Embed Size (px)
Citation preview



i
PENÉLOPE GIACOMITTI
EFEITOS do HERBICIDA “TORDON 2,4-D 64/240 TRIETANOLAMINA BR”
em FILHOTES de MÃES CONTAMINADAS durante a GESTAÇÃO e LACTAÇÃO
Curitiba – Paraná 1995
Dissertação apresentada ao Curso de Pós-Graduação
em Morfologia – área de concentração em
Biologia Celular,
da Universidade Federal do Paraná,
visando à obtenção do título de
mestre em Ciências Morfológicas

ii
Orientadora: Profª Drª Maria Teresa Barros Schütz
Co-orientadora: Profª Dirnei Seli F. Baroni

iii
À DEUS que em Seu infinito Amor me deu condições para desenvolver
e terminar mais este trabalho.
Aos meus pais, Joâni Giacomitti e Alba Ceccon Giacomitti
(sem palavras).
Aos meus irmãos : Cícero Ceccon Giacomitti Lúcia Ceccon Giacomitti Joâni Giacomitti Júnior e
Vitor Hugo Giacomitti, pela paciência.
Ao meu namorado :Leônidas Alves Rosa,
com carinho, pela compreensão.
À minha “nonna”: Gema Catarina Simioni Ceccon
(sem palavras).

iv
Todas as figuras deste trabalho são,
exceção feita a capa inicial,
Clip-art windons 2008
permitida reprodução
não comercializável

v
AGRADECIMENTOS
Agradeço a orientação precisa, a paciência e a presença amiga da Profª Drª Maria Teresa Barros Schütz, que tornou possível levar a bom termo este trabalho. A co-orientação da Profª Dirnei Seli F. Baroni, que soube com maestria diremir todas as dúvidas surgidas a partir da metodologia e do material histológico. Ao Prof. Romeu Afonso Schütz pelo apoio e amizade. A todos os professores e funcionários do Departamento de Fisiologia, de Farmacologia e do Biotério, que de algum modo auxiliaram, aliviando tarefas ou facilitando o acesso ao material necessário, e muito contribuíram para o término deste trabalho. Em especial, Prof. Cláudio da Cunha, pelo auxílio no uso do computador, Cândido José Thomaz Pereira, técnico responsável pelo Biotério do Setor de Ciências Biológicas, Terezinha de Jesus do Carmo Andrade (D. Teresa) e Armelinda Silvino dos Santos e todos/as os/as demais funcionárias/os do Biotério. À Profª Drª Orieta Silveira, professora do Departamento de Bioquímica pelos esclarecimentos. Todo a apoio e amizade do Prof. Dr. Waldemiro Gremski, professor de pós-graduação do Departamento de Biologia Celular. A Joâni Giacomitti Júnior, pelo empenho e capricho na elaboração final deste trabalho. Aos amigos e colegas da turma: Cecília, Luís, Hamilton, Ivone Maria, Ivone Cipriano, Sônia e todos os demais colegas, pelo carinho, incentivo e apoio. À Coordenação do Curso de Pós-Graduação, professores, funcionários e amigos do Departamento de Morfologia/Biologia Celular da Universidade Federal do Paraná, pelo incentivo e apoio. Em especial, Elinor, ex-secretária do curso de pós-graduação, Rubens Gaier e auxiliar, técnicos em histologia, Ana Maria Brauza Cunha, funcionária, e Marlene Bonifácio, atual secretária do curso de pós-graduação. A todos aqueles que direta ou indiretamente tornaram possível a execução e término deste trabalho. Ao CNPq pela bolsa de mestrado e à PRPPG pelo apoio financeiro.

vi
ÍNDICE
PÁGINA
AGRADECIMENTOS .............................................................................. V
ORGANOGRAMAS E TABELAS ......................................................... VIII
GRÁFICOS E FIGURAS ......................................................................... X
ABREVIATURAS .................................................................................. XII
RESUMO ............................................................................................. XIV
ABSTRACT .......................................................................................... XVI
1. INTRODUÇÃO .................................................................................................... 1
2. OBJETIVOS E JUSTIFICATIVAS ....................................................................... 7
3. MATERIAL E MÉTODOS .................................................................................... 9
3.1. Local de trabalho
3.2. Animais experimentais
3.3. Herbicida
3.4. Metodologia...............................................................................................................10
3.4.1. Acasalamento
3.4.2. Nascimento dos Filhotes
3.4.3. Peso Corporal
3.4.4. Reflexo de Endireitamento
3.4.5. Abertura Palpebral
3.4.6. Sexagem
3.4.7. Testes comportamentais...............................................................................14
3.4.7.1. Movimentação espontânea
3.4.7.2. Teste do “Rota-Rod”
3.4.7.3. Exposição ao “Open-Field” .............................................................................. 17
3.4.7.4. Teste do “Plus Maze” ou Labirinto em X ................................................... 20
3.4.7.5. Esquiva Ativa ................................................................................................. 21

vii
3.5. Estatística ....................................................................................................24
3.6. Morfologia ....................................................................................................24
3.6.1. Coloração por H.E.
3.6.2. Nitratação
3.6.3. Evidenciação da Bainha de Mielina
4. RESULTADOS .................................................................................................. 27
4.1. Dias de Acasalamento e Gestação
4.2. Proporção Sexual
4.2.1. Organogramas
4.3. Peso Corporal
4.4. Desenvolvimento pós-natal........................................................................28
4.4.1. Reflexo de Endireitamento
4.4.2. Abertura Palpebral
4.5. Testes Comportamentais aos 40 dias de idade....................................28
4.5.1. Movimentação Espontânea
4.5.2. “Rota-Rod”
4.6. Testes Comportamentais aos 90 dias de idade....................................29
4.6.1. Movimentação Espontânea
4.6.2. “Rota-Rod”
4.6.3. Exposição ao “Open-Field”.................................................................29
4.6.4. Teste do “Plus-Maze” ou “Labirinto em X”........................................31
4.6.5. Esquiva Ativa......................................................................................31
4.7. Fêmeas que não se apresentaram prenhas.............................................32
4.8. Número de Filhotes mortos/devorados..................................................33
4.9. Outras Observações...................................................................................33
4.10. Morfologia....................................................................................................34
4.10.1. Coloração com hematoxilina-eosina (H.E.)
4.10.2. Nitratação
4.10.3. Evidenciação da bainha de mielina
5. DISCUSSÃO ..................................................................................................... 66
6. CONCLUSÃO ................................................................................................... 78
7. REFERÊNCIAS BIBLIOGRÁFICAS .................................................................. 81

viii
LISTA DE ORGANOGRAMAS E TABELAS
página
Organograma 1. Representa o número de machos e fêmeas nascidos na geração F1 do grupo controle ................................................................................ 41
Organograma 2. Representa o número de machos e fêmeas nascidos na geração F2 do grupo controle ................................................................................ 42
Organograma 3. Representa o número de machos e fêmeas nascidos na geração F3 do grupo controle ................................................................................ 43
Organograma 4. Representa o número de machos e fêmeas nascidos na geração F1 do grupo Tordon 1 .............................................................................. 44
Organograma 5. Representa o número de machos e fêmeas nascidos na geração F2 do grupo Tordon 1 .............................................................................. 45
Organograma 6. Representa o número de machos e fêmeas nascidos na geração F3 do grupo Tordon 1 .............................................................................. 46
Organograma 7. Representa o número de machos e fêmeas nascidos na geração F1 do grupo Tordon 2 .............................................................................. 47
Organograma 8. Representa o número de machos e fêmeas nascidos na geração F2 do grupo Tordon 2 .............................................................................. 48
Organograma 9. Representa o número de machos e fêmeas nascidos na geração F3 do grupo Tordon 2 .............................................................................. 49
Tabela 1. Descreve a análise estatística pelo método x2, feita para avaliar a proporção sexual observada nos grupos experimentais (C, T, TT) ....................... 50
Tabela 2. Apresenta os resultados obtidos em cada um dos parâmetros observados durante o teste no “Open-Field” ......................................................... 57
Tabela 3. Representa as porcentagens da ocorrência, em cada geração, de fêmeas não prenhas ou com filhotes mortos e/ou devorados ............................... 64

ix

x
LISTA DE GRÁFICOS E FIGURAS
página
Gráfico 1. Representação do número de dias que os animais demoraram a nascer, a partir do primeiro dia de acasalamento dos pais ................................... 40
Gráfico 2. Representação da proporção sexual observada durante as três gerações (F1, F2 e F3) dos três grupos experimentais ........................................... 51
Gráfico 3. Representa a curva de peso corporal apresentada pelos filhotes dos grupos C e T, da primeira geração, do 1º ao 101º dia de vida .............................. 52
Gráfico 4. Representa o número de dias que os filhotes dos dois grupos experimentais (C e T) demoraram a desenvolver o reflexo de endireitamento e para abrir completamente a fenda palpebral ......................................................... 54
Gráfico 5. Representa a motricidade dos filhotes quando jovens (com 40 dias) e depois de adultos (com 90 dias), a partir do número de vezes que interrompem feixes fotoelétricos ................................................................................................ 55
Gráfico 6. Representa o equilíbrio apresentado pelos filhotes, quando jovens (com 40 dias) e depois já adultos (90 dias), no teste do “Rota-Rod” .................... 56
Gráfico 7.1. Representa a motricidade espontânea e a completa imobilidade dos animais experimentais em resposta a situação de exposição ao “Open-Field”, por 5 minutos .............................................................................................................. 58
Gráfico 7.2. Representa o tempo de “grooming”, o número de “rearing” ocorridos e o número de bolos fecais excretados durante a exposição ao “Open-Field”, por 5 minutos ................................................................................................................. 59
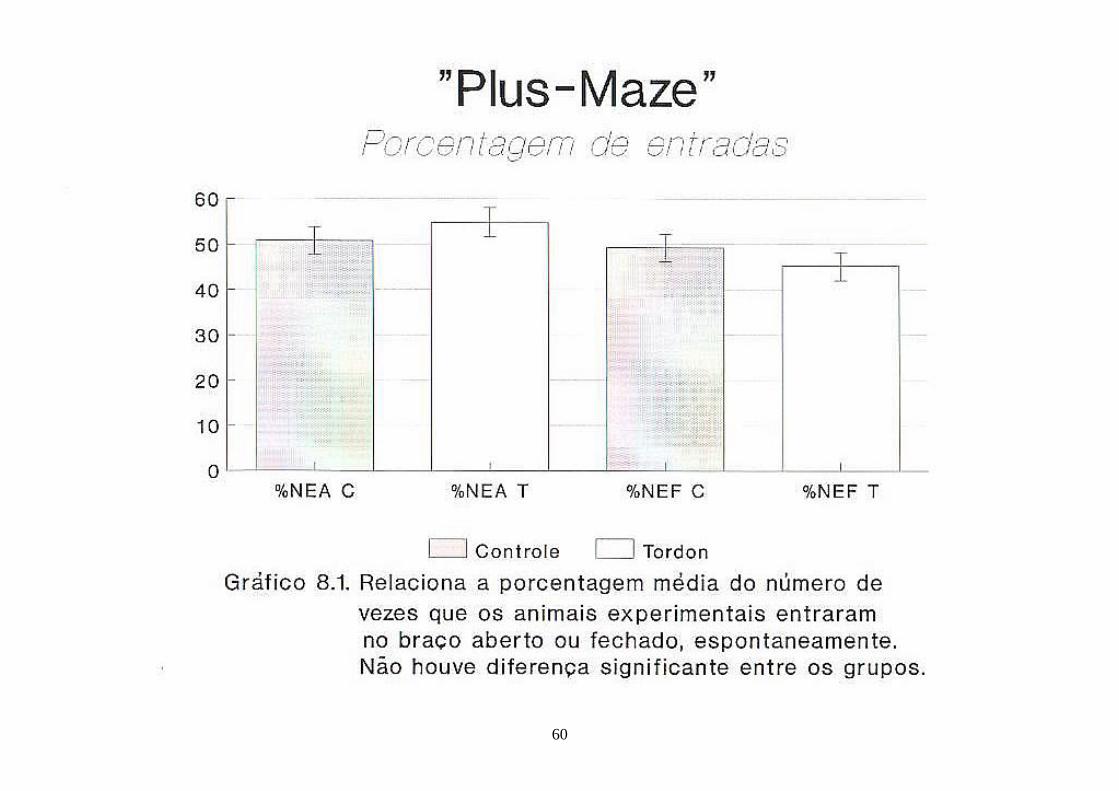
Gráfico 8.1. Representa a porcentagem média do número de vezes que os animais experimentais entraram, espontaneamente, no braço aberto ou fechado durante o teste do “Plus-Maze” ............................................................................. 60
Gráfico 8.2. Representa a porcentagem média de tempo que os animais experimentais permaneceram em cada tipo de braço (aberto ou fechado) durante o teste do “Plus-Maze” .......................................................................................... 61
Gráfico 8.3. Relaciona a proporção de tempo sobre o número de entradas em cada braço (aberto ou fechado), observados durante o teste do “Plus-Maze” ...... 62

xi
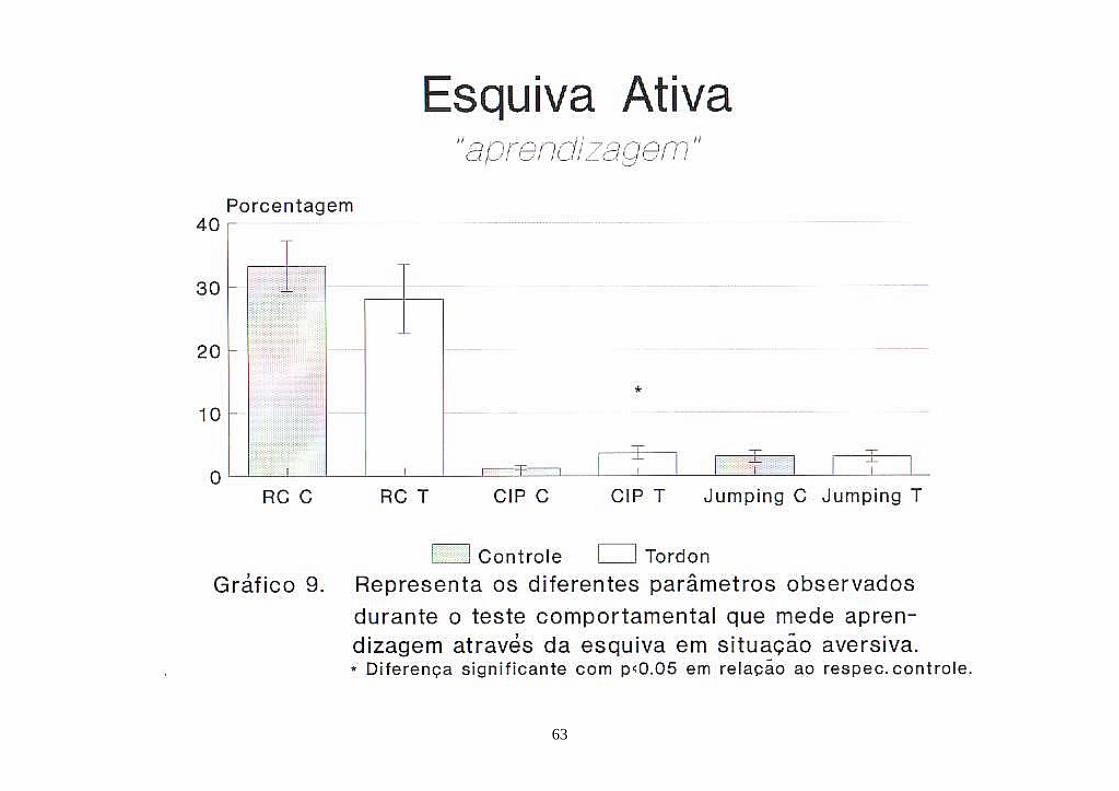
Gráfico 9. Representa os diferentes parâmetros observados durante o teste comportamental que mede aprendizagem através da esquiva em situação aversiva (esquiva ativa) ........................................................................................ 63
Figura 1. Caixa de movimentação espontânea .................................................... 15
Figura 2. Aparelho do “Rota-Rod” ........................................................................ 15
Figura 3. Aparelho do “Open-Field” ...................................................................... 17
Figura 4. Piso do “Open-Field” ............................................................................. 18
Figura 5. Aparelho do “Plus-Maze” ....................................................................... 20
Figura 6. Caixa de esquiva ativa .......................................................................... 22
Figura 7. Epidídimo controle corado por H.E. (200x) ........................................... 35
Figura 8. Epidídimo controle corado por H.E. (400x) ........................................... 35
Figura 9. Testículo controle corado por H.E. (200x) ............................................. 36
Figura 10. Testículo controle corado por H.E. (400x) ........................................... 36
Figura 11. Epidídimo contaminado corado por H.E. (400x) .................................. 37
Figura 12. Testículo contaminado corado por H.E. (400x) ................................... 37
Figura 13. Cérebro controle corado por H.E. (200 e 400x) .................................. 38
Figura 14. Cérebro contaminado corado por H.E. (400x) ..................................... 38
Figura 15. Nitratação de cérebro controle (400x) ................................................. 39
Figura 16. Nitratação de cérebro contaminado (400x) ......................................... 39

