Embed Size (px)
Citation preview

Instituto de Pesquisas Jardim Botânico do Rio de Janeiro Escola Nacional de Botânica Tropical
Programa de Pós-graduação Stricto Sensu
Dissertação de Mestrado
Germinação de Sementes de Espécies da Formação Arbustiva Aberta na Restinga de Massambaba
Aline Cavalcante de Souza
Rio de Janeiro 2010

ii
Instituto de Pesquisas Jardim Botânico do Rio de Janeiro
Escola Nacional de Botânica Tropical Programa de Pós-graduação Stricto Sensu
Germinação de Sementes de Espécies da Formação Arbustiva Aberta na Restinga de Massambaba
Aline Cavalcante de Souza
Dissertação apresentada ao Programa de Pós-Graduação em Botânica, Escola Nacional de Botânica Tropical, do Instituto de Pesquisas Jardim Botânico do Rio de Janeiro, como parte dos requisitos necessários para a obtenção do título de Mestre em Botânica.
Orientador: Dr. Antonio Carlos Silva de Andrade
Rio de Janeiro 2010

iii
Germinação de Sementes de Espécies da Formação Arbustiva Aberta na Restinga de Massambaba
Aline Cavalcante de Souza
Dissertação submetida ao corpo docente da Escola Nacional de Botânica Tropical, Instituto de Pesquisas Jardim Botânico do Rio de Janeiro - JBRJ, como parte dos requisitos necessários para a obtenção do grau de Mestre.
Aprovada por: Prof. Dr. Antonio Carlos Silva de Andrade (Orientador) _________________________________________ Prof.a Dr.a Cláudia Antônia Vieira Rossetto (UFRRJ) ______________________________________ Prof.a Dr.a Dorothy Sue Dunn de Araujo (ENBT) ______________________________________ em _23_/_06_/_2010_/
Rio de Janeiro 2010

iv
Souza, Aline Cavalcante de. S729g Germinação de Sementes de Espécies da Formação Arbustiva
Aberta na Restinga de Massambaba / Aline Cavalcante de Souza. – Rio de Janeiro, 2010.
xvi, 86f. : il. Dissertação (Mestrado) – Instituto de Pesquisas Jardim Botânico
do Rio de Janeiro/Escola Nacional de Botânica Tropical, 2010. Orientadora: Antônio Carlos Silva de Andrade.
Bibliografia.
1. Germinação. 2. Semente. 3. Restinga. 4. Massambaba,
Restinga de (Arraial do Cabo, RJ). 5. Arraial do Cabo (RJ). 6. Rio de Janeiro (Estado). I. Título. II. Escola Nacional de Botânica Tropical.
CDD 582.0467098153

v
“Mas as sementes são invisíveis. Elas dormem nas entranhas da terra até que uma cisme de despertar. Então ela se despreguiça e lança, timidamente, para o sol um inofensivo galhinho”.
Antoine de Saint-Exupéry (O Pequeno Príncipe)
"Hay que endurecer, pero sin perder la ternura jamás."
Ernesto Che Guevara
“A curiosidade é mais importante do que o conhecimento”.
Albert Einstein

vi
AGRADECIMENTOS
Agradeço à Escola Nacional de Botânica Tropical e ao Jardim Botânico do Rio de
Janeiro por tornar possível a realização do meu mestrado.
Ao CNPQ pela concessão da bolsa de mestrado.
Ao meu orientador Antonio Carlos pela orientação durante o curso de mestrado.
À todos do Laboratório de Sementes, Marina de Jesus Fernandes, Ana Paula Cruz,
Charles da Silva, Marly Barbosa, Nilza Urbano, Ricardo Matheus que me ajudaram
sempre.
Aos funcionários da Escola Nacional de Botânica Tropical. Muito obrigada pela atenção e
zelo! Em especial a estimada e querida Janúzia Ferreira, a qual sempre carinhosa, paciente,
e com os melhores conselhos que me ajudaram muito!!
Á todos os estudantes do Laboratório de Sementes, Alba Regina, Thaís Hidalgo,
Amanda Carvalho, Letícia Andrade por tornarem os dias mais agradáveis, e claro pela
ajuda nos experimentos, principalmente nas trocas infindáveis das minhas placas!
À Viviane Stern pela ajuda na identificação das espécies da Restinga de
Massambaba!
À Luiz Roberto Zamith pelas valiosas informações sobre a época de frutificação
das espécies, além de me fornecer informações sobre a germinação de algumas espécies!
Muito obrigada!
Ao meu orientador de Iniciação Científica Eduardo Arcoverde de Mattos, pelo
valioso ensinamento durante minha graduação, o qual foi essencial ao meu mestrado!
Ao Prof. Fábio Scarano pela valiosa ajuda e incentivo na elaboração da minha
dissertação durante a disciplina Seminários I.
Agradeço em especial a Prof.ª Dorothy Araujo e a Prof.ª Alda Lúcia pela enorme
ajuda, respeito e compreensão no momento que eu mais precisei! Muito obrigada! Além

vii
disso, agradeço a Prof. Dorothy pela ajuda na identificação das espécies, e pela revisão do
meu trabalho como banca!
Aos meus professores que me influenciaram muito, os quais me mostraram a beleza
e alegria de ser um biólogo, em especial ao Prof. Ricardo Vieira e a Prof.ª Maria Cristina.
Obrigada pelo exemplo!
É claro aos meus amigos queridos, que me ajudaram nesta caminhada, sempre me
apoiando, me aconselhando e torcendo por mim! Em especial ao Vitor Hugo Maia, grande
amigo e conselheiro, e companheiro de rodízio rs! A querida amiga Danielle Capossoli,
sempre disposta a me ajudar no que fosse preciso! A todos os amigos da turma de
mestrado, em especial as amigas queridíssimas Ana Luiza e Elaine Ribeiro, mais
conhecida com Alzira!
Agradeço em especial a minha querida e sempre companheira Vanessa Rodrigues,
pelas risadas, conselhos, ajuda, compreensão, enfim por ser uma grande pessoa e amiga!
Muito obrigada!
Aos amigos de longa data, Fernanda Celleghin, Juliana Paixão, Flávia Scali, Aline
Leal, Leandro Macedo, Vinícius Galvão, Verônica Araujo, Paloma Loretto, Márcia
Cardoso, Madellon Assis, Mário Bevilaqua, Cyntia Loureiro, Priscilla Anne, por estarem
presente na minha vida e torna-lá mais feliz! Em especial as amigas Tatiana Abreu e
Sabrina Jensen, as quais sempre me passaram a segurança que só um grande amigo pode
dar!
Agradeço muitíssimo a José Fernandes de Santana, por me ajudar sempre que
possível nas saídas de campo, sendo muito importante para a realização desta dissertação!
Sem você não sei o que seria de mim!

viii
Agradeço a minha família por todo apoio, compreensão, incentivo, pelos momentos
felizes, e segurança que sempre me foi dada, em especial a meus pais e irmão, e claro
também ao Billy!
A minha família postiça do Rio Comprido, pelos momentos de alegria!
Ao meu amor e companheiro, Jayme Augusto Prevedello, por todo o incentivo, pela
ajuda em todos os momentos, desde as saídas de campo até na parte intelectual deste
trabalho, pela amizade e segurança! Muito obrigada!
Foram muitas as pessoas que me ajudaram durante este percurso importante na
minha vida, espero não ter esquecido ninguém! Aprendi a dar mais valor às pessoas éticas
e boas, as quais torceram e me apoiaram muito! Esta dissertação é sem dúvida de todos
estes que me ajudaram cada um de uma forma, tornando possível esta dissertação!
Muito Obrigada!!

ix
RESUMO
As exigências germinativas têm um importante papel no padrão de distribuição das plantas. As formações abertas em restingas são compostas por moitas de vegetação intercaladas por areia nua e vegetação esparsa. A superfície da areia nua possui uma baixa capacidade de retenção de água e temperaturas altas são frequentes. Porém, as moitas de vegetação atenuam estas condições, favorecendo a germinação de várias espécies na restinga. Neste tipo de vegetação, é observado que algumas espécies ocorrem somente na areia nua, enquanto outras ocorrem apenas nas moitas de vegetação. O objetivo deste estudo foi avaliar a germinação de sementes em diferentes regimes térmicos e hídricos, visando compreender os nichos preferenciais de germinação das espécies estudadas. Nós selecionamos três espécies associadas às ilhas de vegetação e quatro espécies associadas à areia nua de uma restinga localizada em Arraial do Cabo (RJ, Brasil). Foram testadas temperaturas constantes entre 15 a 40ºC e alternadas de 20/25 a 20/40ºC, com intervalos de 5ºC. Diferentes regimes hídricos também foram simulados com soluções de polietilenoglicol 6000 entre 0 (controle) a -1,2 MPa. Quando comparadas às espécies que ocorrem na areia nua, a germinação das espécies que ocorrem nas moitas de vegetação foi mais inibida por altas temperaturas (constantes e alternadas), e por baixos potenciais hídricos. Porém, algumas exceções foram encontradas, destacando a importância de outros fatores influenciando a distribuição das espécies estudadas, tais como dispersão e o estabelecimento de plântulas. Apenas as espécies que ocorrem na areia nua possuem sementes com dormência primária. Como as condições ambientais na areia nua são mais variáveis e extremas do que nas moitas de vegetação, conclui-se que a dormência é provavelmente uma característica importante para o sucesso de colonização da areia nua, já que aumenta a probabilidade de que a germinação ocorra em condições mais favoráveis ao estabelecimento das plântulas. Palavras-chave: Dormência, alternância de temperaturas, disponibilidade hídrica, moitas de vegetação, umidade do solo.

x
ABSTRACT
The seed germination requirements play an important role in determining de distribution patterns of plants. The open shrub formations in restingas are composed by patches of vegetation surrounded by bare sand with sparse plant cover. The surface of the bare sand has low capability of water retention and high temperatures are frequent. Meanwhile, the patches of vegetation attenuate these conditions, improving germination of many species in the restinga. In this type of vegetation, it is observed that some species occur only at the bare sand, while others species occur only associated with patches of vegetation. The aim of this study was to evaluate seed germination at different temperature and moisture regimes, to understand the preferential niches for germination of studied species. We selected three species associated with patches of vegetation and four species associated with bare sand from restinga located at Arraial do Cabo (RJ, Brazil). Were tested constant temperatures ranged from 15 to 40ºC and alternating temperatures ranged from 20/25 to 20/40ºC with intervals of 5ºC. Different moisture regimes were also simulated with polyethylene glycol 6000 solutions ranging from 0 (control) to -1,2 MPa. When compared to species that occur associated with the bare sand, germination of the species associated with patches of vegetation was more inhibited by high temperatures (constant and alternating), and by low water potentials. However, some exceptions were found, highlighting the importance of other factors influencing the distribution of studied species such as dispersal and establishment of the seedlings. Only species associated with the bare sand had seeds with primary dormancy. As environmental conditions in bare sand are more variable and extreme than in patches of vegetation, it was concluded that dormancy is probably an important feature for successful colonization of the bare sand, since it increases the likelihood that germination occurs in more favorable periods and conditions for establishment of the seedlings. Key words: Dormancy, alternating temperatures, water availability, patches of vegetation, soil moisture.

xi
SUMÁRIO
LISTA DE TABELAS ......................................................................................................XII
LISTA DE FIGURAS ......................................................................................................XIII
RESUMO.............................................................................................................................IX
ABSTRACT ......................................................................................................................... X
1. INTRODUÇÃO ................................................................................................................1
2. OBJETIVOS .....................................................................................................................8
3. HIPÓTESES .....................................................................................................................8
4. MATERIAL E MÉTODOS .............................................................................................9
4.1. Área de Estudo ...............................................................................................................9
4.2. Espécies Estudadas ........................................................................................................13
4.3. Determinação da Umidade Superficial do Solo ............................................................21
4.4. Coleta, Beneficiamento e Armazenamento ..................................................................22
4.5. Determinação do Conteúdo de Água das Sementes .....................................................23
4.6. Procedimentos Gerais para os Experimentos de Germinação ......................................23
4.7. Experimento de Temperatura .......................................................................................25
4.8. Experimento de Disponibilidade Hídrica .....................................................................25
4.9. Análises Estatísticas .....................................................................................................26
5. RESULTADOS ..............................................................................................................27
5.1. Umidade Superficial do Solo ........................................................................................27
5.2. Conteúdo de Água das Sementes .................................................................................29
5.3. Experimentos de Germinação........................................................................................30
5.3.1. Experimento de Temperatura .....................................................................................30
5.3.2. Experimento de Disponibilidade Hídrica ...................................................................47
6. DISCUSSÃO ..................................................................................................................55
6.1. Umidade Superficial do Solo ........................................................................................55
6.2. Conteúdo de Água das Sementes .................................................................................58
6.3. Experimentos de Germinação .......................................................................................58
6.3.1. Experimento de Temperatura .....................................................................................58
6.3.2. Experimento de Disponibilidade Hídrica ...................................................................68
7. CONCLUSÕES...............................................................................................................75
8. REFERÊNCIAS BIBLIOGRÁFICAS .........................................................................76

xii
LISTA DE TABELAS
Tabela 1. Valores de pluviosidade no dia de coleta de amostras do solo em cada mês para o ano de 2009........................................................................................................................29
Tabela 2. Local de ocorrência na APA de Massambaba para as espécies estudadas, e teor de água das sementes. n.a = não foi avaliado .....................................................................30
Tabela 3. Local de ocorrência na APA de Massambaba para as espécies estudadas, temperatura ótima, e tempo em dias para o início e término da germinação em uma das temperaturas ótimas entre parênteses. n.a = não foi avaliado ............................................47

xiii
LISTA DE FIGURAS
Figura 1. Formação arbustiva aberta não inundável na restinga de Massambaba, Arraial do Cabo, RJ. Foto: Aline Cavalcante de Souza. .......................................................................12
Figura 2. Mapa da Restinga de Massambaba, no município de Arraial do Cabo, RJ. A seta indica a localização da área de estudo. (Fonte: Muehe, 1994 apud Dorothy, 2000)............12
Figura 3. Dados de pluviosidade e temperatura média mensal durante o ano de 2009 para a região de Arraial do Cabo, RJ. Fonte: Instituto Nacional de Metereologia (INMET). .......13
Figura 4. Indivíduo de E. ovalifolium na área de estudo (a); detalhe do fruto (b). Foto: Aline Cavalcante de Souza ..................................................................................................14
Figura 5. Indivíduo de C. fluminensis na área de estudo (a); detalhe do fruto (b). Foto: Aline Cavalcante de Souza ..................................................................................................15
Figura 6. Indivíduo de M. obtusifolia na área de estudo (a); detalhe do fruto (b). Foto: Aline Cavalcante de Souza ..................................................................................................16
Figura 7. Indivíduo de L. rigidus na área de estudo (a); detalhe da infrutescência (b). Foto: Aline Cavalcante de Souza. .................................................................................................17
Figura 8. Indivíduo de M. violaceus na área de estudo (a); detalhe do fruto se desprendendo da planta-mãe (b). Foto: Aline Cavalcante de Souza ...................................18
Figura 9. Indivíduo de C. ramosa na área de estudo (a); detalhe do fruto b). Foto: Aline Cavalcante de Souza ............................................................................................................19
Figura 10. Indivíduo de C. flexuosa na área de estudo (a); detalhe do fruto (b). Foto: Aline Cavalcante de Souza ............................................................................................................20
Figura 11. Valores de umidade superficial do solo para amostras da areia nua (■), borda
(○) e interior de moitas (▲) dos meses de Janeiro a Dezembro de 2009. Médias seguidas da mesma letra não indicam diferença significativa entre os ambientes para o mesmo mês. Média ± erro padrão (Anova de um fator, p < 0,05, teste de Tukey). .................................28
Figura 12. Valores de umidade superficial do solo (0-5 cm) para o interior de moitas pequenas (■), interior de moitas médias (●) e para o interior de moitas grandes (∆) entre Janeiro e Dezembro de 2009. Médias seguidas da mesma letra não diferem entre si. Média ± erro padrão (Anova de um fator, p<0,05, teste de Tukey). Média ± erro padrão. ............28
Figura 13. Porcentagem de germinação de sementes de E. ovalifolium em diferentes temperaturas. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p < 0,05, teste de Tukey) .............31
Figura 14. Velocidade média de germinação de sementes de E. ovalifolium em diferentes temperaturas. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p < 0,05, teste de Tukey) .............32
Figura 15. Porcentagem de germinação de sementes de M. obtusifolia em diferentes temperaturas. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p < 0,05, teste de Tukey) .............33
Figura 16. Velocidade média de germinação de sementes de M. obtusifolia em diferentes temperaturas. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p < 0,05, teste de Tukey) .............34

xiv
Figura 17. Porcentagem de germinação de sementes de M. obtusifolia após transferência para temperatura ótima (20ºC). Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anovade um fator, p < 0,05, teste de Tukey). .................................................................................................................................34
Figura 18. Porcentagem de germinação de sementes de C. fluminensis em diferentes temperaturas. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p < 0,05, teste de Tukey). ...........35
Figura 19. Velocidade média de germinação de sementes de C. fluminensis em diferentes temperaturas. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p < 0,05, teste de Tukey). ............36
Figura 20. Porcentagem de germinação de sementes de C. fluminensis após transferência para temperatura ótima (30ºC). Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p < 0,05, teste de Tukey). ................................................................................................................................36
Figura 21. Porcentagem de germinação de sementes de L. rigidus em diferentes temperaturas. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p < 0,05, teste de Tukey). ............38
Figura 22. Velocidade média de germinação de sementes de L. rigidus em diferentes temperaturas. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p < 0,05, teste de Tukey). ............38
Figura 23. Porcentagem de germinação de sementes de L. rigidus após transferência para temperatura ótima (20-30ºC). Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p < 0,05, teste de Tukey). .................................................................................................................................39
Figura 24. Porcentagem de germinação de sementes de M. violaceus em diferentes temperaturas. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p < 0,05, teste de Tukey). ............40
Figura 25. Velocidade média de germinação de sementes de M. violaceus em diferentes temperaturas. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p < 0,05, teste de Tukey). ............41
Figura 26. Porcentagem de germinação de sementes de M. violaceus após a transferência para a temperatura ótima (35ºC). Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p < 0,05, teste de Tukey). .................................................................................................................................41
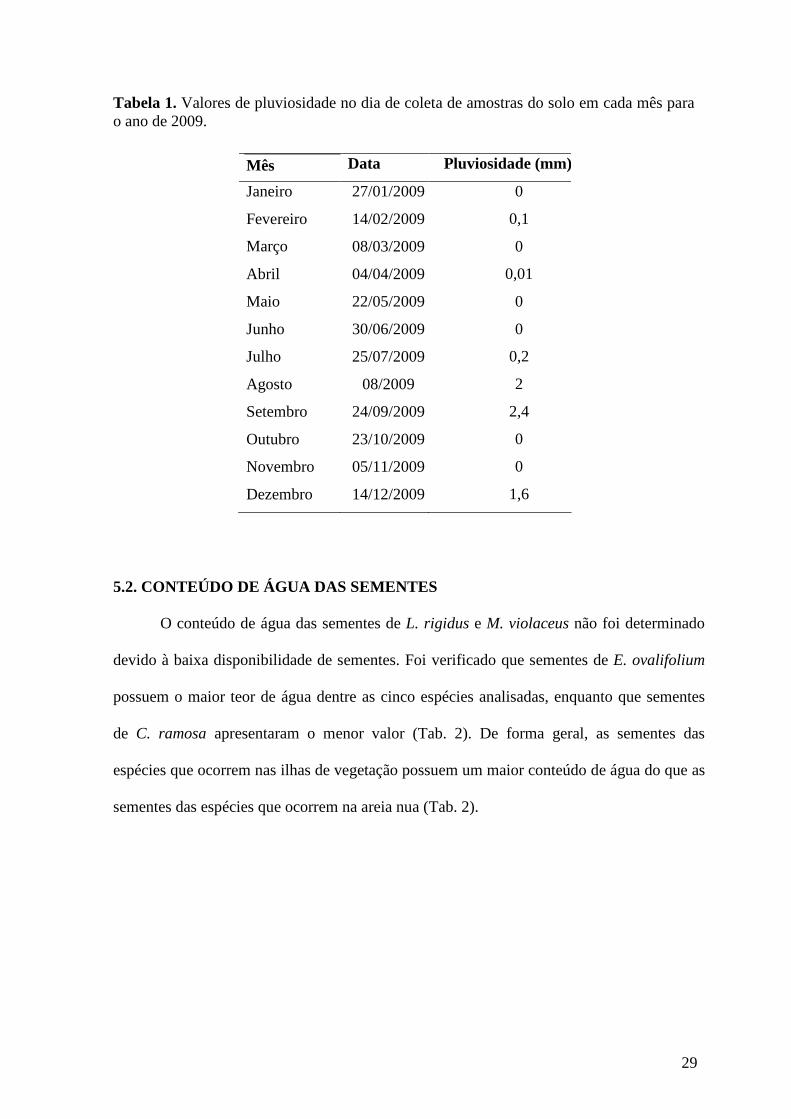
Figura 27. Porcentagem de germinação de sementes de C. ramosa em diferentes temperaturas. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p < 0,05, teste de Tukey). ............43
Figura 28. Velocidade média de germinação de sementes de C. ramosa em diferentes temperaturas. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Kruskal-Wallis, p < 0,05, teste de Dunn). ...................43
Figura 29. Porcentagem de germinação de sementes de C. ramosa após transferência para temperatura ótima (30ºC). Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p < 0,05, teste de Tukey)...................................................................................................................................44

