Embed Size (px)
Citation preview

Universidade Estadual Paulista “Júlio de Mesquita Filho”
Faculdade de Ciências Farmacêuticas
Programa de Pós-Graduação em Alimentos e Nutrição
Área Ciências Nutricionais
JOÃO PAULO BURIAN
Efeito imunomodulatório e controle da infecção fúngica do extrato
de alho (Allium sativum L.) em modelo murino de esporotricose
Araraquara
2016

JOÃO PAULO BURIAN
Efeito imunomodulatório e controle da infecção fúngica do
extrato de alho (Allium sativum L.) em modelo murino de
esporotricose
Dissertação apresentada ao Programa
de Pós-graduação em Alimentos e
Nutrição da Faculdade de Ciências
Farmacêuticas da Universidade Estadual
Paulista “Júlio de Mesquita Filho”,
UNESP, para obtenção do título de estre
em Ciências Nutricionais.
Orientador: Prof. Dr. Luís Vitor Silva do Sacramento
Araraquara
2016


Ficha Catalográfica
Elaborada Por Diretoria Técnica de Biblioteca e Documentação
Faculdade de Ciências Farmacêuticas
UNESP – Campus de Araraquara
Burian, João Paulo
B958e Efeito imunomodulatório e controle da infecção fúngica do extrato de alho (Allium sativum L.)
em modelo murino de esporotricose / João Paulo Burian. – Araraquara, 2016.
58 f.
Dissertação (Mestrado) – Universidade Estadual Paulista. “Júlio de Mesquita Filho”.
Faculdade de Ciências Farmacêuticas. Programa de Pós Graduação em Alimentos e
Nutrição, Área de concentração Ciências Nutricionais.
Orientador: Luís Vitor Silva do Sacramento.
1. Alho. 2. Macrófago. 3. Sporothrix schenckii. 4. Esporotricose. 5. Interleucina. I. Sacramento, Luís Vitor Silva do, orient. II. Título.
CAPES: 50700006

AGRADECIMENTOS
À Prof. Dr.ª Iracilda Zeppone Carlos, que abriu as portas de seu laboratório sem o
qual não seria possível a realização desse trabalho e a toda equipe do laboratório de Imunologia
Clínica, em especial para Marisa e Lucas, pessoas que estiveram sempre me ajudando e
ensinando no decorrer do experimento.
Ao Prof. Dr. Luis Vitor Silva do Sacramento, por toda sua ajuda, dedicação e
paciência em todos os momentos, sendo além de orientador um grande amigo. Meus sinceros
agradecimentos.

SUMÁRIO
1. INTRODUÇÂO. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1
1.1. Justificativa. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2
2. REVISÂO BIBLIOGRÀFICA. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2
2.1. Aspectos gerais do alho. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2
2.1.1. Composição química dos bulbilhos. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3
2.1.2. Compostos bioativos do alho. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4
2.1.3. Ingesta de alho. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5
2.1.4. Biodisponibilidade dos compostos do alho. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 7
2.1.5. Toxicidade. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8
2. 2. Imunidade. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8
2.2.1. Macrófagos. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 9
2.2.2. Óxido nítrico. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10
2.2.3. Interleucinas (IL-1β, IL-10 e IL-12) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10
2.2.4. Alho e imunidade. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12
2.3. Infecções fúngicas. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 13
2.3.1.. Sporothrix schenckii e esporotricose. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 14
3.OBJETIVOS. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15
3.1. Objetivo geral. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15
3.2. Objetivos específicos. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15
4. MATERIAL E MÉTODOS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15
4.1. Obtenção dos extratos. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15
4.2. Ensaio com animais. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15
4.3. Obtenção das células do exsudato peritoneal. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 16
4.4. Avaliação da Viabilidade Celular pelo método de MTT. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 17
4.5. Determinação de NO. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 17
4.6. Determinação de citocinas. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18
4.7. Microrganismo e condições de cultivo. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18
4.8. Infecção experimental. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19
4.9. Determinação da carga fúngica. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19
4.10. Avaliação da capacidade antifúngica por microdiluição. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19
4.11. Análise estatística. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 20
5. RESULTADOS. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 20
5.1. Determinação da viabilidade celular. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 20
5.2. Determinação da liberação de NO. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 20
5.3. Determinação da produção Interleucinas. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 22
5.3.1. Determinação da produção de IL-1β. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 22
5.3.2. Determinação na produção de IL-10. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23
5.3.3. Determinação na produção de IL-12. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 25
5.4. Determinação da carga fúngica (UFC) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 26
5.5. Avaliação da capacidade antifúngica por microdiluição. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 27
6. DISCUSSÃO. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 28
7. CONCLUSÂO. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31
8. REFERÊNCIAS BIBLIOGRÁFICAS. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32
9. ANEXOS. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 40
9.1. Anexo1 - Artigo. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 40
9.2. Anexo 2- Parecer do CEUA. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 55

RESUMO
O alho (Allium sativum L.) é cultivado em todo o mundo como hortaliça condimentar e
medicinal desde 3.000 a. C. A aliina (composto majoritário) e a alinase permanecem em
compartimentos separados nos bulbilhos de alho, e somente quando cortados ou triturados, a
aliina é convertida em alicina pela ação da alinase. A alicina é o principal componente do alho,
sendo atribuída a ela a maior parte das suas atividades biológicas, dentre elas as ações
bactericida, antifúngica e antiviral. Porém, outros compostos do alho apresentam atividade
antioxidante, hipocolesterolemiante, vasodilatadora, ação protetora contra diversos tipos de
câncer e imunomoduladora. Ante a invasão de patógenos, o sistema imunológico desencadeia
respostas que envolvem células como macrófagos e linfócitos. Essa complexa interação entre
células é coordenada pela liberação de citocinas e outros mediadores. A nutrição tem papel
relevante na modulação da resposta imune e inflamatória em diferentes tipos de doenças. As
infecções por fungos são causas importantes de morbidade e mortalidade no homem
principalmente em indivíduos imunossuprimidos. O Sporothrix schenckii, agente causal da
esporotricose (micose subcutânea mais comum na América Latina), é fungo dimórfico, de vida
saprofítica no solo ou em vegetais, infectando homens e os animais principalmente através de
lesões e arranhões na pele. O objetivo desse trabalho foi avaliar a influência do consumo de alho
na imunomodulação de camundongos Swiss saudáveis e infectados por S. schenckii, a partir do
estado funcional dos macrófagos peritoneais desses animais quanto à produção de óxido nítrico
e das citocinas (IL-1β, IL-10 e IL-12) e avaliar o potencial antifúngico do alho frente ao S.
schenckii por meio de teste de concentração inibitória mínima e unidades formadoras de
colônia. Os resultados demonstraram que o alho apresenta potencial antifúngico frente S.
schenckii. A administração oral de extratos de alho influencia a liberação de citocinas por
macrófagos ex vivo, o consumo regular apresenta efeito anti-inflamatório em animais sadios, e
seu uso agudo pode gerar uma resposta inflamatória. Camundongos que consumiram alho
responderam de forma mais efetiva ao combate da infecção.
Palavras-chave: Alho, macrófago, Sporothrix schenckii, esporotricose, interleucina.

ABSTRACT
Garlic (Allium sativum L.) is grown all over the world as seasoning and medicinal food since
3,000 a. C. The aliin (major compound) and alliinase remain in separate compartments in intact
garlic clove, and when the garlic is cut or crushed, the alliin is converted to allicin by the action
of alliinase. Allicin is the main component of garlic is attributed to it most of the biological
activities of garlic, including the bactericide, antifungal and antiviral action. However, other
garlic compounds have antioxidant activity, hypocholesterolemic and vasodilating activities,
guard action against many types of cancer and immunomodulatory. At the invasion of
pathogens, the immune system triggers responses involving cells such as macrophages and
lymphocytes. This complex interaction between cells is coordinated by the release of cytokines
and other mediators. Nutrition plays an important role in modulating the immune and
inflammatory response in different types of diseases. Fungal infections are significant reasons of
human morbidity and mortality mainly in immunocompromised people. The Sporothrix
schenckii, the causal agent of sporotrichosis (most common subcutaneous mycosis in Latin
America), is dimorphic fungus, of saprophytic life in soil or in plant that infects humans and
animals mainly through skin injuries and scratches. The aim of this study was to evaluate the
influence of garlic consumption in immunomodulation of Swiss healthy and infected mice by S.
schenckii from the functioning of peritoneal macrophages of the animals and the production of
nitric oxide and cytokines (IL 1β, IL-10 and IL-12) and evaluating the antifungal potential of
garlic against S. schenckii by minimum inhibitory concentration test and colony forming units.
The results show that garlic has potential antifungal front S. schenckii. Oral administration of
garlic extracts influences the releasing of cytokines by macrophages ex vivo, regular
consumption has anti-inflammatory effects in healthy animals, and its use can generate an acute
inflammatory response. Mices that consumed garlic responded more effectively to combat the
infection.
Keywords: Garlic, macrophage, Sporothrix schenckii, sporotrichosis and interleukin.

1
1. INTRODUÇÃO
O alho (Allium sativum L.) é cultivado em todo o mundo como hortaliça condimentar e
medicinal. Pertence a família Liliaceae, junto com mais de 700 espécies como a cebolinha e o
alho-poró. Seu nome científico é derivado da palavra celta „all‟, que significa ardente ou quente,
e da palavra latina „sativum‟ que significa cultivo ou cultivado. Existem registros do uso de alho
desde 3000 a.C. pelos sumérios e indianos (GARCIA GOMEZ et al.,2014; ABREU MATOS,
2000; HOLUB et al., 2002). O Codex Ebers, papiro médico egípcio traduzido em 1937 contava
com 22 formulações que utilizavam alho (CHARRON et al., 2014 & HARRIS et al., 2001).
Embora muito utilizado na culinária de muitos povos, sua importância nutricional
energética na dieta diária é quase insignificante. Isso é explicado pelo uso como aromatizante
em pequenas quantidades e não como alimento (GARCIA GOMEZ et al., 2014).
A nutrição tem papel relevante na modulação da resposta imune e inflamatória em
diferentes tipos de doenças, uma vez que nutrientes modulam sistemas de defesa celular e
humoral, pela alteração na formação de mediadores ou pela interferência nas vias de transdução
de sinais celulares, podendo alterar o balanço entre citocinas e atenuar a depleção de nutrientes
teciduais (HEPBURN, 2014; PARHAM, 2011).
As infecções por fungos são causas importantes de morbidade e mortalidade no homem
(JANEWAY, et al., 2000). Existem infecções ditas oportunistas, pois os agentes etiológicos
causam doença branda ou nenhuma doença em indivíduos sadios, mas podem infectar e causar
graves doenças em indivíduos imunossuprimidos. Uma simples infecção, como a candidíase,
pode se tornar grave e invasiva, colonizando esôfago, estômago e intestino em pacientes
portadores da síndrome da imunodeficiência adquirida (SIDA) (ABBAS et al., 2012 &
COELHO-CASTELO et al., 2009).
O Sporothrix schenckii, agente causal da esporotricose é fungo dimórfico, de vida
saprofítica no solo ou em vegetais, infectando ocasionalmente o homem e os animais. No
homem a maior parte das infecções é provocada por lesões com farpas, madeiras ou arame
(MARQUES et al., 1993). Conhecida popularmente como a “doença dos jardineiros”, uma vez
que acomete principalmente estes indivíduos pela alta exposição aos espinhos, estes que podem
causar traumas e arranhões, e com isso levando a inoculação do fungo via subcutânea. Pelo
exposto, horticultores, floristas, fazendeiros e pescadores são exemplos de atividades
ocupacionais e de lazer que podem facilitar a exposição ao fungo (ALEGRANCI, 2013).
Trata-se da micose subcutânea mais comum na América Latina, e vem sendo descrita
em diversos países, em animais e humanos, vale ressaltar que na década de 90 no Estado do Rio
de Janeiro a doença assumiu proporções epidêmicas (BARROS et al., 2010).

2
1.1 Justificativa
Na região metropolitana do Estado do Rio de Janeiro, nos últimos anos, vem ocorrendo
uma epidemia zoonótica de esporotricose que se encontra em franco crescimento e
negligenciada pelo poder público. Apesar de se verificar um expressivo aumento no número de
registro de casos, não existem medidas específicas que objetivem o seu controle (SILVA et al.,
2012).
A maioria dos casos de esporotricose no Brasil, transmitidos de forma clássica, ou seja,
pelo solo ou matéria orgânica foram confirmados no Estado de São Paulo, com 235 casos
humanos confirmados até o ano de 1953. Já no Estado do Rio Grande do Sul, um total de 646
casos humanos acumulados em diferentes casuísticas entre os anos 1957 e 2002 (FREITAS,
2014). Nos casos confirmados em humanos no Estado do Rio de Janeiro, a maior parte das
infecções foi originada por ferimentos causados por gatos domésticos (SILVA, et al., 2012).
É preciso considerar também, que o tratamento de infecções fúngicas com agentes
antifúngicos sintéticos pode causar diversos efeitos colaterais em pacientes, especialmente em
pacientes imunocomprometidos ou que estejam recebendo doses repetidas para tratar infecções
persistentes. Dessa forma é preferível prevenir infecções fúngicas em pacientes de alto risco ao
invés de ter que tratá-los (KYM, et al., 2012).
Considerando os agravos a saúde que indivíduos podem apresentar frente a infecções
fúngicas e ao seu tratamento com antifúngicos sintéticos convencionais, o objetivo desse
trabalho foi avaliar a influência do consumo de alho (Allium sativum L.) na imunomodulação de
camundongos Swiss saudáveis e infectados por S. schenckii. uma vez que não existem dados
sobre o alho no controle da esporotricose.
2. REVISÃO BIBLIOGRÁFICA
2.1. Aspectos gerais do alho
A planta do alho pode alcançar até 60 cm de altura, partindo de um bulbo subterrâneo
que é envolvido por uma película esbranquiçada ou rósea, composto de bulbilhos, popularmente
chamados de “dentes”. Suas flores de cor esbranquiçada ou avermelhada são dispostas em
umbela. Pode ser cultivado a partir dos bulbilhos, que devem ser plantados em solo leve, rico
em matéria orgânica e de preferência em locais de clima ameno. A colheita é feita de cinco a
oito meses após o plantio (ABREU MATOS, 2000).
Existem cultivares nacionais de alho com qualidade semelhante aos melhores alhos
importados. As cultivares variam quanto ao aspecto morfológico da planta, aos bulbos, a
suscetibilidade a pragas, a precocidade entre outras variáveis. As cultivares que apresentam

3
melhores características comerciais são: 1 - Grupo comum: Amarante, Gigante Lavínia, Gigante
roxo e Gravatá, e 2- Grupo nobre: Caçador, Quitéria, Jonas e Chonan (SILVA, 2009). O grupo
nobre é caracterizado por bulbos que apresentem até 20 bulbilhos, enquanto os bulbos do grupo
comum possuem mais de 20 bulbilhos (ANAPA, 2016).
O Brasil é um país importador de alho, contabilizando um consumo mensal de 22 mil
toneladas. A produção nacional supre 33% desse mercado, e os estados com maior produção
são Minas Gerais e Goiás. O restante do alho consumido é importado, principalmente da China
e da Argentina (ANAPA, 2015).
2.1.1. Composição química dos bulbilhos
A química dos bulbilhos é bastante complexa e provavelmente desenvolvida como um
mecanismo de autoproteção contra micro-organismos e outras ameaças presentes
principalmente no solo (AMAGASE at al., 2001).
Entre os componentes com atividade biológica, talvez os mais significativos sejam os
compostos organossulfurados, que se apresentam em quantidades de 11 a 35 mg por grama de
alho fresco. O alho contém três vezes mais compostos sulfurados que a cebola, o brócolis e a
couve- flor, além de arginina, oligossacarídeos, flavonóides e selênio (HOLUB et al., 2002;
MILNER & DONATO, 2010). Na Tabela 1 encontra-se a composição centesimal do alho.
Tabela 1 – Composição do alho cru.
Fonte: Tabela Brasileira de Composição de Alimentos (TACO), 2006 – Campinas.
Nutrientes Unidade Valor por 100g
Umidade % 67,5
Energia Kcal 113
Proteína G 7,0
Lipídeos G 0,2
Carboidrato G 23,9
Fibra Alimentar G 4,3
Cinzas G 1,3
Cálcio Mg 14
Magnésio Mg 21
Manganês Mg 0,24
Fósforo Mg 149
Ferro Mg 0,8
Sódio Mg 5
Potássio Mg 535
Cobre Mg 0,15
Zinco Mg 0,8
Retinol Mg NA
Tiamina Mg 0,18
Riboflavina Mg Tr
Piridoxina Mg 0,44
Niacina Mg Tr

4
O sabor e ardor característico do alho é derivado da hidrólise dos compostos S-alk(en)yl
cysteine sulphoxides (ACSOs) pela enzima alinase. Nos bulbilhos de alho foram encontrados
quatro (ACSOs) diferentes: S-metil-L-cisteína sulfóxido (MCSO), S-propil-L-cisteína sulfóxido
(PCSO), trans-S-(1-propenil) – L-cisteína sulfóxido (1-CEPSO) e S-(2-propenil)-L-cisteína
sulfóxido (RANDLE & LANCASTE, 2002). O principal precursor para a caracterização de
compostos voláteis de aroma no alho é o aminoácido cisteína. A dialil tiossulfinato (alicina)
também contribui para seu sabor e aroma característicos (KREST & KEUSGEN, 1999).
O alho tem um elevado teor de enxofre devido a elevadas concentrações de ACSOs e de
seus intermediários metabólicos. O enxofre é um dos seis macronutrientes requerido pelas
plantas e encontra-se nos aminoácidos cisteína e metionina, e em uma variedade de metabólitos.
O enxofre é absorvido pelas raízes como sulfato e transportado pelos vasos para as folhas, onde
a maior parte da assimilação de sulfato e a redução dos compostos orgânicos ocorrem
(RANDLE & LANCASTE, 2002).
2.1.2. Compostos bioativos do alho
A aliina (S-alilcisteína S-óxido) é o principal composto organossulfurado identificado
nos bulbilhos de alho íntegros, pois quando cortados ou triturados, a aliina é convertida pela
ação da alinase, enzima que possui atividade ótima em pH de 5 a 10, podendo ser desnaturada
irreversivelmente em pH de 1,5 a 3,0. A alicina é um composto muito instável e se decompõe
rapidamente (HARRIS et al., 2001; KREST & KEUSGEN, 1999).
Solúvel em solventes orgânicos e pouco solúvel em água, a alicina têm vida média em
água de 30 dias e em ácido cítrico de 60 dias. A transformação de aliina na molécula
biologicamente ativa, alicina, ocorre em poucos segundos após o esmagamento do dente de
alho. A enzima alinase é uma glicoproteína dependente de piridoxal-5-fosfato, constituída de
duas subunidades, e está presente em quantidades elevadas nos dentes de alho, representando
pelo menos 10% do conteúdo de proteína total (LAWSON, 1998; ANKRI & MIRELMAN,
1999; HOLUB et al., 2002; MIRON et al, 2000).
Os dentes de alho possuem baixo odor característico até serem esmagados. A aliina e
alinase permanecem em compartimentos separados no dente de alho intacto, essa organização
funciona como um mecanismo de defesa contra agentes patogênicos. Em uma possível invasão
aos bulbilhos, a membrana que envolve os compartimentos que contêm a enzima e o substrato é
destruída, isso faz com que ocorra a interação entre aliina e alinase o que produz alicina
rapidamente para desativar o invasor. Essa organização também assegura que o mecanismo de
defesa ocorra apenas no local destruído e durante um curto período de tempo, pois a geração

5
massiva de alicina poderia ser tóxica para outros tecidos da planta (ANKRI & MIRELMAN,
1999).
A transformação química da alicina por meio do calor produz outros compostos
sulfurosos, como ajoenos, vinilditiina e trissulfeto de alil metila (ANKRI & MIRELMAN,
1999). Esses compostos estão apresentados na Tabela 2.
Tabela 2 – Compostos sulfurados do alho e possíveis atividades biológicas atribuídas
Fonte: Adaptado de García-Gómez & Sánchez-Muniz (2000).
2.1.3. A ingesta de alho
A ingestão diária de alho é benéfica para a saúde como coadjuvante importante na
vasodilatação, na hipertensão arterial, na dislipidemia e como preventivo do câncer do aparelho
digestivo (JÚNIOR & LEMOS, 2011). Baseado em vários ensaios clínicos em humanos, a
ingesta de alho cru até 10 g/dia (1 a 2 dentes de alho), ou 4 a 6 g de alho em pó/dia, é
considerada segura quando ingerido em uma refeição (SILVA, 2009 & KOCH, 1996).
O uso diário de 4 a 6 g de alho fresco durante 61 dias reduziu em 15% o nível de
triglicérides no sangue e em 12% o de colesterol de adultos com níveis altos (LORENZI &
ABREU MATOS, 2002; SENDL et al. 1992). A redução do colesterol sanguíneo, promovida
por compostos de enxofre do alho solúveis em óleo pode estar relacionada à toxicidade, pois
Composto Possível atividade
Aliina Hipotensor;
Hipoglicemiante;
Ajoeno
(ajocisteína)
Prevenção de
coágulos;
Anti-inflamatório;
Vasodilatador;
Hipotensor;
Antibiótico;
Alicina e
tiosulfinatos
Antibiótica;
Antifúngica;
Antiviral;
Alil maercaptano Hipocolesterolemiante;
S-alil-cisteína e
compostos γ-
Glutâmico
Hipocolesterolemiante;
Antioxidante;
Quimioprotetor
Sulfeto dialil Hipocolesterolemiante.