xii
ABREVIATURAS
AChE---------------------Acetilcolinesterase
ATPase------------------Adenosina tri-fosfatase
2,4-D ---------------------Ácido diclorophenoxiacético
2,4-D éster-------------Éster do ácido diclorophenoxiacético
2,4,5-T-------------------Ácido 2,4,5-triclorophenoxiacético
C ---------------------Grupo controle
Ca++ ---------------------íon Cálcio
14C ---------------------Carbono radioativo com 14 neutrons
5-HTP---------------------5-hidroxitriptofano
5-HIAA-------------------Ácido 5- hidroxiindolacético
5-HT ---------------------Serotonina
CIP ---------------------Cruzamento Inter-Provas
DA ---------------------Dopamina
DL50 ---------------------Metade da dose necessária para matar o animal
DPC ---------------------Situação”esquiva de duas vias”
F1 ---------------------Primeira geração
F2 ---------------------Segunda geração
F3 ---------------------Terceira geração
GABA---------------------Ácido gama aminobutírico
Hz ---------------------Hertz
mA ---------------------mili-ampére
Na+ ----------------------íon sódio
NE ----------------------Norepinifrina
NEA ----------------------Número de Entradas no braço Aberto
NEF ----------------------Número de Entradas no braço Fechado
P ---------------------Geração parental ou pais
P.A. ----------------------produto químico puro Para Análise
PCPA --------------------p-clorofenilalanina

xiii
RC ---------------------Resposta de Cruzamento
“t” ---------------------Teste estatístico “t” de “student”
T ---------------------grupo 1 contaminado com Tordon
TA ---------------------Tempo de permanência no braço Aberto
TBPS----------------------t-butilbiciclofosforotionato
TF ----------------------Tempo de permanência no braço Fechado
T/NEA---------------------Tempo de permanência dividido pelo Número de
Entradas no braço Aberto
T/NEF---------------------Tempo de permanência dividido pelo Número de
Entradas no braço Fechado
TT--------------------------Grupo 2 contaminado com Tordon
S.N.C.---------------------Sistema Nervoso Central
X2---------------------------Teste estatístico – “Qui-Quadrado”

xiv
RESUMO
Ratos das gerações P, F1, F2, foram contaminados com uma solução
do herbicida Tordon 0,1% na água de beber. O processo de contaminação pelo
Tordon foi efetuado sobre os filhotes, através de suas mães, durante o período de
gestação e amamentação.
Nas três gerações foram quantificados: a proporção sexual em cada
ninhada, número de dias para o nascimento dos filhotes, número de filhotes que
morriam após o nascimento em cada ninhada e em cada grupo experimental.
Na geração F1 foram quantificados o reflexo de endireitamento, o
tempo de abertura palpebral, e o peso corporal, bem como, foram feitos testes
comportamentais (movimentação espontânea, teste do “Rota-Rod”, teste do
“Open-Field”, teste do “Plus-Maze” e esquiva ativa). Após todos estes processos,
foi feito um estudo histológico a nível de sistema nervoso central e de testículos.
Nossos resultados mostraram que a proporção sexual entre os
filhotes de mães contaminadas (1 macho : 1 fêmea) é diferente (p<0,05) da
proporção observada entre os filhotes do grupo controle (6 machos : 5 fêmeas).
As fêmeas contaminadas demoraram mais (p<0,05) para dar cria que as fêmeas
do grupo controle. Os animais contaminados nasceram com uma tendência de
peso corporal menor, que se manteve até, aproximadamente, os 49 dias de idade.
Aos 53 dias de idade o peso corporal destes animais tornou-se maior que os dos
controles, permanecendo assim até a idade adulta. O reflexo de endireitamento
levou mais tempo para ser adquirido no grupo Tordon, enquanto que o tempo de
abertura palpebral é igual nos dois grupos.
Nos testes comportamentais, verificamos que a movimentação
espontânea aos 40 e aos 90 dias de idade é maior no grupo Tordon, sendo que
no grupo controle há um decréscimo e no grupo Tordon um aumento da
movimentação espontânea dos 40 para os 90 dias de idade. Por outro lado, no
teste do “Rota-Rod” não se observou diferença entre os grupos experimentais.
No teste do “Open-Field” os animais contaminados ambularam
menos, tiveram um tempo de “freezing” maior, um número de “rearing-up” menor

xv
e um tempo de “grooming” maior. Em relação aos parâmetros de defecação e
micção, não foram observadas diferenças.
No teste de “Plus-Maze”, os parâmetros %TA e T/NEA foram
maiores no grupo Tordon.
No teste de esquiva ativa os animais Tordon mostraram tendência a
um desempenho maior.
Apesar de todos estes resultados, o material histológico corado com
H.E e/ou impregnado por prata não apresentou nenhuma alteração morfológica,
quer seja a nível de S.N.C. ou a nível de testículos.
Assim, devido a todas as alterações comportamentais observadas,
podemos inferir que o herbicida Tordon contamina os filhotes através da barreira
placentária, hemato-encefáfica e mesmo através do leite na amamentação.
Podendo, com isto, causar alterações funcionais irreversíveis no S.N.C.
Alterações estas que foram observadas na idade adulta do animal, quando foram
feitos os testes comportamentais e não havia contato com o herbicida há, pelo
menos, 40 dias.

xvi
ABSTRACT
Three generation of rats P, F1, F2 was contaminated with “Tordon”
0,1% solution in water drink.
The offspring Tordon contamination was done via their mothers
during pregnancy and lactation.
At the three generation was quantified the sexual ratio in each
progeny; number of days for bird offspring and number of dead offspring in each
experimental group.
The F1 was used for quantified the time to respond stand upright
reflex; eyelid opening time; body weight and behavioral tests (spontaneous
movement, “Rota-Rod” time, “Open-Field” test, “Plus-Maze” test and active
avoidance) and histological studies of C.N.S and testicles.
Our experimental results demonstrated a sexual ratio 1 male : 1
female at the contaminated animal (TA) and 6 males : 5 females in control group
(CA); contaminated females had longer gestational period than the control
females; the (TA) were born with non significant lower body weight in comparison
the (CA). However from 53 days old their average body weight become higher
than the mean of non contaminated group; the contaminated rats were able to
responsed to the stand upright reflex later than (CA) in spite of eyelid opening time
being the same for both groups.
At behavioral tests we have found that : Tordon group spontaneous
movement rate was higher than the (CA) at 40 and 90 days old with a increasing
rate in (TA) and decreasing rate in (CA) between first and second measurement.
The results from “Rota-Rod test” were similar to both groups.
Contaminated animals at the Open-Fields test, ambulated less, had
a higher time of freezing, lower number of rearing-up and a higher time of
grooming. Number of boluses and miction equal to control group.
Plus-Maze test showed higher TA% and T/NEA for (TA), while
active-avoidance test demonstrated a tendency of worse of performance for (TA).
In spite of morphological alteration of C.N.S. and testicles were not
observed at histological examinations, the behavioral changes found led us to
condud that probably Tordon herbicide was able to cross the placentary and
hemato-encephalic barrier and were present at the milk, causing irreversible
functional alterations at C.N.S. that stayed until adult age when the behavior test
occurred.

xvii


1
1.INTRODUÇÃO
Segundo as pesquisas da EMATER do Paraná, na região
metropolitana de Curitiba, apenas 13% dos agricultores utilizam agrotóxicos
seguindo as instruções; 75% não respeitam os períodos de carência; 91% não
usam proteção quando fazem aplicação; 67% poluem rios e córregos e 24%
reutilizam as embalagens para guardar ou transportar alimentos (Tararthuch,
1992).
O Tordon é um agrotóxico organoclorado comumente usado em
sítios e fazendas no norte do Paraná para combater ervas daninhas do pasto e
impedir o crescimento de vegetais de folhas largas (dicotiledôneas). O Tordon
nada mais é do que a associação de dois herbicidas : o 2,4-D (ácido 2,4-
diclorofenoxiacético) e o picloram (ácido 4-amino-3,5,6-tricloropicolínico) e seus
sais de potássio (C6H2Cl3KN2O2), que no entanto, recebem "impurezas" durante
a produção, as chamadas dioxinas. Sua composição age exatamente como o
"Agente Laranja" que por sua ação desfolhante foi utilizado, pelos norte-
americanos, como arma química durante a guerra do Vietnã para descamuflar os
soldados. Esta barbárie produziu inúmeras crianças, com má-formações
congênitas, filhas de pais anteriormente expostos ao agrotóxico (Reis, 1991).
O TORDON é composto pelo 2,4-D, pelo picloram e por resíduos
chamados dioxinas. O 2,4-D é prontamente degradado por hidrólise não
representando um risco de longo prazo ao meio. No entanto, o picloram é
altamente persistente, com resíduos detectáveis no solo até três anos após a
aplicação e mantém sua toxicidade por até cinco anos para certas cepas
suceptíveis (Fao, 1964).
A DL50 do 2,4-D ingerido na comida é de 666 mg/kg para ratos e foi
descrita em 1947 por Hill e Carlisle. Os mesmos autores afirmam que a maior
dose, sem efeitos indesejáveis, para macacos é de 214 mg/kg e, que existe
perigo potencial para a saúde humana através da inalação ou ingestão de
alimentos contaminados.
Entretanto, Gorzinski et al. (1987), trabalhando num laboratório da
DOW CHEMICAL COMPANY, observaram que doses de até 20 mg/kg na dieta
de ratos machos e fêmeas não tiveram efeitos tóxicos observáveis.
Goldstein et al.(1959) afirmam que este herbicida se for usado
deverá sê-lo com grande cautela. Segundo o mesmo autor a DL50 oral do Tordon
22K (25% sal potássio do picloram) em ratos é estimada em mais que 10.000

2
mg/kg, enquanto que do ácido picloram é de 8.000 mg/kg (nível igual ao
encontrado por McCollister & Leng, 1969).
Defendendo-se do crescente mal estar criado pelos casos de
contaminação e intoxicação humanas causados pelo Tordon, a empresa DOW
CHEMICAL (que detém a marca registrada do herbicida) pediu revisão das
informações toxicológicas sobre o produto e concluiu-se que a toxidade é baixa e
não haverá risco algum se o produto for usado como recomendado no rótulo
(Lynn, 1965).
Erne (1966) verificou que, quando administrado oralmente, o
composto é prontamente absorvido e distribuído pelo organismo. Os órgãos mais
contaminados são o fígado, rim, pulmão e baço. Dados corroborados por
Guzelian (1982) que descreveu a mesma ordem indicando níveis decrescentes de
intoxicação e incluiu por último os músculos esqueléticos. No mesmo trabalho o
autor observou que a difusão do 2,4-D no tecido adiposo e S.N.C. foi restrito,
enquanto que ocorreu pronta transferência placentária em suínos e foram
encontradas pequenas quantidades de 2,4-D nos ovos de galinhas. Estes
resultados são concordantes com os de Bernard et al. (1985).
Sabe-se que a maior parte do composto herbicida Tordon é
excretada pelos rins por mecanismo aniônico orgânico que, segundo Berndt
(1973), contribui com sua baixa toxicidade.
No entanto, Tararthuch (1992) observou injúria no processo global
de filtração e caracterizou o Tordon como agente nefrotóxico.
Outrossim, Knopp & Shiller (1992) descreveram o aumento
significante do volume de urina após administração oral de dois sais do 2,4-D.
Knopp (1994) observou que os empregados que trabalhavam na produção e
formulação do herbicida excretavam o 2,4-D na urina de maneira cumulativa, o
qual só não era mais detectado após três semanas de interrupção da exposição.
Os autores encontraram aumento das atividades do citocromo hepático p-450.
Cipriano et al. (1991) verificaram que a ação do Tordon em
macrófagos peritoniais intoxicados "in vitro" provoca intensa vacuolização.
Por outro lado, Paulino & Palermo Neto (1991) verificaram que a
intoxicação aguda pelo 2,4-D altera os níveis de algumas enzimas e componentes
dos soro considerados indicadores de tecido injuriado, refletindo principalmente
prejuízo hepático e muscular induzido pelo herbicida.
Opistótonos (contração tetânica em arco) podem aparecer
momentaneamente em animais intoxicados com 2,4-D, através de uma injeção
subcutânea de 150 a 200 mg/kg, e membros posteriores são comumente mais
nocivamente afetados do que membros anteriores (Bucher, 1946).

3
Goldstein et al. (1959) observaram que apenas algumas horas após
o manuseio do 2,4-D para matar ervas daninhas, três pessoas (dois homens com
52 e 65 anos de idade e uma mulher com 50 anos) foram hospitalizadas com
sintomas progressivos de dor, parestesias e paralisias graves. Têm-se poucas
dúvidas de que a contaminação não tenha ocorrido por absorção dérmica do
produto. A prostração foi grande e a recuperação incompleta mesmo após anos.
O diagnóstico de neuropatia periférica foi suportado por exame eletromiográfico.
Pelletier et al. (1990) verificaram que a aplicação dérmica do 2,4-D
provoca níveis altos do herbicida no sangue e na urina 24 horas após a
exposição. Embora Knopp & Schiller (1992) tenham constatado que a absorção
do 2,4-D por aplicação tópica é muito mais baixa do que a observada na dose
oral.
Schulze & Dougherty (1988) verificaram que a administração diária
do 2,4-D inicialmente deprime a atividade locomotora espontânea em ratos e
desenvolve tolerância após 10 dias de exposição repetida.
No entanto, o tratamento crônico com 2,4-D resulta em miopatias
graves e aparentemente irreversíveis em ratos (Dux et al., 1977) e parece que isto
se deve a uma falha na recaptação de Ca++, pela inibição de um passo na
produção de energia mitocondrial (Guzelian, 1982; Pereira, 1991).
Guzelian (1982) observou que a toxicidade neuromuscular e as
anormalidades neuropsiquiátricas em humanos desaparecem após a remoção do
organoclorado. Seu mecanismo de neurotoxicidade tem sido atribuído à inibição
da membrana mitocondrial e sinaptossomal para sódio-potássio ATPase. E, seu
sítio de ação pode ser a formação reticular mesencefálica.
Pereira (1991) observou que o Tordon atua sobre as enzimas da
cadeia respiratória inibindo de maneira significativa as atividades enzimáticas da
NADH-oxidase e NADH citocromo c redutase, alterando a elasticidade da
membrana mitocondrial interna ou a sua permeabilidade para o íon Na+. Verificou
também que o picloram e o 2,4-D tem efeito inferior ao observado com o Tordon
comercial, reforçando a ação de "impurezas" chamadas dioxinas.
Schulze (1988) observou que diferentes formulações do mesmo
herbicida podem produzir efeitos comportamentais diferentes.
Outros autores verificaram, através de cortes histológicos, que o
picloram é altamente carcinogênico em ratos e camundongos (Reuber, 1981).
Resultados contrários aos de John-Greene (1985) que conduziu um estudo sobre
os efeitos teratogênicos do composto para que os produtos Tordon pudessem ter
seu registro continuado.

