Embed Size (px)
Citation preview

i
UNIVERSIDADE ESTADUAL DE CAMPINAS
INSTITUTO DE BIOLOGIA
KELL GRANDJEAN DA COSTA
ADAPTAÇÕES MUSCULARES EM MARCADORES METABÓLICOS
E DE ESTRESSE OXIDATIVO INDUZIDAS EM RATOS PELO
TREINAMENTO RESISTIDO EM ESCADA COM SISTEMA DE
ROLDANAS
Campinas, 2015

ii

iii

iv

v

vi

vii
RESUMO
O objetivo do trabalho foi desenvolver uma escada com sistema de roldanas para aplicação
da sobrecarga, para mimetizar condições semelhantes aos treinos em humanos, uma vez
que o músculo esquelético é muito sensível as variáveis de treinamento (sobrecarga,
repetições, pausas). Outro objetivo foi avaliar os efeitos de oito semanas de treinamento de
força em escada com e sem suplementação antioxidante. Foram analisados marcadores
morfológicos, metabólicos e de estresse oxidativo no sangue e no músculo flexor longo do
hálux (FHL) em ratos divididos em grupo controle e treinado (Protocolo 1), e
posteriormente em ratos submetidos a suplementação antioxidante (óleo de arroz) e
treinamento (Protocolo 2). O sistema de roldanas gerou uma sobrecarga com menor
interferência de atrito, propiciando sessões de treino com contribuição do metabolismo
anaeróbico, e com dano tecidual nas fibras musculares, resultando em aumento de
desempenho ao longo das 24 sessões de treino e adaptações características de um treino
resistido tais como aumentos da AST, dos estoques de glicogênio e atividade da enzima
lactato desidrogenase. Importante salientar que houve hipertrofia das fibras glicolíticas e
oxidativas e aumento da atividade da enzima citrato sintase, demonstrando que além do
aumento na produção de força o treino aumentou a capacidade oxidativa do músculo.
Houve uma maior produção de EROs refletida em aumento significativo da atividade das
enzimas antioxidantes glutationa redutase e catalase. Esse aumento foi suficiente para a
proteção do músculo, uma vez que os valores do marcador de peroxidação lipídica
(TBARs) estavam homogeneamente diminuídos após as 8 semanas de treino quando
comparado ao grupo controle. A possível resposta ergogênica não ocorreu com a
suplementação de óleo de arroz no FHL de ratos treinados divididos em 3 grupos:
suplementados com água, 0.75 ml e 1.5 ml de óleo de arroz. Não encontramos diferenças
significativas no desempenho e na hipertrofia entre os grupos. Houve diminuição
significativa nas concentrações de TBARs nos grupos suplementados com 0,75mL e 1,5mL
de óleo de arroz. No entanto, os efeitos adaptativos na capacidade oxidativa muscular e nas
enzimas antioxidantes foi perdido com a suplementação. Esses dados sugerem cautela na
utilização de antioxidantes com intuito de proteção do aumento de EROs induzido pelo
treinamento, pois vias adaptativas são sinalizadas por EROs.
Palavras-Chaves: Hipertrofia; óleo de arroz, capacidade oxidativa, enzimas antioxidantes,
glicogênio, treinamento de força.

viii

ix
ABSTRACT
The main goal of this study was to develop a ladder for rats with pulley system for the
application of overload, to mimic conditions similar to training in humans, since the
skeletal muscle is very sensitive to the training variables (overload, repetitions, pauses).
Other objective was to evaluate the effect of eight weeks of strength training in ladder in
this system with and without antioxidant supplementation. It was performed morphological
markers, metabolic and oxidative stress analyses in blood and in the flexor hallucis muscle
(FHL) in rats divided into control group and trained group (Protocol 1), and in other group
of rats subjected to antioxidant supplementation (rice oil) and training (Protocol 2). The
pulley system generated an overload with less interference of friction, providing training
sessions with anaerobic metabolism contribution, and tissue damage in the muscle fibers,
resulting in increased performance over the 24 training sessions and adaptations of
resistance training such as increases in (Cross-section-area) CSA, the glycogen stores and
activity of lactate dehydrogenase. It is Important to note that there was hypertrophy of the
glycolytic and oxidative fibers and increased activity of the citrate synthase enzyme,
showing that besides the increase in force production, training also increased muscle
oxidative capacity. There was a higher production of ROS reflected in a significant increase
of antioxidant enzymes catalase and glutathione reductase. This increase was enough for
muscle protection, since the values of lipid peroxidation marker (TBARS) were
homogeneously decreased after 8 weeks of training compared to the control group. We did
not find ergogenic response with the rice oil supplementation in rats trained FHL divided
into 3 groups: supplemented with water, 1.5 ml and 0.75 ml rice bran oil. We found no
significant differences in performance and hypertrophy among groups. There was a
significant decrease in TBARS concentrations in the groups supplemented with 0,75mL
and 1.5 mL of rice oil. However, the adaptive effects in muscle oxidative capacity and the
antioxidant enzymes were lost with the supplementation. These data suggest caution with
the use of antioxidants to protect against the increase of ROS induced by training, because
adaptive pathways of training can be signalize by ROS.
Key Words: Hypertrophy; rice oil, oxidative capacity, antioxidant enzymes, glycogen,
strength training.

x

xi
SUMÁRIO
RESUMO ....................................................................................................................... vii
ABSTRACT ................................................................................................................... ix
LISTA DE FIGURAS ................................................................................................... xv
LISTA DE TABELAS ................................................................................................ xvii
LISTA DE ABREVIATURAS .................................................................................... xix
I. Introdução ................................................................................................................... 1
I.1. Diversidade Fibrilar. ............................................................................................... 3
I.2. Espécies Reativas de O2 (EROs) e Exercício Físico .............................................. 6
I.3. Vias miogênicas para hipertrofia e capacidade oxidativa. ................................... 15
I.4. Modelos animais de treino resistido ..................................................................... 20
II. Objetivos e hipótese do Estudo ............................................................................... 25
III. Material e Métodos ................................................................................................ 26
III.1. Construção da escada com sistema de roldanas. ................................................ 26
III.1.1. Escada.......................................................................................................... 26
III.1.2. Suportes ....................................................................................................... 26
III.1.3. Angulação da escada ................................................................................... 27
III.1.4. Sobrecarga ................................................................................................... 28
III.1.5. Aparato ........................................................................................................ 29
III.2. Protocolo de Treinamento .................................................................................. 29
III.2.1. Animais ....................................................................................................... 29
III.2.2. Semana de Adaptação ................................................................................. 30
III.2.3. Protocolo de Treinamento ........................................................................... 31
III.2.4. Suplementação com óleo de arroz (Protocolo 2) ........................................ 32
III.3. Análises (Protocolo 1 e 2) .................................................................................. 34
III.3.1.Eutanásia, Coleta de Sangue e Tecidos ........................................................ 34
III.3.2. Lactato ......................................................................................................... 35
III.3.3. Creatina Quinase ......................................................................................... 35
III.3.4. Hemograma ................................................................................................. 35
III.3.5. Histologia .................................................................................................... 35

xii
III.3.6. Histoquímica ............................................................................................... 36
III.3.7. Enzimas do Metabolismo Energético (Citrato Sintase e Lactato
desidrogenase) ......................................................................................................... 36
III.3.9. Glicogênio ................................................................................................... 39
III.3.10. Peroxidação Lipídica (TBARs) ................................................................. 39
III.3.11. Análise Estatística e Apresentação dos Resultados................................... 40
IV. Resultados ............................................................................................................... 41
IV.1.Caracterização da escada com sistema de roldanas ............................................ 41
IV.2. Efeitos agudos da sessão de treino resistido ...................................................... 44
IV.3. Efeitos crônicos do protocolo de treinamento (Protocolo 1) ............................. 45
IV.4. A suplementação antioxidante produz efeito ergogênico adicional ao efeito do
treinamento? ................................................................................................................ 51
V. Discussão ................................................................................................................... 55
VI. Considerações Finais .............................................................................................. 66
VII. Conclusões ............................................................................................................. 67
Anexo A ......................................................................................................................... 68
Anexo B .......................................................................................................................... 79
VII. Referências Bibliográficas ................................................................................... 82

xiii
AGRADECIMENTOS
Primeiramente agradeço a orientadora deste estudo e da minha formação
desde iniciação científica, Prof(a). Dra. Denise Vaz de Macedo quem a partir da sala
de aula me recebeu, ensinou e me inseriu no universo da pesquisa e ensino,
propiciando o conhecimento de todas as pessoas que serão citadas. Agradeço ao Prof.
Dr. René Brenzikofer com seu jeito natural para fazer ciência teve papel fundamental
do início ao fim na execução deste estudo. Agradeço ao Prof. Dr. Barreto Martins que
ajudou na construção da escada e me recebeu gentilmente em sua casa com seus
equipamentos de marcenaria. Ao Prof. Dr. Alexandre de Oliveira por me receber em
seu laboratório e Prof. Dr. Gustavo Simoes pelos ensinamentos de bancada. A Prof(a)
Dra. Fernanda Lazarim pela orientação e formação profissional. A banca de
qualificação e defesa pelas discussões. Aos amigos feitos no LABEX: Juliana
Pascuotte parceira deste trabalho e outros, Flaviane Papaléo sempre disposta a ajudar
na bancada. Parceiros de discussões filosóficas, grupos de estudo e bandejão:
Bernardo Ide, Lúcio Muramatsu, Júlia Barreira, Leonardo Carvalho, Pedro Firmino,
Mauro Páscoa. Agradeço ao Prof. Dr. Rodrigo Hohl e Prof. Dr. Eduardo Bodnariuc
pela contribuição na minha formação fora e dentro do ambiente acadêmico. Agradeço
aos Professores Cesar Sartori, Rodrigo Ferraresso, Thiago Lourenço, Fernando
Catanho, Renato Buscariolli, Danilo Ferruci, Mirtes Stancanelli, Carolina Normand
cada um com sua contribuição em minha formação e no estudo. Ao CNPq, HT-Nutri
e FUNCAMP pelo financiamento.
E claro, agradeço pelos conselhos e acolhimento dos pais, irmãos e companheira.

xiv

xv
LISTA DE FIGURAS
Figura 1. A - Corte transversal do músculo Flexor longo do Hálux (FHL) de ratos Wistar corado com
Hematoxilina e Eosina (Objetiva 20x). B – Corte transversal do FHL corado pela técnica da
Nicotinamida Adenina Dinucleotideo tetrazolio Redutase (NADH-TR), que indica a capacidade
oxidativa das fibras de acordo com a intensidade da coloração (mais escura mais oxidativa)
(Objetiva 10x). Em destaque as fibras do tipo I (mais escuras) e fibras do tipo II (mais claras). ........ 3
Figura 2. Mecanismo simplificado dos potenciais indutores da resposta hipertrófica ...................... 6
Figura 3. Distribuição eletrônica de um átomo de Ferro com 4 elétrons desemparelhados na última
camada. ............................................................................................................................................... 7
Figura 4. Mecanismo de redução ímpar da molécula de oxigênio ..................................................... 8
Figura 5. Mecanismo de defesa antioxidante enzimático ................................................................ 10
Figura 6. Estrutura química dos fitoesteróis ferrúlicos (Patel & Naik, 2004) ................................... 14
Figura 7. Resumo das principais vias adaptativas para hipertrofia e capacidade oxidativa: As setas
verdes representam as vias que estão sendo estimulados e em vermelho as vias inibidas. Em azul
claro o princípio básico para resposta hipertrófica ocorrer.............................................................. 16
Figura 8. Rato treinando com sobrecarga aplicada através de tubos falcon (Sukho & Farrar, 2003).
........................................................................................................................................................... 24
Figura 9. Escada, suportes e materiais do sistema de roldanas ....................................................... 26
Figura 10. Triângulo o da angulação da escada ................................................................................ 27
Figura 11. Escada com sistema de roldanas onde pode ser visualizado a roldana 1 embaixo da
plataforma onde o animal chega após uma escalada e a rolda 2 no primeiro degrau da escada
abaixo da plataforma de sustentação da escada .............................................................................. 28
Figura 12. Materiais utilizados na sessão de treino .......................................................................... 29
Figura 13. Design experimental da divisão dos grupos suplementados e submetidos ao protocolo
de treinamento resistido em escada................................................................................................. 33
Figura 14. Figuras representativas de gráficos hipotéticos de Boxplot. A figura A apresenta todos
os dados fornecidos por um gráfico de boxplot. A figura B exemplifica uma condição onde não há
sobreposição dos intervalos de confiança dos boxplot (diferenças significativas entre as amostras
dos boxplots (95%)). Já a figura C exemplifica uma condição onde há a sobreposição dos intervalos
de confiança dos boxplots (ausência de diferenças significativas entre as amostras dos boxplots).
........................................................................................................................................................... 40
Figura 15. Situações do teste de atrito. A - Baldes em equilíbrio apenas com as roldanas. B – Tubos
falcon na escada em equilíbrio com o balde. C – Tubos falcon na escada sem atrito dos degraus em
equilíbrio com o balde....................................................................................................................... 42
Figura 16. Retas de inclinação das cargas de incremento necessárias para tirar o sistema em
equilíbrio em cada carga fixa. Azul – Situação A. Vermelho Situação B. Verde Situação C. CA –
Coeficiente Angular; CL – Coeficiente Linear; R2 – Correlação linear ................................................ 43
Figura 17. Diferentes momentos da concentração (mmol/L) de lactato no sangue após uma sessão
de exercício resistido na escada (A) e da CK (U/L) nos momentos basal, 3 horas após a última
sessão da 4°Semana e 8° Semana (B). .............................................................................................. 44

xvi
Figura 18. Em azul a CM do grupo controle na primeira semana (1°S.C) na quarta semana (4°S.C) e
na oitava semana (8°S.C). Em verde se encontra a CM do grupo treinado na primeira semana
(1°S.Tr) na quarta semana (4°S.Tr) e na oitava semana (8°S.Tr). ...................................................... 45
Figura 19. (A) Erro-padrão da média da AST dos grupos controle (C) e treinado (Tr) e Histograma
da distribuição das AST de 100 fibras individuais de cada animal do grupo treinado (B) e controle
(C). *t-stutent test não pareado : Diferença estatística em relação ao controle (P<0,05) ........... 46
Figura 20. Corte transversal do músculo FHL de um animal do grupo Treinado e outro do controle,
corado com Hematoxilina e Eosina (Objetiva 20x). .......................................................................... 47
Figura 21. Concentrações de Glicogênio no FHL (mmol/kg tecido) do grupo C e Tr. ....................... 47
Figura 22. Atividade (U/g tecido úmido) da enzima Lactato Desidrogenase (A) e da Citrato Sintase
(B) no músculo FHL dos grupos controle (C) e treinado (Tr). ............................................................ 48
Figura 23. Corte transversal do FHL do grupo controle (A) e treinado (B)corado pela técnica de
NADH-TR (Objetiva 10x). ................................................................................................................... 49
Figura 24. Área de Secção Transversa das fibras oxidativas (C vs Tr) e das fibras glicolíticas (C vs Tr).
*, # t-stutent test não pareado: Diferença estatística em relação ao controle (P<0,05) .............. 49
Figura 25. Atividade (U/g tecido úmido) das enzimas antioxidantes SOD, CAT e GR e peroxidação
lipídica (nnamol TEP/g tecido úmido) nos grupos controle (C) e treinado (Tr). ............................... 50
Figura 26. Desempenho (CM) ao longo das semanas de treinamento dos grupos água (azul),
suplementado com 0.75 ml de óleo de arroz (verde) e 1.5 ml de óleo arroz (Vermelho). .............. 51
Figura 27. Área de secção transversa (AST µm2) e peso (gramas) do músculo FHL nos grupos
controle, e suplementados com água, 0.75 ml e 1.5 ml de óleo de arroz. *Kruskal-wallis test com
Dun’s post test: Diferença estatística em relação ao controle (P<0,05). ....................................... 52
Figura 28. Atividade (U/g tecido úmido) das enzimas antioxidantes SOD, CAT e GR e peroxidação
lipídica (nmol TEP/g tecido úmido) nos grupos controle, e suplementados com água e óleo de
arroz. * Kruskal-wallis test com Dun’s post test: Diferença estatística em relação ao controle
(P<0,05). # Kruskal-wallis test com Dun’s post test: Diferença estatística em relação ao grupo
água (P<0,05). .................................................................................................................................. 53
Figura 29. Atividade (U/g tecido úmido) da enzima Citrato Sintase (A), concentração de glicogênio
(mmol/kg tecido) (B) nos grupos controle e suplementados com água, 0.75 ml e 1.5 ml de óleo de
arroz. * Kruskal-wallis test com Dun’s post testDiferença estatística em relação ao controle
(P<0,05). ............................................................................................................................................ 54

xvii
LISTA DE TABELAS
Tabela 1. Composição do óleo de arroz ............................................................................................ 14
Tabela 2. Resumo das vantagens e desvantagens de cada modelo de hipertrofia. ......................... 22
Tabela 3. Semana de Adaptação ....................................................................................................... 30
Tabela 4. Exemplo do protocolo de treino ....................................................................................... 32

xviii

xix
LISTA DE ABREVIATURAS

1
I. Introdução
O sedentarismo ou a falta da prática de atividade física regular tem sido um dos
grandes problemas de saúde encontrado na sociedade moderna. Desde a origem do gênero
Homo a 2,4 milhões de anos atrás, nossos ancestrais passaram aproximadamente 84,000
gerações no período pleistoceno, dos caçadores-coletores. A característica de nômades e a
luta pela sobrevivência na busca de alimentos, água, e fuga de predadores tornavam o
exercício físico uma atividade diária (O’Keefe et al., 2010). As características do genoma
humano evoluíram durante milhares de anos através da seleção natural nesse período. A
taxa de mutação espontânea dos nossos genes é de 0,5% a cada milhão de anos. Ou seja,
nos últimos 10.000 anos nossos genes mudaram 0.005 %. Nossas características inatas
continuam praticamente as mesmas dos nossos ancestrais da idade da pedra, que tinham
uma rotina diária de atividade física (Lieberman, 2013; Trembley et al., 2008).
As revoluções agrícola, industrial e tecnológica mudaram drasticamente o ambiente
em que vivemos com uma maior oferta de alimentos associado a diminuição da necessidade
do homem se movimentar em busca de alimentos. Diversas doenças estão relacionadas à
associação entre hipocinética humana e má alimentação como diabetes, doenças
cardiovasculares (hipertensão, arteriosclerose, infarto do miocárdio) e doenças
neurodegenerativas. Nesse contexto, exercícios resistidos progressivos que induzem
hipertrofia muscular, tem sido utilizados como terapia não medicamentosa contra
osteoporose, acidente vascular cerebral (AVC), doenças cardiovasculares, doenças
metabólicas e musculares. (Layne et al., 1999; Weis et al., 2000; McCartney, 1999;
Tollback et al., 1999).
Já na Grécia antiga os gregos estavam atentos aos benefícios da prática de
atividades físicas. Hipócrates e Galeno foram os primeiros “fisiologistas experimentais” a
pensar no exercício físico de maneira sistematizada. Já utilizavam treinos resistidos e
pensavam em suas consequências no organismo. Exerceram grande influência no
pensamento da civilização ocidental e, consequentemente, na fisiologia do exercício
(Mcardle & Kacth, Kacth 2008).
O exercício físico sob o ponto de vista fisiológico e bioquímico pode ser
caracterizado como um estímulo perturbador da homeostase do organismo, pois gera

2
depleção de substrados energéticos e microtraumas nas fibras musculares dentre vários
outros efeitos, que sinalizam a resposta adaptativa. Principalmente quando o período de
recuperação entre os estímulos é respeitado e adequado, as fibras musculares se regeneram
dos microtraumas e os níveis de atividade enzimática (Ex. lactato desigrogenase e citrato
cintase) e de substratos metabólicos (Ex. glicogênio e foscreatina) são restabelecidos acima
dos limites detectados antes do treinamento, propiciando no próximo estímulo, por
exemplo, uma maior disponibilidade de ATP, nossa moeda energética (Mcardle & Katch,
Katch 2008; Weineck, 2003)
Incrementos nas capacidades de força, potência, e/ou resistência refletem
funcionalmente as adaptações oriundas do treinamento físico direcionado para o
desenvolvimento de determinada capacidade física (Weineck, 2013).
O princípio da sobrecarga é uma das principais variáveis manipuladas no
treinamento resistido para gerar hipertrofia. Um exemplo de sobrecarga pode ser a
intensidade empregada durante o exercício, que deve ser suficiente para provocar alterações
agudas nas estruturas celulares envolvidas com o movimento realizado. Estudos recentes
têm mostrado que o volume ou número de repetições e séries também desempenham um
papel estressor no músculo suficiente para gerar uma resposta hipertrófica semelhante a
intensidade (Burd, et al 2012). Outras variáveis do treinamento como o controle entre as
séries e exercícios, a velocidade de execução do movimento, frequência de treinamento e
tempo de descanso entre os treinos também estão envolvidas com a resposta hipertrófica
(Kramer et al., 2002).
O exercício resistido para ser realizado com alta intensidade deve ser de curta
duração. Isso porque há alta taxa de hidrólise de ATP em um curto período, com grande
contribuição do metabolismo anaeróbico para ressíntese do ATP. A elevada contração
muscular, resulta em pequeno dano tecidual nas fibras musculares. Essas características
metabólicas podem ser evidenciadas, respectivamente, pelo aumento nas concentrações de
lactato muscular e sanguíneo, e pelo aumento das concentrações plasmáticas da enzima
creatina quinase (CK) em diferentes momentos após o exercício (Baird et al., 2012). Para
que o treinamento resistido seja executado por um tempo prolongado há a necessidade dos

3
Figura 1. A - Corte transversal do músculo Flexor longo do Hálux (FHL) de ratos Wistar
corado com Hematoxilina e Eosina (Objetiva 20x). B – Corte transversal do FHL corado
pela técnica da Nicotinamida Adenina Dinucleotideo tetrazolio Redutase (NADH-TR), que
indica a capacidade oxidativa das fibras de acordo com a intensidade da coloração (mais
escura mais oxidativa) (Objetiva 10x). Em destaque as fibras do tipo I (mais escuras) e
fibras do tipo II (mais claras).
esforços serem intercalados por pausas durante as sessões de treino, além da pausa
regenerativa necessária entre as sessões.
Os mecanismos moleculares envolvidos na resposta hipertrófica são de ordem
multifatorial. Além do estresse mecânico gerado durante as contrações e do dano muscular
(microlesões), também fazem parte desse quadro o estresse metabólico, a resposta
hormonal (sinalizações anabólicas) e a disponibilidade de nutrientes. Esse conjunto de
sinais é responsável pelas alterações na plasticidade muscular em resposta ao treinamento
resistido (Schoenfeld, 2013). Refletem as adaptações no nível estrutural (aumento da massa
muscular – que envolve síntese de proteínas contráteis (actina e miosina), citoesqueleto e
aumentos na síntese de enzimas metabólicas, e adaptações no nível neural, com aumento do
recrutamento das fibras musculares acompanhado de maior inervação das fibras musculares
pelos motoneurônios (Fluck, 2003).
I.1. Diversidade Fibrilar.
Apesar das fibras musculares terem uma aparência uniforme com seus núcleos
periféricos e membranas de tecidos conjuntivo em volta (Endomísio e Perimísio) (Figura-
1A) elas são muito heterogêneas em respeito a tamanho, metabolismo e capacidade
contrátil. As fibras musculares foram divididas classicamente em fibras de contração lenta
(Tipo I /escuras) e fibras de contração rápida (Tipo II/claras) (Figura-1b) (Pette & Staron,
2001)

4
Com base na expressão das diferentes isoformas de cadeia pesada da miosina (MHC
(myosin heavy chain)), com a utilização de diferentes modelos de estudos (i.e. animais
(ratos, camundongos, porcos e coelhos) e humanos), e de técnicas de imunohistoquímica
com anticorpos monoclonais, histoquímica pela atividade ATPásica (i.e. velocidade de
hidrólise de ATP na cabeça da miosina) em diferentes pHs (ácido e alcalino), e técnicas de
eletroforese em gel de poliacrilamida (SDS-PAGE) as fibras foram divididas nas chamadas
fibras “puras” do Tipo I, IIa, IId(x) e IIb (Pette & Staron, 2001).
As fibras do tipo I possuem atividade ATPásica mais lenta, e menor velocidade de
contração, assim como uma baixa frequência de disparo de potenciais de ação pelos
motoneurônios que as inervam (10 a 20 Hz). Possuem uma maior concentração de cálcio
intracelular (100 a 300 nM). São mais resistentes a fadiga, conseguindo permanecer por
mais tempo em contração. Por isso são mais vascularizadas e possuem um metabolismo
oxidativo mais elevado, com um maior conteúdo mitocondrial (Rhonda et al. 2006; Wessel
et al. 2010).
As fibras do Tipo IId(x) e IIb possuem alta atividade ATPásica e,
consequentemente, uma maior velocidade de contração. Por isso são denominadas de fibras
de contração rápida. Os motoneurônios que inervam essas fibras possuem uma maior
frequência de disparo dos potenciais de ação (100 – 150 Hz) e menor concentração de
cálcio intracelular (<50nM), por possuírem uma captação de cálcio pelo retículo
sarcoplasmático mais eficiente (Rhonda et al. 2006; Wessel et al. 2010). Estas fibras
necessitam da contribuição do metabolismo anaeróbico para obtenção de ATP, através da
via glicolítica e da fosfocreatina. Essas fibras possuem um tamanho maior (Figura-1B) e
são poucos resistentes a fadiga, justamente por utilizarem uma grande quantidade de ATP
para produzir força em um curto período de tempo. Dessa forma, são utilizadas para
situações onde precisamos de potência (Potência=Trabalho/tempo) para realizar o
movimento (Weineck, 2003).
A fibra do Tipo IIa possui uma particularidade interessante. Ela é considerada uma
fibra de contração rápida, pois seus motoneurônios apresentam alta taxa de disparo de
potencial de ação comparada com os motoneurônio da fibra do tipo I. No entanto possui
tamanho e capacidade oxidativa semelhante a fibra tipo I quando comparada com as fibras

5
do tipo IId e IIb. Ou seja, a fibra do Tipo IIa é considerada uma fibra de característica
intermediária entre as fibras de contração lenta e rápida (Weineck, 2003; Wessel et al.
2010).
Existem ainda as fibras musculares chamadas fibras híbridas, que apresentam duas
ou mais isoformas de MHC (Exemplo: MCHI+MCHIIa (TipoI/IIa); MCHIIa+MCHIId/x
(Tipo IIAD)). Essas fibras permitem a conversão das fibras de características mais
oxidativas em glicolíticas ou vice versa, dependendo do estímulo (Pette & Staron , 2001):
(TipoITipoI/IIATipoIIATipoIIADTipoIIDTipoIIDBTipoIIB)
De acordo com o princípio do tamanho das unidades motoras postulado por
(Henneman, 1965) o recrutamento neural das fibras musculares responde a intensidade do
exercício. Intensidades mais baixas recrutam as fibras do tipo I, sendo necessárias
intensidades mais altas para o recrutamento das fibras do tipo II. Essa proposta vai de
acordo com a frequência de disparos de potenciais de ação e velocidade de contração das
fibras musculares. Ou seja, são necessárias intensidades mais altas para contrações mais
rápidas.
A capacidade do tecido muscular se adaptar aos estímulos produzidos pelo ambiente
já está bem documentada na literatura. O treinamento físico gera alterações fisiológicas,
bioquímicas e morfológicas que alteram a constituição do tecido muscular, fenômeno
conhecido como plasticidade muscular. Muitos dos processos de remodelação do tecido
têm início com a chegada do potencial de ação, que permite alterações nas células
musculares como o influxo de cálcio, a velocidade de contração muscular e o estresse
mecânico, além de consumo de ATP e estresse metabólico. A alteração no fenótipo é
dependente da ativação de vias intracelulares de sinalização de síntese proteica, da secreção
de hormônios anabólicos e fatores de transcrição gênica, levando a alterações fenotípicas na
massa muscular, capacidade contrátil e metabolismo energético otimizado (Pette & Staron
RS, 2001; Rhonda et al. 2006).

