foram adicionados 200 µL do reagente corante BioRad Protein Assay
(BIO-RAD).
As amostras a quantificar, em triplicado, foram diluídas em água, para um
volume total de 800 µL, de modo que a sua quantidade total de proteína se
CH2
N
C2H5
CH3 CH3
NaO3S
NH
OC2H2
N
C2H5
CH2
SO3-
Figura 8 – Molécula do corante CBB G-250, com destaque para o seu grupo carregado negativamente que forma ligações iónicas com os grupos amina carregados positivamente das proteínas;

Materiais e Métodos 39
situasse entre 0 e 20 µg. A estes tubos foram adicionados 200 µL de reagente
corante.
Os tubos foram agitados de modo a homogeneizar a solução e, após
cinco minutos de reacção à temperatura ambiente, foi medida a sua absorvância
a 595nm usando cuvettes de plástico de 1 mL, com 1 cm de comprimento óptico,
e tomando como branco uma solução de 800 µL de água e 200 µL de reagente
corante.
2.2.6 – Cromatografia de Filtração em Gel A expressão filtração em gel, ou exclusão molecular, é utilizada para
descrever a separação de moléculas de diversos tamanhos através grânulos
constituídos por compostos orgânicos poliméricos com a capacidade de
estabelecer uma rede tridimensional de poros que lhes confere propriedade de
crivo molecular (Wilson et al., 2000).
O princípio desta técnica baseia-se no facto de que moléculas com
diferentes tamanhos demoram tempos diferentes a atravessarem uma coluna
constituída por grânulos de gel em equilíbrio com uma fase móvel. Moléculas
grandes, que são excluídas dos poros, passam por fora dos grânulos do gel e
são as primeiras a serem eluídas, enquanto que moléculas mais pequenas, que
podem penetrar nos poros dos grânulos, passam mais devagar, sendo assim
retardado o seu volume de eluição (Wilson et al., 2000). As moléculas de uma
amostra são portanto eluídas seguido uma ordem decrescente do seu tamanho
molecular.
Tendo em consideração as diferenças entre o tamanho dos poros no gel
e os diferentes tamanhos das moléculas, torna-se interessante a observação da
existência de uma relação linear entre os logarítmos das duas variáveis (Glick,
1970), tornando-se possível, por exemplo, com a utilização do mesmo gel,
determinar a massa molecular de moléculas, tendo por base o tempo de eluição
de outras moléculas cuja massa molecular (Mr) seja conhecida, por comparação
0
0
VVtVVeKav −
−=

Materiais e Métodos 40
de um parâmetro de eluição de volume (Kav) da molécula de interesse com Kav
obtido para as moléculas aplicadas na calibração, segundo a fórmula
em que Ve corresponde ao volume de eluição da substância, Vt o volume total
do gel e V0 o “void volume” (volume nulo) da coluna.
2.2.6.1 – Cromatografia de Filtração em Gel em Superose 6
A coluna Superose 6 (AMERSHAM PHARMACIA BIOTECH) é composta
por um gel de polímeros de agarose com partículas na ordem dos 13 µm, e é
específica para separações de moléculas na ordem dos 5 aos 5000 kDa.
Apresenta uma altura de 30 cm e 1 cm de diâmetro, para um volume total (Vt) de
23,56 mL. Tem um V0 de 7,77 mL e o gel garante uma separação funcional para
um fluxo de eluente a exercer uma pressão máxima de 1,5 MPa.
A separação de complexos proteassomais foi realizada de acordo com o
método descrito por Ramos at al. (1998). Extractos proteicos foram fraccionados
com o auxílio da coluna Superose 6 acoplada a um ÄKTA-FPLC (AMERSHAM
PHARMACIA BIOTECH), controlado pelo software Unicorn v3.21. O gel foi
equilibrado em tampão 26S previamente filtrado e desgasificado sob vácuo. A
equilibração do gel foi efectuada pela passagem de, aproximadamente, duas
vezes o Vt da coluna em tampão, sendo o primeiro volume bombeado a 0,2
mL/minuto e o segundo a 0,3 mL/minuto. Injectaram-se, com o auxílio de um
loop apropriado, 200 µL de CE (preparado como mencionado na secção 2.2.4.1)
de cada estirpe a analisar, contendo 1 mg de proteína. A separação foi
executada com um fluxo de eluente (tampão 26S) de 0,3 mL/minuto,
encontrando-se o eluente, a coluna e o colector de fracções numa câmara fria a
aproximadamente 4 ºC. O perfil de eluição das proteínas foi monitorizado a
280nm e foram colhidas fracções de 0,6 mL.
A separação de complexos proteassomais purificados por cromatografia
de metalo-afinidade realizou-se de acordo com o procedimento acima descrito,

Materiais e Métodos 41
mas com a utilização, como tampão de equilibração do gel e eluente, o tampão
Co2+ (20mM de Tris, 5mM de cloreto de magnésio, 100mM de NaCl, 15% (v/v)
de glicerol, pH 8,0).
A calibração da coluna foi realizada utilizando as seguintes proteínas:
citocromo c, aldolase, catalase e tiroglobulina. O “void volume” (V0) foi
determinado com o azul de dextrano. Calculou-se o Kav para cada uma das
proteínas marcadoras, tabela II. O gráfico Kav em função do logarítmo da massa
molecular dos marcadores encontra-se na figura 9.
Tabela II – Proteínas utilizadas para a calibração da coluna SUPEROSE 6;
Proteína Mr (kDa)
log Mr Ve Kav
Citocromo C 13,0 4,11 19,7 0,753
Aldolase 158 5,20 14,9 0,449
Catalase 232 5,37 14,4 0,417
Tiroglobulina 669 5,83 11,3 0,224
2.2.6.2 – Cromatografia de Filtração em Gel para Dessalinização
Uma outra aplicação da filtração em gel, e tirando partido da exclusão
molecular dos géis, é a dessalinização de amostras, nomeadamente de
extractos proteicos. Pode também ser aplicada para trocas de tampão dos
Figura 9 – Calibração da coluna Superose 6, obtida pela regressão linear dos pontos de Kav contra os logarítmos da massa molecular, para cada proteína
y = -0,2998x + 1,9975R2 = 0,9878
0,000
0,200
0,400
0,600
0,800
1,000
4,00 4,50 5,00 5,50 6,00
log Mr
Kav

Materiais e Métodos 42
extractos ou soluções proteicas. As moléculas de elevado Mr são eluídas com o
volume nulo da coluna enquanto moléculas de baixo Mr são distribuídas entre a
fase estacionária, sendo eluídas num volume mais elevado. Na dessalinização
de amostras proteicas, a porosidade do gel é escolhida de modo a excluir as
proteínas sendo eluídas com o volume nulo da coluna, sendo assim separadas
de moléculas pequenas como sais (Janson et al., 1998).
2.2.6.2.1 – Cromatografia de Filtração em Gel em PD-10
As colunas PD-10 (AMERSHAM PHARMACIA BIOTECH) contem como
enchimento o Sephadex G-25 M, um polímero de dextrano, e são especialmente
preparadas para dessalinização e/ou troca de tampão de amostras. Este tipo de
colunas apresenta uma altura de 5 cm e tem um volume de gel de 9,1 mL.
O gel foi equilibrado com 25 mL de tampão Co2+. Foram colocados 2,5 mL
de amostra no topo do gel e desprezou-se o eluente. Depois da saída deste,
adicionaram-se 3,5 mL de tampão Co2+ e recolheu-se o eluente, o extracto
dessalinizado (DCE). O procedimento foi realizado a 4 ºC.
2.2.7 – Electroforese de Biomoléculas As moléculas biológicas, nomeadamente proteínas e ácidos nucleicos,
devido às características individuais dos seus monómeros, são providas de
carga eléctrica. Esta carga é condicionada pelo meio em que as moléculas se
encontram (fisiológico ou não), nomeadamente pela concentração de iões H+
(Plummer, 1987). A electroforese utiliza esta propriedade para promover a
separação das moléculas ao longo de um meio sólido, fazendo uso do princípio
que uma partícula carregada electricamente, sob o efeito de uma diferença de
potencial, se move no sentido do polo de carga contrária à sua.
Esta é uma técnica de grande importância para a separação de moléculas
biológicas uma vez que, para além de ser extremamente sensível a pequenas
diferenças na carga da molécula, normalmente não afecta a estrutura nativa da
mesma (Robyt et al., 1990).

Materiais e Métodos 43
O meio de suporte utilizado na electroforese varia de acordo com as
necessidades de separação e com as características das moléculas a serem
separadas, no entanto, deverá ser o mais quimicamente inerte possível, de
modo a não interferir na migração (Robyt et al., 1990).
2.2.7.1 – Electroforese de Ácidos Nucleicos em Géis de Agarose A agarose, a fracção polissacarídica neutra do agar, é composta por um
polímero linear de D-galactopiranose com uma ligação β-1→4 a 3,6-anidro-L-
galactopiranose que, por sua vez está ligada na forma α-1→3 ao próximo
resíduo de D-galactopiranose (Robyt et al., 1990).
Dependendo da concentração de agarose utilizada, os poros formados
por este polímero são maiores ou menores. A baixas concentrações de agarose,
o gel formado tem a capacidade de separar moléculas de grandes dimensões,
como DNA (Maniatis et al., 1983).
Uma das vantagens no uso deste polímero reside no facto de não
necessitar de qualquer tipo de agentes interligantes ou iniciadores da
polimerização, evitando assim qualquer interferência destes com as moléculas a
separar.
As amostras de DNA foram adicionadas de solução de aplicação
contendo dois corantes (0,042% (m/v) azul de bromofenol (SIGMA) e 0,042%
(m/v) xilenocianol) para verificação da evolução da electroforese e 2,5% (v/v)
Ficoll (tipo 400) para aumentar a densidade das amostras.
Foram preparados géis de 0,8% (m/v) ou 1% (m/v) de agarose, em
tampão TAE (40mM de Tris-HCl, 0,114% (v/v) de ácido acético glacial e 1mM de
EDTA a pH 8,0). A solução de agarose foi aquecida até completa dissolução da
agarose. Após o aquecimento e ainda em estado líquido, foram adicionados
0,05% de brometo de etídio (SIGMA). O brometo de etídio intercala-se nas
bases dos ácidos nucleicos e, neste estado, é fluorescente quando irradiado
com luz Ultravioleta (UV), permitindo assim a visualização dos ácidos nucleicos
no gel. A solução foi colocada numa tina de electroforese horizontal e esperou-

Materiais e Métodos 44
se a completa solidificação da mesma. O gel foi coberto com tampão de
electroforese TAE contendo 0,05% (v/v) de brometo de etídio. As amostras
foram aplicadas e estabeleceu-se uma diferença de potencial de 70 V no
sistema.
2.2.7.2 – Electroforese de Proteínas em Géis de Poliacrilamida Um dos meios sólidos mais usados para a separação de proteínas são
os géis de poliacrilamida. Este tipo de gel, para além de ser quimicamente
inerte, apresenta ainda a vantagem de permitir controlar o tamanho dos seus
poros, levando a que a separação não seja baseada apenas na carga da
proteína mas também na massa molecular e forma da molécula (Voet et al.,
1995). Para além disso, apresenta ainda as vantagens de minimizar a difusão
das proteínas (Robyt et al., 1990), ser facilmente manuseado e ser transparente,
permitindo assim a sua observação sob luz visível e ultravioleta (UV) (Plummer,
1987).
Um gel de poliacrilamida é formado pela polimerização de acrilamida na
presença de um agente de entrecruzamento, geralmente a N’,N’ –
metilenobisacrilamida (bisacrilamida) (Jason et al., 1998). A polimerização é
catalisada pelo persulfato de amónio (PSA) e é iniciada pela N,N,N’,N’-
tetrametiletilenodiamina (TEMED) (figura 10).
Figura 10 – Reacção de polimerização dos monómeros de acrilamida; adaptado de Voet et al., 1995
Acrilamida N,N’-Metilenobisacrilamida

Materiais e Métodos 45
O tamanho dos poros varia consoante a percentagem de acrilamida e
bisacrilamida presentes. Esta percentagem pode ser definida segundo o método
de nomenclatura T e C introduzido por Hjertén (Hjertén, 1962) segundo as
fómulas
em que A e B são as massas, em grama, de acrilamida e bisacrilamida,
respectivamente, e V o volume de água em cm3.
2.2.7.2.1 – Electroforese de Proteínas em Condições Desnaturantes
A separação electroforética realizada em presença de um detergente
aniónico, o dodecilsulfato de sódio (SDS) e de um agente redutor, o 2-
mercaptoetanol permite a separação de polipéptidos de acordo com a sua
massa molecular. O SDS tem a capacidade de formar um forte complexo SDS-
proteína devido às suas características apolares, ligando-se de forma micelar às
regiões apolares da proteína, e ficando a sua zona iónica exposta ao solvente
(Robyt et al., 1990). A ligação do SDS à proteína promove dois efeitos distintos,
a dissociação dos oligómeros proteícos e a disrupção da estrutura secundária da
proteína. Além disso, a saturação da proteína com SDS (1,4 g de SDS por
grama de proteína) leva à camuflagem da carga proteica, passando a migração
no gel a ser totalmente dependente da sua massa molecular (Plummer, 1987). O
uso do 2-mercaptoetanol promove a redução de possíveis pontes dissulfídicas
que ligam resíduos de cisteína inter ou intra subunidades proteicas (Robyt et al.,
1990).
%100)(VBAT ×+
= %)(
100BA
BC+
×=

