Embed Size (px)
Citation preview

MINISTÉRIO DA EDUCAÇÃO UNIVERSIDADE FEDERAL DE PELOTAS PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIA E TECNOLOGIA DE SEMENTES
TÉCNICA DE FOCALIZAÇÃO ISOELÉTRICA NA
DETERMINAÇÃO DE VARIEDADES DE Helianthus
tuberosus L.
Marcelo Marcos Muller
2005

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

ii
MINISTÉRIO DA EDUCAÇÃO UNIVERSIDADE FEDERAL DE PELOTAS FACULDADE DE AGRONOMIA ELISEU MACIEL
TÉCNICA DE FOCALIZAÇÃO ISOELÉTRICA NA DETERMINAÇÃO DE VARIEDADES DE Helianthus
tuberosus L. MARCELO MARCOS MÜLLER
Tese apresentada à Universidade Federal de Pelotas, sob a orientação do Prof. Silmar Teichert Peske, como parte das exigências do Programa de Pós-Graduação em Ciência e Tecnologia de Sementes, para obtenção do título de Doutorado.
PELOTAS Rio Grande do Sul - Brasil
Outubro de 2005

iii
Dados de catalogação na fonte:
(Marlene Cravo Castillo – CRB-10/744)
M958t Muller, Marcelo Marcos Técnica de focalização isoelétrica na determinação de variedades de Helianthus tuberosus / Marcelo Marcos Muller ; orientador Silmar Teichert Peske; co-orientadores Norbert Leist e Paulo Dejalma Zimmer. – Pelotas, 2005. – 37 f. Tese (Doutorado). Ciência e Tecnologia de Sementes. Faculdade de Agronomia Eliseu Maciel. Universidade Federal de Pelotas,. Pelotas, 2005. 1. Helianthus tuberosus 2. Focalização isoelétrica 3. Identificação de genótipos I. Peske, Silmar Teichert (oritentador) II. Título. CDD 633.491

iv
MARCELO MARCOS MÜLLER
UTILIZAÇÃO DA TÉCNICA DE FOCALIZAÇÃO ISOELÉTRICA NA DETERMINAÇÃO DE VARIEDADES DE Helianthus tuberosus L.
Tese apresentada à Universidade Federal de Pelotas, sob a orientação do Prof.Silmar Teichert Peske, como parte das exigências do Programa de Pós-Graduação em Ciência e Tecnologia de Sementes, para obtenção do título de Doutorado.
APROVADA: 11 de outubro de 2005 Prof. Dr. Paulo Dejalma Zimmer Prof. Dr. Antônio Carlos S. Albuquerque Barros Profa. Dra. Beatriz H. da Rocha Gomes Dr. Elbio Treicha Cardoso Prof. Dr. Silmar Teichert Peske (Orientador)

v
Aos meus pais Aos meus amigos

vi
AGRADECIMENTOS À Universidade Federal de Pelotas pela oportunidade de realizar o curso de Pós-Graduação. À Fundação Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) pela concessão da bolsa de estudos, tanto no país quanto no exterior. Ao Professor Silmar Teichert Peske pela valiosa orientação, sobretudo confiança no meu potencial durante o curso e execução do trabalho, o qual não poupou dedicação ao meu amadurecimento e formação profissional
Ao Prof. Paulo Dejalma Zimmer, pela Co-orientação, amizade e ensinamentos transmitidos. Ao Prof. Norbert Leist pela co-orientação na Alemanha e valiosos ensimentos.
Aos colegas da Alemanha, Rainer Knoblauch, Martina Went, Cristina Vogel, Andréa Jonitz, pelo companheirismo e ensinamentos.
Ao doutorando Gaspar Malone pela valiosa contribuição na análise dos resultados do trabalho e pela amizade.
Aos amigos Jackson Giongo, Clóvis, Maribel, Guilherme e Ana Hoffmann por sempre estarem presentes e pelo incentivo e auxílio fundamentais em todos os momentos. Aos colegas, funcionários e estagiários da área de sementes, pelo apoio e dedicação.
E a todos que direta ou indiretamente contribuíram de alguma forma para a realização deste trabalho.

vii
INDICE Página
SUMÁRIO ...................................................................................... vii
SUMMARY..................................................................................... viii
INTRODUÇÃO............................................................................... 01
a – Características botânicas e agrícolas da espécie........... 02
b – Importância econômica e nutricional............................ 05
c – Utilização da técnica de focalização isoelétrica na
determinação de cultivares................................................. 07
MATERIAL E MÉTODOS.............................................................. 09
Material vegetal.................................................................. 09
Preparo das amostras.......................................................... 10
Metodologia....................................................................... 10
RESULTADOS E DISCUSSÃO..................................................... 13
CONCLUSÕES................................................................................ 24
REFERÊNCIAS BIBLIOGRÁFICAS............................................. 25

viii
SUMÁRIO
TÉCNICA DE FOCALIZAÇÃO ISOELÉTRICA NA DETERMINAÇÃO DE
VARIEDADES DE Helianthus tuberosus L. UFPEL 2005. Professor Orientador:
Silmar Teichert Peske; Co-orientadores: Norbert Leist e Paulo Dejalma Zimmer.
Estudou-se a aplicação da técnica de focalização isoelétrica na determinação de 29
cultivares de Helianthus tuberosus. A importância nos estudos relacionados a
Helianthus tuberosus se deve ao aumento da utilização da inulina, uma fibra não
digerível presente em grande quantidade nessa planta, em alimentos dietéticos. A
quantidade de inulina varia conforme a variedade. Para o estudo, foi extraída a proteína
total de 29 cultivares, produzidas pelo instituto de pesquisa agropecuária LAP-
FORCHHEIM, Alemanha. Os resultados apresentados neste trabalho mostram que foi
possível diferenciar, três grupos de cultivares através da similaridade genética média,
sendo que o genótipo 22 (bianka) pode ser separado isoladamente dos demais. A técnica
de focalização isoelétrica aplica-se a uma determinação de cultivar rápida e de baixo
custo de cultivares de Helianthus tuberosus, tornando sua utilização acessível a
qualquer laboratório interessado.
Palavras-chave: Helianthus tuberosus, focalização isoelétrica, identificação de genótipos

ix
SUMMARY
ISOELECTRIC FOCUSING TECHNIQUE IN THE DETERMINATION OF
Helianthus tuberosus L.VARIETIES. UFPEL 2005. Adviser: Silmar Teichert Peske;
Co-advisers: Norbert Leist e Paulo Dejalma Zimmer.
The application of the isoelectric focusing technique in the determination of 29 varieties
of Helianthus tuberosus was study. The importance of studies related with Helianthus
tuberosus is due the rise of use of inulin, a non digerible fiber present in height amount
in this plant, in dietary foods. The inulin amount varies as the varieties. For the study,
protein was extracted of the 29 varieties produced by the institute LAP-FORCHHEIM,
Germany. The results show that it was possible to differentiate three groups of varieties,
through the medium similarity, and that the genotype 22 (bianka) could be separated
alone from the others. The isoelectric focusing can be apply to a fast variety
determination and with low cost of Helianthus tuberosus genotypes, and the use of the
technique can become accessible to any laboratory interested.
Key words: Helianthus tuberosus, isoelectric focusing, variety identification.

1
1
INTRODUÇÃO
A caracterização de cultivares e a pureza genética são importantes requisitos
de qualidade da semente, oferecidos na comercialização e, por essa razão, estão entre os
principais objetivos dos obtentores vegetais, dos institutos de proteção dos direitos dos
produtores de sementes e dos laboratórios de controle de sementes.
Segundo as regras da ISTA (1996), a certificação da pureza genética pode ser
realizada por meio de características morfológicas, fisiológicas, citológicas e
bioquímicas, McDonald et al., (1994).
Dada a importância desse assunto, o Instituto Agrícola de Pesquisa e de
Análises (LUFA – Augustenberg) de Karlsruhe, Alemanha, em parceria com o Instituto
Nacional de Plantas de Lavoura (LAP – Forchheim) de Forchheim, Alemanha, estão
desenvolvendo um projeto de identificação, caracterização e desenvolvimento de
cultivares de Helianthus tuberosus, com o objetivo de definir as cultivares existentes,
para melhorá-las e desenvolver métodos de identificação das mesmas.
O instituto LAP está incumbido de realizar a caracterização morfológica da
espécie, enquanto que o instituto LUFA começou a realizar estudos sobre as técnicas de
identificação de cultivares, que possam produzir resultado satisfatório na análise, além
de serem de fácil implementação em qualquer laboratório.
Os programas de melhoramento de Helianthus tuberosus tiveram no passado
pouco incentivo e eventualmente em intervalos irregulares. Além disso, outro motivo
para a baixa produção dos cultivares de Helianthus tuberosus é a possibilidade do
produtor fazer a propagação vegetativa do material, não comprando sementes