4
No entanto, é de conhecimento da população brasileira alfabetizada
(muito provavelmente não agrícola), através da Folha de São Paulo (Reis,1991)
que estudos intensivos estão sendo feitos sobre a carcinogenicidade dos
herbicidas organoclorados que como o Tordon contêm o 2,4-D. Sendo que esta
patologia é mais comumente associada com hormônios, ou seja, o 2,4-D produz
linfomas não Hodgkin e sarcomas de tecido mole em homens (ou ratos e não
produz em ratas) e produz câncer de fígado somente em ratas que produzam
estrogênio (ou seja, não estéreis).
Dados concordantes com os de Ibrahim et al. (1991) que elaboraram
estudos sobre a carcinogenicidade humana e a opinião predominante entre os
cientistas é que há indicação de que a exposição do 2,4-D pode causar câncer
em humanos como doença de Hodgkin do tecido mole e outros linfomas.
Esta opinião está de acordo com os resultados apresentados por
Reis (1991) que ligam o 2,4-D a tumores cerebrais.
Entretanto, Goetz et al. (1994) estudando os efeitos neurológicos do
Agente Laranja (Tordon) sobre os veteranos da guerra do Vietnã, apesar da
fragilidade metodológica, concluíram que não existe evidência de associação
entre exposição ao herbicida e tumores cerebrais.
Tyynela et al. (1990) descrevem a distribuição dos organoclorados
em diversas áreas cerebrais mostrando que apesar do nível relativamente baixo
(5% relativo ao plasma), de acordo com o conceito corrente de transferência de
droga através de membranas corporais (menor solubilidade lipídica das formas
ionizadas - Bernard et al., 1985), o 2,4-D atravessa a barreira hematoencefálica.
Elo et al. (1988) concluíram que todas as doses tóxicas em ratos
causaram injúria nos vasos sangüíneos cerebrais.
Os resultados de Kim et al. (1983) indicam que o 2,4-D é
transportado do fluído cérebro-espinhal através de um sistema ânion orgânico e
que inibidores deste sistema podem bloquear sua eliminação do cérebro "in vivo".
Dados confirmados por Ylitalo et al. (1990).
Lawrence & Casida (1984) afirmam que os organoclorados são
potentes inibidores competitivos estéreo específicos do t-butilbiciclofosforotionato
(TBPS) que ligando-se ao sítio cerebral específico do ácido gama-aminobutírico
(GABA) regulam o canal de cloro.
Enquanto que, Bernard et al. (1985) concluíram que o 2,4-D causa
um decréscimo da atividade da acetilcolinesterase (AChE) no botão sináptico bem
como mudanças comportamentais e eletrofisiológicas. A diminuição da atividade
da AChE pode ser uma pré-etapa no desenvolvimento de miopatias que ocorrem
após grandes doses de 2,4-D.

5
Elo e MacDonald (1989) endossaram a hipótese de que o 2,4-D
inibe o transporte ácido orgânico para fora do cérebro, o que pode resultar em
níveis cerebrais aumentados de metabólitos acidificados de aminas biogênicas, e
assim pode também ter efeitos sobre a atividade de neurônios serotonérgicos e
dopaminérgicos.
Boyd et al. (1990) determinaram alterações no nível colinérgico do
neostriatum de ratos tratados com inseticidas/pesticidas, bem como observaram
diminuição significante na proporção acetilcolina/colina em seus hipocampos.
Da mesma forma, E. de Duffard et al. (1990 - 28) determinaram o
aumento da serotonina (5-HT) e do ácido 5-hidroxiindolacético (5-HIAA) em ratos
machos e fêmeas adultos expostos ao 2,4-D e, a reversão para níveis mais
baixos que o controle quando os animais foram alimentados sem o herbicida.
Oliveira e Palermo Neto (1993), observando os efeitos do 2,4-D
sobre o comportamento no "open-field", verificaram que a ação do herbicida é
contrária ao 5-hidroxitriptofano (5-HTP). Ratos pré-tratados com 5-HTP e p-
clorofenilalanina (PCPA) tiveram diminuídos e aumentados, respectivamente, os
efeitos do 2,4-D dimetil-lamina. O herbicida administrado oralmente diminuiu os
níveis estriatais do 5-HT ( discordante dos resultados de E. de Duffard et al., 28,
com alimentos) e aumentou os níveis do 5-HIAA, sugerindo, assim, que há
modificação da atividade funcional dentro do S.N.C.
Por outro lado, St. Omer & Mohamed (1987) observaram
neurotoxicidade sutil no desenvolvimento de ratos expostos ao 2,4-D pré-
natalmente. Embora, não se observassem sinais tóxicos claros os resultados
indicaram alterações funcionais.
McCollister & Leng (1969) descreveram os resultados de um estudo
sobre a reprodução durante três gerações de ratos alimentados com picloram na
ração. Não encontraram diferença significante entre os grupos controle e
contaminado com relação a viabilidade, sobrevivência ou crescimento dos jovens.
Não encontraram também resíduos do herbicida no leite de vaca, exceto em uma
das três vacas estudadas que apresentou um resíduo de 0,05 ppm quando em
tratamento. Estes dados não correspondem aos encontrados por Guzelian (1982),
segundo o qual em animais de laboratório a função reprodutora bloqueada ou
enfraquecida foi bem documentada. Lagenbach (1989) afirma que "... os
organoclorados, ... , contaminam intensamente a carne, o leite e seus derivados,
... ". Da mesma forma, Fang et al.(1973) encontraram herbicida organoclorado
marcado com 14C em fetos e no leite.

6
Por outro lado, cortes histológicos indicaram que houve atrofia em
testículos e a produção de esperma foi afetada, com espermatozóides anormais
ou "pacotes" em degeneração, bem como a espermatogênese pareceu ser
suprimida (Eroschenko, 1978).
Evangelista de Duffard et al. (1990) descreveram o papel da
testosterona e/ou sua ausência exercendo influência sobre os efeitos tóxicos do
2,4-D em testes comportamentais.
Vin' et al.(1990) descreveram a ação do 2,4-D sobre a função
reprodutiva de ratas que resultaram em certos distúrbios no ciclo estral, bem
como em um aumento significativo de morte intra-uterina com tratamentos agudos
e crônicos. Várias aberrações cromossomais foram identificadas (resultados estes
que estão de acordo com os dados de Adhikari & Grover, 1988).
Blakley et al. (1989) observaram que a exposição pré-concepcional e
gestacional de ratas ao Tordon 202c acarreta a incidência de má-formações fetais
(palato-fendido, agenesia renal, hidronefrose, agenesia unilateral dos testículos e
hérnia umbilical) e fetos com variantes (especialmente ossificação incompleta do
esqueleto). Chernoff et al., 1.990, citam costelas supernumerárias. Chernoff et al.
(1990) e Blakley et al. (1989) concordam que nas dosagens mais tóxicas o peso
materno foi diminuído e/ou a letalidade materna aumentada. Chernoff et al. (1990)
afirmam ainda que houve diminuição do peso fetal e alargamento da pelve renal e
que o composto 2,4-D foi escolhido porque estudos prévios indicavam baixa ou
nenhuma toxicidade para o desenvolvimento de animais. Blakley et al. (1989)
sugerem que a exposição preconcepcional e gestacional ao Tordon 202c provoca
efeitos teratogênicos e de diminuição do crescimento fetal. Sugere, porém, que
somente exposição no período preconcepcional não é suficiente para somatizar
efeitos embriotóxicos. Este resultado está de acordo com os resultados de
Evangelista de Duffard et al. (1990 - 27) que sugerem que a exposição pré-natal
não tem nenhuma influência sobre a condição pós-natal dos neurotransmissores.
O fator que nos levou a fazer este trabalho foi o fato de que todas as
pessoas (trabalhadores rurais ou não) contaminadas ou intoxicadas pelo herbicida
Tordon apresentaram neuropatias periféricas, confusão mental e até esterilidade
presumida em homens (ou seja, número de espermatozóides zerado).

7
2. OBJETIVOS E JUSTIFICATIVAS
Tendo em vista que os agrotóxicos desenvolvidos para atender as
necessidades dos países desenvolvidos chamados de "primeiro mundo" são
manuseados pelas pessoas menos esclarecidas a seu respeito (mesmo em seus
países de origem) e que por isso são responsáveis pelos inúmeros "acidentes" de
intoxicação e contaminação dos seres vivos e de seu meio ambiente (Lagenbach,
1989), escolhemos um modelo experimental que mimetizasse um nível de
contaminação que não é considerado tóxico e que pode ocorrer normalmente,
principalmente no meio rural.
Este trabalho tem por objetivo estudar os efeitos da contaminação
materna (durante o período de acasalamento, gestação e lactação) com o
herbicida "Tordon 2,4-D 64/240 trietanolamina BR" sobre o desenvolvimento do
Sistema Nervoso Central (S.N.C.) dos filhotes.
Foi escolhido este esquema experimental porque, o mesmo,
compreende a embriogênese e a ontogênese do S.N.C. Os resultados
comportamentais aqui apresentados foram realizados para verificar se ocorre
passagem do herbicida através da placenta e/ou leite e/ou barreira
hematoencefálica.
Neste trabalho também foram avaliados os efeitos do herbicida Tordon sobre a
função reprodutora dos animais experimentais, bem como a proporção de filhotes
machos e fêmeas nascidos em cada ninhada de cada grupo experimental.

8

9
3.MATERIAIS E MÉTODOS
3.1. LOCAL DE TRABALHO
Os Experimentos foram realizados no Laboratório de Fisiologia e
Farmacologia do S.N.C., do Departamento de Fisiologia da UFPr, por apresentar
as condições mínimas de manutenção da temperatura constante para os animais
experimentais, bem como locais adequados para os testes comportamentais.
O material para as lâminas histológicas foi preparado em parte nas
dependências do Departamento de Fisiologia e em parte no Laboratório Central
do Departamento de Morfologia - Biologia Celular da UFPr.
3.2. ANIMAIS EXPERIMENTAIS
Os animais experimentais utilizados foram ratos Wistar (ratos
brancos de laboratório - Ratus ratus, variante albino), obtidos do Biotério do Setor
de Ciências Biológicas da UFPr e/ou de outras instituições através do técnico
responsável - Cândido J. T. Pereira.
3.3. HERBICIDA
O herbicida usado na contaminação dos grupos experimentais é o
agrotóxico comercial "TORDON 2,4-D 64/240 trietanolamina BR"
O TORDON apresenta a seguinte composição: sal trietanolamina do
ácido 4-amino 3,5,6-tricloropicolínico (picloram, sal trietanolamina) 103 g/l.
Equivalente ácido do picloram :64 g/l.
Sal trietanolamina do ácido 2,4-diclorofenoxiacético (2,4-D, sal
trietanolamina) 406 g/l.
Equivalente ácido do 2,4-D: 240 g/l.
O picloram é derivado do ácido picolínico e o 2,4-D do grupo dos
fenoxiacéticos.

10
O picloram é o composto ativo e o 2,4-D se apresenta comumente
como éster no Tordon.
Optou-se pela contaminação crônica oral com uma dose
considerada sem efeito tóxico, propriamente dito, no período reprodutivo do
animal. Em doses maiores o sabor e o odor impedem que o animal beba
normalmente.
A contaminação dos animais ocorre através da água de beber na
proporção de 1 (um) ml do agrotóxico para 999 ml de água (0,1% V/V), durante o
período de acasalamento, gestação e lactação.
Nascidos os filhotes, o trabalho prosseguiu somente com os machos
que, a partir do desmame, receberam água igual a do grupo controle.
3.4. METODOLOGIA
3.4.1. Acasalamento
O período de acasalamento ideal é de dez dias. No entanto, por
medida de precaução, o tempo de acasalamento no nosso experimento foi de
vinte dias.
Acasalaram-se dois grupos experimentais em um total de dez
fêmeas e dois machos. Sendo um macho e cinco fêmeas em cada caixa. Um dos
grupos foi o controle (C) e o outro foi contaminado por Tordon na água de beber
(grupo Tordon 1 ou T).
3.4.2. Nascimento dos filhotes
Verificada a proporção sexual obtida nos dois grupos experimentais
da primeira etapa, propusemo-nos a fazer novos acasalamentos observando-se a
proporção durante três gerações consecutivas (F1, F2 e F3). Onde F1 é a
primeira geração nascida de mães contaminadas, F2 são os filhos de F1 e, F3
são os filhos de F2, sendo que nas três gerações as mães do grupo T foram
contaminadas da mesma maneira com Tordon.

11
Para se chegar a segunda e terceira gerações foram acasalados
machos e fêmeas filhos de mães contaminadas da geração anterior. O modelo
experimental do acasalamento evitou cruzamento consangüíneo.
Observando-se, ainda, a diferença estatística do peso obtido a partir
do 53o dia nos filhotes de mães T em relação aos controles, optou-se por um novo
grupo experimental. Este novo grupo (Tordon 2 ou TT) foi igualmente
contaminado pelo herbicida Tordon diferenciando-se apenas o tempo de
exposição das fêmeas ao agrotóxico. A contaminação começou aos 60 dias de
idade, ou seja, 30 dias antes do período de acasalamento e terminou com o
desmame dos filhotes.
Os grupos C e T foram acasalados quando os animais se
apresentavam em idade de maturidade sexual (entre 90 e 100 dias) nas três
gerações.
Em todos os grupos observou-se também, o número de dias para o
nascimento dos filhotes a partir do primeiro dia de acasalamento; o número de
fêmeas que procriaram, a proporção macho/fêmea de cada grupo e a mortalidade
de animais durante o período de contaminação.
3.4.3. Peso Corporal
Os filhotes machos, apenas da primeira geração dos grupos C e T,
foram pesados individualmente, a partir do primeiro dia após o nascimento até o
101o dia de idade, sendo que a pesagem foi feita de quatro em quatro dias.
Desta maneira os animais são pesados no dia seguinte ao
nascimento (1o dia), no 5o dia, no 9o dia e assim sucessivamente até o 101o.
Cada grupo experimental foi pesado com a mesma idade (em número de dias)
para posterior comparação estatística entre os grupos experimentais.
Outro fator observado, dado o ciclo vital do rato Wistar e como este
animal dorme durante o dia e desperta à noite, foi manter fixo o período de
manuseio e pesagem dos animais das 14 as 18 horas, visando com isso interferir
o mínimo possível em seu comportamento devido ao stress, ou mesmo, para se
ter uma melhor avaliação com relação à proporção idade/peso do animal. Dentro
deste contexto todos os comportamentos quantificados foram verificados no
mesmo período.
3.4.4. Reflexo de Endireitamento

12
O reflexo de endireitamento é um teste que nos dá parâmetros de
como o Sistema Nervoso Central (S.N.C.) está quanto ao seu desenvolvimento
pós-natal.
Consiste em se colocar o animal em decúbito dorsal e verificar
quanto tempo o mesmo demora para pôr-se na posição normal, ou seja, em
decúbito ventral. Se o reflexo for imediato, ou seja, seu tempo de incursão for
menor que um segundo, considera-se reflexo adquirido. Se as ninhadas do grupo
contaminado adquirirem o reflexo no mesmo número de dias que o grupo
controle, pode-se inferir que o desenvolvimento do S.N.C. foi igual nos dois
grupos.
Considera-se que todos os filhotes da mesma ninhada devem
adquirir o reflexo no mesmo dia.
3.4.5. Abertura Palpebral
Outro modo de se verificar o desenvolvimento do animal é pela
observação do número de dias decorridos do nascimento até a abertura palpebral
completa dos filhotes. Como no ítem anterior é importante anotar a idade da
ninhada, para isso todos os filhotes devem estar com as pálpebras
completamente abertas.
3.4.6. Sexagem
Ocorre de 5 a 7 dias após o nascimento. Observa-se o número de
filhotes e a proporção macho/fêmea em cada ninhada. Permanecem com a mãe
até o desmame apenas machos ou apenas fêmeas, mantendo-se no máximo 6
filhotes por mãe. Apenas os machos, por terem um ciclo reprodutivo constante e
contínuo durante toda a vida, serão utilizados nos testes comportamentais e
histológicos. Foram utilizados um número mínimo de 15 animais em cada
experimento comportamental.
Os filhotes fêmeas serão matrizes para um novo acasalamento e
obtenção de uma geração posterior, durante o qual seu processo reprodutivo será
observado.

