Embed Size (px)
Citation preview

1
Marcelo José da Silva de Magalhães
Influência do polimorfismo nos genes tas1r1, tas1r2 e tas1r3 na detecção do
estímulo doce e umami no Homo sapiens.
Belo Horizonte Novembro de 2015

2
Marcelo José da Silva de Magalhães
Influência do polimorfismo nos genes tas1r1, tas1r2 e tas1r3 na detecção do estímulo
doce e umami no Homo sapiens.
Orientadora: Dra. Laila Alves Nahum
Belo Horizonte Novembro de 2015
Dissertação apresentada à Faculdade Infórium
de Tecnologia como parte dos requisitos para a
obtenção do título de Mestre no curso de
Tecnologia da Informação Aplicada à Biologia
Computacional

3
FOLHA DE APROVAÇÃO
Influência do polimorfismo nos genes tas1r1, tas1r2 e tas1r3 na detecção do estímulo doce e
umami no Homo sapiens.
Examinado por:
Prof.a Laila Alves Nahum/ orientadora
Faculdade Infórium de Tecnologia Centro de Pesquisas René Rachou (CPqRR)
Prof. Luiz Alexandre Viana Magno/ Membro titular
Faculdade Infórium de Tecnologia
Prof. Jonas Jardim de Paula/ Membro titular externo
Faculdade de Ciências Médicas de Minas Gerais
Prof. Tadeu Henrique de Lima/ Membro suplente
Universidade do Estado de Minas Gerais (UEMG) Faculdade Infórium de Tecnologia
Belo Horizonte, ----- de ------------------- de 2015.
Dissertação apresentada à Faculdade Infórium
de Tecnologia como parte dos requisitos para a
obtenção do título de Mestre no curso de
Tecnologia da Informação Aplicada à Biologia
Computacional

4
DEDICATÓRIA
A minha esposa Aline, pelo amor incondicional.

5
AGRADECIMENTOS
A Deus que me permitiu escolher o caminho do estudo proporcionando crescimento
intelectual e moral;
Prof.a Laila Alves Nahum, mais que orientadora, um exemplo de dedicação à ciência;
Aos acadêmicos de medicina das faculdades Funorte (Faculdade Unidas do Norte
de Minas) e FipMoc (Faculdades Integradas de Montes Claros). Eles foram mais um
incentivo para o meu aperfeiçoamento;
A Rosângela Silqueira Hickson coordenadora do curso de Tecnologia da Informação
Aplicada à Biologia Computacional por ter proporcionado uma nova oportunidade de
ampliar meus horizonte
A secretária do curso de Tecnologia da Informação Aplicada à Biologia
Computacional Francielle Costa Ramos por todas as orientações prestadas.
Aos meus pacientes, motivo maior para continuar estudando e renovando o
conhecimento;
Aos amigos neurocirurgiões Dr. Virgílio Bueno e Dr. Carlos Antônio que realizaram
inúmeros plantões no Hospital Aroldo Tourinho durante minhas ausências.

6
Influência do polimorfismo nos genes tas1r1, tas1r2 e tas1r3 na detecção do estímulo doce e umami no Homo sapiens.
RESUMO: umami e doce são sabores induzidos pelo glutamato monossódico (GMS)
aspartato e glicose respectivamente, os quais são detectados pelos receptores
heterodiméricos (T1R1, T1R2 e T1R3) acoplados á proteína G presentes nas papilas
gustativas da língua. A detecção do GMS e glicose diferem substancialmente entre
os indivíduos e está associada ao polimorfismo que ocorre nos receptores. Este é
um artigo de revisão das bases de dados PubMed utilizando os seguintes
descritores: “tas1r1” ,”tas1r2”, “tas1r3”, “polimorfismos”, “umami” e “doce”. Esta
revisão aponta que os polimorfismos simples não sinônimos (SNPs) do T1R1
(A372T) e T1R3 (A5T) estão associados ao aumento da percepção dos sabores
umami e doce pelos humanos. Em contraste, os SNPs (R757C) está associado à
redução da percepção do sabor umami pelos humanos.
PALAVRAS CHAVES: humano, polimorfismo, L-glutamato, receptores de sabor,
T1R1, T1R2, T1R3, umami, doce

7
Influence of polymorphisms on tas1r1, tas1r2 e tas1r3 gens in taste detection of sweet and
umami in Homo sapiens
ABSTRACT: Umami and sweet are the typical tastes induced by monosodium
glutamate (MSG), aspartate and glucose, respectively, which are detected by the
heterodimeric G protein–coupled receptor (T1R1, T1R2, and T1R3) presents on taste
bud of tongue. MSG and glucose detection differ substantially among individuals and
are associated with polymorphisms occurring in the T1R1 and T1R3 genes. This
work is a review and the search was conducted on electronic literature databases
(PubMed) using the following descriptors: "tas1r1”, “tas1r2”, “tas1r3” “polimorphisms”,
“umami” and “sweet”. This review points out that SNPs of T1R1 (A372T) and T1R3
(A5T) are associated with the increased perception of umami sensations in humans.
In contrast, the SNP (R757C) of T1R3 is associated with the decreased perception of
umami sensations in humans.
KEYWORDS: human, polymorphism, L-glutamate, taste receptors, T1R1, T1R2,
T1R3, umami, sweet.

8
LISTA DE FIGURAS
FIGURA 1- Corte histológico do tecido epitelial em superfície de papila gustativa exibindo botão gustativo........................................................23 FIGURA 2- Botão gustativo isolado.........................................................................23 FIGURA 3- Inervação gustativa da língua...............................................................25 FIGURA 4- Inervação gustativa da língua...............................................................25 FIGURA 5- Sequência de aminoácidos do receptor T1R1......................................29 FIGURA 6- Sequência de aminoácidos do receptor T1R2......................................31 FIGURA 7- Sequência de aminoácidos do receptor T1R3.................................... 33 FIGURA 8- Polimorfismo de nucleotídeo simples da base nitrogenada citosina pela timina...............................................................................36 FIGURA 9- FIGURA 10- Pequeno trecho de alinhamento das proteínas Q99PG6 (tas1r Mus musculus) e Q7RTX1 (tas1r1 Homo sapiens)..................40 FIGURA 11- Estrutura molecular dos aminoácidos e sua classificação conforme suas propriedades físico-químicas....................................40 FIGURA 12- Alinhamento de sequências dos receptores T1R1, T1,
T1R3 do Homo sapiens......................................................................44
FIGURA 13- Gene tas1r1 e posição do SNPns no éxon III.....................................50
FIGURA 14
Pequeno trecho de alinhamento das seguintes proteínas: Q99PG6 (ts1r1 Mus musculus), A0A088DCW5 (t1r1 Chaetura pelagica), A0A088DBQ4 (t1r1 Gallus gallus) C6L5J3 (t1r1 Vulpes vulpes japonica) e Q7RTX1 (tas1r1 Homo sapiens)
Pequeno trecho de alinhamento realizado com Clustal Omega das proteínas Q7RTX1 na sequência superior (T1R1 Homo sapiens-versão canônica deste receptor) e na sequência inferior a proteína T1R1 com SNPns A372T
....................51
................................................................................38

9
FIGURA 15- FIGURA 16- Gene tas1r3 e posição do SNPns no éxon III FIGURA 17- Gene tas1r3 e posição do SNPns no éxon VI FIGURA 18- FIGURA 19- Pequeno trecho da proteína Q7RTX0-1 (T1R3 Homo sapiens- versão canônica deste receptor) região rica em cisteína FIGURA 20- FIGURA 21- Gene tas1r2 e posição do SNPns I191V no éxon II
FIGURA 22-
FIGURA 23-
Pequeno trecho de alinhamento realizado com Clustal Omega das proteínas Q7RTX1 na sequência superior (T1R3 Homo sapiens-versão canônica deste receptor) e na
sequência inferior a proteína T1R3 com SNPns A537V
Pequeno trecho de alinhamento realizado com Clustal Omega das proteínas Q8TE23 na sequência superior (T1R2 Homo sapiens-versão canônica deste receptor) e na sequência inferior a proteína T1R2 com SNPns I191V A-Receptores T1R1 e T1R3 com o SNPns A110V. B- molécula de glutamato no sítio de ligação do receptor T1R1 e os resíduos de aminoácidos os quais realiza ligações de hidrogênio. C- molécula de glutamato se ligando ao receptor T1R1 portador do SNPns A110V
Pequeno trecho de alinhamento realizado com Clustal Omega das proteínas Q7RTX0 na sequência superior (T1R3 Homo sapiens-versão canônica deste receptor) e na sequência inferior a proteína T1R3 com SNPns R757C
Pequeno trecho de alinhamento realizado com Clustal omega das proteínas Q7RTX0 na sequência superior (T1R3 Homo sapiens-versão canônica deste receptor) e na sequência inferior a proteína T1R3 com SNPns
................................................52
..............................52
..............................53
....................54
.................54
...................57
...................56
....................58
.....................................................60

10
LISTA DE QUADROS
QUADRO 1- Principais periódicos envolvendo polimorfismo e os genes tas1r1, tas1r2 e tas1r3..........................................................................21
QUADRO 2- Resumo dos principais receptores e suas especificidades quanto à modalidade de sabor..............................................................27
QUADRO 3- Domínios do receptor T1R1..................................................................30
QUADRO 4- Domínios do receptor T1R2. ................................................................32
QUADRO 5- Domínios do receptor T1R3. ................................................................34
QUADRO 6- Aminoácidos e suas respectivas representações por siglas e letras..................................................................................................39 QUADRO 7-Polimorfismo de nucleotídeo presentes nos genes tas1r1, tas1r2 e tas1r3.......................................................................................47

11
LISTA DE GRÁFICOS
GRÁFICO 1-
GRÁFICO 2-
GRÁFICO 3-
GRÁFICO 4-
SNPns e SNPs e a probabilidade de desenvolvimento de doença conforme o grau de conservação da região do DNA acometida.
SNPns e SNPs e a probabilidade de desenvolvimento de doença conforme o grau de acessibilidade de um solvente ao sítio da mutação.
Taxas de intensidade para cloreto de sódio 1M, sacarose
1M, ácido cítrico 32 Mm e quinino 1 Mm aplicados
bilateralmente na ponta da língua nas papilas fungiformes
e no dorso da língua nas papilas circunvaladas entre os o
grupo com genótico GG e AA/AG para o SNPns A372T.
Grau de resposta da quimera T1R3+ T1R1 com diferentes
SNPns no aminoácido 537-alanina.
...................................................................38
................................................................41
...................50
.................................................58

12
LISTA DE ABREVIATURAS e SIGLAS
DNA Deoxyribonucleic acid- ácido desoxirribonucleico
GM148 Homônimo do receptor T1R1
GMS Glutamato monossódico
GPCRs G-protein-coupled receptor- receptor acoplado à proteina G
GPR70 Homônimo do receptor T1R1
GPR71 Homônimo do receptor T1R2
HEK293 Human Embryonic Kidney 293-célula embrionária de rim humano 293
HGMD Human Gene Mutation Database-base de dados de mutação dos genes humanos
mGluRs Metabotropic glutamate receptors- receptores metabotrópicos de glutamato
OMIM Online Mendelian Inheritance in Man-
PKD2L1 Receptor do sabor ácido-
RMSD Root-mean-square deviation- desvio da raiz quadrada média
SNP Simple Nucleotide Polymorphism-polimorfismo de nucleotídeo único
SNPns Non-synonymous Simple Nucleotide Polymorphism
polimorfismo de nucleotídeo único não sinônimo
T2R16 Receptor do sabor amargo
T2R38 Receptor do sabor amargo
T2R43 Receptor do sabor amargo
T2R5 Receptor do sabor amargo
T2R8 Receptor do sabor amargo
TAS1R1 Gene do receptor gustativo T1R1
TAS1R2 Gene do receptor gustativo T1R2
TAS1R3 Gene do receptor gustativo T1R3
TR2 Homônimo do receptor T1R2
UniProt Universal Protein Resource-

13
SUMÁRIO
1 INTRODUÇÃO.......................................................................................................15
2 OBJETIVOS ..........................................................................................................16
2.1 Objetivo geral ....................................................................................................16
2.2 Objetivos específicos.......................................................................................16
3 JUSTIFICATIVA ....................................................................................................17
4 OBJETO ................................................................................................................18
4.1 Problema ...........................................................................................................18
4.2 Hipótese básica.................................................................................................18
5 METODOLOGIA ....................................................................................................19
5.1 Critérios de inclusão .........................................................................................19
5.2 Critérios de exclusão........................................................................................19
6 RESULTADOS......................................................................................................20
7 EMBASAMENTO TEÓRICO.................................................................................22
7.1 Aspectos gerais da gustação..........................................................................22
7.2 Aspectos neuroanatômicos da gustação........................................................24
7.3 O sabor doce......................................................................................................26
7.3 Sabor umami......................................................................................................27
7.4 Receptor T1R1....................................................................................................28
7.5 Receptor T1R2....................................................................................................30
7.6 Receptor T1R3....................................................................................................32
7.7 Ativação molecular dos receptores T1R durante a alimentação...................34
7.8 Diferenças nos receptores T1R1 e T1R3 em diferentes espécies................35
7.9 Polimorfismos....................................................................................................35
7.10 Conceitos de homologia, ortologia e paralogia............................................42
7.11 Polimorfismos dos receptores T1R................................................................45
8 DISCUSSÃO...........................................................................................................48
8.1 Influência dos SNPns’s sobre os receptores T1R2 e T1R3 no grau de percepção individual do sabor doce e umami..................................48
8.2 SNPns A372T do receptor T1R1 e aumento da percepção do sabor ............................................................................................................49

14
8.3 SNPns A5T do receptor T1R3 e aumento da percepção do sabor umami..................................................................................................................51
8.4 SNPns R757C do receptor T1R3 e redução da percepção do sabor umami................................................................................................................52 8.5 Influência dos SNPns’s nas regiões ricas em cisteína do receptor T1R3 no grau de percepção individual do sabor doce...................................54 8.6 Polimorfismos I191V e S9C do receptor T1R2 e sua influência na dieta de açúcar..............................................................................................57 8.7 Influência do SNPns A110V na estrutura tridimensional do dímero
T1R1-T1R3 e redução da percepção do sabor umami...................................58
9 CONCLUSÕES.....................................................................................................61
REFERÊNCIAS........................................................................................................62
ANEXO I...................................................................................................................67
ANEXO II .................................................................................................................72

15
1 INTRODUÇÃO
A língua é um órgão que recebe inervação de múltiplos pares de nervos cranianos e
apresenta funções importantes como a fonação, mastigação e a gustação. Por meio
da gustação o indivíduo pode selecionar os alimentos que irá ingerir, procurar
adequadamente fontes de nutrientes ricas em energia, como também, afastar-se de
alimentos potencialmente venenosos, como por exemplo o tanino presente em
diversas plantas.
Sabe-se que os alimentos ricos em carboidratos, açúcares, cetonas, aldeídos e
peptídeos (aspartame) propiciam, em humanos, respostas gustativas do tipo doce, já
os alimentos ricos em L-aminoácidos, especialmente glutamato monossódico e
aspartato, evocam uma sensação denominada umami. A percepção dessas duas
modalidades de sabor é realizada a partir dos receptores T1R1, T1R2 e T1R3
presentes nas papilas gustativas.
Ocorre que alguns polimorfismos presentes nos genes tas1r podem produzir um
receptor com uma sequência peptídica diferente daquela mais frequente, o que por
consequência, em muitos casos, levará a uma modificação no grau de percepção
daquela modalidade de sabor que o receptor desempenha.
O presente trabalho faz uma revisão da literatura a respeito dos polimorfismos
presentes nos genes dos receptores T1R1, T1R2 e T1R3 e sua influência na
percepção dos sabores doce e umami pelo Homo sapiens.

16
2 OBJETIVOS
2.1 Objetivo geral
Revisar a influência dos polimorfismos nos genes dos receptores T1R1, T1R2 e
T1R3 na percepção dos sabores doce e umami pelo Homo sapiens.
2.2 Objetivos específicos
- Descrever quais destes polimorfismos dos genes tas1r1, tas1r2 e tas1r3 são mais
frequentes em humanos.
- Correlacionar os diferentes polimorfismos com a alteração no limiar de percepção
do sabor doce e umami pelo humano.

17
3 JUSTIFICATIVA
Indivíduos de uma mesma população de Homo sapiens apresentam graus distintos
de capacidade de detecção dos sabores doce e umami. Um melhor conhecimento
dos polimorfismos nos genes tas1r1, tas1r2 e tas1r3 no Homo sapiens pode
contribuir para o entendimento do porque alguns indivíduos selecionam
preferencialmente alimentos que proporcionam respostas gustativas do tipo doce e
umami.

18
4 OBJETO
4.1 Problema
Qual a influência do polimorfismo de nucleotídeo único (do inglês, single nucleotide
polymorfism - SNP) na detecção dos sabores doce e umami na espécie Homo
sapiens?
4.2 Hipótese básica orientadora do estudo
A presença de polimorfismo de nucleotídeo único nos genes tas1r1, tas1r2 e tas1r3
pode alterar o limiar da espécie Homo sapiens na detecção dos sabores doce e
umami.

19
5 METODOLOGIA
Trata-se de uma revisão da literatura realizada a partir da reunião de uma base
conceitual e teórica sobre o tema. Foram utilizados para a elaboração desta revisão
apenas periódicos presentes no Pubmed selecionados a partir dos seguintes
descritores: T1R1, T1R2 e T1R3, polymorphims, tas1r1, tas1r2 e tas1r3.
a) critérios de inclusão
Foram incluídos apenas artigos científicos envolvendo Homo sapiens como objeto
de estudo e que apresentem análise de polimorfismos de nucleotídeo simples não
sinônimo dos genes tas1r1, tas1r2 e tas1r3.
b) critérios de exclusão
Foram excluídos artigos que apresentavam o foco do estudo em receptores
provenientes dos genes tas1r1, tas1r2 e tas1r3 localizados fora das papilas
gustativas.

