Embed Size (px)
Citation preview

Instituto de Pesquisas Jardim Botânico do Rio de Janeiro
Escola Nacional de Botânica Tropical
Asteraceae do Parque Estadual do Ibitipoca, Minas Gerais,
Brasil: checklist e taxonomia de Astereae.
Rafael Augusto Xavier Borges
2008


Instituto de Pesquisas Jardim Botânico do Rio de Janeiro
Escola Nacional de Botânica Tropical
Asteraceae do Parque Estadual do Ibitipoca, Minas Gerais, Brasil:
checklist e taxonomia de Astereae.
Rafael Augusto Xavier Borges
Dissertação de Mestrado apresentada ao Programa
de Pós-Graduação em Botânica, Escola Nacional
de Botânica Tropical, do Instituto de Pesquisas
Jardim Botânico do Rio de Janeiro, como parte dos
requisitos necessários à obtenção do título de
Mestre em Botânica.
Orientadora: Rafaela Campostrini Forzza
Rio de Janeiro
Janeiro 2008
I

Asteraceae do Parque Estadual do Ibitipoca, Minas Gerais, Brasil:
checklist e taxonomia de Astereae.
Rafael Augusto Xavier Borges
Dissertação submetida ao corpo docente da Escola Nacional de Botânica
Tropical, Instituto de Pesquisas Jardim Botânico do Rio de Janeiro – JBRJ,
como parte dos requisitos necessários à obtenção do grau de Mestre.
Aprovada por:
Prof._____________________________ - Dr. Jimi Naoki Nakajima
Prof._____________________________ - Dr. Roberto Lourenço Esteves
Prof._____________________________ - Dra. Rafaela Campostrini Forzza
Rio de Janeiro
2008
II

Agradecimentos
Agradeço aos meus pais Renato e Adélia e ao meu irmão Rodolfo, pelo amor sincero,
preocupação, apoio e confiança, sem vocês a vida teria pouco sentido.
À minha amada família pelo carinho e apoio constantes, em especial aos meus queridos
padrinhos Armando e Alcina, pelo exemplo como pessoas maravilhosas e por todo suporte
que têm me oferecido durante esses anos de estudo; aos queridos tios Álvaro e Luiza e aos
primos Guilherme, Bernardo, Bruna e Paula, por toda ajuda, paciência e suporte desde o
processo seletivo e pelos bons momentos que compartilhamos nesses dois anos em que
estivemos mais próximos.
À minha querida amiga e orientadora Rafaela, por todas as oportunidades acadêmicas e
profissionais, pelos ensinamentos, pela confiança, paciência, amizade e apoio. Ao amigo
André Amorim pela oportunidade de trabalho e ensinamentos e ao time do herbário CEPEC:
Márdel, Pedro, Alexandre, Ricardo, Luis Carlos, José Lima, Serginho, Cristiane e Terezinha.
À ENBT e JBRJ pela estrutura e suporte técnico durante o desenvolvimento da dissertação.
Aos professores, pesquisadores e funcionários, principalmente àqueles que participaram
diretamente da minha formação e que tive a oportunidade de conviver mais próximo: Elsie,
Vidal, Cláudio, Leandro, Claudine, Ronaldo, Marli, Luis Fernando, Luzia, Rosângela,
Nilcéia, Denise, Rosana, Carla, Nilton, Joel, Manoel e Edimar.
Aos funcionários do Parque Estadual do Ibitipoca, pela estrutura oferecida e auxílio no
trabalho de campo, principalmente ao Tião e ao senhor Pereira.
Aos sinanterólogos Aristônio Teles, Jimi Nakajima, Roberto Esteves, Mara Magenta, Victoria
Funk, Walter Holmes e Nicholas Hind, que muito me ajudaram nas identificações da flora do
Ibitipoca e que contribuíram com vastas bibliografias para a dissertação.
III

À ilustradora botânica Maria Alice Resende, pelo empenho e dedicação nas belas e bem
trabalhadas ilustrações.
Às amigas e aos amigos da “sala da Rafa” Adriana, Raquel, Valquíria, Ravena, Carol, Bia,
Bruno, Luiz (principalmente nas urgências com a coleção do herbário CESJ) e Eduardo; em
especial à Mariana, que muito contribuiu e auxiliou no meu aprendizado das Asteraceae no
herbário e no campo e ao grande amigo Fabrício, pela amizade, discussões filosóficas e de
“causos” das Minas Gerais.
Aos amigos Alexandre e Rodrigo, pela convivência amistosa na “mansão” e ajuda nos
momentos de dificuldade.
Aos amigos e amigas da pós-graduação e do Jardim Botânico, pelos bons momentos e por
tornarem a vida melhor nas confusões do cotidiano: Nívea, João, Gustavo, Jacira, Vitor,
Jerônimo, Jakeline, Gracialda, Beatriz, Luciano, Fabiana, Izar, Mariana, Marcelinho, Tião,
Pablinho, Rodolfo, Arno, Alba, Leonardo, Ursula, Felipe, Christo e em especial ao “hermano”
Maurício e à Marina pelo carinho e cumplicidade nas etapas finais.
A todos agradeço pela amizade, companhia, ensinamentos, filosofias mundanas, festas e boas
risadas.
Enfim, a todos aqueles que acabei esquecendo (graças a Deus as pessoas queridas são tão
numerosas que não consigo lembrar de todas) as minhas desculpas e o meu muito obrigado.
IV

Resumo
A presente dissertação compreende o estudo da família Asteraceae no Parque Estadual
do Ibitipoca (PEIB), localizado nos municípios de Santa Rita do Ibitipoca e Lima Duarte,
Minas Gerais. O primeiro capítulo apresenta o levantamento florístico das espécies de
Astereae ocorrentes no Parque. A tribo apresenta no local 20 espécies e quatro gêneros:
Baccharis está representado por 17 espécies, enquanto Conyza, Inulopsis e Leptostelma
possuem uma espécie cada. São fornecidas descrições morfológicas, ilustrações, comentários
taxonômicos, dados sobre a distribuição geográfica e uma chave de identificação para os
táxons em questão. O segundo capítulo apresenta a lista de espécies de Asteraceae do PEIB e
uma análise da distribuição geográfica da família em nove localidades de formações serranas
do Brasil. Foram registradas 12 tribos, 50 gêneros e 111 espécies. Eupatorieae, Vernonieae,
Astereae, Baccharis, Mikania e Lessingianthus foram as tribos e gêneros mais diversos,
respectivamente. O PEIB apresenta maior similaridade florística com o Itatiaia e
posteriormente com as Serras da Canastra e do Cipó. O dendrograma obtido pela análise de
agrupamento permite inferir que a vegetação dominante e características geoclimáticas são
fatores importantes na composição local. No entanto, as relações entre as áreas intermediárias
não foram demonstradas, ao contrário do gráfico resultante da análise de ordenação. Nesta
última, foram detectadas conexões entre as floras, representadas por áreas de localização
periférica (Serra do Ibitipoca, Serra da Canastra e Serra de Grão-Mogol.) e central (Serra do
Cipó). Além dos fatores locais, elementos biogeográficos históricos são fundamentais no
entendimento das relações encontradas.
Palavras-chave: campo rupestre, distribuição, diversidade, flora, floresta atlântica, serras.
V

Abstract
This study presents the Asteraceae family in Ibitipoca State Park (PEIB), located in
Santa Rita do Ibitipoca and Lima Duarte, state of Minas Gerais. The first chapter reports the
floristic inventory of Astereae found in PEIB. We found 20 species and four genera: 17
species were scored for Baccharis, while Conyza, Inulopsis and Leptostelma presented a
single species each. Descriptions, illustrations, taxonomic comments, geographic distribution
are given for all taxa, as well an identification key of the species. The second chapter presents
a checklist for Asteraceae of PEIB, with a floristic analyses of nine Brazilian mountain fields.
We found 12 tribes, 50 genera and 111 species. Eupatorieae, Vernonieae, Astereae,
Baccharis, Mikania and Lessingianthus were the most diverse tribes and genera. PEIB flora
was more similar with Itatiaia, and after with Serra da Canastra and Serra do Cipó. Cluster
analyses indicated that vegetation matrix and geoclimatic factors were important for local
composition, nevertheless, relationships among mid areas were shown only in ordination
analyses. Graphics expressed connections among “serras”, represented by remote (Serra do
Ibitipoca, Serra da Canastra and Serra de Grão-Mogol) and central (Serra do Cipó) areas.
Besides local factors, historical biogeographic elements are essential to understand Asteraceae
distribution.
VI

Sumário
Pág.
1. Introdução Geral................................................................................................1
2.Capítulo I: Taxonomia de Astereae (Asteraceae) do Parque Estadual
do Ibitipoca, Minas Gerais, Brasil. ....................................................................3
3.1 Resumo/Abstract.......................................................................................4
3.1 Introdução.................................................................................................5
3.2 Material e Métodos....................................................................................6
3.3 Resultados.................................................................................................7
3.3.1 Chave para a identificação das espécies da tribo Astereae do
Parque Estadual do Ibitipoca.......................................................................8
3.3.2 Baccharis..........................................................................................12
Baccharis aphylla.............................................................................13
Baccharis dracunculifolia.................................................................14
Baccharis itatiaiae............................................................................16
Baccharis ligustrina..........................................................................17
Baccharis lychnophora.....................................................................19
Baccharis microdonta.......................................................................20
Baccharis myriocephala....................................................................22
Baccharis oblongifolia......................................................................23
Baccharis platypoda..........................................................................25
Baccharis reticularia.........................................................................27
Baccharis rufidula.................................………………………....…28
Baccharis salzmanii............…………....……………………..……30
Baccharis schultzii.................................................………….……..32
Baccharis serrula..............................………………………………33
Baccharis serrulata...........................................................................35
Baccharis tarchonanthoides..........................………………………36
VII

Baccharis varians..............................................................................38
3.3.3 Conyza..............................................................................................40
Conyza monorchis.............................................................................40
3.3.4 Inulopsis...........................................................................................41
Inulopsis scaposa..............................................................................42
3.3.5 Leptostelma.......................................................................................43
Leptostelma maxima..........................................................................44
3.4 Referências..............................................................................................46
4.Capítulo II: Análise Florística (Asteraceae) do Parque Estadual do
Ibitipoca, Minas Gerais, Brasil. .......................................................................61
4.1 Resumo/Abstract.....................................................................................62
4.2 Introdução...............................................................................................63
4.3 Material e Métodos..................................................................................64
4.4 Resultados...............................................................................................65
4.5 Discussão................................................................................................67
4.6 Referências..............................................................................................72
2. Conclusões gerais............................................................................................90
3. Referências bibliográficas...............................................................................91
VIII

Introdução Geral
Ao longo dos últimos anos, novos táxons inéditos têm sido descobertos, em proporções
semelhantes as da metade do século XVIII e do final do século XIX (Donoghue & Alverson
2000). Entre os anos de 1989 e 1997 cerca de 21.100 novas espécies de angiospermas
tropicais foram descritas, sendo que 29% das espécies estudadas nas monografias da Flora
Neotropica são novas para a ciência (Prance et al. 2000). Dessa forma, o número de espécies
estimado para as espermatófitas está próximo de 420.000 (Govaerts 2001, 2003).
Tais dados mostram que trabalhos taxonômicos e levantamentos florísticos, mesmo que
atualmente sejam relegados a segundo plano em virtude do “impedimento taxonômico” (Funk
2006), são fundamentais para o desenvolvimento de teorias e padrões em biodiversidade,
assim como representam fontes primárias de informação para ações conservacionistas (Funk
& Richardson 2002, Funk 2006).
Segundo Giulietti et al. (2005), o Brasil possui a flora mais diversa do planeta, com
aproximadamente 56.000 espécies, que correspondem a 19% da flora mundial. Dentre as
famílias com maior riqueza, encontra-se Asteraceae com uma estimativa preliminar de 1.900
espécies. No entanto, esta contagem provavelmente tornar-se-á maior à medida que trabalhos
taxonômicos e levantamentos florísticos forem desenvolvidos (Donoghue & Alverson 2000).
Em Minas Gerais, a família Asteraceae é melhor estudada em áreas da Cadeia do Espinhaço,
onde trabalhos florísticos de longa duração têm se concentrado (e.g. Giulietti et al. 1987, Hind
1995, Roque 1997, Hind 2003 a,b, Hatschbach et al. 2006). Além desta, outras formações
serranas do estado foram inventariadas recentemente (Nakajima 2001) ou os trabalhos ainda
se encontram em desenvolvimento.
O Parque Estadual do Ibitipoca (PEIB) situa-se no sudeste de Minas Gerais nos
municípios de Santa Rita do Ibitipoca e Lima Duarte (21°40’ - 21°44’S e 43°52’ - 43°55’W)
(Fig. 1). Abrange aproximadamente 1.488 ha da Serra de Ibitipoca, formação pertencente ao
1

complexo da Mantiqueira no domínio da floresta atlântica (CETEC 1983). Os campos
rupestres presentes nesta área são disjuntos e diferenciados dos campos encontrados na
Cadeia do Espinhaço de Minas Gerais e Bahia, que são circundados por cerrado e caatinga,
respectivamente (Giulietti & Pirani 1988, Romero 2002). Pela primeira vez todas as espécies
vegetais do PEIB estão sendo catalogadas (Campos 2005, Medeiros 2006, Dias-Melo 2007,
Ferreira 2007, Menini-Neto 2007, Milward-de-Azevedo 2007, Monguilhott 2007, Monteiro &
Forzza 2008, Silva et al. em preparação) e o presente trabalho vem adicionar esforços e
resultados a este projeto.
Fig. 1. Localização do Parque Estadual do Ibitipoca, MG com a indicação das principais localidades (adaptado de Menini-Neto et al 2007a).
2

Capítulo I: Taxonomia de Astereae (Asteraceae) do Parque Estadual de
Ibitipoca, Minas Gerais, Brasil.
3

Abstract - [Astereae (Asteraceae) of Ibitipoca State Park, Brazil]. This work presents the
taxonomy of Astereae found in Ibitipoca State Park, Minas Gerais, located between Santa
Rita do Ibitipoca and Lima Duarte, state of Minas Gerais. We found 20 species and four
genera: 17 species belong to Baccharis, while Conyza, Inulopsis and Leptostelma
presented a single species each. Descriptions, illustrations, taxonomic comments and
geographic distribution are given for all taxa, with an identification key of the species.
Key words: Atlantic forest, campo rupestre, flora, Mantiqueira range.
Resumo – [Taxonomia de Astereae (Asteraceae) do Parque Estadual do Ibitipoca, Minas
Gerais, Brasil]. O presente trabalho compreende o tratamento taxonômico das espécies de
Astereae ocorrentes no Parque Estadual do Ibitipoca, localizado nos municípios de Santa
Rita do Ibitipoca e Lima Duarte, Minas Gerais. A tribo apresenta no local 20 espécies e
quatro gêneros: Baccharis está representado por 17 espécies, enquanto Conyza, Inulopsis
e Leptostelma possuem uma espécie cada. São fornecidas descrições morfológicas,
ilustrações, comentários taxonômicos e dados sobre a distribuição geográfica de todos os
táxons, além de uma chave de identificação das espécies.
Palavras-chave: campo rupestre, flora, floresta atlântica, serra da Mantiqueira.
4

Introdução
Asteraceae abriga cerca de 23.000 espécies organizadas em 1.600-1.700 gêneros, com
distribuição cosmopolita, exceto na Antártica (Funk et al. 2005, Jeffrey 2007). A partir de
análises filogenéticas, foi corroborado que a família é monofilética e tem Calyceraceae como
grupo-irmão (Kim et al. 1992, Gustafsson & Bremer 1995, APG II 2003, Funk et al. 2005,
Jeffrey 2007). As principais sinapomorfias morfológicas que sustentam a família são: flores
dispostas em capítulo circundado por brácteas involucrais, sépalas modificadas em papilho,
anteras singenésicas e fruto cipsela (Bremer 1994, Judd et al. 1999, Funk et al. 2005).
A tribo Astereae é reconhecida como a segunda maior de Asteraceae, constituída por
205 gêneros e cerca de 3.080 espécies (Nesom & Robinson 2007). Possui distribuição
cosmopolita, sendo encontrada geralmente em regiões montanhosas da América do Sul e do
sudoeste da América do Norte, sul do continente africano, Austrália e Nova Zelândia
(Bentham 1873, Grau 1977, Bremer 1994, Nesom 1994a, Funk et al. 2005). No Brasil,
ocorrem 19 gêneros e cerca de 200 espécies sendo Baccharis o maior, com cerca de 140
espécies (Barroso 1976, Bremer 1994, Oliveira et al. 2006, Teles com. pess.).
Determinadas características morfológicas foram inicialmente apontadas como possíveis
sinapomorfias para a tribo: células epidérmicas da corola com espessamentos medianos na
parede externa; ramos do estilete com apêndices agudos, de formato triangular, lanceolado ou
subulado, sem tricomas coletores internamente; superfícies estigmáticas laterais e próximas à
base do estilete (Bremer 1987). Posteriormente, foi reconhecido o monofiletismo de Astereae
(Zhang & Bremer 1993, Bremer 1994), porém há controvérsias quanto à subdivisão e
circunscrição dos gêneros. Até o momento, análises filogenéticas contemplaram parcialmente
a tribo, assim como utilizaram um número insuficiente de caracteres para o melhor
entendimento das relações evolutivas entre os táxons envolvidos (Zanowiak 1991, Nesom
1994a, Cross et al. 2002, Funk et al. 2005).
5

Dentre os estudos que abordaram a taxonomia da tribo Astereae no Brasil, podem ser
destacados os trabalhos de Baker (1882), Luis (1952, 1958), Solbrig (1962), Barroso (1976),
Nesom (1994 a,b), Matzenbacher & Sobral (1996), Oliveira & Marchiori (2005, 2006),
Oliveira et al. (2006).
Levantamentos florísticos e revisões taxonômicas são fundamentais para a conservação,
planos de manejo e catalogação da biodiversidade, principalmente na região tropical (Prance
et al. 2000). No entanto, os dados obtidos para Asteraceae ainda são insuficientes, devido à
grande diversidade da família. No Brasil, Asteraceae tem sido melhor estudada nas serras da
Cadeia do Espinhaço (e.g. Giulietti et al. 1987, Hind 1995, Roque 1997, Hind 2003a,b,
Hatschbach 2006) enquanto que em outras formações montanhosas de Minas Gerais a família
continua pouco conhecida (Nakajima 2001).
Dessa forma, o presente estudo tem por objetivos colaborar para o melhor conhecimento
da família Asteraceae em Minas Gerais, especificamente da tribo Astereae, além de fornecer
informações sobre a flora e para o plano de manejo do Parque Estadual do Ibitipoca.
Material e Métodos
Área de estudo
O Parque Estadual do Ibitipoca (PEIB) situa-se no sudeste de Minas Gerais nos
municípios de Santa Rita do Ibitipoca e Lima Duarte (21°40’ - 21°44’S e 43°52’ - 43°55’W).
Abrange aproximadamente 1.488 ha da Serra de Ibitipoca, que pertence ao complexo da
Mantiqueira no domínio da floresta atlântica (CETEC 1983). Os campos rupestres presentes
nesta área são disjuntos e diferenciados dos campos encontrados nas serras de Goiás e Cadeia
do Espinhaço, que estão inseridas nos biomas cerrado e caatinga (Giulietti & Pirani 1988,
Harley 1995, Romero 2002). Dados e comentários sobre o clima, geomorfologia e vegetação
encontram-se em Menini-Neto et al. (2007 a,b).
6

Trabalho de campo e laboratório
Foram realizadas excursões ao PEIB no período de março de 2004 a maio de 2007.
Todas as coletas foram depositadas no herbário RB. As identificações e os dados sobre
distribuição geográfica foram obtidos das observações de campo, coleções dos herbários
BHCB, CESJ, RB, SP e SPF (acrônimos segundo Holmgren et al. 1990) e de consulta
bibliográfica (De Candolle 1836, Baker 1882, Solbrig 1962, Cuatrecasas 1967, Barroso 1976,
Hind 1995, Munhoz & Proença 1998, Boggan et al. 1997, Hind 2003 a,b, Müller 2006). As
espécies foram descritas e ilustradas a partir de coleções do Parque; quando estas foram
insuficientes, utilizou-se material de outras localidades. Dados como altura das plantas, forma
de vida, coloração das flores e características do habitat, foram obtidos das etiquetas de
herbário ou observados durante as excursões. As descrições seguem a terminologia proposta
por Radford (1974), Stearn (1992) e Weberling (1989) para as sinflorescências (Fig. 1).
Resultados e Discussão
No presente estudo, foram registradas 20 espécies em quatro gêneros. Baccharis é o
mais diverso com 17 espécies, enquanto Conyza, Leptostelma e Inulopsis possuem apenas
uma espécie cada. A maioria dos táxons foi coletada em campo rupestre, geralmente nos
locais de vegetação herbáceo-arbustiva ou próximo de mata ciliar. Ferreira e Magalhães
(1977) publicaram a primeira lista de espécies para a Serra do Ibitipoca, em que não consta
nenhuma espécie de Astereae. Andrade & Sousa (1995) referiram três espécies da tribo
(Baccharis myriocephala, B. platypoda e B. schultzii) em levantamento preliminar para a
mesma área.
Dentre as espécies estudadas, metade tem distribuição apenas no Brasil. Baccharis
itatiaiae distribuí-se pelas serras dos Órgãos, da Mantiqueira e Cadeia do Espinhaço de Minas
Gerais; B. lychnophora está restrita ao Planalto do Itatiaia e também ao Espinhaço de Minas
Gerais; B. reticularia, B. rufidula, B. salzmanii, B. schultzii, B. tarchonanthoides e B.
7

serrulata ocorrem em áreas de cerrado, mata atlântica, campos rupestres, campos de altitude e
restinga, sendo que a última também está presente nos campos sulinos; B. serrula é restrita
aos campos rupestres e campos de altitude de Minas Gerais e São Paulo. Das espécies
encontradas em outros países da América do Sul, B. aphylla e B. platypoda ocorrem nos
páramos da Bolívia e nas serras e cerrados da Região Sudeste do Brasil; B. dracunculifolia e
B. oblongifolia são amplamente distribuídas da Bolívia à Argentina, sendo que a última
ocorre também nas Guianas; B. varians e B. ligustrina são registradas nas montanhas da
Venezuela e Guianas até as serras do sudeste brasileiro, sendo a última encontrada também
nos campos sulinos; Conyza monorchis, Inulopsis scaposa, B. microdonta, B. myriocephala e
Leptostelma maxima ocorrem da Argentina, Paraguai e Uruguai até as serras de Minas Gerais,
sendo que as duas últimas têm registro também para áreas de restinga.
No PEIB, as espécies com maiores populações e encontradas em mais de duas
localidades são B. itatiaiae; B. lychnophora; B. myriocephala; B. platypoda; B. reticularia; B.
salzmanii e B. serrulata. As espécies presentes em populações de tamanho médio em uma ou
duas localidades são: B. oblongifolia; B. rufidula; B. varians; I. scaposa e L. maxima. São
encontradas em pequenas populações: B. aphylla; B. dracunculifolia; B. ligustrina; B.
microdonta; B. schultzii; B. tarchonanthoides; C. monorchis. Apenas B. serrula não foi
recoletada no presente trabalho.
Chave para identificação das espécies da tribo Astereae
do Parque Estadual do Ibitipoca
1. Ervas. Capítulos radiados ou disciformes.
2. Lâmina oboval-elíptica a oboval-lanceolada, completamente híspida. Sinflorescência
escaposa, capítulo solitário .............................................................. 19. Inulopsis
scaposa
8

2’. Lâmina oblonga a oblanceolada, híspida apenas nas margens, ápice ou na nervura
central. Sinflorescência corimbiforme ou paniculiforme, capítulos numerosos.
3. Lâmina membranácea, > 10 cm compr. Capítulos radiados, brácteas involucrais 3-
seriadas; flores do raio ca. 1 cm compr ..................................... 20. Leptostelma maxima
3’. Lâmina papirácea, < 10 cm compr. Capítulos disciformes, brácteas involucrais 2-
seriadas; flores marginais ca. 4 mm compr ................................. 18. Conyza monorchis
1’. Subarbustos, arbustos ou arvoretas. Capítulos discóides
4. Ramos áfilos.
5. Ramos 3-alados. Sinflorescência diplobótrio estaquióide. Capítulo feminino sem
páleas, com mais de 30 flores, corola ligulada. Capítulo masculino com 20-25 flores,
papilho com cerdas de ápice longo-barbelado ................................. 7. B.
myriocephala
5’. Ramos não alados. Sinflorescência espiciforme. Capítulo feminino com páleas, ca. 15
flores, corola 5-lobada. Capítulo masculino ca. 15 flores, papilho com cerdas de ápice
agudo ......................................................................................................... 1. B. aphylla
4’. Ramos folhosos.
6. Folhas com nervação peninérvea.
7. Folhas densamente glanduloso-pontuadas. Plantas femininas e masculinas com
sinflorescências distintas. Capítulo feminino cilíndrico, brácteas involucrais
glandulosas, glabrescentes ............................................................... 9. B. platypoda
7’. Folhas não glanduloso-pontuadas. Plantas masculinas e femininas com o mesmo
tipo de sinflorescência. Capítulo feminino campanulado (cilíndrico em B.
lychnophora e então com brácteas involucrais lanosas).
8. Face abaxial das folhas com indumento lanoso ou tomentoso.
9