xv
Figura 30. Porcentagem de germinação de sementes de C. flexuosa em diferentes temperaturas. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p < 0,05, teste de Tukey) .............45
Figura 31. Velocidade média de germinação de sementes de C. flexuosa em diferentes temperaturas. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Kruskal-Wallis, p < 0,05, teste de Dunn). ...................46
Figura 32. Porcentagem de germinação de sementes de C. flexuosa após transferência para temperatura ótima (30ºC). Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p < 0,05, teste de Tukey). .................................................................................................................................46
Figura 33. Porcentagem de germinação de sementes de E. ovalifolium em diferentes potenciais hídricos. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos no PEG (letras minúsculas) e entre tratamentos após transferência para água destilada (letras maiúsculas). Média ± erro padrão (Anova de um fator, p < 0,05, teste de Tukey). ■ PEG □ Água destilada (0MPa) ......................................................................48
Figura 34. Velocidade média de germinação de sementes de E. ovalifolium em diferentes potenciais hídricos. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Kruskal-Wallis, p<0,05, teste de Dunn). ............49
Figura 35. Porcentagem de germinação de sementes de M. obtusifolia em diferentes potenciais hídricos. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos no PEG (letras minúsculas) e entre tratamentos após transferência para água destilada (letras maiúsculas).. Média ± erro padrão (Anova de um fator, p < 0,05, teste de Tukey). ■ PEG □ Água destilada (0MPa). .............................................................50
Figura 36. Velocidade média de germinação de sementes de M. obtusifolia em diferentes potenciais hídricos. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p < 0,05, teste de Tukey). ....50
Figura 37. Porcentagem de germinação de sementes de C. fluminensis em diferentes potenciais hídricos. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos no PEG (letras minúsculas) e entre tratamentos após transferência para água destilada (letras maiúsculas). Média ± erro padrão (Anova de um fator, p < 0,05, teste de Tukey; Kruskal-Wallis, p<0,05, teste de Dunn). ■ PEG □ Água destilada (0 MPa). ...51
Figura 38. Velocidade média de germinação de sementes de C. fluminensis em diferentes potenciais hídricos. Médias seguidas da mesma letra não diferem entre si. Média ± erro padrão (Anova de um fator, p<0,05, teste de Tukey). .........................................................52
Figura 39. Porcentagem de germinação de sementes de C. ramosa em diferentes potenciais hídricos. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos no PEG (letras minúsculas) e entre tratamentos após transferência para água destilada (letras maiúsculas). Média ± erro padrão (Anova de um fator, p < 0,05, teste de Tukey). ■ PEG □ Água destilada (0 MPa). ....................................................................53
Figura 40. Velocidade média de germinação de sementes de C. ramosa em diferentes potenciais hídricos. Médias seguidas da mesma letra não diferem entre si. Média ± erro padrão (Anova de um fator, p<0,05, teste de Tukey). .........................................................53
Figura 41. Porcentagem de germinação de sementes de C. flexuosa em diferentes potenciais hídricos. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos no PEG (letras minúsculas) e entre tratamentos após transferência para

xvi
água destilada (letras maiúsculas). Média ± erro padrão (Anova de um fator, p < 0,05, teste de Tukey). ■ PEG □ Água destilada (0 MPa). ....................................................................54
Figura 42. Velocidade média de germinação de sementes de C. flexuosa em diferentes potenciais hídricos. Médias seguidas da mesma letra não diferem entre si. Média ± erro padrão (Anova de um fator, p < 0,05, teste de Tukey). .......................................................55

1
1. INTRODUÇÃO
Uma questão central em ecologia se refere à distribuição e abundância das espécies
(Begon et al., 2006). Diversos fatores influenciam esses parâmetros em uma dada
comunidade, os quais atuam como filtros que selecionam as espécies em um determinado
habitat. Estes filtros são divididos em filtros histórico, fisiológico e biótico, os quais
interagem e mudam constantemente no tempo e no espaço (Lambers et al., 1998). O filtro
histórico se refere à capacidade de uma determinada espécie de chegar a certo local. Para
persistir em um determinado habitat, as espécies precisam possuir características
fisiológicas que as capacitem se estabelecer, crescer e reproduzir (filtro fisiológico). As
interações bióticas como competição, predação e parasitismo, representam o filtro biótico,
que podem ou não eliminar uma espécie de uma região (Lambers et al., 1998).
Um dos principais componentes do filtro fisiológico é a capacidade de uma espécie
germinar em determinado local. Este processo é considerado um dos estágios mais críticos
no ciclo de vida da planta, constituindo juntamente com a fase de plântulas, uma das fases
com maior pressão demográfica, e provavelmente a fase com uma maior seleção (Harper,
1977; Whitmore, 1996). O conjunto específico de condições necessárias à germinação
varia de acordo com a espécie, sendo que a existência de lugares adequados que
proporcionam tais condições, chamados de “sítios seguros” (Harper, 1977) ou “nichos de
regeneração” (Grubb, 1977), influenciam a abundância e a distribuição das espécies
(Baskin & Baskin, 1998). Logo, o estudo das exigências necessárias para que o processo
de germinação ocorra é fundamental para a compreensão dos processos como sucessão e
regeneração natural (Khurana & Singh, 2004).
A germinação pode ser definida como a retomada do crescimento do embrião de
uma semente em condições favoráveis de hidratação, temperatura e oxigênio. Este
processo é iniciado pela absorção de água pela semente e terminando com o alongamento

2
do eixo embrionário, geralmente a radícula (Labouriau, 1983). Vários eventos
bioquímicos estão envolvidos neste processo como respiração, hidratação e síntese de
proteínas e alongamento celular. Diversos fatores bióticos e abióticos, como água, luz,
salinidade, temperatura, dispersão, predação atuam em conjunto, regulando o processo de
germinação. Além destes fatores, características da própria semente, como viabilidade,
tamanho, conteúdo de água, tolerância à dessecação, dormência, espessura da testa, entre
outros, também influenciam o processo de germinação (Baskin & Baskin, 1998; Fenner &
Thompson, 2005).
A ausência de germinação de uma semente viável em condições usualmente
consideradas como favoráveis à germinação, ou seja, em condições ideais de hidratação,
temperatura e oxigênio, é definida como dormência. A dormência é causada por um ou
mais bloqueios situados na própria semente que precisam ser superados para que a
germinação ocorra (Baskin & Baskin, 1998). Pode-se dizer que a função básica da
dormência é impedir a germinação em condições desfavoráveis ao estabelecimento das
plântulas, sendo uma característica importante na perpetuação das espécies (Fenner &
Thompson, 2005). A dormência distribui a germinação no tempo, fazendo com que a
emergência de plântulas ocorra em intervalos irregulares, diminuindo a competição e
aumentando a probabilidade de sobrevivência das plântulas (Bewley & Black, 1994).
Sementes dormentes são comuns em espécies que ocorrem em ambientes imprevisíveis e
com condições extremas (Jurado & Flores, 2005).
A água é um fator crucial que regula o processo de germinação, pois somente com
a embebição que se tem o início deste processo. A embebição é regulada principalmente
pela diferença de potencial hídrico do meio circundante e o da semente, com a água se
difundindo do maior potencial hídrico para o menor (Castro & Hilhorst, 2004). Além disso,
o tamanho, forma, permeabilidade do tegumento, a superfície de contato entre a semente e

3
a água, presença de mucilagem, tamanho das partículas do solo, entre outros, também
influenciam a taxa de embebição (Sheldon, 1974). A semente absorve a água a curtas
distâncias, evidenciando a importância da microtopografia do solo onde a mesma se
encontra na relação água semente (Labouriau, 1983).
Uma baixa disponibilidade hídrica, especialmente no começo da embebição,
inviabiliza a seqüência dos eventos germinativos da semente durante a absorção de água,
consequentemente podendo diminuir a porcentagem e a velocidade de germinação ou até
mesmo impedir que este processo ocorra (Castro & Hilhorst, 2004). Além disso, uma baixa
disponibilidade hídrica pode causar a dessecação de sementes, provocando a morte das
mesmas. Porém, é preciso considerar que a quantidade de água exigida para que uma
semente germine, e o grau de tolerância a dessecação varia de acordo com a espécie,
influenciando a distribuição da mesma. É observado que sementes de espécies de
ambientes áridos, geralmente conseguem germinar em uma menor disponibilidade hídrica
e são tolerantes à dessecação (ortodoxas), enquanto que sementes de espécies provenientes
de ambientes úmidos geralmente são sensíveis a dessecação (recalcitrantes) e não
conseguem germinar em condições de deficiência hídrica (Evans & Etherington, 1990;
Facelli & Ladd, 1996; Pammenter & Berjak, 2000; Tweddle et al., 2003).
Cada espécie possui uma faixa de temperatura na qual a germinação ocorre, ou seja,
acima da temperatura máxima e abaixo da temperatura mínima não há germinação. Desta
forma, existem três temperaturas cardinais para a germinação. A temperatura máxima,
mínima e ótima. A faixa ótima de temperatura é aquela na qual há uma maior porcentagem
de germinação em um menor intervalo de tempo (Bewley & Black, 1994). Estas três
temperaturas cardinais variam de acordo com a espécie, tendo relação com sua origem
geográfica, ano da colheita, duração do armazenamento, fatores ambientais durante o
processo de maturação, entre outros. (Labouriau, 1983). De forma geral, espécies tropicais

4
germinam em temperaturas mais elevadas do que espécies de ambientes temperados
(Fenner & Thompson, 2005).
A temperatura influencia a absorção de água pela semente e as reações bioquímicas
que regulam o metabolismo envolvido neste processo, principalmente a entrada de
oxigênio até o embrião, afetando a porcentagem, velocidade de germinação, viabilidade, e
quebra ou indução de dormência (Bewley & Black, 1994; Brändel & Schütz, 2005).
Temperaturas elevadas e flutuações na temperatura são fatores importantes na germinação
de sementes com tegumento duro, uma vez que provocam rachaduras no tegumento,
permitindo a entrada de água e gases (Baskin, 2003). Porém, temperaturas elevadas podem
ser letais, já que elas desnaturam estruturas celulares e causam um rápido envelhecimento
da semente, enquanto que baixas temperaturas reduzem o metabolismo, gerando um atraso
na germinação e uma redução da germinação (Probert, 2000; Zaidan & Barbedo, 2004).
Além disso, altas temperaturas aumentam a evaporação da água no solo, diminuindo a
disponibilidade de água para o processo germinativo (Breshears et al., 1998).
A facilitação é um importante tipo de interação na estruturação de comunidades
formadas por moitas intercaladas por uma matriz, em ambientes com uma baixa
produtividade, principalmente em ambientes áridos (savanas, desertos, e em regiões
costeiras) (Martínez, 2003; Dias et al., 2005; Aerts et al., 2006). Nestes ambientes, o
processo de germinação e estabelecimento de muitas espécies depende completamente das
espécies pioneiras facilitadoras (nurse plant), ou seja, há uma grande limitação da
colonização do solo nu por algumas espécies (Franco & Nobel, 1989; Pugnaire et al.,
1996; Kos & Poschlod, 2007). Este tipo de interação positiva é chamada de nurse plant
syndrome (“síndrome da planta berçário”), na qual a espécie facilitadora aumenta a
disponibilidade hídrica e nutricional, reduz a amplitude térmica e a radiação solar, criando
condições mais favoráveis à germinação, estabelecimento e/ou crescimento de outras

5
espécies de plantas (Callaway, 1995; Pugnaire et al., 2004; Yang et al., 2009).
O processo de facilitação resulta em uma agregação espacial de plântulas e
indivíduos adultos das espécies beneficiadas sob o dossel das espécies facilitadoras (Facelli
& Brock, 2000; Martínez, 2003). Porém, enquanto algumas espécies dependem de plantas
facilitadoras para se estabelecerem em um determinado local, outras espécies não
dependem deste tipo de interação (Went, 1942; Shmida & Whittaker, 1981; Gutiérrez et
al., 1993; Tielbörger & Kadmon, 1997). Desta forma, algumas espécies só ocorrem ou são
mais abundantes em áreas abertas do que sob o dossel de espécies facilitadoras. Tal padrão
de distribuição espacial se deve as condições ambientais distintas entre a região sob o
dossel das plantas facilitadoras e as áreas abertas entre elas, a características germinativas
distintas entre as espécies, diferenças nas exigências durante a fase de estabelecimento das
plântulas, dispersão, entre outros (Franco & Nobel, 1989; Aerts et al., 2006; Kos &
Poschlod, 2007).
Outro exemplo de uma comunidade formada por arbustos os quais são intercalados
por uma vegetação esparsa são as formações abertas de restinga. Este tipo de vegetação é
composta por moitas de vegetação, as quais são intercaladas por areia nua com uma
vegetação esparsa (Araujo et al., 2004). Este padrão de vegetação cria uma variação
espacial na disponibilidade hídrica, radiação solar, temperatura, evaporação, quantidade de
serapilheira e nutrientes, gerando basicamente dois tipos diferentes de microsítios neste
tipo de vegetação, a areia nua e as ilhas de vegetação (Martínez, 2003; Pereira, 2005).
Além da variação espacial há uma variação temporal das condições ambientais. Durante os
meses de verão, a radiação solar é mais intensa, fazendo com que a temperatura da
superfície do solo alcance 70ºC (Scarano, 2002). Além da radiação, a precipitação também
é maior durante esta época do ano, gerando uma maior disponibilidade de água no solo
para os processos de germinação e estabelecimento.

6
Nas ilhas de vegetação, devido ao sombreamento há uma menor exposição à
radiação solar e consequentemente uma menor amplitude térmica e evaporação de água no
solo. Além disso, no interior das moitas a quantidade de serapilheira é maior, fazendo com
que a retenção de água seja mais eficiente, gerando uma maior disponibilidade de água e
nutrientes em comparação com a areia nua (Mantovani & Iglesias, 2008; Mantuano, 2008).
Segundo Mantovani & Iglesias (2008), na restinga de Maricá a temperatura da superfície
do solo no interior das moitas ao longo de um dia de verão fica entre 25 e 29ºC e a
umidade superficial do solo entre 18,2 e 27,3 mg H2O [g peso seco solo] -1, enquanto que
na areia nua a temperatura durante o verão pode alcançar 60ºC ao meio-dia e a umidade
superficial varia ao longo do dia entre 2,6 e 0,9 mg H2O [g peso seco solo] –1. Devido às
condições físicas do substrato arenoso, a percolação de água na areia nua é rápida, fazendo
com que este recurso fique disponível na superfície do solo em um curto intervalo de
tempo, provavelmente apenas após eventos de chuva.
Pelo fato de as restingas serem um ambiente geologicamente mais recente, ser um
ecossistema adjacente a Mata Atlântica sensu stricto, e compartilhar cerca de 80% das
espécies com este ambiente, é considerado que as espécies que colonizaram as restingas
são provenientes da Mata Atlântica (Araujo, 2000). Segundo Scarano (2002), espécies
provenientes da Mata Atlântica com uma ampla plasticidade ecológica conseguiram
colonizar este ambiente com condições extremas como altas temperaturas do solo e do ar,
salinidade, baixa disponibilidade de água e de nutrientes, e dessecação. Outra forma de
colonização da restinga seria a partir do processo de facilitação (Dias et al., 2005; Scarano
et al., 2009). As espécies facilitadoras criam condições favoráveis ao processo de
germinação e estabelecimento de algumas espécies, sendo estas condições similares com as
condições que são encontradas na Mata Atlântica sensu stricto. Ou seja, condições distintas
as encontradas na areia nua, como altas temperaturas e deficiência hídrica, as quais

7
provavelmente atuam como barreiras a colonização de algumas espécies (Scarano et al.,
2009). Logo, a facilitação tem um importante papel nos processos de sucessão e na
formação de moitas destas comunidades, além do aumento da riqueza em espécies (Zaluar
& Scarano, 2000; Scarano, 2002).
A colonização via sementes no solo desnudo em restingas parece ser um processo
restrito a poucas espécies. Allagoptera arenaria (Gomes) Kuntze (Arecaceae) é uma das
poucas espécies capaz de germinar na areia nua (Scarano, 2002). Uma forma freqüente de
colonização da areia nua em restingas se dá por crescimento clonal, sendo observado nas
espécies Clusia hilariana Schltdl. (Clusiaceae) e Achemea nudicaulis (L.) Griseb
(Bromeliaceae), as quais são consideradas espécies facilitadoras (Scarano, 2002). Alguns
estudos verificaram que a germinação e o estabelecimento de determinadas espécies de
restinga são inibidos na areia nua devido à ocorrência de altas temperaturas, baixa
disponibilidade hídrica, e soterramento das sementes (Fialho, 1990; Maun, 1994; Cuzzuol
& Lucas, 1999; Hackbart & Cordazzo, 2003; Pinheiro & Borghetti, 2003; Mantovani &
Iglesias, 2008).
Nas formações abertas de restinga, também é verificado que algumas espécies são
mais abundantes ou só ocorrem na areia nua, enquanto outras espécies são mais abundantes
ou só ocorrem nas moitas, sendo que a areia nua é ocupada principalmente por herbáceas
(Zaluar, 2002; Pereira, 2005). Levando em consideração que na areia nua há uma menor
disponibilidade hídrica e maior amplitude térmica do que nas moitas, é provável que as
espécies que ocorrem em cada microsítio possuam respostas germinativas distintas em
relação à água e à temperatura. Desta forma, o objetivo geral do presente estudo foi
verificar se as respostas germinativas em relação à água e à temperatura influenciam a
distribuição espacial das espécies.

8
2. OBJETIVOS
a) Verificar se a umidade superficial do solo (0-5 cm) varia entre os microsítios areia nua,
borda e interior de moitas e com a redução do tamanho das moitas para o microsítio
interior.
b) Verificar se as espécies que ocorrem em cada ambiente (areia nua ou moita) diferem
quanto às exigências de temperatura para a germinação.
c) Verificar se as sementes das espécies que ocorrem em cada ambiente (areia nua ou
moita) diferem quanto às exigências de água para a germinação.
3. HIPÓTESES
a) Em função do aumento da incidência de radiação luminosa, a umidade superficial do
solo decresce do interior das moitas em direção a areia nua e com a redução do tamanho
das moitas para o microsítio interior.
b) A germinação das sementes das espécies que ocorrem na areia é maior em regimes
térmicos mais similares ao encontrado neste ambiente (maior temperatura e amplitude
térmica), sendo inibida em regimes térmicos mais similares ao encontrado em moitas de
vegetação (menor temperatura e amplitude térmica). O padrão inverso ocorre para as
espécies restritas às moitas de vegetação.
c) As sementes das espécies que ocorrem na areia nua possuem maior capacidade de
germinar em baixos potenciais hídricos do que as sementes das espécies que ocorrem nas
ilhas de vegetação.

