Embed Size (px)
Citation preview

MÁRIO CESAR DO NASCIMENTO BEVILAQUA
LIBERAÇÃO DE GABA INDUZIDA POR AMINOÁCIDOS
EXCITATÓRIOS:
CARACTERIZAÇÃO E EFEITOS DA DESNUTRIÇÃO
EM RETINA DE ROEDORES
DISSERTAÇÃO SUBMETIDA À UNIVERSIDADE FEDERAL
DO RIO DE JANEIRO VISANDO A OBTENÇÃO DO GRAU DE
MESTRE EM CIÊNCIAS BIOLÓGICAS (BIOFÍSICA)
Universidade Federal do Rio de Janeiro
Centro de Ciências da Saúde Instituto de Biofísica Carlos Chagas Filho 2008

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

Bevilaqua, Mário Cesar do Nascimento. LIBERAÇÃO DE GABA INDUZIDA POR AMINOÁCIDOS EXCITATÓRIOS:
CARACTERIZAÇÃO E EFEITOS DA DESNUTRIÇÃO EM RETINA DE RATO /
XI, 98/f Dissertação: Mestrado em Ciências Biológicas (Biofísica)
Universidade Federal do Rio de Janeiro (UFRJ) Instituto de Biofísica Carlos Chagas Filho (IBCCF), 2008.
Orientação: Patrícia Franca Gardino e Karin da Costa Calaza. 1. Retina 2. Glutamato 3. GABA 4. Aminoácidos excitatórios 5. Desnutrição

O presente trabalho foi realizado no Laboratório de Neurobiologia da Retina do programa de Neurobiologia do Instituto de Biofísica Carlos Chagas Filho da UFRJ sob orientação da Professora Patrícia Franca Gardino e co-orientação da professora Karin da Costa Calaza (UFF) na vigência de auxílios concedidos pelo Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Financiadora de Estudos e Projetos (FINEP), Fundação de Amparo a Pesquisa do Estado do Rio de Janeiro (FAPERJ), Conselho de Ensino para Graduados da UFRJ (CEPEG-UFRJ), PRONEX e Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES)

LISTA DE ABREVIAÇÕES
AAE – Aminoácido excitatório
ASP – Aspartato
CCG – Camada de células ganglionares
CNI – Camada nucelar interna
CPE – Camada plexiforme externa
CPI – Camada plexiforme interna
DA – Dopamina
DBR – Dieta básica regional
GABA – Ácido gama-aminobutírico
GABA+ - GABA positivas
GABA-IR – Imunorreatividade para GABA
GAD – Ácido glutâmico descarboxilase
GAT – Transportador de GABA
[3H]-GABA – GABA tritiado
KA – Kainato
NMDA – N-Metil-D-Aspartato
SNC – Sistema nervoso central

AGRADECIMENTOS
Primeiramente agradeço a Deus por ter me concedido a oportunidade de
desenvolver este trabalho.
Agradeço às minhas orientadoras: Patrícia Gardino (Paty) e Karin Calaza! E
aproveito para parabenizá-las por desempenharem tão bem esse papel. Agradeço por serem
tão pacientes comigo, mesmo quando eu perguntava “posso não estar entendendo???”.
Muito obrigado!
Agradeço à minha família. Meus pais: José e Raquel, meus irmãos: Marcos, Márcio
e Cecília. Minhas tias Vilma e Hosana e meus primos. Muito obrigado pelo apoio!
Agradeço a todos os meus amigos! Em especial: Fernanda, Ilduara e Bárbara
(amigas da época da escola), muito obrigado pelos conselhos e companheirismo! Ainda:
Anna Claudia (minha sempre orientadora!), Gisele, Cristiano e Camila (amigos de
laboratório).
Agradeço a todos os componentes dos Laboratórios de Neurobiologia da Retina e
Neuroquímica. A ajuda de vocês foi indispensável!
Agradeço em especial aos professores dos Laboratórios supracitados: Patrícia
Gardino, Jan Nora, Fernando Mello, Maria Cristina Mello e Ricardo Reis. Muito obrigado
por cumprirem tão bem o papel de professor e nos permitirem o acesso ao conhecimento. O
papel de vocês é fundamental!
Agradeço também a Rozilane (Rose) pelo apoio técnico (não só técnico, mas
também nas discussões, experimentos...) Muito obrigado!
Agradeço ainda a todos os professores com os quais cursei disciplinas durante o
mestrado (Ricardo Reis, Edna Yamasaki, Patrícia Gardino, Marcelo Morales, Maira Froes,

Ana Martinez, Mônica Rocha). Esse, sem dúvidas, foi um período de intenso aprendizado!
Sou muito grato por ter tido a oportunidade de aprender com vocês!
Enfim, agradeço a todos os que diretamente ou indiretamente participaram desse
trabalho!

Resumo
Glutamato e GABA são os principais neurotransmissores excitatório e
inibitório, respectivamente, presentes no sistema nervoso central. Na retina de
rato, o GABA é expresso nas células amácrinas e células presentes na camada de
células ganglionares (CCG). Estudos anteriores mostraram que aminoácidos
excitatórios (AAE’s) são capazes provocar a liberação de GABA em células da
retina de aves, de gambás e de macacos. Além disso, o desenvolvimento das
células GABAérgicas apresenta um retardo em ratos desnutridos. Neste estudo, a
liberação de GABA endógeno estimulada pelos AAE’s foi estudada em retinas de
ratos adultos normais e desnutridos, usando a técnica de imunohistoquímica. O
tratamento com NMDA em animais normais resultou na redução do número de
células amácrinas imunorreativas para GABA, não exibindo efeito na CCG.
Diferentemente, NMDA reduziu a imunorreatividade para GABA tanto entre células
amácrinas quanto na CCG em animais desnutridos. Cainato e Aspartato
provocaram liberação de GABA em ambas as populações celulares GABAérgicas
na retina de animais normais e desnutridos. De forma geral, o efeito dos AEE’s foi
mais evidente entre células na CCG de ratos desnutridos. Além disso, dopamina
inibiu parcialmente os efeitos do NMDA e aspartato. A liberação de GABA, em
todas as situações estudadas, ficou caracterizada como sendo causada pela
reversão do transportador de GABA.

Abstract
Glutamate and GABA are respectively the major excitatory and inhibitory
neurotransmitters in the central nervous system. In the rat retina, GABA is
expressed in amacrine cells and cells in the ganglion cell layer (GCL). A number of
studies have demonstrated that excitatory amino acids (EAA) can promote GABA
release from retina cells in different species (avian, opossum and monkeys). Also,
the development of GABAergic cells is retarded in malnourished animals. In this
study, GABA release induced by EAA was studied in retinas from normal and
malnourished adult rats using an immunohistochemistry approach. In normal
animals, NMDA treatment promotes GABA release from amacrine cells, but not
from cells in the GCL. The same treatment showed a different effect in
malnourished rat, where NMDA was able to promote GABA release from cells in
GCL. Cainate and Aspartate promote GABA release from all the cell population
studied. The effect of EAA treatment was more significant on GCL of malnourished
animals. In addition, dopamine partially blocked the reduction of the number of
GABA immunoreactive cells when treated with NMDA or Aspartate. Based on our
data GABA release observed was due to by the reversal of GABA transporter.

SUMÁRIO LISTA DE ABREVIAÇÕES....................................................................................................................i AGRADECIMENTOS........................................................................................................ ii RESUMO..............................................................................................................................iv ABSTRACT...........................................................................................................................v SUMÁRIO............................................................................................................................vi
INTRODUÇÃO .................................................................................................................... 8 1. RETINA ............................................................................................................................ 8
1.1 - Estrutura e principais tipos celulares .................................................................... 8 1.2 – Visão geral do Funcionamento da Retina ........................................................... 11 1.3 – Desenvolvimento da Retina ................................................................................. 13 1.4 – Classificação dos neurônios retinianos............................................................... 14 1.4.1 Fotorreceptores ................................................................................................... 14
1.4.2. Células horizontais ......................................................................................... 15 1.4.3. Células bipolares ............................................................................................ 15 1.4.4. Células Amácrinas.......................................................................................... 17 1.4.5 – Células interplexiformes............................................................................... 18 1.4.6 – Células Ganglionares.................................................................................... 19
2. SISTEMAS DE NEUROTRANSMISSORES NA RETINA.......................................................... 21 2.1. Glutamato .............................................................................................................. 21
2.2. GABA ........................................................................................................................ 27 3. LIBERAÇÃO DE GABA INDUZIDA POR GLUTAMATO E SEUS AGONISTAS......................... 31 4. MATURAÇÃO DO SISTEMA DE NEUROTRANSMISSORES E A DESNUTRIÇÃO ...................... 34
4.1. Caracterização do modelo de desnutrição............................................................ 34 4.2. Os efeitos da DBR ................................................................................................. 35
OBJETIVOS ....................................................................................................................... 39
MATERIAIS E MÉTODOS.............................................................................................. 40 1. SACRIFÍCIO DOS ANIMAIS, DISSECÇÃO E ESTIMULAÇÃO DAS RETINAS ............................ 40
1.1. Fixação e obtenção do tecido................................................................................ 43 1.2. Processamento imunohistoquímico ....................................................................... 44
2. CRIAÇÃO DE ANIMAIS DESNUTRIDOS ............................................................................. 46 2.1. Composição da DBR ............................................................................................. 46
3. ANÁLISE QUANTITATIVA................................................................................................ 48
RESULTADOS ................................................................................................................... 50
1. PADRÃO DE IMUNORREATIVIDADE DE GABA E ANÁLISE DO NÚMERO DE CÉLULAS GABA-POSITIVAS EM RETINAS DE RATOS ADULTOS.......................................................... 50 2. IDENTIFICAÇÃO DOS RECEPTORES DE AAE ENVOLVIDOS NA LIBERAÇÃO DE GABA ..... 52
2.1. Análise do perfil dose-resposta dos Aminoácidos excitatórios ............................. 52 2.1.1. Cainato............................................................................................................ 52 2.1.2. Aspartato......................................................................................................... 52
2.2. O Efeito do tratamento com NMDA e influência da dopamina ............................ 59 2.2.1. Análise qualitativa .......................................................................................... 59

2.2.2. Análise quantitativa ........................................................................................ 60 2.3. Efeito do tratamento com cainato e influência da dopamina................................ 64
2.3.1. Análise qualitativa .......................................................................................... 64 2.3.2. Análise quantitativa ........................................................................................ 65
2.4. Efeito do tratamento com aspartato e influência da dopamina ............................ 68 2.4.1. Análise qualitativa .......................................................................................... 68 2.4.2. Análise quantitativa ........................................................................................ 69
3. EFEITOS DA DESNUTRIÇÃO CRÔNICA SOBRE A LIBERAÇÃO DE GABA INDUZIDA POR AMINOÁCIDOS EXCITATÓRIOS............................................................................................ 72
3.1. padrão de imunoreatividade de GABA e análise do número de células GABA-positivas em retinas de ratos adultos. .......................................................................... 72 3.2. Efeito do tratamento com NMDA em retinas de animais desnutridos .................. 72 3.3. Efeito do tratamento com cainato em retinas de animais desnutridos ................. 77
3.4. EFEITO DO TRATAMENTO COM ASPARTATO EM RETINAS DE ANIMAIS DESNUTRIDOS... 80 4. AVALIAÇÃO DA INTEGRIDADE DO TECIDO...................................................................... 83
4.1. Avaliação morfológica de retinas de animais normais e desnutridos................... 83
DISCUSSÃO ....................................................................................................................... 86 1. CARACTERIZAÇÃO DO PADRÃO DE LIBERAÇÃO DE GABA INDUZIDA POR AMINOÁCIDOS EXCITATÓRIOS EM RETINAS DE RATOS NORMAIS................................................................ 86 2. EFEITO DA DESNUTRIÇÃO CRÔNICA SOBRE A FUNCIONALIDADE DE CIRCUITOS GLUTAMATÉRGICOS\GABAÉRGICOS NA RETINA DE RATOS............................................... 94
CONCLUSÕES................................................................................................................... 98BIBLIOGRAFIA..................................................................................................................99

INTRODUÇÃO
1. Retina
1.1 - Estrutura e principais tipos celulares
A retina é uma estrutura do sistema nervoso central (SNC) que apresenta
elevado grau de segregação celular e organização laminar com diferentes camadas
morfo-funcionais. Muitos estudos se utilizam desta estrutura como modelo
experimental de sistema nervoso central. Isso, devido ao fato da retina se
apresentar fora do neuro-eixo central, o que possibilita sua obtenção sem a
contaminação por outros tecidos nervosos ou conjuntivos. Além disso, existe o
fato desta estrutura apresentar quase todos os neurotransmissores presentes no
restante do sistema nervoso central. A grande diversidade de estudos sobre os
tipos celulares retinianos e moléculas envolvidas na transmissão sináptica,
incluindo aqui os próprios neurotransmissores, são fatores muito importantes para
a utilização deste tecido como modelo de estudo tanto do SNC quanto da
compreensão da circuitaria da retiniana.
A retina é um tecido sensorial especializado que reveste internamente a
porção posterior do olho. É neste tecido que ocorre o primeiro estágio de detecção
do sinal luminoso, é também na retina que se detectam diferentes comprimentos
de onda do espectro da luz visível; já é possível a detecção de contraste
simultâneo e de estímulos em movimento.

Em vertebrados, a retina apresenta três camadas de corpos celulares
(camadas nucleares) alternadas com duas camadas de plexos onde ocorrem
contatos sinápticos (camadas plexiformes). Os principais tipos celulares
encontrados na retina são: células fotorreceptoras, células horizontais, células
bipolares, células interplexiformes, células amácrinas, células ganglionares, e as
células de Müller (principal componente glial). Estes tipos celulares encontram-se
distribuídos em camadas, a saber: a camada nuclear externa (CNE) composta
pelos corpos celulares dos fotorreceptores; a camada plexiforme externa (CPE),
exibindo contatos sinápticos entre os fotorreceptores, células horizontais e células
bipolares; a camada nuclear interna (CNI), onde se encontram os corpos celulares
de células horizontais, bipolares, amácrinas, glia de Muller e células
interplexiformes, além de algumas células ganglionares deslocadas; a camada
plexiforme interna (CPI), onde as células neuronais da CNI fazem contatos
sinápticos com as células ganglionares; a camada de células ganglionares (CCG),
onde estão presentes os corpos celulares das células ganglionares, além de
células amácrinas deslocadas. As células ganglionares emitem projeções para
áreas mais superiores do sistema visual. Tais projeções originam-se logo após a
CCG onde axônios das células ganglionares se fasciculam dando origem à
camada de fibras ópticas (CFO) que compõem as fibras do nervo óptico. (figura 1).

Figura 1. Estrutura da retina. Diagrama esquemático mostrando as
camadas e tipos celulares da retina. Modificado de NEUROSCIENCE, Segunda Edição. Dale Purves et al. 2001.

1.2 – Visão geral do Funcionamento da Retina
O funcionamento do sistema visual depende diretamente do padrão de
organização dos tipos celulares presentes na retina, assim como também do
equilíbrio metabólico entre a síntese e liberação de neurotransmissores e
neuromoduladores produzidos na estrutura.
Brevemente: os raios luminosos atravessam toda a estrutura da retina até
atingirem a porção externa dos fotorreceptores (únicas células da retina sensíveis
à luz) onde ocorrerá a transdução do sinal luminoso em sinal neural, e este
adquirirá natureza eletroquímica. A informação é então transmitida a partir dos
fotorreceptores para as células bipolares e destas para as células ganglionares.
Esta via de transmissão é conhecida como via vertical (figura 1) e ocorre
basicamente por meio de mediação glutamatérgica. A informação luminosa flui
pelos tipos celulares supracitados provocando flutuações excitatórias sublimiares
no potencial elétrico das membranas celulares. O único tipo celular no qual é
produzido efetivamente um potencial de ação é a célula ganglionar.
Células horizontais e amácrinas são componentes da via horizontal ou via
modulatória (figura 1). Nestes tipos celulares, o principal neurotransmissor é o
ácido gamma-amino butírico (GABA).
Existem dois tipos de fotorreceptores: os cones e os bastonetes. Essas
células são responsáveis pela transdução do sinal luminoso que ocorre graças a
presença da rodopsina, nos bastonetes e das opsinas, nos cones. No escuro, as
células fotorreceptoras são despolarizadas (potencial de membrana em torno de -
40mV) por influxo de cátions através de canais iônicos que são mantidos abertos

pelas altas concentrações do segundo mensageiro cGMP. Em resposta à
despolarização, os fotorreceptores liberam glutamato. O aumento progressivo de
intensidade luminosa provoca mudança conformacional nas moléculas de opsinas
e rodopsina o que ativa uma cascata enzimática que, por meio de reações de
hidrólise de cGMP, leva à redução dos níveis de cGMP e o consequente
fechamento dos canais iônicos. Dessa forma, observa-se uma proporcional
redução no potencial de membrana dessas células, o que as torna cada vez mais
hiperpolarizadas saturando-se em torno de -65mV. (Baylor, 1987).
As células fotorreceptoras (cones e bastonetes) fazem contato com células
bipolares e células horizontais. De acordo com o fotorreceptor com quem fazem
contato, as células bipolares, podem ser bipolares de cone ou de bastonete. As
células bipolares de cone estabelecem contato direto com as células ganglionares
e as células bipolares de bastonetes conectam-se a células amácrinas, que
passam a informação às células ganglionares, que através dos seus axônios
projetam-se para os núcleos visuais centrais do sistema nervoso até o córtex
visual primario, onde ocorrerá a percepção do estímulo visual. (Kolb, 1994).
Os axônios das células ganglionares emergem do globo ocular pelo disco
óptico, uma região conhecida como “ponto cego” na retina.
Apesar de se encontrarem astrócitos e microglia na retina, o componente
glial neste tecido, como citado anteriormente, é representado principalmente pelas
células de Müller. As células de Müller, além de exercerem importante papel de
suporte e nutrição, estão envolvidas também na captação de íons de potássio
liberados pelos neurônios durante a despolarização.