9. Folhas com margem denteada na metade superior. Capítulo feminino ca. 40
flores. Flores masculinas com corola hipocrateriforme, papilho caduco
.................................................................................. 16. B. tarchonanthoides
9’. Folhas com margem inteira. Capítulo feminino 10-20 flores. Flores
masculinas com corola infundibuliforme, papilho persistente.
10. Folhas coriáceas. Capítulo feminino cilíndrico, sem páleas
..................................................................................... 5. B. lychnophora
10’. Folhas cartáceas. Capítulo feminino campanulado, com páleas
.......................................................................................... 11. B. rufidula
8’. Face abaxial das folhas glabra.
11. Folha lanceolada a oblanceolada, margem irregular e curtamente
denteada no ápice. Sinflorescência paniculiforme. Cipsela ca. 1 mm
compr. Capítulo masculino 3 mm compr., brácteas involucrais 3-
seriadas ....................................................................... 4. B.
ligustrina
11’. Folha oblonga, margem inteira. Sinflorescência duplo-
paniculiforme. Cipsela ca. 2 mm compr. Capítulo masculino 2 mm
compr., brácteas involucrais 2-seriadas ............... 8. B. oblongifolia
6’. Folhas trinérveas ou uninérveas.
12. Folhas uninérveas.
13. Subarbustos xilopodíferos. Folhas papiráceas, lâmina geralmente tridentada.
Capítulos femininos e masculinos ca. 10 flores. Cipsela convexa, 10-
angulosa ..................................................................................... 17. B. varians
10

13’. Subarbustos sem xilopódio. Folhas coriáceas, lâmina geralmente com
margem superior curto-serreada. Capítulos femininos e masculinos com
menos de 10 flores. Cipsela cilíndrica, 20-angulosa ................ 14. B. serrula
12’. Folhas trinérveas.
14. Capítulo masculino até 2 mm compr., feminino até 3 mm compr. Flores
masculinas com corola hipocrateriforme. Flores femininas ca. 50, corola de
ápice truncado com tricomas adensados ............................... 15. B.
serrulata
14’. Capítulo masculino ≥ 3 mm e feminino ≥ 4 mm. Flores masculinas com
corola infundibuliforme. Flores femininas ≤ 40, corola 5-lobada sem
tricomas adensados.
15. Capítulo feminino campanulado, ca. 40 flores. Capítulo masculino ca.
35 flores .................................................................. 2. B. dracunculifolia
15’. Capítulo feminino cilíndrico, até 15 flores. Capítulo masculino até 20
flores.
16. Folhas elípticas a lanceoladas de margem serreada. Sinflorescência
diplobotrióide de florescências estaquióides, inseridas na axila de
bráctea foliácea. Capítulo feminino ca. 15 flores ..... 6. B. microdonta
16’. Folhas obovais, oblongas ou oblanceoladas de margem inteira a
denteada. Sinflorescência duplo-corimbosa ou diplobotrióide de
florescências não estaquióides. Capítulo feminino até 10 flores.
17. Subarbusto 0,5-1 m. Sinflorescência duplo-corimbosa. Cipsela 5-
angulosa. Capítulo masculino turbinado, flores masculinas com
papilho caduco ......................................................... 3. B.
itatiaiae
11

17’. Arbusto ≥ 1 m. Sinflorescência diplobotrióide. Cipsela 10 a 12-
angulosa. Capítulo masculino campanulado, flores masculinas
com papilho persistente.
18. Receptáculo convexo. Capítulo feminino ca. 5 mm compr. e
masculino ca. 3 mm compr. .......................... 10. B.
reticularia
18’. Receptáculo plano. Capítulo feminino ≥ 8 mm compr. e
masculino ≥ 5 mm compr.
19. Sinflorescência diplobotrióide de florescências
glomeruliformes, homotética. Capítulo feminino ca. 1 cm
compr. Capítulo masculino ca. 10 flores ....... 13. B.
schultzii
19’. Sinflorescência diplobótrio, heterotética. Capítulo
feminino ca. 8 mm compr. Capítulo masculino 10-20
flores ......................................................... 12. B. salzmanii
Baccharis L., Sp. pl. 860. 1753.
Subarbustos, arbustos a arvoretas; dióicos, raramente monóicos. Ramos algumas vezes
áfilos, alados ou não, glabros ou com indumento variável. Folhas alternas, espiraladas a
dísticas, opostas a subopostas, sésseis ou pecioladas; lâmina de forma e tamanho variáveis,
margem inteira, denteada a serreada, membranácea, papirácea ou coriácea, uninérvea,
trinérvea a peninérvea, glabras ou com indumento variável. Sinflorescência de variadas
formas, homotética a heterotética; capítulos discóides ou disciformes, sésseis ou
pedunculados, campanulados ou cilíndricos, com páleas ou não. Brácteas involucrais 3 a 8-
12

seriadas, lanceoladas a ovais, glabras ou com indumento variável. Flores masculinas com
corola infundibuliforme, hipocrateriforme ou campanulada, geralmente 5-lobada, lobos
geralmente revolutos. Anteras com apêndice lanceolado e base obtusa, raro sagitada. Ramos
do estilete em geral triangulares, longo-papilosos. Ovário rudimentar. Flores femininas com
corola tubulosa, de ápice truncado, liguliforme ou 5-lobado. Ramos do estilete lanceolados a
triangulares, papilosos. Papilho persistente a caduco, cerdoso, barbelado, estramíneo ou
creme, cerdas com ápice agudo ou longo-barbelado, espessado ou não. Cipsela cilíndrica a
convexa, 2 a 20-angulosa, glabra ou com indumento variável.
Gênero com 360-400 espécies, que possui distribuição panamericana com elevado
número de espécies nos Andes (Colômbia, Chile e Argentina) e vegetações campestres do
sudeste e sul do Brasil (Bremer 1994, Müller 2006, Nesom 1994a, Nesom & Robinson 2006).
Nomes populares: alecrim, alecrim-do-campo, alecrim-de-vassoura, cambará-de-serra,
carqueja, vassourinha, vassoura-cambará, vassoura-do-campo, vassoura-lageana, vassoura-de-
são-joão (Barroso 1976).
1. Baccharis aphylla (Vell.) DC., Prodr. 5: 424. 1836. Iconografia: Baker (1882); Müller
(2006).
≡ Chrysocoma aphylla Vell., Fl. Flum. 324. 1829.
Fig 2. f-h
Subarbusto, ca. 30 cm alt., ereto, ramificado, xilopodífero. Ramos cilíndricos, estriados,
glabros, resinosos, áfilos; folhas rudimentares raras próximas à base da planta, sésseis, 1-2
mm compr., linear-lanceoladas, ápice agudo, margem inteira, uninérveas, resinosas.
Sinflorescência espiciforme, heterotética, glabra, resinosa, brácteas 0,5 mm compr., ovais,
margem inteira, resinosas. Capítulos numerosos, sésseis; receptáculo plano, alveolado,
fimbriado, glabro. Capítulo masculino campanulado, ca. 5 mm compr., sem páleas; brácteas
13

involucrais 3 a 4-seriadas, externas 1-2,5 x 1-1,3 mm, internas 3-4 x 0,7-1,2 mm, persistentes,
elípticas a lanceoladas, glabras, glandulosas, ápice agudo, ciliado, margem hialina. Flores ca.
15; corola infundibuliforme, ca. 5 mm compr., glabra, 5-lobada, lobos 1,5-2 mm compr.,
glabros, algumas vezes revolutos; estilete 5,5-6,5 mm compr., ramos lanceolados, papilosos.
Capítulo feminino cilíndrico, ca. 7 mm compr., páleas ca. 8 mm compr., glabras, ápice agudo,
ciliado, margem hialina; brácteas involucrais 5 a 7-seriadas, externas 1-3 x 0,8-1,3 mm,
internas 4,5-6 x 1 mm, persistentes, lanceoladas, glabras, glandulosas, ápice agudo, ciliado,
margem hialina. Flores ca. 15; corola ca. 6 mm compr., glabra, 5-lobada, lobos diminutos,
desiguais; estilete ca. 7 mm compr., ramos lanceolados. Papilho 5-8,5 mm compr., persistente,
estramíneo, cerdas com ápice agudo. Cipsela ca. 2 mm compr., cilíndrica, 12 a 15-angulosa,
glabra, glandulosa, ápice truncado.
Material examinado: Minas Gerais, Lima Duarte, Parque Estadual do Ibitipoca: 13.IX.1940,
M. Magalhães 472 (RB); 1.500 m alt., trilha Monjolinho - Gruta dos Viajantes, 19.IX.2006,
R. Borges et al. 259 (RB).
Material adicional: Minas Gerais. Belo Horizonte: Morro das Pedras, 1.000-1.100 m alt.,
11.IX.1945, L.O. Williams & Assis 7529 (RB). São Paulo. São Paulo: 12.XII.1947, A. Betzler
s.n. (RB 61336).
Distribuição geográfica: Bolívia e Brasil: Bahia, Minas Gerais, São Paulo e Paraná.
Encontrada em campo rupestre, campo de altitude e cerrado. No Parque foi observada
em meio a subarbustos próximos a afloramento quartzítico; fértil de maio a dezembro. O
táxon é facilmente reconhecido pela ausência de folhas e alas, e pela sinflorescência multi-
espiciforme. Quando estéril pode ser confundida com B. gracilis DC. ou B. orbignyana Klatt.,
no entanto pode ser diferenciada destas pelo tipo de sinflorescência e número de flores por
capítulo.
14

2. Baccharis dracunculifolia DC., Prodr. 5: 421. 1836. Iconografia: Cabrera (1978).
Fig. 3 h-i
Arbusto, ca. 2 m alt., ereto, ramificado. Ramos cilíndricos, estriados, cicatricosos,
seríceo-glandulosos. Folhas alternas, espiraladas, algumas vezes subopostas, sésseis; lâmina
(0,6) 1-2,5 x 0,3 cm, lanceolada, ápice agudo, base atenuada, margem inteira a tridentada na
porção superior, cartácea, uninérvea, ambas faces glabras, glanduloso-pontuadas.
Sinflorescência diplobótrio heterotético, seríceo-glandulosa. Capítulos numerosos, axilares,
pedúnculos 1-4 mm compr.; receptáculo plano, alveolado, glabro, glanduloso. Capítulo
masculino campanulado, ca. 3 mm compr., brácteas involucrais 4-seriadas, externas 1,5-2 x
0,7-1 mm, internas 2,8-3 x 0,8-1 mm, persistentes, lanceoladas, glabras, glandulosas, ápice
acuminado, ciliado, margem hialina. Flores ca. 35; corola infundibuliforme, ca. 3 mm compr.,
glandulosa, com tricomas esparsos, 5-lobada, lobos ca. 1 mm compr., lanceolados,
glandulosos, revolutos; estilete ca. 3,5 mm compr., ramos triangulares, longo-papilosos.
Capítulo feminino campanulado, ca. 4 mm compr., brácteas involucrais 4-seriadas, externas
1,5-3 x 0,9-1,2 mm, internas 4-4,5 x 0,7-1 mm, persistentes, elípticas a lanceoladas, glabras,
levemente glandulosas; ápice agudo, ciliado, margem hialina. Flores ca. 40; corola ca. 3 mm
compr., glabra, 5-lobada, lobos lanceolados, diminutos; estilete ca. 4,5 mm compr., ramos
lanceolados, curto-papilosos. Papilho 2,5-5 mm compr., persistente, estramíneo, ápice agudo
nas flores femininas e longo-barbelado nas masculinas. Cipsela ca. 1 mm compr., convexa, 10
a 12-angulosa, glandulosa.
Material examinado: Minas Gerais. Lima Duarte, Parque Estadual do Ibitipoca: estrada
centro de visitantes – alojamentos, 1.328 m alt., 31.V. 2007, R. Borges 883 (RB).
Distribuição geográfica: Bolívia, Brasil, Argentina, Paraguai e Uruguai. No Brasil: Bahia,
Mato Grosso, Goiás, Distrito Federal, Minas Gerais, Mato Grosso do Sul, Espírito Santo, Rio
de Janeiro, São Paulo, Paraná, Santa Catarina e Rio Grande do Sul.
15

Encontrada em campo rupestre, campo de altitude, cerrado, mata atlântica, restinga e
campos sulinos. No Parque foi observada em área degradada; fértil durante todo o ano.
Espécie comum em capoeiras e áreas alteradas, ocorrendo geralmente próximo a rodovias e
pastos. B. dracunculifolia é muito utilizada na medicina popular (Freire et al. 2007) e como
produto fitoterápico (Kumazawa et al. 2003, Alencar et al. 2005).
3. Baccharis itatiaiae Wawra, Itin. Prin. S. Coburgi 2: 28. 1888.
Fig. 2 i-n; 8 D
Subarbusto, 0,5-1 m alt., ereto, ramificado. Ramos cilíndricos, estriados, cicatricosos,
glandulosos, glabros. Folhas alternas, espiraladas; pecíolo 3-8 mm compr., glanduloso-
pontuado; lâmina (0,8) 1,5-3,5 (3,8) x (0,8) 1,3-2,5 (3) cm, oboval, ápice obtuso, base
cuneada, decorrente, margem superior denteada, coriácea, trinérvea, ambas faces glabras,
glanduloso-pontuadas. Sinflorescência duplo-corimbosa, heterotética. Capítulos numerosos,
concentrados no ápice das florescências, pedúnculos 0,2-2,6 cm compr.; receptáculo plano
(levemente côncavo nos capítulos masculinos), alveolado, fimbriado, glabro; brácteas 5-10
(14) x 2-4 (7) mm, obovais, ápice truncado, base cuneada, margem denteada na porção
superior, coriáceas, trinérveas, ambas faces glabras, glanduloso-pontuadas. Capítulo
masculino turbinado, 4-5 mm compr.; brácteas involucrais 4-seriadas, externas 2-3 x 1-2 mm,
internas 3,5-4 x 1-1,5 mm, persistentes, lanceoladas, resinosas, ápice agudo, denso-
glanduloso, margem hialina. Flores ca. 15; corola infundibuliforme, 3,5-4 mm compr.,
esparso seríceo-glandulosa, 5-lobada, lobos ca. 1,5 mm compr., revolutos; estilete ca. 4,5 mm
compr., ramos triangulares, curtos, denso-papilosos. Capítulo feminino cilíndrico, 6-7 mm
compr., brácteas involucrais 4-seriadas, externas 5-6 x 1 mm, internas 2-4 x 0,7-1,2 mm,
persistentes, lanceoladas, resinosas; ápice agudo, denso-glanduloso, margem hialina. Flores 4-
7; corola ca. 3 mm compr., 5-lobada, glandulosa, serícea, com tricomas concentrados na base;
16

lobos curtos, triangulares. estilete 3-4 mm compr., ramos lanceolados. Papilho 3-4 mm
compr., caduco, cerdas com ápice agudo nas flores femininas e espesso nas masculinas.
Cipsela 2-2,5 mm compr., cilíndrica a convexa, 5-angulosa, glabra, algumas vezes glandulosa,
ápice rostrado.
Material examinado: Minas Gerais. Lima Duarte, Parque Estadual do Ibitipoca: 30.IX.1970,
U. Confúcio s.n. (CESJ 9425); 8.II.2001, F.S. Araújo at al. 3 (CESJ); 10.II.2001, F.S. Araújo
et al. 32 (CESJ); 19.V.2001, M. Heluey et al. 124 (CESJ); trilha Lombada - Cruzeiro, 1.777 m
alt., 25.VI.2003, R. Borges et al. s.n. (RB 429.699); idem, 26.VII.2004, R.C. Forzza et al.
3508 (RB); idem, 8.III.2006, F.M. Ferreira 1017 (CESJ); idem, 18.V.2006, M.M. Saavedra
et al. 304 (RB); idem, 18.V.2006, M.M. Saavedra et al. 313, 314 (RB).
Distribuição geográfica: Brasil: Cadeia do Espinhaço de Minas Gerais, Serra da Mantiqueira
(MG, RJ) e Serra dos Órgãos no Rio de Janeiro.
Encontrada em campo rupestre e campo de altitude. No Parque foi observada em campo
rupestre, no topo das principais vertentes (Lombada e Pico do Pião); fértil de fevereiro a
setembro. É facilmente reconhecida pelo hábito subarbustivo, estruturas vegetativas e florais
glandulosas e capítulos concentrados no ápice das florescências. Barroso (1976) utiliza a
organização dos capítulos para o posicionamento do táxon no “grupo Platypoda”, junto com
B. platypoda. No entanto, características das folhas e sinflorescência indicam maior
parentesco com as espécies da série Cylindricae, fato observado no presente estudo (Giuliano
2005).
4. Baccharis ligustrina DC., Prodr. 5: 421. 1836. Iconografia: Aristeguieta (1964).
Fig. 4 f-g; 8 B
Arbusto, ca. 1,5 m alt., ereto, ramificado. Ramos cilíndricos, estriados, cicatricosos,
glabros, os mais jovens levemente seríceos. Folhas alternas, espiraladas, sésseis; lâmina (2,5)
17

4-7,5 (8) x 0,8-1 (1,5) cm; lanceolada a oblanceolada, ápice mucronado, base cuneada,
margem revoluta, irregular e curtamente denteada na região superior, cartácea, peninérvea,
face adaxial glabra, face abaxial glabra e esparssamente glanduloso-pontuada. Sinflorescência
paniculiforme, heterotética, levemente serícea. Capítulos numerosos, pedúnculos 0,4-10 cm
compr., receptáculo cônico (levemente côncavo nos capítulos masculinos), alveolado, glabro;
brácteas 2-3 mm compr., lanceoladas, uninérveas, margem ciliada. Capítulo masculino
campanulado, ca. 3 mm compr., sem páleas; brácteas involucrais 3-seriadas, externas 1,5-2 x
0,5-1 mm, internas 2,8-3 x 0,8-1 mm, persistentes, lanceoladas, glabras, ápice agudo, ciliado,
margem hialina. Flores ca. 30; corola infundibuliforme, 2,5-3 mm compr., serícea, 5-lobada,
lobos ca. 1 mm compr., lanceolados, glabros; estilete ca. 3 mm compr., ramos triangulares,
denso-papilosos. Capítulo feminino campanulado, ca. 5 mm compr., páleas ca. 4,5 mm,
glabras, ápice acuminado, ciliado, margem hialina; brácteas involucrais 2-seriadas, externas
1-1,5 x 0,6-0,8 mm, internas 2-3 x 0,6-1 mm, persistentes, lanceoladas, glabras, ápice agudo,
ciliado, margem hialina. Flores ca. 18; corola ca. 3 mm compr., levemente serícea, com
tricomas concentrados na base e próximo ao ápice, 5-lobada, lobos triangulares, diminutos;
estilete ca. 4 mm compr., ramos lanceolados, curto-papilosos. Papilho 2,5-3 mm compr.,
persistente, estramíneo, cerdas com ápice agudo nas flores femininas e longo-barbelado nas
masculinas. Cipsela 1-1,3 mm compr., cilíndrica, achatada, 5 a 7-angulosa, serícea, ápice
truncado, carpopódio desenvolvido.
Material examinado: Minas Gerais: Lima Duarte, Parque Estadual do Ibitipoca: trecho da
praia do Ribeirão à Ponte de Pedra, 1.300-1.500 m alt., 30.IX.1970, D. Sucre 7231 (RB);
idem, 30.VIII.1987, J.R. Stehmann & A.R. Oliveira 1027 (BHCB); Cachoeira dos Macacos,
1.214 m alt., 9.VIII.2005, R.F. Monteiro et al. 57 (RB); idem, 29.VI.2006, R. Borges et al.
145 (RB).
18

Distribuição geográfica: Venezuela, Guiana Inglesa e Brasil: Bahia, Goiás, Distrito Federal,
Minas Gerais, Rio de Janeiro, São Paulo, Paraná, Santa Catarina e Rio Grande do Sul.
Encontrada em campo rupestre, campo de altitude, cerrado, mata atlântica e campos
sulinos. No Parque foi observada na orla de mata ciliar; fértil durante o ano todo. Phillipi
(1891 apud Aristeguieta 1964) e Cabrera (1944) criaram respectivamente os gêneros Psila e
Pseudobaccharis, baseados na presença de páleas nos capítulos femininos, combinando B.
ligustrina nos respectivos gêneros novos. No entanto, o uso desta característica para
segregação do gênero Baccharis é questionável, sendo aqui utilizado o nome mais antigo para
a espécie.
5. Baccharis lychnophora Gardner, J. Bot. (Hooker) 7: 85. 1848.
Fig. 2 a-e, 8 C
Arbusto a arvoreta, 1,5-5 m alt., ereto, ramificado. Ramos cilíndricos, cicatricosos,
pubérulos, alvos a gríseos. Folhas alternas, espiraladas; pecíolo 0,5-2 cm compr., pubérulo;
lâmina (3) 4-7 (13) x (1) 1,5-2,5 (3,8) cm, elíptica a oblanceolada, ápice agudo, base cuneada,
decorrente, margem inteira, coriácea, peninérvea, face adaxial glabra, cerosa, face abaxial
alva a gríseo-lanosa. Sinflorescência duplo-paniculada, heterotética, alvo-lanosa. Capítulos
numerosos, pedúnculos 0,5-1,5 cm compr.; receptáculo plano, alveolado, longo-fimbriado,
glabro; brácteas 0,1-1,5 cm compr., lanceoladas, alvo-lanosas em ambas faces. Capítulo
masculino campanulado, ca. 3 mm compr.; brácteas involucrais 3-seriadas, externas 1,5-2 x 1
mm, internas 2-2,3 x 0,5-0,8 mm, persistentes, lanceoladas, esparso glandulosas, alvo-lanosas,
ápice agudo, ciliado, margem hialina. Flores ca. 23, corola infundibuliforme, ca. 2 mm
compr., 5-lobada, glabra; lobos ca. 1 mm compr., lanceolados, com extremidade curto-
papilosa. estilete ca. 2,5 mm compr., ramos triangulares, denso e longo-papilosos. Capítulo
feminino cilíndrico, 4-5 mm compr.; brácteas involucrais 3-4 seriadas, externas 2-2,5 x 1-1,3
19

mm, internas 3-4 x 1-1,5 mm, persistentes, lanceoladas, esparso glandulosas, alvo-lanosas,
ápice agudo, ciliado, margem hialina. Flores ca. 10-15; corola 2-2,5 mm compr., com poucos
tricomas concentrados no ápice, 5-lobada, lobos triangulares, papilosos. estilete ca. 3 mm
compr., ramos triangulares, curto-papilosos. Papilho 2-3 mm compr., persistente, estramíneo,
cerdas com ápice agudo, longo-barbelado. Cipsela 1,8-2 mm compr., convexa, 5 a 7-angulosa,
glandulosa, ápice truncado.
Material examinado: Minas Gerais. Lima Duarte, Parque Estadual do Ibitipoca: 13.V.1970, L.
Krieger s.n. (CESJ 8.534, RB); 1.X.1970, U. Confúcio s.n. (CESJ 9459); 8.II.2001, F.S.
Araújo 9 (CESJ); 25.III.2001, R.M. Castro 216 (CESJ); Pico do Pião, 1.580 – 1.600 m alt.,
13.V.1970, D. Sucre et al. s.n. (RB 166.050); idem, 16.V.2006, M.M. Saavedra et al. 282,
283 (RB); próximo à Gruta do Cruzeiro, 31.III.2004, R.C. Forzza et al. 3341 (RB); Lombada,
20.I.2005, R.C. Forzza et al. 3961 (RB); idem, 7.III.2006, F.M. Ferreira 968 (CESJ).
Distribuição geográfica: Brasil: Cadeia do Espinhaço (Minas Gerais) e Planalto do Itatiaia
(Minas Gerais e Rio de Janeiro).
Encontrada em campo rupestre e campo de altitude. No Parque foi observada em
vertente inclinada de campo rupestre, junto a vegetação arbustiva e em borda de mata
estacional; fértil durante o ano todo. Com distribuição semelhante a de B. itatiaiae, o táxon é
diferenciado dos demais abordados no estudo pelo indumento pubérulo associado ao capítulo
feminino cilíndrico e flores masculinas com corola infundibuliforme. A espécie foi
classificada como ameaçada de extinção na categoria “criticamente em perigo” para a flora de
Minas Gerais (Mendonça & Lins 2000), porém não consta entre as espécies da flora brasileira
ameaçada (Nakajima 2007).
6. Baccharis microdonta DC., Prodr. 5: 416. 1836. Iconografia: Cabrera (1978).
Fig. 3 f-g
20

