Embed Size (px)
Citation preview

1
Running head: Hexokinase controls ROS formation in plant mitochondria
Dr. Antonio Galina from the Instituto de Bioquímica Médica, Programa de Biofísica e Bioquímica Celular
Universidade Federal do Rio de Janeiro, Cidade Universitária, Rio de Janeiro 21941-590, Brazil, Av.
Brigadeiro Trompowsky, s/n, CCS, Bloco D, sub-solo sala D-013, Laboratório de Bioenergética e
Fisiologia Mitocondrial, Cidade Universitária, Rio de Janeiro, RJ, 21941-590. Tel: (55 21) 22609573
Brazil. E-mail: [email protected]
AREA: Bioenergetics and Photosynthesis
Plant Physiology Preview. Published on December 24, 2008, as DOI:10.1104/pp.108.129247
Copyright 2008 by the American Society of Plant Biologists
www.plantphysiol.orgon April 10, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

2
REACTIVE OXYGEN SPECIES PRODUCTION BY POTATO TUBER MITOCHONDRIA IS
MODULATED BY MITOCHONDRIALLY BOUND HEXOKINASE ACTIVITY.
Juliana Camacho-Pereira*, Laudiene Evangelista Meyer*, Lilia Bender Machado, Marcus Fernandes
Oliveira and Antonio Galina.
From Instituto de Bioquímica Médica, Programa de Biofísica e Bioquímica Celular and Programa de
Biologia Molecular e Biotecnologia, Universidade Federal do Rio de Janeiro, Laboratório de
Bioenergética e Fisiologia Mitochondrial.
Av. Brigadeiro Trompowsky, s/n, CCS, Bloco D, sub-solo sala D-013, Cidade Universitária, , Rio de
Janeiro, RJ, 21941-590. Brazil.
* These authors contributed equally to the results of this work.
www.plantphysiol.orgon April 10, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

3
FOOTNOTES:
This work was supported by grants from the Conselho Nacional de Desenvolvimento Científico e Tecnológico
(CNPq), Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ).
ABBREVIATIONS:
ANT: adenine nucleotide transporter. AOX: alternative oxidase. AUF: arbitrary units of fluorescence. EGTA:
ethylene glycol-bis (-aminoethyl ether)-N,N,N´,N´-tetraacetic acid. ETS: electron transport system. Faf-BSA:
fatty acid free bovine serum albumin. FCCP: carbonyl cyanide p-trifluoromethoxyphenylhydrazone. G6PDH:
glucose-6 phosphate dehydrogenase. LA: linolenic acid. MHP: mannoheptulose. mt-HK: mitochondrial-
associated hexokinase. NAG: N-acetylglucosamine. PEP: phosphoenolpyruvate. PMSF: phenyl methyl
sulfonyl fluoride. PUMP: plant uncoupling protein. ROS: reactive oxygen species. SOD: superoxide dismutase.
St-UCP: Solanum tuberosum uncoupling protein. UCP: uncoupling protein. VDAC: voltage-dependent anion
channel. ΔΨm: mitochondrial membrane potential.
www.plantphysiol.orgon April 10, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

4
ABSTRACT
Potato tuber (Solanum tuberosum) mitochondria (PTM) have a mitochondrially bound hexokinase (mt-
HK) activity that exhibits a pronounced sensitivity to ADP inhibition. Here we investigated the role of
mitochondrial hexokinase activity in potato tuber mitochondria reactive oxygen species generation.
Mitochondrial hexokinase has a ten fold higher affinity for glucose than for fructose (KMGlc = 140 μM
versus KMFrc = 1375 μM). Activation of potato tuber mitochondria respiration by succinate led to an
increase in H2O2 release that was abrogated by mitochondrial hexokinase activation. Mitochondrial
hexokinase activity caused a decrease in the mitochondrial membrane potential (ΔΨm) and an increase
in oxygen consumption by potato tuber mitochondria. Inhibition of glucose phosphorylation by
mannoheptulose or N-acetylglucosamine induced a rapid increase in H2O2 release. The blockage of
H2O2 release sustained by glucose was reverted by oligomycin and atractyloside indicating that ADP re-
cycles through the adenine nucleotide translocator and FoF1ATP synthase is operative during the
mitochondrial hexokinase reaction. Inhibition of mitochondrial hexokinase activity by 60 to 70 % caused
an increase of 50 % in the maximal rate of H2O2 release. Inhibition in H2O2 release by mitochondrial
hexokinase activity was comparable to, or even more potent, than that observed for St-UCP activity. The
inhibition of H2O2 release in potato tuber mitochondria was two orders of magnitude more selective for
the ADP produced from the mitochondrial hexokinase reaction than for that derived from soluble yeast
hexokinase. Modulation of H2O2 release and oxygen consumption by glucose and mitochondrial
hexokinase inhibitors in potato tuber slices shows that hexoses and mitochondrial hexokinase may act
as a potent preventive antioxidant mechanism in potato tubers.
www.plantphysiol.orgon April 10, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

5
INTRODUCTION
Production of reactive oxygen species (ROS) is an unavoidable consequence of aerobic respiration
(Chance et al., 1979). The mitochondrial electron transport system (ETS) is the major site of ROS
production in mammalian and non-photosynthesizing plant cells (Puntarulo et al., 1991; Halliwell and
Gutteridge, 1999). Depending on the mitochondrial respiratory states, a small portion of the consumable
oxygen is partially reduced to generate ROS (Skulachev, 1996; Liu, 1997; Turrens, 1997; Møller, 2001;
Considine et al., 2003; Smith et al., 2004). In plants, the monoelectronic reduction of oxygen by ETS
leads to the production of superoxide radicals (O2˙�) which can be dismutated by SOD, producing
hydrogen peroxide (H2O2), and further decomposed by catalase and/or ascorbate-glutathione peroxidase
cycles (Møller, 2001). An imbalance between the ROS production and antioxidant defenses can lead to
an oxidative stress condition. Increased levels of ROS may be a consequence of the action of plant
hormones, environmental stress, pathogens or high levels of sugars and fatty acids (Bolwell et al., 2002;
Couée et al., 2006; Gechev et al., 2006; Liu et al., 2007; Rhoads and Subbaiah, 2007). These conditions
may lead to storage deterioration or impairment of seedling growth decreasing on crop yield. To avoid
the harmful accumulation of ROS or to fine-tune the steady-state levels of ROS, various enzymatic
systems control the rate of ROS production in mitochondria (Schreck and Baeuerle, 1991; Møller, 2001).
Mitochondrial ROS production is highly dependent on the membrane potential (ΔΨm) generated
by the proton gradient formed across the inner mitochondrial membrane. High ΔΨm was shown to
stimulate ROS production when the ETS is predominantly in a reduced state (i.e. when NADH, FADH2
and O2 are present in abundance but ADP or Pi levels are low). This condition is reached in resting
metabolic states after a full oxidation of glucose or fatty acids. Stimulating electron flow by decreasing
ΔΨm either by the use of uncouplers or by coupling respiration to ATP synthesis, slows the ROS
generation rate (Boveris and Chance, 1973; Korshunov et al., 1997). It has been observed that in
isolated potato tuber mitochondria (PTM) the uncoupling protein (referred to as PUMP in plants, or UCP
in animals) causes a small decrease in ΔΨm when this proton carrier protein is activated by the presence
of anionic fatty acids, a condition that blocks ROS generation (Vercesi et al., 1995; Vercesi et al., 2006).
Nucleotides, such as ATP, antagonize this effect (Considine et al., 2003; Vercesi et al., 2006). On the
other hand, fluctuations in free hexose levels due to environmental or developmental conditions (Morrell
and ap Rees, 1986; Geigenberger and Stitt, 1993; Renz et al., 1993) lead to variations in the oxygen
consumption rate in heterotrophic tissues of plant (Brouquisse et al., 1991, Dieuaide et al., 1992). As a
result, ROS-producing pathways may be either stimulated or repressed (Couée et al., 2006). Unlike
PUMP activity, which is activated by an excess of free fatty acids, a specific mechanism for
mitochondrial ROS production caused by an excess of hexose remains elusive.
The metabolism of free hexoses begins by their phosphorylation in a reaction catalyzed by the
hexokinase:
Hexose + ATP → Hexose 6-P + ADP
www.plantphysiol.orgon April 10, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

6
Hexokinase (HK) is an ubiquitous enzyme found in many organisms. In plants, the binding
mechanism of HK to the outer mitochondrial membrane is not fully established, but some reports indicate
that it may differ considerably from those properties described for mammal cells (Dry et al., 1983;
Miernyk and Dennis, 1983; Rezende et al., 2006). It has been shown that in several mature and
developing plant tissues, multiple HK isoforms are expressed with different kinetic properties and
subcellular localizations. The HK`s are found in cytosol, bound to the mitochondrial membrane or in
stroma of plastids in plant cells (Miernyk and Dennis, 1983; Galina et al., 1995; Damari-Weissler et al.,
2007). Beyond its obvious role in glycolysis regulation, HK activity may also function as a sugar sensor,
triggering a signal transduction pathway in plants (Rolland et al., 2006).
In mammals, HK types I and II are associated with the mitochondrial outer membrane through
the voltage dependent anion channel (VDAC) and adenine nucleotide transporter (ANT). These
associations were found in tissues with a high energy demand, such as heart, brain and tumor cells
(Arora and Pedersen, 1988; BeltrandelRio and Wilson, 1992; Wilson, 2003). In addition, recent evidence
in mammalian cells has shown that binding of HK to VDAC located at the outer mitochondrial membrane
is somehow involved in the protection against pro-apoptotic stimuli (Nakashima et al., 1986; Pastorino et
al., 2002; Cesar and Wilson, 2004). Similar observations were reported for tobacco plant mitochondrial
HK (Kim et al., 2006). However, it has been shown that drugs such as the fungicide clotrimazole and the
anesthetic thiopental, which promptly disrupt the association between mt-HK and VDAC in mammalian
mitochondria, are unable to promote this effect in maize root mitochondria (Rezende et al., 2006). These
observations suggest a different type of association of mt-HK with plant mitochondria. The binding of mt-
HK with mitochondria in many plants involves a common N-terminal hydrophobic membrane anchor
domain of about 24 amino acids which is related to the membrane targeting, but the exact mechanism of
association is unknown (Damari-Weissler et al., 2007).
Recently, our group demonstrated that mt-HK activity plays a key preventive antioxidant role by
reducing mitochondrial ROS generation through a steady-state ADP re-cycling mechanism in rat brain
neurons. The mitochondrial ADP re-cycling leads to a decrease in the ΔΨm coupled to the synthesis of
ATP by oxidative phosphorylation (da-Silva et al., 2004; Meyer et al., 2006).
Although plant HK is recognized to fulfill a catalytic function, the role of mt-HK activity in the
regulation of both mitochondrial respiration and ROS production in plants is unknown. Recently, an
authentic HK activity was detected in PTM (Graham et al., 2007) and its involvement in potato tuber
glycolysis suggested, but its involvement in PTM ROS generation was not explored. We then raise the
hypothesis that HK bound to PTM would contribute to produce a steady-state ADP recycling that
regulates ROS formation. However, whether this association is capable of controlling the rate of ROS
generation in plant mitochondria is unknown. Here, we aim to investigate the role of mt-HK activity in
PTM physiology. The data indicate that mt-HK activity plays a key role as a regulator of ROS levels in
respiring plant tissues exposed to high hexose levels.
www.plantphysiol.orgon April 10, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