2
2
certificadas (Schittenhelm, 1999). Porém o advento das leis de proteção de cultivares e
da adesão dos paises as atas de convenção da UPOV, que passa a proteger o obtentor de
cultivares (UPOV, 1991), vem contribuindo para o incremento de estudos de
melhoramento de cultivares em culturas já desenvolvidas e também em culturas ainda
pouco estudadas até então.
O desenvolvimento de novas cultivares é desejável para incrementar os
sistemas de rotação de culturas.
A escolha da espécie para estudo, que está entre as de interesse mais recente
nos referidos institutos, se deve ao fato da mesma produzir grande quantidade de
inulina, carboidrato utilizado em alimentos dietéticos, como edulcorante natural, e por
não se ter ainda informações precisas sobre os cultivares existentes, além de não se ter
uma metodologia acessível aos laboratórios para a identificação dos cultivares.
a – Características botânicas e agrícolas da espécie
A planta de Helianthus tuberosus é originária da América do Sul e foi se
espalhando pela América Central e do Norte, antes de ser levada para a Europa,
provavelmente junto com a batata no século 17, durante as navegações. Na Europa
recebeu diversas denominações, sendo chamada de Topinambur na Alemanha,
Topinambour na França e Holanda e Jerusalém artichoke na Inglaterra. Com a
concorrência exercida pela batata, o cultivo de Helianthus tuberosus ficou restrito a
pequenas áreas, tomando um pequeno impulso após as guerras mundiais, quando a falta
de alimento e sua rusticidade, incentivaram sua exploração pelos agricultores, uma vez
que seus tubérculos servem como fonte de carboidratos.
Hoje a planta já está espalhada, não só pela América e Europa, como também
pela Ásia, África e Oceania.
A planta pertence à família das Asteraceae (Compositae), sendo muito
semelhante ao girassol (Helianthus annus) em estatura, porém, produzindo flores
menores.
Apresenta a formação de tubérculos enterrados no solo, que armazenam
substâncias de reserva, que serão utilizadas durante o período de germinação (brotação).
A inulina é utilizada em substituição ao amido como carboidrato utilizado como
substância de reserva.

3
3
A planta que eventualmente pode ser considerada como invasora em grandes
culturas, tem sua erradicação dificultada, justamente pela manutenção dos tubérculos no
solo, quando a parte aérea é eliminada (Wyse et al., 1986).
Além do aumento da quantidade de inulina na planta, que é atualmente o
principal interesse do melhoramento, também tem sido conduzidos trabalhos visando o
incremento da matéria seca, assim como uniformização e aumento do tamanho dos
tubérculos, com a opção de serem utilizados na indústria do álcool.
Além da crescente valorização dos seus tubérculos, a planta também pode ser
usada como alimento para ruminantes, com quantidades satisfatórias de todos os
nutrientes, com exceção do fósforo (P) (Seiler, 2004). O mesmo autor também cita a
importância do melhoramento e seleção da planta, como possibilidade de melhorar
também os atributos de qualidade da planta como forragem.
A Figura 1 mostra aspectos morfológicos da planta de Helianthus tuberosus,
destacando a grande produção de massa verde, e a aparência dos tubérculos colhidos e
descascados.
FIGURA 1. Planta de Helianthus tuberosus, tubérculos com casca e descascados.
A propagação de Helianthus tuberosus se faz através dos tubérculos,
semelhante a propagação de batata. A produção média de tubérculos fica entre 16 a 20

4
4
toneladas por hectare, sendo que cada planta produz entre 10 e 17 tubérculos,
dependendo da variedade.
Quanto à exigência de solo, não são aconselháveis solos muito úmidos nem
muito secos, sendo que a planta produz bem em solos mais argilosos, mas
preferencialmente solos arenosos (Schittenhelm, 1994).
A planta de Helianthus tuberosus é considerada como tendo boa resistência a
estresse hídrico (Monti et al., 2005a), o que contrasta com o trabalho de Schittenhelm,
(1999), que diz que a cultura é severamente afetada pelo estresse hídrico, quando
comparado com outros tubérculos como beterraba e chicória, principalmente devido a
sua disposição mais superficial no solo.
Uma vez que as técnicas de plantio e de colheitas de beterraba e de batata
podem ser utilizados para a cultura, praticamente nenhum investimento é necessário
para a sua produção.
O preparo do solo é similar ao dispensado para a batata, sendo bem arado e
com boa profundidade. Após a semeadura, também é feita a elevação do solo ao longo
das fileira, para proporcionar melhor suporte à planta e maior produção.
Devido a enorme estatura que suas plantas podem atingir, a influência de
ervas daninhas é baixa em lavouras de Helianthus tuberosus e representam baixa
percentual de perda de produção (Schittenhelm, 1999).
A forma dos tubérculos pode variar desde redonda até alongada, passando por
oval, cilíndrica e cuneiforme. Além de ser determinado pela cultivar, a forma do
tubérculo também é grandemente influenciado pelo tipo de solo onde o mesmo se
desenvolve.
Os tubérculos de Helianthus tuberosus apresentam 20 a 26% de matéria seca,
da qual 75 a 82% são carboidratos solúveis, sendo 75 a 98% dos açúcares redutores
totais (ART) correspondentes à frutose e o restante, glicose (Mullin et al., 1994).
Helianthus tuberosus é uma espécie que produz tubérculos comestíveis, sendo
seu principal uso na alimentação animal, mas é utilizada também na alimentação
humana, assim como na elaboração de bebidas destiladas (Schittenhelm, 1994).
b – Importância econômica e nutricional

5
5
A utilização de edulcorantes, naturais ou artificiais, na produção de adoçantes
dietéticos em substituição ao açúcar, em função da baixa quantidade de calorias, tem
incentivado a pesquisa e a descoberta de novos produtos que podem ser utilizados para
esse fim. São utilizados como edulcorantes o ciclamato de potássio, o aspartame, e
steviosídeo, a sacarina, a frutose, o manitol, entre outros, porém muitas pesquisas têm
sido realizadas no sentido de avaliar a segurança alimentar desses produtos.
Recentemente pesquisas na Europa mostraram que o uso de certa quantidade
de aspartame diariamente, provocou câncer em ratos, e isso gerou muita dúvida no
mercado consumidor, que já a algum tempo vem buscando alternativas naturais para o
consumo de adoçantes. Na América do Sul, inclusive no Brasil, o steviosídeo, um
edulcorante natural, vem despertando muito interesse da indústria alimentícia.
Juntamente ao steviosídeo, pode se acrescentar a inulina, que também é um
produto natural encontrado em plantas como chicória e Helianthus tuberosus. Isso
aumenta o interesse em pesquisar melhor essas plantas, tanto nos seus aspectos
agronômicos, como na obtenção de técnicas para determinação de cultivares e produção
de sementes.
A inulina é um ingrediente natural dos alimentos, encontrado em percentuais
variados em alimentos dietéticos. Está presente em mais de 36000 espécies de plantas.
(Carpita et al 1989).
A inulina com baixo grau de polimerização pode ser adicionada a alimentos
como adoçante de baixa caloria. Inulina com alto grau de polimerização pode ser usado
como fibra pro-biótica com efeitos benéficos para a saúde (Flamm et al., 2001).
Está presente como carboidrato de reserva em plantas em grande número de
vegetais e plantas, incluindo trigo, cebola, banana, chicória e Helianthus tuberosus.
Devido a não digestibilidade da inulina , ela pode ser recomendada para ser consumida
por diabéticos.
A inulina, considerada o frutano mais comum, é uma fibra alimentar solúvel
que não pode ser digerida pelos humanos. É uma glicofrutana de reserva presente nos
tubérculos de alguns membros da família Compositae, notadamente em Helianthus
tuberosus. É um polissacarídeo linear formado por aproximadamente 35 resíduos ß-D-
frutofuranose unidos por ligações ß-2,1 com uma terminação de α-D glucopiranose
idêntica à da sacarose (Shallenberg, 1982).
Frutanos são os oligo ou polissacarídeos que contém, ao menos, duas
unidades de frutose adjacentes. Uma glucose pode estar presente, mas não é necessário

