Embed Size (px)
Citation preview

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA
MÓDULO
BIOQUÍMICA METABÓLICA
ESCUELA DE CIENCIAS AGRICOLAS PECUARIAS Y DEL MEDIO AMBIENTE.
ECAPMA
AUTOR: JAIRO ENRIQUE GRANADOS MORENO., MSc.
DOCENTE ECAPMA
BOGOTÁ 2010

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
INTRODUCCIÓN
La bioquímica metabólica es la ciencia que estudia todo lo relacionado con la actividad
catalítica en las vías anabólica y catabólica de las diversas biomoléculas que forman parte
de la dinámica celular de organismos animales y vegetales presentes en todo tipo de agro
ecosistemas; por ende, permite comprender la estructura y cinética de los compuestos
que intervienen en las diversas reacciones bioquímicas metabólicas aeróbicas y
anaeróbicas.
Además, teniendo en cuenta que es una ciencia teórico experimental la cual posee
conceptos, leyes, principios y teorías científicas, se constituye en un pilar fundamental
para la interpretación significativa de las múltiples reacciones biomoleculares que
gobiernan los diferentes procesos exergónicos y endergónicos, enfocados a mantener
las funciones vitales de sistemas autótrofos y heterótrofos, los cuales interaccionan
permanentemente garantizando la bioactividad de los componentes de nuestro planeta.
En consecuencia, la intencionalidad del curso se centra en el estudio de la bioquímica
estructural, biotermodinámica , actividad molecular y bioquímica metabólica aplicada,
importantes en lo bioactividad de animales y plantas, de tal forma que puedan ser
interrrelacionados e incorporados por los estudiantes en su dimensión cognitiva, para
lograr un verdadero aprendizaje significativo autónomo de la bioquímica metabólica,
como núcleo esencial en la comprensión de eventos y fenómenos primordiales de las
ciencias agrícolas, pecuarias y del medio ambiente.

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
UNIDAD 1:CINÈTICA Y BIOTERMODINÁMICA METABÓLICA
Capítulo 1: Equilibrio ácido base en monogástricos y rumiantes
Las células animales y vegetales se consideran verdaderos sistemas metabólicos
abiertos, debido a la interacción de un sinnúmero de procesos bioquímicos y
termodinámicos que generan flujos de masa y energía hacia y desde el interior de las
células mencionadas, estos procesos dependen también de factores cinéticos y
moleculares que inciden en la velocidad con que ocurren las reacciones bioquímicas
anabólicas y catabólicas, las cuales determinan las funciones vitales de un organismo
vivo.

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
Lección 1. Soluciones Amortiguadoras
También se les denomina soluciones "Buffer" ó lampón y son aquellas que se
oponen a los cambios de pH, cuando se les adicionan ácidos o álcalis
(hidróxidos). su acción se basa principalmente en la absorción de hidrogeniones
(H+) ó iones hidróxilo (OH').En forma general, una solución amortiguadora está
conformada por una mezcla binaria de un ácido débil y una sal del mismo
ácido proveniente de base fuerte ó también, una base y una sal de esta base
proveniente de un ácido fuerte.
Ejemplo:
Mezcla de ácido acético y acetato de Sodio
Hidróxido de amonio y cloruro de amonio
La aplicación más importante de estas soluciones reside en el estudio de la regulación
del equilibrio ácido=base en los sistemas biológicos, por eso a nivel de experimentos
bioquímicos se utilizan para controlar el pH de reacciones in vitro.
Un amortiguador biológico de vital importancia es el plasma sanguíneo, el cual
regula valores de pH entre 7,2 y 7,3; con variaciones de 0,2 unidades se
presentarían efectos letales para la vida.
1.1 pH de una Solución Amortiguadora
Considerando que la solución amortiguadora es una mezcla de ácido débil con
una sal del mismo ácido proveniente de base fuerte y además que un ácido débil
se ioniza parcialmente, podemos representar la ionización de esta forma:
HA <======> H+ + A-
Aplicando la ley de acción de masas y teniendo en cuenta la constante de disociación se
obtiene la siguiente expresión:

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
pH =pKa + Log HA
A
Donde pka,representa el valor del potencial de la constante de acidez del ácido débil,
A es la concentración del anión común,equivalente a la sal y HA indica la
concentración del ácido débil que forma parte de la solución buffer.En consecuencia,la
anterior ecuación se puede reescribir así:
pH = pKa + Log Ácido
Sal
Esta expresión se conoce como ecuación de Henderson -Hasselbach y sirve para calcular
el pH de mezclas de ácidos débiles y sus sales es decir, soluciones "Buffer", Tampón ó
amortiguadoras
De acuerdo a esta ecuación, se puede deducir, que el pH de una
solución amortiguadora, depende de dos factores:
a)El valor del pKa del ácido débil
b)Las proporciones entre Las concentraciones de sal y ácido
1.2 Capacidad Amortiguadora
Se utiliza para comparar las eficiencias de las soluciones amortiguadoras y se define como:
La cantidad en miliequivalentes(meq) de ácido o base fuerte que puede neutralizar la
solución amortiguadora, sufriendo un cambio de pH en una unidad. Matemáticamente, se
expresa como la relación cociente entre el incremento de ácido o base fuerte con respecto al
incremento del pH, es decir:

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
= B / (pH)
Donde: = Capacidad amortiguadora de la solución
B = Incremento de ácido o base fuerte
meq /(pH) (pH) = incremento en unidades de pH
Lo evidente es que esta capacidad, depende de dos factores:
a) Concentraciones absolutas del sistema
b)Proporción relativa de las formas disociada y sin disociar, siendo máxima
cuando el cociente [sal]/[ácido] es próximo a la unidad.
Lección 2. Sistemas amortiguadores fisiológicos
El equilibrio ácido-base de las células está condicionado por un conjunto de
sistemas amortiguadores, porque estas funcionan dentro de límites estrechos de
pH a causa de su metabolismo.
Los factores de amortiguación más sobresalientes en los organismos vivos, por su
acción rápida y eficiente en la regulación del pH son:
a. Sistema Bicarbonato
b. Sistema Fosfato
c. Hemoglobina

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
d. Proteínas del plasma
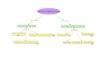
Figura 1. características y tipos de amortiguadores biológicos en animales
La importancia y relevancia de cada uno, depende del tipo de organismo.El
mentefacto mostrado en La figura 1, resume las características, propiedades y
clasificación de las principales soluciones amortiguadoras, presentes en lo
organismos animales.
Soluciones buffer
(amortiguadoras)Soluciones buffer
(amortiguadoras)
Lípidos y
Carbohidratos
Reguladores
de pHReguladores
de pH
Regulan equilibrio
HomeostáticoRegulan equilibrio
Homeostático
ejemplos
Bicarbonatos
(HCO3-)
Bicarbonatos
(HCO3-)
Aminoácidos
(proteínas)Aminoácidos
(proteínas)
Ácidos fuertes
(ácidos
minerales)
ÁCIDO DÉBIL + SALÁCIDO DÉBIL + SAL
Controlan
metabolismo
ácido base
Controlan
metabolismo
ácido base
FosfatosFosfatosHemoglobina
HbHHemoglobina
HbH
Absorben exceso
de iones H+ y OH-Absorben exceso
de iones H+ y OH-
4HPO
propiedades
componentes
excluyen

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
De acuerdo a Miles y Butcher(1995),profesores de la Universidad de Florida, los
amortiguadores (buffers) en los fluidos corporales sirven como una defensa contra el
cambio del PH .Cada compartimiento de fluido contiene tipos y características de
substancias disueltas, algunas que son amortiguadores a un pH fisiológico. Por eso, el
pH es estabilizado por la capacidad amortiguadora de los fluidos corporales.
En animales existen básicamente cuatro principales amortiguadores que se localizan
en los tres diferentes compartimientos fluidos (Cuadro 1).
Cuadro 1.Tipos de amortiguadores fisiológicos y ubicación en los fluídos biológicos
FLUÍDO
SISTEMA AMORTIGUADOR
Sangre
Bicarbonato
Hemoglobina
Proteínas
Fosfatos
Extracelular y cerebroespinal
Bicarbonato
Proteínas
Fosfatos
Intracelular
Proteínas
Fosfatos
Bicarbonato
Orina Fosfato
Amoníaco
Fuente: Miles y Butcher(1995)

