Embed Size (px)
Citation preview

INSTITUTO AGRONÔMICO CURSO DE PÓS-GRADUAÇÃO EM AGRICULTURA
TROPICAL E SUBTROPICAL
TEMPERATURA E DISPONIBILIDADE HÍDRICA E OS SEUS EFEITOS NA FOTOSSÍNTESE E NO
CRESCIMENTO DE LARANJEIRA VALÊNCIA ENXERTADA EM LIMOEIRO CRAVO OU
CITRUMELEIRO SWINGLE
FERNANDA KEILA JUNQUEIRA VILELA
Orientador: Rafael Vasconcelos Ribeiro
Dissertação submetida como requisito parcial para obtenção do grau de Mestre em Agricultura Tropical e Subtropical, Área de Concentração em Tecnologia da Produção Agrícola
Campinas, SP Março 2012

ii

iii

iv
Aos meus pais, Hudson e Wanda
E meus irmãos, Vandison, Fabio e Fabiano,
Por todo apoio, amor e união que
Constituíram a base para esta conquista.
DEDICO
Ao meu noivo Thiago,
Pelo amor, companheirismo, incentivo
E compreensão em todos os momentos.
OFEREÇO

v
AGRADECIMENTOS
À Deus, meu guia, por me proporcionar a vivência ao lado de pessoas tão especiais e
oportunidades únicas;
Ao meu orientador Dr. Rafael Vasconcelos Ribeiro pela excelente orientação, pelos
ensinamentos, entusiasmo contagiante, pela dedicação e incentivos constantes, que
despertaram em mim grande admiração e eterna gratidão;
Ao Dr. Eduardo Caruso pelas idéias para o desenvolvimento do projeto;
Ao IAC e ao programa de Pós-Graduação pela possibilidade de realização deste curso;
À Coordenadoria de Aperfeiçoamento de Ensino Superior (Capes) e à Fundação de Amparo à
Pesquisa do Estado de São Paulo (Fapesp) pela concessão das bolsas de estudos;
Aos amigos José Rodrigues, Paulo Marchiori, Fernanda Castro, Karina Silva e Severino
Nogueira pelo auxílio durante o experimento. Em especial à Ana Carolina Rosa Bueno pela
amizade, pelos conselhos e ajudas incessantes para iniciar o mestrado e durante todo ele.
À amiga Danielle Prudente por acompanhar todo o experimento, auxiliando com grande
responsabilidade, com boas idéias e muitas vezes tornando os momentos difíceis mais leves e
descontraídos.
Ao Dr. Emerson Alves da Silva, ao Dr. Erick Espinoza Núñez, ao Dr. Fernando Alves de
Azevedo, à Dra. Simone Rodrigues da Silva e ao Dr. Eduardo Caruso Machado por
colaborarem com minha banca de defesa.
Aos amigos da pós-graduação, em especial Rafaely Lameira, Patrícia Pessoni e Fernanda
Castro pelos bons momentos de descontração, boas conversas e risadas.
A todos os colegas da Fisiologia e pesquisadores pela boa convivência.

vi
À minha família em Campinas, tia Maurinha, Renata, Maria Laura e Augustinho por todo o
suporte durante minha estadia nesta cidade, pelos bons momentos de descontração e calor
familiar.
Aos meus pais, exemplos de vida, que nunca mediram esforços para que eu pudesse alcançar
meus objetivos, sendo grandes responsáveis por todas as minhas conquistas, uma vez que sem
eles não haveria inspiração e nem força para seguir em frente.
Aos irmãos, cunhadas e lindos sobrinhos, família maravilhosa que em todos os momentos me
fazem mais feliz e motivada por tê-los completando minha vida.
Ao Thiago, meu noivo, por todo companheirismo, apóio e amor. Mais uma motivação na
minha vida, impulsionando meus sonhos a se tornarem realidade. Obrigada por tudo!
A todos aqueles que de uma forma direta ou indireta contribuíram para a realização deste trabalho.

vii
SUMÁRIO LISTA DE ABREVIAÇÕES .................................................................................................viii LISTA DE TABELAS ..............................................................................................................x LISTA DE FIGURAS ..............................................................................................................xi RESUMO ................................................................................................................................xv ABSTRACT ..........................................................................................................................xvii 1 INTRODUÇÃO......................................................................................................................1 2 REVISÃO DE LITERATURA ..............................................................................................3 2.1 Respostas fisiológicas das plantas cítricas ao déficit hídrico ..............................................3 2.2 Respostas fisiológicas das plantas cítricas a variações térmicas .........................................5 2.3 Porta–enxertos: influência no crescimento e metabolismo de plantas cítricas....................7 3 MATERIAL E MÉTODOS....................................................................................................8 3.1 Material vegetal ...................................................................................................................8 3.2 Condições de crescimento ...................................................................................................9 3.3 Avaliações biométricas......................................................................................................10 3.4 Potencial da água na folha, teor relativo de água e condutância hidráulica da planta.......11 3.5 Trocas gasosas e atividade fotoquímica ............................................................................12 3.6 Teores de clorofila e de nitrogênio foliar ..........................................................................13 3.7 Teores de carboidratos nas folhas, caules e raízes.............................................................14 3.8 Teor foliar de prolina.........................................................................................................16 3.9 Análise dos dados ..............................................................................................................16 4 RESULTADOS ....................................................................................................................16 4.1 Ambiente frio (25/10 ºC)...................................................................................................16 4.1.1 Potencial da água na folha, condutância hidráulica, teor relativo de água e prolina......16 4.1.2 Trocas gasosas e atividade fotoquímica .........................................................................18 4.1.3 Teores de clorofila e de nitrogênio foliar .......................................................................22 4.1.4 Teores de carboidratos nas folhas, caules e raízes..........................................................22 4.1.5 Biometria ........................................................................................................................24 4.2 Ambiente quente (35/20 ºC) ..............................................................................................25 4.2.1 Potencial da água na folha, condutância hidráulica, teor relativo de água e prolina......25 4.2.2 Trocas gasosas e atividade fotoquímica .........................................................................27 4.2.3 Teores de clorofila e de nitrogênio foliar .......................................................................30 4.3.4 Teores de carboidratos nas folhas, caules e raízes..........................................................31 4.2.5 Biometria ........................................................................................................................32 5 DISCUSSÃO........................................................................................................................34 5.1 Ambiente frio (25/10 ºC)...................................................................................................34 5.2 Ambiente quente (35/20 ºC) ..............................................................................................38 6 CONCLUSÃO......................................................................................................................42 7 REFERÊNCIAS BIBLIOGRÁFICAS .................................................................................43

viii
LISTA DE ABREVIAÇÕES
ABA Ácido abscísico
AFB Área total das brotações (m2)
AM Amido (mg g-1)
AS Açúcares solúveis (mg g-1)
AT Carboidratos metabolizáveis (mg g-1)
CB Comprimento das brotações (cm)
Cc Concentração de CO2 no cloroplasto (µmol mol-1)
Ci Concentração intercelular de CO2 (µmol mol-1)
CMA Capacidade máxima de armazenamento de água no substrato
DAIT Dias após a imposição do tratamento
DPV Déficit de pressão de vapor do ar (kPa)
DPVf-a Diferença de pressão de vapor entre folha e ar (kPa)
E Transpiração (mmol m-2 s-1)
Ei Transpiração diurna (mol m-2 d-1)
ETR Transporte aparente de elétrons (µmol m-2 s-1)
ETRC Transporte de elétrons destinados à carboxilação da RuBP (µmol m-2 s-1)
ETRO Transporte de elétrons destinados à oxigenação da RuBP (µmol m-2 s-1)
EUA Eficiência do uso da água (mmol mol-1)
FO Fluorescência mínima em tecidos adaptados ao escuro
FO’ Fluorescência mínima após excitação do fotossistema I
FM Fluorescência máxima em tecidos adaptados ao escuro
FM’ Fluorescência máxima em tecidos adaptados à luminosidade
FS Fluorescência instantânea em tecidos adaptados à luminosidade
FSI Fotossistema I
FSII Fotossistema II
FV Fluorescência variável em tecidos adaptados ao escuro
FV/FM Eficiência quântica potencial do fotossistema II
gs Condutância estomática (mol m-2 s-1)
gm Condutância mesofílica (mol m-2 s-1)
Glu Glicose
kL Condutância hidráulica (mmol m-2 s-1 Mpa-1)

ix
MS Matéria seca (g)
MSB Matéria seca das brotações (g)
MSC Matéria seca de caule (g)
MSFM Matéria seca das folhas maduras (g)
MSR Matéria seca das raízes (g)
N Nitrogênio
NFB Número de folhas das brotações (unidade)
NPQ Coeficiente de extinção não-fotoquímica da fluorescência
Pn Assimilação de CO2 (µmol m-2 s-1)
Pni Assimilação diurna de CO2 (mmol m-2 d-1)
Pn/Cc Eficiência de carboxilação (µmol m-2 s-1 Pa-1)
Q Radiação fotossinteticamente ativa (µmol m-2 s-1)
qP Coeficiente de extinção fotoquímica da fluorescência
Rd Respiração (µmol m-2 s-1)
RuBP Ribulose-1,5-bisfosfato
SAC Sacarose (mg g-1)
TRA Teor relativo de água (%)
∆F Fluorescência variável em tecidos adaptados à luminosidade
∆F/FM’ Eficiência quântica efetiva do fotossistema II
Ψ Potencial da água na folha (MPa)

x
LISTA DE TABELAS
Tabela 1 - Características biométricas e assimilação de CO2 das mudas de laranjeira Valência enxertada em limoeiro Cravo ou citrumeleiro Swingle avaliadas antes dos tratamentos térmicos (25/10 ºC e 35/20 ºC), ao término da fase de aclimatação..............................................................................................
9
Tabela 2 - Teores de clorofila a e b, eficiência quântica potencial (FV/FM) e efetiva (∆F/FM’) do fotossistema II, coeficientes de extinção fotoquímica (qp) e não fotoquímica (NPQ) da fluorescência, transporte aparente de elétrons (ETR), transporte de elétrons destinados à carboxilação (ETRC) e à oxigenação (ETRO) da Rubisco em mudas de laranjeira Valência enxertadas em limoeiro Cravo ou citrumeleiro Swingle mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) e crescidas no regime térmico de 25/10 ºC.....................................................
22
Tabela 3 - Teores de clorofila a e b, eficiência quântica potencial (FV/FM) e efetiva (∆F/FM’) do fotossistema II, coeficientes de extinção fotoquímica (qp) e não fotoquímica (NPQ) da fluorescência, transporte aparente de elétrons (ETR), transporte de elétrons destinados à carboxilação (ETRC) e à oxigenação (ETRO) da Rubisco em mudas de laranjeira Valência enxertadas em limoeiro Cravo ou citrumeleiro Swingle mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) e crescidas no regime térmico de 35/20 ºC.....................................................
31

xi
LISTA DE FIGURAS
Figura 1 - Mudas de laranjeiras Valência enxertadas em limoeiro Cravo ou citrumeleiro Swingle utilizadas no experimento. Setas vermelhas indicam as folhas maduras utilizadas nas avaliações do regime térmico 25/10 ºC (a) e as folhas jovens utilizadas nas avaliações do regime térmico 35/20 ºC (b)............................................................................................................
11
Figura 2 - Potencial da água na folha de mudas de laranjeira Valência enxertadas em limoeiro Cravo ou citrumeleiro Swingle mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) no regime térmico de 25/10 ºC (dia/noite). Avaliações realizadas na antemanhã (a) e às 13h00 (b) após 20 dias de déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio padrão. Letras maiúsculas distintas indicam diferenças estatísticas (p<0,05) entre os porta-enxertos na mesma condição hídrica e letras minúsculas distintas representam diferenças estatísticas entre os tratamentos hídricos no mesmo porta-enxerto.............................................
17
Figura 3 - Teor relativo de água (TRA, em a), condutância hidráulica (kL, em b) e teor foliar de prolina (c) em folhas de laranjeiras Valência enxertadas em limoeiro Cravo ou citrumeleiro Swingle mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) no regime térmico de 25/10 ºC (dia/noite). Avaliações realizadas após 20 dias de déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio padrão. Letras maiúsculas distintas indicam diferenças estatísticas (p<0,05) entre os porta-enxertos na mesma condição hídrica e letras minúsculas distintas representam diferenças estatísticas entre os tratamentos hídricos no mesmo porta-enxerto....................................................................................
18
Figura 4 - Assimilação de CO2 (Pn, em a), condutância estomática (gS, em b), condutância mesofílica (gm, em c) e eficiência de carboxilação (Pn/Cc, em d) em laranjeiras Valência enxertadas em limoeiro Cravo ou citrumeleiro Swingle, mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) no regime térmico de 25/10 ºC (dia/noite). Avaliações realizadas às 13h00 após 20 dias de déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio padrão. Letras maiúsculas distintas indicam diferenças estatísticas (p<0,05) entre os porta-enxertos na mesma condição hídrica e letras minúsculas distintas representam diferenças estatísticas entre os tratamentos hídricos no mesmo porta-enxerto..................................................................................
19
Figura 5 - Assimilação diurna de CO2 (Pni, em a), transpiração diurna (Ei, em b) e eficiência do uso da água (EUA, em c) em laranjeiras Valência enxertadas em limoeiro Cravo ou citrumeleiro Swingle, mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) no regime térmico de 25/10 ºC (dia/noite). Avaliações realizadas às 13h00 após 20 dias de déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio padrão. Letras maiúsculas distintas indicam diferenças estatísticas (p<0,05) entre os porta-enxertos na mesma condição hídrica e letras

xii
minúsculas distintas representam diferenças estatísticas entre os tratamentos hídricos no mesmo porta-enxerto.............................................
20
Figura 6 - Respiração foliar (Rd) de laranjeiras Valência enxertadas em limoeiro Cravo (a) ou em citrumeleiro Swingle (b), mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) no regime térmico de 25/10 ºC (dia/noite). Avaliações realizadas após 20 dias de déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio padrão. Letras maiúsculas distintas indicam diferenças estatísticas (p<0,05) entre os períodas na mesma condição hídrica e letras minúsculas distintas representam diferenças estatísticas entre os tratamentos hídricos no mesmo período.............................................................................................
21
Figura 7 - Teores de açúcares solúveis (AS), sacarose (SAC), amido (AM) e carboidratos metabolizáveis (AT) em folhas maduras (a,b), caules (c,d) e raízes (e,f) de mudas de laranjeiras Valência enxertadas em limoeiro Cravo (a,c,e) ou citrumeleiro Swingle (b,d,f) mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) no regime térmico de 25/10 ºC (dia/noite). Avaliações realizadas após 20 dias de déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio padrão. Asteriscos indicam diferença estatística (p<0,05) entre os tratamentos hídricos.........................................................................................................
23
Figura 8 - Crescimento em matéria seca da brotação (MSB, em a), em área foliar (AFB, em b), em comprimento de brotações (CB, em c) e número de folhas das brotações (NFB, em d) de mudas de laranjeira Valência enxertadas em limoeiro Cravo ou citrumeleiro Swingle, mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) no regime térmico de 25/10 ºC (dia/noite). Avaliações realizadas após 20 dias de déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio padrão. Letras maiúsculas distintas indicam diferenças estatísticas (p<0,05) entre os porta-enxertos na mesma condição hídrica e letras minúsculas distintas representam diferenças estatísticas entre os tratamentos hídricos no mesmo porta-enxerto.............................................
24
Figura 9 - Matéria seca de raízes (MSR) de mudas de laranjeira Valência enxertadas em limoeiro Cravo ou citrumeleiro Swingle, mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) no regime térmico de 25/10 ºC (dia/noite). Avaliações realizadas após 20 dias de déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio padrão. Letras maiúsculas distintas indicam diferenças estatísticas (p<0,05) entre os porta-enxertos na mesma condição hídrica e letras minúsculas distintas representam diferenças estatísticas entre os tratamentos hídricos no mesmo porta-enxerto....................................................................................
25
Figura 10 - Potencial da água (Ψ) na folha jovem de mudas de laranjeira Valência enxertadas em limoeiro Cravo ou citrumeleiro Swingle mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) no regime térmico de 35/20 ºC (dia/noite). Avaliações realizadas na antemanhã (a) e às 13h00 (b) no 20º dia após imposição do déficit hídrico. Cada

xiii
histograma representa o valor médio (n=3) ± desvio padrão. Letras maiúsculas distintas indicam diferenças estatísticas (p<0,05) entre os porta-enxertos na mesma condição hídrica e letras minúsculas distintas representam diferenças estatísticas entre os tratamentos hídricos no mesmo porta-enxerto....................................................................................
26
Figura 11 - Teor relativo de água (TRA, em a), condutância hidráulica (kL, em b) e teor foliar de prolina (c) em folhas de laranjeiras Valência enxertadas em limoeiro Cravo ou citrumeleiro Swingle mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) no regime térmico de 35/20 ºC (dia/noite). Avaliações realizadas após 20 dias de déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio padrão. Letras maiúsculas distintas indicam diferenças estatísticas (p<0,05) entre os porta-enxertos na mesma condição hídrica e letras minúsculas distintas representam diferenças estatísticas entre os tratamentos hídricos no mesmo porta-enxerto....................................................................................
27
Figura 12 - Assimilação de CO2 (Pn, em a), condutância estomática (gS, em b), condutância mesofílica (gm, em c) e eficiência de carboxilação (Pn/Cc, em d) em laranjeiras Valência enxertadas em limoeiro Cravo ou citrumeleiro Swingle, mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) no regime térmico de 35/20 ºC (dia/noite). Avaliações realizadas às 13h00 após 20 dias de déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio padrão. Letras maiúsculas distintas indicam diferenças estatísticas (p<0,05) entre os porta-enxertos na mesma condição hídrica e letras minúsculas distintas representam diferenças estatísticas entre os tratamentos hídricos no mesmo porta-enxerto....................................................................................
28
Figura 13 - Assimilação diurna de CO2 (Pni, em a), transpiração diurna (Ei, em b) e eficiência do uso da água (EUA, em c) em laranjeiras Valência enxertadas em limoeiro Cravo ou citrumeleiro Swingle, mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) no regime térmico de 35/20 ºC (dia/noite). Avaliações realizadas às 13h00 após 20 dias de déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio padrão. Letras maiúsculas distintas indicam diferenças estatísticas (p<0,05) entre os porta-enxertos na mesma condição hídrica e letras minúsculas distintas representam diferenças estatísticas entre os tratamentos hídricos no mesmo porta-enxerto.............................................
29
Figura 14 - Respiração foliar (Rd) de laranjeiras Valência enxertadas em limoeiro Cravo (a) ou em citrumeleiro Swingle (b), mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) no regime térmico de 35/20 ºC (dia/noite). Avaliações realizadas após 20 dias de déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio padrão. Letras maiúsculas distintas indicam diferenças estatísticas (p<0,05) entre os períodos na mesma condição hídrica e letras minúsculas distintas representam diferenças estatísticas entre os tratamentos hídricos no mesmo período.............................................................................................
30

xiv
Figura 15 - Teores de açúcares solúveis (AS), sacarose (SAC), amido (AM) e carboidratos metabolizáveis (AT) em folhas maduras (a,b), caules (c,d) e raízes (e,f) de mudas de laranjeiras Valência enxertadas em limoeiro Cravo (a,c,e) ou citrumeleiro Swingle (b,d,f) mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) no regime térmico de 35/20 ºC (dia/noite). Avaliações realizadas após 20 dias de déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio padrão. Asteriscos indicam diferenças estatísticas (p<0,05) entre os tratamentos hídricos.........................................................................................................
32
Figura 16 - Crescimento em matéria seca da brotação (MSB, em a), em área foliar (AFB, em b), em comprimento de brotações (CB, em c) e número de folhas das brotações (NFB, em d) de mudas de laranjeira Valência enxertadas em limoeiro Cravo ou citrumeleiro Swingle, mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) no regime térmico de 35/20 ºC (dia/noite). Avaliações realizadas após 20 dias de déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio padrão. Letras maiúsculas distintas indicam diferenças estatísticas (p<0,05) entre os porta-enxertos na mesma condição hídrica e letras minúsculas distintas representam diferenças estatísticas entre os tratamentos hídricos no mesmo porta-enxerto.............................................
33
Figura 17 - Matéria seca de raízes (MSR) de mudas de laranjeira Valência enxertadas em limoeiro Cravo ou citrumeleiro Swingle, mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) no regime térmico de 35/20 ºC (dia/noite). Avaliações realizadas após 20 dias de déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio padrão. Letras maiúsculas distintas indicam diferenças estatísticas (p<0,05) entre os porta-enxertos na mesma condição hídrica e letras minúsculas distintas representam diferenças estatísticas entre os tratamentos hídricos no mesmo porta-enxerto....................................................................................
34

