Embed Size (px)
Citation preview

i
UNIVERSIDADE FEDERAL DO RIO DE JANEIRO
CENTRO DE CIÊNCIAS DA SAÚDE
INSTITUTO DE BIOLOGIA
PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOGIA
INFLUÊNCIA DA ÁREA E DA HETEROGENEIDADE DE HABITATS NA DIVERSIDADE VEGETAL EM FRAGMENTOS DE FLORESTA ATLÂNTICA
ANA CAROLINA BORGES LINS E SILVA
Tese apresentada ao Programa de Pós-Graduação em Ecologia da Universidade Federal do Rio de Janeiro, como parte dos requisitos necessários à obtenção do grau de Doutor em Ciências Biológicas (Ecologia).
Orientador: Prof. Fabio Rubio Scarano, PhD
RIO DE JANEIRO, RJ - BRASIL
ABRIL DE 2010

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

ii
UNIVERSIDADE FEDERAL DO RIO DE JANEIRO/UFRJ INSTITUTO DE BIOLOGIA
PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOGIA-PPGE
CX.POSTAL 68.020 – ILHA DO FUNDÃO CEP: 21941-590 – RIO DE JANEIRO – RJ – BRASIL
TEL./FAX: (21) 290-3308 TEL.: (21) 562-6320
INFLUÊNCIA DA ÁREA E DA HETEROGENEIDADE DE HABITATS NA DIVERSIDADE VEGETAL EM FRAGMENTOS DE FLORESTA
ATLÂNTICA
ANA CAROLINA BORGES LINS E SILVA
Tese apresentada ao Programa de Pós-Graduação em Ecologia da Universidade Federal do Rio de Janeiro, como parte dos requisitos necessários à obtenção do grau
de Doutor em Ciências Biológicas (Ecologia).
Defendida em 5 de abril de 2010. APROVADA POR:
________________________________________________Dr. Fabio Rubio Scarano (orientador)
________________________________________________Dr. Jean Paul W. Metzger
________________________________________________Dr. Claudio Belmonte de Athayde Bohrer
________________________________________________Dra. Rejan Rodrigues Guedes-Bruni
________________________________________________Dra. Dorothy Sue Dunn de Araújo
________________________________________________Dr. Marcelo Trindade Nascimento (suplente)
________________________________________________Dra. Carla Bernadete Madureira Cruz (suplente)

iii
LINS E SILVA, ANA CAROLINA BORGES Influência da área e da heterogeneidade de habitats na
diversidade vegetal em fragmentos de Floresta Atlântica. [Rio de Janeiro] 2010
XVI. 162 p. 29,7 cm (Instituto de Biologia/UFRJ, D.Sc., Ecologia, 2010)
Tese - Universidade Federal do Rio de Janeiro, PPGE 1. Floresta Atlântica 2. Fragmentos 3. Heterogeneidade de habitats I. IB/UFRJ II. Título ( série )

iv
Da mais alta janela da minha casa
Com um lenço branco digo adeus
Aos meus versos que partem para a Humanidade.
E não estou alegre nem triste.
Esse é o destino dos versos.
Escrevi-os e devo mostrá-los a todos
Porque não posso fazer o contrário
Como a flor não pode esconder a cor,
Nem o rio esconder que corre,
Nem a árvore esconder que dá fruto.
Ei-los que vão já longe como que na diligência
E eu sem querer sinto pena
Como uma dor no corpo.
Quem sabe quem os lerá?
Quem sabe a que mãos irão?
Flor, colheu-me o meu destino para os olhos.
Árvore, arrancaram-me os frutos para as bocas.
Rio, o destino da minha água era não ficar em mim.
Submeto-me e sinto-me quase alegre,
Quase alegre como quem se cansa de estar triste.
Ide, ide de mim!
Passa a árvore e fica dispersa pela Natureza.
Murcha a flor e o seu pó dura sempre.
Corre o rio e entra no mar e a sua água é sempre a que foi sua.
Passo e fico, como o Universo.
Alberto Caeiro (Fernando Pessoa)
O Guardador de Rebanhos, 1914

v
À minha família - meu nicho realizado, meu habitat preferido, minha felicidade não fragmentada,
Ofereço.
À memória de Marlene Farias, cujo ponto final nesta Terra coincidiu com o ponto final desta tese,
Dedico.

vi
AGRADECIMENTOS
Esta tese é fruto de uma conjunção de privilégios recebidos. Minha fé me diz
que, mais que benefícios, uma conjunção de graças me trouxe até aqui. Tudo esteve
ajustado demais, sincrônico demais para ser simplesmente obra do acaso.
Há dez anos, tive o privilégio de ingressar como docente na Universidade
Federal Rural de Pernambuco (UFRPE), que, desde 2006, autorizou que eu me afastasse
das atividades docentes para cursar o Doutorado. Sou grata aos que fazem a Reitoria, à
Pró-Reitoria de Pesquisa e Pós Graduação (PRPPG), à Diretoria do Departamento de
Biologia e à Supervisão da Área de Ecologia, por autorizarem meu afastamento. Aos
colegas de Área Paula Gomes, Ângelo Alves, Carlos Guaraná, Lady Maia, Karine
Magalhães, Fátima Santos, Mônica Botter e Lohengrin Fernandes, que seguraram as
pontas na minha ausência e sempre me deram força e incentivo; aos substitutos que
ministraram minhas aulas; aos colegas de outras áreas acadêmicas que sempre vibraram
com minhas conquistas, em especial a Edson Silva, Emídio Cantídio e Maria José de
Sena. À Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), pela
concessão da bolsa de doutorado através da PRPPG/ UFRPE. A vocês todos, meus
sinceros agradecimentos!
Há seis anos, tenho a honra de integrar a equipe do Projeto Fragmentos, Fases I e
II, desenvolvido no âmbito do Programa de Ciência e Tecnologia para a Mata Atlântica,
com apoio financeiro do CNPq (590039/2006-7) e do Ministério Federal de Educação e
Pesquisa da Alemanha (BMBF, 01 LB 0203 A1). O projeto garantiu a logística sem a
qual este trabalho não seria, nem de longe, possível. Permitiu, também, que uma ótima
equipe se formasse, integrando pessoas das mais diversas áreas em torno de uma
pergunta comum: qual é o presente e o futuro desses fragmentos? Foi um prazer
trabalhar com os colegas da UFRPE, UFPE, IPA e Universidade de Ulm. Obrigada
especialíssimo à coordenadora Mari Rodal, amiga de tantos anos, por acreditar no
projeto, “comprar” minhas idéias e nunca esmorecer nos incentivos. Sou muito grata
também aos colegas do Grupo de Estrutura, Variação Temporal e Florística, cujos
líderes e pesquisadores contribuíram valiosamente para montagem do banco de dados e
identificação botânica. Obrigada a Mari Rodal, Marccus Alves, Ana Du Bocage e
Everardo Sampaio e aos seus extensos grupos de pesquisadores, especialmente
Ladivânia Nascimento, Amanda Menezes, Poliana Mendes, Tassiane Novacosque,

vii
Diego Marcelino, Tânia Costa, Maria Carolina Nunes, Jussara Souza, Fátima Lucena,
Anderson Alves-Araújo, Jefferson Maciel, Diogo Amorim e Aline Melo. Importante
frisar que eu não teria ido muito longe sem o empenho da minha equipe do Laboratório
de Ecologia Vegetal (LEVE), em especial do seu primeiro integrante Henrique Costa,
de “todo mundo” (Juliana Gomes, Kleybiana Dantas e Wilkilane Silva), de Frederico
Brandão e Thiago Campos. Um agradecimento caloroso ao empenho de Marcos
Chagas, nosso fiel escudeiro Marquinhos, que se descobriu mais botânico que mateiro e
cuja ajuda foi essencial no campo. À Michelle Trindade, cuja habilidade com mapas e
geoprocessamento foi fundamental à realização desta tese, com todo apoio logístico do
Geosere, principalmente do Professor Hernande Pereira e dos pesquisadores Sandro
Figueira e Gustavo Sobral. Agradeço também a participação daqueles que deixaram sua
contribuição nas coletas e bancos de dados, em especial a Sarah Freire, Iana Moura,
Clayton Souza, André Lima, Alexandre Gomes, Keila Costa, Fragoso Júnior, Jacemy
Souza, Airton Cavalcanti, Mário Machado e Ana Carla Souto. Meus sinceros
agradecimentos aos colegas alemães que contribuíram para a realização desta tese,
investindo horas na dura tarefa de coordenar acadêmica e financeiramente, além de
dividir conosco as horas de campo e laboratório, muito especialmente a Michael
Schessl, Leonhard Krause, Daniel Piechowski, Holger Teichert, Ute Knörr, Marcus
Braun e Thomas Kimmel. Valeu, turma da Usina!
Há quatro anos, tive o privilégio de ingressar no Programa de Pós Graduação em
Ecologia (PPGE) da Universidade Federal do Rio de Janeiro para realização do
Doutorado. Esta casa deu créditos aos meus anseios de estudar os fragmentos florestais
na minha terra, Pernambuco. Meu agradecimento mais que especial ao professor,
mentor e amigo Fabio Scarano, que desde o primeiro contato acreditou no potencial
desses fragmentos perdidos no canavial, aceitando dividir comigo sua experiência em
conversas sempre produtivas, criticando quando necessário, sugerindo, apoiando e
elogiando (por que não?). Valeu, grande Fabio, por tudo! À coordenação do PPGE, na
sua gestão atual e passadas; à Secretaria do Programa, nas pessoas sempre disponíveis e
prestativas de Sueli Teixeira e Márcia Renauld; aos professores que se empenharam nas
disciplinas, especialmente Ricardo Iglesias, Reinaldo Bozelli e Jean Valentin; a Eduardo
Arcoverde e Pablo Rodrigues pelas valiosas críticas na Qualificação; aos colegas de
curso, em especial a Solange Pessoa, Leonardo Lignani, Miriam Pinto e Erli Costa. Ao
professor Jean Paul Metzger e a Milton César Ribeiro (Miltinho), da Universidade de

viii
São Paulo, com os quais tive o privilégio de cursar Ecologia de Paisagens, trocar muitas
idéias e aprender a pensar nessa escala. Sou grata a vocês, para sempre!
Durante todos esses anos de vigência do Projeto Fragmentos, a Usina São José,
proprietária da área de estudo, virou minha casa e meu laboratório, com o
consentimento e confiança do seu visionário Diretor, Frederico Petribú Vilaça, e do
companheiro biólogo Roberto Siqueira, responsável e grande conhecedor da riqueza
contida naquelas matas. Sou grata pela privilegiada logística e apoio constante da
Diretoria e do Setor Agrícola da Usina, nas suas gestões atual e passada, e pelo apoio
indispensável dos seus funcionários, em especial dos agrônomos Luiz Vital, Antônio
Bandeira e Luiz Lúcio, do nosso sábio auxiliar florestal Lenilson, das secretárias Sueli e
Marluce e do topógrafo João. Minha gratidão a vocês, com os votos que essa parceria
renda produtivos frutos, por muito tempo.
Desde sempre, tenho o privilégio de ser filha de Marieta Borges e Fernando
Lins-e-Silva e neta de Tosca Barbieri, que sempre acreditaram que uma boa educação
seria a melhor herança para mim e para minhas irmãs. Sou eternamente grata a vocês
por pensarem assim e espero ter-lhes dado o orgulho que vocês merecem. Agradeço a
Deus por ter comigo a minha mãe, minha grande família e especiais amigos
testemunhando este momento, enquanto meu pai e minha vó, que trago no coração,
celebram junto ao Pai. A vida me permitiu ainda o privilégio de construir uma família
com Gilmar e termos juntos nossa Laura. Sou a grata a vocês, por todo amor que
recebo, por entenderem minhas ausências, por vibrarem com minhas vitórias e,
principalmente, por me permitirem ser esposa e mãe, minhas maiores conquistas. Amo
vocês, rei e princesa! Sou grata também aos meus especiais amigos e família no Rio de
Janeiro. Sem o apoio de vocês, acredito que eu teria tomado outro destino. Obrigada, de
coração, à família Almeida, dos queridos Álvaro e sua estrela Dorinha (in memoriam),
das primas-irmãs Bia e Zezé (e Fernando); e aos Guedes-Bruni, da querida Rejan e seus
Sérgio, Flora, Tito, Dona Dora (in memoriam) e Cida. Muito obrigada também a Malu
Pires e Mônica Santos, que me abrigaram em São Paulo. A acolhida de todos vocês foi
essencial e incondicional. Dou graças ao privilégio de poder tê-los sempre por perto.
Enfim, por todos que deram as mãos para a realização deste trabalho, sou grata a
Deus. Como diz o poeta, quando “Deus quer, O homem sonha, A obra nasce”.

ix
Resumo Geral
Em fragmentos florestais, o aumento progressivo na riqueza com aumento do tamanho é
explicado como função da área ou da maior heterogeneidade, com base, principalmente,
na Teoria da Biogeografia de Ilhas e nas Teorias de Nicho. Como a pesquisa empírica
aponta que essas hipóteses são relacionadas, comprovadas ou refutadas, em algum grau
e escala, objetivou-se, nesta tese, testar ambas as idéias em comunidades vegetais
fragmentadas numa paisagem de Floresta Atlântica. Inicialmente, analisou-se a relação
entre área, forma e heterogeneidade em 80 fragmentos, nos quais os habitats foram
classificados e quantificados quanto à borda, topografia e presença de água. Utilizando-
se regressões, verificou-se que o número (NH) e a diversidade (DH) de habitats foram
positiva e significativamente relacionados à área dos fragmentos, uma relação menor em
fragmentos < 50 ha. Área e forma juntas explicaram 60% da variância de NH e 53% de
DH. Em seguida, focando em nove fragmentos em três classes de tamanho, assembléias
de dossel (30 parcelas de 100 m2 em cada) e sub-bosque (30 de 25 m2) foram
inventariadas e comparadas entre tamanhos de fragmentos. Amostraram-se 14200
plantas e um total de 300 espécies. A riqueza total variou de 126 ± 25 em fragmentos
grandes a 91 ± 15 nos pequenos, mas apenas o sub-bosque foi significativamente mais
rico nos fragmentos maiores, o que sinaliza um risco de perda de espécies no futuro.
Complementarmente, três fragmentos grandes e três pequenos foram analisados quanto
à heterogeneidade de habitats e especialização de plantas, obtendo-se amostras de dossel
e sub-bosque por habitat. Variáveis estruturais foram comparadas entre habitats e entre
fragmentos, neste caso a partir de estimativas obtidas por rarefação e bootstrapping.
Foram amostradas 308 espécies, sendo a riqueza e diversidade significativamente
maiores em fragmentos grandes. Entre habitats, a similaridade foi mais baixa entre
habitats semelhantes em diferentes fragmentos do que entre habitats distintos em um
mesmo fragmento. Preferências por habitats foram identificadas em 57 (teste do qui-
quadrado) e 49 (análise de espécies indicadoras - ISA) espécies, principalmente por
sítios ripários. Os resultados sugerem que há um efeito da área na riqueza, da
heterogeneidade de habitats na diversidade beta, e da limitação de dispersão na
complementaridade entre fragmentos. Conclui-se, assim, que área e heterogeneidade de
habitats são hipóteses correlacionadas, porém complementares, pois a área per se não é
suficiente para explicar padrões de riqueza com vistas à conservação da diversidade na
paisagem como um todo.

x
General Abstract
In forest fragments, the progressive increase in species richness with increasing size is
explained as a function of either area per se or greater environmental heterogeneity,
based on the Island Biogeography Theory and Niche Theories. Considering that
empirical research has found that these two hypotheses are related, confirmed or
refuted, at some degree or scale, the objective this thesis was to analyze both ideas in
fragmented plant communities in an Atlantic Forest landscape. Initially, the relationship
between area, shape and heterogeneity was analyzed in 80 fragments, in which habitats
were classified according to edge, topography and water presence. Using regression
models, it was found that habitat number (HN) and diversity (HD) were positive and
significantly related to fragment area, with a weaker relationship in fragments smaller
than 50 ha. Area and shape together explained 60% of HN variance and 53% of HD
variance. Secondly, focusing on nine fragments in three size classes, canopy
assemblages (thirty 100m2-plots in each) and understory (thirty 25m2-plots) were
surveyed and compared between fragment size classes. A total of 14,200 plants and 300
species were recorded. Total richness varied between 126 ± 25 in large fragments and
91 ± 15 in small fragments, but only understory assemblages were significantly richer in
large fragments, indicating a risk of future loss of species. Three large and three small
fragments were further analyzed in relation to habitat heterogeneity and plant
specialization, based on canopy and understory samples per habitat. Structural variables
were compared between habitats and fragments; in the latter case richness estimatives
were used applying sample-based rarefaction and bootstrapping. A total of 308 species
were identified, being richness and diversity significantly greater in larger fragments,
independently of habitat diversity. Species similarity was smaller between similar
habitats in different fragments than between different habitats within the same fragment,
for total sample and per stratum. Habitat preferences were identified in 57 (chi-square
test) and 49 (indicative species analyses - ISA) species, mainly for riparian sites. Results
suggest that there is an area effect on species richness, a heterogeneity effect on β-
diversity and an influence of dispersal limitation on species complementarity between
fragments. As a conclusion, although area and heterogeneity are indeed correlated
hypotheses, they are complementary, because area per se alone cannot explain patterns
of richness or guarantee the maintenance of high plant diversity on a landscape level.

xi
SUMÁRIO
Resumo Geral........................................................................................................................ ix
General Abstract ................................................................................................................. x
I. INTRODUÇÃO GERAL ............................................................................................ 1
II. ÁREA DE ESTUDO E METOLOGIA GERAL ................................................... 5
1. Descrição da área ................................................................................................... 6
2. Metodologia geral ................................................................................................. 11
III. CAPÍTULO 1 - Diversidade vegetal em fragmentos florestais: bases teóricas e empíricas ......................................................................................... 14
1. Resumo ..................................................................................................................... 15
2. Introdução ............................................................................................................... 16
3. O estado da arte – ou o que se sabe sobre os determinantes de riqueza e diversidade em comunidades naturais? ............................................... 17
4. Os limites das molduras - ou o que se sabe sobre riqueza e diversidade em comunidades naturais fragmentadas? ..................................... 32
5. Integrando as idéias: heterogeneidade, ilhas e bordas ........................ 42
IV. CAPÍTULO 2 - Integrando área, forma e heterogeneidade de habitats em remanescentes florestais numa paisagem altamente fragmentada da Floresta Atlântica ........................................................................ 46
1. Resumo ..................................................................................................................... 47
2. Introdução ............................................................................................................... 48
3. Material e métodos .............................................................................................. 51
4. Resultados ............................................................................................................... 57
5. Discussão ................................................................................................................. 63
6. Considerações finais e implicações ................................................................ 67
V. CAPÍTULO 3 – Diversidade de assembléias de plantas de dossel e sub-bosque em fragmentos de Floresta Atlântica: a área
importa? ................................................................................................................................. 69
1. Resumo ..................................................................................................................... 70
2. Introdução ............................................................................................................... 71
3. Material e métodos .............................................................................................. 74
4. Resultados ............................................................................................................... 77
5. Discussão ................................................................................................................. 88
6. Considerações finais e implicações ................................................................ 92

xii
VI. CAPÍTULO 4 - Influência da heterogeneidade de habitats na diversidade e especialização de plantas em fragmentos de Floresta Atlântica ................................................................................................................................. 94
1. Resumo ..................................................................................................................... 95
2. Introdução ............................................................................................................... 96
3. Material e métodos .............................................................................................. 99
4. Resultados ............................................................................................................. 103
5. Discussão ............................................................................................................... 116
6. Considerações finais e implicações .............................................................. 119
VII. CONCLUSÃO GERAL ........................................................................................... 121
II. REFERÊNCIAS BIBLIOGRÁFICAS ............................................................... 124
APÊNDICE A ....................................................................................................................... 153

xiii
Índice de Figuras
Figura II.1. Localização da área de estudo, na Zona da Mata Norte de Pernambuco, Nordeste do Brasil. ........................................................................................................... 6
Figura II.2. Paisagem atual na área de estudo, na Zona da Mata Norte de Pernambuco, Nordeste do Brasil. Fonte: Trindade et al. (2008). ........................................................... 7
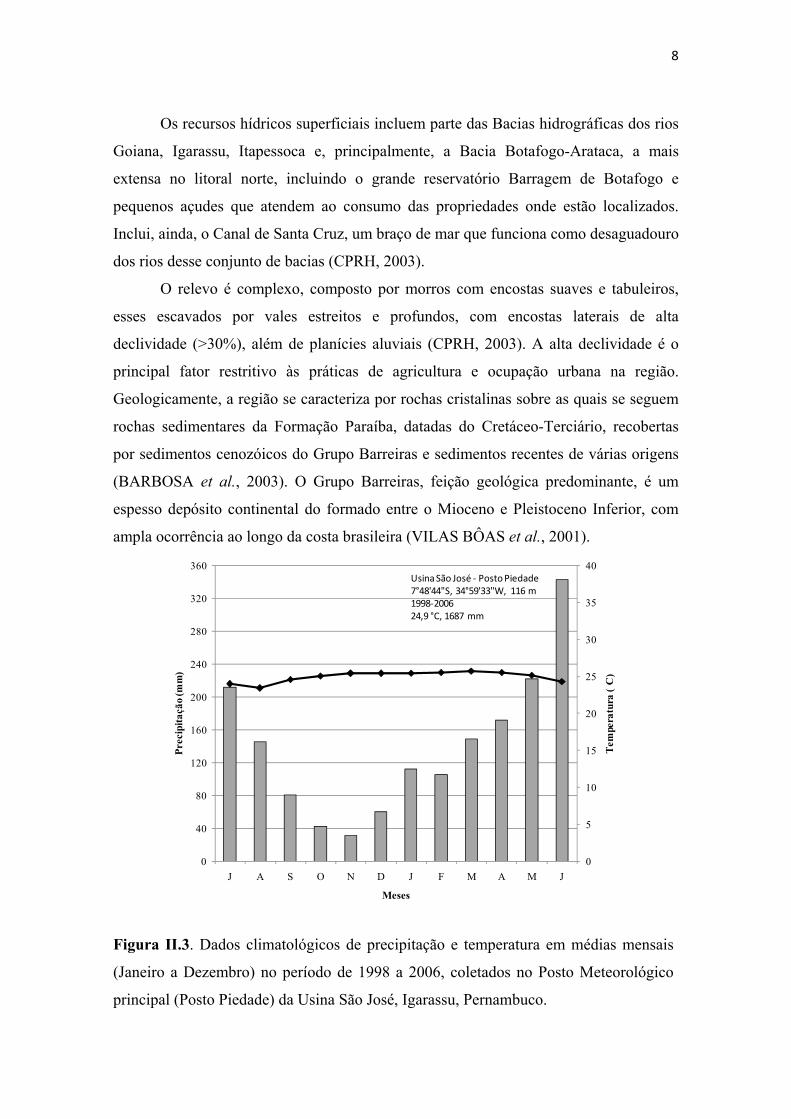
Figura II.3. Dados climatológicos de precipitação e temperatura em médias mensais (Janeiro a Dezembro) no período de 1998 a 2006, coletados no Posto Meteorológico principal (Posto Piedade) da Usina São José, Igarassu, Pernambuco. ............................. 8
Figura II.4. Vista geral da paisagem na Zona da Mata Norte de Pernambuco, com fragmentos florestais dispersos em meio ao canavial (a). Em detalhe, encosta íngreme revestida por floresta (b) e recorte de um fragmento (c). ............................................... 10
Figura IV.1. Localização da área de estudo, no estado de Pernambuco, Nordeste do Brasil e os 80 fragmentos florestais analisados. ............................................................. 53
Figura IV.2. Distribuição dos fragmentos de acordo com o índice de forma (IF) em 80 fragmentos de floresta atlântica na zona da Mata Norte de Pernambuco, Nordeste do Brasil. Valores nas classes correspondem ao limite superior aberto em cada classe. .... 58
Figura IV.3. Regressão logarítmica entre área total do fragmento (AT) e índice de forma (IF) em 80 fragmentos de floresta atlântica na zona da Mata Norte de Pernambuco, Nordeste do Brasil. ......................................................................................................... 58
Figura IV.4. Frequência de ocorrência dos oito tipos potenciais de habitats em 80 fragmentos de floresta atlântica na zona da Mata Norte de Pernambuco, Nordeste do Brasil. .............................................................................................................................. 59
Figura IV.5. Relação entre área total (AT) e proporção de habitats de borda em 80 fragmentos de floresta atlântica na Zona da Mata Norte, Nordeste do Brasil. ............... 60
Figura IV.6. Proporção dos tipos de habitats em relação à topografia em 80 fragmentos de floresta atlântica na zona da Mata Norte de Pernambuco, Brasil. Habitats de fundo de vale estão detalhados em relação à presença de água (permanente ou temporária). ...... 60
Figura IV.7. Relação entre área total (AT) e riqueza de habitats (NH) em 80 fragmentos de floresta atlântica na zona da Mata Norte de Pernambuco, Nordeste do Brasil. ......... 62
Figura IV.8. Relação entre área total (AT) e diversidade de habitats (NH) em 80 fragmentos de floresta atlântica na zona da Mata Norte de Pernambuco, Nordeste do Brasil. .............................................................................................................................. 63
Figura V.1. Localização dos nove fragmentos estudados na paisagem altamente fragmentada na Zona da Mata Norte, Pernambuco, Nordeste do Brasil. ....................... 74

xiv
Figura V.2. Densidades das assembléias de plantas (dossel e sub-bosque) amostradas em nove fragmentos de Floresta Atlântica, agrupados por tamanho, em Pernambuco, Nordeste do Brasil. Barras de erro = erro padrão. .......................................................... 79
Figura V.3. Riqueza de espécies em assembléias de plantas (dossel, sub-bosque e amostra total) em nove fragmentos de Floresta Atlântica, agrupados por tamanho, em Pernambuco, Nordeste do Brasil. Barras de erro = erro padrão. .................................... 80
Figura V.4. Curvas de rarefação baseadas nos indivíduos (médias de 100 re-amostragens) em assembléias de plantas em nove fragmentos de Floresta Atlântica em Pernambuco, Nordeste do Brasil. (a) curvas para amostras totais; (b) curvas para sub-bosque. Códigos alfa-numéricos referem-se aos fragmentos grandes (G), médios (M) e pequenos (P), conforme descrito na Tabela 1................................................................. 81
Figura V.5. Diversidade de espécies em assembléias de plantas (dossel, sub-bosque e amostra total) em nove fragmentos de Floresta Atlântica, agrupados por tamanho, em Pernambuco, Nordeste do Brasil. Barras de erro = erro padrão. .................................... 82
Figura V.6. Densidade relativa de espécies muito frequentes (registradas em todos os fragmentos) e de espécies exclusivas em nove fragmentos de Floresta Atlântica em Pernambuco, Nordeste do Brasil. G = fragmento grande, M = médio e P = pequeno, conforme descrito na Tabela 1........................................................................................ 86
Figura V.7. Distribuição média de espécies em quatro categorias de frequência em fragmentos de Floresta Atlântica em Pernambuco, Nordeste do Brasil, agrupados por tamanho (N=3 fragmentos em cada grupo). Espécies infrequentes (amostradas em 1-2 fragmentos), pouco frequentes (3-5), frequentes (6-7) e muito frequentes (8-9). .......... 88
Figura VI.1. Distribuição percentual dos habitats ocorrentes em seis fragmentos de Floresta Atlântica ao Norte de Pernambuco, Brasil. Linha pontilhada e valores referem-se à diversidade de habitats correspondente (DH, índice de Simpson 1/D). ................ 104
Figura VI.2. Densidade de indivíduos de dossel, sub-bosque e amostra total registrados em seis fragmentos de Floresta Atlântica ao Norte de Pernambuco, Brasil, com indicação do número de habitats amostrados em cada (linha pontilhada), definindo a ordem dos fragmentos no eixo x. G, fragmentos grandes e P, fragmentos pequenos, conforme descrito na Tabela 1...................................................................................... 105
Figura VI.3. Densidade de indivíduos na amostra de dossel e sub-bosque (colunas) e riqueza total (triângulos) nos cinco habitats mais frequentes em seis fragmentos de Floresta Atlântica ao Norte de Pernambuco, Brasil. a) encosta, b) ripário permanente, c) ripário temporário. G – fragmento grande, P – fragmento pequeno (conforme Tabela 1). ...................................................................................................................................... 108
Figura VI.4. Índices de diversidade β calculados para seis fragmentos de Floresta Atlântica ao Norte de Pernambuco, Brasil, para amostra total, dossel e sub-bosque. a) SS – similaridade de Sørensen média; b) βW – Diversidade β de Whittaker; G – fragmento grande, P – fragmento pequeno, conforme descrito na Tabela 1; barras de erro = ± 1 erro padrão. .......................................................................................................................... 110

xv
Figura VI.5. Similaridade de Sørensen média (SSM) entre habitats semelhantes ocorrentes em seis fragmentos de Floresta Atlântica ao Norte de Pernambuco, Brasil, para amostra total e estratos. IRT – Interior ripário temporário (N=4), BE – borda de encosta (6), IE – interior de encosta (6), BRP – borda ripária permanente (5), IT – interior tabuleiro (2), IRP – interior ripário permanente (5). Barras de erro=±1 erro padrão. .......................................................................................................................... 112
Figura VI.6. Correlação entre o número de ocorrências do habitat e riqueza de espécies (amostra total) no habitat, em seis fragmentos de Floresta Atlântica ao Norte de Pernambuco, Brasil. Coeficiente de correlação linear r = 0,8713. Cada ponto no gráfico representa um habitat: IRT – Interior ripário temporário, BE – borda de encosta, IE – interior de encosta, BRP – borda ripária permanente, IT – interior tabuleiro, IRP – interior ripário permanente, BT – borda de tabuleiro, BRT – borda ripária temporária. ...................................................................................................................................... 112

xvi
Índice de Tabelas
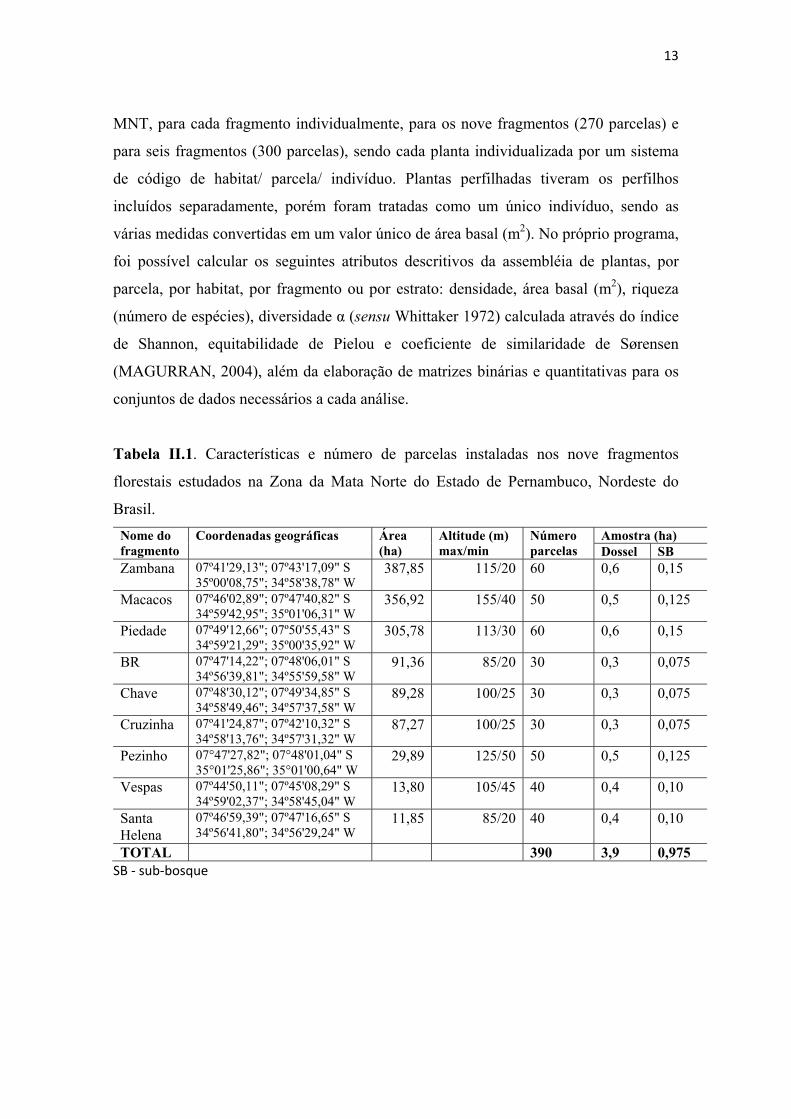
Tabela II.1. Características e número de parcelas instaladas nos nove fragmentos florestais estudados na Zona da Mata Norte do Estado de Pernambuco, Nordeste do Brasil. .............................................................................................................................. 13
Tabela IV.1. Tipos potenciais de habitats com base em três dimensões utilizadas para quantificação da heterogeneidade de habitats em fragmentos de Floresta Atlântica em Pernambuco, Nordeste do Brasil. ................................................................................... 55
Tabela IV.2. Descritores da heterogeneidade de habitats e resultados da regressão logarítmica em fragmentos de floresta atlântica na Zona da Mata Norte de Pernambuco, Nordeste do Brasil, para amostra total e para sub-amostra de fragmentos menores que 50 ha. NH, número de habitats; DH, diversidade de habitats; Min, valor mínimo,; max, valor máximo; N, número de fragmentos; VI, variável independente; AT, área total. .. 61
Tabela V.1. Características dos nove fragmentos florestais estudados na Zona da Mata Norte do estado de Pernambuco, Nordeste do Brasil. .................................................... 77
Tabela V.2. Percentuais de mortos de pé e lianas amostrados no dossel e de mortos de pé no sub-bosque de nove fragmentos de Floresta Atlântica em Pernambuco, Nordeste do Brasil. G, fragmentos grandes; M, médios; e P, pequenos. ....................................... 78
Tabela V.3. Espécies muito abundantes (> 100 indivíduos) e/ou muito frequentes (em 8-9 fragmentos), ordenadas pelo valor absoluto de abundância (amostra total), em nove fragmentos de Floresta Atlântica em Pernambuco, Brasil. Em destaque, densidades das muito abundantes localmente (no fragmento). G, fragmentos grandes; M, médio; P, pequeno, conforme descrito na Tabela 1. ....................................................................... 84
Tabela V.4. Número (± desvio padrão) e proporção média de espécies por categorias de abundância em fragmentos florestais estudados na Zona da Mata Norte, Pernambuco, Nordeste do Brasil. ......................................................................................................... 86
Tabela V.5. Espécies exclusivas de um fragmento e registradas com densidades > 5 indivíduos, amostradas em nove fragmentos de Floresta Atlântica em Pernambuco, Nordeste do Brasil. ......................................................................................................... 87
Tabela VI.1. Características dos seis fragmentos florestais estudados na Região Norte do Estado de Pernambuco, Nordeste do Brasil. N, número; DH, diversidade de habitats (índice de Simpson recíproco). ..................................................................................... 102
Tabela VI.2. Riqueza e diversidade de espécies observados e estimados em seis fragmentos de Floresta Atlântica ao Norte de Pernambuco, Brasil, para amostra total, dossel e sub-bosque. G, fragmentos grandes; P, fragmentos pequenos; S, número de espécies; H', diversidade de Shannon; estimativas obtidas por (a) rarefação (±1 desvio padrão) e (b) bootstrapping........................................................................................... 107
Tabela VI.3. Espécies com associação positiva por habitats (pelo teste do χ2) e/ou indicadoras de habitat (ISA), para oito habitats em seis fragmentos de Floresta Atlântica, Pernambuco, Brasil: IRT – Interior ripário temporário, BE – borda de encosta, IE – interior de encosta, BRP – borda ripária permanente, IT – interior tabuleiro, IRP – interior ripário permanente, BT – borda de tabuleiro, BRT – borda ripária temporária. No resultados da ISA, o valor indicador (INDVAL) marca o habitat de preferência; ns – não significativo. VI – valor de Importância, G – fragmento grande, P – frag. pequeno. ......................................................................................................... 113

1
I. INTRODUÇÃO GERAL

2
No mundo atual, poucas áreas exibem uma cobertura vegetal nativa contínua. A
necessidade de uso da terra, principalmente para exploração agrícola, moldou novas
formas de paisagens ao longo dos últimos séculos. Como resultado, fisionomias
homogêneas vêm sendo transformadas em mosaicos heterogêneos, nos quais a
vegetação remanescente é tipicamente reduzida e fragmentada (SAUNDERS et al.,
1991), especialmente na região tropical, com as maiores perdas de área registradas
(MYERS et al., 2000). Esta interferência humana nas comunidades primitivas
introduziu grandes desafios à ciência ecológica que, se por um lado ainda carece de uma
teoria geral sobre a organização de comunidades (SIMBERLOFF, 2004;
ROUGHGARDEN, 2009), por outro lado, precisa atuar na solução e mitigação dos
problemas gerados na interface natureza-sociedade (KREBS, 2006).
Enquanto termos como fragmentação, efeito de borda, isolamento, mudança e
subdivisão de habitats têm se incorporado rapidamente à Ecologia e Conservação nas
últimas décadas (NOBIS; WOHLGEMUTH, 2004; FAZEY et al., 2005;
LINDENMAYER; FISCHER, 2007), pesquisa empírica e teórica vêm investigando a
importância e o peso de diferentes variáveis na manutenção da diversidade em
paisagens fragmentadas. Embora ainda não se tenha alcançado uma teoria unificadora
(HAILA, 2002; WATSON, 2002), a crescente produção de pesquisa nas últimas
décadas (FISCHER; LINDENMAYER, 2007) vem construindo sólido conhecimento
sobre padrões e processos biológicos ao nível do fragmento e da paisagem (HAILA,
2002). Na escala do fragmento, análises do efeito da área, com base na Teoria da
Biogeografia de Ilhas (MACARTHUR; WILSON, 1967), e do efeito de borda
(SAUNDERS et al., 1991; MURCIA, 1995) têm gerado importante corpo de dados
empíricos. Paralelamente, com o desenvolvimento da Ecologia de Paisagens (TURNER,
2005b), tornou-se mais frequente analisar o efeito da estrutura da paisagem sobre a biota
fragmentada, como, por exemplo, o papel da conectividade (MURPHY; LOVETT-
DOUST, 2004; DAMSCHEN et al., 2006), a influência da matriz (RICKETTS, 2001), e
o tempo de acomodação de populações e comunidades à fragmentação (HELM et al.,
2006; METZGER et al., 2009).
A existência de paisagens altamente fragmentadas tem provocado uma mudança
no paradigma de conservação da biodiversidade, passando da prioridade de se conservar
grandes áreas para uma abordagem de conservação da paisagem (SILVA;
TABARELLI, 2000; LINDENMAYER et al., 2006; METZGER, 2006), na qual

3
também importam os menores fragmentos (TURNER; CORLETT, 1996) e as florestas
secundárias (CHAZDON et al., 2009; DENT; WRIGHT, 2009). Esta é uma tendência
especialmente relevante na região de abrangência da Floresta Atlântica brasileira.
Considerada um dos cinco principais hotspots mundiais de biodiversidade, por
concentrar alto endemismo de espécies (MORELLATO; HADDAD, 2000; MYERS et
al., 2000), esta floresta sofreu intensa redução de área, restando um percentual estimado
máximo de 16% da cobertura original no país (RIBEIRO et al., 2009). Este percentual,
entretanto, não é homogêneo ao longo da região de ocorrência potencial dessas
florestas. Na região ao norte do rio São Francisco, incluindo a faixa costeira de Alagoas
ao Rio Grande do Norte, o chamado Centro de Endemismo Pernambuco sensu Silva e
Casteleti (2005), restam 11,5% da cobertura nativa (RIBEIRO et al., 2009). Mais do
que o total remanescente, chama à atenção a intensa fragmentação nessa região, na qual
o maior fragmento atinge 3500 ha (OLIVEIRA et al., 2004) e a maioria tem tamanho
inferior a 50 ha (RANTA et al., 1998; TRINDADE et al., 2008).
Em fragmentos florestais, as árvores são elementos cruciais que determinam a
arquitetura e as condições microclimáticas (LAURANCE et al., 1998; LAURANCE et
al., 2006) e fisicamente manifestam os efeitos das muitas variáveis ecológicas
(HARPER et al., 2005). Entre os componentes de uma floresta, as árvores parecem ser
os mais resistentes aos efeitos da fragmentação, considerando a longevidade de muitas
espécies. Estudos em florestas fragmentadas amazônicas e atlânticas revelaram,
contrariamente, que a fragmentação provoca mudanças rápidas e profundas nas
comunidades arbóreas, levando a um aumento na mortalidade, declínio de algumas
populações e extinções locais (SILVA; TABARELLI, 2000; LAURANCE et al., 2002;
LAURANCE et al., 2006; OLIVEIRA et al., 2008; SANTOS et al., 2008; LOPES et
al., 2009). Os efeitos marcantes podem também se revelar tardiamente em relação ao
evento de fragmentação, em um atraso de até várias décadas, de modo especial para
árvores e outros grupos biológicos de dispersão limitada (METZGER et al., 2009).
Existindo na forma contínua ou isolada, comunidades florestais são compostas,
em relevantes proporções, por espécies arbóreas com preferências por determinados
habitats (CONDIT et al., 2000; HOFER et al., 2008). Como a fragmentação acontece
com objetivos de uso da terra, alguns ambientes são preferidos por serem mais úteis à
ocupação e agricultura, de forma que os remanescentes, geralmente, cobrem áreas
íngremes, encharcadas e menos produtivas da paisagem (SEABLOOM et al., 2002).

4
Tem-se, assim, que não apenas espécies são perdidas no processo de fragmentação, mas
também se perdem os habitats que provêm condições de ocupação e recursos para várias
espécies (KOLASA; WALTHO, 1998).
Em uma região altamente fragmentada da floresta atlântica no estado de
Pernambuco, Nordeste do Brasil, fragmentos, habitats e comunidades de plantas de
dossel e sub-bosque foram avaliados e deram origem a esta tese, cujo objetivo principal
foi analisar as consequências da perda de área florestal sobre a disponibilidade de
habitats e diversidade vegetal. Os objetivos específicos foram abordados em quatro
capítulos distintos. No primeiro capítulo, objetivou-se revisar as bases teóricas e
empíricas no estudo de comunidades fragmentadas, prioritariamente assembléias
vegetais. São discutidas as teorias e sínteses que formam o arcabouço do tema da tese,
apresentando a relevância de se aplicar tanto teorias espaciais neutras quanto teorias de
nicho (SCARANO; DIAS, 2004) aos estudos de comunidades. No segundo capítulo,
aborda-se a influência da redução da área florestal na perda de habitats. Para tanto, foi
analisada a relação entre área e forma dos fragmentos com a heterogeneidade de
habitats, a partir de oito habitats relevantes para comunidades florestais, mapeados e
quantificados espacialmente em 80 fragmentos. No terceiro capítulo, aborda-se a
influência da redução da área florestal sobre a perda de espécies. Especificamente,
objetivou-se investigar a relação entre o tamanho de fragmentos e assembléias de
plantas de dossel e sub-bosque. A análise foi feita em conjunto e separadamente por
estrato, com objetivo de descrever se esta relação existia para árvores ou apenas no sub-
bosque, devido a sua menor idade e maior suscetibilidade aos efeitos da fragmentação.
No quarto capítulo, foi avaliada a influência da heterogeneidade de habitats na estrutura
e especialização de plantas, o que permitiu distinguir a influência da heterogeneidade de
habitats da influência da área sobre as comunidades de plantas, identificar espécies com
especialização por habitats e discutir a importância da complementaridade na
diversidade de comunidades fragmentadas.
Ao final, são compilados os principais achados desta tese que, em síntese, testa
as seguintes hipóteses: i) a disponibilidade de habitats aumenta com o aumento da área
do fragmento; ii) a riqueza e a diversidade de plantas aumentam com a área do
fragmento; iii) a riqueza e a diversidade vegetal aumentam com o aumento da
heterogeneidade de habitats; e iv) assembléias de dossel e de sub-bosque apresentam
respostas distintas ao efeito da área.

5
II. ÁREA DE ESTUDO E METOLOGIA GERAL

6
1. Descrição da área
Os estudos que formam esta tese foram conduzidos na propriedade da Usina São
José/ Grupo Cavalcanti Petribú – USJ, uma área de aproximadamente 240 km2 na Zona
da Mata Norte do Estado de Pernambuco, Nordeste do Brasil, com sua porção principal
no município de Igarassu e partes nos municípios de Goiana, Itaquitinga, Itapissuma,
Abreu e Lima e Araçoiaba (Figura II.1). A região é uma das mais reduzidas e
fragmentadas da Floresta Atlântica, com percentual remanescente estimado em 11,5%
(RIBEIRO et al., 2009) para a sub-região biogeográfica “Centro de Endemismo
Pernambuco” (SILVA; CASTELETI, 2005), na qual está inserida, e 4,6% da floresta
original no estado de Pernambuco (LIMA, 1998). Na propriedade da USJ, há 202
manchas de vegetação nativa (Floresta Atlântica), sendo 96 áreas de floresta secundária
(capoeiras) e 106 remanescentes de floresta madura (TRINDADE et al., 2008) (Figura
II.2), dos quais subconjuntos de 80 (capítulo 2), nove (capítulo 3) e seis (capítulo 4)
constituíram o material das pesquisas.
Figura II.1. Localização da área de estudo, na Zona da Mata Norte de Pernambuco,
Nordeste do Brasil.
Com base em dados coletados na Estação Meteorológica da Usina São José/
Posto Piedade (7°48'44"S, 34°59'33"W), no período de 1998 a 2006 (Figura II.3), a

7
temperatura média anual é de 24,9ºC, com média de 25,7º C no mês mais quente. A
precipitação média anual é de 1.687 mm, com uma estação seca no verão, sendo
novembro o mês mais seco (31 mm), e uma estação chuvosa de março a agosto,
registrando-se a maior precipitação no mês de junho, quando chovem, em média, 342
mm. A pequena amplitude térmica anual e a sazonalidade da precipitação permitem
caracterizar o clima na área como quente e úmido com verão seco e chuvas de outono-
inverno, característico do As' no sistema de Köppen (REIS, 1970; STRAHLER;
STRAHLER, 1978).
Figura II.2. Paisagem atual na área de estudo, na Zona da Mata Norte de Pernambuco,
Nordeste do Brasil. Fonte: Trindade et al. (2008).
8
Os recursos hídricos superficiais incluem parte das Bacias hidrográficas dos rios
Goiana, Igarassu, Itapessoca e, principalmente, a Bacia Botafogo-Arataca, a mais
extensa no litoral norte, incluindo o grande reservatório Barragem de Botafogo e
pequenos açudes que atendem ao consumo das propriedades onde estão localizados.
Inclui, ainda, o Canal de Santa Cruz, um braço de mar que funciona como desaguadouro
dos rios desse conjunto de bacias (CPRH, 2003).
O relevo é complexo, composto por morros com encostas suaves e tabuleiros,
esses escavados por vales estreitos e profundos, com encostas laterais de alta
declividade (>30%), além de planícies aluviais (CPRH, 2003). A alta declividade é o
principal fator restritivo às práticas de agricultura e ocupação urbana na região.
Geologicamente, a região se caracteriza por rochas cristalinas sobre as quais se seguem
rochas sedimentares da Formação Paraíba, datadas do Cretáceo-Terciário, recobertas
por sedimentos cenozóicos do Grupo Barreiras e sedimentos recentes de várias origens
(BARBOSA et al., 2003). O Grupo Barreiras, feição geológica predominante, é um
espesso depósito continental do formado entre o Mioceno e Pleistoceno Inferior, com
ampla ocorrência ao longo da costa brasileira (VILAS BÔAS et al., 2001).
Figura II.3. Dados climatológicos de precipitação e temperatura em médias mensais
(Janeiro a Dezembro) no período de 1998 a 2006, coletados no Posto Meteorológico
principal (Posto Piedade) da Usina São José, Igarassu, Pernambuco.
0
5
10
15
20
25
30
35
40
0
40
80
120
160
200
240
280
320
360
J A S O N D J F M A M J
Tem
pera
tura
(C
)
Prec
ipita
ção
(mm
)
Meses
Usina São José ‐ Posto Piedade7°48'44"S, 34°59'33"W, 116 m1998‐2006 24,9 °C, 1687 mm

9
A vegetação primária na área é Floresta Ombrófila Densa das Terras Baixas
(VELOSO et al., 1991), atualmente muito reduzida e fragmentada. A floresta original
começou a ser derrubada com a chegada dos colonizadores portugueses no século XVI,
primeiramente para exploração da madeira, e mais tarde para ceder lugar a pastagens e
agricultura, principalmente cana-de-açúcar (CÂMARA, 2003; KIMMEL et al., 2008).
Trindade et al. (2008) descreveram a perda de área florestal nesta paisagem em cerca de
50% entre 1975 e 2005, o que provavelmente ocorreu devido ao incentivo para
produção de biocombustível ocorrido entre meados da década de 1970 a meados dos
anos de 1980, que motivou um pico de desflorestamento para implantação de canaviais
(GOLDEMBERG et al., 2008). A paisagem atual é caracterizada por plantações de
cana-de-açúcar revestindo tabuleiros, encostas e terraços fluviais (CPRH, 2003), com
fragmentos florestais esparsamente distribuídos (Figura II.4), com vários tamanhos e
formas (RANTA et al., 1998; TRINDADE et al., 2008). Remanescentes florestais
cobrem 24% da paisagem, sendo o tamanho médio dos fragmentos de 61 ha, ocupando
as áreas menos úteis para cultivo, que são as íngremes e os fundos de vale, um padrão
frequente em paisagens de floresta atlântica no nordeste (TRINDADE et al., 2008) e
sudeste (SILVA et al., 2007; SILVA et al., 2008d).
Desde 2003, pesquisas científicas vêm sendo desenvolvidas na propriedade USJ,
através do “Projeto Fragmentos”1 (RODAL et al., 2008), cujos resultados revelam que
os remanescentes florestais ainda ostentam uma fisionomia densa, com árvores altas, e
expressivas riqueza de plantas de dossel e sub-bosque (SILVA et al., 2008a; SILVA et
al., 2008c; GOMES et al., 2009). Uma primeira listagem florística catalogou 650
espécies de plantas em seis fragmentos na área, incluindo 280 espécies de ervas e
arbustos, 213 arbóreas e 93 trepadeiras e lianas (ALVES-ARAÚJO et al., 2008). Entre
as espécies lenhosas, destacam-se como mais importantes Eschweilera ovata
(Cambess.) Miers., Pogonophora schomburgkiana Miers ex Benth., Brosimum
guianense (Aubl.) Huber, Tapirira guianensis Aubl., Parkia pendula (Willd.) Benth. ex. 1 Em 2001, foram iniciadas pesquisas científicas na propriedade da Usina São José, através de Convênio de Cooperação Científica com a Universidade Federal Rural de Pernambuco (UFRPE). Em 2003, teve início o Projeto Fragmentos, uma iniciativa da UFRPE, Universidade Federal de Pernambuco, Universidade de Ulm (Alemanha) e Instituto Agronômico de Pernambuco (IPA), viabilizada através da Cooperação Brasil – Alemanha no âmbito do Programa de Ciência e Tecnologia para a Mata Atlântica, com apoio financeiro do Ministério de Ciência e Tecnologia/ CNPq e Ministério Federal de Educação e Pesquisa da Alemanha (BMBF). Na fase I (2003-2006), foram desenvolvidas pesquisas de Florística e Estrutura, Biologia de Solos e etno-ornitologia. Na fase II, na qual se insere este trabalho, o projeto agregou biologia vegetal, animal e de solos, interação planta-animal, ecologia de paisagens e etnobotânica, numa perspectiva interdisciplinar (CNPq 590039/2006-7, BMBF 01 LB 0203 A1).

10
Walp., Schefflera morototoni (Aubl.) Magari, Sleyrm, Inga thibaudiana DC.,
Thyrsodium spruceanum Benth. e Miconia prasina (Sw.) DC. (ROCHA et al., 2008;
BRANDÃO et al., 2009).
A iniciativa de pesquisa conjunta na área representa uma mudança de paradigma
na indústria canavieira na região, que hoje se alia às ações de conservação da Floresta
Atlântica, atuando na preservação de reservas, proteção de mananciais e reflorestamento
(TABARELLI; RODA, 2005). Dada a importância biológica das florestas e seu valor
para pesquisa, ensino, lazer e educação ambiental, além da relevância na proteção dos
recursos hídricos, do relevo e do solo, esta região integra, desde 1993, a área piloto
“Complexo Itamaracá – Itapissuma – Igarassu” da Reserva da Biosfera da Mata
Atlântica no Programa Homem e Biosfera (MAB) da UNESCO (LIMA, 1998).
Figura II.4. Vista geral da paisagem na Zona da Mata Norte de Pernambuco, com
fragmentos florestais dispersos em meio ao canavial (a). Em detalhe, encosta íngreme
revestida por floresta (b) e recorte de um fragmento (c).
a
cb Foto: L. Krause
Foto: A.C.B. Lins e Silva
Foto: A.C.B. Lins e Silva

11
O histórico dos remanescentes florestais foi avaliado através da análise do
material cartográfico disponível, incluindo fotografias aéreas de 1969 elaboradas pelo
Exército Brasileiro/ 3ª Divisão de Levantamento, ortofotocartas de 1975 elaboradas pela
FIDEM (atual Agência Estadual de Planejamento e Pesquisas de Pernambuco), imagens
multiespectrais de alta resolução do satélite IKONOS II, obtidas em 2005, e
documentação de campo e produção agrícola (carta planimétrica) da USJ. Esta análise
permitiu constatar o tempo de isolamento dos fragmentos e ocorrência de alterações
fitofisionômicas nos últimos 40 anos.
2. Metodologia geral
2.1. Amostragem das assembléias de plantas
Para amostragem das assembléias nos fragmentos (Capítulos 3 e 4), adotou-se o
método de parcelas (MUELLER-DOMBOIS; ELLENBERG, 1974), demarcadas com
estacas de PVC e fitilho de polietileno, cada uma medindo 10 x 10 m para amostragem
do dossel. Uma parcela menor para amostragem do estrato de sub-bosque (5 x 5 m) foi
instalada dentro e em um vértice de cada parcela de dossel. No dossel, foram
amostrados todos os indivíduos lenhosos com circunferência à altura do peito – 1,30 m
do solo (CAP) ≥ 15 cm, incluindo árvores, palmeiras, lianas, fetos arborescentes e
indivíduos mortos de pé. No sub-bosque, incluíram-se todas as plantas lenhosas e
mortas de pé com circunferência ao nível do solo (CNS) ≥ 1 cm até CAP < 15 cm.
Optou por definir o critério de amostragem no solo e sem limite de altura, entre as
várias maneiras de se amostrar o sub-bosque, por este ser mais inclusivo para plântulas,
indivíduos jovens, arvoretas e arbustos (GOMES et al., 2009).
O número de parcelas instaladas em cada fragmento variou de acordo com o
objetivo do estudo e hipótese testada. No capítulo 3, são descritos dados obtidos em
nove fragmentos (três pequenos, três médios e três grandes), tendo sido instaladas 30
parcelas de dossel e 30 de sub-bosque em cada, nos habitats mais frequentes,
totalizando 270 parcelas em cada estrato. Nos fragmentos grandes e pequenos, os
habitats existentes foram abordados em mais detalhes (capítulo 4) e, assim, foi
necessária complementação da amostra, instalando-se mais 120 parcelas (dossel e sub-
bosque) distribuídas nesses seis fragmentos. A amostra total envolveu, portanto, 390

12
parcelas em cada estrato (Tabela II.1), totalizando 3,9 ha na amostra de dossel e 0,975
ha na amostra de sub-bosque.
Em cada parcela, as plantas que atenderam ao critério definido foram marcadas
com plaqueta de PVC com numeração sequencial e tiveram suas medidas de
circunferência (CAP ou CNS) e de altura total anotadas. Plantas com diâmetros
múltiplos tiveram todos os perfilhos medidos, desde que pelo menos um atendesse ao
critério estabelecido. Todos os indivíduos marcados foram identificados no campo e
coletados de acordo com normas de coleta botânica (ALVES-ARAÚJO et al., 2008)
para preparação de exsicatas de herbário visando à identificação/ confirmação
taxonômica. A coleta de material botânico foi feita entre 2004 e 2009, e os dados
estruturais obtidos em campo entre 2007 e 2008. No total, o banco de dados incluiu 55
famílias botânicas e 345 espécies/ morfoespécies (Apêndice A). Grande esforço
taxonômico foi empreendido para identificação dos táxons ao nível de família, gênero e
espécie, com auxílio de especialistas e coleções dos Herbários UFP (Geraldo Mariz) e
IPA (Dárdano de Andrade-Lima) e de diversas instituições nacionais e estrangeiras
(PEUFR, HST, JPB, CEPEC, MAC, BHCB, ASE, HUEFS, ALCB, RB, R, NY, MO, G,
P), sob a coordenação taxonômica de M. Alves e A.L du Bocage. Desse esforço, 63%
das espécies foram identificadas totalmente, 64 (19%) até gênero e 52 (15%) foram
identificadas apenas ao nível de família, enquanto 12 (3%) restaram não identificadas,
devido à falta de material fértil, principalmente nas espécies de sub-bosque ou naquelas
registradas em apenas um indivíduo. As plantas indeterminadas foram separadas e
numeradas como morfoespécies, de forma que listas de espécies e estimativas de
riqueza pudessem ser obtidas. Exsicatas foram incluídas na Coleção de Referência da
Flora da Usina São José (Projeto Fragmentos - Fase II), no UFP e IPA. Duplicatas das
coletas férteis foram enviadas aos Herbários ULM, da Universidade de Ulm, Alemanha,
e RB, do Jardim Botânico do Rio de Janeiro. O arranjo das famílias seguiu a proposta
do APG II (AGP-II, 2003) e os nomes das espécies e abreviação dos autores seguiram a
base de dados Tropicos do Missouri Botanical Garden (www.tropicos.org).
2.2. Montagem do banco de dados
O banco de dados geral foi preparado, armazenado e manejado no programa
Mata Nativa 2.0 - MNT (SOUZA et al., 2006). Foram criados projetos no ambiente
13
MNT, para cada fragmento individualmente, para os nove fragmentos (270 parcelas) e
para seis fragmentos (300 parcelas), sendo cada planta individualizada por um sistema
de código de habitat/ parcela/ indivíduo. Plantas perfilhadas tiveram os perfilhos
incluídos separadamente, porém foram tratadas como um único indivíduo, sendo as
várias medidas convertidas em um valor único de área basal (m2). No próprio programa,
foi possível calcular os seguintes atributos descritivos da assembléia de plantas, por
parcela, por habitat, por fragmento ou por estrato: densidade, área basal (m2), riqueza
(número de espécies), diversidade α (sensu Whittaker 1972) calculada através do índice
de Shannon, equitabilidade de Pielou e coeficiente de similaridade de Sørensen
(MAGURRAN, 2004), além da elaboração de matrizes binárias e quantitativas para os
conjuntos de dados necessários a cada análise.
Tabela II.1. Características e número de parcelas instaladas nos nove fragmentos
florestais estudados na Zona da Mata Norte do Estado de Pernambuco, Nordeste do
Brasil. Nome do fragmento
Coordenadas geográficas Área (ha)
Altitude (m) max/min
Número parcelas
Amostra (ha) Dossel SB
Zambana 07º41'29,13"; 07º43'17,09" S 35º00'08,75"; 34º58'38,78" W
387,85 115/20 60 0,6 0,15
Macacos 07º46'02,89"; 07º47'40,82" S 34º59'42,95"; 35º01'06,31" W
356,92 155/40 50 0,5 0,125
Piedade 07º49'12,66"; 07º50'55,43" S 34º59'21,29"; 35º00'35,92" W
305,78 113/30 60 0,6 0,15
BR 07º47'14,22"; 07º48'06,01" S 34º56'39,81"; 34º55'59,58" W
91,36 85/20 30 0,3 0,075
Chave 07º48'30,12"; 07º49'34,85" S 34º58'49,46"; 34º57'37,58" W
89,28 100/25 30 0,3 0,075
Cruzinha 07º41'24,87"; 07º42'10,32" S 34º58'13,76"; 34º57'31,32" W
87,27 100/25 30 0,3 0,075
Pezinho 07°47'27,82"; 07°48'01,04" S 35°01'25,86"; 35°01'00,64" W
29,89 125/50 50 0,5 0,125
Vespas 07º44'50,11"; 07º45'08,29" S 34º59'02,37"; 34º58'45,04" W
13,80 105/45 40 0,4 0,10
Santa Helena
07º46'59,39"; 07º47'16,65" S 34º56'41,80"; 34º56'29,24" W
11,85 85/20 40 0,4 0,10
TOTAL 390 3,9 0,975 SB ‐ sub‐bosque

14
III. CAPÍTULO 1 - Diversidade vegetal em
fragmentos florestais: bases teóricas e empíricas

15
1. Resumo
O estudo de comunidades fragmentadas reúne, como arcabouço, várias teorias e sínteses
ecológicas, cuja revisão constitui o objetivo deste trabalho. Com ênfase nas relações
espécie-espaço em comunidades vegetais florestais, as contribuições teóricas e
empíricas aqui revisadas compõem dois grupos principais. No primeiro grupo, estão as
idéias acerca dos processos determinantes da diversidade, independentemente dos seus
limites espaciais, destacando-se as Teorias de Nicho/ Heterogeneidade de Habitats e a
Teoria neutra da biodiversidade. Nicho e habitat, conceitos desenvolvidos paralelamente
ao longo do século XX, evoluíram para hipóteses explicativas da diversidade biológica,
pois quanto mais nichos e habitats, mais espécies. Frequentemente testadas em
comunidades de plantas, tais teorias se comprovam para pelo menos uma fração das
espécies. Em oposição às idéias de nicho, a teoria neutra assume que as espécies são
ecologicamente equivalentes e seus testes também a corroboram parcialmente. No
segundo grupo, reúnem-se as teorias nas quais as relações espaciais importam. A
primeira delas, a Relação Espécie-Área, ou aumento progressivo no número de espécies
com aumento da área, tem três principais hipóteses explicativas: a riqueza aumenta em
função da área per se, em função da amostra ou devido à maior heterogeneidade de
habitats. Fortemente baseada na primeira, a teoria da biogeografia de ilhas (TBI) soma à
área per se o efeito do isolamento geográfico, que teria uma correlação negativa com a
riqueza. Além das variáveis da TBI, outros aspectos se integram ao estudo de
comunidades fragmentadas, entre eles o efeito de borda. De alta relevância em estudos
de fragmentação, este efeito penetra nos fragmentos, altera a dinâmica e, geralmente,
tem efeito deletério sobre a comunidade de plantas. Considerando a comprovação ou
refutação empírica, em algum grau e escala, de todas as teorias aqui discutidas, ressalta-
se a necessidade de integração das idéias para uma melhor compreensão da diversidade
biológica em comunidades fragmentadas. Despontam como lacunas que precisam ser
preenchidas a investigação da relação entre área e heterogeneidade de habitats em
paisagens fragmentadas e o entendimento da influência de uma e/ ou outra na riqueza e
diversidade de espécies nos remanescentes de vegetação.