6
aumentou a concentração da enzima lactato desidrogenase em contato com os hepatócitos, por
outro lado, a soluções aquosas diminuíram o colesterol sem apresentar hepatotoxicidade
(AMAGASE, et al., 2001). O alho também é usado de maneira tópica em ferimentos infectados,
aplicando-se alho triturado em água sobre o ferimento. Nos casos de corrimento vaginal e mau
odor causado por infecções, coloca-se antes de dormir, um dente de alho perfurado enrolado em
gaze, como se fosse um óvulo vaginal (ABREU MATOS, 2000).
Alho, cebola, alho-poró e cebolinha, têm sido relacionados com proteção contra o
câncer, principalmente no estômago, cólon e reto. O efeito protetor parece estar relacionado
com a presença de compostos organossulfurados, os quais também inibem a carcinogênese no
esôfago, nas glândulas mamárias e no pulmão de animais experimentais (BIANCHINI &
VAINIO, 2001).
Milner (2010) apresenta resultados significativos relacionando o consumo de alho
isoladamente ou associado à cebola na prevenção e combate ao câncer, alguns deles são: A
partir da análise da dieta de 2230 sujeitos, foi possível relacionar o consumo de alho e cebola
com a diminuição de 54% do risco de incidência do câncer de pâncreas; em 51 pacientes com
câncer colorretal tratados com extrato de alho envelhecido por 6 meses, o número de tumores e
seus tamanhos foram menores comparados ao grupo controle; estudo duplo cego com 50
indivíduos japoneses com tipos inoperáveis de câncer de fígado, pâncreas ou cólon receberam
500 mg ao dia de extrato de alho envelhecido por 6 meses. Os tamanhos dos tumores não
diferiram estatisticamente, entretanto, o número de células natural killer e sua atividade
aumentaram; um estudo controle realizado entre 1986 e 1989 na França com 345 pacientes
mostrou que a incidência de câncer de mama foi menor no grupo que consumia regularmente
alho e cebola.
O tratamento com o extrato aquoso de A. sativum em ratos diabéticos, nas doses de 200
e 400 mg kg -1
por via gástrica promoveu melhora no perfil lipídico, especialmente na redução
das taxas de colesterol e de LDL, acentuando seu efeito anti-aterosclerótico. Além disso, o alho
apresentou efeito benéfico na redução da glicemia no grupo que recebeu a maior dose de
extratos, nas condições experimentais analisadas. Houve redução da glicemia só no 29º dia, no
grupo de maior dose (KISS, et al., 2006). O extrato de alho em óleo também apresenta atividade
biológica. Os tiossulfonatos produzidos com a maceração são muito solúveis em óleo e se
transformam em vinilditiinas, sulfidos e ajoenos, compostos a que são atribuídas atividades anti-
trombóticas (SILVA, 2009).
O Extrato de alho apresentou capacidade de neutralizar diretamente um número de
espécies reativas de oxigênio em lesão tecidual em ratos, induzida por queimadura de segundo
grau provocada por água em 90° C, demonstrando um efeito protetor (SENER et al., 2003).
Várias hipóteses têm sido propostas para o mecanismo de ação dos compostos
sulfurosos, entre elas a de que os organossulfurados modulam a atividade de várias enzimas

7
pertencentes aos sistemas citocromo P450s e glutationa S-transferases que desintoxicam os
tecidos de potenciais agentes cancerígenos, protegendo contra danos ao DNA (AMAGASE, et
al., 2001). Contudo, o alho em excesso pode também causar alterações na função plaquetária e
no processo de coagulação. Isso não representa um problema significativo em condições
normais, entretanto deve ser levado em conta em condições de casos pré e pós-operatório
(MILNER & DONATO, 2010).
Análises de expressão gênicas podem levar a descobertas importantes no papel do alho
em respostas biológicas. Essas respostas podem ser estudadas a partir de análises do
transcriptoma (expressão de genes de rotina), epigenéticas (modificação de histonas e metilação
do DNA), proteômica (efeitos sobre as concentrações e atividades de proteínas) e metabolômica
(alterações de metabólitos celulares). Nesse contexto, em células animais, o extrato aquoso de
alho diminui a expressão de RNA mensageiro para a proteína Npc1l1 que possui papel relevante
na absorção de colesterol pelo intestino, por outro lado aumenta a expressão de m-RNA para o
receptor retinóico Rarb, que vem sendo considerado limitador de crescimento tumoral
(CHARRON et al., 2014).
2.1.4. Biodisponibilidade dos compostos do alho
Acredita-se que a biodisponibilidade (quantidade e velocidade de absorção) de
compostos organossulfurados tenha um papel importante em determinadas respostas biológicas
para as diferentes preparações do alho. A S-allil cisteína (SAC) é um dos compostos
organossulfurados solúveis em água e presentes no alho, sua farmacocinética é bem
estabelecida, sendo detectada no plasma sanguíneo, fígado e rim depois da ingestão oral e
rapidamente absorvida no trato gastrointestinal e sua concentração aumenta durante a extração
e/ou envelhecimento. No envelhecimento de longo tempo, pedaços de alho cru são colocados
em etanol (15-20%) por 20 meses ou mais (SILVA, 2009).
Os compostos organossulfurados do alho solúveis em óleo, incluindo alicina, sulfidos,
ajoeno e vinilditinas, não foram encontrados no sangue e urina, mesmo depois do consumo em
grande quantidade (25 g) de alho ou alicina pura (60 mg). O estudo demonstrou a
irreversibilidade da inativação da enzima aliinase em pH baixo, portanto, a enzima aliinase é
inibida pelas condições de acidez do estômago (LAWSON, 1998).
Estudos “in vitro” têm mostrado que quando a alicina é adicionada em sangue fresco é
rapidamente transformada (meia vida menor que 1 minuto) para alil mecarptano (alil-SH), mas a
tentativa de encontrar allil mecarptano no sangue, urina ou fezes depois do consumo de alho ou
de allil mercarptano isolado tem sido falha, indicando que esses compostos são rapidamente
metabolizados. Em resumo, alicina e compostos derivados da alicina são rapidamente

8
metabolizados pelo Alil Metil Sulfido, um composto que estimula a produção de acetona e pode
ser utilizado para medir a biodisponibilidade da alicina (SILVA, 2009).
2.1.5. Toxicidade
A American Dietetic Association (2004) considera o alho um alimento funcional e
recomenda a ingestão de um dente de alho fresco diariamente (3 a 10 gramas) para diminuição
do colesterol total e da fração LDL.
Não está claro se uma resposta ao alho aumenta proporcionalmente com o aumento do
consumo. Revisões fornecem evidências de que apenas 1g de alho cru ao dia pode ser protetor
contra o risco de câncer. Ingestões exageradas podem causar consequências, no entanto, nem
todos os indivíduos que consomem alho em grande quantidade apresentam alguma
complicação. Os raros relatos de caso têm destacado o uso de alho como capaz de causar
reações alérgicas (dermatite de contato alérgica, urticária generalizada, angioedema, pênfigo,
anafilaxia e fotossensibilidade) e queimaduras quando o alho fresco é aplicado sobre a pele
especialmente em curativos. Em um estudo conduzido com 132 crianças alérgicas, o alho estava
ligado com a alergia de apenas 3 casos (MILNER & DONATO, 2010).
Dois artigos japoneses tratam da toxicidade aguda e crônica relacionada ao consumo de
alho. SUMIYOSHI e colaboradores (1984) não observaram sintomas de toxicidade causados
por extrato de alho em ratos Wistar após o consumo oral de 2g kg-1
, 5 vezes por semana durante
6 meses. Também não encontraram diferenças significativas nas análises histológicas e nos
exames urinários e hematológicos comparados ao grupo controle; NAKAGAWA et al. (1984)
estimaram valores de DL50 de extrato de alho para ratos e camundongos por via oral,
intraperitoneal e subcutânea em mais de 30 ml kg-1
. Em 30 ml kg-1
, cinco dos dez ratos machos
e uma de dez em ratas morreram dentro de um dia após a administração intraperitoneal, no
entanto, não houve sinais específicos devido ao extrato de alho nos sobreviventes.
2.2. Imunidade
Ante a invasão de patógenos, o sistema imunológico desencadeia uma série de respostas
que envolvem células como granulócitos, macrófagos e linfócitos. Essa complexa interação
entre células é coordenada pela liberação de citocinas e outros mediadores (COZZOLINO &
COMINETTI, 2013).
As citocinas constituem um grande e variado grupo de proteínas solúveis com funções
diversas. Podem ser imunorreguladoras, sintetizadas quando necessário dentro das células
linforreticulares para desempenhar inúmeros papéis de interação na função do sistema imune e
no controle da hematopoiese (PARHAM, 2011; TREVOR, MASTERS, BERTRAM, 2014).

9
Algumas citocinas são mediadoras e reguladoras da imunidade natural, produzidas por células
dendríticas e macrófagos, por exemplo, e impulsionam o processo da inflamação ou contribuem
para a defesa contra infecções. Outras citocinas produzidas por células T auxiliares contribuem
para a defesa do hospedeiro mediada pela imunidade adaptativa e também regulam a resposta
imune (ABBAS et al., 2012). Na maioria dos casos, as citocinas promovem seus efeitos por
receptores células-alvo relevantes e parecem atuar de uma maneira similar ao mecanismo de
ação dos hormônios. Em outros casos, as citocinas podem ter efeitos antiproliferativos,
antimicrobianos e antitumorais (TREVOR, MASTERS, BERTRAM, 2014).
Uma citocina pode exercer múltiplos efeitos biológicos (pleiotropismo) e por outro lado,
várias citocinas podem exercer a mesma ação (redundantes). Também podem estimular ou inibir
a produção de outras citocinas, antagonizar ou produzir efeito aditivo ou sinérgico entre si
(ALEGRANCI, 2013; ABBAS et al., 2012).
A imunomodulação pode ser exercida mediante a potencialização ou supressão de
elementos do sistema imunológico. A supressão do sistema imunológico pode ter como
consequência a proliferação de microrganismos oportunistas que vivem nos organismos vivos e,
por este motivo, normalmente é tida como um evento prejudicial. No entanto, há circunstâncias
em que a atividade do sistema imune deve ser reduzida ou suprimida. Em casos de transplante,
por exemplo, a tendência natural do sistema imune é rejeitar o órgão enxertado, e potentes
fármacos imunossupressores são necessários para evitar tais rejeições e aumentar a taxa de
sobrevida dos enxertos. Agentes imunossupressores também são usados nas doenças
autoimunes para controlar a resposta imunológica descontrolada contra os próprios tecidos
(FISCHER et al., 2008).
Os macrófagos desempenham um papel fundamental na defesa do organismo, por meio
da fagocitose, geração de radicais livres, mediação de processos inflamatórios e secreção de
uma variedade de substâncias bioquimicamente diferentes, como enzimas, citocinas e
componentes do sistema complemento (FISCHER et al., 2008). Os linfócitos Natural Killer
(NK), que apresentam atividade citotóxica sobre várias células tumorais ou células infectadas
por vírus, são considerados o primeiro mecanismo de defesa do organismo para essas doenças
(FISCHER et al., 2008).
2.2.1. Macrófagos
Macrófagos atuam predominantemente em processos inflamatórios e no controle de
células tumorais. Pertencem ao sistema fagocítico mononuclear e fazem parte, junto com células
NK, da primeira linha de defesa contra patógenos. São células que além de participarem de
eventos regulatórios relacionados com inflamação e ação tumoricida, também atuam no reparo e

10
renovação dos tecidos, na ação microbicida, e na interação da resposta imune inata com a
adquirida (KLIMP et al., 2002).
São classificados como M1 e M2, sendo os M1 ativados por sinalizadores como IFN-y e
LPS, levando a produção de mediadores envolvidos na resposta proinflamatória e os M2, que
podem ser ativados pela IL-4, IL-13, complexos imunes, entre outros, sendo sua principal
característica a produção de altos níveis de IL-10 que atua diminuindo a inflamação e promove
a reparação tecidual. Ambos os tipos de macrófagos produzem enzimas que degradam a L-
arginina, sendo a NOS (óxido nítrico sintase) produzida pelos M1 e a arginase pelos M2, no
entanto os produtos finais são funcionalmente distintos (ALEGRANCI, 2013).
A destruição de invasores por macrófagos é significativamente aumentada com as
propriedades do extrato do alho, uma vez que o número de macrófagos peritoneais é aumentado
frente estímulo parasitário ou infeccioso, e a expressão de receptores nas membranas de
macrófagos também é aumentada (GHAZANFARI et al., 2006).
2.2.2. Óxido nítrico
De acordo com o meio em que está, o óxido nítrico pode ser um oxidante ou um
redutor. Sua meia vida é curta, variando de 3 a 60 segundos, e a especificidade de suas reações é
mínima. É citotóxico e vasodilatador, modulando reações inflamatórias ou anti-inflamatórias,
dependendo do tipo celular e do estímulo. É sintetizado pelas enzimas óxido nítrico sintases
(NOSs) por células do sistema cardiovascular e outras células como as endoteliais, os
macrófagos, os fibroblastos, as plaquetas e as musculares lisas, a partir da L-arginina. Tem
papel importante no controle fisiológico do sistema cardiovascular, atuando como mensageiro
inter e intracelular (CERQUEIRA & YOSHIDA, 2002; MORIHARA et al., 2002).
Especificamente no sistema imune, o NO interfere em vários processos, como na
diferenciação, proliferação e apoptose de células imunológicas, na produção de citocinas e
outros mediadores solúveis e na síntese de componentes da matriz extracelular (FERREIRA, et
al.; 2014).
2.2.3. Interleucinas (IL-1 β, IL-10 e IL12)
Diversas citocinas produzidas por leucócitos são denominadas de interleucinas. A
Interleucina-1, por exemplo, media a inflamação e é produzida principalmente por macrófagos
ativados. Seus efeitos incluem a ativação de células endoteliais, estímulo na produção de
quimiocinas, aumento da produção de neutrófilos pela medula óssea e aumento da atividade
citotóxica das células NK e linfócitos (ABBAS et al., 2012).
Assim como a IL-1, o fator de necrose tumoral alfa é uma citocina inflamatória. Os
principais alvos celulares do TNF-α são os neutrófilos, as células endoteliais, o fígado, o

11
hipotálamo e o músculo esquelético. Entre seus efeitos biológicos estão: ativação da inflamação
e coagulação, febre, catabolismo e apoptose de diversas células (ABBAS et al., 2012).
A IL-1 participa do desencadeamento de uma resposta imune específica que gera
citocinas participantes da resposta Th1 como a IL-12 e o IFN-γ. Todavia, esse processo pode ser
altamente regulado pela ação de antagonistas como a IL-10. A resposta Th1 está relacionada
com a defesa contra protozoários, bactérias intracelulares e vírus, enquanto a resposta Th2 é
mais efetiva contra os helmintos e bactérias extracelulares. Essas respostas são também
antagônicas, desde que o IFN-γ modula negativamente a resposta Th2, e a IL-4 e a IL-10
modulam negativamente a resposta Th1, o que permite uma homeostasia no sistema imune e
uma resposta imunológica balanceada (CORREA-CAMACHO et al., 2007).
Há dois tipos de IL-1: IL-1α e IL-1β, com 31 a 33 kDa cada, respectivamente. Estes
atuam sobre os mesmos receptores, IL-1RI e IL-1RII. A IL-1α é marcadamente associada a
membranas celulares e age através de contatos celulares. Já a IL-1β é sintetizada como uma
proteína precursora (Pro-IL-1β), que não é secretada na forma ativa até ser metabolizada pela
enzima caspase-1. As primeiras citocinas formadas após lesão tecidual ou infecção são IL-1β
(OLIVEIRA et al., 2011).
A interleucina-10 (IL-10) atua como imunossupressor, exercendo atividade anti-
inflamatória. Suprime a atividade de células T, NK e macrófagos ativados, e está, portanto,
envolvida no controle de reações da imunidade natural e da imunidade mediada por células. A
IL-10 em macrófagos causa inibição da produção de IL-12 que consiste na principal forma de
ativação das células NK. Também atua suprimindo a produção de diversas outras citocinas (IL-
1α, IL-1β, IL-6, IL-8, IL, IL-18 e TNF -a). Também suprime a atividade de células dendríticas e
diminui a expressão do gene para síntese de macrófagos e linfócitos. Estudos in vitro
demonstraram que a IL-10 é um forte estimulador da diferenciação de células B e da secreção
de imunoglobulinas. Pode ser produzida por inúmeras células imunológicas, entre elas
macrófagos ativados, linfócitos e células dendríticas, como também pode ser produzida por
células não imunes, apresentando como exemplo os queratinócitos (ABBAS et al., 2012).
Sugere-se que a IL-10 seja importante para o controle de reações inflamatórias na
mucosa, teoria sustentada pela existência de uma rara doença autoimune herdada, geradora de
uma mutação no receptor IL-10 que causa colite severa em seus portadores, destacando a
possível ativação descontrolada de macrófagos reagindo a microbiota intestinal (ABBAS et al.,
2012). Por outro lado, antagonistas de IL-10 podem ter efeito satisfatório durante a ativação de
células-B policlonal e hiperglobulinemia em pacientes com Aids (OLIVEIRA et al., 2011).
Os efeitos supressivos da IL -10 sobre as células Th1 podem ser clinicamente úteis em
prevenir a rejeição de transplantes e tratar doenças autoimunes mediadas por células-T, como
esclerose múltipla e diabetes mellitus tipo I. Efeito benéfico também pode ser observado em
sepse, artrite reumatoide e psoríase (LEE et al., 2006 & OLIVEIRA et al., 2011).