6
6
(Lewis, 1993). São também reservas alternativas ao amido que ocorrem em 15% das
plantas superiores (Hendry, 1993). Podem também ser encontrado em órgãos
especializados, sendo utilizados durante o crescimento após a desfolha, que pode ser
observado pela queda do teor de frutanos antes do crescimento. Em Helianthus
tuberosus, essa queda é iniciada ainda durante a dormência (Martinez, 2000).
A produção de frutano pelas plantas é desejável para indústria de alimentos,
que pode transformá-lo em xarope de frutose por hidrólise, ou para utilização da inulina,
já mencionada acima. Tanto que genes de Helianthus tuberosus responsáveis pela
produção de frutanos foram inseridos em beterrabas, para que as mesmas também
passem a produzir esse carboidrato (Sevenier et al., 1998). Esse exemplo mostra que
através do rápido avanço da engenharia genética, os conceitos de classificação atual, de
acordo com os principais carboidratos de armazenamento, amido, frutano e sacarose,
começam a se tornar rapidamente inapropriados (Schittenhelm, 1999).
Em plantas, os frutanos podem ter outras funções, além de armazenar
reservas. Eles têm sido relacionados à proteção da planta contra déficit hídrico, causado
pela seca ou baixas temperaturas (Hendry and Wallace, 1993; Pilon-Smits et al., 1995).
A presença de frutanos nas plantas pode aumentar a tolerância ao frio e a seca. O
mecanismo por trás do aumento da resistência a vários estresses ainda não é claro, mas
tem sido proposto que os frutanos podem prevenir a danificação da membrana. Além da
importância ecológica, os frutanos são de grande interesse econômico, devido às suas
propriedades benéficas à saúde, que despertaram a atenção da indústria mundial de
alimento (Tomomatsu, 1994).
Os frutanos podem ser utilizados devido as suas características físicas e
fisiológicas, que os torna não somente interessante para o uso na alimentação humana e
animal, mas também em algumas aplicações não alimentares, como por exemplo, em
biopolímeros industriais (Fuchs, 1993). Essa classe de carboidratos tem sido empregada
com sucesso nos casos de constipação, prevenção de doenças do intestino, por estimular
a proliferação das bifidobactérias, e no controle da osteoporose, por estimular absorção
de cálcio (Coussement & Frank, 1998).
O aumento da temperatura durante a produção dos frutanos, se mostrou
prejudicial para a qualidade e quantidade do mesmo (Monti et al., 2005b)
Os teores de frutanos e conseqüentemente de inulina em Helianthus tuberosus
são distintos entre as variedades (Stonzenburg, 2002). Portanto a obtenção de técnicas

7
7
rápidas e de baixo custo para a determinação de variedades desta espécie se torna
importante.
Apesar da principal função dos tubérculos ser a de armazenar carboidratos
que serão utilizados posteriormente para o desenvolvimento de uma nova planta,
também podem ser encontradas proteínas de armazenamento nos tubérculos, porém em
menor quantidade, variando de 1 a 2 % na maioria dos tubérculos, até 10 % na
mandioca (Shewry, 2003). As proteínas de armazenamento classificam-se em quatro
tipos principais: 7S Globulinas; 11S Globulinas, 2S Albuminas e as Prolaminas.
c – Utilização da técnica de focalização isoelétrica na determinação de
cultivares
A técnica de focalização isoelétrica tem sido utilizada para a separação
varietal de diferentes espécies, como milho, trigo, tomate, entre outras (Wang et al.,
2000). Ela consiste na separação das frações protéicas através dos seus pontos
isoelétricos. Esta técnica é baseada na formação de um gradiente de pH, obtido pelo uso
de substâncias conhecidas como anfólitos, que são geralmente ácidos poliméricos
sintéticos (e.g., poliaminoácidos, ácidos policarboxílicos ou ácidos polissulfônicos).
A aplicação de um campo elétrico na mistura de anfólitos gera a formação de
um gradiente de pH. Os anfólitos são mantidos em um meio onde se usa um catodo com
pH alto, tipicamente hidróxido de sódio e um ânodo com pH baixo, tipicamente ácido
fosfórico. As proteínas, que são anfóteras, irão se comportar de maneira idêntica e serão
focalizadas em uma dada posição ditada pelo seu ponto isoelétrico (PI) (Hames e
Rickwood, 1990).
A utilização da focalização isoelétrica, quando possível, resulta em menor
custo para montar o laboratório, comparado a outras técnicas, além de ser uma técnica
relativamente simples e com boa qualidade dos resultados obtidos.
Wang et al (2004), utilizaram a técnica para determinar a pureza genética em
F1 de duas linhas de arroz híbrido, podendo ser usado em substituição ao teste a campo.
Lucchese et al. (1998) utilizou a técnica para identificação de 20 cultivares de
pimenta e encontrou correlação entre os resultados obtidos e a forma da semente. Além
disso, conseguiu separar isoladamente 16 cultivares somente pelo uso da técnica de
focalização isoelétrica, sendo que as pequenas diferenças das quatro restantes foram
separadas por análises de densiometria.

8
8
Lucchese et al. (1997) demonstrou ser possível a utilização da técnica para a
diferenciação de espécies de sementes do gênero Vicia e para a identificação de
cultivares de Vicia sativa e Vicia villosa.
Vieira et. al (2001) trabalhando com seis cultivares de feijão, conseguiu
separá-los em quatro grupos distintos utilizando a focalização isoelétrica.
Na Alemanha, o instituto LAP Forchheim é o responsável oficial, desde 1999
pela pesquisa e melhoramento da planta, trabalhando atualmente com 65 cultivares, com
o objetivo de caracterizar melhor a origem das cultivares, preservá-las e promover a
regulação das mesmas, (Stonzenburg, 2002).
O presente trabalho teve por objetivo avaliar a qualidade e eficiência da
técnica de focalização isoelétrica na determinação de variedades de Helianthus
tuberosus.

9
9
MATERIAL E MÉTODOS
Material Vegetal
Foram utilizados 29 genótipos de Helianthus tuberosus fornecidos pelo LAP -
Forchheim. O experimento foi conduzido no laboratório de eletroforese do instituto
LUFA, em Karlsruhe, Alemanha. A Tabela 1 apresenta os genótipos utilizados no
trabalho.
TABELA 1. Genótipos de Helianthus tuberosus utilizados nas análises. Os números se referem à
identificação dos genótipos no laboratório.
TOPIANKA 4 COLUMBIA 20 TOPSTAR 34
TOPIANKA 34 DUBO 21 GIGANT 35
MEDIUS 7 RICO 23 HENRIETTE 36
MEDIUS 31 MONTEO 24 DLO 40 37
DORNBURGER 13 WALDSPIEL 26 FUSEAU 60 38
DORNBURGER 41 ROTE ZONENKUGEL 27 LANDSORTE ROTE 39
BIANKA 22 BELA 29 LANDSORTE WEISS 40
BIANKA 25 2071-63 30 GUTE GELBE 42
VIOLET DE RENNES 12 BT-3 32 LOLA 67
BOYNARD 19 VÖLKENRODER 33

10
10
Preparo das amostras
Os tubérculos foram mantidos congelados até o momento do processamento
dos mesmos, sendo então descascados e fatiados sobre uma bandeja e levados para secar
durante a noite à temperatura do ar de 60ºC. Utilizou-se 10 tubérculos por genótipo.
Após secas, as amostras foram moídas e peneiradas. Para fazer a extração das
proteínas, foram pesadas 150 mg de cada amostra e colocadas dentro de eppendorfs,
adicionando-se, 0,9mL de água, por uma hora e depois centrifugadas, retirando-se
0,3mL do sobrenadante, para posterior aplicação no gel.
Metodologia
A metodologia aplicada no trabalho foi baseada na metodologia utilizada pelo
instituto, em função de sua vasta experiência na utilização da técnica de IEF em cereais,
porém como no presente trabalho foram utilizados tubérculos para a extração das
proteínas, as técnicas tiveram de ser adaptadas, no que se refere à quantidade e tipo de
anfólito utilizado, voltagem aplicada na corrida do gel, quantidade de amostra aplicada
no gel, entre outros, para que os géis pudessem apresentar bandas mais nítidas e
possíveis de serem avaliadas.
A focalização isoelétrica foi realizada em lâmina gel de 0,5mm de
espessura (17 X 13cm). Todas as análises foram feitas em cubas de eletroforese,
utilizando géis de poliacrilamida, preparados a partir de uma solução pronta de 7,33%
(p/V) de acrilamida, 0,23% (p/V) de N, N-metileno-bisacrilamida, sendo em seguida
acrescentados à uréia, os anfólitos, que produzem o gradiente de pH no gel, APS e
TEMED, nas proporções de 50mL de solução Acrilamida/Bis, (32:1); 16g de uréia;
6,2mL de anfólito, pH 6-7; 2,6mL de anfólito, pH 2-11; 350 µl APS e 50 µl TEMED.
Para o trabalho foi utilizado uma mistura com anfólitos de faixa de pH de 6–7
e 2-11. Essa mistura foi definida após estudos preliminares com todos os anfólitos
utilizados no laboratório, por apresentar melhor padrão de bandas nas faixas de maior
polimorfismo.
São aplicados 7 a 8mL da solução pronta em oito filmes plásticos, colocados
sobre placas de vidro, para dar suporte e para facilitar o manuseio, colocando-se então
outra placa de vidro sobre a anterior para que a solução pudesse se espalhar por toda a
placa antes de polimerizar.
11
11
O processo de polimerização se completa em aproximadamente uma hora,
quando então as duas placas de vidro que moldam o gel são abertas para retirada do
filme, contendo o gel com espessura de aproximadamente 0,5 mm.
O gel então é levado para as cubas de eletroforese, que estão ligadas a um
sistema de refrigeração para impedir o superaquecimento e possível queima do gel,
durante a passagem de alta corrente elétrica. Sobre a chapa de cada cuba é aplicado um
pouco de água, com isso, o filme contendo o gel se adere melhor a chapa, facilitando a
dissipação do calor.
Sobre o gel foram aplicados 15µl de amostra, com auxílio de um pente de
silicone, com espaço para aplicação de 50 amostras. O pente foi colocado no ânodo.
Para gerar a corrente elétrica, foi usada solução catódica de hidróxido de sódio 20
mmoL/L-1 e solução anódica de ácido fosfórico 10 mmoL/L-1., e o programa da corrida
do gel tinha as seguintes etapas:
TABELA 2. voltagem utilizada nas diferentes etapas do processo de focalização isoelétrica
Etapa Voltagem máxima Tempo (min)
1 300 10
2 800 10
3 1000 10
4 1500 10
5 2000 10
6 2500 10
7 3000 10
8 1000 ---
Após 70 minutos de corrida, o gel foi retirado das câmaras e colocado em uma
solução de Tricloroacético (TCA), para fixação das bandas de proteína, por 20 minutos,
seguido do tingimento em Comassie Blue, por 45 minutos e destingimento em solução
de ácido acético e etanol por 20 minutos.
Na avaliação do gel de isofocalização, a ausência e presença de bandas foi
designada por 1 e 0, respectivamente. Foi construída uma matriz binária e a estimativa
de similaridade genética entre cada par de genótipos foi calculada pelo coeficiente de