Arvoreta, 4 m alt., ereta, ramificada. Ramos cilíndricos, estriados, resinosos. Folhas,
alternas, dísticas, algumas vezes opostas; pecíolo 4-7 mm, resinoso, glanduloso-pontuado;
lâmina (1) 2,5-3,5 (4) x 0,5-1 cm, elíptica a lanceolada, ápice agudo, base cuneada,
decorrente, margem curta e irregularmente serreada, papirácea, trinérvea, face adaxial glabra,
resinosa, face abaxial glabra, glanduloso-pontuada. Sinflorescência diplobotrióide homotética,
de florescências estaquióides inseridas na axila de bráctea foliácea. Capítulos numerosos,
sésseis, receptáculo plano, alveolado, fimbriado, glabro; bráctea 0,5-1 cm compr., linear-
lanceolada, margem inteira, algumas vezes denteada, resinosa. Capítulo masculino
campanulado, ca. 3 mm compr.; brácteas involucrais 4-seriadas, externas 1-2 x 0,5-1 mm,
internas 2,5-3,5 x 1 mm, persistentes, elípticas a lanceoladas, glabras, levemente glandulosas;
ápice agudo, ciliado, margem hialina. Flores 10-12; corola infundibuliforme, ca. 4-4,5 mm
compr., tomentosa, glandulosa, 5-lobada, lobos ca. 1 mm compr., levemente tomentosos,
revolutos; estilete ca. 5 mm compr., ramos triangulares, denso-papilosos. Capítulo feminino
cilíndrico, ca. 5 mm compr.; brácteas involucrais 4 a 5-seriadas, externas 1-2,5 x 1 mm,
internas 3-4,5 x 1 mm, persistentes, lanceoladas, glabras, glandulosas; ápice agudo, ciliado,
margem hialina. Flores ca. 15; corola ca. 3 mm compr., glabra, com tricomas concentrados na
base e ápice; ápice irregularmente 5-lobado; estilete 4-4,5 mm compr., ramos lanceolados,
curto-papilosos. Papilho ca. 4 mm compr., caduco, estramíneo, cerdas com ápice agudo nas
flores femininas e espessado nas masculinas. Cipsela ca. 1 mm compr., cilíndrica a convexa,
10 a 12-angulosa, glabra, glandulosa.
Material examinado: Minas Gerais: Lima Duarte, Parque Estadual do Ibitipoca: trilha
interditada próximo ao alojamento novo, 1.326 m alt., 18.V.2006, M.M. Saavedra et al. 316
(RB).
Material adicional: Minas Gerais. Poços de Caldas: Véu das Noivas, 24.II.1965, O. Roppa
587 (RB).
21

Distribuição geográfica: Brasil, Argentina, Paraguai e Uruguai. No Brasil: Minas Gerais, Rio
de Janeiro, São Paulo, Paraná, Santa Catarina e Rio Grande do Sul.
Encontrada em campo rupestre, campo de altitude, mata atlântica, restinga e campos
sulinos. No Parque foi observada em borda de mata estacional, local de drenagem pluvial;
fértil durante o ano todo. Posicionada na série Cylindricae, se distingue das demais espécies
estudadas pelas características da sinflorescência e margem foliar.
7. Baccharis myriocephala DC., Prodr. 5: 426. 1836.
Fig. 5 j-n; 8 E
Subarbusto, ca. 30 cm alt., ereto, ramificado. Ramos áfilos, 3-alados; alas 0,1-1,2 cm,
planas, levemente onduladas, interrompidas. Sinflorescência diplobótrio estaquióide,
heterotética. Capítulos numerosos, em grupos de 3-5, sésseis; receptáculo plano, alveolado,
fimbriado, glabro. Capítulo masculino campanulado, ca. 3-4 mm compr.; brácteas involucrais
3 a 4-seriadas, externas 1,5-2,5 x 0,8-1 mm, internas 3-4,3 x 0,7-1 mm, persistentes, elípticas
a lanceoladas, glandulosas, ápice agudo, margem hialina. Flores 20-25; corola
infundibuliforme, ca. 3,5 mm compr., glandulosa, 5-lobada, lobos ca. 1,5 mm compr.,
lanceolados, glabros, levemente seríceo na base; estilete ca. 4 mm compr., ramos lanceolados,
papilosos. Capítulo feminino cilíndrico, 4-5 mm compr.; brácteas involucrais 3 a 4-seriadas,
externas 2-3 x 0,7-1 mm, internas 3,8-5 x 0,4-0,7 mm, persistentes, elípticas a lanceoladas,
glandulosas, ápice agudo a acuminado, margem hialina. Flores 32-114; corola ligulada, ca.
3,5 mm compr., glandulosa; estilete 4,5-5 mm compr., ramos lanceolados, curto-papilosos.
Papilho 3,5-4 mm compr., persistente, cerdoso, barbelado, estramíneo, cerdas com ápice
agudo nas flores femininas e longo-barbelado nas masculinas. Cipsela ca. 1 mm compr.,
convexa, 15 a 17-angulosa, hirsuta, com tricomas curtos, carpopódio desenvolvido.
Material examinado: Minas Gerais. Lima Duarte, Parque Estadual do Ibitipoca: 14.V.1970, L.
Krieger s.n. (CESJ 8.525, RB); 20.IX.1971, L. Krieger s.n. (CESJ 10.779); 18.VI.1994, M.
22

Garcia et al. 5 (CESJ); VII.1998, L.G. Rodela s.n. (CESJ 40622); 8.II.2001, M. Heluey et al.
33 (CESJ); 24.III.2001, M. Heluey 80 (CESJ); 19.V.2001, M. Heluey 127 (CESJ);
22.VI.2001, M. Heluey 142 (CESJ); Pico do Pião, 15.V.1970, D. Sucre et al. 6864 (RB);
trilha para o Monjolinho, 25.VII.2004, R.C. Forzza et al. 3474 (RB); idem, 19.IX.2006, R.
Borges et al. 257 (RB); trilha Monjolinho - Pico do Pião, 31.V.2007, R. Borges 873 (RB).
Distribuição geográfica: Paraguai e Brasil. No Brasil: Bahia, Minas Gerais, Rio de Janeiro,
São Paulo, Paraná, Santa Catarina, Rio Grande do Sul.
Encontrada em campo rupestre, campo de altitude, restinga, cerrado e nos campos
sulinos. No Parque está geralmente associada à vegetação herbácea densa; fértil durante o ano
todo. B. myriocephala é semelhante a B. trimera e geralmente confundido com este nas
coleções, devido ao considerável número de exsicatas apresentarem apenas florescências
parciais, uma vez que a principal diferença entre os dois táxons está no tamanho e conjunto da
sinflorescência. Müller (2006) considera ambos como sinônimos nomenclaturais de B.
genistelloides subsp. crispa (Sprengel) Joch. Müller, propondo dessa forma a existência de
uma única espécie com distribuição do Peru até a Argentina. Tal critério não foi adotado no
presente trabalho devido à presença de características distintivas entre as duas espécies, como
o número de flores por capítulo. Além disso, grande variação morfológica observada nas
coleções de herbário e no campo, resultantes provavelmente de plasticidade fenotípica aos
diferentes ambientes onde ocorre (Silvertown 1998) e/ou à formação de híbridos (Knobloch
1959, Hellwig 1990, Zanowiak 1991, Müller 2006) com espécies simpátricas da seção
Caulopterae. A espécie também possui propriedades medicinais e é utilizada na medicina
popular (Castro et al. 1999).
8. Baccharis oblongifolia (Ruiz & Pav.) Pers., Syn. pl. 2: 424.1807. Iconografia: Cuatrecasas
(1967), Müller (2006).
23

≡ Molina oblongifolia Ruiz & Pav., Syst. veg. fl. peruv. chil. 203. 1798.
= Baccharis brachylaenoides DC., Prodr. 5: 421. 1836.
Fig. 4 m-n
Arbusto, ca. 2 m alt., ereto, ramificado. Ramos cilíndricos, cicatricosos, hirsutos. Folhas
alternas, espiraladas, pecíolo ca. 6 mm compr., glabro; lâmina (4,5) 5,5-6,5 (7) x (1,5) 2-2,5
cm, oblonga, ápice curtamente mucronado, base atenuada, decorrente, margem inteira,
revoluta, papirácea, peninérvea, ambas faces glabras, face abaxial brilhante. Sinflorescência
duplo-paniculada, heterotética. Capítulos numerosos, pedúnculos 0,4-1,4 mm compr.;
receptáculo cônico, alveolado, glanduloso, fimbriado; brácteas 0,4-2,8 cm compr.,
semelhantes às folhas, bractéolas 2-3 mm compr., lanceoladas, glandulosas, margem ciliada.
Capítulo masculino campanulado, ca. 2 mm compr., sem páleas; brácteas involucrais 2-
seriadas, externas 1-2 x 0,5-1 mm, internas 2,2-3 x 0,5-1 mm, persistentes, elípticas,
glandulosas; ápice agudo, ciliado, margem hialina. Flores 20-25; corola infundibuliforme, ca.
2,5 mm compr., glandulosa, 5-lobada, lobos ca. 0,5 mm compr., lanceolados, glandulosos,
tomentosos na base; estilete 2,5-3 mm compr., ramos triangulares, denso-papilosos. Capítulo
feminino campanulado, ca. 5 mm compr., páleas ca. 5 mm compr., lanceoladas, glandulosas,
ápice acuminado, ciliado, margem hialina; brácteas involucrais 3-seriadas, externas 2-3,5 x 1-
1,3 mm, internas 4-5 x 0,7-1,2 mm, persistentes, elípticas a lanceoladas, glabras, glandulosas;
ápice agudo, ciliado, margem hialina. Flores 15-20; corola 3-3,5 mm compr., glanduloso-
tomentosa na região médio-superior, 5-lobada, lobos triangulares, diminutos; estilete 4-4,5
mm compr., ramos lanceolados, papilosos. Papilho 2-4 mm compr., persistente, estramíneo,
cerdas com ápice agudo. Cipsela ca. 2 mm compr., convexa, levemente achatada, 6-angulosa,
tomentosa, glandulosa, ápice truncado.
Material examinado: Minas Gerais. Lima Duarte, Parque Estadual do Ibitipoca: 13.IX.1940,
M. Magalhães 449 (BHCB); 30.IX.1970, U. Confúcio s.n. (CESJ 9425); 30.IX.1970, 1.550-
24

1.630 m alt., D. Sucre et al. 7210 (RB); 6.III.2006, F.R.G. Salimena 1327 (CESJ); Gruta dos
Viajantes, 25.VII.2004, R.C. Forzza et al. 3483 (RB).
Material adicional: Rio de Janeiro. Nova Friburgo: morro da Caledônia, 15.VI.2004, R.
Mello-Silva et al. 2608 (RB, SPF).
Distribuição geográfica: Venezuela, Guiana Inglesa, Colômbia, Bolívia, Brasil e Argentina.
No Brasil: Bahia, Goiás, Distrito Federal, Mato Grosso, Minas Gerais, Espírito Santo, Rio de
Janeiro, São Paulo, Paraná, Santa Catarina e Rio Grande do Sul.
Encontrada em campo rupestre, campo de altitude, cerrado, mata atlântica, campos
sulinos e restinga. No Parque foi observada em borda de matas nebulares e capões arbustivos
em pequenos afloramentos; fértil de fevereiro a novembro. Diferencia-se de B. ligustrina
pelas folhas mais largas, e pelas sinflorescências e cipselas maiores. Devido sua ampla
distribuição, vários táxons específicos e infra-específicos foram criados para esta espécie; nas
exsicatas provenientes do Brasil, Argentina, Colômbia e Guiana Inglesa, o sinônimo
normalmente utilizado é B. brachylaenoides e os epítetos infra-específicos deste. Apenas
recentemente o conceito mais amplo de B. oblongifolia tem sido adotado (Müller 2006,
Oliveira et al. 2006).
9. Baccharis platypoda DC., Prodr. 5: 409. 1836. Iconografia: Baker (1882), Müller (2006).
Fig. 2 o-p; 8 G-G’’
Arbusto, 1-2 m alt., ereto, ramificado. Ramos cilíndricos, estriados, cicatricosos,
resinosos. Folhas alternas, espiraladas, algumas vezes opostas; pecíolo ca. 6 mm compr.,
glanduloso-pontuado; lâmina (3) 5-7 (8) x (0,8) 2-4 (5) cm, obovada a oblongo-elíptica, ápice
agudo a obtuso, base cuneada, decorrente, margem superior denteada, coriácea, peninérvea,
ambas faces densamente glanduloso-pontuadas. Plantas femininas com sinflorescência
glomerular, homotética e plantas masculinas com sinflorescência corimbiforme de
25

florescências glomerulares, heterotética. Capítulos numerosos, sésseis; receptáculo plano,
alveolado, glabro; brácteas 1-3 mm compr., oblongas, resinosas. Capítulo masculino
campanulado, 4-5 mm compr.; brácteas involucrais 3 a 4-seriadas, externas 2-3 x 0,8-1 mm,
internas 3-4 x 0,8-1 mm, persistentes, lanceoladas, resinosas; ápice agudo, ciliado, margem
hialina. Flores 20-25; corola infundibuliforme, ca. 4 mm compr., levemente serícea; 5-lobada,
lobos ca. 1 mm compr., lanceolados; estilete ca. 4,5-5 mm compr., ramos triangulares, denso-
papilosos. Capítulo feminino cilíndrico, ca. 6 mm compr.; brácteas involucrais 4 a 5-seriadas,
externas 3-4 x 1,5-2 mm, internas 5-7 x 1-2 mm, persistentes, lanceoladas, glandulosas; ápice
agudo, ciliado, margem hialina. Flores ca. 25; corola 4-4,5 mm compr., levemente serícea; 5-
lobada, lobos ca. 0,5 mm compr., lanceolados; estilete 5-6 mm compr., ramos lanceolados,
curto-papilosos. Papilho ca. 4-5 mm compr., creme, caduco e com ápice agudo nas flores
femininas, persistente e com ápice longo-barbelado nas masculinas. Cipsela 2,5-3 mm compr.,
cilíndrica a convexa, 10 a 12-angulosa, glabra, costelas glandulosas.
Material examinado: Minas Gerais. Lima Duarte, Parque Estadual do Ibitipoca: 11.V.1970, L.
Krieger & D. Sucre (CESJ 8571); 11.V.1970, D. Sucre 6675 (RB); 5.V.1987, H.C. Souza s.n.
(BHCB 9816); 27.IV.1988, P.M. Andrade 1173 (BHCB); 11.II.1996, L.G. Rodela s.n. (CESJ
29534); 24.III.2001, M. Heluey et al. 87 (CESJ); Pico do Pião, 1.580-1.600 m alt., 12.V.1970,
D. Sucre & L. Krieger 6705 (RB, SP), 6717 (RB); trilha Monjolinho - Gruta dos Viajantes,
25.VII.2004, R.C. Forzza et al. 3491 (RB); idem, 16.V.2006, M.M. Saavedra et al. 273 (RB);
Gruta dos Três Arcos, 1.670 m alt., 26.VII.2004, R.C. Forzza et al. 3525 (RB); trilha para
Ponte de Pedra, 26.V.2005, M.M. Saavedra et al. 222 (RB).
Distribuição geográfica: Peru, Bolívia e Brasil: Bahia, Minas Gerais, Espírito Santo, Rio de
Janeiro, São Paulo.
Encontrada em campo rupestre, campo de altitude e restinga. No Parque foi observada
em afloramentos rochosos, campos hidromórficos e borda de matas nebulares; fértil durante o
26

ano todo. O táxon é facilmente reconhecido por apresentar folhas peninérveas densamente
glandulosas e sinflorescências distintas nas plantas masculinas e femininas. Giuliano (2005)
considera B. platypoda e B. condensata Rusby distintas e exclusivas da seção Agglomeratae.
Por sua vez, Müller (2006) sinonimiza B. condensata em B. platypoda, desconsidera a seção
proposta anteriormente e posiciona o último táxon no subgênero Baccharis.
10. Baccharis reticularia DC., Prodr. 5: 429. 1836.
Fig. 6 a-g; 8 J
Arbusto, 1-2 m alt., ereto, ramificado. Ramos cilíndricos, estriados, cicatricosos,
glabros, resinosos. Folhas alternas, espiraladas; pecíolo 1-4 mm compr., resinoso; lâmina (1)
1,5-3 (3,5) x 0,5-1 (1,5) cm, oboval a oblanceolada, ápice agudo a obtuso, base cuneada,
decorrente, margem inteira a denteada na porção superior, papirácea, trinérvea, ambas faces
glanduloso-pontuadas. Sinflorescência diplobótrio, homotética. Capítulos numerosos, os
femininos sésseis e os masculinos com pedúnculos 2-5 mm compr.; receptáculo convexo,
alveolado, glabro; brácteas 0,5-1,5 cm compr., oblanceoladas, semelhantes às folhas. Capítulo
feminino cilíndrico, ca. 5 mm compr.; brácteas involucrais 4 a 5-seriadas, externas 1-3 x 1
mm, internas 4-5 x 1-1,5 mm, persistentes, elípticas a lanceoladas, glabras, glandulosas;
ápice agudo, ciliado, margem hialina. Flores ca. 4; corola ca. 2,5 mm compr., levemente
serícea, com tricomas esparsos, 5-lobada, lobos triangulares, glabros; estilete ca. 3 mm
compr., ramos lanceolados, curto-papilosos. Capítulo masculino campanulado, ca. 3 mm
compr.; brácteas involucrais 4 a 5-seriadas, externas 1-2 x 0,7-1 mm, internas 2,5-3 x 1 mm,
persistentes, elípticas a lanceoladas, glabras, glandulosas, ápice agudo, ciliado, margem
hialina. Flores 10-15; corola infundibuliforme, ca. 2,5 mm compr., glandulosa, 5-lobada,
lobos ca. 1 mm compr., lanceolados, glandulosos, com tricomas curtos na base; estilete 2,5
mm compr., ramos triangulares, papilosos. Papilho 2-3 mm compr., estramíneo, caduco e com
27

cerdas de ápice agudo nas flores femininas, persistente e longo barbelado nas masculinas.
Cipsela 1,5-2 mm compr., convexa, 12-angulosa, glandulosa.
Material examinado: Minas Gerais. Lima Duarte, Parque Estadual do Ibitipoca: campo
rupestre ao lado da portaria, 4.II.2004, B.R. Silva et al. 1326 (RB); trilha restaurante - Prainha,
9.III.2004, R.C. Forzza et al. 3080 (RB); idem, 17.V.2006, M.M. Saavedra et al. 300, 301
(RB); idem, 29.VI.2006, R. Borges et al. 149, 150, 151, 152 (RB); trilha Gruta do Fugitivo -
Cascatinha, 11.III.2004, R.C. Forzza et al. 3195 (RB); trilha Lombada - Cruzeiro, 17.V.2006,
M.M. Saavedra et al. 307, 308 (RB); trilha Prainha - Lago dos Espelhos, 29.VI.2006, R.
Borges et al. 139, 140 (RB).
Distribuição geográfica: Brasil: Bahia, Goiás, Distrito Federal, Minas Gerais, Espírito Santo,
Rio de Janeiro, São Paulo, Paraná e Santa Catarina.
Encontrada em campo rupestre, campo de altitude, cerrado, mata atlântica e restinga. No
Parque foi observada em afloramentos rochosos, borda de matas nebulares e de galeria, e em
campo com vegetação arbustiva; fértil durante o ano todo. Foi observada uma grande variação
morfológica foliar nas populações do PEIB, assim como nas coleções de outras localidades.
No entanto, a morfologia das sinflorescências, flores e cipsela, e a composição dos capítulos
apresentam-se estáveis, o que é confirmado em outros estudos em que o táxon foi estudado
(Barroso 1976, Hind 1995, Oliveira & Marchiori 2006). Barroso (1976) posiciona B.
reticularia no “grupo Axilaris”. No entanto a classificação proposta por Giuliano (2005), que
considera B. reticularia na série Cylindricae, confere melhor relação morfológica com as
demais espécies presentes na seção.
11. Baccharis rufidula (Spreng.) Joch. Müller, Syst. Bot. Monogr. 76: 306. 2006.
≡ Conyza rufidula Spreng., Neue Entdeck. Pflanzenk. 2: 141. 1821.
= Baccharis vernonioides DC., Prodr. 5: 422. 1836.
28

Fig. 4 a-e; 8 H
Arbusto a arvoreta, 1,5-3 m alt., ereto, ramificado. Ramos cilíndricos, estriados,
ferrugíneo-tomentosos, glandulosos. Folhas alternas, espiraladas, pecíolo ca. 4 mm compr.;
lâmina (2) 4,5-7 (8) x (1) 1,5-2,3 (3) cm, elíptica a lanceolada, ápice agudo, base obtusa,
margem inteira, cartácea, peninérvea, ambas faces ferrugíneo-tomentosas. Sinflorescência
duplo-paniculada, heterotética. Capítulos numerosos, pedúnculos 1,5-2 cm compr.;
receptáculo cônico, alveolado, fimbriado, glanduloso; brácteas 3-6 mm, lanceoladas,
ferrugíneo-tomentosas. Capítulo masculino campanulado, ca. 2,5 mm compr., sem páleas;
brácteas involucrais 2 a 3-seriadas, externas 1-1,5 x 1 mm, internas 2-2,3 x 0,5-1 mm,
persistentes, elípticas a lanceoladas, glandulosas, glabras, algumas vezes as externas
tomentosas; ápice agudo, ciliado, margem hialina. Flores ca. 30; corola infundibuliforme, ca.
2,5 mm compr., serícea, 5-lobada, lobos ca. 0,5 mm compr., lanceolados, glabros; estilete 2,5-
3 mm compr., ramos triangulares, papilosos; ovário rudimentar ca. 0,5 mm compr., seríceo,
glanduloso. Capítulo feminino campanulado, ca. 5 mm compr., páleas ca. 5 mm compr.,
glabras, ápice acuminado, ciliado, margem hialina; brácteas involucrais 3 a 4-seriadas,
externas 2-3 x 1-1,5 mm, persistentes, internas 4-5 x 0,7-1 mm, caducas, elípticas a
lanceoladas, glandulosas, glabras, algumas vezes as externas tomentosas, ápice agudo, ciliado,
margem hialina. Flores ca. 20; corola ca. 4 mm compr., com tricomas formando um anel
acima da metade superior, 5-lobada, lobos diminutos, lanceolados; estilete ca. 5 mm compr.,
ramos lanceolados, curto-papilosos. Papilho 2-5 mm compr., persistente, estramíneo, cerdas
com ápice agudo. Cipsela ca. 2 mm compr., convexa, 5 a 7-angulosa, tomentosa, glandulosa.
Material examinado: Minas Gerais. Lima Duarte, Parque Estadual do Ibitipoca: trilha para a
Ponte de Pedra, 10.VIII.2005, R.C. Forzza et al. 4144 (RB); borda da mata, entre Mata
Grande e Matinha, 25.VII.2004, R.C. Forzza et al. 3465 (RB); trilha para o Monjolinho, 1.460
29

m alt., 25.VII.2004, R.C. Forzza et al. 3483 (RB); idem, 1.VII.2006, R. Borges et al. 162, 163
(RB); trilha interditada próxima ao alojamento novo, 29.VI.2006, R. Borges et al. 148 (RB);
Distribuição geográfica: Brasil: Minas Gerais, Espírito Santo e Rio de Janeiro.
Encontrada em campo rupestre, campo de altitude e mata atlântica. No Parque foi
observada em campo rupestre, mata de candeia, em borda de mata nebular e de galeria; fértil
de junho a setembro. Espécie facilmente reconhecida pelo indumento ferrugíneo-tomentoso e
sinflorescência duplo-paniculada. Baker (1882) indica Conyza rufidula Spreng. como
sinônimo de B. vernonioides DC., apesar do primeiro táxon ter sido publicado anteriormente.
Barroso (1976) não faz essa observação, nem comenta a categorização feita por Baker (1882),
utilizando também o nome proposto por De Candolle. Somente a partir de Müller (2006), a
prioridade de C. rufidula foi reconhecida e a nova combinação para o gênero Baccharis
realizada.
12. Baccharis salzmanii DC., Prodr. 5: 409. 1836.
Fig. 7 a-e; 8 K
Arbusto, 1,5-2,5 m alt., ereto, ramificado. Ramos cilíndricos, estriados, cicatricosos,
glabros, resinosos. Folhas alternas, espiraladas algumas vezes subopostas; pecíolo 0,5-0,8 mm
compr., glabro, resinoso; lâmina 1,5-5 (5,5) x 0,5-2,7 (3) cm, oboval, oblanceolada ou
elíptica, ápice agudo a obtuso, base atenuada, decorrente, margem irregularmente denteada na
porção superior, coriácea, trinérvea, ambas faces glabras, resinosas, face abaxial glanduloso-
pontuada. Sinflorescência diplobótrio, heterotética. Capítulos numerosos, sésseis ou com
pedúnculos 0,4-1,7 mm compr., inseridos na axila das folhas; receptáculo plano, alveolado,
glabro; brácteas 0,7-1,5 cm compr., oblanceoladas, ambas faces glabras, resinosas. Capítulo
masculino campanulado, ca. 5 mm compr.; brácteas involucrais 5 a 6-seriadas, externas 2-3 x
1-1,5 mm, internas 4-5,5 x 1-1,5 mm, persistentes, elípticas a lanceoladas, glabras,
30

