Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DE SANTA MARIA CENTRO DE CIÊNCIAS RURAIS
PROGRAMA DE PÓS-GRADUAÇÃO EM ENGENHARIA FLORESTAL
FITOSSOCIOLOGIA DE ESPÉCIES NATIVAS DE MATA CILIAR NO ENTORNO DO RESERVATÓRIO
DIVISA, SÃO FRANCISCO DE PAULA, RS
DISSERTAÇÃO DE MESTRADO
DANIELE GUARIENTI RORATO
Santa Maria, RS, Brasil 2012

FITOSSOCIOLOGIA DE ESPÉCIES NATIVAS DE MATA
CILIAR NO ENTORNO DO RESERVATÓRIO DIVISA, SÃO
FRANCISCO DE PAULA, RS
Daniele Guarienti Rorato
Dissertação apresentada ao Curso de Mestrado do Programa de Pós-Graduação em Engenharia Florestal, Área de Concentração em
Silvicultura, da Universidade Federal de Santa Maria (UFSM, RS) como requisito parcial para obtenção do grau de
Mestre em Engenharia Florestal
Orientadora: Prof. Dra. Maristela Machado Araujo
Santa Maria, RS, Brasil. 2012


Universidade Federal de Santa Maria Centro de Ciências Rurais
Programa de Pós-Graduação em Engenharia Florestal
A Comissão Examinadora, abaixo assinada, aprova a Dissertação de Mestrado
FITOSSOCIOLOGIA DE ESPÉCIES NATIVAS DE MATA CILIAR NO ENTORNO DO RESERVATÓRIO DIVISA, SÃO FRANCISCO DE
PAULA, RS
elaborada por Daniele Guarienti Rorato
como requisito parcial para obtenção do grau de Mestre em Engenharia Florestal
COMISÃO EXAMINADORA:
Maristela Machado Araujo, Dra.
(Presidente/Orientadora)
João André Jarenkow, Dr. (UFRGS)
Luciane Almeri Tabaldi, Dra. (UFSM)
Santa Maria, 03 de maio de 2012.

Dedico este trabalho aos meus pais Mário (in memorium) e Gelsa, aos meus irmãos Fabrício e Rodrigo, ao meu namorado Ezequiel e à minha
querida amiga Adriana, que sempre me deram todo o amor e suporte para seguir em frente.

AGRADECIMENTOS
Agradeço primeiramente a Deus pela vida e por inúmeras graças alcançadas.
Aos meus pais, Mário (in memorium) e Gelsa, pelo apoio incondicional, amor, carinho
e dedicação recebido.
Aos meus irmãos Fabrício e Rodrigo e demais familiares, que mesmo distantes,
sempre me deram apoio e incentivo ao longo dessa jornada.
Ao meu grande amor Ezequiel, por toda a ajuda, compreensão e companheirismo.
À minha orientadora, Profª Maristela Machado Araujo, pela dedicação, amizade,
orientação e ensinamentos transmitidos.
Aos professores João André Jarenkow e Luciane Almeri Tabaldi pela disponibilidade
em participar da banca de avaliação.
A Agência Nacional de Energia Elétrica (ANEEL) e a Companhia Estadual de
Energia Elétrica (CEEE) por meio do projeto “Fitossociologia de espécies nativas de mata
ciliar na Barragem do Divisa”, pela oportunidade, auxilio logístico e financeiro para
elaboração desse trabalho.
Ao Alexandre Fanfa Bordin e César Augusto Albrech (CEEE) por todo apoio e
suporte ao longo de todo o projeto.
Ao Sr. Carlos Schneider e demais proprietários de terras adjacentes ao reservatório,
pela hospitalidade e generosidade com que receberam a equipe, liberando parte da área
para a pesquisa e colocando-se sempre a disposição para o que fosse preciso;
À minha grande amiga Adriana, por todo o aprendizado, convivência, dedicação e
amizade incondicional ao longo dessa caminhada.
À equipe do projeto, Fernando, Suelen, Cristina, Luciane, Drica e Thaíse pela ajuda
incansável na realização das coletas a campo. Tenho certeza que guardaremos lembranças
desses momentos para sempre.
Aos colegas Adriana, Ezequiel, Gisele, Suzane, Maurício e Marta, pela troca de
informações ao longo desse período.
Aos funcionários Gervásio, Élio, Maria, João e toda equipe do Viveiro Florestal pelo
convívio e amizade durante todos esses anos.
Aos professores e funcionários do Programa de Pós-Graduação em Engenharia
Florestal, pelos conhecimentos transmitidos e pela oportunidade de realizar esse trabalho.
A Universidade Federal de Santa Maria pela formação profissional.
A todos que de uma forma ou de outra contribuíram para a realização dessa
importante conquista em minha vida.
. A todos vocês meu muito obrigada!

RESUMO
Dissertação de Mestrado Programa de Pós-Graduação em Engenharia Florestal
Universidade Federal de Santa Maria
FITOSSOCIOLOGIA DE ESPÉCIES NATIVAS DE MATA CILIAR NO ENTORNO DO RESERVATÓRIO DIVISA, SÃO FRANCISCO DE
PAULA, RS
AUTORA: DANIELE GUARIENTI RORATO ORIENTADORA: MARISTELA MACHADO ARAUJO
Data e Local da Defesa: Santa Maria, 03 de maio de 2012. O trabalho tem como objetivo caracterizar a estrutura da floresta e composição de espécies presentes na área, identificar a formação de agrupamentos no componente arbóreo e determinar a influência das variáveis do solo nos grupos e espécies. A área de estudo localiza-se no entorno do Reservatório Divisa, município de São Francisco de Paula, RS. O levantamento da vegetação arbóreo-arbustiva foi realizado em quatro fragmentos denominados de F1 (9 ha), F2 (9 ha), F4 (2 ha) e F5 (9 ha). Em cada parcela foi realizada a identificação e medição da circunferência à altura do peito (CAP), conforme as seguintes classes de tamanho: Classe I (10 x 20 m) - indivíduos com CAP ≥ 30 cm; Classe II (10 x 10 m) - indivíduos com 15 ≤ CAP < 30 cm; Classe III (5 x 5 m), os indivíduos com 3 ≤ CAP < 15 cm e a Classe IV (2 x 2 m), os indivíduos com altura ≥ 30 cm e CAP < 3 cm. A estrutura vertical foi avaliada mediante a classificação dos indivíduos inventariados em classes sociológicas, pertencendo à posição sociológica 1 (PS1) os indivíduos do estrato superior, posição sociológica 2 (PS2) os do estrato médio e posição sociológica 3 (PS3) os do estrato inferior. Os dados do inventário florestal constituíram uma matriz parcela x espécie, a qual foi utilizada em análise multivariada, pelo método TWINSPAN, permitindo a classificação de grupos de parcelas, a partir da densidade dos indivíduos. Adicionalmente ao levantamento da vegetação, foram obtidas as variáveis ambientais. A correlação entre os dados de vegetação e dados ambientais foi realizada por meio da Análise de Correspondência Canônica, com uso do programa PC-ORDTM for Windows versão 5.10. Os resultados demonstram a formação de três grupos distintos. O Grupo 1 (GR1) representa a situação própria de mata ciliar, o Grupo 2 (GR2) representa um remanescente de Floresta Ombrófila Mista, sob influência do uso intensivo dos campos na pecuária, e o Grupo 3 (GR3) demonstra uma situação atípica, com a presença de espécies fortemente influenciadas pela profundidade do solo, bem como pela saturação hídrica em maior período de tempo. No GR1, a mistura de espécies de diferentes grupos ecológicos, com predomínio de espécies pioneiras e secundárias, demonstra que a floresta apresenta-se em processo dinâmico de sucessão. O GR2 apresenta influência negativa do gado na sua estrutura e composição. Destacam-se nas matas ciliares (GR1) Eugenia uruguayensis e Sebastiania commersoniana e nos capões (GR2), o predomínio é de Araucaria angustifolia, Podocarpus lambertii e Myrceugenia cucullata, como principais responsáveis pelas formações existentes. Além disso, a lenta decomposição da matéria orgânica potencializada pela maior umidade altera a disponibilidade de nutrientes para as espécies no local. Nos capões, a constante presença do gado altera negativamente a estrutura do sub-bosque, influenciando nas características químicas do solo. Espécies como Araucaria angustifolia, Eugenia uruguayensis, Blepharocalyx salicifolius, Calyptranthes concinna, Lithraea brasiliensis, Myrsine coriacea, Ocotea pulchella e Sebastiania commersoniana podem ser utilizadas na recuperação e enriquecimento de mata ciliar. Palavras-chave: Análise de Agrupamentos; Análise de Correspondência Canônica; Floresta de Araucária; Campos de Cima da Serra.

ABSTRACT
Master Dissertation Post-Graduation Program in Forest Engineering
Federal University of Santa Maria
PHYTOSOCIOLOGY OF RIPARIAN NATIVE SPECIES IN THE ENVIRONMENT DIVISA RESERVOIR, SÃO FRANCISCO DE PAULA,
RS
AUTHOR: DANIELE GUARIENTI RORATO ADVISER: MARISTELA MACHADO ARAUJO
Place and Date of Presentation: Santa Maria, 03rd of May, 2012.
The work aims to characterize the forest structure and species composition in the area, identify the formation of clusters in the tree component and to determine the influence of soil variables in species and in groups. The study area is located at Divisa Reservoir, São Francisco de Paula, RS. The survey of the trees and shrubs vegetation was conducted in four fragments: F1 (9 ha), F2 (9 ha), F4 (2 ha) and F5 (9 ha). In each plot was performed the identification and measurement of the circumference at breast height (CAP) as the following size classes: Class I (10 x 20 m) – individuals with CAP ≥ 30 cm; Class II (10 x 10 m) – individuals with 15 ≤ CAP < 30 cm; Class III (5 x 5 m) – individuals with 3 ≤ CAP < 15 cm and Class IV (2 x 2 m) the individuals with < 3 cm and CAP < 3 cm. The vertical structure was evaluated by the classification of the individuals surveyed on sociological classes, belonging to the sociological position 1 (PS1) individuals in the upper stratum, sociological position 2 (PS2) the stratum medium and sociological position 3 (PS3) in the lower stratum. The forest inventory data constituted a matrix parcel x species, which was used in multivariate analysis, by the method TWINSPAN, allowing the classification groups of parcels, based on the density of individuals. In addition to the vegetation, the environmental variables were obtained. The correlation between vegetation data and environmental data was performed by means of Canonical Correspondence Analysis, using the program PC-ORDTM for Windows version 5.10. The results demonstrate the formation of three distinct groups. Group 1 (GR1) represents the specific situation of riparian vegetation, Group 2 (GR2) represents a remnant of Araucaria Forest, under the influence of intensive livestock farming in the fields, and Group 3 (GR3) demonstrates an atypical situation, with the presence of species strongly influenced by the depth of the soil, and water saturation in the longest period of time. In GR1, the mixture of species of different ecological groups, with predominance of pioneer and secondary species, shows that the forest is presented in the dynamic process of succession. The GR2 has a negative influence of cattle on their structure and composition. Stand out in the riparian (GR1) Eugenia uruguayensis and Sebastiania commersoniana and clumps (GR2), the predominance are Araucaria angustifolia, Podocarpus lambertii and Myrceugenia cucullata, as main responsible for the existing formations. Furthermore, the slow decomposition of organic matter enhanced by higher humidity change the availability of nutrients for the species in place. In the woods, the constant presence of cattle negatively changes the structure of the understory, thus influencing the soil chemical characteristics. Species such as Araucaria angustifolia, Eugenia uruguayensis, Blepharocalyx salicifolius, Calyptranthes concinna, Lithraea brasiliensis, Myrsine coriacea, Ocotea pulchella e Sebastiania commersoniana can be used in the recovery and enrichment of riparian vegetation. Keywords: Cluster Analysis; Canonical Correspondence Analysis; Araucaria Forest; Campos de Cima da Serra.

LISTA DE TABELAS
Tabela 1 - Parâmetros fitossociológicos das espécies pertencentes ao Grupo 1, considerando a Classe I (CAP ≥ 30 cm), Classe II (15 ≤ CAP < 30 cm), Classe III (3 ≤ CAP < 15 cm) e a Classe IV (altura ≥ 30 cm e CAP < 3 cm). ................................. 46 Tabela 2 - Parâmetros fitossociológicos das espécies pertencentes ao Grupo 2, considerando a Classe I (CAP ≥ 30 cm), Classe II (15 ≤ CAP < 30 cm), Classe III (3 ≤ CAP < 15 cm) e a Classe IV (altura ≥ 30 cm e CAP < 3 cm). ................................. 51 Tabela 3 - Parâmetros fitossociológicos das espécies pertencentes ao Grupo 3, considerando a Classe I (CAP ≥ 30 cm), Classe II (15 ≤ CAP < 30 cm), Classe III (3 ≤ CAP < 15 cm) e a Classe IV (altura ≥ 30 cm e CAP < 3 cm). ................................. 56 Tabela 4 - Valores médios relativos aos atributos químicos do solo e interpretação, conforme a SBCS/CQFS (2004) para os grupos fitossociológicos, Reservatório Divisa, São Francisco de Paula, RS .......................................................................... 65 Tabela 5 - Coeficientes de correlação entre as variáveis ambientais nos dois primeiros eixos de ordenação para os dois grupos de vegetação no entorno do Reservatório Divisa, São Francisco de Paula, RS. ................................................... 66 Tabela 6 - Características populacionais dos grupos fitossociológicos, Reservatório Divisa, São Francisco de Paula, RS. ......................................................................... 73

LISTA DE FIGURAS
Figura 1 - Curva espécie-área considerando os indivíduos da Classe I (CAP ≥ 30 cm), em uma área amostral composta de 57 parcelas de 200 m2, totalizando 11,400 m2 (1,14 hectares), em quatro fragmentos de Floresta Ombrófila Mista, São Francisco de Paula, RS. Sendo: Linha contínua: curva de acumulação de espécies; Linha tracejada: amplitude de variação no acúmulo de espécies . ............................................. 39 Figura 2 - Classificação das unidades amostrais em três grupos florísticos, para a Classe I (CAP ≥ 30 cm), em fragmento de Floresta Ombrófila Mista, no entorno do Reservatório Divisa, São Francisco de Paula, RS. ................................................... 44 Figura 3 – Estrutura vertical da população pertencente a três grupos florísticos, considerando PS 1 (Posição sociológica 1, estrato superior), PS 2 (Posição sociológica 2, estrato médio) e PS 3 (Posição sociológica 3, estrato inferior), São Francisco de Paula, RS. ............................................................................................ 58 Figura 4 - Diagrama de ordenação dos grupos analisados com as variáveis ambientais para os dois primeiros eixos produzidos pela Análise de Correspondência Canônica, Reservatório Divisa, São Francisco de Paula, RS. Sendo GR: Grupo; Plot: Parcelas; Cu: Cobre; S: Enxofre; Al: Alumínio; m: Saturação por alumínio; Ca: Cálcio. ....................................................................................................................... 68 Figura 5 - Localização das parcelas com os respectivos grupos fitossociológicos nos quatro fragmentos inventariados, Reservatório Divisa, São Francisco de Paula, RS. .................................................................................................................................. 69 Figura 6 - Diagrama de ordenação das espécies com as variáveis ambientais para os dois primeiros eixos produzidos pela Análise de Correspondência Canônica, Reservatório Divisa, São Francisco de Paula, RS. Sendo Cu: Cobre; S: Enxofre; Al: Alumínio; m: Saturação por alumínio; Ca: Cálcio; Arauca: Araucaria angustifolia; Blepharo: Blepharocalyx salicifolius; Calyp: Calyptranthes concinna; Case.dec: Casearia decandra; Cinnamo: Cinnamomum amoenum; Drimys: Drimys brasiliensis; E.urug: Eugenia uruguayensis; Handroa: Handroanthus albus; Ilex.bre: Ilex brevicuspis; Lithraea: Lithraea brasiliensis; Myrc.cuc: Myrceugenia cucullata; Myrcia p: Myrcia palustris; Myrciant: Myrcianthes gigantea; Myrc.del: Myrciaria delicatula; M.coriac: Myrsine coriacea; O.pul: Ocotea pulchella; Podocar: Podocarpus lambertii; Sebas.com: Sebastiania commersoniana; Symp.uni: Symplocos uniflora e Zanthoxy: Zanthoxylum rhoifolium. ............................................................................................ 71 Figura 7 - Diagrama de ordenação de Araucaria angustifolia (A) e Podocarpus lambertii (B) para os dois primeiros eixos produzidos pela Análise de Correspondência Canônica, Reservatório Divisa, São Francisco de Paula, RS. Sendo GR: Grupo; Plot: Parcelas. ............................................................................. 74
Figura 8 - Diagrama de ordenação de Eugenia uruguayensis para os dois primeiros eixos produzidos pela Análise de Correspondência Canônica, Reservatório Divisa, São Francisco de Paula, RS. Sendo GR: Grupo; Plot: Parcelas. .............................. 76

Figura 9 - Diagrama de ordenação de Myrsine coriacea para os dois primeiros eixos produzidos pela Análise de Correspondência Canônica, Reservatório Divisa, São Francisco de Paula, RS. Sendo GR: Grupo; Plot: Parcelas. ..................................... 76 Figura 10 - Diagrama de ordenação de Sebastiania commersoniana para os dois primeiros eixos produzidos pela Análise de Correspondência Canônica, Reservatório Divisa, São Francisco de Paula, RS. Sendo GR: Grupo; Plot: Parcelas. .................. 77 Figura 11 - Diagrama de ordenação de Myrceugenia cucullata para os dois primeiros eixos produzidos pela Análise de Correspondência Canônica, Reservatório Divisa, São Francisco de Paula, RS. Sendo GR: Grupo; Plot: Parcelas. .............................. 78

LISTA DE ANEXOS
Anexo A - Mapa de uso e ocupação do solo no entorno do Reservatório Divisa, São Francisco de Paula, RS (F1: Fragmento 1; F2: Fragmento 2; F3: Fragmento 3; F4: Fragmento 4; F5: Fragmento 5; F6: Fragmento 6). ................................................... 95 Anexo B - Características autoecológicas das espécies identificadas no Reservatório Divisa, São Francisco de Paula, RS. ......................................................................... 96 Anexo C – Resultados da classificação realizada pelo TWINSPAN, para a vegetação pertencente à Classe I (CAP ≥ 30 cm), no Reservatório Divisa, São Francisco de Paula, RS. ................................................................................................................. 99 Anexo D – Resultados da ordenação utilizando Análise de Correspondência Canônica , para a vegetação pertencente à Classe I (CAP ≥ 30 cm), no Reservatório Divisa, São Francisco de Paula, RS. ....................................................................... 105

SUMÁRIO
1 CONSIDERAÇÕES GERAIS SOBRE O ESTUDO ...................................... 14 1.1 Introdução ................................................................................................... 14
1.1.1 Justificativa ................................................................................................... 15 1.2 Objetivos Geral e Específicos ................................................................... 15 1.3 Revisão bibliográfica .................................................................................. 16 1.3.1 Floresta Ombrófila Mista e Campos de Cima da Serra ................................ 16 1.3.2 Análise de Agrupamentos ............................................................................. 19 1.3.3 Parâmetros Fitossociológicos ....................................................................... 21 1.3.3.1 Estrutura Horizontal ...................................................................................... 22 1.3.3.2 Estrutura Vertical .......................................................................................... 23 1.3.4 Análise de Correspondência Canônica ......................................................... 25 1.4 Material e métodos ..................................................................................... 26
1.4.1 Caracterização geral da área de estudo ....................................................... 26 1.4.2 Definição das áreas a serem inventariadas .................................................. 27 1.4.3 Levantamento da vegetação ........................................................................ 28 1.4.4 Variáveis ambientais ..................................................................................... 29 1.4.4.1 Características topográficas ......................................................................... 29 1.4.4.2 Características químicas e físicas do solo .................................................... 30 1.4.5 Análise estatística ......................................................................................... 32 2 CAPÍTULO I – ANÁLISE DE AGRUPAMENTO EM FRAGMENTOS DE FLORESTA OMBRÓFILA MISTA NA REGIÃO DOS CAMPOS DE CIMA DA SERRA ..................................................................................................................... 35 2.1 Resumo ....................................................................................................... 35 2.2 Introdução ................................................................................................... 36 2.3 Material e métodos ..................................................................................... 38 2.4 Resultados e discussão ............................................................................. 39 2.5 Conclusão ................................................................................................... 59 3 CAPÍTULO II - ANÁLISE DE CORRESPONDÊNCIA CANÔNICA EM FRAGMENTOS DE FLORESTA OMBRÓFILA MISTA SOBRE OS CAMPOS DE CIMA DA SERRA ..................................................................................................... 60 3.1 Resumo ....................................................................................................... 60 3.2 Introdução ................................................................................................... 61 3.3 Material e métodos ..................................................................................... 63 3.4 Resultados e discussão ............................................................................. 64 3.5 Conclusão ................................................................................................... 79 4 CONSIDERAÇÕES FINAIS ......................................................................... 80 REFERÊNCIAS BIBLIOGRÁFICAS ......................................................................... 81 ANEXOS ................................................................................................................... 94

14
1 CONSIDERAÇÕES GERAIS SOBRE O ESTUDO
1.1 Introdução
Os ecossistemas naturais, em especial as florestas, sofrem constante pressão
e degradação a partir da ação antrópica, mediante o desenvolvimento de atividades
agrícolas, ocupação urbana e construção de obras de utilidade pública. Nesse
contexto, dados da FAO (2011) indicam que a América do Sul e a África são as
regiões que estão perdendo suas florestas mais rapidamente, registrando perdas
líquidas anuais de cobertura florestal equivalente a 4 e 3,4 milhões de hectares no
período de 2000-2010, respectivamente.
No art. 2º do Código Florestal brasileiro fica estabelecida a criação das Áreas
de Preservação Permanente (APP) (BRASIL, 1965). Nessas áreas não se pode
retirar a cobertura vegetal original, exceto sob licença do órgão ambiental
competente para a construção de obras de interesse público, como por exemplo,
barragem para a geração de energia elétrica. Mais recentemente, o Estado do Rio
Grande do Sul, por meio do decreto estadual nº 47.137/2010, instituiu o Programa
Estadual de Recuperação de Áreas de Preservação Permanente e Reserva Legal,
demonstrando, com isso, a preocupação do governo perante a questão ambiental
(RIO GRANDE DO SUL, 2010).
A mata ciliar, caracterizada pela vegetação florestal nas nascentes,
reservatórios de água naturais e artificiais, e ao longo dos rios, funciona como um
sistema tampão entre o terreno mais elevado e a linha de drenagem. Entre suas
principais funções cita-se a redução do processo erosivo, a perda de nutrientes e o
fato de constituir corredor de fluxo gênico (LIMA; ZAKIA, 1998).
Uma das atividades essenciais para o estabelecimento de metodologias de
recuperação de áreas alteradas e/ou degradadas é o conhecimento da vegetação
quanto à composição florística e estrutural dos ecossistemas naturais da região,
assim como, peculiaridades ambientais, como o tipo de solo. Essas informações são
subsídios importantes na recomposição da vegetação, tendo como base a

15
ocorrência e o comportamento de espécies vegetais em condições naturais.
Sequencialmente, é necessário identificar o grau de alteração do local e as espécies
potenciais para recuperação, de forma a adequar arranjos de espécies nativas,
produzidas a partir de material genético superior.
1.1.1 Justificativa
O reflorestamento depende do conhecimento prévio da vegetação nativa na
área foco de estudo, assim como das espécies de ocorrência natural e condição
ambiental onde ocorrem. Nesse contexto, o conhecimento da composição florística e
o comportamento fitossociológico das espécies locais são fundamentais para o
entendimento dos ecossistemas, considerando que subsidiam informações para a
elaboração de propostas de restabelecimento e enriquecimento de áreas alteradas.
1.2 Objetivos Geral e Específicos
A pesquisa tem como objetivo geral estabelecer parâmetros fitossociológicos
nos ecossistemas ripários, a partir de um estudo de caso no Reservatório Divisa,
localizado em São Francisco de Paula, RS.
Como objetivos específicos pretende-se:
• Caracterizar a estrutura da floresta e a composição de espécies presentes na
área;
• Identificar a formação de agrupamentos no componente arbóreo;
• Identificar a influência das variáveis do solo nos grupos e espécies, a partir da
Análise de Correspondência Canônica.

16
1.3 Revisão bibliográfica
1.3.1 Floresta Ombrófila Mista e Campos de Cima da Serra
O bioma Mata Atlântica ocupa 15% do território brasileiro e apresenta uma
variedade de formações, dentre elas, a Floresta Ombrófila Mista. O conceito de
Floresta Ombrófila Mista (Floresta de Araucária) se refere à mistura de floras
Austral-Antártica e Andina (RAMBO, 1950) com ingresso recente de espécies da
corrente tropical atlântica e amazônica (BEHLING; PILLAR, 2007), com predomínio
em regiões de clima tipicamente pluvial, principalmente em latitudes maiores a 23º
S, altitudes superiores de 500 m, e em situações afastadas das influências
marítimas (LEITE; KLEIN, 1990).
Essa formação, originalmente distribuída em uma superfície aproximada de
200.000 km², possui remanescentes florestais nos Estados do Paraná, Santa
Catarina, Rio Grande do Sul, São Paulo, Minas Gerais e Rio de Janeiro (INOUE;
RODERJAN; KUNIYOSHI, 1984). Além disso, ocorre com disjunções em países
vizinhos, atingindo a Argentina na província de Misiones, estendendo-se até o oeste
do Paraguai (KLEIN, 1960).
Segundo Reitz; Klein; Reis (1983), esse tipo fitoecológico apresenta como
característica marcante a presença dominante de Araucaria angustifolia no estrato
superior, acompanhada de diversas espécies pertencentes às famílias Lauraceae,
Aquifoliaceae, Sapindaceae, entre outras. Quadros; Pillar (2002) destacaram
também a presença de um denso sub-bosque, com predomínio de espécies da
família Myrtaceae.
A Araucaria angustifolia forma uma cobertura emergente característica,
semelhante a uma formação uniestratificada. No entanto, sob a cobertura das copas
das araucárias, encontram-se outras espécies de árvores, arbustos, ervas, epífitos e
lianas, que contribuem para a diversidade ecológica desses ambientes (KLEIN,
1960).

17
De acordo com Backes (1983), a estratificação da vegetação da Floresta
Ombrófila Mista é muito evidente e composta por quatro níveis ou estratos, sendo o
estrato superior formado exclusivamente por araucárias; o segundo estrato formado,
predominantemente, por Podocarpus lambertii e diversas espécies latifoliadas, como
Ocotea pulchella, Sloanea monosperma e Symplocos uniflora; a vegetação arbustiva
que não apresenta forma contínua, e sim com interrupções, dependendo do estrato
superior; e o estrato herbáceo, quase ausente, em virtude da grande quantidade de
ramos caídos, juntamente com a baixa luminosidade provocada pelos estratos
superiores.
Segundo dados do Inventário Florestal Contínuo do Rio Grande do Sul, a
Floresta Ombrófila Mista, em seus estágios iniciais, médios e avançados de
sucessão, ocupa uma área de 919.565 hectares, o que representa 3,25% da
cobertura florestal do Estado e 18,64% da área total coberta com florestas naturais
(RIO GRANDE DO SUL, 2002).
Atualmente, existem poucos remanescentes de Floresta de Araucária, devido,
principalmente, ao avanço das áreas agrícolas (culturas cíclicas e pastagens),
cultivo de culturas permanentes (Pinus spp. e Eucalyptus spp.) ou vegetação
secundária (LEITE, 2002).
A diminuição da área de cobertura florestal é apenas uma das consequências
negativas evidenciadas nesse tipo de formação. Segundo Hack (2007), a diminuição
da diversidade de espécies, o empobrecimento ecológico e econômico são outros
fatores a serem considerados, pois espécies de maior valor são retiradas desses
locais. Atualmente, a Floresta Ombrófila Mista se encontra em diferentes estágios
sucessionais, possuindo locais em que ocorreram apenas pequenas alterações,
especialmente, por causa de restrições de relevo ou exploração das espécies de
valor comercial (NARVAES, 2004).
Os Campos de Altitude são encontrados na região do Planalto Sul-Brasileiro,
especialmente nos estados do Rio Grande do Sul e Santa Catarina, onde também
são conhecidos como Campos de Cima da Serra (PILLAR et al., 2009). Segundo
esses autores, os campos formam, com frequência, mosaicos com a Floresta de
Araucária, sendo fortemente influenciados pelas atividades humanas. Esses
mosaicos caracterizam um sistema de transição campo-floresta ou áreas de tensão

18
ecológica, no qual duas áreas disjuntas se contatam, mas onde cada uma preserva
suas características ecológicas (IBGE, 1992). Baseando-se em evidências
fitogeográficas, Rambo (1956) descreveu que os campos são o tipo de vegetação
mais antigo e que a expansão da floresta é um processo recente, decorrente das
mudanças no clima.
Por outro lado, esses locais, assim como a Floresta de Araucária, vêm sendo
substituídos pela expansão da agricultura, pecuária e silvicultura. Além disso, a
construção de obras de utilidade pública é outra atividade impactante, acarretando
perda da biodiversidade local (BOND-BUCKUP, 2008).
Considerado como obras de utilidade pública, a construção de reservatórios
artificiais de água, como o Reservatório Divisa, necessita de legislação específica
vigente. No Brasil, a questão ambiental é abordada pela Constituição Federal de
1988, a Lei Federal n° 6.938 de 31 de agosto de 1981, que dispõe sobre a Política
Nacional do Meio Ambiente e a Lei Federal n° 4.771, de 15 de setembro de 1965, a
qual institui o Código Florestal.
O Código Florestal, no artigo 2°, considera de preservação permanente as
florestas e demais formas de vegetação natural situadas ao longo dos rios ou de
qualquer curso d’água ao redor de lagoas, lagos ou reservatórios de água naturais
ou artificiais, nas nascentes, no topo de morros, encostas, restingas, borda de
tabuleiros ou chapadas e altitudes superiores a 1.800 metros. Nesses casos, a
supressão total ou parcial da vegetação só será autorizada quando for necessário a
execução de obras, planos, atividades ou projetos de utilidade pública ou interesse
social (BRASIL, 1965).
Os parâmetros, definições e limites de Áreas de Preservação Permanente de
reservatórios artificiais foram estabelecidos pela Resolução do CONAMA n°
302/2002. No artigo 3° dessa resolução, fica estabelecida a área com largura
mínima, em projeção horizontal, no entorno dos reservatórios artificiais, medida a
partir do nível máximo normal de quinze metros, no mínimo, para os reservatórios
artificiais de geração de energia elétrica com até dez hectares, sem prejuízo da
compensação ambiental. Entretanto, as obras executadas antes dessa data seguem
normas incluídas no Plano de uso e ocupação do solo no entorno do Reservatório

19
Divisa, em que são mantidos 100 metros da vegetação natural local (CEEE-GT,
2009).
1.3.2 Análise de Agrupamentos
Montoya-Maquin; Matos (1967) definiram a floresta como uma estrutura
complexa que pode ser quantificada por diversos parâmetros utilizados para sua
descrição e classificação. Além disso, Austin; Greig Smith (1968) relatam que
nesses ambientes existe grande variação na composição florística, riqueza de
espécies, produtividade (estrutura) e grau de predominância de cada espécie.
De acordo com Braun-Blanquet (1979), a fitossociologia estuda o
agrupamento das plantas, suas inter-relações e dependência dos fatores bióticos e
abióticos, permitindo-se deduzir o aspecto, grau de desenvolvimento, relações de
competição, área de distribuição e outras características da comunidade.
O estudo fitossociológico analisa os diferentes métodos de reconhecimento e
definição de comunidades vegetais no que se refere à origem, estrutura,
classificação e relações com o meio (FELFILI; REZENDE, 2003).
A análise de agrupamentos é um conjunto de procedimentos que busca reunir
objetos em grupos homogêneos (FREI, 2006). Essa técnica multivariada é utilizada
na ciência florestal para agrupar parcelas e espécies com características
semelhantes, a partir de cálculos matemáticos (KENT; COKER, 1992), a qual é
desenvolvida com informações contidas na matriz de dados multivariados (SOUZA
et al., 1990). Segundo Felfili et al. (2011), a utilização da análise multivariada, na
ecologia, permite a detecção dos padrões de similaridade, associação e correlação
entre as variáveis.
Para Kent; Coker (1992), o número de grupos formados depende do tamanho
e da característica do conjunto de dados. Contudo, os autores afirmam que nem
todos os agrupamentos formados devem ser considerados, exceto aqueles que
podem ser verificados in loco e explicados, sendo os demais descartados.