12
12
DICE. Estimadas as similaridades genéticas, os genótipos foram agrupados pelo método
UPGMA (unweighted pair-group method), obtendo-se desta forma, um dendrograma
(Rohlf, 1992).
13
13
RESULTADOS E DISCUSSÃO
Considerando a inexistência de uma metodologia de IEF para Helianthus
tuberosus, os resultados e discussão serão apresentados em duas partes, a primeira,
sobre o processo em si e a segunda sobre a identificação dos acessos, utilizando o
processo ajustado.
1. O processo de ajuste da metodologia de IEF para Helianthus tuberosus.
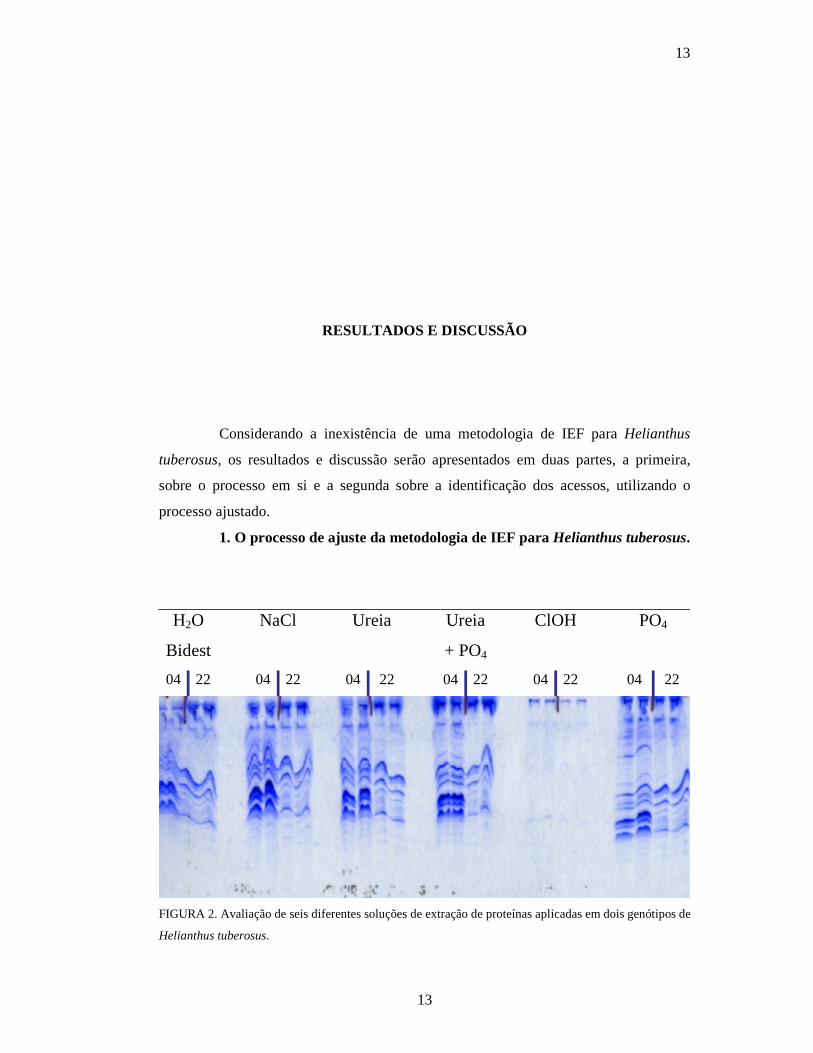
FIGURA 2. Avaliação de seis diferentes soluções de extração de proteínas aplicadas em dois genótipos de
Helianthus tuberosus.
H2O
Bidest
NaCl Ureia Ureia
+ PO4
ClOH PO4
04 22 04 22 04 22 04 22 04 22 04 22

14
14
A Figura 2 apresenta o resultado de estudos preliminares com seis diferentes
tipos de solução extratora. Cada solução age sobre um grupo de proteínas e pode mudar
o número de bandas polimórficas encontradas e ou a qualidade das mesmas.
Pode-se observar na Figura 2 que todas as soluções de extração apresentaram
padrões de bandas similares para extração de proteínas de Helianthus tuberosus, com
exceção do hidróxido de cloro (ClOH), que não extraiu proteínas de maneira eficiente.
Portanto, qualquer uma das soluções extratoras pode ser usada com eficiência, sendo
que no trabalho, foi utilizada somente água bidestilada, por ser a mesma de mais fácil
utilização em qualquer laboratório de análises.
Para se obter os melhores resultados nas corridas dos géis, foi necessário,
além de se testar as soluções extratoras, também testar os anfólitos que poderiam ser
usados no trabalho. Uma vez que ainda não se havia utilizado a técnica de focalização
isoelétrica em Helianthus tuberosus, iniciaram-se os testes utilizando as quantidades de
anfólito e faixas de pH já utilizadas no laboratório para testes rotineiros de análise em
trigo, porém os mesmos não se mostraram eficientes, como pode ser comprovada na
Figura 3, que apresenta os padrões de banda para doze amostras de Helianthus
tuberosus, submetidas à corrida em géis preparados para amostras de trigo, com a
presença de anfólitos na faixa pH entre 4 e 9.
Pode-se observar no gel da Figura 3, a baixa qualidade das bandas
encontradas, sendo também difícil de distinguir as bandas polimórficas, o que foi
dificultado também pela pequena faixa de pH que o anfólito utilizado cobriu,
proporcionando, portanto, uma condensação das bandas.
31 23 07 04 30 22 25 34 35 36 35 36
FIGURA 3. Eletroforegrama de 10 genótipos de Helianthus tuberosus, utilizando géis feitos seguindo
metodologia aplicada para avaliação de amostras de trigo.
Durante todo o experimento, foram feitos mais de 300 géis de poliacrilamida
e realizadas as corridas com as amostras, com o objetivo de testar o maior número de

15
15
variações possíveis dos fatores que poderiam influenciar no resultado final e
proporcionar um melhor padrão de gel para Helianthus tuberosus.
Uma vez que a metodologia de preparação da amostra foi estabelecida,
conforme citado em Material e Métodos e a solução de extração escolhida, em função
dos resultados preliminares apresentados, as principais mudanças realizadas na técnica
de focalização isoelétrica foram quanto ao tipo e quantidade de anfólito utilizado;
relação entre as quantidades de Urea, APS e TEMED utilizadas; relação da proporção
entre acrilamida e bisacrilamida; quantidade de amostra aplicada no gel; programação
da intensidade e do tempo de aplicação da corrente elétrica sobre o gel; distância entre
os pólos negativos e positivos e a posição da colocação da amostra.
Os anfólitos utilizados nas soluções de preparação dos géis de poliacrilamida
para a focalização isoelétrica têm a função de criar um gradiente de pH ao longo do gel,
depois de se passar a corrente elétrica, fazendo com que as proteínas migrem para o pH
correspondente ao seu ponto isoelétrico (pI).
Para cada espécie analisada pela técnica, existem bandas de pH onde se
concentram as maiores quantidades de bandas de proteínas polimórficas. É necessário,
portanto, testar diferentes anfólitos para determinar o que apresenta os melhores
resultados.
No trabalho foram testados mais de 20 anfólitos, sendo que se encontrou a
maior quantidade de bandas polimórficas em pH abaixo de sete.
A quantidade de anfólito utilizado foi dobrada em relação ao que vem sendo
utilizado para análises de milho e trigo, pois as bandas produzidas inicialmente não se
mostravam muito claras, nem uniformes, sendo que com a utilização de maior
quantidade de anfólito, elas ficaram mais definidas.
Para o trabalho, foi definida a utilização de uma mistura de anfólitos na
quantidade de 6,2 mL com pH entre 6-7 e 2,8mL com pH entre 2-11. O primeiro tem a
função de melhorar a focalização das bandas de proteínas nessa faixa de pH e próximas,
enquanto que a utilização do segundo, proporcionou uma melhor disposição das demais
bandas.
A presença ou não da uréia na solução e a proporção de APS e TEMED
também foram analisadas. A uréia também proporcionou melhor definição das bandas
de proteínas e melhor conservação da qualidade do gel, quando o mesmo não era usado
nos primeiros dias. Porém as variações nas quantidades já utilizadas de APS e TEMED,

