Embed Size (px)
Citation preview

UNIVERSIDADE ESTADUAL DE FEIRA DE SANTANA
PROGRAMA DE PÓS-GRADUAÇÃO EM BIOTECNOLOGIA
CAROLINA OLIVEIRA DE CERQUEIRA LIMA
MICROPROPAGAÇÃO E CRESCIMENTO IN VITRO DE
BROTOS DE Orthophytum mucugense WAND. E
CONCEIÇÃO, BROMELIACEAE ENDÊMICA DE
MUCUGÊ-BAHIA
Feira de Santana, BA
2008

ii
CAROLINA OLIVEIRA DE CERQUEIRA LIMA
MICROPROPAGAÇÃO E CRESCIMENTO IN VITRO DE
BROTOS DE Orthophytum mucugense WAND. E
CONCEIÇÃO, BROMELIACEAE ENDÊMICA DE
MUCUGÊ-BAHIA
Dissertação apresentada ao Programa de Pós-graduação
em Biotecnologia, da Universidade Estadual de Feira de
Santana como requisito parcial para obtenção do título de
Mestre em Biotecnologia.
Orientador: Prof. Dr. José Raniere Ferreira de Santana
Co-Orientadora: Dra. Moema Cortizo Bellintani
Feira de Santana, BA
2008

iii
Dedicatória
A minha família, que me acompanhou por
toda a minha vida e que amo acima de tudo.

iv
AGRADECIMENTOS
Ao Professor José Raniere Ferreira de Santana pela experiência, atenção
e orientação;
A Moema Bellintani, pela orientação, atenção, companheirismo, amizade
e carinho. Pelas noites doadas para a realização dessa dissertação;
A Alone Lima principalmente pela amizade e zelo, por todas as ajudas e
atenções que foram fundamentais para a finalização desse trabalho;
A professora Cláudia Helena (Claudinha) pelas orientações e ajuda nos
trabalhos de anatomia realizados nessa dissertação, mas acima de tudo pela
amizade;
Sheila pela atenção e amizade;
A meus Pais pelo carinho e amor mútuo, e pela compreensão das minhas
faltas por causa do trabalho; e principalmente ao meu irmão o qual esta sempre
ao meu lado, seja como biólogo ou simplesmente como irmão e que amo MUITO;
Aos meus irmãos de republica: Lio pelo companheirismo em todas as
horas e momentos da minha vida deste 2001; VV pela atenção; Deni pela
companhia nos finais de semana; Rafa por conseguir dividir o quarto comigo; Lo,
apesar da distância, pelo maior presente que iremos ganhar esse ano: a nossa
princesinha; Dan-Dan pela alegria, Mel pelo sorriso constante, Babi pelas risadas
e Yve pelo zelo;
As minhas amigas da antiga republica: Dani por fazer parte da minha vida
por um longo período, Cel pelo retorno as origens;
Aos meus amigos da turma Bio 2001.1 que mesmo após formatura
continuaram ao meu lado nesses dois anos de mestrado, e espero que pela vida
inteira: Juninho, Gegê, Lulu, Suiki, Dêa, Rose, Paulinho, Elívia, Livan, Geo, Guto,
Rodrigo.
A Carlinha pela amizade e companheirismo;
A Tai, Line e Pri amigas que amo muito;
A Cindy pela amizade e confidências;
Aos meus vizinhos Aigara e Cyrio, pela amizade e pela boa vizinhança, e
um obrigado em especial para Cyrio pela ajuda na hora das coletas;

v
Aos meus amigos do colegiado de Biologia: Giza, Jú, Pri, e
Principalmente Binho pela amizade, carinho e atenção;
Aos meus amigos da Pós: Irailde, Lidiane, Emília, Bruno, Sidnei, Helna,
Lucymeire e Elinalva.
Ao meu amigo Helton por toda atenção nesses dois anos, MUITO
obrigada;
Ao coordenador do curso, o professor Aristóteles Góes Neto pela
atenção;
Aos professores da UEFS que participaram da minha formação
acadêmica, seja na graduação ou na pós: Caio, Glória, Carlos, Dalva, Walter,
Nora, Marilene, Dêa, Ari, Ana Paula, Milton, Áurea, Celeste.
A meus amigos virtuais, que acompanharam todo o meu trabalho,
obrigada pela companhia. Principalmente Gui pela atenção e pelas brincadeiras;
Aos amigos da “ex” casa rosada que estão sempre presentes alegrando
os meus dias: Grandão, Paloma, Moço, Ale e Rodrigo
Aos amigos do Horto pela atenção ajuda: Jane, Cris, Paulinho (Peixinho),
Nando, Tina, Bruninho, Flavinha, Sandrinha, Tecla, Renata e Cimili.
Aos professores do Horto;
Ao projeto Sempre-Viva, pela atenção a auxilio nos períodos de coleta;
A FAPESB pela bolsa;
A todos muito obrigada.

vi
RESUMO
Orthophytum mucugense é uma bromélia endêmica da Chapada Diamantina, Bahia, Brasil, com grande potencial ornamental e relevante importância ecológica. É classificada como vulnerável em decorrência do endemismo restrito ao município de Mucugê, demandando portanto ações que resultem na conservação da espécie. O cultivo in vitro tem sido utilizado para fins comerciais e para preservação de espécies raras ou ameaçadas de extinção podendo se constituir numa alternativa para a espécie em estudo. O presente trabalho avaliou a multiplicação e o crescimento in vitro de O. mucugense em um período curto, com o intuito de otimizar protocolos de micropropagação para produção comercial e conseqüentemente reduzir o seu extrativismo. Para tanto, foram avaliados os efeitos de diferentes fatores que afetam a morfogênese in vitro: idade e tipos de explante; orientação do explante no meio de cultura e dominância apical; fontes de carbono; presença de carvão ativado associado a diferentes concentrações de AIB; tipos de selamento dos frascos de cultura e balanço hormonal. A maior taxa de multiplicação foi obtida com a utilização de explante caulinar com a excisão do ápice na ausência de regulador de crescimento quando se utilizou caules com 40 dias como fonte de explante (média de 2,25 brotos/explante). Para o crescimento in vitro a utilização do açúcar cristal como fonte de carbono diminui os custos do meio sem perder a qualidade dos brotos. A utilização de tampas permeáveis às trocas gasosas durante a fase de enraizamento in vitro permitiu a transferência direta dos brotos para o ambiente ex vitro.
Palavras-chave: Bromeliaceae. Plantas ornamentais. Tipos de explante. Fonte de carbono. Auxina. Citocinina. Selamento.

vii
ABSTRACT
Orthophytum mucugense is a endemic bromeliad to the Chapada Diamantina, Bahia State, Brazil. It has significant potential as an ornamental plant as well as ecological relevance, as it is an endemic species restricted to the municipality of Mucugê. As this species is classified as vulnerable to extinction, proactive measures should be taken to guarantee its conservation. In vitro culture is widely used both commercially and as a technique for preserving rare or threatened species, and represents a viable option for conserving this bromeliad. The present study evaluated the rapid in vitro growth and multiplication of O. mucugense with the intention of optimizing production protocols for micro-propagating these plants in order to reduce pressure from wild harvesting. To that end, different factors that affect in vitro morphogenesis were examined: age and types of explants; explant orientation in the culture medium and apical dominance; carbon sources; presence of activated charcoal associated with different concentrations of IBA; and the types of vessel seals and hormonal balance. A good multiplication rate was obtained only by the excision of the stem apex in the absence of growth regulator and using 40 day-old stems as the explant source (average of 2.25 shoots / explant). The use of crystallized sugar as a carbon source decreases the costs of the in vitro growth medium without compromising shoot quality. The use of permeable vessel caps that allow gas exchange during the in vitro rooting phase permited direct transfer of the shoots to the ex vitro environment. Key words – Bromeliaceae. Ornamental plant. Explants. Carbon sources. Auxin. Cytokinins.

viii
LISTA DE TABELAS
Tabela 1.1 – Percentual de explante responsivos, número de brotos por explante e comprimento dos brotos em Orthophytum mucugense obtidos a partir de plantas com diferentes idades, em meio suplementado com 2,22 µM de BAP associado a duas concentrações de ANA após 60 dias de cultivo. Feira de Santana, BA, 2007.
24
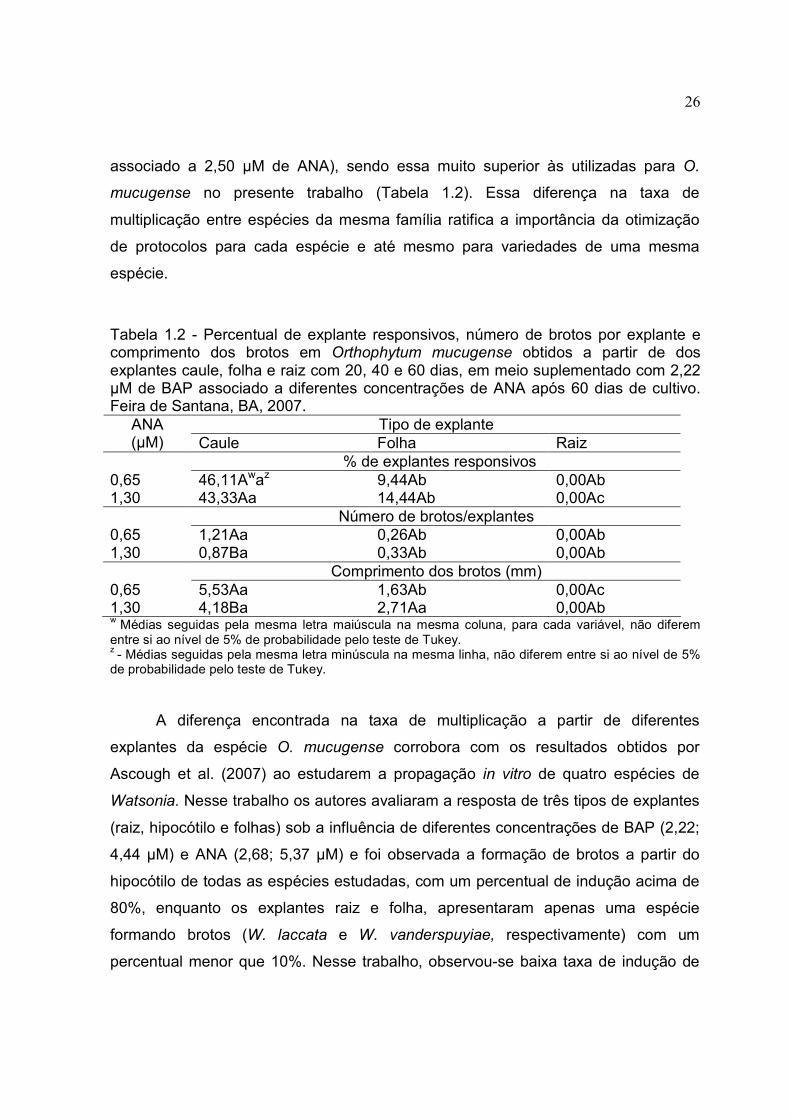
Tabela 1.2 - Percentual de explante responsivos, número de brotos por explante e comprimento dos brotos em Orthophytum mucugense obtidos a partir de dos explantes caule, folha e raiz com 20, 40 e 60 dias, em meio suplementado com 2,22 µM de BAP associado a diferentes concentrações de ANA após 60 dias de cultivo. Feira de Santana, BA, 2007.
26
Tabela 1.3 - Percentual de explante responsivos, número de brotos por explante e comprimento dos brotos em Orthophytum mucugense obtidos a partir do explante caule em diferentes idades de desenvolvimento em meio suplementado com 2,22 µM de BAP associado a duas concentrações de ANA após 60 dias de cultivo. Feira de Santana, BA, 2007.
28
Tabela 1.4 - Percentual de explante responsivos, número de brotos por explante e comprimento dos brotos em Orthophytum mucugense obtidos em função da posição do explante (caule com 40 dias) no meio de cultura MS½ suplementado com 2,22µM de BAP associado a 0,65 µM de ANA após 60 dias de cultivo. Feira de Santana, BA, 2007.
31
Tabela 1.5 - Número de brotos por explante obtidos em plantas de Orthophytum mucugense com e sem ápice caulinar cultivado em meio MS½ na presença ou ausência dos reguladores de crescimento ANA (0,65µM) e BAP (2,22µM ) após 60 dias de cultivo. Feira de Santana, BA, 2007.
33
Tabela 2.1 – Valores médios para comprimento da parte aérea (CPA) e das raízes (CR), e matéria seca das raízes (MSR) e parte aérea de brotos (MSPA) da espécie Orthophytum mucugense cultivados em meio MS ½ suplementado com diferentes fontes de carbono, após 60 dias de cultivo. Feira de Santana-BA, 2007.
46
Tabela 2.2 - Média do comprimento da parte aérea e das raízes e matéria seca da parte aérea e das raízes de brotos de Orthophytum mucugense desenvolvidos em meio MS ½ suplementado com carvão ativado e AIB, após 60 dias de cultivo. Feira de Santana, 2007.
49
Tabela 3.1 – Percentual de explante responsivos, número de brotos por explante, comprimento dos brotos, taxa de sobrevivência e emissão de raiz Orthophytum mucugense a partir do explante caule em diferentes concentrações e tipos de citocininas, após 60 dias de cultivo. Feira de Santana, BA, 2007.
64

ix
Tabela 3.2 - Percentual de explante responsivos, número de brotos por explante, comprimento dos brotos, taxa de sobrevivência e emissão de raiz em Orthophytum mucugense a partir do explante caule em diferentes tipos de citocininas associado a dois tipos de selamentos, após 60 dias de cultivo. Feira de Santana, BA, 2007.
67
Tabela 3.3 Média do comprimento da parte aérea e das raízes e matéria seca da parte aérea e das raízes de brotos de Orthophytum mucugense desenvolvidos em meio MS ½ suplementado com carvão ativado e AIB, após 60 dias de cultivo. Feira de Santana, 2007.
69

x
LISTA DE FIGURAS
Figura 1. Orthophytum mucugense em seu habitat natural, Mucugê-BA, (a) espécie florida; (b) populações em paredões rochosos. Feira de Santana, 2007.
4
Figura 1.1 – Brotações obtidas via organogênese direta em plantas de Orthophytum mucugense em explante com diferentes idades e submetidos a duas concentrações de ANA, após 60 dias de cultivo. A seta mostra a origem dos brotos. A barra representa 0,5 cm. Feira de Santana, 2007.
23
Figura 1.2 - Brotos de Orthophytum mucugense micropropagados a partir dos explantes caule e folha, após 60 dias de cultivo. A barra representa 0,5 cm. Feira de Santana, 2007.
25
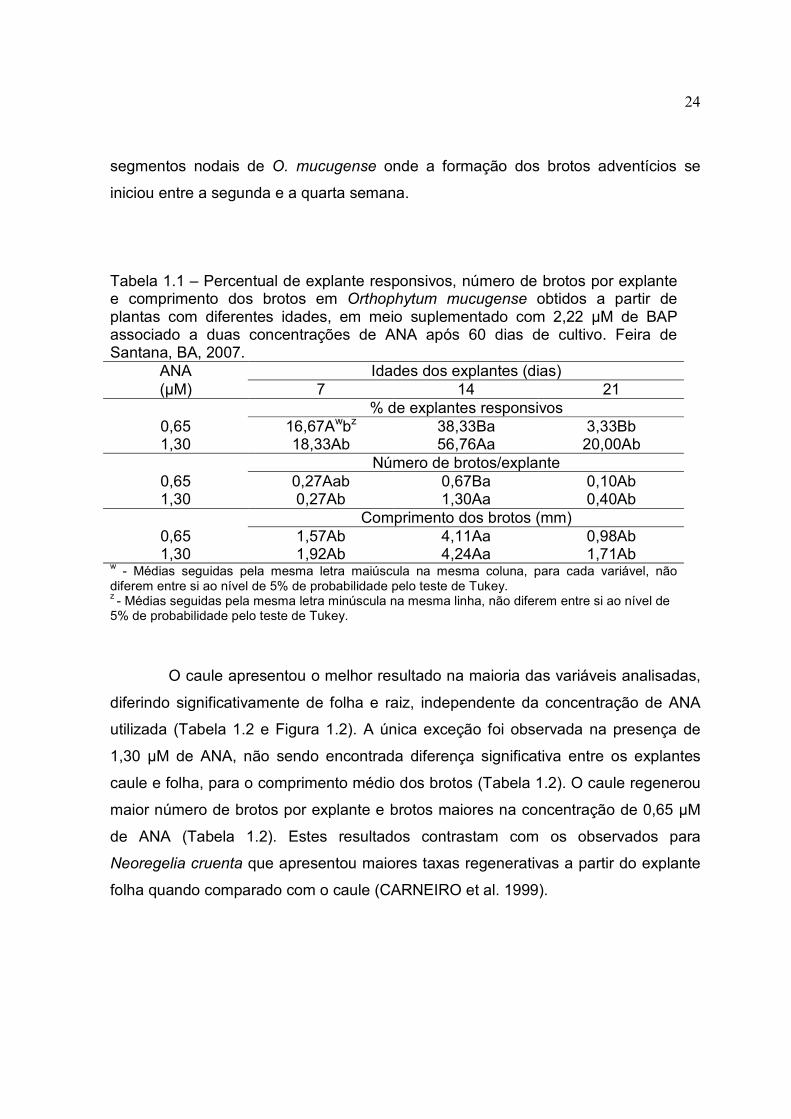
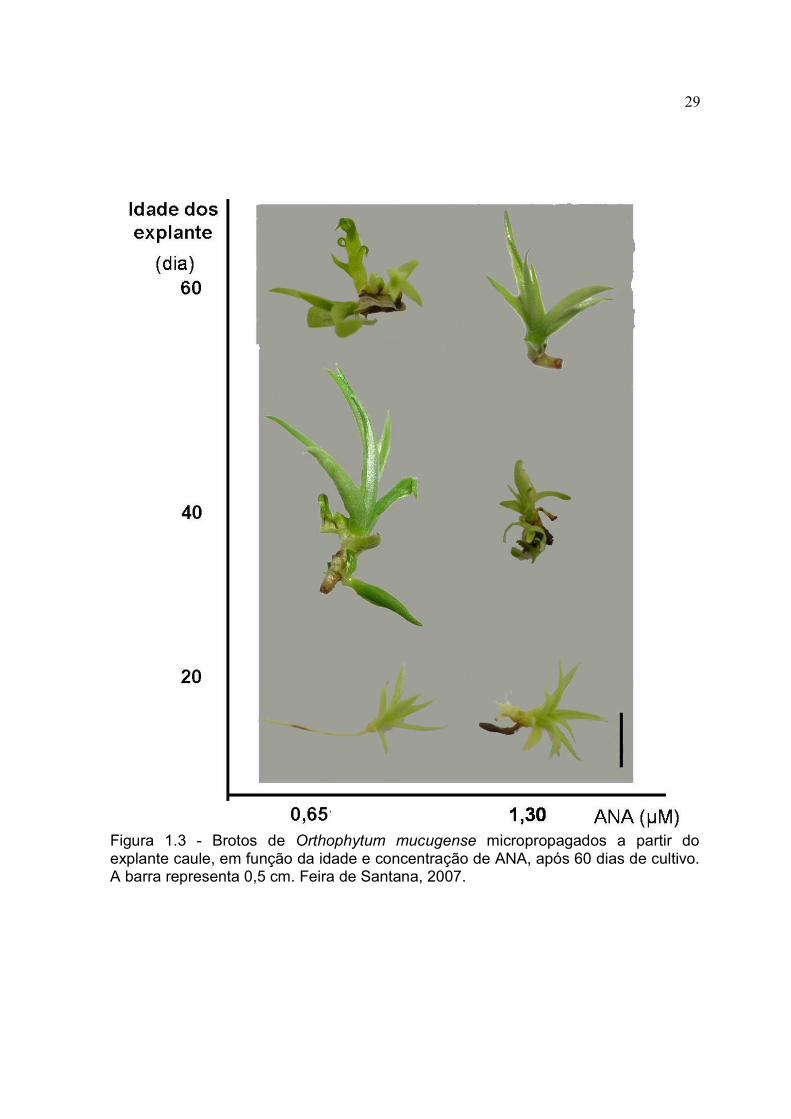
Figura 1.3 - Brotos de Orthophytum mucugense micropropagados a partir do explante caule, em função da idade e concentração de ANA, após 60 dias de cultivo. A barra representa 0,5 cm. Feira de Santana, 2007.
29
Figura 1.4 - Brotos de Orthophytum mucugense, micropropagados a partir do explante caule com 40 dias, em função da orientação do explante no meio de cultura, após 60 dias de cultivo. A barra representa 0,5 cm. Feira de Santana, 2007.
32
Figura 1.5 – Anatomia de Orthophytum mucugense micropropagados a partir dos explantes plantas (d), caule (a-c) e folha(e-f), após 60 dias de cultivo. (a) corte transversal do caule mostrando o inicio da diferenciação a partir do cambio vascular; (b) zona meristematica em um estagio mais avançado de desenvolvimento, (c) o inicio da vascularização do novo broto (d) corte longitudinal da planta mostrando a ligação de dois brotos com a planta a partir dos vasos condutores; (e) corte longitudinal da lamina foliar mostrando nova zona meristematica e a conexão dos vasos condutores; (f) zona meristematica em destaque. (cv = cambio vascular); (m = zona meristematica); (v = vaso condutor); (LF = lâmina foliar); a barra representa 5µm, Feira de Santana, 2007.
34
Figura 2.1. Brotos da espécie Orthophytum mucugense procedentes de meio de cultura MS ½ suplementado com diferentes fontes de carbono (87,64 mM), após 60 dias de cultivo. A barra representa 1,0 cm. Feira de santana, 2007.
47
Figura 3.1 Brotos de Orthophytum mucugense micropropagados a partir do explante caule, em função de diferentes concentrações de CIN, BAP e TDZ sob a influência de dois tipos de selamento, após 60 dias de cultivo. A barra representa 0,5 cm. Feira de Santana, 2007.
68

xi
Figura 3.2 Taxa de sobrevivência de plantas micropropagadas de Orthophytum mucugense em diferentes períodos de aclimatização, após 30 dias. Feira de Santana, 2007.
70
Figura 3.3 Aclimatização de plantas micropropagadas de Orthophytum mucugense em diferentes períodos de adaptação. (A e D) plantas transferidas diretamente após 5 e 10 dias de cultivo respectivamente; (B e E) plantas com cinco dias com garrafa PET após 5 e 10 dias de cultivo respectivamente; (C e F) plantas com dez dias com garrafa PET após 5 e 10 dias de cultivo. Feira de Santana, 2007.
72
Figura 3.4 Anatomia foliar de brotos micropropagados de Orthophytum mucugense sobre a influência de diferentes tipos e concentrações de citocininas e TDZ, sobre a influência de dois selamentos, após 60 dias de cultivo. (A) epiderme adaxial; (B) folhas com ausência de lignina; (C) folhas cultivadas em tubos fechados com algodão com presença de lignina; (D) epiderme adaxial com células epidérmicas desorganizadas; (E) folha sem diferenciação de Parênquima paliçádico e lacunoso; (F) presença de estômato na epiderme adaxial; (G) presença de lignina na epiderme adaxial; (H) presença de lignina na epiderme adaxial; (I) presença de parede espessada entre as células da epiderme; (J) extremidade da folha; (L) epiderme abaxial; (M) folha com diferenciação de Parênquima paliçádico e lacunoso. (seta = parede celular); (F.V = feixe vascular); (est. = estômato); (P.P = parênquima paliçádico); (P.L= parênquima lacunoso). A barra representa 5 µm. Feira de Santana, 2007.
74

xii
SUMÁRIO
INTRODUÇÃO GERAL 1
REFERÊNCIAS 11
CAPÍTULO 1 Otimização do protocolo de multiplicação in vitro de Orthophytum mucugense WAND e CONCEIÇÃO.
15
RESUMO 15
ABSTRACT 15
1.1 INTRODUÇÃO 16
1.2 MATERIAL E MÉTODOS 18
1.2.1 MATERIAL VEGETAL, ASSEPSIA E GERMINAÇÃO 18
1.2.2 CONDIÇÕES DE CULTIVO, pH E ESTERILIZAÇÃO DO MEIO DE CULTURA
18
1.2.3 MULTIPLICAÇÃO 19
1.2.3.1 Idade e tipo do explante em duas concentração de ANA 19
1.2.3.2 Posição do explante 20
1.2.3.3 Remoção do ápice caulinar 20
1.2.4 ANÁLISE HISTOLÓGICA 21
1.2.5 ANÁLISE ESTATÍSTICA 21
1.3 RESULTADOS E DISCUSSÃO 22
REFERÊNCIAS 36

xiii
CAPÍTULO 2 Crescimento in vitro de brotos de Orthophytum mucugense WAND e CONCEIÇÃO obtidos por organogênese direta.
39
RESUMO 39
ABSTRACT 39
2.1 INTRODUÇÃO 40
2.2 MATERIAL E MÉTODOS 42
2.2.1 MATERIAL VEGETAL 42
2.2.2 CRESCIMENTO IN VITRO 42
2.2.2.1 Fonte de carbono 42
2.2.2.2 AIB e carvão ativado 43
2.2.3 CONDIÇÃO DE CULTIVO IN VITRO 43
2.2.4 ANÁLISE ESTATÍSTICA 44
2.3 RESULTADOS E DISCUSSÃO 45
REFERÊNCIAS 50
CAPÍTULO 3 Impacto do selamento dos recipientes na micropropagação de Orthophytum mucugense WAND e CONCEIÇÃO sob a influência de diferentes citocininas.
54
RESUMO 54
ABSTRACT 54
3.1 INTRODUÇÃO 56
3.2 MATERIAL E MÉTODOS 59
3.2.1 MATERIAL VEGETAL 59
3.2.2 MULTIPLICAÇÃO 59
3.2.3 ENRAIZAMENTO 60
3.2.4 ACLIMATIZAÇÃO 60

xiv
3.2.5 ANÁLISE HISTOLÓGICA 61
3.2.6 CONDIÇÃO DE CULTIVO IN VITRO 61
3.2.7 ANÁLISE ESTATÍSTICA 62
3.3 RESULTADO E DISCUSSÃO 63
REFERÊNCIAS 76
CONCLUSÃO GERAL 80
APÊNDICES 81

