Embed Size (px)
Citation preview

Expresión de las isoformas de la cadena pesada de la miosina en los músculos elevadores de la
mandíbula en Pan troglodytes
Neus Ciurana Maynegre


TESIS DOCTORAL
FACULTAD DE MEDICINA Y CIENCIAS DE LA SALUD
PROGRAMA DE DOCTORADO
MEDICINA
Expresión de las isoformas de la cadena pesada de la miosina en los músculos elevadores de la mandíbula en Pan troglodytes
Directores de tesis
Dr. Josep Mª Potau Ginés
Dra. Rosa Artells Prats
Neus Ciurana Maynegre
Barcelona, 2018


A Montse,


Agradecimientos
Recuerdo perfectamente el primer día en la Facultad de Medicina. Todos los
alumnos estábamos en el aula magna y el Decano nos comentó que allí no sólo se
formaban médicos, sino que también existía la opción de realizar un doctorado y
dedicarse a la investigación. En aquel momento pensé que yo nunca haría un doctorado.
Con la licenciatura y el MIR era suficiente. Me equivoqué. Me equivoqué como otras
tantas veces, pero por suerte siempre he tenido personas a mi lado que me han ayudado,
empezando por mi familia. Quiero dar las gracias a mis padres, a mi abuela, a mi
hermano y a mi pareja por su apoyo incondicional y por su ayuda, tanto en los buenos
como en los malos momentos.
Quiero agradecer a todos los compañeros y compañeras de la Unidad de
Anatomía y Embriología Humanas, Anna, Carme, Joan, Marc, Sandra y Ruth, por
haberme dedicado horas de su tiempo a enseñarme todo lo necesario para trabajar en un
laboratorio. Sin vuestra colaboración hubiera sido mucho más complicado aprender a
realizar RT-qPCR, geles de agarosa, geles de poliacrilamida, extracción de proteínas y
un largo etcétera de técnicas y procedimientos.
También agradecer al Dr. Navarro sus consejos y su ayuda, siempre que me he
encontrado en un callejón sin salida. Sin su orientación la realización de la tesis hubiera
sido más difícil.
Por último, un agradecimiento muy especial a mis directores de tesis, la Dra.
Artells y el Dr. Potau. En primer lugar, a la Dra. Artells por su apoyo y todos sus
comentarios que han ayudado a mejorar el trabajo realizado. En segundo lugar, al Dr.
Potau por su dedicación, paciencia y sinceridad entre muchas otras cosas. Han sido
muchos años con momentos compartidos, disecciones, charlas, comidas..., incluso

salidas en bicicleta. Gracias por tratarme como una más de tu familia. Sabes que sin ti
nunca hubiera hecho un doctorado.

Abreviaturas
AA Aminoácido
Ac Anticuerpo
ADP Adenosín difosfato
ADN Ácido desoxirribonucleico
ADNc Ácido desoxirribonucleico complementario
ARN Ácido ribonucleico
ARNm Ácido ribonucleico mensajero
ARNnp Ácido ribonucleico nuclear pequeño
ARNr Ácido ribonucleico ribosómico
ARNt Ácido ribonucleico de transferencia
ASTF Área de sección transversal fisiológica
ATP Adenosín trifosfato
BSA Bovine serum albumine
DTT Dithiothreitol
dNTP Deoxyribose nucleoside triphosphate
DE Desviación estándar
EDTA Ethylenediaminetetraacetic acid
kDa Kilodalton

MHC Myosin heavy chain
MLC Myosin light chain
RNasa Ribonucleasa
RT-qPCR Real time quantitative polymerase chain reaction
SDS-PAGE Sodium dodecyl sulfate polyacrylamide gel electrophoresis
TEMED Tetrametiletilendiamina

9
Índice
1. Introducción ............................................................................................................ 13
1.1. Filogenia de los primates ................................................................................. 13
1.2. Evolución del aparato masticador .................................................................... 18
1.3. Anatomía de los músculos masticadores ......................................................... 20
1.3.1. Morfología externa ................................................................................... 20
1.3.2. Morfología interna .................................................................................... 23
1.4. Características electromiográficas de los músculos masticadores ................... 28
1.5. Características moleculares de los músculos masticadores ............................. 31
1.5.1. Isoformas de la MHC ............................................................................... 31
1.5.2. Métodos de análisis .................................................................................. 36
1.5.3. Composición y distribución de los diferentes tipos de fibras ................... 40
1.6. Expresión genética ........................................................................................... 45
1.6.1. RT-qPCR .................................................................................................. 47
2. Hipótesis y objetivos ................................................................................................ 51
2.1. Hipótesis .......................................................................................................... 51
2.2. Objetivos .......................................................................................................... 52
3. Artículos ................................................................................................................... 57
3.1. Artículo primero: “Expression of MyHC isoforms mRNA transcripts in
different regions of the masseter and medial pterygoid muscles in chimpanzees” .... 57
3.2. Artículo segundo: “Expression of myosin heavy chain isoforms mRNA
transcripts in the temporalis muscle of common chimpanzees (Pan troglodytes) ...... 67
4. Discusión .................................................................................................................. 79
5. Conclusiones ............................................................................................................. 87
6. Anexos ...................................................................................................................... 91
6.1. Anexo 1: peso muscular y porcentajes de expresión del ARNm en Pan
troglodytes .................................................................................................................. 91

10
6.2. Anexo 2: protocolo de la RT-qPCR ................................................................. 92
6.2.1. Extracción del ARN mediante el protocolo TriPure de Roche ................ 92
6.2.2. Síntesis del ADNc .................................................................................... 93
6.2.3. RT-qPCR .................................................................................................. 94
6.3. Anexo 3: resultados no publicados .................................................................. 95
6.4. Anexo 4: objetivos de futuro ......................................................................... 100
6.4.1. Extracción y cuantificación de proteínas ................................................ 101
6.4.2. Electroforesis en gel de poliacrilamida .................................................. 102
7. Bibliografía............................................................................................................ 111

Introducción


13
1. Introducción
1.1. Filogenia de los primates
Los primates han evolucionado de un ancestro común que los estudios
moleculares sitúan en el período Cretácico, hace 85 millones de años (Perelman et al.,
2011). Posteriormente, durante el Eoceno temprano aparecieron los euprimates1, hace
unos 55 millones de años, y tuvo lugar su gran expansión por el mundo que dio lugar a
una gran variedad de especies (Martin, 1990; Ni et al., 2004; Stringer & Andrews,
2005). Actualmente hay unas 400 especies que conforman el orden de los primates
(Groves, 2005; Fleagle, 2013), que se divide en dos subórdenes (Strepsirrhini y
Haplorrhini) (Figura 1).
Figura 1. Árbol filogenético del orden de los primates.
Los Strepsirrhini incluyen a todos los prosimios, excepto a los társidos, y se
dividen en dos superfamilias, la Lemuroidea (lémures) y la Lorisoidea (loris y gálagos)
1 Primates con aspecto moderno

14
(Goodman et al., 1990) (Figura 2). Estos primates presentan un tamaño cerebral
pequeño en relación al tamaño corporal, un hocico puntiagudo y rinario, una
almohadilla de piel húmeda y desprovista de pelo situada alrededor de la nariz con una
función olfativa (Ankel-Simons, 2010). Hoy en día, los lémures se localizan en
Madagascar y constan de varios géneros, entre ellos el género Microcebus que es el
primate vivo más pequeño, con un peso de 30 a 60 gr (Campbell et al., 2007; Ankel-
Simons, 2010). Los loris viven en África y en el sudeste asiático, mientras que los
gálagos viven en África (Campbell et al., 2007; Swindler, 1998). En general, los
Strepsirrhini se alimentan de insectos, caracoles, frutas y, en algunos casos, de lagartos
y resina de los árboles (Campbell et al., 2007; Swindler, 1998).
Figura 2. a) Fotografía de un lémur de la especie Propithecus verreauxi. b) Fotografía de un loris del género Nycticebus (Imágenes realizadas por Michael Melford y Frans Lanting
publicadas en The Guardian).
Los Haplorrhini se subdividen en el infraorden Tarsiiformes (társidos) y el
infraorden Simiiformes (o Anthropoidea). En general, los társidos presentan las mismas
características que los otros prosimios (Martin, 1990), pero carecen de rinario (Figura
3). Estos primates se localizan en el sudeste asiático y son básicamente insectívoros
(Ankel-Simons, 2010; Swindler, 1998). El infraorden Anthropoidea se diferencia de los
prosimios por presentar un tamaño cerebral mayor, por tener un menor número de
a) b)

15
dientes y por la presencia de un tabique postorbital (Campbell et al., 2007). Los
primates Anthropoidea se clasifican en el parvorden Platyrrhini (New World monkeys)
y el parvorden Catarrhini, que se divide a su vez en dos superfamilias, la
Cercopithecoidea (Old World monkeys) y la Hominoidea (Perelman et al., 2011)
(Figura 1).
Figura 3. Fotografía de Tarsius tumpara (Imagen realizada por Geoff Deehan, publicada en The Guardian).
Los Platyrrhini se diferencian de los Catarrhini por presentar algunas
características primitivas, como tener tres premolares superiores e inferiores, pero sobre
todo por la forma de la nariz que es más ancha y plana (Ankel-Simons, 2010; Swindler,
1998) (Figura 4). Estos primates son relativamente pequeños, con un peso corporal de
100 gr (Cebuella) a 7-10 kg (Atelines), y viven en bosques de centroamérica,
sudamérica y México (Campbell et al., 2007; Swindler, 1998). En general, habitan en
los árboles y su dieta consta principalmente de frutas, insectos y raíces (Campbell et al.,
2007). La superfamilia Cercopithecoidea es la más abundante y diversa, incluyendo a
más de 100 especies de primates que viven en bosques tropicales y en las sabanas
africana y asiática, tanto en el hábitat terrestre como en el arbóreo (Lewin, 2000;
Campbell et al., 2007). Presentan una dieta rica en fruta, hojas y raíces y,

16
ocasionalmente, algunas especies también se alimentan de resina de los árboles e
insectos (Cachel, 1979).
Figura 4. Diferencias anatómicas de la región nasal entre un primate platirrino (a) y un primate catarrino (b) (Imagen adaptada de Primates, John P. Rafferty, 2011).
La superfamilia Hominoidea se diferencia de los otros primates, entre otras
cosas, por presentar un gran desarrollo encefálico, una región nasal ancha y una cola
muy reducida o ausente (Fleagle, 2013). Se divide en dos familias, la Hylobatidae y la
Hominidae (Stringer & Andrews, 2005). La familia Hylobatidae está formada por los
gibones que habitan únicamente en el medio arbóreo y presentan algunas características
ancestrales que comparten con los cercopitécidos, como por ejemplo, los caninos en
forma de daga (Stringer & Andrews, 2005; Campbell et al., 2007). Los gibones son los
Hominoidea más pequeños, tienen un peso corporal de 5 a 15 kg y viven en los bosques
tropicales del sudeste asiático (Fleagle, 2013). Estos simios son frugívoros, pero algunas
especies también se alimentan de raíces y de invertebrados como las termitas (Swindler,
1998; Fleagle, 2013) (Figura 5). La familia Hominidae comprende a los grandes simios
(“great apes”) que son principalmente arbóreos, pero que habitan en el hábito terrestre
en diferentes grados. Incluyen los orangutanes (género Pongo), los gorilas (género
Gorilla), los chimpancés (género Pan) y los humanos (género Homo) (Figura 6).
a) b)

17
Figura 5. Imagen de un gibón de la especie Hoolock tianxing (Imagen realizada por Fan Pengfei y publicada en The Guardian).
Figura 6. Imágenes de un orangután (a), de un gorila (b) y de un chimpancé común (c) (Imágenes realizadas por Ian Sample y Alexandra Rosati y publicadas en The Guardian).
Los orangutanes están representados actualmente por dos especies que viven en
las islas Borneo (Pongo pygmaeus) y Sumatra (Pongo abelii) y su peso varía entre 39
kg en las hembras y hasta 80 kg en los machos (Campbell et al., 2007; Swindler, 1998).
El principal componente de la dieta de los orangutanes es la fruta madura, pero también
pueden comer raíces y, en algunos casos, termitas (Taylor, 2006). Los gorilas también
están representados por dos especies (Gorilla gorilla y Gorilla beringei) y se localizan
en África central, presentando como los orangutanes, un marcado dimorfismo sexual. El
peso corporal máximo de las hembras es de 90 kg y el de los machos de hasta 200 kg
a) b) c)

18
(Fleagle, 2013). Los gorilas son principalmente vegetarianos y su dieta consta de una
elevada proporción de hojas, pudiendo añadir también hormigas y termitas (Campbell et
al., 2007). Los últimos grandes simios no humanos son los chimpancés que viven en
África occidental y se alimentan de grandes cantidades de fruta, pequeños invertebrados
y, ocasionalmente, raíces y semillas (Martin, 1990; Campbell et al., 2007; Swindler,
1998). Están representados por dos especies, los chimpancés comunes (Pan troglodytes)
y los bonobos (Pan paniscus) y se consideran los parientes más próximos a los
humanos, en la escala filética. Se calcula que esta divergencia tuvo lugar hace entre 5 y
7 millones de años (Schrago & Russo, 2003; Patterson et al., 2006).
1.2. Evolución del aparato masticador
A lo largo de la evolución de los primates se han sucedido modificaciones
morfológicas en el aparato masticador que se relacionan con el tipo de dieta, estilo de
vida y hábitos alimentarios (Cachel, 1979; Hoh, 2002; Taylor & Vinyard, 2004; Stringer
& Andrews, 2005; Lieberman, 2011). Así, los Strepsirrhini y los társidos presentan una
mandíbula primitiva, con forma de V y con falta de fusión de sus dos mitades a nivel de
la línea media, lo que permite un cierto grado de movilidad durante la masticación
(Lieberman, 2011). Estos prosimios se caracterizan por presentar unos dientes
puntiagudos, con un incisivo medial inclinado hacia delante (excepto en los társidos),
que realizan una función prensil. Estas adaptaciones son idóneas para capturar y matar
insectos, pero también para excavar agujeros en la corteza de los árboles con el objetivo
de obtener resina (Martin, 1990; Stringer & Andrews, 2005; Rosenberger, 2010) (Figura
7).

19
Figura 7. Mandíbula de Galago senegalensis (prosimio).
Los primates antropoideos presentan una mandíbula con forma de U, excepto
algunos Platyrrhini, con sus dos mitades fusionadas a nivel de la línea media
(Lieberman, 2011). La fusión de la mandíbula se relaciona con la capacidad de resistir
una mayor fuerza transversal, generada durante la masticación debido a la acción del
músculo masetero profundo y permite una coordinación más efectiva de los músculos
masticadores (Hylander, 1979; Hylander et al., 2000; Vinyard et al., 2006). Otras
diferencias existentes entre antropoides y prosimios son los incisivos con forma de
espátula, adaptados para cortar el alimento, y la mayor área de superficie de los molares
para aumentar la eficacia del molido (Martin, 1990; Stringer & Andrews, 2005). En los
primates hominoideos, la principal diferencia respecto al aparato masticador se
encuentra entre los humanos y el resto de los grandes simios. Los grandes simios no
humanos ingieren alimentos mecánicamente resistentes que requieren una mayor fuerza
oclusiva y un mayor número de ciclos masticatorios. Por ejemplo, los chimpancés
comunes dedican aproximadamente la mitad del día a la masticación (Lieberman,
2011). En cambio, los humanos presentan un aparato masticador más grácil que genera
una menor fuerza oclusiva en comparación con la de los grandes simios. Estas
diferencias funcionales están relacionadas con las características anatómicas del aparato

20
masticador y con los tipos de fibras que presentan los músculos masticadores
(Lieberman, 2011) (Figura 8).
Figura 8. Mandíbula de un chimpanzé común (a) y de un humano (b).
1.3. Anatomía de los músculos masticadores
Los músculos que actúan sobre la mandíbula se subdividen en dos grupos, los
músculos cuya contracción provoca el descenso de la mandíbula (“jaw-open muscles”)
y los músculos cuya contracción eleva la mandíbula (“jaw-close musles”) (Van Eijden
et al., 1997). Los músculos elevadores de la mandíbula son el masetero, el pterigoideo
medial y el temporal. Estos músculos son los responsables de generar la fuerza oclusiva
y los movimientos de la mandíbula asociados a la masticación y a la mordedura (Taylor
& Vinyard, 2013).
1.3.1. Morfología externa
El masetero es un músculo multipeniforme que está constituido por dos
porciones, la superficial y la profunda (Williams & Warwick, 1980; Lieberman, 2011).
La porción superficial se origina en el arco zigomático y sus fibras se dirigen en sentido
posteroinferior para insertarse en el ángulo de la mandíbula. La porción profunda se
origina en el tercio posterior del arco zigomático y se inserta en la rama de la
a) b)

21
mandíbula, presentado sus fibras una orientación prácticamente vertical (Figura 9). En
los primates no humanos estas dos porciones se pueden separar fácilmente gracias a la
presencia de una lámina aponeurótica entre ellas (Aiello & Dean, 1990).
Figura 9. Imágenes de la porción superficial (a) y profunda (b) del músculo masetero en Pan troglodytes y Homo sapiens (Imágenes cedidas por JM Potau).
El músculo pterigoideo medial se origina en la fosa pterigoidea del esfenoides y
se inserta en la cara interna de la rama y del ángulo de la mandíbula (Williams &
Warwick, 1980) (Figura 10). La orientación de sus fibras es oblicua, similar a la
orientación de las fibras de la porción superficial del masetero y, al igual que éste, se
trata de un músculo multipeniforme (Aiello & Dean, 1990).
a)
b)