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
Como se puede observar, con excepción del amoníaco en la orina y la hemoglobina
en la sangre, los amortiguadores en los compartimientos son idénticos. En
consecuencia, el conocer como estas soluciones disueltas tienen la capacidad de
amortiguar es esencial para poder entender al equilibrio ácido-base.
Sistema Bicarbonato (anhídrido carbónico/bicarbonato) :Este el buffer amortiguador
principal en el fluído extraceular, dentro de.la célula roja de la sangre y en el plasma.
En este sistema el CO2 se comporta como ácido volátil y su concentración puede ser
controlada por medio de la tasa de respiración del animal.
La Siguiente ecuación muestra La formación de iones de hidrógeno en las células rojas de
sangre como resultado del transporte del gas carbónico del tejido a los pulmones
CO2 + H2O H2CO3 HCO3- + H+
Cuando la célula roja de sangre está dentro de los tejidos corporales esta reacción va hacia la
derecha. En los pulmones la reacción va hacia la izquierda. Además, la presión parcial del
CO, es más alta dentro del los tejidos y más baja en los pulmones.
La reducción en la tasa de respiración permite la acumulación de CO2 y mueve la
ecuación hacia la derecha,la concentración de hidrogeniones se incrementa y el pH del
fluído se reduce, lo que produce una condición conocida corno acidosis respiratoria. Si la
tasa de respiración es más rápida que lo normal, la ecuación se mueve hacia la
izquierda y resulta la alcalosis respiratoria. Esto ocurre comúnmente en aves como
resultado del jadeo debido al estrés por calor. Se puede controlar estos disturbios
metabólicos, por medio de aumentar o reducir la tasa respiratoria.
Sistema Fosfato: Todos los fosfatos en el animal vienen de la dieta, a un pH de 7.40,
la mayoría del fosfato en los compartimientos fluídos existe en la forma de las especies
iónicas H2PO4-1 y HPO4
-2 , cuando el pH en los fluídos corporales comienza a decaer, la
especie HPO4-2 se vuelve importante corno un aceptante de protones y se convierte en la

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
especie H2PO4-1,así cuando el pH se eleva por encima de 7.40, la especie H2PO4
-1 dona un
protón al fluído y se convierte de nuevo en la especie HPO42. El sistema fosfatos es el
amortiguador más importante en la orina,debido a que los protones excretados en la orina
son principalmente en la forma de la especie H2PO4-1.
Durante la acidosis prolongada, la amortiguación por fosfato es muy importante, lo cual
se relaciona con los huesos,debido a que son una buena reserva de amortiguadores
como el fosfato cálcico que se presenta en forma de hidroxiapatita,el cual no es muy
soluble, pero su solubilidad es mayor durante la acidosis y algo de fosfato cálcico en los
huesos se convierte en solución. Esto ocurre comúnmente en las ponedoras cuando los
huesos están suministrando calcio para la calcificación del cascarón de huevo,entonces,el
fosfato cálcico se disocia y se convierte en Ca+2 y PO4-3, inmediatamente la especie PO4
-3
acepta un protón y se convierte en la especie HPO4-2. Durante la acidosis esta reacción
continúa y la especie HPO4-2 acepta otro protón y se convierte a H2PO4
-1. Así pues, durante
la acidosis tos huesos...pueden ayudar a mantener el e.quilibrio ácido-base por medio
proporcionar la especie de fosfato que acepta protones, incrementando el pH al nivel desead
7,4
Hemoglobina : La hemoglobina es un amortiguador muy importante y sólo so encuentra
en la célula roja de la sangre. Sirve como un amortiguador excelente por varias razones.
Las dos razones principales son su alta concentración en la sangre y su altísimo
contenido del aminoácido histidina. Este aminoácido tiene una cadena lateral única
llamada imidazol. Esta cadena , puede atraer a los protones y sacarlos de los fluidos
corporales o puede donar protones dichos fluidos en el intento de mantener el pH cerca
de 7.40. Las otras proteínas en los compartimentos de fluído, también le deben su
capacidad de amortiguar a esta cadena lateral. La albúmina es la proteína del plasma
más abundante y contribuye en forma significativa a la amortiguación de la sangre. El
fluido intracelular está lleno de proteínas que funcionan como el sistema más
importante de amortiguación dentro de la célula.En condiciones metabólicas la Hb se
comporta como un ácido débil y la oxihemoglobina como un ácido más fuerte que la
Hb reducida (es decir aquella que lleva un hidrogenión —> HHb).

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
Es importante anotar, que la Hb incide sobre el transporte del CO2 por la sangre,
veamos como lo hace:En las células por efecto de la respiración celular se produce
gas carbónico que pasa a la sangre penetrando los hematíes, quienes contienen la
enzima anhidrasa carbónica y convierten al CO2 en ácido carbónico (H2CO3), este se
disocia en iones bicarbonato e hidrógeno, que harían descender el pH, de no ser
capturados rápidamente por la HbO2-, que se transforma en oxihemoglobina
reducida(HHbO2).

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
Lección 3. Metabolismo ácido base en animales.
El equilibrio ácido-base del organismo animal está localizado en los compartimentos
líquidos. El agua representa aproximadamente el 60% del peso vivo de un animal
adulto y se distribuye en el líquido intracelular (alrededor del 60% del agua total) y el
líquido intersticial, con un 7 a 8% del agua total formando el agua plasmática (Meschy
F.,2000)
El potasio, sodio, cloro y bicarbonato tienen un papel esencial en el mantenimiento
del equilibrio iónico y por ende del metabolismo ácido-base, ya que el proceso
bioquímico de su regulación pasa por los sistemas tampón o de intercambio iónico.Por
lo tanto, la ingestión de agua o de electrolitos desplaza este equilibrio y puede
traducirse en cambios temporales del tamaño de los compartimentos líquidos.
La relación que existe entre electrolitos y equilibrio ácido-base se basa en los
mecanismos de absorción digestiva y los intercambios iónicos entre los
compartimentos digestivos y sanguíneos. La absorción de cationes se hace “en contra”
de los iones H+ y tiene, por tanto, un efecto alcalinizante a nivel sanguíneo, mientras
que la absorción de aniones tiene un efecto inverso debido a la salida de iones
bicarbonato de célula sanguínea. El mantenimiento del equilibrio ácido-base dentro de
los valores fisiológicos pone en juego un sistema principalmente localizado a nivel
sanguíneo (poder tampón de los hematíes y del plasma) y renal.
La hipótesis clásica de compensación de la acidosis metabólica (Pitts, 1948)
considera que el pH plasmático y la concentración en bicarbonato sanguíneo se
mantienen en los valores normales por dos vías complementarias a nivel renal:
• Reabsorción del bicarbonato en el tubo proximal del riñón ; en situación de acidosis la
casi totalidad del bicarbonato filtrado a nivel glomerular es rápidamente reabsorbido;

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
• Salida de protones por acidificación intensa en el tubo distal .
Esta acidificación puede hacerse por dos vías: la del fosfato, generalmente admitida, y
la del catabolismo de la glutamina, hoy en día cuestionada (Atkinson y Bourke, 1995),
que no conduciría a la producción de amoníaco sino directamente ión amonio, sin
acción sobre el equilibrio ácido-base en esa zona de pH. Igualmente, es preciso tener
en cuenta la importante producción iónica derivada del metabolismo de los
aminoácidos o la utilización del bicarbonato en la ureogénesis. El pH de la orina es
ciertamente un indicador fiable (Patience, 1990) y sobre todo de más fácil
determinación que el pH sanguíneo o la concentración de bicarbonatos en plasma.
Lección 4: Balance electrolítico dietario (BED)
Un electrólito es una substancia que cuando se disuelve agua produce una solución de átomos
disueltos o iones con cargas eléctricas. Por ejemplo NaCl se disocia en solución y se vuelve a Na+1
y Cl-1 y cuando KHCO3 se disocia en solución se convierte en K+ y HCO3-1. . Las dietas de
animales no tienen ninguna carga eléctrica neta,por eso, todos los aniones con carga eléctrica
negativa están balanceados con los cationes de carga positiva. De la misma manera, en
todos los fluidos biológicos la suma de la carga positiva tiene que ser igual a la suma de la carga
negativa. Los electrólitos pueden tener diferentes efectos sobre el metabolismo y la fisiología y
por eso pueden afectar al equilibrio eléctrico y consecuentemente al equilibrio ácido-base y al
desempeño del animal. Enfermedades tales como enteritis resultan en una pérdida de
electrólitos,en consecuencia,se tienen que tomar medidas, como la suplementación de
electrólitos, para compensar los perdidos.El equilibrio de electrólitos es la diferencia entre la
concentración total de aniones y cationes dietéticos,así, los elementos minerales en la
dieta que tienen una carga eléctrica negativa se consideran elementos que forman ácidos y
recíprocamente los elementos que forman bases tienen una carga positiva .
La idea de manipular las concentraciones iónicas de la ración a fin de evitar las
consecuencias patológicas de la acidosis (o de la alcalosis) es bastante antigua y
encontró en los años 70 un primer campo de aplicación en avicultura. En rumiantes, la