xv
Temperatura e disponibilidade hídrica e os seus efeitos na fotossíntese e no crescimento de laranjeira Valência enxertada em limoeiro Cravo ou citrumeleiro Swingle
RESUMO
Variações nos regimes térmico e hídrico geram modificações no metabolismo da planta
resultando em mudanças no padrão de crescimento e desenvolvimento. Considerando-se a
informação existente, este estudo foi desenvolvido com o objetivo de testar as seguintes
hipóteses: 1) em ambiente quente, maior fotossíntese e maior crescimento seriam observados
em plantas enxertadas em limoeiro Cravo quando comparadas às enxertadas em citrumeleiro
Swingle, independente da condição hídrica; 2) em ambiente frio com boa disponibilidade
hídrica, o melhor desempenho fotossintético e crescimento ocorreriam em laranjeiras
enxertadas em ‘Swingle’ quando comparadas às enxertadas em ‘Cravo’. Quando há restrição
de água em ambiente frio, não foi possível a elaboração de uma hipótese sobre o desempenho
de laranjeiras, uma vez que são escassos os trabalhos que tratem da interação entre porta-
enxertos, regime hídrico e térmico. Desta forma, este estudo também objetivou avaliar qual
combinação copa/porta-enxerto teria melhor desempenho fisiológico em condições de
restrição hídrica em ambiente frio. As plantas foram submetidas aos regimes de temperatura
(dia/noite) de 25/10 ºC e 35/20 ºC, por 20 dias. Em cada regime térmico, as mudas foram
agrupadas em dois lotes: um submetido à restrição hídrica (40% da capacidade máxima de
armazenamento de água do solo) e outro mantido sob disponibilidade hídrica adequada (80%
da capacidade máxima de armazenamento de água do solo). Foram avaliadas as trocas
gasosas, a atividade fotoquímica, as relações hídricas, a variação de carboidratos nas folhas,
ramos e raízes; teores de prolina, de clorofilas a e b e de nitrogênio foliar e algumas
características biométricas. Os resultados foram submetidos à análise de variância e quando
detectada diferença estatística, as médias foram comparadas pelo teste de Tukey a 5% de
probabilidade. Em ambos os regimes térmicos foi possível observar que o déficit hídrico
causou acúmulo foliar de prolina, redução do potencial da água na folha, reduções na
condutância estomática, na respiração, redução na assimilação de CO2 e consequentemente no
acúmulo de fitomassa em ambos os porta-enxertos, sendo, entretanto, os efeitos da seca mais
severos em plantas enxertadas em ‘Swingle’. Independente do porta-enxerto ou regime
térmico, não houve alterações nos teores de clorofila, na atividade fotoquímica e no teor foliar
de nitrogênio devido ao déficit hídrico. Em ambiente frio e independente da condição hídrica,
o melhor desempenho fotossintético das plantas enxertadas em ‘Swingle’ foi ocasionado pela
maior condutância estomática, maior condutância mesofílica e maior eficiência de

xvi
carboxilação. Como conseqüência, as plantas enxertadas em ‘Swingle’ apresentaram maior
crescimento em relação às enxertadas em ‘Cravo’. Mesmo sendo mais afetadas pelo déficit
hídrico do que as enxertadas em ‘Cravo’, as plantas enxertadas em ‘Swingle’ apresentaram
melhor desenvolvimento. Em ambiente quente e sob déficit hídrico, as plantas enxertadas em
‘Cravo’ mantiveram maior condutância estomática o que resultou em maior fotossíntese e
crescimento quando comparadas às enxertadas em ‘Swingle’. Sob boa disponibilidade hídrica,
a hipótese de melhor crescimento das plantas enxertadas em ‘Cravo’ não foi comprovada,
mesmo com estas apresentando maior desempenho fotossintético quando comparadas às
enxertadas em ‘Swingle’ no regime de 35/20 ºC.
Palavras-chave: Citrus spp., déficit hídrico, frio, porta-enxertos, trocas gasosas.

xvii
Temperature and water availability and their effects on photosynthesis and growth of Valencia sweet orange plants grafted on Rangpur lime or Swingle citrumelo rootstocks
ABSTRACT
Variations in temperature and water regimes affect plant metabolism, changing the pattern of
growth and development. Considering the available information, this study was developed to
test the following hypotheses: 1) under warm condition, higher photosynthesis and higher
growth would be observed in plants grafted on Rangpur lime compared to those grafted on
Swingle citrumelo, regardless of water availability; 2) under cold condition with water
supplying, higher photosynthetic rates and higher growth would occur in orange plants
grafted on ‘Swingle’ compared to ones grafted on ‘Rangpur’. When there is water stress under
cold condition, any assumption regarding the performance of orange plants is not possible as
there are no studies that address the interaction between rootstocks, thermal and water
regimes. Thus, this study also aimed to evaluate which combination scion/rootstock would
have better physiological performance under water stress in a cold environment. The plants
were subjected to temperature regimes (day/night) of 25/10 °C and 35/20 °C for 20 days. In
each thermal regime, the plants were grouped into two lots: one that was submitted to water
restriction (40% of the maximum soil water storage capacity) and one that was maintained at
80% of the maximum soil water storage capacity. We evaluated leaf gas exchange,
photochemical activity, water relations, the concentrations of carbohydrates in leaves, stem
and roots, the leaf concentrations of proline, chlorophylls a and b, and nitrogen. Biometric
characteristics were also evaluated. The results were subjected to the analysis of variance and
when statistical differences were detected, mean values were compared by Tukey test at 5%
probability level. In both thermal regimes the water deficit caused leaf accumulation of
proline and reductions in leaf water potential, stomatal conductance, leaf respiration, and low
CO2 assimilation. As a consequence, the production of phytomass in both rootstocks was
decreased. However, the most severe effects of drought were found in plants grafted on
‘Swingle’. Regardless of the rootstock or thermal regime, there were no changes in
chlorophyll concentration, photochemical activity and leaf nitrogen due to drought. Under
cold condition and regardless water availability, the higher photosynthetic performance of
plants grafted on ‘Swingle’ was caused by higher stomatal conductance, higher mesophyll
conductance and higher carboxylation efficiency. As a result, the plants grafted on ‘Swingle’
showed higher growth compared to ones grafted on ‘Rangpur’. Although most affected by
drought, plants grafted on ‘Swingle’ rootstock exhibited better shoot development as

xviii
compared to ones grafted on ‘Rangpur’ rootstock. In a warm environment and under water
deficit, plants grafted on ‘Rangpur’ rootstock maintained higher stomatal conductance that
resulted in increased photosynthesis and growth when compared to those grafted on
‘Swingle’. Under adequate water supplying and 30/20 ºC, the hypothesis of better growth of
plants grafted on ‘Rangpur’ rootstock was not confirmed, even with these plants showing
higher photosynthetic performance as compared to ones grafted on ‘Swingle’.
Key-words: Citrus spp., drought, cold, rootstock, gas exchange.

1
1 INTRODUÇÃO
Oscilações nos regimes térmico e hídrico geram modificações no metabolismo das
plantas resultando em variações no padrão de crescimento e desenvolvimento. Sabe-se que o
crescimento das plantas é conseqüência de uma complexa cadeia de eventos relacionados à
produção fotossintética, consumo de fotoassimilados e eficiência de utilização de recursos
como água e nutrientes (BUENO et al., 2011; PRADO et al., 2007; RIBEIRO &
MACHADO, 2007).
Entre os fatores limitantes da produção vegetal, o déficit hídrico é tido como o
principal, visto que um dos seus primeiros impactos fisiológicos é a redução da fotossíntese
(FLEXAS et al., 2004a). Sabendo-se que a fotossíntese é a fonte primária dos carboidratos
utilizados no crescimento e na manutenção da planta (RIBEIRO & MACHADO, 2007) e que
o déficit hídrico é um fenômeno que ocorre em grandes extensões de áreas cultiváveis
(PASSIOURA, 2007) torna-se necessário entender como as plantas respondem a esta
condição de estresse, possibilitando-se assim, o desenvolvimento de técnicas de manejo que
impeçam prejuízos ao crescimento e desenvolvimento da planta, garantindo a manutenção
e/ou o aumento da produtividade (CHAVES et al., 2009).
As respostas fotossintéticas ao estresse hídrico são complexas (FLEXAS et al., 2004a;
CHAVES et al., 2009) e os decréscimos da fotossíntese em citros são ocasionados
primariamente pelo fechamento parcial dos estômatos (MAGALHÃES FILHO et al., 2008).
Em condições mais severas de estresse, fatores relacionados ao metabolismo do carbono e à
atividade bioquímica são afetados (DAMOUR et al., 2008), tais como a atividade da enzima
ribulose-1,5-bifosfato carboxilase/oxigenase (Rubisco), a regeneração da ribulose-1,5-
bifosfato (RuBP) e a distribuição de fotoassimilados na planta (FLEXAS et al., 2004b; VU,
1999). O estresse hídrico também pode causar comprometimento dos eventos fotoquímicos
nos tilacóides e conseqüente redução da eficiência quântica da fotossíntese, com inativação do
fotossistema II e da cadeia de transporte de elétrons (FLEXAS et al., 2002; GARCÍA-
SANCHÉZ et al., 2007).
A temperatura também tem papel decisivo no padrão de crescimento dos citros
(BUENO et al., 2011; RAMOS, 2009). Como um dos fatores ambientais mais variáveis, a
temperatura pode suprimir a fotossíntese em condições extremas, fora da faixa ótima para o
crescimento de espécies cítricas, entre 22 e 30 ºC (SUN & MA, 1999).

2
A baixa temperatura pode afetar praticamente todas as principais reações da
fotossíntese, incluindo o transporte de elétrons, o ciclo de Calvin-Benson e a condutância
estomática (ALLEN & ORT, 2001). Por outro lado, a exposição à alta temperatura também
pode levar a reduções na assimilação de CO2 devido ao fechamento estomático, a menor
eficiência de carboxilação da Rubisco e redução da atividade fotoquímica (MACHADO et al.,
2005; RIBEIRO et al., 2006; RIBEIRO et al., 2009b).
Vários estudos têm demonstrado os efeitos da variação da temperatura na fisiologia
dos citros, sendo tratada a variação sazonal da fotossíntese (MACHADO et al., 2002;
RIBEIRO & MACHADO, 2007; RIBEIRO et al., 2009a,b), a temperatura de crescimento
(RAMOS, 2009), os efeitos da baixa temperatura noturna e do substrato na fotossíntese
(MAGALHÃES FILHO et al., 2009; MACHADO et al., 2010; SANTOS et al., 2011) e da
amplitude térmica diária no metabolismo de carbono de plantas cítricas (BUENO et al.,
2011). Embora a literatura seja rica em relação aos efeitos das condições ambientais na
fotossíntese, poucos são os trabalhos que relacionam o padrão de variação fotossintética com
o crescimento dos citros (RAMOS, 2009; BUENO et al., 2011), o que limita o alcance dos
resultados quando se considera o processo produtivo.
A resposta da fotossíntese das laranjeiras à variação da temperatura e da condição
hídrica também é afetada pelo porta-enxerto. Em função do porta-enxerto, há alterações no
crescimento das plantas e na utilização de nutrientes (FALLAHI et al., 2002; MAGALHÃES
FILHO et al., 2008, 2009; MATTOS Jr. et al., 2008; POMPEU Jr., 2005; SETIN et al., 2009).
As diferentes combinações de porta-enxerto e copa causam ainda variação nas trocas gasosas
e afetam o grau de resistência à seca, a resposta às oscilações na temperatura (BUENO et al.,
2011; FALLAHI et al., 2002; MACHADO et al., 2010) e outros estresses abióticos
(SYVERTSEN & LEVY, 2005) e bióticos (MEDINA FILHO et al., 2003). O limoeiro Cravo
é o principal porta-enxerto utilizado na citricultura brasileira por propiciar alta produtividade
e precocidade de produção, além de apresentar resistência à seca (AMARO &
BAPTISTELLA, 2010; DONATO et al., 2007; POMPEU Jr., 2005; MAGALHÃES FILHO
et al., 2008). Quanto ao citrumeleiro Swingle, o segundo porta-enxerto mais utilizado na
citricultura, tem-se a marcante resistência à Morte Súbita dos Citros e ao Declínio, além de
proporcionar bom desenvolvimento das plantas em ambientes frios (POMPEU Jr., 2005;
MULLER et al., 2005; MACHADO et al., 2010).
Considerando-se a informação existente, este estudo foi desenvolvido para testar as
seguintes hipóteses: 1) em ambiente quente (regime 35/20 ºC), maior fotossíntese e maior
crescimento seriam observados em plantas enxertadas em limoeiro Cravo quando comparadas

3
às enxertadas em citrumeleiro Swingle, independente da condição hídrica; 2) em ambiente
frio (regime 25/10 ºC) com boa disponibilidade hídrica, o melhor desempenho fotossintético e
crescimento ocorreriam em laranjeiras enxertadas em citrumeleiro Swingle quando
comparadas às enxertadas em limoeiro Cravo. Quando há deficiência de água em ambiente
frio (regime 25/10 ºC), não foi possível a elaboração de uma hipótese sobre o desempenho de
laranjeiras, uma vez que os trabalhos com a interação entre porta-enxertos, regime hídrico e
térmico são escassos. Desta forma, este estudo também objetivou avaliar qual combinação
copa/porta-enxerto teria melhor desempenho fisiológico em condições de restrição hídrica em
ambiente frio (regime 25/10 ºC).
2 REVISÃO DE LITERATURA
Os citros, por seu amplo cultivo, são submetidos a grande variação sazonal das
condições ambientais (MACHADO et al., 2002, 2010; RIBEIRO & MACHADO, 2007) e
qualquer fator ambiental que afete a fotossíntese também afetará o acúmulo de fitomassa pela
planta (MACHADO et al., 2005). Em condições subtropicais de cultivo dos citros,
temperatura e disponibilidade hídrica são variáveis determinantes do crescimento e
produtividade (ALLEN & ORT, 2001; CERQUEIRA et al., 2004; COELHO et al., 2006;
RIBEIRO & MACHADO, 2007) sendo necessários à planta, mecanismos fisiológicos que se
ajustem às condições climáticas a que estão expostas e que permitam seu desenvolvimento
mesmo em condições limitantes (MACHADO et al., 2010).
2.1 Respostas fisiológicas das plantas cítricas ao déficit hídrico
Plantas cítricas, por serem perenes, são freqüentemente expostas a condições de baixa
umidade no solo e na atmosfera (CAMPOS et al., 2011; ORTOLANI et al., 1991). Apesar de
apresentarem características morfológicas e mecanismos fisiológicos que proporcionam
considerável tolerância ao estresse hídrico quando comparadas a outras plantas perenes
(COELHO et al., 2006; PEREIRA et al., 2009; SILVA et al., 2006a), a produtividade está
diretamente relacionada à disponibilidade de água. Uma vez que a capacidade fotossintética é
bastante afetada pela seca, há decréscimos nos teores de carboidratos acumulados e de
fitomassa (CERQUEIRA et al., 2004; GARCIA-SÁNCHEZ et al., 2007; MAGALHÃES
FILHO et al., 2008).

4
Entre os principais processos afetados pelo déficit hídrico, destacam-se a fotossíntese e
o crescimento celular (BOTA et al., 2004; CHAVES et al., 2009; MAGALHÃES FILHO et
al., 2008). Condições de seca afetam predominantemente a difusão de CO2 através das folhas
devido a uma queda da condutância estomática e mesofílica (FLEXAS et al., 2004a). Os
estômatos se fecham em resposta ao declínio da turgescência das células, ao alto déficit de
pressão de vapor na atmosfera ou a sinais químicos gerados nas raízes (CHAVES et al., 2009;
MEDRANO et al., 2002; SILVA et al., 2006a). Já a redução da condutância mesofílica pode
ser causada por alterações físicas na estrutura foliar ou por alterações na bioquímica e/ou
permeabilidade das membranas (CHAVES et al., 2009).
Além do aspecto difusivo, o estresse hídrico também pode reduzir a atividade
fotoquímica e o teor das clorofilas (GARCÍA–SÁNCHEZ et al., 2007), a síntese de ATP
(FLEXAS et al., 2004b), limitar as reações de carboxilação e a regeneração da RuBP
(FLEXAS & MEDRANO, 2002; FLEXAS et al., 2004b; MEDRANO et al., 2002) e a síntese
e distribuição de fotoassimilados na planta (FLEXAS et al., 2004a; GARCÍA–SÁNCHEZ et
al., 2007).
A utilização de carboidratos pelas plantas depende de vários fatores, como alterações
das condições ambientais e ocorrência de estresses (ITO et al., 2003). Em geral, há uma
tendência de aumento no teor foliar de açúcares solúveis enquanto o teor de amido é reduzido
sob estresse hídrico (CHAVES et al., 2009). Em condições muito severas de seca, o teor de
açúcares solúveis pode também ser reduzido. Essas respostas estão relacionadas à menor
assimilação de CO2 e à mobilização de reservas para o crescimento e manutenção das plantas
sob estresse (MAGALHÃES FILHO et al., 2008).
Em resposta ao déficit hídrico as plantas têm também diversos mecanismos para
amenizar a condição estressante, tais como: aumento da relação raiz:parte aérea, folhas
menores e em menor número e acúmulo de solutos (GARCÍA-SÁNCHEZ et al., 2007). Estas
respostas limitam o consumo de água pela planta e ajudam a manter o status da água e,
portanto, o metabolismo da planta (CHAVES et al., 2009).
O acúmulo de solutos para diminuir o potencial osmótico da folha permite à planta
manter a hidratação da parte aérea quando o solo se torna mais seco. Logo, mantêm-se o
turgor celular e por conseqüência os estômatos permanecem abertos, as trocas gasosas
ocorrem e o efeito do déficit hídrico no crescimento da planta é reduzido (CAMPOS et al.,
2011; CHAVES et al., 2009; GUIMARÃES et al., 2008). O acúmulo de prolina em plantas
em resposta à seca pode influenciar a resistência aos estresses, ajudando na manutenção da
atividade metabólica dos tecidos (SZABADOS & SAVOURÉ, 2009). No entanto, há

5
indicação de que o acúmulo de prolina em condições ambientais adversas está mais
relacionado a distúrbios metabólicos do que ao ajuste osmótico (LUTTS et al., 1999), sendo
um indicador bioquímico de estresse. Em algumas espécies vegetais, o acúmulo de prolina é
resultante de alterações no metabolismo de aminoácidos associados com o aumento do
catabolismo de proteínas e a senescência vegetal (LUTTS et al., 1999; SILVEIRA et al.,
2003).
2.2 Respostas fisiológicas das plantas cítricas a variações térmicas
A temperatura é um fator determinante da produtividade e da distribuição geográfica
de muitas espécies de importância agrícola (ALLEN & ORT, 2001). Plantas cítricas são
cultivadas em extensas áreas com variáveis condições climáticas. No entanto, a faixa térmica
ideal para a atividade fotossintética dos citros está entre 25 e 30 ºC, com as plantas
apresentando maior abertura estomática e maior eficiência fotoquímica e bioquímica da
fotossíntese (GUO et al., 2006; MACHADO et al., 2005; RIBEIRO et al., 2004; SPIEGEL &
GOLDSCHMIDT, 1996). RAMOS (2009) observou que o metabolismo de laranjeiras é tão
fortemente afetado pelo regime térmico de crescimento que mesmo tecidos completamente
expandidos e maduros apresentaram modificações fisiológicas relacionadas à fotossíntese,
exportação de fotoassimilados e aos teores de carboidratos e pigmentos fotossintéticos.
Durante o inverno, as plantas cítricas apresentam menor desempenho fotossintético,
principalmente em decorrência de baixas temperaturas (ALLEN & ORT, 2001; RIBEIRO et
al., 2009a,b; SANTOS et al., 2011). A temperatura basal dos citros está em torno de 13 ºC, ou
seja, abaixo desta o metabolismo das plantas começa a ser seriamente reduzido (SPIEGEL-
ROY & GOLDSCHMIDT, 1996). A baixa temperatura reduz a eficiência fotossintética
devido à menor condutância estomática e a menor eficiência aparente de carboxilação,
resultante de limitações difusivas e metabólicas (MACHADO et al., 2002; 2010; MEDINA et
al., 2002; RIBEIRO et al., 2009a,b; SANTOS et al., 2011).
O frio, além de afetar o metabolismo fotossintético, causa alterações no estado hídrico
da parte aérea das plantas (RIBEIRO et al., 2009a). De acordo com ALLEN & ORT (2001),
reduções da temperatura do ar e da folha normalmente diminuem a demanda evaporativa, no
entanto, com a redução da temperatura no solo as raízes têm menor condutância hidráulica,
com inibição substancial da captação de água do solo. Após o frio, o solo se aquece muito
mais lentamente do que as folhas e o ar circundante, conseqüentemente, a demanda

6
evaporativa aumenta mais rápido do que o abastecimento de água e assim as plantas
permanecem em estado de déficit hídrico mesmo com umidade no solo.
O metabolismo de carboidratos é também modificado em resposta às baixas
temperaturas. Segundo RIBEIRO & MACHADO (2007), menores taxas fotossintéticas e,
portanto, menor fornecimento de fotoassimilados ocorre no inverno. As reações metabólicas,
incluindo as de síntese de sacarose, são mais lentas sob baixas temperaturas (RUELLAND &
ZACHOWSKY, 2010). De fato, ALLEN & ORT (2001) relatam em seu estudo menor
acúmulo de sacarose enquanto os teores de açúcares solúveis aumentaram em plantas em
ambiente frio.
Há evidências de aclimatação térmica da fotossíntese, que desempenha papel crucial
na determinação da produtividade vegetal pelo ajuste da oferta e a demanda de carboidratos,
otimizando a capacidade de crescimento (CHU et al., 2011). Estes ajustes podem ocorrer em
escala de tempo variável e também em resposta a outras mudanças no meio ambiente, tais
como luminosidade, duração do dia e disponibilidade de água (SMITH & STITT, 2007).
A atividade respiratória, processo pelo qual os compostos energéticos e redutores são
formados a partir de carboidratos ou outros substratos, também é influenciada pela
temperatura (ATKIN et al., 2007; LLOYD & FARQUHAR, 2008). A respiração é um
processo metabólico chave, diretamente relacionado com o crescimento das culturas, sendo a
resposta da respiração a temperaturas abaixo ou acima da faixa ótima pouco conhecida
(LOKA & OOSTERHUIS, 2010). A respiração foliar é bastante sensível a mudanças de curto
prazo da temperatura. No entanto, também há evidências de um processo de aclimatação da
respiração foliar a alterações na temperatura de crescimento em longo prazo (CROUS et al.,
2011). Esse processo de aclimatação da respiração ao ambiente tem grande importância na
manutenção da produtividade vegetal, uma vez que pode ajudar na manutenção de um
balanço positivo de carbono, seja pela redução de perda de carbono em condições quentes ou
manutenção de taxas basais de respiração em períodos frios (CHU et al., 2011; CROUS et al.,
2011).
A exposição de plantas cítricas às altas temperaturas, fora da faixa ótima, também
reduz a fotossíntese e o metabolismo de carboidratos. Há decréscimos significativos na
assimilação de CO2 devido ao fechamento dos estômatos e a danos fotoquímicos causados por
injúrias nas membranas fotossintéticas (GUO et al., 2006). O fechamento estomático é uma
estratégia para evitar a perda excessiva de água pelas plantas e ocorre mesmo com boa
disponibilidade hídrica no solo (MACHADO et al., 2002; RIBEIRO et al., 2004; RIBEIRO et
al., 2009a,b).