1.3 – Desenvolvimento da Retina
A retina é um tecido sensorial bastante complexo cuja correta formação
necessita de uma exata sincronização e coordenação de diversos fatores durante
o desenvolvimento. Dentre estes podemos citar: o aporte nutricional necessário
(Almeida et al., 2001; Silveira et al., 2007), a geração dos diversos tipos celulares
de forma apropriada e proporcional, a expressão dos diversos neurotransmissores
(Martins et al., 2007) e aferências de informações luminosas (Landi et al., 2007).
Assim como todas as outras regiões do sistema nervoso central, a retina
origina-se do tubo neural. Durante o desenvolvimento, os olhos emergem de
evaginações do diencéfalo. Tais evaginações são conhecidas como vesículas
ópticas. Cada vesícula óptica se invagina formando um cálice onde a sua parte
interna dá origem ao epitélio neural e a porção externa, ao epitélio pigmentado. A
histogênese da retina segue um padrão especial onde todos os tipos celulares são
gerados e começam a se diferenciar a partir da região central e interna para a as
regiões mais periféricas e externas da retina. A camada nuclear interna (CNI), a
camada plexiforme interna (CPI) e a camada de células ganglioanres (CCG)
desevolvem-se mais precocemente do que as camadas plexiforme externa (CPE)
e nuclear externa (CNE).

1.4 – Classificação dos neurônios retinianos
Como já mencionado, são encontrados basicamente nove tipos celulares na
retina: seis tipos de neurônios – fotorreceptores, células horizontais, células
bipolares, células amácrinas, células ganglionares e células interplexiformes – e
três tipos de células gliais – astrócitos, microglia e células de Müller.
1.4.1 Fotorreceptores
Existem basicamente dois tipos de fotorreceptores que transduzem o
estímulo luminoso em sinal elétrico. Os cones são responsáveis pela visão diurna.
O sistema de cones possui melhor resolução e é responsável pela visão de cores.
Dessa forma, existem três tipos de cones, os azuis, os verdes e os vermelhos que
absorvem comprimentos de luz na faixa do azul, verde e vermelho,
respectivamente. Os cones encontram-se concentrados na região da fóvea.
Os bastonetes encontram-se em maior quantidade que os cones. Eles
detectam luz em baixas intensidades. Possuem grandes quantidades de pigmento
(rodopsina) o que permite aproveitar pequenas quantidades de luz e ampliar este
sinal. Conseqüentemente o sistema de bastonetes é responsável pela visão
noturna.

1.4.2. Células horizontais
Seus corpos celulares estão presentes na porção mais externa da CNI.
Possuem abundantes ramificações que estabelecem contato sináptico com células
bipolares e fotorreceptores. Sua função é de modular o sinal transmitido pelos
fotorreceptores (modulação lateral). Acredita-se que o GABA seja o principal
neurotransmissor utilizado por essas células. Ao receberem input de células
fotorreceptoras, essas células são despolarizadas pelo glutamato liberado no
escuro, pois expressam receptores ionotrópicos. Como resultado da
despolarização, as células horizontais liberam GABA o que leva à hiperpolarização
dos fotorreceptores, estabelecendo, dessa forma, um mecanismo de
retroalimentação negativa (Wu, 1992).
Todos os cones e bastonetes recebem feedback de células horizontais,
mas a proporção numérica dessas células, em relação aos outros tipos celulares é
bastante pequena, geralmente, menos de 5% das células na CNI (Masland, 1986).
1.4.3. Células bipolares
Células bipolares são responsáveis pelo transporte do sinal luminoso (agora
com natureza elétrica) a partir dos fotorreceptores para as células ganglionares.
Interessantemente, as respostas registradas de células ganglionares possuem
natureza bastante diversa, quando comparada às produzidas pelos
fotorreceptores. As células bipolares são implicadas como produtores de tal
diversidade de resposta.

Fotorreceptores, de forma geral, liberam glutamato, mas as células
bipolares respondem de forma diferente às aferências glutamatérgicas. Dessa
forma, distinguem-se dois tipos de células bipolares: as que possuem receptores
de glutamato do tipo ionotrópico, despolarizando-se na presença do
neurotransmissor – bipolares OFF; e as que possuem receptores de glutamato do
tipo metabotrópico, hiperpolarizando-se em resposta ao glutamato – bipolares ON
(Werblin et al., 1991).
As células bipolares de bastonete possuem sempre resposta do tipo ON,
enquanto que as células bipolares de cones podem ser tanto do tipo ON quanto do
tipo OFF. Células bipolares do tipo ON e OFF existem praticamente na mesma
proporção na retina. Essa distinção, criada nesta primeira sinapse retiniana, é
propagada através de todo o sistema visual.
Os campos receptores das células bipolares são circulares e contêm dois
elementos concêntricos e de resposta antagônica. Dessa forma, células bipolares
de centro ON são estimuladas na presença de luz se localizadas no centro do
campo receptor, caso estejam na periferia, são inibidas. Existem ainda as células
bipolares de centro OFF, exibindo comportamento inverso.
Morfologicamente, verifica-se na CPE a formação das chamadas tríades
sinápticas (contato sináptico entre cones ou bastonetes e células bipolares onde
há a participação de duas células horizontais), exibindo as chamadas sinapses em
fita .Os terminais axonais de células bipolares levam a informação a partir da CPE
para a CPI. Nesta camada, as células bipolares estabelecem contato com células
amácrinas e células ganglionares, formando, novamente as sinapses em fita.

Ainda na CPI é possível verificar que as células bipolares do tipo ON e OFF
se estratificam em níveis diferentes: células bipolares do tipo OFF fazem contato
com células ganglionares na superfície externa da CPI, enquanto que as células
bipolares do tipo ON estratificam-se na superfície interna da CPI (Nelson e
Famiglietti, 1978).
1.4.4. Células Amácrinas
As células amácrinas estão localizadas na camada nuclear interna. Seus
prolongamentos fazem contato sináptico com células ganglionares, bipolares,
interplexiforme e outras amácrinas, formando a camada plexiforme interna
(Dowling, 1987). Existem vinte e nove tipos morfológicos de células amácrinas
(Masland, 2001) e provavelmente todos os tipos de neurotransmissores do
sistema nervoso central podem ser encontrados entre os diversos tipos de células
amácrinas (Wagner, 1992). Em contraste com as células horizontais que possuem
um único papel (relacionado com aumento de contraste) as células amácrinas
desempenham várias funções que visam o controle e a forma de respostas das
células ganglionares (Kidd, 1962; Dowling e Boycott, 1966; Kolb e Nelson, 1993).
De acordo com sua localização na retina, podem-se distinguir dois tipos de
células amácrinas: as que têm o corpo celular localizado na camada nuclear
interna e as que apresentam o corpo celular na camada de células ganglionares –
ditas deslocadas (figura 2).

O nome “célula amácrina” (introduzido por Cajal, 1894) significa “célula sem
axônio”. Hoje existem indícios de que alguns tipos de células amácrinas possuem
prolongamentos que funcionam como axônios (Dacey, 1989).
Todas as células ganglionares recebem aferências de células bipolares de
cone, mas sinapses diretas de células bipolares são a minoria de todas as
sinapses nas células ganglionares. A maioria é de células amácrinas. As células
amácrinas fazem sinapses inibitórias com os terminais axonais de células
bipolares, controlando, dessa forma, o sinal de saída para as células ganglionares
(Sterling, 1998).
As células amácrinas tem sido implicadas, além de na modulação do sinal
das células bipolares para as células ganglionares, na sincronização dos disparos
de células ganglionares. Isso se daria com a participação de neurotransmissores
ou de acoplamento – junções comunicantes – entre células ganglionares e células
amácrinas (Vaney, 1994).
1.4.5 – Células interplexiformes
As células interplexiformes possuem como principal característica a
presença de prolongamentos que unem a camada plexiforme externa à interna.
Estas células apresentam seus corpos celulares localizados na camada nuclear
interna. Em comparação aos outros tipos celulares encontrados nessa camada, as
células interplexiforme aparecem em número bastante reduzido.

As células interplexiformes levam informação da camada plexiforme interna
para a camada plexiforme externa, onde fazem contado com células horizontais e
bipolares. (Downling, 1987).
1.4.6 – Células Ganglionares
As células ganglionares, como citado anteriormente, são o único tipo celular
na retina a produzirem efetivamente potenciais de ação. Além disso, essas células
compõem a única via de saída da informação para centros visuais do sistema
nervoso. Essas células estão localizadas na porção mais interna da retina. Seus
axônios se unem, formando a camada de fibras ópticas. Os axônios das células
ganglionares são muito grande e seguem pelo nervo óptico até estruturas mais
centrais, passando pelo quiasma óptico.
As células ganglionares fazem contato com as células bipolares e células
amácrinas na camada plexiforme interna. Existem dois níveis de estratificação
dessas células na CPI. O local de estratificação na CPI e o tipo de célula bipolar
com a qual fazem contato determinam a identidade dessas células como sendo
ganglionares – ON (estratificando-se na porção mais interna da CPI – sublâmina
a), ganglionares – OFF (estratificando-se na porção mais externa da CPI –
sublâmina b) e ganglionares ON-OFF, possuindo dois níveis de estratificação
(figura 2).

Figura 2. Desenho esquemático da retina interna. A figura aponta as células bipolares ON e OFF; células
amácrinas tópicas e deslocadas e diversidade de células ganglionares (ON, OFF e ON-OFF). Bevilaqua, M.C.N. 2008.

2. Sistemas de Neurotransmissores na retina
A transmissão e\ou modulação da informação luminosa entre os neurônios
da retina se dá através liberação de neurotransmissores (na maioria dos casos) ou
por sinapses elétricas (alguns tipos celulares da retina apresentam acoplamento
elétrico – junções comunicantes). A transmissão através da via vertical, como já
mencionado, ocorre principalmente pela liberação de glutamato. Já a modulação e
processamento horizontal ocorre principalmente por meio de transmissão
GABAérgica ou por acoplamento elétrico.
Dentre os neurotransmissores presentes na retina serão destacados no
presente trabalho o glutamato (principal neurotransmissor excitatório no SNC), o
GABA (principal neurotransmissor inibitório no SNC), o aspartato e sua ação como
aminoácido excitatório candidato a neurotransmissor e a dopamina, moduladora
da ação de diversos neurotransmissores.
2.1. Glutamato
O glutamato é considerado o principal neurotransmissor excitatório do SNC,
incluindo a retina. Fotorreceptores, células bipolares e ganglionares liberam
glutamato para mediar a transferência da informação visual para centros visuais
no cérebro (Massey e Redburn, 1987).
A ação pós-sinaptica do glutamato pode ser extraordinariamente diversa.
Isso, graças à extensa de uma extensiva família de receptores de para este

neurotransmissor. Estes receptores podem ser de duas categorias: ionotrópicos
(iGLUr’s) ou metabotrópicos (mGLUr’s) (figura 3).
Os receptores ionotrópicos de glutamato são responsáveis pela
transmissão rápida no SNC. Eles podem ser complexos protéicos homoméricos ou
heteroméricos que se ligam ao glutamato e formam um canal de cátions não
seletivo (Cotman et al., 1989). Os receptores ionotrópicos podem ser classificados
de acordo com sua afinidade a agonistas glutamatérgicos em: N-metil-D-aspartato
(NMDA) e não-NMDA [ácido caínico (KA) e ácido α-amino-3-hidroxi-5-metil-4-
isoxazolepropiônico (AMPA)]. Estes subtipos de receptores ionotrópicos exibem
diferenças na farmacologia, seletividade iônica e propriedades cinéticas.
Receptores metabotrópicos de glutamato possuem habilidade de, por meio
de ativação de proteína G, acionar vias de segundos mensageiros (fosfolipase C
ou adenilil ciclase) que irão provocar respostas diversas na célula pós-sinaptica.
Os receptores do tipo NMDA (NMDAR) constituem uma classe de
receptores com muitas peculiaridades. Estes receptores apresentam alta
permeabilidade ao Ca++ e só são completamente ativados quando estão ligados
ao glutamato e glicina simultaneamente. Os NMDAR formam um canal iônico cujo
poro está constitutivamente bloqueado por Mg++ (quando a membrana apresenta-
se em potencial de repouso – bloqueio voltagem-dependente), dessa forma, é
necessária uma pré despolarizarão para que haja a liberação do canal e sua
ativação pelos agonistas glutamatérgicos. Estes receptores podem ser bloqueados
completamente por antagonistas não-competitivos tais como PCP, quetamina e
MK801 (Nakanishi et al., 1994).

Receptores NMDA são na maioria dos casos hetero-oligômeros compostos
de uma combinação de subunidades que podem ser de dois tipos: NMDAR1
(NR1) ou NMDAR2 (NR2). Existem oito isoformas de NR1 (NR1a-h) que são
geradas por splicing alternativo e quatro isoformas de NR2 (NR2A-D). A
subunidade NR2D possui ainda duas variantes resultantes de splicing alternativo,
NR2D1 e NR2D2. Receptores homoméricos formados por NR2 não são
funcionais, enquanto os formados por NR1, embora possuam correntes iônicas
relativamente baixas, apresentam-se funcionais. Entretanto, formas heteroméricas
formadas por subunidades NR2 e NR1 formam canais com correntes
significativamente mais altas e grande variedade funcional (Witkovsky, 1999;
Wässle, 1998).
Os receptores não-NMDA (AMPA e KA) formam canais iônicos voltagem-
independentes que, quando ativados, apresentam alta condutância a íons Na+.
Os receptores AMPA são canais iônicos pentaméricos formados por uma
ou mais das quatro subunidades constituintes destes receptores. Estas são
conhecidas como: GluR(1-4). possuem alta afinidade ao AMPA. O cainato também
é um importante agonista desses receptores.
Os receptores do tipo cainato são subdivididos em duas categorias:
receptores de baixa afinidade, formados pelas subunidades GluR5, GluR6 e
GluR7 e receptores de alta afinidade, formados pelas subunidades KA1 e KA2.
A ação do glutamato em receptores NMDA e não NMDA pode ser
verificada pelo uso de antagonistas. Assim, NMDAR são bloqueados pelo
antagonista não competitivo MK-801 e os receptores AMPA/KA pelo
CNQX/DNQX.

Vários estudos têm revelado a localização de receptores de glutamato na
retina de rato. Esses estudos geralmente lançam mão de técnicas como
hibridização in sito e imunohistoquímica.
No animal adulto, todas as subunidades de NMDAR podem ser detectadas
(Brandstatter et al., 1994). Análises de hibridização in situ revelam que RNAm de
subunidade NR1 encontra-se largamente distribuída através da retina. Estudos
imunohistoquímicos mostram que todas as subunidades de NMDAR são
expressas na CCG. O padrão de imunomarcação revela ainda que a subunidade
NR1 é expressa em células horizontais, amácrinas, além da CPI, onde é possível
verificar duas bandas de marcação. Ainda na CNI é possível verificar a expressão
de NR1 em células de Muller.(Brandstatter et al., 1994; Blanco et al., 1999;
Gründer et al., 2000). A subunidade NR2A é encontrada em células horizontais.
Duas bandas são formadas na CPI tanto na imunomarcação de NR2A quanto
NR2B. Verifica-se ainda a expressão de NR2B na camada plexiforme externa.
NR2C e D são encontradas em células amácrinas, ganglionares e amácrinas
deslocadas (Flecher et al., 1999).
As subunidades de receptores do tipo AMPA e KA são diferencialmente
distribuídas através da CNI e CCG. Possivelmente, podem ser encontradas em
células horizontais, bipolares, amácrinas e ganglionares. Receptores AMPA
provavelmente possuem envolvimento na transmissão em sinapses em fita da
CPE e CPI (Branstätter et al., 1998).
Finalmente, a ação do glutamato em seus receptores é regulada por um
processo de recaptação citoplasmática que é mediado por transportadores de alta
afinidade (figura 3). Uma importante característica do transporte de glutamato é o

fato de este ser eletrogênico, isto é, existe uma corrente iônica (Na+) que
acompanha o transporte. Existem cinco tipos de transportadores de glutamato
(EAAT 1-5) em ratos. Na retina, estão localizados em células de Müller e alguns
neurônios pós-sinapticos. Embora estas proteínas transportem preferencialmente
o L-glutamato, são capazes de transportar igualmente o L- e D-aspartato (Nicholls
et al., 1990).

Figura 3 – desenho esquemático de uma sinapse glutamatérgica. A figura ilustra a diversidade de receptores glutamatérgicos, assim como os transportadores de aminoácidos excitatórios. Modificado de NEUROSCIENCE, Segunda Edição. Dale Purves et al. 2001.

