Embed Size (px)
Citation preview

Metabolismo dos Carboidratos
Carboidratosprincipal substrato oxidável para a maioria dos seres vivos essencial para o funcionamento do cérebro e das hemácias
a maioria dos monossacarídeos são encontrados na forma de anelgrupo aldeído/cetona reagiu com o grupo álcool da mesma molécula (ligação glicosídica)carbono anomérico: carbono envolvido por uma ligação glicosídica configuração α do C anomérico: quando ele fica para baixo do plano do anelconfiguração β do C anomérico: quando ele fica para cima do plano do anel
digestão dos carboidratosboca: α-amilase salivar atua sobre o amido hidrolisando ligações α(1→4)*humanos não são capazes de quebrar ligações β(1→4), portanto não digerem celuloseestômago: a digestão cessa temporariamente, pois o pH ácido inativa a α-amilase salivarintestino delgado: conteúdo ácido neutralizado pelo bicabornato (secretado pelo pâncreas) e a α-amilase retorna a digestão maltase → maltose → glicose + glicosesacarase → sacarose → glicose + frutose galactase → galactose + glicose
oxidação total da glicose: 36 ATPsprincipais vias do metabolismo de carboidratos: glicólise, desvio das pentoses, glicogênese e glicogenólise, gliconeogênese
Introdução ao metabolismovias: sequência de múltiplos passos as vias podem ser classificadas em catabólicas (degradação) ou anabólicas (síntese)- as vias catabólicas capturam energia química obtida da degradação de moléculas ricas em energia formando ATP, o catabolismo permite que moléculas da dieta sejam convertidas em blocos construtivos a energia gerada da degradação de moléculas ocorre em 3 estágios: hidrólise de moléculas complexas (moléculas complexas são quebradas em seus blocos constitutivos), conversão dos blocos constitutivos em intermediários mais simples (esses blocos constitutivos são posteriormente convertidos em acetil-CoA e outras moléculas simples, parte da energia é capturada como ATP) e oxidação da acetil-CoA (grandes quantidades de ATP são geradas na fosforilação oxidativa à medida que elétrons fluem do NADH e do FADH₂ para o oxigênio)- as reações anabólicas reúnem moléculas pequenas para formar moléculas complexas, estas reações necessitam de energia que é fornecida pela quebra do ATP, frequentemente estas reações envolvem reduções químicas onde o redutor geralmente é fornecido pelo NADH
metabolismo: é a soma de todas as mudanças química que ocorrem em uma célula/tecido/organismoregulação do metabolismo: sinais regulatórios que informam uma determinada célula sobre o estado metabólico do organismo como um todo incluem hormônios, neurotransmissores e a disponibilidade de alimentos - sinais de dentro da célula (intracelular): a velocidade de uma via pode ser influenciada pela disponibilidade de substratos, pela inibição ocasionada pelos produtos ou por alterações nos níveis de ativadores ou inativadores alostéricos, esses sinais intracelulares normalmente determinam respostas rápidas