20
Os métodos de agrupamento podem ser classificados em hierárquicos e não
hierárquicos (FELFILI et al., 2011). Conforme esses autores, o método hierárquico é
de fácil compreensão, pois apresenta os resultados na forma de dendrogramas que
informam os diferentes níveis de similaridade ou dissimilaridade observada no
conjunto de dados, úteis para interpretações ecológicas. No método não hierárquico,
os dados são plotados em gráficos que indicam os grupos formados, no qual os
indivíduos são agrupados em constelações, não sendo demonstradas suas relações
hierárquicas (FELFILI, 2007).
Além disso, Felfili et al. (2011), reforçam que os métodos de agrupamento
podem apresentar características de divisivos, aglomerativos, monotéticos ou
politéticos. O método divisivo inicia com a população total composta por todas as
parcelas, dividindo em grupos menores, e o aglomerativo inicia com cada
“elemento”, ligando-os até que formem grandes grupos; monotético aloca os
elementos baseado na presença e ausência de uma variável, como por exemplo, de
determinada espécie e, no politético, o processo de classificação é fundamentado no
uso dos dados encontrados, como a densidade de indivíduos de cada espécie.
Frei (2006) caracteriza que o método de agrupamento hierárquico consiste
em sucessivos agrupamentos, em que os elementos são agregados ou
desagregados, enquanto o método não hierárquico agrupa elementos em “n” grupos,
definidos previamente. O resultado dos agrupamentos pode ser influenciado pela
escolha da medida de similaridade, bem como pela definição do número de grupos
(DUARTE; SANTOS; MELO, 1999).
Na fitossociologia, pode ser utilizado o conceito de espécies diferenciais, as
quais são responsáveis pela divisão de um conjunto de dados, em grupos menores
de maior similaridade (KENT; COKER, 1992).
Siegmund; Laird; Lairdoffringa (2004) ressaltam a ampla utilização dos
métodos hierárquicos para diversas finalidades. Entre eles, o TWINSPAN (TWo-way
INdicator SPecies ANalysis), utiliza um procedimento multivariado que classifica as
unidades em grupos relativamente homogêneos, sendo amplamente utilizado em
análises fitossociológicas (BRUCE, 1994). Felfili (2007) classifica esse método como
hierárquico, divisível e politético.

21
O método pressupõe a formação de uma matriz baseada nos dados do
número de indivíduos obtidos no inventário da vegetação, na qual as colunas devem
conter as espécies e nas linhas as parcelas. Gauch (1982) desconsidera as
espécies com baixo número de indivíduos devido a pouca influência sobre os
resultados. De acordo com McCune; Mefford (1997), esse método de classificação
pode ser analisado pelo programa PC-ORDTM.
Conforme Hill (1979), no método TWINSPAN, as espécies representadas em
diferentes densidades ficam implícitas nas pseudoespécies, reduzindo os dados
quantitativos. Isso é possível por meio da conversão da densidade em classes que
representam intervalos do número de indivíduos da espécie. Além disso, o resultado
final do TWINSPAN é expresso por uma tabela de duas entradas, com as espécies
distribuídas nas linhas e as parcelas na coluna, conferindo assim, conforme Felfili et
al. (2001), a tabela dicotômica.
Complementando, Araujo et al. (2004) sintetizam o resultado final desse
procedimento como uma matriz organizada contendo a classificação hierárquica das
amostras e espécies conjuntamente.
A formação de agrupamentos na vegetação está relacionada às condições
ambientais associadas, que favorecem a ocorrência, ou não, de determinadas
espécies (FELFILI; REZENDE, 2003).
1.3.3 Parâmetros Fitossociológicos
Estudos de fitossociologia podem ser realizados por meio da análise da
estrutura horizontal da floresta, obtida mediante parâmetros fitossociológicos como
densidade, frequência e dominância das espécies (CAIN; CASTRO, 1959 apud
LONGHI et al., 2000), bem como o índice de valor de importância (FINOL, 1971).
Tais valores fornecem informações sobre a distribuição espacial das populações e
sua participação no contexto do ecossistema.
A análise da estrutura vertical, considerando a posição sociológica que os
indivíduos ocupam na estrutura da floresta, bem como a presença de regeneração

22
natural, confere subsídios importantes para analisar a ordem ecológica existente no
ambiente (FINOL, 1971).
Segundo Queiroz et al. (2006), o estudo dos parâmetros fitossociológicos
contribui para a obtenção de importantes informações acerca de processos
ecológicos de uma determinada fitocenose, no que se refere às características
básicas e fundamentais e todas as transformações permanentes e contínuas que
ocorrem em diversos ambientes ecológicos.
1.3.3.1 Estrutura Horizontal
Felfili; Venturoli (2000) e Felfili; Rezende (2003) descreveram a densidade
absoluta (DA) como o número de indivíduos de uma determinada área, expresso
geralmente em hectares, e a densidade relativa (DR) como a relação entre o número
de indivíduos de uma espécie e o número de indivíduos de todas as espécies,
expresso em percentagem.
De acordo com esses autores, a frequência absoluta (FA) é a relação entre o
número de parcelas em que determinada espécie ocorre e o número total de
parcelas amostradas, ou seja, a ocorrência de cada espécie no total de unidades
amostradas e frequência relativa (FR) como a relação entre a frequência absoluta de
determinada espécie com a soma das frequências absolutas de todas as espécies.
Conforme Longhi et al. (2000), a frequência absoluta é um parâmetro utilizado para
dar uma visão de como as espécies se distribuem na área.
A dominância absoluta (DoA) expressa a cobertura de cada espécie em
relação ao espaço. Além disso, esse dado mostra o grau de influência que cada
espécie exerce sobre o ecossistema (MARTINS, 1991). Com relação à dominância
relativa (DoR), a mesma expressa o espaço horizontal que uma espécie está
ocupando em relação às outras (LONGHI et al., 2000).
O Índice de Valor de Importância (IVI) ou Valor de Importância (VI) hierarquiza
as espécies na comunidade analisada, sendo obtido por meio do somatório dos
parâmetros relativos à densidade (DR), frequência (FR) e dominância (DoR) de cada

23
espécie, podendo apresentar como valor máximo, 300%. Geralmente, esse
parâmetro é utilizado de forma percentual, dividindo-se o valor por três, obtendo
assim o Valor de Importância Relativo (FELFILI; VENTUROLI, 2000).
De acordo com Mueller-Dombois; Ellenberg (1974), qualquer um dos
parâmetros de densidade, frequência ou dominância de uma espécie pode ser
interpretado como seu valor de importância, dependendo do que o pesquisador
pretende abordar.
O mesmo é ressaltado por Araujo (1998), que salienta que a inferência sobre
a hierarquia das espécies por meio do valor de importância, pode ser realizada de
forma mais precisa quando cada parâmetro é analisado de forma associada ao Valor
de Importância, pois algumas espécies têm importância ressaltada em razão,
apenas, de um parâmetro.
Além disso, Mueller-Dombois; Ellenberg (1974) destacaram que o uso de
parâmetros relativos, ao invés dos absolutos, tem informação limitada, pois
ambientes com vegetação densa ou esparsa podem apresentar os mesmos valores
de densidade, frequência e dominância relativas. Contudo, Martins (1991) ressalta
que, apesar das críticas, os valores de importância tem se mostrado satisfatórios,
tanto para separar diferentes tipos de florestas, como para relacioná-los a fatores
ambientais, ou para relacionar a distribuição de espécies aos fatores abióticos.
1.3.3.2 Estrutura Vertical
Finol (1971) sugeriu a inclusão da estrutura vertical na análise estrutural das
florestas, considerando como parâmetros a posição sociológica e a regeneração
natural. Segundo esse autor, somente os parâmetros da estrutura horizontal, em
muitos casos, não permitem uma caracterização real da importância ecológica das
espécies.
Além disso, o estudo da estrutura vertical em florestas naturais gera
informações relativas à dominância, em termos de competição por luz, que algumas

24
espécies exercem sobre outras, ou seja, sobre a exigência lumínica das espécies
(KOHYAMA, 1989).
Moscovich (2006) definiu que a formação de estratos está relacionada com a
diferenciação de crescimento causada pela existência de nichos ecológicos
condicionados às seções horizontais de disponibilidade de luz ao longo do perfil
vertical da floresta. De acordo com Sanquetta (1995), existem diversas razões para
formação de estratos bem definidos em uma floresta natural, como a existência de
aspectos arquitetônicos das árvores de diferentes espécies e em vários estágios de
desenvolvimento.
Conforme esse mesmo autor, a disposição e a altura dos indivíduos na
floresta determinam à penetração da luz, sendo os principais fatores responsáveis
pela formação de grupos de plantas com hábitos similares que ocupam os mesmos
nichos ecológicos e desempenham funções semelhantes.
A formação da regeneração natural, ou das fases iniciais de estabelecimento
e desenvolvimento da vegetação, implica no restabelecimento do ecossistema
florestal, sendo parte do ciclo de crescimento da floresta (HACK, 2007). Por meio
desse estudo, podem-se realizar previsões sobre o comportamento e
desenvolvimento futuro da floresta, considerando a relação e a quantidade de
espécies que constituem o seu estoque, bem como suas dimensões e distribuição
na área (CARVALHO, 1982). Além disso, permite diagnosticar o estado de
conservação do fragmento e a resposta ao manejo (DANIEL; JANKAUSKIS, 1989;
AMADOR; VIANA, 2000), bem como informações referentes à autoecologia, estágio
sucessional e efeitos da exploração florestal, norteando possíveis intervenções
silviculturais (HIGUCHI et al., 1985).
O processo de regeneração natural pode ser limitado pela baixa produção de
sementes, ausência de dispersores de sementes, predação de plântulas,
competição radicular e mudanças nas condições ambientais (STEVENS, 1999).
Na ecologia de florestas, o termo banco de plântulas, é utilizado para
caracterizar a regeneração natural de menor tamanho, representando o conjunto de
indivíduos em desenvolvimento no sub-bosque da floresta. Por outro lado, o conceito
fisiológico determina que um vegetal seja considerado plântula enquanto depender
das reservas da semente (MELO et al., 2004).

25
De acordo com Harper (1977), na ecologia, a presença ou ausência do banco
de plântulas, bem como a sua densidade, dependem da disponibilidade de
sementes e ambientes que apresentem condições adequadas para a sua
germinação e recrutamento, assim como do grupo ecológico ao qual as espécies
pertencem.
Além do banco de plântulas, estudos referentes à regeneração avançada,
considerando os indivíduos de maior tamanho, são de grande importância, pois
ajudam a evidenciar falhas na perpetuação das espécies a médio e longo prazo na
floresta. Entretanto, não existe definição clara e padronizada nas classes de
tamanho utilizadas, dificultando comparações (CHAMI, 2008).
1.3.4 Análise de Correspondência Canônica
A Análise de Correspondência Canônica (CCA) é um método que realiza
análise direta de gradientes, pressupondo respostas unimodais, baseadas na média
ponderada dos dados (TER BRAAK, 1987). Segundo Kent; Coker (1992), a CCA
caracteriza-se como um método de ordenação direta com análise de
correspondência, que permite a ordenação conjunta e simultânea das espécies com
as variáveis ambientais, cujo resultado expressa a variabilidade dos dados
ambientais juntamente com as espécies. Além disso, Felfili (2007) descreveu que,
com base nos resultados pode-se determinar as relações entre a vegetação e o
ambiente.
O método utiliza duas matrizes para a obtenção de suas correlações, sendo
que a matriz principal contém os dados das parcelas e espécies, respectivamente
nas linhas e colunas, enquanto a matriz secundária contém dados de variáveis
ambientais obtidos nas mesmas parcelas amostradas (FELFILI et al., 2011).
Conforme Van Den Berg; Oliveira-Filho (2000), o resultado final da análise de
correspondência canônica é fornecido com base em eixos de ordenação
perpendiculares entre si que demonstram a variação multidimensional dos dados da
vegetação. Os pontos plotados representam as espécies ou parcelas e as setas as

26
variáveis ambientais explicativas. O comprimento e a posição da seta indicam o grau
de correlação de determinado fator com determinado eixo, respectivamente
(FELFILI, 2007). As setas também podem ser estendidas desde a origem,
permitindo explicar o lado oposto de sua direção (KENT; COKER, 1992).
A análise de correspondência canônica desenvolve uma combinação linear de
cada conjunto de variáveis visando maximizar a correlação existente entre as
mesmas (HAIR et al., 1998). Assim, Kent; Coker (1992) descreveram que essa
análise é a mais indicada quando o objetivo é obter uma relação estreita das
variáveis ambientais com abundância de espécies.
1.4 Material e métodos
1.4.1 Caracterização geral da área de estudo
A área de estudo localiza-se no entorno do Reservatório Divisa, município de
São Francisco de Paula, RS (29° 26' 52" S de latitude e 50° 35' 02" W de longitude),
altitude média de 907 m, o qual é gerenciado pela Companhia Estadual de Geração
e Transmissão de Energia Elétrica (CEEE-GT), que é responsável por todas as
atividades executadas nesse perímetro. Além dessa área, foi estudado um
remanescente em propriedade particular do Sr. Carlos Schneider. No entorno do
Reservatório, considerando uma Área de Preservação Permanente (APP) de 15
metros, a partir da lâmina d’água, foram encontrados diferentes usos e ocupação do
solo, dos quais a vegetação de campo nativo predominou com 72,6%, seguido por
plantio de Eucalyptus spp. (8,5%), Pinus spp. (8,2%), vegetação florestal nativa
(4,3%), área de banhado (4,2%) e edificações (1,4%) (Anexo A).
A área está situada na porção nordeste do estado, no sul do município de São
Francisco de Paula. O município é limitado ao norte por Bom Jesus, ao sul por
Taquara, a leste por Cambará do Sul e a oeste por São Marcos.
O município possui área de unidade territorial de 3.274 km2. Climaticamente,
constitui-se na região mais fria do estado e também com as maiores precipitações, a

27
qual, segundo a classificação de Köppen, apresenta clima subtropical do tipo
fundamental Cfb, com verões brandos e úmidos e invernos relativamente úmidos e
frios (MORENO, 1961).
Os solos dominantes na região foram classificados como Cambissolos
Húmicos Alumínicos típicos, Cambissolos Háplicos Alumínicos organossólicos,
Neossolos Regolíticos Húmicos lépticos e Neossolos Litólicos Húmicos típicos, e em
menor frequência, aparecem sob a forma de inclusões nas classes predominantes,
Gleissolos Melânicos e Organossolos Háplicos (STRECK et al., 2008).
A região é denominada como Campos de Cima da Serra, no qual os campos
formam mosaicos com a Floresta Ombrófila Mista, caracterizando sistemas de
transição campo-floresta denominados encrave, fortemente influenciados pela ação
antrópica (PILLAR et al., 2009). Esses encraves são representados pela porção
inicial de um vale remanescente de matas ciliares existentes na área, antes da
inundação da barragem.
Tonetto (2011) analisando a variabilidade dos solos em área de tensão
ecológica entre campo e Floresta Ombrófila Mista, no entorno da Barragem do
Divisa, São Francisco de Paula, RS, verificou que a profundidade do solo variou em
função da declividade do terreno e em relação à altura do lençol freático. Os
Neossolos predominaram nos fragmentos florestais e os Cambissolos na tipologia
campo nativo, esses quando observados na floresta estavam associados a áreas
mais baixas, com influência do lençol freático.
1.4.2 Definição das áreas a serem inventariadas
Com base preliminar na imagem Landsat e classificação de uso e ocupação
do solo, assim como a reambulação, em dezembro de 2010, após plotagem de
todas as formações florestais no entorno do reservatório, definiu-se dentre seis
fragmentos florestais nativos, a amostragem de quatro fragmentos, denominados de
Fragmento 1 (F1), Fragmento 2 (F2), Fragmento 4 (F4) e Fragmento 5 (F5). A
princípio, buscou-se caracterizar todas as situações com base na avaliação

28
preliminar de aspectos como fitofisionomia, declividade, presença de nascentes e
córregos. Assim, dada a similaridade entre o F6 e o F2 e de F3 e F1, selecionou-se
F1, F2, F4 e F5 com base na acessibilidade às áreas por meio de estradas (Anexo
A).
Nesses fragmentos foi utilizada amostragem sistemática, metodologia cuja
seleção de amostras segue processo probabilístico, caracterizado pela seleção
aleatória da primeira unidade amostral, sendo que, a partir da primeira, todas as
demais unidades da amostra são automaticamente selecionadas e sistematicamente
distribuídas na população (PÉLLICO NETO; BRENA, 1997).
Na sistematização dos fragmentos F1, F2 e F5, foram utilizadas faixas
locadas paralelamente a distância de 100 m, contendo parcelas de 10 x 20 m, que
por sua vez mantiveram distância de 20 m. No F4, a distância entre faixas foi
reduzida para 50 m, visando uma intensidade amostral satisfatória, considerando a
pequena área do fragmento.
Em todos os fragmentos buscou-se traçar faixas perpendiculares à linha de
drenagem, obtendo-se assim, um gradiente de vegetação. No entanto, no F5
somente uma das quatro faixas alcançou a linha de drenagem, as demais
terminaram em área de campo ou construção rural. Nos demais fragmentos, todas
as faixas foram locadas no sentido da borda para o leito do rio.
Na demarcação das faixas, o ponto inicial de cada uma foi localizado com
GPS Garmin® 12, e o direcionamento por meio de bússola, modelo A-40 Suunto®.
1.4.3 Levantamento da vegetação
A avaliação da vegetação do componente arbóreo-arbustivo nos fragmentos
nativos foi realizada visando conhecer a composição florística e diversidade da área
de estudo. Para tal, foram inventariados os quatro fragmentos previamente
mencionados, denominados de F1 (9 ha), F2 (9 ha), F4 (2 ha) e F5 (9 ha).
Em cada parcela foi realizada a identificação dos indivíduos e a medição da
circunferência à altura do peito (CAP) das árvores e arbustos, conforme as seguintes

29
classes de tamanho: Classe I (10 x 20 m) - indivíduos com CAP ≥ 30 cm; Classe II
(10 x 10 m) - indivíduos com 15 ≤ CAP < 30 cm. Adicionalmente, para o estudo das
populações, considerou-se a Classe III (5 x 5 m), também denominada de
Regeneração Natural Estabelecida (RNE), observando-se os indivíduos lenhosos
com 3 ≤ CAP < 15 cm e na Classe IV (2 x 2 m) ou Banco de Plântulas (BP), os
indivíduos com altura ≥ 30 cm e CAP < 3 cm. Além disso, a estrutura vertical foi
avaliada mediante a classificação dos indivíduos inventariados em classes
sociológicas, pertencendo à posição sociológica 1 (PS1) os indivíduos do estrato
superior, posição sociológica 2 (PS2) os do estrato médio e posição sociológica 3
(PS3) os do estrato inferior.
No F1, com faixas perpendiculares a linha de drenagem, foram demarcadas 17
parcelas de 10 x 20 m, distribuídas em 4 faixas, enquanto que para o F2, 14 parcelas
em 8 faixas. No F4, utilizou-se 7 parcelas, distribuídas sistematicamente em 3 faixas,
e no F5, 19 parcelas, as quais foram distribuídas em 4 faixas. Em todas as áreas, a
amostragem foi superior a 3% da área considerada do fragmento.
O material botânico de todas as espécies observadas foi coletado e
herborizado no Herbário Florestal do Departamento de Ciências Florestais, da
Universidade Federal de Santa Maria (UFSM), sendo o material catalogado e
disponibilizado para consulta das espécies em trabalhos sequenciais. A identificação
das espécies foi efetuada mediante consulta a especialista na área e literatura
(BACKES; IRGANG, 2002; RIO GRANDE DO SUL, 2002; CARVALHO, 2003, 2006a;
SOBRAL et al., 2006; CARVALHO, 2008).
1.4.4 Variáveis ambientais
1.4.4.1 Características topográficas
Adicionalmente ao levantamento da vegetação, foi obtida a variável
declividade nas parcelas inventariadas, seguindo uma linha principal, interligada
entre e dentro das parcelas, totalizando seis pontos externos (quatro vértices e dois

30
pontos intermediários entre a linha dos 20 m), além da declividade da borda a 6 m
da primeira parcela. Para obtenção dessa variável foi utilizado clinômetro Suunto®.
Com base nesses dados foi obtida a variação de declividade entre as
parcelas, dentro de cada faixa.
1.4.4.2 Características químicas e físicas do solo
Para a caracterização química e física do solo florestal, assim como para
Análise de Correspondência Canônica, foram coletadas 3 amostras simples de solo,
na camada 0-20 cm, formando a amostra composta, em 34 parcelas inventariadas,
as quais foram destorroadas e homogeneizadas. Para tal, em cada ponto de
amostragem foi coletada uma amostra de solo com auxílio de uma pá de 25 cm de
altura. Após a coleta, as folhas, raízes e pedaços de minerais foram retirados e o
solo foi destorroado e homogeneizado em recipiente limpo. Todas as amostras
foram devidamente etiquetadas e armazenadas em sacos plásticos para análises
posteriores.
Do material coletado, foram separadas 500 gramas de cada amostra para a
análise química e 100 gramas para a física. As amostras químicas foram
encaminhadas ao Laboratório de Análise de Solos da UFSM, onde foram realizadas
análises completas, conforme metodologia proposta por Tedesco et al. (1995).
Nessa ocasião, foram determinados pH em água e em KCl, V% (saturação por
bases), m% (saturação por alumínio), H+Al, CTC efetiva, CTC pH7, além dos
conteúdos de matéria orgânica (MO), Al, Ca, Mg, K, S, P, Cu, B e Zn.
As análises físicas foram realizadas no Laboratório de Física do Solo, da
UFSM. A análise granulométrica foi realizada pelo Método da Pipeta (GEE;
BAUDER, 1986). Outro parâmetro analisado foi à umidade gravimétrica e a
densidade aparente do solo, em 34 parcelas inventariadas. A umidade gravimétrica
é definida como a quantidade de água retida por unidade de massa de sólido e a
densidade aparente é o peso seco de um volume determinado de solo, levando em
consideração os poros presentes, sendo por isso, utilizada para avaliar o impacto de

31
modificações no ambiente sobre o solo. Qualquer alteração da estrutura do solo leva
a um aumento da densidade, a qual serve como indicativo de degradação
(AZEVEDO; DALMOLIN, 2006).
A umidade gravimétrica e a densidade aparente do solo foram obtidas por
meio da utilização de um anel de metal, comumente utilizado em coletas de campo,
denominado de Anel de Koppek, que possui 3 cm de altura e diâmetro aproximado
de 6 cm (EMBRAPA, 1997). O anel foi enterrado no solo, e removido
cuidadosamente, eliminando-se os excessos de material até igualar as bordas do
anel. Após, o anel foi armazenado em embalagem impermeável, para não absorver
umidade, evitando pressões sobre a massa de solo durante o transporte. As
análises de umidade gravimétrica e de densidade aparente foram realizadas no
Laboratório de Silvicultura, DCFL, UFSM.
O teor de umidade presente em uma amostra de solo foi obtido pela diferença
entre o peso úmido e peso seco ou massa seca, dividido pelo peso seco e
multiplicado por 100. Esse parâmetro foi determinado pesando-se uma alíquota da
amostra (peso úmido) existente no anel volumétrico de alumínio numerado e de
peso conhecido. Em laboratório, os anéis foram colocados em estufa a 105°C até
peso constante, obtendo-se o peso seco, que descontado do peso do recipiente
fornece a massa ou peso do solo seco. A densidade do solo foi calculada pela razão
da massa seca pelo volume de solo, indicando o grau de alteração do solo.
Segundo Reichardt; Timm (2004), a Densidade do Solo ou Densidade
Aparente ou Densidade Global (ds) é obtida pela fórmula:
ds= ms/V (g.cm-3),
Sendo: ds= densidade aparente; ms=massa seca da amostra solo; V= volume total
da amostra.

32
1.4.5 Análise estatística
A avaliação da intensidade amostral da vegetação arbórea, pertencente à
Classe I (CAP ≥ 30 cm), foi realizada por meio da curva espécie-área. Essa curva é
obtida pela aleatorização da entrada das parcelas, pelo procedimento bootstrap por
permutação (EFRON, 1982).
A ordenada (y) contém o número cumulativo de espécies em função da área
amostrada, representada pela abcissa (x). De acordo com Mueller-Dombois;
Ellenberg (1974), a suficiência amostral é detectada quando a curva, após
apresentar crescimento abrupto, torna-se horizontal. Além disso, a curva espécie-
área permite identificar se a amostragem foi suficiente para representar a ocorrência
das espécies na comunidade (McCUNE; MEFFORD, 1997).
Os dados do inventário florestal foram organizados no programa Microsoft
Excel®, onde foi construída uma matriz parcela x espécie, formada por 57 parcelas
(linhas) e 34 espécies (colunas), obtidas nos quatro fragmentos inventariados
(fragmento 1, 2, 4 e 5) para as espécies com CAP ≥ 30 cm, excluindo as raras,
consideradas com menos de 5 indivíduos. De acordo com Gauch (1982), espécies
raras ou espécies com baixa densidade apresentam pouca ou nenhuma influência
sobre os resultados, podendo ser removidas da análise.
Posteriormente, a matriz foi utilizada em análise multivariada pelo método
TWINSPAN (TWo-way INdicator SPecies ANalysis) (GAUCH, 1982), utilizando o
programa PC-ORDTM for Windows versão 5.10. A utilização desse método permite a
classificação de grupos de parcelas que caracterizam microhábitats, a partir da
densidade dos indivíduos das diferentes espécies presentes. Em cada local são
identificadas as espécies indicadoras e preferenciais do ambiente (HILL, 1979;
FELFILI et al., 2001; ARAUJO et al., 2004). O método fornece a variância explicada,
indicada pelo autovalor da divisão, que acima de 0,3 é considerado de forte
relevância na determinação da variação dos dados (FELFILI et al., 2001). Além
disso, visando atenuar os efeitos dos desvios provocados por valores de abundância
muito elevados, houve necessidade de transformação desses, aplicando ln (a+1),

33
sendo que “a” representa o número de indivíduos em cada parcela (CARVALHO et
al., 2005).
Os níveis de corte para caracterizar as pseudoespécies foram determinados
com base nas densidades observadas para as espécies, em cada parcela, sendo 0
(0 ou 1 indivíduos), 2 (2 a 4), 5 (5 a 7), 8 (8 a 9) e 10 (10 ou mais indivíduos),
representando as pseudoespécies 1, 2, 3, 4 e 5, respectivamente.
Os grupos formados foram analisados em relação à composição florística e
estrutura, utilizando parâmetros como densidade, dominância e frequência absoluta,
além do valor de importância percentual, buscando-se definir diferenças biológicas
presentes em cada local, com base nas classes de tamanho avaliadas.
Adicionalmente, foi obtida a diversidade de espécies em nível de comunidade pelo
Índice de Diversidade de Shannon (H’) e a equabilidade (J) (FELFILI; REZENDE,
2003).
A correlação entre os dados de vegetação e dados ambientais foi realizada
por meio da Análise de Correspondência Canônica (CCA), com uso do programa
PC-ORDTM for Windows versão 5.10. Essa análise partiu da formação de duas
matrizes, a primeira referente à densidade de indivíduos de cada espécie por
parcela, e a segunda, com as características químicas e físicas do solo, bem como
características do relevo, além de conter o grupo como variável categórica. Para
efeito dessa análise, as parcelas pertencentes ao Grupo 3 (19 e 33) foram
removidas por serem consideradas “outliers”.
Para a construção da matriz das variáveis ambientais, foi realizada uma
análise preliminar para identificar variáveis semelhantes entre parcelas, as quais não
apresentaram influencia, podendo-se assim, retirá-las. Tal procedimento torna viável
o processamento, que deve apresentar número de variáveis ambientais menor que o
número de parcelas com dados de vegetação. Desse modo, as características
químicas foram representadas pelo teor de carbono orgânico; pH em água; alumínio
extraível (Al3+); acidez total (H++Al3+), além da concentração de macronutrientes
(Fósforo, Potássio, Cálcio, Enxofre e Magnésio) e micronutrientes (Zinco, Cobre e
Boro), soma de bases, capacidade de troca de cátions (CTC), percentagem de
saturação por bases (V%), percentagem de saturação por alumínio (m%) e teor de
argila.

34
As características físicas analisadas no solo foram textura, representada pela
proporção de areia grossa, areia fina, silte e argila, além da densidade aparente do
solo. Com relação à topografia, foi considerada a variável declividade média em
cada parcela.
Assim, os dados de entrada consistiram em uma matriz principal de
densidade das espécies, pertencentes à Classe I (CAP ≥ 30 cm), composta por 34
parcelas (linhas) e 22 espécies (colunas), e uma matriz secundária com 34 parcelas
(linhas) e 24 variáveis ambientais (colunas). Nessa análise, foram desprezadas as
espécies presentes com densidade inferior a 5 indivíduos. Além disso, as variáveis
ambientais analisadas em modelos unimodais como a CCA, foram homogeneizadas
pela transformação logarítmica (ln (a+1)), especialmente recomendada para dados
de nutrientes do solo (PALMER, 1993).
Com base na análise preliminar dos resultados, foram removidas variáveis
fracamente correlacionadas, considerando valor de correlação inferior a 0,4. Após a
remoção dessas variáveis, os dados foram novamente processados com cinco
variáveis ambientais, com significância das correlações entre as matrizes analisadas
pelo teste de permutação de Monte Carlo, utilizando 5000 interações para aumentar
a precisão no cálculo da probabilidade (McCUNE; GRACE, 2002).
A análise final dos resultados permitiu avaliar quais as variáveis de solo
apresentaram maior influência sobre os grupos, procurando-se identificar a
necessidade nutricional das espécies.