1
INTRODUÇÃO GERAL
A produção de plantas ornamentais é uma atividade agrícola exigente em
relação à qualidade do produto, principalmente se este for para exportação. Nesse
grupo de plantas a aplicação da micropropagação teve uma expressão significativa
no mundo científico, com repercussão direta na economia, devido ao alto valor
agregado ao produto final (TOMBOLATO e COSTA, 1998; BOSA et al., 2003). Nos
laboratórios de pesquisa do País que desenvolvem e adaptam protocolos para
inúmeras espécies ornamentais, a maior freqüência de trabalho são com espécies
das famílias: Araceae, Asteraceae, Bromeliaceae, Cariofilaceae e Zingiberales
(TOMBOLATO e COSTA, 1998).
As bromélias há décadas são apreciadas como plantas ornamentais,
especialmente nos EUA, na Europa e na Austrália, onde seu cultivo movimenta uma
economia considerável, absorvendo, direta ou indiretamente, grande contingente de
mão-de-obra. O Brasil, detentor da maior diversidade de espécies dessa família, só
recentemente começou a desenvolver uma produção comercial expressiva (PAULA,
2004). Sendo essa produção de grande importância, pois grande parte das plantas
utilizadas para ornamentação são obtidas de forma extrativista, o que vem
acarretando uma perda genética significativa, não apenas da espécie a ser
comercializada, como também da flora e fauna associada, pois as Bromélias têm
grande importância na manutenção da biodiversidade, fornecendo água e alimento
além de ser utilizada como abrigo e local de acasalamento para inúmeras espécies
animais (ROCHA et al., 2004).
Além da utilização como plantas ornamentais, espécies da família
Bromeliaceae são utilizadas como fonte de fibras e na alimentação por populações
nativas das três Américas a séculos. O primeiro registro de bromélia da nossa
civilização data de 1493, na segunda viagem de Cristóvão Colombo à América. De
acordo com documentos, os nativos da ilha de Guadalupe, nas Antilhas, utilizavam
uma planta muito saborosa como alimento, denominada por eles “karatas” – hoje o

2
tão comum abacaxi (Ananas comosus). Desde então o abacaxi foi levado para a
Europa e disseminado por todo o mundo (PAULA, 2004).
A família Bromeliaceae esta dividida em três subfamílias: Pitcairnioideae
Harms, Bromelioideae Reichenbach e Tillandsioideae Harms. O número de gêneros
diverge entre os autores. Smith e Downs (1983) na Flora Neotrópica apresentam 46
gêneros. Entretanto a descrição de novas espécies e o crescente estudo da família,
tem ampliado este número para 51 (LEME e MARIGO, 1993), 50 (BRUMMITT, 1992)
e 56 (LEME, 1997).
Schultz (1985) descreve as bromélias como uma grande família de plantas
herbáceas, terrestres ou epifíticas, com caule reduzido. As folhas lanceoladas se
inserem em espiral no caule ou formam uma roseta basal capaz de acumular grande
quantidade de água. Característica que proporciona a essa família uma grande
importância na manutenção da biodiversidade animal e vegetal, a partir do
fornecimento de água e alimento, além de ser utilizada como abrigo e local de
acasalamento para muitas espécies (ROCHA et al., 2004).
Com cerca de 3.000 espécies, Bromeliaceae é considerada a maior família
de plantas essencialmente americana, distribuindo-se do sul dos Estados Unidos até
o Chile. A única exceção é Pitcairnia feliciana (A. Chev.) Harms J. Mildbraed,
encontrada no Golfo da Guiné, África (LEME e MARIGO, 1993).
Dados da biogeografia da família sugerem que as primeiras espécies devem
ter se originado e iniciado a sua diversificação numa época bastante próxima à
separação dos continentes que originou o Oceano Atlântico, há cerca de 200 milhões
de anos (LEME e MARIGO, 1993). São conhecidos três grandes centros de origem e
dispersão de espécies: os Andes, o planalto das Guianas e o leste brasileiro (SMITH,
1934).
As bromélias podem ocorrer em locais diversificados desde o nível do mar,
até em altitudes acima de 3.000m, como na região andina. São encontradas tanto
em regiões secas e quentes, como o sertão nordestino, quanto em regiões frias,
como os Andes e podem crescer em locais muito ensolarados, como em rochas à
beira-mar (PAULA, 2004). No Brasil, encontram-se quase metade das espécies de
bromélias conhecidas, distribuem-se de norte a sul, especialmente na região

3
costeira, na Mata Atlântica e na restinga, podendo aparecer também na Amazônia,
na Caatinga, nos Cerrados e nos campos de altitude, em todos os ecossistemas,
(LEME e MARIGO, 1993; PAULA, 2004). O longo período de estabilidade geológica
ocorrido na região que envolve o território brasileiro proporcionou a sobrevivência e
desenvolvimento de grupos que hoje ocorrem na forma de vários gêneros endêmicos
tais como Canistrum, Cryptanthus, Encholirium, Nidularium, Orthophytum, Quesnelia
e Wittrockia (LEME e MARIGO, 1993; LEME, 1998).
Orthophytum Beer é um gênero composto por espécies terrestres ou
saxícolas. As folhas serreadas são completamente recobertas por escamas, que
refletem parte da luz solar que incide nas plantas. Esse gênero pode ser dividido em
dois grupos: o primeiro reúne maior número de espécies, estas apresentam
inflorescência provida de escapo e folhas verdes; as espécies do outro grupo
apresentam inflorescência séssil e durante a floração, tendem a apresentar folhas
coloridas na parte central da roseta foliar (SMITH e DOWNS, 1983; WANDERLEY,
1990).
Segundo Wanderley e Conceição (2006) as espécies de Orthophytum com
inflorescência séssil, ocorrentes na Chapada Diamantina têm grande valor
ornamental devido à intensa coloração vermelha que suas brácteas e folhas atingem
na antese.
Entre essas espécies ocorrentes na Chapada Diamantina encontramos a
Orthophytum mucugense, uma espécie endêmica de Mucugê-BA. Destaca-se dentro
do grupo de espécies de inflorescência séssil pelas rosetas pequenas, folhas
estreitas, atingindo aproximadamente 4mm de largura, e presença de estolões. A
roseta foliar em O. mucugense é típica para o gênero (ortho= reto; phytum= folha),
ou seja, com presença de folhas geralmente patentes, formando um ângulo reto com
o eixo da planta. As folhas são verdes e, na época de floração, tornam-se parcial ou
completamente vermelhas, conferindo notável valor ornamental a esta espécie
(Figura 1- a) (WANDERLEY e CONCENIÇÃO, 2006).

4
Figura 1. Orthophytum mucugense em seu habitat natural no município de Mucugê-BA, (a) espécie florida; (b) populações em paredões rochosos. Feira de Santana, 2007.

5
Ocorre em paredões rochosos verticais, em geral com mais de 30m de
altura, em beira de rios e cachoeiras, constituindo locais sombreados e úmidos,
muitas vezes em pontos onde há drenagem de água vinda da serra (Figura 1-b). O
substrato é raso e formado principalmente por emaranhado de raízes (WANDERLEY
e CONCEIÇÃO, 2006).
Apesar de formar densas populações e ocorrer em uma Unidade de
Conservação, O. mucugense pode ser classificada como vulnerável em decorrência
da coleta predatória, e do endemismo restrito ao município de Mucugê
(WANDERLEY e CONCEIÇÃO, 2006). Ressaltando a importância de trabalhos de
conservação que protejam esta espécie e que permitam a produção em escala
comercial, reduzindo o extrativismo.
A utilização de técnicas biotecnologias, como a cultura de tecidos, permitem
a propagação rápida e em massa de genótipos superiores estáveis, isentos de
doenças, produzindo mudas de alta qualidade, com características homogêneas, em
espaço e tempo reduzidos, em qualquer época do ano (BOSA et al., 2003;
CARNEIRO e MANSUR, 2004; DROSTE et al., 2005).
O sucesso da cultura in vitro depende da produção de protocolos eficientes,
pois a variabilidade da resposta morfogenética existe não apenas entre espécies do
mesmo gênero, mas também entre genótipos da mesma espécie (GRATTAPAGLIA e
MACHADO, 1998). A micropropagação ou propagação vegetativa in vitro tem
viabilizado a clonagem de várias espécies, a partir de células, órgãos ou pequenos
fragmentos de uma planta matriz (SOUZA et al., 2006).
A atividade comercial da micropropagação, hoje, concentra-se
principalmente na limpeza clonal e na multiplicação de espécies ornamentais
herbáceas e arbustivas e é realizada em diversos países do mundo com destaque
para os países da Europa Ocidental e para os Estados Unidos. No Brasil, onde a
aplicação comercial da micropropagação é relativamente recente, existem diversos
grupos trabalhando em instituições públicas de pesquisa e universidades, entretanto,
poucas são as empresas que atuam na área (GRATTAPAGLIA e MACHADO, 1998).
Com o domínio da técnica de micropropagação é possível utilizá-la para
acelerar o desenvolvimento de plantas que possuem crescimento lento, como

6
algumas espécies de bromélias. A micropropagação pode ser obtida por meio da
proliferação de gemas axilares, multiplicação mediante indução de gemas
adventícias por organogênese direta ou indireta ou via embriogênese somática
(GRATTAPAGLIA e MACHADO, 1998).
A organogênese normalmente envolve a regeneração de gemas a partir de
grupos de células meristemáticas. Processo semelhante ao observado na
embriogênese. Logo, em alguns casos, é difícil saber por qual via ocorreu a
regeneração. O estudo da anatomia auxilia na determinação da via de regeneração,
pois embriões somáticos possuem sistema vascular fechado, sem conexão com o
sistema vascular do explante inicial, e a estrutura formada é bipolar, com a presença
do eixo caulinar e radicular (PERES, 2002).
A opção pelo método e via a ser utilizada depende do produto que se
deseja. Se a pretensão for obter mudas com integridade clonal, com o intuito de
conservação, a multiplicação a partir de gemas axilares é desejável, desde que a
formação de calos seja mínima ou nula (GRATTAPAGLIA e MACHADO, 1998).
A organogênese direta é um dos processos que permite a multiplicação em
larga escala mantendo a identidade genética do individuo propagado, através do
surgimento direto de gemas a partir de tecidos que apresentem potencial
morfogenético sem a formação de calo (GRATTAPAGLIA e MACHADO, 1998).
Devido à complexidade existente na organogênese direta e à resposta
específica de cada genótipo à regeneração, é necessário a avaliação dos fatores que
afetam a morfogênese em cada espécie, tais como o explante, a composição do
meio de cultura e as condições de cultivo (COSTA et al., 2006).
A diferenciação do tecido utilizado como explante tem implicação não
apenas na fidelidade genética, mas também na dificuldade de se obter a
regeneração de plantas. De forma geral, explantes oriundos de tecidos jovens e com
maior atividade metabólica são os mais competentes e possuem maior
potencialidade morfogenética (PINTO et al., 1994). Ao se utilizar explantes de folhas,
partes reprodutivas ou outros tecidos num estádio avançado de diferenciação há, em
geral, a necessidade de se induzir uma volta ao estado meristemático para, em
seguida, iniciar o processo de multiplicação. A desdiferenciação e indução de

7
regeneração de plantas a partir de calos são, muitas vezes, processos difíceis e
demandam tempo até a obtenção de um protocolo eficiente, além de ocasionar o
aparecimento de variação somaclonal (GRATTAPAGLIA e MACHADO, 1998).
De acordo com Grattapaglia e Machado (1998) um dos mecanismo de
controle da resposta dos explantes à morfogênese está no tipo e concentração dos
reguladores de crescimento utilizados no meio de cultura. Assim o sucesso da
micropropagação em qualquer espécie depende também da sensibilidade do tecido
ao regulador de crescimento.
O efeito de cada regulador no processo de crescimento e desenvolvimento
da planta depende: da espécie utilizada, da parte da planta em que é aplicado, do
estágio de desenvolvimento da planta e das condições fisiológicas da mesma, do
estágio de desenvolvimento do tecido, da concentração hormonal utilizada e da
interação entre os hormônios; está também relacionada ao número de receptores,
afinidade do regulador com o receptor, natureza da transdução, estrutura química do
regulador e aos vários fatores ambientais (CID, 2000; TAGLIACOZZO, 1998).
Para ocorrer a resposta da planta ao regulador três fatores são
fundamentais: o regulador deve estar presente na célula correta, deve reconhecer e
se ligar ao grupo de células que respondem a ele e a ligação hormônio-receptor
protéico deve desencadear mudanças metabólicas que amplificam o sinal do
hormônio. Portanto, diferentes partes das plantas nas várias espécies podem
responder por caminhos diversos a um determinado hormônio. A concentração e o
tipo de regulador ideal dependem do tecido utilizado como explante e da espécie
(TAGLIACOZZO, 1998).
Para o sucesso da produção in vitro há a necessidade da transferência dos
brotos micropropagados para o ambiente ex vitro. Porém, para que isso ocorra esses
brotos tem que se desenvolver primeiro in vitro, passando por processo de
crescimento e enraizamento, e depois pelo processo de aclimatização.
Em ambiente in vitro é necessário o desenvolvimento de meios que auxiliem
no crescimento e enraizamento desses brotos. O crescimento da planta é um
processo complexo e dinâmico que está submetido a dois tipos de controle: genético

8
e ambiental. Ambos permitem que as plantas respondam e se adaptem às condições
externas dentro de múltiplas interações (CID, 2000).
As células, tecidos e plântulas cultivadas in vitro não encontram condições
adequadas de iluminação e concentração de gás carbônico e, às vezes, não
apresentam teores de clorofila suficientes para realizar fotossíntese que sustente o
seu crescimento (CALDAS et al., 1998). Plantas cultivadas in vitro necessitam
portanto, de energia independente do processo fotossintético. Com este intuito são
utilizadas fontes exógenas de carbono, que influenciam na fisiologia da planta, na
diferenciação celular, no crescimento heterotrófico do vegetal e na emissão de raízes
(CALVETE et al., 2002; KOZAI, 1991).
O enraizamento in vitro normalmente é obtido com a utilização de auxinas,
sendo o ácido indolbutírico (AIB) a mais utilizada atualmente com esse objetivo
(GRATTAPAGLIA e MACHADO, 1998).
O enraizamento também tem sido favorecido pelo carvão ativado. Esta
substância escurece o meio de cultura impedindo a incidência de luz na base do
explante, reduz a oxidação e adsorve compostos tóxicos presentes no ágar ou
formados durante a oxidação dos compostos fenólicos (GRATTAPAGLIA e
MACHADO, 1998; PAN e STADEN, 1998).
Mesmo com um bom estabelecimento e desenvolvimento dos brotos in vitro,
esses podem apresentar problemas na adaptação ao ambiente ex vitro. Além do
desenvolvimento dos brotos formados, um aspecto importante a ser considerado na
cultura de tecidos é o microclima dentro dos frascos da cultura. Alterações no
ambiente in vitro, possibilitando o menor acúmulo de gases como etileno, e
principalmente a redução da umidade presente no recipiente utilizado para o cultivo
in vitro permitem uma melhor adaptação da planta ao ambiente ex vitro
(BUDDENDORF-JOOTEN e WOLTERING, 1996; DE PROFT et al., 1985; ZOBAYED
et al., 1999).
O que parece ser um ambiente bastante homogêneo tem mostrado ser, na
verdade, o responsável pela variabilidade no comportamento das culturas, que pode
comprometer todo o sistema de micropropagação. Os principais fatores que
determinam a qualidade do microambiente no frasco de cultura são os tipos de

9
tampas e frascos utilizados e a quantidade de meio presente no frasco
(GRATTAPAGLIA e MACHADO, 1998).
Uma vedação total do frasco, além de causar o acumulo de gases liberados
pela cultura, leva à saturação de vapor de água, o que diminui o fluxo transpiratório
nas culturas, podendo causar deficiências minerais de elementos como o cálcio,
ocasionando a necrose dos ápices (GRATTAPAGLIA e MACHADO, 1998). A
utilização de uma tampa que permita trocas gasosas possibilita um melhor controle
dos brotos ao serem transferidos para o ambiente ex vitro. Processo esse
denominado de aclimatização que finaliza o protocolo da micropropagação.
Com o domínio dos fatores que influenciam a micropropagação é possível o
estabelecimento de protocolos que proporcionam a produção em larga escala de
espécies de interesse comercial, além de reduzir problemas ambientais como o
extrativismo (FIALHO, 1990; FIALHO e FURTADO, 1993; LEME e MARIGO, 1993;
OLIVEIRA et al., 1994; OLIVEIRA e ROCHA, 1997). Com essa finalidade métodos de
cultura in vitro têm sido aplicado com sucesso em genótipos específicos de bromélias
(ARRABAL et al., 2002; BELLINTANI, 2006; CARNEIRO et al., 1998, 1999; DROSTE
et al., 2005; MERCIER e KERBAUY, 1997; MERCIER e NIEVOLA, 2003; RECH
FILHO et al., 2005; RODRIGUES et al., 2004).
Trabalhos com micropropagação de O. mucugense e de crescimento in vitro
dessa espécie, indicam que a cultura de tecidos é uma ferramenta viável para a
produção de mudas dessa espécie. De acordo com Bellintani et al. (2006, a) as
concentrações de 0,65 ou 1,30 µM de ANA com 2,22 µM de BAP apresentaram as
maiores médias de brotos por explante. Porem os autores realizaram o experimento
por um período de seis meses, e a partir do quarto mês ocorreu o aparecimento de
calos, o que pode ocasionar o surgimento de variantes somaclonais, que são
indesejáveis quando se deseja produzir protocolos com intuído de conservação.
Necessitando portanto de trabalhos que viabilize uma boa taxa de multiplicação, sem
o aparecimento de calos.
Em relação ao crescimento in vitro dessa espécie, Bellintani et al. (2007)
realizaram experimentos onde foi observado o maior crescimento, em relação ao
comprimento da parte aérea, em plantas germinadas in vitro na presença de 2,0

10
mg.L-1 de giberelina. Em estudos de aclimatização Bellintani et al (2006,b) obtiveram
uma ótima taxa de sobrevivência (90%) ao utilizarem tampas de algodão nos frascos
de cultura no período de enraizamento. Todos os trabalhos realizados de
micropropagação dessa espécie, mostram a produção em um período de cerca de
dez meses. A produção de protocolos que acelerem essa produção proporcionaria
um atrativo para a produção dessa espécies, evitando o extrativismo.
Sendo a espécie em estudo uma bromélia ameaçada de extinção, e os
trabalhos de micropropagação realizados até o momento ocorreram o aparecimento
de calos, o presente estudo avaliou a multiplicação e o crescimento in vitro de
Orthophytum mucugense em um período de seis meses, da germinação até a
aclimatização, com o intuito de acelerar o protocolo de produção de mudas
micropropagadas, reduzindo o extrativismo.

11
REFERÊNCIA
ARRABAL, R.; AMANCIO, F.; CARNEIRO, L.A.; NEVES, L.J.; MANSUR, E. Micropropagation of endangered endemic Brazilian bromeliad Cryptanthus sinuosus (L.B. Smith) for in vitro preservation. Biodiversity and Conservation, Netherlands, v. 11, p.1081-1089, 2002.
BELLINTANI, Moema Cortizo; LIMA, Carolina Oliveira de Cerqueira; BRITO, Alone Lima; SANTANA, José Raniere Ferreira de; DORNELLES, Ana Lúcia Cunha. Estabelecimento in vitro de Orthophytum mucugense e Neoregelia mucugensis, bromélias endêmicas da Chapada Diamantina, Bahia - Brasil.. In: LVII Congresso Nacional de Botânica, 2006, Gramado. Revista Brasileira de Biociências. Porto Alegre : Universidade Federal do Rio Grande do Sul, 2006 (a).
BELLINTANI, M. C. ; LIMA, Carolina Oliveira de Cerqueira ; BRITO, Alone Lima ; SANTANA, José Raniere Ferreira de ; DORNELLES, Ana Lúcia Cunha . Efeito da Ventilação in vitro na Aclimatização de Plantas Micropropagadas de Orthophytum mucugense Wand e Conceição.. In: LVII Congresso Nacional de Botânica, 2006, Gramado. Revista Brasileira de Biociências. Porto Alegre : Universidade Federal do Rio Grande do Sul, 2006 (b).
BELLINTANI, Moema Cortizo; LIMA, Carolina Oliveira de Cerqueira; SANTANA, José Raniere Ferreira de. Avaliação do crescimento in vitro de Orthophytum mucugense na presença de diferentes concentrações de giberelina.. In: 16º Congresso Brasileiro de Floricultura e Plantas Ornamentais e 3º Congresso Brasileiro de Cultura de Tecidos vegetais, 2007, Goiânia GO. Revista Brasileira de Horticultura Ornamental, São Paulo : Sociedade Brasileira de Floricultura e Plantas Ornamentais, v. 13. p. 792-795, 2007.
BOSA, Nair; CALVETE, Eunice O.; NIENOW, Alexandre Augusto; SUZIN, Marilei. Enraizamento e aclimatização de plantas micropropagadas de gipsofila. Horticultura Brasileira, Brasília, v. 21, n. 2, p.207-210, abril-junho 2003.
BRUMMITT, R.K. Vascular Plant Families and Genera. Royal Botanic Gardens, Kew, 804p., 1992.
BUDDENDORF-JOOSTEN, J.M.C.; WOLTERING, E.J. Controlling the gaseous composition in vitro growth of potato plantlets. Scientia Horticulturae, Amsterdam, v.65, p.11-23, 1996.
CALDAS, L. S.; HARIDASAM, P.; FERREIRA, M. E. Meios nutritivos. In: TORRES, A. C.; CALDAS, L. S.; BUSO, J. A. Cultura de tecidos e transformação genética de plantas. Brasília: Embrapa/SPI, v.1, p. 87-132, 1998.

12
CALVETE, Eunice Oliveira; KÄMPF , Atelene Normann; SUZIN Marilei. Concentração de sacarose no enraizamento in vitro de morangueiro. Horticultura Brasileira, Brasília, v. 20, n. 2, p. 186-191, junho 2002
CARNEIRO, L. A.; CÂNDIDO, M. S. D.; ARAUJO, R.F.G.; FONSECA, M.H.P.B.; CROCOMO, O.J.; MANSUR, E. Clonal propagation of Cryptanthus sinuosus L.B. Smith, an endemic stoloniferous Bromeliaceae species from Rio de Janeiro, Brazil. Plant Tissue Culture and Biotechnology, Bangladesh, v. 4, n.3-4, p. 152-158 nov. 1998.
CARNEIRO, L.A.; ARÁUJO, R.F.G.; BRITO, G.J.M.; FONSECA, M.H.P.B.; COSTA, A.; CROCOMO, O.J.; MANSUR, E. In vitro regeneration from leaf explants of Neoregelia cruenta (R. Graham) L.B. Smith, an endemic bromeliad from Eastern Brazil. Plant Cell, Tissue and Organ Culture, Netherlands v. 55, p. 79–83, 1999.
CARNEIRO, L. A.; MANSUR, E. Contribuição de metodologias in vitro para a conservação de Bromeliaceae. Vidalia, Lavras, v.2, n.1 p. 12- 20, 2004.
CID, Luis Pedro Barrueto. Introdução aos hormônios vegetais, Embrapa Recursos Genéticos e Biotecnologia, Brasília, 2000.
COSTA, Maria Angélica P. de C.; SOUZA, Antônio da S. e ALMEIDA, Weliton Antonio B. de. Morfogênese in vitro. In:SOUZA, Antônio da Silva; JUNGHANS, Tatiana Góes. Introdução à micropropagação de plantas. Cruz das Almas: Embrapa, p. 115-130, 2006.
DE PROFT, M.P.; MAENE, L.J.; DEBERGH, P.C. Carbon dioxide and ethylene evolution in the culture atmosphere of Magnolia cultured in vitro. Physiologia Plantarum, Denmark, v.65, p. 375-379, 1985.
DROSTE, A.; SILVA, A.M.; MATOS, A. V.; ALMEIDA, J. W. In vitro culture of Vriesea gigantea and Vriesia philippocoburgii: two vulnerable bromeliads native to southern Brazil. Brazilian Archives of biology and Technology, Curitiba, v. 48, n. 5, p. 717-722, 2005.
FIALHO, R.F. Seed dispersal by a lizard and a treefrog – Effect of dispersal site on seed survivorship. Biotropica, Washington, v. 22, p. 423-424, 1990.
FIALHO, R. F.; FURTADO, A.L.S. Germination of Erythroxylum ovalifolium (Erythroxylaceae) seeds within the terrestrial bromeliad Neoregelia cruenta. Biotropica, Washington, v. 25, p. 359-362, 1993.
GRATTAPAGLIA, Dario; MACHADO, Marcos A. Micropropagação In: TORRES, A. C.; CALDAS, L. S.; BUSO, J. A. Cultura de tecidos e transformação genética de plantas. Brasília: Embrapa/SPI, v.1, p. 183-260,1998.
KOZAI, T. Micropropagation under photoautotrophic conditions. In Micropropagation, technology and application (DEBERGH, P.; ZIMMERMAN, R. eds) Dordrecht, The Netherlands: Kluwer Academic Publishers, p. 444-469, 1991.