7
RESULTS
An Authentic Hexokinase Activity is Associated with Potato Tuber Mitochondria.
Previous studies have shown that HK is bound to mitochondria in mammalian tissues, in different
plant species (Galina et al., 1995; da-Silva et al., 2001; Wilson, 2003; Meyer et al., 2006; Claeysen and
Rivoal, 2007), and even in PTM (Graham et al., 2007). To check whether a particulate HK activity from
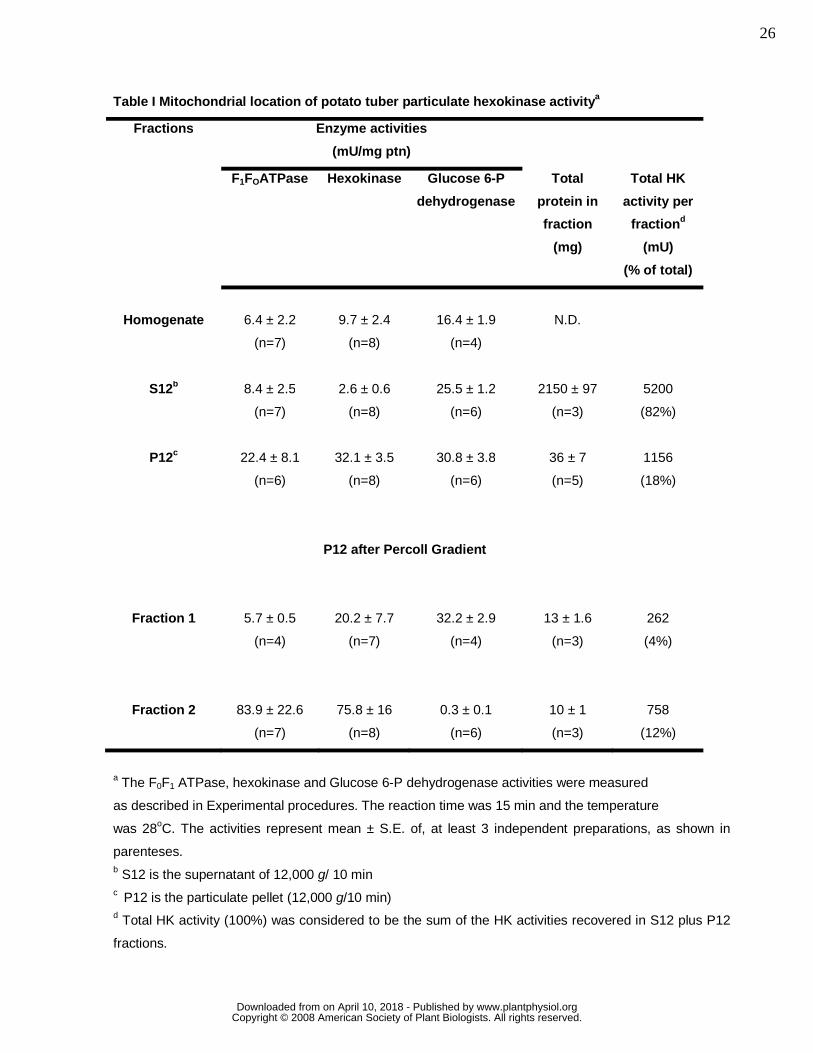
potato tuber is also associated with this organelle, in Table I we determined the activities of HK,
oligomycin-sensitive FoF1ATPase, a mitochondrial enzyme, and glucose-6-phosphate dehydrogenase
(G6PDH), a cytosolic and plastid enzyme (Dennis and Green, 1975; Miernyk and Dennis, 1983) in
different fractions of potato tuber homogenates. Clearly, the majority of HK activity is concentrated in the
particulate fraction (P12) of potato tuber (Table I). As the FoF1ATPase and the G6PDH activities were
also substantial in P12 fraction, a further separation step was carried out on the P12 fraction by using a
self-generated Percoll gradient method, producing two main fractions. One was enriched in plastids
(fraction 1) and the other was enriched in mitochondria (fraction 2) (Neuburger, et al, 1982). Accordingly,
the FoF1ATPase activity was enriched 13 fold in fraction 2 of the Percoll gradient as compared to the
homogenate (Table I). The HK activity was present in both plastid and mitochondrial fractions from the
Percoll gradient, being enriched 8 fold in this fraction and only 2 fold in the plastid fraction (Table I). The
low G6PDH activity detected in the mitochondrial fraction indicates a very low degree of cross-
contamination between these fractions. Almost all particulate G6PDH activity was associated with the
plastid fraction (Table I). These data show that an authentic HK activity is associated with PTM.
Kinetic Properties of Potato Tuber mt-HK and Dependence of Activity on Oxidative
Phosphorylation
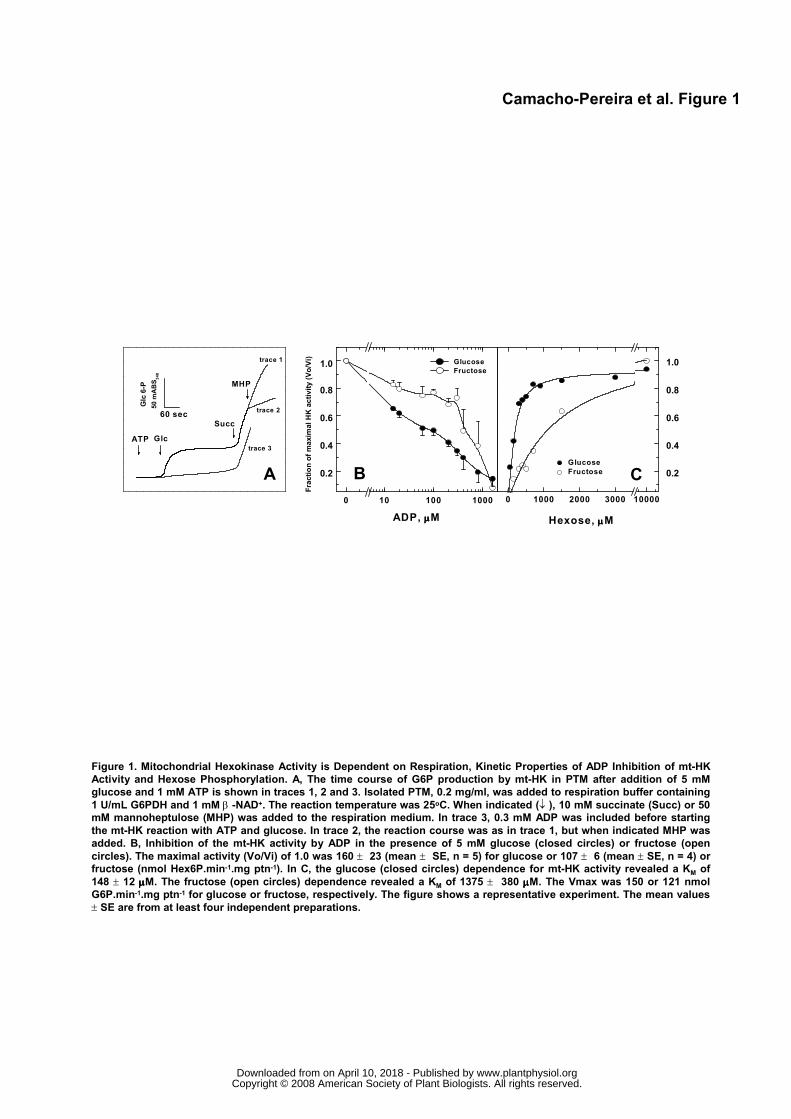
The next step was to evaluate the activity of mt-HK after addition of ATP and glucose using
intact, Percoll-purified PTM (Figure 1). The mt-HK activity was detected immediately after addition of 1
mM ATP and 5 mM glucose (Fig. 1A, traces 1 and 2), but was progressively inhibited by more than 90 %
after 2 minutes. However, the mt-HK activity could be restored by the inclusion of succinate, an
oxidisable substrate that supports the conversion of ADP to ATP during oxidative phosphorylation,
indicating that inhibition observed for mt-HK was due to the ADP accumulated by the mt-HK reaction
(Fig. 1A, traces 1 and 2). The mt-HK sensitivity to ADP inhibition was confirmed when 0.3 mM ADP was
added to the reaction medium prior to starting the mt-HK reaction with ATP plus glucose (Fig. 1A, trace
3). The mt-HK reaction rate in this case was much lower than that observed in the first 2 minutes in the
absence of ADP (Fig. 1A, trace 1). The addition of succinate re-established the maximal rate of mt-HK
activity (Fig. 1A, trace 3). The authenticity of mt-HK activity was confirmed when its specific competitive
inhibitor, 50 mM mannoheptulose (MHP), was added to the reaction medium (Fig. 1A, trace 2). No
oxidative phosphorylation-supported mt-HK activity was observed in the presence of oligomycin (a
specific inhibitor of FoF1ATPase), atractyloside (a specific inhibitor of adenine-nucleotide translocator-
www.plantphysiol.orgon April 10, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