20
6 RESULTADOS
A investigação realizada em agosto de 2015 no Pubmed utilizando os seguintes
descritores: T1R1, T1R2 e T1R3, polymorphims, tas1r1, tas1r2 e tas1r3 e com
restrição de buscas somente para artigos envolvendo Homo sapiens obteve os
seguintes resultados:
-descritores “T1R1” e polymorphims- obtidos três artigos, os quais preenchiam os
critérios de inclusão.
-descritores “tas1r1” e “polymorphims”- obtidos oito artigos, os quais apresentaram
os critérios de inclusão.
-descritores “T1R2” e “polymorphims”- obtido um artigo. Após verificar que esse
resultado de busca apresentou critério de exclusão seu conteúdo não foi utilizado na
confecção deste trabalho.
-descritores “tas1r2” e “polymorphims”- obtidos sete artigos, os quais apresentaram
os critérios de inclusão.
-descritores “T1R3” e “polymorphims”- obtidos cinco artigos. Após verificar que um
dos resultados apresentou critério de exclusão, apenas quatro artigos foram
utilizados na confecção deste trabalho.
-descritores “tas1r3” e “polymorphims”- obtidos 10 artigos. Todos os artigos foram
utilizados na confecção deste trabalho por apresentarem os critérios de inclusão.
Nota-se que durante as sucessivas pesquisas realizadas no Pubmed utilizando os
descritores supracitados, as buscas apresentaram artigos em comum (QUADRO 1).

21
Autor Periódico N° de indivíduos e
nacionalidade CHEN et al., 2009 Am J Clin Nutr N=242, EUA
ENY et al., 2010 Am J Clin Nutr N=1443, Canada
FUSHAN et al., 2009 Curr Biol. N=144, diferentes nacionalidades
JIANG et al., 2004 The Journal of
Biological Chemistry
In vitro
RALIOU et al., 2011 Chem. Senses. In vitro
RAWAL et al., 2013 Chem. Senses. N=92, EUA
SHIGEMURA et al.,
2009
PLoS ONE. In vitro

22
7 EMBASAMENTO TEÓRICO
7.1 Aspectos gerais da gustação
A língua é um órgão que recebe inervação de múltiplos pares de nervos cranianos
dificultando o seu estudo. Além de auxiliar da fonação e mastigação, este órgão
apresenta em sua superfície botões gustativos que produzem estímulos nervosos
conforme a natureza química dos alimentos que, conjuntamente às informações
sobre sua textura, levam o indivíduo a selecionar os alimentos que irá ingerir.
Através da gustação, o organismo cria uma percepção de um estímulo químico
podendo assim procurar adequadamente fontes de nutrientes ricas em energia,
como também, se afastar de alimentos potencialmente venenosos. (LINDEMANN,
2001).
As leves sensações dolorosas na superfície da língua induzidas pelos condimentos
(principalmente a pimenta) também modificam o sabor final dos alimentos através da
estimulação das terminações nervosas livres do nervo trigêmeo presentes neste
órgão. Curiosamente o cheiro age em conjunto com o sabor dos alimentos trazendo
mais informações sobre sua constituição através do nervo olfatório. Corroborando
esses fatos, durante o período em que uma pessoa apresenta um resfriado nota-se
que a anosmia ou hiposmia acarreta uma redução da percepção do sabor do
alimento que aparentemente se torna mais insípido. (LINDEMANN, 2001).
Quanto à gustação, pode-se dizer que os alimentos poderiam ser agrupados em
cinco grandes grupos ou sensações gustativas primárias: ácido, doce, azedo,
amargo e umami. Através da mistura destas diferentes modalidades primárias todos
os diferentes sabores são formados e captados através dos receptores gustativos
(MATSUNAMI, 2000).
As células receptoras localizam-se nos botões gustativos que ficam nas papilas
sobre o dorso da língua. Cada botão gustativo possui cerca de 50-150 células
receptoras (Figuras 1 e 2). No ápice de cada célula gustativa encontramos

23
microvilosidades que permanecem em contato com o poro gustativo e com isso
entram em contato direto com os alimentos durante o processo de alimentação. As
papilas circunvaladas localizadas na região posterior da língua albergam cada uma
cerca de centenas de botões gustativos. As papilas foliadas predominantes na
região postero-lateral da língua contêm dezenas a centenas de botões gustativos e
as papilas fungiformes predominantes nos 2/3 anteriores da língua possuem poucos
botões gustativos. Além da língua, encontramos botões gustativos no palato,
esôfago proximal e pilares das tonsilas (MATSUNAMI, 2000).
As células receptoras apresentam uma vida muito curta sendo substituídas a cada
duas semanas. Um fato ainda não bem elucidado é como se dá a nova ligação da
fibra nervosa para transmitir as informações gustativas após a substituição dessas
células receptoras por outras novas (LINDEMANN, 2001).
Os botões gustativos são mais numerosos nas crianças e após os 45 anos de idade
esse número se torna progressivamente menor, modificando a capacidade gustativa.
Contrariando a crença antiga da existência na língua de locais específicos para
percepção de cada modalidade primária de sabor, nota-se que a percepção de cada
sabor se faz em todo do dorso da língua (GUYTON, 2006).
FIGURA 1- Corte histológico do
tecido epitelial em superfície de
papila gustativa exibindo botão
gustativo. Note seu aspecto esférico
com inúmeras células em seu
interior. FONTE MAGALHÃES, M .
J.S. Capítulo pares cranianos. in.
Neuroanatomia humana aplicada à
clínica médica. Editora Medbook.
FIGURA 2- Botão gustativo
isolado. Cada uma destas
unidades possui cerca de 50 a 150
células receptoras gustativas.
Localizam-se sobre a superfície
das papilas gustativas na língua.
FONTE- MAGALHÃES, M . J.S.
Capítulo pares cranianos. in.
Neuroanatomia humana aplicada
à clínica médica. Editora
Medbook.

24
A gustação fornece ao animal a capacidade de selecionar o alimento e modificar seu
instinto conforme as necessidades e alterações metabólicas do organismo
(fenômeno da preferência gustativa). Quando se come um alimento doce, surge a
sede. Posteriormente, surge um comportamento de repulsa a alimentos com níveis
elevados de açúcar. Indivíduos com insuficiência adrenal apresentam baixas
concentrações de mineralocorticoides, levando a redução dos níveis de sódio no
organismo, aumentando com isso a preferência pelos alimentos salgados
(GUYTON, 2006).
7.2 Aspectos neuroanatômicos da gustação:
Os 2/3 anteriores da língua e mucosa adjacente possuem receptores gustativos com
axônios que alcançam o nervo lingual (V3). Este nervo apresenta anastomose com o
nervo corda do tímpano (ramo do nervo facial) que por sua vez apresenta o corpo
neuronal localizado no gânglio geniculado com o prolongamento central realizando
sinapse no núcleo do trato solitário. O 1/3 posterior da língua e região da orofaringe
possuem receptores gustativos com fibras que percorrem o nervo glossofaríngeo
que possui o corpo neuronal das fibras localizado no gânglio sensorial do nervo
glossofaríngeo (NC IX) com o processo central deslocando-se para o tronco
encefálico chegando ao núcleo do trato solitário. A região da epiglote e parede da
hipofaringe apresenta receptores gustativos que apresentam fibras nervosas que
percorrem o nervo vago. Estas fibras possuem o corpo neuronal localizado no
gânglio sensitivo do nervo vago com o prolongamento central dos axônios
penetrando no tronco encefálico pelo mesmo nervo craniano e chegando ao núcleo
do trato solitário. A mucosa localizada no palato e nasofaringe apresenta receptores
gustativos com fibras que inicialmente passam através de ramos do nervo maxilar
(V2). Essas fibras nervosas levando a informação gustativa atravessam a fossa
pterigopalatina até alcançar o gânglio pterigopalatino. A partir deste gânglio as fibras
alcançam o nervo petroso superficial maior (ramo do nervo facial) chegando até o
gânglio geniculado (gânglio do nervo facial) local onde se encontram os corpos
neuronais desta via gustativa. A via gustativa a partir do núcleo do trato solitário
produz projeções em direção ao núcleo posteromedial do tálamo. As fibras oriundas

25
deste núcleo talâmico direcionam-se para o córtex gustativo localizado na região
anterior da ínsula e opérculo frontal (IVAN, 2003) (Figura 3 e 4).
FIGURA 3- Inervação
gustativa da língua. A
inervação gustativa dos
2/3 anteriores da língua é
realizada pelo nervo facial.
O nervo glossofaríngeo é o
responsável pela
inervação gustativa do 1/3
posterior FONTE-
MAGALHÃES, M. J.S.
Capítulo pares cranianos.
in. Neuroanatomia humana
aplicada à clínica médica.
Editora Medbook
FIGURA 4- Inervação gustativa da língua. Note que as fibras gustativas alcançam o
núcleo do trato solitário através de diferentes nervos cranianos: trigêmeo, facial,
glossofaríngeo e vago. FONTE- MAGALHÃES, M . J.S. Capítulo pares cranianos. in.
Neuroanatomia humana aplicada à clínica médica. Editora Medbook

26
7.3 O sabor doce
Alimentos ricos em carboidratos, proteínas (taumatina e aspartame), açúcares,
cetonas, aldeídos e aminoácidos (alanina, glicina e serina) propiciam em humanos,
respostas gustativas do tipo doce em um grau maior ou menor. Essa atrativa
modalidade de sabor está associada à presença de diferentes sítios de ligação aos
receptores acoplados á proteína G (G-protein-coupled receptors-GPCRs), T1R1,
T1R2 e T1R3 (Quadro 2). Tais receptores são caracterizados pela presença de
domínios amino-terminais longos extracelulares, cuja função está associada ao
reconhecimento e ligação a moléculas presentes nos alimentos que conferem esse
tipo de sabor. Estudos como o de Li (2009) em que a expressão dos receptores
T1R2 e T1R3 foi bloqueada em ratos por alterações genéticas artificiais, as
sensações dessa modalidade de sabor ficaram abolidas nestes roedores. Os felinos
apresentam naturalmente a deleção do gene dos receptores T1R2 explicando o já
conhecido comportamento indiferente destes animais frente a um alimento doce,
fenômeno também conhecido como pseudogenização de gene (LI, 2009).
27
7.4 Sabor umami
A maioria dos mamíferos apresenta uma atração pelos alimentos ricos em L-
aminoácidos, especialmente glutamato monossódico e aspartato. Esses
aminoácidos evocam uma sensação denominada recentemente como umami, que
em japonês significa “sabor delicioso”. Houve um direcionamento ecológico de cada
grupo de animais para determinados tipos de alimentos. Nota-se que somente o L-
glutamato e L-aspartato estimulam fortemente o sabor umami entre humanos e entre
ratos os demais L-aminoácidos estimulam este tipo de resposta (IWASAKI, 1985).
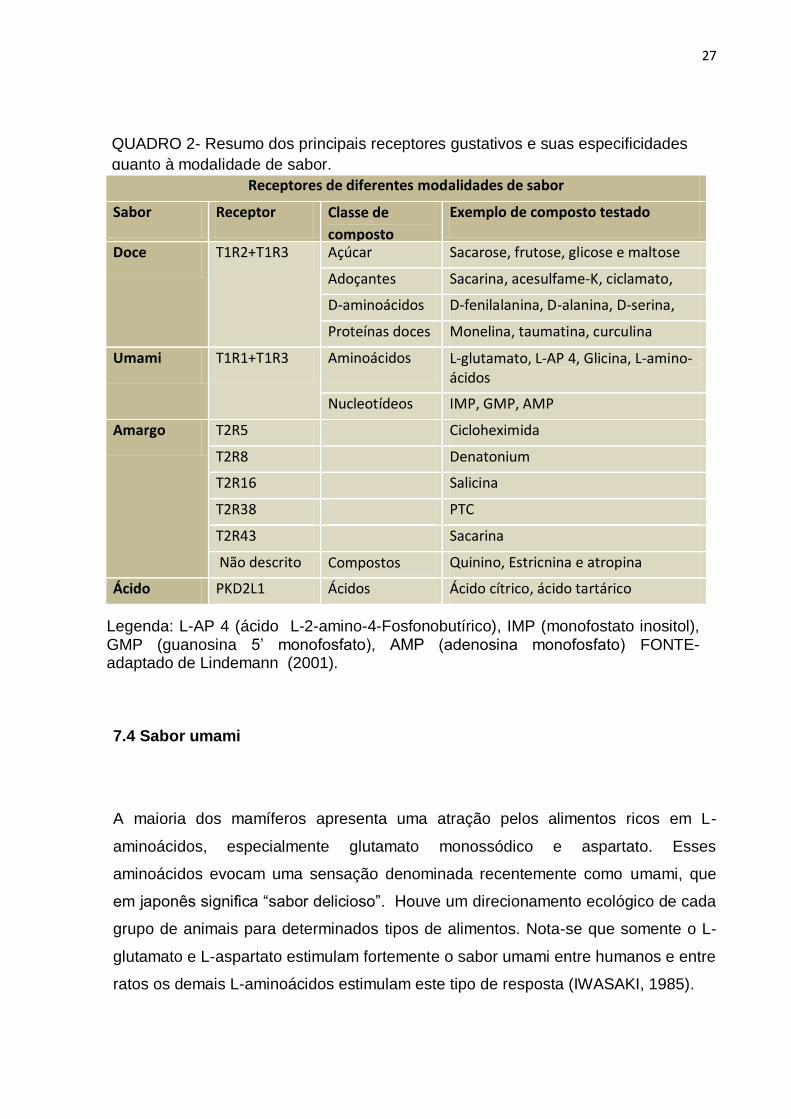
Receptores de diferentes modalidades de sabor
Sabor Receptor Classe de
composto
Exemplo de composto testado
Doce T1R2+T1R3 Açúcar Sacarose, frutose, glicose e maltose
Adoçantes
artificiais
Sacarina, acesulfame-K, ciclamato,
Aspartame D-aminoácidos D-fenilalanina, D-alanina, D-serina,
Proteínas doces Monelina, taumatina, curculina
Umami T1R1+T1R3 Aminoácidos L-glutamato, L-AP 4, Glicina, L-amino- ácidos
Nucleotídeos IMP, GMP, AMP
Amargo T2R5 Cicloheximida
T2R8
T2R44
T2R4
Denatonium
T2R16 Salicina
T2R38 PTC
T2R43
T2R44
Sacarina
Não descrito Compostos
tóxicos
Quinino, Estricnina e atropina
Ácido PKD2L1 Ácidos Ácido cítrico, ácido tartárico
QUADRO 2- Resumo dos principais receptores gustativos e suas especificidades
quanto à modalidade de sabor.
Legenda: L-AP 4 (ácido L-2-amino-4-Fosfonobutírico), IMP (monofostato inositol), GMP (guanosina 5’ monofosfato), AMP (adenosina monofosfato) FONTE- adaptado de Lindemann (2001).

28
Os receptores envolvidos na percepção da modalidade umami são T1R1 e T1R3,
apesar do conhecimento da existência do receptor gustativo acoplado à proteína G
denominado receptor de glutamato metabotrópico truncado 4 (taste-mGluR4)
(CHAUDHARI , 2000) (Quadro 2). Estudos como o de Iwasaki (1985) envolvendo
uma mutação homozigótica em ratos levando a não expressão destes receptores
acarretou a perda da atração por alimentos ricos em glutamato monossódico por
esses animais (IWASAKI, 1985).
7.5 Receptor T1R1
O receptor T1R1 também recebe outras denominações como: TR1, GM148 e
GPR70 (MAX, 2001). A nomenclatura utilizada para o gene do receptor T1R1 é
tas1r1. O gene tas1r1 localiza-se no cromossomo 1 em humanos, mais
precisamente no segmento 1p36.23. Este gene apresenta 22577 pares de bases em
sua extensão apresentando como genes adjacentes nol9 e zbtb48. Após a
transcrição do gene tas1r1 é formado um RNA que quando traduzido leva à
formação de uma proteína de 841 aminoácidos (Figura 5). O receptor T1R1 como os
outros receptores acoplados a proteína G apresenta em sua composição vários
domínios citoplasmáticos, extracelulares e transmembranas sendo este último
também conhecido com helicoidal (UniProt, 2015) (Quadro 3).

29
FIGURA 5- Sequência de aminoácidos do receptor T1R1 (identificador Q7RTX1).
FONTE- Disponível em:< http://www.uniprot.org/uniprot/Q7RTX1>. Acesso em:
20 de setembro de 2015.

30
Domínios do receptor T1R1
Descrição Posição Comprimento
Extracelular 20-567
547
Helicoidal 1 568-588 21
Citoplasmático 589-603 15
Helicoidal 2 604-624 21
Extracelular 625-639 15
Helicoidal 3 640-660 21
Citoplasmático 661-680 20
Helicoidal 4 681-701 21
Extracelular 702-725 24
Helicoidal 5 726-746 21
Citoplasmático 747-761 13
Helicoidal 6 762-782 21
Extracelular 783-795 13
Helicoidal 7 796-816 21
Citoplasmático 817-841 25
7.6 Receptor T1R2
O receptor T1R2 também recebe outras denominações aceitas na literatura como
TR2 e GPR71. A nomenclatura utilizada para gene do receptor T1R2 é tas1r2. O
gene tas1r2 localiza-se no cromossomo 1 no Homo sapiens, mais precisamente no
segmento 1p36.13. Trata-se de um gene que apresenta seis éxons em sua
estrutura. Esse gene tas1r2 apresenta genes adjacentes o aldh4a1 e pax. Depois de
traduzido, o RNA transcrito pelo gene tas1r2 produz uma proteína constituída por
839 aminoácidos (Figura 6) (UniProt, 2015). O receptor T1R2 como outros
receptores acoplados à proteína G apresenta em sua composição vários domínios
citoplasmáticos, extracelulares e transmembranas sendo este último também
conhecido como helicoidal (Quadro 3) (UniProt, 2015).
FONTE-Disponível em:<
http://www.uniprot.org/uniprot/Q7RT
X1>. Acesso em: 20 de setembro
de 2015.
QUADRO 3- Domínios do receptor T1R1.