16
2. Introdução
As teorias e sínteses que mais têm contribuído para esclarecer os padrões de
diversidade vegetal em fragmentos florestais derivam de dois grupos, cuja integração
teórica é complexa e continua a ser um desafio (GARDNER; ENGELHARDT, 2008).
De um lado, situam-se as idéias acerca dos processos determinantes da diversidade,
independentemente dos limites espaciais do ecossistema estudado (WRIGHT, 2002),
aqui se inserindo duas abordagens em lados bem opostos para explicar a organização de
comunidades (KELLY et al., 2008): as Teorias de Nicho/ Heterogeneidade de Habitats
e a Teoria Neutra Unificada da Biodiversidade e Biogeografia. No outro grupo, reúnem-
se as idéias que abordam ecossistemas com limites definidos, quando o recorte espacial
importa e define um novo paradigma (WU; LOUCKS, 1995; CANTRELL; COSNER,
2001), entre elas a Relação Espécie-Área, a Teoria da Biogeografia de Ilhas e o Efeito
de Borda.
Neste texto, objetiva-se sintetizar as teorias e pesquisas nos dois grupos expostos
acima, que formam o arcabouço do estudo de comunidades fragmentadas. Para tanto,
foi feito um amplo trabalho de revisão bibliográfica, seguindo dois eixos principais:
sobre a pesquisa em fragmentação e sobre as teorias de comunidades, listadas a partir de
textos em Ecologia (BEGON et al., 1996; CRAWLEY, 1997; SCARANO; DIAS,
2004). Em ambos os casos, foi utilizada a base de dados Web of Science do Institute for
Scientific Information (ISI, 2010) para localização de trabalhos relevantes, empregando-
se combinações de palavras chave como ‘fragmentation’, ‘theory’, ‘community’, “island
biogeography’, ‘niche theory’, ‘heterogeneity’ e ‘neutral theory’, com foco inicial em
documentos de revisão. Nesses, as primeiras citações, trabalhos teóricos e pioneiros
foram localizados, incluindo artigos e livros ou capítulos de livros. Em um segundo
momento, foram pesquisados trabalhos empíricos sobre os temas, a partir da literatura já
levantada e em novas buscas na base Web of Science. Para localização de pesquisa
original no Brasil, utilizou-se a base eletrônica Scielo (FAPESP/BIREME, 2010).
Ressalta-se que a revisão aqui apresentada enfatiza as relações espécie-espaço e
não as relações espécie-espécie, acreditando que as últimas só podem ser realizadas no
contexto das primeiras (KOLASA; WALTHO, 1998). Também é nossa intenção
investigar, na literatura, se a heterogeneidade de habitats é abordada ou considerada
uma determinante importante da diversidade em ecossistemas fragmentados e priorizar,

17
nesses temas, exemplos advindos da Ecologia vegetal, principalmente comunidades
arbóreas. Portanto, com permissão da metáfora, para entendimento da informação
contida num fragmento de ecossistema é necessário conhecer a ‘arte’ e o recorte a ela
imposto pelas ‘molduras’. E nossa arte é tendenciosamente espacial e heterogênea, com
padrões fundamentalmente vegetais.
3. O estado da arte – ou o que se sabe sobre os
determinantes de riqueza e diversidade em
comunidades naturais?
3.1. Teoria de nicho e heterogeneidade de habitats
Há muito se reconhece, em Ecologia, que cada espécie ocupa um ambiente e
desempenha uma função. Os termos referentes a essas propriedades formalizaram-se no
início no século XX, um referente ao espaço e outro ao papel das espécies no espaço,
respectivamente, habitat e nicho (WHITTAKER et al., 1973). Os dois conceitos, por
serem relacionados, promoveram muita ambiguidade e debates, de forma que várias
sínteses e revisões foram feitas (WHITTAKER et al., 1973; LEIBOLD, 1995; HALL et
al., 1997; KOLASA; WALTHO, 1998; PETREN, 2001; KEARNEY, 2006), buscando
uniformizar as definições e, assim, melhor operacionalizar as pesquisas no tema.
Habitat e nicho começaram como conceitos, mas rapidamente evoluíram para
hipóteses explicativas da diversidade biológica, a partir de uma dedução simples:
quanto mais nichos e habitats, mais espécies (SHMIDA; WILSON, 1985). Esta
dedução, descrita inicialmente com referência ao nicho por MacArthur e Levins (1967),
pode ser sintetizada como: o número total de espécies competindo que podem coexistir
é proporcional à amplitude ambiental dividida pela amplitude de nicho das espécies.
Roughgarden (2009) utiliza a metáfora da ‘prateleira de livros’ para explicar a hipótese:
as espécies são livros, com suas larguras variadas (nichos) e a riqueza resultante
depende do número de livros que a prateleira (ambiente) pode acomodar. Dito de outra
forma, em referência aos habitats, tem-se: habitats são distribuídos de forma
heterogênea no espaço, e porque diferentes espécies necessitam de diferentes habitats,
quanto mais habitats, mais espécies (TURNER; TJØRVE, 2005). Cabe destacar a

18
importância da competição na estruturação das comunidades, cujos conceitos e
mecanismos - não discutidos neste texto - evoluíram paralelamente ao conceito de nicho
e ditaram muito do que foi pesquisado a partir da década de 1920, com o
desenvolvimento dos modelos de V. Volterra, A.J. Lotka e G.F. Gause
(HUTCHINSON, 1957; PETREN, 2001).
A partir das décadas de 1960 e 1970, a relação entre a co-existência de espécies
e a diversidade de nichos e habitats, além da habilidade das espécies na partição de tais
nichos (LEIBOLD, 2008), passaram a ser aspectos fundamentais para o entendimento
da biodiversidade (PETREN, 2001; ROUGHGARDEN, 2009). Como os sistemas
naturais ocupam espaço e quase sempre exibem variabilidade no espaço que ocupam
(CANTRELL; COSNER, 2001), todo um corpo teórico fundamentou-se na
heterogeneidade e partição de recursos, distinguindo-se como teoria de nicho (CHASE;
LEIBOLD, 2003; KELLY et al., 2008; ROUGHGARDEN, 2009).
O que é nicho?
A definição de nicho engloba todas as interações possíveis de uma espécie com
o ambiente em que vive (PETREN, 2001). O termo foi criado e aplicado (mas não
definido) inicialmente por R.H. Johnson, em 1910 (GAFFNEY, 1975) e,
posteriormente, por Grinnel (1917), que detalhou as condições de vida de uma espécie
de ave e seus fatores limitantes, incluindo distribuição geográfica, condições climáticas,
relações tróficas, e o comportamento da espécie. Mais tarde, o termo nicho consagrou-
se no vocabulário de Ecologia através da obra Animal Ecology de Elton (1927), na qual
se cunhou a expressão de nicho como o ‘papel’ da espécie na comunidade (LEIBOLD,
1995). Numa reunião de biologia quantitativa, Hutchinson (1957) formalizou a
definição mais clássica de nicho, redefinida por várias outras vezes por Whittaker et al.
(1973), Leibold (1995), Kolasa e Waltho (1998), Petren (2001) e Chase e Leibold
(2003), para citar algumas, cada uma trazendo uma ênfase especial no enfoque espacial,
funcional ou em ambos.
Classicamente, define-se nicho como um espaço n-dimensional, de n variáveis
físicas e biológicas, que completamente definem as propriedades ecológicas da espécie,
constituindo um hiper-volume ou nicho fundamental para uma espécie S, sendo que as
dimensões efetivamente ocupadas por S definem seu nicho realizado (HUTCHINSON,
1957). Por definição, portanto, o nicho é um atributo de uma espécie e sua população.

19
Em 1973, Whittaker et al. lamentaram que os termos nicho e habitat eram utilizados de
forma confusa, pois o aspecto n-dimensional vinha sendo aplicado aos dois conceitos,
indiscriminadamente. Ao revisarem os termos, definiram que o conceito clássico de
nicho (HUTCHINSON, 1957) envolve um componente funcional e um espacial, e esse
espacial seria o habitat, embora Hutchinson não tenha definido habitat propriamente.
Para Whittaker et al. (1973), habitat pode ser definido para uma espécie ou comunidade,
porém nicho é propriedade da espécie, e abrange as variáveis que descrevem o seu papel
na comunidade. Leibold (1995) identificou duas vertentes nas definições: a que envolve
requisitos ambientais e a que envolve o impacto dos organismos nos recursos
disponíveis, chamando-as, respectivamente, de nicho de requisitos (esse englobando o
habitat) e nicho de impacto, que juntas formariam o nicho total. Para Petren (2001), os
conceitos de nicho e habitat se desenvolveram simultaneamente desde os trabalhos de
Grinnell (1917) e convergiram ao longo do tempo, de forma que habitat e seus
gradientes associados se tornaram um subconjunto de nicho.
Em síntese, as n-dimensões que definem as condições de uma espécie pertencem
a dois componentes: um espacial e outro funcional. Nicho pode se referir apenas ao
componente funcional (WHITTAKER et al., 1973) ou à soma dos dois (GRINNELL,
1917; HUTCHINSON, 1957; LEIBOLD, 1995; PETREN, 2001). As dimensões
espaciais, se integradas ao nicho, pertencem ao subconjunto habitat (PETREN, 2001) ou
nicho de requisitos (LEIBOLD, 1995); se complementares ao nicho, formam o habitat
propriamente dito (WHITTAKER et al., 1973, 1975).
Analisando o desenvolvimento do conceito de nicho, extensamente revisado por
Chase e Leibold (2003), chama à atenção o fato que à medida que a teoria foi sendo
desenvolvida, mais clara ficou a dificuldade em se descrever o nicho realizado ou total
de uma espécie no mundo real. Hutchinson (1957) já esclarecia que a sua definição era
claramente uma abstração e que a ordenação de todas as variáveis era impossível. Para
lidar com esta limitação à pesquisa empírica, muitos ecólogos enveredaram pela criação
de modelos matemáticos que representassem simplificações do mundo real e
permitissem testar a teoria, baseando-se nos mecanismos de competição por nicho (p.
ex. MacArthur e Levins 1967) ou na heterogeneidade ambiental (AUSTIN et al., 1990;
KOLASA; WALTHO, 1998) como fatores explicativos da coexistência de espécies. Os
modelos de nicho são hoje abundantes na literatura (AUSTIN, 2007) e intensamente
utilizados para desenvolver hipóteses sobre os limites de distribuição e prever o

20
comportamento futuro de espécies a partir de algumas dimensões de nicho conhecidas,
tais como recursos, variáveis espaciais ou fisiológicas (KEARNEY, 2006; KEARNEY;
PORTER, 2009).
Visando a operacionalização da pesquisa empírica, Hutchinson (1957) propôs
definir um nicho restrito contendo apenas variáveis nas quais as espécies competem,
enquanto outros recomendaram a construção do nicho básico para uma espécie, a partir
de alguns fatores, como alimento, micro-habitat, características físicas e requerimentos
reprodutivos para animais (PETREN, 2001), ou topografia, luz e variáveis edáficas para
espécies vegetais (WHITTAKER et al., 1973). Em alguns casos, os modelos propostos
foram testados com dados empíricos, o que permitiu confrontar curvas de abundância de
espécies entre si (MACARTHUR, 1957; KELLY et al., 2008; LEVINE;
HILLERISLAMBERS, 2009) ou com as dimensões de nicho escolhidas. Para aplicar
este teste às plantas, o grupo de M.P. Austin, na Austrália, escolheu cinco variáveis
ambientais (precipitação, temperatura, radiação, litologia e topografia) testando-as como
nicho realizado de duas a cinco espécies de Eucalyptus, o que permitiu determinar a
largura e sobreposição de nicho das espécies testadas (AUSTIN et al., 1990; AUSTIN et
al., 2009). Note-se que as variáveis espaciais (habitat) estão, via de regra, incluídas na
lista reduzida de dimensões a serem descritas. Esta abordagem reducionista está
provavelmente apoiada no entendimento que as variáveis abióticas fornecem as
condições para as variáveis bióticas (por exemplo, espaço limita competição), na
opinião crítica de Mitchell (2005).
Apesar de considerada em crescente desuso para alguns, principalmente após o
surgimento da Teoria Neutra Unificada da Biodiversidade e Biogeografia (HUBBELL,
2001), discutida adiante, e seus seguidores “neutralistas” (LEIBOLD, 2008), a
existência do nicho foi recentemente testada em comunidades de plantas anuais, através
da utilização da predição de modelos e pesquisa empírica (LEVINE;
HILLERISLAMBERS, 2009). No experimento, registrou-se, após duas gerações, uma
diversidade 50% maior em comunidades não manejadas, nas quais as diferenças de
nicho aconteciam, do que nas comunidades controladas para essas diferenças,
fornecendo fortes evidências da importância do nicho na manutenção da diversidade.
O que é habitat?
Após descrever nicho, aceitamos que, independentemente do conceito espacial,
funcional ou uma mistura de ambas (WHITTAKER et al., 1973; LEIBOLD, 1995),

21
habitat é parte fundamental ou complementar do nicho, fisicamente possível de
descrever na forma de variáveis ambientais espaciais, e típico de uma população ou de
uma comunidade, cujas populações tenham necessidades semelhantes. Apesar de
parecer simples de definir, constituindo um paradigma fundamental da ecologia básica e
aplicada (MITCHELL, 2005), a terminologia relacionada ao conceito de habitat é
imprecisa e ambígua (HALL et al., 1997), principalmente quando se considera o grupo
biológico ao qual o conceito se aplica. Enquanto alguns autores definem habitat para um
organismo específico, habitat pode significar, para outros, cobertura vegetal nativa ou
selvagem (LINDENMAYER; FISCHER, 2007) ou simplesmente onde um organismo
pode ser encontrado na natureza; e sua abrangência, em maior escala, já foi comparada
aos limites de bioma (PETREN, 2001).
Numa tentativa de unificar esta terminologia, Hall et al. (1997) definiram habitat
como “as condições e recursos presentes em uma área, que produzem ocupação por um
dado organismo – incluindo sobrevivência e reprodução; habitat é específico do
organismo (animal ou planta), e relaciona a presença de uma espécie, população ou
indivíduo às características físicas e biológicas de uma área”. Petren (2001) e Kolasa e
Waltho (1998), tal qual Whittaker et al. (1973), estenderam o conceito à comunidade,
no chamado habitat de comunidade (KOLASA; WALTHO, 1998), já que esta partilha
condições gerais que se aplicam a muitas espécies, coincidindo aproximadamente com
porções homogêneas do ambiente (HUTCHINSON, 1957).
Na sua obra, Kolasa e Waltho (1998) apresentaram uma definição objetiva de
habitat: um conjunto de dimensões descritivas do mundo físico, relevantes para a
ecologia, cujos valores interagem com uma espécie e variam no espaço e no tempo.
Propuseram, ainda, que a caracterização de habitat seja feita hierarquicamente, partindo
de unidades maiores (o habitat de comunidade) para subunidades, essas representando a
‘percepção’ das espécies. Os passos mais importantes em um estudo de habitats e
comunidades seriam (KOLASA; WALTHO, 1998; MITCHELL, 2005): 1) a
identificação de dimensões de habitats relevantes para as espécies presentes, 2) a
identificação de atributos das populações que quantifiquem a influência dos habitats,
tais como abundância e crescimento e 3) a quantificação e correlação adequadas dessas
variáveis.
As idéias integrando heterogeneidade de habitats e diversidade biológica têm seu
nascimento na biogeografia do século XIX, que reconheceu ser o mundo heterogêneo; a

22
expansão dessas idéias, porém, só aconteceu na década de 1960, como uma oposição ao
paradigma de equilíbrio e homogeneidade reinante na ecologia daquela época (WIENS,
2000). Na escala de comunidades, Williams (1964) propôs que a heterogeneidade de
habitats explica a correlação entre área e riqueza específica, criando o que viria a se
chamar “Hipótese da Diversidade de Habitats”. A partir daí, muitas pesquisas
procuraram analisar esta relação, que se mostra positiva na maioria das vezes
(TURNER; TJØRVE, 2005).
A heterogeneidade de habitats existe e, em teoria, pode ser mensurada em várias
escalas. Objetivando viabilizar as pesquisas, especialmente na Ecologia Vegetal, R.H.
Whittaker desenvolveu, na década de 1960, a “Análise de gradientes” (WHITTAKER;
NIERING, 1965; WHITTAKER, 1967), que consiste no estudo das relações de
populações e comunidades ao longo de gradientes ambientais, reduzindo o espaço n-
dimensional a poucas dimensões em um sistema de coordenadas, no qual a distribuição
das espécies será representada (WHITTAKER et al., 1973). Em um gradiente de
habitats, considerando que as espécies evoluem para ocupar diferentes posições, é
possível medir a influência do gradiente na comunidade através de descritores de
diversidade, as chamadas diversidade alfa, tomada em um habitat no gradiente, e a
diversidade beta, medida entre habitats (WHITTAKER, 1972; HALVORSEN;
EDVARDSEN, 2009).
Heterogeneidade de habitats e comunidades vegetais
O desenvolvimento da teoria de nicho e habitats se deu basicamente por
pesquisadores da Ecologia Animal (GRINNELL, 1917; HUTCHINSON, 1957;
WILLIAMS, 1964; MACARTHUR; LEVINS, 1967; LEIBOLD, 1995; HALL et al.,
1997; PETREN, 2001). Esses ecólogos, mesmo quando explicitamente intencionado,
não estenderam com clareza a sistematização de habitat para as espécies vegetais. Por
exemplo, Hall et al. (1997) assumiram um olhar fixamente tendencioso do “ponto de
vista animal” (aves e mamíferos, basicamente) para as definições das diversas variáveis
relacionadas ao habitat, tais como uso (“a forma que um animal usa...”), disponibilidade
(“acessibilidade por animais...”) e seleção (“decisões comportamentais feitas por um
animal...”).
Para plantas, dada a sua natureza modular, imóvel e a ausência de
comportamento, Bazzaz (1991) discutiu se a “seleção”, “escolha” e “especialização” de

23
habitat não seriam primeiramente desempenhadas pelo próprio habitat e em seguida
fixadas pelas plantas através de seleção natural. A plasticidade vegetal, que seria
equivalente ao comportamento nos animais, e os mecanismos de reprodução ampliariam
essa capacidade de escolha das espécies e atuariam na diferenciação de comunidades
(BAZZAZ, 1991). Com esses argumentos, lançou-se luz sobre o fato que as plantas
também selecionam (ou são selecionadas) por diferentes habitats e, mesmo sendo
indivíduos sésseis, a elas também se aplica a idéia de coexistência dependente da
variação espaço-temporal (WRIGHT, 2002) e da interação e competição espacial entre
organismos vizinhos (TILMAN, 1994).
Sob um ‘ponto de vista vegetal’, quais seriam, então, as dimensões mais
relevantes nas pesquisas que integram heterogeneidade de habitats e comunidades?
Historicamente, a análise de gradientes de Whittaker (WHITTAKER; NIERING, 1965;
WHITTAKER, 1967) representou uma exceção à predominância da Ecologia Animal
na formação da teoria de nicho. As dimensões escolhidas por Whittaker e seu grupo
para estudo da vegetação em vários gradientes topográficos norte-americanos foram
altitude, inclinação, material parental (rocha), solo e grau de perturbação. Embora visto
com críticas, principalmente devido à subjetividade dos métodos empregados para
caracterizar as dimensões de habitat (WILSON et al., 2004), a análise se destaca pelo
pioneirismo e influencia pesquisas até os dias de hoje, com crescente refinamento dos
métodos empregados (LOOKINGBILL; URBAN, 2005).
Nas duas últimas décadas, as dimensões de habitats para comunidades de plantas
não mudaram muito em relação àquelas utilizadas por Whittaker, ou seja, variáveis de
relevo e substrato, de forma que habitats são frequentemente definidos com base em
diferenças topográficas e edáficas. De fato, a topografia é considerada o fator abiótico
mais importante causando variação espacial e influenciando a estrutura e diversidade de
comunidades (OLIVEIRA-FILHO et al., 1994b; JOHNSON et al., 2003;
LUNDHOLM, 2009). Os padrões horizontais e verticais dos solos reconhecidamente
também criam oportunidades potenciais para diferenciação de nichos e coexistência
(FITTER et al., 2000; JOHN et al., 2007).
Em florestas tropicais, já há fortes evidências que as plantas lenhosas respondem
às diferenças de nicho associadas à topografia (WRIGHT, 2002), porém para um
percentual variável na comunidade, de pelo menos um terço das espécies. Clark et al.
(1999), examinando a relação entre diversidade de árvore e as dimensões topografia e

24
solo em uma floresta na Costa Rica, encontraram gradientes significativos e sugeriram
que 30% das espécies têm distribuição relacionada ao substrato. Uma parcela de 42%
das espécies mostrou preferência por pelo menos um habitat na comunidade florestal do
Barro Colorado, com destaque para as especialistas por habitats de encosta (HARMS et
al., 2001), enquanto 25% de espécies mostraram preferência por nichos topográficos na
Amazônia equatoriana (VALENCIA et al., 2004). Estudando a diversidade de
pteridófitas, Kessler e Lehnert (2009) também encontraram alta preferência das espécies
por habitats de encosta, nos quais há maior diversidade de microhabitats típicos de
pteridófitas, em contraste com a baixa riqueza de espécies e maior homogeneidade
ambiental no habitat de topo.
A natureza imóvel das plantas faz com que o seu conjunto assuma uma
fisionomia, que aparece como a representação física mais evidente do ecossistema. Por
este motivo, é comum na literatura se descrever habitat como sinônimo de vegetação ou
fitofisionomia, o que é, para alguns, inapropriado (HALL et al., 1997; MILLER, 2000;
FISCHER; LINDENMAYER, 2007; LINDENMAYER; FISCHER, 2007). De fato,
para animais, a percepção e uso dos habitats podem envolver múltiplas fisionomias ou
uma parte restrita de uma fisionomia. Para plantas, uma fisionomia homogênea pode ser
formada por grupos distintos de espécies ocupando habitats específicos com relação às
dimensões físicas, tais quais disponibilidade de água e topografia. É comum, porém,
usar a heterogeneidade fisionômica, resultante de perturbação, sucessão e idade, como
indicadora da heterogeneidade de habitats. Neste caso, as fitofisionomias atuam como
substitutas (surrogates) de idade, variáveis microclimáticas, níveis de perturbação ou
disponibilidade de água (PITMAN et al., 1999). Como resultado, aceitando-se que a
medida direta de uma dimensão de habitat (primária) fornece uma medida indireta de
outra (secundária), numa relação de substituição nem sempre quantitativa, distinguem-
se dois tipos de estudos: 1) aquele nos quais dimensões primárias substituem dimensões
secundárias (PITMAN et al., 1999) e 2) aquele que envolve dois componentes: a
relação dimensão primária - dimensão secundária e a relação habitat - comunidade
vegetal (WEBB; FA'AUMU, 1999; KUBOTA et al., 2004; ZANNE; CHAPMAN,
2005).
Exemplo típico do primeiro grupo, Pitman et al. (1999) estudaram a distribuição
de árvores na Amazônia peruana em quatro fitofisionomias (ou quatro habitats, na
definição dos autores), registrando um percentual de até 26% das espécies com

25
especificidade por habitat. Webb e Fa'aumu (1999) também consideraram floresta
madura e em regeneração como habitats distintos ao estudarem a comunidade arbórea
na ilha de Samoa. O estudo conjugou a fitofisionomia ao substrato (sem o amostrar),
escolhendo dois tipos de floresta madura de acordo com o tipo de solo, e permitiu
registrar 29% das espécies com distribuição agregada em um habitat específico e pelo
menos 25% com distribuição preferencial em floresta madura ou secundária. Zanne e
Chapman (2005) consideraram habitats quatro tipos de cobertura de copa ao estudarem
plantas lenhosas em Uganda. A sua caracterização, entretanto, exemplifica o segundo
grupo, pois, partindo da fisionomia, os autores amostraram variáveis de solo e luz em
cada sítio. Como resultados, encontraram alta similaridade microclimática entre as
fisionomias florestais e uma forte preferência de espécies (de até 72%) por habitats
específicos. Kubota et al. (2004) quantificaram e confirmaram o efeito da topografia nas
variáveis de solo em floresta no Japão, e da topografia e solo nas plantas, encontrando
que a heterogeneidade topográfica, mais que a edáfica, foi fortemente correlacionada à
riqueza e distribuição espacial da comunidade vegetal estudada.
A disponibilidade hídrica é também uma dimensão amplamente considerada,
mensurada de diversas formas, e para a qual as espécies demonstram seletividade. Por
vezes, pode ser substituída por uma descrição qualitativa do sítio, como por exemplo,
“áreas inundadas e áreas bem drenadas” (CLARK et al., 1999) ou “microhabitats
úmidos ao longo de riachos” (KESSLER; LEHNERT, 2009). A variação hídrica
temporal é, da mesma forma, uma medida de heterogeneidade, como descrito por Brose
(2001), que identificou quatro tipos de habitat em áreas úmidas na Alemanha, a
depender do período do ano e abrangência espacial da inundação, revelando ser a
heterogeneidade hídrica espaço-temporal altamente correlacionada aos padrões de
riqueza vegetal encontrados. Topografia também é usada como substituta de
disponibilidade de água, descrevendo-se sítios mais baixos e mais altos, ou adjacente ou
não adjacente aos rios, que revelam, geralmente, correlação com a riqueza e estrutura
vegetal sobrejacente (OLIVEIRA-FILHO et al., 1994a; 1994b; 1994c). Da mesma
forma, variáveis edáficas como classe de drenagem e saturação por bases quantificam
variações no regime de água, produzindo significativas relações espécie-habitat
(BOTREL et al., 2002).
O estudo da heterogeneidade de habitats tem seus problemas intrínsecos. Do
ponto de vista operacional, há o reducionismo obrigatório, a seleção de variáveis por

26
inferência ou conveniência e a frequente conversão de variáveis ambientais contínuas
em discretas (MITCHELL, 2005). Para Wiens (2000), considerando a necessidade de
medição e quantificação, podem-se listar até quatro formas de heterogeneidade, desde a
mais simples, que quantifica as dimensões ambientais em pontos numa área
qualitativamente semelhante até a espacialmente mais integradora, quando pontos são
convertidos numa área com referência espacial. A estrutura das comunidades também
impõe seus percalços, pois as interações entre variáveis atuam em diferentes escalas
espaço temporais (KOLASA; WALTHO, 1998; WRIGHT, 2002; CUSHMAN;
MCGARIGAL, 2004) e as medidas de abundância das populações podem ser resposta a
uma condição ambiental anterior ao momento das pesquisas (MITCHELL, 2005).
Apesar dessas limitações, muitos estudos bem conduzidos apontam para especialização
por habitats e correlação à heterogeneidade ambiental em várias comunidades,
demonstrando que os mecanismos subjacentes à teoria de habitats de fato operam em
comunidades vegetais (WRIGHT, 2002) em diferentes escalas espaciais (JOHNSON et
al., 2003). Ao revisar 52 estudos que compararam a diversidade vegetal e tiveram a
heterogeneidade espacial medida em pelo menos duas áreas, Lundholm (2009)
confirmou os padrões conhecidos: 78% dos estudos utilizaram topografia, variáveis
edáficas ou ambas como dimensões da heterogeneidade de habitats; houve grande
variabilidade de escala nos estudos; e, na maioria, houve relação positiva entre
heterogeneidade de habitats e riqueza ou diversidade vegetal.
Qual o papel das perturbações na criação de novos habitats?
Lamentando a ausência de boas teorias que explicassem a diversidade em
florestas tropicais, Ricklefs (1977) propôs a “Hipótese da Heterogeneidade Ambiental”,
na qual sugeriu que a principal força causadora da heterogeneidade nessas florestas seria
a perturbação e formação de clareiras. A hipótese relaciona a formação de clareiras às
alterações de propriedades microclimáticas e edáficas e, consequentemente, à
regeneração florestal nestas condições. Esta relação entre clareiras, novos habitats e
diversidade foi explicada por Connell (1978) como causada pelo não-equilíbrio
ocasionado por perturbações de frequência e intensidade variáveis, na chamada
“Hipótese do Equilíbrio Intermediário”. Denslow (1987) progrediu ainda mais em
direção a uma síntese teórica da relação entre clareiras e diversidade em florestas
tropicais. A partir de criteriosa revisão, observou que a maioria das árvores tropicais

27
teria alguma dependência de clareiras para crescimento e reprodução, e o
estabelecimento de plântulas e juvenis dessas espécies estaria limitado à quantidade e
duração da radiação. Com base nesta propriedade, as espécies poderiam ser organizadas
em um gradiente de disponibilidade de luz ou em grupos típicos de clareiras com
diferentes tamanhos e regimes de radiação. As clareiras seriam, então, responsáveis pelo
aumento da heterogeneidade de habitats para espécies dependentes de luz e temperatura.
Alternativamente, a maior diversidade de plantas em clareiras estaria ligada à maior
densidade de indivíduos recrutados nesses ambientes (DENSLOW, 1995).
Ainda não há um consenso se a abertura de clareiras atua criando uma
heterogeneidade suficientemente explicativa da diversidade vegetal. Hubbell et al.
(1999) testaram o papel da perturbação por mais de 13 anos em uma floresta tropical no
Panamá e não registraram relação entre a riqueza de espécies lenhosas e presença ou
densidade de clareiras. Já Molino e Sabatier (2001) aplicaram o teste em uma floresta
secundária na Guiana Francesa e encontraram forte relação entre a presença de clareiras
e diversidade, afirmando que os novos habitats criados pela alteração nos regimes de luz
em clareiras não são efêmeros, mas permanecem por décadas após a abertura inicial.
Em estudo recente, Bongers et al. (2009) analisaram o papel das perturbações
em florestas úmidas e secas em Gana, a partir de um extenso banco de dados. Apesar de
terem confirmado uma influência das perturbações na riqueza de espécies, concluíram
que este papel seria mais importante em florestas mais secas. Na úmida floresta
amazônica, porém, a importância das clareiras naturais como geradoras de habitats de
regeneração foi recentemente comprovada. Comparando sítios de floresta primária a
áreas contendo clareiras de diversos tamanhos, Chambers et al. (2009) encontraram
fortes diferenças estruturais e maior diversidade florística nas áreas naturalmente
perturbadas, diferenças essas que seriam melhor percebidas em maiores escalas. A
perturbação seria, então, uma dimensão temporal criando padrões de heterogeneidade
espacial em algum grau (WIENS, 2000; PEREIRA et al., 2007), através de eventos
estocásticos percebíveis em maiores escalas espaciais, que formam gradientes
ecológicos e influenciam a estrutura de comunidades (CHAMBERS et al., 2009).
Um esclarecimento: habitat na Ecologia de Paisagens
A heterogeneidade espacial sempre foi um tema presente nos estudos de
sistemas ecológicos. Na década de 1980, surgiu um novo ramo da Ecologia dedicado ao

28
entendimento das causas e consequências da heterogeneidade, integrando escala espaço-
temporal e ferramentas de geoprocessamento ao estudo de padrões e processos
ecológicos - a Ecologia de Paisagens (TURNER, 2005b). Com os atributos de ser uma
disciplina espacial e visual, a Ecologia de Paisagens agrega elementos da Ecologia e da
Geografia, com abordagens ligeiramente distintas em cada uma (METZGER, 2001). A
abordagem ecológica de paisagens define as pesquisas através da “perspectiva do
organismo estudado”, de forma que a paisagem passa a ser uma abstração da
heterogeneidade em qualquer escala, assumindo uma configuração diferente a depender
da espécie (PICKETT; CADENASSO, 1995; HAILA, 2002).
Neste contexto ecológico e na escala da espécie, o termo habitat naturalmente se
incorporou ao vocabulário e pesquisas em paisagens. Fragmentação, configuração,
abundância, perda, conexão, disponibilidade e qualidade de habitats, por exemplo, são
expressões abundantes nos textos básicos da disciplina (PICKETT; CADENASSO,
1995; METZGER, 2001; TURNER, 2005a) ou em artigos de conservação (HAILA,
2002; VILLARD, 2002; LINDENMAYER; FISCHER, 2007; LAURANCE, 2008).
Seus significados, porém, correspondem ao habitat na Ecologia de Populações ou
Comunidades?
Considerando as origens da Ecologia de Paisagens, o termo habitat ainda tem, na
disciplina, um uso confuso, em função da escala e abordagem do estudo. Quando
trabalhando na escala de espécie (abordagem ecológica), habitat é usado como sinônimo
de território ou área de uso, configurando uma unidade de paisagem no mosaico
descrito. Esta aplicação é semelhante ao habitat revisado neste texto, porém inclui
apenas dimensões espaciais e horizontais, tais como tamanho, perímetro, forma e
contexto, traçando-se a área de sítios com diferentes condições de exploração para
diferentes espécies (GARDNER; ENGELHARDT, 2008). A interação espécie - habitat
nesta abordagem refere-se ao modo como a espécie interage com a área ocupada pelo
habitat e seu contexto na configuração da paisagem. Para descrição desta relação, foi
criado um índice de “especificidade por habitat”, que é uma expressão quantitativa da
contribuição das manchas de habitat à riqueza específica na escala da paisagem
(HALVORSEN; EDVARDSEN, 2009).
Na visão mais geográfica, particularmente usada em estudos de fragmentação e
conservação, definem-se na paisagem unidades de habitat e não-habitat para uma
determinada fitofisionomia (METZGER, 2001), sendo habitat um sinônimo de

29
cobertura vegetal ou forma de uso da terra, que é determinado pela área e contexto de
sítios habitáveis em uma matriz inabitável (GARDNER; ENGELHARDT, 2008). É
necessário cautela, então, ao inventariar pesquisas sobre heterogeneidade de habitats,
pois, em estudos de paisagem, o termo pode ter um sentido distinto, significando
cobertura do terreno e não exatamente relacionado à população ou comunidade
(LINDENMAYER; FISCHER, 2007). Neste caso, está-se descrevendo a
heterogeneidade de manchas na paisagem e não a heterogeneidade de habitats
propriamente ditos.
3.2. A neutralidade
Em oposição ao paradigma clássico de diversidade biológica baseado nas idéias
de nicho, uma nova escola de pensamento fundamentada na idéia que as espécies são
ecologicamente idênticas surgiu, já no século XXI, a partir da publicação da Teoria
Neutra Unificada da Biodiversidade e Biogeografia ou, simplesmente, Teoria Neutra
(HUBBELL, 2001). Neutralidade, nas palavras de Hubbell (2001), significa
equivalência ecológica de todos os indivíduos numa comunidade, ou seja, todos têm a
mesma probabilidade de nascer, morrer, migrar ou até de sofrer especiação.
Para estruturar a teoria, Hubbell partiu da sua própria experiência em florestas
tropicais, da proposta de neutralidade construída na Teoria da Biogeografia de Ilhas
(MACARTHUR; WILSON, 1967), da observação cautelosa de vários modelos
descritivos da abundância de espécies em comunidades e vislumbrou uma possível
analogia ecológica com os modelos de deriva em genética de populações (CHASE;
LEIBOLD, 2003; CASSEMIRO; PADIAL, 2008). Observou, então, faltar uma teoria
explicativa (não descritiva) de comunidades, que tivesse valor preditivo e que incluísse
processos de dinâmica populacional. Sua teoria propôs, assim, uma nova distribuição de
abundâncias de espécies, baseada no tamanho da comunidade, nos padrões
populacionais e nas taxas de migração, extinção e especiação, aplicada em várias
escalas espaciais e temporais.
A teoria e seus modelos neutros trazem uma visão radicalmente diferente das
comunidades ecológicas e uma nova forma de explicar a diversidade de espécies
(BELL, 2001; HUBBELL, 2001; CHAVE, 2004), o que motivou uma busca por
padrões empíricos que confirmassem a neutralidade em comunidades e uma reavaliação

30
das interpretações de diversidade, atraindo seguidores e críticos. Muitas críticas advêm
do fato da teoria: (i) afirmar que as idéias de nicho não são essenciais à coexistência, (ii)
defender que não há equilíbrio de espécies, mas sim de indivíduos, nas comunidades e
(iii) por valorizar a estruturação de comunidades por eventos aleatórios de dispersão e
extinção (HUBBELL, 2001, 2006; CASSEMIRO; PADIAL, 2008). Contra essas
premissas, Chase e Leibold (2003) argumentam que há sim demonstrações empíricas de
que existe equilíbrio nas comunidades, como um retorno após perturbação, e que muitos
padrões empíricos de diversidade não se ajustam às predições do modelo de
neutralidade. Ricklefs (2006a) critica ainda que a aplicação da teoria a amplas escalas
ignora que os ambientes não são uniformes e que as espécies respondem a gradientes
ambientais através de ajustes populacionais e, assim, não são ecologicamente
equivalentes.
Desde a publicação da teoria, vários testes vêm comparando os modelos de
nicho e neutros na estruturação de comunidades, gerando um caloroso debate na
literatura, motivado principalmente pela controversa idéia da equivalência ecológica
entre espécies (HUBBELL, 2006). Na floresta amazônica, Gilbert et al. (2006) testaram
a teoria com base em um banco de dados de vários censos de indivíduos arbóreos,
coletado em 30 parcelas permanentes de 1 ha, em fragmentos de vários tamanhos e
floresta contínua. No teste, empregaram-se as premissas básicas da teoria, entre elas:
todas as árvores são fundamentalmente equivalentes, a densidade da comunidade é
constante, e o ambiente é uniforme. Aplicando um modelo neutro derivado da genética
de populações, Gilbert et al. (2006) obtiveram predições de vários parâmetros da
comunidade e compararam a dados reais. Como resultados, obtiveram ajustes acurados
para os parâmetros de extinção e mortalidade, porém não precisos para mudanças na
composição de espécies, indicando a atuação de processos de nicho, ou seja, que as
espécies respondem diferentemente às mudanças ambientais.
Gilbert e Lechowicz (2004) testaram a teoria neutra e de nichos ambientais na
comunidade vegetal de sub-bosque em uma floresta no Canadá. O estudo partiu da
seguinte hipótese: havendo equivalência ecológica das espécies, as predições do modelo
neutro serão semelhantes à estrutura da comunidade encontrada na natureza,
independentemente da heterogeneidade ambiental, sendo o único fator de influência a
distância de dispersão entre sítios. Os resultados mostraram fortes evidências da
influência de nichos (variáveis edáficas, topográficas e microclimáticas) na estruturação

31
das comunidades e não confirmaram a neutralidade, demonstrando que testes
apropriados da teoria neutra requerem a incorporação (e contestação da influência) de
parâmetros de heterogeneidade ambiental, dispersão e história evolutiva das espécies.
Os resultados obtidos nos exemplos acima corroboram parcialmente ou não
corroboram a teoria neutra, demonstrando que são pertinentes as críticas existentes à
equivalência ecológica (CHASE; LEIBOLD, 2003; LEIBOLD; MCPEEK, 2006;
RICKLEFS, 2006b). Kelly et al. (2008) testaram a equivalência ecológica versus o
papel do nicho na coexistência de espécies e levaram em consideração a história
evolutiva das espécies. Para tanto, escolheram pares de espécies arbóreas em uma
floresta tropical no México, de acordo com similaridades evolutivas (26 pares
pertencentes ao mesmo gênero e 20 à mesma família), o que indicaria possível
similaridade de nichos. Compararam, então, os dois grupos entre si quanto à abundância
relativa nos pares, e compararam cada grupo à distribuição de uma seleção aleatória de
espécies na comunidade. Na hipótese da neutralidade, as distribuições de abundâncias
dos pares relacionados seriam semelhantes nos dois grupos e com a amostra aleatória,
mas isso não ocorreu. Apenas foram semelhantes as distribuições entre pares de uma
mesma família e amostra aleatória, demonstrando haver alguma separação de nicho
entre espécies de um mesmo gênero.
Ao revisar evidências empíricas que confirmassem ou não a teoria, com ênfase
em comunidades vegetais, Chave (2004) encontrou que a teoria responde a alguns
padrões básicos de diversidade, enquanto mostra pouco suporte para outros. Isso o
levou a afirmar que o desenvolvimento ideal de uma teoria de comunidades buscaria a
fusão da teoria neutra com os processos de nicho. Esse ponto é também defendido por
Leibold e McPeek (2006), para quem equivalência total entre espécies é uma abstração
e os modelos neutros e de nicho não são mutuamente exclusivos. Para Leibold (2008),
resultados de estudos como o de Kelly et al. (2008) indicam que ambas as teorias
contribuem, de alguma forma, para explicar a estrutura de comunidades, que seria
dependente da separação de nichos pelo menos em uma fração das suas espécies.

32
4. Os limites das molduras - ou o que se sabe sobre
riqueza e diversidade em comunidades naturais
fragmentadas?
4.1. A relação espécie-área
A relação entre área e número de espécies, ou relação espécie-área, foi descrita
inicialmente no século XIX pelo biogeógrafo De Candolle (SCHEINER, 2004). Apenas
na década de 1920, a partir de estudo de comunidades de plantas, Arrhenius (1921) e
Gleason (1922; 1925) formalizaram a existência da curva espécie-área e sua formulação
matemática. Ambos buscaram, embora com formulações diferentes e em escalas
espaciais distintas, descrever o aumento progressivo no número de espécies com
aumento da área amostral (SCHEINER, 2003).
Segundo Gardner e Engelhardt (2008), os modelos relação espécie-área são
fáceis de formular, porém difíceis de explicar, não por falta de hipóteses, mas por falta
de testes que permitam distinguir os seus diversos fatores causais. Connor e McCoy
(1979) listam que três principais idéias explicam o aumento da riqueza com a área. A
primeira delas, a “hipótese da área per se”, é neutra com relação às espécies e
diversidade de nichos e habitats, explicando a riqueza como uma função da área
(PRESTON, 1960). A teoria da biogeografia de ilhas, discutida adiante, baseou-se nesta
hipótese, vinculando à área a atuação dos processos de imigração e extinção
(MACARTHUR; WILSON, 1967). O processo de imigração seria dependente da
distância da área em questão à fonte de espécies, enquanto a extinção seria diretamente
proporcional à área e ao tamanho das populações ali residentes – quanto maiores,
menores as probabilidades de extinção (CONNOR; MCCOY, 1979). Assim sendo, pela
hipótese da área per se o número de espécies aumentará com a área mesmo em
ambientes homogêneos (KALLIMANIS et al., 2008).
A segunda suposição explicativa da relação espécie-área, a “hipótese da
diversidade de habitats” formulada por Williams (1964), é baseada nas idéias de nicho e
relaciona o aumento da riqueza à maior diversidade de habitats amostrada em áreas
maiores. Por esta hipótese, o número de espécies aumenta com a área apenas se mais
habitats forem amostrados. Enquanto a área per se é mais facilmente comprovada,

33
considerando a sua neutralidade, um menor número de estudos experimentais
demonstrou a influência da diversidade de habitats na relação espécie-área, devido à
própria correlação da diversidade de habitats com área, e à dificuldade em definir
dimensões de habitats relevantes que produzam resposta pela comunidade estudada
(RICKLEFS; LOVETTE, 1999).
A terceira hipótese, proposta por Connor e McCoy (1979) como alternativa às
outras duas, é a chamada “hipótese da amostra passiva”, que explica que mais espécies
são amostradas em maiores áreas porque nessas são feitas maiores amostras. Esta
hipótese, diferentemente das outras duas que se baseiam em processos biológicos, é
baseada puramente na amostragem e, assim sendo, também é neutra com relação ao
nicho ou aos processos populacionais. A idéia da relação espécie-área como resultante
da amostragem foi abordada por Turner e Tjørve (2005) como “hipótese da amostragem
incompleta”, relacionando o aumento da riqueza não só à área, mas também ao número
de indivíduos amostrados e ao posicionamento aleatório das espécies na área.
Schoereder et al. (2004) testaram o efeito amostral em comunidades arbóreas e não
encontraram comprovação desta hipótese, pois a relação espécie-área foi independente
do número de amostras empregadas em áreas com diferentes tamanhos. Ao registrarem
também um aumento da diversidade beta entre amostras numa mesma área, inferiram a
atuação da diversidade de habitats sobre a riqueza (SCHOEREDER et al., 2004). Turner
e Tjørve (2005) somam ainda às três hipóteses apresentadas, a “hipótese da
independência evolutiva”, perceptível quando a relação espécie-área é investigada em
escalas espaciais maiores, tais como províncias biogeográficas. Por não se aplicar à
escala temporal da fragmentação, esta hipótese não é considerada aqui como um quarto
fator explicativo da relação espécie-área.
Apesar das explicações da relação espécie-área ainda causarem considerável
debate (HOLT et al., 1999), sua existência já está comprovada na Ecologia, tanto
empiricamente quanto através de descrições matemáticas (ARRHENIUS, 1921;
GLEASON, 1925; PRESTON, 1960; CONNOR; MCCOY, 1979). Vários tipos de
relação já foram descritos na literatura, em número de três (HOLT et al., 1999) a seis
tipos diferentes de curvas (SCHEINER, 2003), a depender da característica da amostra e
da análise empregada. Em linhas gerais, as curvas são construídas a partir da riqueza
versus área amostral, dando origem a: curva tipo I, obtida de uma amostra única em um
único habitat; curvas tipo II e III, estimadas a partir de várias amostras contíguas (tipo

34
II) ou não contíguas (tipo III) em uma área; e tipo IV, baseada em uma amostra única
que cubra uma área inteira (HOLT et al., 1999; SCHEINER, 2003, 2004). De acordo
com Scheiner (2003; 2004), as curvas que descrevem a relação espécie-área são função
da diversidade alfa e beta (WHITTAKER, 1972), e têm várias aplicações em ecologia e
conservação, de forma que os modelos existentes são úteis tanto para descrição da
riqueza de uma amostra quanto para estimativa do número de espécies em uma área
maior que a amostra (KALLIMANIS et al., 2008).
4.2. A teoria da biogeografia de ilhas e sua adaptação aos
estudos de fragmentação florestal
Em 1967, MacArthur e Wilson publicaram seu célebre livro “A Teoria da
Biogeografia de Ilhas” (TBI), no qual reconheceram o valor da insularidade para a
biologia, tais como seu impacto na evolução e biogeografia. A teoria, considerada
neutra no sentido de equivalência ecológica das espécies (sensu HUBBELL, 2001),
assume que as espécies são essencialmente idênticas, e que as ilhas são idênticas nas
suas características, com exceção de área e isolamento (BROWN; LOMOLINO, 2000).
Assim, relaciona a riqueza de espécies residentes ao tamanho e grau de isolamento da
ilha, e à atuação de processos de dispersão, dependentes de parâmetros demográficos
das diferentes populações no contexto regional. A TBI trouxe muito da relação espécie-
área, mas incorporou o contexto insular e os processos de dispersão a esta relação
(HUBBELL, 2001).
No seu modelo clássico, MacArthur e Wilson (1967) se baseiam em cinco
premissas: há uma relação positiva entre a área de uma ilha e o número de espécies ali
encontrado; há um balanço entre os processos de imigração e extinção em ilhas; a
diversidade das biotas insulares representa um equilíbrio entre esses processos; há um
limite no número de espécies persistindo numa ilha; e os processos de imigração e
extinção são influenciados pelo tamanho e grau de isolamento das ilhas. A integração
dessas premissas levou ao modelo teórico proposto que, em síntese, prevê que ilhas
menores e mais isoladas têm menos espécies que ilhas maiores e mais próximas ao
continente (DEBINSKI; HOLT, 2000), o que foi exemplificado com dados empíricos na
própria obra e em várias pesquisas subsequentes.

35
Mais do que os processos, o padrão insular da relação riqueza-área-isolamento é
frequentemente comprovado na literatura. Visando testar a TBI em floras insulares,
destacamos alguns exemplos em variados números de ilhas e situações geográficas. Nos
Estados Unidos e Canadá, em ilhas com diferentes áreas, McMaster (2005) examinou a
riqueza de plantas e relacionou às variáveis de área e isolamento, confirmando as
predições da teoria: a riqueza foi positivamente relacionada ao tamanho da ilha e
negativamente relacionada à distância da ilha mais próxima. Da mesma forma, Panitsa
et al. (2006) examinou a composição florística em pequenas ilhas na Grécia e encontrou
ser a área o fator que mais explicou a riqueza insular, enquanto Kreft et al. (2008)
examinaram a diversidade de plantas em ilhas e áreas continentais e encontraram que a
variável área foi a determinante mais forte da riqueza de espécies vegetais insulares,
seguida por isolamento. Os princípios gerais da TBI se ajustaram também ao padrão
registrado em ilhas no Mar Báltico, por Hannus e von Numers (2008), que encontraram
a riqueza da flora vascular significantemente associada ao tamanho da ilha.
Como exceção ao padrão área-riqueza, MacArthur e Wilson (1967) notaram a
ocorrência frequente de um fato curioso: em ilhas pequenas, aquém de um certo
tamanho relativo às demais, não há variação da riqueza relacionada à área. Este padrão,
o chamado “Efeito da Pequena Ilha” (SIE na sigla em inglês) aparece comumente em
bancos de dados para vários taxa e o limiar abaixo do qual o efeito é detectável é menor
para plantas (LOMOLINO; WEISER, 2001), o que significa dizer que as variáveis da
TBI são explicativas até ilhas bem pequenas para comunidades vegetais.
Na obra original, MacArthur e Wilson (1967) já previam a aplicabilidade das
idéias por eles formuladas à acelerada quebra de habitats previamente contínuos. De
fato, uma das consequências da remoção da cobertura vegetal natural é que a vegetação
remanescente fica fragmentada na paisagem, o que lhes dá um caráter insular de
isolamento ou de habitat-ilha (SAUNDERS et al., 1991), habitat aqui empregado no
contexto de paisagem. A partir da década de 1970, enquanto o tema fragmentação ia se
tornando um dos principais e mais adotados para a ecologia e conservação (HAILA,
2002), a TBI inspirava muito dos estudos desenvolvidos sobre o tema nos anos
subsequentes à teoria, estimulando uma avalanche de pesquisas em ecossistemas
fragmentados (LAURANCE, 2008) e contribuindo para o desenvolvimento da Ecologia
de Paisagens (METZGER, 2001).