6
Figura 2. Mecanismo simplificado dos potenciais indutores da resposta hipertrófica
A hipertrofia muscular deve ser considerada distinta e diferente da hiperplasia.
Durante a hipertrofia ocorre o aumento de estruturas contráteis e da matriz celular para
suportar este crescimento. Na hiperplasia ocorre o aumento no número de fibras musculares
por inserção de novos mionúcleos (células satélite) e posterior diferenciação (Vierck et al.
2000). A hiperplasia tem sido demonstrada em modelos animais (Kelley, 1996) e em
humanos. No entanto, a real contribuição da hiperplasia para o aumento da área de secção
transversa (AST) do músculo em humanos carece de mais estudos (Schoenfeld, 2010). A
Figura 2 apresenta os potenciais indutores da resposta hipertrófica.
I.2. Espécies Reativas de O2 (EROs) e Exercício Físico
Toda a matéria do universo é composta por átomos constituídos basicamente de um
núcleo (prótons e nêutrons) e elétrons orbitando em volta deste núcleo, segundo o modelo
proposto por Niels Bohr e aprimorado por Erwin Shӧrdinger, com o conceito de orbital
atômico como uma região do espaço onde a probabilidade de encontrar o elétron é maior
(Ohara, 2006).
Para o átomo se manter estável o número de elétrons (cargas negativas) em um
átomo é igual ao número de prótons (cargas positivas). Em cada orbital atômico se
encontram apenas 2 elétrons com spins opostos que são chamados de emparelhados. Spin
representa uma propriedade do elétron de contínua rotação da partícula sobre o seu próprio
eixo, semelhante ao um giro de pião. Essa rotação gera um campo magnético onde sua
força de atração é que permite duas partículas carregadas negativamente se encontrarem no
mesmo orbital (Halliwell, & Gutteridge, 2007). Os orbitais atômicos são divididos em
subcamadas de energia, definido pela distribuição eletrônica de Linus Pauling, que aumenta

7
Figura 3. Distribuição eletrônica de um átomo de Ferro com 4 elétrons desemparelhados
na última camada.
o nível de energia a medida que se distancia do núcleo. Cada subcamada designadas pelas
letras s,p,d, e f, comportam 1,3,5, e 7 orbitais respectivamente (Loewenstrein, 2013).
Os radicais livres são átomos ou moléculas que possuem elétrons com spins no
mesmo sentido na última subcamada, e por isso ocupam orbitais diferentes e são chamados
de desemparelhados. Esta situação faz com que a molécula seja instável. Para se tornar
estável irá roubar elétrons (oxidar) de outras moléculas para completar seus orbitais,
promovendo a oxidação de diferentes macromoléculas como proteínas, lipídios de
membrana e bases nitrogenadas da dupla hélice do DNA. Dependendo da concentração de
oxidantes pode alterar muito o estado redox celular e levar a perda da função. Importante
salientar que existem outros átomos como o Fe, Fe2+,Fe+3 que possuem elétrons
desemparelhados, mas não se comportam como radicais livres (Halliwell, & Gutteridge,
2007, Ohara, 2006). A Figura 3 abaixo representa a distribuição eletrônica do átomo de Fe,
onde ele apresenta 4 elétrons desemparelhados na última subcamada d.
No início da formação da Terra a cerca de 2 bilhões de anos atrás a evolução dos
seres fotossintetizantes permitiu o aparecimento do oxigênio na atmosfera terrestre. O O2
possui uma particularidade interessante. Apesar de estável, a molécula de O2 no seu estado
fundamental se encontra em uma situação denominada oxigênio triplete: com dois elétrons
desemparelhados na última camada com spins no mesmo sentido. Essa condição, conhecida
como restrição de spin impede que a redução da molécula de O2 seja através do

8
recebimento de um par de elétrons, como acontece na maioria das outras moléculas, mas
sim recebendo um elétron de cada vez (Pryor et al, 2006). A consequência disso é que até a
completa redução do O2 a H2O é possível formar as Espécies Reativas de Oxigênio (EROs).
A redução monoeletrónica da molécula de oxigênio (radical ânion superóxido) foi
demonstrada pela primeira vez pela cientista argentina Rebeca Gerschman em 1954. A
produção do radical ânion superóxido leva a produção de outras espécies reativas,
coletivamente chamadas de EROS, de acordo com as reações apresentadas na Figura 4.
Quando o oxigênio recebe um elétron forma o radical ânion superóxido (O-.). Este quando
recebe mais um elétron e 2 H+ forma o peróxido de hidrogênio (H2O2), que pode receber
outro elétron e formar o radical hidroxila (.OH), que finalmente quando recebe um elétron e
um próton é reduzido a água (Ohara, 2006).
Figura 4. Mecanismo de redução ímpar da molécula de oxigênio
Apesar do peróxido de hidrogênio (H2O2) não entrar na definição de um radical livre
ele é considerado da família das EROs, pois tem facilidade para atravessar membranas
lipídicas. Quando recebe um elétron geralmente proveniente do Fe2+ ou Cu+, através da
reação de Fenton gera o radical hidroxila, que possui uma meia vida muito curta sendo,
portanto, altamente reativo. Outro problema é que não há defesa enzimática contra essa
espécie (Balaban, 2005).
Fe2+/Cu+ + H2O2 Fe3+/Cu2+ + OH- + OH
.
Na produção mitocondrial de ATP o O2 molecular é o aceptor final de elétrons das
reações de reoxidação das coenzimas reduzidas NADH e FADH2 produzidas pelo
funcionamento do ciclo de Krebs. A reoxidação das coenzimas ocorre na cadeia de
transporte de elétrons (CTE), com produção de H2O além das coenzimas NAD+ e FAD
como produto final. A energia gerada pelo funcionamento da CTE (reações de óxido
redução) alimenta o bombeamento de prótons pelos complexos mitocondriais I, III e IV, da

9
matriz para o espaço intermembranas, gerando um gradiente eletroquímico de H+ na
membrana mitocondrial. A volta dos H+ através da porção Fo da enzima ATP sintetase
favorece a fosforilação do ADP em ATP.
O O2 se comporta como aceptor final dos elétrons no complexo IV mitocondrial,
mas pode ser reduzido monoeletronicamente nos complexos I e III, produzindo O2.- (Liu et
al, 2002). Há muito tempo foi demonstrado que cerca de 2 a 5 % do oxigênio consumido na
respiração celular são reduzidos a EROs (Boveris, 1972).
Durante o exercício físico o consumo de O2 intramuscular aumenta cerca de 100
vezes, elevando a produção EROs (Davies et al, 1982) demonstrada através de mensuração
direta da produção de EROs por ressonância paramagnética, que detecta os elétrons
desemparelhados através de seu comportamento sob um campo magnético; ou através de
sondas fluoroscentes e fluorógrafos (Bloomer, 2008; Silveira, 2004).
Sistema de Defesa Antioxidante
Nosso organismo possui um elaborado sistema de defesa antioxidante enzimático
(ex. catalase (CAT), superóxido dismutase (SOD), sistema glutationa peroxidase
(GPx)/glutationa redutase (GR), e não enzimático (ex. glutationa reduzida, ácido úrico,
vitaminas antioxidantes (A, C, E), polifenóis, ácidos graxos poliinsaturados), que atuam no
combate ao excesso das EROs produzidas (Halliwell, & Gutteridge, 2007)
A primeira defesa contra o ânion superóxido (O2-) é a reação catalisada pela enzima
superóxido dismutase (SOD), que dismuta o radical superóxido a H2O2 e H2O.
Posteriormente a enzima catalase (CAT) catalisa a reação de redução do H2O2 em H2O e O2
(Ferreira e Matsubara, 1997). A enzima glutationa peroxidase (GPx) também tem
capacidade de detoxificar o H2O2 com a participação da glutationa reduzida (GSH), que
funciona como doador de elétrons na reação. Quando a GSH é oxidada nesta reação, ocorre
a formação da glutationa oxidada (GSSG). Diminuição nos níveis de GSH podem
prejudicar o sistema de defesa antioxidante, e consequentemente a defesa celular contra a
ação das EROs. Para que isso não ocorra a enzima glutationa redutase (GR) as custas de
NADPH reduz a GSSG formada para sua forma reduzida GSH, mantendo desta forma a

10
razão GSH/GSSG alta, preservando as defesas celulares (Smith, 1988). A Figura 5
apresenta um esquema da defesa antioxidante enzimática.
Figura 5. Mecanismo de defesa antioxidante enzimático
Estresse Oxidativo
Estresse oxidativo foi definido pela primeira vez pelo alemão Helmut Sies (1991)
como uma situação de maior produção de oxidantes em relação a capacidade da defesa
antioxidante.
Diversas são as doenças que parecem estar relacionadas com situações de estresse
oxidativo como câncer, doenças cardiovasculares, diabetes, alzheimer, e envelhecimento
(Vollard et al. 2005). O cientista Denham Harman foi um dos primeiros a propor que o
envelhecimento estava relacionado com maiores ataques oxidativos as moléculas orgânicas
do nosso organismo, e que isso poderia ser prevenido com suplementação de antioxidantes
(Harman, 1956).
Os estudos relacionados ao estresse oxidativo e exercício físico na sua maioria
utilizam exercícios de característica aeróbia, ou músculos isolados em modelos animais e
seres humanos, onde a mitocôndria é apontada como a principal fonte de EROs. Estudos
realizados de maneira aguda (logo após a sessão de exercício) mostraram aumento em
marcadores de ataque oxidativo, contribuindo para o conceito que o exercício físico poderia
gerar um quadro de estresse oxidativo (Powers e Jackson 2008).
No entanto, os estudos sobre o efeito crônico do exercício físico demonstraram que
o exercício protege de doenças relacionadas ao estresse oxidativo (Hamilton, 2003;

11
Hawkins, 2003). Nesse sentido, já foi demonstrado que as EROs são sinalizadores das vias
da AMPK (Mitogen-activated protein kinase) e da NFKB (fator nuclear kappa B), que
controlam a expressão gênica de enzimas antioxidantes, proteínas de reparo de DNA,
proteínas de estresse e proteínas da cadeia de transporte de elétrons (Ji et al. 2002; Pattwell
& Jackson, 2007).
Como parece necessário haver um equilíbrio sutil entre a produção de EROs e a
atividade da defesa antioxidante para a resposta adaptativa, surgiu uma teoria para os
efeitos do treinamento baseada no conceito da Hormesis, que discute a necessidade de
exposição a pequenas doses de agentes tóxicos para gerar uma adaptação. Esta teoria
transposta para os radicais livres propõe que a exposição a doses ligeiramente aumentadas
de EROs podem contribuir para o aumento da atividade das enzimas antioxidantes e outras
adaptações benéficas. Se ultrapassada essa produção ótima de EROs instaura-se o estresse
oxidativo, levando a perda de função (Reid, 1992; Radak et al, 2005).
Um dos danos celulares que ocorre com frequência em virtude do estresse oxidativo
é a oxidação de lipídeos das membranas, processo conhecido como peroxidação lipídica,
através de uma série de reações em cadeia, com participação das EROs e oxigênio. Essas
reações incorporam quantidades anormais de oxigênio nos lipídios, aumentando a
vulnerabilidade da célula e seus componentes, podendo levá-la a morte (Halliwell &
Chirico, 1993).
Fontes de EROs no treinamento resistido
A principal fonte de EROs em exercícios resistidos parece estar relacionada com
sua produção, principalmente no pós-exercício (Fisher-Wellman & Bloomer, 2009). Como
já mencionado o treinamento resistido gera um estresse mecânico e dano nas fibras
musculares, cujo reparo e regeneração demandam a instalação de um processo inflamatório.
Neste processo são secretados eicosonóides oriundos do ácido aracdônico, que sinalizam o
aumento da permeabilidade do endotélio vascular na região da inflamação, facilitando o
influxo de células inflamatórias (Tidball, 2005).
Uma das primeiras células inflamatórias a migrar para o tecido lesionado são os
neutrófilos polimorfonucleados, que possuem função de remoção por fagocitose de

12
elementos indesejáveis relacionados a lesão tecidual através da formação de EROs pela
ação da enzima mieloperoxidase e através da ação da enzima NADPH oxidase, em um
processo conhecido como “Burst Respiratório” (Tidball, 2005).
Outra situação relacionada ao dano tecidual promovido principalmente pela fase
excêntrica do movimento durante a contração muscular é a desregulação da homeostase do
cálcio e proteólise de proteínas com ferro na sua estrutura. Com o aumento da
disponibilidade desses elementos nas células aumenta a probabilidade do O2 receber um
elétron, produzindo O2-. e H2O2 (Hellsten, 1999).
A produção de EROs durante o exercício resistido também pode aumentar pela via
da xantina oxidase, enzima responsável pela degradação do AMP, produto da reação
catalisada pela enzima mioquinase (MK) para produção de ATP. A característica de
isquemia/reperfusão promovida pelas contrações intensas intercaladas por pausas durante o
treino resistido; as altas concentrações intracelulares de cálcio e a baixa concentração de O2
fazem com que a enzima xantina desidrogenase sofra conversão para a forma oxidase.
Dessa forma, durante a reperfusão a xantina oxidase utiliza o oxigênio ao invés de NAD+
como aceptor de elétrons, produzindo o O2-. (Hellsten, 2000; Sjödin & Hellsten, 1990).
Outra via que contribui para produção de O2-., é a via da enzima NADPH oxidase,
uma enzima localizada na membrana das células. Em situações de estresse metabólico essa
enzima doa os elétrons do NADPH para o oxigênio produzindo O2-.. Essa via está ativa
quando as concentrações intracelulares de AMP estão aumentadas e parece ser uma via de
produção de EROs fundamental para a sinalização celular (Jiang, & Zhang, 2011).
Estresse Oxidativo e Treino Resistido
Resultados apresentados na literatura têm mostrado aumento em marcadores de
ataque oxidativo em resposta a um protocolo agudo de treinamento resistido (Demici, et al
2010; Zembron-Lacny et al 2006; Hudson et al, 2008), principalmente em protocolos que
enfatizam a fase excêntrica do movimento (Paschalis et al, 2007). Também existem estudos
que não demonstraram alterações (Hellsten, 1997; Ramel et al, 2004) e estudos com
treinamento resistido feito de maneira crônica que mostraram proteção contra o estresse
oxidativo (Parise et al, 2005; 2005; Cakir-Atabek et al, 2010).

13
Essa variação de respostas pode ser devida principalmente a variedade de
protocolos, tipo de contração muscular enfatizado (ações dinâmicas
(concêntrica/excêntrica), isométrica, ou excêntrica), técnicas de análises, tecidos e
momentos de coletas utilizados. Alguns protocolos podem não ter promovido um estímulo
estressor suficiente para gerar uma resposta, ou a técnica utilizada não ser tão específica.
Também o momento de coleta pode não ter sido o melhor, fazendo com que o tecido
(sangue, músculo, saliva) não refletisse o ataque oxidativo ocorrido. Ou seja, quando não
são detectadas alterações em marcadores de ataque oxidativo não significa que elas não
ocorreram, mas por essas limitações não foram mensuradas. Outra situação que pode
interferir nos resultados realizados com humanos é a dieta do indivíduo, uma vez que
diversos alimentos apresentam compostos com propriedades antioxidantes (Fisher-
Wellman & Bloomer, 2009).
Suplementação antioxidante
A suplementação esportiva na atividade física tem o objetivo de contribuir para uma
melhora adicional de desempenho devido a melhor adaptação aos estímulos do treino e
proteção contra lesões indesejadas. A principal teoria por trás da suplementação
antioxidante no esporte é a capacidade de determinadas moléculas sequestrar ou neutralizar
radicais livres, o que preveniria o estresse oxidativo. Há uma considerável propaganda a
respeito da suplementação com antioxidantes na comunidade esportiva, fazendo com que
diversos atletas e treinadores, profissionais ou amadores busquem evitar os danos causados
pelo estresse oxidativo através de suplementação seja através de dietas ricas em nutrientes
com função antioxidante ou produtos industrializados (Yanfat et al., 2009).
O óleo de arroz possui em sua composição antioxidantes da família dos tocoferóis e
tocotrienóis (Vitamina E), ácidos graxos poli-insaturados (PUFAs) (ômegas-6 e 3) e ácidos
graxos saturados (SAFA) e monoinsaturados (MUFA) (Cheruvansky & Thumala,1991),
conforme apresentado na Tabela 1.

14
Figura 6. Estrutura química dos fitoesteróis ferrúlicos (Patel & Naik, 2004)
Tabela 1. Composição do óleo de arroz
Compostos Óleo de arroz
SAFA 20 a 22,6 %
MUFA 37 a 41,0%
PUFA-n6 34,0 a 34,5%
PUFA-n3 1,8% a 2,2 %
Antioxidantes 3 a 5 %
A propriedade antioxidante da Vitamina E e ácidos graxos poli-instaturados está
bem documentada na literatura (Ristow, 2009; Richard et al, 2008). Outro componente
importante dentre os antioxidantes presentes no óleo de arroz é o gamma-oryzanol, que
inicialmente foi considerado o componente único presente no óleo de arroz. Posteriormente
foi caracterizado como sendo um misto de éster ferrúlico de vários esteróis álcoois
triterpenos chamados α, β, γ-oryzanol. Sendo este último o nome mais utilizado. O γ-
oryzanol possui 3 fitoesteróis ferrúlicos em maior concentração: cycloartenyl ferulate
(presente em maior quantidade), 24-methylenei cycloartanyl ferulate e campesteryl
ferulate, todos com uma unidade de ácido ferrúlico em sua estrutura (Islam et al, 2011)
(Figura 6).
A propriedade antioxidante do γ-oryzanol foi demonstrada in vitro com uma
atividade antioxidante quatros vezes maior que a da vitamina E (Kim et al 1995; Hiramitsu
& Armstrong, 1991). Sua capacidade antioxidante parece estar relacionada com a
capacidade do grupamento hidroxila presente na unidade do ácido ferrúlico de doar
hidrogênio para radicais livres, semelhante à ação dos flavonoides e polifenóis (Patel &

15
Naik, 2004). Estudos demonstraram também o γ-oryzanol age na inibição do NF-KB tanto
in vitro (Nagasaka et al 2007) quanto ex vivo (Islam et al, 2008). Um estudo mostrou
diminuição de colesterol plasmático, propondo sua utilização em tratamento de
hiperlipidêmicos (Gerhardt & Gallo, 1998). Outros estudos sugerem ainda a utilização do γ-
oryzanol em treinamento resistidos (Fry et al, 1997; Eslami et al, 2014), sendo que existe
um crescente mercado de suplementação de γ-oryzanol para praticantes do fisiculturismo
(Grunewald & Bailey, 1993).
No entanto, embora exista uma quantidade significativa de componentes
antioxidantes no óleo de arroz, e um mercado crescente da utilização deste óleo como
produto alimentício, principalmente nos países asiáticos (Japão, China, India) (Islam et al,
2011), não encontramos nenhum estudo reportando os efeitos da suplementação de óleo de
arroz associado ao treinamento resistido.
I.3. Vias miogênicas para hipertrofia e capacidade oxidativa.
Para ocorrer o processo hipertrófico é necessário um balanço positivo na razão entre
síntese e degradação de proteínas, no sentido de aumento na síntese de proteínas do
citoesqueleto (microfilamentos e microtubos) e das proteínas contráteis (actina, miosina),
aumentando consequentemente a área de secção transversa, associado a uma diminuição
nas vias de degradação proteica (Toigo & Boutellier, 2006).
A Figura 7 resume as principais vias de sinalização para gerar a resposta
hipertrófica, assim como a melhora da capacidade oxidativa muscular. Destaque para a
participação dos hormônios IGF-1, sinalizado pelo Hormônio do Crescimento (GH), da
Insulina e Testosterona, das EROs, do íon cálcio citoplasmático (Ca2+) e da Adenosina
Monofosfato (AMP).
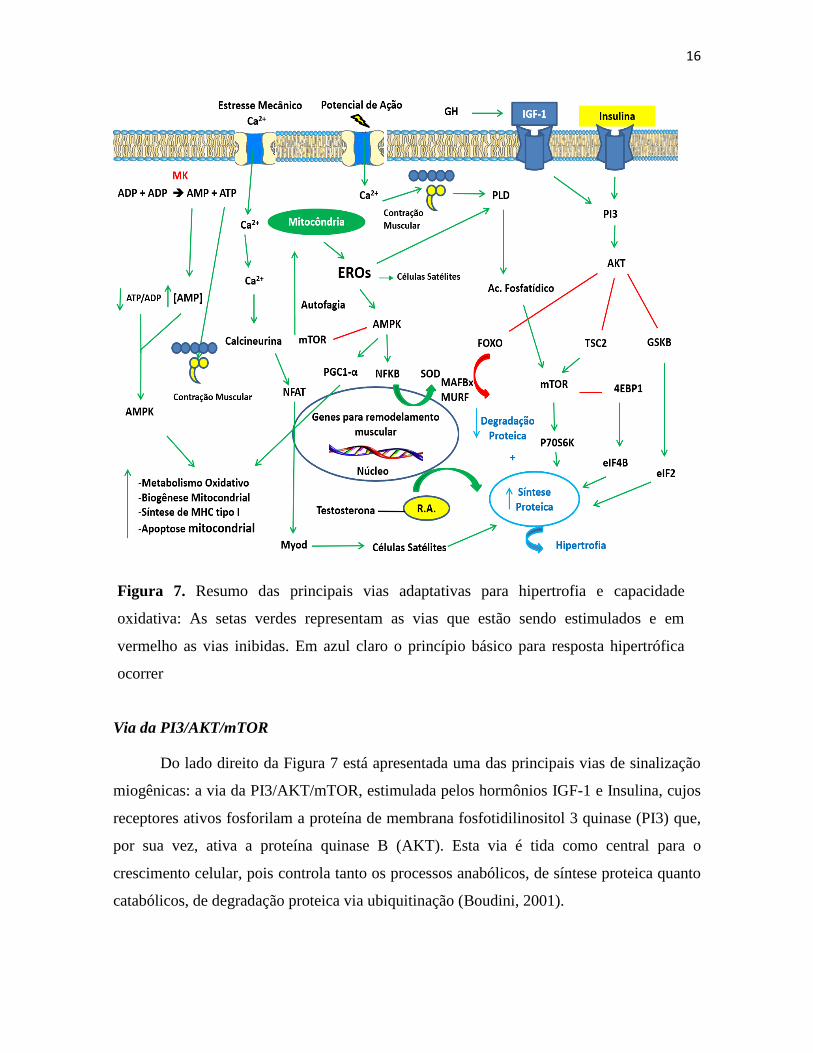
16
Figura 7. Resumo das principais vias adaptativas para hipertrofia e capacidade
oxidativa: As setas verdes representam as vias que estão sendo estimulados e em
vermelho as vias inibidas. Em azul claro o princípio básico para resposta hipertrófica
ocorrer
Via da PI3/AKT/mTOR
Do lado direito da Figura 7 está apresentada uma das principais vias de sinalização
miogênicas: a via da PI3/AKT/mTOR, estimulada pelos hormônios IGF-1 e Insulina, cujos
receptores ativos fosforilam a proteína de membrana fosfotidilinositol 3 quinase (PI3) que,
por sua vez, ativa a proteína quinase B (AKT). Esta via é tida como central para o
crescimento celular, pois controla tanto os processos anabólicos, de síntese proteica quanto
catabólicos, de degradação proteica via ubiquitinação (Boudini, 2001).

17
A ativação da síntese proteica e concomitante inibição da degradação proteica
sinalizada através da via PI3/AKT é regulada pela fosforilação e consequente inativação
das enzimas GSK3β (glicogen Synthase Kinase 3β), TSC2 (Tuberin) e FOXO (forkhead
transcription factor) (Hornberger et al, 2007).
A GSK3β quando inativa permite a ativação do fator eIF2 (eucarotic initiation
factor 2), envolvido na ligação do RNA transportador a subunidade 40s do ribossomo, que
favorece a tradução de proteínas. A proteína TSC2 inativada permite a ativação da mTOR.
E a FOXO quando inativada sai do núcleo da célula, impedindo a ativação de fatores de
transcrição MAFBx (Muscle Atrophy F-box) e MURF (Muscle ring finger), que sinalizam a
síntese de proteínas relacionadas a atrofia muscular via sistema de ubiquitinação e
proteassomos (Welsh et al, 1998, Philippou et al, 2007).
A proteína mTOR (mammalian target of rapamycin) é uma proteína com atividade
serina/treonina quinase de 300 kDa de massa. Foi descoberta em 1991 como uma proteína
alvo do antifungo rapamicina, produzido por uma bactéria de solo descoberta em 1970 na
ilha da Páscoa (Heitman, et al 1991). A mTOR possui duas subunidades (mTORC1 e
mTORC2). A mTOR é responsável pelo controle do crescimento celular através da inibição
do mecanismo de autofagia mitocondrial, e concomitante ativação da transcrição e tradução
gênica em resposta a oferta de nutrientes (Ex. Leucina). Nesse sentido, tem sido muito
estudada em mecanismos relacionados ao câncer, longevidade e doenças neurogenerativas
(Jung, et al 2010).
A estimulação da síntese proteica ocorre quando as enzimas 4E-BP1 e p70s6k estão
fosforiladas. A responsável por isso é a proteína mTOR. A fosforilação da p70s6k leva a
uma hiperfosforilação da S6K, que favorece aumento da tradução do RNA de proteínas
ribossomais e fatores de alongamento. Já a 4E-BP1 quando em repouso se encontra
associada com um fator de iniciação eucariótico denominado eIF4B. Ao ser fosforilada se
dissocia, liberando o fator de iniciação, permitindo com isso o início da tradução (Bodini,
2006).
Estudos mostraram que a inibição da mTOR por rapamicina leva a inibição de até
95% da hipertrofia, demonstrando o papel fundamental da mTOR e das proteínas 4E-BP1 e
p70S6K no processo de hipertrofia muscular (Hornberger, et al., 2007).