31
FIGURA 6- Sequência de aminoácidos do receptor T1R2. (identificador Q8TE23).
FONTE- Disponível em:< http://www.uniprot.org/uniprot/Q8TE23>. Acesso em: 27 de
setembro de 2015.
32
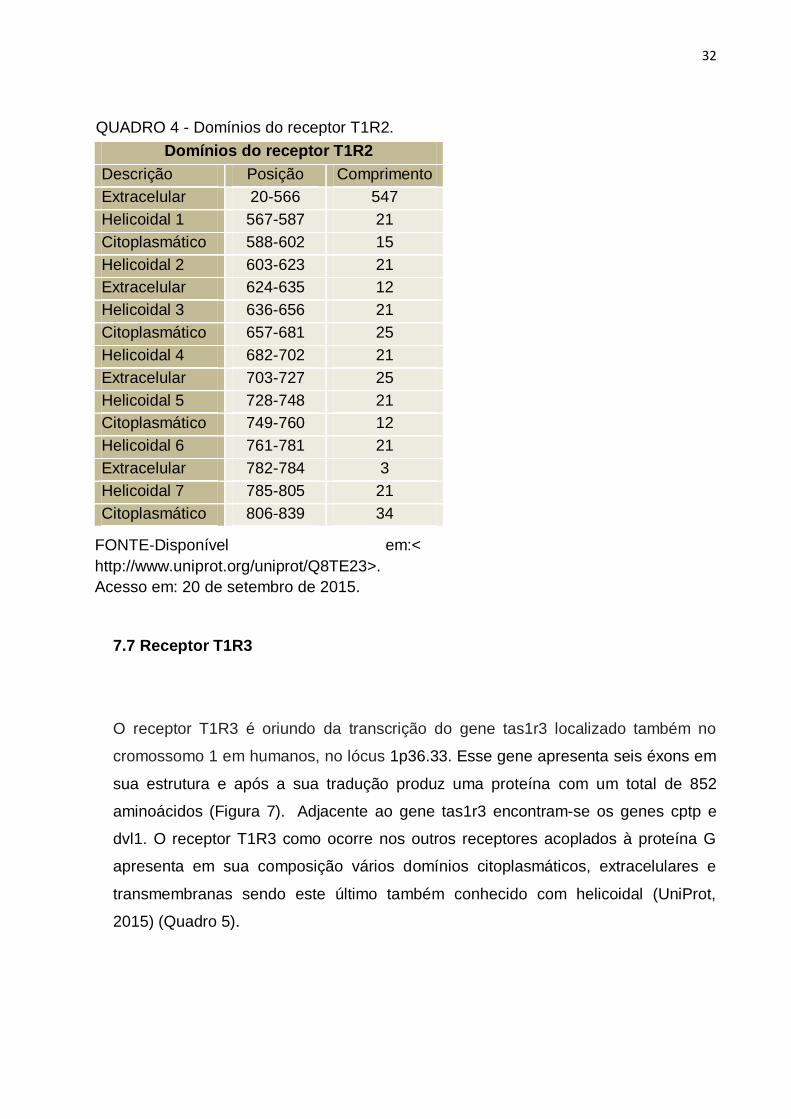
Domínios do receptor T1R2
Descrição Posição Comprimento
Extracelular 20-566
547
Helicoidal 1 567-587 21
Citoplasmático 588-602 15
Helicoidal 2 603-623 21
Extracelular 624-635 12
Helicoidal 3 636-656 21
Citoplasmático 657-681 25
Helicoidal 4 682-702 21
Extracelular 703-727 25
Helicoidal 5 728-748 21
Citoplasmático 749-760 12
Helicoidal 6 761-781 21
Extracelular 782-784 3
Helicoidal 7 785-805 21
Citoplasmático 806-839 34
7.7 Receptor T1R3
O receptor T1R3 é oriundo da transcrição do gene tas1r3 localizado também no
cromossomo 1 em humanos, no lócus 1p36.33. Esse gene apresenta seis éxons em
sua estrutura e após a sua tradução produz uma proteína com um total de 852
aminoácidos (Figura 7). Adjacente ao gene tas1r3 encontram-se os genes cptp e
dvl1. O receptor T1R3 como ocorre nos outros receptores acoplados à proteína G
apresenta em sua composição vários domínios citoplasmáticos, extracelulares e
transmembranas sendo este último também conhecido com helicoidal (UniProt,
2015) (Quadro 5).
QUADRO 4 - Domínios do receptor T1R2.
FONTE-Disponível em:<
http://www.uniprot.org/uniprot/Q8TE23>.
Acesso em: 20 de setembro de 2015.

33
FIGURA 7- Sequência de aminoácidos do receptor T1R3. (identificador Q7RTX0). FONTE- Disponível em:< http://www.uniprot.org/uniprot/Q7RTX0 >. Acesso em: 27 de setembro de 2015.

34
Domínios do receptor T1R3
Descrição Posição Comprimento
Extracelular 21-570
550
Helicoidal 1 571-591 21
Citoplasmático 592-603 12
Helicoidal 2 604-624 21
Extracelular 625-639 15
Helicoidal 3 640-660 21
Citoplasmático 661-682 22
Helicoidal 4 683-703 21
Extracelular 704-729 26
Helicoidal 5 730-750 21
Citoplasmático 751-762 12
Helicoidal 6 763-783 21
Extracelular 784-789 6
Helicoidal 7 790-810 21
Citoplasmático 811-852 42
7.8 Ativação dos receptores T1R durante a alimentação
Durante a ingestão do alimento, alguns açúcares ao entrarem em contato com as
papilas gustativas presentes na língua irão ativar os receptores T1R2 e T1R3
presentes nos botões gustativos ai presentes. Estes dois distintos receptores
gustativos sofrerão inicialmente uma heterodimerização para promover sua ativação.
Adoçantes e sacarose são capazes de ativar o heterodímero T1R2+T1R3 nos
humanos, sendo que nos roedores nota-se que os adoçantes artificiais não
promovem a ativação desses receptores, logo não são capazes de perceber quando
adoçantes estão presentes nos alimentos (WAUSON, 2013). Alimentos ricos em L-
aminoácidos e glutamato monossódico levam a heterodimerização dos receptores
T1R1+T1R3 promovendo a ativação destes. Este sabor recebeu o nome de umami
sendo não promovido por alimentos ricos em D-aminoácidos.
FONTE-Disponível em:<
http://www.uniprot.org/uniprot/Q7RTX0>.
Acesso em: 20 de setembro de 2015.
QUADRO 5 - Domínios do receptor T1R3

35
A heterodimerização dos receptores T1R1+T1R3 e/ou T1R2+T1R3 leva à ativação
da proteína intracitoplasmática α-gustiducina que, por sua vez, libera a subunidade
Gβᵞ acarretando uma elevação da atividade da fosfolipase C β-2. Após essa fase,
nota-se que o inositol trifosfato induz a liberação do Ca2+ intracitoplasmático ativando
o canal iônico TRPM5 desencadeando um potencial de ação através do nervo corda
do tímpano (CHANDRASHEKAR, 2006).
7.8 Diferenças nos receptores T1R1 e T1R3 em diferentes espécies
Humanos e roedores exibem notáveis diferenças na habilidade em detectar certos
adoçantes artificiais. Os roedores, por exemplo, não sentem o sabor do Aspartame,
Monelina ou Taumatina, enquanto os humanos tem sensibilidade submilimolar (10-3
mol/L) a micromolar (10-6 mol/L) para estes compostos (DANILOVA, 1998). Nota-se
que entre humanos, ratos e camundongos os receptores T1R apresentam apenas
70% de identidade em suas sequências de aminoácidos (BROWN, 1993). Por outro
lado, receptores de outras famílias tais como GPCR compartilham mais de 90% de
identidade no nível das sequências de aminoácidos (GENBANK).
7.9 Polimorfismos
Um dos principais mecanismos evolutivos das espécies está associado a mutações
no material genético que podem resultar em divergências entre as proteínas
traduzidas. Essas mutações podem ocorrer através de deleções, substituições,
duplicações e inserções. Polimorfismo genético foi definido em 1971 por Cavalli-
Sforza e Bodmer como a ocorrência em uma mesma população de dois ou mais
alelos em um mesmo lócus com apreciável frequência (HEDRICK, 2011). O
polimorfismo pode contribuir para traços do fenótipo tais como cor da pele,
suscetibilidade a agentes farmacológicos e doenças. Dentre os mais de 30.000
genes presentes no genoma humano, com 3,12 bilhões de nucleotídeos, encontram-
36
se mais de 2 milhões de polimorfismos (MONÇORES, 2008). Esse tipo de alteração
gênica no DNA ocorre aproximadamente a cada 1.250 bases. Sabe-se que o
polimorfismo genético pode acarretar um resultado clínico ou não (DANIELI, 2002;
BIOLO, 2004).
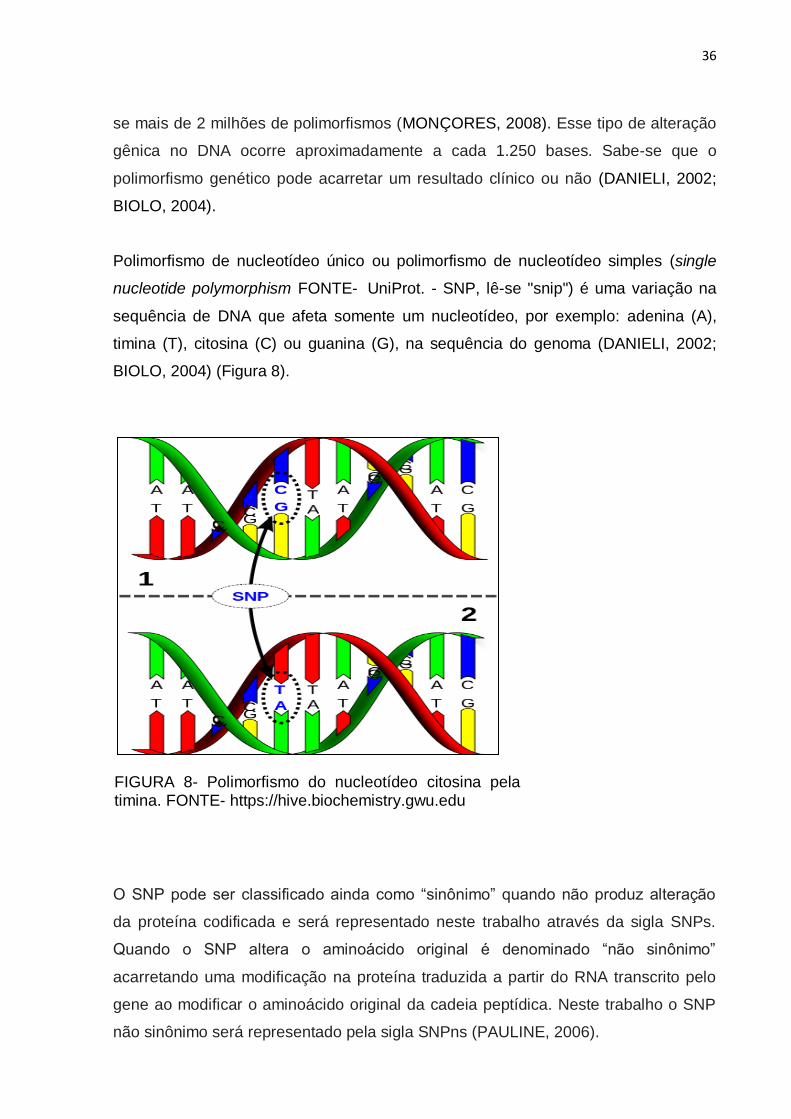
Polimorfismo de nucleotídeo único ou polimorfismo de nucleotídeo simples (single
nucleotide polymorphism FONTE- UniProt. - SNP, lê-se "snip") é uma variação na
sequência de DNA que afeta somente um nucleotídeo, por exemplo: adenina (A),
timina (T), citosina (C) ou guanina (G), na sequência do genoma (DANIELI, 2002;
BIOLO, 2004) (Figura 8).
O SNP pode ser classificado ainda como “sinônimo” quando não produz alteração
da proteína codificada e será representado neste trabalho através da sigla SNPs.
Quando o SNP altera o aminoácido original é denominado “não sinônimo”
acarretando uma modificação na proteína traduzida a partir do RNA transcrito pelo
gene ao modificar o aminoácido original da cadeia peptídica. Neste trabalho o SNP
não sinônimo será representado pela sigla SNPns (PAULINE, 2006).
FIGURA 8- Polimorfismo do nucleotídeo citosina pela timina. FONTE- https://hive.biochemistry.gwu.edu

37
Os SNPns são variantes do DNA que acarretam uma alteração nos aminoácidos
presentes nas proteínas podendo como consequência apresentar grande impacto na
saúde humana. Por isso é de fundamental importância distinguir o SNPs dos SNPns,
já que o primeiro grupo não afeta a sequência final de aminoácidos da proteína
(PAULINE, 2006).
Na população humana estima-se que 67.000 a 200.000 SNPns estejam presentes
no genoma e que cada indivíduo seja heterozigoto para 24.000 a 40.000 SNPns
(MONÇORES, 2008). Através de dois bancos de dados específicos para mutações
no genoma humano (Online Mendelian Inheritance in Man - OMIM e Human Gene
Mutation Database - HGMD) foi constatado que aproximadamente metade das
alterações genéticas por SNPns descritas no genoma já foram encontradas em uma
doença (PAULINE, 2006). Outro achado também associado ao surgimento de
doença com a presença de SNPns é a alteração de aminoácido na sequência
proteica que leve a modificação das características estruturais de sítios de ligação
quando comparado a estrutura da proteína oriunda de DNA sem o SNPns no gene
codificador (STITZIEL, 2004).
Dentre as observações descritas sobre SNPns e manifestação de doenças tem se
observado que o surgimento destas apresentam associação com alterações do DNA
em regiões conservadas (MILLER, 2001) (Figura 9) (Gráfico 1) (Quadro 5).
Usualmente nos alinhamentos de sequências homólogas de proteínas revelam-se as
regiões conservadas ao longo da evolução entre as diferentes espécies. Regiões
conservadas entre proteínas homólogas usualmente desempenham funções
similares (PAULINE, 2006).

38
FIGURA 9- Pequeno trecho de alinhamento das seguintes proteínas: Q99PG6 (tas1r1 Mus musculus), A0A088DCW5 (tas1r1 Chaetura pelagica), A0A088DBQ4 (tas1r1 Gallus gallus ) C6L5J3 (tas1r1 Vulpes vulpes japonica) e Q7RTX1 (tas1r1 Homo sapiens). O alinhamento revela em cinza regiões de sequências conservadas dos aminoácidos. FONTE- Sequências recuperadas do UniProt e alinhadas utilizando o programa Clustal Omega.
FONTE- Adaptado de PAULINE, C.N.G.; HENIKOFF, S. Predicting the Effects of Amino Acid Substitutions on Protein Function. Annu. Rev. Genomics Hum. Genet. 7:61–80, 2006.
GRÁFICO 1- SNPns e SNPs e a probabilidade de desenvolvimento de doença conforme o grau de conservação da região do DNA acometida. Nota-se que a linha verde representa SNPns os quais são menos frequentes em sítios conservados. A linha vermelha revela como as mutações ocorridas nos sítios conservados apresentam probabilidade maior de estarem associados à doença. Por último, a linha azul representa o SNPs onde a falta de modificação da cadeia peptídica não resulta modificações.

39
Alanina Ala A
Cisteína Cys C
Glutamato Glu E
Fenilalanina Phe F
Glicina Gly G
Histidina His H
Isoleucina Ile I
Lisina Lys K
Leucina Leu L
Metionina Met M
Asparagina Asn N
Prolina Pro P
Glutamina Gln Q
Arginina Arg R
Serina Ser S
Treonina Thr T
Selenocisteína Sec U
Valina Val V
Triptofano Trp W
Tirosina Tyr Y
Através deste conhecimento prévio foi possível o desenvolvimento de softwares que
por meio de algoritmos criam um ranqueamento com pontos para cada SNP
identificado na sequência, acusando presença ou não de similaridade entre os
aminoácidos envolvidos (Figura 10 e 11). Para se exemplificar esta ideia, quando um
alinhamento de aminoácidos apresenta tirosina e triptofano em determinada região
da sequência, é provável que outro aminoácido aromático seja também bem
tolerado naquele sítio e desempenhe pouca alteração na função original daquele
sítio já que estes apresentam características físico-químicas parecidas. (PAULINE,
2006).
FONTE- Adaptado de: LEHNINGER, A.L.; NELSON, D.L.; COX, M.M. - Princípios de Bioquímica. São Paulo, Sarvier, 1995.
QUADRO 6- Aminoácidos e suas respectivas representações por siglas e letras respectivamente.