9
4. MATERIAL E MÉTODOS
4.1. ÁREA DE ESTUDO
Restingas são planícies arenosas costeiras associadas a um complexo de
comunidades vegetais que ocorrem em cerca de 79% da costa brasileira (Lacerda et al.,
1993). São originadas a partir de depósitos marinhos durante as transgressões e regressões
marinhas no quaternário, as quais formaram uma sucessão de cordões arenosos (Suguio &
Tessler, 1984). As restingas, assim como os brejos, pântanos, campos de altitude,
constituem ambientes marginais ao Bioma Mata Atlântica (Scarano, 2002). Comparado
com a Mata Atlântica sensu stricto, a diversidade em restingas é menor, além da baixa
ocorrência de espécies endêmicas, evidenciando que não houve tempo suficiente para o
processo de especiação (Scarano, 2002). Segundo Araujo (2000), as restingas que ocorrem
no estado do Rio de Janeiro podem ser divididas de acordo com a sua flora e fisionomia
em dez tipos de formações vegetacionais, sendo influenciadas pela história geológica e
pelo clima.
O presente estudo foi desenvolvido na APA de Massamababa, a leste da cidade do
Rio de Janeiro, no município de Arraial do Cabo, próximo a localidade de Figueira, na
formação arbustiva aberta não inundável (Fig. 1). Esta formação é caracterizada pela
ocorrência de moitas formadas por uma vegetação arbustiva com até 5m de altura,
intercaladas por areia nua e uma vegetação esparsa composta por espécies herbáceas ou
subarbustivas (Araujo et al., 2009). Nesta formação há a presença de dunas fixas altas e
baixas, tornando o relevo bastante diversificado. Erythroxylum ovalifolium e Maytenus
obtusifolia estão entre as espécies lenhosas mais comuns nesta formação, ficando a espécie
herbácea Panicum trinii como uma das mais predominantes no espaço entre moitas
(Araujo et al., 2009).

10
A Área de Proteção Ambiental de Massambaba (APA) está localizada no Centro de
Diversidade Vegetal de Cabo Frio, nos municípios de Saquarema, Araruama e Arraial do
Cabo (aproximadamente 22º56´S). A APA de Massambaba possui uma área de 76,3 Km2 e
inclui a Reserva Ecológica de Jacarepiá e a Reserva Ecológica de Massambaba (Araujo,
1997). Comparando com outras restingas fluminenses, o Centro de Diversidade Vegetal de
Cabo Frio apresenta a maioria das espécies endêmicas às restingas, sendo vinte e seis
encontradas em um total de trinta e seis, além de ser a mais rica em espécies. Esta alta
diversidade em espécies é influenciada pelo clima seco, pela grande variedade de habitats e
pela história geomorfológica (Araujo, 1997; Araujo, et al., 2009).
A restinga de Massambaba possui 48 km de extensão, ocorrendo a partir do
município de Saquarema, mais precisamente da barra da Lagoa de Saquarema, até o Morro
da Atalaia, no município de Arraial do Cabo (Fig. 2). A restinga é composta por um
sistema de dois cordões arenosos paralelos de idades diferentes, os quais são separados por
uma faixa de relevo mais baixo onde há um sistema de lagunas. Estes dois cordões
arenosos separam o mar da lagoa de Araruama, provavelmente a maior lagoa hipersalina
do mundo, com ca. 200 km2 de superfície (Muehe, 1994 apud Dorothy, 2000). Três
esporões penetram esta lagoa, os quais foram formados pelo movimento de sedimentos no
interior da mesma. Na restinga de Massambaba existe uma diferença de relevo entre áreas
com (e.g. Reserva Ecológica Estadual de Massambaba) e sem dunas (e.g. Reserva
Ecológica Estadual de Jacarepiá), sendo que as dunas podem ultrapassar 20m de altura
(Araujo, 2000).
O clima da região de Cabo Frio é sui generis para a região sudeste do Brasil, sendo
classificado como Bsh, uma variação do clima semi-árido quente de Köppen (Barbiére,
1984), o que representa um contraste em relação ao resto do estado. Nesta região há a
presença de ventos predominantes de nordeste durante todo o ano (Barbiére, 1975). A

11
pluviosidade é reduzida em relação ao restante da costa do Rio de Janeiro devido ao
fenômeno da ressurgência, aos ventos alísios constantes secos e quentes ao longo de todo o
ano, à situação de cabo, a uma maior distância da Serra do Mar em relação à costa, entre
outros (Barbiére, 1975; Barbiére & Coe Neto, 1999). Há uma irregularidade no total
pluviométrico e na distribuição das chuvas entre os anos, sendo geralmente os meses mais
chuvosos durante a primavera e o verão (Barbiére, 1975). A média anual de precipitação da
região de Cabo Frio é de cerca de 800 mm, com pluviosidade mensal de 40 mm no
inverno, sendo que o balanço hídrico do solo é negativo ao longo do ano. A média anual da
temperatura é de 25ºC, com mínima de 12ºC e máxima de 36oC (Araujo, 1997).
Os dados climáticos referentes às medias mensais de pluviosidade e temperatura para
o período de 2009, na região de Arraial do Cabo- RJ foram disponibilizados pelo INMET
(Fig. 3).

12
Figura 1. Formação arbustiva aberta não inundável na restinga de Massambaba, Arraial do Cabo, RJ. Foto: Aline Cavalcante de Souza.
Figura 2. Mapa da Restinga de Massambaba, no município de Arraial do Cabo, RJ. A seta indica a localização da área de estudo. (Fonte: Muehe, 1994 apud Dorothy, 2000).

13
0
50
100
150
200
250
300
J F M A M J J A S O N D
Plu
vios
idad
e (m
m)
0,0
5,0
10,0
15,0
20,0
25,0
30,0
Tem
pera
tura
(ºC
)
Figura 3. Dados de pluviosidade e temperatura média mensal durante o ano de 2009 para a região de Arraial do Cabo, RJ. Fonte: Instituto Nacional de Pesquisas (INMET).
4.2. ESPÉCIES ESTUDADAS
As espécies estudadas foram divididas em dois grupos, conforme a distribuição das
mesmas na área de estudo e de acordo com a disponibilidade de sementes (Zaluar, 2002;
Pereira 2005), sendo o total de sete espécies estudadas. O primeiro grupo é formado pelas
espécies que não foram observadas isoladamente no solo desnudo ou foram observadas em
uma baixa freqüência, sendo, portanto o sítio mais favorável à germinação de suas
sementes provavelmente nas moitas de vegetação. O segundo grupo é composto pelas
espécies que foram observadas de forma isolada na areia nua, constituindo o grupo das
espécies que provavelmente possuem a areia nua como sítio mais favorável à germinação.
Para o primeiro grupo as espécies são: Erythroxylum ovalifolium Peyr.
(Erythroxylaceae), Clusia fluminensis Planch. & Triana (Clusiaceae) e Maytenus
obtusifolia Mart. (Celastraceae). Para o segundo grupo as espécies são: Lagenocarpus

14
rigidus Nees (Cyperaceae), Melocactus violaceus Pfeiff (Cactaceae), Chamaecrista
ramosa (Vogel) H. S. Irwin & R. C. Barneby (Fabaceae) e Chamaecrista flexuosa L.
(Greene) (Fabaceae). A seguir uma breve descrição das espécies estudadas:
Erythroxylum ovalifolium
A espécie E. ovalifolium pertence a família Erythroxylaceae, sendo conhecida
popularmente como fruta-de-pomba (Zamith & Scarano, 2004). É uma planta lenhosa de
hábito arbustivo, subarbustivo e arbóreo (Bieras & Sajo, 2004). No estado do Rio de
Janeiro, esta espécie tem ocorrência nas restingas de São João da Barra, Macaé, Maricá,
Cabo Frio, Jacarepaguá, Grumari, Marambaia, Barra de São João e Reserva Biológica da
Praia do Sul (Araujo, 2000). A frutificação é concentrada durante o verão (Fialho, 1990).
Na restinga de Massambaba, esta espécie ocorre nas ilhas de vegetação (Fig. 4) (obs.
pess.), sendo uma das espécies lenhosas mais comuns na formação arbustiva aberta não
inundável (Araujo et al., 2009).
Figura 4. Indivíduo de E. ovalifolium na área de estudo (a); detalhe do fruto (b). Foto: Aline Cavalcante de Souza.
b
a b

15
Clusia fluminensis
Clusia fluminensis pertence à família Clusiaceae, conhecida popularmente como
abaneiro (Zamith & Scarano, 2004). No estado do Rio de Janeiro, C. fluminensis ocorre nas
restingas nas regiões de Cabo Frio, Jacarepaguá, Maricá e Grumari (Araujo, 2000). Sua
frutificação ocorre entre os meses de Julho a Novembro (Zamith & Scarano, 2004). NA
APA da Massambaba, este arbusto ocorre na formação arbustiva aberta não inundável (fig.
5) (Araujo et al., 2009), mais especificamente nas moitas de vegetação (obs.pess.).
Figura 5. Indivíduo de C. fluminensis na área de estudo (a); detalhe do fruto (b). Foto: Aline Cavalcante de Souza.
b
a b

16
Maytenus obtusifolia
Maytenus obtusifolia é uma planta lenhosa arbustiva que pertence à família
Celastraceae, conhecida popularmente como carne-de-anta (Zamith & Scarano, 2004). Para
as restingas do Estado do Rio de Janeiro, esta espécie ocorre em São João da Barra, Macaé,
Cabo Frio, Maricá, Jacarepaguá, Grumari e Marambaia (Araujo, 2000). Sua frutificação é
concentrada no verão (Zamith & Scarano, 2004). Na restinga de Massambaba, é umas das
espécies mais comuns na formação arbustiva aberta não inundável (Fig. 6) (Araujo et al.,
2009), ocorrendo nas ilhas de vegetação (obs. pess).
Figura 6. Indivíduo de M. obtusifolia na área de estudo (a); detalhe do fruto (b). Foto: Aline Cavalcante de Souza.
b
ba
b
a b

17
Lagenocarpus rigidus
Lagenocarpus rigidus pertence à família Cyperaceae, conhecida vulgarmente como
capim-arroz (Araújo Junior et al., 2007). No Rio de Janeiro, ocorre nas restingas de
Macaé, Cabo Frio, Jacarepaguá e Marambaia (Araujo, 2000). Sua frutificação ocorre no
verão (obs. pess.). Na APA da Massambaba esta espécie herbácea ocorre na região entre
moitas na formação arbustiva aberta não inundável (obs.pess) (Fig.7).
Figura 7. Indivíduo de L. rigidus na área de estudo (a); detalhe da infrutescência (b). Foto: Aline Cavalcante.
ab

18
Melocactus violaceus
Melocactus violaceus pertence à família Cactaceae, conhecida vulgarmente como
coroa-de frade (Zamith & Scarano, 2004). É uma planta herbácea globosa, com 12-15 cm
de altura (Freitas, 1992). Esta espécie ocorre nas restingas da Marambaia, Araruama,
Arraial do Cabo, Macaé, Maricá e Barra de São João no estado do Rio de janeiro (Freitas,
1992; Araujo, 2000). Possui frutificação contínua ao longo do ano, e é considerada uma
espécie rara (Zamith & Scarano, 2004). Ocorre na formação arbustiva aberta não inundável
(Araujo et al., 2009), mais especificamente na areia nua (obs. pess.) (Fig.8).
Figura 8. Indivíduo de M. violaceus na área de estudo (a); detalhe do fruto se desprendendo da planta-mãe (b). Foto: Aline Cavalcante de Souza.
a b

19
Chamaecrista ramosa
Chamaecrista ramosa pertence à família Fabaceae, de porte subarbustivo (Madeira
& Fernandes, 1999). Ocorre no estado do Rio de Janeiro nas restingas de Macaé, Maricá,
Marambaia, Jacarepaguá e Cabo Frio (Araujo, 2000). Possui frutificação concentrada
durante a época chuvosa (Madeira & Fernandes, 1999). Para a área de estudo, ocorre na
formação arbustiva aberta não inundável (Araujo et al., 2009), mais especificamente na
areia nua obs. pess.) (Fig.9).
Figura 9. Indivíduo de C. ramosa na área de estudo (a); detalhe do fruto (b). Foto: Aline Cavalcante de Souza.
a b

20
Chamaecrista flexuosa
Chamaecrista flexuosa pertence à família Fabaceae, conhecida popularmente como
peninha (Miotto et al., 2008). Ocorre em quase todo o Brasil, sendo muito comum em
vegetações pioneiras (Bechara et al., 2007). Possui frutificação principalmente entre os
meses de outubro a Abril (Camargo & Miotto, 2004). Na restinga de Massambaba, esta
erva ocorre na formação arbustiva aberta não-inundável (Araujo et al., 2009), sendo
observada na areia nua (obs. pess.) (Fig. 10).
Figura 10. Indivíduo de C. flexuosa na área de estudo (a); detalhe do fruto (b). Foto: Aline Cavalcante de Souza.
a b

21
4.3. DETERMINAÇÃO DA UMIDADE SUPERFICIAL DO SOLO
Foi determinada mensalmente de Janeiro a Dezembro de 2009 a umidade superficial
do solo em três microsítios na formação arbustiva aberta não inundável na restinga de
Massambaba, RJ, a fim de verificar se há uma variação espaço-temporal na umidade
superficial do solo. Os três microsítios amostrados foram a areia nua, a borda e o interior
das moitas. A borda da moita foi classificada como o limite do dossel das espécies
arbustivas presentes na moita.
Um total de quinze moitas foram dividas em três classes de tamanho, sendo cinco
moitas pequenas, cinco médias e cinco grandes, a fim de verificar também se há diferença
na umidade do solo entre os diferentes tamanhos de moita. Uma moita foi definida como
um conjunto ou agrupamento de diferentes formas de vida com pelo menos uma espécie
arbustiva formando um dossel. Este conjunto de formas de vida estará isolado de outro
conjunto por solo arenoso desnudo ou vegetação herbácea, constituindo uma ilha de
vegetação (Zaluar, 2002).
Por ser a forma mais próxima possível de uma moita, uma vez que moitas possuem
um formato irregular, foi selecionada a área da elipse como medida de tamanho para cada
moita (Zaluar, 2002). A área das quinze moitas foi calculada através da fórmula A = D1/2 x
D2/2 x 3,14, onde D1 é o diâmetro maior da moita, e D2 é o diâmetro menor. A área das
quinze moitas amostradas foram 6,74m2, 7,91m2, 9,50m2, 11,04m2, 17,22m2, 30,02m2,
37,96m2, 47,48m2, 50,71m2, 50,94m2, 73,36m2, 87,60m2, 90,99m2, 138,47m2, 199,19m2. O
tamanho médio e erro padrão para a área das cinco moitas pequenas foi de 10,48m2 ± 1,83,
para as cinco moitas médias foi de 43,43m2 ± 4,10, e para as cinco moitas grandes foi de
117,93m2 ± 23,08.
No interior de cada moita foram coletadas quatro amostras, sendo duas na borda e
duas no interior. Os pontos de coleta foram amostrados no quadrante norte e sul. Para

22
verificar se há diferença na umidade do solo entre moitas e areia nua, a uma distância de
1m de cada moita foram coletadas duas amostras espaçadas por 1m na areia nua.
O solo foi coletado na profundidade de 0-5 cm, transportado em um em sacos
plásticos vedados para impedir a perda de umidade para o ambiente. No total foram
coletadas para as quinze moitas, trinta (15 x 2) amostras de solo no interior das moitas,
trinta (15 x 2) amostras na borda das moitas e trinta (15 x 2) na areia nua, totalizando
noventa amostras por mês. No laboratório, o material foi colocado em estufa a 103ºC
durante 24 horas, e a umidade foi calculada a partir do método gravimétrico, através da
fórmula: umidade (g/g) = 100 x (peso da amostra úmida – peso da amostra seca / peso da
amostra seca) (Embrapa, 1997).
4.4. COLETA, BENEFICIAMENTO E ARMAZENAMENTO DE SEMENTES
Frutos maduros de no mínimo sete indivíduos de cada espécie foram coletados em
2008 e 2009 de acordo com a época de frutificação para cada espécie na formação arbustiva
aberta não inundável na APA da Massambaba, no município de Arraial do Cabo, Rio de
Janeiro. Os frutos foram levados em sacos plásticos ou de papel para o Laboratório de
Sementes do Jardim Botânico do Rio de Janeiro. No laboratório, as sementes foram
extraídas dos frutos manualmente ou com o auxílio de peneiras. As sementes foram
homogeneizadas para a posterior montagem dos experimentos. Dependendo da espécie
analisada, as sementes foram armazenadas na câmara de armazenamento a 10ºC em
recipientes plásticos ou de vidro vedados hermeticamente até o início do experimento. O
intervalo entre a coleta e a montagem dos experimentos foi de no máximo 15 dias.

23
4.5. DETERMINAÇÃO DO CONTEÚDO DE ÁGUA DAS SEMENTES
O teor de água foi determinado pelo método da estufa (103 ºC/ 17h; ISTA, 1999)
logo após a coleta dos frutos. Dependendo da disponibilidade de sementes para cada
espécie, de três a quatro amostras de sementes foram pesadas individualmente em balança
com quatro casas decimais antes e após serem colocadas em estufa. Após este procedimento
o teor de água foi calculado através da fórmula:
% Upf = Pu – Ps / Pu. 100, onde:
% Upf = porcentagem de água, base peso fresco
Pu = peso fresco das sementes (g)
Ps = peso seco das sementes (g)
4.6. PROCEDIMENTOS GERAIS PARA OS EXPERIMENTOS DE GERMINAÇÃO
Sementes foram utilizadas para os experimentos em todas as espécies, exceto em E.
ovalifolium e L. rigidus, onde os diásporos eram o pirênio e aquênio, respectivamente.
Antes da montagem dos experimentos, as sementes foram esterilizadas com hipoclorito de
sódio 5% durante um minuto em duas repetições, e depois lavadas em água destilada. Em
seguida, lotes de 40, 30 ou 25 sementes dependendo da disponibilidade das mesmas foram
dispostas em placas de Petri (5 ou 9cm de diâmetro) sobre duas folhas de papel filtro
umedecido com água destilada ou uma solução de PEG 6000. Para diminuir a perda de
água por evaporação, as placas foram cobertas com filme PVC. A posição das placas
dentro dos germinadores foi alterada aleatoriamente todos os dias. O fotoperíodo das
câmaras foi de 8 horas. Para os experimentos de disponibilidade hídrica, as sementes foram
colocadas na temperatura ótima previamente observada no experimento de temperatura,
exceto para a espécie C. fluminensis. A temperatura ótima para esta espécie foi a de 30ºC,
porém a temperatura usada no experimento de disponibilidade hídrica foi a de 20ºC devido

24
à grande infestação de fungos na temperatura ótima. Página: 24
Diante de sua dormência tegumentar, as sementes das espécies C. ramosa e C. flexuosa
foram escarificadas com ácido sulfúrico concentrado (98%) por três minutos, antes da
montagem do experimento de disponibilidade hídrica.
As placas foram vistoriadas diariamente, e o critério de germinação utilizado foi o
de Labouriau (1983), no qual a semente é considerada como germinada quando ocorre a
emergência de uma das partes do embrião de dentro dos envoltórios. Mais precisamente a
emissão da raiz primária com curvatura geotrópica positiva, para eliminar uma possível
“falsa germinação” de um embrião morto que emitiu uma radícula devido à pressão de
embebição (Labouriau, 1983). Não foi utilizado o critério agronômico, no qual a
germinação ocorre quando há emergência da plântula, já que neste critério está incluído
além da fase de germinação, o crescimento inicial da plântula. As sementes germinadas
foram contadas diariamente e eliminadas das placas de Petri para facilitar a contagem. O
experimento foi finalizado quando não ocorreu germinação durante cinco dias
consecutivos.
Para cada tratamento foram calculadas a porcentagem e velocidade de germinação,
além do início e término da germinação. A porcentagem foi calculada pela razão entre o
número de sementes germinadas e o número total de sementes, e a velocidade média de
germinação (V) pela fórmula:
V = 1/t, onde:
t = tempo médio de germinação. Ou seja, o inverso do tempo médio de germinação,
V = ∑ni / ∑ ni.ti, onde:
ti = tempo entre o início do experimento e a i-ésima observação (dia)
ni = número de sementes germinadas no intervalo ti, em dias (Labouriau, 1983).