8
ANT) or in the absence of succinate (data not shown). In addition, these oxidative phosphorylation
inhibitors had no effect on PTM mt-HK activity (data not shown).
Figure 1B shows the ADP dependence for mt-HK inhibition using glucose or fructose as
substrate. ADP caused a half-inhibition of mt-HK activity in the micromolar range with glucose (IC50 ≅ 40
to 70 μM), but much more ADP was needed to cause a similar degree of inhibition (IC50 ≅ 400 to 700
μM) with fructose as substrate. This result suggests that the affinity of mt-HK for glucose and fructose is
different and the inhibitory activity of ADP depends on the substrate used. In fact, the affinity of mt-HK
from PTM was higher for glucose (KM 0.14 mM) than for fructose (KM 1.4 mM). The mt-HK activity
measured with saturating concentrations of hexoses was practically the same (Fig. 1C).
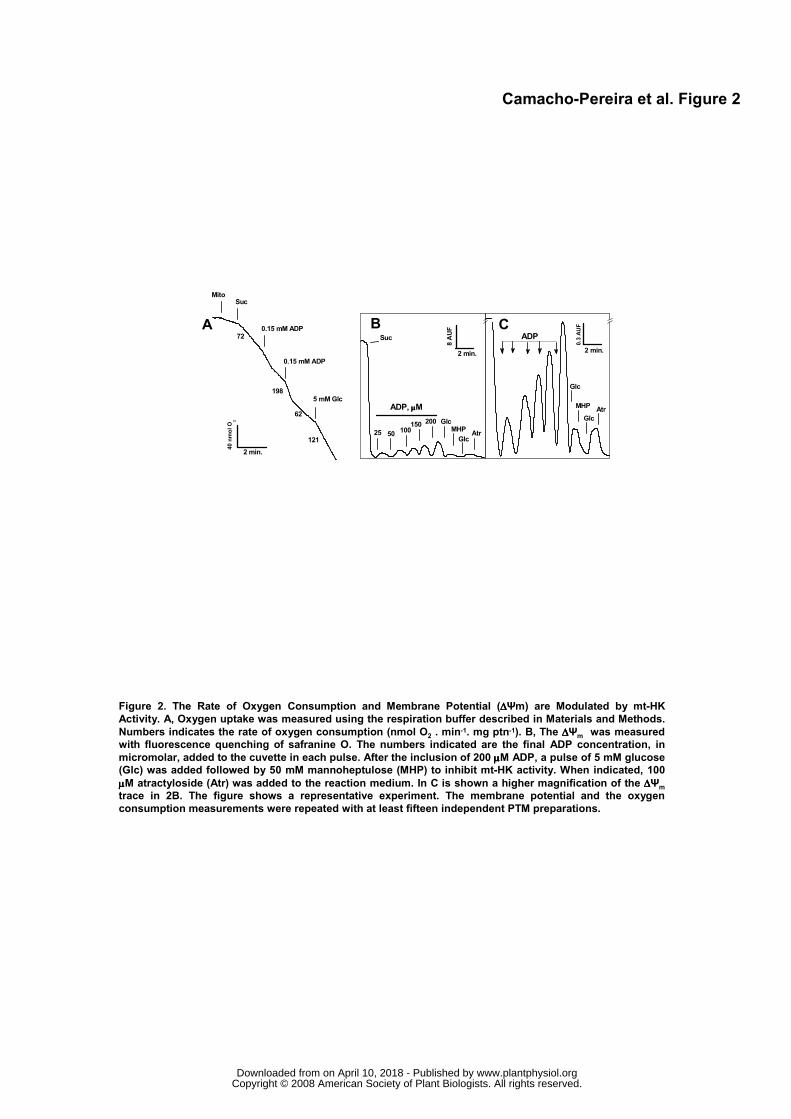
Potato Tuber mt-HK Activity Modulates Oxygen Consumption, ΔΨm and H2O2 Release
The experiments shown in Figure 1 indicated that PTM respiration is important to maintain the
mt-HK activity free of the ADP inhibition. In the next set of experiments, we evaluated the effect of the
mt-HK activity on several mitochondrial functional parameters (Figs. 2, 3 and 4). During PTM respiration,
the addition of small amounts of ADP (0.15 mM) accelerated the rate of O2 consumption (state 3) (Fig.
2A). When all the ADP was converted to ATP, the respiration rate decreased, returning to the resting
state (state 4), but a further addition of glucose permanently stimulated the O2 consumption (Fig. 2A).
Based on previous data from our group (da-Silva et al., 2004; Meyer et al., 2006), glucose-induced O2
consumption in PTM is a result of ADP generated by the mt-HK reaction being transported by the ANT to
the mitochondrial matrix and then being utilized as substrate by the FoF1ATP synthase, decreasing the
ΔΨm and leading to an acceleration of the electron flux in the respiratory electron chain. This can be
confirmed by ΔΨm measurements showing that sequential ADP additions transiently dissipated the ΔΨm
(Figs. 2B and 2C), reaching an estimated ATP concentration of 545 μM. Inclusion of glucose after all
ADP pulses promoted a small, but consistent decrease in ΔΨm (about 3 % of maximal value, Figs. 2B,
2C and 4A), which was reversed by the inclusion of 50 mM MHP (Figs. 2B and 2C). An increase in
glucose concentration to 10 mM further decreased the ΔΨm (Fig. 2C), and interruption of the ADP re-
cycling mechanism by ANT using atractyloside promoted an increase in the ΔΨm (Fig. 2C). These data
show that the mt-HK activity is able to modulate the proton motive force in PTM.
It has been established that mitochondrial H2O2 formation is strongly dependent on high ΔΨm
values (Korshunov et al., 1997). In Figure 3 we observed substantial H2O2 release after induction of
respiration by succinate and ATP. The addition of 1 mM ADP strongly reduced H2O2 release, showing
that a small decrease in ΔΨm due to activation of state 3 respiration, is the main mechanism by which
mitochondrial H2O2 release is prevented (Fig. 3, left trace). A similar blockage in H2O2 release was
observed when ADP was replaced by 5 mM glucose (Fig. 3, right trace). This was reversed by either mt-
HK inhibitors such as MHP (Fig. 3) or NAG (Fig. 4C) by oxidative phosphorylation poisons such as
oligomycin or atractyloside, two inhibitors that promote an increase in ΔΨm in the presence of oxidizable
substrates (Figs. 2, 4A and 4B). These experiments demonstrate that mt-HK activity is able to modulate
www.plantphysiol.orgon April 10, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

9
the ΔΨm and H2O2 release in PTM by an ADP recycling mechanism through ANT/ FoF1ATP synthase
activities. The regulation of H2O2 release from PTM by mt-HK is attained regardless the source of ATP
for the mt-HK is external (Fig. 4B) or from oxidative phosphorylation (Fig. 4A).
Glucose Dependence for mt-HK Activity is Inversely Related to the Rate of H2O2 Release by PTM
The mt-HK activity and the rate of H2O2 release by PTM were measured simultaneously with
increasing amounts of glucose (Fig. 5A). We observed an inverse relation between mt-HK activity and
mitochondrial H2O2 release. The IC50 to inhibit H2O2 release was approximately 20 μM glucose (Fig. 5A).
The dependence on mt-HK activity to abrogate the H2O2 release was confirmed in Figure 5B, when 25
mM NAG, a competitive inhibitor of mt-HK was included in the assay medium. Under this condition, the
IC50 for glucose to inhibit H2O2 release rose to about 800 μM. Importantly, a half maximum inhibition of
the mt-HK activity was able to maintain the rate of H2O2 release at about 15 % of its maximal value (Fig.
5C).
Comparison of the Preventive Antioxidant Role of mt-HK and PUMP Activities in PTM
The rate of H2O2 release, oxygen consumption and ΔΨm levels were titrated with increasing
amounts of a proton ionophore, FCCP, or with the PUMP activator linolenic acid (LA) (Vercesi et al.,
2006) (Fig. 6). In the titration with FCCP (Fig. 6A), it became apparent that a small decrease in the
maximal ΔΨm (about 3 % of AUF of safranine O) by 700 nM FCCP (Fig. 6A, open circles) promoted a
two-fold stimulation of oxygen consumption (Fig. 6A, open stars) and a complete inhibition of H2O2
release (Fig. 6A, open triangles). The effect of maximal mt-HK activity compared with these parameters
was determined and the set point gives a 3 % decrease in ΔΨm, expressed as AUF of safranine O
(closed circle), an increase of 1.8 fold in oxygen consumption (closed stars) and a reduction of at least
95 % in H2O2 release (closed triangle) (see Fig. 6A, vertical dotted line for set point).
In an attempt to compare the impact of mt-HK activity with, that of PUMP activation on the
bioenergetic parameters evaluated above, we next measured the effect of increasing amounts of LA,
either with (Fig. 6B) or without faf-BSA (a protein that sequesters LA) (Fig. 6C). The IC50 for LA to
inhibit H2O2 release was about 30 μM in the presence of faf-BSA (Fig. 6B, open triangles). Under this
specific condition mt-HK activation exerted an even more powerful effect on H2O2 release than did
PUMP activation (Fig. 6C, closed triangle). The IC50 for LA to inhibit H2O2 release when faf-BSA was
omitted from the reaction medium (Fig. 6C) was about 4 μM, confirming the ability of this fatty acid to
activate PUMP. In this condition, 30 μM LA almost completely blocked H2O2 release, similar to mt-HK
activation (Fig. 6C, open and closed triangles).
www.plantphysiol.orgon April 10, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

10
Prevention of H2O2 Release by PTM mt-HK Requires Specific Enzyme Association to
Mitochondria
In order to evaluate whether a specific location of mt-HK on mitochondria is required to prevent
H2O2 release we next measured the rate of H2O2 release in PTM using two alternative hexose
substrates, glucose or fructose at 0.5 mM (Fig. 7A). Mt-HK activity using fructose as substrate is much
lower when compared with glucose, as expected from the apparent affinities of these two substrates
(Fig. 1C). In fact, when 0.5 mM fructose was added to the reaction mixture, a negligible effect was
observed on the rate of H2O2 release, contrasting with the strong reduction in the rate of H2O2 release
when 0.5 mM glucose was used as substrate (Fig. 7A). Using fructose as a substrate, we only observed
a decrease in the rate of H2O2 release when the hexose concentration was increased to 10 mM (Fig.
7A). Regardless of the substrate used, oligomycin promptly increased the H2O2 release by impairing the
ADP recycling activity mediated by mt-HK.
In contrast to PTM mt-HK activity, soluble yeast HK exhibits a high affinity for fructose with an
apparent Km of 0.7 mM (Avigad and Englard, 1968; Bernard 1975). Taking advantage of this difference
in kinetic properties between the two enzymes, we next compared the effect of soluble yeast HK
activation, with that of activating the endogenous PTM mt-HK by using fructose as substrate and
measuring the rate of H2O2 release. The rationale was that at 0.5 mM fructose, PTM mt-HK would not be
active, thus not producing ADP, but addition of soluble yeast HK, which has a higher affinity for fructose,
would promptly catalyze this reaction. Thus, under this condition, the ADP produced from the HK
reaction comes only from the exogenous HK activity. Clearly, increasing amounts of the soluble yeast-
HK decreased the rate of H2O2 release in PTM (Fig. 7A). On comparing at the levels of soluble yeast HK
activity required to impair H2O2 release in PTM with the activity of endogenous mt-HK, we observed a
difference of at least two orders of magnitude (Fig. 7B). This result led us to conclude that specific
location of mt-HK at the outer PTM membrane is absolutely essential to improve ADP delivery for the
oxidative phosphorylation, decreasing PTM H2O2 release.
Glucose Impairs H2O2 Release in Potato Tuber Slices
Potato tuber slices incubated with glucose showed a rate of oxygen consumption 1.6-fold higher
than those without glucose or with sucrose (Fig. 8A). The potato tuber slices treated with FCCP showed
oxygen consumption 2.2-fold higher than that observed for the control (Fig. 8A). To ascertain whether
these increased rates in O2 consumption were related to the rate of H2O2 release, we measured the
accumulation of H2O2 in potato tuber slices medium. The response was the opposite to that observed for
the rate of respiration by potato tuber slices in each condition (Figs. 8B and C). The rate H2O2 release in
disk slices was three-fold lower when the slices were incubated with 5 mM glucose compared to the rate
measured in the absence or in the presence of sucrose, a non-phosphorylatable sugar for mt-HK (Fig.
8B, closed circle and open triangle against open circle). When H2O2 release by potato tuber slices was
measured in the presence of glucose and 50 mM NAG, an inhibitor of mt-HK, the rate of H2O2 release
tended to be higher than that measured only in the presence of glucose (Fig. 8B, closed triangles). This
www.plantphysiol.orgon April 10, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