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
primera aproximación ha sido para la prevención de la fiebre de la leche; más
recientemente han aparecido un cierto número de trabajos relacionados con la especie
porcina. La base es bastante simple: los aniones, a excepción del fosfato y del
bicarbonato, son acidógenos y los cationes son alcalógenos. Según el esquema de
Monguin(1981),el comportamiento de los electrolitos y equilibrio ácido-base es:
(An-Cat)in = acidez ingerida, donde: An:aniones y Cat:cationes
(An-Cat)or = acidez excretada
AEndo = acidez endógena
En estado de equilibrio :
(An-Cat)in + AEndo - (An-Cat)or = 0
Sobre esta base, se han propuesto varias ecuaciones, para monogástricos, se
mantiene generalmente el Balance Electrolítico (BED) expresado en mEq/kg de MS (ó
por 100 g) de alimento.Esto se puede representar de la siguiente forma:
BED =meq (Na+ + K+ - Cl-)/Kg dieta
También:
BED = (Na/23 + K/39 - Cl/35,5) x 1000

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
Con todo rigor, sería preciso probablemente tener en cuenta otros aniones y cationes
con la condición de que no sean metabolizados, es decir iones exclusivamente
destinados a la homoestasis ácido-base, y debería ser tenida en cuenta su eficacia de
absorción digestiva. Así, otras ecuaciones para rumiantes (Horst et al., 1997) y
porcinos han sido propuestas (Patience y Wolynetz, 1990), sin que por el momento
supongan una mejora sensible
.
Lección 5. Aplicaciones del balance electrolítico(BED) en animales.
Los primeros estudios sobre los efectos del equilibrio electrolítico de la ración sobre los
rendimientos fueron realizados con aves en los años 70. Sauveur y Mongin(1978)
encontraron una respuesta curvilínea de la velocidad de crecimiento cuando el BED
aumentaba, siendo el crecimiento máximo para un BED de alrededor de 250
mEq/kgEstos mismos autores también demostraron la existencia de una relación
estrecha entre la acidosis metabólica, caracterizada por un bajo contenido deL anión
bicarbonato: HCO3- en el plasma, y la mayor frecuencia de discondroplasia tibial
Después de una docena de años, estudios similares han sido realizados con la
especie porcina. El destete se de un cambio brutal de la naturaleza y del modo de
presentación del alimento consumido por los lechones, los cuales necesitan una rápida
adaptación de su función digestiva. En particular, la acidificación del alimento en el
estómago debe ser suficiente para controlar el desarrollo de la flora bacteriana (E.
Coli) y permitir una actividad óptima de las enzimas digestivas. En el destete, esta
acidificación es difícil debido a la que la actividad secretora es pequeña y a que el
poder tampón de los alimentos (resistencia a la acidificación) es mucho más
importante que el de la leche (Bolduan et al., 1988). El poder tampón de los alimentos,
definido como la cantidad de HCl necesaria para bajar el pH a 3 (Boltshauser et al.,
1993) ó 4 (Bolduan et al., 1988)y depende principalmente de su contenido en
proteínas y minerales. Paralelamente, también depende del balance electrolítico de la
dieta. Un poder tampón pequeño del alimento corresponde a un valor bajo de BED. En
la práctica, se recomienda utilizar durante este período (0-2 semanas postdestete)
alimentos que presenten un bajo poder tampón (Bolduan et al., 1988). Si bien el efecto
sobre el pH estomacal no ha sido claramente demostrado, la utilización de ácidos

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
orgánicos ha resultado ser interesante durante este período, sobre todo cuando la
dieta no contiene productos lácteos (Easter, 1988 ; Giesting et al., 1991). Sin embargo,
es preciso destacar que estas dietas con bajo poder tampón son igualmente
acidógenas a nivel metabólico, lo que podría tener una incidencia negativa sobre los
rendimientos.
Algunos trabajos han estudiado las posibles interacciones entre la adición de ácidos
orgánicos y de bicarbonato sódico (Krause et al., 1994, Giesting et al., 1991). En
dichos trabajos, los rendimientos óptimos generalmente se obtienen cuando la adición
de ácidos orgánicos está asociada a la incorporación de bicarbonato de sodio
El ejercicio propuesto a continuación, aplica los conceptos del balance electrolítico

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
dietario en la alimentación de aves.
Capítulo 2 : BIOACTIVIDAD DE ENZIMAS EN NUTRICIÓN ANIMAL
Leccción 6: Características de las enzimas
Son proteínas y por lo tanto, cualquier proceso que altere su estructura, como por
ejemplo: El calentamiento excesivo, adición de ácidos o bases fuertes, las
desnaturalizan, perdiendo su actividad biológica y catalítica.
Las enzimas son biocatalizadores de origen proteíco, altamente especializados, esto
significa que actúan casi siempre sobre una sola reacción o determinado tipo de
sustancias llamadas sustratos.
Así existen enzimas que hidrolizan únicamente almidones y se llaman amilasas,
otras hidrolizan grasas y se denominan lipasas. En general, el nombre de la enzima
depende de la sustancia sobre la que actúa y que se llama sustrato.Sin las enzimas
no podrían ocurrir actividades metabólicas como la respiración, la contracción
muscular, el crecimiento, la actividad cerebral o la digestión.
Al igual que los catalizadores inorgánicos, las enzimas no se destruyen durante el
proceso y porque su acción se limita a activar el sustrato, disminuyendo así la
energía de activación que necesita éste para transformarse en un producto
determinado.
1.1 Clasificación de las enzimas
Como se afirmó anteriormente, las enzimas son catalizadores altamente
especializados, esto significa que actúan casi siempre sobre una sola reacción o
determinado tipo de sustrato; así existen biocatalizadores que hidrolizan únicamente
almidones y se llaman amilasas, otras hidrolizan grasas y se denominan lipasas. En
general, el nombre de la enzima depende de la sustancia sobre la que actúa, es decir,

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
el sustrato, su clasificación se da con base en el tipo de reacción catalizada, como se
muestra en el siguiente cuadro:
Cuadro 1. Clasificación de los diversos tipos de enzimas
GRUPO TIPO DE REACCIÓN
CATALIZADA EJEMPLOS
Òxido -reductasas Reacciones de òxido-
reducción
Deshidrogenasa
succínica
Catalasas
Transferasas Transferencia de grupos
funcionales orgánicos
Transaldolasas
Aciltransferasas
Hidrolasas Hidrólisis en presencia
de agua.
Carbohidrasas
Celulasas
Liasas Adiciòn al doble enlace Descarboxilasas
Pirùvico descarboxilasa
Ligasas Unión de moléculas
utilizando ATP.
Glutamina sintetasa
polimerasas
Isomerasas Reacciones de
isomerización
Triosa isomerasa
Fructosa isomerasa
EJEMPLOS:
GLUCOSA-6-P aisomerasafosfohexos FRUCTOSA-6-ISOMERASA
ÀCIDO PIRÚVICO acarboxilas Pirùvico ÀCIDO OXALOACÈTICO