2.2. GABA
O GABA é considerado o principal neurotransmissor inibitório de diversas
estruturas do SNC, incluindo a retina em diferentes espécies (Mosinger et al., 1986;
Yazulla, 1986). GABA é sintetizado pela enzima descarboxilase do ácido glutâmico
(GAD), embora, em estágios precoces do desenvolvimento, uma via de síntese
alternativa a partir de putrescina seja responsável por sua síntese (De Mello, 1976;
Hokoç et al., 1990; Yamasaki et al., 1999). Essa via alternativa parece exercer um
papel menos significativo em animais adultos. Após sua síntese, o GABA é
armazenado em vesículas até a sua liberação (dependente de cálcio). Nem todas
as células que possuem GABA, possuem a sua enzima de síntese. Isso porque
algumas células captam GABA do meio extracelular, via transportadores específicos
em suas membranas. Portanto, estas células possuem GABA muito embora não
sejam capazes de sintetizá-lo (Agardh et al., 1987a, b).
Os receptores para GABA podem ser ionotrópicos (GABAA e GABAC) e
metabotrópicos (GABAB) (figura 4). O GABA pode inibir neurônios por aumentar a
condutância de íons cloreto, pela ativação dos receptores GABA
B
A e GABAC, ou de
íons potássio, via ação em receptores GABABB, causando, em ambos os casos,
hiperpolarização. De fato, é verdade que em alguns estágios do desenvolvimento, o
GABA é responsável por eventos despolarizantes (Yamashita e Fukuda, 1993). Isso
está relacionado à concentração intracelular de íons Cl- que no indivíduo neonato
mostra-se bastante aumentada quando se compara com o adulto. A concentração
intracelular de Cl- é determinada pela expressão de transportadores para estes íons
(NKCC e KCC2). Dessa forma, em estágios precoces do desenvolvimento, estão

presentes os transportadores NKCC, que mantêm alta a concentração intracelular do
ânion. Em estágios mais tardios, há a substituição de NKCC por KCC2, que
transporta o Cl- para o meio extracelular (Ben-Ari et al., 1997). Quando receptores
ionotrópicos de GABA são ativados em indivíduos adultos a resposta verificada é
hiperpolarização, pois o Cl- flui, seguindo o gradiente eletroquímico, para dentro da
célula.
Na retina de rato o GABA é expresso pelas células horizontais e células
amácrinas – tópicas e deslocadas (Mosinger et al., 1986; Yamasaki et al., 1999; Lee
et al., 1999). Os sistemas GABAérgicos e glutamatérgicos interagem na retina.
Como mencionado anteriormente, a via vertical (glutamatérgica) sofre ação
modulatória da via horizontal (GABAérgica). Esta modulação é mediada pela ação
das células horizontais na camada plexiforme externa e pelas células amácrinas
na camada plexiforme interna. A maioria das células amácrinas (92%) são
GABAérgicas e recebem aferência de células bipolares glutamatérgicas (Marc e
Liu, 2000).
Alguns estudos levantam a possibilidade de GABA ter um papel neurotrófico
em períodos precoces do desenvolvimento em função da modulação dos níveis de
cálcio. De fato, tem sido demonstrada ação neurotrófica de GABA (Spoerri, 1988;
Ikeda et al., 1997) assim como glutamato, via NMDAR (Vecino et al., 1999).
A ação sináptica do GABA é terminada pela sua captação seletiva para o
interior do terminal pré-sinaptico e de células gliais e/ou degradação pela enzima
GABA-2-oxoglutarato-transaminase. A captação de GABA se dá por carreadores de
alta afinidade e é observada em células amácrinas, horizontais e células na camada
de células ganglionares. Em retinas de mamíferos, ao contrário do observado em

retina de aves, GABA pode ser também captado por células de Müller (Moran et al.,
1986). A recaptação de GABA é mediado por um transportador que carrega GABA e
sódio para dentro da célula. Quatro tipos de transportadores para GABA foram
descritos no SNC são eles: GAT-1, GAT-2, GAT-3 e GAT-4 (Clark e Amara, 1993;
Liu et al., 1993) muito embora até o momento somente três (GAT-1-GAT-3) tenham
sido localizados na retina (Johnson et al., 1996; Hu et al., 1999). Em função do
aumento de sódio intracelular, próximo destes transportadores, ocorre a reversão da
direção de fluxo e consequentemente a liberação de GABA. De fato, tem sido
observado que reversão do transportador parece mediar grande parte da liberação
de GABA induzida por aminoácidos excitatórios (Tapias e Arias, 1982; Morán e
Pasantes-Morales, 1983; Yazulla, 1983; Yazulla e Kleinschmidt, 1983; Do
Nascimento e De Mello, 1985; Schwartz, 1987; Duarte et al., 1993; Ferreira et al.,
1994; Carvalho et al., 1995).

Figura 4 - Desenho esquemático ilustrando uma sinapse GABAérgica. A figura identifica a diversidade de receptores e transportadores de GABA. Adaptado de Owens e Kriegstein, 2002.

3. Liberação de GABA induzida por glutamato e seus agonistas
É sabido que o glutamato é um potente agente estimulador de atividade
neuronal. A estimulação com L-glutamato é responsável pela liberação de vários
agentes neuroativos, incluindo o GABA na retina de peixe, pinto, gambá e macaco
(Yazulla et al., 1985; Andrade da Costa et al., 2001; Calaza et al., 2001, 2003 e
2005). Muitos estudos sugerem que a liberação de GABA estimulada por glutamato
ou agonistas na retina de diversas espécies se dá de forma independente de cálcio
externo (Duarte et al., 1993; Ferreira et al., 1994; Carvalho et al., 1995) e ocorre via
ativação de receptores ionotrópicos NMDA e não NMDA (do Nascimento et al., 1998;
Castro et al., 1999). Em vários estudos, a liberação de GABA induzida por
aminoácidos excitatórios (AAE’s) foi analisada utilizando-se um protocolo de
liberação de [3H] GABA. Nesses trabalhos, células em cultura eram expostas ao [3H]
GABA que após sua captação, eram estimuladas com os AAE’s. Tal protocolo
experimental, entretanto, não possibilita a identificação das populações neuronais
responsáveis por tal liberação.
Os efeitos dos AAE’s na liberação de GABA podem ser parcialmente
revertidos pela aplicação de MK-801 (antagonista de receptor NMDA) e totalmente
bloqueados pela adição de DNQX (antagonista de receptores não-NMDA) e MK-801.
Isso mostra que ambos os subtipos de receptores ionotrópicos estão envolvidos
neste fenômeno (Duarte et al., 1993; Kubrusly et al., 1998; Calaza et al., 2001).
L- e D-aspartato também têm sido apontados como responsáveis pela
liberação de GABA em vários estudos utilizando-se retina de diversas espécies (Do

Nascimento e De Mello, 1985; Bloomfield e Dowling, 1985 a e b; Cha et al., 1986;
Kubrusly et al., 1998; Calaza, 2001, 2003). Ao contrário do D-glutamato, o D-
aspartato também foi eficaz em provocar a liberação de GABA em células de retina
de pinto em cultura (Kubrusly et al., 1998). Fortes evidências sustentam a hipótese
de que L- e D-aspartato ativam receptores NMDA com alta eficiência. Hashimoto e
Oka (1997) descobriram que as formas isômeras de aspartato ligam-se a sítios de
glutamato com alta afinidade. Além disso, foi observado que L-aspartato induz a
liberação de [3H] GABA na retina pela ativação específica de receptores do tipo
NMDA (Yazulla et al., 1983; Kubrusly et al., 1998; Calaza et al., 2001). Kubrusly e
colaboraderes (1998) demosntraram que tanto L- quanto D-aspartato são capazes
de provocar a liberação de [3H] GABA em culturas de células de retina de pinto via
ativação de receptores NMDA nas mesmas concentrações que glutamato e NMDA.
Poucos são os estudos que identificam os tipos celulares envolvidos na
liberação de GABA induzida por AAE’S. Andrade da Costa e colaboradores (2000)
verificaram que na retina de Cebus Apella glutamato e cainato são responsáveis pela
liberação de GABA de células da retina interna, células amácrinas e interplexiformes.
Nesse animal, a ativação de receptores NMDA não tem nenhum efeito sobre a
liberação de GABA. Em estudos recentes, Calaza e colaboradores (2001, 2003 e
2006 a e b) identificaram os tipos celulares envolvidos na liberação de GABA
induzida por AAE’s, inclusive o aspartato, na retina de pinto e gambá. Esses estudos
mostraram que na retina de pinto NMDA e aspartato não provocaram a liberação de
GABA de células horizontais, entretanto, induziram a redução de, respectivamente,
50 e 60% da imunoreatividade para GABA entre as células amácrinas GABA+. A
exposição das retinas ao cainato provocou liberação de GABA de forma significativa

nas células amácrinas e horizontais. Na retina de gambá, NMDA e cainato
provocaram a redução do número de células amácrinas tópicas e deslocadas
imunoreativas para GABA. Aspartato somente provocou a liberação de GABA a
partir de células amácrinas tópicas, não exercendo nenhum efeito nas deslocadas.
Até o presente momento não existe nenhum dado publicado em relação aos tipos
celulares envolvidos na liberação de GABA induzida por AAE’s na retina de rato. Isso
torna bastante conveniente o estudo do padrão de liberação de GABA em roedores,
mamíferos não marsupiais.
Vários indícios sugerem que a liberação de GABA induzida por glutamato e
seus agonistas, incluindo o aspartato se dê de forma não vesicular. O mais aceito é
que o GABA seja liberado via reversão do transportador. As principais evidências
que sustentam isso são: a liberação de GABA é independente de cálcio externo,
excluindo a possibilidade de liberação vesicular (Tapia e Arias, 1982; Yazulla e
Kleinschmidt, 1983; Do Nascimento e De Mello, 1985); O processo é dependente de
sódio tal como a atividade do transportador (Tapia e Arias, 1982); a exposição do
tecido/cultura de células estimulado(a) ao inibidor específico deste transportador
(NNC-711) abole completamente o efluxo de GABA induzido por AAE’S.
A Dopamina (importante neurotransmissor no SNC, incluindo a retina) tem
sido implicada na modulação da ação glutamatérgica na liberação de GABA. O que
se verifica é que a aplicação de dopamina durante a estimulação com AAE’S é
responsável pela inibição parcial na liberação de GABA. Estudos com retina de
peixes datados da década de 80 (Yazulla e Kleinschmidt, 1982; O’Brien e Dowling,
1985; Kato et al., 1985) mostraram que talvez a dopamina agisse de forma a diminuir
a concentração intracelular de sódio (via ativação de bomba de sódio) o que inibiria o

transporte reverso de GABA. Esses trabalhos propuseram ainda que a dopamina,
via ativação de receptore D1, provocaria aumento nos níveis de AMPc.
Em experimentos com cultura de células de retina de galinha, foi verificado
que somente a liberação de [3H]-GABA induzida por ativação de receptores NMDA é
sensível à modulação dopaminérgica. (Do Nascimento et al., 1998). Nesse trabalho,
constatou-se ainda que a inibição da dopamina não era mediada por receptores
dopaminérgicos clássicos – antagonistas para estes receptores não foram capazes
de bloquear o efeito inibitório. Fortes evidências sugerem a existência de um sítio
específico para a dopamina nos receptores NMDA, no qual a ligação da dopamina
teria um efeito inibitório nas correntes induzidas pela ativação do receptor (Castro et
al., 1999).
4. Maturação do sistema de neurotransmissores e a desnutrição
4.1. Caracterização do modelo de desnutrição
A desnutrição crônica (ingesta insuficiente de nutrientes) tem sido apontada
como importante agente deletério em diversos tecidos, incluindo estruturas do SNC.
As lesões decorrentes da desnutrição sobre o sistema nervoso são bastante
extensas. A desnutrição pode reduzir o número de neurônios, células gliais, e suas
conexões; pode alterar o crescimento dendrítico, causar retardo ou déficit
permanente dificultando a transmissão do sinal entre as células nervosas (Jacobson,
1991).

O modelo de desnutrição a ser utilizado no presente trabalho se caracteriza
como sendo uma grave e crônica restrição nutricional (multideficiente). Tal
desnutrição é provocada pela administração da Dieta Básica Regional (DBR).
A composição da DBR foi determinada de acordo com um inquérito alimentar
realizado na população da Zona da Mata de Pernambuco pela Universidade Federal
de Pernambuco – UFPE (Batista Filho, 1968). O efeito dessa dieta em ratos pode se
comparar ao padrão de desnutrição encontrado nessa população, principalmente
entre as crianças, com sintomatologia comparada ao Marasmo (Teodósio et al.,
1990). Dentre as principais características dessa dieta destaca-se o fato desta ser
pobre em certos nutrientes, principalmente em relação às proteínas. A dieta
comercial apresenta cerca de 22% de proteína (caseína) enquanto a DBR possui
cerca de 7,8% deste nutriente (Guedes et al., 1996).
4.2. Os efeitos da DBR
Os animais tratados com a DBR mostram-se, desde o nascimento, menores
que os animais normais. A curva de crescimentos desses animais é extremamente
débil e não exibem diferenças, em relação ao peso corporal, entre os sexos. O peso
dos cérebros desses animais mostra-se 20% menor enquanto que a taxa de
mortalidade entre eles é de cerca de 24% (Lago et al., 1997).
Dados obtidos a partir da administração da DBR revelam que a desnutrição
provocada é responsável por alterações prejudiciais no aprendizado e
desenvolvimento comportamental. Tais alterações são diretamente proporcionais ao

período e intensidade da carência alimentar. Animais alimentados com a DBR
tendem a demonstrar maior capacidade exploratória, um importante indicador de
carência nutricional. (Teodósio et al., 1979).
Almeida e colaboradores (2001) demonstraram que a expressão de
neurotransmissores como GABA, acetilcolina e proteínas ligadoras de cálcio
(calbidina e calretinina) na retina de ratos desnutridos pela administração da DBR
mostra-se bastante alterada. Tais alterações se caracterizam principalmente como
um possível retardo na maturação dos sistemas metabólicos das substâncias
envolvidas. Surpreendentemente, com a maturidade, verifica-se a compensação do
quadro de retardo de modo que aos 30 dias pós-natal (P30) os animais desnutridos
apresentam o mesmo padrão de expressão de neurotransmissores e proteínas
ligadoras de cálcio que o animal normal. Ainda nesse estudo, verificou-se que o
período de morte celular na retina de ratos desnutridos também se encontra
alterado. O pico de morte celular em animais desnutridos apresentou-se aos 8 dias
pós-natal (P8) (verificado pela presença de núcleos picnóticos), enquanto que nos
animais normais, a morte celular é verificada entre os dias 6 e 7 pós-natal (P6-7).
Algumas alterações causadas pela desnutrição via DBR mostram-se,
aparentemente reversíveis ou seja, possivelmente há uma reorganização do sistema
visando suprir tal carência nutricional. Outras, no entanto, mostram-se irreversíveis,
como, por exemplo o processo de mielinização de fibras centrais (nervo óptico) e
periféricas (nervo sural). Verifica-se, em animais desnutridos a diminuição da bainha
de mielina e o aumento do número de fibras não mielinizadas. Tais características
são verificadas durante o desenvolvimento até a fase adulta (Almeida et al., 2001;
Almeida et al., dados não publicados).

Silveira e colaboradores (2007) mostraram que a neurogênese de células com
fenótipo GABAérgico também se encontra alterada em animais alimentados com a
DBR. Em ratos desnutridos, o pico de geração de células GABAérgicas na retina
interna ocorre no 20º dia embrionário (E20) enquanto que nos animais controle, em
E18. O período de geração de células GABAérgicas na retina interna desses animais
mostrou-se bastante atrasado. Em P4 – idade que caracteriza o fim do período
mitótico de células GABA+ em animais normais (Lee et al., 1999) – há ainda grande
atividade mitótica na retina interna dos animais desnutridos. A desnutrição, nesse
caso, mostra-se responsável por alterar o período de neurogênese de células
GABAérgicas na retina.
Embora, algumas características, principalmente as que sugerem atraso na
formação/maturação da retina, sejam aparentemente reversíveis, ao ponto que na
idade adulta, não se verifica mais diferenças entre os animais normais e os
desnutridos, não se sabe quais seriam as implicações desse atraso no
desenvolvimento funcional da estrutura. Como citado anteriormente, o
desenvolvimento adequado da retina depende diretamente de uma série de eventos
perfeitamente orquestrados que incluem geração de tipos celulares e expressão de
neurotransmissores e substancias neuroativas. A literatura carece de dados que
mostrem os efeitos deste tipo de desnutrição sobre a funcionalidade (em nível
celular) de circuitos neurais e interação entre sistemas de neurotransmissores.
Tendo em vista que a liberação de GABA induzida por AAE’s é um excelente
modelo para o estudo de interações entre sistemas de neurotransmissores, torna-se
bastante conveniente verificar os possíveis efeitos da desnutrição sobre esta

característica funcional em retinas de animais adultos que, pelo menos nos aspectos
já estudados, mostra-se “recuperada” nessa idade.

OBJETIVOS O presente estudo tem como objetivo geral estudar os circuitos
glutamatérgicos/GABAérgicos na retina interna de ratos, bem como os efeitos da
desnutrição sobre tais circuitos.
Os objetivos específicos são:
1) Estabelecer o padrão de dose-resposta de AAE’s como cainato e aspartato
através de imunocitoquímica para GABA.
2) Verificar os efeitos de AAE’s na liberação de GABA em retinas intactas através
de imunocitoquímica para GABA, identificando na estrutura retiniana os tipos
celulares que respondem aos diferentes agonistas glutamatérgicos, incluindo o
aspartato. Estabelecer a contribuição de cada receptor glutamatérgico na
liberação de GABA.
3) Através de análises quantitativas, estabelecer a porcentagem de diferentes
tipos de células que respondem ao tratamento com AAE’s na estrutura da retina.
4) Verificar a modulação dopaminérgica sobre os receptores glutramatérgicos na
liberação de GABA. Identificar as populações celulares que são sensíveis à ação
dopaminérgica (se houver).
5) Através de análises quantitativas, estabelecer a porcentagem de diferentes
tipos de células que respondem ao tratamento com AAE’s e que possuem
modulação dopaminérgica na estrutura da retina.
6) Estabelecer uma análise inicial dos efeitos da desnutrição crônica sobre a
liberação de GABA induzida por AAE’s, incluindo o aspartato.