11
result is in accordance with those shown in Figures 3, 4 and 5 using isolated PTM. The H2O2 release
measured in the presence of FCCP and without glucose was as low as that recorded for the potato
slices incubated with 5 mM glucose without FCCP (Fig. 8B, open square). These results indicate that the
preventive effects of glucose on H2O2 release mediated by the mt-HK activity observed in isolated PTM
are working in a similar manner in the potato tuber slices and are associated with increased O2
consumption.
DISCUSSION
Changes in free hexose levels by carbohydrate starvation and reverse feeding lead to variations
in the respiration rate in heterotrophic plant tissues (Brouquisse et al., 1991; Dieuaide et al., 1992).
These free hexose fluctuations may correspond to certain environmental or developmental conditions
where sugar availability may be limiting (Morrell and ap Rees, 1986; Geigenberger and Stitt, 1993; Renz
et al., 1993; Couée et al., 2006) and are associated with ROS generation.
High glucose levels are toxic in several models because of increased release of ROS by glucose
auto-oxidation and metabolism (Couée et al., 2006). The leakage of ROS from mitochondria is as
important in non-photosynthesizing plant cells as it is in mammalian cells (Skulachev, 1996; Skulachev,
1997; Møller, 2001). In animal cells, high glucose concentrations can lead to activation of NADPH
oxidase, which is one of the sites responsible for glucose toxicity (Bonnefont-Rousselot, 2002). However,
it has been proposed that ROS release by mitochondria via the respiratory chain is a causal link between
high glucose levels and the main pathways responsible for cell damage (Russell et al., 1999; Nishikawa
et al., 2000; Russell et al., 2002). In this regard, our group recently demonstrated that mt-HK activity
plays a central role in preventing mitochondrial ROS generation through a steady-state ADP re-cycling in
rat brain (da-Silva et al., 2004). High glucose levels in plants may not trigger mitochondrial ROS
formation as in mammals for the following reasons: (1) plant mt-HK activities are not strongly inhibited by
hexose-6-phosphate as they are in mammals (Renz and Stitt, 1993; Galina et al., 1995; da-Silva et al.,
2001; da-Silva et al., 2004; Meyer et al., 2006); (2) plant mt-HK activities are strongly inhibited by ADP
(Fig. 1B and Galina et al., 1995; da-Silva et a., 2001). These observations may explain the plant cell
tolerance to mitochondrial ROS formation due to high sugar concentration availability compared to
mammalian cells (Nishikawa et al., 2000; Brownlee, 2001; Couée et al., 2006). Thus, the ADP re-cycling
mt-HK activity would play a key preventive antioxidant role by keeping lower ΔΨm and ROS generation
levels in mitochondria.
In the present work we observed mt-HK activity (Table I) in isolated PTM, in agreement with a
recent report (Graham et al., 2007). The mt-HK activity found in PTM is highly dependent on oxygen
consumption, because the activation of mitochondrial respiration by succinate restores the activity levels
of the enzyme (Fig. 1A). On the other hand, the activity of mt-HK is able to promote an acceleration of
www.plantphysiol.orgon April 10, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

12
oxygen consumption after the conversion of ADP to ATP (Fig. 2A). This would ensure that the
hexokinase could respond rapidly to changes in the cellular demand for glucose-6-P, which is known to
be a key intermediate in several metabolic pathways sensitive to ADP/ATP ratio, including glycolysis,
sucrose synthesis, pentose phosphate pathway, and cellulose biosynthesis.
As previously observed for maize mt-HK, mt-HK activity from PTM is also much more sensitive
to ADP inhibition in the micromolar range when the substrate is glucose than with fructose (Fig. 1B)
(Galina et al., 1995; Galina et al., 1999; da-Silva et al., 2001). This result suggests that the affinity of mt-
HK for glucose and fructose is different. In fact this was shown in Figure 1 and is in accordance with the
kinetic behavior of mt-HK from maize seedling roots (Galina et al., 1999).
Because atractyloside and oligomycin impair the decrease of ΔΨm and the preventive role in
H2O2 formation sustained by the mt-HK reaction, we can conclude that this mt-HK activity is able to
promote modulation of electron flux in ETS via a mechanism of ADP re-cycling through ANT:FoF1ATP
synthase complex (Fig. 2 and Fig. 4). This mechanism of control of H2O2 formation was previously
observed in rat brain mitochondria which contains large amounts (more than 80% of total tissue content)
(Wilson, 2003; da-Silva et al., 2004) of mt-HK activity, but it had not been described so far for plant
mitochondria.
Besides the classical antioxidant enzymes (SOD, catalase, and ascorbate-glutathione
peroxidase), two systems have been identified as participants in controlling pro- and anti-oxidant
balance in plants, the alternative oxidase (AOX) and the uncoupling protein (PUMP). Ultimately, these
two systems work to decrease the long-lived ubisemiquinone (UQH●) concentration which, in turn, is
capable of directly reducing O2 (Skulachev, 1996; Skulachev, 1997). In isolated PTM, the mt-HK activity
achieves the same effect on the ubiquinone pool by decreasing the ΔΨm and increasing the O2
consumption after mt-HK activation (Figs. 2, 4 and 5). These observations are in accordance with the
respiratory control theory in which the ΔΨm is decreased by the presence of ADP (Figs. 2B and C). It is
known that a small decrease in the ΔΨm leads to a large reduction in the rate of H2O2 generation
(Korshunov et al., 1997). Our findings are in line with these data because the activation of mt-HK activity
by glucose reduces ΔΨm and practically abolished the rate of H2O2 generation (Figs. 2 and 3). In
addition, the mt-HK system reduces H2O2 generation by 50 % in PTM at glucose concentrations as low
as 20 μM (Fig. 5A). At saturating glucose concentration, the mt-HK activity level of 30 % (about 20 mU)
of the total amount recovered in PTM is sufficient to reduce the rate of H2O2 generation by 50 % (Figs.
5C and 7B). The half-maximal glucose concentration needed to activate mt-HK (about 100 μM) causes
an almost complete inhibition of H2O2 generation (Figs. 1C and 5A). This indicates that the mt-HK does
not need to be saturated with glucose to cause a significant impairment in ROS generation. That
saturation with glucose is not required is confirmed by comparing the rate of H2O2 generation and the
activity levels of mt-HK treated with its competitive inhibitor, NAG, which promotes a displacement of
glucose concentration required for inhibition to a higher values (i.e. a decrease by more than 80% in
H2O2 generation is achieved only at about 1 mM glucose) (Fig. 5B). A threshold plot of the activity levels
www.plantphysiol.orgon April 10, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

13
of mt-HK and the rate of H2O2 generation reveals that 50 % inhibition in ROS generation requires only
30 % of the total mt-HK activity bound to the PTM membrane (which corresponds to approximately 12 %
of total potato tuber HK activity) (Table I). This estimate leads us to propose that only 6 % of total
glucose phosphorylation occurring at the surface of the PTM membrane would lead to almost total
blockage of H2O2 generation. According, the data on rate of H2O2 release observed in potato tuber slices
are in agreement with the expectation that a small activation of mt-HK by glucose is sufficient to inhibit its
release in slices (Figs. 8B and C). Interestingly, the decrease in ROS generation was accompanied by a
corresponding increase in the oxygen consumption when glucose was added to the incubation medium
of potato tuber slices (Fig 8A).
The effect of glucose in decreasing the H2O2 release by potato tuber slices and in isolated PTM
could be due to a tightly bound mitochondrial HK that guides the ADP delivery to FoF1ATP synthase via
ANT in an efficient channeling to the mitochondrial matrix (Fig 7). Almost two orders of magnitude more
activity from an unbound HK form than from mt-HK is needed to reduce the rate of H2O2 release in
isolated PTM (Fig. 7B). These data suggest that the access to ADP is substantially increased by mt-HK
in PTM and in potato tuber slices. Several lines of evidence indicate that, in addition to its activity, the
localization of mt-HK is relevant for mitochondrial respiration (Moore and Jöbsis, 1970; BeltrandelRio
and Wilson, 1991; Galina et al., 1995). This possibility would be explained in mammalian mitochondria
by the mt-HK binding site in the VDAC-ANT complex. According to our previous study, mt-HK
localization seems to be critical for performing its preventive antioxidant activity, as the ADP would be
rapidly delivered through the VDAC-ANT complex to the FoF1ATP synthase, which phosphorylates it at
the expense of ΔΨm (da-Silva et al., 2004). Although the binding mechanism of mt-HK to plant
mitochondrial outer membranes is not yet fully established (Rezende et al., 2006), the data presented in
Figure 7 is similar to data from previous study in rat brain mitochondria which demonstrated that the mt-
HK activity is more effective in stimulating respiration in the presence of glucose than its nonbindable
chymotrypsin-treated mt-HK form (Moore and Jöbsis, 1970; BeltrandelRio and Wilson, 1991).
Comparison of ADP-recycling activity of mt-HK with the activity of St-UCP as preventive
antioxidant systems in PTM (Fig. 6) shows that mt-HK is able to decrease the ΔΨm, accelerate oxygen
consumption and block H2O2 release to the same extent as observed with St-UCP activated by 30 μM
linolenic acid (Fig. 6C). In contrast, when the St-UCP activity is inhibited by the presence of 1 mg/mL faf-
BSA and 1 mM ATP, the mt-HK activity is even more potent than St-UCP in preventing ROS generation
(Fig. 6B).
These similarities between mt-HK and St-UCP in the response with regard to ROS formation
may indicate that potato tubers have complementary mechanisms against oxidative damage induced by
respiration in heterotrophic plant tissues. In conditions of higher oxidative metabolism fueled by hexoses
may lead to increased ROS formation, the mt-HK activity would play a predominant role as a preventive
mechanism. On the other hand, when the rate of fatty acid β-oxidation is increased, the UCP becomes
the main mechanism to prevent the accumulation of ROS. In plant tissues, we cannot exclude the
www.plantphysiol.orgon April 10, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