glandulosas, ápice agudo, margem hialina. Flores 10-20; corola infundibuliforme, 4-4,5 mm
compr., glandulosa, levemente serícea, 5-lobada, lobos 1-1,3 mm compr., lanceolados,
glabros, estilete ca. 5,5 mm compr., ramos lanceolados, papilosos. Capítulo feminino
cilíndrico, 8-9 mm compr.; brácteas involucrais 5 a 6-seriadas, externas 2,5-3,5 x 1,5 mm,
internas 5-7,5 x 1,5 mm, persistentes, lanceoladas, glabras, glandulosas, ápice agudo, margem
hialina. Flores 5-7; corola ca. 4 mm compr., levemente serícea, com tricomas esparsos, 5-
lobada, lobos triangulares, diminutos, estilete ca. 5,5 mm compr., ramos lanceolados, curto-
papilosos. Papilho 4-4,5 mm compr., estramíneo, caduco e com cerdas de ápice agudo nas
flores femininas, persistente e com cerdas de ápice longo-barbelado nas masculinas. Cipsela
2-3 mm compr., cilíndrica a levemente convexa, 10 a 12-angulosa, glabra.
Material examinado: Minas Gerais. Lima Duarte, Parque Estadual do Ibitipoca: 25.V.1988,
P.M. Andrade 1201 (BHCB); 6.II.1996, L.G. Rodela s.n. (CESJ 28981); Pico do Pião, 1.450
m alt., 14.V.1970, D. Sucre 6830 (RB); idem, L. Krieger s.n. (CESJ 8586); trilha para o
Monjolinho, 25.VII.2004, R.C. Forzza et al. 3473 (RB); idem, 9.VIII.2005, R.C. Forzza et al.
4123 (RB); idem, 30.VI.2006, R. Borges et al. 168, 169 (RB); mata de candeia na trilha para
Ponte de Pedra, 17.V.2006, M.M. Saavedra et al. 298, 299 (RB); trilha Prainha - Lago dos
Espelhos, 16.V.2006, M.M. Saavedra et al. 254, 255 (RB); idem, R. Borges et al. 137, 138
(RB); trilha interditada próxima ao alojamento novo, 18.V.2006, M.M. Saavedra et al. 317,
318 (RB); idem, 29.VI.2006, R. Borges et al. 146, 147 (RB).
Distribuição geográfica: Brasil: Bahia, Goiás, Distrito Federal, Minas Gerais, São Paulo e
Paraná.
Encontrada em campo rupestre, campo de altitude, cerrado e restinga. No Parque
apresenta grandes populações, sendo observada em campo rupestre graminoso ou com
vegetação arbustiva densa, em meio a mata de candeia e em borda de mata estacional e
ombrófila; fértil durante o ano todo. Assim como B. reticularia, a espécie apresenta folhas
31

com grande variação morfológica, porém a morfologia dos capítulos, flores e cipsela permite
reconhecê-la dentre as demais espécies (Barroso 1976, Hind 1995). Além desta variação, foi
também observada a presença de capítulos masculinos e femininos na mesma planta, fato que
será abordado em estudos futuros. O táxon é morfologicamente próximo de B.
pseudomyriocephala Teodoro, diferenciando-se este último pelos capítulos menores e folhas
obovais, de margem curto-denteada e base longo-atenuada. Barroso (1976) identificou coletas
provenientes do PEIB como B. pseudomyriocephala, no entanto, apesar da semelhança foliar
presente no material estudado pela autora, a análise das estruturas reprodutivas permitiu a
identificação correta de B. salzmanii. Devido a difícil distinção entre os dois táxons e a grande
variação morfológica observada, constatamos a necessidade de um estudo que aborde mais
profundamente a identidade de ambos.
13. Baccharis schultzii Baker, in Mart. & Eichler, Fl. bras. 6(3): 78. 1882.
Fig. 7 f-k
Arbusto, ca. 2 m alt., ereto, ramificado. Ramos cilíndricos, estriados, cicatricosos,
glandulosos. Folhas alternas, espiraladas pecíolo 7-8 mm compr., glabro, glanduloso; lâmina
(2,5) 3,5-4 (4,5) x (0,5) 1 cm, elíptica a oblonga, ápice agudo a obtuso, base longamente
cuneada, decorrente, margem denteada, papirácea, trinérvea, ambas faces glabras,
glandulosas. Sinflorescência diplobotrióide, homotética, de florescências glomeruliformes.
Capítulos numerosos, pedúnculos 0,5-1,2 cm compr., originados na axila das folhas e não
ultrapassando o comprimento destas; receptáculo plano, alveolado, glabro, glanduloso;
brácteas 0,5-2,2 cm, oblanceoladas, semelhante às folhas. Capítulo masculino campanulado,
ca. 6 mm compr.; brácteas involucrais 4-seriadas, externas 3-4 x 1,5-2 mm, internas 5-5,5 x
1,5-2 mm, persistentes, lanceoladas, glabras, glandulosas; ápice agudo, margem hialina.
Flores ca. 10; corola infundibuliforme, ca. 4,5-5 mm compr., levemente serícea, 5-lobada,
32

lobos ca. 1 mm compr., lanceolados, revolutos; estilete ca. 6 mm compr., ramos lanceolados,
longo-papilosos. Capítulo feminino cilíndrico, ca. 1 cm compr.; brácteas involucrais 5 a 6-
seriadas, externas 2-5 x 1-1,5 mm, internas 6-7,5 x 1-1,5 mm, persistentes, lanceoladas,
glabras, glandulosas, ápice agudo, margem hialina. Flores 5-7; corola 4,5-5,5 mm compr.,
levemente serícea, 5-lobada, lobos ca. 0,5 mm compr., lanceolados, estilete 6-7 mm compr.,
ramos lanceolados, papilosos. Papilho 4-5 mm compr., estramíneo, caduco e com cerdas de
ápice agudo nas flores femininas, persistente e cerdas com ápice longo-barbelado nas
masculinas. Cipsela ca. 2,5 mm compr., cilíndrica, 10 a 12-angulosa, glabra, ápice truncado.
Material examinado: Minas Gerais. Lima Duarte, Parque Estadual do Ibitipoca: 21.III.1996,
L.G. Rodela s.n. (CESJ 36535); mata ao lado da Potaria do Parque, 23.I.2007, M.M. Saavedra
et al. 340 (RB).
Material adicional: Rio de Janeiro. Teresópolis: Posse, morro das antenas de televisão,
10.II.1968, D. Sucre 2335 (RB).
Distribuição geográfica: Brasil: Minas Gerais, Rio de Janeiro e São Paulo.
Encontrada em campo rupestre, campo de altitude e mata atlântica. No Parque, foi
observada em campo rupestre e borda de mata estacional; fértil de janeiro a junho. Espécie
próxima de B. salzmanii, diferenciando-se desta principalmente pela sinflorescência
composta, formada por florescências glomeruliformes. Barroso (1976) posicionou o táxon no
“grupo Cassiniaefolia”, baseada na morfologia das florescências. Entretanto, a classificação
proposta por Giuliano (2005) situa B. schultzii na série Cylindricae, relacionando melhor a
espécie com as outras do gênero.
14. Baccharis serrula Sch. Bip. ex Baker, in Mart. & Eichler, Fl. bras. 6(3): 70. 1882.
Fig. 3 a-e
33

Subarbusto, 0,5-1 m alt., ereto, ramificado. Ramos cilíndricos, estriados, cicatricosos,
glabros. Folhas alternas, espiraladas, sésseis; lâmina (0,5) 1-1,5 x 0,3-0,4 cm, lanceolada a
oblanceolada, ápice agudo a obtuso, base atenuada, margem curto-serreada na metade
superior, coriácea, uninérvea, ambas faces glabras, glandulosas. Sinflorescência botrióide de
florescências glomerulares, homotética. Capítulos numerosos, sésseis, desenvolvidos na axila
das folhas; receptáculo plano, alveolado, glabro. Capítulo masculino campanulado, 3-4 mm
compr.; brácteas involucrais 3-seriadas, externas 1,5-2,3 x 1-1,5 mm, internas 3-3,5 x 1-1,5
mm, persistentes, elípticas a lanceoladas, glandulosas, ápice agudo, margem hialina. Flores 5-
6; corola infundibuliforme, ca. 3 mm compr., com tricomas curtos concentrados abaixo dos
lobos, 5-lobada, lobos ca. 1 mm compr., lanceolados, glabros; estilete ca. 3,5 mm compr.,
ramos triangulares, papilosos. Capítulo feminino cilíndrico, ca. 0,5 mm compr.; brácteas
involucrais 3 a 4-seriadas, externas 1,5-2,8 x 0,7-1,3 mm, internas 4-5 x 0,5-1,3 mm,
persistentes, lanceoladas, glabras, glandulosas, ápice agudo, margem hialina. Flores
geralmente 1; corola 4-4,8 mm compr., levemente serícea, com tricomas concentrados na
base, 5-lobada, lobos ca. 0,5 mm compr., lanceolados, glabros, estilete ca. 5 mm compr.,
ramos lanceolados, curto-papilosos. Papilho ca. 3,5 mm compr., creme, caduco e com cerdas
de ápice agudo nas flores femininas, persistente e com cerdas de ápice longo-barbelado nas
masculinas. Cipsela 1,5-2 mm compr., cilíndrica, ca. 20-angulosa, glabra, ápice truncado.
Material examinado: Minas Gerais. Lima Duarte, Parque Estadual do Ibitipoca: Pico do Pião,
1.580-1.600 m alt., 12.V.1970, D. Sucre 6693 (RB).
Material adicional: Minas Gerais. Serro: Alto Pico do Itambé, 5.V.1942, M. Magalhães 1622
(RB).
Distribuição geográfica: Brasil: Minas Gerais e São Paulo.
Encontrada em campo rupestre e campo de altitude; fértil de maio a julho. B. serrula é
reconhecido principalmente pela morfologia das folhas, capítulos e sinflorescência. Apresenta
34

semelhança com B. parvidentata Teodoro, diferenciando-se desta pelos capítulos menores,
número de flores menor e cipsela com mais ângulos. O táxon também foi situado no “grupo
Axilaris” por Barroso (1976), apesar da proximidade e melhor posicionamento desta com as
espécies da série Axillares (Giuliano 2005). Durante o trabalho de campo a espécie não foi
observada em nenhuma localidade do PEIB.
15. Baccharis serrulata (Lam.) Pers., Syn. Plant. 2: 423. 1807.
≡ Conyza serrulata Lam., Encycl. 2(1): 85. 1786.
Fig. 3 j-n; 8 F
Arbusto, 1-1,5 m alt., ereto, ramificado. Ramos cilíndricos, estriados, seríceos. Folhas
alternas, espiraladas; pecíolo 0,7-1,3 mm compr., glanduloso, denso-seríceo; lâmina (1) 3,5-
6,5 (8) x 0,7-1,5 (2) cm, lanceolada, ápice agudo, base levemente cuneada, decorrente,
margem serreada, papirácea, trinérvea, face adaxial seríceo-glandulosa, face abaxial
glandulosa, brilhante, com tricomas dispostos nas nervuras principais. Sinflorescência
corimbosa de florescências paniculares, heterotética. Capítulos numerosos, pedúnculos 3-5
mm compr.; receptáculo plano, alveolado, glabro; brácteas 5-8 mm compr., lineares, seríceo-
glandulosas. Capítulo masculino campanulado, ca. 2 mm compr.; brácteas involucrais 3-
seriadas, externas 1-1,5 x 0,5 mm, internas 2,0 x 0,5 mm, persistentes, lanceoladas, glabras;
ápice agudo, ciliado, margem hialina. Flores ca. 20; corola hipocrateriforme, ca. 2 mm
compr., 5-lobada, levemente serícea, lobos ca. 0,5 mm compr., lanceolados, patentes; estilete
2-2,3 mm compr., ramos lanceolados, curtos, denso-papilosos. Capítulo feminino
campanulado, ca. 3 mm compr.; brácteas involucrais 3-4 seriadas, externas 1,5-2 x 0,5-0,7
mm, internas 2-2,5 x 0,3-0,5 mm, persistentes, lanceoladas, glabras, ápice agudo, ciliado,
margem hialina. Flores ca. 50; corola 1-1,5 mm compr., glandulosa, truncada, com tricomas
concentrados no ápice e base; estilete ca. 2 mm compr., ramos lanceolados, curto-papilosos.
35

Papilho ca. 2 mm compr., persistente, estramíneo, cerdas com ápice agudo nas flores
femininas e longo-barbelado nas masculinas. Cipsela 0,5-1 mm compr., convexa, 8 a 10-
angulosa, hirsuta, com tricomas curtos, ápice truncado.
Material examinado: Minas Gerais. Lima Duarte, Parque Estadual do Ibitipoca: 28.IX.1970,
P.I.S. Braga 1881 (RB); 30.IX.1970, P.I.S. Braga 1904 (RB); 3.XI.2000, M.A. Heluey et al. 3
(CESJ); Pico do Pião, 1.580-1.600 m alt., 12.V.1970, D. Sucre et al. 6693 (RB); próximo ao
Rio do Salto, 1.200 m alt., 9.III.2004, R.C. Forzza et al. 3037 (RB); trilha Monjolinho - Gruta
dos Viajantes, 25.VII. 2004, R.C. Forzza et al. 3485 (RB); Gruta dos Três Arcos, 1.IX.2004,
M.M. Saavedra et al. 215 (RB); mata do Cruzeiro, 27.V.2005, M.M. Saavedra et al. 233
(RB); 27.V.2005, M.M. Saavedra et al. 238 (RB); trilha restaurante - Prainha, 16.V.2006,
M.M. Saavedra et al. 249, 250 (RB); mata atrás do alojamento, 20.XI.2006, M.M. Saavedra
et al. 327 (RB); mata ao lado da portaria do Parque, 23.I.2007, M.M. Saavedra et al. 342
(RB).
Distribuição geográfica: Paraguai e Brasil: Pernambuco, Bahia, Minas Gerais, Espírito Santo,
Rio de Janeiro, São Paulo, Paraná, Santa Catarina e Rio Grande do Sul.
Encontrada em campo rupestre, campo de altitude, cerrado, mata atlântica, restinga e
campos sulinos. No Parque foi observada em afloramento rochoso, orla de mata estacional,
nebular e de galeria e em área perturbada; fértil durante o ano todo. Apresenta grandes
populações no PEIB, característica que também é mencionada em estudos anteriores (Barroso
1976, Hind 1995, 2003a). É facilmente reconhecida pelas folhas lanceoladas, trinérveas e de
margem serreada, e pela sinflorescência corimbosa.
16. Baccharis tarchonanthoides DC., Prodr. 5: 414. 1836. Iconografia: Baker (1882).
Fig. 6 h-i
36

Arbusto, 1-2 m alt., ereto, ramificado. Ramos cilíndricos, cicatricosos, pubérulos, alvos.
Folhas alternas, espiraladas; pecíolo ca. 5 mm compr., pubérulo; lâmina (2,6) 3-7 (8,5) x (0,5)
1,5-2 (2) cm, oblanceolada, ápice agudo, base atenuada, decorrente, margem denteada na
metade superior, coriácea, peninérvea, face adaxial glandulosa, face abaxial alvo-lanosa.
Sinflorescência duplo-paniculada heterotética, eixo alvo-lanoso. Capítulos numerosos,
pedúnculo 3-8 mm compr.; receptáculo plano, alveolado, glabro; brácteas 2-5 mm compr.,
lineares a lanceoladas, alvo-lanosas abaxialmente. Capítulo masculino campanulado, 3-4 mm
compr.; brácteas involucrais 2-seriadas, externas 2-3 x 0,6-1 mm, internas 3-3,2 x 0,7-1 mm,
persistentes, lanceoladas, alvo-lanosas, resinosas, ápice agudo, ciliado, margem hialina. Flores
ca. 30; corola hipocrateriforme, ca. 3 mm compr., levemente serícea, 5-lobada, lobos ca. 1,5
mm compr., lanceolados, glabros, revolutos; estilete ca. 3-3,3 mm compr., ramos lanceolados,
papilosos. Capítulo feminino campanulado, 4-5 mm compr.; brácteas involucrais 3-seriadas,
externas 2-3,5 x 0,8-1 mm, internas 4-4,2 x 0,6-1 mm, persistentes, lanceoladas, alvo-lanosas,
resinosas, ápice agudo, ciliado, margem hialina. Flores ca. 40; corola 2,5-3,8 mm compr.,
levemente serícea; 5-lobada, lobos triangulares, papilosos, com tricomas longos, esparsos;
estilete 3-3,5 mm compr., ramos lanceolados, curto-papilosos. Papilho 2-3,3 mm compr.,
estramíneo, persistente e com cerdas de ápice agudo nas flores femininas, caduco e com
cerdas de ápice longo-barbelado nas masculinas. Cipsela 1-1,2 mm compr., cilíndrica a
levemente convexa, 5 a 7-angulosa, glandulosa, coberta por espessa secreção.
Material examinado: Minas Gerais. Lima Duarte, Parque Estadual do Ibitipoca: 13.IX.1940,
M. Magalhães 517 (BHCB, SP, RB); 27.IX.1970, L. Krieger s.n. (CESJ 9284, RB 151.303);
1.X.1970, U. Confúcio s.n. (CESJ 9459, RB 151.294); IX.1987, M. Sobral 5594 (BHCB);
trilha do Monjolinho, 19.IX.2006, R. Borges et al. 259 (RB).
Distribuição geográfica: Brasil: Minas Gerais, Rio de Janeiro, São Paulo e Paraná.
37

Encontrada em campo rupestre, campo de altitude, cerrado e mata atlântica. No Parque
foi observada em campo rupestre junto a vegetação arbustiva e próximo a campo
hidromórfico; fértil durante o ano todo. Semelhante a B. lychnophora pelo indumento, porém
distingui-se facilmente pelas folhas denteadas, capítulo feminino campanulado, flores
masculinas com corola hipocrateriforme e papilho caduco.
17. Baccharis varians Gardner, J. Bot. (Hooker) 7: 84. 1848.
Fig. 4 h-l; 8 L
Subarbusto, 0,5-1 m alt., ereto, ramificado, xilopodífero. Ramos cilíndricos, estriados,
cicatricosos, glabros, glandulosos. Folhas alternas, espiraladas, sésseis; lâmina (0,5) 1-3 x 0,2-
0,5 cm; oblanceolada a linear, ápice agudo, base cuneada a atenuada, margem geralmente
tridentada na metade superior, papirácea, uninérvea, ambas faces glabras. Sinflorescência
diplobótrio, heterotética. Capítulos numerosos, pedúnculo 0,5-2,5 cm compr., desenvolvido
na axila das folhas; receptáculo plano, alveolado, fimbriado, glanduloso; brácteas 5-7 mm
compr., lineares, glabras, glandulosas. Capítulo masculino cilíndrico, ca. 3 mm compr.;
brácteas involucrais 4 a 5-seriadas, externas 1-2 x 0,8-1 mm, internas 2,5-3 x 0,5-1 mm,
persistentes, elípticas a lanceoladas, glandulosas; ápice agudo, margem hialina. Flores ca. 10;
corola infundibuliforme, ca. 3,5 mm compr., glabra, glandulosa, 5-lobada, lobos ca. 1 mm
compr., lanceolados, glabros; estilete ca. 4 mm compr., ramos triangulares, longo-papilosos.
Capítulo feminino cilíndrico, ca. 3,5 mm compr.; brácteas involucrais 3 a 4-seriadas, externas
1-2,5 x 0,5-1 mm, internas 3-3,5 x 0,5-0,7 mm, persistentes, lanceoladas, glabras,
glandulosas, ápice agudo, ciliado, margem hialina. Flores 8-10; corola 2,5 mm compr.,
levemente serícea, com tricomas próximos aos lobos, 5-lobada, com dois lobos maiores que
os demais, lanceolados, glabros, estilete ca. 3,5 mm compr., ramos lanceolados, curto-
papilosos. Papilho ca. 3 mm compr., estramíneo, caduco e com cerdas de ápice agudo nas
38

flores femininas, persistente e com cerdas de ápice longo-barbelado nas masculinas. Cipsela
ca. 1 mm compr., convexa, ca. 10-angulosa, glabra, glandulosa.
Material examinado: Minas Gerais. Lima Duarte, Parque Estadual do Ibitipoca: Pico do Pião,
1.350-1.450 m alt., 14.V.1970, D. Sucre 6841 (RB); L. Krieger s.n. (CESJ 8551); trilha Lago
dos Espelhos - Monjolinho, 9.VIII.2005, R.C. Forzza et al. 4116 (RB); idem, 16.V.2006,
M.M. Saavedra et al. 271, 272 (RB); idem, 1.VII.2006, R.Borges et al. 166, 167 (RB).
Distribuição geográfica: Guiana Inglesa, Suriname, Brasil: Bahia, Goiás, Distrito Federal,
Mato Grosso, Minas Gerais, Rio de Janeiro e São Paulo.
Encontrada em campo rupestre, campo de altitude e cerrado. No Parque foi observada
em campo rupestre com vegetação arbustiva esparsa; fértil durante o ano todo. A espécie
possui características marcantes como hábito subarbustivo, xilopódio, capítulos com ca. 10
flores e corola das flores femininas com 2 lobos maiores que os demais. Posicionado no
“grupo Rufescens” por Barroso (1976), na série Cylindricae por Giuliano (2005) e no “grupo
Baccharis linearifolia” por Müller (2006), o táxon apresenta maior semelhança com as
espécies posicionadas no agrupamento de Barroso (1976). Segundo Hind (1995), a espécie faz
parte do complexo B. linearifolia, no entanto Müller (2006) adota um conceito amplo para as
espécies morfologicamente relacionadas a esta, propondo a sinonímia de B. varians e demais
espécies relacionadas em B. linearifolia. No presente trabalho não adotamos o conceito
proposto pelo último autor devido à complexidade taxonômica que envolve o grupo.
Acreditamos que sejam necessários estudos específicos das espécies envolvidas, que
compreendam hipóteses relativas à história evolutiva destas (e.g. análises filogeográficas),
não se restringindo apenas à análise de coleções de herbário para a proposição de extensas
sinonímias.
Em sua revisão Barroso (1976) reconhece através da análise dos síntipos, que Gardner
indica duas espécies distintas no protólogo de B. varians. O síntipo Gardner 3839 é
39

identificado pela autora como B. leptocephala DC., porém corresponde a B. linearifolia
(Lam.) Pers. E o síntipo Gardner 4913 foi então reconhecido como o lectótipo (designado
como holótipo na obra) do táxon e indicado no emendavit feito pela autora, que agiu
corretamente segundo os artigos 9.2, 9.9 e 9.10 do CINB (McNeill et al. 2007).
Conyza Less., Syn. Gen. Compos. 203-204. 1832
Gênero com 60-100 espécies, que possui distribuição pantropical (América do Sul e
África) (Nesom & Robinson 2007). A divisão infragenérica de Conyza reflete a semelhança
entre as características morfológicas e biogeográficas das espécies (Nesom 1994a). No
entanto, a sistemática de Conyza apresenta problemas quanto às relações e aos limites
taxonômicos com o gênero Erigeron L. Noyes (2000) indica que as espécies ocorrentes na
América são polifiléticas e possuem os processos de especiação vinculados a Erigeron.
Nome-popular: buva, cauda-de-raposa, erva-lanceta, voadeira (Pio-Corrêa 1984a,b,
Neto & Morais 2003).
18. Conyza monorchis (Griseb.) Cabrera, Man. Fl. Alrededores Buenos Aires, 480. 1953.
Iconografia: Baker (1882).
≡ Erigeron monorchis Griseb., Symb. Fl. Argent. 175. 1879.
Fig. 5 h-i
Erva, 0,5 – 1,5 m alt., perene, acaule, tuberosa. Folhas da base rosuladas, sésseis; lâmina
(3,5) 4-6 (7) x (0,6) 0,8-1 cm, oblanceolada, ápice agudo, mucronado, base atenuada, margem
denteada, papirácea, peninérvea, face adaxial glandulosa, ápice e margens híspidas, face
abaxial glanduloso-pontuada; folhas do eixo sésseis, (1,8) 3-3,5 (4) cm, oblanceoladas, ápice
agudo, mucronado, base atenuada, margem denteada, papiráceas, peninérveas, híspidas.
40