MATERIAIS E MÉTODOS
Foram utilizadas, no presente estudo, retinas de ratos albinos de linhagem
Wistar de ambos os sexos provenientes do biotério central do Instituto de
Bioquímica Médica da UFRJ. Os animais eram mantidos sob condições
constantes de temperatura (cerca de 25ºC) e ciclo de claro/escuro (12 horas no
claro e 12 horas no escuro). A manipulação e sacrifício dos animais seguiram as
normas estabelecidas pela Sociedade Brasileira de Neurociências e
Comportamento (SBNeC) e aprovadas pela comissão de uso de animais
experimentais do Instituto de Biofísica Carlos Chagas Filho da Universidade
Federal do Rio de Janeiro.
1. Sacrifício dos animais, dissecção e estimulação das retinas
Animais adultos (com idades de 30 a 45 dias) foram sacrificados por
inalação de clorofórmio. Os olhos foram enucleados e seccionados ao nível da
córnea e gentilmente separado do humor vítreo. A calota posterior do olho,
contendo a retina, foi utilizada em diferentes protocolos experimentais (ver
desenho esquemático de um olho de rato e suas principais estruturas na figura 5).
As retinas eram então imersas em solução de Ringer que tinha a seguinte
composição: NaCl 125mM, KCl 205mM, CaCl2 2,0mM, NaHCO3 25mM, NaH2PO4
1,25mM, glicose 25mM, MgCl2 10mM. Após o preparo, a solução tinha o seu pH

ajustado para 7,2-7,4. A solução de Ringer se caracteriza por ser isosmótica ao
tecido, com osmolaridade entre 300-315mOsM.
Plano de corte
Figura 5 – Desenho esquemático de vista longituninal de um olho de rato evidenciando as principais estruturas e o plano de corte utilizado para a obtenção do tecido retiniano. Bevilaqua, M.C.N. 2008.
Os experimentos de liberação de GABA consistiam na estimulação do
tecido retiniano em solução de Ringer contendo um determinado agonista
glutamatérgico (NMDA – Research Biochemical Inc.; KA – Research Biochemical
Inc.; ou ASP – Sigma). Os dados obtidos dessas retinas estimuladas eram
comparados com a retina controle (retina que foi mantida durante todo tempo em
solução de Ringer livre de agonistas de glutamato). Detalhadamente: durante a

estimulação, as retinas eram mantidas em temperatura constante de 37ºC e
constantemente aeradas com uma mistura carbogênica de 95%O2 e 5%CO2 em
um tubo de ensaio contento a solução de Ringer e um agonista de glutamato
(NMDA - 100µM; KA - 10µM/50µM/100µM; ASP - 100µM/500µM/2mM),
completando um volume final de 2mL durante 45 minutos.
Para verificar o efeito de antagonistas de receptores ionotrópicos de
glutamato, a ação modulatória da dopamina e a participação do transportador
neuronal de GABA, foram utilizados, em uma pré-incubação, respectivamente:
MK-801 (10µM), antagonista não competitivo de receptores do tipo NMDA e
DNQX(100µM), antagonista seletivo de receptores do tipo não-NMDA; dopamina
(2mM) e NNC-711 (100µM), inibidor específico do transportador neuronal de
GABA. A pré-incubação era realizada na ausência dos agonistas glutamatérgicos
e tinha a duração de 30 minutos. Em seguida as retinas eram tratadas com os
agonistas glutamatérgicos ainda na presença das drogas utilizadas na pré-
incubação por 45 minutos (ver tabela 1).
A solução de dopamina era preparada sempre no dia do experimento na
presença de ácido ascórbico (Sigma - 200µM) para prevenir sua oxidação e
pargilina (Sigma - 200µM) para prevenir sua degradação. Pargilina e ácido
ascórbico estavam presentes também na solução de Ringer do experimento em
questão.
Em experimentos com NMDA, o Ringer utilizado na incubação era livre de
magnésio e continha 2mM de glicina, co-agonista de receptores do tipo NMDA.

Tabela 1 – exemplo esquemático de experimento de estimulação de retina com aminoácidos excitatórios
1.1. Fixação e obtenção do tecido
Após o período de incubação, as retinas eram rapidamente lavadas em
tampão fosfato (0,1M - pH 7,2) e fixadas por imersão em solução de
paraformaldeído 4% (PA 4%) e tampão fosfato por duas horas. Em seguida, o
tecido era lavado por três vezes com tampão fosfato e mantido no mesmo por uma
noite. As análises foram realizadas com processamento imunohistoquímico em
cortes transversais de retina. Para obtenção dos cortes, as retinas foram
crioprotegidas sendo colocadas em tampão fosfato com concentrações crescentes
de sacarose (15% e 30%). Após uma noite em tampão fosfato + sacarose 30%, as
retinas eram seccionadas ao meio e, na presença de meio de inclusão aquoso

(OCT – Sakura Finetek, Torrance, CA) eram congeladas em uma placa de
alumínio em nitrogênio líquido (-196ºC) o bloco congelado era então posicionado
em um suporte para cortes em criostato. Cortes transversais de 14µm eram
obtidos em criostato sob temperatura entre -15ºC e -20ºC e colhidos em lâminas
de vidro. As retinas de um mesmo experimento – por exemplo: retina controle;
estimulada com NMDA e estimulada com NMDA na presença de MK-801 – eram
montadas em um mesmo bloco, cortadas e colhidas em uma mesma lâmina. As
lâminas eram gelatinizadas previamente em solução de gelatina (0,5%) e alúmem
de cromo (0,002%) em água destilada. Cada lâmina era gelatinizada por duas
vezes e secavam por, no mínimo, oito horas. Os cortes colhidos em lâminas
gelatinizadas eram mantidos em temperatura ambiente até secarem e após isso,
armazenados em freezer na temperatura de -20ºC.
1.2. Processamento imunohistoquímico
O método de imunohistoquímica foi utilizado para avaliar, indiretamente, por
meio de marcação de GABA remanescente, a quantidade e a origem, em nível
celular, do GABA liberado com a estimulação das retinas.
Para tal, as lâminas contendo cortes obtidos em criostato foram
descongeladas a temperatura ambiente. Como o método imunohistoquímico
utilizado baseia-se na ação de uma enzima peroxidase, foi necessário uma pré-
incubação (por 15 minutos) das lâminas contendo os cortes em peróxido de
hidrogênio – 3% (H2O2) a fim de exaurir a peroxidase endógena. Após isso, as

lâminas eram lavadas em solução tampão fosfato salina (PBS) por três vezes com
intervalo de cinco minutos entre as lavagens – todas as lavagens com PBS eram
feitas dessa forma – e seguia-se a incubação com uma solução de 5% de soro de
albumina bovino (BSA) em PBS adicionado de 0,25% de Triton (PBST) por 45
minutos. Em seguida, as lâminas foram incubadas por 24 horas em solução de
PBST contendo anticorpo anti-GABA (anticorpo primário) obtido a partir de coelho
(Vector) na diluição de 1:5000. Após o período de incubação com anticorpo
primário, as lâminas eram lavadas com PBS e incubadas com o anticorpo
secundário (anti-Rabbit biotinilado, Vector) na diluição de 1:200 por uma hora. Os
cortes eram novamente lavados em PBS e incubados com ABC (complexo
avidina-biotina; Kit Elite Vectastain, Vector – 1:50) por uma hora. Finalmente, após
lavagem em PBS, a reação era revelada com o kit para peroxidase com
cromógeno SG (Vector) por 10 minutos e então eram lavados com PBS e
montados com meio de glicerol tamponado (40% em tampão fosfato 0,2M).
2. Criação de animais desnutridos
Os animais utilizados nesse trabalho foram divididos em dois grupos:
controle (alimentados com dieta comercial - purina®) e desnutridos (alimentados
com a Dieta Básica Regional – DBR). Para a formação do grupo de animais
desnutridos, foi utilizada a substituição da dieta comercial pela Dieta Básica
Regional (DBR) a ser oferecida às ratas durante toda lactação, a partir do primeiro

dia de vida pós-natal (P0) dos filhotes, e posteriormente, diretamente aos filhotes
após o desmame (aos vinte e um dias).
2.1. Composição da DBR
A DBR se caracteriza por ser uma dieta desbalanceada que contém apenas
8% de proteína (a maioria de origem vegetal), baixo teor lipídico (0,8%) e excesso
de glicídios (69%). A dieta é composta por: feijão mulatinho (Phaseolus vulgaris),
Mandioca (Manioc esculenta), batata doce (Ipomaea batatas) e charque (carne
bovina salgada e prensada). Como já mencionado, tal composição foi obtida a
partir de um inquérito alimentar realizado em 1968 por Batista Filho na região de
Zona da Mata em Pernambuco. Nesse inquérito, foram investigados os principais
componentes da dieta alimentar dessa população. Teodósio e colaboradores, em
1990, apresentaram a DBR como um modelo experimental para desnutrição e
disponibilizaram a composição centesimal dessa dieta.
Os alimentos são cozidos e ressecados na estufa (cerca de 65ºC). Após
isso, os alimentos são moídos até serem obtidas farinhas de cada alimento (com
exceção da farinha de mandioca, que pode ser comprada pronta). Os alimentos
triturados são misturados na mesma proporção encontrada nos inquéritos
alimentares e com um pouco de água morna, forma-se uma massa a partir da qual
a ração é confeccionada. As tabelas 2 e 3 mostram a composição centesimal da
dieta comercial e da DBR.

Componentes g% Proteína Carboidrato Lipídio Vitaminas Sais Minerais Fibra Kcal/Kg
Caseína Comercial
27,08 22,0 88,0
Amido de Milho
57,12 0,58 50,44 0,12 0,09 205,16
Óleo Vegetal 7,5 7,5 67,5 Vitamina Hidrossolúvel
1,0 1,0
Vitamina Lipossolúvel
1,0 1,0 1,0 9,0
Sais Minerais 4,0 4,0 Metionina 0,3 Papel de Filtro 2,0 2,0 Total 100,0 22,58 50,44 8,62 2,0 4,09 2,0 369,66
Tabela 2 – Composição centesimal dos alimentos da ração comercial Purina®
Componentes g% Proteína Carboidrato Lipídio Vitaminas Sais Minerais
Fibra Kcal/Kg
Feijão Mulatinho
18,34 3,99 10,66 0,24 0,57 1,09 60,76
Mandioca 64,81 0,84 48,59 0,12 0,43 5,64 198,80 Charque 3,74 2,74 0,43 0,06 0,06 14,57 Gordura 0,35 0,35 3,15 Batata Doce 12,76 0,30 9,99 0,03 0,20 0,48 58,87 Total 100,00 7,87 69,67 0,95 1,02 7,21 336,15
Tabela 3 – Composição centesimal dos alimentos da Dieta Básica Regional. Observe, comparando as tabelas 2 e 3 a porcentagem total de proteína das duas dietas.

3. Análise quantitativa
O número de células imunorreativas ao GABA foi quantificado em secções
radiais de retina de 14µm de espessura. As contagens foram realizadas em
secções radiais alternadas, a fim de evitar a contagem de uma mesma célula
duplamente. Utilizando-se um microscópio Axioskop (Zeiss) de campo claro,
obtinha-se o valor do numero de células GABA+, assim como os perfis de células
não marcadas na CCG e nas três fileiras mais internas da CNI (posição onde se
localizam as células amácrinas tópicas). Calculava-se, em seguida, a
porcentagem de células marcadas em relação ao total de células (marcadas e
não-marcadas). Tais contagens foram realizadas com uma lente objetiva de 40X
de aumento e uma ocular de 10X munida de gratícula (figura 6).
Cada dado referente à contagem de células provém de um experimento
com N=3. Isso quer dizer que três animais foram utilizados. Para cada animal, no
mínimo três lâminas foram analisadas. Logo, para N=3, foi necessário a análise
de, no mínimo, nove lâminas. Cabe lembrar que os cortes das retinas de
diferentes grupos de um mesmo experimento eram colhidas na mesma lâmina,
sofrendo, dessa forma, o mesmo processamento histológico e imunohistoquímico.
Os dados foram analisados estatisticamente usando o one-way ANOVA e o
pós-teste de Bonferroni.

Figura 6 – Esquema de como foi feita a análise quantitativa em cortes de retina. A figura mostra o desenho de gratícula sobreposta à foto de um corte de retina.

RESULTADOS
1. Padrão de imunorreatividade de GABA e análise do número de células GABA-positivas em retinas de ratos adultos.
As retinas de ratos adultos, que não sofreram nenhum tratamento, foram
fixadas logo após a enucleação do olho e processadas imunohistoquímicamente
para GABA. Isso permitiu a análise do padrão de expressão do neurotransmissor.
Nesse estágio, o GABA mostra-se presente em células amácrinas (três fileiras de
células mais internas na CNI) e células na camada de células ganglionares. Além
disso, foi possível verificar marcação para GABA por toda a CPI (figura 7). Os
dados estão de acordo com os expostos na literatura (Mosinger et al., 1986).
A análise quantitativa foi realizada com o intuito de identificar de forma
numérica o padrão de imunoreatividade para GABA nas populações de células
estudadas. A análise quantitativa dos corpos celulares marcados (valores tomados
como porcentagem do total de células presentes na região relativa a células
amácrinas da CNI) revelou que cerca de 47% das células amácrinas eram GABA+
e 51% das células na camada de células ganglionares expressavam GABA na
retina de ratos adultos (figura 7).

CNE CPE CNI CPI CCG
Figura 7. A. Fotomicrografia de corte transversal de retina de rato imunoreagida para GABA. Barra= 10µm. B. Gráfico do número de células GABA+ na CNI (células amácrinas) e células na CCG em retinas não tratadas.

2. Identificação dos receptores de AAE envolvidos na liberação de GABA
2.1. Análise do perfil dose-resposta dos Aminoácidos excitatórios
2.1.1. Cainato Foi necessário o estabelecimento de uma curva dose-resposta para
cainato e aspartato para escolher a concentração ótima a ser utilizada. A dose
utilizada na estimulação com NMDA (100µM) foi obtida a partir de dados da
literatura (Calaza et al., 2001, 2003 e 2006) e mostrou-se eficaz em induzir
diminuição na imunorreatividade para GABA em retinas tratadas. Estudos
referentes ao uso do cainato mostram que este é capaz de estimular o tecido
retiniano de aves e gambá (Calaza et al., 2001, 2003 e 2006) na concentração de
100µM, porém, ao utilizar esta concentração na retina de ratos, observa-se a
destruição parcial do tecido (dado não mostrado). Dessa forma, tornou-se
necessário o estabelecimento de doses ótimas com as quais o tecido seria
estimulado, porém não danificado.
KA, em todas as concentrações utilizadas por 45 minutos, induziu
significativa diminuição na imunorreatividade e no número de células GABA+ tanto
na CNI quanto na CCG sem danificação do tecido (Figuras 8 e 9).
A partir de então, a dose estabelecida para a estimulação com KA foi a de
25µM, por se tratar de um valor intermediário, onde é verificada a estimulação do
tecido sem a sua danificação (figura 23).

CNE CPE CNI CPI CCG
Figura 8 – Fotomicrografias de cortes transversais de retinas imunoreagidas para GABA A. retina controle e após tratamento com: B. KA 10µM. C. KA 25µM. D. KA 50µM. Barra=10µm.
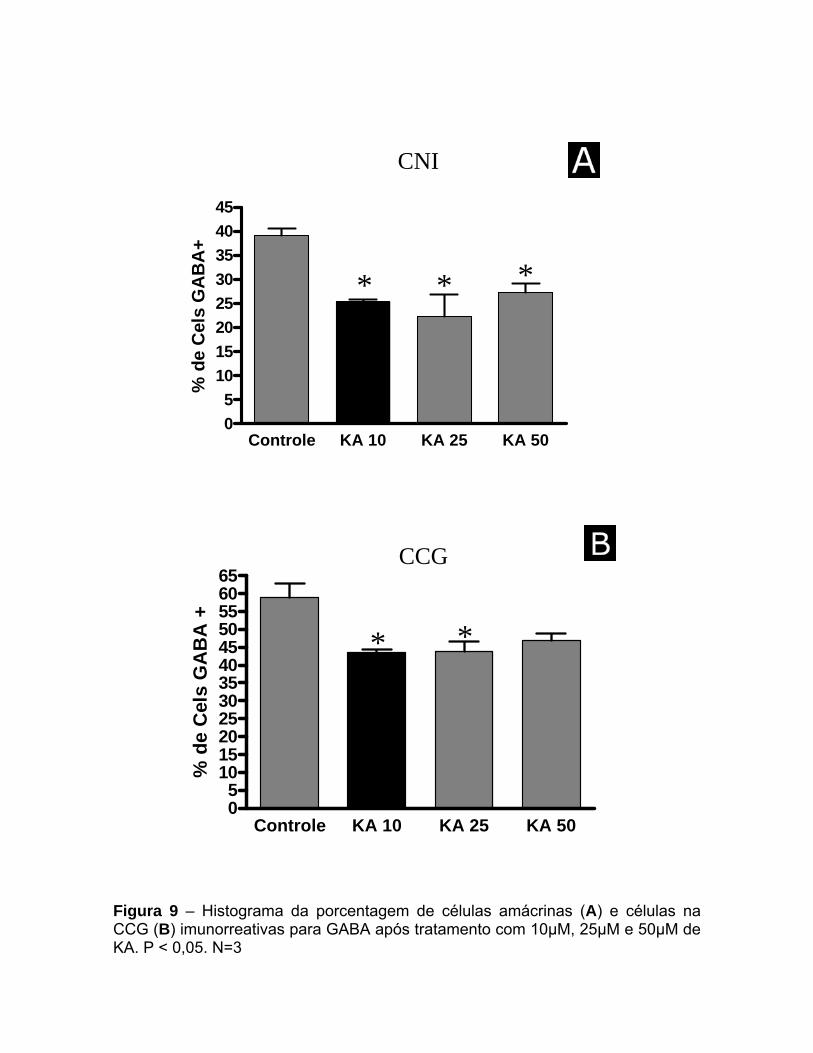
Amácrinas Figura 9 – Histograma da porcentagem de células amácrinas (A) e células na CCG (B) imunorreativas para GABA após tratamento com 10µM, 25µM e 50µM de KA. P < 0,05. N=3
* * *
* *
Controle KA 10 KA 25 KA 5005
1015202530354045
% d
e C
els
GA
BA
+
Ganglionares
CNI
CCG
Controle KA 10 KA 25 KA 5005
101520253035404550556065
% d
e C
els
GA
BA
+

2.1.2. Aspartato
A dose inicial de ASP testada em experimento-piloto foi de 100µM – dose
utilizada por Calaza e colaboradores em trabalhos com retinas de pinto e gambá
(2001, 2003, 2006). Tal dose não se mostrou eficiente em provocar qualquer
liberação de GABA em retinas de ratos. Dessa forma, novamente foi necessário o
estabelecimento de uma curva dose-resposta para verificar se o ASP exerce
algum efeito na retina de rato e, se afirmativo, identificar a concentração
necessária de ASP na qual fosse possível verificar seu efeito na diminuição de
imunorreatividade para GABA em cortes de retina.
As retinas foram tratadas com ASP nas concentrações de 100µM, 2mM e
5mM por 45 minutos. A análise qualitativa mostrou que o ASP foi capaz de
provocar redução significativa no número de células GABA+ quando administrado
em concentrações iguais ou superiores a 2mM (figura 10). Em retinas tratadas
com essa concentração de ASP verificou-se significativa redução na
imunorreatividade para GABA em células amácrinas e células na CCG (figura 10).
A análise quantitativa mostrou que em retinas tratadas com 2mM de ASP a
redução no número de corpos celulares imunorreativos para GABA entre as
células amácrinas na CNI foi de cerca de 30%, enquanto que esta mesma
concentração de ASP foi capaz de reduzir a marcação para GABA em 35% na
CCG. O tratamento com 5mM de ASP provocou efeito mais notável na CNI,
reduzindo a imunorreatividade para GABA entre essas células em 60% (figura 11).