13

14
3.4.7. Testes Comportamentais
3.4.7.1. MOVIMENTAÇÃO ESPONTÂNEA
Este experimento nos dá uma idéia da tendência natural do animal
para a atividade exploratória. Consiste em quantificar a atividade motora do
animal dentro de uma caixa de movimentação espontânea durante cinco minutos.
A contagem é feita por um aparelho eletrônico, acoplado à caixa, que marca
quantas vezes o animal interrompe feixes fotoelétricos quando cruza o assoalho.
Estes feixes são emitidos por três pares de células fotoelétricas dispostas
paralelamente e distantes 15 mm do nível do assoalho da caixa. Localizam-se nas
paredes anterior e posterior.
Esta caixa possui 50,0 cm de largura, 20,5 cm de profundidade e
22,3 cm de altura (medidas internas). As paredes laterais e de fundo são
formadas por alumínio. O teto da caixa é feito do mesmo material e possui
dobradiças para que se abra como porta. O chão é formado por barras paralelas
de quatro milímetros de diâmetro que servem de apoio ao animal não permitindo
que suas excreções se acumulem no interior da caixa. A parede anterior é
formada por acrílico de cor fumê que permite a observação do animal. O ambiente
é mantido o mais silencioso possível e a luz externa não é intensa.
3.4.7.2. TESTE DO "ROTA-ROD"
Este experimento determina a capacidade de equilíbrio do animal.
Rota-rod é um aparelho que consiste em uma barra giratória de 72
cm de comprimento e 2,8 cm de diâmetro, estando elevada a 37 cm da base e
que possui uma rotação de 12 rpm.
Liga-se o aparelho e coloca-se o animal, pela cauda, de modo que
suas patas toquem a barra. Solta-se o animal e cronometra-se o tempo que
conseguir permanecer na barra. O experimento é repetido por três vezes,
somando-se o tempo total de permanência do animal na barra.
Os testes comportamentais de Movimentação Espontânea e no
"Rota-Rod" são feitos aos 40 e aos 90 dias de idade. Os aparelhos utilizados
nestes testes podem ser visualizados na figura 1 e figura 2, respectivamente.

15
FIGURA 1. Mostra um animal experimental dentro da caixa de
Movimentação Espontânea. O reflexo, proporcionado pelo "flash" da máquina
fotográfica, no piso da bancada revela a estrutura do assoalho da caixa deste
modelo experimental.
FIGURA 2. Ilustra o aparelho do "Rota-Rod" com um animal
experimental.

16

17
3.4.7.3. EXPOSIÇÃO AO "OPEN FIELD"
Este teste avalia a reatividade emocional do animal quando exposto
a aversividade do campo aberto.
Sobre o "Open-Field" encontram-se 4 lâmpadas de 60 wats cada e
um auto falante que produz um ruído contínuo e monótono de 78 decibéis que
deixa camuflados outros possíveis ruídos estranhos ao experimento.
O ambiente do "Open-Field" é separado do observador através de
uma cortina, conforme figura 3, que permite a quantificação dos comportamentos
do animal em experimentação.
FIGURA 3. Ilustra o aparelho do "Open-Field".

18
O campo aberto "Open-Field" consiste em uma arena de fundo
redondo com 80 cm de diâmetro e parede lateral de 30 cm de altura. O piso do
aparelho é dividido em raios e círculos concêntricos de maneira a formar figuras
geométricas semelhantes que possuem relativamente a mesma área, conforme
fig.4. Cada figura geométrica é considerada um "quadrado".
FIGURA 4. Ilustra o piso da arena do "Open-Field" com um animal
experimental em posição de "rearing-up´.
Este teste comportamental compreende a observação de vários
parâmetros, tais como ambulação, "freezing", "rearing-up", "grooming", defecação
e micção, durante cinco minutos de experimentação.

19
a. Ambulação
Observa-se quantos "quadrados" foram "invadidos" pelo animal com
pelo menos as duas patas anteriores.
b. Tempo de "Freezing"
Observa-se o tempo de imobilização total do animal, com pelos
eriçados, olhos fixos e orelhas orientadas em dada direção.
c. "Rearing-up"
Observa-se a atitude exploratória do animal contando-se o número
de vezes que explorou o ambiente levantando-se e permanecendo apoiado
apenas sobre as patas posteriores, conforme figura 3.
d. Tempo de "Grooming"
Observa-se o comportamento deslocado do animal. Em ratos
cronometra-se o tempo em segundos que o animal passa se "limpando".
e. Número de bolos fecais
Conta-se, neste tipo de observação, o número de bolos fecais
excretados pelo animal durante os cinco minutos de teste no Open-Field.
f. Micção
Neste parâmetro foi observada a presença ou ausência de urina na
arena após completado o teste de Campo Aberto.

20
3.4.7.4. TESTE DO "PLUS-MAZE" OU LABIRINTO EM "X":
Teste experimental que quantifica medo ou ansiedade, através da
exposição do animal a espaços abertos e fechados.
O "Plus-Maze" é um labirinto em X elevado 50 cm do chão, que
possui dois braços abertos (como prancha de navio pirata) e dois braços fechados
(com "paredes" em suas laterais e em uma de suas extremidades). Os braços se
intercomunicam por uma das extremidades de maneira a formarem um X quando
observados de cima. Cada braço mede 50 cm de comprimento por 10 cm de
largura, e as "paredes" do braço fechado medem 40 cm de altura. Não há parede
nas laterais do quadrado central (10x10 cm) que faz a intercomunicação entre os
braços. Os braços fechados não possuem "teto", permitindo assim a observação
do animal, conforme ilustrado na figura 5.
FIGURA 5. Mostra os quatro braços do "Plus-Maze" e um animal
experimental explorando um deles.

21
Este experimento é realizado em um ambiente com isolamento
acústico da mesma maneira que os outros dois testes comportamentais: "Open-
field" e Esquiva Ativa. Utiliza-se luz vermelha para que o observador possa
observar sem ser observado.
O animal é colocado no quadrado central e observa-se (durante
cinco minutos) sua atividade exploratória pelos braços. Conta-se uma entrada em
cada braço cada vez que o animal fizer uma "invasão" com, no mínimo, as duas
patas dianteiras e, cronometra-se o tempo de permanência durante cada
"invasão". O quadrado central é considerado neutro e o tempo de permanência
neste lugar não é computado.
Segundo Sharon Pellow mede-se a porcentagem do número de
entradas (% de NEA ou % de NEF) e a porcentagem do tempo de permanência
do animal em cada braço (% de TA ou % de TF).
Numa tentativa de melhor evidenciar o medo ou ansiedade dos
animais em relação ao braço aberto, os nossos dados foram computados
verificando-se a relação T/N (tempo de permanência sobre número de entradas)
que nos mostra como o animal entra no braço aberto ou fechado e não apenas o
quanto entra.
3.4.7.5. ESQUIVA ATIVA
Teste experimental que mede o aprendizado do animal. Aprendizado
este que consiste em o animal associar o estímulo incondicionado do som da
campainha com o comportamento de cruzar a caixa (uma "shüttle-box") para se
esquivar do choque elétrico nas patas.
Todas as observações comportamentais deste teste foram feitas
numa "shüttle-box" de alumínio de 50x25x25 cm, pintada com esmalte cinza,
exceto em sua parte anterior que é de vidro transparente. Isto possibilita a
observação do animal durante o experimento. Presa ao teto da linha média da
caixa há uma lâmpada de 10 w e, em situação oposta à lâmpada, do lado externo
da caixa, encontra-se uma campainha. O piso da caixa é formado por uma grelha
metálica, constituída por barras de bronze de 2 mm de diâmetro, separadas entre
si por uma distância de 7 mm. Na linha média do piso, separando o lado direito do
esquerdo da caixa, há uma trave de alumínio de 0,5x0,5x25 cm pintada com
esmalte cinza e colocada no mesmo nível das barras de bronze. Esta "shüttle-
box" está ilustrada na figura 6.

22
FIGURA 6. Mostra a atividade exploratória de um animal
experimental dentro de uma "shüttle-box" utilizada no teste de Esquiva Ativa. Ao
lado visualiza-se o aparelho que produz a estimulação sonora e elétrica.
No teste de Esquiva Ativa o animal experimental é colocado dentro
da caixa de condicionamento onde permanece de 5 a 10 minutos para que possa
explorar livremente a caixa e se habituar a ela. Após este período de tempo inicial
na "shüttle-box", os animais ouvem toques de campainha com 5 seg de duração e
recebem choques elétricos de 1,5 mA, 60 Hz, durante 2 seg, nas patas. Os
intervalos entre os toques de campainha variam aleatoriamente entre 10 e 40
segundos.
Esta situação experimental que constitui o teste DPC, ou situação de
"esquiva ativa de duas vias" típica, consiste de 50 toques de campainha de 5 seg
de duração pareados ao choque nas patas. Caracterizando, assim, contigüidade
de estímulos ou "pareamento", como na situação experimental do paradigma
clássico ou pavloviano. Nas provas em que a campainha provoca uma resposta
de cruzamento (de um lado ao outro da caixa), o animal não recebe choque nas
patas. "Prêmio" por haver aprendido a se esquivar.

23
Na esquiva ativa observam-se os seguintes parâmetros:
a. Resposta de Cruzamento (RC)
Indica o desempenho do animal na "shüttle-box" em termos de
comportamento aprendido. Consiste em quantificar o número de cruzamentos que
ocorrem na "shüttle-box" (de um lado ao outro) dentro do intervalo de cinco
segundos de duração da campainha.
b. Cruzamentos Inter Provas (CIP)
Índice que indica ansiedade. Consiste no número de cruzamentos
que ocorrem na "shüttle-box" sem estimulação externa, ou seja, o animal cruza a
"shüttle-box" no período entre as provas.
c. "Jumping"
Número de saltos que ocorrem durante o tempo de estimulação
sonora (no caso a campainha). O "jumping" nos fornece um índice de reatividade
emocional do animal.
d. Orientação
Neste parâmetro observa-se se o animal procura identificar a origem
do som, ou seja, verifica-se em cada prova se ele está escutando a campainha.

24
3.5. ESTATÍSTICA
Todos os resultados foram expressos através da média ± erro
padrão da média.
Nas observações comportamentais que foram quantificadas com
grandezas paramétricas foi feita a avaliação estatística pelo teste "t" de "student",
tais como na avaliação da variação do peso corporal, número de dias para o
reflexo de endireitamento, número de dias para a abertura da fenda palpebral e
nos parâmetros observados nos testes de Movimentação Espontânea, "Rota-
Rod", "Open-Field" (exceto micção), "Plus-Maze" e Esquiva Ativa. Foi aplicado ,
também, o teste "t" para avaliar o número de dias que os filhotes levaram para
nascer a partir do primeiro dia de acasalamento dos pais.
Os dados não paramétricos foram analisados pelo teste do "Qui-
Quadrado", como a proporção sexual observada, outras observações (fêmeas
estéreis, filhotes mortos/devorados e mães mortas) e o parâmetro micção do teste
do "Open-Field", que também foi analisado pelo teste de FISHER.
3.6. MORFOLOGIA
Foram feitos cortes histológicos do S.N.C. e testículos de 10 filhotes
machos da geração F1 do grupo C e 15 do grupo T, que já contavam com 110
dias de idade e que eram filhos de mães diferentes em ambos os grupos.
Utilizou-se o médoto de perfusão para a retirada dos órgãos, da
seguinte maneira:
Os animais foram anestesiados com éter etílico, via inalação, e
imobilizados em mesas cirúrgicas próprias. Os tecidos abdominais foram
rebatidos e presos por pinças de maneira a permitir a invasão da caixa toráxica e
exposição do coração. Imediatamente o ventrículo esquerdo recebeu cerca de
200 ml de solução salina, (através de uma agulha butterfly, cânula e seringa)
aproveitando as contrações do músculo cardíaco para "limpar" os vasos
sangüíneos dos órgãos em questão. Evitando-se, assim, material desnecessário
na lâmina histológica. A aurícula direita recebeu um pique para que o sangue que
vinha da grande circulação extravasasse. Quando o líquido que jorrava da
aurícula direita tornou-se transparente considerou-se que os vasos sangüíneos
estavam "limpos".

25
Em seguida, através do mesmo processo com outra seringa, os
tecidos foram perfundidos com cerca de 200 ml de formalina (solução de formol a
40%) para que os tecidos fossem sendo fixados o mais breve possível. Como a
partir da invasão toráxica o animal já é considerado morto, não se utiliza mais
anestésico e o tremor observado, bem como a rigidez da cauda e da nuca, é sinal
característico da ação da formalina nos tecidos.
Sendo assim procedeu-se a retirada dos testículos, por simples
desalojamento da bolsa escrotal, e do cérebro, por decaptação da cabeça e
abertura - extremamente cuidadosa - da caixa craniana e das meninges. Os
órgãos foram, então, colocados em um recipiente contendo formalina e após uma
hora de fixação foram seccionados. O cérebro foi dividido em três partes com
cortes frontais: cérebro anterior, diencéfalo e tronco cerebral. Os testículos foram
divididos em três partes por cortes transversais ou duas partes por um corte
longitudinal.
Prosseguiu-se com a metodologia corrente de fixação, emblocagem,
corte, desidratação, diafanização e coloração dos tecidos com hematoxilina-
eosina em algumas das peças, e nitratação em outras.
3.6.1.Coloração por H.E.:
Fixação em formol 40%.
Emblocagem em parafina com 3% de cera de abelha.
Cortes histológicos - A única diferença metodológica foi o corte
seriado do cérebro, do qual aproveitavam-se dez fatias e desprezavam-se vinte,
totalizando, de cinco peças, cerca de quinhentas lâminas aproveitáveis.
Desidratação em soluções com concentração alcóolica crescente -
álcool 70%, álcool 90%, álcool I PA., álcool II PA., ácool III PA.
Diafanização com xilol.
Método corrente de coloração por H.E.
Montagem em lâmina e lamínula com bálsamo do Canadá.
3.6.2. Nitratação
Utilizou-se o método do nitrato de cobalto de DA FANO, descrito por
Beçak & Paulete (1970).