- comunicação entre as células (intercelular): a sinalização entra as células fornece uma integração mais ampla do metabolismo e normalmente resulta em uma resposta que é mais lenta, a comunicação pode ser mediada pelo contato entre suas superfícies, pela formação de junções comunicantes e pela sinalização química entre as células (hormônios e NT)- sistemas de segundos mensageiros: os segundos mensageiros interveem entre o mensageiro original (hormônio ou NT) e o efeito final dentro da célula, são parte de uma cascata de eventos que traduz a ligação do hormônio ou NT em uma resposta celular adenilato ciclase: é uma enzima ligada à membrana que converte ATP em 3’,5’-monofosfato de adenosina (AMPc), o reconhecimento de um sinal químico por alguns receptores de membrana irá disparar um aumento ou redução na atividade dessa enzimaos receptores tem um domínio extracelular (onde se liga o ligante), 7 helices transmembranas e um domínio intracelular (que interage com a proteína G)proteínas reguladoras dependentes de GTP: as proteínas G (se ligam a nucleotídeos de guanosina = GTP/GDP) formam um elo de comunicação entre o receptor e a adenilato ciclase a forma inativa da proteína G liga-se ao GDP, o receptor ativado interage com a proteína G disparando uma troca de GDP por GTP, a proteína G então dissocia-se em uma subunidade α e um dímero βγ, a subunidade α ligada ao GTP move-se do receptor para a adenilato-ciclase que é então ativadaa capacidade de um hormônio ou NT de estimular ou inibir a adenilato-ciclase depende do tipo de proteína G ligada ao receptor as ações do complexo proteína G-GTP são de curta duração por causa da rápida hidrólise de GTP em GDP causando inativação da proteína Gproteína-cinases: o AMPc ativa enzimas (proteínas-cinases dependentes de AMPc), essas enzimas adicionam grupos fosfato às proteínas, esses grupos são removidos pela proteína-fosfatases causando mudanças na atividade enzimática resumindo: o receptor, quando desocupado, não interage com a proteína G → o receptor, quando ocupado, sofre uma alteração conformacional e interage com a proteína G, a qual libera GDP e liga GTP → a subunidade α da proteína G se dissocia e ativa a adenilato-ciclase → quando o hormônio/neurotransmissor não mais está presente, o receptor reverte para o estado basal, o GTP ligado à subunidade α é hidrolisado a GDP e a adenilato-ciclase é desativada
Visão geral da glicólise a via glicolítica é utilizada em todos os tecidos para a quebra de glicose, com o objetivo de fornecer energia (na forma de ATP) e intermediários para outras vias metabólicas praticamente todos os glicídeos podem ser, no final, convertidos em glicose o piruvato é o produto final da glicólise nas células que possuem mitocôndrias e fornecimento adequado de oxigênio glicose aeróbica: o oxigênio é necessário para reoxidação do NADH formado durante a oxidação do gliceraldeído-3-fosfato glicólise anaeróbica: glicose convertida em piruvato, permite a produção contínua de ATP em tecidos que não apresentam mitocôndrias (eritrócitos) ou células em que o oxigênio esteja em quantidade insuficiente
Transporte da glicose para dentro das células- transporte por difusão facilitada, independe de Na⁺: GLUT-1 até GLUT-14 (transportadores de glicose)transportadores na membrana → glicose extracelular liga-se ao transportador → mudança conformacional → transporta a glicose através da membrana os GLUTs possuem especificidade tecidual GLUT-1, GLUT-3 e GLUT-4: envolvidos principalmente na captação de glicose a partir do sangue

GLUT-2: encontrado no fígado/rim/células β do pâncreas, transporta glicose do sangue para as células estando a concentração de glicose alta/baixa no sangue - sistema de co-transporte monossacarídeo-Na⁺: requer energia e transporá a glicose “contra” o gradiente de concentração, o Na⁺ é transportado concomitantemente à glicose para o interior da célula
Reações da glicólise fase de investimento de energia: formas fosforiladas dos intermediários são sintetizadas, à custa de gasto de ATP-fosforilação da glicose: a fosforilação irreversível da glicose retém efetivamente o glicídio na forma de glicose-6-fosfato, utilização de 1 molécula de ATPna maioria dos tecidos é fosforilada pela hexocinase (baixo Km para a glicose, portanto alta afinidade, inibida pela glicose-6-fosfato) e nos hepatócitos/células das ilhotas do pâncreas pela glicocinase (Km muito menor, portanto pouca afinidade, só funciona quando a concentração de glicose nos hepatócitos for alta, impede que grandes quantidades de glicose cheguem à circulação sistêmica após uma refeição rica em carboidratos, não é inibida pela glicose-6-fosfato, sua atividade é aumenta pela insulina)- isomerização da glicose-6-fosfato: reação reversível, conversão da glicose-6-fosfato em frutose-6-fosfato pela fosfoglicose-isomerase- fosforilação da frutose-6-fosfato: reação irreversível, conversão da frutose-6-fosfate em frutose-1,6-diP (bifosfato) pela fosfofrutocinase-1 (PFK-1) que é inibida por ↑ATP e ↓AMP- clivagem da frutose-1,6-bifosfato: reação reversível, conversão da frutose-1,6-bifosfato em diidroxiacetona-fosfato + gliceraldeído-3-fosfato pela aldolase- isomerização da diidroxiacetona-fosfato: reação reversível, conversão da diidroxiacetona-fosfato em gliceraldeído-3-fosfato pela isomerase
resumo: 1 glicose (6 C) → 2 gliceraldeído 3-P (3C)5 reações sendo 2 irreversíveis e 3 reversíveisgasta dois ATPreações de fosforilação (1ª e 3ª)reações de isomerização (2ª e 5ª)reação de quebra (4ª)
fase de produção de energia: produção líquida de 2 moléculas de ATP- oxidação do gliceraldeído-3-fosfato: primeira reação de óxido-redução da glicólise, conversão de gliceraldeído-3-fosfato em 1,3-bifosfoglicerato pela gliceraldeído-3-fosfatoo NAD⁺ é oxidado em NADHsíntese do 1,3-bifosfofosfoglicerato e síntese de 2,3-bifosfoglicertato nos eritrócitos - síntese do 3-fosfoglicerato: reversível, conversão do 1,3-BPG em 3-fosfoglicerato pela fosfoglicerato-cinase, produz ATP - troca do grupo fosfato do carbono 3 para o carbono 2: reversível, feito pela fosfoglicerato-mutase- desidratação do 2-fosfoglicerato: reversível, conversão de 2-fosfoglicerato em fosfoenolpiruvato (PEP) pela enolase- formação do piruvato: irreversível, conversão do PEP em piruvato pela piruvato-cinase, produz ATP
resumo: 2 gliceraldeído 3-P (3C) → 2 piruvatos (3C)4 reações reversíveis e 1 irreversível (10ª )produção de 2 NADH e 4 ATP1 reação de oxidação (6ª ) → produz NADH2 reações de desfosforilação (7ª e 10ª ) → produz ATP