18
Estresse Mecânico
A proteína mTOR também pode ser ativada independente da sinalização pela via da
AKT, através do ácido fosfatídico produzido pela ação da enzima fosfolipase-D (PLD). O
estresse mecânico provocado pela elevada contração muscular em um treino resistido leva a
alterações na estrutura do sarcômero e rompimento de proteínas do citoesqueleto. Na
situação de repouso a fosfolipase D se encontra associada, e inibida pela proteína α-actinina
na linha Z (ponto crítico para transmissão de força) do sarcômero. O rompimento da linha Z
pelo estresse mecânico ativa a enzima PLD, que utiliza os fosfolipídios de membrana para
produção do ácido fosfatídico. Este sinaliza a ativação da mTOR (Hornberger et al, 2006).
É sabido que o estresse mecânico gerado pelo alongamento crônico das fibras
musculares também estimula canais sensíveis ao alongamento, e tem capacidade de
produzir novos sarcômeros em série e contribuir para a hipertrofia, assim como aumentar a
permeabilidade da membrana a entrada do Ca2+ (Zollner et al. 2012). Diversas moléculas
mecano-sensíveis como proteínas de citoesqueleto, integrinas, canais iônicos e fatores de
transcrição gênica estão relacionadas com o remodelamento do tecido muscular. Além
disso, a tensão provocada durante a contração muscular em treinos resistidos causa um
distúrbio na integridade celular que sinaliza para fatores de crescimento, células satélites e
proteínas quinases (Schoenfeld, 2010).
Estresse Metabólico
O estresse metabólico e sua sinalização para vias de hipertrofia no músculo são
oriundos principalmente da contração muscular e da necessidade de ressíntese de ATP
durante um período maior de estímulo. Isto leva a um acúmulo de metabólitos
intracelulares que sinalizam para diferentes mecanismos celulares como recrutamento
neural, resposta hormonal, ativação de proteínas quinases locais, inchaço celular e produção
de EROs (Schoenfeld, 2013).
No centro da Figura 7 está representada a mitocôndria, como uma das principais
fontes de EROs (Balaban et al, 2005). As EROs estimulam a AMPK, uma proteína com
atividade serina-treonina quinase constituída de uma subunidade catalítica (α) e duas
subunidades reguladoras (β, γ) (Cheung et al. 2000). A proteína AMPK é considerada um

19
sensor metabólico da célula, sendo ativada por alterações no estado energético (↓
ATP/AMP) e estado redox (reações de óxido- redução) da célula (Kyriakis & Avruch ,
2001).
Com o acúmulo de ADP intracelular devido ao estresse metabólico a enzima
mioquinase se torna ativa e a tem capacidade de utilizar duas moléculas de ADP para gerar
um ATP e um AMP (adenosina monofosfato). Quando as concentrações intracelulares de
AMP aumentam ocorre sua ligação na subunidade gama da AMPK, permitindo a
fosforilação de proteínas histonas de DNA, expondo os genes e permitindo a atuação dos
fatores de transcrição gênica que também são fosforilados pela AMPK (Cheung et al 2000).
Dentre os fatores de transcrição gênica que são estimulados pela AMPK se
encontram o PGC1-α (Peroxisome proliferator-activated receptor-gamma) e o NFKB. O
PGC1-α aumenta a expressão de enzimas do metabolismo oxidativo, promovendo também
a biogênese mitocondrial. A proteína NFKB estimula a expressão da enzima antioxidante
SOD, além dos genes de atrofia MAFBx e MURF (Powers & Jackson, 2009).
Como já foi discutido o AMP também pode contribuir para o aumento na formação
de radical anion superóxido quando é degradado a ácido úrico, através da via da Xantina
Oxidase. Por sua vez, o radical ânion superóxido também tem capacidade para sinalizar
outras vias adaptativas (Sjödin, 1990). Foi proposto que as EROs ativam a enzima
fosfolipase D, contribuindo para gerar hipertrofia em músculo cardíaco (Tapia et al, 2006).
Também foi demonstrado a participação das EROs na ativação de células satélites (CS)
(Vierck, 2001).
Células Satélites (CS)
As CS são caracterizadas como uma população de células com alta atividade
mitogênica, que contribuem para o processo de crescimento celular pós-natal, reparo de
fibras musculares danificadas e manutenção da integridade músculo-esquelética de adultos
(Hawke & Garry, 2001). O princípio básico de atuação das CS é que elas promovem a
inserção de novos mionúcleos no citosol das fibras musculares, aumentando,
consequentemente a síntese proteica e o domínio nuclear (relação entre volume do citosol e
quantidade de núcleos) (Petrella et al, 2006). No repouso as CS se encontram naturalmente

20
no estado de quienescência (inativas). A contribuição das CS para o processo de hipertrofia
depende de dano tecidual para a sua ativação. Diferentes fatores têm sido relacionados com
a ativação das CS após o dano tecidual provocado pelo estresse mecânico, como a elevação
de Ca2+ intracelular e ativação da calcineurina, o MGF (muscle Growth factor), HGF
(heptocyte growth factor) uma glicoproteína liberada pelo músculo através de uma via
sinalizada por óxido nítrico, e a interleucina-6 (IL-6) produzida pela infiltração de
neutrófilos e macrófagos no tecido danificado durante o processo de inflamação (Silva &
Macedo, 2011).
Após a ativação as CS passam a se chamar de mioblastos. Esses irão passar ainda
pelos estados de proliferação (aumento do número) e diferenciação aonde podem se inserir
nas fibras danificadas gerando novos núcleos, contribuindo para a síntese de proteínas para
regeneração do tecido. Ou podem se fundir com outras CS e se diferenciar em novas fibras,
contribuindo para o fenômeno de hiperplasia (Hawke & Garry, 2001).
Via da Calcineurina/Calmodulina
A calcineurina é uma proteína heterodímera com atividade fosfatase composta por
duas subunidades, uma catalítica ligada a calmodulina, e outra subunidade reguladora que
se liga ao cálcio. Ela se comporta como um sensor da atividade contrátil sensível a
flutuações de Ca2+ (Rhonda et al. 2006). Quando Ca2+ se liga na calmodulina ativa a
calcineurina, que irá desfosforilar o NFAT (Nuclear factor of activated T cells). O NFAT
desfosforilado migra para o núcleo da célula onde irá ativar outros fatores de transcrição
gênica como o MEF2 (Myocite ehancer factor-2), Mygenin e MyoD relacionados com
expressão de proteínas para o remodelamento muscular e com a ativação das células
satélites (Sakuma & Yamaguchi, 2010).
I.4. Modelos animais de treino resistido
Os modelos animais de investigação científica têm sido fundamentais para a
compreensão dos mecanismos moleculares, celulares e metabólicos da resposta hipertrófica
(Carson, 1997; Booth et al., 1998). Assim como para programas específicos de atividade
física na prevenção e tratamento de doenças como osteoporose, AVC, doenças

21
cardiovasculares, doenças metabólicas (Ex. Diabetes) e musculares (sarcopenia)
(Hornberger & Farrar, 2004).
Alguns questionamentos são feitos se é possível transpor as interpretações feitas
nestes modelos para humanos. Não podemos esquecer que a evolução científica da área de
biológicas ocorreu muito por conta da utilização de modelos animais. Dentro de uma
perspectiva evolutiva somos todos parentes dos seres vivos presentes no planeta, alguns
mais próximos e outros separados por milhões de anos da escala da árvore genealógica.
Hipócrates, Galeno, Aristóteles e outros cientistas aprenderam muito sobre o
funcionamento dos órgãos humanos estudando por comparação em modelos animais os
fenômenos biológicos como circulação, respiração e nutrição. No campo da medicina
diversos fármacos foram e continuam a ser descobertos se utilizando de modelos animais
para intepretação dos mecanismos básicos por trás de fenômenos biológicos, assim como
teste destes fármacos que contribuíram para erradicação de diversas doenças (Foster et al,
1983).
Diferentes modelos de treinamento em modelo animal com o objetivo de gerar
hipertrofia muscular esquelética estão descritos na literatura. Dentre eles encontramos
modelos de agachamento (Tamaki et al 1992), eletroestimulação (Baar & Esser, 1999),
mecanismo compensatório de sobrecarga (Degens et al, 1995), estiramento crônico
(Goldspink et al 1995), e treinamento em escada (Hornberger & Farrar, 2004). A Tabela 2
apresenta sucintamente alguns desses modelos, resumindo suas vantagens e desvantagens
para a interpretação de resultados mais próximos da hipertrofia induzida em humanos.

22
Tabela 2. Resumo das vantagens e desvantagens de cada modelo de hipertrofia.
Modelos de treino Vantagens Desvantagens
Agachamento
(Ho et al 1980; Tamaki et al
1992; Klitgaard 1988)
Simula movimento e permite
controle fino das variáveis de treino
semelhante a humanos.
Tempo para resposta hipertrófica
semelhante a humanos.
Condicionamento do animal com
choque ou sistema de recompensa.
Não tem músculo contralateral
Mão-de-obra do pesquisador
Treinamento em escada
(Lee & Farrar, 2003;
Hornberger & Farrar 2004;
Duncan et al, 1998;
Yarasheski et al 1990)
Simula movimento e permite
controle fino das variáveis de treino
semelhante a humanos.
Tempo para resposta hipertrófica
semelhante a humanos
Condicionamento do animal com
choque ou sistema de recompensa.
Não tem músculo contralateral
Mão-de-obra do pesquisador
Eletroestimulação
(Baar & Esser, 1999; Walters
et al, 1999; Wong et al, 1988)
Músculo contralateral disponível
Independente da motivação do
animal
Controle fino das variáveis de
treino.
Repetidas anestesias
Princípio do tamanho não é seguido.
Mudanças fenotípicas na fibra
muscular (IIaIIb)
Compensatório de sobrecarga
(Goldberg, 1967; Degens et al,
1995; Baldwin et al, 1982)
Músculo contralateral disponível
Pouca mão-de-obra para o
pesquisador
Resposta rápida de hipertrofia
Estímulo para induzir hipertrofia
diferente de humanos.
Tempo para hipertrofia diferente de
humanos
Interferência cirúrgica
Estiramento Crônico
(Antonio & Gonyea, 1993;
Goldspink et al 1995)
Músculo contralateral disponível
Pouca mão-de-obra para o
pesquisador
Não cirúrgico
Estímulo para induzir hipertrofia
diferente de humanos.
Tempo para hipertrofia diferente de
humanos

23
Treinamento em escada
Este modelo de treinamento consiste em submeter os animais (ratos) a uma escalada
carregando uma sobrecarga presa na base da cauda, utilizando uma escada apoiada,
geralmente em uma angulação de 80°. Em cada escalada o animal realiza de 8 a 12
movimentos, correspondentes a repetições em seres humanos. Diferentes protocolos de
treino em escada demonstraram hipertrofia muscular (Sukho & Farrar, 2003; Hornberger &
Farrar 2004; Duncan et al, 1998; Yarasheski et al 1990). Os estudos diferem entre os
músculos analisados, as análises realizadas, características do protocolo de treinamentos e
nos métodos de condicionamento para o animal realizar a escalada.
A vantagem deste modelo de treinamento é que os animais realizam as escaladas
conscientes, o número de séries, repetições, intensidade e pausa podem ser controlados
rigorosamente. O movimento realizado pelo animal se assemelha funcionalmente aos
treinos resistidos realizados por humanos, com as ações dinâmicas musculares (contrações
concêntricas e excêntricas), inclusive ações isométricas durante a determinação de carga
máxima para prescrição de intensidade de exercício. Os estudos com esse modelo animal
têm mostrado que a magnitude e o tempo de resposta para hipertrofia (8 semanas) dos
ganhos hipertróficos e de força são semelhantes aos treinos resistidos realizados por
humanos (Sukho & Farrar, 2003; Hornberger & Farrar 2004).
Uma crítica a este modelo é a metodologia utilizada para condicionamento do
animal para realizar a escalada. Alguns autores utilizam choque no período de adaptação
(Sukho & Farrar, 2003; Hornberger & Farrar 2004). Isso pode induzir um estresse adicional
nos animais e acarretar flutuações hormonais que podem confundir os resultados (Lowe &
Always, 2002). Outros autores utilizam comida como recompensa (Yarasheski et al 1990),
o que obviamente não ocorre em um treinamento com humanos. Com os animais fazendo o
exercício motivados com sistemas de recompensa (comida) ou punição (choque) eles irão
fazer o mínimo de exercício possível para satisfazer sua necessidade de comida ou evitar o
choque, que usualmente faz com que eles consumam menos comida (Lowe & Always,
2002). Isso pode refletir em um menor ganho de peso e crescimento relacionado ao grupo
controle, justificando alguns trabalhos em relativizar a massa do músculo pela massa do
animal (Duncan et al, 1998). Outra desvantagem desse modelo de treino resistido é que não

24
Figura 8. Rato treinando com sobrecarga aplicada através de tubos
falcon (Sukho & Farrar, 2003).
permite a análise do músculo contralateral, impondo a utilização de grupos controle para as
comparações.
Em nossa opinião, outra limitação do modelo em escada é o método de aplicação da
sobrecarga feita através de tubos falcon presos na cauda do rato. A carga é colocada dentro
dos tubos, e o animal realiza a escalada carregando esses tubos em atrito com os degraus da
escada (Figura 8). Esse fato pode aumentar a variabilidade das respostas adaptativas ao
treino.

25
II. Objetivos e hipótese do Estudo
Objetivo Geral:
O objetivo da presente dissertação de mestrado foi fabricar a escada com um sistema de
roldanas para aplicação da sobrecarga, para mimetizar condições semelhantes aos treinos
em humanos, uma vez que o músculo esquelético é muito sensível as variáveis de
treinamento (sobrecarga, repetições, pausas). Outro objetivo foi avaliar os efeitos de oito
semanas de treinamento em escada com e sem suplementação antioxidante.
Objetivos específicos:
- Caracterizar resposta do atrito em um experimento com a adição de carga na roldana em
comparação com a adição de cargas nos tubos falcon.
- Não utilizar sistemas de recompensa como comida ou choque para condicionamento do
animal.
- Analisar o desempenho (carga máxima) e marcadores morfológicos (área de secção
transversa), metabólicos (lactato, CK. Glicogênio, enzimas lactato desidrogenase e citrato
sintase) e de estresse oxidativo (atividade de enzimas antioxidantes SOD, CAT e GR e
(TBARs) no sangue e no músculo flexor longo do hálux (FHL) em ratos divididos em
grupo controle e treinado (Protocolo 1).
- Investigar a possível resposta ergogênica da suplementação antioxidante (óleo de arroz)
nos mesmos marcadores apresentados acima (Protocolo 2).
- Testar a reprodutilidade do protocolo de treino resistido em escada com sistema de
roldanas com outro grupo de animais.
Nossa hipótese foi que o sistema de roldanas permitiria gerar uma sobrecarga mais
homogênea, sem o atrito provocado pelos sistemas com tubos normalmente utilizados,
capaz de gerar respostas semelhantes ao um treino resistido. Assim como uma possível
resposta ergogênica com a suplementação de óleo de arroz.

26
III. Material e Métodos
III.1. Construção da escada com sistema de roldanas.
A escada foi construída nas dimensões propostas pelo modelo de treinamento
resistido proposto por Hornerberg Jr. e Farrar (2004), com alteração no modo de aplicação
da sobrecarga e algumas adaptações nos materiais e condições da sala de treinamento
descritas abaixo.
III.1.1. Escada
Escada vertical com 1,1 metros de comprimento e 0,18 metros de largura, e com
2cm entre os degraus. Foi construída utilizando madeira envernizada (laterais) e fios de
cobre encapados (degraus). Os cortes na madeira foram mensurados com uma régua e feitos
com um serra Tico-Tico 450W – Black&Decker KS455.
III.1.2. Suportes
A escada se encontra apoiada em uma angulação de 80 graus em duas plataformas
de madeira envernizada: uma fixada em um banco e outra na parede da sala, através de
suportes para estante. Em ambas as plataformas foram feitos cortes para o encaixe da
escada, onde na plataforma inferior um suporte de madeira suporta a escada através do
degrau. Duas caixas de plástico de 20x20x20 cm foram colocadas sob as plataformas
(Figura 9).
Figura 9. Escada, suportes e materiais do sistema de roldanas

27
Os ratos são colocados na base da escada e ao completar a escalada ficam pelo
tempo proposto para a pausa na caixa superior. Na caixa superior foi adicionado um piso
emborrachado (Figura 9), pois verificamos em estudos pilotos que os animais escorregavam
na caixa ao chegar ao final de escalada, o que prejudicava a sua última repetição.
Importante destacar que nos estudos com o modelo de escada apresentados na literatura a
caixa superior é fechada e escura para diminuir o estresse do animal e facilitar que o animal
realize a escalada e fique na caixa. No entanto em nosso estudo não vedamos a caixa, pois
os animais eram mantidos em ciclo invertido no biotério e o treino era realizado em
ambiente escuro.
III.1.3. Angulação da escada
Foi utilizada uma angulação de 80° como proposto na literatura, que era regulada a
cada sessão de treino. A angulação da escada é um procedimento importante neste
protocolo de treinamento, pois interfere diretamente na força que o animal irá desempenhar
durante sua escalada. Em angulações menores o animal irá produzir menos força. Para
regular a angulação utilizamos uma régua presa na escada e uma linha na forma de pêndulo
(Figura 10) para formar um triângulo, conforme mostrado na figura 10:
Figura 10. Triângulo o da angulação da escada
Neste esquema de um triângulo a escada representa a hipotenusa (x), o pêndulo o
cateto oposto ao ângulo, e o cateto adjacente corresponde a régua. Fixamos um valor de 11
cm para o cateto adjacente e calculamos o cosseno de 80° para achar o valor de x=63,36
cm. Esta foi a distância marcada na escada entre a fixação da régua e do pêndulo. Desta
forma, toda sessão de treino o pêndulo era acertado em cima dos 11 cm na régua e
garantíamos que escada se encontrava em 80°.

28
III.1.4. Sobrecarga
A aplicação da sobrecarga foi feita através de um sistema de roldanas. Para isso
foram utilizadas duas roldanas de plástico com raio de 3,3 cm, sulco (onde passa a linha) de
0,5 cm e orifício central (0,7cm) onde passa a haste (22cm) (Figura 9). Sendo uma haste
com a roldana colocada abaixo de uma plataforma aonde o rato chega ao realizar a escalada
(roldana 1), e outra na base da escada (roldana 2), conforme mostrado na figura 11.
Uma linha (barbante de nylon) passa pelas duas roldanas, sendo uma presa a base do
rabo do rato por um aparato, e outra em um balde suspenso, onde são colocados os
chumbos (Carga) para produzir sobrecarga. Ao realizar uma escalada o rato levanta o balde
por uma altura de 0,9 metros, correspondente ao seu deslocamento na escada e realiza de 8
a 12 movimentos, que são equivalentes as repetições em seres humanos, como pode ser
visualizado na figura 11.
Figura 11. Escada com sistema de roldanas onde pode ser visualizado a roldana 1
embaixo da plataforma onde o animal chega após uma escalada e a rolda 2 no primeiro
degrau da escada abaixo da plataforma de sustentação da escada

29
Figura 12. Materiais utilizados na sessão de treino
III.1.5. Aparato
Consiste em um velcro com uma argola presa, que é utilizada para fixar o gancho
preso na linha que passa pelas roldanas. Um pedaço de esparadrapo era utilizado toda
sessão de treino para fixar o aparato na cauda do rato (3 cm da parte proximal). Uma
tesoura era utilizada com cuidado para tirar o esparadrapo após a sessão de treino. Os
materiais utilizados no treinamento e o aparato podem ser visualizados na figura 12.
III.2. Protocolo de Treinamento
III.2.1. Animais
Foram utilizados 30 ratos (Protocolo 1) e 20 ratos (Protocolo 2) da linhagem Wistar
com 21 dias de vida, adquiridos do Centro de Bioterismo da Unicamp. Todos os ratos
foram alimentados com ração e água ad libitum, sendo mantidos em ambiente com a
temperatura controlada entre 18 e 22ºC e fotoperíodo de 12 horas com ciclo de luz claro-
escuro invertido. Os ratos com massa = 362.6±20.7 g (Protocolo 1) e massa = 343,3±24
(Protocolo 2) iniciaram a semana de adaptação com 60 dias de idade, correspondente a
idade de maturação (Andreollo et al, 2012). Todos os procedimentos adotados foram
aprovados pelo Comitê de Ética em Experimentação Animal (2625-1 Protocolo 1) e (3037-
1 Protocolo 2).

30
III.2.2. Semana de Adaptação
Esta semana consistiu em 5 dias consecutivos de treinamento, no qual os animais
eram colocados na base da escada e estimulados com leves pressões com a pinça na base da
cauda a realizar 4 escaladas, conforme protocolo apresentado na Tabela 3.
Tabela 3. Semana de Adaptação
Para facilitar a familiarização dos animais com a escada, nos três primeiros dias os
animais foram colocados em diferentes pontos da escada durante as escaladas: no meio, na
base e próximo do ponto de chegada, representando tentativas de média e fácil dificuldade.
A semana de adaptação teve o objetivo de separar os animais que sobem a escada
voluntariamente e realizar o Teste de Carga Máxima (TCM) na última sessão da semana.
Assim, foram escolhidos para fazer parte do estudo somente aqueles animais que
realizaram o 4º dia de treino voluntariamente (n=28). O TCM iniciou com a primeira
escalada com um peso correspondente a 75% da massa do animal, após análise subjetiva do
desempenho do animal na última escalada classificadas em: Fácil quando o animal
realizava a escalada sem parar na escada, média quando o animal realizava escala parando
algumas vezes, ou difícil quando o animal parava e precisava de um estímulo com a mão
para completar a escalada. Desta forma eram adicionados 10 gramas se escalada foi difícil,
20 gramas para média e 30 gramas para a difícil, a cada escalada até que o animal não
conseguisse completar a escalada após três ajudas com a mão ou não suportasse se manter
na escada. Os animais realizavam de 2 a 4 repetições para atingir a carga máxima (CM).
Isso é muito importante para que o início do protocolo de treinamento seja realizado com as
intensidades prescritas baseadas em percentuais da CM atingida no TCM. Após esta última
sessão os animais foram separados aleatoriamente em grupo controle (C) (n=10) e treinado
(Tr) (n=18) e separados nas caixas alocadas no biotério em controle e treinado.
1° Dia 2° Dia 3°Dia 4°Dia 5°Dia
4 escaladas
sem carga e
com pausa de
30 segundos
4 escaladas
sem carga e
com pausa de
30 segundos
4 escaladas
apenas com o
balde preso
4 escaladas
com 150g e 2
minutos de
pausa
Teste de carga
máxima (TCM)

31
III.2.3. Protocolo de Treinamento
O protocolo de treinamento consistiu de 8 semanas de treinamento com 3 sessões de
treino por semana (Segunda, Quarta e Sexta-feiras), totalizando 24 sessões de treino. Os
animais realizavam quatro escaladas com 50%, 75%, 90% e 100% da carga máxima
atingida no teste de carga máxima, com 2 minutos de pausa entre as escaladas em cada
sessão de treino. Toda sexta-feira era realizado um novo teste de carga máxima para
compor uma nova carga máxima a ser utilizada no treino da semana seguinte. Somente na
7° e 8° semanas de treino foi realizado teste de carga máxima também na segunda-feira,
visto que os animais apresentavam sinais qualitativos (realizando o treino de maneira fácil)
de adaptação ao treino. Os treinos foram realizados com dois animais por vez, onde cada
animal possuía seu balde com a carga e tempo de pausa individualizados, sendo necessário
apenas trocar o balde no suporte das roldanas e marcar no esparadrapo do aparato o rato
para diferencia-los.
O TCM iniciava após o animal cumprir as 4 escaladas previstas na sessão de treino,
com a adição de 10 a 30 gramas até o animal atingir a sua nova CM, verificada quando o
animal se recusava a subir após 3 ajudas com a mão, ou não conseguia se manter na escada.
A maioria dos animais realizou em média de 6 a 8 escaladas na 3° sessão da semana.
É importante salientar que todos os treinos foram realizados no período da manhã
com a luz da sala apagada, onde apenas um abajur estava ligado para realizar as anotações
no caderno de treino. Além disso, embora o estímulo de ajuda com a mão fosse o método
prioritário para estimular o animal a subir a escada, a pinça sempre esteve presente na sala e
foi utilizada em situações esporádicas quando alguns animais se recusavam a treinar.
No caderno de anotações eram anotadas as cargas individuais de cada animal e
observações decorrentes do treino, como por exemplo, um animal que precisou do estímulo
da pinça na sessão, não conseguiu completar a última escalada sem ajuda, ou não estava
acompanhando as cargas de treino e vinha repetidamente realizando sessões de treino ruins.
Com base nestas anotações e comportamento repetidos como estes, seis animais que
iniciaram o treinamento foram excluídos por não realizarem o treinamento voluntariamente
após os incrementos de carga, ou porque se recusavam a treinar mesmo com estímulo.
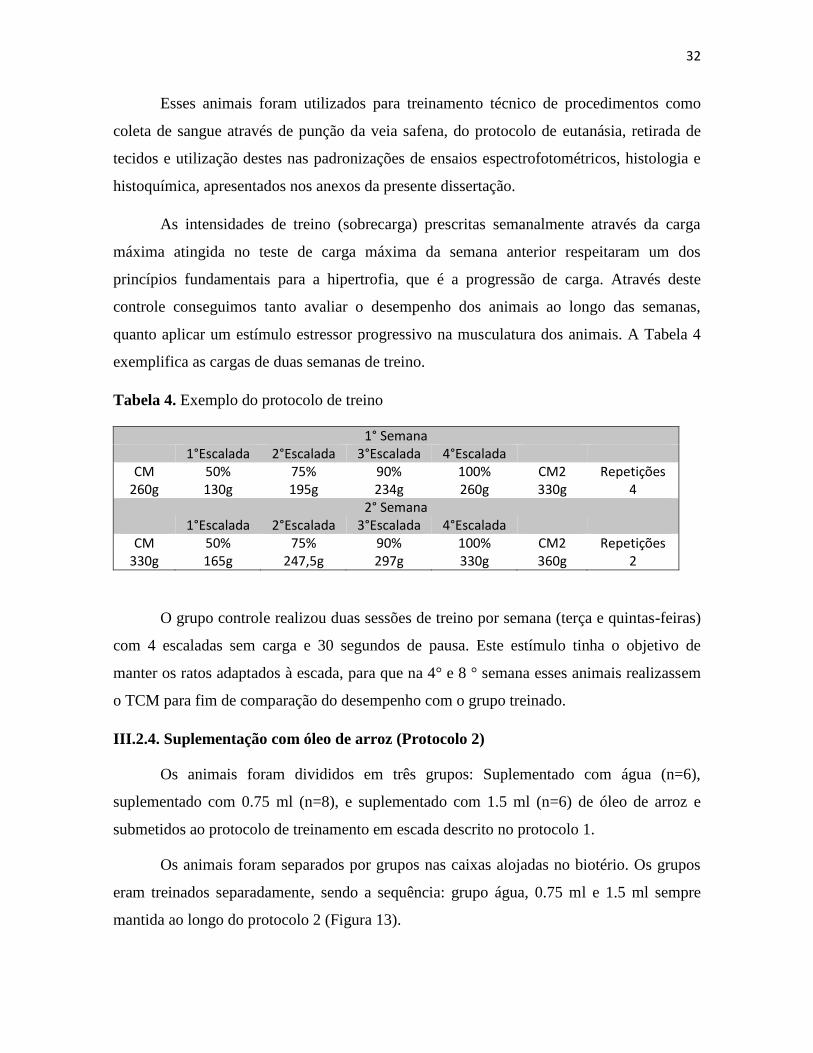
32
Esses animais foram utilizados para treinamento técnico de procedimentos como
coleta de sangue através de punção da veia safena, do protocolo de eutanásia, retirada de
tecidos e utilização destes nas padronizações de ensaios espectrofotométricos, histologia e
histoquímica, apresentados nos anexos da presente dissertação.
As intensidades de treino (sobrecarga) prescritas semanalmente através da carga
máxima atingida no teste de carga máxima da semana anterior respeitaram um dos
princípios fundamentais para a hipertrofia, que é a progressão de carga. Através deste
controle conseguimos tanto avaliar o desempenho dos animais ao longo das semanas,
quanto aplicar um estímulo estressor progressivo na musculatura dos animais. A Tabela 4
exemplifica as cargas de duas semanas de treino.
Tabela 4. Exemplo do protocolo de treino
1° Semana 1°Escalada 2°Escalada 3°Escalada 4°Escalada
CM 50% 75% 90% 100% CM2 Repetições 260g 130g 195g 234g 260g 330g 4
2° Semana 1°Escalada 2°Escalada 3°Escalada 4°Escalada
CM 50% 75% 90% 100% CM2 Repetições 330g 165g 247,5g 297g 330g 360g 2
O grupo controle realizou duas sessões de treino por semana (terça e quintas-feiras)
com 4 escaladas sem carga e 30 segundos de pausa. Este estímulo tinha o objetivo de
manter os ratos adaptados à escada, para que na 4° e 8 ° semana esses animais realizassem
o TCM para fim de comparação do desempenho com o grupo treinado.
III.2.4. Suplementação com óleo de arroz (Protocolo 2)
Os animais foram divididos em três grupos: Suplementado com água (n=6),
suplementado com 0.75 ml (n=8), e suplementado com 1.5 ml (n=6) de óleo de arroz e
submetidos ao protocolo de treinamento em escada descrito no protocolo 1.
Os animais foram separados por grupos nas caixas alojadas no biotério. Os grupos
eram treinados separadamente, sendo a sequência: grupo água, 0.75 ml e 1.5 ml sempre
mantida ao longo do protocolo 2 (Figura 13).