A partir deste experimento, a dose escolhida de ASP para experimentos
futuros foi de 2mM, pois a partir desse valor nota-se o efeito do aminoácido em
induzir a liberação de GABA, sem, no entanto, danificar o tecido (Figura 25).
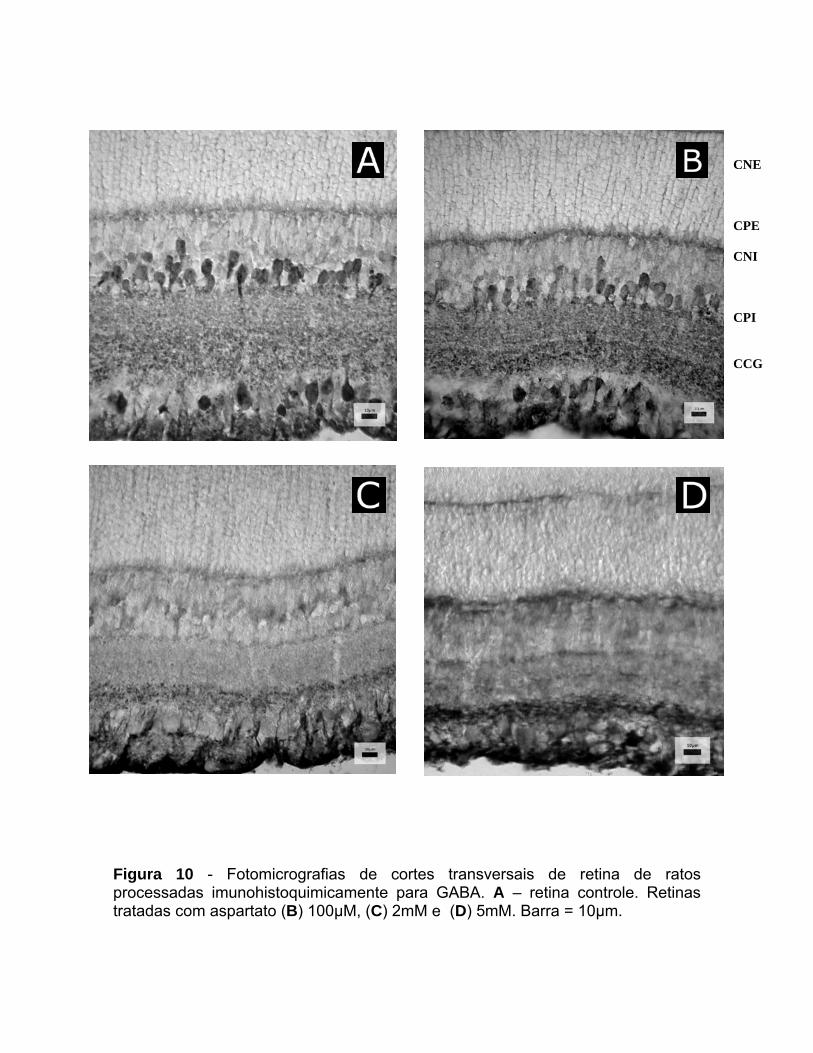
CNE CPE CNI CPI CCG
Figura 10 - Fotomicrografias de cortes transversais de retina de ratos processadas imunohistoquimicamente para GABA. A – retina controle. Retinas tratadas com aspartato (B) 100µM, (C) 2mM e (D) 5mM. Barra = 10µm.

**
* *
Controle 100µM 2mM 5mM0
10
20
30
40
Grupos
% d
e cé
ls. G
AB
A +
/to
tal d
e cé
lula
s am
ácri
nas
Controle 100µM 2mM 5mM05
10152025303540455055
Grupos
% d
e cé
ls. G
AB
A +
/to
tal d
e cé
lula
s na
CC
G
CNI
CCG
Figura 11 – Histogramas da porcentagem de células amácrinas (A) e células na CCG (B) imunorreativas para GABA após tratamento com 100µM, 2mM e 5mM de aspartato. P < 0,05. N=3.

2.2. O Efeito do tratamento com NMDA e influência da dopamina
2.2.1. Análise qualitativa
Qualitativamente o NMDA não parece induzir a diminuição de
imunorreatividade para GABA na CCG.
Retinas tratadas com 100µM de NMDA por 45 minutos sofreram redução na
imunoreatividade para GABA e no número de células GABA+ quando comparada
com retinas controle. A redução foi observada somente na CNI (figura 12).
O antagonista não competitivo do receptor NMDA, MK801, foi utilizado para
confirmar a ação específica de NMDA sobre seus receptores. Como esperado,
MK801 bloqueou completamente o efeito do NMDA (figura 12).
Como já citado, a dopamina tem sido mostrada como agente modulador da
ação do glutamato em receptores do tipo NMDA (Castro et al., 1999) e na
liberação de GABA induzida por NMDA e ASP (Calaza et al., 2001). Retinas
incubadas com NMDA na presença de dopamina (2µM) apresentaram um
bloqueio parcial na ação estimulatória do NMDA, apresentando, dessa forma, uma
menor redução no número de células GABA+ na CNI (figura 12).
Em retinas tratadas com NMDA na presença de NNC-711 foi observado o
completo bloqueio dos efeitos do NMDA.
Estes dados juntos sugerem que de fato os receptores do tipo NMDA estão
sendo ativados durante o tratamento com o NMDA, pois o efeito é completamente
bloqueado com o uso do MK-801. Além disso, a ação do NMDA sobre seu
receptor aparentemente é modulada pela dopamina. Os dados mostram ainda que

o processo de liberação de GABA é provocado pela reversão do transportados de
GABA, pois é completamente bloqueado pelo NNC-711.
2.2.2. Análise quantitativa
A análise quantitativa revelou que a porcentagem de células amácrinas
GABA+ na retina controle é cerca de 35%, mostrando-se reduzida em relação à
retinas intactas, não submetidas à incubação em Ringer. Em acordo com os dados
qualitativos, o tratamento com NMDA por 45 minutos foi eficiente em induzir a
diminuição de cerca de 62,5% do número de células amácrinas GABA+ na CNI.
Entre as células na CCG, o NMDA não exerceu efeito significativo (diminuição da
imunorreatividade para GABA de 15%). Além disso, as retinas que foram tratadas
com NMDA mais dopamina apresentaram uma porcentagem maior de células
amácrinas GABA+ na CPI, reduzindo o efeito do NMDA de 25% em média.
Confirmando a inibição parcial do efeito do NMDA pela dopamina (figura 13)
observada anteriormente em neurônios de hipocampo de rato (Castro et al., 1999).
Tanto o MK801 quanto o NNC-711 bloquearam completamente o efeito do
NMDA (figura 13) Estes dados corroboram a idéia de que a redução do número de
células GABA+ induzida por NMDA ocorre via estimulação direta do receptor
NMDA e reversão do transportador de GABA, caracterizando um processo de
liberação de GABA de forma não vesicular.
Como o NMDA não alterou a imunorreatividade para GABA nem o número
de células GABA+ na CCG, sugere-se que o NMDA não induz a liberação de

GABA nessa população celular ou que a dose utilizada tenha sido insuficiente em
provocar tal efeito.

CNE CPE CNI CPI CCG
Figura 12 - Fotomicrografia de cortes transversais de retina de ratos processadas imunohistoquimicamente para GABA. A – retina controle. Retinas tratadas com (B) NMDA 100µM, (C) 100µM de NMDA + DA e (D) 100µM de NMDA + NNC-711. Barra = 10µm.

*
Amácrinas
Controle
NMDA
NMDA+DA
NMDA+NNC
NMDA + MK
0
10
20
30
40
% d
e cé
lula
s G
AB
A+
Ganglionares
Controle
NMDA
NMDA+DA
NMDA+NNC
05
1015202530354045
% d
e cé
lula
s G
AB
A+
CNI
CCG
Figura 13 – Histograma da porcentagem de células amácrinas (A) e células na CCG (B) imunorreativas para GABA após tratamento com 100µM de NMDA, 100µM de NMDA + DA; 100µM de NMDA + NNC e 100µM de NMDA + MK801. P < 0,05. N=3.

2.3. Efeito do tratamento com cainato e influência da dopamina
2.3.1. Análise qualitativa
Retinas que foram tratadas com 25µM de cainato sofreram redução na
imunorreatividade para GABA tanto na CNI quanto na CCG. Diferentemente do
observado com o NMDA, a dopamina não foi capaz de modular o efeito do cainato
em nenhuma das retinas tratadas. Retinas tratadas com cainato na presença de
dopamina (2µM) apresentaram o mesmo padrão de imunorreatividade que as
retinas tratadas somente com cainato. O DNQX, antagonista específico de
receptores não-NMDA, bloqueou completamente a ação do KA, confirmando a
participação de receptores não-NMDA no processo de liberação de GABA
observado. O efeito do cainato foi completamente bloqueado quando a
estimulação ocorria também na presença de NNC-711. Em conjunto, esses dados
sugerem que o KA atua ativando receptores não-NMDA que não são modulados
pela dopamina e que o efluxo de GABA provocado ocorre via transportador de
GABA (figura 14).
2.3.2. Análise quantitativa
O cainato reduziu em cerca de 50% o número de células amácrinas na CNI
e diferentemente do NMDA, foi capaz de reduzir também o número de células na
CCG em torno de 20%, como verificado também na análise qualitativa. Os

resultados da análise quantitativa confirmam que a dopamina não foi capaz de
modular o efeito da estimulação com KA em nenhuma população celular em
análise. E, finalmente, a quantificação revelou que, como observado de forma
qualitativa, tanto o DNQX quanto o NNC-711 bloquearam totalmente a ação do
cainato, indicando, mais uma vez, que o efluxo de GABA induzido por ativação de
receptores do tipo não-NMDA ocorre via transportador membranar de GABA
(figura 15).
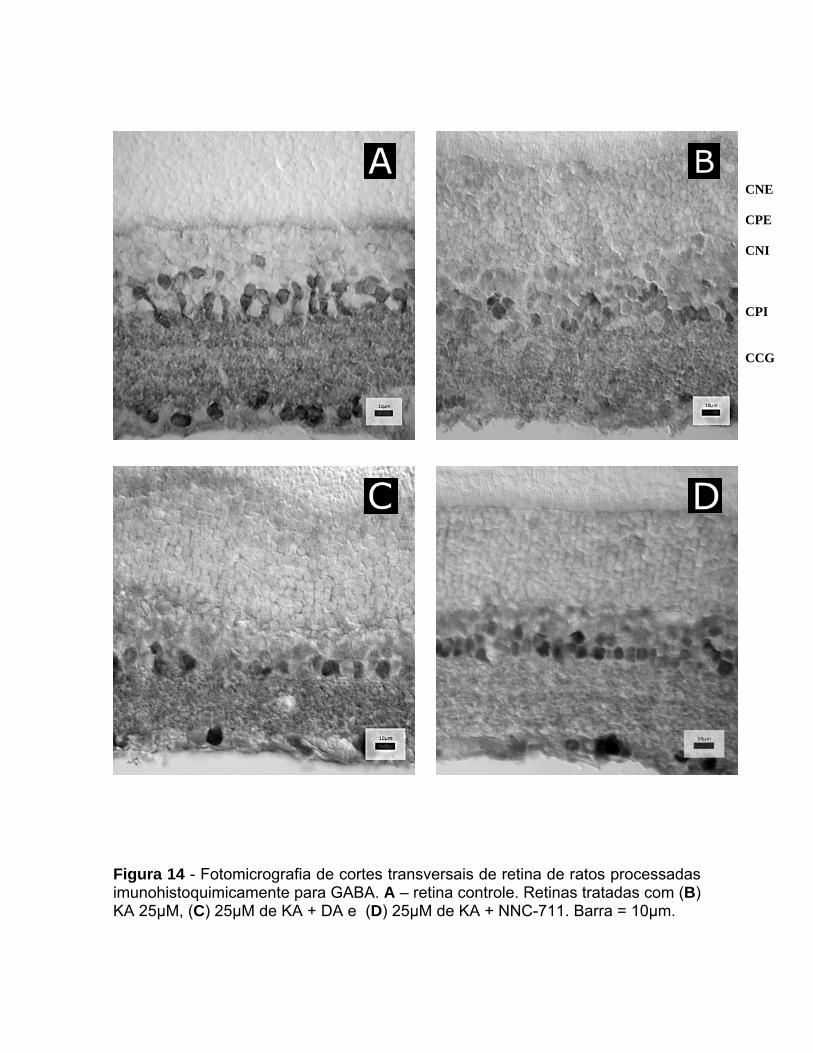
CNE CPE CNI CPI CCG
Figura 14 - Fotomicrografia de cortes transversais de retina de ratos processadas imunohistoquimicamente para GABA. A – retina controle. Retinas tratadas com (B) KA 25µM, (C) 25µM de KA + DA e (D) 25µM de KA + NNC-711. Barra = 10µm.
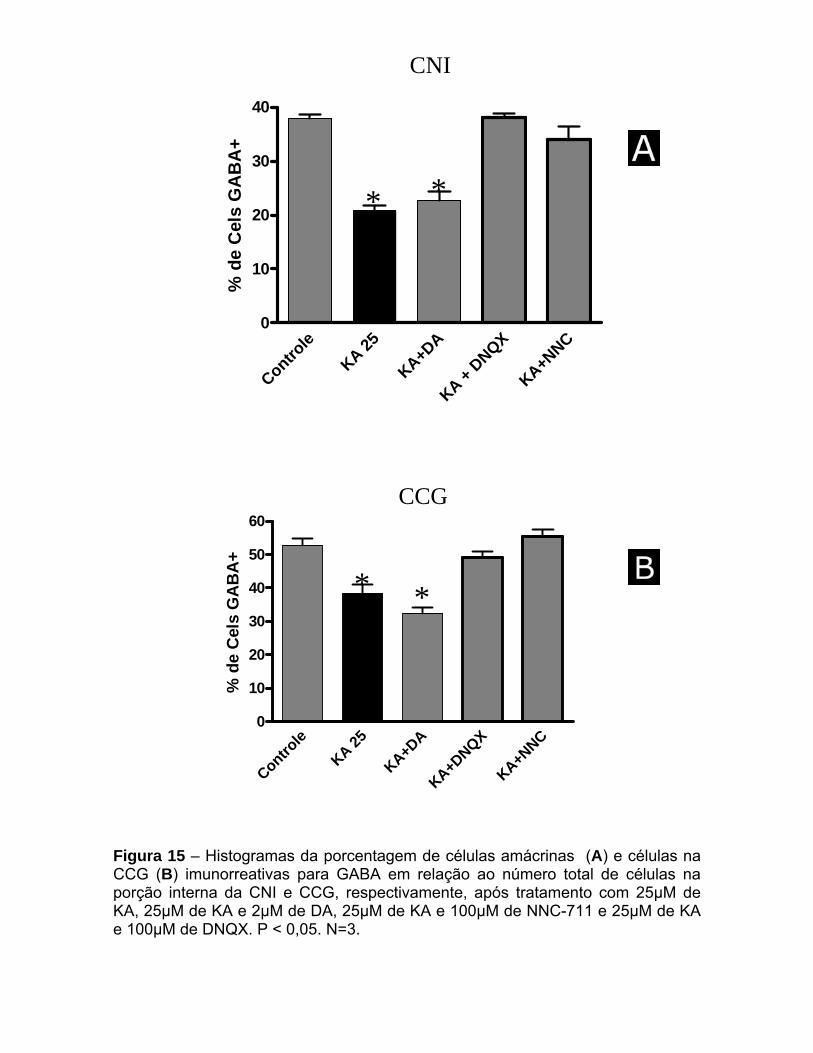
Figura 15 – Histogramas da porcentagem de células amácrinas (A) e células na CCG (B) imunorreativas para GABA em relação ao número total de células na porção interna da CNI e CCG, respectivamente, após tratamento com 25µM de KA, 25µM de KA e 2µM de DA, 25µM de KA e 100µM de NNC-711 e 25µM de KA e 100µM de DNQX. P < 0,05. N=3.
**
**
Amácrinas
Controle
KA 25
KA+DA
KA + DNQX
KA+NNC
0
10
20
30
40
% d
e C
els
GA
BA
+
Ganglionares
Controle
KA 25
KA+DA
KA+DNQX
KA+NNC
0
10
20
30
40
50
60
% d
e C
els
GA
BA
+
CNI
CCG

2.4. Efeito do tratamento com aspartato e influência da dopamina
2.4.1. Análise qualitativa
O tratamento com aspartato (2mM), mesmo na presença de magnésio, foi
eficaz em provocar redução no padrão de imunorreatividade para GABA entre as
células amácrinas e também entre as células na CCG. Da mesma forma que
observado para o NMDA, a dopamina também inibiu parcialmente a redução da
imunorreatividade para GABA induzida por aspartato em ambas as populações
celulares. Ainda, o efeito do tratamento com aspartato foi totalmente bloqueado
quando na presença de MK801, um antagonista clássico de receptor NMDA, o que
confirma a ação do aspartato sobre este tipo de receptor. Quando na presença de
NNC-711, assim como ocorre com o NMDA e com o KA, verifica-se o bloqueio
total do efluxo de GABA induzido por aspartato (figura 16).
Esses dados mostram que provavelmente a ação do aspartato ocorra via
ativação de receptores NMDA, pois o efeito é bloqueado por MK801, apesar de
ocorre mesmo na presença de magnésio. O fato de o efeito ser modulado pela
dopamina reforça as suposições de que haja ativação de receptores NMDA, uma
vez que, como mostrado anteriormente, a dopamina parece não modular
receptores do tipo não NMDA. E como ocorre com os outros AAE’s estudados, o
efluxo de GABA parece ser proveniente de uma reversão no transportador de
GABA, sendo completamente bloqueado pelo NNC-711.