7
A redução da atividade fotoquímica em plantas expostas à alta temperatura (GUO et
al., 2006; MACHADO et al., 2005; RIBEIRO et al., 2006) pode estar associada à fotoinibição
e limitar a regeneração de RuBP dependente do transporte de elétrons (RIBEIRO &
MACHADO, 2007). A alta densidade de fluxo de fótons pode induzir a fotoinibição por uma
superexcitação do fotossistema II quando o fechamento dos estômatos limita a entrada de CO2
(VESTE et al., 2000). Além disso, degradação de clorofilas também já foi relatada sob alta
temperatura (GUO et al., 2006), situação em que também ocorre estímulo da atividade
oxigenase da Rubisco (fotorrespiração) e redução da condutância mesofílica (GUO et al.,
2006; PIMENTEL et al., 2007; RIBEIRO et al., 2009b).
Com o aumento da temperatura do ar e conseqüente aumento da temperatura foliar há
redução da taxa fotossintética, resultante de aumento na taxa respiratória da planta (LLOYD
& FARQUHAR, 2008). LOKA & OOSTERHUIS (2010) observaram em algodão que o
aumento da temperatura noturna provocou aumento significativo da respiração e nos teores de
carboidratos foliares.
2.3 Porta–enxertos: influência no crescimento e metabolismo de plantas cítricas
A resistência a muitos estresses abióticos e as relações hídricas dos citros variam com
o porta-enxerto utilizado de tal forma que eficientes combinações copa/porta-enxerto podem
garantir a manutenção do pomar mesmo sob condições ambientais desfavoráveis (DONATO
et al., 2007; ERISMANN et al., 2008; RIBEIRO & MACHADO, 2007; SYVERSTEN &
LEVY, 2005). Os porta-enxertos de plantas cítricas afetam características da copa e de seus
frutos, como o desenvolvimento vegetativo (ESPINOZA-NÚÑEZ et al., 2011; FOCHESATO
et al., 2006), a eficiência de uso do nitrogênio (SORGONÀ et al., 2011) e a resposta à
disponibilidade de outros nutrientes (MATTOS Jr. et al., 2006; 2007; 2010), a resistência a
doenças (MEDINA FILHO et al., 2003) e a produção e a qualidade dos frutos
(CANTUARIAS et al., 2009; SILVA et al., 2006b,c), tornando-se essenciais na citricultura
(NOGUEIRA et al., 2001; SCHÄFER et al., 2001)
O limoeiro Cravo é o principal porta-enxerto utilizado na citricultura brasileira
(PRUDENTE et al., 2004; STUCHI et al., 2004). Há relatos de que nos pomares do estado de
São Paulo, o seu emprego já era predominante desde a década de 1960 (POMPEU Jr., 2005).
Entre as várias qualidades que justificam seu expressivo uso, destaca-se a resistência à seca
(DONATO et al., 2007; MEDINA et al., 2005). Levando-se em conta que os pomares
brasileiros são na maioria manejados sem o uso de irrigação e que também estão expostos a

8
períodos de déficit hídrico severo durante o ano, o uso de porta-enxertos resistentes à seca
torna-se muitas vezes indispensável (ERISMANN et al., 2008; SCHÄFER et al., 2001). O
limoeiro Cravo também apresenta boa compatibilidade com todas as copas de maior uso no
Brasil, sua produção é precoce bem como a maturação de seus frutos e apresenta alto
rendimento de frutos (AMARO & BAPTISTELLA, 2010; GIRARDI et al., 2010). Os
motivos pelo qual o limoeiro Cravo apresenta boa adaptação ao déficit hídrico ainda não são
totalmente claros, mas há fortes evidências de que isto esteja relacionado à arquitetura e
volume de seu sistema radicular e à capacidade de suas raízes em explorar melhor a água
disponível no solo (MAGALHÃES FILHO et al., 2008).
Já o citrumeleiro Swingle é o segundo porta-enxerto de maior expressão no Brasil.
Atualmente, além das suas vantagens fitossanitárias, ele também tem sido bastante utilizado
em regiões com temperaturas mais baixas devido ao seu bom desempenho produtivo
(MACHADO et al., 2010; POMPEU Jr., 2005). Entretanto, ainda são pouco conhecidos os
aspectos fisiológicos relacionados à resistência do ‘Swingle’ ao frio. MACHADO et al.
(2010) compararam ‘Swingle’ e ‘Cravo’ como porta-enxertos da laranjeira Valência em
condições de frio noturno (8 ºC) e chegaram à conclusão de que as laranjeiras enxertadas
sobre ‘Swingle’ apresentaram menor redução da eficiência fotossintética do que as enxertadas
sobre ‘Cravo’.
O aumento do uso do porta-enxerto ‘Swingle’ se deu a partir dos anos 80, motivado
pela sua resistência à Gomose e tolerância ao Declínio (POMPEU Jr., 2005). Além destas,
também apresenta resistência à Tristeza, Morte Súbita dos Citros e a nematóides; confere
melhor qualidade aos frutos e também proporciona produção mais precoce (SCHÄFER et al.,
2001; GIRARDI et al., 2010). Em contrapartida, o ‘Swingle’ é um porta-enxerto sensível ao
déficit hídrico (MULLER et al., 2005; POMPEU Jr., 2005).
3 MATERIAL E MÉTODOS
3.1 Material vegetal
Mudas de laranjeira Valência [Citrus sinensis (L.) Osbeck] enxertada em limoeiro
Cravo (Citrus limonia Osbeck) ou citrumeleiro Swingle [Citrus paradisi Macf x Poncirus
trifoliata (L.) Raf.] com aproximadamente 5 meses de idade foram cultivadas em recipientes
plásticos com capacidade de 7,0 L contendo substrato orgânico comercial constituído de 80%
de casca de pinus, 5% de carvão e 15% de vermiculita média (Multicitrus, Terra do Paraíso
9
Ltda., SP, Brasil). As mudas foram produzidas pelo sistema de formação de mudas
certificadas em viveiro comercial e permaneceram em casa de vegetação até o surgimento de
brotações.
As plantas foram irrigadas e adubadas freqüentemente até o início do período
experimental. A nutrição das plantas foi realizada utilizando-se solução nutritiva comercial
(Brennfeed®, Brenntag, Brasil) constituída de 94,8 g N L-1, sendo 14,0 g L-1 na forma
amoniacal (N-NH4) e 80,8 g L-1 na forma nítrica (N-NO3); 24 g P L-1 (P2O5), 80 g K L-1
(K2O), 81,5 g Ca L-1 (CaO), 9,2 g Mg L-1 (MgO), 0,28 g Mn L-1, 0,2 g Zn L-1, 1,5 g Cu L-1,
0,72 g Fe L-1 e 0,12 g B L-1. Para a irrigação das mudas, usou-se 2,5 mL da solução comercial
por litro de água destilada. A cada dois dias, as mudas foram irrigadas com 300 mL da
solução diluída.
3.2 Condições de crescimento
Mudas apresentando porte similar (altura, área foliar e brotações) foram selecionadas e
transferidas da casa-de-vegetação para uma câmara de crescimento modelo PGR15
(Conviron, Canadá) onde permaneceram por um período de aclimatação de cinco dias a 25/20
ºC (dia/noite), com 12 horas de fotoperíodo. Após este período, três mudas de cada porta-
enxerto foram avaliadas quanto às características biométricas e de assimilação de CO2 (Tabela
1) para caracterização inicial do lote.
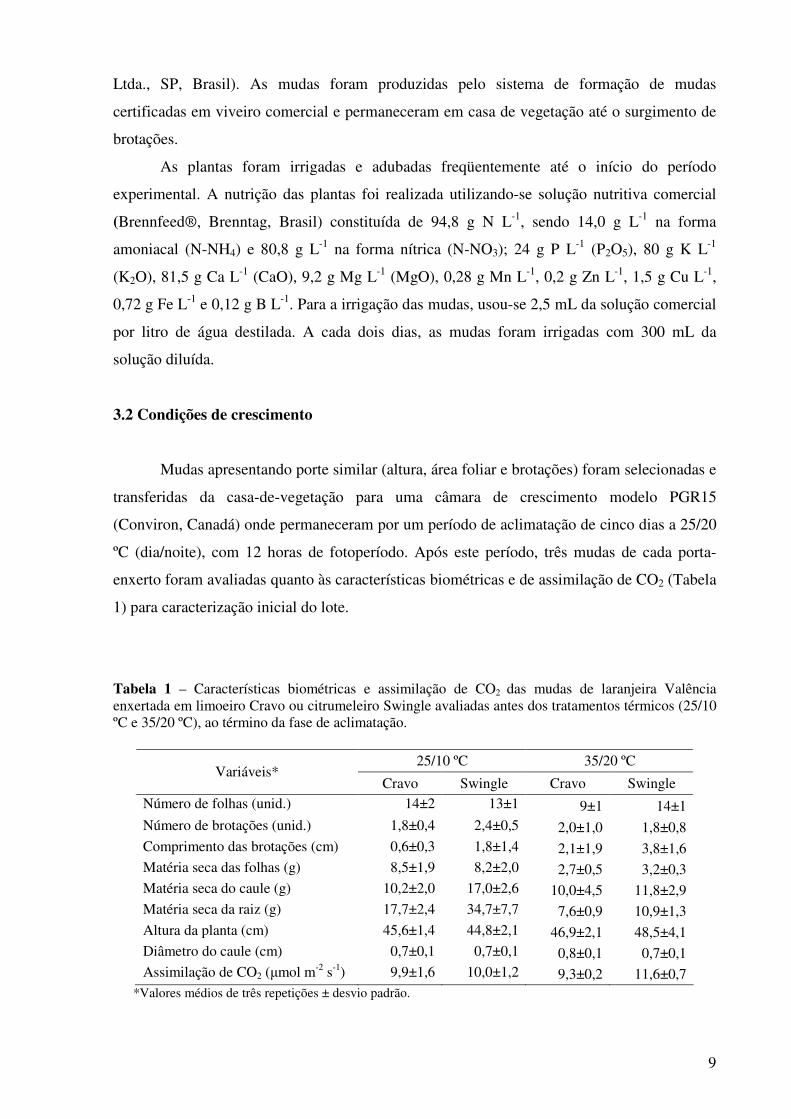
Tabela 1 – Características biométricas e assimilação de CO2 das mudas de laranjeira Valência enxertada em limoeiro Cravo ou citrumeleiro Swingle avaliadas antes dos tratamentos térmicos (25/10 ºC e 35/20 ºC), ao término da fase de aclimatação.
25/10 ºC 35/20 ºC Variáveis*
Cravo Swingle Cravo Swingle Número de folhas (unid.) 14±2 13±1 9±1 14±1 Número de brotações (unid.) 1,8±0,4 2,4±0,5 2,0±1,0 1,8±0,8 Comprimento das brotações (cm) 0,6±0,3 1,8±1,4 2,1±1,9 3,8±1,6 Matéria seca das folhas (g) 8,5±1,9 8,2±2,0 2,7±0,5 3,2±0,3 Matéria seca do caule (g) 10,2±2,0 17,0±2,6 10,0±4,5 11,8±2,9 Matéria seca da raiz (g) 17,7±2,4 34,7±7,7 7,6±0,9 10,9±1,3 Altura da planta (cm) 45,6±1,4 44,8±2,1 46,9±2,1 48,5±4,1 Diâmetro do caule (cm) 0,7±0,1 0,7±0,1 0,8±0,1 0,7±0,1 Assimilação de CO2 (µmol m-2 s-1) 9,9±1,6 10,0±1,2 9,3±0,2 11,6±0,7
*Valores médios de três repetições ± desvio padrão.

10
Após a aclimatação, em cada regime de temperatura as mudas foram agrupadas em
dois lotes: um foi submetido à restrição hídrica, ou seja, umidade do substrato mantida
próxima a 40% da capacidade máxima de armazenamento de água do substrato (CMA) e no
outro a umidade foi mantida próxima a 80% de CMA durante todo o período experimental. A
rega foi suspendida até o conjunto planta+pote atingir o peso correspondente a 80% e 40% da
CMA. Quando atingiram os pesos desejados, considerado dia 0, foram submetidas ao
tratamento térmico. A umidade do substrato foi monitorada a cada dois dias pela pesagem de
três potes de cada tratamento, com uma balança eletrônica. A reposição da água durante o
período experimental foi realizada de modo a manter a umidade do substrato entre 35% e 45%
de CMA (tratamento de restrição hídrica) e entre 75% e 85% de CMA (tratamento controle).
As mudas permaneceram por vinte dias em cada regime de temperatura, com a
amplitude térmica diária constante de 15 ºC (BUENO et al., 2011): 25/10 ºC e 35/20 ºC. Esses
regimes simularam ambientes frios (temperatura média de 17,5 oC) e quentes (temperatura
média de 27,5 oC), nos quais foram avaliados o desempenho fotossintético e o acúmulo de
fitomassa.
As demais condições ambientais foram constantes: radiação fotossinteticamente ativa
(Q) de 800 µmol m-2 s-1, déficit de pressão de vapor do ar (DPV) menor que 1,5 kPa e
fotoperíodo de 12 h (7h00 às 19h00).
3.3 Avaliações biométricas
As avaliações biométricas e morfológicas foram realizadas no início (fim do período
de aclimatação) e após 20 dias da imposição dos tratamentos (DAIT). As frações de folhas
maduras e jovens, de caule e de raiz de cada planta foram coletadas e desidratadas em estufa
de circulação forçada (modelo MA032, Marconi, Brasil) a 60 ºC até obtenção de peso
constante. Nesse momento, as matérias secas de raízes (MSR), de caule (MSC), de folhas
maduras (MSFM) e de brotações (MSB) foram determinadas em balança eletrônica, sendo MSB
composto pela matéria seca dos ramos jovens e das folhas jovens.
A área foliar das brotações (AFB) foi avaliada com o uso de um planímetro digital
modelo Li-3000 (Licor, EUA). O número de folhas e de brotações foi determinado por
contagem e o comprimento das brotações e a altura das plantas foram determinados com o
auxílio de uma trena. O diâmetro do caule (a 3 cm do ponto de enxertia) foi determinado com
um paquímetro digital.

11
3.4 Potencial da água na folha, teor relativo de água e condutância hidráulica da planta
O potencial da água na folha (Ψ) foi medido com uma câmara de pressão modelo 3005
(SoilMoisture, EUA) em folhas localizadas próximas àquelas utilizadas nas medidas de trocas
gasosas. Como no regime de temperatura de 35/20 ºC observou-se crescimento mais rápido e
uniforme das brotações foi possível a avaliação do potencial da água nas folhas jovens.
Figura 1 – Mudas de laranjeiras Valência enxertadas em limoeiro Cravo ou citrumeleiro Swingle utilizadas no experimento. Setas vermelhas indicam as folhas maduras utilizadas nas avaliações do regime térmico 25/10 ºC (a) e as folhas jovens utilizadas nas avaliações do regime térmico 35/20 ºC (b).
Nas medidas de teor relativo de água (TRA), discos foliares foram excisados e
imediatamente pesados em balança analítica de precisão para obtenção do peso fresco (Mf).
Após a pesagem, os discos ficaram por três horas imersos em água destilada para obtenção do
peso túrgido (Mt) e depois foram secos em estufa de ventilação forçada a 60 °C por 48 h para
obtenção do peso seco (Ms). Os valores obtidos nestas pesagens foram utilizados na seguinte
expressão: TRA(%)=[(Mf-Ms)/(Mt-Ms)]x100. Da mesma maneira foi calculado o teor
relativo de água das folhas jovens no regime térmico de 35/20 ºC.
Tanto as medidas de potencial da água como de teor relativo de água foram realizadas
no 20º DAIT. Medidas de TRA foram realizadas às 6h00 e as medidas de Ψ foram realizadas
às 6h00 (antes do início do fotoperíodo) e às 13h00 nos dias de avaliação.
A condutância hidráulica (kL) foi estimada de acordo com a seguinte equação:
kL=E/∆Ψ, em que E é a transpiração avaliada às 13h00 e ∆Ψ=Ψ6-Ψ13 ,sendo que Ψ6 e Ψ13 são
respectivamente o potencial da água na folha medido às 6h00 e às 13h00 (RIBEIRO et al.,
2009a).
(a) (b)

12
3.5 Trocas gasosas e atividade fotoquímica
As trocas gasosas foliares foram avaliadas com um analisador de gases por radiação
infravermelha modelo Li-6400F (Licor, EUA). As variáveis estudadas foram: assimilação de
CO2 (Pn); condutância estomática (gs); transpiração (E) e concentração intercelular de CO2
(Ci). A respiração no escuro (Rd) também foi avaliada nas mesmas folhas. A partir dos
valores de Pn, gs, Ci e Rd foram calculadas a concentração de CO2 no cloroplasto (Cc) e a
condutância mesofílica (gm) de acordo com EPRON et al. (2005).
Medidas de Pn, gs, E e Ci foram realizadas em intervalos de 2 h após o início do
fotoperíodo (7h00) e as avaliações de Rd foram realizadas às 6h00, 13h00 e 19h00, no 20º
DAIT. As avaliações foram realizadas com concentração de CO2 no ar constante (380 µmol
mol-1) e Q de 800 µmol m-2 s-1 [radiação saturante de acordo com MACHADO et al. (2005)],
controlados respectivamente pelo misturador de gases e pela fonte de luz do Li-6400F. As
avaliações foram registradas quando o coeficiente de variação (CV) total foi inferior a 0,5% e
sob estabilidade temporal.
Aos 20 DAIT, as folhas consideradas nas avaliações fisiológicas no regime de 25/10
ºC foram as formadas no período anterior ao experimento, denominadas de maduras. Já aos
20 DAIT do regime de 35/20 ºC (ambiente quente) foram avaliadas as folhas na base das
brotações formadas durante o tratamento térmico, denominadas de jovens. Essa medida foi
possível uma vez que as mudas do regime 35/20 ºC apresentaram crescimento mais vigoroso.
A partir dos dados de Rd, estimou-se a respiração diurna e a noturna para folhas
jovens e maduras, considerando-se Rd constante durante o período diurno (medidas das
13h00) e noturno (medida das 6h00). Calculou-se a assimilação diurna de CO2 (Pni) pela
integração de Pn e a transpiração diurna (Ei) pela integração de E ao longo do 20º DAIT, entre
7h00 e 19h00. Com os dados de Pni e Ei calculou-se a eficiência do uso da água (EUA) pela
relação Pni/Ei.
Medidas da emissão de fluorescência da clorofila a foram realizadas com um
fluorômetro modulado (6400-40 LCF) integrado ao Li-6400F. As folhas foram previamente
adaptadas ao escuro por 30 minutos para a determinação da fluorescência mínima em tecidos
adaptados ao escuro (FO), em seguida, um pulso de luz saturante (λ= 630 nm, Q ~ 6000 µmol
m-2 s-1, 0,8 s) foi aplicado para determinação da fluorescência máxima em tecidos adaptados
ao escuro (FM). A partir dos valores de FO e de FM, calculou-se a fluorescência variável no
escuro (FV=FM-FO), determinando-se, então, a eficiência quântica potencial do fotossistema II

