Embed Size (px)
Citation preview

Introdução à Neurociência Computacional (Graduação)– Antonio Roque – Aula 14
1
Modelagem de Sinapses
Há dois tipos de sinapses, químicas e elétricas. Vamos começar considerando apenas a
sinapse química, que é tida como a mais importante segundo a maioria dos neurocientistas.
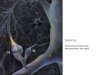
Uma sinapse química padrão conecta o axônio do neurônio que envia o estímulo, chamado
de neurônio pré-sináptico, a um dendrito do neurônio que recebe o estímulo, chamado de
neurônio pós-sináptico (veja a figura abaixo).
Quando um potencial de ação chega ao terminal do axônio do neurônio pré-sináptico, uma
série de eventos acontece:
- Canais de cálcio na membrana do terminal pré-sináptico se abrem e íons de Ca2+
entram na célula pré-sináptica.
- Os íons de Ca2+ provocam a fusão de vesículas que contêm neurotransmissores com
a membrana pré-sináptica, liberando esses neurotransmissores na fenda sináptica.
- Depois que as vesículas liberam seu conteúdo, elas retornam ao meio intracelular do
neurônio pré-sináptico e são “recarregadas” com neurotransmissores para ser usadas
novamente no futuro.

Introdução à Neurociência Computacional (Graduação)– Antonio Roque – Aula 14
2
- Os neurotransmissores liberados na região do meio extracelular entre os dois
terminais sinápticos – chamada de fenda sináptica – se difundem pela fenda
sináptica e se ligam a receptores na membrana do dendrito do neurônio pós-
sináptico.
- Há dois tipos básicos de receptores na membrana do neurônios pós-sináptico:
receptores ionotrópicos e receptores metabotrópicos. Os receptores ionotrópicos são
canais iônicos que se abrem quando se ligam a um neurotransmissor e deixam
passar íons para dentro ou para fora do neurônio pós-sináptico. Os receptores
metabotrópicos serão descritos mais adiante.
- Dependendo do tipo de neurotransmissor, quando ele se liga a um receptor
ionotrópico isso pode provocar uma pequena despolarização local na membrana
(pela entrada de carga positiva) ou uma pequena hiperpolarização local na
membrana (pela entrada de carga negativa).
- Uma despolarização local na membrana é chamada de potencial pós-sináptico
excitatório e uma hiperpolarização local é chamada de potencial pós-sináptico
inibitório. A figura abaixo mostra um exemplo de um potencial pós-sináptico
excitatório.
- Um neurônio pré-sináptico sempre libera o mesmo tipo de neurotransmissor: quando
ele provoca uma despolarização local, o neurônio pré-sináptico é chamado de
excitatório e a sinapse é dita excitatória; quando ele provoca uma hiperpolarização
local, o neurônio pré-sináptico é chamado de inibitório e a sinapse é dita inibitória.
Introdução à Neurociência Computacional (Graduação)– Antonio Roque – Aula 14