22
Figura 10. Imágenes del músculo pterigoideo medial en Pan troglodytes (a) y Homo sapiens (b)
(Imágenes cedidas por JM Potau).
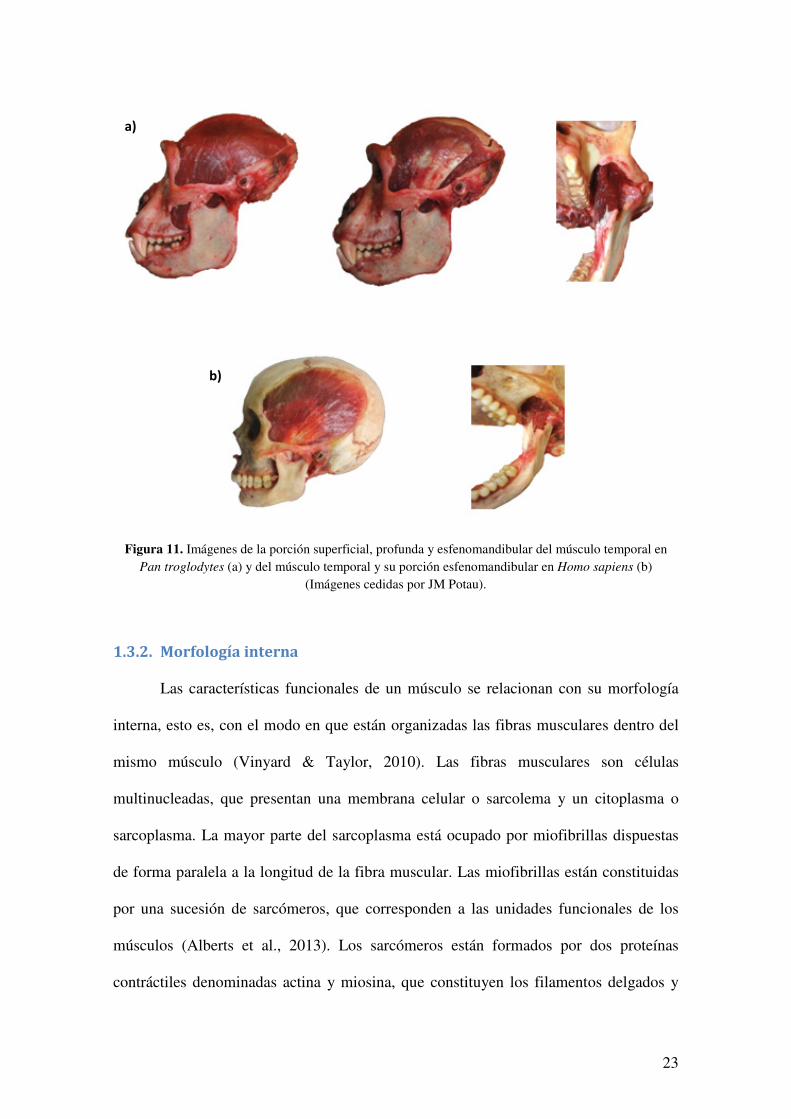
Finalmente, el temporal es un músculo bipeniforme que se origina en la fosa
temporal del cráneo y en la fascia temporal, insertándose en la apófisis coronoides de la
mandíbula y en el borde anterior de la rama mandibular (Aiello & Dean, 1990) (Figura
11). En Pan troglodytes y en los primates no humanos, el músculo temporal está
dividido por un tendón intramuscular en una porción superficial y una porción profunda
(Ross, 1995; Oxnard & Franklin, 2008). En los humanos, la porción superficial es
prácticamente inexistente o está representada por una fina capa muscular con un
impacto funcional mínimo, mientras que la porción profunda constituye la porción
principal del músculo temporal (Oxnard & Franklin, 2008; Lee et al., 2012). La porción
más profunda del temporal ha sido diferenciada por Dunn et al. (1996) con el nombre de
músculo esfenomandibular, que se extiende desde la superficie infratemporal del ala
mayor del esfenoides a la cresta temporal de la mandíbula. Sin embargo, estudios
posteriores consideran el músculo esfenomandibular como una parte del músculo
temporal y no como un músculo independiente, debido a la falta de una inervación
propia y a la ausencia de tejido fascial que delimite los dos vientres musculares (Türp et
al., 1997; Akita et al., 2000; Schön-Ybarra & Bauer, 2001; Skinner & Aziz, 2003; Geers
et al., 2005).
a) b)
23
Figura 11. Imágenes de la porción superficial, profunda y esfenomandibular del músculo temporal en Pan troglodytes (a) y del músculo temporal y su porción esfenomandibular en Homo sapiens (b)
(Imágenes cedidas por JM Potau).
1.3.2. Morfología interna
Las características funcionales de un músculo se relacionan con su morfología
interna, esto es, con el modo en que están organizadas las fibras musculares dentro del
mismo músculo (Vinyard & Taylor, 2010). Las fibras musculares son células
multinucleadas, que presentan una membrana celular o sarcolema y un citoplasma o
sarcoplasma. La mayor parte del sarcoplasma está ocupado por miofibrillas dispuestas
de forma paralela a la longitud de la fibra muscular. Las miofibrillas están constituidas
por una sucesión de sarcómeros, que corresponden a las unidades funcionales de los
músculos (Alberts et al., 2013). Los sarcómeros están formados por dos proteínas
contráctiles denominadas actina y miosina, que constituyen los filamentos delgados y
a)
b)

24
gruesos del sarcómero, respectivamente (Schiaffino & Reggiani, 2011). La miosina, a
su vez, está compuesta por cuatro cadenas ligeras y dos cadenas pesadas, presentando
estas últimas una cabeza que contiene la enzima ATPasa, encargada de proporcionar la
energía para la contracción muscular (Korfage et al., 2005a) (Figura 12).
Figura 12. Esquema que muestra el nivel de organización dentro de la fibra muscular, desde la miofibrilla hasta la miosina, que está compuesta por dos cadenas pesadas (MHC) y cuatro ligeras (MLC)
(Imagen adaptada de Korfage et al., 2005a).
La contracción muscular se debe al deslizamiento de los filamentos de actina
respecto a los filamentos de miosina y el grado de interacción entre estos dos filamentos
determina la tensión que un músculo puede generar. Según la teoría de los filamentos
deslizantes, a medida que el sarcómero disminuye su longitud aumenta el grado de

25
solapamiento entre la actina y la miosina hasta llegar al punto óptimo de interacción,
donde la fuerza muscular generada es máxima. A partir de este punto, si el sarcómero
continúa disminuyendo su longitud, también disminuye la tensión muscular generada
(Sciote & Morris, 2000) (Figura 13).
Figura 13. Relación entre la tensión y la longitud del sarcómero (Imagen adaptada de Sciote & Morris, 2000).
La suma de las tensiones generadas por todos los sarcómeros resulta en la fuerza
generada por todo el músculo. Dependiendo de la organización de las fibras musculares
dentro del músculo, éste es propicio para la generación de una elevada fuerza muscular
o para la generación de una rápida velocidad de contracción (Van Eijden et al., 1997).
Así, los músculos con fibras oblicuas (unipeniformes, bipeniformes y multipeniformes)
son óptimos para la generación de una fuerza muscular elevada. En estos músculos, las
fibras se sitúan formando un ángulo en relación al eje de acción del músculo (ángulo de
penación). Esta disposición permite agrupar a un mayor número de fibras por unidad de
área de sección y, por lo tanto, agrupa a un mayor número de sarcómeros en paralelo,
que ejercen una fuerza muscular mayor (Van Eijden et al., 1997). El área de sección
transerval fisiológica (ASTF) es un parámetro que se relaciona con el número de
Longitud sarcómero (µm)
Ten
sión
rel
ativ
a

26
sarcómeros en paralelo que presenta un músculo y, por lo tanto, se relaciona
directamente con la fuerza muscular que puede desarrollar dicho músculo (Van Eijden
et al., 1997). El ASTF se calcula mediante la seguiente fórmula:
Masa muscular (gr) x cos Ø
ASTF (cm2) =
Longitud fibra muscular x 1.0564 (gr/cm3)
Donde Ø es el ángulo de penación de las fibras musculares y 1.0564 (gm/cm3)
es la densidad del músculo esquelético (Méndez, 1960; Taylor & Vinyard, 2013). Con
esta fórmula se deduce que el ASTF está directamente relacionada con la masa
muscular e inversamente relacionada con la longitud de las fibras musculares (Powell et
al., 1984; Taylor & Vinyard, 2004). Por otro lado, los músculos con fibras largas y
paralelas a su eje longitudinal son óptimos para generar una elevada velocidad de
contracción. Estas fibras presentan un mayor número de sarcómeros en serie cuyo
acortamiento es sumatorio, es decir, la contracción total de la fibra muscular equivale a
la suma de acortamientos de sus sarcómeros (Van Eijden et al., 1997). Por lo tanto, la
longitud de la fibra muscular es proporcional a la velocidad de contracción ya que las
fibras más largas presentan un mayor número de sarcómeros en serie y pueden producir
contracciones más rápidas (Sciote & Morris, 2000; Grünheid et al., 2009). En resumen,
un músculo adaptado a una elevada velocidad de contracción presenta una menor
capacidad de generar tensión muscular y viceversa (Lieber & Friden, 2000).
En Homo sapiens se han descrito diferencias intra e intermusculares en los
elevadores de la mandíbula en relación a la longitud de sus fibras, área de sección
transerval fisiológica y ángulo de penación (Van Eijden et al., 1997). En primer lugar,
se han observado diferencias entre las dos porciones del músculo masetero. La porción

27
superficial presenta una mayor ASTF (6.82 cm2), longitud de fibras (24.6 mm) y ángulo
de penación (16.5º) respecto a la porción profunda, que presenta una ASTF de 3.49 cm2,
una longitud de fibras de 18.0 mm y un ángulo de pennación de 6.7º (Van Eijden et al.,
1997). En segundo lugar, la región anterior del músculo temporal tiene una ASTF de 7.7
cm2 y una longitud de fibras de 27.1 mm, mientras que la región posterior presenta una
ASTF de 5.55 cm2 y una longitud de fibras de fibras de 25.7 mm (Van Eijden et al.,
1997). Finalmente, a nivel intermuscular, el temporal es el que presenta una mayor
ASTF y longitud de fibras entre los músculos elevadores de la mandíbula, mientras que
el pterigoideo medial es el músculo con una menor ASTF (6.0 cm2) y longitud de fibras
(12.9 mm) (Van Eijden et al., 1997).
También se han observado diferencias morfológicas internas en los músculos
masetero y temporal entre Homo sapiens y Pan troglodytes que se relacionan con sus
diferentes requerimientos funcionales (Taylor & Vinyard, 2013). El masetero
superficial y el temporal de Homo sapiens presentan, respectivamente, una ASTF de
4.96 cm2 y 10.39 cm2 (Taylor & Vinyard, 2013), valores muy parecidos a los
observados por otros autores (Van Eijden et al., 1997). En cambio, en Pan troglodytes
el ASTF del masetero superficial es de 16.78 cm2 y en el temporal de 19.95 cm2. Es
decir, Homo sapiens presenta una menor ASTF de los músculos elevadores de la
mandíbula en comparación con Pan troglodytes, pero la longitud de las fibras
musculares en relación a la longitud de la mandíbula es muy similar o superior en los
humanos. Así, en Homo sapiens la relación de la longitud de las fibras musculares con
la longitud de la mandíbula es de 0.248 para el músculo masetero superficial y de 0.282
para el temporal, mientras que en Pan troglodytes esta relación es de 0.168 para el
masetero superficial y de 0.334 para el temporal (Taylor & Vinyard, 2013). Estos

28
resultados sugieren una disminución de la fuerza máxima a expensas de una mayor
velocidad de contracción en Homo sapiens en comparación con Pan troglodytes.
1.4. Características electromiográficas de los músculos masticadores
Una forma de valorar si las diferencias estructurales de los músculos se traducen en
diferencias funcionales es mediante la electromiografía. Esta técnica permite registrar la
señal bioeléctrica generada durante la contracción muscular, cuantificarla y compararla
entre diferentes músculos, individuos o especies mediante el uso de electrodos
intramusculares o de superficie (Grünheid et al., 2009). Esto nos permite determinar
cuánto tiempo ha estado activo un músculo, cuando ha llegado al pico máximo de
actividad y el orden en el que se han activado los diferentes músculos involucrados en
una misma acción o conducta, como la masticación (Vinyard & Taylor, 2010). La
masticación presenta dos fases, la apertura y la clausura (Lieberman, 2011). A su vez, la
fase de clausura se divide en dos partes: una primera parte rápida que finaliza cuando
los dientes contactan con la comida y una segunda parte lenta en la que tiene lugar la
oclusión dentaria (“power stroke”), momento en el que se produce la rotura de los
alimentos (Wall et al., 2006; Ross & Iriarte-Diaz, 2014).
Los estudios electromiográficos realizados en humanos y otras especies de primates
antropoideos muestran una gran variabilidad en el reclutamiento de las fibras de los
músculos elevadores de la mandíbula y de sus partes. Esta variabilidad depende del
movimiento realizado, del punto de oclusión dentaria (o "power stroke") y de la dieta
(Cachel, 1979; Hylander et al., 1987; Wood, 1987; Schindler et al., 2007; Vinyard et al.,
2008). En primer lugar, según el movimiento realizado se han descrito diferencias
funcionales entre las dos porciones del músculo masetero y entre la región anterior,

29
posterior y esfenomandibular del músculo temporal (Zenker, 1955; Wood, 1986;
Hylander et al., 1987; Wood, 1987; Hylander & Johnson, 1994; Blanksma et al., 1997;
Hylander et al., 2005; Fuentes et al., 2012). En relación al músculo masetero, la porción
superficial actúa principalmente en los desplazamientos verticales de la mandíbula y
presenta una actividad sinérgica con el pterigoideo medial, mientras que la porción
profunda actúa en los movimientos laterales y de retrusión de la mandíbula (Wood,
1987; Hylander & Johnson, 1994; Blanksma et al., 1997). En primates antropoideos
como Papio anubis, Macaca fuscata y Aotus trivirgatus se ha observado que la porción
profunda del masetero llega al pico máximo de actividad antes que la porción
superficial (Hylander et al., 1987; Hylander & Johnson, 1994; Hylander et al., 2005).
Este modelo de actividad electromiográfico se relaciona con un desplazamiento lateral
de la mandíbula que ayuda a posicionar los molares justo antes de la oclusión dentaria
(Hylander et al., 1987; Hylander & Johnson, 1994; Vinyard et al., 2008; Ross & Iriarte-
Diaz, 2014). En los humanos se han descrito diferencias funcionales entre las regiones
anterior y posterior del masetero superficial y profundo (Blanksma et al., 1992;
Guzmán-Venegas et al., 2015). Estos estudios muestran que la región anterior del
masetero superficial tiene un mayor reclutamiento que la región posterior en acciones
submaximales, pero no en acciones de fuerza máxima donde presentan el mismo nivel
de actividad (Guzmán-Venegas et al., 2015). Por otro lado, la región anterior del
masetero profundo presenta una mayor actividad en las mordidas en dirección
anteromedial, mientras que la región posterior del masetero profundo actúa en las
mordidas en dirección posterolateral (Blanksma et al., 1992). En relación al músculo
temporal, la región anterior presenta un mayor reclutamiento de sus fibras en los
movimientos verticales de la mandíbula, mientras que la región posterior actúa
principalmente en los movimientos de retrusión (Cachel, 1979; Wood, 1987; Blankma

30
et al., 1997). De la porción esfenomandibular sólo se han realizados estudios
electromiográficos en humanos, en los que se ha observado que participa en los
movimientos laterales de la mandíbula, presentando una actividad sinérgica con el
músculo pterigoideo lateral, y en el mantenimiento de la postura y estabilización de la
mandíbula (Zenker, 1955; Wood, 1986; Fuentes et al., 2012).
En segundo lugar, la variación en el punto de oclusión dentaria provoca un cambio
en el sistema de palancas del aparato masticador que se traduce en variaciones en el
patrón de reclutamiento de los músculos masticadores (Vinyard et al., 2008). Así, en
Homo sapiens se ha observado una menor actividad electromiográfica de los músculos
masetero y temporal cuando el punto de mordida es anterior a la posición de
intercuspidación máxima2, siendo la porción anterior del temporal la que presenta un
mayor reclutamiento de sus fibras en comparación a la región posterior. En cambio, si el
punto de mordida es posterior a la posición de intercuspidación máxima el
reclutamiento de fibras es mayor en la región posterior del temporal en relación a la
región anterior (Ahlgren, 1986; Blanksma & Van Eijden, 1995; Schindler et al., 2007).
Finalmente, en tercer lugar, según el tipo de dieta, los estudios electromiográficos
realizados en el músculo masetero de Papio anubis, Macaca fuscata, Aotus trivirgatus y
Lemur catta muestran una gran variabilidad en el reclutamiento de las fibras de la
porción superficial en comparación con la porción profunda (Hylander et al., 2000;
Vinyard et al., 2006). De hecho, la porción profunda presenta prácticamente la misma
actividad electromiográfica en alimentos duros y blandos, mientras que la porción
superficial disminuye la actividad electromiográfica en alimentos blandos y la aumenta
2 Posición oclusal de la mandíbula en la que las cúspides de los dientes de ambos arcos se interponen
completamente.

31
en alimentos duros (Hylander et al., 2000; Vinyard et al., 2006). En relación al músculo
temporal, Wall et al. (2008) describen en Papio anubis un mayor reclutamiento de las
fibras del temporal superficial anterior en comparación con las fibras del temporal
profundo anterior en alimentos mecánicamente resistentes. Este modelo de
reclutamiento apoya la hipótesis de una fuerza oclusiva elevada (“high occlusal force
hypothesis”), según la cual las partes superficiales de los músculos masticadores son las
encargadas de generar la elevada fuerza muscular necesaria para procesar los alimentos
más resistentes.
1.5. Características moleculares de los músculos masticadores
1.5.1. Isoformas de la MHC
Otra forma de valorar la función de un músculo es analizando la expresión de las
isoformas de la cadena pesada de la miosina (MHC). La MHC es una proteína con
actividad ATPasa que determina la velocidad y la fuerza de contracción de la fibra
muscular (Bottinelli et al., 2006). En los mamíferos, las diferentes isoformas de la MHC
están codificadas por un grupo de genes localizados en los cromosomas 14, 17 y 7
(Tabla 1) (Schiaffino & Reggiani, 1996; Weiss & Leinwand, 1996; Weiss et al., 1999).
En el cromosoma 14 se localizan los genes MYH6 y MYH7 que codifican para las
isoformas cardíacas, la MHC-α cardíaca y la MHC-ß, respectivamente. La isoforma
MHC-α cardíaca se expresa en los atrios del corazón, mientras que la isoforma MHC-ß
se expresa en los ventrículos. Esta última isoforma también se expresa en el músculo
esquelético, donde se denomina MHC-I (Weiss & Leinwand, 1996).