14
possible operation of AOX in detoxifying the formation of superoxide anions. However, in potato tuber
under no abiotic stress, the AOX activity levels are low (Calegario et al., 2003; Considine et al., 2003). In
our experimental conditions the cyanide-resistant respiration is less than 3% when using succinate or
NADH as oxidizable substrates, and about 10% when using pyruvate plus malate as substrates (data not
shown).
CONCLUSION
Thus, besides its involvement in general plant sugar metabolism, in glucose sensing (Rolland et
al., 2006) and in the regulation of programmed cell death (Kim et al., 2006), we propose that mt-HK
plays a specific role in generating ADP to support oxidative phosphorylation, thereby avoiding an ATP
synthesis–related limitation of respiration and subsequent H2O2 release in plants.
MATERIALS AND METHODS
Chemicals and Biological Materials
ADP, ATP, FCCP, horseradish peroxidase, rotenone, safranine O, MHP, NAG, yeast
hexokinase, fatty acid-free bovine serum albumin, β-NAD+, linolenic acid, oligomycin and G6PDH from
Leuconostoc mesenteroides, were purchased from Sigma-Aldrich (St. Louis, MO, USA). Percoll was
from Amersham Biosciences (USA). Amplex Red was purchased from Invitrogen. All other reagents
were analytical grade. Potato tubers (Solanum tuberosum) were purchased from a local supermarket.
Isolation of Potato Tuber Mitochondria by Self-generated Percoll Gradient (PTM)
PTM were obtained as previously described (Neuburger et al., 1982) using a cold extraction
buffer containing: 10 mM Hepes/Tris pH 7.4; 0.3 M manitol; 2 mM EGTA ; 5 mM EDTA; 0.3 mM PMSF;
20 mM β-mercaptoethanol and 0.1 g % (w/v) faf-BSA. The homogenate was strained through eight
layers of cheesecloth and centrifuged at 3,000 x g at 4 °C for 3 min. The supernatant was centrifuged at
12,000 x g at 4 °C for 10 min. The supernatant (S12) was used to determine enzymatic activities. The
mitochondrial pellet (P12) was resuspended in 5 mL of ice-cold extraction buffer and layered in Hitachi
P50 centrifuge tubes containing 35 mL of cold extraction buffer containing 28 % (v/v) Percoll and
centrifuged at 40,000 x g at 4 °C for 30 min. After centrifugation, three major bands were obtained in
sequence; the first band (fraction 1) at the top and the middle band (fraction 2) were used for further
PTM isolation or for enzymatic activity determinations. Mitochondrial activity markers were found to be
enriched in fraction 2. The mitochondrial fraction was removed and diluted with extraction buffer without
Percoll and centrifuged twice at 12,000 x g at 4 °C for 10 min. The final pellet was resuspended in 0.6
mL of extraction buffer and kept in an ice-water bath. The final protein concentration varied from 10 to 20
mg/mL.
www.plantphysiol.orgon April 10, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

15
Enzyme Assays and Kinetic Parameters
The activity of mt-HK was determined by a coupled assay according to Galina et al., (1995).
Briefly, mt-HK activity was determined by NADH formation following the absorbance at 340 nm at 28oC.
The assay medium contained: 20 mM Tris-HCl pH 7.4, 5 mM glucose, 6 mM MgCl2, 1 mM β-NAD+, 1
U/mL G6PDH, 2 mM PEP, 0.1% (v/v) Triton X-100 and 10 U/ml pyruvate kinase. The reaction was
started by adding 1 mM ATP. The final mitochondrial protein concentration varied from 0.05 to 0.1
mg/mL. When mt-HK activity was measured in intact PTM, Triton X-100 was omitted from the reaction
medium which was in this case the same as the respiration buffer used to measure oxygen consumption.
Glucose 6-P dehydrogenase (G6PDH) activity was assayed in a reaction medium containing 50
mM Tris-HCl buffer, pH 7.4, 6 mM MgCl2, 0.1% (v/v) Triton X-100 and 0.5 mM β-NADP+. The reaction
was started by adding 1 mM glucose 6-P, and G6PDH activity was determined by measuring the
absorption of β-NADPH formation at 340 nm.
FoF1 ATPase activity was determined by measuring the release of Pi from ATP in two different
reaction media: (1) in the absence or (2) in the presence of 5 mM NaN3. Both contained 20 mM Tris-HCl
pH 8.0, 5 mM MgCl2, 2 mM ATP and 1 μM FCCP. The reaction was started by the addition of PTM
protein and the difference between the activities in media (1) and (2) was considered as an authentic
FoF1 ATPase. The PTM protein concentration varied from 0.06 to 0.1 mg/mL.
The kinetic parameters were estimated by non-linear regression analysis applied to the
Michaelis-Menten equation using the program package supplied by Origin software (Galina et al., 1999).
Oxygen Uptake Measurements
Oxygen uptake was measured in an oximeter fitted with a water-jacket Clark-type electrode
(Yellow Springs Instruments Co., Model 5300, USA) or in Oxytherm System for Photosynthesis &
Respiration Measurements in Liquid-Phase – Hansatech Inst. (England).The PTM (0.2 mg/mL) were
incubated with 1 to 1.5 mL of the standard respiration buffer containing 0.3 M mannitol, 10 mM Tris-HCl
pH 7.2, 3 mM MgSO4, 10 mM NaCl, 5 mM KH2PO4, 0.3 mM β-NAD+ and 0.1% (v/v) faf-BSA. The cuvette
was closed immediately before starting the experiments. Respiratory control ratio (RCR) values were
obtained with isolated PTM, after complex I inhibition by 1 μM rotenone and complex II activation by 10
mM succinate. Other additions are indicated in the figure legends.
ΔΨm Determination
The mitochondrial membrane potential was measured by using the fluorescence signal of the
cationic dye safranine O, which is accumulated and quenched inside energized mitochondria (Akerman
and Wikströn, 1976). PTM (0.2 mg protein/mL) were incubated in the standard respiration buffer
supplemented with 15 µM safranine. FCCP (2 µM) was used as a positive control to collapse ΔΨm.
Fluorescence was detected with an excitation wavelength of 495 nm (slit 5 nm) and an emission
www.plantphysiol.orgon April 10, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

16
wavelength of 586 nm (slit 5 nm) using a Hitachi Model F-3010 spectrofluorimeter (Tokyo). Data were
reported as arbitrary fluorescence units (AFUs). Other additions are indicated in the figure legends.
Determination of Mitochondrial H2O2 Release
The H2O2 released from PTM was determined by the Amplex Red oxidation method, as previously
described (Smith et al., 2004). Briefly, mitochondria (0.2 mg protein/ml) were incubated in the standard
respiration buffer supplemented with 10 µM Amplex Red and 5 U/mL horseradish peroxidase. Fluorescence
was monitored at excitation and emission wavelengths of 563 nm (slit 5 nm) and 587 nm (slit 5 nm),
respectively. Calibration was performed by the addition of known quantities of H2O2.
Potato Tuber Slice Assays
Small square pieces were cut (8 mm diameter, 2 mm thickness) perpendicular to the stolon-apex
axis of potato tuber (Tiessen et al., 2002). The tuber slices were taken from the middle of the tuber,
avoiding the outer 3 mm and the tuber skin. The slices were incubated for 2 hours in 10 mM MES-KOH,
pH 6.5, in different conditions as shown in figure legend. In oxygen consumption measurements we used
16 slices (0.3 to 0.45 g total slices) in a Hansatech oxygraph chamber of 1 mL and the oxygen
consumption rate was monitored. For H2O2 generation 20 slices were placed in 10 mL of the incubation
buffer in an orbital shaker at 28oC. At different times, aliquots of 0.5 mL were removed and added to 1.5
mL respiration buffer containing 10 μM Amplex Red and 5 U/mL horseradish peroxidase and the
fluorescence measured was taken as the amount of H2O2 accumulated in the incubation buffer of potato
tuber slices.
Protein determination
The protein concentration was determined as described by Lowry (1951), using BSA as standard.
Statistical Analysis
Data were plotted with Origin 7.0 and analyzed by one-way ANOVA and a posteriori Tukey´s test. P
values < 0.05 were considered statistically different.
ACKNOWLEDGMENTS
We are grateful for Dr Martha Sorenson for kind and valuable comments in improving the paper. This
work is dedicated to Leopoldo de Meis in honor of his 70th birthday.
www.plantphysiol.orgon April 10, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

17
LITERATURE CITED
Akerman KE, Wikströn MK (1976) Safranine as a probe of the mitochondrial membrane potential.
FEBS Lett 68: 191-197
Arora KK, Pedersen PL (1988) Functional significance of mitochondrial bound hexokinase in tumor cell
metabolism. Evidence for preferential phosphorylation of glucose by intramitochondrially generated ATP.
J Biol Chem 263: 17422-17428
Avigad G, Englard S (1968) 5-Keto-D-fructose. V. Phosphorylation by yeast hexokinase. J Biol Chem
243: 1511-1513
BeltrandelRio H, Wilson JE (1991) Hexokinase of rat brain mitochondria: Relative importance of
adenylate kinase and oxidative phosphorylation as sources of substrate ATP, and interaction with
intramitochondrial compartments of ATP and ADP. Arch Biochem Biophys 286: 183-194
BeltrandelRio H, Wilson JE (1992) Interaction of mitochondrially bound rat brain hexokinase with
intramitochondrial compartments of ATP generated by oxidative phosphorylation and creatine kinase.
Arch Biochem Biophys 299: 116-124
Bernard EA (1975) Hexokinases from yeast. Methods Enzymol 42:6–20
Bolwell GP, Bindschedler LV, Blee KA, Butt VS, Davies DR, Gardner SL, Gerrish C, Minibayeva F
(2002) The apoplastic oxidative burst in response to biotic stress in plants: a three-component system.
J Exp Bot 53:1367-1376
Bonnefont-Rousselot D (2002) Glucose and reactive oxygen species. Curr Opin Clin Nutr Metab Care
5: 561-568
Boveris A, Chance B (1973) The mitochondrial generation of hydrogen peroxide. General properties
and effect of hyperbaric oxygen. Biochem J 134: 707-716
Brouquisse R, James F, Raymond P, Pradet A (1991) Study of glucose starvation in excised maize
root tips. Plant Physiol 96: 619-626
Brownlee M (2001) Biochemistry and molecular cell biology of diabetic complications. Nature 414: 813-
820
www.plantphysiol.orgon April 10, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

18
Calegario FF, Cosso RG, Fagian MM, Almeida FV, Jardim WF, Jezek P, Arruda P, Vercesi AE
(2003) Stimulation of potato tuber respiration by cold stress is associated with an increased capacity of
both plant uncoupling mitochondrial protein (PUMP) and alternative oxidase. J Bioenerg Biomembr 35:
211-220
Cesar MDC, Wilson JE (2004) All three isoforms of the voltage-dependent anion channel (VDAC1,
VDAC2, and VDAC3) are present in mitochondria from bovine, rabbit, and rat brain. Arch Biochem
Biophys 422: 191-196
Chance B, Sies H, Boveris A (1979) Hydroperoxide metabolism in mammalian organs. Physiol Rev 59:
527–605
Claeyssen E, Rivoal J (2007) Isozymes of plant hexokinase: occurrence, properties and functions.
Phytochemistry 68: 709-731 .
Considine MJ, Goodman M, Echtay KS, Laloi M, Whelan J, Brand MD, Sweetlove LJ (2003)
Superoxide stimulates a proton leak in potato mitochondria that is related to the activity of uncoupling
protein. J Biol Chem 278: 22298-22302
Couée I, Sulmon C, Gouesbet G, El Amrani (2006) An involvement of soluble sugars in reactive
oxygen species balance and responses to oxidative stress in plants. J Exp Bot 57: 449-459
da-Silva WS, Rezende GL, Galina A (2001) Subcellular distribution and kinetic properties of cytosolic
and on-cytosolic hexokinases in maize seedling roots: implications for hexose phosphorylation. J Exp
Bot 52: 1191-1201
da-Silva WS, Gómez-Puyou A, de Gómez-Puyou MT, Moreno-Sanchez R, De Felice FG, de Meis L,
Oliveira MF, Galina A (2004) Mitochondrial bound hexokinase activity as a preventive antioxidant
defense: steady-state ADP formation as a regulatory mechanism of membrane potential and reactive
oxygen species generation in mitochondria. J Biol Chem 279: 39846-39855
Damari-Weissler H, Ginzburg A, Gidoni D, Mett A, Krassovskaya I, Weber AP, Belausov E, Granot
D (2007) Spinach SoHXK1 is a mitochondria-associated hexokinase. Planta 226: 1053-1058
Dennis DT, Green TR (1975) Soluble and particulate glycolysis in developing castor bean endosperm.
Biochem Biophys Res Commun 64: 970-975
www.plantphysiol.orgon April 10, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