3
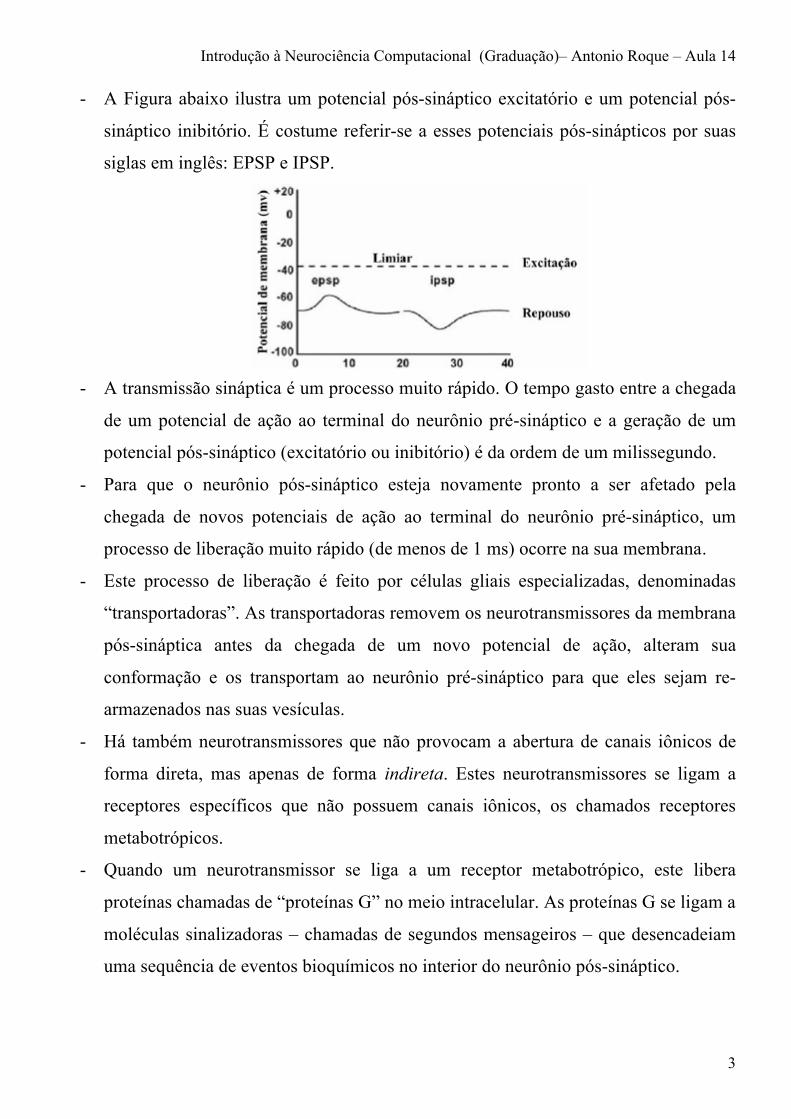
- A Figura abaixo ilustra um potencial pós-sináptico excitatório e um potencial pós-
sináptico inibitório. É costume referir-se a esses potenciais pós-sinápticos por suas
siglas em inglês: EPSP e IPSP.
- A transmissão sináptica é um processo muito rápido. O tempo gasto entre a chegada
de um potencial de ação ao terminal do neurônio pré-sináptico e a geração de um
potencial pós-sináptico (excitatório ou inibitório) é da ordem de um milissegundo.
- Para que o neurônio pós-sináptico esteja novamente pronto a ser afetado pela
chegada de novos potenciais de ação ao terminal do neurônio pré-sináptico, um
processo de liberação muito rápido (de menos de 1 ms) ocorre na sua membrana.
- Este processo de liberação é feito por células gliais especializadas, denominadas
“transportadoras”. As transportadoras removem os neurotransmissores da membrana
pós-sináptica antes da chegada de um novo potencial de ação, alteram sua
conformação e os transportam ao neurônio pré-sináptico para que eles sejam re-
armazenados nas suas vesículas.
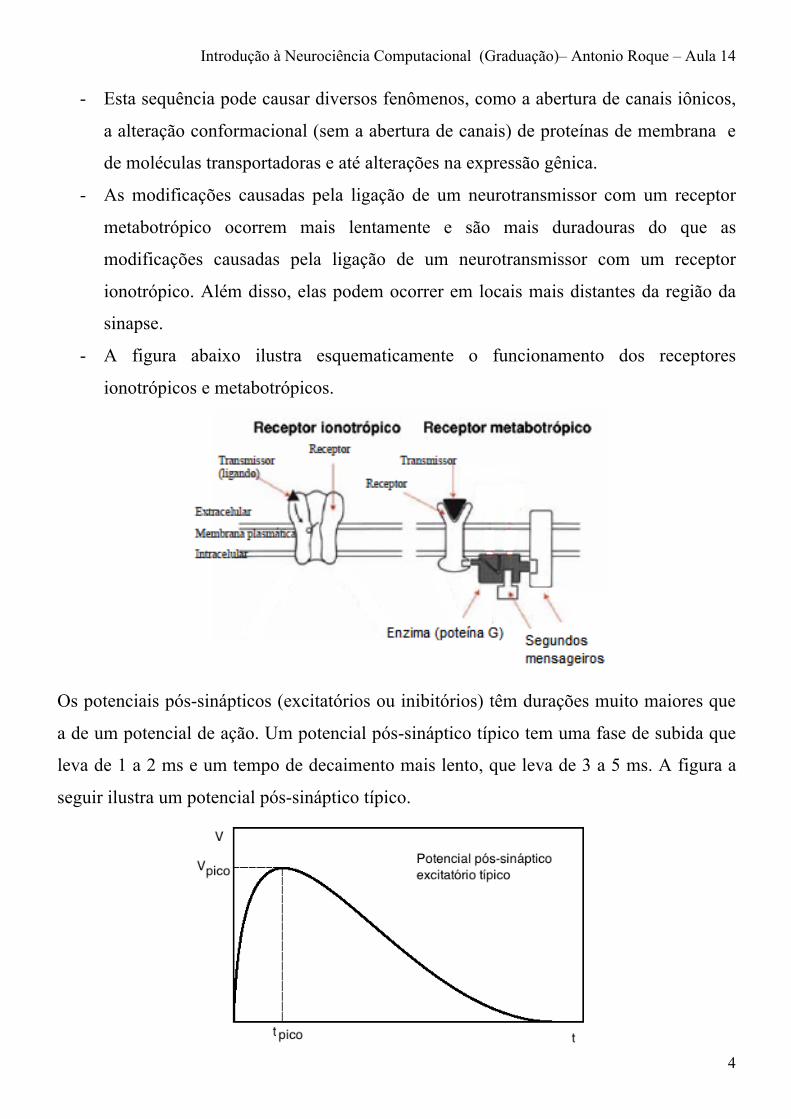
- Há também neurotransmissores que não provocam a abertura de canais iônicos de
forma direta, mas apenas de forma indireta. Estes neurotransmissores se ligam a
receptores específicos que não possuem canais iônicos, os chamados receptores
metabotrópicos.
- Quando um neurotransmissor se liga a um receptor metabotrópico, este libera
proteínas chamadas de “proteínas G” no meio intracelular. As proteínas G se ligam a
moléculas sinalizadoras – chamadas de segundos mensageiros – que desencadeiam
uma sequência de eventos bioquímicos no interior do neurônio pós-sináptico.
Introdução à Neurociência Computacional (Graduação)– Antonio Roque – Aula 14
4
- Esta sequência pode causar diversos fenômenos, como a abertura de canais iônicos,
a alteração conformacional (sem a abertura de canais) de proteínas de membrana e
de moléculas transportadoras e até alterações na expressão gênica.
- As modificações causadas pela ligação de um neurotransmissor com um receptor
metabotrópico ocorrem mais lentamente e são mais duradouras do que as
modificações causadas pela ligação de um neurotransmissor com um receptor
ionotrópico. Além disso, elas podem ocorrer em locais mais distantes da região da
sinapse.
- A figura abaixo ilustra esquematicamente o funcionamento dos receptores
ionotrópicos e metabotrópicos.
Os potenciais pós-sinápticos (excitatórios ou inibitórios) têm durações muito maiores que
a de um potencial de ação. Um potencial pós-sináptico típico tem uma fase de subida que
leva de 1 a 2 ms e um tempo de decaimento mais lento, que leva de 3 a 5 ms. A figura a
seguir ilustra um potencial pós-sináptico típico.