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
H2N-CO-NH2 +3 H2O ureasa 1CO2 + 2NH4OH
2H2O2 catalasa 1O2 + 2H2O
Otras reacciones bioquímicas enzimáticas(RBE) que son importantes en el
metabolismo animal son:
• En la glucólisis o ruptura de la glucosa sanguínea, la primera reacción
es catalizada por una ligasa denominante Hexoquinasa.
GLUCOSA + ATP GLUCOSA-6-FOSFATO +ADP
+ H
• La enzima alcohol deshidrogenase (una óxido-reductasa)
convierte en el hígado, el alcohol etílico, antes de que llegue al
cerebro, en acetaldehído, el cual es usado por la célula para
sintetizar grasas.
Lección 7. Mecanismo de reacción de las enzimas
El mecanismo de una reacción bioquímica enzimática(RBE),está relacionado con
las diferentes etapas que sigue el sustrato, para transformarse en producto. Dicho
mecanismo, fue propuesto por los doctores Leonor Michaelis y Maud
Menten(1913).y se muestra en la siguiente ecuación:
Hexoquinasa

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
Esto implica que el enzima libre (E) puede captar en su centro activo una
molécula de sustrato (S), transformándose en el complejo activado enzima -
sustrato (ES)*, este complejo puede evolucionar de dos formas:
a)Libera el sutrato y regenera la enzima libre, cuando no alcanza la
suficiente energía de activación,es decir, invierte su reacción.
b) Transforma el sustrato en un producto final y regenera la enzima libre de
acuerdo a la cinética K3, cuando supera la barrera energética inicial.
Lección 3. Factores que afectan la cinética enzimática
Las reacciones químicas catalizadas por enzimas, dependen del pH, la
temperatura, el efecto de la concentración de la enzima y de la concentración del
sustrato.
E + S (E--S)*K1
K2
K3 P + E

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
A continuación, se analiza cada factor:
a) Efecto del pH
Debido a que las enzimas son de naturaleza proteica, las alteraciones en el pH
del medio modificarán profundamente el carácter iónico de los grupos aminos y
carboxilos de los aminoácidos que constituyen la proteína y en consecuencia
afectarán su poder catalítico. Es decir, a valores extremos de pH se produce una
inactivación de la enzima. Por lo tanto, es importante establecer m pH óptimo en
estudios enzimáticos, en el cual, la actividad catalítica de la enzima presenta un
rendimiento alto, conociendo este valor, la reacción se debe mantener en este rango
de pH por medio de un buffer o solución amortiguadora, lo mismo ocurre en la
célula, ya que un pequeño cambio en el valor de pH produce también graves
efectos sobre la actividad enzimática, incidiendo como es obvio en el metabolismo
del animal.
pH
V
Vmáx
Ph óptimo
La gráfica anterior nos muestra que cuando se modifica el pH en el transcurso de una
reacción enzimática, aparece un valor en el cual la velocidad es máxima, este punto se
denomina pH óptimo y por encima o debajo de dicho valor, la velocidad disminuye.

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
b) Efecto de la Temperatura
Al igual que el pH, las temperaturas extremas, afectan las reacciones enzimáticas,
dada su naturaleza proteica. Debido a esto, se observa que a diversas temperaturas, la
velocidad de la reacción cambia, tal como lo demuestra la ecuación de Arrhenius. Sin
embargo, a un cierto valor la velocidad de la reacción es máxima, lo cual nos indica una
TEMPERATURA ÓPTIMA de la misma, por encima o por debajo de esta temperatura la
enzima pierde actividad y la velocidad disminuye, tal como lo indica la figura 2
C) Efecto de la Concentración de la Enzima
La velocidad de una reacción enzimática varía, directamente proporcional a la
concentración de la enzima, por lo tanto a mayor [E] se incremente la velocidad, esto es
válido en presencia de un exceso de sustrato.
d) Efecto de la Concentración del Sustrato
Sí mantenemos la concentración de la enzima constante y variamos la
concentración del sustrato, podemos observar que cuando se aumenta la
concentración de éste, inicialmente hay un incremento notable en la velocidad de la
reacción, hasta alcanzar una velocidad máxima estable, después de la cual permanece
invariable por más sustrato que se le agregue. La curva hiperbólica de la figura nos
muestra que a baja concentración de sustrato la relación entre V y [S] es
prácticamente lineal y por lo tanto obedece a una cinética de primer orden con respecto
al sustrato, es decir: V =[ K]
A medida que se incrementa la concentración de sustrato se llega a una zona donde se
presenta una mezcla cinética de primer y segundo orden, finalmente, llega a una
región donde la velocidad es máxima, constante e independiente la concentración del
sustrato, aquí la cinética es de orden cero y no se modifica, poque la enzima ya está
saturada en sus centros activos con el sustrato, también observamos que en el punto
medio de la velocidad máxima aparece siempre una concentración de sustrato fija,
denominada constante de Michaelis, la cual se relaciona con la afinidad de la enzima
por el sustrato.El anterior análisis, nos indica que la cinética enzimática está

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
caracterizada por dos factores fundamentales: La velocidad máxima (Vmáx.) de la
enzima y la constante de Michaelis (Km)
L a ecuación cinética de esta gráfica hiperbólica es:
V
Sus parámetros cinéticos, se pueden explicar así:

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
a.LA VELOCIDAD MÁXIMA (Vmáx) de una reacción enzimática es el
límite de la velocidad cuando la concentración del sustrato tiende al
infinito y es causada por la saturación de los centros activos de la enzima
por el sustrato.
b. LA CONSTANTE DE MICHAELIS (Km) es la concentración del sustrato
en el cual la velocidad de la reacción enzimática es la mitad de su
velocidad máxima.
Lección 4. Cinética enzimática de Michaelis.
Los estudios sistemáticos del efecto de la concentración inical del sustrato sobre
la actividad enzimática comenzaron a realizarse a finales del siglo XIX. Ya en
1882 se introdujo el concepto del complejo enzima-sustrato como intermediario
del proceso de catálisis enzimática. En 1913, Leonor Michaelis y Maud Menten ,
desarrollaron esta teoría y propusieron una ecuación de velocidad que explica el
comportamiento cinético de los enzimas.
Para explicar la relación observada entre la velocidad inicial (v0) y la
concentración inicial de sustrato ([S]0, Michaelis y Menten propusieron que las
reacciones catalizadas enzimáticamente ocurren en dos etapas: En la primera
etapa se forma el complejo enzima-sustrato y en la segunda, el complejo enzima-
sustrato da lugar a la formación del producto, liberando el enzima libre:

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
En este esquema, k1, k2 y k3 son las constantes cinéticas individuales de cada
proceso y también reciben el nombre de constantes microscópicas de velocidad.
Según esto, podemos afirmar que:
v1 = k1 [E] [S]
v2 = k2 [ES]
v3 = k3 [ES]
Despejando [ES], queda que:
, en donde la expresión (k2+k3)/k1 se ha sustituído por KM, o constante de
Michaelis-Menten. Este enlace nos aporta una explicación sobre las razones que
hacen de la KM un parámetro cinético importante.
Para cualquier reacción enzimática, [ET], k3 y KM son constantes. Vamos a
considerar dos casos extremos:
A concentraciones de sustrato pequeñas ([S] << KM) v = (k3 [ET]/KM) [S]. Como
los términos entre paréntesis son constantes, pueden englobarse en una nueva
constante, kobs, de forma que la expresión queda reducida a: v = kobs [S], con lo
cual la reacción es un proceso cinético de primer orden.
A concentraciones de sustrato elevadas ([S] >> KM), v = k3 [ET]. La velocidad de
reacción es independiente de la concentración del sustrato, y por tanto, la
reacción es un proceso cinético de orden cero. Además, tanto k3 como [ET] son
constantes, y nos permite definir un nuevo parámetro, la velocidad máxima de la
reacción (Vmax): Vmax = k3 [ET], que es la velocidad que se alcanzaría cuando todo
el enzima disponible se encuentra unido al sustrato.
Si introducimos el parámetro Vmax en la ecuación general de la velocidad,
obtenemos la expresión más conocida de la ecuación de Michaelis-Menten:

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
V
Hay enzimas que no obedecen la ecuación de Michaelis-Menten. Se dice que su
cinética no es Michaeliana. Esto ocurre con los enzimas alostéricos, cuya
gráfica v frente a [S] no es una hipérbola, sino una sigmoide . En la cinética
sigmoidea, pequeñas variaciones en la [S] en una zona crítica (cercana a la KM)
se traduce en grandes variaciones en la velocidad de reacción.
El siguiente esquema, resume los conceptos básicos de la cinética enzimática,
iniciando el análisis en el comportamiento de las enzimas como biocatalizadores
de origen proteíco