40
FIGURA 10- Pequeno trecho de alinhamento das proteínas Q99PG6 (tas1r1 Mus musculus) e Q7RTX1 (tas1r1 Homo sapiens). Destacado em cinza as similaridades dos aminoácidos com mesma característica química. FONTE- Sequências recuperadas do UniProt e alinhadas utilizando o programa Clustal Omega.
FIGURA 11- Estrutura molecular dos aminoácidos e sua classificação conforme suas propriedades físico-químicas. FONTE- Adaptado de LEHNINGER, A.L.; NELSON, D.L.; COX, M.M. Princípios de Bioquímica. São Paulo, Sarvier, 1995.
41
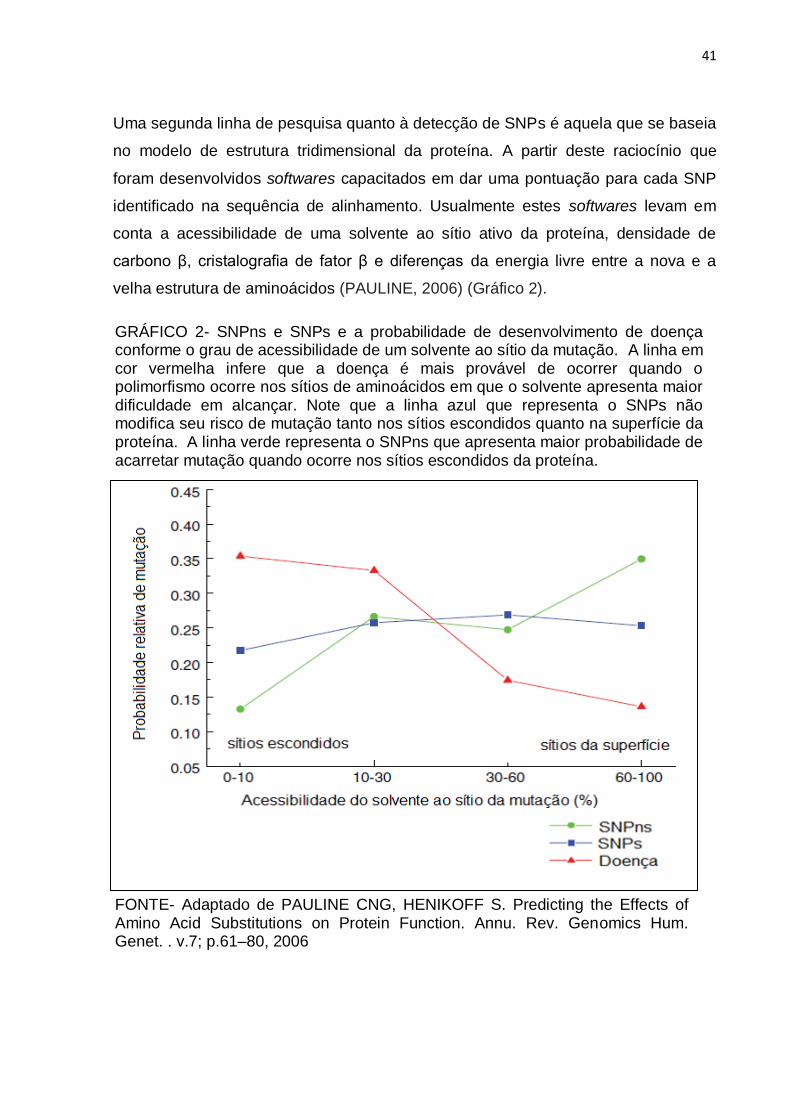
Uma segunda linha de pesquisa quanto à detecção de SNPs é aquela que se baseia
no modelo de estrutura tridimensional da proteína. A partir deste raciocínio que
foram desenvolvidos softwares capacitados em dar uma pontuação para cada SNP
identificado na sequência de alinhamento. Usualmente estes softwares levam em
conta a acessibilidade de uma solvente ao sítio ativo da proteína, densidade de
carbono β, cristalografia de fator β e diferenças da energia livre entre a nova e a
velha estrutura de aminoácidos (PAULINE, 2006) (Gráfico 2).
FONTE- Adaptado de PAULINE CNG, HENIKOFF S. Predicting the Effects of Amino Acid Substitutions on Protein Function. Annu. Rev. Genomics Hum. Genet. . v.7; p.61–80, 2006
GRÁFICO 2- SNPns e SNPs e a probabilidade de desenvolvimento de doença conforme o grau de acessibilidade de um solvente ao sítio da mutação. A linha em cor vermelha infere que a doença é mais provável de ocorrer quando o polimorfismo ocorre nos sítios de aminoácidos em que o solvente apresenta maior dificuldade em alcançar. Note que a linha azul que representa o SNPs não modifica seu risco de mutação tanto nos sítios escondidos quanto na superfície da proteína. A linha verde representa o SNPns que apresenta maior probabilidade de acarretar mutação quando ocorre nos sítios escondidos da proteína.

42
7.10 Conceitos de homologia, ortologia e paralogia
Em biologia evolutiva a homologia se refere aquelas estruturas que possuem a
mesma origem embrionária e desenvolvimento semelhante em diferentes espécies
que descendem de um ancestral em comum. Em alguns casos estas estruturas
podem exercer funções diferentes em diferentes espécies. O conceito de homologia
também pode ser estendido para o estudo das proteínas frequentemente realizado
através do uso de alinhamento das sequências de aminoácidos avaliando desta
forma o grau de similaridade entre si (FITCH, 1970; HÖLTJE, 2003).
As mutações ao longo do tempo evolutivo resultam na origem das diferentes famílias
de proteínas que apresentam algumas diferenças em relação às suas sequências de
aminoácidos, mas as quais conservam alto grau de similaridade estrutural. As
proteínas que evoluem a partir de um ancestral comum são conhecidas como
homólogas (FITCH, 1970). Duas sequências homólogas podem ser praticamente
idênticas, similares em vários aspectos ou até muito diferentes devido a várias
mutações (HÖLTJE, 2003).
Apesar dos alinhamentos serem amplamente difundidos como uma ferramenta para
comparar o grau de similaridade entre duas proteínas avaliando assim se são
homólogas, pesquisas comparando a estrutura tridimensional das proteínas têm
comprovado que essa variável poderia completar uma lacuna na detecção de
proteínas homólogas. A comparação convencional de sequências em alguns casos
é insuficiente para detectar homologia entre proteínas. Duas enzimas como a D-
alanina ligase e a glutationa sintetase são similares tridimensionalmente apesar de
que o alinhamento das sequências primárias falha em confirmar a homologia
(BRENNER, 1998) (Figura 12).
As estruturas tridimensionais de proteínas homólogas são conservadas durante o
processo de evolução, sobretudo no que diz respeito aos resíduos funcionais, pois a
conservação da estrutura é crucial para a manutenção e desempenho de funções
específicas. Usualmente, quando essas regiões conservadas sofrem mutações
podem modificar a funcionalidade. As maiores divergências entre proteínas

43
homólogas ocorrem com mais frequência em regiões próximas da superfície. Nessas
regiões, até mesmo as propriedades físico-químicas dos resíduos que sofreram
mutações podem ser diferentes dos resíduos anteriores ao processo de mutação.
Em geral, os resíduos localizados no interior das proteínas variam com menor
frequência e, quando o fazem, ocorrem normalmente com menor distinção de
propriedades físico-químicas. Um SNPns que apresenta um aminoácido que
compartilha as mesmas características físico-químicas do aminoácido nativo se
enquadra neste caso. Habitualmente, certo conjunto de resíduos de aminoácidos
que compreendem o core da proteína e os principais elementos da estrutura
secundária permanecem mais conservados dentro de uma família de proteínas
homólogas (HÖLTJE, 2003).
No processo de especiação, uma espécie ancestral se diferencia em duas novas
espécies que podem apresentar genes homólogos que desempenham função similar
nestes organismos. Mas à medida que as espécies seguem evoluindo ao longo do
tempo, esses genes acumulam novas mutações e se tornam progressivamente
menos semelhantes. A essas proteínas provenientes da transcrição e tradução
destes genes originados a partir do processo de especiação dá-se o nome de
proteínas ortólogas.
O termo paralogia se refere a genes oriundos da duplicação de um gene ancestral.
Como exemplo pode-se encontrar a cadeia β da hemoglobina que é parálogo do
gene da mioglobina e cadeia α da hemoglobina (GOGARTEN,1999).

44
Fonte: disponível em: http://www.ebi.ac.uk/Tools/services/web/toolresult.ebi?jobId=clustalo-I20150929-024249-0924-16454937-pg&tool=clustalo&showColors=true.
Acesso em: 27 de setembro de 2015.
FIGURA 12- Alinhamento de sequências dos receptores T1R1, T1, T1R3
do Homo sapiens obtidas do UniProt utilizando o programa Clustal
Omega

45
7.11 Polimorfismos dos receptores T1R
Os SNPs presentes no gene tas1r1 podem ocorrer em qualquer parte da sequência
de DNA. Quando presente o SNP nos íntrons não modifica o códon, por outro lado,
quando o SNP acomete os éxons pode resultar em um SNPns, em que a proteína
constituída apresentará uma sequência peptídica diferente daquela original.
Outro ponto relevante a ser considerado é que a frequência dos SNPs presentes nas
sequências do DNA destes receptores é inconstante (RALIOU, 2009; ENY, 2010 e
CHEN, 2009). Para alguns SNPs a frequência se aproxima de 1% e em alguns
casos até superior a 10%. Dentre genes tas1r, o maior número de SPNs foi descrito
no tas1r3 e o menor número no tas1r2. (RALIOU, 2009; ENY, 2010 e CHEN, 2009)
(Quadro 7).
Existe uma variação também da frequência de acordo com os diferentes grupos
étnicos notado nos estudos envolvendo diferentes grupos populacionais em todo
mundo. A população de Camarões apresentou um número maior de SNPns do que
outras sete outras populações estudadas. O SNPns A110V, A372T do gene TAS1R1
têm sido detectado mais frequentemente em populações leucodermas
(SHIGEMURA, 2009 e KIM, 2006).
É descrita a associação entre polimorfismos dos receptores T1R2 e cárie em
humanos (WENDELL, 2010 e KULKARNI, 2013). Também é descrita a associação
do polimorfismo do receptor T1R2 e obesidade (ENY, 2010). A associação da
seleção de alimento e bebida pelos humanos e polimorfismos presentes no receptor
T1R3 também tem sido descrita na literatura. (PIRASTU, 2012)
Os SNPns descritos no tas1r1 apresentaram a frequência compreendida entre 0,5%
até 20% nas populações de diferentes estudos. Importante ressaltar que os SNPns
apresentaram uma distribuição homogênea ao longo dos diferentes éxons deste
gene. (RALIOU, 2011; SHIGEMURA 2009; ENY 2010 e CHEN, 2009).

46
Os SNPns descritos no tas1r2 apresentaram uma frequência compreendida entre
21% e 25% nas populações de diversos estudos. (RALIOU, 2011; SHIGEMURA
2009; ENY 2010 e CHEN, 2009)
Por fim, os SNPns encontrados no tas1r3 foram mais numerosos quando
comparados aos outros dois genes descritos. Sua frequência oscilou entre 0,3 % e
13% nas populações de diferentes estudos. Apresentou como o tas1r1, uma
distribuição homogênea ao longo dos diferentes éxons. (RALIOU, 2011;
SHIGEMURA 2009; ENY 2010 e CHEN, 2009)
As mudanças de aminoácidos envolvidos nos SNPns resultaram em respostas
díspares nos estudos in vitro e clínicos. Muitas vezes a mudança de aminoácido por
outro de mesma propriedade físico-química, como é o caso do A537V presente no
tas1r3, resultou em perda in vitro da resposta a adoçantes artificiais em células
HEK293 portadoras do receptor T1R3 (JIANG, 2004). Outro exemplo seguindo essa
mesma linha de pensamento é o SNPns I191V presente no gene tas1r2. Nesse
caso, a substituição de um aminoácido não polar com radical alifático por outro de
mesma característica fisico-química (isoleucina por valina) foi associada a alterações
na ingestão de carboidratos pela população estudada (ENY, 2010).
Por outro lado, SNPns como o A5T presente no gene tas1r3 acarretou a substituição
do aminoácido alanina por treonina que apresenta característica físico-química
diferente e resultou em um menor limiar para a percepção do glutamato
monossódico por humanos em estudos clínicos. (CHEN, 2009).
Vale ressaltar que o fenômeno do desequilíbrio de ligação (DL) pode levar um
mesmo indivíduo a apresentar diferentes SNPns simultaneamente, não
necessariamente no mesmo cromossomo (FALCONER, 1996). O DL poderia
interferir em estudos clínicos do limiar de percepção dos sabores umami e doce,
especialmente quando um mesmo indivíduo apresentasse simultaneamente
múltiplos SNPns que acarretassem alteração do limiar de percepção dos sabores
umami e doce (FALCONER, 1996). Trata-se então de um possível viés nos estudos
clínicos dos SNPns nos genes tas1r1, tas1r2 e tas1r3 (FALCONER, 1996).

47
Polimorfismos simples do receptor T1R1 Gene Identifica
ção do SNP
Posição do SNP
Nucleotídeo
Alelo Éxon Posição do aminoácido
Aminoácido Frequência do alelo
na população
tas1r1
3537539
2
11 A 1 4 Cisteína >Tirosina raro - 36 C 1 12 Glutamina
>Histidina
6,3% - 255 A - 85 Glutamina >
Glutamina
0,5% 4127802
0
329 T 2 110 Alanina >Valina 2% - 416 T - 139 Treonina >Metionina 1,5% - 519 T - 173 Serina >Serina 0,5%
6174470
0
572 G - 191 Asparagina> Serina 0,5% 1086462
8
1039 A 3 347 Glutamina >Lisina 1% 3416096
7
1114 A 3 372 Alanina >Treonina 20% 4130774
9
1067 G 3 356 Serina >Cisteína raro 3511845
8
1520 A 5 507 Arginina >Glutamina 2,5% tas1r2 9701796 - - - 9 Serina >Cisteína 21%
3587411
6
- - - 191 Isoleucina >Valina 25%
tas1r3
- 13 A 1 5 Alanina >Treonina 13% - 15 A 1 5 Alanina >Alanina 13% - 220 T - 74 Leucina >Leucina
4,9% - 329 C - 110 Metionina >Treonina 0,3% - 459 T 2 153 Treonina >Treonina 3% - 612 T 3 204 Fenilalanina
>Fenilalanina
raro - 740 A 3 247 Arginina>Histidina 7% - 1078 A 3 360 Glicina>Serina raro
3813210 1248 T 3 416 Prolina>Prolina 8% - 1251 A 3 417 Alanina>Alanina raro - 1323 A 4 441 Prolina>Prolina 1% - 1652 A - 551 Serina>Asparagina 1% - 1719 T - 573 Leucina >Leucina 4,2% - 1977 T - 659 Fenilalanina
>Fenilalanina
1% - 2070 A - 690 Leucina >Leucina 0,5%
7914807
3
2246 C 6 749 Fenilalanina >Serina 1% - 2253 A 6 751 Valina >Valina 0,4%
307377 2269 T 6 757 Arginina >Cisteína 9,0% - 2407 T 6 803 Leucina>Leucina raro
3481082
8
2438 T 6 813 Arginina>Lisina 0,5% - 2517 T 6 839 Glicina> Glicina raro
FONTE- adaptado de (RALIOU, 2011; SHIGEMURA 2009; ENY 2010 e CHEN, 2009)
QUADRO 7- Polimorfismo de nucleotídeo único presentes nos genes tas1r1, tas1r2 e
tas1r3.

48
8 DISCUSSÃO
8.1 Influência dos SNPns’s sobre os receptores T1R1, T1R2 e T1R3 no grau de
percepção individual do sabor doce e umami
Polimorfismos presentes nos genes TAS1R que acometem regiões do DNA
responsáveis pela transcrição dos éxons e que levam à alteração da base
nitrogenada no códon podem produzir um receptor com uma sequência peptídica
diferente daquela original conforme já foi supracitado. De forma geral, nota-se que
em muitos casos, a alteração de apenas um aminoácido na sequência peptídica dos
receptores levará a uma modificação no grau de percepção daquela modalidade de
sabor que o receptor desempenha (FUSHAN, 2009). A presença deste SNPns
poderá aumentar ou reduzir a capacidade do indivíduo em detectar através da
gustação a modalidade de sabor umami e doce.
Mesmo quando o SNPns apresente um aminoácido com as mesmas propriedades
fisico-químicas daquele aminoácido substituído não garante a manutenção do grau
de ativação do receptor pela molécula ligante. Alguns estudos têm revelado que
além das propriedades físico-químicas, o tamanho da cadeia lateral do aminoácido
parece interferir na flexibilidade da cadeia polipeptídica interferindo na função desta
(JIANG, 2004 e PAULINE, 2006).
Alguns animais, como os ratos, naturalmente são incapazes de perceber o sabor de
adoçantes artificiais devido a alterações adquiridas em aminoácidos na região rica
em cisteína do receptor T1R3 (trecho 536-545). Experiências in vitro com células
HEK293 onde estes receptores foram modificados geneticamente e que receberam
SNPns com aminoácidos encontrados naturalmente nos humanos propiciou
respostas positivas elevando a concentração de cálcio intracelular mediante a
estímulos químicos produzidos por adoçantes artificiais (JIANG, 2004).
SNPns’s do tas1r1 e tas1r3 apresentam a capacidade de aumentar e reduzir
respectivamente a capacidade do indivíduo em perceber o sabor umami, como é o