12
Secretada predominantemente por macrófagos e células dendríticas, a IL-12 estimula a
produção de IFN-γ por células NK e linfócitos T, aumenta a citotoxicidade mediada por células
NK e estimula a diferenciação de células T. Embora muitas outras células pareçam poder
secretar IL-12, apenas macrófagos e células dendríticas produzem a citocina biologicamente
ativa. Pacientes com mutações no receptor de IL-12, por exemplo, são altamente suscetíveis a
infecções por bactérias intracelulares, fato que denota a importância da IL-12 na diferenciação
de linfócitos, importantes células de defesa contra a infecções intracelulares (ABBAS et al.,
2012). Tem papel fundamental no desenvolvimento de um tipo de célula T auxiliar, sendo um
co-fator de crescimento potente, além de estimulante da adesão de células Th1 já diferenciadas,
uma citocina chave da imunorregulação, está intimamente envolvida no desenvolvimento de
doenças auto-imunes mediadas por células Th1 (JANEWAY, et al., 2000).
2.2.4. Alho e imunidade
TENDE at al. (2014) observaram que o extrato aquoso de alho aumentou
significativamente a contagem de leucócitos totais no fluido peritoneal de ratos Wistars tratados
com a solução, sendo o efeito dose dependente. Por outro lado, as concentrações de células
brancas de forma isolada não apresentaram diferenças estatísticas significantes.
Existem poucos estudos sobre a ação antiviral do alho, comparados aos estudos sobre
sua ação antibacteriana. Entre os estudos, relata-se atividade in vitro contra a influenza A e B,
citomegalovírus, rinovirus, pneumonia viral, rotavirus e HIV. A atividade antiviral é atribuída
somente à alicina e seus metabólitos (HARRIS et al., 2001).
O alho é uma das várias plantas com componentes bioativos que podem servir de
agentes antimicrobianos e tem sido historicamente reconhecido por esta atividade contra vários
agentes de deterioração e bactérias patogênicas (MILNER & DONATO, 2010). A alicina
possui atividade contra uma vasta gama de bactérias Gram - negativas e positivas, incluindo as
cepas de Escherichia coli multirresistentes. O principal efeito antimicrobiano de alicina é
devido à sua reação química com grupos tiol de várias enzimas. Também possui atividade
antifúngica, particularmente contra Cândida albicans e atividade antiparasitária incluindo
alguns dos principais protozoários parasitas intestinais humanos, tais como Entamoeba
histolytica e Giardia lamblia (ANKRI & MIRELMAN, 1999).
BAKRI & DOUGLAS (2005), comprovaram a ação bactericida do extrato aquoso de
alho em bactérias orais patogênicas. A concentração de extrato necessária foi de 35,7 mg mL-1
para bactérias Gram-negativas e de 142 mg mL-1
para Gram-positivas, o que indica que o
extracto de alho inibe o crescimento de agentes patogênicos orais e assim podem ter valor
terapêutico, particularmente para periodontite.

13
No entanto, nem todos os microrganismos são igualmente sensíveis a esses compostos,
ou seja, as taxas de absorção e seu metabolismo podem determinar o grau de resposta global dos
microrganismos para os compostos de enxofre. A adição de alho na manteiga, por exemplo,
aumentou as taxas de inativação de três patógenos (Salmonella spp., Escherichia coli e Listeria
monocytogeneses) quando incubadas a 21 e 37ºC (MILNER & DONATO, 2010). A atividade
antibiótica de 1 mg de alicina tem sido igualada à de 15 UI de penicilina (HOLUB et al., 2002;
SILVA, 2009).
Também é demonstrado na literatura, que suplementos de alho contendo alicina foram
eficientes em diminuir a incidência de resfriados (p>0,001) na população estudada, além disso,
a intensidade dos sintomas e a duração do resfriado foram menores no grupo que consumiu os
produtos de alho (JOSLING, 2001; NANTZ et al., 2012).
2.3. Infecções fúngicas
Fungos são organismos eucarióticos que ocorrem em forma filamentosa ou de levedura.
As leveduras são unicelulares e se reproduzem por brotamento, processo que as células filhas
apresentam-se em tamanhos diferentes, por outro lado, fungos filamentosos reproduzem-se por
divisão celular e possuem longos filamentos de células denominados hifas. Alguns fungos são
dimórficos, podendo existir nas duas formas, por exemplo, são filamentosos em temperatura
ambiente e leveduriforme na temperatura corporal de humanos (LEVINSON, 2014).
Difundidos por todos os ambientes, os fungos possuem um papel ecológico importante
da decomposição do material orgânico. Podem afetar a saúde humana de 3 formas: alergia,
infecção e toxicidade (OLSON, 2013). Diferentes fungos infectam o homem e podem viver em
tecidos extracelulares ou dentro de fagócitos, então as respostas imunológicas a esses
microrganismos combinam respostas a bactérias celulares e intracelures (ALEGRANCI, 2013;
FERREIRA, 2014; ABBAS, 2012).
Os fagócitos, principalmente macrófagos, respondem contra fungos ou micoses
utilizando a enzima NO sintase induzida e outros mediadores secretados para a destruição
desses patógenos. A participação de citocinas é imprescindível, pois aumenta a capacidade
fungicida dos fagócitos, favorecendo a destruição do fungo (COELHO-CASTELO et al., 2009).
Os antifúngicos de maneira geral são classificados como: orais, parentais ou tópicos.
Esses fármacos atuam diretamente nas diversas fases dos fungos, ou seja, desde a germinação de
esporos, tubo germinativo, hifa de penetração, crescimento miceliar e esporulação. O efeito
fungicida de determinado antifúngico está baseado exclusivamente na diferença da composição
lipídica das membranas celulares dos fungos e das células (MARTINEZ, 2006).
A imunidade antifúngica é menos conhecida quando comparada à imunidade contra
bactérias e vírus. Essa falta de conhecimento deve-se em parte à escassez de modelos animais

14
para micoses e, ao mesmo tempo, ao fato de que essas infecções normalmente afligem
indivíduos incapazes de apresentar respostas imunes efetivas (ABBAS et al., 2012).
2.3.1 Sporothrix schenckii e esporotricose
Em Baltimore, EUA no ano de 1898, Benjamin Schenck descreveu pela primeira vez a
esporotricose, isolando o microrganismo responsável pela doença em amostras de tecido
subcutâneo no Hospital John Hopkins (SCHENCK, 1898). Anos mais tarde esse microrganismo
foi identificado pelo micologista Erwin F. Smith como pertencente ao gênero Sporotrichum.
Outros relatos foram surgindo e o agente responsável pela doença foi denominado Sporothrix
schenckii (HEKTOEN &PERKINS, 1900). No Brasil, Lutz e Splendore descreveram em 1907,
os primeiros casos de esporotricose em seres humanos e ratos (BARROS et al., 2010).
A forma miceliar saprofítica do fungo encontrada no meio ambiente cresce em
temperaturas que variam de 25ºC a 30ºC. Esta forma é caracterizada por hifas hialinas e
septadas de 1 a 2 µm de diâmetro, com seus conidióforos e conídios organizados em grupos
assemelhando-se a pétalas de uma flor. A forma leveduriforme é encontrada in vivo, a 37°C,
onde se reproduz por brotamento e não forma conídios. Tipicamente a forma de levedura tem de
4 a 6 µm de diâmetro e frequentemente apresenta forma de charuto. Em laboratório, in vitro a
37°C, o fungo é convertido à forma de levedura em meios suplementados, como o caldo BHI,
cuja colônia apresenta coloração creme (ALEGRANCI, 2013).
Diversos fatores determinam a forma que a infecção se apresenta, como o tamanho do
inóculo, a profundidade da inoculação, a tolerância térmica da cepa e o estado imunológico do
hospedeiro. As lesões costumam ser restritas à pele, tecido celular subcutâneo e vasos linfáticos
adjacentes. Raramente pode disseminar-se para outros órgãos, ou ainda ser sistêmica, resultante
da inalação de esporos. A infecção pode ser classificada em cutânea (fixa ou localizada),
cutâneo-linfática, cutânea disseminada, mucosa e sistêmica (BARROS et al., 2010).
Dessa forma, estudos como o nosso são importantes, uma vez que produtos naturais
podem ser de grande valia no combate a infecções por exemplo, devido suas propriedades
químicas com efeitos biológicos.

15
3. OBJETIVOS
3.1. Objetivo geral
Avaliar a influência do consumo de alho (Allium sativum L.) na imunomodulação de
camundongos Swiss saudáveis e infectados por S. schenckii.
3.2. Objetivos específicos
Avaliar o estado funcional dos macrófagos peritoneais de animais sadios e infectados
quanto à produção de óxido nítrico e das citocinas (IL-1β, IL-10 e IL-12) produzidas em
sobrenadantes de cultura de macrófagos de animais tratados ou não com extrato de alho.
Avaliar o potencial antifúngico do alho frente ao S. schenckii por meio de teste de
concentração inibitória mínima e unidades formadoras de colônia.
4. MATERIAL E MÉTODO
4.1. Obtenção dos extratos
Extratos aquoso e oleoso foram preparados utilizando bulbilhos de alho (Allium sativum
L.) adquiridos em mercado de hortifrutigranjeiros de Araraquara. Depois de descascados, 32
gramas de bulbilhos in natura foram triturados em 200 mL de água destilada em liquidificador
durante um minuto, permanecendo em repouso por meia hora e sendo posteriormente filtrado e
dividido em alíquotas que foram armazenadas em refrigerador durante o experimento. O extrato
oleoso foi obtido utilizando-se o mesmo processo, substituindo-se a água por óleo de soja. O
óleo de soja foi escolhido por ser um óleo bastante consumido na alimentação humana, e por se
tratar de uma substância inócua, quando aplicado em doses baixas como a do estudo.
4.2. Ensaio com animais
O estudo foi conduzido no Laboratório de Imunologia Clínica do Departamento de
Análises Clínicas da FCF- UNESP, campus de Araraquara-SP, sob a supervisão da Prof.ª Dr.ª
Iracilda Zeppone Carlos. Os animais foram mantidos durante todo o período experimental em
condições de biotério, onde foram obedecidos os princípios éticos em experimentação animal
preconizados pelo Colégio Brasileiro de Experimentação Animal, COBEA. O estudo foi

16
aprovado pelo Comitê de Ética em Pesquisa da UNESP - FCF - Araraquara conforme protocolo
de aprovação nº 15/2015 que consta no Anexo1.
Foram utilizados camundongos Swiss, machos, adultos (oito semanas de idade),
provenientes do Biotério da UNESP – Botucatu e mantidos no Biotério da FCF - UNESP -
Araraquara - SP. Os animais foram alimentados com ração balanceada padrão para roedores e
água "ad libitum". Os camundongos foram mantidos em gaiolas coletivas a temperatura
ambiente de 25° C e fotoperíodo de 12 horas de claro/12horas de escuro.
O animais foram divididos em 2 grupos principais, os sadios e os infectados por S.
schenckii, e cada grupo foi dividido em 3 subgrupos, de acordo com o extrato recebido: grupo
controle que recebeu o tampão PBS (GC), grupo que recebeu extrato aquoso (GA) e grupo que
recebeu extrato oleoso (GO). Foram realizados 2 experimentos independentes com 4 animais
por grupo. Todos os grupos receberam no mesmo horário seus respectivos extratos e solução
controle. A dose dos extratos (0,1 mL) utilizada no tratamento correspondeu a 800 mg kg -1
,
equivalente a 16 mg por animal e administrada por via intragástrica (gavagem) uma vez ao dia
durante 15 dias. Considerando o peso médio do camundongo adulto (20 g) a dose administrada
corresponde aproximadamente em ingerir 4 a 5 bulbilhos médios ao dia para um indivíduo de
75 kg. Sabe-se que até 2g de alho kg -1
não causam efeitos adversos.
A Figura A mostra um esquema do cronograma do experimento:
Figura A. Cronograma do experimento.
4.3. Obtenção das células do exsudato peritoneal
Foram obtidas células do exsudato peritoneal (CEPs), após os animais serem
previamente estimulados com 3,0 mL de tioglicolato de sódio 3% através de inoculação
intraperitoneal três dias antes de serem eutanasiados em câmara de CO2. Os animais tiveram a
pele da região abdominal retirada assepticamente em câmara de fluxo laminar e o peritônio
exposto. Foram inoculados 5 mL de solução PBS gelado a pH 7,2 na cavidade abdominal. O
líquido peritoneal resultante foi coletado e transferido para um tubo cônico estéril e centrifugado
Semana 1
Chegada dos animais.
4 semanada de idade.
Divisão dos grupos.
Semana 4
Administração dos
extratos
8 semanas de idade.
Infecção dos animais no 5º
dia após administração
dos extratos.
Semana 5 Semana 6
Eutanásia
15 dias de extratos
10º dia de infecção

17
a 2500 rpm durante 5 min. O sedimento celular foi lavado três vezes com 3 mL de PBS. Após a
última lavagem as células foram ressuspendidas em 1 mL de meio de cultura RPMI-1640
completo para a contagem de células. O número de células foi determinado pela contagem em
câmara hemocitométrica tipo Neubauer e a suspensão celular ajustada à concentração final de
5x106 células para realização dos testes propostos, sendo que as placas com as CEPs foram
incubadas por uma hora em estufa de CO2 a 37ºC para a formação do “tapete” e após este
período desprezou-se o sobrenadante e foram acrescentados os extratos e soluções
correspondentes aos testes.
4.4. Avaliação da Viabilidade Celular pelo método de MTT
Para o ensaio de viabilidade celular, utilizou-se o método baseado na capacidade que
têm as células viáveis de clivar o anel tetrazólico presente no MTT (brometo de 3-4,5-
dimetiltiazol-2-il)-2,5-difenil-tetrazólio) pela ação de enzimas desidrogenases presentes na
mitocôndria ativa, formando cristais de formazana (MOSMANN, 1983). A uma placa de
microtitulação de 96 cavidades de fundo plano foram distribuídos 100 µL por cavidade das
suspensões de células do exsudato peritoneal de animais infectados e não infectados, ajustadas à
concentração de 5x106
células mL-1
em meio de cultura RPMI-1640-C em presença de 100 µL
de cada extrato testado (1 mg mL -1
), LPS a 1 mg mL -1
ou somente meio de cultura RPMI-
1640-C, em triplicata. As placas foram incubadas por 24 h, a 37º C, em estufa contendo tensão
constante de 5% de CO2. As células aderentes foram tratadas com 100 µL de uma solução de
MTT (Across Organics) a 1 mg mL -1
em RPMI-1640. A placa foi incubada por mais 3 horas
nas mesmas condições anteriores. Após esta incubação os sobrenadantes foram descartados e as
células aderentes foram tratadas com 100 µL de isopropanol para solubilizar os cristais de
formazana formados. A leitura da densidade ótica foi determinada em espectrofotômetro de
UV/visível, a 540 nm, com filtro de referência de 620 nm.
4.5. Determinação de NO
O NO foi quantificado espectrofotometricamente pela Reação de Griess (GREEN et al.,
1982). A partir da cultura de células aderentes do exsudato peritoneal ajustadas como descrito
anteriormente no item acima. Foram retirados 100 µL e colocados em placa de cultura de
células de 96 cavidades. Em seguida foram adicionados os extratos a serem testados e suas
respectivas soluções controle. A placa foi incubada por 24 horas em estufa incubadora de CO2 a
37º C. Alíquotas de 50 μL do sobrenadante de cada amostra em triplicata foram transferidas
para uma placa de cultura de células contendo 96 cavidades de fundo plano e acrescidas de igual

18
volume de reagente de Griess. Após 10 minutos de incubação em temperatura ambiente e ao
abrigo da luz, a absorbância foi determinada com filtro de 540 nm em espectrofotômetro. A
concentração de NO liberado no sobrenadante das culturas celulares foi calculada a partir de
uma curva padrão, previamente estabelecida e os valores foram expressos em μmols de
nitrito/5x106 células.
4.6. Determinação de citocinas
As citocinas IL- 1β, IL-10 e IL-12 foram quantificadas nos sobrenadantes de (CEPs)
obtidas e ajustadas como descrito acima. A quantificação foi realizada através do teste
imunoenzimático ELISA de captura para cada citocina, de acordo com as instruções do
fabricante. As absorbâncias foram lidas a 450 nm em espectrofotômetro UV/visível para
microplacas (Multiskan Ascent, Labsystems), e as concentrações de cada citocina foram
determinadas através da utilização de curva padrão previamente estabelecida com concentrações
conhecidas dos padrões de cada citocina. Os resultados foram expressos em pg mL -1
. Os limites
de detecção das citocinas analisadas podem ser consultados na Tabela 3.
Tabela 3. Limite de detecção das citocinas analisadas.
Interleucina pg mL -1
IL-1β 8 - 1000
IL-10 30 - 4000
IL-12 15 - 2000
4.7. Microrganismo e condições de cultivo
Foi utilizada a cepa 16345 de Sporothrix schenckii isolada de um caso de infecção
pulmonar humano de esporotricose (Baltimore, MD), cedida gentilmente pelo Laboratório de
Materiais de Referências da Fundação Oswaldo Cruz, Departamento de Microbiologia, Instituto
Nacional de Controle de Qualidade em Saúde, Rio de Janeiro. Atualmente, este isolado é
mantido no Laboratório de Imunologia Clínica do Departamento de Análises Clínicas da
Faculdade de Ciências Farmacêuticas de Araraquara - UNESP, na fase miceliar em meio
Sabouraud, a temperatura ambiente (± 25°C). A forma leveduriforme é obtida em meio BHI
(Brain Heart Infusion, Difco) líquido a 37°C sob agitação constante de 150 rpm por sete dias.
Em seguida, uma alíquota contendo 107 unidades fúngicas foi transferida para um Erlenmeyer
de 250 mL, contendo 100 mL de caldo BHI e cultivada por cinco dias nas mesmas condições
anteriores, que corresponde com a fase de crescimento logarítmica previamente descrita por
Ferreira et al. (2015).