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
Con el propósito de profundizar este tema, se propone un ejercicio aplicado que
se muestra en el diagrama UVE desarrollado en el esquema superior.
Buscando desarrollar competencias interpretativas y procedimentales en esta
temática,se propone el próximo ejercicio ,de tal forma que lo desarrolle
adecuadamente en el formato propuesto:
La actividad de la enzima L-aspartato-4-carboxilasa (aspartato-
Betadescarboxilasa) se puede seguir midiendo la producción de gas
carbónico: CO2 (g), a partir del aminoácido ácido aspártico(AA), mediante la
RBE:
3 CONCEPTOS
1. ¿Se cumplió la Ley de Michaellis
en este experimento? Porqué?
2. ¿Cuál es el valor de la cons-
tante de Michaellis, la velo-
cidad Máxima y el coeficien-
te de correlación ( r ), de la
cinética estudiada?
3 ¿Cual es la ecuación
de Michaellis de este
experimento?
Caracterización enzimática
de una fosfatasa, que actúa
sobre el glicero-fosfato.
DIMENSIÓN METODOLOGICA
1. ACONTECIMIENTO
Interacción
2. PREGUNTA CENTRALDIMENSIÓN CONCEPTUAL
TCA y colisiones5 TEORIAS
4. PRINCIPIOS
Describen el comportamiento ciné-
ticomolecular de la enzima fosfatasa
sobre los enlaces qcos del
glicerofosfato (sustrato).
Michaellis-Menten
Establece relación entre V de reacción
y la Concentración del sustrato.
10. CONCLUSIÓN
La cinética de la fosfatasa cumplió la Ley
de M-M, debido a la alta correlación lineal
entre los recíprocos de [s | y la V.
8.TABLA de RESULTADOS7. GRAFICAS
VARIABLE VALOR
Km 0.0103 M
V máx 1.012 umol/min
r 0.999
V = 1.012 [S|/0.0103 + [S|
1/V
3.03
200 1/[S|
V
[S|
0.5
0.01
8.TABLA de RESULTADOS7. GRAFICAS
VARIABLE VALOR
Km 0.0103 M
V máx 1.012 umol/min
r 0.999
V = 1.012 [S|/0.0103 + [S|
1/V
3.03
200 1/[S|
V
[S|
0.5
0.01
6. REGISTRO DE DATOS
0.100 0.910
0.050 0.830
0.020 0.670
0.010 0.500
0.005 0.330
[S| (mol/lt) V (umol/min)
pH = 7.0
T = 37 º
6. REGISTRO DE DATOS
0.100 0.910
0.050 0.830
0.020 0.670
0.010 0.500
0.005 0.330
[S| (mol/lt) V (umol/min)
pH = 7.0
T = 37 º
7. TRANSFORMACION DE DATOS (Cálculos)
1/[S| 1/V
10 1.09
20 1.20
50 1.49
100 2.00
200 3.03
Hallar: 1/[S| y 1/V (doble recíproca
Por Regresión lineal se obtiene
b = 0.9873 min/umol; m= 0.0102; R = 0.999
Se calcula KM y V max; así
K M = m / b = 0.0102/0.9873 = 0.0103
V max = 1 / b = 1 / 0.9873 = 1.012 umol/min
7. TRANSFORMACION DE DATOS (Cálculos)
1/[S| 1/V
10 1.09
20 1.20
50 1.49
100 2.00
200 3.03
Hallar: 1/[S| y 1/V (doble recíproca
Por Regresión lineal se obtiene
b = 0.9873 min/umol; m= 0.0102; R = 0.999
Se calcula KM y V max; así
K M = m / b = 0.0102/0.9873 = 0.0103
V max = 1 / b = 1 / 0.9873 = 1.012 umol/min
ENZIMA FOSFATASA
V max [S|
V = --------------
K M + [S|
cumple
CINETICA MICHAELLIANA
Actúa
sobre el
GLICERO
FOSFATO
produce
GLICEROL H3PO4
V [ s |
relaciona
V
[S|
Puede
ser es
Donde
aparece
así
V maxK M
1 / b m / b
es es
ENZIMA FOSFATASA
V max [S|
V = --------------
K M + [S|
cumple
CINETICA MICHAELLIANA
Actúa
sobre el
GLICERO
FOSFATO
produce
GLICEROL H3PO4
V [ s |
relaciona
V
[S|
Puede
ser es
Donde
aparece
así
V maxK M
1 / b m / b
es es
9. INTERPRETACION y DISCUSION:
La Km equivale a una concent. De 0.0103 mol de
glicerofosfato por cada litro de solución.
La gráfica de la doble recíproca presentó una
correlación lineal altamente significativa (p<0.01).

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
ÁCIDO ASPÁRTICO L-ALANINA + CO2(g)
Se realizó un experimento en el laboratorio, para verificar la potencia del
Treo-βetahidroxiaspartato(TBHA), como inhibidor de una fuente microbiana,
que contenía el ácido aspártico y se obtuvieron los siguientes resultados
Tabla 1:Valores de velocidad de la RBE,con y sin inhibidor TBHA, en función de la concentración de L -aspartato
(C)[L-aspartato]
(µmol/L)
(V)VELOCIDAD (µmolCO2/min)
SIN INHIBIDOR (RBE) CON INHIBIDOR(TBHA)
25.0 27.0 12.0
33.3 31.3 15.7
50.0 47.8 18.1
100.0 51.8 24.7
200.0 52.6 25.0
Con base en estos valores de la tabla 1,completar la siguiente tabla 2:
Tabla 2:Valores de la doble recíproca de Linneweaver and Burk.
1/C
(L/μmol)
1/ V (min/μmol)
SIN INHIBIDOR (RBE) CON INHIBIDOR(TBHA)
L-apartato-4-carboxilasa
T=37°C ; pH =
7.0

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
Adicionalmente:
1.Trazar las gráficas de:
1.2 1/V contra 1/C (gráfica de la doble recíproca, para las dos Reacciones
bioquímicas)
1.3 Ajustar las gráficas anteriores(1.2),por el método estadístico de los
mínimos cuadrados, encontrar :Pendiente(m),intercepto(b),ecuación de
regresión y coeficiente de correlación de Pearson(r2).Completar la siguiente
tabla(Deben realizar una por cada RBE)
TENER EN CUENTA: 1/C =X ; 1/ V = Y
Tabla 3.Datos para realizar la regresión lineal por mínimos cuadrados.
X Y X.Y X2 Y2 Ycorregida
X Y X.Y X2 Y2

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
2.Con base en las pendientes e interceptos, hallados en el ajuste de la
gráfica de la doble recíproca, Hallar: Constante de Michaelis-Menten( Km) y
Velocidad máxima (Vmáx),para las dos reacciones bioquímicas.(Completar
la tabla)
Tabla 4.Resultados de los parámetros cinéticos de las RBE.
TIPO DE RBE Vmáx (µmol/min) Km (µmol/L)
Sin inhibidor
Con inhibidot
2.1Encontrar la ecuación de Michaellis-Menten y la ecuación de la doble
reciproca, para ambas reacciones bioquímicas.
Tabla 5.Ecuaciones cinéticas de las RBE
TIPO DE RBE Ecuación de Michaelis
Ecuación de la doble
recíproca
Sin inhibidor
Con inhibidor
Con base en las ecuaciones indicadas en la tabla 5:
2.2 Trazar las gráficas corregidas de:
2.2.1 V contra C (para ambas reacciones)
2.2.2 Doble Recíproca (para ambas reacciones)
2.3Calcular la [L-aspartato] en la reacción bioquímica, con inhibidor y sin
inhibidor, cuando la actividad enzimática (V) es de :15 µmol CO2/min y 125
µmol CO2/min.