49
caso do A372T e G1114A presentes no tas1r1 e R757C presente no tas1r3
(FEENEY, 2011).
8.2 SNPns A372T do receptor T1R1- aumento da percepção do sabor umami
Trata-se de um SNPns (rs34160967) localizado no éxon 3 devido a substituição na
posição 372 do aminoácido alanina pela treonina (Figura 14). O alelo deste SNPns é
encontrado em 20% da população. Estudos in vitro utilizando células HEK293
contendo receptores T1R1 com esse SNPns revelaram uma maior resposta (maior
presença de cálcio intracitoplasmático) ao glutamato monossódico que seu alelo
com alanina. Mesmo em concentrações mais diluídas como 0,3 mM e concentrações
maiores como 5 mM o SNPns A372T desencadeou respostas maiores nestes
experimentos in vitro (SHIGEMURA, 2009).
Um estudo envolvendo 92 americanos adultos com ancestrais europeus realizou
testes clínicos para avaliar o grau de percepção de diferentes modalidades de sabor
em soluções e comprovou que a presença do SNPns na forma heterozigótica (GA) e
homozigótica (AA) reduziu o limiar de percepção dos indivíduos para soluções de
cloreto de sódio (RAWAL, 2013). Os participantes portadores do alelo GG
apresentaram respostas maiores para uma solução salina 1 M quando comparada
aos indivíduos portadores do alelo AA e GA (RAWAL, 2013) (Gráfico 3). Pode-se
inferir que a substituição do aminoácido alanina que é apolar com radical alifático por
treonina que apresenta radical polar e não apresenta carga tenha conferido a este
receptor maior capacidade de formar interações intermoleculares com o ligante
permitindo desta forma uma maior percepção destas moléculas no alimento mesmo
em uma concentração menor (Figura 13).
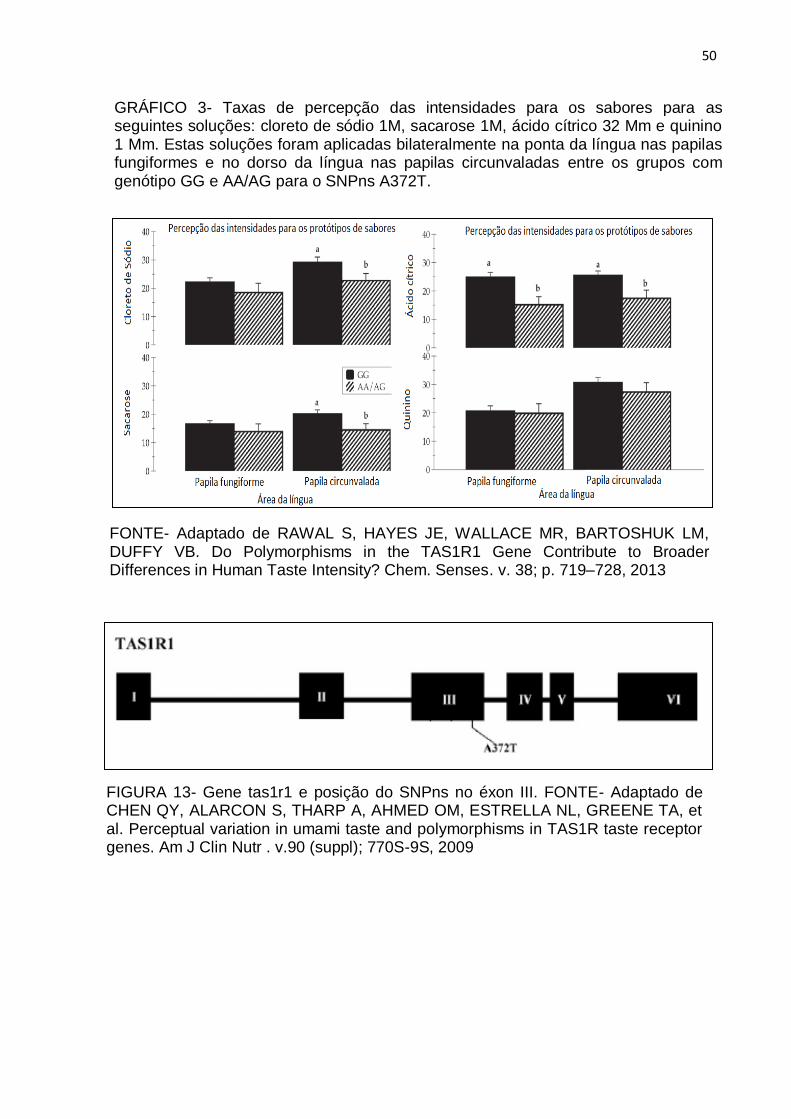
50
FONTE- Adaptado de RAWAL S, HAYES JE, WALLACE MR, BARTOSHUK LM, DUFFY VB. Do Polymorphisms in the TAS1R1 Gene Contribute to Broader Differences in Human Taste Intensity? Chem. Senses. v. 38; p. 719–728, 2013
GRÁFICO 3- Taxas de percepção das intensidades para os sabores para as seguintes soluções: cloreto de sódio 1M, sacarose 1M, ácido cítrico 32 Mm e quinino 1 Mm. Estas soluções foram aplicadas bilateralmente na ponta da língua nas papilas fungiformes e no dorso da língua nas papilas circunvaladas entre os grupos com genótipo GG e AA/AG para o SNPns A372T.
FIGURA 13- Gene tas1r1 e posição do SNPns no éxon III. FONTE- Adaptado de CHEN QY, ALARCON S, THARP A, AHMED OM, ESTRELLA NL, GREENE TA, et al. Perceptual variation in umami taste and polymorphisms in TAS1R taste receptor genes. Am J Clin Nutr . v.90 (suppl); 770S-9S, 2009

51
8.3 SNPns A5T do receptor T1R3 e aumento da percepção do sabor umami.
Trata-se de um SNPns localizado no éxon 1 do gene tas1r3 devido a substituição de
guanina por adenina no nucleotídeo 13, que acarretou a substituição do aminoácido
5 de alanina por treonina. Este alelo está presente em 13% da população de
americanos com ancestrais de origem holandesa (CHEN, 2009) (Figura 15). O
aminoácido encontra-se no domínio extracelular deste receptor.
Um estudo clínico envolvendo 242 indivíduos norte americanos avaliou como a
presença deste alelo treonina interferia na percepção da modalidade de sabor
umami. Foi possível constatar que o alelo treonina promovia nestes indivíduos uma
maior sensibilidade para detecção de glutamato monossódico em soluções quando
comparado ao grupo de indivíduos portadores do alelo alanina. (CHEN, 2009).
Pode-se inferir que a substituição do aminoácido alanina que é apolar com radical
alifático por treonina que apresenta radical polar e não apresenta carga tenha
conferido a este receptor gustativo maior capacidade de se formar interações
intermoleculares com o ligante permitindo desta forma uma maior percepção por
parte do indivíduo destas moléculas no alimento mesmo em uma concentração
menor (ZHANG, 2010) (Figura 16). A partir do estudo de CHEN (2009) infere-se que
a alteração deste aminoácido localizado no domínio extracelular modificou a
FIGURA 14- Pequeno trecho de alinhamento realizado com Clustal Omega das proteínas Q7RTX1 com e sem a presença do SNPns A372T. Note que foi destacado no retângulo vermelho o aminoácido substituído. Note que houve alteração da coloração dos aminoácidos comprovando que houve alteração nas propriedades físico-químicas dos mesmos. FONTE- UniProt.

52
afinidade do receptor pelo ligante. Alguns estudos corroboram que o domínio
extracelular dos receptores T1R1 e T1R3 é o sítio de ligação do glutamato (XU,
2004 e LI, 2007).
8.4 SNPns R757C do receptor T1R3 e a redução da percepção do sabor umami.
Trata-se do SNPns envolvendo o éxon 6 da proteína T1R3 no domínio
transmembrana (terceira alça intracitoplasmática) em que houve uma substituição do
aminoácido arginina pela cisteína (Figura 17). Este alelo T está presente em
aproximadamente 2% da população (CHEN, 2009). Fisiologicamente este domínio
FIGURA 15- Pequeno trecho de alinhamento realizado com Clustal Omega das proteínas Q7RTX0 na sequência superior (T1R3 Homo sapiens-versão canônica deste receptor) e na sequência inferior a proteína T1R3 com SNPns A5T. Note que foi destacado no retângulo vermelho o aminoácido substituído. Observe ainda que houve alteração da coloração dos aminoácidos comprovando que houve alteração nas propriedades físico-químicas dos mesmos. FONTE- UniProt.
FIGURA 16- Gene tas1r3 e posição do SNPns no éxon III. FONTE- Adaptado de CHEN QY, ALARCON, THARP A, AHMED OM, ESTRELLA NL, GREENE TA, et al. Perceptual variation in umami taste and polymorphisms in TAS1R taste receptor genes. Am J Clin Nutr. v. 90 (suppl): 770S-9S, 2009.

53
citoplasmático está envolvido na ativação da proteína G ou ligação a um inibidor
alostérico lactisole do receptor T1R1 (XU, 2004). Embora o receptor T1R3 não
apresente ligação relevante à proteína G, a necessidade de ligação do T1R3 ao
T1R1 para ativar a sinalização interna após acoplar com ligante é fundamental
(GASPARINI, 2002 e PIN, 2005). Ligantes na região transmembrana do receptor
T1R3 como o fármaco lactisole podem acarretar a supressão da percepção do sabor
umami por indivíduos que antes eram capazes de perceber essa modalidade de
sabor (XU ,2004).
Estudos in vitro utilizando células HEK293 contendo receptores T1R3 com esse
SNPns revelaram uma menor sensibilidade ao glutamato monossódico (para
concentrações de 5 e 20 mM) e monofosfato de adenosina que seu alelo com
arginina (para concentrações de 0,3 e 0,5 mM). (SHIGEMURA 2009).
Estudo clínico envolvendo a avaliação da percepção humana de indivíduos
portadores do alelo C revelou uma menor capacidade de percepção do sabor umami
em soluções que indivíduos com o alelo T (CHEN, 2009).
O SNPns R757C ocorreu através da substituição de um aminoácido por outro com
características físico-químicas diferentes, sendo o primeiro (arginina) portador de
uma radical com carga positiva e o segundo (cisteína) polar sem carga, o que
sugere uma possível modificação da função deste domínio transmembrana do T1R3
(Figura 18).
FIGURA 17- Gene tas1r3 e posição do SNPns no éxon VI. FONTE- Adaptado de CHEN QY, ALARCON S, THARP A, AHMED OM, ESTRELLA NL, GREENE TA, et al. Perceptual variation in umami taste and polymorphisms in TAS1R taste receptor genes. Am J Clin Nutr. 90 (suppl): 770S-9S, 2009.

54
8.5 Influência dos SNPns’s nas regiões ricas em cisteína do receptor T1R3 no
grau de percepção individual do sabor doce.
Localizado no receptor T1R3 nota-se que a presença de um segmento rico em
cisteína (intervalo 509-575) é necessário para à ativação do receptor por alguns
tipos de adoçantes artificiais utilizados na alimentação pelos humanos tais como a
Brazina, Monomelina e Aspartame (Figura 19) (MUTO, 2007). Diferentemente dos
humanos, sabe-se que alguns símios e roedores naturalmente não são capazes de
perceber adoçantes artificiais, apesar de serem capazes de perceber o sabor da
sacarose no alimento. Importante salientar que para a percepção do sabor doce se
faz necessário à perfeita integração dos receptores T1R2 e T1R3 (JIANG, 2004).
FIGURA 18- Pequeno trecho de alinhamento realizado com Clustal Omega das proteínas Q7RTX0 na sequência superior (T1R3 Homo sapiens-versão canônica deste receptor) e na sequência inferior a proteína T1R3 com SNPns R757C. Observe que foi no retângulo vermelho o aminoácido substituído. Note que houve alteração da coloração dos aminoácidos comprovando que houve alteração nas
propriedades físico-químicas dos mesmos. FONTE- UniProt
FIGURA 19- Pequeno trecho da proteína Q7RTX0-1 (T1R3 Homo sapiens-versão canônica deste receptor). Destacado em retângulo a região rica em cisteína associada à percepção dos adoçantes artificiais. FONTE-
http://www.uniprot.org/uniprot/Q7RTX0
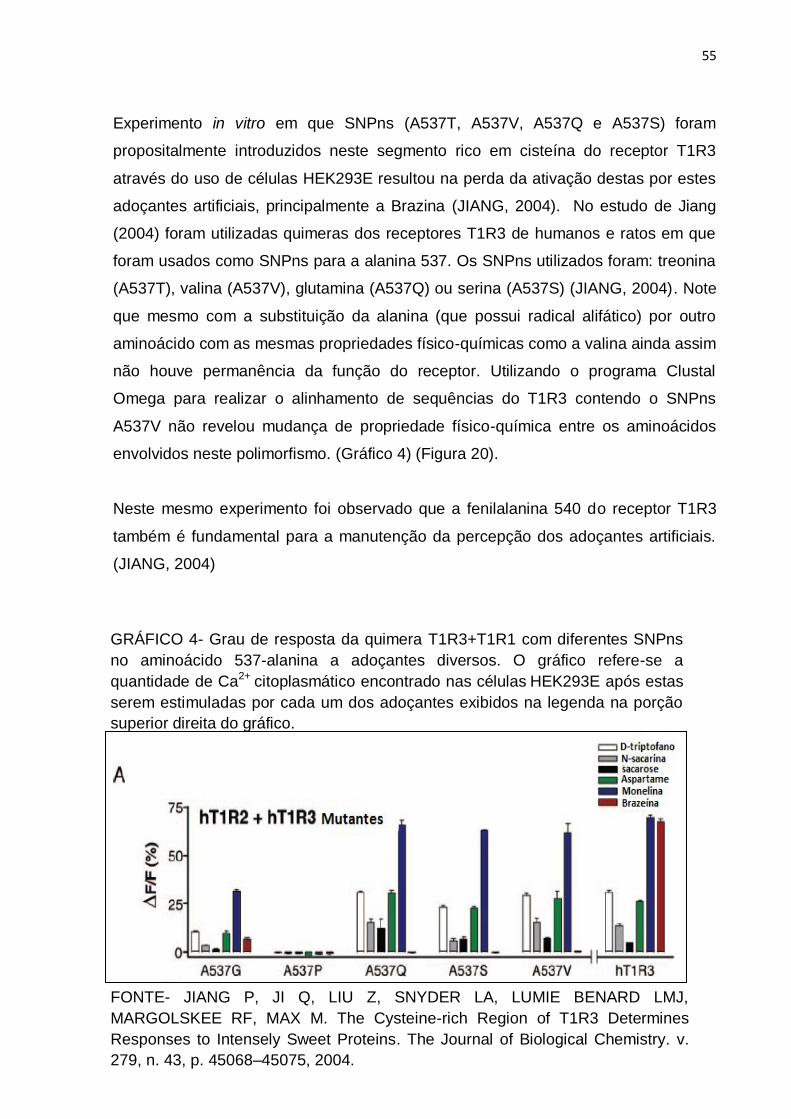
55
Experimento in vitro em que SNPns (A537T, A537V, A537Q e A537S) foram
propositalmente introduzidos neste segmento rico em cisteína do receptor T1R3
através do uso de células HEK293E resultou na perda da ativação destas por estes
adoçantes artificiais, principalmente a Brazina (JIANG, 2004). No estudo de Jiang
(2004) foram utilizadas quimeras dos receptores T1R3 de humanos e ratos em que
foram usados como SNPns para a alanina 537. Os SNPns utilizados foram: treonina
(A537T), valina (A537V), glutamina (A537Q) ou serina (A537S) (JIANG, 2004). Note
que mesmo com a substituição da alanina (que possui radical alifático) por outro
aminoácido com as mesmas propriedades físico-químicas como a valina ainda assim
não houve permanência da função do receptor. Utilizando o programa Clustal
Omega para realizar o alinhamento de sequências do T1R3 contendo o SNPns
A537V não revelou mudança de propriedade físico-química entre os aminoácidos
envolvidos neste polimorfismo. (Gráfico 4) (Figura 20).
Neste mesmo experimento foi observado que a fenilalanina 540 do receptor T1R3
também é fundamental para a manutenção da percepção dos adoçantes artificiais.
(JIANG, 2004)
FONTE- JIANG P, JI Q, LIU Z, SNYDER LA, LUMIE BENARD LMJ,
MARGOLSKEE RF, MAX M. The Cysteine-rich Region of T1R3 Determines
Responses to Intensely Sweet Proteins. The Journal of Biological Chemistry. v.
279, n. 43, p. 45068–45075, 2004.
GRÁFICO 4- Grau de resposta da quimera T1R3+T1R1 com diferentes SNPns
no aminoácido 537-alanina a adoçantes diversos. O gráfico refere-se a
quantidade de Ca2+ citoplasmático encontrado nas células HEK293E após estas
serem estimuladas por cada um dos adoçantes exibidos na legenda na porção
superior direita do gráfico.

56
A alanina 537 apresenta um radical pequeno constituído por grupo (CH3) que
permite maior flexibilidade da cadeia peptídica naquele ponto para que ocorra a
formação da estrutura folha-β. A cisteína 538 forma uma ponte dissulfeto com outra
cisteína nesta mesma região da cadeia polipeptídica (CUFF, 1998). Possivelmente
os SNPns para a alanina 537 (A537T, A537V, A537Q e A537S) promoveriam uma
menor flexibilidade da cadeia peptídica naquele ponto pelo fato da cadeia lateral dos
aminoácidos substituídos apresentarem um maior volume molecular. A alanina
possui o segundo menor agrupamento para cadeia lateral, o CH3. Essa falta de
flexibilidade da cadeia polipeptídica com o SNPns levaria a alteração de sua
estrutura alfa-hélice resultando em uma menor ativação deste receptor pelas
moléculas de adoçantes artificiais (JIANG, 2004).
Por outro lado, o receptor T1R3 de ratos modificados através da introdução de
SNPns na fenilalanina 540 do segmento rico em cisteína pode resultar em resposta
a adoçantes artificiais (JIANG, 2004). Nos humanos, o receptor T1R3 apresenta
naturalmente uma prolina na posição 540. Quando este mesmo receptor de ratos em
experimento in vitro recebe a modificação pelo SNPns F540P resulta em ativação
das células HEK293E pelos adoçantes artificiais. Esses fatos inferem que este
trecho rico em cisteína do receptor T1R3 é apresenta relevância para a percepção
do sabor de adoçantes artificiais (JIANG, 2004).
FIGURA 20- Pequeno trecho de alinhamento realizado com Clustal Omega das proteínas Q7RTX1 na sequência superior (T1R3 Homo sapiens-versão canônica deste receptor) e na sequência inferior a proteína T1R3 com SNPns A537V. Observe que foi destacado no retângulo vermelho o aminoácido substituído. Note que não houve alteração da coloração do aminoácido comprovando a manutenção das propriedades físico-químicas do aminoácido. O programa Clustal Omega considerou os SNPns similares. FONTE- UniProt.