13
LEME, E.M.C. Canistrum – Bromélias da Mata Atlântica. Rio de Janeiro, Salamandra, 107p., 1997.
LEME, E.M.C. Canistropsis – Bromélias da Mata Atlântica. Rio de Janeiro, Salamandra, 107p., 1998
LEME, E.M.C.; MARIGO, L.C. Bromeliads in the Brazilian wilderness. Rio de Janeiro, Marigo Comunicação visual, 183p., 1993.
MERCIER, H.; KERBAUY, G.B. Micropropagation of ornamental bromeliads (Bromeliaceae). In: Biotecchnology in agriculture and forestry – serie high-tech and micropropagation (BAJAJ, Y.P.S. org.) Berlim: Springer-Verlag, p. 43-57, 1997.
MERCIER, Helenice; NIEVOLA, Catarina C. Obtenção de bromélias in vitro como estratégia de preservação. Vidalia, Lavras, v. 1, n. 1, p. 57-62, 2003.
OLIVEIRA, M.G.N; ROCHA, C.F.D.; BAGNAL, T.A. A comunidade animal associada à bromélia tanque Neoregelia cruenta (R. Graham) L. B. Smith. Bromélia, Rio de Janeiro, v.1, n.1, p. 22-29, 1994.
OLIVEIRA, M.G.N.; ROCHA, C.F.D. O efeito da complexidade da bromélia-tanque Neoregelia cruenta (R. Graham) L. B. Smith sobre a comunidade animal associada. Bromélia, Rio de Janeiro, v.4, p.13-22, 1997.
PAN, M.J.; STADEN, J. van. The use of charcoal in in vitro culture – A review. Plant Growth Regulation, New York, v. 26, p. 155–163, 1998.
PAULA, Cláudio Coelho de; SILVA, Helena M. Peregrino da. Cultivo prático de bromélias. Viçosa: UFV, 3ed., 2004.
PERES, Lazaro E. P. Bases fisiológicas e genéticas da regeneração de plantas in vitro. Biotecnologia Ciências e Desenvolvimento, Brasília, v.23, p.44-48, mar./abr., 2002.
PINTO, J.E.B.P., ARELLO, E.F., PINTO, C. A. B. P., BARBOSA, M.H.P. Uso de Explantes e Concentrações de Benzilaminopurina na Multiplicação in vitro de brotos de Kielmeyera coriacea. Pesquisa Agropecuaria Brasileira, Brasília, v 29, n.6, p.867-873. Jun. 1994.
RECH FILHO, A.; DAL VESCO, L.L.; NODARI, R.O.; LISCHKA, R.W.; MULLER, C.V.; GUERRA, M.P. Tissue culture for the conservation and mass propagation of Vriesea reitzii Leme and Costa, a bromeliad threatened of extinction from the Brazilian Atlantic Forest. Biodiversity and Conservation, Netherlands, v. 14, n. 8, p.1799-1808, 2005.
ROCHA, Carlos Frederico Dutra da; COGLIATTI-CARVALHO, Luciana; NUNES-FREITAS, André Felippe; ROCHA-PESSOA, Thereza Christina; DIAS, Aline dos Santos; ARIANI, Cristina Valente; MORGADO, Leila Nunes. Conservando uma larga porção da diversidade biológica através da conservação de Bromeliaceae. Vidalia, Lavras, v.2, n.1, p.52-68, 2004.

14
RODRIGUES, Tatiana Michlovská; PAIVA, Patrícia Duarte de Oliveira; RODRIGUES, Carlos Ribeiro; CARVALHO, Janice Guedes de; FERREIRA, Clarissa Alves & PAIVA, Renato. Desenvolvimento de mudas de bromélia-imperial (Alcantarea imperialis) em diferentes substratos. Ciência e agrotecnologia, Lavras, v. 28, n. 4, p. 757-763, jul./ago., 2004.
SCHULTZ, A. Introdução à Botânica Sistemática. 5 ed., v. II, Porto Alegre, Editora da Universidade de Porto Alegre, 285p. 1985.
SMITH, L.B. Geographical evidence on the lines of evolution in the Bromeliaceae. Botanische Jahrbucher fur Systematik, Stuttgart, v. 66, p. 446-468, 1934.
SMITH, L. B.; DOWNS, R. Monograph Bromelioideae (Bromeliaceae), 2 edição. Flora neotropica, Nova York, v.14, n.3, p. 1493-2142, 1983.
SOUZA, Fernanda Vidigal Duarte; JUNGHANS, Tatiana Góes; SOUZA, Antônio da Silva; SANTOS-SEREJO, Janay Almeida dos; COSTA, Maria Angélica Pereira de Carvalho. Micropropagação, In: SOUZA, Antônio da Silva; JUNGHANS, Tatiana Góes. Introdução à micropropagação de plantas, Cruz das Almas: Embrapa, p.38-50, 2006.
TAGLIACOZZO, Gláucia Moraes Dias. Fitormônios e seus efeitos biológicos in vivo e in vitro. In: TOMBOLATO, Antonio Fernando Caetano; COSTA, Ana Maria Molini. Micropropagação em plantas ornamentais. Campinas, Instituto Agronômico, 1998.
TOMBOLATO, Antonio Fernando Caetano; COSTA, Ana Maria Molini. Micropropagação em plantas ornamentais. Campinas, Instituto Agronômico, 1998.
WANDERLEY, Maria das Graças Lapa. Diversidade e distribuição geográfica das espécies de Orthophytum (Bromeliaceae). Acta botânica Brasílica, São Paulo, v.4, n.1, p. 169-175, 1990.
WANDERLEY, Maria das Graças Lapa; CONCEIÇÃO, Abel Augusto. Notas Taxonômicas e uma nova espécie do gênero Orthophytum Beer (Bromeliaceae) da Chapada Diamantina, Bahia, Brasil. Sitientibus (Série Ciências Biológicas), Feira de Santana, v. 6, n. 1, p.3-8, 2006.
ZOBAYED, S.M.A.; ARMSTRONG, J.; ARMSTRONG,W. Evaluation of a closed system, diffuse and humidity-induced convective through flow ventilation on the growth and physiology of cauliflower in vitro. Plant Cell, Tissue and Organ Culture, Netherlands, v.59, p.113-123, 1999.

15
CAPÍTULO 1
Multiplicação in vitro de Orthophytum mucugense WAND e CONCEIÇÃO.
RESUMO
Orthophytum mucugense é uma bromélia, possui folhas verdes que na época de floração tornam-se parcial ou completamente vermelhas, conferindo-lhe notável valor ornamental. É uma espécie considerada vulnerável devido a sua ocorrência restrita ao município de Mucugê, Chapada Diamantina-BA. Métodos de cultura in vitro têm sido aplicados na conservação e multiplicação de genótipos específicos de bromélias. O presente estudo visa avaliar a multiplicação de O. mucugense com a analise de diferentes concentrações de ANA, tipos e idades do explante e a influência da dominância apical a partir da abscisão do ápice caulinar e da posição do explante no meio. A utilização de caules sem ápice com 40 dias como fonte de explante, cultivado em meio de cultura MS½ sem a presença de reguladores de crescimento proporcionou maior taxa de regeneração com baixo custo de produção. Palavras-chave: Bromeliaceae. Plantas ornamentais. Tipo de explante. Idade do
explante
ABSTRACT
Orthophytum mucugense is a bromeliad that has green leaves that become partially or completely red during flowering, giving the plant significant ornamental value. This specie is considered vulnerable as its occurrence is restricted to the municipality of Mucugê, Chapada Diamantina-BA. In vitro culturing has been used for the conservation and multiplication of specific bromeliad genotypes. The present study examined techniques to optimize the multiplication of O. mucugense by evaluating the effects of varying concentrations of NAA, types and ages of explants, the influence of apical dominance on the stem, and the position of the explant in the culture medium. The highest regeneration rates and lowest production costs were obtained using 40 day-old stems without their apices as the explant source and MS½ medium culture without any growth regulators. Keywords: Bromeliaceae. Ornamental. Explant type. Explant age

16
1.1 INTRODUÇÃO
Diversas espécies de bromélias têm sido amplamente utilizadas na
alimentação ou como fonte de fibras por populações nativas das três Américas e são
também apreciadas como plantas ornamentais, especialmente nos EUA, na Europa
e na Austrália, onde seu cultivo movimenta uma economia considerável, gerando
emprego e renda (LEME, 1998; PAULA e SILVA, 2004). No Brasil, a utilização de
bromélias no mercado ornamental está em expansão o que gera uma demanda de
tecnologia voltada para a produção de mudas em escala comercial (DROSTE et al.,
2005).
Orthophytum mucugense possui roseta foliar típica do gênero com presença
de folhas geralmente patentes, formando um ângulo reto com o eixo da planta. As
suas folhas são verdes e, na época de floração, tornam-se parcial ou completamente
vermelhas, conferindo notável valor ornamental a esta espécie (WANDERLEY e
CONCEIÇÃO, 2006). O. mucugense é considerada vulnerável devido a sua
ocorrência restrita ao município de Mucugê, Chapada Diamantina-BA. Assim, a
realização de trabalhos visando a produção de mudas em grande escala contribuirá
para a redução do extrativismo vegetal.
Métodos de cultura in vitro têm sido aplicados na conservação e
multiplicação de genótipos específicos de bromélias. Esta técnica é muito importante
para a indústria agrícola por proporcionar a rápida propagação, e disponibilizar uma
grande quantidade de mudas com características homogêneas, independente da
estação do ano (CARNEIRO e MANSUR, 2004; DROSTE et al., 2005). Contudo o
sucesso dessa técnica depende da produção de protocolos eficientes, pois a
variabilidade da resposta morfogenética existe até entre genótipos da mesma
espécie (GRATTAPAGLIA e MACHADO, 1998).
O sucesso da micropropagação de qualquer espécie depende do
entendimento dos fatores que controlam a morfogênese. Entre esses, á identificação
dos tecidos mais adequados para essa técnica, o tipo e concentração do regulador
de crescimento utilizado. A resposta do tecido ao regulador de crescimento deve ser

17
atribuída a mudanças na sua sensibilidade que relacionada ao número de
receptores, afinidade do regulador com o receptor, natureza da transdução e
estrutura química do regulador. Pode variar também com o tipo de tecido, estado de
desenvolvimento e condições fisiológicas da planta (HINOJOSA, 2000). De forma
geral, explantes oriundos de tecidos jovens e com maior atividade metabólica são os
mais competentes e possuem maior potencialidade morfogenética (PINTO et al.,
1994).
Outro fator que altera a resposta morfogenética na micropropagação é a
ação da dominância apical. Ela pode ser quebrada pela remoção da gema apical da
planta em crescimento e/ou aplicação de ácido giberélico ou citocinina na planta e
pela alteração da polaridade do vegetal, invertendo sua posição no meio de cultura
(CALDAS et al., 1998; GRATTAPAGLIA e MACHADO, 1998). Sendo múltiplos os
fatores que influenciam na micropropagação e a respostar ser dependente da
espécie em estudo, a necessidade de trabalhos que avaliem esses fatores em
genótipo desejados, principalmente em espécies ameaçadas de extinção.
Em O. mucugense Bellintani (2006) avaliou diferentes concentrações de
ácido naftalenoacético (ANA) e 6-benzilaminopurina (BAP), obtendo uma taxa de
multiplicação (média de 5 brotos por explante), porém foi observado o aparecimento
de calos, após quatro meses de cultivo. Sendo o calo uma estrutura que indica um
possível surgimento de variantes somaclonais, houve a necessidade de se reavaliar
o protocolo de multiplicação dessa espécie e de avaliar outros fatores ainda não
estudos para esta espécie.
O presente estudo visa portanto, melhorar o protocolo de multiplicação em
Orthophytum mucugense avaliando concentrações de ANA, tipos e idades do
explante e a influência da dominância apical a partir da abscisão do ápice caulinar e
da posição do explante do meio.

18
1.2 MATERIAL E MÉTODOS
1.2.1 MATERIAL VEGETAL, ASSEPSIA E GERMINAÇÃO
Sementes de Orthophytum mucugense foram coletadas no Parque Municipal
Sempre-Viva localizado no município de Mucugê-BA. Os experimentos de
multiplicação e as análises histológicas foram realizadas no Laboratório de Cultura
de Tecidos Vegetais (LCTV) e no Laboratório de Micromorfologia Vegetal (LAMIV),
respectivamente, na Universidade Estadual de Feira de Santana (UEFS).
As sementes foram retiradas de frutos maduros e colocadas para secar
sobre papel filtro durante três dias a temperatura ambiente. Sementes secas foram
desinfestadas em álcool 70% durante um minuto e hipoclorito de sódio com 2,5-3%
de cloro ativo por 15 minutos e posteriormente foram lavadas em água destilada
autoclavada duas vezes e inoculadas em frascos de 250 mL contendo 80 mL de
meio de cultura gelificado com 7g.L-1ágar.
Após a emissão dos primórdios foliares as plântulas foram transferidas para
o meio MS (MURASHIGE e SKOOG, 1962) com metade da concentração salina
(MS½) suplementado com 87,64 mM de sacarose e 7g.L-1 de ágar (BELLINTANI,
2006).
1.2.2 CONDIÇÕES DE CULTIVO, pH E ESTERILIZAÇÃO DO MEIO DE CULTURA
Durante todos os experimentos os frascos foram mantidos em sala de
crescimento sob temperatura de 25+ 2ºC, fotoperíodo de 16h e densidade de fluxo
de fótons fotossínteticamente ativos de 60 µmol. µm-2.s-1.
Os meios utilizados tiveram o pH ajustado para 5,8 antes da autoclavagem,
que foi realizada a 121ºC por 15 minutos.

19
1.2.3 MULTIPLICAÇÃO
Nos experimentos de multiplicação foi utilizado como meio básico o MS½
suplementado com 87,64 mM de sacarose e gelificado com 7 g.L-1 de agar, de
acordo com metodologia descrita por Bellintani (2006) para essa espécie.
1.2.3.1 Idade e tipo do explante em duas concentrações de ANA
Para avaliar a influência da idade e do tipo de explante na multiplicação in
vitro de O. mucugense foram utilizados como explante: a) plantas inteiras com idade
de 7, 14 e 21 dias; b) raízes, caules e folhas extraídos de plantas com idade de 20,
40 e 60 dias. As plantas utilizadas para a retirada dos explantes foram colocadas no
escuro 15 dias antes da utilização para o estiolamento e assim auxiliar na retirada
das fontes de explante.
Na multiplicação foi utilizado o meio básico suplementado com 2,22µM de
BAP associado a 0,65 ou 1,30µM de ANA, conforme recomendado por Bellintani
(2006). Foram utilizados como recipientes frascos de 250 mL contendo 80 mL de
meio de cultura.
Foi utilizado o delineamento inteiramente casualizado em esquema fatorial
2x3x4 (concentração de ANA x idade x tipo de explante), com 12 repetições, cada
uma composta de cinco explantes.
Decorridos 60 dias da inoculação foram avaliados: número de brotos
formados por explante, percentual de explantes responsivos e tamanho dos brotos
formados.

20
1.2.3.2 Posição do explante
Para determinar a influência da orientação do explante na regeneração in
vitro foi analisado o efeito da posição do explante no meio de cultura. Caules de
plantas com 40 dias foram inoculados no meio básico suplementado com 2,22 µM de
BAP e 0,65 µM ANA (melhores resultados do experimento anterior) em três posições:
vertical, horizontal e com polaridade invertida. Cada explante foi colocado em um
tubo de ensaio (25x150 mm) com 15 mL de meio de cultura. Os tratamentos
consistiram de dez repetições com cinco explantes cada.
Decorridos 60 dias da inoculação foram avaliados: número de brotos
formados por explante, porcentagem de explantes responsivos e tamanho dos
brotos. Foi avaliada ainda a taxa de sobrevivência dos brotos 30 dias após a
transferência para meio MS½.
1.2.3.3 Remoção do ápice caulinar
A influência da dominância apical na micropropagação foi avaliada pela
excisão do ápice caulinar de plantas estioladas. Plantas com 10 dias de idades foram
mantidas no escuro por 30 dias para induzir o estiolamento. Após esse período
avaliou-se a capacidade regenerativa de plantas estioladas com e sem ápice,
cultivadas no meio básico suplementado com ANA (0,65 µM) e BAP (2,22 µM) ou
sem regulador de crescimento (controle).
O delineamento estatístico foi inteiramente casualizado em esquema fatorial
2x2 (presença ou ausência de regulador de crescimento x tipo de explante), com 20
repetições, cada uma com cinco explantes. Após 60 dias da inoculação foi avaliado o
número de brotos formados por explante.

21
1.2.4 ANÁLISE HISTOLÓGICA
De cada tratamento do experimento 1.2.3.1 foram retiradas cinco amostras
de plantas, caules e folhas micropropagadas para a análise histológica do processo
de organogênese, a fim de confirmar a origem dos brotos produzidos. Essas foram
fixadas em álcool 70% e com auxílio de lâminas de barbear foram realizados cortes
longitudinais e transversais a mão livre. Os cortes foram clarificados com hipoclorito
de sódio a 3% por 15 minutos, lavados em água destilada, corados com safrablau,
analisados em microscópio óptico e fotografados (BUKATSCH, 1972).
1.2.5 ANÁLISE ESTATÍSTICA
Os dados foram submetidos à análise de variância (ANOVA) e as médias
comparadas pelo teste Tukey (0,05), utilizando o programa estatístico SISVAR (v.
4.4, UFLA) (FERREIRA, 2000).

22
1.3 RESULTADOS E DISCUSSÃO
Os resultados obtidos nesse trabalho demonstram que a organogênese
direta em Orthophytum mucugense é influenciada pelo tipo e idade do explante além
de ser potencializada pelo balanço auxina/citocinina e pela remoção do ápice
caulinar.
Os melhores resultados foram obtidos de plantas (explante) com 14 dias de
idade nas três variáveis analisadas, exceto para o número de brotos na presença de
0,65 µM de ANA, situação na qual não foram observadas diferenças significativas
entre as plantas com 7 e 14 dias (Tabela 1.1, Figura 1.1). Resultado diferente do
obtido na bromélia Tillandsia eizii onde não foi observada a influência da idade dos
explantes na taxa de multiplicação in vitro na presença de diferentes concentrações
de ANA (0,54; 2,69; 5,37 µM) e BAP (8,88; 4,44; 2,22 µM). Porém houve diferença
em relação à qualidade dos brotos produzidos, sendo os brotos mais viáveis
formados a partir de explantes que tinham a maior idade avaliada (12 semanas),
explantes com idade intermediaria (3 semanas) levaram à formação de calos e nos
explantes mais novos (0 semanas) não ocorreu o crescimento dos brotos (PICKENS
et al., 2006), resultado não observado nos brotos de O. mucugense os quais
apresentaram boa qualidade independente do tratamento como pode ser observado
na Figura 1.1.
A formação dos primeiros brotos ocorreu na terceira semana após a
inoculação em quase todos os tratamentos, com exceção das plantas com 7 e 21
dias cultivadas na presença de 0,65 µM e 1,30 µM de ANA respectivamente. Nestes
tratamentos os primeiros brotos foram formados na quarta semana após inoculação.
Observou-se também que a formação dos brotos ocorreu na base do explante
(Figura 1.1), divergindo dos resultados obtido em Passiflora cincinnata por Lombardi
et al. (2007) que ao utilizar plantas como fonte de explante, observou a formação dos
brotos a partir da raiz.

23
Figura 1.1 – Brotações obtidas via organogênese direta em plantas de Orthophytum mucugense em explante com diferentes idades e submetidos a duas concentrações de ANA, após 60 dias de cultivo. A seta mostra a origem dos brotos. A barra representa 0,5 cm. Feira de Santana, 2007.
Entre as concentrações de reguladores de crescimento utilizadas, a
combinação de 1,30 µM de ANA com 2,22 µM de BAP proporcionou os melhores
resultados para o número de brotos formados em plantas com 14 dias de idade e
para o percentual de explantes responsivos, em plantas com 14 e 21 dias de idade
(Tabela 1.1).
Considerando os tipos de explantes avaliados, as plantas inteiras formaram
brotos a partir da terceira e quarta semana, o caule apresentou formação de brotos já
na primeira semana após a inoculação, a folha na segunda semana e a raiz não
apresentou competência organogenética nas condições as quais foram submetidos.
Estes dados concordam com os obtidos por Bellintani (2006) ao trabalhar com
24
segmentos nodais de O. mucugense onde a formação dos brotos adventícios se
iniciou entre a segunda e a quarta semana.
Tabela 1.1 – Percentual de explante responsivos, número de brotos por explante e comprimento dos brotos em Orthophytum mucugense obtidos a partir de plantas com diferentes idades, em meio suplementado com 2,22 µM de BAP associado a duas concentrações de ANA após 60 dias de cultivo. Feira de Santana, BA, 2007.
ANA Idades dos explantes (dias) (µM) 7 14 21
% de explantes responsivos 0,65 16,67Awbz 38,33Ba 3,33Bb 1,30 18,33Ab 56,76Aa 20,00Ab Número de brotos/explante
0,65 0,27Aab 0,67Ba 0,10Ab 1,30 0,27Ab 1,30Aa 0,40Ab Comprimento dos brotos (mm)
0,65 1,57Ab 4,11Aa 0,98Ab 1,30 1,92Ab 4,24Aa 1,71Ab
w - Médias seguidas pela mesma letra maiúscula na mesma coluna, para cada variável, não diferem entre si ao nível de 5% de probabilidade pelo teste de Tukey. z - Médias seguidas pela mesma letra minúscula na mesma linha, não diferem entre si ao nível de 5% de probabilidade pelo teste de Tukey.
O caule apresentou o melhor resultado na maioria das variáveis analisadas,
diferindo significativamente de folha e raiz, independente da concentração de ANA
utilizada (Tabela 1.2 e Figura 1.2). A única exceção foi observada na presença de
1,30 µM de ANA, não sendo encontrada diferença significativa entre os explantes
caule e folha, para o comprimento médio dos brotos (Tabela 1.2). O caule regenerou
maior número de brotos por explante e brotos maiores na concentração de 0,65 µM
de ANA (Tabela 1.2). Estes resultados contrastam com os observados para
Neoregelia cruenta que apresentou maiores taxas regenerativas a partir do explante
folha quando comparado com o caule (CARNEIRO et al. 1999).

25
Figura 1.2 - Brotos de Orthophytum mucugense micropropagados a partir dos explantes caule e folha, após 60 dias de cultivo. A barra representa 0,5 cm. Feira de Santana, 2007.
Observou-se formação de brotos ao utilizar o explante folha nas duas
concentrações de ANA avaliadas (Tabela 1.2). A formação direta de gemas
adventícias na base foliar é atribuída à presença de meristemas intercalares nesse
local (MERCIER e NIEVOLA, 2003). Mercier et al.(2003) estudaram a influencia de
auxinas/citocininas endógenas na regeneração de brotos a partir de folhas de
abacaxi cultivadas na presença de 5,37 µM de ANA e 8,87 µM de BA (N6-
benziladenina), tratamento no qual foi observado um aumento na concentração
endógena de iP (2-isopenteniladenina) o que levou os autores a sugerir que a
concentração de iP desencadeia a sinalização indutora da organogênese a partir da
base foliar, atribuindo a produção dessa citocinina à presença de reguladores de
crescimento no meio de cultura.
Carneiro et al. (1998 e 1999) trabalhando com Neoregelia cruenta e
Cryptanthus sinuosus também observaram a formação de brotos a partir de folhas.
Nessas duas espécies de bromélias foram observadas altas taxas de multiplicação
para essa fonte de explante. A alta taxa pode estar relacionada com as elevadas
concentrações dos reguladores usados por esses autores (22,00 µM de BAP
26
associado a 2,50 µM de ANA), sendo essa muito superior às utilizadas para O.
mucugense no presente trabalho (Tabela 1.2). Essa diferença na taxa de
multiplicação entre espécies da mesma família ratifica a importância da otimização
de protocolos para cada espécie e até mesmo para variedades de uma mesma
espécie.
Tabela 1.2 - Percentual de explante responsivos, número de brotos por explante e comprimento dos brotos em Orthophytum mucugense obtidos a partir de dos explantes caule, folha e raiz com 20, 40 e 60 dias, em meio suplementado com 2,22 µM de BAP associado a diferentes concentrações de ANA após 60 dias de cultivo. Feira de Santana, BA, 2007.
Tipo de explante ANA (µM) Caule Folha Raiz
% de explantes responsivos 0,65 46,11Awaz 9,44Ab 0,00Ab 1,30 43,33Aa 14,44Ab 0,00Ac Número de brotos/explantes 0,65 1,21Aa 0,26Ab 0,00Ab 1,30 0,87Ba 0,33Ab 0,00Ab Comprimento dos brotos (mm) 0,65 5,53Aa 1,63Ab 0,00Ac 1,30 4,18Ba 2,71Aa 0,00Ab w Médias seguidas pela mesma letra maiúscula na mesma coluna, para cada variável, não diferem entre si ao nível de 5% de probabilidade pelo teste de Tukey. z - Médias seguidas pela mesma letra minúscula na mesma linha, não diferem entre si ao nível de 5% de probabilidade pelo teste de Tukey.
A diferença encontrada na taxa de multiplicação a partir de diferentes
explantes da espécie O. mucugense corrobora com os resultados obtidos por
Ascough et al. (2007) ao estudarem a propagação in vitro de quatro espécies de
Watsonia. Nesse trabalho os autores avaliaram a resposta de três tipos de explantes
(raiz, hipocótilo e folhas) sob a influência de diferentes concentrações de BAP (2,22;
4,44 µM) e ANA (2,68; 5,37 µM) e foi observada a formação de brotos a partir do
hipocótilo de todas as espécies estudadas, com um percentual de indução acima de
80%, enquanto os explantes raiz e folha, apresentaram apenas uma espécie
formando brotos (W. laccata e W. vanderspuyiae, respectivamente) com um
percentual menor que 10%. Nesse trabalho, observou-se baixa taxa de indução de

27
gemas quando foi utilizado a folha como fonte de explante (média menor de um broto
por explante, Tabela 1.2), corroborando resultados encontrados por Souza et al.
(2006) quando utilizou folhas para indução de gemas em Cucumis melo.
Raízes de O. mucugense não apresentaram competência organogenética
nos níveis de regulador de crescimento aqui utilizados (Tabela 1.2). Apesar desse
órgão possuir tecidos meristemáticos nos ápices e no periciclo, em algumas espécies
as raízes apresentam extrema determinação, sendo difícil a indução de gemas
caulinares (PERES, 2002). Os dados obtidos para O. mucugense diferem do
encontrado por Biasi et al. (2002) para a espécie lenhosa Diospyros kaki na qual foi
observada um maior potencial organogenético em tecidos radiculares quando
comparados com os caulinares na presença de BAP ou TDZ (10,00 µM) ambos
associados ao AIA (0,01 µM), concentrações de BAP superiores ao utilizadas no
presente experimento.
Tendo em vista que o caule foi considerado o melhor explante na
micropropagação de O. mucugense, foi avaliada a interação da idade deste explante
com diferentes concentrações de ANA (Tabela 1.3).
A idade do explante influenciou no percentual de explantes responsivos, no
número de brotos formados por explante e no comprimento dos brotos regenerados
na presença de BAP (2,22 µM) e ANA (0,65 ou 1,30 µM) (Tabela 1.3 e Figura 1.3).
Os explantes mais jovens, com 20 ou 40 dias, em geral apresentaram maior
responsividade que os explantes com 60 dias, exceto quando foram comparados os
explantes com 40 e 60 dias cultivados na presença de 2,22 µM de BAP associado a
0,65 µM de ANA, os quais não apresentaram diferenças significantes quanto ao
percentual de explantes responsivos (Tabela 1.3). Resultado similar ao encontrado
por Carneiro et al. (1999) ao estudar a micropropagação em folhas de Neoregelia
cruenta, onde com o aumento da idade do explante ocorreu a diminuição na
porcentagem de explante responsivo e na média de brotos por explante.
As médias de explantes responsivos obtidas nos caules com 20 e 40 dias
foram semelhantes as observadas em Vriesea gigantea, mas inferior a encontrada
em V. philippocoburgii, ambas espécies cultivadas em meio com concentrações mais
altas de reguladores (8,87 µM de BAP e ANA-2,68 µM) (DROSTE et al., 2005).