36
A TBI reúne, por um lado, elementos importantes de sistemas biológicos e
constitui uma contribuição fundamental à teoria ecológica, mas por outro lado ignora
vários aspectos cruciais para o estudo de biotas continentais. Muitos trabalhos têm sido
produzidos visando à sistematização de conceitos em fragmentação, incluindo a
adaptação dos estudos de ilhas aos fragmentos e paisagens fragmentadas (SAUNDERS
et al., 1991; LAURANCE, 1999a; DEBINSKI; HOLT, 2000; VILLARD, 2002;
FISCHER; LINDENMAYER, 2007; LINDENMAYER; FISCHER, 2007).
Considerando as diferenças entre ilhas e fragmentos, Watson (2002) propôs uma
classificação de ecossistemas tipo-ilha para melhor operacionalizar a aplicação da TBI
aos estudos de fragmentação e permitir comparações e sínteses entre estudos. Foram
criadas, então, oito classes, de acordo com três aspectos: origem (ilha real versus
fragmento de ecossistema), contraste entre mancha e matriz (alto versus baixo) e tempo
de formação (até 200 anos versus mais de 200 anos). Testes da TBI em arquipélagos de
fragmentos florestais (manchas recentes e de baixo contraste com a matriz, de acordo
com a classificação de Watson 2002), embora geralmente testem mais o efeito da área
do que isolamento (ZUIDEMA et al., 1996), na maioria das vezes têm confirmado o
padrão de relação riqueza-área em diversas paisagens fragmentadas, como por exemplo,
em fragmentos florestais na Amazônia (LAURANCE et al., 2002), no estudo da
diversidade de árvores em 21 fragmentos florestais em Gana (HILL; CURRAN, 2003)
ou em 51 remanescentes florestais no Chile (ECHEVERRÍA et al., 2007). A forma dos
fragmentos, variável não considerada na TBI, também tem sido testada e exibe
correlação positiva com a riqueza, principalmente em áreas relativamente menores
(SAUNDERS et al., 1991), de forma que quanto mais regular o fragmento, maior a
riqueza (HILL; CURRAN, 2003). Há exemplos, porém, da não confirmação da relação
riqueza-área (DEBINSKI; HOLT, 2000), demonstrando que paisagens fragmentadas
exibem novos padrões e uma dinâmica típica, indo muito além das relações clássicas da
TBI (EWERS; DIDHAM, 2006).
Haila (2002) e Laurance (2008) discutiram a proporção em que a pesquisa em
fragmentação transcendeu a TBI, listando tópicos a serem considerados para o avanço
da pesquisa empírica em ecossistemas fragmentados, que seriam tão ou mais
importantes que as variáveis insulares básicas. Entre esses pontos, citam a relevância da
heterogeneidade da paisagem e a qualidade da matriz, termo típico do estudo de
paisagens continentais, análogo de água na TBI (CADENASSO et al., 2003a), mas nem

37
sempre inóspita como a água para ilhas reais. As qualidades da matriz, juntamente com
padrões espaciais da paisagem, incluindo o arranjo dos fragmentos e conectividade,
podem compensar o efeito da área e isolamento e facilitar os fluxos biológicos na
paisagem (METZGER, 1997; LAURANCE et al., 2002; DAMSCHEN et al., 2006),
sendo tão importantes para comunidades de plantas quanto as propriedades dos
fragmentos isoladamente. Isso porque, como explicam Murphy e Lovett-Doust (2004),
as plantas não têm uma percepção discreta da paisagem e respondem mais a gradientes
ambientais do que ao binário fragmento e matriz.
Para Laurance (2008), um dos aspectos mais importantes a ser considerado ao se
empregar a teoria insular é que a fragmentação é um processo não aleatório e os habitats
remanescentes ocupam, intencionalmente, as áreas menos privilegiadas para uso
humano e menos úteis na paisagem, tais como encostas íngremes e terras inundadas.
Adicionalmente, a TBI, por ser neutra com relação à ecologia das espécies e dinâmica
de suas populações (CANTRELL; COSNER, 2001; KADMON; ALLOUCHE, 2007),
considera apenas a riqueza e não a composição das espécies. Para Saunders et al.
(1991), a identidade das espécies é fundamental em estudos de fragmentação, já que
número de espécies não reflete a qualidade da biota, oportunamente ‘enriquecida’ por
espécies da matriz adjacente (DEBINSKI; HOLT, 2000). Quando a relação riqueza-área
não se confirma, por exemplo, a riqueza em fragmentos menores pode estar sendo
influenciada pela presença de espécies pioneiras, ruderais ou invasoras, principalmente
na fase inicial pós-fragmentação (JANZEN, 1983) ou em fragmentos sob forte
perturbação (SANTOS et al., 2007b). Uma mudança na composição de espécies e sua
relação com área do fragmento tem sido, de fato, reportada por vários estudos, que
demonstram haver, em menores remanescentes, alteração na estrutura de guildas de
plantas lenhosas, aumentando a importância de espécies ruderais (TABARELLI et al.,
1999), empobrecimento da comunidade vegetal, incorporação de exóticas e
desaparecimento de espécies raras (CAGNOLO et al., 2006) e declínio ou extinção
local de populações devido à alteração de processos de polinização e fecundidade
(LAURANCE et al., 2002).
Outro tópico importante a ser ponderado na aplicação da TBI à fragmentação diz
respeito à origem das biotas em ilhas e em fragmentos. As ilhas têm uma origem
geológica seguida de colonização primária e história evolutiva (processos
biogeográficos), podendo resultar em biotas muito distintas, enquanto fragmentos de

38
ecossistemas, por outro lado, são áreas remanescentes, nas quais persiste uma amostra
de espécies que já existia na área contínua (HAILA, 2002; WATSON, 2002). A biota
resultante em um fragmento, assim, não depende de história evolutiva biológica, mas
sim da história da paisagem e do tempo de acomodação da biota e esta paisagem
(SAUNDERS et al., 1991). Estudos de fato têm mostrado que a riqueza atual de
espécies exibe maior correlação com a estrutura de paisagens passadas do que com a
paisagem presente, como comprovado para a comunidade de árvores numa paisagem de
Mata Atlântica em São Paulo, Brasil (METZGER et al., 2009), o que leva à dedução
que pode ocorrer uma maior perda de espécies no futuro mesmo se a paisagem atual for
mantida (LINDBORG; ERIKSSON, 2004).
Em termos conceituais, o vocábulo fragmentação tem sido usado
indiscriminadamente para se referir a dois processos distintos, que geralmente atuam
juntos: a perda de área e fragmentação propriamente dita (FAHRIG, 2003;
LINDENMAYER; FISCHER, 2007). O modelo de ilhas se adapta aos estudos da
fragmentação propriamente dita (FISCHER; LINDENMAYER, 2007; LAURANCE,
2008), que diz respeito à subdivisão de áreas previamente contínuas, aumenta o número
de fragmentos e produz isolamento. Porém, muitas pesquisas não diferenciam esses dois
processos, dificultando a compreensão do declínio na biodiversidade em função da área
e isolamento como preconizado na TBI. Ao revisar vários trabalhos sobre fragmentação,
Fahrig (2003) observou que a perda de área, medida como o percentual de cobertura
remanescente na paisagem, teria um efeito mais forte na biodiversidade do que a
fragmentação propriamente dita. A separação desses dois processos ainda é um desafio
na pesquisa em fragmentação (LAURANCE, 2008), e a esses se soma ainda outro
processo chave ao entendimento dos efeitos da fragmentação na biodiversidade, o efeito
de borda (RIES et al., 2004).
4.3. O efeito de borda
Todo limite físico cria zonas de contato entre duas situações adjacentes distintas,
na forma de áreas de transição ou bordas abruptas (CADENASSO et al., 2003a). No
processo de fragmentação de ecossistemas terrestres, os novos limites dos fragmentos
passam a exibir condições diferentes da antiga condição contínua, surgindo uma faixa
de transição onde há interações entre o ecossistema fragmentado e o entorno, e o

39
resultado desta interação é o que tem se chamado “efeito de borda” (MURCIA, 1995).
Com a fragmentação acelerada de ecossistemas no último século e o consequente
aumento de habitats de borda (LAURANCE, 2000), o estudo dos padrões e processos
de influência da borda se tornou um tópico fundamental em pesquisas de ecologia e
paisagens (CADENASSO et al., 2003a; HARPER et al., 2005).
Considerando a importância das bordas nas paisagens atuais, várias sínteses têm
sido feitas, visando à sistematização de padrões gerais (MURCIA, 1995; LAURANCE
et al., 2001; CADENASSO et al., 2003a; STRAYER et al., 2003; RIES et al., 2004) e
construção de uma teoria unificadora (CADENASSO et al., 2003b). O que as pesquisas
e sínteses têm buscado responder pode-se resumir em: i) é possível uma teoria geral de
bordas ecológicas?; ii) é possível uma classificação de bordas e unificação de
conceitos?; iii) em que sistemas biológicos as bordas são relevantes?; iv) que grupos
biológicos e em que escala o efeito de borda exerce influência? v) qual a espessura da
zona de borda, ou qual a dimensão espacial e temporal da influência da borda? e vi) o
efeito de borda é deletério para a diversidade biológica?
Entre os avanços na resposta ao primeiro desafio, Cadenasso et al. (2003b)
evoluíram em direção a uma teoria unificadora de bordas, baseando-se numa estrutura
hierárquica entre componentes, fluxos e contrastes. Numa primeira aproximação de
classificação dos componentes, Strayer et al. (2003) observaram que as bordas na
natureza diferem em origem, estrutura espacial, função e dinâmica temporal. Ries et al.
(2004) avançaram ainda mais na proposição de um modelo de respostas ecológicas às
bordas, no qual reconheceram a multiplicidade do efeito de borda e identificaram quatro
mecanismos principais que determinam a abundância dos organismos e dinâmica nessas
condições, dois relacionados ao espaço e ambiente (distribuição espacial dos recursos, e
acesso dos organismos a recursos distintos nos habitats adjacentes) e dois relacionados
aos organismos, fluxos, e suas interações (movimentos de matéria, energia e
organismos, e interações entre espécies).
Quanto aos sistemas biológicos nos quais as bordas exercem influência,
fragmentos florestais são de inegável relevância ecológica e de compreensão
extremamente desafiadora, devido ao contraste microclimático entre floresta e entorno
aberto (HARPER et al., 2005; LAURANCE, 2008). A criação de uma borda numa
floresta gera, assim, novo habitat com condições microclimáticas e edáficas distintas
(DAVIES-COLLEY et al., 2000), definindo novos nichos no qual a comunidade se

40
reestruturará. Bordas florestais são classificadas, sensu Strayer et al. (2003) como
antropogênicas em origem, com espessura variável, sendo permeáveis ou semi-
permeáveis e dinâmicas. Para caracterização da estrutura e dinâmica nessas bordas, têm-
se utilizado como objetos de estudo variáveis bióticas diretas (características
demográficas e estruturais de plantas e animais), bióticas indiretas (interações
biológicas e reprodução) e condições abióticas (variáveis microclimáticas e edáficas,
substâncias químicas) (MURCIA, 1995). Essas variáveis podem ser empregadas para
descrever e avaliar o contraste entre ambientes de borda e interior ou, se medidas ao
longo de um gradiente, para avaliar a espessura ou distância de influência da borda.
Harper et al. (2005) revisaram uma coletânea de artigos sobre efeito de borda
amostrando comunidades de plantas em fragmentos florestais, e formularam uma
síntese fundamental à compreensão da influência das bordas em florestas.
Primeiramente, foi descrita a dinâmica da influência, distinguindo-se as respostas
primárias, incluindo os danos físicos e alterações microclimáticas imediatas após a
criação das bordas, e secundárias, incluindo os processos subsequentes de regeneração,
crescimento, reprodução e mortalidade. Em segundo lugar, foi possível distinguir que a
espessura da zona de borda em florestas no geral atinge até 50m, valor que já tinha sido
referido por Murcia (1995) e Ries et al. (2004) como recorrentes para plantas e
variáveis abióticas, podendo atingir distâncias maiores, a depender da variável testada e
da qualidade da matriz. Excepcionalmente, já foi registrada uma largura de borda de
500m, medidos através da distância de penetração de espécies indicadoras de
perturbação em florestas australianas (LAURANCE, 1991). As evidências mostram que
a distância de influência pode ser maior: nas respostas secundárias do que primárias
(HARPER; MACDONALD, 2002), em bordas mais antigas do que bordas recentes
(WILLIAMS-LINERA et al., 1998), para indivíduos de sub-bosque do que arbóreos
(HARPER; MACDONALD, 2001) e em bordas com vegetação adjacente aberta/ baixa
do que lenhosa (MESQUITA et al., 1999). Isso indica haver uma dinâmica de
desenvolvimento da zona de borda, que exibe, logo após criação do limite físico, grande
intensidade do efeito penetrando uma menor distância e, numa fase posterior de
expansão, um efeito menos intenso penetrando maiores distâncias (HARPER et al.,
2005).
A magnitude e a distância de influência da borda em um fragmento dependente
da sua área e forma (LAURANCE et al., 1998), aumentando a proporção de bordas à

41
medida que a área do remanescente diminui, levando a uma maior razão borda/ interior
e à redução do habitat interior, que pode até desaparecer (BÁLDI, 1999). Fragmentos
pequenos, desta forma, podem estar totalmente sob influência da borda, não contendo as
condições microclimáticas de interior que garantem a sobrevivência de espécies típicas
de floresta madura (MATLACK, 1993; YOUNG; MITCHELL, 1994). Porém, a
magnitude da influência da borda não é função apenas da distância de penetração e área
do fragmento, mas também da forma e qualidade das bordas (homogênea versus
heterogênea). Fernández et al. (2002) modelaram o efeito de borda considerando essas
variáveis e encontraram uma complexa interação, sugerindo que as variáveis forma do
fragmento e qualidade das bordas sejam levadas em consideração para estimar a
influência no remanescente como um todo.
Quanto às bordas terem efeito deletério na diversidade, especialmente de
plantas, o modelo de Ries et al. (2004) confirmou com consistente previsibilidade um
efeito negativo nas bordas de ecossistemas limitados por habitats externos de qualidade
inferior, com baixa disponibilidade ou desprovido de recursos. No nível do fragmento
isoladamente, o efeito negativo ocorre na forma alteração da estrutura da comunidade,
havendo diminuição na diversidade, perda de espécies climácicas e aumento de espécies
secundárias nas bordas (OOSTERHOORN; KAPPELLE, 2000; OLIVEIRA et al.,
2004; SANTOS et al., 2008; GOMES et al., 2009), além de alteração em processos de
regeneração, como chuva de sementes, que apresentam menor riqueza de espécies e
menor número de sementes grandes na borda (MELO et al., 2006). O efeito na
diversidade ocorre concomitantemente às alterações fisionômicas na borda,
representadas pelo aumento da densidade do estrato de sub-bosque (JOSE et al., 1996;
NASCIMENTO; LAURANCE, 2006; SCHEDLBAUER et al., 2007; SILVA et al.,
2008c; GOMES et al., 2009), diminuição da cobertura da copa (LAURANCE, 1991) e
altura do dossel (DIDHAM; LAWTON, 1999), bem como aumento na produção de
serrapilheira, principalmente pelo efeito de dessecação pelo vento (FEELEY, 2004;
SCHESSL et al., 2008).
Além da influência por fragmento, é importante analisar a influência das bordas
na paisagem como um todo. Estudos apontam, por um lado, que o efeito de borda pode
atuar em amplas escalas, produzindo vulnerabilidade nos ecossistemas, principalmente à
invasão de espécies exóticas (LAURANCE, 2000). Por outro lado, a manutenção de
habitats apropriados de interior em vários fragmentos pode ajudar na manutenção da

42
biodiversidade no nível da paisagem (WILLIAMS-LINERA et al., 1998; NELSON;
HALPERN, 2005), indicando que o contexto da paisagem importa para garantir a
cicatrização e enriquecimento de espécies nas bordas, na sequência da dinâmica de
respostas secundárias sensu Harper et al. (2005). Os componentes da paisagem que
podem auxiliar na manutenção da diversidade em longo prazo são plantações florestais,
proximidade a outros fragmentos e até mesmo árvores isoladas (KOLLMANN;
SCHNEIDER, 1999).
5. Conclusão: é possível integrar as idéias de
heterogeneidade, ilhas e bordas?
A pesquisa em comunidades fragmentadas reúne, como pano de fundo,
elementos de todas as teorias e sínteses aqui apresentadas. Porém, a maioria dos
estudos, sob influência da TBI, aborda número de espécies nos fragmentos como função
da área, independentemente dos tipos de habitats presentes nesta área (HAILA, 2002)
ou evitando as zonas de borda para minimizar a influência dessa variável (SANTOS et
al., 2007b; METZGER et al., 2009). A idéia de variabilidade espacial, apesar de
demonstrada e conhecida desde antes dos anos de 1970 (WILLIAMS, 1964), foi
negligenciada por algumas décadas e vista como um fator complicador, em favor da
homogeneidade assumida por conveniência ou simplificação (PICKETT;
CADENASSO, 1995).
Nos primórdios da descrição da relação espécie-área, Arrhenius (1921) já
reconhecia a influência da heterogeneidade e gradientes de habitats na relação, ao citar
que o número de espécies aumenta continuamente à medida que a área aumenta e são
amostradas diferentes associações de plantas, que penetram umas nas outras
continuamente, sem limites visíveis. Da mesma forma, Preston (1960) constatou que
áreas pequenas de tamanho semelhante têm aproximadamente o mesmo número de
espécies, incluindo uma ressalva: desde que seja evitada a escolha de áreas que são, por
exemplo, metade floresta e metade pântano. Reconheceu, assim, que a curva espécie-
área podia ser influenciada pelas mudanças ecológicas inevitáveis e diversidade de
habitats encontrada em grandes áreas. Entre essas mudanças, citou a existência de um
efeito de borda ou ecótono, consequente de um encontro de dois habitats distintos. Na

43
própria obra original da TBI, MacArthur e Wilson (1967) também afirmaram que a área
de uma ilha é correlacionada à diversidade de ambientes presentes, e que esta última
teria um efeito mais direto sobre o número de espécies do que área apenas. Porém,
embora tenham reconhecido que em menores ilhas reduz-se a variedade de habitats,
constataram que não havia boa informação disponível sobre diversidade de habitats e
que esta era, à época, uma variável que apenas começava a ser descrita e medida (pp. 8
e 65).
Os estudos de fragmentação requerem, assim, uma abordagem mais ampla e
integrada das teorias, mas que continue mantendo uma simplicidade sintetizadora. Para
tanto, o efeito de borda e as perturbações se integram à heterogeneidade por gerarem
habitats microclimaticamente diferenciados nos fragmentos; a heterogeneidade de
habitats representa a disponibilidade de recursos e nichos para a comunidade no
fragmento; e a relação espécie-área, para área total do fragmento, é relacionada à
heterogeneidade de habitats presentes. Desta forma, área, isolamento e heterogeneidade
de habitats formam os parâmetros básicos para o entendimento da estrutura de
comunidades isoladas. Só recentemente, a integração dessas variáveis tem sido
considerada, tanto na criação de modelos teóricos (KADMON; ALLOUCHE, 2007)
quanto em estudos empíricos em comunidades insulares e fragmentadas.
Em ambientes insulares, Ricklefs e Lovette (1999) conseguiram isolar os efeitos
da área per se versus diversidade de habitat, a partir de bancos de dados de fauna,
influenciando, nos anos subsequentes, muito da pesquisa desenvolvida e testes de novos
métodos (TRIANTIS et al., 2005). No estudo, analisaram a relação entre área e
diversidade de habitats e riqueza de quatro taxa, em 19 ilhas no Caribe, escolhendo tipos
vegetacionais e elevação como dimensões de habitats, que se mostraram fortemente
correlacionadas. Área e diversidade de habitats apresentaram estreita correlação, porém
uma ou outra mostrou relação significativa para grupos distintos de fauna, a depender da
biologia do grupo. A diversidade de habitats foi, assim, mais influente para grupos não
voadores e com reconhecida especialização por habitats, como anfíbios, por exemplo.
Em ilhas e com enfoque em plantas, as pesquisas empíricas têm de fato
confirmado haver uma correlação positiva entre área e diversidade de habitats, tal qual
descrito por Ricklefs e Lovette (1999). Porém, os estudos têm revelado também que
essas duas variáveis desempenham papéis independentes, principalmente para a flora
nativa. Tal resultado foi encontrado por McMaster (2005), que somou elevação e tipos

44
de solo - variáveis de habitat - às variáveis típicas da TBI para explicar a riqueza
florística em 22 ilhas e encontrou importante poder explicativo da riqueza vegetal na
variável edáfica. Hannus e von Numers (2008), utilizando vegetação como dimensão de
habitat, observaram uma maior correlação entre a riqueza de plantas e um índice obtido
pela multiplicação da área de cada uma das 160 ilhas estudadas pelo seu número de
habitats, do que com área ou número de habitats separadamente. Kreft et al. (2008)
examinaram a diversidade vegetal global em um número maior de ilhas (488) e em mais
970 amostras em continentes e também introduziram, além das variáveis da TBI, uma
variável topográfica, a amplitude de elevação, usada pelos autores como substituta
(surrogate) da complexidade e diversidade de habitats. Como resultado, mostraram uma
relação forte e positiva entre a complexidade topográfica e a riqueza de plantas, além da
correlação área- diversidade de habitats. A frequente covariância entre área e
diversidade de habitats quando esta é descrita através variável topográfica levou
Johnson et al. (2003) a questionar se área e topografia não seriam correlacionadas
simplesmente porque a complexidade topográfica aumenta a superfície e, assim, reflete
um aumento da área per se. Aplicando este teste em ilhas britânicas, encontraram um
efeito da topografia independente da área em um grupo de ilhas, e um efeito dependente
da área em outro, sugerindo a existência de gradientes mais complexos do que o de
topográfico atuando nas comunidades.
Em arquipélagos de fragmentos sobre o continente, há maior versatilidade na
definição das dimensões de habitat a serem relacionadas à riqueza, e geralmente o
número de fragmentos não excede algumas dezenas. Brose (2001), por exemplo,
pesquisou 58 fragmentos de áreas úmidas temporárias, correlacionando a riqueza de
plantas vasculares à área do fragmento, isolamento e diversidade de habitats, definidos
quanto à dinâmica hidrológica. Gignac e Dale (2005) mediram variáveis meteorológicas
e a variação nos sítios propícios ao estabelecimento de criptógamos em 44 fragmentos
florestais de diferentes tamanhos. Já Pereira et al. (2007) estudaram 20 fragmentos
florestais nos quais a heterogeneidade foi medida através de variáveis de solo,
topografia, e índices de borda e perturbação. Nesses estudos, os resultados confirmaram
a hipótese de heterogeneidade, que foi a principal variável explicativa da riqueza,
seguida pela borda, que exerceu efeito negativo na riqueza nos estudos em que o efeito
de borda foi considerado. Porém, mesmo quando utilizando variáveis não topográficas,

45
esses estudos também revelaram a existência de correlação entre tamanho dos
fragmentos e diversidade de habitats.
Se área e diversidade de habitats são correlacionadas, alguns defendem que a
área per se seria suficiente para explicar padrões de riqueza, atuando como substituta da
heterogeneidade. Três estudos recentes fizeram um importante avanço na compreensão
desta relação. Do lado teórico, Kadmon e Allouche (2007) apresentaram um modelo
para unificação da TBI e teoria da heterogeneidade de habitats, incorporando as
variáveis e os processos insulares, e a diversidade de habitats e nichos, confirmando
alguns padrões, porém obtendo alguns resultados inesperados. Entre as confirmações,
estão a correlação positiva entre riqueza e área e negativa entre riqueza e isolamento.
Inesperadamente, porém, o modelo gerado revelou que a heterogeneidade de habitats
pode ter efeito positivo ou negativo na riqueza. Relações positivas são esperadas em
grandes áreas, enquanto em pequenas áreas a heterogeneidade leva a uma redução na
riqueza, devido à diminuição relativa da disponibilidade de cada habitat, aumentando o
risco de extinção estocástica de espécies especialistas. O estudo de Kadmon e Allouche
(2007) revelou que, então, que a heterogeneidade de habitats afeta comunidades
biológicas de forma mais complexa do que tem se assumido. Do lado empírico,
Kallimanis et al. (2008), analisaram a integração de área e heterogeneidade de habitats a
um banco de dados da flora em 237 sítios na Grécia. Considerando o grande número de
áreas amostradas e o rigor na definição dos habitats, a pesquisa permitiu descrever um
padrão interessante: mantendo-se área constante, a riqueza foi correlacionada à
diversidade de habitats e mantendo-se heterogeneidade constante, a riqueza foi
correlacionada à área. Já Báldi (2008) reforçou a defesa da hipótese da heterogeneidade
por não encontrar correlação entre área e heterogeneidade, e registrar forte relação da
riqueza com a heterogeneidade ao estudar 16 reservas na Hungria.
Os resultados de Kadmon e Allouche (2007), Kallimanis et al. (2008) e Báldi
(2008) sinalizam, assim, que as hipóteses de área e heterogeneidade têm papéis
independentes e não são mutuamente exclusivas, e sugerem o abandono da dicotomia e
uma crítica reavaliação e redirecionamento das pesquisas em comunidades
fragmentadas, à luz de um arcabouço teórico mais integrado. Despontam como lacunas
que precisam ser preenchidas a investigação da relação entre área e heterogeneidade de
habitats em paisagens fragmentadas e o entendimento da influência de uma e/ ou outra
na riqueza e diversidade de espécies nos remanescentes de vegetação.

46
IV. CAPÍTULO 2 - Integrando área, forma e
heterogeneidade de habitats em remanescentes
florestais numa paisagem altamente fragmentada da
Floresta Atlântica

47
1. Resumo
Duas são as hipóteses principais que explicam a relação espécie-área: a da área per se e
da heterogeneidade de habitats, esta última menos pesquisada, devido à sua frequente
relação positiva com a área e à dificuldade em se definirem variáveis relevantes de
habitats. Este estudo objetivou descrever a heterogeneidade de habitats e investigar sua
relação com a área e forma de fragmentos florestais numa paisagem altamente
fragmentada (19.200 ha) da Floresta Atlântica. Com base em imagens de satélite de alta
resolução (4 m) e mapas topográficos (curvas de nível a cada 5 m), 80 fragmentos foram
mapeados, nos quais os habitats foram classificados e quantificados de acordo a posição
em relação à borda, relevo e presença de água, resultando em oito habitats possíveis.
Para cada fragmento, calculou-se área total (AT), índice de forma (IF), número (NH),
diversidade de Simpson (DH) e equitabilidade de habitats (EH). Nas análises,
utilizaram-se regressões lineares e logarítmicas, sendo AT e IF as variáveis
independentes, cujo efeito conjunto também foi analisado através de regressão múltipla
(stepwise). AT variou de 1,2 a 790 ha, sendo 77% <50 ha; IF variou de 1,11 a 5,03 e foi
positivamente relacionado à área. Habitats de borda cobriram 40% da área florestal,
sendo borda de encosta o habitat mais frequente, seguido pelo interior de encosta, que
foi o mais importante em cobertura (44%). Quanto à topografia, 77% da área são de
encosta, apenas 2% tabuleiros, e 21% são fundos de vale. Nesses, 72% são áreas ripárias
permanentes. NH variou de 1 a 7, e DH de 1 a 3,32, sendo as duas variáveis positiva e
significativamente relacionadas à TA, uma relação menor em fragmentos < 50 ha. AT e
IF juntas explicaram 60% da variância de NH e 53% de DH, restando uma importante
fração de NH e DH não explicadas. Os resultados reforçam as idéias que: área e
heterogeneidade são hipóteses mutuamente complementares e não exclusivas, com
relação mais sincrônica em áreas maiores e menos previsível em áreas menores. Este
comportamento imprevisível tem implicações cruciais para conservação. Habitats
ripários, por exemplo, altamente vulneráveis às modificações humanas, ocorrem em
fragmentos a partir de 5 ha, perfazendo uma relevante soma desses sítios na paisagem.
Portanto, embora em pequenos fragmentos haja um aumento na proporção de habitats
perturbados ou de relevo mais acidentado, a configuração espacial dos habitats
remanescentes na paisagem deve ser considerada, principalmente no caso de fragmentos
próximos entre si, com vistas à preservação dos habitats em proporções adequadas para
manutenção da diversidade biológica na escala da paisagem.

48
2. Introdução
A heterogeneidade de habitats é uma importante hipótese explicativa da relação
espécie-área – REA (WILLIAMS, 1964; CONNOR; MCCOY, 1979) e,
consequentemente, da diversidade de espécies, o que é amplamente reconhecido na
teoria ecológica e demonstrado em simulações e pesquisa empírica (DUFOUR et al.,
2006). A hipótese prevê maior riqueza de espécies em função da maior heterogeneidade
ambiental em áreas maiores (BÁLDI, 2008), uma relação aparentemente mais forte em
menores escalas, considerando que a heterogeneidade normalmente é dissipada em
maiores escalas (WILLIAMSON et al., 2001). Outra importante explicação para a REA,
a hipótese da área per se, explica o aumento na riqueza de espécies com aumento na
área (PRESTON, 1960; BROSE, 2001) e processos relacionados de dinâmica de
colonização e extinção, como aplicado pela Teoria da Biogeografia de Ilhas
(MACARTHUR; WILSON, 1967). Como alternativa, uma hipótese não biológica
explica a riqueza de espécies como um efeito passivo da amostragem, que seria
incompleta em áreas menores (CONNOR; MCCOY, 1979; TURNER; TJØRVE, 2005).
A existência de diferentes habitats explica a riqueza de espécies porque mais
espécies se beneficiam dos vários nichos ou tipos de vida que os habitats fornecem
(MACARTHUR; MACARTHUR, 1961). A adaptação de uma espécie a um habitat
pode resultar em inferioridade competitiva em outro, levando diferenças na composição
e abundância de espécies entre diferentes habitats (CRAMER; WILLIG, 2005). Como
consequência, considerando que cada tipo de habitat seja grande o suficiente para
suportar populações viáveis, a diversidade de habitats aumenta a diversidade geral inter-
habitats, conhecida como diversidade beta (WHITTAKER, 1972). Por outro ponto de
vista, a hipótese da área per se prevê que o número de espécies aumentará com a área
mesmo se o ambiente permanecer homogêneo (KALLIMANIS et al., 2008).
As duas hipóteses mais importantes que explicam a REA são frequentemente
testadas em comunidades vivendo em ilhas ou em áreas insulares sobre o continente,
tais como fragmentos florestas (WATSON, 2002; LAURANCE, 2008). Nessas áreas
isoladas, o número de espécies residentes, as densidades populacionais e diversidade
resultante são reconhecidas como consequentes da área per se, da heterogeneidade de
habitats ou da integração de ambos (KADMON; ALLOUCHE, 2007). Apesar da
importância da heterogeneidade ambiental ser amplamente reconhecida (WILLIAMS,

49
1964), mesmo pelos defensores da área per se (PRESTON, 1960; MACARTHUR;
WILSON, 1967) sua relação à riqueza de espécies é bem menos estudada do que o
efeito da área. Há duas razões básicas para isso. Primeiramente, essas variáveis são
normalmente correlacionadas (ROSENZWEIG, 1995; KALLIMANIS et al., 2008) e,
assim, a área apenas seria a hipótese principal explicando a riqueza de espécies. Além
disso, a heterogeneidade de habitats é difícil de ser estudada, devido à falta de
consistência e unificação na definição de habitats e suas variáveis relevantes
(TRIANTIS et al., 2003; MITCHELL, 2005).
O termo habitat corresponde ao componente ambiental do nicho em seus
aspectos físicos e químicos, sendo típico de uma população (HALL et al., 1997;
MITCHELL, 2005) ou de comunidades (WHITTAKER et al., 1973; KOLASA;
WALTHO, 1998; PETREN, 2001). As definições de habitat vão da mais simples
“lugar onde um organismo habita” (BEGON et al., 1996), a conceitos mais descritivos
de um local físico onde uma espécie vive ou poderia potencialmente viver, em uma
escala (espacial e temporal) particular, de natureza simplesmente abiótica (KEARNEY,
2006) ou, eventualmente, incluindo componentes bióticos (MITCHELL, 2005). Habitat,
portanto, inclui os recursos e condições que produzem ocupação, incluindo
sobrevivência e reprodução, por um organismo (HALL et al., 1997). Essas condições
podem ser quantificadas como uma série de dimensões físicas descritivas e relevantes,
cujos valores variem espacial e temporalmente, e interagem com uma espécie ou
comunidade (KOLASA; WALTHO, 1998).
Além do problema conceitual, uma peça chave no estudo de habitats está na
identificação de dimensões que sejam, ao mesmo tempo, possíveis de quantificar e
relevantes para a comunidade biológica estudada. Considerando a natureza
multidimensional do espaço ecológico, pode-se tornar muito difícil escolher um fator
ambiental que capture a heterogeneidade, criando diferentes condições ao longo das
quais haja um efeito na partição de espécies (DUFOUR et al., 2006). Além disso, apesar
de, geralmente, as dimensões escolhidas serem muito simplificadas (MITCHELL,
2005), elas devem ainda assim reter uma informação pertinente para os organismos. Em
estudos empíricos, as variáveis frequentemente escolhidas como dimensões de habitat
relevantes para diferentes comunidades são: fatores topográficos (JOHNSON et al.,
2003; HOFER et al., 2008; LUNDHOLM, 2009), edáficos (FITTER et al., 2000;
KUBOTA et al., 2004; JOHN et al., 2007), hídricos (CLARK et al., 1999; BROSE,

50
2001; DRUCKER et al., 2008; KELLER et al., 2009; KESSLER; LEHNERT, 2009;
SFENTHOURAKIS; TRIANTIS, 2009), microclimáticos (CLARK et al., 1999;
DUFOUR et al., 2006; PEREIRA et al., 2007) e cobertura vegetal (TEWS et al., 2004).
Antes de se entender o efeito da área per se e da heterogeneidade de habitats na
diversidade de espécies, a relação entre essas duas hipóteses precisa ser mais bem
investigada. Esta avaliação requer, idealmente, a definição e quantificação apropriada
de dimensões de habitat em um número o quanto maior de áreas isoladas, estimativa da
disponibilidade de habitats em cada área e avaliação de uma eventual relação. Seguindo
este procedimento, área e diversidade de habitats mostraram ser correlacionadas em
ilhas de vários tamanhos e diferentes posições geográficas (RICKLEFS; LOVETTE,
1999; MCMASTER, 2005; TRIANTIS et al., 2005; PANITSA et al., 2006; HANNUS;
VON NUMERS, 2008; KREFT et al., 2008; SFENTHOURAKIS; TRIANTIS, 2009). O
padrão de correlação entre área e heterogeneidade também foi confirmado em
fragmentos continentais de várias origens, tais como áreas úmidas (BROSE, 2001),
florestas manejadas (GIGNAC; DALE, 2005), áreas protegidas (KALLIMANIS et al.,
2008) e minas calcárias (KRAUSS et al., 2009). Há exceções ao padrão, porém, como
demonstrado por Báldi (2008), que não encontrou correspondência entre área e
heterogeneidade em 16 reservas na Hungria, e por Benedick et al. (2006), quando
estudando borboletas em fragmentos florestais em Bornéu. Quando a correlação
positiva entre área e heterogeneidade foi violada, Báldi (2008) viu em seu estudo de
campo um sistema excelente para examinar a contribuição relativa da área versus
heterogeneidade à riqueza de espécies.
Neste estudo, objetivou-se investigar a relação entre área e heterogeneidade de
habitats em fragmentos florestais numa paisagem altamente fragmentada no Centro de
Endemismo Pernambuco (SILVA; CASTELETI, 2005), na Floresta Atlântica brasileira.
Os objetivos principais foram descrever a disponibilidade de habitats e testar sua
positiva relação com área, baseado na quantificação de dimensões de habitats relevantes
para comunidades florestais em 80 fragmentos. Além da área, a forma dos fragmentos
também foi avaliada como uma variável explicativa da heterogeneidade. Intenciona-se
discorrer sobre esta nova faceta na aplicação da teoria de biogeografia de ilhas aos
estudos de fragmentação e na importância e representação de habitats para a
conservação de comunidades florestais fragmentadas neste bioma extremamente
ameaçado.

51
3. Material e métodos
3.1. Área de Estudo
O estudo foi desenvolvido em uma região de 19.200 ha, contida na propriedade
da Usina São José, Pernambuco, Nordeste do Brasil (07º40'21,25", 07º55'50,92"S e
34º54'14,25", 35º05'21,08"W, Figura IV.1). Para descrição da geologia, relevo, clima e
histórico da área, ver item “Área de Estudo”, p. 6. O local escolhido é uma amostra
típica da paisagem na região costeira do Nordeste, caracterizada pela predominância de
plantações de cana-de-açúcar e fragmentos esparsos de floresta ombrófila densa das
terras baixas (RANTA et al., 1998; TRINDADE et al., 2008), vegetação potencial na
área (VELOSO et al., 1991).
3.2. Métodos
Aquisição de dados
Este estudo teve como base duas fontes de dados espaciais disponíveis para todo o
polígono analisado, contendo informações sobre os fragmentos e relevo. A informação
espacial sobre os fragmentos foi obtida a partir de um mosaico digital de imagens
multi-espectrais do satélite IKONOS II, obtidas em janeiro de 2005, com resolução
espacial de 4 m, projeção UTM e Datum WGS 84, georreferenciadas com base em um
levantamento de reconhecimento de campo utilizando GPS com receptores geodésicos
e método diferencial. Com base nesta imagem, todos os fragmentos com área maior
que 1 ha foram vetorizados em tela, seguido de checagem de campo. Atributos de
relevo foram obtidos a partir de mapas topográficos na escala de 1:10.000, elaborados
com base em fotografias aéreas pela atual Agência Estadual de Planejamento e
Pesquisas de Pernambuco (antiga FIDEM), em 1975. Curvas de nível com intervalos
equidistantes de 5 m foram digitalizadas para, a partir delas, gerar-se a altimetria.
Ambas as fontes de dados foram geoprocessadas e integradas utilizando o software
Arcview GIS 9.2 (ESRI, 2006).

52
Classificação dos habitats
Em duas recentes revisões sobre a relação área-habitat e diversidade de espécies,
percebe-se que, para animais, é mais frequente a escolha de cobertura vegetal como
dimensão de habitats (TEWS et al., 2004), enquanto para plantas, variáveis topográficas
e de solo são mais comuns (LUNDHOLM, 2009). Além disso, a escala especial das
dimensões varia muito, desde amplas categorias de cobertura (BÁLDI, 2008) a micro-
habitats relevantes para um táxon em particular como, por exemplo, moluscos
(TRIANTIS et al., 2005) terrestres ou criptógamos (GIGNAC; DALE, 2005). No
primeiro caso, o número de categorias de habitats pode ser simplificado demais,
enquanto dimensões de micro-habitats podem levar a um número excessivo de habitats
possíveis. Numa abordagem ideal, os tipos de habitat devem refletir dimensões que são
proeminentes em paisagens fragmentadas e relevantes para uma ampla gama de
espécies em vários grupos taxonômicos, atuando como estruturas-chave sensu Tews et
al. (2004), isto é, uma estrutura espacial distinta que provê recursos e abrigo para
grupos cruciais de espécies. Habitats foram classificados de acordo com três dimensões:
(i) posição em relação à borda florestal, (ii) relevo e (iii) presença de água. Em todos os
fragmentos, os habitats foram quantificados através de áreas (polígonos). A
heterogeneidade pode ser medida de várias formas, de pontos amostrais a polígonos
espacialmente explícitos (WIENS, 2000). Esta abordagem de área, apesar de arbitrária,
considerando que porções homogêneas de habitats com bordas abruptas constituem
apenas uma aproximação de habitats reais, é espacialmente mais integradora e mais
facilmente aplicável a iniciativas de conservação (WIENS, 2000; HOFER et al., 2008)
e, por este motivo, adotada neste estudo. Definições de cada dimensão e tipos de
habitats resultantes estão descritos a seguir.
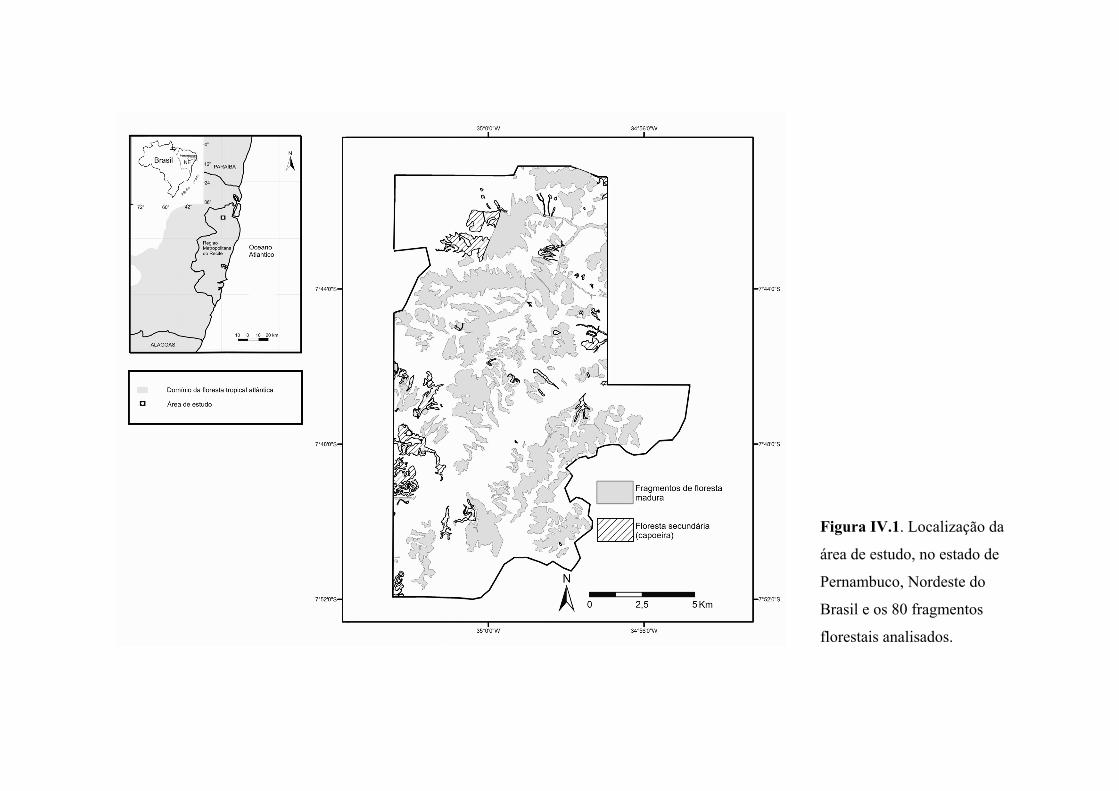
53
Figura IV.1. Localização da
área de estudo, no estado de
Pernambuco, Nordeste do
Brasil e os 80 fragmentos
florestais analisados.

54
Posição em relação à borda: para definição do habitat sob influência da borda, buffers
de 50 m foram gerados a partir do limite do fragmento em direção ao interior, em um
plano de informações contendo todos os polígonos dos fragmentos. Cada buffer
corresponde a um cinturão de influência ao logo do perímetro do fragmento. As áreas
restantes, isto é, áreas além dos 50 m de influência da borda, foram consideradas como
interior. A largura de 50 m para definição da influência da borda foi escolhida com base
na literatura disponível e revisões (MURCIA, 1995; RIES et al., 2004; HARPER et al.,
2005; PEREIRA et al., 2007) e também na pesquisa sobre influência da borda em
fragmentos florestais na área de estudo, desenvolvidos por Silva et al. (2008a), Silva et
al.(2008c) e Gomes et al. (2009). Nesta dimensão, dois habitats são possíveis: borda e
interior.
Relevo: para classificação dos habitats em relação aos atributos de relevo, foi utilizada a
extensão 3D-Analyst no ArcGIS 9.2 (ESRI, 2006). Inicialmente, foram gerados clips
combinando altimetria e todos os fragmentos vetorizados. A partir destes, modelos 3D
(tins) foram criados, com informação sobre a declividade, em percentagem. Após
análises preliminares sobre possíveis classes de declividade, foram escolhidas duas
classes, aplicando-se como divisor a declividade de 10%. Áreas com declividade igual
ou menor que 10% foram classificadas como superfícies plano-suave inclinadas,
enquanto aquelas com declividade > 10% foram classificadas como encostas. Nas
análises, superfícies plano-suave inclinadas foram geradas para altitudes altas e baixas,
essas últimas correspondendo aos terraços fluviais. Foram incluídas como superfícies
planas apenas as de maiores altitudes, correspondendo aos tabuleiros; áreas planas
baixas foram incluídas nos habitats ripários. Como resultado, três habitats possíveis
foram identificados de acordo com o relevo, todos quantificados em projeção
horizontal: tabuleiros altos, encostas e fundos de vale. Os últimos foram classificados
em mais detalhes de acordo com a presença de água, descritos a seguir.
Presença de água: para determinação dos sítios sob influência de corpos d´água, as
características hidrológicas da área foram analisadas, procurando-se identificar água
permanente ou temporária. Um complexo sistema de drenagem existe na área, incluindo
superfícies suaves e muito inclinadas, terraços fluviais, canais horizontais e profundos
ou fundos de vale baixos, escavados nos sedimentos Barreiras. Considerando esta

55
complexidade, uma metodologia foi definida para classificação dos sítios ripários com
base em três fontes de informação. Primeiramente, foram consideradas as áreas
identificadas como terraços na análise de declividade. Como segunda fonte, na ausência
de um terraço plano, mas havendo confirmação da presença de água, através de
checagem de campo ou imagem de satélite, foram usadas as curvas de nível para
demarcar uma área definida entre a cota altitudinal mais baixa no fragmento e a curva
10 a 15m acima desta. Como um último recurso, a ferramenta Hydrology no ArcGIS 9.2
foi utilizada para gerar linhas de drenagem, a partir das quais buffers de 30 m foram
criados para quantificação da influência ripária ao longo dos canais de drenagem.
Habitats ripários foram definidos através da soma dos terraços, fundos de vale baixos e
fundos de vale inclinados. Nesta dimensão, habitats possíveis foram definidos como:
ripários permanentes (com rios, riachos, reservatórios ou nascentes), e ripários
temporários (com drenagem ativa apenas durante chuvas pesadas na estação úmida).
Através da combinação das três dimensões de habitat, foi possível definir oito
tipos de habitat potenciais nos fragmentos estudados, descritos na Tabela IV.1.
Tabela IV.1. Tipos potenciais de habitats com base em três dimensões utilizadas para
quantificação da heterogeneidade de habitats em fragmentos de Floresta Atlântica em
Pernambuco, Nordeste do Brasil.
Dimensão de habitat Tipo de habitat potencial Influência de
borda Relevo Presença de água
Borda
Encosta Não-ripário Borda de encosta Tabuleiro alto Borda de tabuleiro Fundo de vale Ripário permanente Borda ripária permanente
Ripário temporário Borda ripária temporária
Interior
Encosta Não-ripário
Interior de encosta Tabuleiro alto Interior de tabuleiro Fundo de vale Ripário permanente Interior ripário permanente
Ripário temporário Interior ripário temporário
3.3. Análise dos dados
Para cada fragmento, foi calculada a área total (AT), em hectares, e a área
ocupada por cada tipo de habitat, todas em projeção planimétrica. Trindade et al.
(2008), ao analisarem 110 fragmentos em um polígono de 280 km2 circunscrito a esta
paisagem em estudo, encontraram uma diferença pronunciada no número de fragmentos

56
por classe de tamanho. Considerando este padrão, a distribuição dos fragmentos por
classe de tamanho foi avaliada e testada quanto à normalidade, para definição do melhor
modelo de regressão a ser utilizado nas análises, se linear ou logarítmica.
A forma dos fragmentos foi descrita utilizando-se um índice de forma (IF),
calculado para cada fragmento, de acordo com Jorge e Garcia (1997). O índice é
baseado na razão do perímetro pela raiz quadrada da área, descrito como
IF = P/(2√(πA)
Onde P é o perímetro do fragmento (em metros) e A é a área do fragmento (em m2). IF
foi proposto como uma alternativa para reduzir a dependência de escala que existe
quando o índice é baseado na razão entre o perímetro e a área do fragmento (JORGE;
GARCIA, 1997; COMBER et al., 2003), e assume um valor de 1,0 para áreas
perfeitamente circulares, aumentado a medida que a forma se torna mais complexa ou
irregular.
A heterogeneidade de habitats (aqui também chamada disponibilidade de
habitats ou de ambientes) em cada fragmento foi definida através da riqueza de habitats,
isto é, número de habitats presentes (NH) e diversidade de habitats (DH). A diversidade
foi estimada utilizando-se o índice de Simpson inverso ou recíproco (SIMPSON, 1949;
RICKLEFS; LOVETTE, 1999; MAGURRAN, 2004):
DH= 1/ Σ ni=1 pi
2
Onde p é a proporção da área total ocupada pelo habitat i de n habitats. Este índice não
resulta em zero, como seria esperado quando apenas um habitat tivesse sido encontrado
em um fragmento particular, aplicando-se índices baseados na distribuição logarítmica,
como índice de Shannon. Além disso, o índice escolhido é mais sensível e enfatiza o
grau de equitabilidade nas proporções entre áreas de habitats. No presente estudo, o
índice poderia variar de 1, quando apenas um habitat for encontrado em um fragmento,
a 8, se todos os habitats possíveis estiverem representados com proporções iguais numa
área (RICKLEFS; LOVETTE, 1999; NAGENDRA, 2002). A equitabilidade de habitat
(EH) foi também avaliada utilizando-se o índice de equitabilidade de Simpson,
calculado como o valor observado de DH em um fragmento dividido pelo valor máximo
possível de diversidade (NAGENDRA, 2002).
Para análises estatísticas, utilizaram-se técnicas padrões de regressão linear e não
linear, sendo área (AT) e forma (IF) colocadas como variáveis independentes,
objetivando-se analisar possíveis relações entre essas e as variáveis dependentes riqueza

57
de habitats (NH) e diversidade (DH), separadamente. Regressões simples foram
utilizadas para avaliar a relação entre proporções de tipos específicos de habitats e área
(AT). Adicionalmente, uma análise de regressão múltipla tipo stepwise foi utilizada para
avaliar a contribuição conjunta de área e forma para disponibilidade de habitats (NH e
DH). O coeficiente de determinação (R2) ajustado foi escolhido em todos os resultados
significativos (P>0,05) nas análises de regressão. Todas as análises foram feitas no
software STATISTICA versão 9.0 (STATSOFT, 2009).
4. Resultados
4.1. Padrão geral da paisagem
Na área de estudo, há 80 fragmentos de vários tamanhos, com AT variando de
1,2 ha a 790 ha, cobrindo uma área de 4970 ha (26%) na paisagem estudada. Analisando
a distribuição dos fragmentos por classes de tamanho, constatou-se uma distribuição
irregular, não se ajustando à curva de distribuição normal (p<0,001), com
predominância de pequenos fragmentos. Apesar do maior fragmento na área ser mais de
600 vezes maior que o menor fragmento, 62 (77%) remanescentes são menores que 50
ha e apenas 9% são maiores que 250 ha. Uma distribuição normal foi ajustada quando
as áreas foram transformadas (log10), e, por isso, a regressão logarítmica foi utilizada
nas análises.
O índice de forma dos fragmentos (IF) teve ampla variação, de 1,11 a 5,03, com
aproximadamente metade dos fragmentos apresentando índices com valor até 2,0
(Figura IV.2). Isto demonstra que nenhum fragmento na área tem forma regular e que
metade do total tem forma complexa. AT e IF foram positivamente relacionados
(r2=0,525, p<0,001), indicando a tendência de formas mais complexas em fragmentos
maiores (Figura IV.3). O maior valor de IF foi, de fato, obtido para o maior fragmento.
4.2. Frequência e abundância dos tipos de habitats
Uma proporção substancial de habitats de borda foi documentada no
levantamento, sendo a cobertura relativa de habitats de borda e interior de 40% e 60%
da área florestal total, respectivamente. A borda de encosta foi o tipo de habitat mais
frequente, ocorrendo em 78 dos 80 fragmentos (Figura IV.4), nos quais tiveram
proporções de área variando de 15 a 100%. Entretanto, a proporção de habitats de
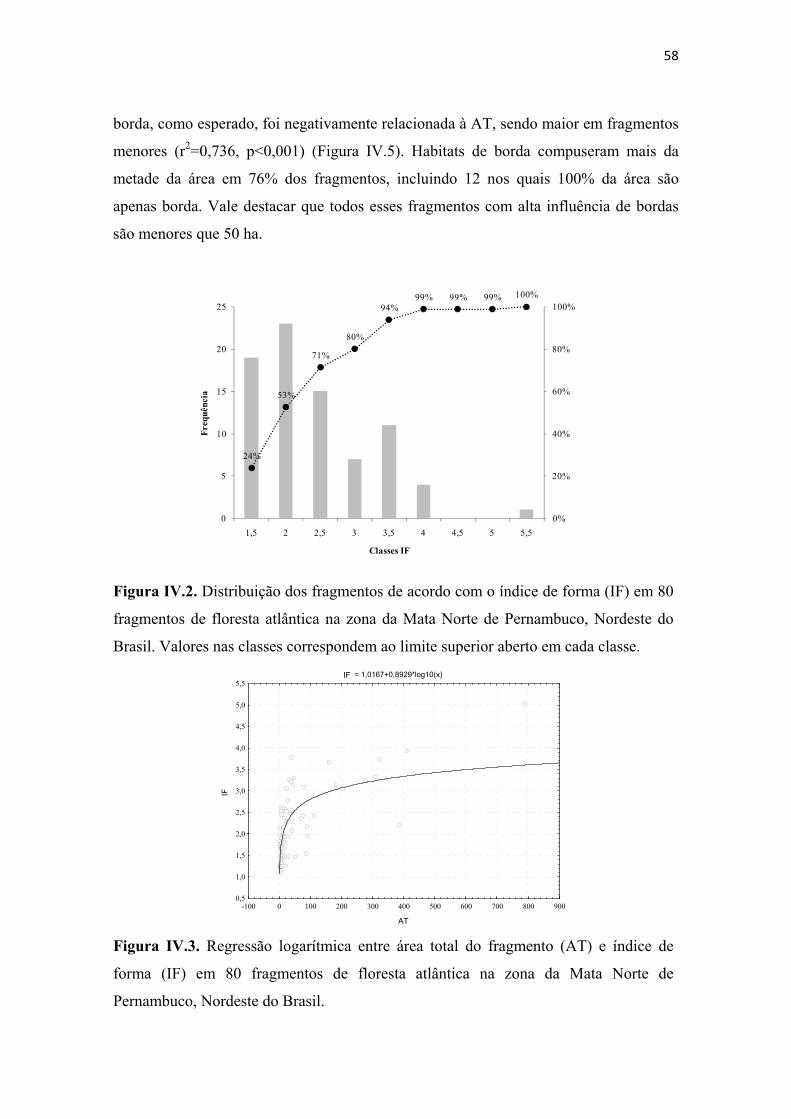
58
borda, como esperado, foi negativamente relacionada à AT, sendo maior em fragmentos
menores (r2=0,736, p<0,001) (Figura IV.5). Habitats de borda compuseram mais da
metade da área em 76% dos fragmentos, incluindo 12 nos quais 100% da área são
apenas borda. Vale destacar que todos esses fragmentos com alta influência de bordas
são menores que 50 ha.
Figura IV.2. Distribuição dos fragmentos de acordo com o índice de forma (IF) em 80
fragmentos de floresta atlântica na zona da Mata Norte de Pernambuco, Nordeste do
Brasil. Valores nas classes correspondem ao limite superior aberto em cada classe.
Figura IV.3. Regressão logarítmica entre área total do fragmento (AT) e índice de
forma (IF) em 80 fragmentos de floresta atlântica na zona da Mata Norte de
Pernambuco, Nordeste do Brasil.
24%
53%
71%
80%
94%99% 99% 99% 100%
0%
20%
40%
60%
80%
100%
0
5
10
15
20
25
1,5 2 2,5 3 3,5 4 4,5 5 5,5
Freq
uênc
ia
Classes IF
SI = 1,0167+0,8929*log10(x)
-100 0 100 200 300 400 500 600 700 800 900
TA
0,5
1,0
1,5
2,0
2,5
3,0
3,5
4,0
4,5
5,0
5,5
SI
IF
IF
AT
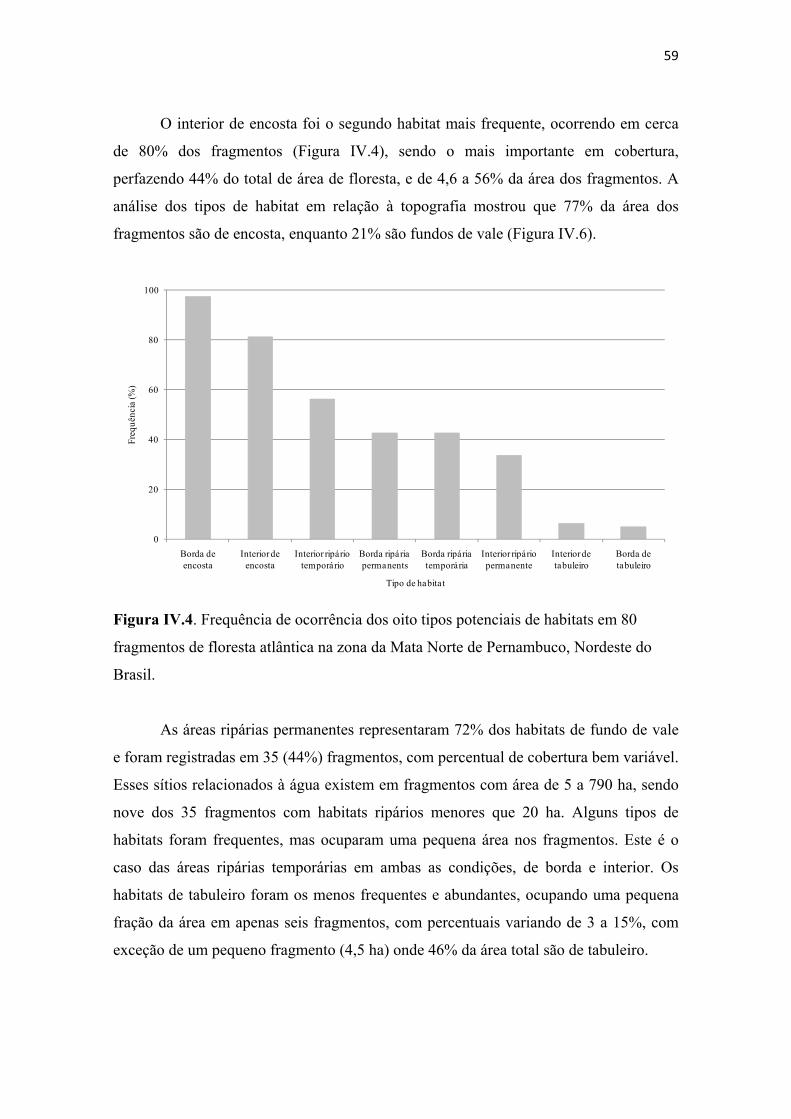
59
O interior de encosta foi o segundo habitat mais frequente, ocorrendo em cerca
de 80% dos fragmentos (Figura IV.4), sendo o mais importante em cobertura,
perfazendo 44% do total de área de floresta, e de 4,6 a 56% da área dos fragmentos. A
análise dos tipos de habitat em relação à topografia mostrou que 77% da área dos
fragmentos são de encosta, enquanto 21% são fundos de vale (Figura IV.6).
Figura IV.4. Frequência de ocorrência dos oito tipos potenciais de habitats em 80
fragmentos de floresta atlântica na zona da Mata Norte de Pernambuco, Nordeste do
Brasil.
As áreas ripárias permanentes representaram 72% dos habitats de fundo de vale
e foram registradas em 35 (44%) fragmentos, com percentual de cobertura bem variável.
Esses sítios relacionados à água existem em fragmentos com área de 5 a 790 ha, sendo
nove dos 35 fragmentos com habitats ripários menores que 20 ha. Alguns tipos de
habitats foram frequentes, mas ocuparam uma pequena área nos fragmentos. Este é o
caso das áreas ripárias temporárias em ambas as condições, de borda e interior. Os
habitats de tabuleiro foram os menos frequentes e abundantes, ocupando uma pequena
fração da área em apenas seis fragmentos, com percentuais variando de 3 a 15%, com
exceção de um pequeno fragmento (4,5 ha) onde 46% da área total são de tabuleiro.
0
20
40
60
80
100
Borda de encosta
Interior de encosta
Interior ripário temporário
Borda ripária permanents
Borda ripária temporária
Interior ripário permanente
Interior de tabuleiro
Borda de tabuleiro
Freq
uênc
ia (%
)
Tipo de habita t

60
Figura IV.5. Relação entre área total (AT) e proporção de habitats de borda em 80
fragmentos de floresta atlântica na Zona da Mata Norte, Nordeste do Brasil.
Figura IV.6. Proporção dos tipos de habitats em relação à topografia em 80
fragmentos de floresta atlântica na zona da Mata Norte de Pernambuco, Brasil. Habitats
de fundo de vale estão detalhados em relação à presença de água (permanente ou
temporária).
Edge % = 107,3749-29,9989*log10(x)
-100 0 100 200 300 400 500 600 700 800 900
TA
10
20
30
40
50
60
70
80
90
100
110
Edge
pro
porti
onPr
opor
ção
de b
orda
AT
Borda %
77.3
1.6
15.1
6.0
21.1
Encostas
Tabuleiros
Ripários permanentes
Ripários temporários

61
4.3. Disponibilidade de habitats em relação à área e forma
dos fragmentos
O número de habitats (NH) registrado nos fragmentos variou de 1 a 7, o que
significa que nenhum fragmentos teve os oito tipos potenciais definidos. NH foi positiva
e significativamente relacionado à TA (Tabela IV.2, Figura IV.7), isto é, um maior
número de habitats é esperado em maiores fragmentos. O índice de diversidade de
habitats (DH) variou de 1 a 3,32 e os valores foram também significativamente
relacionados à TA (Tabela IV.2, Figura IV.8). Índice igual a 1,0 para fragmentos com
apenas um tipo de habitat foi encontrado em oito áreas, todas elas compostas por
habitats de borda, seja de encosta ou ripária. Um alto DH, acima de 2,5, foi registrado
em 41% dos fragmentos, mas em apenas 12 fragmentos DH foi maior que 3,0, a maioria
em áreas maiores que 50 ha. O maior fragmento teve DH de 2,82, enquanto o maior DH
(3,32) foi registrado em um fragmento de 112 ha. Considerando a distribuição irregular
dos fragmentos quanto ao tamanho, foi testada a relação entre AT e NH ou DH em uma
sub-amostra contendo os fragmentos menores que 50 ha, obtendo-se relações
significativas, porém mais fracas: TA explicou 21,1% de NH e apenas 6% de DH
(Tabela IV.2).
Tabela IV.2. Descritores da heterogeneidade de habitats e resultados da regressão
logarítmica em fragmentos de floresta atlântica na Zona da Mata Norte de Pernambuco,
Nordeste do Brasil, para amostra total e para sub-amostra de fragmentos menores que
50 ha. NH, número de habitats; DH, diversidade de habitats; Min, valor mínimo,; max,
valor máximo; N, número de fragmentos; VI, variável independente; AT, área total.
Var
iáve
l
Amostra total Sub-amostra < 50 ha
Valores Resultados
regressão
VI: AT
Valores Resultados
regressão
VI: AT
Média Min Max N r2 Média Min Max N r2
NH 3,65 1 7 80 0,418 ** 3,20 1 6 62 0,211 *
DH 2,21 1 3,32 80 0,324 ** 1,99 1 3,10 62 0,060 *
EH 0,28 0,125 0,415 80 0,316 ** 0,24 0,125 0,388 62 0,076 *
*P < 0,05; **P < 0,001
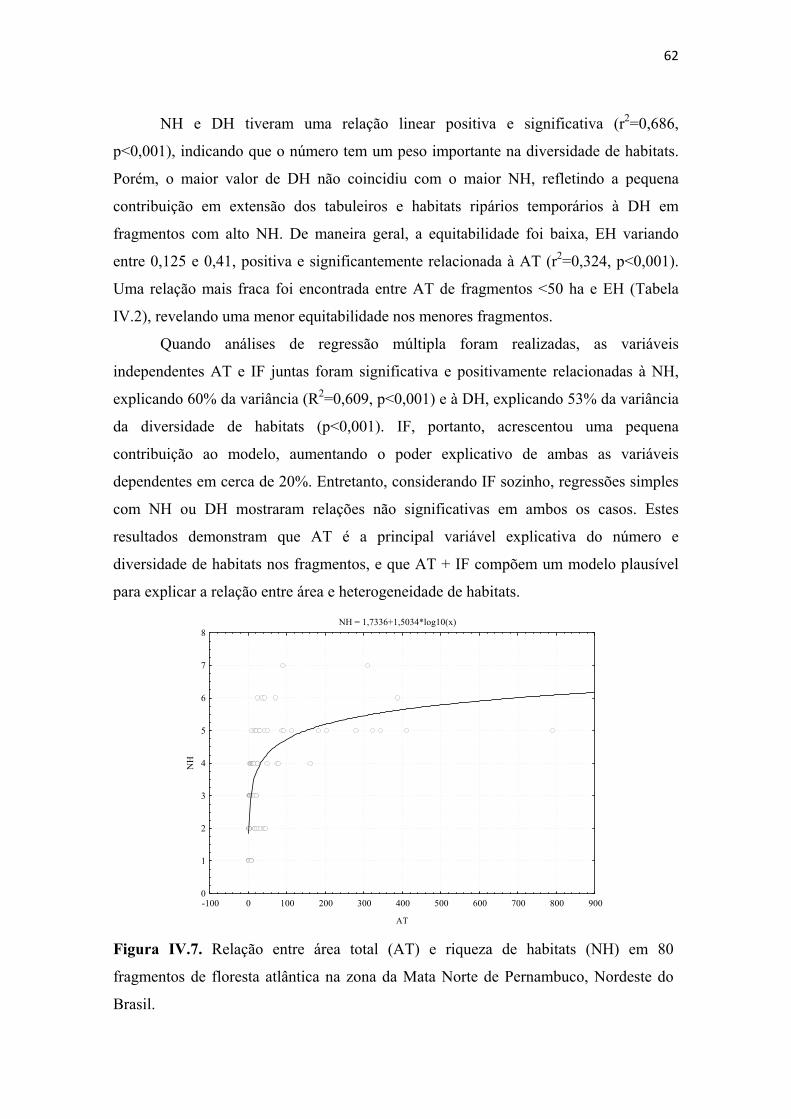
62
NH e DH tiveram uma relação linear positiva e significativa (r2=0,686,
p<0,001), indicando que o número tem um peso importante na diversidade de habitats.
Porém, o maior valor de DH não coincidiu com o maior NH, refletindo a pequena
contribuição em extensão dos tabuleiros e habitats ripários temporários à DH em
fragmentos com alto NH. De maneira geral, a equitabilidade foi baixa, EH variando
entre 0,125 e 0,41, positiva e significantemente relacionada à AT (r2=0,324, p<0,001).
Uma relação mais fraca foi encontrada entre AT de fragmentos <50 ha e EH (Tabela
IV.2), revelando uma menor equitabilidade nos menores fragmentos.
Quando análises de regressão múltipla foram realizadas, as variáveis
independentes AT e IF juntas foram significativa e positivamente relacionadas à NH,
explicando 60% da variância (R2=0,609, p<0,001) e à DH, explicando 53% da variância
da diversidade de habitats (p<0,001). IF, portanto, acrescentou uma pequena
contribuição ao modelo, aumentando o poder explicativo de ambas as variáveis
dependentes em cerca de 20%. Entretanto, considerando IF sozinho, regressões simples
com NH ou DH mostraram relações não significativas em ambos os casos. Estes
resultados demonstram que AT é a principal variável explicativa do número e
diversidade de habitats nos fragmentos, e que AT + IF compõem um modelo plausível
para explicar a relação entre área e heterogeneidade de habitats.
Figura IV.7. Relação entre área total (AT) e riqueza de habitats (NH) em 80
fragmentos de floresta atlântica na zona da Mata Norte de Pernambuco, Nordeste do
Brasil.
NH = 1,7336+1,5034*log10(x)
-100 0 100 200 300 400 500 600 700 800 900
TA
0
1
2
3
4
5
6
7
8
NH
AT

63
Figura IV.8. Relação entre área total (AT) e diversidade de habitats (NH) em 80
fragmentos de floresta atlântica na zona da Mata Norte de Pernambuco, Nordeste do
Brasil.
5. Discussão
Neste estudo, área e forma atuaram na previsão da heterogeneidade de habitats
em fragmentos florestais, integrando as duas hipóteses explicativas principais da relação
espécie-área. Entretanto, apesar da relação existente, uma importante parte da variância
na riqueza e diversidade de habitats (38 a 46%, respectivamente) permanece não
explicada. Relações positivas e significativas têm sido encontradas para diferentes taxa
em ilhas e em fragmentos tipo-ilha (RICKLEFS; LOVETTE, 1999; BROSE, 2001;
MCMASTER, 2005; TRIANTIS et al., 2005; PANITSA et al., 2006; BÁLDI, 2008;
HANNUS; VON NUMERS, 2008; KALLIMANIS et al., 2008; KREFT et al., 2008;
KRAUSS et al., 2009; SFENTHOURAKIS; TRIANTIS, 2009). Em pesquisas nas quais
são informadas as regressões entre área e riqueza ou diversidade de habitats,
coeficientes de determinação significativos variam de valores tão baixos quanto 0,269
(KALLIMANIS et al., 2008) até cerca de 0,8 (MCMASTER, 2005; TRIANTIS et al.,
2005; SFENTHOURAKIS; TRIANTIS, 2009). As evidências apontam, portanto, que a
área explica uma percentagem bem variável da heterogeneidade de habitats, considerada
HD = 1,4333+0,6071*log10(x)
-100 0 100 200 300 400 500 600 700 800 900
TA
0,8
1,0
1,2
1,4
1,6
1,8
2,0
2,2
2,4
2,6
2,8
3,0
3,2
3,4
3,6
HD
AT
DH
DH

64
fraca por certos autores (RICKLEFS; LOVETTE, 1999; PANITSA et al., 2006). Por
este motivo, apesar do constante debate, área per se e heterogeneidade de habitats são
consideradas hipóteses mutuamente complementares e não mutuamente exclusivas
(TRIANTIS et al., 2003; HANNUS; VON NUMERS, 2008), com diferentes respostas
para cada uma de acordo com o grupo taxonômico analisado (RICKLEFS; LOVETTE,
1999).
A forma dos fragmentos, uma variável não avaliada com frequência (HILL;
CURRAN, 2003; GIGNAC; DALE, 2005), acrescentou uma pequena contribuição à
explicação da heterogeneidade de habitats, quando integrada à área. Formas irregulares
são uma característica eminente nos fragmentos nesta paisagem, como revelado por
Trindade et al, (2008) que registraram 98% de todos os fragmentos irregulares,
incluindo todos os fragmentos com área maior que 30 ha. Nossos resultados mostraram
uma relação entre forma e habitats mais fraca do que em outras paisagens de florestas
fragmentadas (HILL; CURRAN, 2003; GIGNAC; DALE, 2005), sendo também aqui
mais fraca a relação entre forma e área. Porém, diferentes índices de forma estão
disponíveis na literatura e são adotados em diferentes pesquisas, variando desde uma
medida de circularidade (MARTINS et al., 2002; TRINDADE et al., 2008), à razão
entre perímetro e área (HILL; CURRAN, 2003) ou à razão entre perímetro e a raiz
quadrada da área (COMBER et al., 2003). Sem dúvida, o entendimento do papel da
forma na diversidade de habitats depende da compreensão da sua relação com a área
(EWERS; DIDHAM, 2006) e da adoção de um único índice de forma nos estudos desta
variável, para o que seria recomendável o índice baseado na razão entre perímetro e raiz
quadrada da área, como sugerido por Jorge e Garcia (1997), Comber et al. (2003) e
Ochoa-Gaona et al. (2004).
Hipoteticamente, a ausência de uma relação forte entre área e disponibilidade de
habitats poderia ser explicada pela alta concentração de fragmentos pequenos, uma
característica comum e restritiva nas paisagens atuais em “arquipélagos” de fragmentos,
especialmente na floresta tropical (TURNER; CORLETT, 1996), tornando a
transformação logarítmica da variável área uma escolha frequente (BROSE, 2001;
HILL; CURRAN, 2003; BENEDICK et al., 2006; BÁLDI, 2008; KALLIMANIS et al.,
2008; KRAUSS et al., 2009). Ao analisarmos uma subamostra de fragmentos mais
homogêneos em tamanho, reunindo aqueles menores que 50 ha, os resultados obtidos
revelaram que o poder explicativo da área foi ainda menor do que para a amostra total.