35
2 CAPÍTULO I – ANÁLISE DE AGRUPAMENTO EM FRAGMENTOS DE FLORESTA OMBRÓFILA MISTA NA REGIÃO DOS CAMPOS DE CIMA DA SERRA 2.1 Resumo
O presente trabalho teve como objetivo verificar se os fragmentos de floresta estudados formam agrupamentos no componente arbóreo, caracterizando a situação da vegetação nos fragmentos estudados, além de determinar quais espécies são as responsáveis pelas principais formações existentes. A área de estudo localiza-se no entorno do Reservatório Divisa, município de São Francisco de Paula, RS. O levantamento da vegetação arbóreo-arbustiva foi realizado em quatro fragmentos denominados de F1 (9 ha), F2 (9 ha), F4 (2 ha) e F5 (9 ha). Em cada parcela foi realizada a identificação dos indivíduos e medição da circunferência à altura do peito (CAP) conforme as seguintes classes de tamanho: Classe I (10 x 20 m) - indivíduos com CAP ≥ 30 cm; Classe II (10 x 10 m) - indivíduos com 15 ≤ CAP < 30 cm; Classe III (5 x 5 m), os indivíduos com 3 ≤ CAP < 15 cm e a Classe IV (2 x 2 m), os indivíduos com altura ≥ 30 cm e CAP < 3 cm. Além disso, a estrutura vertical foi avaliada mediante a classificação dos indivíduos inventariados em classes sociológicas, pertencendo à posição sociológica 1 (PS1) os indivíduos do estrato superior, posição sociológica 2 (PS2) os do estrato médio e posição sociológica 3 (PS3) os do estrato inferior. Os dados do inventário florestal constituíram uma matriz parcela x espécie, a qual foi utilizada em análise multivariada, pelo método TWINSPAN, permitindo a classificação de grupos de parcelas que caracterizam microhábitats, a partir da densidade dos indivíduos de cada espécie. Os grupos formados foram analisados em relação à composição florística e estrutura, em todas as classes de tamanho avaliadas. Os resultados indicam a formação de três grupos distintos. O Grupo 1 (GR1) representa a situação própria de mata ciliar, o Grupo 2 (GR2) representa um remanescente de Floresta Ombrófila Mista, sob influência do uso intensivo dos campos na pecuária, e o Grupo 3 (GR3) demonstra uma situação atípica, com a presença de espécies fortemente influenciadas pela profundidade do solo, bem como pela saturação hídrica em maior período de tempo. Desse modo, a formação de grupos indica a necessidade de manejar a floresta considerando as características dos diferentes ambientes. No GR1, a mistura de espécies de diferentes grupos ecológicos, com predomínio de espécies pioneiras e secundárias, demonstra que a floresta apresenta-se em processo dinâmico de sucessão. O GR2 apresenta influência negativa do gado na sua composição e estrutura. Além disso, destacam-se nas matas ciliares (GR1) Eugenia uruguayensis e Sebastiania commersoniana e nos capões (GR2), o predomínio é de Araucaria angustifolia, Podocarpus lambertii e Myrceugenia cucullata, como principais responsáveis pelas formações existentes. Palavras-chave: Encrave de floresta nativa; Campos de Altitude; Fitossociologia; TWINSPAN.

36
2.2 Introdução
O bioma Mata Atlântica ocupa 15% do território brasileiro e apresenta uma
variedade de formações, dentre elas, a Floresta Ombrófila Mista ou Floresta de
Araucária. Essa tipologia florestal apresenta como característica marcante a
presença dominante de Araucaria angustifolia no estrato superior acompanhada de
diversas espécies pertencentes às famílias Lauraceae, Aquifoliaceae, Sapindaceae,
entre outras (REITZ; KLEIN; REIS, 1983).
A constante ação antrópica nesses ambientes mediante expansão das áreas
cultivadas com agricultura e pecuária, bem como cultivo de culturas permanentes
(Pinus spp. e Eucalyptus spp.), levaram a degradação desses ecossistemas,
restando atualmente poucos remanescentes da Floresta Ombrófila Mista.
Os Campos de Altitude, também conhecidos como Campos de Cima da
Serra, são encontrados na região do Planalto Sul-Brasileiro, especialmente nos
estados do Rio Grande do Sul e Santa Catarina. Essa vegetação forma,
frequentemente, mosaicos com a Floresta de Araucária, caracterizando um sistema
de transição campo-floresta denominado encrave (IBGE, 1992), fortemente
influenciados pelas atividades humanas (PILLAR et al., 2009).
Nesses locais são necessários estudos que levem em consideração aspectos
fitossociológicos da comunidade avaliada. De acordo com Felfili; Rezende (2003),
esses estudos permitem analisar os diferentes métodos de reconhecimento e
definição de comunidades vegetais no que se refere à origem, estrutura,
classificação e relações com o meio. Isso pode ser realizado por meio da utilização
de técnicas multivariadas como a análise de agrupamentos, procedimento que
busca reunir objetos em grupos homogêneos.
Desse modo, a formação de agrupamentos na vegetação está relacionada às
condições ambientais associadas, que favorecem a ocorrência, ou não, de
determinadas espécies (FELFILI; REZENDE, 2003).
Além disso, Queiroz et al. (2006), consideram que o estudo dos parâmetros
fitossociológicos contribui para a obtenção de importantes informações acerca de
processos ecológicos de uma determinada fitocenose, no que se refere às suas

37
características e todas as transformações permanentes e contínuas que ocorrem em
diversos ambientes ecológicos.
Com isso, o conhecimento do comportamento fitossociológico das populações
naturais, associado às informações sobre estrutura e dinâmica, permite que sejam
feitas inferências sobre o manejo e estratégias de conservação das espécies, assim
como subsidiar informações para recuperação de áreas alteradas.
Diante do exposto, o objetivo do trabalho foi caracterizar o componente
arbóreo-arbustivo dos fragmentos de Floresta Ombrófila Mista, por meio dos
seguintes questionamentos: a) os fragmentos de floresta estudados formam
agrupamentos no componente arbóreo da vegetação? b) quais espécies são as
responsáveis pela (s) principal (is) formação (ões) existente (s)? c) qual a situação
atual da vegetação nos fragmentos estudados?

38
2.3 Material e métodos
A caracterização geral da área de estudo, definição das áreas a serem
inventariadas e o levantamento da vegetação, encontram-se descritos no item
“Considerações Gerais sobre o Estudo”.
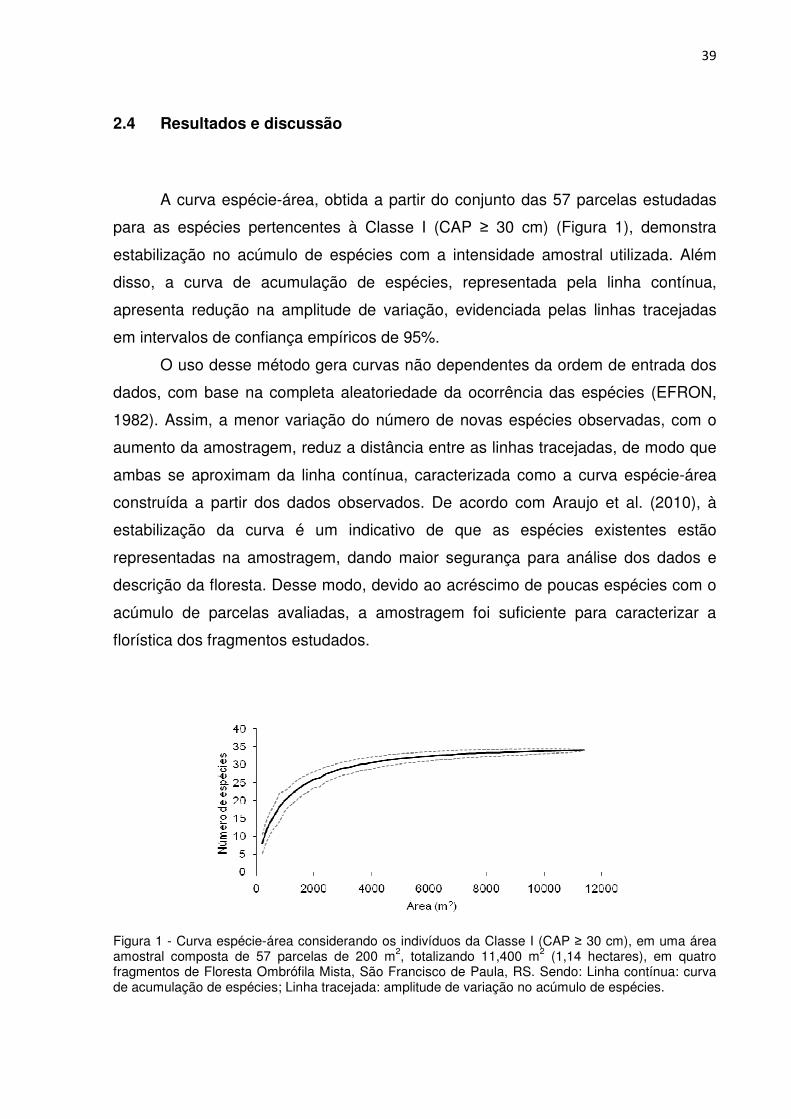
39
2.4 Resultados e discussão
A curva espécie-área, obtida a partir do conjunto das 57 parcelas estudadas
para as espécies pertencentes à Classe I (CAP ≥ 30 cm) (Figura 1), demonstra
estabilização no acúmulo de espécies com a intensidade amostral utilizada. Além
disso, a curva de acumulação de espécies, representada pela linha contínua,
apresenta redução na amplitude de variação, evidenciada pelas linhas tracejadas
em intervalos de confiança empíricos de 95%.
O uso desse método gera curvas não dependentes da ordem de entrada dos
dados, com base na completa aleatoriedade da ocorrência das espécies (EFRON,
1982). Assim, a menor variação do número de novas espécies observadas, com o
aumento da amostragem, reduz a distância entre as linhas tracejadas, de modo que
ambas se aproximam da linha contínua, caracterizada como a curva espécie-área
construída a partir dos dados observados. De acordo com Araujo et al. (2010), à
estabilização da curva é um indicativo de que as espécies existentes estão
representadas na amostragem, dando maior segurança para análise dos dados e
descrição da floresta. Desse modo, devido ao acréscimo de poucas espécies com o
acúmulo de parcelas avaliadas, a amostragem foi suficiente para caracterizar a
florística dos fragmentos estudados.
Figura 1 - Curva espécie-área considerando os indivíduos da Classe I (CAP ≥ 30 cm), em uma área amostral composta de 57 parcelas de 200 m2, totalizando 11,400 m2 (1,14 hectares), em quatro fragmentos de Floresta Ombrófila Mista, São Francisco de Paula, RS. Sendo: Linha contínua: curva de acumulação de espécies; Linha tracejada: amplitude de variação no acúmulo de espécies.

40
Valério et al. (2008), também verificaram estabilização da curva espécie-área
ao analisar a composição florística e estrutural de uma Floresta Ombrófila Mista
Montana, localizada no município de Irati, PR. Nessa situação, as 14 unidades
amostrais utilizadas no trabalho foram suficientes para representar à composição
florística do estrato arbóreo, dada a estabilização da curva em torno de 4.400 m2.
Considerando todas as classes estudadas, foram amostradas 65 espécies,
pertencentes a 49 gêneros e 30 famílias botânicas (Anexo B). Entretanto, analisando
as espécies pertencentes à Classe I (CAP ≥ 30 cm), foram encontrados 755
indivíduos.ha-1 pertencentes a 51 espécies, 43 gêneros e 29 famílias. O índice de
Shannon (H’) gerou um valor de 2,97, indicando baixa diversidade associada à
reduzida equabilidade (J) (0,44), em resposta ao predomínio de poucas espécies.
Araujo et al. (2010), avaliando a fitossociologia de um remanescente de
Floresta Ombrófila Mista em seis conglomerados, com 16 parcelas cada, ao
identificar e medir todos os indivíduos arbóreos e arbustivos com CAP ≥ 30 cm,
encontraram 86 espécies pertencentes a 60 gêneros e 34 famílias. Além disso,
segundo os autores, a vegetação apresentou diversidade (H’= 3,61) e equabilidade
(0,81). As diferenças encontradas podem ter ocorrido devido o estudo ter sido
realizado na FLONA (Floresta Nacional de São Francisco de Paula) sobre a qual
incide baixa intensidade de alteração, assim como representa a típica Floresta
Ombrófila Mista, e não o sistema de transição campo-floresta. Tal constatação pode
ser evidenciada pela maior riqueza florística.
Visando realizar o diagnóstico da flora arbórea de um trecho de Floresta
Ombrófila Mista, inserida no Parque Municipal da Ronda, no município de São
Francisco de Paula, RS, Cappelatti; Schmitt (2011) amostraram 5 ha de floresta, a
fim de caracterizar a flora arbórea, encontrando um total de 67 espécies,
pertencentes a 32 famílias.
Desse modo, embora realizadas no mesmo município, as pesquisas
demonstram diferenças, que provavelmente ocorreram devido à formação de
mosaicos, considerando os fragmentos de Floresta Ombrófila Mista entre os campos
(PILLAR et al., 2009), que caracterizam o sistema de transição campo-floresta,
fortemente influenciado pelas atividades humanas, com reduzido número de

41
espécies que se adaptam a essas condições, enquanto que nos demais estudos a
vegetação representa a típica Floresta Ombrófila Mista.
Na análise da vegetação, a classificação pelo método TWINSPAN, dividiu a
área em três grupos florísticos (Anexo C). De acordo com Felfili et al. (2001) e Araujo
et al. (2004), a utilização desse método permite a classificação de grupos de
parcelas que caracterizam microhábitats, a partir da densidade de indivíduos de
determinado número de espécies. Desse modo, a primeira divisão apresentou
autovalor de 0,3338, dando origem ao Grupo 1 (GR1), e a segunda divisão, com
autovalor de 0,3456, classificou os Grupos 2 (GR2) e 3 (GR3). Os autovalores
fornecem a variância explicada e quando maiores de 0,3 são considerados
relevantes na classificação dos dados (FELFILI et al., 2001).
Conforme observações a campo, o GR1 embora pertencente à Floresta
Ombrófila Mista, apresenta grande número de espécies características de mata
ciliar. A composição e estrutura desse grupo podem ser caracterizadas,
geograficamente, por ocorrer no F2, a jusante do barramento, com menor
declividade (12%), a montante do barramento no F1 (27%) e em encraves,
representados pela porção inicial de um vale, remanescente de matas ciliares
existentes na área, antes da inundação da barragem. Os encraves são encontrados
entre a vegetação herbácea, tipicamente dos Campos de Cima da Serra,
apresentando declividade média de 11%, em relação à linha de drenagem.
O GR2 representa um remanescente de Floresta Ombrófila Mista sob
influência do uso intensivo dos campos na pecuária (capões), enquanto o GR3,
caracterizado apenas por duas parcelas, representa uma situação atípica da região,
em que uma das parcelas (parcela 33) apresentava o predomínio de Eucalyptus
spp., utilizado na recuperação da área. Essa parcela, juntamente com a 19, ocorreu
em Cambissolo (TONETTO, 2011), com a presença de espécies fortemente
influenciadas pela profundidade do solo, bem como pela saturação hídrica em maior
período de tempo.
De forma geral, mesmo que disjuntos, os fragmentos no entorno do
Reservatório Divisa (GR1) apresentam vegetação bastante similar, onde se observa
a presença, apenas, de remanescentes de florestas nativas associados a córregos,
entremeados pelos Campos de Cima da Serra e Capões (GR2).

42
Contudo, constata-se que a floresta ocorre associada à rede de drenagem
natural, composta principalmente por nascentes, o que é confirmado por Behling et
al. (2004), enquanto, quando associada aos campos, é formada por divisores
d’água, que estabelecem apenas canais intermitentes. Dessa forma, os fragmentos
com nascentes são ecossistemas que devem ser preservados de igual forma que os
campos naturais no entorno do Reservatório, pois ambas as tipologias representam
ambientes naturais da região, ou seja, a transição campo-floresta, já descrita por
Pillar et al. (2009).
Uma hipótese para tais circunstâncias é que a região apresenta menor
temperatura e maior umidade, resultando na menor evapotranspiração. Essa
condição favorece a manutenção dos campos, que apresentam vegetação original
composta por herbáceas, essas, por sua vez, possuem sistema radicular superficial
e realizam baixas taxas de evapotranspiração em função da reduzida área foliar.
Desse modo, somente a umidade local é suficiente para a manutenção dessa
vegetação. Em contrapartida, as florestas ocorrem associadas aos córregos
permanentes, pois necessitam de maior umidade em função de seu maior porte.
Longhi et al. (2005), caracterizando os estágios sucessionais em
remanescentes de Floresta Ombrófila Mista na FLONA de São Francisco de Paula,
definiu a presença de três grupos florísticos distintos entre as parcelas avaliadas. Os
grupos se diferenciaram em relação às suas características ecológicas, sendo
denominados com base nas características sucessionais das espécies indicadoras e
preferenciais, de Grupo 1 (Floresta de locais úmidos), Grupo 2 (Floresta secundária)
e Grupo 3 (Floresta primária).
Os grupos formados nesse estudo, com as respectivas parcelas e espécies
indicadoras e preferenciais, podem ser visualizados na Figura 2. As espécies
indicadoras do GR1, formado por 34 parcelas, foram Eugenia uruguayensis e
Myrsine coriacea, ambas representadas pela pseudoespécie 1, ocorrendo em 29 e
15 parcelas, respectivamente.
Conforme Felfili (2007), as espécies indicadoras são aquelas que ocorrem
com maior peso que as demais espécies, em apenas um lado da divisão. Com
relação às espécies preferenciais, essas apresentam duas vezes mais chances de
ocorrer em um lado da divisão que do outro, permitindo fazer inferências sobre a

43
adaptação das espécies ao ambiente, bem como a detecção de espécies chaves de
determinadas condições ambientais. Entre as espécies preferenciais do GR1 (Figura
2), destacam-se Drimys brasiliensis, Eugenia uruguayensis, Ilex brevicuspis, Myrcia
palustris, Myrcianthes gigantea, Myrciaria delicatula, Myrsine coriacea, Ocotea
pulchella, Sebastiania commersoniana, Styrax leprosus, Symplocos uniflora e
Zanthoxylum rhoifolium representadas pela pseudoespécie 1, que indica baixa
densidade por parcela.
Além dessas, destacam-se Drimys brasiliensis, Eugenia uruguayensis,
Lithraea brasiliensis, Myrciaria delicatula, Myrsine coriacea e Sebastiania
commersoniana, pertencentes à pseudoespécie 2, as quais apresentaram de 2 a 4
indivíduos. Longhi et al. (2005), encontraram como espécie indicadora do GR1 a
Araucaria angustifolia, tendo Casearia decandra e Luehea divaricata como espécies
preferenciais. Esse grupo é considerado pelos autores como o mais característico da
Floresta Ombrófila Mista na área.
No GR2, não houve espécies indicadoras, sendo o grupo definido pelas
espécies preferenciais (Figura 2). O grupo foi formado por 21 parcelas,
apresentando como espécies preferenciais Blepharocalyx salicifolius, Calyptranthes
concinna e Myrciaria delicatula para pseudoespécie 1. Myrceugenia cucullata para a
pseudoespécie 2, Araucaria angustifolia e Myrceugenia cucullata para a
pseudoespécie 3 e Araucaria angustifolia para a pseudoespécie 4, constatando-se a
elevada densidade do pinheiro-brasileiro na área.
A espécie indicadora do GR3 (2 parcelas) foi Symplocos uniflora representada
pela pseudoespécie 2 (Figura 2). Entre as preferenciais destacam-se Allophylus
edulis, Eucalyptus grandis, Eucalyptus saligna, Sebastiania commersoniana e
Symplocos uniflora representadas por 1 indivíduo (pseudoespécie 1). Eucalyptus
grandis, Eucalyptus saligna, Sebastiania commersoniana e Symplocos uniflora
representando de 2 a 4 indivíduos (pseudoespécie 2), enquanto Eucalyptus grandis,
Eucalyptus saligna, Podocarpus lambertii, Sebastiania commersoniana, são
representadas por 5 a 7 indivíduos (pseudoespécie 3). Já Eucalyptus saligna e
Sebastiania commersoniana representaram a pseudoespécie 4 (8 a 9 indivíduos).

44
Figura 2 - Classificação das unidades amostrais em três grupos florísticos, para a Classe I (CAP ≥ 30 cm), em fragmento de Floresta Ombrófila Mista, no entorno do Reservatório Divisa, São Francisco de Paula, RS.
Na estrutura do GR1, representado pela Floresta Ombrófila Mista com grande
número de espécies características de mata ciliar, para a Classe I (CAP ≥ 30 cm),
foram encontrados, aproximadamente, 815 indivíduos.ha-1 pertencentes a 42
espécies, 36 gêneros e 24 famílias botânicas. Em estudos na FLONA de São
Abreviaturas: Arauca: Araucaria angustifolia; Blepharo: Blepharocalyx salicifolius; Calyp: Calyptranthes concinna; Drymys: Drimys brasiliensis; E.gran: Eucalyptus grandis; E.salig: Eucalyptus saligna; E.urug: Eugenia uruguayensis; Ile.brev: Ilex brevicuspis; Lithraea: Lithraea brasiliensis; Myrcia p: Myrcia palustris ; Myrciant: Myrcianthes gigantea; Myrc.cuc: Myrceugenia cucullata; Myrc.del: Myrciaria delicatula; M.coriac: Myrsine coriacea; O.pul: Ocotea pulchella; Podocar: Podocarpus lambertii; Sebas.co: Sebastiania commersoniana; Styrax: Styrax leprosus; Symp.uni: Symplocos uniflora; Zanthoxy: Zanthoxylum rhoifolium; Observação: O número depois da abreviatura refere-se à pseudoespécie (nível de corte). O número entre parênteses refere-se às parcelas onde estão contidas as pseudoespécies, em ambos os lados da divisão.

45
Francisco de Paula, Araujo et al. (2010), encontraram para o Grupo 1, também
caracterizado pela presença de espécies típicas de mata ciliar, a presença 877
indivíduos.ha-1 pertencente a 55 espécies arbóreas e arbustivas, 42 gêneros e 23
famílias botânicas. Esses resultados demonstram a similaridade desses grupos
quanto à estrutura (densidade), pois conforme Pagano; Durigan (2000), em solos
úmidos, verifica-se elevada densidade da vegetação arbórea.
Entre as espécies pertencentes a esse grupo, considerando a Classe I (CAP
≥ 30 cm), as que apresentaram o maior valor de importância, foram Araucaria
angustifolia (15,7%), Sebastiania commersoniana (10,6%), Eugenia uruguayensis
(9,3%), Podocarpus lambertii (8,1%) e Lithraea brasiliensis (7,9%), representando
51,6% da estrutura horizontal do grupo (Tabela 1). Em termos fitossociológicos,
Araujo et al. (2010), observaram que, em ambiente similar, Siphoneugena reitzii
apresentou maior valor de importância (12,85%), seguida por Ilex brevicuspis
(8,85%), Podocarpus lambertii (8,29%), Araucaria angustifolia (6,67%) e Vernonia
discolor (5,94%). Quanto à estrutura horizontal desse grupo, as cinco espécies
representaram 42,60%, provavelmente, devido à área estudada pelos autores sofrer
menor alteração, dada sua ocorrência em uma Floresta Nacional.
Nesse estudo, Araucaria angustifolia obteve maior hierarquização devido aos
elevados valores de densidade (DA=119,1 indivíduos.ha-1), dominância
(DoA=6,9155 m2.ha-1) e frequência (FA= 88,2%) (Tabela 1). Enquanto Sebastiania
commersoniana e Eugenia uruguayensis, apesar dos elevados valores de densidade
de 110,3 e 100 indivíduos.ha-1 e frequência de 64,7 e 73,5%, apresentaram baixa
dominância (3,3116 e 2,1124 m2.ha-1), respectivamente.
Podocarpus lambertii e Lithraea brasiliensis, entretanto, apresentam outro
comportamento, dado o valor intermediário para todos os parâmetros (DA= 64,7 e
55,9 indivíduos.ha-1; DoA= 3,4893 e 2,9019 m2.ha-1; FA= 44,1 e 64,7%). Apesar da
dominância semelhante em relação às duas espécies anteriormente citadas,
Podocarpus lambertii e Lithraea brasiliensis possuem pouco mais da metade do
número de indivíduos, comprovando seu maior porte diamétrico na área de estudo.