57
8.6 Polimorfismos I191V e S9C do receptor T1R2 e sua influência na dieta de
açúcar.
Dentre os SNPns mais relevantes descritos para o receptor T1R2 encontram-se
I191V e S9C (Figura 21). Em um estudo realizado em Toronto, Canadá, foi possível
constatar que os alelos serina (S9C) e valina (I191V) estiveram presentes
respectivamente em 21% e 25% dos indivíduos envolvidos naquela pesquisa (ENY,
2010). Estes indivíduos foram examinados quanto à influência da presença destes
alelos no consumo de alimentos contendo açúcar. A metodologia desse estudo se
embasou em questionários nutricionais que revelaram que indivíduos portadores de
um alelo valina (I191V) estiveram associados a um IMC (índice de massa corporal) <
25 kg/m2 mais adequado que seus pares sem este alelo, apesar do valor p não
significativo (ENY, 2010). Ainda nesta mesma pesquisa, os indivíduos que
apresentavam IMC ≥ 25kg/m2, a presença de um alelo valina (I191V) esteve
associada significativamente a um consumo menor de açúcares (frutose, glicose e
sacarose) (ENY, 2010).
A presença do alelo correspondente ao SNPns S9C não acarretou modificações nas
taxas de consumo de açúcares por essa amostra de indivíduos nesta pesquisa
realizada em Toronto (ENY, 2010).
O SNPns I191V localiza-se no domínio extracelular do receptor T1R2 e acarreta a
substituição de um aminoácido não polar com radical alifático por outro de mesmas
características fisico-químicas (isoleucina por valina) (ENY, 2010) (Figura 22).
FIGURA 21- Gene TAS1R2 e posição do SNPns no éxon II. FONTE- Adaptado de CHEN QY, Alarcon S, Tharp A, Ahmed OM, Estrella NL, Greene TA, et al. Perceptual variation in umami taste and polymorphisms in TAS1R taste receptor genes.Am J Clin Nutr 2009; 90 (suppl): 770S-9S

58
8.7 Influência do SNPns A110V na estrutura tridimensional do dímero T1R1-
T1R3 e redução da percepção do sabor umami
Estudos in vitro demonstraram que a presença do SNPns A110V esteve associado a
menores respostas celulares ao glutamato monossódico (RALIOU, 2011).
Experimentos que consideraram similaridade do receptor mGluRs ao receptor T1R1
foram utilizados para melhor compreensão das alterações induzidas por este
polimorfismo no dímero T1R1-T1R3 (ZHANG, 2008).
A sequência de aminoácidos do receptor T1R1 apresenta aproximadamente 30% de
similaridade àquela presente na estrutura primária do receptor mGluRs, de tal forma
que suas estruturas secundárias compartilham alto grau de similaridade. Um estudo
envolvendo as características moleculares da interação do receptor mGluRs à
molécula de glutamato foram utilizadas como analogia para o estudo da ligação do
receptor T1R1 a esse mesmo ligante (ZHANG, 2008). Este mesmo autor constatou
que alguns resíduos de aminoácidos específicos realizavam ligações de hidrogênio
com a molécula do glutamato, mais especificamente R54, S148, T149, R151, S172,
R249 e E301 (ZHANG, 2008).
O trabalho in silico com estruturas proteicas veio facilitar o processo de estudo
abrindo desta forma novas possibilidades para a realização da comparação
tridimensional de estruturas proteicas. A partir do gráfico de Ramachandran indicou
FIGURA 22- Pequeno trecho de alinhamento realizado com Clustal Omega das proteínas Q8TE23 na sequência superior (T1R2 Homo sapiens-versão canônica deste receptor) e na sequência inferior a proteína T1R2 com SNPns I191V. Foi destacado no retângulo vermelho o aminoácido substituído. Note que não houve alteração da coloração dos aminoácidos no SNPns revelando similaridade entre os mesmos. FONTE- FONTE- UniProt.

59
que 98% dos resíduos do receptor mGluRs apresentaram ângulos phi e psi dentro
das zonas permitidas, portando constatando grande possibilidade de sua estrutura in
silico ser próxima daquela encontrada na natureza. Nota-se que após uma
comparação das estruturas tridimensionais das duas subunidades T1R1-T1R3 e
T1R1-110V/T1R3, sendo esta segunda com a presença do SNPns A110V, foi
possível verificar alteração das distâncias entre os resíduos de aminoácidos
presentes nestes receptores. Para a realização do cálculo das distâncias entre
resíduos de aminoácidos presentes em estruturas proteicas pode-se utilizar o erro
médio quadrático (RMSD-root-mean-square deviation). O RMSD desta sobreposição
foi de 0.707 Å (angstrom) para o esqueleto de aminoácidos do receptor T1R1 e 2.55
Å para T1R3. Esse erro médio quadrático para este SNPns não foi significativo na
região onde o mesmo se encontra, mas apresentando por outro lado, modificações
significativas nos resíduos de aminoácidos K155 e R54 localizados no receptor
T1R3 e nos resíduos 110 e 180 do receptor T1R1 (RALIOU, 2011). A presença do
SNPns A110V no T1R1 acarretou uma redução de 8 para 2 no número de resíduos
de aminoácidos do heterodímero T1R1/T1R3 que realizam ligações de hidrogênio à
molécula de glutamato. A redução no número de pontes de hidrogênio entre o
ligante e o heterodímero T1R1/T1R3 poderiam explicar o motivo de suas menores
respostas celulares ao glutamato monossódico nos estudos in vitro. (RALIOU, 2011)
(Figura 23).

60
* Como cada estrutura de aminoácido é provida de um carbono quiral no centro de
sua estrutura isso permite que a cadeia peptídica forme um número muito grande de distintas estruturas tridimensionais. Apesar desta informação, estudos matemáticos comprovaram que somente algumas destas formações tridimensionais são factíveis de serem encontradas na natureza por se tratar de formas mais estáveis energeticamente. O gráfico de Ramachandran é utilizado para encontrar todas as possíveis estruturas proteicas a partir de uma mesma sequência de aminoácidos.
FIGURA 23. A- Receptores T1R1 e T1R3 com o SNPns A110V. B- molécula de glutamato no sítio de ligação do receptor T1R1 e os resíduos de aminoácidos os quais realizam ligações de hidrogênio. C- molécula de glutamato se ligando ao receptor T1R1 portador do SNPns A110V. Observe que apenas dois resíduos de aminoácidos do receptor T1R1 realizam suas ligações de hidrogênio. FONTE- adaptado de RALIOU M, GRAUSO M, HOFFMANN B, LE-POUPON CS, NESPOULOUS C, De´bat HL, Christine Belloir C. and col. Human Genetic Polymorphisms in T1R1 and T1R3 Taste Receptor Subunits Affect Their Function Chem. Senses. v.36 (6); p. 527-537, 2011

61
9 CONCLUSÕES Alguns estudos clínicos demonstraram que a presença de SNPns nos genes tas1r1,
TAS1R2 e TAS1R3 são capazes de alterar o limiar de percepção dos sabores doce e
umami em humanos, podendo inclusive influenciar na decisão da seleção dos
alimentos. É importante ressaltar que estudos in vitro apresentam resultados que
corroboram esses resultados. Nota-se que tanto SNPns localizados em regiões de
domínio transmembrana e extracelular foram capazes de alterar o limiar de percepção
destas modalidades de sabor pelos humanos. Dentre as alterações observadas nos
estudos sobre SNPns foi possível constatar que estes podem reduzir, abolir ou
aumentar o limiar de percepção do sabor doce e umami pelos humanos portadores
destes SNPns. Várias características destes SNPns foram levantadas como hipótese
para explicar as alterações no grau de ativação destes receptores pelos alimentos.
Dentre elas estão: ocorrência do SNPns em região conservada ou rica em cisteína,
mudança de características físico-químicas do aminoácido substituído pelo SNPns e
ocorrência do SNPns em região próxima a aminoácidos que realizam pontes de
dissulfeto. Esta revisão infere que somente a propriedade físico-química (similaridade)
do aminoácido substituído pelo SNPns nos receptores T1R1, T1R2 e T1R3 não
representa argumento científico suficiente para explicar a mudança do limiar de
percepção dos sabores doce e umami presente nos alimentos pelos humanos. A
realização de estudos com uma amostragem maior ou que considere as estruturas
secundárias e terciárias destas proteínas com o SNPns e não somente a estrutura
primária dos receptores T1R1, T1R2 e T1R3 poderiam trazer novos dados sobre a
influência destes na ativação destes receptores.

62
REFERÊNCIAS
ADLER, E.; et al. A novel family of mammalian taste receptors. Cell. v.100, p. 693–
702, 2000. ARAUJO, I.E. Representações gustativas no córtex humano, e o controle central do apetite. Rev. Bras. Psiquiatr. v. 25, supl. 2, 2003.
BIOLO A.; ROHDE L.E. O impacto dos polimorfismos genéticos e da farmacogenética na avaliação e manejo da insuficiência cardíaca. Rev Soc Cardiol RS. v. 3, p. 1-5, 2004.
BRENNER, S.E;, CHOTHIA, C.; HUBBARD, T.J. Assessing sequence comparison methods with reliable structurally identified distant evolutionary relationships. Proc Natl Acad Sci USA. v.95, p. 6073-6078,1998.
BROWN, E.M.; GAMBA, G.; RICCARDI, D.; Lombardi, M.; BUTTERS, R.; KIFOR, O.; SUN, A.; HEDIGER, M.A.; LYTTON, J.; HEBERT, S.C. Cloning and characterization of an extracellular Ca(2_)-sensing re ceptor from bovine parathyroid. Nature. v.366, p. 575–580,1993. CARGILL, M.; ALTSHULER, D.; IRELAND, J.; SKLAR, P.; ARDLIE, K.; et al. Characterization of singlenucleotide polymorphisms in coding regions of human genes. Nat. Genet. v.22, p.231–38, 1999. CHANDRASHEKAR ,J.; HOON, M.A.; RYBA, N.J.; ZUKER, C.S. The receptors and cells for mammalian taste. Nature. v.444, p.288–294, 2006.
CHAUDHARI, N.; LANDIN, A.M.; ROPER, S.D. A metabotropic glutamate receptor variant functions as a taste receptor. Nat Neurosci 2000;3:113–9. CHEN, Q.Y.; ALARCON, S.; THARP, A.; AHMED, O.M.; ESTRELLA, N.L.; GREENE, T.A.; et al. Perceptual variation in umami taste and polymorphisms in TAS1R taste receptor genes. Am J Clin Nutr. v.90 (suppl): 770S-9S, 2009. CUFF, J.A.; CLAMP, M.E.; SIDDIQUI, A.S.; FINLAY, M.; BARTON, G.J. Bioinformatics (Oxf.) v.14, p. 892–893, 1998.
DAMAK, S.; et al. Detection of sweet and umami taste in the absence of taste receptor T1r3. Science. v.301, p. 850–853, 2003. DANIELI, G.A. Genetics and genomics for the cardiologist. Dordrecht (The Netherlands): Kluwer, 2002.
DANILOVA, V.; HELLEKANT, G.; TINTI, J. M.; NOFRE, C. Gustatory responses of the hamster Mesocricetus auratus to various com pounds considered sweet by humans. J. Neurophysiol. v.80, p. 2102– 2112, 1998.

63
ENY, K.M.; WOLEVER, T.M.S. COREY, P.N.; EL-SOHEMY, A. Genetic variation in TAS1R2 (Ile191Val) is associated with consumption of sugars in overweight and obese individuals in 2 distinct populations. Am J Clin Nutr. v.92, p. 1501–10, 2010.
FALCONER, D.S.; MACKAY, T.F.C. Introduction to Quantitative Genetics. 4th ed. Harlow, Essex, UK: Addison Wesley Longman, 1996. FEENEY, E.; O’BRIEN, S.; SCANNELL, A.; MARKEY, A.; GIBNEY, E.R. Genetic variation in taste perception: does it have a role in healthy eating? Proc Nutr Soc. v.70(1), p.135-43, 2011. doi: 10.1017/S0029665110003976. FITCH, W.M. Distinguishing homologous from analogous proteins. Syst Zool.
v.19(2), p.99-113, 1970. FUSHAN, A.A.; SIMONS, C.T.; SLACK, J.P.; MANICHAIKUL, A.; DRAYNA, D. Allelic polymorphism within the TAS1R3 promoter is associated with human taste sensitivity to sucrose. Curr Biol. 11;19(15):1288-93, 2009 GASPARINI, F.; KUHN, R. PIN, J.P. Allosteric modulators of group I metabotropic glutamate receptors: novel subtype-selective ligands and therapeutic perspectives. Curr Opin Pharmacol. v.2, p.43–9, 2002. GenBank accession numbers: AY032620– taste buds, and to study the impact of genetic ablation, AY032623). GOGARTEN, J.P.; OLENDZENSKI, L. Orthologs, paralogs and genome comparisons. Current Opinion in Genetics & Development. v. 9, p.630–636,1999. GUYTON, A.C.; HALL, J.E. Tratado de Fisiologia Médica. 11ª ed. Rio de Janeiro,
Elsevier Ed., 2006.
HECK, G. L.; MIERSON, S.; DESIMONE, J. A. Salt taste transduction occurs through an amiloride-sensitive sodium transport pathway. Science. v. 223, p.403–405, 1984.
HEDRICK, P. Genetics of Populations. Jones & Bartlett Learning. pp. 104–.
ISBN 978-0-7637-5737-3. 2011
HÖLTJE, H.D.; SIPPL, W.; ROGNAN, D.; FOLKERS, G. Introduction to comparative protein modeling. In Molecular Modeling: Basic Principles and Applications.
Weinheim: Wiley-VCH, 2003. HOON, M. A.; et al. Putative mammalian taste receptors: a class of taste-specific GPCRs with distinct topographic selectivity. Cell. v.96, p. 541–551,1999.
HUANG, A.L.; et al. The cells and logic for mammalian sour taste detection. Nature.
v.442, p.934–938, 2006.

64
IWASAKI, K.; KASAHARA, T.; SATO, M. Gustatory effectiveness of amino acids in mice: behavioral and neurophysiological studies. Physiol. Behav. v. 34, p. 531–542, 1985. JIANG, P.; JI, Q.; LIU, Z.; SNYDER, L.A.; LUMIE BENARD, L.M.J.; MARGOLSKEE, R.F.; MAX, M. The Cysteine-rich Region of T1R3 Determines Responses to Intensely Sweet Proteins. The Journal of Biological Chemistry. V. 279, No. 43, p. 45068–45075, 2004.
KIM, U.K.; WOODING, S.; RIAZ, N.; JORDE LB, DRAYNA D. Variation in the human
TAS1R taste receptor genes. Chem Senses. 31(7):599-611,2006.
KULKARNI, G.V.; CHNG, T.; ENY, K.M.; NIELSEN, D.; WESSMAN, C.; EL-
SOHEMY, A. Association of GLUT2 and TAS1R2 genotypes with risk for dental
caries. Caries Res. 47(3):219-25, 2013.
LI, X.; ZHANG, F.; XU, H.; LI, Q. PCT/US patent 2006/041158. 2007. LI, X.; et al. Pseudogenization of a sweet-receptor gene accounts for cats’ indifference toward sugar. PLoS Genet. v.1, p.27–35, 2005. LINDEMANN, B. Receptors and transduction in taste. Nature. v. 413, p. 219–225, 2001. LUGAZ, O.; PILLIAS, A.M.; FAURION A. 2002. A new specific ageusia: some humans cannot taste L-glutamate. Chem Senses. 27:105–115.
LYALL, V. et al. ;The mammalian amiloride-insensitive non-specific salt taste receptor is a vanilloid receptor-1 variant. J. Physiol. (Lond.). v. 558, p.147–159,2004. MATSUNAMI, H.; MONTMAYEUR, J.P.; BUCK, L.B. A family of candidate taste receptors in human and mouse. Nature. v. 404, p. 601–604, 2000.
MAX, M.;SHANKER, Y.G.; HUANG, L.; RONG; M.; LIU, Z.; CAMPAGNE, F.;
WEINSTEIN, H.; DAMAK, S.; MARGOLSKEE, R.F. Tas1r3, encoding a new
candidate taste receptor, is allelic to the sweet responsiveness locus Sac. Nat
Genet. 28(1):58-63, 2001.
MAGALHÃES, M . J.S. Capítulo pares cranianos. in. Neuroanatomia humana
aplicado à clínica médica. Editora Medbook. (Não publicado)
MILLER, M.P.; KUMAR, S. Understanding human disease mutations through the use of interspecific genetic variation. Hum. Mol. Genet. v.10, p. 2319–28, 2001.

65
MONÇORES, M.W.; PEREIRA, S.B.; et. al. Personalized Medicine Applied to
Cardiology. Rev SOCERJ. 21(3):184-193, 2008.
MUELLER, K.L.; et al. The receptors and coding logic for bitter taste. Nature. v. 434,
p. 225–229, 2005. MUTO, T.; TSUCHIYA, D.; MORIKAWA, K.; JINGAMI, H. Structures of the extracellular regions of the group II/III metabotropic glutamate receptors Proc Natl Acad Sci U S A. ;104(10): 3759–3764, 2007 NELSON, G.; et al. An amino-acid taste receptor. Nature. v.416, p.199–202, 2002. NELSON, G. et al. Mammalian sweet taste receptors. Cell. v.106, p. 381–390, 2001. NELSON, G.; CHANDRASHEKAR, J.; HOON, M.A.; Feng, L.; Zhao, G.; Ryba, N.J.; ZUKER, C.S. An amino-acid taste receptor. Nature. v.416, p.199–202, 2002.
PAULINE, C.N.G.; HENIKOFF, S. Predicting the Effects of Amino Acid Substitutions on Protein Function. Annu. Rev. Genomics Hum. Genet, 2006.
PIN, J.P.; KNIAZEFF, J.; LIU J.; et al. Allosteric functioning of dimeric class C G-protein-coupled receptors. FEBS J. v.272, p. 2947–55, 2005.
PIRASTU, N.; ROBINO, A,; LANZARA, C.; ATHANASAKIS, E.; ESPOSITO, L.;
TEPPER, B.J.; GASPARINI , P. Genetics of food preferences: a first view from silk
road populations. J Food Sci. 77(12):S413-8, 2012.
Proc Natl Acad Sci USA.;99(26):16899-903. Proceedings of the Nutrition Society,
v.70(1), p. 135–43, 2002 doi:10.1017/S0029665110003976 RALIOU, M.; GRAUSO, M.; HOFFMANN, B.; LE-POUPON, C.S.; NESPOULOUS, C.; De´BAT, H.L.; BELLOIR C. C.; et al. Human Genetic Polymorphisms in T1R1 and T1R3 Taste Receptor Subunits Affect Their Function. Chem. Senses. v. 36 (6):
p. 527-537, 2011
RAWAL, S.; HAYES, J.E.; WALLACE, M.R.; BARTOSHUK, L.M.; DUFFY, V.B. Do
Polymorphisms in the TAS1R1 Gene Contribute to Broader Differences in human
taste intensity? Chem Senses. v.38(8), p. 719-28, 2013. doi: 10.1093/chemse/bjt040
SHIGEMURA, N.; SHIROSAKI, S.; SANEMATSU, K.; YOSHIDA, R.; NINOMIYA, Y. Genetic and Molecular Basis of Individual Differences in Human Umami Taste Perception. PLoS ONE. v.4(8): e6717, 2009. doi:10.1371/journal.pone.0006717

66
STITZIEL, N.O.; BINKOWSKI, T.A.; TSENG, Y.Y.; KASIF, S.; LIANG, J. TopoSNP: a topographic database of non-synonymous single nucleotide polymorphisms with and without known disease association. Nucleic Acids Res. v.32, p. D520–22, 2004.
The DNA sequence and biological annotation of human chromosome 1. JOURNAL Nature. v.441 (7091), p. 315-321. UniProt. Q8TE23 - TS1R2_HUMAN, 2006. Disponível em: < http://www.uniprot.org/uniprot/Q8TE23>. Acesso em: 21 Abr. 2015. WAUSON, E.M.; LORENTE-RODRÍGUEZ, A.; COBB, M.H. Minireview: Nutrient Sensing by G Protein-Coupled Receptors. Mol Endocrinol. v.27(8), p. 1188–1197, 2013.
WENDELL, S.; WANG, X.; BROWN, M.; COOPER, M.E.; DESENSI, R.S.; WEYANT,
R.J.; CROUT, R.; MCNEIL, D.W.; MARAZITA ,M.L. Taste genes associated with
dental caries. J Dent Res. 89(11):1198-202, 2010.
XU H.; STASZEWSKI, L.; TANG, H.; ADLER, E.; ZOLLER, M.; LI, X. Different functional roles of T1R subunits in the heteromeric taste receptors. Proc Natl Acad Sci USA. v.101, p. 14258–63, 2004.
ZHANG, F.; KLEBANSKY, B.; FINE, R.M.; LIU, H.; XU, H.; SERVANT, G.; ZOLLER, N. M.; TACHDJIAN, C.; LI, X. Molecular mechanism of the sweet taste enhancers. Proc Natl Acad Sci USA. v. 107, p.4752–4757, 2010.
ZHAO, G.Q.; ZHANG, Y.; HOON, M.A., CHANDRASHEKAR, J.; ERLENBACH, I.; RYBA.; et al. The receptors for mammalian sweet and umami taste. Cell. v. 115, p. 255–266, 2003.