13
(FV/FM). A fluorescência instantânea (FS) e máxima (FM’) em tecidos adaptados à
luminosidade também foram avaliadas com o método do pulso de saturação. A fluorescência
mínima em tecidos adaptados à luz (FO’) foi avaliada após a excitação do fotossistema I com
radiação vermelha distante (λ= 740 nm, Q ~ 5 µmol m-2 s-1, 2,0 s). A partir dos valores de FS e
FM’ calculou-se a fluorescência variável no claro (∆F= FM’-FS) e determinou a eficiência
quântica efetiva do fotossitema II (∆F/FM’). Foram também avaliados os coeficientes de
extinção fotoquímica [qP=(FM’-FS)/(FM’-FO’)] e não-fotoquímica [NPQ=(FM-FM’)/FM’] da
fluorescência e o transporte aparente de elétrons [ETR=Qx∆F/FM’x0,5x0,84] (ROHACEK,
2002). No cálculo de ETR foi adotado que as folhas absorvem 0,84 de Q (DEMMIG &
BJÖRKMAN, 1987) e que há partição idêntica de energia entre os fotossistemas, i.e., 0,5. O
índice ETR foi separado em dois componentes, que correspondem ao transporte de elétrons
destinados à carboxilação (ETRC) e à oxigenação (ETRO) da ribulose-1,5-bisfosfato (GENTY
et al., 1989):
ETR = ETRC + ETRO
ETRC=1/3[ETR+8(Pn+Rd)]
ETRO=2/3[ETR-4(Pn+Rd)]
em que: ETRC = transporte de elétrons destinados a carboxilação da RuBP; ETRO = transporte
de elétrons destinados a oxigenação da RuBP, ETR = transporte aparente de elétrons; Pn =
assimilação de CO2; e Rd = respiração no escuro.
3.6 Teores de clorofila e de nitrogênio foliar
Determinações indiretas dos teores de clorofila a e b foram realizadas com um
clorofilômetro modelo CFL1030 (Falker, Brasil). As medidas foram realizadas no 21º DAIT
nas mesmas folhas utilizadas nas medidas de trocas gasosas, sendo os valores expressos como
índice de clorofila (adimensional).
A avaliação do teor de nitrogênio (N) foi realizada em folhas maduras coletadas no 21º
DAIT. Após a coleta, as folhas foram armazenadas em ultrafreezer (-80 ºC), liofilizadas e o
teor de N determinado pelo método micro-Kjeldhal (BATAGLIA et al., 1983). As folhas
foram maceradas em almofariz e em um pequeno pedaço de lenço de papel foi pesado 0,1 g
do material vegetal. Esta amostra foi transferida para um tubo de digestão (tipo Folin-Wu)
onde foi adicionado 1 g de mistura digestora (composta por 100 g de sulfato de potássio, 100
g de sulfato de cobre e 1 g de selênio), 1 mL de H2O2 e 3 mL de ácido sulfúrico p.a.. Esses
tubos foram cobertos com pequenos funis de vidro, colocados em um bloco digestor e

14
aquecidos por 15 minutos a 60 ºC, 30 minutos a 180 ºC e 2 horas a 333 ºC. As amostras foram
digeridas até o clareamento do líquido. Os tubos foram removidos do bloco digestor e
resfriados à temperatura ambiente. Após o resfriamento adicionou-se 5 mL de água
deionizada para evitar que o líquido se solidificasse. Posteriormente seguiu-se a destilação das
amostras: acrescentou-se ao tubo de digestão, através da torneira colocada sobre o conjunto de
destilação, 15 mL de NaOH 10N. O volume destilado de 20 mL foi recolhido em um
erlenmeyer contendo 20 mL de solução de ácido bórico-indicador. O conteúdo do erlenmeyer
foi titulado com solução padronizada de H2SO4. O ponto de viragem foi lilás claro. Junto às
amostras também foram processados dois tubos brancos e um padrão. Posteriormente
realizou-se o cálculo do teor de nitrogênio pela fórmula: N (mg g-1) = (amostra – branco) x
F/MS, em que: amostra = volume gasto na titulação da amostra; branco = volume gasto na
titulação do branco; F = fator de correção do ácido (igual a 1) e MS = massa seca da amostra
utilizada.
3.7 Teores de carboidratos nas folhas, caules e raízes
A quantificação de carboidratos foi realizada em amostras de matéria seca obtidas de
folhas, ramos e raízes coletadas no 21º DAIT. As amostras foram coletadas e posteriormente
secas em estufa (modelo MA032, Marconi, Brasil) de circulação forçada a 60 oC até obtenção
de peso constante, sendo posteriormente armazenadas até o momento das análises. As
amostras foram então maceradas em almofariz e armazenadas em recipientes plásticos.
Nas determinações dos teores de carboidratos foram considerados os açúcares solúveis
(AS), sacarose (SAC), amido (AM) e carboidratos metabolizáveis (AT), sendo AT
determinado indiretamente (AT=AS+AM).
Para extração dos carboidratos foliares utilizou-se uma solução composta por metanol,
clorofórmio e água (MCW) na proporção de 15:5:3 v/v (BIELESKI & TURNER, 1966). Em
um tubo de rosca colocaram-se aproximadamente 75 mg de matéria seca com 3 mL de MCW.
Após 48 h sob refrigeração adicionou-se 1,2 mL de água e 1,8 mL de clorofórmio. Esta
solução foi novamente armazenada sob refrigeração por 24 h. Após esta etapa, o sobrenadante
foi coletado e concentrado a 55 ºC em banho-maria (MA184, Marconi, Brasil). O volume
obtido foi mensurado e as amostras utilizadas para determinação de AS e SAC.
A fração AS foi determinada pelo método fenol-sulfúrico (DUBOIS et al., 1956). Em
10 µL de amostra, foram adicionados 490 µL de água destilada, 500 µL de fenol 5% e 2 mL
de ácido sulfúrico p.a., agitando-se a solução em vórtex. Após o resfriamento da solução,

15
realizou-se a leitura da absorbância a 490 nm, em espectrofotômetro modelo B342II
(Micronal, Brasil), em duplicata. Os valores foram transformados em teor de AS com o
auxílio de uma reta-padrão obtida com concentrações variadas de glicose (0, 5, 10, 20, 30, 40
e 50 µg).
A fração SAC foi determinada pelo método proposto por VAN HANDEL (1968). Em
tubos de ensaio, foram adicionados 15 µL de amostra, 485 µL de água e 500 µL de solução de
hidróxido de potássio 30%. Os tubos foram vedados e incubados a 95 ºC por 10 minutos em
banho-maria. Posteriormente, foram adicionados 500 µL de fenol 5% e 2 mL de ácido
sulfúrico p.a. A solução foi agitada em vórtex e após o resfriamento procedeu-se a leitura da
absorbância a 490 nm, em espectrofotômetro, em duplicata. Os valores foram transformados
em teor de SAC com o auxílio de uma reta-padrão obtida com concentrações variadas de
sacarose (0, 5, 10, 20, 30, 40 e 50 µg).
Para determinação de AM utilizou-se o método enzimático proposto por AMARAL et
al. (2007). Amostras de 10 mg de matéria seca foram depositadas em microtubos de 2 mL.
Para obtenção do precipitado foram feitas quatro extrações com 500 µL de etanol 80% com
incubação em banho-maria por 20 minutos a 80 ºC. O precipitado foi seco durante 72 horas a
temperatura ambiente. Adicionou-se ao precipitado 500 µL (110 U mL-1) de α-amilase (EC
2.3.1.1) termoestável de Bacillus licheniformis (cód. E-ANAAM, Megazyme, Irlanda),
diluída em tampão MOPS 10 mM e pH 6,5. As amostras foram incubadas a 75 ºC (em banho-
maria) por 30 minutos. Este passo foi repetido mais uma vez, totalizando 120 unidades da
enzima. Em seguida adicionou-se 500 µL (30 U mL-1) de amiloglucosidade (EC 3.2.1.3) de
Aspergillus niger (cód. E-AMGPU, Megazyme, Irlanda) em tampão acetato de sódio 100 mM
e pH 4,5. As amostras foram incubadas a 50 ºC por 30 minutos. Este passo foi repetido mais
uma vez, totalizando 30 unidades da enzima. Após, foram acrescentados 100 µL de ácido
perclórico 0,8 M. A quantificação de amido foi realizada em 50 µL da amostra adicionados a
750 µL de glicose PAP Liquiform (Labtest Diagnóstica S.A., Brasil), com posterior incubação
a 30 ºC por 15 minutos. A absorbância foi avaliada com um leitor de microplacas modelo
EL307C (Bio-Tek Instruments, EUA) a 490 nm, em triplicata. Os valores foram
transformados em teor de amido a partir da reta-padrão obtida com as leituras de soluções
contendo 5, 10, 15, 20, 25 e 30 µg de glicose.

16
3.8 Teor foliar de prolina
A quantificação de prolina foi feita segundo os procedimentos descritos por RENA &
MASCIOTTI (1976). Utilizou-se o mesmo extrato purificado das análises de AS e SAC.
Alíquotas de 100 µL do extrato purificado foram colocadas em um tubo de ensaio,
adicionando-se 100 µL de glicina 0,13 M e completando-se o volume de 3 mL com água
destilada. Adicionou-se posteriormente 2 mL de solução de ninhidrina (600 mg de ninhidrina,
15 mL de ácido acético p.a. e 10 mL de ácido fosfórico 6 M). A solução foi agitada em vórtex
para homogeneização e incubada em banho-maria a 95 ºC por 35 minutos. Após a incubação
os tubos foram colocados em banho de gelo, adicionando-se 4 mL de tolueno p.a. e
novamente procedendo-se a agitação em vórtex. A absorbância da solução foi determinada a
515 nm em espectrofotômetro, em duplicata. Os valores foram transformados em teor de
prolina com o auxílio de uma reta-padrão obtida com concentrações variadas de prolina (0,2,
5, 10, 15, 20, 25, 30, 35 e 40 µg). Os resultados foram expressos em µmol g-1.
3.9 Análise dos dados
O delineamento experimental foi em blocos casualizados, em esquema fatorial. As
causas de variação foram combinação copa/porta-enxerto e disponibilidade hídrica, em cada
regime térmico. Os dados obtidos foram submetidos à análise de variância (teste F) e quando
houve diferença significativa, as médias (três repetições) foram comparadas pelo teste de
Tukey a 5% de probabilidade.
4 RESULTADOS
4.1 Ambiente frio (25/10 ºC)
4.1.1 Potencial da água na folha, condutância hidráulica, teor relativo de água e prolina
Tanto em medidas realizadas na antemanhã quanto em medidas realizadas no início da
tarde, observou-se redução significativa do potencial da água na folha (Ψ) devido ao déficit
hídrico em ambos os porta-enxertos (Figura 2). No período da tarde, plantas enxertadas em
‘Swingle’ apresentaram menor Ψ quando comparadas às enxertadas em ‘Cravo’,

17
independente da condição hídrica. Em condições de adequada disponibilidade hídrica os
valores de Ψ em ‘Swingle’ foram 25% mais baixos quando comparadas às plantas enxertadas
em ‘Cravo’. Em plantas submetidas ao déficit hídrico, Ψ foi 18% menor em ‘Swingle’ quando
comparado a ‘Cravo’ (Figura 2b).
Figura 2 - Potencial da água na folha de mudas de laranjeira Valência enxertadas em limoeiro Cravo ou citrumeleiro Swingle mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) no regime térmico de 25/10 ºC (dia/noite). Avaliações realizadas na antemanhã (a) e às 13h00 (b) após 20 dias de déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio padrão. Letras maiúsculas distintas indicam diferenças estatísticas (p<0,05) entre os porta-enxertos na mesma condição hídrica e letras minúsculas distintas representam diferenças estatísticas entre os tratamentos hídricos no mesmo porta-enxerto.
Em plantas enxertadas em ‘Cravo’, o teor relativo de água (TRA) foi reduzido pelo
déficit hídrico, o que não ocorreu em plantas enxertadas em ‘Swingle’ (Figura 3a). A
condutância hidráulica (kL) foi reduzida pelo déficit hídrico independente do porta-enxerto
considerado. Todavia, a redução foi mais acentuada em plantas enxertadas em ‘Cravo’ (-
74%). As plantas dessa combinação apresentaram valores de kL bem superiores (+69%) aos
das plantas enxertadas em ‘Swingle’ em condição de adequada disponibilidade hídrica (Figura
3b).
Houve aumento no teor de prolina foliar em plantas submetidas ao déficit hídrico,
independente do porta-enxerto utilizado. O teor de prolina em plantas sob déficit hídrico foi
maior nas enxertadas em ‘Swingle’ quando comparadas às enxertadas em ‘Cravo’ (Figura 3c).
Aa
BaAbBb
-1,8
-1,2
-0,6
0,0
Cravo Swingle
Aa AaAb Ab
-1,8
-1,2
-0,6
0,0
Cravo Swingle
Ψ (M
Pa)
(a) Antemanhã Irrigado Déficit hídrico (b) 13h00
Porta-enxerto Porta-enxerto

18
Ab Ab
Ba
Aa
0
15
30
45
60
Cravo Swingle
Pro
lina
(µ
mol
g -1)
Porta-enxerto
Figura 3 - Teor relativo de água (TRA, em a), condutância hidráulica (kL, em b) e teor foliar de prolina (c) em folhas de laranjeiras Valência enxertadas em limoeiro Cravo ou citrumeleiro Swingle mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) no regime térmico de 25/10 ºC (dia/noite). Avaliações realizadas após 20 dias de déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio padrão. Letras maiúsculas distintas indicam diferenças estatísticas (p<0,05) entre os porta-enxertos na mesma condição hídrica e letras minúsculas distintas representam diferenças estatísticas entre os tratamentos hídricos no mesmo porta-enxerto.
4.1.2 Trocas gasosas e atividade fotoquímica
Nas medidas de trocas gasosas realizadas no 20º DAIT, observou-se que o déficit
hídrico causou redução na assimilação de CO2 (Pn) nas plantas de ambos os porta-enxertos
(Figura 4a). Indiferente à condição hídrica, plantas enxertadas em ‘Swingle’ apresentaram
maior Pn quando comparadas às enxertadas em ‘Cravo’ na mesma condição hídrica. Plantas
enxertadas em ‘Swingle’ não apresentaram redução da condutância estomática devido ao
déficit hídrico, no entanto, em plantas sobre ‘Cravo’ houve redução (Figura 4b). Tanto em
Aa
BaAb
Ab
0
3
6
9
12
Cravo Swingle
k L
(m
mol
m-2
s-1
MP
a-1)
AaBa
AbAa
0
25
50
75
100
Cravo Swingle
TR
A (
%)
(a)
(b)
Irrigado Déficit hídrico
(c)

19
plantas enxertadas em ‘Cravo’ quanto em ‘Swingle’, o déficit hídrico causou redução também
da condutância mesofílica (gm) e da eficiência de carboxilação (Pn/Cc). Entretanto, as plantas
enxertadas em ‘Swingle’ apresentaram sempre maiores valores de gm e Pn/Cc quando
comparadas às enxertadas em ‘Cravo’, na mesma condição hídrica (Figura 4c,d).
Figura 4 - Assimilação de CO2 (Pn, em a), condutância estomática (gS, em b), condutância mesofílica (gm, em c) e eficiência de carboxilação (Pn/Cc, em d) em laranjeiras Valência enxertadas em limoeiro Cravo ou citrumeleiro Swingle, mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) no regime térmico de 25/10 ºC (dia/noite). Avaliações realizadas às 13h00 após 20 dias de déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio padrão. Letras maiúsculas distintas indicam diferenças estatísticas (p<0,05) entre os porta-enxertos na mesma condição hídrica e letras minúsculas distintas representam diferenças estatísticas entre os tratamentos hídricos no mesmo porta-enxerto.
Plantas enxertadas sobre ambos os porta enxertos apresentaram redução da
assimilação diurna de CO2 (Pni) devido ao déficit hídrico, no entanto, essa redução foi mais
acentuada em plantas enxertadas em ‘Swingle’ (Figura 5a). No tratamento com
disponibilidade hídrica, plantas enxertadas em ‘Swingle’ apresentaram maior Pni quando
comparadas às enxertadas em ‘Cravo’ (Figura 5a). Houve redução da transpiração diurna (Ei)
devido ao déficit hídrico tanto em plantas enxertadas em ‘Cravo’ quanto em ‘Swingle’,
porém, as plantas enxertadas em ‘Cravo’ apresentaram menor Ei quando comparadas às
enxertadas em ‘Swingle’ em condições de déficit hídrico (Figura 5b). A redução da eficiência
do uso da água (EUA) devido ao déficit hídrico ocorreu somente em plantas enxertadas em
AaBa
Ab
Bb
0
2
4
6
Cravo Swingle
Pn
(µm
ol m
-2 s
-1)
AaAa
Ab
Aa
0,000,020,040,060,08
Cravo Swingle
g S (
mol
m-2
s-1
)
Ba
Aa
Bb
Ab
0,00
0,02
0,04
0,06
0,08
Cravo Swingle
g m (
mol
m-2
s-1
)
BaAa
Ab
Bb
0,0
0,2
0,4
0,6
0,8
Cravo Swingle
Pn
/Cc
(µm
ol m
-2 s
-1 P
a-1)
(a)
(c)
Irrigado Déficit hídrico
Porta-enxerto Porta-enxerto
(d)
(b)

20
‘Swingle’. Nesta condição hídrica, as plantas enxertadas em ‘Cravo’ apresentaram maior
EUA quando comparadas às enxertadas em ‘Swingle’. No entanto, estas apresentaram maior
EUA quando comparadas às enxertadas em ‘Cravo’ em condições de adequada
disponibilidade hídrica (Figura 5c).
Figura 5 - Assimilação diurna de CO2 (Pni, em a), transpiração diurna (Ei, em b) e eficiência do uso da água (EUA, em c) em laranjeiras Valência enxertadas em limoeiro Cravo ou citrumeleiro Swingle, mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) no regime térmico de 25/10 ºC (dia/noite). Avaliações realizadas às 13h00 após 20 dias de déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio padrão. Letras maiúsculas distintas indicam diferenças estatísticas (p<0,05) entre os porta-enxertos na mesma condição hídrica e letras minúsculas distintas representam diferenças estatísticas entre os tratamentos hídricos no mesmo porta-enxerto.
Em plantas enxertadas em ‘Cravo’ houve redução tanto da respiração diurna (-48%)
quanto da respiração noturna (-75%) devido ao déficit hídrico (Figura 6a). Em plantas
enxertadas em ‘Swingle’ a redução ocorreu somente na respiração noturna (Figura 6b).
Ba
Aa
Ab Ab
0
50
100
150
200
Cravo Swingle
Pn i
(m
mol
m-2
d-1
)
Aa Aa
BbAb
0
10
20
30
40
Cravo Swingle
Ei (
mol
m-2
d-1
)
Aa
Ba Aa Bb
0,0
1,5
3,0
4,5
6,0
Cravo Swingle
EU
A (
mm
ol m
ol -1)
(a)
(b)
(c)
Porta-enxerto
Irrigado Déficit hídrico