46
(continua) Tabela 1 - Parâmetros fitossociológicos das espécies pertencentes ao Grupo 1, considerando a Classe I (CAP ≥ 30 cm), Classe II (15 ≤ CAP < 30 cm), Classe III (3 ≤ CAP < 15 cm) e a Classe IV (altura ≥ 30 cm e CAP < 3 cm).
NOME CIENTÍFICO
Classe I Classe II Classe III Classe IV DA DoA FA VI DA DoA FA VI DA DoA FA VI DA FA
Araucaria angustifolia 119,1 6,9155 88,2 15,7 32,4 0,1187 20,6 4,9 11,8 0,0006 2,9 0,4 Sebastiania commersoniana 110,3 3,3116 64,7 10,6 70,6 0,3011 14,7 8,7 235,3 0,0071 23,5 4,4 882,4 20,6 Eugenia uruguayensis 100,0 2,1124 73,5 9,3 79,4 0,3750 52,9 13,4 82,4 0,0022 14,7 1,9 220,6 8,8 Podocarpus lambertii 64,7 3,4893 44,1 8,1 11,8 0,0531 11,8 2,3 11,8 0,0002 2,9 0,3 Lithraea brasiliensis 55,9 2,9019 64,7 7,9 11,8 0,0495 11,8 2,2 Myrsine coriácea 35,3 1,7443 44,1 5,1 11,8 0,0539 11,8 2,3 23,5 0,0002 5,9 0,6 Ilex brevicuspis 29,4 2,1769 32,4 4,8 2,9 0,0124 2,9 0,6 73,5 2,9 Myrciaria delicatula 36,8 0,7298 47,1 4,2 108,8 0,4067 55,9 15,6 741,2 0,0329 55,9 14,7 1176,5 32,4 Ocotea pulchella 17,6 2,0519 26,5 3,9 8,8 0,0451 8,8 1,8 70,6 0,0008 14,7 1,5 220,6 8,8 Drimys brasiliensis 35,3 0,5207 29,4 3,2 47,1 0,1873 26,5 7,1 58,8 0,0052 11,8 2,1 73,5 2,9 Blepharocalyx salicifolius 19,1 0,3633 26,5 2,2 26,5 0,0982 20,6 4,4 58,8 0,0021 14,7 1,7 220,6 8,8 Symplocos uniflora 20,6 0,3640 23,5 2,2 20,6 0,1429 14,7 4,1 147,1 5,9 Cinnamomum amoenum 13,2 0,4383 20,6 1,8 8,8 0,0306 8,8 1,6 70,6 0,0011 11,8 1,4 367,6 11,8 Handroanthus albus 10,3 0,7468 14,7 1,8 2,9 0,0096 2,9 0,5 Styrax leprosus 11,8 0,3403 20,6 1,7 2,9 0,0096 2,9 0,5 11,8 0,0001 2,9 0,3 147,1 2,9 Myrceugenia cucullata 13,2 0,3617 17,6 1,6 35,3 0,2018 20,6 6,1 529,4 0,0247 35,3 10,4 294,1 11,8 Calyptranthes concinna 13,2 0,1639 14,7 1,3 23,5 0,1435 17,6 4,5 258,8 0,0087 38,2 5,8 367,6 11,8 Myrcia palustris 8,8 0,2077 17,6 1,3 17,6 0,0706 14,7 3,1 82,4 0,0037 8,8 1,8 73,5 2,9 Zanthoxylum rhoifolium 8,8 0,1878 17,6 1,3 11,8 0,0479 11,8 2,2 23,5 0,0008 5,9 0,7 73,5 2,9 Dicksonia sellowiana 8,8 0,3656 11,8 1,2 Roupala brasiliensis 8,8 0,1665 14,7 1,1 5,9 0,0211 2,9 0,8 23,5 0,0016 2,9 0,6 220,6 8,8
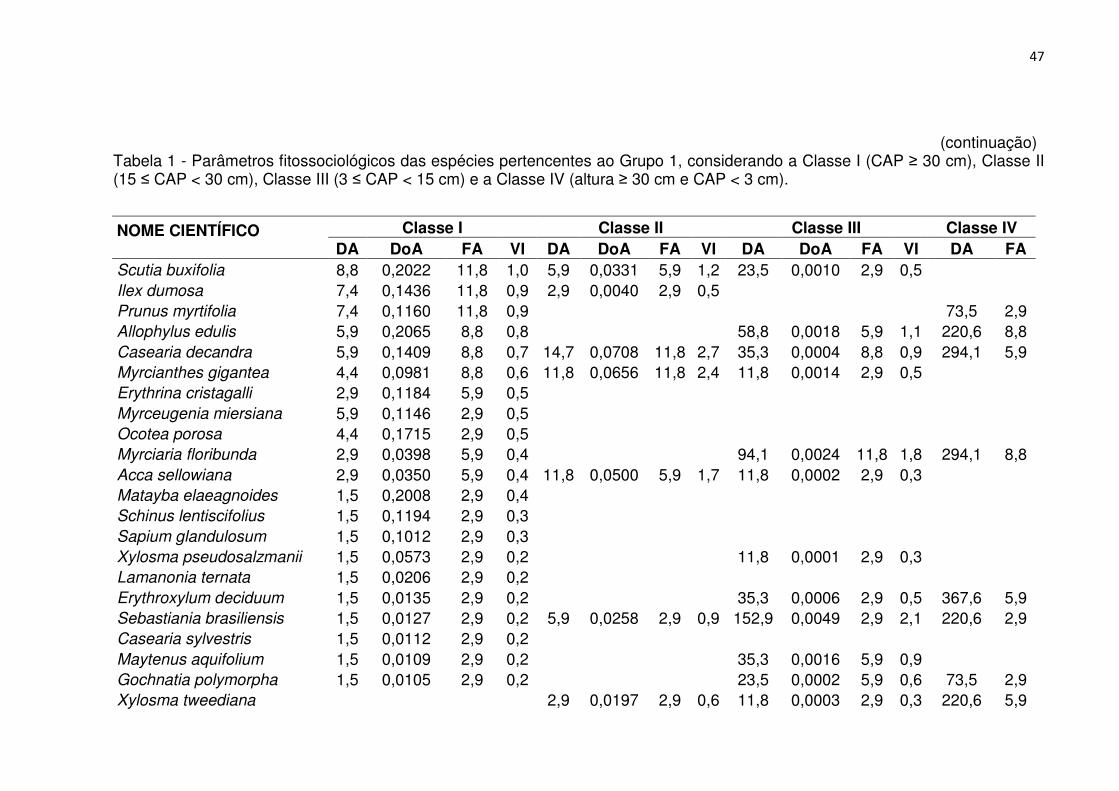
47
(continuação)
Tabela 1 - Parâmetros fitossociológicos das espécies pertencentes ao Grupo 1, considerando a Classe I (CAP ≥ 30 cm), Classe II (15 ≤ CAP < 30 cm), Classe III (3 ≤ CAP < 15 cm) e a Classe IV (altura ≥ 30 cm e CAP < 3 cm).
NOME CIENTÍFICO
Classe I Classe II Classe III Classe IV DA DoA FA VI DA DoA FA VI DA DoA FA VI DA FA
Scutia buxifolia 8,8 0,2022 11,8 1,0 5,9 0,0331 5,9 1,2 23,5 0,0010 2,9 0,5 Ilex dumosa 7,4 0,1436 11,8 0,9 2,9 0,0040 2,9 0,5 Prunus myrtifolia 7,4 0,1160 11,8 0,9 73,5 2,9 Allophylus edulis 5,9 0,2065 8,8 0,8 58,8 0,0018 5,9 1,1 220,6 8,8 Casearia decandra 5,9 0,1409 8,8 0,7 14,7 0,0708 11,8 2,7 35,3 0,0004 8,8 0,9 294,1 5,9 Myrcianthes gigantea 4,4 0,0981 8,8 0,6 11,8 0,0656 11,8 2,4 11,8 0,0014 2,9 0,5 Erythrina cristagalli 2,9 0,1184 5,9 0,5 Myrceugenia miersiana 5,9 0,1146 2,9 0,5 Ocotea porosa 4,4 0,1715 2,9 0,5 Myrciaria floribunda 2,9 0,0398 5,9 0,4 94,1 0,0024 11,8 1,8 294,1 8,8 Acca sellowiana 2,9 0,0350 5,9 0,4 11,8 0,0500 5,9 1,7 11,8 0,0002 2,9 0,3 Matayba elaeagnoides 1,5 0,2008 2,9 0,4 Schinus lentiscifolius 1,5 0,1194 2,9 0,3 Sapium glandulosum 1,5 0,1012 2,9 0,3 Xylosma pseudosalzmanii 1,5 0,0573 2,9 0,2 11,8 0,0001 2,9 0,3 Lamanonia ternata 1,5 0,0206 2,9 0,2 Erythroxylum deciduum 1,5 0,0135 2,9 0,2 35,3 0,0006 2,9 0,5 367,6 5,9 Sebastiania brasiliensis 1,5 0,0127 2,9 0,2 5,9 0,0258 2,9 0,9 152,9 0,0049 2,9 2,1 220,6 2,9 Casearia sylvestris 1,5 0,0112 2,9 0,2 Maytenus aquifolium 1,5 0,0109 2,9 0,2 35,3 0,0016 5,9 0,9 Gochnatia polymorpha 1,5 0,0105 2,9 0,2 23,5 0,0002 5,9 0,6 73,5 2,9 Xylosma tweediana 2,9 0,0197 2,9 0,6 11,8 0,0003 2,9 0,3 220,6 5,9
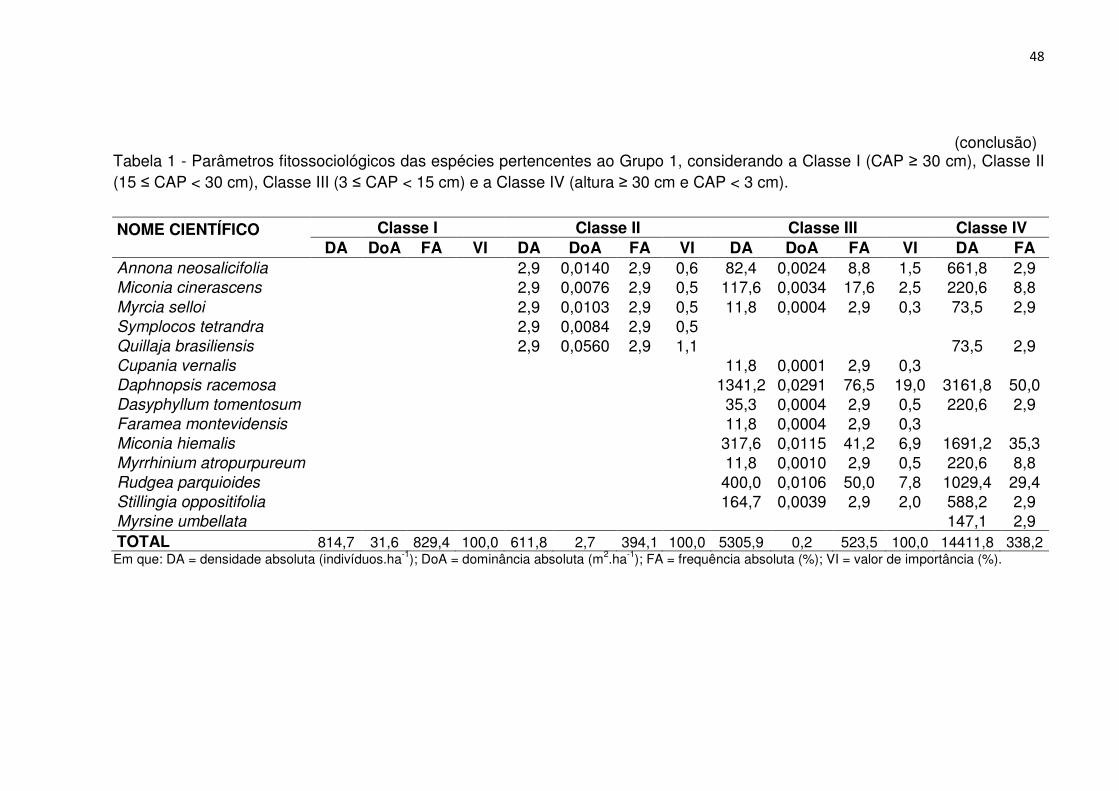
48
(conclusão)
Tabela 1 - Parâmetros fitossociológicos das espécies pertencentes ao Grupo 1, considerando a Classe I (CAP ≥ 30 cm), Classe II (15 ≤ CAP < 30 cm), Classe III (3 ≤ CAP < 15 cm) e a Classe IV (altura ≥ 30 cm e CAP < 3 cm).
NOME CIENTÍFICO
Classe I Classe II Classe III Classe IV DA DoA FA VI DA DoA FA VI DA DoA FA VI DA FA
Annona neosalicifolia 2,9 0,0140 2,9 0,6 82,4 0,0024 8,8 1,5 661,8 2,9 Miconia cinerascens 2,9 0,0076 2,9 0,5 117,6 0,0034 17,6 2,5 220,6 8,8 Myrcia selloi 2,9 0,0103 2,9 0,5 11,8 0,0004 2,9 0,3 73,5 2,9 Symplocos tetrandra 2,9 0,0084 2,9 0,5 Quillaja brasiliensis 2,9 0,0560 2,9 1,1 73,5 2,9 Cupania vernalis 11,8 0,0001 2,9 0,3 Daphnopsis racemosa 1341,2 0,0291 76,5 19,0 3161,8 50,0 Dasyphyllum tomentosum 35,3 0,0004 2,9 0,5 220,6 2,9 Faramea montevidensis 11,8 0,0004 2,9 0,3 Miconia hiemalis 317,6 0,0115 41,2 6,9 1691,2 35,3 Myrrhinium atropurpureum 11,8 0,0010 2,9 0,5 220,6 8,8 Rudgea parquioides 400,0 0,0106 50,0 7,8 1029,4 29,4 Stillingia oppositifolia 164,7 0,0039 2,9 2,0 588,2 2,9 Myrsine umbellata 147,1 2,9 TOTAL 814,7 31,6 829,4 100,0 611,8 2,7 394,1 100,0 5305,9 0,2 523,5 100,0 14411,8 338,2 Em que: DA = densidade absoluta (indivíduos.ha-1); DoA = dominância absoluta (m2.ha-1); FA = frequência absoluta (%); VI = valor de importância (%).

49
No GR1, para as classes que representam os indivíduos de menores
dimensões (Classe II (15 ≤ CAP < 30 cm); Classe III (3 ≤ CAP < 15 cm); Classe IV
(altura ≥ 30 cm e CAP < 3 cm) (Tabela 1), Araucaria angustifolia não está presente
no banco de plântulas (Classe IV), além de pouco representada na regeneração
natural estabelecida (Classe III). Esses dados divergem dos encontrados por Duarte;
Dillenburg (2000) que observaram a ocorrência de Araucaria angustifolia em
diferentes níveis de irradiância. Essa espécie também foi observada desde pioneira
até secundária tardia (RIO GRANDE DO SUL, 2007). Isso evidencia a plasticidade
no desenvolvimento dessa espécie quanto às condições de luminosidade.
Sebastiania commersoniana e Eugenia uruguayensis encontram-se bem
representadas em todas as classes estudadas, sugerindo sua maior possibilidade de
conservação no ambiente (GR1). Isso ocorreu provavelmente, devido à maior
plasticidade em relação às condições de luminosidade, pois apresentam
comportamento de espécie heliófila, mas também são consideradas de pioneira à
secundária tardia. Além disso, cabe destacar que ambas as espécies apresentam
características para ocorrer em florestas aluviais e matas de galeria (REITZ; KLEIN;
REIS, 1983; LORENZI, 2008), ambiente esse semelhante ao do GR1. Tais
características fitossociológicas observadas para as espécies permitem indicá-las
tanto para o enriquecimento de florestas quanto para a recuperação de áreas a
pleno sol, ao longo dos cursos d’água na região.
Podocarpus lambertii, espécie classificada como secundária inicial a
secundária tardia (VACCARO, 2002), apresenta o mesmo comportamento de
Araucaria angustifolia, nas classes de menor tamanho (Tabela 1). Isso confirma o
que descreveu Backes (1983), ao caracterizar a estratificação da vegetação da
Floresta Ombrófila Mista em quatro estratos, sendo o segundo estrato formado
predominantemente por Podocarpus lambertii, entre outras espécies.
De forma semelhante, Lithraea brasiliensis (Tabela 1) não se encontra
representada nas classes de menor tamanho (Classe III e IV). De acordo com
Moscovich (2006), a espécie é classificada como pioneira, e em condições de baixa
intensidade luminosa, não consegue se estabelecer no ambiente. Espécies pioneiras
surgem após perturbações que expõem o solo a luz, pois suas sementes
apresentam maior longevidade (SWAINE; WHITMORE, 1988; OLIVEIRA FILHO,

50
1994). Desse modo, a ausência de Lithraea brasiliensis nas menores classes indica
que sem a abertura do dossel, essa espécie tem menor chance de permanecer na
floresta. Já Araucaria angustifolia, embora pertença ao grupo das pioneiras
(MOSCOVICH, 2006; RIO GRANDE DO SUL, 2007) está presente na Classe III,
demonstrando a perpetuação da espécie característica da tipologia estudada
(REITZ; KLEIN; REIS, 1983), por um maior período.
Analisando o GR1, além de Sebastiania commersoniana e Eugenia
uruguayensis, que estão entre as cinco melhor hierarquizadas e presentes em todas
as classes, cabe destacar Myrciaria delicatula, Ocotea pulchella, Drimys brasiliensis,
Blepharocalyx salicifolius, Cinnamomum amoenum, Styrax leprosus, Myrceugenia
cucullata, Calyptranthes concinna, Myrcia palustris, Zanthoxylum rhoifolium, Roupala
brasiliensis, Casearia decandra e Sebastiania brasiliensis, as quais também
apresentam situação favorável para a sua perpetuação nessa condição ambiental
(GR1). Para esse grupo, destaca-se ainda Daphnopsis racemosa, Rudgea
parquioides, Miconia hiemalis e Miconia cinerascens como espécies arbustivas,
consequentemente, não encontradas no dossel da floresta. O elevado valor de
importância obtido na Classe III (Tabela 1) é a melhor condição para as espécies,
considerando que utilizam sinúsias diferentes.
Para as espécies pertencentes ao Grupo 2 (GR2), representando um
remanescente de Floresta Ombrófila Mista sob influência do uso intensivo dos
campos na pecuária, considerando a Classe I (CAP ≥ 30 cm), foram encontrados
667 indivíduos.ha-1, pertencentes a 30 espécies, 26 gêneros e 17 famílias botânicas
(Tabela 2). O GR2, embora apresente menor número de indivíduos (DA= 666,7 ind.
ha-1) em relação do GR1 (DA= 814,7 ind. ha-1) possui área basal, expressa pela
dominância absoluta total, superior (DoA= 44,2 m2.ha-1) quando comparado com
GR1 (DoA= 31,6 m2.ha-1). Esse fato indica a presença de indivíduos com diâmetro
expressivamente maior no GR2, possivelmente devido à menor influência da água e
menor competição dos indivíduos regenerantes, caracterizando uma diferença
estrutural desses dois ambientes. Entre as que apresentaram o maior valor de
importância, constam Araucaria angustifolia (29,1%), Podocarpus lambertii (23,4%),
Myrceugenia cucullata (8,0%), Lithraea brasiliensis (5,9%) e Sebastiania
commersoniana (3,8%), representando 70,2% da estrutura da floresta (Tabela 2).

51
(continua) Tabela 2 - Parâmetros fitossociológicos das espécies pertencentes ao Grupo 2, considerando a Classe I (CAP ≥ 30 cm), Classe II (15 ≤ CAP < 30 cm), Classe III (3 ≤ CAP < 15 cm) e a Classe IV (altura ≥ 30 cm e CAP < 3 cm).
NOME CIENTÍFICO
Classe I Classe II Classe III Classe IV DA DoA FA VI DA DoA FA VI DA DoA FA VI DA FA
Araucaria angustifolia 185,7 19,1209 95,2 29,1 109,5 0,4657 52,4 23,5 133,3 0,0035 33,3 2,4 595,2 23,8 Podocarpus lambertii 190,5 11,2322 95,2 23,4 61,9 0,2681 28,6 13,3 238,1 9,5 Myrceugenia cucullata 59,5 1,5828 66,7 8,0 161,9 0,5198 76,2 31,5 476,2 0,0522 119,0 12,3 1309,5 52,4 Lithraea brasiliensis 31,0 2,2396 47,6 5,9 4,8 0,0243 4,8 1,4 Sebastiania commersoniana 19,0 1,9705 23,8 3,8 285,7 0,0265 71,4 6,9 238,1 9,5 Blepharocalyx salicifolius 21,4 0,4836 23,8 2,8 119,0 4,8 Eugenia uruguayensis 21,4 0,4730 19,0 2,5 14,3 0,1222 9,5 4,6 19,0 0,0002 4,8 0,3 Ocotea pulchella 7,1 1,3467 14,3 2,2 4,8 0,0185 4,8 1,3 19,0 0,0002 4,8 0,3 595,2 23,8 Annona neosalicifolia 9,5 1,0497 14,3 2,1 19,0 0,0634 14,3 4,4 133,3 0,0468 33,3 6,3 238,1 9,5 Calyptranthes concinna 16,7 0,3793 14,3 1,9 9,5 0,0374 9,5 2,6 190,5 0,0279 47,6 5,5 952,4 38,1 Cinnamomum amoenum 7,1 1,0074 14,3 1,9 304,8 0,0058 76,2 5,4 714,3 28,6 Maytenus aquifolium 11,9 0,5368 14,3 1,8 76,2 0,0266 19,0 3,6 119,0 4,8 Myrciaria delicatula 11,9 0,1587 19,0 1,8 19,0 0,1023 14,3 5,2 190,5 0,0061 47,6 3,6 1190,5 47,6 Campomanesia xanthocarpa 9,5 0,3061 14,3 1,5 133,3 0,0249 33,3 4,4 Ocotea corymbosa 7,1 0,0877 14,3 1,2 19,0 0,0003 4,8 0,3 Ocotea porosa 7,1 0,3622 9,5 1,2 Quillaja brasiliensis 4,8 0,4861 9,5 1,1 Myrcianthes gigantea 9,5 0,1000 9,5 1,1 Ilex brevicuspis 4,8 0,3084 9,5 1,0 Xylosma pseudosalzmanii 7,1 0,1083 9,5 1,0 19,0 0,0003 4,8 0,3 Matayba elaeagnoides 2,4 0,3392 4,8 0,6

52
(continuação)
Tabela 2 - Parâmetros fitossociológicos das espécies pertencentes ao Grupo 2, considerando a Classe I (CAP ≥ 30 cm), Classe II (15 ≤ CAP < 30 cm), Classe III (3 ≤ CAP < 15 cm) e a Classe IV (altura ≥ 30 cm e CAP < 3 cm).
NOME CIENTÍFICO
Classe I Classe II Classe III Classe IV DA DoA FA VI DA DoA FA VI DA DoA FA VI DA FA
Styrax leprosus 2,4 0,1484 4,8 0,5 19,0 0,0002 4,8 0,3 238,1 9,5 Myrceugenia miersiana 2,4 0,0881 4,8 0,5 4,8 0,0272 4,8 1,5 19,0 0,0025 4,8 0,5 Faramea montevidensis 2,4 0,0835 4,8 0,5 Allophylus edulis 2,4 0,0455 4,8 0,4 19,0 0,0003 4,8 0,3 595,2 23,8 Schinus lentiscifolius 2,4 0,0315 4,8 0,4 4,8 0,0248 4,8 1,4 Celtis iguanaea 2,4 0,0303 4,8 0,4 38,1 0,0005 9,5 0,6 Myrciaria floribunda 2,4 0,0194 4,8 0,4 4,8 0,0110 4,8 1,2 57,1 0,0370 14,3 4,3 714,3 28,6 Dasyphyllum spinescens 2,4 0,0171 4,8 0,4 238,1 9,5 Zanthoxylum rhoifolium 2,4 0,0171 4,8 0,4 19,0 0,0002 4,8 0,3 Casearia decandra 9,5 0,0462 9,5 2,8 114,3 0,0012 28,6 1,9 833,3 33,3 Miconia cinerascens 14,3 0,0361 9,5 3,0 19,0 0,0019 4,8 0,5 238,1 9,5 Roupala brasiliensis 4,8 0,0130 4,8 1,2 Symplocos uniflora 4,8 0,0109 4,8 1,2 19,0 0,0003 4,8 0,3 Daphnopsis racemosa 857,1 0,0198 214,3 15,4 1547,6 61,9 Dasyphyllum tomentosum 19,0 0,0002 4,8 0,3 Eugenia involucrata 57,1 0,0006 14,3 1,0 Gochnatia polymorpha 19,0 0,0028 4,8 0,6 238,1 9,5 Miconia hiemalis 19,0 0,0003 4,8 0,3 119,0 4,8 Myrcia palustris 19,0 0,0002 4,8 0,3 Myrrhinium atropurpureum 76,2 0,0008 19,0 1,3 357,1 14,3 Rudgea jasminoides 19,0 0,0249 4,8 2,6 Rudgea parquioides 247,6 0,0122 61,9 5,0

53
(conclusão)
Tabela 2 - Parâmetros fitossociológicos das espécies pertencentes ao Grupo 2, considerando a Classe I (CAP ≥ 30 cm), Classe II (15 ≤ CAP < 30 cm), Classe III (3 ≤ CAP < 15 cm) e a Classe IV (altura ≥ 30 cm e CAP < 3 cm).
NOME CIENTÍFICO Classe I Classe II Classe III Classe IV
DA DoA FA VI DA DoA FA VI DA DoA FA VI DA FA Scutia buxifolia 19,0 0,0003 4,8 0,3 Sebastiania brasiliensis 57,1 0,0009 14,3 1,0 238,1 9,5 Stillingia oppositifolia 457,1 0,0182 114,3 8,9 1190,5 47,6 Xylosma tweediana 19,0 0,0218 4,8 2,3 119,0 4,8 Myrsine coriacea 357,1 14,3 TOTAL 666,7 44,2 585,7 100,0 452,4 1,8 257,1 100,0 4209,5 0,4 1052,4 100,0 13333,3 533,3 Em que: DA = densidade absoluta (indivíduos.ha-1); DoA = dominância absoluta (m2.ha-1); FA = frequência absoluta (%); VI = valor de importância (%).

54
No Grupo 2, Araucaria angustifolia (DA=185,7 indivíduos.ha-1; DoA=19,1209
m2.ha-1; FA= 95,2%) e Podocarpus lambertii (DA=190,5 indivíduos.ha-1;
DoA=11,2322 m2.ha-1; FA= 95,2%) apresentaram semelhança entre os parâmetros
avaliados, sendo a dominância absoluta o principal parâmetro que definiu a
hierarquia dessas espécies (Tabela 2). Conforme Carvalho (2003), os indivíduos de
araucária atingem maior porte, considerando, além da altura, o diâmetro. Além
disso, destaca-se o fato dessa espécie estar presentes em todas as classes de
tamanho avaliadas para esse grupo, o que não é verificado ao observar Podocarpus
lambertii, que está ausente na Classe III. Isso ocorreu, provavelmente, devido a
eventos passados não explicados pela análise pontual, podendo apenas serem
comprovados em estudos de dinâmica propriamente dita.
Com relação à Myrceugenia cucullata (DA=59,5 indivíduos.ha-1; DoA=1,5828
m2.ha-1; FA= 66,7%), evidencia-se sua presença em todas as classes de tamanho
(Tabela 2). A espécie pertence ao grupo ecológico das secundárias iniciais a
secundárias tardias (RIO GRANDE DO SUL, 2007), comprovado pelos parâmetros
fitossociológicos, que na Classe II apresentaram os maiores valores em relação à
Classe I, indicando o melhor desenvolvimento desses indivíduos em ambientes com
certo grau de sombreamento.
Semelhantemente ao observado no Grupo 1, Lithraea brasiliensis (DA=31,0
indivíduos.ha-1; DoA=2,2396 m2.ha-1; FA= 47,6%) (Tabela 2) não se encontra
representada nas classes de menor tamanho (Classe III e IV), confirmando o
descrito por Moscovich (2006), que atribuiu à ausência nas classes de menores
dimensões devido ao caráter pioneiro da espécie. Sebastiania commersoniana
(DA=19,0 indivíduos.ha-1; DoA=1,9705 m2.ha-1; FA= 23,8%), está ausente na Classe
II, devido a extração de madeira, visto que a espécie é uma excelente produtora de
lenha e carvão, com elevado poder calorífico (CARVALHO, 2003), confirmado por
relato de moradores locais, que afirmaram a remoção de indivíduos dessa espécie
para essa finalidade.
No GR2, além de Araucaria angustifolia e Myrceugenia cucullata, cabe
destacar Ocotea pulchella, Annona neosalicifolia, Calyptranthes concinna, Myrciaria
delicatula e Myrciaria floribunda encontram-se bem representadas em todas as
classes, provavelmente dando suporte para a sua perpetuação ao longo do tempo.
Destaca-se também Daphnopsis racemosa, Stillingia oppositifolia, Rudgea

55
parquioides e Rudgea jasminoides como espécies arbustivas, não encontradas no
dossel da floresta.
Além disso, ao comparar as espécies com maior valor de importância
pertencente à Classe I, para GR1 e GR2 observa-se que com exceção de Eugenia
uruguayensis e Myrceugenia cucullata, todas as demais espécies estão entre as
mais representativas em ambos os grupos. Tal fato indica que os grupos formados,
embora apresentem características estruturais bastante peculiares, a composição
florística é semelhante. Corroborando, Pillar et al. (2009) descreveram que
fragmentos de floresta nos Campos de Cima da Serra representam à típica transição
campo-floresta.
Por outro lado, observa-se no GR1, além do maior número de indivíduos total,
maior proporção de espécies ocorrendo em todas as classes de tamanho (8,7%), em
relação ao GR2 (3,4%), o que indica a influência negativa da presença do gado nas
classes de menor tamanho para esse grupo.
No Grupo 3 (GR3), considerando a Classe I (CAP ≥ 30 cm), foram
encontrados 675 indivíduos.ha-1 pertencentes a 6 espécies, 5 gêneros e 4 famílias
botânicas (Tabela 3). As espécies melhor hierarquizadas foram Podocarpus lambertii
(21,9%), Eucalyptus saligna (21,8%), Araucaria angustifolia (18,3%), Sebastiania
commersoniana (16,6) e Eucalyptus grandis (15,5%). Essas espécies juntas somam
94,1% da estrutura horizontal, caracterizando assim, o grupo em estudo. A presença
de espécies exóticas nesse grupo o diferencia dos demais, além da presença de
espécies fortemente influenciadas pela profundidade do solo, caracterizado
exclusivamente nessas parcelas como Cambissolo (TONETTO, 2011), bem como
pela saturação hídrica.
Outro aspecto importante é que as espécies exóticas não se encontram
representadas nas classes de menor tamanho, assim não possuem descendentes
para permanecer na área. Dessa forma, a erradicação dessas espécies pode ser
uma estratégia silvicultural importante, considerando a obrigatoriedade de manter
apenas espécies nativas na mata ciliar. Porém, ressalta-se a necessidade de uma
prévia análise da luminosidade no sub-bosque e de corte direcionado desses
indivíduos, a fim de impedir a germinação de Eucalyptus spp., no banco de
sementes do solo e reduzir o impacto sobre a vegetação existente, respectivamente.
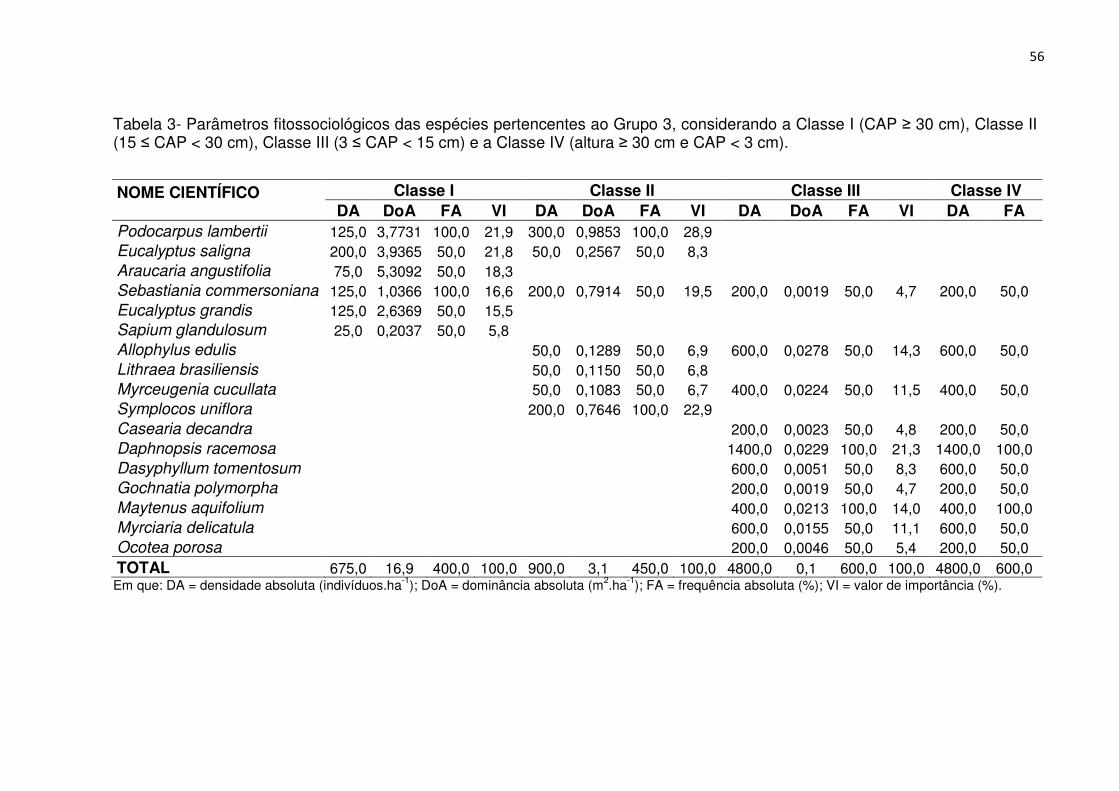
56
Tabela 3- Parâmetros fitossociológicos das espécies pertencentes ao Grupo 3, considerando a Classe I (CAP ≥ 30 cm), Classe II (15 ≤ CAP < 30 cm), Classe III (3 ≤ CAP < 15 cm) e a Classe IV (altura ≥ 30 cm e CAP < 3 cm).
NOME CIENTÍFICO
Classe I Classe II Classe III Classe IV DA DoA FA VI DA DoA FA VI DA DoA FA VI DA FA
Podocarpus lambertii 125,0 3,7731 100,0 21,9 300,0 0,9853 100,0 28,9 Eucalyptus saligna 200,0 3,9365 50,0 21,8 50,0 0,2567 50,0 8,3 Araucaria angustifolia 75,0 5,3092 50,0 18,3 Sebastiania commersoniana 125,0 1,0366 100,0 16,6 200,0 0,7914 50,0 19,5 200,0 0,0019 50,0 4,7 200,0 50,0 Eucalyptus grandis 125,0 2,6369 50,0 15,5 Sapium glandulosum 25,0 0,2037 50,0 5,8 Allophylus edulis 50,0 0,1289 50,0 6,9 600,0 0,0278 50,0 14,3 600,0 50,0 Lithraea brasiliensis 50,0 0,1150 50,0 6,8 Myrceugenia cucullata 50,0 0,1083 50,0 6,7 400,0 0,0224 50,0 11,5 400,0 50,0 Symplocos uniflora 200,0 0,7646 100,0 22,9 Casearia decandra 200,0 0,0023 50,0 4,8 200,0 50,0 Daphnopsis racemosa 1400,0 0,0229 100,0 21,3 1400,0 100,0 Dasyphyllum tomentosum 600,0 0,0051 50,0 8,3 600,0 50,0 Gochnatia polymorpha 200,0 0,0019 50,0 4,7 200,0 50,0 Maytenus aquifolium 400,0 0,0213 100,0 14,0 400,0 100,0 Myrciaria delicatula 600,0 0,0155 50,0 11,1 600,0 50,0 Ocotea porosa 200,0 0,0046 50,0 5,4 200,0 50,0 TOTAL 675,0 16,9 400,0 100,0 900,0 3,1 450,0 100,0 4800,0 0,1 600,0 100,0 4800,0 600,0 Em que: DA = densidade absoluta (indivíduos.ha-1); DoA = dominância absoluta (m2.ha-1); FA = frequência absoluta (%); VI = valor de importância (%).