Materiais e Métodos 46
As amostras foram preparadas para a electroforese desnaturante pela
adição de um tampão amostra desnaturante (SB) 4x concentrado, contendo
320mM de Tris-HCl a pH 6,8, 0,4M de 2-mercaptoetanol, 8% (m/v) de SDS, 15%
(v/v) de glicerol (para aumentar a densidade das amostras) e 0,024% (m/v) de
roxo de m-cresol (como corante de frente de migração). Após a
homogeneização da amostra em tampão SB, as amostras foram colocadas num
banho a 100 ºC durante 5 minutos e aplicadas directamente num gel
desnaturante ou guardadas a –20 ºC, até uso posterior.
Foram utilizados os aparelhos de electroforese vertical Hoefer SE600
(AMERSHAM PHARMACIA BIOTECH) e Mini-Protean II (BIO-RAD),
dependendo do número de amostras a analisar e necessidade de resolução.
Foi utilizado um sistema descontínuo de géis, formado por um gel de
concentração e um de resolução. O primeiro, de menor concentração de
acrilamida, visa acumular as proteínas na interface entre os dois géis de modo a
que estas entrem no gel de resolução ao mesmo tempo. O segundo, de maior
concentração de acrilamida, promove a separação de acordo com a massa
molecular das moléculas.
A partir de uma solução de acrilamida/bisacrilamida 30% T, 2,7% C,
preparou-se um gel de resolução 12%T, 1%C contendo 375mM de Tris-HCl a
pH 8,8, 0,1% (m/v) de SDS, 0,1% (m/v) de PSA e 0,033% (v/v) de TEMED. O gel
de resolução foi coberto com água de modo a minimizar a inibição da reacção
de polimerização pelo oxigénio e para facilitar a formação de uma superfície
regular no topo do gel. A polimerização do gel tornou-se evidente pela
observação de uma interface gel-água. Por cima do gel de resolução
polimerizado foi adicionada a solução do gel de concentração com 4% T, 0,36%
C em 125 mM de Tris-HCl a pH 6,8 juntamente com 0,1% (m/v) de SDS, 0,1%
(m/v) de PSA e 0,033% de TEMED. Imediatamente após a adição da solução do
gel de concentração introduziu-se no molde o pente para a formação dos
“poços” para posterior aplicação das amostras. No final da polimerização do gel
de concentração removeu-se o pente e lavaram-se os “poços” para remover
resíduos de acrilamida não polimerizada. As amostras foram aplicadas com uma

Materiais e Métodos 47
seringa de vidro graduada (Hamilton) e o conjunto foi colocado na tina de
electroforese. O tampão de eléctrodo catódico (25mM de Tris, 192mM de glicina
e 0,1% de SDS) foi colocado na tina superior. O tampão anódico, idêntico ao
anterior, mas contendo 100mM acetato de sódio para aumentar a resolução de
polipéptidos de baixa massa molecular (Christy et al., 1989), foi colocado na tina
inferior.
A diferença de potencial aplicada foi de 60 V ou 130 V, conforme se
pretendeu que a separação ocorresse durante a noite ou ao longo do dia,
respectivamente. A corrente foi interrompida quando a frente de migração atingiu
o final do gel.
2.2.7.2.2 – Electroforese de Proteínas em Condições Nativas
As amostras foram preparadas para a electroforese em condições não
desnaturantes, não sendo por isso usado qualquer reagente que pudesse por
em causa a estrutura natural da proteína. O tampão amostra para condições
nativas usado (SN) estava 4x concentrado e era constituído por 200mM de Tris -
HCl pH 6,8, contendo 15% (v/v) de glicerol e 0,25% (m/v) de azul de bromofenol
como corante da frente de migração. Os tubos contendo as fracções a analisar
em SN foram agitados para homogeneizar a solução e de seguida, as amostras
foram aplicadas no gel para separação, uma vez que, se guardados, poderia
haver disrupção das interacções.
A electroforese em condições nativas foi efectuada com sistema Mini-
Protean II (BIO-RAD).
Assim como para a separação em condições desnaturantes, as amostras
foram separadas num sistema descontínuo de géis. No entanto, devido às
dimensões e formas dos complexos proteicos a separar, o poro do gel
resolvente e de concentração foi maior sendo, portanto, utilizada uma menor
concentração de acrilamida/bisacrilamida. Assim, preparou-se um gel de
resolução com 4% T, 0,4% C contendo 2,5% (m/v) de sacarose, em tampão TBE
(90mM de Tris-HCl com pH 8,3 contendo 80mM de ácido bórico e 100mM de

Materiais e Métodos 48
EDTA ), 0,2% (m/v) de PSA e 0,066% (v/v) de TEMED. A solução do gel de
resolução, depois de inserida nos moldes de vidro, foi coberta com água. Após a
polimerização deste gel, foi adicionada a solução do gel de concentração
composta por 2,5% T, 0,23%C, 2,5% (m/v) de sacarose, 50mM de Tris-HCl a pH
6,8, 0,2% (m/v) de PSA e 0,066% (v/v) de TEMED. Foi introduzido o pente para
a formação dos “poços” e as amostras foram aplicadas com a ajuda de uma
seringa de vidro graduada. O conjunto foi montado na tina de electroforese
contendo tampão TBE previamente arrefecido a 4 ºC. O sistema foi colocado em
gelo de modo a manter a baixa temperatura e foi aplicada uma diferença de
potencial de 120V, interrompida quando a frente de migração chegou ao fundo
do gel.
2.2.8 – Detecção de Polipéptidos em Géis de Poliacrilamida As proteínas ou polipéptidos separados electroforeticamente podem ser
visualizados no gel recorrendo a diferentes métodos. Géis contendo amostras
proteicas relativamente concentradas foram analisados pelo método de
coloração que envolve o corante orgânico azul de Coomassie R-250 (CBB R-
250), cujo limite inferior de detecção corresponde a cerca de 50 a 100 ng de
proteína concentrada numa banda (Neuhoff et al., 1985). A detecção de
quantidades inferiores de proteína foi realizada pelo método do nitrato de prata,
que permite uma detecção de níveis de proteína na ordem de 1 a 10 ng (Blum et
al., 1987).
2.2.8.1 – Coloracão de Proteínas com Azul de Coomassie O azul de Coomassie R-250, devido à sua carga negativa, liga-se aos
resíduos básicos das proteínas “aprisionadas” no gel, não exercendo qualquer
interacção com os polímeros do mesmo, tornando-se assim possível remover o
corante não ligado (Vesterberg et al., 1977).

Materiais e Métodos 49
O gel contendo os polipéptidos a analisar foi mergulhado numa solução
de 0,1% (m/v) de CBB R-250, 45% (v/v) de metanol (PRONALAB) e 10% (v/v)
de ácido acético glacial durante aproximadamente duas horas. O corante não
ligado à proteína foi removido através de uma solução de descoloração com
10% (v/v) de metanol e 10% (v/v) de ácido acético glacial, que foi substituída
diversas vezes, até o fundo do gel se apresentar transparente.
2.2.8.2 – Coloração de Proteínas com Prata
Na coloração por prata, os géis são impregnados com o ião solúvel Ag+ e
revelados por tratamento com um agente redutor. As biomoléculas presentes no
gel, neste caso proteínas, promovem a redução do ião a Ag0, insolúvel e visível,
permitindo a visualização das bandas presentes. A deposição inicial de prata
promove a continuação da reacção num processo autocatalítico (PHARMACIA
BIOTECH, 1996) (figura 11).
Durante o processo de coloração por prata, o gel sofre diversos
tratamentos, com funções específicas: fixação, em que o gel é tratado com
ácido, tornando as moléculas no gel insolúveis e impedindo a sua difusão do gel
durante os tratamentos seguintes; pré-tratamento, através da acção de
reagentes que tornam as proteínas mais reactivas à prata e aceleram a sua
redução a Ag0, aumentando a sensibilidade do método; coloração, por um
Figura 11 – Reacção de redução do ião solúvel Ag+ a prata metálica Ag0 através do contacto com biomoléculas, e posterior reacção autocatalítica; adaptado de PHARMACIA BIOTECH, 1996.

Materiais e Métodos 50
tratamento com nitrato de prata, sob condições ácidas para impedir a redução
do ião solúvel; revelação, através da redução do ião de prata a prata metálica, a
pH elevado; paragem, num passo que impede a continuação da redução do ião
ião de prata solúvel ainda existente.
O protocolo utilizado baseou-se no método desenvolvido por Blum et al.
(1987). Os polipéptidos foram fixados com uma solução constituída por 50%
(v/v) de metanol, 12% (v/v) de ácido acético glacial e 0,05% (v/v) de formaldeído
durante 20 minutos. Seguiram-se três lavagens de 10 minutos cada com 50%
(v/v) de etanol e um pré-tratamento, durante 1 minuto, com uma solução de
tiosulfato de sódio (200 mg.L-1). Após três lavagens de 20 segundos cada, com
água destilada, o gel foi incubado com solução corante de nitrato de prata (2
gr.L-1) contendo 0,075% (v/v) de formaldeído, durante 10 minutos. O excesso de
corante foi removido por lavagem do gel com água destilada, duas vezes
durante 20 segundos e as bandas proteicas reveladas até à intensidade
desejada, com uma solução de carbonato de sódio (60 gr.L-1) contendo 0,05%
(v/v) de formaldeído e tiosulfato de sódio (4 mg.L-1). Após atingida a coloração
desejada dos polipéptidos, o gel foi lavado com água destilada, duas vezes,
durante 2 minutos, e mergulhado numa solução de paragem da reacção
contendo 50% (v/v) de metanol e 12% (v/v) de ácido acético glacial, durante 5
minutos. A lavagem final realizou-se durante 20 minutos, com uma solução de
50% (v/v) de metanol. A totalidade do procedimento foi executada à temperatura
ambiente e com agitação.
2.2.9 – Immunoblot A técnica de immunobloting é extremamente poderosa na identificação de
proteínas separadas numa electroforese em gel de poliacrilamida, sendo mais
sensível que qualquer tipo de corante utilizado directamente nos géis e, para
além disso, é uma técnica que pode ser muito específica na identificação de
determinadas proteínas contra as quais existam anticorpos.

Materiais e Métodos 51
Neste estudo foi aplicada uma técnica de immunoblot que utilizou um
sistema de anticorpos (Abs) primários e anticorpos secundários, ambos
monoclonais, ou seja, com a capacidade de reconhecerem o mesmo antigénio
(Ag). Os primeiros têm a capacidade de reconhecer e formar um complexo Ab-
Ag com um epítopo presente na proteína de interesse, que por sua vez são
reconhecidos pelos Abs secundários (Anti-IgG). Acoplada aos Abs secundários
está uma peroxidase de rábano que tem a capacidade de oxidar certos
substratos, como o luminol, que ficam num estado excitado e decaem por
fosforescência. Esta fosforescência, com emissão de luz visível, pode ser
detectada recorrendo ao uso de uma chapa fotográfica (figura 12).
2.2.9.1 – Electrotransferência de Proteínas Para se proceder a uma correcta análise e tirar completo partido da
capacidade dos anticorpos, as proteínas separadas num gel de poliacrilamida
têm de ser transferidas para um suporte sólido de modo a torná-las mais
acessíveis aos anticorpos.
O processo de transferência das proteínas do gel para a membrana
porosa de fluoreto de polivinilideno (PVDF) (IMMOBILON-P, MILLIPORE) é
efectuado tendo por base a carga intrínseca das proteínas, no caso de
luminol e
amplificador
Figura 12 – Representação esquemática da técnica de immunoblot utilizada. Reconhecimento do epítopo pelo Ab primário (1), seguido do reconhecimento do Ab primário pelo Ab secundário acoplado a uma peroxidase (2) e oxidação de um substrato para emissão de luz (3); adaptado de BOEHRINGER MANNHEIM.
luz
1 2 3

Materiais e Métodos 52
transferência de proteínas separadas por electroforese em condições nativas, ou
na carga negativa dos complexos SDS-proteína, no caso de transferência de
proteínas separadas por electroforese em condições desnaturantes. A aplicação
de um campo eléctrico ao sistema promove a migração das proteínas do gel
para a membrana.
A membrana de PVDF utilizada tem características hidrofóbicas, sendo
por isso necessário, antes da transferência, submergi-la em metanol e equilibrá-
-la em tampão de transferência (25 mM de Tris, 190 mM de glicina, 0,1% (m/v)
de SDS, 20% (v/v) de metanol, pH 8,3). Foi montado um sistema como o
representado na figura 13, tendo o cuidado de eliminar bolhas de ar existentes
entre as partes constituintes, já que estas afectam o sucesso da transferência.
Depois de montado, o sistema foi mergulhado numa tina de transferência,
tendo-se o cuidado de colocar a membrana virada para o ânodo, e o gel para o
cátodo. Aplicou-se uma diferença de potencial de 70V entre os eléctrodos. No
caso de géis de poliacrilamida com 0,75 mm de espessura, a diferença de
potencial foi mantida por 1 hora, enquanto que para géis de poliacrilamida com
1,5 mm de espessura, a transferência foi prolongada até 2 horas.
2.2.9.2 – Fixação das Proteínas à Membrana de PVDF
Placa de suporte Esponja Membrana de PVDF Gel de poliacrilamida 2 folhas de papel 3MM
Figura 13 – Montagem do sistema de transferência

Materiais e Métodos 53
Após a transferência, foi necessário fixar as proteínas à membrana de
PVDF para que estas não se difundissem nos passos seguintes do immunoblot.
Para isso foram testadas três condições de fixação da membrana:
solução de fixação (10% de ácido acético glacial, 25% de isopropanol), 30
minutos num banho a 100 ºC e autoclavagem da membrana durante 20 minutos
(Swerdlow et al., 1986). Amostras correspondentes às fracções 22, 23 e 24
obtidas por filtração em gel (ver secção 2.2.6.1) de um extracto de JD183 foram
separadas por electroforese em gel desnaturante de acrilamida (ver secção
2.2.7.2.1). Fixaram-se as proteínas recorrendo aos métodos acima descritos,
bloqueou-se a membrana, incubou-se com os anticorpos primário e secundário e
revelou-se o immunoblot como descrito, em detalhe, nas secções seguintes.
Pela observação da figura 14 concluiu-se que o melhor método de fixação foi a
fervura durante 30 minutos, uma vez que fornece um sinal mais forte do que o
obtido com a solução de fixação e, no entanto, não leva à perda de sinal como
aconteceu na fixação usando a autoclave.
A B C
22 23 24 22 23 24 22 23 24
Figura 14 – Teste às diferentes condições de fixação das proteínas à membrana. A – Solução de fixação, B – 30 minutos a 100 ºC, C – autoclavagem durante 20 minutos. 22, 23, 24 – fracções colhidas por filtração em gel de um extracto de JD183 e aplicadas para este teste.