65
O padrão observado nos nossos dados mostrou-se semelhante ao reportado em ilhas por
Sfenthourakis e Triantis (2009): em geral, para áreas maiores, a diversidade de habitats
aumenta com o aumento do tamanho de uma forma bem sincrônica, enquanto em áreas
menores esta relação se torna mais variável. Nessas manchas pequenas e mais
frequentes, a disponibilidade de habitats é, portanto, menos uniforme, sendo a riqueza e
diversidade de habitats menos previsíveis com base na área.
Se a relação área-habitat foi menor em manchas menores, o que estaria
contribuindo para uma maior relação em áreas maiores? Por certo, a presença de alguns
tipos de habitats cuja ocorrência é esperada apenas em áreas grandes, como os habitats
de tabuleiro e os ripários. Tabuleiros ocorreram em baixíssima proporção nos
remanescentes florestais. Como áreas úteis para cultivo da cana-de-açúcar, a exclusão
desses habitats é claramente resultante de um processo não aleatório no qual a complexa
topografia da região, aliada às restrições impostas ao uso da terra, atuam na condução
do padrão de fragmentação e nas posições e formas finais dos remanescentes, que
guardam uma amostra tendenciosa da paisagem original (LAURANCE, 2008). Na área
de estudo, a maioria dos fragmentos está em superfícies inclinadas, caracteristicamente
menos úteis para objetivos econômicos como agricultura (SILVA et al., 2007),
principalmente quando se observam os altos níveis de desflorestamento e antropização
nas superfícies mais planas, tanto em baixas (RANTA et al., 1998) quanto altas
altitudes (TRINDADE et al., 2008). Apesar de a heterogeneidade topográfica ser
amplamente reconhecida por influenciar a diversidade de espécies na escala da
paisagem, ao longo de um gradiente altitudinal climático ou edáfico (VALENCIA et al.,
2004), sendo um estimador apropriado da riqueza biológica relativa em paisagens
topograficamente heterogêneas (HOFER et al., 2008), nesses sítios a influência do
relevo é mais indireta por guiar a exclusão de habitats planos, do que um efeito direto
natural (SILVA et al., 2008d).
Habitats ripários, por serem extremamente vulneráveis às modificações humanas
(SFENTHOURAKIS; TRIANTIS, 2009), também seriam uma categoria esperada em
maiores áreas. Na presente análise, áreas ripárias estiveram presentes em fragmentos a
partir de 5 ha, perfazendo uma relevante soma total de sítios ripários na paisagem. Isto
aumenta o valor de pequenos fragmentos para conservação, tanto para proteção de
recursos naturais quanto para a manutenção de espécies ripárias. Estes sítios são
importantes “estruturas-chave” para várias espécies de plantas hidrofílicas, com

66
adaptações morfológicas e fisiológicas (NAIMAN; DÉCAMPS, 1997), garantindo
refúgio, forragem e sítios de reprodução para vertebrados e invertebrados (BOUTIN et
al., 2003) e contendo um número maior de especialistas em relação a outros
ecossistemas (MOUW; ALABACK, 2003). Reconhecidos como um dos mais
dinâmicos e complexos habitats biofísicos entre os ecossistemas terrestres (NAIMAN;
DÉCAMPS, 1997), sítios ripários são considerados de importância central para a
conservação da biodiversidade. Uma vez que habitats ripários são identificados em
vários fragmentos, iniciativas de manejo considerando a rede de drenagem como um
todo podem vir a restaurar a integridade de rios e planícies de inundação (NAIMAN;
DÉCAMPS, 1997). Áreas ripárias podem atuar como um eixo ou área nuclear para
guiar a regeneração florestal (WILLIAMS-LINERA et al., 1998), pois os fluxos
biológicos entre fragmentos podem ser restabelecidos através da criação de corredores
ripários (PUTH; WILSON, 2001).
Nesta paisagem, quase metade da área florestal remanescente está sob efeito de
borda, o que é mais pronunciado em menores fragmentos. Apesar da reconhecida
influência da borda e relevância nos estudos de fragmentação (RIES et al., 2004;
HARPER et al., 2005; LAURANCE, 2008), principalmente devido aos seus efeitos
deletérios, raramente se inclui uma dimensão de habitat relacionada à borda na pesquisa
de heterogeneidade que avalia mais habitats do que o par interior-borda. Uma alta
proporção de bordas tem múltiplas consequências (RANTA et al., 1998). No nível do
fragmento, são esperadas mudanças nas variáveis microclimáticas, estrutura da
vegetação, processos ecológicos e um consequente declínio na riqueza de espécies
(DAVIES-COLLEY et al., 2000; FLETCHER JR, 2005; SANTOS et al., 2008),
enquanto no nível da paisagem, ecossistemas sob efeito de borda são mais vulneráveis a
vários processos externos, tais como invasão biológica (LAURANCE, 2000). Na
paisagem estudada, a manutenção de uma alta proporção de habitats de interior é crucial
para conservação da biodiversidade (WILLIAMS-LINERA et al., 1998; KOLLMANN;
SCHNEIDER, 1999), bem como a indução da recuperação das bordas com vistas a um
enriquecimento progressivo em espécies, como definido por Harper et al. (2005).

67
6. Considerações finais e implicações
Entre os resultados encontrados, uma maior diversidade de habitats em
fragmentos maiores encaixa no padrão esperado. O comportamento imprevisível da
disponibilidade de habitats em pequenos fragmentos, porém, é a exceção que tem
implicações cruciais para conservação. Por um lado, em fragmentos pequenos há um
aumento na proporção de habitats perturbados, tais como ambientes de borda, ou de
habitats de relevo mais acidentado e, consequentemente, mais suscetíveis a perturbações
naturais como deslizamentos, movimentos de matéria e queda de árvores (LINS-E-
SILVA; RODAL, 2008). Além disso, em pequenos fragmentos, a presença de sítios que
são mais frágeis e sensíveis a variações microclimáticas pode ser reduzida. Por outro
ângulo, há casos em que um pequeno fragmento conserva muitos habitats. Nossos dados
revelaram que, entre fragmentos pequenos (<50 ha), 45% têm quatro a seis habitats.
Porém, no mesmo grupo, apenas 25% têm uma diversidade de habitats maior que 2,5.
Isto revela que mesmo quando um pequeno fragmento tem alta riqueza, a equitabilidade
entre tipos de habitats não é alta, levando a uma baixa representação de habitats
particulares. Considerando que a representação de habitats afeta a diversidade através da
alteração significativa das proporções relativas de espécies em habitats contrastantes
(CRAMER; WILLIG, 2005), a pergunta chave é: até que ponto a presença de um
habitat em particular garante uma área adequada para manutenção de populações
viáveis? O modelo unificado de Kadmon e Allouche (2007), integrando área e
heterogeneidade, demonstrou que um efeito negativo da heterogeneidade de habitats na
diversidade pode ocorrer quando áreas pequenas são ricas em habitats. O contexto da
paisagem, porém, poderia auxiliar na compensação deste feito. No caso particular da
paisagem no Centro de Endemismo Pernambuco (SILVA; CASTELETI, 2005), no qual
os fragmentos são próximos entre si (RANTA et al., 1998; TRINDADE et al., 2008),
onde pequenos fragmentos são particularmente importantes para reduzir o isolamento
(RIBEIRO et al., 2009) e onde ainda há fluxo gênico entre indivíduos de populações,
por exemplo, da espécie arbórea Tapirira guianensis (SILVA et al., 2008b), pequenas
porções de habitats em vários fragmentos poderia garantir a manutenção de populações
viáveis (EWERS; DIDHAM, 2006). Atenção especial deve ser dada aos habitats
ripários, devido às suas características antagônicas de serem pequenos em área, mas de
alta importância para conservação.

68
Consideremos as evidências de que, para uma fração considerável de espécies
nas comunidades tropicais, há partição por habitats (CLARK et al., 1999; HARMS et
al., 2001; BOTREL et al., 2002; WRIGHT, 2002; VALENCIA et al., 2004; KESSLER;
LEHNERT, 2009). Se os fragmentos são vistos como mosaicos, cujas partes restringem
a área efetiva de ocupação dessas espécies, a verificação do papel dos habitats
heterogêneos na estrutura das comunidades é essencial à conservação, pois a simples
medida de área do fragmento não é suficiente para assegurar a manutenção de espécies
que dependam de habitats específicos. As três dimensões de habitat utilizadas aqui
proporcionam uma relevante caracterização da disponibilidade de habitats, que pode
guiar a definição de prioridades para futuros inventários e iniciativas de conservação. É
importante ter em mente, ainda, que a relação entre área e heterogeneidade de habitats
não é apenas uma relação numérica, mas a qualidade e configuração espacial dos
habitats remanescentes devem ser consideradas. Desta forma, todos os esforços devem
ser feitos para garantir que todos os habitats sejam preservados em proporções
adequadas para manutenção da diversidade biológica na escala da paisagem.

69
V. CAPÍTULO 3 – Diversidade de assembléias de
plantas de dossel e sub-bosque em fragmentos de
Floresta Atlântica: a área importa?

70
1. Resumo
Neste estudo, objetivou-se inventariar assembléias de plantas de dossel e sub-bosque em
nove fragmentos de floresta atlântica em Pernambuco, Brasil, isolados há pelo menos
25 anos. Utilizando-se três réplicas por tamanhos (grandes, ≈300 ha; médios, ≈100 ha; e
pequenos, ≈ 15 ha), analisou-se a influência da área por estrato, hipoteticamente maior
no sub-bosque, separando-se o efeito da área de um possível efeito de densidade. Em
cada fragmento, instalaram-se 30 parcelas de 0,01 ha para amostra do dossel (CAP ≥ 15
cm); e dentro de cada, uma parcela de 5 x 5 m para amostragem do sub-bosque (CNS ≥
1 cm até CAP < 15 cm), sendo as plantas amostradas marcadas, medidas e identificadas.
Amostras de dossel, sub-bosque e totais foram comparadas entre classes de tamanho de
fragmentos utilizando-se ANOVA de um fator e teste de Tukey, testando-se: percentual
de mortos e lianas, densidade, riqueza, diversidade de Shannon, classes de abundância e
frequência. Estimativas de riqueza também foram obtidas através de curvas de rarefação
com 100 re-amostragens. Os percentuais de mortos de pé e de lianas foram muito
variáveis nos estratos, sem diferenças significativas entre tamanhos. Amostraram-se
4502 plantas no dossel e 9698 no sub-bosque, com médias de densidade
significativamente mais altas no sub-bosque de fragmentos médios. Nos nove
fragmentos, ocorreram 55 famílias e 300 espécies, 192 no dossel e 249 no sub-bosque.
No total, a média de espécies variou de 126 ± 25 em fragmentos grandes a 91 ± 15 nos
pequenos. Nas estimativas por rarefação, o sub-bosque foi significativamente mais rico
nos fragmentos maiores (93,3 ± 13,2). A diversidade do sub-bosque também foi
significativamente maior nos fragmentos grandes, fornecendo evidências à relação
espécie-área. Diferentes respostas em amostras de diferentes idades sinalizaram a
existência de um débito ecológico. Cerca de metade das espécies foram raras, com
proporções por fragmento entre 50 e 60%. Apenas 12% das espécies foram muito
frequentes, cada fragmento exibindo algumas muito abundantes localmente. Quase 60%
das espécies foram infrequentes (120 exclusivas de um fragmento em particular), com
decréscimo nas suas proporções dos fragmentos grandes (29,7%) para os pequenos
(19,5%). Em conjunto, os resultados encontrados descrevem um cenário de alta riqueza,
com riscos de perda de espécies no futuro. Neste contexto, medidas urgentes são
necessárias para minimizar futuras perdas, principalmente a manutenção de todas as
florestas maduras, independentemente do tamanho, nesta paisagem onde pequenas áreas
atuam como importantes complementos para as grandes áreas.

71
2. Introdução
A pesquisa em fragmentação, embora ainda por alcançar uma estrutura
conceitual unificadora (HAILA, 2002; WATSON, 2002), tem como embasamento
teórico principal a relação espécie-área (ARRHENIUS, 1921; GLEASON, 1925), que
descreve o aumento progressivo no número de espécies com aumento da área amostral
(SCHEINER, 2003). MacArthur e Wilson (1967) aplicaram esta relação a ecossistemas
insulares na Teoria de Biogeografia de Ilhas, que prevê um aumento na riqueza em ilhas
maiores. Na teoria, vincularam à área a atuação do processo de extinção local – quanto
maior a ilha, menores as probabilidades de extinção – e vincularam ao isolamento as
probabilidades de migração – menores em ilhas mais isoladas (CONNOR; MCCOY,
1979). A teoria da biogeografia de ilhas explicou o aumento de espécies com o aumento
da área de forma neutra com relação à diversidade de nichos e habitats, assumindo que
as espécies são essencialmente idênticas, e que as ilhas são idênticas nas suas
características, com exceção das variáveis área e isolamento (BROWN; LOMOLINO,
2000). Esta interpretação neutra do efeito da área configura a primeira proposição
explicativa da relação espécie-área, a chamada hipótese da área per se (PRESTON,
1960; RICKLEFS; LOVETTE, 1999; TRIANTIS et al., 2003).
A transposição das idéias da biogeografia de ilhas aos estudos sobre as
consequências da crescente fragmentação de ecossistemas ocorreu de forma acelerada
nos anos subsequentes à publicação da teoria (HAILA, 2002; LAURANCE, 2008),
principalmente com o desenvolvimento da Ecologia de Paisagens (METZGER, 2001).
De fato, mosaicos heterogêneos, nos quais a vegetação remanescente é tipicamente
reduzida e fragmentada para ceder espaço à agricultura, são a forma predominante de
paisagens atuais (SAUNDERS et al., 1991). Esse cenário é mais crítico nos trópicos
úmidos, região de potencial ocorrência das florestas tropicais (WRIGHT; MULLER-
LANDAU, 2006), que, embora reconhecidamente possuam a maior diversidade de
plantas no mundo (GIVNISH, 1999), estão criticamente ameaçadas pela fragmentação e
redução de área (MYERS et al., 2000). A percepção de que a chance de conservação da
diversidade nessas florestas está na manutenção dos remanescentes estimulou o
crescente interesse pela pesquisa em fragmentos florestais tropicais (SAUNDERS et al.,
1991; LAURANCE; BIERREGAARD JR, 1997; LAURANCE, 1999a).

72
Testes da teoria de ilhas em arquipélagos de fragmentos geralmente investigam
mais o efeito da área do que isolamento (ZUIDEMA et al., 1996) e, na maioria das
vezes têm confirmado a relação espécie-área para plantas, em diversas paisagens
(LAURANCE et al., 2002; HILL; CURRAN, 2003; CAGNOLO et al., 2006;
ECHEVERRÍA et al., 2007). Há exemplos, porém, da não confirmação da relação
espécie-área (DEBINSKI; HOLT, 2000; CAYUELA et al., 2006; SANTOS et al.,
2007b), demonstrando que paisagens fragmentadas exibem uma dinâmica típica e
multifacetada, indo além das relações clássicas da teoria (HAILA, 2002; EWERS;
DIDHAM, 2006; LAURANCE, 2008). Novos aspectos tais como a qualidade da matriz
(GASCON et al., 1999), o grau de conectividade (METZGER, 1997; OCHOA-GAONA
et al., 2004; METZGER et al., 2009), o efeito de borda e perturbação antropogênica
(CAYUELA et al., 2006; PEREIRA et al., 2007; TABARELLI et al., 2008) e invasão
de espécies (JANZEN, 1983; TABARELLI et al., 1999; CAGNOLO et al., 2006)
exercem também influência na biota de fragmentos tipo-ilha sobre o continente.
Além dos vários aspectos típicos da fragmentação, remanescentes de áreas
anteriormente contínuas constituem uma amostra da riqueza dessas áreas (WATSON,
2002), cuja dinâmica pós-fragmentação depende da história da paisagem, sendo
esperado um tempo de acomodação da biota nos fragmentos (LAURANCE, 2008).
Desta forma, a riqueza em um fragmento pode estar mais relacionada à estrutura de
paisagens passadas do que a do presente, o que tem sido comprovado em fragmentos de
comunidades vegetais herbáceas (LINDBORG; ERIKSSON, 2004; ERNOULT et al.,
2006; HELM et al., 2006) e arbóreas (PALTTO et al., 2006; METZGER et al., 2009).
Este tempo de acomodação seria especialmente importante para indivíduos de maior
longevidade, como árvores (PALTTO et al., 2006), o que foi comprovado por Metzger
et al. (2009), ao revelar que a estrutura de paisagens passadas, principalmente o
tamanho dos fragmentos, significativamente explica a riqueza da comunidade arbórea
em florestas. Assim, é provável que os efeitos da paisagem atual sejam mais atuantes e
perceptíveis sobre a comunidade de “árvores do futuro” (METZGER, 1998) ou no sub-
bosque (OCHOA-GAONA et al., 2004), que inclui o estrato de regeneração do dossel e
espécies de ciclo de vida mais curto, tais como arvoretas, arbustos, palmeiras e fetos
arborescentes, plântulas e ervas (GOMES et al., 2009).
Entre as florestas tropicais, a floresta atlântica apresenta uma das maiores taxas
de redução de área e fragmentação no mundo (RIBEIRO et al., 2009), sendo

73
considerada um hotspot para conservação da biodiversidade (MYERS et al., 2000). Na
sua porção mais a nordeste, estão os menores percentuais de florestas remanescentes,
especialmente na sub-região biogeográfica “Centro de Endemismo Pernambuco”, sensu
Silva e Casteleti (2005), na qual restam apenas 12% das florestas (RIBEIRO et al.,
2009) e não há nenhum fragmento maior que 3500 ha (OLIVEIRA et al., 2004). Na
paisagem típica desta região, predominam arquipélagos de fragmentos pequenos e
recentes, formados a partir da década de 1970, em sua maioria, e distribuídos com baixa
densidade (≈0,7 fragmento/ km2) em uma matriz de cana-de-açúcar (RANTA et al.,
1998; TRINDADE et al., 2008). Quanto ao isolamento, os fragmentos geralmente
ocorrem em grupos, com uma distância entre manchas de um mesmo grupo inferior a
350 m (RANTA et al., 1998).
Este cenário configura, portanto, um sistema apropriado para analisar a relação
entre área e riqueza de espécies, constituindo um “experimento pronto”, ideal para
estudos de insularização (SAUNDERS et al., 1991; CAGNOLO et al., 2006),
fornecendo réplicas de tamanho em um mesmo contexto de paisagem, com a vantagem
da neutralidade da variável isolamento como possível fonte de influência. Esta análise,
objeto do presente trabalho, permitirá compreender o impacto da redução de área na
diversidade de plantas e, assim, auxiliar na definição de estratégias mais realísticas de
conservação e manejo da floresta atlântica, numa região carente de tais estudos e de
grandes remanescentes, geralmente prioritários para conservação (TABARELLI;
GASCON, 2005). Os nossos objetivos foram: inventariar assembléias de plantas
lenhosas de dossel e sub-bosque em fragmentos de diferentes tamanhos, investigar a
influência da área na diversidade total, analisar a influência da área por estrato,
hipoteticamente maior no sub-bosque, considerando-se sua menor idade e respostas
mais imediatas às mudanças na paisagem, e separar o efeito da área de um possível
efeito de densidade, através da rarefação das amostras.

74
3. Material e métodos
3.1. Área de estudo
Os fragmentos florestais estudados estão localizados nos municípios de Igarassu
e Goiana, Zona da Mata Norte do estado de Pernambuco, Nordeste do Brasil, na
propriedade da Usina São José/ Grupo Cavalcanti Petribú. Para descrição da geologia,
relevo, clima e histórico da área, ver item “Área de Estudo”, p. 6.
Na propriedade, há 106 fragmentos de floresta madura, dos quais nove foram
selecionados para estudo (Figura V.1, Tabela V.1), com base no estado de conservação,
tempo de isolamento e tamanho, sendo três grandes (G), três médios (M) e três
pequenos (P). Todos os fragmentos estudados foram isolados há pelo menos 25 anos.
Figura V.1. Localização dos nove fragmentos estudados na paisagem altamente
fragmentada na Zona da Mata Norte, Pernambuco, Nordeste do Brasil.
G1
G2
G3
M1
M2
M3
P1
P2
P3
Fragmentosestudados

75
Desenho amostral
As assembléias de plantas foram amostradas em dois estratos: dossel e sub-
bosque. O estrato de dossel incluiu todos os indivíduos lenhosos com circunferência a
altura do peito (CAP) ≥ 15 cm, compreendendo árvores, lianas e plantas mortas de pé. A
amostra de sub-bosque incluiu todas as plantas lenhosas vivas e mortas de pé com
circunferência ao nível do solo (CNS) a partir de 1 cm até CAP < 15 cm. Optou-se por
definir o critério de inclusão do sub-bosque ao nível do solo e sem limite de altura, por
ser este mais inclusivo para indivíduos jovens, arvoretas e arbustos (GOMES et al.,
2009).
Nos habitats mais frequentes nos fragmentos, ou seja, borda (até 50 m da borda
florestal), interior de encosta e habitats não ripários de interior (para descrição completa
dos habitats, ver capítulo 2), 30 parcelas de 0,01 ha (10 x 10 m) foram instaladas por
fragmento, para amostragem do dossel, sistematicamente estabelecidas,
georreferenciadas e distribuídas em grupos de 10 parcelas, contíguas em pelo menos um
lado nesses grupos. No interior de cada parcela de dossel, foi instalada parcela menor
para amostragem do sub-bosque (5 x 5 m). As plantas incluídas nas amostras de dossel
e sub-bosque foram marcadas e medidas no campo, coletadas e identificadas, seguindo
procedimentos gerais descritos na “Área de Estudo/ Metodologia Geral”, p. 11.
Análise dos dados
O banco de dados foi preparado e manejado no software Mata Nativa 2.0
(SOUZA et al., 2006). Nas análises, várias questões aplicadas a conjuntos particulares
do banco de dados. Primeiramente, foi explorada a participação dos indivíduos mortos
de pé e lianas nas amostras em fragmentos de tamanhos distintos. Para tanto, totais e
percentuais de indivíduos mortos e lianas foram calculados para cada fragmento e
comparados através de ANOVA de um fator (ZAR, 1999). Em seguida, considerando o
conjunto de indivíduos vivos, os seguintes atributos descritivos de assembléias de
plantas foram obtidos: densidade, riqueza (número de espécies) e diversidade de
Shannon H’ (MAGURRAN, 2004), tabulados para cada estrato e para amostra total por
fragmento. Apesar de a área amostral ser equivalente em todos os fragmentos, a
densidade de indivíduos variou consideravelmente entre fragmentos e estratos, o que
poderia influenciar a riqueza e diversidade devido a um “efeito de densidade”
(DENSLOW, 1995) e não devido à área per se. Para comparar número de espécies com

76
base no número de indivíduos, a riqueza foi estimada utilizando curvas de rarefação.
Estas curvas foram produzidas através de repetidas re-amostragens aleatórias (100
randomizações) das amostras de indivíduos, obtendo-se três curvas para cada fragmento
(amostra total, dossel e sub-bosque) e 27 curvas no total. O resultado de rarefação
corresponde à expectativa estatística de uma curva de acumulação de espécies obtida a
partir de diferentes ordens de amostragem dos indivíduos (GOTELLI; COLWELL,
2001). As curvas de rarefação foram computadas no software EstimateS 7.5.2
(COLWELL, 2006), sem reposição em cada re-amostragem. Para comparação, a riqueza
em cada fragmento, para amostra total, dossel e sub-bosque, foi obtida de um número
equivalente de indivíduos, sendo este número fornecido pela menor densidade da
amostra em questão entre os nove fragmentos.
A relação entre área do fragmento e os atributos vegetais das assembléias foi
analisada através de comparações de médias obtidas de grupos de três fragmentos de
tamanhos similares: grandes, médios e pequenos (G, M, e P, respectivamente; Tabela
V.1). As médias foram comparadas utilizando-se ANOVA de um fator e teste de Tukey
para comparações a posteriori. Adicionalmente, foi testada a relação entre área do
fragmento e diversidade utilizando-se regressão linear simples. Área dos fragmentos, a
variável independente, foi transformada (log10) para esta análise.
Números e percentuais de espécies amostradas em cada estrato e comuns a
ambos foram calculados para cada fragmento. Número de espécies raras (até cinco
indivíduos), comuns (6 a 100 plantas) e espécies muito abundantes (mais de 100
indivíduos) foram obtidos por estrato e por fragmento, assim como números de espécies
exclusivas. Considerando que o limite para definição de espécies raras varia
amplamente, dependendo da escala de investigação (MAGURRAN, 2004), o ponto de
corte em cinco indivíduos foi escolhido, devido à amostra por fragmento ser inferior a 1
ha, não permitindo a definição de raras como aquelas com densidade ≤ 1 planta/ ha
(TABANEZ; VIANA, 2000; GUEDES-BRUNI et al., 2009). As espécies foram
classificadas de acordo com a frequência, como aplicado por Metzger et al. (1997) e
Santos et al. (2007b), com modificações, em: infrequente (registrada em 1-2
fragmentos), pouco frequente (2-5), frequente (6-7) e muito frequente (presente em 8 a
9 fragmentos), para amostra total e por fragmento. Diferenças significativas nas
proporções de espécies com relação ao estrato, abundância e frequência foram
analisadas entre pares de fragmentos utilizando qui-quadrado, enquanto diferenças nas

77
proporções numa mesma classe de cada variável, entre fragmentos de diferentes
tamanhos, foram testadas através de ANOVA de um fator. Todas as análises foram
conduzidas no software BioEstat 5.0 (AYRES et al., 2007).
Tabela V.1. Características dos nove fragmentos florestais estudados na Zona da Mata
Norte do estado de Pernambuco, Nordeste do Brasil.
Nome do fragmento
Código Coordenadas geográficas Área (ha) Altitude max/min (m)
Zambana G1 07º41'29,13"; 07º43'17,09" S 35º00'08,75"; 34º58'38,78" W
387,85 115/20
Macacos G2 07º46'02,89"; 07º47'40,82" S 34º59'42,95"; 35º01'06,31" W
356,92 155/40
Piedade G3 07º49'12,66"; 07º50'55,43" S 34º59'21,29"; 35º00'35,92" W
305,78 113/30
BR M1 07º47'14,22"; 07º48'06,01" S 34º56'39,81"; 34º55'59,58" W
91,36 85/20
Chave M2 07º48'30,12"; 07º49'34,85" S 34º58'49,46"; 34º57'37,58" W
89,28 100/25
Cruzinha M3 07º41'24,87"; 07º42'10,32" S 34º58'13,76"; 34º57'31,32" W
87,27 100/25
Pezinho P1 07°47'27,82"; 07°48'01,04" S 35°01'25,86"; 35°01'00,64" W
29,89 125/50
Vespas P2 07º44'50,11"; 07º45'08,29" S 34º59'02,37"; 34º58'45,04" W
13,80 105/45
Santa Helena P3 07º46'59,39"; 07º47'16,65" S 34º56'41,80"; 34º56'29,24" W
11,85 85/20
4. Resultados
4.1. Descrição geral das assembléias de plantas
Foram amostradas 15.360 plantas, sendo 4995 no dossel e 10.365 no sub-
bosque. No dossel, o percentual de mortos de pé foi muito variável entre fragmentos
(Tabela V.2), sendo mais baixo em M1 (1,8%), seguido por proporções entre 3 e 8% na
maioria dos fragmentos e chegando a mais de 10% em G3, M3 e P2. No sub-bosque,
registrou-se um padrão semelhante, com percentuais de mortos variando entre 1,5 (M1)
e 13,3% (M3). Não houve diferenças significativas entre as médias dos fragmentos de
diferentes tamanhos. No dossel, a contribuição de lianas variou pouco e sem diferenças
significativas, com percentuais entre 0,3 em M2 e 4,7% em P2.
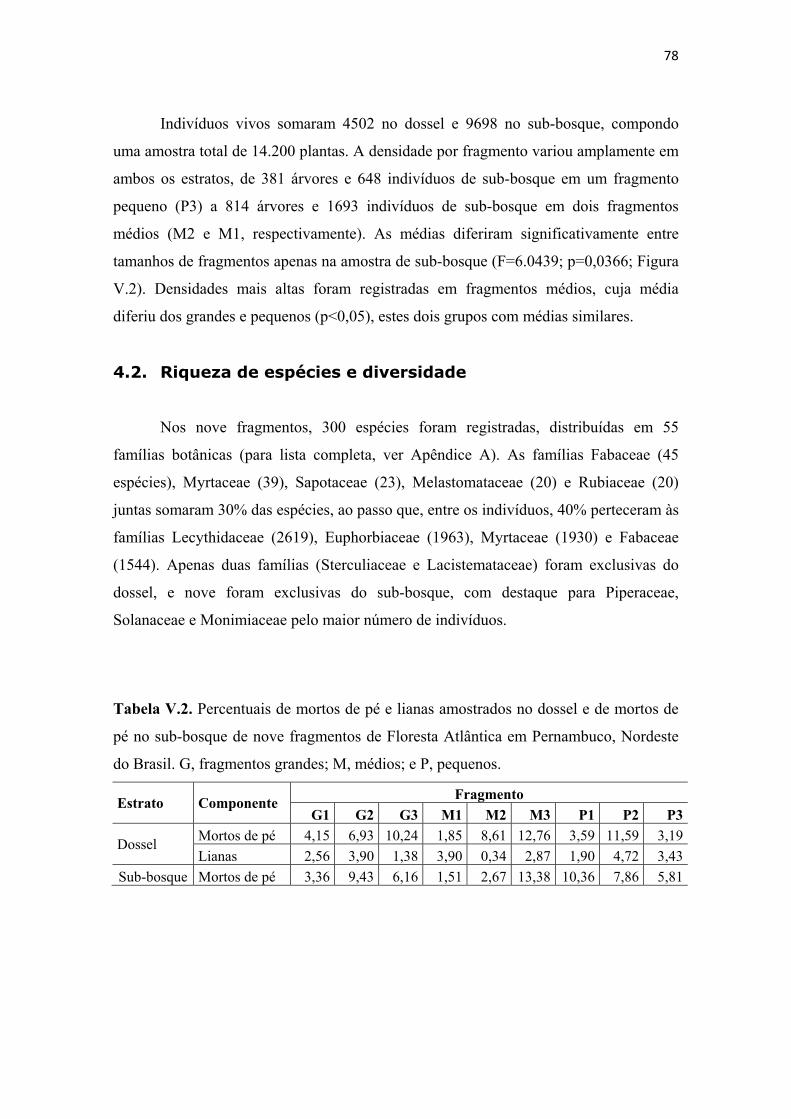
78
Indivíduos vivos somaram 4502 no dossel e 9698 no sub-bosque, compondo
uma amostra total de 14.200 plantas. A densidade por fragmento variou amplamente em
ambos os estratos, de 381 árvores e 648 indivíduos de sub-bosque em um fragmento
pequeno (P3) a 814 árvores e 1693 indivíduos de sub-bosque em dois fragmentos
médios (M2 e M1, respectivamente). As médias diferiram significativamente entre
tamanhos de fragmentos apenas na amostra de sub-bosque (F=6.0439; p=0,0366; Figura
V.2). Densidades mais altas foram registradas em fragmentos médios, cuja média
diferiu dos grandes e pequenos (p<0,05), estes dois grupos com médias similares.
4.2. Riqueza de espécies e diversidade
Nos nove fragmentos, 300 espécies foram registradas, distribuídas em 55
famílias botânicas (para lista completa, ver Apêndice A). As famílias Fabaceae (45
espécies), Myrtaceae (39), Sapotaceae (23), Melastomataceae (20) e Rubiaceae (20)
juntas somaram 30% das espécies, ao passo que, entre os indivíduos, 40% perteceram às
famílias Lecythidaceae (2619), Euphorbiaceae (1963), Myrtaceae (1930) e Fabaceae
(1544). Apenas duas famílias (Sterculiaceae e Lacistemataceae) foram exclusivas do
dossel, e nove foram exclusivas do sub-bosque, com destaque para Piperaceae,
Solanaceae e Monimiaceae pelo maior número de indivíduos.
Tabela V.2. Percentuais de mortos de pé e lianas amostrados no dossel e de mortos de
pé no sub-bosque de nove fragmentos de Floresta Atlântica em Pernambuco, Nordeste
do Brasil. G, fragmentos grandes; M, médios; e P, pequenos.
Estrato Componente Fragmento G1 G2 G3 M1 M2 M3 P1 P2 P3
Dossel Mortos de pé 4,15 6,93 10,24 1,85 8,61 12,76 3,59 11,59 3,19Lianas 2,56 3,90 1,38 3,90 0,34 2,87 1,90 4,72 3,43
Sub-bosque Mortos de pé 3,36 9,43 6,16 1,51 2,67 13,38 10,36 7,86 5,81
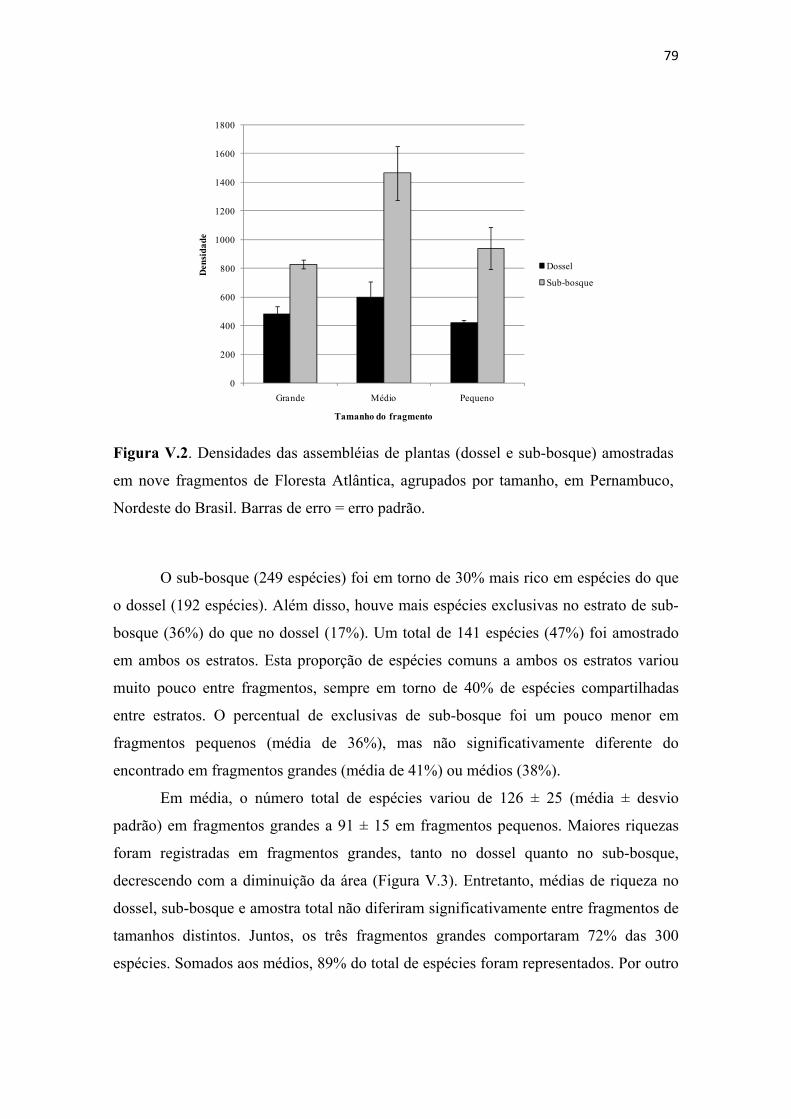
79
Figura V.2. Densidades das assembléias de plantas (dossel e sub-bosque) amostradas
em nove fragmentos de Floresta Atlântica, agrupados por tamanho, em Pernambuco,
Nordeste do Brasil. Barras de erro = erro padrão.
O sub-bosque (249 espécies) foi em torno de 30% mais rico em espécies do que
o dossel (192 espécies). Além disso, houve mais espécies exclusivas no estrato de sub-
bosque (36%) do que no dossel (17%). Um total de 141 espécies (47%) foi amostrado
em ambos os estratos. Esta proporção de espécies comuns a ambos os estratos variou
muito pouco entre fragmentos, sempre em torno de 40% de espécies compartilhadas
entre estratos. O percentual de exclusivas de sub-bosque foi um pouco menor em
fragmentos pequenos (média de 36%), mas não significativamente diferente do
encontrado em fragmentos grandes (média de 41%) ou médios (38%).
Em média, o número total de espécies variou de 126 ± 25 (média ± desvio
padrão) em fragmentos grandes a 91 ± 15 em fragmentos pequenos. Maiores riquezas
foram registradas em fragmentos grandes, tanto no dossel quanto no sub-bosque,
decrescendo com a diminuição da área (Figura V.3). Entretanto, médias de riqueza no
dossel, sub-bosque e amostra total não diferiram significativamente entre fragmentos de
tamanhos distintos. Juntos, os três fragmentos grandes comportaram 72% das 300
espécies. Somados aos médios, 89% do total de espécies foram representados. Por outro
0
200
400
600
800
1000
1200
1400
1600
1800
Grande Médio Pequeno
Den
sida
de
Tamanho do fragmento
Dossel
Sub-bosque
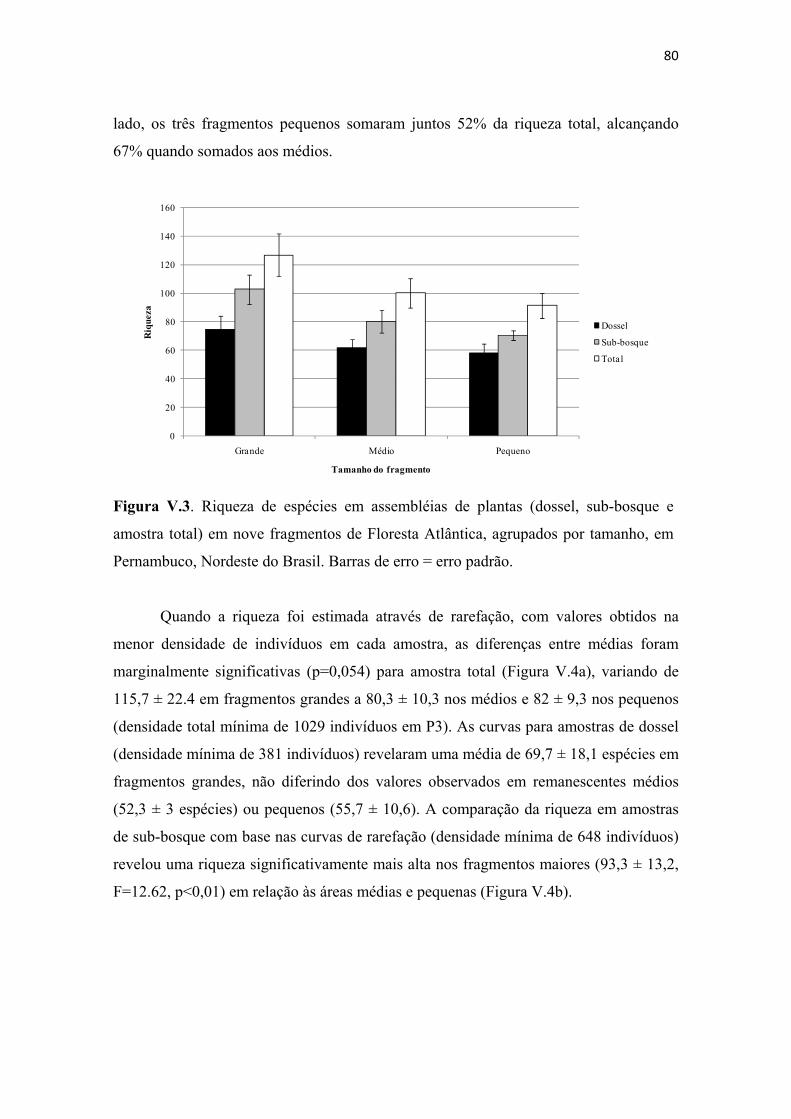
80
lado, os três fragmentos pequenos somaram juntos 52% da riqueza total, alcançando
67% quando somados aos médios.
Figura V.3. Riqueza de espécies em assembléias de plantas (dossel, sub-bosque e
amostra total) em nove fragmentos de Floresta Atlântica, agrupados por tamanho, em
Pernambuco, Nordeste do Brasil. Barras de erro = erro padrão.
Quando a riqueza foi estimada através de rarefação, com valores obtidos na
menor densidade de indivíduos em cada amostra, as diferenças entre médias foram
marginalmente significativas (p=0,054) para amostra total (Figura V.4a), variando de
115,7 ± 22.4 em fragmentos grandes a 80,3 ± 10,3 nos médios e 82 ± 9,3 nos pequenos
(densidade total mínima de 1029 indivíduos em P3). As curvas para amostras de dossel
(densidade mínima de 381 indivíduos) revelaram uma média de 69,7 ± 18,1 espécies em
fragmentos grandes, não diferindo dos valores observados em remanescentes médios
(52,3 ± 3 espécies) ou pequenos (55,7 ± 10,6). A comparação da riqueza em amostras
de sub-bosque com base nas curvas de rarefação (densidade mínima de 648 indivíduos)
revelou uma riqueza significativamente mais alta nos fragmentos maiores (93,3 ± 13,2,
F=12.62, p<0,01) em relação às áreas médias e pequenas (Figura V.4b).
0
20
40
60
80
100
120
140
160
Grande Médio Pequeno
Riq
ueza
Tamanho do fragmento
Dossel
Sub-bosque
Total
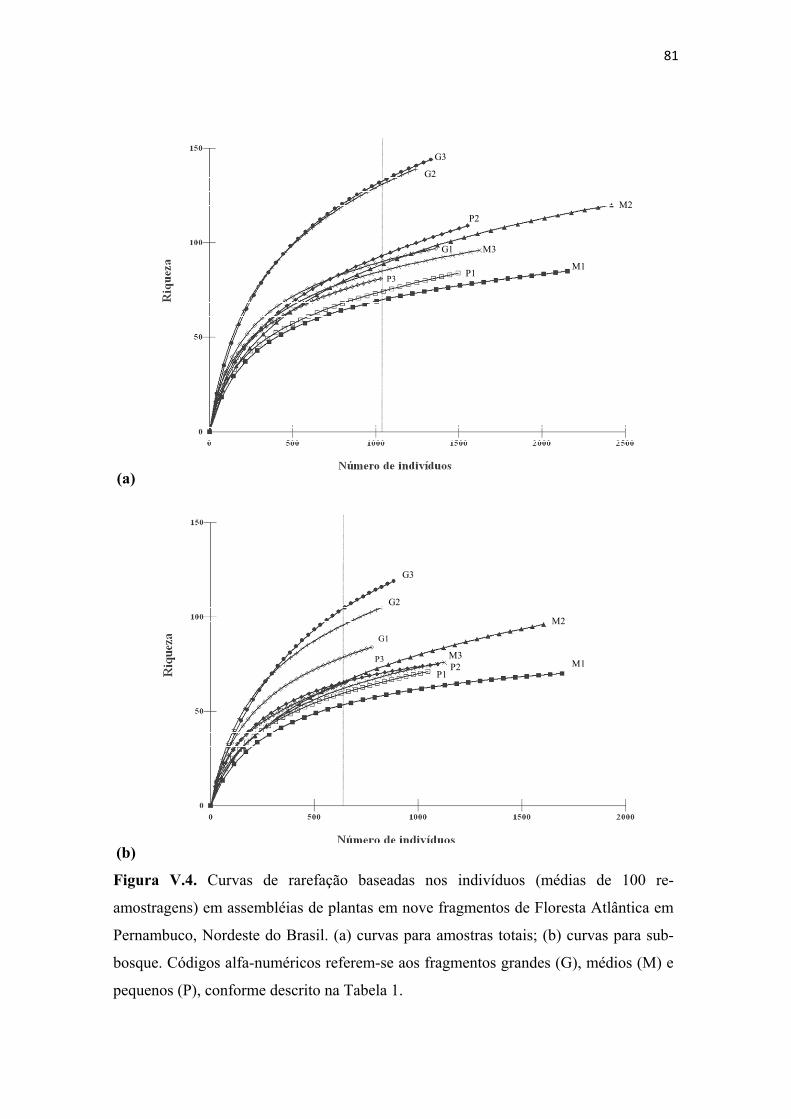
81
(a)
(b)
Figura V.4. Curvas de rarefação baseadas nos indivíduos (médias de 100 re-
amostragens) em assembléias de plantas em nove fragmentos de Floresta Atlântica em
Pernambuco, Nordeste do Brasil. (a) curvas para amostras totais; (b) curvas para sub-
bosque. Códigos alfa-numéricos referem-se aos fragmentos grandes (G), médios (M) e
pequenos (P), conforme descrito na Tabela 1.
G3
G2
M2
M1
M3
P1
P2
P3
G1
G3
G2
M3M1
M2
P1P2
P3
G1
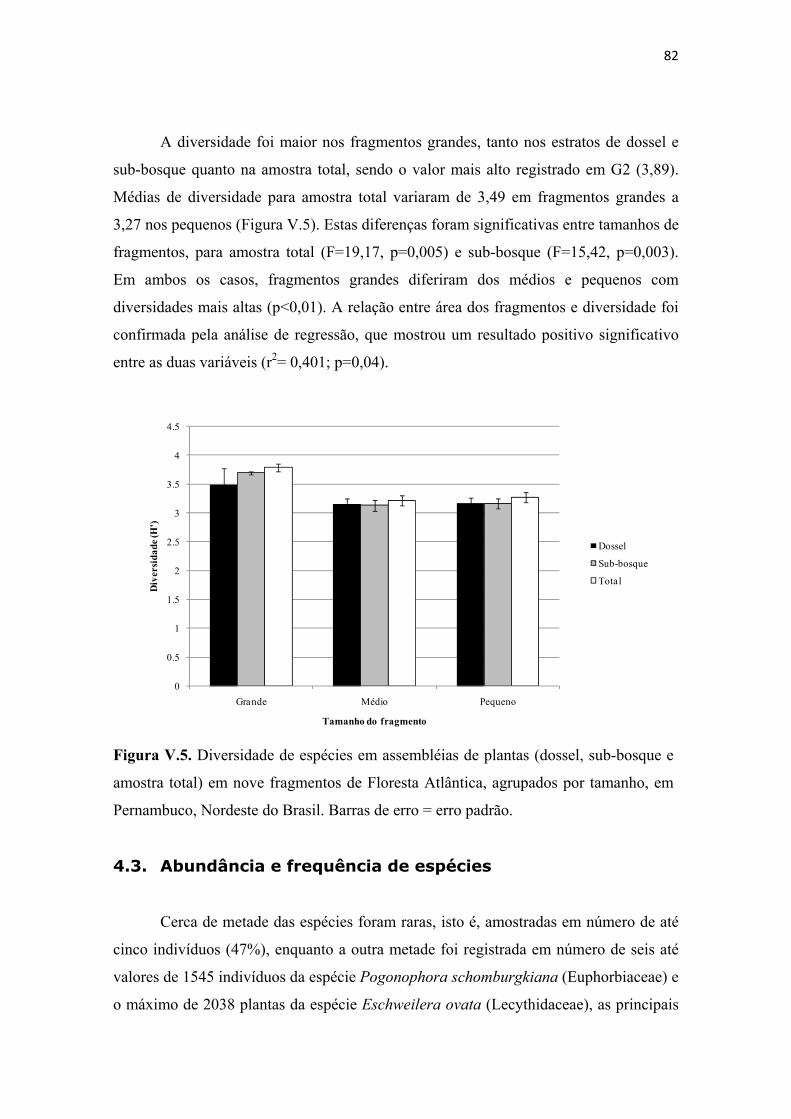
82
A diversidade foi maior nos fragmentos grandes, tanto nos estratos de dossel e
sub-bosque quanto na amostra total, sendo o valor mais alto registrado em G2 (3,89).
Médias de diversidade para amostra total variaram de 3,49 em fragmentos grandes a
3,27 nos pequenos (Figura V.5). Estas diferenças foram significativas entre tamanhos de
fragmentos, para amostra total (F=19,17, p=0,005) e sub-bosque (F=15,42, p=0,003).
Em ambos os casos, fragmentos grandes diferiram dos médios e pequenos com
diversidades mais altas (p<0,01). A relação entre área dos fragmentos e diversidade foi
confirmada pela análise de regressão, que mostrou um resultado positivo significativo
entre as duas variáveis (r2= 0,401; p=0,04).
Figura V.5. Diversidade de espécies em assembléias de plantas (dossel, sub-bosque e
amostra total) em nove fragmentos de Floresta Atlântica, agrupados por tamanho, em
Pernambuco, Nordeste do Brasil. Barras de erro = erro padrão.
4.3. Abundância e frequência de espécies
Cerca de metade das espécies foram raras, isto é, amostradas em número de até
cinco indivíduos (47%), enquanto a outra metade foi registrada em número de seis até
valores de 1545 indivíduos da espécie Pogonophora schomburgkiana (Euphorbiaceae) e
o máximo de 2038 plantas da espécie Eschweilera ovata (Lecythidaceae), as principais
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
Grande Médio Pequeno
Div
ersi
dade
(H')
Tamanho do fragmento
Dossel
Sub-bosque
Total