21
Plantas enxertadas em ‘Cravo’ apresentaram maior respiração durante o dia quando
comparada à respiração noturna, independente da condição hídrica (Figura 6a). Nas plantas
enxertadas em ‘Swingle’ não houve diferença na respiração entre os períodos diurno e
noturno nas plantas mantidas sob adequada disponibilidade hídrica, o que não foi observado
nas plantas sob déficit hídrico (Figura 6b). Nestas, a respiração foi 50% maior durante o dia.
Figura 6 - Respiração foliar (Rd) de laranjeiras Valência enxertadas em limoeiro Cravo (a) ou em citrumeleiro Swingle (b), mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) no regime térmico de 25/10 ºC (dia/noite). Avaliações realizadas após 20 dias de déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio padrão. Letras maiúsculas distintas indicam diferenças estatísticas (p<0,05) entre os períodos na mesma condição hídrica e letras minúsculas distintas representam diferenças estatísticas entre os tratamentos hídricos no mesmo período.
Os teores de clorofila não foram influenciados pelos porta-enxertos ou pelo déficit
hídrico. Quanto à atividade fotoquímica, não foram observadas alterações significativas na
eficiência quântica potencial (FV/FM) e efetiva (∆F/FM’) do fotossistema II, nos coeficientes
de extinção fotoquímica (qp) e não fotoquímica (NPQ) da fluorescência, no transporte
aparente de elétrons (ETR) e no transporte de elétrons para a oxigenação da Rubisco (ETRO)
devido ao déficit hídrico ou aos porta-enxertos. Com relação ao transporte de elétrons para a
carboxilação da Rubisco (ETRC), observou-se redução devido ao déficit hídrico em plantas
enxertadas em ‘Cravo’ (Tabela 2).
Ba
Aa
Bb
Ab
0
1
2
3
Noite Dia
Rd
(µm
ol m
-2 s
-1)
Aa Ab
Bb
Aa
0
1
2
3
Noite Dia
(a) Cravo (b) Swingle
Período Período
Irrigado Déficit hídrico
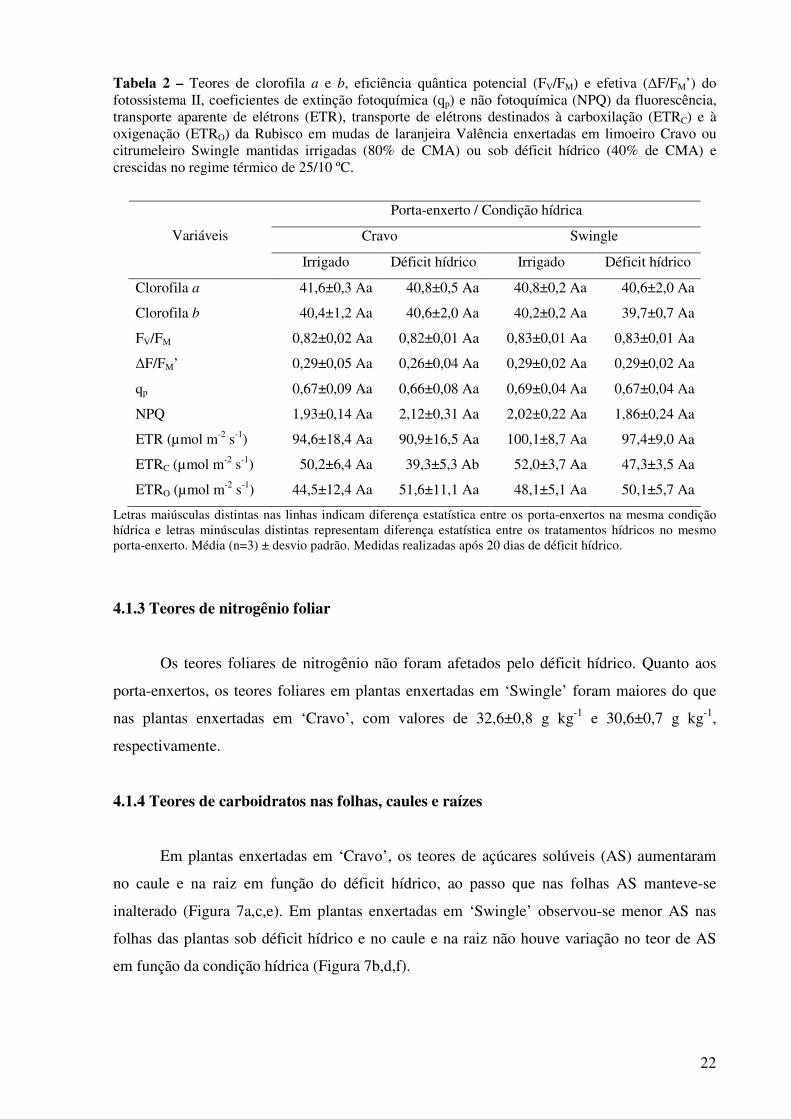
22
Tabela 2 – Teores de clorofila a e b, eficiência quântica potencial (FV/FM) e efetiva (∆F/FM’) do fotossistema II, coeficientes de extinção fotoquímica (qp) e não fotoquímica (NPQ) da fluorescência, transporte aparente de elétrons (ETR), transporte de elétrons destinados à carboxilação (ETRC) e à oxigenação (ETRO) da Rubisco em mudas de laranjeira Valência enxertadas em limoeiro Cravo ou citrumeleiro Swingle mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) e crescidas no regime térmico de 25/10 ºC.
Porta-enxerto / Condição hídrica
Cravo Swingle Variáveis
Irrigado Déficit hídrico Irrigado Déficit hídrico
Clorofila a 41,6±0,3 Aa 40,8±0,5 Aa 40,8±0,2 Aa 40,6±2,0 Aa
Clorofila b 40,4±1,2 Aa 40,6±2,0 Aa 40,2±0,2 Aa 39,7±0,7 Aa
FV/FM 0,82±0,02 Aa 0,82±0,01 Aa 0,83±0,01 Aa 0,83±0,01 Aa
∆F/FM’ 0,29±0,05 Aa 0,26±0,04 Aa 0,29±0,02 Aa 0,29±0,02 Aa
qp 0,67±0,09 Aa 0,66±0,08 Aa 0,69±0,04 Aa 0,67±0,04 Aa
NPQ 1,93±0,14 Aa 2,12±0,31 Aa 2,02±0,22 Aa 1,86±0,24 Aa
ETR (µmol m-2 s-1) 94,6±18,4 Aa 90,9±16,5 Aa 100,1±8,7 Aa 97,4±9,0 Aa
ETRC (µmol m-2 s-1) 50,2±6,4 Aa 39,3±5,3 Ab 52,0±3,7 Aa 47,3±3,5 Aa
ETRO (µmol m-2 s-1) 44,5±12,4 Aa 51,6±11,1 Aa 48,1±5,1 Aa 50,1±5,7 Aa
Letras maiúsculas distintas nas linhas indicam diferença estatística entre os porta-enxertos na mesma condição hídrica e letras minúsculas distintas representam diferença estatística entre os tratamentos hídricos no mesmo porta-enxerto. Média (n=3) ± desvio padrão. Medidas realizadas após 20 dias de déficit hídrico.
4.1.3 Teores de nitrogênio foliar
Os teores foliares de nitrogênio não foram afetados pelo déficit hídrico. Quanto aos
porta-enxertos, os teores foliares em plantas enxertadas em ‘Swingle’ foram maiores do que
nas plantas enxertadas em ‘Cravo’, com valores de 32,6±0,8 g kg-1 e 30,6±0,7 g kg-1,
respectivamente.
4.1.4 Teores de carboidratos nas folhas, caules e raízes
Em plantas enxertadas em ‘Cravo’, os teores de açúcares solúveis (AS) aumentaram
no caule e na raiz em função do déficit hídrico, ao passo que nas folhas AS manteve-se
inalterado (Figura 7a,c,e). Em plantas enxertadas em ‘Swingle’ observou-se menor AS nas
folhas das plantas sob déficit hídrico e no caule e na raiz não houve variação no teor de AS
em função da condição hídrica (Figura 7b,d,f).

23
Figura 7 – Teores de açúcares solúveis (AS), sacarose (SAC), amido (AM) e carboidratos metabolizáveis (AT) em folhas maduras (a,b), caules (c,d) e raízes (e,f) de mudas de laranjeiras Valência enxertadas em limoeiro Cravo (a,c,e) ou citrumeleiro Swingle (b,d,f) mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) no regime térmico de 25/10 ºC (dia/noite). Avaliações realizadas após 20 dias de déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio padrão. Asteriscos indicam diferença estatística (p<0,05) entre os tratamentos hídricos no mesmo porta-enxerto.
O teor de sacarose (SAC) em plantas enxertadas em ‘Cravo’ também aumentou no
caule e na raiz devido ao déficit hídrico, ao passo que nas folhas não houve alteração (Figura
7a,c,e). Já em plantas enxertadas em ‘Swingle’, a única alteração em função do tratamento
hídrico ocorreu no caule, com aumento de SAC devido ao déficit hídrico (Figura 7d).
O déficit hídrico não causou alteração no teor de amido (AM) nas folhas, caule ou
raízes de plantas enxertadas em ‘Cravo’ (Figura 7a,c,e). Já em plantas enxertadas em
‘Swingle’, AM aumentou nas folhas (+52%) e nas raízes (+78%) em plantas sob restrição
hídrica (Figura 7b,f).
0
25
50
75
100
AS SAC AM AT
Teo
r na
fol
ha (
mg
g -1)
0
25
50
75
100
AS SAC AM AT
0
25
50
75
100
AS SAC AM ATTeo
r no
cau
le (
mg
g -1)
0
25
50
75
100
AS SAC AM AT
0
25
50
75
100
AS SAC AM ATTeo
r na
rai
z (m
g g
-1)
0
25
50
75
100
AS SAC AM AT
(c)
(e)
*
*
*
* *
*
*
*
*
*
*
Irrigado Déficit hídrico
Carboidratos Carboidratos
(f)
(d)
(b) (a) Cravo Swingle

24
Quanto aos carboidratos metabolizáveis (AT), houve aumento de AT no caule e nas
raízes de plantas enxertadas em ‘Cravo’ em resposta ao déficit hídrico (Figura 7c,e). Em
plantas enxertadas em ‘Swingle’ houve alteração em função da condição hídrica somente nas
raízes, com aumento de AT em plantas submetidas ao déficit hídrico (Figura 7f).
4.1.5 Biometria
O déficit hídrico causou uma redução de 54% na produção de fitomassa das brotações
(tecidos jovens) das plantas enxertadas em ‘Swingle’, enquanto que nas enxertadas em
‘Cravo’ essa variável não foi afetada (Figura 8a). Tanto em plantas enxertadas em ‘Cravo’
quanto em ‘Swingle’, o déficit hídrico reduziu o crescimento em área foliar (Figura 8b) e em
comprimento da brotação (Figura 8c), e o número de folhas das brotações (Figura 8d), sendo
esse efeito mais severo nas plantas sobre ‘Swingle’.
Figura 8 - Crescimento em matéria seca da brotação (MSB, em a), em área foliar (AFB, em b), em comprimento de brotações (CB, em c) e número de folhas das brotações (NFB, em d) de mudas de laranjeira Valência enxertadas em limoeiro Cravo ou citrumeleiro Swingle, mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) no regime térmico de 25/10 ºC (dia/noite). Avaliações realizadas após 20 dias de déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio padrão. Letras maiúsculas distintas indicam diferenças estatísticas (p<0,05) entre os porta-enxertos na mesma condição hídrica e letras minúsculas distintas representam diferenças estatísticas entre os tratamentos hídricos no mesmo porta-enxerto.
AaAa
Ab
Ab
0
20
40
60
80
Cravo Swingle
NF B
(un
id.)
Ba
Aa
Ba
Ab
0
200
400
600
800
Cravo Swingle
MS
B (
mg
d -1
)
Ba
Aa
Ab
Ab
0
20
40
60
80
Cravo Swingle
AFB
(cm
2 d -1
)
Ba
Aa
Bb
Ab
0,0
0,5
1,0
1,5
2,0
Cravo Swingle
CB (
cm d
-1)
(b)
(c)
(d)
(a)
Porta-enxerto Porta-enxerto
Irrigado Déficit hídrico

25
Além das alterações na parte aérea (brotações), observou-se que a matéria seca das
raízes (MSR) das plantas enxertadas em ‘Cravo’ aumentou em 24% sob déficit hídrico, não
havendo alterações significativas nas plantas enxertadas em ‘Swingle’ (Figura 9). Quanto à
matéria seca do caule (MSC) e das folhas maduras (MSFM), não foram observadas
modificações devido aos regimes hídricos.
Aa
Ab
AaAa
0
10
20
30
40
Cravo Swingle
MS R
(g)
Figura 9 – Matéria seca de raízes (MSR) de mudas de laranjeira Valência enxertadas em limoeiro Cravo ou citrumeleiro Swingle, mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) no regime térmico de 25/10 ºC (dia/noite). Avaliações realizadas após 20 dias de déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio padrão. Letras maiúsculas distintas indicam diferenças estatísticas (p<0,05) entre os porta-enxertos na mesma condição hídrica e letras minúsculas distintas representam diferenças estatísticas entre os tratamentos hídricos no mesmo porta-enxerto.
4.2 Ambiente quente (35/20 ºC)
4.2.1 Potencial da água na folha, condutância hidráulica, teor relativo de água e prolina
Tanto em medidas realizadas na antemanhã quanto em medidas realizadas no início da
tarde, observou-se redução significativa de Ψ devido ao déficit hídrico em ambos os porta-
enxertos (Figura 10). Na antemanhã, o Ψ de plantas sob déficit hídrico enxertadas em ‘Cravo’
foi 17% maior do que o observado nas plantas enxertadas em ‘Swingle’ na mesma condição
hídrica.
Porta-enxerto
Irrigado Déficit hídrico

26
Figura 10 - Potencial da água (Ψ) na folha jovem de mudas de laranjeira Valência enxertadas em limoeiro Cravo ou citrumeleiro Swingle mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) no regime térmico de 35/20 ºC (dia/noite). Avaliações realizadas na antemanhã (a) e às 13h00 (b) no 20º dia após imposição do déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio padrão. Letras maiúsculas distintas indicam diferenças estatísticas (p<0,05) entre os porta-enxertos na mesma condição hídrica e letras minúsculas distintas representam diferenças estatísticas entre os tratamentos hídricos no mesmo porta-enxerto.
O TRA foi maior em plantas sob déficit hídrico em ambos os porta-enxertos (Figura
11a), ao passo que kL foi reduzido em 40% devido ao déficit hídrico apenas em plantas
enxertadas em ‘Cravo’ (Figura 11b). Houve aumento do teor foliar de prolina devido à
restrição hídrica em plantas enxertadas sobre ambos os porta-enxertos. No entanto, o teor de
prolina foi 62% maior em plantas enxertadas em ‘Swingle’ quando comparadas às enxertadas
em ‘Cravo’ com adequada disponibilidade hídrica e 57% maior em plantas sob déficit hídrico
(Figura 11c).
Aa Aa
Ab
Bb
-2,0
-1,5
-1,0
-0,5
0,0
Cravo Swingle
Ψ (
MP
a)
Ba Aa
Ab Ab-2,0
-1,5
-1,0
-0,5
0,0
Cravo Swingle
Irrigado Déficit hídrico
Porta-enxerto Porta-enxerto
(b) 13h00 (a) Antemanhã

27
Ab AbAa
Ba
50
63
75
88
100
Cravo SwingleT
RA
(%
)
AaAa
Aa
Ab
0
1
2
3
4
Cravo Swingle
k L
(mm
ol m
-2 s
-1 M
Pa-1
)
Ab
Bb
Aa
Ba
0
25
50
75
100
Cravo Swingle
Pro
lina
(µm
ol g
-1)
Porta-enxerto
Figura 11 - Teor relativo de água (TRA, em a), condutância hidráulica (kL, em b) e teor foliar de prolina (c) em folhas de laranjeiras Valência enxertadas em limoeiro Cravo ou citrumeleiro Swingle mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) no regime térmico de 35/20 ºC (dia/noite). Avaliações realizadas após 20 dias de déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio padrão. Letras maiúsculas distintas indicam diferenças estatísticas (p<0,05) entre os porta-enxertos na mesma condição hídrica e letras minúsculas distintas representam diferenças estatísticas entre os tratamentos hídricos no mesmo porta-enxerto.
4.2.2 Trocas gasosas e atividade fotoquímica
Nas medidas de trocas gasosas realizadas no 20º DAIT observou-se que plantas em
ambos os porta-enxertos apresentaram redução de Pn devido ao déficit hídrico. Indiferente à
condição hídrica, plantas enxertadas em ‘Cravo’ apresentaram maior Pn quando comparadas
Irrigado Déficit hídrico
(c)
(a)
(b)

28
às enxertadas em ‘Swingle’ na mesma condição hídrica (Figura 12a). Tanto em plantas
enxertadas em ‘Cravo’ quanto em plantas enxertadas em ‘Swingle’ houve redução de gs
devido ao déficit hídrico. Todavia, plantas enxertadas em ‘Cravo’ apresentaram maior gs
quando comparadas às enxertadas em ‘Swingle’ na mesma condição hídrica (Figura 12b).
Somente em plantas enxertadas em ‘Cravo’, gm foi reduzido devido ao déficit hídrico (Figura
12c). Quanto à Pn/Cc não houve diferenças devido aos porta-enxertos ou ao tratamento
hídrico (Figura 12d).
Porta-enxerto Porta-enxerto
Figura 12 - Assimilação de CO2 (Pn, em a), condutância estomática (gS, em b), condutância mesofílica (gm, em c) e eficiência de carboxilação (Pn/Cc, em d) em laranjeiras Valência enxertadas em limoeiro Cravo ou citrumeleiro Swingle, mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) no regime térmico de 35/20 ºC (dia/noite). Avaliações realizadas às 13h00 após 20 dias de déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio padrão. Letras maiúsculas distintas indicam diferenças estatísticas (p<0,05) entre os porta-enxertos na mesma condição hídrica e letras minúsculas distintas representam diferenças estatísticas entre os tratamentos hídricos no mesmo porta-enxerto.
Em plantas enxertadas sobre ambos os porta enxertos houve redução de Pni e de Ei
devido ao déficit hídrico (Figura 13a,b). Plantas enxertadas em ‘Cravo’ apresentaram maior
Pni e Ei quando comparadas às enxertadas em ‘Swingle’ na mesma condição hídrica, tanto no
tratamento irrigado quanto sob déficit hídrico. EUA não foi afetado pelo déficit hídrico nos
porta-enxertos utilizados, porém, plantas enxertadas em ‘Swingle’ apresentaram maior EUA
quando comparadas às enxertadas em ‘Cravo’ na mesma condição hídrica (Figura 13c).
Ba
Aa
BbAb
0
2
4
6
8
Cravo Swingle
Pn
(µ
mol
m-2
s-1
)
Aa
BaAb
Bb
0,00
0,04
0,08
0,12
0,16
Cravo Swingle
g S (
mol
m-2
s-1
)
Aa
Aa
AaAb
0,00
0,03
0,06
0,09
0,12
Cravo Swingle
g m (m
ol m
-2 s-1
) AaAa Aa Aa
0,0
0,2
0,3
0,5
0,6
Cravo SwinglePn/
Cc
(µm
ol m
-2 s
-1 P
a-1)
(a)
(c)
(b)
(d)
Irrigado Déficit hídrico
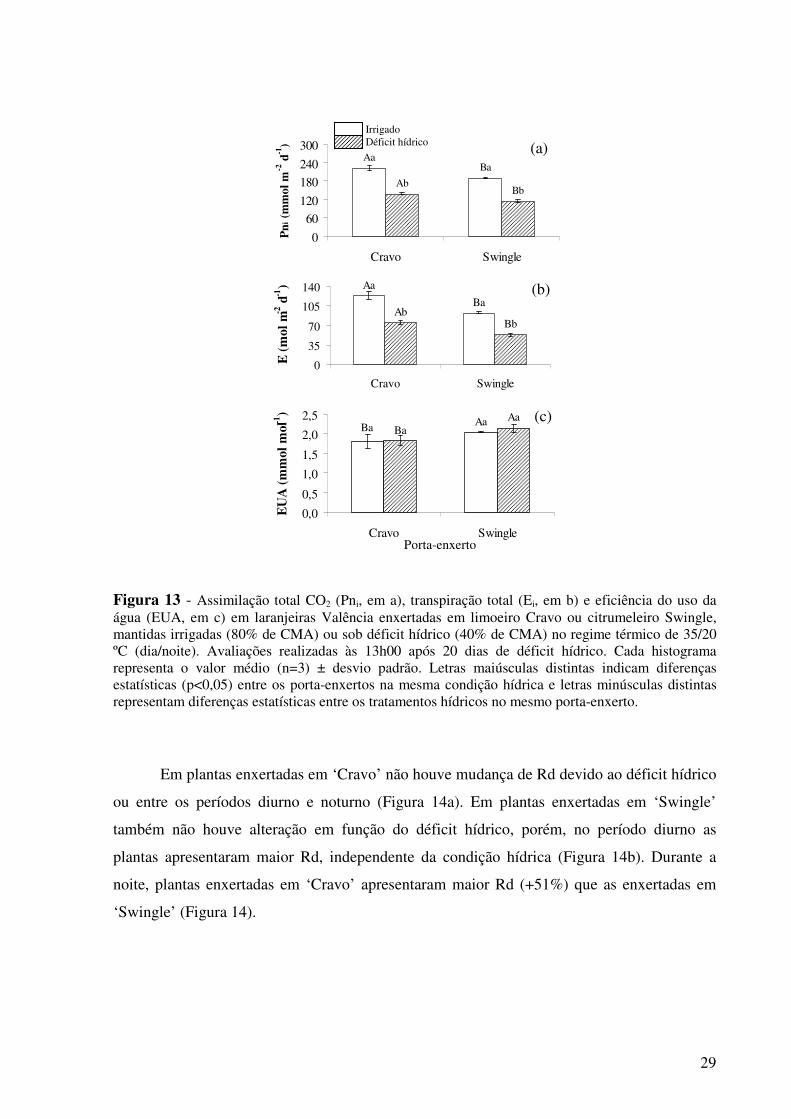
29
Figura 13 - Assimilação total CO2 (Pni, em a), transpiração total (Ei, em b) e eficiência do uso da água (EUA, em c) em laranjeiras Valência enxertadas em limoeiro Cravo ou citrumeleiro Swingle, mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) no regime térmico de 35/20 ºC (dia/noite). Avaliações realizadas às 13h00 após 20 dias de déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio padrão. Letras maiúsculas distintas indicam diferenças estatísticas (p<0,05) entre os porta-enxertos na mesma condição hídrica e letras minúsculas distintas representam diferenças estatísticas entre os tratamentos hídricos no mesmo porta-enxerto.
Em plantas enxertadas em ‘Cravo’ não houve mudança de Rd devido ao déficit hídrico
ou entre os períodos diurno e noturno (Figura 14a). Em plantas enxertadas em ‘Swingle’
também não houve alteração em função do déficit hídrico, porém, no período diurno as
plantas apresentaram maior Rd, independente da condição hídrica (Figura 14b). Durante a
noite, plantas enxertadas em ‘Cravo’ apresentaram maior Rd (+51%) que as enxertadas em
‘Swingle’ (Figura 14).
BaAa
BbAb
0
60
120
180
240
300
Cravo Swingle
Pni
(m
mol
m-2
d-1
)Aa
BaAb
Bb
0
35
70
105
140
Cravo Swingle
E (
mol
m-2
d-1
)
Ba AaBa
Aa
0,0
0,5
1,0
1,5
2,0
2,5
Cravo Swingle
EU
A (
mm
ol m
ol -1)
Porta-enxerto
(c)
(b)
(a) Irrigado Déficit hídrico