Introdução à Neurociência Computacional (Graduação)– Antonio Roque – Aula 14
5
O principal neurotransmissor excitatório do cérebro é o glutamato e o principal
neurotransmissor inibitório é o GABA (ácido γ-aminobutírico). Os dois podem atuar tanto
sobre receptores ionotrópicos como metabotrópicos.
Os principais receptores ionotrópicos para o glutamato são os chamados receptores AMPA
e NMDA. Eles recebem estes nomes por causa das drogas agonistas que os ativam (ácido
α-amino-3-hidroxi-metil-4-isoxazolepropriônico, ou AMPA; e N-metil-D-aspartato, ou
NMDA). Tanto os receptores AMPA como NMDA, quando ligados ao glutamato, atuam
como canais iônicos para cátions em geral (Na+, Ca2+, etc), embora o receptor NMDA seja
mais permeável ao Ca2+ do que o receptor AMPA. As correntes resultantes (para dentro da
célula) têm potenciais de reversão em torno de 0 mV.
A corrente iônica associada ao receptor AMPA é ativada e inativada muito rapidamente. Já
a corrente associada ao receptor NMDA é ativada mais lentamente e a sua inativação é
muito mais lenta. Além disso, a condutância dos receptores NMDA tem uma dependência
com a voltagem mais complicada e o seu comportamento não pode ser reproduzido pelo
modelo padrão de Hodgkin-Huxley. Para modelá-la, é necessário modificar um pouco o
modelo de Hodgkin-Huxley.
O neurotransmissor inibitório GABA ativa dois tipos de receptores, chamados de GABAA
e GABAB. O receptor GABAA é ionotrópico e constitui um canal iônico para o Cl− cuja
condutância se ativa e inativa de maneira relativamente rápida. Já o receptor GABAB é
metabotrópico e produz um aumento mais lento e duradouro da condutância ao K+. As
correntes resultantes (para dentro ou para fora da célula) têm potenciais de reversão em
torno de −75 mV.
Estudos experimentais com o uso da técnica de patch-clamp (ver aula 10), que permite o
registro da atividade de um único canal, mostram que os potenciais pós-sinápticos são
eventos macroscópicos resultantes do comportamento de uma população de canais iônicos
que transitam rapidamente entre os seus estados condutor e o não condutor.