19
4.8. Infecção Experimental
A infecção dos animais foi realizada com a forma leveduriforme de S. schenckii, obtida
pela transferência de uma parte da massa de crescimento fúngico em forma miceliar para meio
caldo BHI (Difco), do qual, após 7 dias de cultivo a 37ºC sob agitação constante de 150 rpm,
uma alíquota contendo 2 x 10 7
leveduras foi transferida para um novo caldo BHI e cultivada
por mais 5 dias nas mesmas condições. Este procedimento garante que a cultura usada para as
infecções esteja em fase logarítmica de crescimento, segundo Ferreira et al. (2015).
A infecção experimental pode ser facilmente induzida em camundongos pela injeção
intraperitoneal de células leveduriformes de S. schenckii, e aproximadamente em 10 dias, os
animais podem desenvolver peritonite ou orquite grave. Para os experimentos os animais foram
inoculados com 200 µL de uma suspensão contendo 10 6
leveduras de S. schenckii em PBS ou
com igual volume de PBS apenas, por via intraperitoneal.
4.9. Determinação da carga fúngica
A avaliação da carga fúngica sistêmica para acompanhamento das infecções foi feita ao
se considerar uma determinação das unidades formadoras de colônia (UFC) no baço dos
animais após sua retirada. Após exposição do peritônio dos animais e coleta das CEPs, o baço
foi assepticamente removido em câmara de fluxo laminar e passado através de uma malha de
nylon com poros de 100μm em 2mL de PBS usando o êmbolo de uma seringa. Uma alíquota do
macerado do baço em PBS foi inoculada em placas de meio sólido MycoselTM
em diluição pré-
determinada para permitir a contagem adequada das colônias. As UFC foram contadas após 3
dias e confirmadas após 5 dias de incubação a temperatura ambiente.
4.10. Avaliação da capacidade antifúngica por microdiluição
A avaliação da atividade antifúngica foi realizada através do teste de microdiluição de
acordo com a Norma M27-A2 do National Committee for Clinical Laboratory Standards. Foi
utilizada a cultura de S. schenckii preparando um inóculo em PBS estéril. Foram realizadas
diluições seriadas em meio RPMI-1640 com MOPS (3-(N-morpholino) propanesulfonic acid),
ajustando a suspensão fúngica em uma concentração de 2,5×103 UFC mL
-1. A concentração
mais elevada dos extratos foi de (160mg mL-1
), a utilizada no tratamento dos animais
(gavagem), seguidas de diluições 1:2. A placa foi mantida em estufa a 37ºC contendo tensão
constante de 5% de CO2. Após 4 dias a leitura foi realizada em espectrofotômetro com
comprimento de onda ajustado para 570nm.

20
4.11. Análise estatística
Os resultados obtidos foram expressos como média ± desvio padrão e submetidos ao
teste de Lilliefors para verificação da normalidade, ao teste de Levene para verificação da
homocedasticidade e à análise de variância (ANOVA one-way – pós teste de Tukey). O valor de
significância adotado foi de 5% (p<0,05).
5. RESULTADOS
5.1. Determinação da viabilidade celular
A determinação da viabilidade celular em células do exsudato peritoneal (CEPs) após
estímulo prévio com tioglicolato de sódio 3% em camundongos Swiss não infectados e
infectados com o fungo S. schenckii foi previamente realizada a fim de observar se a infecção
ou os extratos utilizados como estímulo poderiam ser tóxicos para as culturas celulares. Os
resultados obtidos demonstraram que a viabilidade celular nos camundongos Swiss não
infectados manteve-se acima de 90% e nos infectados acima de 85% mediante os desafios com
os extratos de alho, controle positivo e negativo. O teste de viabilidade celular também mostrou
que os extratos, assim como as condições de cultura e a infecção não foram tóxicos às culturas
celulares, uma vez que a viabilidade dos grupos manteve-se acima de 85%.
5.2. Determinação da liberação de NO
A determinação da liberação de óxido nítrico (NO) foi realizada em sobrenadantes de
culturas de células do exsudato peritoneal (CEPs) de camundongos infectados e não infectados
com o fungo S. schenckii, mediante o desafio com os extratos de alho (1mg mL-1
), e os controles
LPS (controle positivo) ou meio RPMI- 1640-C (controle negativo). O grupo GC não recebeu
extrato de alho via gavagem, no entanto, as CEPs desse grupo foram desafiadas com os dois
extratos diferentes (oleoso e aquoso).
A produção de óxido nítrico (NO) pelas CEPs revelou que animais sadios do grupo GA
têm uma produção significativamente menor (p<0,05) de NO em comparação aos grupos GO e
GC. Não obstante, animais que não foram expostos previamente aos extratos (GC), ao serem
estimulados pelos extratos, apresentam maior produção de NO, comparados ao controle
negativo, o que não ocorre com animais que receberam os extratos de alho via oral (GA). Isso
demonstra que a atividade imunomoduladora do alho pode ser usada de formas diferentes, ou
seja, a resposta muda se ingerido regularmente ou de forma aguda (Figura 1A).

21
No grupo GA infectados com o fungo S. schenckii, houve aumento significativo
(p<0,05) na produção de NO por CEPs desafiadas com extrato aquoso frente ao controle
negativo. Esse resultado mostra que o consumo de alho pode aumentar a produção de NO,
mesmo em condições que a produção já está aumentada, como nesse caso pela infecção
(Figura 1B).
A infecção aumentou a produção de NO em todos os grupos analisados, isso pode ser
observado nas Figuras 1C, 1D e 1E.
A n im a is s a d io s - Ó x id o N ítr ic o
µm
ols
m
L-1
de
nit
rito
GA
(a
qu
os
o)
GA
+G
A -
GO
(o
leo
so
)G
O+
GO
-G
C (
aq
uo
so
)
GC
(o
leo
so
)G
C +
GC
-
0
5
3 0
6 0
9 0
1 2 0
A
a
bcc
**
A n im a is in fe c ta d o s - Ó x id o N ítr ic o
µm
ols
m
L-1
de
nit
rito
GA
(a
qu
os
o)
GA
+G
A -
GO
(o
leo
so
)G
O+
GO
-G
C (
aq
uo
so
)
GC
(o
leo
so
)G
C +
GC
-
0
3 0
6 0
9 0
1 2 0
B
b
a
b c
c
*
G A - Ó x id o N ítr ic o
µm
ols
m
L-1
de
nit
rito
Extr
ato
Co
ntr
ole
+
Co
ntr
ole
-
0
1
3 0
6 0
9 0
1 2 0
C
**
**
**
G O - Ó x id o N ítr ic o
µm
ols
m
L-1
de
nit
rito
Extr
ato
Co
ntr
ole
+
Co
ntr
ole
-
0
4
3 0
6 0
9 0
1 2 0
D
** **
**
G C - Ó x id o N ítr ic o
µm
ols
m
L-1
de
nit
rito
Ext.
Aq
uo
so
Ext.
Ole
oso
Co
ntr
ole
+
Co
ntr
ole
-
0
3 0
6 0
9 0
1 2 0
S a u d á v e is
E
In fe c ta d o s
**
**
****
Figura 1. Produção de NO em cultura de células do exsudato peritoneal (CEPs) de camundongos
Swiss sadios e infectados com o fungo S. schenckii no 10º dia após infecção. GA (grupo que recebeu
extrato aquoso), GO (grupo que recebeu extrato oleoso) e GC (grupo que recebeu tampão PBS) via
gavagem. Os macrófagos dos camundongos foram cultivados em presença dos extratos aquoso e oleoso
(1 mg mL -1
) de alho, LPS (controle +) e meio RPMI-1640-C (controle -). Cada animal estudado foi
avaliado em triplicata no momento de execução do teste. As concentrações de nitrito foram obtidas
através de uma reta padrão previamente estabelecida com concentrações molares conhecidas de nitrito de
sódio (NaNO2) e os resultados expressos em µmols de nitrito mL -1
de 5x106 células, como média ±
desvio padrão de 4 animais por grupo de dois experimentos independentes. Letras iguais não apresentam
diferença significativa. Letras diferentes são estatisticamente diferentes. ANOVA - pós-teste de Tukey
(p<0,05). Comparação entre estímulos do extrato em A e B. Comparação entre sadios e infectados em C,
D e E. (* p<0,05) (**p<0,01).

22
5.3. Determinação da produção Interleucinas
Foi avaliada a produção de citocinas pelas CEPs dos animais dos diferentes grupos
(GA, GO e GC) tratados via intragástrica com diferentes extratos de alho (aquoso e oleoso)
frente aos desafios LPS, RPMI-1640-C e do próprio extrato recebido por gavagem (1 mg mL -
1). O grupo GC não recebeu extratos de alho via gavagem, no entanto, as CEPs desse grupo
foram desafiadas com os extratos (oleoso e aquoso).
5.3.1. Determinação da produção de IL-1β
A determinação da produção de IL-1β foi realizada em sobrenadantes de culturas de
células do exsudato peritoneal (CEPs) de camundongos sadios e infectados, mediante o desafio
com o extrato aquoso e oleoso de alho, e os controles: LPS (controle positivo) ou meio RPMI-
1640-C (controle negativo). Os resultados obtidos foram calculados por meio de curva analítica
previamente estabelecida com concentrações molares conhecidas de IL-1β, como média ±
desvio padrão de quatro animais.
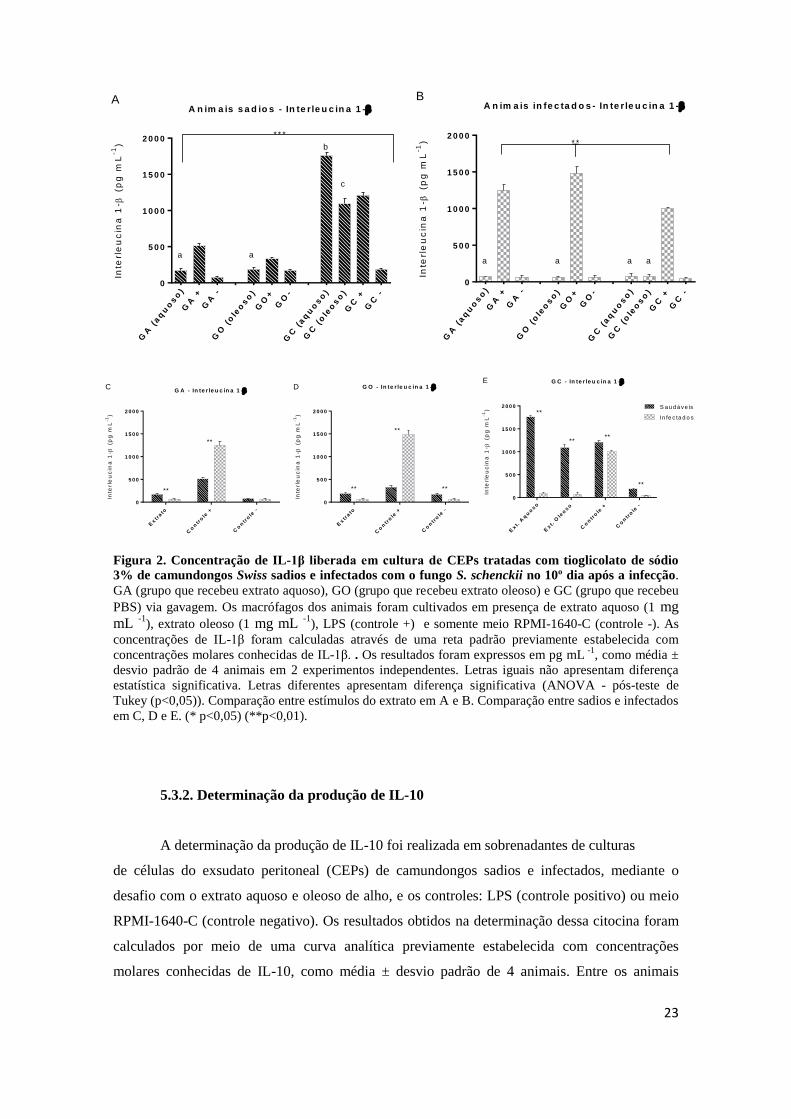
A produção de IL-1β em animais sadios de diferentes grupos foi estatisticamente
diferente entre si (p<0,01), e o grupo dos animais tratados com os extratos apresentou menor
produção da citocina (Figura 2A). Também foi diferente a produção desta citocina entre
animais sadios de um mesmo grupo, comparando o estímulo extrato com seus respectivos
controles (LPS e RPMI) (***p<0,001), o que não ocorreu com os animais infectados (Figura
2B). A produção de IL-1β em animais infectados não diferiu entre os grupos estudados frente
aos desafios com extratos, apenas houve diferença entre os controles positivos, onde todos
foram diferentes entre si, com maior produção para o grupo GO. Nitidamente os extratos de
alho mostraram atividade anti-inflamatória quando administrados de maneira crônica (15 dias),
contudo, CEPs de animais que não tiveram contato prévio com os extratos (GC), quando
desafiados com os extratos de alho, apresentaram valores superiores ao desafio LPS (controle
positivo), indicando que o uso agudo do alho pode ser um potente estimulador da produção de
IL-1β, principalmente em indivíduos sadios. A Figura 2 apresenta o gráfico da produção de
interleucina IL-1β.
23
A n im a is s a d io s - In te rle u c in a 1 -In
terl
eu
cin
a 1
- (
pg
mL
-1)
GA
(a
qu
os
o)
GA
+G
A -
GO
(o
leo
so
)G
O+
GO
-G
C (
aq
uo
so
)
GC
(o
leo
so
)G
C +
GC
-
0
5 0 0
1 0 0 0
1 5 0 0
2 0 0 0
A
a a
b
c
* * *
A n im a is in fe c ta d o s - In te r le u c in a 1 -
Inte
rle
uc
ina
1-
(p
g m
L-1
)
GA
(a
qu
os
o)
GA
+G
A -
GO
(o
leo
so
)G
O+
GO
-G
C (
aq
uo
so
)
GC
(o
leo
so
)G
C +
GC
-
0
5 0 0
1 0 0 0
1 5 0 0
2 0 0 0
B
a a aa
* *
G A - In te r le u c in a 1 -
Inte
rle
uc
ina
1-
(p
g m
L-1
)
Extr
ato
Co
ntr
ole
+
Co
ntr
ole
-
0
5 0 0
1 0 0 0
1 5 0 0
2 0 0 0
C
**
**
G O - In te r le u c in a 1 -
Inte
rle
uc
ina
1-
(p
g m
L-1
)
Extr
ato
Co
ntr
ole
+
Co
ntr
ole
-
0
5 0 0
1 0 0 0
1 5 0 0
2 0 0 0
D
**
**
**
G C - In te r le u c in a 1 -
Inte
rle
uc
ina
1-
(p
g m
L-1
)
Ext.
Aq
uo
so
Ext.
Ole
oso
Co
ntr
ole
+
Co
ntr
ole
-
0
5 0 0
1 0 0 0
1 5 0 0
2 0 0 0 S a u d á v e is
In fe c ta d o s
E
**
****
**
Figura 2. Concentração de IL-1β liberada em cultura de CEPs tratadas com tioglicolato de sódio
3% de camundongos Swiss sadios e infectados com o fungo S. schenckii no 10º dia após a infecção.
GA (grupo que recebeu extrato aquoso), GO (grupo que recebeu extrato oleoso) e GC (grupo que recebeu
PBS) via gavagem. Os macrófagos dos animais foram cultivados em presença de extrato aquoso (1 mg
mL -1
), extrato oleoso (1 mg mL -1
), LPS (controle +) e somente meio RPMI-1640-C (controle -). As
concentrações de IL-1β foram calculadas através de uma reta padrão previamente estabelecida com
concentrações molares conhecidas de IL-1β. . Os resultados foram expressos em pg mL -1
, como média ±
desvio padrão de 4 animais em 2 experimentos independentes. Letras iguais não apresentam diferença
estatística significativa. Letras diferentes apresentam diferença significativa (ANOVA - pós-teste de
Tukey (p<0,05)). Comparação entre estímulos do extrato em A e B. Comparação entre sadios e infectados
em C, D e E. (* p<0,05) (**p<0,01).
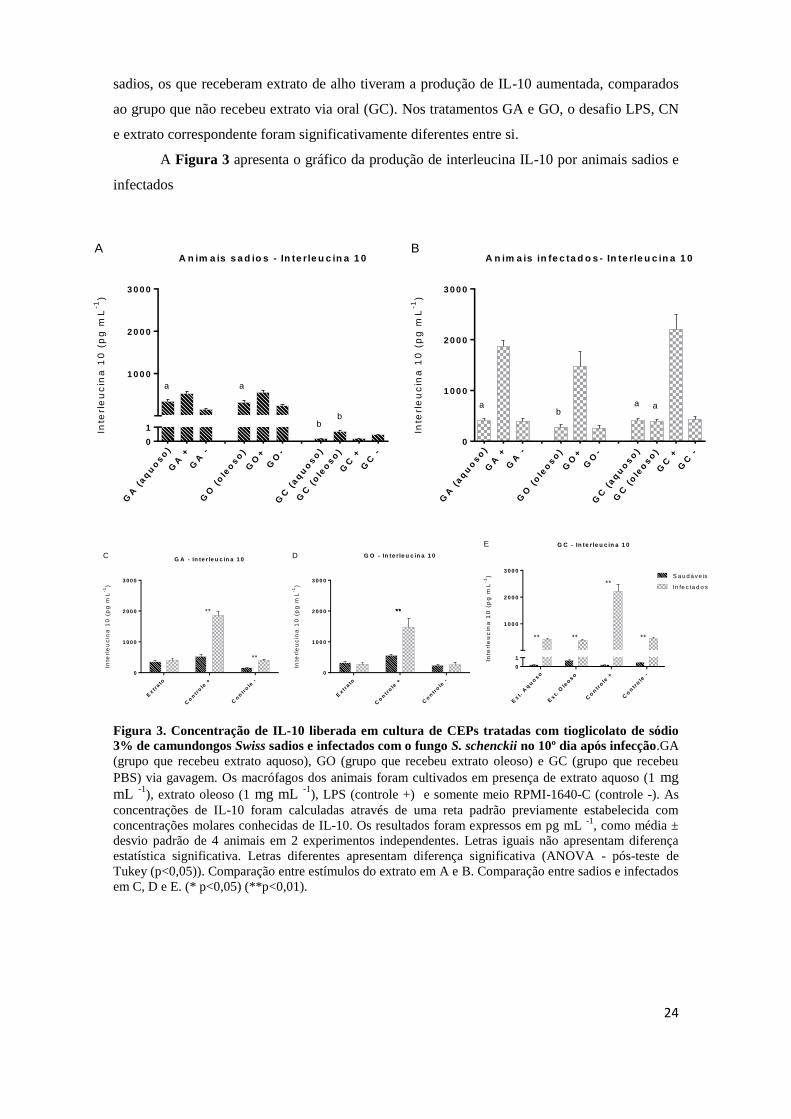
5.3.2. Determinação da produção de IL-10
A determinação da produção de IL-10 foi realizada em sobrenadantes de culturas
de células do exsudato peritoneal (CEPs) de camundongos sadios e infectados, mediante o
desafio com o extrato aquoso e oleoso de alho, e os controles: LPS (controle positivo) ou meio
RPMI-1640-C (controle negativo). Os resultados obtidos na determinação dessa citocina foram
calculados por meio de uma curva analítica previamente estabelecida com concentrações
molares conhecidas de IL-10, como média ± desvio padrão de 4 animais. Entre os animais
24
sadios, os que receberam extrato de alho tiveram a produção de IL-10 aumentada, comparados
ao grupo que não recebeu extrato via oral (GC). Nos tratamentos GA e GO, o desafio LPS, CN
e extrato correspondente foram significativamente diferentes entre si.
A Figura 3 apresenta o gráfico da produção de interleucina IL-10 por animais sadios e
infectados
A n im a is s a d io s - In te rle u c in a 1 0
Inte
rle
uc
ina
10
(p
g m
L-1
)
GA
(a
qu
os
o)
GA
+G
A -
GO
(o
leo
so
)G
O+
GO
-G
C (
aq
uo
so
)
GC
(o
leo
so
)G
C +
GC
-
0
1
1 0 0 0
2 0 0 0
3 0 0 0
A
bb
aa
A n im a is in fe c ta d o s - In te rle u c in a 1 0
Inte
rle
uc
ina
10
(p
g m
L-1
)
GA
(a
qu
os
o)
GA
+G
A -
GO
(o
leo
so
)G
O+
GO
-G
C (
aq
uo
so
)
GC
(o
leo
so
)G
C +
GC
-
0
1 0 0 0
2 0 0 0
3 0 0 0
B
baaa
G A - In te rle u c in a 1 0
Inte
rle
uc
ina
10
(p
g m
L-1
)
Extr
ato
Co
ntr
ole
+
Co
ntr
ole
-
0
1 0 0 0
2 0 0 0
3 0 0 0
C
**
**
G O - In te r le u c in a 1 0
Inte
rle
uc
ina
10
(p
g m
L-1
)
Extr
ato
Co
ntr
ole
+
Co
ntr
ole
-
0
1 0 0 0
2 0 0 0
3 0 0 0
D
******
G C - In te r le u c in a 1 0
Inte
rle
uc
ina
10
(p
g m
L-1
)
Ext.
Aq
uo
so
Ext.
Ole
oso
Co
ntr
ole
+
Co
ntr
ole
-
0
1
1 0 0 0
2 0 0 0
3 0 0 0S a u d á v e is
In fe c ta d o s
E
**
**** **
Figura 3. Concentração de IL-10 liberada em cultura de CEPs tratadas com tioglicolato de sódio
3% de camundongos Swiss sadios e infectados com o fungo S. schenckii no 10º dia após infecção.GA
(grupo que recebeu extrato aquoso), GO (grupo que recebeu extrato oleoso) e GC (grupo que recebeu
PBS) via gavagem. Os macrófagos dos animais foram cultivados em presença de extrato aquoso (1 mg
mL -1
), extrato oleoso (1 mg mL -1
), LPS (controle +) e somente meio RPMI-1640-C (controle -). As
concentrações de IL-10 foram calculadas através de uma reta padrão previamente estabelecida com
concentrações molares conhecidas de IL-10. Os resultados foram expressos em pg mL -1
, como média ±
desvio padrão de 4 animais em 2 experimentos independentes. Letras iguais não apresentam diferença
estatística significativa. Letras diferentes apresentam diferença significativa (ANOVA - pós-teste de
Tukey (p<0,05)). Comparação entre estímulos do extrato em A e B. Comparação entre sadios e infectados
em C, D e E. (* p<0,05) (**p<0,01).