26
Fixação em nitrato de cobalto, impregnação em nitrato de prata
(1,5%), redução na mistura de Ramón y Cajal, desidratação, inclusão e corte.
Virar os cortes por meio de cloreto de ouro (0,2%), fixar em
hipossulfito de Na+ (5%) e montar em lâmina e lamínula com bálsamo da Canadá.
Além de evidenciar os corpos dos neurônios, esta técnica cora em
marrom o Complexo de Golgi.
3.6.3. Evidenciação da bainha de mielina
A técnica LUXOL FAST BLUE - LEVAFIX RED VIOLET para
evidenciação das células gliais e da bainha de mielina, descrita por Vacca (1985),
resulta na mielina corada em azul, grânulos em neurócitos (parecidos com
corpúsculo de Nissl), proteínas e corpúsculos de Nissl em vermelho, musculatura
lisa de vasos sangüíneos normais em vermelho violeta e musculatura lisa de
vasos sangüíneos com fibrose média ou espessamento da íntima, bem como o
colágeno, não se coram.
O método de PTAH modificado para demonstração da glia em tecido
do S.N.C., descrito pela mesma autora acima, tem como resultado a coloração da
glia e mielina em azul, das fibras musculares normais em vários tons de azul e do
músculo degenerado em tons do púrpura ao vermelho. Esta técnica usa uma
grama de IODINE.
A demonstração da mielina degenerada, para animais que não têm
contato com o herbicida a mais de 60 dias como no nosso experimento, pode ser
feita por algumas técnicas.
Como o tempo após o término da exposição é grande (mais de 60
dias), as células fagocitárias provavelmente terão removido a mielina degenerada.
A demonstração positiva pelo método clássico de Marchi - de 1895 (descrito no
Cel. Pat. Tech.) ou de 1935 (descrito por Conn, 1962),ou pelo método de Swank-
Davenport de 1935 (descrito por Conn, 1962 e no Cel. Path. Tech.), resultam na
precipitação do tetróxido de ósmio apresentando mielina degenerada e gorduras
neutras (ou seja, tecido adiposo) na cor preta.
Outro método de demonstração positiva é a tecnica do LUXOL FAST
BLUE-OIL RED, que resulta na coloração azul-esverdeada da mielina normal e
coloração vermelha da mielina degenerada. A coloração por OIL RED não é tão
intensa como quando se faz a contra-coloração com CRESIL VIOLET (Cel. Path.
Tech.).

27
4.RESULTADOS
4.1. DIAS DE ACASALAMENTO E GESTAÇÃO
Um ciclo estral completo varia de 3 a 5 dias numa fêmea adulta (90
dias de idade) de ratos Wistar (ratos brancos de laboratório - Ratus ratus variante
albino), portanto um período de dez dias seria o ideal para o acasalamento. No
entanto, verificamos que enquanto para animais controle este tempo é suficiente,
para animais contaminados pelo Tordon chegou a dobrar (26 dias) e mesmo
assim houveram fêmeas que não tiveram filhotes. O período de gestação normal,
reconhecido por todos os que trabalham com reprodução de ratos, é de 21 dias.
Conforme gráfico 1.
4.2. PROPORÇÃO SEXUAL
As porcentagens da proporção sexual observada em filhotes de
ratas controle (C) é de 56,18±2,039 machos (n = 249) para 43,82±2,039 fêmeas
(n = 196) na geração F1 (p<0,05), 57,02±2,381 machos (n = 269) para
42,93±2,384 fêmeas (n = 202) na geração F2 (p<0,05) e 57,16±2,232 machos (n
= 334) para 42,84±2,232 fêmeas (n = 251) na geração F3 (p<0,01).
Nas ninhadas das mães contaminadas por Tordon (T) verificou-se,
em três gerações consecutivas, a proporção de um macho para uma fêmea.
Conforme demonstrado nos organogramas 4-9 e no gráfico 2.
O teste do "X2" ("qui-quadrado") confirmou a diferença significante
observada no grupo controle, conforme tabela 1.
4.3. PESO CORPORAL
Os animais filhos de mães T nasceram com peso levemente inferior
ao observado no grupo C. Esta tendência permaneceu até o 53o dia, quando
então aumentaram significativamente seu peso corporal (p<0,05) em relação aos
controles. Observou-se que a diferença de peso corporal entre os dois grupos
continuou aumentando na idade adulta (p<0,01).
Estes resultados estão expressos no gráfico 3.

28
4.4. DESENVOLVIMENTO PÓS-NATAL
4.4.1. Reflexo de Endireitamento
Nos animais controle o tempo médio de endireitamento foi de
4,22±0,15 dias que é significantemente menor (p<0,05) que o tempo médio de
endireitamento dos animais filhos de mães contaminadas (T): 4,65±0,07 dias,
descritos no gráfico 4.
4.4.2. Abertura Palpebral
Os animais controle levaram em média 14,51±0,08 dias para a
abertura completa das pálpebras e os animais filhos de mães T levaram
14,6±0,07 dias, com p>0,05. Dados descritos no gráfico 4.
4.5. TESTES COMPORTAMENTAIS AOS 40 DIAS DE IDADE
4.5.1. Movimentação Espontânea
Animais controle apresentaram em média 129,911,96 cruzamentos
na caixa de movimentação espontânea, enquanto que os animais filhos de mães
T apresentaram em média 158,2617,27 cruzamentos, com p>0,05.
4.5.2. "Rota-Rod"
Os animais controle conseguiram permanecer na barra giratória do
"rota-rod" em média por 26,6±3,46 segundos, e os animais filhos de mães T
permaneceram 29,78±2,92 segundos. Tempos estatisticamente iguais (p>0,05).

29
4.6. TESTES COMPORTAMENTAIS AOS 90 DIAS DE IDADE
4.6.1. Movimentaçao Espontânea
Neste teste verificou-se que os animais controle apresentaram um
número de cruzamentos médio de 119,8612,5, enquanto que os filhos de mães
T apresentaram um número de cruzamentos de 169,3818,9, estatisticamente
significante com p<0,05.
Os resultados da movimentação espontânea observada estão
descritos no gráfico 5.
4.6.2. "Rota-Rod"
Da mesma maneira que aos 40 dias o equilíbrio dos animais foi
quantificado na idade adulta.
Os animais controle ficaram em média 50,577,39 seg na barra
giratória do "rota-rod", enquanto que os animais filhos de mães T permaneceram
45,813,79 seg em média, com p>0,05.
Não houve diferença significante entre os grupos, conforme gráfico
6.
4.6.3.Exposição ao "Open-Field"
a. Ambulação
Os animais controle apresentaram ambulação média de 57,055,87
"quadrados" invadidos, enquanto que os filhos de mães T apresentaram
43,003,99 "quadrados" invadidos. Diferença estatisticamente significante com
p<0,05, conforme tabela 2 e gráfico 7.1.
b Tempo de "Freezing"
Os animais controle permaneceram em média 91,5212,00 seg
"congelados", enquanto que os filhos de mães contaminadas permaneceram

30
119,339,53 seg "congelados", embora, de um modo geral, não se
apresentassem com os pêlos eriçados. Dados com uma significância estatística
de p<0,05, conforme tabela 2 e gráfico 7.1.
c. Número de "Rearings"
Os animais controle apresentaram uma média de 16,812,04
"rearings", enquanto que os filhos de mãe T apenas 12,671,47 "rearings",
diferença estatisticamente significante com p<0,05. Conforme tabela 2 e gráfico
7.2.
d. Tempo de "Grooming"
Nos animais controle observou-se que os mesmos apresentaram um
tempo de "grooming" de 11,831,86 segundos e os animais filhos de mães T
apresentaram 19,251,74 segundos, com p<0,05. Conforme tabela 2 e gráfico
7.2.
e. Número de bolos fecais
Para os animais controle obteve-se uma média de 5,710,64 bolos
fecais e para os filhos de mães contaminadas 5,051,96 bolos fecais em média,
com p>0,05. Conforme tabela 2 e gráfico 7.2.
f. Micção
Nos animais controle verificou-se que 90,4% do total apresentaram
micção e nos filhos de mães T 85,7% do total . Resultado considerado
semelhante estatisticamente, pelo teste de FISHER e do "qui-quadrado".
Conforme tabela 2.

31
4.6.4. Teste do "Plus-Maze" ou Labirinto em "X"
No braço aberto do "Plus-Maze", os animais controle apresentaram
uma média de 50,763,04% de NEA (número de entradas no braço aberto) e
12,182,08% de TA (tempo de permanência neste braço). Por sua vez, os
animais filhos de mães T apresentaram uma média de porcentagem de
54,783,21 entradas no braço aberto (% de NEA) e ali permaneceram
22,944,5% do tempo total em segundos. Para % de TA houve diferença
significante com p<0,05.
Por sua vez, no braço fechado os animais C apresentaram
49,243,04% do total de entradas (% de NEF) e os filhos de mães T entraram
45,223,21% do total. Os animais controle permaneceram 754,02% do tempo
total em segundos no braço fechado (% de TF), enquanto que os filhos de mães
contaminadas permaneceram 65,764,96% do tempo total em segundos. Para %
de TF houve diferença significante com p<0,05.
Os animais C tiveram T/NEA igual a 5,510,95 e os filhotes do grupo
T 8,641,44, com p<0,05.
Os animais C apresentaram T/NEF igual a 48,048,26 e os animais
filhos de mães T 71,0820,46. Diferença não significante.
Os parâmetros % de NEA, % de NEF e T/NEF não apresentaram
diferença significante, com p>0,05.
Enquanto que, os parâmetros % de TA, % de TF e T/NEA
apresentaram significância estatística com p<0,05.
Todos os dados do Plus-Maze estão representados nos gráficos 8.1,
8.2 e 8.3.
4.6.5. Esquiva Ativa
a. Resposta de Cruzamento (RC)
A resposta de cruzamento dos animais controle foi de 33,24±4,01%
do total de testes e dos animais filhos de mães T foi de 28,00±5,46%, com
p>0,05.

32
b. Cruzamento Inter-Provas (CIP)
Os animais controle apresentaram 1,22±0,34% de CIP e os
descendentes de mães T 3,63±1,04%. Dados considerados não diferentes
estatisticamente, com p<0,05.
c. "Jumping"
Animais controle e filhos de mães Tordon 1 apresentaram a mesma
porcentagem de saltos, a saber 3,10±0,98% para os controle e 3,07±0,96% para
o segundo grupo. Houveram animais do grupo contaminado que saltaram durante
e após a aplicação do estímulo sonoro.
d. Orientação
Foi 100% positiva para os dois grupos.
Todos os resultados do teste de esquiva ativa estão representados
no gráfico 9.
4.7. FÊMEAS QUE NÃO SE APRESENTARAM PRENHAS
No grupo controle foram observadas 138 fêmeas, das quais
nenhuma deixou de dar cria.
No grupo Tordon 1 observou-se que de 55 fêmeas 10 não tiveram
filhotes. Originando uma média de 18,2% de nuliparidade nas três gerações
consecutivas.
No grupo Tordon 2 a média observada foi de 19,7% de nuliparidade,
ou seja, de 61 fêmeas 12 não tiveram filhotes. Estas fêmeas foram acasaladas
novamente por igual período e mesmo assim 4 delas (acasaladas com machos
diferentes) permaneceram sem filhotes.
Dados observados na tabela 3.

33
4.8. FILHOTES MORTOS/DEVORADOS
Nenhum dos 1506 filhotes do grupo controle foi encontrado morto,
semi-devorado ou dado por desaparecido.
Entretanto, dos 469 filhotes do grupo T 71 (15%) morreram, dos
quais 34 (63%) desapareceram (devorados pela mãe).
E, no grupo TT observou-se que dos 498 filhotes 29 (6%) morreram,
dos quais 16 (55%) desapareceram (devorados pela mãe ).
Provavelmente a mãe devora ou deixa morrer os filhotes não aptos,
como normalmente ocorre em outra espécies de mamíferos (gato e outros felinos,
cão,coelho etc.)
4.9. OUTRAS OBSERVAÇÕES
A contaminação crônica com Tordon foi o suficiente para fazer com
que algumas ninhadas perdessem mais de 70% da pelagem, da mesma forma
que suas mães. Duas dessas mães apresentaram emagrecimento grave e dorso
exageradamente arqueado, deixando seus filhotes com aparente desnutrição. A
partir do desmame, quando não receberam mais o Tordon, os filhotes foram
gradativamente recuperando a pelagem até sua aparente totalidade (em torno do
65o dia).
Quase metade das fêmeas contaminadas, com ou sem filhotes após
o período de gestação, apresentaram-se agressivas ao manuseio.
Comportamento não observado nas fêmeas controle.
Em nossos experimentos, observamos ainda que os ossos dos
crânios dos filhotes do grupo contaminado apresentaram menor resistência ao
manuseio que os do grupo controle. Da mesma forma, Guzelian (1982) afirma que
a firmeza e espessura da casca dos ovos de aves se apresentaram reduzidas.
Das 37 mães do grupo Tordon 1, observou-se que uma da primeira
geração, sem filhotes, e outra da segunda geração, com 11 filhotes, não
conseguiam se levantar nem se alimentar e acabaram morrendo.
Estes animais não passaram pelos testes comportamentais.
Os dados dos ítens 4.8 e 4.9 estão descritos na tabela 3.

34
4.10. MORFOLOGIA
4.10.1. Coloração com Hematoxilina-Eosina (H.E.):
Não foram encontradas diferenças morfológicas nos cérebros e na
espermatogênese (testículos e epidídimos), dos animais dos dois grupos (C e T),
preparados com esta técnica. Conforme figuras 7 a 14.
4.10.2. Nitratação:
Não foram encontradas diferenças morfológicas nos cérebros, dos
animais dos dois grupos (C e T), preparados com esta técnica. Conforme figuras
15 e 16.
4.10.3. Evidenciação da bainha de mielina degenerada:
Não foi possível evidenciar se houve degeneração na bainha de
mielina pela dificuldade de encontrar os corantes CRESIL VIOLET, IODINE,
LUXOL FAST BLUE, OIL RED específico e RED VIOLET e, quando encontrados,
pela dificuldade financeira de aquisição, bem como pela demora da chegada dos
produtos.
Os métodos de simples evidenciação da bainha de mielina com
ácido ósmico, nitratação e outros não são capazes de diferenciar a mielina normal
da degenerada.

35
FIGURA 7. Ilustra epidídimo de animal normal com epitélio íntegro, em aumento
de 200 vezes ao M.O.
FIGURA 8. Ilustra epidídimo de animal normal com epitélio íntregro e estereocílios
(seta menor), em aumento de 400 vezes ao M.O.

36
FIGURA 9. Ilustra túbulo seminífero de testículo normal com núcleo de fibroblasto
(seta menor) e tecido intersticial (seta maior) bem evidentes
(aumento de 200 X aoM.O.).
FIGURA 10. Ilustra a formação de espermatozóides em túbulo seminífero normal
com espermatócito primário (seta curta curta) e espermátide (seta mais
longa) indicados(aumento de 400 X ao M.O.)
A letra "c" indica Célula de Sertoli.

37
FIGURA 11. Ilustra epidídimo de animal contaminado por Tordon evidenciando
estereocílios (seta menor) em epitélio íntegro.
Aumento de 400 vezes observado ao microscópio de luz.
FIGURA 12. Ilustra túbulo seminífero de animal contaminado por Tordon com
núcleo de fibroblasto (seta menor) e espermatogônia (seta maior)
evidentes (400 X ao M.O.).
A espermatogênese não foi alterada de maneira quantitativa.

38
FIGURA 13. Ilustra cérebro de animal controle corado por H.E. em aumento de
400 vezes (13.a) e de 200 X (13.b).
FIGURA 14. Ilustra cérebro de animal contaminado por Tordon corado por H.E.
(400 X). Com esta técnica não foi possível evidenciar nenhuma alteração
morfológica.

39
FIGURA 15. Ilustra cérebro de animal controle impregnado pelo nitrato de
cobalto, com oligodendrócitos bem evidentes, em aumento de 400 vezes.
FIGURA 16. Ilustra cérebro de animal contaminado por Tordon, com
oligodendrócitos bem evidentes, impregnado pelo nitrato de cobalto
(400 X). Não se observou alteração morfológica com esta técnica.