83
entre 32 espécies muito abundantes (Tabela V.3). Considerando a amostra total por
fragmento, proporções de espécies raras variaram de uma média de 59,45% em
fragmentos grandes a 52,88% em fragmentos médios (Tabela V.4). Analisando o dossel,
proporções médias mais altas de espécies raras foram encontradas: 72,17% em
fragmentos grandes, e médias similares em torno de 67% em remanescentes médios e
pequenos. No sub-bosque, proporções médias foram 67,87% em grandes, 56,44% em
médios e 59,96% em fragmentos pequenos. Essas proporções de espécies raras não
diferiram entre fragmentos de tamanhos distintos em nenhum dos estratos.
Apenas 7% de todas as espécies foram registradas em nove fragmentos, e 5% em
oito, juntas perfazendo 12% de espécies muito frequentes (Tabela V.3), que
representaram 38 a 87% da densidade total nos nove fragmentos estudados (Figura
V.6). Apesar de uma clara tendência a uma maior densidade de espécies muito
frequentes em fragmentos médios e pequenos, essas diferenças não foram significativas.
Entre as muito frequentes, a maioria foi também muito abundante, destacando-se,
principalmente, espécies das famílias Fabaceae e Myrtaceae. Excetuando-se as espécies
E. ovata e P. schomburgkiana, as mais frequentes e abundantes, cada fragmento teve
uma ou algumas espécies muito abundantes localmente, como Thyrsodium spruceanum
em G1, Tovomita mangle e Anaxagorea dolichocarpa em G3, Pouteria gardneri e
Albizia pedicellaris em M1, Maytenus distichophyla em M2, Myrcia racemosa em M3,
Myrcia guianensis em P1 e Brosimum guianense em P3 (Tabela V.3).
Quase 60% foram registradas em apenas um ou dois fragmentos, sendo
classificadas como infrequentes, entre as quais 120 espécies (40% do total) foram
exclusivas de um fragmento em particular. O número de exclusivas por fragmento
variou bastante, independentemente da área, em valores tão distintos quanto seis
espécies (menos de 1% dos indivíduos) em G1 a 33 espécies em G3 (7% dos
indivíduos) e 23 espécies (10% dos indivíduos) em M2 (Figura V.6). Entre aquelas 120
exclusivas de um fragmento, 90% foram também raras, isto é, amostradas em números
de até cinco indivíduos, restando apenas 12 espécies exclusivas e, ao mesmo tempo,
com densidade > 5 (Tabela V.5), destacando-se espécies das famílias Myrtaceae e
Euphorbiaceae.
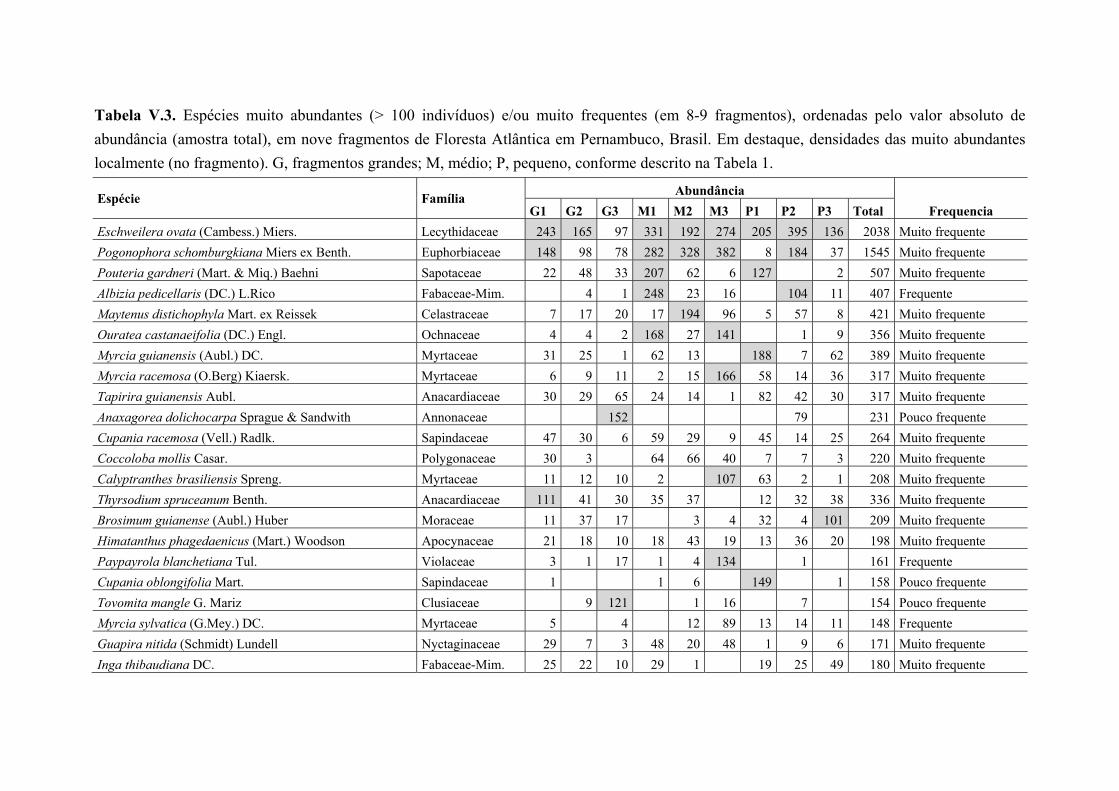
84
Tabela V.3. Espécies muito abundantes (> 100 indivíduos) e/ou muito frequentes (em 8-9 fragmentos), ordenadas pelo valor absoluto de abundância (amostra total), em nove fragmentos de Floresta Atlântica em Pernambuco, Brasil. Em destaque, densidades das muito abundantes localmente (no fragmento). G, fragmentos grandes; M, médio; P, pequeno, conforme descrito na Tabela 1.
Espécie Família Abundância Frequencia G1 G2 G3 M1 M2 M3 P1 P2 P3 Total
Eschweilera ovata (Cambess.) Miers. Lecythidaceae 243 165 97 331 192 274 205 395 136 2038 Muito frequente Pogonophora schomburgkiana Miers ex Benth. Euphorbiaceae 148 98 78 282 328 382 8 184 37 1545 Muito frequente Pouteria gardneri (Mart. & Miq.) Baehni Sapotaceae 22 48 33 207 62 6 127 2 507 Muito frequente Albizia pedicellaris (DC.) L.Rico Fabaceae-Mim. 4 1 248 23 16 104 11 407 Frequente Maytenus distichophyla Mart. ex Reissek Celastraceae 7 17 20 17 194 96 5 57 8 421 Muito frequente Ouratea castanaeifolia (DC.) Engl. Ochnaceae 4 4 2 168 27 141 1 9 356 Muito frequente Myrcia guianensis (Aubl.) DC. Myrtaceae 31 25 1 62 13 188 7 62 389 Muito frequente Myrcia racemosa (O.Berg) Kiaersk. Myrtaceae 6 9 11 2 15 166 58 14 36 317 Muito frequente Tapirira guianensis Aubl. Anacardiaceae 30 29 65 24 14 1 82 42 30 317 Muito frequente Anaxagorea dolichocarpa Sprague & Sandwith Annonaceae 152 79 231 Pouco frequente Cupania racemosa (Vell.) Radlk. Sapindaceae 47 30 6 59 29 9 45 14 25 264 Muito frequente Coccoloba mollis Casar. Polygonaceae 30 3 64 66 40 7 7 3 220 Muito frequente Calyptranthes brasiliensis Spreng. Myrtaceae 11 12 10 2 107 63 2 1 208 Muito frequente Thyrsodium spruceanum Benth. Anacardiaceae 111 41 30 35 37 12 32 38 336 Muito frequente Brosimum guianense (Aubl.) Huber Moraceae 11 37 17 3 4 32 4 101 209 Muito frequente Himatanthus phagedaenicus (Mart.) Woodson Apocynaceae 21 18 10 18 43 19 13 36 20 198 Muito frequente Paypayrola blanchetiana Tul. Violaceae 3 1 17 1 4 134 1 161 Frequente Cupania oblongifolia Mart. Sapindaceae 1 1 6 149 1 158 Pouco frequente Tovomita mangle G. Mariz Clusiaceae 9 121 1 16 7 154 Pouco frequente Myrcia sylvatica (G.Mey.) DC. Myrtaceae 5 4 12 89 13 14 11 148 Frequente Guapira nitida (Schmidt) Lundell Nyctaginaceae 29 7 3 48 20 48 1 9 6 171 Muito frequente Inga thibaudiana DC. Fabaceae-Mim. 25 22 10 29 1 19 25 49 180 Muito frequente
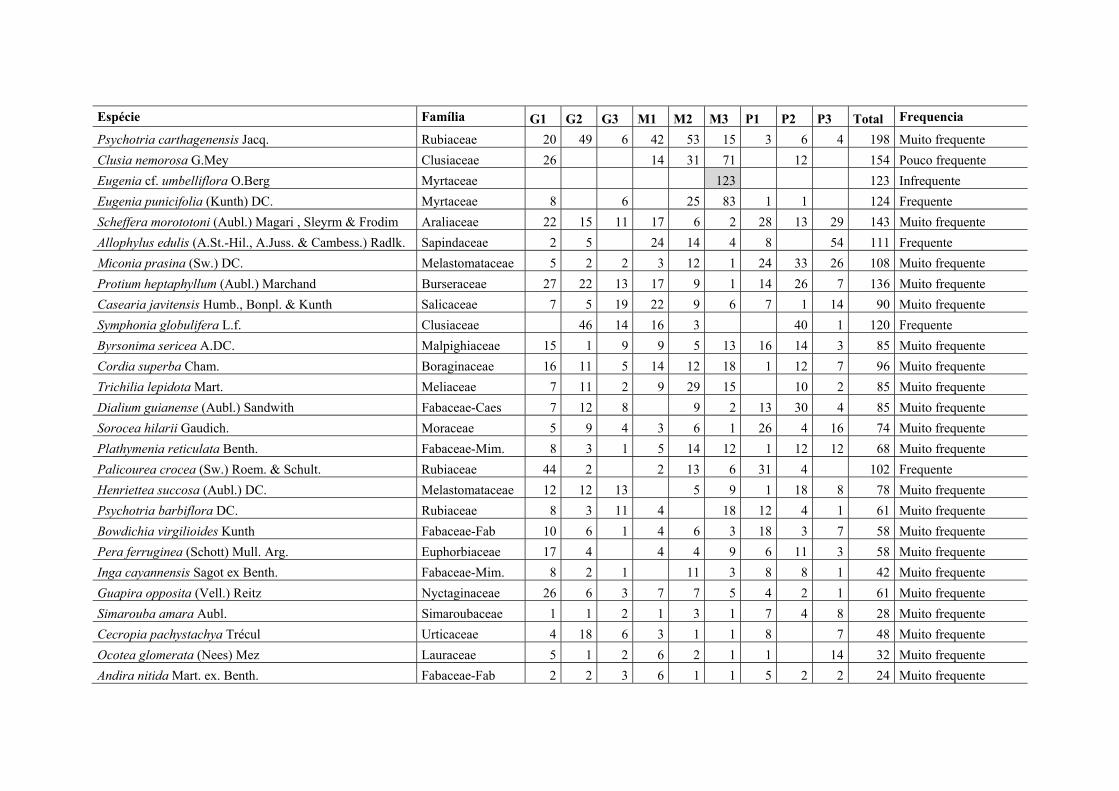
85
Espécie Família G1 G2 G3 M1 M2 M3 P1 P2 P3 Total Frequencia
Psychotria carthagenensis Jacq. Rubiaceae 20 49 6 42 53 15 3 6 4 198 Muito frequente Clusia nemorosa G.Mey Clusiaceae 26 14 31 71 12 154 Pouco frequente Eugenia cf. umbelliflora O.Berg Myrtaceae 123 123 Infrequente Eugenia punicifolia (Kunth) DC. Myrtaceae 8 6 25 83 1 1 124 Frequente Scheffera morototoni (Aubl.) Magari , Sleyrm & Frodim Araliaceae 22 15 11 17 6 2 28 13 29 143 Muito frequente Allophylus edulis (A.St.-Hil., A.Juss. & Cambess.) Radlk. Sapindaceae 2 5 24 14 4 8 54 111 Frequente Miconia prasina (Sw.) DC. Melastomataceae 5 2 2 3 12 1 24 33 26 108 Muito frequente Protium heptaphyllum (Aubl.) Marchand Burseraceae 27 22 13 17 9 1 14 26 7 136 Muito frequente Casearia javitensis Humb., Bonpl. & Kunth Salicaceae 7 5 19 22 9 6 7 1 14 90 Muito frequente Symphonia globulifera L.f. Clusiaceae 46 14 16 3 40 1 120 Frequente Byrsonima sericea A.DC. Malpighiaceae 15 1 9 9 5 13 16 14 3 85 Muito frequente Cordia superba Cham. Boraginaceae 16 11 5 14 12 18 1 12 7 96 Muito frequente Trichilia lepidota Mart. Meliaceae 7 11 2 9 29 15 10 2 85 Muito frequente Dialium guianense (Aubl.) Sandwith Fabaceae-Caes 7 12 8 9 2 13 30 4 85 Muito frequente Sorocea hilarii Gaudich. Moraceae 5 9 4 3 6 1 26 4 16 74 Muito frequente Plathymenia reticulata Benth. Fabaceae-Mim. 8 3 1 5 14 12 1 12 12 68 Muito frequente Palicourea crocea (Sw.) Roem. & Schult. Rubiaceae 44 2 2 13 6 31 4 102 Frequente Henriettea succosa (Aubl.) DC. Melastomataceae 12 12 13 5 9 1 18 8 78 Muito frequente Psychotria barbiflora DC. Rubiaceae 8 3 11 4 18 12 4 1 61 Muito frequente Bowdichia virgilioides Kunth Fabaceae-Fab 10 6 1 4 6 3 18 3 7 58 Muito frequente Pera ferruginea (Schott) Mull. Arg. Euphorbiaceae 17 4 4 4 9 6 11 3 58 Muito frequente Inga cayannensis Sagot ex Benth. Fabaceae-Mim. 8 2 1 11 3 8 8 1 42 Muito frequente Guapira opposita (Vell.) Reitz Nyctaginaceae 26 6 3 7 7 5 4 2 1 61 Muito frequente Simarouba amara Aubl. Simaroubaceae 1 1 2 1 3 1 7 4 8 28 Muito frequente Cecropia pachystachya Trécul Urticaceae 4 18 6 3 1 1 8 7 48 Muito frequente Ocotea glomerata (Nees) Mez Lauraceae 5 1 2 6 2 1 1 14 32 Muito frequente Andira nitida Mart. ex. Benth. Fabaceae-Fab 2 2 3 6 1 1 5 2 2 24 Muito frequente
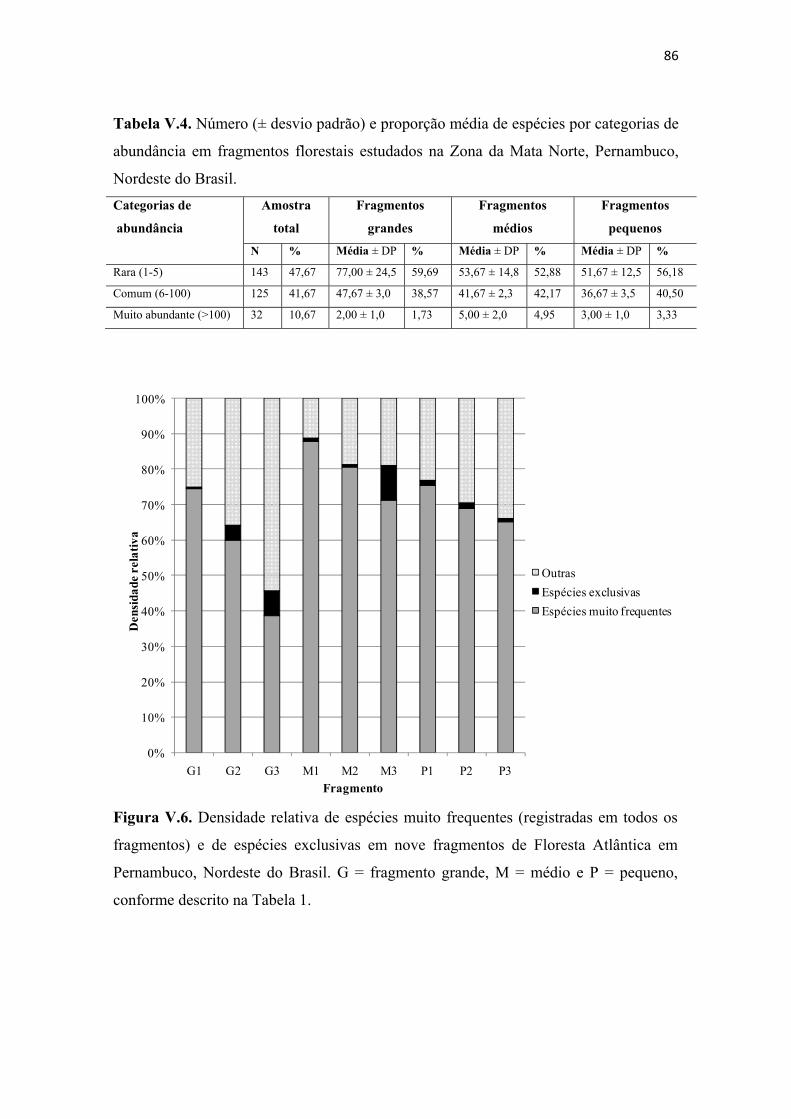
86
Tabela V.4. Número (± desvio padrão) e proporção média de espécies por categorias de
abundância em fragmentos florestais estudados na Zona da Mata Norte, Pernambuco,
Nordeste do Brasil. Categorias de
abundância
Amostra
total
Fragmentos
grandes
Fragmentos
médios
Fragmentos
pequenos
N % Média ± DP % Média ± DP % Média ± DP %
Rara (1-5) 143 47,67 77,00 ± 24,5 59,69 53,67 ± 14,8 52,88 51,67 ± 12,5 56,18
Comum (6-100) 125 41,67 47,67 ± 3,0 38,57 41,67 ± 2,3 42,17 36,67 ± 3,5 40,50
Muito abundante (>100) 32 10,67 2,00 ± 1,0 1,73 5,00 ± 2,0 4,95 3,00 ± 1,0 3,33
Figura V.6. Densidade relativa de espécies muito frequentes (registradas em todos os
fragmentos) e de espécies exclusivas em nove fragmentos de Floresta Atlântica em
Pernambuco, Nordeste do Brasil. G = fragmento grande, M = médio e P = pequeno,
conforme descrito na Tabela 1.
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
G1 G2 G3 M1 M2 M3 P1 P2 P3
Den
sida
de r
elat
iva
Fragmento
OutrasEspécies exclusivasEspécies muito frequentes
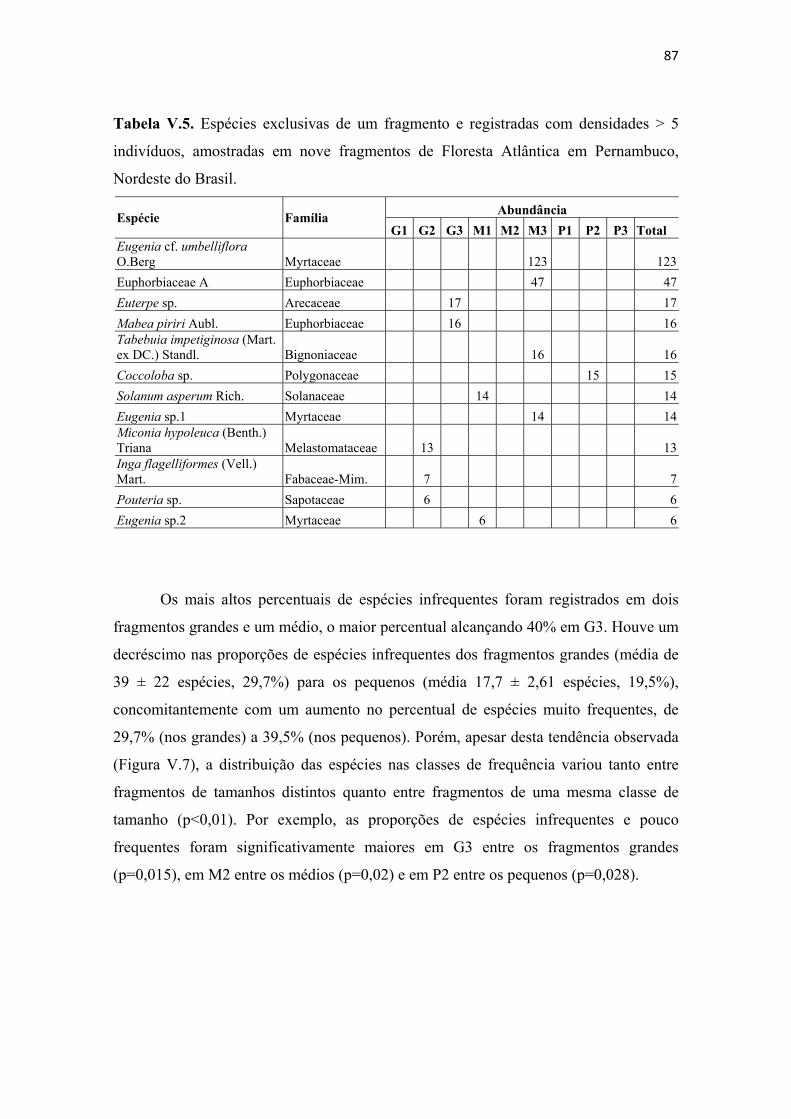
87
Tabela V.5. Espécies exclusivas de um fragmento e registradas com densidades > 5
indivíduos, amostradas em nove fragmentos de Floresta Atlântica em Pernambuco,
Nordeste do Brasil.
Espécie Família Abundância G1 G2 G3 M1 M2 M3 P1 P2 P3 Total
Eugenia cf. umbelliflora O.Berg Myrtaceae 123 123Euphorbiaceae A Euphorbiaceae 47 47Euterpe sp. Arecaceae 17 17Mabea piriri Aubl. Euphorbiaceae 16 16Tabebuia impetiginosa (Mart. ex DC.) Standl. Bignoniaceae 16 16Coccoloba sp. Polygonaceae 15 15Solanum asperum Rich. Solanaceae 14 14Eugenia sp.1 Myrtaceae 14 14Miconia hypoleuca (Benth.) Triana Melastomataceae 13 13Inga flagelliformes (Vell.) Mart. Fabaceae-Mim. 7 7Pouteria sp. Sapotaceae 6 6Eugenia sp.2 Myrtaceae 6 6
Os mais altos percentuais de espécies infrequentes foram registrados em dois
fragmentos grandes e um médio, o maior percentual alcançando 40% em G3. Houve um
decréscimo nas proporções de espécies infrequentes dos fragmentos grandes (média de
39 ± 22 espécies, 29,7%) para os pequenos (média 17,7 ± 2,61 espécies, 19,5%),
concomitantemente com um aumento no percentual de espécies muito frequentes, de
29,7% (nos grandes) a 39,5% (nos pequenos). Porém, apesar desta tendência observada
(Figura V.7), a distribuição das espécies nas classes de frequência variou tanto entre
fragmentos de tamanhos distintos quanto entre fragmentos de uma mesma classe de
tamanho (p<0,01). Por exemplo, as proporções de espécies infrequentes e pouco
frequentes foram significativamente maiores em G3 entre os fragmentos grandes
(p=0,015), em M2 entre os médios (p=0,02) e em P2 entre os pequenos (p=0,028).
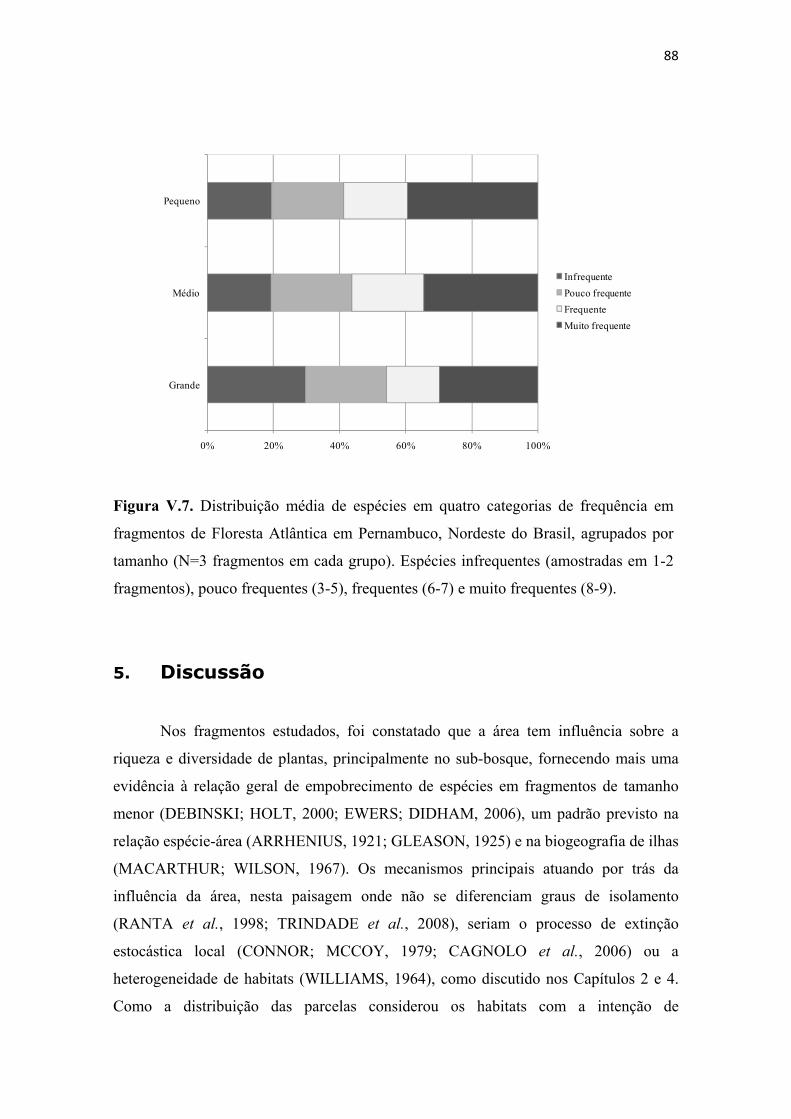
88
Figura V.7. Distribuição média de espécies em quatro categorias de frequência em
fragmentos de Floresta Atlântica em Pernambuco, Nordeste do Brasil, agrupados por
tamanho (N=3 fragmentos em cada grupo). Espécies infrequentes (amostradas em 1-2
fragmentos), pouco frequentes (3-5), frequentes (6-7) e muito frequentes (8-9).
5. Discussão
Nos fragmentos estudados, foi constatado que a área tem influência sobre a
riqueza e diversidade de plantas, principalmente no sub-bosque, fornecendo mais uma
evidência à relação geral de empobrecimento de espécies em fragmentos de tamanho
menor (DEBINSKI; HOLT, 2000; EWERS; DIDHAM, 2006), um padrão previsto na
relação espécie-área (ARRHENIUS, 1921; GLEASON, 1925) e na biogeografia de ilhas
(MACARTHUR; WILSON, 1967). Os mecanismos principais atuando por trás da
influência da área, nesta paisagem onde não se diferenciam graus de isolamento
(RANTA et al., 1998; TRINDADE et al., 2008), seriam o processo de extinção
estocástica local (CONNOR; MCCOY, 1979; CAGNOLO et al., 2006) ou a
heterogeneidade de habitats (WILLIAMS, 1964), como discutido nos Capítulos 2 e 4.
Como a distribuição das parcelas considerou os habitats com a intenção de
0% 20% 40% 60% 80% 100%
Grande
Médio
Pequeno
InfrequentePouco frequenteFrequenteMuito frequente

89
homogeneizar a amostra nos fragmentos, a área figura como a variável explicativa
principal da diversidade vegetal, o que já foi confirmado em diversas paisagens com
alto grau de fragmentação florestal, como na floresta atlântica brasileira (TABARELLI
et al., 1999), na floresta amazônica (LAURANCE et al., 2002), na floresta tropical em
Gana (HILL; CURRAN, 2003), no Chaco argentino (CAGNOLO et al., 2006) e em
florestas temperadas chilenas (ECHEVERRÍA et al., 2007).
A riqueza global do dossel foi mais alta que o esperado nesta região
biogeográfica da floresta atlântica, ao norte do Rio São Francisco, onde têm sido
reportados valores de riqueza de plantas arbóreas e arbustivas, compilados a partir de
estudos em florestas de terras baixas, de 282 (SIQUEIRA et al., 2001), 427 (SILVA;
TABARELLI, 2000) e 452 espécies (SANTOS et al., 2007a). Principalmente quando se
considera que o inventário foi desenvolvido em fragmentos numa mesma paisagem, as
192 espécies arbóreas aqui encontradas correspondem a um percentual entre 40% e 70%
do número estimado na região. Pontualmente, apesar de uma pequena variação
metodológica, quando é amostrado o dossel (DAP > 5 cm) em área de até 1 ha, o
número de espécies por fragmento nesta região tem variado desde 40-50 espécies em
áreas menores, perturbadas ou urbanas (BARBOSA, 1996; LINS-E-SILVA; RODAL,
2008) até 80-120 espécies em fragmentos maiores ou mais preservados (SIQUEIRA et
al., 2001; ROCHA et al., 2008; SILVA et al., 2008a; SILVA JÚNIOR et al., 2008;
BRANDÃO et al., 2009). Em áreas amostrais maiores, incluindo-se árvores com DAP >
10 cm, há registros de 146 espécies em 3,8 ha (OLIVEIRA, 2007) e 219 espécies em
7,5 ha (SANTOS et al., 2008).
A amostra de sub-bosque trouxe significativo acréscimo à riqueza global,
permitindo inventariar tanto espécies típicas deste estrato, tais como as várias
Myrtaceae, Melastomataceae e Rubiaceae, quanto espécies arbóreas em regeneração, a
exemplo das espécies da família Sapotaceae, importante componente do dossel nessas
florestas (OLIVEIRA et al., 2004; SILVA et al., 2008c). Este incremento na riqueza foi,
da mesma forma, demonstrado por Silva et al. (2008a) e Silva et al. (2008c), revelando
ser o sub-bosque essencial para o entendimento da ecologia e dinâmica de comunidades
fragmentadas ou perturbadas (LINS-E-SILVA; RODAL, 2008).
As assembléias amostradas foram dominadas por espécies de baixa frequência
(60%) e raras (47%). Este padrão é relatado em outras paisagens fragmentadas da
floresta atlântica (METZGER et al., 1997; OCHOA-GAONA et al., 2004; SANTOS et

90
al., 2007b) e parece ser típico de florestas tropicais (MARTINS, 1993), nas quais
espécies regionalmente raras aparecem como exclusivas de uma comunidade
(GUEDES-BRUNI et al., 2009), de um fragmento ou de um tipo de paisagem
(METZGER et al., 1997). Entretanto, a presença de espécies raras e infrequentes em
fragmentos florestais não deve ser interpretada – de imediato – como um sinal de bom
estado de conservação do fragmento ou das espécies (SANTOS et al., 2007b). Isto
porque o tempo de resposta biológica a um evento de fragmentação – o tempo de
latência – cria um retardo entre o evento e a acomodação da comunidade, representando
um débito ecológico, que é um custo ecológico futuro de uma situação presente
(TILMAN et al., 1994). Este processo de acomodação pode ser positivo para algumas
populações com altas taxas de colonização (TILMAN et al., 1994) ou negativo, na
forma de extinções – neste caso as raras possivelmente levam desvantagem (HENLE et
al., 2004), principalmente na regeneração do estrato de dossel, onde as raras são mais
importantes (GUEDES-BRUNI et al., 2009). No grupo das lianas, que normalmente
apresenta alta colonização em ambientes alterados (SCHNITZER; BONGERS, 2002),
não parece estar havendo acomodação em direção a um incremento da abundância em
fragmentos menores. Por outro lado, em pelo menos cinco fragmentos foram registrados
percentuais > 5% de plantas mortas de pé, no dossel e sub-bosque. Estes valores,
considerados altos em relação a outros levantamentos de floresta atlântica na região
(SIQUEIRA et al., 2001), representam um potencial efeito negativo na produção de
sementes, dispersão e regeneração florestal (BENITEZ-MALVIDO, 1998).
A comunidade vegetal estudada, separada em estratos – como dublês da idade –
ressaltou novos aspectos da aplicação da teoria de ilhas às comunidades fragmentadas.
Como fragmentos são remanescentes de áreas contínuas, isolados pela imposição de
uma matriz contrastante (WATSON, 2002), nossos dados confirmaram que amostras de
diferentes idades podem apresentar diferentes respostas à fragmentação e redução da
área, numa mistura de efeitos de curto prazo e débitos de extinção de longo prazo
(EWERS; DIDHAM, 2006). Isto porque árvores mais antigas no dossel já estavam ali
antes da fragmentação e, portanto, não refletem uma resposta a este evento, enquanto o
estrato de regeneração já exibe uma resposta à nova cobertura vegetal remanescente. Há
duas maneiras de se testar esta idéia: confrontando uma comunidade atual com a
paisagem atual e passada (LINDBORG; ERIKSSON, 2004; ERNOULT et al., 2006;
HELM et al., 2006; METZGER et al., 2009), ou confrontando uma paisagem atual com

91
estratos de diferentes idades na comunidade. Metzger (1998) investigou a influência da
paisagem na riqueza de diferentes grupos etários em 11 fragmentos florestais ripários
(Bacia do rio Jacaré-Pepira, São Paulo). Distinguindo “árvores do presente” e “árvores
do futuro”, a partir de relações alométricas em plantas com DAP ≥ 3 cm, o estudo
constatou serem as árvores mais jovens (“do futuro”) mais sensíveis à estrutura atual da
paisagem. Ochoa-Gaona et al. (2004) amostraram dossel e sub-bosque de florestas no
México e encontraram relação apenas entre as métricas da paisagem atual (basicamente
forma e isolamento) e o componente arbustivo e de sub-bosque. Nossos dados, embora
distinguindo estruturas etárias de forma diferente, corroboram esta idéia: houve baixa
taxa de renovação nos fragmentos (percentuais < 50% de similaridade entre estratos), e
houve diferenças significativas na riqueza e diversidade do sub-bosque entre fragmentos
de diferentes tamanhos. Neste estrato, no qual ocorrem regenerantes do dossel e
espécies de ciclo de vida mais curto (OLIVEIRA et al., 2001; OLIVEIRA; AMARAL,
2005; GOMES et al., 2009), a riqueza foi menor nos fragmentos menores, o que não
aconteceu com o estrato de dossel, que muito provavelmente ainda responde à cobertura
vegetal anterior à fragmentação.
Cabe ressaltar que a alta regeneração no sub-bosque, principalmente nos
fragmentos médios, causou o chamado “efeito de densidade” (DENSLOW, 1995),
quando um maior número de indivíduos suporta um maior número de espécies. Este
efeito foi revelado ao aplicarmos rarefação às amostras: sem rarefação, não houve
diferenças na riqueza entre tamanhos de fragmentos; após rarefação, a riqueza se
revelou maior nos fragmentos maiores. Este incremento na riqueza, porém, foi de
espécies cujas abundâncias não influenciaram a diversidade nos fragmentos médios, que
foi significativamente menor que os grandes mesmo sem rarefação das amostras.
Segundo Gotelli e Colwell (2001), a comparação da riqueza com base na área amostral
pode esconder uma “armadilha”, pois, mesmo em áreas semelhantes, diferentes
densidades podem surgir como reflexo de diferentes – e aleatórios – regimes locais de
perturbação, densidade de clareiras ou estágio de sucessão.

92
6. Considerações finais e implicações
Em conjunto, os resultados encontrados descrevem um cenário de alta riqueza e
diversidade numa paisagem altamente fragmentada da Floresta Atlântica. Porém, há
riscos de perda de espécies no futuro, mesmo se mantendo a paisagem atual, a depender
do tempo de acomodação da comunidade (LINDBORG; ERIKSSON, 2004). De fato,
predições baseadas em modelos têm mostrado que a própria redução de área já levou à
extinção uma importante parcela da biodiversidade nesta região da floresta atlântica
(TABARELLI et al., 2004). Algumas implicações para conservação podem ser
delineadas a partir deste cenário. Em primeiro lugar, é preciso considerar que, muito
embora a maior parte da riqueza esteja contida nos maiores fragmentos, fragmentos
pequenos sozinhos contêm mais de 50% da riqueza amostrada. Além disso, há um alto
percentual de espécies raras e de espécies de alta frequência, porém com abundâncias
distintas nos vários fragmentos. Neste contexto, pequenas áreas atuam como
importantes complementos para as grandes áreas, suportando diferentes amostras de
espécies (FISCHER; LINDENMAYER, 2007; RODRIGUES et al., 2009), podendo vir
a garantir o balanço entre recrutamento e mortalidade entre fragmentos (MANNING et
al., 2006). Deve-se considerar, ainda, que outros habitats, além dos aqui amostrados,
estão presentes nos vários fragmentos (capítulo 2), a exemplo dos ambientes ripários.
Considerando a dinâmica própria e alto número de especialistas esperado nesses sítios
(NAIMAN; DÉCAMPS, 1997; MOUW; ALABACK, 2003) é possível que as espécies
ali presentes representem um aumento na riqueza dos fragmentos (capítulo 4).
O valor de fragmentos pequenos para manutenção da diversidade em paisagens
fragmentadas é inegavelmente reconhecido (SANTOS et al., 2007b; RICHART;
HEWITT, 2008; RODRIGUES et al., 2009). O conceito de “pequeno”, porém, é muito
relativo. Qual seria o limiar de tamanho para um fragmento ser considerado pequeno?
Turner e Corlett (1996) afirmaram que nas terras baixas tropicais a floresta tropical está
restrita a fragmentos menores que 400 ha, “pequenos e isolados”. Muito do
conhecimento dos efeitos da fragmentação foi gerado em estudos de fragmentos com
menos de 10 ha, considerados por Zuidema et al. (1996) como inviáveis para várias
populações. Todos os fragmentos aqui estudados, mesmo os localmente grandes,
seriam, assim, pequenos e, portanto, extremamente vulneráveis a fatores externos.
Medidas urgentes são necessárias, portanto, para minimizar futuras perdas de espécies.

93
Como iniciativas de conservação numa paisagem fragmentada, Metzger et al.
(2009) sugerem a manutenção das florestas maduras e aumento da área de florestas e
conectividade, através da restauração florestal. Iniciativas de manutenção das florestas
maduras já estão em prática na região, lideradas pelo setor sucroalcooleiro em parceria
com centros de pesquisa (TABARELLI; RODA, 2005). Faz-se necessário, porém, um
controle mais eficiente da exploração dos recursos madeireiros pelas populações locais,
que tem relevante impacto nas comunidades arbóreas (ALBUQUERQUE et al., 2008;
ALMEIDA et al., 2008). Por outro lado, já foram iniciadas e fazem parte da agenda do
setor práticas de restauração florestal e abandono de áreas de cultura para regeneração
natural, formando “capoeiras” com várias idades, que já apresentam aumento
progressivo na riqueza e diversidade (NASCIMENTO, 2010).
Saunders et al. (1991), ao discorrerem sobre as consequências da fragmentação,
relembraram a questão: “qual a área mínima de um ecossistema para preservar sua
diversidade e composição?”. Embora existam modelos propostos para área mínima, com
base em regimes de perturbação natural (LEROUX et al., 2007), a questão nesta região
da floresta atlântica é: mesmo havendo um efeito de área e uma tendência a perda de
espécies, não é possível abrir mão de pequenos fragmentos. Ao contrário, nossos
resultados sinalizam que todos os fragmentos são importantes como refúgio e banco de
espécies (AERTS et al., 2006), para conservação da diversidade atual nesta paisagem.

94
VI. CAPÍTULO 4 - Influência da heterogeneidade de
habitats na diversidade e especialização de plantas
em fragmentos de Floresta Atlântica

95
1. Resumo
Neste trabalho, analisou-se a influência da heterogeneidade de habitats e
especialização de plantas sobre a diversidade de seis fragmentos (3 grandes, ≈ 300 ha; 3
pequenos, ≈15 ha) de Floresta Atlântica. Foram obtidas amostras em parcelas de dossel
(10 x 10 m) e sub-bosque (5 x 5 m) por habitat, em uma área amostral total de 3 ha. Em
cada habitat, calculou-se a densidade, área basal (m2), riqueza e diversidade de
Shannon, comparadas através de análise de variância. Como variou o número de
habitats por fragmento (4 a 6), foram estimadas a riqueza e diversidade totais por
fragmentos através de rarefação e bootstrapping, comparadas pelo teste t. Como
medidas de complementaridade, calcularam-se a similaridade de Sørensen e diversidade
β de Whittaker. Para identificar preferência por habitats, foi aplicado o teste do qui-
quadrado e análise de espécies indicadoras (ISA) às espécies com ≥ 30 indivíduos.
Foram amostrados 13808 indivíduos (4389 no dossel, 9419 no sub-bosque),
pertencentes a 308 espécies. A riqueza e diversidade estimadas foram
significativamente maiores em fragmentos grandes, por estrato e na amostra total. Nos
habitats de maior frequência, borda e interior de encosta, apesar da densidade e área
basal terem sido semelhantes, a riqueza foi significativamente inferior nos fragmentos
pequenos. Nos habitats ripários, registraram-se maiores densidade e área basal, e as
mais baixas diversidades, enquanto o interior de tabuleiro teve a maior riqueza e
diversidade entre todas as áreas. A similaridade média por fragmento (amostra total)
variou de 0,44 e 0,62 e a diversidade βW, de 1,80 a 2,55, sendo maior no sub-bosque,
com complementaridade significativamente maior nos fragmentos grandes. Entre
habitats, a similaridade foi mais baixa entre habitats semelhantes em diferentes
fragmentos do que entre habitats distintos em um mesmo fragmento, tanto na amostra
total, quanto nos estratos. Entre as espécies, 86 foram testadas quanto à preferência por
habitat, resultando em 57 com associação por um a três ambientes e 49 espécies
indicadoras (39 em comum). A maioria teve especialização por habitats ripários
permanentes e temporários. Os resultados sugerem, nesses fragmentos, a atuação de três
processos estruturadores de comunidades fragmentadas: o efeito da área na riqueza, da
heterogeneidade de habitats na diversidade beta, e da limitação de dispersão na
complementaridade entre fragmentos. Conclui-se, assim, que a diversidade só será
preservada com base na manutenção de todos os fragmentos e da soma de porções de
seus habitats.

96
2. Introdução
A crescente fragmentação de áreas naturais tem levado importantes ecossistemas
continentais a existirem sob a forma de fragmentos, com consequente perda de espécies
(HENLE et al., 2004; LINDENMAYER; FISCHER, 2006; FISCHER;
LINDENMAYER, 2007). São notadamente marcantes as modificações na paisagem em
regiões tropicais (LAURANCE, 1999b; WRIGHT; MULLER-LANDAU, 2006),
principalmente na floresta atlântica brasileira, onde os remanescentes são, na maioria,
pequenos e isolados (RIBEIRO et al., 2009). Quando se quer investigar a diversidade
remanescente nessas manchas - questão-chave em qualquer iniciativa de conservação de
comunidades (GROVES et al., 2002; METZGER, 2006) - normalmente se abordam os
efeitos inerentes à condição de fragmentação antropogênica: o de área e o de borda
(LAURANCE et al., 2007). O primeiro, guiado pela teoria da biogeografia de ilhas
(MACARTHUR; WILSON, 1967), prediz que fragmentos menores têm menos
espécies; o segundo prevê uma condição alterada nos novos limites do ecossistema que
estão em contato com o entorno distinto (LAURANCE et al., 2001; RIES et al., 2004).
Outro efeito, mais timidamente investigado (KADMON; ALLOUCHE, 2007), pode ser
ainda somado ao entendimento de comunidades fragmentadas: o da heterogeneidade
interna nos remanescentes.
A variação espacial nas condições e recursos presentes em uma área causa
heterogeneidade. Em estudos de ecologia, a integração entre heterogeneidade e
diversidade biológica teve forte expansão a partir da década de 1960, erguidas sobre a
base sólida de um dos principais paradigmas da ecologia básica e aplicada
(MITCHELL, 2005): o conceito de habitat. Definido como um conjunto de dimensões
do mundo físico, cujos valores variam no espaço e no tempo (KOLASA; WALTHO,
1998), no habitat se produzem as condições de ocupação, sobrevivência e reprodução
por uma espécie (HALL et al., 1997) ou para toda uma comunidade que partilha
condições homogêneas do ambiente (WHITTAKER et al., 1973; KOLASA; WALTHO,
1998; PETREN, 2001). Williams (1964) foi pioneiro ao propor que a correlação entre
área e riqueza específica pode ser explicada pela heterogeneidade de habitats, e, por este
motivo, é considerado o criador da “Hipótese da Diversidade de Habitats”. Se os
habitats são distribuídos de forma heterogênea no espaço, e porque diferentes espécies
necessitam de diferentes habitats, quanto mais habitats, mais espécies (TURNER;

97
TJØRVE, 2005). Por esta hipótese, com o aumento da área, o número de espécies
aumenta apenas se mais habitats forem amostrados (RICKLEFS; LOVETTE, 1999).
As plantas, mesmo sendo indivíduos sésseis, também respondem à variação
espacial dos recursos através de diversos mecanismos (WRIGHT, 2002). A
especialização por habitat em plantas ocorre quando diferentes espécies são mais
adaptadas a diferentes ambientes, nos quais são relativamente mais abundantes,
consistindo numa manifestação da diferenciação por nichos de recursos (HARMS et al.,
2001). Segundo Bazzaz (1991), os mecanismos que explicam a preferência de espécies
vegetais por habitats são passivos, sendo a seleção, escolha e especialização de habitat
primeiramente desempenhada pelo próprio habitat e em seguida fixada pelas plantas
através de seleção natural.
Como resultado de processos complexos da dinâmica das espécies, o padrão de
distribuição de plantas numa comunidade pode ser usado para inferir preferência ou
especialização por habitats (HARMS, 1997). Nos estudos pioneiros de R. Whittaker em
gradientes ambientais (WHITTAKER; NIERING, 1965; WHITTAKER, 1967), a
representação das distribuições das populações revelou tendências à formação de
assembléias de espécies com preferências por alguma zona do gradiente, composto por
poucas dimensões em um sistema de coordenadas (WHITTAKER et al., 1973). A
análise de gradientes levou à formulação de descritores de diversidade que capturassem
a relação entre heterogeneidade e diversidade a partir de parâmetros da própria
assembléia (WHITTAKER, 1972): a diversidade alfa, tomada em um habitat, e a
diversidade beta, medida como a desigualdade ou a taxa de substituição entre dois
habitats (WHITTAKER, 1972; CONDIT et al., 2002; HALVORSEN; EDVARDSEN,
2009).
A distribuição não aleatória de espécies com relação a gradientes ambientais tem
sido documentada em várias escalas e em várias comunidades vegetais, demonstrando
que os mecanismos subjacentes à teoria de habitats de fato operam em comunidades
vegetais (WRIGHT, 2002) e são uma explicação plausível para manutenção da
diversidade (COMITA et al., 2007). Em seis grandes áreas de florestas tropicais, Condit
et al. (2000) demonstraram que a distribuição de indivíduos de populações de plantas
lenhosas raramente é aleatória, e que tanto indivíduos jovens quanto grandes árvores
ocorrem em padrões agregados correspondentes às variações de habitats. Ao revisar 52
estudos que compararam a diversidade vegetal e tiveram a heterogeneidade espacial

98
medida em pelo menos duas áreas, Lundholm (2009) confirmou que a maioria mostrou
relação positiva entre heterogeneidade de habitats e riqueza ou diversidade vegetal,
apesar da grande variabilidade de escala nos estudos, e que as principais dimensões
utilizadas para descrição de habitats para plantas são variáveis de relevo e substrato.
Entre esses, a topografia é considerada o fator abiótico mais importante causando
variação espacial e influenciando a estrutura e diversidade de comunidades
(OLIVEIRA-FILHO et al., 1994b; JOHNSON et al., 2003).
Em florestas tropicais e subtropicais, há fortes evidências que as plantas
lenhosas respondem às diferenças de nicho associadas à topografia (WRIGHT, 2002;
HOFER et al., 2008), para um percentual de pelo menos um terço das espécies
(HUBBELL; FOSTER, 1983; CLARK et al., 1999; HARMS et al., 2001; KUBOTA et
al., 2004; VALENCIA et al., 2004). A disponibilidade hídrica é também uma dimensão
amplamente considerada e para a qual as espécies demonstram seletividade espacial
(OLIVEIRA-FILHO et al., 1994a; 1994b; 1994c; BOTREL et al., 2002; DRUCKER et
al., 2008; KESSLER; LEHNERT, 2009; SFENTHOURAKIS; TRIANTIS, 2009) ou
temporal (BROSE, 2001). Recentemente, variáveis de habitat relacionadas a
perturbações têm sido escolhidas (PEREIRA et al., 2007; SANTOS et al., 2008), o que
é uma tendência natural em ecossistemas fragmentados, nos quais proporções de
habitats perturbados têm aumentado, principalmente sob influência da bordas
(LAURANCE, 2000; TABARELLI et al., 2008). Nesses habitats, alterações
microclimáticas atuam sobre os processos de dinâmica da comunidade (HARPER et al.,
2005; LOPES et al., 2009), levando à formação de uma assembléia distinta, geralmente
por efeito deletério na diversidade (RIES et al., 2004).
A fragmentação é um processo antropogênico e, portanto, não aleatório
(SEABLOOM et al., 2002), de forma que os fragmentos de áreas naturais são, com
frequência, uma amostra tendenciosamente mais representativa de áreas com menor
potencial de uso econômico, tais como encostas íngremes ou áreas inundadas
(TABARELLI et al., 2004; LAURANCE, 2008). Esta seleção determinística pode fazer
com que, numa mesma paisagem, fragmentos com diferentes tamanhos comportem
diferentes números e tipos de habitats. A análise da influência desta heterogeneidade na
estrutura e especialização de plantas em comunidades vegetais fragmentadas foi o
objetivo deste estudo. Para tanto, foram avaliados seis fragmentos na floresta atlântica
do Nordeste do Brasil, nos quais se estratificou a amostra, considerando os habitats

99
existentes, avaliando-se o dossel e o sub-bosque. Foram metas específicas: descrever a
estrutura das assembléias por habitat e por fragmento, distinguir a influência da
heterogeneidade de habitats da influência da área do fragmento, e identificar se as
espécies encontradas apresentam especialização por habitats.
3. Material e métodos
3.1. Área de Estudo
O estudo foi conduzido em seis remanescentes de Floresta Atlântica localizados
na Zona da Mata Norte do Estado de Pernambuco, Nordeste do Brasil, em terras de
propriedade da Usina São José/ Grupo Cavalcanti Petribú - USJ (Tabela VI.1). Para
descrição da geologia, relevo, clima e histórico da área, ver item “Área de Estudo”, pp.
6-11. A vegetação primária na área é Floresta Ombrófila Densa das Terras Baixas,
atualmente muito reduzida e fragmentada. Na propriedade USJ, há 106 fragmentos
florestais, dos quais seis foram escolhidos para estudo, com base no seu tamanho, sendo
três grandes e três pequenos (Tabela VI.1), todos com bom estado de conservação e
tempo de isolamento semelhante, de pelo menos 25 anos.
3.2. Desenho amostral
As assembléias de plantas foram amostradas em dois diferentes estratos: dossel
(plantas lenhosas com circunferência à altura do peito - CAP ≥ 15 cm) e sub-bosque
(plantas lenhosas com circunferência ao nível do solo ≥ 1 cm a CAP < 15 cm).
Procedimentos de amostragem e marcação dos indivíduos em cada estrato, coleta e
identificação botânica estão descritos “Área de Estudo/ Metodologia Geral”, p. 11.
Em cada fragmento, os habitats existentes foram identificados e quantificados e
foi calculada a diversidade de habitats (Tabela VI.1) através do índice de diversidade de
Simpson recíproco – 1/D (SIMPSON, 1949), seguindo metodologia descrita no
Capítulo 2. Em todos os habitats determinados, dez parcelas de 0,01 ha (10 x 10 m)
foram instaladas para amostragem do dossel, sistematicamente localizadas com auxílio
de GPS na área mais central possível do habitat, formando módulos de 20 x 50m. Uma
parcela menor para amostragem de sub-bosque (5 x 5 m) foi instalada em cada uma das

100
parcelas de dossel. A distância entre habitats nos fragmentos foi calculada a partir do
centro da área instalação das parcelas (Tabela VI.1).
3.3. Análise de dados
O banco de dados foi preparado, armazenado e manejado no programa Mata
Nativa 2.0 (SOUZA et al., 2006). Em cada fragmento, foram calculados os seguintes
atributos descritivos da assembléia de plantas, por parcela e por habitat: densidade, área
basal (m2), riqueza (número de espécies) e diversidade α (referente a uma série de
unidades amostrais em um mesmo habitat, sensu Whittaker 1972), calculada através do
índice de Shannon (MAGURRAN, 2004). Em cada situação, foram gerados valores
para amostra total (dossel + sub-bosque) e separadamente por estrato.
Os valores de atributos obtidos foram comparados entre fragmentos ou entre
habitats. Para comparações entre grupos de fragmentos (1 fragmento = 1 amostra),
utilizou-se teste t para amostras independentes e, para comparações entre habitats (1
parcela = 1 amostra), utilizou-se o teste não paramétrico de análise de variância
Kruskal–Wallis, no programa BioEstat 5.0 (AYRES et al., 2007). Em cada fragmento, a
similaridade florística entre habitats foi calculada através do coeficiente de similaridade
de Sørensen (1948), SS. O mesmo coeficiente foi aplicado para calcular similaridade
entre habitats semelhantes em diferentes fragmentos. A riqueza total de cada habitat,
somando-se as amostras nos vários fragmentos, também foi obtida e correlacionada ao
número de fragmentos em que o habitat ocorreu, através do coeficiente de correlação
linear de Pearson.
Devido à variação no número de habitats, a quantidade de parcelas variou entre
fragmentos (Tabela VI.1). Por este motivo, para permitir comparação da riqueza e
diversidade, foi obtida uma curva suavizada de amostragem de espécies para amostra
total de cada fragmento, elaborada através de 100 re-amostragens aleatórias de todas as
parcelas e indivíduos no fragmento, sem substituição, obtendo-se médias para cada
ponto na curva, sendo que cada indivíduo ou amostra teve a mesma probabilidade de ser
incluído na média de riqueza em qualquer ponto da curva (GOTELLI; COLWELL,
2001). Por este procedimento, anula-se o efeito da ordem da amostragem (COLWELL;
CODDINGTON, 1994), permitindo a comparação entre amostras por interpolação, ou
seja, no ponto em que se assemelham no eixo x, como por exemplo, comparando-se

101
todos os fragmentos pelo menor número de parcelas ou indivíduos amostrados.
Também foi utilizado o método não-paramétrico “bootstrapping” (SMITH; VAN
BELLE, 1984) para obter estimativas da riqueza comparáveis entre fragmentos, a partir
de sub-amostras retiradas por 100 vezes, aleatoriamente, da amostra total de cada
fragmento, com substituição, ou seja, cada indivíduo pode aparecer mais de uma vez
numa mesma sub-amostra. As duas estimativas de riqueza foram obtidas no programa
EstimateS 7.5.2 (COLWELL, 2006). Os valores observados (todos os indivíduos
amostrados) e obtidos por rarefação ou bootstrapping para cada fragmento foram
comparados entre si através de teste G (AYRES et al., 2007).
Visando ao entendimento da contribuição dos vários habitats à diversidade
vegetal nos fragmentos, para cada fragmento foram calculados dois índices: a
similaridade média e a diversidade beta (βW). A similaridade média (SSM) foi obtida a
partir da média do índice de similaridade de Sørensen obtido para pares de habitats no
fragmento (VELLEND, 2001). A diversidade βW (WHITTAKER, 1972), que mede a
taxa de substituição de espécies entre áreas, sendo maior quanto menor for a
similaridade entre habitats, foi calculada através da fórmula:
βW = SC/αM
sendo SC a riqueza total combinada registrada no fragmento e αM a média da riqueza por
habitat no fragmento.
Para identificar preferência por habitats entre as espécies, foi aplicado o teste do
qui-quadrado (partição) e análise de espécies indicadoras (ISA). O uso do qui-quadrado
para análise de associações por habitats (HUBBELL; FOSTER, 1983) é visto com
alguma reserva (HARMS et al., 2001; ZANNE; CHAPMAN, 2005), pois o teste
assume independência entre amostras (parcelas ou indivíduos), premissa
frequentemente violada em comunidades vegetais devido aos padrões de dispersão e
recrutamento. Entretanto, nas nossas análises, a distribuição das espécies foi avaliada
por habitats, e as amostras por habitat foram suficientemente afastadas daquelas nos
habitats vizinhos, diminuindo uma possível autocorrelação entre amostras. No teste,
comparou-se a densidade observada para cada espécie em cada um dos habitats
(amostra total) à proporção esperada, obtida com base na área amostral, através da
multiplicação da densidade total da espécie pela proporção da área amostrada naquele
habitat em relação ao total (NP/300), onde NP é o número de parcelas no habitat em
questão e 300 o número total de parcelas amostradas. Como a proporção esperada em

102
cada habitat não pode ser menor que 1 (SIEGEL, 1975), a densidade mínima para
aplicação do teste foi obtida através da multiplicação de 1 por 0,033 (10 parcelas/ 300
parcelas), obtendo-se valor de 30 indivíduos. O número de habitats variou para cada
espécie, pois foram excluídos aqueles no qual a espécie não ocorreu (habitats não úteis
para a espécie). Assim, os grau de liberdade foram ajustados caso a caso, variando entre
1 (dois habitats disponíveis) e 7 (oito habitats disponíveis), como aplicado por Harms
(1997) e Harms et al. (2001). Havendo diferenças significativas para a distribuição das
espécies, complementava-se o teste com a análise de resíduos do qui-quadrado, para
verificação da significância de cada associação (AYRES et al., 2007).
A análise de espécies indicadoras (ISA) foi aplicada utilizando-se o programa
PC-ORD 4.0 (MCCUNE; MELFORD, 1999), a partir de uma matriz quantitativa com
todas as parcelas em todos os habitats, incluindo as espécies com pelo menos 30
indivíduos, para permitir comparações com o teste do qui-quadrado. Na análise, valores
indicadores (INDVAL) foram calculados para cada espécie, combinando a abundância
relativa da espécie e frequência nos habitats, após 1000 permutações (Monte Carlo) para
testar a significância da distribuição da espécie comparada a uma distribuição aleatória
(DUFRÊNE; LEGENDRE, 1997). Espécies com associação e/ ou indicadoras de habitat
foram analisadas quanto à sua posição na assembléia nos diferentes fragmentos, a partir
do seu valor de importância, obtido por meio da soma dos atributos relativos de
densidade, dominância e frequência da espécie em cada habitat.
Tabela VI.1. Características dos seis fragmentos florestais estudados na Região Norte
do Estado de Pernambuco, Nordeste do Brasil. N, número; DH, diversidade de habitats
(índice de Simpson recíproco); distância entre, distância aos outros cinco fragmentos
estudados; distância intra, distância entre habitats no fragmento (média ± IC). Nome Área
(ha) Código área
N habitats
N parcelas
DH Código DH
Distância média entre (km)
Distância média intra (m)
Zambana 387,85 G1 6 60 3.03
A1 7,28 ± 2,51
736,86 ± 356
Macacos 356,92 G2 5 50 2.57
B1 4,63 ± 1,70
428,00 ± 283
Piedade 305,78 G3 6 60 3.08
A2 6,63 ± 2,77 696,38 ± 299
Pezinho 29,89 P1 5 50 2.57
B2 6,30 ± 2,34
119,60 ± 79
Vespas 13,80 P2 4 40 2.41
B3 5,45 ± 1,12
95,00 ± 51
Sta Helena
11,85 P3 4 40 2.69 A3 6,03 ± 2,02 125,00 ± 108