28
A idade do caule só interferiu no número de brotos formados quando os
explantes foram cultivados na presença de 2,22 µM de BAP associado a 1,30 µM de
ANA, concentração na qual explantes com 20 e 40 dias regeneraram maior número
de brotos que explantes com 60 dias, o que confirma que os tecidos mais jovens
apresentam maior potencial morfogenético (PINTO et al. 1994). Nos explantes com
20 e 40 dias foi observado um aumento significativo no número de brotos na maior
concentração de ANA (Tabela 1.3). Bellintani (2006) em estudo com essa mesma
espécie obteve uma média de 4 a 6 brotos por explante em mesma condição de
cultivo. Essa diferença pode estar relacionada com o tempo de permanência dos
explantes no meio de cultura por um período de seis meses, o que mostra que não
apenas a idade do explante como também o tempo de cultivo interfere na taxa de
multiplicação.
Tabela 1.3 - Percentual de explante responsivos, número de brotos por explante e comprimento dos brotos em Orthophytum mucugense obtidos a partir do explante caule em diferentes idades de desenvolvimento em meio suplementado com 2,22 µM de BAP associado a duas concentrações de ANA após 60 dias de cultivo. Feira de Santana, BA, 2007.
Idade do explante (dias) ANA (µM) 20 40 60
% de explantes responsivos 0,65 55,00Awaz 50,00Aab 33,33Ab 1,30 48,33Aa 56,67Aa 25,00Ab Número de brotos/explante 0,65 1,02Aa 1,25Aa 1,37Aa 1,30 0,95Aab 1,30Aa 0,37Bb Comprimento dos brotos (mm) 0,65 4,62Ab 7,80Aa 4,15Ab 1,30 4,15Aab 5,99Aa 2,40Ab w Médias seguidas pela mesma letra maiúscula na mesma coluna, para cada variável, não diferem entre si ao nível de 5% de probabilidade pelo teste de Tukey. z - Médias seguidas pela mesma letra minúscula na mesma linha, não diferem entre si ao nível de 5% de probabilidade pelo teste de Tukey.
29
Figura 1.3 - Brotos de Orthophytum mucugense micropropagados a partir do explante caule, em função da idade e concentração de ANA, após 60 dias de cultivo. A barra representa 0,5 cm. Feira de Santana, 2007.

30
A permanência dos explantes no meio de cultura com reguladores de
crescimento por longo período pode ocasionar a formação de calos, corroborando
observações feitas por Bellintani (2006) em O. mucugense após quatro meses de
cultura, e com isso ocasionar o aparecimento de variantes somaclonais.
Em O. mucugense as diferenças nas taxas de multiplicação de explantes
com 20 e 40 dias, podem decorrer do tamanho reduzido dos explantes com 20 dias
(Figura 1.3). Explantes com pequeno porte normalmente demoram em iniciar a fase
de crescimento e podem apresentar poucas gemas diminuindo assim a taxa de
multiplicação (GRATTAPAGLIA e MACHADO, 1998).
O número de brotos por explante encontrada em caules com 40 dias (Tabela
1.3) corrobora aqueles encontrados por Droste et al. (2005) em Vriesea gigantea
quando utilizou o mesmo meio de cultura (MS ½) e os mesmos reguladores mas em
concentrações maiores (8,87 µM de BAP e 2,68 µM de ANA).
Para a variável comprimento dos brotos, explantes com 40 dias
apresentaram médias superiores às outras idades, independente da concentração do
regulador utilizada, não diferindo significativamente apenas do caule com 20 dias na
concentração de 1,3 µM de ANA (Tabela 1.3).
Ocorreu a diminuição no comprimento dos brotos com o aumento da
concentração do regulador de crescimento ANA. Esse resultado corrobora os obtidos
por Chen et al. (2006) ao avaliar diferentes concentrações de BAP e ANA na espécie
Lychnis senno, ambos sem diferença significativa.
Em relação à orientação do explante no meio de cultura, a posição
horizontal apresentou as maiores médias nas três variáveis analisadas, mas não
diferiu significativamente dos outros dois tratamentos (Tabela 1.4, Figura 1.4). A
produção de AIA no ápice caulinar e o transporte de caráter polar, com requerimento
de energia, estão vinculada à diferenciação vascular e tropismo (HINOJOSA, 2000).
Dessa forma, a produção e transporte da auxina inibe a formação de brotos laterais,
e com a alteração no eixo da planta ao posicionar o explante na posição horizontal,
ocorre a quebra dessa dominância apical, proporcionando o aumento da taxa de
multiplicação, sendo uma possível explicação para os resultados obtidos nesse
trabalho.

31
A taxa de sobrevivência dos brotos formados a partir de explantes na
posição vertical e com a polaridade invertida foi de 95% e na posição horizontal
96,67%. Ocorreu 100% de enraizamento em todos os tratamentos utilizados.
Altas porcentagens de enraizamento de brotos em meio sem regulador de
crescimento foram observadas também em outras espécies de bromélias
(CARNEIRO et al. 1998, 1999; MACÊDO et al. 2003). De acordo com Bosa et al.
(2003) isso ocorre porque algumas espécies vegetais possuem níveis endógenos de
auxina suficientes para promover o enraizamento.
Erig e Schuch (2002) ao estudarem o efeito da orientação, de segmentos
caulinares da espécie lenhosa Malus prunifolia, observaram também a formação de
elevado número de brotos ao utilizarem a posição horizontal em meio MS
suplementado com 4,44 µM de BAP.
Tabela 1.4 - Percentual de explante responsivos, número de brotos por explante e comprimento dos brotos em Orthophytum mucugense obtidos em função da posição do explante (caule com 40 dias) no meio de cultura MS½ suplementado com 2,22µM de BAP associado a 0,65 µM de ANA após 60 dias de cultivo. Feira de Santana, BA, 2007.
Orientação % de explante responsivos
No de brotos/explante Comprimento dos brotos (mm)
Polaridade invertida 27,27a 1,34a 5,27a Vertical 34,54a 1,85a 5,07a Horizontal 45,45a 3,53a 5,85a Médias seguidas pela mesma letra, não diferem entre si ao nível de 5% de probabilidade pelo teste de Tukey.

32
Figura 1.4 - Brotos de Orthophytum mucugense, micropropagados a partir do explante caule com 40 dias, em função da orientação do explante no meio de cultura, após 60 dias de cultivo. A barra representa 0,5 cm. Feira de Santana, 2007.
Os resultados obtidos no experimento que avaliou a influencia da excisão do
ápice caulinar na micropropagação de O. mucugense mostraram uma diferença
significativa na produção de brotos a partir de plantas com e sem ápice (Tabela 1.5).
Plantas sem o ápice caulinar apresentaram maior capacidade regenerativa que as
plantas com ápice. Plantas com ápice não apresentaram diferença significativa para
o número de brotos formados por explante entre meios com e sem regulador,
entretanto, plantas sem o ápice caulinar formaram maior número de brotos na
ausência de reguladores de crescimento. Esses resultados contrastam com os
observados em abacaxi onde ocorreu uma diminuição na taxa de multiplicação com
a retirada do ápice em meio sem regulador de crescimento (MOREIRA et al., 2003).
A produção de AIA nas plantas superiores ocorre no ápice do vegetal, e o
seu transporte assume caráter polar com requerimento de energia (HINOJOSA,
2000). Entre as ações fisiológicas do AIA no vegetal encontramos a dominância
apical que inibe o crescimento das gemas laterais (TAIZ e ZEIGER, 2004). A
remoção do ápice caulinar resulta na quebra da dominância apical, possibilitando o
desenvolvimento das gemas laterais, o que explica a formação de brotos mesmo na
ausência de regulador de crescimento exógeno (Tabela 1.5).

33
A menor média de brotos observada nos explante sem ápice cultivados em
meio com regulador de crescimento, pode ser devido a presença de ANA, que sendo
uma fonte de auxina, poderia ter restabelecido a dominância apical.
Tabela 1.5 - Número de brotos por explante obtidos em plantas de Orthophytum mucugense com e sem ápice caulinar cultivado em meio MS½ na presença ou ausência dos reguladores de crescimento ANA (0,65µM) e BAP (2,22µM ) após 60 dias de cultivo. Feira de Santana, BA, 2007. Tratamento Número de brotos/explante Com ápice Sem ápice ANA (0 µM) + BAP (0 µM) 0,78Awbz 2,25Aa ANA (0,65 µM) + BAP (1,3 µM) 0,60Ab 1,26Ba W – Médias seguidas pela mesma letra maiúscula em cada coluna, não diferem entre si ao nível de 5% de probabilidade pelo teste de Tukey. z – Médias seguidas pela letra minúscula em cada linha, para cada variável, não diferem entre si ao nível de 5% de probabilidade pelo teste de Tukey
A partir da análise histológica foi possível constatar que a exposição dos
explantes em meio com regulador de crescimento proporcionou regiões de intensa
divisão celular (Figura 1.5). Nos explantes plantas e caules, a origem das regiões
com atividade mitótica ocorreu no cambio vascular com a formação de células com
características meristemóides, sendo essas pequenas, com núcleo proeminente e
divisões celulares periclinais e anticlinais (Figura 1.5 a-c). Resultado diferente do
observado em Tillandsia eizii onde foram encontradas células sub-epidérmicas em
divisões mitóticas (PICKENS et al., 2006). A formação dos brotos ocorreu por
organogênese direta nas três fontes de explante utilizadas (plantas, folha e caule)
devido à formação de um tecido apenas com o eixo caulinar (monopolar) e a junção
observada entre os feixes vasculares dos brotos com o explante (Figura 1.5 c-f).
Nas folhas a diferenciação ocorreu no parênquima clorofiliano onde foi
possível observar células em divisões periclinais e anticlinais (Figura 1.5 e-f).
Resultado semelhante ao encontrado por Pedraza-Santos et al. (2006) ao utilizarem
inflorescência imatura de Alstroemeria como fonte de explante, encontrando no
parênquima destas a formação de células com características meristematicas.

34
Figura 1.5 – Anatomia de Orthophytum mucugense micropropagados a partir dos explantes plantas (d), caule (a-c) e folha(e-f). (a) corte transversal do caule mostrando o inicio da diferenciação a partir do cambio vascular; (b) zona meristematica em um estagio mais avançado de desenvolvimento, (c) o inicio da vascularização do novo broto (d) corte longitudinal da planta mostrando a ligação de dois brotos com a planta a partir dos vasos condutores; (e) corte longitudinal da lamina foliar mostrando nova zona meristematica e a conexão dos vasos condutores; (f) zona meristematica em destaque. (cv = cambio vascular); (m = zona meristematica); (v = vaso condutor); (LF = lâmina foliar); a barra representa 5µm, Feira de Santana, 2007.

35
A organogênese foi caracterizada como direta pela presença da ligação
entre os vasos condutores do explante e dos brotos (Figura 1.5 c-e), e a ausência da
desdiferenciação do tecido com a passagem pela fase de calo antes da formação do
broto.
A partir dos resultados obtidos no presente estudo é possível assegurar que
a maior taxa de regeneração de Orthophytum mucugense é obtida utilizando caule
estilados sem ápice, com 40 dias de cultivo como fonte de explante, cultivado em
meio de cultura MS½ sem a necessidade de reguladores de crescimento, o que
possibilita uma diminuição considerável no custo da produção de mudas.

36
REFERÊNCIAS
ASCOUGH, Glendon D.; ERWIN, John; STADEN, Johannes van. In vitro propagation of four Watsonia species. Plant Cell Tissue Organ Culture, Netherlands, v. 88, p. 135–145, 2007
BELLINTANI, Moema Cortizo. Estudos da propagação in vitro e ex vitro de Neoregelia mucugensis Leme, Orthophytum mucugense Wand e Conceição e Orthophytum albopictum Philcox, espécies de Bromeliaceae endêmicas da Bahia. 2006. 115p. Tese (Doutorado em Botânica) – Universidade Estadual de Feira de Santana, Feira de Santana.
BIASI, Luiz Antonio; CARVALHO, Cristina de; WOLF, Geferson Davi; ZANETTE, Flávio. Potencial organogenético de tecidos caulinares e radiculares de caquizeiro. Revista Brasileira de Fruticultura, Jaboticabal, v. 24, n. 1, p. 29-34, abr. 2002.
BOSA, Nair; CALVETE, Eunice O.; NIENOW, Alexandre Augusto; SUZIN, Marilei. Enraizamento e aclimatização de plantas micropropagadas de gipsofila. Horticultura Brasileira, Brasília, v. 21, n. 2, p.207-210, abr.-jun. 2003.
BUKATSCH, F. Bermerkunger zur Doppelfarbung Astrablau-Safranin. Mikrokosmos, v. 61, p.255, 1972.
CALDAS, L. S.; HARIDASAM, P.; FERREIRA, M. E. Meios nutritivos. In: TORRES, A. C.; CALDAS, L. S.; BUSO, J. A. Cultura de tecidos e transformação genética de plantas. Brasília: Embrapa/SPI,. v.1, p. 87-132, 1998.
CARNEIRO, L.A.; CÂNDIDO, M.S.D.; ARAUJO, R.F.G.; FONSECA, M.H.P.B.; CROCOMO, O.J.; MANSUR, E. Clonal propagation of Cryptanthus sinuosus L.B. Smith, na endemic stoloniferous Bromeliaceae species from Rio de Janeiro, Brazil. Plant Tissue Culture and Biotechnology, Bangladesh, v.4, n. 3-4, p.152-158, nov. 1998.
CARNEIRO, L.A.; ARÁUJO, R.F.G.; BRITO, G.J.M.; FONSECA, M.H.P.B.; COSTA, A.; CROCOMO, O.J.; MANSUR, E. In vitro regeneration from leaf explants of Neoregelia cruenta (R. Graham) L.B. Smith, an endemic bromeliad from Eastern Brazil. Plant Cell, Tissue and Organ Culture, Netherlands, v. 55, p. 79–83, 1999.
CARNEIRO, L. A.; MANSUR, E. Contribuição de metodologias in vitro para a conservação de Bromeliaceae. Vidalia, Viçosa, v. 2, n.1, p. 12- 20, 2004.
CHEN, Liping; WANG, Yanju; XU, Chunxia; ZHAO, Mingshui; WU, Jianguo. In vitro propagation of Lychnis senno Siebold et Zucc.,a rare plant with potential ornamental value. Scientia Horticulturae, Amsterdam, v.107, p.183–186, 2006.
DROSTE, A.; SILVA, A.M.; MATOS,A. V.; ALMEIDA, J. W. In vitro culture of Vriesea gigantea and Vriesia philippocoburgii: two vulnerable bromeliads native to southern

37
Brazil. Brazilian Archives of biology and Technology, Curitiba, v. 48, n. 5, p. 717-722, 2005.
ERIG, Alan Cristiano; SCHUCH, Márcio Wulff. Multiplicação in vitro do porta-enxerto de macieira CV. Marubakaido: efeito da orientação do explante no meio de cultura. Revista Brasileira de Fruticultura, Jaboticabal, v. 24, n. 2, p. 293-295, ago., 2002.
FERREIRA, D. F. Análise estatística por meio do SISVAR para Windows: Versão 4.0. In: REUNIÃO ANUAL DA REGIÃO BRASILEIRA DA SOCIEDADE INTERNACIONAL DE BIOMETRIA, 45, 2000, São Carlos. Anais... São Carlos: SSP/SIB, p. 255-268, 2000.
GRATTAPAGLIA, Dario; MACHADO, Marcos A. Micropropagação. In: TORRES, A. C.; CALDAS, L. S.; BUSO, J. A. Cultura de tecidos e transformação genética de plantas. Brasília: Embrapa/SPI, v.1, p. 183-260,1998.
HAMASAKI, Regina M; PURGATTO, Eduardo; MERCIER, Helenice. Glutamine enhances competence for organogenesis in pineapple leaves cultivated in vitro. Brazilian Journal of Plant Physiology, Viçosa, v.17, n. 4, p.383-389, 2005.
HINOJOSA, Gladys Fernández. Auxinas. In: CID, Luis Pedro Barrueto. Introdução aos hormônios vegetais, Brasília: Embrapa Recursos Genéticos e Biotecnologia, p. 15-54, 2000.
LEME, E.M.C., Canistropsis - Bromélias da Mata Atlântica. Rio de Janeiro: Salamandra, 1998. 143p.
LOMBARDI, Simone Pacheco; PASSOS, Ilene Ribeiro da Silva; NOGUEIRA, Maria Cristina Stolf; APPEZZATO-DA-GLÓRIA, Beatriz. In vitro Shoot Regeneration from Roots and Leaf Discs of Passiflora cincinnata Mast. Brazilian Archives of Biology and Technology, Curitiba, v.50, n. 2, p.239-247, Mar. 2007.
MACÊDO, Cristiane Elizabeth Costa de; SILVA, Marcela Gomes da, NÓBREGA, Fabiana Silva da; MARTINS, Camila Pimentel; BARROSO, Paulo Augusto Vianna; ALLOUFA, Magdy Ahmed Ibrahim. Concentrações de ANA e BAP na micropropagação de abacaxizeiro L.Merrill (Ananas comosus) e no cultivo hidropônico das plântulas obtidas in vitro. Revista Brasileira de Fruticultura, Jaboticabal - SP, v. 25, n. 3, p. 501-504, dez. 2003.
MERCIER, Helenice; SOUZA, Beatriz Maia; KRAUS, Jane Elizabeth; HAMASAKI, Regina Mayumi; SOTTA, Bruno. Endogenous auxin and cytokinin contents associated with shoot formation in leaves of pineapple cultured in vitro. Brazilian Journal of Plant Physiology, Viçosa, v.15, n.2, p.107-112, 2003.
MERCIER, Helenice e NIEVOLA, Catarina C. Obtenção de bromélias in vitro como estratégias de preservação. Vidalia, Viçosa, v.1, n.1, p.57-62, 2003.
MOREIRA, Maria Aparecida; PASQUAL, Moacir; CARVALHO, Janice Guedes de; FRÁGUAS, Chrystiane Borges. Estiolamento na micropropagação do abacaxizeiro

38
CV pérola. Ciência e Agrotecnologia., Lavras, v.27, n.5, p.1002-1006, set./out., 2003.
MURASHIGE, T.; SKOOG, F. A revised médium for rapid growth and bio assays with tobacco tissue culture. Physiologia Plantarum, v. 15, p. 473-497, 1962.
PAULA, Cláudio Coelho de; SILVA, Helena M. Peregrino da. Cultivo prático de bromélias. Viçosa: UFV, 3ed., 2004.
PEDRAZA-SANTOS, Martha E.; LÓPEZ-PERALTA, Ma. Cristina; GONZÁLEZ-HERNÁNDEZ, Vitor; ENGLEMAN-CLARK, E. Mark e SÁNCHEZ-GARCÍA, Prometeo. In vitro regeneration of Alstroemeria cv. ‘Yellow King’ by direct organogenesis. Plant Cell, Tissue and Organ Culture, Netherlands, v.84, p.189-198, 2006.
PERES, L. E. P. Bases Fisiológicas e Genéticas da regeneração de plantas in vitro. Biotecnologia Ciências e Desenvolvimento, Brasília, v.25, p. 44-48, 2002.
PICKENS, Kimberly A.; WOLF, Jan; AFFOLTER, James M.; WETZSTEIN, Hazel Y. Adventitious bud development and regeneration in Tillandsia eizii. In Vitro Cellular and Developmental Biology—Plant, Berlin, v. 42, p. 348–353, July–Aug., 2006.
PINTO, J.E.B.P., ARELLO, E.F., PINTO, C. A. B. P., BARBOSA, M.H.P. Uso de Explantes e Concentrações de Benzilaminopurina na Multiplicação in Vitro de brotos de Kielmeyera coriacea. Pesquisa Agropecuária Brasileira, Brasília, v. 29, n.6, p.867-873, Jun., 1994.
SOUZA, Fernanda Vidigal Duarte; GARCIA-SOGO, Begoña; SOUZA,Antonio da Silva; SAN-JUÁN, Amparo Pérez; MORENO, Vicente. Morphogenetic Response of Cotyledon and Leaf Explants of Melon (Cucumis melo L.) cv. Amarillo Oro. Brazilian Archives of Biology and Technology, Curitiba, v. 49, n. 1, p. 21-27, Jan., 2006.
TAIZ, Lincoln; ZEIGER, Eduardo. Fisiologia vegetal. Trad. Eliane Romanato Santarém et al., 3ed. Porto Alegre: Artmed, 2004. 719p.
WANDERLEY, Maria das Graças Lapa; CONCEIÇÃO, Abel Augusto. Notas Taxonômicas e uma nova espécie do gênero Orthophytum Beer (Bromeliaceae) da Chapada Diamantina, Bahia, Brasil. Sitientibus (Série Ciências Biológicas), Feira de Santana, v. 6, n. 1, p.3-8, 2006.

39
CAPÍTULO 2
Influência de diferentes fontes de carbono, AIB e carvão ativado, no crescimento in vitro de brotos de Orthophytum mucugense WAND e CONCEIÇÃO obtidos por organogênese direta.
RESUMO
A espécie Orthophytum mucugense é considerada vulnerável devido ao seu endemismo restrito ao município de Mucugê-BA, na Chapada Diamantina, o que salienta a importância de trabalhos que visem a conservação. O cultivo in vitro proporciona uma alta taxa de multiplicação, produção de plantas saudáveis, além de assegurar o armazenamento de germoplasma. Sendo o crescimento da planta um processo complexo e dinâmico sujeito ao controle genético e ambiental, o presente trabalho avaliou o crescimento de brotos de O. mucugense regenerados in vitro sob a influencia de diferentes fontes de carbono, e a presença de carvão ativado associado a diferentes concentrações de ácido indolbutírico. O uso de sorbitol, sacarose e açúcar cristal apresentaram os melhores desempenhos para o comprimento da parte aérea. A presença de carvão ativado interferiu no desenvolvimento das plântulas, enquanto o ácido indolbutírico influenciou apenas o desenvolvimento da raiz. Palavras-chave: Bromeliaceae. Plantas ornamentais. Fonte de carbono. Carvão
ativado. Ácido indolbutírico.
ABSTRACT
Orthophytum mucugense is considered vulnerable because of its strict endemism in the municipality Mucugê-BA, in the Chapada Diamantina, and has been the object of important conservation work. The in vitro culture provides a high rate of multiplication of healthy plants and ensures germplasm storage. As plant growth is a complex and dynamic process subject to both genetic and environmental control, the present study evaluated the in vitro growth of shoots and the propagation of O. mucugense under the influence of different carbon sources, and in the presence of activated charcoal associated with different concentrations of indole-3-butyric acid (IBA). Sorbitol, sucrose and crystal sugar showed the best results in terms of shoot length; activated charcoal interfered with seedling development; and IBA only influenced root development. Keywords: Bromeliaceae. Ornamental. Carbon source. Activated charcoal. Indole-3-butyric acid.