25
5.3.3. Determinação na produção de IL-12
A determinação da produção de IL-12 foi realizada em sobrenadantes de culturas
de células do exsudato peritoneal (CEPs) de camundongos sadios e infectados, mediante o
desafio com os extrato aquoso e oleoso de alho, e os controles: LPS (controle positivo) ou meio
RPMI-1640-C (controle negativo). Os resultados obtidos foram calculados por meio de uma
curva analítica previamente estabelecida com concentrações molares conhecidas de IL-12, como
média ± desvio padrão de 4 animais por grupo estudado.
A produção de IL-12 por animais sadios, que receberam extrato de alho via oral (GA e
GO) e foram estimulados pelos extratos, foi menor quando comparada aos que não receberam
(GC), entretanto, em animais infectados a resposta foi inversa, animais do grupo GA e GO
estimulados pelos seus respectivos extratos tiveram uma produção elevada, quando comparados
aos animais do grupo controle estimulados igualmente (Figuras 4A e 4B). CEPs do grupo GC
ao serem desafiadas com os extratos apresentaram valores de produção de IL-12 acima do
controle positivo (Figura 4A).

26
A n im a is s a d io s - In te rle u c in a 1 2In
terl
eu
cin
a 1
2 (
pg
mL
-1)
GA
(a
qu
os
o)
GA
+G
A -
GO
(o
leo
so
)G
O+
GO
-G
C (
aq
uo
so
)
GC
(o
leo
so
)G
C +
GC
-
0
5 0
1 0 0
1 5 0
2 0 0
A
b d
c
a
* *
* *
A n im a is in fe c ta d o s - In te rle u c in a 1 2
Inte
rle
uc
ina
12
(p
g m
L-1
)
GA
(a
qu
os
o)
GA
+G
A -
GO
(o
leo
so
)G
O+
GO
-G
C (
aq
uo
so
)
GC
(o
leo
so
)G
C +
GC
-
0
5 0
1 0 0
1 5 0
2 0 0
B
bb
aa
G A - In te rle u c in a 1 2
Inte
rle
uc
ina
12
(p
g m
L-1
)
Extr
ato
Co
ntr
ole
+
Co
ntr
ole
-
0
5 0
1 0 0
1 5 0
2 0 0
C
**
*
**
G O - In te r le u c in a 1 2
Inte
rle
uc
ina
12
(p
g m
L-1
)
Extr
ato
Co
ntr
ole
+
Co
ntr
ole
-
0
5 0
1 0 0
1 5 0
2 0 0
D
**
**
G C - In te r le u c in a 1 2
Inte
rle
uc
ina
12
(p
g m
L-1
)
Ext.
Aq
uo
so
Ext.
Ole
oso
Co
ntr
ole
+
Co
ntr
ole
-
0
5 0
1 0 0
1 5 0
2 0 0S a u d á v e is
In fe c ta d o s
E
***
**
Figura 4. Concentração de IL-12 liberada em cultura de CEPs tratadas com tioglicolato de sódio
3% de camundongos Swiss sadios e infectados com o fungo S. schenckii no 10º dia após infecção.GA
(grupo que recebeu extrato aquoso), GO (grupo que recebeu extrato oleoso) e GC (grupo que recebeu
PBS) via gavagem. Os macrófagos dos animais foram cultivados em presença de extrato aquoso (1 mg
mL -1
), extrato oleoso (1 mg mL -1
), LPS (controle +) e somente meio RPMI-1640-C (controle -). As
concentrações de IL-12 foram calculadas através de uma reta padrão previamente estabelecida com
concentrações molares conhecidas de IL-12. Os resultados foram expressos em pg mL -1
, como média ±
desvio padrão de 4 animais em 2 experimentos independentes. Letras iguais não apresentam diferença
significativa. Letras diferentes apresentam diferença significativa (ANOVA - pós-teste de Tukey
(p<0,05)). Comparação entre estímulos do extrato em A e B. Comparação entre sadios e infectados em C,
D e E. (* p<0,05) (**p<0,01).
5.4. Determinação da carga fúngica (UFC)
Os resultados obtidos estão como média ± desvio padrão de quatro animais por
tratamento em dois experimentos independentes. Os grupos tratados com alho apresentaram
uma capacidade aumentada de controlar a infecção, como pode ser observado na Figura 5.

27
U F C p o r b a ç o
Tra
tam
en
tos
0250
500
750
1000
1250
1500
G A
G O
G C a
b
b
Figura 5. Determinação da carga fúngica (UFC). GA (grupo que recebeu extrato aquoso), GO (grupo
que recebeu extrato oleoso) e GC (grupo que recebeu PBS) via gavagem. Os animais dos grupos GA, GO
e GC foram inoculados por via intraperitoneal com 10 6 leveduras de S. schenckii em PBS. No dia 10 pós-
inoculação, os camundongos foram eutanasiados e os baços removidos para determinação da carga
fúngica sistêmica. Os resultados obtidos estão como média ± desvio padrão de quatro animais por
tratamento em dois experimentos independentes. Letras diferentes representam diferença significativa
(ANOVA - pós teste de Tukey (p<0,01)) .
5.5. Avaliação da capacidade antifúngica por microdiluição
Este teste foi realizado utilizando-se diluições seriadas 1:2 de cada um dos extratos de
alho (aquoso e oleoso). Iniciou-se com a concentração recebida pelos animais via gavagem (160
mg mL -1
). Após isso, foi feita a diluição das amostras com a colocação da suspensão de fungo e
incubado por 4 dias. O ponto de viragem foi observado de maneira visual e lido em
espectrofotômetro com comprimento de onda de 570 nm. Com a leitura da absorbância em
espectrofotômetro, é possível comprovar o que já foi observado no teste visual.
A Tabela 4 apresenta a ação antifúngica dos extratos em sua concentração mais baixa.
Tabela 4. Determinação da Concentração Inibitória Mínima (MIC)
Extrato MIC (mg mL-1
)
Aquoso 0,62
Oleoso 2,5 MIC dos extratos de alho (aquoso e oleoso). Não houve inibição no
controle positivo (apenas fungo) e não houve crescimento no controle
negativo (apenas meio).

28
6. DISCUSSÃO
A atividade antifúngica do alho foi descrita pela primeira vez em 1936 por Schmidt e
Marquardt ao estudarem micoses (HARRIS et al., 2001). Atualmente essa ação encontra-se
bem descrita, e muitos fungos mostraram-se ser susectíveis ao alho, incluindo Candida
albicans, Torulopsis glabrata, Trichophyton, Cryptococcus, Aspergillus niger, Trichosporon e
Rhodotorula. Sabe- se hoje, que o alho in natura ou na forma de extratos reduz o crescimento
de fungos, causa danos à membrana e inibe a síntese de lípidos, proteínas e ácidos nucleicos
(HARRIS et al., 2001; BAYAN, et al., 2014).
Deve-se dar importância à ação antifúngica do alho frente à Candida albicans,onde in
vitro, o alho in natura foi capaz de inibir o crescimento do fungo de maneira mais eficiente que
o Miconazol (50µg), usualmente empregado no combate ao fungo (FONSECA et al., 2014).
Além de inibir o crescimento do fungo, o extrato hidroalcoólico do alho apresenta ação biocida,
ou nesse caso, fungicida (OTA, et al., 2010). Estudos com extrato de alho envelhecido não
demonstraram atividade antifúngica in vitro, no entanto, quando administrados oralmente em
ratos infectados, o número de organismos que foram vistos foi reduzido em até 80% (BAYAN,
et al., 2014), fato que pode ser explicado pela complexa interação entre os compostos do alho e
células do sistema imune. O mesmo ocorreu no presente estudo. Nas duas formas como o
extrato foi oferecido, tanto em meio aquoso, quanto oleoso, o alho foi capaz de reduzir a carga
fúngica nos animais tratados e infectados (Figura 5). O tratamento com alho, então, ocasionou
uma resposta mais efetiva contra a infecção, demonstrando efetividade contra o fungo in vivo.
Dentre os compostos do alho, a alicina isolada mostrou-se eficiente como antifúngico,
diminuindo o crescimento de Candida albicans, Aspergillus niger e Paracoccidiodes sp.,
embora uma combinação entre alicina e ajoeno seja mais eficaz na inibição do crescimento
desses fungos, comparada a alicina pura (BAYAN, et al., 2014). Essa diminuição de
crescimento do fungo é justificada, em parte, pela inibição do complexo da succinato
desidrogenase pela alicina e outros compostos derivados dela (HARRIS et al., 2001; BAYAN,
et al., 2014).
Na imunidade contra fungos, destaca-se a importância da via Th1, onde há uma
liberação de citocinas como IFN-γ, IL-1 e IL-12. Por outro lado, a via Th2 que libera citocinas
estimulantes da produção de anticorpos (IL-4 e IL-5) apresenta valor limitado na defesa do
hospedeiro contra estes agentes. A IL-10 é a citocina chave para regulação desses processos
(GIRALDO & WITKIN, 1998; ALEGRANCI, 2013). Especificamente na resposta imune frente
a esporotricose, macrófagos quando ativados em cultura com componentes da parede celular do
fungo produzem citocinas pro-inflamotórias como IL-1, IL-6, TNF-α, que estimulam a resposta

29
fagocítica (CARLOS et al., 2009) e liberam compostos intermediários de nitrogênio e do
oxigênio com função fungicida (CARLOS et al., 2003).
Em modelos murinos, seis dias após a infecção por S. schenckii atinge-se o pico da
carga fúngica, o qual passa a diminuir até a eliminação total após 28 dias de infecção, embora
experimento de sobrevivência tenha mostrado que alguns animais podem levar mais de 28 dias
para eliminar a infecção (FERREIRA, 2014). No presente estudo, no 10º dia pós-infecção,
período de eliminação ativa do fungo, animais que foram tratados com extratos de alho
apresentaram carga fúngica muito inferior quando comparados aos animais não tratados (Figura
5), demonstrando maior controle da infecção. Ademais, os extratos aquoso e oleoso
demonstraram capacidade em inibir o crescimento de S. schenckii, respectivamente, em doses
de 0,62 e 2,5 mg mL-1
. Assim, a ingestão de extratos de alho de forma crônica, pode representar
uma redução na manifestação da esporotricose, e das lesões, principalmente nas regiões em que
a população está mais exposta ao fungo, e não somente em situações de tratamento da doença,
como descrito na maioria dos trabalhos na literatura.
SOUZA & SOARES (2013), encontraram valores semelhantes aos observados no
presente estudo, onde o extrato aquoso de alho apresentou efeito inibitório no crescimento do
fungo Aspergillus niger. A partir da concentração de 3 mg mL-1
do extrato, ocorreu inibição
total do crescimento miceliar e da esporulação do fungo.
Atualmente discute-se muito sobre a ação antifúngica de determinados fármacos,
considerando a segurança, a eficiência e os custos destas terapias. Uma questão de suma
importância a se considerar é a toxicidade das formulações e sua eficácia. A anfotericina B,
antifúngico muito usado, quando ligada ao colesterol de células humanas ocasiona toxicidade
celular e tecidual nas quais em suas reações há a liberação de prostaglandinas e interleucinas.
Seus efeitos colaterais apresentam-se como febre, taquicardia, náuseas, entre outros
(MARTINEZ, 2006). O alho então pode ser indicado como forma alternativa e complementar
de terapia, diminuindo assim a dose de medicamento usada, uma vez demonstrado que a
ingestão de extratos de alho de forma crônica, pode reduzir a carga fúngica dos animais, além de
favorecer a resposta imunológica sem os prejuízos causados por fármacos sintéticos.
Quanto à produção de NO, CEPs de animais sadios expostos aos extratos (GA e GO)
apresentaram baixa produção frente ao desafio com o próprio extrato, no entanto, em animais do
grupo GC, o estímulo com os extratos propiciou um aumento na produção, ou seja, houve
aumento na produção de NO quando os extratos foram usados de forma aguda. As CEPs de
animais infectados apresentaram valores mais elevados de NO devido à infecção, porém, em
animais que consumiram extratos via oral, sua produção foi maior comparada ao grupo que não
recebeu, além disso, CEPs desse grupo (GA e GO) foram estimuladas a produzir mais NO ex
vivo quando desafiadas com seu respectivo extrato, evidenciando assim o papel

30
imunomodulador do alho, onde animais que consumiram os extratos apresentaram respostas
imunológicas mais efetivas, uma vez que o aumento na produção de NO auxilia na eliminação
do fungo ex vivo.
A habilidade de macrófagos ativos liberarem NO é de suma importância na defesa do
hospedeiro, uma vez que permite o ataque a diversos tipos de patógenos. Porém deve-se levar
em conta que isso só é possível à custa de dano tissular. Apesar de serem extremamente
eficientes na destruição dos agentes infecciosos essas células não são mantidas em estado de
ativação pelo grande consumo de energia que demanda esse estado, além de estarem
intimamente relacionadas à destruição tissular localizada (ALEGRANCI, 2013).
Considerando que a inflamação excessiva é considerada um fator crítico em muitas
doenças humanas, incluindo o câncer, a obesidade, as doenças cardiovasculares,
neurodegenerativas e intestinais, o diabetes tipo II e até mesmo no envelhecimento
(SANTANGELO et al., 2007), conhecer e utilizar alimentos capazes de diminuir a resposta
inflamatória, como exemplo o alho, torna-se muito importante, permitindo modular a secreção
de NO e citocinas por meio da alimentação, oferecendo uma abordagem de tratamento natural
para essas doenças.
Além dos compostos derivados do enxofre, outras moléculas podem apresentar
respostas imunológicas, apresentando resultados em conformidade com os resultados do
presente estudo. De acordo com CHANDRASHEKAR et al. (2001), os frutooligossacarídeos do
alho também provocam ativação de macrófagos, além disso, sabe- se que os frutanos estimulam
seletivamente algumas bactérias benéficas no cólon, modulando a resposta imunológica.
Nessa mesma ótica, considerando que outros compostos presentes no alho podem ter
atividade biológica, LOUIS et al. (2012) verificaram que o extrato da casca de bulbilhos
contribui da mesma maneira que o extrato do bulbo de alho na prevenção de hipertrofia de
cardiomiócitos murinos e na diminuição do estresse oxidativo e apoptose. O mecanismo celular
da ação protetora do alho em células cardíacas pode ser em parte mediado pela molécula de NO.
Extratos aquosos com quantidade iguais de cascas e bulbos descascados foram comparados. O
extrato dos bulbilhos apresentou altos níveis de alicina e allina (0,233 mg mL -1
e 0,04 mg mL -
1) e o extrato da casca do alho apresentou (<0.002 mg mL
-1 e < 0.004 mg mL
-1 ), entretanto,
apesar da discrepância de valores, ambos apresentaram atividade biológica.
Considerando os resultados apresentados pode- se dizer que o alho apresenta atividade
anti-inflamatória quando consumido de forma regular, pois inibindo a produção de citocinas
como a IL-12, por macrófagos e estimulando a produção de IL-10, consequentemente haverá
diminuição da ativação de macrófagos (ALEGRANCI, 2013). A administração oral de alho em
pó em humanos adultos e saudáveis, em dose única de 1g e 3g aumentou significativamente a
excreção urinária de IL-12 em ambas as doses (ALMA et al., 2014). O alho também inibiu a
liberação de citocinas inflamatórias e aumentou a produção de citocinas anti-inflamatórias por