25
O início e término da germinação foram calculados na temperatura ótima observada
para cada espécie como o intervalo em dias para a primeira e última germinação,
respectivamente.
4.7. EXPERIMENTO DE TEMPERATURA
As sementes foram expostas às temperaturas constantes de 15, 20, 25, 30, 35 e 40oC
e alternadas de 20/25, 20/30, 20/35, 20/40oC (16/8h, respectivamente). O regime alternado
de temperatura foi promovido através da troca manual diária das placas de Petri. Durante o
período da manhã as sementes que estavam na câmara de germinação com a temperatura
de 20ºC foram transferidas para uma das câmaras de germinação com umas das
temperaturas usadas (25, 30, 35 e 40ºC), e o inverso durante o período da tarde. Estes
regimes térmicos foram usados para tentar simular condições de temperatura que ocorrem
em restingas, mais precisamente nas moitas de vegetação e na areia nua, sendo que nas
moitas de vegetação a temperatura e amplitude térmica são menores do que na areia nua
(Mantuano, 2008). Para os tratamentos com germinação significativamente inferior a
temperatura ótima ou nula, as sementes foram transferidas para a temperatura ótima
visando avaliar se estes tratamentos promoveram a perda de viabilidade.
4.8. EXPERIMENTO DE DISPONIBILIDADE HÍDRICA
As sementes foram expostas aos potenciais hídricos de 0; -0,2; -0,4; -0,6; -0,8; -1,0
e -1,2 MPa sob temperatura constante ótima, definida previamente pelo experimento de
temperatura. Para a obtenção de potenciais hídricos negativos foram produzidas soluções
com polietilenoglicol 6000 através da fórmula de Michel & Kaufmann (1973). As soluções
de PEG eram trocadas em um intervalo de cinco dias, a fim de minimizar uma variação no
potencial hídrico das soluções. Para minimizar a perda de água das soluções por evaporação,

26
as bandejas contendo as placas de Petri eram envoltas por sacos plásticos. Após a exposição
ao PEG 6000, as sementes que não germinaram foram transferidas para água destilada para
verificar se houve perda da viabilidade durante o período de exposição às soluções.
4.9. ANÁLISES ESTATÍSTICAS
Os dados utilizados na análise estatística nos experimentos de germinação foram a
porcentagem de germinação e a velocidade média de germinação para as réplicas de cada
tratamento. Para verificar diferenças significativas entre os tratamentos para uma dada
espécie foi utilizada a Análise de Variância (ANOVA) de um fator. Os dados relativos à
porcentagem de germinação que não atenderam aos pressupostos de normalidade e
homogeneidade das variâncias foram transformados para arco seno da raiz quadrada de
(x/100) (Zar, 1999), e posteriormente foram submetidos à análise de variância. Para a
velocidade média de germinação foi usada a transformação log (x +1), ou raiz cúbica
quando os dois pressupostos exigidos não foram atendidos. A normalidade foi verificada
através do teste de Shapiro - Wilk, e a homocedasticidade através do teste de Levene (Zar,
1999). Uma vez constatada diferenças significativas entre os tratamentos, foi realizado o
teste de Tukey com nível de significância de 5% para verificar entre quais tratamentos
houve diferença significativa. Quando os dados não atenderam os pré-requisitos exigidos
para as análises paramétricas mesmo após transformação dos dados foi utilizada a análise
de variância de Kruskal-Wallis, e para verificar entre quais tratamentos ocorreram
diferenças significativas foi usado o teste de Dunn com nível de significância de 5% (Zar,
1999).
Os dados de umidade superficial foram analisados através da Anova de um fator para
verificar diferenças significativas entre os microsítios, e entre os diferentes tamanhos de

27
moitas em um determinado mês. Uma vez constatada diferenças significativas entre os
tratamentos, foi realizado o teste de Tukey com nível de significância de 5% (Zar, 1999).
5. RESULTADOS
5.1. UMIDADE SUPERFICIAL DO SOLO
Foi verificado que a umidade superficial do solo (0-5cm) de amostras provenientes
do interior das moitas apresentaram valores significativamente superiores do que amostras
provenientes da borda e/ou da areia nua durante os meses de 2009 (Anova de um fator) ,
exceto em Agosto, onde a umidade do solo de amostras provenientes da areia nua foram
estatisticamente superiores em relação a borda e ao interior (Fig. 11). A umidade superficial
do microsítio borda na maior parte do ano apresentou valores estatisticamente iguais a
umidade superficial da areia nua, exceto em Janeiro, Abril e Junho, onde a umidade do solo
na borda foi maior do que na areia nua (Fig. 11). Desta forma, pode-se observar um
gradiente de umidade durante a maior parte do ano, com amostras do interior das moitas
alcançando os maiores valores de umidade, posteriormente as amostras retiradas da borda, e
por último com os menores valores de umidade do solo as amostras provenientes da areia
nua.
Foram observados valores inferiores de umidade superficial do solo para amostras
provenientes do interior de moitas pequenas em relação ao interior de moitas médias e/ou
grandes ao longo dos meses, exceto para os meses de Julho, Agosto e Dezembro (Fig. 12).
Ao contrário, não foi observada diferença significativa entre o interior de moitas médias e
grandes para todos os meses (Anova de um fator).
Foi verificado que a pluviosidade, no dia de coleta das amostras de solo, foi maior
nos meses de Agosto e Setembro, sendo nula ou baixa na maior parte do ano (Tab. 1).

28
J F M A M J J A S O N D
0
1
2
3
4
5
6
7
8
bb
a
bb
a
b
b
a
b
a
a
c
b
a
b
a
ab
c
b
a
bb
a
c
b
a
b
b
ab
b
a
c
b
a
Um
idad
e do
sol
o (%
)
Meses
Figura 11. Valores de umidade superficial do solo para a areia nua (■), borda (○) e interior
de moitas (▲) de Janeiro a Dezembro de 2009. Médias seguidas da mesma letra não indicam diferença significativa entre os ambientes para o mesmo mês. Média ± erro padrão (Anova de um fator, p<0,05, teste de Tukey).
J F M A M J J A S O N D0
1
2
3
4
5
6
7
8
9
a
a
a
ab
b
a
b
a
a
b
ab
a
a
a
a
a
a
a
b
ab
a
ab
b
a
ab
b
a
ab
b
a
ab
b
a
b
aa
Um
idad
e do
sol
o (%
)
Meses Figura 12. Valores de umidade superficial do solo (0-5 cm) para o interior de moitas pequenas (■), interior de moitas médias (●) e para o interior de moitas grandes (∆) entre Janeiro e Dezembro de 2009. Médias seguidas da mesma letra não diferem entre si. Média ± erro padrão (Anova de um fator, p<0,05, teste de Tukey). Média ± erro padrão.
29
Tabela 1. Valores de pluviosidade no dia de coleta de amostras do solo em cada mês para o ano de 2009.
Mês Data Pluviosidade (mm)
Janeiro 27/01/2009 0
Fevereiro 14/02/2009 0,1
Março 08/03/2009 0
Abril 04/04/2009 0,01
Maio 22/05/2009 0
Junho 30/06/2009 0
Julho 25/07/2009 0,2
Agosto 08/2009 2
Setembro 24/09/2009 2,4
Outubro 23/10/2009 0
Novembro 05/11/2009 0
Dezembro 14/12/2009 1,6
5.2. CONTEÚDO DE ÁGUA DAS SEMENTES
O conteúdo de água das sementes de L. rigidus e M. violaceus não foi determinado
devido à baixa disponibilidade de sementes. Foi verificado que sementes de E. ovalifolium
possuem o maior teor de água dentre as cinco espécies analisadas, enquanto que sementes
de C. ramosa apresentaram o menor valor (Tab. 2). De forma geral, as sementes das
espécies que ocorrem nas ilhas de vegetação possuem um maior conteúdo de água do que as
sementes das espécies que ocorrem na areia nua (Tab. 2).

30
Tabela 2. Local de ocorrência na APA de Massambaba para as espécies estudadas, teor de água das sementes. n.a = não foi avaliado
Espécies Local de ocorrência Teor de água
E. ovalifolium
Moita 52,72 ± 0,50
M. obtusifolia
Moita 40,12 ± 0,76
C. fluminensis
Moita 40,27 ± 3,31
M. violaceus
Areia nua n.a.
L. rigidus
Areia nua n.a.
C. ramosa
Areia nua 21,55 ± 1,98
C. flexuosa
Areia nua 23,15 ± 2,95
5.3. EXPERIMENTOS DE GERMINAÇÃO
5.3.1 EXPERIMENTO DE TEMPERATURA
Erythroxylum ovalifolium
O aumento da temperatura provocou um decréscimo significativo da porcentagem de
germinação, sendo significativamente inferior nas temperaturas de 25, 30, 20-30 e 20-35ºC.
A porcentagem de germinação foi inferior a 50% na temperatura constante de 30ºC e na
alternada de 20-35ºC, e nula nas temperaturas de 35, 40 e 20-40°C (Fig. 13). A velocidade
média de germinação na temperatura de 20-30ºC foi estatisticamente inferior do que a
velocidade nas temperaturas de 20ºC e 30ºC, não ocorrendo diferença significativa entre
todas as demais temperaturas (Fig. 14). A faixa de temperatura ótima para esta espécie foi
obtida nas temperaturas de 15, 20 e 20-25ºC, uma vez que nestas temperaturas foi verificada
uma maior porcentagem e velocidade média de germinação (Fig. 13 e 14). Após a
transferência das sementes não germinadas ou com germinação estatisticamente inferior a

31
temperatura ótima (25, 30, 35, 40, 20-30, 20-35 e 20-40°C) para 20ºC, que corresponde a
uma das temperaturas dentro da faixa de temperatura ótima, foi verificada em todas as
temperaturas uma germinação nula. Através do corte longitudinal das sementes que não
germinaram após a transferência para a temperatura ótima, foi verificado que todas as
sementes estavam inviáveis, uma vez que foi constatada a liberação de fluidos das sementes,
as mesmas possuíam consistência mole, com embriões com coloração amarelada.
15C 20C 25C 30C 35C 40C 20-25 20-30 20-35 20-400
20
40
60
80
100
ab
dd
c
bc
aa
Ger
min
ação
(%
)
Temperatura (ºC)
(Erythroxylum ovalifolium)
Figura 13. Porcentagem de germinação de sementes de E. ovalifolium em diferentes temperaturas. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p<0,05, teste de Tukey).

32
15 20 25 30 35 40 20-25 20-30 20-35 20-400,00
0,01
0,02
0,03
0,04
0,05
0,06
ab ababab
a
b
a
IVG
(di
a-1)
Temperatura (ºC)
Erythroxylum ovalifolium
Figura 14. Velocidade média de germinação de sementes de E. ovalifolium em diferentes temperaturas. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p<0,05, teste de Tukey). Maytenus obtusifolia
A porcentagem e a velocidade de germinação de sementes de M. obtusifolia foram
significativamente afetadas pela temperatura. Foi observado um decréscimo significativo da
porcentagem de germinação em 35ºC e 20-40ºC. A germinação foi nula em 40ºC, e baixa na
temperatura constante de 35ºC, sendo inferior a 50% na temperatura alternada de 20-40ºC
(Fig. 15). Foi observado um decréscimo significativo da velocidade de germinação com o
aumento da temperatura em 30ºC, 35ºC, 20-35ºC e 20-40ºC (Fig. 16). Além disso, a
velocidade na menor temperatura constante (15ºC) foi significativamente inferior a
velocidade nas temperaturas de 20, 25, 20-25 e 20-30ºC. A faixa de temperatura ótima para
esta espécie ocorreu nas temperaturas de 20, 25, 20-25 e 20-30ºC, uma vez que nestas
temperaturas a porcentagem de germinação foi significativamente maior em um menor
intervalo de tempo (Fig. 15 e 16). Após a transferência das sementes dos tratamentos que
obtiveram uma porcentagem de germinação significativamente inferior (35, 40, e 20-40ºC)

33
para a temperatura ótima (20ºC), para as sementes que foram expostas a temperatura de
40ºC a germinação foi nula, sendo que para as temperaturas de 35 e 20-40ºC foi observado
um acréscimo após a transferência (Fig. 17). Porém, em ambos os tratamentos a germinação
foi significativamente inferior em relação ao que foi obtido na temperatura ótima de 20ºC.
Através do corte longitudinal das sementes que não germinaram após a transferência para a
temperatura ótima, foi observado que as sementes expostas a 40ºC apresentavam liberação
de fluidos, consistência mole, e embriões com coloração amarelada, estando, portanto todas
inviáveis. As sementes que foram expostas as temperaturas de 35ºC e 20-40ºC, 84%
estavam inviáveis e 16% viáveis, 88% estavam inviáveis e 12% viáveis, respectivamente.
15 20 25 30 35 40 20-25 20-30 20-35 20-400
20
40
60
80
100 a a aa a
c
b
a
aab
Ger
min
ação
(%
)
Temperatura (ºC)
Maytenus obtusifolia
Figura 15. Porcentagem de germinação de sementes de M. obtusifolia em diferentes temperaturas. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p<0,05, teste de Tukey).

34
15 20 25 30 35 40 20-25 20-30 20-35 20-400,00
0,02
0,04
0,06
0,08
0,10
0,12
0,14
abcbc
cd
ab
a
ee
d
ab
IVG
(di
a -1)
Temperatura (ºC)
Maytenus obtusifolia
Figura 16. Velocidade média de germinação de sementes de M. obtusifolia em diferentes temperaturas. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p<0,05, teste de Tukey).
20 35 40 20-400
20
40
60
80
100
b
b
a
Ger
min
ação
(%
)
Temperatura (ºC)
Maytenus obtusifolia
Figura 17. Porcentagem de germinação de sementes de M. obtusifolia após transferência para temperatura ótima (20ºC). Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p<0,05, teste de Tukey).

35
Clusia fluminensis
Com o aumento da temperatura, foi observado um decréscimo significativo da
porcentagem de germinação em 35ºC, sendo nula nas maiores temperaturas no regime
constante e alternado (40ºC e 20-40ºC) (Fig. 18). A velocidade média de germinação foi
significativamente maior em 30ºC do que em todas as outras temperaturas (Fig. 19). Logo,
a temperatura ótima para esta espécie foi obtida em 30°C, uma vez que nesta temperatura
foi verificada uma maior porcentagem de germinação e um maior valor para a velocidade
de germinação (Fig. 18 e 19). Após a transferência das sementes dos tratamentos com uma
porcentagem de germinação significativamente inferior (35, 40 e 20-40ºC) para a
temperatura ótima (30ºC), foi verificada uma germinação nula para as sementes que foram
expostas a 40ºC. Porém, para as temperaturas de 35ºC e 20-40ºC foi verificado um
aumento significativo na porcentagem de germinação (Fig. 20), uma vez que foi
estatisticamente igual ao obtido na temperatura ótima de 30ºC.
15 20 25 30 35 40 20-25 20-30 20-35 20-400
20
40
60
80
100 a
b
aaaaaa
Ger
min
ação
(%
)
Temperatura (ºC)
Clusia fluminensis
Figura 18. Porcentagem de germinação de sementes de C. fluminensis em diferentes temperaturas. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p<0,05, teste de Tukey).

36
15 20 25 30 35 20-25 20-30 20-350,00
0,02
0,04
0,06
0,08
0,10
0,12
0,14
0,16
0,18
cd
b
bc
cd
a
bb
d
IVG
(di
a -1)
Temperatura (ºC)
Clusia fluminensis
Figura 19. Velocidade média de germinação de sementes de C. fluminensis em diferentes temperaturas. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p < 0,05, teste de Tukey).
30 35 40 20-400
20
40
60
80
100
a
aa
Ger
min
ação
(%
)
Temperatura (ºC)
Clusia fluminensis
Figura 20. Porcentagem de germinação de sementes de C. fluminensis após transferência para temperatura ótima (30ºC). Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p<0,05, teste de Tukey).

37
Lagenocarpus rigidus
Com o aumento da amplitude térmica no regime alternado, foi observado um
acréscimo significativo da porcentagem de germinação, sendo a maior porcentagem de
germinação obtida nas temperaturas de 20-35ºC e 20-40ºC (Fig. 21). Nas temperaturas
constantes de 35ºC e 40ºC a germinação foi nula, e para as demais temperaturas constantes
foi verificada uma baixa porcentagem germinação, sendo estes valores estatisticamente
iguais somente a temperatura alternada de 20-25ºC. A velocidade média de germinação foi
significativamente inferior nas temperaturas de 20ºC e 20-40ºC (Fig. 22). A faixa de
temperatura ótima foi obtida em 20-35ºC, uma vez que apresentou a maior porcentagem de
germinação em um menor intervalo de tempo (Fig. 21 e 22). Os tratamentos com
porcentagem de germinação significativamente inferior foram transferidos (20, 25, 30, 35,
40 e 20-25 ºC) para a temperatura de 20-30ºC. Apesar de 20-30ºC não ser uma das
temperaturas ótimas, após a transferência foi observado um acréscimo significativo da
porcentagem de germinação para as temperaturas de 35ºC e 40ºC (Fig. 23). Não foi
possível realizar o corte longitudinal das sementes para verificar a viabilidade das mesmas
devido ao seu pequeno tamanho.

38
20 25 30 35 40 20-25 20-30 20-35 20-400
20
40
60
80
100
ab
d ddcd
bc
a
Ger
min
ação
(%
)
Temperatura (ºC)
(Lagenocarpus rigidus)
Figura 21. Porcentagem de germinação de sementes de L. rigidus em diferentes temperaturas. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p<0,05, teste de Tukey).
20 25 30 20-25 20-30 20-35 20-400,00
0,01
0,02
0,03
0,04
0,05
0,06
0,07
0,08
b
abab
abab
b
a
IVG
(di
a-1)
Temperatura (ºC)
Lagenocarpus rigidus
Figura 22. Velocidade média de germinação de sementes de L. rigidus em diferentes temperaturas. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p<0,05, teste de Tukey).

39
20 25 30 35 40 20-25 20-350
20
40
60
80
100
c
ab
ab
bc
bcbc
a
Ger
min
ação
(%
)
Temperatura (ºC)
Lagenocarpus rigidus
Figura 23. Porcentagem de germinação de sementes de L. rigidus após transferência para temperatura ótima (20-30ºC). Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p<0,05, teste de Tukey). Melocactus violaceus
Foi verificada a ausência de germinação em todas as temperaturas constantes
testadas, ocorrendo germinação superior a 20% apenas nas temperaturas alternadas. O
aumento da amplitude térmica nas temperaturas alternadas promoveu o acréscimo
significativo da porcentagem de germinação (Fig.24). As temperaturas de 20-35ºC e 20-
40ºC promoveram os maiores valores de porcentagem de germinação, sendo
significativamente superiores aos valores de germinação nas temperaturas de 20-25ºC e 20-
30ºC (fig. 24). Houve diferença significativa para a velocidade média de germinação
apenas entre a temperatura de 20-25ºC e 20-35ºC (Fig. 25). Portanto, a faixa de
temperatura ótima para esta espécie foi obtida nas temperaturas de 20-35ºC e 20-40ºC.
Após a transferência das sementes das temperaturas que apresentaram uma porcentagem de

40
germinação significativamente inferior (20, 25, 30, 35, 40, 20-25 e 20-30ºC) para a
temperatura ótima de 20-35ºC, foi observado um aumento significativo da porcentagem de
germinação apenas nas temperaturas alternadas de 20-25ºC e 20-30ºC. Nas temperaturas
de 20ºC e 30ºC a germinação continuou a ser igual a zero (Fig. 26). Não foi realizado o
corte longitudinal das sementes que não germinaram após a transferência para a
temperatura ótima devido ao pequeno tamanho das mesmas. Porém, foi observada a
liberação de fluidos e uma infestação de fungos em sementes que foram expostas a 40ºC.
20 25 30 35 40 20-25 20-30 20-35 20-400
20
40
60
80
100
c
bc
ab
a
Ger
min
ação
(%
)
Temperatura (ºC)
Melocactus violaceus
Figura 24. Porcentagem de germinação de sementes de M. violaceus em diferentes temperaturas. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p<0,05, teste de Tukey).

41
20-25 20-30 20-35 20-400,000
0,005
0,010
0,015
0,020
0,025
0,030
0,035
0,040
ab
a
ab
b
IVG
(di
a-1)
Temperatura (ºC)
Melocactus violaceus
Figura 25. Velocidade média de germinação de sementes de M. violaceus em diferentes temperaturas. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p<0,05, teste de Tukey).
20 25 30 35 40 20-25 20-30 20-350
20
40
60
80
100
d
cd
bc
ab
aba
Ger
min
ação
(%
)
Temperatura (ºC)
Melocactus violaceous
Figura 26. Porcentagem de germinação de sementes de M. violaceus após a transferência para a temperatura ótima (35ºC). Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p<0,05, teste de Tukey).