67
Anexo I (Manuscrito em preparação)
Do Polymorphisms in the Extracellular Domain of T1R1 and T1R3 Receptors Contribute to Reduce
Human Taste Perception?
Polimorfismos no domínio extracelular dos receptores T1R1 e T1R3 contribuem para
reduzir a percepção do sabor em humanos? Marcelo José da Silva de Magalhães1, Fabiano Sviatopolk-Mirsky Pais2,3 Laila Alves
Nahum2,3
1- Faculdades Unidas do Norte de Minas (FUNORTE) Medical School. Department of Neurosurgery, Aroldo Tourinho Hospital, Montes Claros, MG 39400-162, Brazil.
2- Grupo de Genômica e Biologia Computacional, Centro de Pesquisas René Rachou, Fundação Oswaldo Cruz -FIOCRUZ, Belo Horizonte, MG 30190-002, Brazil. 3- Faculdade Infórium de Tecnologia, Belo Horizonte, MG 30140-061, Brazil
Abstract
The tas1r genes encode heterodimeric receptors that mediate umami (tas1r1 + tas1r3) and sweet (tas1r2 + tas1r3) sensations. The question of interest for this study is whether or not tas1r1 nonsynonymous single polymorphisms (SNPns) in the extracellular domain (ECD) are associated with the reduced perception of taste intensity in humans. Nontaster and hypotaster subjects are associated with SNPs occurring in the tas1r genes. This article points out that SNPs of the extracellular domain of T1R1 and T1R3 are associated with reduced perception of umami and sweet sensations in humans. In rare cases SNPs could increase the taste sensation intensity in humans. The majority of studies described elsewhere suggested that SNPs are located in the ECD reducing the activation of T1R3 and T1R3 taste receptors. Keywords: tas1r1, tas1r3, sweet, polymorphisms, umami.
Resumo
Os genes TAS1R codificam receptores heterodiméricos que mediam as sensações umami e doce. A questão de interesse neste estudo é se os polimorfismos não sinônimos (SNPns) nos domínios extracelulares (DEC) dos receptores T1R1 e T1R3 estão associados a uma redução da percepção destas modalidades de sabor em humanos. Pessoas que não percebem ou percebem pouco estes sabores têm apresentado SNPs no gene tas1r. Este artigo indica que vários SNPs estão associados a uma redução da percepção da sensação umami e doce por estes receptores em humanos. Os SNPs raramente acarretam um aumento na percepção da sensação de sabor em humanos. A maioria dos estudos tem sugerido que os SNPs localizados no DEC reduzem a ativação dos receptores T1R1 e T1R3. Palavras-chave: tas1r1, tas1r3, sabor doce, polimorfismo, sabor umami.

68
Introduction
The neurological ability to perceive food taste seems very import to select more appropriate food for good nutrition among animals.1 Sugar-rich food when selected to nutrition will provide a good amount of calories. In the carnivorous group the capacity to recognize protein-rich foods is relevant for these animals, which have more preference for meat in their diet.1 The taste associated to protein-rich food receives the designation of umami taste. This term comes from the Japanese language and means delicious food.2. The animal ability to recognize sweet and umami tastes depends on the receptors T1R1 and T1R3.2 These receptors are encoded by the tas1r gene, which product is a G protein coupled receptor constituted by seven domains.3 In humans the tas1r is found on chromosome 1, but in rats on chromosome 6.3 These receptors are located on the tongue surface in gustative buds.1,2 The T1R1 and T1R3 receptors on gustative buds are responsible to turn chemical stimuli present in foods in synapse that will stimulate the facial nerve until pons and from there to the brain gustative cortex. This gustative receptor is able to respond to sweet and umami tastes, but not for D-glucose and L-amino acids.1 Umami taste is found in several protein-rich foods with meat providing a good amount of L-amino acids.2 It is worth mentioning that the receptor T1R1+T1R3 is functional only with the heterodimer formation.3 When separated T1R1 and T1R3 are not able to convert chemical stimulus into synapses on the facial nerve.3 Recent studies have shown that nonsynonymous single polymorphisms (SNPs) are usually associated with reduced perception of these taste intensity in humans.4 Very rare SNPs are associated with enhancing these modalities of taste perceptions.4 The main goal of this article is to show that SNPs in the extracellular domain are involved on reducing the perception of taste modalities, which dependents on T1R1 and T1R3.
Methods
The present work discusses articles indexed on PubMed at NCBI. These articles were selected to include only studies involving the polymorphism on tas1r. Studies included on this article were performed in vitro and in vivo. The keywords used on the search were T1R1, T1R3, tas1r1, tas1r3 polymorphism, sweet taste, umami taste, and taste receptor. By using this approach, it was possible find a suitable number of articles and discuss some of them in the present work. It was not our objective to perform an exhaustive search of all articles about polymorphism on tas1r for this article.
Results
Some studies involving taste perception in humans evaluate the influence of each SNP in the subject ability to perceive different solution concentrations. In contrast, there are in vitro studies that mimics the responses to monosodium glutamate (MSG) or glucose using functional expression SNPs of T1R1 or T1R3 in HEK293 or different cells. The latter, usually selected SNPs in the aforementioned studies.
In the present work, we focused on selected three articles that discuss the most common SNPs found in humans.4-6

69
The first study selected was the research that involved a sample population of 242 subjects characterized by the general sensitivity to glutamate through tests repeated with MSG solutions.4 After the characterization, 87 subjects were underwent by sequencing the coding regions of the tas1r1 and tas1r3 genes. Five SNPs in tas1r1 (all nonsynonymous) and 12 SNPs in tas1r3 (four nonsynonymous and eight synonymous) were found.4 tas1r1 presents the SNPs A110V and A372T and tas1r3 the SNPs A5T, A5A, T153T, R247H, P416P, and R757C.4
The second study selected was a research performed in vitro using functional expression of SNPs of T1R1 and T1R3 in HEK293 cells, which was able to quantify the receptor activity when stimulated with MSG.5 The research involved the SNPs A110V and R507Q from the N-terminal ligand-binding domain of T1R1, and F749S and R757C from the transmembrane domain of T1R3.5
The last study chosen to be discussed in the present work was a research performed with 92 adults who were underwent to tastants (NaCl, sucrose, citric acid, and quinine) applied locally to fungiform and circumvallate loci. The whole mouth was sampled and evaluated for the influence of SNPs A372T and intronic rs17492553 C/T.6
Discussion
The tas1r gene encodes a large N-terminal extracellular domain (ECD), seven transmembrane domains (TM1-TM7) that are separated by alternating intracellular and extracellular loops (il-i3 and el-e3, respectively), and an intracellular C-terminal domain. Studies have suggested that the ECD of tas1r1 contains binding sites of umami stimuli and ECD of tas1r3 is also required for receptor-ligand binding activity.8
Studies involving genetic analysis of mouse strains showed differences in preference for sweeteners and glutamate that suggest that SNPs may result in modification on receptor threshold. 9-12 Corroborating these results, studies with humans have shown that the SNPs are also able to modify the receptor threshold. 13
Although the binding role of tas1r ECDs for human taste perception has not been determined, these studies suggested that the majority of polymorphisms occur in the ECDs of tas1rs, thereby compromising the sensitivity to umami stimuli.4 An in vitro study investigated the capacity of R507Q and A110V SNPs to respond to MSG solution activated to approximately 25-50% of the T1R1/T1R3. This means that these two SNPs when present decrease the activation of these specific receptors. On the other hand, the A372T SNP in an in vitro study expressed in HEK293 cells showed comparable sensitivity to MPG to the normal T1R1+T1R3 dimer.
Transmembrane domains (TMD) may be involved in the activation of the G protein or binding to an allosteric modulator.4 Different from the SNPs in the ECD, the TMD R757C SNPs resulted in greater umami ratings of low concentrations of MSG in human experiments suggesting that humans with this SNP are more sensitive to recognize low concentration solutions of MSG.4 Amino terminal SNPs like A5T and R247H affected the human capacity to recognize differences in the MSG solutions only with high solution concentrations.4 Molecular tools such as 3D modeling have helped to explain how the SNP on ECD modify the receptor sensibility to chemical stimuli. Information from the 3D modeling used as a guide to explore the impact of the 110 amino acid replacement due to SNP T1R1/T1R3 receptor activity revealed the introduction of hydrophobic interactions with the T1R3 residue K155. The substitution of alanine by valine in this

70
position suggested the induction of a large conformational change of the T1R1 monomer backbone.5
The cystein-rich region (CRR) seems to be very important for the T1R1 function. The residue 507 of T1R1 is localized in the CRR and fits with the conserved basic residue 519.5,7 This SNP suggests that the substitution leads to the loss of a conserved negative charged residue. Furthermore, this substitution is likely to lead a conformational change with a novel pairing of the neighboring disulphide bridges. These events may explain the loss of the receptor activity observed using a functional assay.7
The SNP A372T localizes in ECD of T1R1. This SNP is alternatively spliced during transcription, suggesting that part of the peptide encoded by this exon is less critical for receptor functioning and instead is involved in other ways to influence taste functioning.6
Conclusion
Polymorphisms in tas1r are associated with modifications of perception of sweet and umami tastes in humans. Nontaster and hypotaster subjects to these types of taste were a more common form of alteration found in recent studies. SNPs that enhanced the ability to identify the sweet or umami tastes are rare. Studies suggested the ECD SNPs of TAS1R3 were usually associated with the decrease of perception of umami sensation.
References 1- Lindemann B. Receptors and transduction in taste. Nature. 2001 Sep 13;413(6852):219-25. Review. PubMed PMID: 11557991. 2- Hoon MA, Adler E, Lindemeier J, Battey JF, Ryba NJ, Zuker CS. Putative mammalian taste receptors: a class of taste-specific GPCRs with distinct topographic selectivity. Cell. 1999 Feb 19;96(4):541-51. PubMed PMID: 10052456. 3- Nelson G, Hoon MA, Chandrashekar J, Zhang Y, Ryba NJ, Zuker CS. Mammalian sweet taste receptors. Cell. 2001 Aug 10;106(3):381-90. PubMed PMID: 11509186. 4- Chen QY, Alarcon S, Tharp A, Ahmed OM, Estrella NL, Greene TA, Rucker J, Breslin PA. Perceptual variation in umami taste and polymorphisms in TAS1R taste receptor genes. Am J Clin Nutr. 2009 Sep;90(3):770S-779S. doi: 10.3945/ajcn.2009.27462N. Epub 2009 Jul 8. PubMed PMID: 19587085; PubMed Central PMCID: PMC3136006.. 5- Raliou M, Grauso M, Hoffmann B, Schlegel-Le-Poupon C, Nespoulous C, Débat H, Belloir C, Wiencis A, Sigoillot M, Bano SP, Trotier D, Pernollet JC, Montmayeur JP, Faurion A, Briand L. Human genetic polymorphisms in T1R1 and T1R3 taste receptor subunits affect their function. Chem Senses. 2011 Jul;36(6):527-37. doi: 10.1093/chemse/bjr014. Epub 2011 Mar 21. PubMed PMID: 21422378.

71
6- Rawal S, Hayes JE, Wallace MR, Bartoshuk LM, Duffy VB. Do polymorphisms in the TAS1R1 gene contribute to broader differences in human taste intensity? Chem Senses. 2013 Oct;38(8):719-28. doi: 10.1093/chemse/bjt040. Epub 2013 Sep 2. PubMed PMID: 24000232; PubMed Central PMCID: PMC3777563. 7- Muto T, Tsuchiya D, Morikawa K, Jingami H. Structures of the extracellular regions of the group II/III metabotropic glutamate receptors. Proc Natl Acad Sci U S A. 2007 Mar 6;104(10):3759-64. Epub 2007 Feb 26. PubMed PMID: 17360426; PubMed Central PMCID: PMC1820657. 8- Li X, Zhang F, Xu H, Li Q. PCT/US patent 2006/041158. 2007. 9- Bachmanov AA, Tordoff MG, Beauchamp GK. Intake of umami-tasting solutions by mice: a genetic analysis. J Nutr. 2000 Apr;130(4S Suppl):935S-41S. PubMed PMID: 10736356; PubMed Central PMCID: PMC3644809. 10- Max M, Shanker YG, Huang L, Rong M, Liu Z, Campagne F, Weinstein H, Damak S, Margolskee RF. Tas1r3, encoding a new candidate taste receptor, is allelic to the sweet responsiveness locus Sac. Nat Genet. 2001 May;28(1):58-63. PubMed PMID: 11326277. 11- Reed DR, Li S, Li X, Huang L, Tordoff MG, Starling-Roney R, Taniguchi K, West DB, Ohmen JD, Beauchamp GK, Bachmanov AA. Polymorphisms in the taste receptor gene (Tas1r3) region are associated with saccharin preference in 30 mouse strains. J Neurosci. 2004 Jan 28;24(4):938-46. PubMed PMID: 14749438; PubMed Central PMCID: PMC1397915. 12- Winnig M, Bufe B, Meyerhof W. Valine 738 and lysine 735 in the fifth transmembrane domain of rTas1r3 mediate insensitivity towards lactisole of the rat sweet taste receptor. BMC Neurosci. 2005 Apr 7;6:22. PubMed PMID: 15817126; PubMed Central PMCID: PMC1084349. 13- Kim UK, Wooding S, Riaz N, Jorde LB, Drayna D. Variation in the human TAS1R taste receptor genes. Chem Senses. 2006 Sep;31(7):599-611. Epub 2006 Jun 26. PubMed PMID: 16801379.