16
16
não surtiram efeitos positivos nos géis. Ficou então definida a utilização de 16g de uréia
para cada 50mL de solução de bisacrilamida.
Além da utilização de uma solução pronta, com proporção definida de
acrilamida e bisacrilamida, foram feitas misturas das duas substâncias em proporções
diferentes das mesmas. Os resultados, todavia, não mostraram melhoras sensíveis na
definição das bandas de proteínas.
A quantidade da amostra aplicada sobre o gel mostrou sensíveis diferenças
nas bandas. Pouca quantidade de amostra produziu bandas muito finas e de difícil
diferenciação, ao passo que quantidades maiores de amostra fizeram com que as
proteínas tivessem dificuldade de atingir seu ponto ideal no gel, diminuindo a distância
percorrida pelas mesmas. Os melhores resultados foram obtidos com aplicação de 15
microlitros de amostra em cada célula.
A utilização de uma maior carga elétrica e por um tempo maior, vem se somar
à necessidade de se aumentar à quantidade de anfólito e de não se usar aplicações
elevadas de amostra, devido a maior dificuldade de migração das proteínas de
Helianthus tuberosus, quando comparadas a outras espécies. Porém, não se conseguiu
estabelecer nesse trabalho, as possíveis causas da maior dificuldade de migração dessas
proteínas.
Novamente, quando se partiu dos valores já padronizados para outras espécies
da programação da voltagem, as bandas se apresentavam condensadas e borradas, não
se tendo facilidade para distingui-las. Foi necessário se chegar à voltagem máxima
permitida pelo aparelho, para atingir os resultados desejáveis. Porém, essa maior
voltagem, que chegou a 3000 V na última fase, também aumentou o risco de
incineração do gel, que ocorria com mais freqüência em géis com falhas, ou seja,
espaços vazios no filme plástico e quando o gel não ficava bem aderido à chapa
dissipadora de calor da câmara.
Porém, a aplicação de alta voltagem desde o início da focalização, assim
como a pré-focalização, ou seja, a passagem de corrente elétrica sem a presença da
amostra, não se mostrou efetivos. No caso da utilização de alta voltagem no início da
corrida, as bandas se apresentaram menos nítidas, com maior dispersão, provavelmente
devido à energia inicial muito alta aplicada sobre as proteínas, dificultando a sua
migração mais coordenada.
Segundo Hames e Rickwood, (1990), a posição de colocação do pente de
silicone para aplicação das amostras no gel, seja mais próximo do catodo, do ânodo ou

17
17
no meio, não tem influência sobre os resultados obtidos. Essa afirmação pode valer para
outras espécies, mas não se aplica a Helianthus tuberosus, pois a colocação do pente de
silicone em qualquer posição que não seja próxima do ânodo, não produziu resultados
satisfatórios, além disso, a distância ideal do pente se dá a 0,5 cm da posição do
eletrodo.
As alterações realizadas permitiram que a técnica de focalização isoelétrica,
aplicada com eficiência em outras espécies, pudesse também ser utilizada para avaliação
de genótipos de Helianthus tuberosus, apresentando resultados reprodutíveis e com a
possibilidade de aplicação prática.
2. Identificação dos acessos por IEF ajustada para Helianthus tuberosus.
Na Figura 4 são apresentados os eletroforegramas de 24 amostras de
Helianthus tuberosus, correspondendo cada amostra, a duas colunas adjacentes, ou seja,
duas repetições.
19 39 23 41 27 32 38 37 21 36 34 22 13 40 42 33 30 07 43 12 35 31 29 20
FIGURA 4. Eletroforegrama de 24 amostras de Helianthus tuberosus, pela técnica de focalização
isoelétrica. Cada duas colunas representa um genótipo, identificado pelo número na linha superior e
identificado na Tabela 1. Os setas dentro da figura indicam diferenças entre os genótipos devido a
presença ou ausência de bandas de proteínas.
Pode-se observar que um grupo de genótipos, que correspondem às colunas
do lado direito do gel, possui uma banda que é inexistente no outro grupo, encontrado a
esquerda do gel. Essa banda, quando presente, situa-se na posição mais próxima do
ânodo, ou seja, em pH baixo. Portanto, as amostras podem ser divididas em dois