2.4.2. Análise quantitativa
A análise quantitativa revelou que o tratamento com aspartato a 2mM por
45 minutos foi responsável pela diminuição do número de células GABA+ de cerca
de 50% na CNI quando comparando com a retina controle. A incubação com
aspartato provocou ainda a diminuição do número de células GABA+ na CCG
(cerca de 23% do controle). O efeito do tratamento com aspartato foi modulado
pela incubação conjunta com dopamina tanto na CNI quanto na CCG, ainda que
de forma não significativa, mas é possível observar uma tendência de que a
dopamina está provocando redução no efeito do aspartato. A redução na
imunorreatividade para GABA provocada pelo tratamento com aspartato foi
completamente bloqueada pela administração de MK-801 durante a estimulação
com o aminoácido excitatório. Verificou-se o bloqueio do efeito do aspartato
também quando a retina foi estimulada na presença de NNC-711 (figura 17).
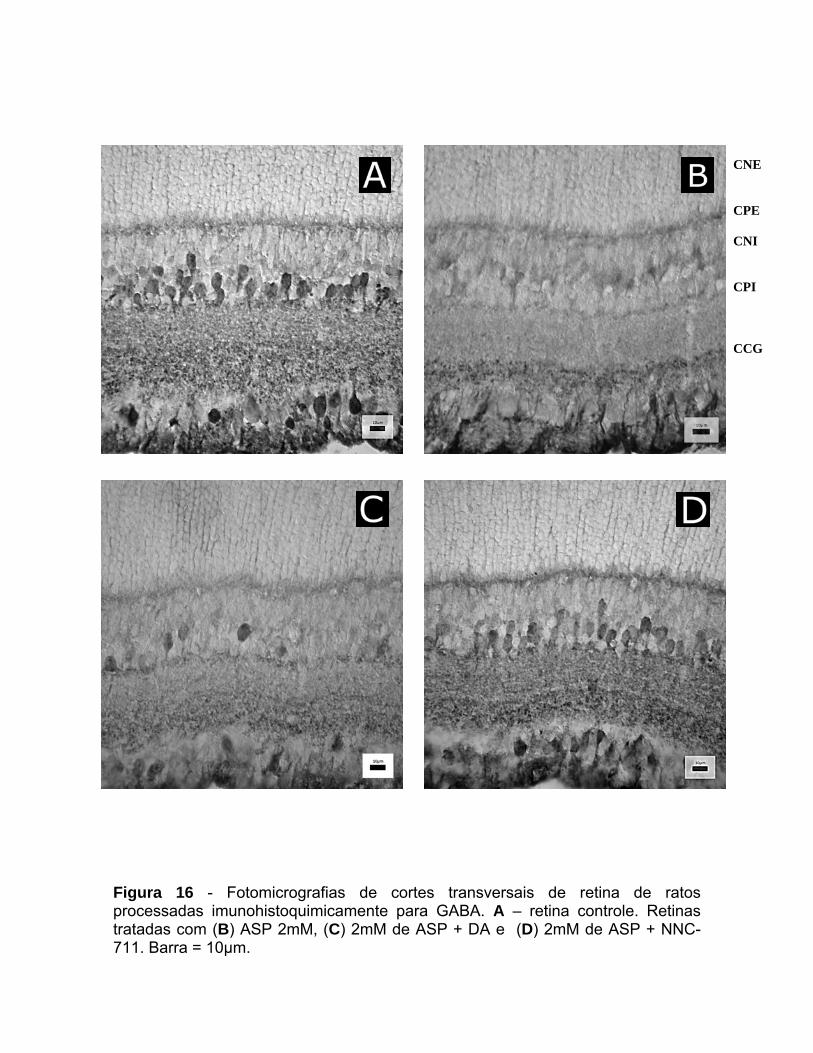
CNE CPE CNI CPI CCG
Figura 16 - Fotomicrografias de cortes transversais de retina de ratos processadas imunohistoquimicamente para GABA. A – retina controle. Retinas tratadas com (B) ASP 2mM, (C) 2mM de ASP + DA e (D) 2mM de ASP + NNC-711. Barra = 10µm.
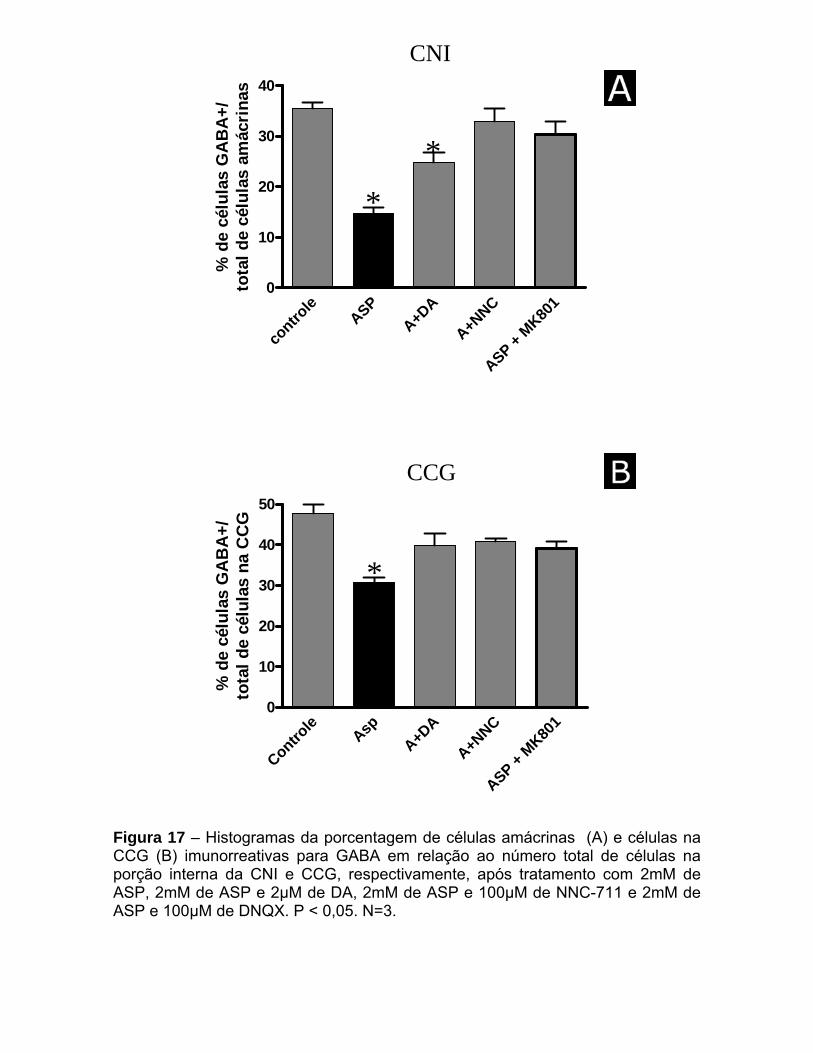
* Figura 17 – Histogramas da porcentagem de células amácrinas (A) e células na CCG (B) imunorreativas para GABA em relação ao número total de células na porção interna da CNI e CCG, respectivamente, após tratamento com 2mM de ASP, 2mM de ASP e 2µM de DA, 2mM de ASP e 100µM de NNC-711 e 2mM de ASP e 100µM de DNQX. P < 0,05. N=3.
*
*
Células Amácrinas
contro
leASP
A+DA
A+NNC
ASP + MK80
10
10
20
30
40
% d
e cé
lula
s G
AB
A+/
tota
l de
célu
las
amác
rina
s
Células Ganglionares
Controle
AspA+D
A
A+NNC
ASP + MK80
10
10
20
30
40
50
% d
e cé
lula
s G
AB
A+/
tota
l de
célu
las
na C
CG
CNI
CCG

3. Efeitos da desnutrição crônica sobre a liberação de GABA induzida por aminoácidos excitatórios
3.1. padrão de imunoreatividade de GABA e análise do número de células GABA-positivas em retinas de ratos adultos.
O padrão de imunorreatividade para GABA foi investigado em retinas de
animais desnutridos aos trinta dias pós natal. Para isso, foi utilizada a mesma
metodologia já descrita para animais alimentados com dieta padrão. Dessa forma,
a população de células GABA+ na CNI de animais desnutridos é de cerca de 50%.
Enquanto que na CCG a porcentagem de células GABA+ é de cerca de 49%.
(figura 18).Como já descrito por Almeida e colaboradores (2001 e 2002), em
retinas de ratos tratados com a DBR, a detecção de GABA aos 30 dias não se
diferencia do encontrado nas retinas de animais controle (alimentados com dieta
comercial) com a mesma idade.

CNE CPE CNI CPI CCG
Figura 18. A. Fotomicrografia de corte transversal de retina de rato desnutrido imunorreagida para GABA. Barra= 10µm. B. Gráfico do número de células amácrinas e células na CCG GABA+ em retinas não incubadas.

3.2. Efeito do tratamento com NMDA em retinas de animais desnutridos
Retinas de animais de 30 dias desnutridos, incubadas com 100µM de
NMDA, apresentaram uma redução significativa da imunorreatividade para GABA
quando comparadas com as retinas controle (também proveniente de animais
desnutridos, porém não estimuladas com NMDA mantidas em solução de Ringer).
Surpreendentemente, a redução podia ser observada, além de na CNI, também na
CCG, onde com a mesma concentração de NMDA, não foram observados
qualquer efeito em retinas de animais alimentados com dieta comercial (comparar
figuras 12 e 19).
A análise quantitativa mostrou que o NMDA provocou uma diminuição na
porcentagem de células GABA+ na CNI de cerca de 25% enquanto que na CCG a
redução foi de 35% (figura 20).
A confirmação de que se tratava da ativação do receptor de NMDA clássico
foi obtida com o tratamento das retinas com NMDA e MK801. Este antagonista de
receptores NMDA bloqueou completamente a ação do NMDA em reduzir o
número de células GABA+ tanto na CNI quanto na CCG (figura 20).

CNE CPE CNI CPI CCG
Figura 19. Retinas de ratos desnutridos imunorreagidas para GABA. A. retina controle. B. retina estimulada com NMDA a 100µM. C. Retina estimulada com NMDA na presença de MK-801. Barra = 10µm.

*
*
CNI
Figura 20 – Histogramas da porcentagem de células amácrinas (A) e células na CCG (B) imunorreativas para GABA em relação ao número total de células na porção interna da CNI e CCG, respectivamente, após tratamento com 100µM de NMDA e 100µM de NMDA na presença de MK801. P < 0,05. N=3.

3.3. Efeito do tratamento com cainato em retinas de animais desnutridos
Em retinas de animais desnutridos, o cainato reduziu significativamente o
número de células amácrinas GABA+ em cerca de 50% e na CCG também em
cerca de 50%. A redução na imunorreatividade provocada por cainato em retinas
de animais desnutridos mostrou-se mais evidente do que em animais alimentados
com dieta comercial (comparar figuras 14 e 21).
O efluxo de GABA provocado pelo tratamento com cainato foi
completamente bloqueado pelo tratamento conjunto de cainato com DNQX,
antagonista de receptores não-NMDA. Tal fato sugere que a liberação de GABA
induzida por cainato observada em animais desnutridos ocorre via ativação de
receptores não-NMDA (figura 22).

CNE CPE CNI CPI CCG
Figura 21. Retinas de ratos desnutridos imunorreagidas para GABA. A. retina controle. B. retina estimulada com KA a 25µM. C. Retina estimulada com KA na presença de DNQX. Barra = 10µm.

*
*
CNI Figura 22 – Histogramas da porcentagem de células amácrinas (A) e células na CCG (B) imunorreativas para GABA em relação ao número total de células na porção interna da CNI e CCG, respectivamente, após tratamento com 25µM de KA e 25µM deKA na presença de DNQX. P < 0,05. N=3.

3.4. Efeito do tratamento com aspartato em retinas de animais desnutridos
O tratamento com aspartato em retinas de animais desnutridos provocou
redução geral na imunorreatividade para GABA. Assim como em animais
alimentados com dieta comercial, nos animais tratados com a DBR o aspartato
exerceu efeito tanto em células amácrinas quanto em células GABAérgicas na
CCG (figura 23). Entre as células amácrinas na CNI o aspartato provocou redução
no número de células GABA+ em cerca de 35%. Na CCG, a redução foi de cerca
de 50%.
O tratamento conjunto das retinas com aspartato e MK-801 impediu
completamente o efluxo de GABA provocado pelo aspartato, sugerindo haver
ativação de receptores do tipo NMDA durante a estimulação com aspartato
também em animais desnutridos (figura 24).

CNE CPE CNI CPI CCG
Figura 23. Retinas de ratos desnutridos imunorreagidas para GABA. A. retina controle. B. retina estimulada com ASP a 2mM. C. Retina estimulada com ASP na presença de MK801. Barra = 10µm.

*
*
CNI
CCG
Figura 24 – Histogramas da porcentagem de células amácrinas (A) e células na CCG (B) imunorreativas para GABA em relação ao número total de células na porção interna da CNI e CCG, respectivamente, após tratamento com 2mM de ASP e 2mM de ASP na presença de MK801. P < 0,05. N=3.

4. Avaliação da integridade do tecido
4.1. Avaliação morfológica de retinas de animais normais e desnutridos
A histologia geral das retinas dos diferentes grupos experimentais foi
avaliada pela observação com a coloração de DAPI, que conhecidamente marca
núcleos celulares.
A estrutura das retinas não foi alterada pelos tratamentos com NMDA, KA e
ASP. Não houve, no entanto, nenhuma indicação morfológica de lesão ou edema
nas camadas nucleares, ou perfil de rompimento celular. As concentrações
utilizadas, bem como o tempo de exposição do tecido aos reagentes excitatórios
não encontraram-se dentro das faixas que provocam excitotoxicidade (figuras 25 e
26).

CNE CPE CNI CPI CCG
Figura 25 – Fotomicrografias de retinas de animais alimentados com dieta comercial coradas com DAPI. A – retina de animal não-estimulado. B – retina de animal tratado com 100µM de NMDA. C – retina de animal tratado com 25µM de cainato. D – retina de animal tratada com 2mM de aspartato. Barra=10µm.

CNE CPE CNI CPI CCG
Figura 26 – Fotomicrografias de retinas de animais desnutridos alimentados com dieta básica regional (DBR) coradas com DAPI. A – retina de animal não-estimulado. B – retina de animal tratada com 100µM de NMDA. C – retina de animal tratada com 25µM de cainato. D – Retina de animal tratada com 2mM de aspartato. Barra = 10µm.

DISCUSSÃO
1. Caracterização do padrão de liberação de GABA induzida por aminoácidos excitatórios em retinas de ratos normais
A liberação de GABA induzida por AAE’s é um fenômeno que tem sido
investigado por diversos autores nos últimos 27 anos. Os primeiros estudos
utilizavam a metodologia que aproveitava a capacidade de células GABAérgicas
da retina de captarem o [3H]-GABA, após a captação, um estímulo excitatório
(gutamatérgico) era oferecido e a liberação do [3H]-GABA analisada (Tapia e
Arias, 1982; Morán e Pasantes-Morales, 1983; Yazulla, 1983; Yazulla e
Kleinschmidt, 1983; Do Nascimento e de Mello, 1985; Schwartz, 1987; Duarte et
al., 1993; Ferreira et al., 1994; Carvalho et al., 1995). Essa abordagem bioquímica
possibilitou importantes descobertas em relação aos mecanismos de liberação de
GABA. Entretanto, tal metodologia não permitia a identificação dos tipos celulares
ou circuitos envolvidos na interação glutamatérgica/GABAérgica na retina.
Andrade da Costa e colaboradores (2000, 2001) e Calaza e colaboradores (2001,
2003, 2006) utilizando-se da técnica de imunohistoquímica identificaram
populações de células envolvidas na liberação de GABA em retinas numa
abordagem in situ.
É sabido que além do L-Glu, aspartato também é capaz de induzir a
liberação de [3H]-GABA (Yazulla et al., 1985; Yaqub e Eldred, 1993) e que esta
liberação ocorre via ativação de receptores do tipo NMDA (Yazulla, 1983; Kubrusly
et al., 1998).