Introdução à Neurociência Computacional (Graduação)– Antonio Roque – Aula 14
6
Por causa disso, a geração dos potenciais pós-sinápticos pode ser bem modelada pelo
formalismo de Hodgkin e Huxley, que descreve o comportamento temporal de voltagens e
correntes em termos de alterações em condutâncias (a única exceção é condutância dos
receptores NMDA). Como se trata de sinapses, essas condutâncias são chamadas de
condutâncias sinápticas.
Segundo o formalismo de Hodgkin-Huxley, a corrente sináptica pela membrana do
neurônio pós-sináptico produzida após a chegada de um potencial de ação ao terminal do
neurônio pré-sináptico é dada por
(1)
onde gsin(t) é a condutância sináptica (que descreve a variação na condutância dos canais
sinápticos na membrana do neurônio pós-sináptico), Vpós(t) é a voltagem de membrana do
neurônio pós-sináptico e Esin é o potencial de reversão (equilíbrio) da sinapse.
Existem várias maneiras de modelar a condutância sináptica gsin(t). Uma maneira muito
popular em modelos de redes de neurônios é assumir que gsin(t) é uma função pré-
determinada de t que vale 0 para t < t0, onde t0 é o instante da chegada do potencial de ação
ao terminal pré-sináptico, e é positiva para t > t0:
(2)
onde é uma constante escolhida de tal forma que gsin(tpico) = gpico, onde gpico é o
valor de pico da curva descrita pela condutância sináptica a partir de t0. As formas
funcionais pré-determinadas mais usadas para a função z(t) são:
a) Decaimento exponencial simples:
(3)
ó

Introdução à Neurociência Computacional (Graduação)– Antonio Roque – Aula 14
7
b) Função alfa (assim chamada porque Rall lhe deu este nome em 1967):
(4)
c) Duas funções exponenciais:
(3)
para a modelagem de neurônios, a condutância de uma sinapse é representada pela
condutância máxima da sinapse (quando todos os canais sinápticos estão abertos), g ,
multiplicada pela fração de canais sinápticos abertos, r(t), onde r é uma variável com valor
entre 0 e 1.
Inserir aqui o material sobre funções alfa (ver curso SAIFR2)
A dependência temporal de r é governada pela dinâmica de liberação de
neurotransmissores do neurônio pré-sináptico após a chegada de um potencial de ação ao
seu terminal e pela dinâmica de ligação desses neurotransmissores com os receptores
localizados no neurônio pós-sináptico.
Essa dinâmica pode ser modelada por um modelo cinético de dois estados (ver, por
exemplo, Destexhe et al., 1998)
,TRTR
β
α
←
→+ (1)
onde R representa a quantidade de receptores não-ligados, T representa a quantidade de
transmissores liberada, TR representa a quantidade de receptores ligados a transmissores e

Introdução à Neurociência Computacional (Graduação)– Antonio Roque – Aula 14
8
α e β representam as taxas de transição entre os dois estados do sistema. As concentrações
de R e de TR devem obedecer à seguinte equação de conservação:
[ ] [ ] .1=+ TRR
Este modelo implica que a fração de receptores ligados (ou de canais abertos) r(t) obedece
à seguinte equação diferencial (considerando que existe um grande número de canais
iônicos e desprezando flutuações estatísticas):
( ) ).()(1)( trtrTdttdr
βα −−= (2)
Esta equação pode ser resolvida exatamente a partir da hipótese de que a liberação de
neurotransmissores ocorre em pulsos quadrados e idênticos disparados sempre que o
potencial de ação pré-sináptico ultrapassa um dado limiar (por exemplo, 0 mV).
Considerando que um pulso se inicia em t0 e vai até t1 (duração igual a t1 − t0) e que a sua
amplitude é constante e igual a Tmax, podemos dividir o pulso em duas fases distintas: (a)
durante um pulso; e (b) após um pulso. A equação (2) pode ser resolvida analiticamente
para cada uma dessas duas fases. Seja:
a) Durante um pulso (t0 < t < t1): T = Tmax;
b) Após um pulso (t > t1): T = 0.
Durante o pulso – caso (a) –, a equação (2) fica,
( ) ( ) .)()(1)(maxmaxmax rTTtrtrT
dttdr
βααβα +−=−−=
Dividindo-se ambos os lados por (αTmax+β),
( ) ( ).)(1
max
max
max
rTT
dttdr
T−
+=
+ βαα
βα
Definindo-se,
( ),
max
max
βαα
+≡∞ T
Tr (3)