32
Tabla 1. Cromosomas y genes de la MHC y la isoforma para la que codifican.
CROMOSOMA GEN MHC
17 MYH1 IIX
17 MYH2 IIA
17 MYH3 Embriónica
17 MYH4 IIB
14 MYH6 α-cardíaca
14 MYH7 Beta/I
17 MYH8 Neonatal
17 MYH13 Extraocular
7 MYH16 Masticadora
En el cromosoma 17 se localizan seis genes que codifican las isoformas MHC-
neonatal o MHC-fetal, MHC-embriónica, MHC-IIA, MHC-IIX, MHC-IIB y MHC-
extraocular (Weiss & Leinwand, 1996). Las isoformas MHC-neonatal y MHC-
embriónica están codificadas por los genes MYH8 y MYH3, respectivamente, y se
expresan durante el desarrollo fetal. Después del nacimiento, son reemplazadas de
forma progresiva por las isoformas MHC-I, MHC-IIA, MHC-IIX y MHC-IIB
(Schiaffino et al., 2015). Las isoformas MHC-I, MHC-IIA, MHC-IIX y MHC-IIB son
las más abundantes en el músculo esquelético de los mamíferos adultos (Pette & Staron,
2000). Estas isoformas difieren en la actividad ATPasa y en la velocidad de liberación
del ADP durante el ciclo de contracción, características que determinan la fuerza y la
velocidad de contracción de la fibra muscular (Bottinelli et al., 1996; Schiaffino &
Reggiani, 2011). El ciclo de contracción, o de puentes cruzados, se inicia con la unión

33
del ATP a la cabeza de la miosina, que se separa de la actina. La cabeza de la miosina
tiene actividad ATPasa e hidroliza el ATP en ADP y Pi, provocando la formación de un
puente cruzado entre la actina y la miosina. A continuación, se libera el Pi y se
desencadena el golpe de potencia que produce el desplazamiento de la actina en relación
a la miosina y genera la fuerza de contracción. Finalmente, se libera el ADP y la
miosina puede volver a unirse al ATP y a separarse de la actina (Schiaffino & Reggiani,
2011; Walklate et al., 2016) (Figura 14).
Figura 14. Representación del ciclo de contracción muscular.
Las isoformas de la MHC con mayor actividad ATPasa hidrolizan el ATP más
rápidamente y, por lo tanto, presentan una mayor potencia muscular3 (Walklate et al.,
2016). Por otro lado, las isoformas de la MHC con mayor velocidad de liberación del
ADP pueden iniciar el ciclo de interacción de la actina con la miosina con mayor
3 La potencia muscular es la energía liberada por unidad de tiempo.

34
rapidez, hecho que confiere a estas isoformas una mayor velocidad de acortamiento de
la fibra muscular y, por lo tanto, una mayor velocidad de contracción.
La isoforma MHC-I presenta una baja velocidad de liberación del ADP y una baja
actividad ATPasa, pero tiene un coste tensional bajo4 (Schiaffino & Reggiani, 2011).
Estas características hacen que la isoforma MHC-I sea apropiada para acciones
musculares prolongadas y de baja intensidad, como el mantenimiento de la postura. De
hecho, esta isoforma se expresa en fibras de oxidación lenta (o "slow-oxidative") que
obtienen el ATP necesario para la contracción por medio de la vía oxidativa (glucólisis
aeróbica) (Korfage et al., 2005a). Estas fibras presentan una elevada resistencia a la
fatiga y son las predominantes en los músculos tónicos y posturales (Schiaffino &
Reggiani, 2011).
Las isoformas MHC-II son más rápidas y potentes que la MHC-I, presentando una
velocidad de liberación del ADP y una actividad ATPasa que aumenta en el orden
MHC-IIA<MHC-IIX<MHC-IIB. La isoforma MHC-IIB, que es la isoforma con mayor
velocidad de contracción y potencia muscular de las tres, no se ha detectado en los
mamíferos de gran tamaño, incluido Homo sapiens (Pette & Staron, 2000). Esto se debe
a una disminución de la actividad transcripcional en la región promotora del gen MYH4
que permite la transcripción de parte del gen, pero no la traducción a proteína (Snow et
al., 1982; Horton et al., 2001; Harrison et al., 2011). La isoforma MHC-IIX presenta un
coste tensional elevado y se expresa en fibras con metabolismo glucolítico de
contracción rápida (o "fast-twitch glycolitic"). Estas fibras obtienen el ATP por medio
de la vía glucolítica (glucólisis anaeróbica) y presentan una baja resistencia a la fatiga
(Korfage et al., 2005a). Finalmente, la isoforma MHC-IIA presenta una actividad
ATPasa y una velocidad de liberación del ADP que se sitúa entre las isoformas MHC-I
4 El coste tensional equivale al consumo de ATP por unidad de tiempo.

35
y MHC-IIX. La isoforma MHC-IIA se expresa en fibras con metabolismo oxidativo
glucolítico de contracción rápida (o "fast-twitch oxidative glycolitic"), es decir, fibras
que obtienen el ATP necesario para la contracción de la vía oxidativa y también de la
vía glucolítica (glucólisis aeróbica y anaeróbica) y que presentan una resistencia a la
fatiga y un coste tensional moderados (Korfage et al., 2005a). Las fibras "fast-twitch
oxidative glycolitic" junto a las fibras "fast-twitch glycolitic" son el tipo de fibras más
abundantes en los músculos fásicos (Schiaffino & Reggiani, 2011).
La isoforma MHC-extraocular está codificada por el gen MYH13 y es específica de
los músculos extraoculares (Weiss et al., 1999). Esta isoforma presenta una elevada
velocidad de contracción y un bajo coste tensional que la hacen apropiada para los
requerimientos funcionales de los músculos extraoculares, los cuales presentan
movimientos sacádicos rápidos pero también movimientos lentos para poder centrar la
mirada en la retina y así enfocar los objetos (Briggs & Schachat, 2000; Sciote & Morris,
2000).
Finalmente, en el cromosoma 7 se localiza el gen MYH16 que codifica la isoforma
MHC-IIM (Desjardins et al., 2002). Esta isoforma se caracteriza por una elevada
actividad ATPasa, capaz de generar una elevada tensión muscular, y por una velocidad
de contracción que se sitúa entre las isoformas MHC-I y MHC-IIA (Hoh, 2002; Toniolo
et al., 2008). La isoforma MHC-IIM solamente se ha descrito en músculos derivados del
primer arco branquial de diferentes especies de mamíferos, incluidos los primates
(Rowlerson et al., 1983; Hoh, 2002). El gen de la isoforma MHC-IIM es el primero en
divergir durante el proceso evolutivo (Qin et al., 2002), hace aproximadamente 400
millones de años, dando a los animales unos músculos masticadores potentes, útiles
para la captura de presas y/o para procesar alimentos mecánicamente resistentes (Figura
15). En los humanos, el gen MYH16 se transcribe solamente a una porción inestable de

36
la cabeza de la miosina, de 76 kDa, debido a una mutación que provoca la aparición de
un codón de stop en la secuencia del ARNm (Stedman et al., 2004). Por lo tanto, en los
humanos el gen MYH16 se transforma en un pseudogen, capaz de transcribir una parte
de la secuencia del gen a ARNm pero sin capacidad para la expresión de la proteína,
hecho que se relaciona con la disminución del tamaño de las fibras de tipo II en los
músculos elevadores de la mandíbula en Homo sapiens (Stedman et al., 2004). Según
Perry et al. (2005), la inactivación del gen MYH16 tuvo lugar hace 5.7 millones de
años, coincidiendo con la divergencia de los géneros Homo y Pan.
GEN PROTEÍNA
MYH8 MHC-neonatal
MYH4 MHC-IIB
MYH1 MHC-IIX
MYH2 MHC-IIA
MYH3 MHC-embriónica
MYH13 MHC-extraocular
MYH6 MHC-α cardíaca
MYH7 MHC-β/I
MYH16 MHC-masticadora
Figura 15. Reconstrucción filogenética de la MHC.
1.5.2. Métodos de análisis
La variedad de isoformas de la MHC da un marco funcional muy amplio al músculo
esquelético. Para valorar estas características funcionales desde un punto de vista
molecular existen diferentes métodos histoquímicos, inmunohistoquímicos o por medio
de electroforesis en gel (SDS-PAGE).

37
Tinción ATPasa
Mediante este método las fibras musculares se clasifican en I, IIA, IIX y IIM
dependiendo de su actividad ATPasa en diferentes medios de pH (Brooke & Kaiser,
1970; Rowlerson et al., 1981). Las fibras de tipo I tienen una elevada actividad ATPasa
en pH ácido, mientras que las fibras de tipo II presentan una elevada actividad ATPasa
en pH básico. Para diferenciar los subtipos de fibras II, la muestra muscular se incuba
previamente en un tampón ácido (fibras IIX) o en un tampón básico (fibras IIA)
(Brooke & Kaiser, 1974; Sciote & Morris, 2000). Las fibras de tipo IIM presentan una
elevada actividad ATPasa en todos los rangos de pH (Rowlerson et al., 1981).
El inconveniente de la tinción ATPasa es que no puede diferenciar fibras puras, que
expresan una sola isoforma de la MHC, de las fibras híbridas, que expresan más de una
isoforma de la MHC (Pette & Staron, 2000). Tampoco puede diferenciar las isoformas
MHC-neonatal y MHC-α cardíaca (Korfage et al., 2005b).
Inmunohistoquímica
Esta técnica consiste en utilizar anticuerpos (Ac) específicos para cada isoforma
de la MHC. Los Ac más utilizados en humanos y primates no humanos se resumen en la
Tabla 2 y en la Tabla 3, respectivamente (Rowlerson et al., 1983; Rowlerson et al.,
1988; Sciote et al., 1994; Korfage & Van Eijden, 1999; Korfage et al., 2000; Wall et al.,
2013).
Uno de los inconvenientes de la inmunohistoquímica es la posibilidad de que un
Ac específico para una isoforma de la MHC reaccione con otra isoforma. Esto es debido
a que las isoformas de la MHC presentan una cadena de aminoácidos muy similar. Por
ejemplo, la isoforma MHC-IIA y la MHC-IIX tienen un 94.8% de aminoácidos
idénticos (Österlund et al., 2012). Otro inconveniente de este método es la dificultad

38
para diferenciar fibras híbridas. Las fibras híbridas pueden variar el contenido de las
diferentes isoformas de la MHC a lo largo de la fibra muscular (Korfage et al., 2016;
Zhang & Gould, 2017). Esto favorece que solamente se observe el Ac de la isoforma de
la MHC con mayor proporción y se subestimen las otras isoformas que componen la
fibra muscular. Por ejemplo, el Ac 332-3D4, utilizado en humanos, no puede diferenciar
fibras de tipo IIA de fibras híbridas que co-expresan las isoformas MHC-IIA y MHC-
IIX (Korfage & Van Eijden, 1999; Korfage & Van Eijden, 2003).
Tabla 2. Anticuerpos más utilizados en los músculos masticadores de Homo sapiens (Sciote et al., 1994; Korfage & Van Eijden, 1999; Korfage et al., 2000).
Anticuerpo MHC
I IIA IIB IIX α neo IIM
219-1D1 + - - - - - -
249-5A4 - - - - + - -
333-7H1 - + - - - - -
340-3B5 - + + + - - -
332-3D4 - + - + - - -
Antifetal - - - - - + -
Anti-IIM - - - - - - +
MY-32 - + - + - + -

39
Tabla 3. Anticuerpos más utilizados en los músculos masticadores de primates no humanos (Rowlerson et al., 1983; Rowlerson et al., 1988; Wall et al., 2013).
Anticuerpo MHC
I IIA IIB IIX α Emb IIM
Anti-I + - - - - - -
Anti-IIA - + - - - - -
Anti-IIM - - - - - - +
Anti-emb - - - - - + -
Anti-IIB - + + - - - -
NOQ7.5.4D + - - - - - -
MY32 - + - + - - +*
* no reacciona a la MHC-IIM en algunos primates del Nuevo Mundo.
SDS-PAGE
Se llama SDS-PAGE a la electroforesis en gel de poliacrilamida con dodecil
sulfato sódico. La electroforesis en gel permite separar las proteínas aplicando un
campo eléctrico. Las proteínas se cargan en una membrana, en este caso de
poliacrilamida, por la que se desplazan en dirección al electrodo con carga opuesta a
una velocidad que depende del tamaño y de la forma de la proteína (Alberts et al.,
2013). Es decir, este método nos permite separar las proteínas de diferente tamaño
molecular y diferente forma. En el caso de las isoformas de la MHC, donde tenemos un
tamaño y una forma prácticamente idénticos, se requiere un gel especial, diseñado por
Talmadge & Roy (1993), que permite separar proteínas con un peso molecular de 1-2
kDa de diferencia. Posteriormente, este protocolo ha sido modificado para mejorar la

40
resolución de las bandas y disminuir el tiempo requerido para realizar la técnica
(Bamman et al., 1999; Roberts et al., 2012).
Uno de los inconvenientes del SDS-PAGE es la imposibilidad de detectar
isoformas de la MHC con una baja proporción o que co-emigran con otra banda de la
MHC (Korfage & Van Eijden, 2003).
1.5.3. Composición y distribución de los diferentes tipos de fibras
Los músculos elevadores de la mandíbula presentan una composición y
distribución muy heterogenia de los diferentes tipos de fibras, tanto en los humanos
como en el resto de primates (Ringqvist, 1974; Maxwell et al., 1979; Vignon et al.,
1980; Eriksson & Thornell, 1983; Rowlerson et al., 1983; Miller & Farias, 1988;
Shaughnessy et al., 1989; Sciote et al., 1994; Korfage & Van Eijden, 1999; Korfage et
al., 2000; Andreo et al., 2002; Wall et al., 2013).
Primates no humanos
En uno de los primeros estudios realizados en primates no humanos, concretamente
en Macaca mulatta, se observaron diferencias en la distribución de las fibras I y II entre
las regiones anterior y posterior de los músculos masetero superficial y temporal
mediante la tinción por ATPasa (Maxwell et al., 1979). Según este estudio, la región
anterior de los dos músculos presenta una mayor proporción de fibras I y una menor
proporción de fibras II que la región posterior. Concretamente, la región anterior del
masetero superficial presenta un 78.93-92.14% de fibras I y un 7.29-21.08% de fibras
II, mientras que la porción posterior del masetero superficial presenta un 27.50-40.14%
de fibras I y un 60-74.25% de fibras II. En relación al músculo temporal, las fibras de
tipo I representan entre el 20.65-49.95% en la región anterior y entre el 9.43-9.92% en
la región posterior, siendo el resto fibras de tipo II. El inconveniente de este estudio es

41
que clasifica a las fibras con una intensidad de tinción intermedia como fibras de tipo I,
hecho que puede sobreestimar la proporción de estas fibras. Posteriormente, Rowlerson
et al. (1983) realizaron un estudio donde se analizaba la composición de los músculos
elevadores de la mandíbula en diferentes especies de primates mediante tinción ATPasa
y tinción indirecta de inmunoperoxidasa (Tabla 4).
Tabla 4. Primates estudiados por Rowlerson et al. (1983).
Parvorden Especie Músculo analizado
Platyrrhini Callithrix penicillata Masetero y temporal
Saimiri sciureus Masetero, temporal y pterigoideo medial
Catarrhini Cercopithecus sp. Masetero y temporal
Macaca fascicularis Masetero, temporal y pterigoideo medial
Macaca irus Masetero y temporal
Pan troglodytes Masetero
En todos los primates estudiados se observaron dos características comunes. La
primera es que las fibras de tipo IIM son las más abundantes en los músculos elevadores
de la mandíbula siendo el resto fibras de tipo I y, en algunos casos, de tipo IIC
(equivalentes a fibras que expresan las isoformas MHC-I y MHC-IIA). Así, en el
masetero de Pan troglodytes las fibras de tipo IIM representan entre un 43-69% y las
fibras de tipo I entre un 31-57% (Rowlerson et al., 1983). La segunda característica es el
aumento gradual de fibras de tipo I a medida que profundizamos en el músculo, en
decremento de las fibras IIM (Rowlerson et al., 1983; Rowlerson et al., 1988).