30
Figura 14 - Respiração foliar (Rd) de laranjeiras Valência enxertadas em limoeiro Cravo (a) ou em citrumeleiro Swingle (b), mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) no regime térmico de 35/20 ºC (dia/noite). Avaliações realizadas após 20 dias de déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio padrão. Letras maiúsculas distintas indicam diferenças estatísticas (p<0,05) entre os períodos na mesma condição hídrica e letras minúsculas distintas representam diferenças estatísticas entre os tratamentos hídricos no mesmo período.
Os teores de clorofila não foram influenciados pelo déficit hídrico em plantas
enxertadas em ‘Swingle’ ou em ‘Cravo’. Quanto à atividade fotoquímica, não foram
observadas alterações significativas em FV/FM, ∆F/FM’, qp, NPQ, ETR e ETRO devido ao
déficit hídrico ou aos porta-enxertos. Quanto a ETRC, observou-se redução devido ao déficit
hídrico em plantas enxertadas tanto em ‘Cravo’ quanto em ‘Swingle’ (Tabela 3).
4.2.3 Teores de nitrogênio foliar
Os teores foliares de N também não foram afetados pelo déficit hídrico. Quanto aos
porta-enxertos, estes também não influenciaram significativamente os teores foliares de N.
Em ‘Cravo’ o teor foi de 26,2±1,3 g kg-1 e em ‘Swingle’ foi de 28,1±0,3 g kg-1.
Aa AaAaAa
0
1
2
3
4
Noite Dia
Rd
(µ
mol
m-2
s-1
)
Ba
Aa Aa
Ba
0
1
2
3
4
Noite Dia
(b) Swingle
Período Período
(a) Cravo Irrigado Déficit hídrico

31
Tabela 3 – Teores de clorofila a e b, eficiência quântica potencial (FV/FM) e efetiva (∆F/FM’) do fotossistema II, coeficientes de extinção fotoquímica (qp) e não fotoquímica (NPQ) da fluorescência, transporte aparente de elétrons (ETR), transporte de elétrons destinados à carboxilação (ETRC) e à oxigenação (ETRO) da Rubisco em mudas de laranjeira Valência enxertadas em limoeiro Cravo ou citrumeleiro Swingle mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) e crescidas no regime térmico 35/20 ºC.
Porta-enxerto / Condição hídrica
Cravo Swingle Variáveis
Irrigado Déficit hídrico Irrigado Déficit hídrico
Clorofila a 38,6±0,2 Aa 37,7±0,2 Aa 39,6±0,3 Aa 39,1±0,1 Aa
Clorofila b 17,7±0,5 Ba 19,6±0,9 Ba 39,1±1,0 Aa 40,1±0,6 Aa
FV/FM 0,79±0,01 Aa 0,78±0,02 Aa 0,80±0,02 Aa 0,81±0,01 Aa
∆F/FM’ 0,22±0,02 Aa 0,21±0,01 Aa 0,25±0,02 Aa 0,21±0,01 Ab
qp 0,54±0,04 Ba 0,54±0,02 Aa 0,67±0,02 Aa 0,60±0,06 Aa
NPQ 2,22±0,14 Aa 2,25±0,26 Aa 2,43±0,20 Aa 2,47±0,12 Aa
ETR (µmol m-2 s-1) 76,2±6,2 Aa 71,1±4,5 Aa 84,6±7,2 Aa 72,7±4,8 Aa
ETRC (µmol m-2 s-1) 51,8±2,5 Aa 44,3±0,9 Ab 51,5±2,2 Aa 43,1±2,0 Ab
ETRO (µmol m-2 s-1) 24,3±3,8 Aa 28,8±3,6 Aa 33,1±5,6 Aa 29,5±3,0 Aa
Letras maiúsculas distintas nas linhas indicam diferença estatística entre os porta-enxertos na mesma condição hídrica e letras minúsculas distintas representam diferença estatística entre os tratamentos hídricos no mesmo porta-enxerto. Média (n=3) ± desvio padrão. Medidas realizadas após 20 dias de déficit hídrico.
4.3.4 Teores de carboidratos nas folhas, caules e raízes
Em plantas enxertadas em ‘Cravo’, não houve alteração nos teores de AS, SAC, AM
ou de AT em folhas em função do déficit hídrico (Figura 15a). Já em plantas enxertadas em
‘Swingle’, maiores teores AS e AT foram observados em folhas de plantas submetidas à
restrição hídrica (Figura 15b).
No caule, observou-se redução dos teores de SAC, AM e AT devido ao déficit hídrico
em plantas enxertadas em ‘Cravo’ (Figura 15c). Já em plantas enxertadas em ‘Swingle’ e
submetidas à restrição hídrica, houve redução dos teores de AS, AM e AT (Figura 15d).
Observou-se nas raízes de plantas enxertadas em ‘Cravo’ um aumento do teor de AS,
SAC e AT em plantas sob déficit hídrico (Figura 15e), ao passo que em plantas enxertadas em
‘Swingle’ houve redução nos teores de todos os carboidratos analisados.

32
Figura 15 – Teores de açúcares solúveis (AS), sacarose (SAC), amido (AM) e carboidratos metabolizáveis (AT) em folhas maduras (a,b), caules (c,d) e raízes (e,f) de mudas de laranjeiras Valência enxertadas em limoeiro Cravo (a,c,e) ou citrumeleiro Swingle (b,d,f) mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) no regime térmico de 35/20 ºC (dia/noite). Avaliações realizadas após 20 dias de déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio padrão. Asteriscos indicam diferenças estatísticas (p<0,05) entre os tratamentos hídricos no mesmo porta-enxerto.
4.2.5 Biometria
O déficit hídrico causou redução no crescimento em MSB, AFB, CB e NFB tanto de
plantas enxertadas em ‘Cravo’ quanto em ‘Swingle’ (Figura 16). A redução em MSB e AFB
em plantas enxertadas em ‘Swingle’ foi mais acentuada, em torno de 71% e 66%,
respectivamente, enquanto que em ‘Cravo’ essa redução foi de 46% e 44%, respectivamente.
Em condições de déficit hídrico, plantas enxertadas em ‘Cravo’ apresentaram maior
crescimento em MSB e em AFB e maior NFB quando comparadas às enxertadas em ‘Swingle’
na mesma condição hídrica (Figura 16a,b,d).
0
25
50
75
100
AS SAC AM AT
Teo
r n
a fo
lha
(mg
g-1)
0
25
50
75
100
AS SAC AM AT
0
25
50
75
100
AS SAC AM AT
Teo
r n
o ca
ule
(m
g g-1
)
0
25
50
75
100
AS SAC AM AT
0
25
50
75
100
AS SAC AM AT
Teo
r n
a ra
iz (
mg
g-1)
0
25
50
75
100
AS SAC AM AT
(a)
(c)
(e)
* *
*
* *
*
*
*
*
*
*
*
* *
*
(b)
(f)
Irrigado Déficit hídrico
Carboidratos
(d)
Carboidratos
Cravo Swingle

33
Figura 16 - Crescimento em matéria seca da brotação (MSB, em a), em área foliar (AFB, em b), em comprimento de brotações (CB, em c) e número de folhas das brotações (NFB, em d) de mudas de laranjeira Valência enxertadas em limoeiro Cravo ou citrumeleiro Swingle, mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) no regime térmico de 35/20 ºC (dia/noite). Avaliações realizadas após 20 dias de déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio padrão. Letras maiúsculas distintas indicam diferenças estatísticas (p<0,05) entre os porta-enxertos na mesma condição hídrica e letras minúsculas distintas representam diferenças estatísticas entre os tratamentos hídricos no mesmo porta-enxerto.
Além das alterações na parte aérea (brotações), observou-se que MSR das plantas
enxertadas em ‘Cravo’ aumentou sob déficit hídrico (+44%), não havendo alterações
significativas nas plantas enxertadas em ‘Swingle’ (Figura 17). Quanto à MSC e MSF não
foram observadas modificações devido aos regimes hídricos.
Aa
BaAbBb
0
15
30
45
60
Cravo SwingleN
FB (
un
id.)
BaAa
Ab
Bb
0
100
200
300
400
Cravo Swingle
MS
B (
mg
d-1) Aa
Ba
BbAb
0
13
25
38
50
Cravo Swingle
AF
B (
cm2 d
-1)
Aa
Aa
AbAb
0,0
0,4
0,8
1,2
1,6
Cravo Swingle
CB (
cm d
-1)
(a)
(c)
(d)
Porta-enxerto Porta-enxerto
(b)
Irrigado Déficit hídrico

34
Aa
Bb
AaBa
0
6
12
18
Cravo Swingle
MS R
(g)
Figura 17 – Matéria seca de raízes (MSR) de mudas de laranjeira Valência enxertadas em limoeiro Cravo ou citrumeleiro Swingle, mantidas irrigadas (80% de CMA) ou sob déficit hídrico (40% de CMA) no regime térmico de 35/20 ºC (dia/noite). Avaliações realizadas após 20 dias de déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio padrão. Letras maiúsculas distintas indicam diferenças estatísticas (p<0,05) entre os porta-enxertos na mesma condição hídrica e letras minúsculas distintas representam diferenças estatísticas entre os tratamentos hídricos no mesmo porta-enxerto.
5 DISCUSSÃO
5.1 Ambiente frio (25/10 ºC)
Os dois parâmetros mais utilizados para medir a intensidade do estresse hídrico em
plantas são Ψ e TRA (FLEXAS et al., 2004b). A partir do estudo desses parâmetros (Figuras
2 e 3b) foi possível verificar que o déficit hídrico foi efetivo e modificou o estado hídrico de
plantas enxertadas em ambos os porta-enxertos, no entanto, as enxertadas em ‘Swingle’ não
apresentaram redução do TRA devido à restrição hídrica.
A capacidade de manter a atividade fisiológica à medida que a água se torna menos
disponível implica em alguns custos. Esses custos, por exemplo, podem se expressar na forma
de acúmulo de solutos para manter a pressão de turgor (GARCÍA-SÁNCHEZ et al., 2007).
De acordo com CAMPOS et al. (2011), a prolina desempenha vários papéis importantes
durante a aclimatação ao estresse, como mediador de ajuste osmótico e de armazenamento de
carbono, nitrogênio e energia. De fato, foi possível verificar que os níveis de prolina
aumentaram nas plantas submetidas à seca (Figura 3c). As plantas enxertadas em ‘Swingle’
apresentaram teores mais elevados de prolina sob estresse hídrico quando comparadas às
enxertadas em ‘Cravo’. Levando-se em conta a ação no ajuste osmótico (CAMPOS et al.
2011), o aumento do teor de prolina em ‘Swingle’ pode ter contribuído para uma melhor
manutenção da água nos tecidos, levando à manutenção de TRA nessas plantas e
Porta-enxerto
Irrigado Déficit hídrico

35
conseqüentemente menor queda de gs e de Pn (Figura 4a,b) (CHAVES et al., 2009;
SZABADOS & SAVOURÉ, 2009).
A redução de TRA em plantas enxertadas em ‘Cravo’ foi provavelmente ocasionada
pelo decréscimo acentuado de kL (Figura 3). Além dos efeitos da falta de água reduzindo kL
(Figura 3a), há a influência da baixa temperatura, aumentando a viscosidade da água e
reduzindo a permeabilidade do protoplasma e o fluxo de água na planta (SELLIN &
KUPPER, 2007).
O menor Ψ nas plantas sob estresse hídrico, como também verificado por
MAGALHÃES FILHO et al. (2008), causa redução de gs, resultando em menor E e Pn
(Figura 4a; 5a,b). A deficiência hídrica diminui a capacidade fotossintética das plantas e isso é
atribuído em grande parte ao decréscimo de gs (FLEXAS et al., 2004a; MAGALHÃES
FILHO et al., 2008). O controle da abertura e fechamento dos estômatos é essencial para
regulação do equilíbrio entre a perda de água pela transpiração e a assimilação de CO2 pela
planta (PIMENTEL, 2004). Um componente importante na regulação estomática é o balanço
hormonal, e dentre os hormônios, o ácido abscísico (ABA) assume destacada função no
controle estomático em condições de déficit hídrico (POPKO et al., 2010). O ABA funciona
como mensageiro químico entre a raiz e a parte aérea, induzindo o fechamento estomático e a
economia de água antes mesmo de variações no Ψ da folha. Sob estresse hídrico, há estímulo
da biossíntese e distribuição de ABA entre os diferentes compartimentos da folha,
promovendo-se a regulação estomática por mudanças do potencial osmótico da célula guarda.
Qualquer redução do estado hídrico na folha aumenta a resposta estomática ao ABA,
diminuindo assim a transpiração e retornando o estado hídrico da planta a valores adequados
(POSPÍSILOVÁ & DODD, 2005).
Em adição a redução da difusão do CO2 através dos estômatos, o estresse hídrico
também resulta em redução da difusão de CO2 pelo mesofilo das folhas (CHAVES et al.,
2009). Plantas enxertadas sobre ambos os porta enxertos apresentaram redução de gm (Figura
4c) o que leva a conclusão de que menor Pn (Figura 4a e 5a) em plantas submetidas a déficit
hídrico foi causado por limitação difusiva. Plantas enxertadas em ‘Swingle’ apresentaram
maior gm e Pn quando comparado às enxertadas em ‘Cravo’, indiferente à condição hídrica.
Esse fato pode estar relacionado ao melhor desempenho das plantas enxertadas em ‘Swingle’
em ambientes frios (MACHADO et al., 2010).
Em ambos os porta-enxertos houve redução de Pn/Cc (Figura 4d) devido ao déficit
hídrico, ou seja, menor Pn teve como provável causa não somente aspectos difusivos como
também bioquímicos. A taxa fotossintética depende da atividade carboxilase da Rubisco e da

36
síntese/regeneração de RuBP (FLEXAS et al., 2004b). Vários estudos sugerem que reduções
na capacidade fotossintética em condições de déficit hídrico resultam de decrécimos no
conteúdo e atividade da Rubisco, bem como de comprometimentos da regeneração da RuBP e
menor fosforilização (MEDRANO et al., 2002). Entretanto, a quantidade de Rubisco é
geralmente pouco afetada pelo estresse hídrico, mesmo prolongado. No caso, a atividade da
proteína é mais afetada pela falta de água do que sua concentração. Já reduções no conteúdo
de RuBP, substrato da Rubisco, em tecidos com baixo teor relativo de água pode resultar de
limitações em uma ou mais enzimas do ciclo de Calvin-Benson (HUCHZERMEYER &
KOYRO, 2005).
Com relação à atividade fotoquímica (Tabela 2), esta se mostrou bastante resistente ao
estresse hídrico em ambos os porta-enxertos, o que também tem sido observado em outros
estudos (FLEXAS et al., 2004a). O único parâmetro afetado foi o ETRC, que foi reduzido em
plantas enxertadas em ‘Cravo’ sob déficit hídrico, este fato pode ter contribuído para a menor
eficiência de carboxilação nessas plantas. Os elétrons produzidos pelos processos
fotoquímicos são utilizados para fixação do carbono. Porém, em condições em que a fixação
de carbono é limitada, como sob déficit hídrico ou baixas temperaturas, outros processos
como a fotorrespiração, reação de Mehler (ciclo da água-água) e metabolismo do nitrogênio
passam a ser importantes drenos desses elétrons (BAKER et al., 2007). Incremento na relação
ETR/Pn é indício de utilização de elétrons em drenos alternativos de elétrons, i.e., rotas
alternativas à redução fotossintética do CO2 (ALLEN & ORT, 2001). Em plantas enxertadas
sobre ambos os porta-enxertos houve aumento na relação ETR/Pn, no entanto em ‘Cravo’
esse incremento foi maior (21,6 para 45,1) do que em ‘Swingle’ (18,7 para 27,0). Como
Pn/Cc e ETRC foram reduzidos e ETR/Pn aumentou em plantas sob ‘Cravo’, podemos
considerar que a menor eficiência de carboxilação está associada à menor regeneração de
RuBP dependente do transporte de elétrons que ocorre em paralelo ao aumento do fluxo de
elétrons para processos não-assimilatórios (drenos alternativos). Em plantas sob ‘Swingle’,
houve apenas o aumento no direcionamento dos elétrons para drenos alternativos, mecanismo
que protegeu as plantas de uma possível fotoinibição induzida pela falta de água ou baixa
temperatura (KOZAKI & TAKEBA, 1996; OSMOND et al., 1997).
Não houve variação dos teores de clorofila em função do déficit hídrico (Tabela 2).
Este fato pode ser justificado pelo estado nutricional das plantas, sabendo-se que N é um dos
componentes da molécula de clorofila (EPSTEIN & BLOOM, 2006) e que os teores de
nitrogênio não foram limitantes e não diferiram entre os tratamentos hídricos em nenhum dos
porta-enxertos. De acordo com FAQUIN (2002), para citros a faixa crítica de N está entre 25