40

41

42

43

44

45

46

47

48

49

50

51

52

53

54

55

56

57

58

59
60

61

62
63

64

65

66
5. DISCUSSÃO
Nos últimos anos uma quantidade considerável de trabalhos têm
sido publicada na imprensa científica a respeito do TORDON e/ou seus
componentes : o 2,4-D (normalmente encontrado na forma de éster neste
composto), o picloram e as dioxinas.
As dioxinas e o picloram já foram muito bem estudados, sendo que o
picloram (2,4,5-T) está proibido nos E.U.A. desde 1985 (Reis, 1991) e no Brasil
desde 1984. A legislação brasileira proibiu os organoclorados (compostos
orgânicos que contém moléculas de cloro e atuam no fator de crescimento dos
vegetais - Ferri, 1986) desde 1979, liberando-os de forma restrita em 1984. Por
isso, o 2,4-D continua sendo empregado sem restrições em quase todos os
países. Mas o Tordon que contém todos os componentes acima, a rigor estaria
proibido por lei, mas continua sendo comercializado.
Apesar dos registros de doses letais, inclusive em seres humanos,
todos os autores pesquisados relataram reversibilidade dos efeitos tóxicos
causados pelo herbicida. Em alguns casos há regeneração total dos tecidos
afetados, porém, em outros tais como no trabalho de Goldstein et al (1959) que
relata neuropatia periférica (sistema nervoso e tecido muscular) ocorrida por
absorção dérmica, o tecido muscular esquelético foi regenerado, mas a
neuropatia (tremores e estados alucinatórios) permaneceu mesmo após anos de
tratamento; no trabalho de Eroschenko (1978) que relata atrofia de testículos,
alguns animais conseguiram regenerar todo o tecido do aparelho reprodutor,
enquanto que outros permaneceram com os testículos e seus dutos severamente
afetados, prejudicando ou abolindo completamente sua capacidade reprodutiva;
no trabalho de Lerda & Rizzi (1991) que relatam astenospermia, necrospermia e
teratospermia em trabalhadores rurais, o número e quantidade dos
espermatozóides voltou ao normal e não se encontraram mais espermatozóides
mortos, mas a grande quantidade de espermatozóides anormais permaneceu
mesmo após anos de não contato com o herbicida; e ainda, no trabalho de E. de
Duffard et al (1990-27) que observaram efeitos permanentes nos níveis de
neurotransmissores cerebrais, ocorridos durante o desenvolvimento cerebral de
ratos nascidos de mães contaminadas durante gestação e lactação.
Tomando por base estes dados da literatura podemos inferir que as
alterações somáticas e comportamentais verificadas em nossos experimentos

67
estão relacionadas com alterações a nível de sistema nervoso, de sistema
reprodutor e, possivelmente a nível cromossomal.
Os efeitos do Tordon em nossos animais experimentais nos mostrou
que este herbicida tem efeito tóxico sobre a fertilidade dos ratos, tanto em fêmeas
como em machos. De tal maneira que filhotes de mães contaminadas nascem
mais tardiamente (p<0,01) que os animais controle. Estes dados concordam com
os trabalhos de Guzelian (1982) e Vin' et al (1990) que dizem "que a
administração crônica do 2,4-D resulta em certos distúrbios no ciclo estral,
manifestado como um prolongamento da fase diestro e mudanças na duração do
estro e meta-estro, bem como em um aumento na proporção de ciclos
anuvolatórios". Deste modo, os autores acima concluem que herbicidas
organoclorados têm "efeito tóxico sobre os sistemas endócrinos e reprodutivos",
tais como número de espermatozóides baixo ou ausente, baixa porcentagem de
espermatozóides móveis, efeito indireto no sistema pituitário hipotalâmico, pela
liberação do hormônio folículo estimulante (FSH) e prevenindo a liberação de
hormônios luteinizantes, com fortes evidências de efeitos estrogênicos diretos.
Por outro lado, Reuber (1981) observou atrofia de testículos de ratos
e camundongos tratados com picloram.
Entre os autores pesquisados acima é consenso geral que os efeitos
dos herbicidas organoclorados mimetizam a situação de excesso de estrogênio.
O tempo que os filhotes do grupo TT levaram para nascer (tempo
entre o acasalamento e o nascimento) diminuiu significativamente (p<0,05) no
decorrer das três gerações, tendendo a voltar ao mesmo tempo do grupo controle,
indicando que houve tolerância ao Tordon. Isto provavelmente é devido a um
processo de seleção natural, onde aqueles que se reproduzem conseguem
eliminar mais rapidamente os compostos herbicidas de seu organismo,
diminuindo seus efeitos tóxicos.
Esta nossa observação está de acordo com o trabalho de Schulze &
Dougherty (1988) que afirmam a ocorrência de desenvolvimento de tolerância ao
herbicida 2,4-D éster após dez dias de exposição.
Por outro lado, observou-se que cerca de 20% das fêmeas
acasaladas, nos dois grupos contaminados, não tiveram filhotes mesmo depois de
um longo período de acasalamento (26 dias) com vários machos. Estes dados
estão de acordo com os resultados de Guzelian (1982). Sendo que nenhuma das
fêmeas do grupo controle, acasalada por dez dias, deixou de ter filhotes.
Além disso, observou-se que a proporção sexual das ninhadas do
grupo controle foi diferente (p<0,05) da proporção obtida nas ninhadas dos grupos
contaminados.

68
Eroschenko (1978) apresentou resultados expressivos da ação do
herbicida organoclorado sobre a reprodução de pombas japonesas. Tais como
mudanças estruturais nos testículos e seus dutos seminíferos (testículos
aumentados com túbulos seminíferos dilatados e epitélio germinativo
severamente dilacerado e/ou testículos atrofiados com túbulos seminíferos
reduzidos e espermatogênese suprimida, onde o epitélio germinativo apresentava
espermatozóides anormais). Conclue o autor que, embora se observassem
reparações estruturais na maioria dos testículos e dutos de alguns animais,
"prejuízos permanentes verificados em testículos e dutos seminíferos" de outros
animais limitavam severamente, se não aboliam por completo, sua capacidade
reprodutiva.
Da mesma forma Lerda e Rizzi (1991), que usaram o teste YFF -
que identifica espermatozóides com dois cromossomos Y devido a não disjunção
meiótica, observaram níveis significantes de astenospermia (redução do número e
motilidade dos espermatozóides), necrospermia (espermatozóides mortos) e
teratospermia (espermatozóides anormais) em trabalhadores rurais que usavam o
herbicida. Embora a astenospermia e a necrospermia diminuíssem com o passar
do tempo, a quantidade de espermatozóides anormais permaneceu. Lembrando
que "a motilidade e a quantidade de espermatozóides normais (número de
espermatozóides por ml de ejaculado) são os dois melhores indicadores de
fertilidade, o último sendo um indicador de toxicidade espermatogênica em
humanos". As autoras demonstraram, assim, que o 2,4-D influencia nocivamente
a fertilidade em machos a partir de um efeito tóxico sobre o epitélio germinativo
que causa alterações na espermatogênese.
Guzelian (1982) estudando os efeitos do 2,4-D afirma que "em
animais de laboratório a função reprodutora bloqueada ou enfraquecida foi bem
documentada". Afirma , também, que em aves, o número e o tamanho dos ovos
se apresentou reduzido.
Vin' et al. (1990) afirmam que a administração crônica do 2,4-D em
ratas "aumenta a proporção de morte intra-uterina, principalmente por causa de
morte pós-implantacional de embriões".
Os dados observados no nosso grupo controle nos levaram a
proporção de 6 machos para cada 5 fêmeas, proporcionalmente diferentes
(p<0,05) entre si, enquanto que, nos grupos contaminados (T e TT) o número de
machos é estatisticamente igual ao número de fêmeas. Todos estes dados
levantados em relação à contaminação pelo Tordon nos leva a inferir que este
herbicida provavelmente ativa ou inativa algum fator seletivo em detrimento do
número de machos, ou a favor do número de fêmeas, nascidos vivos. Estes

69
resultados concordam com Cavalli-Sforza & Bodmer (1971) que descrevem que
as proporções sexuais obtidas em pássaros é diferente da proporção esperada de
1 macho para 1 fêmea, assim como observado também na espécie humana, onde
o número de meninos nascidos é maior do que o número de meninas.
Observando-se que morrem mais meninos durante o primeiro ano de vida e, não
havendo nenhuma guerra ou catástrofe, a proporção sexual tenda a se igualar em
idade adulta.
Em nossos experimentos também observamos que, nos grupos
contaminados, entre 6% e 15% dos filhotes morreram logo após o nascimento.
Dos quais mais da metade foram devorados por suas mães. Provavelmente
repetindo o gesto instintivo, facilmente observado em outras espécies, de devorar
ou deixar morrer os filhotes doentes ou não aptos.
Neste mesmo sentido, Guzelian (1982) observou que os animais
tratados com 2,4-D nascem com peso fetal menor e sua sobrevivência pós-parto
é reduzida e Blakley et al. (1989) observaram aumento da incidência de fetos
mortos/reabsorvidos e fetos mal formados.
Duas das fêmeas contaminadas (com dose de 0,1%) do grupo T
morreram. A primeira sem filhotes, a segunda uma semana após dar cria. Blakley
et al. (1989) e Chernoff et al. (1990) tiveram resultados semelhantes quando
investigaram a toxicidade maternal induzida quimicamente, pois verificaram que o
2,4-D reduz de maneira significante o peso maternal e produz mortalidade das
fêmeas que receberam o composto do 6o ao 15o dia de gestação.
Isto pode explicar porque algumas das fêmeas perderam peso a
ponto de deixar à mostra a coluna vertebral bem arqueada, como se estivessem
em constante posição de ataque, apresentando certa dificuldade de locomoção.
Phyllis et al. (1985) descrevem a diminuição "dramática" da atividade
locomotora espontânea causada pelo tratamento agudo com 2,4-D que diminui a
atividade da acetilcolinesterase (AChE) no botão sináptico, causando "latência
motora distal prolongada".
Outros autores (Goldstein et al., 1959; Dux et al., 1977; Guzelian,
1982; Pereira, 1991) documentam bem a ação do 2,4-D causando miopatias
graves a partir da inibição do potencial elétrico da membrana, diminuindo a
recaptação de Ca++ e aumentando sua concentração intracelular. Ou seja, o
músculo se contrai e, devido a ação direta do 2,4-D sobre a cadeia respiratória
(Pereira, 1991), a mitocôndria sem capacidade de manter os níveis normais de
recaptação de Ca++ (Guzelian, 1982) tem uma diminuição dos níveis de ATP, o
que impede a descontração das fibras musculares. Este processo é dependente
de Ca++ e ATP. Isto pode levar ao extremo de uma paralisia muscular (tetania)

70
seguida de morte (Berwick, 1970) ou apenas opistótonos (contração tetânica em
arco) momentâneos (Bucher, 1946).
Logo após o nascimento dos filhotes verificamos que o
desenvolvimento do S.N.C. foi prejudicado nos recém-nascidos do grupo
contaminado. Evidenciando que houve algum tipo de injúria a nível neural ou
muscular ocasionada pela contaminação com Tordon através do leite e/ou
placenta. Uma vez que os filhotes não têm contato direto com o herbicida em
nenhuma etapa de sua vida.
Estes resultados estão de acordo com Lagenbach (1989) que afirma
que os organoclorados "contaminam intensamente a carne, o leite e seus
derivados", bem como as gorduras animais que concentram alto teor de
organoclorados. Tudo isto poderia servir de contaminação para o ser humano no
período de gestação e amamentação, tal como em nosso experimento.
Muito embora, McCollister & Leng (1969) e Erne (1966) afirmem o
contrário quando dizem que nenhum resíduo do fenoxiherbicida Tordon pôde ser
detectado no leite, o próprio Erne (1966) conclue que houve pronta transferência
placentária em porcas.
Além disso, com um método de marcação radioativa (com 14C) Fang
et al. (1973) puderam detectar o picloram em fetos e no leite, concluindo que a
meia vida do herbicida em ratos adultos é de 3,4 hs e no recém-nascido é de 97
horas.
Com base neste dados, podemos afirmar que os filhotes são
realmente contaminados pela mãe através da placenta e do leite.
Embora a pronta transferência placentária tenha sido comprovada
(Erne, 1966; Guzelian, 1982), apenas a exposição pré-natal ao Tordon (Blakley et
al., 1989) ou a um de seus componentes (E. de Duffard et al., 1990 - 28) não é
considerada suficiente para causar algum prejuízo neurotóxico.
No entanto, efeitos permanentes do herbicida foram estabelecidos
durante a exposição pré- e pós-natal (E de Duffard et al., 1990 - 27). Por isso,
apesar de se considerar que o 2,4-D éster é lentamente absorvido pelo organismo
(Gering, 1973),seus efeitos fetotóxicos evidenciaram a vulnerabilidade do período
de desenvolvimento pré- e pós-natal, incluindo proliferação e desenvolvimento
das células que formam a mielina, pela redução desta bainha na medula espinhal,
cerebelo e tronco cerebral em pintainhos recém-eclodidos de ovos pré-tratados
com o herbicida (Duffard et al., 1987).
Nossos animais experimentais apresentaram uma curva de peso
corporal diferente da observada nos animais controle. Embora não significante,
esses animais nasceram e permaneceram com o peso levemente abaixo do

71
verificado no grupo controle até o 53o dia de idade. Daí em diante, os filhotes
machos do grupo contaminado apresentaram um peso corporal maior (p<0,05)
que os filhotes machos do grupo controle que tinham a mesma idade.
Considerando-se que a função gonadal se torna mais ativa ao redor
deste período, que seria o período de puberdade para o animal, estes resultados
sugerem uma deficiência gonadal nos animais contaminados durante gestação e
lactação. Ou seja, estes animais seriam hipogonádicos ou pseudo
gonadectomizados. O que justificaria o peso corporal aumentado, sendo que, com
o passar dos dias a diferença entre os dois grupos tornou-se maior (p<0,01).
Entretanto, Bucher (1946) já tinha observado um leve retardo no
crescimento de ratos jovens e associou este fato a redução da ingestão de
alimento.
Ratos tratados com 2,4-D apresentam um decréscimo na
capacidade reprodutiva quando adultos (Guzelian, 1982) e a presença de
testosterona é importante para que se evidenciem os efeitos tóxicos do 2,4-D
éster (E. de Duffard et al., 1990 - 28).
Há também relatos na literatura ligando o 2,4-D a tumores cerebrais
somente em ratos e não em ratas (Reis, 1991). Isto mostra um efeito associativo
da testosterona e a toxicidade do Tordon.
Outros efeitos tóxicos foram observados no nível dos
neurotransmissores. St. Omer & Mohammad (1987) observaram níveis
aumentados de dopamina (DA) e nor-epinifrina (NE) no período pós-natal de ratos
contaminados, em relação aos controles.
Do mesmo modo, foram encontradas altas concentrações cerebrais
de serotonina (5-HT) e do ácido 5-hidroxi-indolacético (5-HIAA) em ratos adultos
nascidos de mães tratadas com 2,4-D éster durante gestação e lactação (E. de
Duffard et al., 1990 - 27). Sabe-se que este período de contaminação produz,
segundo os autores, efeitos permanentes nos animais.
Um dos efeitos permanentes observados em nossos animais
experimentais foi o aumento da motricidade. Sabe-se que todo animal novo
movimenta-se mais do que o adulto da mesma espécie. Entretanto, nosso
animais experimentais não só movimentavam-se mais quando jovens, em relação
aos controles da mesma idade, como movimentavam-se mais quando adultos do
que quando jovens. Dado este sustentado, também, pelo maior número de
cruzamentos inter-provas observados do teste de esquiva ativa.
Mohammad & St. Omer (1988) descrevem a ação do Tordon
mudando os níveis cerebrais de glutamato, proteínas, ácidos nucleicos e GABA
(ácido gama-aminobutírico). Estes autores afirmam que os herbicidas