18
18
grandes grupos, em função da presença ou ausência de uma banda localizada em pH
mais ácido do gel (Figura 4 – seta 1).
Tomando-se o grupo sem a presença da banda mencionada acima, pode-se
fazer uma subdivisão em dois subgrupos, em função da presença ou ausência de duas
bandas na região intermediária do gel (Figura 4 – seta 2). As amostras com a presença
das duas bandas, correspondentes às variedades 19, 39, 23, 41, 27, 32 e 38 não puderam
mais ser subdivididas, por não apresentarem diferenças visíveis entre as mesmas no gel.
Isso indica que o número de bandas polimórficas que se consegue, utilizando esta
técnica para essas variedades, é insuficiente para conseguir separar todas, ou que essas
amostras devam ser agrupadas em uma única variedade, uma vez que não se tem
informações precisas ainda sobre todas as cultivares de Helianthus tuberosus.
Eventualmente, tubérculos de uma determinada cultivar, podem ser levada para locais
diferentes, onde também passou a ser cultivada, por receber nome diferente, causando a
confusão.
Ainda do grupo sem a presença da banda próxima ao ânodo e com a presença
da banda intermediária, diferencia-se a variedade 37 das demais, pelo fato da mesma
apresentar duas bandas na região mais próxima do catodo no gel, e que não estão
presentes em nenhum outro genótipo (Figura 4 – seta 3).
A variedade 22 também pode ser diferenciada das demais por apresentar
poucas bandas (Figura 4 – seta 4), principalmente na região básica do gel, onde se
concentram as principais bandas da maioria das variedades.
A variedade 34 se distingue das demais pela ausência de uma banda forte na
região básica, que está presente nas demais variedades deste grupo, com exceção da
variedade 22 (Figura 4 – seta 5).
Por fim, as variedades 21 e 36 podem ser diferenciadas entre si, pelo fato da
variedade 21 possuir duas bandas duplas, na região intermediária, ao passo que a
variedade 36 possui duas bandas simples no mesmo local (Figura 4 – seta 6).
Observando o grupo das amostras com a presença da banda na região ácida
(Figura 4 – seta 1), podem-se agrupar as variedades 13, 40 e 42 e diferenciá-las das
demais, devido à presença de bandas fortes na região intermediária do gel, um pouco
mais abaixo em relação às demais (Figura 4 – seta 7).
A variedade 33 pode ser diferenciada das demais por apresentar uma banda
das demais do mesmo subgrupo analisado. Essa banda se localiza já mais próxima do
catodo (Figura 4 – seta 8).
19
19
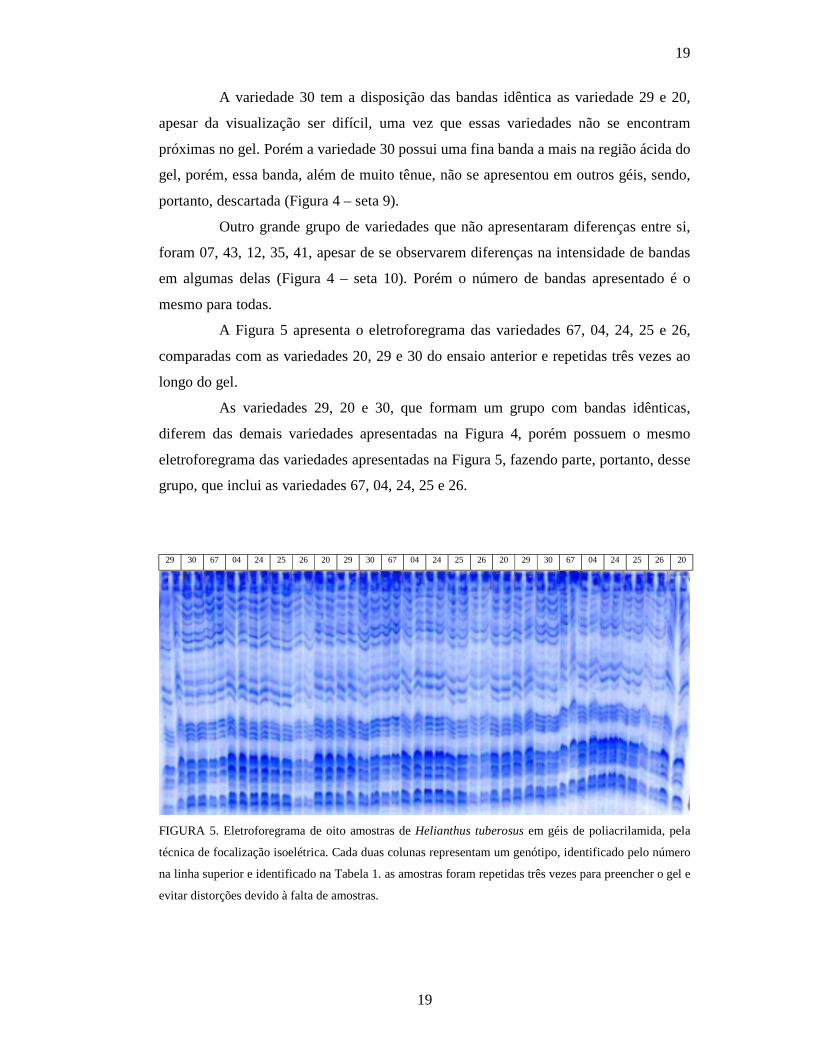
A variedade 30 tem a disposição das bandas idêntica as variedade 29 e 20,
apesar da visualização ser difícil, uma vez que essas variedades não se encontram
próximas no gel. Porém a variedade 30 possui uma fina banda a mais na região ácida do
gel, porém, essa banda, além de muito tênue, não se apresentou em outros géis, sendo,
portanto, descartada (Figura 4 – seta 9).
Outro grande grupo de variedades que não apresentaram diferenças entre si,
foram 07, 43, 12, 35, 41, apesar de se observarem diferenças na intensidade de bandas
em algumas delas (Figura 4 – seta 10). Porém o número de bandas apresentado é o
mesmo para todas.
A Figura 5 apresenta o eletroforegrama das variedades 67, 04, 24, 25 e 26,
comparadas com as variedades 20, 29 e 30 do ensaio anterior e repetidas três vezes ao
longo do gel.
As variedades 29, 20 e 30, que formam um grupo com bandas idênticas,
diferem das demais variedades apresentadas na Figura 4, porém possuem o mesmo
eletroforegrama das variedades apresentadas na Figura 5, fazendo parte, portanto, desse
grupo, que inclui as variedades 67, 04, 24, 25 e 26.
29 30 67 04 24 25 26 20 29 30 67 04 24 25 26 20 29 30 67 04 24 25 26 20
FIGURA 5. Eletroforegrama de oito amostras de Helianthus tuberosus em géis de poliacrilamida, pela
técnica de focalização isoelétrica. Cada duas colunas representam um genótipo, identificado pelo número
na linha superior e identificado na Tabela 1. as amostras foram repetidas três vezes para preencher o gel e
evitar distorções devido à falta de amostras.
20
20
Portanto, pela visualização direta, pode-se separar os 29 genótipos estudados
em 11 grupos, sendo que seis puderam ser caracterizadas isoladamente.
A metodologia desenvolvida neste trabalho foi testada também em outras
espécies para verificar o comportamento e apresentou excelentes resultados, quando
utilizada em amostras de trigo, com grande quantidade de bandas, com boa nitidez e
bem distribuídas.
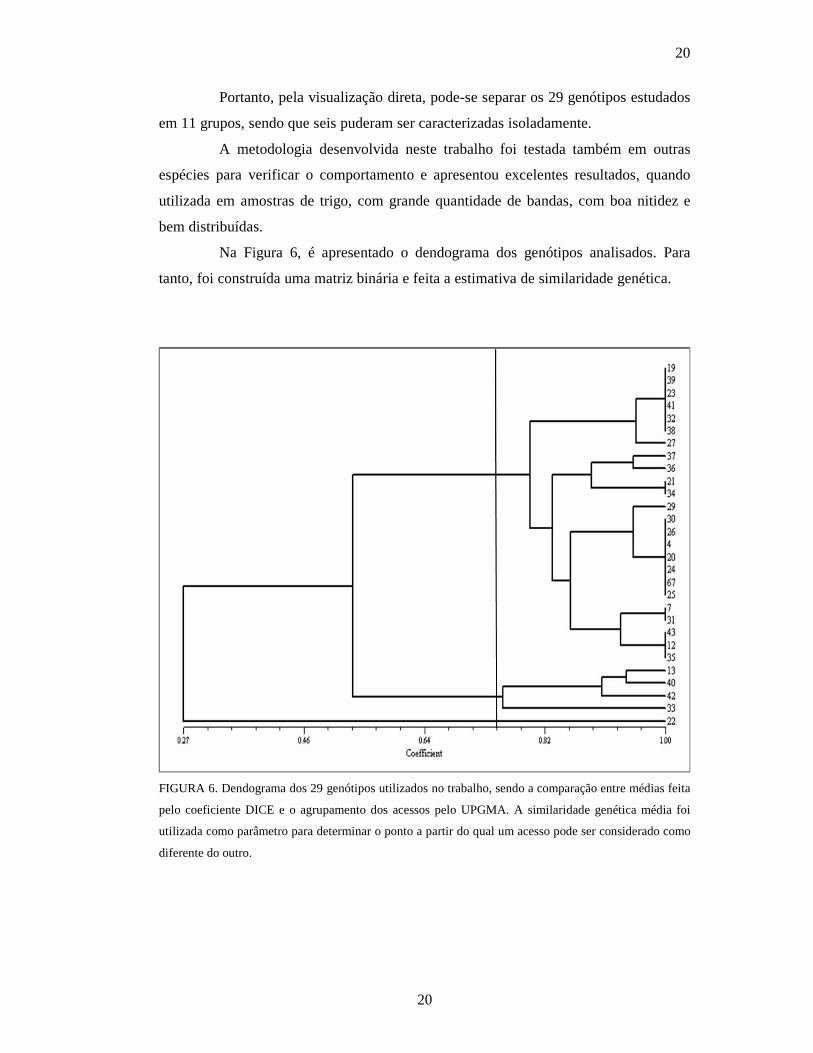
Na Figura 6, é apresentado o dendograma dos genótipos analisados. Para
tanto, foi construída uma matriz binária e feita a estimativa de similaridade genética.
FIGURA 6. Dendograma dos 29 genótipos utilizados no trabalho, sendo a comparação entre médias feita
pelo coeficiente DICE e o agrupamento dos acessos pelo UPGMA. A similaridade genética média foi
utilizada como parâmetro para determinar o ponto a partir do qual um acesso pode ser considerado como
diferente do outro.
1
1
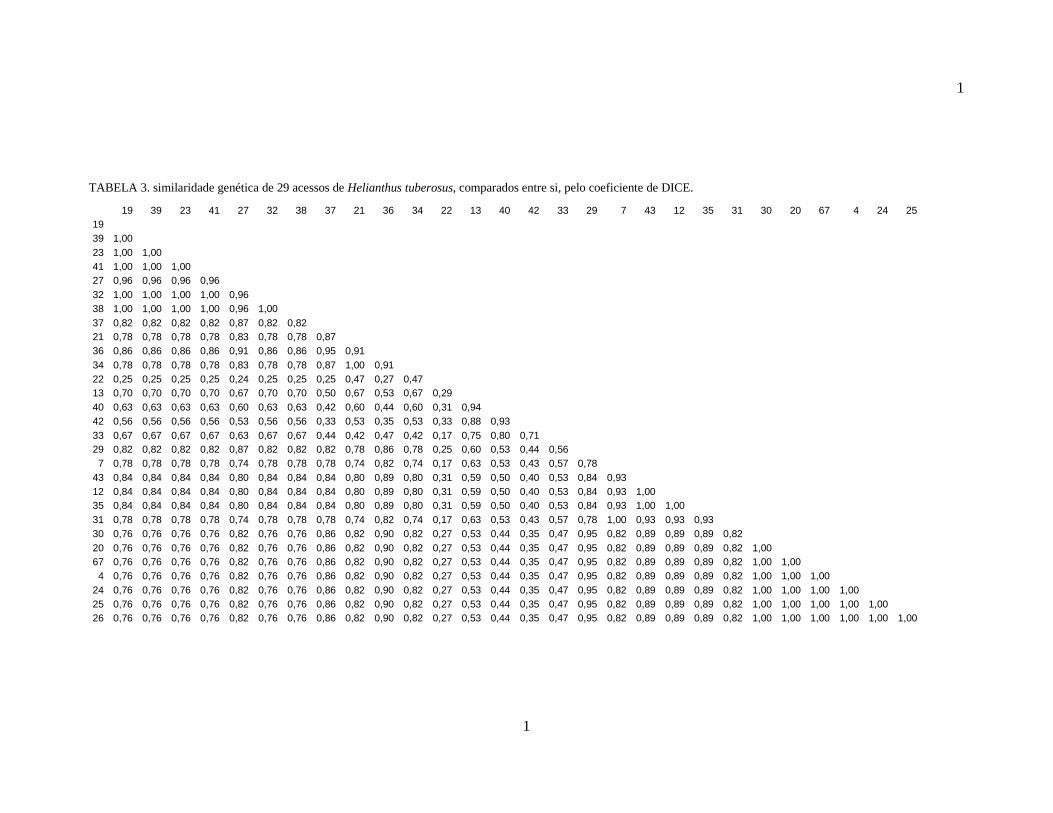
TABELA 3. similaridade genética de 29 acessos de Helianthus tuberosus, comparados entre si, pelo coeficiente de DICE.
19 39 23 41 27 32 38 37 21 36 34 22 13 40 42 33 29 7 43 12 35 31 30 20 67 4 24 25 19 39 1,00 23 1,00 1,00 41 1,00 1,00 1,00 27 0,96 0,96 0,96 0,96 32 1,00 1,00 1,00 1,00 0,96 38 1,00 1,00 1,00 1,00 0,96 1,00 37 0,82 0,82 0,82 0,82 0,87 0,82 0,82 21 0,78 0,78 0,78 0,78 0,83 0,78 0,78 0,87 36 0,86 0,86 0,86 0,86 0,91 0,86 0,86 0,95 0,91 34 0,78 0,78 0,78 0,78 0,83 0,78 0,78 0,87 1,00 0,91 22 0,25 0,25 0,25 0,25 0,24 0,25 0,25 0,25 0,47 0,27 0,47 13 0,70 0,70 0,70 0,70 0,67 0,70 0,70 0,50 0,67 0,53 0,67 0,29 40 0,63 0,63 0,63 0,63 0,60 0,63 0,63 0,42 0,60 0,44 0,60 0,31 0,94 42 0,56 0,56 0,56 0,56 0,53 0,56 0,56 0,33 0,53 0,35 0,53 0,33 0,88 0,93 33 0,67 0,67 0,67 0,67 0,63 0,67 0,67 0,44 0,42 0,47 0,42 0,17 0,75 0,80 0,71 29 0,82 0,82 0,82 0,82 0,87 0,82 0,82 0,82 0,78 0,86 0,78 0,25 0,60 0,53 0,44 0,56 7 0,78 0,78 0,78 0,78 0,74 0,78 0,78 0,78 0,74 0,82 0,74 0,17 0,63 0,53 0,43 0,57 0,78
43 0,84 0,84 0,84 0,84 0,80 0,84 0,84 0,84 0,80 0,89 0,80 0,31 0,59 0,50 0,40 0,53 0,84 0,93 12 0,84 0,84 0,84 0,84 0,80 0,84 0,84 0,84 0,80 0,89 0,80 0,31 0,59 0,50 0,40 0,53 0,84 0,93 1,00 35 0,84 0,84 0,84 0,84 0,80 0,84 0,84 0,84 0,80 0,89 0,80 0,31 0,59 0,50 0,40 0,53 0,84 0,93 1,00 1,00 31 0,78 0,78 0,78 0,78 0,74 0,78 0,78 0,78 0,74 0,82 0,74 0,17 0,63 0,53 0,43 0,57 0,78 1,00 0,93 0,93 0,93 30 0,76 0,76 0,76 0,76 0,82 0,76 0,76 0,86 0,82 0,90 0,82 0,27 0,53 0,44 0,35 0,47 0,95 0,82 0,89 0,89 0,89 0,82 20 0,76 0,76 0,76 0,76 0,82 0,76 0,76 0,86 0,82 0,90 0,82 0,27 0,53 0,44 0,35 0,47 0,95 0,82 0,89 0,89 0,89 0,82 1,00 67 0,76 0,76 0,76 0,76 0,82 0,76 0,76 0,86 0,82 0,90 0,82 0,27 0,53 0,44 0,35 0,47 0,95 0,82 0,89 0,89 0,89 0,82 1,00 1,00 4 0,76 0,76 0,76 0,76 0,82 0,76 0,76 0,86 0,82 0,90 0,82 0,27 0,53 0,44 0,35 0,47 0,95 0,82 0,89 0,89 0,89 0,82 1,00 1,00 1,00
24 0,76 0,76 0,76 0,76 0,82 0,76 0,76 0,86 0,82 0,90 0,82 0,27 0,53 0,44 0,35 0,47 0,95 0,82 0,89 0,89 0,89 0,82 1,00 1,00 1,00 1,00 25 0,76 0,76 0,76 0,76 0,82 0,76 0,76 0,86 0,82 0,90 0,82 0,27 0,53 0,44 0,35 0,47 0,95 0,82 0,89 0,89 0,89 0,82 1,00 1,00 1,00 1,00 1,00 26 0,76 0,76 0,76 0,76 0,82 0,76 0,76 0,86 0,82 0,90 0,82 0,27 0,53 0,44 0,35 0,47 0,95 0,82 0,89 0,89 0,89 0,82 1,00 1,00 1,00 1,00 1,00 1,00