Materiais e Métodos 54
2.2.9.3 – Bloqueamento da Membrana de PVDF
Sendo a membrana de PVDF utilizada, especialmente desenhada para a
retenção de proteínas, tornou-se necessário bloquear toda a área da membrana
não ocupada pelas proteínas transferidas uma vez que, sendo os anticorpos
proteínas, poderia haver ligação dos mesmos à membrana e impedir assim a
análise dos resultados.
Para testar as melhores condições de bloqueamento foram empregues
três soluções diferentes para o efeito: 3% (m/v) de BSA em PBST (8,1 mM de
hidrogeno fosfato de sódio, 1,5 mM dihidrogeno fosfato de potássio, 137 mM de
cloreto de sódio, 2,7 mM cloreto de sódio e 0,1% (v/v) Tween-20), 3% (m/v) leite
em pó (RÉGILAIT) em PBST e 3% (m/v) BSA e 3% (m/v) leite em pó em PBST.
Foram aplicadas três séries de 5, 10 e 17 µg de proteína provenientes de um
extracto proteico de JD183 num gel desnaturante de 12%. Após a electroforese,
os polipéptidos foram transferidos para uma membrana PVDF nas condições
definidas em 2.2.9.1. A membrana foi cortada em três e os péptidos fixados à
membrana por fervura, a 100 ºC, durante 30 minutos. Cada membrana foi
incubada com uma solução de bloqueamento diferente com agitação, durante
uma hora. Processou-se o immunoblot como descrito nas secções seguintes
(figura 15).
A B C
5 10 17 5 10 17 5 10 17
Figura 15 – Teste a diferentes condições de bloqueamento da membrana. A – 3% BSA, B – 3% leite em pó, C – 3% BSA e 3% leite em pó. 5, 10, 17 – quantidade de proteína (em µg) aplicada no SDS-PAGE.

Materiais e Métodos 55
Verifica-se que em A o bloqueamento não foi muito eficaz devido à
obtenção de uma grande coloração do “background”. Em C, o bloqueamento foi
eficaz. No entanto, a perda de sinal relativamente às outras condições de
bloqueamento pode indicar que algumas proteínas ficaram inacessíveis aos
anticorpos por este método de bloqueamento. B representa uma situação
intermédia entre A e C, havendo um bloqueio mais eficaz que em A e não se
verificar perda de sinal como em C. Em todo o trabalho apresentado utilizou-se o
bloqueamento com 3% de leite em pó em PBST.
2.2.9.4 – Incubação com Anticorpos Neste estudo foram usados três tipos de anticorpos primários para a
identificação de dois epítopos diferentes. Utilizaram-se dois anticorpos primários
anti-ha, o 16B12 (BABCO) produzido em ratinho, e o anti-ha 3F10 (ROCHE)
produzido em rato, para a detecção do epítopo 2x YPYDVPDYA, existente na
hemaglutinina do vírus da influenza que se encontra presente nas proteínas de
interesse das estirpes JD183, JD127, JD125, JD129 e AM1 e o anticorpo
primário M2 (SIGMA) (anti-FLAG) para a detecção do epítopo DYKDDDDK
presente nas proteínas de interesse das estirpes JD184 e JD228.
Os anticorpos primários foram especificamente detectados com os
seguintes secundários, conjugados com uma peroxidase de rábano: anti-IgG de
ratinho (AM) (SIGMA) específico para os anticorpos produzidos nesta espécie,
conjugado com uma peroxidase de rábano e anti-IgG de rato (AR) (ROCHE)
específico para os anticorpos 3F10.
A incubação da membrana com o anticorpo primário 16B12 foi feita numa
diluição deste 1:2000 enquanto que a diluição utilizada para o 3F10 foi de
1:8000. O anticorpo anti-FLAG foi utilizado numa diluição de 1:1000, em solução
3% (m/v) de leite em pó em PBST. A membrana foi selada juntamente com a
solução contendo o anticorpo e colocada sob agitação, a 4 ºC, durante a noite.
Após duas lavagens de 5 minutos com PBST, duas lavagens de
semelhante tempo com SAIS (1 M de NaCl, 10 mM de hidrogeno fosfato
disódico e 0,5% (v/v) de Tween-20) e novo bloquamento de 15 minutos,

Materiais e Métodos 56
incubou-se com o anticorpo secundário diluído em 3% (m/v) leite em pó em
PBST. O anticorpo secundário anti-rato foi diluído 1:2000. A diluição a usar para
o anticorpo secundário anti-ratinho foi previamente testada em quatro séries de
amostras provenientes de uma separação por filtração em gel de um extracto de
JD127. Como se pode observar na figura 16 a diluição 1:6000 do anticorpo
apresenta uma razão sinal/ruído de fundo maior do que as outras diluições
testadas revelando-se, portanto, a mais adequada.
Após a incubação com o anticorpo secundário durante 4 horas em saco
selado, com agitação, à temperatura ambiente, procedeu-se a duas lavagens
com PBST durante cinco minutos cada, uma lavagem de 10 minutos com SAIS,
três lavagens de cinco minutos com PBST e uma lavagem de 1 minuto em PBS
(8,1 mM de hidrogeno fosfato de sódio, 1,5 mM de dihidrogeno fosfato de
potássio, 137 mM de cloreto de sódio e 2,7 mM de cloreto de potássio).
16 22 26 30 16 22 26 30 16 22 26 30 16 22 26 30
1:2000 1:3000 1:6000 1:12000 Figura 16 – Teste da diluição do anticorpo secundário anti-IgG de ratinho. As frações 16, 22, 26 e 30, obtidas por separação em filtração em gel de extracto de células JD127 foram aplicadas num gel SDS-PAGE de 12%, transferidas para uma membrana PVDF. O immunoblot processou-se como descrito, mas utilizando as diluições de anticorpo secundário mencionadas.

Materiais e Métodos 57
2.2.9.5 – Revelação do Immunoblot Após a incubação da membrana com os anticorpos primário e secundário,
foi possível detectar a localização destes, e consequentemente das proteínas de
interesse, na membrana, através de luminescência, pela adição de um substrato
da peroxidase que, ao ser oxidado (figura 17) e na presença de um activador,
sobe ao extado excitado, decaindo por fosforescência.
Numa câmara escura retirou-se a membrana da solução de lavagem PBS
onde se encontrava e adicionou-se a solução de reacção (100:1 Solução
A:Solução B (BOEHRINGER MANNHEIM)), cerca de 0,125 mL por cm2 de
membrana. Esperou-se 1 minuto e colocou-se a membrana sobre um vidro,
numa cassete de revelação (AMERSHAM PHARMACIA BIOTECH). Cobriu-se a
membrana com uma folha de papel de acetato, tendo o cuidado de eliminar
bolhas de ar. Posicionou-se uma chapa fotográfica (FUJI) sobre a folha de
acetato de modo a ser impressionada pela luz emitida pela oxidação do luminol.
Após o tempo de exposição do filme, este foi removido da cassete de revelação
e colocado numa solução de revelador (SIGMA) até à intensidade desejada,
passado por água e colocado numa solução de fixador (SIGMA) até à
transparência da chapa. Foi novamente passado por água e colocado a secar.
2.2.10 – Cromatografia de Metalo-Afinidade Na cromatografia de metalo-afinidade, um ião metálico, como o Cu2+,
Zn2+, Cd2+, Co2+ ou Ni2+, ligado a um suporte sólido, é utilizado para
selectivamente ligar proteínas através dos grupos imidazole de resíduos de
luz
Figura 17 – Reacão de oxidação do luminol (diacil-hidrazida) pela peroxidase. Adaptado de BOEHRINGER MANNHEIM

Materiais e Métodos 58
histidina, dos grupos tiol de cisteínas ou grupos indol de triptofanos (Porath,
1985). A imobilização da proteína envolve ligações de coordenação com o
metal que têm de ser suficientemente estáveis para reterem as proteínas a
purificar durante os passos de eluição do material não ligado.
A eluição das proteínas ligadas pode ser feita diminuindo o pH, que
destabiliza o complexo metal-proteína, ou através da adição de grupos
competidores no tampão de eluição.
Neste estudo foi utilizada a resina TALON (CLONTECH) para purificar
proteínas com a tag de 6 histidinas (6His) provenientes de extractos proteicos
das estirpes JD184 e JD228. Esta resina possui um ião de cobalto complexado
através de um agente quelante (TC) ligado a um polímero de agarose (figura
18). A eluição das proteínas ligadas ao metal foi efectuada com o auxílio de
imidazole.
Partiu-se de um extracto de células JD184 ou JD228 em tampão de
extracção MA, e utilizou-se a coluna PD-10 para o trocar por tampão Co2+,
obtendo-se assim, como acima descrito o DCE.
Retiraram-se 2 mL do stock de resina em suspensão (equivalente a 1 mL
de resina), centrifugou-se a 700 g e ressuspendeu-se a resina em 20 mL de
tampão Co2+. Este passo de centrifugação e ressuspensão foi repetido duas
vezes, para assim obter a resina equilibrada no tampão de interesse. Adicionou-
se o DCE à resina equilibrada e colocou-se a 4 ºC, durante 1 hora, com
Figura 18 – Diagrama representativo do sistema de metalo-afinidade utilizado, adaptado de CLONTECH, 1999.

Materiais e Métodos 59
agitação. Transferiu-se a suspensão para uma coluna de vidro vazia, de 10 mL,
e deixou-se empacotar pela acção da gravidade. Abriu-se a coluna e colheu-se a
fracção não ligada (FT), tendo o cuidado de não deixar secar o topo da resina.
Efectuou-se uma lavagem com 20 mL de tampão Co2+ e colheu-se o material
eluído (W1). Repetiu-se este passo com o mesmo volume de tampão e colheu-
se de novo o material eluído (W2). Procedeu-se à eluição do material ligado
através de um gradiente de imidazole. Aplicaram-se 2 mL de cada solução de
imidazole (50 mM, 100 mM, 200 mM, 300 mM, 400 mM) em tampão Co2+, em
sentido crescente de concentração, tendo o cuidado de colher o material eluído
de cada concentração (E1, E2, E3, E4, E5).
2.2.1.1 – Medição da Actividade Quimotríptica do Proteassoma A medição da actividade quimotríptica do proteassoma foi feita com
recurso a um substrato sintético, Suc-Leu-Leu-Val-Tyr-AMC (BACHEM). Na
presença deste substrato, o proteassoma quebra a ligação Tyr-AMC, deixando a
coumarina na sua forma livre.
A 7-amido-4-metilcoumarina, após libertação do péptido, é excitável a 380
nm, decaindo por fluorescência, com o pico máximo de emissão a situar-se nos
440 nm. Deste modo é possível detectar a presença de proteassoma funcional
recorrendo à utilização de um fluorímetro.
A 56 µL de uma solução de substrato a 13mM foram adicionados 1144 µL
de água e 300 µL de um tampão de actividade 5 vezes concentrado (50 mM
Tris, 2 mM ATP, 5 mM cloreto de magnésio, 1 mM DTT, pH 7,8) para uma
concentração final de substrato de 0,5 mM. A 20 µL da amostra a analisar foram
adicionados 20 µL desta solução. Após a homogeneização da mistura, esta foi
incubada num banho a 37 ºC durante 1 hora. A reacção foi interrompida pela
adição de 960 mL de etanol absoluto a –20 ºC e as amostras guardadas a –20
ºC até serem analisadas. Os valores de actividade foram obtidos pela medição
da fluorescência a 440 nm após excitação da amostra a 380 nm num fluorímetro
(FP-777 Spectrofluorimeter – JASCO).

Resultados 60
3 – Resultados 3.1 – Efeito do Cádmio e do Choque Térmico no Crescimento de
S. cerevisiae e na Expressão da Subunidade Proteassomal α7/Prs1p
O trabalho realizado por Jungmann et al. (1993) revela que a presença de
cádmio induz um aumento na expressão dos genes codificantes para várias
proteínas envolvidas na via proteolítica dependente de Ubiquitina/Proteassoma,
como sejam: o UBC7, o UBC5 e o UBI4. A resposta a este stress é diferente da
apresentada pelas células expostas a choque térmico em que, apesar de haver
aumento na expressão de UBC5 e UBI4, não há aumento significativo na
expressão de UBC7 (Seufert e Jentsch, 1990; Finley et al., 1987).
Qualquer acréscimo na indução da expressão de subunidades
proteassomais, em princípio, iria aumentar a quantidade de complexos
precursores do proteassoma na célula, facilitando o seu isolamento e
caracterização.
Na tentativa de observar se a presença de cádmio e exposição a um
choque térmico de 37 ºC, seriam condições de stress capazes de aumentar os
níveis de expressão de subunidades proteassomais e, consequentemente, a
quantidade de proteassoma funcional nas células de levedura de padeiro,
realizou-se a experiência que, de seguida, se descreve.
Células da estirpe JD183 foram crescidas em SD-Ura contendo 50 mM de
Cd2+ e, como controlo, cresceu-se a mesma estirpe na ausência de cádmio. O
crescimento foi monitorizado pela medição da OD600 nm ao longo do tempo. Após
30, 60 e 120 minutos de exposição ao cádmio, retiraram-se 10 µL da cultura e
colocaram-se a recuperar em meio sólido (figura 19A).