103
4. Resultados
4.1. Heterogeneidade de habitats nos fragmentos
O número de habitats por fragmento variou de 4 a 6, de forma que o estudo
envolveu 30 áreas nos seis fragmentos, perfazendo uma amostra total de 3 ha para
dossel e 0,75 ha para sub-bosque. Os distintos percentuais de cobertura de cada habitat
(Figura VI.1) geraram índices de diversidade de habitats (DH) entre 2,41 e 3,08. Para
permitir comparações, os fragmentos foram classificados com base no tamanho
(pequenos versus grandes) e na DH (maior DH versus menor DH), utilizando-se o valor
de 2,60 para separar fragmentos de alta ou baixa DH (Tabela VI.1). Agrupando-se
quanto ao tamanho, os pequenos fragmentos apresentaram média de DH inferior, mas
não significativamente diferente dos grandes (t=1,84; p=0,06). Quanto à DH, um
fragmento pequeno (P3) teve alta DH, enquanto um grande (G2) teve baixa DH,
gerando grupos distintos daqueles formados por tamanho.
Borda e interior, ambos de encosta, foram os dois únicos habitats presentes em
todos os fragmentos estudados, com percentuais de cobertura bem distintos em
fragmentos pequenos e grandes (Figura VI.1). Os demais habitats ocuparam de 12,5%
(P2) a 33% (G1) da área dos fragmentos. Habitats ripários ocorreram em todos os
fragmentos, ocupando entre 11 e 27% das áreas, sendo que ripários permanentes
ocorreram em cinco e temporários, em quatro. Ambientes de tabuleiro estiveram
restritos aos fragmentos grandes, ocupando 6% de G1 e 15% de G3.
4.2. Assembléias de plantas nos fragmentos e habitats
No total, foram amostrados 13808 indivíduos, 4389 no dossel e 9419 no sub-
bosque, pertencentes a 308 espécies/ morfoespécies em 58 famílias botânicas. A
densidade de indivíduos por fragmento variou amplamente, de forma sincrônica com o
número de habitats amostrados (Figura VI.2). A amostra de dossel, para os seis
fragmentos, forneceu uma lista de 206 espécies, enquanto a amostra de sub-bosque
acrescentou mais 102 espécies à lista total, além das 163 espécies em comum com o
dossel (53% do total). Considerando a amostra total, 28% das espécies ocorreram com
um ou dois indivíduos e 46% foram amostradas em número maior que 10 plantas. Por
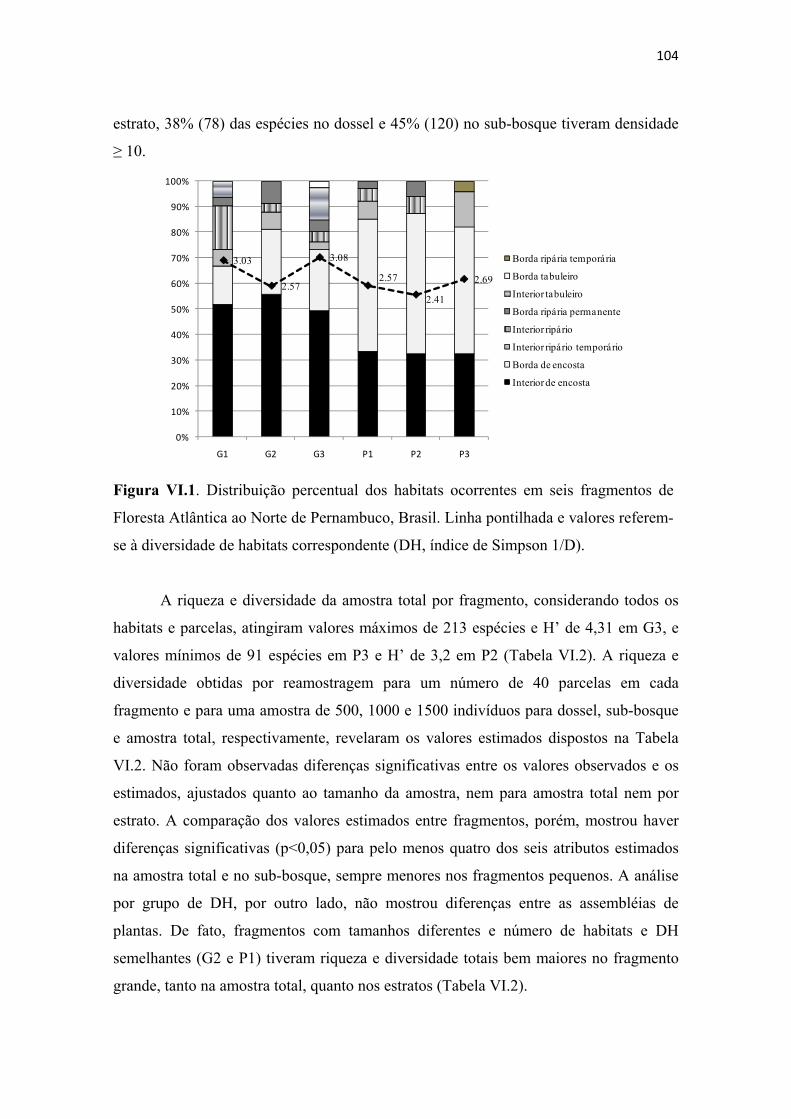
104
estrato, 38% (78) das espécies no dossel e 45% (120) no sub-bosque tiveram densidade
≥ 10.
Figura VI.1. Distribuição percentual dos habitats ocorrentes em seis fragmentos de
Floresta Atlântica ao Norte de Pernambuco, Brasil. Linha pontilhada e valores referem-
se à diversidade de habitats correspondente (DH, índice de Simpson 1/D).
A riqueza e diversidade da amostra total por fragmento, considerando todos os
habitats e parcelas, atingiram valores máximos de 213 espécies e H’ de 4,31 em G3, e
valores mínimos de 91 espécies em P3 e H’ de 3,2 em P2 (Tabela VI.2). A riqueza e
diversidade obtidas por reamostragem para um número de 40 parcelas em cada
fragmento e para uma amostra de 500, 1000 e 1500 indivíduos para dossel, sub-bosque
e amostra total, respectivamente, revelaram os valores estimados dispostos na Tabela
VI.2. Não foram observadas diferenças significativas entre os valores observados e os
estimados, ajustados quanto ao tamanho da amostra, nem para amostra total nem por
estrato. A comparação dos valores estimados entre fragmentos, porém, mostrou haver
diferenças significativas (p<0,05) para pelo menos quatro dos seis atributos estimados
na amostra total e no sub-bosque, sempre menores nos fragmentos pequenos. A análise
por grupo de DH, por outro lado, não mostrou diferenças entre as assembléias de
plantas. De fato, fragmentos com tamanhos diferentes e número de habitats e DH
semelhantes (G2 e P1) tiveram riqueza e diversidade totais bem maiores no fragmento
grande, tanto na amostra total, quanto nos estratos (Tabela VI.2).
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
G1 G2 G3 P1 P2 P3
Borda ripária temporária
Borda tabuleiro
Interior tabuleiro
Borda ripária permanente
Interior ripário
Interior ripário temporário
Borda de encosta
Interior de encosta
3.03
2.57
3.08
2.57
2.41
2.69
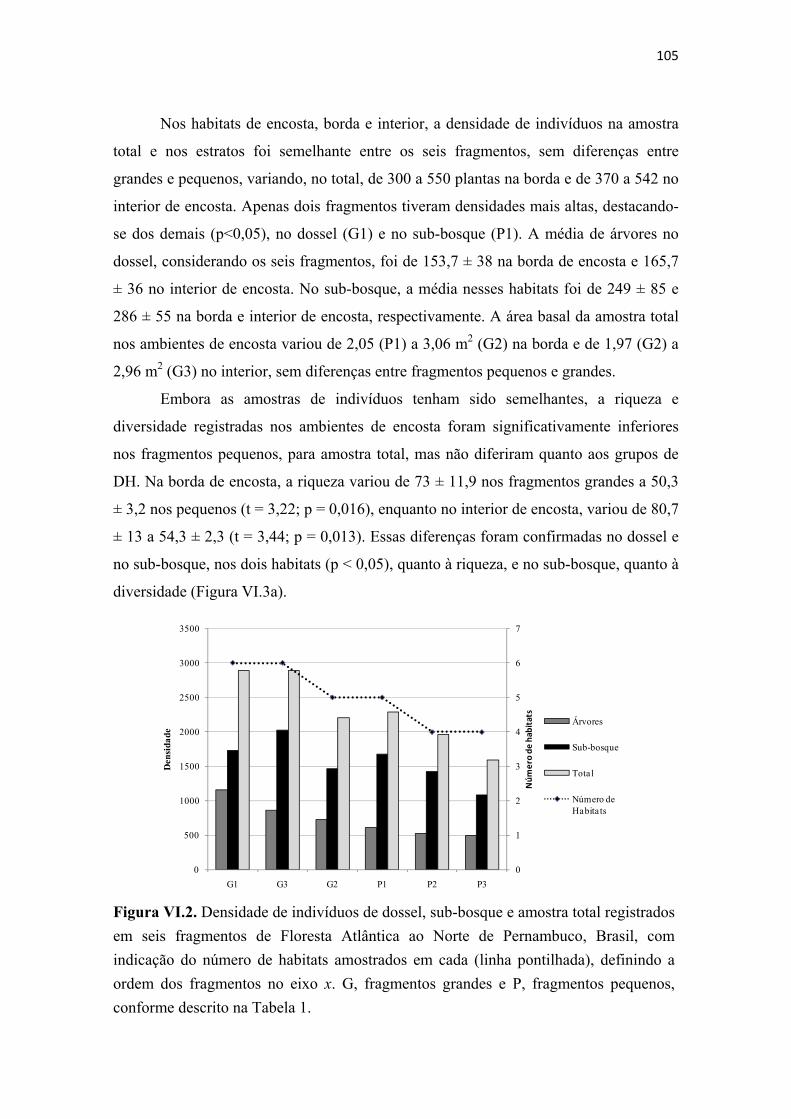
105
Nos habitats de encosta, borda e interior, a densidade de indivíduos na amostra
total e nos estratos foi semelhante entre os seis fragmentos, sem diferenças entre
grandes e pequenos, variando, no total, de 300 a 550 plantas na borda e de 370 a 542 no
interior de encosta. Apenas dois fragmentos tiveram densidades mais altas, destacando-
se dos demais (p<0,05), no dossel (G1) e no sub-bosque (P1). A média de árvores no
dossel, considerando os seis fragmentos, foi de 153,7 ± 38 na borda de encosta e 165,7
± 36 no interior de encosta. No sub-bosque, a média nesses habitats foi de 249 ± 85 e
286 ± 55 na borda e interior de encosta, respectivamente. A área basal da amostra total
nos ambientes de encosta variou de 2,05 (P1) a 3,06 m2 (G2) na borda e de 1,97 (G2) a
2,96 m2 (G3) no interior, sem diferenças entre fragmentos pequenos e grandes.
Embora as amostras de indivíduos tenham sido semelhantes, a riqueza e
diversidade registradas nos ambientes de encosta foram significativamente inferiores
nos fragmentos pequenos, para amostra total, mas não diferiram quanto aos grupos de
DH. Na borda de encosta, a riqueza variou de 73 ± 11,9 nos fragmentos grandes a 50,3
± 3,2 nos pequenos (t = 3,22; p = 0,016), enquanto no interior de encosta, variou de 80,7
± 13 a 54,3 ± 2,3 (t = 3,44; p = 0,013). Essas diferenças foram confirmadas no dossel e
no sub-bosque, nos dois habitats (p < 0,05), quanto à riqueza, e no sub-bosque, quanto à
diversidade (Figura VI.3a).
Figura VI.2. Densidade de indivíduos de dossel, sub-bosque e amostra total registrados em seis fragmentos de Floresta Atlântica ao Norte de Pernambuco, Brasil, com indicação do número de habitats amostrados em cada (linha pontilhada), definindo a ordem dos fragmentos no eixo x. G, fragmentos grandes e P, fragmentos pequenos, conforme descrito na Tabela 1.
0
1
2
3
4
5
6
7
0
500
1000
1500
2000
2500
3000
3500
G1 G3 G2 P1 P2 P3
Núm
ero de
habitats
Den
sida
de
Árvores
Sub-bosque
Total
Número de Habitats

106
Nos ambientes ripários permanentes, foram registrados altos valores de
densidade e área basal para amostra total, tanto em ambientes de borda quanto de
interior (Figura VI.3b). No dossel, a densidade registrada nos ambientes ripários foi
inferior nos fragmentos pequenos (p = 0,01). No sub-bosque, estas diferenças não foram
observadas, tendo-se registrado densidades tão distintas quanto 679 indivíduos no
interior ripário de P2 e 271 em habitat semelhante em G3. Diferenças em área basal com
relação ao tamanho dos fragmentos também não ocorreram. Os valores de diversidade
foram mais baixos em ambientes ripários permanentes que nos demais habitats, com H'
geralmente inferior a 3,0 nos vários fragmentos, independentemente do tamanho, tanto
na amostra total, quanto por estrato. Nos ambientes ripários temporários (Figura VI.3c),
os atributos vegetais foram mais semelhantes aos ambientes de encosta do que aos
ripários permanentes, sem diferenças entre tamanhos de fragmentos.
As duas únicas áreas de interior sobre tabuleiros existentes nos fragmentos
tiveram alta densidade na amostra total (> 520 indivíduos), pela grande contribuição do
sub-bosque, com mais de 400 indivíduos na amostra em cada área, significativamente
maior em G3 (p<0,05). Nesse habitat, registrou-se o mais alto valor de riqueza e
diversidade entre todas as áreas, no fragmento G3, onde habitats de tabuleiro atingem a
maior proporção entre os fragmentos estudados: 119 espécies, sendo 55 no dossel e 102
no sub-bosque, e H' = 3,91.
4.3. Similaridade entre fragmentos e habitats
Valores de similaridade média (SSM) obtidos por fragmento variaram entre 0,44
e 0,62 para amostra total, entre 0,35 a 0,55 para amostra de sub-bosque e de 0,33 a 0,49
no dossel (Figura VI.4a). Claramente, fragmentos menores, nos quais os habitats são
mais próximos (Tabela VI.1), tiveram maior SSM entre habitats, o que foi confirmado
pela análise entre grupos de tamanho, tanto na amostra total (t = 2,70; p = 0,02) quanto
no sub-bosque (p = 0,01), não se registrando diferenças entre grupos de DH.
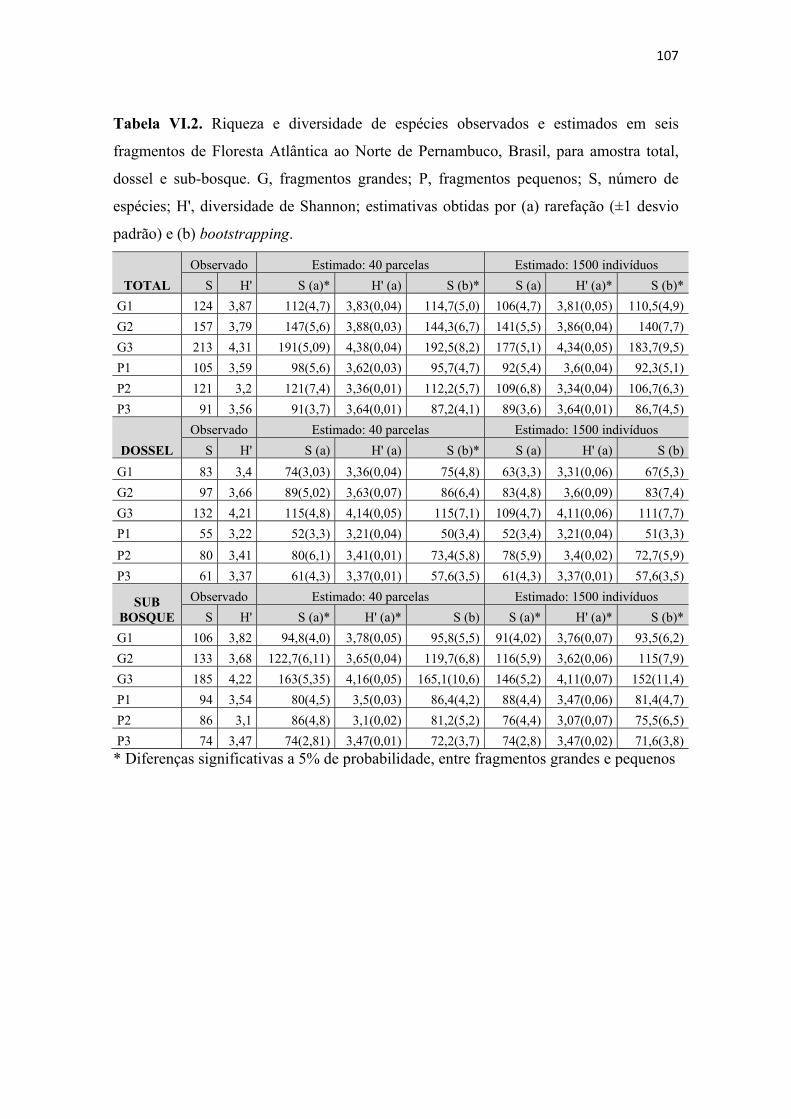
107
Tabela VI.2. Riqueza e diversidade de espécies observados e estimados em seis
fragmentos de Floresta Atlântica ao Norte de Pernambuco, Brasil, para amostra total,
dossel e sub-bosque. G, fragmentos grandes; P, fragmentos pequenos; S, número de
espécies; H', diversidade de Shannon; estimativas obtidas por (a) rarefação (±1 desvio
padrão) e (b) bootstrapping.
TOTAL Observado Estimado: 40 parcelas Estimado: 1500 indivíduos
S H' S (a)* H' (a) S (b)* S (a) H' (a)* S (b)*G1 124 3,87 112(4,7) 3,83(0,04) 114,7(5,0) 106(4,7) 3,81(0,05) 110,5(4,9)G2 157 3,79 147(5,6) 3,88(0,03) 144,3(6,7) 141(5,5) 3,86(0,04) 140(7,7)G3 213 4,31 191(5,09) 4,38(0,04) 192,5(8,2) 177(5,1) 4,34(0,05) 183,7(9,5)P1 105 3,59 98(5,6) 3,62(0,03) 95,7(4,7) 92(5,4) 3,6(0,04) 92,3(5,1)P2 121 3,2 121(7,4) 3,36(0,01) 112,2(5,7) 109(6,8) 3,34(0,04) 106,7(6,3)P3 91 3,56 91(3,7) 3,64(0,01) 87,2(4,1) 89(3,6) 3,64(0,01) 86,7(4,5)
DOSSEL Observado Estimado: 40 parcelas Estimado: 1500 indivíduos
S H' S (a) H' (a) S (b)* S (a) H' (a) S (b)G1 83 3,4 74(3,03) 3,36(0,04) 75(4,8) 63(3,3) 3,31(0,06) 67(5,3)G2 97 3,66 89(5,02) 3,63(0,07) 86(6,4) 83(4,8) 3,6(0,09) 83(7,4)G3 132 4,21 115(4,8) 4,14(0,05) 115(7,1) 109(4,7) 4,11(0,06) 111(7,7)P1 55 3,22 52(3,3) 3,21(0,04) 50(3,4) 52(3,4) 3,21(0,04) 51(3,3)P2 80 3,41 80(6,1) 3,41(0,01) 73,4(5,8) 78(5,9) 3,4(0,02) 72,7(5,9)P3 61 3,37 61(4,3) 3,37(0,01) 57,6(3,5) 61(4,3) 3,37(0,01) 57,6(3,5)
SUB BOSQUE
Observado Estimado: 40 parcelas Estimado: 1500 indivíduos S H' S (a)* H' (a)* S (b) S (a)* H' (a)* S (b)*
G1 106 3,82 94,8(4,0) 3,78(0,05) 95,8(5,5) 91(4,02) 3,76(0,07) 93,5(6,2)G2 133 3,68 122,7(6,11) 3,65(0,04) 119,7(6,8) 116(5,9) 3,62(0,06) 115(7,9)G3 185 4,22 163(5,35) 4,16(0,05) 165,1(10,6) 146(5,2) 4,11(0,07) 152(11,4)P1 94 3,54 80(4,5) 3,5(0,03) 86,4(4,2) 88(4,4) 3,47(0,06) 81,4(4,7)P2 86 3,1 86(4,8) 3,1(0,02) 81,2(5,2) 76(4,4) 3,07(0,07) 75,5(6,5)P3 74 3,47 74(2,81) 3,47(0,01) 72,2(3,7) 74(2,8) 3,47(0,02) 71,6(3,8)* Diferenças significativas a 5% de probabilidade, entre fragmentos grandes e pequenos

108
a)
Borda
Interior
b)
Borda
Interior
c)
Interior
Figura VI.3. Densidade de indivíduos na amostra de dossel e sub-bosque (colunas) e
riqueza total (triângulos) nos cinco habitats mais frequentes em seis fragmentos de
Floresta Atlântica ao Norte de Pernambuco, Brasil. a) encosta, b) ripário permanente, c)
ripário temporário. G – fragmento grande, P – fragmento pequeno (conforme Tabela 1).
0
10
20
30
40
50
60
70
80
90
100
0
100
200
300
400
500
600
700
800
900
G1 G2 G3 P1 P2 P30
10
20
30
40
50
60
70
80
90
100
0
100
200
300
400
500
600
700
800
900
G1 G2 G3 P1 P2 P3
0
10
20
30
40
50
60
70
80
90
100
0
100
200
300
400
500
600
700
800
900
G1 G2 G3 P1 P2 P30
10
20
30
40
50
60
70
80
90
100
0
100
200
300
400
500
600
700
800
900
G1 G2 G3 P1 P2 P3
0
10
20
30
40
50
60
70
80
90
100
0
100
200
300
400
500
600
700
800
900
G1 G2 G3 P1 P2 P3
Sub-bosque
Dossel
Riqueza

109
A diversidade βW calculada por fragmento, para amostra total, variou de 1,80 no
menor fragmento (P3) a 2,55 em G3 (Figura VI.4b). Entre grupos de tamanho, βW da
amostra total foi significativamente maior em fragmentos maiores (t = 3,40; p = 0,01),
não havendo diferenças entre grupos de DH. De fato, o maior e o menor valor de βW
ocorreram em fragmentos com alto DH. Diferenças entre fragmentos pequenos e
grandes mantiveram-se ao serem analisados os valores de βW obtidos de amostras de
dossel (p = 0,01) e sub-bosque (p < 0,01).
Entre os habitats, seis ocorreram em pelo menos dois fragmentos, permitindo
calcular a similaridade entre eles, através da SSM. Esta análise revelou valores de
similaridade mais baixos entre habitats semelhantes em diferentes fragmentos do que
entre habitats distintos em um mesmo fragmento, tanto na amostra total, quanto nos
estratos (p < 0,05). Habitats de interior ripário temporário (N = 4) foram os mais
semelhantes entre si, com mais de 50% de similaridade, seguidos por habitats de borda
e interior de encosta (Figura VI.5). Menores valores de similaridade foram registrados
entre habitats ripários permanentes (N = 5). Entre fragmentos, considerando todas as
parcelas amostradas, a similaridade florística variou entre 0,47 e 0,65 para amostra total,
entre 0,4 e 0,66 para dossel e 0,46 e 0,64 no sub-bosque. Quando se consideraram
apenas as amostras obtidas nos habitats de maior frequência, borda e interior de encosta,
a similaridade entre fragmentos manteve as mesmas proporções, entre 0,42 e 0,66.
Menores similaridades foram registradas entre o fragmento G3 (de maior riqueza) e P1
ou P3 (de menores riquezas).
Analisando por fragmento, registraram-se percentuais de 33 a 49% das espécies
exclusivas em um habitat, e baixas proporções de espécies comuns a todos os habitats
(de 3,7 a 8% nos fragmentos grandes e de 14 a 24% nos pequenos). Analisando-se por
habitat, houve uma forte correlação (r=0,871) entre o número total de espécies no
habitat, somando todas as suas amostras, e o número de fragmentos em que esse habitat
ocorreu (Figura VI.6), demonstrando a contribuição dos vários fragmentos à riqueza
total em cada habitat. De fato, nos cinco habitats que ocorreram em pelo menos quatro
fragmentos, entre 42 e 57% das espécies foram registradas em apenas um fragmento.

110
a)
b)
Figura VI.4. Índices de diversidade β calculados para seis fragmentos de Floresta
Atlântica ao Norte de Pernambuco, Brasil, para amostra total, dossel e sub-bosque. a)
SS – similaridade de Sørensen média; b) βW – Diversidade β de Whittaker; G –
fragmento grande, P – fragmento pequeno, conforme descrito na Tabela 1; barras de
erro = ± 1 erro padrão.
4.4. Associações entre espécies e habitats
Apenas duas espécies ocorreram em todos os fragmentos e habitats, Eschweilera
ovata (Lecythidaceae) e Tapirira guianensis (Anacardiaceae). Entre as demais, 86
espécies com número maior que 30 indivíduos foram testadas quanto à preferência por
habitat. Para este número de espécies, com oito possibilidades de associação de habitat
0.00
0.10
0.20
0.30
0.40
0.50
0.60
0.70
G1 G2 G3 P1 P2 P3
Søre
nsen
méd
io
Amostra total
Dossel
Sub-bosque
0
0.5
1
1.5
2
2.5
3
3.5
G1 G2 G3 P1 P2 P3
Div
ersi
dade
β d
e W
hitta
ker
Amostra total
Dossel
Sub-bosque

111
para cada, havia 688 associações possíveis. Dessas, foram registradas 186 (27%)
associações positivas ou negativas, em 60 espécies. Excluindo-se as espécies sem
associação positiva (3), obteve-se uma lista de 57 espécies, com 80 associações
positivas, sendo 37 espécies com associação por um habitat, 17 por dois habitats e 3 por
três habitats (Tabela VI.3). Entre as espécies com associação por habitat, 46 são de
dossel e 11 são típicas de sub-bosque. Habitats ripários permanentes (borda e interior)
apresentaram maior número de associações positivas, 31 no total, seguidos pelos
ripários temporários (25) e interior de tabuleiro (10).
A análise de espécies indicadoras (ISA) apontou 49 espécies indicadoras, entre
as quais sete foram indicadas apenas pela ISA, 39 concordaram com o teste do qui-
quadrado, com associação pelos mesmos habitats, duas foram apontadas para habitats
distintos nos dois testes e uma (Dialium guianense, Fabaceae) teve, pelo qui-quadrado,
apenas associações negativas (Tabela VI.3). Semelhantemente, os habitats com maior
número de espécies indicadoras foram os ripários (temporário e permanente, borda e
interior) e interior de tabuleiro, que juntos somaram 44 espécies.
Entre as espécies com associação, observou-se variação na posição de
importância (VI) na assembléia do habitat nos vários fragmentos. Como indicado na
Tabela VI.3, cerca de 60% das espécies com associação por habitats foram mais
importantes nos fragmentos grandes, enquanto 40% se destacaram nos pequenos. As
espécies associadas à borda de encosta foram todas arbóreas, figurando, em sua maioria,
entre as mais importantes nos fragmentos grandes. Entre as seis espécies associadas ao
interior de encosta, três são típicas de sub-bosque e três de dossel, com destaque para a
arbórea Eschweilera ovata, sempre na posição de primeira ou segunda em importância
nesse habitat em todos os fragmentos. Nos habitats ripários temporários e permanentes,
as espécies associadas foram, na maioria, do dossel, com importância bem distinta entre
os fragmentos, a exemplo das Annonaceae Anaxagorea dolichocarpa em G3, e Xylopia
frutescens em P1, e da Myrtaceae Calyptranthes brasiliensis em G1. Espécies de dossel
também predominaram como típicas nos ambientes de tabuleiro.
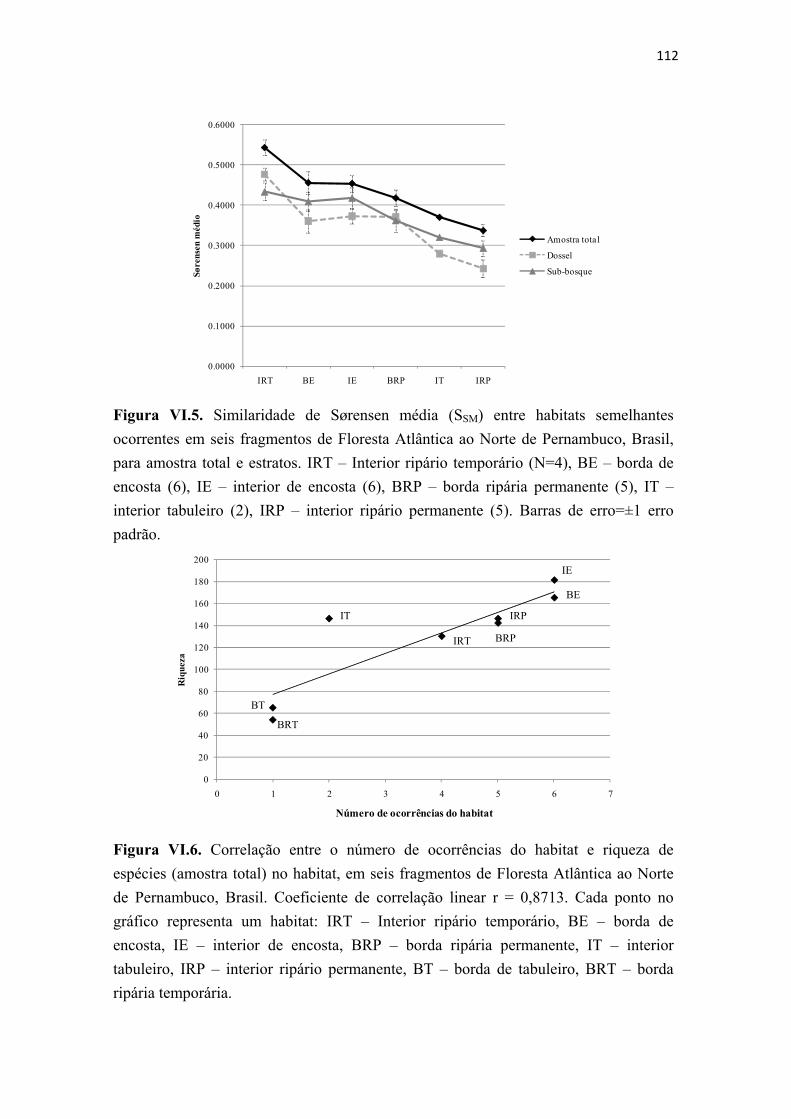
112
Figura VI.5. Similaridade de Sørensen média (SSM) entre habitats semelhantes ocorrentes em seis fragmentos de Floresta Atlântica ao Norte de Pernambuco, Brasil, para amostra total e estratos. IRT – Interior ripário temporário (N=4), BE – borda de encosta (6), IE – interior de encosta (6), BRP – borda ripária permanente (5), IT – interior tabuleiro (2), IRP – interior ripário permanente (5). Barras de erro=±1 erro padrão.
Figura VI.6. Correlação entre o número de ocorrências do habitat e riqueza de espécies (amostra total) no habitat, em seis fragmentos de Floresta Atlântica ao Norte de Pernambuco, Brasil. Coeficiente de correlação linear r = 0,8713. Cada ponto no gráfico representa um habitat: IRT – Interior ripário temporário, BE – borda de encosta, IE – interior de encosta, BRP – borda ripária permanente, IT – interior tabuleiro, IRP – interior ripário permanente, BT – borda de tabuleiro, BRT – borda ripária temporária.
0.0000
0.1000
0.2000
0.3000
0.4000
0.5000
0.6000
IRT BE IE BRP IT IRP
Søre
nsen
méd
io
Amostra total
Dossel
Sub-bosque
0
20
40
60
80
100
120
140
160
180
200
0 1 2 3 4 5 6 7
Riq
ueza
Número de ocorrências do habitat
BRT
BT
IRT
IT
BRP
IRP
BE
IE
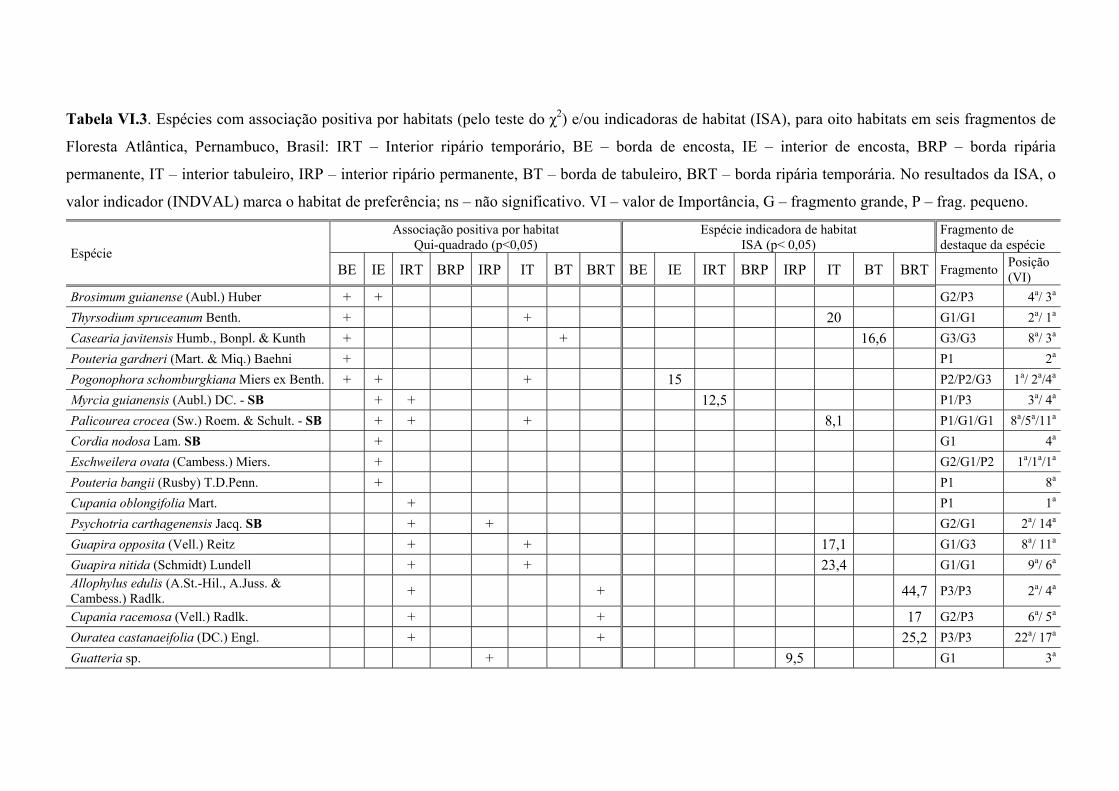
113
Tabela VI.3. Espécies com associação positiva por habitats (pelo teste do χ2) e/ou indicadoras de habitat (ISA), para oito habitats em seis fragmentos de
Floresta Atlântica, Pernambuco, Brasil: IRT – Interior ripário temporário, BE – borda de encosta, IE – interior de encosta, BRP – borda ripária
permanente, IT – interior tabuleiro, IRP – interior ripário permanente, BT – borda de tabuleiro, BRT – borda ripária temporária. No resultados da ISA, o
valor indicador (INDVAL) marca o habitat de preferência; ns – não significativo. VI – valor de Importância, G – fragmento grande, P – frag. pequeno.
Espécie
Associação positiva por habitat Qui-quadrado (p<0,05)
Espécie indicadora de habitat ISA (p< 0,05)
Fragmento de destaque da espécie
BE IE IRT BRP IRP IT BT BRT BE IE IRT BRP IRP IT BT BRT Fragmento Posição (VI)
Brosimum guianense (Aubl.) Huber + + G2/P3 4a/ 3a Thyrsodium spruceanum Benth. + + 20 G1/G1 2a/ 1a Casearia javitensis Humb., Bonpl. & Kunth + + 16,6 G3/G3 8a/ 3a Pouteria gardneri (Mart. & Miq.) Baehni + P1 2a Pogonophora schomburgkiana Miers ex Benth. + + + 15 P2/P2/G3 1a/ 2a/4a Myrcia guianensis (Aubl.) DC. - SB + + 12,5 P1/P3 3a/ 4a Palicourea crocea (Sw.) Roem. & Schult. - SB + + + 8,1 P1/G1/G1 8a/5a/11a Cordia nodosa Lam. SB + G1 4a Eschweilera ovata (Cambess.) Miers. + G2/G1/P2 1a/1a/1a Pouteria bangii (Rusby) T.D.Penn. + P1 8a Cupania oblongifolia Mart. + P1 1a Psychotria carthagenensis Jacq. SB + + G2/G1 2a/ 14a Guapira opposita (Vell.) Reitz + + 17,1 G1/G3 8a/ 11a Guapira nitida (Schmidt) Lundell + + 23,4 G1/G1 9a/ 6a Allophylus edulis (A.St.-Hil., A.Juss. & Cambess.) Radlk. + + 44,7 P3/P3 2a/ 4a
Cupania racemosa (Vell.) Radlk. + + 17 G2/P3 6a/ 5a Ouratea castanaeifolia (DC.) Engl. + + 25,2 P3/P3 22a/ 17a Guatteria sp. + 9,5 G1 3a
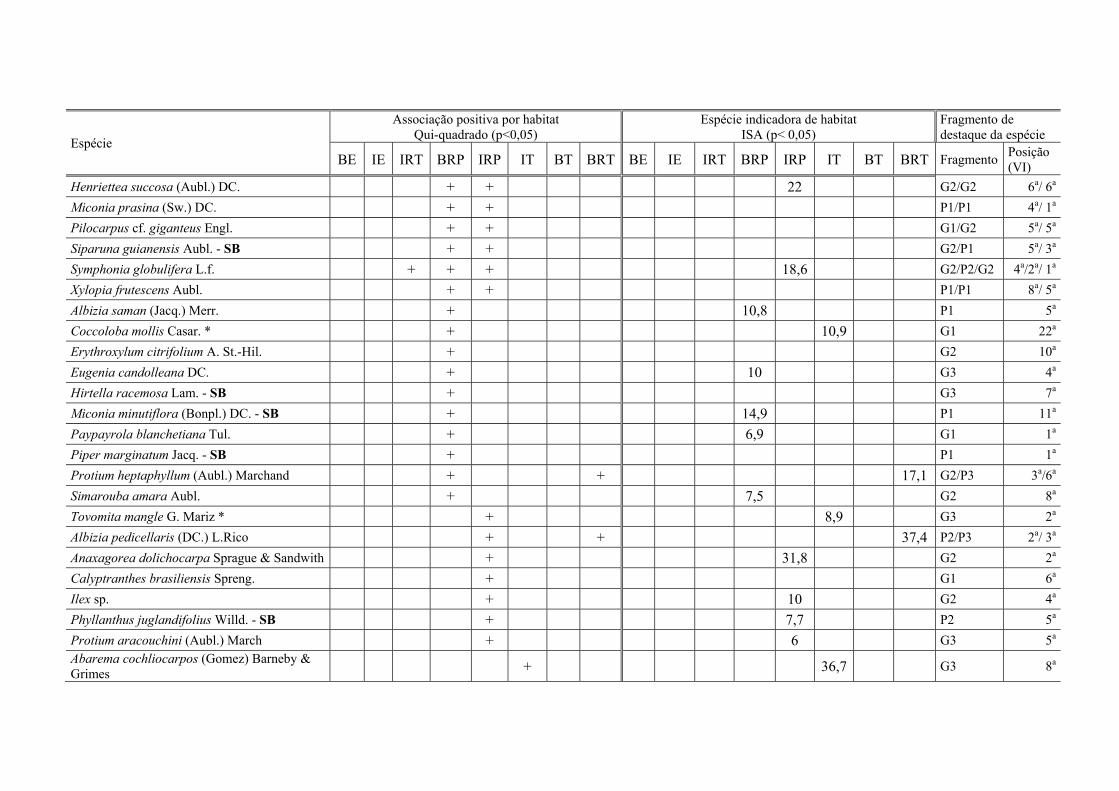
114
Espécie
Associação positiva por habitat Qui-quadrado (p<0,05)
Espécie indicadora de habitat ISA (p< 0,05)
Fragmento de destaque da espécie
BE IE IRT BRP IRP IT BT BRT BE IE IRT BRP IRP IT BT BRT Fragmento Posição (VI)
Henriettea succosa (Aubl.) DC. + + 22 G2/G2 6a/ 6a Miconia prasina (Sw.) DC. + + P1/P1 4a/ 1a Pilocarpus cf. giganteus Engl. + + G1/G2 5a/ 5a Siparuna guianensis Aubl. - SB + + G2/P1 5a/ 3a Symphonia globulifera L.f. + + + 18,6 G2/P2/G2 4a/2a/ 1a Xylopia frutescens Aubl. + + P1/P1 8a/ 5a Albizia saman (Jacq.) Merr. + 10,8 P1 5a Coccoloba mollis Casar. * + 10,9 G1 22a Erythroxylum citrifolium A. St.-Hil. + G2 10a Eugenia candolleana DC. + 10 G3 4a Hirtella racemosa Lam. - SB + G3 7a Miconia minutiflora (Bonpl.) DC. - SB + 14,9 P1 11a Paypayrola blanchetiana Tul. + 6,9 G1 1a Piper marginatum Jacq. - SB + P1 1a Protium heptaphyllum (Aubl.) Marchand + + 17,1 G2/P3 3a/6a Simarouba amara Aubl. + 7,5 G2 8a Tovomita mangle G. Mariz * + 8,9 G3 2a Albizia pedicellaris (DC.) L.Rico + + 37,4 P2/P3 2a/ 3a Anaxagorea dolichocarpa Sprague & Sandwith + 31,8 G2 2a Calyptranthes brasiliensis Spreng. + G1 6a Ilex sp. + 10 G2 4a Phyllanthus juglandifolius Willd. - SB + 7,7 P2 5a Protium aracouchini (Aubl.) March + 6 G3 5a Abarema cochliocarpos (Gomez) Barneby & Grimes + 36,7 G3 8a
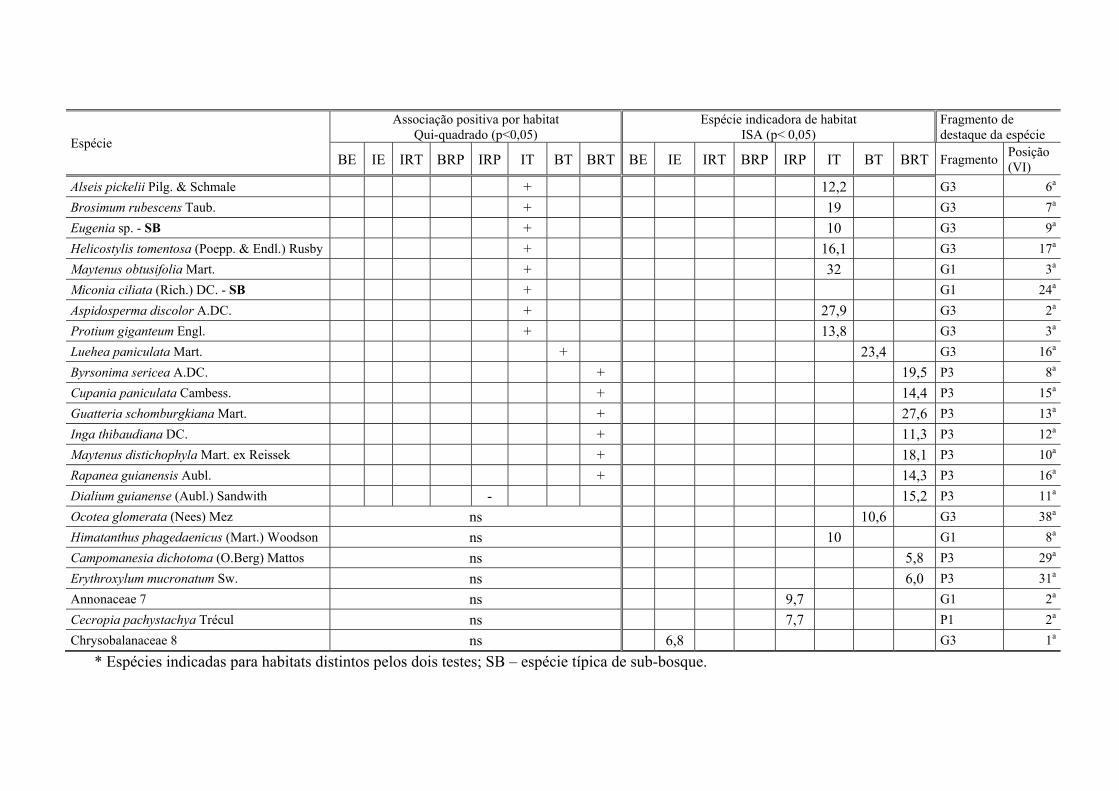
115
Espécie
Associação positiva por habitat Qui-quadrado (p<0,05)
Espécie indicadora de habitat ISA (p< 0,05)
Fragmento de destaque da espécie
BE IE IRT BRP IRP IT BT BRT BE IE IRT BRP IRP IT BT BRT Fragmento Posição (VI)
Alseis pickelii Pilg. & Schmale + 12,2 G3 6a Brosimum rubescens Taub. + 19 G3 7a Eugenia sp. - SB + 10 G3 9a Helicostylis tomentosa (Poepp. & Endl.) Rusby + 16,1 G3 17a Maytenus obtusifolia Mart. + 32 G1 3a Miconia ciliata (Rich.) DC. - SB + G1 24a Aspidosperma discolor A.DC. + 27,9 G3 2a Protium giganteum Engl. + 13,8 G3 3a Luehea paniculata Mart. + 23,4 G3 16a Byrsonima sericea A.DC. + 19,5 P3 8a Cupania paniculata Cambess. + 14,4 P3 15a Guatteria schomburgkiana Mart. + 27,6 P3 13a Inga thibaudiana DC. + 11,3 P3 12a Maytenus distichophyla Mart. ex Reissek + 18,1 P3 10a Rapanea guianensis Aubl. + 14,3 P3 16a Dialium guianense (Aubl.) Sandwith - 15,2 P3 11a Ocotea glomerata (Nees) Mez ns 10,6 G3 38a Himatanthus phagedaenicus (Mart.) Woodson ns 10 G1 8a Campomanesia dichotoma (O.Berg) Mattos ns 5,8 P3 29a Erythroxylum mucronatum Sw. ns 6,0 P3 31a Annonaceae 7 ns 9,7 G1 2a Cecropia pachystachya Trécul ns 7,7 P1 2a Chrysobalanaceae 8 ns 6,8 G3 1a
* Espécies indicadas para habitats distintos pelos dois testes; SB – espécie típica de sub-bosque.

116
5. Discussão
5.1. Heterogeneidade de habitats e diversidade
Neste estudo, área teve maior influência na riqueza e diversidade de plantas, na
escala do fragmento, somando evidências à aplicação da teoria de ilhas a fragmentos
florestais (LAURANCE et al., 2002; HILL; CURRAN, 2003; CAGNOLO et al., 2006;
ECHEVERRÍA et al., 2007), porém divergindo da hipótese da heterogeneidade de
habitats como explicativa da relação espécie-área (WILLIAMS, 1964; CONNOR;
MCCOY, 1979; BROSE, 2001; DUFOUR et al., 2006; BÁLDI, 2008).
Independentemente do número e diversidade de habitats, maiores riquezas ocorreram
em fragmentos grandes, e menores riquezas nos pequenos, mesmo quando um pequeno
(P1) teve número de habitats semelhante a um grande (G2). Este padrão de maior
riqueza em fragmentos maiores foi também independente da densidade de indivíduos,
portanto não influenciado pelo efeito de densidade ou de massa (SHMIDA; WILSON,
1985; DENSLOW, 1995), e da ordem de amostragem seguindo a distribuição dos
habitats.
Apesar da influência da heterogeneidade de habitats na relação espécie-área não
ter sido confirmada, a influência da área na diversidade de habitats se revelou decisiva
para a riqueza de plantas, pois apenas em fragmentos maiores registraram-se habitats de
tabuleiro, detentores da maior riqueza florística neste estudo. Este dado fornece mais
uma evidência às sérias consequências da natureza não aleatória da fragmentação
(SEABLOOM et al., 2002) e à necessidade de se avaliar os habitats remanescentes na
paisagem como um todo, independentemente do tamanho dos fragmentos, buscando
identificar porções significativas de habitats que atuem provendo recursos e abrigo para
grupos cruciais de espécies (TEWS et al., 2004; EWERS; DIDHAM, 2006).
A heterogeneidade de habitats teve influência marcante na diversidade beta.
Mesmo em fragmentos pequenos, nos quais as distâncias entre sítios são menores,
valores de similaridade entre habitats foram, em média, inferiores a 50%, ou seja, um
alto percentual de espécies ocorreu em habitats restritos. Este dado reforça a
necessidade de se considerar a escala do habitat em estudos de diversidade (KOLASA;
WALTHO, 1998), a partir de dimensões relevantes para a assembléia em foco
(LOOKINGBILL; URBAN, 2005; LUNDHOLM, 2009). Neste caso, uma amostra mais

117
completa da riqueza vegetal (308 espécies) foi obtida partindo do habitat para o
fragmento e deste à paisagem. Da mesma forma, se nossa abordagem enfocasse área do
fragmento apenas, teríamos uma medida numérica e dissociada da complementaridade,
que mede a distinção na composição de espécies entre habitats diferentes (COLWELL;
CODDINGTON, 1994). Altos graus de complementaridade vêm sendo registrados em
ecossistemas florestais no nível do fragmento (METZGER et al., 1997; CHUST et al.,
2006; PEREIRA et al., 2007; SANTOS et al., 2007b), mostrando ser esta uma medida
imprescindível ao entendimento de padrões de diversidade em paisagens fragmentadas,
principalmente se obtida ao nível do habitat (CRAMER; WILLIG, 2005).
Entre os fragmentos, habitats de borda predominaram nos menores. Nesses
ambientes, pesquisas têm revelado uma tendência à redução da diversidade alfa
(OLIVEIRA et al., 2004; PEREIRA et al., 2007; LAURANCE, 2008; SILVA et al.,
2008c; GOMES et al., 2009) e simplificação das assembléias de plantas, devido à
proliferação de espécies pioneiras e invasoras (LAURANCE et al., 2007; TABARELLI
et al., 2008), alteração nos processos reprodutivos (LOPES et al., 2009) e aumento nas
taxas de mortalidade (LAURANCE et al., 2007). Os estudos de Santos et al. (2008) e
Oliveira et al. (2008), numa paisagem altamente fragmentada, permitiram descrever a
influência da borda como agente de erosão florística e de colapso estrutural das espécies
de dossel. Além de somar mais uma evidência a esses processos, nossos dados
revelaram que esta influência é dependente da proporção do habitat de borda no
fragmento, já que bordas em fragmentos menores tiveram menor riqueza do que em
fragmentos maiores, principalmente no estrato de dossel.
Habitats de encosta, que ocorreram em maiores proporções, foram os que mais
contribuíram para a riqueza total nos fragmentos, especialmente o interior de encosta.
Porém, em fragmentos menores, a proporção relativamente menor de interior de encosta
coincidiu com uma diminuição da riqueza nesses habitats. Isto pode ser reflexo de um
processo de simplificação da estrutura da assembléia, devido à maior proporção de
bordas, que favoreceria uma homogeneização por extinção seletiva de espécies
secundárias de dossel (LAURANCE et al., 2007) ou uma influência do tamanho no
desaparecimento de espécies mais sensíveis típicas de floresta madura (TILMAN et al.,
1994).

118
5.2. Heterogeneidade de habitats e especialização de plantas
Considerando a alta proporção de raras na amostra e as restrições das análises,
um número relativamente pequeno de espécies (28%) pôde ser testado quanto à
preferência por habitats. Dessas, 66% das espécies analisadas (19% do total) mostraram
associação positiva ou indicação por algum habitat. Este percentual é compatível com
os mais altos registrados em outras comunidades florestais, como, por exemplo: 76% de
34 espécies dominantes no sudoeste da Amazônia (PHILLIPS et al., 2003); 42% de 171
espécies (HARMS et al., 2001) ou 37% de 80 espécies (COMITA et al., 2007), no
Panamá; 36% de 199 espécies na Colômbia (JOHN et al., 2007); 30% de 132 espécies
em La Selva, Costa Rica (CLARK et al., 1999); e 25% em 1104 espécies na Amazônia
(VALENCIA et al., 2004).
A maioria das espécies teve associação ou foi indicadora de habitats ripários
permanentes ou temporários. Estas áreas, embora com menor diversidade alfa entre os
habitats estudados, contribuem com a riqueza total por abrigarem espécies exclusivas,
de distribuição restrita e com adaptações a inundações e intolerância à seca, um padrão
comum em sítios ripários (HARMS et al., 2001; GUNATILLEKE et al., 2006;
DRUCKER et al., 2008). São exemplos particularmente interessantes as espécies
ripárias restritas Symphonia globulifera, de alta frequência nos fragmentos, e Tovomita
mangle e Anaxagorea dolichocarpa, de ocorrência exclusiva em um fragmento. Muitas
espécies tiveram associação com o habitat de tabuleiro, que também apresentou a maior
diversidade alfa, somando mais de 140 espécies, embora ocorrendo em apenas dois
fragmentos. Lamentavelmente, este habitat está entre os menos frequentes e com menor
cobertura nesta paisagem (capítulo 2) e provavelmente sofreu maior desflorestamento
nas décadas passadas (TRINDADE et al., 2008).
Apesar das assembléias terem sido amostradas separadamente em dossel e sub-
bosque, não foram observadas, no geral, grandes diferenças nas respostas dos estratos
nas análises realizadas. O sub-bosque, porém, teve valores de similaridade entre habitats
ligeiramente maiores do que o dossel tanto dentro quanto entre fragmentos. Embora não
tenhamos testado a associação espécie-habitat por estrato, este dado pode estar
sinalizando que as preferências por habitats se definem em diferentes estágios de plantas
lenhosas (WEBB; PEART, 2000), o que leva a distintos valores de diversidade beta em
cada estrato. Comita et al. (2007), em floresta panamenha, só encontraram associações

119
por habitats em grandes árvores e não no estrato de regeneração, enquanto Duque et al.
(2002) registraram padrão contrário na Amazônia. Uma maior similaridade no estrato de
sub-bosque pode apontar, por outro lado, para uma convergência das assembléias por
homogeneização, representando uma resposta tardia à fragmentação (METZGER et al.,
2009), como discutido no capítulo 3.
6. Considerações finais e implicações
A integração entre área e heterogeneidade em comunidades fragmentadas ainda
é um desafio, com um problema implícito de escala. John et al. (2007) argumentam que
há poucos trabalhos avaliando a correspondência entre dimensões de habitat e
distribuição de espécies em escalas em torno de 1 km2, na qual se enquadram os
fragmentos avaliados no nosso estudo. Além disso, tem-se, por um lado, poucas
pesquisas que avaliam a relação entre riqueza, área e alguma medida de
heterogeneidade em um grande número de fragmentos de variados tamanhos, testando o
poder explicativo de cada variável (BROSE, 2001; GIGNAC; DALE, 2005; PEREIRA
et al., 2007). Por outro lado, estudos desenvolvidos em áreas contínuas ou parcelas
permanentes fornecem as principais evidências à existência de preferências por habitats,
seja na Amazônia (PITMAN et al., 1999; PITMAN et al., 2002; PHILLIPS et al.,
2003), Costa Rica (CLARK et al., 1999), Panamá (HUBBELL; FOSTER, 1983;
CONDIT et al., 2000; HARMS et al., 2001; PYKE et al., 2001; CONDIT et al., 2002;
COMITA et al., 2007; ZILLIO; CONDIT, 2007) ou Malásia (WILLS; CONDIT, 1999;
CONDIT et al., 2002; POTTS et al., 2002). Nesses, geralmente investiga-se a qual
mecanismo de estruturação de comunidades as espécies respondem: preferências por
habitats ou limitações na dispersão, esta última explicando a diversidade de forma
neutra (HUBBELL, 2001), por dispersão aleatória limitada pelo aumento da distância
geográfica (PHILLIPS et al., 2003; CHUST et al., 2006).
Nos dados aqui apresentados, foram registradas baixas similaridades (i.e. alta
complementaridade), alto número de espécies exclusivas por habitat e baixa proporção
de espécies comuns entre habitats, mesmo em fragmentos pequenos, o que suscita uma
atuação mais provável, ao nível do fragmento, do processo de preferência por habitats
ou ‘segregação ambiental’ (CHUST et al., 2006). Porém, a alta complementaridade
entre habitats semelhantes em fragmentos distintos sugere a influência da limitação de

120
dispersão, imposta pela distância geográfica ou inospitalidade da matriz, para explicar
este padrão. Na literatura, há evidências, de fato, que os processos de segregação
ambiental e dispersão provavelmente atuam em diferentes escalas espaciais e temporais
(SHMIDA; WILSON, 1985; CONDIT et al., 2002; PHILLIPS et al., 2003; CHUST et
al., 2006). Padrões similares, quando mais de um processo é evidente, foram discutidos
por Condit et al. (2002) para florestas no Panamá, Equador e Peru, por Valencia et al.
(2004) na Amazônia e Hardy e Sonke (2004) em Camarões. No nosso caso, fica clara a
necessidade de mais pesquisas nesta direção, baseadas na estrutura de habitats e
amostrando-se um maior número de fragmentos.
Dos resultados aqui apresentados, portanto, podem-se derivar algumas
implicações para teoria ecológica e conservação: são fornecidas evidências à atuação de
pelo menos três processos estruturadores de comunidades fragmentadas: o efeito da
área, heterogeneidade de habitats e limitação de dispersão; ressalta-se ser a diversidade
beta tão importante quanto à alfa para conservação, como postulado por Condit et al.
(2002); constata-se que grande perda de espécies deve ter ocorrido a partir da perda de
habitats de tabuleiro e ripários, preservados em menores proporções, porém,
respectivamente, de maior diversidade e riqueza em espécies exclusivas; e corrobora,
junto a vários outros autores (TURNER; CORLETT, 1996; BENEDICK et al., 2006;
SANTOS et al., 2007b), que em florestas altamente fragmentadas e reduzidas, a
diversidade só será preservada com base na manutenção de todos os fragmentos e da
soma de porções de seus habitats, com enfoque na complementaridade entre
assembléias.

121
VII. CONCLUSÃO GERAL

122
Nesta tese, transitou-se em uma área de sobreposição de disciplinas e escalas
ecológicas. De um lado, há a pesquisa em fragmentação - no seu senso amplo que
aborda redução de área e quebra de continuidade - na qual a maioria dos estudos
investiga a riqueza como função da área apenas ou testa o efeito de borda ou evita as
zonas de borda e a heterogeneidade ambiental. Do outro lado, há os estudos
sinecológicos que explicam a estruturação de comunidades vegetais a partir da
preferência por nichos de recursos (ou habitats) ou de forma neutra, quando as espécies
são equivalentes. Na interface, há fragmentos de comunidades anteriormente contínuas,
há previsão de perda de habitats e de perda de espécies. A partir dos resultados, foi
possível somar evidências às teorias e listar várias implicações para conservação.
Para integrar área, heterogeneidade e riqueza, a abordagem adotada partiu da
paisagem e terminou no habitat e nas espécies. Numa prospecção inicial, na escala da
paisagem, somaram-se evidências à relação entre área e heterogeneidade de habitats:
são variáveis positiva e significativamente relacionadas. Esta relação, entretanto, se
mostrou menos previsível nos fragmentos menores, que são exatamente a maioria. Em
áreas maiores, a relação foi influenciada por certos tipos de habitat eliminados,
principalmente, dos fragmentos menores no processo de fragmentação, tais como os
sítios ripários e as áreas planas altas (tabuleiro). Na escala do habitat, nas áreas ripárias
perenes ocorreu maior número de espécies exclusivas, exibindo preferências pelo
habitat; e o tabuleiro foi a área com maior riqueza e diversidade.
Habitats ripários resistem em 44% dos fragmentos, mas somam juntos apenas
15% das áreas florestais. Os vales de rios foram os primeiros habitats a serem
explorados na região canavieira de Pernambuco, por apresentarem os solos mais férteis
(KIMMEL et al., 2008). Assim, habitats ripários podem estar isolados uns dos outros há
pelo menos um século, considerando o acelerado desenvolvimento da cultura da cana-
de-açúcar até o final do século XIX (ANDRADE, 1989). É provável que, por isso, as
áreas ripárias amostradas compartilhem tão poucas espécies. Em florestas ripárias, é
uma característica comum a variação da riqueza ao longo do curso dos rios,
contribuindo para uma alta diversidade biótica total. Porém, uma vez isoladas,
comunidades ripárias tendem a diminuir em riqueza (NILSSON; SVEDMARK, 2002).
Antagonicamente à alta riqueza, é alarmante constatar que apenas 2% da área dos
fragmentos são habitats de tabuleiro. Provavelmente, um considerável número de
espécies já foi perdido nessas áreas, cujo desmatamento foi mais recente e impulsionado

123
pela mecanização no cultivo da cana-de-açúcar e adoção de novos métodos de irrigação
na segunda metade do século XX (KIMMEL et al., 2008).
No nível do fragmento, constatou-se que a relação espécie-área é explicada pela
área per se e não pela heterogeneidade de habitats. Em outras palavras, confirmou-se
um aumento do número de espécies com aumento da área, mesmo quando se estimou a
riqueza para amostras equivalentes. A heterogeneidade, por outro lado, foi importante
para explicar a diversidade beta nos fragmentos e a complementaridade da riqueza total
na paisagem. Isso implica que cada porção de habitat em cada fragmento, mesmo nos
pequenos, importa para a riqueza no nível da paisagem. Entretanto, a análise do sub-
bosque, tanto em relação à área quanto à similaridade entre fragmentos, mostra que já se
delineia uma tendência à simplificação das assembléias, provavelmente como uma
resposta secundária à influência das bordas (HARPER et al., 2005; SANTOS et al.,
2008), que dominam 40% da área florestal, e redução da área. As decorrências deste
fato são: apesar da riqueza de habitats e espécies nos pequenos fragmentos contribuir de
forma relevante para a riqueza total, o número de espécies pode ainda diminuir
progressivamente em direção a assembléias mais homogêneas e simplificadas. A alta
complementaridade encontrada soma, ainda, evidências da atuação do processo de
limitação de dispersão, imposta pela distância entre fragmentos ou inospitalidade da
matriz.
Do ponto de vista conservacionista, a sustentação da riqueza e diversidade na
paisagem, em longo prazo, depende i) de medidas efetivas de manutenção dos
fragmentos existentes, quase exclusivamente formados por habitats de encostas, ii) de
ações de recuperação florestal para aumentar a conectividade entre fragmentos de
diferentes tamanhos, iii) da criação de corredores ciliares que reintegrem os habitats
ripários, iv) da proteção das bordas florestais e v) acompanhamento das variações
temporais das assembléias nos diferentes habitats para verificar a regeneração no sub-
bosque.
À luz das teorias de comunidades, se área e diversidade de habitats são
correlacionadas, área per se seria suficiente para explicar padrões de riqueza vegetal,
atuando como substituta da heterogeneidade? A resposta é sim para uma relação
espécie-área simplesmente. Mas a resposta é não quando se deseja a conservação da
riqueza remanescente como um todo, nesta paisagem de Floresta Atlântica.