1
1
Os resultados mostram que a técnica foi capaz de separar os acessos em vários
grupos.
A similaridade genética variou de 0,16 até 1, sendo que a maior distância
genética entre os genótipos analisados individualmente ficou entre o 22 e o 31 . No
entanto, alguns grupos foram formados por mais de um acesso geneticamente igual,
impossibilitando a separação dos mesmos.
Fazendo-se a comparação dos dados do dendograma com a separação visual
dos genótipos, pode-se observar uma correlação entre as duas análises, uma vez que
também se observou no dendograma a formação dos mesmos grupos sem diferenciação
genética. Além disso, a maioria dos genótipos segue o mesmo padrão de separação da
análise visual, porém, um grande número de separações apresentadas no dendograma
não se mostram significativas, em função da pequena quantidade de bandas
polimórficas encontradas. Esse fato é exemplificado pelos quatro genótipos
diferenciados isoladamente na análise visual e que no dendograma formam um grupo
com 90% de similaridade genética.
Quando os agrupamentos são avaliados a partir da similaridade genética
média dos genótipos, que ficou em 0,75, formam-se somente três grupos geneticamente
distintos, um grupo representado unicamente pelo genótipo 22, outro incluindo os
genótipos 13, 40, 42 e 33 e o terceiro grupo formado pelos demais 24 acessos
estudados. Esse resultado é muito diferente do encontrado na avaliação visual e
evidencia a importância do uso de parâmetros estatísticos para a análise dos resultados.
Porém pode-se observar que se a similaridade genética média fosse um pouco maior,
um novo grupo se formaria, separando o genótipo 33 dos genótipos 13, 40 e 42, por esse
estar muito próximo da linha que forma os grupos geneticamente distintos.
O número reduzido de grupos formados pela análise do dendograma em
relação à análise visual pode se explicar pelo baixo número de bandas polimórficas que
separou cada genótipo dos agrupamentos, em relação ao número total de bandas
encontradas. Portanto, a presença de uma ou mais bandas polimórficas encontradas, por
mais que tenha gerado um valor de similaridade diferente, não foi suficiente para
caracterizar um referido genótipo diferente dos demais.
A análise dos perfis isoelétricos de outras categorias de proteínas poderá
contribuir para a separação dos referidos genótipos.

2
2
Os grupos que apresentaram similaridade genética alta, podem na verdade ser
formados por uma mesma variedade que, pelo fato de ser produzida em locais
diferentes, aparece com mais de um nome.
Na Figura 7 são apresentados alguns tubérculos de Helianthus tuberosus e as
formas que os mesmos podem apresentar.
Figura 7: diferentes formas dos tubérculos de Helianthus tuberosus.
Fazendo-se a comparação entre alguns genótipos agrupados pela análise do
dendograma, por não apresentarem distância genética entre os mesmos, e a forma de
seus tubérculos (Figura 7), seguindo caracterização descrita por Stonzenburg (2002),
encontrou-se uma correlação direta, onde um agrupamento formado pelos genótipos 19,
39, 23, 41, 32 e 38 possuem forma arredonda. De forma semelhante, o grupo formado
por 4, 19, 29, 30 e 67 tem a forma dos seus tubérculos arredondada. Esse resultado
também é um indicativo de que a forma dos tubérculos pode ser usado como descritor
morfológico para a diferenciação de cultivares.