Resultados 61
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
4,0
0 50 100 150 200 250 300 350 400 450
Tempo (min)
OD 60
0
Controlo50 microM Cd++
Controlo
60 minutos
30 m
inut
os
120 minutos
0
500
1000
1500
2000
2500
3000
3500
4000
4500
0’ 60’ 100’ 130’ 0’ 120’ 240’
Cd2+ Choque térmico
tempo
Cd2+ 37 ºC
130’ 60’ 30’ 30’ 60’ 0’ 120’0’ 240’
50 µM 100 µM
A
B
Ctempo
Figura 19 – Efeito de cádmio e choque térmico a 37 ºC no crescimento de células de S. cerevisiae e na expressão da subunidade proteassomal α7/Prs1p. Curva de crescimento e diferenças no crescimento de células JD183 sob efeito de 50 µM de cádmio (adicionado no tempo zero) (A). Actividade quimotríptica do proteassoma em extractos proteicos de células expostas a diversos tempos a 50 µM de cádmio e choque térmico a 37 ºC (B). Expressão da subunidade proteassomal α7/Prs1p em células expostas durante os tempos indicados a 50 e 100 µM de cádmio e choque térmico a 37 ºC, analisada por immunoblot após separação por SDS-PAGE (C).
OD
600

Resultados 62
As células controlo apresentam um crescimento superior ao observado
para as células expostas a cádmio. Esta diferença no crescimento é mais notória
a partir dos 165 minutos de monitorização em que se nota um claro afastamento
das linhas de crescimento. A partir de 400 minutos de exposição, a densidade
óptica da cultura não exposta a cádmio chega quase a duplicar a densidade
óptica da cultura exposta a este metal tóxico.
Na recuperação em meio sólido após diversos tempos de exposição ao
cádmio observa-se que o tempo de exposição é inversamente proporcional ao
tamanho das colónias isoladas obtidas, ou seja, quanto mais tempo a cultura
esteve exposta ao agente de stress, menor foi o tamanho das colónias isoladas
obtidas.
De modo a verificar se a exposição a 50 µM de cádmio e o choque
térmico a 37 ºC afectavam, de algum modo, a actividade quimotríptica do
proteassoma, foram feitas culturas de células JD183 expostas a estes indutores
de stress (60, 100 e 130 minutos e 120 e 240 minutos, respectivamente).
Prepararam-se extractos proteicos de acordo com o método descrito em 2.2.4.1
e após a normalização da concentração de proteína, a actividade quimotríptica
foi analisada recorrendo a um substrato sintético do proteassoma, cuja
degradação origina um produto fluorogénico (figura 19 B).
A actividade quimotríptica do proteassoma não sofreu alterações
significativas ao longo do tempo de exposição a cádmio relativamente ao
controlo. Este resultado foi verificado também para a indução de stress por
choque térmico, apesar de se notar um ligeiro aumento da actividade
quimotríptica após 120 minutos de exposição a uma temperatura de 37 ºC.
O efeito da exposição de células ao cádmio e a choque térmico sobre a
expressão de subunidades proteassomais, foi estudado em culturas de células
JD183 expostas durante 30, 60 e 130 minutos a 50 µM de cádmio, 30 e 60
minutos a 100 µM de cádmio e 120 e 240 minutos à temperatura de 37 ºC.
Foram preparados extractos como descrito anteriormente e a concentração de
proteína foi normalizada entre eles. As proteínas foram separadas num SDS-
PAGE (ver secção 2.2.7.2.1). Os polipéptidos separados foram
electrotransferidos para uma membrana de PVDF e a presença da subunidade

Resultados 63
proteassomal marcada nesta estirpe, Prs1p-ha foi monitorizada por immunoblot,
como descrito em 2.2.9, com anticorpos anti-ha (figura 19 C).
As bandas obtidas, correspondentes à subunidade Prs1p-ha, em relação
às culturas expostas às duas concentrações de cádmio e a choque térmico, não
apresentam diferenças significativas de intensidade relativamente aos contolos.
3.2 – Marcação da Subunidade α7/Prs1p com o Epítopo ha em JD305
As estirpes de S. cerevisiae utilizadas neste estudo possuem diversas
subunidades marcadas no seu terminal C com epítopos (tabela I) para que
possam ser detectadas imunoquimicamente com anticorpos monoclonais contra
esses epítopos. Nesta secção são apresentados os resultados obtidos nos
vários passos para a marcação da subunidade proteassomal α7/Prs1p na
estirpe JD305 com o epítopo ha.
3.2.1 – Estratégia Usada para a Marcação da Subunidade α7/Prs1p com ha
Plasmídios de DNA contendo genes de interesse com alterações
desejadas podem ser introduzidos em células competentes de levedura. No
caso de existir homologia entre o gene introduzido e o material genético da
levedura, pode ocorrer integração desse gene modificado por recombinação
homóloga (Heinemann e Sprague, 1991) (figura 20 A). Antes da transformação
das células, o plasmídio deverá ser linearizado, com um enzima de restrição,
cujo único local de corte se encontre no gene de interesse. Neste caso foi
utilizado o enzima Cla I, com um sítio de corte único localizado no interior de um
fragmento 5’∆ do gene PRS1 da levedura contendo os codões que codificam
para o epítopo 2xha antes do codão STOP. A integração é efectuada pela DNA
polimerase da levedura durante a replicação do material genético (caminho da
polimerase indicado pelas setas). O resultado final será a introdução do
plasmídio no genoma da levedura, com o gene de interesse modificado de forma
que a sua expressão origine a proteína marcada com o epítopo 2xha.

Resultados 64
5 148 / 4 973 bp 4 268 bp
M + -
pCR8
Cla I
A B
M - + 1 2 4 5 3 6 7 8 C
1375 bp
947 bp
Prs1p-ha
Gel de agarose
Immunoblot
JD305
YPD
SD-Ura
D JD47-13C JD305
JD183 AM1
pCR8

Resultados 65
3.2.2 – Análise da Linearização do Plasmídio pCR8 O plasmídio pCR8 foi incubado com o enzima de restrição Cla I. Num gel
de agarose foram aplicadas as seguintes amostras: Marcador (DNA λ digerido
com EcoR I e Hind III), plasmídio digerido e plasmídio não digerido. Após a
electroforese o resultado foi observado sob luz UV (figura 20 B).
A separação electroforética do plasmídio digerido deu origem a uma só
banda visível (DNA linear). No caso do plasmídio não digerido, são visíveis duas
bandas características do DNA supercoiled (migração rápida) e circular
(migração lenta).
3.2.3 – Análise da Integração por PCR e Immunoblot Uma vez obtido o plasmídio linearizado, procedeu-se à integração do mesmo no
genoma da estirpe JD305. Obtiveram-se cerca de trinta colónias de células
transformadas em meio SD-Ura. Seleccionaram-se oito colónias e a integração
do plasmídio foi analisada por PCR e por Immunoblot, nestas células (figura 20
C).
A análise por PCR foi efectuada com a utilização de primers desenhados
para a amplificação do material integrado desde o início do gene PRS1 até ao
terminador do citocromo c (ver secção 2.2.3.6). Após a amplificação, os produtos
de PCR foram aplicados num gel de agarose juntamente com um marcador. Na
electroforese em gel de agarose, na linha correspondente ao controlo negativo
não foi observada nenhuma banda, contrariamente, nas linhas referentes ao
controlo positivo e às colónias 4, 5 e 6 observou-se uma banda distinta com um
tamanho entre 1375 e 947 bp, que correspondia ao tamanho esperado para o
fragmento a amplificar. Por immunoblot, em todas as amostras analisadas, à
excepção do controlo negativo, foram observadas bandas únicas e distintas.
JD183 Y13∆Prs1p-ha

Resultados 66
3.2.4 – Observação do Fenótipo da Estirpe Mutante (Y13∆) sob Choque Térmico
Para análise do fenótipo da estirpe JD305 marcada com ha (AM1) e não
marcada (JD305), foram incubadas placas a 37 ºC inoculadas com JD305 e AM1
e as estirpes selvagens correspondentes, JD47-13C e JD183. Após a incubação
de 3 dias, a placa foi analisada e o tamanho das colónias isoladas comparado.
Observou-se que as colónias isoladas nas estirpes selvagens são maiores do
que colónias isoladas nas estirpes mutadas, independentemente da marcação
(figura 20 D).
3.3 – Presença de Várias Subunidades Proteassomais ao Longo das Fracções Obtidas por Filtração em Gel
A presença de várias subunidades proteassomais e do factor Ump1 foi
monitorizada por immunoblot ao longo das fracções obtidas pela separação de
extractos proteicos em Superose 6, após uma electroforese desnaturante. A
actividade catalítica destas fracções foi também observada para as estirpe
selvagem JD183 e estirpe mutante AM1.
3.3.1 – Análise da Actividade Quimotríptica em JD183 e AM1 Após separação dos extractos proteicos provenientes das estirpes JD183
e AM1 por exclusão molecular em Superose 6, procedeu-se à análise da
actividade quimotríptica.
Como apresentado na figura 21 A, para a estirpe selvagem, a actividade
catalítica inicía-se na fracção 13, sofre um aumento intenso até atingir o seu
máximo na fracção 16 e decresce até à fracção 20, a partir da qual deixa de
existir actividade catalítica significativa.

Resultados 67
Figura 21 – Análise da actividade quimotríptica e da presença das subunidades proteassomais Pre1p-ha e Prs1p-ha e do factor Ump1p-ha nos complexos separados por filtração em gel. Os extractos de células JD47-13C (sem marcação), AM1 (Prs1p-ha), JD125 (Pre1p-ha), JD129 (Ump1p-ha) e JD183 (Prs1p-ha) foram separados por filtração em gel numa coluna de Superose 6. A presença dos polipéptidos marcados foi analisada ao longo das fracções colhidas, por immunoblot (B) e a actividade catalítica entre a estirpe selvagem e a estirpe mutada foi comparada (A). As subunidades proteassomais marcadas foram salientadas na estrutura cristalina do proteassoma (C).
0
500
1000
1500
2000
2500
3000
12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30fr acco e s
JD47-13CA M1
CE
Pre1p-ha
Ump1p-ha
Prs1p-ha
Prs1p-ha
158
kDa
232
kDa
669
kDa
A
B
C
*
(JD125)
(JD129)
(AM1)
(JD183)

Resultados 68
A estirpe mutante, à semelhança da selvagem, apresenta o início da
actividade catalítica na fracção 13, com uma subida acentuada da actividade até
à fracção 15 onde atinge valores semelhantes à estirpe selvagem decrescendo a
partir desta até à fracção 19, onde deixa de se observar actividade significativa.
Os valores de actividade monitorizados entre as fracções 16 e 19 da estirpe
mutante são muito inferiores aos observados nas mesmas fracções para a
estirpe selvagem. Um padrão semelhante ao apresentado foi obtido sempre que
se repetiu a experiência.
3.3.2 – Presença de Várias Subunidades Marcadas ao Longo do Padrão de Separação por Filtração em Gel
Extractos de células JD125, JD129, JD183 e AM1 foram passados numa
coluna de Superose 6 e colhidas as fracções daí resultantes. Amostras destas
fracções foram separadas por electroforese em gel desnaturante e a presença
das várias subunidades proteassomais marcadas com ha analisada por
immunoblot com anticorpos contra este epítopo (figura 21 B).
Seguindo uma ordem decrescente de massa molecular, a subunidade
Pre1p (marcada a verde na figura 21 C) está presente nas fracções com
complexos de maior massa molecular até à fracção 23, após a qual se segue
um desaparecimento desta subunidade, voltando a aparecer a partir da fracção
27 e estando presente daí até à fracção 30.
O factor Ump1 (não representado na figura 21 C) encontra-se apenas
presente em complexos colhidos entre as fracções 21 e 24 (figura 21 B)
Curiosamente, a subunidade α7/Prs1p (sinalizada a azul na figura 21 C),
nas duas estirpes analisadas contendo esta subunidade marcada com o epítopo
ha, está presente desde as fracções contendo complexos, de massa molecular
elevada até às fracções contendo os complexos mais leves.