redução de piruvato a lactato: o lactato é formado pela ação da lactato-desidrogenase e é produto final da glicólise anaeróbicaa formação do lactato é o principal destino do piruvato nos eritrócitos, no cristalino e na córnea do olho, na medula renal, nos testículos e nos leucócitos formação de lactato no músculo esquelético: no músculo em exercício, a produção de NADH excede a capacidade oxidativa da cadeia respiratória resultando num aumento da razão NADH/NAD⁺ favorecendo a redução de piruvato a lactato, este se acumula no músculo causando diminuição do pH intracelular podendo levar a cãibras, grande parte do lacato é difundido para a corrente sanguínea podendo ser utilizado pelo fígado para produzir glicose consumo de lactato: o fígado e o coração oxidam lactato produzindo piruvatono fígado o piruvato pode ser convertido em glicose através da gliconeogênese ou oxidado no ciclo do ácido cítrico acidose láctica: altas [lactato] no plasma ocorrem quando há um colapso no sistema circulatório, a falha em levar quantidades adequadas de oxigênio aos tecidos resulta em prejuízo na fosforilação oxidativa e diminuição na síntese de ATP, então as células utilizam a glicose anaeróbica para a produção de ATP produzindo ácido láctico como produto final
os produtos finais (piruvato ou lactato) ainda retém a maior parte da energia originalmente obtida na glicose, o ciclo do ácido cítrico é necessário para liberar completamente essa energiaglicólise anaeróbica: 2 moléculas de ATP são geradas para cada molécula de glicose convertida em 2 moléculas de lactato, não há produção ou consumo líquido de NADHglicólise aeróbica: 2 moléculas de ATP são geradas para cada molécula de glicose convertida em 2 moléculas de piruvato, 2 moléculas de NADH são produzidas, a glicólise aeróbica requer a oxidação da maior parte de NADH pela CTE produzindo aproximadamente 3 ATPs por molécula de NADH que chega à cadeia respiratória
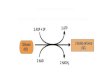
o piruvato pode ser descarboxilado oxidativamente pela piruvato-desidrogenase no músculo cardíaco sendo convertido em acetil-CoA (principal combustível do ácido cítrico), carboxilado a oxalacetato (um intermediário do ciclo do ácido cítrico) fornecendo substrato para gliconeogênese e reduzido a etanol em fungos

![6 - Bio4 [Somente leitura] - agraria.com.br · Acetato de etila Acetato de feniletanol Acetato de isoamila glicólise fermentação Catabolismo aminoácidos ramificados Biosentese](https://img.document.onl/doc/110x75/5c4bed5593f3c34aee54dd76/6-bio4-somente-leitura-acetato-de-etila-acetato-de-feniletanol-acetato.jpg)