3
3
CONCLUSÕES
• Os resultados apresentados neste trabalho indicam que a
técnica de focalização isoelétrica aplica-se a uma determinação rápida e de
baixo custo de cultivares de Helianthus tuberosus, tornando sua utilização
acessível a qualquer laboratório.
• A técnica de focalização isoelétrica foi eficiente na
organização dos acessos em diferentes grupos.
• Através da técnica de IEF, foi possível diferenciar, nas 29
amostras trabalhadas, três grupos distintos

4
4
REFERÊNCIAS BIBLIOGRÁFICAS
BOTT, R. R.; NAVIASO, M. A.; SMITH, J. L. Improving the Quality of
Protein Crystals through Purification by Isoelectric Focusing. The Journal of
Biologicachle Mistry Vol. 257, No. 17, Issue of September 10. pp. 9883-9886, 1982
CARPITA, N. C.; KANABUS, J.; HOUSLEY, T. L. Linkage structure of
fructans and fructan oligomers from Triticum aestivum and Festuca arundinacea
leaves. Journal of Plant Physiology. 134: 162–168. 1989.
COUSSEMENT, P; FRANCK, A. New food applications for inulin. Agro-
food-Industry Hi-Tech 9:26-28. 1998.
FLAMM G., GLINSMANN W., KRITCHEVSKY D., PROSKY L. &
ROBERFROID M. Inulin and oligofructose as dietary fiber: a review of the
evidence. Critical Reviews in Food Science and Nutrition 41, 353–362. 2001.
FUCHS, A. Inulin and inulin-containing crops. Studies in plant science.
Elsevier: Amsterdam, p. 355-357, 1993.
HAMES, B. D.; RICKWOOD D. Gel electrophoresis of proteins, a
practical approach, 2ª ed. IRL Press at Oxford University Press, Oxford. New York.
1990.

5
5
HAMES, B. D.; RICKWOOD, D. Gel electrophoresis of proteins. In:
Practical approach. IRL Press, Washington DC. p 290. 1981.
HENDRY G. Evolutionary origins and natural functions of fructans. A
climatological, biogeographic and mechanistic appraisal. New Phytologist 123, 3–
14. 1993.
HENDRY GAF, WALLACE RK The origin, distribution, and
evolutionary significance of fructans. In M Suzuki, NJ Chatterton, eds, Science and
Technology of Fructans. CRC Press, Boca Raton, FL, pp 119–139. 1993.
ISTA - INTERNATIONAL SEED TESTING ASSOCIATION. Rules for
seed testing. Zürich, 1996. 44p.
LEWIS, D.H. Nomenclature and diagrammatic representation of
oligomeric fructans - a paper for discussion. New Phytologist. (124) 583-593. 1993
LUCHESE, C.; DINELLI, G.; BONETTI, A.; LOVATO, A. Specific and
varietal differentiation in Vicia sativa L. and Vicia villosa Roth by means of IEF-
PAGE and capillary electrophoresis. Seed Science and Technology, Zürich, v. 25, p.
253-262. 1997.
LUCCHESE, C.; DINELLI, G.; MIGGIANO, A.; LOVATO, A.
Identification of pepper (Capsicum spp.) cultivars by field and electrophoresis tests.
Seed Science Technology, 27. p. 37-47. 1998.
McDONALD, M.B.; ELLIOT, L.J. & SEENEY, P.M. DNA extration from
seeds for RAPD analysis in varietal identification studies. Seed Science and
Technology, Zürich, v.22, n.1, p.171-176. 1994.

6
6
MARTINEZ NOE¨ L G, PONTIS HG: Involvement of sucrose synthase in
sucrose synthesis during mobilization of fructans in dormant Jerusalem artichoke
tubers. Plant Science. 159:191-195. 2000.
(A) MONTI, A.; AMADUCCI, M. T.; VENTURI, G. Growth response, leaf
gas exchange and fructans accumulation of Jerusalem artichoke (Helianthus
tuberosus, L.) as affected by different water regimes. European Journal of Agronomy
(available on line).
(B) MONTI, A.; AMADUCCI, M. T.; PRITONI, G.; VENTURI, G. Growth,
fructan yield, and quality of chicory (Cichorium intybus L.) as related to
photosynthetic capacity, harvest time, and water regime. Journal of Experimental
Botany. 56(415):1389-1395. 2005.
MULLIN, W. J.; MODLER, H. W.; FARNWORTH, E. R.; PAYNE, A. The
macronutrient content of fractions from Jerusalem artichoke tubers (Helianthus
tuberosus). Food Chemistry 51(3): 263-269. 1994.
MUSSIGMANN, C.; LEDOIGT, G. Major storage proteins in Jerusalem
artichoke tubers. Plant Physiology and Biochemistry. 27: 81 – 86. 1989.
PILON-SMITS EAH, EBSKAMP MJM, PAUL MJ, JEUKEN MJW,
WEISBEEK PJ, SMEEKENS SCM Improved performance of transgenic fructan-
accumulating tobacco under drought stress. Plant Physiology. 107: 125–130. 1995.
RITSEMA, T.; SMEEKENS, S. Fructans: beneficial for plants and
humans. Current Opinion in Plant Biology. 6, 223-30. 2003.
ROHLF, F.J. Numerical taxonomy and multivariate analysis sistem.
version 1.70. New York: Plenum Press, 1992. 470p.
SCHITTENHELM, S. Influence of genotype and soil tillage on regrowth in
Jerusalem artichoke (Helianthus tuberosus L.). Archieve of Agronomic Soil Science.
38:89–96. 1994.

7
7
SCHITTENHELM, S. Agronomic Performance of Root Chicory,
Jerusalem Artichoke, and Sugarbeet in Stress and Nonstress Environments. Crop
Science. 39:1815–1823. 1999.
SHALLENBERGER, R.S. Advanced sugar chemistry: principles of sugar
stereochemistry. Connecticut: AVI Publishing, 323p. 1982.
SEVENIER, R.; HALL R. D.; VAN DER MEER, I. M.; HAKKERT, H. J. C.;
VAN KOOPS, A. J. High level fructan accumulation in a transgenic sugar beet.
National Biotechnology. 16:843–846. 1998.
SEILER, G. J.; CAMPBELL, L. G. Genetic Variability for Mineral
Element Concentrations of Wild Jerusalem Artichoke Forage. Crop Science.
44:289–292 (2004).
SHEWRY, P. R. Tuber Storage Proteins. Annals of Botany. 91: 755 – 769,
2003.
STONZENBURG, K. Anbau und verwertung von Topinambur
(Helianthus tuberosus L.). Informationen für die Pflanzenproduktion. LAP – Forcheim.
Deutschland. p. 174. 2002.
TOMOMATSU, H. Health effects of oligosaccharides. Food Technology
48:61-65. 1994.
UPOV (Union for the Protection of New Varieties of Plants). International
Convention for the Protection of New Varieties of Plants. UPOV, Geneva,
Switzerland. 1991.
VANHAASTRECHT, J. Promising performers; oligosaccharides present
new product development opportunities for a wide range of processed foods. Int.
Food Ingredients. No. 1: 23–27. 1995.

8
8
VIEIRA, E. S. N.; VON PINHO, E. V. R.; VIEIRA, M. G. G. C.; MANN, R.
S. Similaridade genética entre cultivares de feijão do grupo carioca por meio de
marcadores moleculares de proteínas e enzimas visando a certificação da pureza
genética. Revista Brasileira de Sementes, vol. 23, nº 2, p.35-42, 2001.
WANG, X. F.; KNOBLAUCH, R.; LEIST, N. Varietal descrimination of
tomato (Lycopersicum esculentum L.) by ultrathin-layer isoelectric focusing of
seed protein. Seed Science & Tecnology. 28, 521–526. 2000.
WANG; XIAOFENG; ZHAO; TING; YAN; MIN & LU; YANPENG.
Genetic purity testing of two-line hybrid rice seeds by grow-out of embryo-
containing half-seeds and ultrathin-layer isoelectric focusing of proteins from
corresponding embryoless half-seeds Abstracts 27
ISTA Congress Seed Symposium
Budapest, Hungary May 17
– 19. 42p. 2004.
WYSE, D. L.; YOUNG, F. L.; JONES, R.J. Influence of Jerusalem
artichoke (Helianthus tuberosus) in density and duration of interference on
soybean (Glycine max) growth and yield. Weed Science. 34:243–247. 1986.
Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas
Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo






![TABELA NUTRICIONAL DE - Nutritotal (Abertura)...Tabela nutricional de suplementos para uso em inquéritos dietéticos / Natália Koren Simoni ... [et al.]. ‐‐ São Paulo : Faculdade](https://img.document.onl/doc/110x75/5f6c3681fab810044a20f5c3/tabela-nutricional-de-nutritotal-abertura-tabela-nutricional-de-suplementos.jpg)





