31
macrófagos e linfócitos in vitro (HODGE et al., 2002), evidenciando sua atividade
imunomoduladora.
Como encontrado no presente estudo que em situações normais (sem infecção), os
extratos de alho diminuem a produção de IL-1, o que indica e confirma que o consumo de alho
pode colaborar no controle da aterosclerose, doença responsável pelo maior índice de
morbidade e mortalidade no mundo, e que pode ser agravada pela IL-1. A função principal
atribuída a IL-1 na aterogênese, é a sua capacidade de aumentar a expressão de moléculas de
adesão quando secretada em baixas doses. Em doses mais elevadas, passa a ser liberada na
circulação sanguínea e exerce efeitos endócrinos, estimulando a produção de proteínas
inflamatórias de fase aguda pelo fígado (CORREA-CAMACHO et al., 2007).
A IL-10 participa nesse processo, e o seu papel regulador da IL-10 na aterosclerose
consiste na inibição da liberação de IL-12 induzida por LDL. Essa citocina pode atuar também
inibindo a ação do fator nuclear kappa Β, com consequente diminuição da produção de citocinas
pró-inflamatórias, da inibição da apoptose de macrófagos, da expressão do fator tecidual, do
fibrinogênio entre outros fatores (CORREA-CAMACHO et al., 2007)
Também de acordo com os resultados encontrados neste estudo, o alho pode ser usado
no combate à dor, visto que seu consumo diminui a produção de IL-1β, citocina que produz
inflamação sistêmica através da ativação da ciclooxigenase-2, com a formação de PGE2 no
hipotálamo anterior, tendo importante função no desenvolvimento da dor (OLIVEIRA et al.,
2011).
FABIAN e colaboradores (2013) demonstraram que o tratamento com a alicina em
cultura de adipócitos 3T3-L1, foi capaz de conter a indução inflamatória pelo LPS.
Considerando o fato de que em nosso estudo o alho também teve ação modulatória positiva
sobre o sistema imune, induzida por administração oral, talvez o alho possa ser um alimento
funcional a ser altamente recomendado no tratamento das patologias crônicas contemporâneas,
com estado inflamatório, como é o caso da obesidade.
Considerando os resultados obtidos nesse trabalho, a importância do alho deve ser
considerada no tratamento de infecções fúngicas, uma vez que os extratos se mostraram
eficientes no controle da carga fúngica no baço dos animais (ex vivo) e impediu o crescimento
do fungo em teste de concentração inibitória. O estudo mostrou pela primeira o efeito do alho
sobre a esporotricose.
7. CONCLUSÂO
Extrato de alho apresenta potencial antifúngico frente S. schenckii. e a administração
oral de extratos de alho influencia a liberação de citocinas por macrófagos ex vivo, o consumo
regular apresenta efeito anti-inflamatório em animais sadios, sendo que o uso agudo pode gerar

32
uma resposta inflamatória. Animais que consumiram alho responderam de forma mais efetiva
no combate da infecção.
Contudo mais estudos são necessários para entender completamente o mecanismo de
ação do alho no combate as infecções, e com isso elucidar alternativas para o combate a
infecções fúngicas e possíveis tratamentos menos tóxicos ao hospedeiro.
8. REFERÊNCIAS BIBLIOGRÁFICAS
ABBAS, K. A.; ANDREW H. H. LICHTMAN, A. H. H.; PILLAI, S. Imunologia Celular e
Molecular. Elsevier Brasil, 24 de ago de 2012 - 560p.
ABREU MATOS, F. J. Plantas Medicinais. Guia de seleção e emprego de plantas usadas em
fitoterapia no nordeste do Brasil. Doenças evitáveis. Fortaleza: Imprensa Universitária-UFC, 2000.
346p.
[ADA] American Dietetic Association. Position of the American Dietetic Association: Functional
Foods. Journal of The American Dietetic Association. 2004. v. 104, n 5.
ALEGRANCI, P. Análise dos macrófagos M1 e M2 durante a infecção por Sporothrix schenckii em
modelo murino. Tese de doutorado – UNESP - Araraquara, 2013.97 f.
ALMA, E.; EKEN, A.; ERCIL, H.; YELSEL, K.; DAGLIOGLU, N.The Effect of Garlic Powder on
Human Urinary Cytokine Excretion. Urology Journal, Vol 11, n 1 2014.
AMAGASE, H.; PETESCH, B. L.; MATSUURA, H.; KASUGA, S.; ITAKURA, Y. Intake of Garlic
and Its Bioactive Components. The Journal of Nutrition - American Society for Nutritional Sciences,
2001.
[ANAPA] Associação Nacional de Produtores de Alho.2016.
ANKRI, S.; MIRELMAN, D. Antimicrobial properties of allicin from garlic.Microbes and Infection
1999, 125-129.
ARREOLA, R.; FABIÁN, S. Q.; ROA, R. I. L.; GUTIÉRREZ, E. O. F.; GRAJEDA, J. P. R.;
QUINTANAR, L. C.; SAHAGÚN, D. O. Immunomodulation and Anti-Inflammatory Effects of
Garlic Compounds. Journal of Immunology Research Volume 2015, Article ID 401630, 13 pages
http://dx.doi.org/10.1155/2015/401630.

33
ASADULLAH K, SABAT R, FRIEDRICH M. Interleukin-10: an important immunoregulatory
cytokine with major impact on psoriasis. Curr Drug Targets Inflamm Allergy, 2004;3:185-192.
BAKRI, I. M.; DOUGLAS, C. W. I. Inhibitory effect of garlic extract on oral bactéria. July
2005.Volume 50, Issue 7.
BARROS, M. B. L.; SCHUBACH, T. P.; COLL, J. O.; GREMIÃO, I. D.; WANKE, B.;
SCHUBACH, A. Esporotricose: a evolução e os desafios de uma epidemia. Rev. Panam Salud
Publica 27(6), 2010.
BAYAN, L.; KOULIVAND, P. H.; , ALI GORJI, A. Garlic: a review of potential therapeutic
effects. AJP, Vol. 4, No. 1, Jan-Fev 2014.
BIANCHINI, F.; VAINIO, H. Allium Vegetables and Organosulfur Compounds: Do They Help
Prevent Cancer? Environmental Health Perspectives Vol. 109. n 9. September 2001.
CARLOS, I. Z., SGARBI, D. B.; SANTOS, G. C.; PLACERES, M. C. Sporothrix schenckii lipid
inhibits macrophage phagocytosis: involvement of nitric oxide and tumour necrosis factor-alpha.
Scand. J. Immunol. 2003. v.57, p.214–220.
CARLOS, I. Z.; SASSA, M. F.; SGARBI, D. B.; PLACERES, M. C.; MAIA, D. C. Current research
on the immune response to experimental sporotrichosis. Mycopathologia. 2009. v.168, p.1–10.
CARRADA-BRAVO, T.; OLVERA-MACÍAS, M. I. New observations on the ecology and
epidemiology of Sporothrix schenckii and sporotrichosis. Rev Latinoamer Patol Clin, 2013, Vol. 60.
CERQUEIRA, N. F.; YOSHIDA, W. B. Óxido nítrico. Revisão. Acta Cir. Bras. vol.17. n.6. São
Paulo, 2002.
CHANDRASHEKAR, P. M.; PRASHANTH, K. V. H.; AND VENKATESH, Y. P. Isolation,
structural elucidation and immunomodulatory activity of fructans from aged garlic extract,”
Phytochemistry, vol. 72, no. 2-3, pp. 255–264, 2011.
CHARRON, C. S.; DAWSON, D. D.; NOVOTNY, J. A. Garlic Influences Gene Expression In
Vivo and In Vitro. International Garlic Symposium: Role of Garlic in Cardiovascular Disease

34
Prevention, Metabolic Syndrome, and Immunology. The Journal of Nutrition Supplement (2014).
doi: 10.3945/jn.114.202481.
COELHO-CASTELO, A. A. M.; TROMBONE, A. P. F.; ROCHA, C. D.; LORENZI, J. C. C.
Immune response to infectious diseases. Medicina (Ribeirão Preto) 2009;42(2): 127-42.
CORRÊA-CAMACHO, C. R.; DIAS-MELICIO, L. A.; SOARES, A. M. V. C. Atherosclerosis, an
inflammatory response. Arq Ciênc Saúde 2007 jan-mar;14(1):41-48.
COZZOLINO, S. M. F.; COMINETTI, C. Bases bioquímicas e fisiológicas da nutrição. Nas
diferentes fases da vida, na saúde e na doença. Barueri, SP: Manole, 2013.
FABIÁN, S. Q.; SAHAGÚN, D. O.; CARRERA, M. V.; ROA, R. L. Alliin, a Garlic (Allium
sativum ) compounds, prevents LPS-induced Inflammation in 3T3-L1 adipocytes. HPC. Mediators
of Inflammations. Vol 2013. ID 381815.
FERREIRA, L. S. Avaliação da apoptose e resposta Th17 em modelo murino de esporotricose.
Araraquara, 2014. Tese (Doutorado) – UNESP - FCF. Biociências e Biotecnologia aplicadas à
Farmácia.
FERREIRA, L. S.; GONÇALVES, A. C.; PORTUONDO, D. L.; MAIA, D. C.; PLACERES, M.C.;
BATISTA-DUHARTE, A.; CARLOS, I. Z. Optimal clearance of Sporothrix schenckii requires an
intact Th17 response in a mouse model of systemic infection. Immunobiology 28/02/2015, S0171-
2985.
FONSECA, G. M.; PASSOS, T. C.; NINAHUAMAN, M. F. M. L.; CAROCI, A. S.; COSTA, L. S.
Avaliação da atividade antimicrobiana do alho (Allium sativum Liliaceae) e de seu extrato aquoso.
Rev. Bras. Pl. Med., Campinas, v.16, n.3, supl. I, p.679-684, 2014.
FRESHNEY, R. I. Culture of animal cells: a manual of basic technique. New York: Wiley-Liss,
1994. 486p.
FISCHER, G.; S. HUBNER, O. S.; VARGAS, G. D.; VIDOR, T. Imunomodulação pela própolis.
Arq. Inst. Biol., São Paulo, v.75, n.2, p.247-253, abr./jun., 2008.
FREITAS, D. F. S. Avaliação de fatores epidemiológicos, micológicos, clínicos e terapêuticos
associados à esporotricose. Tese de doutorado em Medicina Tropical. INSTITUTO OSWALDO

35
CRUZ. Rio de Janeiro, 2014.
GARCIA GOMEZ, L. J.; SANCHEZ-MUNIZ, F. J. Revisión: Efectos cardiovasculares del ajo
(Allium sativum). ALAN, Caracas, v. 50, n. 3, set. 2000.
GHAZANFARI, T. Z.; HASSAN, Z. M.; KHAMESIPOUR, A. Enhancement of peritoneal
macrophage phagocytic activity against Leishmania major by garlic (Allium Sativum) treatment.
Journal of Ethnopharmacology 103 (2006) 333–337.
GIRALDO, P.C; WITKIN, S. S. Vaginal Candidiasis: An incomprehensible Challege. DST - Jornal
Bras. doenças sex. transm; 10(5):31-6, 1998.
GREEN, L.C.; WAGNER, D.A.; GLOGOWSKI, J.; SKIPPER, P.L.; WISHNOK, J.S.;
TANNENBAUM, S.R. Analysis of nitrate, nitrite, and [15N]nitrate in biological fluids. Anal
Biochem, v.126, p.131-8, 1982.
GREG HODGE, G.; SANDY HODGE, S.; HAN, P. Allium sativum (garlic) suppresses leukocyte
inflammatory cytokine production in vitro: Potential therapeutic use in the treatment of inflammatory
bowel disease. Cytometry. Vol 48, Issue 4, pag. 209–215, ago. 2002.
HARRIS, J. C.; COTTRELL, S. L.; PLUMMER, S.; LLOYD, D. Antimicrobial properties of
Allium sativum (garlic). Appl Microbiol Biotechnol (2001) 57:282–286.
HEKTOEN, L.; PERKINS, C.F. Refractory subcutaneous abscess caused by Sporothrix schenckii: a
new pathogenic fungus. J. Exp. Med., v.5, p.77–91, 1900.
N.J. HEPBURN, J. N., GARAIOVA, I., WILLIAMS, E. A., MICHAEL, D. R., PLUMMER, S.
Probiotic supplement consumption alters cytokine production from peripheral blood mononuclear
cells: a preliminary study using healthy individuals. Beneficial Microbes: 4 (4) - Pages: 313 – 317.
Dez/ 2013. DOI: http://dx.doi.org/10.3920/BM2013.0012
HOLUB B.J, ARNOTT K, DAVIS J-P, NAGPURKAR A, PESCHELL J. Organosulfur compounds
from garlic. In: Shi J, Mazza G, Maguer M L (ed) Functional Foods. Washington; CRC,
2002;(2):213-231.
JANEWAY, C. A.; TRAVERS, P.; WALPORT, M.; DONALD CAPRA, J. Imunobiologia. O
sistema imune na saúde e na doença. 4 ed. Porto Alegre: Artes Médicas Sul, 2000.

36
JOSLING, P. Preventing the common cold with a garlic supplement: A double-blind, placebo-
controlled survey. Advances in Therapy July 2001, Volume 18, Issue 4, pp 189-193.
JÚNIOR, H. P. L.; LEMOS, A. L. A. Alho. Nutrologia.Diagn Tratamento. 2011;16(1):14-6.
KLIMP, A.H.; DE VRIES, E.G.; SCHERPHOF, G.L.; DAEMEN, T. A potential role of
macrophage activation in the treatment of cancer. Critical Reviews in Oncology/Hematology. v. 44,
n. 2, p. 143-161, 2002.
KISS, A.C.I.; TAKAKU, M.; DAMASCENO, D.C.; CAMPOS, K.E.; SINZATO, Y.K.; LIMA,
P.O.; VOLPATO, G.T. Efeito do extrato aquoso de Allium sativum L. sobre parâmetros bioquímicos
de ratas com diabete induzido por Streptozotocin. Rev. Bras. Pl. Med., Botucatu, v.8, n.3, p.24-30,
2006.
KOCH, H. P.; Toxicology, side effects and unwanted effects of garlic. Garlic: the science and
therapeutic application of Allium sativum L. and related species. Baltimore, 1996. p.221-228.
KREST I, KEUSGEN M. Qualitity of herbal remedies from Allium sativum; differences between
garlic power and fresh garlic. Plant Med 1999;65(2):139-43.
KIM, Y. S.; KIM, K. S.; HAN, I.; KIM, M. H.; JUNG, M. H. Quantitative and Qualitative Analysis
of the Antifungal Activity of Allicin Alone and in Combination with Antifungal Drugs. 2012. PLoS
ONE 7(6): e38242. doi:10.1371/journal.pone.0038242.
LAWSON, L. D.; Garlic: a rewien of this medicinal effects and indicated active compounds.
Phytochemicals of Europe: chemistry and biological activity. Washington, 1998. n 691, p,176-209.
LEVINSON, Warren. Microbiologia Médica E Imunologia. 12.ed. Porto Alegre: Artmed, 2010.Pag
377.
LIMA, D. M. Tabela brasileira de composição de alimentos / NEPA – UNICAMP.- 4. Ed. Campinas:
NEPA-UNICAMP, 2011. 161 p.
LORENZI, H.; ABREU-MATOS, J.F. Plantas medicinais no Brasil, nativas e exóticas. Nova
Odessa, SP: Instituto Plantaum, 2002.

37
LEE M, PARK H, YOUN, J. Interleukin-10 plasmid construction and delivery for the prevention of
type 1 diabetes. Ann N Y Acad Sci 2006;1079:313-319.
LOUIS, X. L.; MURPHY, R.; THANDAPILLY, S. J.; YU, L.; NETTICADAN, T. Garlic extracts
prevent oxidative stress, hypertrophy and apoptosis in cardiomyocytes: a role for nitric oxide and
hydrogen sulfide. BMC Complementary and Alternative Medicine 2012, 12:140.
MARQUES, S. A.; FRANCO, S. R. V. S.; CAMARGO, R. P.; DIAS, L. D. F.; HADDAD JÚNIOR,
V.; FABRIS, V. E. Esporotricose do gato doméstico (Felis catus): transmissão humana. Revista do
Instituto de Medicina Tropical de São Paulo. Instituto de Medicina Tropical, v. 35, n. 4, p. 327-330,
1993.
MARTINEZ, R. Atualização no uso de agentes antifúngicos. J. bras. Pneumol.vol.32 no.5 São
Paulo Sept./Oct. 2006.
MILNER, J. A.; DONATO, F. R. Bioactive Compounds and Cancer. Springer Science & Business
Media. Humana Press. 2010. 896p.
MIRON, T.; RABINKOV, A.; KONSTANTINOVSKI, L.; WILCHEK, M.; MIRELMAN, D. The
mode of action of allicin: trapping of radicals and interaction with thiol containing proteins.
Biochimica et Biophysica Acta 2000, 1379, 233–244.
MORIHARA, N.; SUMIOKA, I.; MORIGUCHI, T.; UDA, N.; KYO, E. Aged garlic extract
enhances production of nitric oxide. Vol. 71. n.5. 21/Jun/2002.pag. 509–517.
MOSMANN, T. Rapid colorimetric assay for cellular growth and survival: application and
citotoxicity assays. J. Immunol. Methods, v.65, n.1-2, p.55-63, 1983.
NAKAGAWA, S.; MASAMOTO, K.; SUMIYOSHI, H.; HARADA, H. Acute toxicity test of garlic
extract. The Journal of Toxicological Sciences. 1984, 9(1):57-60.
NANTZ, M. P.; ROWE, C. A.; MULLER, C. E.; CREASY, R. A.; STANILKA, J. M.; PERCIVAL,
S. S. Supplementation with aged garlic extract improves both NK and γδ-T cell function and reduces
the severity of cold and flu symptoms: A randomized, double-blind, placebo-controlled nutrition
intervention. Clinical Nutrition. Vol 31, Issue 3, June 2012, Pages 337–344.

38
OLIVEIRA, C. M. B.; TSA, R. K. S.; ISSY, A. M.; GEROLA, L. R.; SALOMÃO, R. Citocinas e
dor. Rev. Bras. Anestesiol. vol.61 no.2 Campinas Mar./Abril. 2011. http://dx.doi.org/10.1590/S0034-
70942011000200014.
OLSON, KENT. Manual de toxicologia clínica. 6 ed. Artmed, 2013.Pag 324.
OTA, C. C.;A, SILVA, D. V. G.; JACON, K. C.; BAURA, V.; NUNES, S. Avaliação da atividade
antimicrobiana e anti-inflamatória do extrato hidroalcoólico do allium sativum (alho) Tuiuti: Ciência
e Cultura, n. 43, p. 37-49, Curitiba, 2010.
PARHAM, P. O sistema imune. 3. ed. Porto Alegre: ArtMed, 2011. 608 p.
RANDLE, W. M.; LANCASTER, J. E. Sulphur Compounds in Alliums in Relation to Flavour
Quality..Allium Crop Science: Recent Advances. CAB International 2002. pp. 329-356.
SANTANGELO, C.; VARI, R.; SCAZZOCCHIO, B.; DI BENEDETTO, R.; FILESI, C;
MASELLA, R. Polyphenols, intracellular signalling and inflammation. Ann. Ist. Super Sanita., v.43,
n.4, p.394-405, 2007.
PERCIVAL, S. S. Aged Garlic Extract Modifies Human Immunity. J Nutr. Jan/2016.
SENDL, A.; SCHLIACK, R. L. et al. Inhibition of cholesterol synthesis in vitro by extracts and
isolated compounds prepared from garlic and wild garlic. Aherosclerosis, v.94, n1, p. 79-85. 1992.
SENER, G.; SATIROGLU, H.; SEHIRLI, A. O.; KAÇMAZ, A. Protective effect of aqueous garlic
extract against oxidative organ damage in a rat model of thermal injury. Life Sciences 73 (2003) 81 –
91.
SILVA, E. Y. Y. Propriedades funcionais " in vitro" e "in vivo" de alhos frescos e processados. Tese
de doutorado. Universidade de Brasília, 2009.
SILVA, M. B. T.; MATTOS- COSTA, M. M.; TORRES, C. C. S.; GALHARDO, M. C. G.;
VALLE, A. C. F.; MAGALHÃES, M. A. F. M.; SABROZA, P. C,; ROSELY MAGALHÃES DE
OLIVEIRA, R. M. Urban sporotrichosis: a neglected epidemic in Rio de Janeiro, Brazil. Cad. Saúde
Pública vol.28 no.10 Rio de Janeiro out. 2012.