33
Figura 13. Design experimental da divisão dos grupos suplementados e
submetidos ao protocolo de treinamento resistido em escada
A suplementação foi realizada através de gavagem 5 dias da semana (segunda à
sexta). Nos dias de treino a suplementação era realizada imediatamente após a sessão de
treino, e nos dias sem treino a suplementação era realizada no período da manhã no mesmo
horário das sessões de treino. Importante ressaltar que os animais foram pesados toda
segunda e sextas-feiras, e a dose de suplementação era ajustada de acordo com o peso do
animal para evitar que o animal quando ganhasse massa tomasse uma dose menor relativa a
sua massa que no início do protocolo.
O grupo suplementado com água teve o objetivo servir como grupo controle
treinado que também passou pelo estresse do estímulo da gavagem. As doses de 0,75ml e
1,5 ml óleo de arroz correspondem aproximadamente a 2 e 4 colheres de sopa para
humanos.
Cálculo da dose equivalente em humanos
A dose equivalente em humanos (HED) foi calculada segundo a equação abaixo
que leva em consideração a dose ingerida pelo animal relativizada pela sua massa (animal
dose) e as constantes de fator de conversão relativo área de superfície corporal do animal
(Animal Km factor) e humano (Human Km factor) (FDA, 2005).
HED (mg/kg) = animal dose (mg/kg) x Animal K m factor / Human K m factor
Exemplo do cálculo da dose e transformação para o número de colheres:
A densidade (m/v) do óleo de arroz é de 920 mg/ml. Portanto em 1,5 ml temos 1380
mg. Ao dividir 1380 pelo valor médio do peso dos ratos (0,3356 kg) encontramos a
quantidade administrada aos animais normalizado pelo peso (4112,03 mg/kg) que

34
corresponde a dose ingerida pelo animal relativa ao seu peso (Animal dose). Utilizando as
constantes do Km factor para ratos e humanos, 6 e 37 respectivamente (FDA, 2005), e
colocando na equação temos:
HED = 4112,03 x 6/ 37
HED = 666,81 mg/kg
Este valor corresponde a dose em mg de óleo por kg de peso humano.
Considerando um homem de adulto de 70 kg, ele irá ingerir 666,81 x 70 = 4667,67 mg ou
46 gramas de óleo de arroz nesta dose que também correspondem a 46 ml de óleo. Em uma
colher de sopa cabem 13 ml de óleo de arroz, portanto se dividirmos a dose ingerida pelo
número de colheres, temos: 46/13= 3,5 colheres correspondente a dose de 1,5 ml
suplementada nos animais. Se realizarmos o mesmo cálculo para a dose de 0,75 ml
chegaremos a um valor de 1,8 colheres correspondentes para humanos.
III.3. Análises (Protocolo 1 e 2)
III.3.1.Eutanásia, Coleta de Sangue e Tecidos
Os animais foram sacrificados 72 h após a última sessão de treino com o intuito de
garantir os efeitos crônicos do treinamento. Os mesmos foram anestesiados via
intramuscular na pata dianteira com Zoletil® (50mg/kg peso) em associação com o
relaxante muscular xilazina (10un/kg peso) (Machado et al, 2009). Após a anestesia e perda
dos reflexos de cauda, pálpebra e flexão plantar, 8-10 mL de sangue foram coletados do
ventrículo esquerdo em tubos heparinizados (Vaccuette Greiner Bio-one) através de punção
cardíaca.
O sangue foi imediatamente centrifugado (Centrífuga CT – 6000R) a 1800 g por 10
minutos a 4°C para separação do plasma e células sanguíneas. As hemácias foram lavadas
com solução gelada de Tampão Fosfato 0,1M e NaCl 0,9%, pH 7,4 e centrifugadas a 700g
por 10 minutos a 4°C. Em seguida o sobrenadante foi dispensado, e o procedimento
repetido por três vezes. A papa de hemácias foi hemolisada com água miliQ na proporção
1:1 (v/v). O plasma e o hemolisado foram armazenados a -80ºC para posterior análises.
O músculo Flexor Longo do Hálux (FHL) foi cuidadosamente dissecado das patas
traseiras direita para análise histológica e histoquímica, e da esquerda congelados em

35
nitrogênio líquido e armazenados a -80ºC para posterior análises de ensaios
espectrofotométricos utilizando o espectrofotômetro UV–VIS (Beckman DU-640
Spectrophotometer).
III.3.2. Lactato
Para análise do metabolismo utilizado durante a sessão de treino o sangue foi
coletado na oitava semana com os animais vivos, através de punção da veia safena da pata
traseira dos animais, nos momentos basal (anterior ao treino), logo após, 5min, e 10min
após o treino. O sangue coletado em capilares heparinizados era colocadoFoi fitas reagentes
para mensuração de Lactato da marca ROCHE em lactímetro Accusport Roche.
III.3.3. Creatina Quinase
A coleta de sangue para análise da enzima CK foi feita através da veia safena da
pata traseira dos animais nos momentos basal e 3 horas após a sessão de treino na 4° e 8°
semanas. O sangue foi posteriormente centrifugado em micro centrífuga CT-14000R
Cientex durante 5 minutos a 635 g. O sobrenadante foi congelado em N2 e armazenado. As
análises da CK foram realizadas em soro através de aparelho point of care Reflotron Plus®
(ROCHE®) através de fitas reativas específicas.
III.3.4. Hemograma
Uma alíquota de 200μl de sangue armazenada em eppendorfs com EDTA (20%) foi
analisada no aparelho automatizado Sysmex pocH-100iV-Diff, selecionando no software do
aparelho a calibração para ratos Wistar. Foram analisados os seguintes parâmetros:
quantidade de células vermelhas, leucócitos totais e plaquetas. Esses dados estão
apresentados no Anexo A.
III.3.5. Histologia
Para análise da área de secção transversa (AST-µm2) foram fixadas amostras do
ventre do FHL com um volume aproximadamente de 5mm3 em um pedaço de madeira de
cortiça com Optimal Cutting temperature - Tissue Teck Compound, e congelados por 30
segundos em n-hexano resfriado a -70°C. Posteriormente a esse procedimento foram
armazenados em nitrogênio liquído até serem levados para corte no criostato. 10 Cortes
histológicos seriados de cada músculo com 12 µm de espessura foram feitos em criostato a

36
-25°C e 5 cortes corados com Hematoxilina e Eosina. As imagens foram digitalizadas na
magnitude de 20x no Microscópio Nikon TS100 equipado com câmera digital (Nikon,
Tokyo, Japan, DXM1200i). Para o cálculo de aumento da AST do músculo, foram
mensuradas através do programa Image Pró-Plus 6.0 a AST individual de 100 fibras
musculares aleatórias de cada animal através de 6 fotos de campos aleatórios de um único
corte do músculo.
III.3.6. Histoquímica
Os 5 cortes seriados realizados no criostato restantes foram utilizados para medir a
capacidade oxidativa das fibras musculares através da técnica da Nicotinamida adenina
dinucleotideo tetrazolio redutase (NADH-TR), conforme (Novikoff, 1961). As lâminas
foram retiradas do biofreezer e deixadas em temperatura ambiente por 20 min. As lâminas
foram então colocadas em uma solução contendo NADH 0,45 mM, Nitro Blue Tetrazolium
(NBT) 1mM e tampão MOPs 10 mM por 30 minutos em banho maria 37°C. Nesta técnica
o NBT gera uma coloração escura nas fibras que consumirem mais o NADH, medindo sua
capacidade oxidativa e indiretamente o conteúdo mitocondrial. As imagens foram
posteriormente digitalizadas na magnitude de 10x no Microscópio Nikon TS100 equipado
com câmera digital (Nikon, Tokyo, Japan, DXM1200i). Foram mensuradas a AST de 100
fibras aleatórias oxidativas (escuras) e glicolíticas (claras) utilizando o programa image
pró-plus através de 4 fotos de campos aleatórios do corte transverso do músculo em um
grupo de animais Controle (n=3) vs Treinado (n=3).
III.3.7. Enzimas do Metabolismo Energético (Citrato Sintase e Lactato desidrogenase)
Ressaltamos que o cálculo das atividades enzimáticas, curva-padrão de todas as
enzimas e analitos e análise de confiança das técnicas estão apresentadas no Anexo A da
presente dissertação.
Preparação do Homogenato Muscular: As amostras do FHL (30 mg) foram preparadas
como originalmente descrito por Zerbetto et al (1997), com as modificações de Molnar et al
(2004). As mesmas foram homogeneizadas em 1ml de uma solução gelada contendo 20
mM de MOPS, 440 mM de sacarose, 1 mM de EDTA e 5 mM de PMSF em pH 7,2. Tais
amostras foram posteriormente centrifugadas em ultra-centrífuga a 20.000g por 20 minutos,

37
sendo o sobrenadante congelado em nitrogênio líquido e armazenado a -70°C para posterior
análises em ensaio enzimático espectrofotométrico.
Citrato Sintase
A Citrato Sintase foi quantificada através do método enzimático descrito por Srere
(1969). Utilizamos no ensaio 0,1 mL de DTNB (1mM), 0,03 mL de acetil Coa (10mM) e
um volume de amostra correspondente a 50 µL. A reação foi iniciada acrescentando-se 0,05
mL de oxaloacetato (OAA) (10mM), completando o volume final na cubeta (1mL) com
água. Mensurou-se a atividade da CS pela formação de TNB em comprimento de onda, no
espectrofotômetro, igual a 412 nm, conforme mostrado na reação abaixo:
Acetil-CoA + Oxaloacetato Citrato + Coa-SH
Coa-SH + DTNB Coa + TNB
Lactato Desidrogenase (LDH)
O cálculo da atividade da LDH foi realizado espectrofotometricamente com
alterações segundo (Ralph, 1965; Decker & Lohmann-Matthes, 1988). O princípio do
método consiste em acompanhar espectrofotometricamente a diminuição de absorbância do
NADH em comprimento de onda fixo de 340 nm segundo o esquema abaixo:
Piruvato +NADH Lactato + NAD+
No ensaio foi utilizado 990 uL de uma solução de trabalho com 50mM de Tampão-
Tris, 4mM de EDTA, 120 uM de NADH e 2 mM de piruvato, onde eram acrescentados 10
uL de amostra. O tempo de leitura foi de 1 minuto, determinado pela curva-padrão (Anexo
A).
III.3.8. Enzimas Antioxidantes (SOD, CAT, GR)
Preparação do homogenato de musculo para análises enzimáticas: O FHL (120 mg) foi
homogeneizado sobre banho de gelo com um homogeinezador Politron (Politron PT-MR
2100, Kinematica, Switzerland) durante 30s em velocidade máxima em 3 ml de solução
contendo K2HPO4 (39 mM), EDTA ácido (1 mM), sacarose (440 mM), pH 7,4 e 1 µL
LDH

38
PMSF (phenylmethylsulfonilfluoride) [200mM] na razão 1:25 (tecido/volume) (wt/vol).
Depois foi centrifugado a 790 g por 10 min a 4 C°. O sobrenadante foi retirado transferido
para micro tubos e congelados em nitrogênio líquido para posterior análises. Importante
ressaltar que durante as padronizações verificou-se a necessidade de retirar uma camada de
tecido conjuntivo intramuscular espessa do FHL antes da homogeneização. Caso contrário,
a amostra fica “suja” com muitos pedaços de tecido conjuntivo, que prejudica o caminho
ótico durante os ensaios enzimáticos e deixa o coeficiente de variação da absorbância
elevado.
Superóxido Dismutase. (SOD)
A SOD foi quantificada através do kit de análise da Trevigen ®. Nele os ânions
superóxido originários da conversão da xantina a ácido úrico e H2O2 pela xantina oxidase
(XOD) convertem NBT a NBT-diformazam, o qual absorve luz a 560nm. A SOD reduz a
concentração de O2- e desta forma inibe a taxa de formação de NBT-diformazam, sendo
este o parâmetro de avaliação da atividade da SOD na amostra, conforme pode ser
observado no esquema abaixo.
Cada leitura requer cerca de 5 minutos, sendo posteriormente realizado o cálculo da
atividade da SOD a partir do percentual de inibição da formação de NBT-diformazan
utilizando-se de uma curva padrão de inibição indicada pelo kit. A solução com a SOD
isolada com atividade conhecida é fornecida pelo fabricante do kit.
Catalase (CAT)
A CAT foi quantificada por meio do método enzimático descrito por Aebi (1984).
No ensaio a atividade enzimática é avaliada a partir da decomposição do peróxido de

39
hidrogênio a água, podendo essa ser determinada diretamente pela diminuição da
absorbância, em um comprimento de onda de 240nm:
H2O2 H20 + O2
Glutationa Redutase. (GR)
A GR foi quantificada através do método enzimático descrito por Smith et al
(1988). O procedimento padrão adotado para quantificação da GR foi seguir
espectrofotometricamente a formação de TNB a 412nm a partir do ácido 5,5’-
ditiobinitrobenzóico (DTNB) conforme esquema abaixo.
III.3.9. Glicogênio
A quantificação da concentração de glicogênio no FHL foi feita pelo método
colorimétrico proposto por Lo et al. (1970). Nele a glicose reage com o ácido fenol-
sulfúrico e no final do processo é obtida uma solução laranja, e a variação da intensidade da
cor é analisada por espectrofotômetro a 490 nm. O cálculo da concentração de glicogênio
(mmol/kg tecido) foi realizado partir de uma curva-padrão com quantidades conhecidas de
glicogênio, e está apresentado no Anexo A.
III.3.10. Peroxidação Lipídica (TBARs)
Homogenato de músculo: Amostras do FHL foram homogeneizadas sobre banho de gelo
com um homogeinezador Politron (Politron PT-MR 2100, Kinematica, Switzerland) na
razão 1:10 (tecido/vol) em solução de KCl (1,15%). Posteriormente as amostras foram
divididas em diferentes alíquotas, congeladas em N2 líquido e armazenadas em biofreezer -
70 °C.
Substâncias que reagem ao ácido tiobarbitúrico (TBARs)
O TBARs foi quantificado conforme os métodos descritos por (Ohkawa, 1978 e
Uchiyama & Mihara, 1977). No ensaio os produtos resultantes do ataque oxidativo às
estruturas celulares (peróxidos lipídicos), reagem com o ácido 1-tiobarbitúrico (TBARs),
NADPH + H+ + GSSGGR
NADPH+ + 2GSHNADPH + H+ + GSSGGR
NADPH+ + 2GSH
GSH + DTNB GSTNB + TNBGSH + DTNB GSTNB + TNB

40
formando bases de Schiff. A interação entre os peróxidos lipídicos e o TBARs forma um
complexo colorido, mensurado em espectrofotômetro a um comprimento de onda de 532
nm. No ensaio são acrescentados ácido fosfórico, KCl, ácido tiobarbitúrico e butanol. O
cálculo da concentração de TBARS foi feito através da equação da reta de uma curva-
padrão de tetraetoxipropano (TEP), e relativizado pela quantidade de tecido.
III.3.11. Análise Estatística e Apresentação dos Resultados
Para apresentação dos dados foram utilizados gráfico de erro padrão da média e
gráficos do tipo boxplot utilizando-se o programa MATLAB 7.0 e GraphPad Prism versão
5.00 para Windows (GraphPad Software, San Diego California USA). A apresentação em
gráficos do tipo boxplot possibilita uma melhor visualização da dispersão dos valores
obtidos. Além disto, segundo McGill et al (1978), caso os intervalos de confiança das
medianas não estejam sobrepostos podemos inferir que a diferença entre as amostras é
significativa. A Figura 14 ilustra a forma de apresentação e interpretação do gráfico tipo
boxplot.
Figura 14. Figuras representativas de gráficos hipotéticos de Boxplot. A figura A apresenta
todos os dados fornecidos por um gráfico de boxplot. A figura B exemplifica uma condição
onde não há sobreposição dos intervalos de confiança dos boxplot (diferenças significativas
entre as amostras dos boxplots (95%)). Já a figura C exemplifica uma condição onde há a
sobreposição dos intervalos de confiança dos boxplots (ausência de diferenças
significativas entre as amostras dos boxplots).

41
Foram feiras ainda análises estatísticas utilizando-se o programa GraphPad Prism
versão 5.00 para Windows (GraphPad Software, San Diego California USA). Para verificar
se as distribuições dos dados eram paramétricas ou não paramétricas foi realizado o teste de
normalidade Shapiro-Wilk test. Para amostras paramétricas foi aplicado o teste t-student
não pareado, e com pós-teste de Mann-Whitney para amostras não paramétricas. Foram
consideradas significativas alterações com p<0,05.
Apresentação dos resultados do protocolo 2 e análise estatística.
Em um processo de construção do conhecimento científico a reprodutibilidade é um
fator importante para confiança dos resultados e interpretações obtidas. Nesse sentido na
apresentação dos resultados deste protocolo colocamos juntos os dados obtidos do grupo
controle do protocolo 1 para verificar a reprodutibilidade das análises realizadas, exceto na
análise de desempenho onde julgamos o aumento de carga ao longo das semanas como um
fator de reprodutibilidade.
Para apresentação dos dados foram utilizados gráficos de erro padrão da média e na
análise estatística para verificar se as distribuições dos dados eram paramétricos ou não
paramétricos foi realizado o teste de normalidade Shapiro-Wilk test. Para dados
paramétricos foi aplicado teste de one-way ANOVA com pós-teste de Tukey’s e para dados
não paramétricos Kruskal-wallis com pós-teste de Dun’s foram considerados significativos
valores com p<0,05.
Foi utilizado o programa GraphPad Prism para elaboração dos gráficos e análises
estatísticas. Para análise do desempenho dos animais foi construído um gráfico com a
média das CM ao longo das semanas e uma linha de tendência utilizando o programa
MATLAB 7.0.
IV. Resultados
IV.1.Caracterização da escada com sistema de roldanas
Homogeneidade do Sistema de Roldanas
Comparamos nosso sistema de roldanas com o método de sobrecarga utilizado na
literatura, com os tubos falcon preso na cauda dos ratos (Lee & Farrar, 2003; Hornberger &
Farrar 2004). A principal diferença entre os dois métodos é a ausência ou presença de atrito

42
com os nos degraus da escada. Para isso realizamos um experimento que consistiu em
comparar a carga necessária para tirar um sistema em equilíbrio em 3 situações distintas:
Situação A (Representa o rato treinando em nosso sistema). Dois baldes foram
colocados em equilíbrio (Figura 15 - A), sendo adicionada carga (7 gramas) no balde até a
perda do equilíbrio, e o balde descer em velocidade constante até o chão. O total de carga
adicionada era mensurada.
Situação B (Representa o rato treinando com os tubos). Tubos falcon foram colocados
na escada em equilíbrio com o balde, e o procedimento de acréscimo de carga foi repetido
(Figura 15 - B).
Situação C (Representa a situação B sem atrito). Tubos falcon foram colocados na
escada em cima de uma chapa lisa de aço, para diminuir o atrito com os degraus, em
equilíbrio com o balde (Figura 15 – C), e o procedimento de acréscimo de carga foi
repetido.
Em cada situação foi testado a perda do equilíbrio com a adição de carga no balde e
ou tubos com cargas fixas de 100, 200, 300, 400, 500, 600 e 700 gramas. Mensurados o
incremento de cargas necessárias para tirar o equilíbrio nas três situações com diferentes
Figura 15. Situações do teste de atrito. A - Baldes em equilíbrio apenas com as
roldanas. B – Tubos falcon na escada em equilíbrio com o balde. C – Tubos falcon na
escada sem atrito dos degraus em equilíbrio com o balde.

43
Figura 16. Retas de inclinação das cargas de incremento necessárias para tirar o sistema
em equilíbrio em cada carga fixa. Azul – Situação A. Vermelho Situação B. Verde
Situação C. CA – Coeficiente Angular; CL – Coeficiente Linear; R2 – Correlação linear
cargas em equilíbrio realizamos uma reta (7 pontos) e calculamos a diferença em percentual
da inclinação de cada reta através do coeficiente angular. Os dados estão apresentados na
figura 16.
O
R2 demonstra que o experimento apresentou boa linearidade em todas as situações.
No entanto, a situação A apresentou uma inclinação 40% menor que a situação B, e a
situação C apresentou uma inclinação 11% menor que a situação B. Isso demonstra que a
situação B demandou uma carga de incremento maior para tirar o sistema do equilíbrio
devido ao maior atrito dos tubos com os degraus da escada. O sistema de roldanas permite
gerar uma tração de carga mais homogênea devido a menor interferência de atrito.
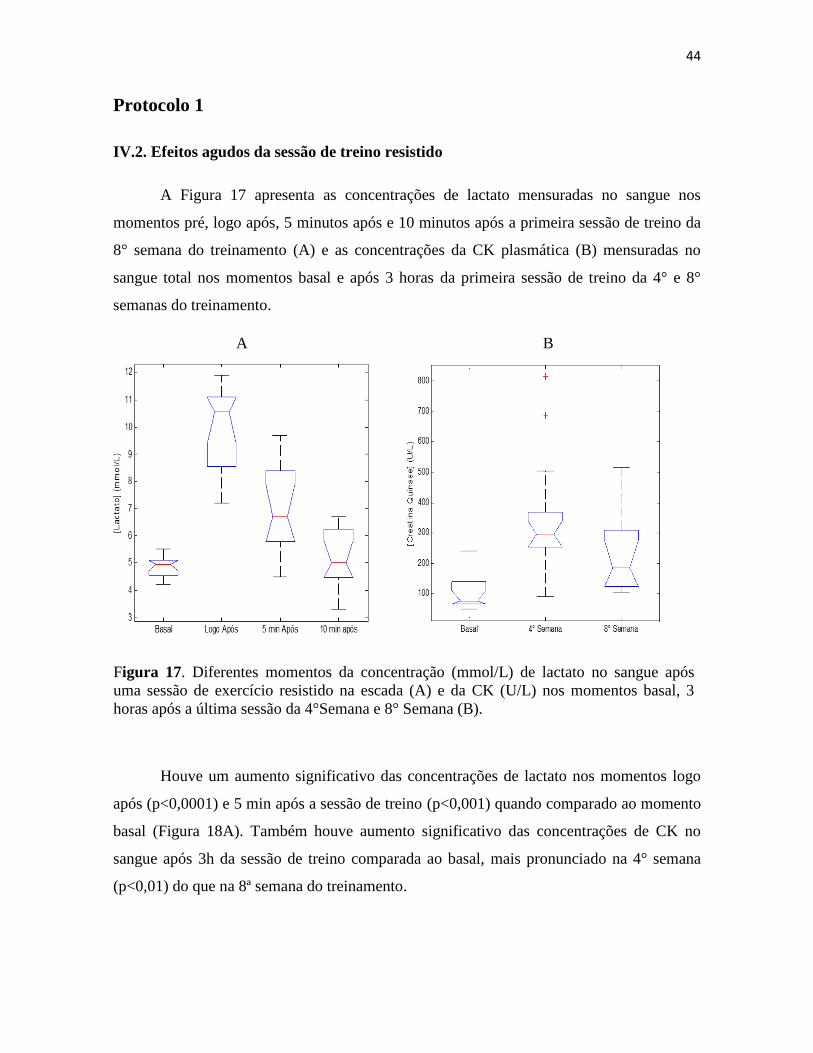
44
Figura 17. Diferentes momentos da concentração (mmol/L) de lactato no sangue após
uma sessão de exercício resistido na escada (A) e da CK (U/L) nos momentos basal, 3
horas após a última sessão da 4°Semana e 8° Semana (B).
Protocolo 1
IV.2. Efeitos agudos da sessão de treino resistido
A Figura 17 apresenta as concentrações de lactato mensuradas no sangue nos
momentos pré, logo após, 5 minutos após e 10 minutos após a primeira sessão de treino da
8° semana do treinamento (A) e as concentrações da CK plasmática (B) mensuradas no
sangue total nos momentos basal e após 3 horas da primeira sessão de treino da 4° e 8°
semanas do treinamento.
A B
Houve um aumento significativo das concentrações de lactato nos momentos logo
após (p<0,0001) e 5 min após a sessão de treino (p<0,001) quando comparado ao momento
basal (Figura 18A). Também houve aumento significativo das concentrações de CK no
sangue após 3h da sessão de treino comparada ao basal, mais pronunciado na 4° semana
(p<0,01) do que na 8ª semana do treinamento.