Resultados 69
3.4. – Análise de Complexos Contendo a Subunidade β4/Pre1p e o factor de maturação Ump1p
Foi analisada uma estirpe contendo a subunidade β4/Pre1p e o factor
Ump1p ambos marcados com o epítopo ha (células JD127).
O extracto foi separado por filtração em gel e as fracções analisadas por
immunoblot após electroforese desnaturante (figura 22 A). Foi observado a
presença de uma banda com início na fracção 14, correpondente a complexos
de maior massa molecular, até à fracção 23. Este sinal é interrompido, não
sendo observável sinal nas fracções 24 e 25, voltando a ser visível na fracção 26
e até à ultima fracção analisada.
É também visível o sinal de um polipéptido de menor massa molecular
nas fracções 21 a 23, não observável em nenhuma das outras fracções
analisadas, referente ao factor de maturação Ump1p.
As proteínas foram transferidas para membrana PVDF e as subunidades
marcadas com ha analisadas por immunoblot com dois conjuntos de anticorpos
anti-ha: (16B12/AM e 3F10/AR). Para verificar se algumas das bandas
observadas correspondiam a ligação não específica por parte dos anticorpos
utilizados, o mesmo procedimento foi efectuado com células JD47-13C (estirpe
não marcada). Os filmes foram expostos à luz emitida pelo decaimento do
luminol durante 15 e 70 minutos.
As fracções resultantes da separação de um extracto de células JD127
foram submetidas a electroforese em géis nativos com o objectivo de observar
complexos com diferentes razões carga/massa que contenham os polipéptidos
marcados. Na figura 22 B podem observar-se os resultados obtidos para as
duas estirpes. Quando fracções correspondentes ao fraccionamento do extracto
proteico de células JD127 foram analisados com o conjunto de anticorpos
16B12/AR, foi visível um sinal repetitivo desde a fracção 16 até à 22. Para além
deste sinal observou-se um complexo distinto que se estende da fracção 21 à
25, e um outro conjunto de sinais cujos polipéptidos mostram uma migração no
gel menor que a do conjunto referido anteriormente, nas fracções 25 a 27.
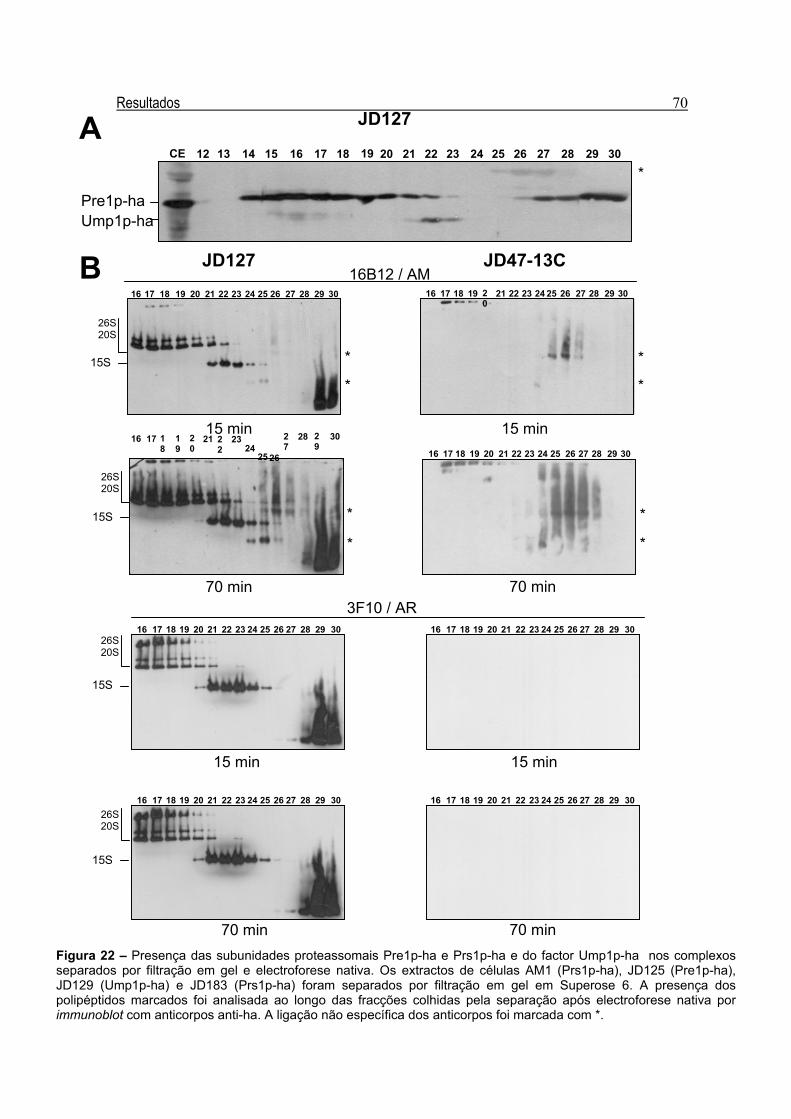
Resultados 70
JD127
A
B
15 min
70 min
CE 13 12 15 14 16 18 17 20 19 21 23 22 24 25 26 28 27 29 30
Pre1p-ha Ump1p-ha
*
JD127
JD47-13C
15 min
70 min
16 18 17 20 19 21 23 22 24 25 26 28 27 29 30
15 min
16 18 17 20 19 21 23 22 24 25 26 28 27 29 30
15 min
16 18 17 20 19 21 23 22 24 25 26 28 27 29 30
70 min
16 18 17 20 19 21 23 22 24 25 26 28 27 29 30
70 min
16 18 17 20 19 21 23 22 24 25 26 28 27 29 30
26S 20S
15S *
*
16 18 17 20
19 21 23 22 24 25 26 28 27 29 30
*
*
16 18
17 20
19
21 23 22 24
25 26
28 27
29
30
26S 20S
15S *
*
16 18 17 20 19 21 23 22 24 25 26 28 27 29 30
*
*
16B12 / AM
3F10 / AR
26S 20S
15S
26S 20S
15S
Figura 22 – Presença das subunidades proteassomais Pre1p-ha e Prs1p-ha e do factor Ump1p-ha nos complexos separados por filtração em gel e electroforese nativa. Os extractos de células AM1 (Prs1p-ha), JD125 (Pre1p-ha), JD129 (Ump1p-ha) e JD183 (Prs1p-ha) foram separados por filtração em gel em Superose 6. A presença dos polipéptidos marcados foi analisada ao longo das fracções colhidas pela separação após electroforese nativa por immunoblot com anticorpos anti-ha. A ligação não específica dos anticorpos foi marcada com *.

Resultados 71
Nas fracções 24, 25 e 26 é também visível um sinal na zona inferior do gel.
Estes dois últimos conjuntos foram salientados e são mais facilmente
observados com 70 minutos de exposição. É também visível um arrastamento
nas três últimas fracções que termina quase no final do gel.
Quanto aos resultados obtidos com este conjunto de anticorpos na estirpe
JD47-13C, observam-se dois conjuntos de sinais. O primeiro estende-se da
fracção 25 à 27, enquanto o segundo aparece com uma maior migração no gel,
nas fracções 24, 25 e 26.
Pela análise com o conjunto de anticorpos 3F10/AR, observa-se existirem
muitas semelhanças com os resultados obtidos com os anticorpos 16B12/AM.
Nomeadamente, são semelhantes os sinais visíveis nas fracções 16 a 21,
apesar de, com estes anticorpos, o sinal na zona superior seja mais intenso,
também, são vísiveis as bandas nas fracções 20 a 25 correspondentes a
complexos com maior migração no gel e ainda o arrastamento nas três últimas
fracções até à frente de migração do gel. Realça-se o desaparecimento de dois
conjuntos de sinais, mencionados na análise dos resultados obtidos com os
anticorpos 16B12/AR. Na estirpe JD47-13C, com a utilização do segundo
conjunto de anticorpos, não existe qualquer sinal visível.
3.5 – Análise de complexos proteassomais contendo β4/Pre1p, α7/Prs1p e Ump1p
Os resultados apresentados na figura 21 B, obtidos por análise das
fracções recolhidas na separação dos extractos das respectivas estirpes por
filtração em gel, após electroforese desnaturante foram comparados com a
análise das referidas fracções separadas por electroforese em condições nativas
(figura 23).

Resultados 72
Na estirpe JD125 é visível a presença da β4/Pre1p num complexo
presente desde as fracções a que corresponde uma massa molecular mais
elevada até à fracção 23 e de novo nas três últimas fracções analisadas, no final
da migração. É ainda observável um conjunto de bandas nas fracções 24 a 26
correspondente a ligação não específica dos anticorpos, como se constatou
pelas análises realizadas e apresentadas na figura anterior.
O complexo contendo Ump1p, analisado na estirpe JD129, é visível nas
fracções 21 a 24.
Quanto à subunidade α7/Prs1p analisada na estirpe JD183, é visível em
complexos de elevada massa molecular nas fracções 16 a 23. Está também
presente nas fracções 20 a 25 em complexos com uma maior migração.
Curiosamente, nestas mesmas fracções observa-se um outro complexo com
uma menor intensidade de sinal e uma migração mais lenta. É, também
claramente, visível a presença desta subunidade em complexos com uma
16 18 17 20 19 21 23 22 24 25 26 28 27 29 30 16 18 17 20 19 21 23 22 24 25 26 28 27 29 30
16 18 17 20 19 21 23 22 24 25 26 28 27 29 30
*
16 18 17 20 19 21 23 22 24 25 26 28 27 29 30
*
JD125 JD129
JD183 AM1 Pre1p-ha Ump1p-ha
Prs1p-ha Prs1p-ha
26S 20S
15S
26S 20S
26S 20S
15S
Figura 23 – Presença das subunidades proteassomais Pre1p-ha e Prs1p-ha e do factor Ump1p-ha ao longo dos complexos separados por filtração em gel e electroforese nativa. Os extractos de células AM1 (Prs1p-ha), JD125 (Pre1p-ha), JD129 (Ump1p-ha) e JD183 (Prs1p-ha) foram separados por filtração em gel em Superose 6. A presença dos polipéptidos marcados foi analisada nas fracções colhidas pela separação após electroforese nativa por immunoblot com anticorpos anti-ha 16B12/AM para as estirpes JD125 e JD129 e 3F10/AR para as estirpes JD183 e AM1. A ligação não específica dos anticorpos foi marcada com *.

Resultados 73
elevada mobilidade, que se estende da fracção 21 à 30. Menos distintamente,
são ainda visíveis dois conjuntos de bandas nas fracções 27, 28 e 29, com uma
mobilidade intermédia entre o complexo 15S e a forma livre, que se parecem
repetir, um ligeiramente acima do outro.
A análise da subunidade α7/Prs1p na estirpe com a delecção da
subunidade α3/Y13, mostrou-nos que esta está presente em complexos de
elevada massa molecular (fracções 16 a 21) e apareceu distribuída ao longo das
fracções 19 a 30, numa forma com elevada mobilidade no gel nativo. Na posição
onde habitualmente se encontra o complexo 15S, nesta estirpe são apenas
visíveis bandas com uma fraca intensidade de sinal, quase indistintas do
background. Também com um sinal muito fraco, aparecem bandas nas fracções
27 a 29, semelhantes às observadas na análise desta subunidade na estirpe
selvagem JD183.
3.6 – Purificação de Complexos contendo α7/Prs1pFLAG-6His Para uma melhor caracterização dos complexos precursores envolvidos
na montagem do proteassoma 20S, testou-se um processo de purificação por
cromatografia de metalo-afinidade, usando a estirpe JD228 com a subunidade
α7/Prs1p marcada com o epítopo FLAG e com 6His. A cromatografia foi
efectuada como descrito em 2.2.10 e o material recolhido foi separado por
electroforese desnaturante e analisado por coloração com prata e por
immunoblot com anticorpos anti-FLAG (figura 24).
No gel submetido a análise pelo método da prata observam-se muitos
polipéptidos contaminantes nas fracções eluídas com imidazole.
No immunoblot observa-se uma banda intensa, correspondente à
subunidade marcada, nas amostras ce, dce, ft e w1, juntamente com outras
bandas não específicas. Nas amostras w10 e w11 não é visível nenhum sinal.
Nas amostras e1 – e5, observa-se a presença da subunidade marcada com uma
intensidade decrescente de sinal.

Resultados 74
ce dce ft w1 w10 w11 e1 e2 e3 e4 e5 ce dce ft w1 w10 w11 e1 e2 e3 e4 e5
JD228
Coloração por prata Immunoblot
Prs1pFLAG6His
Figura 24 – Análise das fracções obtidas por cromatografia de metalo-afinidade através de coloração dos péptidos por nitrato de prata e immunoblot com anticorpos anti-FLAG, após separação por electroforese desnaturante. ce – extracto cru, dce – extracto cru dessalinizado, ft – eluição do material não ligado à resina, w1 - primeiro mililitro da lavagem com tampão de eluição, w10 – décimo mililitro da lavagem com tampão de eluição, w11 – décimo primeiro mililitro da lavagem com tampão de eluição, e1, e2, e3, e4, e5 – eluições com concentrações crescentes de imidazole (50 mM, 100 mM, 200 mM, 300 mM, 400 mM)

Discussão 75
4 – Discussão 4.1 - Efeito do Cádmio e do Choque Térmico no Crescimento de
S. cerevisiae e na Expressão da Subunidade Proteassomal α7/Prs1p
O efeito de vários tipos de stress como o cádmio e choque térmico leva a
mudanças na regulação celular e pode alterar a expressão de várias proteínas
cuja competência é a de a proteger a célula. Algumas das proteínas cuja
expressão é alterada pela presença de cádmio fazem parte da via proteolítica
dependente de ubiquitina/proteassoma (Jungmann et al.,1993).
Sendo o proteassoma um dos complexos proteicos presentes neste
mecanismo de degradação proteica, havia a hipótese de stress induzido por
cádmio e/ou choque térmico provocar efeitos na expressão das subunidades
que formam este complexo. Em caso de aumento de expressão, a quantidade
intracelular disponível para análise seria maior, podendo facilitar o isolamento e
caracterização de complexos precursores na montagem do proteassoma 20S.
4.1.1 – O Cádmio Influencia o Crescimento Celular em Levedura Como o observável na figura 19 A, relativamente ao controlo, a cultura
sujeita ao tratamento com cádmio teve um crescimento menos acentuado, o que
é visível pelo afastamento das curvas de crescimento. Esta diferença é mais
salientada para elevados tempos de exposição. O efeito nocivo do cádmio foi,
também, verificado nas células em recuperação em meio sólido, uma vez que
células mais tempo expostas ao indutor de stress tiveram uma taxa de
recuperação menor, observável pelos diferentes tamanhos das colónias
isoladas.
A diminuição no crescimento das células expostas a cádmio poderá
dever-se ao efeito deste metal nas proteínas, que origina anormalidades na
formação das mesmas (Vallee e Ulmer, 1972). As proteínas malformadas podem
tornar-se tóxicas e formar agregados que podem levar à morte celular.