57
Entre essas espécies, a presença de Podocarpus lambertii (DA=125,0
indivíduos.ha-1; DoA=3,7731 m2.ha-1; FA= 100%) e Araucaria angustifolia (DA=75,0
indivíduos.ha-1; DoA=5,3092 m2.ha-1; FA= 50%) (Tabela 3) é típica e característica
da formação em estudo. Aliado a isso, a presença de Sebastiania commersoniana
em todas as classes, confirma sua preferência ambiental por ambientes ripários,
com maior saturação hídrica, como a situação encontrada nesse grupo.
Contudo, a presença de espécies exóticas, entre as mais representativas,
merece destaque. Conforme Ziller; Zalba; Zenni (2007), o processo de dispersão de
uma espécie exótica pode resultar na ampliação da área de ocorrência da população
de determinada espécie. Nesse caso, quando a espécie exótica passa a dominar o
ambiente e causar a homogeneização da área, provoca efeitos negativos no local. A
homogeneização biótica acarreta em uma condição ambiental, ecologicamente
inferior a original, com perda de biodiversidade no nível de espécies e alteração de
processos ecológicos e evolutivos (CARROLL; DINGLE, 1996).
Embora os Campos de Altitude e os poucos remanescentes de Floresta
Ombrófila Mista estarem sendo fortemente pressionados devido principalmente, ao
avanço das culturas cíclicas, pastagens, pecuária e cultivo de culturas permanentes,
como Pinus spp. e Eucalyptus spp. (PILLAR et al., 2009), o inventário não amostrou
espécies de Pinus spp. em nenhuma classe inventariada, indicando que a espécie,
mesmo sendo considerada invasora, não compromete a vegetação das áreas em
estudo. Contudo, por se tratar de uma espécie heliófila que necessita alta
intensidade lumínica para sobreviver (EMER; FONSECA, 2007), a remoção do
componente arbóreo no local pode determinar seu estabelecimento na área, uma
vez que as sementes possuem dispersão anemocórica e mantém-se viável no banco
de sementes.
Além disso, a presença de espécies do gênero Eucalyptus, evidenciada no
GR3, caracteriza uma situação atípica, relacionada com a utilização de espécies
exóticas na recuperação de área alterada pela construção da barragem, conforme
comunicação pessoal com o Sr. Carlos Schneider, antigo proprietário da área.
Na estrutura vertical, tomando como base os indivíduos arbóreo-arbustivos
(CAP ≥ 15 cm), observa-se para todos os grupos formados, a predominância das
espécies na posição sociológica 2 (PS2), ou estrato médio, representando 82%,
67% e 67%, respectivamente, para os Grupos 1 (Figura 3A), Grupo 2 (Figura 3B) e
Grupo 3 (Figura 3C). De acordo com Sanquetta (1995), existem diversas razões

58
para formação de estratos bem definidos em uma floresta natural, como à existência
de aspectos arquitetônicos das árvores de diferentes espécies e em vários estágios
de desenvolvimento. No presente estudo, a maior densidade de indivíduos com
menor diâmetro no GR1 reduz o crescimento em altura como forma de compensar a
relação altura/diâmetro, proporcionando a menor densidade no estrato superior.
Por outro lado, o maior sombreamento no sub-bosque reduz a densidade no
estrato inferior. Além disso, Watzlawick et al. (2002), analisando o aspecto florístico
e estrutural de um fragmento de Floresta de Araucária, com acesso de bovinos,
verificou que 39% dos indivíduos amostrados foram classificados como dominados e
suprimidos, fato esse atribuído à intensa presença do gado na vegetação. Desse
modo, verifica-se que a presença de bovinos em floresta pode causar danos físicos
e mecânicos aos indivíduos de menor porte. Além disso, Schneider; Galvão; Longhi
(1978) relataram que o pisoteio do gado causa exposição da superfície do solo,
danifica raízes superficiais responsáveis pela absorção nutricional e prejudica a
regeneração natural.
Figura 3 – Estrutura vertical da população pertencente a três grupos florísticos, considerando PS 1 (Posição sociológica 1, estrato superior), PS 2 (Posição sociológica 2, estrato médio) e PS 3 (Posição sociológica 3, estrato inferior), São Francisco de Paula, RS.

59
2.5 Conclusão
Os fragmentos de Floresta Ombrófila Mista sobre os Campos de Cima da
Serra formam agrupamentos no componente arbóreo da vegetação definidos por
matas ciliares e capões, no entorno do Reservatório Divisa, conforme sua
localização.
No entorno do Reservatório a estrutura da vegetação remanescente é
tipicamente ciliar da região, representando encraves na vegetação de campos.
Nas matas ciliares (GR1), destacam-se Eugenia uruguayensis e Sebastiania
commersoniana.
Nos capões (GR2), as espécies predominantes foram Araucaria angustifolia,
Podocarpus lambertii e Myrceugenia cucullata.
No GR1, a mistura de espécies de diferentes grupos ecológicos, com
predomínio de espécies pioneiras e secundárias, demonstra que a floresta
apresenta-se em processo dinâmico de sucessão.
O GR2 apresenta influência negativa do gado na sua composição e estrutura.
A formação de grupos distintos indica a necessidade de manejar a floresta
considerando as características dos diferentes ambientes.

60
3 CAPÍTULO II - ANÁLISE DE CORRESPONDÊNCIA CANÔNICA EM FRAGMENTOS DE FLORESTA OMBRÓFILA MISTA SOBRE OS CAMPOS DE CIMA DA SERRA
3.1 Resumo O presente trabalho teve como objetivo caracterizar a influência do solo no componente arbóreo dos fragmentos, determinar a influência de variáveis do solo nas espécies predominantes com ênfase nas de mata ciliar, além de identificar espécies que apresentam potencial para recuperação e enriquecimento de mata ciliar. A área de estudo localiza-se no entorno do Reservatório Divisa, município de São Francisco de Paula, RS. O levantamento da vegetação arbóreo-arbustiva foi realizado em quatro fragmentos (F1, F2, F4 e F5), em cada parcela foi realizada a identificação e medição da circunferência à altura do peito (CAP) conforme as seguintes classes de tamanho: Classe I (10 x 20 m) - indivíduos com CAP ≥ 30 cm; Classe II (10 x 10 m) - indivíduos com 15 ≤ CAP < 30 cm; Classe III (5 x 5 m), os indivíduos com 3 ≤ CAP < 15 cm e a Classe IV (2 x 2 m), os indivíduos com altura ≥ 30 cm e CAP < 3 m. Adicionalmente ao levantamento da vegetação, foram obtidas as variáveis ambientais como declividade média nas parcelas inventariadas e coleta de amostras de solo, em 34 parcelas, caracterizando aspectos químicos e físicos do solo. A correlação entre os dados de vegetação e dados ambientais para a Classe I (CAP ≥ 30 cm) foi realizada por meio da Análise de Correspondência Canônica. As características físicas do solo não apresentam influência nos grupos formados e nas espécies. No ambiente ciliar, a lenta decomposição da matéria orgânica potencializada, pela maior umidade característica desse ambiente, é responsável pela elevada acidez do solo, influenciando negativamente a disponibilidade de nutrientes para as espécies no local. Entretanto, as espécies florestais parecem adaptadas a tal condição. Nos capões, a constante presença do gado altera negativamente a estrutura do sub-bosque, assim como nas características químicas do solo. Espécies como Araucaria angustifolia, Eugenia uruguayensis, Blepharocalyx salicifolius, Calyptranthes concinna, Lithraea brasiliensis, Myrsine coriacea, Ocotea pulchella e Sebastiania commersoniana podem ser utilizadas na recuperação e enriquecimento de mata ciliar. Palavras-chave: Análise multivariada; Floresta de Araucária; Relação solo-vegetação; Campos de Altitude.

61
3.2 Introdução
A Floresta Ombrófila Mista ou Floresta de Araucária, como também é
conhecida, se refere à mistura de floras. Nessa formação, a Araucaria angustifolia,
espécie típica, forma uma cobertura emergente característica. O segundo estrato é
formado predominantemente por Podocarpus lambertii e diversas espécies
latifoliadas, seguida da vegetação arbustiva.
A Floresta Ombrófila Mista encontra-se entre as tipologias florestais mais
ameaçadas, visto que, inicialmente, a formação ocupava uma superfície aproximada
de 200.000 km², passando a representar 0,7% da área original, considerando
remanescentes em estágios primários e avançados de sucessão (BRASIL, 2002).
Kanieski (2010) relata a necessidade de estudos nessa tipologia, devido à
importância para subsidiar o estabelecimento de planos de conservação para esses
ecossistemas.
Sendo a floresta um ambiente com grande variação na composição florística,
riqueza de espécies e estrutura, diversos parâmetros podem ser utilizados para sua
descrição, classificação e ordenação da vegetação, correlacionada a fatores
ambientais. De acordo com Braun-Blanquet (1979), a fitossociologia estuda o
agrupamento das plantas, suas inter-relações e dependência dos fatores bióticos e
abióticos, visando inferir sobre a origem, estrutura, classificação e outras
características da comunidade. Além disso, a formação de agrupamentos na
vegetação está relacionada às condições ambientais associadas, que favorecem a
ocorrência, ou não, de determinadas espécies (FELFILI; REZENDE, 2003).
Métodos de ordenação são empregados para determinar as relações
existentes entre a vegetação e o ambiente (MATTEUCCI; COLMA, 1982). Dentre
esses métodos, destaca-se a Análise de Correspondência Canônica (CCA), a qual
permite a ordenação conjunta e simultânea das espécies com as variáveis
ambientais, cujo resultado expressa a variabilidade dos dados. Aliado a isso, Felfili
(2007) descreve que, com base nos resultados, pode-se determinar as relações
entre a vegetação e o ambiente.
Assim, para a compreensão das florestas, é fundamental correlacionar as
informações da vegetação com as variáveis ambientais, permitindo-se inferir sobre a
conservação biológica desses ecossistemas e ter subsídio para sua recuperação.
Nesse sentido, foi desenvolvido um estudo em fragmentos de Floresta Ombrófila

62
Mista, no entorno do Reservatório Divisa, São Francisco de Paula, RS, visando: a)
caracterizar a influência do solo no componente arbóreo dos fragmentos; b)
determinar a influência de variáveis do solo nas espécies predominantes, com
ênfase nas de mata ciliar; c) identificar espécies que apresentam potencial para
recuperação e enriquecimento de mata ciliar.

63
3.3 Material e métodos
A caracterização geral da área de estudo e áreas inventariadas; obtenção dos
dados de vegetação e das variáveis ambientais, incluindo as características
morfológicas, químico e físicas do solo, encontram-se descritas no item
“Considerações gerais sobre o estudo”.
A interpretação dos teores nutricionais foi estabelecida com base em curvas
de calibração que definem os teores críticos para uso do solo, e acima dos quais a
probabilidade de resposta das culturas à adição de fertilizantes e corretivos é
pequena ou nula. O teor crítico é o limite inferior da faixa "Alto" (rendimentos
próximos a 90%). As faixas "Muito baixo", "Baixo" e "Médio" correspondem a
rendimentos menores, que são aproximadamente, 40%, de 40 a 75% e de 75 a 90%
do rendimento máximo, respectivamente, enquanto a faixa "Alto" varia entre o teor
crítico até duas vezes esse valor (90 a 180% de rendimento máximo) (SBCS/CQFS,
2004). Essa interpretação, apesar de ser referencial para a agricultura e silvicultura
de espécies comerciais, serve como parâmetro para classificar a tolerância ou
necessidade de espécies florestais nativas, ainda pouco estudadas quanto a esse
aspecto.
Para a Análise de Correspondência Canônica, foi removido da matriz,
contendo os dados da vegetação, as parcelas do Grupo 3 (parcela 19 e parcela 33),
considerando que em análise preliminar, foram observados como “outliers” na
ordenação dos dados.
De forma complementar, foram selecionadas seis espécies entre as melhor
hierarquizadas do GR1 e GR2, tomando-se GR2 como comparativo. Além disso,
foram utilizados aspectos relacionados com o grupo ecológico ao qual pertencem
essas espécies, visando detalhar essas populações na comunidade arbóreo-
arbustiva.

64
3.4 Resultados e discussão
As características físicas do solo analisadas como textura, densidade
aparente (0,71 g.cm-3) e umidade gravimétrica (38,1%) não apresentaram correlação
com os grupos fitossociológicos nem com as espécies.
Em ambos os grupos (GR1 e GR2), com relação à acidez do solo (pH água;
H+Al; Al), pode-se observar os elevados valores de acidez potencial (H+Al) e
alumínio livre no solo (Al) devido às altas concentrações de matéria orgânica,
refletindo no pH baixo ou ácido (Tabela 4). Nessa situação, devido à matéria
orgânica estar sobre o solo, os cátions (H+ e Al+3) não se encontram aderidos às
cargas negativas do solo (SILVA; CAMARGO; CERETTA, 2000), em função da lenta
decomposição da matéria orgânica, decorrente das menores temperaturas nessa
região, comparativamente às outras do RS. Isso confirma o descrito por Lepsch;
Silva; Espironelo (1982) os quais relataram que a taxa de decomposição da matéria
orgânica varia em função das condições de temperatura, umidade, relevo, material
de origem, entre outros.
Observou-se também que no GR1 e GR2, os teores dos macronutrientes (Ca,
Mg, K, P e S), apresentaram variação na interpretação quando comparados entre si,
em relação às concentrações dos micronutrientes (Cu, Zn e B), que segundo a
SBCS/CQFS (2004) apresentaram valores interpretados como altos (Tabela 4). No
caso dos macronutrientes, os baixos teores de K e P, provavelmente se devem, a
elevada mobilidade e baixos teores desses elementos, comumente observados em
solos brasileiros (RAIJ, 1991; TROEH; THOMPSON, 2007).
A variação na concentração dos macronutrientes pode estar associada às
baixas taxas de ciclagem de nutrientes em regiões de clima frio, como a estudada.
Nessas regiões, grande parcela da matéria orgânica e dos nutrientes permanece no
solo e em sedimentos, ao invés de ser reciclada dentro das estruturas orgânicas do
sistema (ODUM, 1988).
Além disso, a elevada acidez indica tolerância das espécies arbóreas
predominantes nessa situação, o que conforme Gonçalves; Benedetti (2005), afeta
consideravelmente o balanço de Ca, Mg e P para a planta. Assim, identifica-se
também a baixa exigência nutricional dessas espécies para esses elementos.
No caso dos micronutrientes, as elevadas concentrações são características
dos solos da região, desenvolvidos a partir de rochas efusivas básicas,

65
intermediárias e ácidas (basalto, riodacitos ou riolitos), apresentando baixa
fertilidade natural (MOSER, 1990).
Tabela 4- Valores médios relativos aos atributos químicos do solo e interpretação, conforme a SBCS/CQFS (2004) para os grupos fitossociológicos, Reservatório Divisa, São Francisco de Paula, RS.
Atributos Químicos Grupos
GR1 Interpretação GR2 Interpretação
pH água 4,7 Baixo 4,8 Baixo H+Al 21,7 19,1 Al 4,9 3,1 Ca 3,0 Médio 5,8 Alto Mg 1,1 Alto 1,5 Alto K 0,2 Muito Baixo 0,3 Muito Baixo P 3,7 Muito Baixo 2,5 Muito Baixo S 10,0 Alto 14,8 Alto Cu 2,8 Alto 6,5 Alto Zn 2,6 Alto 4,1 Alto B 0,5 Alto 0,5 Alto MO 6,4 Alto 5,9 Alto m% 56,9 Baixo 34,2 Médio V 19,9 Muito Baixo 34,0 Muito Baixo CTC efet 9,2 Baixo 10,7 Médio CTC pH7 26,0 Muito Baixo 26,0 Muito Baixo Sendo: pH água: acidez ativa; H+Al: acidez potencial (cmolc.dm3); Al: alumínio (cmolc.dm3); Ca: cálcio (cmolc.dm3); Mg: magnésio (cmolc.dm3); K: potássio (mg.dm³); P: fósforo extraído pelo método Mehlich (mg.dm³); S: enxofre (mg.dm³); Cu: cobre (mg.dm³); Zn: zinco (mg.dm³); B: boro (mg.dm³); MO: matéria orgânica (%); m%: saturação por alumínio (%); V: saturação por bases (%); CTC efet: capacidade de troca de cátions efetiva (cmolc.dm3); CTC pH7: capacidade de troca de cátions potencial (cmolc.dm3); GR1: Grupo1; GR2: Grupo 2.
Por meio da Análise de Correspondência Canônica, pode-se verificar a
influência das variáveis ambientais (Al, Ca, S, Cu, m) na formação dos grupos. A
análise da distribuição das parcelas e espécies, com relação aos fatores ambientais,
para a vegetação arbórea (CAP ≥ 30 cm) indicou autovalores de 0,209 e 0,067, para
o 1° e 2° eixos, respectivamente (Anexo D). Conforme Carvalho et al. (2005),
autovalores baixos indicam que a maior parte das espécies encontram-se
distribuídas em toda a área, variando apenas em sua abundância.
A porcentagem acumulada de variância explicada foi de 16,3%, sendo 12,3%
explicado pelo eixo 1 e 4,0%, no eixo 2. Com base nesses valores, pode-se
considerar que as variáveis ambientais de solo utilizadas explicam, parcialmente, as

66
variações existentes na distribuição das espécies. Desse modo, a variação restante
não explicada pelas variáveis ambientais utilizadas nesse estudo, provavelmente
está associada a variáveis ambientais não registradas, como luminosidade,
pedregosidade, entre outras (NAPPO; OLIVEIRA-FILHO; MARTINS, 2000) e
incluindo as aleatórias e de maior complexidade, como dispersão, polinização e
outras de difícil mensuração. Contudo, a significância da correlação entre a
abundância das espécies e as variáveis ambientais estudadas não foi prejudicada,
considerando que o teste de permutação de Monte Carlo foi significativo, a 2% de
probabilidade de erro (Anexo D).
Autovalores e percentagem de variância explicada, semelhantes, foram
observados em estudos realizados por Avila et al. (2011), considerando a formação
de agrupamentos florísticos na regeneração natural em remanescente de Floresta
Ombrófila Mista, RS, Brasil e, também, por Budke; Jarenkow; Oliveira-Filho (2007),
considerando a estrutura do componente arbóreo e suas relações com a topografia
e o solo, em mata ciliar no Rio Botucaraí, RS, Brasil.
As variáveis mais correlacionadas com o primeiro eixo de ordenação foram o
enxofre (S) e cobre (Cu), enquanto o alumínio (Al), cálcio (Ca) e a saturação por
alumínio (m%) apresentaram maior correlação com o segundo eixo (Tabela 5).
Tabela 5 - Coeficientes de correlação entre as variáveis ambientais nos dois primeiros eixos de ordenação para os dois grupos de vegetação no entorno do Reservatório Divisa, São Francisco de Paula, RS.
Variáveis
Correlação com grupos
Eixo 1 Eixo 2
Alumínio (Al+3) 0,429 0,484
Cálcio (Ca) -0,530 -0,578
Enxofre (S) -0,635 0,063
Cobre (Cu) -0,672 0,390
Saturação por Alumínio (m%) 0,370 0,500
Os resultados da Análise de Correspondência Canônica encontram-se
apresentados na forma de diagrama de ordenação (“biplot”) na Figura 4. Esse

67
apresenta a distribuição das variáveis ambientais (Al, Ca, S, Cu, m) em relação aos
grupos fitossociológicos (GR1 e GR2).
Nesse diagrama, verifica-se que as variáveis enxofre e cobre apresentam
correlação com o Grupo 2 (GR2), ambiente caracterizado como um remanescente
de Floresta Ombrófila Mista, comumente denominados de capões, que encontram-
se sob influência do uso intensivo dos campos na pecuária. Os dejetos produzidos
pelo gado, juntamente com a deposição da biomassa sob o dossel da floresta,
compõem a matéria orgânica do solo, contribuindo para a situação evidenciada.
Observa-se que apesar de S e Cu apresentarem altos teores para ambos os
grupos (Tabela 1), os maiores valores no GR2 influenciaram a classificação desse
grupo (Figura 4). O fato de o enxofre ter, entre as principais fontes, resíduos
orgânicos vegetais e animais (ALVAREZ et al., 2007), sendo encontrado,
principalmente, na forma orgânica em aminoácidos (cistina, cisteína e metionina) e
proteínas (BISSANI; ANGHINONI, 2004), permite inferir que a presença do gado no
GR2, além de alterar a estrutura do sub-bosque, influencia nas características
químicas do solo.
Para esse mesmo grupo, o cobre (Cu+2) apresentou valor médio de 6,5
mg.dm3, superior ao observado no GR1 (2,8 mg.dm3) (Tabela 4). Conforme Abreu;
Lopes; Santos (2007), esse é o micronutriente menos móvel no solo, devido a sua
forte adsorção aos colóides orgânicos e inorgânicos do solo, ficando retido na
matéria orgânica, em especial pelos ácidos húmicos e fúlvicos, formando complexos
estáveis que exercem papel importante na mobilidade e disponibilidade para as
plantas.

68
PLOT 1
PLOT 2
PLOT 3
PLOT 4
PLOT 5
PLOT 6
PLOT 9
PLOT 10
PLOT 13
PLOT 15
PLOT 20
PLOT 23
PLOT 25
PLOT 26
PLOT 28
PLOT 29
PLOT 30PLOT 31
PLOT 34PLOT 36
PLOT 38PLOT 40
PLOT 42
PLOT 44
PLOT 46
PLOT 48
PLOT 50
PLOT 51
PLOT 53
PLOT 54PLOT 55
PLOT 57
Al
Ca
S
Cum
Axis 1
Axi
s 2
GR
12
Figura 4 - Diagrama de ordenação dos grupos analisados com as variáveis ambientais para os dois primeiros eixos produzidos pela Análise de Correspondência Canônica, Reservatório Divisa, São Francisco de Paula, RS. Sendo GR: Grupo; Plot: Parcelas; Cu: Cobre; S: Enxofre; Al: Alumínio; m: Saturação por alumínio; Ca: Cálcio.
Tonetto (2011), estudando a variabilidade de solos em área de tensão
ecológica entre campo e Floresta Ombrófila Mista na Barragem do Divisa, São
Francisco de Paula, RS, destacou que os valores de Cu encontrados, interpretados
como altos, de acordo com a Sociedade Brasileira de Ciência do Solo (SBCS/CQFS,
2004), podem atingir níveis tóxicos às plantas. No entanto, Silva et al. (2011),
observaram que algumas espécies florestais apresentaram tolerância aos elevados
teores de Cu no solo, o que sugere a utilização dessas na revegetação desses
ambientes.
Contudo, a presença de S e Cu, influenciando o Grupo 2, fica evidenciada ao
analisar os seus teores nutricionais, 14,8 e 6,5 mg.dm3, respectivamente (Tabela 4),
e o comprimento das setas em direção à maioria das parcelas desse grupo (Figura
4). Nesse caso, o GR2 apresenta as maiores concentrações desses elementos, o
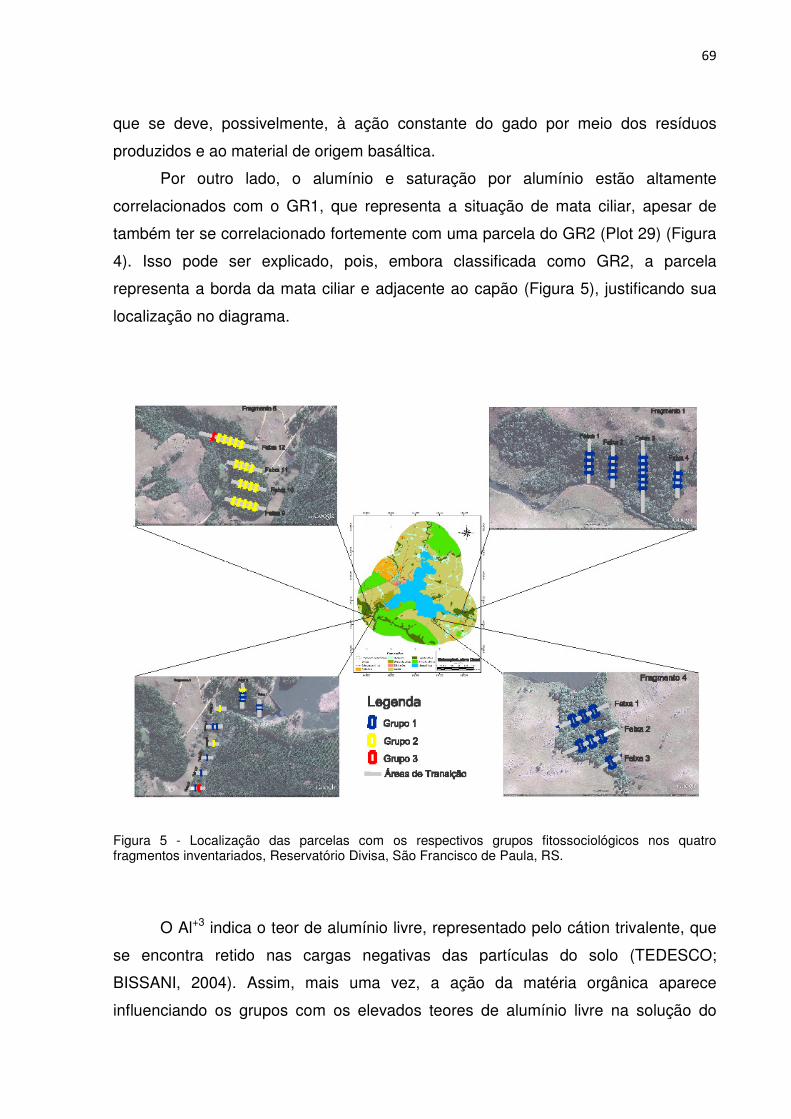
69
que se deve, possivelmente, à ação constante do gado por meio dos resíduos
produzidos e ao material de origem basáltica.
Por outro lado, o alumínio e saturação por alumínio estão altamente
correlacionados com o GR1, que representa a situação de mata ciliar, apesar de
também ter se correlacionado fortemente com uma parcela do GR2 (Plot 29) (Figura
4). Isso pode ser explicado, pois, embora classificada como GR2, a parcela
representa a borda da mata ciliar e adjacente ao capão (Figura 5), justificando sua
localização no diagrama.
Figura 5 - Localização das parcelas com os respectivos grupos fitossociológicos nos quatro fragmentos inventariados, Reservatório Divisa, São Francisco de Paula, RS.
O Al+3 indica o teor de alumínio livre, representado pelo cátion trivalente, que
se encontra retido nas cargas negativas das partículas do solo (TEDESCO;
BISSANI, 2004). Assim, mais uma vez, a ação da matéria orgânica aparece
influenciando os grupos com os elevados teores de alumínio livre na solução do

70
solo, devido ao maior teor de matéria orgânica (MO%), em função das lentas taxas
de intemperismo na região, condicionadas pela altitude, clima frio, material de
origem basáltico e relevo (SILVA; CAMARGO; CERETTA, 2000). Todos esses
aspectos, associados à maior umidade, que além de ser própria da região, é
potencializada na mata ciliar (GR1), acarreta menor saída do Al do complexo do
solo.
Esse resultado pode ser confirmado ao analisar os valores médios de Al, no
GR1 (4,9 cmolc.dm3) em comparação ao GR2 (3,1 cmolc.dm3). A presença de Al em
concentrações elevadas em relação à fertilidade dos solos indica toxidade às
plantas, podendo interferir na disponibilidade de outros elementos (SOUSA;
MIRANDA; OLIVEIRA, 2007).
Outro atributo a ser analisado é a saturação por alumínio (m%), que
apresentou valores de 56,9% (GR1) e 34,2% (GR2). Conforme Troeh; Thompson
(2007), a saturação por alumínio expressa o teor de alumínio livre em relação à CTC
ocupada por alumínio trocável, fornecendo indicativo de toxidez desse elemento às
espécies vegetais. Os efeitos do Al em concentrações tóxicas manifestam-se na
parte aérea e radicular, por meio de sintomas anatômicos, morfológicos e redução
no crescimento, devido à diminuição no alongamento das raízes e a consequente
redução na possibilidade de absorção de nutrientes (SOUSA; MIRANDA; OLIVEIRA,
2007). Desse modo, as elevadas concentrações de alumínio e cobre, possivelmente,
influenciam de forma negativa o desenvolvimento e expansão das florestas no local.
A variável cálcio (Ca) explica a vegetação local, porém, não está direcionada
diretamente para nenhum grupo. Para Larcher (2000), esse nutriente possui a
função de regulação da hidratação, ativação de enzimas e regulador do crescimento,
sendo acumulado, principalmente, nas folhas e na casca das plantas.
Tendo em vista que o comprimento da seta nos biplot (Figuras 5 e 7) é
proporcional à magnitude de mudança na direção indicada (KENT; COKER, 1992),
observa-se a menor influência das variáveis ambientais sobre as espécies (Figura 7)
do que às parcelas (grupos) (Figura 5), o que não permite definir possíveis
exigências nutricionais. Por outro lado, há indicativos que Eugenia uruguayensis e
Myrsine coriacea, ambas espécies indicadoras do GR1, apresentam correlação com
o aumento do alumínio livre no solo e com a saturação por alumínio (Figura 7). Isso
pode ser confirmado pela expressiva densidade de indivíduos dessas espécies, na
maioria das classes de tamanho estudadas na vegetação (Tabela 6).