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
2.3.1Determinar la actividad enzimática de la enzima (V), cuando la [L-
aspartato] es de 75 µmol/L ,para la reacción sin inhibidor y con inhibidor.
2.3.2¿Cómo afectó los parámetros cinéticos (Km y Vmax), la adicion del
inhibidor Treo-βetahidroxiaspartato, la reacción enzimática?
3.Elaborar un diagrama UVE HEURÍSTICO de este experimento
Lección 5. Enzimas exógenas en la nutrición animal(Granados et al.,2000)
Las enzimas son proteínas de estructura tridimensional sumamente comple ja.
Actúan sólo en condiciones definidas de temperatura, pH y humedad y únicamente con
substratos específicos (Btihler y colaboradores, 1998). El uso comercial de enzimas en
nutrición avícola, empezó hace algunos arios con la aplicación de Beta-glucanasas en
dietas a base de cebada, debido a su bajo contenido energético y pobre digestibilidad, por
la presencia de Beta-Glucanos, los cuales forman soluciones de elevada viscosidad en
el intestino de las aves, interfiriendo así con la correcta digestión y difusión de los
nutrientes de la ración alimenticia (Sorensen y Nielsen., 1998).
La utilización de enzimas en dietas balanceadas para aves, ha demostrado contribuir a]
mejoramiento de su comportamiento productivo en factores como: conversión alimenticia,
ganancia de peso y eficiencia en razón al aumento de la degradación de los componentes
antinutricionales de los piensos, basados principalmente en cereales como: cebada
centeno y trigo. El efecto antinutritivo de la cebada se atribuye principalmente al 1,3 - 1,4
Betaglucano, porque su porción soluble es mucho más elevada que la de los
pentosanos,(Buhler y colaboradores., 1998). Estos Betaglucanos son parecidos a la
celulosa y están conformados por moléculas de glucosa unidas entre sí por enlaces
Beta-glucosídicos 1, 4 y 1, 3. Estos son los que originan la intensa ramificación y la
posibilidad de acumulación de agua, que tiene un efecto antinutritivo por aumento de la
viscosidad. El contenido de betaglucanos en la cebada oscila entre 15 y 107 g/kg de
materia seca (Bühler et al.,1998). Otros componentes de la cebada, difíciles de digerir,
son el ácido tífico y sus sales denominadas Fitatos.
Dichas sales son ésteres del ácido hexafosfórico del inositol y pueden formar complejos
insolubles con proteínas y cationes divalentes como calcio, magnesio, Zinc y cobre. Esto
reduce la biodisponibilidad de estos nutrientes, haciéndolos difícilmente digeribles. En

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
consecuencia, el fitato es un factor antinutritivo en la dieta (Basf., 1997). El contenido
de fósforo tífico en la cebada tiene un rango de 2.2 gramos a 2.9 gramos por cada
kilogramo de materia seca (Bubler et al.,1998). Las sales del ácido fitico ó fitatos, sólo
pueden descomponerse por acción de las fitasas que no están presentes de forma
natural en el tubo digestivo de las aves de corral. Estas enzimas., son producidas en el
laboratorio por fermentación microbiana a partir del Aspergillus niger y son utilizadas en
animales monogástricos para degradar los fitatos, incrementando con ello la
biodisponibilidad del fósforo y otros componentes nutricionales de la dieta (Vahan.,
1997). La fitasa cataliza una reacción de defosforilación del fitato a través de un
mecanismo no definido (Hurtado y Resende., 1997).
El producto enzimático más utilizado en la producción de pavos es la fitasa. Fancon
(1997), utilizó esta enzima con 13 mil 280 pavos de las variedad But Big; observó que
se produjeron algunos efectos beneficiosos en los parámetros productivos especialmente,
en la ganancia de púa y el índice de conversión. Vahan (1997), encontró que esta
enzima además de mejorar la digestíbilidad del fósforo, incrementaba también la
digestibilidad del nitrógeno de los componentes de la dieta, aumentando los
rendimientos de carne y la velocidad de crecimiento.
Teniendo en cuenta que las enzimas permiten hacer uso de materias primas poco
utilizadas en la alimentación aviar, se decidió evaluar el efecto de la suplementación
de las enzimas: carbohidrasas, complejo (arabanasa + celulosa + betaglucanasa +
hemicelulosa xilanasa) y fitasa, en una ración alimenticia enriquecida con cebada y su
incidencia en el comportamiento productivo de gallopavos.
Capítulo 3: Biotermodinámica
LECCIÓN 1: Introducción a la termodinámica
La termodinámica es la ciencia que estudia las relaciones entre calor y las
demás formas de transferencias de energía, puesto que toda
transformación biofisicoquímica implica, generalmente, un cambio
energético.

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
Para abordar el estudio de la termodinámica es necesario revisar los conceptos de
Temperatura y Calor.
1.1 Temperatura (T)
La temperatura puede definirse como la propiedad termodinámica que cuantifica la
energía cinética molecular promedio de las moléculas existentes en cualquier
sistema,además, determina el flujo de calor y el equilibrio térmico,así, dos cuerpos
están a la misma temperatura si no hay transferencia de calor cuando se colocan
juntos.
1.1.1 Escalas de Temperatura
La más importantes son: La Celsius o centígrado (°C), kelvin(K), Farenheit
(°F) y la Rankine(°R)
La siguientes ecuaciones muestran la correspondencia matemática entre las
escalas:
K= °C + 273
°F=1,8°C +32
°R = °F + 460
1.2 Calor (Q)
El calor como propiedad termodinámica es una forma de transferencia de energía
debida a la diferencia de temperatura(ΔT), entre dos sistemas,es decir, es energía en
tránsito que depende de T1 y T2.
Teniendo en cuenta el concepto de calor específico, como la cantidad de calor(Q)
que se debe suministrar a unidad de masa (m) para elevar la temperatura en 1
grado (ΔT),se tiene que la ecuación fundamental del calor es:

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
Q= mCP ΔT
Donde:
Q = calor ganado o perdido por el sistema
m = masa de la sustancia en gramos
C p= calor específico de la sustancia,medido a presión constante, en cal/g°C
ΔT= T2 -T1
T2 = Temperatura final del sistema
T1= Temperatura inicial del sistema
Para efectos de resolver algunos ejercicios,se debe tener en cuenta los siguientes
factores de conversión:
Cuadro 1:Tipo de unidades y factores de conversión para masa y energía.
UNIDADES CONVERSIONES
MASA
1g=1000mg , 1kg=1000g ;
1lb=454g
1kg=2,2 lb ; 1tonelada(t)=1000kg
CALOR , TRABAJO Y
ENERGÍA
1cal=4,18J ; 1kcal=4,18KJ
1kcal=1000cal ; 1KJ=1000J
1MJ =106J; 1Mcal =106 cal
1BTU=252 cal
g=gramos ; mg=miligramos ; lb=libras ; cal=calorías ; Kcal=kilocalorías ; J=Joule

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
KJ=kilojoule ; MJ=Megajoule ; BTU=British thermal unit
1.2.1 Calor de Combustión (Energía Bruta)
Cuando una sustancia orgánica se quema por completo hasta sus últimos
productos de oxidación, gas carbónico (CO2), agua(H2O) y otros gases, el calor
liberado se denomina energía bruta o calor de combustión y se expresa como la
variación de la entalpÍa (ΔH) en Kcalorías por mol.
Ejemplo:
Para la Glucosa : ΔH = 673 Kcal/mol, lo cual indica que cuando 1 mol de glucosa, es
decir 180g, se oxidan completamente hasta CO2 y H2O, se generan 673 kilocalorías.
Esta medida es importante para determinar el contenido energético de los
alimentos, que el organismo utiliza y su determinación se efectúa en una
bomba calorimétrica. A nivel de nutrición animal es útil expresar la energía bruta en
Kcal/ g.
Ejemplo:
Para el Salvado de trigo : ΔH = 4,54 cal/g, esto significa que cuando 1 g de salvado
de trigo se oxida completamente, existe una liberación de 4,54 calorías o 19 Joules.
LECCIÓN 2: Postulados fundamentales de la termodinámica
2.1 Primera Ley
Enunciada por Robert Meyer en 1841, es el principio de la conservación de la energía y
puede definirse así: La energía total de un sistema aislado permanece constante, es
decir, la energía no se crea ni se destruye, sólo se transforma de un tipo a otro. Por lo
tanto, cuando desaparece una clase de energía debe producirse una cantidad
equivalente de otra clase.