37
e 27 g kg-1, ou seja, detectados níveis abaixo dessa faixa, o nutriente passa a ser limitante à
produção.
A respiração também foi afetada pela deficiência hídrica (Figura 6), com exceção às
plantas enxertadas em ‘Swingle’ que não apresentaram redução de Rd durante o dia. Este fato
pode ter contribuído, como será apresentado a diante, para o maior crescimento das plantas
enxertadas em ‘Swingle’ sob déficit hídrico, uma vez que altas taxas de respiração estão
associadas ao maior crescimento (AMTHOR, 2000).
O metabolismo de carboidratos na planta é outro fator diretamente relacionado ao
crescimento e sua dinâmica também é alterada em condições de estresses (CHAVES et al.,
2004; ITO et al., 2003). Houve aumento de teores de carboidratos no caule e raízes tanto de
plantas enxertadas em ‘Cravo’ quanto em ‘Swingle’ (Figura 7). Essa estratégia de acúmulo de
reservas pode ter importância na retomada do crescimento quando as condições ambientais se
tornarem adequadas, i.e., houver disponibilidade hídrica. De acordo com SMITH & STITT
(2007), os produtos imediatos da assimilação de CO2 durante o dia são divididos entre
sacarose e amido. Sacarose deve estar imediatamente disponível para o crescimento, enquanto
o amido é acumulado na folha durante o dia e degradado durante a noite para produzir
sacarose. Em situações de severa redução na disponibilidade de carbono, como sob estresse
hídrico, há um ajuste no balanço diurno de carbono e uma redução da taxa de crescimento
(CHAVES et al., 2009). Em folhas de plantas enxertadas em ‘Cravo’, não houve alteração nos
teores de carboidratos devido ao déficit hídrico (Figura 7a). Já nas plantas enxertadas em
‘Swingle’, o aumento no teor foliar de amido concomitante à redução no teor de sacarose
indica redução do crescimento (Figuras 7b e 8).
A matéria orgânica acumulada em uma planta durante o seu crescimento tem origem
no processo fotossintético de fixação de carbono atmosférico, o que representa 95% de toda
sua fitomassa seca (MACHADO et al., 2005). Desta fitomassa, em torno de 60% está nas
raízes e 40% na parte aérea em mudas de laranjeira (SANTOS et al., 2011). O crescimento
observado no regime térmico 25/10 ºC (Figura 8) respondeu ao padrão de assimilação de CO2
das plantas, ou seja, maior Pn em plantas enxertadas em ‘Swingle’ resultou em maior
crescimento em MSB, CB e em AFB em plantas com adequada disponibilidade hídrica quando
comparadas às plantas enxertadas em ‘Cravo’. Sob déficit hídrico, o maior Pn também esteve
associado ao maior crescimento em MSB e CB.
Com a redução do suprimento hídrico, muitas plantas apresentam inibição do
crescimento do caule e da expansão foliar e estímulo do crescimento das raízes (GARCÍA-
SÁNCHEZ et al., 2007). Essas são respostas de aclimatação que limitam o gasto de água

38
pelos tecidos, ajudando a manter o estado hídrico da planta (CHAVES et al., 2009). Ambos os
porta-enxertos apresentaram redução do NFB, da AFB e do CB em condições de deficiência
hídrica (Figura 8). Com relação ao sistema radicular (Figura 9), plantas enxertadas em
‘Swingle’ não apresentaram modificações devido ao déficit hídrico. Já plantas enxertadas em
‘Cravo’, apresentaram aumento significativo do sistema radicular em resposta à baixa
disponibilidade de água no solo, sendo essa uma evidência de que a característica de
tolerância à seca atribuída a este porta-enxerto se relaciona ao vigor de seu sistema radicular
(MAGALHÃES FILHO et al., 2008).
Avaliando a redução relativa de Pn e Rd (controle vs. déficit hídrico) em ambos os
porta-enxertos, pode-se constatar que as plantas enxertadas em ‘Cravo’ foram mais afetadas
se comparadas às enxertadas em ‘Swingle’ (Figuras 4 e 6). No entanto, a redução relativa no
crescimento foi maior nas plantas enxertadas em ‘Swingle’ (Figura 8). Esses resultados
sugerem que a produção de fitomassa foi regulada por outro fator, com provável influência do
balanço hormonal.
Em condições de boa disponibilidade hídrica, o maior crescimento das plantas
enxertadas em ‘Swingle’ justifica-se pelo melhor desempenho fisiológico em ambiente frio se
comparadas às plantas enxertadas em ‘Cravo’. Embora plantas enxertadas em ‘Swingle’
sejam mais suscetíveis à restrição hídrica (POMPEU Jr., 2005), o crescimento observado nas
plantas estressadas foi similar ao crescimento das enxertadas em ‘Cravo’ com boa
disponibilidade hídrica.
5.2 Ambiente quente (35/20 ºC)
As medidas de Ψ evidenciam o fato de que a imposição do déficit hídrico também
afetou as relações hídricas das plantas no regime térmico 35/20 ºC (Figura 10). Foi possível
constatar que plantas enxertadas em ‘Cravo’ sob déficit hídrico amanheceram mais hidratadas
quando comparadas às enxertadas em ‘Swingle’, o que provavelmente favoreceu as trocas
gasosas em ‘Cravo’. O TRA, ao contrário do que é esperado em condições de déficit hídrico,
foi mais elevado nas plantas submetidas à restrição hídrica em ambos os porta-enxertos
(Figura 11a). Essa resposta é um indício de possível ajuste osmótico e pode estar relacionada
aos altos teores de prolina em plantas submetidas ao déficit hídrico (CAMPOS et al., 2011).
Em plantas enxertadas em ‘Swingle’ o teor de prolina foi maior quando comparado ao teor de
plantas enxertadas em ‘Cravo’ (Figura 11c), o que também foi observado no regime térmico

39
25/10 ºC. A produção mais elevada desse aminoácido tem origem genética e plantas mais
sensíveis ao déficit hídrico podem acumular mais prolina (PIMENTEL, 2004).
O baixo Ψ nas plantas sob déficit hídrico ocasionou limitação na difusão de gases
pelos estômatos, havendo redução de gs em ambos os porta-enxertos (Figuras 12b). Porém,
essa redução foi maior em plantas enxertadas em ‘Swingle’, o que está de acordo com o
menor Ψ nas plantas sobre esse porta-enxerto quando comparado ao ‘Cravo’. A redução da gs
determinou menor Pn (Figuras 12a), com menores valores de Pn e gs em plantas enxertadas
em ‘Swingle’ em ambos os regimes hídricos. Em plantas enxertadas em ‘Cravo’, além da
redução de gs, também houve redução de gm (Figura 12c).
Durante a fotossíntese, o CO2 tem que se mover da atmosfera em torno da folha para a
cavidade interna dos estômatos e de lá para o local de carboxilação no estroma do cloroplasto,
através do mesofilo da folha (TAIZ & ZEIGER, 2009). Trabalhos mostram que há diferenças
na concentração de CO2 entre os espaços intercelulares (Ci) e os cloroplastos (Cc), ou seja, a
condutância estomática se difere da condutância mesofílica, com evidências de que Cc é
geralmente menor do que Ci (FLEXAS et al., 2008). Essa diferença em condições de estresse
hídrico aumenta uma vez que há redução de gm, uma resposta de aclimatação importante na
regulação da fotossíntese frente às mudanças ambientais (CHAVES et al., 2009).
Em ambiente quente, o menor Pn devido ao déficit hídrico teve como provável causa
somente fatores difusivos, uma vez que não houve redução na eficiência de carboxilação
(Pn/Cc) em plantas submetidas ao déficit hídrico (Figura 12d). Nesse regime térmico também
foi possível observar a resistência fotoquímica ao estresse hídrico (Tabela 3), como também
relatado em outros trabalhos (FLEXAS et al., 2004a). Com exceção de ETRC, que apresentou
redução devido ao déficit hídrico em plantas enxertadas sobre ambos os porta-enxertos, as
demais variáveis fotoquímicas não foram alteradas. Mesmo ocorrendo redução de ETRC, a
eficiência de carboxilação não foi prejudicada conforme já discutido, o que leva a conclusão
que os elétrons eram suficientes para a fixação de CO2 sob estresse.
Em relação ao teor de carboidratos, enquanto em plantas enxertadas em ‘Cravo’ houve
aumento do teor de carboidratos nas raízes devido ao déficit hídrico, em ‘Swingle’ o padrão
foi inverso, ou seja, houve redução (Figura 15). Se a maior disponibilidade de reservas for
considerada uma estratégia fisiológica para a retomada do crescimento após eventos
estressantes (SMITH & STITT, 2007), pode-se considerar que as plantas enxertadas em
‘Cravo’ serão favorecidas. Com a redução de Pn devido ao déficit hídrico, seria esperada
redução no teor de carboidratos nas folhas (MAGALHÃES FILHO et al., 2008), o que não foi
observado (Figura 15a). Aparentemente, a manutenção ou mesmo aumento dos teores de

40
carboidratos nas folhas das plantas sob déficit hídrico ocorreu às custas do consumo de
reservas do caule (Figura 15b).
O maior Rd noturno em plantas enxertadas em ‘Cravo’ sob déficit hídrico pode estar
relacionado ao maior crescimento dessas plantas se comparadas às enxertadas em ‘Swingle’
na mesma condição hídrica (Figura 14). Quando comparadas às plantas enxertadas em
‘Swingle’, pode-se sugerir que os maiores Pn e Rd observados nas plantas enxertadas em
‘Cravo’ tiveram relação com o maior acúmulo de fitomassa em condição de déficit hídrico, ou
seja, maior crescimento em MSB, em AFB e também maior NFB (Figura 16). Esses resultados
indicam o melhor desempenho das plantas sobre o porta-enxerto ‘Cravo’ em condições de
baixa disponibilidade hídrica no solo, se comparadas às plantas enxertadas em ‘Swingle’.
Enquanto em ‘Swingle’ não houve alterações em MSR sob déficit hídrico, em ‘Cravo’ o
aumento de 44% em MSR evidencia o crescimento do sistema radicular sob seca (Figura 17),
uma estratégia de resistência à seca.
5.3 Fotossíntese e crescimento vs. regime de temperatura
Os regimes de temperatura de 25/10 ºC e 35/20 ºC induziram efeito marcante na
assimilação de CO2 e no crescimento das brotações de laranjeiras. Ao comparar os regimes
térmicos é possível observar que Ψ em ambiente frio é mais baixo, o que possivelmente
causou menor gs e gm, e consequentemente, menor Pn. No regime de 35/20 ºC, os valores de
gm foram maiores do que os observados no regime de 25/10 ºC, especialmente nas plantas
enxertadas em ‘Cravo’. Baixa temperatura do ar noturno tem sido apontada como causadora
do fechamento estomático devido a uma disfunção das células guardas ou por uma disfunção
no estado hídrico da planta, visto que o frio no solo diminui kL e gs (RIBEIRO et al., 2009a).
A redução observada em todas essas variáveis foi menos drástica em plantas enxertadas em
‘Swingle’, o que comprova a maior tolerância da laranjeira a ambientes com temperaturas
mais baixas quando enxertada em ‘Swingle’. Esses resultados estão de acordo com o trabalho
de MACHADO et al. (2010), que testando os mesmos porta-enxertos em condições de apenas
frio noturno (8 ºC), observaram que as plantas apresentaram redução da eficiência
fotossintética em conseqüência de menor gs e menor eficiência aparente de carboxilação.
Além de menor Pn, houve também menor Rd no ambiente frio (com temperatura média de
17,5 ºC), estando ambos os processos relacionados ao menor crescimento de plantas nesse
ambiente.

41
As brotações cresceram de forma vigorosa e uniforme no regime de maior temperatura
média (27,5 ºC), havendo folhas totalmente expandidas aos 20 DAIT. No regime de menor
temperatura, embora ainda tenha havido produção de fitomassa, principalmente em plantas
enxertadas em citrumeleiro Swingle, o crescimento entre as plantas foi bastante desuniforme e
aos 20 DAIT mesmo com muitas folhas jovens, a maioria ainda não se apresentava
completamente expandida, o que inviabilizou a avaliação de tecidos jovens no regime de
25/10 ºC. Essa alteração no crescimento entre os regimes térmicos é justificada pela maior Pni
no regime 35/20 ºC em relação ao regime 25/10 ºC em plantas enxertadas em ‘Cravo’ e em
‘Swingle’ (Figuras 5 e 13). Em relação ao regime de 25/10 ºC, o crescimento em MSB das
plantas enxertadas em ‘Swingle’ foi praticamente o dobro do crescimento observado no
ambiente com 35/20 ºC, revelando que (i) o crescimento das laranjeiras é sensivelmente
alterado pelo porta-enxerto e (ii) que o uso do ‘Swingle’ deve ser priorizado em ambientes
frios.
Independente do porta-enxerto e da condição hídrica, as plantas apresentaram maior
número de folhas em ambiente frio, porém, estas eram menores em tamanho se comparadas às
folhas produzidas no ambiente quente, que eram em menor número (Figuras 8d e 16d). A
relação parte aérea/raiz também diferiu entre os regimes térmicos. No regime de 35/20 ºC,
plantas enxertadas em ‘Cravo’ apresentaram uma redução de 51% da relação parte aérea/raiz
sob déficit hídrico e plantas enxertadas em ‘Swingle’ uma redução em 34%. Esta mudança na
partição de massa seca em ‘Cravo’ foi ocasionada pelo significativo incremento em MSR e
pela redução de MSB. Em ‘Swingle’, porém, a mudança da partição ocorreu somente em
função de menor MSB, uma vez que não houve modificação significativa no sistema radicular
(Figuras 16a e 17). Já no regime de 25/10 ºC a redução da relação parte aérea/raiz foi de 21%
das plantas enxertadas em ‘Cravo’ e de 30% das plantas enxertadas em ‘Swingle’, pelos
mesmos motivos observados no regime de 35/20 ºC (Figuras 8a e 9). A redução da relação
parte aérea/raiz em ‘Cravo’ devido ao déficit foi menor em 25/10 oC quando comparado ao
regime de 35/20 ºC (21% vs. 51%), já em ‘Swingle’ os valores foram bem próximos. Isto se
justifica pelo fato de que no regime 25/10 ºC a redução de MSB não foi significativa e o
aumento de MSR de plantas enxertadas em ‘Cravo’ sob déficit hídrico foi menor no regime de
25/10 ºC quando comparado ao de 35/20 ºC. Essas mudanças morfológicas são mecanismos
de escape à seca e variam entre os porta-enxertos (GARCÍA-SÁNCHEZ, 2007; SILVA et al.,
2006a), permitindo a maior interceptação da água no substrato (sistema radicular mais
desenvolvido) e reduzindo a perda de água pela transpiração (menor área transpirante devido
à redução da massa da parte aérea).

42
6 CONCLUSÃO
Em ambiente frio e independente da condição hídrica, o melhor desempenho
fotossintético das plantas enxertadas em ‘Swingle’ foi ocasionado pela maior condutância
estomática, maior condutância mesofílica e maior eficiência de carboxilação. Como
conseqüência, as plantas enxertadas em ‘Swingle’ apresentaram maior crescimento em
relação às enxertadas em ‘Cravo’. Mesmo sendo mais afetadas pelo déficit hídrico do que as
enxertadas em ‘Cravo’, as plantas enxertadas em ‘Swingle’ apresentaram melhor
desenvolvimento.
Em ambiente quente e sob déficit hídrico, as plantas enxertadas em ‘Cravo’
mantiveram maior condutância estomática o que resultou em maior fotossíntese e crescimento
quando comparadas às enxertadas em ‘Swingle’. Sob boa disponibilidade hídrica, a hipótese
de melhor crescimento das plantas enxertadas em ‘Cravo’ não foi comprovada, mesmo com
estas apresentando maior desempenho fotossintético.

43
7 REFERÊNCIAS BIBLIOGRÁFICAS
ALLEN, D.J.; ORT, D.R. Impacts of chilling temperatures on photosynthesis in warm-climate plants. Trends in Plant Science, London, v.6, p. 36-41, 2001. AMARO, A.A.; BAPTISTELLA, C.S.L. Viveiro de citros - uma visão econômica. Instituto de Economia agrícola – IEA, Texto para discussão, São Paulo, v.23, p. 1-13, 2010. AMARAL, L.I.V.; GASPAR, M.; COSTA, P.M.F.; AIDAR, M.P.M.; BUCKERIDGE, M.S. Novo método enzimático rápido e sensível de extração e dosagem de amido em materiais vegetais. Hoehnea, São Paulo, v. 34, p. 425-431, 2007. AMTHOR, J.S. The McCree-de Wit-Penning de Vries-Thornley Respiration Paradigms: 30 Years Later. Annals of Botany, Oxford, v. 86, p.1-20, 2000. ATKIN, O.K.; SCHEURWATER, I.; PONS, T.L. Respiration as a percentage of daily photosynthesis in whole plants is homeostatic at moderate, but not high, growth temperatures. New Phytologist, London, v.174, p. 367–380, 2007. BAKER, N.R.; HARBINSON, J.; KRAMER, D.M. Determining the limitations and regulation of photosynthetic energy transduction in leaves. Plant, Cell and Environment, Oxford, v. 30, p. 1107-1125, 2007. BATAGLIA, O.C.; FURLANI, A.M.C.; TEIXEIRA, J.P.F.; FURLANI, P.R.; GALLO, J.P. Métodos de análise química de plantas. Campinas: Instituto Agronômico, 1983, 48p. (Boletim Técnico 78). BIELESKI, R.L.; TURNER, A. Separation and estimation of amino acids in crude plant extracts by thin-layer electrophoresis and chromatography. Analytical Biochemistry, New York, v.17, p. 278-293, 1966. BOTA, J.; MEDRANO, H.; FLEXAS, J. Is photosynthesis limited by decreased Rubisco activity and RuBP content under progressive water stress? New Phytologist, London, v.162, p. 671-681, 2004. BUENO, A.C.R.; PRUDENTE, D.A.; MACHADO, E.C.; RIBEIRO, R.V. Daily temperature amplitude affects the vegetative growth and carbon metabolism of orange trees in a rootstock-dependent manner. Journal of Plant Growth Regulation, 2011 (doi: 10.1007/s00344-011-9240-x). CAMPOS, M.K.F.; CARVALHO, K.; SOUZA, F.S.; MARUR, C.J.; PEREIRA, L.F. P.; BESPALHOK FILHO, J.C.; VIEIRA, L.G.E. Drought tolerance and antioxidant enzymatic activity in transgenic ‘Swingle’ citrumelo plants over-accumulating proline. Environmental and Experimental Botany, Oxford, v.72, p. 242–250, 2011. CANTUARIAS, T.; MOURÃO FILHO, F.A.A.; STUCHI, E.S.; SILVA, S.R. ; ESPINOZA, E. Tree performance and fruit yield and quality of Okitsu Satsuma mandarin grafted on 12 rootstocks. Scientia Horticulturae, Amsterdam, v.123, p. 318-322, 2009.

44
CERQUEIRA, E.C.; CASTRO NETO, M.T.; PEIXOTO, C.P.; SOARES FILHO, W.S.; LEDO, C.A.S.; OLIVEIRA, J.G. Resposta de porta-enxertos de citros ao déficit hídrico. Revista Brasileira de Fruticultura, Jaboticabal, v.26, p. 515-519, 2004. CHAVES, M.M., OLIVEIRA, M.M. Mechanisms underlying plant resilience to water deficits: prospects for water-saving agriculture. Journal of Experimental Botany, Oxford, v.55, p. 2365–2384, 2004. CHAVES, M.M.; FLEXAS, J.; PINHEIRO, C. Photosynthesis under drought and salt stress: regulation mechanisms from whole plant to cell. Annals of Botany, Oxford, v.103, p. 551-560, 2009. CHU, Z.; LU, Y.; CHANG, J.; WANG, M.; JIANG, H.; HE, J.; PENG, C.; GE, Y. Leaf respiration/photosynthesis relationship and variation: an investigation of 39 woody and herbaceous species in east subtropical China. Trees, Heidelberg, v. 25, p. 301–310, 2011.
COELHO, E.F.; COELHO FILHO, M.A.; SIMÕES. W.L; COELHO, Y.S. Irrigação em citros nas condições do nordeste do Brasil. Laranja, Cordeirópolis, v.27, p. 297-320, 2006. CROUS, K.; ARAGOZA-CASTELLS, J.; LOW, M.; ELLSWORTH, D.S.; TISSUE, D.T.; TJOELKER, M.G.; BARTON, C.V.M.; GIMENO, T.E.; ATKIN, O. Seasonal acclimation of leaf respiration in Eucalyptus saligna trees: impacts of elevated atmospheric CO2 and summer drought. Global Change Biology, Urbana, v.17, p. 1560–1576, 2011. DAMOUR, G.; VANDAME, M.; URBAN, L. Long-term drought modifies the fundamental relationships between light exposure, leaf nitrogen content and photosynthetic capacity in leaves of the lychee tree (Litchi chinensis). Journal of Plant Physiology, Stuttgart, v.165, p. 1370-1378, 2008. DEMMIG, B.; BJÖRKMAN, O. Comparison of the effect of excessive light on chlorophyll fluorescence (77 K) and photon yield of O2 evolution in leaves of higher plants. Planta, New York, v.171, p. 171-184, 1987. DONATO, S.L.R.; PEREIRA, C.S.; BARROS, Z.J.; SIQUEIRA, D.L.; SALOMÃO, L.C.C. Respostas de combinações de variedades copa e porta-enxerto de citros à deficiência hídrica. Pesquisa Agropecuária Brasileira, Brasília, v.42, p. 1507-1510, 2007. DUBOIS, M.; GILLES, K.A.; HAMILTON, J.K.; REBERS, P.A.; SMITH, F. Colorimetric method for determination of sugars and related substances. Analytical Chemistry, London, v.28, p. 350-356, 1956. EPRON, D.; GODARD, D.; CORNIC, G.; GENTY, B. Limitation of net CO2 assimilation rate by internal resistances to CO2 transfer in the leaves of two tree species (Fagus sylvatica
L. and Castanea sativa Mill.). Plant, Cell and Environment, Oxford, v.18, p. 43-51, 2005. ERISMANN, N.S., MACHADO, E.C., TUCCI, M.L. S. Photosynthetic limitation by CO2 diffusion in drought stressed orange leaves on three rootstocks. Photosynthesis Research, Dordrecht, v.96, p.163-172, 2008.