42
Chamaecrista ramosa
Nenhuma temperatura propiciou uma porcentagem de germinação superior a 50%
para as sementes desta espécie (Fig. 27). A maior porcentagem de germinação (49%) foi
observada na temperatura de 30ºC, sendo significativa a diferença na porcentagem de
germinação neste tratamento em relação às temperaturas de 35, 40, 20-25, 20-30 e 20-40ºC.
A menor velocidade média de germinação foi observada para a temperatura de 40ºC, não
ocorrendo diferença significativa entre as demais temperaturas (Fig. 28). A maior
porcentagem de germinação em um menor intervalo de tempo foi obtida nas temperaturas
de 20, 25, 30 e 20-35ºC, sendo, portanto a faixa de temperatura ótima para esta espécie (Fig.
27 e 28). Após transferência das sementes nos tratamentos com germinação
significativamente inferior (35, 40, 20-25, 20-30 e 20-40ºC) para a temperatura ótima de
30ºC, foi observada a ausência de um acréscimo significativo da porcentagem germinação
em todos os tratamentos (Fig. 29). Não foi realizado corte longitudinal das sementes que
não germinaram após a transferência para a temperatura ótima devido ao pequeno tamanho
das sementes. Porém, foi verificado que as sementes que foram expostas a temperaturas de
40ºC, apresentaram-se com liberação de fluidos e infestação de fungos.
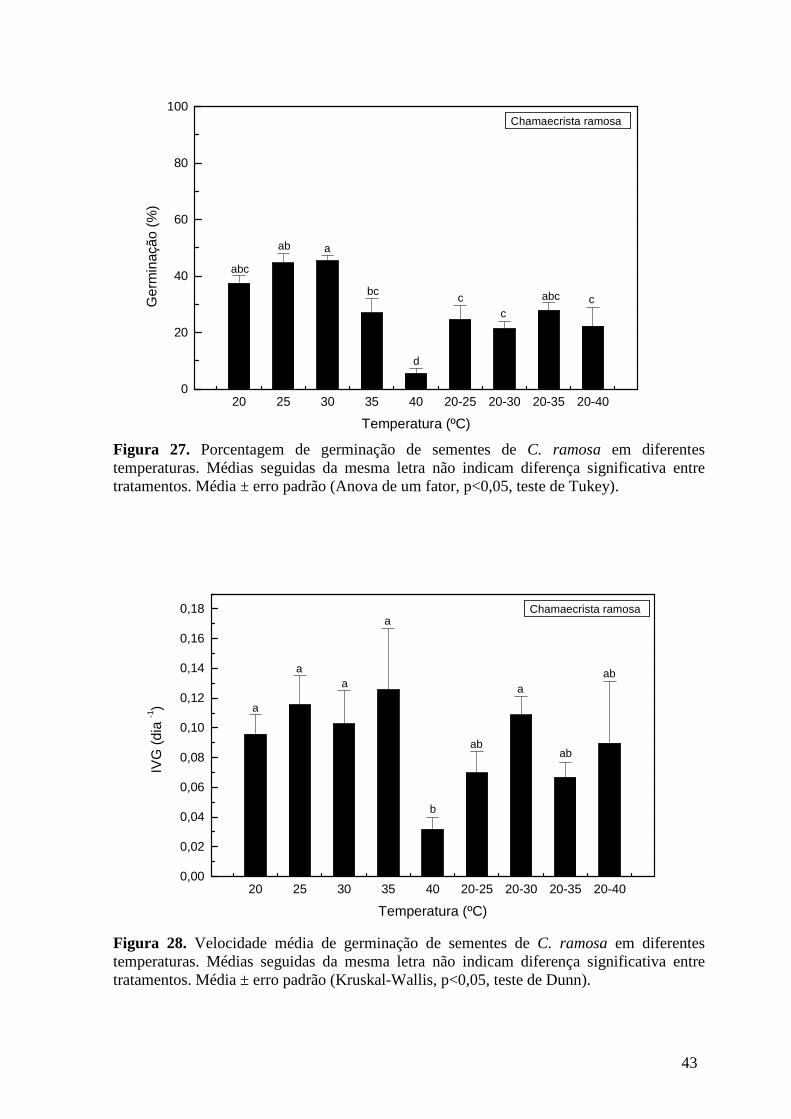
43
20 25 30 35 40 20-25 20-30 20-35 20-400
20
40
60
80
100
cabc
cc
d
bc
aab
abc
Ger
min
ação
(%
)
Temperatura (ºC)
Chamaecrista ramosa
Figura 27. Porcentagem de germinação de sementes de C. ramosa em diferentes temperaturas. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p<0,05, teste de Tukey).
20 25 30 35 40 20-25 20-30 20-35 20-400,00
0,02
0,04
0,06
0,08
0,10
0,12
0,14
0,16
0,18
ab
ab
a
ab
b
a
aa
a
IVG
(di
a -1)
Temperatura (ºC)
Chamaecrista ramosa
Figura 28. Velocidade média de germinação de sementes de C. ramosa em diferentes temperaturas. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Kruskal-Wallis, p<0,05, teste de Dunn).

44
30 35 40 20-25 20-30 20-400
20
40
60
80
100
c
bb
bb
a
Ger
min
ação
(%
)
Temperatura (ºC)
Chamaecrista ramosa
Figura 29. Porcentagem de germinação de sementes de C. ramosa após transferência para temperatura ótima (30ºC). Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p<0,05, teste de Tukey).
Chamaecrista flexuosa
A temperatura influenciou significativamente apenas a porcentagem de germinação
de sementes de C. flexuosa. As temperaturas que proporcionaram uma maior porcentagem
de germinação para C. flexuosa foram as temperaturas de 25, 30 e 35ºC, sendo
significativamente diferentes em relação a todas as outras temperaturas (Fig. 30). As
temperaturas de 20ºC e 40ºC apresentaram um valor significativamente inferior para a
porcentagem de germinação. A velocidade média de germinação não apresentou diferença
significativa entre os tratamentos (Fig. 31). Desta forma, a faixa de temperatura ótima para
esta espécie ocorreu entre as temperaturas de 25, 30 e 35ºC. Após a transferência das
sementes que apresentaram uma porcentagem de germinação significativamente inferior
(20, 40, 20-30, 20-35 e 20-40ºC) para uma das temperaturas ótimas (30ºC), foi observado
um aumento não significativo da porcentagem de germinação em todos os tratamentos (Fig.
32). Não foi realizado corte longitudinal das sementes que não germinaram após a

45
transferência para a temperatura ótima devido ao pequeno tamanho das mesmas. Porém, foi
verificado que as sementes que foram expostas as temperaturas de 35ºC e 40ºC tinham a
consistência mole, liberando fluidos e com infestação de fungos.
20 25 30 35 40 20-30 20-35 20-400
20
40
60
80
100
bcc
c
c
ab
a
a
c
Ger
min
ação
(%
)
Temperatura (ºC)
Chamaecrista flexuosa
Figura 30. Porcentagem de germinação de sementes de C. flexuosa em diferentes temperaturas. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p<0,05, teste de Tukey).

46
20 25 30 35 40 20-30 20-35 20-400,00
0,02
0,04
0,06
0,08
0,10
0,12
0,14
0,16
0,18
a
aaa
a
a
aa
IVG
(di
a-1)
Temperatura (ºC)
Chamaecrista flexuosa
Figura 31. Velocidade média de germinação de sementes de C. flexuosa em diferentes temperaturas. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Kruskal-Wallis, p<0,05, teste de Dunn).
20 30 40 20-30 20-35 20-400
20
40
60
80
100
bb
b
c
bc
a
Ger
min
ação
(%
)
Temperatura (ºC)
Chamaecrista flexuosa
Figura 32. Porcentagem de germinação de sementes de C. flexuosa após transferência para temperatura ótima (30ºC). Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p<0,05, teste de Tukey).

47
Tabela 3. Local de ocorrência na APA de Massambaba para as espécies estudadas, temperatura ótima, e tempo em dias para o início e término da germinação em uma das temperaturas ótimas entre parênteses. n.a = não foi avaliado
Espécies Local de ocorrência
Temperatura ótima (ºC) Início e Término da germinação (dias)
E. ovalifolium Moita 15, 20, 20-25 7- 56 (20ºC)
M. obtusifolia Moita 20, 25, 20-25, 20-30 5 – 23 (20ºC)
C. fluminensis Moita 30 2 – 19 (30ºC)
M. violaceus Areia nua 20-35, 20-40 13 – 65 (20-35ºC)
L. rigidus Areia nua 20-35 9 – 43 (20-35ºC)
C. ramosa Areia nua 20, 25, 30, 20-35 2 – 56 (30ºC)
C. flexuosa Areia nua 25, 30, 35 3 – 38 (30ºC)
5.3.2. EXPERIMENTO DE DISPONIBILIDADE HÍDRICA
Erythroxylum ovalifolium
Foi observada redução significativa da porcentagem e velocidade de germinação
com a diminuição do potencial hídrico. O tratamento que apresentou a maior porcentagem
de germinação (82%) foi o controle (0 MPa). Com a redução do potencial hídrico foi
verificada uma redução significativa da porcentagem de germinação em todos os
tratamentos (Fig. 33). No potencial hídrico de -0,4 MPa a porcentagem de germinação foi
inferior a 50%, sendo baixa em -0,8 MPa, e nula no tratamento com potencial hídrico de -
1,0 MPa. (Fig 33). A velocidade de germinação não foi significativamente afetada pela
redução do potencial hídrico até o tratamento de -0,6 MPa, sendo significativamente inferior
apenas no tratamento de -0,8 MPa (Fig. 34). Foi verificado um aumento na porcentagem de
germinação quando as sementes não germinadas nos potenciais hídricos negativos foram

48
transferidas para água destilada (Fig. 33). Porém, este aumento foi decrescente com a
diminuição do potencial hídrico, sendo a porcentagem final de germinação igual
estatisticamente ao controle apenas para os potenciais hídricos de -0,2 MPa e -0,4 MPa.
Para os demais tratamentos, a germinação após a transferência para água destilada foi
inferior ao controle, sendo o menor valor de germinação obtido no tratamento com menor
potencial hídrico (-1,0 MPa).
0,0 0,2 0,4 0,6 0,8 1,00
20
40
60
80
100
C
BCBC
AB
A
cc
b
b
Aa
Ger
min
ação
(%
)
Potencial Hídrico (-MPa)
Erythroxylum ovalifolium
Figura 33. Porcentagem de germinação de sementes de E. ovalifolium em diferentes potenciais hídricos. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos no PEG (letras minúsculas) e entre tratamentos após transferência para água destilada (letras maiúsculas). Média ± erro padrão (Anova de um fator, p<0,05, teste de Tukey). ■ PEG □ Água destilada (0 MPa).

49
0,0 0,2 0,4 0,6 0,8 1,00,000
0,005
0,010
0,015
0,020
0,025
0,030
0,035
ab
abab
b
a
IVG
(di
a-1)
Potencial Hídrico (-MPa)
Erythroxylum ovalifolium
Figura 34. Velocidade média de germinação de sementes de E. ovalifolium em diferentes potenciais hídricos. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Kruskal-Wallis, p<0,05, teste de Dunn). Maytenus obtusifolia
Com a diminuição do potencial hídrico observou-se uma redução significativa da
porcentagem e velocidade de germinação (Fig. 35 e 36). A maior porcentagem de
germinação (98%) foi verificada no tratamento controle (0 MPa). Com a redução do
potencial hídrico foi observada uma redução significativa da porcentagem de germinação
em todos os tratamentos, exceto em -0,2 MPa. A germinação foi inferior a 50% no potencial
hídrico de -0,8 MPa, e baixa em -1,2 MPa (Fig. 35). A velocidade média de germinação foi
significativamente reduzida em -0,8, -1,0 e -1,2 MPa (Fig. 36). Após a transferência das
sementes não germinadas nas soluções de PEG para a água destilada, foi verificado um
aumento na porcentagem de germinação (Fig. 35). Porém, para o tratamento de -1,2 MPa
não foi observado um acréscimo significativo da porcentagem de germinação, uma vez que
ao contrário de todos os outros tratamentos, a germinação foi estatisticamente inferior ao
controle.

50
0,0 0,2 0,4 0,6 0,8 1,0 1,20
20
40
60
80
100
B
A
A
A
A
A
c
b
b
bb
a
Aa
Ger
min
ação
(%
)
Potencial Hídrico (-Mpa)
Maytenus obtusifolia
Figura 35. Porcentagem de germinação de sementes de M. obtusifolia em diferentes potenciais hídricos. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos no PEG (letras minúsculas) e entre tratamentos após transferência para água destilada (letras maiúsculas).. Média ± erro padrão (Anova de um fator, p < 0,05, teste de Tukey). ■ PEG □ Água destilada (0 MPa).
0,0 0,2 0,4 0,6 0,8 1,0 1,20,00
0,01
0,02
0,03
0,04
0,05
c
bb
aa
aa
IVG
(di
a -1)
Potencial Hídrico (-MPa)
Maytenus obtusifolia
Figura 36. Velocidade média de germinação de sementes de M. obtusifolia em diferentes potenciais hídricos. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos. Média ± erro padrão (Anova de um fator, p < 0,05, teste de Tukey).

51
Clusia fluminensis
Foi observado um decréscimo significativo da porcentagem e velocidade de
germinação com a redução do potencial hídrico. Foi observada maior porcentagem de
germinação no controle (0 MPa), sendo este valor significativamente maior do que todos
os outros tratamentos (Fig. 37). Em -0,4 MPa a porcentagem de germinação foi inferior a
50%, sendo nula nos potenciais hídricos de -0,8 e -1,0 MPa. A velocidade média de
germinação foi maior no controle, apresentando diferença significativa apenas em relação
ao tratamento de -0,6 MPa (Fig. 38). Com a transferência das sementes dos tratamentos
com potenciais hídricos negativos para a água destilada, observou-se um acréscimo não
significativo da porcentagem de germinação em todos os tratamentos (Fig. 37).
0 0,2 0,4 0,6 0,8 10
20
40
60
80
100
CCC
BC
B
d
c
b
Aa
Ger
min
ação
(%
)
Potencial Hídrico (-MPa)
Clusia fluminensis
Figura 37. Porcentagem de germinação de sementes de C. fluminensis em diferentes potenciais hídricos. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos no PEG (letras minúsculas) e entre tratamentos após transferência para água destilada (letras maiúsculas). Média ± erro padrão (Anova de um fator, p<0,05, teste de Tukey; Kruskal-Wallis, p<0,05, teste de Dunn). ■ PEG □ Água destilada (0 MPa).

52
0,0 0,2 0,4 0,6 0,8 1,00,00
0,02
0,04
0,06
0,08
0,10
0,12
0,14
0,16
a
a
b
a
IVG
(di
a-1)
Potencial Hídrico (-MPa)
(Clusia fluminensis)
Figura 38. Velocidade média de germinação de sementes de C. fluminensis em diferentes potenciais hídricos. Médias seguidas da mesma letra não diferem entre si. Média ± erro padrão (Anova de um fator, p<0,05, teste de tukey). Chamaecrista ramosa
Foi observado um decréscimo significativo da porcentagem de germinação com a
redução do potencial hídrico apenas em -0,8 MPa e -1,0 MPa. A maior porcentagem de
germinação foi verificada no controle (82%), e a menor porcentagem de germinação (4%)
foi observada no menor potencial hídrico (-1,0 MPa), ficando abaixo de 50% no potencial
hídrico de -0,8 MPa (Fig. 39). A velocidade média de germinação foi maior no controle,
ocorrendo uma queda significativa na velocidade no tratamento com potencial hídrico de -
0,6, -08 e -1,0 MPa (Fig. 40). Após a transferência das sementes não germinadas nos
tratamentos com soluções de PEG para a água destilada, foi observado um acréscimo da
porcentagem de germinação. Porém no tratamento de menor potencial hídrico (-1,0 MPa)
este acréscimo foi baixo, sendo estatisticamente inferior a porcentagem de germinação no
controle (Fig. 39).

53
0 0,2 0,4 0,6 0,8 10
20
40
60
80
100
B
AB
AAA
c
b
ab
aaAa
Chamaecrista ramosa
Ger
min
ação
(%
)
Potencial Hídrico (- MPa)
Figura 39. Porcentagem de germinação de sementes de C. ramosa em diferentes potenciais hídricos. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos no PEG (letras minúsculas) e entre tratamentos após transferência para água destilada (letras maiúsculas). Média ± erro padrão (Anova de um fator, p<0,05, teste de Tukey). ■ PEG □ Água destilada (0 MPa).
0 0,2 0,4 0,6 0,8 1,00,00
0,05
0,10
0,15
0,20
0,25
0,30
0,35
0,40
b
c
bcb
aa
IVG
(di
a-1)
Potencial Hídrico (-MPa)
Chamaecrista ramosa
Figura 40. Velocidade média de germinação de sementes de C. ramosa em diferentes potenciais hídricos. Médias seguidas da mesma letra não diferem entre si. Média ± erro padrão (Anova de um fator, p<0,05, teste de Tukey).

54
Chamaecrista flexuosa
Foi observado um decréscimo significativo da porcentagem de germinação com a
redução da disponibilidade hídrica em -0,8 e -1,0 MPa (Fig. 41). O decréscimo do potencial
hídrico foi acompanhado de uma redução significativa da velocidade média de germinação.
A maior velocidade média de germinação foi observada no controle (0 MPa) e no potencial
hídrico de -0,2 MPa, sendo estatisticamente inferior nos outros tratamentos (Fig. 42). Foi
observado um acréscimo significativo em todos os tratamentos na porcentagem de
germinação após a transferência das sementes não germinadas nas soluções de PEG para a
água destilada, exceto para o tratamento com potencial hídrico de -1,0 MPa (Fig. 41).
0 0,2 0,4 0,6 0,8 1,00
20
40
60
80
100
B
AB
AA
AB
c
b
aa
a
Aa
Chamaecrista flexuosa
Ger
min
ação
(%
)
Potencial Hídrico (- MPa)
Figura 41. Porcentagem de germinação de sementes de C. flexuosa em diferentes potenciais hídricos. Médias seguidas da mesma letra não indicam diferença significativa entre tratamentos no PEG (letras minúsculas) e entre tratamentos após transferência para água destilada (letras maiúsculas). Média ± erro padrão (Anova de um fator, p<0,05, teste de Tukey). ■ PEG □ Água destilada (0 MPa).

55
0 0,2 0,4 0,6 0,8 10,00
0,05
0,10
0,15
0,20
0,25
0,30
0,35
0,40
c
c
bc
bc
ab
a Chamaecrista flexuosa
IVG
(di
a-1)
Potencial Hídrico (- MPa)
Figura 42. Velocidade média de germinação de sementes de C. flexuosa em diferentes potenciais hídricos. Médias seguidas da mesma letra não diferem entre si. Média ± erro padrão (Anova de um fator, p<0,05, teste de Tukey). 6. DISCUSSÃO
6.1. UMIDADE SUPERFICIAL DO SOLO
Os valores significativamente superiores de umidade superficial do solo para o
microsítio interior das moitas em relação a borda e a areia nua observados na maioria dos
meses do ano, provavelmente é devido ao sombreamento feito pela copa das espécies
lenhosas arbustivas presentes nas moitas. Mantuano (2008) verificou maiores valores de
radiação para a areia nua, menores no interior das moitas, e valores intermediários para a
borda na restinga de Jurubatiba (Macaé-RJ). Portanto o sombreamento feito pela copa
diminui a radiação luminosa e a temperatura no solo no interior da moita, consequentemente
reduzindo a evaporação de água no solo neste microsítio (Breshears et al.,1998; Mantovani
& Iglesias, 2008). Além disso, o interior das moitas por ser um ambiente mais abrigado,

56
recebe menor influência dos ventos comuns nesta região (Barbiére, 1975), os quais
intensificam a evaporação de água no solo.
Ao contrário, o microsítio areia nua por estar totalmente exposto, recebe uma maior
incidência de ventos e de radiação luminosa. Esta por sua vez aumenta a temperatura do
solo, consequentemente favorecendo uma alta taxa de evaporação de água no solo.
Mantuano (2008) verificou que devido à maior quantidade de matéria particulada nas
moitas, a capacidade de campo no solo, ou seja, a retenção de água máxima após a saturação
do solo é de 15% (g H2O retida em 100 g de solo) sob a moita de Clusia hilariana, enquanto
que na areia nua é de apenas 1%. Logo, como a areia nua tem uma menor quantidade de
serapilheira, a percolação de água no solo é maior, consequentemente reduzindo mais
intensamente a umidade nas camadas mais superficiais do solo para este microsítio do que
para o interior da moita, onde a quantidade de serapilheira é maior. A serapilheira também
reduz a desidratação do solo, uma vez que protege a camada superficial da ação do vento e
da radiação solar. Em uma situação intermediária de umidade encontra-se a borda das
moitas, provavelmente por também estar em uma situação intermediária para os valores de
sombreamento, ventos, radiação, temperatura, e consequentemente evaporação de água.
Os valores significativamente superiores de umidade superficial para a areia nua em
relação ao interior e/ou borda das moitas observados durantes os meses de Agosto e
Setembro deve-se provavelmente porque para estes dois meses a pluviosidade no dia de
coleta foram as mais altas (Tab. 1). Juntando-se a isso, deve ser considerado que ao mesmo
tempo em que a copa dos arbustos das moitas de vegetação sombreia o interior da moita,
diminuindo a evaporação de água, a copa intercepta a chuva, diminuindo a quantidade de
água que atinge o interior das moitas em relação à quantidade que atinge a areia nua. Logo,
como a coleta se deu em dias de chuva, provavelmente não houve tempo suficiente para que
ocorresse a evaporação de água no solo nos dois microsítios, ficando apenas o fator

57
interceptação de água pela copa como o prevalente, favorecendo maiores valores para a
areia nua.
Foi observado que a umidade superficial do interior das moitas pequenas era
significativamente inferior do que a umidade superficial do interior de moitas grandes e/ou
médias ao longo do ano. Moitas são como ilhas de vegetação em uma matriz de areia nua,
podendo-se considerar que há, portanto uma borda natural entre dois ambientes muito
distintos. Desta forma, ocorrem mudanças abióticas e bióticas devido à interação entre estes
dois ambientes, sendo este conjunto de alterações chamado efeito de borda (Murcia, 1995).
Logo, é de se esperar que quanto mais próximo da margem de uma moita, maior será a
intensidade do efeito de borda. Consequentemente pode-se supor que a umidade do solo
decresça do interior em direção a margem da moita, uma vez que o efeito de borda leva a
um aumento da radiação luminosa e da temperatura, consequentemente da evaporação,
diminuindo mais intensamente a umidade nas margens das moitas, região adjacente a areia
nua.
Na mesma linha de raciocínio, pode ser esperado que em moitas pequenas o efeito de
borda é mais intenso do que em moitas maiores, uma vez que a intensidade do efeito de
borda é inversamente proporcional ao tamanho do fragmento, neste caso o tamanho das
moitas (Ranta, 1998). Aumentando-se o tamanho da moita, ou seja, aumentando-se a
distância entre o interior e a borda, diminui-se a radiação que chega ao interior,
consequentemente diminuindo a evaporação de água no solo neste microsítio. Desta forma,
a proporção de alteração do ambiente devido ao efeito de borda é maior em moitas menores,
podendo ser considerados ambientes menos heterogêneos em relação à umidade superficial
do solo em relação a moitas médias e grandes.