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
Cualquier cambio en el estado del sistema, incluye un cambio en la energía interna
(ΔE) igual a la cantidad de calor (Q) absorbida por el sistema menos la cantidad de
trabajo (W) realizado por el mismo, esto se puede escribir así:
ΔE = Q - W
Teniendo en cuenta, que el calor es una forma de energía, fácilmente
cuantificable, la mayor parte de las investigaciones sobre las equivalencias e
intercambios de energía que tienen lugar en los procesos fisicoquímicos fueron
analizados con base en los cambios caloríficos de un sistema termodinámico.
La energía Interna (ΔE), se define como la capacidad intrínseca de un sistema para
producir trabajo, incluye todas las formas de energía y resulta del movimiento de las
moléculas, la atracción intermolecular y otros factores fisicoquímicos.
ΔE es una función termodinámica de estado, porque depende únicamente de los
estados inicial y final del sistema, esto significa que es independiente de la tayectoria
de una transformación, así el calor de combustión de la glucosa se puede determinar
por incineración ó a nivel metabólico.
En un organismo animal, ocurren un múltiples reacciones bioquímicas que constituyen
el metabolismo bioenergético. Esta serie de reacciones, se subdividen en 2
categorías básicas: el catabolismo, que implica la desintegración de macromoléculas
en otra más sencillas y el anabolismo, que se refiere al proceso inverso, es decir, a la
síntesis o formación de productos bioquímicos destinados a la estructura del
protoplasma celular.
En toda esta cadena de procesos químicos, se requiere en mayor ó menor
cantidad la energía, por lo tanto, es importante pensar ¿De dónde se pro
duce?, ¿Cómo se produce?, ¿en dónde se almacena?, ¿cómo se utiliza?. Para dar
respuesta a esta serie de interrogantes, comenzaremos por afirmar que la energía
utilizada, es extraída gradualmente de los alimentos (carbohidratos, lípidos y

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
proteínas) a través de tres etapas fundamentalmente oxidativas: Hidrólisis,
formación de Acetil CoA y ciclo de Krebbs con la fosforilación oxidativa, los
productos finales de estas etapas son el gas carbónico, agua y lo más importante:
ENERGÍA, La cual es almacenada en una molécula extraordinaria: El ATP ó
Adenosín Trifosfato.
2.2 Segunda ley de la Termodinámica
La primera ley de la termodinámica, no establece la dirección de flujo del calor, ni
tampoco clarifica sus efectos de éste sobre el sistema y sus entorr Debido a esto,
los eminentes científicos Rudolf J.E. Clausius, lord Kelvin y > Planck, trataron de
generalizar estos conceptos, para así, darle el sentido a las transformaciones
termodinámicas. Con base en sus trabajos experimentales llegaron a los
siguientes enunciados.
a. Primer enunciado (Según kelvin-Planck): No es posible diseñar una
máquina térmica capaz de convertir todo el calor absorbido en trabaje.
Esto significa que la eficiencia o rendimiento de las máquimas térmicas
en menor del 100%.
b. Segundo enunciado (Según Clausius): El calor fluye espontáneamente de
un foco más caliente a un foco más frío y no viceversa.
De estos enunciado, puede deducirse lo siguiente:
- Todos los procesos de la naturaleza tienden a cambiar espontáneamente
en una dirección que conduzca al equilibrio.
- El calor no se transforma en trabajo, sin producir cambios permanentes
en los sistemas o sus proximidades.
- La energía se degrada.
Ejemplo, sí calentamos previamente una barra de hierro y luego la aislamos el calor
no se concentrará únicamente en un extremo, sino que se distribuirá

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
uniformemente en toda la barra, marcando la dirección de flujo de calor desde el
punto más caliente hasta el extremo más frío.
2.2.1 Características de la Entropía(ΔS)
a. La entropía puede considerarse como la medida del desorden de un
sistema termodinámico.
b. En un proceso reversible la entropía global no cambia, es decir ΔS = O,
porque ésta depende únicamente de los estados inicial y final del sistema.
c. En todo proceso irreversible la entropía total aumenta, luego ΔS > O
d. A una temperatura dada, los sólidos tienen una entropía relativamente
baja, los líquidos una cantidad intermedia y los gases la entropía más alta.
e. Al aumentar la temperatura, la entropía y el desorden crecen.
f. En general, se puede afirmar que la entropía del universo va en aumento
ya que la mayoría de los procesos termodinámicos son irreversibles.
g. Cuando los organismos vivos se desarrollan, disminuyen su entropía
(ΔS<0), por causa del orden estructurado de la materia viva. Pero el
descenso se produce a expensas de un incremento entrópico del medio
ambiente.
h. En un sistema, la energía útil está organizada y cuando se utiliza en realización de un
trabajo se convierte en calor, que es una forma de energía, basada en el movimiento
caótico de átomos y moléculas, por lo tanto, incrementa el desorden del sistema y por
consiguiente la entropía.
En síntesis, podemos afirmar que todo proceso natural se realiza con un incremento
entrópico y que la dirección del cambio es aquella que conduce a tal aumento. Este

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
enunciado es la forma más general de la segunda ley de la termodinámica, luego los
otros casos indicados son formas particulares de esta ley.
El ejercicio que se describe a continuación es una aplicación directa de las leyes
biotermodinámicas en la termo-regulación de los animales
Un animal de 480 Kg de masa ,temperatura basal :39°C y calor específico corporal:
4KJ/Kg°C, disipa 0,80Mcal, por causa de un estado febril.
Preguntas centrales
1.¿Cuántos Kilojoules disipa el animal por causa del estado febril?
2.¿Cuál es el aumento de la temperatura(°C) corporal del animal?
3.¿Cuál es la temperatura corporal final del animal, en °F?
4.¿Qué efectos sobre el metabolismo del animal , causará este incremento térmico?
5.¿Cómo se termorregulará el animal en esta situación?
Listado de conceptos para elaborar el mapa conceptual
CONCEPTO CONCEPTO
Calor Basal
Metabolismo Catabolismo
Temperatura Transferencia
Energía Pirógenos
Estado de febril Hipotálamo
Q= mCΔT Corporal
Termorregulación ATP

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
A continuación, se muestra el ejercicio desarrollado y resumido en un diagrama UVE

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
LECCIÓN 3. Energía libre metabólica (ΔG) y equilibrio
La ecuación básica es la siguiente:
ΔG =(ΔH) - T(ΔS)
Esta ecuación de gran aplicación en bioquímica, puesto que combina la primera y
segunda leyes de la termodinámica, proporcionando información valiosa
acerca de la espontaneidad, sentido y estado de equilibrio de una reacción
bioquímica.
3.1 Características de ΔG
a. La energía libre de Gibbs mide la energía necesaria que necesita un sistema
para realizar un trabajo útil.
b. La energía libre del Gibbs es la fuerza impulsora de las reacciones bioquímicas
enzimáticas(RBE)
c. Sí el cambio en la energía libre es negativo la RBE se produce
espontáneamente, por lo tanto, se dice que es exergónica, catabólica libera energía.
Debido a que el cambio entrópico, no es fácilmente medible en una reacción
bioquímica, el profesor Josiah.W. Gibbs , propone una función termodinámica de
estado, que relaciona el cambio de entalpía (ΔH) y el cambio entrópico del sistema
a presión y temperatura constantes, la cual se denomina Energía Libre de Gibbs (ΔG)

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
d. Sí el cambio en la energía libre es positivo la RBE no se produce
espontáneamente, por lo tanto, se dice que es endergónica, anabólica y requiere el
suministro de energía para que ocurra.
e. Sí el cambio en. la energía libre es cero (0), el sistema está en equilibrio, y
no se produce ninguna reacción(RBE)
De esto se puede concluir, que una reacción química RBE, se produce, siempre y cuando
disminuya la energía libre de Gibbs, es decir ΔG<0.
El mentefacto ,resume las bases conceptuales y características biotermodinámicas de la energía
libre de Gibbs.
Lección 4. El adenosín trifosfato (ATP),biomolécula energética
La biomolécula del El ATP es considerada como un nucleótido energético ,porque
contiene :una base nitrogenada que corresponde a la Adenina,un monosacárido pentosa

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
denominado:ribosa y un grupo trifosfato,el cual almacena la energía libre metabólica en
sus enlaces químicos fosfoanhídridos de éster,como lo muestra la figura 4.
Figura 4.Fórmula estructural del adenosín trifosfato (Lehninger.,2000)
Es así como el enlace del último radical fosfato contiene unas 7300 calorías por mol, lo
que significa que cuando éste se rompe por hidrólisis, se deben liberar 7300 calorías
por mol, es decir:
ATP + H2O <======> ADP + Fosfato + 7300 calorías
Esta energía química del ATP la utiliza la célula en su trabajo biológico, transformándola
en energía mecánica, eléctrica, térmica y por último en calor
La reacción anterior es reversible, lo cual indica, que para formar una molécula de ATP,
debe presentarse la reacción entre el ADP(Adenosín difosfato) y una molécula de
fosfato, con un suministro de 7300 calorías por mol, por lo tanto, este proceso consume
energía.
Es importante el análisis cuantitativo de esta energía producida en un organismo
animal, por eso, su valoración se fundamenta principalmente en el calor desprendido o

UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD
Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
liberado por éste, en una bomba calorimétrica o calorímetro de respiración, con este
aparato, se lleva cuenta del ingreso de alimentos, agua y oxígeno, de la excreción de
sólidos, líquidos, gases y la producción de calor.
Esta medición se denomina calorimetría directa y sirve para efectuar un balance
energético en la nutrición animal, es decir, el análisis de energía metabolizable,
energía neta y energía nutritiva total, esenciales en el estado nutricional del animal.
La energía utilizada, es extraída gradualmente de los alimentos (carbohidratos, lípidos y
proteínas) a través de tres etapas fundamentalmente oxidativas: Hidrólisis,
formación de Acetil CoA y ciclo de Krebbs con la fosforilación oxidativa, los
productos finales de estas etapas son el gas carbónico, agua y lo más importante:
ENERGÍA. La cual es almacenada en una molécula extraordinaria: El ATP

46
UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
Lección 5 Flujo y tipos de energía en los animales.
La fuente primaria de toda la energía utilizada por los animales, las plantas y todos
los organismos vivos del planeta está en la energía liberada por el sol. Esta es
captada por las plantas en el proceso de la fotosíntesis, con la formación de
cadenas carbonadas (glucosa, ácidos grasos, aminoácidos, etc.) que conservan
dicha energía y de donde posteriormente los animales la obtienen para suplir sus
necesidades.
Todas las complejas funciones del organismo animal son realizadas por medio de
la energía. Así, el trabajo celular, la biosíntesis, el trabajo osmótico, el trabajo
mecánico y demás funciones orgánicas, son posibles gracias a la energía, la cual
toma de los productos ingeridos, que al ser oxidados en el organismo animal,
liberan la energía contenida en ellos.
En los animales superiores la temperatura se mantiene constante a 37° Celsius,
esto hace que no se pueda utilizar el calor como fuente de energía para realizar el
trabajo. Sin embargo, los animales realizan trabajo; la razón de ello es que la
energía producida en las reacciones química a nivel celular es captada en forma
de energía química y utilizada posteriormente para el trabajo celular.
En este aspecto el ATP juega un importantísimo papel como transportador de toda
la energía química requerida en todas las reacciones del metabolismo. Su
formación, a partir del ADP y el fósforo inorgánico, está acoplada a la degradación
de las moléculas que actúan como combustibles y que liberan la energía requerida
para ello. Posteriormente el ATP libera su energía la cual es usada para todo el
trabajo celular. Tal es el ciclo que se establece entre las plantas y los animales y el
papel del ATP como intermediario en los intercambios de energía a nivel celular,
tanto en las plantas como en los animales.
El ciclo energético en su conjunto incluye los siguientes aspectos:

47
UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
1. La fotofosforilación. Es decir, la captación de la energía de las radiaciones
solares en los cloroplastos de las plantas verdes y su transformación en energía
química en forma de ATP.
2. La utilización de la energía química del ATP para la formación de sustancias
orgánicas tales como glúcidos, lípidos, aminoácidos, etc., por los vegetales, donde
a partir del CO2, y del H2O, se forman las cadenas carbonadas que serán
posteriormente utilizadas por los animales.
3. La respiración celular en las mitocondrias de las células de los animales, donde
estos productos son oxidados a CO2, y H2O liberando su energía que es utilizada
para la síntesis del ATP (fosforilación oxidativa).
4. La utilización de la energía del ATP formado para realizar todo el trabajo celular,
que incluye el trabajo químico o biosintético (síntesis de proteínas, glúcidos,
lípidos, y otras biomacromoléculas ).
5.1Tipos de energía en los animales.
A partir de los aspectos antes analizados, sobre todo el flujo de energía en la
naturaleza y el sistema representado por la cadena respiratoria y la fosfori lación
oxidativa, se comprende el papel de la energía dentro del metabolismo.
Es importante señalar que toda la energía requerida por un sistema metabólico
debe estar presente y por supuesto suministrada por el entorno. Es decir, los
animales requieren del suministro constante de energía la cual obtienen de los
productos alimenticios ingeridos y los vegetales del sol.
La energía contenida en los alimentos ingeridos recibe el nombre de energía bruta
(EB) y se obtiene por combustión completa del alimento en base a materia seca.
La energía bruta de un alimento está dada por la relación que contenga de
carbohidratos, proteínas, grasas y otros compuestos orgánicos. Un gramo de

48
UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
carbohidrato produce por combustión 4.10 kcal, un gramos de proteínas 5,65 kcal
y un gramo de grasas 9,45 kcal. Como es lógico el incremento de proteínas y
sobre todo de grasas en la composición del alimento aumenta el valor energético
de los mismos.
Estos conceptos son muy aplicados en nutrición animal para establecer diferentes
tipos de dietas.
Si a la energía bruta (EB) le descontamos la energía perdida por las heces (EF)
debido a los alimentos sin digerir, así como, a las secreciones del aparato
digestivo, restos celulares, microbios entéricos, etcétera, se obtiene la energía
digestible (ED). La energía digestible depende del coeficiente de digestibilidad de
la ele la dieta lo cual se debe fundamentalmente a la composición química su
solubilidad y posibilidades de hidrólisis, por las enzimas el aparato digestivo de
cada especie animal. La energía digestible varía mucho en dependencia del
alimento y es un concepto de mayor utilidad que el de energía bruta.
Si de la energía digestible descontamos la energía urinaria, debido a productos
absorbidos no oxidados y la energía perdida por los productos gaseosos de la
digestión, sobre todo del metano en los rumiantes, obtenemos la energía
metabolizable (EM). La energía metabolizable representa la suma de la energía
de todos los productos asimilados una vez descontadas las pérdidas anteriores.
Es un índice de gran valor pues a partir de ella, deducido la energía por el
incremento del calor, se obtiene la energía neta (EN) del metabolismo.
La energía neta responde a la energía usada directamente en las funciones
celulares tanto la de mantenimiento (EN de mantenimiento) es decir la energía del
metabolismo basal, actividad corporal, etc. Y la energía de producción (EN de
producción) referida a la energía para crecimiento, trabajo, producción de leche,
lana, reproducción, etc.

49
UNIVERSIDAD NACIONAL ABIERTA Y A DISTANCIA – UNAD Escuela de Ciencias Agrícolas,Pecuarias y del Medio Ambiente.,ECAPMA. Contenido didáctico del curso Bioquímica Metabólica Elaboró: Jairo Granados.,MSc.
Por supuesto todos estos conceptos tienen una utilidad práctica en los sistemas
de alimentación sobre todo el concepto de energía digestible. A nivel metabólico
como ya habíamos expresado se une el término de energía libre para ver la
tendencia de las reacciones. Se refiere a la energía capaz de realizar trabajo
biosintético, osmótico o mecánico y representada por las Kcal captadas en los
enlaces macroenergéticos del ATP a partir de la oxidación de un mol de glucosa,
de ácido graso o de aminoácido. Es decir son conceptos diferentes pues cuando
hablamos de energía bruta o energía metabolizable se refiere a energía de
combustión.
Por ejemplo:
1 gramo de glucosa produce 3,76 Kcal de EB
1 gramo de ácido palmítico produce 9.35 Kcal de EB
Mientras la energía química obtenida en forma de enlaces macroenergéticos del
ATP es:
1 gramo de glucosa: 1.48 Kcal
1 gramo de ácido palmítico 3,58 Kcal
Es decir hay una captación de un 39% para la glucosa de la energía total de estos
compuestos y de un 38% para el ácido palmítico.
Estos conceptos son muy útiles a la hora de entender el flujo energético en la
naturaleza pues permite comprender que en el paso de los compuestos por todos
los procesos metabólicos, por ejemplo de la glucosa al CO2 hay unas 21
reacciones, se va liberando energía en forma de calor e incrementando la
entropía, en definitiva