45
IV.3. Efeitos crônicos do protocolo de treinamento (Protocolo 1)
Desempenho (Carga máxima)
O desempenho dos animais do grupo treinado foi mensurado através do teste de
carga máxima ao longo do protocolo de treino. A Figura 18 apresenta a comparação dos
resultados da carga máxima do grupo Controle (C) e treinado (Tr) após a 1°, 4° e 8°
semanas de treino.
Figura 18. Em azul a CM do grupo controle na primeira semana (1°S.C) na quarta semana
(4°S.C) e na oitava semana (8°S.C). Em verde se encontra a CM do grupo treinado na
primeira semana (1°S.Tr) na quarta semana (4°S.Tr) e na oitava semana (8°S.Tr).
Podemos observar que no início do treinamento a carga máxima dos animais era
praticamente a mesma. Houve aumento na carga máxima após a 4° e 8° para ambos os
grupos. No entanto, houve aumento significativo da carga máxima do grupo treinado em
relação ao controle na 4° semana (p<0,0001) e 8° semanas de treino (p<0,0001).
1°S.Tr 1°S.C 4°S.Tr 4°S.C 8°S.Tr 8°S.C.
250
300
350
400
450
500
550
600
650
Desem
penho -
CM
(G
ram
as)
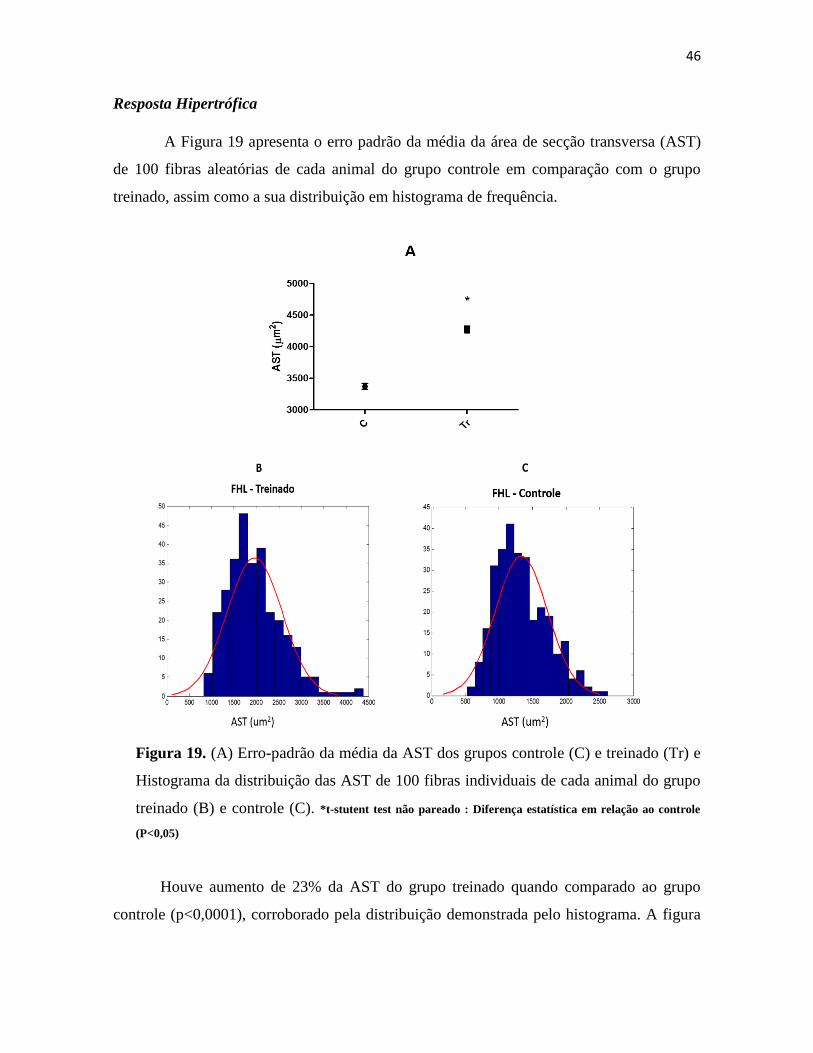
46
Figura 19. (A) Erro-padrão da média da AST dos grupos controle (C) e treinado (Tr) e
Histograma da distribuição das AST de 100 fibras individuais de cada animal do grupo
treinado (B) e controle (C). *t-stutent test não pareado : Diferença estatística em relação ao controle
(P<0,05)
Resposta Hipertrófica
A Figura 19 apresenta o erro padrão da média da área de secção transversa (AST)
de 100 fibras aleatórias de cada animal do grupo controle em comparação com o grupo
treinado, assim como a sua distribuição em histograma de frequência.
Houve aumento de 23% da AST do grupo treinado quando comparado ao grupo
controle (p<0,0001), corroborado pela distribuição demonstrada pelo histograma. A figura

47
20 apresenta uma imagem qualitativa da hipertrofia no músculo de animal submetido ao
protocolo de treinamento resistido comparado com um animal do grupo controle.
Efeito sobre as reservas de Glicogênio (Substrato Energético)
A Figura 21 apresenta as concentrações de glicogênio do FHL dos animais do grupo
controle e treinado após 8 semanas de treinamento resistido em escada.
Podemos observar que o treinamento promoveu um aumento significativo (p<0,01)
dos estoques de glicogênio muscular no grupo treinado comparado ao grupo controle.
C Tr
20
25
30
35
40
45
50
55
[]G
licogênio
(m
mol/kg t
ecid
o)
Figura 20. Corte transversal do músculo FHL de um animal do grupo Treinado e outro do
controle, corado com Hematoxilina e Eosina (Objetiva 20x).
Figura 21. Concentrações de Glicogênio no FHL (mmol/kg tecido) do grupo C e Tr.
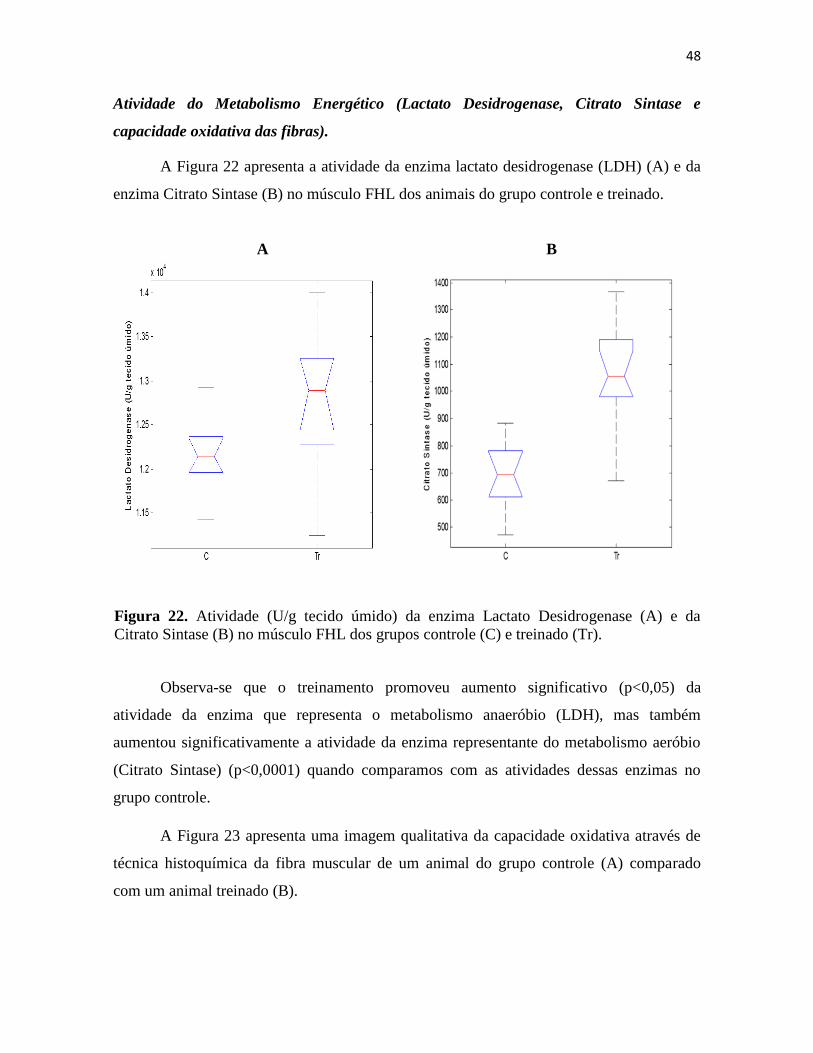
48
Atividade do Metabolismo Energético (Lactato Desidrogenase, Citrato Sintase e
capacidade oxidativa das fibras).
A Figura 22 apresenta a atividade da enzima lactato desidrogenase (LDH) (A) e da
enzima Citrato Sintase (B) no músculo FHL dos animais do grupo controle e treinado.
A B
Observa-se que o treinamento promoveu aumento significativo (p<0,05) da
atividade da enzima que representa o metabolismo anaeróbio (LDH), mas também
aumentou significativamente a atividade da enzima representante do metabolismo aeróbio
(Citrato Sintase) (p<0,0001) quando comparamos com as atividades dessas enzimas no
grupo controle.
A Figura 23 apresenta uma imagem qualitativa da capacidade oxidativa através de
técnica histoquímica da fibra muscular de um animal do grupo controle (A) comparado
com um animal treinado (B).
Figura 22. Atividade (U/g tecido úmido) da enzima Lactato Desidrogenase (A) e da
Citrato Sintase (B) no músculo FHL dos grupos controle (C) e treinado (Tr).

49
Figura 24. Área de Secção Transversa das fibras oxidativas (C vs Tr) e das fibras
glicolíticas (C vs Tr). *, # t-stutent test não pareado: Diferença estatística em relação ao controle
(P<0,05)
A B
Em escuro estão as fibras com maior capacidade oxidativa (Tipo I) e em claro as
fibras menos oxidativas (Tipo II). A imagem indica que houve hipertrofia das fibras II e
também das fibras mais oxidativas, Tipo I. Dessa forma, mensuramos a AST de 100 fibras
oxidativas (escuras) e glicolíticas (Claras) aleatórias de cada animal para verificar a
resposta hipertrófica. Os dados estão apresentados na Figura 24.
Os dados mostraram através do aumento da AST em relação ao controle que as
fibras oxidativas apresentaram hipertrofia (p<0,0001) e as fibras glicolíticas também
Figura 23. Corte transversal do FHL do grupo controle (A) e treinado (B)corado pela
técnica de NADH-TR (Objetiva 10x).

50
(p<0,0001). Mostraram ainda a maior AST das fibras glicolíticas em relação as oxidativas
no grupo treinado quando comparado ao controle.
Marcadores de Estresse oxidativo: Atividade das Enzimas Antioxidantes (SOD, CAT,
GR) e peroxidação lipídica (TBARs).
A Figura 25 apresenta a atividade das enzimas antioxidantes Superóxido dismutase
(SOD - A), Catalase (CAT - B) e Glutationa Redutase (GR - C) no FHL dos grupos
controle e treinado. A Figura 25 - D apresenta as concentrações de TBARs no músculo
FLH.
Figura 25. Atividade (U/g tecido úmido) das enzimas antioxidantes SOD, CAT e GR e
peroxidação lipídica (nnamol TEP/g tecido úmido) nos grupos controle (C) e treinado (Tr).

51
Figura 26. Desempenho (CM) ao longo das semanas de treinamento dos grupos água
(azul), suplementado com 0.75 ml de óleo de arroz (verde) e 1.5 ml de óleo arroz
(Vermelho).
Podemos observar uma tendência de up-regulation na atividade das enzimas
antioxidantes no grupo treinado, sendo estatisticamente significativo apenas para as
enzimas CAT e GR (p<0,05). Por outro lado, é visível que o grupo Tr apresentou menores
concentrações de TBARs e uma resposta muito mais homogênea do que o grupo controle
(p<0,05).
Protocolo 2
IV.4. A suplementação antioxidante produz efeito ergogênico adicional ao efeito do
treinamento?
Desempenho
A Figura 26 apresenta a média das cargas máximas e uma linha de tendência
atingidas pelos grupos ao longo das 8 semanas de treinamento, incluindo a semana de
adaptação. Assim como uma linha tracejada de comparação entre a carga máxima atingida
pelos grupos água e 1.5 ml na última semana de treino.
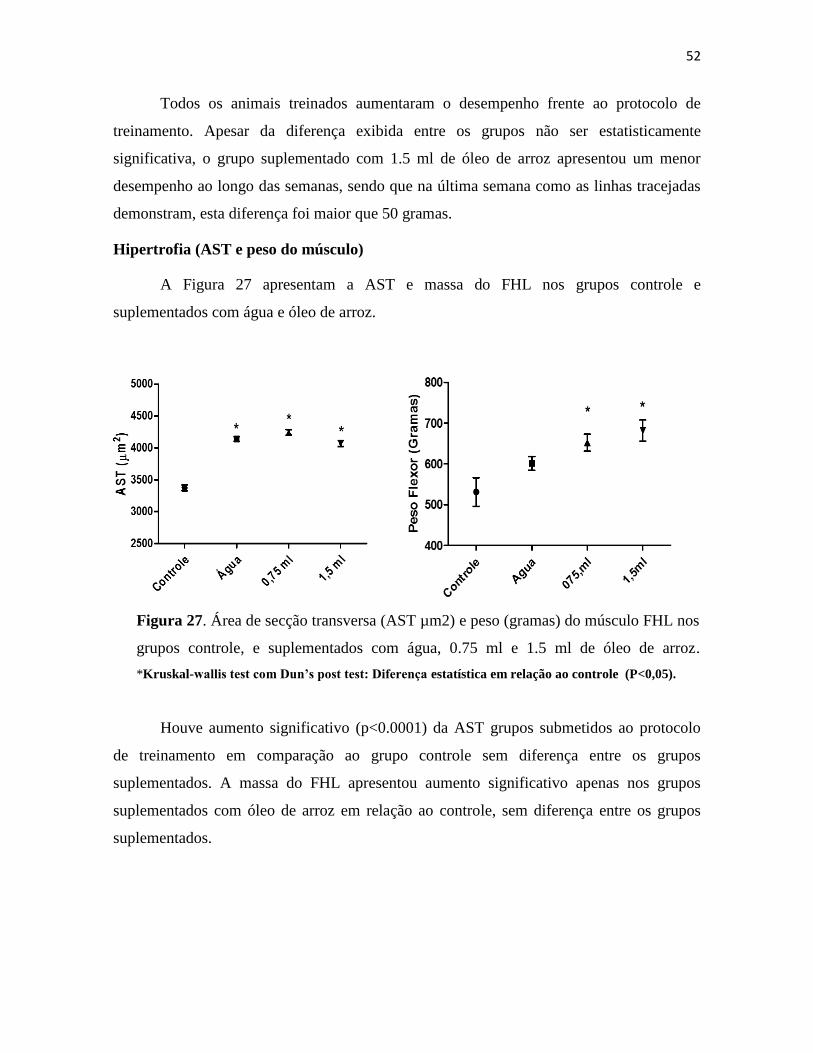
52
Figura 27. Área de secção transversa (AST µm2) e peso (gramas) do músculo FHL nos
grupos controle, e suplementados com água, 0.75 ml e 1.5 ml de óleo de arroz.
*Kruskal-wallis test com Dun’s post test: Diferença estatística em relação ao controle (P<0,05).
Todos os animais treinados aumentaram o desempenho frente ao protocolo de
treinamento. Apesar da diferença exibida entre os grupos não ser estatisticamente
significativa, o grupo suplementado com 1.5 ml de óleo de arroz apresentou um menor
desempenho ao longo das semanas, sendo que na última semana como as linhas tracejadas
demonstram, esta diferença foi maior que 50 gramas.
Hipertrofia (AST e peso do músculo)
A Figura 27 apresentam a AST e massa do FHL nos grupos controle e
suplementados com água e óleo de arroz.
Houve aumento significativo (p<0.0001) da AST grupos submetidos ao protocolo
de treinamento em comparação ao grupo controle sem diferença entre os grupos
suplementados. A massa do FHL apresentou aumento significativo apenas nos grupos
suplementados com óleo de arroz em relação ao controle, sem diferença entre os grupos
suplementados.
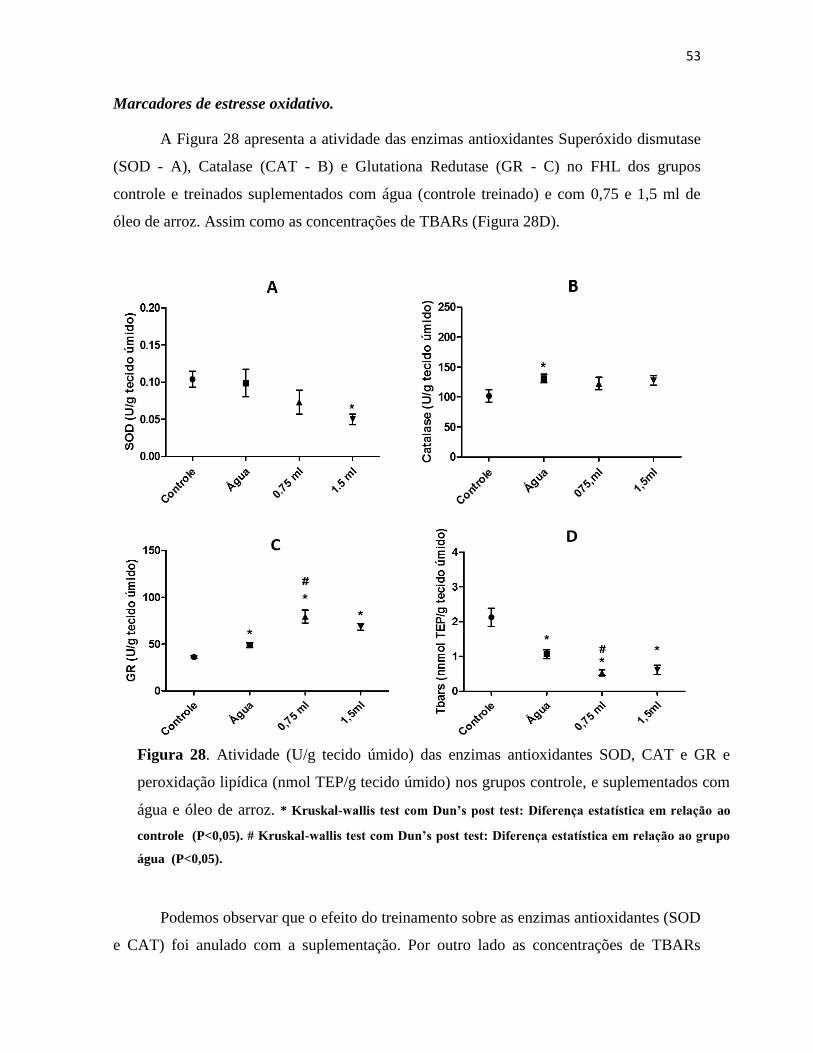
53
Marcadores de estresse oxidativo.
A Figura 28 apresenta a atividade das enzimas antioxidantes Superóxido dismutase
(SOD - A), Catalase (CAT - B) e Glutationa Redutase (GR - C) no FHL dos grupos
controle e treinados suplementados com água (controle treinado) e com 0,75 e 1,5 ml de
óleo de arroz. Assim como as concentrações de TBARs (Figura 28D).
Podemos observar que o efeito do treinamento sobre as enzimas antioxidantes (SOD
e CAT) foi anulado com a suplementação. Por outro lado as concentrações de TBARs
Figura 28. Atividade (U/g tecido úmido) das enzimas antioxidantes SOD, CAT e GR e
peroxidação lipídica (nmol TEP/g tecido úmido) nos grupos controle, e suplementados com
água e óleo de arroz. * Kruskal-wallis test com Dun’s post test: Diferença estatística em relação ao
controle (P<0,05). # Kruskal-wallis test com Dun’s post test: Diferença estatística em relação ao grupo
água (P<0,05).
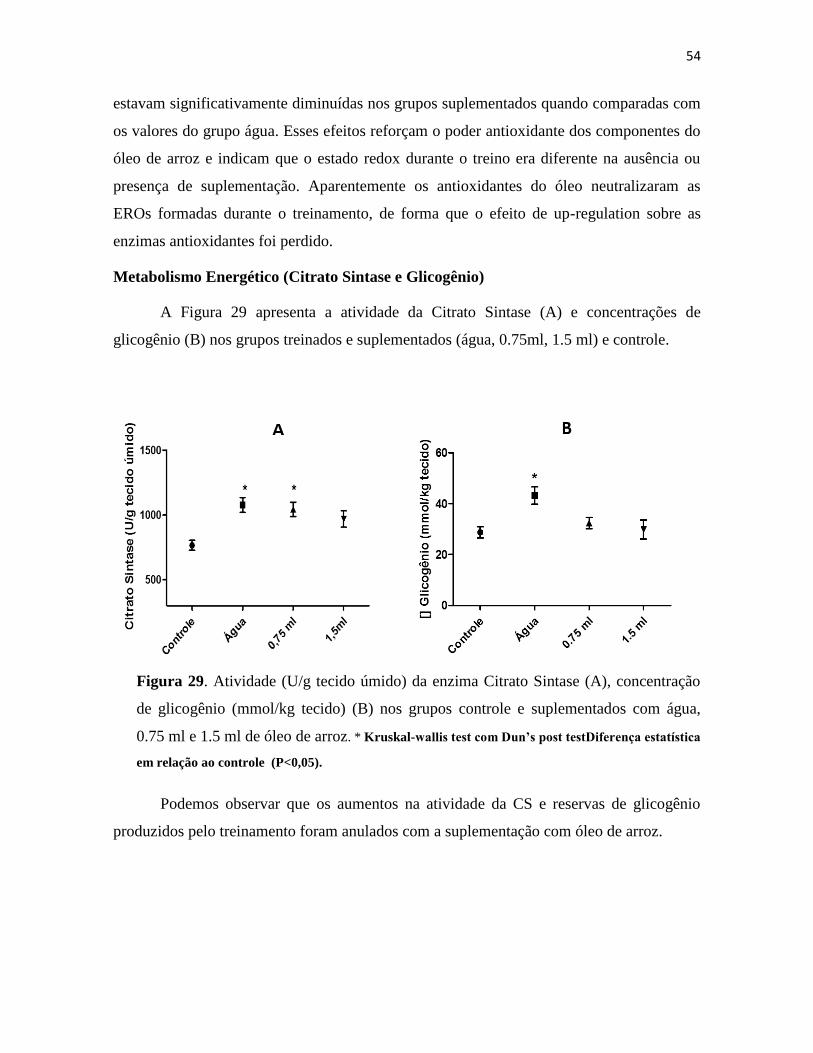
54
Figura 29. Atividade (U/g tecido úmido) da enzima Citrato Sintase (A), concentração
de glicogênio (mmol/kg tecido) (B) nos grupos controle e suplementados com água,
0.75 ml e 1.5 ml de óleo de arroz. * Kruskal-wallis test com Dun’s post testDiferença estatística
em relação ao controle (P<0,05).
estavam significativamente diminuídas nos grupos suplementados quando comparadas com
os valores do grupo água. Esses efeitos reforçam o poder antioxidante dos componentes do
óleo de arroz e indicam que o estado redox durante o treino era diferente na ausência ou
presença de suplementação. Aparentemente os antioxidantes do óleo neutralizaram as
EROs formadas durante o treinamento, de forma que o efeito de up-regulation sobre as
enzimas antioxidantes foi perdido.
Metabolismo Energético (Citrato Sintase e Glicogênio)
A Figura 29 apresenta a atividade da Citrato Sintase (A) e concentrações de
glicogênio (B) nos grupos treinados e suplementados (água, 0.75ml, 1.5 ml) e controle.
Podemos observar que os aumentos na atividade da CS e reservas de glicogênio
produzidos pelo treinamento foram anulados com a suplementação com óleo de arroz.