45
ESPINOZA-NÚÑEZ, E.; MOURÃO FILHO, F.A.A.; STUCHI, E.S.; CANTUARIAS-AVILÉS, T.; DIAS, C.T.S. Performance of ‘Tahiti’ lime on twelve rootstocks under irrigated and non-irrigated conditions. Scientia Horticulturae, Amsterdam, v.129, p. 227–231, 2011. EPSTEIN, E.; BLOOM, A.J. Nutrição mineral de plantas: princípios e perspectivas. Londrina: Editora Planta, 2006. 403p. FALLAHI, E.; COLT, W.M; FALLAHI, B.; CHUN, I. The importance of apple rootstocks on tree growth, yield, fruit quality, leaf nutrition, and photosynthesis with an emphasis on ‘Fuji’. HortTechnology, Alexandria, v.12, p. 38-44, 2002. FAQUIN, V. Diagnose do estado nutricional de plantas. Textos acadêmicos, UFLA/FAEPE, 2002. 77p. FLEXAS, J; MEDRANO, H. Drought-Inhibition of photosynthesis in C3 plants: stomatal and non-stomatal limitation revisited. Annals of Botany, Oxford, v.89, p. 183-189, 2002. FLEXAS, J.; BOTA, J.; LORETO, F.; CORNIC, G.; SHARKEY, T.D. Difusive and metabolic limitations on photosynthesis under dhought and salinity in C3 plants. Plant Biology, Malden, v.6, p.1-11, 2004a. FLEXAS, J.; BOTA, J.; CIFRE, J.; ESCALONA, J. M.; GALMÉS, J.; GULIÁS, J.; LEFI, E.; MARTÍNEZ-CANELLAS, S.F; MORENO, M.T.; RIBAS-CARBÓ, M.; RIERA, D.; SAMPOL, B.; MEDRANO, H. Understanding down-regulation of photosynthesis under water stress: future prospects and searching for physiological tools for irrigation management. Annals of Applied Biology, Warwick, v.144, p. 273-283, 2004b. FLEXAS, J.; RIBAS-CARBÓ, M. DIAZ-ESPEJO, A.; GALMÉS, J; MEDRANO, H. Mesophyll conductance to CO2: current knowledge and future prospects. Plant, Cell and Environment, Oxford, v.31, p. 602–621, 2008. FOCHESATO, M.L.; SOUZA, P.V.D.; SCHÄFER, G.; MACIEL, H.S. Produção de mudas cítricas em diferentes porta-enxertos e substratos comerciais. Ciência Rural, Santa Maria, v.36, p.1397-1403, 2006. GARCÍA-SÁNCHEZ, F.; SYVERTSEN, J.P.; GIMENO, V.; BOTÍA, P.; PEREZ- PEREZ, J.G. Responses to flooding and drought stress by two citrus rootstock seedlings with different water-use efficiency. Physiologia Plantarum, Copenhagen, v.130, p. 532-542, 2007. GENTY, B.; BRIANTAIS, J.M.; BAKER, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochimica et Biophysica Acta, Amsterdam, v.990, p. 87-92, 1989. GIRARDI, E.A.; MOURÃO FILHO, F.A.A.; ALVES, A.S.R. Mudas de laranjeira Valência sobre dois porta-enxertos e sob diferentes manejos de adubação. Revista Brasileira de Fruticultura, Jaboticabal, v. 32, p. 855-864, 2010. GUIMARÃES, E.R.; MUTTON, M.A.; MUTTON, M.J.R.; FERRO, M.I.T.; RAVANELI, G.C.; SILVA, J.A. Free proline accumulation in sugarcane under water restriction and spittlebug infestation. Scientia Agricola, Piracicaba, v.65, p. 628-633, 2008.

46
GUO, Y.P.; ZHOU, H.F.; ZHANG, L.C. Photosynthetic characteristics and protective mechanisms against photooxidation during high temperature stress in two citrus species. Scientia Horticulturae, Amsterdam, v.108, p. 260-267, 2006. HUCHZERMEYER, B; KOIRO, H.W. Enzime cohesion and high turnover metabolite shutting, essential for functioning of pathways, is impaired by changes in cytosolic water potential. In: PESSARAKLI, M. Handbook of Photosynthesis. 2.ed. Arizona, 2005. 928p. ITO, A.; HAYAMA, H.; KASHIMURA, Y. Sugar metabolism in bud during flower bud formation: a comparison of the japanese pear [Pyrus pyrifolia (Burm.) Nak.] cultivars possessing different flowering habitats. Scientia Horticulturae, Amsterdam, v.96, p. 163-175, 2003.
KOZAKI, A.; TAKEBA, G. Photorespiration protects C3 plants from photooxidation. Nature, London, v. 384, p. 557-560, 1996.
LLOYD, J.; FARQUHAR, G.D. Effects of rising temperatures and [CO2] on the physiology of tropical forest trees. Philosophical Transactions of the Royal Society, London, v.363, p. 1811-1817, 2008. LUTTS, S.; MAJERUS, V.; KINETET, J.M. NaCl effects on proline metabolism in rice (Oryza sativa) seedlings. Physiologia Plantarum, Copenhagen, v.105, p. 450-458, 1999. LOKA, D.A.; OOSTERHUIS, D.M. Effect of high night temperatures on cotton respiration, ATP levels and carbohydrate content. Environmental And Experimental Botany, Oxford, v.68, p. 258-263, 2010. MACHADO, E.C.; MEDINA, C.L.; GOMES, M.M.A.; HABERMANN, G. Variação sazonal da fotossíntese, condutância estomática e potencial da água na folha de laranjeira “Valência”. Scientia Agricola, Piracicaba, v.59, p. 53-58, 2002. MACHADO, E.C.; SCHMIDT, P.T.; MEDINA, C.L.; RIBEIRO, R.V. Respostas da fotossíntese de três espécies de citros a fatores ambientais. Pesquisa Agropecuária Brasileira, Brasília, v.40, p. 1161-1170, 2005. MACHADO, D.F.S.P.; MACHADO, E.C.; MACHADO, R.S.; RIBEIRO, R.V. Efeito da baixa temperatura noturna e do porta-enxerto na variação diurna das trocas gasosas e na atividade fotoquímica de laranjeira ‘Valencia’. Revista Brasileira de Fruticultura, Jaboticabal, v.32, p. 351-359, 2010. MAGALHÃES FILHO, J.R.; AMARAL, L.R.; MACHADO, D.F.S.P.; MEDINA, C.L.; MACHADO, E.C. Deficiência hídrica, trocas gasosas e crescimento de raízes em laranjeira ‘Valência’ sobre dois tipos de porta-enxerto. Bragantia, Campinas, v.67, p. 693-699, 2008. MAGALHÃES FILHO, J.R.; MACHADO, E.C.; MACHADO, D.F.S.P.; RAMOS, R.A.; RIBEIRO, R. V. Variação da temperatura do substrato e fotossíntese em mudas de laranjeira ‘Valência’. Pesquisa Agropecuária Brasileira, Brasília, v.44, p. 1118-1126, 2009.

47
MATTOS Jr, D.; QUAGGIO, J.A.; CANTARELLA, H.; ALVA, A.K.; GRAETZ, D.A. Response of young citrus trees on selected rootstocks to nitrogen, phosphorus, and potassium fertilization. Journal of Plant Nutrition, Philadelphia, v.29, p. 1371-1385, 2006. MATTOS Jr, D.; QUAGGIO, J.A.; CANTARELLA, H.; BOARETTO, A.E. Nitrogênio e enxofre na cultura dos citros. In: YAMADA, T.; ABDALLA, S.R.S.; VITTI, G.S. (Org. ). Nitrogênio e enxofre na agricultura brasileira. 1 ed. Piracicaba: International Plant Nutrition Institute - IPNI, 2007, v.1, p. 413-443. MATTOS Jr, D.; BOARETTO, R.M.; ABREU, M.F.; CARVALHO, S.A. Disponibilidade de boro em substrato para produção de porta-enxertos de citros em fase de sementeira. Bragantia, Campinas, v.67, p. 983-989, 2008. MATTOS Jr, D.; RAMOS, U.M.; QUAGGIO, J.A.; FURLANI, P.R. Nitrogênio e cobre na produção de mudas de citros em diferentes porta-enxertos. Bragantia, Campinas, v.69, p. 135-147, 2010. MEDINA, C.L.; SOUZA, R.P.; MACHADO, E.C.; RIBEIRO, R.V.; SILVA, J.A.B. Photosynthetic response of citrus grown under reflective aluminized polypropylene shading nets. Scientia Horticulturae, Amsterdam, v.96, p.115-125, 2002. MEDINA, C.L.; MACHADO, E.C.; RENA, A.B.; SIQUEIRA, D.L. de. Fisiologia dos citros. In: MATTOS JÚNIOR, D.; DE NEGRI, J. D.; PIO, R. M.; POMPEU Jr, J. (Ed.). Citros. Campinas: IAC, 2005. p. 147-195. MEDINA FILHO, H.P.; BORDIGNON, R.; SIQUEIRA, W.J.; FEICHTENBERGER, E.; CARVALHO, M.R.T.; TEÓFILO SOBRINHO, J. Resistência de clones e híbridos de porta-enxertos de citros à gomose de tronco causada por Phytophthora parasitica. Fitopatologia Brasileira, Brasília, v.28, p. 534-540. 2003. MEDRANO, H.; ESCALONA, J.M.; BOTA, J.; GULÍAS, J.; FLEXAS, J. Regulation of photosynthesis of C3 plants in response to progressive drought: the interest of stomatal conductance as a reference parameter. Annals of Botany, Oxford, v.89, p. 895–905, 2002. MÜLLER, G.W.; TARGON, M.L.; CARVALHO, S. A.; SOUZA, A. A.; RODRIGUES, J.C.V. Doenças de citros causadas por vírus e viróides. In: MATTOS JUNIOR, D. et al. Citros, Cap.19, p. 567-604, 2005. NOGUEIRA, R.J.M.C.; MORAES, J.A.P.V.; BURITY, H. A.; BEZERRA NETO, E. Alterações na resistência à difusão de vapor das folhas e relações hídricas em aceroleiras submetidas a déficit de água. Revista Brasileira de Fisiologia Vegetal, Campinas, v.13, p. 75-87, 2001. ORTOLANI, A.; PEDRO JUNIOR, M.J.; ALFONSI, R.R. Agroclimatologia e o cultivo de citros. In: RODRIGUES, O.; VIÉGAS, F.; POMPEU JUNIOR, J.; AMARO, A. A. (ed). Citricultura brasileira. Campinas, Fundação Cargill, p.153-195, 1991. OSMOND, B.; BADGER, M.; MAXWELL, K.; BJÖRKMAN, O.; LEEGOOD, R. Too many photons: photorespiration, photoinhibition and photooxidation. Trends in Plant Science, London, v.2, p.119-120, 1997.

48
PASSIOURA, J. The drought environment: physical, biological and agricultural perspectives. Journal of Experimental Botany, Oxford, v.58, p. 113-117, 2007. PEREIRA, A.B.; VILA NOVA, N.A.; ALFARO, A.T. Necessidades hídricas dos citros e macieiras a partir da área foliar e da energia solar. Revista Brasileira de Fruticultura, Jaboticabal, v.31, p. 671-679, 2009. PIMENTEL, C. A relação da planta com a água. Seropédica, RJ: Edur, 2004. 191p. PIMENTEL, C.; BERNACCHI, C.; LONG, S. Limitations to photosynthesis at different temperatures in the leaves of Citrus limon. Brazilian Journal of Plant Physiology, Londrina, v.19, p. 141-147, 2007. POMPEU Jr., J. Porta-enxertos. In: MATTOS JUNIOR, D.; DENIGRI, J.D.; PIO, R.M.; POMPEU Jr, J. (Eds). Citros. Campinas, v.61, p. 104, 2005. POPKO, J.; HÄNSCH, R.; MENDEL, R.R.; POLLE , A.; TEICHMANN, T. The role of abscisic acid and auxin in the response of poplar to abiotic stress. Plant Biology, Malden, v.12, p. 242–258, 2010. POSPÍSILOVÁ, J.; DODD; I. C. Role of plant growth regulators in stomatal limitations to photosynthesis during water stress. In: PESSARAKLI, M. Handbook of Photosynthesis. 2.ed. Arizona, 2005. 928p. PRADO, A.K.S.; MACHADO, E.C.; MEDINA, C.L.; MACHADO, D.F.S.P.; MAZZAFERA, P. Florescimento e frutificação em laranjeiras ‘Valência’ com diferentes cargas de frutos e submetidas ou não à irrigação. Bragantia, Campinas, v. 66, p. 173-182, 2007. PRUDENTE, R.M.; SILVA, L.M.S.; CUNHA SOBRINHO, A.P. Comportamento da laranjeira ‘Pêra’ sobre cinco porta-enxertos em ecossistema de Tabuleiros Costeiros, Umbaúba-SE. Revista Brasileira de Fruticultura, Jaboticabal, v. 26, p. 110-112, 2004. RAMOS, R.A. Regime térmico e relação fonte-dreno em mudas de laranjeiras: dinâmica de carboidratos, fotossíntese e crescimento. 2009. 52p. Dissertação (Mestrado em Tecnologia da Produção Agrícola) - Instituto Agronômico/IAC, Campinas. RENA, A.B.; MASCIOTTI, G.Z. The effect of dehydration on nitrogen metabolism and growth of bean cultivars (Phaseolus vulgaris L.). Revista Ceres, Viçosa, v.23, p. 288-301, 1976. RIBEIRO, R.V.; MACHADO, E.C.; OLIVEIRA, R.F. Growth- and leaf-temperature effects on photosynthesis of sweet orange seedlings infected with Xylella fastidiosa. Plant Pathology, Oxford, v.53, p. 334-340, 2004. RIBEIRO, R.V.; MACHADO, E.C.; OLIVEIRA, R.F. Temperature response of photosynthesis and its interaction with light intensity in sweet orange leaf discs under non-photorespiratory condition. Ciência e Agrotecnologia, Lavras, v.30, p. 670-678, 2006.

49
RIBEIRO, R.V.; MACHADO, E.C. Some aspects of citrus ecophysiology in subtropical climates: re-visiting photosynthesis under natural conditions. Brazilian Journal of Plant Physiology, Londrina, v.19, p. 393-411, 2007. RIBEIRO, R.V.; MACHADO, E.C.; SANTOS, M.G.; OLIVEIRA, R.F. Photosynthesis and water relations of well-watered orange plants as affected by winter and summer conditions. Photosynthetica, Pragha, v. 47, p. 215-222, 2009a. RIBEIRO, R.V.; MACHADO, E.C.; SANTOS, M.G.; OLIVEIRA, R.F. Seasonal and diurnal changes in photosynthetic limitation of young sweet orange trees. Environmental and Experimental Botany, Oxford, v.66, p. 203-211, 2009b. ROHACEK, K. Chlorophyll fluorescence parameters: the definitions, photosynthetic meaning, and mutual relationships. Photosynthetica, Pragha, v.40, p. 13-29, 2002. RUELLANDA, E.; ZACHOWSKIA, A. How plants sense temperature. Environmental and Experimental Botany, Oxford, v.69, p. 225-232, 2010. SANTOS, C.M.A.; RIBEIRO, R.V.; MAGALHÃES FILHO, J.R.; MACHADO, D.F.S.P.; MACHADO, E.C. Low substrate temperature imposes higher limitation to photosynthesis of orange plants as compared to atmospheric chilling. Photosynthetica, Pragha, v.49, p. 546-554, 2011. SCHÄFER, G.; BASTIANEL, M.; DORNELLS, A.L.C. Porta-enxertos utilizados na citricultura. Ciência Rural, Santa Maria, v.31, p. 723-733, 2001. SELLIN, A.; KUPPER, P. Temperature, light and leaf hydraulic conductance of little-leaf linden (Tilia cordata) in a mixed forest canopy. Tree Physiology, Victoria, v.27, p. 679-688, 2007. SETIN, D.W.; CARVALHO, S.A.; MATTOS Jr., D. Crescimento inicial e estado nutricional da laranjeira ‘Valência’ sobre porta-enxertos múltiplos de limoeiro ‘Cravo’ e citrumeleiro ‘Swingle’. Bragantia, Campinas, v.68, p. 397-406, 2009. SILVA, C.R.; ALVES Jr., J.; SILVA, T.J.A.; FOLEGATTI, M.V.; SANTOS R.A.; SOUZA, L.B. Déficit hídrico em citros: informações para o manejo da irrigação. Laranja, Cordeirópolis, v.27, p. 119-130, 2006a. SILVA, S.R.; STUCHI, E.S.; FRANCO, D.; REIFF, E.T.; SEMPIONATO, O.R. Produção inicial e qualidade dos frutos da laranjeira ‘Moro’ em 16 porta-enxertos em Bebedouro-SP. Laranja, Cordeirópolis, v.27, p. 83-90, 2006b. SILVA, S.R.; STUCHI, E.S.; FRANCO, D.; SEMPIONATO, O.R.; REIFF, E.T. Características da produção de frutos das laranjeiras ‘Natal’ e ‘Valência’ em 13 porta-enxertos em Bebedouro-SP. Laranja, Cordeirópolis, v.27, p. 91-100, 2006c. SILVEIRA, J.A.G.; VIÉGAS, R.A.; ROCHA, I.M.A.; MOREIRA, A.C.O.M.; MOREIRA, R.A.; OLIVEIRA, J.T.A. Proline accumulation and glutamine synthetase are increased by salt-induced proteolysis in cashew leaves. Journal of Plant Physiology, Stuttgart, v.160, p. 115-123, 2003.

50
SMITH, A.M.; STITT, M. Coordination of carbon supply and plant growth. Plant, Cell and Environment, Oxford, v.30, p. 1126-1149, 2007. SORGONÀ, A.; LUPINI, A.; ABENAVOLI, M.R.; Nitrate use-efficiency: a morphological analysis of the above and below-ground functional traits in two citrus rootstocks. Global Journal of Plant Ecophysiology, Jordan, v.1, p.26-37, 2011. SPIEGEL-ROY, P.; GOLDSCHMIDT, E.E. Biology of citrus. Cambridge: Cambridge University Press, 1996. 230 p. STUCHI, E.S.; DONADIO, L.C.; SEMPIONATO, O.R.; PERECIN, D. Yield and fruit quality of ‘Pêra’ sweet orange clone iac on 16 rootstocks in Bebedouro region, State of São Paulo, Brazil. Revista Brasileira de Fruticultura, Jaboticabal, v.26, p. 359-362, 2004. SUN, Z.H., MA, X.T. Thermostability of plasma membrane in citrus leaves. Journal of Huazhong Agricultural University, China, v.18, p. 375–377, 1999. SYVERTSEN, J.P., LEVY, Y. Salinity interactions with other abiotic and biotic stresses in citrus. HortTechnology, Alexandria, v.15, p. 100–103, 2005.
SZABADOS, L.; SAVOURÉ, A. Proline: a multifunctional amino acid. Trends in Plant Science, Amsterdam, v.15, p. 89-97, 2009. TAIZ, L.; ZEIGER, E. Fisiologia vegetal. 2.ed. Porto Alegre: Artmed, 2009. 819p. VAN HANDEL, E. Direct microdetermination of sucrose. Analytical Biochemistry, New York, v.22, p. 280-283, 1968. VESTE, M.; BEN-GAL, A.; SHANI, U. Impact of thermal stress and high VPD on gas exchange and chlorophyll fluorescence of Citrus grandis under desert conditions. Acta Horticulturae, The Hague, n.531, p. 143-149, 2000. VU, J.C.V. Photosynthetic responses of citrus to environmental changes. In: PESSARAKLI, M. Handbook of plant and crop stress. New York: Marcel Dekker, 1999. p. 947-961.



















![Artigo Limoeiro[1]](https://img.document.onl/doc/110x75/54938845b479595b4d8b4881/artigo-limoeiro1.jpg)