39
SOUZA, L. S. S.; SOARES, A. C. F. Extrato aquoso de alho (Allium Sativum l.) no controle de
Aspergillus Niger causador da podridão vermelha em sisal. TECNO-LÓGICA, Santa Cruz do Sul, v.
17, n. 2, p. 124-128, jul./dez. 2013
SUIYOSHI, H.; KANEZAWA, A.; MASAMOTO, K.; HARADA, H.; NAKAGAMI, S.;
YOKOTA, A.; NISHIKAWA, M.; NAKAGAWA, S. Chronic toxicity test of garlic extract in rats. J
Toxicol Sci. 1984 Feb;9(1):61-75.
TENDE, J. A.; EZE, E.D.; MUHAMMAD, Z.; DAIKWO, O. A. Immunomodulatory activity of
Garlic (Allium sativum) in Wistar Rats. Annals of Experimental Biology 2014, 2 (3):74-76.
TREVOR, A. J.; MASTERS, S. B.; BERTRAM, K. G. Farmacologia Básica & Clínica. 12ª Ed. 2014

40
9. ANEXOS
9.1. Anexo1. Artigo
Imunomodulação de camundongos Swiss saudáveis e infectados por indução com Sporothrix
schenckii tratados com extratos de alho (Allium sativum L.)
João Paulo Burian, Luis Vitor Silva do Sacramento, Iracilda Zeppone Carlos.
Burian, JP I
Sacramento, LVS I
Carlos, IZ II
I
Universidade Estadual Paulista “Júlio de Mesquita Filho”
Faculdade de Ciências Farmacêuticas - Departamento de Alimentos e Nutrição
Rodovia Araraquara–Jaú, Km 1, Araraquara/SP - CEP 14801-902
II
Faculdade de Ciências Farmacêuticas da UNESP de Araraquara.
Departamento de Biociências e Biotecnologia Aplicadas à Farmácia
Rodovia Araraquara–Jaú, Km 1, Araraquara/SP - CEP 14801-902

41
RESUMO
O alho (Allium sativum L.) é cultivado em todo o mundo como hortaliça condimentar e
medicinal desde 3.000 a. C. A alicina é o principal componente do alho, sendo atribuída a ela a
maior parte das suas atividades biológicas, dentre elas as ações bactericida, antifúngica e
antiviral. Porém, outros compostos do alho apresentam atividade antioxidante,
hipocolesterolemiante, vasodilatadora, ação protetora contra diversos tipos de câncer e
imunomoduladora. As infecções por fungos são causas importantes de morbidade e mortalidade
no homem principalmente em indivíduos imunossuprimidos. O Sporothrix schenckii, agente
causal da esporotricose (micose subcutânea mais comum na América Latina), é fungo
dimórfico, de vida saprofítica no solo ou em vegetais, infectando homens e os animais
principalmente através de lesões e arranhões na pele. O objetivo desse trabalho foi avaliar a
influência do consumo de alho na imunomodulação de camundongos Swiss saudáveis e
infectados por S. schenckii, a partir do estado funcional dos macrófagos peritoneais desses
animais quanto à produção de óxido nítrico e das citocinas (IL-1β, IL-10 e IL-12) e avaliar o
potencial antifúngico do alho frente ao S. schenckii por meio de teste de concentração inibitória
mínima e unidades formadoras de colônia. Os resultados demonstraram que o alho apresenta
potencial antifúngico frente S. schenckii. A administração oral de extratos de alho influencia a
liberação de citocinas por macrófagos, o consumo regular apresenta efeito anti-inflamatório, e
seu uso agudo pode gerar uma resposta inflamatória. Camundongos que consumiram alho
responderam de forma mais efetiva ao combate da infecção.
Palavras-chave: Alho, macrófago, Sporothrix schenckii, esporotricose, interleucina.

42
ABSTRACT
Garlic (Allium sativum L.) is grown all over the world as seasoning and medicinal food since
3,000 BC. Allicin is the main component of garlic is attributed to it most of the biological
activities of garlic, including the bactericide, antifungal and antiviral action. However, other
garlic compounds have antioxidant activity, hypocholesterolemic and vasodilating activities,
guard action against many types of cancer and immunomodulatory. Fungal infections are
significant reasons of human morbidity and mortality mainly in immunocompromised people.
The Sporothrix schenckii, the causal agent of sporotrichosis (most common subcutaneous
mycosis in Latin America), is dimorphic fungus, of saprophytic life in soil or in plant that
infects humans and animals mainly through skin injuries and scratches. The aim of this study
was to evaluate the influence of garlic consumption in immunomodulation of Swiss infected and
healthy mice by S. schenckii from the functioning of peritoneal macrophages of the animals and
the production of nitric oxide and cytokines (IL 1β, IL-10 and IL-12) and evaluating the
antifungal potential of garlic against S. schenckii by minimum inhibitory concentration test and
colony forming units. The results show that garlic has potential antifungal front S. schenckii.
Oral administration of garlic extracts influences the releasing of cytokines by macrophages,
regular consumption has anti-inflammatory effects, and its use can generate an acute
inflammatory response. Mices that consumed garlic responded more effectively to combat the
infection.
Keywords: Garlic, macrophage, Sporothrix schenckii, sporotrichosis and interleukin.

43
Introdução
O alho (Allium sativum L.) é cultivado em todo o mundo como hortaliça condimentar e
medicinal, desde 3000 a.C. pelos sumérios e indianos (Garcia-Gomez et al.,2014; Abreu-Matos,
2000; Holub et al., 2002). Sua química é bastante complexa e provavelmente desenvolvida como
um mecanismo de autoproteção contra micro-organismos e outras ameaças presentes
principalmente no solo (Amagase et al., 2001).
Entre seus componentes com atividade biológica, talvez os mais significativos sejam os
compostos organossulfurados, que se apresentam em quantidades de 11 a 35 mg por grama de alho
fresco. O alho contém três vezes mais compostos sulfurados que a cebola, além de arginina,
oligossacarídeos, flavonóides e selênio (Holub et al., 2002; Milner and Donato, 2010). Seu sabor
e ardor característico é derivado da hidrólise dos compostos S-alk(en)yl cysteine sulphoxides
(ACSOs) pela enzima alinase (Randle and Lancaste, 2002). A aliina (S-alilcisteína S-óxido) é o
principal composto organossulfurado identificado nos bulbilhos de alho íntegros, pois quando
cortados ou triturtados, a aliina é convertida em alicina pela ação da alinase. A transformação
química da alicina por meio do calor produz outros compostos sulfurosos, como ajoenos,
vinilditiina e trissulfeto de alil metila (Ankri and Mirelman, 1999).
A nutrição tem papel relevante na modulação da resposta imune e inflamatória, uma vez
que nutrientes modulam sistemas de defesa celular e humoral, pela alteração na formação de
mediadores ou pela interferência nas vias de transdução de sinais celulares, podendo alterar o
balanço entre citocinas e atenuar a depleção de nutrientes teciduais (Hepburn, 2014; Parham,
2011).
Diversas citocinas produzidas por leucócitos são denominadas interleucinas. A
Interleucina-1 (IL-1), por exemplo, media a inflamação e é produzida principalmente por
macrófagos ativados. Seus efeitos incluem o estímulo na produção de quimiocinas, aumento da
produção de neutrófilos pela medula óssea e aumento da atividade inflamatória. Todavia, esse
processo pode ser altamente regulado pela ação de antagonistas como a interleucina (IL-10)
(Correa-Camacho et al., 2007; Trevor et al., 2014).

44
A IL-10 atua suprimindo a atividade de macrófagos ativados, células T e células Natural
killer (NK), e também causa inibição da produção de interleucina 12 (IL-12) em macrófagos, o que
consiste na principal forma de ativação das células NK. A IL-12 estimula a produção de IFN-y por
células NK e linfócitos T, aumenta a citotoxicidade mediada por células NK e estimula a
diferenciação de células T. Embora muitas outras células pareçam poder secretar IL-12, apenas
macrófagos e células dendríticas produzem a citocina biologicamente ativa (Oliveira et al., 2011).
Os macrófagos desempenham um papel fundamental na defesa do organismo, pela da
fagocitose, geração de radicais livres, mediação de processos inflamatórios e secreção de várias
substâncias (Fischer et al., 2008). Pertencem ao sistema fagocítico mononuclear e fazem parte,
junto com células NK, da primeira linha de defesa contra patógenos (Klimp et al., 2002). São
classificados como M1 e M2, sendo os M1produtores da enzima óxido nítrico sintase, que degrada
a L-arginina, produzindo o óxido nítrico (NO), substância vasodilatadora que também modula
reações inflamatórias ou anti-inflamatórias, dependendo do tipo celular e do estímulo (Alegranci,
2013; Cerqueira and Yoshida, 2002; Morihara et al., 2002).
As infecções por fungos são causas importantes de morbidade e mortalidade no homem.
São ditas oportunistas, pois os agentes etiológicos causam doença branda ou nenhuma doença em
indivíduos sadios, mas podem infectar e causar graves doenças em indivíduos imunossuprimidos.
Uma simples infecção, como a candidíase, pode se tornar grave e invasiva, colonizando esôfago,
estômago e intestino em pacientes portadores da síndrome da imunodeficiência adquirida
(Alegranci, 2013; Coelho-Castelo et al., 2009).
Os fagócitos, principalmente macrófagos, respondem contra fungos ou micoses utilizando a
enzima NO sintase induzida e outros mediadores secretados para a destruição desses patógenos. A
participação de citocinas é imprescindível, pois aumenta a capacidade fungicida dos fagócitos,
favorecendo a destruição do fungo (Coelho-Castelo et al., 2009).
O Sporothrix schenckii, agente causal da esporotricose é fungo dimórfico, de vida
saprofítica no solo ou em vegetais, infectando ocasionalmente o homem e os animais. No homem a
maior parte das infecções é provocada por lesões com farpas, madeiras ou arame (Marques et al.,
1993). A forma miceliar saprofítica do fungo encontrada no meio ambiente cresce em temperaturas

45
que variam de 25ºC a 30ºC. A forma leveduriforme é encontrada in vivo, a 37°C, onde se reproduz
por brotamento (Alegranci, 2013).
As lesões da esporotricose costumam ser restritas à pele, tecido celular subcutâneo e vasos
linfáticos adjacentes. Raramente pode disseminar-se para outros órgãos, ou ainda ser sistêmica,
resultante da inalação de esporos. A infecção pode ser classificada em cutânea, cutâneo-linfática,
cutânea disseminada, mucosa e sistêmica (Barros et al., 2010). Trata-se da micose subcutânea mais
comum na América Latina, e vem sendo descrita em diversos países, em animais e humanos, vale
ressaltar que na década de 90 no Estado do Rio de Janeiro a doença assumiu proporções epidêmicas
(Barros et al., 2010).
O objetivo desse trabalho foi avaliar a influência do consumo de alho (Allium sativum L.)
na imunomodulação de camundongos Swiss saudáveis e infectados por S. schenckii.
Metodologia
Obtenção e administração dos extratos
Extratos aquoso e oleoso foram preparados utilizando 32g de bulbilhos de alho (Allium
sativum L.) descascados e triturados em 200 mL de água destilada ou óleo de soja, permanecendo
em repouso por meia hora e sendo posteriormente filtrado e refrigerado. A dose dos extratos (0,1
mL) utilizada no tratamento correspondeu a 800 mg kg -1
, administrada por gavagem uma vez ao
dia durante 15 dias.
Obtenção das células do exsudato peritoneal
Células do exsudato peritoneal (CEPs) de camundongos Swiis, machos, adultos, sadios ou
infectados com fungo S. schenckii foram obtidas após os animais serem previamente estimulados
com 3,0 mL de tioglicolato de sódio 3% através de inoculação intraperitoneal três dias antes de
serem eutanasiados em câmara de CO2. A suspensão celular ajustada à concentração final de 5x106
células para realização dos testes propostos.

46
Determinação de NO
O NO foi quantificado espectrofotometricamente pela Reação de Griess (Green et al.,
1982). A determinação da liberação de óxido nítrico (NO) foi realizada em sobrenadantes de
(CEPs) de camundongos infectados e não infectados com o fungo S. schenckii, mediante o desafio
com os extratos de alho (1mg mL-1
), e os controles LPS (controle positivo) ou meio RPMI- 1640-C
(controle negativo). O grupo GC não recebeu extrato de alho via gavagem, no entanto, as CEPs
desse grupo foram desafiadas com os dois extratos diferentes (oleoso e aquoso).
Determinação de citocinas
As citocinas IL- 1β, IL-10 e IL-12 foram quantificadas nos sobrenadantes de (CEPs)
obtidas e desafiadas como descrito acima. A quantificação foi realizada através do teste
imunoenzimático ELISA de captura para cada citocina.
Microrganismo, condições de cultivo e infecção experimental
Foi utilizada a cepa 16345 de Sporothrix schenckii isolada de um caso de infecção
pulmonar humano de esporotricose (Baltimore, MD), cedida gentilmente pelo Laboratório de
Materiais de Referências da Fundação Oswaldo Cruz. Atualmente, este isolado é mantido no
Laboratório de Imunologia Clínica do Departamento de Análises Clínicas da Faculdade de Ciências
Farmacêuticas de Araraquara - UNESP, na fase miceliar em meio Sabouraud, a temperatura
ambiente (± 25°C). A forma leveduriforme é obtida em meio BHI (Brain Heart Infusion, Difco)
líquido a 37°C sob agitação constante de 150 rpm por sete dias. Em seguida, uma alíquota contendo
107 unidades fúngicas foi transferida para um Erlenmeyer de 250 mL, contendo 100 mL de caldo
BHI e cultivada por cinco dias nas mesmas condições anteriores, que corresponde com a fase de
crescimento logarítmica previamente descrita por Ferreira et al. (2015). Os animais foram
inoculados com 200 µL de uma suspensão contendo 10 6
leveduras de S. schenckii em PBS ou com
igual volume de PBS apenas, por via intraperitoneal.

47
Determinação da carga fúngica
A avaliação da carga fúngica sistêmica para acompanhamento das infecções foi feita ao se
considerar uma determinação das unidades formadoras de colônia (UFC) no baço dos animais após
sua retirada. As UFC foram contadas após 3 dias e confirmadas após 5 dias de incubação a
temperatura ambiente.
Avaliação da capacidade antifúngica por microdiluição
A avaliação da atividade antifúngica foi realizada através do teste de microdiluição de
acordo com a Norma M27-A2 do National Committee for Clinical Laboratory Standards. Foi
utilizada a cultura de S. schenckii preparando um inóculo em PBS estéril. Foram realizadas
diluições seriadas em meio RPMI-1640 com MOPS, ajustando a suspensão fúngica em uma
concentração de 2,5×103 UFC mL
-1.
Análise estatística
Os resultados obtidos foram expressos como média ± desvio padrão e submetidos ao teste
de Lilliefors para verificação da normalidade, ao teste de Levene para verificação da
homocedasticidade e à análise de variância (ANOVA one-way – pós teste de Tukey). O valor de
significância adotado foi de 5% (p<0,05).
Resultados
A viabilidade celular nos testes manteve-se acima de 90%, mediante todos os estímulos,
indicando que as análises não foram tóxicas para as células. A produção de óxido nítrico (NO)
pelas CEPs revelou que animais sadios do grupo GA têm uma produção significativamente menor
(p<0,05) de NO em comparação aos grupos GO e GC. Não obstante, animais que não foram
expostos previamente aos extratos (GC), ao serem estimulados pelos extratos, apresentam maior
produção de NO, comparados ao controle negativo, o que não ocorre com animais que receberam
os extratos de alho via oral (GA). Isso demonstra que a atividade imunomoduladora do alho pode