124
II. REFERÊNCIAS BIBLIOGRÁFICAS2
AERTS, R.; VAN OVERTVELD, K.; HAILE, M.; HERMY, M.; DECKERS, J.;
MUYS, B. Species composition and diversity of small Afromontane forest fragments in
northern Ethiopia. Plant Ecology, v.187, p.127-142, 2006.
AGP-II. An update of the Angiosperm Phylogeny Group classification for the orders
and families of flowering plants: APG II. Botanical Journal of the Linnean Society,
v.141, p.399-436, 2003.
ALBUQUERQUE, U. P.; MEDEIROS, P. M.; ARAÚJO, T. A. S.; SILVA, T. C.;
CUNHA, L. V. F. C.; OLIVEIRA JÚNIOR, G. J.; ALMEIDA, C. F. C. B. R. The role
of ethnobotany and environmental perception in the conservation of Atlantic Forest
fragments in northeastern Brazil. Bioremediation, Biodiversity and Bioavailability,
v.2, n.1, p.27-34, 2008.
ALMEIDA, A. L. S.; MEDEIROS, P. M.; SILVA, T. C.; RAMOS, M. A.; SIEBER, S.
S.; ALBUQUERQUE, U. P. Does the June tradition impact the use of wood resources
from an area of Atlantic Forest in Northeastern Brazil? Functional Ecosystems and
Communities, v.2, p.32-44, 2008.
ALVES-ARAÚJO, A.; ARAÚJO, D.; MARQUES, J.; MELO, A.; MACIEL, J. R.;
IRAPUAN, J.; PONTES, T.; LUCENA, M. D. F. D. A.; DU BOCAGE, A. L.; ALVES,
M. Diversity of angiosperms in fragments of Atlantic Forest in the state of Pernambuco,
northeastern Brazil. Bioremediation, Biodiversity and Bioavailability, v.2, n.1, p.14-
26, 2008.
ANDRADE, M. C. História das usinas de açúcar de Pernambuco. 1a ed. Recife:
Fundação Joaquim Nabuco/ Massangana/ MCT-CNPq, 1989. 114p.
ARRHENIUS, O. Species and area. Journal of Ecology, v.9, n.1, p.95-99, 1921.
AUSTIN, M. P. Species distribution models and ecological theory: a critical assessment
and some possible new approaches. Ecological Modelling, v.200, n.1-2, p.1-19, Jan.
2007.
2 Baseada na NBR‐6023/2002 da Associação Brasileira de Normas Técnicas (ABNT)

125
AUSTIN, M. P.; NICHOLLS, A. O.; MARGULES, C. R. Measurement of the realized
qualitative niche: environmental niches of five Eucalyptus Species. Ecological
Monographs, v.60, n.2, p.161-177, 1990.
AUSTIN, M. P.; SMITH, T. M.; VAN NIEL, K. P.; WELLINGTON, A. B.
Physiological responses and statistical models of the environmental niche: a
comparative study of two co-occurring Eucalyptus species. Journal of Ecology, v.97,
n.3, p.496-507, 2009.
AYRES, M.; AYRES JR., M.; AYRES, D. L.; SANTOS, A. A. S. BioEstat: Aplicações
estatísticas nas áreas das Ciências Biomédicas. Belém, Brasília: Instituto Mamirauá,
MCT/ CNPq, 2007.
BÁLDI, A. Microclimate and vegetation edge effects in a reedbed in Hungary.
Biodiversity and Conservation, v.8, n.12, p.1697-1706, 1999.
BÁLDI, A. Habitat heterogeneity overrides the species-area relationship. Journal of
Biogeography, v.35, n.4, p.675-681, 2008.
BARBOSA, J. A.; SOUZA, E. M.; LIMA FILHO, M. F.; NEUMANN, V. H. A
estratigrafia da Bacia Paraíba: uma reconsideração. Estudos Geológicos, v.13, p.89-
108, 2003.
BARBOSA, M. R. V. Estudo florístico e fitossociológico da Mata do Buraquinho,
remanescente de Mata Atlântica em João Pessoa, PB. 1996. Tese (Doutorado em
Biologia Vegetal) - Universidade Estadual de Campinas, 1996, Campinas. 135p.
BAZZAZ, F. A. Habitat selection in plants. The American Naturalist, v.137, p.S116-
S130, 1991.
BEGON, M.; HARPER, J. L.; TOWNSEND, C. R. Ecology: Individuals, populations
and communities. 3rd ed. London/ Oxford: Blackwell Scientific Publications, 1996.
1068p.
BELL, G. Neutral macroecology. Science, v.293, n.5539, p.2413-2418, Sep. 2001.
BENEDICK, S.; HILL, J. K.; MUSTAFFA, N.; CHEY, V. K.; MARYATI, M.;
SEARLE, J. B.; SCHILTHUIZEN, M.; HAMER, K. C. Impacts of rain forest
fragmentation on butterflies in northern Borneo: species richness, turnover and the
value of small fragments. Journal of Applied Ecology, v.43, n.5, p.967-977, Oct. 2006.

126
BENITEZ-MALVIDO, J. Impact of forest fragmentation on seedling abundance in a
Tropical Rain Forest. Conservation Biology, v.12, n.2, p.380-389, 1998.
BONGERS, F.; POORTER, L.; HAWTHORNE, W. D.; SHEIL, D. The intermediate
disturbance hypothesis applies to tropical forests, but disturbance contributes little to
tree diversity. Ecology Letters, v.12, n.8, p.798-805, 2009.
BOTREL, R. T.; OLIVEIRA-FILHO, A. T. D.; RODRIGUES, L. A.; CURI, N.
Influência do solo e topografia sobre as variações da composição florística e estrutura
da comunidade arbóreo-arbustiva de uma floresta estacional semidecidual em Igaí, MG.
Revista Brasileira de Botânica, v.25, n.2, p.195-213, 2002.
BOUTIN, C.; JOBIN, B.; BÉLANGER, L. Importance of riparian habitats to flora
conservation in farming landscapes of southern Québec, Canada. Agriculture,
Ecosystems and Environment, v.94, p.73-87, 2003.
BRANDÃO, C. F. L. S.; MARANGON, L. C.; FERREIRA, R. L. C.; LINS-E-SILVA,
A. C. B. Estrutura fitossociológica e classificação sucessional do componente arbóreo
em um fragmento de floresta atlântica em Igarassu – Pernambuco. Revista Brasileira
de Ciências Agrárias, v.4, n.1, p.55-61, 2009.
BROSE, U. Relative importance of isolation, area and habitat heterogeneity for vascular
plant species richness of temporary wetlands in east-German farmland. Ecography,
v.24, n.6, p.722-730, 2001.
BROWN, J. H.; LOMOLINO, M. V. Concluding remarks: historical perspective and the
future of island biogeography theory. Global Ecology and Biogeography, v.9, n.1,
p.87-92, Jan. 2000.
CADENASSO, M. L.; PICKETT, S. T. A.; WEATHERS, K. C.; BELL, S. S.;
BENNING, T. L.; CARREIRO, M. M.; DAWSON, T. E. An interdisciplinary and
synthetic approach to ecological boundaries. BioScience, v.53, n.8, p.717-722, 2003a.
CADENASSO, M. L.; PICKETT, S. T. A.; WEATHERS, K. C.; JONES, C. G. A
framework for a theory of ecological boundaries. BioScience, v.53, n.8, p.750-758,
2003b.

127
CAGNOLO, L.; CABIDOB, M.; VALLADARESA, G. Plant species richness in the
Chaco Serrano Woodland from central Argentina: Ecological traits and habitat
fragmentation effects. Biological Conservation, v.132, p.510-519, 2006.
CÂMARA, I. G. Brief history of conservation in the Atlantic Forest. In: GALINDO-
LEAL, C.; CÂMARA, I. G. (Eds.) The Atlantic Forest of South America:
Biodiversity Status, Threats and Outlook. Washington DC: Island Press, 2003. Cap. 4,
p. 31-42.
CANTRELL, R. S.; COSNER, C. Spatial heterogeneity and critical patch size: area
effects via diffusion in closed environments. Journal of Theoretical Biology, v.209,
n.2, p.161-171, 2001.
CASSEMIRO, F. A. S.; PADIAL, A. A. Teoria Neutra da Biodiversidade e
Biogeografia: aspectos teóricos, impactos na literatura e perspectivas. Oecologia
Brasiliensis, v.12, n.4, p.706-719, 2008.
CAYUELA, L.; GOLICHER, D. J.; BENAYAS, J. M. R.; GONZALEZ-ESPINOSA,
M.; RAMIREZ-MARCIAL, N. Fragmentation, disturbance and tree diversity
conservation in tropical montane forests. Journal of Applied Ecology, v.43, n.6,
p.1172-1181, Dec. 2006.
CHAMBERS, J.; ROBERTSON, A.; CARNEIRO, V.; LIMA, A.; SMITH, M.-L.;
PLOURDE, L.; HIGUCHI, N. Hyperspectral remote detection of niche partitioning
among canopy trees driven by blowdown gap disturbances in the Central Amazon.
Oecologia, v.160, n.1, p.107-117, 2009.
CHASE, J. M.; LEIBOLD, M. A. Ecological niches: linking classical and
contemporary approaches. 1st ed. Chicago: Chicago University Press, 2003. 212p.
CHAVE, J. Neutral theory and community ecology. Ecology Letters, v.7, n.3, p.241-
253, 2004.
CHAZDON, R. L.; PERES, C. A.; DENT, D.; SHEIL, D.; LUGO, A. E.; LAMB, D.;
STORK, N. E.; MILLER, S. E. The potential for species conservation in Tropical
Secondary Forests. Conservation Biology, v.23, n.6, p.1406-1417, Dec. 2009.

128
CHUST, G.; CHAVE, J.; CONDIT, R.; AGUILAR, S.; LAO, S.; PEREZ, R.
Determinants and spatial modeling of tree beta-diversity in a tropical forest landscape in
Panama. Journal of Vegetation Science, v.17, n.1, p.83-92, Feb. 2006.
CLARK, D. B.; PALMER, M. W.; CLARK, D. A. Edaphic factors and the landscape-
scale distributions of Tropical Rain Forest trees. Ecology, v.80, n.8, p.2662-2675, 1999.
COLWELL, R. K. EstimateS: Statistical estimation of species richness and shared
species from samples. Connecticut: University of Connecticut, 2006.
COLWELL, R. K.; CODDINGTON, J. A. Estimating terrestrial biodiversity through
extrapolation. Philosophical Transactions of the Royal Society (Series B), v.345,
p.101-118, 1994.
COMBER, A. J.; BIRNIE, R. V.; HODGSON, M. A retrospective analysis of land
cover change using a polygon shape index. Global Ecology and Biogeography, v.12,
p.207-215, 2003.
COMITA, L. S.; CONDIT, R.; HUBBELL, S. P. Developmental changes in habitat
associations of tropical trees. Journal of Ecology, v.95, n.3, p.482-492, May. 2007.
CONDIT, R.; ASHTON, P. S.; BAKER, P.; BUNYAVEJCHEWIN, S.;
GUNATILLEKE, S.; GUNATILLEKE, N.; HUBBELL, S. P.; FOSTER, R. B.; ITOH,
A.; LAFRANKIE, J. V.; LEE, H. S.; LOSOS, E.; MANOKARAN, N.; SUKUMAR, R.;
YAMAKURA, T. Spatial patterns in the distribution of tropical tree species. Science,
v.288, n.5470, p.1414-1418, 2000.
CONDIT, R.; PITMAN, N.; LEIGH, E. G., JR.; CHAVE, J.; TERBORGH, J.;
FOSTER, R. B.; NUNEZ, P.; AGUILAR, S.; VALENCIA, R.; VILLA, G.; MULLER-
LANDAU, H. C.; LOSOS, E.; HUBBELL, S. P. Beta-diversity in Tropical Forest trees.
Science, v.295, n.5555, p.666-669, Jan. 2002.
CONNELL, J. H. Diversity in Tropical Rain Forests and coral reefs. Science, v.199,
n.4335, p.1302-1310, Mar. 1978.
CONNOR, E. F.; MCCOY, E. D. The statistics and biology of the species-area
relationship. The American Naturalist, v.113, n.6, p.791-833, 1979.
CPRH. Diagnóstico socioambiental do litoral norte de Pernambuco. Recife:
Companhia Pernambucana do Meio Ambiente, 2003, 214p.

129
CRAMER, M. J.; WILLIG, M. R. Habitat heterogeneity, species diversity and null
models. Oikos, v.108, n.2, p.209-218, 2005.
CRAWLEY, M. J. The Structure of Plant Communities. In: CRAWLEY, M. J. (Ed.)
Plant Ecology. Oxford: Blackwell Science, 1997. Cap. 14, p. 475-531.
CUSHMAN, S. A.; MCGARIGAL, K. Patterns in the species-environment relationship
depend on both scale and choice of response variables. Oikos, v.105, p.117-124, 2004.
DAMSCHEN, E. I.; HADDAD, N. M.; ORROCK, J. L.; TEWKSBURY, J. J.; LEVEY,
D. J. Corridors increase plant species richness at large scales. Science, v.313, p.1284-
1286, 2006.
DAVIES-COLLEY, R. J.; PAYNE, G. W.; VAN ELSWIJK, M. Microclimate gradients
across a forest edge. New Zealand Journal of Ecology, v.24, n.2, p.111-121, 2000.
DEBINSKI, D. M.; HOLT, R. D. A survey and overview of habitat fragmentation
experiments. Conservation Biology, v.14, n.2, p.342-355, 2000.
DENSLOW, J. S. Tropical rain-forest gaps and tree species-diversity. Annual Review
of Ecology and Systematics, v.18, p.431-451, 1987.
DENSLOW, J. S. Disturbance and diversity in Tropical Rain Forests: the density effect.
Ecological Applications, v.5, n.4, p.962-968, 1995.
DENT, D. H.; WRIGHT, S. J. The future of tropical species in secondary forests: A
quantitative review. Biological Conservation, v.142, n.12, p.2833-2843, Dec. 2009.
DIDHAM, R.; LAWTON, J. H. Edge structure determines the magnitude of changes in
microclimate and vegetation structure in tropical forest fragments. Biotropica, v.31,
n.1, p.17-30, 1999.
DRUCKER, D. P.; COSTA, F. V. R. C.; MAGNUSSON, W. E. How wide is the
riparian zone of small streams in tropical forests? A test with terrestrial herbs. Journal
of Tropical Ecology, v.24, n.01, p.65-74, 2008.
DUFOUR, A.; GADALLAH, F.; WAGNER, H. H.; GUISAN, A.; BUTTLER, A. Plant
species richness and environmental heterogeneity in a mountain landscape: effects of
variability and spatial configuration. Ecography, v.29, p.573-584, 2006.

130
DUFRÊNE, M.; LEGENDRE, P. Species assemblages and indicator species: the need
for a flexible asymmetrical approach. Ecological Monographs, v.67, n.3, p.345-366,
1997.
DUQUE, A.; SANCHEZ, M.; CAVELIER, J.; DUIVENVOORDEN, J. F. Different
floristic patterns of woody understorey and canopy plants in Colombian Amazonia.
Journal of Tropical Ecology, v.18, n.4, p.499-525, 2002.
ECHEVERRÍA, C.; NEWTON, A.; LARA, A.; REY-BENAYAS, J. M.; COOMES, D.
A. Impacts of forest fragmentation on species composition and forest structure in the
temperate landscape of southern Chile. Global Ecology and Biogeography, v.16,
p.426-439, 2007.
ELTON, C. S. Animal Ecology. 1st ed. Londres: Sidgwick and Jackson 1927. 207p.
ERNOULT, A.; TREMAUVILLE, Y.; CELLIER, D.; MARGERIE, P.; LANGLOIS,
E.; ALARD, D. Potential landscape drivers of biodiversity components in a flood plain:
Past or present patterns? Biological Conservation, v.127, n.1, p.1-17, 2006.
ESRI. ArcGIS Professional GIS for the desktop: Redlands: Environmental Systems
Research Institute (ESRI), 2006.
EWERS, R. M.; DIDHAM, R. K. Confounding factors in the detection of species
responses to habitat fragmentation. Biological Reviews, v.81, n.1, p.117-142, Feb.
2006.
FAHRIG, L. Effects of habitat fragmentation on biodiversity. Annual Review of
Ecology, Evolution and Systematics, v.34, p.487-515, 2003.
FAPESP/BIREME. SciELO - Scientific Electronic Library Online: FAPESP/
BIREME, 2010.
FAZEY, I.; FISCHER, J.; LINDENMAYER, D. B. What do conservation biologists
publish? Biological Conservation, v.124, n.1, p.63-73, 2005.
FEELEY, K. J. The effects of forest fragmentation and increased edge exposure on leaf
litter accumulation. Journal of Tropical Ecology, v.20, p.709–712, 2004.
FERNÁNDEZ, C.; ACOSTA, F. J.; ABELLÁ, G.; LÓPEZ, F.; DÍAZ, M. Complex
edge effect fields as additive processes in patches of ecological systems. Ecological
Modelling, v.149, n.3, p.273-283, 2002.

131
FISCHER, J.; LINDENMAYER, D. B. Landscape modification and habitat
fragmentation: a synthesis. Global Ecology and Biogeography, v.16, p.265-280, 2007.
FITTER, A.; HODGE, A.; ROBINSON, D. Plant response to patchy soils. In:
HUTCHINGS, M. J.; JOHN, E. A.; STEWART, A. J. A. (Eds.) The ecological
consequences of environmental heterogeneity: the 40th symposium of the British
Ecological Society. Oxford: Blackwell Science, 2000. Cap. 5, p. 71-90.
FLETCHER JR, R. J. Multiple edge effects and their implications in fragmented
landscapes. Journal of Animal Ecology, v.74, p.342-352, 2005.
GAFFNEY, P. M. Roots of the niche concept. The American Naturalist, v.109, n.968,
p.490, 1975.
GARDNER, R. H.; ENGELHARDT, K. A. M. Spatial processes that maintain
biodiversity in plant communities. Perspectives in Plant Ecology Evolution and
Systematics, v.9, n.3-4, p.211-228, 2008.
GASCON, C.; LOVEJOY, T. E.; BIERREGAARD JR, R. O.; MALCOLM, J. R.;
STOUFFER, P. C.; VASCONCELOS, H. L.; LAURANCE, W. F.; ZIMMERMAN, B.;
TOCHER, M.; BORGES, S. Matrix habitat and species richness in tropical forest
remnants. Biological Conservation, v.91, n.2-3, p.223-229, 1999.
GIGNAC, L. D.; DALE, M. R. T. Effects of fragment size and habitat heterogeneity on
cryptogam diversity in the Low-boreal Forest of Western Canada. The Bryologist,
v.108, n.1, p.50-66, 2005.
GILBERT, B.; LAURANCE, W. F.; LEIGH JR, E. G.; NASCIMENTO, H. E. M. Can
neutral theory predict the responses of Amazonian tree communities to forest
fragmentation? The American Naturalist, v.168, n.3, p.304-317, 2006.
GILBERT, B.; LECHOWICZ, M. J. Neutrality, niches, and dispersal in a temperate
forest understory. Proceedings of the National Academy of Sciences, v.101, n.20,
p.7651-7656, 2004.
GIVNISH, T. J. On the causes of gradients in tropical tree diversity. Journal of
Ecology, v.87, n.2, p.193-210, 1999.
GLEASON, H. A. On the relation between species and area. Ecology, v.3, n.2, p.158-
162, 1922.

132
GLEASON, H. A. Species and area. Ecology, v.6, n.1, p.66-74, 1925.
GOLDEMBERG, J.; COELHO, S. T.; GUARDABASSI, P. The sustainability of
ethanol production from sugarcane. Energy Policy, v.36, p.2086-2097, 2008.
GOMES, J. S.; LINS-E-SILVA, A. C. B.; RODAL, M. J. N.; SILVA, H. C. H.
Estrutura do sub-bosque lenhoso em ambientes de borda e interior de dois fragmentos
de floresta atlântica em Igarassu, Pernambuco, Brasil. Rodriguésia, v.60, n.2, p.295-
310, 2009.
GOTELLI, N. J.; COLWELL, R. K. Quantifying biodiversity: procedures and pitfalls in
the measurement and comparison of species richness. Ecology Letters, v.4, n.4, p.379-
391, 2001.
GRINNELL, J. The niche-relationships of the California Thrasher. The Auk, v.34, n.4,
p.427-433, 1917.
GROVES, C. R.; JENSEN, D. B.; VALUTIS, L. L.; REDFORD, K. H.; SHAFFER, M.
L.; MICHAEL, J.; SCOTT, J. V. B.; HIGGINS, J. V.; BECK, M. W.; ANDERSON, M.
G. Planning for biodiversity conservation: putting conservation science into practice.
BioScience, v.52, n.6, p.499-512, 2002.
GUEDES-BRUNI, R.; SILVA, A.; MANTOVANI, W. Rare canopy species in
communities within the Atlantic Coastal Forest in Rio de Janeiro State, Brazil.
Biodiversity and Conservation, v.18, n.2, p.387-403, 2009.
GUNATILLEKE, C. V. S.; GUNATILLEKE, I. A. U. N.; ESUFALI, S.; HARMS, K.
E.; ASHTON, P. M. S.; BURSLEM, D. F. R. P.; ASHTON, P. S. Species-habitat
associations in a Sri Lankan dipterocarp forest. Journal of Tropical Ecology, v.22,
p.371-384, 2006.
HAILA, Y. A conceptual genealogy of fragmentation research: from island
biogeography to landscape ecology. Ecological Applications, v.12, n.2, p.321-334,
2002.
HALL, L. S.; KRAUSMAN, P. R.; MORRISON, M. L. The habitat concept and a plea
for standard terminology. Wildlife Society Bulletin, v.25, n.1, p.173-182, 1997.
HALVORSEN, R.; EDVARDSEN, A. The concept of habitat specificity revisited.
Landscape Ecology, v.24, n.7, p.851-861, Aug. 2009.

133
HANNUS, J. J.; VON NUMERS, M. Vascular plant species richness in relation to
habitat diversity and island area in the Finnish Archipelago. Journal of Biogeography,
v.35, n.6, p.1077-1086, Jun. 2008.
HARDY, O. J.; SONKE, B. Spatial pattern analysis of tree species distribution in a
tropical rain forest of Cameroon: assessing the role of limited dispersal and niche
differentiation. Forest Ecology and Management, v.197, p.191-202, 2004.
HARMS, K. E. Habitat-specialization and seed-dispersal limitation in a Neotropical
forest. 1997. Thesis (PhD in Ecology & Evolutionary Biology) - Princeton University,
1997, Princeton. 225p.
HARMS, K. E.; CONDIT, R.; HUBBELL, S. P.; FOSTER, R. B. Habitat associations
of trees and shrubs in a 50-ha neotropical forest plot. Journal of Ecology, v.89, p.947-
959, 2001.
HARPER, K. A.; MACDONALD, S. E. Structure and composition of riparian boreal
forest: new methods for analyzing edge influence. Ecology, v.82, n.3, p.649-659, 2001.
HARPER, K. A.; MACDONALD, S. E. Structure and composition of edges next to
regenerating clear-cuts in mixed-wood boreal forest. Journal of Vegetation Science,
v.13, p.535-546, 2002.
HARPER, K. A.; MACDONALD, S. E.; BURTON, P. J.; CHEN, J.; BROSOFSKE, K.
D.; SAUNDERS, S. C.; EUSKIRCHEN, E. S.; ROBERTS, D.; JAITEH, M. S.;
ESSEEN, P.-A. Edge influence on forest structure and composition in fragmented
landscapes. Conservation Biology, v.19, n.3, p.768-782, 2005.
HELM, A.; HANSKI, I.; PÄRTEL, M.; NAEEM, S. Slow response of plant species
richness to habitat loss and fragmentation. Ecology Letters, v.9, n.1, p.72-77, 2006.
HENLE, K.; DAVIES, K. F.; KLEYER, M.; MARGULES, C.; SETTELE, J. Predictors
of species sensitivity to fragmentation. Biodiversity and Conservation, v.13, p.207-
251, 2004.
HILL, J. L.; CURRAN, P. J. Area, shape and isolation of tropical forest fragments:
effects on tree species diversity and implications for conservation. Journal of
Biogeography, v.30, p.1391-1403, 2003.

134
HOFER, G.; WAGNER, H. H.; HERZOG, F.; EDWARDS, P. J. Effects of topographic
variability on the scaling of plant species richness in gradient dominated landscapes.
Ecography, v.31, n.1, p.131-139, Feb. 2008.
HOLT, R. D.; LAWTON, J. H.; POLIS, G. A.; MARTINEZ, N. D. Trophic rank and
the species-area relationship. Ecology, v.80, n.5, p.1495-1504, 1999.
HUBBELL, S. P. The unified Neutral Theory of Biodiversity and Biogeography. 1st
ed. Princeton: Princeton University Press, 2001. 375p.
HUBBELL, S. P. Neutral theory and the evolution of ecological equivalence. Ecology,
v.87, n.6, p.1387-1398, 2006.
HUBBELL, S. P.; FOSTER, R. B. Diversity of canopy trees in a neotropical forest and
implications for conservation. In: SUTTON, S. J.; WHITMORE, T. C.; CHADWICK,
A. C. (Eds.) Tropical Rain Forest: Ecology and Management. Oxford: Blackwell
Science, 1983. Cap. 3, p. 25-41.
HUBBELL, S. P.; FOSTER, R. B.; O’BRIEN, S. T.; HARMS, K. E.; CONDIT, R.;
WECHSLER, B.; WRIGHT, S. J.; LOO DE LAO, S. Light gap disturbances,
recruitment limitation, and tree diversity in a Neotropical forest. Science, v.283, p.554-
557, 1999.
HUTCHINSON, G. E. Concluding remarks In: Cold Spring Harbor Symposia on
Quantitative Biology. p. 415-427. 1957.
ISI. Web of Science/ Institute for Scientific Information: Thomson Reuters, 2010.
JANZEN, D. H. No park is an island: increase in interference from outside as park size
decreases. Oikos, v.41, p.402-410, 1983.
JOHN, R.; DALLING, J. W.; HARMS, K. E.; YAVITT, J. B.; STALLARD, R. F.;
MIRABELLO, M.; HUBBELL, S. P.; VALENCIA, R.; NAVARRETE, H.; VALLEJO,
M.; FOSTER, R. B. Soil nutrients influence spatial distributions of tropical tree species.
Proceedings of the National Academy of Sciences, v.104, n.3, p.864-869, 2007.
JOHNSON, M. P.; FROST, N. J.; MOSLEY, M. W. J.; ROBERTS, M. F.; HAWKINS,
S. J. The area-independent effects of habitat complexity on biodiversity vary between
regions. Ecology Letters, v.6, n.2, p.126-132, 2003.

135
JORGE, L. A. B.; GARCIA, G. J. A study of habitat fragmentation in Southeastern
Brazil using remote sensing and geographic information systems (GIS). Forest Ecology
and Management, v.98, n.1, p.35-47, 1997.
JOSE, S.; GILLESPIE, A. R.; GEORGE, S. J.; KUMAR, B. M. Vegetation responses
along edge-to-interior gradients in a high altitude tropical forest in peninsular India.
Forest Ecology and Management, v.87, n.1-3, p.51-62, 1996.
KADMON, R.; ALLOUCHE, O. Integrating the effects of area, isolation, and habitat
heterogeneity on species diversity: a unification of island biogeography and niche
theory. The American Naturalist, v.170, n.3, p.443-454, 2007.
KALLIMANIS, A. S.; MAZARIS, A. D.; TZANOPOULOS, J.; HALLEY, J. M.;
PANTIS, J. D.; SGARDELIS, S. P. How does habitat diversity affect the species-area
relationship? Global Ecology and Biogeography, v.17, n.4, p.532-538, 2008.
KEARNEY, M. Habitat, environment and niche: what are we modelling? Oikos, v.115,
n.1, p.186-191, 2006.
KEARNEY, M.; PORTER, W. Mechanistic niche modelling: combining physiological
and spatial data to predict species' ranges. Ecology Letters, v.12, n.4, p.334-350, Apr.
2009.
KELLER, A.; RODEL, M. O.; LINSENMAIR, K. E.; GRAFE, T. U. The importance of
environmental heterogeneity for species diversity and assemblage structure in Bornean
stream frogs. Journal of Animal Ecology, v.78, n.2, p.305-314, Mar. 2009.
KELLY, C. K.; BOWLER, M. G.; PYBUS, O.; HARVEY, P. H. Phylogeny, niches,
and relative abundance in natural communities. Ecology, v.89, n.4, p.962-970, 2008.
KESSLER, M.; LEHNERT, M. Do ridge habitats contribute to pteridophyte diversity in
tropical montane forests? A case study from southeastern Ecuador. Journal of Plant
Research, v.122, n.4, p.421-428, 2009.
KIMMEL, T.; PIECHOWSKI, D.; GOTTSBERGER, G. The history of fragmentation
of the Lowland Atlantic Forest of Pernambuco, Brazil Bioremediation, Biodiversity
and Bioavailability, v.2, n.1, p.1-4, 2008.

136
KOLASA, J.; WALTHO, N. A hierarchical view of habitat and its relationship to
species abundance. In: PETERSON, D. L.; PARKER, V. T. (Eds.) Ecological Scale:
theory and applications. New York: Columbia University Press, 1998. Cap. 4, p. 55-76.
KOLLMANN, J.; SCHNEIDER, B. Landscape structure and diversity of fleshy-fruited
species at forest edges. Plant Ecology, v.144, p.37-48, 1999.
KRAUSS, J.; ALFERT, T.; STEFFAN-DEWENTER, I. Habitat area but not habitat age
determines wild bee richness in limestone quarries. Journal of Applied Ecology, v.46,
n.1, p.194-202, Feb. 2009.
KREBS, C. J. Ecology after 100 years: progress and pseudo-progress. New Zealand
Journal of Ecology, v.30, n.1, p.3-11, 2006.
KREFT, H.; JETZ, W.; MUTKE, J.; KIER, G.; BARTHLOTT, W. Global diversity of
island floras from a macroecological perspective. Ecology Letters, v.11, n.2, p.116-
127, 2008.
KUBOTA, Y.; MURATA, H.; KIKUZAWA, K. Effects of topographic heterogeneity
on tree species richness and stand dynamics in a subtropical forest in Okinawa Island,
southern Japan. Journal of Ecology, v.92, n.2, p.230-240, 2004.
LAURANCE, W. F. Edge effects in Tropical Forest Fragments: application of a model
for the design of Nature Reserves. Biological Conservation, v.57, p.205-219, 1991.
LAURANCE, W. F. Introduction and synthesis. Special Issue: Ecology and
Management of Fragmented Tropical Landscapes. Biological Conservation, v.91, n.2-
3, p.101-107, 1999a.
LAURANCE, W. F. Reflections on the tropical deforestation crisis. Biological
Conservation, v.91, p.109-117, 1999b.
LAURANCE, W. F. Do edge effects occur over large spatial scales? Trends in
Ecology & Evolution, v.15, n.4, p.134-135, 2000.
LAURANCE, W. F. Theory meets reality: How habitat fragmentation research has
transcended island biogeographic theory. Biological Conservation, v.141, n.7, p.1731-
1744, 2008.

137
LAURANCE, W. F.; BIERREGAARD JR, R. O. (Eds). Tropical Forest Remnants:
ecology, management and conservation of fragmented communities. 1st ed. Chicago:
University of Chicago Press, 1997. 632p.
LAURANCE, W. F.; DIDHAM, R. K.; POWER, M. E. Ecological boundaries: a search
for synthesis. Trends in Ecology & Evolution, v.16, n.2, p.70-71, 2001.
LAURANCE, W. F.; FERREIRA , L. V.; RANKIN-DE-MERONA, J. M.;
LAURANCE, S. G.; HUTCHINGS, R. W.; LOVEJOY, T. E. Effects of Forest
Fragmentation on Recruitment Patterns in Amazonian Tree Communities.
Conservation Biology, v.12, n.2, p.460-464, 1998.
LAURANCE, W. F.; LOVEJOY, T. E.; VASCONCELOS, H. L.; BRUNA, E. M.;
DIDHAM, R. K.; STOUFFER, P. C.; GASCON, C.; BIERREGAARD JR, R. O.;
LAURANCE, S. G.; SAMPAIO, E. Ecosystem decay of Amazonian forest fragments: a
22-year investigation. Conservation Biology, v.16, n.3, p.605-618, 2002.
LAURANCE, W. F.; NASCIMENTO, H. E. M.; LAURANCE, S. G.; ANDRADE, A.;
EWERS, R. M.; HARMS, K. E.; LUIZÃO, R. C. C.; RIBEIRO, J. E. Habitat
fragmentation, variable edge effects, and the Landscape-Divergence Hypothesis. PLoS
ONE. , v.2, n.10, p.e1017, 2007.
LAURANCE, W. F.; NASCIMENTO, H. E. M.; LAURANCE, S. G.; ANDRADE, A.;
RIBEIRO, J. E. L. S.; GIRALDO, J. P.; LOVEJOY, T. E.; CONDIT, R.; CHAVE, J.;
HARMS, K. E.; D’ANGELO, S. Rapid decay of tree-community composition in
Amazonian forest fragments. Proceedings of the National Academy of Sciences,
v.103, n.50, p.19010-19014, 2006.
LEIBOLD, M. A. The niche concept revisited: mechanistic models and community
context. Ecology, v.76, n.5, p.1371-1382, 1995.
LEIBOLD, M. A. Ecology: return of the niche. Nature, v.454, n.7200, p.39-41, 2008.
LEIBOLD, M. A.; MCPEEK, M. A. Coexistence of the niche and neutral perspectives
in community ecology. Ecology, v.87, n.6, p.1399-1410, 2006.
LEROUX, S. J.; SCHMIEGELOW, F. K. A.; LESSARD, R. B.; CUMMING, S. G.
Minimum dynamic reserves: A framework for determining reserve size in ecosystems

138
structured by large disturbances. Biological Conservation, v.138, n.3-4, p.464-473,
2007.
LEVINE, J. M.; HILLERISLAMBERS, J. The importance of niches for the
maintenance of species diversity. Nature, v.461, p.254-257, 2009.
LIMA, M. L. F. C. A Reserva da Biosfera da Mata atlântica em Pernambuco:
situação atual, ações e perspectivas. 1a ed. São Paulo: Conselho Nacional RBMA/
Governo do Estado de São Paulo/ Instituto Florestal, v.12, 1998. 41p.
LINDBORG, R.; ERIKSSON, O. Historical landscape connectivity affects present plant
species diversity. Ecology, v.85, n.7, p.1840-1845, 2004.
LINDENMAYER, D. B.; FISCHER, J. Habitat Fragmentation and Landscape
Change: An ecological and conservation synthesis. 1a ed. Washington, DC: Island
Press, 2006. 344p.
LINDENMAYER, D. B.; FISCHER, J. Tackling the habitat fragmentation panchreston.
Trends in Ecology & Evolution, v.22, n.3, p.127-132, 2007.
LINDENMAYER, D. B.; FRANKLIN, J. F.; FISCHER, J. General management
principles and a checklist of strategies to guide forest biodiversity conservation.
Biological Conservation, v.131, p.433-445, 2006.
LINS-E-SILVA, A. C. B.; RODAL, M. J. N. Tree community structure in an urban
remnant of Atlantic Coastal Forest in Pernambuco, Brazil. Memoirs of the New York
Botanical Garden, v.100, p.511-534, 2008.
LOMOLINO, M. V.; WEISER, M. D. Towards a more general species-area
relationship: diversity on all islands, great and small. Journal of Biogeography, v.28,
n.4, p.431-445, Apr. 2001.
LOOKINGBILL, T. R.; URBAN, D. L. Gradient analysis, the next generation: towards
more plant-relevant explanatory variables. Canadian Journal of Forest Research,
v.35, n.7, p.1744-1753, 2005.
LOPES, A. V.; GIRAO, L. C.; SANTOS, B. A.; PERES, C. A.; TABARELLI, M.
Long-term erosion of tree reproductive trait diversity in edge-dominated Atlantic forest
fragments. Biological Conservation, v.142, n.6, p.1154-1165, Jun. 2009.

139
LUNDHOLM, J. T. Plant species diversity and environmental heterogeneity: spatial
scale and competing hypotheses. Journal of Vegetation Science, v.20, n.3, p.377-391,
Jun. 2009.
MACARTHUR, R. H. On the relative abundance of bird species. Proceedings of the
National Academy of Sciences, v.43, p.293-295, 1957.
MACARTHUR, R. H.; LEVINS, R. The limiting similarity, convergence, and
divergence of coexisting species. The American Naturalist, v.101, n.921, p.377-385,
1967.
MACARTHUR, R. H.; MACARTHUR, J. W. On bird species diversity. Ecology, v.42,
n.3, p.594-598, 1961.
MACARTHUR, R. H.; WILSON, E. O. The Theory of Island Biogeography. 13th ed.
Princeton/ Oxford: Princeton University Press, 1967. 203p.
MAGURRAN, A. E. Measuring Biological Diversity. 1st ed. Malden, Oxford:
Blackwell Science, 2004. 256p.
MANNING, A. D.; FISCHER, J.; LINDENMAYER, D. B. Scattered trees are keystone
structures - Implications for conservation. Biological Conservation, v.132, p.311-321,
2006.
MARTINS, F. R. Estrutura de uma Floresta Mesófila. 2a ed. Campinas: Ed.
Unicamp, 1993. 246p.
MARTINS, I. C. D. M.; SOARES, V. P.; SILVA, E.; BRITES, R. S. Diagnóstico
ambiental no contexto da paisagem de fragmentos florestais naturais “ipucas” no
município de Lagoa da Confusão, Tocantins. Revista Árvore, v.26, n.3, p.299-309,
2002.
MATLACK, G. R. Microenvironmen variation within and among forest edge sites in
the Eastern United States. Biological Conservation, v.66, p.185-194, 1993.
MCCUNE, B.; MELFORD, M. J. PC-ORD: Multivariate analysis of ecological data
Gleneden Beach: MjM Software Design, 1999.
MCMASTER, R. T. Factors influencing vascular plant diversity on 22 islands off the
coast of eastern North America. Journal of Biogeography, v.32, n.3, p.475-492, 2005.

140
MELO, F. P. L.; DIRZO, R.; TABARELLI, M. Biased seed rain in forest edges:
Evidence from the Brazilian Atlantic forest. Biological Conservation, v.132, n.1, p.50-
60, Sep. 2006.
MESQUITA, R. C. G.; DELAMÔNICA, P.; LAURANCE, W. F. Effect of surrounding
vegetation on edge-related tree mortality in Amazonian forest fragments. Biological
Conservation, v.91, p.129-134, 1999.
METZGER, J. P. Relationships between landscape structure and tree species diversity
in tropical forests of South-East Brazil Landscape and Urban Planning, v.37, n.1-2,
p.29-35, 1997.
METZGER, J. P. Changements de la structure du paysage et richesse spécifique des
fragments forestiers dans le Sud-Est du Brésil. Comptes Rendus de L´Académie des
Sciences, Serie III - Sciences de La Vie, v. 321, p.319-333, 1998.
METZGER, J. P. O que é Ecologia de Paisagens? Biota Neotropica, v.1, n.1/2, p.1-9,
2001.
METZGER, J. P. Como lidar com regras pouco óbvias para conservação da
biodiversidade em paisagens fragmentadas. Natureza & Conservação, v.4, n.2, p.11-
23, 2006.
METZGER, J. P.; BERNACCI, L. C.; GOLDENBERG, R. Pattern of tree species
diversity in riparian forest fragments of different widths (SE Brazil). Plant Ecology,
v.133, p.135-152, 1997.
METZGER, J. P.; MARTENSEN, A. C.; DIXO, M.; BERNACCI, L. C.; RIBEIRO, M.
C.; TEIXEIRA, A. M. G.; PARDINI, R. Time-lag in biological responses to landscape
changes in a highly dynamic Atlantic forest region. Biological Conservation, v.142,
n.6, p.1166-1177, 2009.
MILLER, C. J. Vegetation and habitat are not synonyms. Ecological Management &
Restoration, v.1, n.2, p.102-104, Aug. 2000.
MITCHELL, S. C. How useful is the concept of habitat? - a critique. Oikos, v.110, n.3,
p.634-638, 2005.

141
MOLINO, J.-F.; SABATIER, D. Tree diversity in Tropical Rain Forests: a validation of
the Intermediate Disturbance Hypothesis. Science, v.294, n.5547, p.1702-1704, Nov.
2001.
MORELLATO, P. C.; HADDAD, C. F. B. Introduction: The Brazilian Atlantic Forest.
Biotropica, v.32 n.4b, p.786-792, 2000.
MOUW, J. E. B.; ALABACK, P. B. Putting floodplain hyperdiversity in a regional
context: an assessment of terrestrial-floodplain connectivity in a montane environment.
Journal of Biogeography, v.30, p.87-103, 2003.
MUELLER-DOMBOIS, D.; ELLENBERG, H. Aims and Methods of Vegetation
Ecology. 1st ed. New York: John Wiley & Sons, 1974. 547p.
MURCIA, C. Edge effects in fragmented forests: implications for conservation. Trends
in Ecology & Evolution, v.10, n.2, p.58-62, 1995.
MURPHY, H. T.; LOVETT-DOUST, J. Context and connectivity in plant
metapopulations and landscape mosaics: does the matrix matter? Oikos, v.105, p.3-14,
2004.
MYERS, N.; MITTERMEIER, R. A.; MITTERMEIER, C. G.; DA FONSECA, G. A.
B.; KENT, J. Biodiversity hotspots for conservation priorities. Nature, v.403, n.6772,
p.853-858, 2000.
NAGENDRA, H. Opposite trends in response for the Shannon and Simpson indices of
landscape diversity. Applied Geography, v.22, p.175-186, 2002.
NAIMAN, R. J.; DÉCAMPS, H. The ecology of interfaces: Riparian Zones. Annual
Review of Ecology & Systematics v.28, p.621-658, 1997.
NASCIMENTO, H. E. M.; LAURANCE, W. F. Efeitos de área e de borda sobre a
estrutura florestal em fragmentos de floresta de terra-firme após 13-17 anos de
isolamento. Acta Amazonica, v.36, n.2, p.183-192, 2006.
NASCIMENTO, L. M. Sucessão secundária em áreas de Floresta Atlântica de
Pernambuco: mudanças florísticas e estruturais. 2010. Tese (Doutorado em
Botânica) - Universidade Federal Rural de Pernambuco, 2010, Recife. 115p.

142
NELSON, C. R.; HALPERN, C. B. Edge-related responses of understory plants to
aggregated retention harvest in the Pacific northwest. Ecological Applications, v.15,
n.1, p.196-209, 2005.
NILSSON, C.; SVEDMARK, M. Basic principles and ecological consequences of
changing water regimes: riparian plant communities. Environmental Management,
v.30, n.4, p.468-480, 2002.
NOBIS, M.; WOHLGEMUTH, T. Trend words in ecological core journals over the last
25 years (1978-2002). Oikos, v.106, n.2, p.411-421, 2004.
OCHOA-GAONA, S.; GONZALEZ-ESPINOSA, M.; MEAVE, J. A.; BON, V. S. D.
Effect of forest fragmentation on the woody flora of the highlands of Chiapas, Mexico.
Biodiversity and Conservation, v.13, n.5, p.867-884, May. 2004.
OLIVEIRA-FILHO, A. T.; ALMEIDA, R. J. D.; MELLO, J. M. D.; GAVILANES, M.
L. Estrutura fitossociológica e variáveis ambientais em um trecho da mata ciliar do
córrego dos Vilas Boas, Reserva Biológica do POço Bonito, Lavras (MG). Revista
Brasileira de Botânica, v.17, n.1, p.67-85, 1994a.
OLIVEIRA-FILHO, A. T.; VILELA, E. A.; CARVALHO, D. A.; GAVILANES, M. L.
Effects of soils and topography on the distribution of tree species in a tropical riverine
forest in south-eastern Brazil. Journal of Tropical Ecology, v.10, n.04, p.483-508,
1994b.
OLIVEIRA-FILHO, A. T.; VILELA, E. A.; GAVILANES, M. L.; CARVALHO, D. A.
Effect of floading regime and understory bamboos on the physiognomy and tree species
composition of a tropical semideciduous forest in Southeastern Brazil. Vegetation,
v.113, p.99-124, 1994c.
OLIVEIRA, A. N. D.; AMARAL, I. L. D. Aspectos florísticos, fitossociológicos e
ecológicos de um sub-bosque de terra firme na Amazônia Central, Amazonas, Brasil.
Acta Amazônica, v.35, p.1-16, 2005.
OLIVEIRA, M. A. Fragmentação e a riqueza de árvores em escala local e regional
na floresta Atlântica nordestina: implicações para a conservação. 2007. Tese
(Doutorado em Biologia Vegetal) - Universidade Federal de Pernambuco, 2007, Recife.
122p.

143
OLIVEIRA, M. A.; GRILLO, A. S.; TABARELLI, M. Forest edge in the Brazilian
Atlantic forest: drastic changes in tree species assemblages. Oryx, v.38, n.4, p.389-394,
Oct. 2004.
OLIVEIRA, M. A.; SANTOS, A. M. M.; TABARELLI, M. Profound impoverishment
of the large-tree stand in a hyper-fragmented landscape of the Atlantic forest. Forest
Ecology and Management, v.256, n.11, p.1910-1917, Nov. 2008.
OLIVEIRA, R. D. J.; MANTOVANI, W.; MELO, M. M. D. R. F. D. Estrutura do
componente arbustivo-arbóreo da floresta atlântica de encosta, Peruíbe, SP. Acta
Botanica Brasilica, v.15, p.391-412, 2001.
OOSTERHOORN, M.; KAPPELLE, M. Vegetation structure and composition along an
interior-edge-exterior gradient in a Costa Rican montane cloud forest. Forest Ecology
and Management, v.126, n.3, p.291-307, 2000.
PALTTO, H.; NORDÉN, B.; GÖTMARK, F.; FRANC, N. At which spatial and
temporal scales does landscape context affect local density of Red Data Book and
Indicator species? Biological Conservation, v.133, n.4, p.442-454, 2006.
PANITSA, M.; TZANOUDAKIS, D.; TRIANTIS, K. A.; SFENTHOURAKIS, S.
Patterns of species richness on very small islands: the plants of the Aegean archipelago.
Journal of Biogeography, v.33, n.7, p.1223-1234, 2006.
PEREIRA, J. A. A.; OLIVEIRA-FILHO, A. T.; LEMOS-FILHO, J. P. Environmental
heterogeneity and disturbance by humans control much of the tree species diversity of
Atlantic montane forest fragments in SE Brazil. Biodiversity and Conservation, v.16,
n.6, p.1761-1784, 2007.
PETREN, K. Concept of the habitat and the niche. The Encyclopedia of Biodiversity,
v.3, p.303-315, 2001.
PHILLIPS, O. L.; VARGAS, P. N.; MONTEAGUDO, A. L.; CRUZ, A. P.;
CHUSPEZANS, M.-E.; WASHINGTON GALIANO, S.; YLI-HALLA, M.; ROSE, S.
Habitat association among Amazonian Tree Species: a landscape-scale approach.
Journal of Ecology, v.91, n.5, p.757-775, 2003.
PICKETT, S. T. A.; CADENASSO, M. L. Landscape Ecology: spatial heterogeneity in
ecological systems. Science, v.269, n.5222, p.331-334, Jul. 1995.

144
PITMAN, N. C. A.; TERBORGH, J.; SILMAN, M. R.; NUNEZ V, P. Tree species
distributions in an upper amazonian forest. Ecology, v.80, n.8, p.2651-2661, 1999.
PITMAN, N. C. A.; TERBORGH, J. W.; SILMAN, M. R.; V, P. N.; NEILL, D. A.;
CERON, C. E.; PALACIOS, W. A.; MILTON, A. A Comparison of tree species
diversity in two Upper Amazonian forests. Ecology, v.83, n.11, p.3210-3224, 2002.
POTTS, M. D.; ASHTON, P. S.; KAUFMAN, L. S.; PLOTKIN, J. B. Habitat patterns
in Tropical Rain Forests: a comparison of 105 plots in Northwest Borneo. Ecology,
v.83, n.10, p.2782-2797, 2002.
PRESTON, F. W. Time and space and the variation of species. Ecology, v.41, n.4,
p.611-627, 1960.
PUTH, L. M.; WILSON, K. A. Boundaries and corridors as a continuum of ecological
flow control: lessons from rivers and streams. Conservation Biology, v.15, n.1, p.21-
30, 2001.
PYKE, C. R.; CONDIT, R.; AGUILAR, S.; LAO, S. Floristic composition across a
climatic gradient in a neotropical lowland forest. Journal of Vegetation Science, v.12,
n.4, p.553-566, Aug. 2001.
RANTA, P.; BLOM, T. O. M.; NIEMELÄ, J.; JOENSUU, E.; SIITONEN, M. The
fragmented Atlantic rain forest of Brazil: size, shape and distribution of forest
fragments. Biodiversity and Conservation, v.7, n.3, p.385-403, 1998.
REIS, A. C. S. Contribuição ao estudo do clima de Pernambuco. Recife:
CONDEPE, 1970, 41p.
RIBEIRO, M. C.; METZGER, J. P.; MARTENSEN, A. C.; PONZONI, F. J.; HIROTA,
M. M. The Brazilian Atlantic Forest: How much is left, and how is the remaining forest
distributed? Implications for conservation. Biological Conservation, v.142, n.6,
p.1141-1153, 2009.
RICHART, M.; HEWITT, N. Forest remnants in the Long Point region, Southern
Ontario: tree species diversity and size structure. Landscape and Urban Planning,
v.86, p.25-37, 2008.
RICKETTS, T. H. The matrix matters: effective isolation in fragmented landscapes.
The American Naturalist, v.158, n.1, p.87-99, 2001.

145
RICKLEFS, R. E. Environmental heterogeneity and plant species diversity: a
hipothesis. The American Naturalist, v.111, n.978, p.376-381, 1977.
RICKLEFS, R. E. Evolutionary diversification and the origin of the diversity-
environment relationship. Ecology, v.87, n.7, p.S3-S13, Jul. 2006a.
RICKLEFS, R. E. The unified neutral theory of biodiversity: do the numbers add up?
Ecology, v.87, n.6, p.1424-1431, 2006b.
RICKLEFS, R. E.; LOVETTE, I. J. The roles of island area per se and habitat diversity
in the species-area relationships of four Lesser Antillean faunal groups. Journal of
Animal Ecology, v.68, n.6, p.1142-1160, 1999.
RIES, L.; FLETCHER, R. J.; BATTIN, J.; SISK, T. D. Ecological responses to habitat
edges: Mechanisms, Models, and Variability Explained. Annual Review of Ecology,
Evolution and Systematics, v.35, n.1, p.491-522, 2004.
ROCHA, K. D.; CHAVES, L. F. C.; MARANGON, L. C.; LINS-E-SILVA, A. C. B.
Caracterização da vegetação arbórea adulta em um fragmento de floresta atlântica,
Igarassu, PE. Revista Brasileira de Ciências Agrárias, v.31, n.1, p.35-41, 2008.
RODAL, M. J. N.; LINS-E-SILVA, A. C. B.; GOTTSBERGER, G. Foreword: special
issue The Fragments Project. Bioremediation, Biodiversity and Bioavailability, v.2,
n.1, p.1, 2008.
RODRIGUES, P. J. F. P.; ABREU, R. C. R.; BARCELLOS, E. M. B.; LIMA, H. C.;
SCARANO, F. R. Population structure and one-year dynamics of the endangered
tropical tree species Caesalpinia echinata Lam. (Brazilian red-wood): the potential
importance of small fragments for conservation. Rodriguésia, v.60, n.1, p.211-220,
2009.
ROSENZWEIG, M. L. Species diversity in space and time. 1st ed. Cambridge:
Cambridge University Press, 1995. 460p.
ROUGHGARDEN, J. Is there a general theory of community ecology? Biology and
Philosophy, v.24, n.4, p.521-529, 2009.
SANTOS, A. M. M.; CAVALCANTI, D. R.; SILVA, J. M. C. D.; TABARELLI, M.
Biogeographical relationships among tropical forests in north-eastern Brazil. Journal of
Biogeography, v.34, p.437-446, 2007a.

146
SANTOS, B. A.; PERES, C. A.; OLIVEIRA, M. A.; GRILLO, A.; ALVES-COSTA, C.
P.; TABARELLI, M. Drastic erosion in functional attributes of tree assemblages in
Atlantic forest fragments of northeastern Brazil. Biological Conservation, v.141, n.1,
p.249-260, 2008.
SANTOS, K.; KINOSHITA, L. S.; SANTOS, F. A. M. D. Tree species composition and
similarity in semideciduous forest fragments of southeastern Brazil. Biological
Conservation, v.135, n.2, p.268-277, 2007b.
SAUNDERS, D. A.; HOBBS, R. J.; MARGULES, C. R. Biological consequences of
ecosystem fragmentation: a review. Conservation Biology, v.5, n.1, p.18-32, 1991.
SCARANO, F. R.; DIAS, A. T. C. A importância de espécies no funcionamento de
comunidades e ecossistemas. In: COELHO, A. S.; LOYOLA, R. D.; SOUZA, M. B. G.
(Eds.) Ecologia Teórica: desafios para o aperfeiçoamento da Ecologia no Brasil. Belo
Horizonte: O Lutador, 2004. Cap. 3, p. 43-60.
SCHEDLBAUER, J. L.; FINEGAN, B.; KAVANAGH, K. L. Rain forest structure at
forest-pasture edges in Northeastern Costa Rica. Biotropica, v.39, n.5, p.578-584 2007.
SCHEINER, S. M. Six types of species-area curves. Global Ecology and
Biogeography, v.12, n.6, p.441-447, 2003.
SCHEINER, S. M. A mélange of curves - further dialogue about species-area
relationships. Global Ecology and Biogeography, v.13, n.6, p.479-484, 2004.
SCHESSL, M.; SILVA, W. L.; GOTTSBERGER, G. Effects of fragmentation on forest
structure and litter dynamics in Atlantic rainforest in Pernambuco, Brazil. Flora, v.203,
n.3, p.215-228, 2008.
SCHNITZER, S. A.; BONGERS, F. The ecology of lianas and their role in forests.
Trends in Ecology & Evolution, v.17, p.223-230, 2002.
SCHOEREDER, J. H.; GALBIATI, C.; RIBAS, C. R.; SOBRINHO, T. G.; SPERBER,
C. F.; DESOUZA, O.; LOPES-ANDRADE, C. Should we use proportional sampling
for species-area studies? Journal of Biogeography, v.31, n.8, p.1219-1226, 2004.
SEABLOOM, E. W.; DOBSON, A. P.; STOMS, D. M. Extinction rates under
nonrandom patterns of habitat loss. Proceedings of the National Academy of
Sciences, v.99, n.17, p.11229-11234, August 20, 2002. 2002.

147
SFENTHOURAKIS, S.; TRIANTIS, K. A. Habitat diversity, ecological requirements of
species and the Small Island Effect. Diversity and Distributions, v.15, n.1, p.131-140,
2009.
SHMIDA, A.; WILSON, M. V. Biological determinants of species diversity. Journal of
Biogeography, v.12, n.1, p.1-20, 1985.
SIEGEL, S. Estatística não-paramétrica. 1a ed. São Paulo: McGraw-Hill, 1975. 350p.
SILVA, A. G.; SÁ-E-SILVA, I. M. M.; RODAL, M. J. N.; LINS-E-SILVA, A. C. B.
Influence of edge and topography on canopy and sub-canopy structure of an Atlantic
Forest fragment in Igarassu, Pernambuco State, Brazil. Bioremediation, Biodiversity
and Bioavailability v.2, n.1, p.41-46, 2008a.
SILVA, E. F.; OLIVEIRA, C. A. M.; LINS-E-SILVA, A. C. B.; RODAL, M. J. N.
Diversity and genetic structure of natural fragmented populations of Tapiriria
guianensis Aubl. in Northeastern Brazil. Bioremediation, Biodiversity and
Bioavailability, v.2, n.1, p.35-40, 2008b.
SILVA, H. C. H.; LINS-E-SILVA, A. C. B.; GOMES, J. S.; RODAL, M. J. N. The
effect of internal and external edges on vegetation physiognomy and structure in a
remnant of Atlantic Lowland Rainforest in Brazil. Bioremediation, Biodiversity and
Bioavailability, v.2, n.1, p.47-55, 2008c.
SILVA, J. M. C. D.; CASTELETI, C. H. M. Status of the biodiversity of the Atlantic
Forest of Brazil. In: GALINDO-LEAL, C.; CÂMARA, I. G. (Eds.) The Atlantic
Forest of South America: Biodiversity Status, Threats, and Outlook. Washington DC:
Island Press, 2005. Cap. 4, p. 43-59.
SILVA, J. M. C. D.; TABARELLI, M. Tree species impoverishment and the future
flora of the Atlantic forest of northeast Brazil. Nature, v.404, p.72-74, 2000.
SILVA JÚNIOR, J. F.; MARANGON, L. C.; FERREIRA, R. L. C.; FELICIANO, A. L.
P.; BRANDÃO, C. F. L. S.; ALVES JÚNIOR, F. T. Estudo fitossociológico do
componente arbóreo em um remanescente de Floresta Atlântica no Município do Cabo
de Santo Agostinho, PE. Revista Brasileira de Ciências Agrárias, v.3, n.3, p.276-282,
2008.

148
SILVA, W. G.; METZGER, J. P.; BERNACCI, L. C.; CATHARINO, E. L. M.;
DURIGAN, G.; SIMÕES, S. Relief influence on tree species richness in secondary
forest fragments of Atlantic Forest, SE, Brazil. Acta Botanica Brasilica, v.22, p.589-
598, 2008d.
SILVA, W. G.; METZGER, J. P.; SIMÕES, S.; SIMONETTI, C. Relief influence on
the spatial distribution of the Atlantic Forest cover on the Ibiúna Plateau, SP. Brazilian
Journal of Biology, v.67, n.3, p.403-411, 2007.
SIMBERLOFF, D. Community Ecology: is it time to move on? (An American Society
of Naturalists Presidential Address). The American Naturalist, v.163, n.6, p.787-799,
2004.
SIMPSON, E. H. Measurement of diversity. Nature, v.163, p.688, 1949.
SIQUEIRA, S. R.; RODAL, M. J. N.; LINS-E-SILVA, A. C. B.; MELO, A. L.
Physiognomy, structure, and floristics in an area of Atlantic Forest in Northeast Brazil.
Dissertationes Botanicae, v.346, p.11-17, 2001.
SMITH, E. P.; VAN BELLE, G. Nonparametric estimation of species richness.
Biometrics, v.40, p.119-129, 1984.
SØRENSEN, T. A method of establishing groups of equal amplitude in plant sociology
based on similarity of species and its application to analyses of the vegetation on Danish
commons. Biologiske Skrifter / Kongelige Danske Videnskabernes Selskab, v.5 n.4,
p.1-34, 1948.
SOUZA, A. L. D.; SILVA, G. F. D.; CHICHORRO, J. F.; FERREIRA, R. L. C. Mata
nativa 2: Manual do usuário. Viçosa: Cientec, 2006.
STATSOFT. STATISTICA: data analysis software system. Tulsa: StatSoft
Incorporation, 2009.
STRAHLER, A. N.; STRAHLER, A. H. Modern Physical Geography. 1st ed. New
York: John Wiley & Sons, 1978. 502 p.
STRAYER, D. L.; POWER, M. E.; FAGAN, W. F.; PICKETT, S. T. A.; BELNAP, J.
A classification of ecological boundaries. Bioscience, v.53, n.8, p.723, 2003.
TABANEZ, A. A. J.; VIANA, V. M. Patch structure within Brazilian Atlantic Forest
Fragments and implications for conservation. Biotropica, v.32, n.4, p.925-933, 2000.

149
TABARELLI, M.; GASCON, C. Lessons from fragmentation research: Improving
management and policy guidelines for biodiversity conservation. Conservation
Biology, v.19, n.3, p.734-739, Jun. 2005.
TABARELLI, M.; LOPES, A. V.; PERES, C. A. Edge-effects drive Tropical Forest
fragments towards an early-successional system. Biotropica, v.40, n.6, p.657-661,
2008.
TABARELLI, M.; MANTOVANI, W.; PERES, C. A. Effects of habitat fragmentation
on plant guild structure in the montane Atlantic forest of southeastern Brazil. Biological
Conservation, v.91, n.2-3, p.119-127, Dec. 1999.
TABARELLI, M.; RODA, S. A. An opportunity for the Pernambuco Endemism Center.
Natureza & Conservação, v.3, n.2, p.128-134, 2005.
TABARELLI, M.; SILVA, J. M. C.; GASCON, C. Forest fragmentation, synergisms
and the impoverishment of neotropical forests. Biodiversity and Conservation, v.13,
n.7, p.1419-1425, 2004.
TEWS, J.; BROSE, U.; GRIMM, V.; TIELBÖRGER, K.; WICHMANN, M. C.;
SCHWAGER, M.; JELTSCH, F. Animal species diversity driven by habitat
heterogeneity/diversity: the importance of keystone structures. Journal of
Biogeography, v.31, n.1, p.79-92, 2004.
TILMAN, D. Competition and biodiversity in spatially structured habitats. Ecology,
v.75, n.1, p.2-16, 1994.
TILMAN, D.; MAY, R. M.; LEHMAN, C. L.; NOWAK, M. A. Habitat destruction and
the extinction debt. Nature, v.371, n.6492, p.65-66, 1994.
TRIANTIS, K. A.; MYLONAS, M.; LIKA, K.; VARDINOYANNIS, K. A model for
the species-area-habitat relationship. Journal of Biogeography, v.30, n.1, p.19-27, Jan.
2003.
TRIANTIS, K. A.; MYLONAS, M.; WEISER, M. D.; LIKA, K.; VARDINOYANNIS,
K. Species richness, environmental heterogeneity and area: a case study based on land
snails in Skyros archipelago (Aegean Sea, Greece). Journal of Biogeography, v.32,
n.10, p.1727-1735, 2005.