19
Dieuaide M, Brouquisse R, Pradet A, Raymond P (1992) Increased fatty acid beta-oxidation after
glucose starvation in maize root tips. Plant Physiol 99: 595-600
Dry IB, Nash D, Wiskich JT (1983) The mitochondrial localization of hexokinase in pea leaves. Planta
158: 152-156
Galina A, Reis M, Albuquerque MC, Puyou AG, Puyou MT, de Meis L (1995) Different properties of
the mitochondrial and cytosolic hexokinases in maize roots. Biochem J 1: 105-112
Galina A, Logullo C, Souza EF, Rezende GL, da-Silva W (1999) Sugar phosphorylation modulates
ADP inhibition of maize mitochondrial hexokinase. Physiol Plantarum 105: 17-23
Gechev TS, Van Breusegem F, Stone JM, Denev I, Laloi C (2006) Reactive oxygen species as
signals that modulate plant stress responses and programmed cell death. Bioessays 28: 1091-1101
Geigenberger R, Stitt M (1993) Sucrose synthase catalyses a readily reversible reaction in vivo in
developing potato tubers and other plant tissues. Planta 189: 329-339
Gottlob K, Majewski N, Kennedy S, Kandel E, Robey RB, Hay N (2001) Inhibition of early apoptotic
events by Akt/PKB is dependent on the first committed step of glycolysis and mitochondrial hexokinase.
Genes Dev 15: 1406-1418
Graham JW, Williams TC, Morgan M, Fernie AR, Ratcliffe RG, Sweetlove LJ (2007) Glycolytic
enzymes associate dynamically with mitochondria in response to respiratory demand and support
substrate channeling. Plant Cell 19: 3723-3738
Halliwell B, Gutteridge JMC (2007) Free radicals in biology and medicine. Oxford: Oxford University
Press
Kim M, Lim JH, Ahn CS, Park K, Kim GT, Kim WT, Pai HS (2006) Mitochondria-associated
hexokinases play a role in the control of programmed cell death in Nicotiana benthamiana. Plant Cell 18:
2341-2355
Korshunov SS, Skulachev VP, Starkov AA (1997) High protonic potential actuates a mechanism of
production of reactive oxygen species in mitochondria. FEBS Lett 416: 15-18
www.plantphysiol.orgon April 10, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

20
Liu, SS (1997) Generating, partitioning, targeting and functioning of superoxide in mitochondria. Biosci
Rep 17: 259-272
Liu Y, Ren D, Pike S, Pallardy S, Gassmann W, Zhang S (2007) Chloroplast-generated reactive
oxygen species are involved in hypersensitive response-like cell death mediated by a mitogen-activated
protein kinase cascade. Plant J 51: 941-954
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ (1951) Protein measurement with the Folin phenol
reagent. J Biol Chem 193: 265-275
Meyer LE, Machado LB, Santiago AP, da-Silva WS, De Felice FG, Holub O, Oliveira MF, Galina A
(2006) Mitochondrial creatine kinase activity prevents reactive oxygen species generation: antioxidant
role of mitochondrial kinase-dependent ADP re-cycling activity. J Biol Chem 281: 37361-37371
Miernyk JA, Dennis DT (1983) Mitochondrial, plastid, and cytosolic isozymes of hexokinase from
developing endosperm of Ricinus communis. Arch Biochem Biophys 226: 458-468
Moore CL, Jöbsis FF (1970) Some studies on the control of respiration in rat brain mitochondrial
preparations. Arch Biochem Biophys 138: 295–305
Morrell S, ap Rees T (1986) Sugar metabolism in developing tubers of Solanum tuberosum.
Phytochemistry 25: 1579-1585
Møller IM (2001) Plant mitochondria and oxidative stress: electron transport, NADPH turnover, and
metabolism of reactive oxygen species. Annu Rev Plant Physiol Plant Mol Biol 52: 561–591
Nakashima RA, Mangan PS, Colombini M, Pedersen PL(1986) Hexokinase receptor complex in
hepatoma mitochondria: evidence from N,N'-dicyclohexylcarbodiimide-labeling studies for the
involvement of the pore-forming protein VDAC . Biochemistry 25: 1015-1021
Neuburger M, Journet EP, Bligny R, Carde JP, Douce R (1982) Purification of plant mitochondria by
isopycnic centrifugation in density gradients of Percoll. Arch Biochem Biophys 217: 312-323
Nishikawa T, Edelstein D, Du XL, Yamagishi S, Matsumura T, Kaneda Y, Yorek MA, Beebe D,
Oates PJ, Hammes HP, Giardino I, Brownlee M (2000) Normalizing mitochondrial superoxide
production blocks three pathways of hyperglycaemic damage. Nature 13: 787-790
www.plantphysiol.orgon April 10, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

21
Pastorino JG, Shulga N, Hoek JB (2002) Mitochondrial binding of hexokinase II inhibits Bax-induced
cytochrome c release and apoptosis. J Biol Chem 277: 7610-7618
Puntarulo S, Galleano M, Sanchez RA, Boveris A (1991) Superoxide anion and hydrogen peroxide
metabolism in soybean embryonic axes during germination. Biochim Biophys Acta 1074: 277–283
Renz A, Stitt M (1993) Substrate specificity and product inhibition of different forms of fructokinases and
hexokinases in developing potato tubers. Planta 190: 166–175
Rezende GL, Logullo C, Meyer L, Machado LB, Oliveira-Carvalho AL, Zingali RB, Cifuentes D,
Galina A (2006) Partial purification of tightly bound mitochondrial hexokinase from maize (Zea mays L.)
root membranes. Braz J Med Biol Res 39: 1159-1169
Rhoads DM, Subbaiah CC (2007) Mitochondrial retrograde regulation in plants. Mitochondrion 7: 177-
194
Rolland F, Baena-Gonzalez E, Sheen J (2006) Sugar sensing and signaling in plants: conserved and
novel mechanisms. Annu Rev Plant Biol 57: 675-709
Russell, JW, Sullivan, KA, Windebank, AJ, Herrmann, DN, and Feldman, EL (1999) Neurons
undergo apoptosis in animal and cell culture models of diabetes. Neurobiol Dis 6: 347–363
Russell, JW, Golovoy D, Vincent, AM, Mahendru P, Olzmann, JA, Mentzer A, Feldman EL (2002)
High glucose-induced oxidative stress and mitochondrial dysfunction in neurons. FASEB J 16: 1738–
1748
Schreck R, Baeuerle PA (1991) A role for oxygen radicals as second messengers. Trends Cell Biol 1:
39-42
Smith AM, Ratcliffe RG, Sweetlove LJ (2004) Activation and function of mitochondrial uncoupling
protein in plants. J Biol Chem 279: 51944–51952
Skulachev VP (1996) Role of uncoupled and non-coupled oxidations in maintenance of safely low levels
of oxygen and its one-electron reductants. Q Rev Biophys 29: 169-202
Skulachev VP (1997) Membrane-linked systems preventing superoxide formation. Biosci Rep 17: 347-
366
www.plantphysiol.orgon April 10, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

22
Tiessen A, Hendriks JH, Stitt M, Branscheid A, Gibon Y, Farré EM, Geigenberger P (2002) Starch
synthesis in potato tubers is regulated by post-translational redox modification of ADP-glucose
pyrophosphorylase: a novel regulatory mechanism linking starch synthesis to the sucrose supply. Plant
Cell 14: 2191-2213
Turrens, JF (1997) Superoxide production by the mitochondrial respiratory chain. Biosci Rep 17: 3-8
Vander Heiden MG, Plas DR, Rathmell JC, Fox CJ, Harris MH, Thompson CB (2001) Growth factors
can influence cell growth and survival through effects on glucose metabolism. Mol Cell Biol 21: 5899-
5912
Vercesi AE, Martins IS, Silva MAP, Leite HMF, Cuccovia IM, Chaimovich H (1995) PUMPing plants.
Nature 375:24
Vercesi AE, Borecký J, Maia Ide G, Arruda P, Cuccovia IM, Chaimovich H (2006) Plant uncoupling
mitochondrial proteins. Annu Rev Plant Biol 57: 383-404
Wiese A, Gröner F, Sonnewald U, Deppner H, Lerchl J, Hebbeker U, Flügge U, Weber A (1999)
Spinach hexokinase I is located in the outer envelope membrane of plastids. FEBS Lett 461: 13-18
Wilson JE (2003) Isozymes of mammalian hexokinase: structure, subcellular localization and metabolic
function. J Exp Biol 206: 2049-2057
www.plantphysiol.orgon April 10, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

23
FIGURE LEGENDS
Figure 1. Mitochondrial Hexokinase Activity is Dependent on Respiration, Kinetic Properties of ADP
Inhibition of mt-HK Activity and Hexose Phosphorylation. A, The time course of G6P production by mt-
HK in PTM after addition of 5 mM glucose and 1 mM ATP is shown in traces 1, 2 and 3. Isolated PTM,
0.2 mg/ml, was added to respiration buffer containing 1 U/mL G6PDH and 1 mM β-NAD+. The reaction
temperature was 25oC. When indicated (↓), 10 mM succinate (Succ) or 50 mM mannoheptulose (MHP)
was added to the respiration medium. In trace 3, 0.3 mM ADP was included before starting the mt-HK
reaction with ATP and glucose. In trace 2, the reaction course was as in trace 1, but when indicated
MHP was added. B, Inhibition of the mt-HK activity by ADP in the presence of 5 mM glucose (closed
circles) or fructose (open circles). The maximal activity (Vo/Vi) of 1.0 was 160 ± 23 (mean ± SE, n = 5)
for glucose or 107 ± 6 (mean ± SE, n = 4) for fructose (nmol Hex6P.min-1.mg ptn-1). In C, the glucose
(closed circles) dependence for mt-HK activity revealed a KM of 148 ± 12 μM. The fructose (open circles)
dependence revealed a KM of 1375 ± 380 μM. The Vmax was 150 or 121 nmol G6P.min-1.mg ptn-1 for
glucose or fructose, respectively. The figure shows a representative experiment. The mean values ± SE
are from at least four independent preparations.
Figure 2. The Rate of Oxygen Consumption and Membrane Potential (ΔΨm) are Modulated by mt-HK
Activity. A, Oxygen uptake was measured using the respiration buffer described in Materials and
Methods. Numbers indicates the rate of oxygen consumption (nmol O2. min-1. mg ptn-1). B, The ΔΨm was
measured with fluorescence quenching of safranine O. The numbers indicated are the final ADP
concentration, in micromolar, added to the cuvette in each pulse. After the inclusion of 200 μM ADP, a
pulse of 5 mM glucose (Glc) was added followed by 50 mM mannoheptulose (MHP) to inhibit mt-HK
activity. When indicated, 100 μM atractyloside (Atr) was added to the reaction medium. In C is shown a
higher magnification of the ΔΨm trace in 2B. The figure shows a representative experiment. The
membrane potential and the oxygen consumption measurements were repeated with at least fifteen
independent PTM preparations.
Figure 3. Effect of mt-HK Activity on H2O2 Release by PTM. H2O2 release in two independent traces
shows the effect of 1mM ADP (left trace) or 5 mM glucose (Glc) (right trace) during a time course of
H2O2 release. The H2O2 release was detected by Amplex Red and peroxidase, after induction of
respiration by 10 mM succinate and 1 mM ATP when indicated by vertical lines. In the right trace, 50 mM
mannoheptulose (MHP) was added when indicated. The PTM was 0.2 mg/mL and the reaction was
performed at 25oC. The figure shows a representative experiment. The H2O2 release by PTM was
measured with at least ten independent PTM preparations.
www.plantphysiol.orgon April 10, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