42
Otros estudios han valorado si existen o no diferencias intramusculares en la
composición y distribución de los diferentes tipos de fibras en los músculos masetero y
temporal (Miller & Farias, 1988; Andreo et al., 2002; Wall et al., 2013). En un estudio
realizado en Cebus apella por medio de la tinción ATPasa no se describen diferencias
entre la región anterior y la posterior de los músculos temporal superficial y masetero
superficial (Andreo et al., 2002). En cambio, en otro estudio realizado en Macaca
mulatta (“rhesus monkey”), con el mismo método, se observó que la región anterior del
temporal superficial presentaba una mayor proporción de fibras de tipo I (13.1%) que la
región posterior del temporal, tanto en el temporal superficial (6.6%) como en el
temporal profundo (9.5%) (Miller & Farias, 1988). En este mismo estudio, la región
posterior del temporal profundo presentaba un mayor porcentaje de fibras I (9.5%) y IIA
(42.6%) que la región posterior del temporal superficial (6.6% de fibras tipo I y 34.9%
de fibras tipo IIA), pero no se observaron diferencias entre la región anterior del
temporal superficial y la región anterior del temporal profundo (Miller & Farias, 1988).
Estas diferencias sí que fueron descritas por Wall et al. (2013) en Papio anubis por
medio de inmunohistoquímica y SDS-PAGE. Según Wall et al. (2013) el temporal
superficial presenta una mayor proporción de fibras IIM (87.0-88.7%) que la región
anterior del temporal profundo (14.4-60.7%), donde hay una mayor presencia de fibras I
(38.1-83.3%) y algunas fibras de tipo IIA (<5%). El amplio rango del porcentaje de las
fibras IIM y I en la región anterior del temporal profundo se debe al dimorfismo sexual,
presentando las hembras una mayor proporción de fibras I (83.3%) y los machos una
mayor proporción de fibras IIM (60.7%) (Wall et al., 2013).
Homo sapiens
Los músculos elevadores de la mandíbula en Homo sapiens se han analizado por
medio de métodos histoquímicos, inmunohistoquímicos y mediante SDS-PAGE

43
(Ringqvist, 1974; Vignon et al., 1980; Eriksson & Thornell, 1983; Sciote et al., 1994;
Korfage & Van Eijden, 1999; Korfage et al., 2000). Estos músculos muestran
proporciones variables de fibras I, IIA, IIX y fibras híbridas, pero en ningún caso se
describe la presencia de fibras de tipo IIM (Sciote et al., 1994; Korfage & Van Eijden,
1999; Korfage et al., 2000). A diferencia de los músculos de las extremidades, las fibras
híbridas de los músculos elevadores de la mandíbula pueden presentar las isoformas
MHC-α cardíaca y MHC-fetal (Butler-Browne et al., 1988; Bredman et al., 1991; Sciote
et al., 1994; Korfage & Van Eijden, 1999; Weiss et al., 1999). Se cree que la presencia
de las isoformas MHC-α cardíaca y MHC-fetal proporciona un mayor rango funcional a
los músculos elevadores de la mandíbula para así poder optimizar al máximo la fuerza y
la velocidad de contracción en cada acción realizada, ya que las características
contráctiles de las fibras híbridas se sitúan entre las isoformas que co-expresan (Korfage
et al., 2005b). Otra diferencia en relación a los músculos de las extremidades es que las
fibras de tipo I de los músculos elevadores de la mandíbula presentan un diámetro
mayor que las fibras de tipo II (Sciote et al., 2003).
Existe una gran variabilidad en las proporciones de las fibras I, IIA, IIX e
híbridas en los músculos elevadores de la mandíbula en los humanos (Ringqvist, 1974;
Vignon et al., 1980; Eriksson & Thornell, 1983; Shaughnessy et al., 1989; Korfage &
Van Eijden, 1999; Korfage et al., 2000). En general, las fibras puras más frecuentes son
las de tipo I seguidas por proporciones variables de fibras de tipo IIA y IIX. Por otro
lado, las fibras híbridas representan aproximadamente el 40% del total de las fibras,
siendo el tipo de fibra híbrida más habitual la que contiene las isoformas MHC-α
cardíaca + MHC-I + MHC-IIA (Korfage et al., 2000; Korfage et al., 2005b).
Los diferentes estudios realizados por tinción ATPasa muestran unos rangos
muy amplios en las proporciones de las diferentes fibras en los músculos masetero,

44
pterigoideo medial y temporal (Ringqvist, 1974; Vignon et al., 1980; Eriksson &
Thornell, 1983; Shaughnessy et al., 1989). Así, en el músculo masetero la proporción de
fibras de tipo I se sitúa entre el 28.6% y el 62.5%, en el pterigoideo medial entre el 37%
y el 54% y en el temporal entre el 30.9% y el 53.4% (Ringqvist, 1974; Vignon et al.,
1980; Eriksson & Thornell, 1983; Shaughnessy et al., 1989). Estos rangos tan amplios
pueden ser debidos al diferente criterio a la hora de clasificar las fibras híbridas en
fibras de tipo I o fibras de tipo II (apartado 1.5.2.). Otro dato a tener en cuenta es la edad
de los sujetos estudiados. Se ha observado que con el envejecimiento hay una
disminución de fibras de tipo I y un aumento de fibras de tipo II e híbridas en los
músculos elevadores de la mandíbula (Monemi et al., 1999). De hecho, coincide que los
estudios por tinción ATPasa, con menor proporción de fibras tipo I, utilizan muestras
musculares de sujetos con una edad más avanzada que los estudios con mayor
proporción de fibras tipo I, que utilizan sujetos más jóvenes (Ringqvist, 1974; Vignon et
al., 1980; Eriksson & Thornell, 1983; Shaughnessy et al., 1989).
Los estudios inmunohistoquímicos, a diferencia de la tinción por ATPasa,
permiten identificar las diferentes isoformas de la MHC mediante el uso de anticuerpos
específicos. Estos estudios muestran que la isoforma más frecuente en los músculos
elevadores de la mandíbula es la MHC-I, que se expresa en el 70% de las fibras, seguida
de la isoforma MHC-IIA, que se expresa en el 30% de las fibras (Korfage et al., 2000;
Korfage et al., 2005b). Estas dos isoformas de la MHC, junto a la isoforma MHC-IIX,
se pueden expresar en fibras puras o híbridas. En el músculo masetero, las fibras puras
que expresan la isoforma MHC-I representan un 35% de todas las fibras, las fibras puras
que expresan la isoforma MHC-IIA un 8.3% y las fibras puras para la isoforma MHC-
IIX un 14.4%, siendo el 42.4% restante fibras híbridas (Korfage et al., 2000). Entre las
dos porciones del masetero, la superficial presenta una menor proporción de fibras puras

45
que expresan la isoforma MHC-I que la porción profunda (26.8% vs. 47.8%) (Korfage
et al., 2000). En el músculo pterigoideo medial se han descrito unos porcentajes muy
similares a los observados en el masetero para las fibras puras, concretamente un 32.3%
de fibras expresan solamente la isoforma MHC-I, un 5.4% la isoforma MHC-IIA y un
10.6% la isoforma MHC-IIX, mientras que el 51.5% de las fibras restantes son híbridas
(Korfage & Van Eijden, 2000). En el músculo temporal, las isoformas MHC-I y MHC-
IIA que se expresan en fibras puras son más abundantes en comparación con los
músculos masetero y pterigoideo medial, pero presentan una menor proporción de fibras
híbridas (Korfage et al., 2000). Concretamente, un 45.5% de las fibras expresan
únicamente la isoforma MHC-I y un 13.7% la isoforma MHC-IIA, mientras que las
fibras híbridas representan un 30% (Korfage et al., 2000). Entre la región anterior y
posterior del músculo temporal se observan diferencias en la proporción de la isoforma
MHC-I que se expresa en fibras puras, un 51.2% en la región anterior y un 37.9% en la
región posterior (Korfage & Van Eijden, 1999; Korfage et al., 2000).
1.6. Expresión genética
La expresión genética es el proceso en el que los genes se transforman en
proteínas y consta, básicamente, de dos fases: la transcripción y la traducción (Lodish et
al., 2000). La transcripción se realiza dentro del núcleo de la célula y consiste en
sintetizar una cadena de ARN utilizando como molde una de las dos cadenas de ADN
que contienen el gen (Borukhov & Nudler, 2008). Las enzimas encargadas de realizar
esta acción son las ARN polimerasas que desenrollan la doble hélice de ADN y van
agregando uno a uno los nucleótidos de ARN complementarios a los nucleótidos de la

46
cadena de ADN (Kerppola & Kane, 1991)5 (Tabla 5). La cadena de ARN que se obtiene
se denomina ARN mensajero (ARNm) y presenta regiones codificadoras, denominadas
exones, y regiones no codificadoras, denominadas intrones (Alberts et al., 2013).
Durante la transcripción, los intrones son eliminados por medio de los ARN nucleares
pequeños (ARNnp) y los exones se unen entre sí para formar el ARNm maduro, proceso
denominado maduración del ARN por corte y empalme o “RNA splicing” (Crick,
1979).
Tabla 5. Nucleótidos complementarios entre el ADN y el ARN.
ADN ARN
Adenina Uracilo
Timina Adenina
Guanina Citosina
Citosina Guanina
El ARNm maduro es transportado hacia el citoplasma donde tiene lugar la segunda
fase de la expresión genética, la traducción. La traducción es la síntesis de la proteína a
partir del ARNm maduro y se lleva a cabo en los ribosomas (Lodish et al., 2000). Los
ribosomas están constituidos por proteínas ribosomales y por ARN ribosómico (ARNr),
que acomoda el ARNm dentro del ribosoma para que interactúe con el ARN de
transferencia (ARNt) (Lodish et al., 2000). El ARNt es un tipo de ARN con forma de
cruz que en un extremo transporta uno de los veinte aminoácidos necesarios para formar
la proteína y en el otro extremo presenta una secuencia de tres nucleótidos, denominada
anticodón, que es complementaria a una secuencia de tres nucleótidos del ARNm,
5 El ADN presenta los nucleótidos adenina, guanina, citosina y timina, mientras que el ARN presenta los
nucleótidos adenina, guanina, citosina y uracilo.

47
denominada codón. Así, el ARNt convierte la secuencia de nucleótidos del ARNm
maduro en una secuencia de aminoácidos que constituirán la proteína (Alberts et al.,
2013) (Figura 16).
Figura 16. Representación de la síntesis proteica.
1.6.1. RT-qPCR
La RT-qPCR (real time quantitative polymerase chain reaction) sirve para
identificar y cuantificar la expresión de diferentes genes a nivel de su ARNm o
“transcript”. Esta técnica permite amplificar una secuencia definida de ARNm y
discriminarla de otra estrechamente relacionada, con lo que presenta una elevada
especificidad (Bustin, 2000). También nos permite trabajar con material genético de
muestras de tejido muscular muy pequeñas debido a que tiene una elevada sensibilidad
y puede amplificar niveles muy bajos de ARNm (Bustin, 2000). Además, nos permite
trabajar con músculos criopreservados durante largos períodos de tiempo, puesto que
cualquier degradación post-mortem potencial del ARNm puede compensarse

48
normalizando los valores de expresión del gen estudiado con los de un gen endógeno
resistente (Bahar et al., 2007).
Estudios realizados en tejido muscular muestran una correlación entre el ARNm
específico de una isoforma de la MHC y su proteína sugiriendo una regulación
transcripcional de la expresión de los genes de la MHC (Cox & Buckingham, 1992;
Wright et al., 1997; Eizema et al., 2005; Hemmings et al., 2009; Zurmanova & Soukup,
2013; Men et al., 2016). Estos resultados indican que el estudio de la expresión genética
de las isoformas de la MHC puede ser válido para analizar las características
funcionales de un músculo o de sus partes.

Hipótesis y objetivos


51
2. Hipótesis y objetivos
2.1. Hipótesis
La hipótesis que se plantea en el presente proyecto de Tesis Doctoral es que las
diferencias funcionales y ultraestructurales que se han descrito entre los diferentes
músculos elevadores de la mandíbula o entre sus diferentes porciones en los humanos
modernos y en otras especies de primates no hominoideos, mediante el uso de diferentes
técnicas como la electromiografía, la tinción ATPasa o mediante técnicas hitoquímicas
o inmunohistoquímicas, se verán también reflejadas en los patrones de expresión de las
diferentes isoformas de la MHC de estos músculos en Pan troglodytes.
A pesar de que los chimpancés comunes y los bonobos son las especies de
primates más emparentadas con los humanos modernos desde el punto de vista
filogenético, las características funcionales y moleculares de sus músculos masticadores
han sido poco estudiadas hasta la actualidad, posiblemente debido en parte a la
dificultad de acceder al estudio de estos primates afectados por un elevado riesgo de
extinción. En este sentido, el uso de la RT-qPCR para determinar los patrones de
expresión de las isoformas de la MHC puede ser de gran utilidad al permitir analizar
músculos pertenecientes a primates criopreservados durante largos períodos de tiempo
que dificultan el estudio mediante técnicas histoquímicas o inmunohistoquímicas.
De esta forma, pensamos que se podrán detectar diferencias significativas en los
patrones de expresión de las isoformas de la MHC entre los diferentes músculos
elevadores de la mandíbula (masetero, pterigoideo medial y temporal) y entre sus
diferentes porciones en Pan troglodytes que se podrán relacionar con sus respectivos

52
patrones funcionales que han sido estudiados en los humanos modernos y en otras
especies de primates no hominoideos mediante técnicas electromiográficas.
2.2. Objetivos
El objetivo principal de nuestro proyecto de Tesis Doctoral es analizar mediante
RT-qPCR el patrón de expresión de las diferentes isoformas de la MHC en los músculos
elevadores de la mandíbula en Pan troglodytes para valorar si existen diferencias
significativas que puedan relacionarse con las características funcionales de estos
músculos observadas mediante electromiografía en los humanos modernos y otras
especies de primates no hominoideos. Con ello, se pretende mejorar el conocimiento
que se tiene sobre la anatomía, función y composición de los músculos masticadores de
los chimpancés comunes, una especie relativamente poco estudiada a pesar del elevado
grado de parentesco que presenta con los humanos modernos. Pensamos que nuestros
resultados podrán ser aplicados a diferentes campos de estudio como la anatomía
comparada, la anatomía evolutiva o la antropología.
La consecución de este objetivo principal se ha basado en el desarrollo de los
siguientes objetivos específicos:
1- Diseccionar los músculos elevadores de la mandíbula (masetero, pterigoideo
medial y temporal) en diferentes individuos de las especies Homo sapiens y Pan
troglodytes para obtener información sobre su anatomía y su masa muscular
relativa.
2- Recoger muestras de los diferentes músculos diseccionados y de sus porciones
(regiones anterior y posterior del masetero superficial, regiones anterior y
posterior del masetero profundo, regiones anterior y posterior del temporal

53
superficial, regiones anterior y posterior del temporal profundo y región
esfenomandibular del temporal) para el estudio de los patrones de expresión de
las isoformas de la MHC mediante RT-qPCR.
3- Obtener y comparar los patrones de expresión de las isoformas de la MHC entre
las regiones anterior y posterior del masetero superficial.
4- Obtener y comparar los patrones de expresión de las isoformas de la MHC entre
las regiones anterior y posterior del masetero profundo.
5- Obtener y comparar los patrones de expresión de las isoformas de la MHC entre
las porciones superficial y profunda del masetero.
6- Obtener y comparar los patrones de expresión de las isoformas de la MHC entre
la porción superficial del masetero y el pterigoideo medial.
7- Obtener y comparar los patrones de expresión de las isoformas de la MHC entre
la porción profunda del masetero y el pterigoideo medial.
8- Obtener y comparar los patrones de expresión de las isoformas de la MHC entre
las regiones anterior y posterior del temporal superficial.
9- Obtener y comparar los patrones de expresión de las isoformas de la MHC entre
las regiones anterior y posterior del temporal profundo.
10- Obtener y comparar los patrones de expresión de las isoformas de la MHC entre
las regiones anteriores de los temporales superficial y profundo.
11- Obtener y comparar los patrones de expresión de las isoformas de la MHC entre
las regiones posteriores de los temporales superficial y profundo.
12- Obtener y comparar los patrones de expresión de las isoformas de la MHC entre
la porción esfenomandibular del temporal y las otras regiones de este músculo.
13- Comparar los resultados obtenidos con otros estudios que se han realizado en
los músculos masticadores de los humanos y otras especies de primates

54
mediante técnicas electromiográficas, tinciones ATPasa y técnicas histoquímicas
e inmunohistoquímicas.

Artículos


57
3. Artículos
3.1. Artículo primero: “Expression of MyHC isoforms mRNA transcripts in different regions of the masseter and medial pterygoid muscles in chimpanzees”
Resumen
Los músculos masetero y pterigoideo medial de Pan troglodytes actúan
conjuntamente elevando la mandíbula, pero los estudios electromiográficos realizados
en otras especies de primates, incluidos los humanos, muestran diferencias funcionales
entre los dos músculos y entre las dos porciones del masetero (Basmajian & De Luca,
1985; Hylander et al., 1987; Blanksma et al., 1997; Hylander et al., 2000; Wall et al.,
2006; Schindler et al., 2007; Vinyard et al., 2008; Williams et al., 2011; Guzmán-
Venegas et al., 2015). Una forma alternativa a la electromiografía para valorar las
características funcionales de un músculo es el análisis de la expresión de las diferentes
isoformas de la MHC (Bottinelli & Reggiani, 2000).
En este artículo se obtuvieron la masa muscular del masetero y del pterigoideo
medial en cinco Pan troglodytes y se analizaron la expresión del ARNm de las
isoformas MHC-I, MHC-IIA, MHC-IIX y MHC-IIM en diferentes porciones de estos
dos músculos mediante RT-qPCR.
Los resultados obtenidos indican que el masetero presenta una mayor masa
muscular en comparación con el pterigoideo medial (peso medio del masetero de 62 gr
y peso medio del pterigoideo medial de 24 gr) y que la masa de la porción superficial
del masetero es superior a la de la porción profunda (peso medio de la porción
superficial de 45.5 gr y peso medio de la porción profunda de 16.5 gr). El patrón de

58
expresión de las isoformas de la MHC obtenido es similar en todas las regiones
analizadas, siguiendo el patrón MHC-IIM>MHC-IIA>MHC-I>MHC-IIX. No se
identificaron diferencias significativas de los patrones de expresión de las isoformas de
la MHC entre las regiones anterior y posterior del masetero superficial, entre las
regiones anterior y posterior del masetero profundo, entre la porción superficial del
masetero y el pterigoideo medial ni entre la porción profunda del masetero y el
pterigoideo medial. Sin embargo, sí que se observaron diferencias significativas entre
las porciones superficial y profunda del masetero, presentando la porción superficial un
mayor porcentaje de expresión de la isoforma MHC-IIM (40.2% vs. 34.2%).
El patrón de expresión de las isoformas de la MHC obtenido en el masetero y el
pterigoideo medial es típico de músculos fásicos, por presentar un porcentaje de
expresión de la isoforma MHC-I inferior al 50% (21.5% en el masetero y 22.2% en el
pterigoideo medial), y de gran potencia, por presentar un porcentaje de expresión
elevado de la isoforma MHC-IIM (37.2% en el masetero y 37.7% en el pterigoideo
medial). Estos resultados concuerdan bien con el gran desarrollo que presenta la
musculatura masticadora en Pan troglodytes. Por otro lado, las diferencias observadas
entre las porciones superficial y profunda del músculo masetero están en la línea de las
observaciones realizadas por otros autores con el uso de la electromiografía. Estas
observaciones indican un patrón de reclutamiento diferente para las dos porciones del
masetero, presentando la porción superficial una mayor actividad en movimientos
verticales, cuando se requiere una mayor fuerza muscular, y presentando la porción
profunda una mayor actividad en los movimientos laterales de la mandíbula, que sirven
para colocar los molares antes de la oclusión dentaria (Hylander et al., 1987; Hylander
& Johnson, 1994).