72
Anexo II
CONHECIMENTO DA POPULAÇÃO UNIVERSITÁRIA DE MONTES CLAROS-MG SOBRE
O ACIDENTE VASCULAR ENCEFÁLICO.
Knowledge of university population about stroke in Montes Claros-MG
Marcelo José da Silva de Magalhães ¹
Rafael Moreno Ribeiro do Nascimento2,
Alcino Franco de Moura Junior3,
Zuíla Maria de Jesus Rametta4,
Diego Vinícius de Castro Pereira5
1- Neurocirurgião do Hospital Aroldo Tourinho. Mestrando pela Inforium. Professor de medicina da Funorte-Faculdade Unidas do Norte de
Minas e FipMoc-Faculdades Integradas de Montes Claros.
2-Analista administrador de redes. Mestrando pela Inforium. Professor de programação e redes de computadores na UNIMONTES -
Universidade Estadual de Montes Claros e FACIT-Faculdade de Ciências e Tecnologia.
3-Jornalista. Mestrando pela Inforium. Professor do Departamento de Ciências da Computação da UNIMONTES-Universidade Estadual de
Montes Claros , Funorte-Faculdade Unidas do Norte de Minas e FipMoc-Faculdades Integradas de Montes Claros.
4-Matemática- Mestrando pela Inforium. Pós-graduanda em Estatística pela UFMG. Professora da disciplina de matemática na
UNIMONTES-Universidade Estadual de Montes Claros e FASI- Faculdade de Saúde Ibituruna.
5-Analista de Sistemas. Mestrando pela Inforium. Professor da disciplina de Informática na Funorte-Faculdade Unidas do Norte de Minas
RESUMO: O acidente vascular encefálico (AVE) é uma doença com prevalência elevada. Apresenta índices
elevados de morbidade e mortalidade, estando entre as principais causas de incapacidade permanente em
diferentes partes do mundo. O objetivo deste trabalho é ter informações sobre o grau de conhecimento sobre
os fatores de risco sobre acidente vascular encefálico entre os acadêmicos de faculdades localizadas no
município em Montes Claros, Brasil. Estudo epidemiológico realizado com 128 acadêmicos provenientes da
cidade de Montes Claros. Dados foram coletados entre maio e julho de 2014, por meio de entrevista
semiestruturada. Conclui-se que pelo menos 38% da amostra entrevistada conhecia um fator de risco relevante
sobre AVE.
Palavras-chave: Acidente vascular encefálico; epidemiologia; fatores de risco.
ABSTRACT: Stroke is a disease with high prevalence. This disease presents high morbidities and mortality rates
and it is among the main disabilities in different parts of the world. The goal of this reseach is show the
university knowledge about stroke. It is characterized as descriptive, it was made with 128 university students

73
in the Montes Claros city. Data was collected between May and July 2014, through semistructured interviews.
It is concluded at least 38% of universitaries know one main risk factor to stroke.
Keywords: Stroke;epidemiology; risk factors.
Introdução
Cerca de 58 milhões de mortes ocorrem
anualmente em todo o mundo por diferentes
etiologias e cerca de 5,7 milhões são decorrentes
do acidente vascular encefálico (AVE).1,2
Atualmente, o AVE é a segunda causa de morte e
responsável por 10% de todas as mortes no
mundo.1,2 Entretanto 85% das mortes decorrentes
desta doença ocorrem nos países em
desenvolvimento e um terço destes pertencem a
faixa economicamente ativa da população.3,4 Sabe-
se que ocorrerá um aumento de 300% na
incidência do AVE na população idosa latino
americana e asiática nos próximos 30 anos. 3,4 No
Brasil esta doença foi a responsável por 10% dos
óbitos em 2005 e por 10% de todas as admissões
nos hospitais públicos. 3,4 Em um estudo brasileiro
envolvendo 814 pessoas em quatro cidades
apenas 22% da população entrevistada sabia
reconhecer adequadamente um sinal físico
sugestivo de AVE.5 Estudos realizados em países
em desenvolvimento sobre a ativação do serviço
de emergência têm revelado de forma sistemática
que intervenções na população são necessárias
para melhorar o reconhecimento do AVE. Isso
diretamente elevará o número de pacientes
atendidos na fase aguda da doença o que
acarretará uma redução na morbidade e
mortalidade destes. 6
O objetivo deste trabalho é avaliar o nível de
conhecimento de uma pequena amostra de
estudantes universitários em algumas
universidades em Montes Claros sobre o AVE.
Os entrevistados participaram da pesquisa após
assinarem o termo de consentimento livre e
esclarecido.
Os integrantes desta pesquisa não apresentam
conflito de interesse.
Metodologia
Trata-se de um estudo descritivo, de cunho
epidemiológico realizado com acadêmicos dos
cursos de matemática, informática, estatística e
programação da UNIMONTES - Universidade
Estadual de Montes Claros, FipMoc - Faculdades
Integradas de Montes Claros, FACIT - Faculdade de
Ciências e Tecnologia e FASI - Faculdade de Saúde
Ibituruna. Todas estas faculdades localizam-se na
cidade de Montes Claros - MG. A amostragem
aleatória envolveu 128 acadêmicos dos cursos
supracitados durante o período de Maio à Junho
de 2014. O cálculo utilizou um erro amostral
máximo tolerável de 9% como valor de erro
amostral com amostra inicial de 123 pessoas.
Critérios de inclusão foram: idade igual ou superior
à 18 anos, estar matriculados nos cursos
supracitados, assinar o termo de consentimento
livre-esclarecido. Critérios de exclusão: idade
inferior a 18 anos, não estar devidamente
matriculado nas disciplinas de matemática e
programação. Para coleta dos dados trabalhou-se
com entrevista semiestruturada composta por
questões fechadas sobre fatores de risco,
manifestações na fase aguda e conduta frente ao
paciente na fase aguda. As informações coletadas
dos pacientes foram organizadas em banco de
dados no programa Microsoft Office Excel®, 2010.
Posteriormente, procedeu-se à análise estatística
com o programa Minitab 16. Entre as limitações
deste estudo está a amostragem aleatória e
amostra envolvendo somente acadêmicos destas
faculdades acarretando um viés na metologia.

74
Resultados
Nota-se que a amostra foi constituída por 53% de mulheres e 47% de homens, com 57% dos entrevistados com idade entre 17 e 25 anos. Observa-se que a grande maioria dos estudantes, 72% concluíram o ensino médio em escolas públicas e 17% em escolas públicas e privadas. Maioria da amostra, 57% concluiu o ensino médio entre 2010 e 2013. Houve um predomínio de solteiros nesta amostra, constituindo 88% dos entrevistados. (Tabela 01) Pelo menos 38% dos entrevistados conhecia pelo menos um fator de risco para AVE. Entre os fatores de risco para esta doença os mais conhecidos pela população entrevistada foram a hipertensão arterial sistêmica (HAS) 83%, obesidade 75% e o sedentarismo com 74%. O fator de risco menos conhecido pela população foi o ataque isquêmico transitório (AIT) que contou com apenas 41% dos entrevistados. (Tabela 02). Notou-se uma baixa taxa de reconhecimento pelos entrevistados dos fatores protetores para AVE. Menos de 10% destes conhecia que o controle adequado da HAS, baixas taxas de colesterol e atividade física contribuíam para reduzir as taxas de AVE. (Tabela 02) Quanto à capacidade de reconhecer um AVE na fase aguda, 65% dos entrevistados sabia que alterações da fala, dormência ou fraqueza motora poderiam ser sinais da doença em uma fase aguda. Quanto ao manejo correto do paciente na fase aguda do AVE, 85% realizaria a conduta correta frente a um paciente com esta patologia. (Tabela 02)
Características demográficas dos
entrevistados
N %
Sexo
Masculino
Feminino
60
68
47%
53%
Idade Menos de 17 anos
Entre 17 a 18
Entre 19 a 25
Entre 26 a 33
Entre 34 a 41
Entre 42 e 49 anos
1
23
73
21
4
4
1%
18%
57%
17%
3%
3%
Estado civil
Solteiro
Casado/mora com um companheiro
Separado/ divorciado/ esquitado.
Viúvo
112
14
1
0
88%
11%
1%
0%
Conclusão do ensino médio
Entre 2013 e 2010
Entre 2009 e 2005
Entre 2004 e 2000
Entre 1999 e 1995
Entre 1994 e 1990
Antes de 1990
72
31
14
5
4
1
57%
24%
11%
4%
3%
1%
Tipo de escola
Somente em escola pública.
Parte em escola pública e parte em
escola particular.
Somente em escola particular.
Somente em escola indígena ou em
escola situada em comunidade
quilombola.
Parte na escola indígena e parte em
escola não-indígena.
Parte em escola situada em
comunidade quilombola e parte em
escola fora de área quilombola.
91
22
14
0
0
0
72%
17%
11%
0%
0%
0%
Tabela 1 – Dados socioculturais da população universitária sobre o AVE
na cidade de Montes Claros.

75
Grau de conhecimento do
entrevistado sobre diversas os
fatores de risco para AVE
N %
Diabetes
Fibrilação atrial
Sedentarismo
Obesidade
Hipertensão arterial sistêmica
Tabagismo
Etilismo
AIT
76
53
75
76
86
79
72
48
59%
41%
74%
75%
83%
61%
56%
38%
Capacidade de reconhecer um fator
protetor para AVE
Atividade Física
Controle de HAS
Controle de taxa de colesterol
11
9
5
9%
7%
4%
Manejo correto doente na fase aguda
do AVE
109 85%
Capacidade de reconhecer um AVE na
fase aguda
Presença de pessoas no círculo social
que apresentaram AVE
Familiar próximos
Familiar distante
Amigo
Colega de trabalho
Vizinho
76
33
16
14
2
13
58%
26%
13%
11%
2%
10%
Discussão
O acidente vascular encefálico (AVE) é
definido como “sinais de distúrbio focal (por vezes
global) da função cerebral de evolução rápida
durando mais de 1 hora.” 1 Esse distúrbio
neurológico é provocado pela redução do fluxo
sanguíneo numa artéria de pequeno ou médio
calibre que irriga uma parte do encéfalo,
ocasionando um infarto ou quando ocorre uma
hemorragia decorrente da ruptura de uma
artéria.1 Quando o déficit neurológico induzido
pela isquemia apresenta duração inferior a 1 hora,
é definido como ataque isquêmico transitório
(AIT). Reconhecer precocemente os sintomas e
sinais do AVE favorece um tratamento mais rápido
reduzindo a morbi-mortalidade dos pacientes
acometidos.2 Estudos realizados em diferentes
partes do mundo têm mostrado divergentes graus
de conhecimentos da população quanto às
informações referentes a esta doença.2
Um estudo realizado em um hospital de
nível terciário de Karachi, cidade mais populosa do
Paquistão, envolveu 398 pacientes.10 Nesta
pesquisa somente metade dos pacientes sabia que
o órgão envolvido nesta patologia é o cérebro e
cerca de 20% acreditava que o coração seria o
órgão afetado durante o AVE. Apesar desta
informação, coerentemente, 70% e 60% dos
entrevistados respectivamente, reconheciam que
a fraqueza e dormência em face e corpo poderia
representar os sintomas iniciais de um AVE. 10
Nota-se que mesmo em nações localizadas
na Europa, com é o caso da Irlanda, apresentam
taxas preocupantes de desconhecimento das
manifestações do AVE.9 Em um grande estudo
realizado na Irlanda envolvendo 2033 pessoas com
idade superior a 65 anos, menos da metade sabia
reconhecer sintomas das manifestações iniciais de
um AVE, exceto quanto à alteração da linguagem,
em que 54% destes reconhecia esta alteração
como manifestação de um AVE.9
No Brasil, um estudo realizado no
município de Pelotas-RS envolvendo 483
entrevistados foram respondidos como sinal,
sintomas ou sequela de AVE o déficit motor em
79,0% e os distúrbios de linguagem em 82,5%.7
Em nosso estudo, realizado em Montes
Claros-MG, nota-se que apenas 58% dos
entrevistados conseguiram reconhecer que o AVE
pode se apresentar como uma cefaleia de forte
intensidade, dificuldade súbita para falar ou
Tabela 2 – Grau de conhecimento da população universitária sobre o
AVE na cidade de Montes Claros.

76
paralisia de um lado do corpo. Somente 19%
sabem que os sintomas do AIT apresentam
duração de 1 hora. Nota-se que somente 67% dos
entrevistados sabiam que a obstrução de uma
artéria cerebral poderia levar a um quadro de AVE
e 18% sabia que a hemorragia no cérebro poderia
acarretar um AVE. (Tabela 2)
A hipertensão arterial sistêmica (HAS) é o
principal fator de risco modificável para AVE.12
Quanto maior a pressão arterial, maior o risco de
desenvolver esta patologia. Outra doença crônica
envolvida nesta doença está o diabetes melito
(DM), que é o segundo fator de risco modificável
mais importante.12 O diabetes predispõe AVE por
mecanismos aterogênicos diretos e por interagir
com outros fatores de risco, como HAS e
dislipidemia.12 Alguns hábitos também elevam o
risco para AVE, sendo o tabagismo um fator de
risco bem conhecido. Todo fumante reduzirá o
risco de AVE ao parar de fumar, independente de
quanto já tenha fumado. A arritmia mais envolvida
no AVE é fibrilação atrial, especialmente se
associada à lesão valvar.12 Existem alguns fatores
de risco não modificáveis para o AVE, tais como a
idade que é o principal deles.12 Outros fatores de
risco conhecidos são: sedentarismo, síndrome do
anticorpo antifosfolipídeo, consumo de cocaína,
meningite bacteriana ou tuberculosa, neurossífilis,
lúpus eritematoso sistêmico e vasculites
necrosantes.12
O desenvolvimento socioeconômico do
país nem sempre apresenta influência no grau de
conhecimento de sua população quantos aos
fatores de risco para o AVE.11 Corroborando com
estas ideias vejam os resultados destes próximos
estudos.
Um grande estudo realizado em Uganda
envolvendo 5481 participantes oriundos de
cidades e zona rural revelou que somente 27%
destes conhecia um fator de risco para o AVE.11
Somente 56% deste subgrupo conhecia que a HAS
era um fator de risco, com diferença estatística
significativa para os residentes da zona urbana. 11
(Tabela 3)
Um estudo realizado na Austrália com 822
participantes através de uma pesquisa realizada
por meio de telefonemas mostrou que o grau de
conhecimento para todos os fatores de risco para
AVE dentre os entrevistados esteve no patamar
inferior a 40%.12 Somente 31% da população
envolvida nesta pesquisa sabia que HAS era um
fator de risco para AVE.12 (Tabela 3)
No Brasil, em um estudo realizado na
cidade de Vassouras-RJ evolvendo 356
entrevistados, 31,38% da população residente na
zona urbana conhecia um fator de risco para o AVE
e 41,07% conheciam dois ou mais fatores de
risco.8 Por outro lado, entre a população
entrevistada residente na zona rural, 29,16%
conheciam um fator de risco e 29,76% mais de um
fator de risco8. No estudo da cidade de Pelotas-RS,
48,7% dos entrevistados conhecia que a idade
acima de 40 anos era fator de risco, 95,3% a HAS,
88,1% o sedentarismo e 96,5% o consumo de
drogas.7 Em nosso estudo cerca de 83%, 59% e
61%, conheciam respectivamente que a
hipertensão arterial sistêmica,d melitus e
tabagismo são fatores de risco para o AVE. (Tabela
2)
Grau de conhecimento do
entrevistado sobre diversas os
fatores de risco para AVE
N %
Uganda 2013- 1180 indivíduos
Idade avançada
Diabetes Melitus
Cardiopatia
Obesidade
Hipertensão Arterial Sistêmica Tabagismo
Etilismo
Hereditariedade
55
65
18
24
244
3
15
3
12,6%
14,9%
4,1%
5,5%
56%
0,7%
3,4%
0,7%
Irlanda 2009-2033 indivíduos
Hipertensão Arterial Sistêmica
Dislipidemia
Obesidade
Diabetes Melitus
Etilismo
1523
823
608
217
201
75%
40%
30%
11%
10%

77
Sedentarismo
Hereditariedade
363
333
18%
16%
Austrália 2001
Tabagismo
Estresse
Dieta inadequada
Hipertensão Arterial Sistêmica
Obesidade
Hereditariedade
Sedentarismo
Estilo de vida não saudável
Dislipidemia
Etilismo
324
277
265
261
220
182
181
128
111
171
39,4%
33,7%
32,2%
31,8%
26,8%
22,1%
22%
15,6%
13,5%
20,8%
Conclusão
O rápido reconhecimento dos sintomas do
acidente vascular encefálico é a chave para
maximizar os resultados do tratamento desta
patologia acarretando desta forma melhores
resultados funcionais dentre estes pacientes. O
presente estudo revela uma lacuna importante no
conhecimento da população universitária de
Montes Claros investigada quanto aos fatores
protetores do AVE e reconhecimento de alguns
fatores de risco para a esta patologia.
Referências
1. The WHO stepwise approach to stroke
surveillance. Overview and Manual (version2.0).
Noncommunicable Diseases and Mental Health.
World Health Organization.
Disponível em:
<www.who.int/entity/ncd_surveillance/steps/en>.
Acesso em novembro de 2005.
2. Yach D, Hawkes C, Gould CL, Hofman KJ. The global burden of chronic diseases: overcoming impediments to prevention and control. JAMA, 291:2616-2622, 2004.
3. Organização Pan Americana da Saúde. Rede interagencial de informações para a Saúde. Disponível em: <www.opas.org.br/ripsa/produto_final_det.cfm?id=9>. Acessado em 2007. 4. Strong K, Mathers C, Bonita R. Preventing stroke: saving lives around the world. Lancet Neurol, 6: 182-187, 2007. 5. Pontes-Neto OM, Silva GS, Feitosa MR, et al. Stroke awareness in Brazil: alarming results in a community-based study. Stroke, 39:292-296, 2008. 6. Alberts MJ, Perry A, Dawson DV, Bertels C. Effects of public and professional education on reducing the delay in presentation and referral of stroke patients. Stroke, 23: 352-356, 1992. 7. Costa F, Oliveira S, Magalhães P, Costa B, Papini R, Silveira M, Lang M - Nível de conhecimento da população adulta sobre acidente vascular cerebral (AVC) em Pelotas – RS. J Bras Neurocirurg 19 (1): 31-37, 2008.
8. Silva IHB, Junqueira CM, Cortes PC Cardoso
RF et al. Conhecimento Sobre o Acidente
Vascular Cerebral em Vassouras-RJ: Análise
Entre População da Zona Urbana e Zona Rural.
Neurobiologia, 75(3-4), 2012.
9. Hickey A, O'Hanlon A, McGee H, Donnellan C, Shelley E, Horgan F, O'Neill D.
Tabela 3 – Grau de conhecimento da população em diferentes partes do
mundo sobre os fatores de risco para o AVE.

78
Stroke awareness in the general population: knowledge of stroke risk factors and warning signs in older adults. BMC Geriatrics, 9: 35, 2009.
10. Aly Z, Abbas K, Kazim SF, Taj F, Aziz F, Irfan
A, Sheik R et al. Awareness of stroke risk
factors, signs and treatment in a Pakistani
population. J Pak Med Assoc, Vol. 59, No. 7,
July 2009.
11. Jane Nakibuuka, Martha Sajatovic, Elly Katabira, Edward Ddumba, Jayne Byakika-Tusiime, and Anthony J. Furlan, “Knowledge and Perception of Stroke: A Population-Based Survey in Uganda,” ISRN Stroke, vol. 2014, Article ID 309106, 7 pages, 2014. doi:10.1155/2014/309106
12. S. Sug Yoon, R. F. Heller, C. Levi, J. Wiggers,
P. E. Fitzgerald. Knowledge of stroke risk
factors, warning symptoms, and treatment
among an Australian urban population.
Stroke,32(8): 1926–1930, 2001.

0
0







![FRESAMENTO DE ACABAMENTO EM ALTAS VELOCIDADES … · Figura 2.6 - Forças de usinagem e suas componentes para o torneamento [16].....32 Figura 2.7 - Posição geométrica da quina](https://img.document.onl/doc/110x75/5c35aa3909d3f221298ce6db/fresamento-de-acabamento-em-altas-velocidades-figura-26-forcas-de-usinagem.jpg)





