55
V. Discussão
V1. Protocolo 1.
O primeiro desafio proposto no início do projeto de mestrado foi realizado com
êxito. Produzimos uma escada com um sistema de roldanas para a aplicação de sobrecargas
ao invés da sobrecarga aplicada com tubos falcon presos na cauda do animal como os
protocolos descritos na literatura, (Sukho & Farrar, 2003; Hornberger & Farrar 2004;
Duncan et al, 1998; Yarasheski et al 1990). Nossos resultados mostraram que o sistema de
roldanas apresenta um menor atrito quando comparado com a carga colocada nos tubos
falcon arrastados no degrau da escada. Como o músculo esquelético é sensível ao estresse
mecânico, conseguimos gerar uma tensão mais homogênea com o sistema de roldanas, que
gerou respostas adaptativas características de um treino resistido. É importante ressaltar que
o sistema de roldanas facilita a mão-de-obra do pesquisador, uma vez que dois ratos podem
treinar ao mesmo tempo, cada um com seu balde, sendo ainda que no balde também é mais
fácil colocar os chumbos para a sobrecarga.
Outra modificação do protocolo foi a introdução de uma semana de adaptação dos
animais a escada sem a utilização de sistema de recompensa (comida) ou punição
(Choque), que podem causar alterações hormonais e de peso nos animais (Lowe et al,
2002). Na semana de adaptação utilizamos apenas o estímulo da pinça, que proporcionou
que nas semanas de treino quando necessário o estímulo fosse apenas com ajuda da mão.
Outra modificação dos estudos anteriores foi a realização do teste de carga máxima
somente na última sessão de treino da semana ao invés de toda sessão de treino, e a
frequência de treino três vezes na semana, ao invés de a cada dois dias (Hornberger &
Farrar 2004).
As adaptações observadas após 8 semanas de treinamento resistido demonstraram
que as modificações realizadas no modelo animal de treinamento resistido (Hornberger &
Farrar 2004) foram eficazes em induzir aumento de carga no teste de carga máxima
associado a hipertrofia muscular.
O aumento de carga é o primeiro sinal que as adaptações decorrentes de um
treinamento resistido estão ocorrendo. Note que os animais do grupo Tr exibiram aumento

56
significativo na carga comparado ao grupo C, embora o grupo C tenha apresentado uma
pequena adaptação nas semanas 4 e 8, com aumento de carga em relação a semana inicial
(Figura 17). Visto que o grupo controle realizou 2 sessões de treino (sem sobrecarga) toda
semana, além de três testes de carga máxima ao longo das oito semanas ele não pode ser
considerado um grupo sedentário, mas fisicamente ativo. Além disso, não podemos
desconsiderar o crescimento natural dos animais.
A comparação com um grupo controle ativo não prejudicou as comparações com o
grupo Tr. Muito pelo contrário, valorizou, pois mostrou que as adaptações promovidas pelo
protocolo de treinamento resistido foram de fato significativas. Não encontramos na
literatura estudo com um grupo controle com essas características. Normalmente os
estudos comparam os resultados de um grupo treinado contra um grupo de animais
sedentários (Lee & Farrar, 2003; Hornberger & Farrar 2004; Duncan et al, 1998;
Yarasheski et al 1990).
O aumento de carga é decorrente principalmente de um maior recrutamento de
unidades motoras e do aumento da área de secção transversa do músculo (Kraemer et al,
2002; Mcardle & Katch, Katch, 2008). Observamos que o aumento de desempenho no
grupo Tr parece diretamente associado a maior AST no FHL e, possivelmente ao maior
recrutamento neural, apesar deste último não ter sido mensurado.
A originalidade dos resultados apresentados no músculo FHL dos animais treinados
foi mostrar que o treinamento aumentou a capacidade oxidativa do músculo, com aumento
da atividade da citrato sintase e hipertrofia das fibras do tipo I, além de aumentar as
concentrações de glicogênio. Esse quadro adaptativo provavelmente ocorreu em resposta ao
aumento nas EROs com o treinamento, uma vez que detectamos aumento de atividade das
enzimas antioxidantes associado a uma menor peroxidação lipídica. Esse panorama ainda
não tinha sido descrito na literatura neste modelo de treinamento e músculo analisado.
A voluntariedade do treinamento em escada tem sido discutida com ênfase no
aprendizado do animal (Casilhas et al, 2013). Este modelo de treinamento tem sido
utilizado em estudos que investigam alterações na neuroplasticidade, com a justificativa de
gerar um menor estresse ao animal (Casilhas et al, 2013; Casilhas et al, 2012). Diferente do
estudo de Casilhas et al 2013, onde nenhum animal abandonou o treino, reforçando o

57
conceito de aprendizado no nosso estudo 6 animais foram excluídos por não acompanharem
o incremento de carga dos outros animais, e não realizarem o treino voluntariamente,
mesmo que a pinça tenha sido utilizada esporadicamente ao longo das semanas de
treinamento, o que deixa a questão porque esses animais não se adaptaram ao incremento
de carga. No entanto, nenhum animal do grupo controle foi excluído. Todos realizaram as
sessões de treino e testes de carga máxima voluntariamente. Vale ressaltar que essa
diminuição de animais no grupo Tr não prejudicou a interpretação dos resultados, pois
continuamos com um “n” robusto após essa perda (10 C x 12 Tr).
Treinos que geram hipertrofia são caracterizados por exercícios que promovem um
estresse mecânico, com alterações na estrutura do sarcômero e rompimento de proteínas do
citoesqueleto (Ingber, 2006). Aumento das concentrações de CK no sangue tem sido
relacionado com aumento das microlesões musculares (Liu et al, 2005). Foi proposto que 5
a 10 % da CK se encontra ligada a linha M do sarcômero que, quando rompido ativa a
enzima PLD. Essa enzima, como já vimos promove a formação de ácido fosfatídico que
estimula diretamente a mTOR, e consequentemente, a resposta hipertrófica (Hornberger et
al, 2006).
As concentrações de CK no sangue 3h após a sessão de treino confirmam que a
sobrecarga aplicada pelo sistema de roldanas apresentou potencial lesivo necessário para a
resposta hipertrófica no FHL. Interessante que na 8° semana o aumento das concentrações
de CK frente ao basal foi relativamente menor que o aumento observado na 4° semana.
Esses dados confirmam que os animais estavam se adaptando ao protocolo, e de fato, na 7ª
e 8° semanas tivemos que acrescentar mais um teste de carga máxima na segunda-feira
além da sexta-feira devido a observação durante o protocolo que os animais estavam
realizando o treino de maneira mais fácil.
O aumento das concentrações de lactato após a última sessão de exercício
confirmou que as sessões de treino utilizaram o metabolismo glicolítico anaeróbio,
característico de um treinamento de força com caráter hipertrófico. Pudemos observar
também uma cinética de remoção de lactato via MCT4 (monotransportador de carboxilato)
do músculo para o sangue e MCT1 do sangue para outros tecidos. A remoção adequada do
lactato é importante porque ocorre sempre em simporte com um H+, favorecendo com isso

58
o controle do pH intramuscular durante as pausas nas sessões de treino. O lactato também
sinaliza a hipófise para liberação de GH, com a consequente produção de IGF-1 pelo
fígado, e estimulação da PI3, mTOr e síntese proteica (Kraemer & Rogol, 2005).
Efeitos crônicos do treinamento resistido
Nossos dados mostraram que as 24 sessões de treinamento ao longo das 8 semanas
que compreenderam o protocolo de treinamento resistido foram suficientes para gerar
hipertrofia do músculo FHL, da mesma forma que o observado com o modelo de
treinamento resistido apresentado por (Hornberger & Farrar, 2004). Esses autores
encontraram aumento do tempo para atingir a fadiga em contrações in situ, sem alterações
na concentração de ATP e atividade da CK no FHL de ratos da linhagem Sprague Dawley.
Propuseram que o aumento do tempo de resistência a fadiga poderia estar relacionado com
adaptações na atividade da LDH e nos estoques de glicogênio. Nossos resultados
corroboraram essa hipótese.
A reação catalisada pela LDH é fundamental durante contrações intensas, pois
permite a regeneração do NAD+ para a reação catalisada pela enzima gliceraldeído
desidrogenase, permitindo a continuidade da via glicolítica e fornecimento de ATP durante
a contração muscular. O lactato produzido pela atividade dessa enzima, por sua vez,
favorece a retirada de H+ em simporte com lactato através do funcionamento dos MCTs
(Torres & Marzzoco, 2007; Mcardle, Katch, Katch 2008).
Correlacionado com o aumento da atividade da LDH encontramos aumento das
concentrações de glicogênio, favorecendo o funcionamento da via glicolítica. O glicogênio
também se relaciona com a AST do músculo, principalmente por favorecer o inchaço
celular. Para cada grama de glicogênio formado são absorvidas 3 gramas de água. Tem sido
demonstrado também que o inchaço celular promove ativação de proteínas quinases
relacionadas a ativação da síntese proteica (Chan et al 1985 ; Lang et al, 2007 ). O aumento
da concentração de lactato também favorece o inchaço celular alterando o equilíbrio
osmótico da célula (Schoenfeld, 2010).
A depleção dos estoques de glicogênio muscular tem sido um dos fatores que
contribuem para o processo de fadiga muscular durante o exercício (Ørtenblad et al 2013),

59
assim como a produção exacerbada de EROs (Powers & Jackson,2008). O aumento dos
estoques de glicogênio contribui para evitar a fadiga por falta de substrato energético, mas
também pode contribuir para defesa antioxidante contra EROs, pois a glicose-6-fosfato
também é substrato da via das pentoses, uma via que produz a coenzima NADPH que
participa da reação catalisada pela enzima antioxidante glutationa redutase, para
regeneração da GSSG em GSH (Silveira, et al 2008).
Os resultados apresentados no nosso estudo mostraram aumento da atividade da
Citrato Sintase, utilizada como marcador do metabolismo aeróbio (Spina et al 1996). Essa
enzima mais ativa indica que o ciclo de Krebs está mais ativo e produzindo coenzimas
reduzidas para serem oxidadas para produção de ATP na CTE. Isso consequentemente
favorece uma maior oxidação de ácidos graxos, e a utilização de glicose-6-fostato pela via
das pentoses. Hipotetizamos que o aumento da Citrato Sintase neste protocolo é oriundo de
uma necessidade de ressíntese de ATP durante as pausas entre as repetições (escaladas) nas
sessões de treino, mas principalmente na pausa entre as sessões de treino. A instalação de
um processo inflamatório para regeneração do tecido lesionado e síntese de novas proteínas
miofibrilares para o processo de hipertrofia requerem uma alta quantidade de ATP, que é
disponibilizado em situações de repouso pelo metabolismo aeróbio (Børsheim & Bahr,
2003; Tidball, 2005).
Apesar de durante o estímulo do exercício resistido a fonte energética predominante
não ser o ácido graxo, e de ser discutido uma certa “diluição” da quantidade de
mitocôndrias com a hipertrofia, diminuindo a capacidade oxidativa (Chilibeck, 1999). O
aumento da atividade da Citrato Sintase justifica porque treinos resistidos favorecem o
emagrecimento e controle de peso (Kirk et al, 2009; Ormsbee, 1985).
Nesse contexto, treinos resistidos têm sido utilizados principalmente pelas respostas
induzidas após o exercício, tais como o excesso de consumo de oxigênio pós exercício
(EPOC), aumento da massa magra com consequente aumento da taxa de metabolismo
basal, favorecendo a oxidação de ácidos graxos (Børsheim & Bahr, 2003). Braith et al
(2005) encontraram aumento da atividade da Citrato Sintase em músculo cardíaco de
pacientes transplantados após 6 meses de treinamento resistido. Assim como Tang et al
(2006) encontraram aumento da atividade da enzima Citrato sintase em músculo de

60
humanos submetidos a 12 semanas de treinamento resistido. Nossos dados, de aumento da
atividade da Citrato sintase corrobora com esses estudos.
O aumentou da capacidade oxidativa provavelmente foi devido a um maior
conteúdo mitocondrial. Também observamos hipertrofia das fibras oxidativas, que refletem
indiretamente a atividade mitocondrial. As fibras oxidativas possuem uma menor AST
quando comparada com as fibras glicolíticas. Este fenômeno é explicado provavelmente
pelo metabolismo das fibras oxidativas, que apesar de ter um maior potencial de transcrição
e uma alta taxa de síntese proteica com um maior número de células satélites, domínio
mionuclear, mitocôndrias e concentrações de RNAm (Wessel et al, 2010), possuem
também uma alta taxa de degradação proteica via mecanismos de ubiquitinação, e
mecanismos lisossomais com atuação das caspases, sinalizado por uma concentração mais
alta de cálcio intracelular, e também pela produção de EROs. A maior produção de EROs
sinalizam para o NFBK para transcrição de genes de atrofia MuRF e MAFbx (Powers &
Jackson, 2009). Isso explica porque as fibras oxidativas possuem um alto turnover proteico,
que contribui para sua menor AST em relação as fibras glicolíticas.
A literatura aponta que treinos de endurance, de característica aeróbica não induzem
grande magnitude de hipertrofia, mas aumentam a expressão de enzimas oxidativas,
conteúdo mitocondrial, densidade capilar e inclusive a posição da mitocôndria no
sarcolema para favorecer a troca de oxigênio. Já os treinos resistidos apresentam maiores
graus de hipertrofia e aumento da atividade de enzimas do metabolismo anaeróbio
(Holloszy & Coyle, 1984; Wessel et al, 2010). Os dados apresentados no nosso estudo
demonstraram adaptações características de um treino resistido no FHL, com aumento da
capacidade oxidativa e hipertrofia em ambos os tipos de fibras, corroborada com as lâminas
coradas por HE. Ao contrário de outros estudos que não encontraram alterações na
capacidade oxidativa muscular após um treinamento resistido (Chilibeck et al, 1999; Green
et al 1999; MacDougall, 1979).
Para ocorrer o processo de hipertrofia precisamos de um balanço positivo na razão
síntese/degradação de proteínas, principalmente nas fibras oxidativas que possuem sua alta
taxa de degradação proteica. Treinamentos que induzem aumento nos níveis de estresse
oxidativo tem sido relacionados com aumento da atividade dos sistemas de degradação

61
proteica (Powers & Jackson, 2009). Foi proposto que a oxidação dos grupos tióis por EROs
dos canais de rianodina e dihidropiridina aumentam a liberação de cálcio para o citosol da
célula, que ativam as caspases para degradação proteica (Xia et al, 2000). Mas se a
mitocôndria é uma das principais fontes de EROs no músculo (Balaban et al, 2005), e
nossos dados indicam que a capacidade oxidativa do músculo aumentou, como foi possível
ocorrer o processo hipertrófico?
O aumento da atividade das enzimas antioxidantes (SOD, CAT e GR) demonstrou
que houve aumento de EROs. No entanto, esse aumento não prejudicou o processo de
hipertrofia provavelmente porque as células estavam em um estado redox intracelular ótimo
para gerar as adaptações ao treinamento. O aumento da defesa antioxidante correlacionou
com o menor ataque oxidativo as membranas celulares no grupo treinado, evidenciado
pelas menores concentrações de TBARs. Esses dados mostram claramente que o protocolo
de treino aumentou a produção de EROs mas não induziu um quadro de estresse oxidativo.
O aumento não significativo da atividade da SOD no grupo Tr em relação ao grupo
controle corrobora com essa interpretação, visto que o NFKB deve ser estimulado por
EROs para aumentar a expressão da SOD (Gomez-Cabrera, 2008). Nesse sentido,
indiretamente também podemos inferir que não houve a transcrição de genes relacionados a
atrofia muscular (MuRF e MAFbx) (Powers & Jackson, 2009). Já as enzimas CAT e GR
aumentaram significativamente suas atividades. A GR provavelmente se favoreceu das
maiores concentrações de glicogênio e disponibilidade de NADPH para produzir GSH e
este atuar via GPx na proteção contra peróxidos orgânicos.
A enzima CAT aumentou sua atividade provavelmente para manter as
concentrações de H2O2 em níveis ótimos, visto que este sinaliza para o fator de transcrição
PGC-1 alpha relacionado a biogênese mitocondrial. Concentrações baixas de H2O2 não
estimulam a transcrição e altas concentrações estimulam indiretamente através da AMPK
(Kang et al, 2009). No entanto a AMPK estimulada inibe a mTOR e, consequentemente, a
síntese proteica (Gwinn et al, 2008). Portanto a CAT mais ativa e a hipertrofia indicam que
as concentrações de H2O2 estavam ideais para ocorrer o processo de hipertrofia. Foi
demonstrado que a isoforma da PGC-1 alpha (PGC-1alpha4) é estimulada em treinamentos
resistidos, favorecendo a hipertrofia através do aumento da expressão de genes do IGF-1 e

62
de inibição da miostatina (Ruas et al, 2012). Um estudo (Pimenta et al, 2007) realizado com
músculo isolado com contrações intensas, características de um treino resistido, encontrou
aumento agudo da atividade da Citrato sintase, com aumento da expressão de enzimas
antioxidantes SOD, GPx e CAT, sendo que apenas a atividade enzimática da CAT
apresentou aumento, justificando o aumento em decorrência de uma maior capacidade
oxidativa da fibra e consequente produção de EROs.
Outros resultados que se correlacionam com os nossos em treinamentos resistidos.
Parise et al 2005 encontraram aumento da atividade das enzimas antioxidantes SOD e CAT
em músculo de idosos submetidos a 12 semanas de treinamento resistido. Enquanto em
outro estudo Parise et al 2005 não mostraram alterações na atividade da Citrato sintase,
CAT e SOD, mas aumento da atividade do complexo IV da CTE e da razão do complexo
IV com I, assim como menor dano oxidativo ao DNA mitocondrial em músculo de idosos
submetidos a 14 semanas de treinamento. Já Vincent et al 2006 encontraram valores
menores de TBARs em sangue de adultos obesos e com sobrepeso submetidos a 6 meses de
treinamento resistido. Já outro estudo sobre treinamento resistido em escada com ratas
ovariotecmizadas demonstrou que o treino não foi capaz de amenizar o estresse oxidativo
induzido no fígado pela intervenção cirúrgica (Rodrigues et al, 2013).
O único estudo que encontramos com modelo animal, treinamento resistido, e
análise em marcadores de estresse oxidativo em músculo (Scheffer et al, 2012) aplicou um
treinamento resistido em escada com ratos wistar durante 12 semanas divididos em 3
grupos. Cada grupo foi treinado com diferentes intensidades, denominadas pelo autor de
resistência (cargas com 10 a 50 % da massa), hipertrófico (25% a 100% da massa) e de
força (25 a 200 % da massa). Os autores mostraram aumento da produção de ânion
superóxido, da atividade da SOD e GPx e diminuição da CAT no músculo brachioradial do
membro anterior, e maiores concentrações de TBARs e proteínas carboniladas no grupo
submetido ao treino chamado de hipertrófico, sugerindo a instalação de um quadro de
estresse oxidativo neste grupo. No entanto, este estudo não apresentou nenhum dado de
desempenho dos animais ou AST do músculo que pudessem ser correlacionados com o
estresse oxidativo mensurado, ou que a intensidade de treino gerou realmente mais
hipertrofia no grupo hipertrófico. As concentrações de glicogênio aumentaram em todos os

63
grupos em relação ao grupo sedentário, demonstrando que esta adaptação não foi
prejudicada.
Já outro estudo (Cakir-Atabek, 2010) foi feito em seres humanos mas com desenho
experimental semelhante. Adultos jovens foram submetidos a 6 semanas de treinamento
resistido dividido em dois grupos chamados de hipertrófico e de força. Os autores
demonstraram menores valores de malondealdeído e aumento de GSH em ambos os
grupos, sem diferença entre os grupos, justificando que o treinamento resistido crônico
melhora a defesa antioxidante independente da intensidade.
Outro efeito terapêutico importante do treinamento resistido mostrado aqui foi o
aumento da capacidade oxidativa do FHL, demonstrando que é possível com treinamentos
resistidos melhorar o metabolismo aeróbio do músculo. Diversas adaptações estão
relacionadas com aumentado da capacidade oxidativa, como angiogênese, maior oxidação
de ácidos graxos, transportadores de membrana de glicose (GLUT 4) e aumento de
receptores de insulina na membrana e consequentemente contribuindo para o combate a
doenças cardiovasculares, diabetes, e doenças relacionadas a síndrome metabólica da
obesidade (Chibalin et al, 2000; Warburton et al, 2006). Nesse sentido treinos resistidos
podem ser incluídos em periodizações de treinamento também para populações específicas
que necessitam destas adaptações para combater doenças relacionadas ao sedentarismo e
obesidade.
Em resumo, nosso modelo de treino resistido em escada está bem caracterizado.
Gerou adaptações musculares semelhantes a treinos resistidos em humanos, podendo ser
utilizado em investigações futuras com a utilização de diferentes intervenções.
V.2. Protocolo 2
A suplementação antioxidante produz efeito ergogênico adicional ao do treinamento
resistido?
Nossa hipótese não foi confirmada. A suplementação antioxidante com óleo de
arroz não promoveu efeito ergogênico frente ao protocolo de treinamento resistido, pelo
contrário. Na verdade, os antioxidantes presentes no óleo podem ter sido os responsáveis
pela perda de algumas adaptações induzidas pelo treinamento tais como menores

64
concentrações de glicogênio, perda do aumento da atividade da citrato sintase e SOD no
grupo 1.5 ml e sem alterações da atividade da CAT em ambos os grupos suplementados
com óleo de arroz. Esses dados foram acompanhados por menores concentrações de
TBARs em ambos os grupos suplementados, onde o grupo 0,75 ml apresentou diminuição
significativa em relação ao grupo treinado.
Provavelmente o estado mais reduzido evidenciado pelas menores concentrações de
TBARs nos grupos suplementados são resultados das ações antioxidantes do gamma-
oryzanol, Vitamina E e ácidos graxos poliinsaturados presentes no óleo de arroz e
prejudicaram adaptações sinalizadas por EROs.
Detectamos um menor desempenho do grupo suplementado 1.5 ml ao longo das
semanas de treinamento, com uma diferença superior a 50 gramas na última semana de
treino. Essa diferença quando pensamos um treinamento resistido onde a variável de
sobrecarga é uma das principais variáveis manipuladas, e o dano tecidual causado por ela
um dos fatores para gerar uma resposta adaptativa pode fazer diferença. No entanto em
relação a hipertrofia não encontramos diferença entre os grupos, onde todos aumentaram a
AST no FHL em relação ao controle assim como aumentaram a massa. O menor
desempenho pode ter sido devido a perda do efeito do treinamento nas concentrações de
glicogênio e atividade da enzima citrato sintase.
Também tem sido discutido na literatura o efeito pró-oxidante da suplementação
com antioxidantes. Childs, et al 2001 encontraram maiores concentrações de peróxidos
lipícos e de 8-isoprostanos no grupo suplementado com Vitamina C e n-acetyl-cisteína
submetido a um treinamento resistido com ênfase na fase excêntrica. Outro estudo (Ristow
et al, 2009) com humanos diabéticos submetidos a 4 semanas de exercício físico e
suplementados com Vitamina C e E, demonstrou que a suplementação antioxidante inibiu
adaptações benéficas do exercício físico nesta população com menor expressão das enzimas
antioxidantes SOD e GPx, do PGC1-α e PPARγ, assim como menores concentrações de
plasmáticas de adiponectina e de GLUT4 na membrana, que refletiu na perda do aumento
da sensibilidade pela insulina promovido pelo exercício.
Mostramos nesse estudo que a produção de EROs durante o exercício e após o
exercício é necessária uma vez que o protocolo de exercício realizado protegeu do estresse

65
oxidativo com aumento da atividade das enzimas antioxidantes, exibindo uma menor
peroxidação lipídica. Esses dados foram confirmados com outro grupo de animais (grupo
água), mostrando que treinos resistidos realizados de maneira crônica protegem contra o
estresse oxidativo, principalmente pela upregulação do sistema de defesa antioxidante. Isso
se relaciona com o conceito da hormesis que propõe pequenas exposições de EROs para
gerar uma adaptação.
É importante ressaltar que as adaptações observadas no grupo treinado e
suplementado com água apresentou uma boa reprodutibilidade com os resultados obtidos
no protocolo 1, onde encontramos aumento do desempenho e aumento da AST, da enzima
citrato sintase, da massa do músculo, da atividade das enzimas CAT e GR associada a uma
menor concentração de TBARs quando comparado ao grupo controle. Destacamos ainda
que nenhum animal neste protocolo foi excluído por não realizar os treinos
voluntariamente, sendo que foi utilizada a mesma metodologia de condicionamento do
animal com pinça na semana de adaptação, e esporadicamente ao longo das semanas de
treinamento.
Nossos dados reforçam o treinamento resistido como uma terapia antioxidante não
medicamentosa (Gomez-Cabrera, et al 2008). De fato este tipo de treinamento já é
orientado para populações idosas que sofrem com processo de sarcopenia e
consequentemente com quedas por perda da força muscular, sendo os efeitos colaterais
destas quedas um dos maiores causadores de morte nesta população (Hunter et al, 2004).
Nesse sentido além de atenuar a perda de massa magra, treinos resistidos podem ser
indicados para esta população com o intuito de combater a diminuição da capacidade
oxidativa e antioxidante e proteger do estresse oxidativo. Ficou nítido pelos nossos
resultados que a suplementação é no mínimo desnecessária, sendo que doses maiores
prejudicam adaptações induzidas pelo treinamento e consequentemente o desempenho.
Aplicações práticas do Protocolo 2: Esta parte do estudo teve o patrocínio de uma empresa
de nutrição (HT Nutri), que tinha o intuito de comercializar o óleo de arroz para animais
(cavalos de corrida) e humanos, devido ao interesse no gamma-oryzanol para treinos
resistidos. Esses dados foram apresentados para a empressa em conjunto com outros
resultados de outra aluna de mestrado no nosso laboratório. Compuseram um quadro que

66
descarta a suplementação de óleo de arroz como agente ergogênico no exercício e, portanto,
sua comercialização para esse fim. Nesse sentido, nosso estudo contribui para área de
suplementação esportiva com antioxidantes, com a mensagem que é preciso ter cautela na
utilização de antioxidantes com intuito de proteção do estresse oxidativo, pois vias
adaptativas são sinalizadas por EROs. Assim como a suplementação de gamma-oryzanol
por fisiculturistas talvez seja desnecessária.
VI. Considerações Finais
Protocolo 1
O protocolo de treinamento induziu aumento da capacidade oxidativa do músculo
FHL e hipertrofia das fibras do tipo I. Esses dados corroboram com a eficiência de
treinamentos resistidos em estratégias de emagrecimento.
O protocolo de treino protegeu do estresse oxidativo, refletido por uma menor
peroxidação lipídica, associada ao aumento da atividade das enzimas antioxidantes.
Treinos resistidos que induzem hipertrofia podem ser utilizados para melhorar a
capacidade oxidativa do músculo e proteger do estresse oxidativo.
Protocolo 2
Os grupos suplementados com óleo de arroz apresentaram perda de adaptações no
FHL induzidas pelo treinamento no protocolo 1. Com menor desempenho, menor
atividade das enzimas, CS e SOD no grupo 1,5 ml. Assim como menor
concentração de glicogênio e sem alteração da atividade da CAT em ambos os
grupos suplementados.
Os grupos suplementados com óleo apresentaram menores concentrações de
TBARs, que refletem um estado redox mais reduzido.
Os antioxidantes presentes no óleo podem ser os responsáveis pela inibição de vias
adaptativas ao treinamento.

67
A suplementação de óleo de arroz não parece ser uma alternativa interessante para
melhora de desempenho no exercício físico.
VII. Conclusões
As roldanas se mostraram uma alternativa eficiente para gerar uma sobrecarga com
tensão mais homogênea, com menor interferência do atrito que os modelos da
literatura apresentam, assim como é possível realizar o condicionamento do animal
sem utilizar sistema de punição (choque) ou de recompensa (comida).
O modelo de treinamento e a sobrecarga aplicada através do sistema de roldanas
geraram respostas adaptativas esperadas com o treinamento resistido tais como
aumento da área secção transversa, aumento da carga em teste máximo, aumento
das reservas de glicogênio muscular e atividade da LDH.
O aumento da sobrecarga e as adaptações apresentadas no FHL do grupo água no
protocolo 2 demonstraram que o modelo de treino resistido é reprodutível.
A suplementação com óleo de arroz não promoveu efeito ergogênico no
treinamento resistido. Bem como as concentrações de gamma-oryzanol presentes no
óleo.
Quando as cargas de treino, descanso e alimentação são bem planejados a
suplementação pode não ser necessária.