24
Figure 4. Effects of Substrates, Uncouplers and Inhibitors on PTM Membrane Potential (ΔΨm) and H2O2
Release. After addition of 10 mM succinate, the agents were added sequentially in the order indicated
below each bar, separated by slashes (/), to evaluate ΔΨm (A and B) and H2O2 release (C). The
concentrations were: 5 μM FCCP; 150 – 200 μM ADP; 1 mM ATP; 1 - 4 mM glucose; 150 μM
atractyloside; 2 μg/mL oligomycin; 50 mM mannoheptulose; 50 mM N-acetylglucosamine. The
concentration of mitochondrial protein varied between 0.2 and 0.3 mg / mL. A Decrease in membrane
potential is represented as percent of depolarization related to the maximal polarization observed after
addition of succinate. The 100% rate of mitochondrial H2O2 release (225.1 ± 14 pmol.min -1.mg ptn-1)
was that following the addition of 10 mM succinate and 1 mM ATP in at least 6 independent potato tuber
mitochondrial preparations. The addition of glucose was performed after the ΔΨm return to the
hyperpolarization state from state 3 to state 4. Asterisks represent statistically significant differences
between groups, determined by one-way ANOVA and Tukey´s test. (*) p < 0.0001 relative to
Suc/ADP/glucose. (**) p < 0.0001 relative to Suc/ATP/glucose. The values are mean ± SE of percent
values of the numbers (n) of experiments.
Figure 5. Glucose Concentration Dependence for Activate the mt-HK Activity and Inhibition of H2O2
Release by PTM. In A is shown the rate of H2O2 release (open circles) and mt-HK activity (closed circles)
in the absence of NAG, an inhibitor of mt-HK activity. In B is shown the rate of H2O2 release (open
triangles) and mt-HK activity (closed triangles) in the presence of 25 mM NAG. The PTM H2O2 release
was induced by the addition of 10 mM succinate and 1 mM ATP. The final protein concentration of PTM
was 0.2 mg/mL in the respiration buffer. In C is shown a plot of mt-HK activity against the rate of H2O2
release from the data in A and B. The maximal rate of H2O2 release was 210 pmol. min-1. mg ptn-1
measured in the absence of added glucose; the maximal rate of mt-HK activity was 105 nmol. min-1. mg
ptn-1 measured at 10 mM glucose. The values shown in C are means ± SE of at least three independent
PTM preparations.
Figure 6. Comparison of the Preventive Antioxidant Role of mt-HK and PUMP Activation in PTM. A
shows the FCCP titration the rate of H2O2 release (open triangles), rate of oxygen consumption (open
stars) and membrane potential (open circles). The closed symbols plotted on a dotted vertical line
represent the values of these parameters as percentage of maximum, when mt-HK is fully active in the
absence of uncoupler. This activation is equivalent to the uncoupling effect of 700 nM FCCP. In B is
shown the effect of linolenic acid titration, an activator of PUMP, on the same parameters measured in
the presence of 1 mg/mL faf-BSA (which sequesters free fatty acids) and 1 mM ATP (an antagonist of
plant UCP). In C is shown the same experiment as in B, but without faf-BSA and ATP. The mt-HK
activity was induced by 5 mM glucose in the presence of 1 mM ATP. The symbols plotted once dotted
vertical line represent the values of these parameters as percentage of maximum, when mt-HK is fully
active and PUMP is inhibited. The values shown are means ± SE of at least three independent PTM
www.plantphysiol.orgon April 10, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

25
preparations. The error bars for bioenergetics parameters when mt-HK is active shown in A were similar
in B and C, but were omitted for clarity.
Figure 7. Specific Localization of mt-HK Is Important to Prevent H2O2 Release in PTM. In A, left and
middle traces, show different fructose or glucose concentrations being added during a time course of
H2O2 release in PTM to induce the mt-HK activity to different activity levels (the final concentrations are
indicated on the traces). Oligomycin was added to a final concentration of 2 μg/mL. In the right graph,
0.5 mM fructose was added as a saturating substrate for exogenously added soluble yeast-HK. Different
amounts of yeast-HK were added to the respiration buffer which contained 0.2 mg/mL PTM. The
numbers in different traces indicate the final mU attained in the 2 mL reaction mixture. In B is plotted the
percentage of maximum rate of H2O2 release by PTM against the activity level of mt-HK obtained with
different glucose concentrations (Figure 4A) (closed circles) or yeast-HK (open circles). The mt-HK
activity was induced by 0.5 mM (closed star) or 20 mM (open star) fructose in PTM to measure the H2O2
release without yeast-HK. The figure shows a representative experiment. The measurements of H2O2
release by PTM were repeated at least with three independent PTM preparations.
Figure 8. H2O2 Release and Oxygen Consumption in Potato Tuber Slices. A shows the rate of O2
consumption in potato tuber slices. B shows the time course of H2O2 release in potato tuber slices.
Twenty slices were maintained in 10 mM MES-KOH, pH 6.5 medium with different media that contained:
no added sugars (closed circles); 5 mM glucose (open circles); 5 mM glucose plus 50 mM NAG (closed
triangles); 5 mM sucrose (open triangles) and no added glucose plus 1 μM FCCP (open square). C
shows the rate of H2O2 release from potato tuber slices under these same conditions. The values
represent means ± S.E. of four independent experiments.
* At the 0.05 level, the population means are significantly different.
www.plantphysiol.orgon April 10, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.
26
Table I Mitochondrial location of potato tuber particulate hexokinase activitya
Fractions Enzyme activities
(mU/mg ptn)
F1FOATPase Hexokinase Glucose 6-P
dehydrogenase
Total
protein in
fraction
(mg)
Total HK
activity per
fractiond
(mU)
(% of total)
Homogenate
6.4 ± 2.2
(n=7)
9.7 ± 2.4
(n=8)
16.4 ± 1.9
(n=4)
N.D.
S12b 8.4 ± 2.5
(n=7)
2.6 ± 0.6
(n=8)
25.5 ± 1.2
(n=6)
2150 ± 97
(n=3)
5200
(82%)
P12c 22.4 ± 8.1
(n=6)
32.1 ± 3.5
(n=8)
30.8 ± 3.8
(n=6)
36 ± 7
(n=5)
1156
(18%)
P12 after Percoll Gradient
Fraction 1
5.7 ± 0.5
(n=4)
20.2 ± 7.7
(n=7)
32.2 ± 2.9
(n=4)
13 ± 1.6
(n=3)
262
(4%)
Fraction 2
83.9 ± 22.6
(n=7)
75.8 ± 16
(n=8)
0.3 ± 0.1
(n=6)
10 ± 1
(n=3)
758
(12%)
a The F0F1 ATPase, hexokinase and Glucose 6-P dehydrogenase activities were measured
as described in Experimental procedures. The reaction time was 15 min and the temperature
was 28oC. The activities represent mean ± S.E. of, at least 3 independent preparations, as shown in
parenteses. b S12 is the supernatant of 12,000 g/ 10 min c P12 is the particulate pellet (12,000 g/10 min) d Total HK activity (100%) was considered to be the sum of the HK activities recovered in S12 plus P12
fractions.
www.plantphysiol.orgon April 10, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.
Camacho-Pereira et al. Figure 1
Figure 1. Mitochondrial Hexokinase Activity is Dependent on Respiration, Kinetic Properties of ADP Inhibition of mt-HK
Activity and Hexose Phosphorylation. A, The time course of G6P production by mt-HK in PTM after addition of 5 mM
glucose and 1 mM ATP is shown in traces 1, 2 and 3. Isolated PTM, 0.2 mg/ml, was added to respiration buffer containing
1 U/mL G6PDH and 1 mM β -NAD+. The reaction temperature was 25oC. When indicated (↓ ), 10 mM succinate (Succ) or 50 mM mannoheptulose (MHP) was added to the respiration medium. In trace 3, 0.3 mM ADP was included before starting
the mt-HK reaction with ATP and glucose. In trace 2, the reaction course was as in trace 1, but when indicated MHP was
added. B, Inhibition of the mt-HK activity by ADP in the presence of 5 mM glucose (closed circles) or fructose (open
circles). The maximal activity (Vo/Vi) of 1.0 was 160 ± 23 (mean ± SE, n = 5) for glucose or 107 ± 6 (mean ± SE, n = 4) or fructose (nmol Hex6P.min-1.mg ptn-1). In C, the glucose (closed circles) dependence for mt-HK activity revealed a KM of
148 ± 12 µµµµM. The fructose (open circles) dependence revealed a KM of 1375 ± 380 µµµµM. The Vmax was 150 or 121 nmol G6P.min-1.mg ptn-1 for glucose or fructose, respectively. The figure shows a representative experiment. The mean values
± SE are from at least four independent preparations.
MHP
Succ
Glc
trace 1
trace 2
trace 3
60 sec
ATP
50 mABS340
0 10 100 1000
0,2
0,4
0,6
0,8
1,0
Fraction of maximal HK activity (Vo/Vi)
ADP, µµµµM
Glucose
Fructose
0 1000 2000 3000 10000
0,2
0,4
0,6
0,8
1,0
CB
Vmax 0.93489 0.02047
k 148.13775 12.07055
Vmax 1.13023 0.10836
k 1375.22003 381.95563
Hexose, µµµµM
Glucose
FructoseA
Glc 6-P
1.0
0.8
0.6
0.4
0.2
1.0
0.6
0.8
0.4
0.2
www.plantphysiol.orgon April 10, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.
Camacho-Pereira et al. Figure 2
ADP
Glc
MHPGlc
Suc
0.15 mM ADP
Suc
0.15 mM ADP
5 mM Glc
B C
0.3
AU
F
8 A
UF
2 min. 2 min.
A
Mito
ADP, µµµµM AtrMHP
Glc
Glc
Atr50
150100
200
121
62
198
40 n
mo
l O
2
2 min.
72
25
Figure 2. The Rate of Oxygen Consumption and Membrane Potential (∆∆∆∆Ψm) are Modulated by mt-HK
Activity. A, Oxygen uptake was measured using the respiration buffer described in Materials and Methods.
Numbers indicates the rate of oxygen consumption (nmol O2 . min-1. mg ptn-1). B, The ∆∆∆∆Ψm was measured
with fluorescence quenching of safranine O. The numbers indicated are the final ADP concentration, in
micromolar, added to the cuvette in each pulse. After the inclusion of 200 µµµµM ADP, a pulse of 5 mM glucose
(Glc) was added followed by 50 mM mannoheptulose (MHP) to inhibit mt-HK activity. When indicated, 100
µµµµM atractyloside (Atr) was added to the reaction medium. In C is shown a higher magnification of the ∆∆∆∆Ψm
trace in 2B. The figure shows a representative experiment. The membrane potential and the oxygen
consumption measurements were repeated with at least fifteen independent PTM preparations.
www.plantphysiol.orgon April 10, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