59
Como conclusión, el análisis de los patrones de expresión de las diferentes
isoformas de la MHC puede aportar información novedosa relacionada con las
características anatómicas y funcionales de los músculos masticadores de Pan
troglodytes, que se puede complementar con la obtenida por otros autores con el uso de
otras metodologías en otras especies de primates.


61

62

63

64

65


67
3.2. Artículo segundo: “Expression of myosin heavy chain isoforms mRNA transcripts in the temporalis muscle of common chimpanzees (Pan troglodytes)
Resumen
El temporal es uno de los músculos elevadores de la mandíbula junto con el
masetero y el pterigoideo medial. En Pan troglodytes, el músculo temporal está
formado por una porción superficial y una porción profunda (Aiello & Dean, 1990).
Además, algunos estudios realizados en humanos y en Macaca mulatta (Türp et al.,
1997; Schön-Ybarra & Bauer, 2001; Skinner & Aziz, 2003) han descrito una porción
profunda del temporal, denominada porción esfenomandibular, que algunos autores
identifican como un músculo independiente (Dunn et al., 1996). Funcionalmente, el
músculo temporal se puede dividir en una región anterior y una región posterior. Los
estudios electromiográficos realizados en humanos y en otras especies de primates no
hominoideos muestran que la región anterior actúa principalmente en la elevación de la
mandíbula, mientras que la región posterior participa sobretodo en su desplazamiento
posterior y lateral (Blanksma et al., 1997; Hylander et al., 2005; Vinyard et al., 2008).
Por su parte, la porción esfenomandibular actúa principalmente en el mantenimiento de
la postura de la mandíbula (Wood, 1986; Fuentes et al., 2012). En el presente estudio se
analizaron los patrones de expresión de las isoformas de la MHC en las diferentes
porciones del músculo temporal en cinco Pan troglodytes, con la finalidad de observar
diferencias significativas que puedan relacionarse con las diferencias funcionales
observadas por otros autores en los humanos y otras especies de primates no
hominoideos con el uso de la electromiografía.

68
En los cinco chimpancés comunes diseccionados se pudo identificar una porción
esfenomandibular claramente separada del resto del músculo temporal. Sin embargo, su
inervación no estaba diferenciada de la inervación de las regiones anteriores del resto
del temporal (nervio temporal profundo anterior), con lo que no consideramos
pertinente la identificación de la porción esfenomandibular como un músculo
independiente del temporal (Türp et al., 1997; Schön-Ybarra & Bauer, 2001).
Los principales resultados obtenidos indican que la expresión de las isoformas
de la MHC sigue un patrón similar en todas las regiones del músculo temporal de Pan
troglodytes (MHC-IIM>MHC-IIA>MHC-I>MHC-IIX), excepto en la región posterior
del temporal superficial donde la isoforma MHC-I presenta un mayor porcentaje de
expresión que la isoforma MHC-IIA. No se observaron diferencias significativas de los
patrones de expresión cuando se compararon las regiones anterior y posterior del
temporal superficial, las regiones anterior y posterior del temporal profundo, las
regiones anteriores de los temporales superficial y profundo y las regiones posteriores
de los temporales superficial y profundo. Cabe destacar, pero, que se observó una
diferencia no significativa en el porcentaje de expresión de la isoforma MHC-IIM entre
las regiones anterior (47%) y posterior (40.5%) del temporal superficial que podría
relacionarse con las diferencias funcionales observadas entre las dos regiones mediante
el uso de la electromiografía (Blanksma et al., 1997; Hylander et al., 2005; Vinyard et
al., 2008).
Los patrones de expresión de las isoformas de la MHC sí que presentaron
diferencias significativas cuando se compararon las regiones anteriores de los
temporales superficial y profundo con la porción esfenomandibular. En este caso, la
porción esfenomandibular presentó un menor porcentaje de expresión de la isoforma
MHC-IIM en la porción esfenomandibular (33.6%) respecto a la región anterior del

69
temporal superficial (47%) y a la región anterior del temporal profundo (43%). Esta
observación implica una menor capacidad de la porción esfenomandibular de desarrollar
potencia muscular, lo que se relaciona con su función lateralizadora y estabilizadora de
la mandíbula que se ha descrito en diferentes estudios electromiográficos realizados en
Homo sapiens (Wood, 1986; Fuentes et al., 2012).
Como conclusión, las diferencias observadas en los patrones de expresión de las
isoformas de la MHC en las diferentes porciones del músculo temporal de Pan
troglodytes, pueden relacionarse con algunas de las diferencias funcionales que se han
identificado electromiograficamente en otras especies de primates.


71

72

73

74

75

76

Discusión


79
4. Discusión
Los músculos elevadores de la mandíbula de Pan troglodytes presentan un patrón de
expresión característico de los músculos fásicos, con un elevado porcentaje de ARNm
de las isoformas MHC-II (>73%). Este patrón de expresión es el esperado para unos
músculos que actúan durante la fase de clausura del ciclo de la masticación, al final de
la cual tiene lugar la rotura de los alimentos (Lieberman, 2011). La gran potencia de los
músculos elevadores de la mandíbula en Pan troglodytes queda reflejada en la mayor
expresión de ARNm de la isoforma MHC-IIM. Esta isoforma es la más potente debido a
su elevada actividad ATPasa y presenta una velocidad de contracción moderada, que se
sitúa entre las isoformas MHC-I y MHC-IIA (Hoh, 2002; Toniolo et al., 2008). Este
modelo de expresión de los músculos elevadores de la mandíbula se corresponde con
sus características morfológicas, propicias para la generación de una elevada fuerza
muscular (Taylor & Vinyard, 2013). Los tres músculos son peniformes y, por lo tanto,
pueden agrupar un mayor número de sarcómeros en paralelo que les permiten generar
contracciones musculares más potentes (Aiello & Dean, 1990; Van Eijden et al., 1997).
Un parámetro que refleja el número de sarcómeros en paralelo es el ASTF que está
directamente relacionada con el peso muscular (Powell et al., 1984). Nuestros resultados
muestran que los músculos con mayor masa muscular presentan una mayor expresión
de la isoforma MHC-IIM. Así, el músculo temporal tiene un peso de 120.5 gr y una
expresión de ARNm de la isoforma MHC-IIM que representa el 41.5%, el músculo
masetero superficial tiene un peso de 45.5 gr y una expresión de la isoforma MHC-IIM
del 40.2%, el músculo masetero profundo tiene un peso de 16.5 gr y una expresión de la
isoforma MHC-IIM del 34.2% y el músculo pterigoideo medial tiene un peso de 24 gr y
una expresión de la isoforma MHC-IIM del 37.7%. Esta relación, pero, no se cumple en

80
el caso de las dos porciones del músculo temporal, donde la porción superficial presenta
un peso muscular de 39.9 gr y una expresión de ARNm de la isoforma MHC-IIM del
43.8%, mientras que la porción profunda presenta un peso muscular de 68.7 gr y una
expresión de ARNm de la isoforma MHC-IIM del 43.2%.
En general, los músculos elevadores de la mandíbula muestran el mismo patrón de
expresión: MHC-IIM>MHC-IIA>MHC-I>MHC-IIX (Anexo 1). Concretamente, el
músculo masetero muestra unos porcentajes para la isoforma MHC-IIM del 37.2%, para
la isoforma MHC-IIA del 28.6%, para la isoforma MHC-I del 21.5% y para la isoforma
MHC-IIX del 12.8%. Estos porcentajes son muy parecidos a los obtenidos en el
músculo pterigoideo medial, que presenta un porcentaje de ARNm de la isoforma
MHC-IIM del 37.7%, de la isoforma MHC-IIA del 28.3%, de la isoforma MHC-I del
22.2% y de la isoforma MHC-IIX del 11.8%. En cambio, el temporal presenta una
mayor proporción de ARNm de la isoforma MHC-IIM (41.5%) y una menor proporción
de la isoforma MHC-IIA (23.5%). Estos resultados sugieren que el masetero y el
pterigoideo medial presentan unas características funcionales parecidas y están en la
línea de las observaciones electromiográficas. Según los estudios electromiográficos
realizados en diferentes especies de primates antropoideos, incluidos los humanos, el
masetero y el pterigoideo medial actúan de forma sinérgica durante los movimientos de
elevación de la mandíbula, llegando al pico máximo de actividad durante la oclusión
dentaria (Basmajian & De Luca, 1985; Hylander et al., 1987; Aiello & Dean, 1990; Van
Eijden et al., 1997; Schindler et al., 2007; Williams et al., 2011).
A nivel intramuscular, existen diferencias significativas en la expresión de la
isoforma MHC-IIM entre las dos porciones del músculo masetero. El masetero
superficial presenta una expresión de ARNm de la MHC-IIM del 40.2%, mientras que
la porción profunda presenta una expresión del 34.2% (p=0.032). Además, el masetero

81
superficial presenta una menor proporción de ARNm de las isoformas MHC-I y MHC-
IIA. Estas dos isoformas son las menos potentes, pero son las más resistentes a la fatiga
(Schiaffino & Reggiani, 2011). Estas diferencias en el patrón de expresión de las
isoformas de la MHC entre el masetero superficial y el masetero profundo se
correlacionan con los estudios electromiográficos realizados en Papio anubis, Macaca
fuscata, Aotus trivirgatus y Homo sapiens (Hylander et al., 1987; Wood, 1987;
Hylander & Johnson, 1994; Blanksma et al., 1997; Hylander et al., 2005). Estos
estudios muestran que el masetero superficial actúa en los movimientos verticales de la
mandíbula y aumenta el reclutamiento de sus fibras durante la masticación de alimentos
mecánicamente resistentes (Hylander & Johnson, 1994; Blanksma et al., 1997). En
cambio, el masetero profundo desplaza la mandíbula lateralmente y ayuda a colocar los
molares justo antes de la oclusión dentaria, por lo que requiere de movimientos más
precisos y menos potentes (Hylander et al., 1987; Hylander & Johnson, 1994; Vinyard
et al., 2008; Ross & Iriarte-Diaz, 2014). Algunos estudios electromiográficos realizados
en humanos también describen diferencias en el modelo de reclutamiento de las fibras
musculares entre las regiones anterior y posterior del músculo masetero, tanto en su
porción superficial como en la profunda (Blanksma et al., 1992; Guzmán-Venegas et al.,
2015). Estas diferencias no se han visto reflejadas en nuestro estudio en el patrón de
expresión de las isoformas de la MHC, con lo que posiblemente se deban a diferencias
en las características morfológicas de las dos regiones como, por ejemplo, la orientación
de sus fibras musculares.
El músculo temporal se divide funcionalmente en una región anterior y una región
posterior. La región anterior actúa durante la fase de clausura de la mandíbula y
aumenta su actividad durante el procesamiento de alimentos resistentes, en cambio la
región posterior actúa en los movimientos de retrusión de la mandíbula (Cachel, 1979;

82
Wood, 1987; Blanksma et al., 1997; Hylander et al., 2005). Estas dos regiones del
temporal también presentan diferencias en su morfología interna. Así, en Homo sapiens
la porción anterior presenta una ASTF de 7.7 cm2 y la porción posterior de 5.55 cm2
(Van Eijden et al., 1997). Todas estas diferencias, funcionales y estructurales, entre las
regiones anterior y posterior del músculo temporal no se traducen en diferencias
significativas en la expresión de las isoformas de la MHC en Pan troglodytes, pero
hemos observado una tendencia a una mayor expresión de la isoforma MHC-IIM y a
una menor expresión de la isoforma MHC-I en la región anterior del temporal
superficial respecto a su región posterior. Concretamente, la región anterior del
temporal superficial presenta un porcentaje de expresión de la isoforma MHC-IIM del
47% y un porcentaje de expresión de la isoforma MHC-I del 19.9, mientras que la
región posterior del temporal superficial presenta un porcentaje de expresión de la
isoforma MHC-IIM del 40.5 y un porcentaje de expresión de la isoforma MHC-I del
26.9%. La isoforma MHC-I presenta una baja actividad ATPasa y es la isoforma de la
MHC menos potente (Schiaffino & Reggiani, 2011). Esta isoforma se expresa en fibras
de contracción lenta, que son idóneas para las actividades posturales y los movimientos
de precisión (Korfage et al., 2005a). Esto sugiere que la región anterior del temporal
superficial es más potente, mientras que su región posterior tiene una mayor
participación en los movimientos de precisión y posturales de la mandíbula, tal y como
indican los estudios morfológicos y electromiográficos. Posiblemente, una ampliación
de la muestra muscular se traduzca en diferencias significativas en el modelo de
expresión de las isoformas de la MHC entre las regiones anterior y posterior del
músculo temporal.
Donde sí hemos obtenido diferencias significativas es en la expresión de ARNm de
la isoforma MHC-IIM entre las regiones anteriores del temporal superficial y del

83
temporal profundo y la porción esfenomandibular. Algunos autores consideran la
porción esfenomandibular como un músculo independiente debido a que se puede
diferenciar anatómicamente de la porción profunda del temporal (Dunn et al., 1996). En
las disecciones que hemos realizado en cinco Pan troglodytes hemos podido identificar
la porción esfenomandibular como una porción separada del temporal profundo pero su
inervación depende del nervio temporal profundo anterior, igual que el resto de la
región más anterior del temporal, con lo que descartamos que se trate de un músculo
independiente. La porción esfenomandibular presenta una menor proporción de ARNm
de la isoforma MHC-IIM (33.6%) y una mayor proporción de la isoforma MHC-IIA
(31.5%) en comparación con la región anterior del temporal superficial (MHC-IIM 47%
y MHC-IIA 22.5%) y profundo (MHC-IIM 43% y MHC-IIA 22.3%), siendo estas
diferencias significativas en el caso de la isoforma MHC-IIM (temporal superficial vs.
esfenomandibular p=0.032 y temporal profundo vs. esfenomandibular p=0.016) pero
no en el caso de la isoforma MHC-IIA. La isoforma MHC-IIA es la más lenta y menos
potente de las isoformas MHC-II (Schiaffino & Reggiani, 2011). Esta isoforma se
expresa en fibras con metabolismo oxidativo glucolítico de contracción rápida con una
resistencia a la fatiga moderada (Korfage et al., 2005a). Este modelo de expresión se
corresponde con las observaciones realizadas mediante electromiografía en los
humanos, que demuestran una mayor participación de la porción esfenomandibular en el
control postural y en la estabilización de la mandíbula (Zenker, 1955; Wood, 1986;
Fuentes et al., 2012). Además, nuestros resultados apoyan la hipótesis de una fuerza
oclusiva elevada (“high occlusal force hypothesis”), según la cual las partes
superficiales de los músculos de la masticación son las encargadas de generar la fuerza
muscular necesaria para procesar los alimentos más resistentes.

84
Los porcentajes de expresión del ARNm de las isoformas de la MHC que hemos
obtenidos mediante RT-qPCR son diferentes a los porcentajes de tipos de fibras
observados por otros autores mediante otras técnicas en los músculos elevadores de la
mandíbula de diferentes especies de primates no humanos, como Pan troglodytes
(Rowlerson et al., 1983), Macaca mulatta (Maxwell et al., 1979; Miller & Farias, 1988),
Cebus apella (Andreo et al., 2002) y Papio anubis (Wall et al., 2013). Estas diferencias
son debidas, posiblemente, a la presencia de fibras híbridas, que en los músculos
elevadores de la mandíbula de Homo sapiens representan entre el 30% y el 51.5% de las
fibras, y a la capacidad de cada técnica para detectarlas y cuantificarlas (Korfage & Van
Eijden, 1999; Korfage & Van Eijden, 2000; Korfage et al., 2000; Korfage & Van
Eijden, 2003; Österlund et al., 2012; Wall et al., 2013). Las fibras híbridas expresan más
de una isoforma de la MHC y pueden variar su contenido a lo largo de la fibra muscular
(Pette & Staron, 2000; Korfage et al., 2016; Zhang & Gould, 2017). Además, los
estudios realizados por tinción ATPasa en Macaca mulatta (Maxwell et al., 1979;
Miller & Farias, 1988) y Cebus apella (Andreo et al., 2002) no describen la presencia
de fibras IIM, que es el tipo de fibra más habitual en los músculos masticadores de los
primates (Rowlerson et al., 1983).
En resumen, el patrón de expresión de las isoformas de la MHC en los músculos
elevadores de mandíbula en Pan troglodytes nos aporta nueva información a nivel
molecular que se puede relacionar con las características morfológicas y funcionales de
estos músculos observadas mediante otras técnicas.

Conclusiones


87
5. Conclusiones
1- La masa media del masetero en Pan troglodytes es 2.6 veces mayor que la del
pterigoideo medial y la masa media del masetero superficial es 2.8 veces mayor
que la del masetero profundo.
2- El patrón de expresión de las isoformas de la MHC es similar en el masetero
superficial, en el masetero profundo y en el pterigoideo medial, presentando la
secuencia MHC-IIM>MHC-IIA>MHC-I>MHC-IIX.
3- No hemos identificado diferencias significativas en los patrones de expresión de
las isoformas de la MHC entre las porciones anterior y posterior del masetero
superficial y del masetero profundo.
4- No hemos identificado diferencias significativas en los patrones de expresión de
las isoformas de la MHC entre el pterigoideo medial y las porciones superficial
y profunda del masetero.
5- Existen diferencias significativas en el patrón de expresión de la isoforma MHC-
IIM entre la porción superficial y profunda del masetero, presentando la porción
superficial un porcentaje de expresión mayor que la porción profunda.
6- No hemos identificado diferencias significativas en los porcentajes de expresión
de las isoformas MHC-I, MHC-IIA y MHC-IIX entre las porciones superficial y
profunda del masetero.
7- La porción superficial del temporal en Pan troglodytes representa el 33.1% de la
masa total del músculo temporal, la porción profunda representa el 57% y la
porción esfenomandibular el 9.9%.