Sinflorescência paniculiforme de florescências botrióides, heterotética; eixo fistuloso,
estriado, glabro; bractéolas 3-7 mm compr., oblanceoladas, híspidas. Capítulos disciformes,
numerosos, pedúnculos 1,2-4,8 cm compr., campanulados, ca. 5 mm compr.; brácteas
involucrais 2-seriadas, externas 2-3 x 0,3-0,7 mm, internas 4-5 x 0,4-0,9 mm, persistentes,
lanceoladas, seríceas, glandulosas, ápice agudo, ciliado, margem hialina; receptáculo
convexo, alveolado. Flores centrais hermafroditas, ca. 10; corola infundibuliforme, ca. 4,5
mm compr., glandulosa, levemente serícea abaixo dos lobos, 5-lobada; lobos ca. 0,5 mm,
glandulosos, levemente seríceos; anteras com apêndice lanceolado, base obtusa; estilete ca. 5
mm compr., com ramos lanceolados, papilosos. Flores marginais femininas, ca. 35; corola
curto-ligulada, ca. 4 mm compr., glandulosa, levemente serícea abaixo do ápice; limbo ca. 0,5
mm compr., 2-lobado, glanduloso; estilete ca. 4 mm compr., ramos lanceolados, curto-
papilosos. Papilho 4-4,5 mm compr., caduco, cerdoso, barbelado, estramíneo, cerdas com
ápice agudo. Cipsela ca. 2 mm compr., oblonga, achatada, com 2-angulosa, serícea, ápice
truncado, carpopódio desenvolvido.
Material examinado: Minas Gerais. Lima Duarte, Parque Estadual do Ibitipoca: trilha
Monjolinho - Pico do Peão, 1.500 m alt., 31.V.2007, R. Borges 878 (RB).
Material adicional: São Paulo. Bocaina, campo a oeste do acampamento, 1.600 m alt.,
7.XII.1952, Markgraf. & App. 10292 (RB).
Distribuição geográfica: Brasil, Argentina e Uruguai. No Brasil: Minas Gerais, Rio de
Janeiro, São Paulo e Rio Grande do Sul.
Encontrada em campo rupestre, campo de altitude, cerrado e campos sulinos. No Parque
foi observada em campo rupestre próximo a mata nebular; fértil de novembro a março. A
espécie pode ser distinguida das demais estudadas principalmente pela presença de pequeno
tubérculo e capítulos disciformes com flores femininas dispostas em várias séries e poucas
flores hermafroditas no centro. C. monorchis é próxima de C. blanchetii Baker pelo tipo de
41

sinflorescência e morfologia do capítulo, porém a última não possui tubérculos e as folhas são
estreito-lanceoladas e de margem inteira.
Inulopsis (DC.) O. Hoffman, in Engler & Prantl, Nat. Pflanzenfam. 4(5): 145, 149. 1890.
Gênero composto de quatro espécies, distribuídas em vegetação campestre do Brasil,
Bolívia e Paraguai (Nesom 1994b, Nesom & Robinson 2007). No presente estudo
reconhecemos apenas três espécies, pois I. phoenix G.L. Nesom é considerado sinônimo de I.
camporum (Gardner) G.L. Nesom (Teles com. pess.).
Nome-popular: quebra-frasco (Pio-Corrêa 1984c).
19. Inulopsis scaposa (DC.) O.Hoffm., in Engler & Prantl, Nat. Pflanzenfam. 4(5): 149.
1894.
≡ Haplopappus scaposus DC., Prodr. 5: 350. 1836.
Fig. 5 a-e; 8 I
Erva, 15- 50 cm alt., perene, acaule. Folhas rosuladas, sésseis; lâmina (2) 3,5 - 10,5 (20)
x (0,7) 1,1 - 1,7 cm, oboval-elíptica a oboval-lanceolada, ápice agudo a obtuso, mucronado,
base longo-cuneada, margem curto-denteada, papirácea, peninérvea, ambas as faces
glanduloso-pontuadas, híspidas, com tricomas esparsos ou densos nas folhas mais jovens.
Sinflorescência escaposa; escapo 9-40 cm compr., estriado, glabro; brácteas ca. 2 mm compr.,
lineares, uninérveas, ciliadas. Capítulo radiado, solitário, campanulado, 6-7 mm compr.;
brácteas involucrais 3-seriadas, externas 5-5,5 x 0,4-1 mm, internas 4-5,1 x 0,7-1 mm,
persistentes, linear-lanceoladas, glandulosas, ápice agudo, margem hialina; receptáculo
convexo, alveolado, fimbriado, glabro. Flores do raio femininas, ca. 70; corola ligulada,
creme, 6,7-7,1 mm compr., tubo 1,8-2,3 mm compr., seríceo, limbo curtamente 2 a 3-lobado,
glabro; estilete 3,3-4,7 mm compr., ramos triangulares, curto-papilosos. Flores do disco
42

funcionalmente masculinas, ca. 95; corola infundibuliforme, ca. 4 mm compr., 5-lobada,
glandulosa, serícea, tubo 1,5-1,8 mm compr., limbo 2,2-2,5 mm compr., lobos triangulares;
anteras com apêndice lanceolado, base obtusa; estilete 5-5,2 mm compr., ramos triangulares,
longo-papilosos. Papilho 2,7-3,5 mm compr., persistente, cerdoso, barbelado, creme, cerdas
com ápice agudo. Cipsela 1,6-2 mm compr., oboval-oblonga, achatada, com 2 costelas
laterais, tomentosa.
Material examinado: Minas Gerais. Lima Duarte, Parque Estadual do Ibitipoca: 26.IX.1970,
D. Sucre 7110 (RB); 10.II.2001, M.C. Heluey et al. 48 (CESJ, BHCB); trilha Monjolinho -
Lagoa Seca, 30.III.2004, F.M. Ferreira et al. 707 (RB); idem, 16.V.2006, M.M. Saavedra et
al. 266 (RB); base do Pico do Pião, 18.I.2005, R.C. Forzza et al. 3936 (RB).
Distribuição geográfica: Paraguai e Brasil: Goiás, Minas Gerais, Rio de Janeiro, São Paulo,
Paraná e Santa Catarina.
Encontrada em campo rupestre, campo de altitude, cerrado e campos sulinos. No Parque
foi observada em campo rupestre com vegetação herbácea densa; fértil durante o ano todo. I.
scaposa é facilmente distinta das demais espécies estudadas pela sinflorescência escaposa e
capítulo solitário. Dentre as espécies do gênero se assemelha a I. stenophylla Dúsen, porém a
última se distingue prontamente pela presença de folhas lineares e sem glândulas.
Leptostelma D. Don, in Sweet, Brit. Fl. Gard. Ser. 2. 1831.
Gênero que apresenta seis espécies encontradas no Brasil, Argentina, Paraguai e
Uruguai (Teles com. pess.). O gênero foi descrito em 1831 e posteriormente tratado como
seção de Erigeron (Bentham & Hooker 1873), sendo mantido nesta categoria nos estudos
taxonômicos subseqüentes (Baker 1882, Solbrig 1960, 1962, Nesom 1989). Apenas
recentemente Hind & Nesom (2002) reconheceram novamente Leptostelma na categoria
genérica.
43

Nomes populares: margarida-do-banhado, margarida-do-brejo (Solbrig 1962, Pio-
Corrêa 1984c).
20. Leptostelma maxima D. Don, in Sweet, Brit. Fl. Gard. Ser. 2. t. 38. 1831. Iconografia:
Baker (1882), Solbrig (1962).
≡ Erigeron maximus (D.Don) Otto ex DC, Prodr. 5: 284. 1836.
Fig. 5 f-g; 8 A
Erva, 0,6 - 1,5 m alt., perene, acaule. Folhas da base rosuladas, sésseis; lâmina 13- 35 x
(1,2) 2,5-6 cm, oblongas a lanceoladas, ápice agudo, base longo-cuneada, margem curto-
serreada, irregular, denso-serícea, membranácea, peninérvea, face adaxial glanduloso-
pontuada, híspida, com tricomas concentrados próximo às margens e ápice, face abaxial
glabra, com tricomas esparsos sobre a nervura central, densamente serícea nas folhas jovens.
Folhas do eixo sésseis e amplexas, (2,4) 3,5-17 (21,2) x (0,5) 0,8-2 (3) cm, lanceoladas, ápice
agudo, base levemente arredondada, margem curto-serreada, irregular, denso-serícea,
membranácea, peninérvea, nervuras secundárias fracamente impressas, face adaxial seríceo-
glandulosa, face abaxial glandulosa. Sinflorescência corimbiforme; eixo lenhoso, fistuloso,
estriado-sulcado, seríceo-glanduloso; bractéolas 0,4-1 mm compr., oblongas, ápice
acuminado, margem inteira, face adaxial glabra, face abaxial glandulosa. Capítulos radiados,
numerosos, terminais, campanulados, 6-7 mm compr., pedúnculos 2,4-12,5 cm; brácteas
involucrais 3-seriadas, externas 5,5-7,5 x 1,1-1,8 mm, internas 7,2-7,8 x 1,3-1,8 mm,
persistentes, elíptico-lanceoladas, glandulosas, ápice acuminado, margem hialina.
Receptáculo plano a convexo, alveolado, fimbriado, glabro. Flores do raio femininas, ca. 110;
corola ligulada, alva, ca. 11 mm compr., tubo 2,7-3 mm compr., esparssamente seríceo, limbo
44

curtamente 2 a 3-lobado, glabro; estilete 3-3,4 mm compr., ramos triangulares, obtusos,
papilosos. Flores do disco hermafroditas, ca. 240; corola infundibuliforme, 4,7-5,2 mm
compr., 5-lobada, esparssamente serícea, tubo 1,5-1,8 mm compr., lobos 3,2-4 mm compr.,
triangulares, papilosos. Anteras com apêndice lanceolado, base obtusa; estilete 4-4,5 mm
compr., ramos triangulares, longo-papilosos, alargados, base espessa. Papilho 4,2-4,5 mm
compr.; persistente, cerdoso, barbelado, estramíneo, cerdas com ápice agudo. Cipsela 2-2,2
mm compr., oblongo-elíptica, achatada, com 2-angulosa, glabra, ápice levemente atenuado.
Material examinado: Minas Gerais. Lima Duarte, Parque Estadual do Ibitipoca: 14.V.1970, L.
Krieger s.n. (CESJ 8558); 12.XI.1987, H.C. Souza s.n. (BHCB 13907); 10.II.2001, F.S.
Araújo 25 (CESJ); trilha Monjolinho - Lagoa Seca, 30.III.2004, F.M. Ferreira et al. 725
(RB); base do Pico do Pião, 16.V.2006, M.M. Saavedra et al. 287 (RB); trilha Lombada -
Gruta dos Três Arcos, 18.V.2006, M.M. Saavedra et al. 310 (RB); trilha Gruta dos Viajantes -
Pico do Pião, 10.III.2004, R.C. Forzza et al. 3119 (RB).
Distribuição geográfica: Brasil, Argentina, Paraguai. No Brasil: Mato Grosso, Goiás, Minas
Gerais, Espírito Santo, Rio de Janeiro, São Paulo, Paraná, Santa Catarina e Rio Grande do
Sul.
Encontrada em campo rupestre, campo de altitude, cerrado e campos sulinos. No Parque
foi observada em campo rupestre com afloramento rochoso e próximo a mata nebular, nos
locais com solo encharcado ou úmido; fértil durante o ano todo. Os capítulos radiados e a
corola das flores femininas com lígula bem desenvolvida tornam a espécie distinta das demais
abordadas no trabalho. L. maxima apresenta a maior distribuição geográfica entre as espécies
do gênero, sendo observada também uma grande variação morfológica das folhas e do eixo da
sinflorescência.
45

Referências
ALENCAR, S.M., AGUIAR, C.L., PAREDES-GUZMÁN, J., PARK, Y.K. 2005.
Composição química de Baccharis dracunculifolia, fonte botânica das própolis dos
estados de São Paulo e Minas Gerais. Ciência Rural 35(4): 909-915.
ANDRADE, P. M. & SOUSA, H. C. 1995. Contribuição ao conhecimento da vegetação do
Parque Estadual do Ibitipoca, Lima Duarte, Minas Gerais. Rev. Árvore 19(2): 249-261.
ANGIOSPERM PHYLOGENY GROUP II. 2003. An update of the Angiosperm Phylogeny
Group classification for the orders and families of flowering plants. Bot. Journ. Linn.
Soc. 141: 399-436.
ARISTEGUIETA, L. 1964. Flora de Venezuela – Compositae, Instituto Botanico, Ministerio
de Agricultura y Cria, Caracas, vol. 10, parte1, p. 319-331.
BAKER, J.G. 1882. Compositae III Asteroideae, Inuloideae. In: C.F.P. Martius & A.W.
Eichler (eds), Flora brasiliensis, Frid. Fleischer. Leipzig, vol 6, pars 3, p. 1-134.
BARROSO, G. M. 1976. Compositae – Subtribo Baccharidinae Hoffman – Estudo das
espécies ocorrentes no Brasil. Rodriguésia 27(40): 3-273.
BENTHAM, G. 1873. Notes on the classification, history and geographical distribution of the
Compositae. Bot. Journ. Linn. Soc. 13: 335-577.
BENTHAM, G. & HOOKER, J.D. 1873. Genera Plantarum. Lovell Reeve, London, vol.
2(1), p.163-533.
BOGGAN, J., CREMERS, G., FEUILLET, C., FUNK, V.A., HOFF, M. & KELLOFF, C.
1997. Checklist of the Plants of the Guianas (Guyana, Surinam and French Guyana).
Smithsonian Institution,Washington, 2 ed., p. 58-61.
BREMER, K. 1987. Tribal interrelationships of Asteraceae. Cladistics 3: 210-253.
BREMER, K. 1994. Asteraceae Cladistics and Classification. Timber Press, Portland.
46

CABRERA, A.L. 1944. Compuestas Sudamericanas nuevas o criticas. Notas Mus. La Plata,
Bot. 9(46): 249-254.
CABRERA, A. L. 1978. Flora de la Provincia de Jujuy – Compositae. INTA, Buenos Aires,
vol. 10, p. 207-247.
CASTRO, H.G., CASALI, V.W.D., BARBOSA, L.C.A. & CECON, P.R. 1999. Rendimento
de tanino em dois acessos de carqueja (Baccharis myriocephala DC.), em diferentes
épocas de colheita em Viçosa – MG. Rev. Bras. Pl. Med. 1(2): 29-33.
CETEC. 1983. Diagnóstico ambiental de Minas Gerais. Belo Horizonte.
CROSS, E.W., QUINN, C.J. & WAGSTAFF, S.J. 2002. Molecular evidence for the
polyphyly of Olearia (Astereae: Asteraceae). Pl. Syst. Evol. 235:99-120.
CUATRECASAS, J. 1967. Revision de las espécies colombianas del gênero Baccharis. Rev.
Acad. Colomb. Cienc. Exact. Fis. Nat. 13(49): 5-102.
De CANDOLLE, A. P. 1836. Compositae: Baccharis. Prodromus Systematis Naturalis Regni
Vegetabilis. Treuttel & Wurtz, Paris, vol 5, p. 398-430.
FERREIRA, M.B. & MAGALHÃES, G.M. 1977. Contribuição para o conhecimento da
vegetação da Serra do Espinhaço em Minas Gerais (Serras de Grão Mogol e da
Ibitipoca). Anais do XXVI Congresso Nacional de Botânica. Rio de Janeiro, p. 189-202.
FREIRE, S.E., URTUBEY, E., GIULIANO, D. 2007. Epidermal characters of Baccharis
(Asteraceae) species used in traditional medicine. Caldasia 29(1): 23-38.
FUNK, V.A., BAYER, R.J., KEELEY, S., CHAN, R., WATSON, L., GEMEINHOLZER, B.,
SCHILLING, E., PANERO, J.L., BALDWIN, B.G., GARCIA-JACAS, N., SUSANNA,
A. & JANSEN, R.K. 2005. Everywhere but Antarctica: Using a supertree to understand
the diversity and distribution of the Compositae. Biol. Skr. 55: 343-374.
GIULIANO, D.A. 2005. New Infragenera in Baccharis (Asteraceae, Astereae). Novon 15:
534-541.
47

GIULIETTI, A.M; MENEZES, N.L; PIRANI, J.R; MEGURO, M.L. & WANDERLEY, M.G
L. 1987. Flora da Serra do Cipó: Caracterização e lista de espécies. Bol. Bot. Univ. São
Paulo 9: 1-152.
GIULIETTI, A.M. & PIRANI, J.R. 1988. Patterns of Geographic Distribution of some Plant
Species from the Espinhaço Range, Minas Gerais, Brazil. Proceedings of a Workshop
on Neotropical Biodiversity Distribution Patterns. Academia Brasileira de Ciências, Rio
de Janeiro, p. 39-69.
GRAU, J. 1977. Astereae-systematic review. In V.H Heywood & J.B. Harborne (eds) The
Biology and Chemistry of the Compositae. London Academic Press, p. 539-565.
GUSTAFSSON, M.H. & BREMER, K. 1995. Morphology and phylogenetic
interrelationships of the Asteraceae, Calyceraceae, Campanulaceae, Goodeniaceae and
related families (Asterales). Amer. J. Bot. 82(2): 250-265.
HARLEY, R.M. 1995. Introdução. In: Stannard, B.L. Flora of the Pico das Almas, Chapada
Diamantina, Brazil. Royal Botanical Gardens, Kew. pp. 1-42.
HATSCHBACH, G., GUARÇONI, E.A.E., SARTORI, M.A., RIBAS, O.S. 2006. Aspectos
fisionômicos da vegetação da Serra do Cabral – Minas Gerais – Brasil. Bol. Mus. Bot.
Mun., Curitiba 67: 1-33.
HELLWIG, F. H. 1990. Die Gattung Baccharis L. (Compositae-Astereae) in Chile. Mitt. Bot.
Staatssamml. München 29: 1-456.
HIND, D.J.N. 1995. Compositae. In B.L. Stannard (ed) Flora of the Pico das Almas,
Chapada Diamantina, Brazil. Royal Botanical Gardens, Kew. p. 175-278.
HIND, D.J.N. 2003a. Flora of Grão-Mogol, Minas Gerais: Compositae (Asteraceae). Bol.
Bot. Univ. São Paulo 21(1): 179-234.
HIND, D.J.N. 2003b. Compositae. In D.C. Zappi, E. Lucas, B.L. Stannard, E.N. Lughadha,
J.R. Pirani, L.P. Queiroz, S. Atkins, D.J.N. Hind, A.M. Giulietti, R.M. Harley & A.M.
48

Carvalho (eds). Lista das Plantas Vasculares de Catolés, Chapada Diamantina, Bahia,
Brasil. Bol. Bot. Univ. São Paulo. 21(2): 345- 398.
HIND, D.J.N. & NESOM, G.L. 2002. A new combination in Leptostelma D.Don
(Compositae: Astereae). Kew Bull. 57: 478.
HOLMGREN P.K, HOLMGREN, N.H. & BARNETT, L.C. 1990. Index Herbariorum. Part
I: The Herbaria Of The World. New York Botanical Garden, New York.
JEFFREY, C. 2007. Compositae. Introduction with key to tribes. In J.W. Kadereit, C. Jeffrey
(eds) The Families and Genera of Vascular Plants. K. Kubitzki (series editor)
Flowering Plants. Eudicots: Asterales. Springer-Verlag. Berlin, vol. 8, p. 61-87.
JUDD, W.S., CAMPBELL, C.S., KELLOG, E.A., STEVENS, P.F. 1999. Plant Systematics:
A phylogenetic approach. Sinauer Associates, Sunderland, p. 396-401.
KIM, K.J., JANSEN, R.K., WALLACE, R.S., MICHAELS, H.J. & PALMER, J.D. 1992.
Phylogenetic implications of rbcL sequence variation in the Asteraceae. Ann. Miss. Bot.
Gard. 79(2): 428-445.
KNOBLOCH, I.W. 1959. A preliminary estimate of the importance of hybridization in
speciation. Bul. Torrey Bot. Club 86(5): 296-299.
KUMAZAWA, S., YONEDA, M., SHIBATA, I., KANAEDA, J., HAMASAKA, T. &
NAKAYAMA, T. 2003. Direct evidence for the plant origin of brazilian propolis by the
observation of honeybee behavior and phytochemical analysis. Chem. Pharm. Bull.
51(6): 740-742.
LUIS, I.T. 1952. Index Baccharidinarum (Compositae). Contr. Inst. Geobiol. 2: 1-55.
LUIS, I.T. 1958. Novum Index Baccharidinarum (Compositae). Contr. Inst. Geobiol. 9: 1-35.
MATZENBACHER N.I. & SOBRAL, M. 1996. Duas novas espécies de Hysterionica Willd.
(Asteraceae – Astereae) no sul do Brasil. Comun. Mus. Ciênc. Tecnol. – PUCRS. Sér.
Bot. 2(1): 3-14.
49

McNEILL, J., BARRIE, F.R., BURDET, H.M., DEMOULIN, V., HAWKSWORTH, D.L.,
MARHOLD, K., NICOLSON, D.H., PRADO, J. SILVA, P.C., SKOG, J.E.,
WIERSEMA, J.H., TURLAND, N.J. (eds.) 2006. Código Internacional de
Nomenclatura Botânica (Código de Viena). Gantner Verlag, Liechtenstein, p. 14-16.
MENDONÇA, M.P. & LINS, L.V. (org.). 2000. Lista Vermelha das espécies ameaçadas de
extinção da flora de Minas Gerais. Fundação Biodiversitas, Fundação Zôo-Botânica de
Belo Horizonte.
MENINI NETO, L., ALVES, R.J.V., BARROS, F. & FORZZA, R.C. 2007a. Orchidaceae do
Parque Estadual de Ibitipoca, MG, Brasil. Acta bot. bras. 21(3): 687-696.
MENINI NETO, L., ALVES, R.J.V. & FORZZA, R.C. 2007b. A subtribo Pleurothallidinae
(Orchidaceae) no Parque Estadual de Ibitipoca, Minas Gerais, Brasil. Bol. Bot. Univ.
São Paulo 25(2): 253-278.
MUNHOZ, C.B.R. & PROENÇA, C.E.B. 1998. Composição florística do município de Alto
Paraíso de Goiás na Chapada dos Veadeiros. Bol. Herb. Ezechias Paulo Heringer 3:102-
150.
MÜLLER, J. 2006. Systematics of Baccharis (Compositae-Astereae) in Bolívia, including an
overview of the genus. Systematic Botany Monographs. The American Society of Plant
Taxonomists. vol. 76, p. 1-341.
NAKAJIMA, J. N. 2001. A família Asteraceae no Parque Nacional da Serra da Canastra,
Minas Gerais, Brasil. Revista Bras. Bot. 24(4): 471-478.
NAKAJIMA, J.N. (coord) 2007. Euasterídeas II. In: C.S. Martins, G.M. Drummond, M.P.
Mendonça (orgs). Lista das espécies da flora brasileira ameaçadas de extinção segundo
os critérios da IUCN, versão 3.1. Fundação Biodiversitas, Belo Horizonte.
NESOM, G.L. 1989. Infrageneric taxonomy of new world Erigeron (Compositae: Astereae).
Phytologia 67(1): 67-93.
50