Introdução à Neurociência Computacional (Graduação)– Antonio Roque – Aula 14
9
( )βατ
+≡
max
1T , (4)
pode-se reescrever a equação (2) como,
.)( rrdttdr
−= ∞τ (5)
Esta equação tem como solução (lembre-se que ela é válida para t0 < t < t1):
( ) τ)(
00
0
)()(tt
ertrrttr−
−
∞∞ −+=− . (6)
Após o pulso – caso (b) –, a equação (2) torna-se,
),()( trdttdr
β−= (7)
cuja solução (lembre-se que ela é válida para t > t1) é:
.)()( )(11
1ttetrttr −−=− β (8)
A condutância sináptica é modelada pela equação,
).()( trgtgs = (9)
Para cada fase de um pulso (durante ou depois dele), r(t) obedece, ou à equação (6), ou à
equação (8).
Da mesma forma, a corrente sináptica associada à condutância acima é dada por:
( )ss EVtgI −= )(sin , (10)
onde V é o potencial de membrana, Es é o potencial de reversão da corrente sináptica e
gs(t) é a condutância da sinapse dada por (9). Para cada fase de um pulso, ela é modelada
pelas equações (6) ou (8).
A aproximação de tratar as quantidades de neurotransmissores liberadas como pulsos
quadrados idênticos simplifica bastante a resolução numérica de um sistema de neurônios

Introdução à Neurociência Computacional (Graduação)– Antonio Roque – Aula 14
10
acoplados sinapticamente. Ao invés de ter que resolver uma equação diferencial como a
(2) para cada sinapse, basta calcular numericamente a equação (6) ou a (8), dependendo da
fase do pulso em que se esteja, e isto envolve basicamente o cálculo de exponenciais (que
podem ser pré-calculadas e tabeladas para facilitar a eficiência computacional).
Para maiores detalhes sobre como implementar numericamente o modelo descrito pelas
equações (6) e (8), sugere-se o artigo de Destexhe et al. (1998).
Uma simulação de uma sinapse excitatória entre dois neurônios modelada pelas equações
(6) e (8) foi feita por Giugliano e Arsiero (2006). Uma figura desse artigo é reproduzida
abaixo, mostrando que o modelo captura as propriedades de saturação e somação temporal
quando ocorrem múltiplos eventos pré-sinápticos.
Foi mencionado acima que a condutância do receptor NMDA tem uma dependência com a
voltagem diferente da observada nas outras condutâncias. Para modelar essa dependência,
a corrente sináptica por um receptor NMDA costuma ser descrita da forma usual (equação
10) multiplicada por um termo a mais que depende de V:
( )sNMDAsNMDA EVVgtrgI −= )()(NMDA . (11)
Nesta equação, os termos NMDAg , rs(t) e Es têm as interpretações usuais descritas acima.
O termos gNMDA(V), no entanto, descreve uma dependência a mais da condutância do
receptor NMDA em relação à voltagem V devido ao fato de que quando o potencial de
membrana do neurônio pós-sináptico está próximo do seu valor de repouso os receptores
NMDA são bloqueados por íons de magnésio (Mg2+). Quando o neurônio pós-sináptico é
despolarizado, esses íons de magnésio são removidos dos receptores e permitem a
passagem de carga elétrica.