71
Arauca Blepharo
Calyp
Case.dec
Cinnamo
Drimys
E.urug
Handroa
Ilex.bre
Lithraea
M yrc.cuc
M yrcia p
M yrciant
M yrc.del
M .coriac
O.pul
Podocar
Sebas.co
Symp.uni
Zanthoxy
Al
Ca
SCu m Axis 1
Axi
s 2
Figura 6 - Diagrama de ordenação das espécies com as variáveis ambientais para os dois primeiros eixos produzidos pela Análise de Correspondência Canônica, Reservatório Divisa, São Francisco de Paula, RS. Sendo Cu: Cobre; S: Enxofre; Al: Alumínio; m: Saturação por alumínio; Ca: Cálcio; Arauca: Araucaria angustifolia; Blepharo: Blepharocalyx salicifolius; Calyp: Calyptranthes concinna; Case.dec: Casearia decandra; Cinnamo: Cinnamomum amoenum; Drimys: Drimys brasiliensis; E.urug: Eugenia uruguayensis; Handroa: Handroanthus albus; Ilex.bre: Ilex brevicuspis; Lithraea: Lithraea brasiliensis; Myrc.cuc: Myrceugenia cucullata; Myrcia p: Myrcia palustris; Myrciant: Myrcianthes gigantea; Myrc.del: Myrciaria delicatula; M.coriac: Myrsine coriacea; O.pul: Ocotea pulchella; Podocar: Podocarpus lambertii; Sebas.com: Sebastiania commersoniana; Symp.uni: Symplocos uniflora e Zanthoxy: Zanthoxylum rhoifolium.
Por outro lado, Araucaria angustifolia, Blepharocalyx salicifolius e
Calyptranthes concinna apresentaram maior correlação com a variável Ca (Figura 6).
Essas espécies, principalmente na Classe I (CAP ≥ 30 cm), estão representadas
proporcionalmente em ambos os grupos. No caso da Araucaria angustifolia, a casca

72
desprende-se em lâmina da parte superior do fuste (CARVALHO, 2003),
aumentando, assim, a deposição desse material no solo e, consequentemente,
aumentando os teores do cálcio no sítio. Considerando o médio e alto teor (Tabela
4), respectivamente, no GR1 e GR2, possivelmente, Blepharocalyx salicifolius e
Calyptranthes concinna, além de Araucaria angustifolia, são mais tolerantes a esse
elemento. Dessa forma, essas espécies também podem ser observadas como
potenciais para utilização em mata ciliar.
Myrceugenia cucullata e, principalmente, Podocarpus lambertii, que se
encontram entre as espécies melhor hierarquizadas do GR2, para as classes I e II
(Tabela 6), apresentam-se melhor correlacionadas com enxofre e cobre (Figura 6).
Verifica-se que esse grupo apresentou os maiores teores de S e Cu (14,8 e 6,5
mg.dm³), quando comparado ao GR1 (10,0 e 2,8 mg.dm³) (Tabela 4), justificando a
influência dessas variáveis sobre as duas espécies mencionadas. Além disso,
apresentam menor potencial para uso na mata ciliar, considerando reduzida
representatividade nesse ambiente, quando comparado ao GR2.
Sebastiania commersoniana, Lithraea brasiliensis e Ocotea pulchella, apesar
de estarem representadas entre as dez melhor hierarquizadas em GR1 e GR2,
podem ser enfatizadas na mata ciliar, dada sua maior densidade e ocorrência em
solos com menores teores de cobre e enxofre. Isso é demonstrado na Figura 6,
considerando que a linha que indica a variável explicativa (ambiental) pode ser
estendida, no sentido contrário de sua origem, indicando que a menor influência da
variável favorece os elementos do lado oposto (KENT; COKER, 1992).
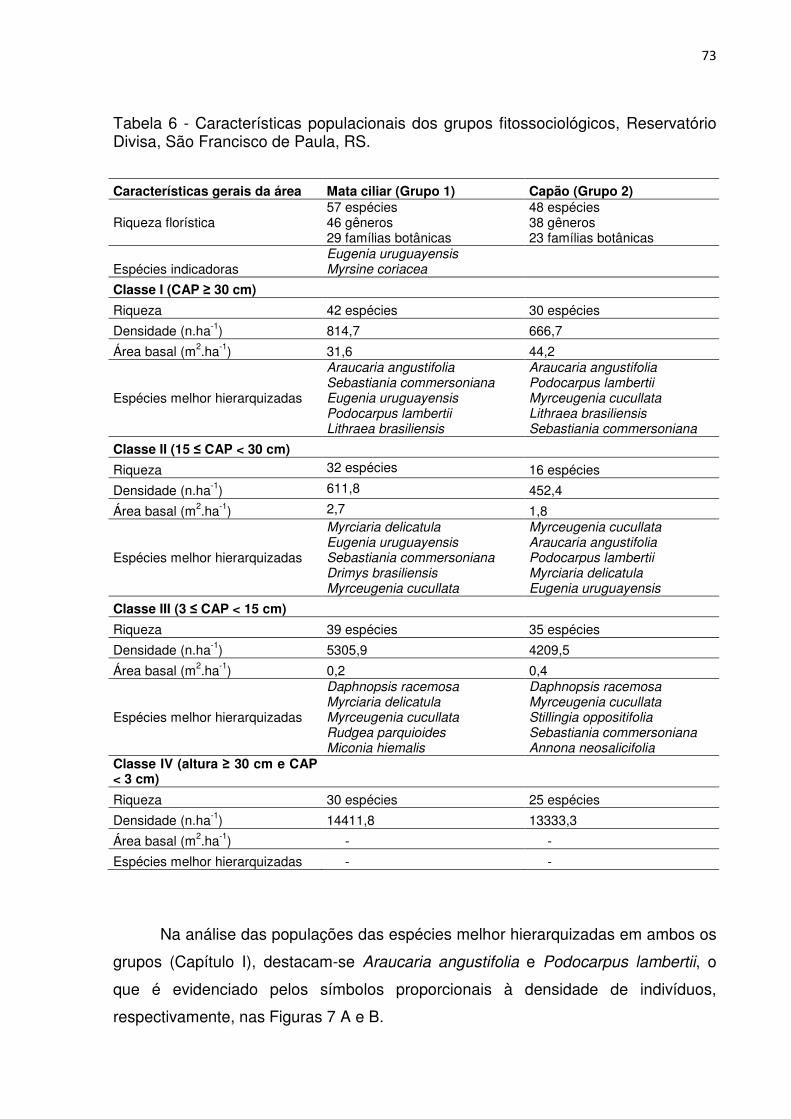
73
Tabela 6 - Características populacionais dos grupos fitossociológicos, Reservatório Divisa, São Francisco de Paula, RS.
Características gerais da área Mata ciliar (Grupo 1) Capão (Grupo 2)
Riqueza florística
57 espécies 46 gêneros 29 famílias botânicas
48 espécies 38 gêneros 23 famílias botânicas
Espécies indicadoras Eugenia uruguayensis Myrsine coriacea
Classe I (CAP ≥ 30 cm)
Riqueza 42 espécies 30 espécies
Densidade (n.ha-1) 814,7 666,7
Área basal (m2.ha-1) 31,6 44,2
Espécies melhor hierarquizadas
Araucaria angustifolia Sebastiania commersoniana Eugenia uruguayensis Podocarpus lambertii Lithraea brasiliensis
Araucaria angustifolia Podocarpus lambertii Myrceugenia cucullata Lithraea brasiliensis Sebastiania commersoniana
Classe II (15 ≤ CAP < 30 cm)
Riqueza 32 espécies 16 espécies
Densidade (n.ha-1) 611,8 452,4
Área basal (m2.ha-1) 2,7 1,8
Espécies melhor hierarquizadas
Myrciaria delicatula Eugenia uruguayensis Sebastiania commersoniana Drimys brasiliensis Myrceugenia cucullata
Myrceugenia cucullata Araucaria angustifolia Podocarpus lambertii Myrciaria delicatula Eugenia uruguayensis
Classe III (3 ≤ CAP < 15 cm)
Riqueza 39 espécies 35 espécies
Densidade (n.ha-1) 5305,9 4209,5
Área basal (m2.ha-1) 0,2 0,4
Espécies melhor hierarquizadas
Daphnopsis racemosa Myrciaria delicatula Myrceugenia cucullata Rudgea parquioides Miconia hiemalis
Daphnopsis racemosa Myrceugenia cucullata Stillingia oppositifolia Sebastiania commersoniana Annona neosalicifolia
Classe IV (altura ≥ 30 cm e CAP < 3 cm)
Riqueza 30 espécies 25 espécies
Densidade (n.ha-1) 14411,8 13333,3
Área basal (m2.ha-1) - -
Espécies melhor hierarquizadas - -
Na análise das populações das espécies melhor hierarquizadas em ambos os
grupos (Capítulo I), destacam-se Araucaria angustifolia e Podocarpus lambertii, o
que é evidenciado pelos símbolos proporcionais à densidade de indivíduos,
respectivamente, nas Figuras 7 A e B.

74
PLOT 1
PLOT 2PLOT 3
PLOT 4
PLOT 5
PLOT 6
PLOT 9
PLOT 10
PLOT 13
PLOT 15
PLOT 20
PLOT 23
PLOT 25
PLOT 26
PLOT 28
PLOT 29
PLOT 30
PLOT 31
PLOT 34PLOT 36
PLOT 38 PLOT 40
PLOT 42
PLOT 44
PLOT 46
PLOT 48
PLOT 50
PLOT 51
PLOT 53
PLOT 54PLOT 55
PLOT 57
Axis 1
Axi
s 2
GR
12
PLOT 1
PLOT 2
PLOT 3
PLOT 4
PLOT 5
PLOT 6
PLOT 9
PLOT 10
PLOT 13
PLOT 15
PLOT 20
PLOT 23
PLOT 25
PLOT 26
PLOT 28
PLOT 29
PLOT 30
PLOT 31
PLOT 34PLOT 36
PLOT 38PLOT 40
PLOT 42
PLOT 44
PLOT 46
PLOT 48
PLOT 50
PLOT 51
PLOT 53
PLOT 54PLOT 55
PLOT 57
Axis 1
Axi
s 2
GR
12
Figura 7 - Diagrama de ordenação de Araucaria angustifolia (A) e Podocarpus lambertii (B) para os dois primeiros eixos produzidos pela Análise de Correspondência Canônica, Reservatório Divisa, São Francisco de Paula, RS. Sendo GR: Grupo; Plot: Parcelas.
A
B

75
A presença de Araucaria angustifolia em elevada densidade e frequência em
todos os grupos analisados é justificada, pois a região representa a Floresta
Ombrófila Mista. Essa vegetação original apresenta como característica marcante a
presença dominante dessa espécie no estrato superior, acompanhada de diversas
espécies pertencentes às famílias Lauraceae, Aquifoliaceae, Sapindaceae, entre
outras (REITZ; KLEIN; REIS, 1983). Além disso, segundo Klein (1960), a espécie
caracteriza-se por formar uma cobertura emergente, onde sob as copas encontram-
se outras espécies de vegetais que contribuem para a diversidade ecológica desses
ambientes.
Assim, a Floresta Ombrófila Mista possui estratificação bem definida e
composta por quatro níveis, sendo o estrato superior formado exclusivamente por
araucárias; o segundo estrato formado predominantemente por Podocarpus lambertii
e diversas espécies latifoliadas, seguido de vegetação arbustiva e do estrato
herbáceo, quase ausente em virtude da grande quantidade de ramos caídos,
juntamente com a baixa luminosidade provocada pelos estratos superiores
(BACKES, 1983). Desse modo, a representatividade de Araucaria angustifolia e
Podocarpus lambertii nos grupos formados é atribuída à formação florestal a que
pertencem.
Na Figura 8, 9 e 10, evidenciam-se as populações de Eugenia uruguayensis,
Myrsine coriacea e Sebastiania commersoniana, respectivamente. Para essas
espécies, observa-se maior densidade de indivíduos nas parcelas representadas
pelo GR1, caracterizado pela Floresta Ombrófila Mista com grande número de
espécies de mata ciliar, com grande expressividade de Myrsine coriacea.
Eugenia uruguayensis, Myrsine coriacea e Sebastiania commersoniana
encontram-se bem representadas no GR1, em relação a todos os parâmetros
fitossociológicos observados (Capítulo I). Essas espécies apresentam plasticidade
em relação às condições de luminosidade, com comportamento de espécie heliófila,
sendo considerada pioneira a secundária tardia (CARVALHO, 2003; LORENZI,
2008). Além disso, Carvalho (2003) destaca que Myrsine coriacea, apesar de ocorrer
em solos secos, muito rasos e de baixa fertilidade química, ocorre também em
várzeas e banhados, o que indica a maior adaptação da espécie em ambientes
úmidos.

76
PLOT 1
PLOT 2PLOT 3
PLOT 4
PLOT 5
PLOT 6
PLOT 9
PLOT 10
PLOT 13
PLOT 15
PLOT 20
PLOT 23
PLOT 25
PLOT 26
PLOT 28
PLOT 29
PLOT 30
PLOT 31
PLOT 34PLOT 36
PLOT 38PLOT 40
PLOT 42
PLOT 44
PLOT 46
PLOT 48
PLOT 50
PLOT 51PLOT 53
PLOT 54PLOT 55
PLOT 57
Axis 1
Axi
s 2
GR
12
Figura 8 - Diagrama de ordenação de Eugenia uruguayensis para os dois primeiros eixos produzidos pela Análise de Correspondência Canônica, Reservatório Divisa, São Francisco de Paula, RS. Sendo GR: Grupo; Plot: Parcelas.
PLOT 1
PLOT 2PLOT 3
PLOT 4
PLOT 5
PLOT 6
PLOT 9
PLOT 10
PLOT 13
PLOT 15
PLOT 20
PLOT 23
PLOT 25
PLOT 26
PLOT 28
PLOT 29
PLOT 30PLOT 31
PLOT 34PLOT 36
PLOT 38PLOT 40
PLOT 42
PLOT 44
PLOT 46
PLOT 48
PLOT 50
PLOT 51PLOT 53
PLOT 54PLOT 55
PLOT 57
Axis 1
Axi
s 2
GR
12
Figura 9 - Diagrama de ordenação de Myrsine coriacea para os dois primeiros eixos produzidos pela Análise de Correspondência Canônica, Reservatório Divisa, São Francisco de Paula, RS. Sendo GR: Grupo; Plot: Parcelas.

77
PLOT 1
PLOT 2
PLOT 3
PLOT 4
PLOT 5
PLOT 6
PLOT 9
PLOT 10
PLOT 13
PLOT 15
PLOT 20
PLOT 23
PLOT 25
PLOT 26
PLOT 28
PLOT 29
PLOT 30
PLOT 31
PLOT 34PLOT 36
PLOT 38 PLOT 40
PLOT 42
PLOT 44
PLOT 46
PLOT 48
PLOT 50
PLOT 51PLOT 53
PLOT 54PLOT 55
PLOT 57
Axis 1
Axi
s 2
GR
12
Figura 10 - Diagrama de ordenação de Sebastiania commersoniana para os dois primeiros eixos produzidos pela Análise de Correspondência Canônica, Reservatório Divisa, São Francisco de Paula, RS. Sendo GR: Grupo; Plot: Parcelas.
A espécie Myrceugenia cucullata apresentou distribuição, principalmente, no
GR2, representado por um remanescente de Floresta Ombrófila Mista sob influência
do uso intensivo dos campos na pecuária (Figura 11). Essa espécie, pertencente ao
grupo ecológico das secundárias iniciais a secundárias tardias (RIO GRANDE DO
SUL, 2007), encontra-se melhor distribuída nesse grupo, pois para se desenvolver,
os indivíduos necessitam de ambientes com certo grau de sombreamento.

78
PLOT 1
PLOT 2
PLOT 3
PLOT 4
PLOT 5
PLOT 6
PLOT 9
PLOT 10
PLOT 13
PLOT 15
PLOT 20
PLOT 23
PLOT 25
PLOT 26
PLOT 28
PLOT 29
PLOT 30
PLOT 31
PLOT 34PLOT 36
PLOT 38PLOT 40
PLOT 42
PLOT 44
PLOT 46
PLOT 48
PLOT 50
PLOT 51PLOT 53
PLOT 54PLOT 55
PLOT 57
Axis 1
Axi
s 2
GR
12
Figura 11 - Diagrama de ordenação de Myrceugenia cucullata para os dois primeiros eixos produzidos pela Análise de Correspondência Canônica, Reservatório Divisa, São Francisco de Paula, RS. Sendo GR: Grupo; Plot: Parcelas.
Considerando a representatividade das espécies, grupo ecológico a que
pertencem, e correlação das espécies com as variáveis ambientais, observa-se que
Araucaria angustifolia, Eugenia uruguayensis, Blepharocalyx salicifolius,
Calyptranthes concinna, Lithraea brasiliensis, Myrsine coriacea, Ocotea pulchella e
Sebastiania commersoniana apresentam maior potencial para utilização no
enriquecimento de mata ciliar, enquanto Myrceugenia cucullata e, principalmente,
Podocarpus lambertii podem ser sugeridas para recuperação de áreas abertas,
considerando seu bom desenvolvimento sob luz direta.

79
3.5 Conclusão
As características físicas do solo não apresentam influência nos grupos
formados e nas espécies.
Na mata ciliar, a lenta decomposição da matéria orgânica potencializada pela
maior umidade é responsável pela elevada acidez do solo, influenciando
negativamente a disponibilidade de nutrientes para as espécies no local. Entretanto,
as espécies florestais parecem adaptadas a tal condição.
Nos capões, a constante presença do gado altera negativamente a estrutura
do sub-bosque, assim como nas características químicas do solo.
A variação na concentração de macronutrientes está associada às baixas
taxas de ciclagem de nutrientes, enquanto as elevadas concentrações de
micronutrientes são típicas de solos derivados do basalto.
Alumínio e saturação por alumínio apresentam maior influência nas espécies
predominantes da mata ciliar enquanto, cobre e enxofre influenciaram as espécies
presentes nos capões.
A elevada densidade de Eugenia uruguayensis e Myrsine coriacea em
ambiente ciliar ocorre devido à tolerância das espécies às elevadas concentrações
de alumínio e saturação por alumínio.
Araucaria angustifolia, Blepharocalyx salicifolius e Calyptranthes concinna são
espécies que requerem maiores teores de cálcio.
Myrceugenia cucullata e, principalmente, Podocarpus lambertii são altamente
correlacionados com cobre e enxofre.
Araucaria angustifolia, Eugenia uruguayensis, Blepharocalyx salicifolius,
Calyptranthes concinna, Lithraea brasiliensis, Myrsine coriacea, Ocotea pulchella e
Sebastiania commersoniana podem ser utilizadas na recuperação e enriquecimento
de mata ciliar.

80
4 CONSIDERAÇÕES FINAIS
As matas ciliares são vegetações típicas da região, com estrutura
diferenciada inclusive dos capões, ocorrendo entremeadas aos campos, e pouco
frequentes no entorno do Reservatório.
A vegetação predominante de campos naturais na região está associada à
rede de drenagem intermitente, enquanto os fragmentos de floresta ocorrem na
presença de córregos formados por nascentes ou continum de matas ciliares,
geralmente contribuindo na manutenção da água no local.
Os fragmentos que se encontram isolados no entorno da barragem (F3 e F4)
são os mais susceptíveis a perda de estrutura e diversidade devido à reduzida fonte
de sementes, merecendo serem avaliados quanto à restrição ao acesso do gado.
Diante da necessidade de restauração e/ou enriquecimento de matas ciliares
na região, os dados obtidos podem ser utilizados na formação de arranjos de
espécies adaptadas, as quais, provavelmente, são pouco influenciadas pelas
características do solo.

81
REFERÊNCIAS BIBLIOGRÁFICAS
ABREU, C. A.; LOPES, A. S.; SANTOS, G. C. G. Micronutrientes. In: NOVAIS, R. F. et al. Fertilidade do solo. Viçosa: Sociedade Brasileira de Ciência do Solo, 2007, p. 645-736. ALVAREZ, V. H. et al. Enxofre. In: NOVAIS, R. F. et al. Fertilidade do solo. Viçosa: Sociedade Brasileira de Ciência do Solo, 2007, p. 595-644. AMADOR, D. B; VIANA, V. M. Dinâmica de “capoeiras baixas” na restauração de um fragmento florestal. Scientia Forestalis, Piracicaba, n. 57, p. 69-85. 2000. ARAUJO, M. M. Vegetação e banco de sementes do solo de florestas sucessionais na região do baixo rio Guamá, Benevides, Pará, Brasil. 1998. 86 f. Dissertação (Mestrado em Ciências Florestais)–Faculdade de Ciências Agrárias do Pará, Belém, 1998. ARAUJO, M. M. et al. Análise de agrupamento da vegetação em fragmento de floresta Estacional Decidual Aluvial, Cachoeira do Sul, RS, Brasil. Ciência Florestal, Santa Maria, v. 14, n. 1, p. 133-147. 2004. ARAUJO, M. M. et al. Análise de agrupamento em remanescente de Floresta Ombrófila Mista. Ciência Florestal, Santa Maria, v. 20, n. 1, p. 1-18. 2010. AUSTIN, M. P.; GREIG-SMITH, P. The application of quantitative methods to vegetation survey. The Journal of Ecology, v. 56, n. 3, p. 827-844. 1968. AVILA, A. L. et al. Agrupamentos florísticos na regeneração natural em remanescente de Floresta Ombrófila Mista, RS, Brasil. Sci. For., Piracicaba, v. 39, n. 91, p. 331-342. 2011. AZEVEDO, A. C; DALMOLIN, R. S. D. Solos e ambiente: uma introdução. Santa Maria: Ed. Pallotti, 2006. 100p. BACKES, A. Dinâmica do pinheiro brasileiro. Iheringia, Porto Alegre, n. 30, p. 49-84. 1983.

82
BACKES, P.; IRGANG, B., Árvores do Sul: guia de identificação & Interesse Ecológico. 1. ed. Santa Cruz do Sul: Instituto Souza Cruz; Porto Alegre: Pallotti 2002. 326p. BEHLING, H. et al. Late Quaternary Araucaria forest, grassland (Campos), fire and climate dynamics, studied by high-resolution pollen, charcoal and multivariate analysis of the Cambará do Sul core in southern Brazil. Palaeogeography, Palaeoclimatology, Palaeoecology, v. 203, p. 277-297. 2004. BEHLING, H.; PILLAR, V. D. Late Quaternary vegetation, biodiversity and fire dynamics on the southern Brazilian highland and their implication for conservation and management of modern Araucaria forest and grassland ecosystems. Philosophical Transactions of the Royal, 2007, 362, p. 243-251. BISSANI, C. A.; ANGHINONI, I. Enxofre, Cálcio e Magnésio. In: BISSANI, C. A. et al. Fertilidade dos solos e manejo da adubação de culturas. Porto Alegre: Genesis, 2004. 207-220p. BOND-BUCKUP, G. Biodiversidade dos Campos de Cima da Serra. Porto Alegre: Libretos, 2008. 146 p. BRASIL. Lei nº 4.771, de 15 de setembro de 1965. Institui o novo Código Florestal. Diário Oficial da União, Brasília, DF, 16 set. 1965. Disponível em:<http://www.planalto.gov.br/ccivil_03/leis/L4771.htm>. Acesso em 10 Agost. 2011. BRASIL. Proposta do grupo de trabalho preservação e recuperação da Floresta Ombrófila Mista no Estado de Santa Catarina: portaria ministerial 49 de 06 de fevereiro de 2002. Ministério do Meio Ambiente, Brasília, 2002. 77p. BRAUN-BLANQUET, J. Fitosociologia: bases para el estudio de las comunidades vegetales. 3. ed. Madrid: H. Blume, 1979. 820p. BRUCE, R. G. H. The application of quantitative method of classification to strategic ecological survey in Britain. In: KLIJN, F. (Ed.). Ecossistem classification for environmental management. Netherlands: Klwer Academic Publishers, p. 173-182. 1994. BUDKE, J. C.; JARENKOW, J. A.; OLIVEIRA-FILHO, A. T. Relationships between tree component structure, topography and soils of a riverside forest, Rio Botucaraí, Southern Brazil. Plant Ecology, Dordrecht, v. 189, p. 187-200. 2007.

83
CAPPELATTI, L.; SCHMITT, J. L. Flora arbórea de área de Floresta Ombrófila Mista em São Francisco de Paula, RS, Brasil. Pesquisas Botânica, São Leopoldo, n. 62, p. 253-261. 2011. CARROLL, S. P.; DINGLE, H. The biology of post-invasion events. Biological Conservation, v. 78, p. 207-214. 1996. CARVALHO, J. O. P. Análise estrutural da regeneração natural em floresta tropical densa na região do Tapajós no Estado do Pará. 1982. 128f. Dissertação (Mestrado em Ciências Florestais)–Universidade Federal do Paraná, Curitiba, 1982. CARVALHO, D. A. et al. Distribuição de espécies arbóreo-arbustivas ao longo de um gradiente de solos e topografia em um trecho de floresta ripária do Rio São Francisco em Três Marias, MG, Brasil. Revista Brasileira de Botânica, São Paulo, v. 28, n. 2, p. 329-345. 2005. CARVALHO, P. E. R. Espécies arbóreas brasileiras. Brasília: Embrapa Informações Tecnológicas, Colombo: Embrapa Florestas, v. 1, 2003. 1039p. CARVALHO, P. E. R. Espécies arbóreas brasileiras. Brasília: Embrapa Informação Tecnológica, Colombo: Embrapa Florestas, v. 2, 2006a. 627p. CARVALHO, P. E. R. Espécies arbóreas brasileiras. Brasília: Embrapa Informação Tecnológica; Colombo, PR: Embrapa Florestas, v. 3, 2008. 593p. CARVALHO, P. E. R. Saboneteira. Colombo: Embrapa Florestas, 2006b. 6p. (Circular técnica, n. 116). Disponível em: < (http://ainfo.cnptia.embrapa.br/digital/bitstream/CNPF-2009-09/40889/1/circ tec116.pdf) >. Acesso em: 06 Mar. 2012. CEEE-GT- COMPANHIA ESTADUAL DE GERAÇÃO E TRANSMISSÃO DE ENERGIA ELÉTRICA. Plano de uso e ocupação do solo no entorno do Reservatório Divisa - UHE Bugres, 2009. 228p. CHAMI, L. B. Vegetação e mecanismos de regeneração natural em diferentes ambientes da Floresta Ombrófila Mista na Flona de São Francisco de Paula, RS. 2008. 125f. Dissertação (Mestrado em Engenharia Florestal)-Universidade Federal de Santa Maria, Santa Maria, 2008.

84
CRESTANA, M. S. M. et al. Florestas-sistemas de recuperação com essências nativas, produção de mudas e legislações. 2. ed. Campinas: CATI, 2004. 216p. DANIEL, O.; JANKAUSKIS, J. Avaliação de metodologia para o estudo do estoque de sementes do solo, em floresta de terra firme na Amazônia brasileira. IPEF: v. 41-42, p. 18-26. 1989. DUARTE, L. S.; DILLENBURG, L. R. Ecophysiological responses of Araucaria angustifolia (Araucariaceae) seedlings to different irradiance levels. Australian Journal of Botany, v.48, n.4, p.531-537, 2000.
DUARTE, M. C.; SANTOS, J. B.; MELO, L. C. Comparison of similarity coefficients based on RAPD markers in the common bean. Genetics and Molecular Biology,Ribeirão Preto, v. 22, n. 3, p. 427-432. 1999. EBLING, A. A. Dinâmica e projeção de um remanescente de floresta com araucária em São Francisco de Paula, RS. 2012. 142f. Dissertação (Mestrado em Ciências Florestais)-Universidade Estadual do Centro-Oeste, Paraná, 2012. EFRON, B. The jackknife, the bootstrap and other resampling plans. Philadelphia: Society for Industrial and applied mathematics, 1982. 92p. EMBRAPA- EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA. Manual de métodos de análises de solo. 2. ed. Rio de Janeiro: Ministério da Agricultura e do Abastecimento, 1997. 212p. EMER, C.; FONSECA, C. R. Floresta com araucária resiste à invasão por árvores exóticas. In: CONGRESSO DE ECOLOGIA DO BRASIL, 2007, Caxambu. Anais... Caxambu, MG, 2007. 2p. FAO. State of the World’s Forest. Rome: Food and Agriculture of the United Nations, 2011. FELFILI, J. M. Análise multivariada em estudos de vegetação. Brasília: Universidade de Brasília, 2007. 60p. FELFILI, J. M.; REZENDE, R. P. Conceitos e métodos em fitossociologia. Comunicações Técnicas Florestais, Brasília, v. 5, n. 1, 68p. 2003.

85
FELFILI, J. M.; VENTUROLI, F. Tópicos em análise de vegetação. Comunicações Técnicas Florestais, Brasília, v. 2, n. 2, 34p. 2000. FELFILI, J. M. et al., Comparação entre as unidades fisiográficas Chapada Pratinha, Veadeiros e Espigão Mestre do São Francisco. In: FELFILI, J. M.; SILVA JR., M. C. da (Orgs.). Biogeografia do bioma Cerrado: estudo fitofisiônomico na Chapada do Espigão Mestre do São Francisco. Brasília: UnB, 2001. p. 80-94. FELFILI, J. M. et al. Análise Multivariada: princípios e métodos em estudos de vegetação. In: ________. Fitossociologia no Brasil: métodos e estudos de casos. Viçosa: Ed. UFV, v. 1, 2011. 556p. FINOL, U. H. Nuevos parámetros a considerarse en el análisis estructural de las sevas virgens tropicales. Revista Forestal Venezoelana, Merida, v.14, n. 21, p. 29-42. 1971. FREI, F. Introdução à análise de agrupamentos: teoria e prática. São Paulo: Editora UNESP, 2006. 109p. GAUCH, H. G. Multivariate analysis in community ecology. Cambridge: Cambridge University Press, 1982. 180p. GEE, G. W.; BAUDER, J. W. Particle size analysis. In: KLUTE, A., ed. Methods of Soil Analysis: part I. 2. ed. Madison, American Society of Agronomy, 1986. p. 383-411. GOMES, M. Reavaliação taxonômica de algumas espécies dos gêneros Coussarea Aubl. e Faramea Aubl. (Rubiaceae, tribo Coussareae). Acta Botânica, v. 17, p. 449-466. 2003. GONÇALVES, J. L. M.; BENEDETTI, V. Nutrição e fertilização florestal. Piracicaba: IPEF, 2005. 427 p. HACK, C. Respostas da vegetação remanescente e da regeneração natural em Floresta Ombrófila Mista cinco anos após intervenções de manejo. 2007. 96 f. Dissertação (Mestrado em Engenharia Florestal)-Universidade Federal de Santa Maria, Santa Maria, 2007. HAIR, J. F. et al. Multivariate Data Analysis. Rio de Janeiro: Editora Prentice-Hall do Brasil, 1998. 730p.

86
HARPER, J. L. Population biology of plants. London: Academic Press, 1977. 892p. HILL, M. O. TWINSPAN: a fortran program for arranging multivariate data in an ordered two way table by classification of individual and attributes. Ithaca, NY: Cornell University, 1979. 60p. HIGUCHI, N. et al. Bacia 3 – Inventário diagnóstico da regeneração natural. Acta Amazonica, Manaus, v. 15, n. 1/2, p. 199- 233. 1985. IBGE- INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA. Manual técnico da vegetação brasileira. Rio de Janeiro, 1992. 91p. INOUE, M. T.; RODERJAN, C. V.; KUNIYOSHI, S. Y. Projeto madeira do Paraná. Curitiba: FUPEF, 1984. 260p. KANIESKI, M. R. Caracterização florística, diversidade e correlação ambiental na Floresta Nacional de São Francisco de Paula, RS. 2010. 101f. Dissertação (Mestrado em Engenharia Florestal) - Universidade Federal de Santa Maria, Santa Maria, 2010. KENT, M.; COKER, P. Vegetation description analyses. London: Behaven Press, 1992. 363p. KLEIN, R. M. O aspecto dinâmico do pinheiro brasileiro. Sellowia, Itajaí, v. 12, n. 12, p. 17- 48. 1960. KOYAMA, T. Simulation of the structural development of warm-temperature rain forest stands. Annals of Botany, London, v. 63, n.6, p. 625-634, 1989. LANDRUM, L. R. Campomanesia, Pimenta, Blepharocalyx, Legrandia, AACC, Myrrhinium and Lumma (Myrtaceae). Flora Neotropica, New York, n. 45, p. 1-178. 1986. LARCHER, W. Ecofisiologia vegetal. São Carlos: Rima, 2000. 531p. LEGRAND, C. D.; KLEIN, R. M. Mirtáceas: Gomidesia. In: Reitz, R. (Ed.). Flora Ilustrada Catarinense. Itajaí: Herbário Barbosa Rodrigues, 1967. p. 1-44.

87
LEGRAND, C. D.; KLEIN, R. M. Mirtáceas: Eugenia. In: Reitz, R. (Ed.). Flora Ilustrada Catarinense. Itajaí: Herbário Barbosa Rodrigues, 1969. p. 45-216. LEITE, P. F. Contribuição ao conhecimento fitoecológico do Sul do Brasil. Ciência & Ambiente, Santa Maria, v. 1, n. 1, p. 51-73. 2002. LEITE, P. F; KLEIN, R. M. Vegetação. In: IBGE. Geografia do Brasil: Região Sul. Rio de Janeiro, 1990. p. 115-150. LEPSCH, I. F.; SILVA, N. M ; ESPIRONELO, A. Relação entre matéria orgânica e textura de solos sob cultivo de algodão e cana-de-açúcar, no Estado de São Paulo. Bragantia, Campinas, v. 41, p. 231-236. 1982. LIMA, W. P.; ZAKIA, M. J. B. Indicadores hidrológicos em áreas florestais. Série Técnica IPEF, Piracicaba, v.12, n.31, p.53-64, 1998. LONGHI, R. A. Livro das árvores: árvores e arvoretas do Sul. 2. ed. Porto Alegre: L&PM, 1995. 176p. LONGHI, S. J. Comunicação pessoal. 2012. LONGHI, S. J. et al. Aspectos fitossociológicos de fragmento de floresta estacional decidual, Santa Maria, RS. Ciência Florestal, v. 10, n. 2, p. 59-74. 2000. LONGHI, S. J. et al. Classificação e caracterização de estágios sucessionais em remanescentes de Floresta Ombrófila Mista na FLONA de São Francisco de Paula, RS, Brasil. Ciência Florestal, Santa Maria, v. 16, n. 2, p. 113-125. 2005. LOPES, S. B.; GONÇALVES, L. Elementos Para Aplicação Prática das Árvores Nativas do Sul do Brasil na Conservação da Biodiversidade. Rio Grande do Sul: Fundação Zoobotânica do Rio Grande do Sul, 2006. 18p. Disponível em: <http://www.fzb.rs.gov.br/jardimbotanico/downloads/paper_tabela_aplicacao_arvores_rs.pdf>. Acesso em: 12 fev. 2012. LORENZI, H. Árvores brasileiras: manual de identificação e cultivo de plantas arbóreas nativas do Brasil. Nova Odessa: Ed. Plantarum, v. 1, 1992. 352p. LORENZI, H. Árvores brasileiras: manual de identificação e cultivo de plantas arbóreas nativas do Brasil. 2. ed. Nova Odessa: Plantarum, v. 2, 2002a. 381p.