40
2.1 INTRODUÇÃO
A família Bromeliaceae possui espécies de grande importância econômica
no mercado alimentício e ornamental. Com cerca de 3.000 espécies constitui a maior
família de plantas essencialmente americana, distribuindo-se do sul dos Estados
Unidos até o Chile, com uma única exceção encontrada no Golfo da Guiné na África
(LEME e MARIGO, 1993; KUNHA, 1995).
Segundo Wanderley e Conceição (2006) as espécies de Orthophytum com
inflorescência séssil, ocorrentes na Chapada Diamantina são muito valorizadas sob a
perspectiva ornamental devido à coloração de suas brácteas e folhas, que atingem
uma intensa cor vermelha na antese. Esta característica torna estas espécies
também muito importantes por atrair beija-flores, desempenhando um importante
papel ecológico.
A espécie Orthophytum mucugense Wand. e Conceição possui
inflorescência séssil, roseta foliar pequena e folhas estreitas que atingem
aproximadamente 4 mm de largura. Apresenta estolões e forma densas populações
em paredões rochosos verticais, na beira de rios e cachoeiras (WANDERLEY e
CONCEIÇÃO, 2006). Apesar de formar densas populações essa espécie é
considerada vulnerável devido ao seu endemismo, tornando necessários trabalhos
que visem a sua conservação.
O cultivo in vitro de bromélias é muito utilizado para fins comerciais e tem
contribuído para a rápida propagação e a preservação de espécies raras ou
ameaçadas de extinção (CARNEIRO e MANSUR, 2004; RECH FILHO et al., 2005;
SILVA et al., 2007). A manutenção in vitro assegura o armazenamento do
germoplasma e é vantajosa inclusive por proporcionar uma alta taxa de
multiplicação, com produção de plantas saudáveis e a conservação de grande
variedade de genótipos em espaços reduzidos (ENGELMANN, 1991).
Para o sucesso do cultivo in vitro há a necessidade de estudos para cada
espécie, que avaliem as respostas morfofisiológicas nesse ambiente, visto que o
crescimento e desenvolvimento da planta são processo complexos e dinâmicos que

41
estão submetidos a dois tipos de controle: genético e ambiental. Ambos permitem
que as plantas respondam e se adaptem às condições externas dentro de múltiplas
interações (HINOJOSA, 2000).
O ambiente in vitro não proporciona condições adequadas de iluminação,
concentração de gás carbônico e, às vezes, as plantas desenvolvidas nesse
ambiente não apresentam teores de clorofila suficientes para realizar fotossíntese
que sustente o crescimento do vegetal (CALDAS et al., 1998). Plantas cultivadas in
vitro necessitam de suplementos de carbono, independente do processo
fotossintético, influenciando na fisiologia, na diferenciação celular e no crescimento
vegetal (CALVETE et al., 2002).
Algumas espécies de bromélias têm o crescimento lento quando
comparadas com outras espécies vegetais, devido ao mecanismo de fixação do
carbono durante a fotossíntese (RAVEN et al., 2001). Espécies com mecanismo
CAM de fixação de carbono normalmente são plantas de crescimento lento,
capacidade reprodutiva e condições de floração limitada, sendo a propagação
convencional um processo desvantajoso (MALDA et al., 1999).
O estímulo a um desenvolvimento vegetativo mais rápido aumentaria a
eficiência do processo de propagação e aceleraria a produção das bromélias em
escala comercial, reduzindo o extrativismo. Entre os fatores que determinam o cultivo
in vitro estão a fonte de carbono, o carvão ativado e o tratamento com reguladores
vegetais, os quais tem interferido na velocidade de crescimento de muitas espécies
(CHAGAS et al., 2005; LEITE et al., 2003).
O presente estudo avaliou a influência de diferentes fontes de carbono e a
presença de carvão ativado associado a diferentes concentrações de ácido
indolbutírico (AIB) no crescimento in vitro de Orthophytum mucugense.

42
2.2 MATERIAL E MÉTODOS
2.2.1 MATERIAL VEGETAL
Foram utilizados brotos com cerca de 5 mm de comprimento oriundos de
cultivo em meio de cultura MS com metade da concentração salina (MS½)
suplementado com 0,65µM de ANA e 2,22µM de BAP, conforme trabalhos
desenvolvidos por Bellintani (2006).
2.2.2 CRESCIMENTO IN VITRO
O crescimento in vitro foi analisado através de dois experimentos: o primeiro
avaliou o efeito de diferentes fontes de carbono e o segundo a influência do ácido
indolbutírico (AIB) associado com carvão ativado.
2.2.2.1 Fonte de carbono
Para avaliar a influência de diferentes fontes de carbono, foram utilizados
sacarose, glicose, maltose, sorbitol, açúcar cristal, açúcar mascavo e manitol na
concentração de 87,64 mM, tomando como base a concentração de sacarose
estabelecida por Bellintani (2006). Para a concentração de açúcar cristal e o
mascavo foi tomada como referência a utilizada para sacarose em grama, que
corresponde a 30 g.L-1. Foi utilizado 80 mL de meio de cultura MS½ por frascos de
250 mL, tampados com filme PVC.

43
O delineamento estatístico utilizado foi inteiramente casualizado com seis
repetições de dez brotos cada. Após 60 dias de cultivo, avaliou-se o comprimento da
parte aérea e da raiz e a matéria seca da parte aérea e das raízes.
2.2.2.2 AIB e carvão ativado
Para avaliar o efeito do AIB e do carvão ativado, plantas foram cultivadas em
meio MS½ suplementado com sacarose (87,64 mM) na presença de ácido
indolbutírico-AIB (0,00; 1,11 e 2,22 µM) combinado com dois níveis de carvão ativado
(0,00 e 83,2 mM) em arranjo fatorial 3x2 (concentração de AIB x níveis de carvão
ativado).
O delineamento estatístico utilizado foi inteiramente casualizado com 12
repetições de cinco brotos cada. Após 60 dias, avaliou-se o comprimento da parte
aérea e da raiz, e o peso seco da parte aérea e das raízes.
2.2.3 CONDIÇÃO DE CULTIVO IN VITRO
Os experimentos foram conduzidos em sala de crescimento com
temperatura de 25 ± 2ºC, fotoperíodo de 16h e densidade de fluxo de fótons
fotossinteticamente ativos de 60 µmol.m-2.s-1.
Todos os meios utilizados tiveram o pH ajustado para 5,8 antes da
autoclavagem, que foi realizada sob temperatura de 121ºC por 15 minutos.

44
2.2.4 ANÁLISE ESTATÍSTICA
Os dados foram submetidos a análise de variância (ANOVA) e as médias
comparadas pelo teste Tukey (0,05), utilizando o programa estatístico SISVAR v. 4.4,
desenvolvido pela UFLA (FERREIRA, 2000).

45
2.3 RESULTADOS E DISCUSSÃO
Os resultados obtidos nesse trabalho demonstram efeito significativo da
fonte de carbono no desenvolvimento de brotos micropropagados via organogênese
direta (Tabela 2.1) e que o carvão ativado influenciou o crescimento da parte aérea
e, quando associado ao AIB estimula o enraizamento (Tabela 2.2).
Sorbitol, sacarose e açúcar cristal demonstraram ser as melhores fontes de
carbono para estimular o aumento do comprimento da parte aérea em O.
mucugense. Rego-Oliveira et al. (2003) observaram que a sacarose também
estimula o crescimento de brotos de Oncidium varicosum, mas apenas até a
concentração de 175,28 mM pois acima disso ocorreu a diminuição na altura da
parte aérea da planta.
Brotos cultivados em meio suplementado com maltose apresentaram uma
baixa taxa de crescimento. Resultado semelhante foram obtido para brotos de
Oncidium varicosum cultivados em 27,75 mM de maltose (REGO-OLIVEIRA et al.,
2003).
Os melhores resultados para variável comprimento das raízes foram obtidos
quando se utilizou o sorbitol e o manitol como fonte de carbono. A glicose promoveu
uma das menores taxas de crescimento radicular (Tabela 2.1). Estes resultados
contrastam com os observados para Quercus suber, espécie lenhosa que obteve
excelente enraizamento na presença de glicose, e que não formou raízes na
presença de sorbitol (ROMANO et al., 1995).
Os brotos desenvolvidos em meio suplementado com açúcar mascavo
apresentaram pequeno crescimento das raízes e alteração na coloração das folhas
novas em relação aos outros tratamentos (Figura 2.1). As folhas desenvolvidas após
a exposição dos brotos ao meio de cultura com açúcar mascavo apresentaram
coloração verde clara. Em bananeira também foi observada uma alteração na
coloração das plantas desenvolvidas na presença de açúcar mascavo quando
comparadas com plantas procedentes de meio com açúcar cristal ou sacarose
(BERNARDI et al., 2004).

46
O amarelecimento das folhas pode ocorrer pela deficiência em algum
mineral e é denominada de clorose. Um dos minerais que em concentrações
deficientes promove essa característica em folhas novas é o cálcio, que além da
clorose pode induzir a formação de folhas curvadas para baixo e raízes
acastanhadas, curtas e altamente ramificadas, com redução severa no crescimento
(TAIZ e ZEIGER, 2004), características semelhantes às encontradas nos brotos de
O. mucugense desenvolvidos em meio com açúcar mascavo (Figura 2.1).
A diferença na coloração pode ter ocorrido também pela falta de carbono, já
que o açúcar mascavo apresenta um maior grau de impureza o que pode ter refletido
diretamente no reduzido tamanho da parte aérea e das raízes (Tabela 2.1; Figura
2.1).
Tabela 2.1 – Valores médios para comprimento da parte aérea (CPA) e das raízes (CR), e matéria seca das raízes (MSR) e parte aérea de brotos (MSPA) da espécie Orthophytum mucugense cultivados em meio MS ½ suplementado com diferentes fontes de carbono, após 60 dias de cultivo. Feira de Santana-BA, 2007.
Fonte de carbono
CPA (mm) CR (mm) MSPA (mg/planta)
MSR (mg/planta)
Sacarose 22,05ab 16,65b 6,47a 1,40a
Glicose 19,03b 8,65c 6,20a 0,80a
Maltose 18,50b 8,77bc 6,40a 1,20a
Sorbitol 26,82a 25,23a 8,80a 1,00a
Açúcar cristal 21,84ab 13,83bc 6,80a 1,00a
Açúcar mascavo 16,84b 9,42bc 4,60a 1,20a
Manitol 18,22b 24,62a 5,80a 0,60a
Médias seguidas pela mesma letra, em cada coluna, não diferem entre si ao nível de 5% de probabilidade pelo teste de Tukey.

47
Figura 2.1. Brotos da espécie Orthophytum mucugense procedentes de meio de cultura MS ½ suplementado com diferentes fontes de carbono (87,64mM ), após 60 dias de cultivo. A barra representa 1,0cm. Feira de Santana, 2007.

48
Na cultura de tecidos vegetais tem se observado cada vez mais o interesse
na diminuição de custos para a produção de mudas micropropagadas (KOZAI e
KUBOTA, 2001).
O bom crescimento observado nos brotos cultivados em meio com açúcar
cristal indica que essa fonte de carbono, comercialmente mais acessível, pode ser
utilizada com sucesso na propagação in vitro de O. mucugense. A substituição de
sacarose P.A. por açúcar cristal na cultura de tecidos, no Brasil, permite uma
redução de até 50,2% no custo de produção de um litro de meio de cultura, como foi
observado por Bernardi et al. (2004). Os autores calcularam que o custo de um litro
de meio ao utilizar a sacarose P.A. era de R$ 4,16 e com açúcar cristal R$ 2,07.
Em relação a matéria seca das raízes e da parte aérea, não foram
observadas diferenças significativas para as fontes de carbono utilizadas (Tabela
2.1). Estes resultados diferem dos encontrados por Nicoloso et al. (2003) para Pfaffia
glomerata. O autor avaliou cinco fontes de carboidratos (sacarose, frutose, glicose,
lactose e maltose) em quatro concentrações e obteve um maior ganho de matéria
seca da parte aérea e das raízes quando utilizou a sacarose, sendo essa fonte a
melhor em todas as variáveis analisadas, enquanto a lactose apresentou o menor
rendimento para quase todas as variáveis com exceção apenas no número de
brotações, que na concentração de 83,28 mM se igualou à sacarose.
Observou-se que a interação “carvão ativado x AIB” interfere no
desenvolvimento da parte aérea e das raízes (Tabela 2.2). O comprimento da parte
aérea de brotos cultivados em meio suplementado com 1,11 µM de AIB foi
beneficiado pela presença de carvão ativado. No entanto, a matéria seca da parte
aérea não foi influenciada pelo carvão ativado ou pelo AIB, demonstrando que
apesar de ocorrer um alongamento, não foi observado desenvolvimento da parte
aérea de brotos sob a influencia destas substâncias (Tabela 2.2).
As concentrações de AIB utilizadas nesse experimento não influenciaram no
comprimento da parte aérea de O. mucugense. Esse resultado concorda com os
obtidos por Rios (2004) que, ao estudar o efeito de diferentes concentrações de AIB
(0,00; 1,48; 2,46; 4,90 µM) em Gypsophila paniculata, não observou alterações no
crescimento da parte aérea.
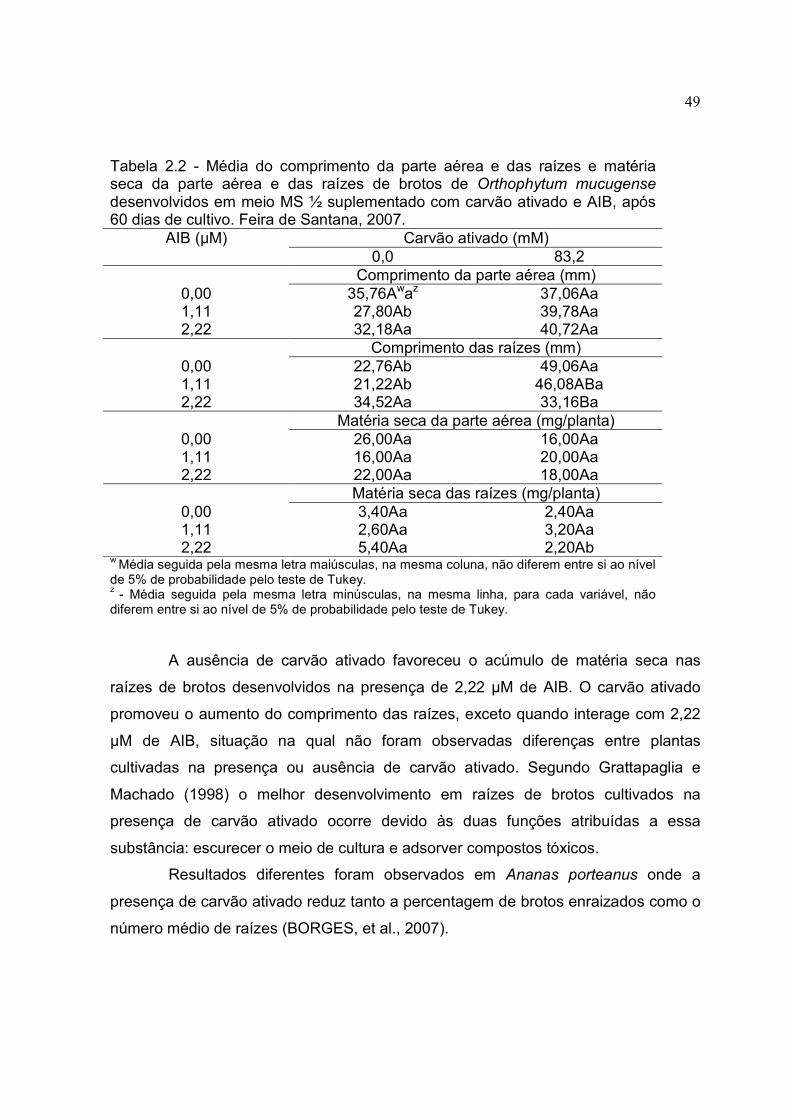
49
Tabela 2.2 - Média do comprimento da parte aérea e das raízes e matéria seca da parte aérea e das raízes de brotos de Orthophytum mucugense desenvolvidos em meio MS ½ suplementado com carvão ativado e AIB, após 60 dias de cultivo. Feira de Santana, 2007.
AIB (µM) Carvão ativado (mM) 0,0 83,2 Comprimento da parte aérea (mm)
0,00 35,76Awaz 37,06Aa 1,11 27,80Ab 39,78Aa 2,22 32,18Aa 40,72Aa Comprimento das raízes (mm)
0,00 22,76Ab 49,06Aa 1,11 21,22Ab 46,08ABa 2,22 34,52Aa 33,16Ba Matéria seca da parte aérea (mg/planta)
0,00 26,00Aa 16,00Aa 1,11 16,00Aa 20,00Aa 2,22 22,00Aa 18,00Aa Matéria seca das raízes (mg/planta)
0,00 3,40Aa 2,40Aa 1,11 2,60Aa 3,20Aa 2,22 5,40Aa 2,20Ab
w Média seguida pela mesma letra maiúsculas, na mesma coluna, não diferem entre si ao nível de 5% de probabilidade pelo teste de Tukey. z - Média seguida pela mesma letra minúsculas, na mesma linha, para cada variável, não diferem entre si ao nível de 5% de probabilidade pelo teste de Tukey.
A ausência de carvão ativado favoreceu o acúmulo de matéria seca nas
raízes de brotos desenvolvidos na presença de 2,22 µM de AIB. O carvão ativado
promoveu o aumento do comprimento das raízes, exceto quando interage com 2,22
µM de AIB, situação na qual não foram observadas diferenças entre plantas
cultivadas na presença ou ausência de carvão ativado. Segundo Grattapaglia e
Machado (1998) o melhor desenvolvimento em raízes de brotos cultivados na
presença de carvão ativado ocorre devido às duas funções atribuídas a essa
substância: escurecer o meio de cultura e adsorver compostos tóxicos.
Resultados diferentes foram observados em Ananas porteanus onde a
presença de carvão ativado reduz tanto a percentagem de brotos enraizados como o
número médio de raízes (BORGES, et al., 2007).

50
Foi possível observar também que na presença de carvão ativado, ocorreu
uma redução no comprimento das raízes com o aumento da concentração de AIB
(Tabela 2.2).
Apesar do carvão ativado ter aumentado o comprimento das raízes de O.
mucugense, os brotos formaram raízes mesmo na ausência do carvão ativado e do
regulador de crescimento. Estes resultados confirmam os obtidos por Bellintani
(2006) que analisou o enraizamento da mesma espécie sob a influência de diferentes
concentrações de sacarose (0; 43,82 e 87,64 mM) e carvão ativado (0; 83,2 mM).
Umas das funções das auxinas é estimular a iniciação e formação de tecidos
meristemáticos e atuar no alongamento dos tecidos que darão origem aos primórdios
radiculares (BOSA et al., 2003). Algumas espécies apresentam níveis endógenos de
auxina suficientes para promover o enraizamento, sem necessitar de reguladores
externos, como foi observado em O. mucugense no presente trabalho e por Bellintani
(2006) nessa mesma espécie e em brotos de outras espécies da família
Bromeliaceae (MACÊDO et al., 2003; SILVA et al., 2007) e em Gypsophila paniculata
(BOSA et al., 2003).
De acordo com os resultados obtidos neste trabalho, sugere-se a utilização
de açúcar cristal como fonte de carbono, a fim de diminuir custos de produção e o
uso do carvão ativado para otimizar a fase de enraizamento.

51
REFERÊNCIAS
BELLINTANI, Moema Cortizo. Estudos da propagação in vitro e ex vitro de Neoregelia mucugensis Leme, Orthophytum mucugense Wand e Conceição e Orthophytum albopictum Philcox, espécies de Bromeliaceae endêmicas da Bahia. 2006. 115p. Tese (Doutorado em Botânica) – Universidade Estadual de Feira de Santana, Feira de Santana.
BERNARDI, Walter Fernando; RODRIGUES, B. I.; CASSIERE NETO, P.; ANDO, A.; TULMANN NETO, A.; CERAVOLO, L. C.; MONTES, S. M. N. M. Micropropagação de baixo custo em bananeira C.V. Maça em meios com diferentes fontes de carbono. Revista Brasileira de Fruticultura, Jaboticabal, v. 26, n. 3, 2004.
BORGES, Neiliane Santiago Sombra; CORREIA, Diva, LIMA, Raimundo Nonato. Multiplicação e enraizamento in vitro de brotos de abacaxi ornamental Ananás porteanus Hort Veitch ex C.Koch. EMBRAPA. Disponível em: http://www.redbio.org/portal/encuentros/enc_2001/posters/01/posterpdf/01-004.pdf. Acesso em: 10 de dez. 2007.
BOSA, Nair; CALVETE, Eunice O.; NIENOW, Alexandre Augusto; SUZIN, Marilei. Enraizamento e aclimatização de plantas micropropagadas de gipsofila. Horticultura Brasileira, Brasília, v. 21, n. 2, p.207-210, abr.-jun., 2003.
CALDAS, L. S.; HARIDASAM, P.; FERREIRA, M. E. Meios nutritivos. In: TORRES, A. C.; CALDAS, L. S.; BUSO, J. A. Cultura de tecidos e transformação genética de plantas. Brasília: Embrapa/SPI, v. 1, p. 87-132, 1998.
CALVETE, Eunice Oliveira; KÄMPF , Atelene Normann; SUZIN Marilei. Concentração de sacarose no enraizamento in vitro de morangueiro. Horticultura Brasileira, Brasília, v. 20, n. 2, p. 186-191, jun., 2002.
CARNEIRO, L.A.; MANSUR, E. Contribuição de metodologias in vitro para a conservação de Bromeliaceae. Vidalia, Viçosa, v. 2, n.1, p. 12-20, 2004.
CHAGAS, E. A.; PASQUAL, M.; RAMOS, J. D.; PIO, L. A. S.; DUTRA, L. F.; CAZETTA, J. O. Cultivo de embriões imaturos de citros em diferentes concentrações de carvão ativado e ácido giberélico. Ciência e Agrotecnologia, Lavras, v. 29, n. 6, p. 1125-1131, 2005.
ENGELMANN, F. In vitro conservation of tropical plant germplasm – a review. Euphytica, Netherlands, v. 57, p. 227-243, 1991.
FERREIRA, D. F. Análise estatística por meio do SISVAR para Windows: Versão 4.0. In: REUNIÃO ANUAL DA REGIÃO BRASILEIRA DA SOCIEDADE INTERNACIONAL DE BIOMETRIA, 45., 2000, São Carlos. Anais... São Carlos: SSP/SIB, p. 255-268, 2000.

52
GRATTAPAGLIA, Dario; MACHADO, Marcos A. Micropropagação In: TORRES, A. C.; CALDAS, L. S.; BUSO, J. A. Cultura de tecidos e transformação genética de plantas. Brasília: Embrapa/SPI, v.1, p. 183-260. 1998.
HINOJOSA, Gladys Fernández. Auxinas. In: CID, Luis Pedro Barrueto. Introdução aos hormônios vegetais, Brasília: Embrapa Recursos Genéticos e Biotecnologia, p. 15-54, 2000.
KOZAI, T; KUBOTA, C. Developing a photoautotrophic micropropagation system for woody plants. Journal of Plant Research, Tokyo, v.114, p.525-537, 2001.
KUNHA, N. América das Bromélias. Nova Ciência, São Paulo, v. 26, p. 32-39, 1995.
LEITE, V. M.; ROSOLEM, C. A.; RODRIGUES, J. D. Gibberellin and cytokinin effects on soybean growth. Scientia Agricola, Piracicaba, v. 60, n. 3, p. 537-541, 2003.
LEME, E.M.C.; MARIGO, L.C., Bromeliads in the Brazilian wilderness. Rio de Janeiro: Marigo Comunicação visual, 1993. 183p.
MACÊDO, Cristiane Elizabeth Costa de; SILVA, Marcela Gomes da, NÓBREGA, Fabiana Silva da; MARTINS, Camila Pimentel; BARROSO, Paulo Augusto Vianna; ALLOUFA, Magdy Ahmed Ibrahim. Concentração de ANA e BAP na micropropagação de abacaxizeiro L.Merrill (Ananas comosus) e no cultivo hidropônico das plântulas obtidas in vitro. Revista Brasileira de Fruticultura, Jaboticabal - SP, v. 25, n. 3, p. 501-504, dez. 2003.
MALDA, Guadalupe; SUZÁN, Humberto; BACKHAUS, Ralph. In vitro culture as a potential method for the conservation of endangered plants possessing crassulacean acid metabolism. Scientia Horticulturae, Amsterdam, v. 81, p.71-87, 1999.
NICOLOSO, Fernando Teixeira; ERIG, Alan Cristiano; RUSSOWSKI, Denise; MARTINS, Cristiane Flores. Efeito de doses e fontes de carboidratos no crescimento de plantas de ginseng brasileiro [Pfaffia glomerata (spreng.) pedersen] cultivadas in vitro. Ciência e Agrotecnologia, Lavras, v.27, n.1, p.84-90, jan./fev., 2003.
RAVEN, Peter H.; EVERT, Ray F.; EICHHORN, Susan E. Biologia Vegetal. 6 ed. Rio de Janeiro:Guanabara, 2001. 906p.
RECH FILHO, A.; DAL VESCO, L. L.; NODARI, R. O.; LISCHKA, R. W.; MÜLLER, C. V.; GUERRA, M. P. Tissue culture for the conservation and mass propagation of Vriesea reitzii Leme and Costa, a bromeliad threatened of extinction from the Brazilian Atlantic Forest. Biodiversity and Conservation, Netherlands, v. 14, n. 8, p. 1799 -1808, 2005.
REGO-OLIVEIRA, Luciana do Valle; FARIA, Ricardo Tadeu de; FONSECA, Inês Cristina de Batista; SACONATO, Charlen. Influence of source and concentration of carbohydrate on shoot growth and rooting of Oncidium varicosum Lindl. (Orchidaceae). Semina: Ciências Agrárias, Londrina, v.24, n.2, p.265-272, jul./dez., 2003.