Discussão 76
4.1.2 – O Cádmio e Choque Térmico não Afectam os Níveis de Expressão nem a Actividade Quimotríptica do Proteassoma
Pela observação da figura 19 B, verifica-se que não houve diferenças
significativas na actividade quimotríptica do proteassoma de células expostas a
cádmio e choque térmico, por comparação com os controlos, apesar dos vários
tempos de exposição testados. Este facto poderá ser explicado em conjunto com
os resultados apresentados na figura 19 C, que mostra não haver aumento na
expressão da subunidade proteassomal α7/Prs1p. É provável que este tipo de
stress, apesar de aumentar a expressão de determinadas proteínas
relacionadas com o sistema de conjugação com Ub, não o faça com os
polipéptidos que constituem o proteassoma. Assim, a actividade quimotríptica
não é afectada uma vez que a concentração de proteassoma presente a nível
intracelular pode não ter sofrido alterações por exposição aos agentes de stress
testados.
4.2 – Marcação da Subunidade α7/Prs1p com o Epítopo ha
A detecção específica de subunidades proteassomais pode realizar-se
com o auxílio de anticorpos. Utilizou-se um epítopo sintético, ha, para a
marcação das subunidades no seu terminal carboxílico, de modo a não interferir
com o tamanho e estrutura do propeptídio apresentado por algumas delas que,
como já mencionado é importante para a montagem correcta do proteassoma
(Chen et al., 1996).
4.2.1 – Verificação da Linearização do pCR8 por Cla I Como observável no gel de agarose (figura 20 B), no poço onde foi
aplicado o plasmídio incubado com o enzima de restrição Cla I é visível uma
única banda, correspondente à forma linearizada do plasmídio de 4612 bp, por

Discussão 77
comparação com as posições relativas das bandas do marcador. No poço
correspondente ao plasmídio não digerido são visíveis duas bandas distintas
cuja migração não é comparável com o marcador, visto tratar-se das formas
características de um plasmídio não linearizado: a forma circular e a forma
supercoiled.
4.2.2 – Verificação da Integração por PCR e Immunoblot A correcta integração do pCR8 no genoma da levedura foi verificada
recorrendo à técnica de PCR e Immunoblotting. Para a primeira foram
desenhados primers para a amplificação de modo a obter um produto com início
na origem do gene PRS1 da levedura até ao terminador do citocromo c
introduzido.
Como é visível na figura 20 C, o controlo negativo não originou produto,
como esperado, uma vez que a estirpe utilizada contém no seu genoma o início
do gene PRS1, mas não o terminador do citocromo c. As colónias 4, 5 e 6 deram
origem a um produto com um padrão de migração semelhante ao obtido no
controlo positivo, com aproximadamente o tamanho esperado (1288 bp). As
restantes colónias não deram origem a produtos de PCR. No entanto, existe a
possibilidade de ter havido problemas na reacção de amplificação, uma vez que
estas colónias, à semelhança do controlo positivo e colónias 4, 5 e 6, deram
origem a uma banda pela reacção com anticorpos anti-ha, com um padrão de
migração semelhante ao destas, como observável no immunoblot (figura 20 C),
banda esta que está ausente no controlo negativo.
4.2.3 – A Estirpe Mutante JD305 (Y13∆) é Sensível a Stress Provocado por Choque Térmico
Como é visível na figura 20 D, quando colocadas a 37 ºC, o crescimento
da estirpe com a delecção da subunidade α3/Y13 é severamente afectado
apresentando um menor nível de multiplicação, por comparação do tamanho das

Discussão 78
colónias isoladas relativamente às estirpes selvagens, o que mostra um menor
nível de resistência a este stress por parte desta estirpe. Este efeito é
observável na a estirpe marcada com Prs1p-ha e na estirpe não marcada,
demonstrando assim que a marcação não influencia o fenótipo desta estirpe em
relação ao stress induzido por choque térmico.
Provavelmente, a delecção da subunidade levará a que haja um menor
nível de proteassomas funcionais dentro da célula, ou um abaixamento na
funcionalidade dos mesmos, que permite a acumulação de agregados proteicos
e proteínas anormais a nível intracelular formados por indução do choque
térmico e que, normalmente, são eliminados por acção da via proteolítica
dependente de Ub/proteassoma.
4.3 – Existem Diferenças na Distribuição das Várias Subunidades ao Longo de Complexos Separados por Exclusão Molecular
A passagem dos extractos por uma coluna de exclusão molecular permite
separar complexos distintos, com diferentes massas moleculares, envolvidos na
formação do proteassoma 20S e proteassoma 26S.
A análise das actividades quimotrípticas dos complexos presentes nas
fracções colhidas dá uma ideia precisa das fracções onde é eluído o
proteassoma completamente montado e activo (figura 21 A), uma vez que, só
depois da montagem e da degradação do factor Ump1, o proteassoma fica
funcional para a degradação de outros substratos (Ramos et al., 1998).
A análise das fracções por immunoblot, após separação por electroforese
desnaturante, permite associar a presença da subunidade numa determinada
fracção a complexos com uma massa molecular correspondente ao volume de
eluição da fracção em causa.

Discussão 79
4.3.1 – A Estirpe Mutante AM1 Apresenta uma Actividade Catalítica Semelhante à Estirpe Selvagem no Proteassoma 26S, mas mais Reduzida no Proteassoma 20S
A estirpe mutante tem um padrão de actividade quimotríptica semelhante
à estirpe selvagem até à fracção 15 (correspondente a uma massa molecular
extrapolada de cerca de 1050 kDa), a partir da qual sofre um decréscimo. Os
valores de actividade da fracção 16 à 19, a partir da qual deixa de ter actividade
catalítica significativa, são inferiores aos da estirpe selvagem (figura 21 A).
Parece claro que a mutação (delecção da subunidade α3/Y13) afecta a
actividade catalítica do proteassoma 20S. É no entanto desconhecida a razão
deste efeito. Curiosamente, as fracções correspondentes a massa molecular
mais elevada que o proteassoma 20S não mostram grandes diferenças em
relação à estirpe selvagem.
Estes resultados sugerem que a ligação dos complexos regulatórios ao
proteassoma 20S venha, de algum modo, minimizar o efeito inibitório que possa
estar a ser induzido por uma alteração estrutural devido à falta da subunidade
α3/Y13.
4.3.2 – A Subunidade α7/Prs1p Está Presente ao Longo de Todas as Fracções Obtidas por Filtração em Gel
A análise da presença das subunidades proteassomais β4/Pre1p e
α7/Prs1p e do factor Ump1p nas fracções obtidas por filtração em gel dos
extractos das estirpes correspondentes (figura 21 B) mostrou-nos que a
α7/Prs1p está presente ao longo de todas as fracções obtidas, e deste modo,
presente em toda a gama de complexos proteassomais separados, na estirpe
selvagem JD183 e mutante AM1. Em contraste está o resultado obtido pela
análise da subunidade β4/Pre1p que apenas se encontra presente nas fracções
correspondentes a massas moleculares baixas e nas fracções correspondentes
a massas moleculares acima dos 200 kDa.

Discussão 80
O factor Ump1, em concordância com os resultados apresentados por
Ramos et al. (1998), é apenas detectado nas fracções 21 a 24, o que poderá
significar que este factor é integrado no complexo 15S e degradado após o
acoplamento de dois destes complexos, para dar origem ao proteassoma 20S,
segundo modelo apresentado por Ramos et al. (1998).
Estes resultados levam a crer que a subunidade α7/Prs1p está presente
desde os estágios iniciais da montagem do proteassoma, sendo integrada muito
cedo ou mesmo sendo ela a iniciadora da montagem do proteassoma 20S. O
“desaparecimento” da subunidade β4/Pre1p nas fracções 24 a 26 pode significar
que esta subunidade é separada na sua forma livre (fracções 27 a 30) e que
posteriormente só é de novo encontrada integrada num complexo pré-existente.
Estas observações foram confirmadas através da análise por immunoblot das
fracções obtidas por filtração em gel de extractos de JD127 (β4/Pre1p-
ha/Ump1p-ha). Como observável na figura 22 A, os padrões são semelhantes
aos obtidos anteriormente com estes polipéptidos marcados em estirpes
separadas, sendo mais uma vez notório o intervalo no padrão da β4/Pre1p-ha
nas fracções 24 e 25 que, como descrito acima, poderá indicar que esta
subunidade é incorporada num nível avançado da montagem do proteassoma, e
está ausente dos passos iniciais.
Os resultados obtidos com o factor Ump1p marcado anteriormente (figura
21 B) foram também observados com esta estirpe (figura 22 A).
As fracções colhidas foram também analisadas por electroforese em
condições nativas, tendo como controlo de especificidade de anticorpo as
fracções obtidas a partir da estirpe não marcada JD47-13C. A primeira grande
observação provém do resultado obtido com esta estirpe em que se verifica a
existência de dois conjuntos de sinais correspondentes a ligação não específica
por parte do conjunto de anticorpos 16B12/AM (assinalados com * na figura 22
B) uma vez que esta estirpe não possui qualquer marcação para ser detectada
por estes anticorpos. Comprova-se que estas bandas são ligação não específica
pela observação do resultado obtido, com esta estirpe, recorrendo à utilização

Discussão 81
do conjunto de anticorpos 3F10/AR, em que não houve qualquer reacção
observável, nem mesmo após 70 minutos de exposição (figura 22 B).
Nos resultados obtidos por immunoblot, a partir da separação
electroforética em condições nativas das amostras colhidas por fraccionamento
em coluna de Superose 6 de extractos proteicos de células JD127, observou-se
o conjunto proteassoma 26S/20S, determinado pela baixa mobilidade dos
complexos e pela massa molecular correspondente às fracções em que são
identificados (figura 22 B). É também visível o complexo 15S (sinal visível da
fracção 20 à 26), cujo sinal é proveniente não só do factor Ump1p marcado mas
também da subunidade β4/Pre1p, também marcada nesta estirpe e que, como
descrito por Ramos et al. (1998), está presente neste complexo. A partir da
fracção 28 é visível um arrastamento que deverá corresponder à migração da
subunidade β4/Pre1p na forma livre.
Estes resultados estão de acordo com os obtidos após separação das
fracções em gel desnaturante (figura 22 A), apesar de nos géis nativos, o
intervalo de sinal não ser tão evidente como no desnaturante.
A análise da subunidade β4/Pre1p na estirpe JD125 e do Ump1p na
estirpe JD129, recorrendo ao uso de géis nativos (figura 23), está também de
acordo com o observado com os géis desnaturantes (figura 21 B). Apesar do
blot obtido para JD125 apresentar uma intensidade de sinal fraca, possivelmente
devido a qualquer anormalidade no decorrer do procedimento, é visível a
presença da subunidade β4/Pre1p em complexos correspondentes ao
proteassoma 26S/20S e na sua forma livre nas fracções 28 a 30. Ao contrário do
esperado, não é detectada a presença desta subunidade no complexo 15S. No
entanto, isto poderá dever-se à degradação do complexo ou à possível
irregularidade na detecção, uma vez que esta subunidade foi detectada neste
complexo com o uso de um gel de acrilamida nativo a 6% (resultado não
apresentado). O Ump1p é detectado nas fracções 21 a 24 (figura 23) como
esperado por comparação com o gel desnaturante (figura 21 B).

Discussão 82
4.3.3 – A Delecção da Subunidade α3/Y13 Provoca Alterações na
Distribuição da Subunidade α7/Prs1p
Apesar de a subunidade α7/Prs1p ser detectada na estirpe selvagem
(JD183) e na estirpe mutante com delecção da subunidade α3/Y13 (AM1),
existem grandes diferenças no padrão de distribuição desta subunidade na
montagem do proteassoma 20S.
Na estirpe JD183 (figura 23), observaram-se claramente os complexos de
massa molecular elevada correspondentes ao proteassoma 26S/20S
demonstrando que esta subunidade está presente no proteassoma montado,
como descrito por Groll et al. (1997). A presença desta subunidade no complexo
15S, nesta estirpe, é também evidente. No entanto, o aparecimento de um
conjunto de sinais extremamente semelhantes aos revelados pelo complexo
15S, numa posição imediatamente acima deste, apesar de com uma intensidade
de sinal mais baixa, poderá significar a existência de alterações estruturais ou a
existência de uma modificação num dos constituintes do complexo que altere a
sua razão carga/massa e consequentemente a mobilidade deste complexo.
Como esperado, observou-se a presença da subunidade na forma livre nas
últimas frações analisadas. No entanto, inesperadamente, este sinal é também
visível em fracções correspondentes à separação de massa molecular mais
elevada, e é observado desde a fracção 19 à 30. Como o extracto foi separado
numa coluna de exclusão molecular e, o que se observa, é o sinal
correspondente à forma livre, admite-se que tenha havido dissociação desta
subunidade a partir de complexos precursores na formação do proteassoma,
uma vez que pensamos que esta é uma das subunidades iniciais na formação
de complexos, como acima descrito. A dissociação pode ter sido causada pelo
tratamento sofrido com a electroforese, após a colheita das fracções, uma vez
que, provavelmente, estes complexos terão um tempo de semi-vida bastante
curto e serão extremamente instáveis. As bandas visíveis, se bem que com fraca
intensidade, nas fracções 27, 28 e 29 poderão ter origem na detecção desses
complexos precursores (figura 23).