58
6.2. CONTEÚDO DE ÁGUA DAS SEMENTES
O conteúdo de água das sementes das espécies que ocorrem em moitas de vegetação
foi maior do que nas sementes que ocorrem na areia nua. O conteúdo de água das sementes
de L. rigidus e M. violaceus (areia nua) não foi analisado devido à baixa disponibilidade de
sementes. De forma geral, sementes recalcitrantes apresentam um alto conteúdo de água em
relação às sementes ortodoxas, as quais possuem uma alta tolerância à dessecação (Tweddle
et al., 2003). Braz e de Mattos (2009) observaram que as sementes da espécie E. ovalifolium
são sensíveis à dessecação. Logo, possivelmente as sementes das outras duas espécies que
ocorrem em moitas de vegetação apresentariam sementes recalcitrantes, uma vez que
possuem um alto conteúdo de água, enquanto que as sementes das quatro espécies que
ocorrem na areia nua apresentariam uma maior chance de serem ortodoxas. A tolerância à
dessecação pode favorecer a colonização da areia nua, já que como neste ambiente há uma
menor disponibilidade hídrica, as sementes são expostas a uma maior taxa de dessecação do
que nas moitas de vegetação. Ou seja, a chance de perda de viabilidade de uma semente
recalcitrante é maior na areia nua. Desta forma, o nível de tolerância à dessecação pode ser
uma característica que influencia a ocorrência de determinada espécie na areia nua ou nas
moitas de vegetação, necessitando de estudos mais detalhados.
6.3. EXPERIMENTOS DE GERMINAÇÃO
6.3.1. EXPERIMENTO DE TEMPERATURA
A germinação de sementes das espécies E. ovalifolium, M. obtusifolia e C.
fluminensis que ocorrem nas moitas de vegetação não ocorreu nas temperaturas mais
elevadas através da observação numérica dos resultados. O aumento da temperatura
propiciou uma redução significativa da porcentagem de germinação, sendo nula nas

59
temperaturas mais elevadas, com a conseqüente perda da viabilidade da maioria das
sementes, porém com uma diferença no grau de tolerância a temperaturas elevadas entre as
espécies. Dentre as três espécies que ocorrem em moitas, através da observação numérica
dos resultados, E. ovalifolium apresentou menor tolerância a temperaturas elevadas no
regime constante e alternado, uma vez que a redução significativa da germinação ocorreu
em temperaturas mais baixas (25ºC e 20-30ºC) do que em M. obtusifolia e C. fluminensis
(35ºC e 20-40ºC). Além disso, através do corte longitudinal das sementes após recuperação
na temperatura ótima, foi verificado que E. ovalifolium apresentou perda total de viabilidade
das sementes nas temperaturas mais elevadas, enquanto que as outras duas espécies de
moitas não. A redução da germinação das sementes de E. ovalifolium em temperaturas
elevadas já tinha sido observada por Fialho (1990), que verificou maior mortalidade das
sementes que estavam na borda das moitas do que no interior da bromélia Neoregelia
cruenta.
Apesar de inferior a 50% de germinação, M. obtusifolia foi a única dentre as três
espécies que ocorrem em moitas que conseguiu germinar na maior temperatura alternada
(20-40ºC), apresentando maior tolerância no regime alternado. Já no regime constante, C.
fluminensis foi a mais tolerante, uma vez que apresentou a maior germinação em 35ºC.
Considerando que em condições naturais a temperatura ao longo do dia varia, é mais
provável que no campo a germinação de C. fluminensis seja mais inibida na areia nua do
que em M. obtusifolia. Após a recuperação na temperatura ótima, foi verificado que C.
fluminensis foi a única dentre as três espécies que ocorrem em moitas que não apresentou
perda de viabilidade das sementes em 35ºC e 20-40ºC. Este resultado sugere que as
sementes desta espécie têm maior capacidade de sobreviver a temperaturas elevadas,
germinando somente quando condições mais amenas se instalam. Já em M. obtusifolia, foi
observado um efeito deletério causado por altas temperaturas sobre as sementes como em E.

60
ovalifolium, uma vez que quando expostas as temperaturas de 35, 40 e 20-40ºC, a maioria
das sementes perderam a viabilidade.
Portanto, os resultados sugerem que a germinação de sementes das espécies E.
ovalifolium, M. obtusifolia e C. fluminensis deve ser maior sob as moitas de vegetação, onde
a temperatura máxima e amplitude térmica são menores, ocorrendo raramente na areia nua
em função da exposição das sementes a altas temperaturas. Porém, devido à diferença no
grau de tolerância a temperaturas elevadas, a inibição da germinação na areia nua deve ser
maior em sementes de E. ovalifolium, em seguida em C. fluminensis, e por último em
sementes de M. obtusifolia. A redução da germinação em temperaturas elevadas também foi
observada em outros estudos realizados com espécies de restinga (Fialho, 1990; Garcia &
Lucas, 1994; Cuzzuol & Lucas, 1999; Pinheiro & Borghetti, 2003; Mantovani & Iglesias,
2008).
A velocidade de germinação também foi reduzida nas temperaturas mais altas para
as três espécies que ocorrem em moitas, exceto para E. ovalifolium que não houve efeito
significativo da temperatura sobre a velocidade de germinação em praticamente todas as
temperaturas. Dentre as três espécies que ocorrem em moitas, C. fluminensis foi a espécie
que germinou mais rápido. A velocidade média de germinação na temperatura ótima (30ºC)
foi maior para esta espécie, com a germinação começando no segundo dia e terminando no
décimo nono. A velocidade de germinação pode ter várias conseqüências ecológicas. Uma
rápida velocidade de germinação pode ser uma estratégia para se estabelecer o mais rápido
possível quando as condições se tornam favoráveis ao estabelecimento da plântula, como
em condições de maior disponibilidade hídrica (Borghetti & Ferreira, 2004). Além disso, a
rápida germinação em C. fluminensis pode diminuir o tempo de exposição das sementes a
ação de predadores, fungos, patógenos, e a dessecação após a dispersão, reduzindo a
probabilidade de morte das sementes (Tweddle et al., 2003). Alguns estudos observaram

61
que os indivíduos que se estabelecem primeiro devido a uma rápida germinação, possuem
maiores chances de ocupar locais sem competição, podendo impedir a germinação e o
estabelecimento de outras espécies, através do aumento da quantidade de serapilheira, de
sombreamento, e redução da disponibilidade de água e nutrientes através de suas raízes
(Grace, 1987; Norden et al., 2009).
Ao contrário, a germinação de E. ovalifolium foi a mais lenta. O maior valor de
velocidade média de germinação desta espécie na temperatura ótima de 20ºC foi duas vezes
menor quando comparada ao maior valor de velocidade em 20ºC para M. obtusifolia, e três
vezes menor em relação a C. fluminensis na temperatura ótima de 30ºC. A germinação foi
iniciada somente no sétimo dia, sendo finalizada somente depois de cinqüenta e seis dias, ou
seja, o período de germinação foi mais amplo em E. ovalifolium. A velocidade média de
germinação em sementes de M. obtusifolia foi intermediária, começando no quinto dia e
terminado no vigésimo terceiro. A menor velocidade de germinação em sementes de E.
ovalifolium distribui a germinação no tempo, aumentando a probabilidade de que algumas
plântulas encontrem condições favoráveis ao estabelecimento. (Harper, 1977). Além disso,
esta maior distribuição da germinação no tempo pode ser benéfica, uma vez que impede
uma germinação uniforme em função de uma chuva isolada na estação de seca, ou em
ambientes onde há uma imprevisibilidade no regime de chuvas, consequentemente evitando
a morte de toda uma coorte de plântulas (Doussi & Thanos, 2002).
A resposta germinativa em relação à temperatura em M. violaceus e L. rigidus que
ocorrem na areia nua foi distinta em comparação com as três espécies de moita E.
ovalifolium, M. obtusifolia e C. fluminensis. O regime alternado promoveu uma alta
porcentagem de germinação em M. violaceus e L. rigidus, enquanto que no regime
constante a germinação foi significativamente reduzida. Ao contrário do que foi observado
nas três espécies que ocorrem em moita, o aumento da amplitude térmica não gerou uma

62
redução da germinação, mas um acréscimo da porcentagem de germinação, com a faixa de
temperatura ótima obtida nas maiores temperaturas alternadas (20-35ºC e 20-40ºC). Ou
seja, as maiores temperaturas alternadas além de não causarem a perda de viabilidade das
sementes de M. violaceus e L. rigidus, promoveram a maior porcentagem de germinação.
Logo, a germinação de sementes destas duas espécies provavelmente é restrita a areia nua
onde a amplitude térmica é maior.
A exposição das sementes de L. rigidus a 35ºC e 40ºC não teve um efeito negativo
como encontrado nas três espécies de moitas. Apesar de a germinação ter sido nula nesta
espécie nas temperaturas constantes de 35ºC e 40ºC, não houve perda de viabilidade das
sementes nestas temperaturas, já que a germinação foi alta (ca. 90%) quando as sementes
foram transferidas para um regime alternado (20-30ºC). Ou seja, a exposição prévia a
temperaturas elevadas promoveu uma alta porcentagem de germinação em L. rigidus.
A germinação em L. rigidus foi similar ao encontrado em espécies de Cyperaceae,
com uma baixa germinação em temperaturas constantes, e maior em temperaturas
alternadas, sendo que quanto maior a amplitude térmica maior a germinação,
principalmente em espécies de ambientes abertos (Schütz, 2000; Leck & Schütz, 2005). A
exigência por flutuações na temperatura é uma dentre as formas de detecção de clareiras
(‘‘gap detection mechanisms’’) e da profundidade na qual a semente está localizada no
solo. Ou seja, a germinação é restrita a clareiras onde a competição é menor, e em menores
profundidades do solo onde a emergência da plântula é mais provável (Thompson &
Grime, 1983; Honda & Katoh, 2007). Esta exigência por temperaturas alternadas pode ser
um mecanismo presente nas sementes de M. violaceus e L. rigidus para evitar a
germinação sob uma vegetação densa onde a competição é mais intensa, no caso as moitas
de vegetação, sendo maior na areia nua onde a variação na temperatura é maior.

63
A germinação de sementes de M. violaceus foi bastante peculiar, já que nenhuma
semente germinou no regime constante, ocorrendo germinação apenas no regime alternado.
Logo, a germinação em M. violaceus ocorreu em uma menor faixa de temperatura dentre
as sete espécies estudadas, podendo ser ampliada após a quebra de dormência. A resposta
germinativa de M. violaceus não vai de acordo com muitos estudos que observaram que a
germinação em cactáceas geralmente ocorre em uma ampla faixa de temperatura, em geral
entre 15ºC e 30ºC (Nolasco et al., 1996; Ortega-Baes & Rojas- Aréchiga, 2007), com faixa
de temperatura ótima entre 20ºC e 30ºC (Rojas-Aréchiga et al., 1998; Rojas-Aréchiga &
Vázquez-Yanes, 2000). Sementes de M. violaceus foram tolerantes a altas temperaturas, já
que a germinação desta espécie foi maior nas temperaturas alternadas mais elevadas (20-
35ºC e 20-40ºC). Dau & Labouriau (1974) também verificaram uma alta tolerância a
temperaturas elevadas em Pereskia aculeata, espécie que também ocorre em restingas.
Este resultado contrasta com outros estudos que verificaram que em geral a germinação e o
estabelecimento de plântulas de algumas cactáceas ocorrem sob o dossel de espécies
facilitadoras, onde há uma menor amplitude térmica (Franco & Nobel, 1989; Valiente-
Banuet & Ezcurra, 1991; Martins, 2007).
Após a transferência das sementes de M. violaceus expostas a temperaturas
constantes para a temperatura ótima de 20-35ºC, não foi observado um acréscimo
significativo da germinação em nenhum tratamento. A ausência de germinação das
sementes que foram expostas a 40ºC após a transferência para a temperatura ótima,
provavelmente se deve a perda de viabilidade das sementes devido à longa exposição em
40ºC (c.a 70 dias), uma vez que foi observada uma infestação de fungos e a liberação de
fluidos das sementes. Para as demais temperaturas constantes a baixa germinação
provavelmente se deve a uma mudança na sensibilidade das sementes à temperatura ótima.
Ou seja, as sementes que foram expostas durante cerca de 60 dias em uma determinada

64
temperatura constante e depois foram submetidas à temperatura ótima de 20-35ºC, não
responderam da mesma forma como as sementes expostas diretamente em 20-35ºC.
A velocidade de germinação em L. rigidus foi significativamente reduzida nas
temperaturas extremas (20ºC e 20-40ºC). Em M. violaceus a menor temperatura alternada
reduziu significativamente a velocidade média de germinação. Temperaturas baixas
diminuem a velocidade das reações enzimáticas na semente, consequentemente reduzindo a
velocidade de germinação (Bewley & Black, 1994). A germinação em M. violaceus e L.
rigidus na temperatura ótima demorou mais tempo para começar dentre as sete espécies
estudadas, sendo que M. violaceus foi a espécie que também levou mais tempo para o
término da germinação, apresentando a menor velocidade média de germinação na
temperatura ótima entre as sete espécies estudadas.
Apesar das sementes de M. violaceus e L. rigidus apresentarem uma testa rígida,
não é possível afirmar a presença de dormência física, já que não foram realizados testes de
embebição devido à baixa disponibilidade de sementes. Levando em consideração que a
dormência fisiológica é mais comum em espécies de cactáceas e ciperáceas, pode ser o tipo
de dormência nas duas espécies estudadas (Schütz, 2000; Leck & Schütz, 2005; Orozco-
Segovia et al., 2007). Sementes de M. violaceus e L. rigidus possuem alta probabilidade de
formar um banco de sementes devido à presença de dormência, o pequeno tamanho das
sementes, e uma baixa germinação em temperaturas constantes (Grime et al., 1981). Além
disso, é observado que algumas sementes de cactáceas e ciperáceas possuem alta
longevidade, contribuindo também para a formação de um banco de sementes (Rojas-
Aréchiga & Vázquez-Yanes, 2000; Schütz, 2000).
A maior porcentagem de germinação em C. flexuosa ocorreu em temperaturas
constantes amenas e mais elevadas (25, 30 e 35ºC). Desta forma, provavelmente a
germinação de C. flexuosa não parece ocorrer de forma preponderante na areia nua como

65
em M. violaceus e L. rigidus, ou nas moitas de vegetação como em E. ovalifolium, M.
obtusifolia e C. fluminensis. Em C. ramosa, a maior germinação foi observada em 20, 25,
30 e 20-35ºC, sendo este resultado similar ao observado nas espécies M. obtusifolia e C.
fluminensis. Logo, a resposta à temperatura não explica a ocorrência destas duas espécies
na areia nua. Porém, como a maior porcentagem de germinação foi ca. de 50% em C.
ramosa e ca. de 80% em C. flexuosa, pode ser que as sementes de ambas as espécies não
foram expostas a melhor temperatura, dificultando a interpretação dos dados. Uma vez que
a superação de dormência em leguminosas se dá em temperaturas elevadas, pode ser que a
germinação destas espécies seja maior em uma temperatura alternada mais elevada típica
da areia nua que não foi testada. Um indício seria a germinação em C. flexuosa no regime
alternado. Apesar de a porcentagem de germinação ser significativamente inferior no
regime alternado em C. flexuosa, foi observado que o aumento da amplitude térmica gerou
um acréscimo da germinação, indicando que possivelmente a germinação poderia ser
maior em uma temperatura alternada mais elevada. Portanto, estudos testando a
germinação em temperaturas alternadas mais elevadas para C. ramosa e C. flexuosa são
necessários.
Chamaecrista flexuosa foi a única dentre as sete espécies estudas que não
apresentou um efeito significativo da temperatura sobre a velocidade média de germinação.
Em C. ramosa apenas em 40ºC houve redução significativa da velocidade de germinação.
Dentre as quatro espécies que ocorrem na areia nua, o período de germinação na
temperatura ótima foi menor em C. flexuosa.
As sementes de C. ramosa e C. flexuosa apresentam a testa rígida, porém não é
possível afirmar a presença de dormência física, porque não foram realizados testes de
embebição devido à baixa disponibilidade de sementes. A dormência física é comum em
sementes de leguminosas (Argel & Paton, 1999). A exposição das sementes a temperaturas

66
constantes elevadas, altas temperaturas alternadas, e a um choque térmico logo após um
incêndio, são os principais fatores responsáveis pela quebra de dormência em sementes
com dormência física no ambiente natural (Baskin, 2003; Van Klinken & Flack, 2005).
Segundo Bechara et al. (2007) a superação de dormência tegumentar em sementes de C.
flexuosa se deu através da imersão em água a 80ºC por 10 segundos. Provavelmente C.
ramosa também possui o mesmo tipo de dormência da sua congenérica, uma vez que suas
sementes também possuem testa rígida e que este tipo de dormência é comum em espécies
de leguminosas. No presente estudo a superação de dormência foi maior em 30ºC, sendo
significativamente superior a todas as temperaturas alternadas em C. flexuosa, e exceto em
relação a 20-35ºC em C. ramosa. Provavelmente isto ocorreu porque o tempo de exposição
à maior temperatura no regime alternado é menor (8h) do que no regime constante. Veasey
& Martins (1991) também verificaram que a superação de dormência em algumas espécies
de leguminosas era maior em uma temperatura constante elevada (40ºC) do que em uma
alternada (25-40ºC).
Após a transferência das sementes para a temperatura ótima não foi verificado um
acréscimo significativo da germinação em ambas as espécies. Provavelmente as sementes
que foram expostas as temperaturas de 35ºC e 40ºC perderam a viabilidade, já que se
encontravam deterioradas e fungadas em C. flexuosa, e em C. ramosa o mesmo foi
observado para a temperatura de 40ºC. Porém, nos outros tratamentos a baixa recuperação
pode ser devido à perda de habilidade de responder a temperatura ótima, como já
mencionada para as sementes de M. violaceus.
Todas as sementes das espécies que ocorrem na areia nua possuem dormência
primária, a qual possivelmente contribuiu para que a porcentagem de germinação das quatro
espécies não superasse 90% em nenhum tratamento, e para um período de germinação mais
amplo em comparação com as três espécies de moita, exceto em relação a E. ovalifolium. A