88
8- En todos los chimpancés comunes diseccionados hemos identificado una porción
esfenomandibular del músculo temporal claramente separada del temporal
profundo. Sin embargo, la inervación de esta porción esfenomandibular se
verifica por el nervio temporal profundo anterior, igual que la porción anterior
del resto del temporal.
9- El patrón de expresión de las isoformas de la MHC en las diferentes porciones
del músculo temporal es similar al observado en los músculos masetero y
pterigoideo medial, siguiendo la secuencia MHC-IIM>MHC-IIA>MHC-
I>MHC-IIX. La única excepción es la región posterior del temporal superficial,
donde se sigue la secuencia MHC-IIM>MHC-I>MHC-IIA>MHC-IIX.
10- No hemos observado diferencias significativas en los patrones de expresión de
las isoformas de la MHC entre las regiones anterior y posterior del temporal
superficial ni entre las regiones anterior y posterior del temporal profundo.
11- El patrón de expresión de las isoformas de la MHC presenta diferencias
significativas entre la porción esfenomandibular y las regiones anteriores del
temporal superficial y del temporal profundo. Concretamente la porción
esfenomandibular presenta una menor expresión de la isoforma MHC-IIM.
12- El patrón de expresión de las isoformas de la MHC en los músculos elevadores
de la mandíbula de Pan troglodytes, obtenido mediante RT-qPCR, se relaciona
con las diferencias funcionales observadas en estos músculos mediante el uso de
la electromiografía en otras especies de primates.

Anexos


91
6. Anexos
6.1. Anexo 1: peso muscular y porcentajes de expresión del ARNm en
Pan troglodytes
La siguiente tabla muestra el peso medio y los porcentajes de expresión del
ARNm en los músculos masetero (porciones superficial y profunda), pterigoideo medial
y temporal (regiones anterior, posterior y esfenomandibular) en Pan troglodytes.
Músculo
Peso medio %MHC-I %MHC-IIA %MHC-IIX %MHC-IIM
(DE) (DE) (DE) (DE) (DE)
Masetero 62 (27.2) 21.5 (4.9) 28.6 (2.2) 12.8 (5.1) 37.2 (3.7)
Porción superficial 45.5 (19.9) 20.3 (5.1) 26.7 (2.5) 12.8 (6.5) 40.2 (5.1)
Región anterior 20.3 (5.6) 27.4 (2.0) 13.1 (6.7) 39.2 (6.0)
Región posterior 20.3 (4.6) 26.0 (2.9) 12.6 (6.7) 41.1 (4.9)
Porción profunda 16.5 (7.5) 22.7 (5.1) 30.4 (2.2) 12.7 (4.0) 34.2 (2.7)
Región anterior 22.6 (5.5) 30.9 (2.3) 12.7 (5.8) 33.8 (1.6)
Región posterior 22.8 (4.8) 30.0 (3.5) 12.6 (4.7) 34.7 (3.9)
Pterigoideo medial 24 (8.6) 22.2 (3.0) 28.3 (1.4) 11.8 (7.4) 37.7 (8.1)
Temporal 120.5 (45.9) 22.0 (2.8) 23.5 (4.8) 13.0 (1.4) 41.5 (5.0)
Superficial 39.9 (11.1)
Región anterior 19.9 (10.1) 22.5 (5.6) 10.6 (6.8) 47 (10.4)
Región posterior 26.9 (7.9) 18.6 (7.5) 14.0 (5.8) 40.5 (16.4)
Profundo 68.7 (7.4)
Región anterior 20.5 (4.6) 22.3 (5.0) 14.2 (7.6) 43.0 (4.9)
Región posterior 20.9 (5.1) 22.5 (7.5) 13.3 (6.5) 43.3 (10.9)
Esfenomandibular 11.9 (4.2) 21.8 (7.2) 31.5 (8.0) 13.1 (7.0) 33.6 (3.2)

92
6.2. Anexo 2: protocolo de la RT-qPCR
La realización de la RT-qPCR se basa en tres procesos diferentes: la extracción
del ARN de las muestras musculares que se van a analizar, la síntesis de ADN
complementario (ADNc) y la RT-qPCR propiamente dicha.
6.2.1. Extracción del ARN mediante el protocolo TriPure de Roche
En primer lugar, se cogen 100 mg de tejido muscular y se colocan en una placa
de Petri. Seguidamente, se añade 1 ml de TriPure y se homogeneiza el tejido trabajando
siempre en frío. Una vez se tiene homogeneizado el tejido, se pipetea en un tubo
Eppendorf de 1.5 ml y se deja incubar durante 5 minutos a temperatura ambiente (15-25
ºC). Pasados los 5 minutos, se añaden 0.2 ml de cloroformo y se agita fuertemente el
tubo Eppendorf durante 15 segundos para que se mezcle bien el contenido. Se vuelve a
incubar el tubo Eppendorf a temperatura ambiente durante 15 minutos. Posteriormente,
se centrifuga la muestra durante 15 minutos a 12000 g y a 2-8 ºC de temperatura.
Después de la centrífuga, el contenido del tubo Eppendorf se separa en una capa
superior que contiene el ARN y una capa inferior que contiene el ADN y las proteínas.
Se aísla el ARN de la fase acuosa incolora de la capa superior en un nuevo tubo
Eppendorf de 1.5 ml y se añaden 0.5 ml de isopropanol. Se invierte manualmente el
tubo varias veces para que se mezclen los distintos componentes y se deja en el
congelador a -20 ºC durante 24 horas.
Una vez pasadas 24 horas, se retira el tubo Eppendorf del congelador y se incuba
a temperatura ambiente durante 20-30 minutos. Pasado este tiempo, se centrifuga la
muestra durante 10 minutos a 12000 g y a 2-8 ºC de temperatura. Se retira el tubo
Eppendorf de la centrifugadora y se descarta el sobrenadante, siendo cuidadosos para no

93
descartar también el pellet6. Se añade 1 ml de etanol al 75% y se realiza un vórtice.
Posteriormente, se centrifuga de nuevo la muestra durante 5 minutos a 7500 g y a 2-8 ºC
de temperatura. Se descarta el sobrenadante y se deja que el exceso de etanol se seque al
aire. Una vez se ha secado el etanol, se vuelve a suspender el pellet de ARN en 50 µl de
agua libre de RNasa, pipeteando repetidas veces para disolver el pellet de ARN y se
incuba durante 15 minutos a 55-60 ºC de temperatura. Pasado este tiempo, se procede a
cuantificar el ARN utilizando el espectrofotómetro NanoDrop 1000 (ThermoFisher) y
se almacena hasta su utilización a -80 ºC.
6.2.2. Síntesis del ADNc
Para realizar la síntesis del ADNc se descongela en primer lugar el ARN
obtenido en el proceso anterior y se realiza un vórtice. Seguidamente, se calcula el
volumen de ARN y de agua libre de RNasa para un volumen final de ADNc de 20 µl.
Es fundamental obtener la misma concentración de ADNc en todas las muestras ya que
posteriormente vamos a cuantificarlas y a analizar sus niveles de expresión. Se pipetean
los volúmenes calculados para cada muestra en su pozo correspondiente de la placa
MicroAmpTM optical 96 (ThermoFisher). Después, se añaden en cada pozo 2 µl de
tampón o buffer, 1.4 µl de MgCl2, 4 µl de dNTP (deoxyribose nucleoside triphosphate),
1 µl de random hexamer, 1 µl de RNA inhibitor y 1 µl de multiscribe. Para mezclar los
diferentes componentes, se tapan los pozos con tiras y se centrifuga la placa durante 1
minuto. Finalmente, se coloca la placa en el termociclador con las condiciones
siguientes: 10 minutos a 25 ºC, 48 minutos a 30 ºC y 5 minutos a 95 ºC para obtener el
ADNc.
6 El pellet es el precipitado de ARN, generalmente invisible, situado en la base del tubo Eppendorf.

94
6.2.3. RT-qPCR
Se realiza la RT-qPCR en un volumen total de 20 µl en ABI Prism 7700
Sequence Detection System (Applied Biosystems). En primer lugar, se pipetean 3 µl de
ADNc de cada muestra en su pozo correspondiente y 3 µl de agua libre de RNasa en un
pozo como control de contaminación. En segundo lugar, se añaden 10 µl de TaqMan, 6
µl de agua libre de RNasa y 1 µl del primer correspondiente (gen objetivo) en cada
pozo. Para finalizar, se centrifuga la placa durante 1 minuto y se realiza la RT-qPCR
con las siguientes condiciones: 2 minutos a 50 ºC, 40 ciclos de 10 minutos a 95 ºC y 15
segundos a 95 ºC y, al final, 1 minuto a 60 ºC.
Al finalizar la RT-qPCR se cuantifica el nivel de expresión de los genes de cada
isoforma de la MHC analizados mediante el método 2-∆∆Ct (Figura 17).
Figura 17. Expresión de las isoformas de la MHC-I, MHC-IIA, MHC-IIX y MHC-IIM en el músculo temporal de Pan troglodytes.

95
6.3. Anexo 3: resultados no publicados
Además de los resultados obtenidos en Pan troglodytes, se ha obtenido una serie
de resultados que aún no se han publicado en relación con la masa muscular y el patrón
de expresión de las isoformas de la MHC en los músculos elevadores de la mandíbula
de Homo sapiens (Tabla 6).
Tabla 6. Porcentajes de expresión del ARNm de las isoformas MHC-I, MHC-IIA, MHC-IIX y MHC-IIM en los músculos elevadores de la mandíbula de Homo sapiens y sus diferentes partes.
Músculo
Peso medio %MHC-I %MHC-IIA %MHC-IIX %MHC-IIM
(DE) (DE) (DE) (DE) (DE)
Masetero 17.8 (5.0) 28,6 (3.8) 34.0 (7.7) 26.4 (5.5) 11.0 (9.5)
Porción superficial 13.2 (3.4) 28.3 (4.4) 31.9 (9.0) 27.8 (7.0) 12.1 (9.4)
Región anterior 29.7 (7.1) 32.7 (9.8) 26.8 (8.8) 10.9 (11.8)
Región posterior 26.9 (3.6) 31.0 (9.8) 28.8 (5.7) 13.3 (8.5)
Porción profunda 4.6 (2.1) 28.9 (4.3) 36.2 (7.9) 25.0 (6.4) 9.9 (9.9)
Región anterior 28.3 (5.0) 37.9 (11.6) 22.9 (9.4) 10.9 (10.5)
Región posterior 29.4 (4.3) 34.6 (7.4) 27.0 (8.3) 9.0 (9.9)
Pterigoideo medial 7.6 (2.7) 29.6 (2.6) 30.4 (5.9) 28.6 (7.3) 11.3 (9.7)
Temporal 24.3 (9.44)
Región anterior 26.3 (11.7) 35.3 (6.0) 25.1 (6.1) 13.3 (12.4)
Región posterior 28.5 (12.1) 39.2 (12.4) 20.6 (7.5) 11.8 (15.1)
Esfenomandibular 37.6 (10.6) 38.5 (7.6) 18.2 (6.1) 5.6 (8.7)
Una de las adaptaciones más características que diferencian a Homo sapiens del
resto de primates hominoideos es la reducción del tamaño de los músculos elevadores
de la mandíbula (Aiello & Dean, 1990). El análisis del peso muscular nos indica que

96
esta reducción de tamaño ha afectado de forma similar a los músculos masetero, a sus
dos porciones, y al pterigoideo medial, y en un orden de magnitud superior al músculo
temporal. Así, si se considera la masa muscular, la relación Pan troglodytes/Homo
sapiens para el músculo masetero es de 3.5, siendo de 3.4 para la porción superficial y
de 3.6 para la porción profunda, y para el músculo pterigoideo medial es de 3.4. En
cambio, la relación existente entre el peso muscular del temporal de Pan troglodytes y
de Homo sapiens es de 5. Esto se debe a la ausencia de la porción superficial del
músculo temporal en Homo sapiens, ya que si eliminamos esta porción en Pan
troglodytes, la relación Pan troglodytes/Homo sapiens en el músculo temporal es de 3.3
(Oxnard & Franklin, 2008).
Los resultados de la RT-qPCR realizada en los músculos masetero, pterigoideo
medial y temporal de Homo sapiens muestran que se expresan todas las isoformas de la
MHC estudiadas, incluida la isoforma MHC-IIM. Posiblemente el ARNm de la
isoforma MHC-IIM detectado se corresponde con la parte del gen que se transcribe en
la porción inestable de la cabeza de la miosina, pero en ningún caso este ARNm llega a
traducirse en una proteína funcional (Stedman et al., 2004). Por este motivo, el ARNm
de la isoforma MHC-IIM no se ha tenido en cuenta para el análisis estadístico y
solamente podemos valorar las diferencias entre Homo sapiens y Pan troglodytes en la
expresión de la isoforma MHC-I y de las isoformas MHC-II en conjunto, es decir, la
expresión de las isoformas MHC-IIA y MHC-IIX en Homo sapiens y la suma de la
expresión de las isoformas MHC-IIA, MHC-IIX y MHC-IIM en Pan troglodytes (Tabla
7).

97
Tabla 7. Porcentajes de expresión del ARNm de las isoformas MHC-I y MHC-II en los músculos elevadores de la mandíbula en Homo sapiens y Pan troglodytes.
Músculo %MHC-I (DE)
Homo sapiens
%MHC-I (DE)
Pan troglodytes
%MHC-II (DE)
Homo sapiens
%MHC-II (DE)
Pan troglodytes
Masetero 32.3 (4.1) 21.5 (4.9) 67.7 (4.1) 78.5 (4.9)
Superficial 32.4 (5.9) 20.3 (5.1) 67.6 (5.9) 79.7 (5.0)
Profundo 32.1 (3.4) 22.7 (5.1) 67.9 (3.4) 77.3 (5.2)
Pterigoideo medial 33.6 (3.1) 22.2 (3.0) 66.4 (3.1) 77.8 (3.0)
Temporal
Región anterior 29.7 (10.7) 20.2 (4.6) 70.3 (10.7) 79.8 (4.6)
Región posterior 31.2 (10.4) 23.9 (5.1) 68.8 (10.4) 76.1 (5.1)
Esfenomandibular 39.6 (9.3) 21.8 (7.2) 60.4 (9.3) 78.2 (7.2)
Los humanos presentan una mayor proporción de ARNm de la isoforma MHC-I.
Como se ha comentado anteriormente, esta isoforma presenta una baja velocidad de
contracción y una baja potencia muscular debido a su baja velocidad de liberación del
ADP y a su baja actividad ATPasa, respectivamente, pero tiene un coste tensional bajo
(Schiaffino & Reggiani, 2011). Esto hace que la isoforma MHC-I sea idónea para
contracciones prolongadas y de baja intensidad. En cambio, Pan troglodytes presenta
una mayor proporción de ARNm de las isoformas MHC-II. Estas isoformas pueden
generar una elevada potencia muscular, en especial la isoforma MHC-IIM, pero se
expresan en fibras con una baja resistencia a la fatiga (Korfage et al., 2005a; Schiaffino
& Reggiani, 2011). Estos resultados sugieren que Homo sapiens ha perdido potencia
muscular a cambio de una mayor resistencia a la fatiga y están en la línea de las
observaciones realizadas por Taylor & Vinyard (2013). Según estos autores, los
músculos elevadores de la mandíbula en Homo sapiens presentan una menor ASTF y,

98
por lo tanto, una menor potencia muscular en comparación con los mismos músculos en
Pan troglodytes. Como se ha comentado anteriormente, el ASTF está directamente
relacionada con la masa muscular y, de hecho, la relación entre el ASTF del masetero
superficial de Pan troglodytes y Homo sapiens es de 3.4 (16.78 cm2 / 4.96 cm2), similar
a la pérdida de masa muscular que hemos observado en nuestra muestra (Powell et al.,
1984; Taylor & Vinyard, 2013).
El patrón de expresión de las isoformas de la MHC obtenido en Homo sapiens es
el mismo para todos los músculos: MHC-IIA>MHC-I>MHC-IIX (Tabla 8). Estos
resultados sugieren que los músculos masetero, pterigoideo medial y temporal presentan
unas características funcionales similares tal y como indican los estudios
electromiográficos, que muestran una activación generalizada de estos músculos durante
la elevación de la mandíbula (Blanksma et al., 1997).
Tabla 8. Porcentajes de expresión del ARNm de las isoformas MHC-I, MHC-IIA y MHC-IIX en los músculos elevadores de la mandíbula de Homo sapiens y sus diferentes partes.
Músculo %MHC-I (DE) %MHC-IIA (DE) %MHC-IIX (DE) %MHC-II (DE)
Masetero 32.3 (4.1) 37.8 (6.4) 30.0 (6.6) 67.7 (4.1)
Superficial 32.4 (5.9) 35.7 (8.6) 31.9 (7.8) 67.6 (5.9)
Profundo 32.1 (3.4) 39.9 (6.2) 28.0 (7.8) 67.9 (3.4)
Pterigoideo medial 32.1 (3.1) 34.3 (5.9) 32.1 (6.0) 67.9 (3.1)
Temporal
Región anterior 29.7 (10.7) 41.2 (7.6) 29.1 (6.2) 70.3 (10.7)
Región posterior 31.2 (10.4) 43.3 (12.1) 25.4 (8.6) 68.7 (10.4)
Esfenomandibular 39.6 (9.3) 41.0 (7.9) 19.4 (6.4) 60.4 (9.3)

99
A nivel intramuscular, no hemos obtenido diferencias significativas entre las dos
porciones del músculo masetero, pero existe una tendencia a la mayor expresión de
ARNm de la isoforma MHC-IIX en el masetero superficial y a la mayor expresión de
ARNm de la isoforma MHC-IIA en el masetero profundo. La isoforma MHC-I presenta
un patrón de expresión prácticamente idéntico en las dos porciones del masetero. Este
modelo de expresión se corresponde con los estudios electromiográficos que muestran
una mayor actividad del masetero superficial durante la fase de clausura de la
mandíbula, cuando se requiere de una elevada fuerza muscular, mientras que la porción
profunda actúa en movimientos laterales y de retrusión de la mandíbula, posiblemente
relacionados con la colocación de los molares (Wood, 1987; Blanksma et al., 1997).
Tampoco existen diferencias significativas a nivel de expresión del ARNm entre las
regiones anterior y posterior del masetero superficial y profundo, aunque el porcentaje
de expresión de la isoforma MHC-IIX es ligeramente menor en la región anterior
(30.2% en el masetero superficial y 26.3% en el masetero profundo) que en la región
posterior (33.5% en el masetero superficial y 29.8% en el masetero profundo). Este
modelo de expresión está en la línea de las observaciones realizadas por Guzmán-
Venegas et al. (2015) que muestran una mayor actividad electromiográfica de la región
anterior del masetero en acciones submaximales, en las cuales se reclutan en primer
lugar las fibras más resistentes a la fatiga.
Por otro lado, el modelo de expresión del ARNm de las isoformas MHC-I,
MHC-IIA y MHC-IIX en el músculo temporal refleja la división funcional descrita en
los estudios morfológicos y electromiográficos (Blanksma & Van Eijden, 1995;
Blanksma et al., 1997; Van Eijden et al., 1997). La región anterior del temporal actúa
durante la elevación de la mandíbula y presenta una mayor proporción de la isoforma
MHC-IIX, mientras que la región posterior participa en el mantenimiento de la postura

100
de la mandíbula y en su movimiento de retrusión, presentando una mayor proporción de
las isoformas más tónicas, la MHC-I y la MHC-IIA. Sin embargo, todas estas
diferencias observadas no son significativas. Las únicas diferencias significativas que
hemos observado respecto a la expresión de las isoformas de la MHC se han
identificado entre la región anterior del músculo temporal y su porción
esfenomandibular. La porción esfenomandibular presenta una mayor expresión de la
isoforma MHC-I y una menor expresión de la isoforma MHC-IIX en comparación con
la región anterior del temporal. Estos resultados se corresponden con las observaciones
realizadas en Pan troglodytes y coinciden con los estudios electromiográficos que
muestran una mayor participación de la porción esfenomandibular del temporal en el
mantenimiento de la postura y la estabilización de la mandíbula (Zenker, 1955; Wood,
1987; Fuentes et al., 2012).
Los porcentajes de expresión de las isoformas de la MHC que hemos obtenido
en nuestro estudio difieren notablemente de los porcentajes de tipos de fibras
musculares obtenidos en estudios histoquímicos e inmunohistoquímicos en los mismos
músculos. Este hecho se debe probablemente al papel que juegan las fibras híbridas en
este tipo de estudios (Ringqvist, 1974; Vignon et al., 1980; Eriksson & Thornell, 1983;
Shaughnessy et al., 1989; Korfage et al., 2000; Korfage et al., 2005b).
6.4. Anexo 4: objetivos de futuro
Actualmente, estamos trabajando en la cuantificación de las isoformas MHC-I y
MHC-II en los músculos elevadores de la mandíbula mediante SDS-PAGE. Este
método nos puede ser de utilidad para valorar si los niveles de ARNm de las isoformas
MHC-I y MHC-II obtenidos por RT-qPCR se correlacionan con los niveles de proteína.