L-Glutamato e, em menor escala, o aspartato são os principais inputs
excitatórios de células GABAérgicas na retina. Trabalhos relativamente recentes
têm, por meio de imunohistoquímica, identificado populações celulares que
respondem ao estímulo por AAE’s com liberação de neurotransmissores. Andrade
da Costa e colaboradores (2000) mostraram que em retina de Cebus Apella o KA
provoca diminuição na imunorreatividade para GABA em células amácrinas e
células interplexiformes, enquanto que o NMDA não se mostra eficiente em induzir
liberação de GABA nas populações de células GABAérgicas investigadas. Em
2001 Calaza e colaboradores mostraram que em retina de pinto o NMDA induz a
liberação de GABA de forma significante entre células amácrinas enquanto que o
KA produz o mesmo efeito nessa população de células e ainda induz a liberação
de GABA entre as células horizontais. Neste mesmo trabalho foi investigado o
efeito do tratamento com aspartato e se descobriu que este aminoácido atua de
forma a provocar a liberação de GABA somente nas células amácrinas, não
exercendo efeito sobre as células horizontais, assim como o NMDA. O efluxo de
GABA induzido por AAE’s também foi verificado em retinas de gambá – mamífero
marsupial. Este estudo mostrou que ambos NMDA e KA provocam decréscimo de
80 e 50%, respectivamente no número de células amácrinas GABA+ e células
GABAérgicas na CCG. Nesse animal, o aspartato foi capaz de induzir liberação de
GABA somente entre células amácrinas (Calaza et al., 2006). A identificação
histológica de subpopulações de células GABAérgicas na retina de rato –
mamífero placentário – que respondem aos diferentes agonistas de glutamato não
havia sido caracterizado até o momento. A identificação dessas populações
celulares torna-se de grande importância, pois acrescentam dados a respeito do

funcionamento do tecido, afirmam a retina como modelo de estudo para interações
de sistemas de neurotransmissores e, somado a dados oriundos de outras
espécies, permite o estabelecimento de relações filogenéticas entre as espécies
estudadas em relação ao funcionamento de circuitos na retina interna.
No presente trabalho ficou estabelecido que todos os agonistas
glutamatérgicos estudados (NMDA, KA e ASP) foram capazes de induzir
diminuição na imunorreatividade para GABA entre as células amácrinas de forma
significativa. Estes dados estão de acordo com o observado em outras espécies –
pinto e gambá – (Calaza et al., 2001, 2003 e 2006), exceto para o Cebus Apella,
onde o NMDA não produz nenhum efeito nessa população de células (Andrade da
Costa et al., 2000).
A dose de NMDA utilizada nos trabalhos de Calaza foi a mesma utilizada no
presente estudo (100µM). Comparando-se o efeito do NMDA em retinas de pinto,
gambá e rato verifica-se que o NMDA mostra-se mais eficiente em provocar a
redução do número de células amácrinas GABA+ na retina de gambá (80% de
redução) e na retina de rato (62,5% de redução) quando se compara com a retina
de pinto, onde a mesma dose de NMDA provocou redução de GABA-IR de 50%
entre as células amácrinas. NMDA a 100µM não foi capaz de induzir liberação de
GABA entre células na CCG de retinas de ratos. Este dado, no entanto, não
permite afirmar a incapacidade deste aminoácido em estimular esta população
celular. Seria necessário investigar o efeito do NMDA em outras doses para
averiguar se células da CCG são menos sensíveis ou completamente insensíveis
ao estímulo por NMDA.

O tratamento com KA provocou diminuição no número de células GABA+
tanto na CNI (50% das células amácrinas) quanto na CCG (20%). Em relação às
outras espécies estudadas (Andrade da Costa et al., 2000, 2001 e Calaza et al.,
2001, 2003 e 2006) pode-se afirmar que todas possuem sub-populações de
células amácrinas sensíveis ao KA na retina. O efeito do KA na retina de rato
mostrou-se quantitativamente semelhante ao encontrado na retina de pinto. A
retina de gambá mostra-se de forma global mais sensível ao KA quando se
compara com a retina de rato. O KA é responsável pela redução de 80 e 50%,
respectivamente no número de células GABA+ entre as células amácrinas e
células na CCG em retinas de gambá (Calaza et al., 2006).
O aspartato tem sido apontado como um possível neurotransmissor
excitatório na retina. Existem evidências de que o ASP é expresso por diversos
tipos celulares na retina (Yaqub e Eldred, 1991; Sun e Crossland, 2000). Ao
contrário do glutamato que não consegue distinguir entre os vários subtipos de
receptores para AAE’s, o aspartato é capaz de ativar seletivamente receptores
NMDA e aparentemente não exerce qualquer efeito sobre receptores não-NMDA.
Até o momento, não existem dados publicados a respeito dos efeitos do aspartato
em induzir liberação de GABA em retina de mamíferos não-marsupiais. Dados do
presente estudo confirmam evidências anteriores de que o ASP pode atuar como
agonista glutamatérgico também na retina de rato. Isso porque este foi capaz de
provocar liberação de GABA tanto entre as células amácrinas quanto em células
da CCG. Na retina de rato, o ASP mostrou efeito bastante semelhante ao
encontrado em retinas de pinto e gambá (Calaza et al., 2001, 2003, 2006),
provocando a diminuição no número de células amácrinas GABA+ em cerca de

50%. Embora o ASP (na concentração utilizada) não seja capaz de provocar
liberação de GABA em células da CCG em retinas de gambá, mostrou-se eficiente
em induzir a diminuição do número de células imunorreativas para GABA na CCG
em 23% na retina de ratos.
(ver tabela 4 para comparação entre as espécies já estudadas).
Cabe lembrar que o aspartato induz a liberação de GABA mesmo na
presença de magnésio (íon que conhecidamente bloqueia o canal de receptores
do tipo NMDA e que existe em concentrações na faixa de 1mM no espaço
extracelular de SNC). Kubrusly e colaboradores (1998), em relação ao efeito do
tratamento com ASP em culturas de células perfundidas com [3H]-GABA,
consideraram a hipótese de que o ASP estaria provocando efluxo de [3H]-GABA
mesmo na presença de Mg++ graças ao transporte eletrogênico de aspartato
(Wadiche et al., 1995). De acordo com essa hipótese, correntes iônicas
associadas ao transporte de ASP seriam responsáveis pela pré-despolarização
necessária para desobstruir receptores adjacentes do tipo NMDA (bloqueados
pelo Mg++) que, após isso, podem ser ativados pelo ASP. Assim, a combinação de
dois fenômenos (ativação de receptores NMDA e transporte eletrogênico de ASP)
seria responsável pelo papel do ASP em induzir liberação de GABA via ativação
de receptores NMDA.
Os efeitos dos diferentes AAE’s nas células GABA+ devem ser decorrentes
da ativação de receptores clássicos já que a liberação de GABA induzida por
NMDA/ASP e KA é completamente abolida por MK801 e DNQX, antagonistas de
receptores NMDA e não-NMDA, respectivamente. A possibilidade de que a
liberação de GABA induzida por AAE’s na retina de rato fosse decorrente de

vazamento por lise celular foi descartada pela análise da integridade do tecido
corado com DAPI (corante de núcleo celular) após a estimulação com os
diferentes agonistas. Tal análise mostrou que, mesmo após a incubação com
AAE’s, a arquitetura do tecido retiniano mostrava-se igual a de uma retina fixada
logo após a enucleação.
A dopamina, uma catecolamina presente na retina, tem sido implicada na
modulação da liberação de GABA induzida por glutamato. De fato, parece haver
grande interação entre células GABAérgicas e dopaminérgicas no processamento
visual em nível de retina (Downling, 1987; Gardino et al., 1993). Na retina de
peixe-dourado, a dopamina é responsável pelo bloqueio da liberação de GABA
provocada pela estimulação com glutamato. Nesse animal, o efeito da dopamina é
aparentemente modulado pela ativação de receptores de dopamina D1 presentes
no tecido (Yazulla e Kleinschmidt, 1982; Kato et al., 1985; O’Brien e Downling,
1985). Parte da liberação de [3H]-GABA na retina de galinha é bloqueada pela
dopamina (do Nascimento et al., 1998). Embora a análise bioquímica realizada na
retina de galinha tenha mostrado o efeito modulatório da dopamina sobre a
liberação de [3H]-GABA, esta não permite a identificação dos tipos celulares que
poderiam estar sendo influenciados por esta amina. O efeito da dopamina na
retina de galinha mostrou-se de diferente natureza em relação aos dados
provenientes de experimentos com peixe-dourado (provavelmente por diferenças
nos tipos celulares envolvidos nesta circuitaria). Na retina de aves, a dopamina
pareceu agir de forma independente de ativação de seu receptor. O efeito
somente foi verificado na estimulação com NMDA, não havendo nenhum efeito
sobre a liberação de [3H]-GABA induzida por KA. Tais dados sugerem que a

dopamina, nessa classe, age de forma direta sobre o receptor NMDA (Do
Nascimento et al., 1998). Ainda não é sabido, até o momento, se a dopamina
exerce seu efeito modulatório em retinas de mamíferos placentários e se seu
efeito nessa sub-classe ocorre via ativação de receptores dopaminérgicos ou
interação com receptores glutamatérgicos.
Na retina de rato, não só foram identificadas as subpopulações de células
GABAérgicas que sofrem ação dos agonistas de glutamato, incluindo o aspartato,
como também foi possível caracterizar o grupo de células influenciado pela
dopamina. Dessa forma, o efeito da dopamina na retina de rato mostrou-se
bastante semelhante ao encontrado na retina de pinto. Tanto o NMDA quanto o
aspartado pareceram ter seus efeitos modulados pela dopamina. A liberação de
GABA induzida por cainato não sofreu qualquer efeito pela incubação conjunta
com a dopamina. Tais dados corroboram as evidências de que o efeito da
dopamina se dê via interação com sítios específicos no receptor NMDA.
Dados anteriores sugeriam a participação do transportador de GABA na
liberação de GABA induzida por AAE’s nas células amácrinas (Hofmann e Möckel,
1991; Duarte et al., 1993; Do Nascimento et al., 1998b; Andrade da Costa et al.,
2000; Calaza et al., 2001, 2003, 2006). Baseando-se nesta evidência, pareceu
importante verificar qual era a natureza da liberação de GABA verificada em
retinas de ratos. Nesse sentido, foi avaliada a contribuição desse transportador na
liberação de GABA induzida por KA, NMDA e ASP. Em todos os casos, NNC-711
inibiu o decréscimo na imunorreatividade para GABA o que reforça a idéia de que
o transportador de GABA atue por meio de sua reversão de modo a permitir – em
grande parte – a liberação de GABA induzida pelos AAE’s.

Tabela 4 – Tabela comparativa dos efeitos de tratamentos com aminoácidos excitatórios, AAE’s na presença de dopamina e AAE’s na presença de NNC em retinas de animais de diferentes grupos taxonômicos: Ave - Gallus domesticus; mamífero marsupial - Didelphis marsupialis; mamífero placentário roedor – Rattus norvegicus albinus (Wistar); mamífero placentário primata - Cebus apella.

2. Efeito da desnutrição crônica sobre a funcionalidade de circuitos glutamatérgicos\GABAérgicos na retina de ratos
A retina (quando comparada a outras estruturas centrais como hipocampo e
córtex cerebral) pode ser considerada um tecido que exibe menos mecanismos de
plasticidade. Porém, estudos recentes têm identificado fatores que alteram a
morfologia e funcionalidade do tecido, como inputs sensoriais e aporte protéico
oferecido pela alimentação durante o desenvolvimento (Almeida et al., 2001;
Silveira et al., 2007; Xu e Tian, 2007; Landi et al., 2007). Nesse sentido, pareceu
bastante importante investigar a funcionalidade dos sistemas de
neurotransmissores em retinas de rato sob condições que conhecidamente
provocam alguma alteração no tecido. Nesse sentido foi realizada a análise dos
efeitos da desnutrição crônica nos parâmetros caracterizados no presente estudo
em animais normais. A escolha da desnutrição como injúria se mostrou bastante
conveniente devido ao fato de se tratar de um modelo baseado em uma dieta
humana, além disso, tal escolha se mostrou bastante conveniente devido ao
volume de informações oriundos de trabalhos do Laboratório de Neurobiologia da
Retina – onde o presente estudo foi realizado. Estudos anteriores utilizando a
DBR, mesma fonte de desnutrição utilizada na presente análise, mostram que a
retina responde ao insulto durante o desenvolvimento como atrasos em todos os
parâmetros investigados: expressão de neurotransmissores como GABA,
acetilcolina, proteínas ligadoras de cálcio e geração de células amácrinas
GABAérgicas. No entanto, em animais adultos (P30) o tecido se mostrou
completamente recuperado (Almeida et al., 2001; Silveira et al., 2007). Como já é

sabido, o desenvolvimento normal do tecido depende de diversos fatores que
devem acontecer de forma orquestrada. Embora morfologicamente recuperada
não se sabia, até o presente momento, se a desnutrição provoca alguma alteração
funcional no tecido retiniano.
Dados caracterizados pelo presente estudo sugerem que de uma forma
geral a desnutrição protéica multideficiente causada pelo uso da DBR provocou
uma maior sensibilidade ao estímulo pelos AAE’s entre as células na CCG. A
estimulação com NMDA provocou uma redução de cerca de 34% no número de
células GABA+ na CNI, comparando-se com retina de animal desnutrido não
estimulada pelo NMDA. Ao passo que a estimulação de retinas de animais
normais com a mesma dose de NMDA provocou redução de cerca de 62% no
número de células GABA+ na mesma população de células. Isso sugere que as
células amácrinas de animais desnutridos são bem menos suscetíveis à
estimulação por NMDA. Os efeitos de KA e ASP entre células amácrinas são
exatamente os mesmos entre animais desnutridos e normais.
Na CCG verifica-se um aumento no efeito dos três agonistas
glutamatérgicos utilizados na indução da liberação de GABA. Nesse sentido, o
efeito do NMDA na CCG foi quase três vezes maior do que o notado em animais
normais (onde o NMDA não foi capaz de induzir liberação de GABA de forma
significativa). O tratamento com cainato provocou redução no número de células
GABA+ de 50% comparando-se com retina não estimulada. O efeito desse
aminoácido foi 2,5 vezes maior do que o observado em animais normais. O
tratamento com ASP também provocou efeito mais notável em retinas de animais
desnutridos. O efeito do ASP foi duas vezes maior na CCG desses animais.

A B
CCG
normal desnutrido05
1015202530354045
*
Red
ução
da
IRca
usad
a pe
lo tr
atam
ento
com
NM
DA
CNI
Normal desnutrido0
10
20
30
40
50
60
70
*
Redu
ção
da IR
caus
ada
pelo
trat
amen
toco
m N
MDA
C D
CNI
normal desnutrido0
10
20
30
40
50
60
Redu
ção
da IR
caus
ada
pelo
trat
amen
toco
m K
A
CCG
normal desnutrido0
10
20
30
40
50
60
*
Redu
ção
da IR
caus
ada
pelo
trat
amen
toco
m K
A
Figura 27 – gráficos comparativos: redução de imunoreatividade entre animais normais e desnutridos após o tratamento com diversos agonistas glutamatérgicos. A e B, efeito do tratamento com NMDA na CNI e CCG. C e D, efeito do tratamento com KA na CNI e CCG. E e F, efeito do tratamento com ASP na CNI e CCG.
CNI
normal desnutrido0
10
20
30
40
50
60
Redu
ção
da IR
caus
ada
pelo
trat
amen
toco
m A
SP
CCG - ASP
normal desnutrido0
10
20
30
40
E F
50
60
Redu
ção
da IR
caus
ada
pelo
trat
amo
com
ASP
ent

Em conjunto, os dados obtidos nos animais desnutridos sugerem que a
injúria da desnutrição durante o desenvolvimento esteja provocando uma
alteração na funcionalidade do tecido (pelo menos nas células GABAérgicas na
CCG). Com base nos dados não é possível identificar o motivo do efeito
diferencial do tratamento com os AA’Es na porção mais interna da retina. O
aumento na expressão de receptores ionotrópicos de glutamato seria uma
explicação plausível para o observado. Embora dados anteriores tenham
mostrado que o tecido retiniano em animais adultos se mostra completamente
recuperado dos efeitos provocados pela desnutrição, resultados obtidos no
presente estudo mostram que a funcionalidade do tecido adulto foi afetada pela
injúria.

CONCLUSÕES
1 – Os aminoácidos excitatórios provocam liberação de GABA na retina de ratos
nas populações celulares estudadas. NMDA na concentração utilizada não
provoca efeitos em células na CCG.
2 – A dopamina age de forma a modular os efeitos provocados pelo NMDA e ASP
em subpopulações de células na CNI e CCG.
3 – Aspartato atua como agonista glutamatérgico endógeno agindo sobre
receptores do tipo NMDA também na retina de ratos, mamíferos placentários.
4 – A liberação de GABA em retinas de ratos é provocada pela reversão de seu
transportador.
5 – A desnutrição crônica provoca maior resposta ao tratamento com todos os
AAE’s estudados principalmente na CCG.

BIBLIOGRAFIA
AGARDH, E., EHINGER, B. e WU, J. Y. GABA and GAD-like immunoreactivity in
the primate retina. Histochemistry, v. 86, p. 485-490, 1987a.
AGARDH, E., BRUUN, A., EHINGER, B., EKSTRÖM, P., VAN VEEN, T. e WU,
J.Y. Gamma-aminobutyric acid- and glutamic acid decarboxylase-
immunoreactive neurons in the retina of different vertebrates. J. Comp.
Neurol., v. 258, p. 622-630, 1987b.
ALMEIDA, M.F.; YAMASAKI, E.N.; SILVEIRA, A.C.; GUEDES, R.C.; HOKOÇ, J.N.
The GAbAergic and cholinergic systems in the retina are differentially affected
by postnatal malnutrition during the suckling period. Nutr Neurosci v. 4(3):p.
223-38, 2001.
ALMEIDA, M.F.; YAMASAKI, E.N.; SILVEIRA, A.C.; GUEDES, R.C.; HOKOÇ, J.N.
MARTINEZ, A.M.B. Quantitative and qualitative ultrastructural analysis of
control and malnourished rat optic nerve. Acta Microscopica Proceedings of
the XVIII Congress of the Brasilian Society for Microscopy and
Microanalysisi. Suplemento B, p. 253-254, 2001.
ASCHER, P. AND L. NOWAK. The role of divalent cations in the N-methyl-D-
aspartate responses of mouse central neurones in culture. J Physiol., v.399, p.
247-66, 1988
ANDRADE da COSTA, B.L.S., DE MELLO, F.G. e HOKOÇ, J.N. Transporter-
mediated GABA release induced by excitatory amino acid agonist is associated
with GAD-67, but not GAD-65 immunoreactive cells of the primate retina.
Brain Res., v. 863, p. 132-42, 2000.