67
dormência é mais comum em ambientes imprevisíveis e com condições severas, como em
regiões semi-áridas (Fenner & Thompson, 2005; Flores et al., 2008). É considerada uma
característica importante porque distribui a germinação no tempo, fazendo com que a
emergência de plântulas ocorra em intervalos irregulares, reduzindo a competição, e
aumentando a probabilidade de que algumas sementes germinem em condições favoráveis
ao estabelecimento (Bewley & Black, 1994). Além disso, em ambientes onde há uma alta
imprevisibilidade no regime de chuvas, a dormência reduz o risco de mortalidade de toda
uma coorte de plântulas, já que impede uma germinação uniforme em função de uma chuva
isolada seguida por condições desfavoráveis (Jurado & Flores, 2005). Ao contrário do que
foi observado para as espécies que ocorrem na areia nua, não foi observado nenhum
mecanismo de dormência primária em sementes das espécies E. ovalifolium, M. obtusifolia
e C. fluminensis que ocorrem em moitas. A ausência de dormência nas sementes das três
espécies permite que a germinação ocorra quando a semente for exposta a condições
favoráveis logo após a dispersão.
A distribuição das espécies estudadas também pode estar sendo influenciada por
outros fatores como a dispersão de sementes. A dispersão zoocórica influencia o sucesso
de recrutamento em plantas, uma vez que o dispersor deposita as sementes em locais
específicos que podem ser adequados ao processo germinativo (Passos & Oliveira, 2003).
Castiglioni et al. (1995) e Gomes (2006) verificaram que a dispersão de sementes de E.
ovalifolium e M. obtusifolia em restingas era feita por aves. Gomes (2006) verificou que
além de não matarem as sementes, as aves as depositavam principalmente nas moitas de
vegetação, onde as condições de temperatura são mais amenas. Provavelmente C.
fluminensis também deve ter como dispersores algumas aves pelo fato de apresentar um
arilo com coloração laranja, que é uma característica atrativa para aves. Outros estudos
também verificaram que a dispersão de sementes por aves era importante na agregação

68
espacial destas espécies sob o dossel de outras plantas, uma vez que a chuva de sementes
era concentrada sob o dossel de árvores que eram usadas como poleiros (Debussche &
Isenmann, 1994; Verdu & Garcia-Fayos, 1996). Da mesma forma, a dispersão pode estar
contribuindo para a ocorrência das espécies C. ramosa e C. flexuosa na areia nua. Os frutos
destas duas espécies não possuem características atrativas, possuindo dispersão do tipo
balística (obs. pess.), a qual impede o alcance de longas distâncias pelas sementes (Wilson,
1993). Ou seja, a deposição de sementes se daria na areia nua e não nas moitas de
vegetação.
6.3.2. EXPERIMENTO DE DISPONIBILIDADE HÍDRICA
Vários estudos verificaram uma relação entre a resposta germinativa das sementes à
água de várias espécies com o habitat, clima e o tipo de solo. De forma geral, as sementes
das espécies que ocorrem em ambientes mais secos conseguem germinar em potenciais
hídricos mais negativos do que as sementes das espécies que ocorrem em ambientes mais
úmidos (Khatri et al., 1991; Facelli & Ladd, 1996; Adams, 1999; Boydak et al., 2003;
Tobe et al., 2005; Daws et al., 2008). Por exemplo, Cluff e Roundy (1988) observaram que
algumas sementes de uma espécie de deserto conseguiam germinar em -2,0 MPa. Evans &
Etherington (1990) verificaram que algumas espécies de ambientes úmidos tinham a
germinação significativamente reduzida em -0,05 MPa, enquanto que algumas espécies de
ambientes secos tinham a germinação reduzida apenas em -1,5 MPa. A maior tolerância a
baixos potenciais hídricos seria uma característica vantajosa, pois permite a germinação em
locais com deficiência hídrica onde as espécies menos tolerantes não conseguem germinar,
permitindo o estabelecimento em locais com uma competição menos intensa (Evans &
Etherington, 1990).

69
Porém, alguns estudos não observaram esta relação entre a resposta germinativa à
água e a distribuição das espécies. Sy et al. (2001) observaram que algumas espécies de
ambientes mais secos não apresentaram maior capacidade de germinar em potenciais
hídricos mais negativos, em comparação com espécies de ambientes mais úmidos. Zeng et
al. (2010) verificaram que a espécie que ocorre no ambiente mais seco tinha a germinação
mais afetada pela redução do potencial hídrico. Schütz et al. (2002) não verificaram
relação entre a tolerância das sementes de quatro eucaliptos a uma baixa disponibilidade
hídrica e o tipo de solo. Esta menor capacidade de germinar em menores potenciais
hídricos seria uma estratégia para evitar que a semente germine em uma condição
desfavorável ao estabelecimento da plântula, já que a habilidade das sementes em germinar
em condições de deficiência hídrica não confere obrigatoriamente em uma maior
capacidade da plântula em sobreviver em uma baixa disponibilidade hídrica. Ou seja, a
germinação em baixos potenciais hídricos nem sempre favorece o estabelecimento (Qi &
Redmann, 1993; Tobe et al., 2005)
O comportamento germinativo de todas as cinco espécies estudadas foi uma
redução da porcentagem e velocidade de germinação com a redução do potencial hídrico,
porém em diferentes níveis. As duas espécies que ocorrem na areia nua C. ramosa e C.
flexuosa apresentaram maior capacidade de germinar em baixos potenciais hídricos do que
as espécies E. ovalifolium e C. fluminensis que ocorrem em moitas através da observação
numérica dos resultados. A redução da porcentagem de germinação foi significativa em -
0,2 MPa em E. ovalifolium e em C. fluminensis, enquanto que em C. ramosa e C. flexuosa
a redução foi significativa apenas em -0,8 MPa. Dentre as cinco espécies, C. fluminensis
foi a espécie que apresentou uma menor tolerância a redução do potencial hídrico, já que a
germinação foi ca. de 20% em -0,4 MPa e baixa em -0,6 MPa. Maytenus obtusifolia
apresentou a maior capacidade de germinar em baixos potenciais hídricos, já que a

70
germinação foi em torno de 40% em -1,0 MPa, enquanto que em todas as espécies a
germinação foi baixa neste potencial hídrico. Outros estudos com espécies que ocorrem em
restingas também verificaram a redução da porcentagem e velocidade de germinação em
função da redução do potencial hídrico (Martins, 2007; Mantovani & Iglesias, 2008; Braz
& de Mattos, no prelo).
A maior capacidade das sementes de C. ramosa e C. flexuosa que ocorrem na areia
nua em germinar em baixos potenciais hídricos pode permitir a germinação na areia nua,
onde a ocorrência de altos potenciais hídricos é rara. Da mesma forma, a baixa tolerância a
baixos potenciais hídricos das sementes de E. ovalifolium e C. fluminensis provavelmente
impede a germinação na areia nua, fazendo com que este processo ocorra de forma
preponderante nas ilhas de vegetação, reduzindo o risco de dessecação das plântulas.
Segundo Scarano et al. (2009), as ilhas de vegetação nas formações abertas, as quais são
formadas pelo processo de facilitação, oferecem nichos similares aos encontrados na Mata
Atlântica sensu stricto, favorecendo a colonização de várias espécies menos tolerantes as
condições extremas da areia nua.
A menor capacidade de germinar no potencial hídrico de -1,0 MPa em C. ramosa e
C. flexuosa do que em M. obtusifolia pode ser uma estratégia para impedir que a
germinação ocorra em uma condição de extrema aridez, evitando a morte das plântulas por
dessecação. A maior tolerância das sementes de M. obtusifolia em relação à diminuição do
potencial hídrico não era esperada, já que esta espécie ocorre nas ilhas de vegetação.
Porém, como já mencionado, a maior capacidade das sementes em germinar em condições
de menor disponibilidade hídrica nem sempre está relacionado com a sobrevivência das
plântulas em condições de seca. Qi & Redmann (1993) verificaram que sementes de
plantas C4 apresentaram maior capacidade de germinar em baixos potenciais hídricos,
porém a sobrevivência das plântulas era baixa em condições de deficiência hídrica. Logo, a

71
maior tolerância a uma baixa disponibilidade hídrica em sementes de M. obtusifolia pode
expor as plântulas à dessecação. Além disso, outros fatores podem estar atuando mais
fortemente para a maior ocorrência de M. obtusifolia nas moitas como dispersão e
predação de sementes.
Além da redução da porcentagem de germinação, foi observado que a velocidade
média de germinação das sementes das cinco espécies estudadas decresceu conforme a
redução da disponibilidade hídrica. A diminuição do potencial hídrico devido ao aumento
da concentração de PEG diminui a entrada de água na semente, reduzindo a porcentagem e
a velocidade de germinação das espécies estudadas. Em potenciais hídricos muito baixos, a
embebição é limitada, permitindo a ativação das atividades metabólicas sem que ocorra a
emissão da radícula (Castro & Hilhorst, 2004). Esta redução da porcentagem e velocidade
de germinação pode ser prejudicial, uma vez que restringe a germinação a locais mais
úmidos, e expõe as sementes a um período maior de tempo a ação de predadores, fungos e
patógenos, e ao risco de dessecação causando a morte das sementes não ortodoxas (Schütz
et al., 2002; Tweddle et al., 2003; Daws et al., 2005).
A germinação das sementes após a transferência para água destilada foi decrescente
com a redução do potencial hídrico em todas as cinco espécies. A baixa recuperação
referente às sementes que foram expostas aos potenciais hídricos mais negativos
provavelmente não é devido a um efeito tóxico do reagente PEG 6000, já que devido ao
seu alto peso molecular ele não é absorvido pelas sementes (Bradford, 1995). Porém,
segundo Mexal et al. (1975) a concentração de oxigênio em soluções de PEG 6000 é
menor do que em água pura, sendo que quanto maior for a concentração de PEG menor é a
concentração de oxigênio na solução. A baixa concentração de oxigênio induz a
anaerobiose, resultando na produção de etanol que é tóxico para as sementes (Kozlowski,
1997). Desta forma, a menor porcentagem de germinação em água destilada das sementes

72
que foram expostas a um baixo potencial hídrico pode ser devido à maior perda de
viabilidade das sementes devido à baixa concentração de oxigênio. Porém, a baixa
recuperação em C. ramosa pode ser devido a uma alta infestação de fungos em -1,0 MPa.
Outros estudos também verificaram um efeito negativo do PEG sobre a porcentagem de
germinação (Capron et al., 2000; Dias et al., 2009).
Apesar da baixa germinação após a transferência para água destilada das sementes
que foram expostas a baixos potenciais hídricos, foi verificado que algumas sementes
permaneceram viáveis. Sementes de M. obtusifolia que foram expostas ao potencial hídrico
de -1,0 MPa apresentaram a maior recuperação em água destilada, enquanto que sementes
de E. ovalifolium e C. ramosa apresentaram a menor recuperação. A ausência de
germinação e a capacidade de permanecer viável em baixos potenciais hídricos pode ser
uma característica vantajosa nas sementes, pois impede que a germinação ocorra em
condições de deficiência hídrica, diminuindo a probabilidade de dessecação das plântulas.
Desta forma, as sementes adiam a germinação para períodos ou locais com uma maior
disponibilidade hídrica, fazendo com que o processo germinativo ocorra em condições
mais favoráveis ao estabelecimento da plântula (Khurana & Singh, 2004; Zeng et al.,
2010).
Apesar de todas as cinco espécies estudadas ocorrerem em uma região de clima
semi-árido, onde há uma irregularidade na distribuição de chuvas e no total pluviométrico
entre os anos (Barbiére, 1975), elas não estão expostas as mesmas condições
microclimáticas. Foi observado no presente estudo que a umidade nas moitas de vegetação
é maior do que na areia nua na maior parte do ano. Além disso, provavelmente a flutuação
hídrica é menor nas moitas do que na areia nua devido à maior quantidade de serapilheira,
já que esta aumenta a capacidade de retenção de água no solo (Pugnaire et al., 2004;
Eckstein & Donath, 2005; Mantuano 2008). Ou seja, após um evento de chuva o

73
suprimento de água no solo nas moitas provavelmente deve ser mantido mais elevado por
mais tempo do que na areia nua, diminuindo o risco de expor as sementes ou plântulas à
dessecação. Ao contrário, na areia nua provavelmente há uma maior flutuação hídrica e a
predominância de baixos potenciais hídricos.
A presença de dormência é comum em sementes que ocorrem em ambientes
imprevisíveis e secos (Rice, 1985; Khurana & Singh, 2001; Jurado & Flores, 2005), e em
clareiras (Roberts 1981 apud Rice 1985; Vázquez-Yanes & Orozco-Segovia, 1993). Freas
& Kemp (1983) estudando três espécies do deserto verificaram que as sementes de duas
espécies que eram dispersas em uma época onde a distribuição das chuvas era mais
irregular e menos abundante eram dormentes, enquanto que as sementes da outra espécie
que dispersava em uma época onde as chuvas eram mais regulares e abundantes não eram
dormentes. A dormência distribui a geminação no tempo e no espaço, aumentando a
probabilidade de que algumas sementes germinem em condições favoráveis ao
estabelecimento da plântula. Além disso, a dormência impede a morte de toda uma coorte
de plântulas por dessecação em função de uma chuva errática (Freas & Kemp, 1983;
Baskin & Baskin, 1998; Jurado & Flores, 2005).
No presente estudo foi observada dormência primária nas sementes de C. ramosa e
C. flexuosa que ocorrem na areia nua, a qual foi superada nos experimentos de
disponibilidade hídrica, enquanto que as sementes das espécies restritas às moitas não
possuem dormência primária. Desta forma, pode-se supor que por ser um ambiente com
uma maior flutuação e deficiência hídrica, a presença de dormência nas sementes das
espécies que ocorrem na areia nua pode evitar a morte de toda uma coorte de plântulas por
dessecação após um evento isolado de chuva, podendo ser essencial ao sucesso da
colonização deste ambiente. Além disso, como a dormência distribui a germinação no
tempo, ela aumenta a probabilidade de que algumas sementes germinem em condições

74
mais favoráveis ao estabelecimento, os quais provavelmente são mais raros na areia nua do
que nas moitas de vegetação. Em contrapartida, como nas moitas de vegetação a umidade
superficial do solo é maior e a flutuação hídrica é menor do que na areia nua, a
probabilidade de expor as plântulas à dessecação é menor. Logo, a ausência de dormência
nas três espécies de moitas permite a pronta germinação assim que a disponibilidade
hídrica se eleve, aumentando o tempo de exposição das sementes a maiores potenciais
hídricos.

75
7. CONCLUSÕES
• A umidade superficial do solo é crescente desde a areia nua até o interior das
moitas, e é maior em moitas médias e grandes para o microsítio interior.
• As espécies que ocorrem na areia nua são capazes de germinar sob temperaturas
elevadas e alternadas, enquanto que a germinação das espécies que ocorrem em
moitas é favorecida sob temperaturas constantes e amenas.
• As espécies que ocorrem na areia nua apresentam maior capacidade de
germinar sob baixos potenciais hídricos (condições típicas deste ambiente), do
que as espécies que ocorrem em moitas de vegetação.

76
8. REFERÊNCIAS BIBLIOGRÁFICAS
Adams, R. 1999. Germination of Callitris seeds in relation to temperature, water stress, priming, and hydration-dehydration cycles. Journal of Arid Environments 43: 437-448.
Aerts, R.; November, E.; Van der Borght, I.; Behailu, M.; Hermy, M. & Muys, B. 2006.
Effects of pioneer shrubs on the recruitment of the fleshy-fruited tree Olea europaea ssp. cuspidata in Afromontane savanna. Applied Vegetation Science 9: 117 – 126.
Araujo, D.S.D. 1997. Cabo Frio Region, south-eastern Brazil. In: Davis, S.D.; Heywood,
V.H.; Herrera-MacBryde, O.; Villa-Lobos, J. & Hamilton, A.C. (eds.). Centres of plant diversity: a guide and strategy for their conservation: The Americas, vol. 3. WWF/IUCN, Oxford. p. 373-375.
_____. 2000. Análise florística e fitogeográfica das restingas do estado do Rio de Janeiro.
Tese de Doutorado, Universidade Federal do Rio de Janeiro, Rio de Janeiro.
_____.; Pereira, M.C.A. & Pimentel, M.C.P. 2004. Flora e estrutura de comunidades no Parque Nacional da Restinga de Jurubatiba – síntese dos conhecimentos com enfoque especial para formação aberta de Clusia. In: Rocha, C.F.D.; Esteves, F. A. & Scarano, F.R. (eds.). Pesquisas de longa duração na Restinga de Jurubatiba: ecologia, história natural e conservação. Editora Rima, São Carlos. p. 59-76.
_____., SÁ, C.F.C.; Fontella-Pereira, J.; Garcia, D.S.; Ferreira, M.V.; Paixão, R.J.;
Schneider, S.M. & Fonseca-Kruel, V.S. 2009. Área de proteção ambiental de Massambaba, Rio de Janeiro: caracterização fitofisionômica e florística. Rodriguésia 60 (1): 67-96.
Araújo Júnior, L.C.; Góes, G.S. & Moreira, E.F. 2007. Dispersão de pólen por vento em
Lagenocarpus rigidus (Kunth) Nees (Cyperaceae), no parque sempre-viva, Mucugê, Bahia. Anais do VIII Congresso de Ecologia do Brasil, p. 1-2.
Argel, P.J. & Paton, C.J. 1999. Overcoming legume hardseededness. In: Loch, D.S. &
Ferguson, J.E. (eds.). Forage seed production: tropical and sub-tropical species. CAB International, Wallingford. 2(14): 247-265.
Barbiére, E.B. 1975. Ritmo climático e extração de sal em Cabo Frio. Revista Brasileira de
Geografia 37(4): 23-109. _____. 1984. Cabo Frio e Iguaba Grande, dois microclimas distintos a um curto intervalo
especial. In: Lacerda, L.D.; Araujo, D.S.D.; Cerqueira, R. & Turcq, B. (orgs). Restingas: origem, estrutura e processos. CEUFF, Niterói, p. 3 -12.
_____. & Coe Neto, R. 1999. Spatial and temporal variation of rainfall of the east
fluminense coast and Atlantic Serra do Mar, state of Rio de Janeiro, Brazil. In: Knoppers, B.; Bidone, E.D. & Abrão, J.J. (eds). Environmental Geochemistry of Coastal Lagoon Systems of Rio de Janeiro, Brazil. Série Geoquímica Ambiental, 6: 47-56.

77
Baskin, C.C. 2003. Breaking physical dormancy in seeds: focussing on the lens. New Phytologist 158(2): 229 – 232.
_____. & Baskin, J.M. 1998. Seeds: ecology, biogeography, and evolution of dormancy
and germination. Academic Press, San Diego, 666 p. Bechara, F.C.; Fernandes, G.G. & Silveira, R.L. 2007. Quebra de dormência de sementes
de Chamaecrista flexuosa (L.) Greene Leguminosae visando a restauração ecológica do Cerrado. Revista de Biologia Neotropical 4(1): 58-63.
Begon, M.; Townsend, C.R. & Harper, J.L. 2006. Ecology: from individuals to ecosystems.
4ed. Blackwell Publishing Press, Oxford, 738 p. Bewley, J.D. & Black, M. 1994. Seeds: physiology of development and germination. 2ed.
Plenum Press, New York, 445 p. Bieras, A.C. & Sajo, M.G. 2004. Ontogenia foliar de três espécies de Erythroxylum P.
Browne (Erythroxylaceae) ocorrentes no Cerrado. Revista Brasileira de Botânica 27(1): 71-77.
Borghetti, F. & Ferreira, A.G. 2004. Interpretação de resultados de germinação. In:
Ferreira, A.G. & Borghetti, F. (orgs.). Germinação: do básico ao aplicado. Editora Artmed, Porto Alegre, p. 209-222.
Boydak, M.; Dirik, H.; Tilki, F. & Çalikoglu, M. 2003. Effects of water stress on
germination in six provenances of Pinus brutia seeds from different bioclimatic zones in Turkey. Turkish Journal of Agriculture and Forestry 27: 91-97.
Bradford, K.J. 1995. Water relations in seed germination. In: Kigel, J. & Galili, G. (eds.).
Seed Development and Germination. Marcel Dekker, New York, p. 351-396. Brändel, M. & Schütz, W. 2005. Temperature effects on dormancy levels and germination
in temperate forest sedges (Carex). Plant Ecology 176: 245-261. Braz, M.I.G. & Mattos, E.A. No prelo. Seed dispersal phenology and germination
characteristics of a drought-prone vegetation in Southeastern Brazil. Biotropica. Breshears, D.D.; Nyhan, J.W.; Heil, C.E. & Wilcoxt, B.P. 1998. Effects of woody plants
on microclimate in a semiarid woodland: soil temperature and evaporation in canopy and intercanopy patches. International Journal of Plant Sciences 159(6): 1010- 1017.
Callaway, R.M. 1995. Positive interactions among plants. Botanical Review 61: 306-349. Camargo, R.A. & Miotto, S.T.S. 2004. O gênero Chamaecrista Moench (Leguminosae-
Caesalpinoideae) no Rio Grande do Sul. Iheringia 59(2): 131-148. Capron, I.; Corbineau, F.; Dacher, F.; Job, C.; Côme, D. & Job, D. 2000. Sugarbeet seed
priming: effects of priming conditions on germination, solubilization of 11-S globulin and accumulation of LEA proteins. Seed Science Research 10:243-254.