72
organoclorados são potentes inibidores competitivos estéreo-específicos do t-
butilbiciclofosforotionato (TBPS), indicando interferência no sistema gabaérgico
que regula o canal de cloro (Lawrence & Casida, 1984).
Os autores acima concluem que as maiores classes de herbicidas
organoclorados agem de maneira similar, "isto é, como antagonistas não
competitivos do GABA-A", sendo esta sua base de ação convulsivante (sistema
neuromuscular) em mamíferos.
A função muscular e o sistema de equilíbrio são bem evidenciados
no teste do "Rota-Rod". Em nossos animais experimentais não se observou
diferença significante no tempo de permanência no "Rota-Rod" em relação aos
controles, aos 40 e aos 90 dias de idade. Os mesmos resultados foram
encontrados por Schulze & Dougherty (1988). Sugerindo com isso que não houve
alteração muscular ou de equilíbrio. No entanto, o teste de movimentação
expontânea mostrou que há exacerbação da movimentação nos animais
contaminados maternalmente. Isto indica que áreas do S.N.C., que não as do
equilíbrio, podem estar alteradas.
Já em ambiente aversivo, no teste do "Open-Field", os animais
contaminados através das mães tiveram a sua atividade locomotora deprimida,
aumentando o tempo parado (que nestes animais pareceu mais ser
catatonia/apatia do que medo ou pavor). Resultados semelhantes aos
encontrados por Schulze (1987 e 1988) e Schulze & Dougherty (1988).
Todos os parâmetros observados no "Open-Field" que tiveram
resultados estatisticamente diferentes (p<0,05) revelaram detrimento da atuação
os animais contaminados. Tais como, ambulação diminuída, "freezing"
aumentado e "rearing" diminuído, indicando que estes animais exploraram menos
o ambiente, bem como, tempo de "grooming" aumentado indicando que estes
animais têm uma reação emocional aumentada. Todos estes resultados sugerem
que a contaminação pelo Tordon produz uma reatividade emocional mais intensa
que o observado nos animais controle, embora, o número de bolos fecais e a
porcentagem de micção dos animais contaminados tenham sido considerados
estatisticamente iguais ao observado nos animais controle.
Oliveira & Pallermo Neto (1993) obtiveram resultados semelhantes
com a administração oral aguda de 2,4-D em ratos. Este tratamento foi suficiente
para diminuir a locomoção e freqüência de "rearings", bem como para aumentar a
duração da imobilidade. Estes resultados são iguais aos encontrados em nosso
experimento, os quais se mantêm na idade adulta dos animais contaminados.
Os autores acima verificaram ainda que o herbicida diminuiu os
níveis estriatais de 5-HT (resultados contrários aos encontrados por E. de Duffard

73
et al., 1990-27, em ratos adultos contaminados pela mãe) e aumentou os níveis
do 5-HIAA estriatal e do tronco cerebral (resultados semelhantes aos encontrados
por E. de Duffard et al., 1990-27, em ratos adultos contaminados pela mãe).
No teste do "Plus-Maze" observou-se que os animais contaminados,
através da mãe, permaneceram mais tempo no braço aberto (%TA), embora
tenham entrado o mesmo número de vezes que os animais controle. Isto nos leva
a levantar duas hipótese : ou os animais contaminados são menos medrosos (ou
ansiosos) na situação do "Plus-Maze" ou estes animais têm um prejuízo da
percepção dos estímulos deste teste. Esta segunda hipótese se baseia no fato de
que o animal nascido de mãe contaminada não só entra no braço aberto e
permanece mais tempo lá, como também chega até a borda do braço aberto, olha
para baixo, continua andando e cai. Isto poderia estar relacionado ao fato deste
mesmo animal apresentar menor atividade exploratória ("rearing") no "Open-
Field".
No teste de esquiva ativa nossos animais experimentais
apresentaram o mesmo desempenho que os controles. Sua orientação também
apresentou-se normal.
Embora, estes resultados indiquem aparente normalidade, nossos
animais experimentais mostraram uma tendência de menor desempenho na
aquisição e retenção da aprendizagem na esquiva ativa. Por outro lado,
apresentavam maior ambulação (CIP), cruzando a caixa na ausência de
estímulos externos, indicando claramente que há prejuízo a nível de S.N.C. (nos
processos de inibição comportamental, que permitiriam ao animal estabelecer
associação entre os estímulos da esquiva ativa). Este prejuízo se expressa na
vida adulta dos animais, embora sua contaminação tenha ocorrido apenas
durante o desenvolvimento embrionário e na lactação.
Tyynela et al. (1990) conseguiram verificar, por marcação do
herbicida com 14C, que os dois compostos que compõem o Tordon (2,4-D e
picloram) realmente atravessam a barreira hemato-encefálica por difusão (e
outros mecanismos ainda não elucidados) e se distribuem de maneira semelhante
no cérebro. Isto reforça a nossa hipótese de prejuízo cerebral nos animais
contaminados.
Sabe-se que o Tordon é lipossolúvel em concentrações baixas e
solubiliza-se em pH 7.0 (Profa. Dra. Orieta Silveira). A concentração utilizada em
nosso experimento é baixa (0,1%).
Baker (1984) observou a interação dos organoclorados com a
proteína heme do sangue. Elo et al. (1988) chegaram a verificar injúria cérebro-
vascular causada pelo 2,4-D.

74
Com isso, e apesar de ser considerada restrita a infiltração do
herbicida no tecido adiposo e dentro do S.N.C. (Erne, 1966), Lagenbach (1989)
encontrou alto teor de organoclorados no tecido adiposo animal. Lembrando, que
a bainha de mielina é formada essencialmente por várias camadas sobrepostas
da bicamada lipídica que forma a membrana celular.
Isto sugere que há saturação do composto dentro dos tecidos e
concorda com os resultados obtidos sobre as mudanças ocorridas na
permeabilidade e fluidez da membrana plasmática (Pritchard et al., 1982 e Videla
et al., 1990), onde a interação dos pesticidas organoclorados (Antunes-Madeira et
al., 1981) com a bicamada lipídica podem formar radicais livres, intermediários
que ligando-se a lipídeos ou proteínas interfeririam na fluidez da membrana
(Baker & Dyke, 1984; Baker et al., 1985 e Videla et al., 1990).
Esta mudança de fluidez e permeabilidade inibe o transporte de
elétrons (Pritchard et al.,1982) e promove a perda da capacidade que a
mitocôndria tem de manter níveis normais de recaptação de Ca++ (Dux et al.,
1977 e Guzelian, 1982) pela inibição de um passo no processo de produção de
energia mitocondrial (Pereira, 1991). Assim, há menos energia disponível para as
atividades comuns a cada tipo de célula.
Um neurônio que apresente um aumento de Ca++ citoplasmático
teria despolarização da membrana neuronal, promovendo desligamento de
neurotransmissores (Guzelian, 1982). Tendo diminuída sua ATPase mitocondrial
haveria menor quantidade de energia disponível para a recaptação de
neurotransmissores. Em conseqüência, a quantidade e freqüência do potencial
sináptico ficariam alteradas, se não bloqueassem o próprio potencial de ação.
A toxicidade neuromuscular do 2,4-D é primariamente pós-juncional
por natureza (Guzelian, 1982). Uma vez que é a recaptação dos
neurotransmissores que está diminuída ou inibida, causando "diminuição da
AChE no botão sináptico" (Bernard et al., 1985) e aumento de sua concentração
na placa motora, torna-se evidente que as mudanças eletrofisiológicas levem a
mudanças comportamentais.
Hrdina et al. (1971) acreditam que os efeitos neurotóxicos dos
herbicidas organoclorados envolvem mecanismos colinérgicos do núcleo estriado.
A acetilcolina (ACh) é secretada entre outros pelos motoneurônios que inervam os
músculos esqueléticos, tendo geralmente efeito excitatório. Isto explicaria a
movimentação exagerada do animal contaminado maternalmente.
Além de afetar o sistema colinérgico, outros autores sugerem
interferência na ação de neurônios dopaminérgicos e serotonérgicos (Elo &
MacDonald, 1989;), bem como modificação na maturação do sistema

75
serotonérgico (E. de Duffard et al., 1990 - 27). Isto está de acordo com os
resultados que obtivemos da observação dos filhotes contaminados que
apresentaram retardo no desenvolvimento pós-natal do S.N.C., atividade
neuromuscular aumentada no correr do seu desenvolvimento e percepção
alterada para vários estímulos em idade adulta.
A recaptação do 2,4-D em neo-natos ocorre contra um gradiente de
concentração (Kim et al., 1983), portanto com gasto de energia (que já está
debilitada), causando saturação do composto nos tecidos fetais incluindo o
cérebro (Kim et al., 1983; Schulze & Dougherty, 1988 e E de Duffard et al., 1990 -
27). O 2,4-D inibe o transporte ácido orgânico para fora do cérebro (Elo &
MacDonald, 1989) e isto pode explicar o aumento da concentração de alguns
neurotransmissores e seus metabólitos.
De acordo com Gonzalez et al.(1994) serotonina e testosterona
têm efeito oposto sobre o comportamento sexual em machos. Se isto realmente
ocorre pode-se inferir que o aumento da 5-HT em ratos adultos contaminados, na
vida intra-uterina e na lactação, diminua a ação da testosterona, e estes animais
passem a apresentar comportamento de animal gonadectomizado (castrado). Isto
explica o aumento de peso iniciado na fase de puberdade nos nossos animais
experimentais, persistindo na idade adulta.
Apesar de todo comportamento alterado, o animal contaminado
apresentou, a nível morfológico, espermatogênese quantitativamente normal. No
entanto, sabemos que, qualitativamente, sua espermatogênese está modificada,
possivelmente apresentando alto nível de teratospermia (produção de
espermatozóide anormal), descrita por Lerda & Rizzi (1991), uma vez que se
verificou alteração no processo reprodutivo destes animais.
Da mesma forma, o tecido nervoso não apresentou alterações
estruturais, quando corado com H.E. e quando impregnado por prata (ao M.O.),
que justificassem as reações do animal contaminado nos testes comportamentais.
Entretanto, se as alterações funcionais observadas a nível de
sistema nervoso e sistema reprodutor não causaram alterações morfológicas que
pudessem ser evidenciadas pelas técnicas por nós utilizadas para microscópio de
luz, outras técnicas mais específicas podem, possivelmente, vir a evidenciar as
alterações funcionais já descritas.
Assim, apesar de alguns autores acharem que o Tordon e/ou seus
componentes apresentam baixa (McCollister & Leng, 1969; Berndt &
Koschier,1973 e Hayes et al., 1986) ou nenhuma (John-Greene et al., 1985)
toxicidade, inclusive embriológica, os nossos dados mostram que os efeitos deste
herbicida podem ser devastadores. Tendo deprimido o medo ou a função do

76
sistema de inibição comportamental, o animal pode colocar-se em situação de
risco diminuindo as possibilidades de sobrevivência no próprio habitat.
O tratamento de extensas terras, ou de campos cortados por
pequenos veios, com dosagens efetivas do herbicida Tordon, além de contaminar
o solo por até 5 anos, polui toda a água que através da chuva, ou outro veículo,
entre em contato com o agrotóxico. E, mesmo doses sub-crônicas (0,1%)
dissolvidas na água que serve aos seres vivos que habitam esta área, ou ao seu
redor, são capazes de provocar efeitos neurotóxicos permanentes em seus
descendentes.
Por isso, sabemos que a dita lei de 1979, como tantas outras, não
impediu que interesses escusos de grandes companhias fizessem, mais uma vez,
com que nosso país servisse de aterro público ao "lixo" proibido em seu país de
origem, bem como não impediu que o não menos aviltante desinteresse das
chamadas "autoridades competentes" se fizesse presente.
Quando há interesse as pessoas são informadas e pela educação
inteligente a população evitaria um número enorme de acidentes de saúde, de
alimentação e outros. Com certeza não envenenaria aquilo que é básico para sua
sobrevivência neste planeta.

77

78
6. CONCLUSÃO
Verificando os efeitos do herbicida Tordon em filhotes de mães
contaminadas, podemos concluir que:
1. Nosso trabalho sugere que o herbicida atravessa a barreira
placentária e a barreira hemato-encefálica.
2. A contaminação reduz o número de ninhadas e o tempo de
sobrevida dos filhotes.
3. Os filhotes nascem significativamente mais tardiamente (p<0,01)
que os controles.
4. Por motivo ainda não elucidado, há um detrimento no número de
machos ou um favorecimento no número de fêmeas que nascem
de mães contaminadas.
5. Os descendentes de mães contaminadas são realmente
contaminados.
6. O desenvolvimento do S.N.C. é significativamente mais lento.
7. A contaminação através da mãe produz efeitos neurotóxicos
observados durante toda a vida do animal.
8. Em ambiente novo, uma situação considerada aversiva, há um
prejuízo da atividade exploratória do animal contaminado.
9. Apesar de se movimentar muito mais do que quando jovem,
contrariamente ao controle, o animal contaminado através da
mãe apresenta um aumento significativo em seu peso corporal,
causado, possivelmente, por uma deficiência no sistema
reprodutor que é reflexo do retardo na maturação ou mau
funcionamento do S.N.C.

79
10. O sistema de equilíbrio, dentro do S.N.C., não parece ter sido
afetado pelo Tordon.
11. Os filhotes, aparentemente, não percebem as situações de
perigo, e podem colocar-se em situação de risco.
12. A falta de "medo" deveria aumentar sua capacidade de
aprendizagem, pois o animal estaria menos "ansioso", no entanto
observou-se que o animal contaminado tem uma capacidade de
retenção de aprendizagem levemente inferior ao observado no
controle. Evidenciando uma tendência de prejuízo em nível de
S.N.C., nos processos envolvidos na aquisição e consolidação da
memória.
13. Aparentemente não houve alteração, a nível morfológico, dos
tecidos que apresentaram alteração funcional, tais como
testículo, epidídimo e cérebro.
Por isso, podemos afirmar que os filhos de mães contaminadas por
TORDON, além de serem realmente contaminados, apresentam prejuízo
permanente em nível de S.N.C. que se reflete no sistema neuromuscular e no
sistema reprodutor do animal.

80

81
7. REFERÊNCIAS BIBLIOGRÁFICAS
1. ADHIKARI, N. & GROVER, I.S. Genotoxic effects of some systemic
pesticides: in vivo chromosomal aberrations in bone marrow cells in rats.
Environmental Molecular Mutagen, 12 (2): 235-42. 1988.
2. ANTUNES-MADEIRA, M.C.; CARVALHO, A.P. & MADEIRA, V.M.C.
Interactions of insectides with erythrocyte membranes. Pesticides
Biochemistry Physiology, 15: 79-89. 1981.
3. ANTUNES-MADEIRA, M.C. & MADEIRA, V.M.C. Partition of lindane in
synthetic and native membranes. Biochemistry Biophysics Acta, 820: 165-
72. 1985.
4. ANTUNES-MADEIRA, M.C. & MADEIRA, V.M.C. Membrane fluidity as
affected by the insecticide lindane. Biochemistry et Biophysics Acta, 982:
161-166. 1989.
5. BAKER, M.T.; NELSON, R.M. & VAN DYKE, R.A. The formation of
chlorobenzene by the reductive metabolism of lindane in rat liver
microsomes. Archives of Biochemistry and Biophysics, 236 (2): 506-514.
Fevereiro de 1985.
6. BAKER, M.T. & VAN DYKE, R.A. Metabolism-dependent binding of the
chlorinated insecticide DDT and its metabolite, DD, to microsomal protein
and lipids. Biochemical Pharmacology, 33 (2): 255-260. 1984.
7. BEÇAK, W. & PAULETE-VANRELL, J. Técnicas de Citologia e Histologia,
116,125-127, 211-212. Livraria Nobel S.A. 1ª edição. 1970.
8. BERNARD, P.A.; TOYOSHIMA, E.; ECCLES, C.U.; MAYER, R.F.;
JOHNSON, K.P. & MAX, S.R. 2,4-Dichlorophenoxyacetic acid (2,4-D)
reduces acetylcholinesterase activity in rat muscle. Experimental
Neurology, 87: 544-556. 1985.