Discussão 83
Na estirpe AM1 é também evidente a presença da subunidade α7/Prs1p
no proteassoma 26S/20S. A presença desta subunidade no complexo 15S,
perfeitamente visível na estirpe selvagem, é quase imperceptível na estirpe
mutante (figura 23). Numa das hipóteses colocadas para explicar este facto
pensou-se que, devido à falta da subunidade α3/Y13, os complexos mais leves
que o complexo 15S poderiam sofrer alterações estruturais que impedissem a
formação deste. No entanto, a acontecer, certamente levaria a uma acumulação
de complexos malformados detectável por um aumento da intensidade das
bandas correspondentes às fracções anteriores ao complexo 15S no gel
desnaturante (figura 21 B), o que não se verificou. Nesta estirpe, à semelhança
do que se observa para a JD183, a forma livre está presente da fracção 19 à 30
(figura 23). Assim como para a estirpe mutante, isto poderá ser consequência da
dissociação desta subunidade de complexos precursores da biogénese do
proteassoma 20S. Na estirpe JD183 são igualmente visíveis bandas nas
fracções 27, 28 e 29 que poderão indicar a presença destes complexos (figura
23).
4.4 – Purificação de Complexo contendo α7Prs1pFLAG-6His
Posteriormente à purificação de complexos precursores da montagem do
proteassoma 20S, poder-se-iam utilizar técnicas avançadas, como por exemplo
a electroforese bi-dimensional para os caracterizar.
Os polipéptidos contidos nas amostras resultantes da cromatografia de
metalo-afinidade foram comparados após separação por electroforese
desnaturante, coloração com prata e immunoblot.
Como se pode observar na figura 24 (immunoblot), a cromatografia de
metalo-afinidade permite purificar especificamente material contendo a
subunidade de interesse, no entanto, as condições utilizadas têm de ser
melhoradas, uma vez que, na coloração por prata, são visíveis contaminantes
nas fracções eluídas com imidazole. A quantidade de resina utilizada foi
insuficiente já que, pela observação da amostra FT (material não ligado),

Discussão 84
verifica-se que é perdido material de interesse, o que pode indicar a saturação
da resina.
A presença de material de interesse ao longo do gradiente de imidazole
pode ser explicado pela razão tamanho/posições de ligação. Isto é, neste tipo de
cromatografia existe uma baixa de eficácia com o aumento do tamanho da
molécula a purificar. No entanto, para complexos proteassomais com tamanhos
elevados, como seja o proteassoma 20S, passam a existir dois locais de ligação
da molécula à resina (uma subunidade marcada com 6His por cada meio
proteassoma) o que torna a ligação mais estável.
As fracções de eluição e1, e2 e e3, foram separadas numa coluna de
filtração em gel (resultados não apresentados) dando origem a resultados não
conclusivos, que podem ser indicadores da necessidade de afinamento de
condições da cromatografia, nomeadamente ao nível do aumento da quantidade
de material utilizado.

Conclusões 85
5 – Conclusões No decorrer deste trabalho, foi conseguido um conjunto de resultados que
permitiu o desenvolvimento das conclusões abaixo descritas.
A nível do efeito dos indutores de stress aplicados em levedura, como o
cádmio e choque térmico, conclui-se que o primeiro influencia o
desenvolvimento normal de levedura de uma forma negativa. No entanto, apesar
do proteassoma ser um dos principais componentes da via de degradação
proteolítica dependente de ubiquitina/proteassoma, e desta via ser a principal
responsável pela eliminação de proteínas malformadas e não funcionais
originadas por indutores de stress como os testados, pelos resultados obtidos
conclui-se que a exposição a cádmio ou a choque térmico não influencia o nível
de expressão do proteassoma nem a actividade quimotríptica do mesmo. Na
estirpe mutante AM1, o crescimento em condições de choque térmico é inibido
em relação à estirpe selvagem.
A subunidade α3/Y13 é essencial para a existência de uma correcta
actividade quimotríptica pelas diferenças observadas entre esta actividade na
estirpe mutante e na estirpe selvagem.
A observação da distribuição das várias subunidades estudadas em
complexos separados por exclusão molecular permite concluir que a ordem de
inserção das subunidades durante da montagem do proteassoma é diferente
para cada uma delas. A subunidade β4/Pre1p é incorporada em complexos com
cerca de 400 kDa contendo a α7/Prs1p, enquanto que esta é incorporada em
complexos com inferior massa molecular.
A delecção da subunidade α3/Y13 provoca alterações ao nível da
distribuição normal da subunidade α7/Prs1p.

Perspectiva Futura 86
6 – Perspectiva Futura No decorrer deste trabalho, e pela observação dos resultados
experimentais, foram encontradas respostas a algumas questões. No entanto,
muitas outras ficaram por responder ou foram levantadas. De futuro, é nossa
intenção tentar responder a algumas dessas questões, nomeadamente no
campo dos passos iniciais da montagem do proteassoma 20S.
Um dos nossos objectivos futuros é conseguir caracterizar complexos que
estejam envolvidos nos passos iniciais de montagem do proteassoma. Uma das
aproximações será a purificação desses complexos recorrendo à utilização de
cromatografias de metalo-afinidade, já iniciadas no presente trabalho,
cromatografias de imuno-afinidade e imuno-precipitação, e posterior
caracterização por electroforese bi-dimensional.
Seria também interessante aprofundar um pouco mais o conhecimento
sobre o efeito da delecção da subunidade α3/Y13 na estrutura e actividade do
proteassoma 20S.

Bibliografia 87
7 – Bibliografia
• AGATEP R, KIRKPARICK R, PARCHALIUK D, WOODS R, GIETZ R (1998)
Transformation of Saccharomyces cerevisiae by the Lithium Acetate / Single-
Stranded Carrier DNA / Polyethylene Glycol (LiAc/ss-DNA/PEG) Protocol.
Technical Tips Online (http://tto.trends.com)
• ACHSTETTER T, EHMANN C, OSAKI A (1984) Proteolysis in Eukaryotic
Cells. Proteinase yscE, a new Yeast Peptidase. J Biol Chem 259, 13344
• ARENDT C, HOCHSTRASSER M (1997) Identification of the Yeast 20S
Proteasome Catalytic Centers and Subunit Interactions Required for Active
Site Formation. Proc Natl Acad Sci USA 94, 7156
• ARENDT C, HOCHSTRASSER M (1999) Eukaryotic 20S Proteasome
Catalytic Sununit Propeptides Prevent Active Site Inactivation by N-terminal
Acetylation and Promote Particle Assembly. EMBO J 18, 3575
• ARRIGO A, TANAKA K, GOLDBERG A (1988) Identity of the 19S Prosome
Particle with the large Multifunctional Protease Complex of Mammalian Cells
(The Proteasome). Nature 331, 192
• BAUMEISTER W, DAHLMANN B, HEGERL R (1988) Electron Microscopy
and Imaging Analysis of the Multicatalytic Proteinase. FEBS Lett 241, 239
• BLUM H, BEIER H, GROSS A (1987) Improved Silver Staining of Plant
Proteins, RNA and DNA in Polyacrylamide Gels. Electrophoresis 8, 93
• BOEHRINGER MANNHEIM BM Chemiluminescence Blotting Substrate
(POD)
• BOHLEY P, SEGLEN P (1992) Proteases and Proteolysis in the Lysossome.
Experientia 48, 151
• BRADFORD M M (1976) A rapid and sensitive method for the quantitation of
microgram quantities of protein utilizing the principle of protein-dye binding.
Anal Biochem 72,248

Bibliografia 88
• BRANNIGAN J, DODSON G, DUGGLEBY H (1995) A Protein Catalytic
Framework with an N-terminal Nucleophile is Capable of Self-activation.
Nature 378, 416
• CHEN P, HOCHSTRASSER M (1996) Autocatalytic Subunit Processing
Couples Active Site Formation in the 20S Proteasome to Completion of
Assembly. Cell 86, 961
• CHRISTY KG JR, LATART DB, OSTERHOUDT HW (1989) Modifications for
SDS-PAGE of proteins. Biotechniques 7, 692
• CHU-PING M, SLAUGHTER C, DeMARTINO G (1992) Identification,
Purification and Characterization of a Protein Activator (PA28) of the 20S
Proteasome (Macropain). J Biol Chem 267, 10515
• CLONTECH (1999). TALON® Metal Affinity Resins – User Manual
• COHEN SN, CHANG AC, HSU L (1972) Nonchromosomal antibiotic
resistance in bacteria: genetic transformation of Escherichia coli by R-factor
DNA. Proc Natl Acad Sci USA 69, 2110
• DAHLMANN B, KUEHN L, REINAUER H (1983) Identification of three high
molecular mass cysteine proteinases from rat skeletal muscle. FEBS Lett
160, 243
• DAHLMANN B, KOPP F, KUEHN L (1989) The Multicatalytic Proteinase
(Prosome) is Ubiquitous from Eukaryotes to Archeobacteria. FEBS Lett 251,
125
• DJABALLAH H, RIVETT A (1992) Peptidylglutamylpeptide Hydrolase Activity
of the Multicatalytic Proteinase Complex: Evidence for a New High-affinity
Site, Analysis of Cooperative Kinetics, and the Effect of Manganese Ions.
Biochemistry 31, 4133
• DISCOLL J, GOLDBERG A (1990) The Proteasome (Multicatalytic Protease)
is a Component of the 1500 kDa Proteolytic Complex which Degrades
Ubiquitin-Conjugated Proteins. J Biol Chem 265, 4789
• DITZEL L, HUBER R, MANN K (1998) Conformational Constrains for Protein
Self-cleavage in the Proteasome. J Mol Biol 279, 1187

Bibliografia 89
• DOHMEN J, STRASSER A, HONER C, HOLLENBERG C (1991) An efficient
transformation procedure enabling long-term storage of competent cells of
various yeast genera. Yeast 7, 691
• DOHMEN RJ, STAPPEN R, MCGRATH JP, FORROVA H, KOLAROV J,
GOFFEAU A, VARSHAVSKY A (1995) An essential yeast gene encoding a
homolog of ubiquitin-activating enzyme. J Biol Chem 270, 18099
• EDMUNDS P, PENNINGTON R (1982) A high molecular weight peptide
hydrolase in erythrocytes. Int J Biochem 14, 701
• EYTAN E, GANOTH D, ARMON T (1989) ATP Dependent Incorporation of
20S Protease into the 26S Complex that Degrades Proteins Conjugated to
Ubiquitin. Proc Natl Acad Sci USA 86, 7751
• FINLEY D, ÖZKAYNAK E, VARSHAVSKY A (1987) The yeast polyubiquitin
gene is essential for resistance to high temperatures, starvation, and other
stresses. Cell 48, 1035
• FRENTZEL S, PESOLD-HURT B, SEELIG A (1994) 20S Proteasomes are
Assembled Via Distinct Precursor Complexes. Processing of LMP2 and LMP7
Proproteins Takes Place in 13-16S Preproteasome Complexes. J Mol Biol
236, 975
• GERARDS W, ENZLIN J, HANER M (1997) The Human alpha-type
Proteasomal Subunit HsC8 Forms a Double Ring-like Structure but does not
Assemble Into Proteasome-like Particles with the beta-type Subunits HsDelta
or HsBPROS26. J Biol Chem 272, 10080
• GERARDS W, de JONG W, BLOEMENDAL H (1998) The Human
Proteasomal Subunit HsC8 Induces Ring Formation of Other alpha-type
Subunits. J Mol Biol 275, 113
• GERARDS W, de JONG W, BOELENS W, BLOEMENDAL H (1998)
Structure and Assembly of the 20S Proteasome. Cell Mol Life Sci 54, 253
• GIETZ R, WOODS R (1994). Molecular Genetics of Yeast: Practical
Approaches. Oxford University Press. U.S.A.
• GLICK D (1970) Methods of Biochemical Analysis, Vol. 18. Wiley-
Interscience. U.S.A.

Bibliografia 90
• GROLL M, DIETZEL L, LÖWE J (1997) Structure of the 20S Proteasome
from Yeast at 2.4 Å Resolution. Nature 386, 463
• GRZIWA A, BAUMEISTER W, DAHLMANN B (1991) Localization of Subunits
in Proteasomes from Thermoplasma acidophilum by Immunoelectron
Microscopy. FEBS Lett 290, 186
• HASE J, KOBASHI K, NAKAI N (1980) The Quaternary Structure of Carp
Muscle Alkaline Protease. Biochim Biophys Acta 661, 205
• HEGERL R, PFEIFER G, PÜHLER G (1991) The Three-dimensional
Structure of Proteasomes from Thermoplasma acidophiulum as Determined
by Electron Microscopy Using Random Conical Tilting. FEBS Lett 283, 117
• HEINEMANN J, SPRAGUE G (1991) Transmission of Plasmid DNA to Yeast
by Conjugation with Bacteria. Guide to Yeast Genetics and Molecular Biology
– Methods in Enzymology 194, 187
• HEINEMEYER W, FISCHER M, KRIMMER T (1997) The Active Sites of the
Eukariotic 20S Proteasome and Their Involvment in Subunit Precursor
Processing. J Biol Chem 272, 25200
• HEINEMEYER W, KLEINSCHMIDT J, SAIDOWSKY J (1991) Proteinase
yscE, the yeast Proteassome/Multicatalytic-Multifunctional Proteinase:
Mutants Unravel its Funtion in Stress Induced Proteolysis and Uncover its
Necessity for Cell Survival. EMBO J 10, 555
• HERSHKO A, CIECHANOVER A (1998) The Ubiquitin System. Ann Rev
Biochem 67, 425
• HJERTÉN S (1962) Archives Biochemical and Biophysical Supplements
1,147
• HILT W, ENENKEL C, GRUHLER A (1993) The PRE4 Gene Codes for a
Subunit of the Yeast Proteassome Necessary for Peptidylglutamylpeptide-
hydrolysing Activity. Mutations Link the Proteasome to Stress- and Ubiquitin-
dependent Proteolysis. J Biol Chem 268, 3479
• HILT W, WOLF D (2000) Proteasomes: The World of Regulatory Proteolysis.
Landes Biosciences / Eurekah.com. U.S.A.