78
Castiglioni, G.D.A.; Cunha, L.S.T. & Gonzaga, L.P. 1995. Ramphocelus bresilius como dispersor das sementes de plantas da restinga de Barra de Maricá, Estado do Rio de Janeiro (Passeriformes: Emberizidae). Ararajuba 3: 94-99.
Castro, R.D. & Hilhorst, H.W.M. 2004. Embebição e reativação do metabolismo. In:
Ferreira, A.G. & Borghetti, F. (orgs). Germinação: do básico ao aplicado. Editora Artmed, Porto Alegre, p. 149-162.
Cluff, G.J. & Roundy, B.A. 1988. Germination responses of desert saltgrass to temperature
amd osmotic potential. Journal of Range Management 41(2): 150-153. Cuzzuol, G.R.F. & Lucas, N.M.C. 1999. Germinação de sementes de Matelea maritima
(Jack.) Woods (Asclepiadaceae). Revista Brasileira de Botânica 22(1): 1-7. Dau, L. 1960. Microclimas das restingas do sudeste do Brasil. I – Restinga interna de Cabo
Frio. Arquivos do Museu Nacional L: 79- 133. _____. & Labouriau, L.G. 1974. Temperatura control of seed germination in Pereskia
aculeata Mill. Anais da Academia Brasileira de Ciências 46(2): 311-322. Daws, M.I.; Crabtree, L.M.; Dalling, J.W.; Mullins, C.E. & Burslem, D.F.R.P. 2008.
Germination responses to water potential in neotropical pioneers suggest large-seeded species take more risks. Annals of Botany 102: 945-951.
_____.; Garwood, N.C. & Pritchard, H.W. 2005. Traits of recalcitrant seeds in a semi-
deciduos tropical forest in Panamá: some ecological implications. Functional Ecology 19: 874-885.
Debussche, M. & Isenmann, P. 1994. Bird-dispersed seed rain and seedling establishment
in patchy Mediterranean vegetation. Oikos 69(3): 414-426. Dias, M.A.; Aquino, L.A.; Dias, D.C.F.S. & Alvarenga, E.M. 2009. Physiological quality
of sugar beet (Beta vulgaris L.) seeds under osmotic conditioning and treatments with fungicide. Revista Brasileira de Sementes 31(2): 118-194.
Dias, A.T.C.; Zaluar, H.L.T.; Ganade, G. & Scarano, F.R. 2005. Canopy composition
influencing plant patch dynamics in a Brazilian sandy coastal plain. Journal of Tropical Ecology 21(3): 343 – 347.
Doussi, M.A. & Thanos, C.A. 2002. Ecophysiology of seed germination in Mediterranean
geophytes. 1. Muscari spp. Seed Science Research 12: 193-201. Eckstein, R.L. & Donath, T.W. 2005. Interactions between litter and water availability
affect seedling emergence in four familial pairs of floodplain species. Journal of Ecology 93: 807-816.
EMBRAPA (Empresa Brasileira de Pesquisa Agropecuária). 1997. Manual de métodos de
análise de solo. 2ed. Centro Nacional de Pesquisa de Solos, Rio de Janeiro. 212 p.

79
Evans, C.E. & Etherington, J.R. 1990. The effect of soil water potential on seed germination of some British plants. New Phytologist 115(3): 539 – 548.
Facelli, J.M. & Brock, D.J. 2000. Patch dynamics in arid lands: localized effects of Acacia
papyrocarpa on soils and vegetation of open woodlands of South Australia. Ecography 23(4): 479 – 491.
_____. & Ladd, B. 1996. Germination requirements and responses to leaf litter of four
species of eucalypt. Oecologia 107(4): 441- 445. Fenner, M. & Thompson, K. 2005. The Ecology of Seeds. Cambridge University Press,
Cambridge, 250 p. Fialho, R.F. 1990. Seed dispersal by a lizard and a treefrog – effect of dispersal site on
seed survivorship. Biotropica 22(4): 423-424. _____.; Jurado, E. & Jiménez-Bremont, J.F. 2008. Breaking seed dormancy in specially
protected Turbinicarpus lophophoroides and Turbinicarpus pseudopectinatus (Cactaceae). Plant Species Biology 23: 43-46.
Franco, A.C. & Nobel, P.S. 1989. Effect of nurse plants on the microhabitat and growth of
cacti. The Journal of Ecology 77(3): 870-886. Freas, K.E. & Kemp, P.R. 1983. Some relationship between environmental reliability and
seed dormancy in desert annual plants. Journal of Ecology 71(1): 211-217. Freitas, M.F. 1992. Cactaceae da Área de Proteção Ambiental da Massambaba, Rio de
Janeiro, Brasil. Rodriguésia 42/44: 67-91. Garcia, Q.S. & Lucas, N.M.C. 1994. Germinative behaviour of Jacquinia brasiliensis
seeds. Revista Brasileira de Botânica 17(1): 13-18. Gomes, V.S.M. 2006. Variação espacial e dieta de aves terrestres na restinga de
Jurubatiba, RJ. Tese de Doutorado em Ecologia, Universidade Federal do Rio de Janeiro, Rio de Janeiro.
Grace, J.B. 1987. The impact of preemption on the zonation of two Typha species along lakeshores. Ecological Monographs 57(4): 283-303.
Grime, J.P.; Mason, G.; Curtis, A.V.; Rodman, J. & Band, S.R. 1981. A comparative study
of germination characteristics in a local flora. The Journal of Ecology 69(3): 1017-1059.
Grubb, P.J. 1977. The maintenance of species-richness in plant communities: the
importance of the regeneration niche. Biological Reviews 52(1): 107-145. Gutiérrez, J.R.; Meserve, P.L.; Contreras, L.C.; Vásquez, H. & Jaksic, F.M. 1993. Spatial
distribution of soil nutrients and ephemeral plants underneath and outside the canopy of

80
Porlieria chilensis shrubs (Zygophyllaceae) in arid coastal Chile. Oecologia 95: 347-352.
Hackbart, V.C.S. & Cordazzo, C.V. 2003. Ecologia das sementes e estabelecimento das
plântulas de Hydrocotyle bonariensis Lam. Revista Atlântica 25(1): 61-65. Harper, J.L. 1997. Population biology of plants. Academic Press, London, 892 p. Honda, Y. & Katoh, K. 2007. Strict requirement of fluctuating temperatures as a reliable
gap signal in Picris hieracioides var. japonica seed germination. Plant Ecology 193: 147-156.
ISTA (International rules for seed testing). 1999. Seed Science and Technology. Zürich:
ISTA. 333 p. (Supplement). Jurado, E. & Flores, J. 2005. Is seed dormancy under environmental control or bound to
plant traits? Journal of Vegetation Science 16: 559-564. Khatri, R.; Sethi, V. & Kauskhik, A. 1991. Inter- population variations of kochia indica
during germination under different stresses. Annals of Botany 67: 413-415. Khurana, E. & Singh, J.S. 2001. Ecology of seed and seedling growth for conservation and
restoration of tropical dry forest: a review. Environmental Conservation 28(1): 39-52. _____. & Singh, J.S. 2004. Germination and seedling growth of five tree species from
tropical dry forest in relation to water stress: impact of seed size. Journal of Tropical Ecology 20: 385-396.
Kos, M. & Poschlod, P. 2007. Seeds use temperature cues to ensure germination under
nurse-plant shade in xeric Kalahari savannah. Annals of Botany 99: 667-675. Kozlowski, T.T. 1997. Responses of Woody plants to flooding and salinity. Tree
Physiology Monograph 1: 1-29. Labouriau, L.G. 1983. A Germinação das sementes. Organização dos Estados Americanos,
Washington, 170 p. Lacerda, L.D.; Araujo, D.S.D. & Maciel, N.C. 1993. Dry coastal ecosystems of the tropical
Brazilian coast. In: van der Maarel, E. (ed.) Dry coastal ecosystems: Africa, America, Asia and Oceania. Elsevier. Amsterdam. p. 477-493.
Lambers, H.; Chapin III, F.S. & Pons, T.L. 1998. Plant physiological ecology. Springer-
Verlag, New York, 540 p. Leck, M.A. & Schütz, W. 2005. Regeneration of Cyperaceae, with particular reference to
seed ecology and seed banks. Perspectives in Plant Ecology, Evolution and Systematics 7: 95–133.

81
Madeira, J.A. & Fernandes, G.W. 1999. Reproductive phenology of sympatric taxa of Chamaecrista (Leguminosae) in Serra do Cipó, Brazil. Journal of Tropical Ecology 15: 463-479.
Mantovani, A. & Iglesias, R.R. 2008. Factors limiting seed germination of terrestrial
bromeliads in the sandy coastal plains (restinga) of Maricá, Rio de Janeiro, Brazil. Rodriguésia 59(1): 135-150.
Mantuano, D.G. 2008. Crescimento clonal em Neoregelia cruenta (R. Graham) L.B. Smith
(Bromeliaceae) na Restinga de Jurubatiba: estrutura populacional, plasticidade morfo-anatômica e integração fisiológica. Tese de Doutorado em Botânica, Escola Nacional de Botânica Tropical, Rio de Janeiro.
Martínez, M.L. 2003 Facilitation of seedling establishment by an endemic shrub in tropical
coastal sand dunes. Plant Ecology 168: 333-345. Martins, L.S.T. 2007. Germinação de sementes de Pilosocereus arrabidae (LEM.) Byl. &
Row. (Cactaceae) de Arraial do Cabo, Rio de Janeiro. Dissertação de Mestrado em Botânica, Escola Nacional de Botânica Tropical, Rio de Janeiro.
Maun, M.A. 1994. Adaptations enhancing survival and establishment of seedlings on
coastal dune systems. Vegetatio 111(1): 59-70. Mexal, J.; Fisher, J.T.; Osteryoung, J. & Reid C.P.P. 1975. Oxygen availability in
polyethylene glycol solutions and its implications in plant-water relations. Plant Physiology 55(1): 20-24.
Michel, B.E. & Kaufmann, M.R. 1973. The osmotic potential of polyethylene glycol 6000.
Plant Physiology 51(5): 914-916. Miotto, S.T.S.; Lüdtke, R. & Oliveira, M.L.A. 2008. A família Leguminosae no Parque
Estadual de Itapuã, Viamão, Rio Grande do Sul, Brasil. Revista Brasileira de Biociências 6(3): 269-290.
Murcia, C. 1995. Edge effects in fragmented forests: implications for conservation. Trends
in Ecology and Evolution 10: 58-62. Nolasco, H.; Vega-Villasante, F.; Romero-Scmidt, H.L. & Diaz-Rondero, A. 1996. The
effects of salinity, acidity, light and temperature on the germination of seeds of cárdon (Pachycereus pringlei (S. Wats.) Britton & Rose, Cactaceae). Journal of Arid Environments 33: 87-94.
Norden, N.; Daws, M.I.; Antoine, C.; Gonzalez, M.A.; Garwood, N.C. & Chave, J. 2009.
The relationship between seed mass and mean time to germination for 1037 tree species across five tropical forests. Functional Ecology 23: 203-210.
Orozco-Segovia, A.; Marqués-Guzmán, J.; Sánchez-Coronado, M.E.; Gamboa de Buen,
A.; Baskin, J.M. & Baskin, C.C. 2007. Seed anatomy and water uptake in relation to seed dormancy in Opuntia tomentosa (Cactaceae, Opuntioideae). Annals of Botany 99: 581-592.

82
Ortega-Baes, P. & Rojas-Aréchiga, M. 2007. Seed germination of Trichocereus terscheckii
(Cactaceae): light, temperature and gibberellic acid effects. Journal of Arid Environments 69: 169-176.
Pammenter, N.W. & Berjak, P. 2000. Evolutionary and ecological aspects of recalcitrant
seed biology. Seed Science Research 10: 301-306. Passos, L. & Oliveira, P.S. 2003. Interactions between ants, fruits, and seeds ina restinga
forest in south-eastern Brazil. Journal of Tropical Ecology 19: 261-270. Pereira, M.C.A. 2005. Fitossociologia da formação aberta de Clusia do Parque Nacional
da Restinga de Jurubatiba-RJ. Tese de Doutorado em Ecologia, Universidade Federal do Rio de Janeiro, Rio de Janeiro.
Pinheiro, F. & Borghetti, F. 2003. Light and temperature requirements for germination of
seeds of Aechmea nudicaulis (L.) Griesebach and Streptocalyx floribundus (Martius ex Schultes F.) Mez (Bromeliaceae). Acta Botanica Brasilica 17(1): 27-35.
Probert, R.J. 2000. The role of temperature in the regulation of seed dormancy and
germination. In: Fenner, M. (ed.). Seeds: Ecology of regeneration of plant communities. CABI Publishing, Wallingford, p. 261 – 292.
Pugnaire, F.I.; Armas, C. & Valladares, F. 2004. Soil as a mediator in plant-plant
interactions in a semi-arid community. Journal of Vegetation Science 15: 85 – 92. _____.; Haase, P.; Puigdefabregas, J.; Cueto, M.; Clark, S.C. & Incoll, L.D. 1996.
Facilitation and sucession under the canopy of a leguminous shrub, Retama sphaerocarpa, in a semi-arid environment in south-east Spain. Oikos 76(3): 455-464.
Qi, M.Q. & Redmann, R.E. 1993. Seed germination and seedling survival of C3 and C4
grasses under water stress. Journal of Arid Environments 24: 277-285. Ranta, P. 1998. The fragmented Atlantic rain forest of Brazil: size, shape and distribution
of forest fragments. Biodiversity and Conservation 7: 385-403. Rice, K.J. 1985. Responses of Erodium to varying microsites: the role of germination
cueing. Ecology 66(5): 1651-1657. Rojas-Aréchiga, M. & Vásquez-Yanes, C. 2000. Cactus seed germination: a review.
Journal of Arid Environments 44: 85-104. _____.; Vázquez-Yanes, C. & Orozco-Segovia, A. 1998. Seed response to temperature of
Mexican cacti species from two life forms: an ecophysiological interpretation. Plant Ecology 135: 207-214.
Scarano, F.R. 2002. Structure, function and floristic relationships of plant communities in
stressful habitats marginal to the Brazilian Atlantic Rainforest. Annals of Botany 90: 517-524.

83
_____.; Barros, C.F.; Loh, R.K.T.; Mattos, E.A. & Wendt, T. 2009. Plant morpho-physiological variation under distinct environmental extremes in restinga vegetation. Rodriguésia 60(1): 221 – 235.
Schütz, W. 2000. Ecology of seed dormancy and germination in sedges (Carex).
Perspectives in Plant Ecology, Evolution and Systematics 3(1): 67-89. _____.; Milberg, P. & Lamont, B.B. 2002. Germination requirements and seedling
responses to water availability and soil type in four eucalypt species. Acta Oecologica 23(1):23-30.
Sheldon, J.C. 1974. The behaviour of seeds in soil: III. The influence of seed morphology
and the behaviour of seedlings on the establishment of plants from surface-lying seeds. The Journal of Ecology 62(1): 47-66.
Shmida, A. & Whittaker, R.H. 1981. Pattern and biological microsite effects in two shrub
communities, southern California. Ecology 62(1): 234-251. Suguio, K. & Tessler, M.G. 1984. Planícies de cordões litorâneos quaternários do Brasil:
origem e nomenclatura. In: Lacerda, L.D.; Araujo, D.S.D.; Cerqueira, R. & Turcq, B. (orgs). Restingas: origem, estrutura e processos. CEUFF, Niterói. p. 15-26.
Sy, A.; Grouzis, M. & Danthu, P. 2001. Seed germination of seven Sahelian legume
species. Journal of Arid Environments 49: 875-882. Thompson, K. & Grime, J. 1983. A comparative study of germination responses to
diurnally-fluctuating temperatures. Journal of Applied Ecology 20:141-156. Tielbörger, K. & Kadmon, R. 1997. Relationships between shrubs and annual communities
in a sandy desert ecosystem: a three-year study. Plant Ecology 130: 191 – 201. Tobe, K.; Zhang, L. & Omasa, K. 2005. Seed germination and seedling emergence of three
annuals growing on desert sand dunes in China. Annals of Botany 95: 649-659. Tweddle, J.C.; Dickie, J.B.; Baskin, C.C. & Baskin, J.M. 2003. Ecological aspects of seed
desiccation sensitivity. Journal of Ecology 91(2): 294-304.
Valiente-Banuet, A. & Ezcurra, E. 1991. Shade as a cause of the association between the
cactus Neobuxbaumia tetezo and the nurse plant Mimosa luisana in the Tehuacan Valley, Mexico. The Journal of Ecology 79(4): 961-971.
Van Klinken, R.D. & Flack, L. 2005. Wet heat as a mechanism for dormancy release and
germination of seeds with physical dormancy. Weed Science 53(5): 663-669. Vázquez, C. & Orozco-Segovia, A. 1993. Patterns of seed longevity and germination in the
tropical rainforest. Annual Review of Ecology and Systematics 24: 69-87.

84
Veasey, E.A. & Martins, P.S. 1991. Variability in seed dormancy and germination potential in Desmodium Desv. (Leguminosae). Revista Brasileira de Genética 14(2): 527- 545.
Verdu, M. & Garcia-Fayos, P. 1996. Nucleation processes in a Mediterranean bird-
dispersed plant. Functional Ecology 10(2): 275-280. Went, F.W. 1942. The dependence of certain annual plants on shrubs in southern
California deserts. Bulletin of the Torrey Botanical Club 69(2): 100 – 114. Whitmore, T.C. 1996. A review of some aspects of tropical rain forest seedling ecology
with suggestions for further inquiry. In: Swaine, M.D. (ed). The ecology of tropical forest tree seedlings. UNESCO and Parthenon Publishing group, Paris, p. 3-39.
Wilson, M.F. 1993. Dispersal mode, seed shadows, and colonization patterns. Vegetatio
107/108: 260-280. Yang, L.; Liu, N.; Ren, H. & Wang, J. 2009. Facilitation by two exotic Acacia: Acacia
auriculiformis and Acacia mangium as nurse plants in south China. Forest Ecology and Management 257: 1786–1793.
Zaidan, L.B.P. & Barbedo, C.J. 2004. Quebra de dormência em sementes. In: Ferreira,
A.G. & Borghetti, F. (orgs.). Germinação: do básico ao aplicado. Editora Artmed, Porto Alegre, p. 135-146.
Zaluar, H.L.T. 2002. Dinâmica da vegetação em restingas abertas fluminenses: uma
aproximação através da interação entre plantas. Tese de Doutorado em Ecologia, Universidade Federal do Rio de Janeiro, Rio de Janeiro.
_____. & Scarano, F.R. 2000. Facilitação em restingas de moitas: um século de buscas por espécies focais. In: Esteves, F.A. & Lacerda, L.D. (Eds.). Ecologia de restingas e lagoas costeiras. Rio de Janeiro: NUPEM-UFRJ, p. 3-23.
Zamith, L.R. & Scarano, F.R. 2004. Produção de mudas de espécies das Restingas do município do Rio de Janeiro, RJ, Brasil. Acta Botânica Brasilica 18(1): 161-176.
Zar, J.H. 1999. Biostatistical analysis. 4ed. Prentice Hall, New Jersey, 663 p.
Zeng, Y.J.; Wang, Y.R. & Zhang, J.M. 2010. Is reduced seed germination due to water limitation a special survival strategy used by xerophytes in arid dunes? Journal of Arid Environments 74: 508-511.

85





