150
TRINDADE, M. B.; LINS-E-SILVA, A. C. B.; SILVA, H. P.; FIGUEIRA, S. B.;
SCHESSL, M. Fragmentation of the Atlantic Rainforest in the Northern coastal region
of Pernambuco, Brazil: recent changes and implications for conservation.
Bioremediation, Biodiversity and Bioavailability, v.2, n.1, p.5-13, 2008.
TURNER, I. M.; CORLETT, R. T. The conservation value of small, isolated fragments
of lowland tropical rain forest. Trends in Ecology & Evolution, v.11, n.8, p.330-333,
1996.
TURNER, M. G. Landscape ecology in North America: past, present, and future.
Ecology, v.86, n.8, p.1967-1974, 2005a.
TURNER, M. G. Landscape ecology: what is the state of the science? Annual Review
of Ecology, Evolution, and Systematics, v.36, n.1, p.319-344, 2005b.
TURNER, W. R.; TJØRVE, E. Scale-dependence in species-area relationships.
Ecography, v.28, n.6, p.721-730, 2005.
VALENCIA, R.; FOSTER, R. B.; VILLA, G.; CONDIT, R.; SVENNING, J.-C.;
HERNANDEZ, C.; ROMOLEROUX, K.; LOSOS, E.; MAGARD, E.; BALSLEV, H.
Tree species distributions and local habitat variation in the Amazon: large forest plot in
eastern Ecuador. Journal of Ecology, v.92, n.2, p.214-229, 2004.
VELLEND, M. Do commonly used indices of β-diversity measure species turnover?
Journal of Vegetation Science, v.12, n.4, p.545-552, 2001.
VELOSO, H. P.; RANGEL FILHO, A. L. R.; LIMA, J. C. A. Classificaçao da
vegetaçao brasileira, adaptada a um sistema universal. 1a ed. Rio de Janeiro: IBGE/
Departamento de Recursos Naturais e Estudos Ambientais, 1991. 124p.
VILAS BÔAS, G. S.; SAMPAIO, F. J.; PEREIRA, A. M. S. The Barreiras Group in the
northeastern coast of the state of Bahia, Brazil: depositional mechanisms and processes.
Anais da Academia Brasileira de Ciências, v.73, n.3, p.417-427, 2001.
VILLARD, M.-A. Habitat fragmentation: major conservation issue or intellectual
attractor? Ecological Applications, v.12, n.2, p.319-320, 2002.
WATSON, D. M. A conceptual framework for studying species composition in
fragments, islands and other patchy ecosystems. Journal of Biogeography, v.29, n.5/6,
p.823, 2002.

151
WEBB, C. O.; PEART, D. R. Habitat associations of trees and seedlings in a Bornean
Rain Forest. Journal of Ecology, v.88, n.3, p.464-478, 2000.
WEBB, E.; FA'AUMU, S. Diversity and structure of tropical rain forest of Tutuila,
American Samoa: effects of site age and substrate. Plant Ecology, v.144, n.2, p.257-
274, 1999.
WHITTAKER, R. H. Gradient analysis of vegetation. Biological Reviews, v.42, p.207-
264, 1967.
WHITTAKER, R. H. Evolution and measurement of species diversity. Taxon, v.21,
n.2/3, p.213-251, 1972.
WHITTAKER, R. H.; LEVIN, S. A.; ROOT, R. B. Niche, habitat, and ecotope. The
American Naturalist, v.107, n.955, p.321-338, 1973.
WHITTAKER, R. H.; LEVIN, S. A.; ROOT, R. B. On the reasons for distinguishing
"Niche, Habitat, and Ecotope". The American Naturalist, v.109, n.968, p.479-482,
1975.
WHITTAKER, R. H.; NIERING, W. A. Vegetation of the Santa Catalina Mountains,
Arizona: a gradient analysis of the south slope. Ecology, v.46, n.4, p.429-452, 1965.
WIENS, J. A. Ecological heterogeneity: an ontogeny of concepts and approaches. In:
HUTCHINGS, M. J.; JOHN, E. A.; STEWART, A. J. A. (Eds.) The ecological
consequences of environmental heterogeneity: the 40th symposium of the British
Ecological Society. Oxford: Blackwell Science, 2000. Cap. 2, p. 9-31.
WILLIAMS-LINERA, G.; DOMÍNGUEZ-GASTELÚ, V.; GARCÍA-ZURITA, M. E.
Microenvironment and floristics of different edges in a fragmented Tropical Rainforest.
Conservation Biology, v.12, n.5, p.1091-1102, 1998.
WILLIAMS, C. B. Patterns in the balance of nature and related problems in
quantitative ecology. 1st ed. London, New York: Academic Press, v.3, 1964. 324p.
WILLIAMSON, M.; GASTON, K. J.; LONSDALE, W. M. The species-area
relationship does not have an asymptote! Journal of Biogeography, v.28, n.7, p.827-
830, 2001.

152
WILLS, C.; CONDIT, R. Similar non-random processes maintain diversity in two
tropical rainforests. Proceedings of the Royal Society of London Series B-Biological
Sciences, v.266, n.1427, p.1445-1452, Jul. 1999.
WILSON, J. B.; AGNEW, A. D. Q.; SYKES, M. T. Ecology or mythology? Are
Whittaker’s “gradient analysis” curves reliable evidence of continuity in vegetation?
Preslia, v.76, p.245-253, 2004.
WRIGHT, J. Plant diversity in tropical forests: a review of mechanisms of species
coexistence. Oecologia, v.130, n.1, p.1-14, 2002.
WRIGHT, S. J.; MULLER-LANDAU, H. C. The future of Tropical Forest species.
Biotropica, v.38, n.3, p.287-301, 2006.
WU, J. G.; LOUCKS, O. L. From balance of nature to hierarchical patch dynamics: A
paradigm shift in ecology. Quarterly Review of Biology, v.70, n.4, p.439-466, Dec.
1995.
YOUNG, A.; MITCHELL, N. Microclimate and vegetation edge effects in a
fragmented podocarp-broadleaf forest in New Zealand. Biological Conservation, v.67,
n.1, p.63-72, 1994.
ZANNE, A. E.; CHAPMAN, C. A. Diversity of woody species in forest, treefall gaps,
and edge in Kibale National Park, Uganda. Plant Ecology, v.178, n.1, p.121-139, 2005.
ZAR, J. H. Biostatistical Analysis. 4th ed. New Jersey: Prentice Hall, 1999. 663p.
ZILLIO, T.; CONDIT, R. The impact of neutrality, niche differentiation and species
input on diversity and abundance distributions. Oikos, v.116, n.6, p.931-940, Jun. 2007.
ZUIDEMA, P. A.; SAYER, J. A.; DIJKMAN, W. Forest fragmentation and
biodiversity: the case for intermediate-sized conservation areas. Environmental
Conservation, v.23, n.4, p.290-297, 1996.

153
APÊNDICE A
Lista florística geral de espécies ocorrentes no dossel e sub-bosque em 390 parcelas de
dossel e 390 de sub-bosque, amostradas em nove fragmentos de Floresta Atlântica em
Pernambuco, Nordeste do Brasil. G - fragmento grande; M - fragmento médio; e P -
fragmento pequeno; IRT - Interior ripário temporário; BE - borda de encosta; IE -
interior de encosta; BRP - borda ripária permanente; IT - interior tabuleiro; IRP -
interior ripário permanente; BT - borda de tabuleiro; BRT - borda ripária temporária.
Em uma célula correspondente a um fragmento/ habitat, D significa que a espécie
ocorreu apenas no dossel; SB, apenas no sub-bosque; e DSB, nos dois estratos.
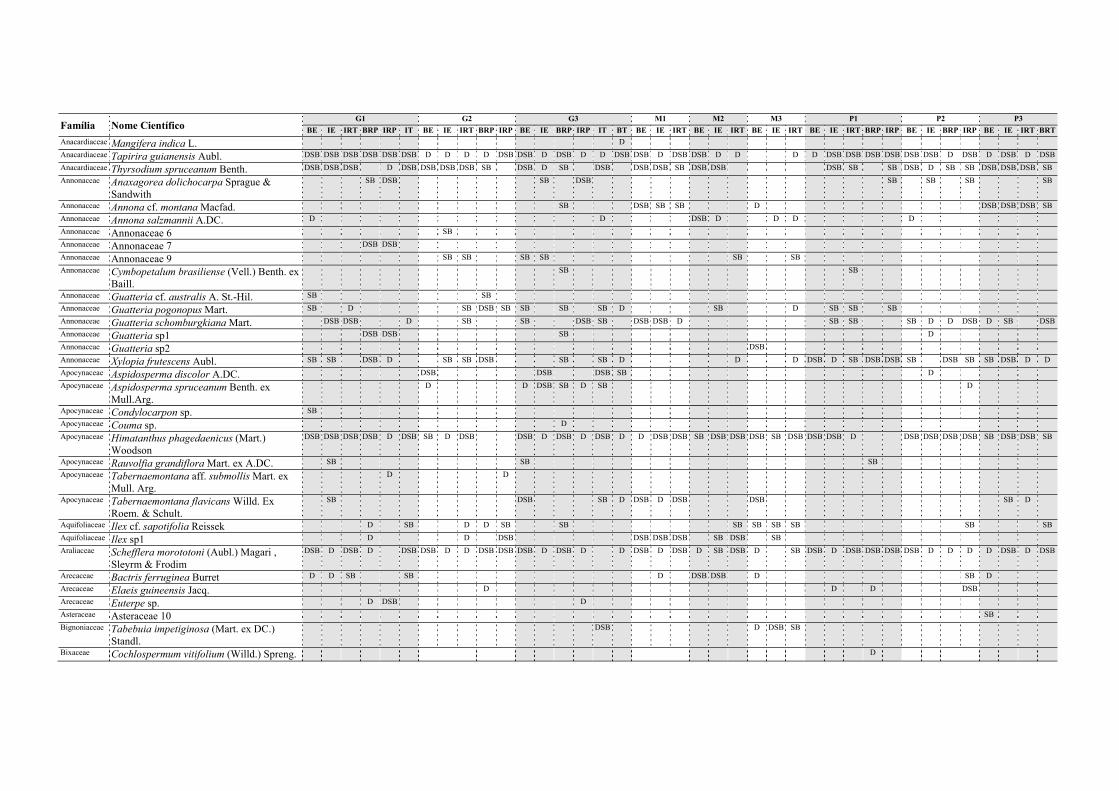
154
Família Nome Científico G1 G2 G3 M1 M2 M3 P1 P2 P3
BE IE IRT BRP IRP IT BE IE IRT BRP IRP BE IE BRP IRP IT BT BE IE IRT BE IE IRT BE IE IRT BE IE IRT BRP IRP BE IE BRP IRP BE IE IRT BRT Anacardiaceae Mangifera indica L. D Anacardiaceae Tapirira guianensis Aubl. DSB DSB DSB DSB DSB DSB D D D D DSB DSB D DSB D D DSB DSB D DSB DSB D D D D DSB DSB DSB DSB DSB DSB D DSB D DSB D DSB Anacardiaceae Thyrsodium spruceanum Benth. DSB DSB DSB D DSB DSB DSB DSB SB DSB D SB DSB DSB DSB SB DSB DSB DSB SB SB DSB D SB SB DSB DSB DSB SB Annonaceae Anaxagorea dolichocarpa Sprague &
Sandwith SB DSB SB DSB SB SB SB SB
Annonaceae Annona cf. montana Macfad. SB DSB SB SB D DSB DSB DSB SB Annonaceae Annona salzmannii A.DC. D D DSB D D D D Annonaceae Annonaceae 6 SB Annonaceae Annonaceae 7 DSB DSB Annonaceae Annonaceae 9 SB SB SB SB SB SB Annonaceae Cymbopetalum brasiliense (Vell.) Benth. ex
Baill. SB SB
Annonaceae Guatteria cf. australis A. St.-Hil. SB SB Annonaceae Guatteria pogonopus Mart. SB D SB DSB SB SB SB SB D SB D SB SB SB Annonaceae Guatteria schomburgkiana Mart. DSB DSB D SB SB DSB SB DSB DSB D SB SB SB D D DSB D SB DSB Annonaceae Guatteria sp1 DSB DSB SB D Annonaceae Guatteria sp2 DSB Annonaceae Xylopia frutescens Aubl. SB SB DSB D SB SB DSB SB SB D D D DSB D SB DSB DSB SB DSB SB SB DSB D D Apocynaceae Aspidosperma discolor A.DC. DSB DSB DSB SB D Apocynaceae Aspidosperma spruceanum Benth. ex
Mull.Arg. D D DSB SB D SB D
Apocynaceae Condylocarpon sp. SB Apocynaceae Couma sp. D Apocynaceae Himatanthus phagedaenicus (Mart.)
Woodson DSB DSB DSB DSB D DSB SB D DSB DSB D DSB D DSB D D DSB DSB SB DSB DSB DSB SB DSB DSB DSB D DSB DSB DSB DSB SB DSB DSB SB
Apocynaceae Rauvolfia grandiflora Mart. ex A.DC. SB SB SB Apocynaceae Tabernaemontana aff. submollis Mart. ex
Mull. Arg. D D
Apocynaceae Tabernaemontana flavicans Willd. Ex Roem. & Schult.
SB DSB SB D DSB D DSB DSB SB D
Aquifoliaceae Ilex cf. sapotifolia Reissek D SB D D SB SB SB SB SB SB SB SB Aquifoliaceae Ilex sp1 D D DSB DSB DSB DSB SB DSB SB Araliaceae Schefflera morototoni (Aubl.) Magari ,
Sleyrm & Frodim DSB D DSB D DSB DSB D D DSB DSB DSB D DSB D D DSB D DSB D SB DSB D SB DSB D DSB DSB DSB DSB D D D D DSB D DSB
Arecaceae Bactris ferruginea Burret D D SB SB D DSB DSB D SB D Arecaceae Elaeis guineensis Jacq. D D D DSB Arecaceae Euterpe sp. D DSB D Asteraceae Asteraceae 10 SB Bignoniaceae Tabebuia impetiginosa (Mart. ex DC.)
Standl. DSB D DSB SB
Bixaceae Cochlospermum vitifolium (Willd.) Spreng. D

155
Família Nome Científico G1 G2 G3 M1 M2 M3 P1 P2 P3
BE IE IRT BRP IRP IT BE IE IRT BRP IRP BE IE BRP IRP IT BT BE IE IRT BE IE IRT BE IE IRT BE IE IRT BRP IRP BE IE BRP IRP BE IE IRT BRT Boraginaceae Cordia nodosa Lam. SB SB SB SB SB SB SB SB SB SB SB SB SB SB SB SB SB SB SB SB SB Boraginaceae Cordia sellowiana Cham. SB Boraginaceae Cordia superba Cham. DSB DSB SB DSB DSB DSB DSB SB SB DSB DSB SB DSB DSB D DSB DSB SB DSB DSB DSB D DSB SB Burseraceae Burseraceae 2 D SB Burseraceae Protium aracouchini (Aubl.) March SB SB D DSB DSB SB SB SB Burseraceae Protium giganteum Engl. DSB DSB D SB D DSB SB DSB DSB DSB SB SB D SB D Burseraceae Protium heptaphyllum (Aubl.) Marchand DSB DSB SB DSB DSB DSB DSB DSB DSB DSB DSB DSB D SB D DSB SB SB DSB SB SB SB SB DSB SB SB DSB DSB SB SB SB SB SB DSB Burseraceae Tetragastris catuaba Cunha D SB SB SB DSB SB D Burseraceae Tetragastris sp. D Celastraceae Celastraceae 1 SB SB SB SB SB Celastraceae Maytenus distichophyla Mart. ex Reissek D DSB SB SB SB DSB SB SB DSB DSB DSB DSB SB D DSB SB DSB DSB DSB DSB DSB DSB SB SB DSB DSB SB SB SB SB Celastraceae Maytenus obtusifolia Mart. D D SB DSB SB D DSB SB Celastraceae Maytenus sp1 SB SB SB Celastraceae Maytenus sp2 SB SB Celastraceae Maytenus sp3 SB SB D D SB Chrysobalanaceae Chrysobalanaceae 5 SB
Chrysobalanaceae Chrysobalanaceae 7 D Chrysobalanaceae Chrysobalanaceae 8 DSB DSB DSB Chrysobalanaceae Couepia rufa Ducke. SB SB SB D D DSB DSB D Chrysobalanaceae Hirtella racemosa Lam. DSB DSB SB SB SB SB SB SB SB SB SB SB SB SB SB SB SB Chrysobalanaceae Hirtella sp1 SB SB Chrysobalanaceae Hirtella sp2 SB SB SB SB SB SB Chrysobalanaceae Hirtella sprucei Benth. D SB Chrysobalanaceae Licania kunthiana Hook. f. DSB SB SB SB SB Chrysobalanaceae Licania octandra (Hoffmanns. ex Roem. &
Schult.) Kuntze SB SB SB
Chrysobalanaceae Licania tomentosa (Benth.) Fritsch D SB SB SB DSB SB DSB SB SB DSB D D D DSB D Clusiaceae Clusia nemorosa G.Mey DSB D DSB D D DSB DSB D D DSB D D D D DSB DSB D D D Clusiaceae Garcinia gardneriana (Planch. & Triana)
Zappi SB SB DSB DSB DSB DSB D
Clusiaceae Symphonia globulifera L.f. DSB DSB SB DSB DSB DSB DSB SB SB SB DSB SB SB SB Clusiaceae Tovomita brasiliensis (Mart.) Walp. DSB Clusiaceae Tovomita brevistaminea Engl. SB SB Clusiaceae Tovomita mangle G. Mariz SB DSB SB SB DSB DSB D DSB DSB SB Combretaceae Buchenavia tetraphylla (Aubl.)
R.A.Howard D D D D D D
Combretaceae Combretaceae 1 D SB D Combretaceae Combretaceae 2 SB D D Connaraceae Rourea sp. SB SB SB Cyatheaceae Cyathea phalerata Mart. DSB D
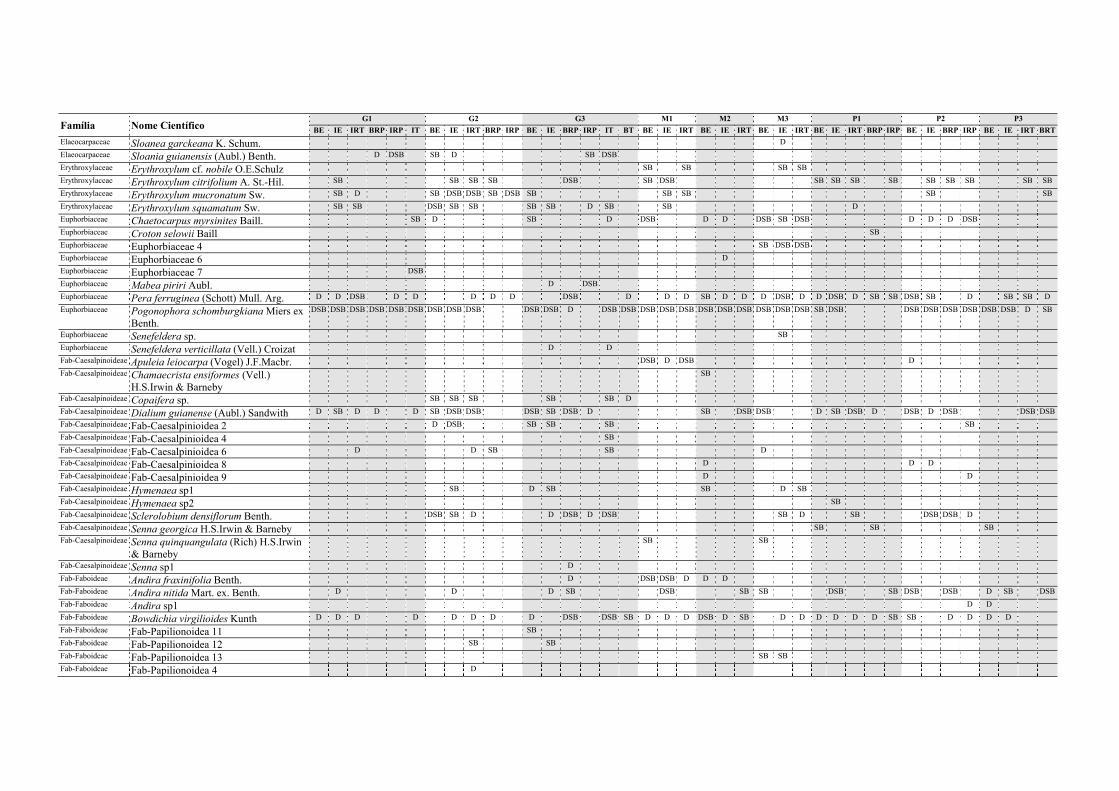
156
Família Nome Científico G1 G2 G3 M1 M2 M3 P1 P2 P3
BE IE IRT BRP IRP IT BE IE IRT BRP IRP BE IE BRP IRP IT BT BE IE IRT BE IE IRT BE IE IRT BE IE IRT BRP IRP BE IE BRP IRP BE IE IRT BRT Elaeocarpaceae Sloanea garckeana K. Schum. D Elaeocarpaceae Sloania guianensis (Aubl.) Benth. D DSB SB D SB DSB Erythroxylaceae Erythroxylum cf. nobile O.E.Schulz SB SB SB SB Erythroxylaceae Erythroxylum citrifolium A. St.-Hil. SB SB SB SB DSB SB DSB SB SB SB SB SB SB SB SB SB Erythroxylaceae Erythroxylum mucronatum Sw. SB D SB DSB DSB SB DSB SB SB SB SB SB Erythroxylaceae Erythroxylum squamatum Sw. SB SB DSB SB SB SB SB D SB SB D Euphorbiaceae Chaetocarpus myrsinites Baill. SB D SB D DSB D D DSB SB DSB D D D DSB Euphorbiaceae Croton selowii Baill SB Euphorbiaceae Euphorbiaceae 4 SB DSB DSB Euphorbiaceae Euphorbiaceae 6 D Euphorbiaceae Euphorbiaceae 7 DSB Euphorbiaceae Mabea piriri Aubl. D DSB Euphorbiaceae Pera ferruginea (Schott) Mull. Arg. D D DSB D D D D D DSB D D D SB D D D DSB D D DSB D SB SB DSB SB D SB SB D Euphorbiaceae Pogonophora schomburgkiana Miers ex
Benth. DSB DSB DSB DSB DSB DSB DSB DSB DSB DSB DSB D DSB DSB DSB DSB DSB DSB DSB DSB DSB DSB DSB SB DSB DSB DSB DSB DSB DSB DSB D SB
Euphorbiaceae Senefeldera sp. SB Euphorbiaceae Senefeldera verticillata (Vell.) Croizat D D Fab-Caesalpinoideae Apuleia leiocarpa (Vogel) J.F.Macbr. DSB D DSB D Fab-Caesalpinoideae Chamaecrista ensiformes (Vell.)
H.S.Irwin & Barneby SB
Fab-Caesalpinoideae Copaifera sp. SB SB SB SB SB D Fab-Caesalpinoideae Dialium guianense (Aubl.) Sandwith D SB D D D SB DSB DSB DSB SB DSB D SB DSB DSB D SB DSB D DSB D DSB DSB DSB Fab-Caesalpinoideae Fab-Caesalpinioidea 2 D DSB SB SB SB SB Fab-Caesalpinoideae Fab-Caesalpinioidea 4 SB Fab-Caesalpinoideae Fab-Caesalpinioidea 6 D D SB SB D Fab-Caesalpinoideae Fab-Caesalpinioidea 8 D D D Fab-Caesalpinoideae Fab-Caesalpinioidea 9 D D Fab-Caesalpinoideae Hymenaea sp1 SB D SB SB D SB Fab-Caesalpinoideae Hymenaea sp2 SB Fab-Caesalpinoideae Sclerolobium densiflorum Benth. DSB SB D D DSB D DSB SB D SB DSB DSB D Fab-Caesalpinoideae Senna georgica H.S.Irwin & Barneby SB SB SB Fab-Caesalpinoideae Senna quinquangulata (Rich) H.S.Irwin
& Barneby SB SB
Fab-Caesalpinoideae Senna sp1 D Fab-Faboideae Andira fraxinifolia Benth. D DSB DSB D D D Fab-Faboideae Andira nitida Mart. ex. Benth. D D D SB DSB SB SB DSB SB DSB DSB D SB DSB Fab-Faboideae Andira sp1 D D Fab-Faboideae Bowdichia virgilioides Kunth D D D D D D D D DSB DSB SB D D D DSB D SB D D D D D D SB SB D D D D Fab-Faboideae Fab-Papilionoidea 11 SB Fab-Faboideae Fab-Papilionoidea 12 SB SB Fab-Faboideae Fab-Papilionoidea 13 SB SB Fab-Faboideae Fab-Papilionoidea 4 D
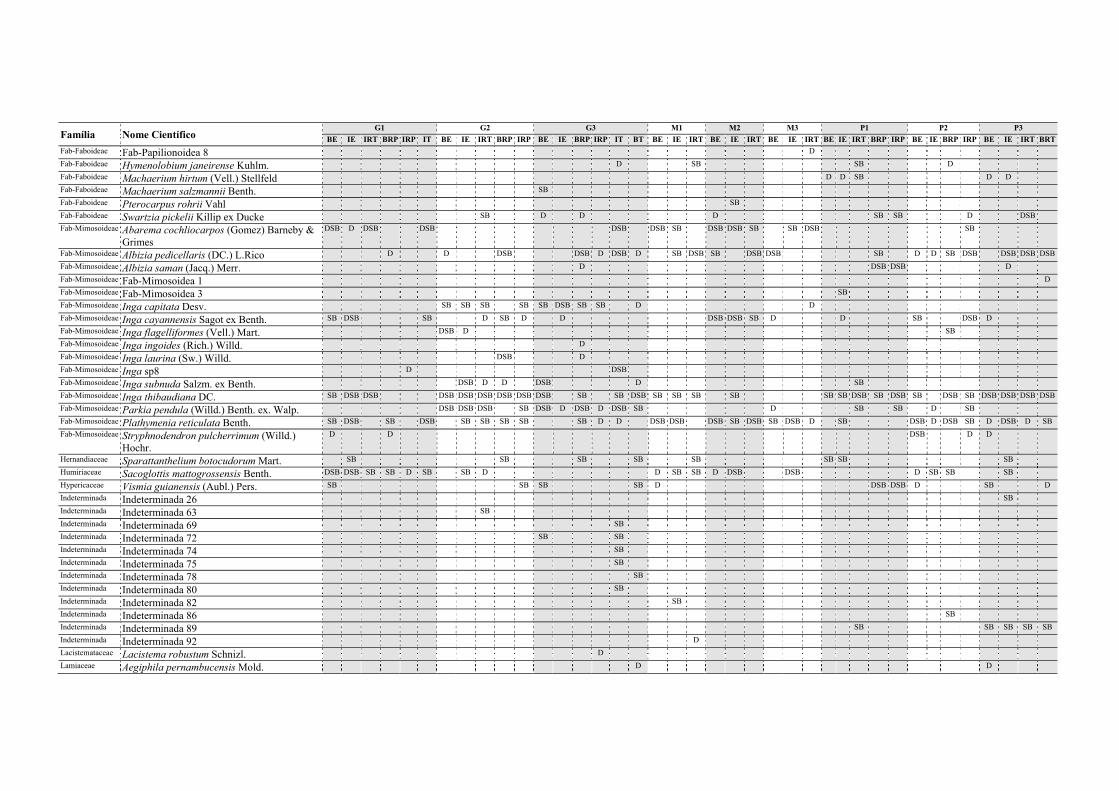
157
Família Nome Científico G1 G2 G3 M1 M2 M3 P1 P2 P3
BE IE IRT BRP IRP IT BE IE IRT BRP IRP BE IE BRP IRP IT BT BE IE IRT BE IE IRT BE IE IRT BE IE IRT BRP IRP BE IE BRP IRP BE IE IRT BRT Fab-Faboideae Fab-Papilionoidea 8 D Fab-Faboideae Hymenolobium janeirense Kuhlm. D SB SB D Fab-Faboideae Machaerium hirtum (Vell.) Stellfeld D D SB D D Fab-Faboideae Machaerium salzmannii Benth. SB Fab-Faboideae Pterocarpus rohrii Vahl SB Fab-Faboideae Swartzia pickelii Killip ex Ducke SB D D D SB SB D DSB Fab-Mimosoideae Abarema cochliocarpos (Gomez) Barneby &
Grimes DSB D DSB DSB DSB DSB SB DSB DSB SB SB DSB SB
Fab-Mimosoideae Albizia pedicellaris (DC.) L.Rico D D DSB DSB D DSB D SB DSB SB DSB DSB SB D D SB DSB DSB DSB DSB Fab-Mimosoideae Albizia saman (Jacq.) Merr. D DSB DSB D Fab-Mimosoideae Fab-Mimosoidea 1 D Fab-Mimosoideae Fab-Mimosoidea 3 SB Fab-Mimosoideae Inga capitata Desv. SB SB SB SB SB DSB SB SB D D Fab-Mimosoideae Inga cayannensis Sagot ex Benth. SB DSB SB D SB D D DSB DSB SB D D SB DSB D Fab-Mimosoideae Inga flagelliformes (Vell.) Mart. DSB D SB Fab-Mimosoideae Inga ingoides (Rich.) Willd. D Fab-Mimosoideae Inga laurina (Sw.) Willd. DSB D Fab-Mimosoideae Inga sp8 D DSB Fab-Mimosoideae Inga subnuda Salzm. ex Benth. DSB D D DSB D SB Fab-Mimosoideae Inga thibaudiana DC. SB DSB DSB DSB DSB DSB DSB DSB DSB SB SB DSB SB SB SB SB SB SB DSB SB DSB SB DSB SB DSB DSB DSB DSB Fab-Mimosoideae Parkia pendula (Willd.) Benth. ex. Walp. DSB DSB DSB SB DSB D DSB D DSB SB D SB SB D SB Fab-Mimosoideae Plathymenia reticulata Benth. SB DSB SB DSB SB SB SB SB SB D D DSB DSB DSB SB DSB SB DSB D SB DSB D DSB SB D DSB D SB Fab-Mimosoideae Stryphnodendron pulcherrimum (Willd.)
Hochr. D D DSB D D
Hernandiaceae Sparattanthelium botocudorum Mart. SB SB SB SB SB SB SB SB Humiriaceae Sacoglottis mattogrossensis Benth. DSB DSB SB SB D SB SB D D SB SB D DSB DSB D SB SB SB Hypericaceae Vismia guianensis (Aubl.) Pers. SB SB SB SB D DSB DSB D SB D Indeterminada Indeterminada 26 SB Indeterminada Indeterminada 63 SB Indeterminada Indeterminada 69 SB Indeterminada Indeterminada 72 SB SB Indeterminada Indeterminada 74 SB Indeterminada Indeterminada 75 SB Indeterminada Indeterminada 78 SB Indeterminada Indeterminada 80 SB Indeterminada Indeterminada 82 SB Indeterminada Indeterminada 86 SB Indeterminada Indeterminada 89 SB SB SB SB SB Indeterminada Indeterminada 92 D Lacistemataceae Lacistema robustum Schnizl. D Lamiaceae Aegiphila pernambucensis Mold. D D
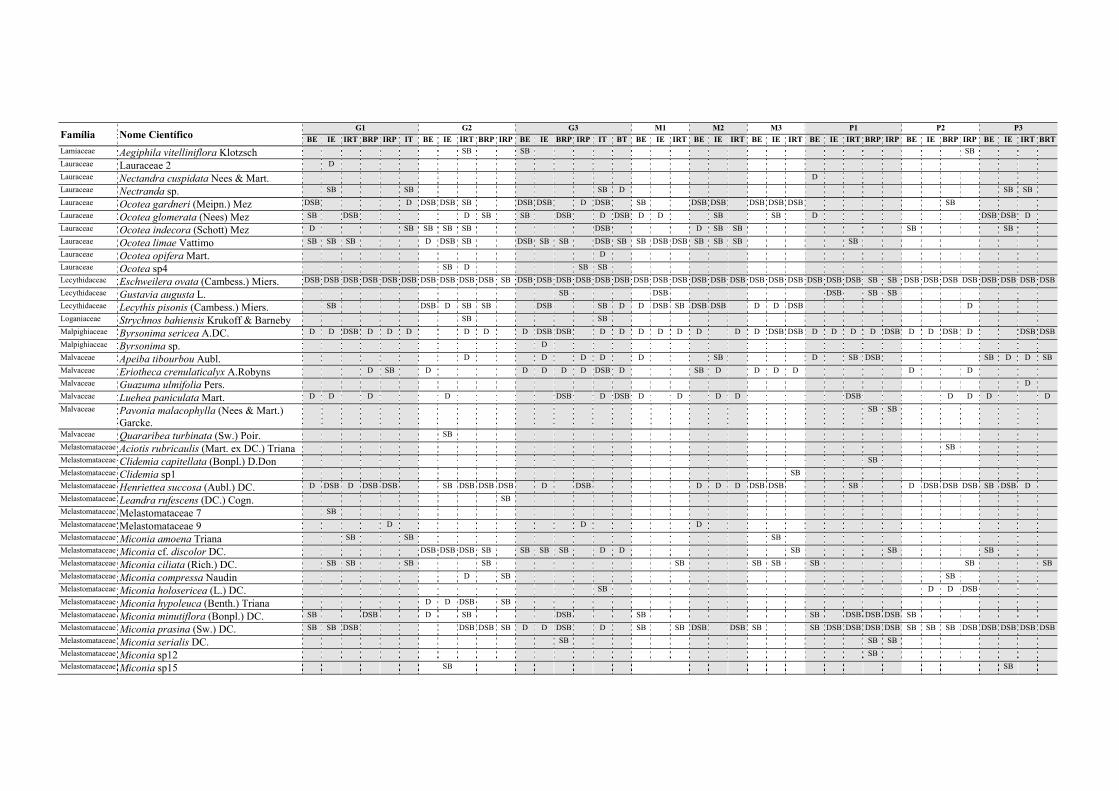
158
Família Nome Científico G1 G2 G3 M1 M2 M3 P1 P2 P3
BE IE IRT BRP IRP IT BE IE IRT BRP IRP BE IE BRP IRP IT BT BE IE IRT BE IE IRT BE IE IRT BE IE IRT BRP IRP BE IE BRP IRP BE IE IRT BRT Lamiaceae Aegiphila vitelliniflora Klotzsch SB SB SB Lauraceae Lauraceae 2 D Lauraceae Nectandra cuspidata Nees & Mart. D Lauraceae Nectranda sp. SB SB SB D SB SB Lauraceae Ocotea gardneri (Meipn.) Mez DSB D DSB DSB SB DSB DSB D DSB SB DSB DSB DSB DSB DSB SB Lauraceae Ocotea glomerata (Nees) Mez SB DSB D SB SB DSB D DSB D D SB SB D DSB DSB D Lauraceae Ocotea indecora (Schott) Mez D SB SB SB SB DSB D SB SB SB SB Lauraceae Ocotea limae Vattimo SB SB SB D DSB SB DSB SB SB DSB SB SB DSB DSB SB SB SB SB Lauraceae Ocotea opifera Mart. D Lauraceae Ocotea sp4 SB D SB SB Lecythidaceae Eschweilera ovata (Cambess.) Miers. DSB DSB DSB DSB DSB DSB DSB DSB DSB DSB SB DSB DSB DSB DSB DSB DSB DSB DSB DSB DSB DSB DSB DSB DSB DSB DSB DSB DSB SB SB DSB DSB DSB DSB DSB DSB DSB DSB Lecythidaceae Gustavia augusta L. SB DSB DSB SB SB Lecythidaceae Lecythis pisonis (Cambess.) Miers. SB DSB D SB SB DSB SB D D DSB SB DSB DSB D D DSB D Loganiaceae Strychnos bahiensis Krukoff & Barneby SB SB Malpighiaceae Byrsonima sericea A.DC. D D DSB D D D D D D DSB DSB D D D D D D D D DSB DSB D D D D DSB D D DSB D DSB DSB Malpighiaceae Byrsonima sp. D Malvaceae Apeiba tibourbou Aubl. D D D D D SB D SB DSB SB D D SB Malvaceae Eriotheca crenulaticalyx A.Robyns D SB D D D D D DSB D SB D D D D D D Malvaceae Guazuma ulmifolia Pers. D Malvaceae Luehea paniculata Mart. D D D D DSB D DSB D D D D DSB D D D D Malvaceae Pavonia malacophylla (Nees & Mart.)
Garcke. SB SB
Malvaceae Quararibea turbinata (Sw.) Poir. SB Melastomataceae Aciotis rubricaulis (Mart. ex DC.) Triana SB Melastomataceae Clidemia capitellata (Bonpl.) D.Don SB Melastomataceae Clidemia sp1 SB Melastomataceae Henriettea succosa (Aubl.) DC. D DSB D DSB DSB SB DSB DSB DSB D DSB D D D DSB DSB SB D DSB DSB DSB SB DSB D Melastomataceae Leandra rufescens (DC.) Cogn. SB Melastomataceae Melastomataceae 7 SB Melastomataceae Melastomataceae 9 D D D Melastomataceae Miconia amoena Triana SB SB SB Melastomataceae Miconia cf. discolor DC. DSB DSB DSB SB SB SB SB D D SB SB SB Melastomataceae Miconia ciliata (Rich.) DC. SB SB SB SB SB SB SB SB SB SB Melastomataceae Miconia compressa Naudin D SB SB Melastomataceae Miconia holosericea (L.) DC. SB D D DSB Melastomataceae Miconia hypoleuca (Benth.) Triana D D DSB SB Melastomataceae Miconia minutiflora (Bonpl.) DC. SB DSB D SB DSB SB SB DSB DSB DSB SB Melastomataceae Miconia prasina (Sw.) DC. SB SB DSB DSB DSB SB D D DSB D SB SB DSB DSB SB SB DSB DSB DSB DSB SB SB SB DSB DSB DSB DSB DSB Melastomataceae Miconia serialis DC. SB SB SB Melastomataceae Miconia sp12 SB Melastomataceae Miconia sp15 SB SB
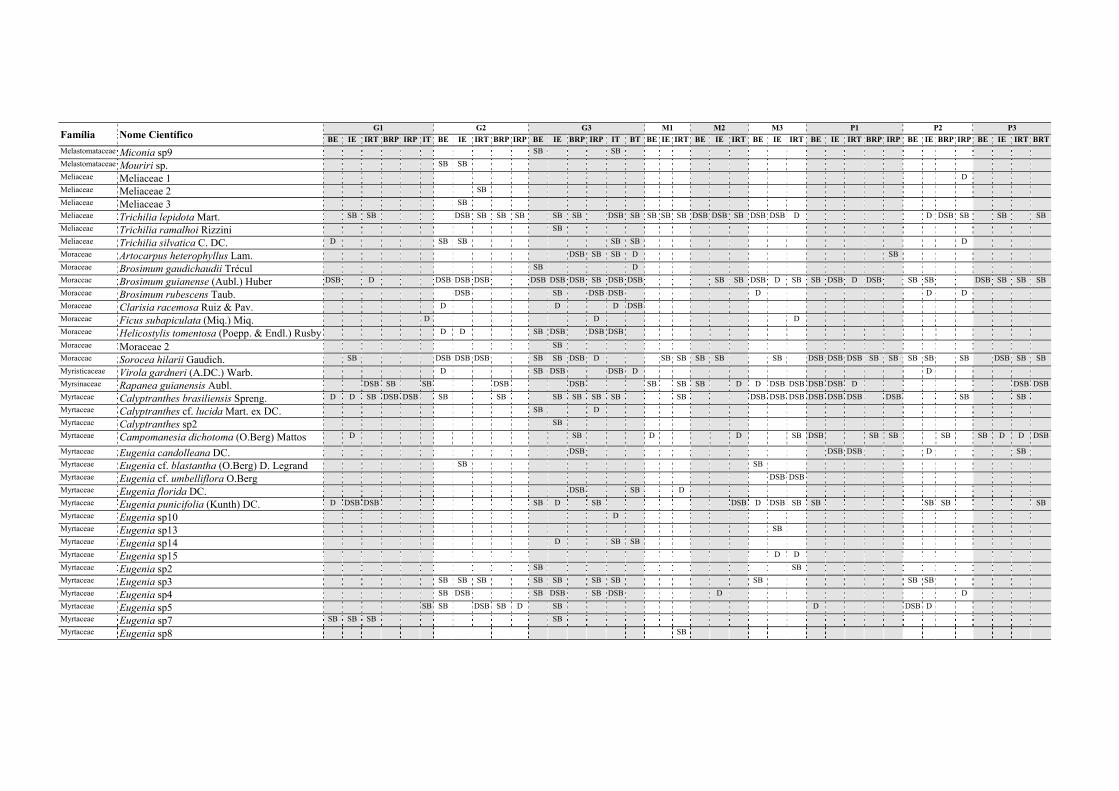
159
Família Nome Científico G1 G2 G3 M1 M2 M3 P1 P2 P3
BE IE IRT BRP IRP IT BE IE IRT BRP IRP BE IE BRP IRP IT BT BE IE IRT BE IE IRT BE IE IRT BE IE IRT BRP IRP BE IE BRP IRP BE IE IRT BRT Melastomataceae Miconia sp9 SB SB Melastomataceae Mouriri sp. SB SB Meliaceae Meliaceae 1 D Meliaceae Meliaceae 2 SB Meliaceae Meliaceae 3 SB Meliaceae Trichilia lepidota Mart. SB SB DSB SB SB SB SB SB DSB SB SB SB SB DSB DSB SB DSB DSB D D DSB SB SB SB Meliaceae Trichilia ramalhoi Rizzini SB Meliaceae Trichilia silvatica C. DC. D SB SB SB SB D Moraceae Artocarpus heterophyllus Lam. DSB SB SB D SB Moraceae Brosimum gaudichaudii Trécul SB D Moraceae Brosimum guianense (Aubl.) Huber DSB D DSB DSB DSB DSB DSB DSB SB DSB DSB SB SB DSB D SB SB DSB D DSB SB SB DSB SB SB SB Moraceae Brosimum rubescens Taub. DSB SB DSB DSB D D D Moraceae Clarisia racemosa Ruiz & Pav. D D D DSB Moraceae Ficus subapiculata (Miq.) Miq. D D D Moraceae Helicostylis tomentosa (Poepp. & Endl.) Rusby D D SB DSB DSB DSB
Moraceae Moraceae 2 SB Moraceae Sorocea hilarii Gaudich. SB DSB DSB DSB SB SB DSB D SB SB SB SB SB DSB DSB DSB SB SB SB SB SB DSB SB SB Myristicaceae Virola gardneri (A.DC.) Warb. D SB DSB DSB D D Myrsinaceae Rapanea guianensis Aubl. DSB SB SB DSB DSB SB SB SB D D DSB DSB DSB DSB D DSB DSB Myrtaceae Calyptranthes brasiliensis Spreng. D D SB DSB DSB SB SB SB SB SB SB SB DSB DSB DSB DSB DSB DSB DSB SB SB Myrtaceae Calyptranthes cf. lucida Mart. ex DC. SB D Myrtaceae Calyptranthes sp2 SB Myrtaceae Campomanesia dichotoma (O.Berg) Mattos D SB D D SB DSB SB SB SB SB D D DSB
Myrtaceae Eugenia candolleana DC. DSB DSB DSB D SB Myrtaceae Eugenia cf. blastantha (O.Berg) D. Legrand SB SB Myrtaceae Eugenia cf. umbelliflora O.Berg DSB DSB Myrtaceae Eugenia florida DC. DSB SB D Myrtaceae Eugenia punicifolia (Kunth) DC. D DSB DSB SB D SB DSB D DSB SB SB SB SB SB Myrtaceae Eugenia sp10 D Myrtaceae Eugenia sp13 SB Myrtaceae Eugenia sp14 D SB SB Myrtaceae Eugenia sp15 D D Myrtaceae Eugenia sp2 SB SB Myrtaceae Eugenia sp3 SB SB SB SB SB SB SB SB SB SB Myrtaceae Eugenia sp4 SB DSB SB DSB SB DSB D D Myrtaceae Eugenia sp5 SB SB DSB SB D SB D DSB D Myrtaceae Eugenia sp7 SB SB SB SB Myrtaceae Eugenia sp8 SB
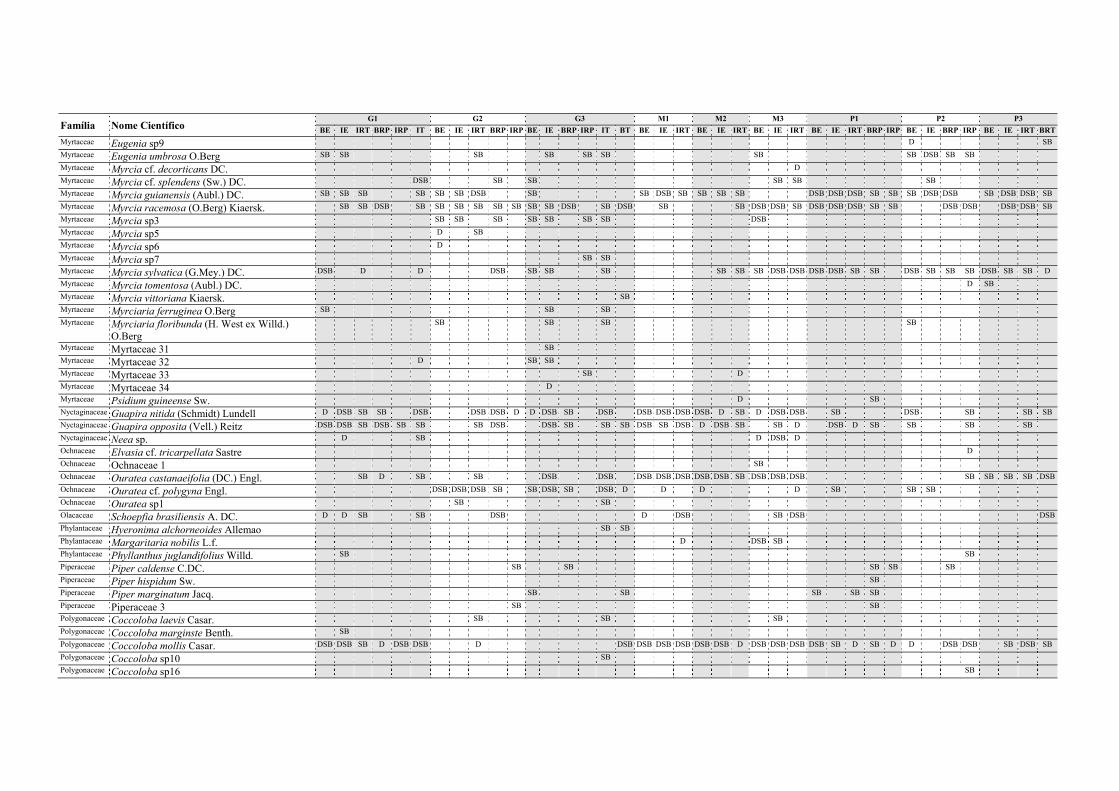
160
Família Nome Científico G1 G2 G3 M1 M2 M3 P1 P2 P3
BE IE IRT BRP IRP IT BE IE IRT BRP IRP BE IE BRP IRP IT BT BE IE IRT BE IE IRT BE IE IRT BE IE IRT BRP IRP BE IE BRP IRP BE IE IRT BRT Myrtaceae Eugenia sp9 D SB Myrtaceae Eugenia umbrosa O.Berg SB SB SB SB SB SB SB SB DSB SB SB Myrtaceae Myrcia cf. decorticans DC. D Myrtaceae Myrcia cf. splendens (Sw.) DC. DSB SB SB SB SB SB Myrtaceae Myrcia guianensis (Aubl.) DC. SB SB SB SB SB SB DSB SB SB DSB SB SB SB SB DSB DSB DSB SB SB SB DSB DSB SB DSB DSB SB Myrtaceae Myrcia racemosa (O.Berg) Kiaersk. SB SB DSB SB SB SB SB SB SB SB SB DSB SB DSB SB SB DSB DSB SB DSB DSB DSB SB SB DSB DSB DSB DSB SB Myrtaceae Myrcia sp3 SB SB SB SB SB SB SB DSB Myrtaceae Myrcia sp5 D SB Myrtaceae Myrcia sp6 D Myrtaceae Myrcia sp7 SB SB Myrtaceae Myrcia sylvatica (G.Mey.) DC. DSB D D DSB SB SB SB SB SB SB DSB DSB DSB DSB SB SB DSB SB SB SB DSB SB SB D Myrtaceae Myrcia tomentosa (Aubl.) DC. D SB Myrtaceae Myrcia vittoriana Kiaersk. SB Myrtaceae Myrciaria ferruginea O.Berg SB SB SB Myrtaceae Myrciaria floribunda (H. West ex Willd.)
O.Berg SB SB SB SB
Myrtaceae Myrtaceae 31 SB Myrtaceae Myrtaceae 32 D SB SB Myrtaceae Myrtaceae 33 SB D Myrtaceae Myrtaceae 34 D Myrtaceae Psidium guineense Sw. D SB Nyctaginaceae Guapira nitida (Schmidt) Lundell D DSB SB SB DSB DSB DSB D D DSB SB DSB DSB DSB DSB DSB D SB D DSB DSB SB DSB SB SB SB Nyctaginaceae Guapira opposita (Vell.) Reitz DSB DSB SB DSB SB SB SB DSB DSB SB SB SB DSB SB DSB D DSB SB SB D DSB D SB SB SB SB Nyctaginaceae Neea sp. D SB D DSB D Ochnaceae Elvasia cf. tricarpellata Sastre D Ochnaceae Ochnaceae 1 SB Ochnaceae Ouratea castanaeifolia (DC.) Engl. SB D SB SB DSB DSB DSB DSB DSB DSB DSB SB DSB DSB DSB SB SB SB SB DSB Ochnaceae Ouratea cf. polygyna Engl. DSB DSB DSB SB SB DSB SB DSB D D D D SB SB SB Ochnaceae Ouratea sp1 SB SB Olacaceae Schoepfia brasiliensis A. DC. D D SB SB DSB D DSB SB DSB DSB Phylantaceae Hyeronima alchorneoides Allemao SB SB Phylantaceae Margaritaria nobilis L.f. D DSB SB Phylantaceae Phyllanthus juglandifolius Willd. SB SB Piperaceae Piper caldense C.DC. SB SB SB SB SB Piperaceae Piper hispidum Sw. SB Piperaceae Piper marginatum Jacq. SB SB SB SB SB Piperaceae Piperaceae 3 SB SB Polygonaceae Coccoloba laevis Casar. SB SB SB Polygonaceae Coccoloba marginste Benth. SB Polygonaceae Coccoloba mollis Casar. DSB DSB SB D DSB DSB D DSB DSB DSB DSB DSB DSB D DSB DSB DSB DSB SB D SB D D DSB DSB SB DSB SB Polygonaceae Coccoloba sp10 SB Polygonaceae Coccoloba sp16 SB

161
Família Nome Científico G1 G2 G3 M1 M2 M3 P1 P2 P3
BE IE IRT BRP IRP IT BE IE IRT BRP IRP BE IE BRP IRP IT BT BE IE IRT BE IE IRT BE IE IRT BE IE IRT BRP IRP BE IE BRP IRP BE IE IRT BRT Polygonaceae Coccoloba sp3 SB SB SB SB SB SB Polygonaceae Coccoloba sp4 D Rubiaceae Alseis pickelii Pilg. & Schmale DSB SB D SB D D DSB SB SB SB SB SB D SB DSB DSB Rubiaceae Amaioua guianensis Aubl. D SB Rubiaceae Gonzalagunia dicocca Cham. & Schltdl. SB SB Rubiaceae Malanea cf. bahiensis Mull. Arg. SB SB Rubiaceae Palicourea crocea (Sw.) Roem. & Schult. SB SB SB SB SB SB SB SB SB SB SB SB SB SB SB SB SB SB Rubiaceae Posoqueria cf. grandiflora Standl. SB Rubiaceae Posoqueria longiflora Aubl. SB SB SB SB SB SB Rubiaceae Psychotria barbiflora DC. SB SB SB SB SB SB SB SB SB SB SB SB SB SB SB SB SB SB SB SB SB SB Rubiaceae Psychotria bracteocardia (DC.) Mull.Arg. SB SB SB SB SB Rubiaceae Psychotria capitata Ruiz & Pav. SB SB SB Rubiaceae Psychotria carthagenensis Jacq. SB SB SB SB SB SB SB SB SB SB SB SB DSB SB SB DSB DSB SB SB SB SB SB SB SB SB SB SB DSB SB Rubiaceae Psychotria erecta (Aubl.) Standl. & Steyerm. SB SB SB SB SB SB SB SB SB Rubiaceae Psychotria hoffmanseggiana (Willd. ex Roem.
& Schult.) Mull.Arg. SB
Rubiaceae Psychotria mapouriodes DC. D Rubiaceae Rubiaceae 12 SB Rubiaceae Rubiaceae 18 SB Rubiaceae Rubiaceae 19 DSB Rubiaceae Rubiaceae 7 SB Rubiaceae Rubiaceae 9 DSB SB SB SB SB Rubiaceae Salzmannia nitida DC. SB SB SB SB SB SB Rutaceae Hortia arborea Engl. DSB Rutaceae Pilocarpus cf. giganteus Engl. DSB SB DSB DSB SB SB Salicaceae Banara cf. brasiliensis (Schott) Benth. SB Salicaceae Casearia arborea (Rich.) Urb. D D D SB Salicaceae Casearia decandra Jacq. D Salicaceae Casearia javitensis Humb., Bonpl. & Kunth DSB DSB D D DSB D SB SB DSB DSB SB DSB DSB DSB SB DSB SB SB SB SB D SB DSB SB DSB DSB Salicaceae Casearia sp2 SB Salicaceae Casearia sp3 D D DSB DSB DSB DSB DSB SB SB D D DSB DSB D SB Salicaceae Casearia sylvestris Sw. SB SB SB SB Sapindaceae Allophylus edulis (A.St.-Hil., A.Juss. &
Cambess.) Radlk. SB SB DSB DSB SB SB DSB DSB SB SB SB SB SB SB SB DSB DSB DSB DSB DSB
Sapindaceae Cupania oblongifolia Mart. D D DSB DSB DSB SB D DSB DSB DSB DSB SB SB DSB Sapindaceae Cupania paniculata Cambess. SB DSB DSB SB SB D DSB DSB DSB SB Sapindaceae Cupania racemosa (Vell.) Radlk. DSB DSB DSB DSB DSB SB DSB DSB SB SB SB DSB DSB DSB DSB DSB SB DSB DSB SB DSB SB DSB SB DSB DSB DSB DSB DSB Sapindaceae Cupania revoluta Rolfe SB Sapindaceae Cupania sp3 SB Sapindaceae Paulinia trigonia Vell. SB SB Sapindaceae Sapindaceae 1 D
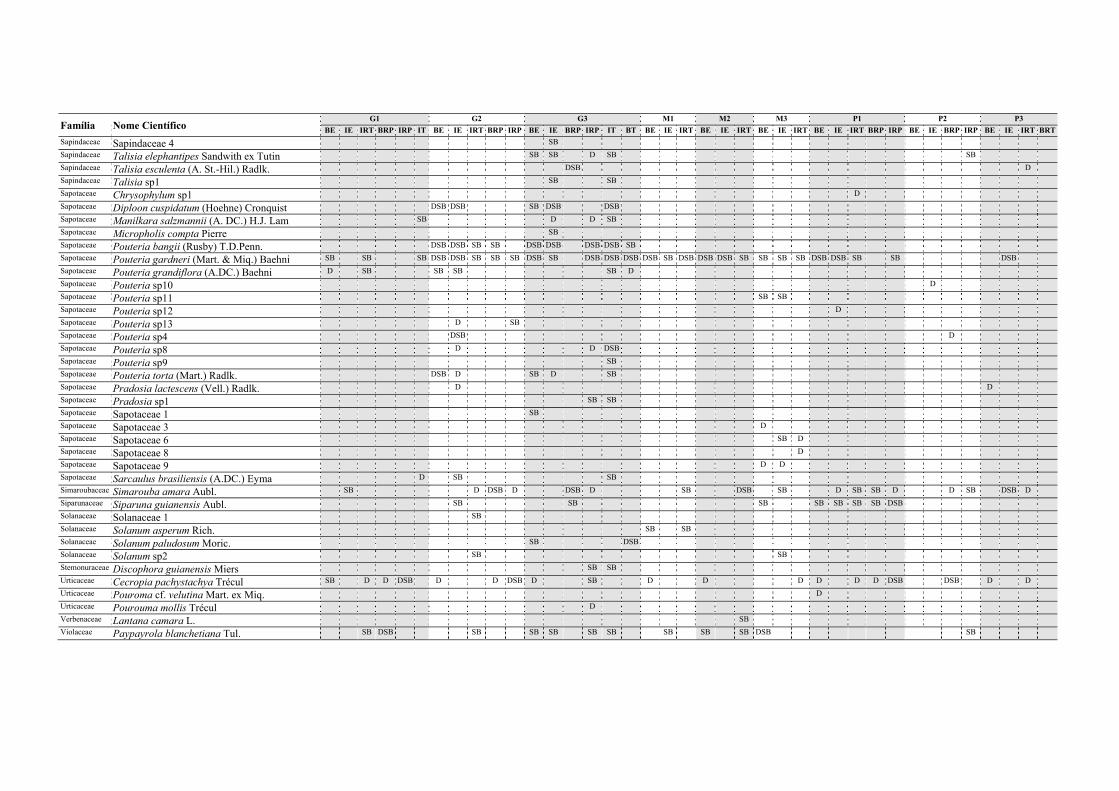
162
Família Nome Científico G1 G2 G3 M1 M2 M3 P1 P2 P3
BE IE IRT BRP IRP IT BE IE IRT BRP IRP BE IE BRP IRP IT BT BE IE IRT BE IE IRT BE IE IRT BE IE IRT BRP IRP BE IE BRP IRP BE IE IRT BRT Sapindaceae Sapindaceae 4 SB Sapindaceae Talisia elephantipes Sandwith ex Tutin SB SB D SB SB Sapindaceae Talisia esculenta (A. St.-Hil.) Radlk. DSB D Sapindaceae Talisia sp1 SB SB Sapotaceae Chrysophylum sp1 D Sapotaceae Diploon cuspidatum (Hoehne) Cronquist DSB DSB SB DSB DSB Sapotaceae Manilkara salzmannii (A. DC.) H.J. Lam SB D D SB Sapotaceae Micropholis compta Pierre SB Sapotaceae Pouteria bangii (Rusby) T.D.Penn. DSB DSB SB SB DSB DSB DSB DSB SB Sapotaceae Pouteria gardneri (Mart. & Miq.) Baehni SB SB SB DSB DSB SB SB SB DSB SB DSB DSB DSB DSB SB DSB DSB DSB SB SB SB SB DSB DSB SB SB DSB Sapotaceae Pouteria grandiflora (A.DC.) Baehni D SB SB SB SB D Sapotaceae Pouteria sp10 D Sapotaceae Pouteria sp11 SB SB Sapotaceae Pouteria sp12 D Sapotaceae Pouteria sp13 D SB Sapotaceae Pouteria sp4 DSB D Sapotaceae Pouteria sp8 D D DSB Sapotaceae Pouteria sp9 SB Sapotaceae Pouteria torta (Mart.) Radlk. DSB D SB D SB Sapotaceae Pradosia lactescens (Vell.) Radlk. D D Sapotaceae Pradosia sp1 SB SB Sapotaceae Sapotaceae 1 SB Sapotaceae Sapotaceae 3 D Sapotaceae Sapotaceae 6 SB D Sapotaceae Sapotaceae 8 D Sapotaceae Sapotaceae 9 D D Sapotaceae Sarcaulus brasiliensis (A.DC.) Eyma D SB SB Simaroubaceae Simarouba amara Aubl. SB D DSB D DSB D SB DSB SB D SB SB D D SB DSB D Siparunaceae Siparuna guianensis Aubl. SB SB SB SB SB SB SB DSB Solanaceae Solanaceae 1 SB Solanaceae Solanum asperum Rich. SB SB Solanaceae Solanum paludosum Moric. SB DSB Solanaceae Solanum sp2 SB SB Stemonuraceae Discophora guianensis Miers SB SB Urticaceae Cecropia pachystachya Trécul SB D D DSB D D DSB D SB D D D D D D DSB DSB D D Urticaceae Pouroma cf. velutina Mart. ex Miq. D Urticaceae Pourouma mollis Trécul D Verbenaceae Lantana camara L. SB Violaceae Paypayrola blanchetiana Tul. SB DSB SB SB SB SB SB SB SB SB DSB SB
Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo





