NESOM, G.L. 1994a. Subtribal classification of the Astereae (Asteraceae) Phytologia 76:
193-274.
NESOM, G.L. 1994b. Inulopsis synopsis (Asteraceae: Astereae). Phytologia 76(2): 115-124.
NESOM, G.L. & ROBINSON, H. 2007. Astereae. In J.W. Kadereit, C. Jeffrey (eds) The
Families and Genera of Vascular Plants. K. Kubitzki (series editor) Flowering Plants.
Eudicots: Asterales. Springer-Verlag. Berlin, vol. 8, p.284-342.
NETO, G.G. & MORAIS, R.G. 2003. Recursos medicinais de espécies do cerrado de Mato
Grosso: um estudo bibliográfico. Acta bot. bras. 17(4): 561-584.
NOYES, R.D. 2000. Biogeographical and evolutionary insights on Erigeron and allies
(Asteraceae) from ITS sequence data. Pl. Syst. Evol. 220: 93-114.
OLIVEIRA, A.S. & MARCHIORI, J.N.C. 2005. O gênero Baccharis L., seção
Oblongifolieae DC. (Asteraceae - Astereae) no sul do Brasil. Balduinia 5: 6-15.
OLIVEIRA, A.S. & MARCHIORI, J.N.C. 2006. O gênero Baccharis L. (Asteraceae -
Astereae) no sul do Brasil 1. Seção Cylindricae Heering, série Cylindricae (Heering)
Giuliano. Balduinia 7: 1-36.
OLIVEIRA, A.S., DEBLE, L.P., SCHNEIDER, A.A. & MARCHIORI, J.N.C. 2006.
Checklist do gênero Baccharis para o Brasil (Asteraceae, Astereae). Balduinia 9: 17-27.
PIO-CORRÊA, M. 1984a. Dicionário das Plantas Úteis do Brasil e das exóticas cultivadas.
Imprensa Nacional, Rio de Janeiro, vol. 2.
PIO-CORRÊA, M. 1984b. Dicionário das Plantas Úteis do Brasil e das exóticas cultivadas.
Imprensa Nacional, Rio de Janeiro, vol. 4.
PIO-CORRÊA, M. 1984c. Dicionário das Plantas Úteis do Brasil e das exóticas cultivadas.
Imprensa Nacional, Rio de Janeiro, vol. 5.
PIRANI, J.R., MELLO-SILVA, R. & GIULIETTI, A.M. 2003. Flora de Grão-Mogol, Minas
Gerais, Brasil. Bol. Bot. Univ. São Paulo 21(1): 1-24.
51

PRANCE, G.T., BEENTJE, H., DRANSFIELD, J. & JOHNS, R. 2000. The tropical flora
remains undercollected. Ann. Miss. Bot. Gard. 87: 67-71.
RADFORD, A. E. 1974. Vascular plants systematics. Harper & Row, New York.
ROQUE, N. & PIRANI, J.R. 1997. Flora da Serra do Cipó, Minas Gerais: Compositae –
Barnadesieae e Mutisieae. Bol. Bot. Univ. São Paulo 16: 151-185.
ROMERO, R. 2002. Diversidade dos campos rupestres de Goiás, sudoeste e sul de Minas
Gerais. In E.L. Araújo, A.N. Moura, E.V. Sampaio, L.M.S. Gestinari, & J.M.T. Carneiro
(eds) Biodiversidade, Conservação e Uso Sustentável da Flora do Brasil. UFRPE -
Imprensa Universitária, Recife, p. 81-86.
SILVERTOWN, J. 1998. Plant phenotypic plasticity and non-cognitive behaviour. Trends
Ecol. Evol. 13: 255–256.
SOLBRIG, O.T. 1960. The South American sections of Erigeron and their relation to
Celmisia. Contr. Gray Herb. 188: 65-85.
SOLBRIG, O.T. 1962. The south american species of Erigeron. Contr. Gray Herb. 191: 3-79.
STEARN, W.T. 1992. Botanical Latin. David & Charles Book, England, p.303-532.
WEBERLING, F. 1989. Morphology of flowers and inflorescences. Cambridge University
Press, Cambridge.
ZANOWIAK, D.J. 1991. An analysis of systematic and phyletic relationships within the
Baccharidinae (Asteraceae: Astereae). PhD thesis, A&M University, Texas.
ZAPPI, D.C., LUCAS, E., STANNARD, B.L., LUGHADHA, E.N., PIRANI, J.R.,
QUEIROZ, L.P., ATKINS, S., HIND, D.J.N., GIULIETTI, A.M., HARLEY, R.M. &
CARVALHO, A.M. (eds) 2003. Lista das Plantas Vasculares de Catolés, Chapada
Diamantina, Bahia, Brasil. Bol. Bot. Univ. São Paulo. 21(2): 345- 398.
ZHANG, X. & BREMER, K. 1993. A cladistic analysis of the tribe Astereae (Asteraceae) with
notes on their evolution and a revised subtribal classification. Pl. Syst. Evol. 184: 259-283.
52

A B C D E1
E2 F G H
I J K
L M N
53

Fig. 1. Sinflorescências presentes nas espécies de Astereae do Parque Estadual do Ibitipoca, MG: a –
espiciforme; b – escaposa; c – corimbifrme heterotética; d – paniculiforme heterotética de florescências
botrióides; e1 – corimbiforme heterotética de florescências glomerulares; e2 – glomerular homotética; f –
paniculiforme heterotética; g – diplobótrio heterotético; h – duplo-corimbosa heterotética; i – diplobótrio
homotético; j – duplo-paniculada heterotética; k - diplobotrióide homotética de florescências estaquióides
inseridas na axila de bráctea foliácea; l – botrióide homotética de florescências glomerulares; m – diplobotrióide
homotética de florescências glomerulares; n – corimbosa homotética de florescências paniculadas.
54

Fig. 2: Baccharis lychnophora: a – ramo florífero, b – capítulo masculino, c – flor masculina d – capítulo
feminino, e – flor feminina (R.C. Forzza et al. 3961); B. aphylla: f – capítulo feminino, g – flor feminina, h -
pálea (R. Borges et al. 259); B. itatiaiae: i – ramo florífero, j - capítulo feminino, k – flor feminina, l – cipsela, m
– capítulo masculino, n – flor masculina (M.M. Saavedra 314); B. platypoda: o – capítulo feminino, p – flor
feminina (R.C. Forzza et al. 3491).
55

Fig. 3: Baccharis serrula: a – ramo florífero, b – capítulo feminino, c – flor feminina, d – capítulo masculino, e –
flor masculina (D. Sucre 6693); B. microdonta: f - capítulo feminino, g – folha (M.M. Saavedra 316); B.
dracunculifolia: h - capítulo feminino, i – folha (R. Borges 883); B. serrulata: j – ramo florífero, k – capítulo
masculino, l – flor masculina, m – capítulo feminino, n – flor feminina (R.C. Forzza 3485, M.M. Saavedra 249).
56

Fig. 4: Baccharis rufidula: a – ramo florífero, b – capítulo feminino, c – flor feminina, d – capítulo masculino, e
– flor masculina (R.C. Forzza et al. 3483, R. Borges et al. 148); B. ligustrina: f - capítulo masculino, g – flor
feminina (R.F. Monteiro 57, R. Borges 145); B. varians: h - hábito, i – capítulo feminino, j – flor feminina, k –
capítulo masculino, l – flor masculina (R. Borges et al. 166, 167); B. oblongifolia: m – capítulo masculino, n –
flor feminina (R.C. Forzza et al. 3492).
57

Fig. 5: Inulopsis scaposa: a – hábito, b – capítulo, c – flor do disco, d – flor do raio, e – receptáculo (F.M.
Ferreira et al. 707); Leptostelma máxima: f – capítulo, g – flor do raio (M.M. Saavedra et al. 287, R.C. Forzza
et al. 3119); Conyza monorchis: h – capítulo, i – flor marginal (R. Borges 878); Baccharis myriocephala: j –
ramo florífero, k – capítulo masculino, l – flor masculina, m - capítulo feminino, n – flor feminina (R. Borges et
al. 257, R. Borges 873).
58

Fig. 6. Baccharis reticularia: a – ramo florífero (planta masculina), b – capítulo masculino, c – flor masculina, d
– ramo florífero (planta feminina), e – capítulo feminino, f - flor feminina, g – receptáculo (R. Borges et al. 149,
150); B. tarchonanthoides: h - capítulo feminino, i – flor masculina (R. Borges et al. 263).
59

Fig. 7: Baccharis salzmanii: a – ramo florífero, b – capítulo feminino, c – flor feminina, d – capítulo masculino,
e – flor masculina (R. Borges et al. 169); B. schultzii: f - ramo florífero, g – capítulo masculino, h - flor
masculina, i - capítulo feminino, j – flor feminina, k - receptáculo (M.M. Saavedra et al. 340).
60

Fig. 8: Espécies da tribo Astereae no Parque Estadual do Ibitipoca, MG. A – Leptostelma maxima; B – Baccharis ligustrina; C – B. lychnophora; D – B. itatiaiae; E – B. myriocephala; F – B. serrulata; G – B. platypoda, G’ sinflorescência feminina, G’’ sinflorescência masculina; H – B. rufidula; I – Inulopsis scaposa; J – B. reticularia; K – B. salzmanii; L – B. varians.
61

Capítulo II: Análise florística de Asteraceae do Parque Estadual do Ibitipoca,
Minas Gerais, Brasil
62

Abstract – [Floristic analyses of the Asteraceae of Ibitipoca State Park, Minas Gerais,
Brazil]. This work presents a checklist for Asteraceae of Ibitipoca State Park (PEIB), with a
geographic distribution analyses among nine Brazilian mountain fields. We found 12 tribes,
50 genera and 111 species. Eupatorieae, Vernonieae, Astereae, Baccharis, Mikania and
Lessingianthus were the most diverse tribes and genera, respectively. PEIB flora was more
similar with Itatiaia, and than with Serra da Canastra and Serra do Cipó. Cluster analyses
indicated that vegetation matrix and geoclimatic factors were very important to determine
local composition. Nevertheless, relationships among mid areas were shown only in
ordination analyses, that expressed strong connections among mountains, represented by
remote (Serra do Ibitipoca, Serra da Canastra and Serra de Grão-Mogol) and central (Serra do
Cipó) campos rupestres. Besides local factors, historical biogeography elements are essential
to understand Asteraceae flora distribution.
Key words: Atlantic forest, campo rupestre, distribution, diversity, flora.
Resumo – [Análise florística de Asteraceae do Parque Estadual do Ibitipoca, Minas Gerais,
Brasil]. O presente estudo apresenta a lista de espécies de Asteraceae do Parque Estadual do
Ibitipoca (PEIB) e uma análise da distribuição geográfica da família em nove localidades de
formações serranas do Brasil. Foram registradas 12 tribos, 50 gêneros e 111 espécies, sendo
uma maior diversidade encontrada em Eupatorieae, Vernonieae e Astereae (tribos);
Baccharis, Mikania e Lessingianthus (gêneros). O PEIB apresenta maior similaridade
florística com o Itatiaia e com as Serras da Canastra e do Cipó, respectivamente. O
dendrograma obtido pela análise de agrupamento permite inferir que a matriz de vegetação
dominante e características geoclimáticas são fatores que explicam a composição local. No
entanto as relações entre as áreas intermediárias não foram demonstradas, ao contrário do
gráfico resultante da análise de ordenação. Nesta última foram detectadas fortes conexões
entre os campos, que são representadas pelas floras de campos rupestres periféricos (Serra do
Ibitipoca, Serra da Canastra e Serra de Grão-Mogol.) e central (Serra do Cipó). Além dos
fatores locais, elementos biogeográficos históricos são fundamentais no entendimento das
relações encontradas.
Palavras-chave: campo rupestre, distribuição, diversidade, flora, floresta atlântica.
63

Introdução
Levantamentos biológicos e revisões taxonômicas são fundamentais para o
desenvolvimento de estudos ecológicos e evolutivos. O conhecimento da flora tem auxiliado
no entendimento dos padrões fitogeográficos das espécies e de organização espacial das
comunidades vegetais (e.g. Gentry 1982, Prance 1982, Crisci et al. 1991, 2006, Nekola &
White 1999, Qian 1999, Sklénar & Jorgensen 1999, Almeida et al. 2004, Kelloff & Funk
2004, Quijano-Abril et al. 2006, Menini-Neto et al. 2007a, Moreira-Muñoz & Muñoz-Schick
2007, Safford 2007). Da mesma forma, hipóteses biogeográficas envolvendo a caracterização
de centros de diversidade e endemismo, assim como estratégias para a conservação de
espécies vegetais, são exemplos de desdobramentos da análise de dados resultante de
trabalhos florísticos (Morin et al. 1988, Clarke & Funk 2005, Funk 2006).
Asteraceae possui ampla distribuição geográfica, sendo melhor representada em
fisionomias campestres e menos freqüentes nos ambientes florestais de baixa altitude (Jeffrey
2006). Nas serras e campos do Brasil, Paraguai, Uruguai e norte da Argentina, assim como
nos Andes e montanhas da Venezuela e Guianas, a família tem um elevado número de
espécies sendo considerável a ocorrência de endemismos e novidades taxonômicas (Giulietti
& Pirani 1988, Martinelli & Bandeira 1989, Hind 1995, 2003a,b, Holmes 1995, Boggan et al.
1997, Romero & Nakajima 1999, Safford 1999, Zuloaga & Morrone 1999, Romero 2002,
Vitta 2002, Young et al. 2002, Giulietti et al. 2005). Aliado a este fato, também foi observada
uma conexão fitogeográfica destas áreas, principalmente devido à grande semelhança entre
táxons, formas de vida e fisionomias (Rambo 1951, 1953, Brade 1956, Smith 1962, Giulietti
& Pirani 1988, Harley 1995, Luteyn 2002, López 2003, Safford 1999, 2007).
Em áreas serranas do Brasil, destacam-se os trabalhos fitogegráficos dos campos
rupestres da Cadeia do Espinhaço e disjunções, presentes em Minas Gerais e Bahia (Giulietti
& Pirani 1988, Harley 1995) e dos campos de altitude da região sudeste (Safford 1999).
64

Estudos que abordaram a distribuição de Asteraceae nas serras da região central e sudeste do
Brasil são praticamente inexistentes (Nakajima 2000, Almeida et al. 2004). Além disso, no
estado de Minas Gerais, os trabalhos taxonômicos relativos à família estão concentrados nas
serras da Cadeia do Espinhaço (e.g. Giulietti et al. 1987, Roque 1997, Hind 2003a) com
exceção do estudo relizado por Nakajima (2001) na Serra da Canastra, havendo dessa forma a
necessidade de se catalogar o restante do estado. Assim, o estudo apresenta o levantamento da
flora de Asteraceae do Parque Estadual do Ibitipoca e uma análise da distribuição da família
em diferentes áreas serranas do Brasil.
Material e Métodos
Área de estudo
O Parque Estadual do Ibitipoca (PEIB) nos municípios de Santa Rita do Ibitipoca e
Lima Duarte, Minas Gerais. A Serra de Ibitipoca, pertence ao complexo da Mantiqueira, com
altitudes compreendidas entre 1.200 a 1.800 m. Segundo classificação de Koppen, o clima da
região é Cwb, mesotérmico úmido, com verões amenos e invernos secos. A precipitação anual
média é de 1.532 mm e a temperatura média é de 18,9ºC (CETEC 1983). A área estudada
caracteriza-se pela presença de diferentes ambientes e tipos vegetacionais, onde ocorrem
variações de formas campestres como os campos rupestres com afloramentos, campos com
predomínio de comunidade arbustiva e campos com predomínio de “candeia” (Eremanthus
erythropappus); e florestais como as florestas semidecíduas, de galeria, nebulares e ombrófila.
Informações completas sobre a caracterização do PEIB encontram-se em Menini-Neto et al.
(2007 a,b) e referências contidas neste.
Levantamento dos dados e análises estatísticas
Foram realizadas excursões ao PEIB no período de março de 2004 a maio de 2007.
Todas as coletas foram depositadas no herbário RB. Os dados sobre composição florística das
demais áreas (Tab.1) foram obtidos através de consulta bibliográfica (Baker 1873, 1876,
65

1882, 1884, Rizzini 1954, Brade 1956, Solbrig 1962, Barroso 1957, 1976, Giulietti et al.
1987, Hind 1995, 2003a,b, Munhoz & Proença 1998, Nakajima 2001) e a partir do acervo dos
herbários BHCB, CESJ, RB, SP e SPF (acrônimos segundo Holmgren et al. 1990) (Tab.1,
Fig. 1).
Os nomes foram uniformizados através da exclusão dos sinônimos nomenclaturais e da
padronização dos gêneros de Eupatorieae, Senecioneae e Vernonieae segundo King &
Robinson (1987), Hind (1993) e Robinson (1999), respectivamente.
A similaridade entre as áreas foi calculada através do índice de Jaccard (McCune &
Grace 2002) e as relações destas foram expressas através de análises de agrupamento e
ordenação. O agrupamento foi obtido pelo método de médias não ponderadas (UPGMA)
através do programa FITOPAC (Shepherd 1996) e a ordenação foi realizada pelo método de
escala multi-dimensional (MDS) (McCune & Grace 2002), desenvolvida no pacote estatístico
XLSTAT do Microsoft® Excel 2002.
Resultados
No PEIB foram registradas 12 tribos, 50 gêneros e 111 espécies de Asteraceae (Tab. 2),
que correspondem a aproximadamente 12% do total estimado de 900 espécies de
angiospermas registradas na área (Forzza com. pess.). Destas, Lessingianthus sp. e Trixis sp.
são espécies novas para a ciência, e Mikania capricorni consta na lista das espécies da flora
brasileira ameaçada de extinção, sendo classificada como vulnerável (Nakajima 2007).
Ageratum fastigiatum, Heterocondylus alatus, Achyrocline satureioides e Baccharis
ligustrina possuem a maior amplitude de distribuição nas áreas amostradas reflexo da
ampla distribuição geográfica destes. Pelo contrário, Austrocritonia angulicaulis, Baccharis
serrula, Graphistylis dichroa, Gyptidium trichobasis, Heterocondylus jaraguensis, H.
pumilus, Lepidaploa helophila, Lessingianthus rubricaulis, Lessingianthus sp., Mikania
capricorni, M. testudinaria, M. vismiaefolia, Stevia lundiana, Trixis sp. e Verbesina
66

floribunda foram registradas somente no PEIB (Tab. 2). Com exceção das duas novas
espécies, esses táxons também ocorrem em outras áreas de campos rupestres da Cadeia do
Espinhaço, como as serras de Ouro Preto, Ouro Branco, Diamantina e Contendas do Sincorá,
além de formações campestres da região Sul e de cerrados de outras regiões do Brasil.
Os gêneros mais diversos no Parque foram Baccharis e Mikania, com 17 espécies cada,
e Lessingianthus com oito espécies (Tab.2). As Serras do Cipó, da Canastra e Catolés
apresentaram maior riqueza para os mesmos gêneros. Dentre as outras áreas de estudo,
Lychnophora (Grão-Mogol), Agrianthus (Pico das Almas), Calea (Chapada dos Veadeiros),
Senecio (Itatiaia) e Piptocarpha (Serra dos Órgãos) possuem maior riqueza do que pelo
menos um dos três gêneros mais diversos no PEIB.
No PEIB, as tribos que apresentaram maior riqueza foram Eupatorieae com 43 espécies
(cerca de 40% do total), Vernonieae e Astereae com 21 e 20 espécies, respectivamente
(aproximadamente 18% cada uma) (Tab.3). Estas porcentagens também foram observadas nos
campos de altitude do Rio de Janeiro. Nos campos rupestres do Espinhaço de Minas Gerais,
as tribos Vernonieae e Eupatorieae apresentaram pequena diferença entre o número de
espécies (30-35% do total), sendo uma maior riqueza encontrada para Vernonieae, exceto em
Grão-Mogol. Na Chapada dos Veadeiros, a tribo Vernonieae também foi a mais diversa (35%
do total), seguida por Eupatorieae e Heliantheae. Já nas serras da Chapada Diamantina, a tribo
Eupatorieae possui o maior número de espécies (34-37% do total), seguida por Vernonieae.
Considerando-se todas as áreas abordadas neste estudo, observamos que Astereae,
Gnaphalieae e Senecioneae são distintamente mais diversas nos campos de altitude do Rio de
Janeiro, enquanto as tribos Heliantheae e Neurolaeneae são mais diversas nos campos
rupestres da Chapada dos Veadeiros e Serra da Canastra. Mutiseae e Barnadesieae por sua vez
são mais diversificadas nos campos de altitude, Serra do Cipó, Canastra e Catolés. Por outro
lado, Venonieae e Eupatorieae são as tribos com alta diversidade em todas as áreas
67

campestres abordadas.
Foi registrado um total de 879 espécies nas nove áreas analisadas, sendo as maiores
similaridades específicas encontradas nos campos rupestres da Bahia (0,39) nos campos de
altitude do Rio de Janeiro (0,30). O PEIB possui maior similaridade com o Itatiaia (0,22) e
menor com a Chapada dos Veadeiros (0,05) (Tab. 4). Tais relações também foram observadas
na análise de ordenação, através dos valores de distância entre as áreas, com exceção às
proximidades encontradas entre Serra dos Órgãos – Catolés, Serra da Canastra – Chapada dos
Veadeiros e Serra de Grão-Mogol – Chapada dos Veadeiros (Tab. 5).
A análise de agrupamento mostrou que a Serra de Ibitipoca está associada à Serra da
Canastra e ambos formam um grupo com a Serra do Cipó. Este grupo está relacionado a um
segundo, formado pela Serra dos Órgãos e Itatiaia, estando os dois grupos ligados a um
terceiro composto pela Serra de Grão-Mogol, Pico das Almas e Catolés. A Chapada dos
Veadeiros forma um quarto grupo disposto externamente às demais áreas (Fig. 2). Na análise
multidimensional, as relações das distâncias entre as localidades foram semelhantes àquelas
observadas para os valores de similaridade (Fig.3).
Discussão
Apesar do PEIB possuir a menor área entre as serras abordadas neste estudo (Tab.1), a
riqueza de espécies apresentada em Asteraceae é elevada, fato também reportado para
Orchidaceae (Menini-Neto et al. 2007a). O grande número de espécies encontrado para a
família no PEIB pode ser explicado pela presença de diferentes geoambientes (Dias et al.
2002), o que também foi verificado na Serra da Canastra (Nakajima 2000). Tal característica
está presente em outras formações montanhosas do Brasil (Harley & Simmons 1986, Giulietti
& Pirani 1988, Harley 1995, Munhoz & Proença 1998, Safford 1999, Vitta 2002, Pirani et al.
2003, Zappi et al. 2003). Conceição & Pirani (2005, 2007) mostraram que os diversos habitats
dos campos rupestres da Chapada Diamantina, em uma escala local, apresentaram
68

composição florística distinta e com distribuição restrita, embora a riqueza de espécies não
seja significativamente diferente entre estes.
Além do conjunto de variados ambientes, a diversidade de Asteraceae também está
ligada ao fato da Serra de Ibitipoca ser um campo rupestre disjunto (Giulietti & Pirani 1988,
Romero 2002), situado sobre um maciço periférico a noroeste da Serra da Mantiqueira e ao
sul da Cadeia do Espinhaço, onde é observada uma confluência vegetacional em que
predomina a floresta atlântica (CETEC 1983). Em sete dos oito geoambientes caracterizados
no PEIB (Dias et al. 2002) desenvolvem-se diversos tipos vegetacionais, sendo predominante
o campo rupestre, que por sua vez difere daquele caracterizado para as áreas centrais da
Cadeia do Espinhaço, a exemplo da Serra da Canastra e Chapada dos Veadeiros (Giulietti &
Pirani 1988, Harley 1995, Romero 2002, Menini-Neto et al. 2007a).
Baccharis, Mikania e Lessingianthus compreendem um elevado número de espécies da
família Asteraceae. Estes gêneros posuem a América do Sul como um dos principais centros
de diversidade, especificamente a região central e o leste do Brasil, razão pela qual se observa
uma elevada riqueza no PEIB (Barroso 1976, King & Robinson 1987, Bremer 1994, Holmes
1995, Robinson 1999, Giuliano 2001, Funk et al. 2005, Dematteis 2006, Jeffrey 2006, Müller
2006).
A presença de formações florestais na área do Parque também condiciona a alta
diversidade de espécies lianescentes de Mikania (Holmes 1995, Ritter & Waechter 2004), que
também conta com espécies arbustivas nos ambientes campestres. Em Baccharis, a presença
conjunta de espécies tipicamente dos campos de altitude e dos campos rupestres confere uma
grande diversidade do gênero na área (Giulietti et al. 1987, Nakajima 2001, Safford 1999,
2007). As espécies de Baccharis predominam em vegetações abertas e grande parte destas
ocupam rapidamente diferentes tipos de habitat (Barroso 1976, Müller 2006). No PEIB, são
encontradas em mata ciliar, bordas de mata estacional e ombrófila, e principalmente nas
69