48
ser usada de formas diferentes, ou seja, a resposta muda se ingerido regularmente ou de forma
aguda (Figura 1A).
No grupo GA infectados com o fungo S.schenckii, houve aumento significativo (p<0,05) na
produção de NO por CEPs desafiadas com extrato aquoso frente ao controle negativo. Esse
resultado mostra que o consumo de alho pode aumentar a produção de NO, mesmo em condições
que a produção já está aumentada, como nesse caso pela infecção (Figura 1B). A infecção
aumentou a produção de NO em todos os grupos analisados, isso pode ser observado nas Figuras
1C, 1D e 1E.
A n im a is s a d io s - Ó x id o N ítr ic o
µm
ols
m
L-1
de
nit
rito
GA
(a
qu
os
o)
GA
+G
A -
GO
(o
leo
so
)G
O+
GO
-G
C (
aq
uo
so
)
GC
(o
leo
so
)G
C +
GC
-
0
5
3 0
6 0
9 0
1 2 0
A
a
bcc
**
A n im a is in fe c ta d o s - Ó x id o N ítr ic oµ
mo
ls
mL
-1 d
e n
itri
to
GA
(a
qu
os
o)
GA
+G
A -
GO
(o
leo
so
)G
O+
GO
-G
C (
aq
uo
so
)
GC
(o
leo
so
)G
C +
GC
-
0
3 0
6 0
9 0
1 2 0
B
b
a
b c
c
*
G A - Ó x id o N ítr ic o
µm
ols
m
L-1
de
nit
rito
Extr
ato
Co
ntr
ole
+
Co
ntr
ole
-
0
1
3 0
6 0
9 0
1 2 0
C
**
**
**
G O - Ó x id o N ítr ic o
µm
ols
m
L-1
de
nit
rito
Extr
ato
Co
ntr
ole
+
Co
ntr
ole
-
0
4
3 0
6 0
9 0
1 2 0
D
** **
**
G C - Ó x id o N ítr ic o
µm
ols
m
L-1
de
nit
rito
Ext.
Aq
uo
so
Ext.
Ole
oso
Co
ntr
ole
+
Co
ntr
ole
-
0
3 0
6 0
9 0
1 2 0
S a u d á v e is
E
In fe c ta d o s
**
**
****
Figura 1. Produção de NO em cultura de células do exsudato peritoneal de camundongos
Swiss sadios e infectados com o fungo S. schenckii no 10º dia após infecção. GA (grupo que
recebeu extrato aquoso), GO (grupo que recebeu extrato oleoso) e GC (grupo que recebeu
tampão PBS) via gavagem. Letras iguais não apresentam diferença significativa. Letras
diferentes são estatisticamente diferentes. ANOVA - pós-teste de Tukey (p<0,05). Comparação
entre estímulos do extrato em A e B. Comparação entre sadios e infectados em C, D e E. (*
p<0,05) (**p<0,01).
A produção de IL-1β em animais sadios de diferentes grupos foi estatisticamente diferente
entre si (p<0,01), e o grupo dos animais tratados com os extratos apresentou menor produção da
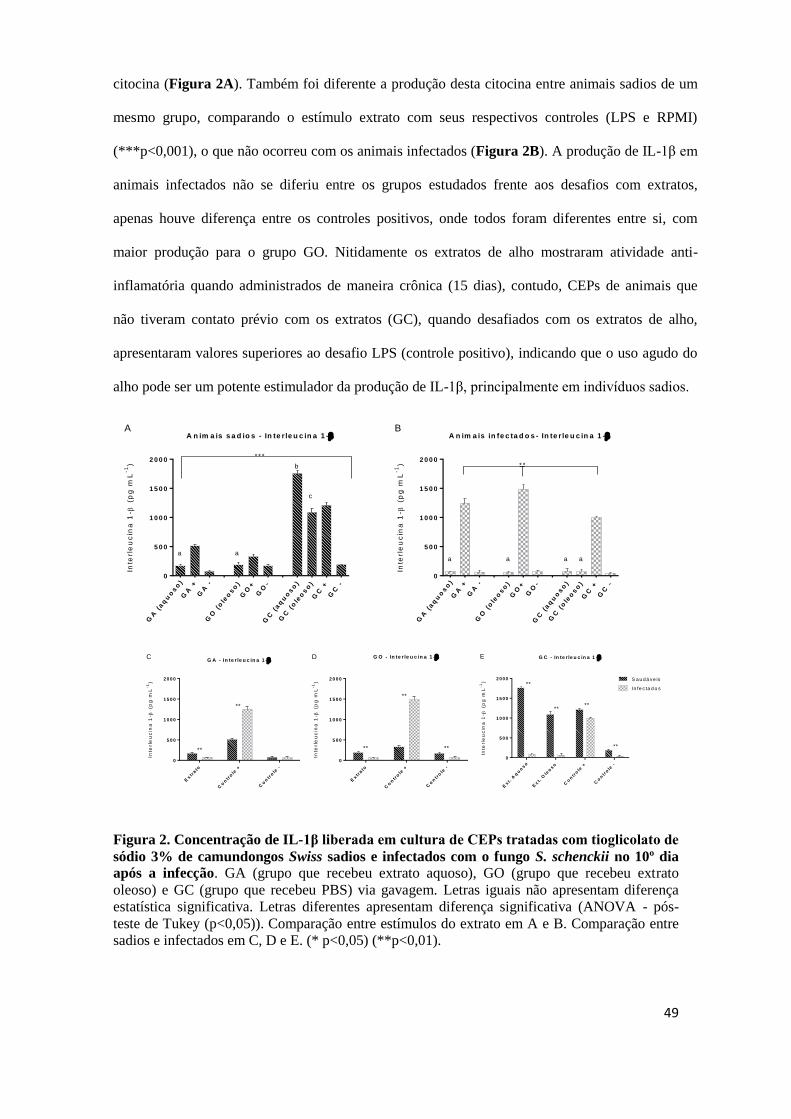
49
citocina (Figura 2A). Também foi diferente a produção desta citocina entre animais sadios de um
mesmo grupo, comparando o estímulo extrato com seus respectivos controles (LPS e RPMI)
(***p<0,001), o que não ocorreu com os animais infectados (Figura 2B). A produção de IL-1β em
animais infectados não se diferiu entre os grupos estudados frente aos desafios com extratos,
apenas houve diferença entre os controles positivos, onde todos foram diferentes entre si, com
maior produção para o grupo GO. Nitidamente os extratos de alho mostraram atividade anti-
inflamatória quando administrados de maneira crônica (15 dias), contudo, CEPs de animais que
não tiveram contato prévio com os extratos (GC), quando desafiados com os extratos de alho,
apresentaram valores superiores ao desafio LPS (controle positivo), indicando que o uso agudo do
alho pode ser um potente estimulador da produção de IL-1β, principalmente em indivíduos sadios.
A n im a is s a d io s - In te rle u c in a 1 -
Inte
rle
uc
ina
1-
(p
g m
L-1
)
GA
(a
qu
os
o)
GA
+G
A -
GO
(o
leo
so
)G
O+
GO
-G
C (
aq
uo
so
)
GC
(o
leo
so
)G
C +
GC
-
0
5 0 0
1 0 0 0
1 5 0 0
2 0 0 0
A
a a
b
c
* * *
A n im a is in fe c ta d o s - In te r le u c in a 1 -
Inte
rle
uc
ina
1-
(p
g m
L-1
)
GA
(a
qu
os
o)
GA
+G
A -
GO
(o
leo
so
)G
O+
GO
-G
C (
aq
uo
so
)
GC
(o
leo
so
)G
C +
GC
-
0
5 0 0
1 0 0 0
1 5 0 0
2 0 0 0
B
a a aa
* *
G A - In te r le u c in a 1 -
Inte
rle
uc
ina
1-
(p
g m
L-1
)
Extr
ato
Co
ntr
ole
+
Co
ntr
ole
-
0
5 0 0
1 0 0 0
1 5 0 0
2 0 0 0
C
**
**
G O - In te r le u c in a 1 -
Inte
rle
uc
ina
1-
(p
g m
L-1
)
Extr
ato
Co
ntr
ole
+
Co
ntr
ole
-
0
5 0 0
1 0 0 0
1 5 0 0
2 0 0 0
D
**
**
**
G C - In te r le u c in a 1 -
Inte
rle
uc
ina
1-
(p
g m
L-1
)
Ext.
Aq
uo
so
Ext.
Ole
oso
Co
ntr
ole
+
Co
ntr
ole
-
0
5 0 0
1 0 0 0
1 5 0 0
2 0 0 0 S a u d á v e is
In fe c ta d o s
E
**
****
**
Figura 2. Concentração de IL-1β liberada em cultura de CEPs tratadas com tioglicolato de
sódio 3% de camundongos Swiss sadios e infectados com o fungo S. schenckii no 10º dia
após a infecção. GA (grupo que recebeu extrato aquoso), GO (grupo que recebeu extrato
oleoso) e GC (grupo que recebeu PBS) via gavagem. Letras iguais não apresentam diferença
estatística significativa. Letras diferentes apresentam diferença significativa (ANOVA - pós-
teste de Tukey (p<0,05)). Comparação entre estímulos do extrato em A e B. Comparação entre
sadios e infectados em C, D e E. (* p<0,05) (**p<0,01).

50
Entre os animais sadios, os que receberam extrato de alho tiveram a produção de IL-10
aumentada, comparados ao grupo que não recebeu extrato via oral (GC). Nos tratamentos GA e
GO, o desafio LPS, CN e extrato correspondente foram significativamente diferentes entre si.
A n im a is s a d io s - In te rle u c in a 1 0
Inte
rle
uc
ina
10
(p
g m
L-1
)
GA
(a
qu
os
o)
GA
+G
A -
GO
(o
leo
so
)G
O+
GO
-G
C (
aq
uo
so
)
GC
(o
leo
so
)G
C +
GC
-
0
1
1 0 0 0
2 0 0 0
3 0 0 0
A
bb
aa
A n im a is in fe c ta d o s - In te rle u c in a 1 0
Inte
rle
uc
ina
10
(p
g m
L-1
)
GA
(a
qu
os
o)
GA
+G
A -
GO
(o
leo
so
)G
O+
GO
-G
C (
aq
uo
so
)
GC
(o
leo
so
)G
C +
GC
-
0
1 0 0 0
2 0 0 0
3 0 0 0
B
baaa
G A - In te rle u c in a 1 0
Inte
rle
uc
ina
10
(p
g m
L-1
)
Extr
ato
Co
ntr
ole
+
Co
ntr
ole
-
0
1 0 0 0
2 0 0 0
3 0 0 0
C
**
**
G O - In te r le u c in a 1 0
Inte
rle
uc
ina
10
(p
g m
L-1
)
Extr
ato
Co
ntr
ole
+
Co
ntr
ole
-
0
1 0 0 0
2 0 0 0
3 0 0 0
D
******
G C - In te r le u c in a 1 0
Inte
rle
uc
ina
10
(p
g m
L-1
)
Ext.
Aq
uo
so
Ext.
Ole
oso
Co
ntr
ole
+
Co
ntr
ole
-
0
1
1 0 0 0
2 0 0 0
3 0 0 0S a u d á v e is
In fe c ta d o s
E
**
**** **
Figura 3. Concentração de IL-10 liberada em cultura de CEPs tratadas com tioglicolato de
sódio 3% de camundongos Swiss sadios e infectados com o fungo S. schenckii no 10º dia
após infecção.GA (grupo que recebeu extrato aquoso), GO (grupo que recebeu extrato oleoso) e
GC (grupo que recebeu PBS) via gavagem. Os macrófagos dos animais foram cultivados em
presença de extrato aquoso (1 mg mL -1
), extrato oleoso (1 mg mL -1
), LPS (controle +) e
somente meio RPMI-1640-C (controle -). Letras iguais não apresentam diferença estatística
significativa. Letras diferentes apresentam diferença significativa (ANOVA - pós-teste de
Tukey (p<0,05)). Comparação entre estímulos do extrato em A e B. Comparação entre sadios e
infectados em C, D e E. (* p<0,05) (**p<0,01).
A produção de IL-12 por animais sadios, que receberam extrato de alho via oral (GA e GO)
e foram estimulados pelos extratos, foi menor quando comparada aos que não receberam (GC),
entretanto, em animais infectados a resposta foi inversa, animais do grupo GA e GO estimulados
pelos seus respectivos extratos tiveram uma produção elevada, quando comparados aos animais do
grupo controle estimulados igualmente (Figuras 4A e 4B). CEPs do grupo GC ao serem desafiadas

51
com os extratos apresentaram valores de produção de IL-12 acima do controle positivo (Figura
4A).
A n im a is s a d io s - In te rle u c in a 1 2
Inte
rle
uc
ina
12
(p
g m
L-1
)
GA
(a
qu
os
o)
GA
+G
A -
GO
(o
leo
so
)G
O+
GO
-G
C (
aq
uo
so
)
GC
(o
leo
so
)G
C +
GC
-
0
5 0
1 0 0
1 5 0
2 0 0
A
b d
c
a
* *
* *
A n im a is in fe c ta d o s - In te rle u c in a 1 2
Inte
rle
uc
ina
12
(p
g m
L-1
)
GA
(a
qu
os
o)
GA
+G
A -
GO
(o
leo
so
)G
O+
GO
-G
C (
aq
uo
so
)
GC
(o
leo
so
)G
C +
GC
-
0
5 0
1 0 0
1 5 0
2 0 0
B
bb
aa
G A - In te rle u c in a 1 2
Inte
rle
uc
ina
12
(p
g m
L-1
)
Extr
ato
Co
ntr
ole
+
Co
ntr
ole
-
0
5 0
1 0 0
1 5 0
2 0 0
C
**
*
**
G O - In te r le u c in a 1 2
Inte
rle
uc
ina
12
(p
g m
L-1
)
Extr
ato
Co
ntr
ole
+
Co
ntr
ole
-
0
5 0
1 0 0
1 5 0
2 0 0
D
**
**
G C - In te r le u c in a 1 2
Inte
rle
uc
ina
12
(p
g m
L-1
)
Ext.
Aq
uo
so
Ext.
Ole
oso
Co
ntr
ole
+
Co
ntr
ole
-
0
5 0
1 0 0
1 5 0
2 0 0S a u d á v e is
In fe c ta d o s
E
***
**
Figura 4. Concentração de IL-12 liberada em cultura de CEPs tratadas com tioglicolato de
sódio 3% de camundongos Swiss sadios e infectados com o fungo S. schenckii no 10º dia
após infecção.GA (grupo que recebeu extrato aquoso), GO (grupo que recebeu extrato oleoso) e
GC (grupo que recebeu PBS) via gavagem. Os macrófagos dos animais foram cultivados em
presença de extrato aquoso (1 mg mL -1
), extrato oleoso (1 mg mL -1
), LPS (controle +) e
somente meio RPMI-1640-C (controle -). Letras iguais não apresentam diferença significativa.
Letras diferentes apresentam diferença significativa (ANOVA - pós-teste de Tukey (p<0,05)).
Comparação entre estímulos do extrato em A e B. Comparação entre sadios e infectados em C,
D e E. (* p<0,05) (**p<0,01).
Determinação da carga fúngica (UFC)
Os grupos tratados com alho apresentaram uma capacidade aumentada de controlar a
infecção, como pode ser observado na Figura 5.

52
U F C p o r b a ç o
Tra
tam
en
tos
0250
500
750
1000
1250
1500
G A
G O
G C a
b
b
Figura 5. Determinação da carga fúngica (UFC). GA (grupo que recebeu extrato aquoso),
GO (grupo que recebeu extrato oleoso) e GC (grupo que recebeu PBS) via gavagem. Os
animais dos grupos GA, GO e GC foram inoculados por via intraperitoneal com 10 6
leveduras
de S. schenckii em PBS. No dia 10 pós-inoculação, os camundongos foram eutanasiados e os
baços removidos para determinação da carga fúngica sistêmica. Os resultados obtidos estão
como média ± desvio padrão de quatro animais por tratamento em dois experimentos
independentes. Letras diferentes representam diferença significativa (ANOVA - pós teste de
Tukey (p<0,01)) .
Avaliação da capacidade antifúngica por microdiluição
A Tabela 1 apresenta a ação antifúngica dos extratos em sua concentração mínima.
Tabela 1. Determinação da Concentração Inibitória Mínima (MIC) Extrato MIC (mg mL
-1)
Aquoso 0,62
Oleoso 2,5
MIC dos extratos de alho (aquoso e oleoso). Não houve
inibição no controle positivo (apenas fungo) e não houve
crescimento no controle negativo (apenas meio).
Discusão
Sabe- se hoje, que o alho in natura ou na forma de extratos reduz crescimento de fungos,
causa danos à membrana e inibe a síntese de lípidos, proteínas e ácidos nucleicos (Harris et al.,
2001; Bayan, et al., 2014). Estudos com extrato de alho envelhecido não demonstraram atividade
antifúngica in vitro, no entanto, quando administrados oralmente em ratos infectados, o número de
organismos que foram vistos foi reduzido em até 80% (Bayan, et al., 2014), fato que pode ser
explicado pela complexa interação entre os compostos do alho e células do sistema imune.

53
Nesse estudo, no 10º dia pós-infecção, período de eliminação ativa do fungo, animais
infectados que foram tratados com extratos de alho apresentaram carga fúngica muito inferior
quando comparados aos animais não tratados, ou seja, ocasionou uma resposta mais efetiva no
controle da infecção demonstrando efetividade contra o fungo in vivo Ademais, os extratos aquoso
e oleoso demonstraram capacidade em inibir o crescimento de S. schenckii, respectivamente, em
doses de 0,62 e 2,5 mg mL-1
, demonstrando efetividade contra o fungo in vitro.
A maior aptidão no controle da infecção pode ser justificada pelo fato de que a destruição
de invasores por macrófagos é significativamente aumentada com as propriedades do extrato do
alho, que aumenta número de macrófagos peritoneais, sua ação e a expressão de receptores em suas
membranas (Ghazanfari et al., 2006). O tratamento com alho, então, pode ser indicado como forma
alternativa e complementar de terapia, diminuindo assim a dose de medicamento usada.
Na imunidade contra fugos, destaca-se a importância da via Th1, onde há uma liberação
de citocinas como IFN-γ, IL-1 e IL-12. Por outro lado, a via Th2 que libera citocinas estimulantes
da produção de anticorpos (IL-4 e IL-5) apresenta valor limitado na defesa do hospedeiro contra
estes agentes. A IL-10 age regulando esse processo (Giraldo and Witkin, 1998; Alegranci, 2013).
Na resposta imune frente à esporotricose, macrófagos quando ativados em cultura com
componentes da parede celular do fungo produzem citocinas pro-inflamotórias como a IL-1, que
estimulam a resposta fagocítica (Carlos et al., 2009) e liberam compostos intermediários de
nitrogênio e do oxigênio com função fungicida (Carlos et al., 2003). A habilidade de macrófagos
ativos liberarem NO é de suma importância na defesa do hospedeiro, uma vez que permite o ataque
a diversos tipos de patógenos (Alegranci, 2013; Ferreira, 2014).
O mesmo pode ser visto nesse trabalho, onde animais que consumiram o alho apresentaram
níveis mais elevados de interleucinas pró-inflamatórias (IL-1β e IL-12) quando infectados, por
outro lado, quando em estado saudável, a citocina anti-inflamatória (IL-10) sofreu acréscimo na
produção.
Considerando que a inflamação excessiva é considerada um fator crítico em muitas
doenças humanas, incluindo o câncer, a obesidade, as doenças cardiovasculares,
neurodegenerativas e intestinais, o diabetes tipo I e até mesmo no envelhecimento (Santangelo et

54
al., 2007), conhecer e utilizar alimentos capazes de diminuir a resposta inflamatória, como exemplo
o alho, torna-se muito importante, permitindo modular a secreção de NO e citocinas por meio da
alimentação, oferecendo uma abordagem de tratamento natural para essas doenças.
Também de acordo com os resultados encontrados neste estudo, o alho pode ser usado no
combate à dor, visto que seu consumo diminui a produção de IL-1β em animais saudáveis. A IL-1β
produz inflamação sistêmica através da ativação da ciclooxigenase-2, com a formação de PGE2 no
hipotálamo anterior, tendo importante função no desenvolvimento da dor (Oliveira et al., 2011).
Fabian et al. (2013) demonstraram que o tratamento com a alicina em cultura de adipócitos
3T3-L1, foi capaz de conter a indução inflamatória pelo LPS. Considerando o fato de que em nosso
estudo o alho também teve ação modulatória positiva sobre o sistema imune, induzida por
administração oral, talvez o alho possa ser um alimento funcional a ser altamente recomendado no
tratamento das patologias crônicas contemporâneas, com estado inflamatório, como é o caso da
obesidade.
Em suma, extrato de alho apresenta potencial antifúngico frente S. schenckii. e a
administração oral de extratos de alho influencia a liberação de citocinas por macrófagos, o
consumo regular apresenta efeito anti-inflamatório, sendo que o uso agudo pode gerar uma resposta
inflamatória. Animais que consumiram alho responderam de forma mais efetiva no combate da
infecção.
Contudo mais estudos são necessários para entender completamente o mecanismo de ação
do alho no combate as infecções, e com isso elucidar alternativas para o combate a infecções
fúngicas e possíveis tratamentos menos tóxicos ao hospedeiro.

55
9.2. Anexo2. Parecer do CEUA
Protocolo CEUA/FCF/CAr nº 15/2015
Interessado: JOÃO PAULO BURIAN
Orientador: Prof. Dr. Luis Vitor Silva do Sacramento
Projeto: “Influência do extrato de alho (Allium sativum L.) na imunomodulação de
camundongos Swiss”
Parecer nº 33/2015 – Comissão de Ética no Uso de Animais
O Coordenador da Comissão de Ética no Uso de Animais, desta
Faculdade, aprovou ad-referendum as adequações feitas no protocolo do
projeto de pesquisa “Influência do extrato de alho (Allium sativum L.) na
imunomodulação de camundongos Swiss”, apresentado pelo graduando JOÃO
PAULO BURIAN, sob orientação da Prof. Dr. Luis Vitor Silva do
Sacramento, do Departamento de Alimentos e Nutrição desta Faculdade.
O relatório final do protocolo de pesquisa deverá ser
entregue em OUTUBRO de 2015, em formulário para este fim.
Araraquara, 24 de abril de 2015.
Prof. Dr. CARLOS CÉSAR CRESTANI
Coordenador da CEUA





