ANDRADE DA COSTA, B.L., DE MELLO, F.G., HOKOÇ, J.N. Comparative study
of glutamate mediated gamma-aminobutyric acid release from nitric oxide
synthase and tyrosine hydroxylase immunoreactive cells of the Cebus apella
retina. Neurosci Lett v. 13;302(1):p. 21-4, 2001.
BATISTA-FILHO, M. Pesquisa nutricional da zona da mata. Recife. UFPE, pp.133,
1968.
BAYLOR, D. A. Photoreceptor signals and vision. Proctor lecture. Invest
Ophthalmol Vis Sci., v.28(1): p. 34-49, 1987.
BLANCO, R. e DE LA VILLA, P. Ionotropic glutamate receptors in isolated
horizontal cells of the rabbit retina. Eur. J. Neurosci., v. 11, p. 867-873, 1999.
BLOOMFIELD, S.A. e DOWLING, J.E. Roles of aspartate and glutamate in
synaptic transmission in rabbit retina. II. Inner plexiform layer. J.
Neurophysiol., v. 53, p. 714-725, 1985a.
BLOOMFIELD, S.A. e DOWLING, J.E. Roles of aspartate and glutamate in
synaptic transmission in rabbit retina. II. Outer plexiform layer. J.
Neurophysiol., v. 53, p. 699-713, 1985b.
BRÄNDSTATTER, J.H., HARTVEIT, E., SASSOE-POGNETO e WÄSSLE, H.
Expression of NMDA and high-affinity kainate receptor subunit mRNAs in the
adult rat retina. Eur. J. Neurosci., v. 6, p. 1100-1112, 1994.
CAJAL, S.R. (1894) In: News Ideas on the Structure of the Nervous System in Man
and Vertebrates (by Swanson, N. & Swanson, L., 1990). The Retina, cap.6,
The MIT Press, London, England.
CALAZA, K. C., DE MELLO, F. G., ET AL. GABA release induced by aspartate-
mediated activation of NMDA receptors is modulated by dopamine in a

selective subpopulation of amacrine cells. J Neurocytol v.30(3): p. 181-93,
2001.
CALAZA, K. C., DE MELLO, M.C., DE MELLO, F.G., GARDINO, P.F. Local
differences in GABA release induced by excitatory amino acids during retina
development: selective activation of NMDA receptors by aspartate in the inner
retina. Neurochem Res v. 28(10): p. 1475-85, 2003
CALAZA, K. C., HOKOC, J. N. ET AL. GABAergic circuitry in the opossum retina: a
GABA release induced by L-aspartate. Exp Brain Res v.172(3): p. 322-30,
2006.
CARVALHO, A.P, FERREIRA, I.L., CARVALHO, A.L. e DUARTE, C.B. Glutamate
receptor modulation of 3H-GABA release and intracelular calcium in chick retina
cells. An. N. Y. Acad. Sci., v. 757, p. 439-456, 1995.
CASTRO, N.G., De MELLO, M.C.F., De MELLO, F.G. e ARACAVA, Y. Direct
inhibition of the N-metyl-D-aspartate receptor channel by dopamine and (+)-
SKF38393. Br. J. Pharmacol., v. 126, p. 1847-1855, 1999.
CHA, J.J., O’BRIEN, D.R. e DOWLING, J.E. Effects of D-aspartate amino acid-
induced release of [3H]GABA from goldfish retina. Brain Res., v. 376, p. 140-148,
1986.
CLARK J.A. e AMARA S.G. Amino acid neurotransmitter transporters: structure,
function, and molecular diversity. Bioessays, v. 15(5), p. 323-332, 1993.
COTMAN, C. AND D. MONAGHAN. Multiple excitatory amino acid receptor
regulation of intracellular Ca2+. Implications for aging and Alzheimer's disease.
Ann N Y Acad Sci v. 568: 138-48. (1989)

DO NASCIMENTO, J.L.M., KUBRUSLY, R.C.C., REIS, R.A.M., DE MELLO,
M.C.F. e DE MELLO, F.G. Atypical effect of dopamine in modulating the
functional inhibition of NMDA receptors of cultured retina cells. Eur. J.
Pharmacol., v. 343, p. 103-110, 1998a.
DO NASCIMENTO, J.L.M, VENTURA, A.L.M. e PAES DE CARVALHO, R.
Veratridine- and glutamate-induced release of [3H]-GABA from cultured chick
retina cells: possible involvement of a GAT-1-like subtype of GABA transporter.
Brain Res., v. 798, p. 217-222, 1998b.
DACEY, D. M. Axon-bearing amacrine cells of the macaque monkey retina. J
Comp Neurol v.284(2): p. 275-93, 1989
DO NASCIMENTO, J.L.M, e DE MELLO, F.G. Induced release of γ-aminobutyric
acid by a carrier-mediated high-affinity uptake of L-glutamate in cultured chick
retinal cells. J. Neurochem., v. 45, p. 1820-1827, 1985.
DOWLING, J. E. AND BOYCOTT, B. B. Organization of the primate retina: electron
microscopy. Proc R Soc Lond B Biol Sci., v.166(2): p. 80-111, 1966
DOWLING J.E. “The Retina: An approachable part of the brain”. Harvard
University Press, Cambridge. 1987.
DUARTE, C.B., FERREIRA, I.L., SANTOS, P.F., OLIVEIRA, C.R. e CARVALHO,
A.P. Glutamate increases the [Ca2+]i but stimulates Ca2+-independent release of
3H-GABA in cultured chick retina cells. Brain Res., v. 611, p. 130-138, 1993.
FERREIRA, I.L., DUARTE, C.B., SANTOS, P.F., CARVALHO, C.M. e CARVALHO,
A.P. Release of 3H-GABA evoked by glutamate receptor agonists in cultured
chick retina cells: effect of Ca2+. Brain Res., v. 664, p. 252-256, 1994.

FLETCHER, E. L., HACK, I., BRANDSTÄTTER, J.H., WÄSSER, H.
Immunocytochemical localization of NMDA receptors in the rat retina. Invest
Ophthalmol Vis Sci v. 40, 1999
GARDINO, P.F., DOS SANTOS, R.M. e HOKOÇ, J.N. Histogenesis and
topographical distribution of tyrosine hydroxylase immunoreactive amacrine
cells in the developing chick retina. Dev. Brain Res., v. 72, p. 226-236, 1993.
GRUNDER, T., K. KOHLER, ET AL. The distribution and developmental regulation
of NMDA receptor subunit proteins in the outer and inner retina of the rat. J
Neurobiol v.44(3): p.333-42, 2000
GUEDES, RC, MONTEIRO, JS, TEODOSIO, NR. Malnutrition and brain function:
experimental studies using the phenomenon of cortical spreading depression.
Rev Bras Biol, v. 56 (2), p.293-301, 1996.
HAMASSAKI-BRITTO, D.E., HERMMANS-BORGMEYER, I., HEINEMANN, S. e
HUGHES, T.E. Expression of glutamate receptor genes in the mammalian
retina: the localization of the GluR1 through GluR7 mRNAs. J. Neurosci., v.
13, p. 1888-1898, 1993.
HASHIMOTO, A. e OKA, T. Free D-aspartate and D-serine in the mammalian
brain and periphery. Prog. Neurobiol., v. 52, p. 325-353, 1997.
HOFMANN, H.D. e MÖCKEL, V. Release of γ-aminobutyric acid from cultured
amacrine-like neurons mediated by different aminoacid receptors. J.
Neurochem., v. 56, p. 923-932, 1991.

HOFFMANN, H., GREMME, T., HATT, H. e GOTMANN, K. Synaptic activity-
dependent developmental regulation of NMDA receptor subunit expression in
cultured neocortical neurons. J. Neurochem., v. 75, p. 1590-1599, 2000.
HOKOÇ, J.N., VENTURA, A.L.M., GARDINO, P.F. e DE MELLO, F.G.
Developmental immunoreactivity for GABA and GAD in the avian retina: possible
alternative pathway for GABA synthesis. Brain Res., v. 532, p. 197-202, 1990.
HU, M., BRUUN, A. e EHINGER, B. Expression of GABA transporter subtypes (GAT-
1, GAT-3) in the developing retina. Acta Ophtalmol. Scand., v. 77, p. 261-265,
1999.
IKEDA Y., NISHIYAMA N., SAITO H. e KATSUKI H. GABAA receptor stimulation
promotes survival of embryonic rat striatal neurons in culture. Dev. Brain Res.,
v. 98, p. 253-258, 1997.
JOHNSON, J., CHEN, T.K., RICKMAN, D.W., EVANS, C. e BRECHA, N.C.
Multiple γ-aminobutyric acid plasma membrana transporters (GAT-1, GAT-2,
GAT-3) in the rat retina. J. Comp. Neurol., v. 375, p. 212-224, 1996.
KATO, S., NEGISHI, K. e TERANISHI, T. Dopamine inhibits calcium-independent
γ-[3H] aminobutyric acid release induced by kainate and high K+ in the fish
retina. J. Neurochem., v. 44, p. 893-899, 1985.
KIDD, M. Electron microscopy of the inner plexiform layer of the retina in the cat
and the pigeon. J Anat v.96: p. 179-87, 1962.
KOLB, H. e NELSON, R. Off-alpha and off-beta ganglion cells in the cat retina: II:
neural circuitry as revealed by electron microscopy of HRP stains. J. Comp.
Neurol., v. 329, p. 85-110, 1993.

KOLB, H. The architecture of functional neural circuits in the vertebrate retina (The
Proctor Lecture). Invest. Ophthalmol. Visual Sci., v. 35, p. 2385-2402, 1994.
KUBRUSLY, R.C.C., DE MELLO, M.C.F. e DE MELLO, F.G. Aspartate as a
selective NMDA receptor agonist in cultured cells from the avian retina.
Neurochem. Int., v. 32, p. 47-52, 1998.
LAGO, ES, TEODOSIO, NR, PESSOA, DC, CABRAL, FILHO JE. Longevity of
malnourished rats fed with the regional basic diet (RBD) of the northeast of
Brazil. Arch Latinoam Nutr, v. 47(4), p. 338-342, 1997.
LANDI, S., A. SALE, ET AL. Retinal functional development is sensitive to
environmental enrichment: a role for BDNF. FASEB J v.21(1): p. 130-9, 2007.
LEE, J.A., HOMMA, H., TASHIRO, K., IWATSUBO, T. e IMAI, K. D-aspartate
localization in the rat pituitary gland and retina. Brain Res., v. 838, p. 193-199,
1999.
LIU, Q.R., LÓPEZ-CORCUERA, B., MANDIYAN, S., NELSON, H., e NELSON, N.
Molecular characterization of four pharmacologically distinct γ-aminobutyric acid
transporters in mouse brain. J. Biol. Chem., v. 268, p. 2106-2112, 1993.
MARC, R.E.e LIU, W.L.S. Fundamental GABAergic amacrine cell circuitries in the
retina: nested feedback, concatenated inhibition, and axosomatic synapses. J.
Comp. Neurol., v. 425, p. 560-582, 2000.
MARTINS, R. A. AND PEARSON, R. A. Control of cell proliferation by
neurotransmitters in the developing vertebrate retina." Brain Res. 2007
MASLAND, R.H. The functional architecture of the retina. Sci Am v. 255(6):102-
11. 1986.

MASLAND, R.H. Neuronal diversity in the retina. v. 11(4):431-6, 2001.
MASSEY, S.C. e REDBURN, D. Transmitter circuits in the vertebrate retina. Prog.
Neurobiol., v. 28, p. 55-96, 1987.
MONAGHAN, D. T., R. J. BRIDGES, ET AL. The excitatory amino acid receptors:
their classes, pharmacology, and distinct properties in the function of the
central nervous system. Annu Rev Pharmacol Toxicol v.29: p. 365-402,
1989.
MORAN, J., PASANTES-MORALES, H. e REDBURN, D.A. Glutamate receptor
agonists release [3H]-GABA preferentially from horizontal cells. Brain Res., v.
398(2), p. 276-287, 1986.
MORAN, J. e PASANTES-MORALES, H. Effects of excitatory amino acids, and of
their agonists and antagonists on the release of neurotransmitters from the
chick retina. J. Neurosci. Res., v. 10, p. 261-271, 1983.
MOSINGER, J.L., YAZULLA, S. e STUDHOLME, K.M. GABA-like immunoreactivity
in the vertebrate retina: a species comparison. Exp. Eye Res., v. 42, p. 631-44,
1986.
NAKANISHI, S., MASU, M., BESSHO, Y., NAKAJIMA, Y., HAYASHI, Y e
SHIGEMOTO, R. Molecular diversity of glutamate receptors and their
physiological functions. EXS., v. 71, p. 71-80, 1992.
NELSON, R., FAMIGLIETTI, E.V. e KOLB, H. Intracelular staining reveals different
levels of stratification for on- and off-center ganglion cells in cat retina. J.
Neurophysiol., v. 41, p. 472-483, 1978.
NICHOLLS, D. AND ATTWELL, D. The release and uptake of excitatory amino acids.
Trends Pharmacol Sci v.11(11): p. 462-8, 1990.

O’BRIEN, D.R. e DOWLING, J.E. Dopaminergic regulation of GABA release from
the intact goldfish retina. Brain Res., v. 360, p. 41-50, 1985.
SCHWARTZ, E.A. Depolarization without calcium can release γ-aminobutyric acid
from a retinal neuron. Science, v. 238, p. 350-355, 1987.
SCHNAPF, J. L. AND BAYLOR, D. A. How photoreceptor cells respond to light.
Sci Am v.256(4): p. 40-7, 1987.
SILVEIRA, A.C.; GARDINO, P.F.; BEVILAQUA, M.C.N.; HOKOÇ, J.N.
Neurogenesis of GABAergic cells in the retina of malnourished rats. Int J Dev
Neurosci. V. 25(5):p. 325-33, 2007.
SPOERRI, P.E. Neurotrophic effects of GABA in cultures of embryonic chick brain
and retina. Synapse, v. 2, p. 11-22, 1988.
STERLING, P. “Knocking out”a neural circuit. Neuron, v. 21, p. 643-652, 1998.
SUN, H. e CROSSLAND, W.J. Quantitative assessment of localization and
colocalization of glutamate, aspartate, glycine and GABA immunoreactivity in
the chick retina. Anat. Rec., v. 260, p. 158-179, 2000.
TAPIA, R. e ARIAS, C. Selective stimulation of neurotransmitter release from chick
retina by kainic and glutamic acids. J. Neurochem., v. 39, p. 1169-1178, 1982.
TEODOSIO, NR, FILHO, JE, GUEDES, RC, COSTA, JA, RAMOS, CFB, DA
SILVA, AT. Learned and emotional behavior in chronically malnourished
rats. Acta Physiol Lat Am,v. 29 (4-5), p. 255-262, 1979.
TEODOSIO, NR, LAGO, ES, ROMANI, SA, GUEDES, RC. A regional basic diet
from northeast Brazil as a dietary model of experimental malnutrition. Arch
Latinoam Nutr., v. 40 (4), p. 533-547, 1990.

VECINO, E., UGARTE, M., NASH, M.S. e OSBORNE, N.N. NMDA induces BDNF
expression in the albino rat retina in vivo. NeuroReport, v. 10, p. 1103-1106,
1999.
WADICHE, J.I., AMARA, S.G., KAVANAUGH, M.P. Ion fluxes associated with
excitatory amino acid transport. Neuron. v. 15(3):p.721-8., 1995.
WAGNER, H. J. AND I. WULLE. Contacts of dopaminergic interplexiform cells in
the outer retina of the blue acara. Vis Neurosci v. 9(3-4): p.325-33. 1992.
WEILER, R., BALDRIDGE, W.H., MANGEL, S.C. e DOWLING, J.E. Modulation of
dopamine release in the fish retina by light and prolonged darkness. Vis.
Neurosci., v. 14, p. 351-356, 1997.
WERBLIN, F. Synaptic connections, receptive fields, and patterns of activity in the
tiger salamander retina. A simulation of patterns of activity formed at each
cellular level from photoreceptors to ganglion cells [the Friendenwald lecture].
Invest Ophthalmol Vis Sci v.32(3): p. 459-83, 1991.
WU, S.M. Feedback connections and operation of the outer plexiform layer of the
retina. Curr. Opin. Neurobiol., v. 2, p. 173-178, 1992.
VANEY, D. I. Territorial organization of direction-selective ganglion cells in rabbit
retina. J Neurosci v.14(11 Pt 1): 6301-16, 1994.
XU, H.P.; TIAN, N. Retinal ganglion cell dendrites undergo a visual activity-
dependent redistribution after eye opening. J Comp Neurol. v. 503(2):p.244-
59, 2007.
YAMASHITA, M. e FUKUDA, Y. Calcium channels and GABA receptors in the
early embryonic chick retina. J. Neurobiol., v. 24, p. 1600-1614, 1993.

YAQUB, A. e ELDRED, W.D. Effects of excitatory amino acids on
immunocytochemically identified populations of neurons in turtle retina. J.
Neurocytol., v. 22, p. 644-662, 1993.
YAZULLA, S. Stimulation of GABA release from retinal horizontal cells by
potassium and acidic amino acid agonists. Brain Res., v. 275, p. 61-74, 1983.
YAZULLA, S. e KLEINSCHMIDT, J. Carrier-mediated release of GABA from retinal
horizontal cells. Brain Res., v. 263, p. 63-75, 1983.
YAZULLA, S., CUNNINGHAM, J.O. e NEAL, M. Stimulated release of GABA and
glycine from goldfish retina. Brain Res., v. 345, p. 384-388, 1985.
YAZULLA, S. GABAergic mechanisms in the retina. Prog. Ret. Res., v. 5, p. 1-52,
1986.

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo





