53
RIOS, José Fernando. Micropropagação de Gypsophila paniculata pela cultura de segmentos nodais e calogênese a partir de segmentos foliares. 2004. Dissertação (Mestrado em Agronomia) Universidade Federal do Paraná, Curitiba.
ROMANO, A.; NORONHA, C.; MARTINS-LOUCAO, M. A. Role of carbohydrates in micropropagation of cork oak. Plant Cell, Tissue and Organ Culture, Netherlands, v. 40, n. 2, p. 159-167, 1995.
SILVA, André Luís Lopes da; DORNELLES, Eduardo Bortoluzzi; BISOGNIN, Dilson Antônio; FRANCO, Elci Terezinha Henz e HORBACH, Micheli Angélica. Micropropagation of Dyckia agudensis Irgang & Sobral – an extinction threatened bromeliad. Iheringia, Série Botânica, Porto Alegre, v. 62, n. 1, p. 39-43, jan./jun. 2007.
TAIZ, Lincoln; ZEIGER, Eduardo. Fisiologia Vegetal. 3 ed., Porto Alegre: Artmed, 2004. 719p.
WANDERLEY, Maria das Graças Lapa; CONCEIÇÃO, Abel Augusto. Notas Taxonômicas e uma nova espécie do gênero Orthophytum Beer (Bromeliaceae) da Chapada Diamantina, Bahia, Brasil. Sitientibus, Série Ciências Biológicas, Feira de Santana, v. 6, n.1, p. 03-08, 2006.

54
CAPÍTULO 3
Fatores que afetam a micropropagação de Orthophytum mucugense WAND e
CONCEIÇÃO: reguladores de crescimento, carvão ativado e fechamento dos
recipientes de cultivo.
RESUMO Orthophytum mucugense é uma espécie endêmica restrita ao município de Mucugê, na Chapada Diamantina-BA. A ocorrência restrita torna importante a realização de trabalhos que visem a conservação. Para o uso da micropropagação com finalidade de conservação é necessário a produção de plantas com estabilidade genética, sendo importante o desenvolvimento de métodos que reduzam a ocorrência de mutações e a variação somaclonal. A regeneração de brotos a partir de organogênese direta e a redução da concentração e do tempo de exposição dos explantes aos reguladores de crescimento diminuem as chances de ocorrência de alterações genéticas e a utilização de tampas que permitam trocas gasosas, nos frascos de culturas, é fundamental para acelerar o processo de aclimatização. O objetivo do presente trabalho foi avaliar o efeito de diferentes tipos e concentrações de reguladores de crescimento sob a formação de brotos de O. mucugense cultivados em tubos sob a influência de dois tipos de selamento e avaliar também o desenvolvimento desses brotos em meio de enraizamento com posterior aclimatização. Os resultados obtidos no presente estudo mostram que a utilização de TDZ promove uma boa taxa regenerativa após 60 dias de cultivo, que o AIB não interferiu na formação de raízes e que o fechamento dos tubos com algodão, no período de enraizamento, acelera a aclimatização da espécie O. mucugense permitindo a transferência direta das plantas para o ambiente ex vitro.
Palavras-chave: Bromeliaceae. BAP. CIN. TDZ. Carvão ativado. AIB. Aclimatização.
ABSTRACT
Orthophytum mucugense is an endemic plant restricted the municipality of Mucugê, in the Chapada Diamantina-BA; and its limited occurrence makes conservation efforts more urgent and important. The use of micropropagation in conservation efforts must consider the necessity of producing genetically stable plants, and it will be important to develop methods that reduce the occurrence of mutations and somatic-clonal variations. The regeneration of shoots from direct organogenesis, the reduction of the concentration of growth regulators, and diminishing explant exposure time to these

55
substances reduces the probability of genetic alterations. Additionally, the use of permeable caps that facilitate gas exchange in culture vessels is essential to accelerate the acclimatization processes. The present study evaluated the effects of different types and concentrations of growth regulators on bud formation in O. mucugense explant grown in culture tubes sealed with two different types of caps sealing, and evaluated the short term development of these shoots in rooting culture medium during later acclimatization. The results indicated that the use of TDZ promotes good regeneration rates after 60 days of cultivation, that IBA did not interfere in root formation, and that sealing the culture tubes with cotton during the rooting phase accelerates acclimatization of O. mucugense and allows the transfer of these plants directly to the ex vitro environment. Keywords: Bromeliaceae. BAP. CIN. TDZ. Activated charcoal. IBA. Acclimatization.

56
3.1 INTRODUÇÃO
Com cerca de 3000 espécies, Bromeliaceae forma a maior família de plantas
essencialmente americana (LEME e MARIGO, 1993). No Brasil, encontram-se quase
metade das espécies de bromélias conhecidas, distribuindo-se de norte a sul,
especialmente na região costeira, na Mata Atlântica e na restinga, podendo aparecer
também na Amazônia, na caatinga, no cerrado e nos campos de altitude, em todos
os ecossistemas (LEME e MARIGO, 1993; PAULA, 2004). No Brasil ocorrem
gêneros endêmicos como o Orthophytum que é composto por espécies de notável
valor ornamental (WANDERLEY, 1990)
Orthophytum mucugense é considerada vulnerável por ter ocorrência restrita
ao município de Mucugê-BA, na Chapada Diamantina. Esta espécie possui
características comerciais devido à coloração vermelha que suas folhas possuem no
período de floração (WANDERLEY e CONCEIÇÃO, 2006). Trabalhos visando a
propagação podem ajudar na preservação desta espécie.
Com esse intuído, a micropropagação vem sendo aplicada em bromélias
como Vriesea reitzii, Cryptanthus sinuosus e Dyckia agundensis (CARNEIRO et al.,
1998; CARNEIRO e MANSUR, 2004; RECH FILHO et al., 2005; SILVA et al., 2007).
Neste processo é necessária a criação de protocolos que permitam a multiplicação
em grande escala, em um período curto de tempo para cada espécie a ser
preservada.
A variabilidade da resposta morfogenética existente entre os vegetais reflete
a necessidade de estabelecer protocolos eficientes para cada espécie
(GRATTAPAGLIA e MACHADO, 1998). Sendo necessário avaliar fatores que afetam
a morfogênese, como os tipos de reguladores utilizados no período de multiplicação
e enraizamento (COSTA et al., 2006).
O tipo e a concentração do regulador de crescimento utilizado é fundamental
para uma resposta morfogenética. Essa resposta é atribuída a mudanças na
sensibilidade do tecido ao regulador, dependendo do número de receptores e
característica do regulador que irá sinalizar a transdução (HINOJOSA, 2000). A

57
importância de pesquisar qual regulador induz a uma resposta morfogenética para
determinada espécie está relacionado às diferenças que existem entre as ações dos
reguladores de crescimento. De modo geral a benzilaminopurina (BAP) promove
uma alta taxa de multiplicação, já a cinetina (CIN), além de induzir brotações, age no
crescimento, enquanto o thidiazuron (TDZ) pode induzir organogênese direta e
indireta, além de estimular também a embriogênese somática, porém já foi relatada a
dificuldade de enraizamento dos brotos induzidos por esse regulador (CARNEIRO et
al., 1998; CALDAS et al., 1998; CID, 2000; ROY et al., 2007; SAINI e JAIWAL, 2002;
YAN et al, 2006).
Brotos regenerados in vitro podem apresentar dificuldade de enraizamento,
tornando necessária a utilização de reguladores com a finalidade de promover a
formação de raízes. As auxinas são os principais reguladores utilizados no
enraizamento, pois têm a capacidade de promover o desenvolvimento de raízes
adventícias no caule, que irá auxiliar na fase de aclimatização (TAGLIACOZZO,
1998).
Altas taxas de sobrevivência na aclimatização de espécies micropropacadas
dependem dos tratamentos utilizados na transição para o ambiente externo. No
período de aclimatização devem ocorrer modificações morfológicas, anatômicas e
fisiológicas que permitam a transição para um novo ambiente (CALVETE, et al.
2002). Nessa fase é necessária a utilização de instrumentos que proporcionem
características semelhantes às encontradas no ambiente ex vitro. Tendo o ambiente
in vitro uma alta taxa de umidade, as plantas não apresentam características
anatômicas e nem fisiológicas que auxiliem no controle da transpiração
(BUDDENDORF-JOOTEN e WOLTERING, 1996; DE PROFT et al., 1985; SEELYE
et al., 2003; ZOBAYED et al., 1999). Assim, um modelo de cultivo que possibilite o
aumento das trocas gasosas tem facilitado e acelerado a fase de aclimatização das
mudas (JACKSON, 2003).
Em trabalhos realizados com O. mucugense, Bellintani (2006) induziu
número considerável de brotações (média de 5 brotos por explante). Todavia, foi
necessário um período longo (seis meses) de exposição dos explantes aos
reguladores de crescimento. Neste mesmo trabalho foi observada a formação de

58
calos após quarto meses de cultura, o que é indesejável devido a possível ocorrência
de alteração genética. Considerando as fases de multiplicação, crescimento,
enraizamento e aclimatização, a produção de mudas de O. mucugense durou 280
dias (BELLINTANI, 2006).
O objetivo do presente trabalho foi avaliar o efeito de diferentes tipos e
concentrações de reguladores de crescimento sobre a formação de brotos de O.
mucugense considerando a influência de dois tipos de selamento dos recipientes de
cultivos, e avaliar também o desenvolvimento desses brotos em meio de
enraizamento com posterior aclimatização.

59
3.2 MATERIAL E MÉTODOS
3.2.1 MATERIAL VEGETAL
Foram utilizados caules, sem a presença do ápice, de plantas com 40 dias
de cultivo (ver capítulo 1), germinadas in vitro em meio contendo 7g.L-1 de ágar e
cultivadas em meio MS com metade da concentração salina (MS½) suplementado
com 87,64 mM de sacarose (BELLINTANI, 2006). As plantas foram colocadas no
escuro 30 dias antes de serem utilizadas. Foi utilizado tubos de ensaio (25x150 mm)
com 15 mL de meio de cultura.
3.2.2 MULTIPLICAÇÃO
No experimento de multiplicação foi utilizado como meio básico o MS½
suplementado com 87,64 mM de sacarose e gelificado com 7 g.L-1 de ágar (Bellintani
2006).
Para a multiplicação foi avaliada a influência de 6-benzilaminopurina (BAP),
cinetina (CIN) e thidiazuron (TDZ), em duas concentrações (2,22 e 4,44 µM). Os
tubos de ensaio com os explantes foram submetidos a dois tipos de selamento (PVC
e algodão) para avaliar a influência das trocas gasosas na multiplicação.
Foi utilizado o delineamento inteiramente casualizado em esquema fatorial
2x3x2 (selamento x tipo de regulador de crescimento x concentração), com seis
repetições, e cinco explantes por parcela.
Decorridos 60 dias da inoculação foram avaliados: o percentual de explantes
responsivos, o número de brotos formados por explante e o tamanho dos brotos
formados. Em seguida os brotos foram transferidos para tubos de ensaio fechados
com algodão contendo meio MS½ sem regulador de crescimento. Após 60 dias foi
avaliada a taxa de enraizamento e sobrevivência das plantas.

60
3.2.3 ENRAIZAMENTO
Foi observada uma baixa taxa de enraizamento nos brotos formados em
meio suplementado com o regulador TDZ independente da concentração e do tipo de
selamento utilizado. Esses brotos foram então transferidos para meio de
enraizamento. No entanto os brotos produzidos em meio com BAP e CIN produziram
raízes e por isso não foram induzidos à formar raízes.
O meio de enraizamento utilizado foi o MS½ suplementado com 1,11 ou
2,22µM de ácido indolbutírico (AIB) combinado com dois níveis de carvão ativado
(0,00 e 83,2 mM).
O delineamento estatístico utilizado foi inteiramente casualizado em arranjo
fatorial 2x2 (concentração de AIB x níveis de carvão ativado), com seis repetições de
cinco brotos cada. Após 60 dias foi avaliado o comprimento da parte aérea e das
raízes, e o peso seco da parte aérea e das raízes.
3.2.4 ACLIMATIZAÇÃO
Plantas enraizadas foram transferidas para substrato composto de
vermiculita + terra (1:1) (BELINTTANI, 2006). A influência da umidade na
aclimatização foi avaliada. Para tanto, as plantas transferidas para o viveiro com 70%
de luminosidade foram cobertas com garrafas PET transparentes por 0, 5 ou 10 dias.
Dois dias antes da retirada das garrafas as tampas foram removidas.
O delineamento estatístico utilizado foi inteiramente casualizado com 20
repetições de três plantas cada. A cada cinco dias foi avaliada a taxa de
sobrevivência dessas plantas.

61
3.2.5 ANÁLISE HISTOLÓGICA
A fim de avaliar a influência de diferentes tipos de selamento na
micropropagação, foi realizada a análise histológicas de cinco amostra de folhas dos
brotos produzidos em cada tratamento. As amostras foram fixadas em álcool 70% e
com auxílio de lâminas de barbear foram realizados cortes transversais à mão livre
nas folhas. Os cortes foram clarificados com hipoclorito de sódio a 3% por 15
minutos, lavados em água destilada e corados com safrablau (BUKATSCH, 1972).
Devido ao pequeno porte das folhas dos brotos (média de 2 mm) algumas
foram cortadas no micrótomo. Essas foram emblocadas em historesinas e passaram
por uma desidratacção etílica. Depois de emblocadas foram realizados cortes de 10
µm, os quais foram analisados em microscópio óptico e fotografados.
3.2.6 CONDIÇÃO DE CULTIVO IN VITRO
Os experimentos foram conduzidos em sala de crescimento com
temperatura de 25 ± 2ºC, fotoperíodo de 16h e densidade de fluxo de fótons
fotossinteticamente ativos de 60 µmol.m-2.s-1.
Todos os meios utilizados tiveram o pH ajustado para 5,8 antes da
autoclavagem, que foi realizada sob temperatura de 121ºC por 15 minutos.

62
3.2.7 ANÁLISE ESTATÍSTICA
Os dados foram submetidos à análise de variância (ANOVA) e as médias
comparadas pelo teste Tukey (0,05), utilizando o programa estatístico SISVAR v. 4.4,
desenvolvido pela UFLA (FERREIRA, 2000).

63
3.3 RESULTADOS E DISCUSSÃO
Os resultados obtidos nesse trabalho demonstram que a organogênese
direta em Orthophytum mucugense é influenciada pelo selamento do recipiente de
cultivo e pelo tipo e concentração de citocinina utilizados. Observou-se também que
o tipo de citocinina interfere no enraizamento dos brotos.
A resposta do explante aos reguladores foi rápida, com a formação de brotos
após uma semana de cultura. Resultado superior ao obtido por Bellintani (2006)
(terceira a quarta semana), com a mesma espécie na presença do ácido
naftalenoacético (ANA) e BAP. Em Cryptanthus sinuosus a formação de brotos foi
considerada lenta (6-7 semanas) na presença de BAP em concentrações iguais ou
superiores (4,44; 13,20 e 22,00 µM) às utilizadas no presente trabalho (CARNEIRO
et al., 1998). Em Cichorium intybus a formação de brotos também foi rápida (9 dias)
ao utilizar 2,32 µM de CIN, 2,22 µM de BAP ou 0,04 µM de TDZ associados com
1,71 µM de AIA ou 1,61 µM de ANA (YUCESAN et al., 2007).
O tipo e as concentrações dos reguladores utilizados não interferiram no
percentual de explantes responsivos (Tabela 3.1), porém a associação do TDZ com
o selamento algodão otimizou a taxa de responsividade dos explantes seguido pelos
tratamentos com CIN e BAP também associados ao selamento algodão (Tabela 3.2).
A média de explantes responsivos foi semelhante à obtida para a mesma espécie por
Bellintani (2006) ao analisar a interação de diferentes concentrações de ANA (0,00;
0,33; 0,65 e 1,30 µM) e BAP (0,00; 1,11; 2,22 e 4,44 µM).
O percentual de explantes responsivos obtido para O. mucugense no
tratamento com TDZ, independente da concentração utilizada, foi superior ao obtido
na micropropagação a partir de folhas de Cryptanthus sinuosus em todas as
concentrações de TDZ avaliadas (0,01; 0,04; 0,07 e 0,10 µM), mas inferior às obtidas
a partir dos caules dessa espécie (CARNEIRO et al., 1998).
Em cotilédones e segmentos foliares de Brassica juncea também foi
observado uma alta taxa de explantes responsivos na presença de TDZ associado
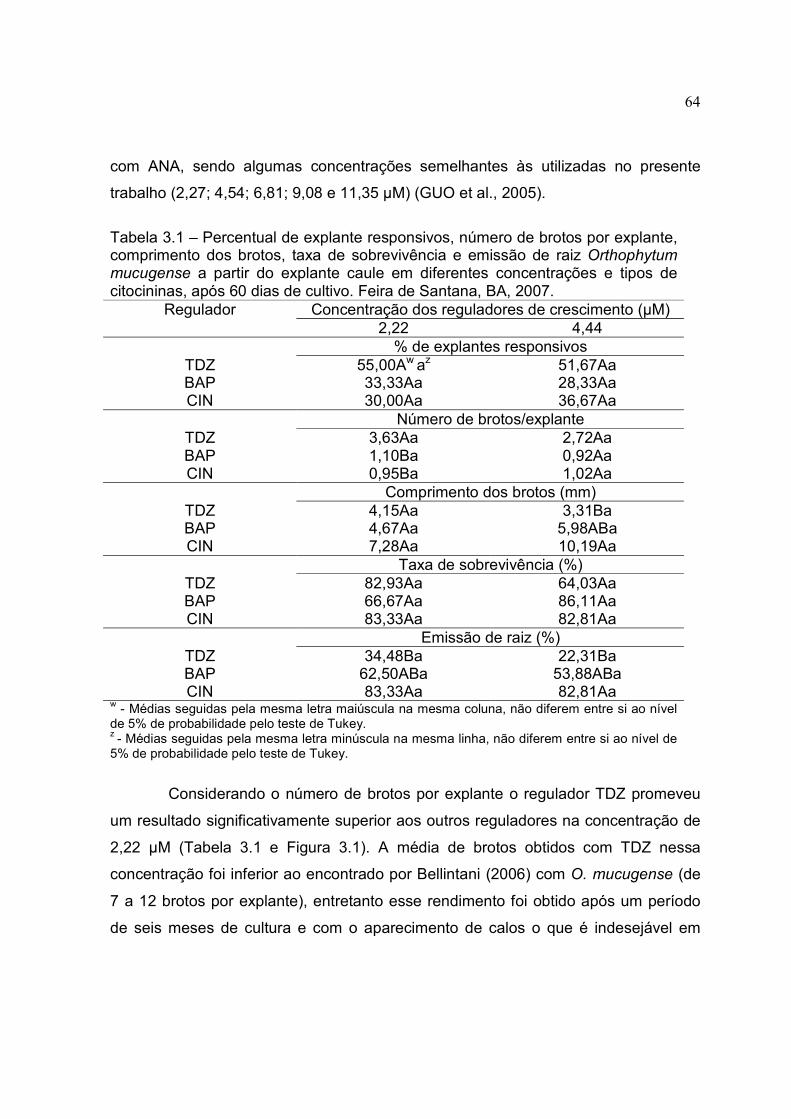
64
com ANA, sendo algumas concentrações semelhantes às utilizadas no presente
trabalho (2,27; 4,54; 6,81; 9,08 e 11,35 µM) (GUO et al., 2005).
Tabela 3.1 – Percentual de explante responsivos, número de brotos por explante, comprimento dos brotos, taxa de sobrevivência e emissão de raiz Orthophytum mucugense a partir do explante caule em diferentes concentrações e tipos de citocininas, após 60 dias de cultivo. Feira de Santana, BA, 2007.
Regulador Concentração dos reguladores de crescimento (µM) 2,22 4,44
% de explantes responsivos TDZ 55,00Aw az 51,67Aa BAP 33,33Aa 28,33Aa CIN 30,00Aa 36,67Aa Número de brotos/explante
TDZ 3,63Aa 2,72Aa BAP 1,10Ba 0,92Aa CIN 0,95Ba 1,02Aa Comprimento dos brotos (mm)
TDZ 4,15Aa 3,31Ba BAP 4,67Aa 5,98ABa CIN 7,28Aa 10,19Aa Taxa de sobrevivência (%)
TDZ 82,93Aa 64,03Aa BAP 66,67Aa 86,11Aa CIN 83,33Aa 82,81Aa Emissão de raiz (%)
TDZ 34,48Ba 22,31Ba BAP 62,50ABa 53,88ABa CIN 83,33Aa 82,81Aa
w - Médias seguidas pela mesma letra maiúscula na mesma coluna, não diferem entre si ao nível de 5% de probabilidade pelo teste de Tukey. z - Médias seguidas pela mesma letra minúscula na mesma linha, não diferem entre si ao nível de 5% de probabilidade pelo teste de Tukey.
Considerando o número de brotos por explante o regulador TDZ promeveu
um resultado significativamente superior aos outros reguladores na concentração de
2,22 µM (Tabela 3.1 e Figura 3.1). A média de brotos obtidos com TDZ nessa
concentração foi inferior ao encontrado por Bellintani (2006) com O. mucugense (de
7 a 12 brotos por explante), entretanto esse rendimento foi obtido após um período
de seis meses de cultura e com o aparecimento de calos o que é indesejável em

65
trabalhos de conservação devido à possibilidade de aparecimento de alterações
somaclonais.
O aumento da concentração de TDZ proporcionou uma diminuição no
percentual de explantes responsivos e no número de brotos formados, sugerindo que
altas concentrações desse regulador podem ter efeito inibitório para a
micropropagação da espécie em estudo. Já em lâminas foliares de Cichorium intybus
foi observado o aumento no número de brotos com o aumento da concentração de
TDZ de 0,02 µM para 0,09 µM (YUCESAN et al., 2007). Na multiplicação a partir de
cotilédones da espécie Selenicereus megalanthus também foi obtido um bom
resultado na presença de TDZ em concentrações superiores (100, 200 e 400 µM) às
utilizadas no presente trabalho, sendo o maior número de brotos por explante (sete)
obtido na concentração de 200 µM de TDZ (PELAH et al., 2002).
A taxa de multiplicação de O. mucugense na presença de TDZ foi melhor
quando se utilizou o selamento PVC (Tabela 3.2).
As concentrações de BAP avaliadas influenciaram de modo semelhante no
número de brotos formados a partir do caule de O. mucugense (Tabela 3.1). Já em
Tillandsia eizii foi observado o aumento no número de brotos com o aumento da
concentração de BAP (8,88 µM) associado com 2,69 µM de ANA (PICKENS et al.,
2006). Porém os resultados obtidos para O. mucugense corroboram os observados
para Prunus salicina quando cultivada em diferentes concentrações de BAP (2,22;
4,44; 8,87 e 13,30 µM) (ROGALSK et al., 2003).
Diferente do observado para O. mucugense, em Gerbera jamesonii foi
observada uma maior produção de brotos em explantes foliares na presença de 8,80
µM de BAP em comparação com outras fontes de citocininas (CIN e TDZ)
(OLIVERA-ORTEGA, et al., 2000).
Os reguladores de crescimento utilizados só interferiram no comprimento
dos brotos quando utilizados na concentração de 4,44 µM, tendo a CIN o melhor
resultado, apesar de não diferir do BAP (Tabela 3.1, Figura 3.1). O efeito da CIN
sobre o comprimento dos brotos foi maior ainda quando se utilizou a tampa de PVC
(Tabela 3.2). O pequeno porte observado nos brotos formados a partir do estímulo
com TDZ está de acordo com o observado em Cryptanthus sinuosus quando