101
El protocolo utilizado es el descrito por Talmadge & Roy (1993) junto con las
modificaciones propuestas por Bamman et al. (1999), Khon & Myburgh (2006) y
Mizunoya et al. (2008). Estos autores proponen aumentar la relación de acrilamida-bis
acrilamida (99:1) y la concentración del tampón superior para mejorar la resolución y la
separación de las bandas de las diferentes proteínas.
6.4.1. Extracción y cuantificación de proteínas
Para la extracción de las proteínas de una muestra muscular de 0.3 cm3 se utiliza un
tampón o buffer de lisis compuesto por: Tris-HCl, NaCl y tritón. Se tritura la muestra
muscular en 100 µl de tampón de lisis con 2 µl de cóctel inhibidor de proteasas y 1 µl
de ortoganato sódico 100X. Una vez triturada la muestra muscular, se pipetea en un
tubo Eppendorf de 1.5 ml y se deja en hielo durante una hora, realizándose vórtices cada
5-10 minutos. Posteriormente, se centrifuga la muestra a 14000 rpm durante 15 minutos
a 4 ºC. Seguidamente, se separa el sobrenadante (proteína) y se coloca en otro tubo
Eppendorf de 1.5 ml.
Una vez extraída la proteína, se cuantifica utilizando el método Bradford. En
primer lugar, se realiza una recta estándar con BSA (albúmina de suero bovino) para
poder conocer la concentración de proteína de nuestras muestras musculares (Tabla 9).

102
Tabla 9. Valores de BSA y de H2O miliQ utilizados para realizar la recta estándar.
Concentración (µg/µl) BSA (µl) H2O miliQ (µl)
0 0 200
1 0.25 199.75
2.5 0.625 199.375
5 1.250 198.75
7.5 1.875 198.125
10 2.5 197.5
15 3.75 196.25
20 5 195
En segundo lugar, se pipetean en un pozo 0.5 µl de muestra muscular con 199.5 µl
de H2O miliQ. Cada muestra muscular se pipetea en dos pozos diferentes de la placa
para cuantificar todas las muestras musculares por duplicado. Finalmente, se pipetean
en un pozo 0.5 µl del tampón de lisis y 199.5 µl de H2O miliQ para utilizarlo como
“blanco” (nos va a servir para quitar el ruido de fondo).
Una vez pipeteada la recta estándar, las muestras musculares y el “blanco” se
añaden 50 µl de Bradford en todos los pozos y se mezcla. Se protege de la luz con papel
de aluminio y se deja reposar a temperatura ambiente durante 30 minutos. Finalmente,
se cuantifican las muestras en el espectrofotómetro con una absorbancia de 595 nm.
6.4.2. Electroforesis en gel de poliacrilamida
Para realizar la electroforesis en gel de poliacrilamida (SDS-PAGE) se utiliza el
sistema de mini-geles de Bio-rad (Mini-Protean Tetra Cell®) (Figura 18).

103
Figura 18. Material utilizado para la preparación del gel de poliacrilamida.
Para preparar un gel de 0.75 mm de grosor, se necesitan los componentes
indicados en la Tabla 10.
Tabla 10. Componentes para la realización de un gel de poliacrilamida de 0.75 mm de grosor.
Componentes
Glicerol 100%
Acrilamida-bis 30% (99:1)
Tris 1.5 M (pH 8.8)
Tris 0.5 (pH 6.7)
Glicina 1M
EDTA 100 mM (pH 7.0)
SDS 10%
H2O destilada
TEMED
Amonio persulfato 10%
Previamente a la preparación del gel separador y del gel compactador se prepara el
marco de fundición (Figura 19).
Soporte de fundición
Placa de vidrio con espaciadores de 0.75mm
Placa de vidrio
Marco de fundición
Junta
Peine de 10 pozos

104
Figura 19. Colocación de los dos vidrios en el marco de fundición (izquierda) y el soporte de fundición preparado para añadir la solución del gel separador y del gel compactador (derecha).
Una vez tenemos el marco de fundición colocado en el soporte de fundición, se
prepara el gel separador (separating gel) con una composición de glicerol del 35% y del
8% acrilamida-bis acrilamida (99:1) (Tabla 11). Los últimos elementos que se añaden a
la solución del gel son el amonio persulfato al 10% y el TEMED
(tetrametiletilendiamina) para que inicien la polimerización. Inmediatamente después,
se mezcla la solución y se vierte en el soporte de fundición, dejando espacio suficiente
en la parte superior para el gel compactador.
Tabla 11. Componentes y volúmenes para la realización del gel separador.
Gel separador Volumen (µl)
Glicerol 100% 1.500
Acrilamida-bis 30% (99:1) 1.334
Tris 1.5 M (pH 8.8) 0.660
Glicina 1M 0.500
SDS 10% 0.200
H2O destilada 0.747
TEMED 0.003
Amonio persulfato 10% 0.050

105
Una vez el gel separador ha polimerizado, se prepara el gel compactador
(stacking gel) con una composición de glicerol del 35% y de 4% acrilamida-bis
acrilamida (99:1) (Tabla 12). Igual que antes, el amonio persulfato al 10% y el TEMED
son los últimos componentes que se añaden a la solución. Inmediatamente después, se
mezcla la solución, se vierte en el soporte de fundición y se coloca el peine para
muestras vigilando que no queden burbujas.
Tabla 12. Componentes y volúmenes para la realización del gel compactador.
Gel compactador Volumen (µl)
Glicerol 100% 1.500
Acrilamida-bis 30% (99:1) 1.334
Tris 0.5 M (pH 6.7) 0.700
EDTA 100 mM (pH 7.0) 0.200
SDS 10% 0.200
H2O destilada 1.680
TEMED 0.003
Amonio persulfato 10% 0.050
Mientras polimeriza el gel compactador se preparan los tampones, el superior y
el inferior. El tampón inferior (lower buffer) consiste en 50 mM de Tris (base), 75 mM
de glicina y 0.05% w/v de SDS, mientras que el tampón superior (upper buffer)
equivale a seis veces la concentración del tampón inferior.
Una vez tenemos el gel polimerizado, se coloca en la cubeta o tanque, se añade el
tampón superior y se preparan las proteínas (Figura 20). En primer lugar, se calcula la
cantidad de proteína para un volumen final de 20 µl. En general, se utiliza una
concentración de proteína de 5 a 15 µg en cada muestra y se utiliza el tampón de lisis
para ajustar el volumen. Se pipetea el volumen de proteína y de tampón de lisis en un

106
tubo Eppendorf de 0.2 ml y se añade 2 µl de DTT (1,4-Dithiothreitol) y 4 µl de laemmli
buffer 5X. Se mezclan bien los componentes y se desnaturalizan las proteínas a una
temperatura de 95 ºC durante 3 minutos.
Figura 20. Tanque con el gel de poliacrilamida.
Antes de colocar las proteínas en los pozos, se quita el peine y se lava el pozo con
un poco de tampón superior para eliminar los residuos no polimerizados. Seguidamente,
se cargan las proteínas en el pozo correspondiente dejando un pozo libre para cargar 10
µl de marcador de peso molecular o ladder. Finalmente, se añade el tampón inferior y
dejamos correr el gel durante 20 horas a 70 voltios a una temperatura aproximada de 4
ºC (Figura 21).
Figura 21. Tanque con el gel de poliacrilamida sin el peine y con los dos tampones, el superior y el inferior (izquierda). Tanque con el gel de poliacrilamida conectado a la fuente de energía (derecha).

107
Pasadas las 20 horas, se saca el gel con cuidado, se lava con agua destilada un par
de veces y se tiñe con azul Coomassie R-250 durante 1 hora. Después, se elimina el
exceso de tinción con uno o dos lavados de agua destilada. Finalmente, se realiza la
cuantificación de las proteínas de las isoformas MHC-I y MHC-II por densitometría
mediante el programa ImageJ (Figura 22).
Figura 22. SDS-PAGE donde vemos las bandas correspondientes a la MHC-I, MHC-II y MHC-IIM.
En el futuro queremos ampliar la muestra de primates y comparar la expresión de
los ARNm de las isoformas MHC-I y MHC-II con la cuantificación de su proteína
mediante SDS-PAGE. Esto puede aportar nueva información en relación con las
características funcionales de los músculos elevadores de la mandíbula que puede
complementar a la obtenida por otros métodos.
MHC-IIM
MHC-II MHC-I


Bibliografía


111
7. Bibliografía
Ahlgren, J. (1986). EMG pattern of temporalis in normal occlusion. The European
Journal of Orthodontics, 8(3), 185-191.
Aiello, L., & Dean, C. (1990). An introduction to human evolutionary anatomy.
London: Academic Press.
Akita, K., Shimokawa, T., & Sato, T. (2000). Positional relationships between the
masticatory muscles and their innervating nerves with special reference to the lateral
pterygoid and the midmedial and discotemporal muscle bundles of temporalis. The
Journal of Anatomy, 197(2), 291-302.
Alberts, B., Bray, D., Hopkin, K., Johnson, A., Lewis, J., Raff, M., Roberts, k., &
Walter, P. (2013). Essential cell biology. Garland Science.
Andreo, J., Oliveira, J., Navarro, J., Roque, D., Roque, J., & Buchain, R. (2002).
Histoenzymology and morphometry of the masticatory muscles of tufted capuchin
monkey (Cebus apella Linnaeus, 1758). Okajimas folia anatomica Japonica, 79(1), 33-
41.
Ankel-Simons, F. (2010). Primate anatomy: an introduction. London: Academic Press.
Bahar, B., Monahan, F. J., Moloney, A. P., Schmidt, O., MacHugh, D. E., & Sweeney,
T. (2007). Long-term stability of RNA in post-mortem bovine skeletal muscle, liver and
subcutaneous adipose tissues. BMC Molecular Biology, 8(1), 108.

112
Bamman, M. M., Clarke, M. S., Talmadge, R. J., & Feeback, D. L. (1999). Enhanced
protein electrophoresis technique for separating human skeletal muscle myosin heavy
chain isoforms. Electrophoresis, 20(3), 466-468.
Basmajian, J. V., & De Luca, C. (1985). Muscles alive. Muscles alive: their functions
revealed by electromyography. Williams & Wilkins.
Blanksma, N., & Van Eijden, T. (1995). Electromyographic heterogeneity in the human
temporalis and masseter muscles during static biting, open\close excursions, and
chewing. Journal of dental research, 74(6), 1318-1327.
Blanksma, N., Van Eijden, T., Van Ruijven, L., & Weijs, W. (1997).
Electromyographic heterogeneity in the human temporalis and masseter muscles during
dynamic tasks guided by visual feedback. Journal of dental research, 76(1), 542-551.
Blanksma, N., Van Eijden, T., & Weijs, W. (1992). Electromyographic heterogeneity in
the human masseter muscle. Journal of dental research, 71(1), 47-52.
Borukhov, S., & Nudler, E. (2008). RNA polymerase: the vehicle of transcription.
Trends in microbiology, 16(3), 126-134.
Bottinelli, R., Canepari, M., Pellegrino, M., & Reggiani, C. (1996). Force‐velocity
properties of human skeletal muscle fibres: myosin heavy chain isoform and
temperature dependence. The Journal of physiology, 495(2), 573-586.
Bottinelli, R., & Reggiani, C. (2000). Human skeletal muscle fibres: molecular and
functional diversity. Progress in biophysics and molecular biology, 73(2-4), 195-262.

113
Bredman, J., Wessels, A., Weijs, W., Korfage, J., Soffers, C., & Moorman, A. (1991).
Demonstration of ‘cardiac-specific’ myosin heavy chain in masticatory muscles of
human and rabbit. The Histochemical journal, 23(4), 160-170.
Briggs, M. M., & Schachat, F. (2000). Early specialization of the superfast myosin in
extraocular and laryngeal muscles. Journal of Experimental Biology, 203(16), 2485-
2494.
Brooke, M. H., & Kaiser, K. K. (1970). Muscle fiber types: how many and what kind?
Archives of neurology, 23(4), 369-379.
Brooke, M. H., & Kaiser, K. K. (1974). The use and abuse of muscle histochemistry.
Annals of the New York Academy of sciences, 228(1), 121-144.
Bustin, S. A. (2000). Absolute quantification of mRNA using real-time reverse
transcription polymerase chain reaction assays. Journal of molecular endocrinology,
25(2), 169-193.
Butler‐Browne, G., Eriksson, P. O., Laurent, C., & Thornell, L. E. (1988). Adult human
masseter muscle fibers express myosin isozymes characteristic of development. Muscle
& nerve, 11(6), 610-620.
Cachel, S. M. (1979). A functional analysis of the primate masticatory system and the
origin of the anthropoid post‐orbital septum. American journal of physical
anthropology, 50(1), 1-17.
Campbell, C. J., Fuentes, A., MacKinnon, K. C., Panger, M., & Bearder, S. K. (2007).
Primates in perspective. New York: Oxford University Press.

114
Cox, R. D., & Buckingham, M. E. (1992). Actin and myosin genes are transcriptionally
regulated during mouse skeletal muscle development. Developmental biology, 149(1),
228-234.
Crick, F. (1979). Split genes and RNA splicing. Science, 204(4390), 264-271.
Desjardins, P. R., Burkman, J. M., Shrager, J. B., Allmond, L. A., & Stedman, H. H.
(2002). Evolutionary implications of three novel members of the human sarcomeric
myosin heavy chain gene family. Molecular Biology and Evolution, 19(4), 375-393.
Dunn, G. F., Hack, G. D., Robinson, W. L., Koritzer, R. T., Sharawy, M., & Tillman, L.
J. (1996). Anatomical observation of a craniomandibular muscle originating from the
skull base: the sphenomandibularis. Cranio, 14(2), 97-105.
Eizema, K., van den Burg, M. M., de Jonge, H. W., Dingboom, E. G., Weijs, W. A., &
Everts, M. E. (2005). Myosin heavy chain isoforms in equine gluteus medius muscle:
comparison of mRNA and protein expression profiles. Journal of Histochemistry &
Cytochemistry, 53(11), 1383-1390.
Eriksson, P. O., & Thornell, L. E. (1983). Histochemical and morphological muscle-
fibre characteristics of the human masseter, the medial pterygoid and the temporal
muscles. Archives of Oral Biology, 28(9), 781-795.
Fleagle, J. G. (2013). Primate adaptation and evolution. London: Academic Press.
Fuentes, E., Frugone, R., Paolinelli, C., Hack, G., & Bittner, V. (2012).
Electromyographic activity of the sphenomandibularis and lateral pterygoid human
muscles during mandibular lateral movements. Chirurgia, 25, 97-99.

115
Geers, C., Nyssen-Behets, C., Cosnard, G., & Lengelé, B. (2005). The deep belly of the
temporalis muscle: an anatomical, histological and MRI study. Surgical and Radiologic
Anatomy, 27(3), 184-191.
Goodman, M., Tagle, D. A., Fitch, D. H., Bailey, W., Czelusniak, J., Koop, B. F.,
Benson, P., & Slightom, J. L. (1990). Primate evolution at the DNA level and a
classification of hominoids. Journal of molecular evolution, 30(3), 260-266.
Groves, C. (2005). Mammal species of the world: A taxonomic and geographic
reference (3rd ed). Baltimore: John Hopkins University Press.
Grünheid, T., Langenbach, G. E., Korfage, J. A., Zentner, A., & Van Eijden, T. M.
(2009). The adaptive response of jaw muscles to varying functional demands. The
European Journal of Orthodontics, 31(6), 596-612.
Guzmán-Venegas, R. A., Picand, J. L. B., & de la Rosa, F. J. B. (2015). Functional
compartmentalization of the human superficial masseter muscle. PloS One, 10(2),
e0116923.
Harrison, B. C., Allen, D. L., & Leinwand, L. A. (2011). IIb or not IIb? Regulation of
myosin heavy chain gene expression in mice and men. Skeletal muscle, 1(1), 5.
Hemmings, K., Parr, T., Daniel, Z., Picard, B., Buttery, P., & Brameld, J. (2009).
Examination of myosin heavy chain isoform expression in ovine skeletal muscles.
Journal of animal science, 87(12), 3915-3922.
Hoh, J. F. (2002). Superfast'or masticatory myosin and the evolution of jaw-closing
muscles of vertebrates. Journal of Experimental Biology, 205(15), 2203-2210.