82
9. BERNDT, W.O. & KOSCHIER, F. In vitro uptake of 2,4-
dichlorophenoxyacetic acid (2,4-D) and 2,4,5-trichlorophenoxyacetic acid
(2,4,5-T) by renal cortical tissue of rabbits and rats. Toxicology and Applied
Pharmacology, 26: 559-70. 1973.
10. BERWICK, P. 2,4-Dichlorophenoxyacetic acid poisoning in man. JAMA,
214 (6): 1114-17. 09 de novembro de 1970.
11. BLAKLEY, P.M.; KIM, J.S. & FIRNEISZ, G.D. Effects of gestational
exposure to Tordon 202c on fetal growth and development in CD-1 mice.
Journal of Toxicology and Environmental Health, 28 (3): 309-16. 1989.
12. BLAKLEY, P.M.; KIM, J.S. & FIRNEISZ, G.D. Effects of preconceptional
and gestational exposure to Tordon 202c on fetal growth and development
in CD-1 mice. Teratology, 39: 547-553. 1989.
13. BOYD, C.A.; WEILER, M.H. & PORTER, W. P. Behavioral and
neurochemical changes associated with chronic exposure to low-level
concentration of pesticide mixtures. Journal of Toxicology and
Environmental Health, 30: 209-21. 1990.
14. BUCHER, N.L.R. Effects of 2,4-dichlorophenoxyacetic acid on experimental
animals. Procedure Society Experimental Biology Medical, 63: 204-205.
1946.
15. CAVALLI/SFORZA, L.L. & BODMER, W.F. The Genetics of Human
Populations. Editora Freeman, 650-651. 1971.
16. CHERNOFF, N.; SETZER, R.W.; MILLER, D.B.; ROSEN, M.B. &
ROGERS, J.M. Effects of chemically induced maternal toxicity on prenatal
development in the rat. Teratology, 42 (6): 651-58. Dezembro de 1990.
17. CIPRIANO, I.M.; QUADROS, J.; TASSINARI, C.M. & BUCHI, D.F. Effects
of Tordon herbicide on mouse peritoneal macrophages. XIII Colóquio da
Sociedade Brasileira de Microscopia Eletrônica, vol.II: 357-58. 1991.

83
18. CLARK, D.E.; YONG, J.E.; YONGER, R.L.; HUNT, L.M. & McLARAN, J.K.
The fate of 2,4-dichlorophenoxiacetic acid in sheep. J. Agric. Food Chem.,
12: 43-45. 1964.
19. CONN, H.J. Staining Procedures, page 148. The Williams & Wilkins
Company. Baltimore. 1962.
20. CULLING, C.F.A.; ALLINSON, R.T. & BARR, W.T. Cellular Pathology
Technique, page. 384-388. Butterworth’s. 4th edition. 1985.
21. DUFFARD, R.O.; MORO, G.B.M. & DE DUFFARD, A.M.E. Vulnerability of
myelin development of the chick to the herbicide 2,4-dichlorophenoxyacetic
butyl ester. Neurochemical Research, 12 (12): 1077-80. 1987.
22. DUX, E.; TÓTH, I.; DUX, L.; JOD, F. & KISZELY, G. The possible cellular
mechanism of 2,4-dichlorophenoxyacetate - induced myopathy. Febs
Letters, 82 (2): 219-222.1977.
23. ELO, H.A.; HERNOVEN, H. & YLITALO, P. Comparative study on
cerebrovascular injuries by three chlorophenoxyacetic acids (2,4-D, 2,4,5-T
and MCPA). Comp. Biochemistry Physiology C., 90(1): 65-68. 1988.
24. ELO, H.A. & MacDONALD, E. Effects of 2,4-dichlorophenoxyacetic acid
(2,4-D) on biogenic amines and their acidic metabolites in brain and
cerebrospinal fluid of rats. Archives Toxicology, 63 (2): 127-30. 1989.
25. ERNE, K. Distribution and elimination of chlorinated phenoxyacetic acids in
animals. Acta Veterinarian Scandinavia, 7: 240-256.1966.
26. EROSCHENKO, V.P. Alterations in the testes of the Japanese quail during
and after the ingestion of the insecticide kepone. Toxicology and Applied
Pharmacology, 43: 535-545. 1978.
27. EVANGELISTA de DUFFARD, A.M.; ALDERETE, M.N. & DUFFARD, R.
Changes in brain serotonin and 5-hydroxyindolacetic acid levels induced by
2,4-dichlorophenoxyacetic butyl ester. Toxicology, 64 (3): 265-70. 03 de
dezembro de 1990.

84
28. EVANGELISTA de DUFFARD, A. M.; ORTA, C. & DUFFARD, R.
Behavioral changes in rats fed a diet containing 2,4-dichlorophenoxyacetic
butyl ester. Neurotoxicology, 11 (4): 563-72. Inverno de 1990.
29. FANG, S.C.; FALLIN, E.; MONTGOMERY, M.L. & FREED, V.H. The
metabolism and distribution of 2,4,5- trichlorophenoxyacetic acid in female
rats. Toxicology and Applied Pharmacology, 24: 555-563. 1973.
30. FAO Meeting Report na.PL/1963/63; Who / food Add/1964/23.
31. GERHART, J.M.; HONG, J.S. & TILSON, H.A. Studies on the possible sites
of chlordecone-induced tremor in rats. Toxicology and Applied
Pharmacology, 70: 382-389. 1983.
32. GERING, P.J.N.; KRAMER, C.G.; SCHURTZ, B.A.; ROSE, J.K. & ROWE,
V.K. The fate of 2,4,5-trichlorophenoxyacetic acid (2,4,5-T) following oral
administration to man. Toxicology and Applied Pharmacology, 26: 352-361.
1973.
33. GOETZ, C.G.; BOLLA, K.I. & ROGERS, S.M. Neurologic health outcomes
and Agent Orange. Neurology, 44 (5): 801-809. Maio de 1994.
34. GOLDSTEIN, N.P.; JONES, P.H.; BROW, J.R. & ROCHESTER, M.
Peripheral neuropathy after exposure to an ester of dichlorophenoxyacetic
acid. JAMA, 7 (10) 1306-09. 1959.
35. GONZALEZ, M.I.; FARABOLLINI, F.; ALBONETTI, E. & WILSON, C.A.
Interactions between 5-hydroxytryptamine (5-HT) and testosterone in male
and female rats. Pharmacology Biochemistry and Behavior, 47 (3): 591-
601. Março de 1994.
36. GORZINSKI, S.J.; JOHNSON, K.A., CAMPBELL, R.A. & LANDRY, T.D.
Dietary toxicity of picloram herbicide. Journal of Toxicology and
Environmental Health, 20: 367-77. 1987.

85
37. GORZINSKI, S.J.; KOCIBA, R.J.; CAMPBELL, R.A.; SMITH, F.A.; NOLAN,
R.J. & EISENBRANDT, D.L. Acute, pharmacokinetic and subchronic
toxicological studies of 2,4-dichlorophenoxyacetic acid. Fundamental and
Applied Toxicology, 9 (3): 423-35. Outubro de 1987.
38. GUZELIAN, P.S. Comparative toxicology of chlordecone (kepone) in
humans and experimental animals. Annual Review Pharmacology
Toxicology, 22: 89-113. 1982.
39. HAYES, J.R.; CONDIE, L.W. & BORZELLECA, J.F. Acute, 14-day repeated
dosing and 90-day subchronic toxicity studies of potassium picloram.
Fundamental and Applied Toxicology, 7: 464-70. 1986.
40. HILL, E.V. & CARLISLE, H. Toxicity of 2,4-dichlorophenoxyacetic acid for
experimental animals. Journal of Industrial Hygiene and Toxicology, 29 (2):
85-95. Março de 1947.
41. HRDINA, P.D.; SINGHAL, R.L.; PETERS, D.A.V. & LING, G.M. Role of
brain acetylcholine and dopamine in acute neurotoxic effects of DDT.
European Journal of Pharmacology, 15: 379-382. 1971.
42. IBRAHIM, M.A.; BOND, G.G.; BURKE, T.A.; COLE, P.; DOST, F.N.;
ENTERLINE, P.E.; GOUGH, M.; GREENBERG, R.S.; HALPERIN, W.E. &
McCONNELL, E. et al. Weight of the evidence on the human
carcinogenicity of 2,4-D. Environmental Health Perspectives, 96: 213-22.
Dezembro de 1991.
43. JOHN-GREENE, J.A.; OUELLETTE, J.H.; JEFFRIES, T.K.; JOHNSON,
K.A. & RAO, K.S. Teratological evaluation of picloram potassium salt in
rabbits. Food Chemistry Toxicology, 23 (8): 753-556. 1985.
44. KIM, C.S.; O'TUAMA, L.A.; MANN, J.D. & ROE, C.R. Saturable
accumulation of the anionic herbicide, 2,4-dichlorophenoxyacetic acid (2,4-
D), by rabbit choroid-plexus : early development origin and interaction with
salicicates. Journal of Pharmacology and Experimental Therapeutics, 225
(3): 699-704. 1983.

86
45. KNOPP, D. Assessment of exposure to 2,4-dichlorophenoxyacetic acid in
the chemical industry - results of a 5-years biological monitoring study.
Occupational and Environmental Medicine, 51 (3): 152-159. Março de
1994.
46. KNOPP, D. & SCHILLER, F. Oral and dermal application of 2,4-
dichlorophenoxyacetic acid sodium and dimethylamine salts to male rats :
investigations on absorption and excretion as well as induction of hepatic
mixed-function oxidase activities. Archives Toxicology, 66 (3): 170-74.
1992.
47. LAGENBACH, T. Pesticidas: Tecnologia sofisticada em mãos
desinformadas. Ciência Hoje, 10 (56): 66-68. 1989.
48. LAWRENCE, L. J. & CASIDA, J.E. Interactions of lindane, toxaphene e
cyclodienes with brain - specific t-butilbicyclophosphorotrionate receptor.
Life Sciences, 35: 171-178. 1984.
49. LERDA, D. & RIZZI, R. Study of reproductive function in persons
occupationally exposed to 2,4-dichlorophenoxyacetic acid (2,4-D). Mutation
Research, 262: 47-50. 1991.
50. LYNN, G.E. A review of toxicological information on Tordon herbicides.
Dow Earth, 20: 6-8. 1965.
51. MAIA, V. Técnicas Citológicas. Atheneu Editora São Paulo. 2ª edição.
1979.
52. McCOLLISTER, D.D. & LENG, M.L. Toxicology of picloram and safety
evaluation of Tordon herbicides. Dow Earth, 25: 5-10. 1969.
53. MOHAMMAD, F.K. & St. OMER, V.E. Effects of prenatal exposure to 2,4-D
/ 2,4,5-T mixture on postnatal changes in rat in brain glutamate, GABA,
protein, and nucleic acid levels. Bulletin Environmental Contamination
Toxicology, 40 (2): 294-300. Fevereiro de 1988.

87
54. OLIVEIRA, G.H. & PALERMO NETO, J. Effects of 2,4-
dichlorophenoxyacetic acid (2,4-D) on open-field behavior and
neurochemical parameters of rats. Pharmacologic Toxicology, 73 (2): 79-
85. Agosto de 1993.
55. PAULINO, C.A. & PALERMO NETO, J. Effects of acute 2,4-
dichlorophenoxyacetic acid intoxication on some rat serum components and
enzyme activities. Brazilian Journal Medical Biology Research, 24 (2): 195-
98. 1991.
56. PELLETIER, O.; RITTER,L. & CARON, J. Effects of skin preapplication
treatments and postapplication cleansing agents on dermal absorption of
2,4-dichlorophenoxyacetic acid dimethylamine by Fischer 344 rats. Journal
Toxicology Environmental Health, 31 (4): 247-60. Dezembro de 1990.
57. PEREIRA, L.F. Efeitos do herbicida Tordon 2,4-D 64/240 trietanolamina Br
sobre o metabolismo energético de mitocôndrias de fígado e córtex renal.
Tese apresentada ao curso de Pós-Graduação em Bioquímica da
Universidade Federal do Paraná, visando à obtenção do título de mestre
em Bioquímica. Curitiba - Paraná. 1991.
58. PRITCHARD, J.B. Toxic substances and cell membrane function. Fed.
Proc., 36: 2220. 1979.
59. PRITCHARD, J.B. Accumulation of anionic pesticides by rabbit choroid
plexus in vitro. Journal Pharmacologic Experimental Therapy, 212 (2): 354-
59. 1980.
60. PRITCHARD, J.B.; KRALL, A.R. & SILVERTHORN, S.U. Effects of anionic
xenobiotics on rat kidney. I. Tissue and mitochondrial respiration.
Biochemistry Pharmacology, 32 (2): 149-55. 1982.
61. REIS, J. Norte-americanos discutem uso do componente do "agente
laranja". Folha de São Paulo, 15 de março de 1991.
62. REUBER, M.D. Carcinogenicity of picloram. Journal of Toxicology and
Environmental Health, 7: 207-222. 1981.

88
63. SCHULZE, G.E. Formulation and food deprivation affects 2,4-D
neurobehavioral toxicity in rats. Neurotoxicology Teratology, 9 (5): 363-67.
Setembro/outubro de 1987.
64. SCHULZE, G.E. 2,4-D-n-butyl ester (2,4-D ester) induced ataxia in rats:
role for n-butanol formation. Neurotoxicology Teratology, 10 (1): 81-84.
Janeiro/fevereiro de 1988.
65. SCHULZE, G.E. & DOUGHERTY, J.A. Neurobehavioral toxicity of 2,4-D-n-
butyl ester (2,4-D ester): tolerance and lack of cross- tolerance.
Neurotoxicology Teratology, 10(1): 75-79. Janeiro/fevereiro de 1988.
66. SCHULZE, G.E. & DOUGHERTY, J.A. Neurobehavioral toxicity and
tolerance to the herbicide 2,4-dichlorophenoxyacetic acid-n-butyl ester (2,4-
D ester). Fundamental Applies Toxicology, 10(3): 413-24. Abril de 1988.
67. SHANTOR, V.F.; KAGAN, N.A.; KARASEVA, A.E.; KREMKO, L.M. &
KRYSANOVA, A.I. Kompleksnoe gigienicheskoe reglamentirovanie
gerbitsida fenagonav vode vodoemov i produktakh pitaniia. [Complex
hygienic standartization of the herbicide fenagon in reservoir water and food
products.] Gigienicheskoe Sanit., (10): 10-12. Outubro de 1988.
68. St. OMER, V.E. & MOHAMMAD, F.K. Ontogeny of swimming behavior and
brain catecholamine turnover in rats prenatally exposed to a mixture of 2,4-
dichlorophenoxyacetic acid. Neuropharmacology, 26 (9): 1351-58.
Setembro de 1987.
69. TARARTHUCH, A.L. Nefrotoxicidade do herbicida Tordon 2,4-D 64/240
trietanolamina BR no rato adulto: avaliação funcional e aspectos
histológicos. Tese apresentada ao curso de Pós-Graduação em Morfologia
do Dep. de Biologia Celular da Universidade Federal do Paraná, para
obtenção do título de mestre em Ciências Morfológicas. Curitiba - Paraná.
1992.
70. TYYNELA, K; ELO, H.A. & YLITALO, P. Distribution of three common
chlorophenoxyacetic acid herbicides into the rat brain. Archives Toxicology,
64 (1):61-65. 1990.

89
71. VACCA, L.L. Myelin. Laboratory Manual of Histochemistry, 251-263.Raven
Press. 1985.
72. VIDELA, L.A.; BARROS, S.B.M. & JUNQUEIRA, V.B.C. Lindane-induced
liver oxidative stress Free Rad. Biol. Med., 9: 169-179. 1990.
73. VIN', V.D.; SLOZINA, N.M. & NIKITIN, A.I. Nature of the estrous cycle,
incidence of anovulation, chromosome gametopathies and fetal death in
female rats with 2,4-dichlorophenoxyacetic acid (2,4-D) before pregnancy.
Arkhives Anatomy Histology Embryology, 99 (9): 69-73. Setembro de 1990.
74. YLITALO, P.; NARHI, U. & ELO, H.A. Increase in the acute toxicity and
brain concentrations of chlorophenoxyacetic acids by probenecid in rats.
Genetic Pharmacology, 21 (5): 811-14. 1990.

0





