66
estimulado com 0,2 e 10 µM de TDZ (CARNEIRO et al., 1998). Em Gerbera
jamesonii também foi observado um melhor desenvolvimento dos brotos produzidos
em meio com 2,20 µM CIN em comparação aos produzidos na presença de BAP e
TDZ, sendo observada a formação de calos na presença de TDZ (OLIVERA-
ORTEGA et al., 2000).
Segundo Caldas et al. (1998) as citocininas atuam de forma diferenciada. O
BAP tem capacidade de induzir grande número de brotos e alta taxa de
multiplicação, enquanto CIN influencia no crescimento mas não tem grande
influência na ocorrência de brotações múltiplas, fato que justifica a baixa taxa de
multiplicação mas o elevado comprimento dos brotos observados ao utilizar a CIN.
Outro tipo de interferência no comprimento dos brotos foi observado em
relação ao tipo de selamento utilizado. Sendo essa a única variável que mostrou
diferença significativa em relação ao tipo de selamento utilizado nos meios com
presença de BAP (Tabela 3.2).
A taxa de sobrevivência dos brotos não foi afetada por nenhum dos
tratamentos testados, apresentando valores maiores que 60% em todas as analises
(Tabelas 3.1 e 3.2).
Já a taxa de emissão de raízes foi maior nos brotos cultivados em meio com
CIN, nas duas concentrações avaliadas, não diferindo dos brotos sob influência do
BAP (Tabela 3.1). O bom enraizamento observado nos brotos formados na presença
de BAP corrobora os resultados observados por Bellintani (2006) para a mesma
espécie. A autora ao avaliar diferentes concentrações de sacarose e carvão ativado
obteve 100% de enraizamento nos brotos formados em meio MS½ suplementado
com sacarose (87,64 mM), BAP (2,22 µM) e ANA (0,65 µM).
Na espécie Dyckia agudensis também foi observado enraizamento na
ausência de reguladores em brotos produzidos em meio com diferentes
concentrações de BAP (2,22; 4,44 e 8,87 µM) associado ao AIB (2,46; 4,90 e 9,8 µM)
(SILVA et al., 2007).
A baixa taxa de enraizamento observada nos brotos de O. mucugense
originários do meio com TDZ contrasta com os resultados obtidos para outras
espécies. Em Cryptanthus sinuosus a utilização de baixas concentrações de TDZ
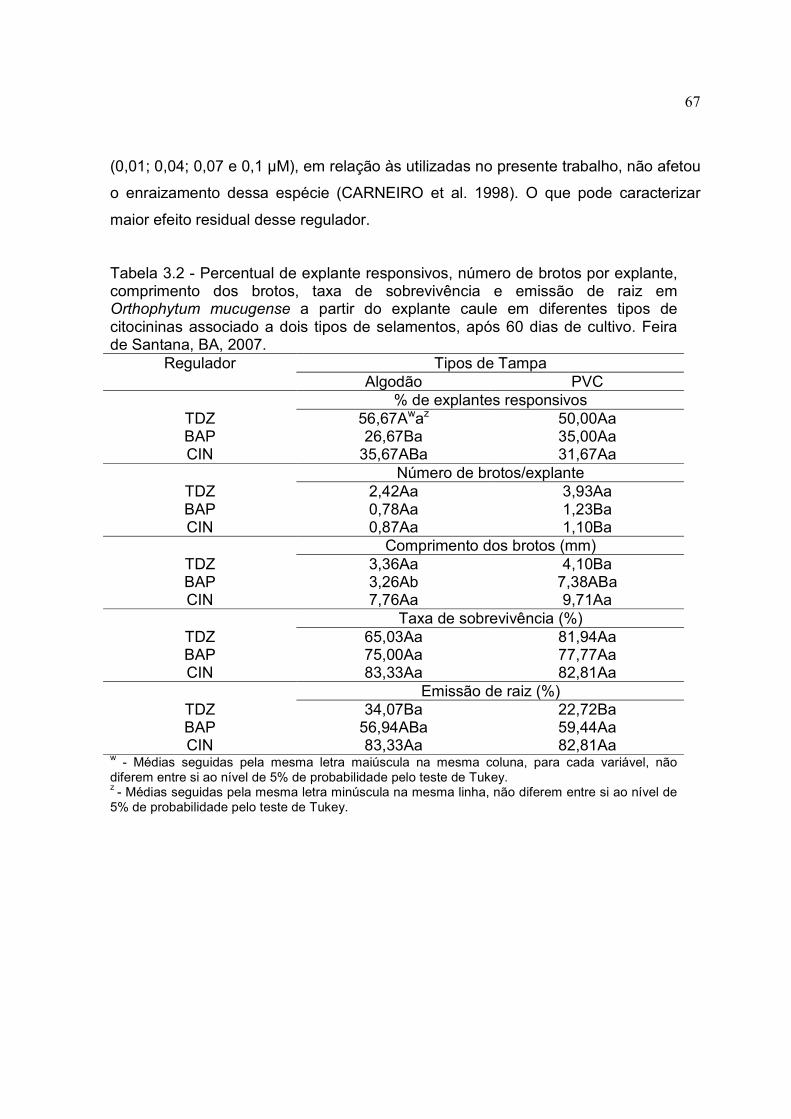
67
(0,01; 0,04; 0,07 e 0,1 µM), em relação às utilizadas no presente trabalho, não afetou
o enraizamento dessa espécie (CARNEIRO et al. 1998). O que pode caracterizar
maior efeito residual desse regulador.
Tabela 3.2 - Percentual de explante responsivos, número de brotos por explante, comprimento dos brotos, taxa de sobrevivência e emissão de raiz em Orthophytum mucugense a partir do explante caule em diferentes tipos de citocininas associado a dois tipos de selamentos, após 60 dias de cultivo. Feira de Santana, BA, 2007.
Regulador Tipos de Tampa Algodão PVC
% de explantes responsivos TDZ 56,67Awaz 50,00Aa BAP 26,67Ba 35,00Aa CIN 35,67ABa 31,67Aa Número de brotos/explante
TDZ 2,42Aa 3,93Aa BAP 0,78Aa 1,23Ba CIN 0,87Aa 1,10Ba Comprimento dos brotos (mm)
TDZ 3,36Aa 4,10Ba BAP 3,26Ab 7,38ABa CIN 7,76Aa 9,71Aa Taxa de sobrevivência (%)
TDZ 65,03Aa 81,94Aa BAP 75,00Aa 77,77Aa CIN 83,33Aa 82,81Aa Emissão de raiz (%)
TDZ 34,07Ba 22,72Ba BAP 56,94ABa 59,44Aa CIN 83,33Aa 82,81Aa
w - Médias seguidas pela mesma letra maiúscula na mesma coluna, para cada variável, não diferem entre si ao nível de 5% de probabilidade pelo teste de Tukey. z - Médias seguidas pela mesma letra minúscula na mesma linha, não diferem entre si ao nível de 5% de probabilidade pelo teste de Tukey.
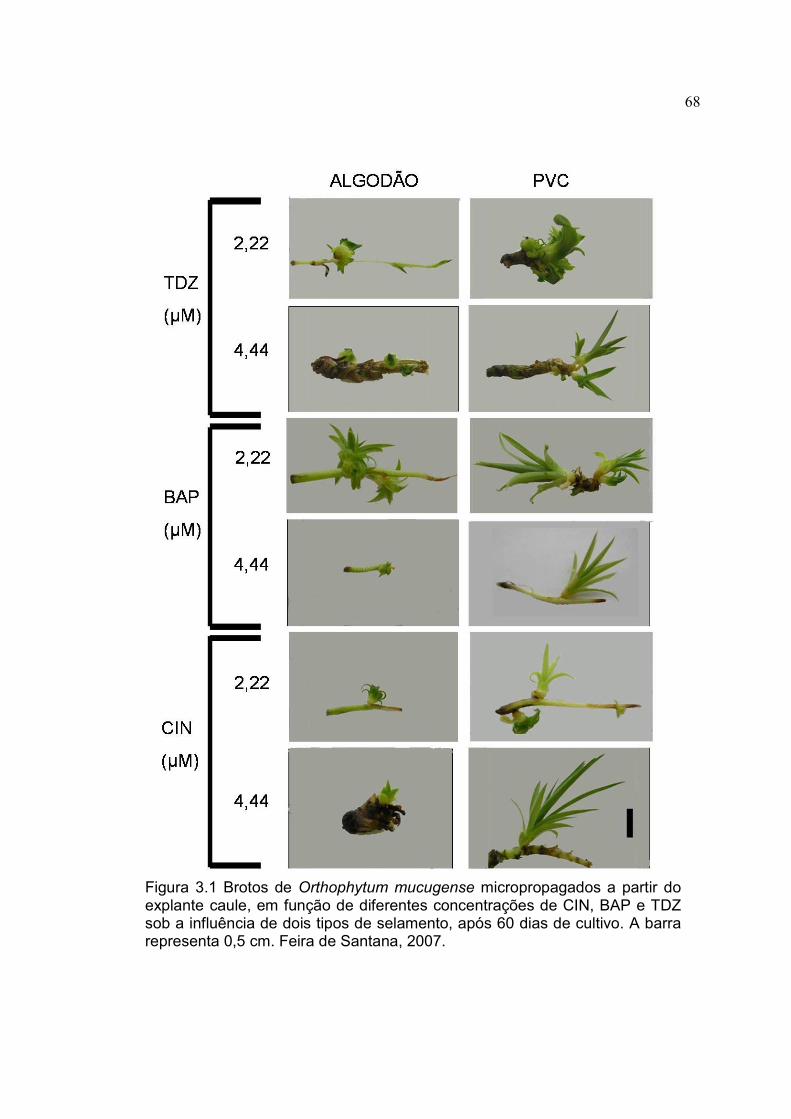
68
Figura 3.1 Brotos de Orthophytum mucugense micropropagados a partir do explante caule, em função de diferentes concentrações de CIN, BAP e TDZ sob a influência de dois tipos de selamento, após 60 dias de cultivo. A barra representa 0,5 cm. Feira de Santana, 2007.
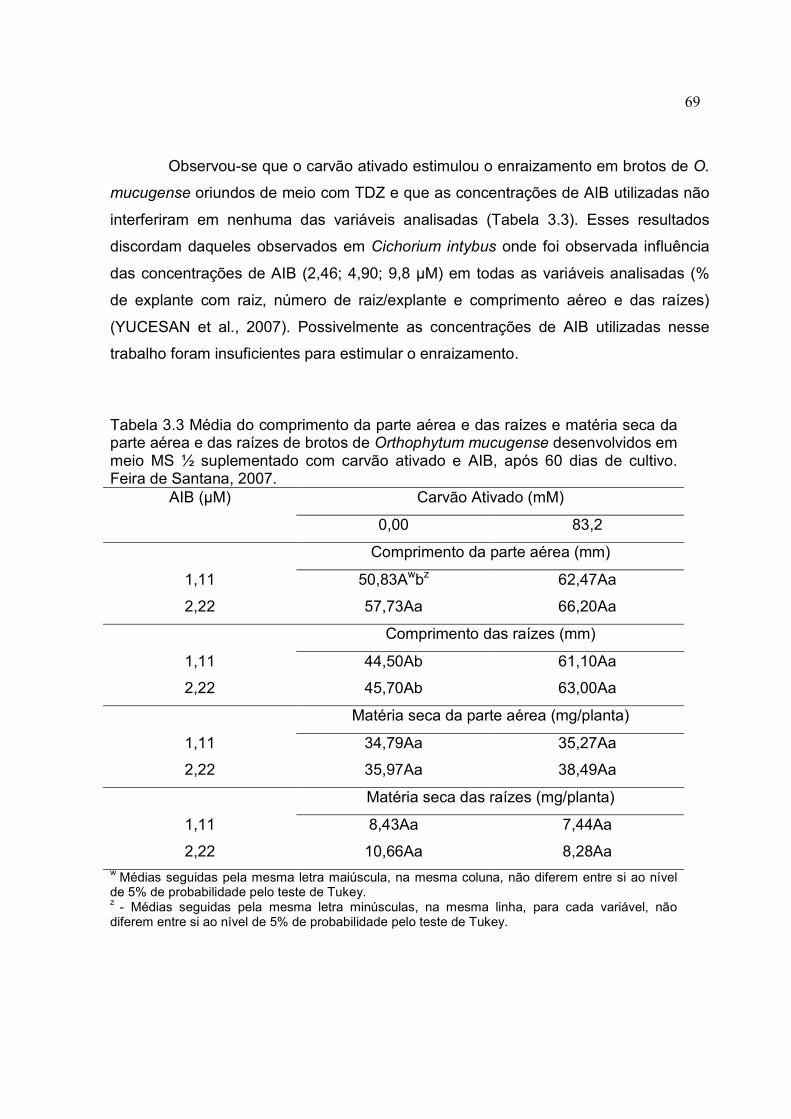
69
Observou-se que o carvão ativado estimulou o enraizamento em brotos de O.
mucugense oriundos de meio com TDZ e que as concentrações de AIB utilizadas não
interferiram em nenhuma das variáveis analisadas (Tabela 3.3). Esses resultados
discordam daqueles observados em Cichorium intybus onde foi observada influência
das concentrações de AIB (2,46; 4,90; 9,8 µM) em todas as variáveis analisadas (%
de explante com raiz, número de raiz/explante e comprimento aéreo e das raízes)
(YUCESAN et al., 2007). Possivelmente as concentrações de AIB utilizadas nesse
trabalho foram insuficientes para estimular o enraizamento.
Tabela 3.3 Média do comprimento da parte aérea e das raízes e matéria seca da parte aérea e das raízes de brotos de Orthophytum mucugense desenvolvidos em meio MS ½ suplementado com carvão ativado e AIB, após 60 dias de cultivo. Feira de Santana, 2007.
AIB (µM) Carvão Ativado (mM)
0,00 83,2
Comprimento da parte aérea (mm)
1,11 50,83Awbz 62,47Aa
2,22 57,73Aa 66,20Aa
Comprimento das raízes (mm)
1,11 44,50Ab 61,10Aa
2,22 45,70Ab 63,00Aa
Matéria seca da parte aérea (mg/planta)
1,11 34,79Aa 35,27Aa
2,22 35,97Aa 38,49Aa
Matéria seca das raízes (mg/planta)
1,11 8,43Aa 7,44Aa
2,22 10,66Aa 8,28Aa w Médias seguidas pela mesma letra maiúscula, na mesma coluna, não diferem entre si ao nível de 5% de probabilidade pelo teste de Tukey. z - Médias seguidas pela mesma letra minúsculas, na mesma linha, para cada variável, não diferem entre si ao nível de 5% de probabilidade pelo teste de Tukey.
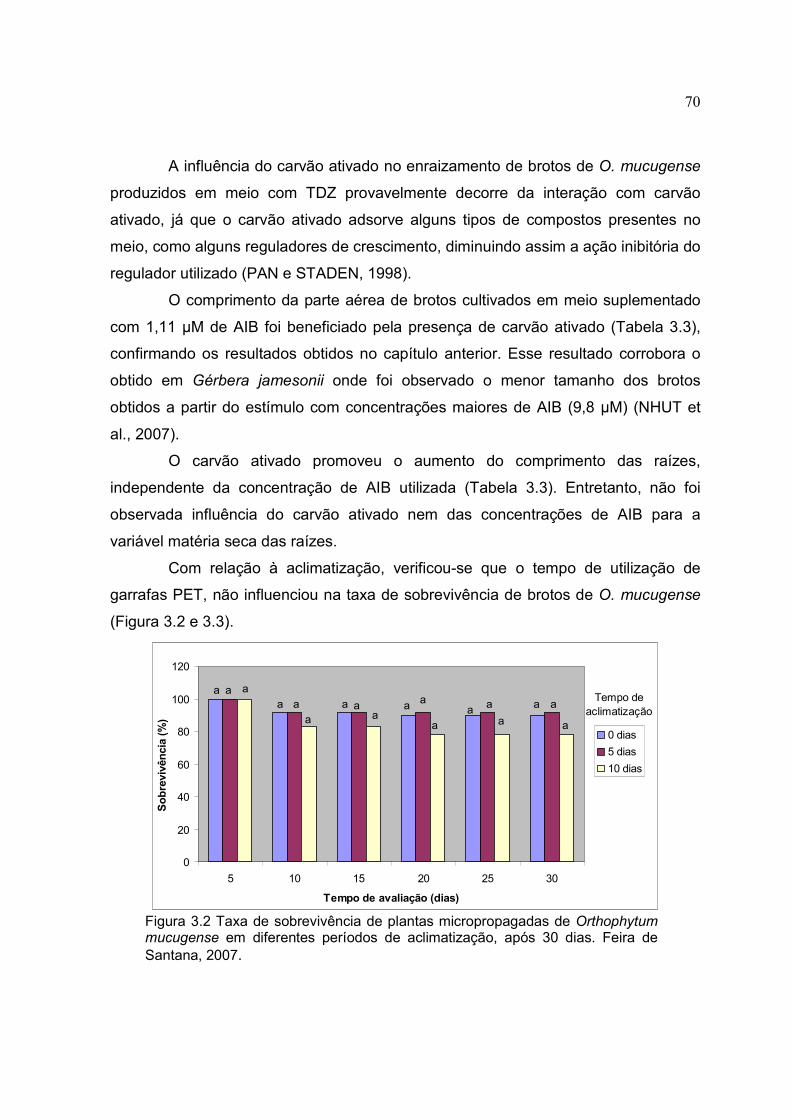
70
A influência do carvão ativado no enraizamento de brotos de O. mucugense
produzidos em meio com TDZ provavelmente decorre da interação com carvão
ativado, já que o carvão ativado adsorve alguns tipos de compostos presentes no
meio, como alguns reguladores de crescimento, diminuindo assim a ação inibitória do
regulador utilizado (PAN e STADEN, 1998).
O comprimento da parte aérea de brotos cultivados em meio suplementado
com 1,11 µM de AIB foi beneficiado pela presença de carvão ativado (Tabela 3.3),
confirmando os resultados obtidos no capítulo anterior. Esse resultado corrobora o
obtido em Gérbera jamesonii onde foi observado o menor tamanho dos brotos
obtidos a partir do estímulo com concentrações maiores de AIB (9,8 µM) (NHUT et
al., 2007).
O carvão ativado promoveu o aumento do comprimento das raízes,
independente da concentração de AIB utilizada (Tabela 3.3). Entretanto, não foi
observada influência do carvão ativado nem das concentrações de AIB para a
variável matéria seca das raízes.
Com relação à aclimatização, verificou-se que o tempo de utilização de
garrafas PET, não influenciou na taxa de sobrevivência de brotos de O. mucugense
(Figura 3.2 e 3.3).
0
20
40
60
80
100
120
5 10 15 20 25 30
Tempo de avaliação (dias)
Sobrevivência (%
)
0 dias
5 dias
10 dias
Tempo de aclimatização
a a
a
a a a
a aa
a a
a
a a
a
a a
a
Figura 3.2 Taxa de sobrevivência de plantas micropropagadas de Orthophytum mucugense em diferentes períodos de aclimatização, após 30 dias. Feira de Santana, 2007.

71
Os resultados obtidos no presente trabalho demonstram que é possível
transferir as plântulas de O. mucugensis produzidas in vitro diretamente para o
ambiente ex vitro, sem passar por uma fase de aclimatização (40 dias), como
sugerido por Bellintani (2006).
Para tanto, sugere-se que durante o período de enraizamento, seja realizada
a rustificação das plântulas utilizando tubos com tampão de algodão. A utilização dos
tampões que permitiram trocas gasosas, no período de multiplicação, possibilitou a
formação de paredes celulares mais espessadas (Figura 3.4).
A menor taxa de sobrevivência observada para o tratamento de 10 dias foi
devido ao desenvolvimento de fungos, por causa da alta umidade observada no
microclima obtido com as garrafas PET. A morte de plantas associada a fungos
representou aproximadamente 30% do total de plantas perdidas.

72
Figura 3.3 Aclimatização de plantas micropropagadas de Orthophytum mucugense em diferentes períodos de rustificação. (A e D) plantas transferidas diretamente após 5 e 10 dias de cultivos respectivamente; (B e E) plantas com cinco dias com garrafa PET após 5 e 10 dias de cultivo respectivamente; (Ce F) plantas com dez dias com garrafa PET após 5 e 10 dias de cultivo respectivamente. Feira de Santana, 2007.
.

73
As análises histológicas demonstraram que o desenvolvimento das folhas de
brotos micropropagados de O. mucugense é influenciado pelo tipo de selamento
(Figura 3.4). As folhas de brotos procedentes dos tubos fechados com algodão
apresentaram células da epiderme lignificadas em quase todos os tratamentos
(Figura 3.4 c, g, i). Já as folhas procedentes dos tubos fechados com PVC não
apresentaram paredes lignificadas em quase todos os tratamentos com exceção do
BAP na concentração de 4,44 µM (Figura 3.4, h). Esse resultado corrobora o obtido
por Bellintani (2006) para a mesma espécie.
A presença de espessamento das paredes das células epidérmicas é uma
característica observada em alguns membros da família Bromeliaceae
(TOMLINSON, 1969; SAJO et al., 1998). Esse espessamento pode estar relacionado
com a redução da evaporação da água dos tecidos, evitando o estresse hídrico e
assegurando a sobrevivência das espécies em condições extremas (ESAÚ, 1977).
Características anatômicas que favorecem a adaptação ao ambiente ex vitro
foram observadas também em Brassica oleracea que apresentou um ótimo
desenvolvimento dos brotos que cresceram em ambiente com trocas gasosas. Em
contrapartida a vedação dos frascos de cultura apresentou efeitos inibidores sobre o
crescimento e desenvolvimento, além de induzir hiperidricidade e redução de clorofila
(ZOBAYED et al., 1999). Resultado observado também para Scrophularia
yoshimurae (LAI et al., 2005).
No cultivo in vitro de morangueiro Calvete et al. (2002) observaram também
uma fina camada de cutícula depositada sobre a epiderme, de plantas cultivadas
sem a presença de trocas gasosas quando comparadas com a camada mais
espessa presente em plantas cultivadas no ambiente ex vitro. Comportamento
semelhante ao observado para O. mucugense (Figura 3.4 h).
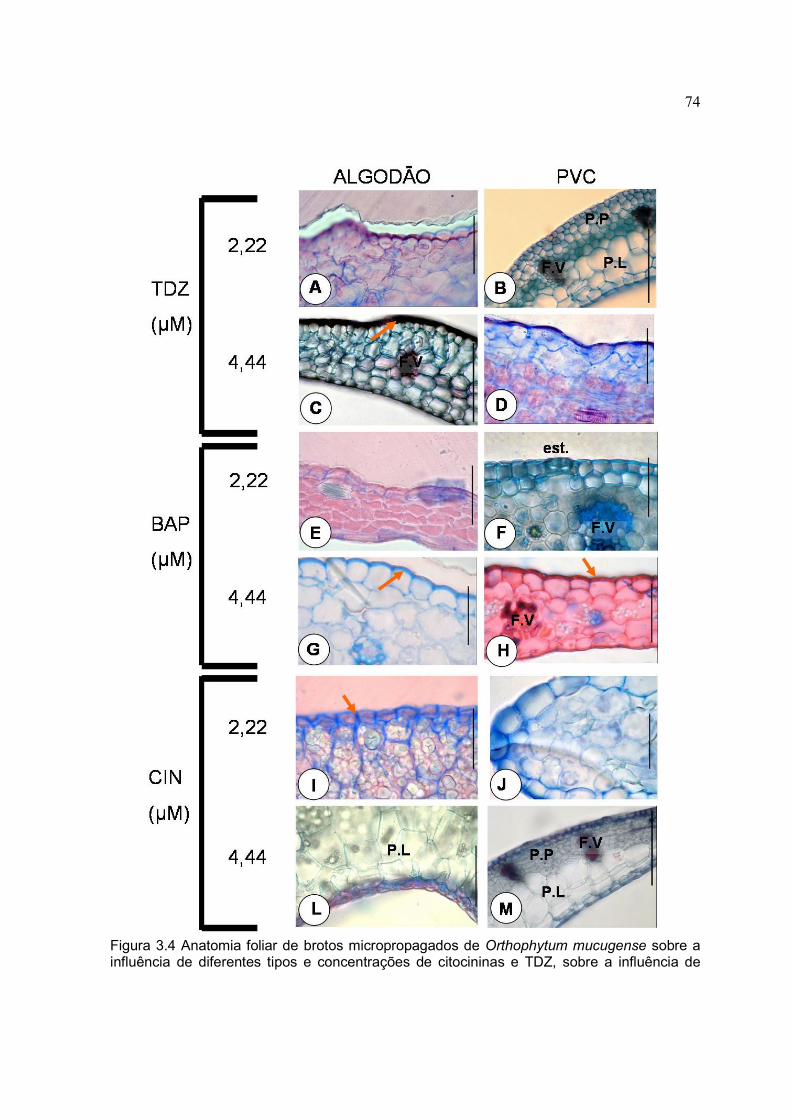
74
Figura 3.4 Anatomia foliar de brotos micropropagados de Orthophytum mucugense sobre a influência de diferentes tipos e concentrações de citocininas e TDZ, sobre a influência de

75
dois selamentos, após 60 dias de cultivo. (A) epiderme adaxial; (B) folhas com ausência de lignina; (C) folhas cultivadas em tubos fechados com algodão com presença de lignina; (D) epiderme adaxial com células epidérmicas desorganizadas; (E) folha sem diferenciação de Parênquima paliçádico e lacunoso; (F) presença de estômato na epiderme adaxial; (G) presença de lignina na epiderme adaxial; (H) presença de lignina na epiderme adaxial; (I) presença de parede espessada entre as células da epiderme; (J) extremidade da folha; (L) epiderme abaxial; (M) folha com diferenciação de Parênquima paliçádico e lacunoso. (seta = parede celular); (F.V = feixe vascular); (est. = estômato); (P.P = parênquima paliçádico); (P.L= parênquima lacunoso). A barra representa 5 µm. Feira de Santana, 2007.
As diferenças encontradas entre as folhas de brotos formados em tubos
fechados por PVC ou algodão justificam as diferenças encontradas no comprimento
dos brotos (Tabela 3.2). A formação de estruturas de proteção requer gasto de
energia impossibilitando assim o crescimento em dimensões das folhas originadas
de tubos fechados com algodão. Além de estruturas de proteção frascos de cultura
de tecidos que apresentem tampas que possibilitem trocas gasosas reduzem a
umidade relativa do ambiente conduzindo ao aumento da transpiração vegetal
associado com o desenvolvimento de estômatos funcionais que controlam a perda
de água pela planta (SEELYE, et al., 2003).
A baixa ocorrência de parede lignificada foi observada também em folhas de
Ananas comosus cultivadas in vitro sem a presença de reguladores. Essas folhas
após seis meses de aclimatização tiveram toda sua epiderme recoberta por cutícula
(BARBOZA et al., 2006). Resultado semelhante também foi obtido para Annona
squamosa onde foi observado um maior desenvolvimento da epiderme nos
tratamentos que permitiram trocas gasosas (ARMSTRONG et al., 1997).
A partir dos resultados obtidos no presente estudo verifica-se a possibilidade
de diminuir o período de exposição do explante a reguladores vegetais, obtendo uma
boa taxa regenerativa utilizando o TDZ, com posterior transferência dos brotos para
meio de enraizamento na presença de carvão ativado. O fechamento dos tubos com
algodão durante a fase de enraizamento, promove a rustificação e facilita a
aclimatização da espécie O. mucugense, permitindo a transferência direta das
plantas para o ambiente ex vitro, sem a necessidade de controlar a umidade relativa
nessa fase.