Bibliografia 91
• HOCHSTRASSER M (1996) Ubiquitin-dependent Protein Degradation. Ann
Rev Gen 30, 405
• HOUGH R, PRATT G, RECHSTEINER M (1986) Ubiquitin-Lysosyme
Conjugates. Identification and Characterization of an ATP-dependent
Protease from Rabbit Reticulocyte Lysates. Jl Biol Chem 261, 2400
• HOUGH R, PRATT G, RECHSTEINER M (1987) Purification of Two High
Molecular Weight Proteases from Rabbit Reticulocyte Lysate. J Biol Chem
262, 8303
• JÄGER S, GROLL M, HUBER R (1999) Proteasome β-type Subunits:
Unequal Roles of Propeptides in Core Particle Maturationand a Hierarchy of
Active Site Funtion. J Mol Biol 297, 997
• JAP B, PÜHLER H, LUCKE H (1993) Preliminary X-ray Crystallographic
Study of the Proteasome from Thermoplasma acidophilum. J Mol Biol 234,
881
• JASON J, RYDÉN L (1998) Protein Purification – Principles, High Resolution
Methods, and Applications. 2nd Ed.. John Wiley and Sons Inc.. U.S.A.
• JENTSCH S, SEUFERT W, SOMMER T (1990) Ubiquitin-conjugating
Enzymes: Novel Regulators of Eukaryotic Cells. Trends Biochem Sci 15, 195
• JENTSCH S (1992) The Ubiquitin-conjugation System. Ann Rev Gen 26, 179
• JUNGMANN J, REINS H, SCHOBERT C, JENTSCH S (1993) Resistance to
Cadmium Mediated by Ubiquitin-dependent Proteolysis. Nature 362, 369
• KOPP F, HENDIL KB, DAHLMANN B, KRISTENSEN P, SOBEK A,
UERKVITZ W (1997) Subunit arrangement in the human 20S Proteasome.
Proc Natl Acad Sci USA 94, 2939
• LÖWE J, STOCK D, JAP B (1995) Crystal Structure of the 20S Proteasome
from the archaeon T. acidophilum at 3.4 Å Resolution. Science 268, 533
• MANIATIS T, FRITSCH E, SAMBROOK J Molecular Cloning, 1989. 2nd Ed..
U.S.A.
• MAYR J, SEEMÜLLER E, MÜLLER S (1998) Late Events in the Assembly of
the 20S Proteasomes. J Struct Biol 124, 179

Bibliografia 92
• McGRATH J, JENTSCH S, VARSHAVSKY A (1991) UBA1: An essential
yeast gene encoding ubiquitin-activating enzyme. EMBO J 10, 227
• NANDI D, WOODWARD E, GINSBURG D (1997) Intermediates in the
Formation of Mouse 20S Proteasomes: Implications for the Assembly of
Precursor β Subunits. EMBO J 17, 5363
• NEUHOFF V, AROLD N, TAUBE D, EHRHARDT W (1988); Improved
staining of proteins in polyacrylamide gels including isoelectric focusing gels
with clear background at nanogram sensitivity using Coomassie Brilliant Blue
G-250 and R-250. Electrophoresis 9, 225
• NIEDERMANN G, GRIMM R, GEIER E, MAURER M, REALINI C,
GARTMANN C, SOLL J, OMURA S, RECHSTEINER MC, BAUMEISTER W,
EICHMANN K. (1997) Potential immunocompetence of proteolytic fragments
produced by proteasomes before evolution of the vertebrate immune system.
J Exp Med 186, 209
• ORLOWSKI M, WILK S (1981) A Multicalatytic Protease Complex from
Pituitary that forms Enkephalin and Enkephalin Containing Peptides. Biochim
Biophys Res Comm 101, 814
• ÖZKAYNAK E, FINLEY D, SOLOMON M, VARSHAVSKY A (1987) The yeast
ubiquitin genes: a family of natural gene fusions. EMBO J 6, 1429
• PETERS J, CEJKA Z, HARRIS J (1993) structural features of the 26S
Proteasome Complex. J Mol Biol 234, 932
• PETERS J, HARRIS J, FINLEY D (1998) Ubiquitin and the Biology of the
Cell. Plenum Press. U.K & U.S.A.
• PHARMACIA BIOTECH (1996) Automated Silver and Coomassie Staining of
Polyacrilamide gels. Protocol Guide Hoefer ® Automated Gel Stainer
• PICKART C (1997) Targeting of Substrates to the 26S Proteasome. FASEB J
11, 1055
• PLUMMER D T (1987) An Introduction to Practical Biochemistry. 3rd Ed..
McGraw-Hill International Limited. U.K.
• PORATH J (1985) Immobilized Metal Ion Affinity Chromatography – A
Powerful Method for Protein Purification. Walter de Gruyter & Co.. U.S.A.

Bibliografia 93
• QIAGEN (1998) QIAprep Miniprep Handbook
• RAMOS P, HÖCKENDORFF J, JOHNSON ES, VARSHAVSKY A, DOHMEN
J (1998) Ump1p is Required for Proper Maturation of the 20S Proteasome
and Becomes its Substrate Upon Completion of the Assembly. Cell 92, 489
• ROBYT J F, WHITE B J (1990) Biochemical Techniques – Theory and
Practice. 2nd Ed.. Waveland Press Inc.. U.S.A.
• SCHMIDT M, KLOETZEL P (1997) Biogenesis of eukaryotic 20S
proteasomes: the complex maturation pathway of a complex enzyme. FASEB
J 11, 1235
• SCHMDTKE G, SCHMIDT M, KLOETZEL P (1997) Maturation of Mammalian
20S Proteasome: Purification and Characterization of 13S and 16S
Proteasome Precursor Complexes. J Mol Biol 268, 95
• SEEGER M, FERRELL K, DUBIEL W (1997) The 26S Proteasome: A
Dynamic Structure. Mol Biol Rep 24, 83
• SEEMÜLLER E, LUPAS A, ZÜHL F (1995) The Proteasome from
Thermoplasma acidophilum is Neither a Cysteine nor a Serine Protease.
FEBS Lett 359, 173
• SEEMÜLLER E, LUPAS A, STOCK D (1995) Proteasome from
Thermoplasma acidophilum: A Threonine Protease. Science 268, 579
• SEUFERT W, JENTSCH S (1990) Ubiquitin-conjugating enzymes UBC4 and
UBC5 mediate selective degradation of short-lived and abnormal proteins.
EMBO J 9, 543
• SMITH S, KOEGL M, JENTSCH S (1996) Role of the Ubiquitin/Proteasome
System in Regulated Protein Degradation in Saccharomyces cerevisiae. Biol
Chem 337, 437
• STEIN R, MELANDRI F, DICK L (1996) Kinetic Characterization of the
Chymotryptic Activity of the 20S Proteasome. Biochemistry 35, 3899
• SWERDLOW PS, FINLEY D, VARSHAVSKY A (1986) Enhancement of
immunoblot sensitivity by heating of hydrated filters. Anal Biochem 156, 147
• SWITZER R, MERRIL C, SHIFRIN S (1979) Anal Biochem 98, 231

Bibliografia 94
• TANAKA K, TSURUMI C (1997) The 26S Proteasome: Subunits and
Functions. Mol Biol Rep 24, 3
• TANAMURA T, NAGY I, LUPAS A, LOTTSPEICH F, CEJKA Z (1995) Curr
Biol 5, 766
• VALLEE B, ULMER D (1972) Biochemical effects of mercury, cadmium, and
lead. Ann Rev Biochem 41, 91
• VESTERBERG O, HANSEN L, SJÖSTEN A (1977) Staining of proteins after
isoelectric focusing in gels by new procedures. Biochim. Biophys. Acta, 160
• VIJAY-KUMAR S, BUGG C, COOK W (1987) Structure of ubiquitin refined at
1.8 A resolution. J Mol Biol 194, 531
• VOET D, VOET J (1995) Biochemistry. 2nd Ed.. John Wiley and Sons Inc..
U.S.A.
• VOGES D, ZWICKL P, BAUMEITER W (1999) The 26S Proteasome: A
Molecular Machine Designed for Controlled Proteolysis. Annual Reviews
Biochemistry 68, 1015
• WENZEL T, ECKERSKORN C, LOTTSPEICH F (1994) Existence of a
Molecular Ruler in Proteasomes Suggested by Analysis of Degradation
Products. FEBS Lett 349, 205
• WILK S, ORLOWSKI M (1980) Cation-sensitive Neutral Endopeptidase:
Isolation and Specificity of the Bovine Pituitary Enzyme. J Neurochem 35,
1172
• WILK S, ORLOWSKI M (1983) Evidence that Pituitary Cation-sensitive
Neutral Endopeptidase is a Multicatalytic Complex. J Neurochem 40, 842
• WILSON K, WALKER J (2000) Principles and Techniques of Practical
Biochemistry. 5th Ed.. Cambridge University Press. U.K.
• WÓJCIK C, SCHROETER D, WILK S (1996) Ubiquitin-mediated Proteolysis
Centers in HeLa Cells: Indication from Studies of an Inhibitor of the
Chymotripsin-like Activity of the Proteasome. Eur J Cell Biol 71, 311
• YANG Y, FRÜH K, AHN K (1995) In vivo Assembly of the Proteasomal
Complexes, Implications for Antigen Processing. J Biol Chem 270, 27687

Bibliografia 95
• YANISCH C, VIERA J, MESSING J (1985) Improved M13 phage cloning
vectors and host strains: nucleotide sequences of the M13mp18 and pUC19
vectors. Gene 33, 103
• ZÜHL F, TAMURA T, DOLENC I, CEJKA Z, NAGY I (1997) Subunit topology
of the Rhodococcus proteasome. FEBS Lett 400, 83
• ZÜHL F, SEEMÜLLER E, GOLBIK R (1997) Dissecting the Assembly
Pathway of the 20S Proteasome. FEBS Lett 418, 189
• ZWICKL P, LOTTSPEICH F, BAUMEISTER W (1992) Expression of
Funtional Thermoplasma acidophilum Proteasomes in Escherichia coli. FEBS
Lett 312, 157
• ZWICKL P, GRZIWA A, PÜHLER G, DAHLMANN B, LOTTSPEICH F,
BAUMEISTER W (1992) Primary structure of the Thermoplasma proteasome
and its implications for the structure, function, and evolution of the
multicatalytic proteinase. Biochemistry 31, 964
• ZWICKL P, KLEINZ J, BAUMEISTER W (1994) Critical Elements in
Proteasome Assembly. Nat Struct Biol 1, 765

Anexos 96
8 – Anexos Em anexo encontra-se uma cópia do trabalho apresentado em forma de
poster no “27th Meeting of the Federation of European Biochemical Societies”, 30
de Junho a 5 de Julho, 2001; Lisboa, Portugal.
António MARQUES†, Jürgen DOHMEN‡, Paula RAMOS†
†ADQuímica, FCT, Universidade do Algarve, PORTUGAL‡Institut für Genetik, Universität zu Köln, DEUTSCHLAND
DISTRIBUTION OF THE α7/Prs1p SUBUNIT (HsC8 HOMOLOG) IN YEAST
PROTEASOMAL COMPLEXES
INTRODUCTIONMany short-lived proteins including various regulatory
proteins in the cell suffer selective ubiquitylation and destruction. Ubiquitin-mediated proteolysis (UMP) therefore serves an important role in a variety of basic cellular functions. In a very simplified view, the UMP system functions in two steps: first, the substrate proteins are modified by the covalent attachment of ubiquitin molecules, second, these polyubiquitylated proteins are recognized by the 26S proteasome and degraded inside its catalytic core, the 20S proteasome 1.
In its mature and active state, the eucaryotic 20S proteasome is characterized by the presence of 14 different subunits, seven of the α-type and seven of the β-type, organized in four heptameric stacked rings with the arrangement α7β7β7α7, as showing in its crystal structure 2 (Fig.1).
Fig. 1 – Crystal structure of the 20S proteasome. Side view (left) and top view (right).
In the past few years much information has been accumulated on many steps of protein degradation, nevertheless our knowledge on the 20S proteasome assembly process is still diffuse with fragments of information stemming from studies donein various species and under different conditions 3.
METHODS
Extracts from a Saccharomyces cerevisiaestrain expressing the α subunit α7/Prs1p tagged with the haepitope, and other with the βsubunit β4/Pre1 and Ump1 tagged with the same epitope, were separated by gel filtration on a Superose 6 column. The fractions were scanned for proteasomal activity (chymotryptic) and analysed by PAGE and SDS-PAGE. The polypeptides were transfered to a PVDF membrane and incubated with anti-ha antbody (Fig. 2)
Fig. 2 – Schematic resuming the methods used;
RESULTS and DISCUSSIONOur results showed that α7/Prs1p is present in all
fractions from the free subunit to the catalytic active 26S proteasome fractions. This observations suggested the presence of distinct complexes containing this subunit. By comparison, the analysis for the presence of β4/Pre1 subunit in proteasomal complexes showed a gap between the Ump1 complex (half proteasome) 4 and what appears, according to its molecular weight, to be the free subunit (Fig. 3).
These results suggest that α7/Prs1p could be one of the initiators or participate in very early events of the proteasomeassembly process, before the two Ump1 complexes assemble together, as shown in a model previously reported 4.
Analysis of the fractions from the α7/Prs1p-ha tagged strain on a native gel (Fig. 4) revealed the presence of this subunit in complexes lighter than the Ump1 complex but heavier than the free subunits, in agreement with the SDS-PAGE analysis.
19 20 21 22 23 24 25 26 27 28 29 30 31 32
26S/20S proteasome
Ump1 complex
**
Fig. 3 – Chymotryptic activity in the fractions obtained after gel filtration on a Superose 6 column and the corresponding SDS-PAGE and immunoblot patterns of Prs1p-ha and Pre1p-ha (blue and green subunits respectively on the proteasome structure) and Ump1p-ha.
Fig. 4 – PAGE analysis followed by anti-ha immunoblot of the fractions collected using the α7/Prs1-ha tagged strain; ** - α7/Prs1p containing complexes.
CHYMOTRYPTIC ACTIVITY
PAGE SDS-PAGE
IMMUNOBLOT (Anti-ha) 1. Hershko A, Ciechanover A; 1998; Annu. Rev. Biochem 67:425-479
2. Groll M, Ditzel L, Löwe J, Stock D, Bochtler M, Bartunik H, Huber R; 1997; Nature 386:463-471
3. Hilt W, Wolf G (ed.); 2001; “Proteasomes: The world of regulatory proteolysis”; Landes & Co.; USA
4. Ramos P, Höckendorff J, Johnson E, Varshavsky A, Dohmen J; 1998; Cell 92:489-499
This work was supported by a grant from Fundação para a Ciência e Tecnologia
0
200
400
600
800
1000
1200
1400
1600
12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30
Fluo
resc
ênci
a
Prs1p-ha
Pre1p-ha
Ump1p-ha
Fluorescence





