Introdução à Neurociência Computacional (Graduação)– Antonio Roque – Aula 14
11
Jahr e Stevens (1990) propuseram, a partir de um ajuste de curvas experimentais, uma
expressão para descrever o comportamento de gNMDA(V) que tornou-se amplamente
utilizado em modelos computacionais. A expressão deles é:
[ ] ( ) .mV 13,16/expmN 57,3
Mg1)(12 −+
⎟⎟⎠
⎞⎜⎜⎝
⎛−+= VVgNMDA (12)
Os canais iônicos formados pelos receptores NMDA conduzem tanto íons de cálcio (Ca2+)
como cátions monovalentes (principalmente Na+). A entrada no neurônio pós-sináptico de
íons de cálcio por seus receptores NMDA é um fenômeno crítico para a modificação de
longa duração da eficácia sináptica.
Note que a abertura dos receptores NMDA depende tanto da despolarização do neurônio
pré-sináptico (pela chegada de um potencial de ação no seu terminal) como da
despolarização do neurônio pós-sináptico (para que haja a retirada dos íons de magnésio).
Portanto, os receptores NMDA atuam como detectores de coincidência de atividade nos
neurônios pré- e pós-sináptico. Eles, portanto, desempenham um importante papel com
relação à chamada regra de Hebb para plasticidade sináptica que será vista na próxima
aula.
O fenômeno conhecido como depressão sináptica pós-ativação refere-se à diminuição da
amplitude do potencial pós-sináptico durante uma ativação sináptica repetitiva. Pode-se
modelar este fenômeno segundo o formalismo proposto por Abbott et al. (1997).
Segundo este formalismo, a condutância de uma sinapse é modelada por
),()()(sin trtzgtg = (13)
onde z(t) é uma variável que controla a eficácia sináptica. O valor de repouso desta
variável é tomado como 1, porém, sempre que ocorrer uma transmissão sináptica esse
valor é reduzido por um fator constante f (f < 1),

Introdução à Neurociência Computacional (Graduação)– Antonio Roque – Aula 14
12
.fzz→
Após a redução, z(t) retorna exponencialmente ao seu valor de repouso com uma constante
de tempo τrec,
).(1)(rec tz
dttdz
−=τ (14)
Caso o intervalo entre dois spikes seja grande o suficiente, a variável de controle da
eficácia sináptica voltará ao seu valor de repouso. Com a diminuição do intervalo entre
spikes, isto é, com o aumento da freqüência dos potenciais de ação pré-sinápticos, a
variável de controle sofre forte redução. Isto implica em potenciais pós-sinápticos menores
e na conseqüente depressão pós-ativação.
Este mesmo modelo pode ser usado para modelar o fenômenos de facilitação sináptica de
curta duração, basta fazer f > 1.
O resultado de uma simulação da mesma sinapse entre dois neurônios do exemplo anterior
(Giugliano e Arsiero, 2006) com o modelo de depressão sináptica pós-ativação está
mostrado na figura abaixo.
Com relação às sinapses elétricas, pode-se modelá-las como resistências elétricas
conectando os citoplasmas de dois neurônios. A figura abaixo ilustra o circuito equivalente
correspondente a esse modelo. A sinapse elétrica é modelada como uma gap junction
conectando dois compartimentos modelados segundo o esquema de Hodgkin-Huxley.

Introdução à Neurociência Computacional (Graduação)– Antonio Roque – Aula 14
13
A conexão entre os citoplasmas das duas células é feita por uma resistência ôhmica RGJ.
As resistências variáveis modelando os canais ativos das duas células não estão mostradas
para não sobrecarregar a figura. A figura também ilustra o processo de estimulação de uma
célula por injeção de corrente externa, Iinj, mostrando que parte dela pode escapar pela gap
junction e ir para a outra célula.
Referências
1. Abbott, L. F., Varela, J. A., Sen, K. and Nelson, S. B., Synaptic depression and cortical
gain control. Science, 275:220-224, 1997.
2. Destexhe, A., Mainen, Z. F. and Sejnowski, T. J., Kinetic Models of Synaptic
Transmission. In: Koch, C. and Segev, I. (eds.), Methods in Neural Modeling: From
Ions to Networks, 2nd. Edition. Cambridge, MA: MIT Press, 1998, pp. 1-25.
3. Giugliano, M. and Arsiero, M., Modeling of biological neuronal networks. In: Akay,
M. (ed.), Wiley Encyclopedia of Biomedical Engineering, New York, Wiley, 2006.
4. Jahr, C. E. and Stevens, C. F., A quantitative description of NMDA receptor channel
kinetic behavior. Journal of Neuroscience, 10:1830-1837, 1990.