formações de campo.
Os campos de altitude possuem dominância de espécies de Astereae, Gnaphalieae e
Senecioneae (Safford 1999) advinda da conexão florística com os Andes através dos campos
sulinos (Mondin & Baptista 1996, Safford 2007). Já a maior riqueza observada em
Heliantheae e Neurolaeneae nos campos rupestres da Chapada dos Veadeiros e Serra da
Canastra deve-se em consequência da alta diversidade destas tribos nas áreas savânicas e
montanas neotropicais, que incluem os cerrados do planalto central brasileiro. Tal relação é
verificada principalmente para as subtribos Ecliptinae e Helianthinae (Robinson 1981,
Schiling & Panero 2002), por exemplo, nos gêneros Calea (Urbatsch et al. 1986)
Dimerostemma (Moraes et al. 2007) e Aspilia (Santos 2001).
De forma geral, a organização do dendrograma pode ser interpretada segundo a
vegetação dominante e características geoclimáticas locais. Porém, os grupos formados não
correspondem aos valores máximos de similaridade para as áreas de composição florística e
localização geográfica intermediária, que é uma distorção própria da análise (Clarke &
Warmick 1994). Já a ordenação dos dados permite inferir processos que corroboram padrões
de distribuição geográfica descritos para campos rupestres e de altitude (Giulietti & Pirani
1988, Harley 1995, Safford 1999).
A análise também permite indicar a Serra do Cipó como uma região geográfica de
conexão central entre as floras dos diferentes campos rupestres, que estão sob influência dos
respectivos biomas em que se encontram (Caatinga, Cerrado ou Mata Atlântica). Da mesma
forma, foram verificadas regiões geográficas de conexão periférica: a Serra de Ibitipoca
estabelece uma passagem para os campos de altitude; a Serra da Canastra ao sul e a Serra de
Grão-Mogol ao norte fazem a comunicação com a flora dos campos rupestres de Goiás e com
os campos rupestres da Bahia, respectivamente.
A maior similaridade florística do PEIB com o Itatiaia pode ser explicada por fatores
70

como condições geoclimáticas similares, distância geográfica (Nekola & White 1999) e
influência da vegetação. A porção central da Mantiqueira surgiu posteriormente à formação
da porção sul e do Caparaó (Safford 1999), ou seja, os maciços do Itatiaia e Passa Quatro
(porção central) apresentam uma dinâmica vegetacional semelhante a que ocorreu nos campos
rupestres do sul do Espinhaço. O registro no PEIB de Baccharis itatiaiae, Chromolaena
ascendens, Dasyphyllum flagellare, Hypochaeris lutea, Inulopsis scaposa, Lessingianthus
linearis, Mikania acuminata, Piptocarpha leprosa, Senecio colpodes e Vernonanthura
westiniana exemplifica a flora compartilhada entre estas duas serras, padrão também
detectado em Almeida et al. (2004) para áreas da Mantiqueira e Espinhaço. Vale destacar que
a relação campo rupestre/campo de altitude não foi expressa nos trabalhos de Giulietti &
Pirani (1988), Harley (1995), Safford (1999, 2007) e Nakajima (2000).
Aliado aos fatores ambientais acima citados, os eventos biogeográficos históricos são
fundamentais para o entendimento da distribuição das espécies em escalas mais amplas
(Nekola & White 1999). As relações verificadas entre a flora de Asteraceae dos campos
rupestres e campos de altitude são também decorrentes de prováveis eventos de dispersão,
vicariância e hibridação, que resultaram em especiações e extinções através do tempo (Vitta
2002). Os campos rupestres da Cadeia do Espinhaço apresentam padrões de distribuição
similares para diferentes famílias, entre as quais supostamente se inclui Asteraceae, o que
indica uma seqüência de eventos históricos em comum a partir da estabilização geológica
desta no Cretáceo (Giulietti & Pirani 1988). Neste mesmo período geológico houve o
soerguimento da região central da Mantiqueira, que se tornou estável no Quaternário (Safford
1999). Segundo Harley (1995), os elementos florísticos dos campos rupestres datam do
Terciário inferior, porém novos elementos chegaram continuamente até estes durante o
Quaternário período em que os campos de altitude ocuparam grandes extensões das serras
do sudeste brasileiro (Safford 1999). A Serra da Canastra e a Chapada dos Veadeiros
71

estiveram supostamente ligadas a estas formações durante as flutuações climáticas deste
último período geológico, quando as vegetações savânicas formaram extensos corredores
ligando o Brasil Central às Guianas e Venezuela (Bigarella et al. 1975). Tal fato é
corroborado pela presença de várias espécies na Serra da Canastra, que antes se acreditava ter
distribuição disjunta entre os campos rupestres de Goiás e Minas Gerais (Nakajima 2000,
Romero 2002).
Finalmente, mesmo apresentando a menor área em relação aos demais parques estaduais
de Minas Gerais (Vitta 2002), o PEIB é a quinta Unidade de Conservação mais visitada do
Brasil recebendo cerca de 40.000 turistas por ano (Menini-Neto et al. 2007a). A visitação
intensa em determinados locais do Parque tem fragilizado os diversos ambientes peculiares a
este. A presença de espécies exóticas e/ou ruderais como Sonchus oleraceus, Galinsoga
parviflora e algumas da família Poaceae (Ferreira 2007, Dias-Melo 2007) são ameaças diretas às
comunidades originais.
Além de Mikania capricorni, seis espécies de Orchidaceae e quatro de Bromeliaceae registradas
no PEIB estão ameaçadas de extinção (Nakajima 2007, Menini-Neto et al. 2007a, Monteiro &
Forzza 2008), havendo dessa forma a necessidade de ações que minimizem os impactos causados
pela visitação. Adicionalmente às 111 espécies de Asteraceae registradas para o PEIB, foram
encontradas no entorno mais 19 diferentes espécies, que estão susceptíveis ao desaparecimento da
região com o desenvolvimento de atividades impactantes no local.
Os resultados obtidos no presente estudo demonstram que os trabalhos taxonômicos e
levantamentos florísticos, mesmo atualmente relegados a segundo plano em virtude do
“impedimento taxonômico” (Funk 2006), são fundamentais para o desenvolvimento de teorias
e padrões em biodiversidade, assim como representam fontes primárias de informação para
ações conservacionistas (Funk & Richardson 2002, Funk 2006). O reduzido número de
inventários de longo prazo e que não contam com a participação de taxonomistas ─ a exemplo
do levantamento de Andrade & Sousa (1995) que indicaram apenas 16 gêneros e 23 espécies
72

de Asteraceae para o PEIB ─ têm dificultado o avanço científico em várias áreas do
conhecimento, principalmente nos estudos biogeográficos históricos e ecológicos.
Referências
ALMEIDA, A.M., PRADO, P.I. & LEWINSOHN, T.M. 2004. Geographical distribution of
Eupatorieae (Asteraceae) in south-eastern and south brazilian mountain ranges. Plant
Ecology 174: 163–181.
ANDRADE, P. M. & SOUSA, H. C. Contribuição ao conhecimento da vegetação do Parque
Estadual do Ibitipoca, Lima Duarte, Minas Gerais. R. Árvore 19(2): 249-261.
BAKER, J.G. 1873. Compositae I Vernoniaceae. In: C.F.P. Martius & A.W. Eichler, Flora
brasiliensis, Frid. Fleischer. Leipzig, vol 6, pars 2, p. 1-179.
BAKER, J.G. 1876. Compositae II Eupatoriaceae. In: C.F.P. Martius & A.W. Eichler, Flora
brasiliensis, Frid. Fleischer. Leipzig, vol 6, pars 2, p. 181-374.
BAKER, J.G. 1882. Compositae III Asteroideae, Inuloideae. In: C.F.P. Martius & A.W.
Eichler, Flora brasiliensis, Frid. Fleischer. Leipzig, vol 6, pars 3, p. 1-134.
BAKER, J.G. 1884. Compositae IV Helianthoideae-Mutisiaceae. In: C.F.P. Martius & A.W.
Eichler, Flora brasiliensis, Frid. Fleischer. Leipzig, vol 6, pars 3, p. 138-398.
BARROSO G.M. 1957. Flora do Itatiaia I – Compositae. Rodriguésia 20: 175–241.
BARROSO, G.M. 1976. Compositae – Subtribo Baccharidinae Hoffman – Estudo das
espécies ocorrentes no Brasil. Rodriguésia 27(40): 3-273.
BIGARELLA, J.J., ANDRADE-LIMA, D. & RIEHS, P.J. 1975. Considerações a respeito das
mudanças paleoambientais na distribuição de algumas espécies vegetais e animais no
Brasil. Anais da Academia Brasileira de Ciências 47: 411-464 (suplemento).
BOGGAN, J., CREMERS, G., FEUILLET, C., FUNK, V.A., HOFF, M. & KELLOFF, C.
1997. Checklist of the Plants of the Guianas (Guyana, Surinam and French Guyana).
Smithsonian Institution,Washington, D.C., 2nd edition, pp. 58-61.
73

BRADE, A.C. 1956. A flora do Parque Nacional do Itatiaia. Boletim do Parque Nacional do
Itatiaia 5: 1–112.
BREMER, K. 1994. Asteraceae Cladistics and Classification. Timber Press, Portland.
CETEC. 1983. Diagnóstico Ambiental de Minas Gerais. Belo Horizonte.
CLARKE, H.D. & FUNK, V.A. 2005. Using checklists and collections data to investigate
plant diversity: II. An analysis of five florulas from northeastern South America. Proc.
Acad. Nat. Sci. Philadelphia 154:29-37.
CLARKE, K.R. & WARWICK, R.M. 1994. Change in marine communities: an approach to
statistical analysis and interpretation. Plymouth Marine Laboratory, Plymouth.
CONCEIÇÃO, A.A. & PIRANI, J. R. 2005. Delimitação de habitats em campos rupestres na
Chapada Diamantina: substratos, composiçãoflorística e aspectos estruturais. Bol. Bot.
Univ. São Paulo 23(1): 85-111.
CONCEIÇÃO, A.A. & PIRANI, J. R. 2007. Diversidade em quatro áreas de campos rupestres
na Chapada Diamantina, Bahia, Brasil: espécies distintas, mas riquezas similares.
Rodriguésia 58 (1): 193-206.
CRISCI, J.V., CIGLIANO, M.M., MORRONE, J.J., ROIG-JUNET, S. 1991. Historical
biogeography of southern South America. Syst. Zool. 40 (2): 152-171.
CRISCI, J.V., SALAS, O.E., KATINAS, L. & POSADAS, P. 2006. Bridging historical and
ecological approaches in biogeography. Australian Systematic Botany 19: 1-10.
DEMATTEIS, M. 2006. Two new species of Lessingianthus (Vernonieae, Asteraceae) from
the Brazilian highlands. Bot. Journ. Linn. Soc. 150, 487–493.
DIAS, H.C.T., FERNANDES FILHO, E.I., SCHAEFER, C.E.G.R., FONTES, L.E.F. &
VENTORIM, L.B. 2002. Geoambientes do Parque Estadual do Ibitipoca, município de
Lima Duarte-MG. R. Árvore 26(6): 777-786.
74

DIAS-MELO, R. 2007. A subfamília Panicoideae (Poaceae) no Parque Estadual do Ibitipoca,
Minas Gerais, Brasil. Dissertação de mestrado, Escola Nacional de Botânica Tropical –
JBRJ, Rio de Janeiro.
FERREIRA, F.M. 2007. Subfamílias Aristidoideae, Bambusoideae, Chloridoideae,
Danthonioideae e Pooideae (Poaceae) no Parque Estadual do Ibitipoca, Minas Gerais,
Brasil. Dissertação de mestrado, Universidade Federal do Rio de Janeiro.
FUNK, V.A. & RICHARDSON, K.S. 2002. Systematic data in biodiversity studies: use it or
lose it. Syst. Biol. 51(2): 303–316.
FUNK, V.A., BAYER, R.J., KEELEY, S., CHAN, R., WATSON, L., GEMEINHOLZER, B.,
SCHILLING, E., PANERO, J.L., BALDWIN, B.G., GARCIA-JACAS, N., SUSANNA,
A. & JANSEN, R.K. 2005. Everywhere but Antarctica: Using a supertree to understand
the diversity and distribution of the Compositae. Biol. Skr. 55: 343-374.
FUNK, V.A. 2006. Floras: a model for biodiversity studies or a thing of the past? Taxon
55(3): 581-588.
GENTRY, A. H. 1982. Neotropical floristic diversity: phytogeographical connections
between Central and South America, Pleistocene climatic fluctuations, or an accident of
the andean orogeny? Ann. Miss. Bot. Gard. 69(3): 557-593.
GIULIANO, D.A. 2001. Clasificación infragenérica de las especies argentinas de Baccharis
(Asteraceae, Astereae). Darwiniana 39(1-2): 131-154.
GIULIETTI, A.M, MENEZES, N.L, PIRANI, J.R, MEGURO, M.L. & WANDERLEY, M.G
L. 1987. Flora da Serra do Cipó: Caracterização e lista de espécies. Bol. Bot. Univ. São
Paulo 9: 1-152.
GIULIETTI, A.M. & PIRANI, J.R. 1988. Patterns of Geographic Distribution of some Plant
Species from the Espinhaço Range, Minas Gerais, Brazil. In: Proceedings of a
Workshop on Neotropical Biodiversity Distribution Patterns. Academia Brasileira de
75

Ciências, Rio de Janeiro, pp. 39-69.
GIULIETTI, A.M., HARLEY, R.M., QUEIROZ, L.P., WANDERLEY, M.G.L. & VAN DEN
BERG, C. 2005. Biodiversity and conservation of plants in Brazil. Conservation
Biology 19(3): 632-639.
HARLEY, R.M. & SIMMONS, N.A. 1986. Florula of Mucugê, Chapada Diamantina, Bahia,
Brazil. Royal Botanical Gardens, Kew.
HARLEY, R.M. 1995. Introdução. In: Stannard, B.L. Flora of the Pico das Almas, Chapada
Diamantina, Brazil. Royal Botanical Gardens, Kew. pp. 1-42.
HIND, D.J.N. 1993. A checklist of the brazilian Senecioneae (Compositae). Kew Bull. 48(2):
279-295.
HIND, D.J.N. 1995. Compositae. In: Stannard, B.L. Flora of the Pico das Almas, Chapada
Diamantina, Brazil. Royal Botanical Gardens, Kew. pp. 175-278.
HIND, D.J.N. 2003a. Flora of Grão-Mogol, Minas Gerais: Compositae (Asteraceae). Bol.
Bot. Univ. São Paulo 21(1): 179-234.
HIND, D.J.N. 2003b. Compositae. In D.C. ZAPPI, E. LUCAS, B.L. STANNARD, E.N.
LUGHADHA, J.R. PIRANI, L.P. QUEIROZ, S. ATKINS, D.J.N. HIND, A.M.
GIULIETTI, R.M. HARLEY & A.M. CARVALHO (eds.). Lista das Plantas Vasculares
de Catolés, Chapada Diamantina, Bahia, Brasil. Bol. Bot. Univ. São Paulo. 21(2): 345-
398.
HOLMES, W.C. 1995. Review preparatory to an infrageneric classification of Mikania
(Tribe: Eupatorieae). In: D.J.N. HIND, C. JEFFREY & G.V. POPE (eds.). Advances in
Compositae Systematics, Royal Botanic Gardens, Kew, pp. 239-254.
HOLMGREN P.K, HOLMGREN, N.H. & BARNETT, L.C. 1990. Index Herbariorum. Part
I. The Herbaria Of The World. 8th ed. The New York Botanical Garden, New York.
JEFFREY, C. 2006. Compositae. Introduction with key to tribes. In: K. KUBITZKI (ed.);
76

J.W. KADEREIT & C. JEFFREY (vol. eds.). The Families and Genera of Vascular
Plants. Flowering Plants. Eudicots: Asterales. Springer-Verlag. Berlin, vol. 8, p. 61-87.
KELLOFF C.L. & FUNK, V.A. 2004. Phytogeography of the Kaieteur Falls, Potaro Plateau,
Guyana: floral distributions and affinities. J. Biogeogr. 31: 501–513.
KING, R.M. & ROBINSON, H. 1987. The genera of the Eupatorieae (Asteraceae).
Monographs in Systematic Botany, vol. 22. Misssouri Botanical Garden. St. Louis.
LÓPEZ, R.P. 2003. Phytogeographical relations of the andean dry valleys of Bolivia. J.
Biogeog. 30: 1659-1668.
LUTEYN, J.L. 2002. Diversity, adaptation and endemism in neotropical Ericaceae:
biogeographical patterns in the Vaccinieae. The Botanical Review 68(1): 55-87.
MARTINELLI, G. & BANDEIRA, J. 1989. Campos de altitude. Editora Index, Rio de
Janeiro.
McCUNE, B. & GRACE, J.B. 2002. Analysis of ecological communities. MjM Software
Design, Oregon.
MENINI-NETO, L., ALVES, R.J.V., BARROS, F. & FORZZA, R.C. 2007a. Orchidaceae do
Parque Estadual de Ibitipoca, MG, Brasil. Acta bot. bras. 21(3): 687-696.
MENINI NETO, L., ALVES, R.J.V. & FORZZA, R.C. 2007b. A subtribo Pleurothallidinae
(Orchidaceae) no Parque Estadual de Ibitipoca, Minas Gerais, Brasil. Bol. Bot. Univ.
São Paulo 25(2): 253-278.
MONDIN, C.A. & BAPTISTA, L.R.M. 1996. Relações biogeográficas da tribo Mutisieae
(Asteraceae), sensu Cabrera, no Rio Grande do Sul. Comun. Mus. Ciênc. Tecnol. –
PUCRS. Sér. Bot. 2(1): 49-152.
MONTEIRO, R.F. & FORZZA, R.C. 2008. A família Bromeliaceae no Parque Estadual de
Ibitipoca, Minas Gerais, Brasil. Bol. Bot. Univ. São Paulo (no prelo).
MORAES, M.D., PANERO, J.L. & SEMIR, J. 2007. New combinations in Dimerostemma
77

(Asteraceae: Heliantheae – Ecliptinae), Phytologia 89(1): 115-120.
MOREIRA-MUÑOZ, A. & MUÑOZ-SCHICK, M. 2007. Classification, diversity, and
distribution of Chilean Asteraceae: implications for biogeography and conservation.
Diversity Distrib. 13(6): 818-828.
MORIN, N.R., WHETSTONE, R.D., WILKEN, D., TOMLINSON, K.L. 1988. Floristics for
the 21st century. Monographs in Systematic Botany, vol. 28. Missouri Botanical Garden.
St. Louis.
MÜLLER, J. 2006. Systematics of Baccharis (Compositae-Astereae) in Bolívia, including an
overview of the genus. Systematic Botany Monographs. The American Society of Plant
Taxonomists. vol. 76, p. 1-341.
MUNHOZ, C.B.R. & PROENÇA, C.E.B. 1998. Composição florística do município de Alto
Paraíso de Goiás na Chapada dos Veadeiros. Bol. Herb. Ezechias Paulo Heringer 3:102-
150.
NAKAJIMA, J.N. 2000. A família Asteraceae no Parque Nacional da Serra da Canastra,
Minas Gerais, Brasil. Tese de doutorado, Universidade Estadual de Campinas.
NAKAJIMA, J.N. 2001. A família Asteraceae no Parque Nacional da Serra da Canastra,
Minas Gerais, Brasil. Revista Bras. Bot. 24(4): 471-478.
NAKAJIMA, J.N. (coord) 2007. Euasterídeas II. In: C.S. Martins, G.M. Drummond, M.P.
Mendonça (orgs). Lista das espécies da flora brasileira ameaçadas de extinção segundo
os critérios da IUCN, versão 3.1. Fundação Biodiversitas, Belo Horizonte.
NEKOLA, J.C. & WHITE, P.S. 1999. The distance decay of similarity in biogeography and
ecology. J. Biogeogr. 26: 867–878.
PIRANI, J.R., MELLO-SILVA, R. & GIULIETTI, A.M. 2003. Flora de Grão-Mogol, Minas
Gerais, Brasil. Bol. Bot. Univ. São Paulo 21(1): 1-24.
PRANCE, G. T. 1982. A review of the phytogeographic evidences for Pleistocene climate
78

changes in the Neotropics. Ann. Miss. Bot. Gard. 69(3): 594-624.
QIAN, H. 1999. Floristic analysis of vascular plant genera of North America north of Mexico:
characteristics of phytogeography. J. Biogeogr. 26: 1307–1321.
QUIJANO-ABRIL, M.A., CALLEJAS-POSADA, R. & MIRANDA-ESQUIVEL, D.R. 2006.
Areas of endemism and distribution patterns for neotropical Piper species (Piperaceae).
J. Biogeogr. 33: 1266–1278.
RAMBO, B. 1951. O elemento andino no pinhal rio-grandense. Anais Botânicos do Herbário
Barbosa Rodriguez 3: 7–39.
RAMBO, B. 1953. A história da flora do planalto rio-grandense. Anais Botânicos do
Herbário Barbosa Rodriguez 5: 185–232.
RITTER, M.R. & WAECHTER, J.L. 2004. Biogeografia do gênero Mikania Willd.
(Asteraceae) no Rio Grande do Sul, Brasil. Acta bot. bras. 18(3): 643-652.
RIZZINI, C.T. 1954. Flora Organensis. Arq. Jardim Bot. Rio de Janeiro, 13: 118-243.
ROBINSON, H. 1981. A revision of the tribal and subtribal limits of the Heliantheae
(Asteraceae). Smithsonian Contrib. Bot. 51: 1–102.
ROBINSON, H., 1999. Generic and subribal classification of American Vernonieae.
Smithsonian Contrib. Bot. 89: 1–116.
ROMERO, R. & NAKAJIMA, J.N. 1999. Espécies endêmicas do Parque Nacional da Serra
da Canastra, Minas Gerais. Revista Brasil. Bot. 22(2): 259-265 (suplemento).
ROMERO, R. 2002. Diversidade dos campos rupestres de Goiás, sudoeste e sul de Minas
Gerais. In: E.L. ARAÚJO, A.N. MOURA, E.V. SAMPAIO, L.M.S. GESTINARI, &
J.M.T. CARNEIRO (eds.) Biodiversidade, Conservação e Uso Sustentável da Flora do
Brasil. UFRPE - Imprensa Universitária, Recife, pp. 81-86.
ROQUE, N. & PIRANI, J.R. 1997. Flora da Serra do Cipó, Minas Gerais: Compositae –
Barnadesieae e Mutisieae. Bol. Bot. Univ. São Paulo 16: 151-185.
79

SAFFORD, H.F. 1999. Brazilian Páramos I. An introduction to the physical environment and
vegetation of the campos de altitude. J. Biogeogr. 26:693-712.
SAFFORD, H.F. 2007. Brazilian Páramos IV. Phytogeography of the campos de altitude. J.
Biogeogr. 34(10): 1701-1722.
SCHILLING, E.E. & PANERO, J.L.. 2002. A revised classification of subtribe Helianthinae
(Asteraceae: Heliantheae). I. Basal lineages. Bot. Journ. Linn. Soc. 140: 65–76.
SANTOS, J.U.M. 2001. O gênero Aspilia Thou. (Asteraceae-Heliantheae) no Brasil. Coleção
Adolpho Ducke, Museu Paraense Emílio Goeldi, Belém.
SHEPHERD, G.J. 1996. FITOPAC I ─ Manual do usuário. Departamento de Botânica,
Universidade Estadual de Campinas, Campinas, São Paulo, 96 p.
SKLENÁR, P. & JØRGENSEN, P.M. 1999. Distribution patterns of páramo plants in
Ecuador. J. Biogeogr. 26: 681–691.
SMITH, L.B. 1962. Origins of the flora of southern Brazil. Contrib.US Nat. Herb. 35: 215-249.
SOLBRIG, O.T. 1962. The south american species of Erigeron. Contr. Gray Herb. 191: 3-79.
URBATSCH, L.E., ZLOTSKY, A., PRUSKI, J.F. 1986. Revision of Calea sect. Lemmatium
(Asteraceae: Heliantheae) from Brazil. Syst. Bot. 11(4): 501-514.
VITTA, F. A. 2002. Diversidade e conservação da flora nos campos rupestres da Cadeia do
Espinhaço em Minas Gerais. In: E.L. ARAÚJO, A.N. MOURA, E.V. SAMPAIO,
L.M.S. GESTINARI, & J.M.T. CARNEIRO (eds.) Biodiversidade, Conservação e Uso
Sustentável da Flora do Brasil. UFRPE - Imprensa Universitária, Recife, pp. 90-94.
YOUNG, K.R., ULLOA, C.U., LUTEYN, J.L. & KNAPP, S. 2002. Plant evolution and
endemism in South America: An introduction. The Botanical Review 68(1): 4-21.
ZAPPI, D.C., LUCAS, E., STANNARD, B.L., LUGHADHA, E.N., PIRANI, J.R.,
QUEIROZ, L.P., ATKINS, S., HIND, D.J.N., GIULIETTI, A.M., HARLEY, R.M. &
CARVALHO, A.M. (eds) 2003. Lista das Plantas Vasculares de Catolés, Chapada
80

Diamantina, Bahia, Brasil. Bol. Bot. Univ. São Paulo. 21(2): 345- 398.
ZULOAGA, F.O. & MORRONE, O. (org.) 1999. Catálogo de plantas vasculares de la
República Argentina II. Acanthaceae – Euphorbiaceae (Dicotyledonae). Monographs in
Systematic Botany. vol. 74. Missouri Botanical Garden. St. Louis.
81