68
Anexo A
Em processo um de investigação científica através de análises laboratoriais que
indiretamente refletem os efeitos induzidos, a padronização dos experimentos e técnicas
utilizadas é uma etapa importante para a confiabilidade dos dados obtidos e das
interpretações realizadas. Nesse sentido a construção de curva-padrão foi o método
escolhido para padronização das técnicas. A curva-padrão apesar do norme “curva” é uma
reta de correlação entre uma variável dependente (eixo x) e uma variável independente
(eixo y). Esta relação permite estudar o quão duas variáveis mudam em conjunto. As curvas
foram utilizadas para medir a linearidade das atividades enzimáticas, concentração de
glicogênio e substâncias que reagem com ácido tiobarbitúrico (TBARs).
Encontrar uma faixa linear com a curva-padrão permite escolher uma situação de
análise para futuras investigações com grupos experimentais distintos, onde aumentos ou
diminuições da atividade enzimática ou concentração refletem apenas a intervenção
realizada. Assim como permite ao pesquisador o aprendizado e manipulação da técnica,
para que variável humana tenha pouca interferência no experimento.
Foram feitas ainda as análises de cada ponto da curva em triplicata o que permitiu
plotar os gráficos com média e desvio-padrão (DP) e calcular o coeficiente de variação
(CV) (DP dividida pela média multiplicado por 100) em percentual, sendo que quanto
menor o CV mais homogênea se encontra a amostra. Calculamos ainda o standart erro
estimate (SEE) que mede o erro (distância) dos valores reais na reta, dos valores estimados
pela equação da Reta (Equação: Raiz quadrada de (Y-^Y)2/(n-2). Onde Y são os valores
reias, ^Y são os valores estimados pela equação da Reta. E n o número de observações)
sendo que pequenos valores significam pouca variação. As figuras abaixo apresentam as
curva-padrão para cada técnica utilizada, assim como os valores de amostra utilizados para
análise, tempo de leitura e CV das triplicatas.
Para análises enzimáticas foram realizadas curvas em diferentes tempos de leitura,
visto que as concentrações de substratos, enzima-substrato e produtos variam em função do
tempo em decorrente da reação enzimática.
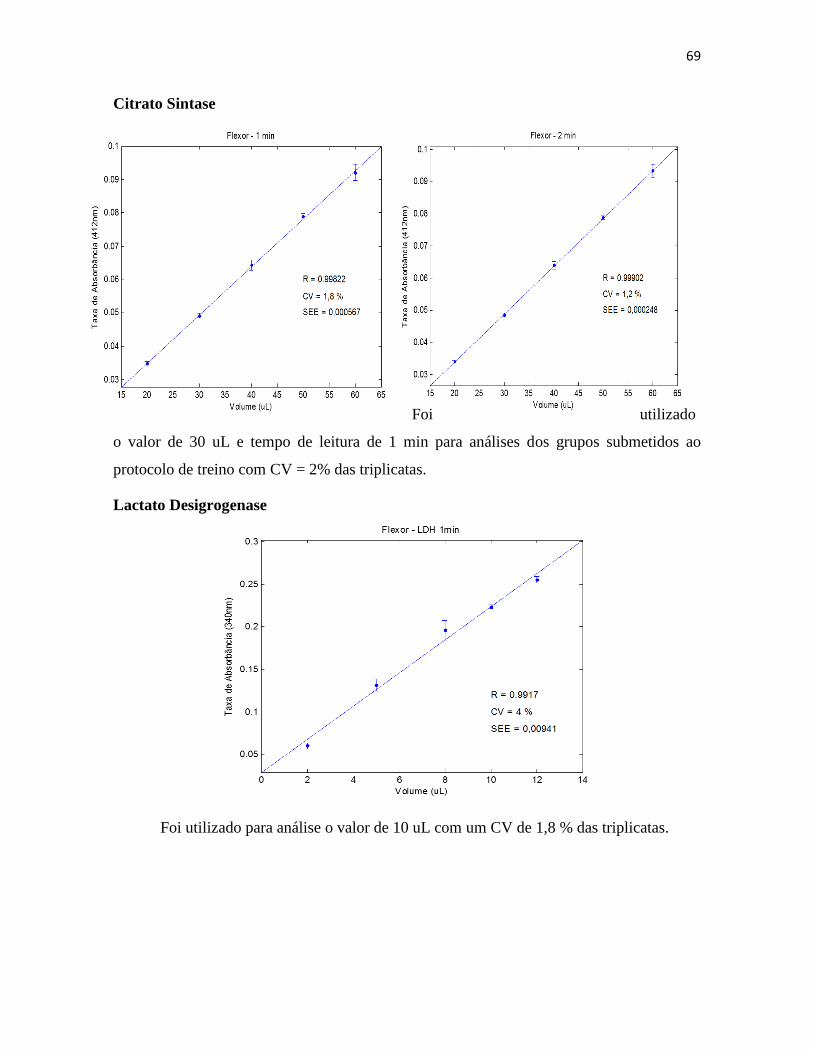
69
Citrato Sintase
Foi utilizado
o valor de 30 uL e tempo de leitura de 1 min para análises dos grupos submetidos ao
protocolo de treino com CV = 2% das triplicatas.
Lactato Desigrogenase
Foi utilizado para análise o valor de 10 uL com um CV de 1,8 % das triplicatas.
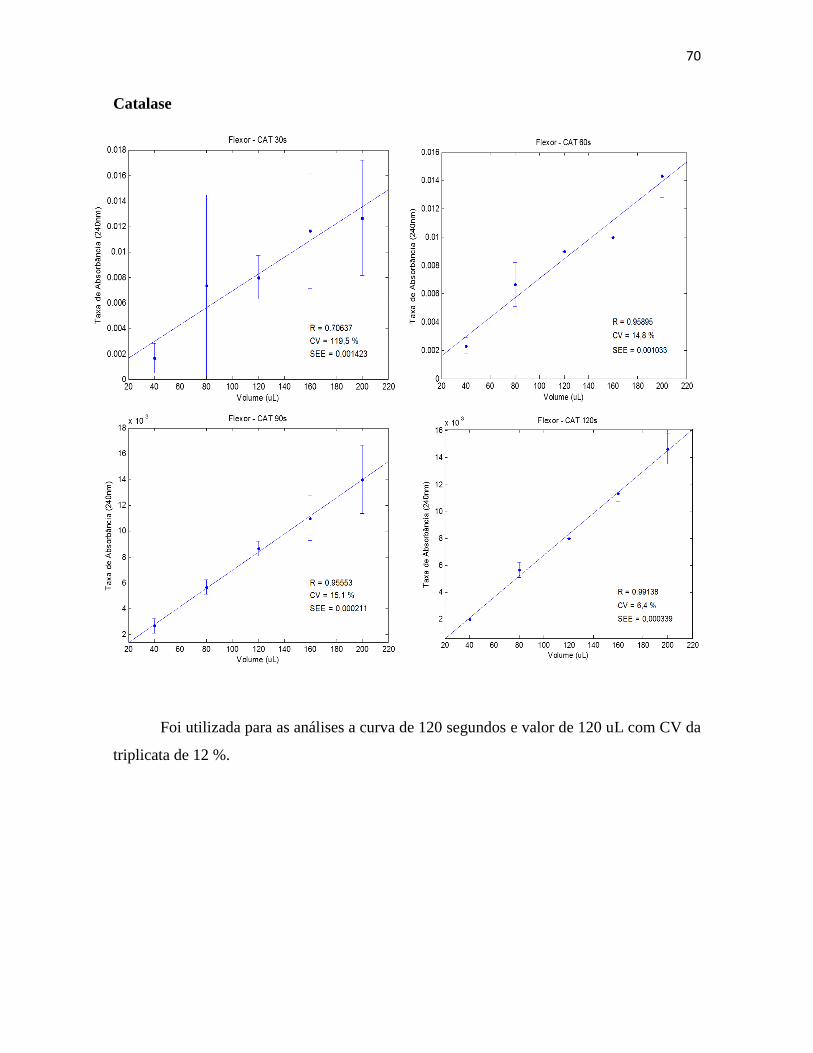
70
Catalase
Foi utilizada para as análises a curva de 120 segundos e valor de 120 uL com CV da
triplicata de 12 %.

71
Glutationa Redutase
Foi utilizada a curva de 4 minutos e valor de 40 uL com CV=12,4 % das triplicatas.

72
Superóxido Dismutase
Foi utilizado o valor de 10 uL para as análises com um CV de 1,8 % entre as
duplicatas.
Curva-Padrão do KIT
Para encontrar melhor linearidade e volumes iguais da curva do FHL a amostra da
SOD do kit foi diluída e o valor obtido multiplicado pela diluição.

73
Cálculo da atividade enzimática
A atividade enzimática é geralmente acompanhada espectrofotometricamente pelo
consumo de um substrato ou formação de um produto pela enzima onde um determinado
comprimento de onda é emitido em uma cubeta contendo o ensaio enzimático. Quando a
luz visível ou ultravioleta incide sobre qualquer corpo ou solução, uma parte é refletida,
outra absorvida e outra transmitida. Essas relações podem ser entendidas a partir da
equação 1 abaixo.
Equação 1: I0= Ir + Ia + I
Onde I0 é a luz incidente e Ir ,Ia e I são respectivamente as intensidades da luz
refletida, absorvida e transmitida. A luz refletida geralmente é muito baixa quando
utilizamos recipientes adequados ao comprimento de onda, as cubetas de quartzo
(λ<340nm) ou de vidro (λ>340nm). Chama-se transmitância a razão entre intensidade da
luz transmitida (I) e a luz incidente (I0).
Do ponto de vista experimental a transmitância também é chamada de Absorbância
(A). Ela é um parâmetro mais adequado porque existe uma relação linear entre ela e a
concentração de uma substância absorvente na solução, esta relação é dada pela lei de
lambert-Beer (equação 2).
Equação 2: C= A/Ɛ x L
Onde C é igual a concentração da substância, A = absorbância, Ɛ = coeficiente de extinção
molar e L = caminho ótico.
O coeficiente de extinção molar é característico da cada substância e pode ser
encontrado em tabelas de livro de bioquímica. Ele representa capacidade de 1 mol da
substância em absorver luz em determinado comprimento de onda. É geralmente expresso
em mol/cm e deve ser levado em conta para calcular a atividade da enzima.
Outros fatores devem ser levados em consideração quando vamos medir a atividade
de uma enzima. Um deles é que sua atividade varia em função do tempo, pois com passar
do tempo as concentrações de substrato e produto também mudam, alterando sua atividade.
Para isso o aparelho espectrofotômetro nos da um valor de rate ou taxa de absorbância em
determinado tempo, geralmente expresso em abs/segundos. Outro fator é a diluição da

74
amostra na cubeta e do tecido analisado no tampão (homogenato). Desta forma a equação 3
leva em conta todos esses fatores:
Equação 3:
Taxa (abs/min) x Volume total na cubeta (ml) x Volume do homogenato (ml)
Atividade = ---------------------------------------------------------------------------------------------------------------------------
Coef. Molar (ml.umol-1.cm-1) x Largura cubeta (cm) x Volume de amostra (ml) x Massa de tecido (g)
Em vermelho temos exatamente a equação da lei de lambert-beer. No caso de
enzimas não estamos interessados na concentração de determinado produto ou substrato,
mas sim a taxa que estas substâncias são formadas ou consumidas. Sendo assim a
concentração no caso foi substituída por atividade e absorbância pelo rate dado no
espectrofotômetro. Em verde temos a diluição da amostra na cubeta e azul a diluição do
homogenato. Cortanto as dimensões:
Temos a atividade expressa em umol/min g de tecido úmido. A massa do tecido no caso foi
pesada antes da homogenização do tecido. Esta notação também é igual U/g de tecido
úmido.
Obs: Em determinadas situações de padronização de técnicas, quando atividade da enzima
é muito alta, devemos diluir a amostra para conseguir encontrar uma faixa linear de leitura
dada pela curva-padrão. Nestas situações é necessário multiplicar o valor do rate obtido
pelo fator de diluição.

75
Exemplo:
Para diluir a amostra deve-se utilizar a solução do tampão homogenato. Se pegar 100 uL de
amostra e diluirmos em 900 uL de tampão, a diluição foi de 1:10. Ou seja o valor de rate
deverá ser multiplicado por 10!
Segue um exemplo do cálculo da atividade enzimática:
Foi realizado o ensaio enzimático em triplicata anotando 3 vezes o valor do rate de
(0.253,0.255,0.257 abs/segundos). É feita a média deste valor e utilizada na equação 3
substituindo a taxa de abs. Outros valores necessários para a equação são:
Ɛ= 3,94 ml.µmol-1.cm-1 para o TNB (produto medido no ensaio enzimático).
Volume da cubeta = 1 ml
Volume da amostra = 0,05 ml
Volume do Homogenato = 1 ml
Peso do tecido = 0,035 gramas
Largura da cubeta = 1 cm
Média do rate = 0.255 abs/segundos*
*A taxa do rate precisa ser multiplicada por 60min pois o U é dado em minutos (umol/min)
Rate = 15,3 abs/min
Desta forma temos:
15,3 x 1 x 1
Atividade = ---------------------------------------------
3,94 x 1 x 0,05 x 0,035
Atividade = 2219 U/ g tecido úmido ou 2219 umol/min/g tecido úmido.

76
Glicogênio
Para a padronização da técnica fizemos experimentos iniciais que constaram da
determinação do espectro de absorção de uma solução de glicogênio em água 100 g/ml
(A) e curva padrão (B) obtida com soluções de glicogênio em concentrações que variaram
de 10 a 100 g/ml.
O cálculo da concentração de glicogênio das amostras foi efetuado utilizando-se a
equação abaixo:
Concentração de Glicogênio (g/100g tecido) = (A490/k) x (V/v) x (10-4/w)
Onde: A490 = absorbância a 490 nm; k = coeficiente angular da curva padrão de
glicogênio; V = volume total da amostra de glicogênio; v = volume da alíquota da amostra
usada no ensaio; w = massa do tecido
A unidade g (glicogênio) / 100 g tecido foi transformada em mmol unidade glicosil / Kg
tecido, uma vez que 1 grama de glicogênio / 100g de tecido equivale a 55,5 mmol unidade
glicosil /Kg tecido (Passonne.Jv e Lauderda.Vr, 1974).
(A) Espectro de absorção de glicogênio em água (100g/ml). (B) Curva padrão de
glicogênio em água (10 a 100 g/ml) dosado pela adição de fenol 5% em meio ácido.
Cada ponto representa a média de três determinações.
0 20 40 60 80 100
0,0
0,2
0,4
0,6
0,8
1,0
1,2
Y=A+Bx
A= -0,009
B= 0,01
R= 0,999
P<0,0001
N=5
A4
90
g /mL Glicogênio400 450 500 550 600
0,0
0,2
0,4
0,6
0,8
1,0
1,2
Ab
s. (u
.a)
(nm)
A B
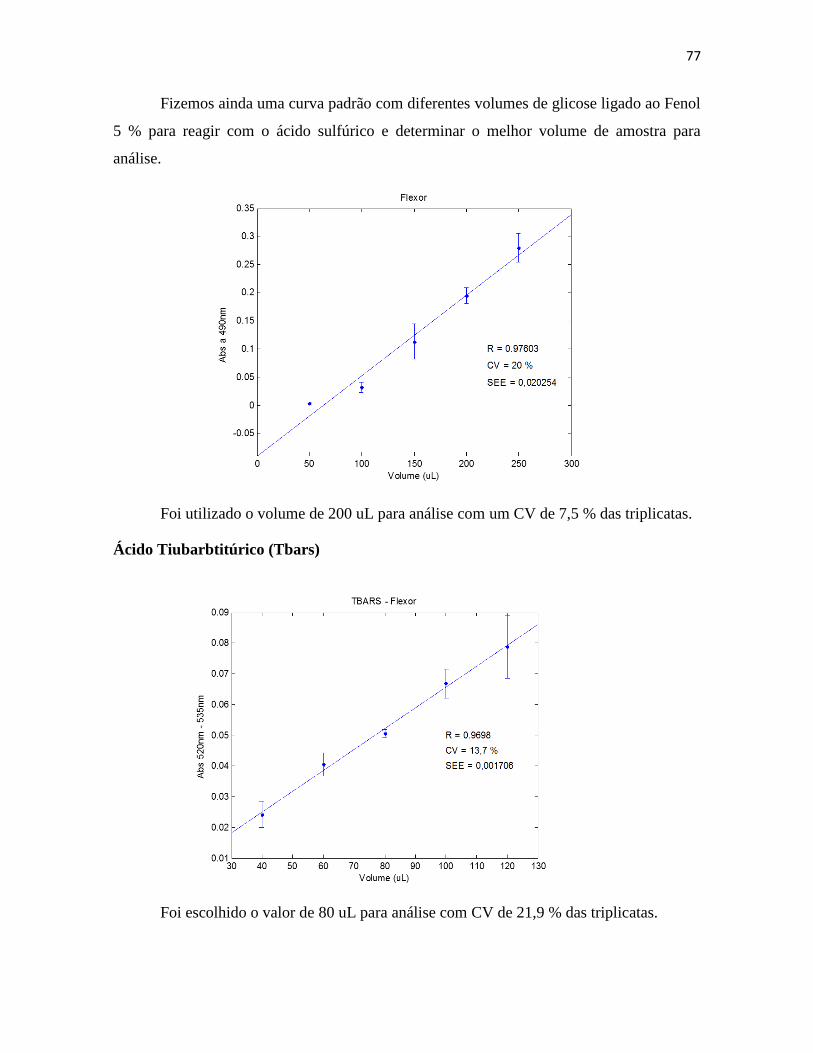
77
Fizemos ainda uma curva padrão com diferentes volumes de glicose ligado ao Fenol
5 % para reagir com o ácido sulfúrico e determinar o melhor volume de amostra para
análise.
Foi utilizado o volume de 200 uL para análise com um CV de 7,5 % das triplicatas.
Ácido Tiubarbtitúrico (Tbars)
Foi escolhido o valor de 80 uL para análise com CV de 21,9 % das triplicatas.

78
Cálculo da concentração de TBARs
A partir curva de TEP foi possível encontrar o coeficiente angular e linear da reta e
com equação da reta (y=ax+b) onde a=coeficiente angular e b= linear, substituímos o valor
de Y pela absorbância encontrada com a leitura da amostra e encontramos X= nnmol de
TEP. Relativizando este valor pela massa de tecido encontramos a concentração de TBARs
em nnmol/ grama de tecido úmido.
0.2 0.25 0.3 0.35 0.4 0.45 0.5 0.55 0.6 0.65
0.06
0.08
0.1
0.12
0.14
0.16
0.18
Curva TEP
Concentração TEP (nnmol/ml)
Abs 5
20nm
- 5
35nm

79
Anexo B

80

81

82
VII. Referências Bibliográficas
Aebi,H. Catalase in vitro. In: Methods in Enzymology. 105:121-126, 1984.
Andersen, M.L, Bignotto M, Machado RB, Tufik S. Different stress modalities result in
distinct steroid hormone responses by male rats. Braz J Med Biol Res. 2004;37(6):791-7.
Andreollo, N.A., Santos E.F, Araújo, M.R., Lopes, L.R. Rat's age versus human's age: what
is the relationship? Arq Bras Cir Dig. 2012 Jan-Mar;25(1):49-51.
Antonio, J., Gonyea, W.J. Role of muscle fiber hypertrophy and hyperplasia in
intermittently stretched avian muscle. J Appl Physiol. 1993;74:1893–1898.
Baar, K,, Esser, K. Phosphorylation of p70S6k correlates with increased skeletal muscle
mass following resistance exercise. Am J Physiol. 1999;276:C120–C127.
Balaban, R.S., Nemoto. S., Finkel, T. Mitochondria, oxidants, and aging Cell. 2005 Feb
25;120(4):483-95.
Baird, M. F., Graham,S.M., Baker, J.S., Bikerstaf, G.,F. Creatine-Kinase-and Exercise-
Related Muscle Damage Implications for Muscle Performance and Recovery. Journal of
Nutrition and Metabolism. Volume 2012.
Baldwin, K.M., Valdez, V., Herrick, R.E., MacIntosh, A.M., Roy, R.R. Biochemical
properties of overloaded fast-twitch skeletal muscle. J Appl Physiol. 1982;52:467–472.
Bloomer, R.J. Efect of exercise on oxidative stress biomarkers. Adv Clin Chem. 46:1–50,
2008.
Bodine, S.C., Stitt, T.N., Gonzalez, M., Kline, W.O., Stover, G.L., Bauerlein, R.,
Zlotchenko, E., Scrimgeour, A., Lawrence, J.C., Glass, D.J., Yancopoulos, G.D.
Akt/mTOR pathway is a crucial regulator of skeletal muscle hypertrophy and can prevent
muscle atrophy in vivo. Nat Cell Biol 3: 1014–1019, 2001.
Bodine, S.C. mTOR signaling and the molecular adaptation to resistance exercise Med Sci
Sports Exerc. 2006 Nov;38(11):1950-7.
Booth FW, Tseng BS, Fluck M, Carson JA. Molecular and cellular adaptation of muscle in
response to physical training. Acta Physiol Scand. 1998;162:343–350.

83
Børsheim, E., Bahr, R. Effect of exercise intensity, duration and mode on post-exercise
oxygen consumption. Sports Med. 2003;33(14):1037-60.
Boveris, A., Oshino, N., Chance, B. The cellular production of hydrogen peroxide.
Biochem J. 1972 Jul;128(3):617-30.
Braith, R.,W., Magyari, P.M., Pierce, G.L., Edwards, D.G., Hill, J.A., White, L.J., Aranda,
J.M. Effect of resistance exercise on skeletal muscle myopathy in heart transplant
recipients. Am J Cardiol. 2005 May 15;95(10):1192-8.
Burd, N.A., Mitchell, C.J., Churchward-Venne, T.A., Phillips, S.M. Bigger weights may
not beget bigger muscles: evidence from acute muscle protein synthetic responses after
resistance exercise Appl. Physiol. Nutr. Metab. 37: 551–554 (2012).
Bucci, L.R., Blackman, G., Defoyd, W., Kaufmann, R., Mandel, T., C., Sparks, W.S. Effect
of ferulate on strength and body composition of weightlifters. J Appl Sport Sci Res. 1990;
4 : 104-9.
Cakir-Atabek, H., Demir, S., Pinarba Şili, R.D,, Gündüz, N. Effects of different resistance
training intensity on indices of oxidative stress. J Strength Cond Res. 2010
Sep;24(9):2491-7.
Cassilhas RC, Lee KS, Fernandes J, Oliveira MG, Tufik S, Meeusen R, de Mello MT.
Spatial memory is improved by aerobic and resistance exercise through divergent
molecular mechanisms. Neuroscience. 2012 Jan 27;202:309-17.
Cassilhas, R.C., Reis, I.T.,Venâncio, D., Fernandes, J., Tufik, S., Marco de Mello, M., T.
Modelo animal de exercício resistido progressivo: descrição detalhada do modelo e a sua
implicação à pesquisa básica com exercício. Motriz: rev. educ. fis. vol.19 no.1 Rio
Claro Jan./Mar. 2013.
Carson, J.A. The regulation of gene expression in hypertrophying skeletal muscle. Exerc
Sport Sci Rev. 1997;25:301–320.
Chan, S.T., Johnson, A.W., Moore, M.H., Kapadia, C.R., Dudley, H.A. Early weight gain
and glycogen-obligated water during nutritional rehabilitation. Hum Nutr Clin Nutr 36:
223–232, 1982.

84
Cheung, P.C., Salt, I.P., Davies, S.P., Hardie, D.G., and Carling, D. 2000. Characterization
of AMP-activated protein kinase gamma subunit isoforms and their role in AMP binding.
Biochem. J. 346(3): 659–669.
Cheruvanky, R., Thummala, R.C. Nutritional and biochemical aspects of the hypolipidemic
action of rice bran oil. Am. Coll. Of Nutrition,10(4): 593-601, 1991.
Chibalin, A.V., Yu, M., Ryder, J.W., Song, X.M., Galuska, D., Krook, A., Wallberg-
Henriksson, H., Zierath, J.R. Exercise-induced changes in expression and activity of
proteins involved in insulin signal transduction in skeletal muscle: differential effects on
insulin-receptor substrates 1 and 2. Proc Natl Acad Sci U S A. 2000 Jan 4;97(1):38-43.
Childs, A., Jacobs, C., Kaminski, T., Halliwell, B., Leeuwenburgh, C. Supplementation
with vitamin C and N-acetyl-cysteine increases oxidative stress in humans after an acute
muscle injury induced by eccentric exercise. Free Radic Biol Med. 2001 Sep
15;31(6):745-53.
Chilibeck P.D., Syrotuik, D.G.,Bell, G.J. The effect of strength training on estimates of
mitochondrial density and distribution throughout muscle fibres. Eur J Appl Physiol
Occup Physiol. 1999. 80, 604–60.
Davies, K.J.A., Quintanilha, A.T., Brooks,G.A., Packer,L. Free radicals and tissue damage
produced by exercise. Biochem. Biophys. Res. Com. 107(4): 1198-1205, 1982.
Decker, T., Lohmann-Matthes, M.L. A quick and simple method for the quantitation of
lactate dehydrogenase release in measurements of cellular cytotoxicity and tumor necrosis
factor (TNF) activity. J Immunol Methods. 1988 Nov 25;115(1):61-9.
Degens, H., Meessen, N.E., Wirtz, P., Binkhorst, R.A. The development of compensatory
hypertrophy in the plantaris muscle of the rat. Anat Anz. 1995;177:285– 289.
Deminice, R., Sicchieri, T., Payão, P.O., Jordão, A.A. Blood and salivary oxidative stress
biomarkers following an acute session of resistance exercise in humans. Int J Sports Med.
2010 Sep;31(9):599-603.
Donatto, F.,F., Neves, R.X., Rosa, F.O., Camargo, R.G., Ribeiro, H., Matos-Neto, E.M.,
Seelaender, M. Resistance exercise modulates lipid plasma profile and cytokine content in
the adipose tissue of tumour-bearing rats. Cytokine. 2013 Feb;61(2):426-32.

85
Duncan, N.D., Williams, D.A., Lynch, G.S. Adaptations in rat skeletal muscle following
long-term resistance exercise training. European Journal of Applied Physiology and
Occupational Physiology, New York, v. 77, n.4, p. 372-378, 1998.
Eslami, S., Esa, N.M., Marandi, S.M., Ghasemi, G., Eslami S. Effects of gamma oryzanol
supplementation on anthropometric measurements & muscular strength in healthy males
following chronic resistance training. Indian J Med Res. 2014 Jun;139(6):857-63.
Ferreira, A.L.A., Matsubara, L.S., Radicais Livres: Conceitos, doenças relacionadas,
sistema de defesa e estresse oxidativo. Rev. Ass. Med. Brasil 43(1): 61-8, 1997.
FDA. Guidance for industry-Estimating the maximum safe dose in initial clinical trials for
therapeutics in adult healthy volunteers. Rockville. 2005.
Fisher-Wellman, K., Bloomer, R.J. Acute exercise and oxidative stress: a 30 year history.
Dyn Med. 2009 Jan 13;8:1.
Fluck, M. Molecular mechanisms in muscle adaptation. Ther Umsch. 60 (7):371-81. 2003.
Foster, J.L., Small, J.D., Fox, J.G. The mouse in biomedical research. New York:
Academic Press. 1983.
Fry, A.C., Bonner, E., Lewis, D.L, Johnson, R.L, Stone, M.H, Kraemer, W.J. The effects of
gamma-oryzanol supplementation during resistance exercise training. Int J Sport Nutr.
1997 Dec;7(4):318-29
Gerhardt, A.L, Gallo, N.B. Full-fat rice bran and oat bran similarly reduce
hypercholesterolemia in humans. Journal Nutrition. 1998 May;128(5):865-9.
Gomez-Cabrera, M.C., Domenech, E., Viña, J. Moderate exercise is an antioxidant:
upregulation of antioxidant genes by training. Free Radic Biol Med. 2008 Jan
15;44(2):126-31.
Gomez-Cabrera, M.C., Domenech, E., Romagnoli, M., Arduini, A., Borras, C., Pallardo,
F.V., Sastre, J., Viña, J. Oral administration of vitamin C decreases muscle mitochondrial
biogenesis and hampers training-induced adaptations in endurance performance. Am J Clin
Nutr. 2008 Jan;87(1):142-9.