Camacho-Pereira et al. Figure 3
Figure 3. Effect of mt-HK Activity on H2O2 Release by PTM. H2O2 release in two independent traces shows
the effect of 1mM ADP (left trace) or 5 mM glucose (Glc) (right trace) during a time course of H2O2 release.
The H2O2 release was detected by Amplex Red and peroxidase, after induction of respiration by 10 mM
succinate and 1 mM ATP when indicated by vertical lines. In the right trace, 50 mM mannoheptulose (MHP)
was added when indicated. The PTM was 0.2 mg/mL and the reaction was performed at 25oC. The figure
shows a representative experiment. The H2O2 release by PTM was measured with at least ten independent
PTM preparations.
ATP
Succ
ADP
60 sec
H
2O
2
350 pmol
MHP
Glc
ATP
Succ
www.plantphysiol.orgon April 10, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

Suc
Suc / FCCP
Suc / ADP
Suc / ADP / Glc At
r
OligoMHPNAG
0
2
4
6
8
10
12
14
50
100
150
* **
Decrease of ∆Ψ
∆Ψ
∆Ψ∆Ψ
mform
ed by succinate.
(% of Maxim
um)
*
Suc/ADP/Glc Suc / ATP
Suc / FCCP
Suc / ATP / Glc At
r
OligoMHP
NAG
(5)(5)(5)(5)
(10)
(5)(5)(5) (7)
(10)
Suc/ATP/Glc
CB
Suc/ATP/Glc
A
(10)
Suc
Suc / ATP
Suc / FCCP
Suc / ATP / Glc A
tr
OligoMHP
NAG
0
20
40
60
80
100 (4)(5)(6)(5)
(11)
(8)
(10)
****** Rate of mitochondrial H
2O
2 release.
(% of Maxim
um)
**
Camacho-Pereira et al. Figure 4
Figure 4. Effects of Substrates, Uncouplers and Inhibitors on PTM Membrane Potential (∆Ψm) and H2O2 Release. After addition of 10 mM
succinate, the agents were added sequentially in the order indicated below each bar, separated by slashes (/), to evaluate ∆Ψm (A and
B) and H2O2 release (C). The concentrations were: 5 µM FCCP; 150 – 200 µM ADP; 1 mM ATP; 1 - 4 mM glucose; 150 µM atractyloside;
2 µg/mL oligomycin; 50 mM mannoheptulose; 50 mM N-acetylglucosamine. The concentration of mitochondrial protein varied between
0.2 and 0.3 mg / mL. A, Decrease in membrane potential is represented as percent of depolarization related to the maximal polarization
observed after addition of succinate. The 100% rate of mitochondrial H2O2 release (225.1 ± 14 pmol.min -1. mg ptn-1) was that following
the addition of 10 mM succinate and 1 mM ATP in at least 6 independent potato tuber mitochondrial preparations. The addition of glucose
was performed after the ∆Ψm return to the hyperpolarization state from state 3 to state 4. Asterisks represent statistically significant
differences between groups, determined by one-way ANOVA and Tukey´s test. (*) p < 0.0001 relative to Suc/ADP/glucose. (**) p <
0.0001 relative to Suc/ATP/glucose. The values are mean ± SE of percent values of the numbers (n) of experiments.
www.plantphysiol.orgon April 10, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

Camacho-Pereira et al. Figure 5
Figure 5. Glucose Concentration Dependence for Activate the mt-HK Activity and Inhibition of H2O2 Release by PTM. In A is shown the
rate of H2O2 release (open circles) and mt-HK activity (closed circles) in the absence of NAG, an inhibitor of mt-HK activity. In B is
shown the rate of H2O2 release (open triangles) and mt-HK activity (closed triangles) in the presence of 25 mM NAG. The PTM H2O2
release was induced by the addition of 10 mM succinate and 1 mM ATP. The final protein concentration of PTM was 0.2 mg/mL in the
respiration buffer. In C is shown a plot of mt-HK activity against the rate of H2O2 release from the data in A and B. The maximal rate of
H2O2 release was 210 pmol. min-1. mg ptn-1 measured in the absence of added glucose; the maximal rate of mt-HK activity was 105
nmol. min-1. mg ptn-1 measured at 10 mM glucose. The values shown in C are means ± SE of at least three independent PTM
preparations.
C
0 10 100 1000 10000
B
Glucose, µµµµMGlucose, µµµµM
% of maxim
al mt-HK activity
A
0
20
40
60
80
100
% of maxim
al rate of ROS form
ation
% m
t-HK activ
ity with
25 m
M NAG
0 10 100 1000 10000
0
20
40
60
80
100
% ra
te of R
OS fo
rmatio
n with
25 m
M NAG
0 20 40 60 80 100
0
20
40
60
80
100
% of maxim
al H
2O
2 release rate
% of maximal mt-HK activity
in Potato Tuber Mitochondria
www.plantphysiol.orgon April 10, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

Figure 6. Comparison of the Preventive Antioxidant Role of mt-HK and PUMP Activation in PTM. A shows the
FCCP titration the rate of H2O2 release (open triangles), rate of oxygen consumption (open stars) and
membrane potential (open circles). The closed symbols plotted on a dotted vertical line represent the values of
these parameters as percentage of maximum, when mt-HK is fully active in the absence of uncoupler. This
activation is equivalent to the uncoupling effect of 700 nM FCCP. In B is shown the effect of linolenic acid
titration, an activator of PUMP, on the same parameters measured in the presence of 1 mg/mL faf-BSA (which
sequesters free fatty acids) and 1 mM ATP (an antagonist of plant UCP). In C is shown the same experiment as
in B, but without faf-BSA and ATP. The mt-HK activity was induced by 5 mM glucose in the presence of 1 mM
ATP. The symbols plotted once dotted vertical line represent the values of these parameters as percentage of
maximum, when mt-HK is fully active and PUMP is inhibited. The values shown are means ± SE of at least
three independent PTM preparations. The error bars for bioenergetics parameters when mt-HK is active shown
in A were similar in B and C, but were omitted for clarity.
Camacho-Pereira et al. Figure 6
A B C
0,00 0,1 1 10
0
20
40
60
80
95
100
Bioenergetic m
itochondrial Parameters
(% of maxim
al)
FCCP, µ µ µ µM
0,0 1 10 1000
20
40
60
80
98
99
100
Linolenic acid, µµµµM
0,0 1 10 1000
20
40
60
80
98
99
100
0.0 0.1 0.0 0.0
www.plantphysiol.orgon April 10, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

0 10 100 1000
0
20
40
60
80
100
Rate of MItochondrial H
2O
2 release
(% of maxim
al rate)
HK activity, mU
Oligo
Oligo
0.5 mM Glc
10 mM
Fruct
0.5 mM
Fruct
Succ +
ATP
B
60 sec
3150
1575
630
378 189
Yeast-HK (mU)
Succ +
ATP
0.5 mM
Fruct
H
2O
2
700 pmol
A
Camacho-Pereira et al. Figure 7
Figure 7. Specific Localization of mt-HK Is Important to Prevent H2O2 Release in PTM. In A, left and middle traces, show different
fructose or glucose concentrations being added during a time course of H2O2 release in PTM to induce the mt-HK activity to different
activity levels (the final concentrations are indicated on the traces). Oligomycin was added to a final concentration of 2 µg/mL. In the
right graph, 0.5 mM fructose was added as a saturating substrate for exogenously added soluble yeast-HK. Different amounts of
yeast-HK were added to the respiration buffer which contained 0.2 mg/mL PTM. The numbers in different traces indicate the final mU
attained in the 2 mL reaction mixture. In B is plotted the percentage of maximum rate of H2O2 release by PTM against the activity
level of mt-HK obtained with different glucose concentrations (Figure 4A) (closed circles) or yeast-HK (open circles). The mt-HK
activity was induced by 0.5 mM (closed star) or 20 mM (open star) fructose in PTM to measure the H2O2 release without yeast-HK.
The figure shows a representative experiment. The measurements of H2O2 release by PTM were repeated at least with three
independent PTM preparations.
www.plantphysiol.orgon April 10, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.

0 30 60 90 120
Potato Tuber Slices
H2O
2 , pmol/ g of fresh weight
35.0
52.5
17.5
CB
minutes
A
0.0
Control G
lc
Glc + NAG
Sucrose
FCCP
0,0
0,4
0,8
1,2
1,6
*
**
Rate of H2O2 release
pmol / 30 m
in . g of fresh weigth
*
Control
Glc
Glc + NAG
Sucrose
FCCP
0
5
10
15
20
25
30
*
Rate of O2 consumption,
nmol/ m
in.g tissue
Figure 8. H2O2 Release and Oxygen Consumption in Potato Tuber Slices. A shows the rate of O2 consumption in potato tuber slices.
B shows the time course of H2O2 release in potato tuber slices. Twenty slices were maintained in 10 mM MES-KOH, pH 6.5 medium
with different media that contained: no added sugars (closed circles); 5 mM glucose (open circles); 5 mM glucose plus 50 mM NAG
(closed triangles); 5 mM sucrose (open triangles) and no added glucose plus 1 µM FCCP (open square). C shows the rate of H2O2
release from potato tuber slices under these same conditions. The values represent means ± S.E. of four independent experiments.
•At the 0.05 level, the population means are significantly different.
Camacho-Pereira et al. Figure 8
1.6
1.2
0.8
0.4
0.0
www.plantphysiol.orgon April 10, 2018 - Published by Downloaded from Copyright © 2008 American Society of Plant Biologists. All rights reserved.












![vermelho Iván Argote...Mais recentemente, Argote apresentou, em 2017, na exposição Future Generation Art Prize @ Venice, o monumento Sweet Potato [Batata doce]. Essa grande escultura](https://img.document.onl/doc/110x75/601d8ca2da194f78a95ce905/vermelho-ivn-argote-mais-recentemente-argote-apresentou-em-2017-na-exposio.jpg)
