Tabela 1. Códigos, coordenadas geográficas e área das localidades analisadas. Estados: MG (Minas Gerais), BA (Bahia), RJ (Rio de Janeiro) e GO (Goiás).C ód igo L oc ali dad e E sta do Mu nic íp io L atitud e (S ) Lo ng itu de (W ) Á rea (h a) Tá xon s R e fe rênc ia sC ip S erra do C ipó M G S an ta na do R iac ho 1 9º 2 5 ’ 43 º 2 7’ 33 80 0 15 5 G iu liet t i et a l. 1 98 7G m G rã o M o gol M G G rão M o go l 1 6º 3 0 ’ 42 º 8 0’ 33 32 4 81 H in d 20 03 aC an S erra da C an as tra M G S ão R oq ue d e M ina s 2 0º 2 0 ’ 46 º 8 5’ 70 52 5 21 5 N a kajima 20 01Ib i S er ra do Ib it ipo ca M G Lim a D u ar te 2 1º 4 2 ’ 43 º 5 3’ 19 23 11 1 pre se nte tra ba lhoP a P ico d as A lma s B A R io d e C on tas 1 3º 3 3 ’ 41 º 5 7’ 17 00 0 13 2 H in d 19 95C at C ato lé s B A C atol és 1 3º 2 0 ’ 41 º 4 5’ 25 10 00 17 9 H in d 20 03 bIta I ta t ia ia R J Ita tia ia 2 2º 2 5 ’ 44 º 3 5’ 28 15 5 20 8 B a rro so 1 95 7O rg S erra do s Ó rgã os R J Te re só po lis 2 2º 3 0 ’ 43 º 0 1’ 10 52 7 15 4 R iz zin i 1 95 4V ea C ha pa da d os V ea de iro sG O A lto P ara ís o 4 7º 3 0 ' 13 º 4 6' 25 94 00 12 0 M un ho z & P roe nç a 19 98
82
Tabela 2. Lista das espécies de Asteraceae registradas no PEIB e suas ocorrências nas demais áreas análisadas. Os materiais testemunho estão sob o número de coleta de Bruno Silva (BS), Dimitri Sucre (DS), Fabrício Ferreira (FF), Luiz Giordano (LG), Mariana Saavedra (MS), Rafael Augusto (RA), Rafaela Forzza (RF) (para as siglas vide Tabela 1).
Táxons Coletor Cip Can Gm Pa Cat Ita Org Vea
ASTEREAE (4 gen.; 20 spp.)
Baccharis aphylla (Vell.) DC. RA 259 X XBaccharis dracunculifolia DC. RA 883 X X XBaccharis itatiaiae Wawra RF 3508 X XBaccharis ligustrina DC. RA 145 X X X X X X X XBaccharis lychnophora Gardner RF 3341 X XBaccharis microdonta DC. MS 316 X XBaccharis myriocephala DC. RA 257 X X X XBaccharis oblongifolia (Lam.) Pers. RF 3492 X X X X XBaccharis platypoda DC. RF 3491 X X X X XBaccharis reticularia DC. RA 149 XBaccharis rufidula (Spreng.) Joch. Mull. RF 3483 X XBaccharis salzmanii DC. RA 169 X XBaccharis schultzii Baker MS 340 X XBaccharis serrula Sch.Bip. ex Baker DS 6693Baccharis serrulata (Lam.) Pers. MS 249 X X X X X XBaccharis tarconanthoides DC. RA 263 X XBaccharis varians Gardner RA 166 X XConyza monorchis (Griseb.) Cabrera RA 878 XInulopsis scaposa (DC.) O. Hoffm. FF 707 X XLeptostelma maxima D.Don MS 287 X X X XBARNADESIEAE (1 gen.; 1 sp.)Dasyphyllum flagellare (Casar.) Cabrera RF 3493 X X XCICHORIEAE (2 gen.; 2 spp.)Hypochaeris lutea (Vell.) Britten MS 352 X X X XSonchus oleraceus L. MS 345 X XCOREOPSIDEAE (1 gen.; 1 sp.)Bidens segetum Mart. ex Colla MS 288 X X XEUPATORIEAE (17 gen.; 43 spp.)Ageratum fastigiatum (Gardner) R.M.King & H.Rob. RF 3472 X X X X X X
Austrocritonia angulicaulis (Sch.Bip. ex Baker) R.M.King & H.Rob. MS 344
Austroeupatorium inulaefolium (Kunth) R.M.King & H.Rob. RA 885 X X X
Ayapana amygdalina (Lam.) R.M.King & H.Rob. MS 200 X X X X X
Barrosoa organensis (Gardner) R.M.King & H.Rob. RF 3111 X
Campuloclinium parvulum (Glaz.) R.M.King & H.Rob. RA 876 X
83

Táxons Coletor Cip Can Gm Pa Cat Ita Org Vea
Chromolaena ascendens (Sch.Bip. ex Baker) R.M.King & H.Rob. MS 253 X X
Chromolaena barbacensis (Hieron.) R.M.King & H.Rob. RF 3471 X X X
Chromolaena decumbens Gardner MS 253 X XChromolaena laevigata (Lam.) R.M.King & H.Rob. MS 326 X X X X
Chromolaena maximiliani (Schrader ex DC.) R.M.King & H.Rob. RA 884 X X X
Chromolaena pedalis (Sch.Bip. ex Baker) R.M.King & H.Rob. RF 3081 X
Grazielia gaudichaudiana (DC.) R.M.King. & H.Rob. RA 877 X X X
Grazielia intermedia (DC.) R.M.King. & H.Rob. MS 302 X X
Gyptidium trichobasis (Baker) R.M.King. & H.Rob. MS 294
Heterocondylus alatus (Vell.) R.M.King & H.Rob. RF 4112 X X X X X
Heterocondylus amphidictyus (DC.) R.M.King. & H.Rob. MS 259 X
Heterocondylus jaraguensis (B.L. Rob.) R. M.King & H.Rob. MS 333
Heterocondylus pumilus (Gardner) R.M.King. & H.Rob. FF 701
Koanophyllon thysanolepis (B.L. Rob.) R.M.King. & H.Rob. MS 332 X
Mikania acuminata DC. RF 4126 X XMikania buddleiaefolia DC. RA 266 XMikania camporum B.L. Rob. FF 715 XMikania capricorni B.L. Rob. MS 216Mikania decumbens Malme MS 274 XMikania hoffmanniana Dúsen ex Malme RF 3475 XMikania lanuginosa DC. RF 4281 XMikania lasiandrae DC. MS 353 X X X XMikania lindbergii Baker RF 3486 X X XMikania microdonta DC. MS 227Mikania nummularia DC. RF 3543 X XMikania obtusata DC. CESJ 29832Mikania sessilifolia DC. MS 262 X X X X XMikania ternata (Vell.) B.L. Robinson RF 3256 X X XMikania testudinaria DC. RF 3545Mikania vismiaefolia DC. MS 189Mikania warmingii Sch. Bip. ex Baker RF 3208 XPraxelis kleinioides (Humb., Bonpl. & Kunth) Sch.Bip.
RF 3045 X X X X
Pseudobrickellia angustissima (Spreng. ex Baker) R.M.King. & H.Rob. RA 256 X X X
Stevia lundiana DC. FF 699
84

Táxons Coletor Cip Can Gm Pa Cat Ita Org Vea
Stevia urticaefolia Billb. RF 3082 XSymphyopappus decussatus Turcz. MS 258 X X XTrichogonia villosa Sch.Bip. ex Baker RF 3917 X X X XGNAPHALIEAE (4 gen.; 4 spp.)Achyrocline satureioides (Lam.) DC. RF 3557 X X X X X XChionolaena arbuscula DC. MS 329 XLucilia lycopodioides (Less.) S.E. Freire RF 4114 X X XStenophalium chionaeum (DC.) Anderberg MS 188 X XHELIANTHEAE s.s. (3 gen.; 3 spp.)Aspilia duarteana Santos RF 3271 XTilesia baccata (L.) Pruski MS 341 X XVerbesina floribunda Gardner RF 3066MILLERIEAE (1 gen.; 1 sp.)Galinsoga parviflora Cav. MS 347 X X X XMUTISIAE (3 gen.; 6 spp.)Gochnatia paniculata (Less.) Cabrera RF 3478 X XRichterago amplexifolia (Gardner) Kuntze RF 3490 XRichterago radiata (Vell.) Roque RF 2643 X XTrixis nobilis (Vell.) Katinas MS 278 XTrixis verbasciformis Less. RF 3319 XTrixis sp. RF 3518NEUROLAENEAE (1 gen.; 1 sp.)Calea clausseniana Baker MS 337 XSENECIONEAE (5 gen.; 8 spp.)Dendrophorbium pellucidinerve (Sch.Bip. ex Baker) C. Jeffrey MS 210 X
Erechtites valerianaefolia DC. CESJ26821 X X XGraphistylis dichroa (Bong.) D.J.N.Hind MS 315Pentacalia desiderabilis (Vell.) Cuatrec. MS 331 X XSenecio adamantinus Bong. RA 157 X X X X XSenecio brasiliensis (Spreng) Less. RF 2671 X X XSenecio colpodes Bong. MS 284 XSenecio emiliopsis C. Jeffrey FF 719 XVERNONIEAE (8 gen.; 21 spp.)Chresta plantaginifolia (Less.) Gardner MS 334 X XChrysolaena simplex (Less.) Dematteis CESJ 13230 X X X XEremanthus erythropappus (DC.) MacLeish MS 297 X X X X
Eremanthus incanus (Less.) Less. RF 4140 X X XHololepis pedunculata (DC. ex Pers.) DC. BS 1313 X XLepidaploa decumbens (Gardner) H.Rob. MS 280 XLepidaploa helophila (Mart. ex DC.) H.Rob. MS 241
Lessingianthus buddleifolius (Mart. ex DC.) H.Rob. MS 291 X X X
Lessingianthus linearis (Spreng) H.Rob. MS 269 X X X X XLessingianthus psilophyllus (DC.) H.Rob. MS 236 X X X
85

Táxons Coletor Cip Can Gm Pa Cat Ita Org Vea
Lessingianthus rosmarinifolius (Less.) H.Rob. LG 2418 X
Lessingianthus rubricaulis (Bonpl.) H.Rob. MS 270 X
Lessingianthus tomentellus (Mart. ex DC.) H.Rob. RF 3306 X X
Lessingianthus venosissimus (Sch.Bip. ex Baker) H.Rob. FF 698
Lessingianthus sp. RF 3962Piptocarpha leprosa (Less.) Baker RF 3709 XPiptocarpha macropoda (DC.) Baker RF 4276 X X X XStenocephalum megapotamicum (Spreng) Sch.Bip. FF 711 X X X
Vernonanthura diffusa (Less.) H.Rob. RF 4297 X XVernonanthura phosphorica (Vell.) H.Rob. RA 879 X X X X
Vernonanthura westiniana (Less.) H.Rob. MS 292 X X
Tabela 3. Número de espécies das nove tribos mais diversas nas áreas estudadas (para as siglas vide Tabela 1).
Tribo Ibi Cip Can Gm Pa Cat Ita Org VeaAstereae 20 20 20 4 17 14 52 33 9
Eupatorieae 43 41 63 28 45 66 53 42 27Gnaphalieae 4 4 3 3 6 3 17 8 1Heliantheae 3 5 17 3 6 10 4 6 16Vernonieae 21 53 67 25 29 42 32 25 42Mutisieae 6 17 12 5 7 9 10 9 3
Barnadesieae 1 4 3 2 1 2 3 4 0Neurolaeneae 1 4 9 4 5 4 1 1 10Senecioneae 9 4 4 5 6 6 20 14 3
Total 108 152 198 79 122 156 192 142 111
86

Tabela 4. Matriz de similaridade florística representada pelos valores do índice de Jaccard (para as siglas vide Tabela 1).Ib i C ip C an G m Pa C at Ita O rg V ea
Ib i 1C ip 0,16 5 1C an 0,18 6 0 ,18 2 1G m 0,07 8 0 ,17 2 0 ,09 8 1Pa 0,08 4 0 ,09 8 0 ,08 8 0 ,1 38 1C at 0,08 5 0 ,10 8 0 ,07 2 0 ,1 31 0 ,3 94 1Ita 0,22 0 0 ,08 9 0 ,11 5 0 ,0 43 0 ,0 65 0 ,0 80 1O rg 0,14 7 0 ,05 1 0 ,07 9 0 ,0 44 0 ,0 59 0 ,0 73 0,3 05 1Vea 0,05 5 0 ,10 4 0 ,11 2 0 ,0 86 0 ,0 63 0 ,0 74 0,0 41 0,0 34 1
Tabela 5. Matriz de distâncias relativas à duas dimensões obtida pela análise MDS (para as siglas vide Tabela 1).Ib i C ip C an G m Pa C at Ita O rg V ea
Ib i 0C ip 0,61 0 0C an 0,44 1 0 ,48 4 0G m 1,13 5 0 ,53 2 0 ,97 3 0Pa 1,11 5 0 ,66 9 1 ,14 6 0 ,4 89 0C at 1,02 8 0 ,73 8 1 ,16 7 0 ,7 38 0 ,2 83 0Ita 0,40 5 0 ,75 2 0 ,80 7 1 ,1 90 0 ,9 97 0 ,8 17 0O rg 0,72 4 0 ,83 6 1 ,05 9 1 ,1 35 0 ,8 08 0 ,5 63 0,3 55 0Vea 0,94 4 0 ,46 8 0 ,58 6 0 ,5 80 0 ,9 74 1 ,1 35 1,1 91 1,3 04 0
87

Figura 1. Localização das nove áreas serranas amostradas no presente estudo (para as siglas
vide Tabela 1).
88

Figura 2. Dendrograma obtido pelo método de UPGMA (Correlação
Cofenética = 0,8990), resultante da análise de similaridade florística
entre as nove áreas (para as siglas vide Tabela 1).
89
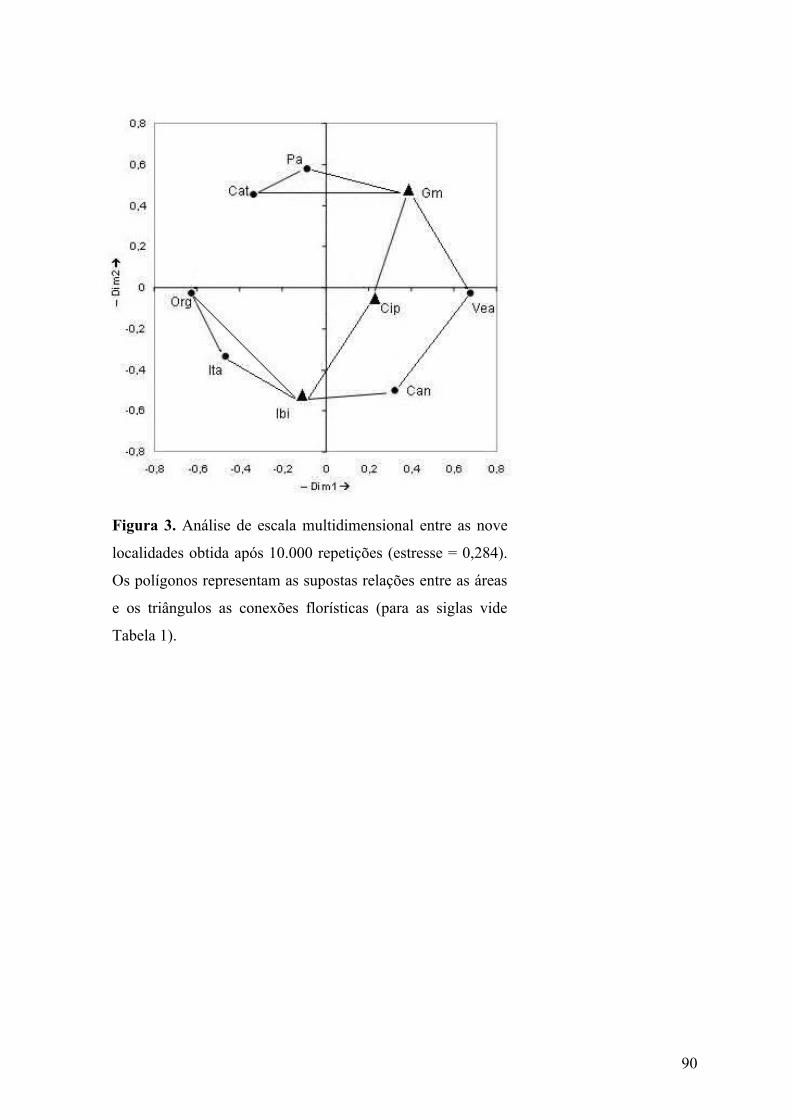
Figura 3. Análise de escala multidimensional entre as nove
localidades obtida após 10.000 repetições (estresse = 0,284).
Os polígonos representam as supostas relações entre as áreas
e os triângulos as conexões florísticas (para as siglas vide
Tabela 1).
90

Conclusões Gerais
• Asteraceae é uma das maiores famílias no Parque Estadual do Ibitipoca (PEIB), sendo
constituída principalmente por táxons com distribuição tropical e encontrados em
formações montanhosas do continente sul-americano. Alguns táxons também ocorrem em
áreas de vegetação campestre, savânica e florestal.
• Da mesma forma, Astereae é uma das maiores tribos do PEIB, sendo Baccharis o gênero
mais representativo. A maioria das espécies da tribo presentes no Parque possui ampla
distribuição pelo Brasil, sendo que metade também ocorre em outros países da América
do Sul.
• Apesar de sua pequena área, o PEIB possui características peculiares que conferem a este
uma elevada riqueza, com a presença de espécies ameaçadas de extinção e outras descritas
recentemente para a Ciência. No entanto, a intensa visitação tem fragilizado os vários
ambientes do Parque, o que têm permitido o estabelecimento de espécies exóticas e/ou
ruderais.
• As floras de Asteraceae dos campos rupestres e de altitude são fortemente influenciadas:
pelo bioma e vegetação dominante em que as serras estão inseridas; pelas características
ambientais e geomorfológicas locais. Além disso, os eventos biogeográficos históricos são
fundamentais no entendimento das conexões existentes entre as áreas estudadas.
• O presente estudo reforça a idéia de que os trabalhos taxonômicos são fundamentais para
o desenvolvimento de teorias e padrões em biodiversidade, assim como são ferramentas
básicas para ações conservacionistas.
91

Referências Bibliográficas
CAMPOS, B.C. 2005. A família Melastomataceae nos campos rupestres e cerrados de altitude
do Parque Estadual do Ibitipoca, Lima Duarte, MG, Brasil. Dissertação de Mestrado,
Escola Nacional de Botânica Tropical - JBRJ, Rio de Janeiro.
CETEC. 1983. Diagnóstico ambiental de Minas Gerais. Belo Horizonte.
DIAS-MELO, R. 2007. A subfamília Panicoideae (Poaceae) no Parque Estadual do Ibitipoca,
Minas Gerais, Brasil. Dissertação de Mestrado, Escola Nacional de Botânica Tropical -
JBRJ. Rio de Janeiro.
DONOGHUE, M.J. & ALVERSON, W.S. 2000. A new age of discovery. Ann. Missouri Bot.
Gard. 87: 110-126.
FERREIRA, F.M. 2007. Subfamílias Aristidoideae, Bambusoideae, Chloridoideae,
Danthonioideae e Pooideae (Poaceae) no Parque Estadual do Ibitipoca, Minas Gerais,
Brasil. Dissertação de Mestrado, Universidade Federal do Rio de Janeiro, Rio de
Janeiro.
FUNK, V.A. & RICHARDSON, K.S. 2002. Systematic data in biodiversity studies: use it or
lose it. Syst. Biol. 51(2): 303–316.
FUNK, V.A. 2006. Floras: a model for biodiversity studies or a thing of the past? Taxon
55(3): 581-588.
GIULIETTI, A.M, MENEZES, N.L, PIRANI, J.R, MEGURO, M.L. & WANDERLEY, M.G
L. 1987. Flora da Serra do Cipó: Caracterização e lista de espécies. Bol. Bot. Univ. São
Paulo 9: 1-152.
GIULIETTI, A.M. & PIRANI, J.R. 1988. Patterns of Geographic Distribution of some Plant
Species from the Espinhaço Range, Minas Gerais, Brazil. In: Proceedings of a
Workshop on Neotropical Biodiversity Distribution Patterns. Academia Brasileira de
Ciências, Rio de Janeiro, pp. 39-69.
92

GIULIETTI, A.M., HARLEY, R.M., QUEIROZ, L.P., WANDERLEY, M.G.L. & VAN DEN
BERG, C. 2005. Biodiversity and conservation of plants in Brazil. Conservation
Biology 19(3): 632-639.
GOVAERTS, R. 2001. How many species of seed plants are there? Taxon 50: 1085–1090.
GOVAERTS, R. 2003. How many species of seed plants are there – a response. Taxon 52:
583–584.
HATSCHBACH, G., GUARÇONI, E.A.E., SARTORI, M.A., RIBAS, O.S. 2006. Aspectos
fisionômicos da vegetação da Serra do Cabral – Minas Gerais – Brasil. Bol. Mus. Bot.
Mun., Curitiba 67: 1-33.
HIND, D.J.N. 1995. Compositae. In: Stannard, B.L. Flora of the Pico das Almas, Chapada
Diamantina, Brazil. Royal Botanical Gardens, Kew. pp. 175-278.
HIND, D.J.N. 2003a. Flora of Grão-Mogol, Minas Gerais: Compositae (Asteraceae). Bol.
Bot. Univ. São Paulo 21(1): 179-234.
HIND, D.J.N. 2003b. Compositae. In D.C. ZAPPI, E. LUCAS, B.L. STANNARD, E.N.
LUGHADHA, J.R. PIRANI, L.P. QUEIROZ, S. ATKINS, D.J.N. HIND, A.M.
GIULIETTI, R.M. HARLEY & A.M. CARVALHO (eds.). Lista das Plantas Vasculares
de Catolés, Chapada Diamantina, Bahia, Brasil. Bol. Bot. Univ. São Paulo. 21(2): 345-
398.
MEDEIROS, E.S.S. 2006. Flora do Parque Estadual do Ibitipoca, Minas Gerais, Brasil –
Família Piperaceae. Dissertação de Mestrado, Escola Nacional de Botânica Tropical -
JBRJ, Rio de Janeiro.
MENINI NETO, L., ALVES, R.J.V., BARROS, F. & FORZZA, R.C. 2007. Orchidaceae do
Parque Estadual de Ibitipoca, MG, Brasil. Acta bot. bras. 21(3): 687-696.
93

MILWARD-DE-AZEVEDO, M.A. 2007. Passifloraceae do Parque Estadual de Ibitipoca,
Minas Gerais. Bol. Bot. Univ. São Paulo 25(1): 71-79.
MONGUILHOTT, L. 2007. Apocynaceae do Parque Estadual de Ibitipoca, Minas Gerais,
Brasil. Dissertação de Mestrado, Universidade de São Paulo, São Paulo.
MONTEIRO, R.F. & FORZZA, R.C. 2008. A família Bromeliaceae no Parque Estadual de
Ibitipoca, Minas Gerais, Brasil. Bol. Bot. Univ. São Paulo (no prelo).
NAKAJIMA, J. N. 2001. A família Asteraceae no Parque Nacional da Serra da Canastra,
Minas Gerais, Brasil. Revista Bras. Bot. 24(4): 471-478.
PRANCE, G.T., BEENTJE, H., DRANSFIELD, J. & JOHNS, R. 2000. The tropical flora
remains undercollected. Ann. Missouri Bot. Gard. 87:67-71.
ROMERO, R. 2002. Diversidade dos campos rupestres de Goiás, sudoeste e sul de Minas
Gerais. In: E.L. ARAÚJO, A.N. MOURA, E.V. SAMPAIO, L.M.S. GESTINARI, &
J.M.T. CARNEIRO (eds.) Biodiversidade, Conservação e Uso Sustentável da Flora do
Brasil. UFRPE - Imprensa Universitária, Recife, pp. 81-86.
ROQUE, N. & PIRANI, J.R. 1997. Flora da Serra do Cipó, Minas Gerais: Compositae –
Barnadesieae e Mutisieae. Bol. Bot. Univ. São Paulo 16: 151-185.
SILVA, D.B., FORZZA, R.C. & ALVES, M. Cyperaceae e Juncaceae no Parque Estadual de
Ibitipoca, Minas Gerais, Brasil. (em preparação).
94







![Restauração - jbrj.gov.br · 2 Manual técnico para a restauração de áreas degradadas no Estado do R io de Janeiro / Luiz Fernando Duarte de Moraes ...[et al.] – Rio de Janeiro](https://img.document.onl/doc/110x75/5bccdd3209d3f218478c51cf/restauracao-jbrjgovbr-2-manual-tecnico-para-a-restauracao-de-areas.jpg)





