86
Gomez-Cabrera, M.C, Ristow, M., Viña, J. Antioxidant supplements in exercise: worse
than useless? Am J Physiol Endocrinol Metab. 2012 Feb 15;302(4):E476-7.
Green H, Goreham C, Ouyang J, Ball-Burnett M & Ranney D (1999). Regulation of fiber
size, oxidative potential, and capillarization in human muscle by resistance exercise. Am J
Physiol Regul Integr Comp Physiol 276, R591–R596
Grunewald, K.K., Bailey, R.S. Commercially marketed supplements for bodybuilding
athletes. Sports Med. 1993 Feb;15(2):90-103.
Gwinn, D.M., Shackelford, D.B., Egan, D.F., Mihaylova, M.M., Mery, A., Vasquez, D.S.,
Turk, B.E., Shaw, R.J. AMPK phosphorylation of raptor mediates a metabolic checkpoint.
Mol Cell. 2008 Apr 25;30(2):214-26.
Goldspink, D.F., Cox, V.M., Smith, S.K., Muscle growth in response to mechanical stimuli.
Am J Physiol. 1995;268:E288–E297.
Goldberg AL. Work-induced growth of skeletal muscle in normal and hypophysectomized
rats. Am J Physiol. 1967;213:1193–1198.
Hamilton K.L., Staib, J.L., Phillips, T., Hess, A., Lennon, S.L., Powers, S.K. Exercise,
antioxidants, and HSP72: protection against myocardial ischemia/reperfusion. Free Radic
Biol Med. 2003. 34: 800–809
Halliwell, B., and Gutteridge, J. 2007. Free Radicals in biology and medicine. 4th ed.
Oxford. Oxford, UK. 851 pp.
Halliwell,B. Chirico,S. Lipid peroxidation: its mechanism, measurement, and significance.
Am J Clin Nutr. 1993 May;57.
Harman, D. (1956) Aging: a theory based on free radical and radiation chemistry. J
Gerontol 11: 298–300.
Hawke, T.J., Garry, D.J. Myogenic satellite cells: physiology to molecular biology. J Appl
Physiol 2001;91(2):534-51.
Hawkins, S.A.,Wiswell, R.A., Marcell, T.J. Exercise and the master athlete–a model of
successful aging? J Gerontol A Biol Sci Med Sci 2003 58: 1009–1011.

87
Sies, H. Oxidative stress: from basic research to clinical application. Am J Med. 1991 Sep
30;91(3C):31S-38S.
Henneman, E., Somjem, G., Carpenter, D. Excitability and inability of motoneurons of
different size. Journal of Neurophysiology. vol. 28, p. 599-620, 1965.
Hellsten, Y., Frandsen, U., Orthenblad, N., Sjødin, B., Richter, E.A. Xanthine oxidase in
human skeletal muscle following eccentric exercise: a role in inflammation. J Physiol.
1997 Jan 1;498 ( Pt 1):239-48.
Hellsten, Y., Richter, E.A., Kiens, B., and Bangsbo, J. AMP deamination and purine
exchange in human skeletal muscle during and after intense exercise. J. Physiol. 1999.
520(3): 909–920.
Hellsten, Y. The role of xanthine oxidase in exercise. In: C.K. Sen, L. Packer, and O.
Hanninen (Eds.), Handbook of Oxidants and Antioxidants in Exercise, pp. 153- 176.
Amsterdam: Elsevier Scienc. 2000.
Hiramitsu, T., Armstrong, D. Preventive effect of antioxidants on lipid peroxidation in the
retina. Ophthalmic Res. 1991;23(4):196-203.
Ho, K.W., Roy, R.R., Tweedle, C.D., Heusner, W.W., Van Huss, W.D., Carrow, E.
Skeletal muscle fiber splitting with weight-lifting exercise in rats. Am J Anat.
1980;157:433–440.
Holloszy, J.O., Coyle, E.F. Adaptations of skeletal muscle to endurance exercise and their
metabolic consequences. J Appl Physiol Respir Environ Exerc Physiol. 1984
Apr;56(4):831-8.
Holvoet, P. Relations between metabolic syndrome, oxidative stress and inflammation and
cardiovascular disease. Verh K Acad Geneeskd Belg. 2008;70(3):193-219
Hornberger, T.A., Farrar, R.P. Physiological hypertrophy of the FHL muscle following 8
weeks of progressive resistance exercise in the rat. Canadian Journal of Applied
Physiology, Champaign, v. 29, p. 16-31, 2004.

88
Hornberger,T.A., W. K. Chu, Y. W. Mak, J. W. Hsiung, S. A. Huang, and S. Chien. The
role of phospholipase D and phosphatidic acid in the mechanical activation of mTOR
signaling in skeletal muscle. PNAS March 21, 2006 vol. 103 no. 12.
Hornberger, T.A., Sukhija, K.B., Wang, X.R, Chien, S. mTOR is the rapamycin-sensitive
kinase that confers mechanically-induced phosphorylation of the hydrophobic motif site
Thr(389) in p70(S6k). FEBS Lett. 2007;581(24):4562-6.
Hudson, M.B., Hosick, P.A., McCaulley, G.O., Schrieber, L., Wrieden, J., McAnulty, S.R.,
Triplett, N.T, McBride, J.M, Quindry, J.C. The effect of resistance exercise on humoral
markers of oxidative stress. Med Sci Sports Exerc. 2008 Mar;40(3):542-8.
Hunter, G.R., McCarthy, J.P., Bamman, M.M. Effects of Resistance training on older
adults. Sports Med. 2004,34(5):329-48.
Islam, M.S., Murata T, Fujisawa M, Nagasaka R, Ushio H, Bari AM, Hori M, Ozaki H.
Anti-inflammatory effects of phytosteryl ferulates in colitis induced by dextran sulphate
sodium in mice. Br J Pharmacol. 2008 Jun;154(4):812-24.
Islam, M.S, Nagasaka, R., Ohara, K., Hosoya, T., Ozaki, H., Ushio, H., Hori, M. Biological
abilities of rice bran-derived antioxidant phytochemicals for medical therapy. Curr Top
Med Chem. 2011;11(14):1847-53.
Jensen, J., Lai, Y.C. Regulation of muscle glycogen synthase phosphorylation and kinetic
properties by insulin, exercise, adrenaline and role in insulin resistance. Arch Physiol
Biochem. 2009 Feb;115(1):13-21.
Ji, L.L. Antioxidant enzyme response to exercise and aging. Med Sci Sports Exerc. 1993
Feb;25(2):225-31.
Ji, L.L.Exercise-induced modulation of antioxidant defense. Ann N Y Acad Sci. 2002
Apr;959:82-92.
Jiang, F., Zhang Y, Dusting GJ. NADPH oxidase-mediated redox signaling: roles in
cellular stress response, stress tolerance, and tissue repair. Pharmacol Rev. 2011
Mar;63(1):218-42
Jung, C.H., Ro, S.H., Cao, J., Otto, N.M., Kim, D.H. mTOR regulation of autophagy.
FEBS Lett. 2010 Apr 2;584(7):1287-95.

89
Kang, C., O'Moore, K.M., Dickman, J.R., Ji, L.,L. Exercise activation of muscle
peroxisome proliferator-activated receptor-gamma coactivator-1alpha signaling is redox
sensitive. Free Radic Biol Med. 2009 Nov 15;47(10):1394-400.
Kelley, G. Mechanical overload and skeletal muscle fiber hyperplasia: A meta-analysis. J
Appl Physiol 81: 1584–1588, 1996.
Kirk, E.P., Donnelly, J.E., Smith, B.K., Honas, J., Lecheminant, J.D., Bailey, B.W.,
Jacobsen, D.J, Washburn, R.A. Minimal resistance training improves daily energy
expenditure and fat oxidation. Med Sci Sports Exerc. 2009 May;41(5):1122-9.
Klitgaard, H. A model for quantitative strength training of hindlimb muscles of the rat. J
Appl Physiol. 1988;64:1740–1745.
Kim, S.,J., Han, D., Moon, K.D., Rhee, J.S.,Measurement of superoxide dismutase-like
activity of natural antioxidants. Biosci Biotechnol Biochem. 1995 May;59(5):822-6.
Kraemer, W,J,, Adams, K,, Cafarelli, E,, Dudley, G,A,, Dooly, C , Feigenbaum, M.S,,
Fleck, S,J,, Franklin, B,, Fry, A,C,, Hoffman, J,R,, Newton, R.U,, Potteiger, J,, Stone,
M,H,, Ratamess, N, A,, and Triplett-McBride, T, (2002), American College of Sports
Medicine position stand. Progression models in resistance training for healthy adults, Med.
Sci. Sports Exerc. 34: 364-380.
Kraemer, W.J., Rogol, A.D. Endocrine System in Sports and Exercise. Volume XI of the
encyclopedia of sports medicine. Blacwell. 2005.
Kyriakis,J.M., Avruch, J. Mammalian mitogen-activated protein kinase signal transduction
pathways activated by stress and inflammation. Physiol Rev 81: 807–869, 2001.
Lang F. Mechanisms and significance of cell volume regulation. J Am Coll Nutr.
2007;26(5 Suppl.):613S–23S.
Layne, J.E., Nelson, M.E. The effects of progressive resistance training on bone density: a
review. Med SciSports Exerc. 1999;31:25–30.
Lieberman, D.E. The Story of the human Body. Evolution, Health and Disease. Editora:
Pantheon Books, New York. 2013.
Liu, Y., Fiskum, G., Schubert, D. Generation of reactive oxygen species by the
mitochondrial electron transport chain. J Neurochem. vol. 80(5):780-7, 2002.

90
Liu, C.Y.,Lai,Y.C.,Wu, Y.C., Tzeng, C.H., Lee, S.D. “Macroenzyme creatine kinase in the
era of modern laboratory medicine,” Journal of the Chinese Medical Association, vol. 73,
no. 1, pp. 35–39, 2010.
Loewenstrein, W.R. Physics in Mind. A quantum view of the brain. Basic Books. New
York. 2013
Low, SY, Rennie, MJ, and Taylor, PM. Signaling elements involved in amino acid
transport responses to altered muscle cell volume. FASEB J 11: 1111–1117, 1995.
Lowe, A.D., Alway, E.S. Animal Models for Inducing Muscle Hypertrophy: Are They
Relevant for Clinical Applications in Humans? Journal of Orthopaedic & Sports
Physical Therapy. 2002.
Machado, E.F; Normand, A.C; Nunes, L.A; Brenzikofer, R; Macedo D.V. Effects of
different general anesthetics on serum hemolysis and hepatic and muscular glycogenolysis
in rats. Braz J Med Biol Res.vol. 42(11): p.1035-8, 2009.
MacDougall JD, Sale DG, Moroz JR, Elder GC, Sutton JR & Howald H (1979).
Mitochondrial volume density in human skeletal muscle following heavy resistance
training. Med Sci Sports 11, 164–166.
McCartney, N. Acute responses to resistance training and safety. Med Sci Sports Exerc.
1999;31:31–37.
Mcardle, D.W., Katch, L.F., Katch, L. V. Fisiologia do Exercício: Energia, Nutrição e
Desempenho Humano.6º. ed. Rio Janeiro, Guanabara Koogan, 2008.
McGill, R., Tukey, J.W., Larsen, W. Variations of box plot. The American Statistician,
32(1): 12-16,1978.
Molnar, A.A., Alves, A.A., Pereira-da-Silva, L., Macedo, D.V., Dabbeni-Sala, F.
Evaluation by blue native polyacrylamide electrophoresis colorimetric staining of the
effects of physical exercise on the activities of mitochondrial complexes in rat muscle.
Braz Journ Med Bio Res. 37: 939-947, 2004.
Musarò, A., Fulle, S., Fanò, G. Oxidative stress and muscle homeostasis. Curr Opin Clin
Nutr Metab Care. 2010 May;13(3):236-42.

91
Nagasaka, R., Chotimarkorn, C., Shafiqul, I.M., Hori, M., Ozaki, H., Ushio H. Anti-
inflammatory effects of hydroxycinnamic acid derivatives. Biochem Biophys Res
Commun. 2007 Jun 29;358(2):615-9. Epub 2007 May 7.
Novikoff, A. B., Shin, W. Y. and Drucker, J. Mitochondrial localization of oxidative
enzymes: staining results with two tetrazolium salts. J. Biophys. Biochem. Cytol.(1961) 9,
47-61.
Ohara, A. Radicais livres: bons, maus e naturais. Ed. Oficina de Textos. São Paulo, 2006.
Okawa, H., Onishi, N., Yagi, K. Assay for lipid peroxides in animal tissues by
thiobarbituric acid reaction. Analytical Biochemistry. 95:351-358, 1979
Ormsbee, M.J., Thyfault, J.P., Johnson, E.A., Kraus, R.M., Choi, M.D., Hickner, R. Fat
metabolism and acute resistance exercise in trained men. J Appl Physiol (1985). 2007
May;102(5):1767-72
Ørtenblad, N., Westerblad, H., Nielsen, J. Muscle glycogen stores and fatigue. J Physiol.
2013 Sep 15;591(Pt 18):440513.
O’Keefe,J., Robert Vogel, Carl J. Lavie, Loren Cordain Achieving Hunter-gatherer Fitness
in the 21st Century: Back to the Future. The American Journal of Medicine, Vol xx, No
x, Month 2010.
Parise, G., Brose, A.N., Tarnopolsky, M.A. Resistance exercise training decreases oxidative
damage to DNA and increases cytochrome oxidase activity in older adults. Exp Gerontol.
2005 Mar;40(3):173-80.
Parise, G., Phillips, S.M., Kaczor, J.J., Tarnopolsky, M.A. Antioxidant enzyme activity is
up-regulated after unilateral resistance exercise training in older adults. Free Radic Biol
Med. 2005 Jul 15;39(2):289-95.
Paschalis, V., Nikolaidis, M.G., Fatouros, I.G., Giakas, G., Koutedakis, Y., Karatzaferi, C.,
Kouretas, D., Jamurtas, A.Z. Uniform and prolonged changes in blood oxidative stress after
muscle-damaging exercise. In Vivo. 2007 Sep-Oct;21(5):877-83.
Patel, M., Naik. S.,N. Gamma-oryzanol from rice bran oil – A review. Journal of scientific
& industrial research. Vol. 63, July 2004. pp 569-578.
Pette. D., Staron, R,S.Transitions of muscle fiber phenotypic profiles. Histochem cell boil.
2001. 115:359-372.
Petrella, J.K., Kim, J., Cross, J.M., Kosek, D.J., Bamman, M.M. Efficacy of myonuclear
addition may explain differential myofiber growth among resistance-trained young and
older men and women. Am J Physiol Endocrinol Metab. 2006;291(5):E937

92
Philippou, A., Halapas, A., Maridaki, M., Koutsilieris, M. Type I insulin-like growth factor
receptor signaling in skeletal muscle regeneration and hypertrophy. J Musculoskelet
Neuronal Interact. 2007;7(3):208-18.
Pimenta, Ada, S., Lambertucci, R.H., Gorjão, R., Silveira, Ldos, R., Curi,R. Effect of a
single session of electrical stimulation on activity and expression of citrate synthase and
antioxidant enzymes in rat soleus muscle. Eur J Appl Physiol. 2007 Dec;102(1):119-26.
Epub 2007 Oct 26.
Powers S.,K., Jackson, M.J. Exercise-induced oxidative stress: cellular mechanisms and
impact on muscle force production. Physiol Rev. 2008 Oct;88(4):1243-76.
Rhonda, Bassel-Duby, & Eric, N. Olson. Signaling Pathways in Skeletal Muscle
Remodeling. Annu. Rev. Biochem. 2006.75:19-37.
Pryor, W.A., Houk, K.N., Foote, C.S., Fukuto, J.M.,Ignarro, L.J., Squadrito, G.L., Davies,
K.J. Free radical biology and medicine: it's a gas, man! Am J Physiol Regul Integr Comp
Physiol. 2006 Sep;291(3):R491-511.
Radak, Z., Chung, H.Y., Goto, S. Exercise and hormesis: oxidative stress-related adaptation
for successful aging. Biogerontology. 2005;6(1):71-5.
Ramel, A., Wagner, K.H., Elmadfa, I. Plasma antioxidants and lipid oxidation after
submaximal resistance exercise in men. Eur J Nutr. 2004 Feb;43(1):2-6. Epub 2004 Jan 6.
Ralph, L., Brinster. Lactate dehydrogenase activity in the preimplanted mouse embryo.
Biochim. Biophvs. Acta, 110 (1965) 439-441.
Reid, M.B., Durham, W.J. Generation of reactive oxygen and nitrogen species in
contracting skeletal muscle: potential impact on aging. Ann N Y Acad Sci.959:108-16,
2002.
Richard, D., Kefi, K., Barbe, U., Bausero, P., Visioli, F. Polyunsaturated fatty acids as
antioxidants. Pharmacol Res. 2008 Jun;57(6):451-5.
Ristow, M., Zarse, K., Oberbach, A., Klöting, N., Birringer, M., Kiehntopf, M., Stumvoll,
M., Kahn, C.R., Blüher, M. Antioxidants prevent health-promoting effects of physical
exercise in humans. Proc Natl Acad Sci U S A. 2009 May 26;106(21):8665-70. doi:
10.1073/pnas.0903485106. Epub 2009 May 11.

93
Rodrigues, M.F., Stotzer, U.S., Domingos, M.M., Deminice, R., Shiguemoto, G.E., Tomaz,
L.M., Sousa, N.M., Ferreira F.C., Leite, R.D., Selistre-de-Araújo, H.S., Jordão-Júnior,
A.A., Baldissera, V., Perez, S.E. Effects of ovariectomy and resistance training on
oxidative stress markers in the rat liver. Clinics (Sao Paulo). 2013 Sep;68(9):1247-54.
Ruas, J.L, White, J.P., Rao, R.R., Kleiner, S., Brannan, K.T., Harrison, B.C., Greene, N.P.,
Wu, J., Estall, J.L., Irving, B.A., Lanza, I.R., Rasbach, K.A., Okutsu, M., Nair, K.S., Yan,
Z., Leinwand,A., Spiegelman, B.M. A PGC-1α isoform induced by resistance training
regulates skeletal muscle hypertrophy. Cell. 2012 Dec 7;151(6):1319-31
Sakuma, K., Yamaguchi, A. The functional role of calcineurin in hypertrophy,
regeneration, and disorders of skeletal muscle. J Biomed Biotechnol. 2010;2010:721219.
Scheffer, D.L., Silva, L.A., Tromm, C.B., da Rosa, G.L., Silveira, P.C., de Souza, C.T.,
Latini, A., Pinho, R.A. Impact of different resistance training protocols on muscular
oxidative stress parameters. Appl Physiol Nutr Metab. 2012 Dec;37(6):1239-46.
Schoenfeld, B.J. The mechanisms of muscle hypertrophy and their application to resistance
training. J Strength Cond Res. 2010 Oct;24(10):2857-72.
Schoenfeld, B.J. Potential mechanisms for a role of metabolic stress in hypertrophic
adaptations to resistance training. Sports Med. 2013 Mar;43(3):179-94.
Sjödin, B., Hellsten-Westing, Y., Apple, F.S. Biochemical mechanisms for oxygen free
radical formation during exercise. Sports Med. vol.10(4): p. 236-54, 1990.
Silva, F. O. C., Macedo, D. V. Exercício físico, processo inflamatório e adaptação: uma
visão geral. Revista Brasileira de Cineantropometria & Desempenho Humano
(Impresso), v. 13, p. 320, 2011
Silveira, L.R., Considerações críticas e metodológicas na determinação de espécies reativas
de oxigênio e nitrogênio em células musculares durante contrações. Arq Bras Endocrinol
Metab vol.48 no.6 São Paulo Dec. 2004.
Silveira, L.R.,Hirabara, S.M., Lambertucci, R.H., Leandro, C.V., Fiamoncini,J.,
Pinheiro,C.H.J, D'Angelo,A.C.A., Bassit,R.A., Pithon-Curi, T.C. Rui Curi. Regulação
metabólica e produção de espécies reativas de oxigênio durante a contração muscular:
efeito do glicogênio na manutenção do estado redox intracelular. Rev Bras Med
Esporte vol.14 no.1 Niterói Jan./Feb. 2008.

94
Smith, I.K. ; Vierheller, T.L. ; Thorne, C.A. Assay of Glutathione Reductase in Crude
Tissue Homogenates using 5,5-Dithiobis(2-nitrobenzoic Acid). Anal Bioch 175: 408-413,
1988.
Spiering, B.A., Kraemer, W.J., Anderson, J.M., Armstrong, L.E., Nindl, B.C., Volek,
J.S., Judelson D.A., Joseph, M., Vingren J.L., Hatfield D.L., Fragala, M.S., Ho,
J.Y, Maresh, C.M. Effects of elevated circulating hormones on resistance exercise-induced
Akt signaling. Med Sci Sports Exerc. 2008 Jun;40(6):1039-48.
Stott, G. H. What is Animal Stress and how is it Measured? Journal of Animal Science
1981, 52:150-153.
Srere, P.A. (1969). Citrate Syntase. Methods in enzymology 13:3-5.
Sukho, Lee, Farrar, R. P., Resistance training induces muscle-specific changes in muscle
mass and function in rat. Journal of Exercise Physiology (online). Volume 6 Number 2
May 2003.
Tang, J.E., Hartman, J.W., Phillips, S.M., Increased muscle oxidative potential following
resistance training induced fibre hypertrophy in young men. Appl Physiol Nutr Metab.
2006 Oct;31(5):495-501.
Tamaki, T., Uchiyama, S., Nakano, S. A weight-lifting exercise model for inducing
hypertrophy in the hindlimb muscles of rats. Med Sci Sports Exerc. 1992; 24:881–886
Tappia, P.S., Dent, M.R., Dhalla, N.S. Oxidative stress and redox regulation of
phospholipase D in myocardial disease. Free Radical Biol Med. 2006;41(3):349–61.
Thomas Decker, Marie-Luise Lohmann-Matthes. A quick and simple method for the
quantitation of lactate dehydrogenase release in measurements of cellular cytotoxicity and
tumor necrosis factor (TNF) activity. Journal of Immunological Methods, 15 (1988) 61-
69.
Tidball, J.G. Inflammatory processes in muscle injury and repair. Am J Physiol Regul
Integr Comp Physiol. 2005 Feb;288(2):R345-53
Tollback, A., Eriksson, S., Wredenberg, A. Effects of high resistance training in patients
with myotonic dystrophy. Scand J Rehabil Med. 1999;31:9–16.

95
Toigo, M.,Boutellier, U. New fundamental resistance exercise determinants of molecular
and cellular muscle adaptations. Eur J Appl Physiol 97: 643–663, 2006.
Torres, Bayardo B.; Marzzoco, Anita. Bioquímica Básica. 3. ed. Rio de Janeiro:
Guanabara Koogan, 2007.
Tremblay,M.S., Esliger, D.W., Copeland, J.L., , J.D., Bassett, D.R. Moving forward by
looking back: lessons learned from long-lost lifestyles. Appl Physiol Nutr Metab.
2008;33:836-842.
Uchiyama, M; Mihara, M. Determination of Malonaldehyde precursor in tissues by
thiobarbituric acid test. Anal Bioch. 86: 271-278, 1978.
Urhausen A, Gabriel H, Kindermann W. Blood hormones as markers of training stress and
overtraining. Sports Med. 1995;20(4):251-76.
Urso, M.L., Clarckson, P.M. Oxidative stress, exercise, and antioxidant supplementation
Toxicology. 189, 41_/54, 2003.
Uttara, B., Singh, A.V., Zamboni, P., Mahajan, R.T. Oxidative stress and
neurodegenerative diseases: a review of upstream and downstream antioxidant therapeutic
options Curr Neuropharmacol. 2009 Mar;7(1):65-74.
Vollaard N.B, Shearman J.P, Cooper C.E. Exercise-induced oxidative stress: myths,
realities and physiological relevance. SportsMed 35:1045–1062, 2005.
Vierck, J., O’Reilly, B., Hossner, K., Antonio, J., Byrne., K, Bucci, L., and Dodson, M.
Satellite cell regulation following myotrauma caused by resistance exercise. Cell Biol Int
24: 263–272, 2000.
Warburton, D.E., Nicol, C.W., Bredin, S.S. Health benefits of physical activity: the
evidence. CMAJ. 2006 Mar 14;174(6):801-9.
Walters, T.J., Sweeney, H.L., Farrar, R.P. Influence of electrical stimulation on a fast-
twitch muscle in aging rats. J Appl Physiol. 1991;71:1921–192.
Welsh, G.I, Miller, C.M., Loughlin, A.J., Price, N.T, Proud, C.G. Regulation of eukaryotic
initiation factor eIF2B: glycogen synthase kinase-3 phosphorylates a conserved serine
which undergoes dephosphorylation in response to insulin. FEBS Lett. 1998;421(2):125-
30.

96
Weineck, J. Treinamento ideal : instruções técnicas sobre o desempenho fisiologico,
incluindo considerações especificas de treinamento infantil e juvenil. 9ª Edição, Editora
Manole, São Paulo, 2003.
Weiss, A., Suzuki, T., Bean, J., Fielding, R.A. High intensity strength training improves
strength and functional performance after stroke. Am J Phys Med Rehabil. 2000;79:369–
376.
Wessel, T.V., Hann, A., Van der Laarse, W.J., Jaspers, W.J. The muscle fiber type–fiber
size paradox: hypertrophy or oxidative metabolism? Eur J Appl Physiol. 2010
Nov;110(4):665-94.
White, J.P., Song Gao, Puppa, M.J., Shuichi Sato, Welle, S.L., Carson, J.A. Testosterone
regulation of Akt/mTORC1/FoxO3a signaling in skeletal muscle. Molecular and Cellular
Endocrinology 365 (2013) 174–186.
Wong, T.S., Booth, F.W. Skeletal muscle enlargement with weight-lifting exercise by rats.
J Appl Physiol. 1988;65:950–954.
Xia R, Stangler T, Abramson JJ. Skeletal muscle ryanodine receptor is a redox sensor with
a well defined redox potential that is sensitive to channel modulators. J Biol Chem. 2000
Nov 24;275(47):36556-61
Yfant, C., Akerstrom, T., Nielsen, S., Nielsen, A. R., Mounier, R., Mortensen, O. H.,
Lykkesfeldt, J., Rose, A. J., Fisher, C.P., Pedersen, B. K. Antioxidant Supplementation
Does Not Alter Endurance Training Adaptation. Medicine & Science in Sports & E x e r
c i s e. 1388 – 4207. 2010.
Yarasheski, K.E., Lemon, P.W., Gilliteaux, J. Effect of heavy-resistance exercise training
on muscle fiber composition in young rats. Journal of Applied Physiology, Bethesda, v.
69, p. 434-437, 1990.
Wang, C.Y., Yang, T.T., Chen, C.L., Lin, W.C., Lin, C.F. Reactive oxygen species-
regulated glycogen synthase kinase-3β activation contributes to all-trans retinoic acid-
induced apoptosis in granulocyte-differentiated HL60 cells. Biochem Pharmacol. 2014
Mar 1;88(1):86-94.

97
Zerbetto, E., Vergani, L., Dabbeni-Sala, F. Quantification of muscle mitochondrial
oxidative phosphorylation via histochemical staining of blue native polyacrylamide gels.
Electrophoresis. 18: 2059-2064, 1997.
Zembron-Lacny, A., Ostapiuk, J., Slowinska-Lisowska, M., Witkowski, K., Szyszka, K.
Pro-antioxidant ratio in healthy men exposed to muscle-damaging resistance exercise. J
Physiol Biochem. 2008 Mar;64(1):27-35.
Zollner, A.M., Abilez, O.J., BOL, M., Kunl, E. Stretching Skeletal Muscle: Chronic Muscle
Lengthening through Sarcomerogenesis.PLOS ONE. October 2012,Volume 7,Issue 10.





