88
LORENZI, H. Árvores brasileiras: manual de identificação e cultivo de plantas arbóreas nativas do Brasil. 4. ed. Nova Odessa: Plantarum, v. 1, 2002b. 381p. LORENZI, H. Árvores brasileiras: manual de identificação e cultivo de plantas arbóreas nativas do Brasil. 5. ed. Nova Odessa: Instituto Plantarum, v. 1, 2008. 368p. LORENZI, H. Árvores brasileiras: manual de identificação e cultivo de plantas arbóreas nativas do Brasil. Nova Odessa: Ed. Plantarum, v. 2, 1998. 368p. LORENZI, H. Árvores brasileiras: manual de identificação e cultivo de plantas arbóreas nativas do Brasil. 1. ed. Nova Odessa: Instituto Plantarum, v. 3, 2009. 384p. MARTINS, F. R. Estrutura de uma Floresta Mesófila. Campinas: UNICAMP, 1991. 246p. MATTEUCCI, S. D.; COLMA, A. Metodologia para el estudio de la vegetacion. Washington: Secretaria General de la Organización de los Estados Americanos – Programa Regional de Desarrollo y Tecnogico, 1982. 169p. McCUNE, B.; GRACE, J. B. Analysis of ecological communities. Oregon: MJM Software, 2002. 300p. McCUNE, B., MEFFORD, M. J. PC-ORD for Windows: multivariate analysis of ecological data - version 3.12. Oregon: MJM Software Design, Gleneden Beach, 1997. MELO, F. P. L. et al. Recrutamento e estabelecimento de plântulas. In: FERREIRA, A. G.; BORGHETTI, F. (Orgs.). Germinação: do básico ao aplicado. Porto Alegre: Artmed, 2004. p. 225-236. MONTOYA-MAQUIN, J. M; MATOS, G. F. Um enfoque fisionômico- estrutural para la descripcion de la vegetacion. Turrialba, Turrialba, v.17, n. 2, p. 197–207. 1967. MORENO, J. A. Clima do Rio Grande do Sul. Porto Alegre: Secretaria da Agricultura, 1961. 73 p.

89
MOSCOVICH, F. A. Dinâmica de crescimento de uma Floresta Ombrófila Mista em Nova Prata, RS. 2006. 130f. Tese (Doutorado em Engenharia Florestal)-Universidade Federal de Santa Maria, Santa Maria, 2006. MOSER, J. M. Solos. In: IBGE. Geografia do Brasil: Região Sul. Rio de Janeiro, 1990. p. 85-111. MUELLER-DOMBOIS, E.; ELLENBERG, F. Aims and methods of vegetation ecology. New York: Wiley & Sons, 1974. 547p. NAPPO, M. E.; OLIVEIRA-FILHO, A. T.; MARTINS, S. V. A estrutura do sub-bosque de povoamento homogêneos de Mimosa scabrella Bentham, em área minerada, em Poços de Caldas, MG. Ciência florestal, Santa Maria, v. 10, n. 2, p. 17-19. 2000. NARVAES, I. S. Classificação e caracterização da regeneração natural em Floresta Ombrófila Mista na Floresta Nacional de São Francisco de Paula, RS. 2004. 143f. Dissertação (Mestrado em Engenharia Florestal)–Universidade Federal de Santa Maria, Santa Maria, 2004. NELVING JUNIOR, L. I.; REITZ, R. Timeleaceas. In: REITZ, R (Org.). Flora ilustrada Catarinense. Itajaí, 1968. 21p. ODUM, E. P. Ecologia. 2. ed. Rio de Janeiro: Guanabara Koogan, 1988. 434p. OLIVEIRA FILHO, A. T. Estudos ecológicos da vegetação como subsídio para programas de revegetação com espécies nativas: uma proposta metodológica. Cerne, v. 1. n. 1, p. 64-72. 1994. PAGANO, S. N.; DURIGAN, G. Aspectos da ciclagem de nutrientes em matas ciliares do Oeste do Estado de São Paulo, Brasil. In: RODRIGUES, R. R.; LEITÃO FILHO, H. F. (Eds.). Matas c i l i a r e s: c o n s e r v a ç ã o e r e c u p e r a ç ã o. São Paulo: EDUSP/FAPESP, 2000. p.109-123. PALMER, M. W. Putting things in even better order: the advantages of canonical correspondence analysis. Ecology, v.74, p. 2215-2230. 1993. PÉLLICO NETO, S.; BRENA, D. A. Inventário Florestal. Curitiba: Editorado pelos autores, 1997. 136p.

90
PILLAR, V. P. et al. Campos Sulinos - conservação e uso sustentável da biodiversidade. Brasília: Ministério do Meio Ambiente, 2009. 409p. QUADROS, F. L. F; PILLAR, V. P. Transições floresta-campo do Rio Grande do Sul. Ciência & Ambiente, Santa Maria, v. 1, n. 1, p. 109-118. 2002. QUEIROZ, J. A. et al. Análise da Estrutura Fitossociológica da Serra do Monte, Boqueirão, Paraíba. Revista de Biologia e Ciências da Terra, João Pessoa, v. 6, n. 1, p. 251-259, 2006. RAIJ, B. Van. Fertilidade do solo e adubação. Piracicaba: Ceres; Potafos, 1991. 343p. RAMBO, B. A porta de Torres: estudo fitogeográfico. Anais Botânicos do Herbário Barbosa Rodrigues 2: 1950, p. 125-136. RAMBO, B. A fisionomia do Rio Grande do Sul. Porto Alegre: Selbach, 1956. 471p. REICHARDT, K.; TIMM, L. C. Solo, Planta e Atmosfera. Conceitos, Processos e Aplicações. Barueri, SP: Manole, 2004. 478p. REITZ, R.; KLEIN R. M.; REIS, A. Projeto Madeira do Rio Grande do Sul. Itajaí: Sellowia, n. 34-35, p. 1- 525. 1983. RIO GRANDE DO SUL. Inventário Florestal Contínuo do Rio Grande do Sul. Secretaria Estadual do Meio Ambiente. Porto Alegre; Santa Maria: SEMA/DEFAP, v. 1, 2002. 706p. RIO GRANDE DO SUL. Diretrizes ambientais para restauração de matas ciliares. Secretaria Estadual do Meio Ambiente. Departamento de Florestas e Áreas Protegidas. Porto Alegre: SEMA, 2007. 33p. RIO GRANDE DO SUL. Decreto Estadual nº 47.137, de 30 de março de 2010. Institui o Programa Estadual de Recuperação de Áreas de Preservação Permanente – APP’s e Reserva Legal, denominado Ambiente Legal, e dá outras providências. Diário Oficial do Estado, Porto Alegre. Disponível em: <http://www.sema.rs.gov.br/sema/html/dec_47137.html>. Acesso em: 10 Nov. 2011.

91
SANQUETTA, C. R. Análise da estrutura vertical de florestas através do diagrama h-M. Revista Ciência Florestal, Santa Maria, v. 1, n. 5, p. 55-68. 1995. SAWCZUK, A. R. Florística e estrutura horizontal no período 2002-2008 de um Fragmento de Floresta Ombrófila Mista no Centro-Sul do Estado do Paraná. 2009. 157f. Dissertação (Mestrado em Ciências Florestais)-Universidade Estadual do Centro-Oeste, Paraná, 2009. SCHNEIDER, P. R., GALVÃO, F., LONGHI, S. L. Influência do pisoteio de bovinos em áreas florestais. Floresta, Curitiba, v. 9, n. 1, p. 19-23. 1978. SCHORN, L. A. et al. Avaliação de técnicas para indução da regeneração natural em área de preservação permanente sob uso anterior do solo com Pinus elliottii. Floresta, Curitiba, v. 40, n. 2, p. 281-294, abr/jun. 2010. SIEGMUND, K. D.; LAIRD, P. W.; LAIRDOFFRINGA, I. A. A. comparison of cluster analysis methods using DNA methylation data. Bioinformatics, v. 20, n. 12, p. 1896-1904. 2004. SILVA, L. S.; CAMARGO, F. A. O.; CERETTA, C. A. Composição da fase sólida orgânica do solo. In: MEURER, E. J. Fundamentos de química do solo. Porto Alegre: Genesis, 2000. 174p. SILVA, R. F. et al. Comportamento de Peltophorum dubium (Spreng.) Taub., Paraptadenia rigida (Benth.) Brenan e Enterolobium contortisiliquum (Vell.) Morong cultivadas em solo contaminado com cobre. Ciência Florestal, Santa Maria, v. 21, n. 1, p. 103-110. 2011. SOBRAL, M. et al. Flora Arbórea e Arborescente do Rio Grande do Sul, Brasil. São Carlo: RiMa: Novo Ambiente, 2006. 350p. SOCIEDADE BRASILEIRA DE CIÊNCIA DO SOLO (SBCS/CQFS). Manual de adubação e calagem para os Estados do Rio Grande do Sul e Santa Catarina. 10. ed. Porto Alegre: Comissão de Química e Fertilidade do Solo, 2004. 400 p. SOUZA, A. L. et al. Análise multivariada para manejo de floresta natural na Reserva Florestal de Linhares, Espírito Santo: análise de agrupamento e discriminante. Revista Árvore, Viçosa, v. 14, n. 2, p. 85-101. 1990.

92
SOUSA, D. M. G.; MIRANDA, L. N.; OLIVEIRA, S. A. Acidez do solo e sua correção. In: NOVAIS, R. F. et al. Fertilidade do solo. Viçosa, MG: Sociedade Brasileira de Ciência do Solo, 2007. p.205-274. STEVENS, A. D. Influência da agricultura itinerante na regeneração da vegetação de pousio no leste da Amazônia. Eschborn: Deutsche Gesellschaft für Technische Zuisammenarbeit (GTZ) GmbH, 1999. 68p. STRECK, E. V. et al. Solos do Rio Grande do Sul. 2. ed. Porto Alegre: EMATER/RS-ASCAR, 2008. 222p. SWAINE, M. D.; WHITMORE, T. C. On the definition of ecological species groups in tropical rain forests. Vegetation, Dordrecht, v. 75, p. 81-86. 1988. TEDESCO, M. J.; BISSANI, C. A. Acidez do solo e seus efeitos nas plantas. In: BISSANI, C. A. et al. Fertilidade dos solos e manejo de adubação de culturas. Porto Alegre: Genesis, 2004. p. 75-92. TEDESCO, M. J. et al. Análises de solos, plantas e outros materiais. 2. ed. Porto Alegre: UFRGS, 1995. 174p. (Boletim técnico, 5). TER BRAAK, C. J. F. The analysis of vegetation environment relationships by canonical correspondence analysis. Vegetatio, v. 69, n. 1, p. 69-77. 1987. TONETTO, T. S. Variabilidade de solos em área de tensão ecológica entre campo e Floresta Ombrófila Mista na Barragem do Divisa – São Francisco de Paula/RS. 65f. (Relatório de Estágio)-Universidade Federal de Santa Maria, Santa Maria, 2011. TROEH, F. R.; THOMPSON, L. M. Solos e fertilidade do solo. São Paulo: Andrei, 2007. 718p. VACCARO, S. Crescimento de uma Floresta Estacional decidual, em três estágios sucessionais, no município de Santa Tereza, RS, Brasil. 2002.137f. Tese (Doutorado em Engenharia Florestal)-Universidade Federal de Santa Maria, Santa Maria, 2002. VALÉRIO, A. F. et al. Análise da composição florística e da estrutura horizontal de uma Floresta Ombrófila Mista Montana, Município de Irati, PR – Brasil. Rev. Acad., Ciênc. Agrár. Ambient., Curitiba, v. 6, n. 2, p. 137-147. 2008.

93
VAN DEN BERG, E.; OLIVEIRA FILHO, A. T. Composição florística e estrutura fitossociológica de uma floresta ripária em Itutinga, MG, e comparação com outras áreas. Revista Brasileira de Botânica, São Paulo, v. 23, n. 3, p.231-253, 2000. WATZLAWICK, L. F. et al. Aspectos florístico e estrutural de um fragmento de Floresta com Araucária utilizado para o pastoreio de bovinos. Ciência & Natura, Santa Maria, v. 24, p. 63-76. 2002. WURDACK, J. J. Melastomataceae of Santa Catarina. Sellowia v. 14, n. 14, p. 109-217. 1962. ZILLER, S. R.; ZALBA, S. M.; ZENNI, R. D. Modelo para o desenvolvimento de uma estratégia nacional para espécies exóticas invasoras. Programa de Espécies Exóticas Invasoras, The Nature Conservancy e Programa Global de Espécies Invasoras – GISP, 2007.

94
ANEXOS

95
Anexo A - Mapa de uso e ocupação do solo no entorno do Reservatório Divisa, São Francisco de Paula, RS (F1: Fragmento 1; F2: Fragmento 2; F3: Fragmento 3; F4: Fragmento 4; F5: Fragmento 5; F6: Fragmento 6).

96
(continua) Anexo B - Características autoecológicas das espécies identificadas no Reservatório Divisa, São Francisco de Paula, RS.
Família Nome científico Nome vulgar Ocorrência GE Fruto S EL
Anacardiaceae
Lithraea brasiliensis Marchand Aroeira-brava MG ao RS11 P1 Drupa; carnoso; indeiscente11 R29 H11 Schinus lentiscifolius Marchand. Aroeira-brava RJ ao RS13 P13 Drupa; seco; globoso13 H13
Annonaceae Annona neosalicifolia H. Rainer Araticum Sul; Sudeste; MS; RS13 P13 Sincarpo; indeiscente13 H13
Aquifoliaceae
Ilex brevicuspis Reissek Caúna MG; SP; RS11 Si1 Baga; indeiscente11 O29 H11 Ilex dumosa Reissek Caúna MG ao RS11 Si1 Baga ou drupa; indeiscente11 O3 H11
Araucariacea Araucaria angustifolia (Bertol.) Kuntze Araucária MG; RJ; RS15 P1, 2 Pseudofruto reunido em pinha3 R3 H15
Asteraceae
Dasyphyllum spinescens (Less.) Cabrera Sucará RJ ao RS13 P1 Aquênio; deiscente13 M13 Dasyphyllum tomentosum (Spreng.) Cabrera Sucará MG; RS13 P6 Aquênio; deiscente13 R29 M13 Gochnatia polymorpha (Less.) Cabr. Cambará RS3 Si3 Aquênio; deiscente3 R3 H3
Bignoniaceae Handroanthus albus (Cham.) Mattos Ipê-da-Serra RJ; MG; RS9 P9 Cápsula; seco; deiscente9 R3 H9
Cannabaceae Celtis iguanaea (Jacq.) Sarg. Esporão-de-galo EUA; UY; RS20 P10 Drupa; carnoso; indeiscente20 R29
Celastraceae Maytenus aquifolium Mart. Cancorosa Sul; Sudeste13 S13 Cápsula; seco; deiscente13 R29 E13
Cunoniaceae Lamanonia ternata Vell. Guaraperê MG; SP; RS15 P8 Cápsula; seco; deiscente15 O3 H15
Dicksoniaceae Dicksonia sellowiana Hook. Xaxim C1
Erythroxylaceae Erythroxylum deciduum A.St.-Hil. Cocão PI; NE; MS; RS11 P11 Drupa; carnoso; indeiscente11 R29 H11
Euphorbiaceae
Sapium glandulosum (L.) Morong Pau-de-leite UY; PY; AR; MG; RS14
P1, 2;
Si2 Cápsula; seco; deiscente14 R3 H3 Sebastiania brasiliensis Marchand Leiteiro St1, 17 Cápsula; seco; deiscente20 R29 E8
Sebastiania commersoniana (Baill.) L.B. Sm. & Downs Branquilho RJ; MG; RS9 P9; Si1 Deiscente9 H9 Stillingia oppositifolia Baill. ex Müll.Arg. Leiterinho MG ao RS20 C17 Cápsula; seco; deiscente9 H9
Fabaceae Erythrina cristagalli L. Cortiçeira-do-banhado RS4 P; Si4 Legume; deiscente4 O4 H4
Lauraceae
Cinnamomum amoenum (Nees) Kosterm. Canela PB; PR20 St8 Drupa20 Ocotea corymbosa (Meisn.) Mez Canela RJ; SP; MG9 Si; C5 Bacáceo5 O5 H9 Ocotea porosa Nees et Martius ex Nees. Imbúia PR; SC; RS9 P9 Carnoso; indeiscente9 R3 H9 Ocotea pulchella (Nees) Mez Canela-lageana ES; MG; MS; RS9 Si1; St2 Baga; carnoso; indeiscente9 R4 H9
Melastomataceae Miconia cinerascens Miq. Pixirica MG ao RS25 P; Si2 Baga20 Miconia hiemalis A.St.-Hil. & Naudin ex Naudin Pixirica SP ao RS20 P32 Cápsula20

97
(continuação) Anexo B - Características autoecológicas das espécies identificadas no Reservatório Divisa, São Francisco de Paula, RS.
Família Nome científico Nome vulgar Ocorrência GE Fruto S EL
Myrtaceae
Acca sellowiana (O.Berg) Burret Goiaba-da-serra RS; PR15 Si2 Carnoso; indeiscente15 H15
Blepharocalyx salicifolius (Kunth) O. Berg Murta GO; BA; MG ao RS11 Si2 Baga; carnoso; indeiscente11 R29 H11
Calyptranthes concinna DC. Guamirim-ferro MG ao RS11 St2 Baga; carnoso; indeiscente11 H11
Campomanesia xanthocarpa O. Berg. Guabiroba MG; SP; MS; RS9 Si1; St2; C2 Carnoso; indeiscente9 R4 H9
Eucalyptus grandis Hill (ex Maiden) Eucalipto Eucalyptus saligna Smith. Eucalipto
Eugenia involucrata DC. Cerejeira MG ao RS9 C1 Carnoso; indeiscente9 R4 H9 Eugenia uruguayensis Cambess. Guamirim PR ao RS24 St1 Drupa24 Myrceugenia cucullata D. Legrand Guamirim St2; C1 E8 Myrceugenia miersiana (Gardner) D. Legrand & Kausel Guamirim-piloso RJ; MG ao RS13 C1 Elipsóide; carnoso13 E13
Myrcia palustris DC. Pitangueira-do-mato RJ ao RS26 St2 Drupa20
Myrcia selloi (Spreng.) N. Silveira Cambuí MG; RS13 St2 Baga globosa13 R29 E13 Myrcianthes gigantea (D. Legrand) D. Legrand Araçá-do-mato UY; RS14 St1 Baga14 Myrciaria delicatula O. Berg Camboim MG ao RS13 C1 Globoso13 H13 Myrciaria floribunda (H.West ex Willd) O.Berg Cambuí MG; RS13 C1 Globoso13 E13 Myrrhinium atropurpureum Schott Murtilho RJ ao RS27 St1 E19
Podocarpaceae Podocarpus lambertii Klotzsch ex Endl. Pinheiro-bravo MG; RJ ao RS9 Si7; St2 Pedúnculo; carnoso9 R3 H9 Primulaceae
Myrsine coriacea (Sw.) R.Br. Capororoca BA; MG ao RS14 P14 Drupa; carnoso14 Myrsine umbellata Mart. Capororoca RJ; RS; SP4 P14 Drupa; esférico; carnoso14 O4
Proteaceae Roupala brasiliensis Klotzsch Carvalho-brasileiro BA; MG ao PR9 C1 Seco; deiscente9 R29 H9
Quillajaceae Quillaja brasiliensis (A.St.-Hil. & Tul.) Mart. Pau-sabão SP ao RS13 P13 Tomentoso13 R18 H13 Rhamnaceae Scutia buxifolia Reissek Coronilha PR ao RS13 P; Si2 Baga; carnoso; indeiscente13 H13

98
(conclusão) Anexo B - Características autoecológicas das espécies identificadas no Reservatório Divisa, São Francisco de Paula, RS.
Em que: GE: Grupo ecológico; P: Pioneira; S: Secundária; Si: Secundária inicial; St: Secundária tardia; C: Clímax; S: Semente; O: Ortodoxa; R: Recalcitrante; EL: Exigência lumínica; E: Esciófila; H: Heliófila; M: Mesófila. 1 MOSCOVICH, 2006; 2 RIO GRANDE DO SUL, 2007; 3 CARVALHO, 2003; 4 CARVALHO , 2006b; 5 CARVALHO, 2008; 6 REITZ; KLEIN; REIS, 1983; 7 VACCARO, 2002; 8 LONGHI, 2012 (comunicação pessoal); 9 LORENZI , 1992; 10 LONGHI, 1995; 11 LORENZI , 1998; 13 LORENZI, 2009; 14 BACKES; IRGANG, 2002; 15 LORENZI, 2002a; 16 LORENZI, 2002b; 17 CHAMI, 2008; 18 CARVALHO, 2006a; 19 LOPES; GONÇALVES, 2006; 20 SOBRAL et al., 2006; 21 NELVING JUNIOR; REITZ, 1968; 24 LEGRAND; KLEIN, 1969; 25 WURDACK, 1962; 26 LEGRAND; KLEIN, 1967; 27 LANDRUM, 1986; 28 GOMES, 2003; 29 CRESTANA et al., 2004; 30 SAWCZUK, 2009; 31 EBLING, 2012; 32 SCHORN et al., 2010.
Família Nome científico Nome vulgar Ocorrência GE Fruto S EL
Rosaceae Prunus myrtifolia (L.) Urb. Pessegueiro-do-mato
MG; RS; RJ; SC; SP5 Si; St2 Drupa5 R5
Rubiaceae
Faramea montevidensis (Cham. & Schltdl.) DC. Café-do-mato SP ao RS28 E8 Rudgea jasminoides (Cham.) Müll. Arg. Café-do-mato Sul; Sudeste20 St30 R29 E8 Rudgea parquioides (Cham.) Müll.Arg. Café-do-mato Sul20 C17 E8
Rutaceae Zanthoxylum rhoifolium Lam. Mamica-de-cadela Em todo o Brasil9 Si1 Seco; deiscente9 O4 H9
Salicaceae
Casearia decandra Jacq. Chá-de-bugre Em todo o Brasil16 Si1 Cápsula; deiscente; glabra16 R3 H16
Casearia sylvestris Swartz. Guaçatunga Em todo o Brasil15 Si1 Cápsula; deiscente15 R4 H15
Xylosma pseudosalzmanii Sleumer Sucará SP ao RS20 Si1 Xylosma tweediana (Clos) Eichler Sucará SC; RS20 Si1
Sapindaceae
Allophylus edulis (A.St.-Hil., Cambess. & A. Juss.) Radlk. Chal-Chal
MS; CE; AM; MG; BA; RJ; RS15 Si2 Drupa; indeiscente15 R4 E15
Cupania vernalis Camb. Camboatá-vermelho
MG; MS; SP ao RS9 St1, 2 Seco; deiscente9 R4 H9
Matayba elaeagnoides Radlk. Camboatá-branco SP; MG ao RS9 St1, 2 Seco; deiscente9 R4 M9 Styracaceae Styrax leprosus Hook. & Arn. Carne-de-vaca MG ao RS13 P13 Drupa escamosa13 R5 H13
Symplocaceae
Symplocos tetrandra (Mart.) Miq. Sete-sangrias MG ao RS20 St31 Drupa20 R4 H13
Symplocos uniflora (Pohl) Benth. Sete-sangrias MG; RS4 Si1,2;
St2 Drupa globosa13 Thymelaeaceae Daphnopsis racemosa Griseb. Embira CE ao RS21 St; C2 Drupa21 H1
Winteraceae Drimys brasiliensis Miers Casca-de-anta Em todo o Brasil9 St; C5 Baga; indeiscente5 R5

99
(continua) Anexo C - Resultados da classificação realizada pelo TWINSPAN, para a vegetação pertencente à Classe I (CAP ≥ 30 cm), no Reservatório Divisa, São Francisco de Paula, RS.
************* Two-way Indicator Species Analysis (TWINSPAN) ************* PC-ORD, 5.10 7 Nov 2011, 10:51 RESUL TWIN_57PLOT 34 SP_SM Number of samples: 57 Number of species: 34 Length of raw data array: 965 non-zero items SPECIES NAMES 1 AACC | 2 Allophy | 3 Annona | 4 Arauca | 5 Blepharo 6 Calyp | 7 Case.dec | 8 Cinnamo | 9 Dickso | 10 Drimys 11 E.gran | 12 E.salig | 13 E.urug | 14 Handroa | 15 Ilex.bre 16 Ilex.du | 17 Lithraea | 18 Maytenus | 19 Myrc.cuc | 20 Myrc.mie 21 Myrcia p | 22 Myrciant | 23 Myrc.del | 24 M.coriac | 25 O.porosa 26 O.pul | 27 Podocar | 28 Prunus.m | 29 Roupala | 30 Scutia 31 Sebas.co | 32 Styrax | 33 Symp.uni | 34 Zanthoxy | SAMPLE NAMES 1 Plot 1 | 2 Plot 2 | 3 Plot 3 | 4 Plot 4 | 5 Plot 5 6 Plot 6 | 7 Plot 7 | 8 Plot 8 | 9 Plot 9 | 10 Plot 10 11 Plot 11 | 12 Plot 12 | 13 Plot 13 | 14 Plot 14 | 15 Plot 15 16 Plot 16 | 17 Plot 17 | 18 Plot 18 | 19 Plot 19 | 20 Plot 20 21 Plot 21 | 22 Plot 22 | 23 Plot 23 | 24 Plot 24 | 25 Plot 25 26 Plot 26 | 27 Plot 27 | 28 Plot 28 | 29 Plot 29 | 30 Plot 30 31 Plot 31 | 32 Plot 32 | 33 Plot 33 | 34 Plot 34 | 35 Plot 35 36 Plot 36 | 37 Plot 37 | 38 Plot 38 | 39 Plot 39 | 40 Plot 40 41 Plot 41 | 42 Plot 42 | 43 Plot 43 | 44 Plot 44 | 45 Plot 45 46 Plot 46 | 47 Plot 47 | 48 Plot 48 | 49 Plot 49 | 50 Plot 50 51 Plot 51 | 52 Plot 52 | 53 Plot 53 | 54 Plot 54 | 55 Plot 55 56 Plot 56 | 57 Plot 57 | Cut levels: 0.0000 2.0000 5.0000 8.0000 10.0000 Options: Minimum group size for division = 5 Maximum number of indicators per division = 5 Maximum number of species in final table = 200 Maximum level of divisions = 6 Length of data array after defining pseudospecies: 833 Total number of species and pseudospecies: 92 Number of species: 34 (excluding pseudospecies and ones with no occurrences) CLASSIFICATION OF SAMPLES **************************************************************************** DIVISION 1 (N= 57) i.e. group * Eigenvalue: 0.3338 at iteration 4 INDICATORS and their signs: E.urug 1(-) Podocar 2(+) Myrc.cuc 1(+) M.coriac 1(-) Maximum indicator score for negative group 0 Minimum indicator score for positive group 1

100
(continuação) Anexo C - Resultados da classificação realizada pelo TWINSPAN, para a vegetação pertencente à Classe I (CAP ≥ 30 cm), no Reservatório Divisa, São Francisco de Paula, RS. ITEMS IN NEGATIVE GROUP 2 (N = 34) i.e. group *0 Plot 20 Plot 21 Plot 23 Plot 24 Plot 25 Plot 27 Plot 28 Plot 30 Plot 31 Plot 32 Plot 34 Plot 35 Plot 36 Plot 37 Plot 38 Plot 39 Plot 40 Plot 41 Plot 42 Plot 43 Plot 44 Plot 45 Plot 46 Plot 47 Plot 48 Plot 49 Plot 50 Plot 51 Plot 52 Plot 53 Plot 54 Plot 55 Plot 56 Plot 57 BORDERLINE NEGATIVES (N = 5) Plot 20 Plot 31 Plot 36 Plot 53 Plot 55 MISCLASSIFIED NEGATIVES (N = 2) Plot 24 Plot 30 ITEMS IN POSITIVE GROUP 3 (N = 23) i.e. group *1 Plot 1 Plot 2 Plot 3 Plot 4 Plot 5 Plot 6 Plot 7 Plot 8 Plot 9 Plot 10 Plot 11 Plot 12 Plot 13 Plot 14 Plot 15 Plot 16 Plot 17 Plot 18 Plot 19 Plot 22 Plot 26 Plot 29 Plot 33 BORDERLINE POSITIVES (N = 1) Plot 26 NEGATIVE PREFERENTIALS Drimys 1( 12, 0) E.urug 1( 29, 4) Ilex.bre1( 12, 2) Myrcia p1( 11, 0) Myrciant1( 7, 2) Myrc.del1( 23, 6) M.coriac1( 15, 0) O.pul 1( 12, 4) Sebas.co1( 22, 7) Styrax 1( 7, 1) Symp.uni1( 11, 3) Zanthoxy1( 8, 1) Drimys 2( 9, 0) E.urug 2( 24, 3) Lithraea2( 13, 2) Myrc.del2( 17, 2) M.coriac2( 7, 0) Sebas.co2( 13, 2) POSITIVE PREFERENTIALS Myrc.cuc1( 10, 21) Podocar 1( 16, 22) Myrc.cuc2( 5, 13) Podocar 2( 9, 19) Myrc.cuc3( 0, 5) Podocar 3( 4, 11) Arauca 4( 0, 5) NON-PREFERENTIALS Arauca 1( 30, 21) Blepharo1( 12, 5) Calyp 1( 10, 5) Cinnamo 1( 7, 3) Lithraea 1 (22, 11) Arauca 2 (23, 20) Arauca 3(8, 7) -------- E N D O F L E V E L 1 -------- **************************************************************************** DIVISION 2 (N= 34) i.e. group *0 Eigenvalue: 0.2583 at iteration 7 INDICATORS and their signs: Podocar 1(+) Drimys 1(-) Blepharo 1(-) Calyp 1(-) Cinnamo 1(-) Maximum indicator score for negative group -1 Minimum indicator score for positive group 0 ITEMS IN NEGATIVE GROUP 4 (N = 18) i.e. group *00 Plot 21 Plot 34 Plot 35 Plot 36 Plot 37 Plot 38 Plot 39 Plot 40 Plot 41 Plot 42 Plot 43 Plot 44 Plot 45 Plot 46 Plot 48 Plot 49 Plot 50 Plot 53 BORDERLINE NEGATIVES (N = 2) Plot 40 Plot 48 ITEMS IN POSITIVE GROUP 5 (N = 16) i.e. group *01 Plot 20 Plot 23 Plot 24 Plot 25 Plot 27 Plot 28 Plot 30 Plot 31 Plot 32 Plot 47 Plot 51 Plot 52 Plot 54 Plot 55 Plot 56 Plot 57 BORDERLINE POSITIVES (N = 2) Plot 52 Plot 56