76
REFERÊNCIAS
ARMSTRONG, J.; LEMOS, E. E. P.; ZOBAYED, S.M. A.; JUSTIN, S. H. F.W.; ARMSTRONG, W. A humidity-induced convective throughflow ventilation system benefits Annona squamosa L. explants and coconut colloid. Annals of Botany, Devon, v. 79, p. 31-40, 1997
BARBOZA, Sarah Brandão Santa Cruz; GRACIANO-RIBEIRO, Dalva; TEIXEIRA, João Batista; PORTES, Tomás Aquino e SOUZA, Luiz Augusto Copati. Anatomia foliar de plantas micropropagadas de abacaxi. Pesquisa Agropecuária Brasileira, Brasília, v. 41, n. 2, 2006.
BELLINTANI, Moema Cortizo. Estudos da propagação in vitro e ex vitro de Neoregelia mucugensis Leme, Orthophytum mucugense Wand e Conceição e Orthophytum albopictum Philcox, espécies de Bromeliaceae endêmicas da Bahia. Tese (Doutorado em Botânica) – Universidade Estadual de Feira de Santana, Feira de Santana, 2006.
BUDDENDORF-JOOSTEN, J.M.C.; WOLTERING, E.J. Controlling the gaseous composition in vitro growth of potato plantlets. Scientia Horticulturae, Amsterdam, v.65, p.11-23, 1996.
BUKATSCH, F. Bermerkunger zur Doppelfarbung Astrablau-Safranin. Mikrokosmos, v. 61, p.255, 1972.
CALDAS, L. S.; HARIDASAM, P.; FERREIRA, M. E. Meios nutritivos. In: TORRES, A. C.; CALDAS, L. S.; BUSO, J. A. Cultura de tecidos e transformação genética de plantas. Brasília: Embrapa/SPI,. v.1, p. 87-132, 1998.
CALVETE, Eunice Oliveira; KÄMPF , Atelene Normann; SUZIN Marilei. Concentração de sacarose no enraizamento in vitro de morangueiro. Horticultura Brasileira, Brasília, v. 20, n. 2, p. 186-191, junho 2002.
CARNEIRO, L.A.; CÂNDIDO, M.S.D.; ARAUJO, R.F.G.; FONSECA, M.H.P.B.; CROCOMO, O.J.; MANSUR, E. Clonal propagation of Cryptanthus sinuosus L.B. Smith, na endemic stoloniferous Bromeliaceae species from Rio de Janeiro, Brazil. Plant Tissue Culture and Biotechnology, Bangladesh, v. 4, n.3-4, p. 152- 158, nov., 1998.
CARNEIRO, L. A.; MANSUR, E. Contribuição de metodologias in vitro para a conservação de Bromeliaceae. Vidalia, Viçosa, n.2, v.1 p. 12- 20, 2004.
CID, Luis Pedro Barrueto. Citocininas. In: CID, Luis Pedro Barrueto. Introdução aos hormônios vegetais, Brasília, Embrapa Recursos Genéticos e Biotecnologia, p. 55-77, 2000.

77
COSTA, Maria Angélica P. de C.; SOUZA, Antônio da S.; ALMEIDA, Weliton Antonio B. de. Morfogênese in vitro. In:SOUZA, Antônio da Silva e JUNGHANS, Tatiana Góes. Introdução à micropropagação de plantas. Cruz das Almas: Embrapa, p. 115-130, 2006.
DE PROFT, M.P.; MAENE, L.J.; DEBERGH, P.C. Carbon dioxide and ethylene evolution in the culture atmosphere of Magnolia cultured in vitro Physiologia Plantarum, Denmark, v.65, p. 375-379, 1985.
ESAÚ, K. Anatomy of seed plants. London: John Wiley and Sons, 1977. 550p.
FERREIRA, D. F. Análise estatística por meio do SISVAR para Windows: Versão 4.0. In: REUNIÃO ANUAL DA REGIÃO BRASILEIRA DA SOCIEDADE INTERNACIONAL DE BIOMETRIA, 45., 2000, São Carlos. Anais... São Carlos: SSP/SIB, p. 255-268, 2000.
GRATTAPAGLIA, Dario; MACHADO, Marcos A. Micropropagação In: TORRES, A. C.; CALDAS, L. S.; BUSO, J. A. Cultura de tecidos e transformação genética de plantas. Brasília: Embrapa/SPI, v.1, p. 183-260. 1998.
GUO, de-Ping; ZHU, Zhu-Jun; HU, Xiao-Xiang; ZHENG, Si-Jun. Effect of cytokinins on shoot regeneration from cotyledon and leaf segment of stem mustard (Brassica juncea var. tsatsai). Plant Cell, Tissue and Organ Culture, Netherlands, v. 83, p.123–127, 2005.
HINOJOSA, Gladys Fernández. Auxinas. In: CID, Luiz Pedro Barrueto. Introdução aos hormônios vegetais. Brasília: Embrapa Recursos Genéticos e Biotecnologia, p. 15-48, 2000.
JACKSON, M. B. Aeration stress in plant tissue cultures. Bulgarian Journal of Plant Physiology, Sofia, v. 28, p. 96-109, 2003.
LAI, Chien-Chou; LIN, Hong-Ming; NALAWADE, Satish Manohar; FANG, Wei; TSAY, Hsin-Sheng. Hyperhydricity in shoot cultures of Scrophularia yoshimurae can be effectively reduced by ventilation of culture vessels. Journal of Plant Physiology, v. 162, p. 355—361, 2005.
LEME, E.M.C.; MARIGO, L.C. Bromeliads in the Brazilian wilderness. Rio de Janeiro, Marigo Comunicação visual, 183p., 1993.
MERCIER, Helenice; NIEVOLA, Catarina C. Obtenção de bromélias in vitro como estratégia de preservação. Vidalia, Viçosa, v. 1, n. 1, p. 57-62, 2003.
NHUT, Duong Tan; THUY AN, Truong Thi; HUONG, Nguyen Thi Dieu; DON, Nguyen Trinh; HAI, Nguyen Thanh; THIEN, Nguyen Quoc; HONG VU, Nguyen. Effect of genotype, explant size, position, and culture medium on shoot generation of Gerbera jamesonii by receptacle transverse thin cell layer culture. Scientia Horticulturae, Amsterdam, v. 111, p. 146-151, 2007.
OLIVERA-ORTEGA, Victoria Z.; GUTIÉRREZ-ESPINOSA, Maria A.; GUTIÉRREZ-ESPINOSA, Jorge A.; ANDRADE-RODRÍGUEZ, Maria. Cultivo in vitro de gérbera

78
(Gérbera jamesonii H. Bolus) y su aclimatación en invernadero. Bioagro, Viçosa, v. 12, n. 3, p. 75-80, 2000.
PAN, M.J.; STADEN, J. van. The use of charcoal in in vitro culture – A review. Plant Growth Regulation, Netherlands, v. 26, p. 155–163, 1998.
PAULA, Cláudio Coelho de; SILVA, Helena M. Peregrino da. Cultivo prático de bromélias. Viçosa: UFV, 3ed., 2004.
PELAH, D.; R. KAUSHIK, A.; MIZRAHI, Y.; SITRIT, Y. Organogenesis in the vine cactus Selenicereus megalanthus using Thidiazuron. Plant Cell, Tissue and Organ Culture, Netherlands, v. 71, p. 81–84, 2002.
PICKENS, Kimberly A.; WOLF, Jan; AFFOLTER, James M.; WETZSTEIN, Hazel Y. Adventitious bud development and regeneration in Tillandsia eizii. In Vitro Cellular and Developmental Biology—Plant, Berlin, v. 42, p. 348–353, July–Aug., 2006.
RECH FILHO, A.; DAL VESCO, L.L.; NODARI, R.O.; LISCHKA, R.W.; MULLER, C.V.; GUERRA, M.P. Tissue culture for the conservation and mass propagation of Vriesea reitzii Leme and Costa, a bromeliad threatened of extinction from the Brazilian Atlantic Forest. Biodiversity and Conservation, Netherlands, n. 14, v. 8, p.1799-1808, 2005.
ROGALSKI, Marcelo; GUERRA, Miguel Pedro; SILVA, Aparecido Lima da. multiplicação in vitro da ameixeira ‘santa rosa’: efeito da citocinina BAP. Revista Brasileira de Fruticultura, Jaboticabal, v. 25, n. 2, p. 365-367, 2003.
ROY, Jonojit; NAHA, Soumi; MAJUMDAR, Madhumita; BANERJEE, Nirmalya. Direct and callus-mediated protocorm-like body induction from shoot-tips of Dendrobium chrysotoxum Lindl. (Orchidaceae). Plant Cell, Tissue Organ Culture, Netherlands, v. 90, p. 31–39, 2007.
SAINI, Raman; JAIWAL, Pawan K. Age, position in mother seedling, orientation, and polarity of the epicotyl segments of blackgram (Vigna mungo L. Hepper) determines its morphogenic response. Plant Science, v. 163, p. 101-109, (2002).
SAJO, M.G.; MACHADO, S.R.; CARMELLO-GUERREIRO, S.M. Aspectos estruturais de folhas de bromélias e suas implicações na agrupamento de espécies. In: LEME, E.M.C. Canistropsis – Bromélias da Mata Atlântica, Rio de Janeiro: Salamandra, p. 102-111, 1998.
SEELYE, John F.; BURGE, Garry K.; MORGAN, Ed R. Acclimatizing Tissue Culture Plants: Reducing the Shock. Combined Proceedings International Plant Propagators’ Society, Mechanicsburg, v. 53, p. 85-90, 2003.
SILVA, André Luís Lopes da; DORNELLES, Eduardo Bortoluzzi; BISOGNIN, Dilson Antônio; FRANCO, Elci Terezinha Henz; HORBACH, Micheli Angélica. Micropropagation of Dyckia agudensis Irgang & Sobral – an extinction threatened bromeliad. Iheringia, Série Botânica, Porto Alegre, v. 62, n. 1, p. 39-43, jan./jun. 2007.

79
TOMLINSON, P.B. Commelinales-Zingiberales. In: METCALFE, C.R. Anatomy of the Monocotyledones, v. 3, Oxford: Clarendon Press, p. 1-446, 1969.
TAGLIACOZZO, Gláucia Moraes Dias. Fitormônios e seus efeitos biológicos in vivo e in vitro. In: TOMBOLATO, Antonio Fernando Caetano; COSTA, Ana Maria Molini. Micropropagação em plantas ornamentais. Campinas, Instituto Agronômico, p. 58-62, 1998.
YAN, Ning; HU, Hong; HUANG, Jia-lin; XU, Kun; WANG, Hua; ZHOU, Zhe-kun. Micropropagation of Cypripedium flavum through multiple shoots of seedlings derived from mature seeds. Plant Cell, Tissue and Organ Culture, Netherlands, v. 84, p, 113–117, 2006.
YUCESAN, Buhara;TURKER, Arzu Ucar; GUREL, Ekrem. TDZ-induced high frequency plant regeneration through multiple shoot formation in witloof chicory (Cichorium intybus L.). Plant Cell Tissue Organ Culture, Netherlands, v. 91, p. 243–250, 2007.
WANDERLEY, Maria das Graças Lapa. Diversidade e distribuição geográfica das espécies de Orthophytum (Bromeliaceae). Acta Botânica Brasílica, São Paulo, v.4, n.1, p. 169-175, 1990.
WANDERLEY, Maria das Graças Lapa; CONCEIÇÃO, Abel Augusto. Notas Taxonômicas e uma nova espécie do gênero Orthophytum Beer (Bromeliaceae) da Chapada Diamantina, Bahia, Brasil. Sitientibus Série Ciências Biológicas, Feira de Santana, v. 6, n. 1, p.3-8, 2006.
ZOBAYED, S.M.A.; ARMSTRONG, J.; ARMSTRONG,W. Evaluation of a closed system, diffuse and humidity-induced convective through flow ventilation on the growth and physiology of cauliflower in vitro. Plant Cell, Tissue and Organ Culture, Netherlands, v.59, p.113-123, 1999.

80
CONSIDERAÇÕES FINAIS
Os resultados obtidos no presente trabalho permitem concluir que:
1 – A indução de brotos in vitro de O. mucugense pode ser otimizada utilizando caule
estiolado com 40 dias de idade, após a remoção do ápice caulinar, sem a
necessidade de reguladores de crescimento e para aumentar a eficiência da
regeneração sugere-se a utilização de PVC no selamento dos frascos de cultura
durante a fase de multiplicação;
2 – Recomenda-se a utilização de açúcar cristal, como fonte de carbono, a fim de
diminuir custos de produção e o uso do carvão ativado para otimizar a fase de
enraizamento de mudas de O. mucugense;
3 – O tempo de aclimatização de mudas micropropagadas de O. mucugense pode
ser reduzido utilizando tampas que permitam as trocas gasosas durante a fase de
enraizamento in vitro.
81
APÊNDICES
QUADROS DE ANÁLISE DE VARIÂNCIAS
QUADROS DE ANÁLISE DE VARIÂNCIAS REFERENTES AO CAPÍTULO 1
Tabela 1.1 – Resumo da análise de variância para média dos números de brotos, comprimento dos brotos e percentagem de explante responsivos em Orthophytum mucugense, obtidos a partir de explante com diferentes idades, em meio suplementado com 2,22 µM de BAP associado a duas concentrações de ANA após 60 dias de cultivo. Feira de Santana, BA, 2007.
Quadrados médios
Causa de variação GL no de brotos/explante
comprimento dos brotos
% de explantes responsivos
Idade do explante 1 1,74** 2,92ns 2688,89*
Conc. de ANA 2 19,17** 56,15** 8872,22**
Idade x ANA 2 0,60ns 0,55ns 505,56ns
Resíduo 66 0,22 3,27 429,30
CV 93,68 74,69 81,08
** - significativo ao nível de 1% de probabilidade pelo teste de Tukey * - significativo ao nível de 5% da probabilidade pelo teste de Tukey ns- não significativo ao nível de 5% de probabilidade pelo teste de Tukey
82
Tabela 1.2 – Resumo da análise de variância para média dos números de brotos, comprimento dos brotos e percentagem de explante responsivos em Orthophytum mucugense, obtidos a partir de diferentes explante com diferentes idades, em meio suplementado com 2,22 µM de BAP associado a duas concentrações de ANA após 60 dias de cultivo. Feira de Santana, BA, 2007. Quadrados médios Causa de variação GL nº de
brotos/explante Comprimento dos brotos
% de explante responsivos
Conc de ANA (A) 1 0,43ns 0,43ns 29,63ns
Tipo de explante (B) 2 20,73** 425,53** 38605,56**
Idade do explante (C) 2 0,06ns 17,59ns 1316,67*
A x B 2 0,87ns 26,45* 279,63ns
A x C 2 0,78ns 5,30ns 190,74ns
BxC 4 0,77ns 34,49** 1797,22**
A x B x C 4 0,61ns 3,98ns 115,74ns
Residuo 198 0,37 7,46 392,26
CV 136,16 116,69 104,85
** - significativo ao nível de 1% de probabilidade pelo teste de Tukey * - significativo ao nível de 5% da probabilidade pelo teste de Tukey ns- não significativo ao nível de 5% de probabilidade pelo teste de Tukey
Tabela 1.3 – Resumo da análise de variância para média dos números de brotos, comprimento dos brotos e percentagem de explante responsivos em Orthophytum mucugense, obtidos em função da posição do explante (caule com 40 dias) no meio de cultura MS½ suplementado com 2,22µM de BAP associado a 0,65 µM de ANA após 60 dias de cultivo. Feira de Santana, BA, 2007.
Quadrados médios Causa da variação
GL N de brotos/explante
comprimento dos brotos
% de explante responsivos
Tratamento 2 14,33ns 1,79ns 921,21ns Resíduo 30 8,86 14,09 652,12
CV 132,78 69,57 71,42 ns- não significativo ao nível de 5% de probabilidade pelo teste de Tukey

83
Tabela 1.4 – Resumo das análises de variância para número de brotos por explente em Orthophytum mucugense formados em função da remoção do ápice de plantas estioladas e da presença de ANA e BAP, Feira de Santana-BA, 2007.
Quadrados médios
Causa de variação GL
Presença do ápice 1 11,34**
ANA e BAP 1 3,42**
Presença do ápice x ANA e BAP
1 1,64*
Resíduo 36 0,42
CV 52,99
** - significativo ao nível de 1% de probabilidade pelo teste de Tukey * - significativo ao nível de 5% da probabilidade pelo teste de Tukey
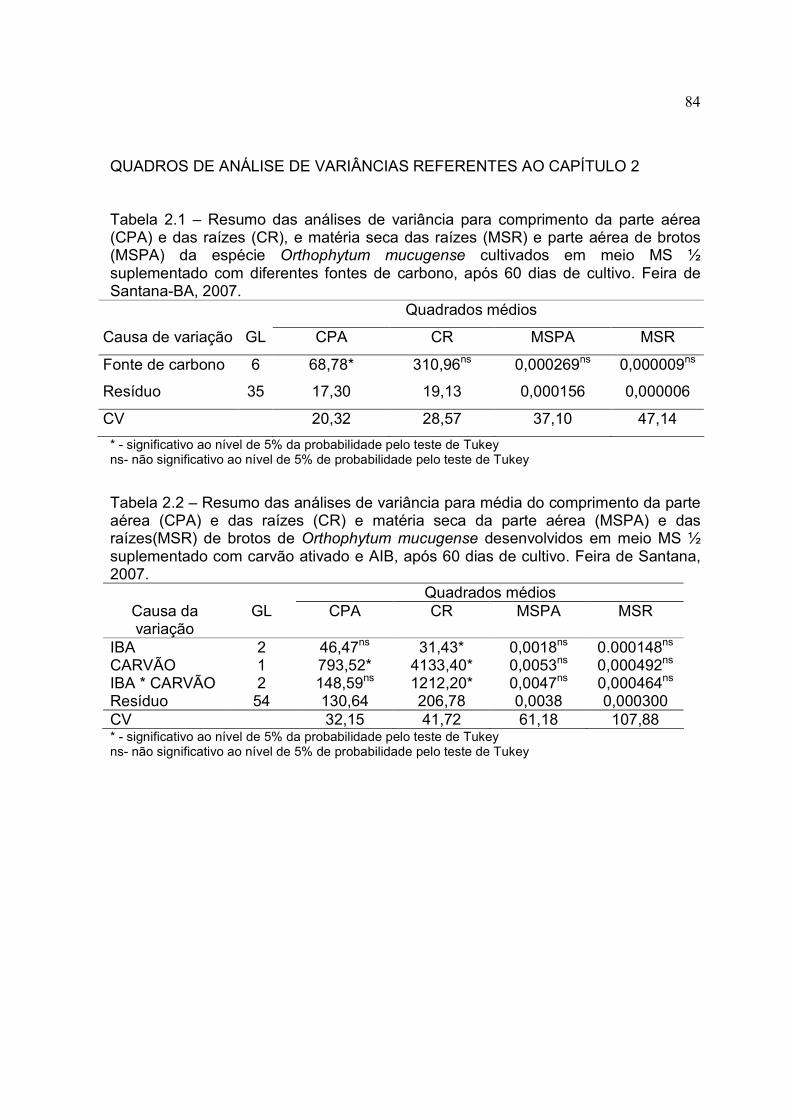
84
QUADROS DE ANÁLISE DE VARIÂNCIAS REFERENTES AO CAPÍTULO 2
Tabela 2.1 – Resumo das análises de variância para comprimento da parte aérea (CPA) e das raízes (CR), e matéria seca das raízes (MSR) e parte aérea de brotos (MSPA) da espécie Orthophytum mucugense cultivados em meio MS ½ suplementado com diferentes fontes de carbono, após 60 dias de cultivo. Feira de Santana-BA, 2007. Quadrados médios
Causa de variação GL CPA CR MSPA MSR
Fonte de carbono 6 68,78* 310,96ns 0,000269ns 0,000009ns
Resíduo 35 17,30 19,13 0,000156 0,000006
CV 20,32 28,57 37,10 47,14
* - significativo ao nível de 5% da probabilidade pelo teste de Tukey ns- não significativo ao nível de 5% de probabilidade pelo teste de Tukey
Tabela 2.2 – Resumo das análises de variância para média do comprimento da parte aérea (CPA) e das raízes (CR) e matéria seca da parte aérea (MSPA) e das raízes(MSR) de brotos de Orthophytum mucugense desenvolvidos em meio MS ½ suplementado com carvão ativado e AIB, após 60 dias de cultivo. Feira de Santana, 2007. Quadrados médios
Causa da variação
GL CPA CR MSPA MSR
IBA 2 46,47ns 31,43* 0,0018ns 0.000148ns CARVÃO 1 793,52* 4133,40* 0,0053ns 0,000492ns IBA * CARVÃO 2 148,59ns 1212,20* 0,0047ns 0,000464ns Resíduo 54 130,64 206,78 0,0038 0,000300 CV 32,15 41,72 61,18 107,88 * - significativo ao nível de 5% da probabilidade pelo teste de Tukey ns- não significativo ao nível de 5% de probabilidade pelo teste de Tukey
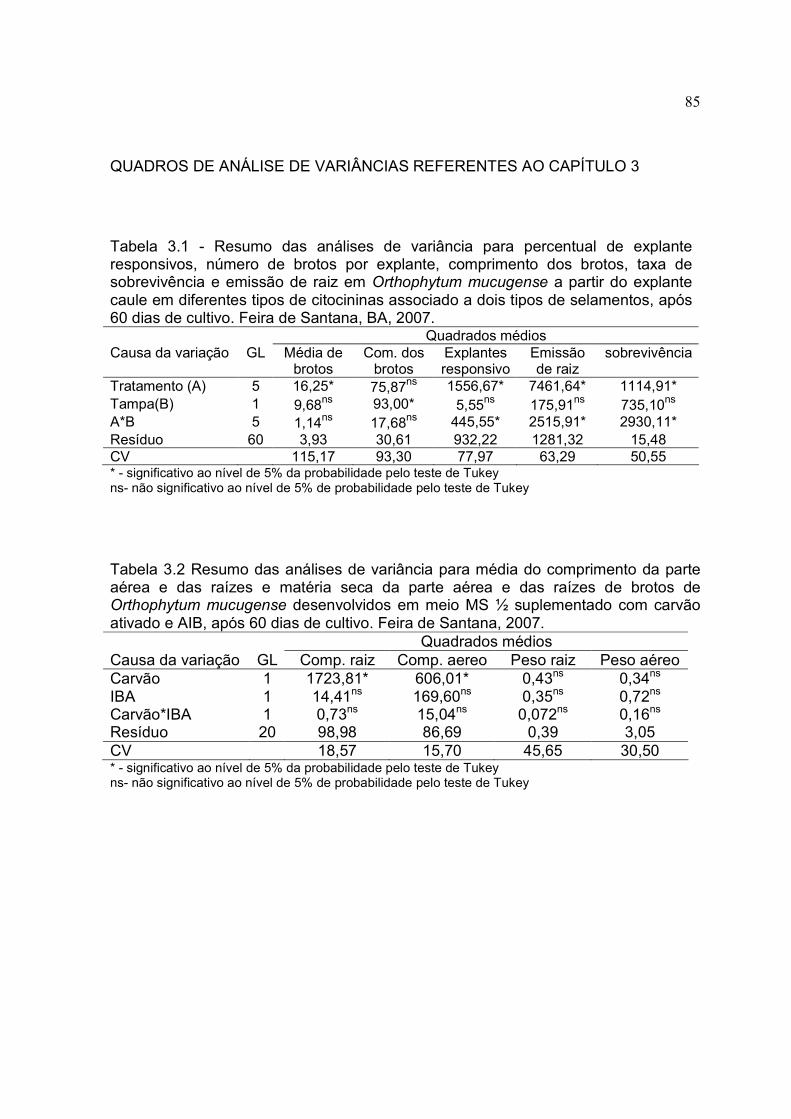
85
QUADROS DE ANÁLISE DE VARIÂNCIAS REFERENTES AO CAPÍTULO 3
Tabela 3.1 - Resumo das análises de variância para percentual de explante responsivos, número de brotos por explante, comprimento dos brotos, taxa de sobrevivência e emissão de raiz em Orthophytum mucugense a partir do explante caule em diferentes tipos de citocininas associado a dois tipos de selamentos, após 60 dias de cultivo. Feira de Santana, BA, 2007. Quadrados médios Causa da variação GL Média de
brotos Com. dos brotos
Explantes responsivo
Emissão de raiz
sobrevivência
Tratamento (A) 5 16,25* 75,87ns 1556,67* 7461,64* 1114,91* Tampa(B) 1 9,68ns 93,00* 5,55ns 175,91ns 735,10ns A*B 5 1,14ns 17,68ns 445,55* 2515,91* 2930,11* Resíduo 60 3,93 30,61 932,22 1281,32 15,48 CV 115,17 93,30 77,97 63,29 50,55 * - significativo ao nível de 5% da probabilidade pelo teste de Tukey ns- não significativo ao nível de 5% de probabilidade pelo teste de Tukey
Tabela 3.2 Resumo das análises de variância para média do comprimento da parte aérea e das raízes e matéria seca da parte aérea e das raízes de brotos de Orthophytum mucugense desenvolvidos em meio MS ½ suplementado com carvão ativado e AIB, após 60 dias de cultivo. Feira de Santana, 2007. Quadrados médios Causa da variação GL Comp. raiz Comp. aereo Peso raiz Peso aéreo Carvão 1 1723,81* 606,01* 0,43ns 0,34ns IBA 1 14,41ns 169,60ns 0,35ns 0,72ns Carvão*IBA 1 0,73ns 15,04ns 0,072ns 0,16ns Resíduo 20 98,98 86,69 0,39 3,05 CV 18,57 15,70 45,65 30,50 * - significativo ao nível de 5% da probabilidade pelo teste de Tukey ns- não significativo ao nível de 5% de probabilidade pelo teste de Tukey





