116
Horton, M. J., Brandon, C. A., Morris, T. J., Braun, T. W., Yaw, K. M., & Sciote, J. J.
(2001). Abundant expression of myosin heavy-chain IIB RNA in a subset of human
masseter muscle fibres. Archives of Oral Biology, 46(11), 1039-1050.
Hylander, W. L. (1979). The functional significance of primate mandibular form.
Journal of Morphology, 160(2), 223-239.
Hylander, W. L., & Johnson, K. R. (1994). Jaw muscle function and wishboning of the
mandible during mastication in macaques and baboons. American journal of physical
anthropology, 94(4), 523-547.
Hylander, W. L., Johnson, K. R., & Crompton, A. (1987). Loading patterns and jaw
movements during mastication in Macaca fascicularis: A bone‐strain,
electromyographic, and cineradiographic analysis. American journal of physical
anthropology, 72(3), 287-314.
Hylander, W. L., Ravosa, M. J., Ross, C. F., Wall, C. E., & Johnson, K. R. (2000).
Symphyseal fusion and jaw-adductor muscle force: an EMG study. American journal of
physical anthropology, 112(4), 469-492.
Hylander, W. L., Wall, C. E., Vinyard, C. J., Ross, C., Ravosa, M. R., Williams, S. H.,
& Johnson, K. R. (2005). Temporalis function in anthropoids and strepsirrhines: an
EMG study. American journal of physical anthropology, 128(1), 35-56.
Kerppola, T. K., & Kane, C. M. (1991). RNA polymerase: regulation of transcript
elongation and termination. The FASEB Journal, 5(13), 2833-2842.
Kohn, T. A., & Myburgh, K. H. (2006). Electrophoretic separation of human skeletal
muscle myosin heavy chain isoforms: the importance of reducing agents. The Journal of
Physiological Sciences, 56(5), 355-360.

117
Korfage, J., Brugman, P., & Van Eijden, T. (2000). Intermuscular and intramuscular
differences in myosin heavy chain composition of the human masticatory muscles.
Journal of the neurological sciences, 178(2), 95-106.
Korfage, J., Koolstra, J., Langenbach, G., & Van Eijden, T. (2005a). Fiber-type
composition of the human jaw muscles-(part 1) origin and functional significance of
fiber-type diversity. Journal of dental research, 84(9), 774-783.
Korfage, J., Koolstra, J., Langenbach, G., & Van Eijden, T. (2005b). Fiber-type
composition of the human jaw muscles-(part 2) role of hybrid fibers and factors
responsible for inter-individual variation. Journal of dental research, 84(9), 784-793.
Korfage, J., Kwee, K., Everts, V., & Langenbach, G. (2016). Myosin heavy chain
expression can vary over the length of jaw and leg muscles. Cells Tissues Organs,
201(2), 130-137.
Korfage, J., & Van Eijden, T. (1999). Regional differences in fibre type composition in
the human temporalis muscle. Journal of anatomy, 194(3), 355-362.
Korfage, J., & Van Eijden, T. (2000). Myosin isoform composition of the human medial
and lateral pterygoid muscles. Journal of dental research, 79(8), 1618-1625.
Korfage, J., & Van Eijden, T. (2003). Myosin heavy chain composition in human
masticatory muscles by immunohistochemistry and gel electrophoresis. Journal of
Histochemistry & Cytochemistry, 51(1), 113-119.
Lee, J. Y., Kim, J. N., Kim, S. H., Choi, H. G., Hu, K. S., Kim, H. J., Song, W. C., &
Koh, K. S. (2012). Anatomical verification and designation of the superficial layer of
the temporalis muscle. Clinical Anatomy, 25(2), 176-181.

118
Lewin, R. (2000). Evolución humana: la más amplia perspectiva acerca de los orígenes
de la humanidad. Salvat.
Lieber, R. L., & Friden, J. (2000). Functional and clinical significance of skeletal
muscle architecture. Muscle & nerve, 23(11), 1647-1666.
Lieberman, D. (2011). The evolution of the human head. Harvard University Press.
Lodish, H., Berk, A., Zipursky, S. L., Matsudaira, P., Baltimore, D., & Darnell, J.
(2000). Molecular cell biology (4th ed). New York: W. H. Freeman.
Martin, R. D. (1990). Primate origins and evolution. A phylogenetic reconstruction.
New Jersey: Princeton University Press.
Maxwell, L. C., Carlson, D. S., McNamara, J. A., & Faulkner, J. A. (1979).
Histochemical characteristics of the masseter and temporalis muscles of the rhesus
monkey (Macaca mulatta). The Anatomical Record, 193(3), 389-401.
Men, X., Deng, B., Tao, X., Qi, K., & Xu, Z. W. (2016). Association analysis of myosin
heavy-chain genes mRNA transcription with the corresponding proteins expression of
Longissimus muscle in growing pigs. Asian-Australasian journal of animal sciences,
29(4), 457-463.
Méndez, J. (1960). Density and composition of mammalian muscle. Metabolism, 9,
184-188.
Miller, A. J., & Farias, M. (1988). Histochemical and electromyographic analysis of
craniomandibular muscles in the rhesus monkey, Macaca mulatta. Journal of Oral and
Maxillofacial Surgery, 46(9), 767-776.

119
Mizunoya, W., Wakamatsu, J. I., Tatsumi, R., & Ikeuchi, Y. (2008). Protocol for high-
resolution separation of rodent myosin heavy chain isoforms in a mini-gel
electrophoresis system. Analytical biochemistry, 377(1), 111-113.
Monemi, M., Kadi, F., Liu, J. X., Thornell, L., & Eriksson, P. (1999). Adverse changes
in fibre type and myosin heavy chain compositions of human jaw muscle vs. limb
muscle during ageing. Acta physiologica scandinavica, 167(4), 339-346.
Ni, X., Wang, Y., Hu, Y., & Li, C. (2004). A euprimate skull from the early Eocene of
China. Nature, 427(6969), 65-68.
Österlund, C., Lindström, M., Thornell, L. E., & Eriksson, P. O. (2012). Remarkable
heterogeneity in myosin heavy-chain composition of the human young masseter
compared with young biceps brachii. Histochemistry and cell biology, 138(4), 669-682.
Oxnard, C., & Franklin, D. (2008). Ghosts of the past I: some muscles and fasciae in the
head domain. Folia Primatologica, 79(6), 429-440.
Patterson, N., Richter, D. J., Gnerre, S., Lander, E. S., & Reich, D. (2006). Genetic
evidence for complex speciation of humans and chimpanzees. Nature, 441(7097), 1103-
1108.
Perelman, P., Johnson, W. E., Roos, C., Seuánez, H. N., Horvath, J. E., Moreira, M. A.,
Kessing, B., Pontius, J., Roelke, M., Rumpler, Y., Schneider, M. P., Silva, A., O’Brien,
S. J., & Pecon-Slattery, J. (2011). A molecular phylogeny of living primates. PLoS
genetics, 7(3), e1001342.
Perry, G. H., Verrelli, B. C., & Stone, A. C. (2005). Comparative analyses reveal a
complex history of molecular evolution for human MYH16. Molecular Biology and
Evolution, 22(3), 379-382.

120
Pette, D., & Staron, R. S. (2000). Myosin isoforms, muscle fiber types, and transitions.
Microscopy research and technique, 50(6), 500-509.
Powell, P. L., Roy, R. R., Kanim, P., Bello, M. A., & Edgerton, V. R. (1984).
Predictability of skeletal muscle tension from architectural determinations in guinea pig
hindlimbs. Journal of Applied Physiology, 57(6), 1715-1721.
Qin, H., Hsu, M. K., Morris, B. J., & Hoh, J. F. (2002). A distinct subclass of
mammalian striated myosins: Structure and molecular evolution of "superfast" or
masticatory myosin heavy chain. Journal of molecular evolution, 55(5), 544-552.
Rafferty, J. P. (2011). Primates (The Britannica guide to predators and prey). New
York: Britannica Educational Publishing.
Ringqvist, M. (1974). Fiber types in human masticatory muscles. Relation to function.
European Journal of Oral Sciences, 82(4), 333-355.
Roberts, M. D., Dalbo, V. J., Sunderland, K. L., & Kerksick, C. M. (2012).
Electrophoretic separation of myosin heavy chain isoforms using a modified mini gel
system. The Journal of Strength & Conditioning Research, 26(12), 3461-3468.
Rosenberger, A. L. (2010). Adaptive profile versus adaptive specialization: fossils and
gummivory in early primate evolution The evolution of exudativory in primates (pp.
273-295). New York: Springer.
Ross, C. (1995). Muscular and osseous anatomy of the primate anterior temporal fossa
and the functions of the postorbital septum. American journal of physical anthropology,
98(3), 275-306.

121
Ross, C. F., & Iriarte‐Diaz, J. (2014). What does feeding system morphology tell us
about feeding? Evolutionary Anthropology: Issues, News, and Reviews, 23(3), 105-120.
Rowlerson, A., Mascarello, F., Barker, D., & Saed, H. (1988). Muscle-spindle
distribution in relation to the fibre-type composition of masseter in mammals. Journal
of anatomy, 161, 37-60.
Rowlerson, A., Mascarello, F., Veggetti, A., & Carpenè, E. (1983). The fibre-type
composition of the first branchial arch muscles in Carnivora and Primates. Journal of
Muscle Research & Cell Motility, 4(4), 443-472.
Rowlerson, A., Pope, B., Murray, J., Whalen, R., & Weeds, A. (1981). A novel myosin
present in cat jaw-closing muscles. Journal of Muscle Research & Cell Motility, 2(4),
415-438.
Sciote, J., Rowlerson, A., Hopper, C., & Hunt, N. (1994). Fibre type classification and
myosin isoforms in the human masseter muscle. Journal of the neurological sciences,
126(1), 15-24.
Sciote, J. J., Horton, M. J., Rowlerson, A. M., & Link, J. (2003). Specialized cranial
muscles: how different are they from limb and abdominal muscles? Cells Tissues
Organs, 174(1-2), 73-86.
Sciote, J. J., & Morris, T. J. (2000). Skeletal muscle function and fibre types: the
relationship between occlusal function and the phenotype of jaw-closing muscles in
human. Journal of orthodontics, 27(1), 15-30.
Schiaffino, S., & Reggiani, C. (1996). Molecular diversity of myofibrillar proteins: gene
regulation and functional significance. Physiological reviews, 76(2), 371-423.

122
Schiaffino, S., & Reggiani, C. (2011). Fiber types in mammalian skeletal muscles.
Physiological reviews, 91(4), 1447-1531.
Schiaffino, S., Rossi, A. C., Smerdu, V., Leinwand, L. A., & Reggiani, C. (2015).
Developmental myosins: expression patterns and functional significance. Skeletal
muscle, 5(1), 22.
Schindler, H., Rues, S., Türp, J., Schweizerhof, K., & Lenz, J. (2007). Jaw clenching:
muscle and joint forces, optimization strategies. Journal of dental research, 86(9), 843-
847.
Schön Ybarra, M. A., & Bauer, B. (2001). Medial portion of M. temporalis and its
potential involvement in facial pain. Clinical Anatomy, 14(1), 25-30.
Schrago, C. G., & Russo, C. A. (2003). Timing the origin of New World monkeys.
Molecular Biology and Evolution, 20(10), 1620-1625.
Shaughnessy, T., Fields, H., & Westbury, J. (1989). Association between craniofacial
morphology and fiber-type distributions in human masseter and medial pterygoid
muscles. The International journal of adult orthodontics and orthognathic surgery,
4(3), 145-155.
Skinner, C. E., & Aziz, M. A. (2003). New methods of dissecting the masticator space
of the rhesus macaque (Macaca mulatta). Primates, 44(4), 311-320.
Snow, D., Billeter, R., Mascarello, F., Carpene, E., Rowlerson, A., & Jenny, E. (1982).
No classical type IIB fibres in dog skeletal muscle. Histochemistry, 75(1), 53-65.
Stedman, H. H., Kozyak, B. W., Nelson, A., Thesier, D. M., Su, L. T., Low, D. W.,
Bridges, C. R., Shrager, J. B., Minugh-Purvis, N., & Mitchell, M. A. (2004). Myosin

123
gene mutation correlates with anatomical changes in the human lineage. Nature,
428(6981), 415-418.
Stringer, C., & Andrews, P. (2005). The complete world of human evolution. London:
Thames & Hudson Ltd.
Swindler, D. R. (1998). Introduction to the Primates. Seattle: University of Washington
Press.
Talmadge, R. J., & Roy, R. R. (1993). Electrophoretic separation of rat skeletal muscle
myosin heavy-chain isoforms. Journal of Applied Physiology, 75(5), 2337-2340.
Taylor, A. B. (2006). Feeding behavior, diet, and the functional consequences of jaw
form in orangutans, with implications for the evolution of Pongo. Journal of Human
Evolution, 50(4), 377-393.
Taylor, A. B., & Vinyard, C. J. (2004). Comparative analysis of masseter fiber
architecture in tree‐gouging (Callithrix jacchus) and nongouging (Saguinus oedipus)
callitrichids. Journal of Morphology, 261(3), 276-285.
Taylor, A. B., & Vinyard, C. J. (2013). The relationships among jaw‐muscle fiber
architecture, jaw morphology, and feeding behavior in extant apes and modern humans.
American journal of physical anthropology, 151(1), 120-134.
Toniolo, L., Cancellara, P., Maccatrozzo, L., Patruno, M., Mascarello, F., & Reggiani,
C. (2008). Masticatory myosin unveiled: first determination of contractile parameters of
muscle fibers from carnivore jaw muscles. American Journal of Physiology-Cell
Physiology, 295(6), C1535-C1542.

124
Türp, J. C., Cowley, T., & Stohler, C. S. (1997). Media hype: musculus
sphenomandibularis. Cells Tissues Organs, 158(2), 150-154.
Van Eijden, T., Korfage, J., & Brugman, P. (1997). Architecture of the human
jaw‐closing and jaw‐opening muscles. The Anatomical Record, 248(3), 464-474.
Vignon, C., Pellissier, J., & Serratrice, G. (1980). Further histochemical studies on
masticatory muscles. Journal of the neurological sciences, 45(2-3), 157-176.
Vinyard, C. J., & Taylor, A. B. (2010). A Preliminary Analysis of the Relationship
Between Jaw‐Muscle Architecture and Jaw‐Muscle Electromyography During Chewing
Across Primates. The Anatomical Record, 293(4), 572-582.
Vinyard, C. J., Wall, C. E., Williams, S. H., & Hylander, W. L. (2008). Patterns of
variation across primates in jaw-muscle electromyography during mastication.
American Zoologist, 48(2), 294-311.
Vinyard, C. J., Wall, C. E., Williams, S. H., Johnson, K. R., & Hylander, W. L. (2006).
Masseter electromyography during chewing in ring‐tailed lemurs (Lemur catta).
American journal of physical anthropology, 130(1), 85-95.
Walklate, J., Ujfalusi, Z., & Geeves, M. A. (2016). Myosin isoforms and the
mechanochemical cross-bridge cycle. Journal of Experimental Biology, 219(2), 168-
174.
Wall, C. E., Briggs, M. M., Huq, E., Hylander, W. L., & Schachat, F. (2013). Regional
variation in IIM myosin heavy chain expression in the temporalis muscle of female and
male baboons (Papio anubis). Archives of Oral Biology, 58(4), 435-443.

125
Wall, C. E., Vinyard, C. J., Johnson, K. R., Williams, S. H., & Hylander, W. L. (2006).
Phase II jaw movements and masseter muscle activity during chewing in Papio anubis.
American journal of physical anthropology, 129(2), 215-224.
Wall, C. E., Vinyard, C. J., Williams, S. H., Johnson, K. R., & Hylander, W. L. (2008).
Specialization of the superficial anterior temporalis in Baboons for mastication of hard
foods. In: Vinyard, C. J., Ravosa, M. J., Wall, C. E. (Eds.), Primate Craniofacial
Function and Biology (pp. 113–124). New York: Springer.
Weiss, A., & Leinwand, L. A. (1996). The mammalian myosin heavy chain gene
family. Annual review of cell and developmental biology, 12(1), 417-439.
Weiss, A., Schiaffino, S., & Leinwand, L. A. (1999). Comparative sequence analysis of
the complete human sarcomeric myosin heavy chain family: implications for functional
diversity. Journal of molecular biology, 290(1), 61-75.
Williams, P., & Warwick R. (1980). Gray’s anatomy. Edinburgh: Churchill
Livingstone.
Williams, S. H., Vinyard, C. J., Wall, C. E., Doherty, A. H., Crompton, A. W., &
Hylander, W. L. (2011). A preliminary analysis of correlated evolution in mammalian
chewing motor patterns. Integrative and comparative biology, 51(2), 247-259.
Wood, W. (1986). A functional comparison of the deep and superficial parts of the
human anterior temporal muscle. Journal of dental research, 65(6), 924-926.
Wood, W. (1987). A review of masticatory muscle function. Journal of Prosthetic
Dentistry, 57(2), 222-232.

126
Wright, C., Haddad, F., Qin, A. X., & Baldwin, K. M. (1997). Analysis of myosin
heavy chain mRNA expression by RT-PCR. Journal of Applied Physiology, 83(4),
1389-1396.
Zenker, W. (1955). New findings in temporal muscle in man. Zeitschrift fur Anatomie
und Entwicklungsgeschichte, 118(4), 355-368.
Zhang, M., & Gould, M. (2017). Segmental distribution of myosin heavy chain
isoforms within single muscle fibers. The Anatomical Record, 300(9), 1636-1642.
Zurmanova, J., & Soukup, T. (2013). Comparison of myosin heavy chain mRNAs,
protein isoforms and fiber type proportions in the rat slow and fast muscles.
Physiological research, 62(4), 445-453.





