101
(continuação) Anexo C - Resultados da classificação realizada pelo TWINSPAN, para a vegetação pertencente à Classe I (CAP ≥ 30 cm), no Reservatório Divisa, São Francisco de Paula, RS. NEGATIVE PREFERENTIALS Blepharo1( 11, 1) Calyp 1( 8, 2) Cinnamo 1( 7, 0) Drimys 1( 12, 0) Styrax 1( 6, 1) Zanthoxy1( 6, 2) Blepharo2( 4, 0) Cinnamo 2( 4, 0) Drimys 2( 9, 0) M.coriac2( 6, 1) Symp.uni2( 4, 1) POSITIVE PREFERENTIALS Myrciant1 (0, 7) Podocar 1( 2, 14) Podocar 2( 0, 9) Podocar 3( 0, 4) Sebas.co3( 2, 4) NON-PREFERENTIALS Arauca 1( 16, 14) E.urug 1( 16, 13) Ilex.bre1( 5, 7) Lithraea1( 11, 11) Myrc.cuc1( 4, 6) Myrcia p1( 6, 5) Myrc.del1( 13, 10) M.coriac1( 9, 6) O.pul 1( 6, 6) Sebas.co1( 10, 12) Symp.uni1( 7, 4) Arauca 2( 15, 8) E.urug 2( 12, 12) Lithraea2( 7, 6) Myrc.del2( 9, 8) Sebas.co2( 5, 8) Arauca 3( 4, 4) **************************************************************************** DIVISION 3 (N= 23) i.e. group *1 Eigenvalue: 0.3456 at iteration 14 INDICATORS and their signs: Symp.uni 2(+) Maximum indicator score for negative group 0 Minimum indicator score for positive group 1 ITEMS IN NEGATIVE GROUP 6 (N = 21) i.e. group *10 Plot 1 Plot 2 Plot 3 Plot 4 Plot 5 Plot 6 Plot 7 Plot 8 Plot 9 Plot 10 Plot 11 Plot 12 Plot 13 Plot 14 Plot 15 Plot 16 Plot 17 Plot 18 Plot 22 Plot 26 Plot 29 ITEMS IN POSITIVE GROUP 7 (N = 2) i.e. group *11 Plot 19 Plot 33 NEGATIVE PREFERENTIALS Blepharo1( 5, 0) Calyp 1( 5, 0) Myrc.del1( 6, 0) Myrc.cuc2( 13, 0) Arauca 3( 7, 0) Myrc.cuc3( 5, 0) Arauca 4( 5, 0) POSITIVE PREFERENTIALS Allophy 1( 1, 1) E.gran 1( 0, 1) E.salig 1( 0, 1) Sebas.co1( 5, 2) Symp.uni1( 1, 2) E.gran 2( 0, 1) E.salig 2( 0, 1) Sebas.co2( 1, 1) Symp.uni2( 0, 2) E.gran 3( 0, 1) E.salig 3( 0, 1) Podocar 3( 9, 2) Sebas.co3( 0, 1) E.salig 4( 0, 1) Sebas.co4( 0, 1) NON-PREFERENTIALS Arauca 1( 20, 1) Lithraea1( 10, 1) Myrc.cuc1( 20, 1) Podocar 1( 20, 2) Arauca 2( 19, 1) Podocar 2( 17, 2) -------- E N D O F L E V E L 2 -------- **************************************************************************** DIVISION 4 (N= 18) i.e. group *00 Eigenvalue: 0.2663 at iteration 11 INDICATORS and their signs: Myrc.del 2(+) Symp.uni 1(+) Sebas.co 1(+) M.coriac 2(+) Maximum indicator score for negative group 2 Minimum indicator score for positive group 3 ITEMS IN NEGATIVE GROUP 8 (N = 11) i.e. group *000 Plot 34 Plot 35 Plot 36 Plot 38 Plot 39 Plot 40 Plot 43 Plot 44 Plot 48 Plot 49 Plot 53 BORDERLINE NEGATIVES (N = 1) Plot 44

102
(continuação) Anexo C - Resultados da classificação realizada pelo TWINSPAN, para a vegetação pertencente à Classe I (CAP ≥ 30 cm), no Reservatório Divisa, São Francisco de Paula, RS. ITEMS IN POSITIVE GROUP 9 (N = 7) i.e. group *001 Plot 21 Plot 37 Plot 41 Plot 42 Plot 45 Plot 46 Plot 50 NEGATIVE PREFERENTIALS Scutia 1( 3, 0) Calyp 2( 3, 0) Arauca 3( 4, 0) POSITIVE PREFERENTIALS Case.dec1( 0, 2) Cinnamo 1( 3, 4) Ilex.du 1( 0, 3) Myrcia p1( 2, 4) Podocar 1( 0, 2) Roupala 1( 0, 3) Sebas.co1( 4, 6) Symp.uni1( 1, 6) Cinnamo 2( 1, 3) Lithraea2( 3, 4) Myrc.del2( 3, 6) M.coriac2( 1, 5) Roupala 2( 0, 2) Symp.uni2( 0, 4) E.urug 3( 0, 3) NON-PREFERENTIALS Arauca 1( 11, 5) Blepharo1( 8, 3) Calyp 1( 4, 4) Drimys 1( 7, 5) E.urug 1( 9, 7) Ilex.bre1( 3, 2) Lithraea1( 7, 4) Myrc.cuc1( 3, 1) Myrc.del1( 7, 6) M.coriac1( 4, 5) O.pul 1( 4, 2) Styrax 1( 4, 2) Zanthoxy1( 4, 2) Arauca 2( 11, 4) Blepharo2( 3, 1) Drimys 2( 5, 4) E.urug 2( 6, 6) Sebas.co2( 3, 2) **************************************************************************** DIVISION 5 (N= 16) i.e. group *01 Eigenvalue: 0.3191 at iteration 7 INDICATORS and their signs: Lithraea 2(+) Ilex.bre 1(+) Sebas.co 2(+) Maximum indicator score for negative group 2 Minimum indicator score for positive group 3 ITEMS IN NEGATIVE GROUP 10 (N = 12) i.e. group *010 Plot 20 Plot 27 Plot 28 Plot 30 Plot 31 Plot 47 Plot 51 Plot 52 Plot 54 Plot 55 Plot 56 Plot 57 BORDERLINE NEGATIVES (N = 1) Plot 20 ITEMS IN POSITIVE GROUP 11 (N = 4) i.e. group *011 Plot 23 Plot 24 Plot 25 Plot 32 NEGATIVE PREFERENTIALS Case.dec1( 3, 0) Dickso 1( 3, 0) Myrcia p1( 5, 0) Scutia 1( 3, 0) Symp.uni1( 4, 0) Myrc.del2( 7, 1) Podocar 2( 8, 1) Podocar 3( 4, 0) POSITIVE PREFERENTIALS AACC 1( 0, 1) Blepharo1( 0, 1) Ilex.bre1( 3, 4) Maytenus1( 0, 1) Myrc.cuc1( 3, 3) Prunus.m1( 1, 1) AACC 2( 0, 1) Lithraea2( 2, 4) Myrc.cuc2( 1, 2) O.pul 2( 1, 1) Prunus.m2( 0, 1) Sebas.co2( 4, 4) Sebas.co3( 1, 3) Sebas.co4( 0, 2) Sebas.co5( 0, 2) NON-PREFERENTIALS Arauca 1( 11, 3) E.urug 1( 11, 2) Lithraea1( 7, 4) Myrciant1( 5, 2) Myrc.del1( 8, 2) M.coriac1( 4, 2) O.pul 1( 4, 2) Podocar 1( 11, 3) Sebas.co1( 8, 4) Arauca 2( 6, 2) E.urug 2( 10, 2) Ilex.bre2( 2, 1) Arauca 3( 3, 1) E.urug 3( 2, 1) **************************************************************************** DIVISION 6 (N= 21) i.e. group *10 Eigenvalue: 0.2965 at iteration 9 INDICATORS and their signs: E.urug 2(+) Lithraea 1(+) Case.dec 1(+) Maximum indicator score for negative group 1 Minimum indicator score for positive group 2

103
(continuação) Anexo C - Resultados da classificação realizada pelo TWINSPAN, para a vegetação pertencente à Classe I (CAP ≥ 30 cm), no Reservatório Divisa, São Francisco de Paula, RS. ITEMS IN NEGATIVE GROUP 12 (N = 17) i.e. group *100 Plot 1 Plot 2 Plot 3 Plot 4 Plot 5 Plot 7 Plot 8 Plot 9 Plot 10 Plot 11 Plot 12 Plot 13 Plot 14 Plot 15 Plot 16 Plot 17 Plot 18 BORDERLINE NEGATIVES (N = 1) Plot 2 ITEMS IN POSITIVE GROUP 13 (N = 4) i.e. group *101 Plot 6 Plot 22 Plot 26 Plot 29 NEGATIVE PREFERENTIALS Blepharo1( 5, 0) Myrc.del1( 6, 0) Sebas.co1( 5, 0) Myrc.cuc3( 5, 0) Arauca 4( 5, 0) Arauca 5( 4, 0) POSITIVE PREFERENTIALS Allophy 1( 0, 1) Case.dec1( 0, 2) Cinnamo 1( 2, 1) E.urug 1( 1, 3) Lithraea1( 6, 4) Myrciant1( 0, 2) Roupala 1( 0, 1) Styrax 1( 0, 1) Symp.uni1( 0, 1) E.urug 2( 0, 3) Lithraea2( 0, 2) Myrciant2( 0, 2) NON-PREFERENTIALS Arauca 1( 17, 3) Calyp 1( 4, 1) Myrc.cuc1( 16, 4) O.pul 1( 3, 1) Podocar 1( 17, 3) Arauca 2( 16, 3) Myrc.cuc2( 11, 2) Podocar 2( 14, 3) Arauca 3( 6, 1) Podocar 3( 7, 2) Podocar 4( 3, 1) **************************************************************************** DIVISION 7 (N= 2) i.e. group *11 Group too small for further division. -------- E N D O F L E V E L 3 -------- Como as demais divisões não apresentaram autovalor considerável, não foram apresentadas nesse anexo. **************** THIS IS THE END OF THE DIVISIONS REQUESTED **************** **************************************************************************** ORDER OF SPECIES INCLUDING RARER ONES 8 Cinnamo | 1 AACC | 10 Drimys | 14 Handroa | 16 Ilex.du 24 M.coriac| 30 Scutia | 21 Myrcia p| 9 Dickso | 15 Ilex.bre 28 Prunus.m| 13 E.urug | 23 Myrc.del| 29 Roupala | 32 Styrax 34 Zanthoxy| 2 Allophy | 17 Lithraea| 26 O.pul | 33 Symp.uni 7 Case.dec| 22 Myrciant| 31 Sebas.co| 5 Blepharo| 6 Calyp 19 Myrc.cuc| 20 Myrc.mie| 25 O.porosa| 27 Podocar | 4 Arauca 3 Annona | 11 E.gran | 12 E.salig | 18 Maytenus| ORDER OF SAMPLES 35 Plot 35 | 36 Plot 36 | 34 Plot 34 | 38 Plot 38 | 48 Plot 48 49 Plot 49 | 53 Plot 53 | 39 Plot 39 | 40 Plot 40 | 43 Plot 43 44 Plot 44 | 37 Plot 37 | 42 Plot 42 | 45 Plot 45 | 46 Plot 46 50 Plot 50 | 21 Plot 21 | 41 Plot 41 | 27 Plot 27 | 28 Plot 28 51 Plot 51 | 20 Plot 20 | 30 Plot 30 | 31 Plot 31 | 47 Plot 47 54 Plot 54 | 55 Plot 55 | 56 Plot 56 | 57 Plot 57 | 52 Plot 52 23 Plot 23 | 24 Plot 24 | 25 Plot 25 | 32 Plot 32 | 1 Plot 1 2 Plot 2 | 10 Plot 10 | 11 Plot 11 | 14 Plot 14 | 7 Plot 7 8 Plot 8 | 9 Plot 9 | 15 Plot 15 | 16 Plot 16 | 17 Plot 17 3 Plot 3 | 5 Plot 5 | 12 Plot 12 | 13 Plot 13 | 18 Plot 18 4 Plot 4 | 6 Plot 6 | 22 Plot 22 | 26 Plot 26 | 29 Plot 29 19 Plot 19 | 33 Plot 33 |

104
(conclusão) Anexo C - Resultados da classificação realizada pelo TWINSPAN, para a vegetação pertencente à Classe I (CAP ≥ 30 cm), no Reservatório Divisa, São Francisco de Paula, RS. TWO-WAY ORDERED TABLE 3333445344434445242252334555552223 111 111 111 22213 564889390347256011781001745672345212014789567352384626993 8 Cinnamo --11---2---2--221--------------------------1---1-----1--- 0000 1 AACC ----1--2------------------------2------------------------ 00010 10 Drimys -1-233-212-23122----------------------------------------- 00010 14 Handroa -2-1-------------1----------12--------------------------- 000110 16 Ilex.du -----------11-2------1--1-------------------------------- 000110 24 M.coriac -1---2-1--12222-2------1---1121-1------------------------ 000110 30 Scutia 1-2--1--------------------11-1--------------------------- 000110 21 Myrcia p -----11-----1111----1---12-11---------------------------- 000111 9 Dickso ----------------1-------1---21--------------------------- 0010 15 Ilex.bre ---11----1-1--2---3---21------1121------------11--------- 0010 28 Prunus.m --------1------1--------1--------2----------------------- 0010 13 E.urug 2-12121222-2332123253122222-223--2----------------122-2-- 00110 23 Myrc.del 2-14-1--11222222-2-12-2222-22--1-21-1-2---1------12------ 00110 29 Roupala ---------------122-1---1------------------------------1-- 00111 32 Styrax -----1-1-12--1--1-2--------------------------------1----- 00111 34 Zanthoxy --2----11-1----1-1--------21--------1-------------------- 00111 2 Allophy 1-----------1----------------2-----------------------1--1 0100 17 Lithraea 1-22211-1---2-222-2121---1-11-2222----11111-------11221-1 0100 26 O.pul 11--1---1-----1-2---111--2----2--1-------1--1-----1--1--- 0100 33 Symp.uni --------1--21-1222----1-1-1-3----------------------1---22 0100 7 Case.dec ------------1---2-221--------------------------------11-- 0101 22 Myrciant ------------------1-111------11--1------------------22--- 0101 31 Sebas.co --1-----3322-21111-2121132---12553-----------21111-----14 0101 5 Blepharo 1--1-112122--1-1-3------------1---2--1-2--1---1---------- 011 6 Calyp 22-1-----2-11--11--1---------1-----1-13---1--------1----- 011 19 Myrc.cuc 12--1------2----------12---1--2-1222121132121423-331212-1 100 20 Myrc.mie -----2-------------------------------------------2------- 100 25 O.porosa ----------------2-------------------------2------1------- 100 27 Podocar --------------1-1-422233-1312112-13211253243531222243-233 100 4 Arauca 223232223322222--13322-1111123321-22222245555221322223-2- 101 3 Annona ------------1---------------------1--2--1---------------- 11 11 E.gran --------------------------------------------------------3 11 12 E.salig --------------------------------------------------------4 11 18 Maytenus ---------------------------------1--12------1------------ 11 000000000000000000000000000000000011111111111111111111111 000000000000000000111111111111111100000000000000000000011 0000000000011111110000000000001111000000000000000001111 001111111110000011000111111111 00000000000111111 00000111100001 000000001 00000111111000001 ********** TWINSPAN completed **********

105
(continua) Anexo D - Resultados da ordenação utilizando Análise de Correspondência Canônica, para a vegetação pertencente à Classe I (CAP ≥ 30 cm), no Reservatório Divisa, São Francisco de Paula, RS.
********************** Canonical Correspondence Analysis ********************** PC-ORD, 5.10; 5 Jan 2012, 16:32 DATA MATRICES Main matrix: 32 PLOTS (rows) 20 ESPÉCIES (columns) Second matrix: 32 PLOTS (rows) 6 ENVIRON (columns) OPTIONS SELECTED Axis scores standardized by Hill`s (1979) method. Axes scaled to optimize representation of rows: PLOTS (Scores for PLOTS are weighted mean scores for ESPÉCIES) Scores for graphing PLOTS are linear combinations of ENVIRON Monte Carlo test: null hypothesis is no relationship between matrices Random number seed: 1157 The following CATEGORICAL VARIABLES ARE EXCLUDED from the analysis: ---------- GR ---------- Consider recoding the variables into binary variables (0/1 dummy variables), then treating them as Q=Quantitative. Note that a categorical variable with n categories should be represented by n-1 dummy variables. RAW CORRELATIONS AMONG VARIABLES IN SECOND MATRIX Al Ca S Cu m Al 1.000 -0.921 -0.278 -0.037 0.964 Ca -0.921 1.000 0.429 0.063 -0.885 S -0.278 0.429 1.000 0.456 -0.267 Cu -0.037 0.063 0.456 1.000 -0.011 m 0.964 -0.885 -0.267 -0.011 1.000 WEIGHTED CORRELATIONS AMONG VARIABLES IN SECOND MATRIX (weighted by row totals in main matrix) Al Ca S Cu m Al 1.000 -0.924 -0.212 -0.082 0.964 Ca -0.924 1.000 0.370 0.083 -0.889 S -0.212 0.370 1.000 0.405 -0.201 Cu -0.082 0.083 0.405 1.000 -0.065 m 0.964 -0.889 -0.201 -0.065 1.000 ITERATION REPORT ----------------------------------------------------------------- Calculating axis 1 Residual = 0.50E+04 at iteration 1 Residual = 0.16E+00 at iteration 2 Residual = 0.10E-01 at iteration 3 Residual = 0.71E-03 at iteration 4 Residual = 0.63E-04 at iteration 5 Residual = 0.61E-05 at iteration 6

106
(continuação) Anexo D - Resultados da ordenação utilizando Análise de Correspondência Canônica, para a vegetação pertencente à Classe I (CAP ≥ 30 cm), no Reservatório Divisa, São Francisco de Paula, RS. Residual = 0.61E-06 at iteration 7 Residual = 0.62E-07 at iteration 8 Residual = 0.64E-08 at iteration 9 Residual = 0.66E-09 at iteration 10 Residual = 0.95E-13 at iteration 14 Solution reached tolerance of 0.100000E-12 after 14 iterations. ----------------------------------------------------------------- Calculating axis 2 Residual = 0.20E+01 at iteration 1 Residual = 0.23E-03 at iteration 2 Residual = 0.98E-04 at iteration 3 Residual = 0.45E-04 at iteration 4 Residual = 0.21E-04 at iteration 5 Residual = 0.98E-05 at iteration 6 Residual = 0.46E-05 at iteration 7 Residual = 0.21E-05 at iteration 8 Residual = 0.98E-06 at iteration 9 Residual = 0.46E-06 at iteration 10 Residual = 0.21E-09 at iteration 20 Residual = 0.68E-13 at iteration 30 Solution reached tolerance of 0.100000E-12 after 30 iterations. ----------------------------------------------------------------- Calculating axis 3 Residual = 0.20E+01 at iteration 1 Residual = 0.19E-06 at iteration 2 Residual = 0.81E-07 at iteration 3 Residual = 0.34E-07 at iteration 4 Residual = 0.14E-07 at iteration 5 Residual = 0.57E-08 at iteration 6 Residual = 0.24E-08 at iteration 7 Residual = 0.99E-09 at iteration 8 Residual = 0.41E-09 at iteration 9 Residual = 0.17E-09 at iteration 10 Residual = 0.52E-13 at iteration 19 Solution reached tolerance of 0.100000E-12 after 19 iterations. ----------------------------------------------------------------------------- AXIS SUMMARY STATISTICS Number of canonical axes: 3 Total variance ("inertia") in the species data: 1.6908 ----------------------------------------------------------- Axis 1 Axis 2 Axis 3 ----------------------------------------------------------- Eigenvalue 0.209 0.067 0.046 Variance in species data % of variance explained 12.3 4.0 2.7 Cumulative % explained 12.3 16.3 19.0 Pearson Correlation, Spp-Envt* 0.870 0.772 0.593 Kendall (Rank) Corr., Spp-Envt 0.677 0.536 0.375 ----------------------------------------------------------- * Correlation between sample scores for an axis derived from the species data and the sample scores that are linear combinations of the environmental variables. Set to 0.000 if axis is not canonical. MULTIPLE REGRESSION RESULTS: Regression of PLOTS in ESPÉCIES space on ENVIRON ---------------------------------------------------------------------------------- Canonical Coefficients -------------------------------------------------------- Standardized Original Units ------------------------ ------------------------------

107
(continuação) Anexo D - Resultados da ordenação utilizando Análise de Correspondência Canônica, para a vegetação pertencente à Classe I (CAP ≥ 30 cm), no Reservatório Divisa, São Francisco de Paula, RS. Variable Axis 1 Axis 2 Axis 3 Axis 1 Axis 2 Axis 3 S.Dev ---------------------------------------------------------------------------------- 1 Al 0.139 -0.340 -0.298 0.199 -0.486 -0.426 0.701E+00 2 Ca -0.354 -0.428 0.319 -0.469 -0.567 0.423 0.756E+00 3 S -0.142 0.091 -0.136 -0.610 0.388 -0.581 0.234E+00 4 Cu -0.316 0.116 0.040 -0.805 0.295 0.102 0.393E+00 5 m -0.280 0.147 0.644 -0.201 0.105 0.462 0.139E+01 -------------------------------------------------------------------------------- Scores that are derived from the scores of ESPÉCIES (WA Scores) FINAL SCORES and raw data totals (weights) for 32 PLOTS -------------------------------------------------------------------- Raw Data Axis 1 Axis 2 Axis 3 Totals -------------------------------------------------------------------- 1 PLOT 1 -1.024883 0.252479 -0.120091 3.2041 2 PLOT 2 -1.227241 -0.008619 0.019082 1.9542 3 PLOT 3 -1.121379 0.071386 0.207154 3.1303 4 PLOT 4 -0.643517 0.363607 -0.283039 3.4874 5 PLOT 5 -0.926165 0.272124 0.063483 2.2833 6 PLOT 6 -0.442172 0.036562 0.248299 3.4594 7 PLOT 9 -0.907887 -0.085280 -0.299451 2.9243 8 PLOT 10 -0.472592 0.231181 0.531231 1.6812 9 PLOT 13 -0.853784 -0.361927 0.227245 1.6812 10 PLOT 15 -0.753422 -0.076118 -0.078166 3.5386 11 PLOT 20 -0.143831 -0.763009 -0.097762 3.0792 12 PLOT 23 -0.203030 -0.205227 -0.276640 5.2587 13 PLOT 25 0.037985 -0.197551 0.340391 3.4294 14 PLOT 26 0.092117 -1.016094 -1.055027 3.0615 15 PLOT 28 0.048970 -0.432591 0.043108 4.6257 16 PLOT 29 -0.381415 0.023867 -0.145149 2.3802 17 PLOT 30 0.065715 0.262789 -0.006998 4.0615 18 PLOT 31 -0.398864 0.569669 0.079319 3.7305 19 PLOT 34 0.413455 -0.288908 0.448198 3.0035 20 PLOT 36 -0.207960 -0.285071 0.272003 3.4082 21 PLOT 38 0.642247 0.273554 -0.167224 4.8854 22 PLOT 40 0.453972 -0.169250 0.327700 4.1406 23 PLOT 42 0.843376 0.284126 -0.312495 4.8608 24 PLOT 44 0.245611 -0.155291 0.442026 3.0792 25 PLOT 46 0.674241 0.462753 -0.167509 5.7658 26 PLOT 48 0.326377 0.354115 -0.668646 3.6124 27 PLOT 50 0.996911 0.272605 0.282761 4.4417 28 PLOT 51 0.335600 -0.430754 -0.374474 4.4417 29 PLOT 53 0.257566 -0.208409 -0.214633 1.6812 30 PLOT 54 0.500769 0.040992 0.012712 3.7324 31 PLOT 55 0.014134 0.052148 1.103910 2.6232 32 PLOT 57 0.438859 0.182784 0.416939 4.3345 -------------------------------------------------------------------- Scores that are linear combinations of ENVIRON (LC Scores) FINAL SCORES and raw data totals (weights) for 32 PLOTS -------------------------------------------------------------------- Raw Data Axis 1 Axis 2 Axis 3 Totals -------------------------------------------------------------------- 1 PLOT 1 -1.114988 -0.050648 -0.026075 3.2041 2 PLOT 2 -0.892897 0.136430 0.164931 1.9542 3 PLOT 3 -0.763238 0.175536 0.053869 3.1303 4 PLOT 4 -0.397682 0.592552 -0.456074 3.4874 5 PLOT 5 -0.612867 0.309716 0.130578 2.2833

108
(continuação) Anexo D - Resultados da ordenação utilizando Análise de Correspondência Canônica, para a vegetação pertencente à Classe I (CAP ≥ 30 cm), no Reservatório Divisa, São Francisco de Paula, RS. 6 PLOT 6 -0.874071 0.022843 0.195839 3.4594 7 PLOT 9 -0.837483 0.300183 0.237683 2.9243 8 PLOT 10 -0.613734 0.086418 0.108969 1.6812 9 PLOT 13 -0.818087 -0.163108 -0.476298 1.6812 10 PLOT 15 -0.345836 -0.330487 -0.400770 3.5386 11 PLOT 20 0.309897 -0.539958 0.169086 3.0792 12 PLOT 23 -0.402023 -0.358793 -0.295109 5.2587 13 PLOT 25 -0.211954 -0.178841 0.161588 3.4294 14 PLOT 26 0.004509 -0.563187 -0.447364 3.0615 15 PLOT 28 -0.117067 -0.295062 0.166598 4.6257 16 PLOT 29 0.237917 0.182423 0.045875 2.3802 17 PLOT 30 0.339979 0.288038 0.052140 4.0615 18 PLOT 31 -0.123617 0.248826 0.144948 3.7305 19 PLOT 34 0.253229 -0.182566 0.225551 3.0035 20 PLOT 36 -0.076458 -0.229367 0.268866 3.4082 21 PLOT 38 0.425815 0.098850 -0.156139 4.8854 22 PLOT 40 0.897937 0.081882 0.003679 4.1406 23 PLOT 42 0.622847 0.214814 -0.371108 4.8608 24 PLOT 44 0.405473 -0.034140 0.154104 3.0792 25 PLOT 46 0.158267 0.460927 -0.008890 5.7658 26 PLOT 48 0.286552 -0.022854 -0.021288 3.6124 27 PLOT 50 0.633384 0.034389 0.139565 4.4417 28 PLOT 51 0.537420 -0.046201 0.045599 4.4417 29 PLOT 53 -0.152087 -0.002560 0.152544 1.6812 30 PLOT 54 0.505193 -0.082650 -0.038073 3.7324 31 PLOT 55 0.071739 -0.114377 0.328805 2.6232 32 PLOT 57 0.227221 -0.163144 0.132540 4.3345 -------------------------------------------------------------------- FINAL SCORES and raw data totals (weights) for 20 ESPÉCIES -------------------------------------------------------------------- Raw Data Axis 1 Axis 2 Axis 3 Totals -------------------------------------------------------------------- 1 Arauca -0.834615 -0.576271 -0.359426 18.1564 2 Blepharo -0.357584 -0.456651 -0.231239 3.4082 3 Calyp -0.373174 -1.038491 0.850167 2.8062 4 Case.dec 0.934041 -1.989000 -1.432143 1.8062 5 Cinnamo 1.506626 0.333079 -0.604178 1.8573 6 Drimys 2.029484 1.541760 -1.464018 3.8396 7 E.urug 0.743228 0.201201 0.359264 11.4762 8 Handroa 0.599626 -1.945042 2.804625 1.2041 9 Ilex.bre 0.013580 1.798419 0.285539 2.9365 10 Lithraea 0.587729 -0.715326 -0.843098 8.7072 11 Myrc.cuc -2.105236 1.210497 -0.377685 8.6000 12 Myrcia p 1.787954 0.720189 0.086034 2.2833 13 Myrciant 0.680456 -4.127118 -2.880763 1.6812 14 Myrc.del 1.022456 1.367117 -0.442224 8.6178 15 M.coriac 0.658056 0.690177 -0.431100 2.8854 16 O.pul 0.504882 -0.426556 -1.134335 3.9645 17 Podocar -1.463290 0.253635 0.445996 12.8977 18 Sebas.co 0.315673 -1.038979 1.306265 8.5086 19 Symp.uni 1.391337 0.844853 1.694929 3.1864 20 Zanthoxy 1.229494 -0.626745 3.910494 2.1584 -------------------------------------------------------------------- CORRELATIONS AND BIPLOT SCORES for 5 ENVIRON -------------------------------------------------------------------- Correlations* Biplot Scores Variable Axis 1 Axis 2 Axis 3 Axis 1 Axis 2 Axis 3

109
(conclusão) Anexo D - Resultados da ordenação utilizando Análise de Correspondência Canônica, para a vegetação pertencente à Classe I (CAP ≥ 30 cm), no Reservatório Divisa, São Francisco de Paula, RS. -------------------------------------------------------------------- 1 Al 0.493 0.627 0.239 0.200 0.157 0.050 2 Ca -0.609 -0.749 -0.110 -0.247 -0.188 -0.023 3 S -0.730 0.081 -0.307 -0.296 0.020 -0.064 4 Cu -0.772 0.505 -0.025 -0.314 0.126 -0.005 5 m 0.425 0.648 0.443 0.172 0.162 0.093 ------------------------------------------------------------------ * Correlations are "intraset correlations" of ter Braak (1986) INTER-SET CORRELATIONS for 5 ENVIRON ----------------------------------------- Correlations Variable Axis 1 Axis 2 Axis 3 ----------------------------------------- 1 Al 0.429 0.484 0.142 2 Ca -0.530 -0.578 -0.065 3 S -0.635 0.063 -0.182 4 Cu -0.672 0.390 -0.015 5 m 0.370 0.500 0.263 ----------------------------------------- Note: Obtain joint plots or biplots by selecting GRAPH, then requesting "Joint plots" from the GRAPH menu. MONTE CARLO TEST RESULTS -- EIGENVALUES ------------------------------------------------------------ Randomized data Real data Monte Carlo test, 4999 runs ------------ --------------------------- Axis Eigenvalue Mean Minimum Maximum p ------------------------------------------------------------ 1 0.209 0.105 0.051 0.193 0.0002 2 0.067 0.070 0.037 0.136 3 0.046 0.049 0.023 0.090 ------------------------------------------------------------ p = proportion of randomized runs with eigenvalue greater than or equal to the observed eigenvalue; i.e., p = (1 + no. permutations >= observed)/(1 + no. permutations) p is not reported for axes 2 and 3 because using a simple randomization test for these axes may bias the p values. MONTE CARLO TEST RESULTS -- SPECIES-ENVIRONMENT CORRELATIONS ------------------------------------------------------------ Randomized data Real data Monte Carlo test, 4999 runs ------------ --------------------------- Axis Spp-Envt Corr. Mean Minimum Maximum p ------------------------------------------------------------ 1 0.870 0.733 0.552 0.922 0.0036 2 0.772 0.682 0.438 0.873 3 0.593 0.630 0.372 0.856 ------------------------------------------------------------ p = proportion of randomized runs with species-environment correlation greater than or equal to the observed species-environment correlation; i.e., p = (1 + no. permutations >= observed)/(1 + no. permutations) p is not reported for axes 2 and 3 because using a simple randomization test for these axes may bias the p values. ***************************** Operation completed *****************************





























